avaliação in silico da afinidade da testosterona e seus ... · 3 resumo abbas, a.r. avaliação...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE CIÊNCIAS FARMACÊUTICAS
Curso de Graduação em Farmácia-Bioquímica
Avaliação in silico da afinidade da testosterona e seus derivados
sintéticos ao receptor androgênico humano genótipo selvagem
ADAM REIAD ABBAS
Trabalho de Conclusão do Curso de
Farmácia-Bioquímica da Faculdade de
Ciências Farmacêuticas da
Universidade de São Paulo.
Orientador(a):
Prof.(a). Dr(a) Gustavo Henrique
Goulart Trossini
São Paulo
2018

SUMÁRIO
Pág.
Lista de Abreviaturas .......................................................................... 1
RESUMO .......................................................................................... 3
1. INTRODUÇÃO 5
1.1 conceitos gerais 5
1.2 – O receptor Androgênico 5
1.3 – Esteroides Anabólico-Androgênicos 10
1.4 – Técnicas computacionais e Química Computacional 12
2. OBJETIVOS 12
3. MATERIAIS E MÉTODOS 13
3.1. Material 13
3.2. Métodos 13
3.2.1. Seleção da proteína alvo 13
3.2.2. Desenho dos ligantes que serão avaliados 14
3.2.3. Estudos de ancoramento (docking) molecular 14
4. RESULTADOS 15
4.1. Validação da metodologia por reedocking 15
4.2. Obtenção das estruturas tridimensionais das moléculas de interesse e
preparo para análise de ancoramento (docking) molecular
16
4.3. Avaliação comparativa entre pontos de interação de cada EAA com o AR 18
5. DISCUSSÃO 22
6. CONCLUSÃO 34
7. BIBLIOGRAFIA 34

1
LISTA DE ABREVIATURAS
AR Receptor Androgênico
ARG Arginina
ASN Asparagina
DBD DNA Binding Domain
DHT Di-hidrotestosterona
DNA Ácido Desoxirribonucleico
EAS Esteroides Anabóbico-Androgênicos
ER Receptor Estrogênico
FDA Food and Drug Adminstration
GLN Glutamina
GR Receptor de glicocorticoides
HIV Vírus da imunodeficiência humana
IGF Fator de Crescimento Semelhante à insulina
LBDD Ligand based drug design
LDB Ligand Binding Domain
LITEC Laboratório de Integração entre Técnicas Experimentais e
Computacionais no Planejamento de Fármacos
NTD N-terminal Domain
PDB Protein data bank
PR Receptor de Progesterona
QSAR Quantitative structure–activity relationship

2
RMSD root-mean square deviation
RNA Ácido ribonucleico
SBDD Structure based drug design
THG Tetra hidro Gestrinona
THR Treonina

3
RESUMO ABBAS, A.R. Avaliação in silico da afinidade da testosterona e seus derivados sintéticos ao receptor androgênico humano genótipo selvagem. 2018. no. f. Trabalho de Conclusão de Curso de Farmácia-Bioquímica – Faculdade de Ciências Farmacêuticas – Universidade de São Paulo, São Paulo, ano. Palavras-chave: Planejamento de fármacos; testosterona; esteroides sintéticos; receptor androgênico INTRODUÇÃO: Andrógenos e o receptor androgênico desempenham um papel crucial na expressão de características fenotípicas masculinas. Estudos atuais buscam elucidar de forma clara as alterações conformacionais que ocorrem com o receptor androgênico quando em ligação com diferentes compostos. Em especial, o estudo da ligação do receptor com análogos esteroides androgênicos tem interesse especial, uma vez que esses possuem relevância clínica, como nos estudos para desenvolvimento de fármacos para controle de tumores androgênio-dependentes, esportiva, devido ao crescente abuso de substancias para aumento de performance e suas implicações éticas, bem como econômica, como o uso de derivados anabólico-androgênicos sintéticos em ramos como a pecuária. OBJETIVO: Avaliar pontos de interação e grau de afinidade do hormônio testosterona e seus derivados sintéticos com o domínio de interação com ligante do receptor androgênico humano, genótipo selvagem, in silico. MATERIAIS E MÉTODOS: Avaliação computacional de ligação entre os compostos e forma selvagem do receptor androgênico humano, a partir da estrutura cristalográfica obtida na base de dados PDB (Protein Data Bank). Para tal, foram utilizados programas específicos bem como as estações de trabalho do LITEC (Laboratório de Integração entre Técnicas Experimentais e Computacionais no Planejamento de Fármacos). RESULTADOS: A validação do método por redocking como base a estrutura cristalográfica do domínio de ligação com o ligante do AR complexado com a molécula de testosterona obtidos através do Protein Data Bank se mostrou eficaz e fidedigna, sendo válida para as análises realizadas nesse trabalho. A avaliação de pontos de interação hidrofílicos e hidrofóbicos dos ligantes propostos ao receptor androgênico se mostrou condizente com os dados da literatura, fornecendo ainda dados mais concretos com relação a potência de esteroides de uso e abuso corrente em performance esportiva. CONCLUSÃO: Os derivados anabólico-androgênicos pertencentes a família 19-nor se mostraram os mais potentes em relação aos demais devido a melhor acomodação ao AR. Contrário a literatura, os membros pertencentes à família de derivados de DHT, tidos como de baixa potência apresentaram diversos pontos de interação importante com o AR. Estudos mais detalhados, bem como dados in vivo, se mostram necessários para confirmação de tais hipóteses. Os achados são promissores para futuros estudos de agentes moduladores do receptor androgênico, visando a dissociação do potencial anabólico dos efeitos androgênicos.

4

5
1. INTRODUÇÃO 1.1 conceitos gerais O desenvolvimento de novos fármacos, atualmente, ocorre de maneira mais
direcionada que no passado. Os avanços nas técnicas de análise molecular
permitiram a elucidação das estruturas de moléculas bioativas, tanto sintéticas
quanto naturais1,2. Tal processo, hoje, pode ser realizado por avaliação
comparativa de moléculas com propriedades físico-químicas e com efeitos
biológicos semelhantes sem conhecimento a fundo das características ultra
estruturais do sitio de ligação onde se acoplam. Tal técnica, conhecida como
LBDD (Ligant Based Drug Design)3,4 se difere justamente por essa característica
em relação à outra metodologia de desenvolvimento, na qual se há conhecimento
da estrutura do sitio de ligação do ligante, podendo ser explorado de forma mais
precisa as propriedades físico-quimicas tanto do ligante quanto do receptor. Tal
técnica, denominada de desenvolvimento baseado na estrutura (structure based
drug design - SBDD) permite, dessa maneira, o desenvolvimento de novos
ligantes de forma a explorar outras regiões do sitio de ligação, caso haja
viabilidade4,5.
Em relação aos esteroides anabólico-androgênicos, são conhecidos hoje mais de
200 compostos, naturais ou sintéticas, capazes de interagir com o chamado
receptor androgênico (AR).
1.2 – O receptor Androgênico O receptor androgênico (AR) (NR3C4, subfamília de receptor nuclear, grupo C,
gene 4) pertence a grupo de receptores responsivos a hormônios esteroides junto
com o receptor estrogênico (ER), receptor de glicocorticoides (GR), receptor
progesterona (PR) e de mineralocorticoides (MR)6,7,8. O AR pode ser classificado
como um fator de transcrição dependente de ligante que é responsável pela
expressão de um conjunto específico de genes, sendo que a sua ligação com
seus ligantes nativos, como a testosterona e a di-hidrotestosterona, dá início ao
processo de diferenciação sexual masculina9. O gene responsável pela
transcrição do AR se localiza no cromossomo X, região Xq 11-12, possuindo 8

6
exons separados por regiões intrônicas de 0,7-26 kb10,11,12. O mecanismo
dinâmico desse processo é retratado na figura 1.
Figura 1. Mecanismo dinâmico de ação do complexo ligante-AR em células prostáticas. No caso, após síntese nos testículos, a testosterona é convertida em seu derivado reduzido, DHT, podendo assim sinalizar vias de crescimento e desenvolvimento em tecidos específicos, como no caso da próstata (adaptado)9
O gene AR codifica uma proteína de 110kDA constituída por 919 aminoácidos
(figura 2). Assim como outros membros da família de receptores nucleares, o AR
é formado por 3 domínios funcionais majoritários:
• Domínio N-terminal (NTD, formado pelos resíduos 1-555), seguido por,
• Domínio de ligação ao DNA (DBD, formado pelos resíduos 555-623) e,
• Domínio C-terminal de associação ao ligante (LBD, formado pelos resíduos
665-919), o qual é conectado ao DBD por uma região de alça flexível
(resíduos 623-665).
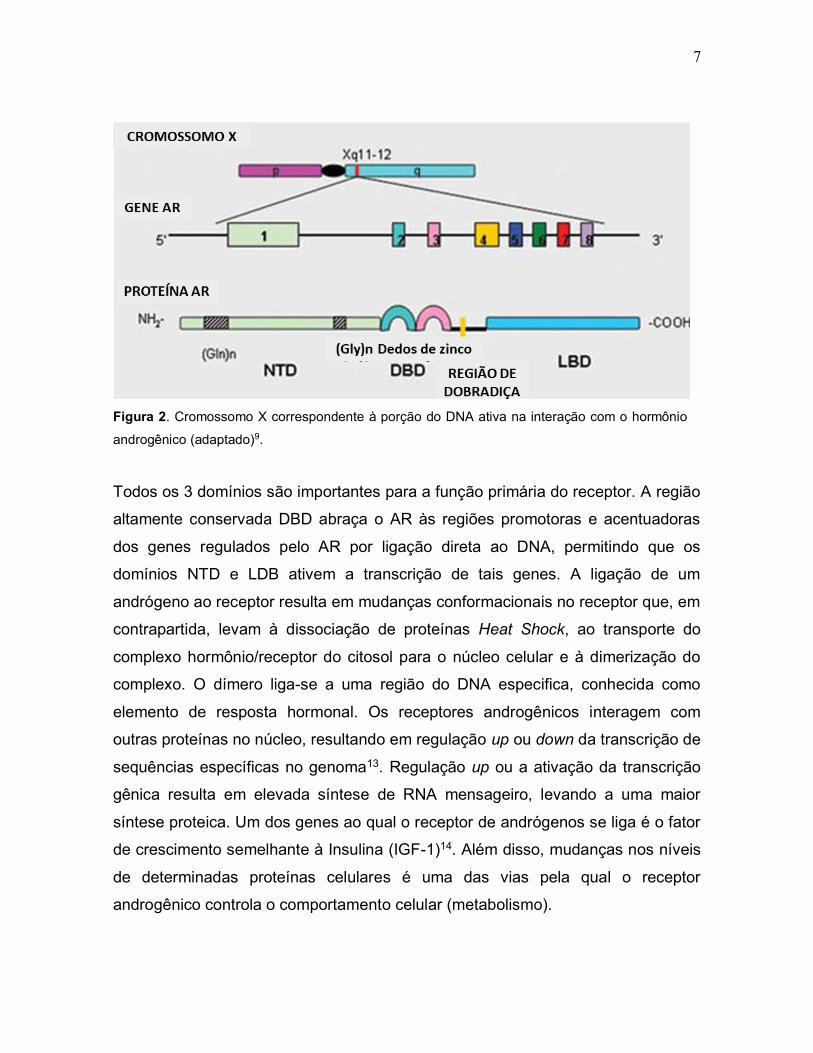
7
Figura 2. Cromossomo X correspondente à porção do DNA ativa na interação com o hormônio
androgênico (adaptado)9.
Todos os 3 domínios são importantes para a função primária do receptor. A região
altamente conservada DBD abraça o AR às regiões promotoras e acentuadoras
dos genes regulados pelo AR por ligação direta ao DNA, permitindo que os
domínios NTD e LDB ativem a transcrição de tais genes. A ligação de um
andrógeno ao receptor resulta em mudanças conformacionais no receptor que, em
contrapartida, levam à dissociação de proteínas Heat Shock, ao transporte do
complexo hormônio/receptor do citosol para o núcleo celular e à dimerização do
complexo. O dímero liga-se a uma região do DNA especifica, conhecida como
elemento de resposta hormonal. Os receptores androgênicos interagem com
outras proteínas no núcleo, resultando em regulação up ou down da transcrição de
sequências específicas no genoma13. Regulação up ou a ativação da transcrição
gênica resulta em elevada síntese de RNA mensageiro, levando a uma maior
síntese proteica. Um dos genes ao qual o receptor de andrógenos se liga é o fator
de crescimento semelhante à Insulina (IGF-1)14. Além disso, mudanças nos níveis
de determinadas proteínas celulares é uma das vias pela qual o receptor
androgênico controla o comportamento celular (metabolismo).

8
Uma função do receptor androgênico, independente da ligação direta ao seu alvo
no DNA, é mediada pelo recrutamento de outras proteínas ligantes ao DNA. Por
exemplo, o fator de resposta ao soro, uma proteína que ativa diversos genes, os
quais levam ao desenvolvimento e crescimento muscular15.
Mais recentemente, receptores androgênicos têm mostrado um segundo
mecanismo de ação. Como já demonstrado para outros receptores hormonais,
como o receptor estrogênico, os receptores androgênicos podem apresentar
atividade que independa de suas interações com o DNA16,17.
Os receptores interagem com determinadas proteínas de transdução no
citoplasma, de modo que a ligação de andrógenos aos seus receptores
citoplasmáticos podem causar rápidas mudanças na função celular, independente
de mudanças na transcrição gênica, como por exemplo, mudanças em transporte
iônico. A regulação em vias de transdução de sinal pode levar a alterações
indiretas na transcrição gênica, por exemplo, pela fosforilação de outros fatores de
transcrição18.
Dado o vasto material disponível em literatura relacionados a estrutura do receptor
além de atividade de esteroides disponíveis em bibliotecas de bancos de dados
foram realizados estudos relacionando dados de QSAR e ancoragem molecular
(docking) entre o receptor e diversos ligantes19,20, buscando relacionar os dados
obtidos a partir dos modelos de QSAR propostos com dados in vivo de potencial
anabólico e androgênico disponíveis na literatura20. Tais dados, ainda que
componham o único conjunto descrito na literatura onde se faz menção ao
potencial anabólico ou androgênico de cada esteroide, já foram extensamente
reavaliados, uma vez que a metodologia de avaliação (desenvolvimento do
músculo eretor do ânus em ratos para efeito anabólico/miotrófico versus
desenvolvimento de tecidos do aparelho genitourinário, como próstata e vesícula
seminal, para efeito androgênico)21. Ao que parece, os dados seriam
inconsistentes devido a diferenças na densidade de receptores androgênicos em
diferentes tecidos, no que tange ao potencial miotrófico, bem como a redução na
atividade na enzima 5a-Redutase em outros, o que seria o fator diferencial para o
potencial androgênico de um esteroide (sua afinidade por tal enzima)22.

9
Há um significativo aumento no interesse na pesquisa de agentes anabólicos no
âmbito clínico para o tratamento de pessoas que sofram com certos tipos de
condições que debilitam suas ações, como por exemplo, aquelas associadas à
perda de tecido muscular devido à idade. Dentre esses compostos, há, por
exemplo, a oxandrolona, que vem sendo empregada em situações relacionadas à
perda muscular ocasionada pelo HIV, queimaduras severas, traumas após
cirurgias de grande porte, doenças neuromusculares, entre outros. Esse composto
é o único aprovado, dentro da classe de esteroides anabólicos androgênicos, para
uso clínico nos EUA, envolvendo o tratamento das distrofias musculares de
Becker e Duchenne. Análises envolvendo o uso deste composto no tratamento de
desordens catabólicas, perdas envolvidas com HIV e problemas musculares
apresentaram uma evidência clínica concisa sobre a sua eficácia farmacológica. A
oxandrolona está envolvida na melhoria da composição corporal, força, função
muscular. Diferentemente de outros esteroides administrados por via oral, a
oxandrolona apresenta uma menor hepatotoxicidade - que envolve possível
ocorrência de icterícia, hepatite colestática, peliose, hiperplasias e neoplasias -,
vinculada aos esteroides do tipo C17-alfa-alquilados. Por outro lado, em doses
elevadas de uso, seus efeitos hepatotóxicos começam a apresentar-se mais
significativos, além de extremamente caros23.
Há questões importantes a serem consideradas no emprego e no
desenvolvimento de um fármaco esteroide, como seu potencial anabólico, que
está vinculada ao crescimento e divisão celulares e desenvolvimento de diversos
tipos de tecidos, especialmente o muscular e ósseo, potencial androgênico,
relacionada ao controle do desenvolvimento e à manutenção das características
sexuais secundárias masculinas, hepatotoxicidade devido aos processos de
metabolização e excreção, bem como potencial de redução e aromatização e
efeitos centrais23.
A aromatização consiste em um processo de conversão de um esteroide
androgênico em seu correspondente estrogênico via atuação da enzima
aromatase, sendo um efeito colateral indesejável em tratamentos de distrofias
musculares. Analogamente, a redução é o processo de conversão de um

10
esteroide androgênico em seu correspondente mais potente (similar ao DHT) via
atuação da enzima 5α-Redutase24.
Dessa maneira, o estudo do potencial anabólico/androgênico de esteroides
sintéticos por ferramentas computacionais, apresenta potencial para, no futuro, o
desenvolvimento racional de novos medicamentos com potencial clinico, assim
como novas rotas de síntese além de aplicabilidade em outras áreas, como
veterinária e agropecuária.
1.3 – Esteroides Anabólico-Androgênicos
Andrógenos exercem diversos efeitos no organismo, incluindo tecidos
reprodutivos, musculo, ossos, entre diversos outros. Os ditos efeitos androgênicos
podem ser classificados como aqueles que causam exacerbação das
características sexuais secundárias masculinas enquanto que os anabólicos
aqueles que aumentam os processos de síntese proteica no tecido muscular.
Os efeitos de andrógenos são modulados em nível celular por conjuntos de
enzimas presentes em cada tecido especifico. Por exemplo, nos tecidos
reprodutivos, a testosterona pode ser considerada como um pró hormônio, uma
vez que é convertida prontamente nos mesmos pela enzima 5α-redutase em seu
derivado reduzido, a di-hidrotestosterona (DHT). Já em outros tecidos, como o
tecido adiposo, a mesma é convertida, agora via ação de outra enzima, em seu
derivado aromático, o estradiol. Já no tecido muscular, não se detecta atividade da
redutase, sendo a testosterona o principal ligante a se associar ao AR nesse
tecido25,26.
Com a adição de modificações estruturais na molécula de testosterona, os efeitos
anabólicos dos esteroides podem ser incrementados. Entretanto, ainda assim não
é possível dissociar totalmente esses derivados de seus efeitos androgênicos
(figura 3).

11
Figura 3: modificações estruturais na molécula de testosterona que aumentam a atividade anabólica e reduzem atividade androgênica (adaptado)27
Por fins didáticos, os EAS utilizados nesse trabalho foram separados em
categorias segundo os critérios de Vida20. A saber:
• Derivados de testosterona: conjunto de moléculas que agrupam os
derivados sintéticos obtidos a partir de modificações na estrutura original da
testosterona. Exemplos são boldenona, metandrostenolona,
fluoximesterona, 4-Cl-dihidrometiltestosterona).
• Derivados 19-nor: conjunto de moléculas agrupadas pela perda do carbono
19 da estrutura original da molécula de testosterona. Tal modificação
molecular impede a metabolização desses compostos pelo citocromo 19A1
(Cyp19A1) Aromatase, responsável pela conversão de andrógenos em
estrógenos no organismo24. Dentre eles, destacam-se a gestrinona, a
nandrolona e a trembolona.
• Derivados de DHT: conjunto de moléculas que agrupam os derivados
sintéticos obtidos a partir de modificações na molécula de DHT (metabólito
obtido a partir da redução da insaturação entre C3-C4 por intermédio da

12
enzima 5ɑ Redutase). Alguns exemplos são o stanozolol, oxandrolona,
metenolona, mesterolona e drostanolona.
1.4 – Técnicas computacionais e Química Computacional
Atualmente, com avanço das pesquisas na área tecnológica, a Química
computacional (QC) se mostra como uma ferramenta importante para
compreensão da dinâmica em sistemas biológicos. Essa potente ferramenta pode,
assim, ser classificada como um conjunto de técnicas que utilizam métodos
computacionais na resolução de problemas químicos (como estrutura e
conformação tridimensional)28, por meio da utilização de recursos matemáticos,
mecânica clássica e quântica, bem como conhecimentos de química teórica, de
modo a entender, sob determinadas condições, os métodos realizados utilizam
cálculos para o processamento, visualização e manipulação de resultados na
busca de moléculas bioativas e/ou no entendimento de seu mecanismo de
ação28,29.
A partir da década de 1980, houve um maior desenvolvimento e melhoria nos
softwares e hardwares, juntamente com avanços no conhecimento bioquímico
sobre as bases moleculares das doenças, o que permitiu um avanço significativo
na química computacional aplicada ao planejamento de fármacos permitindo a
utilização desta tecnologia na academia e nas indústrias químicas e
farmacêuticas30.
2. OBJETIVO(S) Avaliação in silico de pontos de interação de diversos esteroides
anabólico/androgênicos, suas propriedades físico-químicas e comparação de sua
afinidade pelo receptor androgênico

13
3. MATERIAIS E MÉTODOS 3.1. Material Para confecção e elaboração desse trabalho, foram utilizados:
• Estação de trabalho computacional disponível no LITEC/FCF-USP;
• Softwares computacionais:
o Pymol (gratuito);
o Chimera (gratuito);
o GRID (disponível no laboratório do Prof. Dr. Gustavo H. G. Trossini);
o GOLD (disponível no laboratório do Prof. Dr Gustavo H. G. Trossini),
o Autodock Vina e ROCS (disponível no laboratório do Prof. Dr.
Gustavo H. G. Trossini)
3.2. Métodos 3.2.1. Seleção da proteína alvo A escolha da proteína alvo a partir da base de dados PDB depende de diversos
fatores experimentais associados à estrutura cristalográfica selecionada. O critério
base para escolha de uma estrutura é seu valor de resolução. A resolução é uma
medida diretamente relacionada com a distância mínima que dois objetos estão
separados e ainda assim serem vistos como dois objetos independentes. Valores
numéricos pequenos para resolução significam uma pequena incerteza, logo
temos um alto grau de resolução (baixa incerteza). Já valores altos significam
grande incerteza, logo, baixa resolução31. O cristal 2AM9 para a forma selvagem
do AR apresentou alto grau de resolução com valor de 1,64Å, sendo assim a
escolha para esse trabalho. Andrógenos exercem seus efeitos devido a sua
ligação com o receptor androgênico (AR). Adicionalmente aos seus ligantes
naturais androgênicos, o AR pode se ligar a uma variedade imensa de agonistas e
antagonistas sintéticos com diferentes graus de afinidade. Para a identificação dos
determinantes moleculares que levam a essa variabilidade na afinidade de
ligação, em geral são feitas determinações de estrutura cristalográfica da proteína
ligante ou ainda de regiões especificas da mesma. Pereira de Jésus-Tran e

14
colaboradores32 em seu trabalho de 2006 fizeram a determinação da estrutura
cristalográfica do domínio de ligação ao ligante do receptor androgênico humano,
quando complexado com seus dois ligantes naturais, testosterona e di-
hidrotestosterona, além de um esteroide sintético utilizado classificado como
dopping esportivo, a tetrahidrogestrinona (THG), em uma relução de 1,64, 1,90, e
1,75 A˚, respectivamente. A comparação dessas estruturas permitiu demonstrar
uma grande flexibilidade e variedade de resíduos no LBD capazes de acomodar
uma serie de estruturas de ligantes e, como já era de se esperar, a própria
estrutura do ligante é quem determina o número de interações feitas com o LBD
do AR. É com base nessa premissa que foram avaliados os tipos de interações e
número de interações dos compostos propostos nesse trabalho com o AR.
3.2.2. Desenho dos ligantes que serão avaliados
Para preparação das estruturas dos ligantes propostos foi utilizado a ferramenta
ChemBio3D®, parte do pacote software Chemdraw®.
Após a obtenção das estruturas 2 e 3D, foi realizado a obtenção da conformação
mais estável do ligante através do software Chimera®.
3.2.3. Estudos de ancoramento (docking) molecular Para os estudos de docking foram utilizadas as estruturas cristalográficas do
receptor androgênico extraídas do Protein Data Bank – PDB (2AM9) de boa
definição e resolução, sem mutações no sítio ativo (proteína selvagem).
O método de docking é uma ferramenta computacional utilizada para predizer
orientações mais prováveis de uma molécula em relação a outra quando ligadas
entre si, formando um complexo estável33. O conhecimento de orientação
preferencial de uma molécula é um dos pontos chave para prever o grau de
associação ou ligação entre as duas moléculas envolvidas no processo, processo
esse que pode ser convertido em valores numéricos, através de uma função de
ranqueamento ou score. A maior parte dos algoritmos utilizados por softwares de
modelagem molecular e docking são capazes de gerar um número elevado de
confôrmeros possíveis, o que também acaba por exigir algoritmos de forma a
determinar quais dessas estruturas serão de maior interesse para o analisador, o

15
que é preocupante uma vez que deve-se avaliar uma série de estruturas
plausíveis para complexos intermoleculares34,35. Sendo assim, a maneira
encontrada em nosso trabalho para garantir a validação da metodologia foi
confrontar os dados gerados tanto pelo software Gold® quanto por outras
ferramentas, como o software de linha aberta Autodock VINA. Os valores de R²
para análise de regressão para ambos foram significativos para os parâmetros
escolhidos pela equipe de análise. Foram gerados para cada um dos EAS
avaliados, bem como para a própria testosterona 10 possíveis confôrmeros que
satisfizeram os critérios de acoplamento. A área de docking foi definida por caixas
de 5, 10 e 15Å a partir do centro do esqueleto base ciclopentanofenantreno de
andrógenos. Ligações e seus graus de torção foram escolhidos como livres
(flexibilidade completa). Tais resultados demonstram a robustez e solidez da
metodologia empregada frente à proposta desse trabalho.
4. RESULTADOS 4.1. Validação da metodologia por reedocking De forma a melhorar a acurácia e robustez do método, foram realizados estudos
de docking tendo como base a estrutura cristalográfica do domínio de ligação com
o ligante do AR complexado com a molécula de testosterona, obtidos no Protein
Data Bank (código de busca 2AM9 – resolução 1.64 Å). Tal estrutura foi utilizada
como “plano de fundo” para se avaliar os possíveis pontos de interação de cada
um dos esteroides avaliados nesse trabalho. Para tal, foram geradas 10 poses
para a molécula de testosterona, avaliando-se os principais pontos de distância
envolvidos na farmacodinâmica do complexo ligante-receptor, abrangendo uma
área de 5 a 15 Å ao redor do domínio de ligação. Os médias de distância entre os
principais resíduos envolvidos com a atividade hormonal para áreas de 5, 10 e 15
Å, bem como as médias de RMSD e Scores para cada uma das áreas
empregadas se encontram na tabela 1.
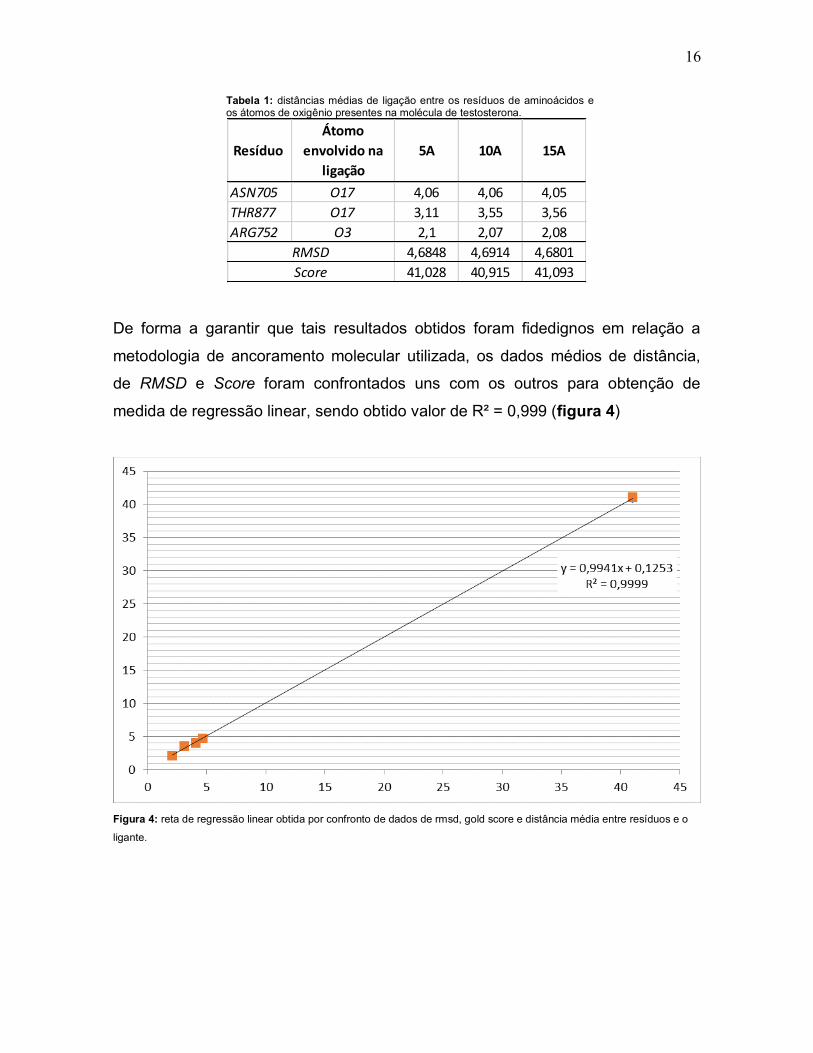
16
Tabela 1: distâncias médias de ligação entre os resíduos de aminoácidos e os átomos de oxigênio presentes na molécula de testosterona.
Resíduo Átomo
envolvido na ligação
5A 10A 15A
ASN705 O17 4,06 4,06 4,05THR877 O17 3,11 3,55 3,56ARG752 O3 2,1 2,07 2,08
4,6848 4,6914 4,680141,028 40,915 41,093
RMSDScore
De forma a garantir que tais resultados obtidos foram fidedignos em relação a
metodologia de ancoramento molecular utilizada, os dados médios de distância,
de RMSD e Score foram confrontados uns com os outros para obtenção de
medida de regressão linear, sendo obtido valor de R² = 0,999 (figura 4)
Figura 4: reta de regressão linear obtida por confronto de dados de rmsd, gold score e distância média entre resíduos e o
ligante.

17
4.2. Obtenção das estruturas tridimensionais das moléculas de interesse e
preparo para análise de ancoramento (docking) molecular Para a obtenção de estruturas tridimensionais visando análise de ancoramento,
docking, é necessário que alguns passos sejam seguidos.
Primeiramente, após desenhadas em programa especifico (Chemdraw® -
ferramenta ChemBio3D®) as mesmas passem por processos de minimização de
energia, para correção de ângulos e distancias de ligação (Chimera®). Uma busca
conformacional foi executada, visando a obtenção de mínimo de energia, local ou
global. Uma única fórmula molecular pode representar um número elevado de
isômeros, cada qual um mínimo local na superfície de energia (superfície de
energia potencial), criada a partir da energia total (energia eletrônica mais a
energia de repulsão entre os núcleos) em função das coordenadas de todos os
núcleos. O ponto estacionário é classificado como a geometria na qual a derivada
da energia com respeito a todos os deslocamentos dos núcleos é igual a zero. Um
mínimo local (de energia) é um ponto estacionário onde todos os deslocamentos
implicam um aumento na energia. Um mínimo local também é chamado de
mínimo global se corresponder ao isômero mais estável, ou seja, se não houver
nenhum outro ponto estacionário com menor energia na superfície de energia
potencial. Se uma determinada mudança de coordenada em particular implicar em
um decréscimo na energia total em duas direções, este ponto estacionário
corresponde a estrutura de transição e a coordenada é chamada de coordenada
de reação. Este processo de determinação de pontos estacionários é chamado de
otimização de geometria33. Entretanto, tal procedimento não foi realizado para os
compostos propostos uma vez que fugiria ao escopo do trabalho.
Para cada uma das 14 moléculas avaliadas foram geradas 10 poses de maior
estabilidade em relação ao domínio de ligação entre o ligante-receptor obtido pela
validação de docking com a molécula de testosterona. A escolha da melhor pose
se baseou-se nos menores valores de distância entre os resíduos base (ASN705,
THR877, ARG752) com os átomos de oxigênio presentes em cada uma das
moléculas. Tais valores podem ser observados na tabela 2. Um dos compostos
sugeridos para análise nesse trabalho foi excluído durante a fase de estudo, a

18
oximetalona, uma vez que os confôrmeros gerados para avaliação se ancoraram
ao sitio de ligação do AR de maneira invertida. Tal dado requer estudos mais
avançados e que fogem ao escopo desse trabalho.
Tabela 2: poses de escolha de cada um dos esteroides anabólico-androgênicos avaliados de acordo
com as distâncias dos resíduos primordiais para ação da testosterona no AR mediante docking de
acordo com os critérios estabelecidos durante a validação do método.
ASN705 THR877 ARG752DHT 5 6,3 5,7 2,1mesterolona 10 6,4 6,3 2,2drostanolona 9 7,3 7,1 2,4metenolona 5 7 6,9 2,1oxandrolona 6 6,9 6,5 2stanozolol 1 5,4 2,3 1,7boldenona 10 5,9 5,3 1,7metandrostenolona 10 6,3 4,8 2,14-Cl-Dihidrometiltestosterona 5 5,7 5,7 1,6Fluoximesterona 4 5,7 5,6 3,3gestrinona 4 6,4 2,4 1,7nandrolona 3 7,7 7,2 1,7trembolona 9 6,7 6 1,7
resíduoPose de escolha
Derivado EAA
4.3. Avaliação comparativa entre pontos de interação de cada EAA com o AR
As tabelas 3, 4 e 5 a seguir indicam os principais pontos de interação de cada um
dos EAS e os aminoácidos do AR, avaliados nesse trabalho, além daqueles
estabelecidos como pontos primordiais para atividade biológica para essa classe
de compostos. Tal indagação surge pelo fato que, apesar de ASN705 e THR877
serem considerados fundamentais para a atividade de testosterona no AR, os
derivados esteroides sintéticos parecem não realizar tal tipo de interação. Logo,
sua elevada potência conhecida na prática clínica pode estar associada com
outros possíveis pontos de interação entre ligantes e receptor.
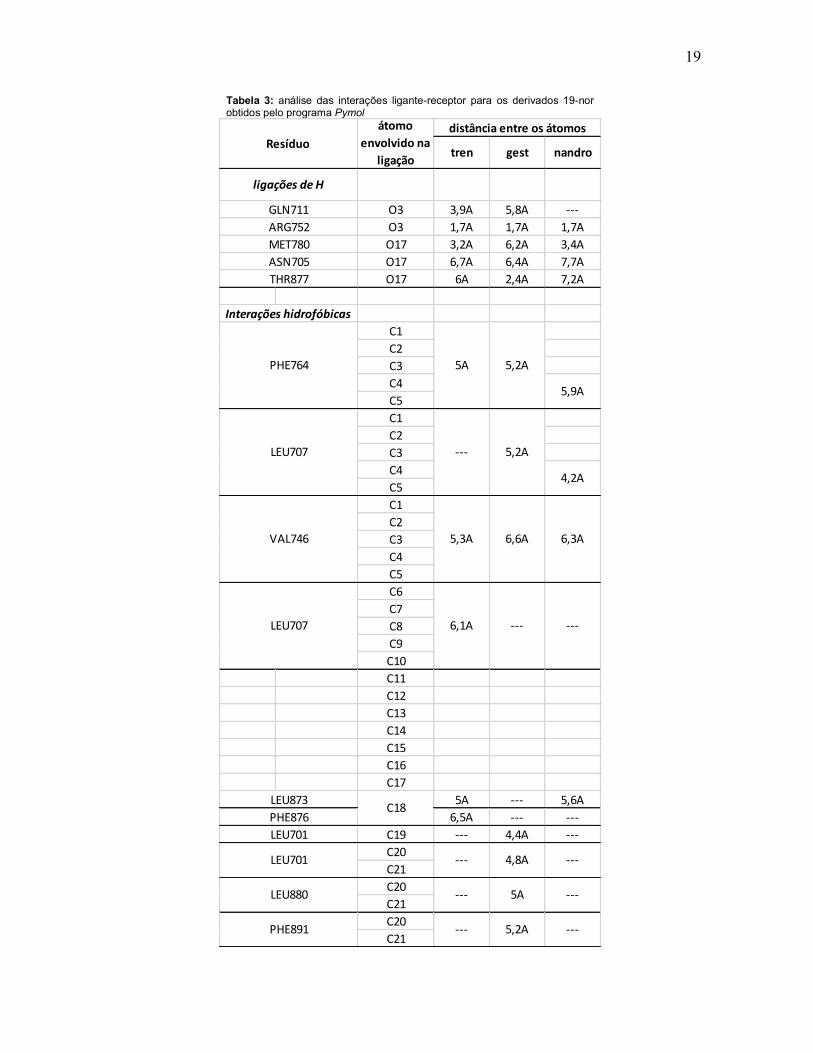
19
Tabela 3: análise das interações ligante-receptor para os derivados 19-nor obtidos pelo programa Pymol
tren gest nandro
O3 3,9A 5,8A ---O3 1,7A 1,7A 1,7AO17 3,2A 6,2A 3,4AO17 6,7A 6,4A 7,7AO17 6A 2,4A 7,2A
C1C2C3C4C5C1C2C3C4C5C1C2C3C4C5C6C7C8C9C10C11C12C13C14C15C16C17
5A --- 5,6A6,5A --- ---
C19 --- 4,4A ---C20C21C20C21C20C21
LEU701
átomo envolvido na
ligaçãoResíduo
distância entre os átomos
ligações de H
Interações hidrofóbicas
ARG752
--- 4,8A ---LEU701
LEU880 --- 5A ---
6,3A6,6A
5,9A
5,2A
PHE891 5,2A ------
LEU707 6,1A
LEU707 5,2A---
--- ---
4,2A
VAL746
PHE876C18
5A
GLN711
MET780
PHE764
LEU873
ASN705THR877
5,3A
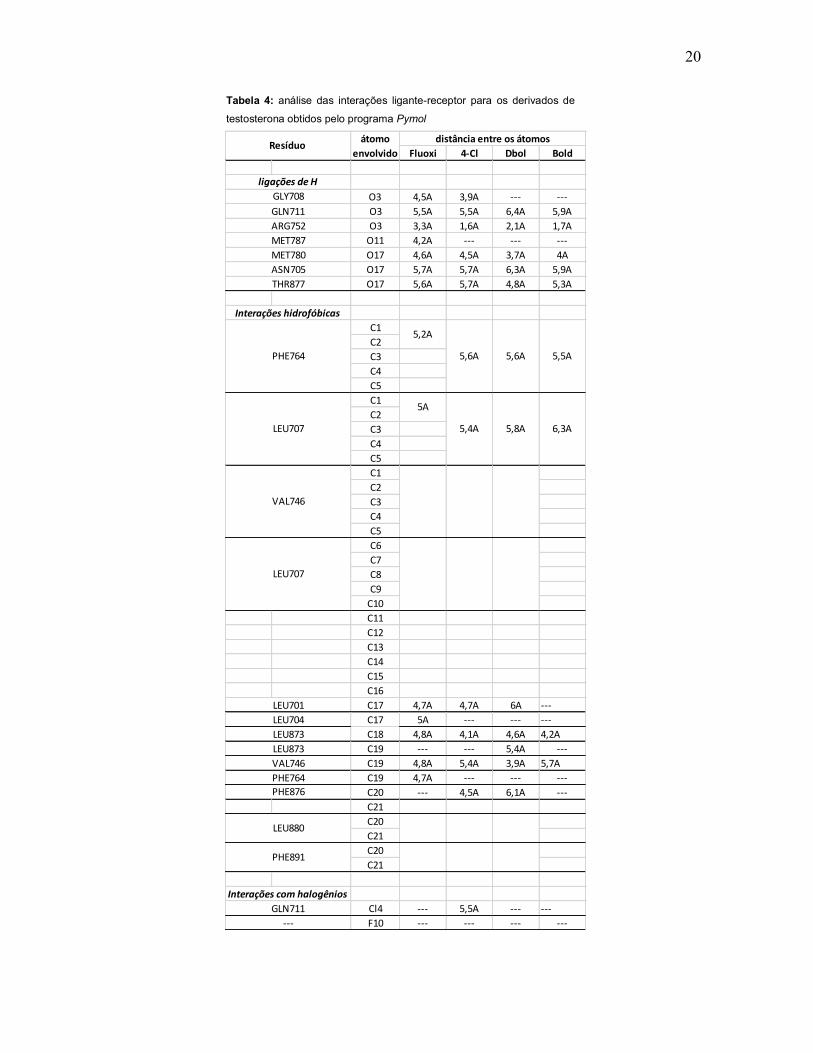
20
Tabela 4: análise das interações ligante-receptor para os derivados de
testosterona obtidos pelo programa Pymol
Fluoxi 4-Cl Dbol Bold
O3 4,5A 3,9A --- ---O3 5,5A 5,5A 6,4A 5,9AO3 3,3A 1,6A 2,1A 1,7AO11 4,2A --- --- ---O17 4,6A 4,5A 3,7A 4AO17 5,7A 5,7A 6,3A 5,9AO17 5,6A 5,7A 4,8A 5,3A
C1C2C3C4C5C1C2C3C4C5C1C2C3C4C5C6C7C8C9C10C11C12C13C14C15C16C17 4,7A 4,7A 6A ---C17 5A --- --- ---C18 4,8A 4,1A 4,6A 4,2AC19 --- --- 5,4A ---C19 4,8A 5,4A 3,9A 5,7AC19 4,7A --- --- ---C20 --- 4,5A 6,1A ---C21C20C21C20C21
Cl4 --- 5,5A --- ---F10 --- --- --- ---
6,3A
PHE764
5,2A
5,5A
MET787
PHE876
GLN711
ligações de H
Interações hidrofóbicas
---
LEU701LEU704
Interações com halogênios
ARG752
PHE891
distância entre os átomos
GLY708
5,6A
5A
5,8A
LEU880
LEU873LEU873VAL746
VAL746
LEU707
THR877
PHE764 5,6A
LEU707 5,4A
Resíduo átomo envolvido
GLN711
MET780ASN705

21
Tabela 5: análise das interações ligante-receptor para os derivados de di-hidrotestosterona obtidos pelo
programa Pymol
stan oxan primo master prov dht
O3 --- 2A 2,1A 2,4A 2,2A 2,1AO3 --- 6,6A 6,4A 6,5A 6,3A 6,2AO17 --- 3,8A 3A 4,6A 3,7A 4,1AO17 5,4A 6,9A 7A 7,3A 6,3A 5,7AO17 2,3A 6,5A 6,9A 7,1A 6,4A 6,3AN2 1,7A --- --- --- --- ---N1 3,4A --- 6,4A --- --- ---N1 4,7A
C1 ---C2C1 --- --- 4,9A --- ---C2 --- 6,7A ---C1 --- 4,6AC2 4,4A ---C1 --- --- --- 4,1A 4,4AC7C8C9C10C11C12C13C14C15C16C17 --- 5A --- --- --- ---C17 4,8A --- --- --- --- ---C17 4,4A 4,6A --- --- --- ---C17 6,8A --- --- --- --- ---C18 --- 4,9A --- --- 4,6A 3,8AC19 6A --- 5,6A --- --- ---C19 5,9A 5A 4,5A --- 4,6A 4,2AC19 --- --- 5,8A --- --- ---C20C21C20C21C20C21
GLN711
LEU873
PHE764
VAL746
---
LEU701LEU873
LEU873TRP741VAL746LEU873
4,8A---
LEU707
Resíduo átomo envolvido
ligações de H
distância entre os átomos
LEU880
MET745
PHE876
ARG752
MET780ASN705THR877
Interações hidrofóbicas
ARG752GLN711
5,1A

22
5. DISCUSSÃO Sabe-se que pontos de interação hidrofílica, através da formação de ligações de
hidrogênio, são de crucial importância para a ligação do composto ligante ao sitio
ativo do AR (LBD). Os resíduos de ARG752, ASN705 E THR877são de crucial
importância para atividade dos ligantes naturais testosterona e di-
hidrotestosterona ao AR. Como regra, a maioria dos andrógenos que se ligam
com alta afinidade ao AR possuem um grupo carbonila no carbono 3 do anel A,
além de um grupo hidroxila ligado ao carbono 17, em posição beta, possibilitando
a formação de interações de hidrogênio com os resíduos acima detalhados27. Uma
vez que ligações de hidrogênio são muito mais potentes que interações
hidrofóbicas (como interações de van der Waals), é de se entender o porque de
tais interações serem o principal agente a explicar o motivo pelo qual andrógenos
se ligam ao AR com tanta afinidade em faixas de concentração da ordem de
nanomolar38. Ainda, ligações de hidrogênio (interações eletrostáticas de forma
geral) são necessárias para promover ligação do ligante ao receptor, enquanto
que as ligações hidrofóbicas (ou interações eletrodinâmicas) teriam por papel
garantir alto grau de seletividade do receptor a determinadas classes de ligantes
além de auxiliar na estabilização do ligante ao receptor.
A maioria dos trabalhos envolvendo análise de interação entre ligantes esteroides
ao AR descreve a interação do átomo de oxigênio da carbonila em C3 como
crucial para atividade do mesmo10,27,33,38,39, sendo que mutações nesse resíduo
(ARG752GLN) afetam a atividade do receptor, produzindo um fenótipo conhecido
como síndrome de insensibilidade androgênica, conduzindo a quadros de pseudo-
hermafroditismo38. É também de conhecimento que a distância de ligação
envolvidos na formação de ligações de hidrogênio é proporcional à força de
interação envolvida entre os mesmos, devido ao vetor força gerado. Os dados
presentes na tabela 2 nos fornecem as distancias de ligação entre a carbonila do
C3 e o resíduo de ARG752 dos compostos avaliados nesse trabalho. Observa-se
que derivados 19-nor apresentam distancias de ligação muito menores que de
outros ligantes, como a própria testosterona e o DHT. Outros esteroides
pertencentes a outras famílias, como os próprios derivados de DHT apresentaram

23
maior grau de interação do que os ligantes naturais. Curiosamente, os membros
derivados dessa família são tidos como de baixa potencia e ligação ao AR20.
Kicman, em revisão24 aponta para a inconsistência dos dados produzidos pelo
trabalho de Vida20, no qual se adotaram índices anabólico-androgênicos para uma
extensa gama de derivados sintéticos incompatíveis com novos trabalhos. As
fundamentações de tais índices são oriundas de metodologia modificada de
Eisenberg e Gordan39 por Hershberger e colaboradores40. Eisenberg e Gordan39
propuseram o uso do desenvolvimento do músculo elevador do ânus (elevator ani)
de ratos como critério para mensuração de anabolismo, uma vez que ratos
castrados tratados com diversos esteroides, como proprionato de testosterona ou
ainda 17-α-metil-testosterona, paralelo a um baixo índice de desenvolvimento das
glândulas seminais (um órgão responsivo a andrógenos). Hershberger e
colaboradores40 utilizaram como critério de avaliação androgênica o
desenvolvimento da parte ventral da próstata de ratos gonadectomizados. O
índice de mensuração proposto foi:
Razão Anabólico-androgênico = (peso do músculo eretor do anus experimental –
peso do musculo eretor do anus controle)/(peso da próstata ventral experimental –
peso da próstata ventral controle)
Kruskemper41 discutiu diversas falhas em tais índices para avaliação anabólico-
androgênico. Por exemplo, as vesículas seminais reagem de forma mais lenta a
certos andrógenos, de forma que a administração por curto período de tempo do
composto poderia levar a um viés de análise. Ainda, diversos outros trabalhos
demonstraram que a administração de curto prazo em ratos gonadectomizados
levaram a desenvolvimento de órgãos associados com o aparelho urogenital em
ratos, mas não levaram ao desenvolvimento de outros músculos como os da
coxa42,43,44. Uma hipótese discutida sobre os mecanismos associados a
anabolismo-androgenicidade é de que determinados tecidos teriam maior
sensibilidade a andrógenos por apresentarem maior densidade da enzima 5α-
Redutase, que poderia tais compostos em derivados mais potentes. Dessa

24
maneira, ainda não se tem claro a razão pela qual determinados esteroides não
aparentarem dissociação entre índices anabólicos androgênicos, ainda que
possuam modificações moleculares que em tese diminuíram tais discrepâncias.
Por exemplo, o derivado 19-nor-testosterona (nandrolona), ainda que possa sofrer
redução, leva a produção de um derivado androgênico de menor afinidade ao AR
que o próprio DHT. Entretanto, sabe-se que derivados de DHT, como estanozolol
e oxandrolona podem apresentar desenvolvimento de órgãos sensíveis a
andrógenos. Isso demonstra que, mesmo compostos com baixo índice
androgênico teórico, podem levar a características androgenizantes (virilizantes)
quando administrados de forma crônica, como observado em mulheres e
crianças24.
Alvarez-Ginarte e colaboradores33 buscaram uma correlação entre técnicas de
Virtual Screening para ligante e receptor com os dados compilados por Vida20 na
busca de compostos esteroides anabólicos líderes, que pudessem apresentar
maior dissociação do índice anabólico-androgênico. O grupo deduziu conclusões
importantes sobre pontos de interação dos ligantes com o AR, baseados nos
dados de interação do mesmo com testosterona e DHT:
• O átomo de oxigênio da carbonila em C3 que contem um par de elétrons
livres que pode interagir com resíduos hidrofílicos da vizinhança do anel A.
• O átomo de oxigênio da hidroxila associada a C17 possui uma carga parcial
positiva que permite que o mesmo interaja com resíduos eletronegativos do
anel D.
• Testosterona e DHT possuem interações hidrofóbicas semelhantes com
outros ligantes e o AR.
O trabalho classificou um composto como sendo um anabólico potente caso
obedeça os modelos QSAR propostos pelo grupo além de apresentar um grupo
carbonil (cetona) e hidroxil nas posições descritas, que favorecem a ligação do
andrógeno ao receptor no sitio de ligação LBD. Em relação a interações
hidrofóbicas, o grupo considerou como boas interações aquelas que apresentaram
distancia menor que 4Å.

25
Ainda que a maior parte dos trabalhos descreve a interação da hidroxila em C17
como um ponto importante para atividade de derivados esteroides sintéticos,
poucos demostraram tal relevância com relação aos derivados sintéticos de uso
corrente para propósitos escusos, como o dopping esportivo. Mesmo o trabalho de
Álvarez-Ginarte e colaboradores33, ao selecionar através de virtual screening uma
coleção de mais de 200 compostos, não parece ter demostrado diferença
relevante desse grupo para maior grau de anabolismo. Mesmo em nosso trabalho,
como pode ser visto na tabela 2, verifica-se que as distancias de ligação da
hidroxila em C17 aos resíduos da vizinhança do anel D são muito maiores do que
aquelas estabelecidas como aceitáveis para ligações de hidrogênio (menores que
3Å). Isso se deve ao arranjo tridimensional adquirido pelos ligantes uma vez
associados ao LDB do AR. A figura 5 aponta as distancias de ligação do derivado
gestrinona aos resíduos de ARG752 e THR877 bem como uma vista geral da
mesma no LDB. De forma geral, sabe-se que os derivados 19-nor (trembolona,
gestrinona e nandrolona) apresentam alto índice de anabolismo e baixo índice
androgênico.

26
Figura 5: a) distância de ligação do derivado gestrinona ao resíduo ARG752 b) distância de ligação do derivado gestrinona ao resíduo THR877 c) vista geral de gestrinona no LDB.
A figura 6 aponta os pontos de interação entre o derivado trembolona no LDB bem
como sua visão geral associada ao receptor e, em conjunto com a gestrinona, e
outros trabalhos que descrevem o uso de outro esteroide, a tetrahidrogestrinona
(THG) por atletas, nos fornece dados importantes acerca de possíveis
mecanismos de sua alta potência e ligação ao receptor. As interações hidrofóbicas
parecem apresentar pouca relevância nos trabalhos descritos anteriormente.
Entretanto a tabela 3 nos fornece dados das interações ligante-receptor para os
derivados 19-nor em relação aos resíduos que circundam o ligante. Notamos que,
apesar das distancias serem maiores que 4Å, como preconizados no trabalho de
Alvarez-Ginarte e colaboradores33, os derivados aparentam distancias de ligação
semelhantes entre si e, para trembolona e gestrinona, que possuem sistemas de
duplas ligações integradas, observamos que tal grau de estabilidade permite que
ambas se acomodem de forma quase espelhada em plano longitudinal no
receptor.

27
Figura 6: a) ponto de interação entre o derivado trembolona ao resíduo ARG 752 b) vista geral do derivado associado ao receptor
Outro dado importante que vale discussão em nosso trabalho é com relação aos
esteroides agrupados como Derivados de testosterona. Ainda que tenham sido
avaliadas possíveis interações hidrofóbicas importantes para interação entre os
derivados e o AR, nenhum dos resíduos avaliados apresentaram interações com o
esqueleto esteroide dos compostos avaliados. Ao que parece, as ligações de
hidrogênio parecem ser os maiores determinantes para atividade desses em
relação ao receptor. Alguns desses derivados, como a fluoximesterona
(halostestin) e o 4-Cl-dihidrometiltestosterona (turinabol) chamam atenção por

28
conterem halogênios em sua estrutura, o que poderia levar a duas possíveis
interações com o AR:
• Por ligações de hidrogênio com resíduos na vizinhança do halogênio
• Pela exploração de bolsões na vizinhança do halogênio, que permitiram
melhor acomodamento dos mesmos no AR.
Na figura 7, observamos os resíduos ao redor do átomo de cloro presente na
molécula de turinabol. É possível que as cargas eletrônicas ao redor do mesmo
possam acomodar de forma harmônica o composto no AR, o que em parte
explicaria seu elevado potencial anabólico. Da mesma forma, podemos usar tal
critério para descrever o potencial anabólico elevado da molécula de
fluoximesterona (halotestin), comumente utilizada de forma abusiva por atletas45.
Figura 7: a) possível bolsão ao redor do grupo Cl da molécula de 4-Cl-di-hidro-metil-testosterona (turinabol) b) possível bolsão ao redor do grupo F da molécula de fluoximesterona (halotestin).

29
Outro ponto importante a ser discutido ainda com relação aos derivados de
testosterona é a mudança de potencia em relação à ligação ao AR em decorrência
da alquilação em alfa do C17. Tal modificação é realizada a fim de que o esteroide
apresente viabilidade quando administrado oralmente, de forma a sobreviver ao
efeito de primeiro passo hepático. Entretanto, sabe-se que a presença desse
grupo alquil (geralmente metil) é responsável por elevar o índice anabólico dos
esteroides. Comparando dois membros dessa mesma família, a saber, boldenona
e metandrostenolona (dianabol). Como é possível verificar na tabela 4, a presença
de um grupo metil na molécula de metandrostenola em C17 permite explorar
interações hidrofóbicas em tal vizinhança, ainda que de forma fraca devido as
distâncias de ligação serem maiores que 4A (figura 8). Nota-se ainda que, a única
diferença estrutural entre os mesmos reside apenas na presença do grupo metil
associado ao C17, demonstrando a importância dessa modificação molecular para
a potência dos EAS (figura 9).
Figura 8: possíveis interações hidrofóbicas entre os resíduos na vizinhança do grupo metil associado em C17 de metandrostenolona (dianabol); a)PHE891 b) PHE876 c) LEU880 d) LEU701.

30
Figura 9: a) conformação da molécula de boldenona no LDB do AR. b) conformação da molécula de metandrostenolona (dianabol) no LDB do AR.
Ainda que não tenha sido explorado nesse trabalho, outro caso onde se verifica
aumento de potencia do esteroide por metilação em C17 ocorre com dois
derivados de DHT, a saber, a drostanolona, e seu derivado 17 alquilado, metil-
drostanolona, conhecido popularmente como SuperDrol, o qual foi popularmente
comercializado nos anos 2000 como um suplemento nutricional na forma de
dimetazina, podendo ser classificado como um pró fármaco, uma vez que o
mesmo é formado por duas moléculas de metil-drostanolona unidos por um átomo
de nitrogênio, levando a sua retirada do mercado 2006 pelo FDA46 (figura 10).

31
Figura 10: molécula de dimetazina, classificada como um pró-hormônio ou pró fármaco, após sofrer metabolização libera 2 moleculas de metil-drostanolona, popularmente conhecido como superdrol.
Os esteroides membros da família dos derivados de DHT são tidos comumente
como compostos de baixa potencia e fraca ligação ao AR. Sua modificação base
consiste na perda da dupla ligação entre C4-C5, sitio de ação da enzima 5α-
Redutase, que leva a formação de derivados reduzidos. Dessa maneira, a
formação de compostos derivados reduzidos em tecidos androgênicos se
reduziria, como preconizado na revisão de Kicman24. Entretanto, sabe-se que o
uso crônico de tais compostos não os dissocia de efeitos virilizantes. Os dados da
tabela 5 se tornam ainda mais interessantes quando analisamos caso a caso
pontos de interação dos mesmos com o AR.
No caso da molécula de estanozolol, verifica-se que a presença do anél pirazol
unido ao anel A do esqueleto esteroide diminui muito a distancia de interação de
hidrogênio entre o par de elétrons do átomo de nitrogênio e o resíduo ARG752.
Ainda, se observa que esse mesmo grupo é capaz de interagir com outros
resíduos em sua vizinhança, como GLN711. Ainda, o oxigênio presenta na
hidroxila associada a C17 apresenta uma distancia de ligação de hidrogênio
reduzida em relação aos ligantes naturais com o resíduo de THR877. Ainda que
existam, com distancias de ligação relativamente altas, as interações hidrofóbicas

32
de estanozolol com o AR parecem desempenhar um papel de menor importância
de interação entre os mesmos (figura 11).
Outra interação que merece destaque e é pouco discutida é a de resíduo MET780
com o oxigênio presenta na hidroxila associada a C17 na molécula de metenolona
(primobolan), de 3A. Tal valor ainda que alto pode ser aceito como uma interação
de hidrogênio fraca.
Outras interações tidas como responsáveis pelo aumento na atividade anabólica
associada as modificações moleculares na molécula de testosterona, como a
alquilação de C1 e/ou C2, propostas por Fragkaki e colaboradores27, não
mostraram interações hidrofóbicas com distancia menor ou igual a 4A entre os
resíduos metil presentes tanto na molécula de drostanolona (masteron) quanto de
metenolona (primobolan) com resíduos de aminoácidos ao redor da vizinhança do
anel A. Possivelmente esses grupos aumentem a atividade anabólica de tais
moléculas devido ao acomodamento do ligante ao receptor de forma mais estável
em decorrência da presença de bolsões hidrofóbicos capazes de acomodar tais
grupos (figura 12)
Figura 11: interações de hidrogênio entre os resíduos ARG752, GLN 711 e THR877 com a molécula de estanozolol. Observe que a presença do anel pirazol diminui a distância de interação do par eletrônico com o resíduo de ARG752.

33
Figura 12: a) possível bolsão hidrofóbico em interação com o grupo metil em C2 da molécula de drostanolona
(masteron). b) possível bolsão hidrofóbico em interação com grupo metil em C1 da molécula de metenolona
(primobolan).

34
6. CONCLUÕES Mesmo com a necessidade de maior detalhamento nas análises para real
compreensão da correlação entre as interações que os ligantes apresentam ao AR
com sua atividade e potência, fica claro nesse trabalho que determinados pontos
de interação são cruciais para atividade dos esteroides anabólico-androgênicos.
Os derivados anabólico-androgênicos pertencentes a família 19-nor se mostraram
os mais potentes em relação aos demais devido a melhor acomodação ao AR.
Contrário a literatura, os membros pertencentes à família de derivados de DHT,
tidos como de baixa potência apresentaram diversos pontos de interação
importante com o AR. Estudos mais detalhados, bem como correlação com dados
in vivo, se mostram necessários para confirmação de tais hipóteses. Os achados
são promissores para futuros estudos de agentes que possam atuar moduladores
do receptor androgênico, visando a dissociação do potencial anabólico dos efeitos
androgênicos, bem como aumento de seletividade de tecido e futura
aplicabilidade, especialmente clínica, como no tratamento de quadros de caquexia
crônica ou ainda agentes com potencial anti tumoral, especialmente em tecidos
sensíveis à andrógenos, como a próstata e o aparelho urogenital de forma geral.
7. BIBLIOGRAFIA
1. SEDDON, G; LOUNMAS, V; Mc GUIRE, R; VAN DEN BERGH, T; BYWATER, R.P;
OLIVEIRA, L. VRIEND, G.; Drug design for ever, from hype to hope. Journal of Computer-Aided Molecular Design. 26;137–150. 2012.
2. BLEICHER, K.H.; BOHM, H.J.; MULLER, K.; A.I. Alanine, Hit and lead generation: beyond high-throughput screening, Nature Reviews Drug Discovery 2;369–378. 2003.
3. BOHACEK, R.S.; Mc MARTIN, C.; GUIDA, W.C.; The art and practice of structure-based drug design: a molecular modeling perspective. Medicinal Research Reviews. 16;13–50. 1996.
4. SONG, C.M.; LIM, S.J.; TONG, J.C. Recent advances in computer-aided drug design. Briefings in Bioinformatics. 10;579–591. 2009.

35
5. GUIDO, R.V.C.; OLIVA, G.; ANDRICOPULO, A.D.; Modern drug discovery technologies: opportunities and challenges in lead discovery. Combinatorial Chemistry &High Throughput Screening. 14;830–839. 2011.
6. MANGELSDORF, D.J.; THUMMEL, C.; BEATO, M.; HERRLICH, P.; SCHULTZ, G.; UMESONO, K.; et. al. The nuclear receptor superfamily: the second decade. Cell. 83; 835–9. 1995
7. TSAI, M.J.; O'MALLEY, B.W. Molecular mechanisms of action of steroid/ thyroid receptor superfamily members. Annu. Rev. Biochem. 63; 451–86. 1994
8. Nuclear Receptors Nomenclature Committee. A unified nomenclature system for the nuclear receptor superfamily. Cell. 97; 161–3.1999.
9. TAN, M.H.E; JUN, L.I.; XU, H.E.; MELCHER, K.; YONG, E-L. Androgen receptor: structure, role in prostate cancer and drug discovery. Acta Pharmacologica Sinica. 36; 3-23. 2015.
10. LUBAHN, D.B.; JOSEPH, D.R.; SULLIVAN, P.M.; WILLARD, H.F.; FRENCH, F.S.;
WILSON, E.M. Cloning of human androgen receptor complementary DNA and localization to the X chromosome. Science. 240; 327–30. 1988.
11. BROWN, C.J.; GOSS, S.J.; LUBAHN, D.B.; JOSEPH, D.R.; WILSON, E.M.;
FRENCH, F.S.; et. al. Androgen receptor locus on the human X chromosome: regional localization to Xq11–12 and description of a DNA polymorphism. Am. J. Hum. Genet. 44; 264–9. 1989.
12. MIGEON, B.R.; BROWN, T.R.; AXELMAN. J.; MIGEON, C.J. Studies of the locus for androgen receptor: localization on the human X chromosome and evidence for homology with the Tfm locus in the mouse. Proc. Natl. Acad. Sci. USA. 78; 6339–43. 1981.
13. HEEMERS, H.V.; TINDALL, D.J. Androgen receptor (AR) coregulators: a diversity of functions converging on and regulating the AR transcriptional complex. Endocr. Rev. 28 (7); 778–808. 2007.
14. PANDINI, G.; MINEO, R.; FRASCA, F.; ROBERTS, C.T.J.; MARCELLI, M.; VIGNERI, R.; BELFIORE, A. Androgens up-regulate the insulin-like growth factor-I receptor in prostate cancer cells. Cancer Res. 65 (5); 1849–57. 2005.
15. VLAHOPOULOS, S.; ZIMMER, W.E.; JENSTER, G.; BELAGULI, N.S.; BALK, S.P.; BRINKMANN, A.O.; LANZ, R.B.; ZOUMPOURLIS, V.C.; SCHWARTZ, R.J. Recruitment of the androgen receptor via serum response factor facilitates expression of a myogenic gene. J. Biol. Chem. 280(9); 7786–92. 2005.

36
16. HEINLEIN, C.A.; CHANG, C. The roles of androgen receptors and androgen-binding proteins in nongenomic androgen actions. Mol. Endocrinol. 16 (10); 2181–7. 2002.
17. FIX, C.; JORDAN, C.; CANO, P.; WALKER, W.H.; Testosterone activates mitogen-activated protein kinase and the cAMP response element binding protein transcription factor in Sertoli cells. Proc Natl Acad Sci USA. 101(30): 10919–24. 2004.
18. PI, M.; PARRILL, A.L.; QUARLES, L.D. GPRC6A Mediates the Non-genomic
Effects of Steroids. The Journal of Biological Chemistry. 285(51);39953-39964. 2010.
19. ÁLVAREZ-GINARTE, Y.M.; MARRERO-PONCE, Y.; RUIZ-GARCIA, A.; MONTERO-CABRERA, L.A.; DE LA VEGA, J.M.G.; NOHEDA-MARIN, P.; CRESPO-OTERO, R.; ZARAGOZA, F.T.; GARCIA-DOMENECH, R. Applying pattern recognition methods plus quantum and physico-chemical molecular descriptors to analyze the anabolic activity of structurally diverse steroids. Journal of Computational Chemistry. 29;317–333. 2008.
20. VIDA, J.A.; Androgens and Anabolic Agents, Chemistry and Pharmacology. New York and London. 1969.
21. NIMNI, M.E.; GEIGER, E. Non-suitability of levator ani method as an index of anabolic effect of steroid. Proc Soc Exp Biol Med. 94:606–610.1957.
22. CELOTTI, F.; CESI, P.N. Anabolic steroids—a review of their effects on the muscles, of their possible mechanisms of action and of their use in athletics. J Steroid Biochem Mol Biol 43;469–477.1992.
23. ORR, R.; FIATARONE-SINGH, M. The anabolic androgenic steroid oxandrolone in the treatment of wasting and catabolic disorders: review of efficacy and safety. Drugs. 64(7);725-50. 2004.
24. KICMAN, A.T.; Pharmacology of anabolic steroids. British Journal of Pharmacology. 154(3);502-521. 2008.
25. ORWOLL, E.S. Androgens as anabolic agents for bone. Trends Endocrinol Metab. 7;77–84.1996.
26. ZITZMANN, M.; NIESCHLAG, E.; Androgens and bone metabolism Testosterone. Cambridge University Press; UK; 233–254 in NIESCHLAG, E.; BEHRE, H.M. (eds).3º ed. 2004.
27. FRAGKAKI, A.G.; ANGELIS, Y.S.; KOUPPARIS, M.; TSANTILI-KAKOULIDOU, A.;
KOKOTOS, G.; GEORGAKOPOULOS, C. Structural characteristics of anabolic

37
androgenic steroids contributing to binding to the androgen receptor and to their anabolic and androgenic activities: Applied modifications in the steroidal structure. Steroids. 74(2);172-197. 2009.
28. GASTEIGER, E.; JUNG, E.; BAIROCH, A. SWISS-PROT: connecting biomolecular
knowledge via a protein database. Curr Issues Mol Biol. 3(3);47-55. 2001.
29. BIRD, A. Il2 transcription unleashed by active DNA demethylation. Nat Immunol. 4(3);208-9. 2003.
30. WERMUTH, C. G. Similarity in drugs: reflections on analogue design. Drug Discov Today. 11(7-8);348-54. 2006.
31. CHEN, S-C.; BAHAR, I. Mining frequent patterns in protein structures: a study of
protease families. Bioinformatic. 20(1):i77-i85.2004.
32. PEREIRA DE JÉSUS-TRAN, K.; CÔTÉ, P-L.; CANTIN, L.; BLANCHET, J.;
LABRIE, F.; BRETON, R. Comparison of crystal structures of human androgen receptor ligand-binding domain complexed with various agonists reveals molecular determinants responsible for binding affinity. Protein Science: A Publication of the Protein Society. 15(5);987-999. 2006.
33. ALVAREZ-GINERTE, Y.M.; MONTERO-CABRERA, L.A.; DE LA Vega, J.M.G.; BENCOMO-MARTÍNEZ, A.; PUPO, A.; AGRAMONTE-DELGADO A.; MARRERO-PONCE, Y.; RUIZ-GARCÍA, J.A.; MIKOSCH, H. Integration of ligand and structure-based virtual screening for identification of leading anabolic steroids. The Journal of Steroid Biochemistry and Molecular Biology. 138;348-358. 2013.
34. LENGAUER, T.; RAREY, M. Computational methods for biomolecular docking. Current Opinion in Structural Biology. 6;402–406. 1996.
35. JORGENSEN, W.L. The many roles of computation in drug discovery, Science. 303;1813–1818. 2004.
36. MOWSZOWICZ, I.; RIAHI, M.; WRIGHT, F.; BOUCHARD, P.; KUTTENN, F.; MAUVAIS-JARVIS, P. Androgen receptor in human skin cytosol. J. Clin. Endocrinol.Metab. 52;338–344. 1981.
37. BOHL, C.E.; CHANG, C.; MOHLER, M.L.; et. al. A Ligand-Based Approach To Identify Quantitative Structure–Activity Relationships for the Androgen Receptor. Journal of medicinal chemistry. 47(15);3765-3776. 2004.
38. SAKAI, N.; YAMADA, T.; ASAO, T.; BABA, M.; YOSHIDA, M., MURAYAMA, T. Bilateral testicular tumors in androgen insensitivity syndrome. Int.J. Urol. 7; 390–392. 2000.

38
39. EISENBERG, E.; GORDAN, G.S. The levator ani muscle of the rat as an index of myotrophic activity of steroidal hormones. J Pharmacol Exp Ther. 99;38–44. 1950.
40. HERSHBERGER, L.G.; SHIPLEY, E.G.; MEYER, R.K. Myotrophic activity of 19-nortestosterone and other steroids determined by modified levator ani muscle method. Proc Soc Exp Biol Med. 83;175–180. 1953.
41. KRUSHKEMPER, H.L. Anabolic Steroids. Academic Press: New York; 1968
42. NIMNI, M.E.; GEIGER, E. Non-suitability of levator ani method as an index of anabolic effect of steroid. Proc Soc Exp Biol Med. 94;606–610. 1957.
43. SCOW, R.O.; HAGAN, S.N. Effect of testosterone propionate on myosin, collagen and other protein fractions in striated muscle of gonadectomized rats. Endocrinology. 60;273–278. 1957.
44. HAYES, K.J. So-called levator ani of rat. Acta Endocrinologica. 48;337–347. 1965.
45. JONES, T.M.; FANG, V.S.; LANDAU, R.L.; ROSENFIELD, R.L. The Effects of Fluoxymesterone Administration on Testicular Function. The Journal of Clinical Endocrinology & Metabolism. 44(1);121–129. 1977.
46. JOSEPH, J.F.; PARR M.K. Synthetic Androgens as Designer Supplements. Current Neuropharmacology. 13(1);89-100. 2015.
_26 de setembro de 2018_______ _26 de setembro de 2018____ Data e assinatura do aluno(a) Data e assinatura do orientador(a)