bioww1.ead.upe.br/nead20201/conteudos/biologia/4... · as bases da hereditariedade foram...
TRANSCRIPT

BIOL O G I A
G e n é t i c a G e r a l
P r o f a . M a r í l i a d e F r a n ç a R o c h a
2a edição | Nead - UPE 2013

Dados Internacionais de Catalogação-na-Publicação (CIP)Núcleo de Educação à Distância - Universidade de Pernambuco - Recife
Rocha, Marília de FrançaBiologia: Genética Geral/Marília de França Rocha. – Recife: UPE/NEAD, 2011.
75 p.
1. Biologia 2. Genética 3. Educação à Distância I. Universidade de Pernambuco,
Núcleo de Educação à Distância II. Título
CDD – 17ed. – 575.1
Claudia Henriques – CRB4/1600BFOP-112/2011
R672b

UNIVERSIDADE DE PERNAMBUCO - UPE
ReitorProf. Carlos Fernando de Araújo Calado Vice-ReitorProf. Rivaldo Mendes de Albuquerque
Pró-Reitor AdministrativoProf. Maria Rozangela Ferreira Silva
Pró-Reitor de PlanejamentoProf. Béda Barkokébas Jr.
Pró-Reitor de GraduaçãoProfa. Izabel Christina de Avelar Silva
Pró-Reitora de Pós-Graduação e Pesquisa Profa. Viviane Colares Soares de Andrade Amorim
Pró-Reitor de Desenvolvimento Institucional e ExtensãoProf. Rivaldo Mendes de Albuquerque
NEAD - NÚCLEO DE ESTUDO EM EDUCAÇÃO A DISTÂNCIA
Coordenador GeralProf. Renato Medeiros de Moraes
Coordenador AdjuntoProf. Walmir Soares da Silva Júnior
Assessora da Coordenação GeralProfa. Waldete Arantes
Coordenação de CursoProf. José Souza Barros
Coordenação PedagógicaProfa. Maria Vitória Ribas de Oliveira Lima
Coordenação de Revisão GramaticalProfa. Angela Maria Borges CavalcantiProfa. Eveline Mendes Costa LopesProfa. Geruza Viana da Silva
Gerente de ProjetosProfa. Patrícia Lídia do Couto Soares Lopes
Administração do AmbienteJosé Alexandro Viana Fonseca
Coordenação de Design e ProduçãoProf. Marcos Leite
Equipe de DesignAnita Sousa/ Gabriela Castro/Renata Moraes/ Rodrigo Sotero
Coordenação de SuporteAfonso Bione/ Wilma SaliProf. José Lopes Ferreira Júnior/ Valquíria de Oliveira Leal
Edição 2013Impresso no Brasil
Av. Agamenon Magalhães, s/n - Santo AmaroRecife / PE - CEP. 50103-010Fone: (81) 3183.3691 - Fax: (81) 3183.3664


GENÉTICA GERAL
Profa. Marília de França Rocha Carga Horária I 15h
APRESENTAÇÃO DA DISCIPLINA
Olá! Sejam bem-vindos a disciplina Genética Geral!
As bases da hereditariedade foram estabe-lecidas por experimentos realizados por um monge chamado Gregor Mendel na metade do século XIX, em Brünn, atual Brno na re-pública Tcheca. Em uma simples definição, a genética estuda a hereditariedade e a variação de características dos organismos. No estudo de uma característica precisamos esclarecer se a mesma é herdável, ou seja, determinada genicamente, e onde o gene ou genes que expressam a característica estão localizados. Envolve conhecer também, as interações en-tre os alelos dos genes que as determinam, e entre genes não-alélicos se a característica for condicionada por mais de um loco gênico. Compreender como os genes determinantes de uma característica se expressam e como são regulados e qual o papel do ambiente na expressão do fenótipo.
Nesta disciplina, Genética Geral, que é a pri-meira do elenco das obrigatórias da área de Genética, vamos trabalhar a compreensão dos mecanismos de transmissão e o entendimento da variação genética nas populações. Nas pró-ximas disciplinas iremos aprofundar os conhe-cimentos sobre os cromossomos (Citogenéti-ca) e sobre a expressão e regulação dos genes (Genética Molecular).
Apesar da genética ser desenvolvida a partir de pesquisas realizadas em laboratórios e áre-

as (microorganismos, vegetal, animal, huma-no, educação) experimentais ou aplicadas de instituições de ensino e pesquisa em todo o globo terrestre, ela está sempre presente no nosso cotidiano. Lendo jornais, livros, revistas ou assistindo a filmes, noticiários e até novelas, dificilmente você não encontrará algum tema relacionado à genética. Os avanços do conhe-cimento e as inovações derivadas da ciência e tecnologia muitas vezes provocam medo e des-confiança na população, o que têm feito com que a mídia procure transmitir, em linguagem acessível, o real significado de tais transforma-ções e sua implicações éticas, sociais e políti-cas. Contudo, a falta de formação específica de jornalistas e profissionais afins muitas vezes provoca interpretações equivocadas sobre a importância da genética na sociedade.
Enquanto você está lendo este texto, pesqui-sadores do mundo inteiro estão produzindo novos conhecimentos. O ritmo de novas infor-mações nesta área é alucinante. Para enfren-tarmos essa tempestade de idéias precisamos nos valer de duas ferramentas: uma boa fun-damentação, só conseguida através da leitura e esforço pessoal e a capacidade de usar os recursos disponíveis na internet. Esta nossa jor-nada será coberta de êxito se vocês ao termi-narem este módulo, estiverem compartilhando o nosso entusiasmo em prosseguir na viagem do conhecimento, desafio lançado pela Fix (a esfinge do conhecimento, a mascote da gené-tica neste curso, que estará sempre fazendo perguntas e lançando desafios).
Apresento Fix,
Oi, sou Fix, leoa alada com cabeça humana e só posso ser vencida pelo intelecto. Meu pri-meiro desafio foi quan-do viajava pela Grécia anos atrás.
Decifra-me ou te devo-ro! “Qual o ser que anda de manhã com quatro patas, ao meio dia com duas e, à tarde, com três e que, contrariamente à lei geral, é mais fraco
quando têm maior número de membros?” Essa é fácil, porém desafio mesmo é o que te-nho para vocês.
O caminho do conhecimento é árduo, mas ex-tremamente gratificante para aquele que nele se aventurar. Convido a quem tiver coragem a en-trar na maravilhosa viagem do conhecimento.

GENÉTICA GERAL
Profa. Marília de França Rocha Carga Horária I 60h
1. EMENTA A disciplina Genética Geral apresenta conceitos fundamentais em genética clássica, baseados em princípios básicos sobre a hereditariedade, mostrando que a unidade, a diversidade e as características adaptativas dos organismos são consequências da história evolutiva. Identifica as principais formas de transmissão de genes e os fatores que alteram os padrões simples de herança, reconhece a manifestação dos genes na distribuição de características qualitativas e quantitativas bem como a interação entre o genótipo e o ambiente na determinação do fenótipo do indivíduo. Além disso, busca fami-liarizar os estudantes com os conhecimentos básicos necessários à compreensão de como os conceitos científicos se desenvolvem a partir de observações e de experimentação, visando ao desenvolvimento do raciocínio estruturado.
2. CONTEÚDO PROGRAMÁTICO E METODOLOGIA
Capítulos sobre os temas básicos em Genética Geral.
1. Mendelismo: experimentos de Mendel, símbolos e terminologias, monoibridismo, dominância e recessividade, retrocruza-mento e cruzamento teste, diibridismo, poliibridismo.
2. Padrões de herança: herança monogêni-
ca, herança de genes organelares, herança multifatorial, penetrância e expressividade, dominância incompleta, codominância, alelos letais, alelos múltiplos, proporções diíbridas modificadas, epistasia.

3. Base cromossômica da herança e mapea-mento gênico: teoria cromossômica, cro-mossomos, não disjunção cromossômica e genes, descoberta dos genes ligados, mecanismos de permuta gênica, mapas de ligação.
4. Genética evolutiva: teorias evolutivas x ge-
nética mendeliana, teoria das frequências alélicas (Equilíbrio de Hardy- Weinberg), processos que influenciam as variações, evolução e diversidade, macroevolução.
As atividades solicitadas nos capítulos terão a finalidade de aprofundar as informações, de redirecionar a aprendizagem e de aferir o co-nhecimento (algumas atividades selecionadas nos capítulos, como exercícios, pesquisas, dis-cussão em fóruns e as avaliações presenciais obrigatórias).

ca
pít
ulo
1
9
MENDELISMO
Profa. Marília de França Rocha Carga Horária I 15hColaboradoras - Profa. Maria Teresa Marquim N. Cornélio Profa. Rita de Cássia de Moura
INTRODUÇÃO
Neste capítulo, vamos iniciar a jornada pelo mundo maravilhoso da genética. Se você es-tiver disposto(a) a vencer os obstáculos, ao tomar contato com uma terminologia mais especializada, o aprendizado ser-lhe-á muito agradável e de grande valia. Além disso, ao se familiarizar com os conhecimentos básicos necessários à compreensão de como os con-ceitos científicos se desenvolvem a partir de observações e de experimentação, você estarádesenvolvendo um raciocínio estruturado e compreendendo a importância da genética nas questões éticas, sociais e políticas.
OBJETIVO ESPECÍFICO
• DiscutiraimportânciadasdescobertasdeMendel e suas implicações fundamentais para o estudo da hereditariedade.
1. MENDELISMO
1.1 QUEM FOI MENDEL
Johann Mendel (1822-1884; o nome Gregor foi acrescentado ao tornar-se religioso) (fig. 1) nasceu na Moravia, Europa Central. Ao ter-minar os estudos secundários, ingressou no monastério católico de St. Thomas, na cida-de de Brünn, atual Brno na república Tcheca. Para saber mais sobre Mendel acesse: http://www.monkinthegarden, ou para obtenção da versão original do artigo de Mendel: http://www.netspace.org/MendelWeb.

ca
pít
ulo
1
10Como o monas-tério agostiniano era dedicado ao ensino de ciên-cias e pesquisas científicas, Men-del foi enviado para a universi-dade de Viena, e como não obteve sucesso nos exa-mes, retornou a Brünn. Lá iniciou um programa de pesquisa de hibridização de plantas e estabe-leceu as regras básicas da herança, o que lhe valeu, após a sua morte, o título de fundador da Genética.
2. AS DESCOBERTAS DE MENDEL2.1 O ORGANISMO EXPERIMENTAL DE MENDEL
Mendel iniciou seus trabalhos com variedades de ervilha de jardim. Utilizou Pisum sativum,
P. quadratum, P. umbellatum e P. saccharatum, cujos resultados foram apresentados em 1865, em um congresso local, e publicados no ano seguinte nos Anais da Sociedade de Pesquisas em Ciências Naturais de Brünn, sob o título de “Experiências sobre híbridos em plantas”. A escolha da espécie foi fundamental para o su-cesso de Mendel. A ervilha apresenta inúmeras características que a tornam apropriada para estudos genéticos. Apresenta várias gerações no ano, é de porte pequeno, tem flor comple-ta, autopoliniza-se, embora permita a realiza-ção de fertilização cruzada, cada vagem forne-ce várias sementes, e, conquanto não fosse do conhecimento de Mendel, todas as espécies são diplóides.
Um outro ponto importante foi a escolha de sete características (propriedades específicas) que permitiam a rápida classificação de indi-víduos. Os geneticistas usam este termo como sinônimo de caráter. A planta só poderia apre-sentar uma forma alternativa para cada carac-terística (fig. 2) e não poderia apresentar am-bas de uma só vez, de maneira a ser capaz de distinguir um padrão de herança, caso existisse algum. As formas são chamadas, atualmente, de fenótipos.
Altura do caule Alto : Baixo:
Morfologia da va-gem
Completa : Constringida:
Cor da vagem Verde : Amarela:
Morfologia da ervi-lha
Lisa : Rugosa:
Cor da ervilha Amarela : Verde:
Localização da flor Axial : Terminal:
Cor da flor Violeta : Branca:
Figura 2- Características contrastantes em ervilha
Características
Figura 1 - Johann MendelFonte: http://history.nih.gov

ca
pít
ulo
1
112.2 O EXPERIMENTO DE MENDEL
Embora não tenha terminado a sua formação universitária, Mendel era professor, e possuía bons conhecimentos na área de Biologia e Físi-ca. Seu sucesso deve-se não apenas à escolha do material de estudo, mas também aos re-gistros e análises cuidadosas dos experimentos realizados, análise estatística, teste das previ-sões de hipóteses e análise populacional. Seu método resume-se em:
• Seleção de sete pares de características,cada uma das quais com duas formas distin-tas (fig. 2);
• Cruzamentodeindivíduoscomcaracterís-ticas contrastantes;
• Realizações de cruzamentos recíprocos(usou cada uma das sete características como progenitor feminino e como proge-nitor masculino, por exemplo, fêmea alta x macho baixo e fêmea baixa x macho alto);
• Estudoinicialdeumacaracterísticaporvez; • Análise de cada característica por várias
gerações; • Classificaçãodeindivíduosemcadaclasse
de expressão do caráter em cada geração; • Proteçãodehíbridosdetodainfluênciade
polens externos; • Utilizaçãodeprincípiosdeprobabilidade; • Abordagem, posteriormente, pares de
características e até três conjuntos de caracteres.
Uma condição era importante: não poderia haver redução marcante da fertilidade dos hí-bridos e dos seus descendentes em sucessivas gerações.
2.3 PRINCÍPIOS MENDELIANOS
Mendel foi capaz de deduzir três princípios fundamentais da Genética (segregação, do-minância e distribuição independente) que contêm as bases subjacentes da herança e da atividade gênica.
Cruzamentos monoíbridos
O primeiro passo de Mendel foi estudar a he-rança de cada par de características. Para isto, ele utilizou cruzamentos recíprocos e a fertili-zação cruzada de plantas com formas alterna-tivas do mesmo caráter, por exemplo, cor da flor (fig 3). Em cada caso, a geração inicial (F1) mostrava, apenas, uma das características da geração parental (P). Então, não faria diferen-ça o modo pelo qual seria feito o cruzamento. Se um genitor puro apresentar flor violeta, e o outro, flor branca, toda a prole da F1 possuirá flores violetas.
Contudo, se indivíduos da F1 fossem autofer-tilizados, a segunda geração de plantas (F2) apresentaria ambas as características. Para este exemplo, violeta x branca, a geração F2 conti-nha 705 flores violetas e 224 flores brancas. O fenótipo branco que não apareceu na F1 rea-pareceu na F2, o que fornece uma proporção de 3,15 de flores violetas para 1 de flor branca. Um padrão começou a se repetir para cada um dos cruzamentos monoíbridos. Os resultados para cada um dos cruzamentos estão resumi-dos no quadro 1. Mendel admitiu que a pro-porção era de 3:1, estando dentro dos limites de erro experimental.
Figura 3 - Cruzamentos recíprocos e prole
Xvioleta branca
Geração parental P
1ª Geração F1
Geração parental P
1ª Geração F1
Xbranca violeta

ca
pít
ulo
1
12
Quadro 1 - Resultados de cruzamentos monoíbridos Geração F2
Cruzamento Geração F1 Números Proporção
Caules altos x baixos Altas 787 altas:277 baixas 2,84 : 1
Vagens completas X Completas 882 comp:299 constring 2,95 : 1
constringidas
Vagens verde x amarelas Verdes 42 8 verdes:152 amarelas 2,82 : 1
Ervilhas lisas x rugosas Lis as 5.474 lisas:1.850 rugosas 2,96 : 1
Ervilhas amarelas x verdes Amarelas 6.022 am: 2.001 verdes 3,01 : 1
Flores axiais x terminais Axiais 651 axiais: 207 terminais 3,15 : 1
Flores violetas x brancas Violetas 705 violetas: 224 brancas 3,15 : 1
Total 14.949 : 5.010 2,98 : 1
As conclusões de Mendel
Mendel propôs que cada característica ou tra-ço (por exemplo, cor da flor) é controlado por um fator (gene) que poderia existir em mais de uma forma (alelo). Mendel, sem nenhum co-nhecimento de citologia, foi capaz de deduzir que os fatores seriam estruturas físicas dentro da célula, e, com isso, propôs a existência dos genes. Sua primeira conclusão foi: cada fator existe aos pares. O que equivale dizer que, para cada gene, existe um par de alelos.
Durante a reprodução sexual, os alelos de cada genitor se separam ou segregam, produzindo gametas com apenas um alelo daquela carac-terística estudada. A isto, chamamos princípio da segregação ou 1ª Lei de Mendel.
Ele, também, usou símbolos na forma de le-tras para denotar as características. Maiúsculas para a versão que aparecia com maior frequ-ência e minúsculas para a de menor frequên-cia, e introduziu as terminologias: dominante para essa predominância de um caráter na primeira geração híbrida; e recessivo para as características alternativas, o que é conhecido como o princípio da dominância.
Introduzindo alguns termos. Cada letra repre-senta um alelo, e indivíduos representados por Aa são ditos heterozigotos apresentando dois alelos diferentes. Indivíduos de linhagens puras AA ou aa são chamados de homozigotos. Des-ta forma, uma planta AA é homozigota domi-nante, Aa é heterozigota, e aa, homozigota re-cessiva. A constituição genética representada por letras, descrevendo os genes herdados porum indivíduo é o genótipo, enquanto o fenóti-po especifica a característica observada.
Um grande desafio para Mendel foi expli-car não a proporção 3:1, mas as proporções 1:2:1, subjacentes a todas as proporções 3:1. Ele cruzou plantas com sementes amarelas e verdes e autofecundou as plantas da F1 (se-mentes amarelas), obtendo uma F2 com ¾ amarelas e ¼ verde. Ele selecionou algumas plantas amarelas da F2 e as autofecundou, obtendo novamente uma mistura de ervilhas amarelas e verdes e uma proporção de três amarelas para uma verde. Ao serem autofe-cundadas, as plantas com ervilhas verdes da F2 apresentaram, apenas, ervilhas verdes (vv). Em conclusão, todas as ervilhas verdes de F2 eram puras. Entretanto, das ervilhas amarelas 2/3 eram iguais às amarelas de F1 (Vv), e 1/3 era como as parentais (VV). Os tipos gaméti-cos femininos serão fertilizados aleatoriamen-te pelos tipos gaméticos masculinos. Uma boa forma de mostrar isso graficamente é utilizar o quadrado de Punnett (fig. 4).

ca
pít
ulo
1
13
ParentaisV V = X v v =
GametasV = v=
F1 Zigoto Vv =
1/2 V1/2 v
1/2 V1/4 V / V 1/4 V v
1/2 v1/4 V / v 1/4 v / v
1 V / V 2 V / v 1 v / v
Figura 4 - Modelo de Mendel para explicar a proporção 1:2:1
Membros de um par de genes são separados por uma barra, que é usada para demonstrar que eles são, de fato, um par.
Quando cruzamos plantas puras com semen-tes amarela x verde, desconhecemos o genó-tipo das plantas de F2 cujo fenótipo é amare-lo, pois este poderá ser homozigoto VV (com probabilidade de 1/3) ou heterozigoto Vv (com probabilidade de 2/3). Uma forma de desco-brirmos o genótipo desconhecido é cruzar esta planta com sementes amarelas com outra de sementes verdes (homozigota para a caracte-rística recessiva). Desta forma, realizamos um cruzamento teste. Aquela que for homozigota dominante apresentará uma prole totalmente amarela, e a heterozigota, por sua vez, mos-trará prole amarela e verde numa proporção de 1:1.
Consideramos cruzamento teste, ao cruzarmos um indivíduo com fenótipo dominante cujo genótipo se deseja determinar com um indiví-duo de fenótipo recessivo. O retrocruzamento, por sua vez, ocorre ao cruzarmos um indiví-duo com fenótipo dominante cujo genótipo se deseja determinar e um genitor recessivo. Ambos têm sido de grande importância no estudo de: identificação de genótipos, ligação
fatorial, melhoramento vegetal e animal. No google acadêmico http://www.scholar.google.com você poderá encontrar diversos artigos sobre estes tipos de cruzamento.
CRUZAMENTOS DIÍBRIDOS
O próximo passo de Mendel foi realizar cru-zamentos envolvendo dois pares de caracterís-ticas, para saber se eram herdados juntos ou independentemente.
Se dois genes estão no mesmo cromossomo, os alelos em um cromossomo são separados dos do outro por uma barra, por exemplo, AB/ab ou Ab/aB. Se estiverem em cromossomos diferentes, separamos os pares de genes por ponto-e-vírgula, como por exemplo, A/a; B/b. Se desconhecemos que os genes estão no mesmo cromossomo ou em cromossomos di-ferentes, iremos separar os genes por um pon-to, por exemplo, A/a . B/b.
Para efetuar um cruzamento diíbrido, Mendel usou uma linhagem com sementes amarelas e lisas e outra, com sementes verdes e rugosas. A prole F1 apresentou sementes amarelas e li-sas. De uma planta autofecundada desta prole, pode-se obter quatro combinações diferentes na F2, duas das quais não existiam nas gera-ções anteriores (sementes amarelas rugosas e sementes verdes lisas). Por exemplo, em certo cruzamento, ele obteve 350 sementes amare-las e lisas, 108 verdes e lisas, 101 amarelas e rugosas e 32 verdes e rugosas; o que o levou a uma proporção 9,06:3,11:2,91:0,92, muito próxima de 9:3:3:1. Mendel realizou inúmeros cruzamentos com a combinação de outras ca-racterísticas, obtendo proporção similar. Mais uma vez, Mendel precisou explicar seus resul-tados. Ele somou algumas classes fenotípicas de F2 para determinar se as proporções mo-noíbridas (3:1) estavam ou não presentes. Ao somar as sementes amarelas (315 + 101) e assementes verdes (108 + 32) das diferentes classes fenotípicas, ele obteve uma proporção muito próxima de 3:1, levando-o à conclusão de que a proporção 9:3:3:1 era a combinação aleatória de duas proporções 3:1 (Fig 5).

ca
pít
ulo
1
14
¾ lisas (R-) ¾ x ¾ = 9/16 amarelas-lisas (V-R-)
¼ rugosas (rr) ¾ x ¼ = 3/16 amarelas-rugosas (v-rr)
¾ lisas (R-) ¼ x ¾ = 3/16 verdes-lisas (vvR-)
¼ rugosas (rr) ¼ x ¼ = 1/16 verdes-rugosas (vvrr)
¾ de F2 são amarelas V-
¼ de F2 são verdes Vv
Fig 5 – Diagrama ramificado apresentando dois genes.
As características de um organismo são deter-minadas por genes. Definindo V e v como os alelos que determinam sementes amarelas e verdes e R e r como sementes lisas e rugosas, respectivamente, os progenitores são VVRR e vvrr. Conseqüentemente a F1 apresenta ge-nótipo VvRr e esperam-se os gametas VR, Vr, vR e vr. Considerando que cada gameta seja produzido com a frequência de ¼, metade dos gametas possuirá V, e a outra v, o mesmo ocorrendo para o segundo gene. Novamente podemos nos valer do quadrado de Punnett para observar as fusões dos gametas e obser-var tanto os fenótipos quanto os genótipos da F2 em um cruzamento diíbrido (fig. 6).
Como há evidência de que indivíduos VvRr produzem o gameta VR em frequência de ¼ ,pode-se concluir que: a probabilidade de um gameta conter os fatores V e R é o produto dasprobabilidades de conter cada gene em sepa-rado (½ x ½ = ¼), o que também é verificado em relação aos outros gametas (Vr, vR, vr ). Mendel , então, chegou ao princípio da com--binação dos diferentes caracteres: Cada cará-ter é herdado independentemente do outro, e a proporção de dominante para recessivo não fica alterada pelo outro caráter.
Parentais V / V ; R / R = x v / v ; r / r =
Gametas VR = vr =
F1 Zigoto V / v ; R / r =
F2 F1 (V / v ; R / r ) x F1 (V / v R / r)
VR Vr
vr
vR
VR
VVRR
VVRr
VvRr
VvRR
Vr VVRr VVrr Vvrr VvRr
vr VvRr Vvrr vvrr vvRr
vR VvRR VvRr vvRr vvRR
9 : 3 : 3 : 1 :
Fig 6 - Resultado de Cruzamento diíbrido

ca
pít
ulo
1
15
3/4 de F2 : sementesamarelas (V-)
1/4 de F2 : sementesverdes (vv)
¾ lisas (R-)
¼ rugosas (rr)
¾ lisas (R -)
¼ rugosas (rr)
¾ flor violeta (B-)27/64 Amarela, lisa,violeta
¼ flor branca (bb)9/64 Amarela, lisa, branca
¾ flor violeta (B-)9/64 Amarela, rugosa, violeta
¼ flor branca (bb)3/64 Amarela, rugosa,branca
¾ flor violeta (B-)9/64 Verde, lisa, violeta
¼ flor branca (bb)3/64 Verde, lisa, branca
¾ flor violeta (B-)3/64 Verde, rugosa,violeta
¼ flor branca (bb)1/64 Verde, rugosa,branca
Fig. 7 - Diagrama ramificado apresentando três genes.
REGRA DO PRODUTO (REGRA DO E)
Probabilidade de ocorrência simultânea de dois ou mais eventos independentes.
Vejamos como poderemos calcular com fa-cilidade a ocorrência de eventos aleatórios. Imaginemos que lancemos ao ar, simultanea-mente, duas moedas. Qual a probabilidade de obtermos cara na primeira e cara na segunda nesta sequência? Perceba que o aparecimento de cara na primeira moeda não influencia o surgimento de cara na segunda moeda. Isso quer dizer que os dois eventos são indepen-dentes. Quando os eventos são independen-tes, a probabilidade da ocorrência de ambos é igual ao produto de suas probabilidades in-dividuais.
A probabilidade de obtermos cara e cara no lançamento simultâneo é dada, então, pelo produto das probabilidades parciais:
P (cara e cara) = ½ x ½ = ¼
Veja a tabela 1 dos possíveis resultados no lan-çamento simultâneo de duas moedas e confira o resultado que acabamos de obter:
Das quatro situações possíveis, apenas a pri-meira realmente satisfaz à condição do pro-blema. Logo, a probabilidade de surgir uma combinação cara e cara é 1/4.
Tabela 1 - Possíveis lançamentos1ª moeda e 2ª moeda
cara e cara
coroa e coroa
cara e coroa
coroa e cara
Passemos, agora, a aplicar esse conhecimento à Genética, analisando dois exemplos:
a) A probabilidade de uma planta F2 ser de genótipo VVRrbb é o produto das proba-bilidades de ser VV (semente amarela), de ser Rr (semente lisa) e de ser bb (flor bran-ca), o que equivale a (1/4) x (2/4) x (1/4) = 2/64. De forma semelhante se quero saber a probabilidade de determinado fenótipo, por exemplo, amarela-lisa-branca calculo o produto das probabilidades de cada fenó-tipo, ou seja, (3/4) x (3/4) x (1/4) = 9/64.

ca
pít
ulo
1
16b) Um casal heterozigoto, para uma dada ca-
racterística, deseja saber qual a probabili-dade de ter duas crianças, sendo a primei-ra heterozigota e a segunda, homozigota dominante.
P Aa x Aa Se P(Aa)= ½ e P(AA)= ¼ ,
Gametas A a A a então:
F1 AA Aa Aa aa P(Aa e AA) = ½ x ¼ = 1/8
Marido Aa X Mulher Aa
Gametas: A a A a
Introdução: Metade das cartas de um bara-lho é formada por cartas de naipes pretos, e a outra metade, por naipes vermelhos. Assim, cada vez que se retira uma carta de um bara-lho completo, a chance de tirar uma carta de naipe vermelho é igual à chance de tirar uma carta de naipe preto, ou seja, ½ (0,5 ou 50%). Da mesma maneira: cada vez que, na fecunda-ção, igual número de espermatozóides com o X ou o Y rodearem um óvulo, a probabilidade de entrar um espermatozóide com o X ou a de entrar um espermatozóide com o Y é a mes-ma: ½.
Objetivo: Entender melhor o uso de probabi-lidades, para aplicá-la a exercícios práticos em genética.
Material: Baralho e tabelas.
Procedimento: As cartas de naipes vermelhos não são todas iguais. Metade é do naipe ouros ( ) e metade é do naipe copas ( ). A mesma coisa acontece com as cartas de naipes pretos: metade é do naipe paus ( ) e metade é do naipe espadas ( ). Suponha agora que as car-tas de naipes vermelhos e as de naipes pretos representem os genes D e d dos gametas; ( )= óvulo D; ( ) = espermatozóide D; ( )= óvulo d e ( ) espermatozóide = d. Onde D- é destro e dd canhoto.
Divida um baralho em dois maços, de acordo com a cor das cartas. Embaralhe cada um de-les e retire uma carta de cada maço, formando um par. Anote na linha da jogada 1 da sua tabela a combinação obtida, os genótipos e os fenótipos. Em seguida, recoloque as cartas em seus maços e embaralhe-os novamente e repi-ta a operação até completar toda a sua tabela.
JOGADAS COMBINAÇÕES GENÓTIPOS( )( )= D( )( )= d
FENÓTIPOS
1.
2.
3.
4.
5.
REGRA DA SOMA (REGRA DO OU)
Probabilidade de ocorrência de dois ou mais eventos mutuamente excludentes.
Quando os eventos são mutuamente excluden-tes, a probabilidade de ocorrência de qualquer um deles é a soma de suas probabilidades in-dividuais. Assim, no nosso exemplo teríamos:
P (ás de copas ou ás de espada) = P (ás de copas) + P (ás de espada)
P (ás de copas) + P (ás de espada) = 1/52 + 1/52 = 2/52 = 1/26
Veja este outro exemplo: qual a probabilidade de um casal, heterozigoto para determinada característica, ter uma criança de genótipo AA ou aa?
Fazendo a separação dos gametas e o quadro de cruzamentos, veremos que:
P(AA) ou P(aa) = P(AA)+P(aa)
P(AA)+P(aa) = ¼ + ¼ = ½
Proposta da Fix: Vamos aprender jogando?
A a
A AA Aa
a Aa aa

ca
pít
ulo
1
17Suponha que esses genótipos e fenótipos se-jam os de cinco filhos de um casal Dd.
1. Qual é a probabilidade de se formar um zigoto DD?
2. Qual é a probabilidade de se formar um zigoto dd?
3. Qual é a probabilidade de se formar um zigoto Dd?
4. Quantos filhos destros e quantos filhos ca-nhotos têm esse casal?
5. Por que a probabilidade de o casal ter um filho canhoto é menor do que a de ter um filho destro?
Envie a sua tabela e as respostas das perguntas de 1 a 5 para os tutores.
Obs: A resposta da questão 4 não poderá ser igual à dos seus colegas, pois cada um montou a sua própria tabela.
3. UTILIZANDO O TESTE DO QUI-QUADRADOComo podemos observar ao longo do texto, Mendel se deparou com resultados que são próximos, mas não, idênticos à proporção esperada. Atualmente aplicamos um teste estatístico muito simples chamado, teste do Qui-quadrado - , e destina-se a comparar pro-porções entre os resultados observados e os previstos por uma hipótese.
A fórmula para -c2 consiste:
c2 = S[(O-E)2 / E]
O= frequência observada para a classeE= frequência esperada para a classe(O-E)= desvio entre cada valorå= somatório
O c2 emprega números absolutos e não fracio-nários. Quanto maior a amostra, mais confiá-vel será o teste.
O c2 resultado do depende do tamanho das amostras e do número de classes da amostra, o que nos fornece os graus de liberdade (GL).Quando um determinado número de classes é fixado, e todos, exceto um, foram arrumados, os demais não estão livres, mas devem preen-cher um determinado nicho vazio. No caso delançamento de um dado (seis lados) o GL=5, de uma moeda (cara ou coroa ) o GL=1. Pro-porções mendelianas, por exemplo, com duas classes (3:1) possuem GL=1, com três classes (1:2:1) GL=2, com quatro classes (9:3:3:1) GL= 3, e assim por diante. Logo, o número de GL é geralmente igual ao número de classes menos um.
Mas ainda estamos no meio do caminho. As hipóteses ainda precisam ser analisadas pelo teste do c2 . Então, após o emprego da fór-mula para o cálculo do c2 (c2 c) e a determi-nação dos GL, devemos consultar a tabela de valores de variáveis com distribuição c2 (tabe-la 2) que mostra os graus de liberdade (GL) nas linhas e o valor das probabilidades (P) nas colunas. O c2 de tabela (c2 t) é função do nível de significância (limites que afirmam se um determinado desvio decorre ou não do acaso) do teste e do número de graus de liberdade. Por exemplo, 3,841 é o c2 t para 1GL em 5% de significância.
Na tabela 2, estão apresentados alguns dos valores de variáveis com distribuição c2 com diferentes graus de liberdade e níveis de sig-nificância.
Para verificar a probabilidade de concordância entre o valor obtido e a hipótese proposta, é necessário consultar a tabela do c2 . Para tal, localize o número de GL à esquerda. Em segui-da, percorra a linha referente ao grau de liber-dade em busca de um valor de c2 tabelado ( c2
t) igual ou aproximado ao c2 calculado (c2 c). O valor do c2 c poderá estar entre dois valores do c2 t. Por fim, verifica-se o valor de P, direta-mente acima, no alto da linha, correspondenteaos c2 tabelados.

ca
pít
ulo
1
18
Tabela 2 – Tabe la parcial do qui-quadrado (c2 )
GL Probabilidade ( P )
0,99 0,95 0,90 0,50 0,20 0,10 0,05 0,01 0,001
1 0,0002 0,004 0,016 0,455 1,642 2,710 3,841 6,635 10,827
2 0,020 0,103 0,211 1,386 3,219 4,605 5,991 9,210 13,815
3 0,115 0,352 0,584 2,366 4,642 6,251 7,815 11,345 16,266
5 0,297 0,711 1,064 3,3571 5,989 7,779 9,488 13,277 18,467Para obtenção da tabela completa da distribuição de qui-quadrado acesse http://www.cultura.ufpa.br/dicas/biometria
Imagine que está sendo testada uma hipótese de nulidade que é verdadeira, embora isto não seja conhecido. Se o valor de c2 c associado a 1GL for 0,0002, verificaremos (ver tabela 2) que a obtenção ao acaso de um valor maior ou igual a ele é bastante provável (0,99), en-tretanto se o valor c2c associado a 1GL for de 10,827, poderemos concluir que é improvável obter ao acaso um valor maior ou igual a este (0,001).
Imagine ainda outras situações nas quais, com base em uma hipótese de 1:1 (GL=1), obtivés-semos, respectivamente, os valores de 9 e 0,9 para os c2c. No primeiro exemplo, c2 c=9, o valor 9 para 1GL se encontra na tabela entre P=0,01 (6,635) e P=0,001 (10,827). Portanto a adequação desse resultado para uma hipóte-se 1:1 não é boa. Por outro lado, no segundo exemplo, o valor 0,9 para 1GL se encontra na tabela entre P=0,50 (0,455) e P=0,20 (1,642).Podemos, então, prever que desvios devidos ao acaso, iguais ou maiores que os observa-dos, ocorreriam em 20 a 50% das tentativas. Os dados se ajustam à proporção 1:1.
O valor calculado pode não estar nos extremos da distribuição da variável, então fixar o nível de significância é importante e deve ser esta-belecido antes de o experimento ser realizado. É o risco que se corre de rejeitar uma hipótese ver-dadeira ou aceitar uma hipótese falsa. O nível de significância de 0,05 é geralmente escolhido como um padrão arbi-trário, definindo desta maneira, que valores me-nores que o tabelado são
devidos ao acaso e valores maiores que o ta-belado são improváveis de terem ocorrido ao acaso. Assim, se o valor calculado é maior que o tabelado, admite-se que os desvios entre os valores observado e esperado não são devidos ao acaso, rejeitando-se a hipótese de nulidade. Convencionaremos, então, que se houver uma probabilidade de menos de 5% de se observar um desvio do esperado, a hipótese será tida como falsa e rejeitada. Entretanto, se a hipóte-se obtiver um valor maior ou igual a 5% (P < 0,05), mesmo assim, não significa que a hipó-tese seja verdadeira e, sim, que os resultados são condizentes com a hipótese.
Algumas vezes o nível de significância de 0,01 é utilizado. Neste caso, há uma probabilida-de muito pequena (1%) de que uma hipótese verdadeira venha a ser rejeitada, em compen-sação existe uma grande chance de que uma falsa hipótese venha a ser aceita.
Vale a pena lembrar: Geralmente usamos o nível de significância 0,05, embora os valores críticos da distribuição do c2 sejam variáveis 0,99-0,001.
Vamos usar os dados de Mendel (tabela 3) para analisar hipóteses com o teste do c2.
Tabela 3 - Números observados e esperados em diferentes classes fenotípicas
Fenótipo dassementes
O E O - E (O - E)2 (O - E)2/E
Lisa e amarela 315 (9/16)x556= 312,75 2,25 5,0625 0,0162
Lisa e verde 108 (3/16)x556= 104,25 3,75 14,0625 0,1349
Rugosa e amarela 101 (3/16)x556= 104,25 -3,25 10,5625 0,1013
Rugosa e verde 32 (1/16)x556= 34,75 -2,75 7,5625 0,2176
Total 556 556 0 0,4 7

ca
pít
ulo
1
19Hipóteses:H0: Os genes de ervilha que determinam a morfologia e a cor das sementes apresentam distribuição independente.
H1: Os genes de ervilha que determinam a morfologia e a cor das sementes não apresen-tam distribuição independente.
1o passo: Calcular o c2
c2 = å [(o-e)2 /e]c2 = 0,0162 + 0,1349 + 0,1013 +0,2176 = 0,47
O valor calculado é próximo de zero. Não ha-vendo motivo para rejeitar a hipótese, e mes-mo que a hipótese seja verdadeira, não iremos obter as mesmas proporções.
Quando os desvios entre os eventos esperados e observados são pequenos, o c2 se aproxima de zero, e o grau de concordância é bom.
2o passo: Definir os graus de liberdade 4 clas-ses fenotípicas – 1 = 3; GL= 3
3o passo: Comparar o valor do 2c com o 2t Considerando o valor de P=0,05 e GL=3 o va-lor tabelado (2t) é de 7,815.2c = 0,47 < 2t 7,815
4o passo: Análise da hipóteseAceita-se Ho. Entretanto, aceita-se que os des-vios entre os valores observados e esperados são devidos, apenas, ao acaso e não, ao fato de os genes não terem distribuição independente.
Fogem ao escopo deste capítulo explicações mais detalhadas sobre testes estatísticos. Para maiores detalhes sobre o teste do 2 e obten-ção da tabela completa da distribuição de 2 acesse http://www.cultura.ufpa.br/dicas/biome/bioqui.htm ou http://www.ufv.br/dgb/gbol/htm/gbol23.htm
4. O IMPACTO DAS IDÉIAS DE MENDEL
4.1 HERANÇA TÊNUE X HERANÇA SÓLIDA
Na época de Mendel, predominava a idéia da herança por mistura dos caracteres dos ances-trais, e o critério aceito era de que a herança não poderia ser estudada de modo científico ou estatístico. Mendel contribuiu de forma efetiva, para a erradicação da herança tênue ou por mistura. Ele enfatizou que, se os fatores paternos e maternos diferiam, eles jamais se fundiriam, mas se separariam sempre de novo, durante a formação da célula germinativa. Era a descoberta de que as partículas existem em conjunto (genes e seus alelos), o que reforçou a idéia da herança sólida ou particulada. Além disso, a constância da proporção 3:1 refutou o postulado de partículas múltiplas e afirmou o de uma única partícula. As conclusões de Mendel lançaram os alicerces para o desenvol-vimento da Genética.
4.2 REDESCOBRIDORES DE MENDELEm 1900, ocorreu um fato curioso na histó-ria da Biologia. Três botânicos, Hugo de Vries, Carl Correns e Tschermack, dentro de um espa-ço curto de meses, publicaram relatos sobre a descoberta de leis de hereditariedade, mas, ao compilar dados da literatura, descobriram que Mendel os havia antecipado 35 anos atrás. Se-ria injusto não acrescentar a esta lista o nome de Bateson, responsável pela tradução do tra-balho de Mendel e publicação do mesmo no Journal of the Royal Horticultural Society em 1900. Bateson foi responsável juntamente com seus colaboradores por importantes con-tribuições para a nossa compreensão atual so-bre hereditariedade, além de descobrir alguns desvios mendelianos, como a ligação incom-pleta e a poligenia.
4.3 TERMINOLOGIA E SIMBOLOGIA
Mendel utilizou símbolos na forma de letras para descrever os fenômenos hereditários e pôde analisar os resultados dos cruzamentos matematicamente. Isto foi uma conquista me-todológica. Os símbolos representam os genes

ca
pít
ulo
1
20e seus alelos. William Bateson aperfeiçoou a escolha de símbolos gênicos e convencionou a notação na qual os alelos para características dominantes e recessivas de um determinado gene eram representados por uma única letra que indicava a característica mutante, como por exemplo, W para flores violetas e w para flores brancas. Bateson criou as palavras ge-nética, alelomorfo (alelo), homozigoto e hete-rozigoto e introduziu a prática de indicar as
gerações em esquemas como P, F1, F2. Entre-tanto, com as novas pesquisas, o número de genes identificados excedeu a capacidade do alfabeto e novas estratégias precisaram ser uti-lizadas, como utilizar duas ou mais letras para representar um gene (tabela 4). Com a desco-berta dos alelos múltiplos, as letras começaram a ser associadas a expoentes, ou até mesmo, a símbolos hifenados associados a números para identificar alelos mutantes.
Organismo Gene selvagem Fenótipo Gene mutante Fenótipo
Bactéria(E. coli)
Str S Sensível aoantibiótico
estreptomicina
StrR Resistente aestreptomicina
Fungo(Aspergillus)
arg-1+ Produz arginina arg-1 Não produz arginina
Milho Wx Endospermaceráceo
wx Endosperma amiloso
Tomate Y Cor vermelha y Cor amarela
Besouro wb Asa azul wg; wt Asa verde, asaturquesa
Camundongo A Pigmentação escura AY Pigmentaçãoamarela
Carneiro(raça Ancon)
a Pernas longas A Pernas curtas
Humano HbA Hemoglobina normal HbS Hemoglobinafalcêmica
Pesquisa da Fix: atualmente exis-tem muitas no-tações represen-tando genes e seus alelos. Pes-quise diferentes tipos de nota-ções em orga-nismos modelos como, em Droso-phila (moscadas- frutas), camundongo, humano, Arabidopsis (planta), Neurospora (fungo), e Escherichia coli (bactéria).
Desafio da Fix: se ao invés de plantas diplóides sexuadas e características qualitativas, Mendel tivesse utilizado plantas com reprodução as-sexuada e características quantitativas, como, por exemplo, altura, ele não poderia ter de-duzido as “leis da herança”. Discuta com seus colegas o porquê.
Nos livros-texto indicados, existe uma significa-tiva quantidade de exercícios que vocês podem resolver, embora estejamos propondo alguns exercícios para resolução imediata e envio das respostas para serem analisadas.
ATIVIDADEREVISANDO CONHECIMENTOS
1. Após completar o diagrama sobre as ter-minologias utilizadas na genética clássica você verá, na coluna em destaque, o nome de um famoso cientista. Comente breve-mente sobre a importância do seu traba-lho para a genética e a repercussão na co-munidade científica.
1. Constituição genética de um organis-
mo ou de uma célula.

ca
pít
ulo
1
212. Indivíduo que apresenta alelos dife-
rentes para um determinado loco cro-mossômico.
3. Alelo que só expressa o seu efeito fe-
notípico, quando se encontra em ho-mozigose.
4. Tipo de textura da semente da ervilha
Pisum sativum.
5. A expressão do genótipo é visivelmente observada.
6. Cruzamento de um híbrido de F1 com um dos tipos parentais.
7. Cromossomos que formam bivalentes na meiose.
8. Cor da semente da ervilha observada como caráter recessivo.
9. Alelo que sempre expressa o seu efeito fenotípico, mesmo que esteja em hete-rozigose.
10. Tipo de cruzamento envolvendo dois
genes. 11. Diferentes formas de um gene que
ocorre em um determinado loco de um cromossomo.
12. Organismo ou célula com dois conjun-
tos de cromossomos (2n).
1-
2-
3-
4-
5-
6-
7-
8-
9-
10-
11-
12-
2. Suponha que você reproduziu alguns dos experimentos de Mendel, utilizando uma outra espécie de planta, na qual foi anali-sado o modo de herança para dois carac-teres: Textura da folha (lisa ou rugosa) e borda da folha (crenada ou lobada).
Partindo da geração parental, foi realiza-do o cruzamento entre duas plantas puras para estas características, sendo uma com folhas rugosas e crenadas e outra com fo-lhas lisas e lobadas, obtendo-se, na gera-ção F1, todas com folhas lisas e lobadas.

ca
pít
ulo
1
22Estas foram cruzadas entre si e você obteve, na geração F2 400 plantas, das quais 200 apresentaram folhas lisas e lobadas, 85, folhas lisas e crenadas, 85, folhas rugosas e lobadas e 30, folhas rugosas e crenadas.
Com base nestes dados, desenvolva as
questões: a) Quais as formas alélicas recessivas?
Justifique sua resposta. b) Através do quadro de Punnett, de-
monstre todos os possíveis genótipos pre-sentes na geração F2.
c) Na geração F2, que proporção fenotí-
pica se espera obter de cada tipo? Aplique o teste do qui-quadrado para verificar se os resultados obtidos estão de acordo com a hipótese de segregação independente.
3. Os genótipos abaixo estão constituídos de
alelos compostos por cinco (5) genes de segregação independente. Através de um diagrama ramificado, demonstre os dife-rentes tipos de gametas que podem ser formados por cada genótipo.
a) AABbCcDdEE b) AABBCcDdEe
REFERÊNCIAS
LEITURA FUNDAMENTAL
GRIFFITHS, A. J. F.; WESSLER, S. R.; LEWONTIN, R. C.;GELBART, W. M.; SUZUKI, D. T.e MILLER, J. H. Introdução à Genética: Rio de Janeiro: Guanabara Koogan S/A, 2006.
LEITURA COMPLEMENTAR
SNUSTAD, D. P. e SIMMONS M. J. Fundamen-tos de Genética: Guanabara Koogan S/A, 2000.
HENING, R. M. O Monge no Jardim: O gênio esquecido e redescoberto de Gregor Mendel, o pai da genética. Rio de mJaneiro: Rocco, 2001.
SITES
Sociedade brasileira de genética: http://www.sbg.org.br
Mendel : http://www.monkinthegarden ehttp://www.netspace.org/MendelWeb
Geral: http://www.scholar.google.com
Testes estatísticos:http://www.cultura.ufpa.br/dicas/biome/bioqui.htm ehttp://www.ufv.br/dgb/gbol/htm/gbol23.htm
Genética Geral: http://www.ufv.br/dgb/gbol/

ca
pít
ulo
2
23
PADRõES DE HERANÇA
INTRODUÇÃO
Neste segundo capítulo, iremos dar conti-nuidade à nossa jornada na construção do conhecimento genético. Convidamos vocês a conhecer os principais padrões de heran-ça dos genes subjacentes às características genéticas. Características qualitativas ou descontínuas são condicionadas pela heran-ça monogênica, enquanto as características quantitativas ou contínuas e semicontínuas ou quase-contínuas fazem parte da heran-ça de características complexas. Vamos nos familiarizar com heredogramas e sua inter-pretação. Aprenderemos, também, sobre padrões não-mendelianos de herança, como o mosaicismo, a herança organelar ou cito-plasmática e a herança epigenética. Além disso, iremos observar as alterações das pro-porções mendelianas e discutir como ocor-rem as interações gênicas.
OBJETIVOS ESPECÍFICOS
• Identificarascaracterísticasqueapresen-tam padrões de herança mendelianos e não-mendelianos;
• Compreendercomoosfatoresgenéticos
e ambientais influenciam a variação fe-notípica.
Profa. Marília de França Rocha Carga Horária I 15hColaboradoras - Profa. Maria Teresa Marquim N. Cornélio Profa. Rita de Cássia de Moura

ca
pít
ulo
2
24
PADRõES DE HERANÇAOs distúrbios genéticos podem ser mendelia-nos (monogênicos) ou não-mendelianos (mul-tifatoriais, mitocôndriais, cromossômicos, epi-genéticos).
1. HERANÇA MONOGÊNICA1.1 CARACTERÍSTICAS QUALITATIVAS
No primeiro capítulo, vimos que os genes encontram-se nos cromossomos, e as caracte-rísticas são ditas dominantes, ao se manifes-tarem no heterozigoto, e recessivas, quando não o fazem. Dominância e recessividade são, então, propriedades do fenótipo e não, do genótipo, embora muitas vezes tenham sido aplicadas aos genes.
Características mendelianas, qualitativas ou descontínuas, estão restritas a um loco gênico, apresentam fenótipos contrastantes e distri-buição populacional descontínua, praticamen-te sem efeito ambiental.
Nos humanos, são conhe-cidas mais de 14 mil ca-racterísticas mendelianas e o principal ponto de busca para maiores informações sobre estas características, seja de ordem patológica ou não, está na base de dados OMIM (Online Men-delian Inheritance in Man), catálogo criado por Victor McKusic continuamente atualizado e acessível em http://www3.ncbi.nlm.nih.gov/omim.
1.2 CONSTRUÇÃO DE HEREDOGRAMAS
Os termos heredograma e árvore genealógica são alternativas para o termo pedigree, mui-to associado a animais. É através do estudo dos genes em genealogias que identificamos os padrões de herança, mas, considerando o tamanho limitado das famílias humanas, fre-qüentemente não temos a certeza do modo de herança de determinada característica, ape-nas, pela análise de uma única família.
Como vocês podem perceber, não é fácil traba-lhar com humanos, pois, além do tamanho da prole (quantidade de filhos) ser limitado, exis-tem outras dificuldades e implicações éticas, tais como: a presença de doenças genéticas de manifestação tardia, a impossibilidade de fa-zer cruzamentos experimentais com humanose, ainda, a forma de averiguação da família.
TERMINOLOGIA E SIMBOLOGIA
Para a construção de heredogramas, vamos precisar aprender a empregar alguns símbolos adotados internacionalmente, como os obser-vados na Figura 1.
= Homem = Número de filhos do sexo indicado
= Mulher = Indivíduos afetados
= Repdorução =Portdora de genes recessivo ligado ao X
= Genitores e filos; 1 menino e 1 menina (em oredem de nas- cimento)
= Morte
= Dizigóticos (gêmeos não idênticos)
= Aborto ou natimorto
=Propósito
= Monozigóticos (gêmeos idênticos)
I = Identificação de pessoas no II heredograma: aqui o propósito é o filho 2 da geração II ou II.2
= Sexo não
especificado
= Casamento consaguíneo
Figura 1- Simbologia internacional utilizada em heredogramas.
2 4
2
2
2

ca
pít
ulo
2
25As gerações são numeradas em algarismos romanos. Os indivíduos da mesma geração (prole e pessoas fora da família com as quais se casam) são identificados por algarismos arábicos. Não é obrigatório representar o cônjuge, quando este não apresentar a ca-racterística em estudo (geralmente destaca-da na cor negra). Os filhos são colocados em ordem de nascimento da esquerda para a direita, e os números, dentro de quadrados (homem) ou círculos (mulher) indicam vá-rios irmãos do sexo representado com igual fenótipo.
Para a montagem de um heredograma, de-vemos contar, inicialmente, com informações obtidas através do probando (propósito ou caso índice), assinalado com uma seta - indiví-duo da família que chama primeiro a atenção dos pesquisadores.
Heredogramas podem ser construídos com características normais (variantes fenotípi-cas), como a capacidade de dobrar ou não a língua, lóbulo da orelha livre ou aderente, dentre outras, onde não há nenhuma impli-cação clínica; ou montados a partir de carac-terísticas patológicas (doenças ou síndromes monogênicas).
Na maioria das vezes, os heredogramas são construídos por motivos médicos e estão associados a distúrbios geralmente raros. Construção e análise de heredogramas são de fundamental importância na determina-ção do padrão de transmissão de um gene e tornam-se indispensáveis no aconselhamento genético.
1.3 TIPOS DE HERANÇA MONOGÊNICA
É do conhecimento de vocês que os carac-teres mendelianos podem ser determinados por locos em cromossomos autossômicos ou alossômicos (sexuais X ou Y). Caracte-rísticas autossômicas (representadas ape-nas por letras) e características ligadas ao X (representadas por letras sobrescritas no X) podem ser dominantes ou recessivas. São
dominantes, quando se manifestam em ho-mozigose ou heterozigose (AA-Aa; X A X A -X A X a ), e recessivas, quando se apresen-tam apenas em homozigose (aa; X a X a ). Entretanto, as características ligadas ao X e ao Y exibem algumas peculiaridades. O ma-cho, ao portar apenas um cromossomo X em suas células diplóides (X A Y ou X a Y), é dito hemizigoto.
Para uma melhor compreensão das caracte-rísticas ligadas aos alossomos, precisamos de alguns conhecimentos básicos da citogenéti-ca, tais como: geralmente, animais e plantas apresentam indivíduos masculinos ou femi-ninos e, na maioria dos casos, o sexo é de-terminado por um par de cromossomos se-xuais. Na nossa espécie, por exemplo, temos 46 cromossomos: 22 pares autossômicos e dois cromossomos sexuais. A fêmea apresen-ta dois cromossomos X e, por formar apenas um tipo de gameta contendo um cromos-somo X é o sexo homogamético. O macho, por sua vez, apresenta um X e um Y (consi-deravelmente menor que o X), sendo capaz de formar dois tipos de gametas, portanto é o sexo heterogamético responsável pela de-terminação do sexo da prole. Em relação ao cromossomo Y, apenas os homens herdam seus genes, com os pais transmitindo genes, diretamente, para os seus filhos.
Os três principais padrões de herança mono-gênica encontrados em humanos são: a he-rança autossômica dominante (AD), a herança autossômica recessiva (AR) e a herança ligada ao X recessiva (LXR). A herança ligada ao X do-minante, por sua vez, é rara.
Na Tabela 1, constam alguns exemplos de ca-racterísticas humanas com herança autossômi-ca ou ligada ao X.

ca
pít
ulo
2
26
Tabela 1: características humanas hereditárias monogênicasCondição clínica Tipo de Herança Efeito mutacional
1 Doença de Waardenburg I AD Alteração no controle da transcrição (genePAX3)
2 Acondroplasia (nanismo) AD Mutação no receptor para o fator de crescimento de fibroblastos (FGFR3)
3 Doença de Huntington AD Expansão repetida do códon CAG no gene para a huntingtina (proteína do sistema nervoso central)
4 Neurofibromatose I AD Alteração no gene supressor de tumor (Nf1)
5 Hipercolesterolemia familiar AD Receptores de LDL (lipoproteína de baixa densida-de) deficientes ou ausentes
6 Fibrose cística AR Defeito em gene (CFTR) para proteínatransmembrana
7 Fenilcetonúria AR Deficiência da enzima fenilalanina-hidroxilase
8 Albinismo oculocutâneo AR Deficiência da enzima tirosinase
9 Doença de Tay-Sachs AR Deficiência da enzima b-hexosaminidase
10 Hipotireoidismo AR Deficiência da enzima tiroxina
11 Distrofia Muscular Duchenne LXR Defeito em gene (DMD) para proteína distrofina
12 Daltonismo LXR Ausência de cones (células da retina) para a cor vermelha (Protanopia) ou verde (deuteranopia)
13 Hemofilia A LXR Deficiência ou ausência do fator VIII da coagula-ção sanguínea
14 Hemofilia B LXR Deficiência ou ausência do fator IX da coagulação sanguínea
15 Síndrome de Lesch-Nyhan LXR Deficiência da enzima hipoxantina-guaninafos-forribosil-transferase
16 X-Frágil LXD Expansão repetida do códon CGG no gene FMR1 para uma proteína de ligação ao RNAm
17 Incontinência pigmentar LXD Alteração na deposição da melanina
18 Raquitismo hipofosfatêmico LXD Defeito relacionado com a absorção intestinal do cálcio ou com a reabsorção do fósforo
Autossômica dominante (AD); Autossômica recessiva (AR); Ligada ao X recessiva (LXR); Ligada ao X dominante (LXD).
Vocês podem obter, na página da Sociedade Brasileira de Genética (SBG) – http://www.sbg.org.br, o E-book do prof. Bernardo Beiguel-man, intitulado: “A interpretação genética da variabilidade humana”. O livro trata de carac-teres, heredogramas e padrões de herança.
1.3.1 AUTOSSÔMICA DOMINANTE - AD
Critérios para o reconhecimento de herança AD:
A característica analisada:
• ocorreemambosossexos;
•étransmitidapelosdoissexos;
• pode ser transmitida diretamente de ho-mem para homem (de pai para filho);
• emgeral,estápresenteemtodasasgera-ções;
• apresenta-seem,pelomenos,umdosge-nitores;
• surge,emmédia,em50%dosfilhosafeta-dos de um afetado.
Na AD, um dos exemplos mais comuns é a po-lidactilia, que se caracteriza pela presença de um ou vários dedos supranumerários. Na he-xadactilia, por exemplo, o sexto dedo extra se encontra freqüentemente situado após o quin-to dedo (mínimo).
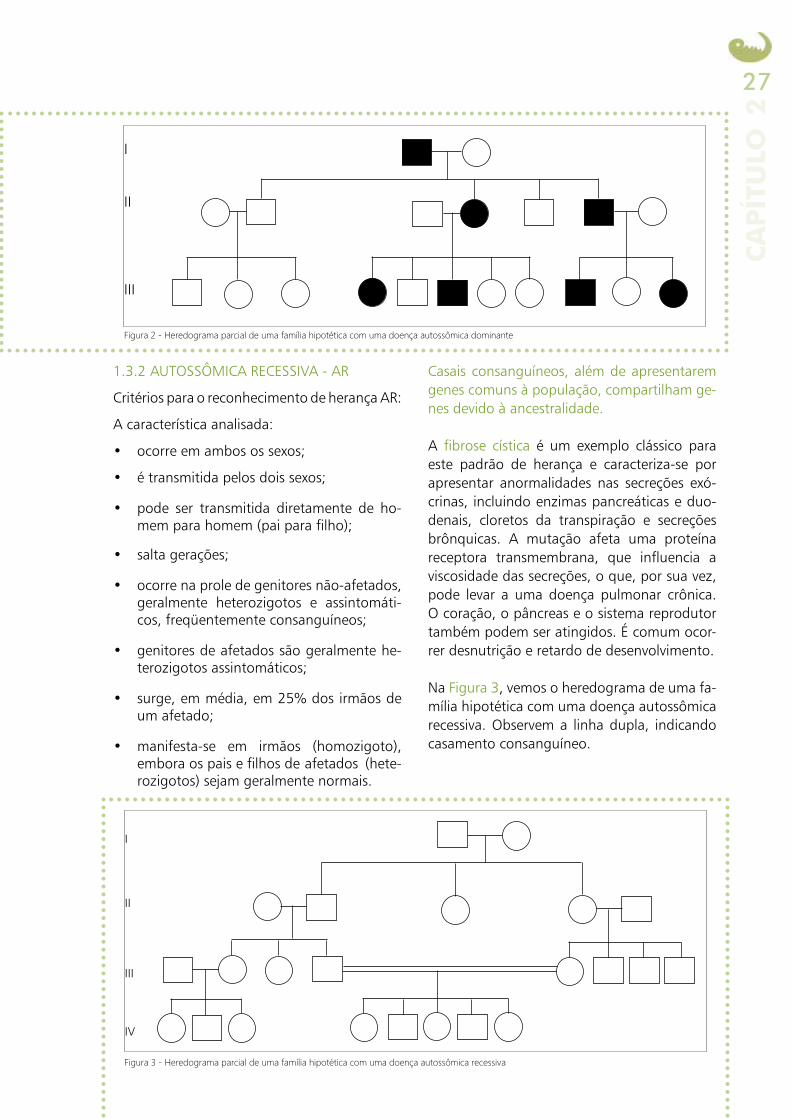
A Figura 2 ilustra o heredograma parcial de uma família hipotética com uma doença au-tossômica dominante.
ca
pít
ulo
2
27
I
II
III
Figura 2 - Heredograma parcial de uma família hipotética com uma doença autossômica dominante
1.3.2 AUTOSSÔMICA RECESSIVA - AR
Critérios para o reconhecimento de herança AR:
A característica analisada:
• ocorreemambosossexos;
• étransmitidapelosdoissexos;
• pode ser transmitida diretamente de ho-mem para homem (pai para filho);
• saltagerações;
• ocorrenaproledegenitoresnão-afetados,geralmente heterozigotos e assintomáti-cos, freqüentemente consanguíneos;
• genitoresdeafetadossãogeralmentehe-terozigotos assintomáticos;
• surge,emmédia,em25%dosirmãosdeum afetado;
• manifesta-se em irmãos (homozigoto),embora os pais e filhos de afetados (hete-rozigotos) sejam geralmente normais.
Casais consanguíneos, além de apresentarem genes comuns à população, compartilham ge-nes devido à ancestralidade.
A fibrose cística é um exemplo clássico para este padrão de herança e caracteriza-se por apresentar anormalidades nas secreções exó-crinas, incluindo enzimas pancreáticas e duo-denais, cloretos da transpiração e secreções brônquicas. A mutação afeta uma proteína receptora transmembrana, que influencia a viscosidade das secreções, o que, por sua vez, pode levar a uma doença pulmonar crônica. O coração, o pâncreas e o sistema reprodutor também podem ser atingidos. É comum ocor-rer desnutrição e retardo de desenvolvimento.
Na Figura 3, vemos o heredograma de uma fa-mília hipotética com uma doença autossômica recessiva. Observem a linha dupla, indicando casamento consanguíneo.
I
II
III
IV
Figura 3 - Heredograma parcial de uma família hipotética com uma doença autossômica recessiva
ca
pít
ulo
2
28Observem o modo de transmissão vertical em que o pai passa o gene para filhos do sexo masculino, nos heredogramas para AD e AR.
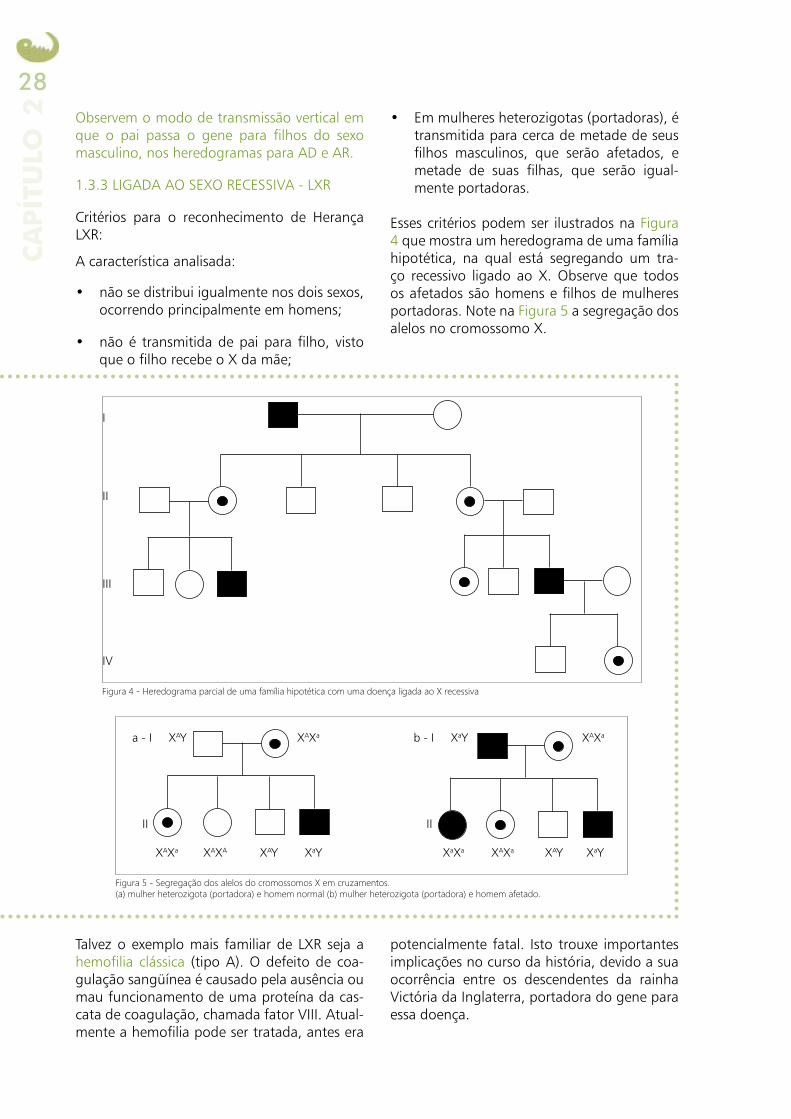
1.3.3 LIGADA AO SEXO RECESSIVA - LXR
Critérios para o reconhecimento de Herança LXR:
A característica analisada:
• nãosedistribuiigualmentenosdoissexos,ocorrendo principalmente em homens;
• nãoé transmitidadepaipara filho, visto
que o filho recebe o X da mãe;
• Emmulheresheterozigotas(portadoras),étransmitida para cerca de metade de seus filhos masculinos, que serão afetados, e metade de suas filhas, que serão igual-mente portadoras.
Esses critérios podem ser ilustrados na Figura 4 que mostra um heredograma de uma família hipotética, na qual está segregando um tra-ço recessivo ligado ao X. Observe que todos os afetados são homens e filhos de mulheres portadoras. Note na Figura 5 a segregação dos alelos no cromossomo X.
I
II
III
IV
Figura 4 - Heredograma parcial de uma família hipotética com uma doença ligada ao X recessiva
a - I XAY XAXa b - I XaY XAXa
II II
XAXa XAXA XAY XaY XaXa XAXa XAY XaY
Figura 5 - Segregação dos alelos do cromossomos X em cruzamentos.(a) mulher heterozigota (portadora) e homem normal (b) mulher heterozigota (portadora) e homem afetado.
Talvez o exemplo mais familiar de LXR seja a hemofilia clássica (tipo A). O defeito de coa-gulação sangüínea é causado pela ausência ou mau funcionamento de uma proteína da cas-cata de coagulação, chamada fator VIII. Atual-mente a hemofilia pode ser tratada, antes era
potencialmente fatal. Isto trouxe importantes implicações no curso da história, devido a sua ocorrência entre os descendentes da rainha Victória da Inglaterra, portadora do gene para essa doença.

ca
pít
ulo
2
29Para saber mais sobre hemofilia, consulte os li-vros indicados nas referências e os sites: http://www.drauziovarella.ig.com.br/entrevistas/hemo-filia_ana.asp e http://www.google.com.
1.3.4 LIGADA AO SEXO DOMINANTE - LXD
Critérios para o reconhecimento de Herança LXD:
A característica analisada:• nãosedistribuiigualmentenosdoissexos,
ocorrendo, principalmente, em mulheres;
• nãoétransmitidadepaiparafilho;
• estápresenteem,pelomenos,umdosge-nitores de um afetado;
• aparecedeformamaisleveevariávelnasmulheres afetadas do que nos homens (por causa da inativação do X);
• nopaiafetadoétransmitidaparatodasasfilhas, que serão afetadas e para nenhum dos filhos masculinos;
• surge, em média, em 50% dos filhos de uma afetada, independentemente do sexo.
A herança LXD pode ser confundida com a DA, entretanto ela se distingue através da descen-dência de homens afetados. Na LXD, todas as filhas de um pai afetado serão afetadas, e ne-nhum filho o será, diferentemente da AD, em que os filhos de ambos os sexos de um pai afe-tado poderão apresentar a característica.
Existem poucos distúrbios conhecidos para esse tipo de herança, um deles é o raquitismo resis-tente à vitamina D, também conhecido como raquitismo hipofosfatêmico, caracterizado, principalmente, por hipofosfatemia, baixa es-tatura e arqueamento dos membros inferiores.
1.3.5 LIGADA AO Y OU HOLÂNDRICA
Critérios para o reconhecimento de Herança ligada ao Y:
A característica analisada:
• sóocorreemindivíduosmasculinos;
• estápresentenopaideumafetado;
• surgeem todosos filhos (masculinos)deum afetado.
Até o momento, não foram demonstrados ca-sos convincentes de variantes fenotípicas não sexuais associadas ao Y, e provavelmente, não existem doenças ligadas ao Y.
1.3.6 CARACTERÍSTICAS LIMITADAS OU INFLUENCIADAS PELO SEXO
As características limitadas ou influenciadas pelo sexo representam formas especiais de he-rança autossômica. Na limitada ao sexo, a ex-pressão restringe-se a, apenas, um dos sexos. Na puberdade precoce, em meninos, as epí-fises ósseas sofrem fusão precoce, resultando em um homem de baixa estatura. Na influen-ciada pelo sexo, por sua vez, a expressão do gene autossômico é influenciada por hormô-nios. Um exemplo clássico seria o da calvície humana, em que o gene se comportaria como dominante para os homens e recessivo para as mulheres.
ANÁLISE DE HEREDOGRAMAS
Desafio da Fix: Você poderá ten-tar construir um heredograma da sua família com uma característi-ca simples, como ter o lóbulo da orelha solto ou pregado. Para isto, você seguirá o roteiro da prática, tente incluir o maior nú-mero possível de indivíduos. Divirta-se.
ROTEIRO DE PRÁTICA - FORMA DE HERANÇA
(Fonte: Material produzido pelos professores Francisco de Assis Maia de Lima e Marília de França Rocha para o Curso de Aperfeiçoamen-to em Biologia para Professores de 2º Grau - Módulo de Genética e Evolução – Natal-RN, (1997) - com modificações e a permissão dos autores).

ca
pít
ulo
2
30INTRODUÇÃO
As pessoas diferem com relação à expressão de muitas características. Uns têm certas ca-racterísticas herdadas tanto do pai quanto da mãe, mas algumas não ocorrem em nenhum dos genitores. São muitos os exemplos fami-liares, tais como cor dos olhos, tipo de cabelo, altura, inteligência, forma do lóbulo da orelha, grupos sangüíneos.
OBJETIVO
Investigar o modo de transmissão da caracte-rística Lóbulo da orelha.
MATERIAL A ser construído: A ser preenchido:
Heredograma TabelaQuestionário
Ajuste a tabela para o número de indivíduos da sua família.
TABELA - Amostra familiar em relação ao tipo de lóbulo da orelhaLÓBULOIndivíduo
Aderente Livre LÓBULOIndivíduo
Aderente Livre
1 Avô paterno 6 Tio(s) paterno(s )
2 Avó pater na 7 Tia(s) paterna(s )
3 Avô mater no 8 Tio(s) materno(s )
4 Avó mater na 9 Tia(s) materna(s )
5 Pai 10 Irmão (ã)
TOTAL TOTAL
a) Qual das características (lóbulo livre ou aderente) pode saltar uma ou mais ge-rações?
( ) lóbulo livre ( ) lóbulo aderente
b) Os indivíduos que apresentam lóbulos livres têm sempre o pai ou a mãe com lóbulos livres?
( ) Sim ( ) Não
c) Os indivíduos que apresentam lóbulos aderentes têm sempre o pai ou a mãe com lóbulos aderentes?
( ) Sim ( ) Não
d) Casais com lóbulos livres podem gerar filhos com lóbulos livres e aderentes?
( ) Sim ( ) Não
e) Casais com lóbulos aderentes podem ge-rar filhos com lóbulos livres e aderentes?
( ) Sim ( ) Não
g) Qual o tipo da herança? ( )AR ( )AD ( )LXR ( )LXD
PROCEDIMENTO
1. Observe as orelhas dos seus pais, irmãos e parentes (se residirem na mesma cidade) e verifique se apresentam lóbulos aderen-te ou livre (figura). Anote os resultados na tabela.
Figura- Orelhas com lóbulos livres (a) e aderentes (b).
2. De posse do levantamento, construa o he-redograma da sua família quanto à caracte-rística, utilizando a simbologia adequada:
3. Analise os dados, preenchendo o ques-tionário.
Vamos fazer uma amostra das genealogias feitas pela turma? Combinem uma exposição com o tutor. Disponibilize o seu heredograma (se for da sua vontade) ou o heredograma de um dos componentes do grupo. Você pode ou não incluir fotos de sua família.
Envie o questionário para que ele possa ser pontuado.

ca
pít
ulo
2
31
2. HERANÇA NÃO-MENDELIANAAbordaremos algumas condições nas quais não são observados os padrões de herança mendeliana.
2.1 HERANÇA DE CARACTERÍSTICAS COMPLEXAS
Os distúrbios multifatoriais são ocasionados pela combinação de efeitos aditivos de genes e fatores ambientais. Abrangem a maioria das malformações congênitas isoladas (não sin-drômicas) e diversas doenças comuns da fase adulta.
As malformações congênitas mais freqüentes envolvem o sistema nervoso central, o car-diovascular, o esqueletal e o urinário. No sexo masculino, predominam o lábio leporino com ou sem palato fendido, a estenose pilórica, ta-lipes equinovarus e malformações cardíacas, enquanto no feminino, as mais comuns são a anencefalia com ou sem espinha bífida, palato fendido isolado, deslocamento congênito do quadril, talipes calcaneus valgus, talipes meta-tarsus varus e pé postural.
Diabetes, hipertensão, doença coronariana, cardiopatia isquêmica e outros distúrbios car-diovasculares, febre reumática, cárie dentária, transtornos do humor (depressivos, bipolares,esquizofrenia), transtornos relacionados ao ál-cool, transtornos de ansiedade (pânico, fobias, transtorno obsessivo compulsivo, doença de Al-zheimer) são exemplos de doenças comuns da meiaidade e de transtornos comuns psíquicos.
Foge ao nosso objetivo dar descrições das doenças relacionadas. Para maiores infor-mações, consultar os livros “Genética Hu-mana” de Borges-Osório e Robinson, 2001, “Thompson e Thompson – Genética Médica” de Nussbaum, McInnes e Willard, 2002. Vo-cês também pode mconsultar os sites: http: / / www.scholar.google.com e http://www.images.google.com.br para o tema malfor-mações: http://www.diabetes.org.br e http://www.sbh.org.br. para as doenças comuns diabetes e hipertensão, respectivamente. Para
transtornospsíquicos, acessar: http://www.neurociencias.org.br/Home.php e clicar em O que é?..
2.1.1 CLASSIFICAÇÃO
Características quantitativas contínuas
• estão distribuídas em diferentes genes eem vários locos gênicos, cada um com pe-quenos efeitos sobre a característica;
• apresentammudançasquantitativasmen-suráveis (efeito aditivo);
• distribuiçãopopulacional contínua ebas-tante influenciada pelo ambiente.
Características quantitativas descontínuas
• estão distribuídas em diferentes genes eem vários locos gênicos, onde alguns locos podem desempenhar papel mais impor-tante que outros;
• apresentamdistribuiçãopopulacionaldes-contínua, com influência ambiental variável.
2.1.2 CRITÉRIOS PARA O RECONHECIMENTO
• As características multifatoriais se distri-buem na população, de acordo com uma curva normal;
• Osgenessofremefeitodoambiente;
• A herdabilidade indica qual o papel dosgenes na determinação de um fenótipo;
• Paisnormaiscomcriançaafetadaapresen-tam risco aumentado de ter uma segunda criança afetada;
• Osriscosderecorrênciaaumentam,quan-do um membro da família é afetado (pa-rentes em primeiro grau).
2.2 HERANÇA DE GENES ORGANELARES OU HERANÇA CITOPLASMÁTICA
A herança de genes organelares envolve o DNA de organelas citoplasmáticas com capa-

ca
pít
ulo
2
32cidade replicativa, mitocôndrias e plastídios. As mitocôndrias são encontradas em animais e vegetais. Os plastídios são exclusivos dos ve-getais, dos quais os mais importantes são os cloroplastos.
O genoma mitocondrial é pequeno em relação ao nuclear, contudo, responsável por ocasio-nar doenças genéticas humanas transmitidas, exclusivamente, por mulheres (herança mito-condrial). Ao ocorrer uma mutação no geno-ma mitocondrial, uma pessoa pode apresentar duas populações mitocondriais distintas, uma com a mutação e outra sem. A isso, chama-mos de heteroplasmia, causa significativa da variabilidade na expressão clínica de doenças mitocondriais. Os distúrbios mitocondriais ca-racterizam-se geralmente por problemas mus-culares e cerebrais. Temos como exemplos: a neuropatia óptica hereditária de Leber (LHON, perda visual progressiva e cegueira) e a síndro-me de Kearns-Sayre (fraqueza dos músculos cardíaco e esquelético da face, tronco e extre-midades), dentre outras..
O programa GBOL disponível no site http://www.ufv.br/dgb/gbol apresenta texto e aplica-tivo com uma série aleatória de exercícios, ten-do como exemplos: o feijão, a Mirabilis jalapa (planta) e a Limnaea peregra (caramujo). O usuário pode avaliar se as respostas apresenta-das estão corretas, clicando no botão Corrigir ou acessar as respostas corretas através do bo-tão Responder.
2.3 MOSAICISMO
Mosaicismo é a presença de duas ou mais li-nhagens celulares geneticamente diferentes, oriundas de um mesmo zigoto, podendo ocor-rer no indivíduo ou em um dos seus tecidos. O mosaicismo pode ser cromossômico ou gênico e acontecer na linhagem somática ou germi-nativa. Se incidir na linhagem somática, só ha-verá prejuízo para o organismo, se a mutação ocasionar um câncer, ou se o evento ocorrer no embrião inicial, afetando uma parte signifi-cativa do organismo como um todo. Assim, o indivíduo poderá apresentar sinais clínicos de uma doença. Mutações na linhagem germina-tiva, por sua vez, podem causar uma doença no filho de um casal normal, sem história fa-miliar prévia para essa mutação. As alterações
cromossômicas também exemplificam a he-rança nãomendeliana, mas serão vistas deta-lhadamente na disciplina de Citogenética.
2.4 HERANÇA EPIGENÉTICA
Este tipo de herança é devido a modificações herdáveis no funcionamento dos genes que não sejam decorrentes de mudanças na sequ-ência de bases do DNA do organismo. Temos como exemplos: o imprinting parental, a ina-tivação do cromossomo X e a paramutação. Estes dois últimos exemplos serão vistos, res-pectivamente, nas disciplinas de Citogenética e Genética Molecular.
2.4.1 DISSOMIA UNIPARENTAL E IMPRESSÃO GENÔMICA
A dissomia uniparental representa uma exce-ção à primeira lei de Mendel, pois apenas um dos genitores contribui com seus alelos para um determinado fenótipo.
A impressão genômica ou imprinting parental é caracterizada por genes homólogos, com di-ferentes expressões fenotípicas, devido à ori-gem parental. Desta forma, alguns genes só se expressam, quando herdados de um dos genitores.
As síndromes de Angelman (pouca coordena-ção muscular, mandíbula grande, risos incon-troláveis, retardo mental grave) e de Prader--Willi (obesidade, ingestão incontrolável de comida, baixa estatura, graus variados de re-tardo mental) ilustram bem estas duas condi-ções. O gene para estas síndromes está situado no braço longo do cromossomo 15 (na mesma região 15q11-q13), porém a forma como ele é herdado ocasiona uma delas. Na síndrome de Angelman, há uma dose dupla paterna (disso-mia uniparental) ou deleção materna; na sín-drome de Prader-Willi, a dose dupla é materna (dissomia uniparental), e a deleção é paterna. Ainda não existe uma boa compreensão do controle genético, envolvendo a impressão genética, embora alterações da estrutura da cromatina e metilação tenham sido propos-tas como possíveis mecanismos. Para saber mais, acesse: http://www.neurociencias.org.br/Home.php e clique em O que é?

ca
pít
ulo
2
33
3. VARIAÇõES NA EXPRESSÃO DOS GENESAlgumas variações na expressão dos genes fre-qüentemente distorcem o padrão mendeliano básico.
3.1 INTERAÇõES ENTRE ALELOS
3.1.1 ALELOS MÚLTIPLOS
Quando uma característica apresenta mais de dois alelos diferentes para o mesmo loco, di-zemos que se trata de alelismo múltiplo. Um exemplo clássico é o do grupo sangüíneo ABO, determinado por três alelos de um gene que interagem, para produzirem os quatro A B ti-pos de fenótipos desse sistema. Os alelos IA e IB determinam, respectivamente, os antígenos A e B, embora Ii seja um alelo nulo e não produ-za proteína antigênica. Dessa forma, IA e IB são dominantes em relação ao alelo Ii.
3.1.3 DOMINÂNCIA INCOMPLETA
Na dominância incompleta, o heterozigoto apresenta um fenótipo quantitativamente inter-mediário aos homozigotos. Nas plantas mara-vilha, também conhecidas como quatro-horas, quando uma linhagem pura com pétalas ver-melhas (c+ c+ ) é cruzada com uma + linhagem pura de pétalas brancas (cc), a F1 (c
+ c) apre-senta um fenótipo de pétalas rosa. Ao haver autofecundação das plantas F1 , surge uma F2 com o seguinte resultado: ¼ pétalas vermelhas : ½ pétalas rosa : ¼ pétalas brancas. Ao invés de uma proporção fenotípica de 1:3, temos a proporção 1:2:1.
3.1.4 ALELOS LETAIS
Alguns alelos mutantes são capazes de cau-sar a morte de um indivíduo e são conhecidos como alelos letais. Provavelmente, temos uma quantidade significativa desse tipo de alelo em nossos genomas, embora a maioria destes seja letal recessivo.
Em humanos, um exemplo de letal dominante é a acondroplasia (AD), tipo de nanismo gené-tico mais freqüente. Pessoas não acondroplá-sicas são de genótipo aa, acondroplásicos sãoheterozigotos (Aa), e, em homozigose AA, o gene é letal, causando morte geralmente nos primeiros meses de idade. A capacidade re-produtiva dos afetados é reduzida em 20% do normal. Dessa forma a maioria dos casos observados é esporádica e causada por muta-ção nova. As síndromes de Rett e Aicardi são alguns exemplos de LXD, descritas, exclusiva-mente, em mulheres, consideradas letais no sexo masculino.
3.1.5 EPISTASIA
A epistasia é uma situação na qual um gene mascara o efeito de outro, ou seja, um gene que exerce sua expressão, cancelando a ex-pressão de alelos de outro gene. Vamos reto-mar o exemplo do sistema ABO. Para formar os antígenos A e B, precisamos da informação dos alelos IA e IB . Mas, para que isso aconteça, antes, fazse necessária a presença de um gene precursor de antígeno, em que o alelo H forma o precursor, e o alelo h não forma o precursor.
Genótipo Fenótipo Antígenos Anticorpos
IAIA, IAIi A A Anti-B
IBIB, IBIi B B Anti-A
IAIB AB AB Não produz
IIII O Falta de proteína antigênica
A nti-A e Anti-B
Outros exemplos conhecidos são os alelos para a cor da pelagem de coelhos (Aguti, Chinchila, Himalaia e Albina) e de ratos (Cinza, Amarela, Preta). Em alguns casos especiais, pode haver mais de 50 alelos para um único gene, embora apenas duas formas alélicas sejam herdadas e se expressem. Isso pode ser observado em cer-tos genes de compatibilidade nas plantas e ge-nes de histocompatibilidade nos vertebrados. Em humanos, temos, por exemplo, o sistema HLA – antígeno leucocitário humano.
3.1.2 CODOMINÂNCIA
Na codominância, ambos os alelos de um par de genes se expressam independentemente no heterozigoto. No exemplo acima, vimos que no genótipo IA IB cada um dos alelos (IA e IB ) produz seu próprio antígeno, logo eles dem.onstram uma relação de codominância. O gru-pamento sangüíneo MN também exemplifica esta situação.

ca
pít
ulo
2
34
Dessa forma, se um indivíduo tiver a informa-ção para um determinado tipo sangüíneo (IA e/ou IB), e não para o precursor (hh), ele não poderá formar o antígeno e será um falso O. Este exemplo é conhecido como fenótipo Bombaim, por ter sido observado pela primei-ra vez, em Bombaim, na Índia.
Um outro exemplo de epistasia recessiva é a cor da pelagem em cães Labrador, determina-da pelos alelos B e b, representando as cores preta e marrom, respectivamente. O alelo e de outro gene é epistático sobre os alelos B e b, conferindo uma pelagem amarela, por impe-dir a deposição de pigmento na pelagem do animal. Assim os genótipos B-ee e bbee pro-duzem cães de pêlo amarelo, B-E cães de pêlo preto e bbE- cães de pelo marrom, produzin-do uma proporção fenotípica de 9 (preto): 3 (marrom): 4 (amarelo).
A epistasia demonstra a interação entre os genes, de alguma via bioquímica ou desenvolvimental.
3.1.6 SUPRESSORES E MODIFICADORES
Genes supressores revertem o efeito de uma mutação de outro gene, restaurando o fenó-tipo selvagem. Nos genes modificadores, uma mutação altera um segundo loco gênico, mu-dando a expressão de um gene mutado ante-riormente, em um primeiro loco. A supressão, às vezes, é confundida com a epistasia. Entre-tanto, a proporção modificada indica o tipo de interação. Na mutação supressora, em ge-ral, apenas dois fenótipos segregam, enquan-to, na epistasia, três fenótipos segregam (ver exemplo do Labrador). Para maiores detalhes de interações de alelos de um gene ou entre genes consultem o capítulo seis (6) do livro “Introdução à Genética” de Grifffiths e cola-boradores, 2006, e acessem o site http://www.ufv.br/dgb/gbol.
3.2 PENETRÂNCIA E EXPRESSIVIDADE
A penetrância de uma característica pode ser definida como: a probabilidade de que uma pessoa com o genótipo estudado manifeste esta característica. Em heranças dominantes, apenetrância reduzida (ou incompleta) pode ser um complicador na avaliação de heredo-gramas, pois a característica salta gerações. A penetrância é como um jogo de tudo ou nada, a característica é ou não expressa.
A característica pode ter expressividade variá-vel, manifestando desde o fenótipo mais leve ao mais grave, em diferentes indivíduos. Na síndrome de Waardenburg, os pacientes apre-sentam alteração do pigmento (pele, cabelo, olho), deficiência auditiva e anomalias discre-tas da face. Diferentes membros da mesma fa-mília podem mostrar distintos aspectos da sín-drome (fig. 6), podendo oscilar desde a perda moderada de audição e mecha branca capilar até a surdez profunda, nariz com ponte alta e larga, heterocromia da íris (olho direito e es-querdo com cores diferentes) e grisalhamento prematuro do cabelo. Esta variabilidade pode ser decorrente de efeitos ambientais, genes modificadores e heterogeneidade genética.
Gene precursor de antígeno
Gene para antí-genos A e B
Fenótipo
HH, Hh IAIA, IAIi A
HH, Hh IBIB, IBIi B
HH, Hh IAIB AB
HH, Hh IiIi O
hh IAIA, IAIi, IBIB, IBIi, IAIB, IiIi
Falso O (Bombaim)
http://www.salud.gob.mx/unidades/pediatria/Imagenes/SW3.jpgFigura 6 - Aspectos variáveis da Síndrome de Waardenburg
http://www.atlas-of-ophthalmology.com/atlasimg/6_106212145428_low_thumb.jpg

ca
pít
ulo
2
353.3 HETEROGENEIDADE GENÉTICA
Na heterogeneidade genética, diferentes tipos de mutação no mesmo loco ou em diferen-tes locos produzem fenótipos semelhantes. A surdez congênita hereditária é decorrente, na maioria dos casos (75%), a genes autossômi-cos recessivos (locos diferentes), e em menor proporção de genes autossômicos dominan-tes (3%) e de genes recessivos ligados ao sexo (2%). O mecanismo de ação dos genes varia, afetando diferentes partes do ouvido interno, todavia, todos resultam no mesmo fenótipo, surdez. Vale destacar que, além dos aspectos genéticos, fatores adquiridos, como meningi-te, rubéola materna, infecções do ouvido, den-tre outros, podem causar a deficiência auditiva.
3.4 ANTECIPAÇÃO E IDADE DE MANIFESTAÇÃO
A antecipação é a tendência de algumas ca-racterísticas surgirem mais cedo e com maior gravidade, a cada geração subseqüente. Este fenômeno parece estar ligado ao aumento de repetições de trinucleotídeos. Estas repetições ocorrem normalmente em vários genes e em número variável. Quando este número ultra-passa a normalidade surgem doenças. Um exemplo é a Distrofia miotônica, doença sis-têmica variável com disfunção muscular carac-terística. Imaginem uma mãe afetada discre-tamente (com 250 repetições CTG) e ao gerar um filho (com mais de 1.000 repetições CTG), esse apresenta graves dificuldades cardio-res-piratórias, chegando a falecer nas primeiras semanas de vida. Todos os indivíduos têm umnúmero adequado de repetições de trincas de nucleotídeos, neste caso CTG, mas algumas pessoas, ao se reproduzirem, podem passar adiante uma repetição maior que a normal, acarretando maior gravidade à doença, de for-ma que a cada geração a doença tende a se manifestar mais precocemente.
Outros exemplos seriam a Síndrome do X--frágil, retardo mental com vários sinais físi-cos, cujo triplete de bases seria o CGG, e a Doença de Huntington, doença degenerativa do sistema nervoso central, com a sequência repetida CAG.
Em relação à idade de manifestação, uma do-ença pode estar presente desde o nascimento (Acondroplasia), manifestar-se no início da in-fância (Distrofia muscular de Duchenne), naadolescência ou idade adulta (Doença de Hun-tington) e na terceira idade (Doença de Al-zheimer). Vamos usar a doença de Alzheimer como exemplo. Essa doença afeta cerca de 5% das pessoas com mais de 65 anos e aproxima-damente 20% daquelas com mais de 80 anos. Há perda progressiva de memória, seguida por distúrbios do comportamento e deterioração cognitiva generalizada. Embora seja uma do-ença de início tardio, em raras ocasiões, essa doença pode ocorrer em idade precoce, sendo geralmente autossômica dominante, enquan-to a de início tardio não o é.
Se você quiser ter uma idéia do que significa uma doença dessas para uma família, assista a um dos filmes lançados nos anos de 2001: Íris, Uma canção para Martin, O filho da noiva e 2004: Diário de uma paixão.
3.5 PLEIOTROPIA
Cada gene apresenta um efeito primário e, a partir deste, podem surgir diversas consequên-cias. A pleiotropia é ocasionada pelos efeitos múltiplos produzidos por um único gene. Al-gumas doenças metabólicas genéticas exem-plificam bem esta situação. Mutações gênicas causadoras de defeitos enzimáticos bloqueiam vias metabólicas específicas e acarretam sub-seqüentemente, acúmulo de substratos e de-ficiências de produtos. O efeito primário da fenilcetonúria é a deficiência da enzima feni-lalanina-hidroxilase, e seus efeitos secundários são: retardo mental, excreção de fenilcetonas na urina, pigmentação clara e odor de mofo. Entretanto, estes efeitos secundários podem ser evitados, se a doença for diagnosticada a tempo. Isto é feito através do conhecido exa-me do pezinho.
Vocês já devem ter percebido que os rótulos de alimentos trazem várias informações. Isto é importante, pois algumas pessoas têm res-trições a determinadas substâncias. Alimentos light reduzem o teor de gordura, alimentos diet reduzem o açúcar, alguns cereais matinais alertam: contém glúten, nos refrigerantes avi-sam: atenção contém fenilalanina e assim por

ca
pít
ulo
2
36diante. No caso do nosso exemplo, a fenilceto-núria, o tratamento com dieta pobre em feni-lalanina deve ser iniciado no primeiro mês de vida e mantido, sem interrupção, pelo menos, ao longo da infância e adolescência. Por ser um aminoácido essencial, a fenilalanina não pode ser removida, totalmente, da dieta.
Pesquisa da Fix: Faça uma pes-quisa na internet sobre o teste do pezinho e sua importância no diagnóstico de doenças e, em especial, a fenil-cetonúria. Dis-cuta com seus colegas.
Desafio da Fix: Assistam ao filme “O óleo de Lorenzo”. O filme pode ser encontrado nas locadoras, sob a forma de fita ou DVD. Bom filme e boa pesquisa!
Em 1984, Michaela e Augusto Odone desco-brem que Lorenzo é portador de uma doença grave, a adrenoleucodistrofia (ALD). Seus pais, então, iniciam uma luta para salvá-lo. Come-çam a estudar genética, fisiologia, bioquímica, promovem simpósios e tentam várias dietas na intenção de descobrir a cura da ALD.
A ALD ocorre devido a uma ß oxidação pobre ou colapso dos ácidos graxos no peroxissoma. A enzima que degrada estes ácidos é produzi-da no citoplasma (ribossomos livres) e, então, é enviada, através da membrana do peroxis-soma, até o lúmen dessa organela. A primeira enzima no caminho da degradação da longa cadeia de ácidos graxos é a Coa (coenzima A). Esta enzima catalisa a adição de acetil-CoA para as cadeias longas de ácidos graxos, e a degradação depende desse primeiro passo. Pesquisas recentes mostraram que a ALD não é devida a uma síntese deficiente de VLCFA--CoA, (VLCFA = very long chain fatty acids, = ácidos graxos de cadeia longa) e, sim, devido a um transporte deficiente de uma proteína que move a VLCFA através da membrana do peroxissoma.
Fórum sobre o filme O óleo de Lorenzo:
No filme, quando Peter Ustinov diz a Michae-la que “ALD é passada só através da mãe”, o médico percebe o impacto da informação e diz a Michaela que ela não deveria se sentir cul-pada pela doença de Lorenzo. Michaela tinha um alelo dominante normal e um alelo reces-sivo com a mutação, mas ela não desenvolvia a doença porque o alelo dominante fazia um transportador que atuava no peroxissoma.
Pesquisem: Sobre outros tipos de ALD. Descu-bram o destino do rapaz. Quem está tomando conta dele, como ele faz para se comunicar, qual dos pais já faleceu, o que a família está fazendo em relação às pesquisas com bainha de mielina?, etc.
Respondam:- Qual o tipo de herança da ALD?- Quais são os outros tipos de ALD?- Como se encontra Lorenzo atualmente?
ATIVIDADEREVISANDO OS CONHECIMENTOS
1. Muitas doenças genéticas têm o padrão de herança já bem estabelecido. Embora nestes casos o fator determinante para a expressão da doença seja genético, mui-tos avanços científicos têm conseguido dar aos indivíduos uma vida melhor e, mesmo, prolongar a longevidade. O aconselha-mento genético e o acompanhamento do paciente são fatores fundamentais para a família.
O texto abaixo fala sobre uma dessas do-
enças: a Distrofia Muscular de Duchenne (DMD).
“Até os três anos, Lucas jogava bola e brin-
cava como qualquer criança saudável. Aos poucos, sua mãe começou a perceber que o menino caía com frequência, apoiava-se nas pernas e na cintura para se levantar e andava nas pontas dos pés. Ela procu-rou os médicos e descobriu o diagnóstico do filho: Distrofia Muscular de Duchenne,

ca
pít
ulo
2
37uma doença genética degenerativa e sem cura...Lucas hoje tem 18 anos. Está vivo e anda com as próprias pernas...Apesar de pouco conhecida, a Distrofia Muscular de Duchenne é a mais comum entre as doen-ças genéticas fatais. O gene defeituoso é transmitido pela mãe. A doença só se ma-nifesta em meninos. Os músculos do corpo se degeneram e progressivamente vão per-dendo a mobilidade, devido à produção insuficiente de uma proteína chamada dis-trofina, componente da fibra muscular...” Modificado e adaptado da revista Época, outubro de 2006.
As informações apresentadas na reporta-
gem podem ser bem entendidas por qual-quer leitor, entretanto você como futuro biólogo poderá compreendê-las e interpre-tá-las com maiores detalhes!
I- Qual o provável padrão de herança desta
doença?
a) Autossômica dominante b) Autossômica recessiva c) Dominante ligada ao sexo d) Recessiva ligada ao sexo e) Ligada ao Y II- Das afirmativas abaixo, qual pode jus-
tificar (ou explicar) a alternativa assinalada por você na questão I?
a) O gene defeituoso é transmitido pela mãe, podendo ela ser portadora e não ex-pressar a doença.
b) Como o gene é transmitido pela mãe, todos os descendentes do sexo masculino manifestarão a doença.
c) A doença somente se manifesta em meninos, pois a mãe transmite este gene apenas para os filhos do sexo masculino.
d) O gene defeituoso tanto pode estar presente na mãe como também no pai, portanto independente do sexo, o gene irá se manifestar igualmente.
e) O gene é responsável por uma proteí-na chamada distrofina, presente nas célu-las somáticas, sendo, portanto, transmiti-do através de um autossomo.
Pesquisa da Fix:
Pesquise, na Net, informa-ções sobre o quadro clínico, p rognós t i co , forma de tra-tamento e gru-pos de pesquisa que estudem a DMD no Brasil.
2. O heredograma a seguir representa o estu-do de uma família em que alguns indivídu-os apresentam polidactilia (dedos extras), característica de herança autossômica do-minante com penetrância reduzida. Anali-se-o e assinale a alternativa correta.
I 1 2
II
1 2 3 4 5 6 7 8 9
III 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
a) O indivíduo II2 deve conter o gene em estudo, por ser do sexo masculino e por sua filha ter expressado a característica.
b) Não há possibilidade de o casal III3 X III4 ter um filho que expresse este caráter.

ca
pít
ulo
2
38 c) O indivíduo II8 deve conter o alelo em questão, porém este não se expressou.
d) Os indivíduos I1, II1 e III14 são homozigotos.
e) O indivíduo II5 é homozigoto para o caráter em estudo.
3.Sobre os tipos de relações entre alelos de um mesmo gene ou genes diferentes, marque a alternativa que faz a correta associação.
a) A 2 V, B 5 IV, C 1 II, D 3 I, E 4 III b) A 2 II , B 1 V, C 5 I, D 3 IV, E 4 III c) A 5 I, B 4 III, C 3 IV, D 1 II, E 2 V d) A 4 V, B 5 II, C 3 IV, D 2 III, E 1 I e) A 4 III, B 1 II, C 2 I, D 5 IV, E 3 V
REFERÊNCIAS
LEITURA FUNDAMENTAL
GRIFFITHS, A. J. F.; WESSLER, S. R.; LEWONTIN, R. C.;GELBART, W. M.; SUZUKI, D. T.e MILLER, J. H. Introdução à Genética: Rio de Janeiro: Guanabara Koogan S/A, 2006.
NUSSBAUM, R. L., McINNES, R. R. e WILLARD, H. F. Thompson e Thompson – Genética Mé-dica. Rio de Janeiro: Guanabara Koogan S. A., 2002.
Tipo de relação Característica / conceito Exemplo
A - Dominância
1- Os dois alelos se expressam, pro-duzindo, no heterozigoto, fenótipos (ou características) que estão presen-tes em ambos os homozigotos (do-minante e recessivo).
I- Alelos relacionados com a cor da pelagem em cães Labra-dor: preta, marrom e amarela.
B - Co-dominância2- Um dos alelos sempre expressa o seu efeito, seja na condição homozi-gota ou heterozigota.
II- lelos para a forma da se-mente da ervilha Pisum sati-vum: lisa e rugosa.
C - Epistasia
3- Mais de dois tipos de alelos dife-rentes para um mesmo loco. Todos os tipos contribuem para um mesmo caráter.
III- Alelos para a cor das péta-lasvermelha, branca ou róse a, na planta Boca-de-Leão –Antir-rhinum majus.
D - Alelos múltiplos
4- Os dois alelos se expressam, pro-duzindo, no heterozigoto, fenótipo intermediário ao presente nos homo-zigotos (dominantes e recessivos).
IV- Série de alelos que codifi-cam os antígenos para o siste-ma sangüíneo ABO.
E - Dominânciaincompleta
5- Alelos de um gene impedem ou inibem a expressão de um ou mais pares de alelos de outros genes.
V- Alelos para o sistema san-güíneo MN.
BORGES-OSÓRIO, M. R. e ROBINSON, W. M. Genética Humana. Porto Alegre: Artmed, 2001.
SNUSTAD, D. P. e SIMMONS M. J. Fundamen-tos de Genética: Guanabara Koogan S/A, 2001.
LEITURA COMPLEMENTAR
E-book - BEIGUELMAN B. A interpretação ge-nética da variabilidade humana. (www.desvir-tual.com/bbeiguel/e-book)
SITES
Gerais: http://www.google.com
OMIM: http://www3.ncbi.nlm.nih.gov/omim
Genética Geral: http://www.ufv.br/dgb/gbol
Sociedade Brasileira de Genética: http://www.sbg.org.br

ca
pít
ulo
2
39Hemofilia: http://www.drauziovarella.ig.com.br/entrevistas/hemofilia_ana.asp
Malformações: http://www.scholar.google.c om e http://www.images.google.com.br
Diabetes: http://www.diabetes.org.br
Hipertensão: http://www.sbh.org.br
Transtornos psíquicos: http://www.neurocien-cias.org.br/Home.php


ca
pít
ulo
3
41
BASE CROMOSSôMICA DA HERANÇA E MAPEAMENTO GÊNICO
INTRODUÇÃO
Neste terceiro capítulo, compreenderemos como os fatores abstratos (genes), postulados por Mendel, foram localizados e mapeados em cromossomos da mosca das frutas (Dro-sophila melanogaster), por Thomas Morgan e seu grupo. Veremos um pouco do desenvol-vimento histórico da teoria cromossômica da herança, da evidência da ligação e da segrega-ção cromossômica anormal (não-disjunção), que ocasiona anomalias da herança. Faremos uso da análise de crossing, para mapear as po-sições dos genes nos cromossomos.
OBJETIVOS ESPECÍFICOS
• Compreender a base cromossômica domendelismo e os procedimentos para o mapeamento gênico por frequência de re-combinação.
BASE CROMOSSôMICA DA HERANÇA EMAPEAMENTO GÊNICONa história da genética, podemos destacar dois períodos: o primeiro, do ano de 1900 até 1909, no qual ocorreu a redescoberta do
Profa. Marília de França Rocha Carga Horária I 15hColaboradoras - Profa. Maria Teresa Marquim N. Cornélio Profa. Rita de Cássia de Moura

ca
pít
ulo
3
42trabalho de Mendel e do início da validação da hereditariedade mendeliana; o segundo, a partir de 1910, quando tiveram início os estu-dos da natureza e arranjo dos genes nos cro-mossomos.
1. BASE CROMOSSôMICA DO MENDELISMONo primeiro capítulo, percebemos que os re-sultados de Mendel ficaram “desconhecidos” para a comunidade científica até 1900, pois muitos pesquisadores não estavam cientes desse trabalho, apesar do interesse na heredi-tariedade. Uma questão importante para essa época era a localização do material hereditário na célula. Os locais mais óbvios para iniciar a investigação seriam óvulos e espermatozóides. O núcleo foi considerado um bom local para abrigar o material hereditário, visto que ocorre uma grande diferença de volume entre os cito-plasmas dessas células (fig.1).
Figura 1- Gametas feminino e masculino. Note a diferença de tamanho ocasionada pelo volume desigual de citoplasma.Fonte: Fhttp://www.reprodusite.hpg.ig.com.br/fertilizacao_1.jpg
Waldeyer tenha compreendido o que estava observando e os nomeou de cromossomos, que significa corpos coloridos.
No ano de 1902, Walter Sutton, um estu-dante da Universidade de Columbia (Estados Unidos), observou que o agrupamento de cro-mossomos paternos e maternos em pares, no momento da formação do zigoto, e sua sub-seqüente separação a cada divisão celular (mi-tose ou meiose) eram a prova da continuidadecromossômica e poderiam constituir a base fí-sica para a lei da hereditariedade de Mendel.
Nesta mesma época, Theodor Boveri, profes-sor de Zoologia e Anatomia comparativa da Universidade de Würzburg (Alemanha), estava trabalhando com uma espécie de ouriço-do--mar e produzindo embriões com números variáveis de cromossomos. Segundo Boveri, apenas aqueles que possuíam 36 cromosso-mos apresentavam desenvolvimento normal, e cada cromossomo possuía, então, uma “quali-dade diferente” ou individualidade. Desta for-ma, o número de pares de cromossomos seria o mesmo para todos os indivíduos da mesma espécie.
A individualidade (Boveri) dos cromossomos nas espécies complementava a continuidade (Sutton) dos cromossomos nas gerações su-cessivas de células. Esses trabalhos uniram es-ses dois pesquisadores em uma das idéias mais revolucionárias da biologia do século XX: “A teoria de Sutton-Boveri da hereditariedade pormeio de cromossomos” ou simplesmente te-oria cromossômica da herança, que forneceu uma estrutura unificadora para todos os estu-dos sobre a herança.
A combinação da evidência citológica com os temas da genética, feita por Montgomery, Correns, Wilson, Sutton e Boveri, originou uma nova disciplina, a citogenética, que lançou uma ponte para o mecanismo da hereditariedade. Entretanto, como vocês podem perceber, con-ceitos novos necessitam de tempo para serem aceitos, e, com essa teoria não foi diferente. A princípio, vários cientistas, em especial, Bate-son e Morgan, não acreditavam na teoria de Sutton- Boveri, pois não havia evidência direta de que os genes localizavamse em cromos-somos. Bateson decidiu aceitar a teoria dos
1.1 A TEORIA CROMOSSôMICA DA HERANÇA
Em 1840, os cromossomos já haviam sido ob-servados por diversos cientistas, embora ape-nas, em 1888, o anatomista alemão W. von

ca
pít
ulo
3
43cromossomos dois anos antes da sua morte, quando as provas se tornaram incontestáveis. Morgan, por sua vez, na tentativa de encon-trar uma alternativa, chegou a revelações que o fizeram mudar de idéia e contribuir com al-gumas das descobertas mais importantes para a nova ciência em desenvolvimento.
Vale a pena lembrar: Edmund Beecher Wilson, embriologista e um dos principais nomes da citogenética, foi o mestre e mentor de toda a equipe de Morgan e, como colega de trabalho e amigo, exerceu grande influência sobre Mor-gan, na aceitação da teoria cromossômica da herança.
1.2 QUEM FOI MORGAN
Thomas Hunt Morgan (fig 2) nasceu em 25 de setembro de 1866, em Lexington, Kentucky, nos Estados Unidos. Considerado um aluno crítico e independente, obteve seu bachare-lado na Universidade de Kentucky e seu dou-torado, na Universidade Johns Hopkins. Em 1904, iniciou seus trabalhos como professor de Zoologia Experimental na Universidade de Columbia, em Nova York, onde permaneceu até 1928. Após essa data, Morgan trabalhou como professor de biologia e diretor do Labo-ratório G. Kerckhoff no Instituto de Tecnologia da Califórnia em Pasadena, onde permaneceu até sua morte, em 04 de dezembro de 1945. Morgan deixou contribuições de grande im-portância na embriologia experimental, no campo da regeneração celular e, em especial, na genética de Drosophila.
O ano de 1910 foi o ano da primeira publi-cação de Mor-gan em Droso-phila.
Para saber mais, acessem:http://nobel-prize.org/no-b e l _ p r i z e s /medicine/lau-reates/1933/ morgan-b io.html
Figura 2- Thomas Hunt Morgan Fonte: http://nobelprize.org/no-bel_prizes/ medicine/laureates/1933/morganbio. html
1.3 O ORGANISMO EXPERIMENTAL DE MORGAN
O gênero Drosophila compreende um grande número de espécies, sendo a D. melanogas-ter (fig 3) a mais intensamente estudada, des-de os dias da Sala das Moscas (laboratório de Morgan na Universidade de Columbia) até os dias atuais. Centenas de laboratórios no mun-do inteiro investigam os genes e cromossomos deste organismo modelo. Vários são os atri-butos que fazem desta mosca um organismo experimental, como por exemplo, a produção de gerações a cada duas ou três semanas, fá-cil manutenção, ocupação restrita de espaço, virtualmente imune a doenças. Uma das pro-priedades mais importantes é a presença de apenas quatro pares de cromossomos distin-tos uns dos outros, sendo um deles o par de cromossomos sexuais, XX para a fêmea e XY para o macho.
As informações sobre Drosophila estão atu-almente disponíveis em um banco de dados eletrônico constantemente atualizado, chama-do FlyBase acessível em http://www.morgan.harvard.edu.
Figura 3- Drosophila melanogasterFonte: http://neofronteras.com/wpcontent/photos/drosophila.jpg
1.4 EVIDÊNCIA DE LIGAÇÃO AO SEXO
O que determina o sexo de um organismo tem sido objeto de especulação desde épocas re-motas. Existem diversas modalidades de deter-minação sexual, envolvendo sistemas cromos-sômicos simples ou múltiplos. Entretanto, em alguns organismos, a determinação é ambien-tal. Essas questões serão abordadas na discipli-na Citogenética.

ca
pít
ulo
3
44Em 1909, Morgan começou a conduzir ex-perimentos genéticos em Drosophila. Ele ha-via visitado o laboratório de Hugo de Vries na Holanda, ficando impressionado com as mu-tações na planta Oenothera. Resolveu, então, expor suas moscas a diferentes tipos de tem-peraturas, substâncias químicas e radiações, no intuito de produzir fenótipos diferentes.
Um certo dia, apareceu um único macho de olhos brancos, em uma população de moscas de olhos vermelhos. Esse macho foi prontamente acasalado com suas irmãs de olhos vermelhos.
Embora a progênie F1 fosse constituída por moscas com olhos vermelhos, reapareceram
machos de olhos brancos na F2, mostrando a recessividade do caráter olho branco. Entre-tanto, todas as moscas de olhos brancos eram machos e havia duas fêmeas para um macho entre aquelas de olhos vermelhos (fig 4). Ele observou que cruzamentos recíprocos davam resultados distintos, e as proporções fenotípi-cas eram diferentes entre machos e fêmeas, o que contrariava os princípios de Mendel. Mor-gan buscou hipóteses para explicar esses des-vios na segregação. Ele propôs que o fator da cor dos olhos estava acoplado com o cromos-somo X (determinante do sexo), mas não com o Y, e que os fenótipos branco e vermelho se deviam a alelos diferentes, representados por w+ (selvagem vermelho) e w (mutante branco).
P-
G-F1-
G-
F2-
Fêmea de olhos vermelhos x Macho de olhos brancos
XW+XW+ XWY
Fêmea de olhos vermelhos Macho de olhos vermelhos
XW+XW XW+Y
½ fêmeas de olhos vermelhos XW+XW+ XW+XW
¼ machos de olhos vermelhos XW+Y
¼ machos de olhos brancos XWY
Figura 4- Resultado do cruzamento entre uma fêmea selvagem de olhos vermelhos (Xw+ Xw+) e um macho de olhos brancos (X wY)
XW+ XW Y
XW+ XW XW+ Y
Outros resultados confirmaram os elementos de suas hipóteses. Em um cruzamento entre uma fêmea heterozigota (XW+ XW) e um ma-cho mutante hemizigoto (XWY), independen-temente do sexo, metade da prole tinha olhos brancos e metade olhos vermelhos (fig 5). Em outro experimento (fig 6), fêmeas de olhos brancos (XWXW) foram cruzadas com machos de olhos vermelhos (XW+Y), e todos os machos apresentaram olhos brancos. Ao intercruzar essa prole, a F2 mostrou uma proporção de 1
(fêmea de olhos brancos): 1 (fêmea de olhos vermelhos): 1 (macho de olhos brancos): 1 (macho de olhos vermelhos). Esse experimento apoiava a idéia de genes situados em cromos-somos, porém não estabelecia firmemente a teoria cromossômica.
Morgan e seu grupo identificaram vários genes (ligados ao cromossomo X e em autossomos) em Drosophila e concluíram que esses estavam situados em cromossomos.

ca
pít
ulo
3
45
1.5 A NÃO DISJUNÇÃO
Calvin B. Bridges, um dos colaboradores de Morgan, cruzou fêmeas de olhos brancos com machos de olhos vermelhos em grande esca-la. Apesar de as moscas da F1 serem em sua maioria fêmeas de olhos vermelhos e machos de olhos brancos, algumas exceções, como fêmeas de olhos brancos e machos de olhos vermelhos, foram observadas. Ele cruzou es-sas fêmeas especiais com machos normais de olhos vermelhos obtendo uma prole normal e uma prole especial. Os machos especiais, por sua vez, eram todos estéreis.
A explicação dada por Bridges era que as fême-as de olhos brancos resultavam do comporta-mento anormal do cromossomo X, originário das fêmeas da geração parental. Assim, fême-as especiais obtinham ambos os cromossomos X de suas mães de olhos brancos (XwXw). Nor-malmente, os cromossomos X deveriam segre-gar um do outro, mas ocasionalmente uma falha na separação, a qual Bridges nomeou de não-disjunção, poderia ocorrer formando um ovócito com dois cromossomos X e outro sem nenhum X. Tais ovócitos anormais, ao serem fecundados por espermatozóides normais, produziriam prole com números de cromos-soos sexuais diferentes do habitual. A Figura
P -
G -
F1-
Fêmea de olhos vermelhos x Macho de olhos brancos
Xw+Xw XwY
¼ fêmeas de olhos vermelhos
Xw+Xw
¼ fêmeas de olhos brancos
XwXw
¼ machos de olhos vermelhos
Xw+Y
¼ machos de olhos brancos
XwY
Figura 5 - Resultado do cruzamento entre uma fêmea heterozigota (Xw+Xw) e um macho mutante (XwY)
Xw+ Xw Xw Y
P -
G -
F1-
Fêmea de olhos brancos x Macho de olhos vermelhos
XWXW XW+Y
½ fêmeas de olhos vermelhos Xw+Xw
½ m achos de olhos brancos XwY
Figura 6 - Resultado do cruzamento entre fêmea de olhos brancos (XwXw) com macho de olhos vermelhos (Xw+Y)
Xw XW+ Y

ca
pít
ulo
3
467 ilustra o processo. Os óvulos com um X w , com dois X w ou sem o X, ao serem fecunda-dos pelos espermatozóides com o Xw+, origi-nam indivíduos de olhos vermelhos, com as seguintes constituições cromossômicas: fêmea (Xw+Xw), metafêmea (Xw+Xw Xw - geralmente morre) e macho (Xw+O), respectivamente. Es-ses mesmos óvulos, ao serem fecundados por espermatozóides com Y, produzem os seguin-tes zigotos: macho (XwY) de olhos brancos, fê-mea especial (XwXwY) de olhos brancos e um indivíduo inviável YO, respectivamente.
1.6 PADRõES DE HERANÇA DE GENES LIGADOS
Algumas das primeiras evidências da ligação vieram de experimentos com ervilhas, realiza-dos por Bateson e Punnett, cujas frequências das quatro classes da F2 desviavam-se, signifi-cativamente, das proporções mendelianas de 9:3:3:1.
Fenótipos Representantes Proporções %
Flores vermelhas e grãos de pólen longos
583 0 ,73
Flores brancas e grãos de pólen curtos
170 0 ,21
Flores vermelhas e grãos de pólen curtos
26 0 ,03
Flores brancas e grãos de pólen longos
24 0 ,03
Total 803 1 ,0
Ao fazer a análise citológica, Bridges concluiu que
• ocromossomoYnãoatuavaparaadeter-minação sexual, mas era importante para a fertilidade do macho, visto que os machos XO eram estéreis.
• os indivíduos especiais eram produzidospor não disjunção do cromossomo X.
Bridges forneceu a prova direta da ligação ao sexo, corroborando definitivamente a teoria cromossômica da hereditariedade.
Havia uma hiper-representação das classes parentais (583+170) contra as não-parentais (26+24). Bateson e Punnett propuseram que a F1 havia produzido mais gametas do tipo pa-rental que o normal e, assim, o acoplamento físico entre os alelos dominantes e recessivos podiam ter evitado a distribuição independen-te, mas eles não sabiam qual a natureza do acoplamento.
O desvio nas proporções ocorria por causa da ligação entre o gene para a cor das flores e o gene para o tamanho do grão de pólen. Umaparte da prole mostrava fenótipo não-parental em baixa frequência, indicando que os alelos dos dois genes se recombinaram na F1.
Qualquer produto meiótico com uma nova combinação de alelos é por definição um re-combinante.
Morgan encontrou um desvio similar na pro-porção de 1:1 entre os tipos parentais e 1:1 entre os tipos não-parentais, estudando dois genes autossômicos em Drosophila. Um dos genes afetava a cor dos olhos (pr=púrpura e pr+ =vermelho), e o outro o tamanho das asas(vg+=vestigial e vg =normal). Morgan reali-zou um cruzamento com diíbridos, seguido de um cruzamento teste (no primeiro capítulo, foi explicada a nomenclatura para genes ligados). Vejamos estes dados.
Figura 7 -Não-disjunção
G
P -XW XW XW+ Y

ca
pít
ulo
3
47P pr/pr - vg/vg x pr+/pr+ . vg+/vg+
púrpura vestigial vermelho normalF1 pr+/pr . vg+/vg vermelho normalCruzamento pr+/pr . vg+/vg x pr/pr . vg/vg teste vermelho normal púrpura vestigial
ResultadosParentais pr+. vg+ 1.339 vermelho normalParentais pr . vg 1.195 púrpura vestigialRecombinantes pr+. vg 151 vermelho vestigialRecombinantes pr . vg+ 154 púrpura normalTotal 2.839
Observe que pr .vg (151) e pr.vg (154) são as classes recombinantes, por estarem em per-centual menor que 50% do total da prole.
Desafio da Fix: vocês devem ter percebido o uso do cruzamento teste para genes autossômicos liga-dos. Por que não há necessidade de cruzamento teste para genes ligados ao X? Discuta com seus colegas.
Agora observemos outro cruzamento realiza-do por Morgan, porém com os alelos em outra associação. Cada genitor é homozigoto para o alelo dominante de um gene e para o alelo recessivo do outro.
P pr+/pr+ - vg/vg x pr/pr . vg+/vg+
F1 pr+/pr . vg+/vgCruzamento pr+/pr . vg+/vg x pr/pr . vg/vg teste
ResultadosRecombinantes pr+. vg+ 157 Recombinantes pr . vg 146Parentais pr+. vg 965Parentais pr . vg+ 1.067Total 2.335
Temos, novamente, um desvio das proporções, constituindo uma exceção ao princípio de Men-del da distribuição independente. Compare os dois cruzamentos e veja que, independente-mente da combinação entre os alelos, as maio-res classes continuam sendo as parentais. O ex-perimento de Morgan mostrou genes ligados em diferentes conformações (fig 8). Na primei-ra, denominada conformação cis ou acoplada, os dois alelos dominantes (selvagens) estão presentes no mesmo homólogo (pr+vg+/prvg). Na segunda, chamada conformação trans ou em repulsão, os alelos se encontram em ho-mólogos diferentes (pr+vg/prvg+).
Figura 8 - Genes ligados nas conformações cis e trans
A B A b
a b a B
Morgan sugeriu que os genes para ambos os fenótipos estão situados no mesmo par de cro-mossomos homólogos. Esta hipótese explica, porque as combinações alélicas dos parentais permanecem juntas. Mas, para explicar o apa-recimento de combinações nãoparentais, Mor-gan propôs que, durante a sinapse, os cromos-somos trocam partes durante o crossing-over (assunto trabalhado anteriormente no capítulo 6 de Citologia). Esse processo, também conhe-cido como permuta, pode ser visualizado cito-logicamente, através dos quiasmas, estrutura em forma de cruz, entre duas cromátides não--irmãs. A Figura 9 ilustra essa troca física de segmentos cromossômicos.
Quiasmas são contados pela análise citológica e cromossomos recombinantes através de aná-lise genética.
pr pr vg+
vg
vg+
pr+
pr+ vg
Figura 9 - Crossing-over

ca
pít
ulo
3
481.7 O CROSSING COMO CAUSA DA RECOMBINAÇÃO GÊNICA
A produção de novas combinações alélicas é conhecida por recombinação. No entanto, en-quanto a recombinação de genes não ligados ocorre por distribuição independente (separa-ção dos cromossomos homólogos durante a anáfase I), a recombinação de genes ligados é devida ao crossing-over (troca de segmentos entre cromátides homólogas na prófase I).
Morgan e seu grupo estudaram mais de mil genes mutantes classificados em quatro gru-pos de ligação, correspondendo de modo no-tável aos quatro cromossomos de Drosophi-la. Entretanto, apenas em 1931, Curt Sterns, estudando alguns cromossomos anormais de Drosophila, demonstrou a associação entre a recombinação genética e a troca de material cromossômico. Os resultados dos experimen-tos de Harriet Creighton e Bárbara McClintock, no cromossomo 9 do milho, foram apresenta-dos no mesmo ano e apoiaram fortemente a troca física entre cromossomos pareados, cau-sando recombinação.
1.8 MENDEL E A LIGAÇÃO
Vimos, anteriormente, que Mendel trabalhou com sete características. Se na ervilha, existem sete pares de cromossomos, seria esperado en-contrar um gene para cada característica, em cada um dos sete cromossomos. Entretanto, Mendel trabalhou com três genes situados no cromossomo 4 (flores axiais e terminais – ale-los T e t, vagem completa ou constringida – alelos C e c, caules altos ou baixos – alelos H e h), dois no cromossomo 1 (sementes amarelasou verdes – alelos V e v, flores brancas ou vio-letas – alelos B e b), um no cromossomo 5 (va-gem verde ou amarela – alelos A e a) e um nocromossomo 7 (sementes lisas ou rugosas – alelos R e r). Então, por que Mendel não des-cobriu a ligação? Uma resposta possível é queas distâncias dos genes V a B, situados no cromossomo 1, e T a C e T a H, situados no cromossomo 4, eram grandes. Todavia, C e H estão relativamente próximos um do outro, todavia Mendel parece não ter realizado esse cruzamento, o que o levaria à quebra da cons-tância da segregação independente. Essa apa-rente sorte em nada diminui o brilhantismo das conclusões de Mendel.
2. MAPEAMENTO GENÉTICOMorgan observou diferentes percentuais na prole recombinante, de acordo com os genes estudados e imaginou que estas variações da frequência de recombinação pudessem indicar as distâncias entre os genes nos cromossomos. Este problema foi resolvido por Alfred H. Stu-tervant, que calculou a posição e a sequência dos genes no cromossomo X de Drosophila melanogaster, construindo o primeiro mapa genético.
Esses mapas são muito úteis, pois permitem compreender o funcionamento gênico, auxiliar no procedimento de isolamento de um gene, prever quais genes poderão vir juntos a uma determinada característica selecionada para estudo, entender mecanismos de doenças, comparar o conteúdo informacional de um cromossomo em espécies diferentes ou corre-latas, auxiliando na compreensão das relações evolutivas.
2.1 CALCULANDO A DISTÂNCIA ENTRE DOIS GENES
Stutervant estimou a distância entre pontos em um cromossomo, contando o número de crossings entre eles. Ele postulou que a distân-cia entre dois pontos no mapa genético de um cromossomo é o número médio de crossings entre eles.
Para facilitar o trabalho, convencionou-se que a distância entre dois locos, acarretando uma frequência de recombinação (FR) de 0,01 (1%) seria dita uma unidade de mapa (u.m.) ou um centimorgan (cM).
Unidade de mapa genético é a distância entre os genes para os quais um produto da meiose em cem é recombinante.
O cálculo da distância entre dois genes é bem simples. Vamos tomar como exemplo o cruza-mento a abaixo.
A/a.B/b X a/a.b/b
Parentais Aa.Bb 440

ca
pít
ulo
3
49Parentais aa.bb 460Recombinantes Aa.bb 48Recombinantes aa.Bb 52Total 1.000
A frequência de recombinantes é dada pela quantidade de recombinantes dividida pelo total da amostra. Neste caso, 48 somado a 52 é igual a 100, dividido pelo total 1.000, ob-temos a frequência de recombinação (FR) de 0,10 u.m ou 10%.
FR = rec/N 100/1.000 = 0,10 u.m.
A maior frequência de recombinação detectá-vel entre dois locos ligados é sempre inferior a 50%. Genes separados por distância igual ou superior a 50 u.m. segregam como genes não--ligados.
2.2 CRUZAMENTO TESTE DE TRÊS PONTOS
Para fazer o mapeamento, é necessário conhe-cer a distância e o posicionamento dos locos no cromossomo. O teste dos três pontos per-mite determinar a distância entre três locos e a ordem seqüencial dos genes. Nesse teste, um triplo heterozigoto é cruzado com um triplo homozigoto recessivo, fornecendo uma des-cendência com números variáveis de crossings e frequências.
Agora vamos analisar os dados:
• Pelasclassesmaisfreqüentes(1e2),defi-nimos a conformação cis.
• As classesmenos freqüentes (7 e 8) nos
mostram que B é o gene do meio, por dife-rir da configuração em cis ABC e abc para, respectivamente, AbC e aBc.
• AdistânciaentreA-Bédadapelasomados
recombinantes simples A-B (3,4) com os duplo-recombinantes (7,8). Desta forma, temos 44 + 37 + 1 + 1 = 83 em um total de 1.000, ou seja, 8,3%, 8,3u.m (unidades de mapa) ou 8,3cM (centimorgan).
• De forma similar, calculamos a distânciaentre B-C. O somatório das classes 5, 6 (re-combinantes simples B-C) com os duplore-combinantes (7 e 8). Nesse caso, obtemos 16 + 19 + 1 + 1 = 37 em um total de 1.000, ou seja, 3,7%, 3,7u.m ou 3,7cM.
O cálculo das distâncias entre cada dois locos é obtido pela soma da frequência de recom-binantes simples para esses dois locos mais a frequência de duplo-recombinantes. Colocando isso em um mapa:
Classe Número de crossings
Frequência
Não-recombinantes (parentais)
0 alta
Recombinantes sim-ples (A-B) e (B-C)
1 intermediária
Duplo-recombinantes 2 menor
Os não-recombinantes evidenciam a confor-mação cis (ABC/abc) ou trans (AbC/aBc; Abc/aBC; ABc/abC). Os duplo-recombinantes per-mitem identificar a ordem correta dos genes (ABC; ACB ou BAC), por indicar o gene do meio.
Vamos fazer uma tentativa passo a passo. Considere o exemplo a seguir.
Em um cruzamento entre um triplo heterozi-goto (ABC/abc) com seu homozigoto recessivo (abc/abc), foram obtidos os seguintes dados.
Classe Genótipo Fenótipo Número
1 Não-recomninante AaBbCc ABC 450
2 Não-recomninante aabbcc abc 432
3 Recombinante simple (A-B)
aaBbCc aBC 44
4 Recombinante simple (A-B)
Aabbcc Abc 37
5 Recombinante simple (B-C)
aabbCc abC 16
6 Recombinante simple (B-C)
AaBbcc ABc 19
7 Duplo-recombi-nante
aaBbcc aBc 1
8 Duplo-recombi-nante
AabbCc AbC 1
Total 1.000
A < 8,3 > B < 3,7 > ou A B C 0 8,3 12

ca
pít
ulo
3
50Vimos um cruzamento na conformação cis, o nível seguinte de complexidade é um cruza-mento na configuração trans.
208/1.000 = 0,208 ou FR de 20,8%). Mas, podemos corrigir o valor, ao somarmos os du-plorecombinantes duas vezes (35 + 30 + 69 + 74 + 1 + 3 + 1 + 3 = 216/1.000 = 0,216), assim obteremos 21,6%.
2.3 FREQUÊNCIA DE RECOMBINAÇÃO E INTENSIDADE DE LIGAÇÃO
A distância entre dois locos é definida pela FR entre eles. Dessa forma, quanto maior a dis-tância entre dois locos, maior a probabilidade de ocorrer um quiasma entre eles.
Como vocês já devem ter percebido, a teoria cromossômica forneceu a base para o desen-volvimento da estrutura hipotética do mapa de ligação, no entanto, entre os primeiros estudos com genes ligados, havia algumas discrepân-cias. Frequências de recombinação menores que 20 a 25% avaliavam diretamente a distân-cia de mapa, entretanto frequências maiores, em geral, subestimavam esta distância. Estas discrepâncias foram prontamente eliminadas por Hermann Muller, outro membro da equipe de Morgan, ao detectar a presença de crossin-gs duplos ao longo do cromossomo.
Além disso, Muller notou que a presença de um quiasma poderia interferir na presença de um segundo crossing na vizinhança, fenôme-no chamado de interferência.
As classes duplo-recombinantes podem ser utilizadas para deduzir a extensão da inter-ferência.
No último exemplo, vimos que a FR de A-C é de 0,147, e o valor de C-B é de 0,069. Des-sa forma, se não ocorre interferência, os du-plorecombinantes podem ser esperados para uma frequência den 0,01014 x 1.000 = 10,14 duplo-recombinantes. Todavia, na amostra, só foi possível observarmos quatro, o que mostra algum tipo de interferência.
A interferência é definida como I. Mas precisa-mos obter inicialmente o coeficiente de coin-cidência (coc), que é a proporção dos recom-binantes duplos observados em relação aos esperados.
Classe Genótipo Fenótipo Número
1 Não-recomninante aaBbCc aBC 418
2 Não-recomninante Aabbcc Abc 370
3 Recombinante simple aabbCc abC 35
4 Recombinante simple AaBbcc ABc 30
5 Recombinante simple aabbcc abc 69
6 Recombinante simple AaBbCc ABC 74
7 Duplo-recombinante aaBbcc aBc 1
8 Duplo-recombinante AabbCc AbC 3
Total 1.000
A< 14,7 >C< 6,9 >B OU A C B 0 14,7 21,6
Agora vamos analisar os dados:
• Pelasclassesmaisfreqüentes(1e2),defi-nimos a conformação trans.
• As classesmenos freqüentes (7 e 8) nosmostram que C é o gene do meio, por dife-rir da configuração trans, de aBC para aBc e de Abc para AbC.
• AdistânciaentreA-Cédadapelasomadosrecombinantes simples A-C (5,6) com os duplo-recombinantes (7,8). Desta forma, temos 69 + 74 + 1 + 3 = 147 em um to-tal de 1.000, ou seja, 14,7%, 14,7u.m (uni-dades de mapa) ou 14,7cM (centimorgan).
• De forma similar, calculamos a distânciaentre C-B. O somatório das classes re-combinantes simples C-B (3 e 4) com os duplorecombinantes (7 e 8). Nesse caso, obtemos 35 + 30 + 1 + 3 = 69 em um total de 1.000, ou seja, 6,9%, 6,9u.m ou 6,9cM.
• Todososlocosestãoligados,poisosvalo-
res de FR são menores que 50%.
Colocando isso em um mapa:
Vamos adicionar mais um aspecto à análise. Observem que as duas distâncias entre A-C (14,7) e C-B (6,9) somam 21,6, maior do que ocalculado para A-B (35 + 30 + 69 + 74 =

ca
pít
ulo
3
51Assim, I = 1 – coc . Logo,
I = 1 - FR observada de recombinantes duplos FR esperada de recombinantes duplos
Em nosso exemplo,
I = 1 - 4 I = 1 - 0 ,39 = 0 ,61% 10,14
A interferência é considerada completa quan-do um crossingover inibe a ocorrência de outro próximo. Nesses casos, coc=0 e I=1, contudo, na maioria das vezes, os valores de I estão en-tre 0 e 1.
Desafio da Fix: Vocês poderão tentar realizar uma prática simples e interessante com peque-nos grupos, intitulada “Entendendo ligação gênica: Uma simulação”. Para isso acessem a página da Revista Genét ica na Escola no si te http://www.sbg.org.br (GeneticaEscola2/Web/vol1Num1.htm).
2.4 O IMPACTO DAS IDÉIAS DE MORGAN E SEU GRUPO
As pesquisas da equipe de Morgan sobre o olho branco da mosca das frutas evidenciaram a linearidade dos cromossomos, posteriormen-te associada à estrutura linear do DNA. Mos-traram, também, que o gene w+ podia mutar para um segundo alelo w, e este, por sua vez num terceiro ou num quarto. Além disso, des-cobriram a reversibilidade dessas mutações. A descoberta das mutações não significou um enfraquecimento da hereditariedade sólida e, sim, uma confirmação do caráter constante do material genético. No exemplo das moscas, só existiam indivíduos de olhos vermelhos, até surgir um novo elemento, um macho de olhosbrancos. A mutação permite a mudança evolu-tiva necessária para a ação da seleção natural.
Vale a pena lembrar que, além da mutação como fonte primordial de variabilidade, a se-gregação e a recombinação gênica produzem novas combinações, fornecendo novos genóti-pos à população.
2.5 MAPEAMENTO NOS DIAS ATUAIS
Mapas genéticos clássicos (mapeamento de li-gação por recombinação) de espécies, como Drosophila melanogaster e Mus musculus (ca-mundongo), são baseados em genes e sofremrefinamento contínuo, sendo obtidos pelos es-forços colaborativos de grupos de pesquisa em todo o mundo. Ao contrário desses, o mapa genético humano não poderia ser baseado em genes, pois a frequência de cruzamentos entre dois indivíduos, sofrendo de diferentes desor-dens genéticas, é extremamente baixa.
A construção dos mapas genéticos humanos esperou pelo advento dos marcadores molecu-lares que apresentassem segregação mende-liana e fossem polimórficos. No ano de 1981, existiam mapas de ligação humanos bastante incompletos e de apenas alguns cromossomos. Para essa época, os marcadores consistiam em proteínas de grupos sangüíneos, proteínas séricas e tipos de HLA tissular, raros e pouco informativos. Apenas em 1987, foi construído o primeiro mapa genético humano global, ba-seado em RFLP (polimorfismo de comprimento de fragmento de restrição). Sete anos depois (1994), mapas de altíssima resolução já haviamsido obtidos, principalmente com a utilização de VNTR (número variável de repetições em tandem) de DNA, na forma de mini ou micros-satélites, e SNPs (polimorfismos de nucleotído único) de DNA. Além dos mapas genéticos (construídos a partir da frequência de recom-binação entre marcadores), foram construídos mapas citológicos (baseados em características citológicas, como o padrão de bandeamento cromossômico observado ao microscópio) e mapas físicos (determinados pelas distâncias de bases que separam os marcadores molecu-lares) do genoma humano, como: de hibridi-zação de células somáticas, citogenético, de sítios de quebra cromossômica, de restrição, com hibridização in situ fluorescente (FISH), de clones contínuos (contigs), de sítios marcados por sequência (STS), de marcadores de sequ-ências expressas (EST), de seqüenciamento de DNA.
Muitas mudanças ocorreram, e os mapas de alta densidade que, atualmente, integram os mapas genético, citológico e físico dos cro-

ca
pít
ulo
3
52mossomos estão sendo construídos, não ape-nas para humanos mas também para vários outros organismos modelo, como Escherichia coli (bactéria), Saccharomyces cerevisiae (leve-dura), Arabdopsis thaliana (planta), permitin-do uma melhor compreensão da evolução dos genomas.
Foge ao nosso objetivo dar descrições detalha-das de outras abordagens para mapeamento, pois você ainda precisa trilhar as disciplinas de Citogenética e Genética Molecular para uma melhor compreensão.
ATIVIDADE
REVISANDO OS CONHECIMENTOS
1. Um experimento de retrocruzamento, en-volvendo um triíbrido, resultou na seguin-te descendência:
nico desse organismo, os genes para cor do corpo (preto ou cinza) e o tipo de asa (longa ou vestigial) estão localizados no cromossomo 2. Suponha que esses genes distam um do outro em 10 unidades de mapa.
Representação dos alelos: b+ - cor cinza vg+ – asa longa b - cor preta vg – asa vestigial
Considerando esses dados, verifique seus conhecimentos sobre genes ligados, anali-sando as proposições e, em seguida, assi-nalando a alternativa correta.
I- Uma fêmea dihíbrida que porta os genes em posição trans, durante a game-togênese, havendo crossing-over, poderá originar dois tipos de gametas recombi-nantes, com as respectivas frequências: b+vg+ 5% e bvg 5%.
II- Uma fêmea de genótipo b+vg/bvg+,
durante a gametogênese, não havendo crossing-over, poderá dar origem a quatro tipos de gametas nas seguintes propor-ções: 25% b+vg+ ; 25% bvg; 25% b+vg e 25% bvg+.
III- A frequência de recombinação entre os
genes b e vg ou b+e vg+ será de 20%. 3. Em certas variedades de plantas, a cor da
flor azul é dominante sobre a cor vermelha e a forma do pólen oblonga é dominante sobre a forma esférica. Os genes para essas características estão ligados e os seus ale-los estão representados a seguir:
V - flor azul E - pólen oblongo v - flor vermelha e - pólen esférico
Suponha que uma dessas plantas, com flor azul e pólen oblongo, foi cruzada com uma planta de flor vermelha e pólen esféri-co. Deste cruzamento obtiveram-se:
• 180plantas com flores azuis e pólen
oblongo; • 190 plantas com flores vermelhas e
pólen esférico;
444 AbC / abc 18 Abc / abc431 aBc / abc 17 aBC / abc42 ABc / abc 2 ABC / abc43 abC / abc 3 abc / abc
1. Com base nesses dados, analise as afirma-tivas, assinalando-as como verdadeiras ou falsas:
( ) Os genes “b” e “c” estão em posição cis no heterozigoto parental.
( ) Os genes “a”, “b” e “c” segregam-se
independentemente. ( ) A distância entre os genes “a” e “b”
pode ser estimada na ordem de 9 unidades de mapa.
( ) Pode-se estimar que esses genes en-
contram-se na sequência b –c –a, no ma-peamento cromossômico do organismo estudado.
2. Você já tem conhecimento de que a Dro-sophila melanogaster tem sido um mode-lo biológico muito utilizado pela genética clássica e molecular para compreensão dos modos de herança. No mapeamento gê-

ca
pít
ulo
3
53 • 50plantascomfloresazuisepólenes-
férico; • 60plantascomfloresvermelhasepó-
len oblongo. Pelos resultados obtidos, é possível deduzir
o genótipo dos parentais e dos descenden-tes. Sendo assim, responda as questões abaixo:
a) Os genótipos dos parentais-planta com
flor azul e pólen oblongo X planta com flor vermelha e pólen esférico podem ser representados por:
( ) VE/ ve e ve / ve, respectivamente. ( ) Ve / vE e VE / VE, respectivamente. ( ) vE / Ve e ve / ve, respectivamente. ( ) Ve / ve e vE / Ve, respectivamente.
b) Dentre os descendentes, o total de plan-tas com genótipos recombinantes é igual a:
( ) 270 ( ) 110 ( ) 240 ( ) 60
REFERÊNCIAS
LEITURA FUNDAMENTAL
GRIFFITHS, A. J. F.; WESSLER, S. R.; LEWONTIN, R. C.;GELBART, W. M.; SUZUKI, D. T.e MILLER, J. H. Introdução à Genética: Rio de Janeiro: Guanabara Koogan S/A, 2006.
SNUSTAD, D. P. e SIMMONS M. J. Fundamen-tos de Genética:Guanabara Koogan S/A, 2001.
SITES
Morgan:http://nobelprize.org/nobel_prizes/medicine/laureates/19 33/morgan-bio.html
Drosophila: http://www.morgan.harvard.edu.
Revista Genética na Escola: http: //www.sbg.org.br (GeneticaEscola2/Web/vol1Num1.htm).


ca
pít
ulo
4
55
GENÉTICA EVOLUTIVA
INTRODUÇÃO
Neste quarto capítulo, vamos compreender a importância da evolução como conteú-do central e unificador dentro da Biologia. Distinguiremos as principais teorias evolu-tivas e enfocaremos processos que influen-ciam as variações genéticas. Faremos uso da análise do princípio de Hardy-Weinberg. Abordaremos a visão atual da seleção na-tural e estenderemos a discussão sobre as restrições ao processo evolutivo. Por fim, trataremos de aspectos da especiação e da macroevolução.
OBJETIVOS ESPECÍFICOS
• Desenvolver as bases para a compre-ensão do pensamento evolutivo e po-pulacional.
GENÉTICA EVOLUTIVA
“Nada em biologia faz sentido exceto à luz da evolução.”(Theodosius Dobzhansky)
Profa. Marília de França Rocha Carga Horária I 15hColaboradoras - Profa. Maria Teresa Marquim N. Cornélio Profa. Rita de Cássia de Moura

ca
pít
ulo
4
56
1. O DESENVOLVIMENTO DO PENSAMENTO EVOLUTIVO
1.1 QUEM FOI DARWIN
Charles Darwin (fig 1) nasceu em 12 de fe-vereiro de 1809, em Shrewsbury, Inglaterra. Darwin obteve seu bacharelado em humani-dades em 1831, e, nesse mesmo ano, embar-cou no H.M.S. Beagle como companheiro de viagem do capitão Robert Fitzroy, tornando-se em pouco tempo o naturalista oficial do na-vio. A viagem durou cinco anos (27/12/1831 a 02/10/1836) e foi o acontecimento mais importante de sua vida, pois oportunizou ob-servações e indagações sobre aspectos geoló-gicos, da vida animal e vegetal, de situações políticas e sociais. Em 1858, no encontro da Sociedade Linneana de Londres, Darwin apre-sentou sua teoria da evolução por meio da se-leção natural, conjuntamente a um manuscrito de Alfred Russel Wallace (1823- 1913) (fig 2) de conteúdo análogo. Darwin publicou, no ano de 1859, A origem das espécies por meio da seleção natural, livro que revolucionou o pensamento ocidental. Publicou inúmeros li-vros e morreu em Down House, no dia 19 de abril de 1882.
Para saber mais, leiam a bibliografia indicada sobre Darwin.
Figura 2 - Alfred Russel WallaceFonte: http://www.ucl.ac.uk/taxome/jim/Mim/Wallace.JPG
Figura 1 - Charles Darwin fonte: http://www.victorianweb.org/science/ da-rwin/darwin_beard.gif
1.2 A VISÃO DE MUNDO NA ÉPOCA DE DARWIN
As visões de mundo comuns na época de Char-les Darwin eram:
(1) um mundo constante e recentemente cria-do (dogma cristão, vitalismo);
(2) um mundo de duração longa ou eterna,
tendendo a melhoramento ou perfeição (visão teleológica);
(3) um mundo eterno, constante ou cíclico,
sem exibir uma direção ou meta, simples-mente devido ao acaso ou à necessidade, como dito por Demócrito e seus seguido-res.
O vitalismo (1) e a teleologia cósmica (2) são duas crenças, atualmente, refutadas. O vitalis-mo era a crença na existência de uma força oculta (mística), responsável pelas manifesta-ções de vida nos organismos. A teleologia li-dava com a explicação de processos naturais que conduziam automaticamente a um fim definido ou a uma meta. Nela estão inclusos os ortogenistas (por acreditarem que havia na natureza viva um anseio intrínseco no sentido da perfeição) e a teoria da evolução de Lamar-ck (segundo a qual formas complexas de vida

ca
pít
ulo
4
57surgiam a partir de múltiplas formas primiti-vas, oriundas de geração espontânea, e carac-teres adquiridos pelo uso seriam transmitidos aos descendentes).
1.3 AS TEORIAS DE DARWIN
O evolucionista e ornitólogo Ernest Mayr con-siderou que o paradigma de Darwin engloba, pelo menos, cinco teorias independentes: evo-lução propriamente dita, ancestralidade co-mum, gradualismo, variação das espécies e se-leção natural. Essas diferentes teorias mostram diferentes aspectos do processo evolutivo.
Na evolução propriamente dita, o mundo e os organismos estão sempre em modificação ao longo do tempo. O conceito de evolução não era novidade e autores como Buffon (Georges Louis Leclerc), Lamarck (Jean-Baptiste Pierre Antoine de Monet) e o próprio avô de Darwin, Erasmus Darwin, já haviam abandonado a visão de espécies fixas e imutáveis muito antes; o pro-blema era explicar o mecanismo de evolução.
Os quatro ramos da Biologia que serviram de base para Darwin, biogeografia, paleontolo-gia, embriologia e morfologia, acumulam, atu-almente, um conjunto imenso de dados, en-dossando a evolução. Além desses, dispomos de dados em outras áreas, como genética, bio-química , biologia molecular. Nos dias atuais, não há dúvida de que os seres vivos tenham evoluído. Esse não é um ponto de debate na comunidade científica, apesar do que pregam os criacionistas.
Quando Darwin embarcou no Beagle, acredi-tava na estabilidade das espécies (essencialis-mo – crença em que a diversidade consistia em um número limitado de essência, delimitadas eimutáveis). Porém a sua visita a Galápagos abalou sua certeza. Nessas ilhas, ele pôde ob-servar tentilhões ou pássaros similares a esses e suspeitou serem variedades (Mayr conside-rava que os (Geospiza) tentilhões na realidade eram (Mimus) tordos-dosremédios). Ao chegar à Inglaterra, Darwin enviou suas coleções para as mãos de vários especialistas. O ornitologista John Gould demonstrou que não existiam va-riedades e, sim, espécies distintas dessas aves. Darwin compreendeu que todas derivavam de um ancestral comum, o que sugeria a imagem
de uma árvore da vida conectando os ances-trais e as espécies atuais (fig 3). Essa teoria de ancestralidade comum obteve aceitação ime-diata. Subitamente, toda a hierarquia de clas-sificação de Lineu tornavase lógica.
Figura 3 -Árvore da vidaFonte: http://www.amnh.org/exhibitions/darwin/images/ e xhi-bit/gallery/lg/darwin_tree_lg.jpg
Com a descoberta da similaridade entre o có-digo genético para procariotos e eucariotos, atualmente, passou-se a ter um consenso de que todos os organismos existentes no planetaTerra possuem uma origem única.
Na terceira teoria, gradualismo, a transforma-ção evolutiva ocorria gradativamente, nunca aos saltos (saltacionismo - originada por mu-tações drásticas). Uma nova espécie evoluiria lentamente, através de espécies preexistentes. Essa teoria encontrou forte oposição, inclusive de amigos que o defendiam, como T. H. Hux-ley, conhecido como o buldogue de Darwin. O gradualismo abria espaço para críticas, como a falta de intermediários que explicassem a tran-sição entre as espécies. Darwin alegou que es-sas lacunas se deviam a falhas do registro fóssil.

ca
pít
ulo
4
58A variação das espécies tratava da problemá-tica da origem da diversidade orgânica. Da-rwin introduz o pensamento populacional, propondo que a variação existente dentro de uma espécie é devida às diferenças entre os indivíduos, ou seja, não são os indivíduos que mudam ao longo do processo evolutivo, e sim, as populações. Os tentilhões (tordos-dos--remédios) ilustravam bem os aspectos da ori-gem da diversidade e a origem da adaptação, porém, mesmo assim, em seu livro, é notória a sua confusão acerca de espécies e especiação. Lembrem que Mendel também possuía esse mesmo problema conceitual. Em biologia, in-clusive no presente, um dos conceitos mais di-fíceis de ser trabalhado é o de espécie.
O livro Ensaio sobre o princípio da população de Thomas Malthus foi crucial para o desenvol-vimento da teoria da seleção natural. Segundo a tese do crescimento exponencial de Malthus: “as populações crescem mais rapidamente do que os recursos dos quais dependem”. De tal modo, uma parcela da variação nas popula-ções ocorre em caracteres que afetam as chan-ces de sobrevivência e reprodução dos orga-nismos. Darwin e Wallace compreenderam esse processo de sobrevivência e reprodução desiguais, independentemente, e uniram uma série de observações e idéias em uma teoria ousada e revolucionária, a teoria da seleção natural.
Dentre as cinco teorias, a da seleção natural foi a mais difícil de ser aceita. Ela representava a rejeição de qualquer pensamento teleológico e determinista (determinismo é a crença de que a maioria dos processos na natureza inanima-da é estritamente determinada por leis naturais – físicas) no mun do orgânico. Além disso, dois problemas se apresentavam: a ausência de um mecanismo convincente de herança e a apa-rente falta de direção do processo evolutivo.
A natureza da herança e a origem da variação nunca ficaram claras para Darwin, apesar de Mendel ter sido seu contemporâneo. Darwin propôs a teoria da pangênese, na qual todas as partes do corpo fornecem material gené-tico para os órgãos reprodutores e, particu-larmente, para os gametas, na forma de gê-mulas. Essa explicação para a hereditariedade mostrou-se totalmente incorreta. A sua crença
parcial na herança por mistura, suas conces-sões ao ambiente como fonte de indução de variação e especulações sobre caracteres ad-quiridos herdáveis tornavam impossível a reso-lução do problema.
1.4 O IMPACTO DAS IDÉIAS DE DARWIN
Em todas as eras, descobertas científicas modi-ficam as concepções das visões de mundo de uma dada época. Lembrem-se de Galileu que refutou o antropocentrismo e serviu de ala-vanca para a mecânica cartesiana e de Newton fornecendo o embasamento para a física mo-derna. A revolução darwiniana abalou as mais arraigadas concepções que possuíamos sobre nós mesmos e sobre a nossa origem. A visão de mundo do homem ocidental jamais seria amesma depois da publicação de A origem das espécies. Darwin refutou a crença na criação individual de cada espécie, ao estabelecer o conceito da ancestralidade comum, através da qual todos os seres vivos descendem de um an-cestral comum. Por extensão, humanos evolui-riam, segundo o mesmo princípio. As noções correntes de um mundo perfeito, constante e em progresso foram profundamente abaladas pela demonstração de que a evolução causa mudança e adaptação, regida de modo contí-nuo pela seleção natural. Ele utilizou um con-ceito inteiramente novo, atualmente chamado de pensamento populacional no lugar do pen-samento tipológico (essencialismo).
A maneira essencialista de pensar é incapaz de comodar a variação, o que originou uma concepção errônea de raças humanas, con-duzindo ao racismo. Os conhecimentos da genética evolutiva e de populações podem ser utilizados para demonstrar, de forma ra-cional, que, em relação à espécie humana, não existem as “raças puras” e, portanto, qualquer filosofia política baseada neste fal-so conceito é insustentável. Existem vários artigos, tratando do tema raça. Vocês podem ler o artigo “Ambigüidades que limitam uma definição de raça” (Scientific American Brasil edição nº 20 – jan 2004) eparaisso,acessem: http://www2.uol.com.br/sciam/conteudo/materia/materia_38.htm.

ca
pít
ulo
4
591.5 TEORIAS EVOLUTIVAS E A GENÉTICA MENDELIANA
A compreensão dos caracteres particulados de Mendel apontava para uma herança sólida, tendo auxiliado na refutação das teorias evo-lutivas que rivalizavam com a seleção natural. Estas teorias eram o neolamarkismo (herança de caracteres adquiridos) e outras formas de herança tênue; teorias autogenéticas baseadas no progresso evolutivo com um fim determi-nado (ortogênese, aristogênese, e outras) e as teorias da evolução saltacional, com rápida emergência de formas de vida (mutacionismo - De Vries).
Não cabe comentar cada teoria; sobre isso, existe uma extensa bibliografia indicada ao fi-nal do capítulo.
Apesar da redescoberta das leis de Mendel em 1900, a aceitação da seleção natural ainda le-varia bastante tempo para se concretizar.
1.6 A SÍNTESE EVOLUTIVA
As idéias de Darwin estavam relegadas a um segundo plano, por volta do final do século XIX. Apenas na década de 1920, a divergência entre geneticistas e naturalistas começou a di-minuir, impulsionando a construção da teoria evolutiva mais influente do século XX, a teoria sintética da evolução.
Na década de 1930, ocorreu a síntese fishe-riana. Os geneticistas populacionais Ronald Aylmer Fisher, John B. S. Haldane (Inglaterra) e Sewall Wright (EUA) mostraram que genes com pequenas vantagens seletivas podiam ser incorporados ao genótipo de populações. Assim, nascia a Genética de Populações. A evolução era definida como a mudança nas frequências gênicas de uma dada população, ocasionada pela seleção lenta de pequenas mutações. Em conjunto, demonstraram que a variação poderia ser explicada pela herança mendeliana e pela seleção natural. Naquela época, lidava-se com a origem das adaptações, entretanto o problema da origem da biodiver-sidade continuava.
O livro de Theodosius Dobzhansky, Genética e a origem das espécies (1937), conseguiu unir
as duas áreas de estudo em biologia evolutiva a evolução filética em populações (anagênese) e a origem da biodiversidade (cladogênese), le-vando ao início da construção da teoria sinté-tica da evolução ao longo da década de 1940.
A síntese evolutiva, conhecida como a segun-da revolução darwiniana, tratou basicamente da origem e do significado da biodiversidade, promovendo um consenso entre naturalistas egeneticistas. Os naturalistas aprenderam com os geneticistas que os caracteres são particula-dos, a herança é sempre sólida e não há influên-cia herdável do ambiente. Os geneticistas, por sua vez, entraram em contato com uma vasta literatura sobre variação geográfica e especia-ção. Havia, ainda, outro ponto de discórdia: o tipo de variação (gradual ou saltacional), atual-mente resolvido através da teoria do equilíbrio pontuado, que apóia a existência de períodos de poucas mudanças (equilíbrio), interrompi-dos (pontuados) por fases de diversificação.
Restou, apenas, um ponto de desacordo entre naturalistas e geneticistas, o objeto de seleção. Atualmente, uma nova visão está sendo cons-truída e concebe-se que a seleção natural pode atuar em diversos níveis de organização bio-lógica: genes, linhagens celulares, organismos individuais, grupos de organismos, populações e talvez, até mesmo espécies. Não é questio-nada a validade da seleção natural, mas os ní-veis de organização em que esta pode atuar e como ocorrem as inter-relações. Os construto-res dessa síntese foram: Bernhard Rensch, Er-nest Mayr, G. G. Simpson, G. L. Stebbins, J. B. S. Haldane, Julian Huxley, R. A. Fisher, Sergei Chetverikov, Sewall Wright, Theodosius Do-bzhansky, dentre outros.
2. O PRINCÍPIO DE HARDY-WEINBERG
Em 1908, G. H. Hardy (matemático britânico) e Wilhelm Weinberg (médico alemão) publi-caram, independentemente, trabalhos descre-vendo uma relação matemática capaz de pre-ver frequências genotípicas de uma população através de suas frequências alélicas (gênicas). Essa relação é conhecida como princípio ou equilíbrio de Hardy-Weinberg (H-W).

ca
pít
ulo
4
60Suponhamos que um determinado gene está segregando dois alelos, A e a, sendo a frequ-ência de A=p e a de a=q. Em termos de meio-se, podemos considerar que a probabilidade de um gameta (óvulo ou espermatozóide) le-var o alelo A ou a é p ou q, respectivamente. Então, a probabilidade de produzir um indiví-duo homozigoto dominante (AA) é p x p = p2 e um homozigoto recessivo (aa) é q x q = q2 . Para os heterozigotos, existem duas possibili-dades A x a ou a x A, assim a probabilidade de formar um zigoto Aa é 2pq. Essas frequências previstas são os termos na 2 expressão do bi-nômio (p + q)2.
(p + q)2 = p2 + 2pq + q2 =1
Em uma população em equilíbrio, as frequ-ências dos genes se mantêm constantes de geração em geração. Cruzamentos ao acaso e população numerosa contribuem para a es-tabilidade da população. Entretanto ocorrem fatores que podem alterar o equilíbrio gené-tico da população, como: reproduções con-sangüíneas ou preferenciais, sobrevida e fer-tilidade desigual (mutação e seleção natural), isolamento de populações e migração. Uma propriedade da segregação mendeliana é o estabelecimento de um novo equilíbrio de ge-nótipos na população, após uma geração de reprodução aleatória.
2.1 CALCULANDO AS FREQUÊNCIAS ALÉLICAS (GÊNICAS) E GENOTÍPICAS
Ao trabalharmos com populações, analisamos uma amostra representativa de indivíduos. Imaginem uma população humana na qual ocorrem os alelos do grupamento sangüíneo MN. Os genótipos possíveis de serem encon-trados em relação a esse loco são: MM, MN e NN. Como não ocorre dominância, há uma perfeita correspondência entre o número de indivíduos das classes genotípicas e fenotípicas correspondentes. Vamos seguir, passo a passo, o cálculo dessas frequências para uma popula-ção imaginária.
Passo 1 – Trabalhamos com números relati-vos e não absolutos. Iremos dividir cada classe pelo N (número total da amostra). Obtemos, dessa maneira, as frequências genotípicas ob-servadas para essa população (Veja a quarta coluna).
Passo 2 – Para calcular as frequências alélicas, somamos a frequência de homozigotos com a metade dos heterozigotos.
Assim, pA = D + ½ H e qa = R + ½ H. M = p = MM + ½ MN = 0,26 + 0,29 = 0,55 e N = q = NN + ½ NN= 0,16 + 0,29 = 0,45
Passo 3 – Podemos estimar agora as frequên-cias genotípicas esperadas.
Aplicando a fórmula binomial (p + q)2 , onde p= 0,55 e q = 0,45. Então, p2 + 2pq + q2 = (0,55)2 + 2 x (0,55) x (0,45) + (0,45)2 = 0,30 + 0,50 + 0,20 = 1
Desafio da Fix: Quando resul-tados observa-dos e esperados são compara-dos, verifica--se a existência de diferenças maiores ou menores entre eles. Aplique o teste do qui--quadrado, para verificar se a população se en-contra ou não em equilíbrio de Hardy- Wein-berg. Lembrem que não utilizamos números relativos nesse teste e, sim, absolutos. Então, é só multiplicar as frequências pelo N (77) da população. Preencham a tabela da Fix e en-viem o resultado.
Fenótipo Genótipo Número de indivíduos
Frequência geno-típica observada
MM LMLM 20 20 / 77 = 0,26
MN LMLN 45 47 / 77 = 0,58
NN LNLN 12 12 / 77 = 0,16
Total 77 1,00

ca
pít
ulo
4
61
2.2 CALCULANDO AS FREQUÊNCIAS, QUANDO NÃO É POSSÍVEL DISCRIMINAR OS HETEROZIGOTOS
Não é admissível usar o procedimento expli-cado anteriormente, quando em um loco um alelo domina outro (A > a) e não é possível discriminar os heterozigotos (Aa) dos homo-zigotos (AA). Vale a pena salientar que este método é menos preciso que o anterior, e só deverá ser empregado, quando não for possí-vel identificar a frequência dos três genótipos.
Suponha que temos conhecimento das fre-quências relacionadas aos fenótipos, mas não, dos genótipos. Por exemplo, A_=0,84 e aa=0,16. Como não sabemos a quantidade exata de AA e Aa, tiramos a raiz quadrada dos homozigotos recessivos, visto que aa=q2 , en-tão q=√aa.Seq2 =0,16 então q=0,4. Sendo p + q = 1, obtemos p através da fórmula p= 1 - q, onde p= 1 - 0,4, p=0,6. De posse das frequências alélicas, podemos calcular as frequências genotípicas esperadas.
Genótipos Frequências
AA p2 (0,6)2= 0,36
Aa 2pq 2 x (0,6) x (0,4) = 0,48
aa q2 (0,4)2= 0,16
Total 1 1,00
O princípio de Hardy-Weinberg também se aplica aos genes ligados ao X e aos genes com alelos múltiplos. Fogem ao escopo des-se capítulo explicações detalhadas sobre es-ses casos. Busquem exemplos na literatura indicada. Vocês também podem obter na página da Sociedade Brasileira de Genética (SBG) – http://www.sbg.org.br, o E-book do prof. Bernardo Beiguelman, intitulado: “Ge-nética de Populações Humanas”. O livro tra-ta de fatores evolutivos e do problema da consanguinidade.
No programa GBOL, disponívelnosite http://www.ufv.br/dgb/gbol, vocês podem ler e aplicar seus conhecimentos em genéti-ca de populações. Existem diversos aplicati-vos que permitem simular a descendência de populações com diferentes constituições ge-notípicas admitindo diferentesacasalamentos, além de uma série de exemplos, para avaliar oequilíbrio de Hardy-Weinberg, incluindo o uso do qui-quadrado.
3. PROCESSOS QUE INFLUENCIAM AS VARIAÇõES GENÉTICAS
As várias forças de estrutura de cruzamen-to, mutação, recombinação, seleção, deriva e fluxo gênico (migração) agem simultane-amente e moldam a composição genética das populações, produzindo variação intra e interpopulacional. A figura 4 apresenta o panorama dos processos que levam à evolu-ção e à interação entre eles na formação de novas espécies.
A variação herdada pode ser estudada pelo confronto de diferenças macroscópicas ( mor-fologia) o u microscópicas, desde a compa-ração de cariótipos (cromossomos) até dife-renças entre sequências d e aminoácidos em proteínas ou de nucleotídeos no DNA.
Tiposangüíneo
Frequência genotípica observada
Frequênciagenotípicaesperada
MM 0,26 0,30
MN 0,58 0,50
NN 0,16 0,20
Total 1,00 1,00
Tabela da Fix: A população encontra-se ou não em equilíbrio de H-W?Qui-quadradocalculado X2
c
Qui-quadradotabelado X2
t
Significância(0,05)
Populaçãoem equilíbrio
X2 c menor X2
t ( ) Sim
X2 c maior X2
t ( ) Não
ca
pít
ulo
4
62
População original
Variação devida a mutação e recombinação
Separação geográfica
Efeito do fundador Efeito do fundador
Seleção no ambiente local e deriva genética
Migração
Divergência (recombinação - seleção - deriva)
Nova espécie 1 Nova espécie 2
/ / / /
/ / / / /
/ / /
/ / /
/ /
/ /
/ / / /
/ / / /
/ / / / /
Figura 4 - Processos evolutivos e sua intereção na formação de novas espécies (Adaotados do Griffiths et al, 2006)
3.1 ESTRUTURA DE CRUZAMENTO
A transmissão da informação genética ocorre de diversas formas, dependendo do tipo de sistema genético: haplóide sexuado (levedu-ras como Neurospora crassa), diplóide sexua-do (maioria das plantas e animais); haplóide assexuado (bactérias) e diplóide assexuado (dinoflagelados, rotíferos). Em assexuados, o genoma é transmitido como uma unidade, enquanto nos sexuados, os descendentes são uma mistura do grupo de genes parentais.
3.2 MUTAÇÃO E RECOMBINAÇÃO
A produção da variação tem início com uma mutação, fonte primordial da mudança. Grande parte da formação de novos genes ocorre por duplicação de genes isolados pré--existentes ou por duplicação total do geno-ma (poliploidia), ampliando a quantidade de DNA disponível, sem o comprometimento de funções préexistentes. Entretanto, após o esta-belecimento de um novo alelo, é a recombina-ção dos genes parentais que irá gerar a grande quantidade de combinações genéticas em no-vos genótipos, que formarão os indivíduos a serem selecionados na próxima geração.

ca
pít
ulo
4
633.3 SELEÇÃO
Quando Darwin usou a palavra seleção, usa-va-a no sentido de “escolha do melhor”, e o ambiente, como a força externa de seleção. Atualmente, a seleção é vista como a elimina-ção dos menos aptos a sobreviverem e numa parcela menor a seleção dos mais aptos. Des-sa forma, sobra uma quantidade enorme de fenótipos variados que enfrentam o ambiente com ou sem sucesso. Assim, as características selecionadas são as mais favoráveis dentro de um elenco de variações disponíveis de uma po-pulação em um dado momento.
Vale a pena lembrar que o ambiente está em processo contínuo de mudança, em consequ-ência das ações dos organismos. O surgimento da clorofila causou profunda modificação na Terra. Algas e plantas lançaram grandes quan-tidades de oxigênio na atmosfera, e surgiu a camada de ozônio. Alguns processos compor-tamentais causados por nossa espécie trazem modificações ao planeta: o uso indiscriminado de antibióticos e venenos causam resistência a bactérias e insetos, respectivamente; o uso inadequado das fontes de energia, desmata-mento e queimadas provocam o efeito estu-fa; a destruição de habitats e o contato com organismos exóticos podem levar à produção de novos patógenos (vírus do HIV - AIDS). Um exemplo clássico é o das mariposas salpicadas ou sarapintadas (Biston betularia) (fig 5), onde, em ambiente livre de poluição, a forma clara constitui a maioria, e, em ambientes poluídos, a forma escura se sobressai.
Figura 5 - Biston betulariaFonte: http://sti.br.inter.net/rafaas/biologia-ar/images/acbi0307.jpg
Reflexão da Fix: A metamorfo-se contínua da Terra resulta da interação dos inúmeros seres existentes em seus ambientes. Se morressem todos os hu-manos, a vida prosseguiria? E se as plantas morressem?
3.3.1 TIPOS DE SELEÇÃO
1- Natural e artificial - o sucesso reprodutivo direciona a seleção natural, de forma tal que a seleção artificial pode ser limitada pela seleção natural. Um bom exemplo é a seleção artificial para tamanho de corpo no besouro da fari-nha, Tribolium castaneum. Nesse experimen-to, Franklin Enfield selecionava as pupas mais pesadas para serem os genitores das próximas gerações, obtendo besouros cada vez maiores, mas, em um dado momento, a seleção natural limitou o aumento corpóreo através da infer-tilidade (leiam o enfoque técnico das páginas 92 e 93 do livro Fundamentos de Genética do Snustad e Simmons, 2001).
A seleção sexual é um componente especial da seleção natural. Esse tipo de seleção consiste em dois processos diferentes. (1) A competi-ção intra-sexual por parceiros, que irá se refle-tir no dimorfismo sexual (machos maiores que fêmeas em mamíferos, em especial, nos ele-fantes marinhos) e no desenvolvimento de ar-mas (como chifres, garras, dentes de combate e habilidade de enganar competidores, como em determinados peixes que imitam fêmeas e nadam junto a essas, sem serem banidos pelos machos maiores). (2) A escolha intersexual de parceiros, traduzida por mecanismos indica-dores (a produção de ornamentos é custosa, e apenas machos com boa resistência a pató-genos são capazes de mantê-los), processo de Fisher (estabilidade de 50:50 no balanço en-tre os sexos, macho e fêmea) e predisposições sensoriais das fêmeas (preferência por deter-minadas cores, cantos).
Acessem e leiam o texto do Futuyma “Evolução, ciência e sociedade“ na página da Sociedade Bra-sileira de Genética (SBG) – http://www.sbg.org.br. O encarte trata de biologia evolutiva.

ca
pít
ulo
4
64Desafio da Fix: Imagine-se em uma excursão no pantanal. Um queixada (porco selva-gem) corre na direção de três pessoas: al-guém do seu núcleo familiar (pai, mãe, ir-mãos), um amigo e um desconhecido. Você tem a possibilidade de resgatar um deles. Quais seriam suas reações? Discuta com seus colegas.
3.4 DERIVA GENÉTICA
A mudança ao acaso na constituição genética de uma população é chamada deriva genética. Ela é maior em populações pequenas do que em grandes, por ser mais provável que uma amostragem pequena apresente maio-res desvios do que uma composição total. No-vas populações podem ser formadas por um pequeno grupo de indivíduos originários de uma população maior. Esse grupo difere das frequências genéticas da população parental, devido ao acaso. Assim, alguns alelos podem estar presentes em frequências altas ou baixas em relação à população inicial. Isso é conheci-do como efeito do fundador. De forma similar, se uma catástrofe (tsunami, enchente, fura-cão, terremoto, erupção vulcânica) eliminassegrande parte de uma população, poucos in-divíduos sobreviveriam para se reproduzir, e a composição genética mudaria, como se pas-sasse por um bottleneck ou gargalo genético, havendo perda de genes e aumento da frequ-ência de outros.
Desafio da Fix: Vo-cês podem tentar realizar a práti-ca sobre deriva. Para isto acessem: www.sbg.org.br/GeneticaEscola2/web/vol1Num2.htm. Maneira lúdi-ca de se entender deriva alélica.
2- Seleção estabilizadora, direcional e disrup-tiva - a Figura 6 representa os três tipos de se-leção em relação ao favorecimento de caracte-rísticas ou alelos. (1) A seleção estabilizadora conserva os valores intermediários originados pelos heterozigotos Aa (peso de bebês, tama-nho de ninhadas), (2) a direcional beneficia valores de uma característica em detrimento da outra, representada pelos alelos A ou a (aumento do tamanho corpóreo de cavalos e do cérebro humano) e (3) a disruptiva elimina heterozigotos e restaura homozigotos AA e aa (gametas de tamanhos diferentes – anisoga-mia), favorecendo valores extremos.
Figura 6 - Tipos de seleção Fonte: Escaneado de Snustad e Simmons(2001).
Freq
uênc
ia
Genitores
Freq
uênc
iaFr
equê
ncia
(a) Seleçãodirecional
Genitores
GenitoresMédia
(b) Seleçãodisruptiva
(c) Seleçãoestabilizadora
3- Dependente da densidade e dependente da frequência - como os próprios nomes indicam, a primeira se refere à densidade populacional na qual a seleção atua. Maiores densidades trazem menores recursos. O exemplo mais co-mum de seleção dependente da frequência é o processo de Fisher. Assim, se há mais machos do que fêmeas, é melhor para os pais produ-zirem mais fêmeas e viceversa. A população tende a um equilíbrio de 50% para cada sexo.
4- Individual, parentesco e de grupo - as ca-racterísticas evoluem, aumentando o sucesso reprodutivo de indivíduos e grupos familiares. A seleção de grupo raramente é efetiva, por ser mais numerosa e apresentar maior variabi-lidade genética.

ca
pít
ulo
4
65No final da década 1960, descobriu-se que havia um grau considerável de variação ge-nética nas espécies, sugerindo a existência de um estoque de variantes genéticas nas popu-lações. O geneticista Motoo Kimura propôs a teoria neutra da evolução molecular, na qual a maior parte das variantes genéticas, cria-da incessantemente pelas mutações, persiste ou não numa população por deriva (acaso) e não, por efeito da seleção natural. Um argu-mento a favor dessa teoria é a taxa relativa-mente constante de substituições de aminoá-cidos em todas as linhagens de descendência de um ancestral comum. Cada proteína pos-suiria seu próprio relógio molecular, o qual marcaria o passo das modificações evolutivas. Entretanto, o relógio não é constante o su-ficiente, ocorre influência do tempo de ge-rações na evolução sinônima, e por fim, há discordância entre níveis de variação genética e taxas evolutivas. Ohta desenvolveu uma ver-são modificada da teoria neutra (teoria apro-ximadamente neutra), de acordo com a qual a maior parte da evolução molecular ocorre à medida que uma mutação quase neutra é substituída por outra.
Apesar de o papel do acaso (processo es-tocástico) ser tema antigo de discussão em evolução, somente nos dias atuais lhe é atri-buído o devido valor. O acaso tem função tanto na geração de novos indivíduos, atra-vés da mutação e recombinação, como tam-bém durante o processo probabilístico de determinação do sucesso reprodutivo desses indivíduos (seleção) e fixação de um alelo por deriva.
O perturbador livro do Monod, O acaso e a necessidade, apresenta a origem e a evolução da vida como um “acidente” dentro do grandeuniverso regido pelas leis físicas.
3.5- FLUXO GÊNICO
Em contraste com a deriva genética, que oca-siona uma variação aleatória nas frequências alélicas de populações pequenas, o fluxo gê-nico é definido como a lenta difusão de genes através de uma barreira. Processo que envolve uma grande população e uma mudança gra-dativa nas frequências gênicas. Dessa manei-ra, os genes das populações migrantes e suas
frequências características vão gradualmente se dispersando no pool gênico da população para a qual migraram.
4. LIMITES E NOVIDADES EVOLUTIVAS
4.1 LIMITES
A variação existente em uma população é mol-dada pela sua história evolutiva, pelo desen-volvimento e pela fisiologia dos organismos. Limites impostos pela química e física não po-dem ser desprezados. Organismos nos quais estão ausentes sistemas respiratório e circu-latório não podem ultrapassar o tamanho de 1mm, limite imposto pela taxa de difusão de oxigênio na água. Animais pesados necessitam de membros mais grossos para a sustentação do peso. Nas brânquias de seres aquáticos cir-culam grandes volumes de água, causando enorme perda de calor, o que inviabilizaria a homeotermia.
A seleção natural pode modificar, apenas, a variação existente em dado genoma. Não há formação de organismos ideais e, sim, mais adaptados para aquele momento (tempo) e ambiente. Características vestigiais constituem evidência morfológica esclarecedora, mos-trando seres repletos de pequenas e toleráveis imperfeições, como: mamilos em machos de mamíferos, serpentes com rudimentos de pel-ve e minúsculas pernas no interior do corpo, besouros dotados de asas sob suas carapaças e incapazes de voar.
Espécies aparentemente diferentes têm simi-laridades subjacentes, devido ao compartilha-mento de atributos de seu ancestral comum. Um bom exemplo disso pode ser lido no artigo “Uma nadadeira é um braço é uma asa” da revista National Geographic de novembro de 2006. Esse artigo poderá ser acessado no site:http://www.nationalgeographic.abril.com.br/ngbonline/edicoes/0611/wings.shtml.
Assim, a evolução de uma espécie é explicada através da história de vida, que é influencia-da pela composição genotípica e conectada às trocas de caracteres ou traços. As pressões

ca
pít
ulo
4
66seletivas atuam nessas variações, dependendo da idade, do tamanho do organismo, da plas-ticidade fenotípica, da capacidade reprodutiva e do tamanho da prole. Dessa forma, o contex-to filogenético, no qual se apresentam os ca-racteres de uma espécie, representa os efeitos históricos de sua construção. A figura 7 repre-senta a forma através da qual um organismo é desenhado.
desenvolvimento envolvem mecanismos com-plexos pelos quais um organismo cresce, até atingir a plenitude de sua forma e tamanho.
5. EVOLUÇÃO E DIVERSIDADE
5.1 O CONCEITO DE ESPÉCIE
Como vimos anteriormente, um dos concei-tos mais polêmicos dentro da biologia é o conceito de espécie. Vários autores, desde os tempos de Platão e Aristóteles até Linneu e início do século XIX, identificaram espécies com base, apenas, em suas diferenças mor-fológicas. Trata-se do conceito tipológico de espécie (CTE). Entretanto, em anos recentes, descobriu-se um número considerável de es-pécies animais que não eram distinguidas pela morfologia, mas, por suas característi-cas não-morfológicas, como por exemplo, o comportamento. Além disso, algumas carac-terísticas intra-específicas podem apresentar notável modificação por sexo, idade, estação do ano ou variação genética comum. Espécies crípticas ou irmãs, por exemplo, podem in-validar um conceito de espécie apoiada, ape-nas, no grau de diferença.
Não existe um conceito que atenda a todas as espécies, pois o sistema de reprodução assexuado ou sexuado é uma barreira para a formação de um conceito geral. O conceito que ancora o reconhecimento de espécies na reprodução é conhecido por conceito bioló-gico de espécies (CBE). A construção desse conceito tem uma longa história que iniciou com Buffon e prosseguiu com Jordan, Poul-ton, Stresemann, Rensch e Dobzhansky. Cou-be a Mayr uma definição concisa e utilizada em larga escala em trabalhos que tratam do CBE: “espécies biológicas são grupos de po-pulações naturais intercruzantes e reproduti-vamente (geneticamente) isoladas de outros grupos similares”. Dessa maneira, o status de espécie é propriedade de populações e não, de indivíduos. De tal forma, o CBE é inaplicá-vel para organismos assexuados que formam clones e não, populações. Outro problema é a hibridização interespecífica. Em vegetais e fungos, híbridos férteis são comuns. Mas fi-
História de vida(limites)
Pressõesseletivas
Trocas
Contextofilogenético
(limites)
Composiçãogenotípica
Figura 7 - Como os organismos são desenhados?
4.2 NOVIDADES EVOLUTIVAS
Uma novidade evolutiva é uma estrutura que não tem antecedente claro em animais ances-trais nem correspondentes afins (homólogos) em parentes contemporâneos. Dedos, mem-bros, olhos e penas são exemplos de estrutu-ras novas. Atualmente, sabemos que as penas antecederam o vôo e já existiam em dinossau-ros, provavelmente para aquecimento térmico. Chamamos de exaptação as estruturas preexis-tentes utilisadas para realizar novas funções.
Novidades evolutivas podem advir de três tipos de mudanças. (1) Substituições de aminoáci-dos, alterando a função protéica, (2) mudan-ças em sequências de DNA que regulam genes, ocasionando alterações na forma e função de partes do organismo e a (3) adição de DNA ex-tra por duplicação ou transposição, liberando o DNA novo para atingir novas funções, en-quanto as antigas são mantidas pelos genes originais.
O campo da biologia do desenvolvimento evolutivo ou evo-devo auxilia na investigação dessas novidades evolutivas através do estudo de um grupo de genes de desenvolvimento conhecidos como genes Hox. Os processos de

ca
pít
ulo
4
67caria a pergunta: quanto fluxo gênico pode existir, para que o significado do CBE seja mantido?
Os paleontólogos estudavam espécies fósseis e necessitavam de um conceito adequado à discriminação de espécies distribuídas na di-mensão do tempo. Simpson criou o conceito filogenético de espécies (CFE): “uma espécie evolutiva é uma linhagem (uma sequência an-cestral-descendente de populações), que está evoluindo, separadamente, das outras, com seu próprio papel e tendências evolutivas uni-formes”. O maior problema seria a divisão ex-trema de espécies. Essa definição só é aplicável para espécies monotípicas.
O conceito ecológico de espécies (CEE), por sua vez, baseado na ocupação de nicho por uma espécie, não é operacional. Populações locais podem diferir em sua ocupação de nicho. As-sim, teríamos que considerar duas populações locais como espécies separadas, mesmo se tra-tando de uma só espécie.
O conceito adotado depende em parte do tipo de pesquisa realizada. Taxonomistas podem preferir o conceito tipológico, pesquisadores que trabalham com populações adotam o CBE, e o paleontólogo, por sua vez, não pode deixar de se ater à dimensão do tempo.
5.2 ESPECIAÇÃO
Cada espécie biológica reúne o seu conjunto de genótipos balanceados e harmoniosos. Um entrecruzamento entre espécies diferentes leva a um colapso dessa harmonia, refletindo-se em prole com reduzida viabilidade e fertilida-de. A aquisição de mecanismos de isolamento previne essas misturas entre diferentes espé-cies, mantendo o acervo gênico.
5.2.1 MECANISMOS DE ISOLAMENTO
Existem dois tipos de mecanismos de isolamen-to reprodutivo, o pré-zigótico e o pós-zigótico. O pré-zigótico impede a união dos gametas de duas espécies e, conseqüentemente, a forma-ção de híbridos na prole de indivíduos perten-centes a espécies diferentes. Esse mecanismo pode ser ocasionado por isolamento ecológico(habitats diferentes), sazonal (estações de re-
produção diferentes), comportamental (fraca atração sexual), mecânico (genitálias ou partes florais incompatíveis), gaméticos (incompati-bilidade fisiológica de gametas), entre outros. Nos vagalumes, por exemplo, o comportamen-to de chamada da fêmea é único. Cada espécieemite luz em diferentes intensidades (fraca ou forte), cores (verde a alaranjado) e vezes de acender e apagar a luz.
O mecanismo pós-zigótico, por outro lado, inclui a inabilidade de desenvolvimento dos embriões híbridos, a esterilidade de híbridos adultos e a degradação de gerações posterio-res de genótipos recombinantes. Na maioria das vezes, isso ocorre devido aos pareamentos inadequados entre cromossomos diferentes, gerando gametas inviáveis. Um exemplo bem conhecido é o da mula, híbrido estéril do cru-zamento de égua e jumento.
A divergência de sequências de DNA pode atuar como uma barreira para a recombinação, ofe-recendo uma base estrutural para o isolamento.
5.2.2 TIPOS DE ESPECIAÇÃO
Existem vários tipos de especiação, entretan-to vamos destacar dois tipos: (1) a especiação geográfica ou alopátrica e (2) a especiação simpátrica. A primeira caracteriza-se pela ocor-rência de modificação gradual das populações isoladas geograficamente de suas espécies pa-rentais. Caso se encontrem, vão estar reprodu-tivamente isoladas, e o fluxo gênico será zero. A segunda é definida como a origem de uma nova espécie através da especialização ecoló-gica dentro dos limites da espécie parental. Dessa forma, envolve fluxo gênico diminuído (diferente de zero) em combinação com a se-leção divergente, entre os grupos diferentes. Um exemplo desse tipo de especiação é a poli-ploidia, causada por um aumento do número de conjuntos de cromossomos, trazendo um isolamento reprodutivo.
Desafio da Fix: Vocês podem tentar realizar uma prática sobre o problema das adap-tações, representan-do o que ocorre na natureza em termos

ca
pít
ulo
4
68de disponibilidade de recursos (quantidade de sementes) e características morfológicas (bicos das aves). Para isto, acessem. www.sbg.org.br/GeneticaEscola2/web/vol1Num1.htm. Os tentilhões de Galápagos: O que Darwin não viu, mas os Grants viram.
6. MACROEVOLUÇÃOA microevolução descreve processos que ocor-rem dentro de espécies e de populações, en-quanto a macroevolução descreve padrões através das comparações entre espécies e grandes grupos, como famílias, ordens e filos.
6.1 HISTÓRIA DA VIDA
Em seu clássico de 1944, O que é vida?, o físico e filósofo austríaco Erwin Schrödinger inspirou toda uma geração de cientistas. A vida, definiu ele, é uma matéria que, tal como um cristal (um estranho “cristal aperiódico”), repete sua estrutura ao crescer. Porém, a vida é muito mais imprevisível do que qualquer mineral. A vida surgiu por volta de 3,5 bilhões de anos atrás, em nosso planeta e poderá terminar em meros cem milhões de anos, quando o sol vier a morrer.
Escutando música com a Fix: existe uma músi-ca chamada Planeta sonho, do grupo 14 bis, que trata da problemática da morte do sol. Escute-a e discuta com seus colegas as conse-quências disso.
Grande parte das pesquisas sobre a origem da vida é realizada em laboratórios, buscan-do compreender os tipos de reações químicas que poderiam ter ocorrido na Terra há 4 bi-lhões de anos. Embora não saibamos ao certo como a vida começou, existem várias hipóte-ses que tratam do tema: (1) a sopa primitiva de Oparin e Haldane; (2) a hipótese de que a vida haveria surgido em um substrato sólido, a pirita; (3) a evolução de ciclos químicos em gotículas de água nas nuvens e (4) a proposta de uma origem vinda do espaço, através da presença de matéria orgânica em meteoritos, dentre outras.
O ácido ribonucléico (RNA) é cotado como a molécula replicável ancestral, pelo fato de ser
mais simples, de fita única, assumir estruturas diferentes e ter papel catalítico (enzimático). O ácido desoxirribonucléico (DNA) o procedeu e, provavelmente, por ser mais estável e ter uma menor taxa de mutação, estabeleceu-se como repositório da informação genética. As células evoluíram como um sistema simples de mem-branas e uma molécula replicadora. A divisão classificatória mais inicial de vida celular é uma árvore trifurcada em arqués, bactérias e euca-riotos. Para saber mais sobre a origem da vida e o mundo de RNA, consulte os capítulos 1 e 2 de Matioli (2001).
O mundo era domínio absoluto dos unicelu-lares, mas, há cerca de 1,5 bilhões de anos, surge a vida pluricelular com tipos celulares es-pecializados. Entretanto, o documentário fós-sil de plantas e animais pluricelulares só teve início no Cambriano (450 milhões de anos atrás). A explosão de vida nesse período é as-sunto de intensa pesquisa. Hipóteses recentes examinam mudanças no ambiente externo ou inovações biológicas internas ou consideram a mistura de ambas, embora não haja consenso.
A saída dos organismos da água para a terra foi um processo lento. Primeiro os micróbios, depois os fungos e as plantas. Os principais eventos da evolução de plantas terrestres fo-ram a formação de esporos resistentes, o de-senvolvimento de tecido vascular, seguido das raízes e folhas. As plantas com sementes cons-tituem os principais grupos atuais de plantas terrestres e estão classificadas em gimnosper-mas (coníferas) e angiospermas (plantas com flores). Essas últimas proliferaram mais tarde, no Cretáceo (200-250 milhões de anos), e sua origem encontra-se ligada à coevolução com insetos polinizadores. Bem mais tarde (60 mi-lhões de anos), surgem as gramíneas que coe-voluiram com os mamíferos herbívoros.
Os fósseis de vertebrados mais antigos são os peixes, datando do Cambriano e Pré-Cambria-no superior. Assim como as plantas, alguns peixes também conquistaram a terra. A maio-ria dos peixes ósseos atuais possui nadadeiras raiadas, embora os tetrápodes (anfíbios, rép-teis, aves e mamíferos) atuais descendam de peixes ancestrais de nadadeiras lobadas (pul-monados e celacantos). Em algum momento da história evolutiva, existiu uma espécie de

ca
pít
ulo
4
69peixe que originou outras, e por meio de ge-rações sucessivas e etapas de ramificação da árvore da vida, surgiram muitas espécies, in-cluindo os peixes atuais e o homem.
O grande passo na evolução dos vertebrados terrestres foi o surgimento do ovo amniótico, libertando répteis, aves e mamíferos da de-pendência da postura na água. Répteis e aves evitam o ataque do sistema imunológico, pro-tegendo o feto com o ovo com casca, expelin-do-o do corpo materno. Dentre os mamíferos, os monotremados (ornitorrinco) põem ovos, marsupiais dão a luz prematuramente a seus embriões, e placentários isolam o feto no úte-ro através da placenta, que o alimenta e o pro-tege do sistema imunológico da mãe.
O predomínio dos dinossauros impedia que os nossos ancestrais mamíferos se tornassem maiores que um gato doméstico; mas uma catástrofe provocada por um asteróide ou co-meta resultou na extinção desses répteis há 65 milhões de anos. Sem a presença desses, os mamíferos puderam conquistar diversos nichos ecológicos, em explosiva diversifica-ção demonstrada pelo registro fóssil. Os ma-míferos viventes são divididos em prototérios, metatérios e eutérios ou placentários. Perten-cemos a esse último grupo e descendemos de ancestrais primatas arborícolas.
A despeito das diferenças morfológicas, o DNA de macacos e humanos é muito semelhante. Ocorre um alto grau de homologia (mais de 95%) entre chimpanzés e humanos. Dessa forma, humanos e macacos são parentes pró-ximos, e esse alto grau de identidade sugere que divergimos de um ancestral comum, bem recentemente no tempo evolutivo, talvez entre 5 a 9 milhões de anos atrás.
6.2 ORIGENS HUMANAS
Os fósseis, apesar de escassos, nos deram im-portantes informações sobre a evolução hu-mana. Todos os primeiros fósseis hominídeos vieram da África. Há 3-4 milhões de anos, sur-giu o Australopithecus afarensis, representado pela famosa Lucy. O gênero Homo apareceu há 2-2,5 milhões de anos. Entre 1,9-1,5 mi-lhões de anos passou a existir um hominídeo de compleição moderna, o Homo ergaster. Há
cerca de 1 milhão de anos, o Homo erectus, deixava vestígios fósseis fora da África, en-contrados na China e Indonésia. Entre 600 a 200 mil anos, vivia na Europa e África o Homo heidelbergensis com 93% da capacidade ce-rebral atual. Entre 230- 30 mil anos surgia o Homo neanderthalensis. De fato, eles podem ter convivido e competido com as populações que evoluíram nos humanos modernos (150 mil anos). Ao comparar fósseis de hominíde-os extintos, pesquisadores desenharam várias árvores evolucionárias humanas. Ao longo de milhões de anos, mais de uma dúzia de espé-cies de hominídeos evoluíram em simpatria. Contudo, há cerca de 30 mil anos, só resta-va uma espécie de hominídeo: Homo sapiens. Atualmente, podemos estar em qualquer lugar do planeta em horas ou dias, não havendo iso-lamento para uma nova espécie de humanos surgir.
A evolução humana foi marcada por cinco grandes transições:
(1) 5 milhões de anos - saída da floresta para a savana;
(2) 2,5 milhões - produção das primeiras fer-ramentas de pedra;
(3) 1,5 milhões - produção dos machados de mão,
(4) 500 mil anos - domínio do fogo;
(5) 50 mil anos - conquista do pensamento simbólico e expressão na forma de pin-turas em cavernas, joalheria (entalhes e conchas), escultura, armas intrincadas e enterros elaborados.
Ocorreram várias mudanças morfológicas de-pois que nossos ancestrais deixaram as flores-tas e passaram a viver em savanas: o aumen-to do cérebro, a mudança nas mandíbulas e nos dentes (nossos ancestrais eram prognatos e possuíam grandes caninos), o bipedalismo (locomoção ereta sobre duas pernas) e suas adaptações (comprimento de braços, posição de crânio sobre a coluna cervical, estreitamen-to da pelve), dentre outras.
A mudança no comportamento social e cultu-ral permitiu que formássemos grupos de indi-

ca
pít
ulo
4
70víduos cada vez maiores. No início, não pas-sávamos de 55 indivíduos (3 milhões de anos atrás - Australopithecus afarensis), todavia com o surgimento do gênero Homo (2 milhões de anos) éramos capazes de formar grupos de 80indivíduos, que foram aumentando para 100 (1 milhão de anos) e 150 (100 mil anos). Atual-mente, formamos grupos enormes, entretanto se estima que o número de 150 indivíduos per-mite um contato social adequado, sendo pos-sível conhecer individualmente cada pessoa. A principal mudança subjacente à cultura huma-na foi a linguagem, que nos permitiu penetrar no mundo simbólico. A linguagem permitiu estreitar laços com uma quantidade maior deindivíduos, o que seria impossível, apenas, com a catação (grumming).
Desafio da Fix: Assistam ao fil-me BBC Worl-dwide Ltd 2003. Homens das cavernas. Su-perinteressante - Editora Abril, 2004. Nesse fil-me, dividido em episódios, vocês irão entrar em contato com os diversos hominídeos que co-existiram na pré-história. Iniciem a jornada, observando nossa ancestral Lucy (Australopi-thecus afarensis) e todas as modificações ao longo da nossa jornada para a humanização. Divirtam-se!
Os genes determinam o curso da seleção na-tural, mas as invenções humanas podem alte-rar as aptidões. A coevolução da cultura e dos genes tornou possível mudanças importantes, como a adoção de novas dietas, a conquista de novos habitats e a adaptação a patógenos. A maioria dos mamíferos, inclusive o homem, pára de produzir a lactase ao se tornar adul-ta, embora algumas pessoas sejam capazes de continuar bebendo leite por ainda produzir a enzima. Essa capacidade tornou-se comum com a domesticação do gado (10 mil anos atrás). Em regiões de malária, doenças, como atalassemia e a anemia falciforme, são comuns, embora raras ou inexistentes em outras partes do mundo. O gene para a anemia falciforme
marca a expansão da agricultura. A floresta devastada dava lugar a plantações. Ao chover, poças eram formadas na terra erodida, auxi-liando na proliferação de mosquitos (Anophe-les), que portam o Plasmodium falciparum, causador da malária.
Fórum: pesquisem sobre a anemia falciforme. Comparem a adaptabilidade de pessoas com genótipos diferentes (HbA HbA, HbA HbS, HbS HbS) em áreas com e sem malária. O que po-deríamos esperar em termos de diminuição de frequência na população para o alelo HbS se a malária for erradicada?
Modificamos o ambiente, e, hoje, temos o controle de muitas doenças que nos matariam antes de alcançarmos a idade reprodutiva. As-sim, à medida que as diferenças no sucesso reprodutivo humano diminuem, a seleção na-tural torna-se mais fraca.
A biologia evolutiva procura explicar, as diver-sas características dos organismos, baseada nos princípios da adaptação, do acaso e da his-tória, ocupando, por isso, uma posição central dentro das Ciências Biológicas. Entendendo a história da vida na Terra, os pesquisadores po-dem criar novos remédios e vacinas, determi-nando como devem ser administrados. Perce-ber como surgiu a AIDS, utilizar fósseis como marcadores em formações ricas em petróleo, evitar o uso indiscriminado de antibióticos e venenos, auxiliar na preservação de espécies, dentre outros exemplos, nos faz compreender que somos parte de um complexo e delicado sistema de intercâmbio com toda a natureza e que nossas ações modificam o planeta como um todo.
No nordeste (Bahia), existe um grupo muito atuante nessa área da biologia, Grupo de Pes-quisa em História, Filosofia e Ensino de Ciên-cias Biológicas, coordenado pelo prof Charbel Niño El-Hani: http://www.gphfecb.ufba.br/. Vale a pena conferir!

ca
pít
ulo
4
71
REFERÊNCIASLEITURA FUNDAMENTAL
GRIFFITHS, A. J. F.; WESSLER, S. R.; LEWONTIN, R. C.;GELBART, W. M.; SUZUKI, D. T.e MILLER, J. H. Introdução à Genética: Rio de Janeiro: Guanabara Koogan S/A, 2006.
MATIOLI, S. R. Biologia Molecular e Evolução. Ribeirão Preto: Holos Editora, 2001.
RIDLEY, M. Evolução. Porto Alegre: Artmed, 2006.
SNUSTAD, D. P. e SIMMONS M. J. Fundamen-tos de Genética: Guanabara Koogan S/A, 2001.
LEITURA COMPLEMENTAR
DARWIN, C. A origem das espécies. Belo Hori-zonte-São Paulo: Itatiaia-EDUSP, 1985.
FUTUYMA, D. Biologia Evolutiva. Ribeirão Pre-to: SBG, 1992.
MAYR, E. Biologia, ciência única - reflexões so-bre a autonomiade uma disciplina científica. São Paulo: Editora Schwarcz Ltda,2005.
MEYER, D. e EL-HANI, C. N. Evolução: o sen-tido da biologia.Coleção paradidáticos. São Paulo: Editora UNESP, 2005.
MONOD, J. O acaso e a necessidade: ensaio sobre a filosofia natural da biologia moderna. Petrópolis: Editora Vozes, 2006.
STEARNS, S. C. e HOEKSTRA, R. F. Evolução: uma introdução. São Paulo: Editora Atheneu, 2003.
FILME
BBC Worldwide Ltd 2003. Homens das cavernas. Superinteressante - Editora Abril, 2004. (solicite pela internet: www.super interessante.com.br/ loja ou por email: [email protected])
SITES
Artigo sobre raça: (Scientific American Brasil edi-ção nº 20 – jan 2004):
http://www2.uol.com.br/sciam/conteudo/ma-teria/materia_38. htm.
Sociedade Brasileira de Genética (SBG): http://www.sbg.org.br
Programa GBOL: http://www.ufv.br/dgb/gbol
Prática sobre deriva:www.sbg.org.br/GeneticaEscola2/web/vol-1Num2.htm
Artigo “Uma nadadeira é um braço é uma asa” da revista National Geographic 2006:http://www.nationalgeographic.abril.com.br/ng-bonline/edicoe s/0611/wings.shtml.
Prática - Os tentilhões de Galápagos: O que Da-rwin não viu, mas os Grants viram:www.sbg.org.br/GeneticaEscola2/web/vol-1Num1.htm
Grupo de Pesquisa em História, Filosofia e Ensino de Ciências Biológicas: http://www.gphfecb.ufba.br/
GLOSSÁRIOAconselhamento genético - fornecimento de informações genéticas para facilitar uma to-mada de decisão informada.
Adaptação - uma particularidade de um in-divíduo que auxilia na sua sobrevivência e re-produção.
Alelismo múltiplo - relação entre dois ou mais alelos.
Alelo - forma alternativa de um mesmo gene.
Alelo letal - gene que em homozigose provoca a morte do indivíduo.
Anagênese - mudança evolutiva em uma linha-gem filogenética única.
Ancestral comum - relativo aos nossos ante-passados, os que nos antecederam e nos dei-xaram informações que são compartilhadas com todos os seres.

ca
pít
ulo
4
72Antecipação - tendência de algumas caracte-rísticas surgirem mais cedo e com maior gravi-dade a cada geração subseqüente.
Autofertilização - união de gametas oriundos de um mesmo indivíduo.
Autofertilizados - ver autofertilização. Características qualitativas - características mo-nogênicas cuja variação entre as classes é des-contínua, permitindo a rápida distinção entre as diferentes classes fenotípicas.
Características quantitativas - características poligênicas cuja variação entre as classes é contínua, permitindo a separação de classes através de divisão arbitrária da variação.
Catação (grumming) - processo de limpeza do pêlo de primatas, onde um indivíduo retira piolhos, pulgas, carrapatos, e restos de comida aderidos. Comportamento que amplia os laços de afetividade e cooperação do grupo.
Cladogênese - origem e multiplicação de es-pécies e de táxons superiores. Classificação fi-logenética.
Código genético - sistema utilizado para rela-cionar as sequências de nucleotídeos do DNA e do RNA mensageiro e as sequências de ami-noácidos nas proteínas formadas na tradução.
Codominância - relação entre alelos, na qual cada um deles se manifesta no fenótipo do he-terozigoto.
Conformação cis - ou acoplada. Dois alelos do-minantes (selvagens) estão presentes no mes-mo homólogo.
Conformação trans - ou em repulsão. Os alelos se encontram em homólogos diferentes.
Crossing - termo inglês para a palavra permuta. Troca de segmentos entre cromátides (não ir-mãs) pertencentes a cromossomos homólogos.
Cruzamento diíbrido - cruzamento em que analisamos dois genes.
Cruzamento monoíbrido - cruzamento em que analisamos apenas um gene.
Deriva genética - variação aleatória na frequ-ência gênica de uma geração para outra em uma população.
Determinismo - é a crença de que a maioria dos processos na natureza inanimada é estrita-mente determinada por leis naturais – físicas.
Determinista - ver determinismo.
Diplóide - célula ou indivíduo que apresenta dois conjuntos de cromossomos homólogos.
Dissomia uniparental - herança de dois alelos homólogos de um dos genitores.
Distúrbios multifatoriais - doenças causadas pela interação de fatores genéticos e ambientais.
Diversidade - existência, numa dada região, de uma grande variedade de espécies, ou de ou-tras categorias taxonômicas.
Doenças comuns - doenças multifatoriais como diabetes e doenças cardiovasculares.
Dominância incompleta - interação entre ale-los em que o fenótipo do heterozigoto é inter-mediário ao dos parentais homozigotos.
Dominante - caráter que se expressa na pre-sença de outro alelo do mesmo loco gênico, mascarando a expressão do outro.
Efeito do fundador - é o fenômeno da perda de variação genética, quando uma nova colô-nia é formada por um número menor de indi-víduos do que o da população original.
Epistasia - alteração na expressão de um gene por outro gene (gene encobre o efeito de outro).
Equilíbrio pontuado - alternância, numa linha-gem filética, de mudanças evolutivas extrema-mente rápidas e outras normais ou lentas.
Especiação - processo evolutivo que acarreta na multiplicação das espécies.
Espécies crípticas - ou irmãs, são populações fenotipicamente muito semelhantes ou idênti-cas que se reproduzem isoladamente uma da outra.

ca
pít
ulo
4
73Essencialismo – crença em que a diversidade consistia em um número limitado de essência, delimitadas e imutáveis. Pensamento tipológico.
Evolução - Darwin definiu-a como “a descen-dência com modificações”. É a mudança, entre as gerações, nas linhagens das populações.
Expressividade variável - amplitude na qual os efeitos gênicos se manifestam e o fenótipo pode variar de leve a severo.
Fenótipo - determinado aspecto exibido por um caráter em função da inter-relação entre a composição genética de um indivíduo e seu ambiente. Inclui não apenas características morfológicas, mas também as fisiológicas e comportamentais.
Fertilização cruzada - cruzamento entre indiví-duos da mesma espécie.
Gargalo genético - perda de genes e aumento da frequência de outros devido a catástrofes, reduzindo a população reprodutiva e alteran-do a composição genética.
Gene - sequência de nucleotídeos que codifi-cam um RNA ou um polipeptídeo.
Genes Hox - grupo de genes que atuam no desen-volvimento. Auxiliam a determinação e diferen-ciação celular nas várias regiões do organismo.
Genes ligados - genes situados em um mesmo cromossomo.
Genes modificadores - gene que atua sobre outro modificando ligeiramente a característi-ca determinada por este.
Genes supressores - genes que suprimem a ati-vidade de outros ou revertem o efeito de uma mutação de outro gene, restaurando o fenóti-po selvagem.
Genótipo - constituição genética de um in-divíduo.
Geração inicial - o mesmo que geração paren-tal, aquela que inicia uma família.
Geração parental - o mesmo que geração ini-cial, aquela que inicia uma família.
Gradualismo - transformação evolutiva origi-nada por mutações lentas e graduais.
Grupo de ligação - conjunto de genes situados no mesmo cromossomo.
Hemizigoto - célula ou indivíduo que só apresen-ta um alelo para um determinado loco gênico.
Herança - capacidade de transmissão de certo caractere de uma geração a outra.
Herança epigenética - alteração na expressão gênica não causada por uma mudança na es-trutura do gene.
Herança mitocondrial - herança onde os genes são oriundos apenas da mãe através das mito-côndrias.
Herança organelar - herança não nuclear, os genes são provenientes das mitocôndrias e/ou cloroplastos.
Herança sólida - ou particulada é aquela trans-mitida por genes.
Herança tênue - ou por mistura é aquela onde se acreditava na mistura de informações gené-ticas. Refutada.
Heredograma - diagrama que apresenta ge-rações de parentes, a relação destes com o probando, e sua situação em relação a uma determinada característica genética.
Heterogeneidade genética - termo que signi-fica variação. Na heterogeneidade alélica as mutações diferentes no mesmo gene causam um fenótipo idêntico. Na heterogeneidade de loco, fenótipos similares são causados por mu-tações em locos diferentes.
Heterozigotos - indivíduo que mostra alelos di-ferentes para o loco gênico considerado.
Hipótese de nulidade - postula não haver dife-rença entre as populações ou grupos em com-paração, com relação ao fator, característica ou condição de interesse. Ver hipótese nula.
Hipótese nula - hipótese principal é aquela que iguala o parâmetro a ser testado a algum valor referencial. Ver hipótese de nulidade.

ca
pít
ulo
4
74Homozigotos - indivíduo com alelos idênticos no mesmo loco gênico.
Imprinting parental - expressão diferencial de um gene dependendo do genitor de origem.
Inativação do X - processo de compensação de dosagem gênica em fêmeas de mamíferos, onde um dos cromossomos X é quase inteiramente heterocromatinizado e silenciado genicamente.
Interferência - processo no qual a presença de um quiasma pode interferir na presença de um segundo crossing na vizinhança.
Letal dominante - gene que mata o indivíduo quando em homozigose dominante.
Letal recessivo - gene que mata o indivíduo quando em homozigose recessiva.
Ligação - situação na qual dois ou mais genes segregam juntos, durante a transmissão para a descendência, em virtude de sua proximidade no mesmo cromossomo.
Ligação incompleta - o mesmo que distribui-ção independente.
Macroevolução - evolução em grande escala que ocorre em nível superior ao de espécie.
Malformações congênitas - defeitos de nasci-mento oriundos de processos desenvolvimen-tais intrinsecamente anormais.
Mapa genético - definição da localização das unidades genéticas em cada cromossomo.
Mapas citológicos - baseados em característi-cas citológicas, como o padrão de bandeamen-to cromossômico observado ao microscópio.
Mapas físicos - determinados pelas distâncias de bases que separam os marcadores moleculares.
Metilação - adição de um grupo metil a citosi-na no DNA, com papel regulatório na expres-são gênica.
Mosaicismo - presença de duas ou mais linha-gens de células originadas de um só zigoto.
Mutação - alteração do material genético.
Não-disjunção - não separação de cromosso-mos homólogos ou cromátides irmãs duran-te a anáfase I e II, respectivamente, levando a uma distribuição desigual destes nos pólos da célula em divisão.
Organismos modelos - organismos utilizados para estudos, como por exemplo, Escherichia coli, Arabdopsis thaliana, Neurospora crassa, Dro-sophila melanogaster. Através deles inferimos processos semelhantes para táxons próximos.
Paradigma - sistema de crenças, valores e ge-neralizações que domina uma ciência ou um ramo da ciência em determinada época.
Paramutação - processo pelo qual o RNA pode influenciar a hereditariedade à semelhança do DNA.
Penetrância - expressão tudo ou nada de um genótipo mutante.
Permuta - troca de segmentos entre cromáti-des (não irmãs) pertencentes a cromossomos homólogos.
Pleiotropia - efeitos fenotípicos múltiplos de um único gene.
Poligenia - efeito aditivo de genes que podem determinar aspectos quantitativos do fenótipo.
Polimórficos - ver polimorfismo.
Polimorfismo - ocorrência simultânea de di-versos fenótipos, onde uma população possui mais do que um alelo para determinado loco.
Poliploidia - condição na qual o número de cromossomos é um múltiplo inteiro do núme-ro haplóide.
Pool gênico - ou acervo gênico é a composição genética total de uma população.
Princípio da distribuição independente - pares de genes em cromossomos diferentes se distri-buem independentemente na meiose.
Princípio da dominância - ocorre predominância de um caráter na F1 (primeira geração híbrida) considerado dominante em relação às carac-terísticas alternativas chamadas de recessivas.

ca
pít
ulo
4
75Princípio da segregação - durante a reprodu-ção sexual os alelos de cada genitor se sepa-ram ou segregam, produzindo gametas comapenas um alelo daquela característica estudada.
Princípio de Hardy-Weinberg - é a razão das frequências genotípicas que se originam quan-do os cruzamentos são aleatórios, a popula-ção é grande e os fatores evolutivos não estão atuando.
Probando - propósito ou indicador, indivíduo portador de uma anomalia que chama a aten-ção médica para o estudo da família.
Quadrado de Punnett - quadro usado para de-terminar os genótipos de um dado cruzamento.
Quiasmas - pontos de trocas entre cromosso-mos homólogos, evidência física da permuta.
Recessivo - caráter que se expressa apenas em dose dupla do mesmo alelo no loco gênico.
Recombinação - evento que ocorre durante a segregação de cromossomos (recombinação cromossômica) ou durante a meiose (recom-binação gênica).
Relógio molecular - é a teoria na qual molé-culas evoluem em uma taxa aproximadamente constante.
Repetições de trinucleotídeos - repetições de trincas de nucleotídeos que podem estar em quantidades normais ou anormais, gerando doenças.
Saltacionismo - transformação evolutiva origi-nada por mutações drásticas.
Seleção natural - processo pelo qual formas de organismos de uma dada população que estão melhores adaptadas ao ambiente presente au-mentam em frequência em relação as demais, ao longo de uma série de gerações.
Sexo heterogamético - indivíduo que produz mais de um tipo de gameta, determinando o sexo da espécie.
Sexo homogamético - indivíduo que produz apenas um tipo de gameta.
Teleologia cósmica - explicação de processos naturais que conduzem automaticamente a um fim definido ou a uma meta.
Teoria cromossômica da herança - teoria que uniu as idéias de continuidade e individualida-de dos cromossomos, de Sutton e Boveri, res-pectivamente.
Teoria neutra - é a teoria em que a maior par-te da evolução em nível molecular ocorre por deriva.
Teste dos três pontos - teste no qual um triplo heterozigoto é cruzado com um triplo homozi-goto recessivo, fornecendo uma descendência com números variáveis de crossings e frequên-cias, permitindo determinar a distância entre três locos e a ordem seqüencial dos genes.
Unidade de mapa - ou centimorgan (cM) é a distância entre dois locos acarretando uma fre-quência de recombinação (FR) de 0,01 (1%).
Vitalismo - crença na existência de uma força oculta e invisível nos organismos vivos, respon-sável pelas manifestações da vida em organis-mos vivos.