1.8 padrÕes e mecanismos genÉticos de...
TRANSCRIPT

Módulo V – Processos Reprodutivos
1.8 PADRÕES E MECANISMOS GENÉTICOS
Rubiani de Cássia PagottoDoutora em Genética. Professora
Universidade Federal de Rondônia
Introdução e Objetivos
Comecemos o estudo entendendo o porquê da
expressão Genética Mendeliana
Na segunda metade do século XX, o monge austríaco Gregor Johann híbridos de plantas e descreveu características hereditárias da ervilha comum num edeste trabalho, Mendel demonstrgenealogia de um casaldescendência. Com is
Mas a sua descoberta só teve validade quando vários cientistas, dentre eles os naturalistas Correns, Tschermak e De Vries chegaram à conclusão de que os estudos do monge austríaco eram corretos e válidos para todos os seres vivos.
Objetivos
Ao final dos seus estudos nest
• Identificar as leis de Mendel.
• Identificar padrões da herança genética.
• Reconhecer como os mecanismos genéticos interferem na vida humana.
Gregor Johann Mendel
Processos Reprodutivos
E MECANISMOS GENÉTICOS DE HERANÇA
Rubiani de Cássia Pagotto Professora Adjunta do Departamento de Biologia da
Federal de Rondônia – UNIR, Porto Velho, RO.
Introdução e Objetivos
Comecemos o estudo entendendo o porquê da
Genética Mendeliana.
Na segunda metade do século XX, o monge austríaco Gregor Johann Mendel desenvolveu experimentos com híbridos de plantas e descreveu características hereditárias da ervilha comum num estudo publicado em 1866. A partir
e trabalho, Mendel demonstrou que, se conhecermos a genealogia de um casal, poderemos prever qual será sua descendência. Com isto, Mendel criou a Genética.
Mas a sua descoberta só teve validade 32 anos depois, quando vários cientistas, dentre eles os naturalistas Correns, Tschermak e De Vries chegaram à conclusão de que os estudos do monge austríaco eram corretos e válidos para todos os seres vivos.
ta unidade, você será capaz de:
Identificar as leis de Mendel.
Identificar padrões da herança genética.
Reconhecer como os mecanismos genéticos interferem na vida
242
Genética Mendeliana é um conjunto de princípios relacionados à transmissão hereditária das características de um organismo a seus filhos. Consiste na base principal da genética clássica. Originou-se dos trabalhos de Mendel publicados em 1865 e 1866, os quais foram considerados controversos inicialmente, e redescobertos em 1900. Somente quando incorporada à teoria do cromossomo de Thomas Hunt Morgan, em 1915, foi que a Genética Mendeliana se tornou a essência da genética clássica (Wikipédia).

Módulo V – Processos Reprodutivos
Breve histórico do
Desde a antiguidade que o homem já carrega dentro de si o que poderíamos chamar de um “conceito intuitivo de genética”. No Talmud, livro sagrado dos que possui alguns textos inseridos no Antigo Testamento da Bíblia Sagrada Cristãos, existem restrições e situaçõessuas razões, utilizando conceitos básicos de genética e padrões genéticos de herança. Um exemplo disto é a circuncisão (remoção do prepúcio, prega cutânea que recobre aglande do pênis), que, na cultura judaica, significa a lembrança da aliança com Deus.
O Rabino Simon Bem Gamaliel proibiu a filhos de três irmãs mais velhas de sua mãe morreram após a mesma. Após o procedimento, os meninos tiveram hemorragia e faleceram. Hoje, com o conhecimento dos padrões de herança mendeliana, podemos explicar esrabino. No antigo Egito temos conhecimento do uso de técnicas de melhoramento vegetal. Os povos do velho mundo, muito antes dos primeiros tratados de genética serem publicados, já tinham uma prática de melhoramento genético animal. Entretanto, a ciência da Genética permaneceu esquecida até meados do século XX, quando três pesquisadores (Hugo De Vries, Karl Correns e E. Tschermak), independentemente, redescobriram os trabalhos de Gregor Mendel,considerado como o “Pai da Genética”.
Genótipo e Fenótipo
Vamos tratar de alguns termos técnicos para um melhor desenvolvimento do tema.
Genótipo = grupo de genes de um indivíduo.
Fenótipo = é o resultado do efeito combinado da ação de genes e fatores ambientais. Pode ser visível a olho nuutilização de técnicas que permitem definisanguíneos ABO.
Quando observamos uma flor, um animal ou uma pessoacaracterísticas físicas que os identificam, como por exemplo
• a cor das pétalas de uma rosa
• a presença ou ausência de espinhos no caule da roseira
• a pelagem lisa ou malhada em um gato ou cachorro
• os olhos claros ou escuros de uma criança.
Todas essas características demonstram FENÓTIPO é o resultado da interação do seu GENÓTIPO com o meio no qual o indivíduo vive.
Processos Reprodutivos
istórico do surgimento da Genética
Desde a antiguidade que o homem já carrega dentro de si o que poderíamos chamar de um “conceito intuitivo de genética”. No Talmud, livro sagrado dos judeus, que possui alguns textos inseridos no Antigo Testamento da Bíblia Sagrada dos Cristãos, existem restrições e situações, que hoje podemos explicar cientificamente suas razões, utilizando conceitos básicos de genética e padrões genéticos de herança.
o é a circuncisão (remoção do prepúcio, prega cutânea que recobre a que, na cultura judaica, significa a lembrança da aliança com Deus.
O Rabino Simon Bem Gamaliel proibiu a circuncisão num menino porque os filhos de três irmãs mais velhas de sua mãe morreram após a mesma. Após o procedimento, os meninos tiveram hemorragia e faleceram. Hoje, com o conhecimento dos padrões de herança mendeliana, podemos explicar esta atitude do
No antigo Egito temos conhecimento do uso de técnicas de melhoramento vegetal. Os povos do velho mundo, muito antes dos primeiros tratados de genética
já tinham uma prática de melhoramento genético animal. a Genética permaneceu esquecida até meados do século XX,
quando três pesquisadores (Hugo De Vries, Karl Correns e E. Tschermak), independentemente, redescobriram os trabalhos de Gregor Mendel, atualmente
ai da Genética”.
Vamos tratar de alguns termos técnicos para um melhor desenvolvimento do tema.
= grupo de genes de um indivíduo.
= é o resultado do efeito combinado da ação de genes e fatores tais. Pode ser visível a olho nu (cor da pelagem dos animais) ou pela
m defini-lo, como por exemplo, no caso dos grupos
Quando observamos uma flor, um animal ou uma pessoa, notamos identificam, como por exemplo:
cor das pétalas de uma rosa;
presença ou ausência de espinhos no caule da roseira;
pelagem lisa ou malhada em um gato ou cachorro;
aros ou escuros de uma criança.
Todas essas características demonstram o fenótipo do indivíduo, sendo que o FENÓTIPO é o resultado da interação do seu GENÓTIPO com o meio no qual o
243
O rabino intuiu que havia uma herança genética nessa família que impedia as crianças de parar de sangrar após ferimento. Provavelmente eram portadores de hemofilia, que é uma alteração genética e hereditária no sangue, caracterizada por um defeito na coagulação.As primeiras referências sobre a hemofilia humana são do século III e estão descritas nos escritos judaicos - Talmud.

Módulo V – Processos Reprodutivos
Algumas vezes notamos características em um in(copiam) um fenótipo com base sabidamente genética, comocoloração de cabelos em humanos. Na espécie humana temos, em algumas civilizações, o hábito de mudar a coloração natural dos cabelos, geralmente simulando uma coloração de ocorrência natural na população. característica, que simula uma coloração geneticamente definida e de ocorrência natural na população, dizemos que o indivíduo é uma FENOCÓPIA.
ATIVIDADE
Observe animais, plantas e pessoas características fenotípicas em cada um deles.
Primeira e Segunda Leis de Mendel
Em 1865, Mendel apresentou tratado sobre “As Leis da Hereditariedade”, hoje chamadas elaborar este precioso trabalho, Mendel dedicou parte de sua vida ao cultivo e observação das ervilhas de jardim (Pisum sativum
Mendel selecionou, cultivou e observou sete característicervilhas de jardim (tabela 1). Os dados obtidos foram analisados matematicamente, o que para a época era um feito bastante inusitado. Com base em tais análises, postulou o que hoje conhecemos como as Leis de Mendel.
Característica Fenótipo ACor da semente Amarela Forma da semente Lisa Cor da vagem Verde Forma da vagem Lisa Altura do pé da ervilha Alta (160 cm)Posição da flor Ao longo dos ramosCor da flor Púrpura
Tabela 1: Listagem das características fenotípicas estudadas por Mendel em seus experimentos de cruzamento entre diversas linhagens de ervilhas.
ATIVIDADE
Mendel foi um homem brilhante, como se costuma dizerde seu tempo”. Sua principal contribuição para a ciência foi lançar as bases para o conhecimento da transmissão das características hereditárias dos seres vivos.
Sua história é emocionante!
Faça uma pesquisa na internet sobre a vida do “Pai da Genética” e descubra como ele se tornou monge e quais outras contribuições ele deixou ao mundo.
Sugestão de leitura: Cruz & Silva (2002) jardins do mosteiro CIÊNCIA HOJE • vol. 31 • nº 184, pags
Processos Reprodutivos
Algumas vezes notamos características em um indivíduo que mimetizam (copiam) um fenótipo com base sabidamente genética, como, por exemplo, a coloração de cabelos em humanos. Na espécie humana temos, em algumas civilizações, o hábito de mudar a coloração natural dos cabelos, geralmente
loração de ocorrência natural na população. Em relação a esta que simula uma coloração geneticamente definida e de ocorrência
dizemos que o indivíduo é uma FENOCÓPIA.
Observe animais, plantas e pessoas à sua volta. Liste no mínimo três características fenotípicas em cada um deles.
egunda Leis de Mendel
Mendel apresentou à Sociedade de História Natural de Berno um tratado sobre “As Leis da Hereditariedade”, hoje chamadas “Leis de Mendel”. Para
e precioso trabalho, Mendel dedicou parte de sua vida ao cultivo e Pisum sativum).
ndel selecionou, cultivou e observou sete características fenotípicas das ervilhas de jardim (tabela 1). Os dados obtidos foram analisados matematicamente, o que para a época era um feito bastante inusitado. Com base em tais análises, postulou
onhecemos como as Leis de Mendel.
tipo A Fenótipo B Verde
Rugosa Amarela Ondulada
Alta (160 cm) Baixa (40 cm) Ao longo dos ramos Terminal
Branca
Listagem das características fenotípicas estudadas por Mendel em seus experimentos de cruzamento entre diversas linhagens de ervilhas.
brilhante, como se costuma dizer: “uma mente à frente de seu tempo”. Sua principal contribuição para a ciência foi lançar as bases para o conhecimento da transmissão das características hereditárias dos seres vivos.
Faça uma pesquisa na internet sobre a vida do “Pai da Genética” e descubra como ele se tornou monge e quais outras contribuições ele deixou ao mundo.
Cruz & Silva (2002) Gregor Mendel: persistência nos CIÊNCIA HOJE • vol. 31 • nº 184, pags. 75-77.
244
Fenocópia = uma característica produzida por um efeito ambiental que mimetiza um fenótipo produzido por um genótipo.
Você sabia que.... Nas experiências com hibridação em ervilhas, acredita-se que Mendel tenha analisado aproximadamente 12 mil plantas e 300 mil sementes?!!! O jardim onde Mendel cultivou suas ervilhas localizava-se no mosteiro agostiniano de São Tomás de Brünn, com extensão aproximada de 7 m de largura por 35 m de comprimento?
Saiba mais: visite o site http://www.mendelWeb.org/ e leia o trabalho de Mendel “Experiments in Plant Hybridization “, lido no dia 8 de março de 1865, durante a Reunião da Sociedade de História Natural da cidade de Brünn.

Módulo V – Processos Reprodutivos
O sucesso de Mendel em seus experimentos deveueles a capacidade de inovação e de discernimento do monge que, ao contrário dos pesquisadores da sua época, formulou hipóteses baseadas em suas observações iniciais e então prosseguiu sua experimentação, realizando cruzamentos adicionais para testar matematicamente suas hipóteses. Em outras palavras, adotou o método científico. Escolheu para seus estudos usendo que essas variedades diferiam entre si em várias características que possuíam formas alternativas bem distintas, por exemplo, semente verde ou amarela. Geneticamente, cada variedade era pura pelo fato das erviautofecundação natural.As flores das ervilhas possuem uma morfologia que propicia a seus grãos de pólen fecundar os óvulos produzidos no gineceu da própria flor. As plantas originárias deste tipo de polinização carregam, desidênticas de cada um de seus genes e são ditas HOMOZIGOTAS.
Para cruzar duas linhagens diferentes, Mendel lançou mão de uma artificial. Antes da flor amadurecer ele abria uma estrutura da flor (quilha) e retirava os estames. Após o amadurecimento da flor, ele colocava sobre o estigma o pólen que havia coletado de outra flor, proveniente de uma linhagem alternativa (Esquema 1 abaixo).
Esquema 1: Procedimento adotado por Mendel para fecundação artificial das flores de ervilhas.
Realizando a fecundação cruzada entre variedades com características contrastantes, Mendel reconheceu que, na primeira geração (F1) obtida, todas as plantas exibiam o mesmo fenótipo de um dos parentais. Entretanto, quando fazia a fecundação entre indivíduos F1, obtinha uma segundindivíduos exibindo o fenótipo idêntico a de F1 e outros iguais ao fenótipo que não havia aparecido em F1.
É muito importante ressaltar quecaracterísticas uma a uma, o que
Planta que
produz vagem
verde
Retirada dos
estames
Flor
madura
homozigotasPlanta que
produz vagem
verde
Retirada dos
estames
Flor
madura
Planta que
produz vagem
verde
Planta que
produz vagem
verde
Retirada dos
estames
Retirada dos
estames
Flor
madura
Flor
madura
homozigotas
Processos Reprodutivos
O sucesso de Mendel em seus experimentos deveu-se a vários fatores, dentre eles a capacidade de inovação e de discernimento do monge que, ao contrário dos
poca, formulou hipóteses baseadas em suas observações iniciais e então prosseguiu sua experimentação, realizando cruzamentos adicionais para testar matematicamente suas hipóteses. Em outras palavras, adotou o método científico. Escolheu para seus estudos uma espécie com uma grande variabilidade, sendo que essas variedades diferiam entre si em várias características que possuíam formas alternativas bem distintas, por exemplo, semente verde ou amarela. Geneticamente, cada variedade era pura pelo fato das ervilhas de jardim realizar
.As flores das ervilhas possuem uma morfologia que propicia a seus grãos de pólen fecundar os óvulos produzidos no gineceu da própria flor. As
e tipo de polinização carregam, desta forma, duas cópias idênticas de cada um de seus genes e são ditas HOMOZIGOTAS.
Para cruzar duas linhagens diferentes, Mendel lançou mão de uma fecundação . Antes da flor amadurecer ele abria uma estrutura da flor (quilha) e retirava
o amadurecimento da flor, ele colocava sobre o estigma o pólen que havia coletado de outra flor, proveniente de uma linhagem alternativa (veja o
Procedimento adotado por Mendel para a realização da fecundação artificial das flores de ervilhas.
cruzada entre variedades com características Mendel reconheceu que, na primeira geração (F1) obtida, todas as
plantas exibiam o mesmo fenótipo de um dos parentais. Entretanto, quando fazia a fecundação entre indivíduos F1, obtinha uma segunda geração (F2) com alguns indivíduos exibindo o fenótipo idêntico a de F1 e outros iguais ao fenótipo que não
É muito importante ressaltar que, inicialmente, Mendel estudava as , o que possibilitou a ele reconhecer o padrão de
Planta que
produz vagem
amarela
Coleta e
transferência do
pólen
Semente
heterozigotafecundação
Plantas
homozigotasPlanta que
produz vagem
amarela
Coleta e
transferência do
pólen
Semente
heterozigotafecundação
Planta que
produz vagem
amarela
Planta que
produz vagem
amarela
Coleta e
transferência do
pólenCo
leta e
transferência do
pólen
Semente
heterozigotafecundação
Plantas
homozigotas
245
Autofecundação. Processo de propagação sexuado que se verifica naturalmente em muitas espécies vegetais, que contam com os aparelhos reprodutores masculino e feminino na mesma planta. Neste tipo de fecundação o gameta masculino fecunda o gameta feminino do próprio organismo.
Algumas definições: GENES ALELOS são aqueles genes localizados em um mesmo locus de cromossomos homólogos. GENÓTIPOS HOMOZIGOTOS são aqueles cujos genes alelos são idênticos (ex. genótipos homozigotos do sistema de grupos sanguíneos ABO em humanos IAIA, IBIB, ii). GENÓTIPOS HETEROZIGOTOS são aqueles cujos genes alelos possuem sequências diferentes um do outro (ex. genótipos heterozigotos do sistema de grupos sanguíneos ABO em humanos IAIB, IAi, IBi).

Módulo V – Processos Reprodutivos
transmissão descrito acima. Ele estudou cada uma das sete características listadas na tabela 1 separadamente e obteve para todo
Dessa forma, quando o monge observou a característica “cor da vagem”, por exemplo, notou que cruzando plantas puras que exibiam vagem verde com plantas puras que apresentavam vagem amarela, obtinha em F1 100% de plantas h(hoje chamadas de heterozigotas), cujas vagens eram de coloração verde. A geração F2, entretanto, exibia aproximadamente 3 plantas que formavam vagens verdes para cada planta que formava vagem amarela. De fato, os dados obtidos por Mendel para a citada característica foram 428 plantas produtoras de vagens verdes e 152 plantas produtoras de vagens amarelas, ou seja, uma proporção de 2,82:1 (lêpara cada amarela).
O “Pai da Genética” então denominou de “característica dominante” aquela que era observada em 100% de F1 e em ¾ de F2. A característica alternativa, que somente reaparecia nas plantas F2Concluiu, pelos resultados obtidos, recessiva deveriam estar presentes em F1, mas de forma “encoberta”foram transmitidos para F2, se manifestarvagens de cor amarela.
Na tentativa de explicar seus resultados, características estudadas eram condicionadas por aquilo que ele chamou de “fatores” (hoje, nomeamos tais fatores de GENEpares nos indivíduos.
Na formação dos gametas, os fatores seriam separados de forma que um gameta possuiria somente uma unidade de cada fator. Após os cruzamentos, os gametas masculinos fecundariam os femininosAssim, os novos organismos seriam novamente portadores de um par de cada fator, sendo uma unidade proveniente do pai e outra da mãe.
Todas essas considerações e explicações elaboradas por Mendel podem ser resumidas no que chamamos de Primeira Lei de Mendel Gametas.
Após analisar as características individualmenteexperimentos realizando cruzamentos Seus resultados demonstravam quequais exibindo duas características distintas, em F1 todos os descendentes exibiam somente as características denominadas dominantescruzamento entre indivíduos F1, existia indivíduos exibindo diferentes combinações das duas características:
Em um dos seus experimentoscomprimento das hastes simultaneamente (sua hipótese estivesse correta, em F2 cada característica seria exibida em 50% das plantas. Os vários caracteres podem ser identificados por “A”, “B”, “a” e “b”“A” simbolizava flores púrpuras, “a” flores brancas, “B” hastes lcurtas. Nos cruzamentos dos parentais ele utilizou a combinação das que apresentavam forma AAbb (flores púrpuras e hastes curtas) com a forma aabb (flores brancas e hastes curtas), e aaBB (flores brancas e hastes longas) foi fertilizadoaabb (flores brancas e hastes curtas). Em F1, como resultado dos cruzamentos obteve plantas híbridas Aabb (flores púrpuras e hastes curtas) do primeiro(flores brancas e hastes longas), do segundo. Para a produção de F2 utilizou como
Processos Reprodutivos
transmissão descrito acima. Ele estudou cada uma das sete características listadas na separadamente e obteve para todos os resultados semelhantes.
Dessa forma, quando o monge observou a característica “cor da vagem”, por exemplo, notou que cruzando plantas puras que exibiam vagem verde com plantas puras que apresentavam vagem amarela, obtinha em F1 100% de plantas híbridas
cujas vagens eram de coloração verde. A geração bia aproximadamente 3 plantas que formavam vagens verdes para
cada planta que formava vagem amarela. De fato, os dados obtidos por Mendel para a citada característica foram 428 plantas produtoras de vagens verdes e 152 plantas
, ou seja, uma proporção de 2,82:1 (lê-se 2,82 verdes
O “Pai da Genética” então denominou de “característica dominante” aquela que era observada em 100% de F1 e em ¾ de F2. A característica alternativa, que
s F2, ele chamou de “característica recessiva”. que os “fatores” responsáveis pela característica
recessiva deveriam estar presentes em F1, mas de forma “encoberta”, e que quando foram transmitidos para F2, se manifestaram apenas nos indivíduos puros, formando
Na tentativa de explicar seus resultados, formou a hipótese de que as características estudadas eram condicionadas por aquilo que ele chamou de “fatores” (hoje, nomeamos tais fatores de GENES), e que estes “fatores” deviam existir aos
os fatores seriam separados de forma que um gameta possuiria somente uma unidade de cada fator. Após os cruzamentos, os gametas masculinos fecundariam os femininos e originariam indivíduos filhos.
, os novos organismos seriam novamente portadores de um par de cada fator, sendo uma unidade proveniente do pai e outra da mãe.
Todas essas considerações e explicações elaboradas por Mendel podem ser Primeira Lei de Mendel ou Lei da Pureza dos
Após analisar as características individualmente, Mendel elaborou experimentos realizando cruzamentos e analisando as características duas a duas. Seus resultados demonstravam que, ao cruzar duas linhagens puras, cada uma das quais exibindo duas características distintas, em F1 todos os descendentes exibiam somente as características denominadas dominantes. Já em F2, originário do cruzamento entre indivíduos F1, existia indivíduos exibindo diferentes combinações
Em um dos seus experimentos, em que observou a cor das flores e o comprimento das hastes simultaneamente (veja Esquema 2), Mendel previu que, se sua hipótese estivesse correta, em F2 cada característica seria exibida em 50% das plantas. Os vários caracteres podem ser identificados por “A”, “B”, “a” e “b”, onde “A” simbolizava flores púrpuras, “a” flores brancas, “B” hastes longas, e “b” hastes curtas. Nos cruzamentos dos parentais ele utilizou a combinação das que apresentavam forma AAbb (flores púrpuras e hastes curtas) com a forma aabb (flores
aaBB (flores brancas e hastes longas) foi fertilizado com aabb (flores brancas e hastes curtas). Em F1, como resultado dos cruzamentos obteve plantas híbridas Aabb (flores púrpuras e hastes curtas) do primeiro, e plantas aaBb (flores brancas e hastes longas), do segundo. Para a produção de F2 utilizou como
246

Módulo V – Processos Reprodutivos
planta doadora do óvulo os híbridos Aabb (o que possibilitava a produção de óvulos Ab e ab) e o pólen produzido pelos híbridos aaBb (os grãos de pólen deveriam portar os fatores aB ou ab). As plantas produzidas em F2, então, deveriam ser AaBb (flores púrpuras e hastes longas) + aaBb (flores brancas e hastes longas) + Aabb (flores púrpuras e hastes curtas)+ aabb (flores brancas e hastes curtas).
Esquema 2: Cruzamento diibrido realizado por Gregor Mendel, considerando as características cor das flores e com
Das 45 fertilizações em F1 que realizou, resultaram 187 sementesproduziram flores em F2. Dentre estflores brancas ou, levando em consideração o tamanho da hascontra 79 de hastes curtas. Considerando a combinação das características47 plantas com hastes longas e flores púrpuras, 40 com hastes longas e flores brancas, 38 de hastes curtas e flores púrpuras e, finalmente, 41 planbrancas. Este experimento podia ser satisfatoriamente explicado por sua teoria!
Outros experimentos considerando pares de caracteres foram realizados e os resultados obtidos estavam em perfeita concordância com sua teoria de qervilhas híbridas formam óvulos e células de pólen as quais, em suas constituições, representam em igual número todas as formas constantes as quais resultam das combinações das características unitárias na fertilização
Tais experimentos levaram àSegregação Independente ou Segunda Lei de Mendel.
X
Gametas Ab
Gametas Ab
47
AAbb aabb
Aabb
AaBb
X
Gametas Ab
Gametas Ab
47
AAbb aabb
Aabb
X
Gametas Ab
Gametas Ab
47
X
Gametas Ab
X
Gametas AbGametas Ab
Gametas Ab
47
Gametas AbGametas Ab
4747
AAbb aabb
Aabb
AaBb
Processos Reprodutivos
lanta doadora do óvulo os híbridos Aabb (o que possibilitava a produção de óvulos Ab e ab) e o pólen produzido pelos híbridos aaBb (os grãos de pólen deveriam portar os fatores aB ou ab). As plantas produzidas em F2, então, deveriam ser AaBb (flores
as e hastes longas) + aaBb (flores brancas e hastes longas) + Aabb (flores púrpuras e hastes curtas)+ aabb (flores brancas e hastes curtas).
Cruzamento diibrido realizado por Gregor Mendel, considerando as características cor das flores e comprimento da haste, em ervilhas.
Das 45 fertilizações em F1 que realizou, resultaram 187 sementes, das quais 166 duziram flores em F2. Dentre estas, 85 plantas apresentavam flores púrpuras e 81
flores brancas ou, levando em consideração o tamanho da haste, 87 com hastes longas contra 79 de hastes curtas. Considerando a combinação das características, observou 47 plantas com hastes longas e flores púrpuras, 40 com hastes longas e flores brancas, 38 de hastes curtas e flores púrpuras e, finalmente, 41 plantas de hastes curtas e flores
e experimento podia ser satisfatoriamente explicado por sua teoria!
Outros experimentos considerando pares de caracteres foram realizados e os resultados obtidos estavam em perfeita concordância com sua teoria de que “as ervilhas híbridas formam óvulos e células de pólen as quais, em suas constituições, representam em igual número todas as formas constantes as quais resultam das combinações das características unitárias na fertilização” (Mendel, 1865).
experimentos levaram à elaboração do que conhecemos como Lei da Segunda Lei de Mendel.
Geração Parental
X
ab abaB
Geração F1
X
ab abaB
Geração F2
40 38 41
aabbaaBB aabb
aaBb
Aabb aabb aaBb
Geração Parental
X
ab abaB
Geração F1
X
ab abaB
Geração F2
40 38 41
aabbaaBB aabb
aaBb
Geração Parental
X
ab abaB
Geração F1
X
ab abaB
Geração F2
40 38 41
Geração Parental
X
ab abaB
Geração Parental
X
abab abaB abaB
Geração F1
X
ab abaB
Geração F2
40 38 41
Geração F1
X
abab abaB abaB
Geração F2
40 38 41
Geração F2
40 38 41
aabbaaBB aabb
aaBb
Aabb aabb aaBb
247

248
Módulo V – Processos Reprodutivos
AS LEIS DE MENDEL
Primeira Lei de Mendel ou Lei da Pureza dos Gametas: “As características dos indivíduos são condicionadas por pares de fatores (genes), que se separam durante a formação dos gametas, indo apenas um fator do par para cada gameta”.
Segunda Lei de Mendel ou Lei da Segregação Independente: “Na herança de duas ou mais características, os fatores, segregados na formação dos gametas, não se fundem no híbrido, mas se distribuem independentemente nos gametas segundo todas as combinações possíveis”.
Meiose e sua relação com os “Fatores Mendelianos”
Como dito anteriormente, atualmente chamamos os fatores mendelianos de genes. Sabemos também que os genes estão situados nos cromossomos e que estes, por sua vez, segregam-se durante a meiose para originar gametas haplóides, ou seja, com metade do número de cromossomos que define a espécie. Vejamos o Esquema 3 relacionando a segregação dos genes durante a meiose.
Esquema 3: Segregação de um par de genes alelos durante a meiose, em um indivíduo heterozigoto (Aa).
Cromossomo portando gene A
Cromossomo portando gene a
Cromossomo portando gene A
Cromossomo portando gene a
Cromossomo portando gene A
Cromossomo portando gene a

Módulo V – Processos Reprodutivos
Inicialmente, vamos observar o que acontece com um par de genes alelos, segregando em um indivíduo heterozigoto para cromossomos se segregam na meiose, junto com eles também são segregados os genes. Se considerarmos dois genes heterozigotos durante a segregaçãoa formação de quatro tipos diferentes de gametas (gametas que possuem exatamente a mesma combinação que o indivíduo recebeu de seus pais são chamados de “parentais”combinação diferente são chamados de “recombinantes”.
Esquema 4: Segregação de dois pares de genes alelos durante a meiose, em um indivíduo duplo heterozigoto (AaBb)
Agora que você sabe como os genes alelos se separam na meiose, para a formação de gametas haplóides, podemos então montar um esquema, tanto para definir os possíveis gametas produzidos pelo indivíduoformados a partir do cruzamento de casais.
Um dos métodos utilizados é conhecido por “quadrado de Punnet”, que permite interpretar o comportamento dos cromossomos durante a meiose, combinando, simultaneamente, com a herança das características. Por convenção, utilizamos letras maiúsculas para designar características dominantesrecessivas são assinaladas por letras minúsculas.
Outra convenção é que a letra utilizada para designar a característica, geralmente, é a primeira letra do nome da caracteríde vagem das ervilhas, usamos “r” de rugoso para designar a característica, sendo que R denomina do caráter dominante (vagens lisas) e rugosa).
Exemplo 1: Suponha um indivíduo heterozigoto pem cromossomos diferentes: AaBb. Utilizando o quadrado de Punnet (veja instruções a seguir), podemos definir os gametas que serão produzidos pelo referido indivíduo.
a) Desenhe uma tabela com igual nque o número de linhas seja igual ao número de genes estudados + 1.
b) Coloque na primeira linha, segunda colunaprimeiro gene e, na terceira coluna, o segundo alelo do mesmo gene
Processos Reprodutivos
Inicialmente, vamos observar o que acontece com um par de genes alelos, segregando em um indivíduo heterozigoto para formação de gametas. Quando os cromossomos se segregam na meiose, junto com eles também são segregados os genes. Se considerarmos dois genes heterozigotos durante a segregação, será possível a formação de quatro tipos diferentes de gametas (veja o Esquema 4). Aqueles dois gametas que possuem exatamente a mesma combinação que o indivíduo recebeu de seus pais são chamados de “parentais”, enquanto aqueles que exibem uma
são chamados de “recombinantes”.
Segregação de dois pares de genes alelos durante a meiose, em um indivíduo duplo heterozigoto (AaBb)
Agora que você sabe como os genes alelos se separam na meiose, para a formação de gametas haplóides, podemos então montar um esquema, tanto para
metas produzidos pelo indivíduo como os indivíduos formados a partir do cruzamento de casais.
Um dos métodos utilizados é conhecido por “quadrado de Punnet”, que permite interpretar o comportamento dos cromossomos durante a meiose,
com a herança das características. Por convenção, utilizamos letras maiúsculas para designar características dominantes, enquanto as
inaladas por letras minúsculas.
Outra convenção é que a letra utilizada para designar a característica, é a primeira letra do nome da característica. Por exemplo, no caso do tipo
de vagem das ervilhas, usamos “r” de rugoso para designar a característica, sendo denomina do caráter dominante (vagens lisas) e r o caráter recessivo (vagem
Suponha um indivíduo heterozigoto para dois genes localizados em cromossomos diferentes: AaBb. Utilizando o quadrado de Punnet (veja instruções a seguir), podemos definir os gametas que serão produzidos pelo referido indivíduo.
Desenhe uma tabela com igual número de linhas e colunas, de forma o número de genes estudados + 1.
b) Coloque na primeira linha, segunda coluna, o primeiro alelo do o segundo alelo do mesmo gene.
OUOU
249
Quadrado de Punnet: Consiste em um quadro, com número de fileiras e de colunas que correspondem, respectivamente, aos tipos de gametas masculinos e femininos formados no cruzamento.

250
Módulo V – Processos Reprodutivos
c) Coloque na segunda linha, primeira coluna, o primeiro alelo do segundo gene e, na terceira linha, o segundo alelo.
d) Complete os quadros colocando o equivalente existente na primeira linha e primeira coluna.
alelos A A B AB aB b Ab Ab
No final do preenchimento, você terá nas casas, exceto da primeira linha da primeira coluna, os gametas possíveis de serem formados por esse indivíduo.
Exemplo 2: Outro método pelo qual podemos igualmente definir os gametas facilmente é descrito a seguir:
a) Coloque em linhas consecutivas os alelos de cada um dos genes.
b) Faça linhas unindo um alelo de cada gene por vez e copie os alelos formando pares, como mostrado no esquema abaixo.
Utilizando os mesmos métodos descritos anteriormente, você pode esquematizar também os possíveis nascimentos provenientes de cruzamentos entre indivíduos.
Exemplo 3: Um casal formado por dois indivíduos heterozigotos para uma dada característica formará a seguinte prole:
a)
Gametas A A A AA AA a Aa AA
Alelos gametas A AB a a B B Ab b
a b
Gametas
Prole

Módulo V – Processos Reprodutivos
Gametas ♀
A
a gametas ♂
A
a
b)
ATIVIDADE
Fixando a aprendizagem:
Exercício 1: Utilizando um dos mindivíduos provenientes dos cruzamentos abaixo:
a) Cruzamento entre um indivíduo duplo heterozigoto (AaBa) e um indivíduo AABb.
b) Cruzamento entre indivíduo triplo heterozigoto e um triplo homozigoto
c) Cruzamento entre indivíduo homozigoto para dois genes e heterozigoto para um terceiro gene e um indivíduo triplo heterozigoto
Exercício 2: Realize um cruzamento entre dois indivíduos puros (homozigotos), sendo um portando o gene dominante e o outro o genApós, realize cruzamentos entre a F1 resultante e determine a proporção de genótipos esperados. Considerando que o alelo A determina o caráter A e o alelo o caráter recessivo “a”, que só será manifestado quando em homozigoseproporção fenotípica esperada em F2?
Exercício 3: Agora refaça o exercício 2, considerando dois pares de genes aleAa e BB, simultaneamente.
Padrões de HerançaVimos até aqui como Gregor Mendel, utilizando
reconheceu a transmissão das características hereditárias em ervilhas de jardim, iniciando assim uma nova era no entendimento de como se dão as das famílias.
Somente por volta do ano de cromossomos dentro do núcleo eram os portadores dos fatores hereditários“estruturas em fila ou cadeia que se autoduplicam quando a célula se dividia”
Processos Reprodutivos
PROLE
AA
Aa
Aa
aa
dos métodos descritos, determine os gametas e os indivíduos provenientes dos cruzamentos abaixo:
a) Cruzamento entre um indivíduo duplo heterozigoto (AaBa) e um indivíduo
b) Cruzamento entre indivíduo triplo heterozigoto e um triplo homozigoto.
Cruzamento entre indivíduo homozigoto para dois genes e heterozigoto para um terceiro gene e um indivíduo triplo heterozigoto.
Realize um cruzamento entre dois indivíduos puros (homozigotos), sendo um portando o gene dominante e o outro o gene recessivo. Após, realize cruzamentos entre a F1 resultante e determine a proporção de genótipos esperados. Considerando que o alelo A determina o caráter A e o alelo “a”, o caráter recessivo “a”, que só será manifestado quando em homozigose, qual a
rção fenotípica esperada em F2?
Agora refaça o exercício 2, considerando dois pares de genes alelos,
Padrões de Herança Vimos até aqui como Gregor Mendel, utilizando-se do método científico,
reconheceu a transmissão das características hereditárias em ervilhas de jardim, iniciando assim uma nova era no entendimento de como se dão as heranças dentro
volta do ano de 1883, Wilhelm Roux postulou que os cromossomos dentro do núcleo eram os portadores dos fatores hereditários -- “estruturas em fila ou cadeia que se autoduplicam quando a célula se dividia”. Em
251
Você sabia que a maioria dos animais e plantas seguem, basicamente, as mesmas regras de transmissão genética que humanos? Pois é, a reprodução envolve a presença de dois sexos e a produção de gametas (reprodução sexuada), temos os mesmos modelos de transmissão de genes.

Módulo V – Processos Reprodutivos
1902, Walter Sutton postula que os genes estatermo “gene” foi utilizado pela primeira vez por botânico dinamarquês, no ano de 1909.
Para a interpretação dos modos de herança exibidos pelas características hereditárias nas famílias precisamos identificar se o gene responsável pela característica está localizado em um um cromossomo alossomo (sexual)se manifesta sempre que um determinado alelo estádeterminado alelo está presente em dos
HEREDOGRAMA é a forma esquemática para se representar uma famíliaforma que podemos identificar o sexo dos indivíduos, suas posições relativas em seu núcleo familiar e se exibem ou não uma característica. Para tanto, existem símbolos utilizados internacionalmente, ou seja, convencionados seus significados. Por exemplo:
Dessa forma, podemos identificar um padrão de herança de um gene que está segregando em um cromossomo autossômico se:
1. A característica é observada em igual proporção tanto noquanto feminino.
2. Homens e mulheres transmitem a cproporção, independente do sexo.
Quando um gene está localizado na região não homóloga do cromossomo X, isto é, não existe um locus alelo no cromossomo Y, dizemos que sua herança é uma Herança Ligada ao X ou Herança Ligada ao Sexo
• A característica é observada em proporção desigual quando consideramos o sexo do indivíduo.
• Homens afetados NUNCA transmitem a característica para seus FILHOS, mas SIM para suas FILHASdependendo se a mesma exibe um padrão de Dominância ou Recessividade
• NETOS de homens afetados, filhos de suas filhas, poderão exibir a característica.
Será observada uma proporção de aproximadamente 50% de FILHOS afetados nascidos de mulheres heterozigotas para a característica em questão.
2ª. geração
1ª geração
Núcleo familiar identificando o probando ou caso índice, ou seja, o indivíduo através do qual se chegou à família em questão.
1ª geração
2ª geração
Probando
Processos Reprodutivos
que os genes estavam localizados nos cromossomos. O termo “gene” foi utilizado pela primeira vez por Wilhelm Ludvig Johannsen, um
no ano de 1909.
Para a interpretação dos modos de herança exibidos pelas características amos identificar se o gene responsável pela
característica está localizado em um cromossomo autossômico (não sexual) ou em ). Após, é necessário identificar se a característica
se manifesta sempre que um determinado alelo está presente ou se somente quando determinado alelo está presente em dose dupla, isto é, em homozigose.
é a forma esquemática para se representar uma família, de forma que podemos identificar o sexo dos indivíduos, suas posições relativas em seu
se exibem ou não uma característica. Para tanto, existem símbolos utilizados internacionalmente, ou seja, convencionados seus significados. Por
Dessa forma, podemos identificar um padrão de herança de um gene que está segregando em um cromossomo autossômico se:
A característica é observada em igual proporção tanto no sexo masculino
Homens e mulheres transmitem a característica para todos os filhos, em igual
Quando um gene está localizado na região não homóloga do cromossomo X, isto é, não existe um locus alelo no cromossomo Y, dizemos que sua herança é uma
Herança Ligada ao Sexo. O padrão observado neste caso é:
A característica é observada em proporção desigual quando
Homens afetados NUNCA transmitem a característica para seus mas SIM para suas FILHAS, que poderão ou não exibir a característica,
dependendo se a mesma exibe um padrão de Dominância ou Recessividade.
NETOS de homens afetados, filhos de suas filhas, poderão exibir a
uma proporção de aproximadamente 50% de FILHOS afetados ascidos de mulheres heterozigotas para a característica em questão.
Mãe falecida
Núcleo familiar identificando o probando ou caso índice, ou seja, do qual se chegou à família em questão.
Mãe falecida
Probando
252

253
Módulo V – Processos Reprodutivos
5b- Herança autossômica recessiva
• Manifesta-se em ambos os sexos com aproximadamente mesma freqüência;
• Pula gerações, ou seja, afetados geralmente nascem de pais não afetados;
• Transmitida igualmente por homens e mulheres;
• Geralmente aparece em prole descendente de casamento consangüíneo.
5 a- Herança autossômica dominante
• Manifesta-se em ambos os
sexos com aproximadamente
mesma freqüência;
• Não pula gerações;
• Transmitida igualmente por homens e mulheres;
• Filhos de casais onde um dos
conjuges é afetado tem 50% de
chances de nascerem afetados.
5c- Herança dominante ligada ao sexo
• Manifesta-se em ambos os sexos com freqüência aumentada no sexo feminino;
• Indivíduos afetados possuem ao menos um dos genitores afetados;
• Afetados do sexo masculino transmitem a característica para 100% de suas filhas mas nunca para seus filhos;
• Afetados do sexo feminino transmitem tanto para seus filhos como filhas, com igual probabilidade;
• Freqüentemente os indivíduos femininos são afetados de forma mais leve e variável do que os masculinos.
5d- Herança recessiva ligada ao sexo
• Afeta principalmente os indivíduos masculinos;
• Os afetados em geral nascem de genitores não-afetados;
• Nos casos em que o pai é afetado e a mãe é portadora indivíduos femininos podem ser afetados;
• Não há transmissão de indivíduo masculino para indivíduo masculino na genealogia.
Tendo determinado a localização do gene em análise em um cromossomo sexual ou não, resta então determinar a relação existente entre os alelos deste gene. Basicamente, podemos identificar três relações entre alelos de um mesmo gene: dominância, recessividade ou ausência de dominância (veja abaixo o Esquema 5).
Esquema 5: Representação exemplificando heredogramas representativos de diferentes padrões de herança mendeliana.
Para uma característica que exibe um Padrão de Herança Autossômica Dominante (veja no Esquema 5 a Fugura 5a), a característica analisada será evidenciada em um heredograma de forma que preencha os requisitos vistos anteriormente para uma herança autossômica e:
1. Manifestar-se em todas as gerações.
2. Uma criança afetada tem, no mínimo, um dos pais também exibindo a característica.
De modo similar, podemos definir um Padrão de Herança Autossômica Recessiva (veja no Esquema 5 a Figura 5b) se, além de preencher os requisitos de uma herança em cromossomo não sexual, ter as seguintes características:
1. Pula gerações, isto é, uma criança exibindo a caraterística possui pais não afetados.
2. Normalmente sua ocorrência é observada em filhos de casais consanguíneos (parentes “de sangue”).

254
Módulo V – Processos Reprodutivos
Se, entretanto, o gene determinante da característica estiver localizado na região não homóloga do cromossomo X, tendo o alelo responsável pela característica em questão uma relação de dominância em relação à característica alternativa, dizemos que seu padrão é de Herança Dominante Ligada ao X ou dominante ligada ao sexo (veja no Esquema 5 a Figura 5c), e serão observadas no heredograma:
1. Homens afetados transmitem a característica para 100 % de suas FILHAS e para nenhum de seus filhos.
2. Mulheres afetadas, quando heterozigotas, transmitem a característica para 50% de seus filhos ou filhas, independente do sexo. Se homozigotas, 100% de sua prole exibirá também a característica.
Em se tratando de uma característica ligada ao X que exiba uma relação de RECESSIVIDADE em relação ao caráter alternativo, teremos uma Herança Recessiva Ligada ao X ou Recessiva Ligada ao Sexo (veja no Esquema 5 a Figura 5d), obedecendo os seguintes critérios:
1. Raras em mulheres, pois nessas, para se manifestar, o gene precisa estar em homozigose;
2. Filhas de homens afetados são, geralmente, portadoras assintomáticas, isto é, possuem o gene alelo determinante da característica em heterozigose e, consequentemente, exibem o fenótipo alternativo.
3. Portadoras assintomáticas transmitem o gene para 50% de sua prole, sendo que, exceto se casada com um afetado, somente 50% dos FILHOS apresentarão a característica.
HERANÇAS RELACIONADAS AO SEXO
Os cromossomos alossomos, em humanos, são representados por dois pares parcialmente homólogos: cromossomo X e cromossomo Y. O cromossomo Y é menor que o X, possui menos genes e, nele, estão localizados os genes determinantes para o desenvolvimento dos caracteres sexuais masculinos. Comparativamente, podemos dizer que existem nestes cromossomos uma região que é compartilhada por ambos (região homóloga) e uma outra região onde os genes presentes não possuem seu respectivo locus no outro cromossomo (região não homóloga). Dessa forma, temos dentre as heranças determinadas por genes localizados em alossomos ou por genes autossômicos, mas cujo efeito sofre influência dos hormônios sexuais. Além das já anteriormente estudadas, as Heranças ligadas ao sexo apresentam-se como:
• Herança restrita ao sexo: localizada na porção não homóloga do cromossomo Y. Tais genes são chamados genes holândricos e são, consequentemente, transmitidos e expressos exclusivamente em homens. Exemplo: hipertricose auricular.
• Herança influenciada pelo sexo: genes localizados em cromossomos autossomos que sofrem influência dos hormônios sexuais. Como exemplo podemos citar o gene da calvicie humana e a produção de leite em bovinos.

Módulo V – Processos Reprodutivos
ATIVIDADE
Observe nas pessoas de sua família se possuem o lóbulo da orelha solto ou pregado. Elabore o heredograma assinalando os indivíduos que possuem lóbulo da orelha solto. Quanto maior o número de gerações sobre as quais você informações, melhor será o resultado de suas observações. Analise o heredograma obtido e determine o padrão de herança da característica “lóbulo da orelha solto”. Repita o procedimento, agora assinalando no heredograma as pessoas que não possuem a capacidade de enrolar a língua.
Padrão de Herança com Ausência de Dominância ou Codominante
Quando a característica exibe uma relação de ausência de dominância entre os alelos do gene responsável pela mesma, podexibir uma característica intermediária entre os fenótipos homozigotos. Esherança é muito comum em vegetais ou quando se analisa a expressão de uma proteína no sangue. Então, se considerarmos a chamada de Maravilha, notaremos que existe uma gama de fenótipos relativos coloração de suas pétalas. Assim, ao realizarmos o cruzamento entre um indivíduo que produz flores cor maravilha com um outro que produz flores cor branca, a prole resultante produzirá flores cor-de-rosa
Em “linguagem genética” podemos definir como A = alelo responsável pela manifestação do caráter flores maravilha, edo caráter flores brancas. Dessa forma, os indivíduos homozigotos AA produzem flores maravilha, enquanto os homozigotos aaheterozigotos, Aa, produzem flores exibindo um fenótipo intermédiario entre os fenótipos apresentados pelos dois homozigotos, ou seja, flores cor
É muito importante notar que a expressão fenotípica de um gene alelo não significa necessáriamente uma variação patológica! A variabilidade genética observada a partir da existência de genes alelos é a responsável, em grande parte, pela identidade fenotípica dos indivíduos. Se esindivíduos seriam idênticos!
O gene da Hemoglobina (Hb) humana, que possui uma vasta gama de tipos alélicos, pode ser utilizado como exemplo de Herança Autossômica Recessivamesmo modo que Herança Autossômica Codominante, dependendo da forma como o fenótipo é analisado. Se levarmos em conta o alelo Hb*S, responsável pela produção de uma hemoglobina pouco eficaz na troca de gases, identificamos o fenótipo Anemia Falciforme em indivíduos homozifatal se o paciente não receber acompanhamento médico adequado. Entretanto, quando em heterozigose, raramente o indivíduo portador do alelo S manifestará fenótipo patológico. Consideramosfenótipo recessivo e, consequentemente, exibindo uma Herança Autossômica Recessiva. Já quando analisamos em laboratório, o sangue de indivíduos falcêmicos e de indivíduos portadores assintomáticos da Hb*S, podemos reconhecer o homozigotoHb*S/Hb*S do heterozigoto Hb*A/Hb*S, bem como do homozigoto normal Hb*A/ Hb*A. Sob este ponto de vista o gene da hemoglobina pode ser considerado exibindo uma Herança Autossômica Codominante ou Herança Autossômica Sem Dominância.
Processos Reprodutivos
Observe nas pessoas de sua família se possuem o lóbulo da orelha solto ou pregado. Elabore o heredograma assinalando os indivíduos que possuem lóbulo da orelha solto. Quanto maior o número de gerações sobre as quais você obtiver informações, melhor será o resultado de suas observações. Analise o heredograma obtido e determine o padrão de herança da característica “lóbulo da orelha solto”. Repita o procedimento, agora assinalando no heredograma as pessoas que não
de de enrolar a língua.
Padrão de Herança com Ausência de Dominância ou Codominante
Quando a característica exibe uma relação de ausência de dominância entre os alelos do gene responsável pela mesma, podemos identificar o heterozigoto que irá
aracterística intermediária entre os fenótipos homozigotos. Este tipo de herança é muito comum em vegetais ou quando se analisa a expressão de uma
, se considerarmos a Mirabillis japala, planta popularmente mos que existe uma gama de fenótipos relativos à
coloração de suas pétalas. Assim, ao realizarmos o cruzamento entre um indivíduo que produz flores cor maravilha com um outro que produz flores cor branca, a prole
rosa.
Em “linguagem genética” podemos definir como A = alelo responsável pela o do caráter flores maravilha, e a = alelo responsável pela manifestação
do caráter flores brancas. Dessa forma, os indivíduos homozigotos AA produzem nquanto os homozigotos aa produzem flores brancas e o
heterozigotos, Aa, produzem flores exibindo um fenótipo intermédiario entre os fenótipos apresentados pelos dois homozigotos, ou seja, flores cor-de-rosa.
É muito importante notar que a expressão fenotípica de um gene alelo não variação patológica! A variabilidade genética
observada a partir da existência de genes alelos é a responsável, em grande parte, ca dos indivíduos. Se esta variação não existisse, todos os
O gene da Hemoglobina (Hb) humana, que possui uma vasta gama de tipos pode ser utilizado como exemplo de Herança Autossômica Recessiva, do
erança Autossômica Codominante, dependendo da forma como o fenótipo é analisado. Se levarmos em conta o alelo Hb*S, responsável pela produção de uma hemoglobina pouco eficaz na troca de gases, identificamos o fenótipo Anemia Falciforme em indivíduos homozigotos para o referido alelo. Tal patologia é fatal se o paciente não receber acompanhamento médico adequado. Entretanto, quando em heterozigose, raramente o indivíduo portador do alelo S manifestará fenótipo patológico. Consideramos, assim, o alelo S como responsável por um fenótipo recessivo e, consequentemente, exibindo uma Herança Autossômica Recessiva. Já quando analisamos em laboratório, o sangue de indivíduos falcêmicos e de indivíduos portadores assintomáticos da Hb*S, podemos reconhecer o homozigoto Hb*S/Hb*S do heterozigoto Hb*A/Hb*S, bem como do homozigoto normal Hb*A/
e ponto de vista o gene da hemoglobina pode ser considerado exibindo uma Herança Autossômica Codominante ou Herança Autossômica Sem Dominância.
255

Módulo V – Processos Reprodutivos
ATIVIDADE
Faça uma pesquisa a respeito dos tipos de heranças em humanos e elabore um quadro onde se possa encontrar, no mínimo, um exemplo de variação normal e um de variação patológica na espécie humana, assinalando o tipo de herança exibida por tal variação. Se preferir, use como base o quadro a seguir, como no exemplo.
T i p o d e H e r a n ç a A u t o s s ô m i c a
Dominan te Reces s iv
Variação n o r m a l Fator Rh
patológica albinismo
Padrões não Mendelianos de herança
Polialelia
Até agora estudamos, basicamente, caracteristicas determinadas por genes que possuem somente dois alelos. Entretanto, um mesmo sob múltiplas formas alélicas, fato esvários alelos de um mesmo gene podem exibir diferentes relações entre seus produtos alélicos, de forma que em um mesmo Sistema Genético podemos encontrar alelos com relação de codominância entre si e, ao mesmo tempo, com relação de dominância e recessividade em relação a um terceiro alelo. Tal é o caso ddeterminantes do grupo sanguíneo ABO em humanos (Quadro 1).
Um outro exemplo clássico de polialelia é o do sistema genético determinante da cor de pelagem em coelhos. Nesse sistemaque exibe, na população, quatro alelos diferentes: aguthimalaia e albino, sendo que aguti enquanto albino é recessivo em relação a todos os outros. Já alelo himalaia.
ATIVIDADE
Considerando as características dos alelos responsáveis pela pelagem de coelhos, faça o esquema de um cruzamento entre macho agutis heterozigoto para chinchila, e uma fêmea himalaia heterozigota para o alelo albino.
1. Qual será a prole resultante em F1?
2. Se for permitido que os indivíduos F1 se acasalem, quais os tipos de acasalamentos possíveis e quais as respectivas proles?
Processos Reprodutivos
pesquisa a respeito dos tipos de heranças em humanos e elabore um quadro onde se possa encontrar, no mínimo, um exemplo de variação normal e um de variação patológica na espécie humana, assinalando o tipo de herança exibida por
se como base o quadro a seguir, como no exemplo.
A u t o s s ô m i c a L i g a d a a o S e x o C o d o m i n a n t e
Reces s iva Dominan te R e c e s s i v a
Fator Rh
albinismo
Padrões não Mendelianos de herança
Até agora estudamos, basicamente, caracteristicas determinadas por genes que possuem somente dois alelos. Entretanto, um mesmo locus gênico pode apresentar-se
ltiplas formas alélicas, fato este designado por polialelia ou multialelia. Esses elos de um mesmo gene podem exibir diferentes relações entre seus
produtos alélicos, de forma que em um mesmo Sistema Genético podemos encontrar alelos com relação de codominância entre si e, ao mesmo tempo, com relação de
ação a um terceiro alelo. Tal é o caso dos alelos determinantes do grupo sanguíneo ABO em humanos (Quadro 1).
Um outro exemplo clássico de polialelia é o do sistema genético determinante da cor de pelagem em coelhos. Nesse sistema, encontramos um gene autossômico que exibe, na população, quatro alelos diferentes: aguti ou selvagem, chinchila,
é dominante em relação a todos os outros três, enquanto albino é recessivo em relação a todos os outros. Já a chinchila domina o
sticas dos alelos responsáveis pela pelagem de coelhos, faça o esquema de um cruzamento entre macho agutis heterozigoto para
e uma fêmea himalaia heterozigota para o alelo albino.
prole resultante em F1?
Se for permitido que os indivíduos F1 se acasalem, quais os tipos de e quais as respectivas proles?
256
Saiba mais sobre a genética dos grupos sanguíneos pesquisando no livro “Os sistemas sanguíneos eritrocitários” (BEIGUELMAN, B, 3a. ed. Ribeirão Preto, Funpec Editora, 2003).
Módulo V – Processos Reprodutivos
Alelos do sistema ABO
Genótipos possíveis
Fenótipos
A ( IA) IA IA IA i
Sangue tipo A
IA IB
Sangue tipo AB
B ( IB) IB IB
IB i Sangue tipo B
O ( i ) I i Sangue tipo O
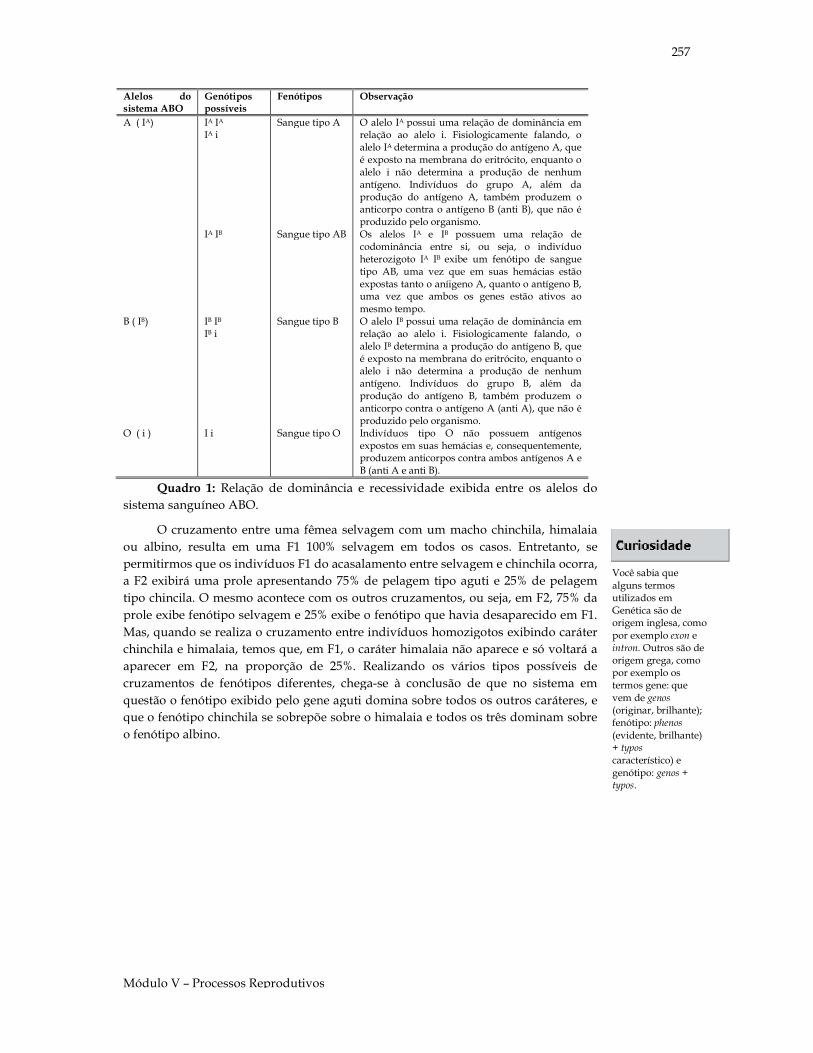
Quadro 1: Relação de dominância e recessividade exibida entre os alelos do sistema sanguíneo ABO.
O cruzamento entre uma fêmea selvagem com um macho chinchila, hiou albino, resulta em uma F1 100% selvagempermitirmos que os indivíduos F1 do acasalamento entre selvagem e chinchila ocorra, a F2 exibirá uma prole apresentando 75% de pelagem tipo aguti e 25% de pelagem tipo chincila. O mesmo acontece com os outros cruzamentos, ou seja, em F2, 75% da prole exibe fenótipo selvagem e 25% exibe o fenótipoMas, quando se realiza o cruzamento entre indivíduos homozigotos exibindo caráter chinchila e himalaia, temos que, em F1, o caráter himalaia não aparece e só voltará a aparecer em F2, na proporção de 25%. Realizando os vários tipos posscruzamentos de fenótipos diferentes, chegaquestão o fenótipo exibido pelo gene aguti domina sobre todos os outros caráteresque o fenótipo chinchila se sobrepõe sobre o himalaia e todos os três dominam sobre o fenótipo albino.
Processos Reprodutivos
Fenótipos Observação
Sangue tipo A O alelo IA possui uma relação de dominância em relação ao alelo i. Fisiologicamente falando, o alelo IA determina a produção do antígeno A, que é exposto na membrana do eritrócito, enquanto o alelo i não determina a produção de nenhum antígeno. Indivíduos do grupo A, além da produção do antígeno A, também produzem o anticorpo contra o antígeno B (anti B), que não é produzido pelo organismo.
Sangue tipo AB Os alelos IA e IB possuem uma relação de codominância entre si, ou seja, o indivíduo heterozigoto IA IB exibe um fenótipo de sangue tipo AB, uma vez que em suas hemácias estão expostas tanto o aníigeno A, quanto o antígeno B, uma vez que ambos os genes estão ativos ao mesmo tempo.
Sangue tipo B O alelo IB possui uma relação de dominância em relação ao alelo i. Fisiologicamente falando, o alelo IB determina a produção do antígeno B, que é exposto na membrana do eritrócito, enquanto o alelo i não determina a produção de nenhum antígeno. Indivíduos do grupo B, além da produção do antígeno B, também produzem o anticorpo contra o antígeno A (anti A), que não é produzido pelo organismo.
Sangue tipo O Indivíduos tipo O não possuem antígenos expostos em suas hemácias e, consequentemente, produzem anticorpos contra ambos antígenos A e B (anti A e anti B).
Relação de dominância e recessividade exibida entre os alelos do
O cruzamento entre uma fêmea selvagem com um macho chinchila, himalaia 1 100% selvagem em todos os casos. Entretanto, se
permitirmos que os indivíduos F1 do acasalamento entre selvagem e chinchila ocorra, a F2 exibirá uma prole apresentando 75% de pelagem tipo aguti e 25% de pelagem
cila. O mesmo acontece com os outros cruzamentos, ou seja, em F2, 75% da prole exibe fenótipo selvagem e 25% exibe o fenótipo que havia desaparecido em F1.
, quando se realiza o cruzamento entre indivíduos homozigotos exibindo caráter em F1, o caráter himalaia não aparece e só voltará a
aparecer em F2, na proporção de 25%. Realizando os vários tipos possíveis de fenótipos diferentes, chega-se à conclusão de que no sistema em
gene aguti domina sobre todos os outros caráteres, e que o fenótipo chinchila se sobrepõe sobre o himalaia e todos os três dominam sobre
257
Você sabia que alguns termos utilizados em Genética são de origem inglesa, como por exemplo exon e intron. Outros são de origem grega, como por exemplo os termos gene: que vem de genos (originar, brilhante); fenótipo: phenos (evidente, brilhante) + typos característico) e genótipo: genos + typos.

Módulo V – Processos Reprodutivos
Interação Gênica
Até agora, mesmo quando tratamos de dois ou mais Mendeliana da Segregação Independente), cada um dos características distintas como, por exemplo, da haste. Entretanto, alguns genes igene não alélico, ou seja, agem conjuntamente na determinação de uma mesma característica fenotípica determinando uma percebe-se uma distorção nas proporções mendelianas.
A forma dos frutos em abóbora é um exemplo desse tipo de fenômeno (1). Para a determinação do fenótipo dois pares de genes não alélicos estão envolvidos, em que a forma de fruto discóide é determinada pela presença, na mesma planta, de no mínimo um AABb, etc. Já frutos esféricos são obtidos toda vez que somente um dos genes possui ao menos um alelo dominante presente. Exemplo: AAbb, Aabb, aaBb,entretanto, o indivíduo apresentar genótipo duplo hoproduzirá frutos alongados. As proporções observadas a partir de um cruzamento entre dois indivíduos duplo heterozigoto será 9/16 de frutos com formas discói6/16 de frutos esféricos e 1/16 de frutos alongados.
Figura 1: Variedades de moranga (Curcubita máxima).http://www.kokopelli-seed-foundation.com/actu/
Existe um tipo especial de interação gênicaoutro não alélico. É a epistasia. Ao gene inibido damos o nome de hipostáticoinibidor, epistático. Um exemplo é a determinação da cor do fruto da amoranga, no qual estão envolvidos os pares de genes Aa e Bb. O gene B produz frutos amarelos, mas na presença do gene A, ele é inibido e produz frutos brancos, como seu recessivo b. O indivíduo duplo homozigoto recessivo, aabb, produzirá frutos verdes. Dizemos que o gene B é hipostático aA
Processos Reprodutivos
quando tratamos de dois ou mais loci simultaneamente (Lei da Segregação Independente), cada um dos loci era responsável por
, por exemplo, a cor da vagem da ervilha e comprimento da haste. Entretanto, alguns genes influenciam no fenótipo determinado por outro gene não alélico, ou seja, agem conjuntamente na determinação de uma mesma característica fenotípica determinando uma interação gênica. Quando isto acontece,
se uma distorção nas proporções mendelianas.
A forma dos frutos em abóbora é um exemplo desse tipo de fenômeno (Figura 1). Para a determinação do fenótipo dois pares de genes não alélicos estão
a forma de fruto discóide é determinada pela presença, na alelo dominante em cada loci. Exemplo: AaBb,
etc. Já frutos esféricos são obtidos toda vez que somente um dos genes possui presente. Exemplo: AAbb, Aabb, aaBb, etc. Se,
o indivíduo apresentar genótipo duplo homozigoto recessivo (aabb), produzirá frutos alongados. As proporções observadas a partir de um cruzamento entre dois indivíduos duplo heterozigoto será 9/16 de frutos com formas discóides,
1/16 de frutos alongados.
Figura 1: Variedades de moranga (Curcubita máxima). Fonte: foundation.com/actu/new_news.cgi?id_news=70
Existe um tipo especial de interação gênica, na qual um gene inibe a ação do Ao gene inibido damos o nome de hipostático, e ao
inibidor, epistático. Um exemplo é a determinação da cor do fruto da abóbora-estão envolvidos os pares de genes Aa e Bb. O gene B produz
frutos amarelos, mas na presença do gene A, ele é inibido e produz frutos brancos, como seu recessivo b. O indivíduo duplo homozigoto recessivo, aabb, produzirá
es. Dizemos que o gene B é hipostático aA , e este é epistático aB.
258
Visite o site: http://www.kokopelli-seed-foundation.com/p/index.html e saiba mais sobre programas de educação e pesquisa em produção de sementes orgânicas, preservação de recursos genéticos, reflorestamento, criação de abelhas e jardins que praticam a agricultura de orgânicos.
O termo locus, do latim “lugar” deve ser destacado no texto, usando fonte em itálico e, quando se referir a dois ou mais locus, utilizar o plural corretamente (em latim, palavras que no singular terminam com “us”, fazem o plural trocando-o por “i”). Ex.: um gene = locus gênico , dois ou mais genes = loci gênicos.
O grupo sanguíneo ABO também pode ser utilizado com um exemplo de epistasia em humanos. O alelo h determina a não produção da substância precursora H. Assim, independente de qual gene A ou B, o indivíduo homozigoto hh não produzirá nenhum antígeno, pois não haverá a produção da substância precursora, exibindo assim o fenótipo O, que manifesta-se da mesma forma que o fenótipo O convencional, ou seja, sem apresentação de antígenos A ou B na

Módulo V – Processos Reprodutivos
VOCÊ SABIA QUE um gene pode influenciar em vários proceao mesmo tempo e determinar vários efeitos fenotípicos ao mesmo tempo?
Esse fenômeno é conhecido como manifestar-se de várias formas. Um exemplo desmanifestações fenotípicas observadas na anemia falciforme, ocasionada pela homozigose do alelo Hb*S. A estrutura primária alterada na proteína produzidapartir deste gene irá conduzir de forma ineficiente os gases no organismo. Ao mesmo tempo, quando em baixa concentração de Opolimeriza-se na hemácia, forçando sua membrana e levando, tomar a forma alongada ou em forma de “foice”, daí o nome da doença. A falcização dos eritrócitos dificulta a passagem dos mesmos nos microcapilconsequentemente levando à não condução de Omesmos e, então, há a trombose e ulceração do desencadeiam um processo doloroso.
ATIVIDADE
Colocando em prática o aprendizado!
1. Com base no que você estudou até aquide variedades de abóbora citados.
2. Em galináceos existem quatro tipose noz. Faça uma pesquisa bibliográfica
a) explique como são determinados tais fenótipos
b) identifique o número de genes e a relação existente entre eles, no referido fenótipo;
c) busque a explicação de como é certas raças de galinhas.
Herança citoplasmática
Você já aprendeu que o DNA, em eucariotos, não se encontra restrito ao núcleo celular, isto é, encontramos também esscitoplasmáticas (mitocôndrias nos animais e cloroplastos nos vegetais).
O DNA mitocondrial caracterizacircular e possuir genes importantes para as funções metabólicas celulares. genes codificam para as proteínas da cadeia reenquanto que os outros demais genes codificam rRNAs e tRNAs para a síntese das proteínas expressas pelo DNA mitocondrial.
As mitocôndrias de um indivíduo são originárias daquelas presentes no óvulo que lhe deu origem. Então, o DNA mitocondrial mesma mitocôndria, encontramos duas ou mais moléculas dcaracterística, origem materna, é de especial interesse para evolução de populações.
Processos Reprodutivos
um gene pode influenciar em vários processos metabólicos vários efeitos fenotípicos ao mesmo tempo?
Esse fenômeno é conhecido como Pleiotropia, e um gene pleiotrópico pode se de várias formas. Um exemplo deste fenômeno são as diversas
manifestações fenotípicas observadas na anemia falciforme, ocasionada pela homozigose do alelo Hb*S. A estrutura primária alterada na proteína produzida a
e gene irá conduzir de forma ineficiente os gases no organismo. Ao mesmo tempo, quando em baixa concentração de O2, tal proteína desestabiliza-se e
se na hemácia, forçando sua membrana e levando, assim, a hemácia a ongada ou em forma de “foice”, daí o nome da doença. A falcização
citos dificulta a passagem dos mesmos nos microcapilares, não condução de O2 até os tecidos banhados pelos
trombose e ulceração do tecido. Todos estes fatores desencadeiam um processo doloroso.
Colocando em prática o aprendizado!
que você estudou até aqui, elabore os esquemas de cruzamento
Em galináceos existem quatro tipos de forma de crista: simples, ervilha, rosa e noz. Faça uma pesquisa bibliográfica, e:
explique como são determinados tais fenótipos;
dentifique o número de genes e a relação existente entre eles, no
usque a explicação de como é determinada a cor das penas em certas raças de galinhas.
Você já aprendeu que o DNA, em eucariotos, não se encontra restrito ao núcleo também essa macromolécula dentro de organelas
animais e cloroplastos nos vegetais).
O DNA mitocondrial caracteriza-se por ser uma molécula de dupla hélice circular e possuir genes importantes para as funções metabólicas celulares. Treze genes codificam para as proteínas da cadeia respiratória e para ATP-sintase, enquanto que os outros demais genes codificam rRNAs e tRNAs para a síntese das
pressas pelo DNA mitocondrial.
um indivíduo são originárias daquelas presentes no óvulo o DNA mitocondrial é de origem materna e, em uma
encontramos duas ou mais moléculas do DNA circular. Sua é de especial interesse para o estudo de dinâmicas e
259

Módulo V – Processos Reprodutivos
Poligenes ou Herança Quantitativa
Como já dito anteriormente, fenótipo é a indivíduo e sua interação com o ambiente. Consideremosem humanos. Tal fenótipo é determinado por dois pares de genes não alélicos com ação aditiva. Sendo assim, quanto mais genes dominantes possuinesses loci não alélicos que manifestam sua ação na produção da melanina, maior será a produção de tal proteína e, consequentemente, mais escura será a pele desindivíduo. De forma inversa, quanto mais genes recessivos possuir um indivíduonessa gama de genes não alélicos que agem determinando a produção de melanina, mais clara (porém não albina) será a pele do indivíduo. quantitativa, e os genes envolvidos na herança são dito
Exemplo: Genes envolvidos Pp
Genótipos poss
PPSS
PPSs
PPss
PpSS
PpSs
Ppss
ppSS
ppSs
ppss
Considere o seguinte:
PPSS: sempre será negro
Mas quando misturados, passam a ser mulatos da seguinte forma:
Para 1 s ou 1 p: escuro (1)
Para 2 s, 2 p ou 1 s + 1 p: médio (2)
Para 1 p + 2 s ou 1 s + 2 p: claro (3)
Altura, peso, cor da pele, etcfenotípica contínua, ou seja, não há uma definiçãoe começa outra. A maioria das características de variação fenotípica contínua são determinadas por poligenes.
Os primeiros conceitos acerca da transmissão das características quantitativas foram desenvolvidos em 1910, por
Processos Reprodutivos
Quantitativa
Como já dito anteriormente, fenótipo é a manifestação do genótipo de um indivíduo e sua interação com o ambiente. Consideremos, por exemplo, a cor da pele em humanos. Tal fenótipo é determinado por dois pares de genes não alélicos com ação aditiva. Sendo assim, quanto mais genes dominantes possuir um indivíduo
não alélicos que manifestam sua ação na produção da melanina, maior será a produção de tal proteína e, consequentemente, mais escura será a pele deste indivíduo. De forma inversa, quanto mais genes recessivos possuir um indivíduo nessa gama de genes não alélicos que agem determinando a produção de melanina,
ina) será a pele do indivíduo. Este tipo de herança é dita e os genes envolvidos na herança são ditos poligenes.
s Pp e Ss
Genótipos possíveis Fenótipos
PPSS Negro
PPSs Mulato escuro
PPss Mulato médio
PpSS Mulato escuro
PpSs Mulato médio
Ppss Mulato claro
ppSS Mulato médio
ppSs Mulato claro
ppss Branco
ppss: sempre será branco
Mas quando misturados, passam a ser mulatos da seguinte forma:
médio (2)
: claro (3)
etc, são heranças que apresentam uma variação fenotípica contínua, ou seja, não há uma definição exata de onde termina uma classe e começa outra. A maioria das características de variação fenotípica contínua são
Os primeiros conceitos acerca da transmissão das características quantitativas s em 1910, por Nilsson-Ehle, quando descreveu a determinação
260
Amplie seus conhecimentos sobre herança poligênica. Visite o site: http://cienciahoje.uol.com.br/121736 e leia o texto “Os muitos genes da estatura humana”, do Prof. Dr. Sérgio Danillo Pena.
Aprofunde seus conhecimentos sobre heranças complexas lendo o artigo da Dra. Mary Feitosa & Dr. Henrique Krieger, publicado no periódico Ciência e Saúde Coletiva, vol. 7 (1): 73-83, (2002).

Módulo V – Processos Reprodutivos
Transcrição
Duplicação
da cor do grão do trigo. O cruzamento de licom linhagens puras de grãos brancosprodutoras de grãos vermelho-médioindivíduos produtores de grãos cujas cores variam do vermelhopassando pelos fenótipos vermelho-
ATIVIDADE
Numa determinada espécie de planta, foi deindivíduo é de 5 cm e a máxima é deplantas de 5 cm originaram, respectivamente, 19, 78, 121, 81 e 21 indivíduos exibindo altura de 5 cm, 10 cm, 15 cm, 20 cm e
Pergunta-se: quantos genes estão envolvidos na determinação despécie vegetal?
Mecanismos Moleculares de Replicação
O DNA é uma molécula fita dupla, antiparalela, responsável pela manutenção da informação genética da célula. Estsemiconservativa, isto é, cada fita dupla dará origem a duas novas fitas compostas por uma fita antiga e uma fita recémperíodo “S” do ciclo celular.
Podemos considerar essa macromolécula como a molécula bocorrências celulares, controlando todo o fluxo de informações na célula. Sem que haja sua replicação (duplicação), não ocorre a divisão celular. Sem que sua informação seja transferida para o formato de RNA (decodificada em aminoácidos (traduçãoprotéica (veja abaixo o Esquema 6).
Esquema 6: Fluxo da informação genética em organismos eucariontes.
O processo de transformação da informação de DNA para RNAproteína, já foi aprendido em unidades anteriores.
Neste momento, vamos olhar um pouco mais de perto o mecanismo pelo qual o DNA se duplica (veja abaixo o Esquema 7).
Processos Reprodutivos
Transcrição Transcrição
da cor do grão do trigo. O cruzamento de linhagens puras de grãos vermelho-escuros inhagens puras de grãos brancos origina uma F1 com 100% de plantas
médio. O cruzamento de indivíduos F1 resulta em F2, indivíduos produtores de grãos cujas cores variam do vermelho-escuro até o branco,
-claros, vermelho-médios e róseos.
Numa determinada espécie de planta, foi definido que a altura mínima de um e a máxima é de 25 cm. Os cruzamentos de plantas de 25 cm com
plantas de 5 cm originaram, respectivamente, 19, 78, 121, 81 e 21 indivíduos exibindo e 25 cm.
ntos genes estão envolvidos na determinação da altura nessa
Mecanismos Moleculares de Replicação
O DNA é uma molécula fita dupla, antiparalela, responsável pela manutenção formação genética da célula. Esta molécula se autoreplica de maneira
ada fita dupla dará origem a duas novas fitas compostas por uma fita antiga e uma fita recém-sintetizada. Sua síntese ocorre durante o
a macromolécula como a molécula base para todas as ocorrências celulares, controlando todo o fluxo de informações na célula. Sem que
), não ocorre a divisão celular. Sem que sua informação seja transferida para o formato de RNA (transcrição), para então ser
tradução), originando as proteínas, não há síntese
: Fluxo da informação genética em organismos eucariontes.
de transformação da informação de DNA para RNA, e deste para rendido em unidades anteriores.
Neste momento, vamos olhar um pouco mais de perto o mecanismo pelo qual quema 7).
261

Módulo V – Processos Reprodutivos
Esquema 7: Representação do processo de duplicação (replicação) do DNAcom identificação das principais proteínas atuantes no mesmo; b) Helicase;Topoisomerase; d) SSB; e) RNA Primase; f) DNA Polimerase III; g) DNA I; h) DNA Ligase.
Ao analisar o Esquema 7, vale lembrarexistam disponíveis enzimas e substrato (nucleotídeo trifosfato)transcorra bem. Tais matérias-primas são obtidas durante a fase G1 do ciclo celular e nas diversas vias metabólicas celulares.
O início da duplicação ocorre com a ligação ao DNA dupla fita de uma enzima, a Helicase, responsável pela quebra das ligaçduas fitas. Esta ligação ocorre em sítios específicos chamados dereplicação). Conforme a Helicase vai quebrando as pontes de hidrogênio, a fita vai sofrendo uma super helicoidização indesejadatopoisomerase.
Para prevenir a reorganização das pontes de hidrogênioHelicase, uma série de proteínas ligantes de DNA (SSB) ligarecém-abertas. Agora o DNA está pronto para ser copiado pela DNA
Entretanto, o DNA Polimeraseterminais 3´OH já existentes. Sendo assimRNA Primase, que tem a capacidade de iniciar uma nova cadeia de ácido nucléico a partir do reconhecimento do terminal 3´OH da cadeiaribonucleotídeos na nova fita, liga-incorporação de 1 a 60 ribonucleotídeos (es
Processos Reprodutivos
Fragmentos de Okazaki
Representação do processo de duplicação (replicação) do DNA: a) ais proteínas atuantes no mesmo; b) Helicase; c)
; d) SSB; e) RNA Primase; f) DNA Polimerase III; g) DNA Polimerase
Ao analisar o Esquema 7, vale lembrar que, na célula, é necessário que já existam disponíveis enzimas e substrato (nucleotídeo trifosfato), para que a síntese
primas são obtidas durante a fase G1 do ciclo celular e nas diversas vias metabólicas celulares.
duplicação ocorre com a ligação ao DNA dupla fita de uma enzima, a Helicase, responsável pela quebra das ligações de ponte de hidrogênio que unem as
a ligação ocorre em sítios específicos chamados de ori (origem de licase vai quebrando as pontes de hidrogênio, a fita vai
sofrendo uma super helicoidização indesejada, que será corrigida pela ação de uma
Para prevenir a reorganização das pontes de hidrogênio, quebradas pela as ligantes de DNA (SSB) liga-se às duas fitas simples
abertas. Agora o DNA está pronto para ser copiado pela DNA Polimerase III.
Polimerase III só consegue incorporar nucleotídeos em terminais 3´OH já existentes. Sendo assim, é necessário que uma outra enzima, a
, que tem a capacidade de iniciar uma nova cadeia de ácido nucléico a partir do reconhecimento do terminal 3´OH da cadeia, molde e, incorporando
-se à extremidade 3ÓH da fita molde, e inicia a incorporação de 1 a 60 ribonucleotídeos (este número depende do organismo em
262
Topoisomerase é uma enzima que introduz clivagens reversíveis em uma dupla hélice de DNA com o objetivo de destorcer ou desenrolar dobramentos ou torções excessivas.
Assista a uma animação sobre a síntese do DNA em: http://objetoseducacionais2.mec.gov.br/bitstream/handle/mec/7633/sintese%20DNA%20(1).swf?sequence=1

Módulo V – Processos Reprodutivos
particular), criando assim uma fita nova de RNA que teste terminal livre, o DNA Polimerase III pode ligarseguimento à duplicação pela incorporação de desoxirribonucleotídeos.
Considerando que o DNA é uma dupla fita antiparalela, teremospontes de hidrogênio são quebradasmodo que uma Primase irá se ligar no início de ORIPrimase só se ligará após certo número de ligações de pontes de H haverem sido quebradas. Sendo assim, uma das fitas seroutra será feita “aos pedaços” de formde Okazaki (veja novamente no Esquema 7
Após a ligação do DNA Polimeraseos ribonucleotídeos localizados no iníciexonucleásica da DNA Polimerase substituirá por nucleotídeos de DNA. Entretanto, como oa capacidade de ligar os terminais 5´existenteinserido, tal ligação é feita pela DNA Ligase, de forma que a nova fita de DNA, ao final do processo, possui somente nucleotídeos de DNAeles.
Em alguns vírus, o material genético é o RNA (retrovquando estes infectam uma célula precisam sintetizar uma molécula de DNA a partir deste RNA molde para, só então, o DNA poder ser incorporado ao DNA do hospedeiro. A síntese de DNA a partir de RNA é realizada pela enzima reversa viral.
Você sabia que o HIV (Vírus da Imunodeficiência Humana) é um retrovírusAssim, quando ele infecta um indivíduoinserir seu genoma no genoma do hospedeiro. Chamamos de o tratamento com fármacos que atquais o vírus toma para se reproduzir dentro da célula, sendo que podemos dividesta terapêutica em três categorias: virucidas, imunomoduladores e antivirais. Um dos modos de ação dos antivirais é pela inibi
Recombinação
A recombinação, com as mutações, é um mecanismo pelo qual a variabilidade genética é inserida em um genoma e, consequentemente, em uma população. Estmecanismo ocorre durante a prófase I da meiosecromossomos homólogos por ocasião do cromátides parentais durante a meiose.
Os Crossing-Over possuem uma característica muito interessanteaumento da sua probabilidade de ocorrência quanto maior for a genes. Desta forma, a frequência de novas combinações gênicas nos diz se dois genes estão mais próximos ou mais distantes um do outro, auxiliandogenes nos cromossomos.
Processos Reprodutivos
criando assim uma fita nova de RNA que tem o terminal 3ÓH livre. Com A Polimerase III pode ligar-se à cadeia nova e dar
uplicação pela incorporação de desoxirribonucleotídeos.
ando que o DNA é uma dupla fita antiparalela, teremos, quando as pontes de hidrogênio são quebradas, duas fitas que terão polarização oposta, de
irá se ligar no início de ORI, enquanto na outra fita, a só se ligará após certo número de ligações de pontes de H haverem sido Sendo assim, uma das fitas será continuamente formada, enquanto que a
de forma descontínua, dando origem aos Fragmentos (veja novamente no Esquema 7, apresentado anteriormente).
Polimerase III e a inserção de desoxirribonucleotídeos, os ribonucleotídeos localizados no início da nova fita serão corrigidos pela ação
I, que removerá os nucleotídeos de RNA e os ídeos de DNA. Entretanto, como o DNA Polimerase I não tem
a capacidade de ligar os terminais 5´existentes ao 3´OH do nucleotídeo recém- ligação é feita pela DNA Ligase, de forma que a nova fita de DNA, ao
possui somente nucleotídeos de DNA sem espaços vazios entre
Em alguns vírus, o material genético é o RNA (retrovírus) e, sendo assim, célula precisam sintetizar uma molécula de DNA a partir
e RNA molde para, só então, o DNA poder ser incorporado ao DNA do síntese de DNA a partir de RNA é realizada pela enzima Transcriptase
o HIV (Vírus da Imunodeficiência Humana) é um retrovírus? ssim, quando ele infecta um indivíduo, ele se utiliza da Transcriptase reversa para
inserir seu genoma no genoma do hospedeiro. Chamamos de “terapêutica antiviral” o tratamento com fármacos que atacam o vírus, interferindo nos caminhos pelos quais o vírus toma para se reproduzir dentro da célula, sendo que podemos dividir
em três categorias: virucidas, imunomoduladores e antivirais. Um dos modos de ação dos antivirais é pela inibição da Transcriptase reversa.
Recombinação
com as mutações, é um mecanismo pelo qual a variabilidade inserida em um genoma e, consequentemente, em uma população. Este
mecanismo ocorre durante a prófase I da meiose, e reorganiza os alelos nos por ocasião do Crossing-Over (CO), uma quebra e união de
cromátides parentais durante a meiose.
possuem uma característica muito interessante, que é o aumento da sua probabilidade de ocorrência quanto maior for a distância entre dois
ência de novas combinações gênicas nos diz se dois genes estão mais próximos ou mais distantes um do outro, auxiliando-nos a mapear os
263
A enzima Transcriptase reversa viral, usada pelo HIV, pode ser vista em seu ciclo numa apresentação neste endereço: http://www.fsc.ufsc.br/~canzian/bau/aids/ciclo-hiv.ppt
Você já se perguntou por que, com a exceção de gêmeos idênticos, duas pessoas não são iguais? Enquanto os genes determinam a maioria das nossas características, a exata combinação de genes que nós herdamos, e nossos traços físicos, é parcialmente definida num processo que nossos cromossomos sofrem, conhecido como recombinação gênica. Consultado em: http://www.cefeteq.br/dna/dnaint/crossingover.htm ).

264
Módulo V – Processos Reprodutivos
Para percebermos se houve ou não a recombinação entre dois genes é necessário que sejam informativos, ou seja, que haja a possibilidade de distinguir seus alelos que se encontrem em heterozigose no indivíduo analisado. Então é possível “seguir o caminho” de um gene através das gerações, identificando a transmissão materna e paterna.
Quando observamos a disposição dos genes em um cromossomo, podemos classificar tal disposição em CIS ou TRANS. Posição CIS é quando dois alelos estão no mesmo cromossomo, e TRANS quando estão em homólogos opostos.
Por exemplo: se analizarmos dois genes, o gene da β globina (Hb) e o gene Transferase S de Glutation classe } (GST-P1) transferase PI, localizados no cromossomo 11 da espécie humana, e considerarmos um indivíduo duplo heterozigoto para ambos os genes, sendo portador dos genes Hb*A e Hb*S e GSTP1*A e GSTP1*B, podemos identificar a fase em que se encontram os alelos, dependendo se estão ou não no mesmo homólogo. Assim, se o genótipo do indivíduo for Hb*A, GSTP1*A/Hb*S, GSTP1*B, dizemos que os genes Hb*A e GSTP1*A e, Hb*S e GSTP1*B, estão em fase CIS, enquanto Hb*A e GSTP1*B e Hb*S e GSTP1*A estão em fase TRANS. Deste modo, se não houver ocorrência de recombinação entre os dois genes, os gametas formados por este indivíduo serão idênticos aos que deram origem ao indivíduo: “Hb*A, GSTP1*A” e “Hb*S e GSTP1*B”. Tais gametas são ditos “tipos parentais”.
Entretanto, se houver Crossing-Over entre estes dois genes, os gametas resultantes serão parte “tipo parental” e parte “recombinante”, ou seja, mais de 50% deles será Hb*A, GSTP1*A e Hb*S, GSTP1*B, enquanto que em uma proporção menor que 50% será Hb*A, GSTP1*B e Hb*S, GSTP1*A. Sabemos que a proporção será diferente de 50%, pois já temos o conhecimento de que ambos os genes encontram-se no mesmo cromossomo, ou seja, estão “ligados” (veja abaixo o Esquema 8).
Genótipo Genótipo
Hb*A,
GSTP1*A
Hb*S,
GSTP1*B
Hb*A,
GSTP1*A
Hb*S,
GSTP1*B
Sem ocorrência de CO Gametas: Com ocorrência de CO
Hb*A, GSTP1*A
tipo
parental Hb*A, GSTP1*A
Hb*S, GSTP1*B Hb*S, GSTP1*B
tipo
recombinant
e
Hb*A, GSTP1*B
Hb*S, GSTP1*A
Esquema 8: Representação dos possíveis gametas resultantes em um indivíduo duplo heterozigoto, quando considerados dois genes no mesmo cromossomo e presença ou ausência de recombinação entre eles.

Módulo V – Processos Reprodutivos
Observe que no Esquema 8 a proporção de gametas parentais/recombinantes nos permite inferir a distância, em centiMorgans (cM), entre dois genes não alelos. Esta medida de unidade de recombinação irá aumentarcromossomo estiverem dois genes não alelosestes genes se localizarem.
Quando realizamos um cruzamento considerando a segregação de dois genes não alélicos e observamospodemos afirmar que estes genes estão se segregando segundo as proporções esperadas na Segunda Lei de Mendel e, consequentemente, são dispostos em cromossomos distintos. Mas se a proporção for menor que esevidências de ambos os genes estarem localizados no mesmo cromossomo. Dizemoentão, que estes dois genes não são independentes
O estudo de ligação entre genes nos permite não só localizaos outros em um determinado cromossomodos alelos, determinar a transmissão de características que são determinadas por genes ligados a genes conhecidos. Estmelhoramento genético animal e vegetal, bem como para aplicações diagnósticas.
ATIVIDADE
Vamos praticar o aprendizado, respondendo às questões:
1. Quais os fatores que podem ocorrer seinformação ser transferida para o formato de RNA?
2. Cite uma característica do Crossing
Mutação Gênica e Reparo
Não existem no mundo dois indivíduos geneticamente idênticos, exceto no caso de gêmeos univitelinos. Entretanto, se o fenótipo é a expressão do genótipo, modulada pelo ambiente, como explicar a variabilidade fenotípica que existe nas espécies?
Existem dois mecanismos moleculares que inserem variabilidade genética em uma espécie: a mutação e a recombinação gênica. Destprimário na criação de variação nos genes. É sofre modificações estruturais, originando novos alelos.
Podemos classificar as mutações em relação ao tipno DNA, alteração que causou no gene ou no ocorreu, etc.
Você sabia que a Alfa-1-Antitripsina (A1AT) é uma glicoproteína altamente polimórfica, sintetizada pelos hepatócitos e macrófagos alveoreações de proteção dos tecidos pulmonares contra o ataque proteolítico pelas proteases? O gene responsável pela sua produção possui 7 região 14q31-32, tendo sido descrito, para o mesmo, mais de 75 variantesquais alguns alelos que determinam uma produção insuficiente da proteína, levando
Processos Reprodutivos
a proporção de gametas parentais/recombinantes nos permite inferir a distância, em centiMorgans (cM), entre dois genes não alelos.
a medida de unidade de recombinação irá aumentar, quanto mais distante no cromossomo estiverem dois genes não alelos, e diminuir, quanto mais próximos
Quando realizamos um cruzamento considerando a segregação de dois genes não alélicos e observamos uma frequência de recombinação (θ = 50%),
es genes estão se segregando segundo as proporções Mendel e, consequentemente, são dispostos em
cromossomos distintos. Mas se a proporção for menor que esta, então temos vidências de ambos os genes estarem localizados no mesmo cromossomo. Dizemos,
es dois genes não são independentes, e sim que estão ligados.
O estudo de ligação entre genes nos permite não só localizá-los uns em relação aos outros em um determinado cromossomo, como também, por meio da segregação dos alelos, determinar a transmissão de características que são determinadas por
es ligados a genes conhecidos. Esta é uma importante ferramenta para o melhoramento genético animal e vegetal, bem como para aplicações diagnósticas.
Vamos praticar o aprendizado, respondendo às questões:
Quais os fatores que podem ocorrer sem que haja replicação e sem a informação ser transferida para o formato de RNA?
Cite uma característica do Crossing-Over.
Mutação Gênica e Reparo
Não existem no mundo dois indivíduos geneticamente idênticos, exceto no caso de gêmeos univitelinos. Entretanto, se o fenótipo é a expressão do genótipo, modulada pelo ambiente, como explicar a variabilidade fenotípica que existe nas
Existem dois mecanismos moleculares que inserem variabilidade genética em ão e a recombinação gênica. Destes, a mutação é o mecanismo
primário na criação de variação nos genes. É por meio dela que um gene selvagem sofre modificações estruturais, originando novos alelos.
Podemos classificar as mutações em relação ao tipo de alteração que aconteceu causou no gene ou no seu produto, ao tipo celular onde
ntitripsina (A1AT) é uma glicoproteína altamente polimórfica, sintetizada pelos hepatócitos e macrófagos alveolares e envolvida nas reações de proteção dos tecidos pulmonares contra o ataque proteolítico pelas
O gene responsável pela sua produção possui 7 exons e está localizado na 32, tendo sido descrito, para o mesmo, mais de 75 variantes, dentre as
quais alguns alelos que determinam uma produção insuficiente da proteína, levando
265
Sobre a utilização das análises de ligação como ferramenta de melhoramento genético, leia o artigo “Marcadores Moleculares e sua Aplicação no Melhoramento Genético de Plantas “, de BARED e Cols (1997).
Uma mutação é uma mudança em uma sequência de nucleotídeos de uma molécula de DNA. É resultado de erros na duplicação do DNA ou de danos ao DNA causados por agentes mutagênicos, tais como radiação e produtos químicos que reagem com o DNA e modificam suas bases. Muitas são simples alterações de base única (mutações de ponto), enquanto outras são caracterizadas pelo aumento (inserção), ou diminuição (deleção), do número de bases de determinada sequência, variando de um a muitos nucleotídeos.

266
Módulo V – Processos Reprodutivos
a um quadro de enfisema pulmonar em adultos ou hepatopatia em crianças e adultos.
O alelo selvagem (alelo PI*M) foi descoberto ser, na realidade, um grupo de alelos resultantes de mutações neutras (PI*M1Val213, PI*M1Ala213, etc), que só foram reconhecidas a partir de estudos moleculares. Já os alelos causadores de deficiência, alelo PI*S (Glu-264-Val*) e alelo PI*Z(Glu-342-Lys), foram originários de mutações de sentido trocado nos exons 3 e 5, respectivamente. O alelo raro PI*Mmalton é o produto de uma deleção de 3 pares de base que retira uma fenilalanina no resíduo 52 da proteína. Uma mutação de ponto do tipo transversão (C→G) leva à criação de um stop códon no alelo PI*Nulo Granite Falls, enquanto uma inserção de um T no códon 353 induz a um erro na matriz de leitura e, consequente criação de um códon de terminalização no alelo PI*Nulo Mattawa.
* Obs.: Lê-se substituição de um ácido glutâmico por uma valina no resíduo 264 da proteína.
Em relação ao tipo de célula de ocorrência, nós podemos subdividir as mutações em duas classes: as mutações somáticas e as mutações germinativas. No primeiro tipo, independente da sua consequência, somente o indivíduo na qual ela ocorreu sofrerá as consequências, isto é, ela não será passada para sua prole. Ao contrário, quando uma mutação ocorre em uma célula germinativa, sua consequência será sentida na prole do indivíduo em que ela ocorreu.
Uma mutação pode ocorrer de duas formas: espontaneamente ou induzida por agentes mutagênicos. As mutações são ditas espontâneas quando surgem a partir de erros na duplicação do DNA, envolvendo a função de “leitura de correção” das DNAs Polimerases que sintetizam a nova fita, ainda na forquilha de replicação. Quando uma mutação ocorre, não naturalmente, mas por indução de agentes externos (físicos, como radiação UV, ou químicos, como brometo de etídio, por exemplo), dizemos que é uma mutação induzida. Neste caso, o agente causal interage com a fita parental do DNA, causando uma mudança estrutural que afeta o pareamento de bases do nucleotídeo.
Quando somente uma base é alterada, essa mutação é dita mutação de ponto. Uma mutação de ponto ocorre quando uma base do DNA é trocada por outra. Em relação ao tipo de base que é trocada, podemos classificar uma mutação de ponto em Transição ou Transversão. Dizemos transição quando uma base é trocada por outra da mesma categoria, ou seja, púrica é trocada por púrica (A trocada por G e vice-versa), ou pirimídica trocada por pirimídica (C trocada por T, e vice-versa). Já no caso de uma mutação do tipo transversão, uma base púrica (A ou G) é trocada por uma pirimídica (C ou T), e vice-versa. Estas mudanças poderão ou não acarretar alteração no funcionamento do genoma. Aquelas que ocorrem em regiões de DNA intrônico e em regiões não codificantes de componentes gênicos ou suas sequências relacionadas, ou seja, em 98,5% dos genomas humanos, ditas mutações silenciosas.
As mutações ocorrentes nas outras, aproximadamente 1,5% de regiões do genoma, ditas regiões codificadoras de genes, são de grande interesse e podem ser classificadas de acordo com seus efeitos (veja Esquema 9):
1. Se a mutação não acarreta nenhuma alteração na proteína, ou seja, o aminoácido codificado pelo códon mutado é o mesmo que o codificado pelo códon
Leia sobre mutação induzida das plantas para combater crise de alimentos em: http://noticias.ambientebrasil.com.br/noticia/?id=40039 Consultado em 23/08/09.
Leia o artigo “Humano é fruto de seleção natural recente” em : http://noticias.terra.com.br/ciencia/interna/0,,OI2415815-EI296,00.html (consultado em 23/08/09).

267
Módulo V – Processos Reprodutivos
antigo, dizemos que é uma mutação sinônima ou silenciosa. Esta mutação não causa nenhum efeito no genoma que ocorreu.
2. Se a mutação for não sinônima, isto é, altera o aminoácido codificado pelo novo códon, ela mesma, assim poderá ou não causar efeitos no genoma. Se não levar a nenhum efeito, dizemos que é uma mutação neutra. Embora altere o aminoácido codificado, ele é funcionalmente semelhante ao aminoácido original e nenhuma mudança é notada na função da proteína. Caso a proteína tenha não somente sua estrutura primária, mas também sua função alterada, então dizemos que é uma mutação de sentido trocado.
3. Um tipo especial de mutação de sentido trocado é aquela capaz de converter um códon de finalização em outro códon que especifica um aminoácido. Neste caso, ao invés do término da tradução, os aminoácidos continuarão sendo colocados no terminal C. A maioria das proteínas pode tolerar pequenos aumentos no seu tamanho sem alterar sua função. Entretanto, grandes inserções de aminoácidos extras irão interferir na estrutura terciária da molécula, e isto poderá reduzir sua atividade.
4. Uma mutação pode converter um códon que especifica um aminoácido em um códon de término de cadeia. Esta mutação sem sentido resulta em uma proteína mais curta que o normal, uma vez que há a tradução do RNAm para o novo stop códon. O efeito na atividade protéica dependerá de quanto pequena é a nova proteína: geralmente, seu efeito é drástico e a proteína é não funcional, entretanto, se o novo códon foi criado na porção terminal 3´ do gene, pode acontecer da proteína não ter seu sítio ativo alterado e funcionar normalmente.
Legenda:
Adenina
Timina
Guanina
Cisina
Transversão
Transição a)
5´ 3´
3´ 5´
Sequência original
Polipeptídeo
5´ 3´
3´ 5´
Mutação neutra
5´ 3´
3´ 5´
Mutação sentido trocado
5´ 3´
3´ 5´
Mutação sem sentido
5´ 3´
3´ 5´
Inserção de base
5´ 3´
3´ 5´
Deleção ou perda de base
c)
b)
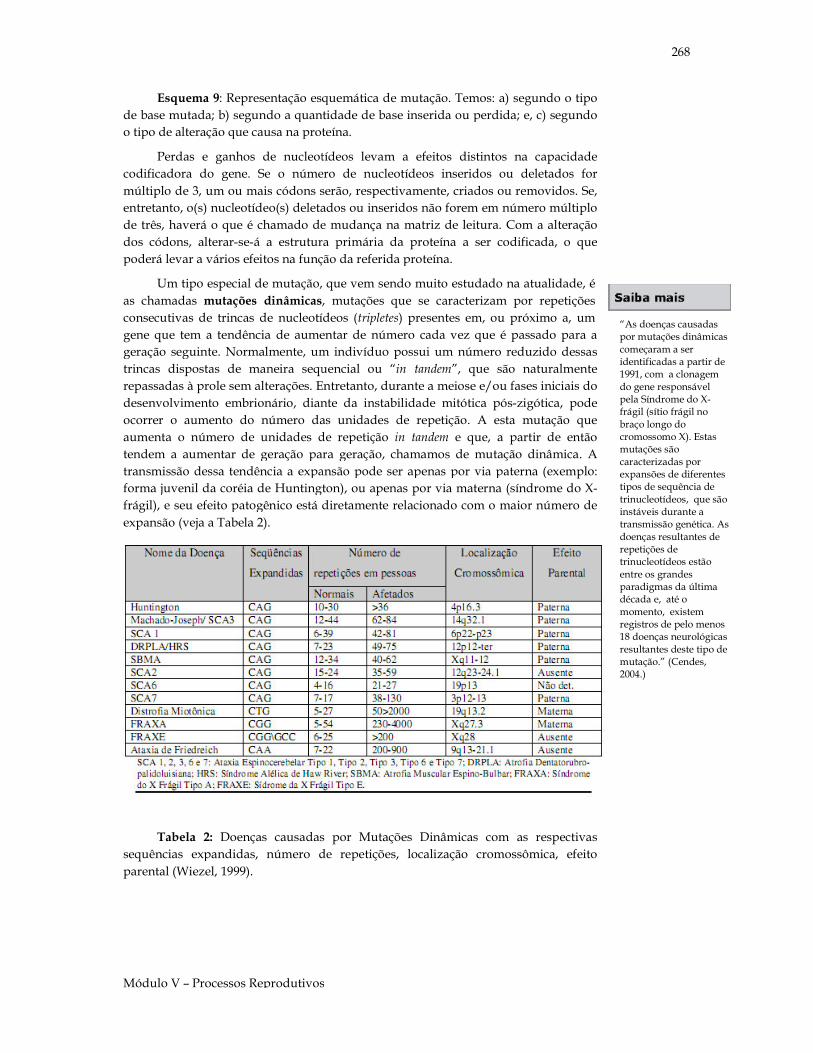
Módulo V – Processos Reprodutivos
Esquema 9: Representação esquemática de mutação. Temos:de base mutada; b) segundo a quantidade de base inserida ou perdida; o tipo de alteração que causa na proteína.
Perdas e ganhos de nucleotídeos levam a efeitos distintos na capacidade codificadora do gene. Se o número de nucleotídeos inseridos ou deletados for múltiplo de 3, um ou mais códons serão, respectivamente, criados oentretanto, o(s) nucleotídeo(s) deletados ou inseridos não forem em número múltiplo de três, haverá o que é chamado de mudança na matriz de leitura.dos códons, alterar-se-á a estrutura primárpoderá levar a vários efeitos na função da referida proteína.
Um tipo especial de mutação,as chamadas mutações dinâmicas,consecutivas de trincas de nucleotídeos (gene que tem a tendência de aumentar de número cada vez que é passado para a geração seguinte. Normalmente, um indivídutrincas dispostas de maneira sequrepassadas à prole sem alterações. Entretanto, durante a meiose e/desenvolvimento embrionário, diante da ocorrer o aumento do número das unidades de repetição. A estaumenta o número de unidades de repetição tendem a aumentar de geração para geraçãotransmissão dessa tendência a expansão pode ser apenas por via paterna (exforma juvenil da coréia de Huntington)frágil), e seu efeito patogênico está diretamente relacionado com o maior número de expansão (veja a Tabela 2).
Tabela 2: Doenças causadas sequências expandidas, número de repetições, localização cromossômica, efeito parental (Wiezel, 1999).
Processos Reprodutivos
sentação esquemática de mutação. Temos: a) segundo o tipo a quantidade de base inserida ou perdida; e, c) segundo
tipo de alteração que causa na proteína.
Perdas e ganhos de nucleotídeos levam a efeitos distintos na capacidade codificadora do gene. Se o número de nucleotídeos inseridos ou deletados for múltiplo de 3, um ou mais códons serão, respectivamente, criados ou removidos. Se, entretanto, o(s) nucleotídeo(s) deletados ou inseridos não forem em número múltiplo
de mudança na matriz de leitura. Com a alteração á a estrutura primária da proteína a ser codificada, o que
poderá levar a vários efeitos na função da referida proteína.
, que vem sendo muito estudado na atualidade, é , mutações que se caracterizam por repetições
consecutivas de trincas de nucleotídeos (tripletes) presentes em, ou próximo a, um m a tendência de aumentar de número cada vez que é passado para a
geração seguinte. Normalmente, um indivíduo possui um número reduzido dessas uencial ou “in tandem”, que são naturalmente Entretanto, durante a meiose e/ou fases iniciais do
diante da instabilidade mitótica pós-zigótica, pode do número das unidades de repetição. A esta mutação que
aumenta o número de unidades de repetição in tandem e que, a partir de então tendem a aumentar de geração para geração, chamamos de mutação dinâmica. A
a tendência a expansão pode ser apenas por via paterna (exemplo: forma juvenil da coréia de Huntington), ou apenas por via materna (síndrome do X-
seu efeito patogênico está diretamente relacionado com o maior número de
Doenças causadas por Mutações Dinâmicas com as respectivas ências expandidas, número de repetições, localização cromossômica, efeito
268
“As doenças causadas por mutações dinâmicas começaram a ser identificadas a partir de 1991, com a clonagem do gene responsável pela Síndrome do X-frágil (sítio frágil no braço longo do cromossomo X). Estas mutações são caracterizadas por expansões de diferentes tipos de sequência de trinucleotídeos, que são instáveis durante a transmissão genética. As doenças resultantes de repetições de trinucleotídeos estão entre os grandes paradigmas da última década e, até o momento, existem registros de pelo menos 18 doenças neurológicas resultantes deste tipo de mutação.” (Cendes, 2004.)

269
Módulo V – Processos Reprodutivos
Embora tenha sido dito anteriormente que as mutações que ocorrem em somente cerca de 1,5% do genoma podem levar a alterações de função, por estarem em regiões codificadoras, mutações em regiões intrônicas ou regiões adjacentes, a ligação intron-exon também será importante se alterarem o nucleotídeo envolvido nas interações RNA–proteína e RNA-RNA que ocorrem durante o processamento dos diferentes tipos de introns. Muitos exemplos neste sentido podem ser dados utilizando a patologia conhecida por β-talassemia, uma vez que algumas mutações no gene da β-globina (cromossomo 11) criam sítios “Splice” alternativos, produzindo transcritos alterados.
Uma mutação, entretanto, só irá modificar o fenótipo do organismo se tiver um efeito sobre o mesmo. Nestes termos, podemos ainda classificar as mutações como mutação de ganho ou perda ou alteração de função. Vamos conhecer cada um destes tipos de mutação a seguir.
Perda de função é o resultado normal de uma mutação que reduz ou inibe a atividade protéica. Geralmente possui característica recessiva, uma vez que um indivíduo que sofreu a mutação é heterozigoto (manteve um gene intacto, enquanto o outro foi mutado) e não apresenta sintomas ou fenótipo que revelem sua presença (veja novamente o Esquema 9 apresentado anteriormente).
Tal fato acontece porque o gene selvagem manteve sua função e a proteína codificada por ele é capaz de suprir as necessidades do organismo. Como exemplo clássico, podemos citar um erro inato do metabolismo, a Fenilcetonúria, doença causada por uma mutação que altera a enzima hidroxilase de fenilalanina, que converte a fenilalanina em tirosina. Sem a enzima de conversão o organismo, que recebe suprimento de fenilalanina pela dieta, não é capaz de degradá-la em tirosina, é desta forma que o aminoácido essencial é armazenado. Uma vez que a fenilalanina é tóxica para as células, estas lançam mão de vias metabólicas alternativas que transformam a fenilalanina em fenilpiruvato, fenilactato e fenilacetato. A fenilalanina e seus catabólitos, também tóxicos ao organismo, se acumulam nos tecidos, em especial nos tecidos neurais, e são eliminados na forma de fenilcetonas.
A criança homozigota para tal deficiência não é identificada até aproximadamente os 3 meses de vida, exceto se realizado teste laboratorial específico (teste do pezinho). A partir desta idade, com o acúmulo das substâncias tóxicas acima relacionadas nos tecidos, a criança começa a apresentar sinais de apatia, atraso no desenvolvimento e, com um ano de idade, apresentará déficit neuropsicomotor irreversível. A realização do teste de triagem neonatal diagnostica a presença da deficiência enzimática em tempo hábil para, por meio de dieta rigorosa de restrição da ingestão do aminoácido fenilalanina, evitar e prevenir o desenvolvimento da patologia.
No caso de indivíduos heterozigotos (dizemos portadores da mutação), não há evolução da doença, pois o gene normal produz enzima suficiente para manter o nível de conversão de fenilalanina em tirosina.
Leia o artigo da Dra. Iscia Lopes Cendes, “Expansão de poliglutaminas e doenças neurodegenerativas”publicado na revista Ciência e Cultura, vol. 56, nº. 1, São Paulo, Jan./Mar. 2004, em: http://cienciaecultura.bvs.br/scielo.php?script=sci_arttext&pid=S0009-67252004000100021&lng=pt&nrm=iso (consultado em 23/08/09).
Leia o texto “Fenilcetonúria”, de Luís Filipe Pedrosa da Costa, em http://projecto.tutorial.googlepages.com/Fenilcetonurialuis.pdf e saiba detalhes sobre mecanismos bioquímicos e genéticos deste erro inato do metabolismo.

Módulo V – Processos Reprodutivos
Assim como as hemoglobinopatias, a fenilcetonúria é um ótimo modelo para estudos genéticos. Podemos estudar padrão de nas quais o gene está segregando. Se considerarmos o efeito do gene sobre oorganismo, podemos utilizar esta patologia como um exemplo de uma única mutação altera a via metabólica do amconsequentemente, altera a produção da partir do metabolismo da tirosina. Destalém de várias alterações neuromotoras e déficit cognitivopigmentação e odor característico na urina, pela aácido homogentísico. Também podemos explicar a utilizando este “modelo”. O gene da fenilcetonúria (PHA) está localizado no cromossomo 12, e várias mutações podem ocorrer em um de seus 13 mutações podem afetar de diferentes maneiras a enzima hidroxilase de fenilalanina, quer deixando a enzima instável, quer alterando sua formafuncione corretamente. Esta doença é, com a galactosemia, as mucopolissacaridoses, entre outras, classicamente utilizada como modelo de
Esse tipo de mutação, excepcionalmente, pode apresentar mecanismo de expressão dominante quando recessivo, insuficiente para o funcionamento normal (haploinsuficiência). Exemplo clássico é o da hipercolesterolemia familiar, na qual compromete a síntese de receptores críticoscomprometendo o transporte intracelular e o catabolismo da LDL, resultando no acumulo de colesterol no plasma. Estaumentando muito o risco de infarto agudo do miocárdio. Indivíduos homozipara o gene mutante possuem um fenótipo ainda mais grave.
Mutações de ganho de função atividade anormal a uma proteína. Geralmente possuem caráter dominante, ou seja, manifestam-se mesmo em heterozigose. O ganho deaumento da dosagem gênica levando a um excesso do produto gênico, aumento da atividade protéica pelo prolongamento da vida média da proteínaatividade reguladora de seu inibidor constitutivo normalexpressão do RNAm , que afetam o local ou tempo da expressão gênica.
Como exemplo de mutação de ganho de função podemos citar vários Oncogenes. O gene RAS é responsável por uma proteína de membrana plasmática homônima, que está envolvida, quando ativada pela ligação com GTP, na regulação da proliferação celular, através da interação com fatores de crescimento. Uma mutação que altere um único par de bases no proteína RAS alterada, promovendotumores. Este oncogene já foi descrito como estando associado a vários tipos diferentes de câncer, como, por exemplomelanoma, leucemia monocítica aguda,
As células possuem enzimas que reparam o DNA,minimizar o número de mutações que ocorrem. Estvias diferentes. Umas na fase pré-replicativa
Processos Reprodutivos
Assim como as hemoglobinopatias, a fenilcetonúria é um ótimo modelo para estudos genéticos. Podemos estudar padrão de herança recessivo analisando famílias
o gene está segregando. Se considerarmos o efeito do gene sobre o a patologia como um exemplo de pleiotropia, pois
uma única mutação altera a via metabólica do aminoácido fenilalanina e, altera a produção de melanina (pigmento da pele), que é formada
do metabolismo da tirosina. Desta forma, os indivíduos com fenilcetonúria, além de várias alterações neuromotoras e déficit cognitivo, apresentam baixa pigmentação e odor característico na urina, pela alteração na via de metabolização do ácido homogentísico. Também podemos explicar a heterogeneidade genética
e “modelo”. O gene da fenilcetonúria (PHA) está localizado no e várias mutações podem ocorrer em um de seus 13 exons. Estas
mutações podem afetar de diferentes maneiras a enzima hidroxilase de fenilalanina, quer alterando sua forma, fazendo com que ela não
a doença é, com a galactosemia, as mucopolissacaridoses, tre outras, classicamente utilizada como modelo de Erros Inatos do Metabolismo.
e tipo de mutação, excepcionalmente, pode apresentar mecanismo de são dominante quando recessivo, pois a quantidade total do produto gênico é
insuficiente para o funcionamento normal (haploinsuficiência). Exemplo clássico é o na qual uma mutação no gene do receptor de LDL
de receptores críticos relacionados à membrana, assim o transporte intracelular e o catabolismo da LDL, resultando no
umulo de colesterol no plasma. Esta alteração dobra o nível do colesterol sérico, aumentando muito o risco de infarto agudo do miocárdio. Indivíduos homozigotos para o gene mutante possuem um fenótipo ainda mais grave.
Mutações de ganho de função são menos comuns. Elas conferem uma atividade anormal a uma proteína. Geralmente possuem caráter dominante, ou seja,
se mesmo em heterozigose. O ganho de função pode ocorrer pelo aumento da dosagem gênica levando a um excesso do produto gênico, aumento da atividade protéica pelo prolongamento da vida média da proteína, ou inibindo a atividade reguladora de seu inibidor constitutivo normal, e por alterações na
que afetam o local ou tempo da expressão gênica.
Como exemplo de mutação de ganho de função podemos citar vários é responsável por uma proteína de membrana plasmática
homônima, que está envolvida, quando ativada pela ligação com GTP, na regulação através da interação com fatores de crescimento. Uma
mutação que altere um único par de bases no RAS é suficiente para produção de uma promovendo o crescimento celular e consequente formação de
tumores. Este oncogene já foi descrito como estando associado a vários tipos por exemplo, o carcinoma de cólon, pulmão, pâncreas, a aguda, etc.
ssuem enzimas que reparam o DNA, numa tentativa de ero de mutações que ocorrem. Estas enzimas trabalham em duas
replicativa, e buscam nucleotídeos com estruturas
270
Oncogenes são genes com mutação que produzem uma proteína aberrante que estimula a divisão, proliferação ou diferenciação celular, mesmo na ausência de sinais de crescimento adequados. A maioria deles deriva de proto-oncogenes, que são genes normais que participam da regulação do crescimento.

Módulo V – Processos Reprodutivos
não usuais no DNA, trocando-os antes da replicação. Outras são enzimas ditas pósreplicativas, uma vez que verificam o DNA recentemente sintetizado em busca de erros, corrigindo-os. Juntas, estas enzimas compõem o
Considerando que diariamente nosso organeventos mutagênicos, quer seja por erros no mecanismo de duplicação e sistema enzimático de revisão de erros, quer seja por ataques de agentes nocivos como raios ultravioletas, agentes químicos e poluentes do ambiente, é fánecessidade de um sistema de reparo eficiente para evitar que o genoma sofra danos excessivos.
No geral, os sistemas de reparo possuem três etapas básicas
• EXCISÃO: primeiramente o DNA mutado é reconhecido e retirado
• POLIMERASE: logo em seguida pela DNA Polimerase.
• LIGASE: então, as extremidades do DNA recémseladas à fita existente pela DNA ligase.
Esses três mecanismos básicos de reparo pósleitura 3´→5´da DNA Polimerase,replicação do DNA.
ATIVIDADE
No quadro a seguir, você poderá obter uma breve revisão das diversas formas de classificação das mutações. Faça uma pesquisa sobre mutações genéticas e complete-a inserindo exemplos dess
Quadro 2- Exercício de Classificação das Mutações
Classificação por
Tipos
Forma de ocorrência
Espontâneas
Induzidas Tipo de alteração
Troca de base
Ganho/perda de base(s)
Efeito no organismo
Sinônima ou silenciosa
Não sinônima
Ação sobre o fenótipo
Perda de função
Ganho de função
Processos Reprodutivos
os antes da replicação. Outras são enzimas ditas pós-replicativas, uma vez que verificam o DNA recentemente sintetizado em busca de
as enzimas compõem o Sistema de Reparo.
Considerando que diariamente nosso organismo está sujeito a uma série de eventos mutagênicos, quer seja por erros no mecanismo de duplicação e sistema
quer seja por ataques de agentes nocivos como raios ultravioletas, agentes químicos e poluentes do ambiente, é fácil entender a necessidade de um sistema de reparo eficiente para evitar que o genoma sofra danos
No geral, os sistemas de reparo possuem três etapas básicas:
primeiramente o DNA mutado é reconhecido e retirado.
logo em seguida é restaurada a sequência original
ntão, as extremidades do DNA recém-substituído são fita existente pela DNA ligase.
es três mecanismos básicos de reparo pós-replicação, associados à prova de Polimerase, reduzem drasticamente a taxa de erro de
No quadro a seguir, você poderá obter uma breve revisão das diversas formas de classificação das mutações. Faça uma pesquisa sobre mutações genéticas e
a inserindo exemplos dessas alterações.
Exercício de Classificação das Mutações
Subtipos Exemplos
Transição
Transversão
Ganho/perda de Deleção
Inserção
Sinônima ou silenciosa
Neutra
Sentido trocado
Sem sentido
Alteração na matriz de leitura
Gene haplosuficiente Gene haploinsuficiente
271

Módulo V – Processos Reprodutivos
Esses mecanismos, normalmente, são utilizados pelas células em quatro diferentes tipos de sistemas de reparo do DNA, a saber:
1. Sistema de reparo direto que age diretamente nos nucleotídeos alterados, convertendo a sua estrutura original
2. Sistema de reparo por excisãocontendo uma alteração é cortado e, apósPolimerase.
3. Sistema de reparo de erros de pareamento que corrige erros de dupela retirada da fita simples de DNA contendo a mutaçãoseguir.
4. Sistema de reparo por recombinaçãoDNA, utilizando enzimas de recombinação e a dupla fita de DNA, por um processo similar à recombinação.
Um grande número de proteínas estde reparo, e mutações nos genes responsáveis pela produção das mesmas podem ocasionar uma falha nos mecanismos de reparo das mutações.
O Xeroderma pigmentoso é um exegene envolvido no sistema de reparo e, consequEsta doença se caracteriza pela apresentação descamativa da pele com pigmentação anormal, e numerosos cânceres de pele (melanomaetc.). Mutações em 7 genes, envolvendo diferentes cromossomos autossomos, estão associadas ao Xeroderma pigmentoso, genética para esta patologia.
RESPOSTA SOS: esse sistema de reparoverdade um complexo regulatório. polinucleotídio seja replicada, com a DNA mais ou menos aleatoriamente.
Atualmente se conhece várias por alterações no sistema de reparo de mutações, comomama, câncer de ovário, telangiectasia atáxica, Síndromes de Bloom e Werner, anemia de Fanconi, etc.
É importante lembrar que, alétambém possuem sistemas que possibilitam regiões alteradas replicarem prévio. Claro que esses mecanismos possibilitam, em alguns casos, o surgimento de doenças. Porém, a criação e manutenção da variabilidmecanismo que permite ao organismo estar apto à evolução.
Se uma mutação persiste em uma célula germinativa, a despeito de todos os mecanismos de reparo, e se mantémaior a 1%, dizemos então que este gene que agora possui duas ou mais formas alternativas, alelo selvagem e alelo(s) mutante(s), é um
Processos Reprodutivos
normalmente, são utilizados pelas células em quatro diferentes tipos de sistemas de reparo do DNA, a saber:
Sistema de reparo direto que age diretamente nos nucleotídeos alterados, estrutura original.
Sistema de reparo por excisão, no qual um segmento de polinucleotídio contendo uma alteração é cortado e, após, resintetizada a sequência correta pela DNA
Sistema de reparo de erros de pareamento que corrige erros de duplicação, pela retirada da fita simples de DNA contendo a mutação, e reparando-a logo a
Sistema de reparo por recombinação, que é usado para corrigir erros no DNA, utilizando enzimas de recombinação e a dupla fita de DNA, por um processo
Um grande número de proteínas está envolvido na realização desses sistemas e mutações nos genes responsáveis pela produção das mesmas podem
anismos de reparo das mutações.
é um exemplo clássico de uma mutação afetando um no sistema de reparo e, consequentemente, definindo uma patologia.
a doença se caracteriza pela apresentação descamativa da pele com pigmentação e numerosos cânceres de pele (melanomas, carcinomas de células basais,
etc.). Mutações em 7 genes, envolvendo diferentes cromossomos autossomos, estão Xeroderma pigmentoso, determinando uma alta heterogeneidade
e sistema de reparo do DNA, caracterizado em E.coli, é na rdade um complexo regulatório. Ele permite que uma região mutada de um
polinucleotídio seja replicada, com a DNA Polimerase selecionando nucleotídeos
Atualmente se conhece várias patologias causadas, direta ou indiretamente, por alterações no sistema de reparo de mutações, como, por exemplo, câncer de mama, câncer de ovário, telangiectasia atáxica, Síndromes de Bloom e Werner,
É importante lembrar que, além de mecanismos de reparo, os organismos também possuem sistemas que possibilitam regiões alteradas replicarem sem reparo
es mecanismos possibilitam, em alguns casos, o surgimento de doenças. Porém, a criação e manutenção da variabilidade genética também é o mecanismo que permite ao organismo estar apto à evolução.
Se uma mutação persiste em uma célula germinativa, a despeito de todos os , e se mantém na população com uma frequência igual ou
que este gene que agora possui duas ou mais formas alternativas, alelo selvagem e alelo(s) mutante(s), é um gene polimórfico. Todos os
272

273
Módulo V – Processos Reprodutivos
alelos deste gene que possuírem frequência igual ou superior a 1% serão também ditos polimórficos, e aqueles que são encontrados na população com frequência inferior a 1%, são ditos alelos raros.
Alterações cromossômicas estruturais
Nesta classe, estão reunidas as inserções, deleções, duplicações e transversões. Todas elas alteram a sequência das bases do DNA que forma o cromossomo. As aberrações estruturais dos cromossomos resultam de quebra cromossômica seguida de reconstituição em uma combinação anormal.
Quando o conjunto cromossômico possui informações a mais ou a menos, são ditos rearranjos não equilibrados. Exemplo: deleções, duplicações, etc. Já se o conjunto cromossômico possui o complemento normal de informações, como, por exemplo, nas inversões, estas alterações são ditas rearranjos equilibrados.
A seguir veremos as várias alterações cromossômicas estruturais que uma célula pode apresentar (Figuras 4 a 6).
• Deleção: perda de um segmento cromossômico. Origina-se por quebra cromossômica e perda do segmento acêntrico ou por Crossing-Over desigual entre cromossomos homólogos desalinhados ou cromátides-irmãs. Dependendo ainda da localização da alteração, as deleções podem ser classificadas em intercalar (intersticial) ou terminal. São exemplos clínicos dos efeitos da deleção cromossômica em um organismo as Síndromes de Angelman e Prader-Willi, Síndrome de Willians, Síndrome de Smith-Magenis, etc.
• Duplicações: podem originar-se por Crossing-Over desigual ou por segregação anormal da meiose num portador de uma translocação ou inversão. Este tipo de anomalia poderá originar indivíduos com trissomias parciais (CID 10- Q92.3 Trissomia parcial minor).
• Cromossomos em anel: as deleções terminais nos dois braços de um cromossomo podem dar origem a um cromossomo em anel, se as extremidades livres fraturadas se soldarem. Já foram descritos casos de formação de anéis em todos os cromossomos humanos. O cromossomo 22 em anel tem mais de 60 casos clínicos descritos na literatura, entretanto não há uma sintomatologia “padrão” que possa definir como Síndrome.
• Isocromossomos: são cromossomos que apresentam deficiência total de um dos braços e duplicação completa do outro. Foram relatadas presenças de isocromossomos em tumores do mediastino (cromossomo 12), na Síndrome de Ullrich-Turner (Xp ), etc.
• Inversão é a ocorrência de duas quebras em um cromossomo unifilamentoso durante a interfase e a soldadura em posição invertida do fragmento ao restante do cromossomo. A inversão é dita paracêntrica se as quebras ocorrerem em um mesmo braço cromossômico, e é denominada pericêntrica se o fragmento cromossômico invertido incluir o centrômero.
Acesse a página http://genetica.ufcspa.edu.br/seminarios%20monitores/alteracoes%20numericas.pdf e leia o texto “Alterações Cromossômicas Numéricas e suas Implicações Clínicas: Dissomia Uniparental e Imprinting Genômico”, da profª. Elizabeth Castro, da Fundação Faculdade Federal de Ciências Médicas de Porto Alegre.
Saiba mais, acesse os sites http://www.swbrasil.org.br/ , http://www.xfragil.org.br/ , http://www.lookfordiagnosis.com/ e conheça alguns exemplos de patologias causadas por alterações cromossômicas estruturais.

274
Módulo V – Processos Reprodutivos
Figura 4: Representação esquemática de alterações cromossômicas estruturais.
• Cromossomos dicêntricos: são cromossomos que apresentam dois centrômeros. Eles tendem a quebrar-se na anáfase se os dois centrômeros estiverem próximos; se um centrômero for inativado, um cromossomo dicêntrico pode ser estável. Na espécie humana geralmente não se observa a existência de síndromes ou patologias ocasionadas por este tipo de alteração.
• Translocação : dois cromossomos sofrem quebras e há a soldadura de um segmento cromossômico a uma região fraturada de outro.
• Translocações recíprocas: resultam de quebra de cromossomos não homólogos com trocas recíprocas de segmentos soltos. Quando os cromossomos de uma translocação recíproca balanceada se pareiam na meiose, forma-se uma figura quadrirradial (em forma de cruz). Na anáfase os cromossomos se segregam a partir desta configuração de três maneiras possíveis (Figura 5).
São exemplos de translocação balanceada a ocorrida entre 22pter e 9pter. O novo cromossomo 22 é chamado de cromossomo Philadelphia. Neste evento, um novo gene (BRC-ABL), proveniente da fusão da sequencia de DNA do cromossomo 22 (ponto de quebra do cluster – BRC) e do gene ABL, localizado no cromossomo 9, é criado. O produto deste novo gene é fundamental para a patogênese da leucemia mielóide crônica (LMC).
• Translocações robertsonianas: Envolvem dois cromossomos acrocêntricos que se fundem próximos à região do centrômero com perda dos braços curtos. Embora o portador de uma translocação robertsoniana seja fenotipicamente normal, há um risco de gametas e de prole não balanceados. No caso de gametas não balanceados, estes carregarão porções a mais ou a menos dos cromossomos que sofreram translocação, dando origem a indivíduos anormais, portadores de trissomias e/ou mononossomias (Figura 6). O exemplo clássico desta translocação é a t(14q;21q). Esta é a responsável por aproximadamente 5% dos casos de Síndrome de Down.

275
Módulo V – Processos Reprodutivos
Figura 5: Representação esquemática de uma translocação equilibrada, com os possíveis erros gaméticos que produzirão.
Figura 6: Representação esquemática de translocação robertsoniana e consequentes gametas a serem formados, pós-segregação na anáfase.
Alterações Cromossômicas Numéricas

Módulo V – Processos Reprodutivos
Quando uma célula apresenta uma alteração no número de cromossomos, dizemos que é poliplóide ou aneuplóide
Chamamos de poliplóide se ncomo, por exemplo, um triplóide (3n) ou um tetraplóide (4n). Entretantopoliploidia é incompatível com a vida, na espécie humana. Já em vegetais, a poliploidia está relacionada com macrossomia, ou seja, plantas mais robustas, como é o caso do tomate tetraplóide.
Uma célula pode ser aneuplóide por possuir um complemento crincompleto ou desequilibrado. Se faltar um cromossomo do par homólogo, chamamos monossomia. Se, ao contrário, existir um cromossomo extra, além do par homólogo, dizemos trissomia. Se um determinado cromossomo estiver quadruplicado, isto é, ocorrência de 4 homólogos na célula, então estetrassômica.
Dentre as trissomias de ocorrência naXX ou XY + 21), ou Síndrome de Down, é a mais freqcromossômico congênito mais comum associado ao retardo mental grave. As características fenotípicas desta síndrome são bem marcantes e discernívenascimento. Entretanto, os traços fenotípicos característicos da Síndrome de Down são expressos variavelmente e não estão presentes em todos os indivíduos afetados. A confirmação do diagnóstico deve ser feita com análise cariotípica, uma vez que estasíndrome é geneticamente caracterizada pela presença de 47 cromossomos.
A incidência dessa patologia está diretamente relacionada com o aumento da idade materna, ou seja, uma mulher com 20 anos de idade, sem antecedentes familiares, tem aproximadamente uuma criança com trissomia do 21. Já aos 45 anos, esperacrianças nascidas. A contribuição paterna para os casos de Síndrome de Down estestimada em cerca de 5 a 15% das crianças.
Estudos indicam que aproximadamente 40 a 50% dos abortos espontâneos do primeiro trimestre de gravidez são trissômicos para um autossômico. Todas as trissomias humanas estão associadatrissomias do 13 (Síndrome de Patau) sobrevivência a termo, porém de sobrevida curta,
Depois da trissomia do 21, as aneuploidias (inteiros isolados) mais frequentes são aquelas envolvendo os cromoss
Sabemos que as mulheres possuem em suas células somáticas duas cópias do cromossomo X, enquanto que os homens possuem um cromossomo X e um cromossomo Y. Indivíduos do sexo masculinoX, são ditos portadores da Síndrome de Klinefeltercomo 47, XXY. Ao contrário, existem mulheres portadoras de uma única cópia do cromossomo X. Elas têm como representação cariotípica a fórmula 45,portadoras da Síndrome de Turner. Outsexuais já foram descritas, como a Síndrome do Triplo X (48,Duplo Y (47, XYY). Entretanto, estas são bem menos frequ
Processos Reprodutivos
Quando uma célula apresenta uma alteração no número de cromossomos, aneuplóide.
nela exibir um ou mais conjuntos haplóides extras, um triplóide (3n) ou um tetraplóide (4n). Entretanto, a
poliploidia é incompatível com a vida, na espécie humana. Já em vegetais, a poliploidia está relacionada com macrossomia, ou seja, plantas mais robustas, como é
Uma célula pode ser aneuplóide por possuir um complemento cromossômico incompleto ou desequilibrado. Se faltar um cromossomo do par homólogo,
. Se, ao contrário, existir um cromossomo extra, além do par . Se um determinado cromossomo estiver
ência de 4 homólogos na célula, então esta célula é dita
Dentre as trissomias de ocorrência na espécie humana, a trissomia do 21 (47, XX ou XY + 21), ou Síndrome de Down, é a mais frequente. É o distúrbio cromossômico congênito mais comum associado ao retardo mental grave. As
índrome são bem marcantes e discerníveis ao nascimento. Entretanto, os traços fenotípicos característicos da Síndrome de Down são expressos variavelmente e não estão presentes em todos os indivíduos afetados. A confirmação do diagnóstico deve ser feita com análise cariotípica, uma vez que esta síndrome é geneticamente caracterizada pela presença de 47 cromossomos.
a patologia está diretamente relacionada com o aumento da idade materna, ou seja, uma mulher com 20 anos de idade, sem antecedentes familiares, tem aproximadamente uma chance em cada 2.000 nascimentos de gerar uma criança com trissomia do 21. Já aos 45 anos, espera-se um bebê Down a cada 40 crianças nascidas. A contribuição paterna para os casos de Síndrome de Down está
as crianças.
os indicam que aproximadamente 40 a 50% dos abortos espontâneos do primeiro trimestre de gravidez são trissômicos para um autossômico. Todas as trissomias humanas estão associadas a déficit de desenvolvimento, exceto para as
atau) e 18 (Síndrome de Edwards), que demonstram porém de sobrevida curta, e são fatais.
Depois da trissomia do 21, as aneuploidias (perda ou adição de cromossomos entes são aquelas envolvendo os cromossomos sexuais.
Sabemos que as mulheres possuem em suas células somáticas duas cópias do cromossomo X, enquanto que os homens possuem um cromossomo X e um cromossomo Y. Indivíduos do sexo masculino, com uma cópia extra do cromossomo
a Síndrome de Klinefelter, e sua fórmula cariotípica é escrita como 47, XXY. Ao contrário, existem mulheres portadoras de uma única cópia do cromossomo X. Elas têm como representação cariotípica a fórmula 45, X0, e são portadoras da Síndrome de Turner. Outras aneuploidias ligadas aos cromossomos sexuais já foram descritas, como a Síndrome do Triplo X (48, XXX), e a Síndrome do
tanto, estas são bem menos frequentes.
276
As manifestações da trissomia do 21 são conhecidas como a Síndrome de Down, uma combinação específica de características fenotípicas que incluem retardo mental e uma face típica, e é causada pela existência de três cromossomos 21 (um a mais do que o normal). Veja o site: http://www.abcdasaude.com.br/artigo.php?393 consultado em 23/08/09.

Módulo V – Processos Reprodutivos
Corpúsculo de Barr e a Hipótese de Lyon
Na década de 40, Murray Barr e Ewart Bertram observaram diferenças na posição de uma massa com coloração interna escura intensa no núcleo das células da interfase. Também notaram que esta massa, conhecida hoje como Corpúsculo de Barr, estava associada somente às células corpúsculos nada mais seriam do que um cromossomo X muito condensado. Daí o termo “cromatina sexual” como uma outra forma de se designar o Corpúsculo de Barr.
Foi observado que em homens portadores da Síndromeencontrado um corpúsculo de Barr, enquanto em mulheres com Síndrome de Turner a cromatina sexual está ausente.
Em 1961, Mary Lyon propôs a hipótese do X inativo para explicar o que acontecia com os genes no corpúsculo de Barr. Segundo a hencontrados no cromossomo X condensado, isto é, no corpúsculo de Barrgeneticamente inativos. Esta inativação ocorreria muito cedo no desenvolvimento, durante o estágio de blastocisto, sendo realizada aleate igualaria a expressão gênica em homens e mulheres, conhecido como “compensação de dose”Lionização.
A hipótese de Lyon explica fenômenos conhecidos como a pelagem de três cores em gatas, a expressão mais amena da deficiência homozigotas, etc.
É interessante notar que as monossomias de cromossomos autossomos são consideradas fatais, enquanto que uma mulher com monossomia do X pode passar a vida toda, ou grande parte dela, sem o diagnóstico. Da mesma forma, as condições trissomicas nos autossomos levam a grandes danos na formação e desenvolvimento do organismo, enquanto a trissomia dos alossomos podepróximo ao normal. Ao que parece,sobreviver com o excesso de genes do que com a falta deles, explicando as raras síndromes de trissomia de autossomos
Já no caso das trissomias e monossomias envolvendo os cromoshoje sabemos que nas células femininas um dos dois cromossomos X permanece inativo, como um mecanismo de compensação de dose. Uma vez que o cromossomo Y é pequeno e possui um número muito restrito de genes, a inativação de um cromossomo X nas mulheres faz com que ambos os organismos, masculino e feminino, exibam a mesma atividade gênica. cromossomo X é conhecido por Hipótese de Lyon.
ATIVIDADE
Fixando o que aprendeu.
1. Faça uma pesquisa na internet e procure informações sobre Síndrome de Down, Síndrome de Edwards, Síndrome de Patau, Síndrome de Turner e Síndrome
Processos Reprodutivos
Corpúsculo de Barr e a Hipótese de Lyon
Na década de 40, Murray Barr e Ewart Bertram observaram diferenças na posição de uma massa com coloração interna escura intensa no núcleo das células da interfase. Também notaram que esta massa, conhecida hoje como Corpúsculo de
nte às células femininas, o que sugeriu que os corpúsculos nada mais seriam do que um cromossomo X muito condensado. Daí o termo “cromatina sexual” como uma outra forma de se designar o Corpúsculo de
Foi observado que em homens portadores da Síndrome de Klinefelter é encontrado um corpúsculo de Barr, enquanto em mulheres com Síndrome de Turner
Em 1961, Mary Lyon propôs a hipótese do X inativo para explicar o que acontecia com os genes no corpúsculo de Barr. Segundo a hipótese de Lyon, os genes encontrados no cromossomo X condensado, isto é, no corpúsculo de Barr, estariam
a inativação ocorreria muito cedo no desenvolvimento, durante o estágio de blastocisto, sendo realizada aleatoriamente em cada blastocélula, igualaria a expressão gênica em homens e mulheres, por meio de um fenômeno
conhecido como “compensação de dose”, sendo a inativação do X conhecida como
A hipótese de Lyon explica fenômenos conhecidos como a pelagem de três ores em gatas, a expressão mais amena da deficiência em G6PD em mulheres
É interessante notar que as monossomias de cromossomos autossomos são consideradas fatais, enquanto que uma mulher com monossomia do X pode passar a
grande parte dela, sem o diagnóstico. Da mesma forma, as condições trissomicas nos autossomos levam a grandes danos na formação e desenvolvimento
a trissomia dos alossomos pode apresentar um fenótipo próximo ao normal. Ao que parece, o organismo está mais bem preparado para sobreviver com o excesso de genes do que com a falta deles, explicando as raras síndromes de trissomia de autossomos, que culminam com gravidez a termo.
Já no caso das trissomias e monossomias envolvendo os cromossomos X e Y, hoje sabemos que nas células femininas um dos dois cromossomos X permanece inativo, como um mecanismo de compensação de dose. Uma vez que o cromossomo Y é pequeno e possui um número muito restrito de genes, a inativação de um
mulheres faz com que ambos os organismos, masculino e feminino, exibam a mesma atividade gênica. O mecanismo de inativação aleatória do cromossomo X é conhecido por Hipótese de Lyon.
Faça uma pesquisa na internet e procure informações sobre Síndrome de Down, Síndrome de Edwards, Síndrome de Patau, Síndrome de Turner e Síndrome
277

Módulo V – Processos Reprodutivos
de Klinefelter. Não deixe de colocar a fórmula cariotípica característica de cada uma dessas aneuploidias, bem como as principais características fenotípicas.
2. Quando tratamos de uma característica apresentando um padrão de ausência de dominância, cujo gene responsável está localizado na região não homóloga do cromossomo X, o fenótipo heterozigoto só se manifestexplicar o por quê?
3) Comente o texto abaixo, transcrito do artigo “Diabetes Insipidus Nefrogênico: Conceitos Atuais de Fisiopatologia e Aspectos Clínicos” (Rocha e 2002, in Arq. Brás. Endocrinol. Metab
“O diabetes insipidus nefrogênico (DIN) é uma rara doença renal, caracterizada pela impossibilidade de concentrar a urina, apesar da concentração sangaumentada de AVP. Em geral, a doença é caracterizada por início na infância, história persistente, poliúria e hipostenúria resistente à administração de vasopressina. É importante salientar que pacientes com outras formas de diabetes insipidus podem apresentar a sintomatologia acima descritaingesta diária de água superior a 18 litros não t(provavelmente têm polidipsia primária). Devido a sua localização no cromossoma X (Xq28), mutações inativadoras no receptor AVPR2 causam um fenótipo caracterizado por episódios de desidratação logo após o nascimento, hipernatremia e hipertermia. Os episódios de desidratação podem ser tão severos que diminuem a pressão de perfusão arterial, levando a uma oxigenação deficiente do cérebro, rins e outros órgãos e, conseqüentemente, retardo mental, insuficiência reAs pacientes heterozigotas (portadoras) apresentam graus variáveis de poliúria e polidipsia... masculino (hemizigotos), não sendo afetados por variações de caráter dominante ou recessivo para uma doença ligada ao X, como as obs
Estrutura e Evolução dos Genes
Atualmente, baseados na tripla função que as moléculas de RNA exercem nas células, é aceita a hipótese de que o RNA precedeu a formação do DNA. A existência de moléculas de RNA com atividade catalítica; a presença de algumas moléculas idênticas ou muito semelhantes aos monômeros de RNA em todos os seres vivos, que atuam como cofatores; o fato do DNA não ser quimicamente tão flexível e os desoxirribononucleotídeos serem derivados dos ribonucleotídeos, entre outras, fortalecem esta chamada “hipótese do mundo de RNA
O DNA fita dupla mantém como se fosse um “backup”genética na fita complementar. É uma molécula extremamente resistente, permitindo maior estabilidade à informação. Essgenomas de DNA em preferência aos de RNA.
Processos Reprodutivos
de Klinefelter. Não deixe de colocar a fórmula cariotípica característica de cada uma como as principais características fenotípicas.
2. Quando tratamos de uma característica apresentando um padrão de ausência de dominância, cujo gene responsável está localizado na região não homóloga do cromossomo X, o fenótipo heterozigoto só se manifesta no sexo feminino. Você sabe
3) Comente o texto abaixo, transcrito do artigo “Diabetes Insipidus Nefrogênico: Conceitos Atuais de Fisiopatologia e Aspectos Clínicos” (Rocha e cols,
Metab., vol. 44, nº. 4):
“O diabetes insipidus nefrogênico (DIN) é uma rara doença renal, caracterizada pela impossibilidade de concentrar a urina, apesar da concentração sanguínea normal ou aumentada de AVP. Em geral, a doença é caracterizada por início na infância, história familiar positiva, sede persistente, poliúria e hipostenúria resistente à administração de vasopressina. É importante salientar que pacientes com outras formas de diabetes insipidus podem apresentar a sintomatologia acima descrita; e pacientes com
diária de água superior a 18 litros não têm DIN (provavelmente têm polidipsia primária). Devido a sua localização no cromossoma X (Xq28), mutações inativadoras no receptor AVPR2 causam um fenótipo caracterizado por episódios de desidratação logo após o
cimento, hipernatremia e hipertermia. Os episódios de desidratação podem ser tão severos que diminuem a pressão de perfusão arterial, levando a uma oxigenação deficiente do cérebro, rins e outros órgãos e, conseqüentemente, retardo mental, insuficiência renal, etc. As pacientes heterozigotas (portadoras) apresentam graus
eis de poliúria e polidipsia... Indivíduos do sexo masculino (hemizigotos), não sendo afetados por variações de caráter dominante ou recessivo para uma doença ligada ao X, como as observadas em mulheres.“
strutura e Evolução dos Genes
Atualmente, baseados na tripla função que as moléculas de RNA exercem nas células, é aceita a hipótese de que o RNA precedeu a formação do DNA. A existência de moléculas de RNA com atividade catalítica; a presença de algumas moléculas dênticas ou muito semelhantes aos monômeros de RNA em todos os seres vivos, que atuam como cofatores; o fato do DNA não ser quimicamente tão flexível e os desoxirribononucleotídeos serem derivados dos ribonucleotídeos, entre outras,
hipótese do mundo de RNA”.
m como se fosse um “backup” (cópia) da informação uma molécula extremamente resistente, permitindo
r estabilidade à informação. Essas características teriam propiciado a seleção dos genomas de DNA em preferência aos de RNA.
278
Hipótese do mundo de RNA A hipótese afirma que as primeiras formas de vida dependiam da atividade catalítica e replicativa do RNA, e que DNA e proteínas teriam assumido suas funções atuais. atuais.ais.posteriorment

279
Módulo V – Processos Reprodutivos
O surgimento dos genomas de DNA e de Polimerases de alta fidelidade possibilitam o aparecimento de genomas maiores, com capacidade de codificação aumentada. Fenômenos como duplicação gênica e o aparecimento de exons e introns, de novas proteínas, com novas funções, culminaram na diversificação e complexidade das formas de vida que hoje conhecemos.
Estudos filogenéticos indicam que eucariotos e arqueobactérias formam um grupo-irmão, excluindo-se as eubactérias. O grupo-irmão possui várias características em comum, como, por exemplo, a ausência de parede celular bacteriana, semelhança de proteínas e vias metabólicas, etc. Atualmente, assume-se que os procariotos são anteriores aos eucariotos modernos, que possuem mitocôndria.
Para explicar vários aspectos do padrão de evolução de sequências de DNA, que codificam para proteínas, existe um amplo conhecimento teórico que, no geral, se reporta a taxas de mutação, de evolução, de restrições funcionais às estruturas protéicas, etc.
Iniciaremos nossa discussão sobre a evolução de sequências gênicas enumerando as principais características dos genes procariotos e eucariotos. A estrutura gênica de ambos os grupos compreendem a região do genoma entre pontos correspondentes às bases dos terminais 5' e 3' do RNA mensageiro maduro. Sequências regulatórias estão presentes em ambos os terminais do gene e são necessárias para assinalar o local de início e, às vezes, de término de transcrição.
Procariotos, de um modo geral, possuem uma estrutura gênica menos complexa que eucariotos. Em ambos os casos, podemos dividir o gene em região codificadora e não codificadora (UTR - do inglês untranslated region). Mas, enquanto um gene procariótico é descrito como contínuo, organizado em sistemas de operons formando RNA policistrônico (poligênico), um gene eucariótico é descontínuo, composto por exons e introns, e seu sistema regulatório é extremamente complexo.
Os genes e proteínas de procariotos são ditos colineares, ou seja, a sequência de nucleotídeos de um gene corresponde exatamente à sequência de aminoácidos da proteína por ele codificada. Tal fato já não acontece entre genes e proteínas eucariontes, em virtude da existência de sequências internas de DNA no gene (introns), que não representam uma proteína. Somente as sequências denominadas como exônicas possuem aminoácidos correspondentes na proteína.
Em procariotos a síntese protéica é simultânea à de RNA mensageiro, enquanto que, em eucariotos, a síntese de RNA mensageiro ocorre no núcleo, sendo que essa molécula, após sofrer uma série e alterações pós-transcricionais será transportada para o citoplasma, onde ocorrerá a tradução. Uma das modificações pós-transcricionais é justamente a retirada dos íntrons, daí não possuírem uma sequência de aminoácidos correspondentes na proteína sintetizada (veja na Figura 7).

Módulo V – Processos Reprodutivos
E I
UTR 5´ 5´
Gene eucarioto
Gene procarioto
5´
UTR
Figura 7: Esquema comparativo da organização dos genes em procariotos e eucariotos.
As várias diferenças entre os genes dos dois tipos de organismos nos indicam que vários mecanismos de diferenciação ocorreram originandoanalisamos a distribuição dos genes, observagenoma formando grupos com características evolutivas peculiares (genes ribossômicos, genes da família das globinas,apresentam sequências nucleotídicas semelhantes, porém diferindo quanto à função.
O mecanismo originário dessde duplicação gênica, via mecanismos de retrApós esse evento, novas cópias podem manter a mesma sequ(exemplo: genes RNA ribossômicos)gênicas. Alguns membros destas famílias são npseudogenes surgem a partir da duplicação gênica genômicomutações, e serem inativados ao longo de gerações sucessivas. No cromossomo 22 foram identificadas 134 dessas sequidentificados neste cromossomo.
Outros mecanismos, tais como o intrônicas, introduzindo nesses sítios de na produção de novos polipeptídios a partir de um mesmo segmento de DNA.
A presença de genes sobrepostos, um mesmo segmento de DNA utilizando quadros de leitura diversos, pode emergir pelo uso da fita complementar de uma sequencia de DNA. Também o fenômeno conhecido por “compartilhamento de genes”, que nada mais é que o recrutamenadicional secundária, sem alteração de sua estrutura, já foi descrito em proteínas responsáveis pela manutenção da transparência e pela difração da luz no cristalino (cristalinas).
ATIVIDADE
Faça uma pesquisa sobre as ações do mecanismo
Processos Reprodutivos
l = intron; E = exon; UTR = região não traduzida (região promotora e reguladora do gene)
E E E I I UTR 3´
3´
UTR
Esquema comparativo da organização dos genes em procariotos e
várias diferenças entre os genes dos dois tipos de organismos nos indicam vários mecanismos de diferenciação ocorreram originando-os. Quando
analisamos a distribuição dos genes, observa-se que alguns deles estão dispostos no genoma formando grupos com características evolutivas peculiares (genes
s globinas, etc.). Famílias gênicas ou multigênicas ências nucleotídicas semelhantes, porém diferindo quanto à função.
ses grupos gênicos parece ser, prioritariamente, o via mecanismos de retrotransposição ou duplicação do DNA.
cópias podem manter a mesma sequência de DNA genes RNA ribossômicos), ou divergirem, dando origem às famílias
as famílias são não funcionais (pseudogenes). Estes pseudogenes surgem a partir da duplicação gênica genômico, seguida de acúmulo de
ao longo de gerações sucessivas. No cromossomo 22 am identificadas 134 dessas sequências, que correspondem a 20,7% dos genes
Outros mecanismos, tais como o Crossing-Over desigual, mutações em regiões s sítios de splicing alternativos, também podem resultar
na produção de novos polipeptídios a partir de um mesmo segmento de DNA.
resença de genes sobrepostos, um mesmo segmento de DNA utilizando quadros de leitura diversos, pode emergir pelo uso da fita complementar de uma
encia de DNA. Também o fenômeno conhecido por “compartilhamento de genes”, que nada mais é que o recrutamento de um gene para exercer uma função adicional secundária, sem alteração de sua estrutura, já foi descrito em proteínas responsáveis pela manutenção da transparência e pela difração da luz no cristalino
s ações do mecanismo Crossing-Over desigual.
280
UTR = região não traduzida (região
3´
A Doença de Alzheimer é uma afecção neurodegenerativa progressiva e irreversível, de aparecimento insidioso, que acarreta perda da memória e diversos distúrbios cognitivos. É uma síndrome progeróide genética com alteração da atividade dos genes ribossômicos.

Módulo V – Processos Reprodutivos
Estratégias de Pesquisa sobre Herança
Quando observamos uma característica hereditária em uma família podemos definir o padrão de herança da mesma, inferir o número de genes que estão atuando para o aparecimento do fenótipo em estudo,abordagem os heredogramas obtidos e as informade uma abordagem familial, entretanto também podemos utilizar outra abordagem clássica, que é estudando os tipos dtipo de prole proveniente de tais cruzamentos. Estabordagem populacional.
Nessa abordagem, levamos em conta quais e em que proporções cada tipo de casal (ambos apresentam a mesma cadiferente) aparece na população e quais as proporções observadas entre suas proles.
Na Tabela 3 estão indicadas as frequporcentagem, na prole de casais, quando se levam em cona, frequentes na população de onde procedem os casais. coleta uma amostra populacional, levando em conta o tipo de genótipo dos casais amostrados e o tipo e proporção observadas nas suas proles, podemos ipadrão de herança que é seguido pela característica.
Genótipo casal Prole AA AA × AA 100 AA x AA 50 AA x AA - Aa x AA 25 Aa x AA - aa x AA -
Tabela 3: Frequências genotípicas esperadas, casais, quando se levam em conta os alelos autossômicos A, a, população de onde procedem os casais ( Beiguelman, B., 2008
ATIVIDADE
Fixando a aprendizagem.
1. Dentre os 120 filhos de casais MN × MNMN e 28 do grupo N (Beiguelman, 2008)de herança você pode sugerir para o sistema sangu
2. Imagine que um casal heterozigoto (MN) tenha tido quatro filhos. Quantos filhos você espera que possuam o mesmo genótipo dos pais? Elabore um heredograma fictício para esta família.
Ambas as estratégias ainda são de uso corrente para o estabelecimento das bases e padrões de herança das características herdáveis.
Processos Reprodutivos
Estratégias de Pesquisa sobre Herança
Quando observamos uma característica hereditária em uma família podemos definir o padrão de herança da mesma, inferir o número de genes que estão atuando
o do fenótipo em estudo, etc. Utilizamos como base para esta abordagem os heredogramas obtidos e as informações neles contidas. Chamamos isto de uma abordagem familial, entretanto também podemos utilizar outra abordagem clássica, que é estudando os tipos de casais que exibem a característica e o número e
eniente de tais cruzamentos. Esta segunda abordagem é a
levamos em conta quais e em que proporções cada tipo de casal (ambos apresentam a mesma característica, ou cada cônjuge exibe um fenótipo diferente) aparece na população e quais as proporções observadas entre suas proles.
3 estão indicadas as frequências genotípicas esperadas, em quando se levam em conta os alelos autossômicos A,
ão de onde procedem os casais. Sendo assim, quando se levando em conta o tipo de genótipo dos casais
amostrados e o tipo e proporção observadas nas suas proles, podemos inferir o padrão de herança que é seguido pela característica.
Prole Aa Prole aa - - 50 - 100 - 50 25 50 50 - 100%
ências genotípicas esperadas, em porcentagem, na prole de quando se levam em conta os alelos autossômicos A, a, frequentes na
população de onde procedem os casais ( Beiguelman, B., 2008).
Dentre os 120 filhos de casais MN × MN, 24 eram do grupo M, 68 do grupo (Beiguelman, 2008). Com base nos dados fornecidos, que padrão
ode sugerir para o sistema sanguíneo MN?
Imagine que um casal heterozigoto (MN) tenha tido quatro filhos. Quantos espera que possuam o mesmo genótipo dos pais? Elabore um
heredograma fictício para esta família.
estratégias ainda são de uso corrente para o estabelecimento das bases e padrões de herança das características herdáveis.
281

282
Módulo V – Processos Reprodutivos
Conclusão
Não devemos esquecer que na atualidade sabemos que o modo como o gene determina um fenótipo está intimamente ligado, não só à sua seqüência de DNA, mas também ao ambiente em que ele se expressa. A partir da década de 50 (século XX), houve um grande avanço nas pesquisas para o entendimento dos padrões de herança e da associação e ligação existentes entre os genes, quando o Dr. Newton Morton publicou seus artigos sobre análise de ligação em heredogramas humanos.
De lá para cá, muitas implementações, estatísticas e computacionais, foram inseridas nas pesquisas genéticas, sendo que estratégias baseadas em coletas de dados bem estruturadas e complexas análises estatísticas estão ajudando a desvendar a importância dos genes em características multifatoriais, tais como diabetes e hipertensão.
Na última década do século XX, a biologia molecular firmou-se como uma ciência forte e influente, principalmente em relação a abordagens de investigação e terapêuticas genéticas. Atualmente, utilizando técnicas moleculares, podemos criar organismos capazes de sintetizar moléculas úteis, tanto nas áreas médicas quanto biológicas e agrárias.
Técnicas de mutagênese propiciam a substituição de um alelo por outro, que foi geneticamente modificado. Em alguns casos, esses genes foram introduzidos num organismo na tentativa de induzir o organismo a sintetizar uma proteína específica. Tal é o exemplo da insulina, na qual bactérias geneticamente modificadas produzem insulina, de forma que possibilitou o barateamento da substância que, agora, é produzida em escala comercial e, consequentemente, a melhora da qualidade de vida de pacientes diabéticos.
Várias outras moléculas são produzidas dessa forma: endorfinas, fator VIII da hemofilia, interferons, etc. Em outros casos, um gene específico é inativado e introduzido no organismo, de forma a se aprender sobre seu papel no organismo. Esta estratégia é utilizada para produzir camundongos com genes inativados (knockout), utilizados em pesquisas em todo o mundo.
Você sabia que o Brasil é um país que se destaca mundialmente em relação às suas produções em pesquisas genéticas? Leia o texto disponível em http://www.fmrp.usp.br/revista/2006/vol39n1/11_o_emprego_camundongos_pesquisa.pdf sobre as vantagens de se usar camundongos geneticamente modificados em pesquisas médicas.

283
Módulo V – Processos Reprodutivos
Referências
Adkison, L.R. & Brown, M.D. Genética – Série Elsevier de formação básica integrada. Rio de Janeiro, Elsevier Ltda., 2008.
BERED, Fernanda; BARBOSA NETO, José Fernandes e CARVALHO, Fernando Irajá Félix de. Marcadores moleculares e sua aplicação no melhoramento genético de plantas. Cienc. Rura, 1997, vol. 27, no. 3, pp. 513-520. **
Beiguelman, B. A Interpretação Genética Da Variabilidade Humana. Ribeirão Preto: SBG, 2008.
Borovik, CL; Muller, R, e cols. Caracterização de um cromossomo 22 em anel por citogenética molecular. Einstein, 2003; 1:113-6
Curiel, D.; Brantly, M.; Curiel, E; Stier, L.; Crystal; R. G. al -Antitrypsin Deficiency Caused by the al -Antitrypsin Null Mattawa Gene- an Insertion Mutation Rendering the al -Antitrypsin Gene Incapable of Producing a-l –Antitrypsin. The Journal of Clinical Investigation, 1989. Vol 83: 1144-1152.
Griffiths, A.J.F., et al. 2006. Introdução à Genética. Rio de Janeiro, Ed. Guanabara Koogan, 8ª ed.
Jorde, L.B., Carey, J.C., Bamshad, M.J. & White, R.L. Genética Médica, Rio de Janeiro, Elsevier Ltda., 2000, 3ª ed.
Kaczor, MP.; Sanak,M.; Szczeklik, A. Rapid and Inexpensive Detection of α1-Antitrypsin Deficiency-Related Alleles S and Z by a Real-Time Polymerase Chain Reaction Suitable for a Large-Scale Population-Based Screening. J. Mol Diagn. 2007. Vol 9: 99–104.
Lewin, B. GENES VIII. Pearson Education Inc., New Jersey. Upper Saddle River, 2004.
Matioli, S.R. Biologia molecular e evolução. Ribeirão Preto, Editora Holos, 2004
Mendel, G. 1865, “EXPERIMENTS IN PLANT HYBRIDIZATION in http://www.mendelWeb.org/
Strachan, T. & READ, A. P. Genética Molecular Humana. Porto Alegre, Editora Artes Médicas, 2002*
Wood, AM.& Stockley, RA. Alpha One Antitrypsin Deficiency: From Gene to Treatment. Respiration. 2007, Vol. 74:481–492.
*disponíveis online em www.ncbi.nih.gov/books
** Disponível em: <http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0103-84781997000300026&lng=pt&nrm=iso>. ISSN 0103-8478. doi: 10.

284
Módulo V – Processos Reprodutivos
Retirado do Google Acadêmico (Faculdades Integradas “Antônio Eufrásio de Toledo” - Faculdade de Direito de Presidente Prudente - Clonagem: O Duelo entre a Engenharia Genética e o direito. Cláudia Regina de Oliveira. Presidente Prudente/SP 2002 - http://intertemas.unitoledo.br/revista/index.php/Juridica/article/viewFile/66/72
Cláudia Emília Veira Wiezel. Segregação e Frequências Alélicas dos Loci SCA1 (Ataxia Espinocerebelar Tipo 1) e MJD (Doença de Machado-Joseph) em indivíduos normais. 1999. Dissertação (Mestrado em Ciências Biológicas (Genética)) - Faculdade de Medicina de Ribeirão Preto. Orientador: Aguinaldo Luiz Simões
Drª. Iscia Lopes Cendes, Expansão de poliglutaminas e doenças neurodegenerativas, publicado na revista Ciência e Cultura, vol.56, no.1, São Paulo Jan./Mar. 2004 em: http://cienciaecultura.bvs.br/scielo.php?script=sci_arttext&pid=S0009-67252004000100021&lng=pt&nrm=iso consultado em 23/08/09