º período faciplac... · importantes funções na regulação local da foliculogênese. diversos...
TRANSCRIPT

1

2

3

4
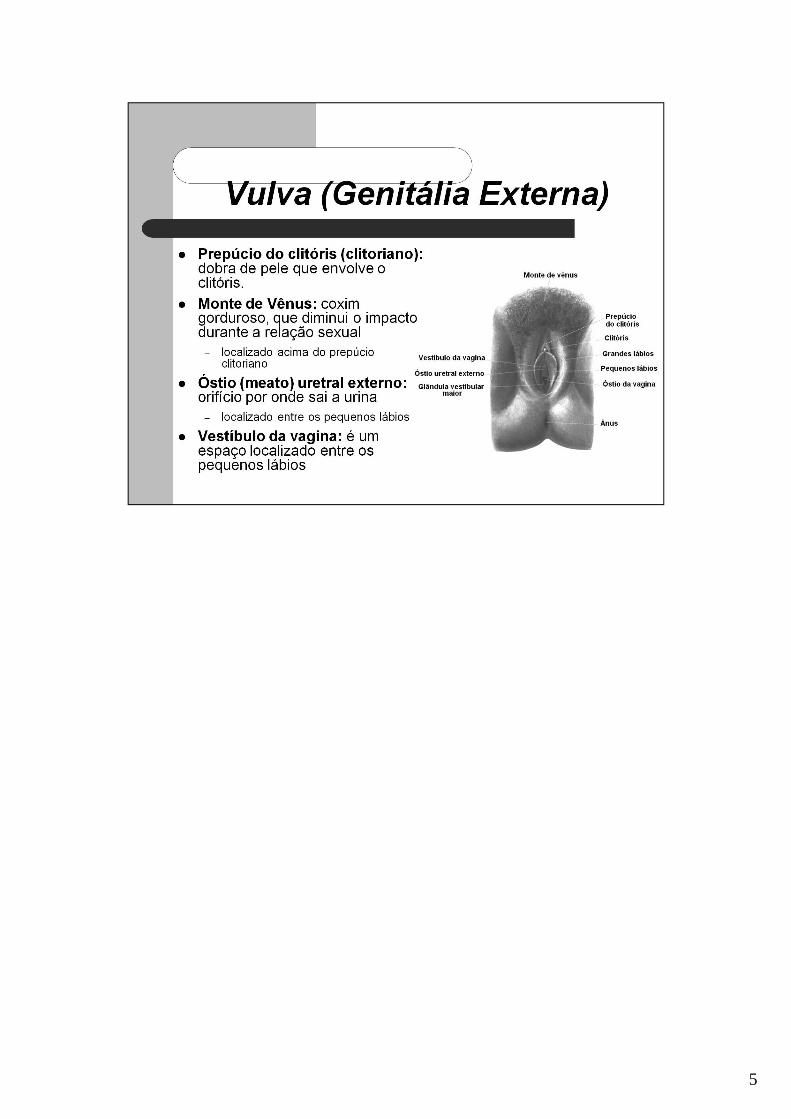
5

6

7

8

9

10

11
O ovário mamífero é um órgão dinâmico encarregado de proporcionar um ambiente ideal para aprodução de hormônios e liberação dos gametas femininos. No ovário, estão contidos milharesde folículos ovarianos, que constituem sua unidade básica estrutural e funcional. Afoliculogênese ovariana é um processo complexo que consiste do desenvolvimento de folículosprimordiais até o estágio de folículo pré-ovulatório, durante o qual ocorre o crescimentooocitário e intensa proliferação das células da granulosa. Vários fatores de crescimentoproduzidos pelas células foliculares, frequentemente, atuam modulando os efeitos dasgonadotrofinas FSH e LH, controlando, assim, a foliculogênese, entre eles, fator de crescimentoepidermal (EGF), fatores de crescimento de fibroblastos (FGF), fator de crescimento semelhanteà insulina-1 (IGF-1), fatores de crescimento transformante- (TGF- ) e Kit ligante (KL). Algunsdesses fatores de crescimento, como EGF, FGF, IGF, TGF- , proteínas morfogenéticas ósseas dostipos 2 (BMP-2), 4 (BMP-4), 6 (BMP-6), 7 (BMP-7) e 15 (BMP-15), fator de diferenciação docrescimento-9 (GDF-9), ativina-A e KL, estimulam o desenvolvimento dos folículos ovarianos,além de estarem envolvidos no controle dos processos de esteroidogênese e atresia folicular. Jáoutras substâncias, como as inibinas, exercem uma ação inibitória sobre a secreção degonadotrofinas e controlam, desta forma, a ação do FSH no ovário.

KL = Kit ligante; GDF-9 = fator de diferenciação do crescimento-9. Família EGF: o próprio EGF, TGF- , HB-EGF,anfiregulina, betacelulina e epiregulina. Família TGF- : o próprio TGF- , proteínas morfogenéticas ósseas dos tipos 2(BMP-2), 4 (BMP-4), 6 (BMP-6), 7 (BMP-7) e 15 (BMP-15), ativina-A, inibina, hormônio anti-Mülleriano (AMH).A foliculogênese ovariana é um processo complexo caracterizado pela formação, crescimento e maturação folicular. Éum processo que consiste no desenvolvimento de folículos primordiais até o estágio de folículos pré-ovulatórios,durante o qual ocorre crescimento oocitário e intensa proliferação das células da granulosa. Este processo é controladopor uma interação entre fatores de crescimento locais e gonadotrofinas de origem hipofisária. Recentemente, tem sidodemonstrado que os membros da superfamília dos fatores de crescimento transformante- (TGF- ) exercemimportantes funções na regulação local da foliculogênese. Diversos estudos e revisões têm mostrado que váriasproteínas pertencentes à família TGF- são expressas em oócitos, células da granulosa e da teca e funcionam comoreguladores intraovarianos dos processos de ativação de folículos primordiais, proliferação de células da granulosa e dateca, esteroidogênese e maturação oocitária, bem como no processo de atresia. Os principais fatores desta família queexercem funções no controle dos processos reprodutivos são as proteínas morfogenéticas ósseas dos tipos 2 (BMP-2), 4(BMP-4), 6 (BMP-6), 7 (BMP-7), 15 (BMP-15), o fator de diferenciação do crescimento-9 (GDF-9), a ativina-A, a inibina, ohormônio anti-Mülleriano (AMH) e o próprio fator de crescimento transformante- (TGF- ).Quando uma criança do sexo feminino nasce, cada ovócito I está circundado por camada única de células da granulosa,sendo denominado folículo primário. Durante toda a infância, acredita-se que as células da granulosa forneçam anutrição necessária para o ovócito I, além de secretar um fator inibidor da maturação dos ovócitos, que mantém osmantém em seu estado primordial, detendo-o na prófase da meiose por todo esse período de tempo. A seguir, após apuberdade, quando o FSH e o LH da adeno-hipófise começam a ser secretados em grandes quantidades, todo o ovário,juntamente com alguns de seus folículos, começa a crescer. A primeira fase do crescimento folicular consiste emaumento moderado do próprio ovócito I, cujo diâmetro aumenta por duas a três vezes. Segue-se, então, o crescimentode camadas adicionais de células da granulosa, e, nesse estágio, o folículo passa a ser conhecido como folículo primário.Parte do processo de desenvolvimento até esse estágio pode ocorrer na ausência de FSH e de LH, mas odesenvolvimento posterior não é possível sem a presença desses dois hormônios.Desenvolvimento dos folículos antrais e vesiculares: Durante os primeiros dias após o início da menstruação, verifica-seaumento leve e moderado nas concentrações de FSH e LH, sendo que a elevação do FSH precede por alguns dias a do LH.Esses hormônios, em particular o FSH, são responsáveis pelo crescimento acelerado de 6 a 12 folículos primários a cadamês. O efeito inicial consiste na rápida proliferação das células da granulosa, dando origem a muito mais camadas dessascélulas. Além disso, muitas células fusiformes derivadas do interstício ovariano também se acumulam, formando váriascamadas por fora das células da granulosa, dando origem à segunda classe de células, denominada teca. A teca édividida em duas subcamadas: na teca interna, as células adquirem características epitelioides, semelhantes às dascélulas da granulosa e adquirem a capacidade de secretar hormônios esteroides, assemelhando-se, nesse aspecto, àcapacidade das células da granulosa de secretar hormônios ligeiramente diferentes. A subcamada externa, denominadateca externa, consiste numa cápsula de tecido conjuntivo altamente vascular, que se transforma na cápsula do folículoem desenvolvimento. Após a fase proliferativa inicial de crescimento, que dura alguns dias, a massa de células dagranulosa secreta o líquido folicular que contém altas concentrações de estrogênio. Em consequência do acúmulo desselíquido, surge um antro no interior da massa de células da granulosa. Uma vez formado o antro, as células da granulosa eas células da teca proliferam ainda mais rapidamente, a velocidade de secreção aumenta, e cada um dos folículos emcrescimento transforma-se em folículo antral. O crescimento inicial do folículo primário até o estágio antral é estimuladoprincipalmente pelo FSH. A seguir, ocorre crescimento acelerado dos folículos antrais, resultando na formação defolículos muito maiores, denominados folículos vesiculares. 12

O crescimento acelerado dos folículos antrais decorre dos seguintes fatores: (1) Oestrogênio é secretado no folículo e induz a formação de números cada vez maiores dereceptores de FSH pelas células da granulosa; esse processo estabelece um efeito defeedback positivo ao tornar as células da granulosa mais sensíveis do que nunca ao FSHda adeno-hipófise. (2) O FSH hipofisário e os estrogênios combinam-se no sentido depromover a formação de receptores de FSH sobre as células da granulosa originais,permitindo, assim, que essas células sejam estimuladas pelo LH além da estimulação doFSH e provocando aumento muito rápido da secreção folicular. (3) O aumento dosestrogênios do folículo e o LH da adenohipófise atuam em conjunto, estimulando aproliferação das células teçais foliculares, bem como sua secreção. Por conseguinte,quando os folículos antrais começam a crescer, seu crescimento posterior ocorre commuita rapidez. Depois de 1 semana ou mais de crescimento — porém antes de ocorrer àovulação —, um dos folículos começa a se destacar dos demais, os quais começam,então, a involuir (processo conhecido como atresia).
13

14

15
Na espécie humana, a ovogênese inicia-se antes do nascimento. As ovogônias começam a se diferenciar em ovócito I(primário) por volta do terceiro mês de vida intra-uterina. No nascimento, os ovócitos primários estão em prófase I, quesó é retomada até a maturidade sexual. A partir da puberdade, mensalmente um ovócito I, sob a ação dos hormôniosgonadotróficos, sofre a 1ª divisão meiótica, originando o ovócito secundário (ovócito II), que recebe quase todo ocitoplasma, e o primeiro corpo polar (célula pequena, não funcional, que logo degenera), que recebe muito poucocitoplasma. Antes da ovulação, o núcleo do ovócito secundário inicia a segunda divisão meiótica, mas progride até ametáfase II, quando a divisão é interrompida. Se um espermatozoide penetra nesse ovócito, a segunda divisão meióticaé completada e novamente maior parte do citoplasma é mantida em uma célula, o óvulo fecundado. A outra célula,o segundo corpo polar, é uma célula também pequena, não funcional, que logo degenera. Essa meiose irregular garantiráum gameta com maior quantidade de nutrientes.Calcula-se que, no nascimento, o número total de ovócitos primários varie de 700.000 a 23 milhões. Durantea infância, a maioria dos ovócitos torna-se atrésico; somente cerca de 400.000 estão presentes no início depuberdade, e menos de 500 serão ovulados.

16

17

18

19

Os estrogênios e a progesterona são transportados no sangue ligados principalmente à albuminaplasmática e a globulinas de ligação específica a estrogênio e progesterona. Todavia, a ligação desseshormônios às proteínas plasmáticas é frouxa o suficiente para que sejam rapidamente liberados nos tecidosdentro de 30 minutos ou mais. O fígado conjuga os estrogênios com a consequente formação deglicuronídios e sulfatos; cerca de um quinto desses produtos conjugados é secretado na bile, enquanto amaior parte do restante é excretada na urina. Além disso, o fígado converte os potentes estrogêniosestradiol e estrona no estrogênio quase totalmente destituído de potência, o estriol. Dentro de poucosminutos após sua secreção quase toda a progesterona é degradada a outros esteroides que não possuemefeito progesterônico. Também neste caso, a exemplo dos estrogênios, o fígado é especialmenteimportante no processo de degradação metabólica.
20

Os esteroides são sintetizados nos ovários a partir do colesterol. A testosterona e aprogesterona (P4) são sintetizadas primeiro e depois são convertidas em estrogêniospelas células da granulosa. Na fase lútea, produz muito mais P4 do que é convertida, porisso tem mais P4 no sangue.As células da teca interna do folículo antral, pelo estímulo do LH, convertem colesterolem testosterona. Essa testosterona é convertida em Estradiol pela enzima Aromatase,nas células da granulosa, pelo estímulo do FSH.Os ovários secretam 1/15 da quantidade de testosterona do homem.
21

Os receptores de estrogênio são em sua maior parte nucleares, mas também ocorrem no citoplasma. Sãomembros da superfamília de receptores de hormônios esteroides, que atuam como fatores de transcrição,alterando a expressão gênica após a sua ativação. A ligação do estrogênio com o receptor promove atransferência para o núcleo, onde o complexo receptor-hormônio liga-se a elementos de resposta dosestrogênios, modulando a transcrição gênica (efeitos genômicos). Alguns efeitos rápidos dos estrogêniosnão podem ser explicados por um mecanismo de transcrição e representam o resultado da açãoestrogênica direta sobre as membranas celulares, mediadas pelas formas dos receptores de estrogênio damembrana (efeitos não-genômicos). Alternativamente, os efeitos não-genômicos podem ser exercidospelos receptores de estrogênio clássicos em interação direta com enzimas intracelulares. Entre os exemplosde alguns dos efeitos não-genômicos rápidos dos estrogênios, destacam-se seus efeitos diretos sobre avasculatura e a ativação das vias de sinalização relacionadas com os fatores de crescimento. Os estrogêniosproduzem vasodilatação a curto prazo e diminuem o tônus do músculo liso vascular pelo aumento naformação e liberação de óxido nítrico e prostaciclinas endoteliais, bem como a abertura de canais de cálciomediada pelo GMPc (monofosfato de guanosina cíclico). Também promovem a rápida ativação das vias desinalização relacionadas com os fatores de crescimento através das cascatas de sinalização das proteínasquinases ativadas pelos mitógenos em diversos tecidos, como os osteoblastos, células endoteliais,neurônios e células do câncer de mama humano.
22

*O desenvolvimento de pêlos é mais pelos andrógenos na puberdade (adrenais).
23

24

A progesterona produzida pelo corpo lúteo exerce feedback negativo nos hormônios dohipotálamo e da hipófise, inibindo a produção de GnRH, LH e FSH. Ela promove odesenvolvimento dos alvéolos mamários e das glândulas uterinas, e também inibe a contraçãodo miométrio.Progesterona: parece exercer a maior parte de seus efeitos pela regulação direta da transcriçãogênica, através de duas proteínas receptoras específicas (A e B). Essas proteínas receptoras deprogesterona originam-se de um único gene e atuam como fatores de transcrição induzíveis porligantes, regulando a expressão dos genes através da ligação a elementos de resposta daprogesterona específicos no DNA, A expressão dos receptores de progesterona é supra-reguladapelos estrogênios e infra-regulada pela progesterona na maioria dos tecidos. A exposição préviaao estrogênio induz à produção dos receptores de progesterona, sendo necessária à ação daprogesterona sobre o trato reprodutor. A expressão dos receptores de progesterona no úteroaumenta durante a segunda metade do ciclo menstrual, quando então, à medida que os níveisséricos de progesterona aumentam, o nível total de receptores de progesterona diminui noútero.
25

26

27

Menopausa = poucos folículos = estrogênios = não exerce mais feedback negativosobre FSH e LH aumento da concentração de gonadotrofinas na urina e redução deestrógeno.
28

29

30

Pico de LH ~16h antes da ovulação
31

32

Antes da puberdade, tanto no macho quanto na fêmea, os centros tônico e cíclico do hipotálamo liberampulsos de baixa frequência e amplitude de GnRH. Após a puberdade, na fêmea, o centro tônico controla osníveis basais de GnRH, mas são mais altos que na fêmea pré-puberal porque a frequência de pulsosaumenta. O centro pulsátil controla o pico pré-ovulatório de GnRH. O macho não desenvolve um centropulsátil.Figura: O hipotálamo libera GnRH no sistema-porta hipotalâmico-hipofisário. O GnRH estimula aadenohipófise a produzir FSH (Hormônio Folículo-estimulante) e LH (Hormônio Luteinizante), queestimulam o folículo ovariano a produzir estradiol (E2). O E2, por sua vez, exerce feedback positivo sobreo GnRH, estimulando sua produção. Ele também estimula a produção de inibina, que juntamente com oestradiol inibem a produção de FSH, mas o E2 continua estimulando a produção de LH. Este terá seu pico,provocando a ovulação.
33

34
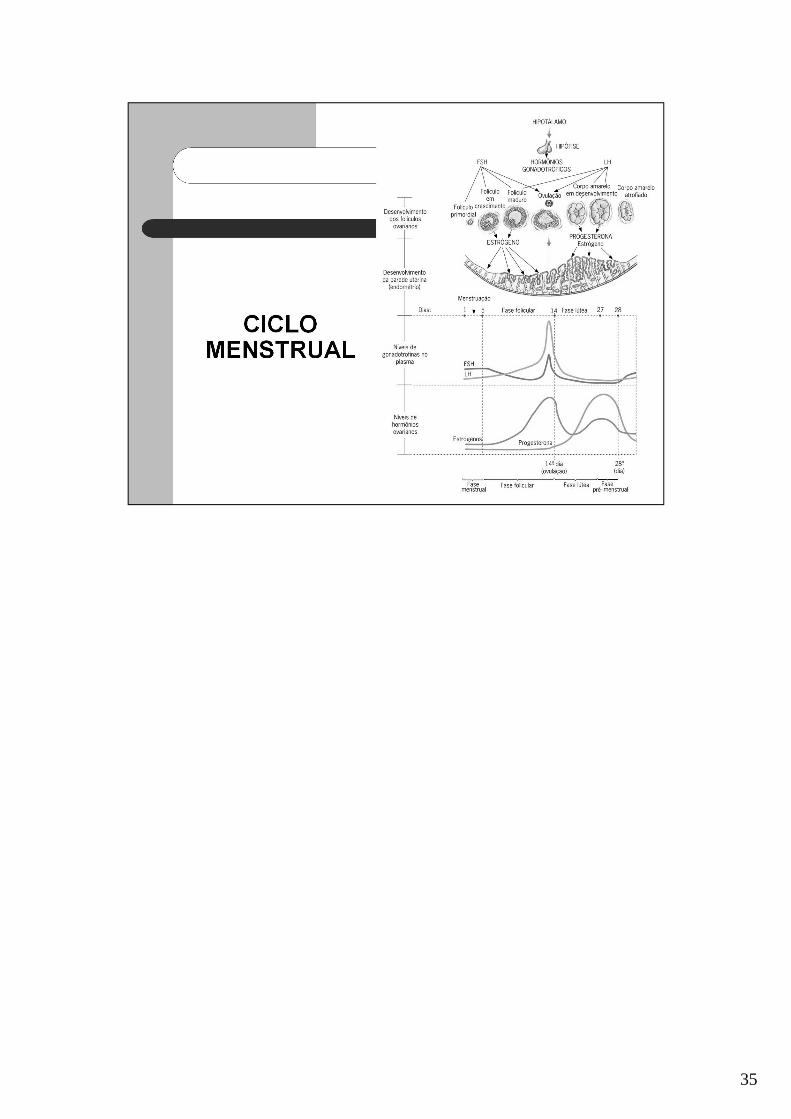
35

O crescimento do ovócito até o estágio antral é independente de gonadotrofinas.Depois ele é influenciado pelo FSH.
Nos primeiros dias do ciclo, as taxas de secreção de FSH aumentam significativamente.Este hormônio induz o crescimento e desenvolvimento de 6 a 12 folículos primordiais acada ciclo.
36

A alta secreção de estrógeno pelo folículo dominante, reduz a secreção de FSH pelahipófise. Assim, as células do folículo dominante que já estao bem sensíveis ao FSHpodem se desenvolver mesmo que ele esteja em baixa concentração.
37

38

Luteinização = tornar amarelo
39

40

41

42

43

44
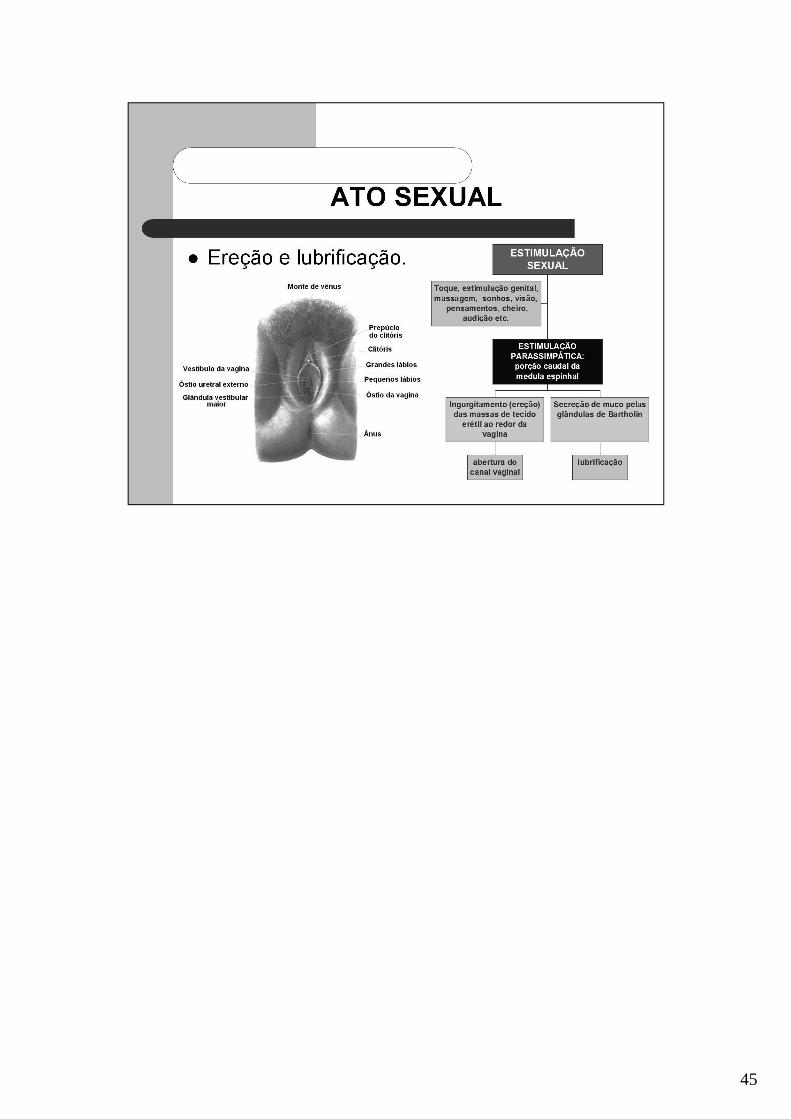
45
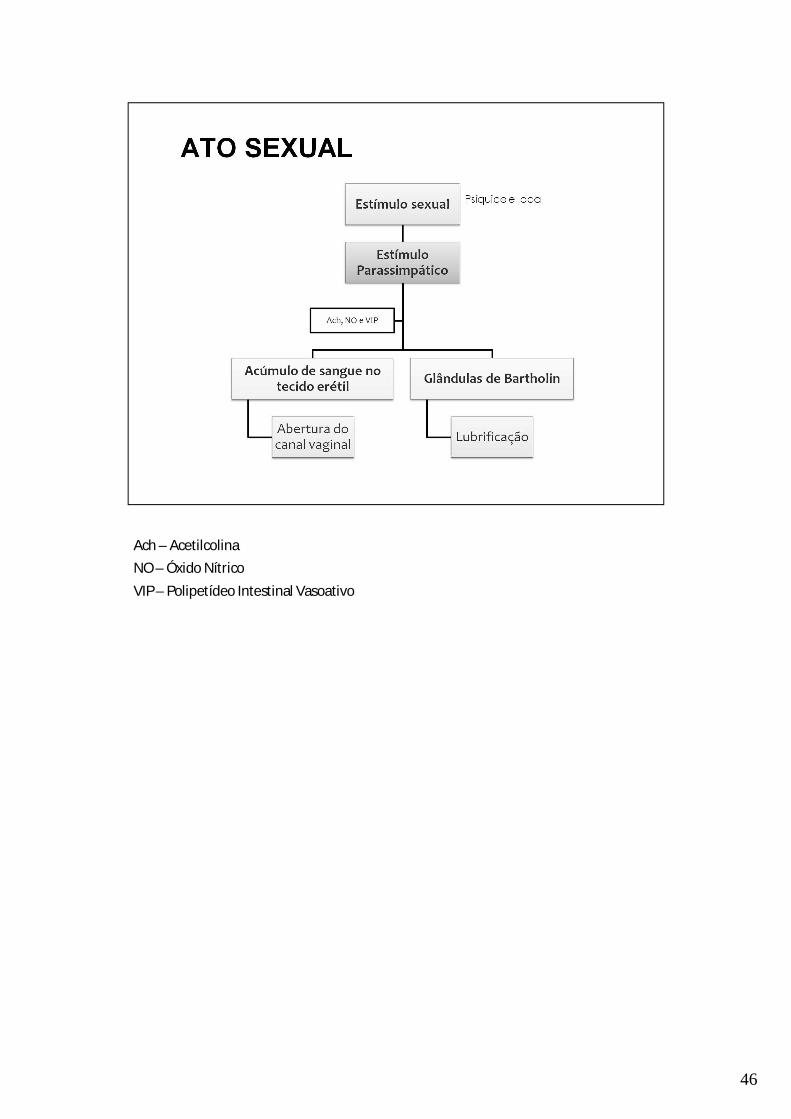
46
Ach – AcetilcolinaNO – Óxido NítricoVIP – Polipetídeo Intestinal Vasoativo

Orgasmo: quando o grau de estimulação sexual (maior na área do clitóris) atingeintensidade suficiente, o útero e as tubas uterinas iniciam contrações peristálticasrítmicas, em direção à cavidade abdominal (orgasmo). Acredita-se que as contraçõesperistálticas impulsionem o sêmen para as tubas uterinas.
47

48

49

50

51

52

53

54

55

56
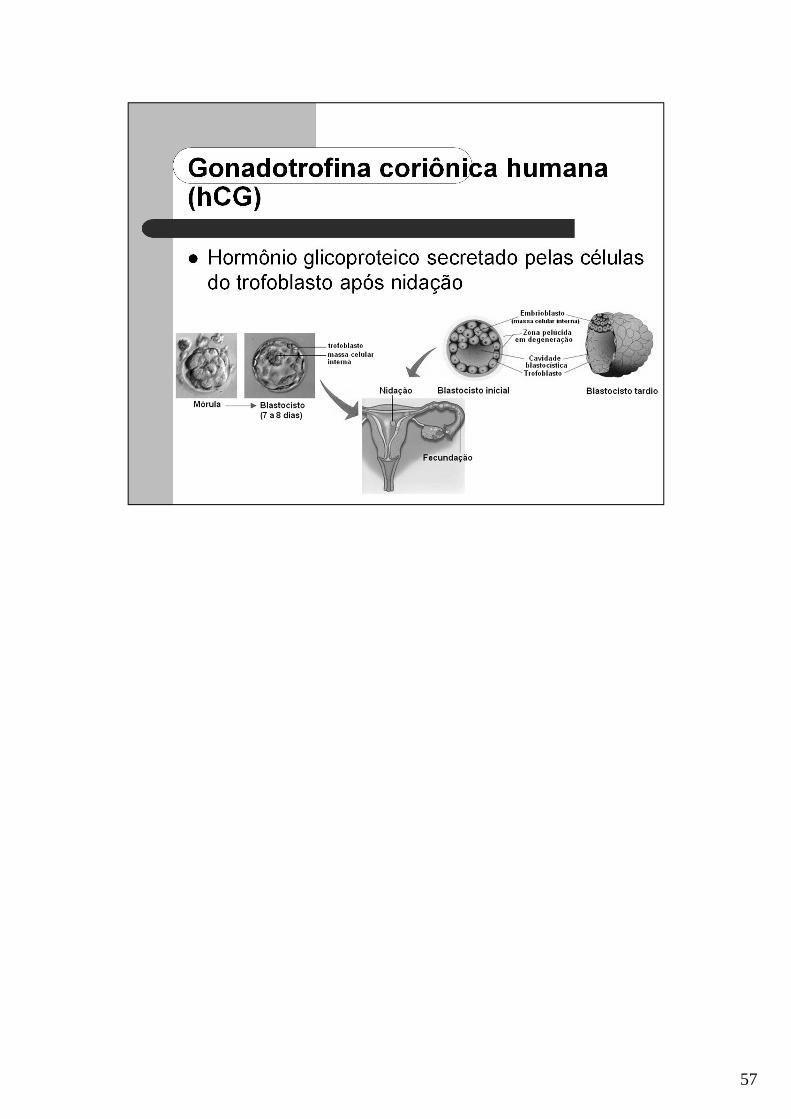
57

58

59
Por volta da 13ª semana de gestação, com a placenta já formada e madura produzindoestrógeno e progesterona, ocorre declínio acentuado na concentração de hCG einvolução do corpo lúteo.

A principal fonte de estrogênio durante a fase inicial da gravidez é o corpo lúteo,posteriormente substituído pela placenta como fonte de produção. O principalestrogênio produzido pelas células sinciciotrofoblásticas da placenta é o estriol, e suaprodução requer uma interação coordenada entre a produção dos hormôniosesteroides pelas glândulas suprarrenais do feto e da mãe. A placenta carece de 17 -hidroxilase e 17,20-desmolase, sendo, por isto, incapaz de converter a progesterona emestrogênio ou de produzir androgênios. Esta falta de produção placentária deandrogênios protege o feto feminino da masculinização. A forte atividade de aromatase,que inativa os androgênios derivados das suprarrenais maternas e fetais tambémcontribui para esta proteção. Os principais efeitos fisiológicos do estrogênio durante agravidez incluem: (1) aumento do útero, (2) aumento das mamas e crescimento dasestruturas dos dutos das mamas, (3) aumento da genitália externa feminina, (4)relaxamento dos ligamentos pélvicos, (5) sensibilização aos efeitos da ocitocina, e (6)inibição da produção de leite.
60

A principal fonte de progesterona durante a fase inicial da gravidez é o corpo lúteo, sob aregulação do hCG. Com cerca de 8 semanas de gestação, a placenta (sinciciotrofoblasto) passa aconstituir a principal fonte de progesterona. O colesterol para a síntese placentária daprogesterona provém das LDL circulantes, que se ligam ao receptor LDL nas células trofoblásticase sofrem endocitose. A progesterona desempenha papel importante na manutenção do úterodurante a gravidez, inibindo a síntese das prostaglandinas e modulando a resposta imune parapreservar a gravidez.A progesterona (P4) exerce efeito sobre as glândulas endometriais e as células contêmglicogênio, proteínas, lipídios e minerais necessários para o desenvolvimento do embrião. Ascélulas do endométrio ficam ainda mais volumosas e formam a decídua. Essa é a forma doembrião se alimentar na primeira semana e até a 8ª semana.
61

A progesterona ainda estimula o centro respiratório, fazendo com que aumente aventilação materna mãe manda mais oxigênio para o feto.
62

Hormônio lactogênico placentário humano (HPL): hormônio proteico que tem efeitolipolítico, aumenta a resistência materna à ação da insulina e estimula o pâncreas nasecreção de insulina, ajudando no crescimento fetal maior quantidade de glicose e denutrientes para o feto em desenvolvimento. É sintetizado pelo sinciciotrofoblasto esecretado na circulação fetal e materna depois de 6 semanas de gestação. No feto, oHPL atua através de receptores lactogênicos e de um receptor específico de HPL paramodular o desenvolvimento embrionário, regular o metabolismo intermediário eestimular a produção de IGF, de insulina, dos hormônios adrenocorticais e dosurfactante pulmonar.
63

Durante a gravidez, o GH-V, uma variante de GH expresso pela placenta, torna-se o GHpredominante da mãe. Esse hormônio possui semelhança estrutural com o GH daadenohipófise e não é liberado no feto. A partir da 15ª a 20ª semana de gestação até otérmino da gravidez, o GH placentário substitui gradualmente o GH da hipófise materna.O GH-V estimula a produção de IGF-1 e modula o metabolismo intermediário materno,aumentando a disponibilidade de glicose e aminoácidos para o feto.Hormônio lactogênio placentário humano (HPL) e somatomamotropina coriônicahumana (GH-V): importantes na regulação do metabolismo materno e fetal, bem comono crescimento e desenvolvimento do feto.
64

Hormônio de liberação da corticotropina (CRH): é produzido pelas células dosinciciotrofoblasto e trofoblasto da placenta. Sua estrutura e função são semelhantes àsdo CRH derivado do hipotálamo. A produção placentária de CRH está associada àduração da gestação, sendo secretado na circulação materna em grandes quantidadesdurante o terceiro trimestre de gestação, e podendo desempenhar um importantepapel no início do trabalho de parto. O CRH exerce várias funções dentro do ambienteuterino, com a indução da produção de prostaglandinas e manutenção do fluxosanguíneo placentário.
65

66
Durante toda a gravidez, as grandes quantidades de estrogênios secretadas pelaplacenta fazem com que o sistema de ductos das mamas cresça e ramifique.Simultaneamente, o estroma das mamas aumenta em quantidade, e grandesquantidades de gordura são depositadas no estroma. Quatro outros hormônios sãoigualmente importantes para o crescimento do sistema de ductos: GH, prolactina,glicocorticoides adrenais e insulina.O desenvolvimento final das mamas em órgãos secretores de leite requer progesterona.Quando o sistema de ductos estiver desenvolvido, a progesterona, agindosinergicamente com o estrogênio e os hormônios mencionados, causará o crescimentoadicional dos lóbulos mamários, com multiplicação dos alvéolos e desenvolvimento decaracterísticas secretórios nas células dos alvéolos.

67

68

69
O ato de mamar promove a inibição da secreção de PIH (hormônio inibidor da prolactina),causando assim a secreção da prolactina, que estimula a produção e a secreção de leite para osalvéolos. Porém, o leite não flui com facilidade dos alvéolos para o sistema de dutos e, portanto,não drena de modo contínuo pelos mamilos. Assim, o leite deve ser ejetado ou "descer" dosalvéolos para os dutos para que o bebê possa obtê-lo. Esse processo é desencadeado por umreflexo combinado neurogênico e hormonal, envolvendo o hormônio neuro-hipofisário ocitocina.Quando o bebê suga, ocorre transmissão de impulsos sensitivos por nervos somáticos dosmamilos até a medula espinhal e, daí, para o hipotálamo, ocasionando também a secreção deocitocina. A ocitocina é, então, transportada pelo sangue até as mamas, onde provoca acontração das células mioepiteliais que circundam as paredes externas dos alvéolos, expelindo,assim, o leite dos alvéolos para os dutos e sua ejeção.

70

71

Alterações no ciclo menstrual:Falhas na menstruação, geralmente com um histórico de se ter um ou mais períodosmenstruais normais durante a puberdade (amenorreia secundária)Períodos menstruais irregulares que podem ser mais ou menos frequentes, variando demuito leves a muito intensosDesenvolvimento de características sexuais masculinas (virilização):Tamanho reduzido das mamasAprofundamento do timbre da vozHipertrofia do clitórisAumento de pelos no peito, abdome e face, assim como ao redor dos mamilos(chamado hirsutismo)Rareamento dos cabelos, chamado calvície de padrão masculinoOutras alterações na pele:Acnes que pioramMarcas escuras ou grossas na pele e dobras ao redor das axilas, nas virilhas, no pescoçoe nas mamas devido à sensibilidade à insulinahttp://www.minhavida.com.br/saude/temas/sindrome-do-ovario-policistico
72

73