o ciclo celular compreende o conjunto de transformações ...biologia.bio.br/curso/2º período...
TRANSCRIPT

1

2

O ciclo celular compreende o conjunto de transformações pelas quais a célulapassa desde a sua formação até sua divisão ou morte.Compreende duas etapas: a intérfase e a mitose.
3

4

5

G0 é um estado quiescente no qual as células adultas maduras podem ficar portempo indeterminado. Nesse estágio, as células permanecem metabolicamenteativas, mas não se dividem ou, então, se dividem apenas quando estimuladas porsinais extracelulares, com a finalidade de renovação tecidual após morte ou lesãocelular (hepatócitos por exemplo, que entram em G0, mas após dano ao órgãopodem voltar a G1 e continuar o ciclo celular). G0 depende da história da célula alongo prazo: em cada tipo celular, cada fase do desenvolvimento do animalobedece a leis distintas, o que reflete as diferenças em sua maquinaria de controleinterno. Por exemplo, no corpo humano algumas células como os neurônios quenão continuam se replicando e sim se mantendo e criando comunicaçõesintercelulares. O estado G0 está muito relacionado com a redução progressiva dassequências teloméricas do DNA durante as sucessivas divisões celulares,sugerindo que G0 pode ser provocado pela incapacidade de manter ocomprimento dos telômeros (ou porque as células são deficientes em telomerase).G1 ("Gap 1") – Intervalo após a mitose durante o qual as células se preparampara iniciar a síntese de DNA. Este período é caracterizado pela transcriçãogênica e tradução, levando à síntese de proteínas necessárias para a síntese deDNA.S – Período no qual ocorre a duplicação do DNA celular.G2 ("Gap 2") – Intervalo após a síntese de DNA, durante o qual as células sepreparam para a divisão.M – Período de divisão celular (mitose ou meiose).
6

Uma vez que tenha passado pelo ponto de restrição (ponto R), a célula estácomprometida a prosseguir até o final da divisão, mesmo na ausência deestímulos adicionais.
7

Caso a célula não receba os estímulos necessários para romper o ponto derestrição, ela entra em G0.
8

Mais animação: http://highered.mcgraw-hill.com/sites/0072495855/student_view0/chapter2/animation__control_of_the_cell_cycle.html
9

Antes que a célula possa se dividir ela tem que crescer até alcançar um tamanhoadequado e constante. Em função disto, cerca de 95% do ciclo são gastos emintérfase, mas o tempo médio total desta fase é variável de tipo celular para tipocelular. A duração varia também com as condições fisiológicas em que a célula seencontra, como idade celular, disponibilidade de hormônios e de fatores decrescimento, temperatura, pressão osmótica, pressão hidrostática e pressão deoxigênio externas, e mesmo com o ritmo circadiano que ocorre em animais.Existem também notáveis diferenças quanto à duração do ciclo celular entre osorganismos. Em geral o ciclo dura 12 horas, em tecidos de mamíferos comcrescimento muito rápido, e 24 horas em tecidos com crescimento mais lento. Emeucariotos unicelulares (leveduras, por exemplo) o tempo de geração é bem maiscurto (aproximadamente 1 hora e meia para a formação de duas células-filhas).Nas células embrionárias, G1 é ausente ou tem duração negligenciável logo apósa fertilização. Neste caso, não ocorre crescimento celular.De todas as fases do ciclo, G1 é a mais variável na maioria das células de animaise plantas. Em geral ocupa muitas horas, durante as quais as células crescem. Esseperíodo pode variar individualmente de célula para célula, pois é o que mais sofreinfluência de fatores extracelulares. Também é o período em que vários inibidorese mutações são capazes de bloquear a proliferação. Depois que as células entramna fase S, fatores extracelulares não mais determinam os eventos do ciclo celular,os quais passam a depender de controles disparados intracelularmente. Portanto,as demais etapas do ciclo, incluindo a mitose, têm tempos de duração maisconstantes. A mitose dura mais ou menos 1 hora. G2 em geral tem duração de 2 a4 horas e o período S dura de 7 a 8 horas. Apesar dessas fases serem maisconstantes, a duração de cada uma varia entre espécies e também entre diferentesestágios do desenvolvimento de um mesmo indivíduo.Nas células embrionárias, G1 é ausente ou tem duração negligenciável logo apósa fertilização. Neste caso, não ocorre crescimento celular. 10

O ciclo pode durar algumas horas em tecidos de renovação rápida e até mesesem outros tipos celulares. Alguns tipos de células (terminalmente diferenciadas)não se dividem e permanecem em G0. Outras entram em G0 e, após um dano aoórgão, voltam a G1 e continuam o ciclo celular (ex: células hepáticas)
11

12

13
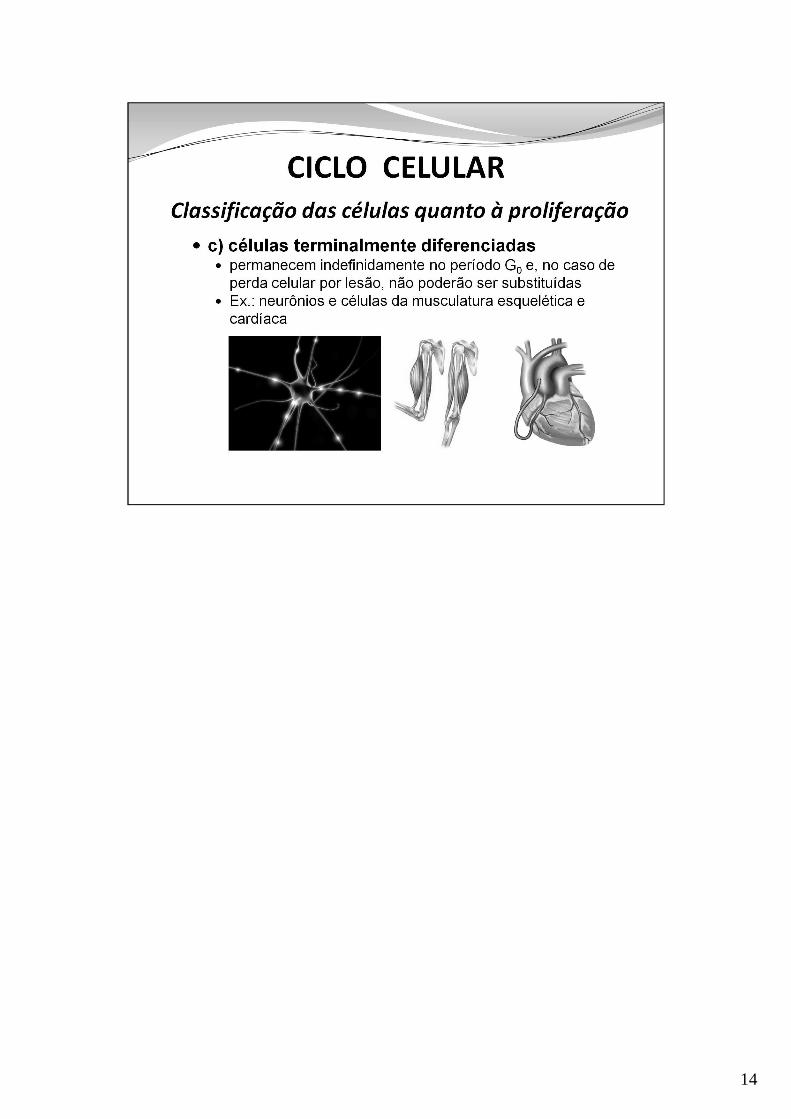
14

Controle Epigenético: qualquer atividade reguladora de genes que não envolvemudanças na seqüência do DNA (código genético) e que pode persistir por umaou mais gerações. As histonas são suscetíveis a uma grande variedade demodificações pós-traducionais, tais como acetilação, fosforilação, metilação eubiquitinação.A maioria dessas modificações acontece no domínio N-terminal das histonas, queé rico nos aminoácidos básicos lisina e arginina (mas também podem ocorrer nosdomínios globulares). Enzimas como a acetiltransferase, as quinases e asmetiltransferases, que depositam marcadores químicos nas histonas (acetil,fosfato e metil, respectivamente), são reguladores importantes da atividadegênica, da dinâmica dos cromossomos, da regulação do ciclo celular e daorganização do genoma.Metilação: adição de radical metil ( CH3) nos resíduos de lisina e/ou argininadas histonas H3 e H4, realizada por metiltransferases: media o silenciamento datranscrição em locais de heterocromatina; afeta a regulação da transcrição emloci de eucromatina.Acetilação e ubiquitinação: reduzem a carga positiva ou o caráter básico dashistonas, reduzindo a interação eletrostática com o DNA favorecem atranscriçãoDesacetilação: aumenta a carga positiva das histonas, aumentando a interaçãoeletrostática com o DNA inibe a transcrição.Fosforilação: o grupo fosfato é fortemente negativo, então sua adição induzforças na cadeia protéica que podem levar a uma radical alteração em suaconformação. Desse modo, uma proteína pode expor os aminoácidos antesescondidos em seu centro e mudar muito suas características. Por exemplo, umaproteína apolar e hidrofóbica pode se tornar polar e hidrofílica. A desfosforilaçãoé realizada pelas fosfatases. 15

Quando ocorrem mutações, proto-oncogenes tornam-se oncogenes, que sãocarcinogênicos e causam multiplicação celular excessiva. Essas mutações levamo proto-oncogene a expressar em excesso sua proteína estimuladora docrescimento ou a produzir uma forma mais ativa. Os genes supressores detumores, em contraste, contribuem para o desenvolvimento de câncer quando sãoinativados por mutações. O resultado é a perda da ação de genes supressoresfuncionais, o que priva a célula de controles cruciais para a inibição decrescimento inapropriado.
16

17

18

19

Substâncias de natureza polipeptídica = polipeptídeos e proteínas. Em geral, ospolipeptídeos com 100 ou mais aminoácidos são chamados de proteínas, eaqueles com mais de 10 e menos de 100 aminoácidos são denominadospolipeptídeos.Antes que a célula possa se dividir ela tem que crescer até alcançar um tamanhoadequado e constante, o que depende, entre outros fatores, da disponibilidade dosfatores de crescimento.Crescimento celular= aumento da massa celular através da síntese de proteínas eoutras macromoléculas.
20

21

Os fatores de crescimento que estimulam a proliferação celular (mitógenos)podem ser divididos em dois grandes grupos: fatores de competência e fatores deprogressão. Os fatores de competência, tais como o fator de crescimento defibroblastos (FGF), fator de crescimento derivado de plaquetas (PDGF), fator decrescimento de hepatócito (HGF) e o fator de crescimento mitógeno de Bischoffpara células precursoras de mioblastos, os quais atuam inicialmente e sãodiretamente responsáveis pela reentrada da célula no ciclo de divisão celular, ouseja, pela sua passagem do estágio G0 para G1. Já os fatores de progressão, comoo fator de crescimento semelhante à insulina 1 e 2 (IGF-1 e IGF-2) e o fator decrescimento epidérmico (EGF), atuam posteriormente no ciclo celular eestimulam a fase de replicação do DNA, que representa a passagem da fase G1para S do ciclo. Assim, os fatores de competência estimulam o crescimento decélulas (aumento de massa celular), promovendo a síntese de proteínas e outrasmacromoléculas e inibindo a sua degradação, enquanto os fatores de progressãocontrolam a taxa de divisão celular atuando na fase G1 do ciclo celular. Oresultado final é a ativação de complexos G1/S-Cdk, que superam as barreirasinibidoras que normalmente bloqueiam a progressão para a fase G1-S (ver aulasobre Controle e regulação do ciclo celular).
22

Receptores de superfície:1- Receptores com atividade tirosina quinase intrínseca autofosforilação.2- Receptores sem atividade catalítica intrínseca ativam proteínas citosólicas(tirosina quinsase) que fosforilam o receptor.3- Receptores associadas a proteína G geram mensageiros secundáriosintracitoplasmáticosSistema de transdução de sinais: processo pelo qual sinais extracelulares sãodetectados e convertidos em sinais intracelulares o que gera uma resposta celularespecífica.A célula estável, após a cascata de fosforilação proteica, entra no ciclo celular(sai de G0 e entra em G1). O sistema de transdução de sinais transfere ainformação para o núcleo, onde ocorre alterações na expressão gênica(transcrição dos genes) controlado for fatores reguladores conhecidos comofatores de transcrição.
23

A ubiquitina é uma proteína não-histônica ácida, que desempenha uma funçãoimportante na regulação de proteínas. Ela marca proteínas indesejadas para que ocomplexo proteico denominado proteassoma possa destrui-las. No ciclo celular, amarcação das ciclinas com ubiquitina leva à sua destruição pelo proteassoma.
24

O ciclo celular é governado por complexos de quinase formados por umasubunidade catalítica, a Cdk (quinase dependente de ciclina) e uma subunidaderegulatória, a ciclina.As ciclinas ativam as CDKs formando complexos moleculares ciclinas-CDKregulam a atividade de outras proteínas cruciais no ciclo celular, fosforilando-asnos seus locais reguladores. A fosforilação funciona como um interruptor para aatividade enzimática e as quinases são os responsáveis por ligar e/ou desligar esteinterruptor, de modo a permitir o retorno ao nível anterior de estimulação quandoo sinal hormonal termina. Desta forma, não é necessário sempre degradarproteínas e transcrever/traduzir novas proteínas toda vez que a célula precisaralterar seu metabolismo: basta apenas ativar ou inibir as proteínas de acordo coma necessidade não havendo nenhuma regra para qual estado é o ligado, ou seja,uma fosforilação pode tanto ativar quanto desativar uma proteína.
25

Proteassomos: grandes máquinas proteolíticas encontradas em todas às célulaseucarióticas.
26

27

Complexos protéicos ubiquitinizam a ciclina.Ubiquitinação: marca a ciclina para ser degradada nos proteassomos.
28

As maiores áreas de controle do ciclo celular incluem a transição G1-S, ondeocorre a maior parte dos defeitos relacionados ao câncer, o ponto de checagemG2/M e sua ativação em resposta a danos no DNA, e o ponto de checagem dofuso mitótico.
29

30

A fase G1 possui um ponto de checagem, G1/S, controlado pela via do pRb(proteína do retinoblastoma). Uma vez que a célula passe este ponto, ela écompelida a replicar seu DNA. Caso ocorra cópia incorreta do DNA durante S oulesão do DNA, a célula não passará o ponto de checagem G2/M, e ocorreráparada de crescimento e apoptose (senescência e morte programada) induzidospelo gene p53.
31

Proteínas da família INK4 ligam e inibem especificamente CDK4 e CDK6 eproteínas CIP/KIP interagem com o complexo ciclina E/CDK2, inibindo o ciclocelular.A proteína do retinoblastoma (pRb, supressor tumoral) regula negativamente ofator de transcrição E2F. Quando a pRb se liga ao E2F, este não pode mais iniciara transcrição dos genes requeridos para a transição de G1 para a fase S. Isso ajudaa assegurar que a replicação do DNA ocorra no momento correto. A proteína Rbfosforilada pelos complexos Cdk 4,6/ciclina D é responsável pela progressão dociclo celular de G1 para S, pois a fosforilação da pRb libera o fator de transcriçãoE2F que inicia a transcrição dos genes cujas proteínas são necessárias para a faseS. Isso permite que a célula continue se desenvolvendo além do ponto derestrição até concluir seu ciclo. A desfosforilação de Rb por fosfatases restringeprogressão do ciclo.
32

O período G1 é o intervalo de tempo que transcorre desde o fim da fase M até oinício da síntese de DNA (intervalo pós-mitótico e pré-sintético).Na fase G1 existe apenas um centrossomo (par de centríolos - centro organizadorde microtúbulos) localizado próximo ao núcleo.
33

34

35

36

A condensação cromossômica é fundamental para evitar o emaranhamento ourompimento do material genético durante sua distribuição às células-filhas.
37

38
Centrossomos + fibras radiais = áster.

Com o rompimento da carioteca, não há mais limites físicos entre o citoplasma eo material nuclear.Esses eventos ocorridos a partir do rompimento da carioteca são considerados pormuitos autores como uma etapa distinta denominada prometáfase.
39

As fibras polares atravessam a região central da célula, sem alcançar o pólooposto.
40

41

42

43

44

45

46

47

48

49

50

Durante a diacinese os quiasmas são mantidos, o que é importante para adistribuição correta dos cromossomos. A falta de quiasmas pode levar a umasegregação incorreta dos cromossomos homólogos na anáfase.
51

52

53
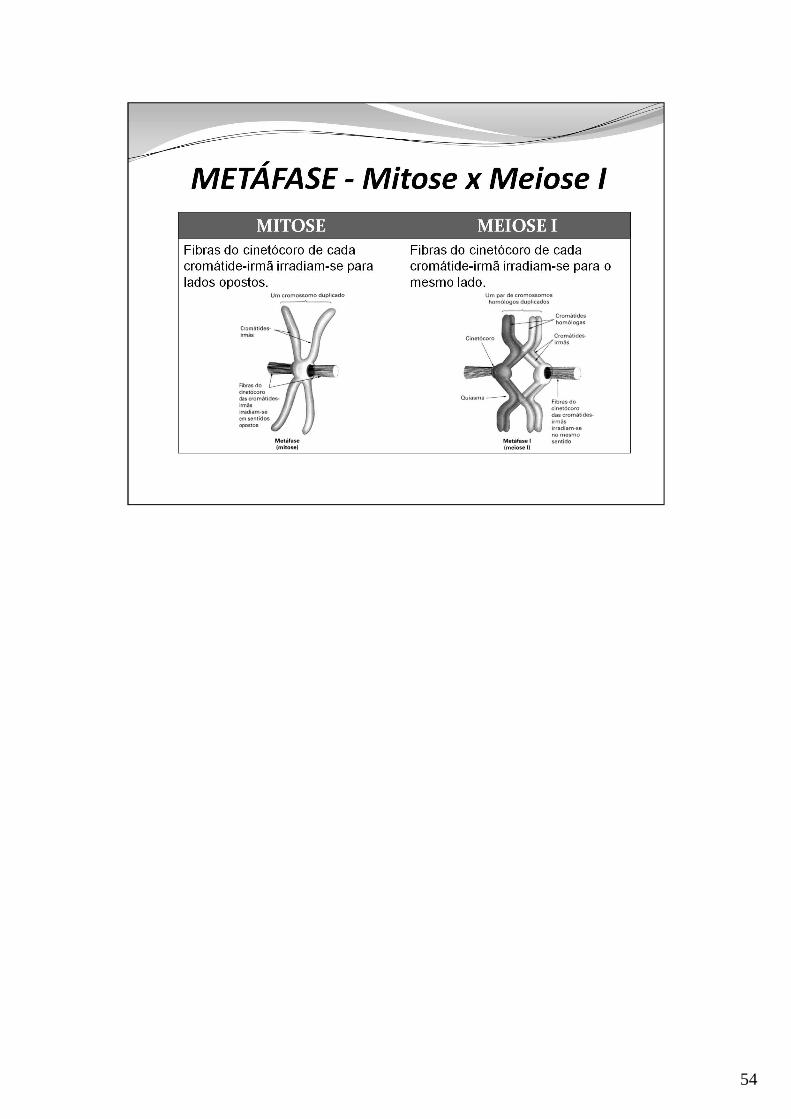
54

55

56

57

58

59

60

61

Intercinese: curto intervalo entre a primeira e a segunda divisões da meiose. Nemsempre existe intercinese entre as duas divisões da meiose, ou esta é tão curta quepode ser considerada como inexistente.
62

63

64

65

66
Na espécie humana, a ovogênese inicia-se antes do nascimento. As ovogôniascomeçam a se diferenciar em ovócito I (primário) por volta do terceiro mês devida intra-uterina. No nascimento, os ovócitos primários estão em prófase I, quesó é retomada até a maturidade sexual.

67
A primeira etapa, ou meiose I, é irregular pela ocorrência de uma citocinesediferencial, que dá origem a um corpúsculo polar e ao ovócito II (secundário). Asegunda etapa, ou meiose II, é iniciada, porém pára em metáfase II. Desta forma,a partir da puberdade, a mulher libera na ovulação um ovócito II por mês. Sehouver fecundação, a meiose II será completada, originando um óvulo poucoantes da fusão dos pronúcleos e formação do zigoto. Isto porque a citocinese dameiose II também é diferencial, e origina um corpúsculo polar e um óvulo. Essameiose irregular garantirá um gameta com maior quantidade de nutrientes.

68

69