versidade estadual do oeste do paranÁ centro de ...tede.unioeste.br/bitstream/tede/1917/1/ariane...
TRANSCRIPT

VERSIDADE ESTADUAL DO OESTE DO PARANÁ
CENTRO DE ENGENHARIAS E CIÊNCIAS EXATAS
PROGRAMA DE PÓS-GRADUAÇÃO EM RECURSOS PESQUEIROS E
ENGENHARIA DE PESCA
ARIANE FURTADO DE LIMA
ALIMENTAÇÃO INICIAL DO “JUNDIÁ”, Rhamdia voulezi Haseman 1911: EFEITOS
DA PRIVAÇÃO ALIMENTAR SOBRE O CRESCIMENTO E PADRÕES NA
SELEÇÃO DE PRESAS AO LONGO DO PERÍODO LARVAL
Toledo
2013

ARIANE FURTADO DE LIMA
ALIMENTAÇÃO INICIAL DO “JUNDIÁ”, Rhamdia voulezi Haseman 1911: EFEITOS
DA PRIVAÇÃO ALIMENTAR SOBRE O CRESCIMENTO E PADRÕES NA
SELEÇÃO DE PRESAS AO LONGO DO PERÍODO LARVAL
Dissertação apresentada ao Programa de Pós-
Graduação Stricto Sensu em Recursos Pesqueiros e
Engenharia de Pesca – Nível de Mestrado, do Centro
de Engenharias e Ciências Exatas, da Universidade
Estadual do Oeste do Paraná, como requisito parcial
para a obtenção do título de Mestre em Recursos
Pesqueiros e Engenharia de Pesca.
Área de concentração: Recursos Pesqueiros e Engenharia
de Pesca.
Orientadora: Profa. Dr
a. Maristela Cavicchioli Makrakis
Toledo
2013

Catalogação na Publicação elaborada pela Biblioteca Universitária
UNIOESTE/Campus de Toledo.
Bibliotecária: Marilene de Fátima Donadel - CRB – 9/924
Lima, Ariane Furtado de
L732a Alimentação inicial do Jundiá, Rhamdia voulezi Haseman
1911 : efeitos da privação alimentar sobre o crescimento e
padrões na seleção de presas ao longo do período larval /
Ariane Furtado de Lima. -- Toledo, PR : [s. n.], 2013.
59 f.
Orientadora: Profa. Dra. Maristela Cavicchioli Makrakis
Dissertação (Mestrado em Recursos Pesqueiros e
Engenharia de Pesca) - Universidade Estadual do Oeste do
Paraná. Campus de Toledo. Centro de Engenharias e Ciências
Exatas.
1. Aqüicultura 2. Jundiá (Peixe) – Larva – Nutrição 3.
Jundiá (Peixe) – Larva – Crescimento 4. Peixe – Tamanho 5.
Alimentação (Zooplâncton) 6. Alimentação (Artêmia) 7.
Seletividade alimentar I. Makrakis, Maristela Cavicchioli,
Orient. II. T
CDD 20. ed. 639.3752

FOLHA DE APROVAÇÃO
ARIANE FURTADO DE LIMA
ALIMENTAÇÃO INICIAL DO “JUNDIÁ”, Rhamdia voulezi Haseman 1911: EFEITOS
DA PRIVAÇÃO ALIMENTAR SOBRE O CRESCIMENTO E PADRÕES NA
SELEÇÃO DE PRESAS AO LONGO DO PERÍODO LARVAL
Dissertação apresentada ao Programa de Pós-graduação Stricto Sensu em Recursos Pesqueiros
e Engenharia de Pesca – Nível de Mestrado, do Centro de Engenharias e Ciências Exatas, da
Universidade Estadual do Oeste do Paraná, como requisito parcial para a obtenção do título de
Mestre em Recursos Pesqueiros e Engenharia de Pesca, pela Comissão Julgadora composta
pelos membros:
COMISSÃO JULGADORA
____________________________________________
Profa. Dr
a. Maristela Cavicchioli Makrakis
Universidade Estadual do Oeste do Paraná
(UNIOESTE)
____________________________________________
Profa. Dr
a. Elaine Antoniassi Luiz Kashiwaqui
Universidade Estadual de Mato Grosso do Sul
(UEMS)
____________________________________________
Prof. Dr. David Augusto Reynalte-Tataje
Universidade Federal de Santa Catarina
(UFSC)
____________________________________________
Prof. Dr. Sérgio Makrakis
Universidade Estadual do Oeste do Paraná
(UNIOESTE)
____________________________________________
Aprovada em:
Local de defesa: Auditório da Unioeste/Campus de Toledo.

DEDICO:
Aos meus pais Eli Lima e Nuceli Furtado de Lima, pela educação, exemplos,
e incentivo na vida.
A minha irmã Neuzeli e meu cunhado Júlio pelo suporte e apoio nos estudos.
A minha irmã Silvia, meu cunhado Neemias, meus sobrinhos André e Filipe e minhas
sobrinhas Camille e Danielle
Por fazerem parte da minha vida.
Ao meu Pai Celestial (Jesus)
por ser o sustento e a
razão da minha vida.
Sempre amarei todos vocês!!
Á meus grandes amigos Cerly e Pr. Werner Maier pelos conselhos, orações, carinho e
suporte em toda trajetória dos meus estudos no Paraná. Muito Obrigada!
“Com Deus conquistaremos a vitória (...)”
Salmos 108:13

AGRADECIMENTOS
A minha orientadora Dra. Maristela Cavicchioli Makrakis por tudo que nos ensinou desde a
graduação, por suas idéias de pesquisa, orientação, confiança e principalmente pelo seu
profissionalismo, honestidade e postura ética que serão espelhos na minha vida profissional. Muito
Obrigada Maris....
Ao meu cunhado Júlio Amilcar Ramos Cabral, por dedicar seu precioso tempo nas correções
ortográficas.
Ao professor Dr. Sérgio Makrakis, por ter aberto as portas do GETECH para o meu estágio e
pesquisa tanto durante o mestrado como também durante toda a trajetória da graduação. Obrigada
também por acreditar no meu potencial em realizar o meu mestrado.
À Dra. Michele de Faveri Gimenes, pela sua preciosa amizade, sugestões de análise, auxílio e apoio
durante o treinamento em Maringá (Nupélia) e durante a fase experimental deste trabalho.
Á Dra. Elaine Antoniassi Luiz Kashiwaqui, pela sua disposição em sanar as frequentes dúvidas em
estatística e pela sua preciosa amizade.
A minha amiga Suelen Pini, pelo companheirismo, dedicação e indispensável ajuda durante a fase
experimental e na tabulação dos dados. Muito Obrigada Sú!!
Ao Luiz Antonio Hesper, assistente do PREP, pela disposição e excelente atendimento ao longo deste
tempo. Muito obrigada Luizinho!!!
As todos os colegas e amigos do GETECH: Tiago; Leandro; Dimas; Fábio; Regis; Carol; Fer,
Dhonatan; Adriano; Bruna, Luciana; Lucileine; Paty e todos aqueles que já passaram pelo Getech,
muito Obrigada pela amizade, apoio nos meus trabalhos de laboratório e memoráveis momentos de
diversão e algumas vezes de contrariedades. Gente sentirei saudades!!! Vocês fizeram parte do
plano de Deus na minha vida para o meu crescimento pessoal e espiritual, acreditem!!
Ás minhas queridas amigas Micheli, Wiviany, Salete, Rúbia, Jurací, Celma e Elieusa pela nossa
amizade.
Ás minhas queridas companheiras do InPAA, Renate e Sília.
Á todos do Nupélia pelo estágio e treinamento na identificação do zooplâncton. Em especial ao Diogo
Castanho de Amaral, Clarisse Leal, Fabiana Palazzo, Ana Paula Fernandes e Louizi Braghin pela
atenção especial. Muito Obrigada!!
Ao InPAA (Instituto de Pesquisa em Aquicultura Ambiental), pelo espaço concedido para a realização
do experimento.
Á equipe do CDT-Iguaçu (Centro de Difusão de Desenvolvimento Tecnológico do Rio Iguaçu), pelo
fornecimento das larvas.
Ao GETECH (Grupo de Pesquisa em Tecnologia de Produção e Conservação dos Recursos
Pesqueiros e Hídricos), pelo apoio logístico para a realização desta pesquisa.
A CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), pela concessão de bolsa
e apoio financeiro para a execução deste trabalho.

ALIMENTAÇÃO INICIAL DO “JUNDIÁ”, Rhamdia voulezi Haseman 1911: EFEITOS
DA PRIVAÇÃO ALIMENTAR SOBRE O CRESCIMENTO E PADRÕES NA
SELEÇÃO DE PRESAS AO LONGO DO PERÍODO LARVAL
RESUMO
Neste estudo foram avaliados os efeitos da privação alimentar sobre o crescimento das larvas
de jundiá Rhamdia voulezi bem como a preferência alimentar durante o período larval. Para
isto foram realizados dois experimentos que tiveram início logo após a absorção completa do
saco vitelínico, que ocorreu no 4o dia após a eclosão (DAE). O primeiro experimento de
privação alimentar foi realizado com 8 tratamentos constituídos por diferentes regimes
alimentares, incluindo o tratamento controle (TC) no qual não houve fornecimento de
alimento. O alimento foi fornecido pela primeira vez no 4o, 5
o, 6
o, 7
o, 8
o, 9
o e 10
o DAE,
correspondendo aos tratamentos: T4; T5, T6, T7, T8; T9 e T10, respectivamente. A
alimentação consistiu de zooplâncton e artêmia recém-eclodida. Para avaliar a preferência
alimentar, foram realizados 3 ensaios de seletividade alimentar ao longo de todo o período
larval, compreendendo diferentes estágios de desenvolvimento e classes de comprimento
padrão. Neste, as larvas foram alimentadas exclusivamente com zooplâncton coletado de
tanques de cultivo. Os resultados demonstraram que as larvas de jundiá que receberam
alimento do 4o ao 6
o DAE tiveram desenvolvimento significativamente maior que aquelas
alimentadas mais tardiamente, a partir do 7o DAE. O ponto de não retorno foi registrado neste
mesmo período, no 7o DAE. Com relação à seletividade alimentar, houve variações nos
padrões de seleção de presas nos diferentes dias de vida analisados. Inicialmente no 5o
DAE,
estágio de pré-flexâo, as larvas selecionaram fortemente os rotiferas (não identificados) e os
cladoceras (Diaphanosoma spinulosum, D. brevireme, Moina sp., M. micrura, M. minuta).
Nos demais dias de vida analisados, os cladoceras continuaram a ser fortemente selecionados.
Entretanto no 8o DAE, estágio de flexão, selecionaram menos intensamente os copepodas
Argyrodiaptomus azevedoi e Termocyclops decipiens e no 10o DAE, estágio de pós-flexão,
consumiram especialmente Argyrodiaptomus furcatus, Notodiaptomus spinuliferus e
Metacyclops mendocinus. Conclui-se que o retarde da primeira alimentação afeta
drasticamente o crescimento das larvas, e que, se a alimentação for iniciada antes do ponto de
não retorno há chances de recuperação e sobrevivência larval. Para isto, é importante que haja
disponibilidade adequada de alimento, com variedade de tipo e de tamanho de modo que
possam compensar as limitações morfológicas das larvas no início do seu desenvolvimento.
Concomitante ao seu crescimento, as larvas tendem a se especializar em determinados itens
presa, existindo uma seleção de presas tanto pelo tipo como também pelo tamanho, onde o
tamanho da presa consumida está relacionado com o tamanho da larva e o tamanho de sua
boca. Inicialmente as larvas de jundiá têm preferência por itens presas menores, os rotíferos e
cladoceras, posteriormente, quando estão mais desenvolvidas, preferem alimentar-se de presas
maiores, os copepodas.
PALAVRAS-CHAVE. Inanição, crescimento, alimentação, seletividade alimentar, seleção
de presas, tamanho de boca.

INITIAL FEEDING OF “JUNDIÁ”, Rhamdia voulezi Haseman 1911: EFFECTS OF
FEEDING PRIVATION ON GROWTH AND PATTERNS IN THE SELECTION OF
PREY ALONG LARVAL PERIOD
ABSTRACT
The effects of feeding privation on growth of larvae of “jundiá” Rhamdia voulezi as well as
the feeding preference during larval period were evaluated. For this purpose two experiments
were conducted which began soon after complete absorption of the yolk sac, the fourth day
after hatching (DAH). The feeding privation experiment was performed in 8 treatments
consisting of different diets, including the control treatment (CT) in which there was no food
supply. The food was provided for the first time in the 4th, 5th, 6th, 7th, 8th, 9th and 10th
DAH corresponding to treatments: T4, T5, T6, T7, T8, T9 and T10, respectively. The food
consisted of zooplankton and newly hatched artemia. We performed 3 trials of selectivity to
analyze the feeding preference throughout the larval period, comprising different larval stages
and classes of standard length. In this, the larvae were fed exclusively on zooplankton
collected from fish ponds. The results demonstrated that the larvae fed on the fourth to sixth
DAH had a significantly higher growth than those fed later, from the seventh DAH. The point
of no return was recorded in the same period in the 7th DAH. With respect to feeding
selectivity, there were variations in the patterns of prey selection in different days of life
analyzed. Initially on the 5th DAH, preflexion stage, larvae strongly selected rotiferas
(unidentified) and cladocerans (Diaphanosoma spinulosum, D. brevireme, Moina sp., M.
micrura, M. minuta). In the remaining days of life examined, the cladocerans continued to be
strongly selected. However on the 8th DAH, flexion stage, they selected less intensely the
copepods Argyrodiaptomus azevedoi and Termocyclops decipiens and more strongly A.
furcatus, Notodiaptomus spinuliferus and Metacyclops mendocinus at 10 DAH, postflexion
stage. The delay of the first feeding drastically affected the growth of the larvae, and that if
feeding is started before the point of no return there are chances of recovery. So, it is
important to have suitable availability of food, with a variety of type and size so that they can
compensate for the morphological limitations of larvae in the early of their development.
Concomitant to its growth, the larvae tend to specialize in certain prey items. The size of prey
consumed is related to the size of the larvae and the gape size. Initially larvae have a
preference for smaller prey, rotifers and cladocerans, and later when they are more developed
prefer to feed on larger prey, the copepods.
KEYWORDS. Starvation, growth, feeding, feeding selectivity, prey selection, gape size.

Dissertação elaborada e formatada conforme as
normas da publicação científica Animal Feed and
Technology e Aquaculture Disponíveis em: <
http://www.scielo.br/revistas/isz/pinstruc.htm> e
http://onlinelibrary.wiley.com/journal/10.1111/%28I
SSN%291365-2109/homepage/ForAuthors.html>

SUMÁRIO
I EFEITOS DA PRIVAÇÃO ALIMENTAR SOBRE O CRESCIMENTO LARVAL DO
“JUNDIÁ” Rhamdia voulezi Haseman 1911
1 INTRODUÇÃO .................................................................................................................................. 12
2 MATERIAIS E MÉTODOS ............................................................................................................... 14
2.1 Análise de dados ............................................................................................................................ 16
3 RESULTADOS ................................................................................................................................... 16
3.1 Eclosão e início da alimentação exógena ...................................................................................... 16
3.2 Período crítico para a sobrevivência larval .................................................................................... 17
3.3 Comprimento padrão e ganho de peso ........................................................................................... 17
3.4 Ponto de não retorno ...................................................................................................................... 21
3.5 Relação peso/comprimento e coeficiente de crescimento ............................................................. 21
4 DISCUSSÕES ..................................................................................................................................... 23
REFERÊNCIAS .................................................................................................................................. 28
II SELETIVIDADE ALIMENTAR DE LARVAS DO “JUNDIÁ” Rhamdia voulezi
Haseman 1911: ALGUMAS IMPLICAÇÕES PARA O SEU CULTIVO
1 INTRODUÇÃO .................................................................................................................................. 34
2 MATERIAIS E MÉTODOS ............................................................................................................... 35
2.1 Análise de dados ............................................................................................................................ 37
3 RESULTADOS ................................................................................................................................... 39
3.1 Seletividade alimentar taxonômica ................................................................................................ 39
3.2 Seletividade por classe de tamanho ao longo do período larval .................................................... 44
3.3 Relação entre tamanho de boca e comprimento padrão ................................................................ 46
3.4 Tamanho da presa x tamanho de boca do predador ....................................................................... 47
3.5 Seletividade por tamanho de boca ................................................................................................ 47
4 DISCUSSÕES ..................................................................................................................................... 48
REFERÊNCIAS ..................................................................................................................................... 54

CAPÍTULO I
EFEITOS DA PRIVAÇÃO ALIMENTAR SOBRE O CRESCIMENTO LARVAL DO
“JUNDIÁ” Rhamdia voulezi Haseman 1911

EFEITOS DA PRIVAÇÃO ALIMENTAR SOBRE O CRESCIMENTO LARVAL DO
“JUNDIÁ” Rhamdia voulezi Haseman 1911
RESUMO
Neste estudo foram avaliados os efeitos da privação alimentar sobre o crescimento das larvas
de jundiá Rhamdia voulezi. O experimento foi iniciado imediatamente após a completa
absorção do saco vitelínico, que ocorreu no 4o dia após a eclosão (DAE). Para tanto, foram
realizados 8 tratamentos constituídos por diferentes regimes alimentares, incluindo o
tratamento controle (TC), no qual não houve fornecimento de alimento. O alimento foi
fornecido pela primeira vez no 4o, 5
o, 6
o, 7
o, 8
o, 9
o e 10
o DAE, correspondendo aos
tratamentos: T4; T5, T6, T7, T8; T9 e T10, respectivamente. Para a alimentação das larvas foi
utilizado zooplâncton coletado de tanques piscícolas previamente adubados e artêmia salina
recém-eclodida. Ao final do experimento, após 14 dias, foram avaliados o comprimento
padrão (CP) e peso de um total de 594 larvas. As larvas que receberam alimento do 4o ao 6
o
DAE tiveram desenvolvimento significativamente maior que aquelas alimentadas mais
tardiamente, a partir do 7o DAE. A partir deste momento, as larvas alcançaram o ponto de não
retorno. Conclui-se que a primeira alimentação deve ser iniciada até o 6o DAE de modo que
seu desenvolvimento e crescimento não sejam afetados.
PALAVRAS-CHAVE. Inanição, primeira alimentação, desenvolvimento larval.

EFFECTS OF FEEDING PRIVATION ON LARVAL GROWTH “JUNDIÁ” Rhamdia
voulezi Haseman 1911
ABSTRACT
The effects of feeding privation were evaluated on growth of larvae of “jundiá” Rhamdia
voulezi. The experiment was started immediately after complete absorption of the yolk sac,
coinciding with the 4th days after hatching (DAH). Then, we conducted seven treatments
including the control treatment (TC) in which food was not provided. The food was provided
for the first time in 4th, 5th, 6th, 7th, 8th, 9th and 10th DAH corresponding to the treatments:
T4, T5, T6, T7, T8, T9 and T10, respectively. Zooplankton collected from previously
fertilized fish ponds and newly newly hatched artemia was used to feed the larvae. At the end
of the experiment, after 14 days, we assessed the standard length (SL) and weight of a total of
594 larvae. Larvae that received food from the 4th to the 6th DAH had a development
significantly greater than those fed later, from the 7th DAH. From this moment, the larvae
reached the point of no return. It was concluded that the first feeding should be initiated on
the 6th DAH so that their development and growth is not affected.
KEYWORDS. Starvation, first feeding, larval development.

12
1. Introdução
Assim que eclodem, muitas larvas de peixes são exclusivamente dependentes das
reservas nutritivas provenientes do saco vitelínico (Geurden et al., 2007). Uma vez que estas
reservas endógenas se esgotam, as larvas precisam estabelecer com sucesso a transição da
alimentação endógena para exógena de modo a garantir a sua sobrevivência e crescimento
nesta fase (Blaxter e Ehrlich, 1974). Para isto alguns fatores são importantes, como a
disponibilidade e composição do alimento (Hunter, 1981; Sánchez-Velasco, 1998),
temperatura, fotoperíodo (Paul, 1983; Porter e Bailey, 2007) desenvolvimento do aparelho de
digestão, acuidade visual e habilidade natatória da larva (Makrakis et al., 2005). Assim sendo,
quando estes fatores não estão em conformidade com o presente estado das larvas, as mesmas
podem não ter êxito na sua primeira alimentação, tornando-as vulneráveis à inanição (Gisbert,
2004; Zhang et al., 2009).
A inanição tem sido uma das maiores causas de mortalidade de larvas de peixes tanto
no ambiente natural como na aquicultura (May, 1971; Leggett e Deblois, 1994; Dou et al.,
2002; Yúfera e Darias, 2007). Isto porque durante o desenvolvimento ontogenético inicial, a
maioria dos peixes apresentam baixa capacidade de natação e de alimentação (Houde, 1974;
Dou et al., 2002). Assim, se forem privados de alimentos adequados nesta fase, os reflexos
mais notórios são o aumento da vulnerabilidade a predadores, mortalidade, deformidades
morfológicas e reduzida taxa de crescimento (Houde, 1974; Hunter, 1981; Dou et al., 2002;
Gisbert et al., 2004; Zhang et al., 2009; Faria et al., 2011). A privação alimentar também afeta
outros parâmetros, em especial os bioquímicos e peso (Hung et al., 1997).
Muitas espécies de peixes adultos podem suportar consideráveis períodos de privação
alimentar, desenvolvendo, portanto, uma habilidade de sobrevivência na ausência ou escassez
do alimento e de se recuperarem completamente se realimentadas (Love, 1970; Salam et al.,

13
2000). O mesmo não é verificado em peixes na fase larval, pois em questão de poucos dias as
larvas chegam a um estado de inanição irreversível, em que mesmo que estejam vivas e haja
disponibilidade de alimento não conseguem mais se alimentar e a mortalidade aumenta
acentuadamente. Blaxter e Hempel (1963) denominaram este estado crítico de “ponto de não
retorno”.
Vários métodos tem sido utilizados para descrever os efeitos da privação alimentar em
adultos e larvas de peixes, tais como: medidas morfométricas e gravimétricas (peso) (Blaxter
e Hempel, 1963; Ehrlich et al., 1976; Powell e Chester, 1985; Bisbal e Bengstson, 1995;
Mookerji e Rao, 1999; Gisbert et al., 2004; Peña e Dumas, 2005; Shan et al., 2009; Faria et
al., 2011); faixa de RNA/DNA (Faria et al., 2011); critérios histológicos (Gisbert et al., 2004);
atividade enzimática (Chen et al., 2007) e atividade natatória (Faria et al., 2011). Tais
informações são importantes porque permitem determinar o tempo máximo no qual os peixes
são capazes de suportar a ausência do alimento até alcançar o ponto de não retorno (Gisbert et
al., 2004; Peña e Dumas, 2005). Além disto, estes estudos podem ser uma ferramenta útil para
entender as causas de mortalidade precoce de peixes em ambiente natural (O'Connell, 1976),
bem como, em condições de cultivo numa das etapas mais críticas, a larvicultura.
Até o momento, existem poucos trabalhos que enfoquem inanição em larvas de peixes.
As pesquisas mais recentes são com espécies marinhas, como os de Gisbert et al. (2004) com
larvas de Paralichthys californicus, Penã e Dumas (2005) para larvas de Paralabrax
maculatofasciatus, Chen et al. (2007) com larvas de Seriola lalandi e Shan et al. (2008, 2009)
para larvas de Oplegnathus fasciatus e Miichthys miiuy, respectivamente. No Brasil, estas
informações são praticamente inexistentes tanto para espécies de água doce como marinhas.
Neste contexto, os efeitos da privação alimentar sobre o crescimento larval do jundiá
Rhamdia voulezi Hanseman 1911 foram avaliados no presente estudo. O jundiá Rhamdia
voulezi Hanseman 1911, é uma espécie de médio porte, endêmica da bacia do rio Iguaçu

14
(Baumgartner et al., 2012), cuja reprodução acontece entre setembro a fevereiro com picos
entre novembro e dezembro (Suzuki e Agostinho, 1997). Seu cultivo em cativeiro ainda está
em fase inicial, provavelmente devido ao endemismo e as poucas informações existentes
sobre a mesma. Especificamente neste estudo foram avaliados: (1) o início da alimentação
exógena; (2) os efeitos do retarde da primeira alimentação no crescimento larval e (3) o ponto
de não retorno.
2. Materiais e métodos
O experimento foi desenvolvido no Instituto de Pesquisa em Aquicultura Ambiental
(InPAA) da Universidade Estadual do Oeste do Paraná, município de Toledo-PR, no mês de
fevereiro de 2012. As larvas de jundiá Rhamdia voulezi foram obtidas por meio de reprodução
induzida e fornecidas pelo Centro de Difusão e Desenvolvimento Tecnológico do rio Iguaçu
(CDT), localizado no município de Boa Vista da Aparecida, PR.
O experimento foi inteiramente casualizado e teve início logo após a absorção do saco
vitelínico, no 4o dia de vida após a eclosão (DAE). Foram realizados 8 tratamentos em
triplicatas constituídos por diferentes regimes alimentares. O alimento foi fornecido pela
primeira vez no 4o, 5
o, 6
o, 7
o, 8
o, 9
o e 10
o DAE, correspondendo aos tratamentos: T4; T5, T6,
T7, T8; T9 e T10, respectivamente. Também foi realizado um tratamento controle (TC), do
qual não recebeu alimento. Os tratamentos foram conduzidos em recipientes plásticos com
volume útil de 10L cada, dotados de sistema de aeração constante, numa densidade de 20
larvas/L. Cada recipiente foram cobertos por lona preta para a ausência total de luz, sendo
descobertos apenas para os manejos diários de limpeza e alimentação.
O alimento foi fornecido duas vezes ao dia, às 09h00min e 16h00min, com
zooplâncton coletado de viveiros aquícolas e artêmias recém-eclodidas. Estes viveiros foram
previamente calados com CaCo3 (250g/m²) e adubados com NPK 4-14-8 (200g/m²) para

15
promover o crescimento da comunidade planctônica. A eclosão das artêmias foi feita em
garrafas pet de 2L, utilizando-se uma salinidade de 10 ppm, luz e aeração constante. Antes de
serem colocadas nas garrafas para a eclosão, era feito a descapsulação com hipoclorito de
sódio (1 mL NaClO/100 mL H2O) durante 15 minutos. A coleta do zooplâncton foi feita
através de rede de plâncton de malhagem de 100 µm. Posteriormente, era feita a
homogeneização e em seguida, distribuídos na mesma proporção para todos os aquários por
meio de um copo de 200 mL. Procedimento semelhante foi adotado para o fornecimento das
artêmias, porém, antes era feita a lavagem para a remoção do sal, e só então eram fornecidas
por meio de pipetas de 5 mL.
Antes da última alimentação, era feita a sifonagem de todos recipientes para remoção
dos detritos. Neste momento, era feito a coleta de três larvas de cada tratamento e a leitura dos
parâmetros físico e químicos da água. As larvas coletadas foram submetidas à eutanásia, em
solução contendo benzocaína (250 mg/L) e em seguida fixadas em formol 4% para posterior
análise em laboratório. As leituras da temperatura, oxigênio dissolvido, pH e condutividade
elétrica foram realizadas com o uso do multiparâmetro da marca YSI modelo professional
Plus, com a finalidade de controlar o experimento, mantendo-os dentro do limite de tolerância
para peixes neotropicais conforme Kubtza (1999) e Sipaúba -Tavares (1994).
Simultaneamente à coleta de larvas, eram feitas observações visuais quanto ao
comportamento das mesmas na coluna de água. Com estas observações foi possível notar os
primeiros reflexos causados pela inanição.
Ao final do experimento, após 14 dias, foram avaliados o comprimento padrão-CP
(mm) e peso (g) de 594 larvas no total. Alguns tratamentos foram finalizados antes que os
demais devido à mortalidade em massa causada pela privação, logo, tiveram menor duração
(9 e 6 dias). Isto também justifica as diferenças do número amostral (N) entre os tratamentos

16
(ver Tabela 1). As medições do CP foram realizadas em um microscópio estereoscópico da
marca Olympus, modelo SZX7 equipado com câmera digital DP-25 com captura de imagem e
software DP2-BSW. Quando as pesagens, foram feitas com balança analítica (0,00001 g) da
marca SHIMADZU, modelo AW 220.
2.1. Análise dos dados
O crescimento das larvas submetidas à privação alimentar por diferentes períodos
(tratamentos) foi avaliado através da Análise não paramétrica de Kruskal-Wallis, sendo o
comprimento padrão (CP) e peso as variáveis dependentes e dias de vida (DAE) a variável
independente. As amostras coletadas nos diferentes dias de vida que não tinham um número
amostral mínimo de 6 larvas, devido à mortalidade, foram desconsideradas da análise.
Também foram observados a relação peso/comprimento e o coeficiente de crescimento
em relação aos diferentes tratamentos. Para isto, foram feitas regressões lineares com dados
transformados em log(x). A relação peso/comprimento foi observada por meio do coeficiente
de determinação (valor de R²) e o coeficiente de crescimento através do valor de b, que é o
coeficiente de regressão extraído da equação da reta: y = a + bx.
O software Statistica v.7.0 foi utilizado para as análises.
3. Resultados
3.1. Eclosão e início da alimentação exógena
A eclosão das larvas de Rhamdia voulezi ocorreu 15 horas após a fertilização numa
temperatura média de 29,28 ºC. No 2o DAE (dias de vida após a eclosão), a boca apresentava-
se aberta e saco vitelínico relativamente grande. Somente no 4o DAE (CP: 4,45±0,38 mm), a
larva apresentou o saco vitelínico completamente absorvido. Neste momento, o alimento foi

17
disponibilizado pela primeira vez para as larvas do tratamento 4 (T4). A ocorrência de
canibalismo foi observada imediatamente após este período.
Convêm ressaltar, que mesmo antes da exaustão completa do saco vitelínico, as larvas
exibiram elementos típicos de comportamento de forrageamento, como boa habilidade
natatória, olhos bem pigmentados, barbilhões formados e tentativas de abocanhar.
3.2. Comprimento padrão e peso
O crescimento em comprimento padrão só foi observado para os tratamentos T4, T5 e
T6 (Kruskal-Wallis, p<0,05), ao contrário dos demais (T7, T8, T9 e T10) (Kruskal-Wallis,
p>0,05) que não apresentaram crescimento (Tabela 1).
Com relação ao peso das larvas, os resultados não foram distantes do crescimento em
comprimento padrão. As larvas dos tratamentos T4, T5 e T6 foram significativamente
maiores em peso do que aquelas dos tratamentos T7, T8, T9 e T10 (Kruskal-Wallis, p<0,05 e
p>0,05, respectivamente) (Tabela 1).
Isto indica que o desenvolvimento das larvas de Rhamdia voulezi só é possível se
alimentadas a partir do 4o, 5
o e 6
o DAE. Se alimentadas após este período, o desenvolvimento
das larvas é interrompido (Fig. 1 e Fig. 2).
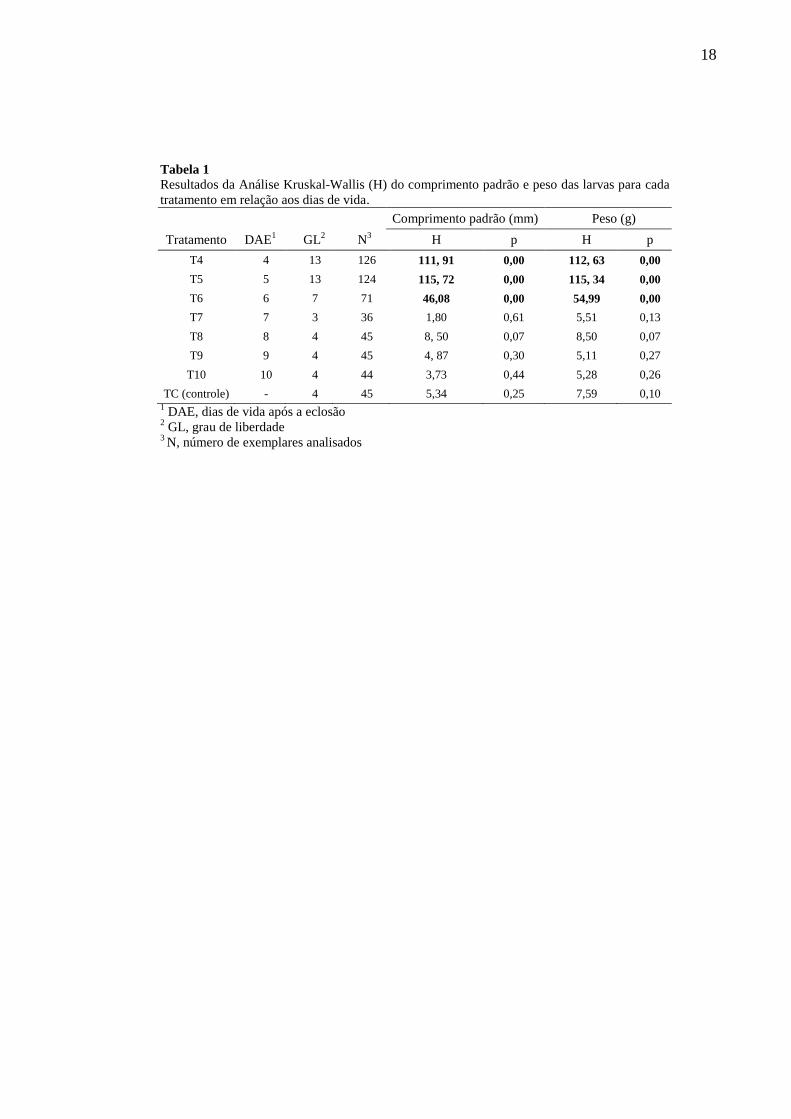
18
Tabela 1
Resultados da Análise Kruskal-Wallis (H) do comprimento padrão e peso das larvas para cada
tratamento em relação aos dias de vida.
Comprimento padrão (mm) Peso (g)
Tratamento DAE1 GL
2 N
3 H p H p
T4 4 13 126 111, 91 0,00 112, 63 0,00
T5 5 13 124 115, 72 0,00 115, 34 0,00
T6 6 7 71 46,08 0,00 54,99 0,00
T7 7 3 36 1,80 0,61 5,51 0,13
T8 8 4 45 8, 50 0,07 8,50 0,07
T9 9 4 45 4, 87 0,30 5,11 0,27
T10 10 4 44 3,73 0,44 5,28 0,26
TC (controle) - 4 45 5,34 0,25 7,59 0,10 1 DAE, dias de vida após a eclosão
2 GL, grau de liberdade
3 N, número de exemplares analisados
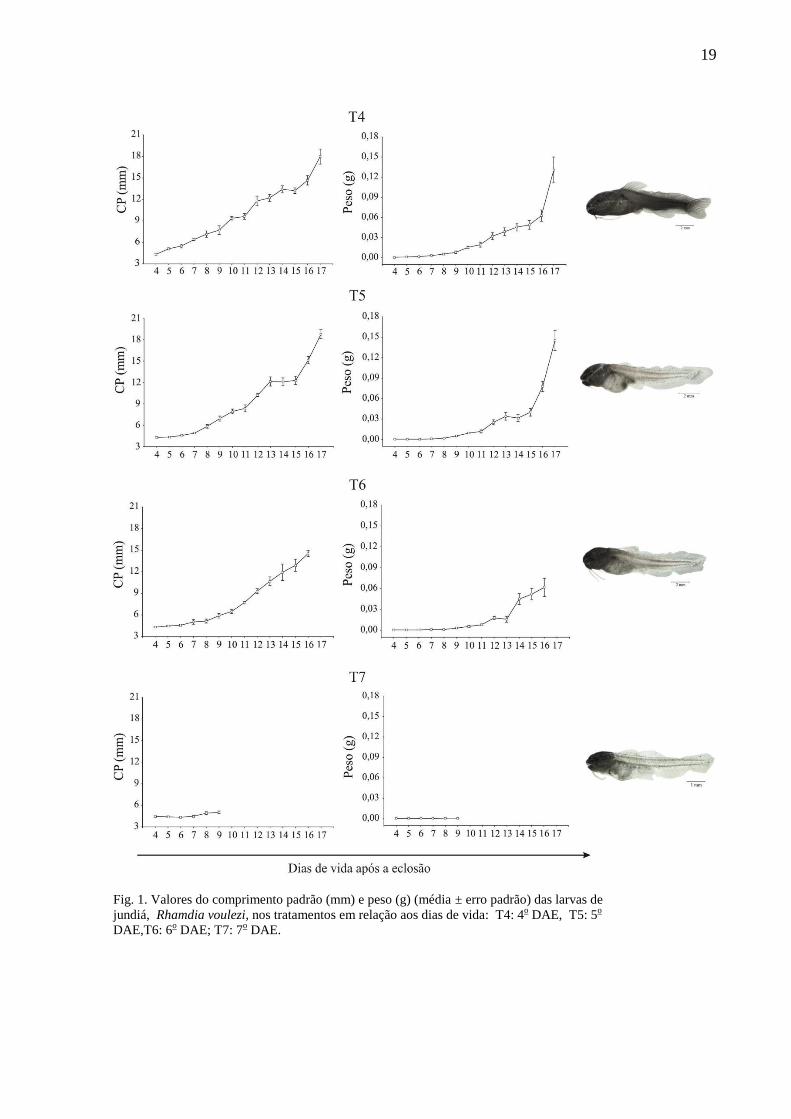
19
Fig. 1. Valores do comprimento padrão (mm) e peso (g) (média ± erro padrão) das larvas de
jundiá, Rhamdia voulezi, nos tratamentos em relação aos dias de vida: T4: 4o DAE, T5: 5
o
DAE,T6: 6o DAE; T7: 7
o DAE.

20
Fig. 2. Valores do comprimento padrão e peso (média ± erro padrão) das larvas de jundiá, Rhamdia voulezi, nos
tratamentos em relação aos dias de vida: T8: 8o DAE; T9: 9
o DAE; T10: 10
o DAE; TC: Tratamento controle.

21
3.4. Ponto de não retorno
O sétimo dia de vida após a eclosão (7o DAE) numa temperatura média de 29,79 ºC,
foi caracterizado como o ponto de não retorno para a larva de jundiá Rhamdia voulezi. O
alimento foi disponibilizado pela primeira vez neste período, porém, as larvas não
conseguiram se recuperar, havendo grande mortalidade (Fig. 1 e Fig. 2).
A partir do 5o DAE, foi observado nas larvas submetidas à privação alimentar a prática
de canibalismo, apresentando sinais de ferimentos e nadadeiras retalhadas. Também foi
possível notar mudanças de comportamento relacionado à natação. Algumas larvas exibiam
natação horizontal e sinais de letargia.
3.5. Relação peso e comprimento
A análise de regressão linear do peso em função do comprimento padrão para os
diferentes tratamentos indicou que apenas nos tratamentos T4, T5 e T6 (4o, 5
o e 6
o DAE) o
peso apresentou-se fortemente associado com a variação do comprimento padrão (Fig. 3). O
T5 apresentou um valor relativamente alto do coeficiente de determinação (R² = 0,72), o que
não se pode afirmar que o peso variou em função do comprimento padrão neste tratamento,
pois se trata de um número reduzido de larvas (7 do total de 52 larvas) que se destacaram em
relação as demais, provavelmente devido ao canibalismo. Assim sendo, para esta relação, tais
larvas canibalistas foram desconsideradas, e o coeficiente de determinação passou a ter um
valor não significativo (R² = 0,14), assim como os demais tratamentos (T7, T8, T9, T10 e TC)
(Fig. 3).
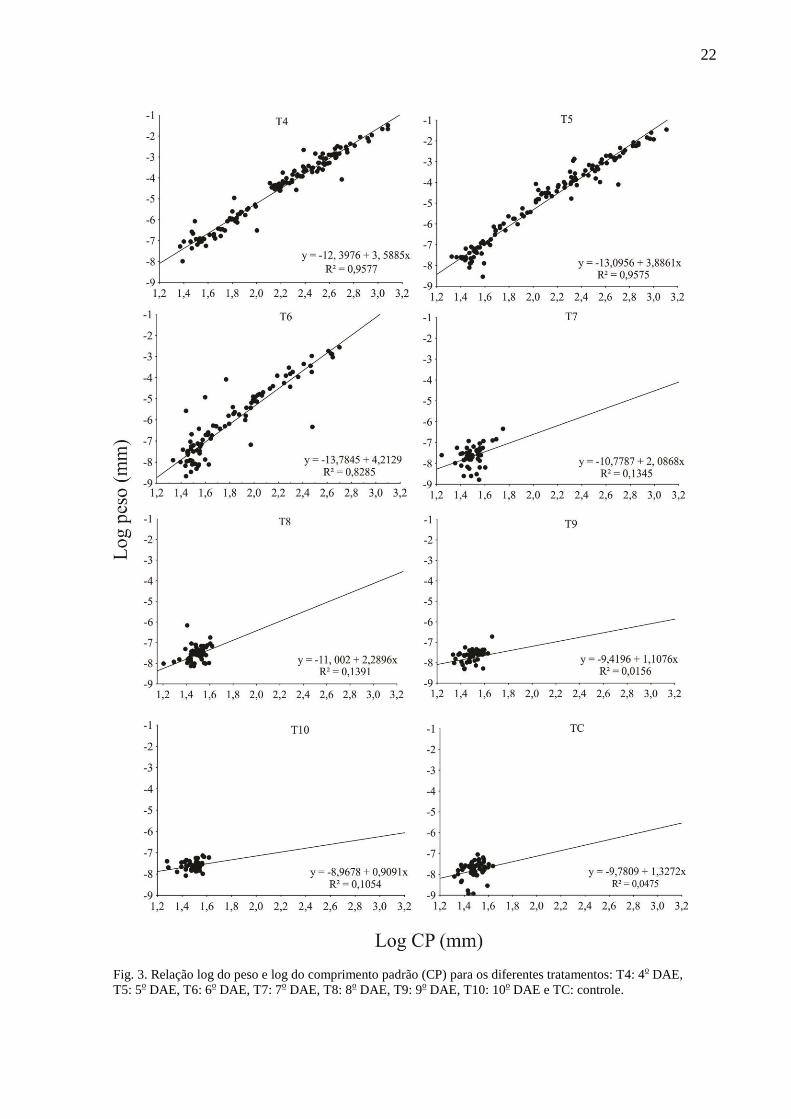
22
Fig. 3. Relação log do peso e log do comprimento padrão (CP) para os diferentes tratamentos: T4: 4o DAE,
T5: 5o DAE, T6: 6
o DAE, T7: 7
o DAE, T8: 8
o DAE, T9: 9
o DAE, T10: 10
o DAE e TC: controle.
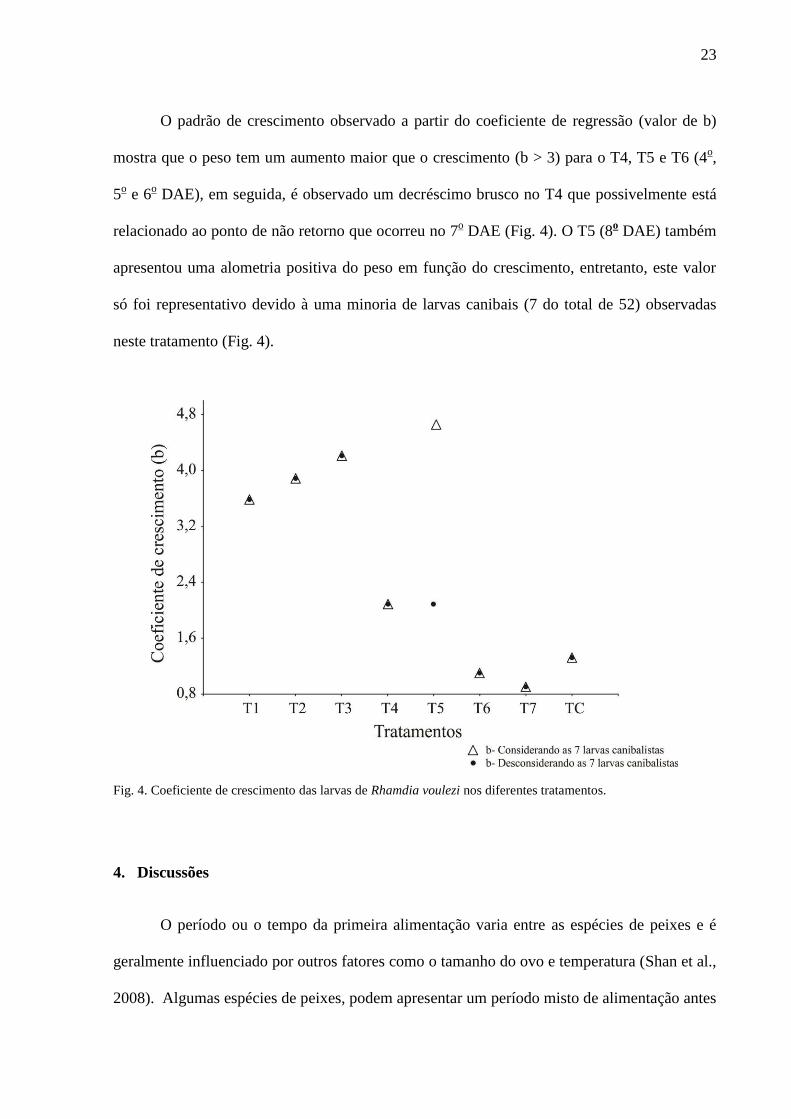
23
O padrão de crescimento observado a partir do coeficiente de regressão (valor de b)
mostra que o peso tem um aumento maior que o crescimento (b > 3) para o T4, T5 e T6 (4o,
5o e 6
o DAE), em seguida, é observado um decréscimo brusco no T4 que possivelmente está
relacionado ao ponto de não retorno que ocorreu no 7o DAE (Fig. 4). O T5 (8
o DAE) também
apresentou uma alometria positiva do peso em função do crescimento, entretanto, este valor
só foi representativo devido à uma minoria de larvas canibais (7 do total de 52) observadas
neste tratamento (Fig. 4).
Fig. 4. Coeficiente de crescimento das larvas de Rhamdia voulezi nos diferentes tratamentos.
4. Discussões
O período ou o tempo da primeira alimentação varia entre as espécies de peixes e é
geralmente influenciado por outros fatores como o tamanho do ovo e temperatura (Shan et al.,
2008). Algumas espécies de peixes, podem apresentar um período misto de alimentação antes

24
da completa exaustão do vitelo (Zhang et al., 2009), porém, outras, só iniciam a primeira
alimentação logo após a total absorção do saco vitelínico (Houde, 1974) ou dias depois
(Gisbert e Williot, 1997; Gisbert et al., 2004). Nakaghi et al. (2011) evidenciaram para larvas
de tilápia Oreochromis niloticus que o início da primeira alimentação antes ou logo após a
depleção total do saco vitelínico não influenciou a sobrevivência das mesmas. Desta forma,
presume-se que o início da primeira alimentação deve ser feita dentro do limite de tolerância
ao jejum de cada espécie.
Sabe-se que o período larval é caracterizado por altas taxas de mortalidade,
principalmente na transição da alimentação endógena para exógena (Blaxter e Ehrlich, 1974;
Fuiman, 2002). Assim, a sobrevivência e o crescimento nesta fase depende, dentre outros
fatores, com o grau de desenvolvimento de órgãos e estruturas necessários a alimentação,
como boca e aparelho digestivo (Makrakis et al., 2005; 2008), capacidade de natação, que está
relacionado com o sucesso de busca pelo alimento, acuidade visual (Yúfera e Darias, 2007;
Makrakis et al., 2008) e disponibilidade de alimento (Sánchez-Velasco, 1998). Larvas que não
conseguem superar essas limitações a tempo, tornam-se vulneráveis a inanição e
consequentemente, à predação (Gisbert et al., 2004; Dou et al., 2005).
A inanição ainda nas primeiras fases de desenvolvimento resulta em graves
deformidades morfológicas, atraso no crescimento e alta mortalidade (Gisbert et al., 2004;
Shan et al., 2009). Para Blaxter e Hempel (1963), as larvas privadas de alimento podem
alcançar um estado de inanição irreversível, tornando-se demasiadamente fracas para se
alimentar e morrem. Estes autores denominou este estado como “ponto de não retorno”. O
ponto de não retorno (PNR) para larvas de Rhamdia voulezi foi verificado no sétimo dia de
vida após a eclosão a uma temperatura de 29,79 ºC. Os tratamentos que receberam alimento
após este período continuaram a apresentar acentuada mortalidade e não tiveram um padrão
de desenvolvimento semelhante àqueles que foram alimentados precocemente (até o 6o DAE).

25
A capacidade de resistência à privação alimentar antes do ponto de não retorno é
dependente do tamanho da larva, temperatura e também da espécie de peixe (Blaxter e
Ehrlich, 1974; Houde, 1974). Como exemplo, uma espécie de água doce, Odontesthes
bonariensis, Strussmann e Takashima (1989) observou o ponto de não retorno em oito dias de
vida após a eclosão a uma temperatura de 26 ºC. Para espécies marinhas, o ponto de não
retorno aconteceu em sete dias após a eclosão para larvas de Paralichthys californicus
(Gisbert et al. 2004) e quatro à cinco dias após a eclosão para larvas de Paralabrax
maculatofasciatus (Peña e Dumas 2005).
Alguns métodos, desde morfológicos até bioquímicos têm sido utilizados para avaliar
as condições das larvas de peixes submetidos a diferentes períodos de privação alimentar.
Neste estudo, o atraso da primeira alimentação foi avaliado através do crescimento
(comprimento padrão) e peso, que foram intensamente afetados em larvas alimentadas mais
tardiamente, após o 6o
DAE. Larvas de outras espécies de água doce, tais como: Later
calcarifer (Kailasam et al., 2007) e Siniperca scherzeri (Zhang et al., 2009), o crescimento
também foi influenciado pelo atraso da primeira alimentação. Vale ressaltar ainda que
mudança de comportamento do peixe como letargia, natação errática (natação horizontal) e
canibalismo, constatados neste estudo, também podem ser os primeiros sinais de inanição.
Para Chen et al. (2007) a habilidade que as larvas tem em suportar determinados
períodos de privação alimentar é crucial para a sobrevivência e crescimento da mesmas,
embora, deve ser evitada antes do seu completo desenvolvimento, pois podem causar danos
em células do intestino que são importantes na digestão e absorção dos alimentos. Não
obstante desta hipótese, Dou et al. (2002) alertam que a alimentação iniciada em torno do
ponto de não retorno reduz a capacidade das larvas de se alimentarem devido ás deformidades
morfológicas e fisiológicas causadas pela inanição.
26
Neste sentido, as informações acerca dos efeitos da privação alimentar sobre o
desenvolvimento dos peixes, bem como, o tempo máximo em que as larvas são capazes de
resistir antes do PNR, são primordiais para a piscicultura, pois permite fornecer um protocolo
do tempo ideal para o inicio da primeira alimentação, e assim, evitar maiores mortalidades na
fase larval. Na ecologia, permite entender as altas taxas de mortalidade precoce de populações
de ambiente natural que levam a variações no tamanho do estoque (O'Connell, 1976).
No geral, nossos resultados indicam que o crescimento larval do jundiá tanto em
tamanho como em peso foram significativamente afetados pelo atraso da primeira
alimentação que deve ser iniciada no quarto dia de vida após a eclosão, sendo tolerável até o
sexto dia de vida após a eclosão. A partir deste período, no sétimo dia após a eclosão, as
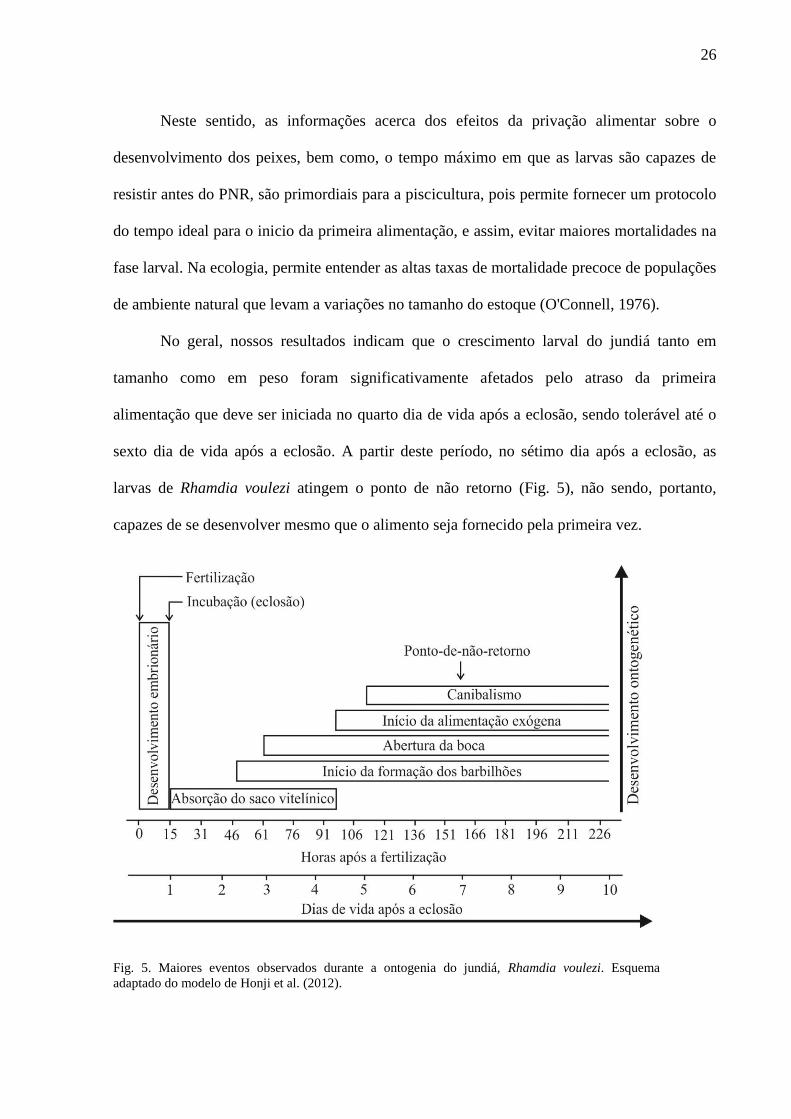
larvas de Rhamdia voulezi atingem o ponto de não retorno (Fig. 5), não sendo, portanto,
capazes de se desenvolver mesmo que o alimento seja fornecido pela primeira vez.
Fig. 5. Maiores eventos observados durante a ontogenia do jundiá, Rhamdia voulezi. Esquema
adaptado do modelo de Honji et al. (2012).

27
Agradecimentos
Agradecemos a Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
pela concessão de bolsa e ao Grupo de Pesquisa em Tecnologia de Produção e Conservação
dos Recursos Pesqueiros e Hídricos (GETECH) pelo apoio logístico e financeiro para a
realização da pesquisa.
Referências
Baumgartner, G., Pavanelli, C.S., Baumgartner, D., Bifi, A.G., Debona, T., Frana, V.A.,
2012. Peixes do Baixo rio Iguaçu. Primeira ed. Eduem, Maringá.
Blaxter, J.H.S., Hempel, G., 1963. The influence of egg size on herring larvae (Clupea
harengus). J. Cons. Int. Explor. Mer. 28, 211-240.
Blaxter, J.H.S., Ehrlich, K.F., 1974. Changes in behavior during starvation of herring and
plaice larvae. In: Blaxter, J.H.S (ed.), The early life history of fish. Springer-Verlag, Berlin,
pp. 575-588.
Bisbal, G., Bengtson, D.A., 1995. Descriptions of the starving condition in summer flounder,
Paralichthys dentatus, early life-history stages. Fish B-NOAA. 93, 217-230.
Chen, B.N., Qin, J.G., Carragher, J.F., Clarke, S.M., Kumar, M.S., Hutchinson, W.G., 2007.
Deleterious effects of food restrictions in yellowtail kingfish Seriola lalandi during early
development. Aquacult. 271, 326-335.
Dou, S.Z., Masuda, R., Tanaka, M., Tsukamoto, K., 2002. Feeding resumption,
morphological changes and mortality during starvation in Japanese flounder larvae. J. Fish
Biol. 60, 1363-1380.
Dou, S., Masuda, R., Tanaka, M., Tsukamoto, K., 2005. Effects of temperature and delayed
first feeding on survival and growth of Japanese flounder larvae. J. Fish Biol. 66, 362-377.
Ehrlich, K.F., Blaxter, J.H.S., Pemberton, R. 1976. Morphological and histological changes
during growth and starvation of herring and plaice larvae. Mar. Biol. 35, 105-118.
Faria, A.M., Muha, T., Morote, E., Chícaro, M.A. 2011. Influence of starvation on the critical
swimming behaviour of the Senegalese sole (Solea senegalensis) and its relationship with
RNA/DNA ratios during ontogeny. Sci. Mar. 75, 87-94.
Fuiman, L.A., 2002. Special considerations of fish eggs and larvae. In: Fuiman, L.A., Werner,
R.G. (eds.), The unique contributions of early life stages. Fishery science, Oxford, Blackwell
Science, pp.1-32.
Geurden, M. Aramendi, J. Zambonino-Infante, Panserat, S., 2007. Early feeding of
carnivorous rainbow trout (Oncorhynchus mykiss) with a hyperglucidic diet during a short

28
period: effect on dietary glucose utilization in juveniles. Am. J. of Physiol. 292, R2275-
R2283.
Gisbert, E., Williot, P., 1997. Larval behavior and effect of the timing of initial feeding on
growth and survival of Siberian sturgeon (Acipenser baeri) larvae under small scale hatchery
production. Aquacult. 156, 63-76.
Gisbert, E., Conklin, D.B., Piedrahita, R.H., 2004. Effects of delayed first feeding on the
nutritional condition andmortality of California halibut larvae. J. Fish Biol. 64, 116-132.
Gomes, L.C., Golombieski, J.I., Gomes, A.R.C., Baldisserotto, B., 2000. Biologia do jundiá
Rhamdia quelen (Teleostei, Pimelodidae). Ciênc. Rural. 30, 179-185.
Hempel, G., Blaxter, J.H.S., 1963. On the condition of herring larvae. Rapports et Proces
Verbaux des Reunions du Conseil International pour l’Exploration de la Mer. 154, 35-40.
Honji, R.M., Tolussi, C.E., Mello, P.H., Caneppele, D., Moreira, R.G., 2012. Embryonic
development and larval stages of Steindachneridion parahybae (Siluriformes: Pimelodidae):
implications for the conservation and rearing of this endangered neotropical species. Neotrop.
Ichthyol. 10, 313-327.
Houde, E.D., 1974. Effects of temperature and delayed feeding on growth and survival of
larvae of three species of subtropical marine fishes. Mar. biol. 26, 271-285.
Hung, S.S.O., Liu,W., Li, H., Storebakken, T., Cui, Y., 1997. Effect of starvation on some
morphological and biochemical parameters in while sturgeon, Acipenser transmontanus.
Aquacult. 151, 357-363.
Hunter, J.R., 1981. Feeding ecology and predation of marine fish larvae. In: Lasker, R. (ed.)
Marine Fish Larvae: Morphology, Ecology and Relation to Fisheries. University of
Washington Press, Seattle, pp. 34-77.
Kailasam, M., Thirunavukkarasu, A. R., Selvaraj, S., Stalin, P., 2007. Effect of delayed initial
feeding on growth and survival of Asian sea bass Lates calcarifer (Bloch) larvae. Aquacult.
271, 298-306.
Kubitza, F., 1999. Qualidade da água na produção de peixes, primeira ed. CIP-USP, Jundiaí,
São Paulo.
Love, R.M., 1970. The chemical Biology of Fishes, Primeira ed. Academic Press, London.
Leggett, W.C, Deblois, E., 1994. Recruitment in marine fishes: is it regulated by starvation
and predation in the egg and larval stages? Neth. J. Sea Res. 32, 119-134.
Makrakis, M.C., Nakatani, K., Bialetzki, A., Sanches, P.V., Baumgartner, G., Gomes, L.C.,
2005. Ontogenetic shifts in digestive tract morphology and diet of fish larvae of the Itaipu
Reservoir, Brazil. Environ. Biol. Fishes. 72, 99-107.

29
Makrakis, M. C., Nakatani, K., Bialetzki, A., Gomes, L. C., Sanches, P. V. & Baumgartner,
G., 2008. Relationship between gape size and feeding selectivity of fish larvae from a
Neotropical Reservoir. J. Fish Biol. 72, 1690-1707.
May, R. C. 1971. Effects of delayed initial feeding on larvae of the grunion, Leuresthes tenuis
(Ayres). Fish B-NOAA. 69, 411-425.
Mookerji, N., Rao, R., 1999. Rates of yolk utilization and effects of delayed initial feeding in
the larvae of the freshwater fishes rohu and singhi. Aquacult. 7, 45-56.
Nakaghi, L.S.O., Batista, W.K., Paes, M.C.F., Makino, L.C., Koberstein, T.C.R.D., 2011.
Influence of the first feeding on absorption of yolk and survival rate of Oreochromis niloticus
larvae. Rev. Acad., Ciênc. Agrár. Ambient. 9, 257-262.
O'Connell, C.P., 1976. Histological criteria for diagnosing the starving condition in early post
yolk sac larvae of the northern anchovy, Engraulis mordax Girard. J. Exp. Mar. Biol. Ecol.
25, 285-312.
Paul, A.J., 1983. Light, temperature, nauplii concentrations, and prey capture by first feeding
pollock larvae Theragra chalcogramma. Mar. Ecol. Prog. Ser. 13, 175-179.
Peña, R.A., Dumas, S., 2005. Effect of delayed first feeding on development and feeding
ability of Paralabrax maculato fasciatus larvae. J. Fish Biol. 67, 640-651.
Porter, S.M., Bailey, K.M., 2007. Optimization of feeding and growth conditions for walleye
pollock Theragra chalcogramma (Pallas) larvae reared in the laboratory. Alaska Fisheries
Science Center (U.S.), United States, National Marine Fisheries Service, Seattle, WA, Alaska
Fisheries Science Center.
Powell, A.B., Chester, A.J., 1985. Morphometric indices of nutritional condition and
sensitivity to starvation of spot larvae. Trans. Am. Fish. Soc. 114, 338-347.
Salam, A., Ali, M., Masud, S., 2000. Effect of various food deprivation regimes on body
composition dynamics of thaila, Catla catla. J. Mater. Sci. Res. 11, 26-32.
Sanchez-Velasco, L. 1998. Diet composition and feeding habits of fish larvae of two
coccurring species (Pisces: Callionymidae and Bothidae) in the North-western Mediterranean.
J. Mar. Sci. 55, 299-308.
Shan, X.J., Quan, H.F., Dou, S.Z., 2008. Effects of delayed first feeding on growth and
survival of rock bream Oplegnathus fasciatus larvae. Aquacult. 277, 14-23.
Shan, X.J., Cao, L., Huang, W., Dou, S.Z., 2009. Feeding, morphological changes and
allometric growth during starvation in miiuy croaker larvae. Environ. Biol. Fishes. 86, 121-
130.
Sipaúba-Tavares, L.H. 1994. Limnologia aplicada à aquicultura. Primeira ed. FUNEP, São
Paulo.
Strussmann, C.A., Takashima, F., 1989. PNR, histology and morphometry of styarved
pejerrey Odontesthes bonariensis larvae. Nippon Suis. Gak. 55, 237-246.

30
Suzuki, H.I., Agostinho, A.A., 1997. Reprodução de peixes do reservatório de Segredo. In:
Agostinho, A.A., Gomes, L.C. (Eds.), Reservatório de Segredo: bases ecológicas para o
manejo. Eduem, Maringá, pp. 163-182.
Yúfera, M., Darias, M.J., 2007. The onset of exogenous feeding in marine fish larvae.
Aquacult. 268, 53-63.
Zhang, L., Wang, Y.J., Hu, M.H., Fan, Q.X., Cheung, P.K.S., Shin, H., Cao, L., 2009. Effects
of the timing of initial feeding on growth and survival of spotted mandarin fish Siniperca
scherzeri larvae. J. Fish Biol. 75, 1158-1172.

31
CAPÍTULO II
SELETIVIDADE ALIMENTAR DE LARVAS DO JUNDIÁ Rhamdia
voulezi Haseman 1911: ALGUMAS IMPLICAÇÕES PARA O SEU
CULTIVO

32
SELETIVIDADE ALIMENTAR DE LARVAS DO JUNDIÁ Rhamdia
voulezi Haseman 1911: ALGUMAS IMPLICAÇÕES PARA O SEU
CULTIVO
RESUMO
A seletividade alimentar nos diferentes dias de vida, bem como o papel da boca na seleção de
presas foi avaliado. Para tanto, foram realizados 3 ensaios de seletividade alimentar nos
diferentes dias de vida após a eclosão (DAE). As larvas foram alimentadas com zooplâncton
coletados em tanques piscícolas. Houve variações nos padrões de seleção de presas nos
diferentes dias de vida analisados. Inicialmente no 5o
DAE, estágio de pré-flexão, as larvas
selecionaram fortemente os rotiferas (não identificado) e os cladoceras (Diaphanosoma
spinulosum, D. brevireme, Moina sp., M. micrura, M. minuta). Nos demais dias de vida
analisados, os cladoceras continuaram a ser fortemente selecionados. Entretanto no 8o DAE,
estágio de flexão, selecionaram menos intensamente os copepodas Argyrodiaptomus azevedoi
e Termocyclops decipiens e no 10o DAE, estágio de pós-flexão, mais fortemente
Argyrodiaptomus furcatus, Notodiaptomus cf. spinuliferus e Metacyclops mendocinus.
Concomitante ao seu crescimento, as larvas tendem a se especializar em determinados itens
presa, existindo uma seleção de presas tanto pelo tipo como também pelo tamanho, onde o
tamanho da presa consumida está relacionado com o tamanho da larva e o tamanho de sua
boca. Inicialmente as larvas têm preferência por itens presas menores, os rotíferos e
cladoceras, posteriormente, quando estão mais desenvolvidas, preferem alimentar-se de presas
maiores, os copepodas.
Palavras-chave: Alimentação, larvas de peixes seleção de presas, tamanho da boca.

33
FEEDING SELECTIVITY OF THE LARVAE JUNDIÁ Rhamdia voulezi Haseman
1911: SOME IMPLICATIONS FOR YOUR CULTURE
ABSTRACT
The feeding selectivity on different days of life, as well as the role of the mouth in prey
selection was evaluated. Thus, we performed 3 trials of feeding selectivity in different days
after hatching (DAH), comprising different larval stages and classes of standard length. The
larvae were fed zooplankton collected in fish ponds. There were variations in the patterns of
prey selection in different days of life analyzed. Initially on the 5th DAH, preflexion stage,
larvae strongly selected rotiferas (unidentified) and cladocerans (Diaphanosoma spinulosum,
Diaphanosoma brevireme, Moina sp., Moina micrura, Moina minuta). In the remaining days
of life examined, the cladocerans continued to be strongly selected. However on the 8th DAH,
flexion stage, they selected less intensely the copepods Argyrodiaptomus azevedoi and
Termocyclops decipiens and more strongly Argyrodiaptomus furcatus, Notodiaptomus cf.
spinuliferus and Metacyclops mendocinus at 10 DAH, postflexion stage. Concomitant to its
growth, the larvae tend to specialize in certain prey items. The size of prey consumed is
related to the size of the larvae and the gape size. Initially larvae have a preference for
cladocerans, and later when they are more developed prefer to feed on copepods.
Keywords: Feeding, larvae fish, prey selection, gape size.

34
1. Introdução
A maior parte da mortalidade em peixes de ambiente natural ou de condições de cultivo,
ocorre em suas primeiras fases de desenvolvimento, principalmente durante a transição da
alimentação endógena para a exógena (Kamler, 1992; Fuiman, 2002; Braaten et al., 2007;
Olivotto et al., 2010; Anto e Turingan, 2010). No entanto, a sobrevivência e o crescimento
nesta fase, estão diretamente relacionado com a quantidade do suprimento alimentar
endógeno e com a disponibilidade de presas zooplanctônicas adequadas à sua primeira
alimentação (Mercier et al., 2004; Makrakis et al., 2008).
O termo “adequadas” é porque muitas larvas de peixes não consomem
indiscriminadamente todos os recursos alimentares em sua volta. Estudos mostram que boa
parte das espécies mesmo em suas primeiras fases de desenvolvimento, apresenta um
comportamento seletivo na escolha do alimento, mantendo sua alimentação associada a tipos
e tamanhos específicos de presas (por exemplo, Sabates e Saiz, 2000; Nip et al., 2003;
Ostergaard et al., 2005; Makrakis et al., 2008; Anto et al., 2009; Morote et al., 2011).
Mas, nem sempre a preferência alimentar por determinados tipos de presas é constante
ao longo do período larval, podem variar simultaneamente com o desenvolvimento (Anneville
et al., 2011). Logo, presume-se que as limitações morfológicas, como tamanho da larva e
tamanho da boca influenciam diretamente na captura do alimento. Além disto, as interações
entre presa e predador também são levadas em consideração. Grande parte destas interações
são determinadas por processos como o número de encontro, ataque, captura, ingestão,
rejeição e escape (Fiksen e MacKenzie, 2002, Sarma et al., 2003).
A compreensão de todos os mecanismos que regem a tomada de alimento pelas larvas,
é essencial para entender alguns aspectos relacionados ao comportamento alimentar (Mikheev

35
e Wanzenböck, 2010), como também, colaborar com alguns “insights” para o seu cultivo,
principalmente no processo de gerir a transição da dieta entre os alimentos.
O jundiá Rhamdia voulezi pertence à família dos Heptapteridade, é uma espécie
nativa, de médio porte, encontrada apenas na bacia do rio Iguaçu (Bockmann e Guazzelli
2003, Baumgartner et al., 2012). O cultivo desta espécie está em fase inicial, pois há carência
de uma literatura especializada que contemple todos os aspectos biológicos e
comportamentais da mesma. Neste contexto, este estudo investigou o comportamento
alimentar das larvas desta espécie, buscando responder as seguintes questões: As larvas
selecionam o alimento ou capturam conforme sua abundância? Existe mudanças na
preferência alimentar durante o desenvolvimento larval? O tamanho do zooplâncton ingerido
acompanha o tamanho da larva e o incremento de sua boca?
2. Materiais e métodos
Este estudo foi conduzido no Instituto de Pesquisa em Aquicultura Ambiental (InPAA) da
Universidade Estadual do Oeste do Paraná, município de Toledo-PR, no mês de fevereiro de
2012. As larvas foram obtidas por meio de reprodução induzida utilizando reprodutores
capturados no rio Iguaçu e fornecidas pelo Centro de Difusão e Desenvolvimento
Tecnológico do rio Iguaçu (CDT), localizado no município de Boa Vista da Aparecida, PR.
Para avaliar a seletividade alimentar ao longo do período larval, foram realizados três
ensaios de seletividade alimentar com larvas em diferentes dias de vida após a eclosão (DAE,
contemplando os diferentes estágios de desenvolvimento/classes de tamanho. O primeiro
ensaio foi feito logo após a absorção completa do saco vitelínico, no 4o DAE, o segundo
ensaio foi realizado no 8o DAE e o terceiro no 10
o DAE, sendo conduzidos aleatoriamente em
4 repetições mais o controle que conteve somente zooplâncton. Para tanto, as larvas foram
previamente submetidas ao jejum num período de 24 horas. Posteriormente, estas foram

36
transferidas para recipientes plásticos com volume útil de 3L cada, dotados de sistema de
aeração constante numa densidade de 10 larvas/L. Estes recipientes foram cobertos por lona
preta, mantendo-se o ambiente interno escuro, sendo descobertos apenas para alimentação.
O alimento foi fornecido duas vezes ao dia, às 8h30min e às 16h30min, com
zooplâncton provenientes de tanques piscícolas. Estes tanques foram previamente calados
com CaCo3 (250 g/m²) e adubados com NPK 4-14-8 (200 g/m²) para promover o crescimento
do plâncton. A coleta do zooplâncton foi feita através de rede de plâncton de malhagem 68
µm. Posteriormente, era feita a homogeneização e em seguida, distribuídos na mesma
proporção para todos os aquários.
Ao final de cada ensaio, após 24 horas, as larvas eram coletadas, submetidas à
eutanásia, em solução contendo benzocaína (250 mg/L) e em seguida fixadas em formol 4%
para posterior análise em laboratório. O mesmo procedimento foi adotado para a coleta do
zooplâncton do ensaio controle.
Para controle dos parâmetros físico e químicos da água, foi realizado para cada
ensaio a leitura, às 16h30min, da temperatura, pH, oxigênio dissolvido e condutividade
elétrica através do multiparâmetro da marca YSI modelo professional Plus, mantendo-os
dentro do limite de tolerância conforme Kubtza (1999) e Sipaúba -Tavares (1994) para peixes
neotropicais.
As larvas coletadas foram classificadas em diferentes estágios de desenvolvimento:
pré-flexão; flexão e pós-flexão conforme Ahlstrom e Ball (1954) e Kendall et al. (1984).
Concomitante as classificações em estágios de desenvolvimento, foram realizadas as
medições do comprimento padrão (CP) e do comprimento da maxila superior, conforme a
metodologia de Shirota (1978), a fim de analisar o tamanho da boca (TB) das larvas. As
medições das mesmas foram feitas em um microscópio estereoscópico Olympus modelo

37
SZX7 equipado com câmera digital DP-25 com captura de imagem e software de medição
DP2-BSW.
Os organismos zooplanctônicos da dieta e do ambiente (ensaio controle) foram
identificados, mensurados e quantificados ao menor nível taxonômico possível com o auxílio
de Matsumura-Tundisi (1976), El-Moor Loureiro (1997), Silva (2003), Previattelli (2006),
Neves (2011) e Joko (2011) através de um microscópio óptico Olympus modelo CX21. Para a
análise do zooplâncton do ambiente foram retiradas três sub-amostras de 2,5mL e
posteriormente extrapolado para o volume total do ambiente (aquário de 3L).
Para a análise dos itens alimentares consumidos, foram analisadas 40 larvas por
ensaio, totalizando 120 larvas. Para larvas em estágio de pré-flexão e flexão analisou-se o
conteúdo total e aquelas em estágio de pós-flexão, apenas 2/3 do tubo digestório foi analisado
devido ao grau de digestão dos itens alimentares na sua porção final. As presas intactas tanto
da dieta quanto do ambiente foram medidas, considerando o comprimento total que inclui
espinhos e elmos.
2.1. Análise dos dados
Para caracterizar a seletividade alimentar pelas larvas através da composição taxonômica e
tamanho do zooplâncton disponível, foram obtidos valores de seletividade alimentar (Chesson
1978, 1983) através da seguinte fórmula:
m
1jjj
iii
n/r
n/r
,i = 1,...m
onde ri e ni são a proporção do tipo de presa i/tamanho i na dieta e no ambiente
respectivamente, e m corresponde ao número de tipos/tamanhos de presas disponíveis. Esse

38
índice varia de 0 a 1, onde valores de seletividade igual a 1/m indicam seleção neutra, valores
acima de 1/m indicam seleção positiva e valores inferiores a 1/m indicam seleção negativa.
Para determinar os valores de seletividade pelo tamanho da presa, os organismos
zooplanctônicos foram divididos em 8 classes de tamanho (incremento 0,19). E para calcular
a seletividade alimentar por tamanho de boca, as larvas foram separadas em quatro classes de
tamanho de boca com incremento de 0,2.
Para analisar se o tamanho do zooplâncton ingerido aumenta conforme o aumento do
tamanho da boca e idade larval foi realizado a MRPP (Procedimento de Permutação Multi-
resposta). Para isto, consideraram-se as diferentes classes de tamanho de boca (classe 1,2, 3 e
4) com os tamanhos de presas na dieta, e diferentes dias de vida (4o, 8o e 10o DAE) em
relação aos diferentes tamanhos de presas na dieta. Antes desta análise, as larvas foram
separadas em 4 classes de tamanho de boca com incremento de 0,2.
O teste de Mantel (Fortin e Gurevitch, 1993) foi usado para examinar se há associação
entre os itens consumidos com os disponíveis no ambiente. A matriz das classes de tamanho
presente na dieta com a matriz das classes de tamanho disponível no ambiente e a matriz das
espécies de zooplâncton ingerido com a matriz das espécies do zooplâncton disponível foi
correlacionada. Todas as análises foram conduzidas no PC-ORD software versão 5.0.
Com o objetivo de verificar se o aumento do tamanho da boca da larva de Rhamdia
voulezi foi significativo com a variação do comprimento padrão, foi feito uma análise de
regressão linear com dados logaritimizados.

39
3. Resultados
3.1. Seletividade alimentar taxonômica
Foi possível verificar diferenças nos padrões de seletividade alimentar nos diferentes dias de
vida das larvas (Fig. 1). As larvas do 4o DAE (4,42 a 5,48 mm de comprimento padrão-CP;
estágio de pré-flexão e início de flexão) selecionaram positivamente os cladoceras,
especialmente Diaphanosoma spinulosum (Dspi), Diaphanosoma brevireme (Dbr), Moina sp.
(Msp), Moina micrura (Mmc) e Moina minuta (Mmi) e também os rotiferas não identificados
(Rotni).
Para as larvas do 8o DAE (6,08 a 7,37 mm de comprimento padrão-CP; estágio de
flexão), observou-se uma forte seleção para os cladoceras: Diaphanosoma spinulosum (Dspi),
Diaphanosoma brevireme (Dbr) e Diaphanosoma birgei (Dbi) e para o copépoda
Metacyplops mendocinus (Mme). Os demais copepodas, Argyrodiaptomus azevedoi (Argz) e
Termocyclops decipiens (Tde), foram fracamente selecionados.
Já as larvas com 10o DAE (7 a 10 mm de comprimento padrão-CP; estágio em início
de pós-flexão e pós-flexão), selecionaram preferencialmente os copepodas da ordem
Calanoida, Argyrodiaptomus furcatus (Argf) e Notociaptomus cf. spinuliferus (Nots) e da
ordem dos Cyclopoidas, Metacyclops mendocinus (Mme). Os cladoceras Diaphanosoma
brevireme (Dbr) continuaram a fazer parte da seleção.
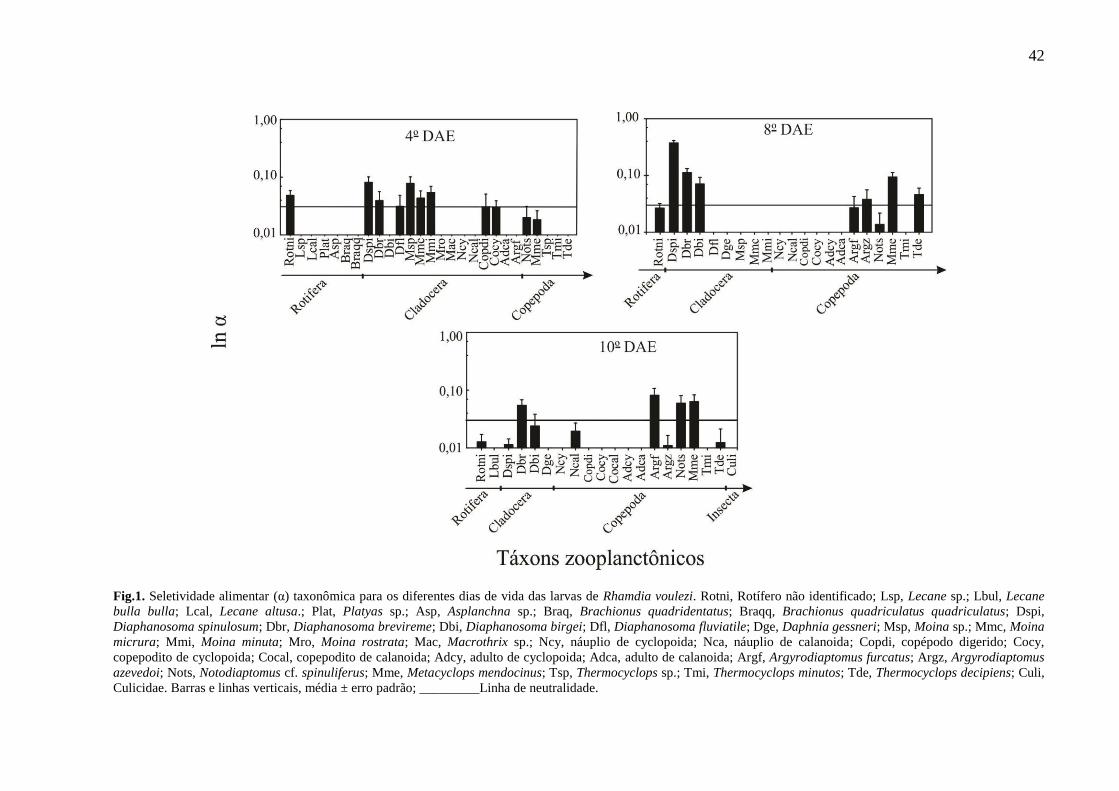
42
Fig.1. Seletividade alimentar (α) taxonômica para os diferentes dias de vida das larvas de Rhamdia voulezi. Rotni, Rotífero não identificado; Lsp, Lecane sp.; Lbul, Lecane
bulla bulla; Lcal, Lecane altusa.; Plat, Platyas sp.; Asp, Asplanchna sp.; Braq, Brachionus quadridentatus; Braqq, Brachionus quadriculatus quadriculatus; Dspi,
Diaphanosoma spinulosum; Dbr, Diaphanosoma brevireme; Dbi, Diaphanosoma birgei; Dfl, Diaphanosoma fluviatile; Dge, Daphnia gessneri; Msp, Moina sp.; Mmc, Moina
micrura; Mmi, Moina minuta; Mro, Moina rostrata; Mac, Macrothrix sp.; Ncy, náuplio de cyclopoida; Nca, náuplio de calanoida; Copdi, copépodo digerido; Cocy,
copepodito de cyclopoida; Cocal, copepodito de calanoida; Adcy, adulto de cyclopoida; Adca, adulto de calanoida; Argf, Argyrodiaptomus furcatus; Argz, Argyrodiaptomus
azevedoi; Nots, Notodiaptomus cf. spinuliferus; Mme, Metacyclops mendocinus; Tsp, Thermocyclops sp.; Tmi, Thermocyclops minutos; Tde, Thermocyclops decipiens; Culi,
Culicidae. Barras e linhas verticais, média ± erro padrão; _________Linha de neutralidade.

43
3.2. Seletividade por classe de tamanho do zooplâncton ao longo do período larval
A seletividade alimentar pelo tamanho do zooplâncton, diferiu entre os dias de vida (Fig. 2).
As larvas no 4o DAE selecionaram preferencialmente presas menores de 0,01 a 0,50 mm. As
larvas do 8o DAE, selecionaram fracamente presas de 30 e 70 mm e mais fortemente presas de
50mm de tamanho. As larvas maiores do 10o DAE, selecionaram presas de todas as classes de
tamanho, sendo positivamente somente para os tamanhos de 0,50, 0,70 e 1,30 mm.
Na MRPP, aplicada com o objetivo de testar tais diferenças de seletividade por
tamanho nos diferentes dias de vida após a eclosão (comparação por pares), as comparações
por pares evidenciaram que o tamanho das presas consumidas no 4o DAE foram
significativamente diferentes daquelas consumidas no 8o
e 10o DAE. Aquelas consumidas no
8o e 10
o DAE foram semelhantes. A Tabela 1 mostra os resultados da análise.
Tabela 1 Comparações pareadas do tamanho do zooplâncton
consumido para os diferentes grupos (dias de vida após a eclosão-
DAE).
Grupos
comparados
T
A
p (Monte Carlo)
1ax2
b -15,2119 0,0862 p<0,01
1ax3
c -8,8562 0,0448 p<0,01
2bx3
c 3,5224 0,0163 p<0,01
a 4o DAE;
b 8
o DAE;
c 10
o DAE.
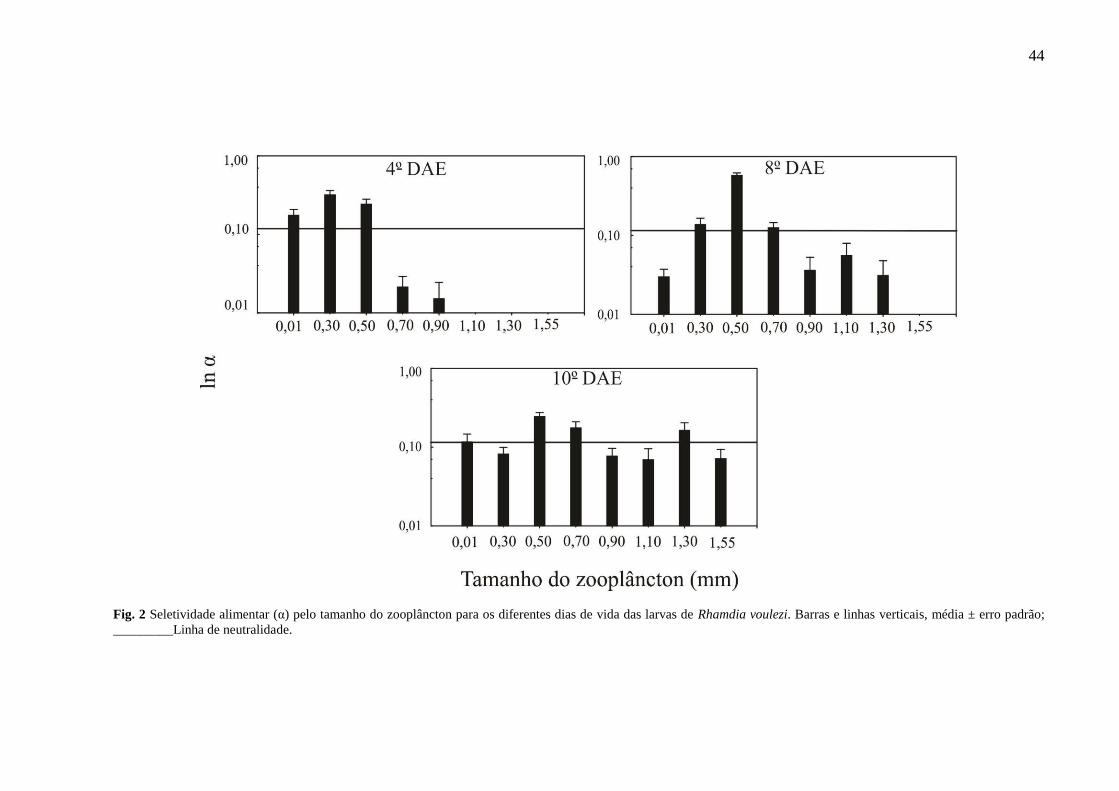
44
Fig. 2 Seletividade alimentar (α) pelo tamanho do zooplâncton para os diferentes dias de vida das larvas de Rhamdia voulezi. Barras e linhas verticais, média ± erro padrão;
_________Linha de neutralidade.
46
O teste de Mantel aplicado para comparar as presas consumidas com as que estavam
disponíveis no ambiente revelou que não há uma relação entre estas variáveis tanto para as
espécies (Mantel teste: N= 114, r= 0,026, Z= 0,184, p<0,05) como para as classes de tamanho
(Mantel teste: N=113, r= 0,026, Z= 0,17, p<0,5). Isto reforça a hipótese de que as larvas
selecionaram o alimento conforme a composição e tamanho do mesmo.
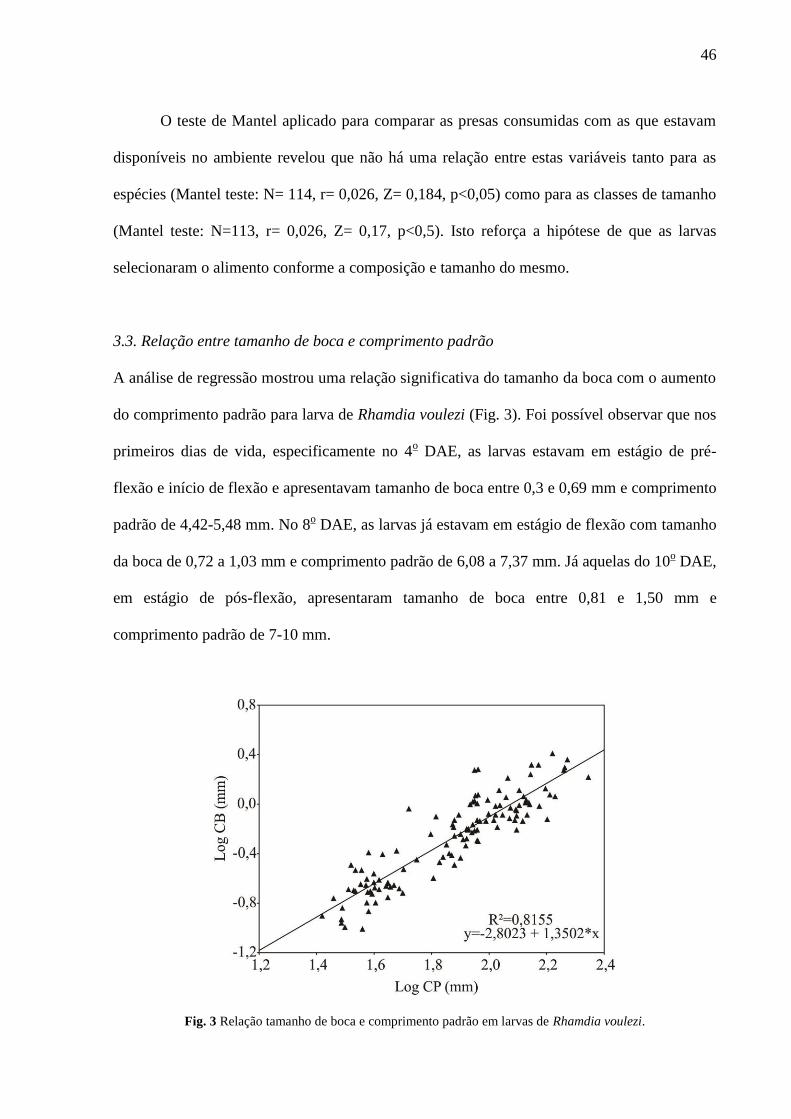
3.3. Relação entre tamanho de boca e comprimento padrão
A análise de regressão mostrou uma relação significativa do tamanho da boca com o aumento
do comprimento padrão para larva de Rhamdia voulezi (Fig. 3). Foi possível observar que nos
primeiros dias de vida, especificamente no 4o DAE, as larvas estavam em estágio de pré-
flexão e início de flexão e apresentavam tamanho de boca entre 0,3 e 0,69 mm e comprimento
padrão de 4,42-5,48 mm. No 8o DAE, as larvas já estavam em estágio de flexão com tamanho
da boca de 0,72 a 1,03 mm e comprimento padrão de 6,08 a 7,37 mm. Já aquelas do 10o DAE,
em estágio de pós-flexão, apresentaram tamanho de boca entre 0,81 e 1,50 mm e
comprimento padrão de 7-10 mm.
Fig. 3 Relação tamanho de boca e comprimento padrão em larvas de Rhamdia voulezi.
47
3.3. Tamanho da presa x tamanho de boca do predador
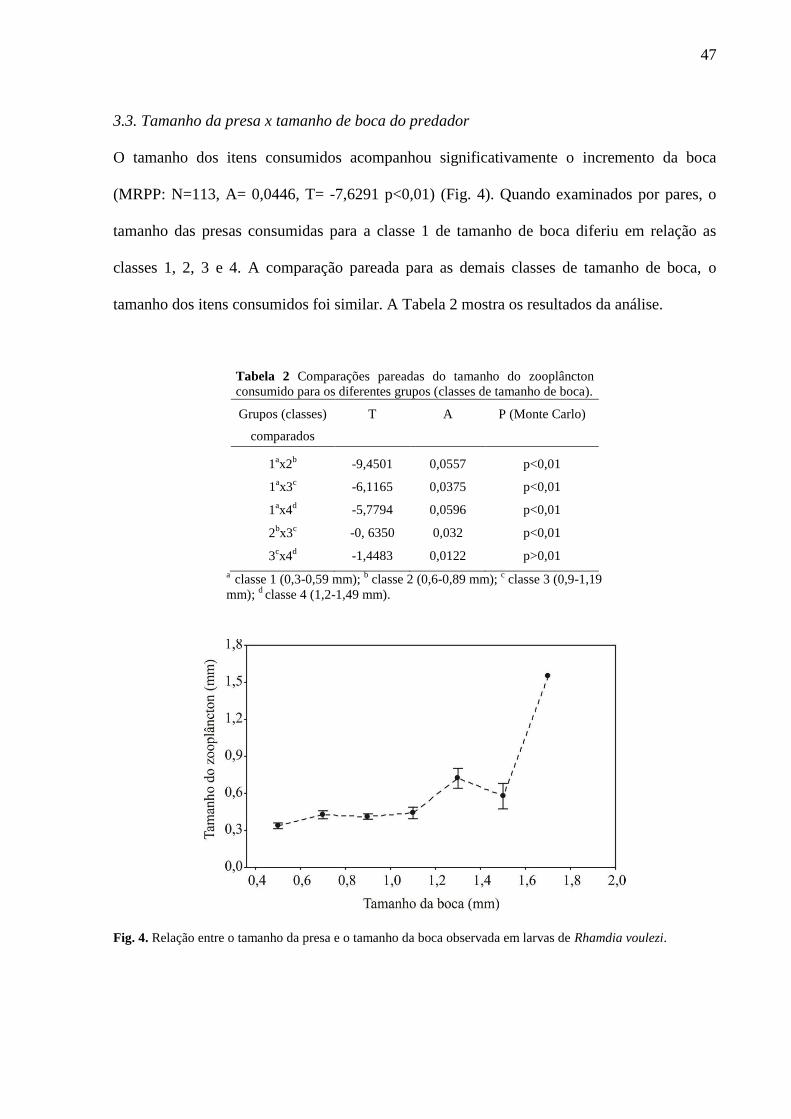
O tamanho dos itens consumidos acompanhou significativamente o incremento da boca
(MRPP: N=113, A= 0,0446, T= -7,6291 p<0,01) (Fig. 4). Quando examinados por pares, o
tamanho das presas consumidas para a classe 1 de tamanho de boca diferiu em relação as
classes 1, 2, 3 e 4. A comparação pareada para as demais classes de tamanho de boca, o
tamanho dos itens consumidos foi similar. A Tabela 2 mostra os resultados da análise.
Tabela 2 Comparações pareadas do tamanho do zooplâncton
consumido para os diferentes grupos (classes de tamanho de boca).
Grupos (classes)
comparados
T
A
P (Monte Carlo)
1ax2
b -9,4501 0,0557 p<0,01
1ax3
c -6,1165 0,0375 p<0,01
1ax4
d -5,7794 0,0596 p<0,01
2bx3
c -0, 6350 0,032 p<0,01
3cx4
d -1,4483 0,0122 p>0,01
a classe 1 (0,3-0,59 mm); b classe 2 (0,6-0,89 mm);
c classe 3 (0,9-1,19
mm); d classe 4 (1,2-1,49 mm).
Fig. 4. Relação entre o tamanho da presa e o tamanho da boca observada em larvas de Rhamdia voulezi.
48
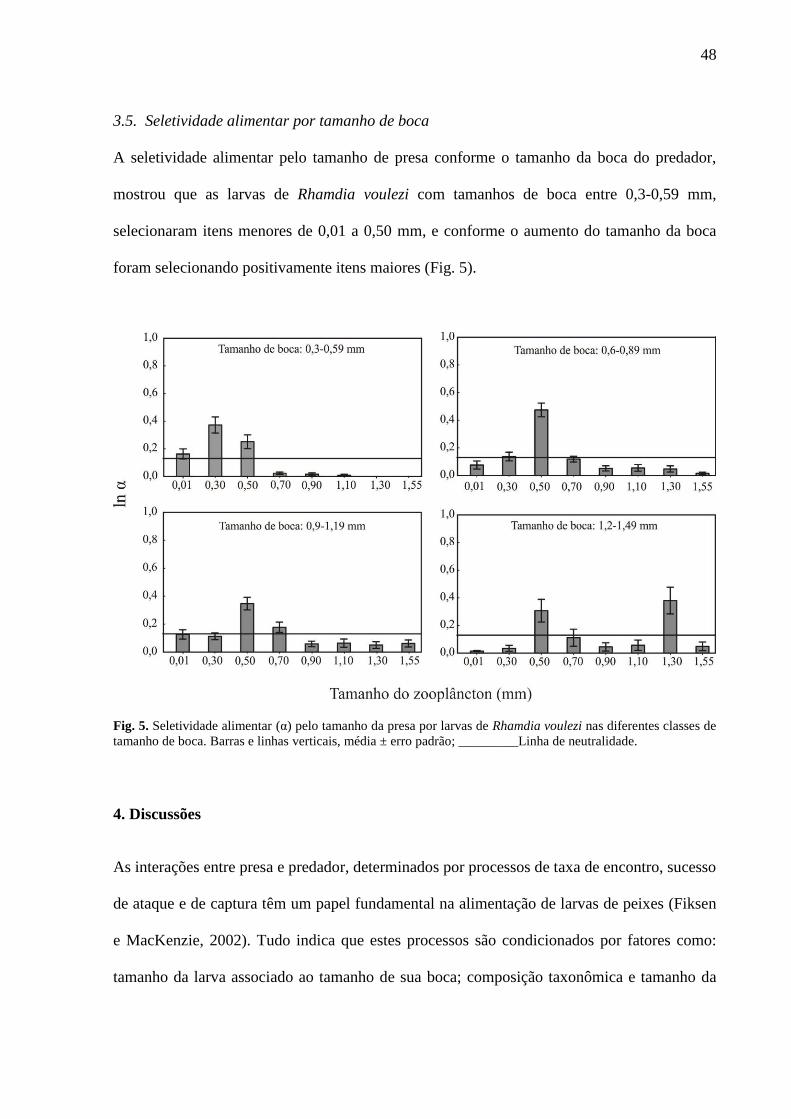
3.5. Seletividade alimentar por tamanho de boca
A seletividade alimentar pelo tamanho de presa conforme o tamanho da boca do predador,
mostrou que as larvas de Rhamdia voulezi com tamanhos de boca entre 0,3-0,59 mm,
selecionaram itens menores de 0,01 a 0,50 mm, e conforme o aumento do tamanho da boca
foram selecionando positivamente itens maiores (Fig. 5).
Fig. 5. Seletividade alimentar (α) pelo tamanho da presa por larvas de Rhamdia voulezi nas diferentes classes de
tamanho de boca. Barras e linhas verticais, média ± erro padrão; _________Linha de neutralidade.
4. Discussões
As interações entre presa e predador, determinados por processos de taxa de encontro, sucesso
de ataque e de captura têm um papel fundamental na alimentação de larvas de peixes (Fiksen
e MacKenzie, 2002). Tudo indica que estes processos são condicionados por fatores como:
tamanho da larva associado ao tamanho de sua boca; composição taxonômica e tamanho da

49
presa, e tais fatores se mostraram determinantes nos padrões de seletividade alimentar pelas
larvas de Rhamdia voulezi.
As larvas em geral, no início da alimentação exógena, apresentam-se pouco
desenvolvidas, com tamanho de boca limitado e sistema digestivo pouco funcional no qual
dificulta a sua alimentação (Wittenrich et al., 2009; Anto et al., 2009; Anto e Turigan, 2010).
Contudo, mesmo limitadas tanto morfologicamente quanto fisiologicamente, as larvas,
conseguem escolher ativamente as presas que preferem se alimentar (Nip et al., 2003;
Ostergaard et al., 2005; Islam e Tanaka, 2006; Anto et al., 2009; Russo et al., 2009).
Entretanto, a escolha pelo alimento é influenciada por diversos fatores que vão desde
os próprios atributos da larva até as condições do ambiente em que vivem. Dentre estes
fatores a preferência alimentar da espécie, grau de desenvolvimento e disponibilidade de
alimento no ambiente tem sido amplamente relatados em outros estudos com larvas, como por
exemplo os de: Shaw et al., 2003; Nunn et al., 2007; Russo et al., 2009; Makrakis et al., 2005;
Hunter, 1981; Sánchez-Velasco, 1998; Mikheev e Pavlov, 2003.
Para Nunn et al. (2007) a disponibilidade de alimento é considerada um fator chave
que determina o padrão da alimentação de larvas. Portanto, deve-se levar em consideração
que o forrageamento larval não depende somente da abundância do alimento, mas também da
qualidade e distribuição do tamanho disponível (Soares, 2003; Makrakis et al., 2008). Uma
das razões, pode ser decorrentes de suas limitações morfológicas, assim como, da estratégia
alimentar da espécie. Estudos mostram que larvas de determinadas espécies, não utilizam todo
o alimento em sua volta, mas, mantem sua preferência associada a tipos e tamanhos
específicos de presas (Checkley, 1982; Govoni et al., 1986; Scharf et al., 2000). Porém, em
outros casos, há espécies (larvas) que apresentam um comportamento mais generalista,
mudando sua preferência conforme a disponibilidade do alimento (Scharf et al., 2000). Como
exemplo, um estudo feito por Makrakis et al., 2008, constatou que larvas Hypophthalmus

50
edentatus alimentam-se de itens presas mais abundantes no ambiente, ao passo que larvas de
Plagioscion squamosissimus e de Iheringichthys ilabrosus mantem sua preferência alimentar
associada a tipos específicos de presas. Neste estudo, a comparação do alimento consumido
com o que estava disponível mostrou que estas duas variáveis não estiveram associadas,
indicando que as larvas de Rhamdia voulezi selecionaram o alimento independente da sua
disponibilidade.
De acordo com Morote et al. (2008) as estratégias alimentares das larvas estão
associadas, além da disponibilidade de alimento, com o grau de desenvolvimento das
mesmas. Não obstante desta hipótese, neste estudo, o padrão de seletividade alimentar variou
gradativamente ao longo de todo o período larval. Os rotiferas e os cladoceras foram as
principais presas selecionadas pelas larvas mais novas do 4o DAE, entretanto, à medida que
foram crescendo, estenderam sua preferência pelos copepodas, principalmente no 10o DAE.
Ribeiro e Nuñer (2008) também verificaram algumas variações na seleção de presas por
larvas de Salminus brasiliensis no decorrer dos dias. A preferência por cladoceras foram
registrados até o 11o DAE, a partir daí, selecionaram preferencialmente larvas de insetos. Para
larvas de Sorubim lima, Rossi (2001) constatou que inicialmente, os rotiferas foram os
principais componentes da dieta, e, à medida que foram crescendo predaram seletivamente os
cladoceras e também as larvas de insetos. Sabatés et al. (2003) ao estudar o padrão de
alimentação de larvas de duas espécies marinhas, constatou mudanças no padrão alimentar ao
longo do desenvolvimento larval, como também verificou diferenças na estratégia de
forrageamento entre as espécies estudadas.
Variações na utilização dos recursos ainda nas primeiras fases de desenvolvimento dos
peixes foram constatados em outros estudos como os de Rossi, 2001; King, 2005; Makrakis et
al., 2005; Tonkin et al., 2006; Anneville et al., 2011; Robert et al., 2011; Santim, 2012.
Provavelmente estas variações podem ser coincidentes com o desenvolvimento da larva,

51
associado com o melhoramento na visão, habilidade natatória (Schiemer, 1989; Bremigan e
Stain, 1994; Machácek e Matena, 1997; Sabatés et al., 2003; Ibrahim et al., 2006; Makrakis et
al., 2008; Nunn et al., 2012) e desenvolvimento do aparelho de alimentação (Turingan et al.,
2005, Makrakis et al., 2005, Beck e Turingan, 2007; Anto et al., 2009).
Entretanto, à medida que as larvas crescem suas limitações morfológicas diminuem e
consequentemente são capazes de explorarem outros tipos de alimento (Santim, 2012). Isto
justifica o fato das larvas de jundiá terem transacionado sua alimentação para copepodas a
partir do 8o DAE.
Makrakis et al. (2005) ao estudar a alimentação inicial de Plagiosciom
squamosissimus, associou o consumo de copepodas, com a maior acuidade visual, capacidade
natatória e protração da boca da larva. O consumo de presas maiores como os copepodas de
calanoida (Argyrodiaptomus azevedoi, Argyrodiaptomus furcatus e Notodiaptomus cf.
spinuliferus), além de estar relacionado com o maior desenvolvimento da larva, pode vir da
suposição de que as mesmas tentam maximizar o ganho de energia disponível a fim de
compensar os gastos de energia com ataques, captura, ingestão e manipulação do alimento
(Beyer, 1980; Pyke, 1984; Adriaens et al., 2001; Nunn et al., 2012). Para Zaret (1980), os
copepodas em geral, são realmente difíceis de serem capturados porque possuem movimentos
erráticos de natação, ao passo que os rotíferos e cladóceros são mais vulneráveis, pois sua
velocidade de natação é menor (Zaret, 1980; Epp e Lewis, 1984). Além do movimento mais
lento, os olhos contrastantes dos cladóceros aumentam sua visibilidade o que pode torná-los
ainda mais vulneráveis aos predadores. Sendo assim, se levar-se em consideração as
características destes organismos, explica-se o fato das larvas mais velhas e mais
desenvolvidas terem continuado a selecionar cladoceras, mesmo sendo capazes de capturar
outros itens (como, por exemplo, os copepodas). Para Drenner et al. (1978 e 1986) e Mayer e

52
Wahl (1997) a preferência alimentar das larvas de peixes geralmente está associado a
determinados tamanhos e tipos específicos de presas e preferem cladócera à copépoda.
Neste estudo o tamanho da larva associado ao tamanho da boca dominou os padrões
de seleção. Zavala-Camin (1996) ressalta que o tamanho da presa tem relação direta com o
tamanho do predador, e que como a maioria dos peixes ingerem suas presas inteiras, existe
um tamanho máximo de presa em relação ao tamanho da boca do predador. O tamanho dos
itens ingeridos por larvas de Rhamdia voulezi aumentou conforme o crescimento das mesmas.
A relação positiva entre tamanho de presa e tamanho da larva de peixes também foi
constatado em outros trabalhos como, por exemplo, Govoni et al. (1983); Kellermann (1990);
Sabate e Saiz (2000). Já com relação ao tamanho de boca, o aumento no tamanho de presas
ingeridas acompanhou o incremento da boca. Resultado semelhante foi verificado por
Makrakis et al. (2008) para larvas de P. squamosissimus, I. labrosus e H. edentatus. Assim,
estes resultados reforçam a hipótese de que para algumas espécies de peixes, o tamanho da
boca da larva muitas vezes limita o tamanho da presa a ser consumida (Gerking, 1994; Rao,
2003; Makrakis et al., 2008; Anto e Turingan, 2010; Nunn et al., 2012).
Em conclusão, os resultados obtidos mostraram que as larvas de Rhamdia voulezi
tendem a se especializar em determinados itens presa ao longo do desenvolvimento larval
(Fig. 6). Inicialmente as larvas têm preferência por itens presas menores, os rotíferos e
cladoceras, e, à medida que foram crescendo, estenderam sua preferência por presas maiores,
os copepodas. Variações no tamanho do zooplâncton ingerido em relação ao tamanho da larva
e de sua boca indicam que estes fatores são determinantes na alimentação inicial desta
espécie. Considerando que a sobrevivência e o crescimento das larvas estão diretamente
relacionados à eficiência na primeira alimentação, tais informações podem ser importantes
para a larvicultura das mesmas, principalmente no processo de gerir a transição entre os
alimentos ao longo do período larval.
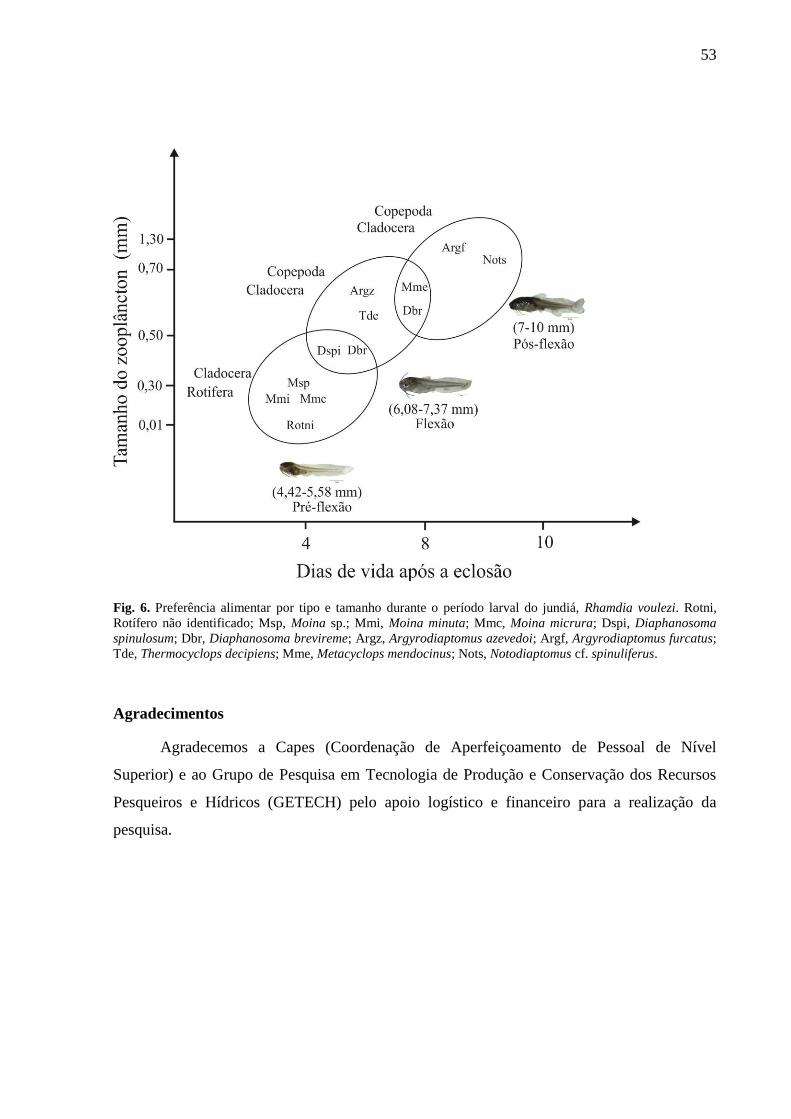
53
Fig. 6. Preferência alimentar por tipo e tamanho durante o período larval do jundiá, Rhamdia voulezi. Rotni,
Rotífero não identificado; Msp, Moina sp.; Mmi, Moina minuta; Mmc, Moina micrura; Dspi, Diaphanosoma
spinulosum; Dbr, Diaphanosoma brevireme; Argz, Argyrodiaptomus azevedoi; Argf, Argyrodiaptomus furcatus;
Tde, Thermocyclops decipiens; Mme, Metacyclops mendocinus; Nots, Notodiaptomus cf. spinuliferus.
Agradecimentos
Agradecemos a Capes (Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior) e ao Grupo de Pesquisa em Tecnologia de Produção e Conservação dos Recursos
Pesqueiros e Hídricos (GETECH) pelo apoio logístico e financeiro para a realização da
pesquisa.

54
Referências
Adriaens, D., Aerts P., Verraes, W., 2001. Ontogenetic shift in mouth opening mechanisms
in catfish (Clariidae, Siluriformes): A response to increasing functional demands. J.
Morphol. 247, 197-216.
Alhstrom, E.H., Ball, O.P., 1954. Description of eggs and larvae of jack mackerel (Trachurus
symetricus) and distribuition and abundance of larvae in 1950 and 1951. Fish. B-NOAA. 56,
209-245.
Anneville, O., Berthon, V., Glippa, O., Mahjoub, M.S., Molineiro, J.C., Souissi, S., 2011.
Ontogenetic dietary changes of whitefish larvae: insights from field and experimental
observations. Environ Biol Fishes. 91, 27-38.
Anto J., Majoris J., Turingan R.G., 2009. Prey selection and functional morphology through
ontogeny of Amphiprion clarkii with a congeneric comparisson. J. Fish Biol. 75, 575-590.
Anto, J., Turingan, R.G., 2010. Relating the Ontogeny of Functional Morphology and Prey
Selection With Larval Mortality in Amphiprion frenatus. J. Morphol. 271, 682-696.
Baumgartner, G., Pavanelli, C.S., Baumgartner, D., Bifi, A.G., Debona, T., Frana, V.A., 2012.
Peixes do Baixo rio Iguaçu, Primeira ed. Eduem, Maringá.
Beck, J.L., Turingan, R.G., 2007. The effects of zooplankton swimming behavior on prey-
capture kinematics of red drum larvae, Sciaenops ocellatus. Mar. Biol. 151,1463-1470.
Beyer, J.E., 1980. Feeding success of clupeoid fish larvae and stochastic thinking. Dana. 1,
65-91.
Boockmann, F.A., Guazzelli, G.M., 2003. Family Heptapteridae. In: Reis, R.E., Kullander,
S.O., Ferraris Junior, C.J. (Eds.), Check list of the freshwater fishes of south and Central
America, EDIPUCRS, Porto Alegre, pp. 406-331.
Braaten, P.J., Fuller, D.B., McClenning, N.D., 2007. Diet composition of larval and young-
of-year shovelnose sturgeon in the Upper Missouri River. J. Appl. Ichthyol. 23, 516-520.
Bremigan, M.T., Stain, R.A., 1994. Gape-dependent larval foraging and zooplankton size:
implications for fish recruitment across systems. Can. J. Fish Aquat. Sci. 51, 913-922.
Checkley, D.M., 1982. Selective feeding by Atlantic herring (Clupea harengus) larvae on
zooplankton in natural assemblages. Mar. Ecol. Prog. Ser. 9, 245-253.
Chesson, J., 1978. Measuring preference in selective predation. Ecol. 59, 211-215.
Chesson, J., 1983. The estimation and analisys of preference and its relationships to foraging
models. Ecol. 64, 1297-1304.
Drenner, R.W., Strickler, J.R., O’Brien, W.J., 1978. Capture probability: the role of
zooplankton escape in the selective feeding of planktivorous fish. J. Fish Res. Board Can. 35,
1370-1373.

55
Drenner, R.W., Threlkled, S.T., McCracken, M.D., 1986. Experimental analysis of direct and
indirect effects of clupeid on plankton community structure. Can. J. Fish Aquat. Sci. 43,
1935-1945.
El-Moor, Loureiro L., 1997. Manual de identificação de cladóceros límnicos do Brasil.
Primeira ed. Editora Universa, UCB, Brasília.
Epp, R.W., Lewis, W.M.Jr., 1984. Cost and speed of locomotion for rotifers. Oecol. 61, 289-
292.
Fiksen, Ø., Mackenzie, B.R., 2002. Process-based models of feeding and prey selection in
larval fish. Mar. Ecol. Prog. Ser. 243, 151-164.
Fortin, M., Gurevitch, J., 1993. Mantel tests: spatial structure in field experiments. In:
Scheiner, S.M., Gurevitch (eds.), Design and Analysis of Ecological Experiments, Chapman
& Hall, New York. pp. 342-359.
Fuiman, L.A., 2002. Special considerations of fish eggs and larvae. In: Fuiman, L.A,
Werner, R.G (Eds.), The unique contributions of early life stages. Fishery science, Blackwell
Science, Oxford. pp. 1-32.
Gerking, S.D., 1994. Feeding Ecology of Fish. Academic Press, San Diego, London.
Gomes, L.C., Golombieski, J.I., Gomes, A.R.C., Baldisserotto B., 2000. Biologia do jundiá
Rhamdia quelen (Teleostei, Pimelodidae). Ciênc. Rural. 30, 179-185.
Govoni, J.J., Hoss, D.E., Chester, A.J., 1983. Comparative feeding of three species of larval
fishes in the northern Gulf of Mexico: Brevoortia patronus, Leiostomus xanthurus, and
Micropogonias undulatus. Mar. Ecol. Prog. Ser. 12, 189-199.
Govoni, J.J., Ortner, P.B., Al-Yamani, F., Hill, L.C., 1986. Selective feeding of spot,
Leiostomus xanthurus, and Atlantic croaker, Micropogonias.undulatus, larvae in the northern
Gulf of Mexico. Mar. Ecol. Prog. Ser. 28,175-183.
Hunter, J.R., 1981. Feeding ecology and predation of marine fish larvae. In: Lasker, R. (Ed.),
Marine Fish Larvae: Morphology, Ecology and Relation to Fisheries. University of
Washington Press, Seattle, pp. 34-77.
Ibrahim, F.S., Rana, K.J., Goddard, J.S., Alamri, I.S., 2006. Morphological development
of post hatch larval goldlined sea bream Rhabdosargus sarba (Forskal, 1775). Aquacult. Res.
37, 1156-1164.
Islam, M.S., Tanaka, M., 2006. Ontogenetic dietary shift of Japenese sea bass during larva -
juvenile transition in Ariake Bay. Mar. Ecol. Prog. Ser. 323, 305-310.
Joko, C.Y., 2011. Taxonomia de rotíferos monogonontas da planície de inundação do alto rio
Paraná (MS/PR). Tese (Doutorado), Programa de Pós Graduação em Ecologia de Ambientes
Aquáticos e Continentais, UEM, Maringá, 187 pp.
Kamler, E., 1992. Early life history of fish: an energetics approach. Chapman e Hall, London.

56
Kellermann, A., 1990. Food and feeding dynamics of the larval Antarctic fish Nototheniops
larseni. Mar. Biol. 106,159-167.
Kendall, A.W.Jr., Alhstrom, E.H., Moser, H.G., 1984. Early life history stages of fishes and
their characters. Special Publication of the American Society of the Ichthyologists and
Hepertologists. 1, 11-22.
King, A.J., 2005. Ontogenetic dietary shifts of fishes in an Australian floodplain river. Mar
Freshwat. Res. 56, 215-225.
Machácek, J., Matena, J., 1997. Diurnal feeding patterns of age-0 perch (Perca fluviatilis) and
roach (Rutilus rutilus) in a steep-sided reservoir. Arch. Hydrobiol. 49, 59-70.
Makrakis, M.C., Nakatani, K., Bialetzki, A., Sanches, P.V., Baumgartner, G., Gomes, L.C.,
2005. Ontogenetic shifts in digestive tract morphology and diet of fish larvae of the Itaipu
Reservoir, Brazil. Environ. Biol. Fishes. 72, 99-107.
Makrakis, M.C., Nakatani, K., Bialetzki, A., Gomes, L.C., Sanches, P.V., Baumgartner, G.,
2008. Relationship between gape size and feeding selectivity of fish larvae from a Neotropical
Reservoir. J. Fish Biol. 72, 1690-1707.
Mayer, M., Wahl, D.H., 1997. The relationship between prey selectivity and growth and
survival in a larval fish. Can. J. Fish Aquat. Sci. 54, 1504-1512.
Matsumura-Tundisi, T., 1976. Latitudinal distribution of Calanoid Copepods in freshwater
aquatic systems of Brazil. Rev. Bras. Biol. 46, 527-553.
Mercier, L., Audet, C., Noue, J., Parent, B., Parrish, C.C., Ross, N.W., 2004. First feeding of
winter flounder (Pseudopleuronectes americanus) larvae: use of Brachionus plicatilis
acclimated at low temperatures as live prey. Aquacult. 229, 361-376.
Mikheev, V.N., Pavlov, D.S., 2003. Nonlethal Interspecific Interactions among Foraging Fish
and the Concept of “Triotroph”. J. Ichthyol. 43, S151-S167.
Mikheev, V.N., Wanzenböck, J., 2010. Diet changes in prey size selectivity in larvae of perch
Perca fluviatilis. J. Ichthyol. 50, 1014-1020.
Morote, E., Olivar, M.P., Pankhurst, P.M., Villate, F., Uriarte, I., 2008. Trophic ecology of
bullet tuna Auxis rocheilarvae and ontogeny of feeding-related organs. Mar. Ecol. Prog. Ser.
353, 243-254.
Morote, E., Olivar, M.P., Bozzano, A., Villate, F., Uriarte, I., 2011. Feeding selectivity in
larvae of the European hake (Merluccius merluccius) in relation to ontogeny and visual
capabilities. Mar. Biol. 158, 1349-1361.
Neves, G.P., 2011. Copépodes planctônicos (Crustacea, Calanoida e Cyclopoida) em
reservatórios e trechos lóticos da bacia do Rio da Prata (Brasil, Paraguai, Argentina e
Uruguai): taxonomia, distribuição geográfica e alguns atributos ecológicos. Tese (Doutorado),
Instituto de Biociências de Botucatu, Unesp, São Paulo, 238 pp.

57
Nip, T.H.M., Ho, W.Y., Wong, K., 2003. Feeding ecology of larval and juvenile black
seabream (Acanthopagrus schlegeli) and Japanese seaperch (Lateolabrax japonicus) in Tolo
Harbour, Hong Kong. Environ. Biol. Fish. 66, 197-209.
Nunn, A.D., Harvey, J.P., Cowx, I.G., 2007. The food and feeding relationships of larval and
0+year juvenile fishes in lowland rivers and connected waterbodies. II. Prey selection and the
influence of gape. J. Fish Biol. 70, 743-757.
Nunn, A.D., Tewson, L.H., Cowx, I.G., 2012. The foraging ecology of larval and juvenile
fishes. Rev. Fish Biol. Fisher. 22, 377-408.
Olivotto, I., Piccinetti, C.C., Avelha, M.A., Rubio, C.M., Carnevali, O., 2010. Feeding
strategies for striped blenny Meiacanthus grammistes larvae. Aquacult. Res. 41, e307-e315.
Ostergaard, P., Munk, P., Janekarn, V., 2005. Contrasting feeding patterns among species of
fish larvae from the tropical Andaman Sea. Mar. Biol. 146, 595-606.
Paul, A. J., 1983. Light, temperature, nauplii concentrations, and prey capture by first feeding
pollock larvae Theragra chalcogramma. Mar. Ecol. Prog. Ser. 13, 175-179.
Paula-Souza, G., 1978. Reprodução de Rhamdia branneri, 1911 (Pisces, Siluriformes) e suas
relações com fatores abióticos. Dissertação (mestrado), Pós-graduação em Zoologia, UFPR.
Porter, S.M., Bailey, K.M., 2007. Optimization of feeding and growth conditions for walleye
pollock Theragra chalcogramma (Pallas) larvae reared in the laboratory. AFSC Processed
Rep. 2007-06, 20 p. Alaska Fish. Sci. Cent., NOAA, Natl.Mar, Fish. Serv.,7600 Sand Point
Way NE, Seattle WA 98115.
Previattelli, D., 2006. Sistemática do gênero Argyrodiaptomus Brehm, 1933 (Crustacea:
Copepoda: Diaptomae). Dissertação (Mestrado), Programa de Pós-Graduação em Biologia
Tropical e Recursos Naturais. Instituto Nacional de Pesquisas da Amazônia, UFAM, Manaus.
Pyke, G.H., 1984. Optimal foraging theory: a critical review. Rev. Ecol. Evol. Systemat. 15,
523-575.
Rao, T.R., 2003. Ecological and ethological perspectives in larval fish feeding. J. Appl.
Aquacult. 13, 145-178.
Ribeiro, D.F.O., Nuñer, A.P.O., 2008. Feed preferences of Salminus brasiliensis (Pisces,
Characidae) larvae in fish ponds. Aquacult. 274, 65-71.
Robert, D., Levesque, K., Gagné, J.A., Fortier, L., 2011. Change in prey selectivity during the
larval life of Atlantic cod in southern Gulf of Lawrence. J. Plankton Res. 33, 195-200.
Rossi, L.M., 2001. Ontogenetic diet shifts in a neotropical catfish, Sorubim lima (Schneider)
from the River Paraná System. Fish. Manag. Ecol. 8, 141-152.
Russo, T., Boglione, C., Marzi, P., Cataudella, S., 2009. Feeding preferences of the dusky
grouper (Epinephelus marginatus, Lowe 1834) larvae reared in semi-intensive conditions: a
contribution addressing the domestication of this species. Aquacult. 289, 289-296.

58
Sabatés, A., Saiz, E., 2000. Intra and interspecific variability in prey size and niche breadth of
myctophiform fish larvae. Mar. Ecol. Prog. Ser. 201, 261-271.
Sabatés, A., Bozzano, A., Vallvey, I., 2003. Feeding pattern and the visual light environment
in myctophid fish larvae. J. Fish Biol. 63, 1476-1490.
Scharf, F.S., Juanes, F., Rountree , R.A., 2000. Predator size–prey size relationships of marine
fish predators: interspecific variation and effects of ontogeny and body size on trophic-niche
breadth. Mar. Ecol. Prog. Ser. 208, 229-248.
Schiemer, F., Keckeis, H., Wanzenböck, J., 1989. Foraging in cyprinids during early
development. Pol. Arch. Hydrobiol. 36, 467- 474.
Sánchez-Velasco, L., 1998. Diet composition and feeding habits of fish larvae of two
coccurring species (Pisces: Callionymidae and Bothidae) in the North-western Mediterranean.
J. Marine Sci. 55, 299-308.
Santin, M., 2012. Ontogenia e seletividade alimentar de larvas e juvenis de três espécies de
peixes Neotropicais de diferentes categorias tróficas. Tese (doutorado), Uem, Maringá,
Paraná.
Sarma, S.S.S., López-Rómulo, J.A., Nandini, S., 2003. Larval feeding behaviour of blind fish
Astyanax fasciatus (Characidae), black tetra Gymnocoryabus tennetzi (Characidae) and angel
fish Pterophyllum scalare (Cichlidae) fed zooplankton. Hydrol. 510, 207-216.
Shaw, G.W., Pankhurst, P.M., Purser, G.J., 2003. Prey selection by greenback Flounder
Rhombosolea tapirina (Günther) larvae. Aquacult. 228, 249-265.
Shirota, A., 1978. Studies on the mouth size of fish larvae. II: Specific characteristics of the
upper jaw length. B. Jpn. Soc. Sci. Fish. 44, 1171-1177.
Silva, W.M., 2003. Diversidade dos Cyclopoida (Copepoda, Crustacea) de água doce do
Estado de São Paulo: taxonomia, ecologia e genética. Tese (doutorado), Centro de Ciências
Biológicas e da Saúde, UFSCAR, São Paulo.
Silva, W.M., 2003. Diversidade dos Cyclopoida (Copepoda, Crustacea) de água doce do
Estado de São Paulo: taxonomia, ecologia e genética. Tese (doutorado), Centro de Ciências
Biológicas e da Saúde, UFSCAR, São Paulo.
Soares, C.M., 2003. Relações ecológicas de quatro espécies de peixes nativos em fase de
desenvolvimento inicial: alterações nas comunidades, alimentação e tamanho da boca. Tese
(doutorado), UEM, Maringá, Paraná.
Tonkin, Z.D., Humphries, P., Pridmore, P.A., 2006. Ontogeny of feeding in two native and
one alien fish species from the Murray-Darling Basin, Australia. J. fish Biol. 76, 303-315.
Turingan, R.G., Beck, J.L., Krebs, J.M., Licamele, J.D., 2005. Development of feeding
mechanics in marine fish larvae and the swimming behavior of zooplankton prey:
Implications for rearing marine fishes. In: Lee, C.S., O’Bryen, P.J., Marcus, N.H. (Eds.),
Copepods in Aquaculture. Blackwell Publishing, Ames, Lowa, pp.119-132.

59
Zaret, T.M., 1980. Predation and Freshwater Communities. Yale University Press, New
Haven.
Zavala-Camin, L.A. 1996. Introdução aos estudos sobre alimentação natural em peixes.
EDUEM, Maringá.