variação temporal de aves insetívoras que utilizam o sub ... · toda vez que eu tinha viagem de...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DE MONTES CLAROS – UNIMONTES
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
Karen Mirele Caldeira
Variação temporal de aves insetívoras que utilizam o sub-
bosque em uma Floresta Estacional Decidual do norte de
Minas Gerais
Montes Claros, Minas Gerais
Maio de 2014

Karen Mirele Caldeira
Variação temporal de aves insetívoras que utilizam o sub-
bosque em uma Floresta Estacional Decidual do norte de
Minas Gerais
Montes Claros, Minas Gerais
Maio de 2014
Dissertação apresentada ao Programa
de Pós-Graduação Stricto Sensu em
Ciências Biológicas da Universidade
Estadual de Montes Claros como pré-
requisito necessário para a conclusão do
curso de Mestrado em Ciências
Biológicas.
Orientador: Prof. Dr. Lemuel Olívio Leite
Co-orientador: Prof. Dr. Rodrigo Oliveira Pessoa

Karen Mirele Caldeira
Variação temporal de aves insetívoras que utilizam o sub-
bosque em uma Floresta Estacional Decidual do norte de
Minas Gerais
______________________________________________
Lemuel Olívio Leite (Orientador/Unimontes)
___________________________________________
Rodrigo Oliveira Pessoa (Unimontes)
____________________________________________
Ronaldo Reis Júnior (Unimontes)
Montes Claros, Minas Gerais
Maio de 2014
Dissertação apresentada ao Programa
de Pós-Graduação Stricto Sensu em
Ciências Biológicas da Universidade
Estadual de Montes Claros como pré-
requisito necessário para a conclusão do
curso de Mestrado em Ciências
Biológicas.

AGRADECIMENTOS
Agradeço a Deus por sempre iluminar meu caminho, por me ajudar nas horas de
dificuldades e por permitir a realização desse trabalho.
Aos meus pais, Jaime e Glória, pelo infinito apoio, dedicação, amor e por sempre
estarem ao meu lado.
Às minhas irmãs, Karine e Kenia, pelos momentos de descontração e por sempre
me ajudar nas horas de sufoco! À minha avó, Dona Ci, pelas orações e por se preocupar
toda vez que eu tinha viagem de campo. Às minhas tias e familiares pelo incentivo. Amo
todos vocês!
Aos colegas de mestrado pela amizade, apoio, conselhos e pelos momentos
divertidos que jamais esquecerei! Em especial, agradeço à Janes, que sempre me
acompanhou e com quem aprendi muito, tanto com relação à vida acadêmica quanto
pessoalmente. Será sempre uma amiga especial pra mim e a quem eu desejo tudo de
melhor, pois merece! Valeu Tchuca linda!
Ao Paulim, por ter tido paciência e me ajudar sempre com as minhas perguntas,
dificuldades, análises, mais análises, até mesmo pela internet, enquanto estava viajando.
Serei eternamente grata! Também não posso esquecer de agradecê-lo pelo companhia até
tarde no lab., na internet, nas passarinhadas, nos perrengues e por torcer sempre e ser tão
atencioso comigo! Grande consideração por você! Te adoro, Picareta!
Ao Hugo, uma pessoa que eu nunca vi igual! Sempre se preocupando comigo, me
fazendo rir e compartilhando algumas experiências. Aprendi muito com você, nem que a
maioria das coisas tenha sido só palhaçadas! Tenho um carinho enorme por você, meu
querido!
À Sarinha da Biotech, Lets e Paulim, terei vocês sempre em minhas lembranças
pelas risadas, companhia, ajuda, torcida e puxões de orelha... os quais não foram poucos!
Vocês têm lugar especial em meu coração meus amigos!
À Nescas, Hugo e Janes pela nossa expedição histórica! Agradeço toda a ajuda,
paciência, descontração, companhia e principalmente bom humor para enfrentar todas as
dificuldades! Ficará sempre em minha memória e coração tudo o que vivemos!
Às amigas de longa data que, mesmo de longe, se faziam presentes em ligações de
preocupação e incentivo, em especial Jane, Thalis, Laudy e Josi!
A todos os colegas do Laboratório de Ornitologia, tanto os que já estão em outra

caminhada, em outro lugar e aos integrantes atuais pelo esforço durante as coletas ao
longos dos anos desse trabalho, digitação de planilhas etc, mas principalmente pela
companhia, aprendizado e divertimento. Cada um de vocês é inesquecível para mim!
Às pessoas que colaboraram com alguns dados como, por exemplo, Giovana, Neto
Sarinha, Joseph, Yule e Mário, por terem paciência com os meus vários pedidos e dúvidas.
Também, à Patrícia (de Fred), Elytania, Vanessinha, Alberto, Dinas, Amanda, Cássia e
Renato, pela boa vontade em compartilhar informações e por se disporem a me ajudar na
ideia inicial... não deu certo, mas ainda sim valeu! A alguns consultores ambientais, à
galera do Wikiaves e curadores de museus com quem entrei em contato.
Ao Augusto Guilherme, que sempre foi muito educado, empenhado e solucionou
diversas problemas de falta de grana para campo, além de nos informar sobre a obtenção e
comprovação dos recursos que conseguíamos do PROAP. Foi extremamente importante!
Aos professores do Programa de Pós-graduação em Ciências Biológicas pelos
ensinamentos, discussões e apoio.
Ao orientador Lemuel pela orientação, conselhos e paciência. Muito obrigada por
tudo!
Ao Ronaldo e a Nescas, por aceitarem participar da banca, pelas correções e
sugestões.
Ao Tropi-Dry, pelo apoio aos projetos de pesquisa em matas secas, incluindo o
trabalho com as aves.
À CAPES pela concessão de bolsa.
A todos que de algum modo colaboraram para a realização dessa dissertação.
Muito obrigada!

SUMÁRIO
1. RESUMO ..................................................................................................
2. INTRODUÇÃO ………………………………………............................
3. OBJETIVOS ………………………………………….............................
3.1 Objetivo Geral …………………………..................................................
3.2 Objetivos Específicos ……………………...............................................
4. HIPÓTESES ……………….....................................................................
4.1 Predições ………………….......................................................................
5. MATERIAIS E MÉTODOS ……………………….................................
5.1 Área de Estudo ……………………..........................................................
5.2 Amostragem das Aves ………………......................................................
5.3 Análises Estatísticas ………………………..............................................
6. RESULTADOS ........................................................................................
7. DISCUSSÃO ............................................................................................
8. REFERÊNCIAS BIBLIOGRÁFICAS .....................................................
9. APÊNDICES ............................................................................................
07
09
12
12
12
13
14
15
15
19
21
23
34
41
47

7
1. RESUMO
A dinâmica e estrutura das comunidades animais podem variar em escala espacial e/ou
temporal, levando a mudanças nos padrões de abundância relativa, riqueza e composição
de espécies. Tais mudanças foram bastante estudadas com relação às aves insetívoras que
utilizam o sub-bosque em ambientes úmidos, no entanto, poucos foram realizados em
ambientes sazonais, como as florestas deciduais, às quais se apresentam geralmente como
um mosaico em diferentes estágios de sucessão ecológica. Em razão desse grupo de aves
ser territorialista, ter baixa mobilidade, apresentar um elevado grau de especialização
ecológica e sensibilidade às mudanças em populações de presas das quais se alimentam,
conhecer como a comunidade de aves insetívoras que utiliza o sub-bosque muda ao longo
do tempo pode mostrar como a sazonalidade e a sucessão ocorre em florestas estacionais
deciduais. Desse modo, o objetivo desse trabalho foi verificar como a riqueza, abundância
e a composição de aves insetívoras que utilizam o sub-bosque variam em relação à
sazonalidade climática e à regeneração natural em três estágios sucessionais - inicial,
intermediário e tardio, em uma Floresta Estacional Decidual localizada no Parque Estadual
da Mata Seca, norte de Minas Gerais. O estudo foi realizado entre outubro de 2007 e junho
de 2013, num total de seis anos. Em cada ano foram realizadas quatro coletas, às quais
correspondem ao final da estação chuvosa (FC), início da estação seca (IS), final da
estação seca (FS) e início da estação chuvosa (IC), sendo utilizadas redes de neblina para a
captura das aves. No estágio inicial, a riqueza diferiu entre as estações e entre os anos,
enquanto a abundância apenas entre as estações. No intermediário, houve diferença
significativa na riqueza e abundância somente entre as estações. No tardio, houve diferença
significativa somente com relação à abundância, a qual variou apenas entre as estações. A
composição de espécies mostrou o mesmo padrão nos três estágios. Houve variação na
composição de aves entre as estações, mas não ao longo dos seis anos de regeneração
natural em cada estágio. Com relação às espécies indicadoras, de modo geral, foi
observado um baixo número de espécies características em relação às estações em cada
estágio, considerando o total de espécies capturadas em cada um. Diante de tais
informações, as aves insetívoras que utilizam o sub-bosque na Floresta Estacional Decidual
estudada, de modo geral, apresentaram variação na riqueza, abundância e composição de
espécies significativamente relacionadas à sazonalidade climática, como era esperado. Tal
observação indica as mudanças nos aspectos estruturais da comunidade de aves insetívoras

8
que utilizam o sub-bosque na FED do PEMS estão mais relacionadas às mudanças em uma
escala temporal menor, ou seja, entre as estações, do que a mudanças estruturais na
vegetação ao longo dos anos. Isso sugere que a sazonalidade climática é o principal fator
que estrutura a comunidade de aves na floresta estudada.

9
2. INTRODUÇÃO
Diversos fatores influenciam a dinâmica e a estrutura das comunidades animais.
Esses fatores podem variar em escala espacial e/ou temporal, levando a mudanças nos
padrões de abundância relativa, riqueza e composição de espécies no tempo e no espaço
(Begon et al., 2007). De acordo com a vasta literatura disponível, há uma grande variedade
de fatores relacionados às tais mudanças nos atributos das comunidades: heterogeneidade
espacial, microclimas e diversidade na estrutura do ambiente (MarcArthur & MacArthur,
1961; Lasky & Keitt, 2010; Willson, 1974; Karr, 1976; Antunes et al., 2009); perturbações
físicas, fragmentação e sucessão (Sekercioglu et al., 2002; Loiselle & Blake, 1994);
condições climáticas (Karr, 1976), abundância de recursos (Loiselle & Blake, 1991; Karr,
1976; Antunes et al., 2009), produtividade (Knapp & Smith, 2001), maturidade e
severidade dos ambientes; interações biológicas, como a competição e predação; e
características intrínsecas dos animais, como comportamento de migração, forrageio e
atividades reprodutivas (Begon et al., 2007; Moermond, 1979).
Com relação à escala espacial, ambientes que apresentam maior heterogeneidade
espacial e são estruturalmente mais diversos, possuem maior diversidade de plantas, níveis
de estratos verticais, fornecendo mais micro-habitats, microclimas, refúgios contra
predadores e variedade de nichos aos organismos, promovem um aumento na diversidade
de animais (Begon et al., 2007 ; MacArthur & MacArthur l961; Willson, 1974).
Na escala temporal, as mudanças nas condições climáticas entre os anos podem
exercer um impacto significativo na disponibilidade de alimento para os animais (Karr,
1976). Da mesma forma, considerável variação tanto entre os anos (Knapp & Smith, 2001)
quanto em relação às estações (Begon et al., 2007) é observada em relação à produtividade,
sendo que uma produtividade mais alta está relacionada a uma maior variedade de recursos

10
disponíveis, o que leva a um aumento da riqueza em espécies (Begon et al., 2007). Essas
variações temporais em condições e recursos, em escalas de tempo distintas, podem ser
previsíveis ou imprevisíveis para os organismos, influenciando profundamente a riqueza
em espécies (Loiselle & Blake, 1994). Em ambientes sazonais e previsíveis, por exemplo,
as espécies estão adaptadas às condições predominantes em épocas distintas do ano, e essas
diferenças permitem a coexistência de um maior número de espécies em comparação com
ambientes mais estáveis (Begon et al, 2007). Nesse sentido, torna-se interessante verificar
como os animais respondem às mudanças ao longo do tempo para melhor entendimento
das dinâmicas e estrutura das diferentes comunidades biológicas.
Estudos sobre comunidades de aves, por exemplo, mostram que esses organismos
também são influenciados pelas características estruturais e variação temporal nos
ambientes. MacArthur & MacArthur (l961) observaram a existência de uma correlação
direta entre diversidade de estratos de uma floresta e a diversidade de espécies de aves.
Willson (1974) encontrou uma correlação com a composição florística. Karr & Roth
(1971) sugeriram que a densidade de folhagem estaria correlacionada positivamente com a
diversidade de espécies de aves, porém, florestas muito densas restringiriam a mobilidade
das aves, reduzindo a diversidade. Karr (1976) verificou os efeitos da variação no ambiente
físico, especificamente o padrão sazonal de precipitação, sobre a estabilidade sazonal da
riqueza em espécies e densidade das comunidades de aves encontrando que a variação
sazonal na estrutura da comunidade diminuiu com o aumento da complexidade da
vegetação. Já alguns trabalhos encontraram que a variação sazonal na disponibilidade de
alimento é o principal fator que influencia as aves (Poulin et al., 1994; Loiselle & Blake,
1991), levando a mudanças na dinâmica e estrutura de comunidades.
Apesar das relações citadas anteriormente, diferenças na riqueza em espécies,
abundância e composição nas comunidades de aves são observadas entre ambientes

11
diferentes e ao longo do tempo, sendo explicadas pelas distintas respostas das espécies
(Loiselle & Blake, 1994; Holmes & Recher, 1986). Essas constituem de diferenças
comportamentais associadas ao forrageio, migração, movimentação, reprodução e
características da história de vida (Loiselle & Blake, 1991; Robinson & Holmes, 1982;
Piratelli & Pereira, 2002). Assim, estudos que reúnam as aves de acordo com algum
aspecto similar entre as mesmas podem mostrar diferenças mais precisas dentro das
comunidades. Nesse sentido, muitos trabalhos foram realizados utilizando as aves
insetívoras que utilizam o sub-bosque, em razão de serem territorialistas (Poulin et al.,
1994), terem baixa mobilidade (Sekercioglu et al., 2002; Martin & Karr, 1986) e por
apresentarem um elevado grau de especialização ecológica e sensibilidade às mudanças em
populações de presas das quais se alimentam (Canaday, 1997). Apesar disso, flutuações
podem ocorrer devido à variação na abundância de recursos e disponibilidade de
artrópodes (Manhães & Dias, 2011; Jaksic, 1981).
Em Florestas Estacionais Deciduais, caracterizadas por apresentar sazonalidade
climática marcante e em diferentes estágios sucessionais, variações na riqueza, abundância
e composição de espécies devem ser observadas não somente entre os ambientes, devido às
diferenças estruturais, mas talvez, principalmente, devido às variações na sazonalidade
climática e em aspectos da vegetação ao longo dos anos. Dessa forma, conhecer como a
comunidade de aves insetívoras que utiliza o sub-bosque muda ao longo do tempo pode
mostrar como a sazonalidade e a sucessão ocorre em florestas estacionais deciduais.

12
3. OBJETIVOS
3.1 Objetivo Geral
Verificar como a riqueza, abundância e a composição de aves insetívoras que
utilizam o sub-bosque variam em relação à sazonalidade climática (coleta) e à regeneração
natural (ano) em cada estágio sucessional (inicial, intermediário e tardio) de uma Floresta
Estacional Decidual localizada no Parque Estadual da Mata Seca, norte de Minas Gerais.
3.2 Objetivos Específicos
1) Verificar se há mudança na riqueza, abundância e composição de aves que utilizam
o sub-bosque entre as estações em cada estágio;
2) Verificar se há mudança na riqueza, abundância e composição de aves que utilizam
o sub-bosque entre os anos em cada estágio;
3) Verificar se existem espécies indicadoras em relação às estações e aos anos de
regeneração natural em cada estágio;

13
4. HIPÓTESES
Hipótese 1: Há variação na riqueza, abundância e composição de aves insetívoras
que utilizam o sub-bosque entre as estações e entre os anos no estágio inicial de
regeneração natural;
Hipótese 2: Há variação na riqueza, abundância e composição de aves insetívoras
que utilizam o sub-bosque entre as estações nos estágios intermediário e tardio de
regeneração natural;
Hipótese 3: Não há variação na riqueza, abundância e composição de aves
insetívoras que utilizam o sub-bosque entre os anos nos estágios intermediário e
tardio de regeneração natural;
Hipótese 4: Há mudança de espécies indicadoras entre as estações e entre os anos
no estágio inicial de regeneração natural;
Hipótese 5: Há mudança de espécies indicadoras entre as estações, mas não entre os
anos nos estágios intermediário e tardio.

14
4.1. Predições
i. A riqueza de espécies e a abundância de aves são maiores na estação chuvosa e
menores na estação seca no estágio inicial;
ii. A riqueza de espécies e a abundância aumentam ao longo dos anos no estágio
inicial;
iii. A riqueza de espécies e a abundância mudam são maiores na estação chuvosa
nos estágios intermediário e tardio;
iv. A riqueza de espécies e a abundância não mudam entre os anos nos estágios
intermediário e tardio;
v. A composição de espécies varia entre as estações e entre os anos no estágio
inicial;
vi. A composição de espécies varia com entre as estações, mas não entre os anos
nos estágios intermediário e tardio;
vii. Há mudança de espécies indicadoras entre as estações e com a regeneração
natural no estágio inicial;
viii. Não há mudança de espécies indicadoras entre as estações e não com a
regeneração natural nos estágios intermediário e tardio.

15
5. MATERIAL E MÉTODOS
5.1 Área de Estudo
Este estudo foi realizado no Parque Estadual da Mata Seca (PEMS) (Figura 1), uma
área de proteção integral criada no ano 2000, que está situada no vale do Médio Rio São
Francisco (Madeira et al., 2009), em uma ampla área de transição entre os biomas Cerrado
e Caatinga (Santos et al., 2007). O parque está localizado na cidade de Manga, norte de
Minas Gerais, Brasil, entre as coordenadas 14°97’02”S- 43°97’02”W e 14°53’08”S-
44°00’05”W e, atualmente, possui uma área de 15.466,44 hectares, sob responsabilidade
do Instituto Estadual de Florestas de Minas Gerais (IEF-MG).
Figura 1. Mapa indicando a distribuição das Florestas Tropicais Secas (FTSs; em vermelho) no
Brasil, com destaque para a localização e delimitação da área do Parque Estadual da Mata Seca
(PEMS), situado na região norte do estado de Minas Gerais (em cinza). Modificado de Espírito-
Santo et al. (2009).

16
Segundo a classificação de Köppen, o clima da região do norte de Minas Gerais é
do tipo Aw, definido como tropical sazonal caracterizado por um verão chuvoso e uma
estação seca bem acentuada no inverno (Figura 2). Neste tipo climático, há pelo menos um
mês com precipitação inferior a 60 mm e a temperatura média do mês mais frio é superior
a 18°C. A temperatura anual média é de 24ºC (Antunes, 1994) e a precipitação anual
média, com base numa estação meteorológica na cidade de Manga, localizada a 10 km da
área de estudo, é de 818 ± 242 mm (Madeira et al., 2009).
O PEMS possui distintas formações vegetais, mas o tipo predominante é a Floresta
Estacional Decidual (FED) (IEF, 2000), conhecida regionalmente como matas secas. Estas
florestas caracterizam-se pelo elevado grau de deciduidade foliar (Scariot & Sevilha, 2005)
e, no PEMS, as árvores perdem cerca de 90-95% das suas folhas durante a estação seca
(maio a outubro) (Madeira et al., 2009), ocorrendo, portanto, em regiões com duas estações
(seca e chuvosa) bem definidas (Murphy & Lugo, 1986).
Em Minas Gerais, o PEMS é a única unidade de conservação que possui Florestas
Secas em solos não-cársticos (chamada também de caatinga arbórea no nordeste) e mata
Figura 2. Aspecto da vegetação da Floresta Estacional Decidual (FED) do Parque Estadual
da Mata Seca (PEMS) durante a estação úmida (à esquerda) e estação seca (à direita). Fotos: Laboratório de Ornitologia da Universidade Estadual de Montes Claros.

17
ciliar ainda preservada (IEF, 2000) à margem esquerda do Rio São Francisco.
Antes da sua formação pela desapropriação de quatro fazendas no ano 2000, as
principais atividades econômicas realizadas na área do parque eram a pecuária intensiva e
plantios (Madeira et al., 2009). Essas atividades levaram à formação de ambientes em
diferentes graus de perturbação antrópica (pastos e fragmentos de FED de diferentes
idades), de acordo com a caracterização da área, com base na estrutura da vegetação, feita
em janeiro de 2006 por Madeira et al. (2009). Nesse estudo, foram selecionadas três
estágios em diferentes estágios de regeneração natural do PEMS (inicial, intermediário e
tardio), cuja descrição, baseada em Madeira et al. (2009), é apresentada à seguir:
Estágio inicial de regeneração natural: esse ambiente foi abandonado desde 2000,
após o uso como pastagem durante vários anos. É caracterizado por manchas
esparsas de vegetação lenhosa, com árvores de pequeno porte, arbustos e
predominância de vegetação herbáceo-arbustiva, compondo um dossel aberto de
cerca de quatro metros de altura. Apesar de abandonado em 2000, gado de
pastagens vizinhas utilizou a área ocasionalmente por algum tempo depois.
Atualmente se encontra com 13 anos em processo de regeneração.
Estágio intermediário de regeneração natural: esta área foi utilizada como pastagem
por um período desconhecido, sendo abandonado no final de 1980, após uso para
criação de gado. Estas matas secundárias são caracterizadas pela presença de dois
estratos verticais de vegetação. O primeiro estrato é composto por árvores de
aproximadamente 10-12 metros de altura, com algumas árvores emergentes de até
15 metros. O segundo estrato é composto por um sub-bosque denso, com muitas
árvores juvenis e alta abundância de lianas (trepadeiras). Atualmente se encontra
com 24-32 anos em regeneração natural.

18
Estágio tardio de regeneração natural: sem histórico de desmatamento nos últimos
50 anos. Assim, como o estágio intermediário, é caracterizado por dois estratos
verticais, entretanto, o primeiro é composto de árvores mais altas, que formam um
dossel fechado de 18-20 metros de altura. O segundo estrato é composto por um
sub-bosque esparso com pouca penetração de luz e baixa densidade de árvores
jovens e de lianas. Atualmente se encontra com 57 anos em regeneração natural.
A fenologia da vegetação no PEMS foi estudada por Pezzini et al. (2014), entre
2007 e 2008. Os autores verificaram que a sazonalidade tem forte influência sobre a
vegetação nos três estágios sucessionais, com maior proporção de indivíduos apresentando
folhas verdes na estação chuvosa e folhas secas na estação seca. Características estruturais
e composição florística da vegetação no parque foram amostradas por Madeira et al.
(2009), cuja amostragem foi realizada em janeiro de 2006, e Nunes et al. (2013), cuja
amostragem foi em março de 2009. Ainda não há nenhum trabalho que tenha comparado
as informações obtidas nos dois trabalhos, dessa forma, não foi possível no presente estudo
verificar como a estrutura da vegetação na área de estudo muda em cada estágio ao longo
dos anos. Apesar disso, são informados aqui os resultados encontrados pelos autores
citados anteriormente quanto a um índice de complexidade da vegetação - índice de
complexidade de Holdridge – CHCI , o qual utiliza os parâmetros quantitativos altura, área
basal, densidade, riqueza de espécies, para indicar o nível de complexidade da vegetação.
Ainda que tenham ocorrido diferenças quanto às amostragens, os valores de CHCI podem
mostrar possíveis diferenças, por inferência, sobre a variação na estrutura da floresta entre
os diferentes anos dos estudos. Os valores de CHCI entre os estudos foram: inicial: 0.6 ± 0.5
e 4.97 ± 3.44; intermediário: 15.0 ± 8.3 e 37.61 ± 15.71; tardio 46.1 ± 25.7 e 111.27 ±
64.05, sendo o primeiro valor referente ao estudo realizado em 2006 (Madeira et al., 2009)
e o segundo ao estudo realizado em 2009 (Nunes et al., 2013) no PEMS. É observado um

19
aumento do CHCI entre os estudos, o que pode indicar um aumento da complexidade da
vegetação, em cada estágio, ao longo do tempo. Contudo, uma organização dos dados
seguida de testes para verificar se há diferenças entre os dados, torna-se necessária para
uma análise segura da variação estrutural em cada estágio entre os anos.
5.2 Amostragem das Aves
As amostragens da avifauna ocorreram em três estágios de regeneração natural de
uma Floresta Estacional Decidual (FED) do Parque Estadual da Mata Seca (PEMS), sendo:
uma área em estágio inicial e dois estágios florestais, denominados intermediário e tardio.
Elas foram realizadas entre outubro de 2007 e junho de 2013, num total de seis anos (ver
relação das coletas no Apêndice 1). Em cada ano foram realizadas quatro coletas, as quais
correspondem ao final da estação chuvosa (FC), início da estação seca (IS), final da
estação seca (FS) e início da estação chuvosa (IC). Em cada coleta foram utilizadas três
parcelas.
Cada coleta teve duração de quatro dias e meio sendo utilizadas, para a captura das
aves, redes de neblina (12m de comprimento X 3m de altura, 20mm de malha) (Figura 4),
que ficaram abertas por um período de 6 horas à partir do amanhecer, e checadas a cada 30
minutos.
As aves capturadas foram identificadas de acordo com Sick (1997), Ridgely &
Tudor (1989), Ridgely & Tudor (1994) e a nomenclatura seguiu o Comitê Brasileiro de
Registros Ornitológicos (CBRO, 2011).

20
Figura 4. Redes de neblina abertas para a captura das aves durante a estação chuvosa (à esquerda);
Indivíduo de Thamnophilus pelzelni (fêmea) sendo retirado da rede após ser capturado. Fotos
retiradas na Floresta Estacional Decidual (FED) do Parque Estadual da Mata Seca (PEMS). Fotos: Laboratório de Ornitologia da Universidade Estadual de Montes Claros
Neste estudo, foram utilizadas as espécies de aves com hábito alimentar insetívoro
e que utilizam frequentemente o sub-bosque. Essas informações foram baseadas em
literatura, como Sick (1997), Martins (2007), Silveira & Machado (2012), Stotz et al.
(1996), em estudos realizados pelos pesquisadores do Laboratório de Ornitologia da
Universidade Estadual de Montes Claros (Unimontes) sobre o conteúdo estomacal das aves
capturadas no PEMS, assim como através de observações pessoais feitas no mesmo
parque. Esse grupo de aves, em específico, foi selecionado porque ocorre ocorrem em
todos os estágios e durante todo o ano no Parque Estadual da Mata Seca, mesmo durante a
estação seca, permitindo realizar comparações sazonais e entre os anos da riqueza,
abundância e composição.

21
5.3 Análises Estatísticas
Para verificar se a riqueza e abundância de aves variam significativamente entre as
coletas (FS: final da estação seca; IC: início da estação chuvosa; FC: final da estação
chuvosa; e IS: início da estação) e entre os seis anos do estudo, foram realizados modelos
lineares generalizados (GLMs), em que a riqueza e abundância de aves foram as variáveis
respostas (quantitativas) e, coletas e anos, variáveis explicativas, ambas qualitativas. Para
ambas as análises, foi utilizada a distribuição Poisson, com as devidas correções em caso
de sub ou sobredispersão. Os modelos foram testados através da ANOVA, com teste Chi-
quadrado, e nível de significância considerando p<0,05. Uma vez que as duas variáveis
explicativas possuem mais de dois níveis (quatro níveis para coleta e seis níveis para anos),
foi realizada uma análise de contraste para verificar quais níveis foram significativamente
diferentes entre si. Essas análises foram realizadas no programa R (R Development Core
Team, 2013).
Para verificar a ordenação da composição de aves entre as coletas (FS, IC, FC e IS)
e entre os seis anos do estudo, em cada estágio, foram realizadas análises de escalonamento
não-métrica (NMDS), utilizando 1000 permutações o índice de similaridade de Bray-
Curtis, uma vez que que é bastante útil na analise de composição de comunidades com
base em dados de abundancia (Clarke, 1993). A significância dos grupos formados na
NMDS foi avaliada através de uma análise de similaridades (ANOSIM), também com
1000 permutações e utilizando a medida de distância de Bray-Curtis. Após verificar
significância entre os grupos, foram realizados testes de comparações múltiplas para
observar entre quais grupos houve diferença significativa. Os valores da significância entre
os grupos foram posteriormente corrigidos com o método BY de Benjamini e Yekutieli
(2001), pois é indicado para controlar as chances de ocorrer o erro Tipo I, que

22
normalmente acontece quando são testadas várias hipóteses através das comparações
múltiplas. Todas essas análises foram realizadas no programa R (R Development Core
Team, 2013).
Para verificar quais as espécies que caracterizaram cada padrão encontrado em
relação às coletas e aos anos na NMDS, foi realizada a análise de espécies indicadoras
(ISA- Indicator Species Analysis), proposta por Dufrêne & Legendre (1997). Esta análise
confere a cada espécie um “valor de indicação”, que varia de 0 a 100, em porcentagem,
através de uma combinação da abundância relativa e a frequência relativa de cada espécie
nos grupos definidos previamente. Nesse estudo, com relação à coleta, foram utilizados os
quatro grupos: FS, IC, FC, IS; e com relação a ano, foram utilizados seis grupos,
correspondentes a cada ano de estudo. O valor 0 (não-indicador) significa que a espécie
está ausente naquele grupo, e o valor 100 significa uma indicação perfeita, em que todos os
indivíduos da espécie ocorrem apenas em um grupo. A significância da indicação de cada
espécie foi testada através do teste de Monte Carlo, com 4999 permutações. As espécies
que apresentaram valores de indicação, entre os grupos, significativamente diferentes
(p<0,05) e valor de indicação superior a 25% foram consideradas indicadoras. Essa análise
foi realizada no programa PC-ORD 6.0 (McCune & Mefford, 1997).

23
6. RESULTADOS
Ao longo dos seis anos de estudo, foi capturado um total de 1998 indivíduos
correspondentes a 51 espécies e a 15 famílias de aves (Apêndice 2) em 19440 horas-rede.
Foram registrados 554 indivíduos e 40 espécies no inicial, 772 indivíduos e 44 espécies no
intermediário e, 672 indivíduos e 36 espécies no tardio. As espécies mais abundantes no
estágio inicial foram Lanio pileatus (N=111), Turdus amaurochalinus (N=88),
Myiodynastes maculatus (N=54), Myiopagis virdicata (N=46); Nos estágios mais
florestais, a espécie insetívora Thamnophilus pelzeni foi a mais abundante, apresentando
abundância igual a 106, no intermediário, e 148 no tardio. Além dessa espécie, também
foram observadas, no intermediário, maiores abundâncias de Lanio pileatus (N=72),
Hemitriccus margaritaceiventer (N=71), Formicivora melanogaster (N=62), Sittasomus
griseicapillus (N=55); e, no tardio, Sittasomus griseicapillus (N=84), Myiopagis viridicata
(N=56), Lathrotriccus euleri (N=47).
No estágio inicial, a riqueza diferiu entre as coletas (p<0,05) e entre os anos
(p<0,01) (Figura 5). A riqueza não foi diferente entre o final da chuva (FC) e final da seca
(FS), assim como entre o início da seca (IS) e o início da chuva (IC), sendo que a riqueza
média foi maior no agrupamento formado pelo FC-FS, igual a 5.4, e IS-IC, igual a 4. Com
relação aos anos, houve diferença significativa apenas entre o último ano do estudo (ano
seis) e os demais (p< 0,01). A riqueza média do ano seis foi igual a 6.5, enquanto no
agrupamento formado pelos cinco primeiros anos (1-2-3-4-5) foi 4,3.

24
No estágio inicial, houve diferença significativa na abundância somente entre as
coletas (p<0,001) (Figura 6), ocorrendo diferença apenas entre o final da seca (FS) e as
demais coletas (IC, FC e IS). A abundância média de FS foi igual a 11,7 e a do
agrupamento IC-FC-IS foi 6,4.
Figura 5. Variação significativa na riqueza de espécies de aves em relação às coletas (FS: final da estação
chuvosa; IC: início da estação chuvosa; FC: final da estação chuvosa; IS: início da estação seca) – à esquerda;
e em relação aos seis anos do estudo – à direita, em um estágio inicial de regeneração natural em uma Floresta
Estaciona Decidual (FED) do Parque Estadual da Mata Seca (PEMS), norte de Minas Gerais. Barras com a
mesma cor não apresentam diferenças significativas entre si.

25
No intermediário, houve diferença significativa na riqueza somente entre as coletas
(p<0,01) (Figura 7). A análise de contraste mostrou diferença na riqueza apenas entre fina
da chuva (FC) e as outras coletas (IS, FS e IC). A riqueza média foi de 8,5 no FC e no
agrupamento IS-FS-IC foi igual a 6,4.
Figura 6. Variação significativa na abundância de aves em relação às coletas (FS: final da estação chuvosa;
IC: início da estação chuvosa; FC: final da estação chuvosa; IS: início da estação seca) – à esquerda; e a
ausência de variação significativa em relação aos seis anos do estudo – à direita, em um estágio inicial de
regeneração natural em uma Floresta Estaciona Decidual (FED) do Parque Estadual da Mata Seca (PEMS),
norte de Minas Gerais. Barras com a mesma cor não apresentam diferenças significativas entre si.

26
No estágio intermediário, com relação à abundância, houve diferença significativa
apenas entre as coletas (p<0,01) (Figura 8). Foi observada diferença na abundância
somente entre início da chuva (IC) e as demais coletas (FC, IS e FS). A abundância média
foi de 11,7 no agrupamento formado por FC-IS-FS e de 7,6 em IC.
Figura 7. Variação significativa na riqueza de espécies de aves em relação às coletas (FS: final da estação
chuvosa; IC: início da estação chuvosa; FC: final da estação chuvosa; IS: início da estação seca) – à esquerda;
e a ausência de variação significativa em relação aos seis anos do estudo – à direita, em um estágio
intermediário de regeneração natural em uma Floresta Estaciona Decidual (FED) do Parque Estadual da Mata
Seca (PEMS), norte de Minas Gerais. Barras com a mesma cor não apresentam diferenças significativas entre
si.

27
No tardio, não houve diferença significativa na riqueza de espécies nem entre as
coletas (p>0,05), nem entre os anos do estudo (p>0,05) (Figura 9).
Figura 8. Variação significativa na abundância de aves em relação às coletas (FS: final da estação chuvosa;
IC: início da estação chuvosa; FC: final da estação chuvosa; IS: início da estação seca) – à esquerda; e a
ausência de variação significativa em relação aos seis anos do estudo – à direita, em um estágio intermediário
de regeneração natural em uma Floresta Estaciona Decidual (FED) do Parque Estadual da Mata Seca (PEMS),
norte de Minas Gerais. Barras com a mesma cor não apresentam diferenças significativas entre si.

28
Figura 9. Ausência de variação significativa na riqueza de espécies de aves tanto em relação às coletas (FS:
final da estação chuvosa; IC: início da estação chuvosa; FC: final da estação chuvosa; IS: início da estação
seca) – à esquerda; quanto em relação aos seis anos do estudo – à direita, em um estágio tardio de regeneração
natural em uma Floresta Estaciona Decidual (FED) do Parque Estadual da Mata Seca (PEMS), norte de Minas
Gerais. Barras com a mesma cor não apresentam diferenças significativas entre si.
No tardio, houve diferença significativa na abundância somente entre as coletas
(p<0,01) (Figura 10). A análise de contraste mostrou que a abundância foi diferente entre
início da chuva (IC) e as demais (FC, IS e FS). A abundância média foi igual a 10,4 no
agrupamento FC-IS-FS, e igual a 6,2 na coleta IC.
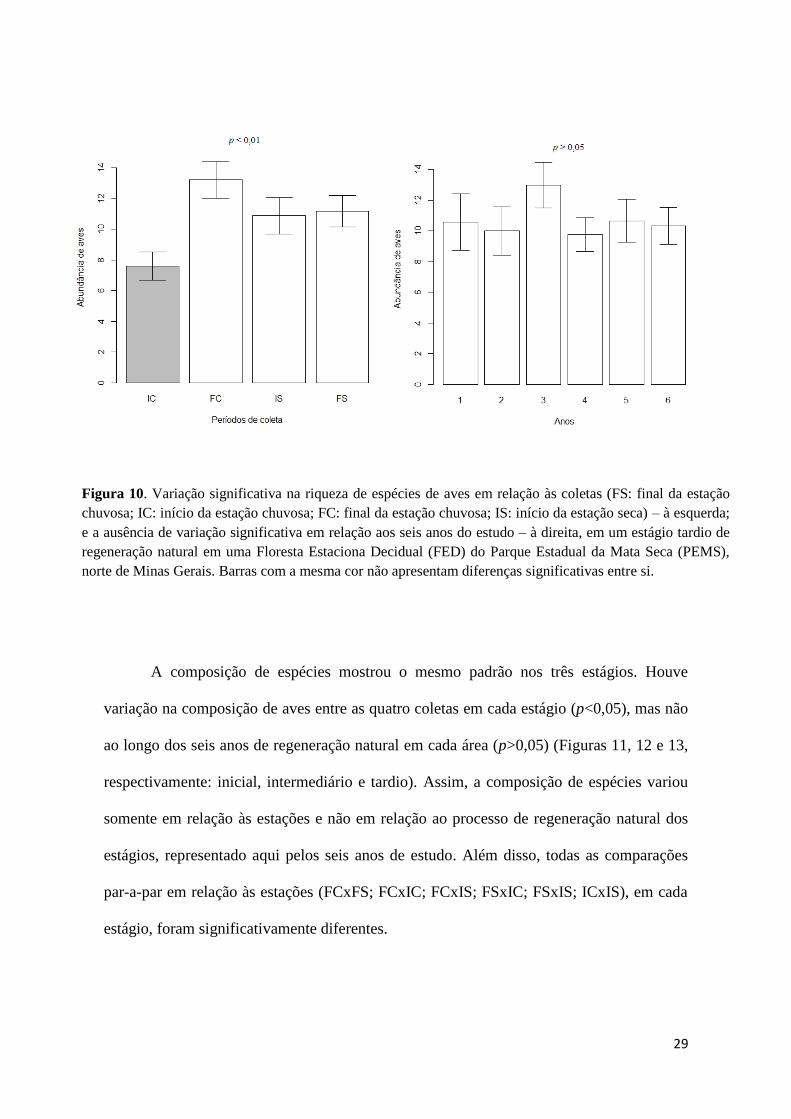
29
Figura 10. Variação significativa na riqueza de espécies de aves em relação às coletas (FS: final da estação
chuvosa; IC: início da estação chuvosa; FC: final da estação chuvosa; IS: início da estação seca) – à esquerda;
e a ausência de variação significativa em relação aos seis anos do estudo – à direita, em um estágio tardio de
regeneração natural em uma Floresta Estaciona Decidual (FED) do Parque Estadual da Mata Seca (PEMS),
norte de Minas Gerais. Barras com a mesma cor não apresentam diferenças significativas entre si.
A composição de espécies mostrou o mesmo padrão nos três estágios. Houve
variação na composição de aves entre as quatro coletas em cada estágio (p<0,05), mas não
ao longo dos seis anos de regeneração natural em cada área (p>0,05) (Figuras 11, 12 e 13,
respectivamente: inicial, intermediário e tardio). Assim, a composição de espécies variou
somente em relação às estações e não em relação ao processo de regeneração natural dos
estágios, representado aqui pelos seis anos de estudo. Além disso, todas as comparações
par-a-par em relação às estações (FCxFS; FCxIC; FCxIS; FSxIC; FSxIS; ICxIS), em cada
estágio, foram significativamente diferentes.

30
Figura 11. Ordenação da composição de aves insetívoras que utilizam o sub-bosque em um estágio inicial de
regeneração natural em relação às coletas (FS: final da estação chuvosa; IC: início da estação chuvosa; FC: final da
estação chuvosa; IS: início da estação seca) – à esquerda; e em relação aos seis anos do estudo – à direita, em
Floresta Estaciona Decidual (FED) do Parque Estadual da Mata Seca (PEMS), norte de Minas Gerais.
Figura 12. Ordenação da composição de aves insetívoras que utilizam o sub-bosque em um estágio intermediário
de regeneração natural em relação às coletas (FS: final da estação chuvosa; IC: início da estação chuvosa; FC: final
da estação chuvosa; IS: início da estação seca) – à esquerda; e em relação aos seis anos do estudo – à direita, em
Floresta Estaciona Decidual (FED) do Parque Estadual da Mata Seca (PEMS), norte de Minas Gerais.

31
Com relação às espécies indicadoras, de modo geral, foi observado um baixo
número de espécies características em relação às estações em cada estágio, considerando o
total de espécies capturadas em cada um. No inicial, do total de 40 espécies amostradas,
apenas três foram indicadoras em três coletas (Tabela 1), só não havendo no início da
estação seca. As espécies características foram: Myiodynastes maculatus (N=54) no final
da seca (FS) e Myiopagis viridicata (N=46), do final da chuva, as quais foram também a
segunda e a terceira espécie com maior abundância entre as estações nesse estágio; e
Pachyramphus polychopterus, no inicio da chuva (IC). As espécies Lanio pileatus (N=111)
e Turdus amaurochalinus (N=88), não foram consideradas indicadoras de nenhuma
estação, pois ocorrem em todas as coletas.
Figura 13. Ordenação da composição de aves insetívoras que utilizam o sub-bosque em um estágio tardio de
regeneração natural em relação às coletas (FS: final da estação chuvosa; IC: início da estação chuvosa; FC: final da
estação chuvosa; IS: início da estação seca) – à esquerda; e em relação aos seis anos do estudo – à direita, em
Floresta Estaciona Decidual (FED) do Parque Estadual da Mata Seca (PEMS), norte de Minas Gerais.

32
Tabela 1. Valores significativos da análise de espécies indicadoras, considerando o
valor de indicação >25% e a significância do Teste de Monte Carlo (p<0,05), de aves
insetívoras que utilizam o sub-bosque em relação às coletas (FS: final da estação
chuvosa; IC: início da estação chuvosa; FC: final da estação chuvosa; IS: início da
estação seca) em um estágio inicial de regeneração natural em uma Floresta
Estacional Decidual do Parque Estadual da Mata Seca, norte de Minas Gerais, Brasil.
Espécie Valor de indicação (%) p Coleta
Myiodynastes maculatus 43 0.0002 FS
Myiopagis viridicata 36 0.0026 FC
Pachyramphus polychopterus 32 0.0026 IC
No intermediário, de um total de 44 espécies, apenas cinco espécies indicadoras
foram encontradas (Tabela 2) e elas estiveram mais relacionadas ao final da estação
chuvosa (FC) e final da estação seca (FS), com mais espécies indicadoras no FC. As
espécies Myiopagis viridicata (N=46) e Sittasomus griseicapillus (N=55) apresentaram os
maiores valores de indicação em relação à coleta FC. Somente uma espécie, Cnemotriccus
fuscatus (N=25), foi indicadora do FS e nenhuma foi para o início da seca e início da
chuva. As espécies Thamnophilus pelzelni (N=106) e Lanio pileatus (N=72) não foram
consideradas indicadoras de nenhuma estação por ocorrerem em todas as coletas.
Tabela 2. Valores significativos da análise de espécies indicadoras, considerando o
valor de indicação >25% e a significância do Teste de Monte Carlo (p<0,05), de
aves insetívoras que utilizam o sub-bosque em relação às coletas (FS: final de seca;
IC: início da chuva; FC: final de chuva e IS: início da seca) em um estágio
intermediário de regeneração natural em uma Floresta Estacional Decidual do
Parque Estadual da Mata Seca, norte de Minas Gerais, Brasil.
Espécie Valor de indicação (%) p Coleta
Cnemotriccus fuscatus 33 0.0002 FC
Myiarchus tyrannulus 36 0.0010 FS
Myiopagis viridicata 46 0.0002 FC
Pachyramphus polychopterus 30 0.0016 FC
Sittasomus griseicapillus 45 0.0004 FC

33
O estágio tardio, no qual foi amostrado um total de 36 espécies, foram encontradas
cinco espécies indicadoras. Este ambiente foi o único que apresentou espécies indicadoras
em todas as coletas, sendo observado maior valor de indicação para a espécie Myiopagis
viridicata (N=56) no final da estação chuvosa (FC). Ocorreram mais espécies indicadoras
no final da estação seca (FS), com valores de indicação baixos como, por exemplo,
Myiodynastes macultaus (N=12) e Tolmolmyias sulphurescens (N=18). As espécies
Sittasomus griseicapillus (N=84) e Thamnophilus pelzelni (N=148) não foram
consideradas indicadoras de nenhuma estação por ocorrerem em todas as coletas.
Tabela 3. Valores significativos da análise de espécies indicadoras, considerando o
valor de indicação >25% e a significância do Teste de Monte Carlo (p<0,05), de
aves insetívoras que utilizam o sub-bosque em relação às coletas (FS: final de seca;
IC: início da chuva; FC: final de chuva e IS: início da seca) em um estágio maduro
de regeneração natural em uma Floresta Estacional Decidual do Parque Estadual da
Mata Seca, norte de Minas Gerais, Brasil.
Espécie Valor de indicação (%) p Coleta
Lathrotriccus euleri 26 0.0416 FS
Myiodynastes maculatus 25 0.0110 FS
Myiopagis viridicata 46 0.0006 FC
Synallaxis scutata 28 0.0028 IS
Tolmomyias sulphurescens 25 0.0212 FS
Com relação aos anos, não foi realizada a análise de espécies indicadoras
para nenhum estágio uma vez que não houve diferença significativa na composição de
espécies entre os anos em nenhum desses ambientes.

34
7. DISCUSSÃO
As aves insetívoras que utilizam o sub-bosque na Floresta Estacional Decidual
(FED) estudada, de modo geral, apresentaram variação na riqueza, abundância e
composição de espécies significativamente relacionadas à sazonalidade climática, como
era esperado. Em florestas deciduais, a marcante variação na precipitação entre as
diferentes épocas do ano (Murphy & Lugo, 1986) limita o recurso hídrico para as plantas
durante a estação seca, fazendo com que grande parte das espécies perca cerca de 90% de
suas folhas nessa época do ano, enquanto a produção de folhas ocorre de forma sincrônica
e restrita à estação chuvosa na região (Pezzini et al., 2014). Tal característica resulta em
mudança na produtividade da vegetação entre as estações, a qual influencia os recursos
disponíveis, como, por exemplo, os insetos, os quais flutuam marcadamente entre as
estações (Neves, 2009). Desse modo, em ambientes sazonais, a variação na disponibilidade
de insetos (Janzen, 1973; Wolda, 1988), pode ser um dos principais fatores que determina
a estrutura de comunidades (Karr, 1971; Loiselle & Blake, 1991).
Mudanças na abundância e distribuição de insetos em relação à sazonalidade
pronunciada podem levar a diferentes respostas das espécies, como, por exemplo,
mudanças comportamento de forrageio das espécies (Murakami, 2002), deslocamentos
entre os habitats (Loiselle & Blake, 1991), migração (Karr, 1971) e o fitness dos
indivíduos, às quais afetam a estrutura da comunidade de aves (Murakami, 2002; Loiselle
& Blake, 1991). Na FED estudada, por exemplo, Gonçalves (2011) encontrou variação
significativa na abundância de artrópodes disponíveis no ambiente para uma ave insetívora
florestal - Choca-do-planalto (Thamnophilus pelzelni), com maior abundância de insetos
disponíveis durante a estação chuvosa, influenciando alguns aspectos do comportamento
de forrageio dessa espécie.

35
No estágio inicial, como era esperado, houve variação na riqueza, abundância e
composição de aves insetívoras que utilizam o sub-bosque entre as estações. Esse resultado
pode ser explicado pelas variações nos recursos nesse estágio mudarem amplamente entre
as estações, devido às mudanças na cobertura foliar e menor estabilidade climática
(Pezzini, 2008; May, 1982) em ambiente ainda não estruturado. Tal característica pode ser
inferida dos trabalhos de Madeira et al. (2009), Nunes et al. (2013) e Pezzini (2008), os
quais encontraram os menores valores dos parâmetros de complexidade estrutural nesse
estágio em comparação aos outros, na área de estudo. De acordo com Madeira et al.
(2009), esse estágio possui machas esparsas de vegetação lenhosa, com árvores de pequeno
porte, arbustos e predominância de vegetação herbáceo-arbustiva, compondo um dossel
aberto de cerca de quatro metros de altura. Stouffer & Bierregaard (1995) estudaram
fragmentos antes e após o isolamento em florestas de terra-firme na Amazônia e seus
efeitos sobre aves insetívoras de sub-bosque, encontrando que a cobertura vegetal pode ser
o principal fator na mudança da diversidade dessas aves. Nesse sentido, deve haver uma
maior variação na disponibilidade de insetos entre as estações, fazendo com que as aves se
desloquem para os estágios mais florestais em busca de recursos, resultando nas mudanças
observadas.
As variações esperadas na estrutura da comunidade de aves do estágio inicial entre
os anos não foram encontradas para a abundância e composição de espécies. Este resultado
contraria ao que era esperado, uma vez que os atributos da comunidade de aves mudam
rapidamente em estágios iniciais, devido à modificação na estrutura do ambiente (Loiselle
& Blake, 1994). No começo da sucessão, espécies de plantas pioneiras colonizam
rapidamente espaços abertos no ambiente, promovendo aumento da heterogeneidade,
microhabitats e alimento para os animais (Begon et al, 2007). Uma provável ausência de
mudanças nesses aspectos pode ser explicada pelo fato de FEDs se recuperarem lentamente

36
(Quesada et al., 2009), em comparação a floresta úmidas, diferentemente do que era
pensado (Murphy & Lugo, 1986). Dessa forma, mudanças na estrutura da vegetação
perceptíveis para as aves insetívoras de sub-bosque ainda não devem ter ocorrido.
Os estágios intermediário e tardio apresentaram variação na abundância e
composição de espécies entre as estações, e não houve variação na riqueza no tardio. As
mudanças esperadas nesses estágios mostram que a sazonalidade climática na área é um
importante fator que influência as aves nesses estágios mais florestais, ainda que estes,
teoricamente, devido à estrutura complexa, apresentassem um tamponamento das variações
climáticas (Karr, 1976) sobre a avifauna. As mudanças esperadas estão relacionadas à
sazonalidade marcante na FED estudada, que influenciam os recursos para as aves mesmo
em ambientes florestais. Por exemplo, Neves (2009) encontrou uma variação nas guildas
de insetos entre o dossel e o sub-bosque no estágio tardio do PEMS entre as estações, com
flutuações ocorrendo entre o início e fim de cada estação. Um aspecto interessante a ser
observado também é que alguns trabalhos na área de estudo observaram que esses estágios
não são diferentes quanto à riqueza e composição de espécies de aves (Espírito-Santo et al.,
2013), sendo observada similaridade na composição entre as estações chuvosas desses
estágios, assim como entre as estações secas. Essa informação foi confirmada para outros
grupos de animais, como para a composição de insetos herbívoros e formigas (Neves,
2009). Dados da vegetação (Nunes et al., 2013; Madeira et al., 2009) no PEMS, no entanto,
mostram que existem diferenças esses estágios, com maior complexidade estrutural no
tardio. Isso indica que para as aves insetívoras que utilizam o sub-bosque as mudanças no
ambiente podem ser mais bem percebidas, tendo em vista que a maioria das espécies é
sensível às alterações no habitat (Stouffer & Bierregaard, 1995). A ausência de variação
riqueza significativa entre as estações no tardio, sugere que pode estar ocorrendo uma
substituição de espécies nesse estágio, com o ambiente se tornando adequado a diferentes

37
espécies entre as estações (Loiselle & Blake, 1992), levando a mudanças na abundância e
composição entre as estações, mas não no número de espécies.
Nesse estudo, não houve variação na estrutura da comunidade de aves insetívoras
de sub-bosque em relação aos anos, tanto no estágio intermediário quanto no tardio, como
esperado. Esse resultado pode ser explicado, provavelmente, por esses estágios
apresentarem estrutura mais complexa e, assim, haver pouca variação no clima entre os
anos (Karr, 1976), ocorrendo apenas a substituição de espécies ao longo do tempo (Blake
& Loiselle, 2001). De acordo com Loiselle e Blake (1992), as variações na abundância de
aves em ambiente florestal pode ser uma consequência de flutuações irregulares na
abundância das espécies não associadas às alterações de habitat ou sucessão. Dessa forma,
mudanças nos atributos da comunidade de aves provavelmente mudam em relação às
estações e não entre os anos na floresta em estudo.
Com relação às variações na riqueza e abundância de aves insetívoras de sub-
bosque entre o início e fim das estações seca e chuvosa, em cada estágio, não foi observado
o padrão esperado na variação dos atributos nos ambientes. Ou seja, maior riqueza e
abundância no período chuvoso e o contrário no período de seca. Uma explicação para isso
pode ser uma variabilidade ampla das respostas das espécies às mudanças entre as estações
na floresta estacional estudada. A ausência de diferença na riqueza e abundância entre as
estações seca e chuvosa indica que esses atributos são influenciados pelas mudanças na
disponibilidade de recursos (insetos) entre as estações devido ao efeito pronunciado da
sazonalidade climática na estrutura da vegetação, como em relação à cobertura foliar
(Pezzini et al., 2009). Ainda que maior abundância de insetos esteja concentrada na estação
chuvosa na área de estudo, esses organismos variam entre o início e o fim de cada estação,
como foi encontrado por Neves (2009). Nesse sentido, essas variações resultam em
mudanças nas atividades das aves entre as estações, como, por exemplo, em relação aos

38
deslocamentos durante as atividades de forrageio, influenciando as taxas de capturas das
espécies e, consequentemente, nos valores de riqueza e abundância obtidos.
Espécies indicadoras de cada estágio foram observadas somente em relação às
estações, pois não houve diferença significativa na composição de aves em cada estágio
durante os anos desse estudo. No inicial ocorreram poucas espécies indicadoras entre as
estações, sendo apenas três: Myiopagis viridicata e Pachyramphus polychopterus na
estação chuvosa e Myiodynastes maculatus na estação seca, indicando que poucas das
espécies consideradas podem ser especialistas em forragear nesse estágio e isso pode estar
relacionado às flutuações mais amplas nas condições ambientais nesse ambiente (May,
1982).
Um baixo número de espécies indicadoras também foi encontrado nos estágios
intermediário e tardio, sendo apenas cinco em cada um. No intermediário, três espécies
foram indicadoras do final da estação chuvosa, enquanto no tardio três foram indicadoras
do final da estação seca, sugerindo que os estágios devem apresentar diferenças na
disponibilidade de recursos ao longo do tempo, como previsto pela sazonalidade. Desse
modo, mesmo nos ambientes florestais, pode haver diferenças não só estruturais, mas
também na disponibilidade de recursos para as espécies, levando a mudanças no uso do
estágio ao longo do tempo.
De modo geral, apesar do baixo número de espécies indicadoras ao longo das
estações em cada estágio, algumas espécies foram indicadoras tanto em estágios diferentes
quanto em estações diferentes, mas também algumas ocorreram em estágios diferentes na
mesma estação. Por exemplo, a espécie Pachyramphus polychopterus foi característica do
inicial e do intermediário, porém no início da estação chuvosa no inicial e final da estação
chuvosa no intermediário. Já M. maculatus foi indicadora do inicial e do tardio no final da

39
estação seca. Tais diferenças sugerem que as aves insetívoras de sub-bosque devem
mostrar uma plasticidade no uso do habitat entre as estações, a qual pode estar relacionada
a diferenças na disponibilidade e uso de recurso entre os indivíduos de uma mesma
espécie. Além disso, algumas espécies podem ser migratórias, como é o caso da espécie
Myiopagis viridicata, que ocorre nessa floresta durante a estação chuvosa, indicando que o
comportamento de migração pode ocorrer devido às mudanças na disponibilidade de
recursos entre as estações (Karr, 1971). A grande maioria de das espécies não ter sido
indicadora, pode-se inferir que o habitat pode ser usado de forma mais generalista pelas
espécies, ocorrendo pouca especificidade por estação/estágio. Com base nessas
informações, pode ser sugerido que, de modo geral, a sazonalidade climática na FED
estudada permite que poucas espécies sejam mais adaptadas a determinadas estações em
cada ambiente, tendo em vista o baixo número de espécies características de cada estação
nos estágios.
As mudanças nos aspectos estruturais da comunidade de aves insetívoras que
utilizam o sub-bosque na FED do PEMS estão mais relacionadas às mudanças em uma
escala temporal menor, ou seja, entre as estações, do que a mudanças estruturais na
vegetação ao longo dos anos. Isso sugere que a sazonalidade climática é o principal fator
que estrutura a comunidade de aves na floresta estudada. No entanto, deve-se ter cautela
em assumir os resultados encontrados como definitivos, tendo em vista que o tempo de
realização desse trabalho pode não ser suficiente para verificar mudanças nos atributos das
aves estudadas em relação ao processo de sucessão. O uso de medidas quantitativas dos
aspectos estruturais da vegetação, assim como estudos sobre o comportamento,
disponibilidade e uso dos recursos por algumas espécies (Loiselle & Blake, 1991) poderão
mostrar com mais precisão a importância da sazonalidade e das diferenças estruturais ao
longo do tempo na dinâmica e estrutura da comunidade de aves insetívoras que usam o

40
sub-bosque nessa floresta estacional decidual. Apesar disso, o presente trabalho mostrou
algumas tendências das variações na estrutura da comunidade de aves insetívoras de sub-
bosque em relação à sazonalidade climática e ao processo de regeneração natural de cada
estágio na FED do PEMS, considerando alguns aspectos estruturais disponíveis na
literatura sobre esses ambientes.

41
8. REFERÊNCIAS BIBLIOGRÁFICAS
Antunes, F. Z. Caracterização climática – Caatinga do estado de Minas Gerais. Informe
Agropecuário. v.17, p.15-19, 1994.
Antunes, A. Z.; Wyrgun, B.; Eston, M. Composição das comunidades de aves em duas
florestas secundárias contíguas no sudeste do Brasil. Revista Instituto Florestal, São
Paulo, v. 21, n. 1, p. 93-106, 2009.
Begon, M.; Townsend, C. R.; Harper, J. L. Ecologia: de indivíduos a ecossistemas. 4°
ed., Editora: Artmed, Porto Alegre - RS, 2007, 752 p.
Benjamini, Y. & Yekutieli, D. (2001). The control of the false discovery rate in multiple
testing under dependency. Annals of Statistics, v. 29, p. 1165–1188.
Blake, J. G. & Loiselle, B. A. Bird assemblages in second-growth and old-growth forests,
Costa Rica: perspectives from mist nets and point counts. The Auk, v. 118, n. 2, p. 304-
326, 2001.
Canaday, C. Loss of insectivorous birds along a gradient of human impact in Amazonia.
Biological Conservation, v.77, p. 63-77, 1997.
CBRO - Comitê Brasileiro de Registros Ornitológicos. 2011. Listas das aves do Brasil.
10ª Edição, Versão 25/01/2011. Disponível em <http://www.cbro.org.br>. Acesso em:
[9/12/2013].
Clarke, K. R. Non-parametric multivariate analyses of changes in community structure.
Australian Journal of Ecology, v. 18, p. 117-143, 1993.
Dufrêne, M. & Legendre, P. Species assemblages and indicator species: the need for a
flexible asymmetrical approach. Ecological Monographs, v. 67, p. 345-366, 1997.
Espírito-Santo, M. M.; Sevilha, A. C.; Anaya, F. C.; Barbosa, R.; Fernandes, G. W.;
Sanchez-Azofeifa, G. A.; Scariot, A.; Noronha, S. E.; Sampaio, C. A. Sustainability of

42
tropical dry forests: two case studies in southeastern and central Brazil. Forest Ecology
and Management, v. 258, p. 922–930, 2009.
Espírito-Santo, M. M.; Leite, L. O.; Neves, F. S.; Nunes, Y. R. F.; Borges, M. A. Z.;
Falcão, L. A. D.; Pezzini, F. F.; Berbara, R. L.; Valério, H. M.; Fernandes, G. W.; Leite,
M. R.; Clemente, C. M. S.; Leite, M. E. Tropical Dry Forests of northern Minas Gerais,
Brazil: diversity, conservation status, and natural regeneration. Em: Sanchéz-Azofeifa, A.;
Powers, J. S.; Fernandes, G. W.; Quesada, M. (Ed). Tropical Dry Forests in the
Americas: ecology, conservation and, management. CRC Press, p. 69-82, 2013.
Gonçalves, R. M. M. Dieta e comportamento de forrageio de (Hellmayr, 1924) (Aves:
Thamnophilidae) em relação à disponibilidade de presas em uma área de Floresta
Estacional Decidual. Dissertação de Mestrado. Universidade Estadual de Montes Claros,
Montes Claros-MG, 2011.
Holmes, R. T. & Recher, H. F. Determinants of guild structure in forest bird communities:
an intercontinental comparison. The Condor, v. 88, n. 4, p. 427-439. 1986.
IEF- Instituto Estadual de Florestas. Parecer técnico para a criação do Parque Estadual da
Mata Seca. Relatório técnico, Belo Horizonte – MG, 2000.
Jaksic, F. M. Abuse and misuse of the term "guild" in ecological studies. Oikos, v. 37,
p.397-400, 1981.
Janzen, D. H. Sweep samples of tropical foliage insects: effects of seasons, vegetation
types, elevation, time of day, and insularity. Ecology, v.54, p.667−701, 1973.
Karr, J. R. & Roth, R. R. Vegetation structure and avian diversity in several new world
areas. The American Naturalist, v. 105, p. 423-435, 1971.
Karr, J. R. Seasonality, resource availability, and community diversity in tropical birds
communities. The American Naturalist, v.110, n.976, p.973-994, 1976.

43
Knapp, A. K. & Smith, M. D. Variation among biomes in temporal dynamics of
abovegroung primary production. Sciense, v. 291, 481-484, 2001.
Lasky, J. R. & Keitt, T. H. Abundance of Panamanian dry-forest birds along gradients of
forest cover at multiple scales. Journal of Tropical Ecology, v. 26, p. 67–78, 2010.
Loiselle, B. A. & Blake, J. G. Resource abundance and temporal variation in fruit eating
birds along a wet forest elevational gradient in Costa Rica. Ecology, v. 72, p. 180-193,
1991.
Loiselle, B. A. & Blake, J. G. Population variation in a tropical bird community.
BioScience, v. 42, n. 11, p. 838- 845, 1992.
Loiselle, B. A. & Blake, J. G. Annual variation in birds and plants of a tropical second-
growth woodland. The Condor, v.96, p.368-380, 1994.
MacArthur, R. H. & MacArthur, J. W. On bird species diversity. Ecology, v. 42, n. 3, p.
594-598, 1961.
Madeira, B. G.; Espírito-Santo, M. M.; D’Ângelo-Neto, S.; Nunes, Y. R. F.; Sánchez-
Azofeifa, G. A.; Fernandes, G. W.; Quesada, M. Changes in tree and liana communities
along a successional gradient in a tropical dry forest in south-eastern Brazil. Plant
Ecology, v. 201, p. 291-304, 2009.
Manhães, M. A. & Dias, M. M. Spatial dynamics of understorey insectivorous birds and
arthropods in a southeastern Brazilian Atlantic woodlot. Brazilian Journal of Biology, v.
71, n. 1, p.1-7. 2011.
Martin, T. E. & Karr, J. Temporal dynamics of Neotropical birds with special reference to
frugivores in second-growth woods. The Wilson Bulletin, v. 98, n. l, p. 38-60, 1986.

44
Martins, F. C. Estrutura de comunidade de aves em remanescentes de Floresta
Estacional Decidual na região do Vale do Rio Paranã- GO e TO. Tese de Doutorado,
Instituto de Ciências Biológicas da Universidade de Brasília, Brasília-DF, 136 p., 2007.
May, P. G. Secondary succession and breeding bird community structure: patterns of
resource utilization. Oecologia, v. 55, n. 2, p. 208-216, 1982.
Moermond, T. C. Habitat constraints on the behavior, morphology, and community
structure of Anolis lizards. Ecology, v. 60, n. 1, p. 152-164, 1979.
McCune, B. & Mefford, M. J. Multivariate Analysis of Ecological Data. MjM Software,
Gleneden Beach, Oregon, version 3.0., 1997.
Murakami, M. Foraging mode shifts of four insectivorous bird species under temporally
varying resource distribution in a Japanese deciduous forest. Ornithologial Sciense, v. 1,
p. 63–69, 2002.
Murphy, P. G. & Lugo, A. E. Ecology of Tropical Dry Forest. Annual Review of Ecology
and Systematics, v.17, p.67-88, 1986.
Nunes, Y. R. F.; Luz, G. R.; Souza, S. R.; Silva, D. L; Veloso, M. D. M; Espírito-Santo,
M. M; Santos, R. M. Floristic, structural, and functional group variations in tree
assemblages in a Brazilian Tropical Dry Forest: effects of successional stage and soil
properties. Em: Sanchéz-Azofeifa, A.; Powers, J. S.; Fernandes, G. W.; Quesada, M. (Ed).
Tropical Dry Forests in the Americas: ecology, conservation and, management. CRC
Press, p. 325-350, 2013.
Neves, F. S. Dinâmica espaço-temporal de insetos associados a uma Floresta
Estacional Decidual. Tese de Doutorado, Universidade Federal de Minas Gerais, Belo
Horizonte-MG, 139 p., 2009.

45
Pezzini, F. F.; Ranieri, B. D.; Brandão, D. O.; Fernandes, G. W.; Quesada, M.; Espírito-
Santo, M. M.; Jacobi, C. M. Changes in tree phenology along natural regeneration in a
seasonally dry tropical forest. Plant Biosystems, p. 1-10, 2014.
Pezzini, F. F. Fenologia e características reprodutivas em comunidades arbóreas de
três estágios sucessionais em Floresta Estacional Decidual do norte de Minas Gerais.
Dissertação de Mestrado. Universidade Estadual de Montes Claros, 130 p., 2008.
Piratelli, A. & Pereira, M. R. Dieta de aves na região leste de Mato Grosso do Sul, Brasil.
Ararajuba, v. 10, p.131–139, 2002.
Poulin, B.; Lefebvre, G.; McNeil, R. Characteristics of feeding guilds and variation in diets
of bird species of three adjacent tropical sites. Biotropica, v. 26, n. 2, p. 187-197, 1994.
Quesada, M.; Sanchez-Azofeifa, G. A.; Alvarez-Añorve, M.; Stoner, K. E.; Avila-
Cabadilla, L.; Calvo-Alvarado, J.; Castillo, A.; Espírito-Santo, M. M.; Fagundes, M.;
Fernandes, G. W.; Gamon, J.; Lopezaraiza-Mikel, M.; Lawrence, D.; Morellato, L. P. C.;
Powers, J. S.; Neves, F. S.; Rosas-Guerrero, V.; Sayago, R.; Sanchez-Montoya, G.
Succession and management of tropical dry forests in the Americas: review and new
perspectives. Forest Ecology and Management, v. 258, p. 1014–1024, 2009.
R Development Core Team (2013). R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria. Disponível em:
<http://www.R-project.org>.
Ridgely, R.S. & Tudor, G. The birds of South America: the oscines passerines. v. I.
University of Texas Press, Austin, 1989, 596 p.
Ridgely, R.S. & Tudor, G. The birds of South America: the suboscine passerines. v. II.
University of Texas Press, Austin, 1994, 940 p.
Robinson, S. K. & Holmes, R. T. Foraging behavior of forest birds: the relationships
among search tactics, diet, and habitat structure. Ecology, v. 63, p.1918-193, 1982.

46
Santos, R. M; Vieira, F. A.; Fagundes, M.; Nunes, Y. R. F.; Gusmão, E. Riqueza e
similaridade florística de oito remanescentes florestais no norte de Minas Gerais, Brasil.
Revista Árvore, v. 31, n. 1, p. 135-144, 2007.
Scariot, A. & Sevilha, A. C. Biodiversidade, estrutura e conservação de florestas
estacionais deciduais no Cerrado. In: Scariot, A.; Souza-Silva, J. C.; Felfili, J. M. (Ed)
Cerrado: ecologia, biodiversidade e conservação. Brasília: Ministério do Meio
Ambiente, p.122-129, 2005.
Sick, H. Ornitologia Brasileira. Rio de Janeiro: Ed. Nova Fronteira, 1997. 999 p.
Silveira, M. H. B. & Machado, C. G. Estrutura da comunidade de aves em áreas de
caatinga arbórea na Bacia do Rio Salitre, Bahia, Brasil. Revista Brasileira de
Ornitologia, v. 20, n. 3, p. 161-172, 2012.
Sekercioglu, C. H.; Ehrlich, P. R.; Daily, G. C.; Aygen, D.; Goehring, D.; Sandi, R. F.
Disappearance of insectivorous birds from tropical forest fragments. Proceedings of the
National Academy of Sciences, v. 99, n. 1, p. 263–267, 2002.
Stotz, D. F.; Fitzpatrick, J. W.; Parker III, T.; Moskovits, D. K.. Neotropical Birds:
Ecology and conservation. Univrsity of Chicago Press, Chicago, 1996.
Stouffer, P. C. & Jr. Bierregaard, R. O. Use of amazonian forest fragments by understory
insectivorous birds. Ecology, v. 76, n. 8, p. 2429-2445, 1995.
Willson, M. F. Avian community organization and habitat structure. Ecology, v. 55, p.
1017-1029, 1974.
Wolda, H. Insect seasonality: why? Annual Review of Ecology and Systematics, v. 19, p.
1−18, 1988.

47
9. APÊNDICES
Apêndice 1. Relação das coletas realizadas em uma Floresta Estacional
Decidual do Parque Estadual da Mata Seca (PEMS), localizado no norte de
Minas Gerais, Brasil, entre os anos de 2007 e 2013. São informados a data
(mês/ano) de cada coleta, o período de realização de cada coleta, em
relação à sazonalidade climática (Final da estação seca - FS; Início da
estação chuvosa - IC; Final da estação chuvosa - FC; e Início da estação
seca - IS), e em relação a ano (N=6; cada ano equivale ao agrupamento de
quatro coletas considerando o início e fim de cada estação).
Data (mês/ano) Período de coleta Ano
Outubro/2007 Final da estação seca - FS
1 Dezembro/2007 Início da estação chuvosa - IC
Abril/2008 Final da estação chuvosa - FC
Junho/2008 Início da estação seca - IS
Outubro/2008 Final da estação seca - FS
2 Dezembro/2008 Início da estação chuvosa - IC
Março/2009 Final da estação chuvosa - FC
Junho/2009 Início da estação seca - IS
Outubro/2009 Final da estação seca - FS
3 Dezembro/2009 Início da estação chuvosa - IC
Abril/2010 Final da estação chuvosa - FC
Junho/2010 Início da estação seca - IS
Outubro/2010 Final da estação seca - FS
4 Dezembro/2010 Início da estação chuvosa - IC
Abril/2011 Final da estação chuvosa - FC
Junho/2011 Início da estação seca - IS
Setembro/2011 Final da estação seca - FS
5 Dezembro-Janeiro/2011 Início da estação chuvosa - IC
Abril/2012 Final da estação chuvosa - FC
Junho/2012 Início da estação seca - IS
Outubro/2012 Final da estação seca - FS
6 Dezembro/2012 Início da estação chuvosa – IC
Abril/2013 Final da estação chuvosa – FC
Junho/2013 Início da estação seca - IS

48
Apêndice 2. Lista das espécies de aves insetívoras que utilizam o sub-bosque em uma Floresta Estacional Decidual do Parque Estadual da Mata
Seca (PEMS), no norte de Minas Gerais, Brasil. São apresentadas as ordens, famílias e nomes científicos das espécies de aves e seus respectivos
valores de abundância em cada estação (FS: final do período seco; IC: início do período chuvoso; FC: final do período chuvoso; e IS: início do
período seco), ano (N= 6) e estágio (Inicial, Intermediário e Tardio).
Ordem/Família/Espécie Estágios Ano 1
Ano 2
Ano 3
Ano 4
Ano 5
Ano 6
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
Galbuliformes
Bucconidae
Nystalus maculatus
Inicial - - - -
- - - -
2 - - 1
2 1 - -
- 1 - -
- - 1 1
Intermediário - - - -
- - 2 1
- - 1 1
1 - 1 1
- - - -
- - - 3
Tardio - - - -
- 1 - -
- - - -
- 1 - -
1 - - 1
- 1 - -
Piciformes
Picidae
Picumnus pygmaeus
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário - - - -
- - - 1
1 - - -
1 - - 2
- - - 1
- - - -
Tardio - - - -
- - - -
- - - -
1 - - 1
1 - - 2
2 - - 1
Picumnus cirratus
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário - - 1 -
- - - -
- - - -
- - - 2
- - - -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Picumnus albosquamatus
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário - - 1 -
- - 1 -
- - - -
1 - - -
- - - -
- - 1 -
Tardio - - 1 -
1 - - -
- - - -
- - - -
- - - -
- - - -
Continua

49
Ordem/Família/Espécie Estágios Ano 1
Ano 2
Ano 3
Ano 4
Ano 5
Ano 6
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
Veniliornis passerinus Inicial - - - - - - - - - - - 1 - - - - 1 - - - - - - -
Intermediário 1 - - - - - - - - - - - - - - - - - 1 - - - - -
Tardio - - - - - - - 1 - - - 1 - - 2 - - - - - - - - -
Piculus chrysochloros
Inicial - - - -
- - - -
- - - 1
- - - -
- - - 1
- - - -
Intermediário - - - -
- - - -
- - - -
- - - -
- - - -
1 - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Colaptes melanochloros
Inicial - - - -
1 - - -
- - - -
1 - - -
- - - -
- - - -
Intermediário - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Celeus flavescens
Inicial - - - -
- - 1 -
- - - -
- - - -
- - - -
- 1 - -
Intermediário - - - 1
- - - 1
- 1 - -
- - - -
- - - -
- 1 - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Passeriformes
Thamnophilidae
Myrmorchilus strigilatus
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário - - - -
- - - 2
- - - -
- - - -
- - 1 -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Formicivora melanogaster
Inicial - - - -
- - - -
- - - -
- - - -
4 - 2 3
3 2 2 2
Intermediário 1 - 2 5
1 2 3 3
6 2 4 2
2 - 1 6
4 1 6 2
4 - 2 3
Tardio - - 1 3
- 1 - 2
2 2 1 1
4 1 3 4
1 1 1 4
2 2 2 6
Continua
50
Continua
Ordem/Família/Espécie Estágios Ano 1
Ano 2
Ano 3
Ano 4
Ano 5
Ano 6
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
Herpsilochmus sellowi Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário - - - -
- - - -
- - - -
1 - - -
2 - - 2
1 - - 1
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Herpsilochmus atricapillus
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário - - - -
1 - 4 -
- - - -
- - - -
1 - 1 -
2 - - -
Tardio - - - -
- - - -
- - - -
- - 1 -
- - - -
- - - -
Sakesphorus cristatus
Inicial - - - -
- - - -
- - - -
- - - -
1 - - -
- - - -
Intermediário - - 1 1
- - - -
- - - -
- - - 1
2 - - 3
1 - - 1
Tardio - - - -
- - - -
- - - -
- - - 1
2 - - 3
- - - 1
Thamnophilus capistratus
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário - - - -
- - - -
- - - 1
- - - -
2 - - -
- - - 1
Tardio - - - -
- - - -
- - - -
- - - 1
- - - 1
- - - 1
Thamnophilus pelzelni
Inicial 2 - - -
- - - 1
- - - -
1 1 - -
3 - 2 2
5 - 1 3
Intermediário 9 4 2 4
6 3 5 1
7 2 4 4
6 5 5 8
6 2 3 4
1 4 4 7
Tardio 10 3 4 10
10 4 6 6
6 1 10 4
8 1 7 8
8 4 3 11
7 3 11 3
Dendrocolaptidae
Sittasomus griseicapillus
Inicial - - - -
- - 1 1
- - 1 -
- - - 1
1 - 1 -
1 - 3 -
Intermediário 1 1 5 5
1 - 4 3
- 1 7 4
- 1 4 -
5 - 4 1
- - 6 2
Tardio 2 1 7 7
1 1 6 6
- 3 4 4
2 4 3 4
2 3 4 8
3 2 5 2

51
Ordem/Família/Espécie Estágios Ano 1
Ano 2
Ano 3
Ano 4
Ano 5
Ano 6
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
Campylorhamphus trochilirostris Inicial - - - -
- - - -
- - - -
- - 1 -
1 - - -
- - - -
Intermediário - - - -
- - - -
- - - -
- - - -
1 - - -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Lepidocolaptes angustirostris
Inicial 1 - 1 3
- - - 2
2 1 1 3
4 1 1 2
1 - 2 2
1 - 3 -
Intermediário - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Lepidocolaptes wagleri
Inicial - - - -
- - - -
- - - -
- - - -
1 - - -
- - - -
Intermediário 1 - - 3
1 - 1 1
- - 1 -
1 - - 1
2 - 1 2
1 1 - -
Tardio 3 - 2 3
1 2 - 1
1 - 1 1
- - - 2
- - 1 2
2 1 2 2
Dendrocolaptes platyrostris
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
4 - - -
Intermediário 1 - - -
1 - - -
- - - -
- 1 1 -
1 - - -
- 1 2 -
Tardio 2 2 1 1
1 1 1 3
- 1 2 2
1 1 1 3
1 - 1 -
3 1 - -
Xiphocolaptes f. franciscanus
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário - 1 - -
- - - -
- - - -
- - - -
- - - -
- - 1 -
Tardio 1 - - -
- 1 - -
1 1 - -
- - - 2
- 1 1 -
1 - - -
Furnariidae
Synallaxis frontalis
Inicial - - - -
- - - -
- - 1 -
- - - -
- - - -
- 2 - -
Intermediário 1 - - -
- - - -
- - - -
- 1 - -
- - 2 -
- - - -
Tardio - - - 1
- - - -
- - - -
- - - -
- - - -
- - - -
Continua

52
Ordem/Família/Espécie Estágios Ano 1
Ano 2
Ano 3
Ano 4
Ano 5
Ano 6
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
Synallaxis scutata Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário - 1 - -
- 1 - -
- - - -
- - - -
- - 1 -
1 - - -
Tardio - - - 2
- - - 1
- - - -
- - - 1
- - - -
- - - 2
Tityridae
Myiobius barbatus
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário - - - -
- - - -
- - - -
- - - -
- - - -
- - - 1
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Pachyramphus viridis
Inicial - - - -
- 1 - -
- - - -
- - - -
- - - -
- - - -
Intermediário - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Pachyramphus polychopterus
Inicial - 5 1 -
- 2 - -
7 4 1 -
- 1 - -
- 1 - -
- 2 2 -
Intermediário - - 2 -
- - 4 -
3 - 1 -
- 1 2 -
- 1 - -
- - 1 -
Tardio - - - -
- - 1 -
- 1 - -
- - - -
- - - -
- - - -
Pachyramphus validus
Inicial - 1 - -
- 1 - -
- - - -
- - - -
- - - -
- - - -
Intermediário - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Rhynchocyclidae
Leptopogon amaurocephalus
Inicial - - - -
- 1 - -
- - - -
- - - -
- - - -
- - 1 -
Intermediário - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Tardio - - - -
- 1 - -
- - - -
- - - -
- - - -
- - - -
Continua

53
Ordem/Família/Espécie Estágios Ano 1
Ano 2
Ano 3
Ano 4
Ano 5
Ano 6
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
Tolmomyias sulphurescens Inicial - - 1 -
- - - -
2 - 1 1
1 1 - -
- - - -
- - - -
Intermediário 3 - 2 1
3 - - -
1 1 1 -
- - - 1
- 1 - -
- - - -
Tardio 3 - - -
1 1 - -
1 - 1 -
4 1 - 1
- 1 - -
1 2 - 1
Tolmomyias flaviventris
Inicial 1 - - -
- - - -
- - 1 -
- - - -
- - - -
- 3 1 -
Intermediário - - 3 3
3 1 1 6
4 4 8 5
4 3 1 2
4 2 5 1
1 2 5 3
Tardio 2 - 1 1
- 1 - -
2 1 2 1
6 1 2 3
1 - 1 7
1 - 2 2
Hemitriccus margaritaceiventer
Inicial - - - -
- - - -
- - - -
- - - -
- - - 1
2 1 2 1
Intermediário - - - 3
- - - 1
- 1 - -
2 1 - -
- 1 4 5
3 2 1 2
Tardio 1 - - -
- - - -
- - - -
1 - - -
- - - -
1 - - 1
Tyrannidae
Camptostoma obsoletum
Inicial 2 - - 1
- - 1 -
- - - -
- - - -
- - 1 -
- - 1 -
Intermediário - - - 1
- - 1 -
- - 1 -
3 - 1 3
1 - - -
- - - -
Tardio - - - 1
- - 1 -
- - 2 -
- - - -
- - - -
1 - - -
Myiopagis viridicata
Inicial - 5 1 -
- 3 6 -
7 3 8 -
- - 3 -
- 4 3 -
- 1 2 -
Intermediário - 1 10 -
- - 2 -
3 3 13 -
- 4 1 -
- 1 - -
- 2 6 -
Tardio - 2 6 -
- - 4 -
9 3 10 -
- 7 1 -
- 2 1 -
- - 11 -
Phaeomyias murina
Inicial - - - -
1 - 1 -
- - - -
- - - -
- - - -
- - - -
Intermediário - - - -
- - 2 -
- - - -
- - - 2
- - 1 -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Continua

54
Ordem/Família/Espécie Estágios Ano 1
Ano 2
Ano 3
Ano 4
Ano 5
Ano 6
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
Myiarchus swainsoni Inicial 1 - - -
- - 2 -
- - - 1
1 - - -
- - - -
- - 1 -
Intermediário - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Myiarchus ferox
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- 1 - -
Intermediário - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Myiarchus tyrannulus
Inicial 3 1 - 2
1 - 2 4
- 3 - 2
- - 1 -
5 - 1 7
2 - 2 -
Intermediário 6 - 2 -
4 - 3 -
3 - - -
1 1 - -
1 - 1 -
- 1 - -
Tardio - - - -
- - - 1
1 - - -
- - - -
1 - - 2
1 - - 1
Casiornis fuscus
Inicial - - - -
- - - -
- - 1 1
- - - -
- - - -
1 - 1 -
Intermediário 1 1 1 2
2 4 - -
3 1 1 -
- 3 - 1
- - 4 -
- 1 - -
Tardio - - - -
- 1 - -
2 1 - -
- - - -
- - - -
1 3 - -
Myiodynastes maculatus
Inicial 14 3 2 2
7 4 - 1
5 3 - -
3 2 - -
- 1 1 -
6 - - -
Intermediário - - - 1
1 - 1 -
1 1 - -
1 1 - -
- 2 3 -
- 4 - -
Tardio - - - -
2 1 - -
3 1 - -
3 1 - -
- - - -
1 - - -
Cnemotriccus fuscatus
Inicial - - - -
- - - -
- - 2 -
- 1 - -
- - - -
2 - 1 2
Intermediário 2 - 1 -
- - 1 -
- - 4 -
- - - -
- - 1 -
2 - 4 -
Tardio - - - -
- - - -
- - - -
- - - -
- - 1 -
- - 2 2
Continua

55
Ordem/Família/Espécie Estágios Ano 1
Ano 2
Ano 3
Ano 4
Ano 5
Ano 6
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
Lathrotriccus euleri Inicial - - - -
- - - -
- - 1 -
- 1 - -
- - 1 -
- - 2 1
Intermediário 2 - 2 -
2 2 - 1
- 2 - -
1 1 1 3
- 1 - -
1 - - -
Tardio 10 - - -
3 - 1 -
4 3 1 -
1 4 1 -
- 6 1 -
6 4 2 -
Vireonidae
Cyclarhis gujanensis
Inicial 1 - - -
- - - -
- - - -
- - - -
1 - - 1
- - - -
Intermediário 2 - - 2
- - - 1
- - - -
- - - 2
- - - -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - 1
Hylophilus amaurocephalus
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
1 - - -
Intermediário - - - -
- - - -
- - - -
1 - - -
1 - - -
- - - -
Tardio - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Corvidae
Cyanocorax cyanopogon
Inicial 3 - 1 -
- - - -
- - 1 1
- - - -
- - - -
3 - 1 -
Intermediário - - - 1
- - 1 -
- - - 1
- - - -
- - - -
- - - -
Tardio - - - -
- - - 1
- - - -
- - - -
- - - 1
- - - -
Troglodytidae
Troglodytes musculus
Inicial - - - -
- - - -
- - - -
- - - -
- - - -
- - - -
Intermediário 2 - 1 1
- 1 4 -
2 10 2 1
2 2 1 -
- 2 2 -
2 - - 1
Tardio 1 - - 3
2 1 1 1
2 - - 3
3 1 3 3
- 1 - 1
3 - 2 1
Polioptilidae
Polioptila plumbea
Inicial - - - -
- - - -
2 - - -
- - - -
2 - - -
- - - -
Intermediário - - - 3
- - - -
- - - -
- - - -
1 - - -
- - - 1
Tardio - - - -
- - - -
- - - -
- 1 - -
- - - -
- - - -
Continua

56
Ordem/Família/Espécie Estágios Ano 1
Ano 2
Ano 3
Ano 4
Ano 5
Ano 6
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
FS IC FC IS
Polioptila dumicola Inicial - - - -
- - - -
- - - 1
- - - -
- - - -
- - - 1
Intermediário 1 - - -
- - - -
- 2 1 -
- - - 1
1 - - 2
- - - -
Tardio - - - -
- - - -
1 - - -
- - - -
- - - -
- - - -
Turdidae
Turdus amaurochalinus
Inicial 5 5 8 -
8 5 1 2
2 5 5 1
15 3 2 6
- 5 2 -
1 5 2 -
Intermediário 1 1 1 -
- 1 4 -
3 1 - -
- 2 - -
- 2 - -
- 2 1 -
Tardio 2 - 1 -
- - - -
- - - -
1 - - -
- - 2 -
1 - - -
Turdus a. crotopezus
Inicial - - 1 -
- - 1 -
- - - 1
- - - -
- - - -
- - - -
Intermediário - - 1 -
- - - -
- - 1 -
- - - -
- - - -
- - - -
Tardio - - - -
- - - -
- - - -
1 - - -
- - - -
- - 1 -
Thraupidae
Lanio pileatus
Inicial 14 4 3 -
4 5 2 -
12 4 - 2
2 4 4 2
6 6 5 10
10 9 - 3
Intermediário 2 2 1 -
1 3 - 8
7 2 1 8
2 1 2 1
4 1 1 4
1 7 2 11
Tardio - - - 3
- 2 - -
4 - 3 3
4 2 4 5
3 1 1 -
2 1 2 3
Parulidae
Basileuterus flaveolus
Inicial 1 - - -
- - - -
- - - 1
- - - -
- - 1 -
- - 1 -
Intermediário 2 - - -
- - 1 -
- - - -
- - - 1
- - 3 -
- - 1 -
Tardio - - 1 -
- - - -
- - 1 -
- - - -
- - - -
- - 1 -

57