universidade vale do rio doce especializaÇÃo...
TRANSCRIPT

UNIVERSIDADE VALE DO RIO DOCE
FACULDADE DE CIENCIAS DA SAÚDE
ESPECIALIZAÇÃO LATU SENSO – ANALISES CLINICAS E GESTÃO DE LABORATORIO
Pedro Henrique Ferreira Marçal
CITOCINAS INTRACITOPLASMATICAS EM PACIENTES COM HANSENIASE E CONTATOS INTRA E EXTRADOMICILIARES: ESTUDO DE CASO
Governador Valadares
2011

2
Pedro Henrique Ferreira Marçal
CITOCINAS INTRACITOPLASMATICAS EM PACIENTES COM HANSENIASE E CONTATOS INTRA E EXTRADOMICILIARES: ESTUDO DE CASO
Trabalho de Conclusao de curso para Obtenção de grau de especialista em analyses clinicas,
Apresentada a faculdade de ciencias da saude da Universidade vale do Rio doce
Oritentador (a): Dra. Lúcia Alves de Oliveira Fraga
Governador Valadares 2011

3
RESUMO
A hanseníase é uma doença infecciosa crônica que afeta principalmente a pele e nervos
periféricos (Lockwood, 2004), com manifestações clinicas que dependem da resposta imune do
hospedeiro ao Mycobacterium leprae (SARNO et al., 1988).
Atualmente não existe nenhum teste laboratorial sensível e específico para o diagnóstico
da infecção assintomática pelo Mycobacterium leprae ou para prever a progressão para
hanseníase entre indivíduos expostos (STEFANI, 2008; ARAOZ et al., 2006). Esses testes,
devem requerer tanto os ensaios baseados em anticorpos, como ensaios de células T (DUTHIE et
al., 2007; DUTHIE et al., 2008). Recentemente vários antígenos inéditos de Mycobacterium
leprae têm sido testados como reagentes diagnósticos para hanseníase (GELUK et al., 2005;
GELUK, OTTENHOFF, 2006; GELUK et al., 2008; REECE et al., 2006; SPENCER et al.,
2005).
Produção de INF-γ induzida por Mycobacterium leprae em células mononucleares do
sangue periférico, tem sido usado para mensurar resposta immune celular (SAMPAIO, 1991).
Produção de IL4 e IL10, levam a ativação de linfócitos B e produção de anticorpos contra
antígenos do Mycobacterium leprae, sendo o bacilo intracelular obrigatório, a imunidade humoral
instalada, conduz ao polo de susceptibilidade à enfermidade (SARNO et al., 1988).
A proposta deste estudo inicial foi avaliar o perfil imunológico de individuos
diagnosticados com hanseníase, e contatos intradomiciliares, residentes em Governador
Valadares – MG, através da análise do padrão de citocinas intracitoplasmáticas IFN-γ, IL-4, IL-
10 em populações de leucócitos do sangue periférico dos mesmos, após incubação com antígenos
específicos do M. leprae, auxiliando na compreensão dos mecanismos imunológicos associados a
variações na susceptibilidade à infecção pelo M. leprae,
Os resultados mostraram que, frente a estímulo com M. leprae, paeientes multibacilares e
um contato de paciente multibacilar apresentaram alta porcentagem de células T CD3+IL4+
(0,98%; 0,86% e 0,77%), respcctivamente, bem como células T CD3+IL10+ (0,72%, 0,81% e
0,7%), respectivamente. Quanto à produção de IFN-γ, pacientes paucibacilares, um contato de
paciente multibacilar e um contato de paciente paucibacilar, apresentaram alta porcentagem de
células produtoras (1,32%; 1,02%, 1,1% e 0,92%), respectivamente. Interressantemente, o
contato extracomiciliar, que apresentara alta frequência de linfócitos T CD3+IL4+IL10+, durante

4
o experimento controle (sem estimulo) e com o estímulo de PHA, quando suas células entraram
em contato com o antigeno M.leprae, houve uma modulação da resposta, ocorrendo uma
diminuição na frequencia de células produtoras de IL4 e IL10 (0,42% e 0,39%), respectivamente,
e um aumento na porcentagem de linfócitos T CD3+ IFN-γ+ (0,78%).
Palavras-chave: Hanseníase, Mycobacterium Leprae, citocinas intracitoplasmáticas.

5
Dedico este trabalho ao Grande Deus
Que em todo tempo permanence com
Suas maos estendidas sobre nós.

6
AGRADECIMENTOS
A Dra. Lúcia, que não mede esforços para o crescimento na pesquisa nesta universidade e
em seus alunos;
Aos amigos e tecnicos do NPqImuno, cujo auxílio foi excencial para a finalização dos
resultados;
Aos familiares amigos, por acreditarem em mim, durante todo o tempo.

7
SUMÁRIO
I INTRODUÇAO…………………………………………………………………………9
II REVISAO DA LITERATURA……………………………………………………...11
2.1 HANSENIASE……………………………………………………………………….11
2.2 ETIOLOGIA………………………………………………………………………….11
2.3 CELULAS DE SHWAN……………………………………………………………..12
2.4 FORMAS CLINICAS………………………………………………………………..13
2.5 IMUNOLOGIA………………………………………………………………………14
2.6 DIAGNOSTICO……………………………………………………………………...15
2.7 TRATAMENTO……………………………………………………………………...16
2.8 CITOMETRIA DE FLUXO APLICADA A HANSENIASE………………………..16
III METODOLOGIA…………………………………………………………………...20
3.1 GRUPO POPULACIONAL DO ESTUDO………………………………………….20
3.2 TERMO DE CONSENTIMENTO LIVRE ESCLARECIDO………………………..21
3.3 COLETA DE SANGUE PARA OS ENSAIOS IMUNOLOGICOS………………...21
3.4 DETECÇAO DE CITOCINAS INTRACITOPLASMATICAS……………………..21
3.5 OBTENÇAO DE DADOS NO CITOMETRO DE FLUXO E ANALISE DOS
RESULTADOS…………………………………………………………………………..21
IV RESULTADOS E DISCUSSAO……………………...………………………….…24
4.1 PRODUÇÃO DE CITOCINAS INTRACELULARES (IL4, IL10 E INF-Γ) EM
LINFÓCITOS T CD3+ DE INDIVÍDUOS MULTIBACILARES, PAUCIBACILARES,
CONTATOS E SADIO APÓS CULTURA IN VITRO SEM
ESTIMULAÇÃO................................................................................................................24
4.2 PRODUÇÃO DE CITOCINAS INTRACELULARES (IL4, IL10 E INF-Γ) EM
LINFÓCITOS T CD3+ DE INDIVÍDUOS MULTIBACILARES, PAUCIBACILARES,
CONTATOS E SADIO APÓS CULTURA IN VITRO COM ESTÍMULO DE
SEB.....................................................................................................................................26
4.3 PRODUÇÃO DE CITOCINAS INTRACELULARES (IL4, IL10 E INF-Γ) EM
LINFÓCITOS T CD3+ DE INDIVÍDUOS MULTIBACILARES, PAUCIBACILARES,

8
CONTATOS E SADIO APÓS CULTURA IN VITRO COM ESTÍMULO DE
MYCOBACTERIUM LEPRAE...........................................................................................29
V CONCLUSAO………………………………………………………………………..33
VI REFERENCIAS BIBLIOGRAFICAS……………………………………………..34

9
I INTRODUÇÃO
A hanseníase, uma das mais antigas doenças humanas, ainda é um importante problema
de saúde pública para muitos países endêmicos, como o Brasil (COLE et al., 2001).
O agente causador da hanseníase, o Mycobacterium leprae foi o primeiro patógeno
bacteriano a ser identificado como causa de uma doença infecciosa humana há mais de 130 anos
(SCOLLARD et al., 2006).
A doença é transmitida de pessoa a pessoa através do convívio de indivíduos susceptíveis
com doentes das formas contagiantes, sem tratamento, com um período médio de incubação, de 2
a 5 anos, tendo como principal via de eliminação dos bacilos, a via aérea superior. É uma
patologia que apresenta um amplo espectro clínico-imunológico, variando dos pólos
imunologicamente estáveis tuberculóide e lepromatoso ao dimorfo, imunologicamente instável,
dependendo da resposta imune do indivíduo (SAPKOTA, et al., 2009).
As características imunológicas das formas espectrais da hanseníase incluem pacientes
tuberculóides que apresentam baixos títulos de anticorpos para o Mycobacterium leprae e forte
resposta imune celular específica (RIC) do tipo Th1 caracterizada por produção de citocinas
como interferon gama (IFN-γ), a doença é localizada e o paciente apresenta poucos bacilos e
lesões limitadas (até cinco lesões na pele). Pacientes com a forma clínica Virchoviana, se
encontram no pólo de extrema suscetibilidade ao M. leprae, no qual a proliferação disseminada
do bacilo resulta em lesões da pele, difusamente distribuídas, associadas a uma potente resposta
imune humoral com produção de níveis elevados de anticorpos anti-M. Leprae (SCOLLARD et
al., 2006; LYON, et al. 2008). Em outras formas clínicas, tais como dimorfos, dimorfos-
tuberculóide, dimorfos-dimorfos e dimorfos-virchovianos, a progressiva redução da resposta
imune mediada por célula é acompanhada por lesões de pele e nervos, aumento da carga bacilar e
dos níveis de anticorpos (OTTENHOFF, 1994a; OTTENHOFF, 1994b).
Os esforços da Organização Mundial da Saúde para eliminar a hanseníase até o ano 2000
foram baseados em importantes avanços na terapia antimicobacteriana na década de 80. Apesar
do grande declínio na prevalência observado na maioria dos países endêmicos na última década, a
detecção de casos novos permanece alta (WHO, 2010). O desenvolvimento de testes diagnósticos
aplicáveis em situação de campo é uma prioridade em pesquisa, visto que o diagnóstico da
hanseníase é baseado principalmente em manifestações clínicas e a escassez de sintomas no

10
início da doença pode contribuir para erros no diagnóstico ou para o sub-diagnóstico (STEFANI,
2008).
Atualmente não existe nenhum teste laboratorial sensível e específico para o diagnóstico
da infecção assintomática pelo Mycobacterium leprae ou para prever a progressão para
hanseníase entre indivíduos expostos (STEFANI, 2008). Esses testes, devem requerer tanto os
ensaios baseados em anticorpos, como ensaios de células T. Recentemente vários antígenos
inéditos de Mycobacterium leprae têm sido testados como reagentes diagnósticos para hanseníase
(ARAOZ et al., 2006; DUTHIE et al., 2007; DUTHIE et al., 2008; GELUK et al., 2005; GELUK,
OTTENHOFF, 2006; GELUK et al., 2008; REECE et al., 2006; SPENCER et al., 2005).
O principal objetivo desse estudo inicial foi avaliar indicadores imunológicos associados à
proteção e/ou adoecimento, em pacientes com hanseníase e seus contatos intradomiciliares,
residentes em Governador Valadares-MG, que possam servir de ferramenta para estratégias de
controle da doença.
Sabendo-se que nas doenças transmissíveis a distribuição de casos não ocorre ao acaso na
população, estando os casos agregados em territórios definidos como consequência da
transmissão; torna-se relevante a busca de novos casos e consequentemente o investimento em
métodos complementares de diagnóstico.
A avaliação de parâmetros imunológicos neste estudo irá auxiliar na compreensão dos
mecanismos imunológicos associados a variações na susceptibilidade à infecção pelo M. leprae,
através da análise do padrão de citocinas intracitoplasmáticas IFN-γ, IL-4, IL-10 em populações
de leucócitos do sangue periférico dos casos e indivíduos comunicantes, após incubação com
antígenos específicos do M. leprae.

11
II REVISAO DA LITERATURA
2.1 HANSENÍASE
A hanseníase é uma doença infecciosa sistêmica de curso crônico, que afeta
principalmente os nervos periféricos, a pele e a mucosa das vias aéreas superiores. Admite-se que
a via de entrada do Mycobacterium leprae é o trato respiratório superior, por onde atinge a
mucosa e dissemina-se pelo organismo. A hipótese da via de transmissão aérea é confirmada pela
grande quantidade de bacilos íntegros isolados de secreções nasais de pacientes. Também é
considerada uma doença incapacitante, uma vez que ocorre dano neural causado pela invasão do
M. leprae e comprometimento do sistema nervoso periférico, irreversível em vários casos
(GALLO et al., 2005).
Dados oficiais da Organização Mundial de Saúde mostram que mais de 213.000 pessoas
estão infectadas, principalmente na Ásia e na África, com aproximadamente 249.000 novos casos
relatados em 2008. Áreas como: Brasil, Angola e Índia apresentam alta endemicidade da doença.
O Brasil encontra-se como o primeiro país com maior taxa de incidência da doença (WHO,
2009).
A resposta imune do hospedeiro frente ao bacilo determina a evolução clínica na
hanseníase. Acredita-se que mais de 99% da população apresenta resposta imune adequada contra
a hanseníase e não desenvolve a doença (SPIERINGS, 2000). Entre os sintomáticos, a doença
pode se manifestar em diferentes formas clínicas.
2.2 ETIOLOGIA
O M. leprae, agente causador da hanseníase, é um patógeno intracelular obrigatório que
infecta predominantemente macrófagos e células de Schwann (BLOOM, 1986). Possui a forma
de um bastonete reto ou ligeiramente encurvado, de 1,5 a 8 micra de comprimento por 0,2 a 0,5
mícron de largura. Cora-se em vermelho pela fucsina e não se descora pela lavagem com álcool
ácido, é, portanto, um bacilo álcool-ácido resistente (BAAR) (REES, 1984). É uma bactéria de
crescimento lento, fazendo uma divisão binária a cada 12-14 dias (WHO, 1987).

12
Compartilha com outras micobactérias características como a abundância de lipídios na
forma de ácidos micólicos (ácidos graxos de elevado peso molecular) e lipoarabinomanana
(LAM) em seu envelope celular. Além disso, mais externamente, estão localizados glicolipídieos,
como o PGL-1 (glicolipídeo fenólico-1), encontrado exclusivamente no M. leprae. Sua estrutura
é composta de um trissacarídeo, fenol, fitiocerol e de ácido micoseossídico (HUNTER &
BRENNAN, 2001). Estudos anteriores já mostraram que o PGL-1 pode ser encontrado em
tecidos, no sangue circulante e na urina de doentes multibacilares. Além disso, ele também foi
detectado em tatus infectados com o M. leprae (HUNTER, STEWART & BRENNAN, 1985).
Este antígeno não apresenta reação cruzada com M. tuberculosis ou outras microbactérias e
estimula uma potente resposta IgM que é proporcional a carga bacteriana nos pacientes (CHO et
al., 1983).
Desde sua descoberta em 1873 por Armauer Hansen todas as tentativas para cultivar este
microorganismo falharam (HUNTER, 2001). Entretanto, em 1962, foi demonstrado por Shepard
que o coxim plantar de camundongos infectados com 103 a 104 bacilos promovia uma lesão
localizada de hanseníase durante 9 a 12 meses (Shepard, 1962). Outra fonte de M. leprae
utilizada para estudos é o tatu nove bandas (Dasypus novencinctus). Ele permite o crescimento do
bacilo de forma disseminada durante 18 a 24 meses comprometendo pele, nervos periféricos,
medula óssea, fígado, baço, linfonodos, pulmões, meninges e olhos. Após a purificação os bacilos
podem ser usados vivos por até uma semana ou letalmente irradiados (KIRCHHEIMER &
STORRS, 1971).
O genoma do M. leprae foi completamente seqüenciado em 2001 e gerou grande
expectativa sobre o conhecimento de sua funcionalidade na patogenia da hanseníase. Apenas
49,5% do genoma do M. leprae (1605 genes) contêm genes que codificam proteínas sendo o
restante constituído de pseudogenes ou genes degenerados (COLE et al., 2001). Quando
comparado ao M. tuberculosis percebe-se uma perda maciça de genes pelo M. leprae, o que
poderia explicar o seu longo tempo de geração e sua incapacidade de multiplicação in vitro
(VISSA & BRENNAN, 2001).
2.3 CÉLULAS DE SHWAN

13
Célula de Schwann é um tipo de célula glial que produz a mielina que envolve os axónios
dos neurónios no sistema nervoso periférico, isolando eletricamente os nervos e assim permitindo
a propagação rápida de potenciais de ação (SAPKOTA, et al., 2009).
Cada célula de Schwann envolve um segmento de um axónio, enrolando-se em volta
deste e de si própria, até cem vezes. A sua membrana celular contém lipídeos e glicoproteínas
produzidas na célula com propriedades especificas para as necessidades de isolamento dos
neurónios (RAMBUKKANA et al., 2002).
As células de Schwann (SC) são um importante alvo para a infecção por M. leprae, o que
induz à lesão do nervo, desmielinização e conseqüente incapacidade física. Experimentos
realizados com culturas de tecido nervoso (in vitro) e modelos animais (in vivo) confirmaram o
tropismo do M. leprae pelos nervos periféricos. A ligação do M. leprae induz a desmielinização
das SCs e perda axonal da condutância (RAMBUKKANA et al., 2002).
2.4 FORMAS CLINICAS
A doença apresenta um amplo espectro, variando do polo tuberculóide ao lepromatoso
com formas no boderline imunologicamente instáveis, dependendo da resposta imune do
indivíduo (SAPKOTA, et al., 2009). O teste de intradermoreação de Mitsuda utilizado no
diagnóstico é positivo para a forma clínica tuberculóide (Paucibacilar-PB), doença localizada,
com vigorosa resposta imune celular, poucos bacilos e lesões limitadas (até cinco lesões no
corpo). O teste é negativo nos pacientes com a forma clínica Virchoviana (Multibacilar-MB), o
pólo de extrema suscetibilidade ao M. leprae, no qual a proliferação disseminada do bacilo
resulta lesões da pele, difusamente distribuídas, associadas a uma potente resposta imune
humoral com produção de níveis elevados de anticorpos anti-M. Leprae (LYON, et al. 2008). Em
outras formas clínicas, tais como dimorfos, dimorfos-tuberculóide, dimorfos-dimorfos e
dimorfos-virchovianos, a progressiva redução da resposta imune mediada por célula é
acompanhada por lesões de pele e nervos, aumento da carga bacilar e dos níveis de anticorpos
(OTTENHOFF, 1994 a; OTTENHOFF, 1994b).
A imunidade humoral presente no pólo virchoviano, exibe altos títulos de anticorpos
contra antígenos do M. Leprae, principalmente o glicolipídeo-fenólico1(PGL-1), sem, contudo
conferir proteção significativa, pois o indivíduo elimina bacilos e contribui para disseminação

14
bacilar (YAMAMURA, et al., 1991; CHIN, et al., 1992; HARBOE, 1985; SIELING, 1992).
Ocorre nesses pacientes, aumento dos níveis de anticorpos dos isotipos IgM, IgG, IgE e IgA,
especialmente contra a proteína PGL-1 (CUNHA, 1998; FOSS et al., 1993; KLATSER, 1994;
BUHRER-SÉKULA et al., 2007; LYON etal., 2008). A imunidade celular, presente no polo
tuberculóide, é caracterizada pela presença de citocinas do tipo 1 (IFN-γ, IL-2 e linfotoxina) e
confere ao indivíduo resistência ao bacilo. Macrófagos e linfócitos ativados por citocinas,
formam granulomas, com a presença de células TCD4+ no centro e, linfócitos TCD8+ na periferia
(MODLIN, et al., 1988; GOULART & GOULART, 2009).
2.5 IMUNOLOGIA
Patógenos intracelulares tais como as micobactérias, são reconhecidos por células
fagocitárias da imunidade inata, direta ou indiretamente via diferentes receptores. A ativação
celular através do receptor Toll pode contribuir para os mecanismos de defesa do hospedeiro
regulando a fagocitose e ativando mecanismos antimicrobianos, tais como, a indução de
linfócitos Th1. Não existe um consenso com relação aos fatores de risco para a indução de uma
maior susceptibilidade ou resistência ao bacilo. Entretanto, alguns autores têm demonstrado que
fatores genéticos, tais como, variações nas regiões promotoras de certos genes (PARK2,
PCARG), poderiam estar associados ao desenvolvimento da forma clínica lepromatosa da
hanseníase (CHER et al., 1987).
A avaliação da imunidade celular através da detecção de citocinas intracitoplasmáticas em
células TCD4+ e TCD8+ de pacientes hansenianos indicou a aplicação testa técnica para
avaliação imunológica de indivíduos contatos de pacientes (ANTAS et al., 2004).
Como descrito anteriormente, a imunidade mediada por célula representa um fator
importante no controle da infecção pelo M. Leprae. As células Th1 produzem IFN-γ que ativa
macrófago e IL-2 que estimula o crescimento de células T antígeno-específicas, resultando na
redução da carga bacilar ou cura. Células Th2 produzem IL-4, IL-5 e IL-10 e ampliam a resposta
humoral. IL-4 estimula a produção de IgE, e ambas IL-4 e IL-10 estimulam células B e inibem a
ativação de macrófago resultando em infecção progressiva (CHER et al., 1987; SIELING et al.,
1994a).

15
Acentuando o fenômeno da imunoregulação na doença em questão, a presença da IL-10
nas lesões suprime a produção de citocinas do tipo 1 por células TCD4, com uma redução
significativa de IFN-γ e IL-2, resultando em uma resposta ineficiente aos antígenos de M. leprae
(MISRA et al., 1995). É importante salientar que, uma produção crônica de IL-10 pode levar a
uma diferenciação de células T CD4, originando uma subpopulação de células T reguladoras
(Tr1) que produzem altos níveis de IL-10 mantendo a supressão da resposta imune antígeno-
específica (ASSEMAN, 1998). Tem sido também investigado o papel imunomodulador do fator
transformador do crescimento-beta1 (TGF-β1) na hanseníase por GOULART et.al (1996). Estes
autores detectaram grande quantidade de células macrofágicas positivas para TGF- β 1 no
infiltrado virchoviano, e ao contrário, constataram ausência dessa citocina no granuloma da
forma tuberculóide. Tem sido demonstrada também, a produção de TGF- β1 ativo em
sobrenadantes de cultura de macrófagos, de portadores de hanseníase com uma produção
significativamente maior naqueles, da forma virchoviana, quando comparados com os pacientes
da forma tuberculóide (GOULART. et al, 1996; GOULART. et., 1999; GOULART. et al., 2000;
GOULART. et al., 2009 ). O TGF- β1 contrapõe os efeitos do IFN-γ na ativação da atividade
antimicrobiana mediada pelo óxido nítrico (NO) e inibe a síntese de TNF-α, dessa forma
contribuindo para a manutenção da infecção (BERMUDEZ, 1993; GRENN et al., 1994,
NELSON et al., 1991).
Recentemente, SAVAGE et al. 2008 apud STEFANI, 2008, demonstrou a presença de
macrófagos no sangue periférico capazes de induzir células T reguladoras do tipo
Foxp3+GITR+CD25+. Estes autores, reforçam a importância da compreensão dos mecanismos
derivados da atuação desses macrófagos, considerando que estas células apresentam um papel
especial no controle e exacerbação das doenças inflamatórias crônicas, como é o caso da
hanseníase. Dentro desse contexto, postula-se que a indução precoce do TGF- β 1 seria essencial
para estabelecer o curso da infecção hansênica na ausência de IFN- γ, levando a proliferação
descontrolada do bacilo dentro do macrófago, o que promoveria o desenvolvimento do padrão de
resposta do tipo 2, que é encontrada na forma virchoviana, com inibição da resposta do tipo 1
(GOULART et. al, 2002).
2.6 DIGNÓSTICO

16
O diagnóstico de hanseníase é essencialmente clínico e baseado em um ou mais dos três
sinais: manchas hipocrômicas ou avermelhada com perda de sensibilidade, nervos
periféricosespessados e bacilo álcool-ácido resistente detectado em esfregaços de pele ou biópsia
de material (WALKER, LOCKWOOD, 2006). Identificação do bacilo é feita através da
coloração do esfregaço de raspado dérmico e biópsias de pele (baciloscopia). O índice
baciloscópico (IB) é uma escala logarítimica (variando 1-6 +) quantificando a densidade de M.
leprae em esfregaços de raspado dérmico e é utilizado para avaliar a carga bacteriana para
classificação e resposta ao tratamento (PINHEIRO, 2011).
É portanto um diagnóstico baseado principalmente em manifestações clínicas e a escassez
de sintomas no início da doença pode contribuir para erros no diagnóstico ou para o
subdiagnóstico. O diagnóstico precoce e a introdução do tratamento específico são importantes
para reduzir fontes de transmissão e para prevenir doença grave com incapacidade e deficiência
física (STEFANI, 2008).
A avaliação da imunidade celular através da detecção de citocinas intracitoplasmáticas em
células TCD4+ e TCD8+ de indivíduos com hanseníase indicou a aplicação desta técnica para
avaliação imunológica de indivíduos contatos de pacientes, uma vez que permite prever a
resposta immune desse indivíduo frente ao estímulo específico com ML (ANTAS et al., 2004).
2.7 TRATAMENTO
A doença é curável e tratada com poliquimioterapia (PQT). Segundo WHO, 2010, a
rifampicina (600mg, uma vez por mês), clofazimina (300mg, uma vez por mês e 50mg / dia) e
dapsona (100mg por dia) são usadas para o tratamento de pacientes MB, por um período de um
ano. Para os pacientes PB, apenas a rifampicina (600mg, mensal) e dapsona (100mg por dia) são
empregados por seis meses.
Em indivíduos impossibilitados de tomar clofazimina ou dapsona, outros agentes como,
fluoroquinolonas, ofloxacina, moxifloxacina ou pefloxacina, ciclinamino e o macrolídeo
claritromicina, são ativos contra o M. leprae e podem potencialmente ser usados como agentes de
segunda linha (BRITTON, LOCKWOOD, 2004).
2.8 CITOMETRIA DE FLUXO APLICADA Á HANSENÍASE

17
De acordo com Shapiro 2003, a citometria de fluxo é um processo no qual são medidas
características físicas e/ou químicas de células ou de outras partículas. Possui utilidade em
diversas áreas como a Imunologia, a Hematologia, a Oncologia, a Anatomia Patológica e a
Biologia Celular. Esta técnica permite a análise de diversas características, tais como o tamanho
celular, a complexidade citoplasmática, a expressão de antígenos celulares, assim como pode ser
realizado o estudo do ciclo celular, na contagem absoluta de células e na separação de populações
celulares com fenótipos específicos (“sorting”), e o estudo de diversos parâmetros celulares como
a concentração do Ca²+ intracelular, o pH intracelular e a apoptose (BROWN e WITTWER
2000; ORFAO e BUITRAGO 1995).
Antes do uso da citometria de fluxo, as células eram visualizadas com um microscópio de
fluorescência. Embora os citômetros de fluxo estivessem disponíveis no comércio desde o início
dos anos 70, o seu uso era em primeiro lugar para a investigação, devido à raridade, ao elevado
custo e à complexidade do seu funcionamento (NICHOLSON, 2002).
É uma tecnologia com uma velocidade de análise superior à outras, visto que se consegue
ler entre 500 a 5.000 células por segundo. O citômetro de fluxo possui um computador acoplado,
o qual recebe os sinais que posteriormente serão convertidos em dados digitais para serem
representados através de gráficos monoparamétricos e biparamétricos.
A técnica consiste em uma suspensão celular que é injetada no citômetro, dirigindo-se
para o núcleo central contido na câmara de fluxo. Nesta câmara existe um líquido de
envolvimento (Sheath fluid) que não se mistura com a suspensão celular (Figura 1). No núcleo
central as células formam um fluxo laminar que mantém hidrodinâmicamente, as células
alinhadas de forma que estas passem uma a uma em frente ao feixe (RIBEIRO, 2010).

18
FIGURA 1 – A focagem hidrodinâmica forma uma fila única de células.
FONTE: RIBEIRO, 2010
A interação das células ou das partículas com o feixe luminoso origina sinais que vão ser
alcançados por detectores adequados. A informação recolhida pode agrupar-se em dois tipos
fundamentais: a que é originada pela dispersão da luz e a que está relacionada com a emissão de
luz por fluorocromos presentes na célula ou na partícula, ao serem excitados pela fonte luminosa
(Figura 2).
A dispersão da luz é um processo físico em que uma célula interage com a luz incidente e
muda a direção da luz. As características celulares que contribuem para a dispersão da luz são o
tamanho celular, a membrana celular, o núcleo e o material granular do interior da célula.
A luz não se dispersa de igual forma em todas as direções. A luz que dispersa para frente
(“forward scatter”, FSC) é uma medida do tamanho celular, enquanto a luz que é dispersa em
ângulo reto (“side scatter”, SSC) depende da complexidade celular (ORFÃO E BUITRAGO
1995).
FIGURA 2 – Esquema de funcionamento de um citômetro de fluxo.
FONTE: RIBEIRO, 2010.

19
A fluorescência é indicada como a emissão de energia luminosa pelos compostos
fluorescentes que se encontram ligados aos anticorpos monoclonais. Estes compostos absorvem
energia, fazendo com que um elétron passe a um nível energético superior. No momento em que
esse elétron regressa ao seu estado de repouso, emite um fóton e libera energia (Orfão e Buitrago
1995).
Os sinais vão depois ser convertidos em dados digitais para serem representados através
de gráficos monoparamétricos e biparamétricos (ORFÃO E BUITRAGO 1995).
Toda esta tecnologia aumenta a sensibilidade e a especificidade da identificação de
moléculas de superfície celular que atuam como marcadores fenotípicos que podem ser
associados às funções celulares.
Dessa forma, a citometria de fluxo apresenta grande aplicabilidade, uma vez que aumenta
o conhecimento ciêntífico, com relação à resposta imune de um grupo específico de indivíduos
frente e estímulos específicos como o Mycobacterium leprae.

20
III. METODOLOGIA
3.1 GRUPO POPULACIONAL DO ESTUDO
Foram incluídos neste estudo 4 casos novos de hanseníase, 4 contatos intradomiciliares, 1
contato extradomiciliar (professional de saúde que trabalha a 31 anos no CREDEN-PES) e 1
indivíduo sadio como controle negativos, residentes no município de Governador Valadares –
MG. Dois (2) dos casos novos de hanseníase eram multibacilares e foram chamados de M1 e M2,
os outros dois (2) casos novos eram paucibacilares, sendo chamados de P1 e P2. A tabela 1,
apresentada informações importantes sobre esses individuos. Dois (2) dos contatos
intradomiciliares eram contatos dos pacientes multibacilares, chamados de CM1 e CM2, já os
outros dois (2), eram contatos dos pacientes paucibacilares, CP1 e CP2. O contato extradomiciliar
foi chamado de CE e o paciente sadio de S.
Tabela 1: Características dos casos novos de hanseníse diagnosticados, participantes
do estudo.
Para fins operacionais, deve-se considerar como contato intradomiciliar toda e qualquer
pessoa que resida ou tenha residido nos últimos cinco anos com o doente. Os dados referentes à
identificação dos casos novos e contatos intradomiciliares que fizaram parte desse estudo foram
fornecidos pelo SINAN (Sistema Nacional de Agravos de Notificação) e prontuários clínicos.
Os contatos intradomiciliares foram submetidos a exame dermato-neurológico por
clínicos experientes para confirmar estar este, sem sintomas de hanseníase.
Pacientes Forma
clínica
Classificação
Operacional
Índice
Baciloscópio
(IB)
Data do
diagnóstico
Tempo de
tratamento
M1 Multibacilar Virchowiano 2,5 27/09/2010 3 meses
M2 Multibacilar Dimorfo 1,5 19/08/2010 4 meses
P1 Paucibacilar Tuberculóide 0 30/08/2010 4 meses
P2 Paucibacilar Tuberculóide 0 08/11/2010 1 mês

21
3.2 TERMO DE CONSENTIMENTO LIVRE ESCLARECIDO
O Consentimento pós-informado (TCLE) foi obtido de todos os participantes. Em caso de
menores, a inclusão só será feita após assinatura do consentimento pelos pais ou guardião.
3.3 COLETA DE SANGUE PARA OS ENSAIOS IMUNOLÓGICOS.
O sangue dos indivíduos participantes do estudo foi coletado por profissionais
qualificados da área de saúde, no CREDEN-PES, respeitando as técnicas de biossegurança, tais
como uso de luvas, material estéril e descartável. Foram coletados cerca de 30 ml de sangue
utilizando tubos heparinizados, estéreis a vácuo e descartáveis (VACUTAINER, Grand Island,
NY, USA), para mensuração de citocinas intracitoplasmáticas.
3.4 DETECÇÃO DE CITOCINAS INTRACELULARES
O sangue dos indivíduos foi colocado em RPMI-suplementado e em contato com o
antígeno ML por tempo variável. Brefeldina A (10ng/ml) foi adicionada 4 horas antes do final da
cultura. As células foram lavadas com PBS e incubadas por 30 minutos, ao abrigo da luz, a
temperatura ambiente, com anticorpos monoclonais de superfície, a saber CD3FITC. Em seguida,
as amostras foram lavadas com PBS por centrifugação (400g, 7 minutos) e o sobrenadante
desprezado. Após lise das hemácias, fixação e permeabilização com solução de lise, formaldeído
(4%) e solução de soponina (0.5%), respectivamente, as células foram incubadas com anticorpos
monoclonais anti-citocinas (IL4, IL10 e INF-γ), marcados com PE por 30 minutos, ao abrigo da
luz, a temperatura ambiente. Após incubação as células foram lavadas com PBS e, em seguida
adicionada solução fixadora (MFF- 10g/L). A leitura foi realizada em Citometro de Fluxo
Beckman Coulter EPICS XL-MCL.
3.5 OBTENÇAO DE DADOS NO CITOMETRO DE FLUXO E ANALISE DOS
RESULTADOS

22
As amostras foram analisadas em citômetro de fluxo Beckman Coulter EPICS XL-MCL.
A aquisição dos dados e análise dos resultados foi realizada utilizando-se o software do
equipamento denominado EXPO 32. Foram coletados 50.000 eventos para a análise após
estímulação in vitro. A identificação das populações celulares de interesse, bem como a
determinação do valor percentual de populações celulares e das citocinas produzidas foi feita
através de um sistema de computador acoplado ao citômetro. A Figura 3 mostra de forma
esquemática a seqüência de procedimentos necessários para a análise dos dados do fenótipo
celular e perfil de citocinas após estimulação in vitro. O primeiro passo consiste na identificação
da população de leucócitos em estudo: os linfócitos (L). Na figura 3A temos um gráfico do tipo
pontual (“dot plot”) onde pode-se avaliar o tamenho celular (FSC) versus a granulosidade (SSC).
Após a seleção da região de interesse, a mesma é analisada utilizando-se a intensidade de
fluorescência apresentada pelas células presentes na região selecionada, em gráficos de
fluorescência 1 (FL1- fluorescência verde obtida pela marcação com isotiocianato de fluoresceína
– FITC) versus fluorescência 2 (fluorescência laranja obtida pela marcação com ficoeritrina –PE).
No exemplo apresentado na Figura 3B temos CD- FITC versus IL- PE.
Figura 3: Análise de leucócitos do sangue periférico por citometria de fluxo. A Figura
1a representa um perfil celular de tamanho versus granulosidade, obtido após cultivo celular com
CD4-FITC
Ant
i IL-
10 P
E
B
4,33 %1,20%
Tamanho
Gra
nulo
sida
de
Linfócitos
A
Tamanho
Gra
nulo
sida
de
Linfócitos
A
CD3-‐FITC

23
o antígeno ML. A Figura 3b representa um perfil celular obtido em um gráfico de fluorescência
1 (CD3-FITC) versus fluorescência 2 (IL-10-PE).

24
IV RESULTADOS E DISCUSSAO
A apresentação dos resultados foi realizada em duas partes: a primeira parte se refere aos
dados obtidos na análise do perfil de citocinas intracelulares de leucócitos do sangue periférico
após cultura in vitro sem estímulo (meio de cultura), a segunda aos dados obtidos em condições
diferentes de estimulação (estimulação com mitógeno inespecífico – SEB, e antígeno específico –
Mycobacterium leprae), também in vitro dos leucócitos do sangue periférico. Os resultados são
expressos sob a forma de média do percentual de células positivas, simultaneamente tanto para o
marcador de superfície celular quanto para a citocina de interesse.
Avaliou-se a população linfócitos T CD3+, para se traçar um perfil geral de citocinas
presentes nos indivíduos participantes do estudo. Para estudar diferenças no padrão imunológico
entre os sujeitos selecionados, analisou-se inicialmente o perfil de células positivas para citocinas
intracelulares do tipo 1 (IFN-g) e do tipo 2 (IL-4 e IL-10) utilizando leucócitos em culturas-
controle (na presença do meio de cultura RPMI). A segunda avaliação foi da cultura de leucócitos
na presença dos antígenos já citados. Essa análise teve como objetivo estudar a resposta imune,
pela produção de citocinas pelos linfócitos T CD3+, dos indivíduos mediante a estimulações
específicas.
4.1 PRODUÇÃO DE CITOCINAS INTRACELULARES (IL4, IL10 E INF-Γ) EM
LINFÓCITOS T CD3+ DE INDIVÍDUOS MULTIBACILARES, PAUCIBACILARES,
CONTATOS E SADIO APÓS CULTURA IN VITRO SEM ESTIMULAÇÃO.
Os resultados a seguir se referem ao perfil total de citocinas (quadrantes 1 e 2 do gráfico
de fluorescência 1 versus fluorescência 2: anticorpos marcados com FITC e PE respectivamente,
representado na figura 3b expresso por linfócitos T CD3+ dos pacientes participantes do estudo.
O principal objetivo foi verificar a porcentagem de linfócitos T CD3+ produtoras das
citocinas de IL4, IL10 e INF-γ. Essa análise torna-se importante no sentido de que permite a
visualização da produção basal das citocinas citadas.

25
Figura 4: Porcentagem de linfócitos T CD3+, produtores de IL4, em cultura controle (sem
estímulo), em pacientes multibacilares, paucilares, contatos intra e extradomiciliares e indivíduo
sadio.
Figura 5: Porcentagem de linfócitos T CD3+, procutores de IL10, em cultura controle
(sem estímulo), em pacientes multibacilares, paucilares, contatos intra e extradomiciliares e
indivíduo sadio.
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
M1 M2 P1 P2 CM1 CM2 CP1 CP2 CE S CD3 -‐ IL4 0.33 0.32 0.15 0.16 0.11 0.3 0.12 0.15 0.24 0.1
% linfócitos T CD3+ IL4+
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
M1 M2 P1 P2 CM1 CM2 CP1 CP2 CE S CD3 -‐ IL10 0.29 0.37 0.1 0.19 0.13 0.21 0.11 0.12 0.21 0.09
% linfócitos T CD3+ IL10+

26
Figura 6: Porcentagem de linfócitos T CD3+, procutores de IFN-gama, em cultura
controle (sem estímulo), em pacientes multibacilares, paucilares, contatos intra e
extradomiciliares e indivíduo sadio.
As figuras 4, 5 e 6, representam a porcentagem de linfócitos T CD3+, produtores de IL4,
IL10 e INF-gamma, respectivamente. Em relação á produção da citocina IL4, observa-se um
maior percentual de células T CD3+ nos pacientes M1 (0,33%), M2 (0,32%), CM2 (0,30%) e CE
(0,24%). O mesmo perfil é observado para a produção de IL10, onde os pacientes M1, M2, CM2
e CE, se destacam na porcetam de células T CD3+ produtoras. Por outro lado, a observação do
gráfico que representa a produção de INF-gamma, permite visualizar que os pacientes que
apresentam maior percentual de células produtoras são P2, P1 e CM1.
4.2 PRODUÇÃO DE CITOCINAS INTRACELULARES (IL4, IL10 E INF-Γ) EM
LINFÓCITOS T CD3+ DE INDIVÍDUOS MULTIBACILARES, PAUCIBACILARES,
CONTATOS E SADIO APÓS CULTURA IN VITRO COM ESTÍMULO DE SEB.
O objetivo desta parte experimental do estudo foi verificar a porcentagem de linfócitos T
CD3+ produtoras das citocinas de IL4, IL10 e INF-γ, frente a um estímulo inespecífico. Utilizou-
se o SEB, que é um superantígeno do Staphilococcos aureus. Essa análise torna-se importante no
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
M1 M2 P1 P2 CM1 CM2 CP1 CP2 CE S CD3 -‐ IFN 0.21 0.22 0.35 0.64 0.3 0.16 0.21 0.19 0.15 0.11
% linfóticos T CD3+ IFN-‐gam
a+
27
sentido de que permite a verificação da positividade dos experimentos, uma vez que o SEB é um
mitógeno inespecífico, ocorre indução da liberação de citocinas de forma exacerbada,
funcionando assim como um controle positivo do experimento.
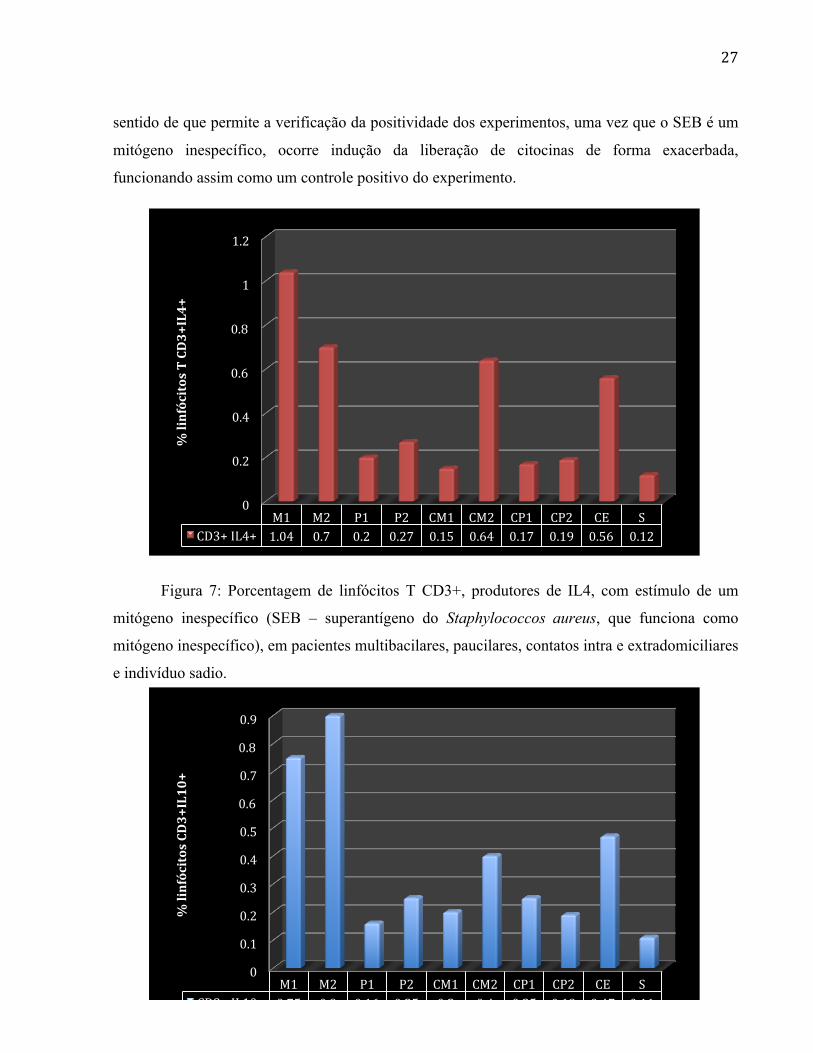
Figura 7: Porcentagem de linfócitos T CD3+, produtores de IL4, com estímulo de um
mitógeno inespecífico (SEB – superantígeno do Staphylococcos aureus, que funciona como
mitógeno inespecífico), em pacientes multibacilares, paucilares, contatos intra e extradomiciliares
e indivíduo sadio.
0
0.2
0.4
0.6
0.8
1
1.2
M1 M2 P1 P2 CM1 CM2 CP1 CP2 CE S CD3+ IL4+ 1.04 0.7 0.2 0.27 0.15 0.64 0.17 0.19 0.56 0.12
% linfócitos T CD3+IL4+
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
M1 M2 P1 P2 CM1 CM2 CP1 CP2 CE S CD3+ IL10+ 0.75 0.9 0.16 0.25 0.2 0.4 0.25 0.19 0.47 0.11
% linfócitos CD3+IL10+

28
Figura 8: Porcentagem de linfócitos T CD3+, produtores de IL10, com estímulo de um
mitógeno inespecífico (SEB – superantígeno do Staphylococcos aureus, que funciona como
mitógeno inespecífico), em pacientes multibacilares, paucilares, contatos intra e extradomiciliares
e indivíduo sadio.
Figura 9: Porcentagem de linfócitos T CD3+, produtores de IFN-gamma, com
estímulo de um mitógeno inespecífico (SEB – superantígeno do Staphylococcos aureus, que
funciona como mitógeno inespecífico), em pacientes multibacilares, paucilares, contatos intra e
extradomiciliares e indivíduo sadio.
Os resultados permitem observar, que o estímulo com o PHA, permitiu um aumento na
porcentagem de células produtoras para todas as citocinas, quando comparado com a cultura sem
estímulo. Apesar disso, os pacientes M1 (1,04%), M2 (0,7%), CM2 (0,64%) e CE (0,56%),
continuam como os maiores produtores de IL4 (1,04%; 0,7%; 0,64%; 0,56%) e IL10 (0.75%;
0,9%; 0,4% e 0,47%). Os pacientes P2 (1,49%), P1 (0,92%) e CM1 (0.95%), continuam como
aqules com maior porcentagem de linfócitos T CD3+IFN-gama+.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
M1 M2 P1 P2 CM1 CM2 CP1 CP2 CE S CD3+IFN+ 0.41 0.52 0.92 1.49 0.95 0.45 0.81 0.34 0.62 0.32
% linfócitos T CD3+ INF-‐gm
a+

29
4.3 PRODUÇÃO DE CITOCINAS INTRACELULARES (IL4, IL10 E INF-Γ) EM
LINFÓCITOS T CD3+ DE INDIVÍDUOS MULTIBACILARES, PAUCIBACILARES,
CONTATOS E SADIO APÓS CULTURA IN VITRO COM ESTÍMULO DE
MYCOBACTERIUM LEPRAE.
Os próximos resultados, referem-se à porcentagem de citocinas produzidas por linfócitos
T CD3+, após estimulo específicos com antígeno bruto do Mycobacterium leprae. Esses
resultados cumprem a finalização de um dos principais objetivos do estudo, que é prever a
resposta imune dos indíduos, na presença do bacilo, o que permite avaliar o perfil de resistência
ou susceptibilidade o bactéria já citada.
Figura 10: Porcentagem de linfócitos T CD3+, produtores de IL4, com estímulo
especifico (Mycobacterium leprae), em pacientes multibacilares, paucilares, contatos intra e
extradomiciliares e indivíduo sadio.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
M1 M2 P1 P2 CM1 CM2 CP1 CP2 CE S CD3+ IL4+ 0.98 0.86 0.15 0.18 0.12 0.77 0.14 0.16 0.42 0.12
% linfócitos T CD3+ IL4+

30
Figura11: Porcentagem de linfócitos T CD3+, produtores de IL10, com estímulo
de um antígeno específico (Mycobacterium leprae), em pacientes multibacilares, paucilares,
contatos intra e extradomiciliares e indivíduo sadio.
Figura 12: Porcentagem de linfócitos T CD3+, produtores de IL4, com estímulo
especifico (Mycobacterium leprae), em pacientes multibacilares, paucilares, contatos intra e
extradomiciliares e indivíduo sadio.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
M1 M2 P1 P2 CM1 CM2 CP1 CP2 CE S CD3+ IL10+ 0.72 0.81 0.12 0.21 0.19 0.7 0.22 0.19 0.39 0.1
% linfócitos T CD3+IL10+
0
0.2
0.4
0.6
0.8
1
1.2
1.4
M1 M2 P1 P2 CM1 CM2 CP1 CP2 CE S CD3+IFN+ 0.21 0.27 1.02 1.32 1.1 0.34 0.92 0.28 0.78 0.24
% linfócitos T CD3+ IFN+

31
Estudos utilizando sangue periférico, tem também demonstrado um perfil de citonas
específico para pacientes com hanseníase. Mutis et al.(32), estudando o padrão de citocinas
produzidas pelos linfócitos T do sangue periférico em hansenianos, encontraram linfócitos com
atividades semelhantes aos Th1 (helper 1) e Th2 (helper 2). Segundo os autores, na forma
Tuberculóide (paucibacilar) os bacilos estimulariam os linfócitos Th1 a produzirem altos níveis
de IFN-y, responsáveis pela manutenção da resposta imune celular. Já na forma Virchowiana
(multibacilar), a estimulação dos linfócitos Th2 levaria e produção das citocinas IL-4, IL-5, IL-6
e IL-10. A IL-4 e IL-10 são supressoras da atividade macrofágica, e a 1L-4, paralelamente,
estimula linfócitos B a produzirem anticorpos. Assim, o aumento dessas citocinas possibilitaria a
persistência e replicação do M. leprae no interior do fagócito.
Pelos resultados apresentados acima, o estudo realizado segue o mesmo padrão de resposta.
Nota-se que 3 dos 10 individuos apresentaram produção de IL4 e IL10 de forma mais
pronunciada. Os pacientes multibacilares e um dos contatos de paciente multibacilar (CM2),
mostraram alta porcentagem de linfócitos T CD3+IL4+ e IL10+. Segundo GOULARD et al,
2002, células T que produzem IL-4, IL-5 e IL-10, chamadas Th2, aumentam a resposta humoral,
IL-4 estimula a produção de IgE e ambas, IL-4 e IL-10 estimulam células B e inibem ativação de
macrófago resultando em infecção progressiva.
Interressantemente o individuo contato extradomiciliar (CE), manteve produção alta de IL4
e IL10 pela cultura sem estímulo e com SEB, mas quando os leucocitos do sangue perifico desse
paciente foram colocados em contato com o antigeno do Mycobacterium leprae, houve uma
modulação da resposta, e a porcentagem de células T CD3+IFN+ aumentou, diminuindo a
porcentagem de células T CD3+IL4+IL10+.
Quanto a produção de IFN, pacientes P2, P1, CM1 e CP1, mantiveram alta porcentagem de
células produtoras em todos os experimentos. Segundo Goulard et al, 2002. Células T que
produzem interleucina 2 (IL-2) e interferon γ (IFN-γ), chamadas Th1,aumentam a imunidade
mediada por células7. IFN-γ ativa macrófago e IL-2 estimula o crescimento de células T
antígeno-específicas, resultando em doença mais branda ou cura.
Num estudo realizado por SARNO et al., 2002, foi feito mensuração de INF-γ no
sobrenadante de cultura de leucócitos de num grupo de contatos intradociliares de pacientes com
hanseníase, com estímulo de M.leprae, esse grupo foi acompanhado durante dois anos, e parte
dos contatos que não respondiam ao antigeno com produção dessa citocina durante os dois anos,

32
ao final desse period desenvolveu hanseníase. Diferentemente de outra parte dos contatos que
desde o inicio do estudo, vinha apresentando alta produção de INF-γ, nesses, em grande parte,
não houve relato de desenvolvimento da doença, e, nos que houve, ela se desenvolveu de maneira
mais branda, com excelente prognóstico.

33
V CONCLUSAO
Diante do exposto, e com relação a outros estudos, conclui-se que a participação das
citocinas esta diretamente envolvida no curso da infecção pelo Mycobacterium leprae. Já esta
muito bem evidenciado na literatura que o desenvolvimento da doença nas formas mais graves,
relacionadas a grande proliferação bacilar, se dá pela ativação de linfícitos Th2, pela
predisposição na produção de IL4, IL10, dentre outras citocinas. O INF-γ, aparece nesse contexto
como fator de proteção ao bacilo, se sentido de que leva a uma ativação de imunidade celular,
capaz de destrui-lo.
Assim, torna-se relevante o uso de tecnicas para mensuração de citocinas, com o objetivo
de prever questões relacionadas a resistência ou susceptibilidade ao Mycobacterium leprae, em
individuos constatemente exposto ao bacilo, chamados de contatos intra ou extradomiciliares.
A tecnica de mensuração de citocinas intracitoplasmáticas, com usa da citomentria de
fluxo, contribui em muito, no entendimento desse processo imunitário, uma vez que responde a
pergunta de qual o tipo celular é capaz de produzir determinada citocina.
São nccessários novos prolongados, que permitem um acompanhamente continuo desses
pacientes, para verificar o desenvolvimento ou não da enfermidade, mas desde já, baseando-se
em dados da literature, pode-se pressupor que os individuos desses estudo, que apresentaram alta
porcentagem de células produtoras de INF-γ, apresentam capacidade em responder ao bacilo com
resposta imune celular (hipersesibilidade do tipo IV), e portando resistencia no densenvolvimento
da doença, e o contrario também é verdadeiro, individuos que responderam ao bacilo com
produção de IL4 e IL10, apresentam susceptibilidade e maior predisposição no denvolvimento de
hanseníase.

34
VI REFERENCIAS BIBLIOGRAFICAS
LOCKWOOD DN, BRITTON WJ. Leprosy. Lancet 363, 1209–1219 (2004).
SARNO EN, ESPINOSA M, Sampaio EP, VIEIRA LM, FIGUEIREIDO AA, MIRANDA
CF, et al. Immunological responsiveness to M. leprae and BCG antigens in 98 leprosy
patients and their household contacts. Braz. J. Med. Biol. Res. 1988;21(3):461-470.
STEFANI MMA. Challenges in the post genomic era for the development of tests for
leprosy diagnosis. Revista da Sociedade Brasileira de Medicina Tropical 41(Suplemento II):89-
94, 2008.
ARAOZ R, HONORE N, BANU S, DEMANGEL C, CISSOKO Y, ARAMA C, UDDIN
MK, HADI SK, MONOT M, CHO SN, JI B, BRENNAN PJ, SOW S, COLE ST. Towards an
immunodiagnostic test for leprosy. Microbes and Infection 8: 2270-2276, 2006.
DUTHIE MS, GOTO W, IRETON GC, REECE ST, CARDOSO LP, MARTELLI CM,
STEFANI MM, NAKATANI M, DE JESUS RC, NETTO EM, BALAGON MV, TAN E,
GELBER RH, MAEDA Y, MAKINO M, HOFT D, REED SG. Use of protein antigens for
early serological diagnosis of leprosy. Clinical and Vaccine Immunology 14: 1400-1408, 2007.
DUTHIE MS, GOTO W, IRETON GC, REECE ST, SAMPAIO LH, GRASSI AB,
SOUSA ALM, MARTELLI CM, BALAGON MVF., TAN E, STEFANI MM, REED SG.
Antigen- specific T cell responses of leprosy patients. Clinical and Vaccine Immunology
15:1659- 1665, 2008.
GELUK A, KLEIN MR, FRANKEN KL, VAN MEIJGAARDEN KE, WIELES B,
PEREIRA KC, BUHRER- SEKULA S, KLATSER PR, BRENNAN PJ, SPENCER JS,
WILLIAMS DL, PESSOLANI MC, SAMPAIO EP, OTTENHOFFTH. Post genomic approach
to identify novel Mycobacterium leprae antigens with potential to improve
immunodiagnosis of infection. Infection and Immunity 73: 5636-5644, 2005.

35
GELUK A, OTTENHOFF TH. HLA and leprosy in the pre and postgenomic eras.
Human Immunology 67: 439-445, 2006.
GELUK A, VAN DER PLOEG J, TELES RO, FRANKEN KL, PRINS C, DRIJFHOUT
JW, SARNO EN, SAMPAIO EP, OTTENHOFF TH. Rational combination of peptides
derived from different Mycobacterium leprae proteins improves sensitivity for
immunodiagnosis of M. leprae infection. Clinical and Vaccine Immunology 15: 522-533, 2008.
REECE ST, IRETON G, MOHAMATH R, GUDERIAN J, GOTO W, GELBER R,
GROATHOUSE N, SPENCER J, BRENNAN P, REED SG. ML0405 and ML2331 are
antigens of Mycobacterium leprae with potential for diagnosis of leprosy. Clinical and
Vaccine Immunology 13: 333-340, 2006.
SPENCER JS, DOCKRELL HM, KIM HJ, MARQUES MA, WILLIAMS DL, MARTINS
MV, MARTINS ML, LIMA MC, SARNO EN, PEREIRA GM, MATOS H, FONSECA LS,
SAMPAIO EP, OTTENHOFF TH, GELUK A, CHO SN, STOKER NG, COLE ST, BRENNAN
PJ, PESSOLANI MC. Identification of specific proteins and peptides in Mycobacterium
leprae suitable for the selective diagnosis of leprosy. Journal of Immunology 175: 7930-7938,
2005.
COLE ST, EIGLMEIER K, PARKHILL J, JAMES KD, THOMSON NR, WHEELER PR,
HONORE N, GARNIER T, CHURCHER C, HARRIS D, MUNGALL K, BASHAM D,
BROWN D, CHILLINGWORTH T, CONNOR R, DAVIES RM, DEVLIN K, DUTHOY S,
FELTWELL T, FRASER A, HAMLIN N, HOLROYD S, HORNSBY T, JAGELS K, LACROIX
C, MACLEAN J, MOULE S, MURPHY L, OLIVER K, QUAIL MA, RAJANDREAM MA,
RUTHERFORD KM, RUTTER S, SEEGER K, SIMON S, SIMMONDS M, SKELTON J,
SQUARES R, SQUARES S, STEVENS K, TAYLOR K, WHITEHEAD S, WOODWARD JR,
BARRELL BG. Massive gene decay in the leprosy bacillus. Nature 409: 1007-1011, 2001.

36
OTTENHOFF THM. Immunology of leprosy. Tropical and Geographical Medicine, v. 46,
p. 72-80, 1994a.
OTTENHOFF THM. Immunology of leprosy: lessons from and for leprosy.
International journal of leprosy, v. 62, p. 108-121, 1994b.
SCOLLARD DM, ADAMS LB, GILLIS TP, KRAHENBUHL JL, TRUMAN RW,
WILLIAMS DL. The continuing challenges of leprosy. CLINICAL MICROBIOLOGY
REVIEWS 19: 338-381, 2006.
SAPKOTA B, NEUPANE K.D., MAHARJAN R.K. Single lesion multibacillary leprosy,
a treatment enigma: a case report. Journal of Medical Case Reports. 3 (8): 2009.
LYON S, LYON AC, DA SILVA RC, GROSSI MA, LYON SH, BÜHRER-SÉKULA S,
ROCHA MO.A comparison of ML Flow serology and slit skin smears to assess the bacterial
load in newly diagnosed leprosy patients in Brazil. Leprosy Rev. 2008 Jun;79(2):162-70.
WHO: Global leprosy situation. Weekly epidemiological record 35, 337–348 (2010).
Gallo, M. E. N., Sampaio, E. P., Nery, J. A. C., Moraes, M. O., Antunes, S. L. G.,
Pessolani, M. C. V., & Sarno, E. N. 2005. Hanseníase: Aspectos Epidemiológicos, Clínicos e
Imunológicos. Coura, J.R. (ed.) Dinâmica das doenças infecciosas e parasitárias: 1383-1394.
Guanabara Koogan.
WHO. Global leprosy situation. Weekly epidemiological record. 25[82], 225-232.
Disponível em: http://www.who.int/wer. Acesso em: 25 mai. 2009.
SPIERINGS E, DE BOER T, ZULIANELO L, OTTENHOFF TH. Novel mechanisms in
the immunopathogenesis of leprosy nerve damage: the role of Schwann cells, T cells and
Mycobacterium leprae. Immunol. Cell Biol. 2000 Ago;78(4):349-355.

37
BLOOM BR. Learning from leprosy: a perspective on immunology and the Third
World. J. Immunol. 1986;137(1):i-x.
ANTAS PRZ, SALES JS, PEREIRA KC, OLIVEIRA EB, CUNHA KC, SARNO EN,
SAMPAIO EP. Patterns of intracellular cytokines in CD4 and CD8 T cells from patients
wits mycobacterial infections. Braz. J. Méd. Biol. Res., v. 37, n. 8, p. 1119-1129, 2004.
REES RJ. LEPRA's elective period student programme, 1973-1983. Lepr Rev. 1984;
55(4):321-325.
WHO, 1987.Laboratory techniques for leprosy.p.62.
HUNTER SW, BRENNAN PJ. A novel phenolic glycolipid from Mycobacterium leprae
possibly involved in immunogenicity and pathogenicity. J. Bacteriol. 1981;147(3):728-735.
HUNTER SW, STEWART C, BRENNAN PJ. Purification of phenolic glycolipid I from
armadillo and human sources. Int. J. Lepr. Other Mycobact. Dis. 1985;53(3):484- 486.
CHO SN, YANAGIHARA DL, HUNTER SW, GELBER RH, BRENNAN PJ. Serological
specificity of phenolic glycolipid I from Mycobacterium leprae and use in serodiagnosis of
leprosy. Infect. Immun. 1983;41(3):1077-1083.
KIRCHHEIMER WF, STORRS EE. Attempts to establish the armadillo
(Dasypusnovemcinctus Linn.) as a model for the study of leprosy. I. Report of lepromatoid
leprosy in an experimentally infected armadillo. Int. J. Lepr. Other Mycobact. Dis.
1971;39(3):693-702.
VISSA VD, BRENNAN PJ. The genome of Mycobacterium leprae: a minimal
mycobacterial gene set. Genome Biol. 2001;2(8): 1023.

38
RAMBUKKANA A, ZANAZZI G, TAPINOS N, SALZER JL. Contact-dependent
demyelination by Mycobacterium leprae in the absence of immune cells. Science 296, 927–
931 (2002).
YAMAMURA M, UYEMURA K, DEANS R.J, WEINBERG K, REA TH, BLOOM BR,
MODLIN RL. Defining protective responses to pathogens: cytokine profiles in leprosy
lesions. Science, v. 254, p. 277-279, 1991.
CHIN- A- LIEN RAM, FABER WR, VAN RENS MM, LEIKER DL, NAAFS B,
KLATSER PR. Follow-up of multibacilary leprosy patients using a phenolic glycolipid-1
based ELISA. Do increasing ELISA- values after discontinuation of treatment indicate
relapse? Leprosy Review, v. 63, p. 21-27, 1992.
HARBOE M. The immunology of leprosy. Hasting RC (ed) Leprosy. 1edition. New York:
Churchill Livingstone Inc, 1985, p.53-87.
SIELING PA, MODLIN RL. Cytokine patterns at the site of mycrobacterial infection. Immunobiology, v. 191, p. 378-3870, 1994a.
SIELING PA, MODLIN RL. T cell and cytokine patterns in leprosy skin lessons.
Springer Seminars in Immunopathology, v. 13, p. 413-426, 1992.
CUNHA MGS. Níveis de anti-PGL-1 no soro de pacientes com hanseníase tratados
com quinolona e poliquimioterapia. Tese de Mestrado. Faculdade de Medicina de Ribeirão
Preto, Universidade de São Paulo, Ribeirão Preto, 1998.
FOSS NT, CALLERA F, ALBERTO FL. Anti- PGL1 levels is leprosy patients and their
contacts. Brazilian Journal of Medical and Biological research, v. 26, p. 43-51, 1993.
KLATSER PR. Serology of Leprosy. Tropical and Geographical Medicine, v. 46, p. 115-
118, 1994.

39
BÜHRER-SÉKULA S, VISSCHEDIJK J, GROSSI MA, DHAKAL KP, NAMADI AU,
KLATSER PR, OSKAM L. The ML flow test as a point of care test for leprosy control
programmes: potential effects on classification of leprosy patients. Leprosy Rev. 2007 Mar;
78 (1): 70-9.
MODLIN RL, MEHARA V, WONG L, FUJIMARA Y, CHANG W-C, HORWITZ DA,
BLOOM BR, REA TH, PATTENGALE PK. Supressor T Lynphocites from lepromatous leprosy
skin lesions. Journal of Immunology, v. 137, p. 2831-2834, 1988.
GOULART L.R, GOULART I.M.B. Leprosy pathogenetic background: a review and
lessons from other mycobacterial diseases. Arch. Dermatol. Res. 301:123–137, 2009.
CHER DJ, MOSMANN TR. Two types of murine helper T cell clone.II, Delayed- type
hypersensitivity is mediated by Th1 clones. Journal of Immunology, v. 138, p. 3688-3694,
1987.
MISRA N, SELVAKUMAR M, SINGH S, BHARADWAJ M, RAMESH V, MISRA RS,
NATH I. Monocyte derived IL-10 and PGE2 are associated with the absence of TH1 cells
and in vitro T cells suppression in lepromatous leprosy. Immunology Letters, v. 48, p. 123-
128, 1995.
ASSEMAN C, POWRIE F. Interleukin 10 is a growth factor for a population pf
regulatory T cells. Gut, v. 42, p. 157-158, 1998.
GOULART IMB, FIGUEIREDO F, COIMBRA T, FOSS NT. Detection transforming
growth factor-b1 in dermal lesions pf different clinical forms of leprosy. American Journal of
Pathology, v. 148, p. 991-917, 1996.
GOULART IMB. Produção de TGF-beta1 por células mononucleares do sangue
periférico de portadores de hanseníase e seu papel na regulação da reposta inflamatória.
Tese de Doutorado, Universidade de São Paulo, Ribeirão Preto, 1999.

40
GOULART IMB, MINEO JR, FOSS NT. Production of transforming growth factor
beta-1 by blood monocytes from patients with different clinical forns of leprosy. Clinical and
Experimental Immunology, v. 122, p. 330-334, 2000.
GOULART IMB, PENNA GO, CUNHA G. Imunopatologia da hanseníase: a
complexidade dos mecanismos da resposta imune do hospedeiro ao Mycobcterium leprae.
Revista da Sociedade Brasileira de Medicina Tropical, v. 35, n; 4, p. 365-375, jul. 2002.
BERMUDEZ LE. Production of transforminf growth factor-beta by Mycobacterium
avium-infected human macrophages is associated with unresponsiveness to INF-gama.
Journal of Immunology, v. 150, p.1838-1845, 1993.
GRENN JS, SCHELLER LF, MARLETTA MA, SEGUIN MC, KLOTZ FW, SALYTER
M, NELSON BJ, NANCY CA. Nitric oxide: cytokine-regulation of nitric oxide in host
resistance to intracellular pathogens. Immunology Letters, v. 43, p. 87-94, 1994.
NELSON BJ, RALPH P, GREEN SJ, NANCY CA. Differential susceptibility of
activated macrophage cytotoxic effector reations to the suppressive effects of transforming
growth factor-beta1. Journal of Immunology, v. 146, p. 1849-1857, 1991.
WALKER SL, LOCKWOOD DN: The clinical and immunological features of leprosy.
Br. Med. Bull. 77–78, 103–121 (2006).
PINHEIRO, R.O.., et al. Mycobacterium leprae-host-cell interations and genetic
determinants in leprosy: an overview. Future Microbiol. 2011.
BRITTON WJ, LOCKWOOD DN. Leprosy. Lancet 363, 1209–1219 (2004).
BROWN, M; Wittwer, C. Flow cytometry: Principles and Clinical Applications in
Hematology. Clinical Chemistry; 46(8B), 2000. 1221-1229p.

41
ORFÃO, A; Buitrago, JMG. Fundamentos de la citometría de flujo. In: Orfao, A; Buitrago,
JMG. La Citometría de flujo en el Laboratório Clínico. Salamanca: SEQC, 1995. 11-30p .
NICHOLSON, JKA. Immunophenotyping of Lymphocytes by Flow Cytometry. In: Rose,
NR; Hamilton, RG; Detrick, B. Manual of Clinical Laboratory Immunology. Washington
DC: ASM Press, 2002. 137-139p.
RIBEIRO, Neuza Filipa da Silva. Relatório em estágio em citometria.2010. 69 f.
Dissertação (Mestrado em Oncologia) – Instituto de Ciências Biomédicas de Abel Salazar,
Universidade do Porto, Portugal, 2010.