universidade federal do cearÁ instituto de … · obrigado pelas oportunidades que você me deu e...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO CEARÁ
INSTITUTO DE CIÊNCIAS DO MAR PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS MARINHAS TROPICAIS
FRANCISCO SYLVÂNIO FERREIRA DA SILVA
EFLUENTE DA AGROINDÚSTRIA DO CAJU E ÓLEO VEGETAL RESIDUAL
COMO MATÉRIAS-PRIMAS PARA PRODUÇÃO DE BIOSSURFACTANTES POR
BACTÉRIAS DE ORIGEM MARINHA
FORTALEZA
2017

FRANCISCO SYLVÂNIO FERREIRA DA SILVA
EFLUENTE DA AGROINDÚSTRIA DO CAJU E ÓLEO VEGETAL RESIDUAL COMO
MATÉRIAS-PRIMAS PARA PRODUÇÃO DE BIOSSURFACTANTES POR
BACTÉRIAS DE ORIGEM MARINHA
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Marinhas Tropicais da Universidade Federal do Ceará, como requisito parcial à obtenção do título de mestre em Ciências Marinhas Tropicais. Área de concentração: Utilização e manejo de ecossistemas marinhos e estuarinos. Orientadora: Profª. Drª. Oscarina Viana de Sousa. Coorientadora:. Drª. Karla Maria Catter.
FORTALEZA
2017


FRANCISCO SYLVÂNIO FERREIRA DA SILVA
EFLUENTE DA AGROINDÚSTRIA DO CAJU E ÓLEO VEGETAL RESIDUAL COMO
MATÉRIAS PRIMAS PARA PRODUÇÃO DE BIOSSURFACTANTES POR
BACTÉRIAS DE ORIGEM MARINHA
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Marinhas Tropicais da Universidade Federal do Ceará, como requisito parcial à obtenção do título de mestre em Ciências Marinhas Tropicais. Área de concentração: Utilização e manejo de ecossistemas marinhos e estuarinos.
Aprovada em: 26/06 /2017
BANCA EXAMINADORA
________________________________________ Profª. Drª. Oscarina Viana de Sousa (Orientadora)
Universidade Federal do Ceará (UFC)
_________________________________________ Profª. Drª. Regine Helena Silva dos Fernandes Vieira
Universidade Federal do Ceará (UFC)
_________________________________________ Profª. Drª. Francisca Gleire Rodrigues de Menezes

À Vânia Maria Ferreira por seu amor
incondicional, dedico.

AGRADECIMENTOS
A Deus por me permitir concluir esta etapa tão importante de minha vida.
À minha mãe Vania Maria por ser meu porto seguro, força e apoio bem
presente em todos os momentos. Dos mais felizes aos mais tristes. Você é a minha
heroína!
Ao Caio César por trazer felicidade aos meus dias quando eles estavam
mais difíceis. Minha gratidão!
Ao meu irmão de alma e amigo do peito Gabriel Neto. Você é um ser
iluminado que veio na terra com o propósito de me guiar nos momentos em que eu
mais precisei de direção. Você tem um lugar enorme em meu coração.
De coração, agradeço à minha orientadora, professora e fonte de
inspiração Oscarina Viana de Sousa. Obrigado por acreditar em mim e me ensinar a
buscar sempre o meu melhor. Obrigado pelas oportunidades que você me deu e por
acreditar no meu potencial. Meus mais sinceros agradecimentos.
À professora Regine Helena Silva dos Fernandes Vieira por sua
contribuição em minha vida acadêmica. Obrigado por me abrir as portas do
laboratório e me permitir conhecer o mundo da microbiologia. Meus sinceros
sentimentos de gratidão. Com carinho, Sinfrônio.
À minha “mãe” de laboratório Karla Catter pela coorientação desse
trabalho, inspiração e companheirismo ao longo de minha caminhada acadêmica.
Sou eternamente grato a você me apresentar o mundo dos biossurfactantes e por
fazer dele um caminho mais tranquilo de ser percorrido. Muito obrigado!
Ao Victor Conde, meu “irmão” de laboratório, pelo companheirismo e por
dividir comigo momentos de verdadeiro aprendizado.
Aos meus amigos Felipe Souza e Patrícia Andrade por serem válvulas de
escape quando os dias estão mais pesados. Guardarei para sempre a alegria e o
carinho de vocês.
Ao grupo Biodançantes, em especial ao João Salmito, por me ensinar a
manter sempre viva a minha criança interior. Vocês possuem uma energia
maravilhosa e que deve ser compartilhada com o mundo todo. Gratidão!
Aos queridos Jéssica Lucinda, Edirsana Carvalho, Adson Queiróz,
Lorrana Santana e Déborah Amarante por acompanharem de perto meus momentos
de bancada e fora dela. Obrigado por dividirem comigo sua amizade e

companheirismo. Vocês são maravilhosos!
Às queridas Rosa Rebouças e Cristiane Teles por todos os ensinamentos,
broncas, risadas e voltas pra casa nesses dois anos e alguns meses de mestrado. O
ensinamento de vocês vai muito além da bancada. Os levarei para a vida. Vocês têm
o meu abraço mais caloroso.
À Gleire Menezes e Rafael Rodrigues, meus gurus na Biologia Molecular.
Agradeço imensamente a ajuda e o companheirismo de vocês. Gleire, obrigado por
me permitir fazer morada nesse coração virginiano. Rafael, você merece todo o
sucesso do mundo! Muito obrigado!
À Marina Rodríguez por dividir comigo seu conhecimento e sabedoria de
vida. Obrigado por toda a sua ajuda. Você é uma inspiração para mim.
Aos demais amigos do Laboratório de Microbiologia Ambiental e do
Pescado: Mariana Franco, Rubson Matheus, Raul Vitor, Maria João, Giselle Cristina,
Rebeca Martins, Graciene Fernandes, Jamille Rabelo, Mariana Lima, Jade Abreu,
Jéssica Costa e Patrícia Carneiro. Agradeço a cada um de vocês por toda ajuda
nesse período. Desejo todo sucesso pra cada um de vocês!
À minha amiga, irmã de alma e companheira de bancada Juliana Moraes
e seu esposo Diego Moraes pelos auxílios, incontáveis horas trabalhadas, finais de
semanas cansativos e ótimos momentos vividos ao longo de nossa caminhada no
mundo dos biossurfactantes. Vocês são anjos de luz que possuem a minha eterna
gratidão.
Ao Centro de Diagnóstico de Enfermidades de Organismos Aquáticos
(CEDECAM), do Instituto de Ciências do Mar (LABOMAR – UFC), em especial o
professor Rodrigo Maggioni pela parceria na etapa de sequenciamento genético das
cepas bacterianas utilizadas no trabalho.
Aos meus colegas de mestrado em especial às queridas Júlia Rabelo e
Thaís Chaves pelas conversas, apoio mútuo e risadas quando estávamos mais
desesperados. Força!
À secretária do Programa de Pós- Graduação em Ciências Marinhas
Tropicais Isabela Agadir Abreu pelas conversas e ensinamentos. Você é um amor!
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
(CAPES) pela concessão da bolsa de pesquisa.

“Mas as árvores não guardam rancor.
Trataram de continuar a viver - e nos
toquinhos surgiram brotos verdes, como
um gesto de perdão.” (Rubem Alves)

RESUMO
Os biossurfactantes são compostos tensoativos produzidos por microrganismos que
apresentam a capacidade de solubilizar substâncias de diferentes polaridades. Tais
compostos podem ser produzidos a partir de diferentes substratos e possuem
aplicação em diferentes setores industriais. O objetivo desta pesquisa foi verificar o
potencial de produção de biossurfactantes por bactérias marinhas utilizando efluente
de uma indústria de beneficiamento da castanha do caju (ECC) e óleo vegetal
residual como matérias-primas. Foram testadas 20 cepas bacterianas pertencentes
ao acervo do Laboratório de Microbiologia Ambiental e do Pescado (LAMAP). As
cepas foram identificadas através de sequenciamento da região do 16S do rDNA.
Foram realizados testes de hemólise e produção de ramnolipídeos, indicativos da
biossíntese de biossurfactantes. Em seguida, foram realizadas fermentações
utilizando ambos os substratos e os metabólitos produzidos foram submetidos aos
testes do colapso da gota, volume de emulsificação, estabilidade da emulsão,
atividade de emulsificação e antimicrobiana. As amostras que apresentaram os
melhores resultados foram submetidas ao teste ecotoxicológico utilizando sementes
de pepino (Cucumis sativus L.) como organismo indicador. As cepas foram
identificadas como espécies dos gêneros: Pseudomonas, Bacillus, Staphylococcus,
Vibrio, Zobelella, Oceanimonas e Brachybacterium. Das 20 cepas testadas, 65%
apresentaram atividade hemolítica enquanto 40% produziram ramnolipídeos. A partir
da metabolização do efluente agroindustrial e óleo residual pelas cepas,
respectivamente, 90% e 70% apresentaram atividade no teste do colapso da gota;
60% e 20% produziram emulsões com qualidade e boa estabilidade. Foi verificado
que 50% dos metabólitos produzidos a partir do ECC e 5% do óleo residual
apresentaram atividade antimicrobiana frente a bactéria Staphylococcus aureus
(ATCC 25923). Os metabólitos produzidos a partir dos dois substratos apresentaram
efeitos tóxicos contra sementes de pepino. Esses resultados evidenciam o potencial
para produção de biossurfactantes de boa qualidade a partir de resíduos poluentes.
Contudo, existe a necessidade de uma melhor caracterização desses bioprodutos,
isolamento e avaliação de seus efeitos tóxicos no ambiente.
Palavras-chave: Poluição. Surfactantes biológicos. LCC.

ABSTRACT
Biosurfactants are tensoative compounds produced by microorganisms that have the
ability to solubilize substances with different polarities. These compounds can be
produced from different substrates and can be applied in different industrial areas.
Thus, the aim of this research was to investigate the potential of marine bacteria to
produce biosurfactants using effluent from processing of cashew nut (CNE) and
residual vegetable oil as a carbon source. Twenty bacterial strains from the
bacteriological collection of the Laboratory of Fish and Environmental Microbiology
(LAMAP) were tested. The strains were identified by 16S rRNA sequencing.
Hemolytic activity and rhamnolipids production were performed as an indicative of
biosurfactant biosynthesis. Then, fermentations were realized using both substrates
and the metabolites produced were submitted to the following tests: droplet collapse,
emulsion index, emulsification activity and antimicrobial activity. The samples that
showed the best results were submitted to the ecotoxicological test using cucumber
seeds (Cucumis sativus L.) as indicator organism. The strains were identified as
species from the genus: Pseudomonas, Bacillus, Staphylococcus, Vibrio, Zobelella,
Oceanimonas e Brachybacterium. Of the 20 strains tested, 65% showed hemolytic
activity while 40% produced rhamnolipids. From the metabolization of the
agroindustrial effluent and the residual oil, respectively, 90% and 70% of bacterial
strains showed activity in drop collapse, while 60% and 20% produced emulsions
with good quality and stability. In addition, 50% of the metabolites produced from
CNE and 5% from residual oil showed antimicrobial activity against standard strain
Staphylococcus aureus (ATCC 25923). The metabolites produced with both
substrates showed toxic effects against cucumber seeds. These results showed the
potential to produce good quality biosurfactant compounds from pollutant residues.
However, a better characterization of these bioproducts, isolation and evaluation of
their toxic effects on the environment is necessary.
Keywords: pollution. biological surfactants. CNSL.

LISTA DE FIGURAS
Figura 1 Estruturas químicas dos componentes do LCC: a) ácido anacárdico, b)
cardol, c) cardanol....................................................................................... 19
Figura 2 Ilustração do processo de produção dos biossurfactantes.......................... 30
Figura 3 Fontes de carbono utilizadas na produção de biossurfactantes pelas
cepas bacterianas: a) efluente do beneficiamento da castanha do caju; b)
óleo vegetal residual.................................................................................... 30
Figura 4 Halos de hemólise em ágar sangue indicativo da produção de
biossurfactantes após 48 horas de incubação............................................ 40
Figura 5 Halos azuis indicativos da produção de ramnolipídeos em meio ágar azul
de metileno................................................................................................. 41
Figura 6 Meio de cultivo após as 72 horas de produção de biossurfactantes
contendo efluente do beneficiamento da castanha do caju (a) e óleo
vegetal residual (b) como fontes de carbono............................................. 45
Figura 7 Líquidos metabólicos extraídos após a produção de biossufactantes........ 45
Figura 8 Atividade antimicrobiana dos líquidos metabólicos (resultado negativo)
extraídos a partir das fermentações com efluente do beneficiamento da
castanha do caju frente à bactéria Escherichia coli ATCC25922............... 55
Figura 9 Atividade antimicrobiana dos líquidos metabólicos (resultado negativo)
extraídos a partir das fermentações com óleo vegetal residual frente à
bactéria Escherichia coli ATCC25922...................................................... 56
Figura 10 Atividade antimicrobiana dos líquidos metabólicos (resultado positivo)
extraídos a partir das fermentações com óleo vegetal residual frente à
bactéria Staphylococcus aureus ATCC 25923......................................... 56
Figura 11 Atividade antimicrobiana dos líquidos metabólicos (resultado positivo)
extraídos a partir das fermentações com efluente do beneficiamento da
castanha do caju frente à bactéria Staphylococcus aureus ATCC 25923.. 57
Figura 12 Teste de fitotoxicidade do líquido metabólico produzido pela cepa
Pseudomonas sp. 9C a partir de óleo vegetal residual utilizando

sementes de pepino Aodai (Cucumis sativus L.)......................................... 65
Figura 13 Teste de toxicidade do efluente do beneficiamento da castanha do caju
utilizando sementes de pepino Aodai (Cucumis sativus L.)........................ 66
Figura 14 Teste de toxicidade do surfactante SDS utilizando sementes de pepino
Aodai (Cucumis sativus L.).......................................................................... 67

LISTA DE GRÁFICOS
Gráfico 1 Estabilidades das emulsões formadas utilizando líquido metabólico
extraído das fermentações com bactérias do gênero Pseudomonas a
partir de efluente do beneficiamento da castanha de caju (EBCC) e
óleo vegetal residual (OVR).................................................................. 51
Gráfico 2 Estabilidades das emulsões formadas utilizando líquido metabólico
extraído das fermentações com as bactérias das espécies Vibrio
fluvialis, Z. taiwanensis e do gênero Bacillus a partir de efluente do
beneficiamento da castanha de caju (EBCC) e óleo vegetal residual
(OVR)................................................................................................... 52
Gráfico 3 Estabilidades das emulsões formadas utilizando SDS e os líquidos
metabólicos extraídos das fermentações com as bactérias O.
baumannii, Brachybacterium sp. 11SEDH, Staphylococcus sp. 26B a
partir de efluente do líquido da castanha de caju (EBCC) e óleo
vegetal residual (OVR)......................................................................... 53
Gráfico 4 Comprimento das raízes das sementes de pepino Aodai (Cucumis
sativus L.) no teste com os líquidos metabólicos extraídos a partir
das fermentações com efluente do beneficiamento da castanha do
caju com as cepas Pseudomonas sp. 2D, Pseudomonas sp. 2E,
Pseudomonas sp. 7A e Pseudomonas sp.7C...................................... 61
Gráfico 5 Comprimento das raízes das sementes de pepino Aodai (Cucumis
sativus L.) no teste com os líquidos metabólicos produzidos a partir
das fermentações com efluente do beneficiamento da castanha do
caju com as cepas Pseudomonas sp. 9A, Pseudomonas sp. 9B,
Pseudomonas sp. 9C e Pseudomonas sp.18D................................ 61
Gráfico 6 Comprimento das raízes das sementes de pepino Aodai (Cucumis
sativus L.) no teste com os líquidos metabólicos extraídos a partir
das fermentações com efluente do beneficiamento da castanha do
caju com as cepas V. fluvialis 21A, V. fluvialis 36C, Z. taiwanensis
11SA, Brachybacterium sp. 11SEDH e Staphylococcus sp. 26B e do

efluente................................................................................................. 62
Gráfico 7 Comprimento das raízes das sementes de pepino Aodai (Cucumis
sativus L.) no teste com os líquidos metabólicos extraídos a partir
das fermentações com óleo vegetal residual com as cepas
Pseudomonas sp. 7C, Pseudomonas sp. 9B, Pseudomonas sp. 9C,
Pseudomonas sp. 18D, O. baumannii 12SC e com o surfactante
sintético SDS........................................................................................ 64

LISTA DE TABELAS
Tabela 1 Reagentes e condições utilizadas na amplificação da
subunidade 16S das bactérias através da Reação em Cadeia de
Polimerase...................................................................................... 27
Tabela 2 Reagentes do Mix de PCR e condições de termociclagem do
sequenciamento.................................................................................. 28
Tabela 3 Codificação, origem e resultado da identificação genética das
bactérias marinhas selecionadas na presente pesquisa.................... 35
Tabela 4 Resultados da atividade hemolítica e produção de ramnolipideos
das cepas isolados de água e sedimento do Mucuripe (MUC),
produtoras de biossurfactantes........................................................... 39
Tabela 5 Resultados do teste qualitativo do colapso da gota (TQCG) e
valores de pH dos líquidos metabólicos extraídos a partir de
efluente do beneficiamento da castanha do caju................................ 43
Tabela 6 Resultados do teste qualitativo do colapso da gota (TQCG) e
valores de pH dos líquidos metabólicos extraídos a partir de óleo
vegetal residual................................................................................... 44
Tabela 7 Resultados dos testes de volume de emulsificação (VE) e
atividade de emulsificação realizados utilizando os líquidos
metabólicos extraído......................................................................... 48
Tabela 8 Resultados da determinação da atividade antimicrobiana dos
líquidos metabólicos extraídos utilizando resíduos orgânicos como
fonte de carbono................................................................................. 55
Tabela 9 Percentuais de germinação (%G) das sementes de pepino Aodai
(Cucumis sativus L.) utilizadas no teste toxicológico com os líquidos
metabólicos produzidos a partir do efluente do beneficiamento da
castanha do caju................................................................................. 59
Tabela 10 Percentuais de inibição (%IP) do crescimento das sementes de
pepino Aodai (Cucumis sativus L.) utilizadas no teste toxicológico

com os líquidos metabólicos produzidos extraídos a partir do
efluente do beneficiamento da castanha de caju................................
60
Tabela 11 Percentuais de germinação (%G) das sementes de pepino Aodai
(Cucumis sativus L.) utilizadas no teste toxicológico com os líquidos
metabólicos produzidos extraídos a partir das fermentações com
óleo vegetal residual........................................................................... 62
Tabela 12 Percentuais de inibição (%IP) do crescimento das sementes de
pepino Aodai (Cucumis sativus L.) utilizadas no teste toxicológico
com os líquidos metabólicos produzidos extraídos a partir das
fermentações com óleo vegetal residual............................................. 63

LISTA DE ABREVIATURAS E SIGLAS
ATCC American Type Culture Collection
BLAST Basic Local Alignment Search Tool
B.O.D Biochemical oxygen demand
CE Ceará
CLSI Clinical and Laboratory Standard Institute
CTAB Brometo de cetiltrimetilamônio
DNA Ácido desoxirribonucleico
DNTP’s Desoxirrimonucleotídeos
DQO Demanda química de oxigênio
EC Estímulo de crescimento
EPS Exopolissacarídeos
ES Estabilidade da emulsão
IP Percentual de inibição
LB Lúria Bertani
LCC Líquido da castanha de caju
LM Líquido metabólico
MgCl2 Cloreto de magnésio
MT Muito tóxico
NaCl Cloreto de sódio
PCR Reação em Cadeira de Polimerase
pH Potencial hidrogeniônico
q.s.p Quantidade suficiente para
rDNA DNA ribossômico
SDS Sodium dodecyl sulfate
TSA Ágar Triptona Soja
T Tóxico
U.A.E. Unidades de atividade de emulsificação
VE Volume de emulsificação
VEt Volume de emulsificação no tempo t
VE0 Volume de emulsificação no tempo 0

LISTA DE SÍMBOLOS
% Porcentagem
H2O Água
°C Graus Célsius
mL Mililitros
® Marca registrada
mM Milimolar
U Unidades
pb Pares de bases
kb Quilobase
V Voltagem
mA Miliampére
pmol Picomoles
µl Microlitro
rpm Rotações por minuto
(v/v) Volume/volume
g Grama
µm Micrômetro
mm Milímetro
M Molar
nm Nanômetro
α Alfa
β Beta
γ Gama

SUMÁRIO
1 INTRODUÇÃO ........................................................................................ 14
2 REVISÃO BIBLIOGRÁFICA................................................................... 16
2.1 Biotecnologia marinha.......................................................................... 16
2.2 Poluição por resíduos........................................................................... 17
2.3 Efluente do beneficiamento da castanha do caju…..…………….... 18
2.4 Óleo vegetal residual……......…….……………………….…………….. 20
2.5 Reaproveitamento de resíduos..…………………………………..…..... 21
2.6 Biossurfactantes................................................................................. 22
3 HIPÓTESE CIENTÍFICA....……….…………………………...………….… 25
4 OBJETIVOS.......................................................................................... 25
4.1 Objetivo Geral........................................................................................ 25
4.2 Objetivos Específicos............................................................................ 25
5 MATERIAL E MÉTODOS....................................................................... 26
5.1 Seleção dos microrganismos.............................................................. 26
5.2 Identificação dos microrganismos produtores de biossurfactantes 26
5.2.1 Extração do DNA total.......................................................................... 26
5.2.2 Amplificação de fragmentos da região 16S rDNA por Reação em
Cadeia de Polimerase (PCR) utilizando iniciadores universais........
26
5.2.3 Visualização dos produtos da extração e dos amplicons gerados.. 27
5.2.4 Sequenciamento do DNA bacteriano................................................. 27
5.2.5 Alinhamento das sequências nucleotídicas obtidas........................ 28
5.3 Caracterização do potencial biotecnológico das estirpes
bacterianas.............................................................................................
29
5.3.1 Atividade hemolítica.............................................................................. 29
5.3.2 Teste de detecção de ramnolipideos................................................. 29
5.3.3 Produção de biossurfactantes a partir de resíduos orgânicos........ 29
5.3.4 Teste qualitativo do colapso da gota................................................. 31
5.3.5 Volume de emulsificação (VE) e estabilidade da emulsão (ES)........ 31
5.3.6 Atividade antimicrobiana..................................................................... 32
5.3.7 Atividade de emulsificação.................................................................. 32
5.3.8 Teste de toxicidade dos subprodutos microbianos........................... 33

6 RESULTADOS E DISCUSSÃO............................................................... 35
6.1 Identificação genotípica dos microrganismos com potencial
produtor de biossurfactantes...............................................................
35
6.2 Atividade hemolítica e produção de ramnolipídeos........................... 39
6.3 Teste qualitativo do colapso da gota e pH das fermentações........... 42
6.4 Volume de emulsificação, estabilidade da emulsão e atividade de
emulsificação.........................................................................................
47
6.5 Atividade antimicrobiana ..................................................................... 54
6.6 Teste de toxicidade com sementes de pepino.................................... 58
7 CONCLUSÕES........................................................................................ 70
REFERÊNCIAS....................................................................................... 71

14
1 INTRODUÇÃO
A vida surgiu nos oceanos há 3,5 bilhões de anos. Sua rica biodiversidade,
ainda desconhecida, desperta interesse pela busca de recursos marinhos com
potencial uso biotecnológico. Dentre os diferentes bioprodutos obtidos de
organismos marinhos destacam-se as drogas utilizadas no combate a diferentes
doenças, cosméticos, proteínas e materiais de interesse (TEIXEIRA et al., 2010).
Como parte da biodiversidade marinha, as bactérias destacam-se, pois
desempenham papel importante na produção de metabólitos com aplicações
distintas e na degradação de poluentes devido a sua ampla adaptação às diferentes
variações ambientais intrínsecas ao ambiente marinho. (THAVASI; SHARMA;
JAYALAKSHMI, 2011; DASH et al., 2013).
A decomposição biológica de poluentes através de microrganismos
(bactérias, bolores e leveduras) é denominada biorremediação. Este processo utiliza
o metabolismo microbiano para transformar essas substâncias no ambiente, onde a
microbiota utiliza os poluentes presentes no meio impactado como fonte de
nutrientes, reduzindo suas concentrações e proporcionando a sua mineralização
(ATLAS; CERNIGLIA, 1995; BRUSSEAU, 2006; JAIN; BAJPAI, 2012).
Inúmeros microrganismos têm sido descritos na literatura como
degradadores de poluentes e produtores de metabólitos de interesse. Dentre eles,
destacam-se os gêneros Pseudomonas, Bacillus, Acinetobacter, Burkolderia,
Rhodococcus, Nocardia, Serratia, Candida e Aspergilus (CASTIGLIONI; BERTOLINI;
COSTA, 2009, SAKALLE; RAJKUMAR, 2009, OBAYORI; SALAM; OMOTOYO, 2012;
HAMZA; MOHAMMED; SALE, 2012; SOBRINHO, et al., 2013,
(VARADAVENKATESAN; MURTY , 2013). Dentre esses, os gêneros Pseudomonas
e Bacillus têm sido amplamente estudados na produção de metabólitos com
propriedades surfactantes.
Biotensoativos ou biossurfactantes são compostos ativos de superfície
produzidos por microrganismos que reduzem a tensão superficial entre substâncias
de diferentes polaridades. Essa propriedade permite a sua aplicação em diferentes
setores industriais, como na remoção e recuperação de óleo na indústria petrolífera,
processos de biorremediação, indústria farmacêutica e alimentícia (NITSCHKE;
PASTORE, 2002). A biorremediação de poluentes hidrofóbicos pode ser
potencializada com o auxílio desses compostos, pois proporcionam a solubilização

15
de poluentes insolúveis no ambiente, aumentando a sua biodisponibilidade para a
comunidade microbiana degradadora (MONTAGNOLLI; LOPES; BIDOI, 2015).
Os biossurfactantes podem ser produzidos a partir de substratos de baixo
custo. O uso de fontes de carbono alternativas na sua produção permitirá uma
redução nos custos, visto que os surfactantes produzidos quimicamente têm como
matéria prima o petróleo (BEZERRA, 2012; PINTO et al., 2009). Dentre as matérias
primas de baixo valor com potencial uso na produção de biossurfactantes estão os
óleos vegetais e o efluente do beneficiamento da castanha do caju.
Os óleos vegetais são amplamente utilizados na produção de alimentos.
Seu descarte inadequado compromete os custos de tratamento de efluentes, assim
como alteram o equilíbrio natural dos corpos hídricos receptores (LOPES; BALDIN,
2009). De forma semelhante, a agroindústria do caju é reconhecida pela geração de
um subproduto tóxico ao ambiente denominado líquido da castanha de caju (LCC). É
necessário o tratamento desse efluente ou a sua adequação aos padrões de
lançamento devido aos efeitos toxicológicos que este apresenta aos corpos hídricos
receptores.
A utilização de resíduos como matéria prima para a elaboração de novos
produtos possibilita a redução dos impactos ambientais causados pelo seu
lançamento no ambiente, além de gerar lucro a partir de substratos de baixo custo
(COSTA-NETO et al., 2000). Nesse contexto, a busca por matérias residuais
domésticas e industriais como base para produção de compostos biossurfactantes é
uma abordagem biotecnológica que agrega valor econômico e é ambientalmente
correta no tratamento de compostos com potencial poluidor.

16
2 REVISÃO BIBLIOGRÁFICA
2.1 Biotecnologia marinha
Os oceanos correspondem a 72% da superfície do planeta, possuindo
representantes de todos os domínios existentes na Terra (VARELA et al., 2014). No
ambiente marinho ocorre intensa variação de temperatura, salinidade, oxigênio,
correntes marítimas e outros fatores abióticos. Entretanto, apesar desta variação
extrema, estes ambientes apresentam uma biodiversidade altamente diversificada,
variando desde microrganismos até animais de grande porte (DASH et al., 2013).
Os ecossistemas marinhos vêm despertando interesse devido a sua rica
biodiversidade que oferece inúmeros bioprodutos de interesse humano (SATPUTE et
al., 2010). De acordo com Teixeira et al. (2010), a utilização dos recursos marinhos
pelo homem se deu historicamente através da pesca, extração de petróleo e
derivados, cultivo de organismos marinhos, lazer e turismo. Ainda segundo os
autores, a grande biodiversidade existente nesse ambiente ainda permanece
desconhecida.
A prospecção de recursos marinhos tem por finalidade a busca por
bioprodutos com uso na indústria biotecnológica. Renduelez e Días (2014) definem
biotecnologia como a utilização de agentes biológicos para uso humano. Segundo os
autores, diferentes bens ou serviços podem ser extraídos a partir do metabolismo de
microrganismos ou de partes de material biológico de animais e plantas.
Teixeira et al. (2010) apresentam o conceito de biotecnologia marinha
como “ o desenvolvimento de drogas viáveis obtidas pela bioprospecção marinha e o
desenvolvimento de organismos geneticamente modificados para propósitos
ambientais ou aquicultura”. Ainda de acordo com os autores, a biotecnologia marinha
atua em diversas áreas, dentre elas: busca por novos fármacos isolados de
diferentes organismos; desenvolvimento de tecnologias de proteção ambiental;
desenvolvimento de métodos moleculares de identificação de doenças em animais
marinhos de interesse na pesca e aquicultura.
As bactérias marinhas produzem metabólitos extracelulares que possuem
características e aplicações distintas (THAVASI; SHARMA; JAYALAKSHMI, 2011).
Dentre os diferentes produtos biológicos obtidos do ambiente marinho podem ser
citados: os inúmeros fármacos e cosméticos isolados de algas, fungos e

17
invertebrados marinhos; produtos químicos de interesse comercial e biomateriais
como ágar, quitinas, proteínas, enzimas fluorescentes, aditivos suplementares em
alimentos, dentre outros (TEIXEIRA et al., 2010; BARRETO et al., 2014).
Mesmo o ambiente marinho oferecendo inúmeros produtos de interesse
humano, esse ecossistema ainda sofre com a poluição (SATPUTE et al., 2010).
Entretanto, sabe-se que as bactérias marinhas possuem a capacidade de formar
biofilme e excretar metabólitos poliméricos que atuarão na captação de poluentes
para as células. Além disso, são capazes de suportar variações ambientais extremas,
típicas deste ambiente. Tais características tornam esses microrganismos aptos para
remediar áreas contaminadas (DASH et al., 2013). A remoção de poluentes no
ambiente por meio de microrganismos ou plantas é possível devido a capacidade
destes organismos em utilizar poluentes como fonte de carbono (MUCHA ; MARISA ;
ALMEIDA, 2014).
2.2 Poluição por resíduos
O crescimento econômico e populacional somado à busca de maior
conforto e qualidade de vida impulsiona a utilização de recursos naturais para
produção de bens de consumo, gerando grandes quantidades de resíduos que irão
modificar o meio ambiente circundante (CANESIN et al., 2014 ; SILVA ; SANTOS ;
GOMES, 2014).
Uma parcela dos resíduos gerados nos processos industriais são
descartados sob condições sanitárias e ambientais inadequadas (ARCHELA et al.,
2003; GOUVEIA, 2012). Segundo Moreira et al., (2009), a agroindústria no Brasil
produz uma grande diversidade de subprodutos e resíduos, como o bagaço da cana-
de-açúcar, coco verde, caju, etc. Há pouco mais de uma década, a produção de
resíduos agroindustriais era de aproximadamente 250 milhões de toneladas por ano
(TAMANINI; HAULY, 2004). Entretanto, esse setor tem gerado, nos últimos anos,
uma quantidade crescente de rejeitos em virtude do incentivo mundial para a
produção de alimentos industrializados (SANTOS, 2014).
O aumento na geração de resíduos da agroindustria motiva a
necessidade de buscar tecnologias viáveis para a sua transformação com o objetivo
de reduzir seus impactos no meio ambiente (FIGUEIREDO; TANAMATI, 2010).
Moreira et al., (2009) destacam que a disposição inadequada de rejeitos

18
agroindustriais apresenta sérios problemas aos ambientes em que são lançados
devido à elevada quantidade e as dificuldades na sua disposição. Entretanto, de
acordo com Sater et al., (2011), apesar das indústrias agrícolas possuírem uma
preocupação em gerenciar e reaproveitar seus subprodutos, muitas ainda descartam
seus rejeitos de forma inadequada causando sérios impactos no meio ambiente
como a contaminação do solo e do lençol freático.
Dentre os diferentes resíduos agroindustriais com potencial poluidor pode-
se destacar o efluente gerado a partir do processamento da castanha de caju e os
resíduos oleosos de origem vegetal (PIMENTEL, 2008; LIMA, 2007). Ambos ao
serem lançados na natureza sem o devido tratamento irão causar impactos
significativos tanto ao meio ambiente quanto a saúde humana (ROSA et al., 2011;
OLIVEIRA, 2014).
2.3 Efluente do beneficiamento da castanha do caju
Conhecido popularmente como cajueiro, o Anacardium occidentale L. é
uma espécie vegetal da família Anacardiacea, nativa do Brasil e encontrada
principalmente na África, Ásia, em áreas costeiras e campos do Norte e Nordeste do
Brasil (CHAVES et al., 2010; SANJEEVA et al., 2014). O fruto do cajueiro, a
castanha, possui localizado na casca um líquido escuro, corrosivo e inflamável
conhecido como líquido da castanha de caju (LCC), enquanto na parte mais interna
encontra-se a amêndoa (MAZZETO; LOMONACO; MELE, 2009; ANDRADE et al.,
2011).
O aproveitamento industrial do caju consiste, majoritariamente, no
beneficiamento da castanha e, em segundo plano, o aproveitamento do pedúnculo.
Neste processo, apenas 15% do pedúnculo são aproveitados na produção de sucos,
geleias, doces, farinhas, fermentados e néctares (MOREIRA et al., 2009). De acordo
com Pimentel et al. (2011), o beneficiamento da castanha do caju consiste na
retirada da amêndoa do interior do fruto e posterior extração do LCC a partir da
casca através do cozimento da castanha ou prensado a frio.
De acordo com Sanjeeva et al., (2014), o LCC é classificado de acordo
com o método de extração empregado em dois tipos: LCC natural e LCC técnico.
Segundo os autores, a extração a partir de solventes orgânicos dá origem ao LCC
natural, constituído por ácido anacárdico (78%), cardol (10-15%) e pequenas frações

19
de 2-metil cardol, cardanol e material polimérico. Por sua vez, a extração por
aquecimento promove a descarboxilação do ácido anacárdido, gerando o LCC
técnico, constituído por 60-65% de cardanol, 20-23% de material polimérico, 10-12%
de cardol, 1-2% de metil-cardol (MAZZETO; LOMONACO; MELE, 2009; SANJEEVA
et al.,2014).
A figura 1 apresenta as estruturas químicas dos compostos presentes no
LCC. O ácido anacárdico é constituído por 15 átomos de carbono e um grupo
carboxila na posição orto e sofre rápida descarboxilação dando origem ao cardanol,
também constituído por 15 átomos de carbono na posição meta contendo ou não
duplas ligações (IUNESCU et al., 2012).
Figura 1 – Estruturas químicas dos componentes do LCC: a) ácido anacárdico, b) cardol, c) cardanol
Fonte: MAZZETO; LOMONACO; MELE, 2009
No processo de beneficiamento da castanha do caju o LCC produzido é
removido para posterior comercialização devido às propriedades de seus
constituintes fenólicos saturados e insaturados que os permitem serem utilizados em
diferentes setores industriais (JERONIMO et al., 2012; MAIA et al., 2012).
O seu principal componente, o cardanol, possui propriedades
antioxidantes, antimicrobianas, antitumorais, antisséptica, antivermífuga,
gastroprotetora e inibidora de enzimas. Tais características possibilitam seu uso na
indústria farmacêutica, formulação de pesticidas e na produção de surfactantes
(MAZZETO; LOMONACO; MELE, 2009; YULIANA, TRAN-TCHI; JU , 2012; RIBEIRO
et al., 2013).
Segundo Santaella et al. (2002), o efluente gerado no fim do processo de
beneficiamento da castanha do caju é produzido, principalmente, durante a lavagem
das castanhas, umidificação das mesmas e lavagem dos gases e equipamentos

20
utilizados na indústria. Há uma preocupação com os efeitos desse efluente nos
corpos hídricos devido à alta quantidade de fenóis e DQO presentes (PIMENTEL,
2008; PIMENTEL et al., 2011). Somado a isso, vale ressaltar a grande quantidade de
água consumida durante o processo de beneficiamento (JERONIMO et al., 2012).
Apesar da remoção do LCC para uso comercial é nítida a necessidade de
tratamento do efluente gerado, pois mesmo após a remoção do subproduto, o
efluente proveniente do processo de beneficiamento da castanha ainda contém
traços do LCC, apresentando sérios riscos aos corpos hídricos receptores
(PIMENTEL, 2008).
2.4 Óleo vegetal residual
Alimentos fritos são amplamente consumidos em escala mundial. De
acordo com dados do Instituto de Economia Agrícola (2011), os óleos vegetais são
produzidos a partir de diferentes oleaginosas, sendo o óleo de palma, também
conhecido como dendê, o mais consumido mundialmente, representando 32% do
consumo. Em segundo lugar está o óleo de soja representando 29% do consumo
mundial. No Brasil, grande parte do óleo vegetal destinado à alimentação humana
(cerca de 95%) é derivado da soja, correspondendo também a matéria prima
principal para a produção de biodiesel no país (OSAKI; BATALHA, 2011).
De acordo com Freire, Mancini-Filho e Ferreira (2013), existem dois tipos
de frituras por imersão: a contínua e a descontínua. Na primeira, amplamente
utilizada em escala industrial, o alimento é frito de uma só vez com aquecimento
contínuo. Na segunda, normalmente empregada em estabelecimentos comerciais e
em residências, o óleo é aquecido para preparo de uma refeição, resfriado e
reutilizado posteriormente. Segundo os autores, em ambos os processos ocorrem
reações físico-químicas diferentes, como hidrólise, oxidação e polimerização,
gerando diferentes subprodutos que podem alterar as características do óleo como
cor (tornando-se mais escuro), viscoso, ácido e com odor desagradável.
São considerados resíduos de óleo de cozinha os óleos ou gorduras
descartados em esgotamento sanitário após utilização no processo de fritura a altas
temperaturas (ZHANG; WANG; MORTIMER, 2012). A utilização de óleos em
processos alimentícios de diferentes setores produz grandes quantidades de
resíduos (RUFINO; SARUBBO; TAKAKI, 2007). Infelizmente, no Brasil, grande parte

21
do óleo vegetal é descartado de forma indevida no solo, nos corpos hídricos, redes
de esgotos ou em processos de incineração (THODE - FILHO et al., 2013).
O descarte de óleo residual doméstico na rede pública de esgoto (quando
existe) apresenta sérios problemas econômicos, pois aumentam os custos de
tratamento de efluentes devido à obstrução dos canos da rede coletora (SEGATTO,
2013). Quando lançado diretamente no solo pode ocasionar a sua
impermeabilização, prejudicando o escoamento das águas pluviais e contribuindo
para eventos de enchentes ou até mesmo migrar para camadas mais profundas
atingindo o lençol freático (FREITAS; BARATA; NETO, 2010). Nos corpos hídricos, o
óleo impede a oxigenação e prejudica fortemente os organismos dependentes de
oxigênio (SOARES; PUPO; MOURÃO, 2013). Por outro lado, este resíduo já
desperta interesse como matéria prima para diferentes produtos, como tintas,
biodiesel, detergentes, sabão etc. (WILDNER; HILLIG, 2012).
2.5 Reaproveitamento de resíduos
A indústria de alimentos gera inúmeros resíduos que servem como fonte
de proteínas, óleos essenciais e enzimas aptas a serem reaproveitadas e
recuperadas (COELHO et al., 2001). O reaproveitamento destes resíduos evita o seu
encaminhamento aos aterros sanitários, gerando novas alternativas de utilização
(COSTA-NETO et al, 2000).
A reutilização de resíduos orgânicos como matéria prima possibilita a
geração de renda, energia e diversos produtos na indústria química, contribuindo
para a minimização dos impactos ambientais e os riscos à saúde causados pelo seu
descarte inapropriado (SANTOS, 2014). A utilização de fontes alternativas de
energia e matéria prima tem sido crescente diante da redução das reservas
petrolíferas e do rigor da legislação ambiental (SEXENA; JAWALE; JOSHIPURA,
2013).
O reuso de resíduos agroindustriais na produção de novos produtos está
relacionado não só com a redução de impactos ambientais ocasionados pela má
disposição desses rejeitos, mas está atrelada ao seu aproveitamento energético
visando à geração do lucro, pois muitos destes rejeitos são considerados matérias
primas de baixo custo (COSTA-NETO et al., 2000).
Dentre os diversos exemplos de utilização de resíduos podem ser citados

22
a utilização de casca de coco oriundas da industrialização da água de coco como
substrato para a produção de enzimas (COELHO et al, 2001), a produção de
biodiesel a partir de óleo de cozinha residual (FREITAS; BARATA; NETO, 2010), e a
utilização de resíduos agroindustriais como fonte de matéria prima na produção de
tensoativos microbianos (RUFINO; SARUBBO; TAKAKI, 2007; BEZERRA, 2012;
KHOPADE et al., 2012).
Os tensoativos de origem microbiológica ou comumente denominados
biossurfactantes são substâncias que apresentam ampla utilização na indústria
devido as suas inúmeras propriedades emulsificantes, tensoativas, antimicrobianas,
umectantes, dentre outras (FRANÇA, 2014). A utilização de resíduos de baixo valor
pode auxiliar na redução dos custos de produção dos surfactantes químicos, além
de auxiliar na redução dos problemas ambientais advindos do descarte inadequado
de efluentes e resíduos industriais (BEZERRA, 2012).
2.6 Biossurfactantes
Agentes ativos de superfície ou surfactantes são substâncias anfipáticas,
que possuem uma porção hidrofílica e outra hidrofóbica na mesma molécula. Tal
configuração permite a formação de um filme molecular que reduz a tensão
superficial e interfacial entre fases distintas como óleo e água (NITSCHKE ;
PASTORE, 2002).
Os surfactantes são produzidos quimicamente a partir de petróleo (PINTO
et al., 2009). Por sua vez, os biossurfactantes são compostos surfactantes de origem
microbiológica que possuem propriedades semelhantes aos surfactantes de origem
sintética (THAVASI; SHARMA; JAYALAKSHMI, 2011; ZHENG et al., 2012; FRANÇA,
2014). Dentre as características dos biossurfactantes está a capacidade de formar e
manter emulsões estáveis (UZOIGWE et al., 2015). Emulsões consistem em
misturas coloidais de dois líquidos imiscíveis pela ação de um emulsificante
(FRANZOL; REZENDE, 2015). Além de possuir atividade tensoativa e emulsificante,
alguns biossurfactantes possuem propriedades umectantes e antimicrobianas
(FRANÇA, 2014). Tais características permitem a sua utilização em diferentes
setores industriais com destaque na recuperação de óleo da indústria petrolífera,
alimentícia, farmacêutica e de cosméticos (UZOIGWE et al., 2015).
Os biossurfactantes são classificados de acordo com a sua natureza

23
química e com o microrganismo produtor (FRANÇA, 2014). Entretanto, Uzoigwe et al.
(2015) os classifica de acordo com o seu peso molecular em dois tipos:
Biossurfactantes: são moléculas de baixo peso molecular,
geralmente constituídos de açúcares, aminoácidos, ácidos pesados
e grupos funcionais, como o ácido carboxílico, por exemplo. Estes
são capazes de reduzir a tensão superficial, assim como produzir
emulsões estáveis;
Bioemulsificantes: são compostos de alto peso molecular
constituídos por uma mistura de heteropolissacarídeos,
lipopolissacarídeos, lipoproteínas e proteínas que apresentam forte
atividade emulsificante, porém não são eficientes em reduzir a
tensão superficial.
Segundo Nitschke e Pastore (2002), os biossurfactantes constituem uma
das principais classes de surfactantes naturais. Dentre os principais microrganismos
produtores desses compostos destacam-se as bactérias, fungos e leveduras
(KHOPADE et al., 2012 ; VARADAVENKATESAN; MURTY , 2013). Diferentes
gêneros bacterianos estão descritos como produtores de biossurfactantes, dentre
eles Pseudomonas, Bacillus, Acinetobacter, Nocardia, Aerobacillus, Nocardiopsis,
Staphylococcus, Klebsiella, Rhodococcus, dentre outros (ROCHA e SILVA et al.,
2014, VARADAVENKATESAN; MURTY, 2013; BAO et al., 2014; VYAS ; DAVE, 2011;
ZHENG et al., 2012; KHOPADE et al., 2012 ; KESKIN et al., 2015; JAIN et al., 2013,
PACHECO et al., 2010).
Em áreas poluídas (solo ou água), os biossurfactantes promovem uma
melhor dispersão do óleo, aumentando sua biodisponibilidade e acesso aos
microrganismos degradadores por meio da solubilização em micelas (ACCORSINI,
2010). O aumento da biodisponibilidade dos poluentes hidrofóbicos promovida pela
ação dos biossurfactantes acelera o processo de degradação microbiana em áreas
contaminadas (VARADAVENKATESAN ; MURTY, 2013 ; MONTAGNOLLI; LOPES;
BIDOI, 2015).
Os biossurfactantes possuem inúmeras vantagens quando comparados
aos surfactantes de origem sintética, como por exemplo: apresentam maior

24
biodegradabilidade, baixa toxicidade, são seletivos e compatíveis com os
surfactantes químicos, são eficientes em condições extremas de temperatura, pH e
salinidade (SOUZA; SALGUEIRO; ALBUQUERQUE, 2012). O longo período de
permanência dos surfactantes químicos no ambiente se dá devido a sua natureza
xenobiótica (VARADAVENKATESAN; MURTY, 2013).
Diferente dos surfactantes químicos, os biossurfactantes podem ser
produzidos a partir de diferentes fontes de carbono, dentre elas: hidrocarbonetos,
carboidratos e resíduos agroindustriais (RAHMAN et al., 2002; VIRAMONTES-
RAMOS et al., 2010; BEZERRA, 2012; DASGUPTA; GHOSH; SENGUPTA, 2013;
CAMPOS; STAMFORD; SARUBBO, 2014). Contudo, sabe-se que a utilização de
diferentes substratos na produção desses compostos pode mudar a sua
configuração molecular e suas propriedades (CAMPOS, STAMFORD; SORUBBO,
2014).
Apesar das vantagens dos biossurfactantes, seu uso na indústria ainda é
limitado, pois são menos competitivos que os surfactantes sintéticos por conta dos
altos custos dos substratos e da recuperação do bioproduto durante a produção
(SOUSA, 2011; SOBRINHO et al., 2013). Portanto, é importante que fontes de
carbono alternativas e de baixo custo sejam utilizadas, visando à redução dos custos
de produção dos biossurfactantes (BEZERRA, 2012).
Tendo como base a problemática da geração de resíduos e a possível
utilização destes na produção compostos na indústria química, este trabalho teve
como objetivo avaliar o potencial biotecnológico de bactérias marinhas produzirem
biossurfactantes utilizando efluente da agroindústria do caju e óleo vegetal residual
como fontes de carbono. O reaproveitamento desses resíduos como matéria prima
na produção de biossurfactantes poderá minimizar os impactos gerados pelo
descarte inadequado desses rejeitos no meio ambiente.

25
3 HIPÓTESE CIENTÍFICA
As bactérias marinhas possuem uma diversidade metabólica que lhes
permite utilizar resíduo da agroindústria e óleo vegetal residual como fontes de
carbono e energia produzindo metabólitos com propriedades surfactantes.
4 OBJETIVOS
4.1 OBJETIVO GERAL
Avaliar o potencial biotecnológico de bactérias isoladas de ambiente
marinho da região portuária de Fortaleza para sintetizarem compostos
biossurfactantes a partir de resíduo da agroindústria do caju e óleo vegetal residual
de uso doméstico.
4.2 OBJETIVOS ESPECÍFICOS
1. Realizar identificação genética de bactérias marinhas produtoras de
biossurfactantes;
2. Verificar o potencial dessas bactérias produzirem compostos
biossurfactantes através dos testes de hemólise e detecção de
ramnolipídeos;
3. Realizar fermentações a partir de efluente da indústria do
beneficiamento do caju e óleo vegetal residual;
4. Avaliar a produção dos biossurfactantes a partir do teste do colapso da
gota, volume de emulsificação, estabilidade da emulsão e atividade
emulsificante utilizando os líquidos metabólicos livres de células;
5. Testar a atividade antimicrobiana dos líquidos metabólicos contra
bactérias patogênicas;
6. Testar a toxicidade dos líquidos metabólicos utilizando sementes de
pepino Cucumis sativus L. como organismo indicador.

26
5 MATERIAL E MÉTODOS
5.1 Seleção dos microrganismos
Foram selecionadas 20 cepas bacterianas provenientes de amostras de
água e sedimento do porto do Mucuripe, Fortaleza- CE, pertencentes à coleção
bacteriológica do Laboratório de Microbiologia Ambiental e do Pescado. As culturas
puras foram renovadas em Ágar Triptona Soja (TSA) e incubadas a 35°C por 24
horas. Após esse período as cepas foram incubadas em estufa B.O.D. para posterior
realização dos testes indicativos da produção de biossurfactantes e das
fermentações. Estas cepas foram selecionadas tendo como base o seu potencial
produtor de biossurfactantes a partir de querosene comercial, verificado por Silva et
al. (2017).
5.2 Identificação dos microrganismos produtores de biossurfactantes
5.2.1 Extração do DNA total
As cepas bacterianas previamente purificadas foram inoculadas em meio
Luria Bertani (LB) a 1% de NaCl e incubadas a 35°C por 48 horas. Após este
período, retirou-se uma alíquota de 1mL do inóculo para a extração do DNA total
utilizando kit comercial Wizard® Genomic DNA Purification Kit (Promega) de acordo
com as instruções do fabricante.
5.2.2 Amplificação de fragmentos da região 16S rDNA por Reação em Cadeia
de Polimerase (PCR) utilizando iniciadores universais
O DNA total extraído foi utilizado como molde na amplificação da região
conservada do DNA ribossomal, subunidade 16S, através da Reação em Cadeia de
Polimerase (PCR) utilizando os iniciadores universais 1401L (5’- CGG TGT GTA CAA
GGC CC - 3’) e 968U (5’-AAC GCG AAG AAC CTT AC- 3’) descritos por Pereira et
al., (2011) e Zhang e Fang (2001). Os volumes e concentrações dos reagentes, as
condições de termociclagem e o tamanho dos amplicons gerados estão descritos na
tabela 1. As amplificações foram realizadas em termociclador AmpliTherm modelo

27
TX96.
Tabela 1 - Reagentes e condições utilizadas na amplificação da subunidade 16S das
bactérias através da Reação em Cadeia de Polimerase
REAGENTES DA REAÇÃO CONDIÇÕES DE TERMOCICLAGEM AMPLICONS
(pb)
Tampão 0,8 X
Desnaturação 94°C / 2min
433
MgCl2 0,4 mM
DNTPs 0,032mM
Iniciador F 0,16 mM Desnaturação/
anelamento
30 ciclos:
94°C/1min.; 52°C/1min.;
72°C /2min.
Iniciador R 0,16 mM
Taq 0,04 U
DNA * Extensão final 72°C / 8 min
H2O ultrapura q.s.p. 25µl
Fonte: Elaborada pelo autor. (*) concentração não padronizada; pb: pares de base.
5.2.3 Visualização dos produtos da extração e dos amplicons gerados
Os produtos da extração de DNA e da reação de PCR foram visualizados
através da eletroforese em gel de agarose (1%) acrescido do corante GelRed para
auxiliar na visualização das bandas com aplicação de luz ultravioleta em um
transluminador (Espectroline-UV). Foi utilizado um marcador molecular de 1kb DNA
ladder (Sigma) para visualizar o tamanho dos amplicons gerados. A migração das
moléculas no gel ocorreu com voltagem de 120V, amperagem de 120mA com tempo
de duração de 60 minutos. Os géis foram fotodocumentados em sistema Kodak
EDAS290.
5.2.4 Sequenciamento do DNA bacteriano
Após a amplificação da subunidade 16S rDNA, as amostras foram
encaminhadas ao Centro de Diagnóstico para Enfermidades de Organismos
Aquáticos (CEDECAM), do Instituto de Ciências do Mar (LABOMAR – UFC) para
sequenciamento do DNA bacteriano utilizando o BigDye® Terminator v3.1 Cycle
Sequencing Kit. Em microplaca de PCR contendo 96 poços foram aplicados os
componentes descritos na tabela 2, seguidos das etapas de desnaturação,
anelamento, extensão e resfriamento.

28
Tabela 2 – Reagentes do Mix de PCR e condições de termociclagem do
sequenciamento
Reagentes Concentração Condições de termociclagem
Big Dye 0,5x Desnaturação 96°C / 15s Primer 1401L 3,2 pmol
Desnaturação/anelamento 40 ciclos: 96°C/15s, 50°C/15s Primer U968 3,2 pmol Tampão 5x 0,5x Extensão final
60°C/4 min;
Água ultra pura -
Amostra de PCR 0,8µL Resfriamento 4°C Volume final 10µL
Fonte: Elaborada pelo autor.
A microplaca foi tratada com isopropanol 65% (40 µL para cada amostra)
para lavagem dos amplicons seguidas de agitação em vórtex e deixadas à
temperatura ambiente por 20 minutos. Após esse período, a mesma foi centrifugada
a 4000 rpm por 30 minutos a 22°C com posterior descarte do sobrenadante. Foi
adicionado etanol 60% (150 µL por amostra) seguido de centrifugação a 4000 rpm
por 10 minutos a 22°C. Após o descarte do álcool a placa contendo as amostras foi
seca através de centrifugação rápida (“spin”). As amostras purificadas foram
ressuspendidas em formamida (20 µL por amostra) seguidas de desnaturação a
95°C por 5 minutos e transferidas para bloco a 4°C. As amostras foram aplicadas e
lidas em sequenciador automático capilar ABI 3500.
5.2.5 Alinhamento das sequências nucleotídicas obtidas
Os fragmentos amplificados foram sequenciados através da análise das
sequências obtidas quanto à qualidade dos nucleotídeos e similaridade, utilizando-
se o programa MEGA (TAMURA et al., 2013) versão 6 com posterior comparação
com as sequências depositadas no GenBank (BENSON et al., 2002) utilizando o
Basic Local Alignment Search Tool (BLAST).

29
5.3 Caracterização do potencial biotecnológico das estirpes bacterianas
5.3.1 Atividade hemolítica
A produção de biossurfactantes está relacionada com a capacidade das
bactérias quebrarem as células vermelhas do sangue. Esta avaliação prévia foi
realizada de acordo com a metodologia descrita por Carrilo et al. (1996), com
modificações. As cepas bacterianas, após 24horas de crescimento, foram inoculadas
em placas de Petri com ágar sangue contendo 5% (v/v) de sangue de carneiro
desfibrilado. Após a inoculação, as placas foram incubadas a 35°C por 48 horas. A
positividade no teste foi evidenciada através da formação de zonas de hemólise ao
redor das colônias.
5.3.2 Teste de detecção de ramnolipídeos
A produção de ramnolipídeos seguiu a metodologia descrita por Schultz
(2010), com adaptações. As cepas com 24 horas de crescimento foram inoculadas
em placas de Petri contendo meio ágar azul de metileno (lactose 10g; fosfato de
potássio 2g; azul de metileno 0,005g; brometo de cetiltrimetilamônio 0,2g; peptona
bacteriológica 10g; ágar 15g; água deionizada q.s.p. 1000 mL). Após a inoculação,
as placas foram incubadas por até 96 horas a 35°C. A produção de ramnolipídeos foi
evidenciada pela formação de um halo azul ao redor da colônia.
5.3.3 Produção de biossurfactantes a partir de resíduos orgânicos
A produção de biossurfactantes foi realizada de acordo com a
metodologia descrita por Lima e Silva et al. (2010) com adaptações (Figura 2). As
cepas com crescimento de 24 horas foram inoculadas em meio LB e incubadas por
16 horas a 35°C. Posteriormente, foi retirado 1mL do meio de cultivo contendo
crescimento bacteriano e inoculado novamente em meio LB acrescido das fontes de
carbono nas concentrações de 2% (v/v) para o efluente do beneficiamento da
castanha de caju (Figura 3a) e 1% para o óleo vegetal residual (Figura 3b). Os
frascos permaneceram sob agitação magnética de 150 rpm a 35°C por 72 horas.
Após o período de agitação foi extraído o líquido metabólico das bactérias através de

30
centrifugação do meio de cultivo a 12000 rpm por 15 minutos a 4°C, seguido de
filtração em membranas de 0,22µm.
Figura 2: Ilustração do processo de produção dos biossurfactantes
Fonte: Adaptado de Silva (2014).
Figura 3 : Fontes de carbono utilizadas na produção de biossurfactantes pelas cepas bacterianas: a) Efluente do beneficiamento da castanha do caju; b) Óleo vegetal residual
Fonte: Autor.
a b
Ágar TSA 35°C por 24
horas Meio LB
50 mL
1 mL
Meio LB
+ substrato
Estufa Shaker a 35°C / 150
rpm por 72 horas
Centrifugação a 12000 rpm
por 15 min / 4°C
Filtragem em membrana
0,22 µm

31
5.3.4 Teste qualitativo do colapso da gota
Este teste foi realizado segundo a metodologia descrita por Bodour e
Miller-Maier (1998). Uma alíquota de 10µL de óleo de motor Castrol®10W20 foi
aplicada nos poços de uma microplaca de 96 poços seguido de descanso por 24
horas. Após o período, foram dispensadas 10µL (em triplicata) dos líquidos testes
nos poços untados: a cultura fermentada (meio com as células bacterianas após 72
horas) e do líquido metabólico resultante dessa fermentação (meio isento de células).
Foi usada água destilada esterilizada como controle negativo. Após 1 minuto, foi
verificado o comportamento da gota em contato com a superfície hidrofóbica. O
espalhamento ou o colapso da gota quando depositada na placa untada com óleo foi
interpretado como positivo, indicativo da possível redução da tensão superficial,
característica dos biossurfactantes. Entretanto, caso a gota permanecesse coesa
(semelhante ao comportamento apresentado pela gota de água) admitia-se
resultado negativo.
5.3.5 Volume de emulsificação (VE) e estabilidade da emulsão (ES)
Foi utilizada a metodologia descrita por Das, Das e Mukheryee (1998),
com modificações. Em tubos de 12mm x 100mm foram adicionados 2mL do líquido
metabólico e 2mL de óleo mineral, em triplicata. Os tubos foram agitados por 2
minutos utilizando-se agitador de tubos do tipo vórtex (Biomixer Q-L 901) e deixados
em repouso pelo mesmo período. Após o descanso, foi medida a altura da camada
emulsificada. A emulsão foi medida após 24 horas para determinação do volume de
emulsificação (VE) utilizando-se a fórmula 1. Foi acompanhado o comportamento
das emulsões através de medições até 168 horas para determinação da sua
estabilidade através da fórmula 2 descrita por Fennema (1985). O surfactante
sintético Dodecil Sulfato de Sódio (SDS) foi utilizado como controle positivo no teste.
Foram consideradas boas emulsões aquelas que após 24 horas de formadas
mantivessem VE maior ou igual a 50% da emulsão formada após os 2 minutos de
agitação (BENTO; CAMARGO; GAYLARDE, 2008).
VE(%)= x 100 (1)

32
%ES = x 100 (2)
5.3.6 Atividade antimicrobiana
A propriedade antimicrobiana dos biossurfactantes produzidos foi testada
através da metodologia descrita pelo Clinical and Laboratory Standard Institute (CLSI,
2010). A ação antimicrobiana dos líquidos metabólicos extraídos foi testada frente às
cepas padrões de Staphylococcus aureus ATCC 25923 e Escherichia coli ATCC
25922. A concentração celular dessas cepas foi padronizada comparando-se à
turvação 0,5 de MacFarland, após leitura em espectrofotômetro (BioMate 3). Após o
ajuste, os inóculos foram dispensados uniformemente em placas de Petri contendo
ágar Mueller Hinton (Difco) utilizando-se cotonetes estéreis. Foram confeccionados
poços nas placas utilizando-se cilindros previamente esterilizados e aplicados 50µl
de água destilada estéril (controle negativo) e dos líquidos metabólicos, em duplicata,
para cada bactéria testada. Como controle positivo foi usado o antimicrobiano
cloranfenicol (30µl). As placas foram incubadas a 35°C por 48 horas. A formação de
halos de inibição ao redor dos poços foi interpretada como resultado positivo no
teste e a ausência negativa.
5.3.7 Atividade de emulsificação
Este teste foi realizado segundo a técnica descrita por Cirigliano e
Carman (1984), com modificações. Em tubos de ensaio com tampa de rosca foram
adicionados 2mL do líquido metabólico, 2mL de tampão de acetato de sódio 0,1 M
(pH 3) e 1mL de óleo mineral, em triplicata. Os tubos foram agitados por 2 minutos e
permaneceram em repouso pelo período de 10 minutos. Após esse tempo, a
emulsão formada foi lida em espectrofotômetro com comprimento de onda de 540
nm. O resultado foi expresso em Unidades de Atividade de Emulsificação (U.A.E.).
Foi utilizado caldo LB acrescido de acetato de sódio como branco.

33
5.3.8 Teste de toxicidade dos subprodutos microbianos
Os líquidos metabólicos livres de células que obtiveram os melhores
resultados nos testes indicativos da produção de biossurfactantes foram
selecionados para o teste ecotoxicológico utilizando-se sementes de pepino
(Cucumis sativus L.) do tipo Aodai segundo a metodologia descrita por Rodríguez e
Perera (2009). Este teste foi realizado com o objetivo de se verificar os efeitos
toxicológicos dos subprodutos microbianos nos processos de germinação e
desenvolvimento das raízes das sementes. As mesmas foram adquiridas em
estabelecimento comercial e selecionadas com base na forma, tamanho e cor. Após
a seleção, foram desinfetadas por imersão em hipoclorito de sódio (Dinâmica) por 20
minutos, seguidas de sucessivas lavagens com água destilada, secagem e
armazenamento a 4°C na ausência de luz. Em placas de Petri contendo papel de
filtro absorvente, em duplicata, foram dispostas 20 sementes e aplicados 3mL de
cada tratamento. Foram utilizados 5 tratamentos para cada amostra selecionada nas
concentrações de 100%, 50%, 25%, 10% e 5% do líquido metabólico. Foi utilizada
água mineral como controle negativo (K-) e solução de NaCl a 0,5% como controle
positivo (K+). Após aplicação dos controles e amostras, as placas foram incubadas a
20°C por 120 horas. Após esse período foi contabilizado o número de sementes
germinadas e medidas as raízes crescidas tendo como critério de medição aquelas
com tamanho maior ou igual a 0,5 centímetros. A toxicidade foi avaliada através da
verificação da inibição do desenvolvimento das raízes (efeitos subletais) e do
percentual de sementes germinadas (efeitos letais). O percentual de inibição (IP%)
do desenvolvimento das raízes foi determinado de acordo com a fórmula 3, onde: IP
positivo indica condição tóxica, ou seja, inibição do crescimento das raízes; IP
negativo representa um estímulo de crescimento das raízes; e IP igual a 0 indica
ausência de toxicidade.
(3)
Os efeitos letais evidenciados pelo percentual de germinação das sementes foram
determinados de acordo com a seguinte classificação:

34
Não tóxicas = tratamentos com percentual de germinação acima de 90% em
relação ao controle;
Tóxicas = tratamentos com percentual de germinação entre 75 e 90% de
germinação em relação ao controle;
Muito tóxicos = tratamentos com percentual de germinação abaixo de 75%
com relação ao controle.
Além dos líquidos metabólitos, foi avaliada a toxicidade do surfactante
SDS e do efluente do beneficiamento da castanha do caju visando um melhor
entendimento dos efeitos desses compostos sobre as sementes de pepino.

35
6 RESULTADOS E DISCUSSÃO
6.1 Identificação genética dos microrganismos com potencial produtor de
biossurfactantes
Na tabela 3 está descrita a origem das cepas e o resultado do
sequenciamento genético. O gênero Pseudomonas foi o que apresentou o maior
número de isolados (8). Além deste grupo, foram identificadas cepas pertencentes
aos gêneros Bacillus (2), Staphylococcus (1) e Brachybacterium (1). Dentre as cepas
de Bacillus, uma foi identificada como pertencente à espécie Bacillus pumilus. Foram
identificadas bactérias pertencentes às espécies bacterianas Víbrio fluvialis (3),
Zobellella taiwanensis (3) e Oceanimonas baumannii (2).
Tabela 3 – Codificação, origem e resultado da identificação genética das bactérias marinhas selecionadas na presente pesquisa
Fonte: Elaborada pelo autor.
A utilização de técnicas moleculares tem possibilitado um melhor
entendimento da dinâmica microbiana de áreas impactadas por petróleo (SANTOS,
Cepa Origem Espécie/ gênero Similaridade (%)
2D Água Pseudomonas sp. 99 2E Água Pseudomonas sp. 99 7A Água Pseudomonas sp. 99 7C Água Pseudomonas sp. 99 9A Água Pseudomonas sp. 99 9B Água Pseudomonas sp. 98 9C Água Pseudomonas sp. 99 18D Água Pseudomonas sp. 99 21A Água Vibrio fluvialis 100 31A Água Vibrio fluvialis 100 36C Água Vibrio fluvialis 100 26B Água Staphylococcus sp. 99 12E Água Bacillus sp. 99
12SEDB Sedimento Bacillus pumilus 100 11SA Sedimento Zobellella taiwanensis 100 11SB Sedimento Zobellella taiwanensis 100 11SD Sedimento Zobellella taiwanensis 100 12SC Sedimento Oceanimonas baumannii 99 12SE Sedimento Oceanimonas baumannii 99
11SEDH Sedimento Brachybacterium sp. 99

36
2012). Além disso, o estudo da comunidade bacteriana degradadora de
hidrocarbonetos em solos impactados por petróleo possibilita um melhor
entendimento do processo de biorremediação dessas áreas, principalmente quando
da sua associação com as raízes de plantas, caracterizando o processo de
fitorremediação (TSUBOI et al., 2015).
O sequenciamento da região 16S rDNA de bactérias tem gerado uma
grande quantidade de informações a respeito da diversidade microbiana. Essas
informações são depositadas em banco de dados genético e permite a identificação
de bactérias cultiváveis e não cultiváveis através da comparação de similaridades
das sequências de DNA (SANTOS, 2012).
De acordo com Satpute et al. (2010), muitos microrganismos produtores
de biossurfactantes têm sido isolados de ambientes impactados por óleo, petróleo e
derivados. No presente estudo, o gênero Pseudomonas foi o que apresentou maior
expressividade, representando um total de oito cepas. Esse grupo bacteriano é
reconhecido por sua capacidade de degradar hidrocarbonetos e utilizá-los como
única fonte de carbono (ZHAO et al., 2009 ; SANTOS, 2012), assim como a sua
capacidade de produzir biossurfactantes tem sido amplamente estudada (LIMA e
SILVA et al., 2010; BEZERRA, 2012; DHAIL; JASUJA, 2012; KACZOREK et al.,
2012).
Semelhante ao gênero Pseudomonas, o gênero Bacillus tem merecido
destaque na produção de biossurfactantes. De acordo com Varadavenkatesan e
Murty (2013), algumas bactérias desse gênero produzem lipopetídeos cíclicos como
a surfactina, presentes nas espécies B. subtilis e B. pumillus (BUGAY, 2009). A
surfactina tem sido um dos biossurfactantes mais estudados e aplicados
comercialmente (OLIVEIRA, 2010; PEREIRA et al., 2013). Em seu estudo de
produção de biossurfactante em caldo de fermentação, Bueno, Silva e Garcia-Cruz
(2010) utilizaram uma espécie de B. pumillus isolada de solo contaminado por
hidrocarboneto, mostrando que essa espécie habita áreas impactadas por petróleo e
derivados.
Foram identificadas três cepas bacterianas pertencentes à espécie Víbrio
fluvialis. Segundo Ramamurthy et al. (2014) esta espécie encontrada em ambientes
costeiros tem sido descrita como um patógeno eminente e de difícil identificação
devido as suas semelhanças fenotípicas com Vibrio cholerae e Aeromonas spp.
Entretanto, técnicas moleculares têm facilitado a distinção entre as espécies. Tonini,

37
Rezende e Grativol (2010) e Santos (2012) citam o gênero Vibrio como degradador
de compostos de petróleo.
Hamdan e Fulmer (2011) verificaram os efeitos de um dispersante
químico sobre a microbiota do óleo depositado em uma área costeira afetada pelo
vazamento da Deepwater Horizon e constataram que o dispersante químico não
inibiu o crescimento de Vibrio spp. identificados no local. A análise genética
identificou quatro cepas desse gênero, dentre elas V. fluvialis. Entretanto, de acordo
com os autores a capacidade desse gênero bacteriano de degradar petróleo e seus
componentes ainda é pouco conhecida. Verugopal e Saramma (2006) verificaram a
produção de proteases com potencial uso como aditivos em detergentes por V.
fluvialis isolados de mangue. Esses trabalhos reforçam o potencial de uso
biotecnológico dos metabólitos produzidos pelas cepas de V. fluvialis identificadas na
presente pesquisa.
Foi identificada uma cepa pertencente ao gênero Staphylococcus.
Pesquisas que avaliam a microbiota de áreas impactadas por petróleo e derivados
têm apontado bactérias pertencentes ao gênero Staphylococcus como potenciais
degradadores dessas substâncias (OLIVEIRA, 2009; LOPES, 2010). Entretanto, são
poucos os estudos que descrevem bactérias desse gênero como produtoras de
biossurfactantes.
Eddouaouda et al. (2012) verificaram que uma bactéria desse gênero foi
capaz de produzir biossurfactantes com boa estabilidade e com ação antimicrobiana
contra bactérias patogênicas (dentre elas, S. aureus e E. coli) a partir de diferentes
fontes de carbono, incluindo óleo vegetal. Keskin et al. (2015) verificaram que os
metabólitos produzidos por Staphylococcus xylosus STF1 isolada de solo
contaminado por petróleo apresentou alta atividade emulsificante e antimicrobiana.
Nesses dois estudos, os biossurfactantes produzidos pertenciam ao grupo dos
lipopeptídeos.
De acordo com Nitschke e Pastore (2002), as células de Staphylococcus
aureus possuem alta hidrofobicidade superficial. Essas células com alta atividade
surfactante pertencem ao grupo dos biossurfactantes particulados. Apesar de
possuir essa capacidade, essa espécie é considerada patogênica e apresenta
multirresistência a antimicrobianos. Esses fatores podem inviabilizar a sua aplicação
em processos de biorremediação e produção de biossurfactantes (CHINELLATO,
2017). Entretanto, de acordo com Eddououda et al. (2012), outras bactérias

38
consideradas patogênicas, como Pseudomonas aeruginosa, por exemplo, têm sido
amplamente descritas como produtoras de biossurfactantes a partir de diferentes
fontes de carbono.
Das cepas originadas de amostras de sedimento três foram identificadas
como pertencentes à espécie Zobellella taiwanensis. O primeiro registro desta
espécie foi feito por Lin e Shieh (2006) ao estudarem duas cepas bacterianas
isoladas de sedimento de manguezal em Taiwan. Segundo os autores, essas se
apresentam em forma de bastonetes móveis Gram negativos, são heterotróficas,
anaeróbicas facultativas, e apresentam teor de CG em seu DNA de 62-64 mol%.
Essa espécie é conhecida por atuar no processo de nitrificação heterotrófica e
desnitrificação aeróbica, removendo altas concentrações de amônio de áreas
contaminadas por amônia (LEI et al., 2016). Porém, existe pouca informação sobre a
produção de biossurfactantes por essa espécie.
Duas cepas isoladas de sedimento foram identificadas como
Oceanimonas baumannii. Esta espécie foi descrita e isolada primeiramente por
Brown, Sutcliffe e Cummings (2001) de estuário do River Wear, Reino Unido. Os
autores verificaram que essa bactéria foi capaz de crescer em meio de cultivo
contendo fenol como substrato a uma concentração de 5% de NaCl. Descrita
originalmente como Oceanomonas baumannii, a nomenclatura foi corrigida
posteriormente para Oceanimonas baumannii.
Segundo Ramezani, Amoozegar e Ventosa (2015), essas bactérias são
halotolerantes e se apresentam como bastonetes Gram negativos. Os autores
isolaram uma bactéria identificada como Oceanimonas sp. GK1 com capacidade de
acumular inclusões de lipídeos intracelulares identificados como poli–β-hidrobutirato
(PHB) em condições limitadas de oxigênio. Bactérias desse gênero habitam áreas
marinhas, locais onde alguns de seus espécimes foram isolados e identificados.
Uma cepa isolada de sedimento marinho foi identificada como
pertencente ao gênero Brachybacterium. As bactérias desse gênero são Gram
positivas com células de formato cocóide e parede celular constituída por ácido
meso-diaminopimélico (tipo D-Glu2) e MK-7 como menaquinona principal. Além disso,
o teor de GC em seu DNA é de 68 a 72%mol (GVOZDYAK; NOGINA; SCHUMANN,
1992; TAKEUCHI; FANG; YOKOT, 1995; HOANG et al., 2014). Segundo Hoang et al.
(2014), bactérias desse gênero têm sido isoladas de superfície de alimentos, órgãos
de animais, plantações de ginseng e sedimento costeiro contaminado por petróleo.
39
Membros desse grupo bacteriano têm sido descrito como degradadoras de
hidrocarbonetos (LILY et al., 2010). Porém, informações acerca do genoma desses
organismos ainda são limitadas (WANG et al., 2014).
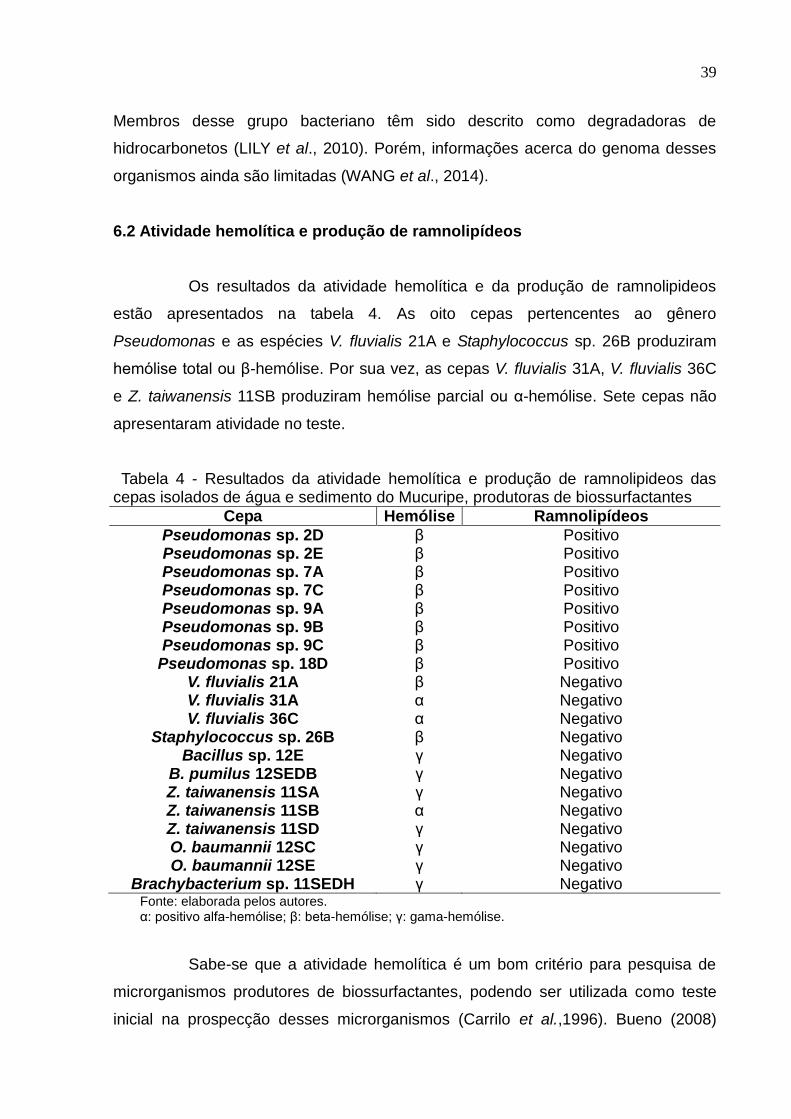
6.2 Atividade hemolítica e produção de ramnolipídeos
Os resultados da atividade hemolítica e da produção de ramnolipideos
estão apresentados na tabela 4. As oito cepas pertencentes ao gênero
Pseudomonas e as espécies V. fluvialis 21A e Staphylococcus sp. 26B produziram
hemólise total ou β-hemólise. Por sua vez, as cepas V. fluvialis 31A, V. fluvialis 36C
e Z. taiwanensis 11SB produziram hemólise parcial ou α-hemólise. Sete cepas não
apresentaram atividade no teste.
Tabela 4 - Resultados da atividade hemolítica e produção de ramnolipideos das cepas isolados de água e sedimento do Mucuripe, produtoras de biossurfactantes
Cepa Hemólise Ramnolipídeos
Pseudomonas sp. 2D β Positivo Pseudomonas sp. 2E β Positivo Pseudomonas sp. 7A β Positivo Pseudomonas sp. 7C β Positivo Pseudomonas sp. 9A β Positivo Pseudomonas sp. 9B β Positivo Pseudomonas sp. 9C β Positivo
Pseudomonas sp. 18D β Positivo V. fluvialis 21A β Negativo V. fluvialis 31A α Negativo V. fluvialis 36C α Negativo
Staphylococcus sp. 26B β Negativo Bacillus sp. 12E γ Negativo
B. pumilus 12SEDB γ Negativo Z. taiwanensis 11SA γ Negativo Z. taiwanensis 11SB α Negativo Z. taiwanensis 11SD γ Negativo O. baumannii 12SC γ Negativo O. baumannii 12SE γ Negativo
Brachybacterium sp. 11SEDH γ Negativo Fonte: elaborada pelos autores. α: positivo alfa-hemólise; β: beta-hemólise; γ: gama-hemólise.
Sabe-se que a atividade hemolítica é um bom critério para pesquisa de
microrganismos produtores de biossurfactantes, podendo ser utilizada como teste
inicial na prospecção desses microrganismos (Carrilo et al.,1996). Bueno (2008)

40
utilizou como critério de seleção prévia de bactérias produtoras de biossurfactantes a
produção de β-hemólise no teste hemolítico.
As cepas isoladas de amostras de água apresentaram melhor
desempenho no teste hemolítico quando comparados às isoladas de sedimento. A
figura 4 mostra os halos de hemólise em ágar sangue produzidos por algumas
bactérias utilizadas na presente pesquisa.
Figura 4 – Halos de hemólise em ágar sangue indicativo da produção de biossurfactantes após 48 horas de incubação
Fonte: Autor.
De acordo com Schultz (2010), o teste de hemólise seleciona
microrganismos capazes de crescer utilizando proteínas do ferro, presentes na
hemoglobina. Além disso, a presença de halos de hemólise no teste pode estar
associada não apenas à produção de biossurfactantes, mas também à presença de
hemolisinas produzidas pelas bactérias. Segundo Thavasi, Sharma e Jayalakshmi
(2011), esse teste seleciona tanto bactérias produtoras como não produtoras de
biossurfactantes.
Sabe-se que algumas espécies dos gêneros Vibrio e Staphylococcus
produzem halos de hemólise no teste devido à produção de hemolisinas
(RAMAMURTHY et al., 2014; DIAS; PINHEIRO; ALVES, 2015). Possivelmente, a
positividade no teste hemolítico das cepas de V. fluvialis e Staplylococcus sp. 26B
pode estar relacionada tanto com a produção de hemolisinas em meio ágar sangue,
quanto à produção de biossurfactantes. Contudo, mesmo a presença de hemolisinas
Bacillus spp.
V. fluvialis
V. fluvialis
Pseudomonas spp.
Pseudomonas spp.
Z. taiwanensis

41
influenciando nos resultados do teste, esta determinação mostra-se como um
indicativo da presença de compostos biossurfactantes produzidos não só por
bactérias, mas também por fungos filamentosos (SPERB et al. 2015).
O teste de hemólise pode ser utilizado para selecionar os organismos
produtores de biossurfactantes. Porém, sempre que possível, deve ser realizado em
conjunto com outros testes para uma melhor análise dos resultados. É difícil realizar
prospecção de microrganismos produtores de biossurfactantes com base em um
único teste específico. O teste de produção de ramnolipídeos torna-se
imprescindível para complementar a identificação de bactérias alvo, pois pode ser
utilizado para detectar os surfactantes aniônicos biológicos e outros glicolipídeos
(SCHULTZ, 2010; SATPUTE et al., 2010; SHOEB et al., 2015).
As oito cepas pertencentes ao gênero Pseudomonas apresentaram
resultado positivo no teste de detecção de ramnolipídeos. Na figura 5 os halos azuis
em volta da colônia confirmam que os biossurfactantes produzidos por essas
bactérias pertencem ao grupo dos glicolídeos. As cepas de Vibrio fluvialis e
Staphylococcus sp. 26B que apresentaram atividade hemolítica não apresentaram
resultados no teste de ramnolipídeos confirmando a especificidade desse teste para
selecionar produtores de glicolipídeos.
Figura 5 - Halos azuis indicativos da produção de ramnolipídeos em meio ágar azul
de metileno
Fonte: Autor.

42
Os biossurfactantes produzidos por bactérias do gênero Pseudomonas
são denominados ramnolipídeos e pertencem à classe dos glicolipídeos. Esses
compostos apresentam uma porção hidrofílica onde predomina uma ramnose e outra
porção hidrofóbica, constituída por ácidos graxos. Esses biossurfactantes podem ser
produzidos a partir de açúcares e outros substratos renováveis (MAGALHÃES, 2012;
SATPUTE et al., 2010).
Varadavenkatesan e Murty (2013) ao realizarem prospecção de bactérias
produtoras de biossurfactantes verificaram que a cepa pertencente ao gênero
Bacillus apresentou melhor desempenho no teste hemolítico. Essa cepa quando
submetida ao teste de detecção de ramnolipideos não apresentou halo no meio de
cultivo, sendo o único teste com resultado negativo desse isolado bacteriano. No
presente estudo, os isolados de Bacillus não apresentaram atividade no teste
hemolítico e nem no de detecção de ramnolipídeos.
6.3 Teste qualitativo do colapso da gota e pH das fermentações
Foram realizadas 39 fermentações, 20 contendo efluente do LCC como
fonte de carbono e 19 contendo óleo vegetal residual. Os resultados do teste
qualitativo do colapso da gota e o pH das fermentações e dos líquidos metabólicos
extraídos estão apresentados nas tabelas 5 e 6.
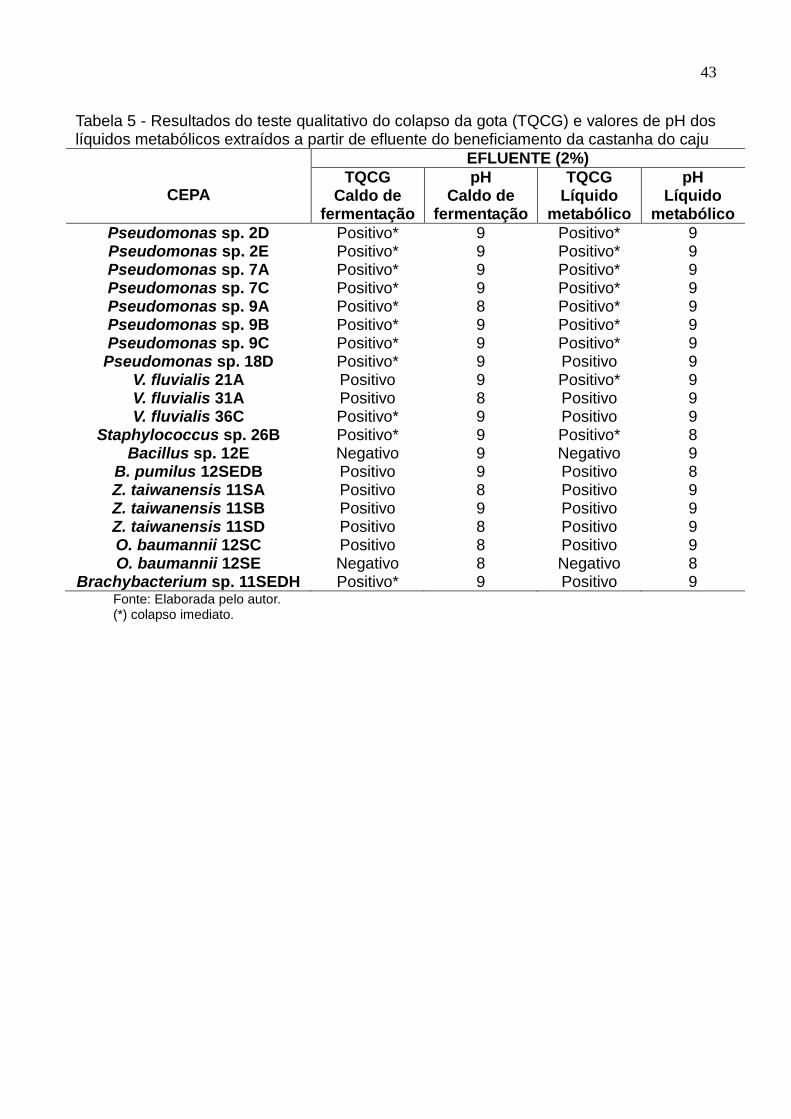
43
Tabela 5 - Resultados do teste qualitativo do colapso da gota (TQCG) e valores de pH dos líquidos metabólicos extraídos a partir de efluente do beneficiamento da castanha do caju
CEPA
EFLUENTE (2%)
TQCG Caldo de
fermentação
pH Caldo de
fermentação
TQCG Líquido
metabólico
pH Líquido
metabólico
Pseudomonas sp. 2D Positivo* 9 Positivo* 9 Pseudomonas sp. 2E Positivo* 9 Positivo* 9 Pseudomonas sp. 7A Positivo* 9 Positivo* 9 Pseudomonas sp. 7C Positivo* 9 Positivo* 9 Pseudomonas sp. 9A Positivo* 8 Positivo* 9 Pseudomonas sp. 9B Positivo* 9 Positivo* 9 Pseudomonas sp. 9C Positivo* 9 Positivo* 9
Pseudomonas sp. 18D Positivo* 9 Positivo 9 V. fluvialis 21A Positivo 9 Positivo* 9 V. fluvialis 31A Positivo 8 Positivo 9 V. fluvialis 36C Positivo* 9 Positivo 9
Staphylococcus sp. 26B Positivo* 9 Positivo* 8 Bacillus sp. 12E Negativo 9 Negativo 9
B. pumilus 12SEDB Positivo 9 Positivo 8 Z. taiwanensis 11SA Positivo 8 Positivo 9 Z. taiwanensis 11SB Positivo 9 Positivo 9 Z. taiwanensis 11SD Positivo 8 Positivo 9 O. baumannii 12SC Positivo 8 Positivo 9 O. baumannii 12SE Negativo 8 Negativo 8
Brachybacterium sp. 11SEDH Positivo* 9 Positivo 9 Fonte: Elaborada pelo autor. (*) colapso imediato.

44
Tabela 6 - Resultados do teste qualitativo do colapso da gota (TQCG) e valores de pH dos líquidos metabólicos extraídos a partir de óleo vegetal residual
CEPA
ÓLEO VEGETAL RESIDUAL (1%)
TQCG Caldo de
fermentação
pH Caldo de
fermentação
TQCG Líquido
metabólico
pH Líquido
metabólico
Pseudomonas sp. 2D Positivo* 7 Positivo* 8 Pseudomonas sp. 2E Positivo* 8 Positivo* 8 Pseudomonas sp. 7A Positivo* 8 Positivo* 8 Pseudomonas sp. 7C Positivo* 8 Positivo* 8 Pseudomonas sp. 9A Positivo* 7 Positivo* 8 Pseudomonas sp. 9B Positivo* 8 Positivo* 8 Pseudomonas sp. 9C Positivo* 7 Positivo* 8
Pseudomonas sp. 18D Positivo* 7 Positivo* 8 V. fluvialis 21A Positivo 7 Negativo 8 V. fluvialis 36C Positivo 6 Positivo 6
Staphylococcus sp.26B Negativo 8 Negativo 9 Bacillus sp. 12E Positivo 8 Positivo 9
B. pumilus 12SEDB Positivo* 8 Positivo* 9 Z. taiwanensis 11SA Positivo 8 Negativo 8 Z. taiwanensis 11SB Negativo 9 Negativo 9 Z. taiwanensis 11SD Negativo 9 Negativo 8 O. baumannii 12SC Positivo 8 Positivo 9 O. baumannii 12SE Negativo 9 Negativo 9
Brachybacterium sp. 11SEDH Negativo 9 Negativo 9 Fonte: Elaborada pelo autor. (*) colapso imediato.
As figuras 6 e 7 mostram os caldos de fermentação após as 72 horas de
produção de biossurfactantes (meios com células bacterianas) e os líquidos
metabólicos extraídos (meio isento de células), respectivamente. Não foi possível
realizar fermentações com a cepa V. fluvialis 31A utilizando o óleo vegetal residual,
pois esta se mostrou inviável para cultivo (sem crescimento em meio de cultura) no
momento da realização do experimento.

45
Figura 6 - Meios de cultivo após as 72 horas de produção de biossurfactantes contendo: (a) efluente do beneficiamento da castanha do caju e (b) óleo vegetal residual como substratos
Fonte: Autor
Figura 7 - Líquidos metabólicos extraídos após a produção de biossurfactantes
Fonte: Autor.
De acordo com Satpute et al. (2010), os biossurfactantes podem
permanecer aderidos às células dos microrganismos produtores ou ser excretados
no meio de cultivo. A realização do teste qualitativo do colapso da gota com o meio
contendo as células bacterianas (meio de cultivo após a fermentação) e com o
líquido metabólico (meio de cultivo isento de células) permite inferir sobre essa
característica.
Dezoito (18) líquidos metabólicos apresentaram resultados positivos no
teste qualitativo do colapso da gota utilizando o efluente como substrato. Das
amostras produzidas a partir da fermentação com efluente, apenas o metabólito da
a b

46
cepa Bacillus sp.12E apresentou resultado negativo em ambos os meios (com
células e isento de células). Durante as fermentações, o pH dos caldos de
fermentação e dos líquidos metabólicos atingiram condições alcalinas (variando de 8
a 9), demonstrando um aumento do pH durante o processo de produção dos
biossurfactantes. Na fermentação com óleo vegetal residual, 14 cepas apresentaram
resultado positivo no teste com as células bacterianas, enquanto apenas 12
apresentaram atividade no meio isento de células. O pH de ambos os meios variou
de condições neutras a alcalinas (variando de 7 a 9).
Em um trabalho anteriormente realizado com as mesmas bactérias, Silva
(2014) verificou esse mesmo comportamento no pH durante fermentações utilizando
querosene como fonte de carbono. Ferreira (2015) verificou que, na produção de
biossurfactantes a partir de hidrocarbonetos de petróleo por bactérias isoladas de
amostras de rizosfera e água de manguezais no Estado do Ceará, o pH das
fermentações variou de condições neutras a alcalinas, semelhantes à presente
pesquisa.
De acordo com a literatura, o teste do colapso da gota tem sido utilizado
na prospecção de bactérias produtoras de biossurfactantes (VIRAMONTES-RAMOS
et al., 2010; DHAIL; JASUJA 2012). No teste, o espalhamento sofrido pela gota ao
entrar em contato com a superfície hidrofóbica está relacionado com a redução da
tensão superficial, característica dos biossurfactantes (SEN et al., 2010). Vale
salientar que grande parte dos metabólitos testados produziram colapso da gota
imediato, sem a necessitar de espera de 1 minuto prescrita pela metodologia,
indicando forte atividade.
O líquido metabólito produzido pela cepa Bacillus sp.12E durante a
fermentação com efluente não apresentou atividade no teste do colapso da gota.
Porém, quando o teste foi realizado com o líquido metabólito, produzido pela
fermentação com óleo, houve atividade. Possivelmente, a diferença nos tipos e nas
concentrações dos substratos utilizados produziram diferentes produtos metabólicos,
visto que o tipo de biossurfactante produzido depende de certas variáveis, dentre
elas, o tipo de substrato utilizado (CAMPOS; STAMFORD; SORUBBO, 2014).
Bactérias do gênero Bacillus têm sido descritas como produtoras de
compostos biossurfactantes a partir de substratos hidrofóbicos (BUENO; SILVA;
GARCIA-CRUZ, 2010). Ao estudar a produção de biossurfactante por uma cepa do
gênero Bacillus utilizando diferentes fontes de carbono, dentre elas óleo residual de

47
fritura, Varadavenkatesan e Murty (2013) utilizaram o teste do colapso da gota como
indicativo da produção de biossurfactante e verificaram que a cepa RT10 que
apresentou melhor desempenho nos testes realizados, apresentou atividade nesse
teste.
As bactérias identificadas como Pseudomonas sp., V. fluvialis 36C, B.
pumillus 12SEDB e O. baumannii 12SC produziram os melhores resultados no teste
com ambos os substratos. Dentre esses grupos bacterianos, os gêneros
Pseudomonas e Bacillus têm sido amplamente descritos como produtores de
biossurfactantes a partir de diferentes substratos (BUENO; SILVA; GARCIA-CRUZ,
2010; BEZERRA, 2012; DECESARO et al., 2013). Entretanto, informações a
respeito da produção de tensoativos microbianos por espécies de V. fluvialis e O.
baumannii ainda são escassas.
De acordo com Burch et al. (2011) a técnica do colapso da gota utilizando
caldo de fermentação (técnica convencional) não é capaz de detectar baixas
concentrações de biossurfactantes. Em estudo anterior, os mesmos autores
compararam o teste do colapso da gota convencional com outro modificado
atomicamente e executado em meio sólido. Como resultados, eles verificaram que o
teste modificado foi mais sensível ao detectar um maior número de bactérias
produtoras de biossurfactantes do que o teste convencional. Segundo eles, o teste
modificado permite detectar baixas quantidades de biossurfactantes e até mesmo
biossurfactantes hidrofóbicos, diferente do teste convencional.
6.4 Volume de emulsificação (VE), estabilidade da emulsão (ES%) e atividade
de emulsificação
Os resultados dos testes de volume e atividade de emulsificação estão
expressos na tabela 7. O volume de emulsão assim como a sua estabilidade foram
determinados para todos os líquidos metabólicos extraídos. Na determinação do
volume de emulsificação com os metabólitos produzidos a partir das fermentações
com o efluente nota-se que as cepas de Pseudomonas sp., V. fluvialis 21A, Z.
taiwanensis 11SA, Brachybacterium sp. 11SEDH e Staphylococcus sp. 26B
produziram subprodutos com propriedades emulsificantes ou seja, formaram
emulsões acima de 50%, com máximos valores de 79% para as cepas
Pseudomonas sp. 2D, Pseudomonas sp. 2E e Pseudomonas sp. 9B. Os

48
subprodutos dessas bactérias apresentaram desempenho semelhante ao produzido
pelo surfactante SDS na concentração de 1mg/mL (78%).
Tabela 7- Resultados dos testes de volume de emulsificação (VE) e atividade de emulsificação realizados utilizando os líquidos metabólicos extraídos
Cepa L.M. Efluente LCC L.M. Óleo vegetal
VE (%) U.A.E. VE (%) U.A.E.
Pseudomonas sp. 2D 79 ± 1,73 5,39 ± 0,47 0 3,86 ± 1,04 Pseudomonas sp. 2E 73 ± 4,58 2,71 ± 0,84 3 ± 3,00 3,65 ± 0,49 Pseudomonas sp. 7A 78 ± 3,00 4,01 ± 0,75 76 ± 2,12 4,51 ± 0,31 Pseudomonas sp. 7C 79 ± 3,46 4,53 ± 0,42 76 ± 1,73 4,64 ± 0,58 Pseudomonas sp. 9A 71 ± 1,73 4,29 ± 0,16 6 ± 5,20 5,10 ± 0,09 Pseudomonas sp. 9B 79 ± 4,58 4,92 ± 0,08 17 ± 13,53 4,03 ± 0,70 Pseudomonas sp. 9C 75 ± 3,00 4,53 ± 0,36 87 ± 3,0 4,51 ± 0,23
Pseudomonas sp. 18D 78 ± 3,00 4,81 ± 0,83 81 ± 0 4,10 ± 0,18 V. fluvialis 21A 54 ± 5,20 4,20 ± 2,44 8 ± 9,17 4,46 ± 0,06 V. fluvialis 31A 29 ± 1,73 4,78 ± 0,49 N.D N.D V. fluvialis 36C 44 ± 22,11 4,48 ± 0,34 45 ± 29,55 2,15 ± 1,44
Staphylococcus sp. 26B 67 ± 1,73 4,90 ± 0,33 5 ± 1,73 3,42 ± 0.20 Bacillus sp. 12E 20 ± 32,08 1,60 ± 1,43 0 2,49 ± 0,70
B. pumilus 12SEDB 47 ± 9,17 3,74 ± 0,19 0 3,64 ± 0,33 Z. taiwanensis 11SA 62 ± 12,12 4,62 ± 0,50 0 1,22 ± 0,11 Z. taiwanensis 11SB 25 ± 24,06 4,23 ± 1,56 0 4,02 ± 0,16 Z. taiwanensis 11SD 33 ± 25,63 4,59 ± 0,76 0 2,28 ± 1,23 O. baumannii 12SC 44 ± 24,25 4,84 ± 0,16 51 ± 39,00 4,08 ± 0,13 O. baumannii12SE 3 ± 5,20 4,8 2± 0,08 53 ± 19,29 0
Brachybacterium sp. 11SEDH 74 ± 7,55 5,22 ± 0,05 0 1,38 ± 0,21 SDS (1mg/ml) 78 ± 3,00 5,17 ± 0,13 78 ± 3,00 5,17 ± 0,13
Fonte: Elaborada pelo autor. ND: Não determinado.
Nos líquidos metabólicos produzidos com resíduo de óleo vegetal, as
cepas Pseudomonas sp. 7A, Pseudomonas sp. 7C, Pseudomonas sp. 9C e
Pseudomonas sp. 18D apresentaram os melhores resultados juntamente com as
cepas O. baumannii 12SC e O. baumannii 12SE. O metabólito da cepa
Pseudomonas sp. 9C apresentou volume de emulsificação de 87%, valor superior ao
determinado para o SDS. A bactéria O. baumannii 12SE produziu metabólito com
bom desempenho no teste do volume de emulsificação, porém não produziu
atividade no teste do colapso da gota. Possivelmente, os biossurfactantes
produzidos por essa cepa possuem alta massa molecular, típico dos
bioemulsificantes (PACWA-PLOCINICZAK et al., 2011).
Segundo Sanches (2016), diferentes variáveis podem alterar o tipo e a
qualidade dos biossurfactantes durante seu processo de produção, dentre elas a

49
fonte de carbono, pH, temperatura, salinidade e nutrientes. Os metabólitos
produzidos a partir das fermentações realizadas com efluente produziram melhores
emulsões do que aqueles produzidos a partir das fermentações com óleo residual.
Possivelmente, essa diferença pode estar associada às diferentes fontes de carbono
utilizadas no processo de produção dos biossurfactantes.
De acordo com a literatura, os biossurfactantes podem ser produzidos a
partir de diferentes matérias primas, sejam elas solúveis ou insolúveis em água
(BENTO; CAMARGO; GAYLARD, 2008). Jain et al. (2013), testaram diferentes
fontes de carbono na produção de biossurfactantes por Klebsiella sp. e verificaram
que substratos solúveis em água foram mais eficientes do que os insolúveis.
Entretanto, substratos hidrofóbicos têm maior efetividade na produção de
ramnolipídeos (SALAZAR-BRYAM, 2016).
Na presente pesquisa, quando comparados os grupos bacterianos
identificados em relação ao volume de emulsificação, as cepas de Pseudomonas sp.
produziram os melhores resultados com ambos os substratos testados. Entretanto,
nas determinações com o óleo residual um número menor de indivíduos desse
gênero produziu bons volumes, ocorrendo variações na produção dos
biossurfactantes entre as bactérias desse grupo.
Os testes que utilizam os líquidos metabólicos na avaliação da produção
de biossurfactantes fornecem informações indiretas a respeito da produção de tais
compostos, pois se assume que os biotensoativos (quando produzidos) são
excretados como metabólitos secundários pelos micro-organismos produtores,
permanecendo livres no líquido metabólico (SHOEB et al., 2015).
Dhail e Jasuja (2012) verificaram que a cepa MW2 identificada como
Pseudomonas sp. apresentou volume de emulsificação de 70% no estudo da
produção de biossurfactantes por bactérias marinhas a partir de hidrocarbonetos.
Essa cepa além de apresentar bons valores de emulsão obteve resultado positivo no
teste do colapso da gota. Em seu estudo na produção de biossurfactante por P.
aeruginosa utilizando manipueira como fonte de carbono, Bezerra (2012) alcançou
índices de emulsificação acima de 65%. Na presente pesquisa, as cepas
pertencentes ao gênero Pseudomonas apresentaram volumes de emulsão
semelhantes aos encontrados pelos autores citados, confirmando o potencial desse
grupo bacteriano em produzir compostos biossurfactantes a partir de diferentes
fontes de carbono.

50
Todos os líquidos metabólicos produzidos a partir do efluente
apresentaram boas U.A.E. na determinação da atividade emulsificante, com mínima
de 1,20 para Bacillus sp.12E e máximas de 5,39 e 5,22 para Pseudomonas sp. 2D e
Brachybacterium sp. 11SEDH, respectivamente. Os metabólitos dessas duas cepas
produziram U.A.E. superiores ao SDS (5,17). Nas determinações com os líquidos
metabólicos produzidos a partir de óleo residual a menor U.A.E. encontrada foi de
1,22 para Z. taiwanensis 11SA. O maior valor encontrado foi de 5,10 para a cepa
Pseudomonas sp. 9A. Entretanto, apesar do subproduto dessa cepa apresentar
unidades maiores, produziu baixo volume de emulsificação (6%). O líquido
metabólico da cepa Pseudomonas sp. 7C produziu o segundo maior valor de U.A.E.,
de 4,64 e produziu alto volume de emulsificação, sendo considerada mais eficiente
do que a cepa Pseudomonas sp. 9A por apresentar melhor emulsão e em maior
quantidade.
Altos valores de U.A.E. estão associados a uma alta dispersão do
componente hidrofóbico no tampão, indicando a presença de biossurfactantes,
enquanto baixos valores estão associados a baixos níveis desses compostos
(BANAT et al. ,1991). Thavasi, Sharma e Jayalakshmi (2011) verificaram que o
biossurfactante produzido por espécies de Pseudomonas sp.e Bacillus sp. a partir de
óleo cru obtiveram altos valores de U.A.E. Os autores encontraram valores de 1,95
U.A.E. para cepa de Pseudomonas sp. enquanto as cepas de Bacillus sp. obtiveram
valores entre 0,5 a 3 unidades. Mulani et al. (2017) obtiveram valores de U.A.E.
entre 1 e 3 para espécies de Bacillus sp. isolados de sedimento marinho. No
presente estudo, as cepas de Pseudomonas sp. e Bacillus sp. produziram
metabólitos, a partir de ambos os substratos, com desempenho similar a esses
autores na determinação da atividade de emulsificação.
De acordo com Willumsen e Karlson (1997), emulsões são consideradas
estáveis quando conseguem manter mais de 50% da emulsão original após 24 horas
de formadas. Na presente pesquisa, todas as cepas do gênero Pseudomonas
produziram emulsões com estabilidade acima de 70% até 168 horas de observação
com os líquidos metabólicos extraídos a partir do efluente como apresentado no
gráfico 1. Entretanto, dos líquidos metabólicos extraídos a partir do óleo vegetal,
apenas as cepas Pseudomonas sp. 7C, Pseudomonas sp. 9C e Pseudomonas sp.
18D obtiveram estabilidades acima do valor de referência (50%). Os metabólitos
dessas três cepas produziram emulsões estáveis utilizando os dois substratos

51
testados.
Gráfico 1 - Estabilidades das emulsões formadas utilizando líquido metabólico extraído das fermentações com bactérias do gênero Pseudomonas a partir de efluente do beneficiamento da castanha de caju (EBCC) e óleo vegetal residual (OVR)
Fonte: Elaborado pelo autor.
As cepas de Pseudomonas identificadas no presente trabalho
apresentaram resultados distintos nos testes realizados quando comparadas as
fontes de carbono utilizadas. Dentre as oito cepas desse grupo, as estirpes
Pseudomonas sp. 7C, Pseudomonas sp. 9C e Pseudomonas sp. 18D produziram
os metabólitos com melhores resultados nos testes indicativos da produção de
biossurfactantes utilizando as fontes de carbono testadas. Uma possível explicação
para esse fato é a diferença de fontes de carbono utilizadas nos processos de
produção dos biotensoativos, pois a utilização de diferentes substratos na produção
desses compostos pode mudar a sua estrutura molecular e suas propriedades
(CAMPOS; STAMFORD; SORUBBO, 2014). Além disso, vale salientar a possível
variedade entre essas cepas, visto que sua identificação não permitiu classifica-las
até espécie. Segundo Bueno, Camargo e Gaylarde (2008), mesmo a produção de
biossurfactantes acontecendo entre os diversos grupos microbianos, em alguns
casos um tipo de biossurfactante é limitado a um gênero e às vezes a uma espécie.
No gráfico 2 estão expressas as estabilidades das emulsões formadas
com os líquidos metabólicos das cepas de Víbrio fluvialis, Bacillus sp. e Zobellella

52
taiwanensis. Dentre as três cepas de V. fluvialis identificadas, apenas a cepa V.
fluvialis 36C produziu emulsão estável ao final das 168 horas de observação
utilizando o líquido metabólico produzido a partir do efluente. Não houve produção
de emulsões estáveis por qualquer cepa dessa espécie usando resíduo oleoso como
fonte de carbono. A cepa V. fluvialis 21A apesar de produzir um volume de
emulsificação de 54% não conseguiu manter-se estável ao final do período de
observação.
Das duas cepas identificadas como pertencentes ao gênero Bacillus,
apenas o metabólito produzido pela cepa de Bacillus sp.12E, a partir do efluente,
produziu boa emulsão. A cepa B. pumillus 11SEDH apresentou estabilidade de 49%,
sendo considerada baixa. Nenhuma das cepas desse gênero apresentou emulsão
estável no teste com os metabólitos obtidos a partir do óleo residual.
A cepa Z. taiwanensis 11SA foi a única, dentre as cepas pertencentes a
essa espécie, capaz de manter boa estabilidade (81%) no final das 168 horas de
observação na determinação com o líquido metabólico produzido a partir do efluente.
As cepas dessa espécie não apresentaram metabólitos com boa estabilidade a partir
das fermentações com óleo vegetal residual.
Gráfico 2 - Estabilidades das emulsões formadas utilizando líquido metabólico extraído das fermentações com as bactérias das espécies Vibrio fluvialis, Z. taiwanensis e do gênero Bacillus a partir de efluente do beneficiamento da castanha de caju (EBCC) e óleo vegetal residual (OVR)
Fonte: Elaborado pelo autor.
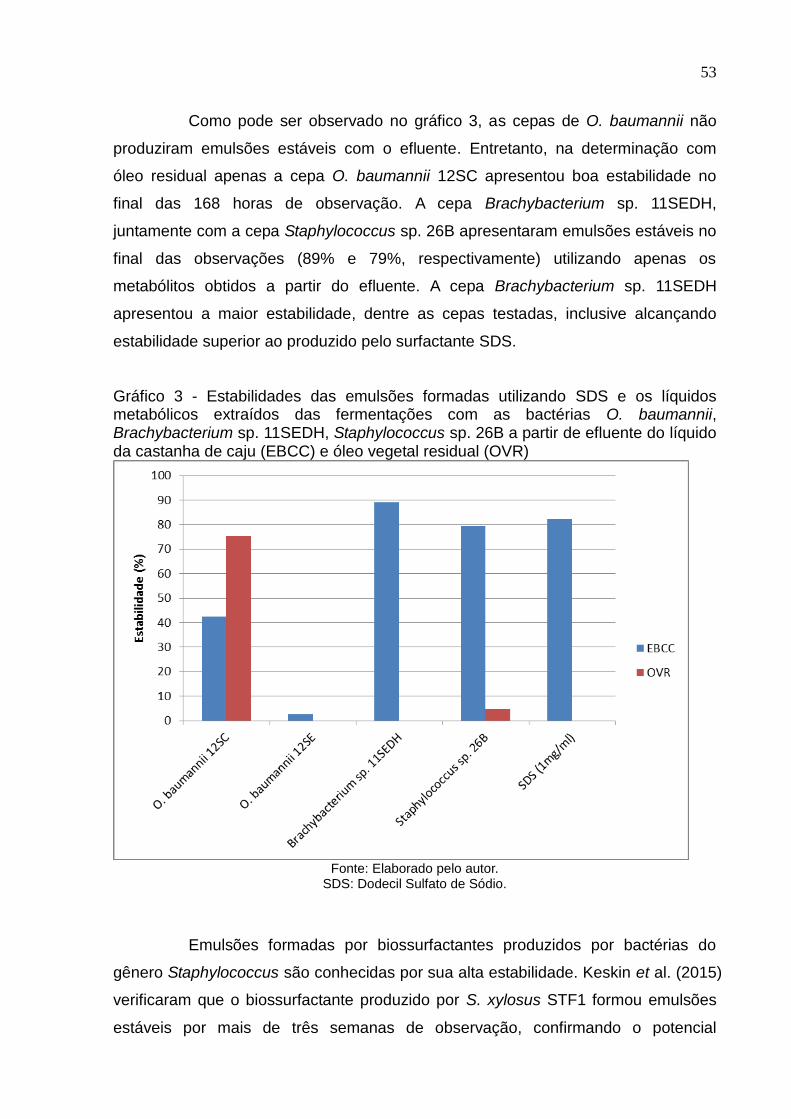
53
Como pode ser observado no gráfico 3, as cepas de O. baumannii não
produziram emulsões estáveis com o efluente. Entretanto, na determinação com
óleo residual apenas a cepa O. baumannii 12SC apresentou boa estabilidade no
final das 168 horas de observação. A cepa Brachybacterium sp. 11SEDH,
juntamente com a cepa Staphylococcus sp. 26B apresentaram emulsões estáveis no
final das observações (89% e 79%, respectivamente) utilizando apenas os
metabólitos obtidos a partir do efluente. A cepa Brachybacterium sp. 11SEDH
apresentou a maior estabilidade, dentre as cepas testadas, inclusive alcançando
estabilidade superior ao produzido pelo surfactante SDS.
Gráfico 3 - Estabilidades das emulsões formadas utilizando SDS e os líquidos metabólicos extraídos das fermentações com as bactérias O. baumannii, Brachybacterium sp. 11SEDH, Staphylococcus sp. 26B a partir de efluente do líquido da castanha de caju (EBCC) e óleo vegetal residual (OVR)
Fonte: Elaborado pelo autor.
SDS: Dodecil Sulfato de Sódio.
Emulsões formadas por biossurfactantes produzidos por bactérias do
gênero Staphylococcus são conhecidas por sua alta estabilidade. Keskin et al. (2015)
verificaram que o biossurfactante produzido por S. xylosus STF1 formou emulsões
estáveis por mais de três semanas de observação, confirmando o potencial

54
emulsificante dos metabólitos de bactérias desse gênero.
Diante dos resultados das estabilidades, nota-se que os líquidos
metabólicos produzidos a partir do efluente produziram emulsões mais estáveis ao
longo do tempo. De acordo com Franzol e Rezende (2015), emulsões que
apresentam elevada estabilidade demoram longos tempos para que ocorra a
separação das fases. As cepas que apresentaram boas emulsões e conseguiram
mantê-las estáveis ao final das 168 horas de estudo apresentam um elevado
potencial biotecnológico, visto que a efetividade de um agente emulsificante está
intimamente relacionada com a sua alta estabilidade. Vale salientar que essas
amostras com elevada estabilidade apresentaram desempenho similar a um
surfactante sintético amplamente utilizado nos diversos setores industriais,
mostrando o potencial de uso desses biossurfactantes na solubilização de
substâncias hidrofóbicas em diferentes áreas da indústria.
6.5 Atividade antimicrobiana
Os resultados do teste de atividade antimicrobiana dos líquidos
metabólicos produzidos a partir das fermentações estão apresentados na tabela 8.
Nenhum dos metabólicos produzidos com ambos os substratos apresentou atividade
frente à cepa de E. coli ATCC 25922 como pode ser visualizado nas figuras 8 e 9.
Entretanto, dos 20 líquidos metabólicos extraídos a partir da fermentação com
efluente, 10 apresentaram ação contra a cepa de S. aureus ATCC 25923 como
mostra a figura 10.

55
Tabela 8 - Resultados da determinação da atividade antimicrobiana dos líquidos metabólicos extraídos utilizando resíduos orgânicos como fonte de carbono
Cepa EFLUENTE DO LCC (2%) ÓLEO VEGETAL (1%)
E. coli
ATCC 25922
S. aureus
ATCC 25923
E. coli
ATCC 25922
S. aureus
ATCC 25923 Pseudomonas sp. 2D Negativo Positivo Negativo Negativo Pseudomonas sp. 2E Negativo Positivo Negativo Negativo Pseudomonas sp. 7A Negativo Positivo Negativo Negativo Pseudomonas sp. 7C Negativo Positivo Negativo Negativo Pseudomonas pp. 9A Negativo Positivo Negativo Negativo Pseudomonas sp. 9B Negativo Positivo Negativo Positivo Pseudomonas sp. 9C Negativo Positivo Negativo Negativo
Pseudomonas sp. 18D Negativo Positivo Negativo Negativo V. fluvialis 21A Negativo Positivo Negativo Negativo V. fluvialis 31A Negativo Negativo - - V. fluvialis 36C Negativo Negativo Negativo Negativo
Staphylococcus sp. 26B Negativo Negativo Negativo Negativo Bacillus sp. 12E Negativo Negativo Negativo Negativo
B. pumilus 12SEDB Negativo Negativo Negativo Negativo Z. taiwanensis 11SA Negativo Negativo Negativo Negativo Z. taiwanensis 11SB Negativo Positivo Negativo Negativo Z. taiwanensis 11SD Negativo Negativo Negativo Negativo O. baumannii 12SC Negativo Negativo Negativo Negativo O. baumannii 12SE Negativo Negativo Negativo Negativo
Brachybacterium sp. 11SEDH
sppspp. 11SEDH
Negativo Negativo Negativo Negativo Fonte: Elaborada pelo autor.
Figura 8 - Atividade antimicrobiana dos líquidos metabólicos (resultado negativo) extraídos a partir das fermentações com efluente do beneficiamento da castanha do caju frente à bactéria Escherichia coli ATCC 25922
Fonte: Autor.
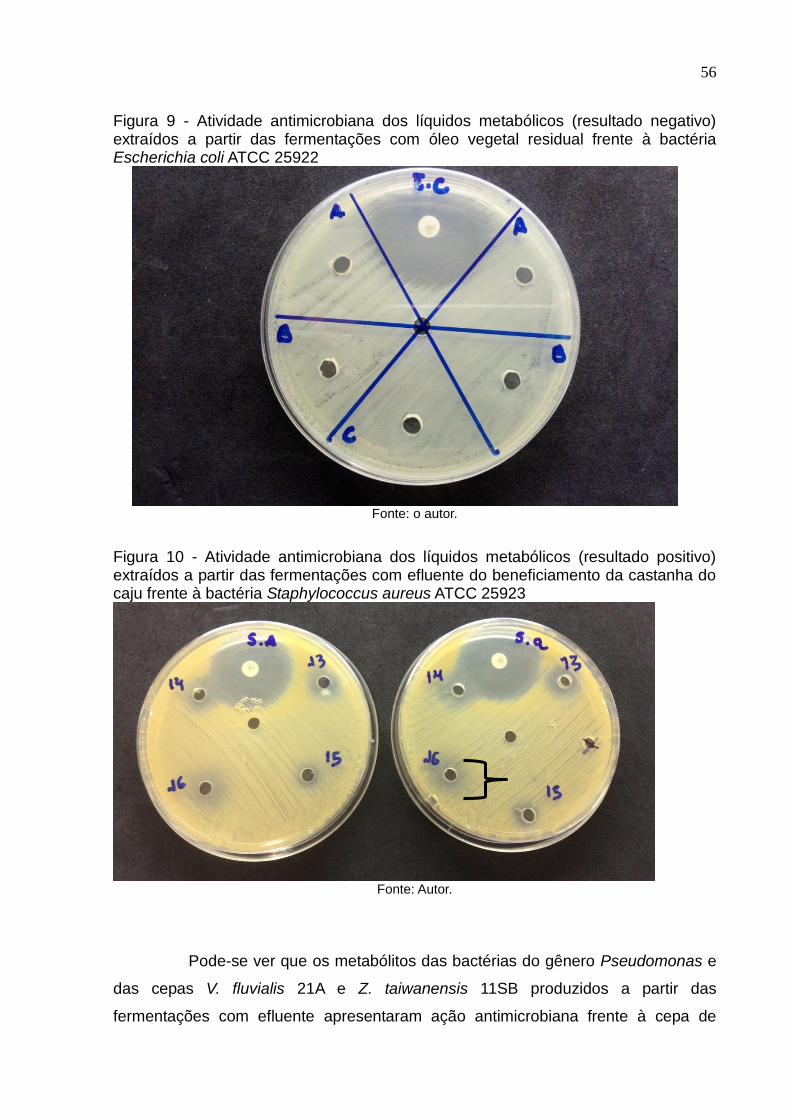
56
Figura 9 - Atividade antimicrobiana dos líquidos metabólicos (resultado negativo) extraídos a partir das fermentações com óleo vegetal residual frente à bactéria Escherichia coli ATCC 25922
Fonte: o autor.
Figura 10 - Atividade antimicrobiana dos líquidos metabólicos (resultado positivo) extraídos a partir das fermentações com efluente do beneficiamento da castanha do caju frente à bactéria Staphylococcus aureus ATCC 25923
Fonte: Autor.
Pode-se ver que os metabólitos das bactérias do gênero Pseudomonas e
das cepas V. fluvialis 21A e Z. taiwanensis 11SB produzidos a partir das
fermentações com efluente apresentaram ação antimicrobiana frente à cepa de

57
Staphylococcus aureus testada. A cepa Pseudomonas sp. 9B foi a única bactéria que
conseguiu produzir metabólitos com propriedades antimicrobianas frente à cepa
Gram positiva testada utilizando os dois substratos testados como pode-se observar
na figura 11. As demais cepas produziram compostos com ação antimicrobiana
apenas quando fermentadas em meio contendo efluente da castanha do caju.
Figura 11 - Atividade antimicrobiana dos líquidos metabólicos (resultado positivo) extraídos a partir das fermentações com óleo vegetal residual frente à bactéria Staphylococcus aureus ATCC 25923
Fonte: Autor.
Sabe-se que os ramnolipídeos produzidos por bactérias do gênero
Pseudomonas possuem ação antimicrobiana e antifúngica já conhecidas
(MAGALHÃES, 2012; EL-SHESHTAWY; DOHEIM, 2014). Esses dados confirmam o
potencial desse grupo bacteriano em produzir metabólicos com ação bioativa frente
às bactérias patogênicas. Alguns lipopeptídeos produzidos por bactérias do gênero
Bacillus também possuem ação antimicrobiana, de acordo com Satpute et al. (2010).
Contudo, na presente pesquisa, não foi detectado nenhum metabólito com atividade
antimicrobiana produzido por bactérias desse gênero.
Estudos que avaliam a produção de biossurfactantes e metabólitos com
ação antimicrobiana produzidos por bactérias dos gêneros Vibrio e Zobelella ainda
são escassos. Contudo, bactérias do gênero Víbrio, assim como o gênero Bacillus

58
são conhecidas por produzirem polímeros de alto peso molecular denominados
exopolissacarídeos (EPS) que envolvem a célula bacteriana e as auxilia a suportar
condições ambientais extremas. Tais bioprodutos têm despertado interesse na
indústria farmacêutica e na recuperação de poluentes no ambiente (SATPUTE et al.,
2010).
De acordo com a literatura, alguns biossurfactantes apresentam ação
contra bactérias, fungos e vírus (BENTO; CAMARGO; GAYLARDE, 2008; EL-
SHESHTAWY; DOHEIM, 2014). Inicialmente, muitos biossurfactantes foram
utilizados como antibióticos devido a sua propriedade antimicrobiana (SATPUTE et
al., 2010). Segundo Bharali e Komwar (2011), bactérias Gram negativas possuem
maior resistência à ação dos surfactantes do que bactérias Gram positivas, pois
além de possuírem lipopolissacarídeos como constituintes da parede celular,
apresentam proteínas de superfície que garantem maior proteção à célula bacteriana
contra ação dos tensoativos. Por sua vez, a susceptibilidade de bactérias Gram
positivas à ação dos surfactantes se dá devido à existência de uma única camada de
lipopolissacarídeos (EL-SHESHTAWY; DOHEIM, 2014).
De acordo com Osmari (2013) o líquido da castanha de caju possui
atividade antimicrobiana conhecida devido à presença de terpenóides e compostos
fenólicos. Dentre os compostos presentes com ação antimicrobiana pode-se citar o
ácido anacárdico (GONÇALVES, 2007). Parasa et al. (2011) verificaram que extratos
acetônicos e metanólicos de LCC apresentaram ação antimicrobiana contra cepas
de Staphylococus aureus resistentes a meticilina (SARM). Os autores explicaram tal
efeito devido à presença de ácido anacárdico nos extratos.
6.6 Teste de toxicidade com sementes de pepino
Os líquidos metabólicos que apresentaram os melhores resultados nos
testes indicativos da produção de biossurfactantes com ambos os substratos foram
selecionados para o teste ecotoxicológico utilizando sementes de pepino. Na tabela
9 estão expressos os resultados do teste tendo como critério de classificação da
toxicidade o percentual de germinação. Como se pode observar na tabela, todos os
líquidos metabólicos produzidos a partir do efluente foram classificados como “muito
tóxicos” em todas as concentrações testadas, com exceção do metabólito extraído
da cepa Pseudomonas sp. 9B na concentração de 5% que foi classificado como
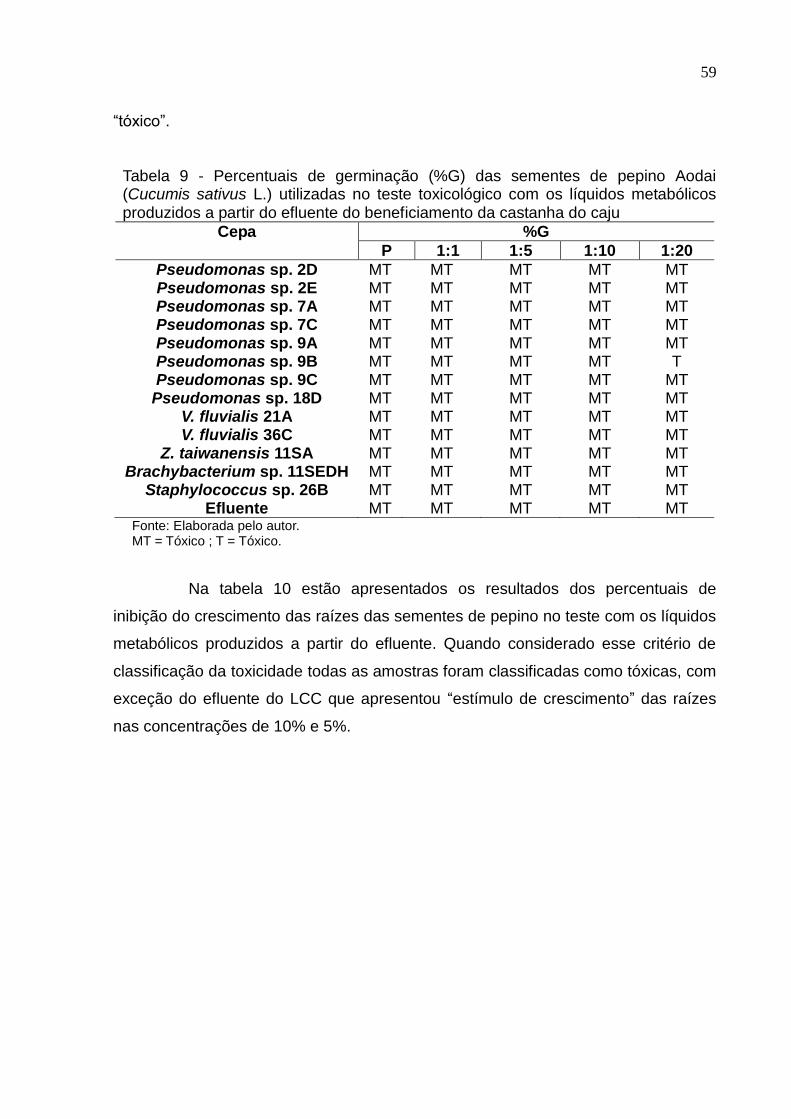
59
“tóxico”.
Tabela 9 - Percentuais de germinação (%G) das sementes de pepino Aodai (Cucumis sativus L.) utilizadas no teste toxicológico com os líquidos metabólicos produzidos a partir do efluente do beneficiamento da castanha do caju
Cepa %G
P 1:1 1:5 1:10 1:20
Pseudomonas sp. 2D MT MT MT MT MT Pseudomonas sp. 2E MT MT MT MT MT Pseudomonas sp. 7A MT MT MT MT MT Pseudomonas sp. 7C MT MT MT MT MT Pseudomonas sp. 9A MT MT MT MT MT Pseudomonas sp. 9B MT MT MT MT T Pseudomonas sp. 9C MT MT MT MT MT
Pseudomonas sp. 18D MT MT MT MT MT V. fluvialis 21A MT MT MT MT MT V. fluvialis 36C MT MT MT MT MT
Z. taiwanensis 11SA MT MT MT MT MT Brachybacterium sp. 11SEDH MT MT MT MT MT
Staphylococcus sp. 26B MT MT MT MT MT Efluente MT MT MT MT MT
Fonte: Elaborada pelo autor. MT = Tóxico ; T = Tóxico.
Na tabela 10 estão apresentados os resultados dos percentuais de
inibição do crescimento das raízes das sementes de pepino no teste com os líquidos
metabólicos produzidos a partir do efluente. Quando considerado esse critério de
classificação da toxicidade todas as amostras foram classificadas como tóxicas, com
exceção do efluente do LCC que apresentou “estímulo de crescimento” das raízes
nas concentrações de 10% e 5%.

60
Tabela 10 - Percentuais de inibição (%IP) do crescimento das sementes de pepino Aodai (Cucumis sativus L.) utilizadas no teste toxicológico com os líquidos metabólicos produzidos extraídos a partir do efluente do beneficiamento da castanha de caju
Cepa %IP
P 1:1 1:5 1:10 1:20
Pseudomonas sp. 2D T T T T T Pseudomonas sp. 2E T T T T T Pseudomonas sp. 7A T T T T T Pseudomonas sp. 7C T T T T T Pseudomonas sp. 9A T T T T T Pseudomonas sp. 9B T T T T T Pseudomonas sp. 9C T T T T T
Pseudomonas sp. 18D T T T T T V. fluvialis 36C T T T T T
Z. taiwanensis 11SA T T T T T Brachybacterium sp. 11SEDH T T T T T
Staphylococcus sp. 26B T T T T T Efluente T T T EC EC
Fonte: Elaborada pelo autor. T = Tóxico; EC: estímulo de crescimento.
Os gráficos 4, 5 e 6 mostram as médias das medidas do elongamento das
raízes após as 120 horas de incubação. Nota-se que os líquidos metabólicos
produzidos pelas cepas Pseudomonas sp. 7C, Pseudomonas sp. 9A, Pseudomonas
sp. 9C,, V. fluvialis 21A, V. fluvialis 36C e Brachybacterium sp. 11SEDH só
favoreceram o crescimento das raízes a partir da concentração de 50%. Os demais
metabólitos só permitiram o desenvolvimento radicular das sementes a partir da
concentração de 25%. Os componentes presentes no efluente favoreceram o
crescimento das raízes em todos os tratamentos.

61
Gráfico 4 – Comprimento das raízes das sementes de pepino Aodai (Cucumis sativus L.) no teste com os líquidos metabólicos extraídos a partir das fermentações com efluente do beneficiamento da castanha do caju com as cepas Pseudomonas sp. 2D, Pseudomonas sp. 2E, Pseudomonas sp. 7A e Pseudomonas sp.7C
Fonte: Elaborado pelo autor. LM: Líquido metabólico.
Gráfico 5 - Comprimento das raízes das sementes de pepino Aodai (Cucumis sativus L) no teste com os líquidos metabólicos produzidos a partir das fermentações com efluente do beneficiamento da castanha do caju com as cepas Pseudomonas sp. 9A, Pseudomonas sp. 9B, Pseudomonas sp. 9C e Pseudomonas sp.18D
Fonte: Elaborado pelo autor. LM: Líquido metabólico.

62
Gráfico 6 - Comprimento das raízes das sementes de pepino Aodai (Cucumis sativus L.) no teste com os líquidos metabólicos extraídos a partir das fermentações com efluente do beneficiamento da castanha do caju com as cepas V. fluvialis 21A, V. fluvialis 36C, Z. taiwanensis 11SA, Brachybacterium sp. 11SEDH e Staphylococcus sp. 26B e do efluente
Fonte: Elaborado pelo autor. LM: Líquido metabólico.
Semelhante aos líquidos metabólicos produzidos a partir do efluente,
aqueles produzidos a partir de óleo vegetal residual também apresentaram efeitos
“muito tóxicos” às sementes, de acordo com a classificação baseada na
porcentagem de germinação como pode ser observado na tabela 11.
Tabela 11– Percentuais de germinação (%G) das sementes de pepino Aodai (Cucumis sativus L.) utilizadas no teste toxicológico com os líquidos metabólicos produzidos extraídos a partir das fermentações com óleo vegetal residual
Cepa %G
P 1:1 1:5 1:10 1:20
Pseudomonas sp. 7C MT MT MT MT MT Pseudomonas sp. 9B MT MT MT MT MT Pseudomonas sp. 9C MT MT MT MT MT
Pseudomonas sp. 18D MT MT MT MT MT O.baumannii12SC MT MT MT MT MT
SDS 1mg/mL MT MT MT MT MT Fonte: Elaborada pelo autor. MT = Muito tóxico.

63
Na tabela 12 estão apresentados os percentuais de inibição das raízes
das sementes de pepino no teste com os metabólitos produzidos a partir do óleo
residual. Nota-se que todas as amostras foram consideradas tóxicas em todos os
tratamentos, com exceção do líquido metabólico produzido pela cepa Pseudomonas
sp. 9C na diluição de 5% e do surfactante SDS que proporcionaram um estímulo de
crescimento das raízes.
Tabela 12 - Percentuais de inibição (%IP) do crescimento das sementes de pepino Aodai (Cucumis sativus L.) utilizadas no teste toxicológico com os líquidos metabólicos produzidos extraídos a partir das fermentações com óleo vegetal residual
Cepa %IP
P 1:1 1:5 1:10 1:20
Pseudomonas sp. 7C T T T T T Pseudomonas sp. 9B T T T T T Pseudomonas sp. 9C T T T T EC
Pseudomonas sp. 18D T T T T T O. baumannii 12SC T T T T T
SDS 1mg/mL T T T EC EC Fonte: Elaborada pelo autor. T= Tóxico; EC = Estimulação de crescimento.
No gráfico 7 estão apresentadas as médias das medições da raízes das
sementes de pepino crescidas até 120 horas de incubação no teste com os
metabólitos produzidos a partir do óleo residual. Os líquidos metabólicos produzidos
pelas cepas Pseudomonas sp. 9B, Pseudomonas sp. 9C e Pseudomonas sp.18D só
permitiram o desenvolvimento radicular das sementes a partir da diluição de 50%,
enquanto que apenas o subproduto da cepa O. baumannii 12SC permitiu o
desenvolvimento das raízes a partir da concentração de 25% do líquido metabólico
testado. O líquido metabólico produzido pela cepa Pseudomonas sp.7C não permitiu
germinação nem crescimento de raízes em nenhum dos tratamentos realizados,
mostrando um efeito altamente tóxico desse subproduto para as sementes vegetais.
No teste com o SDS as sementes só desenvolveram seu sistema radicular a partir
da diluição da amostra pura (concentração de 100%).

64
Gráfico 7 - Comprimento das raízes das sementes de pepino Aodai (Cucumis sativus L.) no teste com os líquidos metabólicos extraídos a partir das fermentações com óleo vegetal residual com as cepas Pseudomonas sp. 7C, Pseudomonas sp. 9B, Pseudomonas sp. 9C, Pseudomonas sp. 18D, O. baumannii 12SC e com o surfactante sintético SDS
Fonte: Elaborado pelo autor. LM: Líquido metabólico.
A avaliação da qualidade ambiental baseada apenas em parâmetros
físico-químicos não oferece informações reais sobre os efeitos de poluentes nos
organismos. Essa lacuna vem sendo preenchida pelos ensaios ecotoxicológicos que
possibilitam um entendimento ecológico da ação desses poluentes no ambiente.
Esses ensaios caracterizam a toxicidade de substâncias expondo-as a organismos
bioindicadores (SOBRINHO et al., 2013).
O período de germinação e desenvolvimento das plantas apresenta maior
sensibilidade a fatores ambientais adversos. Por essa razão, os testes fitotóxicos
baseados no índice de germinação de sementes têm sido comumente utilizados
para avaliar os efeitos de contaminantes no desenvolvimento de plantas graças à
sensibilidade das sementes durante as etapas de germinação e desenvolvimento
das plântulas e ao seu baixo custo para realização (CRUZ, 2013; SOBRINHO et al.,
2013).
Os líquidos metabólicos produzidos pelas cepas Staphylococcus sp. 26B
e Pseudomonas sp. 2E a partir do efluente apresentaram declínio do elongamento
das raízes na última diluição testada. Com exceção dessas amostras e do metabólito
produzido pela cepa Pseudomonas sp. 7C a partir do óleo vegetal, todos os líquidos

65
metabólicos testados (a partir de ambos os substratos) só favoreceram o
desenvolvimento radicular das sementes na medida em que eram diluídas. Diante
disso, pode-se observar que a diluição das amostras permitiu reduzir os efeitos
tóxicos dos subprodutos microbianos no organismo indicador testado.
Segundo o percentual de inibição das sementes, apenas o líquido
metabólico produzido pela cepa Pseudomonas sp. 9C a partir do óleo vegetal
residual apresentou estímulo de crescimento das raízes na concentração de 5%
como pode ser observado na figura 12.
Figura 12 - Teste de toxicidade do líquido metabólico produzido pela cepa Pseudomonas sp. 9C a partir da fermentação com óleo vegetal residual
Fonte: Autor.
Em seu estudo de produção de biossurfactantes por Candida sphaerica a
partir de óleo vegetal utilizando resíduo oleoso e licor de milho, Sobrinho et al. (2013)
verificaram que o líquido metabólico produzido pela estirpe apresentou característica
tóxica para as sementes vegetais testadas, inibindo a germinação e o crescimento
das raízes. Contudo, ao se testar os biossurfactantes brutos extraídos quimicamente,
nenhum deles apresentou toxicidade na concentração de 200mg/L para sementes

66
de Solanum gilo. Os biossurfactantes extraídos nas concentrações de 200mg/L,
400mg/L e 600mg/L não apresentaram efeitos tóxicos para sementes de Brassica
oleracea. De acordo com os autores, os líquidos metabólicos inibiram a germinação
e o crescimento de sementes devido à presença de substâncias extracelulares.
Portanto, possivelmente, os metabólitos produzidos pelas bactérias testadas na
presente pesquisa possuem algum componente extracelular que inibiu o
desenvolvimento das sementes de pepino no teste.
No teste com o efluente houve germinação e desenvolvimento das raízes
das sementes em todos os tratamentos, como mostra a figura 13. Contudo, o fato do
efluente promover um efeito de crescimento das raízes não o caracteriza como
atóxico, pois obteve valor de IP negativo. Possivelmente, os diferentes componentes
do efluente podem ter sido utilizados como fonte de nutrientes e/ou matéria orgânica
para as sementes favorecendo um maior crescimento de suas raízes em relação ao
controle negativo.
Figura 13 – Teste de toxicidade do efluente do beneficiamento da castanha do caju utilizando sementes de pepino Aodai (Cucumis sativus L.)
Fonte: Autor.
O SDS apresentou esse mesmo comportamento como pode ser
observado na figura 14. Contudo, esse surfactante só permitiu o
desenvolvimento do sistema radicular das sementes à medida que houve sua
diluição para concentrações abaixo de 1mg/mL.

67
Figura 14 - Teste de toxicidade do surfactante SDS utilizando sementes de pepino Aodai (Cucumis sativus L.)
Fonte: O autor.
Dos líquidos metabólicos produzidos a partir de óleo vegetal residual,
apenas uma amostra apresentou estímulo de crescimento. É importante salientar
que as interpretações dos percentuais de inibição das raízes definem três possíveis
efeitos: tóxicos, não tóxicos e estímulo de crescimento. Sendo assim, apesar das
amostras de efluente, SDS e do líquido metabólico da cepa Pseudomonas sp. 9C
(produzido a partir de resíduo oleoso) apresentarem um estímulo de crescimento às
raízes, esse resultado não pode ser interpretado como não tóxico, pois promoveu
crescimento radicular maior em relação ao controle negativo utilizado. Esse mesmo
efeito foi encontrado por Rodríguez e Perera (2009), onde avaliaram a toxicidade de
amostras de água de dois rios da cidade de Havana, Cuba. Segundo os autores,
esse efeito teve como causa o fenômeno de hormesis, evidenciado pelo significativo
estímulo de crescimento das raízes. Vale salientar que os rios analisados possuíam
alta carga orgânica o que pode ter favorecido um maior crescimento das raízes.
Além disso, sabe-se que o efluente do LCC também possui elevadas cargas de

68
matéria orgânica (PIMENTEL et al., 2011) o que pode ter favorecido o elevado
crescimento radicular das sementes.
Uma das vantagens dos biossurfactantes em relação aos surfactantes
sintéticos é a sua baixa toxicidade. De acordo com Felix (2012), os biossurfactantes
são atóxicos, pois suas propriedades tensoativas apresentam grande
compatibilidade com as membranas biológicas. A não toxicidade dos
biossurfactantes é essencial para sua aplicação ambiental (SOBRINHO et al., 2013).
Porém, na presente pesquisa, os líquidos metabólicos (de ambos os substratos)
apresentaram efeitos tóxicos frente às sementes de pepino nas concentrações
testadas. Além da presença de compostos extracelulares nos líquidos metabólicos,
deve-se considerar que as fontes de carbono utilizadas na produção dos
biossurfactantes apresentam efeitos deletérios em muitos organismos bioindicadores.
Pimentel et al. (2011) verificaram que o efluente do beneficiamento da
castanha de caju apresentou ação altamente tóxica para Artemia sp. Na presente
pesquisa, o efluente testado estimulou o crescimento das sementes de pepino,
indicando que algum componente do efluente serviu como fonte de carbono e/ou
nutriente para as sementes permitindo um maior elongamento das raízes. A
presença de compostos hidrofóbicos como óleos vegetais podem reduzir ou tornar
mais lento o processo de germinação e desenvolvimento radicular de plantas, pois
reduzem o teor de água necessário para o desenvolvimento dos processos
fisiológicos (REZENDE, 2006).
As cepas testadas produziram metabólitos com características
biossurfactantes a partir do efluente da castanha do caju e óleo vegetal residual
conferindo um potencial uso biotecnológico dessas bactérias na produção de
tensoativos microbianos a partir de resíduos. Vale destacar que os diferentes
resíduos agrícolas e industriais despertam interesse como fonte de carbono na
produção de biossurfactantes devido ao seu potencial nutricional (SARUBBO et al.,
2016).
Na presente pesquisa, apesar dos resultados dos testes de
caracterização dos biossurfactantes indicarem a viabilidade de se produzir
tensoativos a partir dos substratos testados, os produtos metabólicos das bactérias
apresentaram efeitos tóxicos às sementes vegetais utilizadas no teste
ecotoxicológico. Faz-se necessária uma caracterização desses metabólitos e
extração dos biossurfactantes produzidos para realização de estudos mais

69
conclusivos a respeito de seus efeitos toxicológicos nos organismos indicadores,
pois a presença de compostos extracelulares no líquido metabólico pode mascarar o
efeito dos biossurfactantes nos organismos testes.

70
7 CONCLUSÕES
Diante dos resultados, se pode concluir que:
Foi possível produzir compostos biossurfactantes a partir de efluente
do beneficiamento da castanha do caju e óleo vegetal residual
utilizando bactérias marinhas;
Dentre os grupos bacterianos identificados, as cepas Pseudomonas
sp.,Vibrio fluvialis, Zobellella taiwanensis, Oceanimonas baumannii e
Staphylococcus sp. 26B obtiveram os melhores resultados no testes
indicativos da produção de biossurfactantes;
Três cepas de Pseudomonas sp. produziram metabólitos com
propriedades tensoativas utilizando ambos os substratos testados,
confirmando o potencial de uso desses resíduos como matéria prima
na produção de biossurfactantes;
As bactérias do gênero Pseudomonas e as espécie Z. taiwanensis e
Víbrio fluvialis testadas produziram metabólitos com ação
antimicrobiana frente à cepa de Staphylococcus aureus ATCC 25923;
O líquido metabólito produzido pela cepa de Pseudomonas sp. 9C a
partir de óleo residual, o surfactante SDS e o efluente da agroindústria
do caju estimularam o crescimento das raízes das sementes de pepino
no teste toxicológico, enquanto os demais metabólitos testados foram
tóxicos para as sementes;
É necessário investimento em tratamento e adequação dos resíduos
testados para que esses possam ser utilizados como fonte de carbono
na produção de biossurfactantes, visto que os subprodutos
microbianos se mostraram tóxicos ao organismo indicador testado.

71
REFERÊNCIAS
ANDRADE, T. J. et al. Antioxidant properties and chemical composition of technical Cashew Nut Shell Liquid (tCNSL). Food Chemistry., United Kingdom, v. 126, n. 3, p. 1044-1048, June 2011. ACCORSINI, F. R. Isolamento de leveduras de um consórcio especializado e avaliação de seu potencial na prod ução de biossurfactantes em fontes alternativas de carbono. 2010. 76 f. Dissertação ( Mestrado em Microbiologia Agropecuária) – Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista “Júlio de Mesquita Filho”, Jaboticabal, 2010. ARCHELA, E. et al. Considerações sobre a geração de efluentes líquidos em centros urbanos. Geografia., Londrina, v. 12, n.1, p. 517- 525, jan./jun. 2003. ATLAS, R. M. ; CERNIGLIA, C. E. Bioremediation of Petroleum Pollutants. BioScience., Washington, v. 45, n.5, p. 332-338, May 1995. BANAT, I.M. et al. Biosurfactant production and use in oil tank clean-up. World Journal of Microbiology and Biotechnology., England, v. 7, n. 1, p. 80-88, Jan 1991. BAO, M. et al. Lipopeptide biosurfactant production bacteria Acinetobacter sp. D3-2 and its biodegradation of crude oil. Environmental Science: Processes & Impacts., London, v. 16, n. 4, p. 897-903, Apr. 2014. BARRETO, M. C. et al. Potencial biotecnológico do mar dos Açores. Boletim de Biotecnologia., Porto, Série 2, n. 5, p. 14-15, jun. 2014. BENTO, F. M. ; CAMARGO, F.A.O. ; GAYLARDE, C.C. Biossurfactantes. In: MELO, I. S.; AZEVEDO, J. L. (Eds.), Microbiologia ambiental, 2. ed. Embrapa. Jaguariúna, 2008. BENSON, D. A. et al. 2002. GenBank. Nucleic Acids Research., England, v. 30, n. 1, p. 17-20, Jan. 2002. BEZERRA, M. S. Estudo da produção de biossurfactantes sintetizados por Pseudomonas aeruginosas AP029-GVIIA utilizando manipueira como fonte de carbono. 2012. 153 f. Tese (Doutorado em Engenharia Química) – Departamento de Engenharia Química, Universidade Federal do Rio Grande do Norte, Natal, 2012. BHARALI, P. ; KONWAR, B. K. Production and Physico-chemical Characterization of a Biosurfactant Produced by Pseudomonas aeruginosa OBP1 Isolated from Petroleum Sludge. Applied Biochemistry and Biotechnology., United States, v.164, n.8, p. 1444-1460, Aug. 2011. BODOUR, A. A. ; MILLER-MAIER, R. M. Application of a modified drop collapse technique for surfactant quantitation and screening of biosurfactant-producing microorganisms. Journal of Microbiological Methods., Amsterdam, v. 32, n. 3, p. 273-280, May. 1998.

72
BROWN, G. R. ; SUTCLIFFE, I. C. ; CUMMINGS, S. P. Reclassification of [Pseudomonas] doudoroffii (Baumann et al. 1983) into the genus Oceanomonas gen. nov. as Oceanomonas doudoroffii comb. nov., and description of a phenol-degrading bacterium from estuarine water as Oceanomonas baumannii sp. nov. International J ournal of Systematic and Evolutionary Microbiology., v. 51, p. 67–72, Jan 2001. BRUSSEAU, M. L. Soil and Groundwater Remediation. In: PEPPER, I. ; GERBA, C. ; BRUSSEAU, C. Environmental and pollution science, 2. ed. Amsterdam; Boston: Academic Press, 2006. Cap. 19. BUENO, S. M. Bactérias produtoras de biossurfactantes: isolamento, produção, caracterização e comportamento num sistema modelo. 2008. 89 f. Tese (Doutorado em Ciências e Tecnologia de Alimentos) – Instituto de Biociências, Letras e Ciências Exatas , Universidade Estadual Paulista “Júlio de Mesquita Filho”, Ribeirão Preto, 2008. BUENO, S. M. ; SILVA, A. N. ; GARCIA-CRUZ, C. H. Estudo da produção de biossurfactante em caldo de fermentação. Química Nova., São Paulo, v.33, n.7, p.1572-1577, 2010. BUGAY, C. Biossurfactantes produzidos por Bacillus sp.: estudos de produção e caracterização. 2009. 82 p. Dissertação (Mestrado em Química) – Departamento de Química, Universidade Federal do Paraná, Curitiba, 2009. BURCH, A. Y. et al. Comparison of biosurfactant detection methods reveals h ydrophobic surfactants and contact-regulated production. Environmental Microbiology., United Kingdom, v. 13, n. 10, p. 2681-2691, Oct. 2011. CAMPOS, J. M. ; STAMFORD, L. M. ; SARUBBO, L. A. Production of a bioemulsifier with potential application in the food industry. Applied Biochemistry and Biotechnology., United States, v. 172, n. 6, p. 3234-3252, Mar. 2014. CANESIN, E. A. et al. Characterization of residual oils for biodiesel production. Electronic Journal of Biotechnology., Valparaíso, v. 17, p. 39-45, Jan. 2014. CARRILO, P.G. et al. Isolation and selection of biosurfactant-producing bacteria. World Journal of Microbiological and Biotechnology., England, v. 12, n.1, p. 82-84, Jan. 1996. CASTIGLIONI, G. L. ; BERTOLIN, T. E. ; COSTA, J. A.V. Produção de biossurfactante por Aspergillus fumigatus utilizando resíduos agroindustriais como substrato. Química Nova., São Paulo, v. 32, n. 2, p. 292-295, fev. 2009. CHAVES, M. H. et al. Fenóis totais, atividade antioxidante e constintuintes químicos de extratos de Anacardium occidentale L., Anacardiaceae. Revista Brasileira de Farmacognosia., Rio de Janeiro, v. 20, n. 1, p.106-112, jan./mar. 2010. CHINELLATO, R. A. M. Diversidade e sucessão bacteriana em sedimentos contaminados por hidrocarbonetos, derivados e por dispersão de espumas para controle de incência (AFFF), no Estuário de Santos – SP. 2017. 61 p.

73
Dissertação (Mestrado em Biodiversidade Aquática) - Instituto de Biociências, Universidade Estadual Paulista “Júlio de Mesquita Filho”, São Vicente, 2017. CIRIGLIANO, M.C. ; CARMAN, G.M. Isolation of a bioemulsifier from Candida lipolytica. Applied and Environmental Microbiology., Washington, v. 48, n. 4, p. 7 47-750, Oct. 1984. CLINICAL AND LABORATORY STANDARDS INSTITUTE - CLSI/NCCLS. Performance standards for antimicrobial susceptibility testing: nineteenth informational supplement. 19th ed. M100-S19., Wayne, PA. v .29, n. 3, p.149, 2010. COELHO, M. A. Z. et al. Aproveitamento de resíduos agroindustriais: produção de
enzimas a partir da casca de coco verde. Boletim do Centro de Pesquisa de
Processamento de Alimentos., Curitiba, v.19, n.1, p.33-42, jan./jun. 2001.
COSTA-NETO, P.R. C. et al. Produção de biocombustível alternativo ao óleo diesel através da transesterificação de óleo de soja usado em frituras. Química Nova., São Paulo, v. 23, n. 4, p. 531-537, 2000. CRUZ, J. M. Avaliação Ecotoxicológica da Biodegradação Utilizando Inóculo Enriquecido com Bacillus subtilis em Solo Contaminado com Petróleo, Diesel e Biodiesel. 2013. 68 f. Dissertação (Mestrado em Microbiologia Aplicada) – Instituto d e Biociências, Universidade Estadual Paulista, Rio Claro, 2013. DAS, M.; DAS, S. K.; MUKHERJEE, R. K. Surface active properties of the culture filtrates of a Micrococcus species grown on n-alkanes and sugars. Bioresource Technology., New York, v. 63, n. 3, p. 231-235, Mar. 1998. DASGUPTA, D.; GHOSH, R.; SENGUPTA, T. Biofilme mediated enhanced crude oil degradation by newly isolated Pseudomonas species. ISRN Biotechnology., Cairo, v. 2013, Article ID 250749, 13 p., 2013. DASH, H. R. et al. Marine bacteria: potential candidates for enhanced bioremediation. Applied Microbiology and Biotechnology., Berlin, v. 97, n. 2, p. 561–571, Jan. 2013. DECESARO, A. et al. Produção de biossurfactantes por microrganismos isolados de solo contaminado com óleo diesel. Quimica Nova., São Paulo, v. 36, n. 7, p. 947-9 54, 2013. DHAIL, S. ; JASUJA, N. D. Isolation of biosurfactant-producing marine bacteria. African Journal of Environmental Science and Technology.,[S.l], v. 6, n. 6, p. 263-266, June 2012.
DIAS, AL P. M. ; PINHEIRO, M. G. ; ALVES, F. A. Características epidemiológicas e fatores de virulência em Staphylococcus aureus. Acta Scientiae & Technicae.,Rio de Janeiro, v. 3, n. 3, p. 9-23, Jan 2015.

74
EDDOUAOUDA, K. et al. Characterization of a novel biosurfactant produced by Staphylococcus sp. strain 1E with potential application on hydrocarbon bioremediation. Journal of Basic Microbiology., Germany, v. 52, n. 4, p. 408-418, Aug 2012. EL-SHESHTAWY, H. S. ; DOHEIM, M. M. Selection of Pseudomonas aeruginosa for biosurfactant production and studies of its antimicrobial activity. Egyptian Journal of Petroleum., Cairo, v. 23, n.1, 6 p., Mar. 2014. FELIX, A.K. N. Caracterização e estudo da aplicabilidade do biossurfactante produzido por Bacillus subtilis LAM1005 a partir do suco de caju. Dissertação (Mestrado em Engenharia Química) – Centro de Tecnologia, Universidade Federal do Ceará, Fortaleza, 2012. FENNEMA, O. R. Food Chemistry. 2. ed. New York: Food science and technology, 1985. 991p. FERREIRA, V. C. Produção de biossurfactantes por bactérias isoladas do solo,
rizosfera e água do mangue do rio Cocó (Fortaleza – CE). 2015. 71 p.
Dissertação (Mestrado em Engenharia de Pesca) – Departamento de Engenharia de Pesca, Universidade Federal do Ceará, 2015. FIGUEIREDO, P. G. ; TANAMATI, F. Y. Adubação orgânica e contaminação ambiental. Revista Verde de Agroecologia e Desenvolvimento Sustentável., Pombal, v. 5, n. 3, p.01-04, jul./set. 2010. FRANÇA, I. W. L. Estudo da produção e aplicabilidade de biossurfactante bacteriano produzido por cepa isolada de manguezal cearense. 2014. 137 f. Dissertação (Mestrado em Engenharia Química) – Centro de Tecnologia, Universidade Federal do Ceará, 2014, Fortaleza, 2014. FRANZOL, A. ; REZENDE, C. Estabilidade de emulsões: um estudo de caso envolvendo emulsionantes aniônico, catiônico e não-catiônico. Polímeros., São Carlos, v. 25, n. especial, p. 1-9, dez. 2015.
FREIRE, P. C. M. ; MANCINI- FILHO, J. ; FERREIRA, T. A. P. C. Principais alterações físi co-químicas em óleos e gorduras submetidos ao processo de fritura por imersão: re gulamentação e efeitos na saúde. Revista de Nutrição., Campinas, v. 26, n. 3, p. 353-368, maio/jun. 2013. FREITAS, C. F. ; BARATA, R. A. R. ; NETO, L. S. M. Utilização de óleo de cozinha usado como fonte alternativa na produção de energia renovável, buscando reduzir os impcatos ambientais. In: XXX Encontro Nacional de Engenharia de Produção, 2010, São Carlos. Anais... São Carlos, 2010. Disponível em: <http://www.resol.com.br/textos/enegep2010_tn_stp_123_796_16087.pdf>. Acesso em 06 jun. 2016. GONÇALVES, A. L. Estudo da atividade antimicrobiana de algumas árvores medicinais nativas com potencial de conservação/recuperação de florestas tropicais. 2007. 193 p. Tese (Doutorado em Ciências Biológicas) – Instituto de

75
Biociências do Campus de Rio Claro, Universidade Estadual Paulista, Rio Claro, 2007. GOUVEIA, N. Resíduos sólidos urbanos: impactos socioambientais e perspectivas de manejo sustentável com inclusão social. Ciência & Saúde Coletiva., Rio de Janeiro, v . 17, n. 6, 1503-1510, jun. 2012. GVOZDYAK, 0. R. ; T. M. NOGINA, P. ; SCHUMANN. Taxonomic study of the genus Brachybacterium: Brachybacterium nesterenkovii sp. nov. International Journal of Systematic and Evolutionary Microbiology., United Kingdom, v. 42, v. 1, p. 4274-4 278, Jan. 1992. HAMDAN L. J. ; FULMER, P. A. Effects of COREXIT® EC9500A on bacteria from a b each oiled by the Deepwater Horizon spill. Aquatic Microbial Ecology., Germany, v. 63, p. 101-109, Mar 2011. HAMZA, U. D. ; MOHAMMED, I. A. ; SALE, A. Potential of bacterial isolates in bioremediation of petroleum refinery wastewater. Journal of Applied Phytotechnology in Environmental Sanitation.,Surabaya, v. 1, n. 3, p.131-138, Apr. 2012. HOANG, V. A. et al. Brachybacterium ginsengisoli sp. nov., isolated from soil of a ginseng field. International Journal of Systematic and Evolutionary Microbiology., United Kingdom, v. 64, p. 3063-3068, Sept. 2014. INSTITUTO DE ECONOMIA AGRÍCOLA. Óleos vegetais para alimento ou para biodiesel? Análises e Indicadores do Agronegócio, São Paulo, v.6, n. 6, Jun., 8 p. 2011. IUNESCU, M. et al. Polyols and Rigid Polyurethane Foams from Cashew Nut Shell Liquid. Journal of Polymers and the Environment., United States, v. 20, n. 3, p. 647-658, 2012. JAIN, P. K. ; BAJPAI, V. Biotechnology of bioremediation – a review. International J ournal of Environmental Sciences., [S.l], v. 3, n. 1, p. 535-549, 2012. JAIN, R. M. et al. Production and structural characterization of biosurfactant produced by an alkaliphilic bacterium, Klebsiella sp. : Evaluation of different carbon sources. Colloids and Surfaces B: Biointerfaces., [S.l], v. 108, p. 199-204, Aug. 2013.
JERONIMO, C. E. M. et al. Bioaumentação para degradação de efluente do processamento da castnha de caju. Holos., Natal, ano 28, v. 3, p. 47-59, 2012. KACZOREK, E. et al. Cell surface properties of Pseudomonas stutzeri in the process of diesel oil biodegradation. Biotechnology Letters., Netherlands, v. 34, n. 4, p. 857-862, May 2012. KESKIN, N. O. S. et al. Production and structural characterization of biosurfactant produced by newly isolated Staphylococcus xylosus STF1 from petroleum

76
contaminated soil. Journal of Petroleum Science and Engineering., Netherlands, v. 133, p. 689-694, Sept. 2015. KHOPADE, A. et al. Production and stability studies of the biosurfactant isolated from marine Nocardiopsis sp. B4. Desalination., Netherlands, v. 285, p.198-204, Jan. 2012. LEI, Y. et al. A novel heterotrophic nitrifying and aerobic denitrifying bacterium, Zobellella taiwanensis DN-7, can remove high-strength ammonium. Applied Microbiology and Biotechnology., Germany, v. 100, n. 9, p. 4219-4229, May 2016. LILLY, M. K. et al. Degradation of Anthracene by a novel strain Brachybacterium paraconglomeratum BMIT637C (MTCC 9445). International Journal of Environmental Sciences., [S.l], v. 3, n. 4, p. 1242-1252, 2010. LIMA, C. J. B. Produção de biossurfactante por Pseudomonas aeruginosa empregando óleo de soja residual. 2007. 168 f. Tese (Doutorado em Engenharia Química) – Faculdade de Engenharia Química, Universidade Federal de Uberlândia, Uberlândia, 2007. LIMA E SILVA, T. A. et al. Produção de biossurfactante por Pseudomonas fluorescens UCP 1514 utilizando milhocina como substrato. Exacta., São Paulo, v. 8, n.1, p. 19-26, 2010. LIN, Y. ; SHIEH, W. Y. Zobellella denitrificans gen. nov., sp. nov. and Zobellella taiwanensis sp. nov., denitrifying bacteria capable of fermentative metabolism. International Journal of Systematic and Microbiology., United Kingdom, v. 56, p. 1209-1215, 2006. LOPES, P. F. Diversidade taxonômica e potencial de biodegradação de bactérias isoladas de reservatórios de petróleo da Bacia de Campos (RJ). 2010. 68 p. Dissertação ( Mestrado em Biotecnologia) - Instituto de Ciências Biomédicas, Universidade de São Paulo, São Paulo, 2010. LOPES, R. C. ; BALDIN, N. Educação ambiental para a reutilização do óleo de cozinha na produção de sabão – projeto “ECOLIMPO” In: IX CONGRESSO NACIONAL DE EDUCAÇÃO, 2009, Curitiba. Anais... Curitiba, 2009. Disponível em: < http://www.pucpr.br / eventos /educere /educere2009 /anais/ pdf/2078_1012.pdf>. Acesso em 04 jun. 2017. MAGALHÃES, L. Avaliação da atividade antimicrobiana de ramnolipídeos sobre Listeria monocytogenes. 2012. 73 p. Dissertação (Mestrado em Química) – Instituto de Química, Universidade de São Paulo, São Carlos, 2012. MAIA, F. J. et al. Thermo-oxidative evaluation of new cardol derivatives as antioxidants for mineral oils. Journal of Thermal Analysis and Calorimetry., Netherlands, v. 109, p. 1013-1018, Aug. 2012. MAZZETO, S. E. ; LOMONACO, D.; MELE, G. Óleo da castanha de caju: oportunidades e desafios no contexto do desenvolvimento e sustentabilidade

77
industrial. Química Nova, São Paulo, v. 32, n.3, p.732-741, 2009. MONTAGNOLLI, R. N. ; MATOS LOPES, P. R. ; BIDOIA, E. D. 2015. Assessing Bacillus subtilis biosurfactant effects on the biodegradation of petroleum products. Environmental Monitoring and Assessment., Dordrecht, v. 187, n. 1, p. 4116, Jan. 2015. MOREIRA, S. A. et al. Remoção de metais de solução aquosa usando bagaço de caju. Química Nova., São Paulo, v. 32, n. 7, p.1717-1722, ago. 2009. MUCHA, A. ; MARISA, C. ; ALMEIDA, R. Biorremediação de contanimantes em ambientes costeiros e estuarinos. Boletim de Biotecnologia., Porto, Série 2, n. 5, p. 47-49, jun. 2014. MULANI, N. et al. Biodegradation of crude oil using marine Bacillus species from Vadinar coast, Gujarat, India. Current Science., India, v. 112, n. 3, p. 569-567, Feb 2017. NITSCHKE, M. ; PASTORE, G.M. Biossurfactantes: propriedades e aplicações. Química Nova., São Paulo , v. 25, n.5, p. 772-776, 2002. OBAYORI, O. S. ; SALAM, L. B. ; OMOTOYO, I. M. Degradation of weathered crude oil (Escravos Light) by bacterial strains from hydrocarbons-polluted site. African Journal of Microbiology Research., Nigeria, v. 6, n. 26, p. 5426-5432, July 2012. OLIVEIRA, D. W. F. Produção de biossurfactantes por Bacillus subtilis LAM1005 utilizando suco de caju clarificado. 2010. 103 f. Dissertação ( Mestrado em Engenharia Química) - Centro de Tecnologia, Universidade Federal do Ceará, Fortaleza, 2010. OLIVEIRA, E. M. S. Tratamento de efluentes da indústria de benefriciamento da castanha de caju usando processos eletroquímicos de oxidação avançada. 2014. 84 p. Dissertação (Mestrado em Ciências Naturais) – Faculdade de Ciências Exatas e Naturais – FANAT, Universidade do Estado do Rio Grande do Norte, Mossoró, 2014. OLIVEIRA, N. C. Seleção de microrganismos endofíticos com potencialidades para a biorremediação de ambientes contaminados com hidrocarbonetos de petróleo e/ou derivados. 2009. 80 f. Dissertação (Mestrado em Medicina Tropical) – Instituto de Patologia Tropical e Saúde Pública, Universidade Federal de Goiás, Goiânia, 2009. OSAKI, M. ; BATALHA, M. O. Produção de biodiesel e óleo vegetal no Brasil: realidade e desafio. Organizações Rurais & Agroindustriais., Lavras, v. 13, n. 2, p. 227-242, 2011. OSMARI, M.P. Líquido da castanha de cajú associado a fontes de nitrogênio não protéico na alimentação de bovinos. 2013. 56 f. Tese (Doutorado em Zootecnia) – Centro de Ciências Agrárias , Universidade Estadual de Maringá, Maringá, 2013.

78
PACHECO, G. J. et al. Biosurfactant production by Rhodococcus erythropolis and its application to oil removal. Brazilian Journal of Microbiology., São Paulo, v. 41, p. 685-693, 2010.
PACWA-PLOCINICZAK, M. et al. Environmental applications of biosurfactants: recent advances. Internacional Journal of Molecular Sciences., Switzerland, v. 12, n.1, p. 633-654, Jan 2011. PARASA, L. S. et al. In vitro-antimicrobial activity of cashew (Anacardium occidentale, L.) nuts shell liquid against methicillin resistant Staphylococcus aureus (MRSA) clinical isolates. International Journal of Pharmacy and Pharmaceutical Sciences., India, v. 3, n. 4, p. 436-440, Oct./Dez. 2011. PEREIRA , J. F. B. et al. Optimization and characterization of biosurfactant production by Bacillus subtilis isolates towards microbial enhanced oil recovery applications. Fuel., Netherlands, v. 111, p. 259-268, Sept. 2013. PEREIRA, C. et al. Bacteriophages with Potential for Inactivation of Fish Pathogenic Bacteria: Survival, Host Specificity and Effect on Bacterial Community Structure. Marine Drugs., Switzerland, v. 9 , p. 2236-2255, 2011. PIMENTEL, M. F. Análise ecotoxicológica do efluente da indústria de beneficiamento da castanha de cajú antes e após tratamento em reator aeróbio inoculado com fungos. 2008. 68 f. Dissertação (Mestrado em Ciências Marinhas Tropicais – Instituto de Ciências do Mar, Universidade Federal do Ceará, Fortaleza, 2008. PIMENTEL, M. F. et al. O uso de Artemia sp. como organismos-teste para avaliação
da toxicidade das águas residuárias do beneficamento da castnha de cajú antes e
após tratamento em reator biológico experimental.Journal of the Brazilian Society
of Ecotoxicology., Itajaí, v. 6, n.1, p.15-22, 2011.
PINTO, M. H. ; MARTINS, R. G. ; COSTA, J. A. V. Avaliação cinética da produção de biossurfactantes bacterianos. Química Nova, São Paulo, v. 32, n. 8, p. 2104-2108, ago. 2009.
RAHMAN, K. S. et al. Rhamnolipid Biosurfactant Production by Strains of Pseudomonas aeruginosa Using Low-Cost Raw Materials. Biotechnology Progress., United States, v. 18, n. 6, p. 1277-1281, nov./dez. 2002. RAMAMURTHY, T. et al. Vibrio fluvialis: an emerging human pathogen. Frontiers in Microbiology., Switzerland ,v. 5, Article 91, Mar. 2014. RAMEZANI, M. ; AMOOZEGAR, M. A. ; VENTOSA, A. Screening and comparative assay of poly-hydroxyalkanoatesproduced by bacteria isolated from the Gavkhooni Wetlandin Iran and evaluation of poly-β-hydroxybutyrate productionby halotolerant bacterium Oceanimonas sp. GK1. Annals of Microbiology., Germany, v. 65, p. 517-526, Mar. 2015.

79
RENDUELES, M. ; DÍAS, M. Biotecnología Industrial. Arbor., v. 190, n. 768, p. (a155), 2014. REZENDE, I. M. Efeito do solo contaminado por óleo diesel na germinação e nas plantas jovens de Schinus terebinthifolius raddi (Anacardiaceae). 2006.70 p Dissertação (Mestrado em Ciências Biológicas) – Universidade Federal do Paraná, Curitiba, 2006. RIBEIRO, V. G. et al. Magnetic nanoparticles coated with anacardic acid derived from cashew nut shell liquid. Journal of Materisl Science., Netherlands, v. 48, n. 22, p. 7875-7882, Nov. 2013. ROCHA E SILVA, N. M. P. et al. Screening of Pseudomonas species for biosurfactant production using low-cost substrates. Biocatalysis and Agricultural Biotechnology., Netherlands, v. 3, n.2, p. 132-139, Apr. 2014. RODRÍGUEZ, M. T. T. ; PEREIRA, N. M. H. Determinación toxicológica en aguas de río mediante el empleo de un bioensayo con planta. Higiene y Sanidad Ambiental., Granada, v. 9, p. 505-509, 2009. ROSA, M. F. et al. Valorização de resíduos da agroindústria. In: II Simpósio Internacional sobre Gerenciamento de Resíduos Agropecuários e Agroindustriais – II SIGERA, 2011, Foz do Iguaçu. Anais... – Foz do Iguaçu: SBERA, 2011. Disponível em : < http://www.sbera.org.br/2sigera/obras/p12.pdf>. Acesso em 06 jun. 2016. RUFINO, R. D. ; SARUBBO, L. A. ; CAMPOS - TAKAKI, G. M. Enhancement of stability of biosurfactant produced by Candida lipolytica using industrial residue as substrate. World Journal of Microbiological and Biotechnology., Seoul, v. 23, p. 729-734, May 2007.
SALAZAR- BRYAM, A. M. Aproveitamento de glicerol residual como fonte de carbono para produção de ramnolipideos por linhagens de Pseudomonas aeruginosa. 2016. 89 f. Dissertação (Mestrado em Ciências Biológicas – Microbiologia Aplicada) – Instituto de Biociências de Rio Claro, Universidade Estadual Paulista, 2016. SANCHES, M. A. Utilização de resíduos vegetais para a produção de biossurfactantes por fungos isolados de amostras de solo da região amazônica. 2013. 65 f. Dissertação (Mestrado em Ciências Farmacêuticas) – Faculdade de Ciências Farmacêuticas, Universidade Federal do Amazonas, Manaus, 2013. SARUBBO, L. A. et al. Production of a Low-cost Biosurfactant for Application in the Remediation of Sea water Contaminated with Petroleum Derivates. Chemical Engineering Transactions., Italy, v. 49, p. 523-528, 2016. SAKALLE, K. ; RAJKUMAR, S. Isolation of crude oil degrading marine bacteria and assessment for biosurfactant production. The Internet Journal of Microbiology., Texas, v. 7, n. 2 , p. 7, 2009.

80
SANJEEVA, S. K. et al. Distilled technical cashew nut shell liquid (DT-CNSL) as an effective biofuel and additive to stabilize triglyceride biofuels in diesel. Renewable Energy., United Kingdom, v. 71, p. 81-88, Nov. 2014. SANTAELLA, S. T. et al. Emprego de fungos para tratamento de efluentes da indústria de castanha de caju. In: VI Simpósio Ítalo-Brasileiro de Engenharia Sanitária e Ambiental, 2002, Vitória. Anais... Vitória, 2002. Disponível em: <http://www.bvsde.paho.org/bvsacd/sibesa6/ccinq.pdf> . Acesso em 06 jun. 2016. SANTOS, A. C. F. Diversidade e estrutura de comunidades bacterianas em ambientes contaminados com petróleo. 2012. 149 f. Tese (Doutorado em Genética e Biologia Molecular) - Universidade Estadual de Santa Cruz , Ilhéus, 2012. SANTOS, R. M. Produção e caracterização de bio-óleo a partir de resíduo agroindustrial de semente de mangaba. 2014. 82 f. Dissertação (Mestrado em Química) – Univrsidade Federal de Sergipe, São Cristóvão, 2014. SATER, O. et al. Estudo comparativo da carbonização de resíduos agrícolas e florestais visando à substituição da lenha no processo de secagem de grãos de café. Revista Ceres., Viçosa, v. 58, n. 6, p.717-722, nov./dez. 2011. SATPUTE, S. K. et al. Biosurfactants, bioemulsifiers and exopolysaccharides from marine microorganisms. Biotechnology Advances., United States, v. 28, p. 436-450, July/Aug. 2010. SCHULTZ, F. M. Avaliação de microrganismos com potencial de degradação de diesel e biodiesel. 2010. 113 f. Dissertação (Mestrado em Microbiologia Agrícola e do Ambiente) – Instituto de Ciências Básicas da Saúde, Universidade Federal do Rio Grande do Sul, Porto Alegre, 2010. SEGATTO, F. B. B. Conhecendo as formas de descartes do óleo saturado de cozinha para verificar a educação ambiental na escola. Revista Eletrônica em Gestão, Educação e Tecnologia Ambiental - REGET., Santa Maria, v. 10, n. 10, p. 2122-229, jan./abr. 2013. SEN, R.et al. (Edit). Biossurfactantes. USA, Springer, 2010. 359p. SEXENA, P. ; JAWALE, S. ; JOSHIPURA, M. A review on prediction of properties of biodiesel and blands of biodiesel. Procedia Engineering., United Kingdom, v. 51, p. 395-402, 2013. SHOEB, E. et al. Screening and characterization of biosurfactant-producing bacteria isolated from the Arabian Sea coast of Karachi. Turkish Journal of Biology.,Turkey, v. 39, p. 210-2016, 2015. SILVA, F. S. F. Prospecção de cepas bacterianas produtoras de biossurfactantes na região do porto do Mucuripe, Fortaleza-Ceará. 2014. 62 f. Monografia (Graduação em Ciências Ambientais) – Instituto de Ciências do Mar (LABOMAR), Universidade Federal do Ceará, Fortaleza, 2014.

81
SILVA, F. S. F. et al. Produção de biossurfactantes por cepas bacterianas de origem marinha utilizando querosene como fonte de carbono. Revista de Ciências Ambientais – RCA., Canoas, v. 11, n.1, p.7-18, 2017. SILVA, J. S.; SANTOS, S. S. ; GOMES, F. G. G. A biotecnologia como estratégias de reversão de áreas contaminadas por resíduos sólidos. Revista Eletrônica em Gestão, Educação e Tecnologia Ambiental – REGET., Santa Maria, v. 18, n. 4, p. 1361-1370, dez. 2014.
SOARES, V. S. ; PUPO, D. D. ; MOURÃO, R. C. Produção de sabão artesanal no município de Sorriso – MT: alternativa sustentável ao óleo usado. IBEAS – Instituto Brasileiro de Estudos Ambientais. In: IV Congresso Brasileiro de Gestão Ambiental, 2013, Salvador. Anais... Salvador, 2013. Disponível em : < http://www.ibeas.org.br/ congresso/Trabalhos2013/III-035.pdf. Acesso em: 04 jun. 2016. SOBRINHO, H. B. S,. et al. Assessment of toxicity of a biosurfactant from Candida sphaerica UCP 0995 cultivated with indusatrial residues in a bioreactor. Electronic Journal of Biotechnology., Valparaíso, v. 16, n. 4, p. 4-4, July 2013. SOUZA, F. A. S. D. ; SALGUEIRO, A. A. ; ALBUQUERQUE, C. D. C. Production of bioemulsifier by Yarrowia lipolytica in sea water using diesel oil as the carbon source. Brazilian Journal of Chemical Engineering., São Paulo, v. 29, n. 1, p. 61-67, jan./mar. 2012. SOUSA, M. Bioconversão do glicerol para produção de biossurfactantes: aplicação no preparo de emulsões. 2011. 120 p. Dissertação (Mestrado em
Engenharia Química) – Centro de Tecnologia,Universidade Federal do Ceará,
Fortaleza, 2011. SPERB, J. G. C. et al. Avaliação quantitativa da produção de lipases e biossurfactantes por fungos isolados de resíduos oleosos. Engevista., Rio de Janeiro, v. 17, n. 3, p. 385-397, set., 2015. TAMANINI, C. ; HAULY, M. C. O. Resíduos agroindustriais para produção biotecnológica de xilitol. Semina: Ciências Agrárias., Londrina, v. 25, n.4, p.315-330, out./dez. 2004. TAMURA, K. et al. MEGA 6 : Molecular Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution., Chicago, v. 30,n. 12 p. 2725-2729, Oct. 2013. TAKEUCHI, M. ; FANG, C.X. ; YOKOTA, A. Taxonomic Study of the Genus Brachybacterium: Proposal of Brachybacterium conglomeratum sp. Nov., nom. Rev., Brachybacterium paraconglomeratum sp. nov., and Brachybacterium rhamnsorium sp. nov. International Journal of Systematic Bacteriology., United Kingdom, v. 45, n. 1, p.160-168, Jan 1995. TEIXEIRA,V. L. et al.Caracterização do Estado da Arte em Biotecnologia Marinha no Brasil. Brasília: Ministério da Saúde; Ministério de Ciência e Tecnologia; Organização Pan-Americana de Saúde, 2010. 134 p. (Série B. Textos Básicos de

82
Saúde). THAVASI, R. ; SHARMA, S. ; JAYALAKSHMI, S. Evaluation of screening methods for the isolation biosurfactant producing marine bacteria, Journal of Petroleum & Environmental Biotechnology., Los Angeles, S1:001, p. 6, 2011. THODE- FILHO, S. et al. Tecnologia ambiental aplicada ao gerenciamento e processamento do óleo vegetal residual no estado do Rio de Janeiro. Revista Eletrônica em Gestão, Educação e Tecnologia Ambiental – REGET, Santa Maria, v. 15, n.15, p. 3026-3035, out. 2013. TONINI, R.M.C. ; REZENDE, C.E. ; GRATIVO, A.D. Degradação e biorremediação de compostos do petróleo por bactérias: revisão. Oecologia Australis., Rio de Janeiro, v. 14, n. 4, p.1010-1020, dez. 2010. TSUBOI, S. et al. Diversity of alkane hydroxylase genes on the rhizoplane of grasses planted in petroleum-contaminated soils. Springerplus., Germany, v. 4, 10 p., 2015. UZOIGWE, C. et al. Bioemulsifiers are not biosurfactants and require different screening approaches. Frontiers in Microbiology., Switzerland, v. 6, Article 245, 6 p., Apr. 015. VARADAVENKATESAN, T. ; MURTY, V. R. Production of a Lipopeptide Biosurfactant by a Novel Bacillus sp. and Its Applicability to Enhanced Oil Recovery. ISRN Microbiology., United States, v. 2013, Artigo 621519, 8 p., 2013. VARELA, J. et al. Os oceanos e a biotecnologia marinha: um novo desafio para Portugal. Boletim de Biotecnologia., Porto, Série 2, n. 5, p. 8-10, jun. 2014. VENUGOPAL, M. ; SARAMM, A.V. Characterization of alcaline protease from Vibrio fluvialis strain isolated from a mangrove sediment sample and its application as a laundry detergente additive. Process Biochemistry., London, v. 41, p. 1239-1243, 2006. VIRAMONTES-RAMOS, S. et al. Selection of biosurfactan/bioemulsifier-producing bacteria from hydrocarbon-contaminated soil. Brazilian Journal of Microbiology., São Paulo, v. 41, p. 668-675, Oct. 2010. VYAS, T. K ; DAVE, B. P. Production of biosurfactant by Nocardia otitidiscaviarum and its role in biodegradation of crude oil. International Journal of Environmental Science & Technology.,New York, v. 8, n. 2, p. 425-432, Mar. 2011. WANG, X. et al. Draft Genome Sequence of Brachybacterium phenoliresistens Strain W13A50, a Halotolerant Hydrocarbon-Degrading Bacterium. Genome Announcements., Washington DC, v. 2, n. 5, pii: e00878, Sept./Oct. 2014. WILDNER, L. B. A. ; HILLING, C. Reciclagem de óleo comestível e fabricação de sabão como instrumentos de educação ambiental. Revista Eletrônica em Gestão, Educação e Tecnologia Ambiental - REGET., Santa Maria, v. 5, n. 5, p.813-824, 2012.

83
WILLUMSEN, P.A. ; KARLSON, U. Screening of bacteria, isolated from PAH–
contaminated soils, for production of biosurfactants and bioemulsifiers.
Biodegradation., Dordrecht, v. 7, n. 5, p. 415–423, 1997.
YULIANA, M. ; TRAN-THI, N. Y. ; JU, Y. H. Effect of extraction methods on characteristic and composition of Indonesian cashew nut shell liquid. Industrial Crops and Products., Netherlands, v. 35, p. 230-236, Jan. 2012. ZHANG, H. ; WANG, Q. ; MORTIMER, S. R. Waste cooking oil as an energy resource: review of Chinese policies. Renewable and Sustainable Energy Reviews., United Kingdom, v. 16, n. 7, p. 5225-5231, Sept. 2012. ZHANG, T. ; FANG, H. H. P. Phylogenetic diversity of a SRB-rich marine biofilm. Applied Microbiology and Biotechnology., Germany, v. 57, p. 437-440, Oct. 2001. ZHAO et al. Degradation of phenanthrene by bacterial strain isolated from soil in oil refinery fields in Shanghai China. Journal of Hazardous Materials., Amsterdam, v. 164, n. 2-3, p. 863-869, May 2009. ZHENG, C. et al. Characterization and emulsififying property of a novel bioemulsifier by Aeribacillus pallidus YM-1. Journal of Applied Microbiology., United Kingdom, v. 113, p.44-51, July 2012.