universidade federal de mato grosso faculdade de agronomia ... · realizado as análises...
TRANSCRIPT

Universidade Federal de Mato Grosso
Faculdade de Agronomia, Medicina Veterinária e Zootecnia
Departamento de Fitotecnia e Fitossanidade
SEGUNDO RELATÓRIO PARCIAL DE ATIVIDADES
ATRIBUTOS MICROBIOLÓGICOS DO SOLO EM SISTEMAS INTEGRADAS NA
REGIÃO NORTE DE MATO GROSSO
Pesquisador responsável:
Profa. Dra Daniela Tiago da Silva Campos - UFMT/FAMEV
Janeiro - 2014
Cuiabá-MT

2
SEGUNDO RELATÓRIO PARCIAL
Projeto Agrisus No: PA 1045-12
Título do projeto: Atributos microbiológicos do solo em sistemas integradas na
região norte de Mato Grosso
Interessado: Profa. Dra. Daniela Tiago da Silva Campos
Instituição: UNIVERSIDADE FEDERAL DE MATO GROSSO, Avenida Fernando
Correa da Costa, Cuiabá, MT. Fone: (65) 3615-8606. E-mail:
Local da Pesquisa: Experimental – Santa Carmen e Nova Canaã do Norte, MT. Análises
– UFMT/FAMEVZ, Laboratório de Microbiologia do Solo.
Valor financiado pela Fundação Agrisus: R$ 26.800,00
Vigência do Projeto: 04/09/12 a 30/12/14
RELATÓRIO PARCIAL DE ATIVIDADES 2013-2
1. RESUMO DO RELATÓRIO
O presente relatório compila os resultados do projeto de pesquisa cadastrado na
Agrisus pelo número PA 1045/12, onde avalia duas áreas distintas de integração lavoura
pecuária e pecuária floresta, sendo elas a ILP, localizada em Santa Carmem, e a iLPF,
localizada em Nova Canaã do Norte, sendo assim os principais resultados encontrados
nessa primeira fase do projeto são:
- iLP: os sistemas de iLP utilizados surtiram diferenças estatística nos parâmetros
microbiológicos avaliados e entre a mata nativa, lavoura e pastagem para as variáveis
enzima e enumeração de micro-organismos. A atividade da enzima B-glicosidase foi
mais intensa na mata nativa, as configurações de iLP-A onde havia soja na safra,
milheto consorciado com Brachiaria ruziziensis, na entressafra e a iLP-C com soja na
safra, milho consorciado com Brachiaria brizantha, na entressafra foram as que mais se
aproximaram aos teores encontrados na mata nativa. Para a enumeração de micro-
organismos os resultados foram bem variados, com destaque a iLP-C que apresentou os
maiores valores, ultrapassando até mesmo a mata nativa. Até o momento estes
resultados demonstram que os sistemas de integração são capazes de manter a qualidade

3
biológica do solo, mesmo após a sua sucessão.
-iLPF: as áreas de iLPF com as diferentes conformações apesentaram diferença
estatística significativa para as variáveis fosfatase ácida e B-glicosidase, e foi possível
verificar que a distancia do pé da planta perene influencia na atividade bem como na
presença dos micro-organismos do solo; aos três metros do pé da planta apresentaram-
se maior número e maior atividade microbiana.
2. INTRODUÇÃO
Este projeto foi elaborado em virtude da demanda de conhecimentos da
dinâmica da microbiota do solo em sistemas de iLP e iLPF. Os trabalhos com os
sistemas integrados no Laboratório de Microbiologia do Solo da UFMT/FAMEV,
campus de Cuiabá, MT iniciaram-se em 2008. Neste período vários ganhos foram
alcançados com os resultados.
No sistema de iLP tivemos a participação de uma aluna de graduação que
desenvolveu seu trabalho de conclusão de curso e atualmente temos outra aluna de
graduação desenvolvendo trabalhos com fungos micorrízicos arbusculares e uma aluna
de mestrado do curso de Pós Graduação em Agricultura Tropical (PPGAT) que tem
como tema da dissertação avaliar a resiliência da iLP nos atributos químicos e
microbiológicos do solo.
Na iLPF já foi concluída uma dissertação no ano de 2012 que teve como tema a
análise dos atributos microbiológicos nos solos. Atualmente, a aluna que desenvolveu
este trabalho está cursando o doutorado no PPGAT e dará continuidade aos trabalhos
com a microbiota do solo. Porém, a iLPF já está no momento em quem se tem pasto e
animais e objetiva-se avaliar qual a influência destes fatores nos atributos
microbiológicos.
No estado de Mato Grosso, em especial a região Norte e Médio Norte, devido à
exploração agrícola e pecuária, com o uso intensivo do solo associado a manejos
inadequados, levou à degradação do solo, perda do potencial produtivo e elevados
custos de produção, além de gerar sérios problemas tanto econômicos como ambientais,
e a necessidade de abertura de novas áreas na região.
A qualidade do solo tem chamado atenção, e a quantificação de alterações nos
seus atributos, decorrentes da intensificação de sistemas de uso e manejo, têm sido
amplamente realizadas para monitorar a produção sustentável dos solos.

4
Frente a esta realidade, deve-se buscar alternativas viáveis para utilização
sustentável dos recursos naturais, baseadas em conservação de solo e ambiente,
maximizando o uso de recursos e a produção agropecuária. Nesse contexto, podem-se
destacar práticas agropecuárias, tais como o sistema de plantio direto na palha (SPD) e a
diversificação das atividades, por meio da integração Lavoura-Pecuária (iLP) e
integração Lavoura-Pecuária-Floresta (iLPF). A inclusão de pastagens e floresta em
áreas agrícolas pode ser uma ferramenta útil na recuperação de áreas degradadas, bem
como um meio para garantir a sustentabilidade deste sistema.
Esses sistemas de produção que integram agricultura e pecuária (iLP), bem
como agricultura, pecuária e floresta (iLPF) são práticas recentes empregadas no Estado
de Mato Grosso, o qual é responsável por uma das maiores produções agrícolas
nacionais. Esses sistemas de manejo buscam maximizar a utilização da área, integrando
grandes culturas como, milho, soja e feijão, com brachiaria, e floresta, com árvores
geralmente utilizadas para reflorestamento, como o eucalipto, mogno e outras florestais.
Para que os sistemas integrados sejam adotados em escala, a realização de
pesquisas nas áreas já instaladas e a demonstração de resultados práticos aos produtores
por meio de dias de campo, palestras e publicações são indispensáveis para que os
mesmos sejam incentivados e os sistemas ganhem âmbito de áreas comerciais. E desta
forma propõe-se este projeto de pesquisa.
3. MATERIAL e MÉTODOS
O trabalho está sendo realizado com solo coletado em duas fazendas particulares
do norte de Mato Grosso. Nestas áreas estão dispostas unidades de referência
tecnológica (URT) da EMBRAPA Arroz e Feijão, nos quais estão um sistema de
integração lavoura-pecuária (ILP), no município de Santa Carmem, e um sistema de
integração lavoura-pecuária-floresta (iLPF), no município de Nova Canaã do Norte.
3.1 Avaliação microbiológica do solo sob Integração Lavoura-Pecuária (ILP)
O trabalho na iLP está sendo conduzido em uma Unidade de Referência
Tecnológica (URT) da Embrapa Arroz e Feijão, no município de Santa Carmem,
localizada na Fazenda Dona Isabina, onde o sistema iLP foi estabelecido em dezembro
de 2006, atualmente está com rotação de lavoura soja e milho safrinha. Estas áreas

5
foram georeferenciadas, compostas de piquetes de 20 hectares, com a descrição
apresentada na Tabela 1.
Tabela 1. Descrição da configuração dos tratamentos da iLP, URT Santa Carmem, MT.
Trat. Sistema Configuração anterior e atual do campo Área
01 iLP/ A Sucessão de iLP, que havia cultivo de soja na safra, milheto
consorciado com Brachiaria ruziziensis, na entressafra. Atualmente
com lavoura de soja e milho safrinha.
20 ha
02 iLP/ B Sucessão de iLP, cultivo de arroz na safra, seguido de milho, sorgo ou
milheto consorciado com Brachiaria ruziziensis, na entressafra.
Atualmente com lavoura de soja e milho safrinha.
20 ha
03 iLP/ C Sucessão de iLP, cultivo de soja na safra, milho consorciado com
Brachiaria brizantha, na entressafra. Atualmente com lavoura de soja e milho safrinha.
20 ha
04 iLP/ D Sucessão de iLP, cultivo de Brachiaria brizantha por 1 ano mais gado
com rotação de lavoura por 3 anos. Atualmente com lavoura de soja e
milho safrinha.
20 ha
05 iLP/ E Sucessão de iLP, cultivo de Brachiaria brizantha por 1 ano + gado com
rotação de lavoura por 3 anos. Atualmente com lavoura de soja e milho
safrinha.
20 ha
06 Lavoura Monocultivo de soja (Glycine max (L.) sob manejo convencional. 20 ha
07 Pastagem Pastagem com cultivo de Brachiaria brizantha, há 7 anos. 20 ha
08 Mata
nativa
Mata nativa com caracteristica da região. 20 ha
A coleta de solo foi realizada em setembro/2012, correspondente ao período de
seca na região. Amostras de solo deformadas foram coletadas na profundidade de 0-20
cm para as análises microbiológicas. Onde coletou-se 5 amostras compostas de 3
subamostras simples em cada piquete de todas as áreas descritas anteriormente.
O solo coletado foi homogeneizado, adicionado em sacos plásticos,
devidamente identificados, acondicionados em caixas térmicas contendo gelo e
transportados para ao Laboratório de Microbiologia do Solo da Faculdade de
Agronomia e Medicina Veterinária na Universidade Federal de Mato Grosso, onde
foram armazenados em câmara fria à 4 °C até a execução das análises, para todas as
análises laboratoriais foram feitas triplicatas de cada amostra.
3.2 Avaliação microbiológica do solo sob integração lavoura-pecuária-floresta
(iLPF)

6
O estudo está sendo desenvolvido na Fazenda Gamada, localizada no
município de Nova Canaã do Norte – MT, em uma área experimental de integração
lavoura-pecuária-floresta (ILPF) já instalada desde 2009, constituída por 5 piquetes de 5
ha (250 x 200), conforme Tabela 2. Em março de 2011 a pastagem foi introduzida em
meio a lavoura de grãos (mistura de sementes ao fertilizante), a qual está sendo
pastejada até o presente momento.
Tabela 2. Descrição dos tratamentos da iLPF, em Nova Canaã do Norte, MT.
Sistema Configuração Área
iLPF linha simples
de eucalipto
Faixa de 200 m de largura com povoamento de eucalipto
(Eucalyptus urograndis), em linhas únicas de 250 m de
comprimento separadas entre si por 20 m, com plantio de soja (Glycine max) na safra, milho (Zea mays) na safrinha e
Brachiaria ruziziensis na entressafra, nas entrelinhas.
5 ha
iLPF linha dupla de
eucalipto
Faixa de 200m de largura com povoamento de eucalipto
(Eucalyptus urograndis) distribuídos em sub-faixas compostas
por duas linhas de 250 m de comprimento separadas entre si por
3 m, com distância entre plantas de 2 m e distancia entre sub-
faixas de 20m , com plantio de soja (Glycine max) na safra,
milho (Zea mays) na safrinha e Brachiaria ruziziensis na
entressafra, nas entrelinhas.
5 ha
iLPF linha tripla eucalipto
Faixa de 200 m de largura com povoamento de eucalipto (Eucalyptus urograndis) distribuídos em sub-faixas composta
por três linhas de 250 m de comprimento separadas entre si por
3 m, com distância entre plantas de 2 m e distância entre sub-
faixas de 20 m, com plantio de soja (Glycine max) na safra,
milho (Zea mays) na safrinha e Brachiaria ruziziensis na
entressafra, nas entrelinhas.
5 ha
Rotação
soja/pastagem
Área anteriormente com plantio de soja (Glycine max) na safra,
milho (Zea mays) na safrinha e atualmente Brachiaria
ruziziensis mantida por 3 anos.
5 ha
Mata nativa Área com mata nativa característica da região. 5 ha
Foram amostrados em todos os cinco tratamentos, o solo com três repetições por
tratamento (três faixas/pontos dentro do talhão de 5 ha). As amostras foram retiradas no
sentido transversal às linhas das espécies florestais, partindo do meio da área ocupada
pela pastagem (a 10 m das árvores), até o meio da próxima faixa de pastagem (10 m
após as árvores), ficando as linhas florestais no centro da área de amostragem, conforme
figura abaixo.

7
Figura 1. Área desde o início e esquema de pontos de coleta (pontos azuis).
Foram coletados também amostras de solo de mata, localizada próximo à sede
da fazenda, e de lavoura tradicional, sendo 3 pontos por tratamento.
As amostras foram retiradas da camada de 0-20 cm de profundidade nos pontos
a 10 m, 6 m e 3 m florestais para ambos os lados, mais um ponto no sub-bosque
entremeio as florestais, totalizando 7 amostras em cada repetição, com um total de 21
amostras por piquete (resultado das 3 repetições), totalizando 35 amostras de campo.
As análises microbiológicas foram feitas no laboratório de Microbiologia do
Solo, da Universidade Federal de Mato Grosso, Campus Cuiabá – MT.
3.2.1 Procedimento Analítico
No Laboratório de Microbiologia do Solo da Faculdade de Agronomia,
Medicina Veterinária e Zootecnia na Universidade Federal de Mato Grosso, foi
realizado as análises microbiológicas das amostras coletadas na iLP e iLPF.
3.2.1.1 Atividade enzimática
Para a determinação da atividade das enzimas β-glicosidase e fosfatases foram
utilizados os métodos descritos por Tabatabai e Bremner (1969), adaptados como
descrito abaixo.
Enzima β-glicosidase
O substrato utilizado na reação desta enzima foi o p-nitrofenil-β-D-
Glucopiranosídeo 0,05 mol L-1
e uma solução tampão estoque universal modificada
(MUB estoque) preparada para uma solução MUB pH 6 a partir de 200 mL de MUB

8
estoque, para 1000 mL de solução, corrigindo o pH com HCl 0,1 mol L-1
.
Amostras de 0,5 g solo foram colocadas em tubos de ensaio separadamente,
utilizando três repetições para cada tratamento. Em seguida foram adicionados 2 mL de
MUB pH 6 e 0,5 mL de p-nitrofenil 0,05 mol L-1
em todos os frascos. Os tubos de
ensaio foram fechados e incubados a 37 °C por uma hora. Após incubação foram
adicionados 0,5 mL CaCl2, 2 mL solução extratora de Tris-Hydroxymetyl-Amino-
Metano (THAM pH 12) e 1 mL de água destilada. Procedeu-se em seguida à filtragem
em papel filtro Whatman n° 02.
A intensidade da coloração amarela do filtrado foi determinada em
espectrofotômetro a 400 nm de absorbância. A quantidade de p-nitrofenol formada em
cada amostra foi determinada com base em curva padrão preparada com concentrações
conhecidas de p-nitrofenol (0, 1, 2, 3, 4, 5 mg de p-nitrofenol para solução em total de 5
mL, aferidos com água destilada). A atividade enzimática foi expressa em μg de p-
nitrofenol liberado por hora por grama de solo seco (μg p-nitrofenol h-1
g solo seco-1
).
Enzimas fosfatases ácida e alcalina
Na fosfatase ácida, o substrato utilizado na reação da enzima foi o p-nitrofenil
fosfato 0,05 mol L-1
em uma solução tampão universal estoque modificada (MUB
estoque) preparada para uma solução MUB pH 6,5, obtida a partir da titulação de 200
mL MUB estoque, para 1000 mL solução, com adição de HCl 0,1 mol L-1
para
regulação do pH.
Amostras de 0,5 g solo foram colocadas em tubos de ensaio separadamente,
utilizando três repetições e um controle, composto por todos os reagentes exceto o solo.
Em seguida foram adicionados 2 mL de MUB em todos os frascos, e 0,5 mL de p-
nitrofenil 0,05 mol L-1
.
Os tubos de ensaio foram fechados e incubados a 37 °C por uma hora e então
levados a agitação por mais uma hora. Após incubação e agitação, foram adicionados
0,5 mL CaCl2 0,5 mol L-1
, 2 mL NaOH 0,5 mol L-1 e 1 mL de água destilada,
procedendo em seguida à filtragem em papel filtro Whatman n° 02. A intensidade da
coloração amarela do filtrado foi determinada em espectrofotômetro a 400 nm de
absorbância.
A quantidade de p-nitrofenol formada em cada amostra foi determinada com
base em curva padrão preparada com concentrações conhecidas de p-nitrofenol (0, 1, 2,

9
3, 4, 5, mg de p-nitrofenol em total de 5 mL, completos com água destilada). A
atividade enzimática é expressa em μg de p-nitrofenol liberado por hora por grama de
solo seco (μg p-nitrofenol h-1
g solo seco-1
).
Para a fosfatase alcalina seguiu-se o mesmo procedimento descrito na fosfatase
ácida, com exceção do pH do MUB, onde foi alcalinizado para pH 11.
3.2.1.2 Contagem de fungos, bactérias, actinomicetos e celulolíticos totais
Para a determinação da comunidade microbiana foi utilizado o método da
diluição seriada e plaqueamento em meio seletivo (Wollum, 1982). A partir das
amostras de 10 g de solo para execução da metodologia.
A partir dessa amostra foi realizada diluição seriada até 10-5. Após a
homogeneização, com auxílio de pipetador automático, foram distribuídos 200 μL de
cada diluição sobre o meio agarizado e uniformizado com o auxílio da alça de Drigalski.
As placas, de 9 cm de diâmetro, foram incubadas invertidas a 26 ± 2 ºC. A avaliação foi
realizada 48 h, 24 h e 5 dias após incubação, respectivamente, contando-se o número de
unidades formadoras de colônias em cada placa.
Os valores obtidos a partir da contagem direta das colônias foram calculados
para unidades formadoras de colônias por mililitro (UFC mL-1
). Foram utilizadas três
placas para cada diluição, e consideradas para cálculos apenas as diluições cujas placas
tinham entre 30 a 300 colônias.
3.2.1.3 Análise estatística dos dados
Os dados foram submetidos à análise de variância e as médias comparadas pelo
teste de Kruskal Wallise e Tukey a 1% de probabilidade, utilizando-se o programa
Assistat, versão 7.6, beta 2011 (Silva e Azevedo, 2002).
4. RESULTADOS E DISCUSSÃO
Em virtude de o projeto estar em andamento, aqui apresentamos apenas os resultados
das coletas do período de chuvas na região.
10
4.1 Caracterização microbiológica do solo sob sistemas de iLP
4.1.2. CBM, respiração basal e quociente metabólico do solo
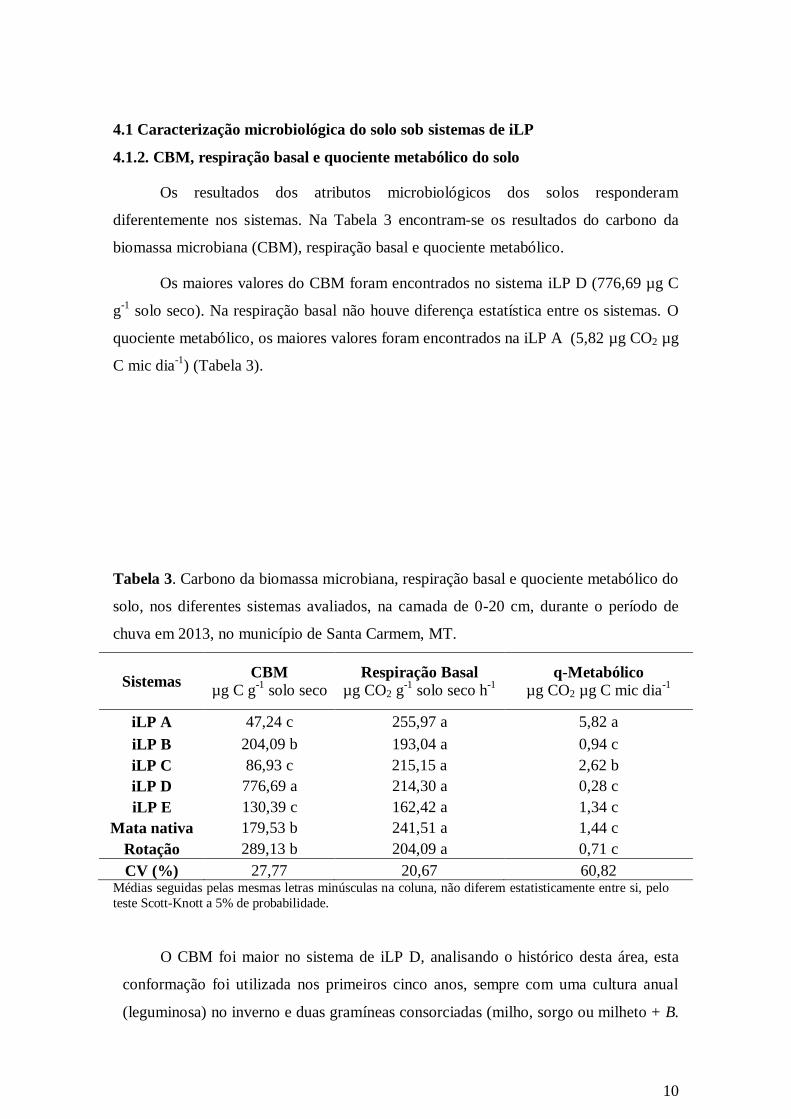
Os resultados dos atributos microbiológicos dos solos responderam
diferentemente nos sistemas. Na Tabela 3 encontram-se os resultados do carbono da
biomassa microbiana (CBM), respiração basal e quociente metabólico.
Os maiores valores do CBM foram encontrados no sistema iLP D (776,69 µg C
g-1
solo seco). Na respiração basal não houve diferença estatística entre os sistemas. O
quociente metabólico, os maiores valores foram encontrados na iLP A (5,82 µg CO2 µg
C mic dia-1
) (Tabela 3).
Tabela 3. Carbono da biomassa microbiana, respiração basal e quociente metabólico do
solo, nos diferentes sistemas avaliados, na camada de 0-20 cm, durante o período de
chuva em 2013, no município de Santa Carmem, MT.
Sistemas CBM
µg C g-1
solo seco
Respiração Basal
µg CO2 g-1
solo seco h-1
q-Metabólico
µg CO2 µg C mic dia-1
iLP A 47,24 c 255,97 a 5,82 a
iLP B 204,09 b 193,04 a 0,94 c
iLP C 86,93 c 215,15 a 2,62 b
iLP D 776,69 a 214,30 a 0,28 c
iLP E 130,39 c 162,42 a 1,34 c
Mata nativa 179,53 b 241,51 a 1,44 c
Rotação 289,13 b 204,09 a 0,71 c
CV (%) 27,77 20,67 60,82 Médias seguidas pelas mesmas letras minúsculas na coluna, não diferem estatisticamente entre si, pelo
teste Scott-Knott a 5% de probabilidade.
O CBM foi maior no sistema de iLP D, analisando o histórico desta área, esta
conformação foi utilizada nos primeiros cinco anos, sempre com uma cultura anual
(leguminosa) no inverno e duas gramíneas consorciadas (milho, sorgo ou milheto + B.

11
ruziziensis) no verão, mudando esse cenário apenas no ano agrícola de 2010/2011 para
uma gramínea (arroz) no inverno e duas gramíneas no verão (piatã + estilosantes).
Assim ao longo deste histórico a iLP D sempre esteve com três culturas diferentes ao
longo de cada ano agrícola, essa diversificação de culturas em uma mesma área
favoreceu o desenvolvimento da biomassa microbiana do solo.
Chavéz et al., (2011) em seu estudo sobre diversidade metabólica e atividade
microbiana de solos sob iLP, confirma o exposto anterior, e explica que, essa
diversidade de espécies vegetais, proporcionam maiores quantidades de diferentes
substratos (exsudatos liberados pelas plantas) que são utilizados pela microbiota do solo
como fonte de carbono.
Por outro lado, Avarenga e Noce (2005) mencionam que, o acúmulo de palhada
e raízes, proporcionado pela pastagem, tende a proporcionar aumento da matéria
orgânica do solo, proporcionando mais fonte de carbono para os micro-organismos.
Para a respiração basal do solo, não diferiu estatisticamente entre os sistemas de
uso do solo. Para Insam e Domsch, (1988), a biomassa mais eficiente, seria aquela que
perde menos carbono na respiração e incorpora mais à suas células, assim, neste estudo
o sistema de iLP D se destaca, por ter incorporado maior quantidade de CBM e ter
perdido menos carbono pela respiração basal do solo.
Esse olhar crítico sobre a BMS mais eficiente, também foi relatado por Caetano
et al., (2013) e Pezarico et al., (2013), que estudaram indicadores de qualidade do solo,
os mesmos enfatizam a importância da utilização da respiração basal para
complementar os resultados da biomassa microbiana, para posterior inferir sobre o
estado da atividade dessa comunidade.
Valores elevados de qCO2, para alguns autores (Carneiro et al., 2008; Silva et
al., 2010; Gomide et al., 2011, Melloni et al., 2013), podem indicar condições de
estresse, uma vez que, os micro-organismos tendem a consumir mais substrato,
ocorrendo um maior gasto de energia para manutenção da comunidade microbiana.
Neste estudo, para o qCO2 os maiores valores foram encontrados na iLP A. Este
resultado, pode estar relacionado a uma condição de estresse, uma vez que, esta área foi
mantida por 2 anos consecutivos sob cultivo de pastagem (marandu), neste período o
aumentando do pisoteio proporcionado pelos bovinos, pode ter aumentado a
compactação do solo. Apesar de não se ter sido feita esta análise, vários autores (Moura
Filho e Buol, 1972; Camargo e Alleoni, 1997; Siqueira Junior, 2005; Araújo et al.,

12
2007), relatam que a compactação modifica os atributos biológicos, limitando as trocas
gasosas, a infiltração e redistribuição de água, e bem como outros atributos, como os
físicos do solo (aeração, temperatura e resistência) e a disponibilidade de nutrientes.
A qualidade do substrato disponível para decomposição é outro fator a ser
considerado, de acordo com sua composição, podendo ser mais fácil ou não a sua
degradação, como exemplo compostos muito lignificados, são mais lentos a sua
degradação, desta forma a microbiota têm um elevado gasto energético para degradar
esse produto, até à sua incorporação.
Em estudos realizados por Pariz e colaboradores (2008), concluíram que as
espécies forrageiras estudadas, sendo uma delas a Brachiária brizanta cv. marandu,
apresentaram relação C/N alta, sendo que o marandu chegou a ter uma relação C/N de
40, com isso, sua decomposição foi mais lenta que as demais forrageiras estudadas.
Assim, além do condicionante de estresse, a qualidade do substrato, no caso a forrageira
marandu, pode ter colaborado para os elevados valores de qCO2 na iLP A.
Os resultados de Santos (2005) confirmam que o qCO2 pode estar relacionado
com a atividade decompositora da biomassa microbiana, altos valores incluem maior
gasto energético e consequentemente menor eficiência para incorporar o carbono na
célula microbiana.
O sistema integrado de produção na conformação D proporcionou estabilidade
ao metabolismo microbiano, tal fato, pode ter sido favorecido pela diversificação de
culturas em uma mesma área, que como já falado anteriormente, sempre esteve com três
culturas diferentes ao longo de cada ano agrícola, esse fator ao longo dos anos
proporcionou essa estabilidade ao sistema solo.
4.1.3. Atividade enzimática do solo
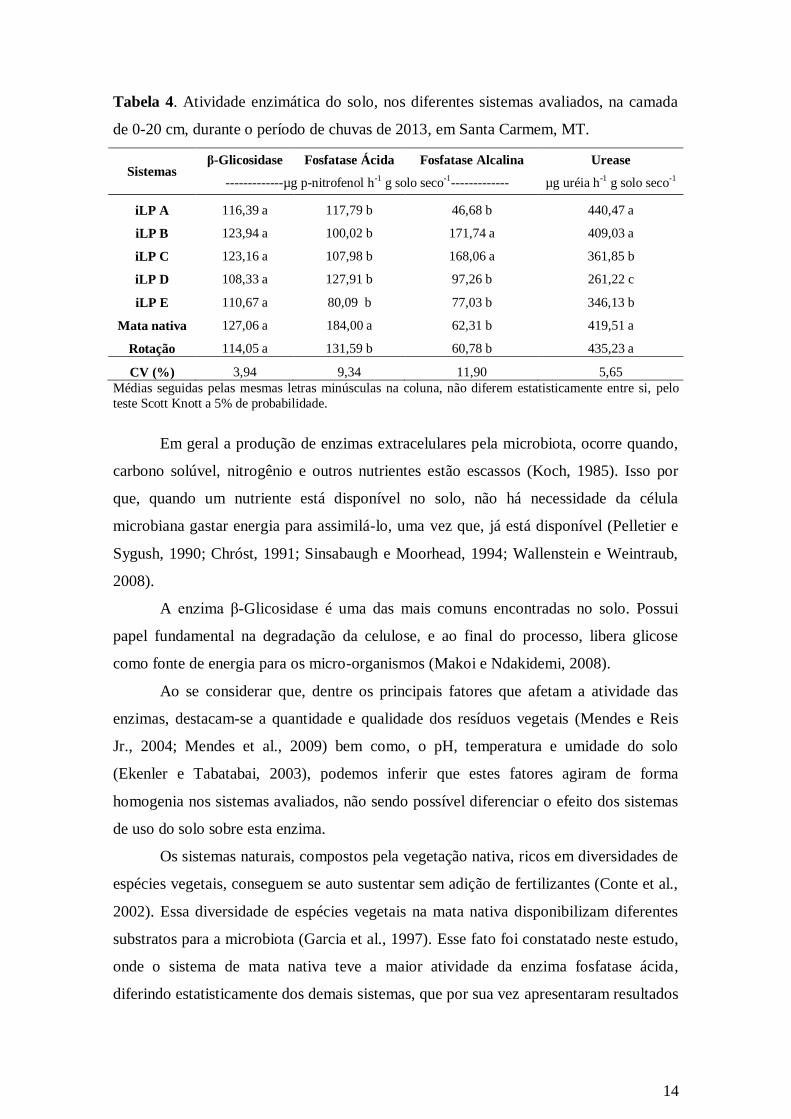
A atividade das enzimas β-glicosidase, fosfatase ácida e alcalina, e urease do
solo tiveram resultados variáveis entre os sistemas. Em todos os sistemas avaliados, em
geral a mata teve a maior atividade destas enzimas (Tabela 4).
A enzima β-Glicosidase, não houve diferença estatística entre os sistemas. Para a
enzima fosfatase ácida, a mata teve a maior atividade (184,00 µg p-nitrofenol h-1
g solo
seco-1
), diferindo estatisticamente dos demais sistemas. Na fosfatase alcalina, observou-
se que, nos sistemas iLP B e iLP C, tiveram maiores valores desta enzima (171,74 e

13
168,06 µg p-nitrofenol h-1
g solo seco-1
, respectivamente) não diferindo estatisticamente
entre si (Tabela 4).
Na urease do solo, os sistemas de iLP A, iLP B, mata e rotação, apresentaram os
maiores valores desta enzima (440,47 ; 409,03; 419,51; 435,23 µg uréia h-1
g solo seco-
1, respectivamente ), não diferindo estatisticamente entre eles (Tabela 4).
14
Tabela 4. Atividade enzimática do solo, nos diferentes sistemas avaliados, na camada
de 0-20 cm, durante o período de chuvas de 2013, em Santa Carmem, MT.
Sistemas β-Glicosidase Fosfatase Ácida Fosfatase Alcalina Urease
-------------µg p-nitrofenol h-1 g solo seco-1------------- µg uréia h-1 g solo seco-1
iLP A 116,39 a 117,79 b 46,68 b 440,47 a
iLP B 123,94 a 100,02 b 171,74 a 409,03 a
iLP C 123,16 a 107,98 b 168,06 a 361,85 b
iLP D 108,33 a 127,91 b 97,26 b 261,22 c
iLP E 110,67 a 80,09 b 77,03 b 346,13 b
Mata nativa 127,06 a 184,00 a 62,31 b 419,51 a
Rotação 114,05 a 131,59 b 60,78 b 435,23 a
CV (%) 3,94 9,34 11,90 5,65
Médias seguidas pelas mesmas letras minúsculas na coluna, não diferem estatisticamente entre si, pelo
teste Scott Knott a 5% de probabilidade.
Em geral a produção de enzimas extracelulares pela microbiota, ocorre quando,
carbono solúvel, nitrogênio e outros nutrientes estão escassos (Koch, 1985). Isso por
que, quando um nutriente está disponível no solo, não há necessidade da célula
microbiana gastar energia para assimilá-lo, uma vez que, já está disponível (Pelletier e
Sygush, 1990; Chróst, 1991; Sinsabaugh e Moorhead, 1994; Wallenstein e Weintraub,
2008).
A enzima β-Glicosidase é uma das mais comuns encontradas no solo. Possui
papel fundamental na degradação da celulose, e ao final do processo, libera glicose
como fonte de energia para os micro-organismos (Makoi e Ndakidemi, 2008).
Ao se considerar que, dentre os principais fatores que afetam a atividade das
enzimas, destacam-se a quantidade e qualidade dos resíduos vegetais (Mendes e Reis
Jr., 2004; Mendes et al., 2009) bem como, o pH, temperatura e umidade do solo
(Ekenler e Tabatabai, 2003), podemos inferir que estes fatores agiram de forma
homogenia nos sistemas avaliados, não sendo possível diferenciar o efeito dos sistemas
de uso do solo sobre esta enzima.
Os sistemas naturais, compostos pela vegetação nativa, ricos em diversidades de
espécies vegetais, conseguem se auto sustentar sem adição de fertilizantes (Conte et al.,
2002). Essa diversidade de espécies vegetais na mata nativa disponibilizam diferentes
substratos para a microbiota (Garcia et al., 1997). Esse fato foi constatado neste estudo,
onde o sistema de mata nativa teve a maior atividade da enzima fosfatase ácida,
diferindo estatisticamente dos demais sistemas, que por sua vez apresentaram resultados

15
homogêneos.
Estes resultados encontram-se de acordo com os vários trabalhos apresentados
em literatura, onde a maior atividade da fosfatase ácida foi apresentada na vegetação
nativa sob diferentes solos (Carneiro et al., 1999; Mendes et al., 2003; Matsuoka, 2001;
Caravaca et al., 2002; Schmitz, 2003; Matsuoka, 2006).
Segundo Gomide et al., (2011), a indução da produção desta enzima pelos
micro-organismos é induzida pela baixa disponibilidade de P no solo. Assim como já
relatado em outros autores que trabalharam com essa e outras enzimas, que não há
necessidade da célula microbiana gastar energia para assimilá-lo, uma vez que, já está
disponível (Pelletier e Sygush, 1990; Chróst, 1991; Sinsabaugh e Moorhead, 1994;
Wallenstein e Weintraub, 2008).
Deve-se destacar também, a contribuição dos altos teores de matéria orgânica,
como contribuinte dos aumentos da atividade da fosfatase ácida na mata. A matéria
orgânica fornece complexos húmicos que mantém a atividade desta enzima no solo
(Dick et al., 1988).
A fosfatase ácida possui extrema importância na mineralização do P orgânico do
solo, especialmente quando se trata de solos do Cerrado, em que a disponibilidade deste
nutriente é restrita (Tabatabai, 1994).
Ainda segundo este autor, a fosfatase alcalina difere da ácida, pelo pH ótimo de
atividade, onde a alcalina age em pH 9 a 10 a ácida em pH 4 a 6,5. Ambas são
secretadas por micro-organismos do solo, e possuem papel importante no ciclo do P.
A fosfatase alcalina teve respostas diferentes quando comparadas com a
fosfatase ácida, onde a iLP B e iLP C tiveram valores mais altos que a mata nativa. O
pH e a qualidade da matéria orgânica nessas iLPs pode ter favorecido este resultado.
Para Gonnety et al., (2012) estudos da atividade das enzimas extracelulares, podem
fornecer informações sobre a disponibilidade temporal e espacial de compostos
orgânicos específicos, como a disponibilidade de C, N e P.
A atividade da urease respondeu de forma homogênea nos sistemas avaliados,
com exceção da iLP C, iLP D e iLP E que tiveram menores valores.
O sistema de uso do solo possui grande influência sobre a atividade da enzima
urease, Marchiori-Júnior (1998) identificou uma redução na atividade da urease,
aproximadamente 33% em solos com pastagens há 10 anos e 72% com algodão, quando
comparados com solos de mata nativa, no Estado de São Paulo.

16
Em estudos com comparações entre sistema de plantio direto (PD) e
convencional (CN), a urease esteve superior no sistema plantio direto. Apresentando
similaridade com os solos sob campo natural em termos da atividade de uma das
enzimas ligadas ao ciclo do N. O resultado foi atribuído aos maiores teores de MO em
PD, o que estimularia a atividade microbiana do solo (Van den Bosshe et al., 2008;
Lisboa et al., 2012).
Sinsabaugh (1994), concluiu que os ensaios enzimáticos oferecem vantagens
quanto à sensibilidade, especificidade e facilidade. Isso por que respondem rapidamente
a perturbações no solo, o que é interessante para o monitoramento da dinâmica espacial
e temporal da atividade microbiana e investigações na ciclagem de nutrientes mediados
por esta.
O plantio direto, sem revolvimento do solo foi relatado como capaz de melhorar
a atividade enzimática do solo (Van den Bossche et al., 2008; Lisboa et al., 2012), e
neste trabalho o plantio direto aliado ao sistema de integração de culturas foi capaz de
se aproximar à mata nativa quanto a produção de enzimas e bem como a produção de
biomassa microbiana, resultados esses positivos para a sustentabilidade do solo.
4.1.4. População de micro-organismos do solo
A população cultivável de bactérias, micro-organismos celulolíticos,
actinomicetos e fungos do solo foram variáveis entre os sistemas e épocas (Tabela 5). A
população de bactérias foi maior na iLP A, iLP B e na mata, estes não diferiram
estatisticamente entre si.
O sistema que favoreceu o desenvolvimento de actinomicetos foi a iLP A, tal
observação também foi constatada neste mesmo sistema para os micro-organismos
celulolíticos. Já a população de fungos do solo foi maior na iLP B (Tabela 5).
Tabela 5. População de micro-organismos cultiváveis do solo, nos diferentes sistemas
avaliados, na camada de 0-20 cm, durante o período de chuvas de 2013, em Santa
Carmem, MT.
Sistemas Bactérias Actinomicetos Mic. Celulolíticos Fungos
----------------Nº UFC x 106 g
-1 solo---------------- Nº UFC x 10
4 g
-1 solo
iLP A 1,42 a 1,33 a 1,38 a 3,69 b
iLP B 1,19 a 1,08 c 1,13 b 4,61 a
iLP C 0,79 b 0,83 e 0,99 c 3,07 c

17
iLP D 0,94 b 0,99 d 1,17 b 2,58 c
iLP E 0,77 b 1,19 b 0,90 c 2,43 c
Mata nativa 1,33 a 1,13 c 0,96 c 2,77 c
Rotação 0,91 b 0,65 f 0,30 d 1,88 d
CV (%) 19,26 8,55 12,78 23,43 Médias seguidas pelas mesmas letras minúsculas na coluna, não diferem estatisticamente entre si,
pelo teste Scott-Knott a 5% de probabilidade.

18
Os sistemas que favoreceram o crescimento bacteriano do solo, foram a iLP A,
iLP B e a mata, provavelmente estimulado pelas culturas, que influenciaram
diretamente sobre as comunidades bacterianas da rizosfera.
Porém, pelo método de diluição seriada, algumas informações são limitadas,
como por exemplo, a diversidade de espécies, que forneceria informações sobre as
comunidades funcionais. Assim, não se sabe se esse número é em quantidade ou
qualidade.
Mas sabe-se que quanto maior o aporte de matéria orgânica, diversificada em
compostos, maior será a diversidade de espécies microbianas (Garcia et al., 1997), o que
seria o ideal ao se pensar em nicho ecológico, onde uma comunidade interage com
outras e atua em partes de determinado processo, como a ciclagem de nutrientes, que
depende de um conjunto de comunidades microbianas funcionais, para exercer tal
função. Neste contexto os sistemas de integração se destacam pela diversificação de
culturas e o não revolvimento do solo, que favorece a diversidade microbiana.
Em geral, a influência do manejo e das culturas sobre as populações de
actinomicetos do solo é menor do que sobre as demais bactérias e os fungos. Devido o
crescimento lento e a baixa capacidade competitiva desses micro-organismos, por não
conseguirem se predominar em substratos orgânicos, onde outros micro-organismos
apresentam capacidade de colonização mais rápida (Siqueira e Franco, 1988; Pereira, et
al., 1999).
Apesar dos actinomicetos apresentarem baixa capacidade competitiva, esses
micro-organismos formam esporos e conídios, essas estruturas, lhes confere resistência
e permite a sua sobrevivência em condições adversas, isso demonstra a importância de
estudos ecológicos das populações de actinomicetos (Williams, et al., 1972; Vobis,
1997).
Na iLP A foram obtidos os maiores valores de micro-organismos celulolíticos.
Tal resultado pode ser atribuído aos resultados de bactérias, onde tiveram resultados
parecidos nos mesmos sistemas. Isso por que, o meio de cultura utilizado para
enumeração de micro-organismos celulolíticos, favorece o crescimento de bactérias,
actinomicetos e fungos, sem distinção. Apenas classifica-os quanto à degradação da
celulose.
Neste sentido, os resultados das populações de bactérias corroboram com os dos
micro-organismos celulolíticos. Podendo inferir, que o tipo de comunidade

19
predominante de micro-organismos celulolíticos está mais associado a bactérias do que
a fungos. Apesar dos fungos também serem notados como positivos a degradação da
celulose, mas não foram tão predominantes neste estudo, quanto às bactérias.
A população de fungos foi favorecida pela iLP B, onde teve o maior número
dessa população. A comunidade de fungos do solo é menor quando comparado com as
de bactérias (Siqueira e Franco, 1988), tais resultados são similares com os obtidos
neste trabalho.
Porém para Perez et al., (2004), a condição da mata, onde há uma grande
deposição de resíduos orgânicos, maior quantidade de raízes e de água retida no solo,
estimulam a comunidade microbiana do solo. Essa comunidade tende a ser maior em
solos em condição de mata do que em solos agrícolas que sofrem perturbações.
Tal resultado não foi observado neste estudo, em geral os solos sob integração
Lavoura-Pecuária, favoreceram o crescimento microbiano. Isso se deve ao aporte de
matéria orgânica, o não revolvimento do solo e a diversificação de culturas em uma
mesma área. O que seria o ideal ao se pensar em sustentabilidade das práticas agrícolas.
Essa preocupação com a sustentabilidade se deve ao fato que, a perda da
biodiversidade gera não somente impactos ecológicos, mas também sociais e
econômicos. Uma vez que, o desequilíbrio ecológico tende a se compensar de alguma
forma, e geralmente mais drástica e dispendiosa.
Os micro-organismos do solo são responsáveis por processos que têm influencia
direta ou indiretamente na produtividade e sustentabilidade dos ecossistemas terrestres.
O método de contagem desses micro-organismos, apesar de ser visto com ressalvas,
ajuda a entender os processos que nele ocorrem, uma vez que, aliando com outros
atributos, ele serve como indicador do impacto de diferentes atividades antrópicas
(Previati et al., 2012).
4.2 Caracterização microbiológica do solo sob sistemas de iLPF, rotação
soja/pastagem e mata nativa
A rotação soja/pastagem apresentou a maior atividade da enzima β-glicosidase,
significativamente maior e diferente dos demais sistemas. Entretanto, o sistema de mata
nativa foi menor e diferente dos demais. Para a atividade da fosfatase ácida, observa-se

20
a mata com maior resultado nesta enzima, enquanto o sistema de iLPF linha tripla de
eucalipto nas distancia 0 e 6 m da planta, foram os menores resultados encontrados.
Resultados da literatura demonstram que a atividade da β-glicosidase é maior em
sistema sem revolvimento do solo, demonstrando o aumento de carbono no solo, devido
à mantença na atividade rizosférica permanente nessa área (Lisboa et al., 2012), porém
esse resultado não vem de encontro ao apresentado na Tabela 5, uma vez que o solo de
mata nativa apresentou os menores resultados.
Porém o relato de Lisboa et al. (2012) vem de encontro com os resultados de
maior atividade na área sob sistema de rotação soja/pastagem e na distância de 3 m da
planta nos três sistemas de iLPF avaliados, uma vez que nessas áreas entra-se a maior
abrangência da atividade rizosférica.
Tabela 6. Atividade das enzimas β-glicosidase e Fosfatase ácida em solo sob
integração Lavoura-Pecuária-Floresta na região Norte de Mato Grosso,
durante o período de chuvas da região, maio de 2013, na profundidade 0-20
cm.
Tratamentos* β-glicosidase Fosfatase ácida
----------- µg p-nitrofenol h-1 g solo seco -1 -----------
LS 0 m 326,54 g 476,05 b
LS 3 m 426,70 b 447,36 ef
LS 6 m 387, 22 e 442,75 ef
LS 10 m 482,76 d 468,68 c
LD 0 m 311,54 fg 461,84 d
LD 3 m 411,69 bcd 410,51 h
LD 6 m 445,52 bc 446,44 de
LD 10 m 207,18 i 426,17 fg
LT 0 m 442,62 bcd 294,70 j
LT 3 m 409,33 cd 388,14 gh
LT 6 m 360,10 ef 278,90 j LT 10 m 371,55 efg 353,66 i
Pasto 706,36 a 243,11 de
Mata 283,90 h 534,22 a *LS – Linha Simples de eucalipto; LD – Linha Dupla de eucalipto; LT – Linha Tripla de eucalipto;
* 0 m, 3 m, 6 m, 10 m distância das coletas em relação ao pé da planta de eucalipto
Médias seguidas pela mesma letra na coluna não diferem entre si pelo teste de Kruskal Wallis, a 1 % de
probabilidade.
Em um estudo realizado por Chaer e Tótola (2007), avaliando atividade
enzimática em solo de eucalipto, obtiveram para a β-glicosidase, valores menores do
que os encontrados no presente trabalho, sendo nas áreas com vegetação natural as
maiores concentrações, o que não corrobora com os resultados da Figura 1, para a
maioria dos sistemas.
Na atividade enzimática da fosfatase ácida observou-se variação entre as

21
distâncias avaliadas, entretanto não apresenta uma tendência de atividades, como a β-
glicosidase apresentou.
Na literatura foram encontrados valores semelhantes, para a maioria dos
tratamentos avaliados; para uma mesma profundidade de avaliação, Matsuoka et al.
(2003) quantificaram a atividade da fosfatase ácida em solos sob Cerradão e
encontraram valores de 383,1 μg p-nitrofenol g de solo h-1
.
A área do presente estudo é uma transição de Cerrado para Floresta Amazônica e
a atividade média encontrada foi superior ao encontrado por Matsuoka et al. (2003), e
menores aos apresentados por Mendes e Vivaldi (2001), os quais quantificaram a
atividade da fosfatase em 2.421 μg p-nitrofenol g-1
de solo h-1
em solo de Cerradão, na
mesma profundidade.
Os autores Chuderova e Zubeta (1969), Dick (1994) e Matsuoka et al. (2003)
destacam que a atividade das fosfatases está diretamente relacionada aos níveis de P do
solo, além do pH, o qual caracteriza a fosfatase em ácida e alcalina.
Estudos demonstram que a atividade das enzimas fosfatase ácida e β-glicosidase
são afetadas pelas práticas de manejo do solo, sendo verificadas reduções acentuadas
em sistema de manejo convencional, em detrimento dos sistemas considerados
conservacionistas, como o plantio direto (Aon et al., 2001). Esse resultado não foi
evidenciado, entretanto destaca-se o resultado das áreas com maior presença rizosférica,
as quais apresentaram maior atividade destas enzimas.
As enzimas apresentam grande importância para os solos, pois grande parte das
transformações bioquímicas que ocorrem neste ambiente é dependente ou relacionada à
presença de enzimas e a avaliação de suas atividades pode ser útil para indicar se um
solo está desempenhando adequadamente os processos que estão intimamente ligados a
sua qualidade (Reis Junior e Mendes, 2009).
O número de bactérias totais presentes nos solos foi determinado por unidades
formadoras de colônias (UFC), utilizando-se os meios de cultura ágar nutriente, extrato
de solo e celulolítico obteve-se o maior crescimento médio para todos os tratamentos
avaliados (Tabela 6).

22
Tabela 7. Número de Unidades Formadoras de Colônias em meios Batata dextrose ágar
(BDA), Ágar nutriente (AN), Extrato de solo (ES) e Meio Celulolítico (CEL),
em solo sob integração Lavoura-Pecuária-Floresta na região Norte de Mato
Grosso, durante o período de chuvas da região, maio de 2013, na profundidade
0-20 cm.
Tratamentos* UFCs BDA UFCs AN UFCs ES UFCs CEL
x 101 UFCs g solo-1 ------------------- x 103 UFCs g solo-1 -------------------
LS 0 m 7 e 373 d 353 j 260 i
LS 3 m 4 f 318 e 872 d 577 f
LS 6 m 9 b 397 d 1141 b 956 c
LS 10 m 10 a 423 d 686 g 734 e
LD 0 m 6 e 243 g 425 i 387 h
LD 3 m 7 e 834 a 1368 a 1372 a
LD 6 m 9 d 814 c 1060 c 1042 b
LD 10 m 7 c 431 c 824 e 1096 b
LT 0 m 6 e 185 i 210 l 542 g LT 3 m 7 c 272 f 681 g 851 d
LT 6 m 4 g 327 e 528 h 608 f
LT 10 m 4 f 477 b 724 f 711 e
Pasto 2 h 158 j 235 l 178 j
Mata 7 d 228 h 316 j 162 j *LS – Linha Simples de eucalipto; LD – Linha Dupla de eucalipto; LT – Linha Tripla de eucalipto;
* 0 m, 3 m, 6 m, 10 m distância das coletas em relação ao pé da planta de eucalipto
Médias seguidas pela mesma letra não diferem entre si pelo teste de Kruskal Wallis, a 1 % de
probabilidade.
As áreas de iLPF apresentaram a maior quantidade de micro-organismo nos
quatro meios avaliados. Para a contagem em meio BDA, no qual obtém-se crescimento
fúngico, a iLPF linha simples de eucalipto com distância de 10 m da planta apresentou o
maior número de UFCs, diferente dos demais.
Para os meios AN, onde há crescimento bacteriano, Extrato de Solo, contagem
de UFC de actinomicetos, e Celulolítico, contagem de micro-organismos que
apresentam halo de degradabilidade do meio, composto por celulose, houve um
sincronismo de crescimento, sendo o solo destaque a área sob iLPF linha dupla de
eucalipto com distância de 3 m da planta.
Segundo Gonçalves et al. (1999) e Vargas e Scholles (2000) áreas com presença
de diferentes espécies vegetais exercem maior influência sobre os micro-organismos e
na quantidade de carbono, devido ao acúmulo de matéria orgânica, o que favorece o
crescimento e atividade microbiana na camada superficial do solo. Esta afirmação pode
justificar o maior número de UFC nos sistemas de iLPF, entretanto a mesma relação
poderia ser considerada para mata nativa, a qual possui ainda mais diversidade, porém
não apresentou maior número de UFC em nenhum dos casos.
Em ecossistemas naturais, a cobertura vegetal permanente proporciona proteção
contínua do solo, além de adicionar grandes quantidades de nutrientes principalmente

23
por meio de resíduos. Seus efeitos sobre a comunidade microbiana podem interagir com
os efeitos provocados pelas flutuações hídricas e térmicas que ocorrem durante o ano,
influenciando em menor ou maior grau essas populações (Tsai, 1992).
A adoção de sistemas de iLPF buscam a mesma relação solo-planta apresentado
pelos ecossistemas naturais, pois a integração com espécies florestais é constante e
permanente na área o que beneficia a atividade microbiana do solo, e como pode ser
visualizado na Tabela 3, neste estágio de desenvolvimento do sistema de iLPF, eles
apresentam mais estabilidade no número de UFC que o sistema de mata nativa.
Melloni et al. (2001) afirmam que o manejo do solo para cultivo geralmente
provoca desequilíbrio pronunciado nas populações da comunidade microbiana. Além
disso, defendem que a ocorrência de UFCs em mata nativa tende a ser maior em relação
à de campo, ambas as afirmações não foram observadas no presente trabalho.
Os habitats apresentam grandes diferenças em suas características, de maneira
que um habitat favorável ao crescimento de uma população de organismo pode ser
extremamente nocivo a outro (Madigan, 2004).
5. CONCLUSÕES
Como conclusões preliminares em virtude do projeto estar em andamento, após
1 ano de avaliação dos sistemas de integração Lavoura-Pecuária, sobre as variáveis
microbiológicas: carbono da biomassa; respiração basal; quociente metabólico;
atividade das enzimas β-glicosidase, fosfatase ácida e alcalina, urease; a população de
micro-organismos celulolíticos, actinomicetos, bactérias e fungos do solo, respondem
sensivelmente as alterações em função do uso do solo.
O sistema integrado de produção, em especial a conformação D, onde houve a
maior diversificação de culturas ao logo de cada ano, como grãos e pastagem na mesma
área, este fato favoreceu a estabilidade na estrutura e função da microbiota do solo.
Para as áreas de iLPF, conclui-se que a as atividades de ambas as enzimas
avaliadas não foram beneficiadas pelo sistema, sendo o pasto destaque para o pasto e
fosfatase ácida para a mata. Para crescimento de micro-organismos, a linha simples de
eucalipto a 10 m do pé da planta apresentou maior crescimento de fungos, enquanto que
o sistema com linha dupla de eucalipto aos 3 m do pé da planta foi destaque para os
demais crescimentos, bactérias totais, celulolíticos e actinomicetos.

24
6. REFERÊNCIAS BIBLIOGRÁFICAS
AON, M.A.; SARENA, D.E.; BURGOS, J.L.; CORTASSA, CS. Microbiological,
chemical and physical properties of soils subjected to conventional or no-till
management: An assessment of their quality status. Soil Tillage Research, v.60, p.173-
1, n. 86, 2001.
BALDANI, J.I.; BALDANI, V.L.D.; XAVIER, D.F.; BODDEY, R.M.;
DÖBEREINER, J. Efeito da calagem no número de actinomicetos e na porcentagem de
bactérias resistentes à estreptomicina na rizosfera de milho, trigo e feijão. Revista de
Microbiologia, São Paulo, v.13, n.3, p.250-263, 1982.
CARNEIRO, M. A. C. ; SIQUEIRA, J. O.; MOREIRA, F. M. de S. Carbono,
nitrogênio, biomassa e atividade microbiana do solo de áreas em cronosseqüências de
reabilitação após a mineração de bauxita. Revista Brasileira de Ciência do Solo, v.
32, p. 621-632, 2008.
CHAER, G.M.; TÓTOLA, M.R. Impacto do manejo de resíduos orgânicos durante a
reforma de plantios de eucalipto sobre indicadores de qualidade do solo. Revista
Brasileira de Ciência do Solo, v.31, p.1381-1396, 2007.
CHUDEROVA, A.I.; ZUBETA, T. Phosphatase activity in dernopodzolic soils.
Pochvovendenie, Moscow, v.11, p.47-53, 1969.
COELHO, R.R.R. Ocorrência de actinomicetos em solos de cerrado, capazes de utilizar
compostos aromáticos. Rio de Janeiro: UFRJ, 1976. 111p. Dissertação (Mestrado).
DICK, R.P.; BREAKWELL, D.P.; TURCO, R.F. Soil enzyme activities and
biodiversity measurements as integrative microbiological indicators. In: DORAN, J.W.
e JONES, A.J., eds. Methods for assessing soil quality. Madison Soil Science, Society
of America, 1996. p.247-272.
EIVAZI, F. e TABATABAI, M.A. Glucosidases and galactosidases in soils. Soil
Biology Biochemistry. Elmsford, v. 20, p. 601-606, 1988.
GONÇALVES, A.S.; MONTEIRO, M.T.; BEZERRA, F.E.A.; GUERRA, J.G.M.; DE-
POLLI, H. Estudo de Variáveis de Solo, Vegetação e Condicionamento de
Amostras de Solo Sobre a Biomassa Microbiana do Solo no Estado do Rio de
Janeiro. Seropédica, RJ: EMBRAPA Agrobiologia, 1999. 18p. (Documento 90).
LISBOA, B. B.; VARGAS, L. K.; SILVEIRIA, A. O. da; MARTINS, A. F.;
SELBACH, P. A. Indicadores microbianos de qualidade do solo em diferentes sistemas
de manejo. Revista Brasileira de Ciência do Solo, 36: 45-55, 2012.
MADIGAN, M.T. 2004. Microbiologia de Brock. Prentice Hall, São Paulo. 608p.
MATSUOKA, M. Atributos Biológicos de solos cultivados com videira na região da
Serra Gaúcha. 2006. 171f. Tese (Doutorado) – Programa de Pós-Graduação em Ciência

25
do Solo, Faculdade de Agronomia, Universidade Federal do Rio Grande do Sul, Porto
Alegre, 2006.
MATSUOKA, M.; MENDES, I.C.; LOUREIRO, M.F. Biomassa microbiana e
atividade enzimática em solos de Cerrado e sistemas agrícolas anuais e perenes na
região de Primavera do Leste/MT. Planaltina, DF: EMBRAPA Cerrados, 2003. 24p.
(Boletim de pesquisa e Desenvolvimento 34)
MELLONI, R., PEREIRA, E.G., TRANNIN, I.C.B., SANTOS, D.R., MOREIRA,
F.M.S. SIQUEIRA, J.O. Características biológicas de solos sob mata ciliar e campo
cerrado no sul de Minas Gerais. Ciência Agrotecnologica, Lavras, v.25, n.1, p.7-13,
2001.
MELO, V.S; ROULAND, C.; DESJARDINS, T.; SERRAZIN, M.; SILVA JÚNIOR,
M.L.; SANTOS, M.M.L.S.; FERREIRA, W.C.; SANTOS, E.R.; ARAUJO, R.C. de
M.; RUIVO, M.L.P. Atividade enzimática de latossolo Amarelo na Amazônia oriental,
sob diferentes tipos de cobertura vegetal. In: FERTBIO, 2006, Bonito, MS. Resumos
Expandidos. Bonito: SBCS, 2006. CDROM.
MENDES, I.C. e VIVALDI, L. Dinâmica da biomassa e atividade microbiana em
uma área sob mata de galeria na região do DF. In: RIBEIRO, J.F.; FONSECA,
C.E.L.; SOUSA-SILA, J.C. (Ed.) Cerrado: caracterização e recuperação de Matas de
Galeria. Planaltina, DF: Embrapa Cerrados, p.664-687, 2001.
RAYMUNDO JÚNIOR, O.R. Fungos filamentosos, actinomicetos e ácidos húmicos,
em diferentes profundidades de Latossolo Vermelho Amarelo textura média em área de
cerrado, no município de Corumbataí, SP. Rio Claro: UNESP, 1992. 149p. Dissertação
(Mestrado).
REIS JUNIOR, F.B. e MENDES, I.C. Atividade enzimática e a qualidade dos solos.
Planaltina, DF: EMBRAPA Cerrados, 2009. (Rede Técnica)
TABATABAI, M.A. Soil enzymes. In WEAVER, R.W.; ANGLE, J.S.; BOTTOMLEY,
P.S. Methods of soil analysis microbiological and biochemical properties. Madison Soil
Science, Society of America, p.775-833, 1994.
TSAI, S.M.; BARAIBAR, A.V.L.; ROMANI, V.L.M. Efeito de fatores do solo. In:
CARDOSO, E.J.B.N.; TSAI, S.M.; NEVES, M.C.P. (Eds.). Microbiologia do solo.
Campinas: Sociedade Brasileira de Ciência do Solo, p.59-72, 1992.
VARGAS, L.K.; SCHOLLES, D. Biomassa microbiana e produção de C-CO2 e N
mineral de um podzólico vermelho-escuro submetido a diferentes sistemas de manejo.
Revista Brasileira de Ciência do Solo, Viçosa, v.24, p.35-42, 2000.

26
7. DESCRIÇÃO DAS DIFICULDADES E MEDIDAS CORRETIVAS
Algumas dificuldades foram encontradas no momento da coleta dos solos, uma
vez que devemos primeiramente realizar os gastos e depois solicitar a restituição dos
mesmos. Mas tudo foi sanado da melhor forma. As secretárias da FEALQ/AGRISUS
são muito eficientes e cooperam para que tudo dê certo no final.
8. RELATÓRIO PRÁTICO
A partir de estudos e avaliações nos sistemas de integração Lavoura-Pecuária
(iLP), esses dados poderão contribuir gerando informações do comportamento da
microbiota do solo ao longo do tempo, e a capacidade de manutenção da qualidade
biológica do solo nestes sistemas mesmo após a sua sucessão.
A avaliação do sistema de iLPF quanto a distância do pé da planta perene irá
fornecer dados consistentes para inferirmos sobre o desenvolvimento dos micro-
organismos e as suas interações, sendo que é conhecida interação de diferentes grupos
com respectivas raízes. Além disso, nos dará subsidio para inferir sobre as condições de
ciclagem de nutrientes e de interferência quanto à relação do tipo de matéria orgânica
disponível no solo, bem como a dinâmica das comunidades microbianas frente ao
pastejo de animais na área.
A pesquisa proposta por este projeto demonstra a preocupação científica para
com os recursos naturais, no interesse de estudar e disponibilizar uma tecnologia
sustentável e inovadora com capacidade de recuperar áreas degradadas, conservar o solo
e ainda potencializar a produção no estado de Mato Grosso.

27
9. COMPENSAÇÕES OFERECIDAS À FUNDAÇÃO AGRISUS
A maior compensação oferecida à Fundação Agrisus vem em forma da produção
científica que é alcançada pelos integrantes do projeto. Dentre elas tem-se a participação
em congressos nacionais e internacionais, dias de campo que são realizados pelos
professores e pesquisadores da UFMT, campus de Cuiabá e Sinop, MT e pela Embrapa.
_______________________________________________________
Profa. Dra. Daniela Tiago da Silva Campos
Cuiabá, 16 de Dezembro de 2013.

28
10. ANEXOS
10.1 TRABALHOS PUBLICADOS
ANEXO 1. Aceite do trabalho proveniente dos dados da integração Laovoura-Pecuária-
Floresta, para apresentação no XXXIV Congresso Brasileiro do Solo,
Florianópolis, Julho-Agosto 2013.

29
ANEXO 2. Certificado de apresentação e publicação de trabalho proveniente de dados
da integração Lavoura-Pecuária, para apresentação no XXXIV Congresso
Brasileiro do Solo, Florianópolis, Julho-Agosto 2013.

30
ANEXO 3. Certificado de apresentação e publicação de trabalho proveniente de dados
da integração Lavoura-Pecuária, para apresentação no XXXIV Congresso
Brasileiro do Solo, Florianópolis, Julho-Agosto 2013.

31
10.2 FOTOS DA ÁREA DE ILPF
ANEXO 4. Sistema de integração Lavoura-Pecuária-Floresta, Maio-2013, Nova Canaã
do Norte, MT.
ANEXO 5. Apresentação dos resultados no Dia de Campo (esquerda) e equipe de
coleta do solo (direita), sistema de integração Lavoura-Pecuária-Floresta,
Maio-2013, Nova Canaã do Norte, MT.