universidade de lisboa -...
TRANSCRIPT

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
EXISTEM LIMITES PARA A DISPERSÃO E COLONIZAÇÃO
DE NOVOS HABITATS PELO LAGOSTIM AMERICANO
PROCAMBARUS CLARKII?
UM ESTUDO A MÉDIO PRAZO NA BACIA DO RIO SADO E
ELABORAÇÃO DE UM PLANO DE CONTENÇÃO
PEDRO HENRIQUE SABINO DE PEREIRA LEITÃO
MESTRADO EM ECOLOGIA E GESTÃO AMBIENTA
2009

2
UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
EXISTEM LIMITES PARA A DISPERSÃO E COLONIZAÇÃO
DE NOVOS HABITATS PELO LAGOSTIM AMERICANO
PROCAMBARUS CLARKII?
UM ESTUDO A MÉDIO PRAZO NA BACIA DO RIO SADO E
ELABORAÇÃO DE UM PLANO DE CONTENÇÃO
PEDRO HENRIQUE SABINO DE PEREIRA LEITÃO
LICENCIADO EM CIÊNCIAS BIOLÓGICAS
DISSERTAÇÃO PARA A OBTENÇÃO DO GRAU DE MESTRE EM ECOLOGIA E
GESTÃO AMBIENTAL
ORIENTADOR CIENTÍFICO: PROF. DOUTOR RUI MIGUEL REBELO
2009

i
AGRADECIMENTOS
Gostaria primeiramente de agradecer a toda minha família, meus irmãos Thiago e
Paulo pelo apoio e motivação e em especial meus pais, Manuel e Idalina, pelo excelente
exemplo de vida e amor que nunca faltou. Me proporcionaram todas as condições
necessárias para chegar até aqui. A tese é para vocês, muito obrigado.
Gostaria de agradecer ao meu orientador Prof. Doutor Rui Miguel Rebelo, pela
excelente condução na elaboração dessa tese e todos os ensinamentos prestados.
Agradeço a FACUL pela oportunidade de realização do mestrado e aos professores
do Departamento de Biologia Animal pelos conhecimentos muito bem ministrados que
tanto colaborou na minha formação.
Agraço muito a todas as pessoas que colaboraram comigo no trabalho de campo.
Agraço a todos meus amigos do mestrado, em especial às amigas Sílvia e Joana pelo
companheirismo em todas as horas.

ii
RESUMO
O lagostim vermelho da Louisiana Procambarus clarkii (Girard, 1852), foi
inicialmente introduzido no início da década de 1970 em Espanha, espalhando-se
rapidamente para Portugal através de colonização natural ou por introdução humana. A
expansão foi rápida: entretanto, estudo inicial conduzido em 2003 e 2004 encontrou
locais na bacia do rio Sado que ainda não haviam sido colonizados. Seleccionamos um
total de 104 pontos do estudo de 2003/04, e foram visitados no ano hidrológico de
2008/09 para constatar alterações na distribuição de lagostim e alguns factores
envolvidos na colonização e erradicação em novos habitats. Ao contrário do esperado,
verificamos que houve uma estagnação e ligeira contracção da distribuição de P. clarkii
comparado com 2003/04. Altitude, velocidade da corrente, vegetação aquática, tipos de
substrato e intervenção humana nas envolvente dos pontos estiveram relacionados com
a presença de lagostim em 2009, presença em ambos anos (2003/04 e 2008/09), perda
de lagostim 2009 e extinção local de lagostim em ambos os anos. Também encontramos
evidências relacionadas com a seca extrema de 2005 e subsequentes anos relativamente
secos na contracção da distribuição de P. clarkii em 2008/09 onde 21% dos pontos eram
considerados permanentes em 2002-04 agora são temporários. A expansão dessa
espécie invasora na área de estudo, aparentemente alcançou seus limites, com excepção
de alguns casos de pontos permanentes isolados que ainda são vulneráveis a introdução.
Palavras-chave: Procambarus clarkii; espécie invasora; distribuição; bacia do rio Sado;
plano de contenção; intervenção no solo

iii
ABSTRACT
The Louisiana red crayfish Procambarus clarkii (Girard, 1852) was introduced in
south-western Spain in the 1970’s, spreading to Portugal either by natural colonization
or by human introductions. Its expansion was fast; however, a study conducted from
2002 to 2004 found several locations in an area in the Sado river basin that had not yet
been colonized. With the propose to refresh his distribution and to find some limiting
factors involved in the colonization and eradication in new habitats, 104 of the 2002-04
study sites were revisited during the hydrological year of 2008/09, in order to check for
changes in the distribution of this crayfish. Contrary to our expectations, we found that
P. clarkii has now a more restricted distribution than in 2002-04. Elevation, current
velocity, water body vegetation, substrate types and human intervention around the sites
had strong relation with the presence of crayfish in 2009, presence of crayfish in both
years (2003/04 and 2008/09), local extinction in 2009 and crayfish absence in both
years. We found some relation between the extreme drought of 2005 and the subsequent
relatively dry years, with the contraction of the distribution whereas 21% of the sites
that were considered permanent in 2002-04 are now temporary. The expansion of this
invasive in our study area has apparently reached its limits, except in the few cases of
isolated permanent water bodies that are still vulnerable to its human introduction.
Keywords: Procambarus clarkii; invasive species; distribution; Sado river basin; control plan; impact land use

iv
ÍNDICE 1 – INTRODUÇÃO ......................................................................................................01
1.2. – Objectivos ...................................................................................................08
2 – MATERIAIS E MÉTODOS ..................................................................................09
2.1 Área de Estudo .................................................................................................09
2.2 Métodos de Amostragem……………………………………………………...12
3 – RESULTADOS …………………………………………………………………..17
3.1 Expansão de P. clarkii em 2003/04 e 2008/09………………………………...17
3.2.Distância a “fontes” de P. clarkii………………………………………………21
3.3 Modelação………………………………………………………………….…..23
3.3.1. Resultados para a distribuição de P. clarkii em 2008/09……………..…23
3.3.2. Comparação entre 2003 e 2009………………………………………….24
3.4 Abundância……………………………………………………………………..26
3.4.1. Estruturas de tamanho corporal dos indivíduos capturados,
agrupadas por tipo de habitat…………………………………………..….……30
3.4.2. Mudas……………………………………………………………………32
3.4.3. Sex ratio………………………………………………………………….34
4 – DISCUSSÃO E CONSIDERAÇÕES FINAIS…………………………..………35
4.1. Modelos…………………………………………………………….………….37
4.2. Abundância…………………………………………………………………….42
4.2.1Estruturas de tamanho corporal dos indivíduos capturados,
agrupadas por tipo de habitat………………………………………………...…43
4.2.2. Mudas……………………………………………………………………44
4.2.3. Sex ratio……………………………………………………………..…...44
5 – REFERÊNCIAS BIBLIOGRÁFICAS .................................................................46
ANEXOS:…………………………………………………………………………..….57
ANEXO I: …………………………………………………………………………….. 57
ANEXO II………………………………………………………………………………59

v
ANEXO III……………………………………………………………………..………60
ANEXO IV…………………………………………………………………….……….62
ANEXO V……………………………………………………………………………...63
ANEXO VI…………………………………………………………………..…………64
ANEXO VII…………………………………………………………………………….66
ANEXO VIII…………………………………………………………………...………68
ANEXO IX ……………………………………………………………………...……..75
ANEXO X ……………………………………………………….…………...………..77

1
INTRODUÇÃO
O reconhecimento dos problemas associados às invasões biológicas tem crescido
exponencialmente e segundo Lockwood et al. 20071 tal é devido a 3 razões: 1-) Os
efeitos negativos de algumas espécies não nativas são demasiado grandes para serem
ignorados; 2-) Tem aumentado o número de espécies transportadas dos seus habitats de
origem para novas localidades; 3-) Com tantas espécies invasoras, é já difícil a
realização de estudos ecológicos de campo sem encontrar nenhuma espécie invasora.
Os termos espécie não-nativa, não-indígena ou exótica são usados para descrever as
espécies que estão fora da sua extensão geográfica natural devido ao transporte
humano1, para áreas onde nunca antes foram registadas.2 Após a introdução, a espécie
exótica pode estabilizar-se e tornar-se auto-suficiente, mas sem capacidade de dispersar
e causar impactos, sendo denominada de espécie naturalizada3. As espécies invasoras,
no entanto, são aquelas exóticas naturalizadas já estabilizadas em ecossistemas naturais
ou semi-naturais4, com capacidade de dispersão1 e que ocupam o território de forma
excessiva,2 podendo constituir ameaças à saúde humana e serem causadoras de
impactos tanto económicos quanto nos ecossistemas nativos.3,5,1,4.
A introdução deliberada de espécies exóticas visa satisfazer algumas necessidades
humanas, tal como a produção de alimentos e o controle biológico de pragas4, satisfazer
a procura de produtos exóticos, ou ainda satisfazer a procura por animais de estimação
ou plantas de jardim3. Há também introduções acidentais que, nos dias de hoje, são
consequência da globalização e aumento do transporte entre países4, o que torna assim
os humanos nos maiores responsáveis pelo aumento massivo da taxa de invasão de
espécies em áreas novas.1
Muitas dessas introduções são benéficas para os humanos6 ou não causam alterações
ambientais.7,8 Apenas uma pequena, mas significante porção de espécies não nativas se
torna invasora9,10e é responsável pelo declínio e extinção de espécies
nativas.11,12,10,13,14,15,16
Actualmente, a problemática relacionada com a introdução de espécies exóticas é
um dos 5 maiores obstáculos para a conservação da natureza,7,4sendo considerada a
segunda maior causas para a redução da biodiversidade12,18em especial para as espécies
mediterrânicas.4Os custos que o mundo paga pela perda da biodiversidade, pelas

2
tentativas de erradicação das espécies invasoras e pela mitigação dos impactos nas
espécies nativas estão na ordem dos biliões de dólares por ano.19
As regiões mediterrânicas, que apresentam alta biodiversidade, são especialmente
susceptíveis à invasão e introdução de espécies exóticas e aos seus efeitos,20,4em
particular os ecossistemas aquáticos de água doce.21,17,22
Introduções e impactos nos sistemas aquáticos
Os sistemas aquáticos de água doce são altamente susceptíveis a colonização de
espécies invasoras como resultado da forte afinidade dos humanos com a água (para
comércio, transportes, recreação ou razões estéticas) e da enorme capacidade de
dispersão das espécies de água doce.23,24,25Os lagos e pontos isolados também estão
particularmente vulneráveis à introdução de novas espécies.17,27
Estudos recentes sugerem que a biodiversidade em águas doces está a declinar tão
rapidamente como nos ecossistemas terrestres mais afectados (Ricciardi & Rasmussen
1999 in27) e a homogeneização biótica28é um fenómeno em aceleração em muitos
sistemas de água doce.29A introdução deliberada ou acidental de espécies não-nativas é
mais negativa para habitats aquáticos que para habitats terrestres,17onde as principais
perturbações são a perda de habitat, poluição e sobre-exploração.4 parei aqui…
As perturbações dos ambientes aquáticos têm facilitado o estabelecimento de muitas
espécies não nativas aquáticas1,tendo como consequência a rápida alteração da
composição da flora e fauna aquáticas.30,31,17,32,33,22
Os crustáceos são um grupo cujos efeitos da introdução em sistemas de água doce
estão melhor documentados, sendo responsáveis pelo declínio e extinção de diversas
espécies nativas.23,34,35
A introdução de lagostins não nativos não é uma prática recente na Europa (27) e
ocorreu na maioria dos países europeus36, sempre motivada pelo seu valor comercial
gastronómico. São, como outros decápodes, omnívoros generalistas,37 alimentando-se
de invertebrados bentónicos, detritos, macrófitas e algas em ambientes lóticos e
lênticos.38São organismos com período de vida relativamente longo e subsistem em
altas densidades. Os seus impactos estendem-se ao longo de toda a cadeia trófica, sendo
geralmente considerados espécies-chave nos ecossistemas que ocupam.

3
Procambarus clarkii
Procambarus clarkii (Girard 1852), popularmente conhecido por Lagostim-
vermelho-da-Louisiana ou Lagostim-vermelho-americano é originário do continente
norte-americano. Juntamente com as outras espécies do género Procambarus (cerca de
107), ocorre naturalmente no Noroeste do México e Centro-Sul dos USA (Hobbs 1972
in39), estendendo-se pelo Oeste do Texas ao Este do Alabama e ao Norte do estado do
Tenessee e Illinois. (Hobbs 1972 in27)
Diferentemente da espécie de lagostim nativa de Portugal (Austropotamobius
pallipes, família Astacidae), que está associada a habitats de águas correntes, límpidas e
oxigenadas, em pequenos cursos de água de montanha frequentemente em zonas de
floresta,40,41 P. clarkii (família Cambaridae) evita corpos de água e áreas com forte
corrente. Ocupa uma variada gama de habitats, sendo tipicamente encontrado em sapais,
rios, cursos de água com pouca corrente, albufeiras e canais de irrigação.39
Em virtude da sua enorme capacidade de adaptação e seu elevado valor comercial,
foi introduzido com sucesso em diversos estados dos EUA e ainda um pouco por todo o
mundo. Actualmente P. clarkii é o lagostim mais cosmopolita, sendo encontrado em
habitats naturais em todos os continentes com excepção da Austrália e Antárctica.42
P. clarkii possui muitas características que lhe conferem sucesso como invasor12 e a
reputação de lagostim de água doce dominante em todo o mundo.43 É uma espécie r-
estrategista, com curto tempo entre gerações,44,45,46 alta fecundidade44,46e alta taxa de
crescimento.47,48 É ainda capaz de se adaptar a variadas condições ambientais,49,50 assim
como a diferentes recursos alimentares44,50,51,52,53e possui grande capacidade de
dispersão e colonização.54,55,27 É uma espécie com elevada plasticidade
ecológica45,56sendo a maior em toda a ordem Decapoda.43São capazes de tolerar baixos
níveis de oxigénio e altas temperaturas,57 água com alguma salinidade e secas
sazonais.43
Quando introduzido, P. clarkii pode tornar-se rapidamente a espécie principal do
ecossistema,58,59causando mudanças dramáticas nas comunidades nativas de animais e
plantas. O lagostim é considerado um consumidor omnívoro e oportunista,60adequando
a sua alimentação consoante o habitat e podendo em algumas circunstâncias adoptar o
canibalismo.61 Segundo Gherardi 2006,27 P. clarkii possui uma plasticidade alimentar
que lhe permitiu encontrar desde material vegetal, animal, sedimentos e até detritos no
seu intestino. Entre os animais que fazem parte da sua dieta incluem-se moluscos,

4
insectos, vermes, ovos e larvas de anfíbios, e mesmo alguns peixes, sendo que os
recursos animais predominam na dieta dos juvenis.
Cruz e Rebelo identificaram a vulnerabilidade de muitas espécies de anfíbios de
Portugal à predação por esta espécie, devido à elevada taxa de predação de ovos e
larvas, podendo levar à diminuição populacional e até à extinção local de espécies
mostrando que o lagostim pode reduzir significantemente a riqueza específica de
anfíbios em vários habitats.14,15,16
Esta espécie também afecta negativamente a cobertura de macrófitas em alguns
sistemas aquáticos;35 devido a destruição das macrófitas não consumidas62,63,64,65 e a
possível alteração da composição da comunidade de invertebrados associadas à elas.66
P. clarkii pode também estar associado à eutrofização de diversos habitats por alterar a
turbidez da água.66
Para além da competição directa por habitats e por recursos, P. clarkii, juntamente
com outro lagostim americano introduzido - Pacifasciatus leniusculus - têm contribuído
para o declínio do lagostim nativo europeu por serem vectores da “peste do lagostim”.
Esta é causada por um fungo, Aphanomyces astaci, que provoca a afanomicose,
responsável pela mortalidade dos lagostins da família Astacidae na ordem de 100% dos
infectados.
O lagostim-vermelho-da-Louisiana também está directamente ligado a problemas
económicos, por diminuir o recrutamento de espécies de peixes com interesse
comercial37 e por reduzir a colheita de arroz (com perdas económicas de 6,3% em
Portugal67,68). Em elevadas densidades os lagostins podem diminuir em até 80% a
eficácia na germinação do arroz por alimentarem-se das sementes e das plantas pós-
germinação. Nos arrozais e nos reservatórios de água, o comportamento escavador do P.
clarkii pode provocar, também, a perda de água.69
História da introdução de Procambarus clarkii na Europa
Em Junho de 1973 um lote contendo 100Kg de P. clarkii vivos foi importado de
New Orleans, Louisiana (EUA) para uma fazenda em Espanha, na província de
Badajoz, e um ano depois um segundo lote, desta vez com 400Kg foi libertado em
tanques no baixo Guadalquivir (Puebla del Rio, Sevilha) (Hasburgo-Lorena 1986 in27).
P. clarkii rapidamente se naturalizou. A ausência de filtros nos tanques onde foram
libertados, combinada com a sua capacidade de escapar, facilitaram a colonização de

5
valas e canais próximos. Na perspectiva de um bom negócio, a expansão do lagostim foi
acelerada pelos pescadores que distribuíram indivíduos por toda a zona. Este foi o
primeiro passo para a expansão para toda a Península Ibérica, incluindo Açores, e para a
sua recente translocação para muitos outros países da Europa.
Existe uma grande procura nos mercados europeus por lagostins, que foram
considerados um prato tradicional e saudável em muitos países. A primeira introdução
em Espanha foi ajudada e solicitada por instituições locais, por constituir uma hipótese
de melhoria económica e de aumento dos rendimentos em regiões carenciadas. Segundo
Hasburgo-Lorena 1986 in27, no momento da introdução todos os procedimentos legais
foram seguidos e respeitados, tendo inclusive sido verificada a ausência de lagostim
nativo Austropotamobius pallipes , nas regiões de introdução do P. clarkii. Pouco se
sabia de seu potencial para vir a tornar-se uma praga e havia confiança, cientificamente
suportada, que o P. clarkii não afectaria a espécie nativa europeia.
Estudos genéticos revelam que muitas das populações da Europa são o resultado de
diversas introduções de lagostins provenientes de diferentes áreas.70 Enquanto a
Espanha é uma importante fonte de P. clarkii, torna-se claro que o comércio de animais
vivos de outras áreas mais distantes tem sido responsável por algumas introduções com
sucesso no ambiente natural43,70,27 em Itália e em alguns países europeus.70
Em conjunto com a introdução para fins comercias, utilizou-se, em algumas
circunstâncias, P. clarkii como um meio de controlo biológico para reduzir as
populações de caracóis envolvidos no ciclo biológico da esquistossomose (doença
causada por Schistosoma sp.), motivo pelo qual foi introduzido no Quénia.
Procambarus clarkii em Portugal
Em Portugal esta espécie foi assinalada pela primeira vez em 1979 no rio Caia, um
afluente do Guadiana71 tendo sofrido uma franca expansão desde essa data.72 Em 1986
ocupava já uma vasta área no Sul de Portugal e em 1987 foi assinalado pela primeira
vez no Baixo Mondego, tendo na década de 90 os primeiros registos da sua presença na
bacia do rio Sado (Rebelo, com. pess.). Actualmente é referida, segundo o Decreto-Lei
nº 565/99 para as bacias hidrográficas dos rios Douro, Leça, Vouga, Mondego, Lis,
Tejo, Sado, Mira e Guadiana e das ribeiras do Oeste e do Algarve, com maior expressão
nas bacias do rio Guadiana e do rio Sado. É comum encontrá-los em reservatórios ou
canais de irrigação, e em campos de arroz73 mostrando-se uma importante praga nos
arrozais em Portugal.51

6
Em Portugal possui um estatuto de espécie introduzida e de espécie de risco
ecológico, de acordo com o Decreto-Lei 565/99 de 21 de Dezembro, entendendo-se
“risco ecológico” como o “impacto negativo potencial, susceptível de causar uma
modificação significativa nos ecossistemas de um dado território”.
Os lagostins são predados por peixes, aves e alguns macroinvertebrados que podem
predar principalmente os seus ovos. Os mamíferos como os ratos de água (Arvicola
sapidus), as ratazanas (Rattus norvegicus) e as lontras (Lutra lutra) também predam o
lagostim. Muitos predadores terrestres têm alterado a sua dieta, passando a consumir
elevadas quantidades de lagostim, desempenhando esta espécie provavelmente um papel
importante na conservação da lontra na Península Ibérica.74 Entre os peixes predadores
estão a perca sol (Lepomis gibbosus), o achigã (Micropterus salmoides), o barbo
(Barbus barbus), a carpa (Cyprinus carpio), o escalo (Leuciscus cephalus), o góbio
(Cottus gobio), a truta (Salmo trutta) e a enguia (Anguilla anguilla). Já entre as aves são
geralmente predados por garças e cegonhas, assim como por algumas aves limícolas.72
O reconhecimento do lagostim como presa já está assim difundido a muitos
predadores autóctones. Entretanto estudos recentes indicam o reconhecimento do
lagostim como possível predador por formas larvares de algumas espécies de anuros.75
Métodos de erradicação
Actualmente muitos esforços têm sido feitos na tentativa de mitigar os impactos de
P. clarkii, apesar de não se conhecer um modo eficaz de erradicar uma população de P.
clarkii.76,34,55
A gestão possível para a eliminação ou redução das populações de lagostim inclui
opções por métodos mecânicos, físicos, químicos ou biológicos, o repovoamento da
população da espécie de lagostim nativo (ameaçado pela praga do fungo e competição
com invasora), a criação de lagostins nativos resistentes ao fungo e o uso de legislação
para proibir o transporte e a libertação do lagostim invasor.
Métodos preventivos: Em Portugal o Decreto-Lei 565/99 de 21/12/99 proíbe a
introdução e disseminação de lagostim. Esta legislação provou ser insuficiente face às
motivações sociais para a introdução. Fiscalizações mais regulares, juntamente com
educação ambiental e maior divulgação da causa podem contribuir para o controlo.
Métodos físicos e mecânicos: A redução pode ser conseguida através dos métodos
físicos de controlo, mas a erradicação é muito difícil, a menos que a população esteja
particularmente restrita em distribuição e tamanho. Todos os métodos físicos têm um

7
custo para o ambiente que deve ser pesado e analisado contra os benefícios do controlo.
Os métodos físicos de controlo incluem a drenagem de charcos, desvios de ribeiras e
construções de barreiras (tanto físicas quanto eléctricas). Já os métodos mecânicos de
controlo do lagostim incluem o uso de armadilhas, redes e pesca eléctrica. A
armadilhagem continuada é preferível à de curto prazo e intensa, que pode provocar
respostas inesperadas na população, tais como a maturação acelerada de jovens e o
aumento da fecundidade nos animais que restam.
Método químico: Os produtos químicos que podem ser usados para o controle de
lagostim incluem os biocidas organofosfatados, organoclorados e piretróides; os
indivíduos são afectados de diferentes formas consoante o seu tamanho, pois quanto
menor mais susceptível é o indivíduo. Os biocidas não são específicos para lagostins, e
outros invertebrados, tal como os artrópodes, provavelmente são eliminados junto com
os lagostins. Outras consequências negativas são a bioacumulação na cadeia alimentar.
Uma possível solução será a utilização de armadilhas iscadas com ferormonas. É um
método direccionado apenas a uma espécie, o que o torna bastante eficaz pois pode ser
usado na presença de outras espécies de lagostins e em populações de baixa densidade.
No entanto, é efectivo apenas no período de reprodução, atraindo somente os machos.
Método biológico: Os possíveis métodos de controlo biológico incluem o uso de
peixes predadores, agentes patogénicos que afectam o lagostim e o uso de
microrganismos que produzem toxinas, por exemplo, a bactéria Bacillus thuringiensis
var. israeliensis (Holdich et al., 1999 in39). Apenas o uso de peixes predadores tem tido
algum sucesso: enguias e percas são exemplos de predadores de lagostins em Portugal.
(Holdich et al., 1999 in39). O sucesso dessa abordagem está dependente do tipo de peixe
predador usado e do local.
O trabalho actual é um estudo comparativo que visa determinar padrões de
distribuição e colonização de novos habitats por P. clarkii. O estudo anterior foi
realizado por Cruz (2006)14 em 2003/04 de modo que poderemos avaliar algumas
alterações na distribuição em 2008/09. A comparação permitir-nos-á identificar os
factores ambientais e de uso do solo que contribuem e/ou limitam a distribuição e a
colonização de P. clarkii.

8
Objectivos
Com o entendimento da problemática que envolve P. clarkii, o principal objectivo
do trabalho é actualizar a distribuição e entender os factores que limitam a colonização
em determinadas zonas; adicionalmente, foi elaborado um plano de contenção de
lagostim para controlar a sua dispersão e proteger pontos importantes para a
biodiversidade. Com esses propósitos definimos esses objectivos:
• Determinar a distribuição actual de Procambarus clarkii na bacia do rio Sado e
compará-la com a sua distribuição em 2003 (último estudo realizado na área);
• Comparar os factores que limitaram em ambos anos a colonização de alguns habitats
e facilitaram a colonização de outros;
• Identificar a influência dos diferentes usos do solo na colonização de novos habitats
por P. clarkii;
• Estimar a abundância de P. clarkii e compará-la entre vários tipos de habitats
ocupados;
• Produzir um modelo de gestão para a contenção da espécie na área de estudo,
determinando assim a melhor época do ano para uma possível acção, os pontos-
chave onde actuar e o modo de acção.

9
2. MATERIAIS E MÉTODOS
2.1 Área de Estudo
A área de estudo localiza-se no Noroeste do Baixo Alentejo, Sudoeste de Portugal e
da Península Ibérica, uma região próxima da área de introdução e de expansão inicial de
P. clarkii na Península.14 Foi a região seleccionada por Cruz & Rebelo (op. cit.) por ser
um local onde nunca terá existido lagostim nativo, e onde as espécies de fauna nativa
não terão nenhuma adaptação referente ao reconhecimento de P. clarkii como potencial
predador e/ou possível presa. A área tem formato triangular a oeste da bacia
hidrográfica do rio Sado, e é naturalmente limitada a Oeste, pelo Oceano Atlântico, e a
Norte e Leste, pelo rio Sado (figura 1), tendo aproximadamente 1500Km2 de extensão.
Figura 1: Mapa da área de estudo, com a localização da bacia do rio Sado em Portugal, mostrando com os
pontos de amostragem em 2008/09: os quadrados representam os pontos em ribeiras e os círculos os
pontos em charcos.

10
A bacia do rio Sado mantém largas áreas de habitats naturais representativos dos
que podem ser encontradas no SO da Península Ibérica, ocorrendo em dois tipos de
substrato geológico: áreas de solo de constituição arenosa e uma pequena cadeia de
montanhas de formação xistosa – Serra de Grândola (ponto mais alto a 325m n.m.m.).
Esta área inclui também diversos corpos de água naturais e artificiais, incluindo charcos,
ribeiras, represas, lagoas e arrozais. A área com substrato arenoso é caracterizada por
declives suaves, com máxima elevação de 120m, contrastando com a cadeia
montanhosa.
O clima da região é Mediterrânico com alguma influência Atlântica com uma
precipitação média de 500mm e temperatura média anual de 15,6 ºC, e um período
xérico de três a quatro meses, de Junho a Setembro77, o que resulta em um regime
hidrológico típico onde a maioria dos corpos de água seca no Verão excepto alguns rios,
represas e lagoas. Recentemente, no ano de 2005, Portugal foi acometido por um
período atípico de seca extrema para a região, onde foram registados os valores mais
baixos de precipitação total desde 193178. O clima é muito semelhante entre a região
montanhosa e a zona arenosa, pois a diferença entre as elevações não é muito
significativa.
A bacia do rio Sado foi colonizada por P. clarkii na década de 1990, estando hoje
expandido por quase todos seus rios constituintes, inclusive com populações viáveis em
ribeiras e charcos de pequenas dimensões com regime de seca anual.26O local de estudo
engloba áreas com alta densidade de lagostim tal como os canais de rega em campos de
arroz, lagoas costeiras, represas, charcos temporários e o rio Sado, e também áreas que
ainda não foram colonizadas, na sua maioria na Serra de Grândola. O rio Sado, as
ribeiras permanentes de médio porte e as lagoas litorais são pontos que pensamos ser
uma fonte de dispersão de lagostim.
O uso florestal ocupa apenas 33,1% da área total da bacia hidrográfica do rio Sado;
a área com ocupação agro-florestal (montados de sobro e azinho, com culturas arvenses
ou pastagens) representa 12,5%.79A vegetação da região consiste em, maioritariamente,
floresta de montado na serra de Grândola e no vale do Sado. Os montados constituem,
de um modo geral, meios abertos em que as árvores apresentam uma disposição
relativamente regular com actividades de explorações agro-florestais de sobro (Quercus
suber) e/ou azinho (Quercus rotundifoliae) com exploração do subcoberto para
agricultura e/ou pastorícia. Pinheiro-Bravo (Pinus pinaster) e Pinheiro-Manso (Pinus
pinea) plantados ao longo da costa, plantações de Eucalipto (Eucalyptus globulus) e

11
campos de arroz (Oryza sativa) encontram-se em áreas menos extensas (o eucalipto
apresenta uma baixa representatividade na totalidade da superfície florestal total da
bacia - 4,9%).
O presente trabalho é focado em três tipos de habitats aquáticos presentes na bacia
do rio Sado: habitats dulçaquícolas lênticos (águas paradas) representado pelas represas,
açudes e os charcos temporários mediterrâneos. Já os habitats lóticos (águas correntes)
podem dividir-se de acordo com o regime hídrico, caudal permanente e temporário; e as
lagoas costeiras. Os charcos temporários mediterrânicos são um habitat prioritário80para
a conservação, juntamente com as lagoas costeiras, que podem conter uma diversidade
florística e faunística muito elevada, e albergar espécies com amplitudes ecológicas
muito específicas, dependentes da ausência dos predadores típicos de sistemas
permanentes.81
A intervenção humana nesta região acontece de várias formas e com diferentes
graus de impacto no ambiente. Em grande parte a intervenção humana é de baixo
impacto, visto que a actividade predominante é de produção agro-florestal com maior
significado na tirada de cortiça. Também são frequentes actividades de pastoreio
geralmente associadas ao montado e pequena agricultura de subsistência. Já as
actividades de maior impacto são a produção intensiva de arroz, cereais e a produção
florestal de eucalipto e pinheiro.
Locais de Amostragem
Foram seleccionados 104 pontos, sendo que 50 são Ribeiras e 54 são Charcos
(Figura 1). 15 pontos de ribeiras são permanentes e 35 temporários. Dentre os charcos
15 são permanentes e 39 são temporários. As amostras foram feitas em rios principais e
seus afluentes pertencentes a bacia do rio Sado e pontos isolados próximos.
Dentre os 104 pontos seleccionados, 69 foram regularmente amostrados durante o
presente trabalho. Os demais pontos ou estiveram secos permanentemente durante o
período do trabalho (visitados, mas não amostrados) ou foram considerados com
lagostim após a primeira visita, devido o cumprimento de alguns critérios tais como: ser
permanente, ter havido lagostim em todas as fases de colecta do trabalho anterior, não
ter sofrido qualquer alteração. Nestes casos, e após a confirmação visual da presença de
lagostim na primeira visita, não foram mais amostrados, visto que uma vez introduzido
em habitats favoráveis, P. clarkii muito dificilmente é eliminado.44

12
Em relação ao ano de 2003/04 foram amostrados 3 novos pontos de amostragem ao
trabalho anterior; por outro lado, houve 2 pontos que foram amostrados por Cruz
2003/04, sobre os quais não obtivemos algumas informações.
2.2 Métodos de Amostragem
O presente trabalho tem como objectivo actualizar a distribuição de P. clarkii e
comparar o seu comportamento de dispersão com um trabalho anterior realizado em
2003/0414 nessa mesma área; portanto, os métodos de amostragem e de caracterização
ambiental agora usados foram os mesmos utilizados e definidos por esses autores.
Em 2003/04 foi aplicado um esquema de amostragem estratificado, procurando
manter um número semelhante de pontos seleccionados para cada tipo diferente de
habitat estudado, tal como recomendado por Hirzel & Guisan (2002).82Para isso foram
usadas algumas informações ambientais como critérios de selecção (sistema lêntico vs
lótico, presença de P. clarkii e tamanho do ponto de amostragem). De um total de 129
pontos amostrados em 2003/04, foram escolhidos para o actual trabalho 104 pontos.
Para a escolha foram prioritários os pontos que tiveram algum tipo de alteração do uso
do solo entre os dois trabalhos, os pontos temporários e os pontos permanentes
localizados próximo aos temporários, por poderem funcionar como fonte de P. clarkii.
Tentámos manter o equilíbrio entre os estratos.
Para cada ponto de amostragem foi feita a caracterização do substrato do fundo e
das margens, a caracterização da vegetação aquática e das margens, e determinada a
presença ou ausência de P. clarkii. Foi ainda caracterizado o uso do solo a 10m e a
100m da área circundante a cada ponto. Características intrínsecas aos pontos, como o
seu hidroperíodo, profundidade, largura, comprimento, área, corrente, altitude, sombra e
visibilidade também foram recolhidas.
Determinação da presença de P. clarkii
Para cada ponto de amostragem foi utilizado um método de armadilhagem, com
armadilhas de funil iscadas (mais eficaz para indivíduos adultos) e passagens de
camaroeiro (mais eficaz para os indivíduos em estádio larvar e juvenil). Usou-se cinco
armadilhas de funil de 20cm de comprimento, sendo três de 5 litros (43mm de abertura)
e duas de 1,5 litros (25mm de abertura), iscadas com comida para gatos com sabor a
peixe e posicionadas com o intuito de amostrar todos os diferentes microhabitats. As
armadilhas foram colocadas num intervalo de 12 horas durante a noite, período que se

13
verifica maior actividade do lagostim. Para o método de passagem de camaroeiro,
utilizou-se um camaroeiro de 30cm de diâmetro, rede de 2mm de malha e cor verde,
realizando 3 passagens (com cerca de 3 metros de extensão cada) por ponto durante o
dia.
Para complementar a amostragem foi utilizada uma pesquisa visual83realizada por
uma única pessoa por todo o transecto, procurando o lagostim e indícios de presença
indirectos, tais como pedaços de carapaças de lagostim na margem do corpo de água ou
até dejectos de lontra com presença de carapaça de lagostim. Para os dejectos de lontra
com lagostim assumimos a presença do lagostim no ponto de amostragem apenas em
casos de ponto isolados, distantes de outro ponto com lagostim, ou se esta foi
confirmada por outros métodos. Em alguns casos foram levados em consideração os
relatos dos proprietários das terras onde se encontravam os pontos.
No próprio local de amostragem, todos os lagostins capturados por qualquer um dos
métodos foram medidos (comprimento de carapaça em mm), determinado o sexo,
verificado o estádio de desenvolvimento (adulto, juvenil, larva) e verificado se o
indivíduo se encontrava em fase de muda, uma indicação de crescimento e de elevada
disponibilidade alimentar. Após a recolha dos dados os indivíduos foram devolvidos aos
seus habitats.
Caracterização dos pontos de amostragem
A caracterização dos pontos de amostragem e do habitat envolvente também foi
feita de acordo com os métodos utilizados por Cruz & Rebelo (2006)15 (ver tabela no
Anexo I). Para a determinação da cobertura da vegetação e do substrato usou-se o
método do quadrado pontual descrito por Barbour et al. (1998)84 com 10 pontos
distantes entre si a 10 cm por transecto. A amostragem foi feita em 8 transectos
distribuídos aleatoriamente ao longo das margens, com vista a amostrar a maior
variedade de microhabitats. A distância mínima entre cada par de transectos, para cada
ponto, foi de 1m. Foi determinada a percentagem de cobertura para o substrato do
fundo, substrato marginal, vegetação marginal, vegetação aquática e sombra no corpo
de água. Em cada local de amostragem foram retiradas as coordenadas geográficas.
A ocupação do solo na região envolvente aos pontos de amostragem foi determinada
aos 10m e aos 100m. Em cada ponto, foi determinada a percentagem de ocupação para
cada tipo de uso do solo de forma visual. Posteriormente, fez-se a correspondência entre
cada tipo de uso do solo e diferentes níveis de intervenção humana, de acordo com a

14
tabela (2) indicada no Anexo II. Para cada ponto foi determinado a classe de impacto
aos 10m e aos 100m (Tabela 3 Anexo II).
Segundo os resultados de 2003/04, a distância entre os pontos isolados e as ribeiras
é um factor importante para a colonização destes habitats por P. clarkii. Assim, foi
medida a distância dos pontos isolados às ribeiras para constatar possíveis alterações
das distâncias de colonizações percorridas. Para esta medição foi utilizado o software
ArcGis 8.9, que permitiu também construir um mapa das zonas de risco.
A amostragem de lagostim foi realizada em três períodos durante o ano hidrológico
de 2008/09. Em Novembro (Outono), em Fevereiro e Março (Inverno) e Abril e Maio
(Primavera).
As caracterizações físicas dos corpos de água foram feitas durante o Inverno no mês
de Fevereiro por ser o período de cheia e a cobertura de vegetação foi caracterizada
durante a Primavera.
Análise estatística
Foi realizada uma regressão logística binária para cada variável dependente (Tabela
4, Anexo III) para identificar os factores relacionados com a distribuição em cada ponto
de amostragem, em cada época do ano, e para as comparações com 2003/04. O
coeficiente de correlação de Pearson foi determinado e nos casos em que r > 0.7, uma
das variáveis (a variável de entre as correlacionadas com o menor valor biológico) foi
retirada e excluída da regressão.
Para a regressão, seguimos os procedimentos estabelecidos por Hosmer &
Lemeshow (2000).85Primeiro conduzimos uma regressão logística univariada para cada
variável independente (Tabela 5, Anexo III) e seleccionadas as varáveis com o p-value
˂ 0.25 no teste Wald. Para essas variáveis não seleccionadas na análise Univariada,
foram realizadas as devidas transformações. Se após as transformações o p-value no
teste de Wald fosse significativo para qualquer das variáveis dependentes, isto é
(˂0.25), tal foi considerado evidência de não linearidade e a variável entrou nos teste
subsequentes.
Após a selecção das variáveis foi realizada, então, uma regressão logística
multivariada no qual o nível de significância de 0.1 foi requerido para as variáveis
entrarem nos modelos.
Nos 69 pontos amostrados calculou-se o índice de abundância relativa a partir dos
valores de captura com cada um dos métodos utilizados a fim de comparar a ocupação

15
nas diferentes épocas do ano, em pontos temporários e permanentes, em ribeiras e
charcos e entre o trabalho actual (2008/09) e o realizado por Cruz (2006).14 Realizamos
o teste de Mann Whitney U para comparar as abundâncias e indivíduos em muda entre
os habitats permanentes e temporário.
Foram também determinadas as classes de tamanho predominantes em cada tipo de
habitat e em cada estação do ano. Foi também comparada a frequência de indivíduos em
muda nos diferentes corpos de água.
Para a identificação dos sexos foi usado o carácter morfológico “presença ou
ausência de gonopódia” (i. e. 1º par de pleópodes).72Os machos possuem esta
característica que está ausente nas fêmeas, entretanto, essa distinção só parece ser
possível a partir dos 20 mm de comprimento total (Huner in Romaire et al, 1977 in72).
A medida usada neste trabalho para diferenciar os adultos foi o comprimento de
aproximadamente 13 a 15mm de tamanho do cefalotórax (incluindo rostro).
O sex-ratio foi calculado a partir dos indivíduos já com o sexo diferenciado, com
tamanho a partir 13 a 15mm72segundo a fórmula Nº Machos / Nº Fêmeas por tipo de
habitat e por estação do ano.
Todos os tratamentos estatísticos dos dados foram realizados no software SPSS
Statistic 17.0 e todas as imagens tratadas no software ArcGis 8.3.
Plano de gestão para a contenção de P. clarkii
Para a elaboração do plano de gestão para a contenção do lagostim, foram levados
em consideração trabalhos sobre o tema na literatura, o anexo 1 da Directiva Habitats,
os resultados obtidos por Cruz (2006)14 e o trabalho actual, visando a integração dos
trabalhos científicos com as problemáticas reais, tendo como resultado uma aplicação
prática do conhecimento gerado na Universidade no campo da Gestão Ambiental e
Conservação de Natureza.
O plano tem como objectivos seleccionar o melhor local para a acção, considerando
os habitats que correm o maior risco com a presença de P. clarkii, a melhor época do
ano, as condições necessárias e os melhores métodos para a realização da acção com o
propósito de mitigar seus impactos. O plano contém uma componente educacional, a
qual envolve a realização do trabalho em conjunto com as escolas locais, visando um
esclarecimento sobre a problemática perante a comunidade e colaboração no trabalho de
campo.
Respostas que o plano de contenção visa fornecer:

16
• Para quê fazer contenção dessa espécie nessa área? Traz benefícios?
• Como fazer visto a dificuldade de erradicação da espécie?
• Onde há maior necessidade de contenção e onde será eficaz?
• Quais os critérios para a selecção dos locais para acção?
Conhecendo as 3 etapas que as espécies exóticas têm que ultrapassar para tornarem-
se invasoras, descritas por Lockwood et al (2007) 1, o plano de contenção visa actuar
nos pontos 1 e 3: consciencializar alunos e comunidade local para a problemática da
introdução de espécies exóticas para evitar futuros acidentes (etapa nº1), e na etapa nº 3
para evitar e dispersão dos indivíduos nas ribeiras e impedir a colonização dos charcos
temporários. Em relação ao ponto nº 2 é complicado obter qualquer alteração visto que
P clarkii já está estabelecido na área.
A produção de plano em formato não científico visa a utilização do protocolo por
pessoas de fora do âmbito universitário e científico, sendo de carácter prático, de fácil
entendimento e utilização.

17
3.RESULTADOS
3.1 Expansão de P. clarkii em 2003/04 e 2008/09:
No decorrer do presente trabalho, constatou-se uma alteração do hidroperíodo em 22
dos pontos amostrados (21% do total) em relação a 2003/04, que foram considerados
permanentes nesse estudo (tendo em conta dois ou três anos completos de trabalho de
campo) e que em 2009 tiveram um carácter temporário.
Em 2008/09 foram encontrados lagostins em 42 dos 104 pontos de amostragem, o
que corresponde a 40% do total de pontos (Figura 2).
104
129
40%46%
0
20
40
60
80
100
120
140
2003/04 2008/09
Nº
de p
onto
s
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Per
cent
agem
de
pont
os c
olon
izad
os
Total de pontos
% Colonizados
Figura 2: Total de pontos amostrados e a percentagem de pontos colonizados por P. clarkii em 2003/04 e
2008/09.
De um total de 74 pontos temporários, 19 (26%) tiveram lagostim e em 30 pontos
permanentes, 23 (77%) tiveram lagostim (Tabela 6).
Verificou-se uma diferença muito grande na expansão do lagostim por período de
amostragem: no Outono apenas 23 pontos apresentaram P. clarkii e no período
seguinte, o Inverno, estes aumentaram para 41 (Figura 3). A sazonalidade foi assim um
factor muito marcante para a presença de lagostim em 2008/09, o que não aconteceu em
2003/04, quando o aumento do Outono para o Inverno foi apenas em 2 locais. No
trabalho actual, houve na Primavera uma redução de pontos colonizados de 41 para 39,
o que foi devido ao balanço entre a seca de 3 charcos temporários e uma nova
colonização numa ribeira temporária. Pelo contrário, em 2003/04 houve um aumento
constante na colonização de novos pontos por período, e na Primavera chegou a haver
46 pontos colonizados, 3 somados ao Inverno desse mesmo ano. Em 2009, a maior

18
expansão da espécie nas ribeiras ocorreu na Primavera, enquanto nos charcos o período
de maior relevância foi o Inverno.

19
Figura 3: Mapas da área de estudo e sua localização em Portugal, com as colonizações comparativas no
Outono (menor extensão) e Inverno (maior extensão) de 2009, respectivamente. Os pontos amostrados
estão representados por: quadrado amarelo para ribeiras colonizadas e quadrado preto para não
colonizadas; círculos amarelos para charcos colonizados e círculos pretos para não colonizados.
Tal como esperado, as ribeiras permanentes apresentaram a maior extensão de
presença de lagostim em 2008/09, com 13 pontos de presença no Outono e 14 no
Inverno e Primavera. No caso das ribeiras temporárias houve colonização ao longo do
ano hídrico, passando de 1 ponto com lagostim no Outono para 8 no Inverno e 9 na
Primavera. Já nos charcos, os temporários foram aqueles onde se registaram mais casos
de colonização, passando de 1 ponto no Outono para 10 no Inverno. O Inverno foi o
período com mais pontos colonizados em todos os tipos de habitat, com excepção das
ribeiras temporárias.
Dos pontos em que o lagostim esteve presente em algum dos períodos de
amostragem em 2009, 45% foram charcos e 55% ribeiras; 55% permanentes e 45%
temporários (Tabela 6). Em 2003/04, do total de 59 pontos com lagostim em algum
período de amostragem, 51% eram charco e 49% ribeiras; 63% permanentes e 37%
temporários (Tabela 6). Ao analisarmos as colonizações por tipos de habitats (Figura 4)
verificamos que, apesar do maior número de pontos amostrados em 2003/04, as ribeiras
temporárias em 2008/09 tiveram maior número de colonizações.
Tabela 6: Sumário de colonização por P. clarkii em 2003/04 e 2008/09. Representado por ++: Percentagem ao total de pontos amostrados; +Percentagem sobre os pontos com presença em alguma vez.
2003/04 2008/09 Total de pontos 129 104 Presença alguma vez 59 (46%) ++ 42 (40%) ++ Permanentes 37 (63%) + 24 (57%) + Temporários 22 (37%) + 18 (43%) + Ribeiras 29 (49%) + 23 (55%) + Charcos 30 (51%) + 19 (45%) + Colonizado no Outono 41 23 Colonizado no Inverno 43 41 Colonizado na Primavera 46 39

20
Colonização 2003/04
16
511 9
41
15
611 11
43
15
711 13
46
05
101520253035404550
Ribeira
/Per
m.
Ribeira
/Tem
p.
Charco
/Per
m.
Charco
/Tem
p.
Total 2
003/0
4
Nº
loca
is c
olon
izad
os
Colonização Outono
Colonização Inverno
Colonização Primavera
Colonização 2008/09
13
1
8
1
23
148 9 10
41
149 9 7
39
05
101520253035404550
Ribeira
/Per
m
Ribeira
/Tem
p
Charco
/Per
m
Charco
/Tem
p
Total 2
008/0
9
Nº
loca
is c
olo
niza
dos
Colonização Outono
Colonização Inverno
ColonizaçãoPrimavera
Figura 4: Colonizações por tipo de habitat em 2003/04 e 2008/09 feitas com os dados presentes na
tabela 7 no Anexo IV.
Uma vez que o número de pontos de amostragem foi inferior em 2008/09 (104,
contra 129, em 2003/04), compara-se de seguida os resultados de 2003/04 com os
actuais, para os mesmos 104 pontos visitados em ambos trabalhos. Considerando apenas
esses 104 pontos de amostragem, em 2003/04 foram colonizados por lagostim 49
pontos, 7 pontos a mais que a distribuição actual. (Figura 5 no Anexo V)
Em 2009 houve 4 novos pontos colonizados, sendo 3 charcos (2 deles temporários),
e uma ribeira permanente. Considerando o balanço entre colonizações e extinções,
houve uma manutenção da situação nos pontos permanentes, com os mesmos números
de pontos com ganho e com perda de lagostim; estes habitats permanentes (charcos e
ribeiras) foram os que menos sofreram extinções locais de lagostim (Tabela 7). Os

21
habitats que obtiveram o maior número de extinções locais de lagostim foram os
charcos temporários, seguidos das ribeiras temporárias, com 5 e 4 pontos
respectivamente.
Tabela 7: Comparação da distribuição entre 2003/04 e 2008/09 por tipos de habitats com o número de pontos que nunca foram colonizados em 2003/04 e 2008/09, que ganhou lagostim em 2008/09, perdeu lagostim em 2008/09 e com presença em ambos anos.
Sem lagostim 2003 e 2009
Ganhou 2009
Perdeu 2009
Presença 2003 e 2009
Ribeiras/Perm. 1 1 0 13 Ribeiras/Temp. 15 0 4 8 Charcos/Perm. 4 1 2 8 Charcos/Temp. 28 2 5 7
Total 48 4 11 36
Entre 2003/04 e 2009, 82% dos pontos mantiveram o mesmo estatuto e apenas 18%
se alteraram, com enfoque nas extinções locais, que contribuíram para 75% das
alterações (Figura 6 Anexo VI). As ribeiras permanentes foram os pontos que menos
sofreram alteração, sendo o habitat mais importante para a manutenção de populações
de lagostim. Do total de 15 pontos em ribeiras permanentes amostrados, apenas 1 nunca
foi colonizado por P. clarkii.
3.2.Distância a “fontes” de P. clarkii
No presente trabalho foi dado um enfoque às distâncias entre que os indivíduos de
P. clarkii teriam de percorrer para colonizar pontos isolados a partir de ribeiras (tabelas
8 e 9 Anexo VII), não levando em consideração as distâncias percorridas dentro das
próprias ribeiras. Obteve-se uma distância de colonização mínima de 20 m, máxima de
1250 m e média de 229 m. Na sua maioria, os pontos colonizados estavam a menos de
270 metros de distância da fonte de lagostim e apenas 3 pontos estavam a uma maior
distância. A média da distância de charcos permanentes colonizados foi de 318 m e para
os charcos temporários foi de 57.3 m.
Os charcos permanentes não colonizados estavam a uma distância média de 617 m
de um ponto fonte, e em apenas 2 casos estavam a uma distância menor que a média dos
pontos colonizados (229m). Os charcos temporários não colonizados estavam a uma
distância média de 433m de um ponto fonte.

22
Em 2003/04 a distância entre um ponto fonte e um ponto isolado colonizado foi em
charcos temporários menor que 250m e para charcos permanente a máxima foi maior
que 1 quilómetro; no entanto, a maioria de dos pontos colonizados estavam a menos de
200 metros da fonte.
Ao contrário do esperado, em 2009 muitos pontos a uma distância menor que a
documentada em 2003/04 não foram colonizados. São 8 os pontos; 4 destes estão nas
proximidades da Ribeira de Castelhanos.
Foi possível a elaboração de uma imagem da área de estudo, utilizando a distância
máxima de colonização, que identifica áreas de risco para a colonização de Lagostim
(Figura 7).
Figura 7: Mapa da área de estudo e sua localização em Portugal, com pontos amostrados e área de risco de colonização. Os quadrados amarelos representam os pontos colonizados, enquanto os pretos, os não colonizados. A linha vermelha construída a partir da distância máxima de colonização em 2008/09 representa área de risco de colonização por P. clarkii.

23
3.3 Modelação
3.3.1. Resultados para a distribuição de P. clarkii em 2008/09 Foi realizada uma regressão logística multivariada com as variáveis anteriormente
seleccionadas para o modelo referente às variáveis dependentes (ver tabela 10, Anexo
VIII).
Presença de Lagostim em 2009
O modelo final incluiu 4 variáveis (Tabela 11, Anexo VIII), 2 referentes a
características dos habitats (gravilha margem e vegetação aquática emergente) e 2
características geográficas (altitude e distâncias dos charcos às ribeiras). A gravilha
condicionou de forma negativa a presença de lagostim, juntamente com a Altitude e a
distância, no entanto a vegetação aquática emergente afectou positivamente a
probabilidade de presença de P. clarkii. O modelo apresentou um muito bom ajuste aos
dados (R2 Nagelkerke = 0.897).
Colonização no Outono de 2009
Apenas duas variáveis foram seleccionadas para a elaboração do modelo (Tabela
11): a altitude afectou negativamente a probabilidade de presença de P. clarkii nesta
época, e a única variável seleccionada positivamente foi o total de vegetação aquática.
O teste do R2 de Nagelkerke indicou um ajuste do modelo aos dados relativamente
baixo (R2 Nagelkerke = 0.548).
Colonização no Inverno de 2009
Neste período no qual foi registada a maior expansão do lagostim e o maior número
de pontos colonizados. Foram seleccionadas para o modelo 5 variáveis, com um óptimo
ajuste (R2 Nagelkerke = 0.907) (Tabela 11). Três das variáveis estão relacionadas com
características do habitat – a vegetação aquática emergente e a profundidade afectaram
positivamente e a variável gravilha margem negativamente a probabilidade de presença
de P. clarkii nesta época. As duas variáveis restantes são altitude e distâncias dos
charcos às ribeiras, ambas com influência negativa.

24
Colonização na Primavera de 2009
O modelo incluiu 4 variáveis com um bom ajustamento aos dados (R2 Nagelkerke =
0.871) (Tabela 11). O período da Primavera teve como característica o começo da
diminuição de área colonizada por P. clarkii, pois houve a perda de alguns pontos
colonizados no Inverno. Afectaram positivamente a probabilidade de presença de P.
clarkii as variáveis ribª (1)/ charco (0) e profundidade. Com influência negativa, foram
seleccionadas as variáveis gravilha margem, altitude e distâncias dos charcos às
ribeiras.
3.3.2. Comparação entre 2003 e 2009
De forma a entender melhor as condicionantes da presença da P. clarkii na Bacia do
rio Sado a médio prazo, realizámos testes comparativos entre os dois anos de
amostragem para identificar possíveis padrões que favorecem ou limitam a presença de
lagostim.
Presença em 2003 e 2009
A regressão logística multivariada seleccionou 3 variáveis com um bom ajuste aos
dados (R2 Nagelkerke = 0.72) (Tabela 11). Estas variáveis foram as que
consistentemente explicam a distribuição do lagostim na área de estudo: ribª (1) /
charco (0) e vegetação aquática emergente afectaram positivamente a probabilidade de
presença, enquanto que a variável altitude afectou negativamente esta probabilidade.
Ganhou lagostim em 2009
Não foi possível produzir nenhum modelo para esta variável dependente, o que quer
dizer que nenhuma das características medidas explicou as novas colonizações.
Perdeu lagostim em 2009
Foram seleccionadas 3 variáveis que afectaram positivamente a probabilidade de um
ponto ter perdido lagostim em comparação com 2003. São elas: solo nu, classe de
impacto a 10m e areia fundo. O ajuste ao modelo é baixo (R2 Nagelkerke = 0.294)
(Tabela 11). O baixo número de pontos com perda de lagostim (11 pontos) pode
explicar a baixa capacidade de explicação do modelo.

25
Sem lagostim em 2003 e 2009
Este é um teste que permite avaliar e identificar os factores e características
ambientais que limitam de forma mais permanente a distribuição de P. clarkii na área.
Após a regressão logística multivariada, foram seleccionadas 11 variáveis significativas
para o modelo que obteve um muito bom ajuste aos dados (R2 Nagelkerke = 0.869)
(Tabela 11). As variáveis que afectaram positivamente a probabilidade de nunca ter
havido lagostim foram: ribª (1) / charco (0), pedra fundo, gravilha fundo, área,
corrente, altitude e distâncias dos charcos às ribeiras. As variáveis total vegetação
aquática, largura, comprimento e cascalho fundo afectam negativamente a
probabilidade. Este foi o modelo que incluiu o maior número de variáveis significativas.
Presença em ambos os Outonos de 2003 e 2009
No ano de 2003/04, a maioria dos pontos já estavam com água no Outono,
permitindo a realização da amostragem. Já em 2009, todos os pontos temporários
estavam secos excepto em 2 pontos em arrozais. Por esse motivo não foi efectuada esta
comparação.
Presença em ambos os Invernos de 2003 e 2009
A regressão logística multivariada seleccionou 6 variáveis significativas para a
presença de P. clarkii em ambos os Invernos. As que afectaram de forma positiva a
probabilidade de presença nesta estação do ano foram: profundidade, cascalho fundo e
sombra (Tabela 11). As variáveis que afectam de maneira negativa foram: permanente
(1) / temporário (0) em 2009, altitude e gravilha margem. O modelo apresentou um
bom ajuste do aos dados (R2 Nagelkerke = 0.795).
Presença em ambas as Primaveras de 2003 e 2009
Para a presença de P. clarkii em ambas as Primaveras foram seleccionadas pela
regressão logística multivariada as variáveis vegetação aquática emergente, altitude e
cascalho fundo, sendo a altitude a única que afectou negativamente a probabilidade de
presença em ambas as Primaveras (Tabela 11). Houve um bom ajuste do modelo aos
dados (R2 Nagelkerke = 0.702).
Em geral, as variáveis que actuaram negativamente na probabilidade de presença de
P. clarkii foram área do ponto, corrente nas ribeiras (características estruturais);

26
gravilha na margem, pedra no fundo, gravilha no fundo, solo nu, areia no fundo
(substrato); altitude, distância dos charcos às ribeiras (características geográficas) e
impacto na envolvente de 10m (intervenção humana). Já a presença de P. clarkii esteve
dependente das seguintes variáveis: ser ribeira, profundidade, largura, comprimento,
sombra (características estruturais); cascalho no fundo (substrato); vegetação
emergente, total de vegetação aquática (vegetação) e hidroperíodo temporário (apenas
para o Inverno).
3.4 Abundâncias
Ao longo de todo o trabalho, foram capturados 343 lagostins em todos os habitats e
estações do ano (103 em ribeiras permanentes, 75 em ribeiras temporárias, 31 em
charcos permanentes e 134 em charcos temporários).
De um modo geral, a maior abundância relativa encontrada foram nos charcos
temporários com 2.4 inds/armadilha, seguida de 2.1 inds/armadilha nos charcos
permanentes, ambos na Primavera (Tabela 12 Anexo XIX). Os menores valores de
captura com armadilhas ocorreram em ribeiras permanentes no Outono (0.2
ind/armadilha) e Inverno (0.22 ind/armadilha), e para o método de passagem de
camaroeiro os menores valores foram relativos às ribeiras temporárias (0.04
ind/passagem) e permanentes (0.08 ind/passagem), ambos na Primavera. O Outono foi a
estação com menores valores de abundância relativa, com capturas apenas nas ribeiras
permanentes.
No Inverno houve uma maior eficácia de captura nas passagens de camaroeiro do
que nas armadilhas, quando foram obtidos valores de 1.85 inds/passagem e 1.66
inds/passagem em ribeiras permanentes e charcos permanentes, respectivamente. Na
Primavera não houve capturas com camaroeiro nos charcos; já nas ribeiras houve
capturas com valores muito baixos, tais como 0.08 e 0.04 ind/passagem em ribeiras
permanentes e ribeiras temporárias, respectivamente. Nessa época do ano os valores de
abundância relativa obtidos por armadilhagem mostraram-se superiores nos charcos
temporários e charcos permanentes.
Charcos
Os valores de captura de indivíduos por armadilha (figura 8) foram superiores em
todas as estações do ano para os charcos temporários, menos no Outono onde os valores
para ambos os charcos foram nulos (quando todos os charcos temporários estiveram
completamente secos). É de salientar que nos charcos permanentes apenas se capturou

27
lagostim em armadilhas na Primavera. Trata-se de corpos de água grandes e de
amostragem complicada e é nesta estação que os indivíduos se encontram em maior
actividade.
Charcos
0
1
2
3
4
5
Outono Inverno Primavera
Nº
Indi
vídu
os/a
rmad
ilha
Charco permanentes 2008/09 Charco temporários 2008/09
Charcos permanentes 2003/04 Charcos temporários 2003/04
Figura 8: Abundância relativa de P. clarkii obtidos através do método de armadilhas em charcos
permanentes e temporários nos dois estudos (2003/04 e 2008/09). As barras verticais representam os erros
padrão.
Em comparação com 2003/04, houve uma diminuição dos números de P. clarkii
capturados por armadilhagem. Salienta-se o facto de os charcos permanentes em
2003/04 não terem registado valores nulos de abundância; nesse ano o Outono
(Nov./Dez.) foi o único período em que os charcos permanentes registaram valores
superiores aos charcos temporários, facto não ocorrido em 2009.
O método de passagem de camaroeiro (figura 9) mostrou-se eficaz para a captura
nos charcos apenas no Inverno, onde os maiores valores foram referentes aos
permanentes; nas outras estações do ano não foram capturados lagostins com passagens
de camaroeiro.

28
Charcos
0
0.5
1
1.5
2
2.5
3
Outono Inverno Primavera
Nº
indi
vídu
os/p
assa
gem
Charcos permanentes 2008/09 Charcos temporários 2008/09Charcos permanentes 2003/04 Charcos temporários 2003/04
Figura 9: Abundância relativa de P. clarkii obtidos através do método passagem de camaroeiro em
charcos permanentes e temporários nos dois estudos (2003/04 e 2008/09). As barras verticais representam
os erros padrão.
Em comparação com 2003/04, houve um aumento dos valores de abundância
relativa de P. clarkii capturados no Inverno. O facto que mais contrasta na comparação
entre os dois trabalhos é um maior valor de abundância nos charcos permanentes no
Inverno de 2009, com 1.66 inds/passagem quando em 2003/04 não passou de 0.2
inds/passagem.
Ribeiras
Não foi possível comparar os valores de abundância relativa entre os dois anos para
os pontos de amostragem localizados em ribeiras. Em 2008/09, os valores de
abundância obtidos por armadilhagem em ribeiras não diferiram muito entre as ribeiras
permanentes e temporárias, sendo maiores nas temporárias no Inverno e na Primavera.
No Outono, as Ribeiras Permanentes obtiveram valores de abundância semelhantes aos
verificados no Inverno (Fig. 10).

29
Ribeiras
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Outono Inverno Primavera
Nº
ind/
arm
adilh
a
Ribeiras permanentes Ribeiras temporárias
Figura 10: Abundância relativa de P. clarkii obtidos através do método de armadilhas em ribeiras
permanentes e temporárias em 2008/09. As barras verticais representam os erros padrão.
Com a utilização do método de passagem de camaroeiro obtiveram-se valores mais
elevados de inds/passagem nas ribeiras permanentes em relação às temporárias em todas
as estações do ano (figura 11). No Inverno o valor de abundância em ribeiras
permanentes foi 4 vezes superior ao valor encontrado nas ribeiras temporárias.
Ribeiras
0
0.5
1
1.5
2
2.5
3
Outono Inverno Primavera
Nº
ind/
pass
agem
Ribeiras permanentes Ribeiras temporárias
Figura 11: Abundância relativa de P. clarkii obtidos através do método de passagem de camaroeiro em
ribeiras permanentes e temporárias em 2008/09. As barras verticais representam os erros padrão.
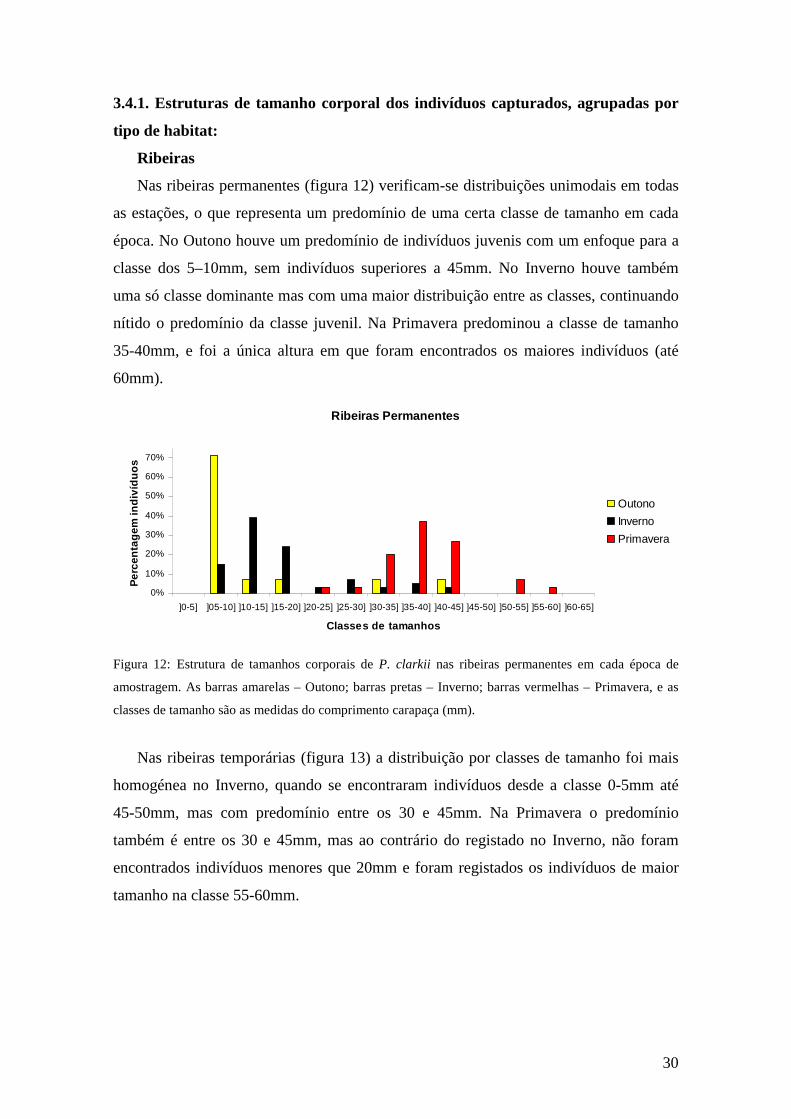
30
3.4.1. Estruturas de tamanho corporal dos indivíduos capturados, agrupadas por
tipo de habitat:
Ribeiras
Nas ribeiras permanentes (figura 12) verificam-se distribuições unimodais em todas
as estações, o que representa um predomínio de uma certa classe de tamanho em cada
época. No Outono houve um predomínio de indivíduos juvenis com um enfoque para a
classe dos 5–10mm, sem indivíduos superiores a 45mm. No Inverno houve também
uma só classe dominante mas com uma maior distribuição entre as classes, continuando
nítido o predomínio da classe juvenil. Na Primavera predominou a classe de tamanho
35-40mm, e foi a única altura em que foram encontrados os maiores indivíduos (até
60mm).
Ribeiras Permanentes
0%
10%
20%
30%
40%
50%
60%
70%
]0-5] ]05-10] ]10-15] ]15-20] ]20-25] ]25-30] ]30-35] ]35-40] ]40-45] ]45-50] ]50-55] ]55-60] ]60-65]
Classes de tamanhos
Per
cent
agem
indi
vídu
os
Outono
Inverno
Primavera
Figura 12: Estrutura de tamanhos corporais de P. clarkii nas ribeiras permanentes em cada época de
amostragem. As barras amarelas – Outono; barras pretas – Inverno; barras vermelhas – Primavera, e as
classes de tamanho são as medidas do comprimento carapaça (mm).
Nas ribeiras temporárias (figura 13) a distribuição por classes de tamanho foi mais
homogénea no Inverno, quando se encontraram indivíduos desde a classe 0-5mm até
45-50mm, mas com predomínio entre os 30 e 45mm. Na Primavera o predomínio
também é entre os 30 e 45mm, mas ao contrário do registado no Inverno, não foram
encontrados indivíduos menores que 20mm e foram registados os indivíduos de maior
tamanho na classe 55-60mm.

31
Ribeiras Temporárias
0%
10%
20%
30%
40%
50%
60%
70%
]0-5] ]05-10] ]10-15] ]15-20] ]20-25] ]25-30] ]30-35] ]35-40] ]40-45] ]45-50] ]50-55] ]55-60] ]60-65]
Classes de tamanho
Per
cent
agem
de
indi
vídu
os
Outono
Inverno
Primavera
Figura 13: Estrutura de tamanhos corporais de P. clarkii nas ribeiras temporárias em cada época de
amostragem. As barras amarelas – Outono; barras pretas – Inverno; barras vermelhas – Primavera, e as
classes de tamanho são as medidas do comprimento carapaça (mm).
Charcos
Nos charcos permanentes (figura 14) verificaram-se distribuições unimodais em
todas as estações, o que representa um predomínio de certa classe de tamanho em cada
época. Assim, no inverno apenas foram capturados indivíduos entre os 5mm e os
20mm, com enfoque na classe de 10-15mm, que representou 50% dos indivíduos. Na
Primavera encontraram-se indivíduos entre as classes 20-25mm até 55-60mm, com
maior variabilidade que no Inverno. As classes com maior representação foram 30-
35mm e 35-40mm, mas é de salientar a alta percentagem (quase 10%) de indivíduos
entre os 55-60mm.
Charcos Permanentes
0%
10%
20%
30%
40%
50%
]0-5] ]05-10] ]10-15] ]15-20] ]20-25] ]25-30] ]30-35] ]35-40] ]40-45] ]45-50] ]50-55] ]55-60] ]60-65]
Classes de tamanhos
Per
cent
agem
de
indi
vídu
os
Outono
Inverno
Primavera
Figura 14: Estrutura de tamanhos corporais de P. clarkii nos charcos permanentes em cada época de
amostragem. As barras amarelas – Outono; barras pretas – Inverno; barras vermelhas – Primavera, e as
classes de tamanho são as medidas do comprimento carapaça (mm).

32
Nos charcos temporários (figura 15), não houve uma moda definida na distribuição
das classes de tamanho, principalmente no Inverno, quando houve um predomínio das
classes 5-10mm à 15-20mm. Foi o único tipo de habitat em que foram registados
indivíduos das maiores classes de tamanho até 55-60mm no Inverno. Na Primavera
houve uma ausência de indivíduos das classes de tamanho inferiores a 20-25mm, sendo
as classes modais de 30-35mm e 40-45mm, que juntas constituem quase 50% dos
indivíduos.
Charcos Temporários
0%
10%
20%
30%
40%
50%
]0-5] ]05-10] ]10-15] ]15-20] ]20-25] ]25-30] ]30-35] ]35-40] ]40-45] ]45-50] ]50-55] ]55-60] ]60-65]
Classes de tamanhos
Per
cent
agem
de in
diví
duos
Outono
Inverno
Primavera
Figura 15: Estrutura de tamanhos corporais de P. clarkii nos charcos temporários em cada época de
amostragem. As barras amarelas – Outono; barras pretas – Inverno; barras vermelhas – Primavera, e as
classes de tamanho são as medidas do comprimento carapaça (mm).
O teste de Mann Whitney para a comparação de abundâncias entre os habitats
permanentes e temporário não verificou resultados significativos. (tabela 13, Anexo
XIX).
3.4.2. Mudas
Nas ribeiras não foram encontrados indivíduos em fase de muda no Outono (Figura
16). Em ribeiras permanentes encontrou-se uma maior percentagem de indivíduos nesta
fase na Primavera (10% do total de indivíduos), enquanto nas ribeiras temporárias foi o
Inverno a estação onde foi encontrada uma maior quantidade de indivíduos em muda
(26% do total). No entanto, mesmo na Primavera 12% dos indivíduos estavam em muda
nas ribeiras temporárias, um valor mais elevado que nas ribeiras permanentes.

33
Indivíduos em Muda
0%
5%
10%
15%
20%
25%
30%
Outono Inverno PrimaveraPer
cent
agem
de
indi
vídu
os
em m
uda
Ribeira/Perm.
Ribeira/Temp.
Figura 16: Percentagem de indivíduos em muda nas ribeiras permanentes e temporárias durante as épocas
de amostragem.
Nos charcos não foram encontrados indivíduos em fase de muda no Outono (Figura
17). A maior percentagem de indivíduos em fase de muda (30%) foi verificada nos
charcos permanentes no Inverno; na Primavera a percentagem caiu para 0,2%. Nos
charcos temporários, e tal como nas ribeiras temporárias, o Inverno manteve-se como o
período com maior percentagem de indivíduos em muda, com 16% do total, caindo na
Primavera para 8%.
Indivíduos em Muda
0%
5%
10%
15%
20%
25%
30%
Outono Inverno PrimaveraPer
cent
agem
de
indi
vídu
os
em m
uda
Charco/Perm.
Charco/Temp.
Figura 17: Percentagem de indivíduos em muda nos charcos permanentes e temporárias durante as épocas
de amostragem.
O teste de Mann Whitney para a comparação dos indivíduos em muda entre os
habitats permanentes e temporário não verificou resultados significativos. (tabela 14
Anexo XIX).
34
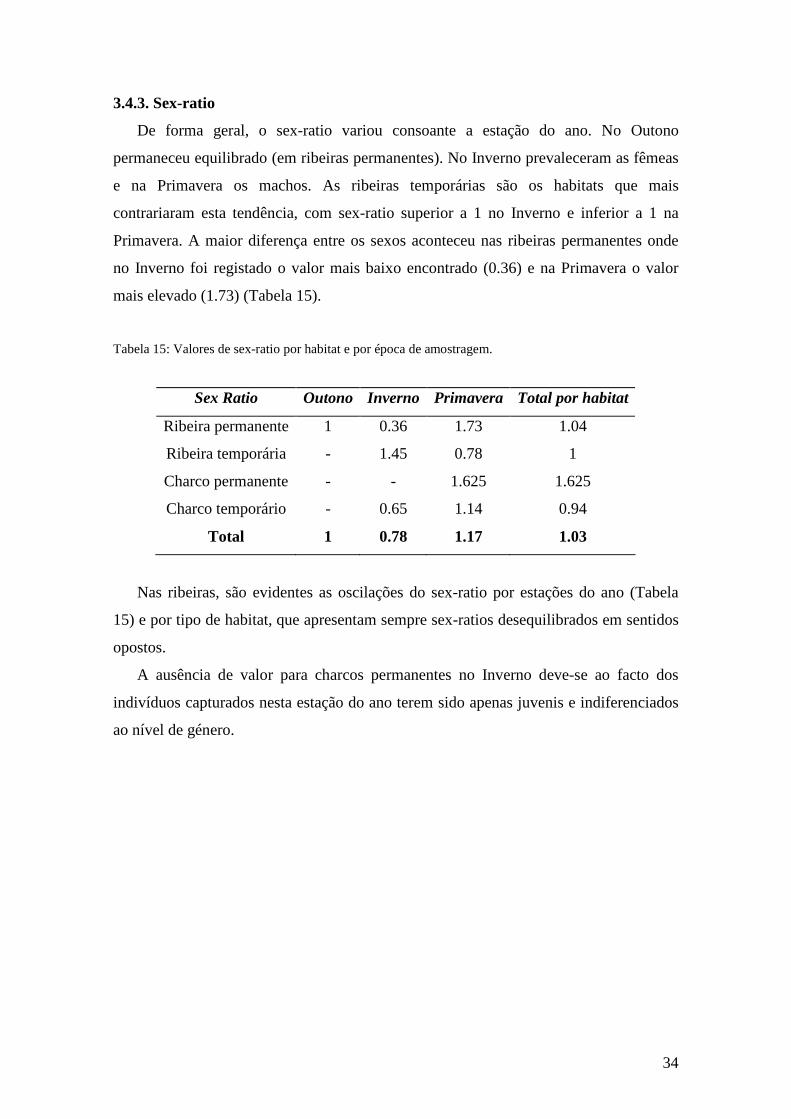
3.4.3. Sex-ratio
De forma geral, o sex-ratio variou consoante a estação do ano. No Outono
permaneceu equilibrado (em ribeiras permanentes). No Inverno prevaleceram as fêmeas
e na Primavera os machos. As ribeiras temporárias são os habitats que mais
contrariaram esta tendência, com sex-ratio superior a 1 no Inverno e inferior a 1 na
Primavera. A maior diferença entre os sexos aconteceu nas ribeiras permanentes onde
no Inverno foi registado o valor mais baixo encontrado (0.36) e na Primavera o valor
mais elevado (1.73) (Tabela 15).
Tabela 15: Valores de sex-ratio por habitat e por época de amostragem.
Nas ribeiras, são evidentes as oscilações do sex-ratio por estações do ano (Tabela
15) e por tipo de habitat, que apresentam sempre sex-ratios desequilibrados em sentidos
opostos.
A ausência de valor para charcos permanentes no Inverno deve-se ao facto dos
indivíduos capturados nesta estação do ano terem sido apenas juvenis e indiferenciados
ao nível de género.
Sex Ratio Outono Inverno Primavera Total por habitat
Ribeira permanente 1 0.36 1.73 1.04
Ribeira temporária - 1.45 0.78 1
Charco permanente - - 1.625 1.625
Charco temporário - 0.65 1.14 0.94
Total 1 0.78 1.17 1.03

35
4. DISCUSSÃO E CONSIDERAÇÕES FINAIS
O presente trabalho indica que houve uma contracção na distribuição de P. clarkii
na bacia do rio Sado entre os anos de 2003 e 2009, e que esta contracção esteve
relacionada com factores geográficos, características do substrato e impactos resultantes
da intervenção humana. Em relação ao estudo realizado em 2003, houve também um
aumento no número de variáveis explicativas da distribuição do lagostim; nesse ano, as
variáveis envolvidas foram apenas a distância entre o ponto e uma eventual “fonte” de
lagostins, a elevação e a velocidade de corrente. Finalmente, os pontos temporários
assumiram uma maior importância em 2009, pois houve alteração do hidroperíodo em
21% dos pontos anteriormente considerados permanentes e que se revelaram de carácter
temporário em 2009.
Procambarus clarkii está largamente distribuído pelo globo44 e encontra-se em
expansão no SO da Península Ibérica.47,90,86No entanto, no presente trabalho foram
documentadas limitações à distribuição e colonização de novos habitats na bacia do rio
Sado, encontrando-se restrito a algumas áreas. O número de pontos que nunca foram
colonizados é superior ao número de pontos que sempre apresentaram lagostim;
adicionalmente, houve mais extinções locais de P. clarkii entre os dois anos de estudo
do que colonizações de novos pontos.
A estagnação e contracção da distribuição de lagostim na área de estudo é explicada
pelo facto de que o carácter colonizado ou não colonizado por lagostim se ter mantido
igual de 2003/04 para 2009 em 82% dos pontos, com apenas 18% dos locais a sofrer
alterações (75% das quais foram extinções locais). Essas flutuações ocorreram
predominantemente nos charcos temporários mediterrânicos, considerados como
habitats peculiares e de dinâmicas próprias.
Os pontos permanentes são os mais estáveis e registaram poucas alterações; em
2009 houve um aumento na distribuição do lagostim nas ribeiras permanentes, o que
constitui uma colonização possivelmente definitiva, visto que nos pontos de ribeiras
permanentes não houve perdas de lagostim. Entretanto os pontos temporários
confirmam-se como habitats mais dinâmicos, sendo os mais alterados, com ênfase na
perda de lagostim ocorrida em 9 desses pontos. No Outono de 2008, que foi muito seco,
mais de 50% dos pontos com lagostim foram em ribeiras permanentes, nas quais nem
sempre se manteve água corrente, muitas vezes com conservação de apenas alguns
pegos. No ano de 2008/09 esses reservatórios foram muito importantes para a

36
manutenção das populações de P. clarkii e serviram como focos para a sua expansão no
Inverno, quando quase duplicou a sua frequência de ocorrência. Estes pontos “fontes de
lagostim” são de extrema importância para qualquer acção de erradicação ou de
contenção de P. clarkii nesta área. Tal como previsto em 2003/04, a invasão em habitats
permanentes tendeu a aumentar, excepto em ribeiras em maior altitude e com corrente
mais forte.
Os habitats temporários estão em menor risco de serem colonizados. Quando se
encontram próximos a uma fonte de lagostim, é provável que tenham sido colonizados
pela dispersão por terra dos próprios indivíduos adultos de P. clarkii,87,54,26 devido a
grande capacidade de dispersão e colonização desta espécie. Em relação aos pontos
mais distantes de uma fonte é possível suspeitar de uma introdução mediada por seres
humanos. Provavelmente a introdução propositada não é frequente na área de estudo.
Visto que esta espécie de lagostim é generalista, pensava-se que a sua expansão era
basicamente limitada por factores geográficos,26,97entretanto o presente trabalho
verificou a importância de períodos climáticos extremos, de alterações no uso do solo e
da vegetação como características que também podem limitar a sua expansão.
Sazonalidade
No presente trabalho, a sazonalidade foi um factor marcante para a distribuição de
P. clarkii na área de estudo e mais importante que em 2003/04. Neste ano, as diferenças
climatéricas entre as estações foram mais acentuadas que em 2003/04. Este aspecto é
importante para uma intervenção de contenção de lagostim, visto que em anos mais
secos como 2008/09 a ocorrência de P. clarkii diminuiu em quase 50% no Outono,
sendo essa a melhor época para esse tipo de trabalho. Em 2003/04 houve sempre um
aumento da área colonizada entre o Outono e a Primavera, o que não se verificou em
2009, quando houve perda entre o Inverno e a Primavera, devido à dessecação de alguns
locais, que aconteceu mais cedo em 2009.
Na bacia do rio Sado, em apenas um local amostrado em ribeira permanente não
havia lagostim. Em comparação, nos charcos temporários houve grandes flutuações
entre as estações, com apenas 1 ponto colonizado no Outono passando para 10 pontos
no Inverno, sendo importante sua identificação para futuras intervenções e contenção.
As ribeiras são ambientes mais estáveis de fácil colonização e recolonização de zonas
que entretanto secaram; já os charcos, por estarem isolados, são ambientes mais difíceis
de colonizar e recolonizar.

37
No período entre os dois estudos houve um ano extremamente seco – 2005 - o que
pode ter contribuído para a erradicação do lagostim em charcos permanentes. De facto,
os 2 charcos permanentes que perderam lagostim secaram completamente em 2005
(relato dos proprietários). Alguns dos charcos temporários que foram colonizados em
2003/04 cumpriam todos os critérios para serem recolonizados, isso é, estarem muito
próximos a ribeiras (entre 15m e 270m), possuírem vegetação aquática, intervenção
mínima entre os charco e a fonte de lagostim. No entanto não foram colonizados em
2008/09; neste caso, a razão para a não colonização pode estar relacionada com a
diminuição da abundância de indivíduos na ribeira.
Distância entre o local “fonte” de lagostim e o local a colonizar
Os trabalhos prévios sobre as distâncias de deslocação do lagostim referem-se às
distâncias percorridas geralmente em arrozais ou em ribeiras,87 ou seja, habitats
aquáticos. Entretanto, Kerby et al. (2005)(55) descreveram a importância da dispersão
por terra. Para a bacia do rio Sado foi verificada uma grande importância da distância
percorrida por terra pelos indivíduos para a sua dispersão.26 Segundo estes últimos
autores, a distância entre os pontos isolados e as ribeiras “fonte” de lagostim é um factor
determinante para o sucesso de colonização de novos pontos e no ano de 2003/04 essa
distância chegou até 250 metros. Em 2009, esta distância continua a ser um factor
determinante para a colonização de novos habitats por P. clarkii na bacia do rio Sado,
verificando-se apenas uma ligeira alteração nas distâncias percorridas, pois grande parte
dos pontos colonizados estavam a uma distância menor que 270m.
A distância máxima de colonização de charcos temporários documentada neste
trabalho foi percorrida entre o rio Sado (fonte para a colonização) e um charco. Entre
eles existe uma floresta de pinhal pouco intervencionada que pode ter fornecido a
sombra e a humidade necessária para a realização do trajecto. Segundo Aquiloni et. al.
(2005)88apenas a temperatura do ar e da água são factores importantes para a
movimentação de lagostim.
4.1. Modelos
Análise para 2009
Verificamos que factores como a vegetação emergente a altitude e a distância dos
charcos às ribeiras e gravilha na margem foram importantes para a presença de P.
clarkii em 2009. A presença em 2009 esteve positivamente dependente da vegetação

38
emergente; já a altitude, distância dos charcos às ribeiras e gravilha na margem
influenciaram a espécie de forma negativa. Cruz et al. (2007)26,97identificaram
características geográficas como sendo importantes para a expansão de lagostim, entre
elas a altitude e a distância dos charcos às ribeiras, factores confirmados para 2009.
A altitude foi um factor limitante para todas as estações do ano em 2009. No
Outono, a presença de vegetação aquática total foi determinante para a manutenção de
lagostins nos pontos. No Inverno destacamos a vegetação aquática emergente para
pontos com presença de lagostim e a distância do ponto isolado à ribeira juntamente
com gravilha na margem como factores limitantes à colonização no Inverno e também
na Primavera, visto que são períodos de dispersão e colonização de novos habitats. O
facto de ser ribeira influenciou positivamente a presença de lagostim na Primavera
período assinalado com o início da contracção da distribuição, quando alguns pontos,
principalmente charcos, já estavam completamente secos.
Análise comparativa 2003/2009
O resultado das análises comparativas permitiu observar padrões, características
essenciais ou limitantes para a presença de P. clarkii.
As características que foram importantes para a presença de lagostim em ambos os
trabalhos foram o facto do ponto de amostragem se encontrar numa ribeira e conter
vegetação aquática emergente. Já o único factor limitante foi a altitude.
As ribeiras das zonas de menor altitude na área de estudo, constituem importantes
locais para a manutenção de populações viáveis de lagostim. As futuras expansões estão
dependentes dessas populações estabelecidas em ribeiras que também apresentaram
vegetação emergente.
Uma questão muito importante para este trabalho é o facto de procurarmos entender
os mecanismos que afectam a distribuição do P. clarkii ao longo de vários anos,
sobretudo os factores relacionados com a extinção local do lagostim em alguns pontos,
dada a necessidade de obter instrumentos para seu controlo e erradicação. Foram 11 os
pontos que perderam lagostim; estão espalhados por toda a área e não estão restritos a
um tipo de habitat, pois apenas as ribeiras permanentes não perderam lagostim.
Podemos assinalar no entanto os habitats temporários como aqueles com maiores
perdas. Os charcos permanentes nos quais houve aparente extinção do lagostim foram
apenas dois; nestes, este facto pode ser explicado devido à sua seca por completo no ano
de 2005, ano em que foram registados os valores mais baixos do total de precipitação
desde 1931 nesta região.78 Assim, a seca, ou períodos mais secos parecem ser

39
importantes para a erradicação de lagostim. A análise dos pontos temporários que
perderam lagostins demonstrou que características como solo nu, classe de impacto a
10m e areia no fundo podem estar relacionadas com esta eliminação. Ao analisarmos as
variáveis seleccionadas podemos considerar que esses factores estão relacionados
principalmente com as necessidades exigidas durante a dispersão por terra para a
colonização de pontos isolados.
O facto de alguns locais nunca terem sido colonizados por P. clarkii pode estar
relacionado com um conjunto de características, onde distinguimos a altitude, a
velocidade da corrente, a distância entre os pontos isolados e as ribeiras, e algumas
características do substrato como pedra no fundo ou gravilha no fundo. No entanto a
vegetação aquática total, largura, comprimento e cascalho no fundo contribuem para a
colonização e manutenção de lagostim.
Os pontos que ganharam lagostim em 2009 foram poucos e a sua colonização não
foi explicada por nenhum modelo.
Análise por variável independente seleccionada pelos modelos
Altitude
A variável altitude esteve presente de forma muito significativa em todos os testes,
em todas as épocas de amostragem, sempre de forma negativa, o que indica que a
altitude influencia de forma negativa a presença de lagostim, actuando assim como um
factor limitante da sua distribuição, o que concorda com resultados obtidos em 2003/04
e com os trabalhos de Gil-Sánchez & Alba-Tercedor (2002).89
Velocidade da corrente
O resultado obtido para a velocidade da corrente está de acordo com outros autores,
pois segundo Kerby et al., (2005),55 a velocidade da corrente pode limitar a expansão do
lagostim para montante no rio e Gil-Sánchez & Alba-Tercedor (2002)89assim como
Cruz & Rebelo97 também encontraram relação negativa entre a velocidade de corrente e
presença de lagostim em cursos de água.
Distância dos charcos as ribeiras
Esta variável é significativa em todas as épocas do ano, menos no Outono, um
resultado esperado pois no Outono a maioria dos charcos estavam secos. Esta variável já
foi discutida acima.

40
Substrato
Correia (1995)90 refere que P. clarkii necessita de substrato silicoso devido à sua
actividade escavadora que é apenas possível em locais com o ratio “sílica/substratos
grosseiros” maior que 0.1 ou 0.2. Gil- Sánchez & Alba-Tercedor (2002)89 e Cruz et al.
200615não encontraram nenhuma associação entre a distribuição do P. clarkii e a
composição do substrato.
O presente trabalho obteve resultados diferentes ao nível do substrato onde uma
diferente composição foi seleccionada positivamente e negativamente: substratos de
pedra e gravilha influenciaram negativamente a presença de lagostim nas análises
comparativas (2003/04 e 2008/09), já a presença de cascalho no fundo beneficiou a
presença de lagostim.
As diferentes composições de substrato podem interferir na capacidade escavadora
do lagostim, e a única característica seleccionada para os modelos que explicam a
presença de lagostim em 2009 foi gravilha na margem. Está variável afecta
negativamente a probabilidade de haver lagostim, assim como gravilha no fundo, pedra
no fundo, areia no fundo e solo nu no resultado comparativo em ambos os anos de
estudo.
Vegetação
Segundo Garvey (1994, in88), os microhabitats caracterizados por uma elevada
biomassa vegetal proporcionam refúgios seguros para as espécies de lagostim. Nesse
estudo as variáveis relacionadas com a vegetação estão sempre positivamente
relacionadas com a presença de lagostim.
A vegetação aquática esteve sempre positivamente relacionada com a presença de P.
clarkii em 2009. Neste caso a vegetação aquática emergente foi seleccionada para os
testes com a Presença em 2009 e Colonização de Inverno 2009.
O total de vegetação aquática apareceu como factor que aumenta a probabilidade de
encontrar P. clarkii no Outono. No Outono verificou-se elevado número de pontos
secos e de ribeiras com alguns reservatórios de água, o que intensifica a predação no
local e a presença de vegetação aquática, por fornecer refúgios seguros (Garvey, 1994
in88), pode ter sido o factor que assegurou a manutenção de lagostim em determinados
pontos.
As macrófitas podem representar até 75% da dieta de um lagostim (Momot 1967;
Prins 1968 in91), entretanto estudos realizados com P. clarkii em Espanha concluíram
que a vegetação representa 58% da sua dieta.92 Ilhéu & Bernardo (1993)69 num estudo

41
sobre preferências alimentares em Portugal verificaram uma maior percentagem de
detritos provenientes de material vegetal.
Para além da alimentação, a vegetação é também usada principalmente pelas fêmeas
como refúgio durante o período de menor actividade88 ao contrário dos machos que têm
preferência pela margem dos corpos de água.
Dimensões do corpo de água
A variável área (medições apenas para charcos) apresentou uma relação positiva
com os locais que nunca contiveram lagostim, resultados contrário ao esperado. Neste
estudo, esta significância refere-se apenas à grande probabilidade dos charcos de
grandes dimensões não terem lagostim na área de estudo. As razões pelas quais os
charcos de grande área não estão colonizados por P. clarkii estão provavelmente
relacionadas com a particularidade de a distância entre eles e as ribeiras ser muito
elevada. Alguns também são locais sem vegetação aquática emergente.
A profundidade assim como o comprimento e a largura foram seleccionadas para as
épocas do ano com maior quantidade de água nas ribeiras e nos charcos e revelaram-se
características importantes para a presença de P. clarkii.
Em ribeiras é menor a probabilidade de introdução humana do que em represas ou
lagoas. Muitos pontos de ribeira onde nunca houve P. clarkii estão localizados na Serra
de Grândola, região de maior altitude e rios de menor profundidade.
Intervenção Humana
A classe de impacto a 10m foi a única variável relacionada com a intervenção
humana seleccionada e esteve positivamente relacionada com a perda lagostim 2009.
Este foi um aspecto inovador do trabalho, relacionando a utilização do solo com a
distribuição e colonização por P. clarkii.
A classe de impacto está relacionada com o somatório de vários tipos de intervenção
humana na envolvente imediata dos corpos de água. A classe de impacto máxima a 10m
é referente a determinados tipos de uso do solo: Estrada; Lavra intensiva; Hortas;
Suinicultura; Vilas; Arrozal; Corte florestal; Cereais, etc.
A interpretação dos impactos que os diferentes tipos de usos do solo têm sobre
dispersão e colonização de novos habitats por P. clarkii, foi feita com base na relação
entre o comportamento que o lagostim apresenta, atravessando terra seca para colonizar
novos pontos e a capacidade de cada tipo de uso de solo para dificultar essa locomoção,
quer por barreiras físicas, quer por aumentar a exposição dos indivíduos à radiação
directa. Aparentemente, apesar de ser uma espécie invasora e generalista, adaptando-se

42
a várias condições, o lagostim necessita de uma certa estabilidade ambiental em redor
dos pontos de água para poder realizar a dispersão, e a perturbação da envolvente de
alguns corpos de água terá impedido a colonização ou recolonização de alguns pontos.
Segundo Lockwood et al. (2007)1 as perturbações causadas no ambiente promovem
e facilitam a invasão de espécies exóticas, e a maioria das espécies não nativas são
encontradas em áreas com perturbação humana. Lindqvist & Huner (1999)93 salientam a
capacidade de P. clarkii colonizar habitats perturbados e em áreas que têm sido
modificadas pelo homem. No entanto, esta não é uma lei fundamental na Ecologia1 e
nossos resultados mostram que o impacto e a perturbação elevadas entre um ponto
“fonte” e charcos isolados podem evitar a colonização dos mesmos por impossibilitar a
dispersão por terra.
4.2. Abundância:
A abundância de lagostim pode ser muito elevada em habitats temporários26 quando
comparada aos habitats permanentes90 onde o P. clarkii é consumido por
peixes.94,90,95Segundo Momot et al. (1978)94 e Huner, (2002)46, a espécie pode atingir
elevadas densidades quando é pouco consumida por peixes e/ou quando existe uma
densa cobertura de vegetação, o que geralmente ocorre nos pontos temporários.
Cruz et al. (2004)26 verificou que após a colonização de um corpo de água, P. clarkii
é capaz de rapidamente alcançar valores de abundância elevados. De modo semelhante,
os charcos temporários colonizados em 2008/09 registaram os maiores valores de
abundância relativa. Entretanto, nas ribeiras o hidroperíodo mais frequente foi o
permanente.
Em 2003/04 os charcos temporários foram locais de grande abundância de lagostins,
característica verificada mesmo logo após a colonização. Em 2008/09 não foi diferente
com abundâncias relativas de lagostim por armadilhas atingindo os valores mais
elevados em comparação com todos os outros habitats.
Houve uma diminuição dos valores de abundância relativa de P. clarkii capturados
por armadilhagem em comparação com 2003/04. No entanto, para o método de
passagem de camaroeiro verificou-se um notável aumento dos valores de abundância
relativa no Inverno. Repetiu-se o pico de indivíduos no Inverno nos charcos
temporários. Houve uma grande diferença nos valores de abundância nos charcos
permanentes no Inverno, com 1.66 inds/passagem em 2008/09 e apenas 0.2

43
ind/passagem em 2003/04. Essa grande diferença pode ser indicadora de alteração do
período de reprodução nos charcos permanentes.
O maior valor de abundância referente ao método de passagem de camaroeiro foi
registado nas ribeiras permanentes (4 vezes maior no Inverno), contrastando assim com
o verificado nos charcos, que apresentam maior abundância nos pontos temporários.
4.2.1Estruturas de tamanho corporal dos indivíduos capturados, agrupadas por
tipo de habitat:
Os resultados obtidos estão em plena concordância com os de Anastácio (1995)72e
Adão (1991)96: os juvenis estão quase completamente ausentes da população na
Primavera, e atingem maiores abundâncias percentuais durante o Outono e Inverno com
o pico mais importante na variação anual de juvenis.
Em charcos permanentes encontramos total domínio de juvenis no Inverno e de
adultos na Primavera. Esse resultado pode indicar que os indivíduos adultos
permanecem em menor actividade no Inverno, enquanto os juvenis estão mais activos
ou pode também indicar falha na amostragem nos charcos permanentes com a
necessidade da utilização de diferentes métodos de captura.
Em charcos temporários a presença de quase todas as classes de tamanho no Inverno
indica que os indivíduos adultos são provenientes da migração e os juvenis da sua
rápida reprodução.
As ribeiras permanentes são o único local onde se registou reprodução do lagostim
no Outono, e podem actuar como “reservatórios” de lagostim durante os períodos mais
secos do ano.
Nas ribeiras temporárias, e ao contrário do que aconteceu nas ribeiras permanentes
encontram-se indivíduos adultos de várias classes de tamanho no Inverno, o que indica
a chegada de indivíduos adultos e o início da reprodução.
Assim como em Anastácio (1995)72 e Adão (1991)96, no Outono e no Inverno
predominaram indivíduos juvenis, independentemente do habitat e do hidroperíodo.
Entretanto na Primavera verificou-se o predomínio de adultos e a ausência de juvenis.

44
4.2.2. Muda
A frequência de indivíduos em muda está directamente relacionada com a taxa de
crescimento (Thorp & Wineriter, 1981 in47) que verificamos ser mais elevada nos
habitats temporários, provavelmente pelo baixo risco de predação por peixes e por
possuirem uma menor densidade de indivíduos adultos predadores de juvenis.26,50Os
nossos resultados concordam com Cruz et al. (2004),26quando a frequência de
indivíduos em muda foi superior nos charcos temporários em relação aos permanentes
em todos os períodos de amostragem. A excepção ocorreu no Inverno de 2009, quando
obtivemos uma maior frequência em charcos permanentes.
Segundo os resultados obtidos para as classes de tamanho em 2009, podemos
verificar o elevado crescimento no período entre a amostragem no Inverno e na
Primavera. De acordo com Thorp & Wineriter, 1981 (in47) o elevado crescimento pode
explicar o alto nº de indivíduos em muda no Inverno de 2009.
4.2.3.“Sex ratio”
A elevada proporção de fêmeas nos charcos temporários no Inverno pode ser um
indicativo de que foram esses os indivíduos migradores e colonizadores de novos
habitats em 2009 na área de estudo. Já no período seguinte, com o crescimento dos
juvenis, o “sex ratio” tendeu para estabilizar com um ligeiro aumento no nº de machos.
Uma das explicações apontadas para uma maior abundância de fêmeas é a de que
estariam sujeitas a uma menor agressividade intraespecífica. Este facto levaria a uma
mortalidade inferior (Sokol in Holdich & Lowery, 1981 in72). Segundo Anastácio a
maioria dos estudos publicados para lagostins de água doce apontam para valores de sex
ratio equilibrados, assim como os resultados obtidos para o total de indivíduos na bacia
do rio Sado em 2009, que também são semelhantes aos resultados de Correia (1990)
(in72), num estudo realizado em Elvas, Portugal.
Actualmente, segundo o grupo de especialistas em espécies invasoras da IUCN
(ISSG), as alterações climáticas podem ainda piorar os problemas relacionados com as
espécies invasoras, pois podem causar distúrbios e alterações no ambiente e possível
eliminação de algumas espécies nativas, dando lugar às espécies oportunistas e
generalistas.
No ano de 2005, um episódio de extrema seca ocorreu em Portugal período no qual
não se registavam valores tão baixos do total de precipitação desde 1931.78 Durante este

45
ano e os subsequentes verificou-se um aumento no período xérico que influencia
directamente a manutenção dos corpos de água e consequentemente as espécies
aquáticas. Se a contracção da distribuição de P. clarkii na área de estudo está
relacionada com este período seco, as futuras alterações climáticas poderão até vir a
dificultar a expansão e manutenção das populações desta espécie, podendo esta ser
erradicada dos pontos isolados.
De modo geral, concluímos que no período entre os anos de 2003 e 2009 houve uma
estagnação e contracção na distribuição de P. clarkii na bacia do rio Sado, explicadas
pela manutenção de colonização da maioria dos pontos. Houve poucas alterações nas
colonizações, sobretudo, foram mais importantes os locais com perda de lagostim. Esta
contracção esteve relacionada com factores geográficos, características do substrato e
impactos resultantes da intervenção humana. Condições climatéricas extremas, como a
seca em 2005, também terão actuado na contracção da distribuição.
As ribeiras permanentes constituíram um habitat de manutenção de populações de
lagostim durante todo o ano e actuam como fonte de colonizadores para os pontos
temporários. A colonização entre 2003 e 2009 de 1 ribeira e 1 charco, ambos
permanentes, e a ausência de extinções locais em ribeiras permanentes evidenciam uma
tendência de expansão dessa espécie que ainda está a decorrer; no entanto, pode ter sido
retardada pelas condições climatéricas verificadas ao longo dos últimos anos.
Com base na identificação dos factores que estão relacionados com a manutenção
desta espécie num local ou com a sua extinção local, é possível definir planos de
contenção da espécie que possam ter alguma probabilidade de sucesso. Apresentamos
assim em seguida uma proposta de plano de contenção (Anexo X) para uma área
localizada na Serra de Grândola, local onde observamos grande alteração, que visa
evitar a expansão e alastramento de P. clarkii em conjunto com uma componente de
educação ambiental.

46
5. REFERÊNCIAS BIBLIOGRÁFICAS Utilizadas as normas da revista científica Nature 1. Lockwood, J. L., Hoopes, M. F., Marchetti, M. P. In Invasion Ecology (eds
Lockwood, et al.) 1-106 (Oxford Blackwell, 2007)
2. Diário da república, 21-12-1999 Decreto-Lei 565/99, p.9000-9114 in world wild web
http://dre.pt/
3. Hulme, P. E. In Biological invasion a Global perspective (eds. Drake, J. A &
Mooney, H. A.) 56-80, 1989
4. Vié, J.-C., Hilton-Taylor, C. & Stuart, S.N. Wildlife in a Changing World – An
Analysis of the 2008 IUCN Red List of Threatened Species. (Gland, Switzerland: IUCN
180 pp, 2009).
5. Invasive Species Advisory Committe Invasive Species Definition Clarification and
Guidance White Paper. www.invasivespecies.gov (2006).
6. Ewel, J. J., et al. Deliberate introductions of species: Research needs. BioScience 49,
619–630 (1999).
7. Williamson, M., & Fitter, A. The varying success of invaders. Ecology 77, 1666–
1670 (1996).
8. Jerscke, J. M., & Strayer, D. L. Invasion success of vertebrates in Europe and North
America. Proc. Nat. Acad. Sci. USA 102, 7198–7202 (2005).
9. Kolar, C. S., & Lodge, D. M. Progress in invasion biology: Predicting invaders.
Trends Ecol. Evol. 16, 199–204 (2001).
10. Crooks, J. A. Characterizing ecosystem-level consequences of biological invasions:
The role of ecosystem engineers. Oikos 97, 153–166 (2002).

47
11. Lodge, D. M. Biological invasions: lessons for ecology. - Trends Ecol. Evol. 8, 133-
137 (1993).
12. Sakai, A. K., et al. The population biology of invasive species. Annual Reviews in
Ecology and Systematics 32, 305-332 (2001).
13. Cruz, M. J., & Rebelo, R. Vulnerability of Southwest Iberian amphibians to an
Introduced Crayfish, Procambarus clarkii. Amphibia-Repitilia 26, 293-304 (2005).
14. Cruz, M. J., Impacts of an exotic species, the red swamp crayfish (Procambarus
clarkii) on amphibian communities of Southwest Iberian Peninsula. Tese doutoramento,
Departamento de Biologia Animal, FCUL Universidade de Lisboa (2006).
15. Cruz, M. J., Rebelo, R. & Crespo, E. G., Effects of an introduced Crayfish,
Procambarus clarkii, on the distribution of SOUTHWEST IBERIAN amphibians in
their Breeding Habitats. Ecography 29, (2006).
16. Cruz, M. J., Pascoal, S., Tejedo, M. & Rebelo, R. Predation by na exotic crayfish,
procambarus clarkii, on Natterjack toad, Bufo calamita, embryos: its role on the
exclusion of this amphibian from its breeding ponds. Copeia in press (2006).
17. Sala, O. E., et al. Biodiversity scenarios for the year 2100. Science 287, 1770–1774
(2000).
18. Baillie, J. E. M., Hilton-Taylor, C. & Stuart, S. N. 2004 UICN red list of threatened
species. A global species assessment. IUCN, Gland, Switzerland and Cambridge (2004).
19. Simons, S. & Poorter, M. 2009 Global Invasive Species Programme (GISP)
Proceedings of an Expert Workshop on Preventing Biological Invasions: Best Practices
in Pre-Import Risk Screening for Species of Live Animals in International Trade.
(University of Notre Dame, Indiana, USA, 2008).

48
20. Macdonald, I. A. W., Graber, D. M., DeBenedetti, S., Groves, R. H. & Fuentes, E.R.
Introduced species in nature reserves in Mediterranean-type climatic regions of the
world. Biol. Conserv. 44, 37-66 (1988).
21. Hobbs, R. J. & Mooney, H. A. Broadening the extinction debate: population
deletions and additions in California and Western Australia. Conservation Biology 12,
271–283 (1998).
22. Garcia-Berthou, E., et al. Introduction pathways and establishment rates of invasive
aquatic sp Journal of Fisheries and Aquatic Sciences 62, 453–463 (2005).
23. Lodge, D. M., et al. Predicting impact of freshwater exotic species on native
biodiversity: Challenges in spatial scaling. Aust. J. Ecol. 23, 53–67 (1998).
24. Gherardi F. Are non-indigenous species ‘ecological malignancies’? Ethol. Ecol.
Evol. 12, 323–325 (2000).
25. Beisel, J. N. The elusive model of a biological invasion process: Time to take
differences among aquatic and terrestrial ecosystems into account? Ethol. Ecol. Evol.
13, 193–195 (2001).
26. Cruz, M. J., Andarde, P., Pascoal, S. & Rebelo, R. Colonization of temporary ponds
by the red swamp crayfish, Procambarus clarkii. Revista Biologia 22, 79-90. (2004).
27. Gherardi F. Crayfish invading Europe: the case study of Procambarus clarkii.
Marine and freshwater behavior and Physiology 39(3), 175-191 (2006).
28. McKinney, M. L. & Lockwood, J. L. Biotic homogenization: A few winners
replacing many losers in the next mass extinction. Trends Ecol. Evol. 14, 450–453
(1999).
29. Rahel, F. J. Homogenization of fish faunas across the United States. Science 288,
854–856 (2000).

49
30. Moyle, P. B. & Light, T. Biological invasions of fresh water: empirical rule and
assembly theory. Biol. Conserv. 78, 149-161 (1996).
31. Hobbs, R. J. & Mooney, H. A. Broadening the extinction debate. Population
deletions and additions in California and Western Australia. Conservation Biology 12,
271-283 (1998).
32.Vila, M., Garcia-Berthou, E., Sol, D. & Pino, J. Survey of the naturalised plants and
vertebrates in peninsular Spain. Eco. Mediterr. 27, 55-67 (2001).
33. Josefsson, M. & Andersson, B. The environmental consequence of alien species in
the Swedish lakes Mälaren, Hjälmaren, Vänern and Vätern. Ambio 30, 514-521 (2002).
34. Lodge, D. M., Taylor, C. A., Holdich, D. M. & Skurdal, J. Nonindigenous
crayfishes threaten North American freshwater biodiversity: Lessons from Europe.
Fisheries 25, 7–20 (2000).
35. Nyström, P., Svensson, O., Lardner, B., Brönmark, C. & Granéli, W. The influence
of multiple introduced predators on a littoral pond community. Ecology 82, 1023–1039
(2001).
36. Gherardi, F. & Holdich, D. M. (eds) Crayfish in Europe as alien species. How to
make the best of a bad situation? Rotterdam: Balkema. A. A. (1999).
37. Nyström, P. In Crayfish in Europe as alien species. How to make the best of a bad
situation? (eds Gherardi, F. & Holdich, D. M.) Rotterdam: A.A. Balkema. pp 63–85
(1999).
38. Lodge, D. M., Kershner, M. W., Aloi, J. E. & Covich, A. P. Effects of an
omnivorous crayfish (Orconectes rusticus) on a freshwater littoral food web. Ecology
75, 1265–1281 (1994).
39. IUCN/SSC Invasive species Specialist Group (ISSG). In world wild web
www.issg.org (2009).

50
40. Bruxelas, S. & Bernardo, J. M. Recuperação das populações de Lagostim-depatas-
brancas Austropotamobius pallipes na área do Centro Rural do Planalto
Mirandês. Relatório final. Direcção Geral das Florestas, Ministério da Agricultura, do
Desenvolvimento Rural e das Pescas. 2000.
41. www.icnb.pt (2009).
42. Huner, J. V. Introductions of the Louisiana red swamp crayfish, Procambarus clarkii
(Girard): An update Freshw. Crayfish 3, 193–202 (1977).
43. Henttonen, P. & Huner, J. V. In Crayfish in Europe as alien species. How to make
the best of a bad situation? (eds Gherardi, F., Holdich, D. M.,) Rotterdam: A.A.
Balkema. pp 13–30 (1999).
44. Hobbs, H. H. et al. A review of global crayfish introductions with particular
emphasis on two north american species. Crustaceana 56, 299-316 (1989).
45. Gutiérrez-Yurrita, P. & Montes, C. Bioenergetics and phenology of reproduction of
the introduced red swamp crayfish, Procambarus clarkii, in Doñana National Park,
Spain, and implications for species management. Freshw. Biol. 42, 561–574 (1999).
46. Huner, J. V. In Biology of freshwater crayfish (eds Holdich, D. M.). Oxford:
Blackwell. pp 541–584 (2002).
47. Anastácio, P. M. & Marques, J. C. Population biology and production of the red
swamp crayfish Procambarusclarkii (Girard) in the lower Mondego valley, Portugal. J.
Crust. Biol. 15, 156-168 (1995).
48. Reynolds, J. D. Growth and reproduction. In Biology of freshwater crayfish(eds
Holdich, D. M.) 152-191. (Blackwell Science, 2002).
49. Bravo, M. A., Duarte, C. M. & Montes, C. Environmental factors controlling the
life history of Procambarus clarkii (Decapoda, Cambaridae) in a temporary marsh of

51
the Doñana National Park (SW Spain). Verh. Internat. Verein. Limnol. 25, 2450-2453
(1994).
50. Gutiérrez-Yurrita, P. J., Sancho, G., Bravo, M. A., Baltanás, A. & Montes, C. Diet
of the red swamp crayfish Procambarus clarkii in natural ecosystems of the Doñana
national park temporary fresh-water marsh (Spain). J. Crustacean Biol. 18, 120-127
(1998).
51. Correia, A. M. Niche breath and trophic diversity: feeding behaviour of the red
swamp crayfish (Procambarus clarkii) towards environmental availability of aquatic
macroinvertebrates in a rice field (Portugal). Acta Oecologica 23, 421-429 (2002).
52. Harper, D. M., et al. Distribution and abundance of the Louisiana red swamp
crayfish Procambarus clarkii Girard at Lake Naivasha, Kenya, between 1987 and 1999.
Hydrobiologia 488: 143-151 (2002).
53. Smart, A. C., et al. Feeding of the exotic Louisiana red swamp crayfish,
Procambarus clarkii (Crustacea, Decapoda), in an African tropical lake: Lake Naivasha,
Kenya. Hydrobiologia 488, 129-142 (2002).
54. Barbaresi, S., Tricarico, E., Santini, G. & Gherardi, F. Ranging behaviour of the
invasive crayfish, Procambarus clarkii. J. Nat. Hist. 38, 2821–2832 (2004).
55. Kerby, J. L., Riley, S. P. D., Kats, L. B. & Wilson, P. Barriers and flow as limiting
factors in the spread of an invasive crayfish (Procambarus clarkii) in southern
California streams. Biological Conservation 126, 402-409 (2005).
56. Gherardi, F., et al. In Crayfish in Europe as alien species. How to make the best of a
bad situation? (eds Gherardi, F., Holdich, D. M.). Rotterdam: A.A. Balkema. 107–128
(1999).
57. Ilhéu, M., Acquistapace, P., Benvenuto C. & Gherardi, F. Shelter use of
the red swamp crayfish (Procamabrus clarkii) in dry-season stream pools. Arch.
Hydrobiol 154(4), 535-546 (2003).

52
58. Nyström, P., Brönmark, C. & Granéli, W. Patterns in benthic food webs: A role for
omnivorous crayfish? Freshwat. Biol. 36, 631–646 (1996).
59. Nyström, P. In Biology of freshwater crayfish, (eds Holdich, D. M.) 192-235
(Blackwell Science, 2002).
60. Hobbs, H. H. Jr. In Contributions in Biology and Geology, Vol 85. 110 (Milwaukee
Public Museum, 1993)
61. Pérez-Bote, J. L. Feeding ecology of exotic red swamp crayfish, procambarus clarkii
(Girard, 1852) in Guadiana river (SW Iberian Península). Crustaceana 77 (11), 1375-
1387 (2005).
62. Lodge, D. M. Biological invasions: lessons for ecology. Trends Ecol. Evol. 8, 133-
137 (1993).
63. Lodge, D. M., Kershner, M. W., Aloi, J. E. & Covich, A. P. Effects of an
omnivorous crayfish (Orconectes rusticus) on a freshwater littoral food web. Ecology
75, 1265–1281 (1994).
64. Momot, W. T. Redefining the role of crayfish in aquatic ecosystems. Reviews in
Fisheries Science 3(1), 33–63 (1995).
65. Nyström, P. & Strand, J. A. Grazing by a native and an exotic crayfish on acquatic
macrophytes. Freshwat. Biol. 36, 673–682 (1996).
66. Nyström, P., Brönmark, C. & Granéli, W. Patterns in benthic food webs: a role for
omnivorous crayfish? Freshwat. Biol. 36, 631-646. (1996).
67. Anastácio, P. M., Correia, A. M. & Menino, J. P. Processes and patterns of plant
destruction by crayfish: Effects of crayfish size and developmental stages of rice. Arch.
Hydrobiol. 162, 37–51 (2005).

53
68. Anastácio, P. M., Parente, V. S. & Correia, A. M. Crayfish effects on seeds and
seedlings: Identification and quantification of damage. Freshwat. Biol. 50, 697–704.
(2005).
69. Ilhéu, M. & Bernardo, J. M. Food preference and predation of red swamp
crayfish Procambarus clarkii - experimental assessment. Freshwater Crayfish 9,
359-364 (1993).
70. Barbaresi, S., Fani, R., Gherardi, F., Mengoni, A. & Souty-Grosset, C. Genetic
variability in European populations of an invasive American crayfish: Preliminary
results. Biol. Invas. 5, 269–274 (2003).
71. Ramos, M. A. & Pereira, T. M. Um novo Astacidae para a fauna portuguesa:
Procambarus clarkii (Girard, 1852). Bol.Inst. Nac. Invest. Pescas, Lisboa, 6, 37-47
(1981).
72. Anastácio, P. M. Ciclo biológico e produção do lagostim vermelho da Lousiana
(Procambarus clarkii, Girard ) na região do baixo Mondego. Tese de Doutoramento,
Departamento de Zoologia, Universidade de Coimbra (1993).
73. Oliveira, J. & Fabião, A. Growth responses of juvenile red swamp crayfish,
Procambarus clarkii Girard, to several diets under controlled conditions. Aquaculture
Research 29, 123-129 (1998).
74. Beja, P.R. Temporal and spatial patters of rest-site use by four female otters Lutra
lutra along the South-West coast of Portugal. J. Zool., Lond. 239, 741-753 (1996).
75. Almeida, E. R. L. & Rebelo, R. Estudo das Respostas Anti-Predatórias de
Discoglossus galganoi (Capula, Nascetti, Lanza, Bullini & Crespo, 1985) e Bufo bufo
(Linnaeus, 1758) na presença de um predador exótico e um autóctone. Tese de
Mestrado, Departamento Biologia Animal, FCUL, Universidade de Lisboa (2008).
76. Barbaresi, S. & Gherardi, F. The invasion of the alien crayfish Procambarus
clarkii in Europe, with particular reference to Italy. Biol. Invas. 2, 259–264 (2000).

54
77. Correia, A. I. & Santos-Reis, M. In Caracterização da flora e fauna do montado da
Herdade da Ribeira Abaixo, Grândola, Baixo- Alentejo. (Centro de Biologia Ambiental,
Universidade de Lisboa, Lisboa, 1999).
78. Instituto de Meteorologia, IP Portugal. In world wild web www.meteo.pt (2005).
79. Plano de Bacia Hidrográfica do Rio Sado Anexo Temático 5 – Análise da Ocupação
do Solo e Ordenamento do Território – - Parte 1 – Análise da Ocupação do Solo in
world wild web www.ccdr-a.gov.pt/
80. Anexo I Directiva Habitats In world wild web http: //portal/icnb.pt
81. Brönmark, C. & Hansson, L. A. In The Biology of Lakes and Ponds. (Oxford:
Oxford University Press, 1998).
82. Hirzel, A. & Guisan, A. Which is the optimal sampling strategy for habitat
suitability modelling. Ecol. Model. 157, 331-341 (2002).
83. Crump, M. L. & Scott, N. J. In. Measuring and monitoring biological diversity:
standard methods for amphibians (eds Heyer, W. R., Donnelly, M. A., McDiarmid, R.
W., Hayek, L.-A. C. & Foster, M. S.) 84-92 (Smithsonian Institution Press, 1994).
84. Barbour, M. G., et al. In Terrestrial plant ecology-3rd (eds. Barbour, M. G., et
al) 229 ( Benjamin/Cumming, Menlo Park, California, 1998).
85 Hosmer, D. W. & Lemeshow, S. Applied logistic regression. Wiley-Interscience
Publication. (2000).
86. Jiménez, A., Cano, E. & Ocete, M. E. Mortality and survival of Procambarus clarkii
Girard, 1852 upon exposure to different insecticide products. Bull. Environ. Contam.
Toxicol. 70, 131-137 (2003).

55
87. Gherardi, F. & S. Barbaresi, Invasive crayfish: activity patterns of Procambarus
clarkii in the rice fields of Lower Guadalquivir (Spain). Archiv fur Hydrobiology 168,
(2000).
88. Aquiloni, L., Ilhéu, M. & Gherardi, F. Habitat use and dispersal of the invasive
crayfish Procambarus clarkii in ephemeral water bodies of Portugal. Mar. Freshw.
Behav. Physiol. 38, 225–236 (2005).
89. Gil-Sánchez, J. M. & Alba-Tercedor, J. Ecology of the native and introduced
crayfishes Austropotamobius pallipes and Procambarus clarkii in southern Spain and
implications for conservation of the native species. Biol. Conserv. 105, 75-80 (2002).
90. Correia, A. M. Biologia populacional e interacções tróficas de Procambarus clarkii
(Crustácea, Decapoda, Cambaridae) no sistema hidrográfico do rio Tejo (Ribatejo,
Portugal). Tese de Doutoramento, Departamento de Zoologia e Antropologia, FCUL,
Lisboa (1995).
91. Geiger, W., Alcorlo, P., Baltanas, A. & Montes, C. Impact of an introduced
Crustacean on the trophic webs of Mediterranean wetlands. Biol. Invas. 7, 49–73(2005).
92. Gherardi, F. & Barbaresi, S. Feeding opportunism of the red swamp crayfish,
Procambarus clarkii, an invasive species. Freshwater Crayfish 16, 77-85 (2008).
93. Lindqvist, O. V. & Huner, J. V. In Crayfish in Europe as alien species. How to
make the best of a bad situation? (eds Gherardi, F., Holdich, D. M.) 23–30 (Rotterdam:
A.A. Balkema, 1999).
94. Momot, W. T., Gowing, H. & Jones, P. D. The dynamics of crayfish and their role
in ecosystems. Am. Mid. Nat. 99, 10-35 (1978).
95. Elvira, B., Nicola, G. G. & Almodovar, A. Pike and red swamp crayfish: a new case
on predator-prey relationship between aliens in central Spain. J. Fish Biol. 48, 437-446
(1996).

56
96. Adão, M. H. S. M. Procambarus clarkii (Girard, 1852) (Decapoda, Cambaridae)
espécie exótica em Portugal. Considerações sobre a ecobiologia e aspectos da sua
biologia populacional na barragem de Monte Novo (Alentejo, Portugal). Provas de
aptidão pedagógica e científica. Universidade de Évora (1991).
97. Cruz, M. J. & Rebelo, R. Colonization of fresh water habitats by na introduced
crayfish, Procambarus clarkii, in Southwest Iberian Península. Hydrobiologia (2007).

57
ANEXO I
Tabela 1: Tabela de campo para caracterização ambiental dos pontos de amostragem. O Nº de pontos representam quantas vezes cada tipo de substrato marginal, ou do fundo, da vegetação marginal ou aquática foram identificados por transecto.
Trans Substrato margs Nº pts Veget margs Nº pts Substrato fundo Nº pts Veg. Aq Nº pts
Rocha Arbórea Rocha Aq flutuante
Pedra > 30 cm Arbustiva Pedra > 30 cm Aq submersa
Casc 7.5 a 30 cm Herbácea Casc 7.5 a 30 cm Emergente
Gravilha 2 a 75 mm Musgo Gravilha 2 a 75 mm
Areia 0.5 a 2 mm Solo nú Areia 0.5 a 2 mm sombra
Argila < 0.5 mm Argila < 0.5 mm
1
Manta morta Manta morta
Rocha Arbórea Rocha Aq flutuante
Pedra > 30 cm Arbustiva Pedra > 30 cm Aq submersa
Casc 7.5 a 30 cm Herbácea Casc 7.5 a 30 cm Emergente
Gravilha 2 a 75 mm musgo Gravilha 2 a 75 mm
Areia 0.5 a 2 mm Solo nú Areia 0.5 a 2 mm sombra
Argila < 0.5 mm Argila < 0.5 mm
2
Manta morta Manta morta
Rocha Arbórea Rocha Aq flutuante
Pedra > 30 cm Arbustiva Pedra > 30 cm Aq submersa
Casc 7.5 a 30 cm Herbácea Casc 7.5 a 30 cm Emergente
Gravilha 2 a 75 mm musgo Gravilha 2 a 75 mm
Areia 0.5 a 2 mm Solo nú Areia 0.5 a 2 mm sombra
Argila < 0.5 mm Argila < 0.5 mm
3
Manta morta Manta morta
Rocha Arbórea Rocha Aq flutuante
Pedra > 30 cm Arbustiva Pedra > 30 cm Aq submersa
Casc 7.5 a 30 cm Herbácea Casc 7.5 a 30 cm Emergente
Gravilha 2 a 75 mm musgo Gravilha 2 a 75 mm
Areia 0.5 a 2 mm Solo nú Areia 0.5 a 2 mm sombra
Argila < 0.5 mm Argila < 0.5 mm
4
Manta morta Manta morta
Rocha Arbórea Rocha Aq flutuante
Pedra > 30 cm Arbustiva Pedra > 30 cm Aq submersa
Casc 7.5 a 30 cm Herbácea Casc 7.5 a 30 cm Emergente
Gravilha 2 a 75 mm musgo Gravilha 2 a 75 mm
Areia 0.5 a 2 mm Solo nú Areia 0.5 a 2 mm sombra
Argila < 0.5 mm Argila < 0.5 mm
5
Manta morta Manta morta
Rocha Arbórea Rocha Aq flutuante 6
Pedra > 30 cm Arbustiva Pedra > 30 cm Aq submersa

58
Casc 7.5 a 30 cm Herbácea Casc 7.5 a 30 cm Emergente
Gravilha 2 a 75 mm musgo Gravilha 2 a 75 mm
Areia 0.5 a 2 mm Solo nú Areia 0.5 a 2 mm sombra
Argila < 0.5 mm Argila < 0.5 mm
Manta morta Manta morta
Rocha Arbórea Rocha Aq flutuante
Pedra > 30 cm Arbustiva Pedra > 30 cm Aq submersa
Casc 7.5 a 30 cm Herbácea Casc 7.5 a 30 cm Emergente
Gravilha 2 a 75 mm musgo Gravilha 2 a 75 mm
Areia 0.5 a 2 mm Solo nú Areia 0.5 a 2 mm sombra
Argila < 0.5 mm Argila < 0.5 mm
7
Manta morta Manta morta
Rocha Arbórea Rocha Aq flutuante
Pedra > 30 cm Arbustiva Pedra > 30 cm Aq submersa
Casc 7.5 a 30 cm Herbácea Casc 7.5 a 30 cm Emergente
Gravilha 2 a 75 mm Gravilha 2 a 75 mm
Areia 0.5 a 2 mm Solo nú Areia 0.5 a 2 mm sombra
Argila < 0.5 mm Argila < 0.5 mm
8
Manta morta Manta morta

59
ANEXO II
Tabela 2: Ocupação do uso do solo e seu respectivo tipo de intervenção.
Intervenção Máxima
Estrada; Lavra intensiva; Hortas; Suinocultura; Vilas; Arrozal; Corte florestal; Solo nu; Cereais e etc.
Intervenção Média
Exploração florestal: Eucalipto; pinheiros; Montado com muito pastoreio; Herbáceas; Olival com prado
Intervenção Mínima
Mato mediterrânico c/ sobreiros; Matos Altos; Galeria Ripícola densa; Montado com lavra pouco Habitual; Caniços, Juncos; Vegetação de
duna secundária
Tabela 3: Classes de impacto determinada para cada ponto de amostragem. Classe de impacto 1 representa menor impacto possível e 7 o máximo. 0 e 1 correspondem a ausência e presença do tipo de intervenção na envolvente a 10m e 100m.
Classe de Impacto Máx. Intervenção Méd Intervenção Min Intervenção
1 0 0 1
2 0 1 1
3 0 1 0
4 1 1 1
5 1 0 1
6 1 1 0
7 1 0 0

60
ANEXO III
Tabela 4: Variáveis dependentes testadas para 2008/09 e comparativo entre 2003/04 e 2008/09.
Variáveis Dependentes 2009
Presença Lagostim 2009
Colonizado Outono 2009
Colonizado Inverno 2009
Colonizado Primavera 2009
Variáveis Dependentes Comparativas 2003/09
Presença Lagostim 2003/09
Ganhou Lagostim 2009
Perdeu Lagostim 2009
Nunca Lagostim 2003/09
Presença Outonos 2003/09
Presença Inverno 2003/09
Presença Primavera 2003/09
Tabela 5: Variáveis medidas em cada ponto de amostragem, unidades e transformações usadas.
Variáveis Unidade Tranformações Habitat (lêntico / lótico) Hidroperíodo 03 Hidroperíodo 09 Sombra Profundidade/100 Profundidade ˂50 (0), ˃ 50cm (1) Percentagem de visibilidade Largura Comprimento Área Corrente Altitude Distâncias dos charcos às ribeiras Variáveis substrato Margem Rocha mãe Pedra ˃30cm Cascalho 7.5 a 30cm Gravilha 2 a 75mm Areia0.5 a 2mm Argila ˂ 0.5mm
Ribeira (1) / Charco (0) Permanente(1)/Temporário(0) Permanente(1)/Temporário(0) % Cobertura cm % Cobertura m m m m/seg m (n m m) m % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura
log (x+1) log (x+1) log (x+1) log (x+1) log (x+1) log (x+1) log (x+1) log (x+1) arcoseno arcoseno arcoseno arcoseno arcoseno arcoseno

61
Manta morta Fundo Rocha mãe Pedra ˃30cm Cascalho 7.5 a 30cm Gravilha 2 a 75mm Areia0.5 a 2mm Argila ˂ 0.5mm Manta Morta Vegetação Margem Vegetação arbórea Vegetação arbustiva Vegetação herbácea Vegetação musgo Solo nu Corpo d’água Veg. Aquática flutuante Veg. Aquática submersa Veg. Aquática emergente Total vegetação aquática Intervenção humana 100m Máxima intervenção Média intervenção Mínima intervenção Classes de impactos
Intervenção comparativa 10m Máxima intervenção Média intervenção Mínima intervenção Classes de impactos
% Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura % Cobertura Presença (1) / Ausência (0) Presença (1) / Ausência (0) Presença (1) / Ausência (0) Ganho(1) / Perda (0) Presença (1) / Ausência (0) Presença (1) / Ausência (0) Presença (1) / Ausência (0)
arcoseno arcoseno arcoseno arcoseno arcoseno arcoseno arcoseno log (x+1) log (x+1) log (x+1) log (x+1) log (x+1) log (x+1) log (x+1) log (x+1) log (x+1)
ANEXO IV

62
Tabela 7: Presença de lagostim por época de amostragem em ambos os estudos (2003/04 e 2008/09).
2003/04 Coloniz. Outono 03/04 Coloniz.Inverno 03/04 Coloniz.Primavera 03/04 Ribeiras 21 (52.5%) 21 (50%) 22 (48%) Charcos 20 (47.5%) 22 (50%) 24 (52%) Ribª/Perm. 16 15 15 Ribª/Temp. 5 6 7 Charco/Perm. 11 11 11 Charco/Temp. 9 11 13 Total 41 43 46
2008/09 Coloniz. Outono 08/09 Coloniz. Inverno 08/09 Coloniz. Primavera 08/09 Ribeiras 14 (60%) 22 (54%) 23 (59%) Charcos 9 (40%) 19 (46%) 16 (41%) Ribª/Perm. 13 14 14 Ribª/Temp. 1 8 9 Charco/Perm. 8 9 9 Charco/Temp. 1 10 7 Total 23 41 39

63
ANEXO V
. Figura 5: Mapas da área de estudo e da colonização máxima em 2003/04 e 2008/09 respectivamente. Os pontos amostrados estão representados por: quadrado amarelo para ribeiras colonizadas e quadrado preto para não colonizadas; círculos amarelos para charcos colonizados e círculos pretos para não colonizados

64
ANEXO VI

65
Figura 6: Mapas da área de estudo e pontos que sofreram alteração entre o anos 2003/04 e 2008/09. O primeiro representa a área total de estudo e sua localização em Portugal e o segundo é uma ampliação da zona amostrada. Os pontos amostrados não alterados estão representados por: quadrado amarelo para ribeiras colonizadas e quadrado preto para não colonizadas; círculos amarelos para charcos colonizados e círculos pretos para não. Já os pontos que sofreram alteração de colonização estão representados por: quadrado vermelho para ribeiras que ganharam lagostins e quadrado azul para as ribeiras que perderam lagostins; círculos vermelhos para charcos que ganharam lagostins e círculos azul para charcos que perderam lagostins.

66
ANEXO VII
DISTÂNCIAS
Tabela 8: Distâncias dos charcos às ribeiras (excepção dos permanentes com lagostim), com o nome dos
pontos amostrados e a distância ao corpo de água mais próxima.
Distâncias dos charcos temporários colonizados Ponto Corpo de água mais próximo Distância (m) M03 Rio Sado 103 M06 Rio Sado arrozal 0 M01 Rio Sado arrozal 0 R02 Rio Sado 270 E04 Barranco Palhota 15 E12 Ribeira Lousal 30 S01b Charco S01 30 Ribeira Corona 700 C09 Ribeira Grândola 20 C10 Ribeira Grândola 100 H07 Ribeira Melides arrozal 20 Distância dos charcos temporários não colonizados Ponto Corpo de água mais próximo Distância (m) C20 Ribeira de Grândola 590 C24 Ribeira Fonte dos Narizes 400 C18 Ribeira Fonte dos Narizes 320 C25 Afluente Fonte dos Narizes 140 C07a Ribeira dos Castelhanos 15 C04 Ribeira dos Castelhanos 107 C03 Ribeira dos Castelhanos 187 C05 Ribeira dos Castelhanos 270 C16 Ribeira dos Castelhanos 600 C14 Afluente Barranco Possancas 290 C17 Ribeira dos Castelhanos 450 E17 Barranco Palhota 310 E05 Barranco Palhota 880 E08 Barranco Palhota 990 E10 Barranco Palhota 1200 F07 Rio Sado 40 F11 Rio Sado 140 I14 Rio Sado 215 I15 Rio Sado 272 T01 Ribeira de Grândola 400 D11 Ribeira de Grândola 100 R01 R02 10 Rio Sado 260 R03 Rio Sado 210 H01b Ribeira de Melides 140 G03 Afluente Ribeira das Fontainhas 240 Ribeira das Fontainhas 843 C21 Barranco Freixeira 390 C22 Ribeira de Grândola 100
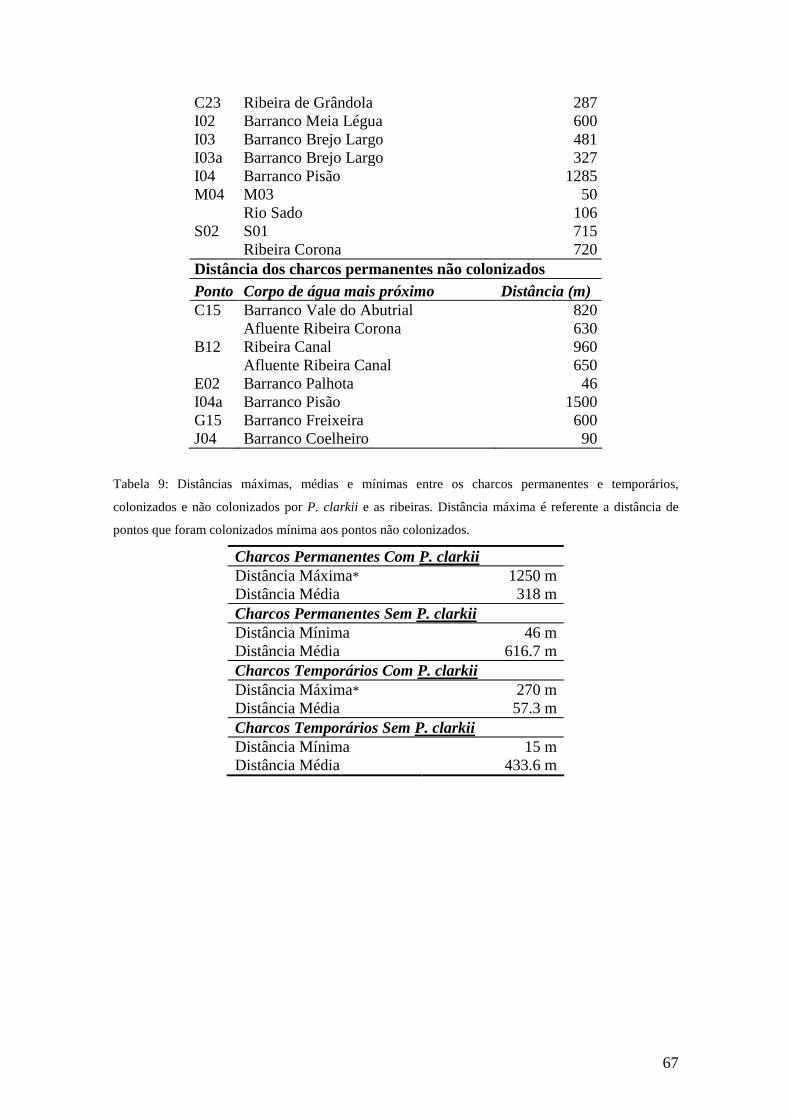
67
C23 Ribeira de Grândola 287 I02 Barranco Meia Légua 600 I03 Barranco Brejo Largo 481 I03a Barranco Brejo Largo 327 I04 Barranco Pisão 1285 M04 M03 50 Rio Sado 106 S02 S01 715 Ribeira Corona 720 Distância dos charcos permanentes não colonizados Ponto Corpo de água mais próximo Distância (m) C15 Barranco Vale do Abutrial 820 Afluente Ribeira Corona 630 B12 Ribeira Canal 960 Afluente Ribeira Canal 650 E02 Barranco Palhota 46 I04a Barranco Pisão 1500 G15 Barranco Freixeira 600 J04 Barranco Coelheiro 90
Tabela 9: Distâncias máximas, médias e mínimas entre os charcos permanentes e temporários,
colonizados e não colonizados por P. clarkii e as ribeiras. Distância máxima é referente a distância de
pontos que foram colonizados mínima aos pontos não colonizados.
Charcos Permanentes Com P. clarkii Distância Máxima* 1250 m Distância Média 318 m Charcos Permanentes Sem P. clarkii Distância Mínima 46 m Distância Média 616.7 m Charcos Temporários Com P. clarkii Distância Máxima* 270 m Distância Média 57.3 m Charcos Temporários Sem P. clarkii Distância Mínima 15 m Distância Média 433.6 m

68
ANEXO VIII
Tabela 10: Variáveis seleccionadas na regressão logística univariadas com coeficiente B. e sig.(significância) onde o P-value requerido foi menor 0.25.
Variáveis
Presença lagostim 2009 B. Sig. Ribª (1) / Charco (0) 0.86 0.036 Permanente (1) / Temporário (0) 2003 2.564 0.000 Permanente (1) / Temporário (0) 2009 2.252 0.000 Gravilha margem 0.028 0.183 Argila margem -0.006 0.254 Manta morta margem 0.013 0.144 Pedra fundo -0.043 0.26 Cascalho fundo arc 0.1940569 0.0795079 Vegetação aquática submersa 0.009 0.241 Vegetação aquática emergente log (x+1) 1.071 0.001 Total vegetação aquática log (x+1) 1.180 0.000 Sombra log (x+1) 0.698 0.011 Profundidade/100 log (x+1) 6.357 0.000 Profundidade˂50 (0), ˃ 50cm (1) 2.036 0.000 Percentagem visibilidade arc 0.3657732 0.0003834 Largura log (x+1) 1.202 0.000 Comprimento 0.011 0.007 Área 0.000201 0.118 Corrente 3.456 0.246 Altitude log (x+1) -1.698 0.001 Mínima intervenção 100m 0.671 0.146 Máxima intervenção10m 1.0766333 0.0218649 Classes de impacto 10m 0.2335092 0.0541878 Distâncias dos charcos às ribeiras log (x+1) -0.612 0.001 Vegetação aquática flutuante log (x+1) 0.715 0.080 Vegetação arbórea log (x+1) -0.013 0.232
Colonizado Outono 2009 B. Sig. Ribª (1) / Charco (0) 0.932 0.055 Permanente (1) / Temporário (0) 2009 4.535 0.000 Areia margem arc 0.260 0.015 Manta morta fundo 0.012 0.200 Vegetação aquática emergente log (x+1) 0.942 0.012 Total vegetação aquática log (x+1) 1.260 0.002 Sombra log (x+1) 1.188 0.000 Profundidade/100 1.804 0.000 Profundidade ˂50 (0), ˃ 50cm (1) 3.045 0.000 Percentagem Visibilidade arc 0.434 0.004 Largura log (x+1) 1.419 0.000 Comprimento 0.010 0.006 Área 0.000 0.022 Altitude -0.016 0.002 Mínima Intervenção 100m 1.827 0.019 Classes de impactos 100m -0.204 0.170
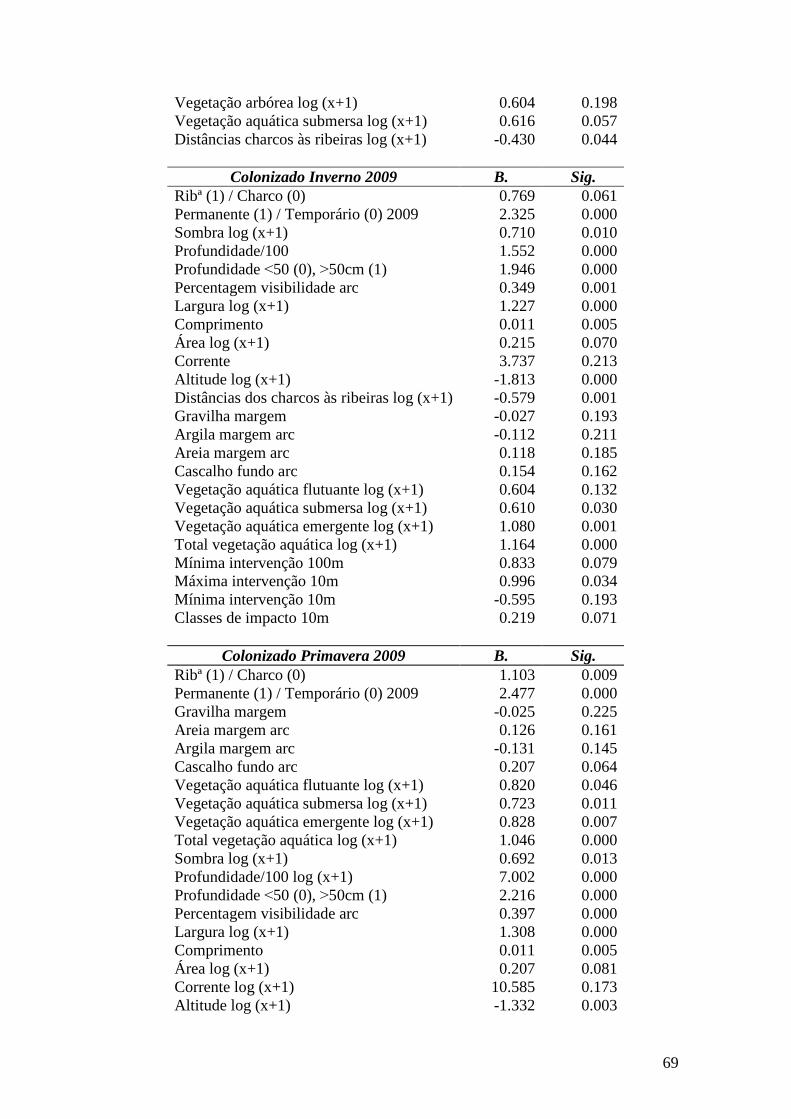
69
Vegetação arbórea log (x+1) 0.604 0.198 Vegetação aquática submersa log (x+1) 0.616 0.057 Distâncias charcos às ribeiras log (x+1) -0.430 0.044
Colonizado Inverno 2009 B. Sig. Ribª (1) / Charco (0) 0.769 0.061 Permanente (1) / Temporário (0) 2009 2.325 0.000 Sombra log (x+1) 0.710 0.010 Profundidade/100 1.552 0.000 Profundidade ˂50 (0), ˃ 50cm (1) 1.946 0.000 Percentagem visibilidade arc 0.349 0.001 Largura log (x+1) 1.227 0.000 Comprimento 0.011 0.005 Área log (x+1) 0.215 0.070 Corrente 3.737 0.213 Altitude log (x+1) -1.813 0.000 Distâncias dos charcos às ribeiras log (x+1) -0.579 0.001 Gravilha margem -0.027 0.193 Argila margem arc -0.112 0.211 Areia margem arc 0.118 0.185 Cascalho fundo arc 0.154 0.162 Vegetação aquática flutuante log (x+1) 0.604 0.132 Vegetação aquática submersa log (x+1) 0.610 0.030 Vegetação aquática emergente log (x+1) 1.080 0.001 Total vegetação aquática log (x+1) 1.164 0.000 Mínima intervenção 100m 0.833 0.079 Máxima intervenção 10m 0.996 0.034 Mínima intervenção 10m -0.595 0.193 Classes de impacto 10m 0.219 0.071
Colonizado Primavera 2009 B. Sig. Ribª (1) / Charco (0) 1.103 0.009 Permanente (1) / Temporário (0) 2009 2.477 0.000 Gravilha margem -0.025 0.225 Areia margem arc 0.126 0.161 Argila margem arc -0.131 0.145 Cascalho fundo arc 0.207 0.064 Vegetação aquática flutuante log (x+1) 0.820 0.046 Vegetação aquática submersa log (x+1) 0.723 0.011 Vegetação aquática emergente log (x+1) 0.828 0.007 Total vegetação aquática log (x+1) 1.046 0.000 Sombra log (x+1) 0.692 0.013 Profundidade/100 log (x+1) 7.002 0.000 Profundidade ˂50 (0), ˃ 50cm (1) 2.216 0.000 Percentagem visibilidade arc 0.397 0.000 Largura log (x+1) 1.308 0.000 Comprimento 0.011 0.005 Área log (x+1) 0.207 0.081 Corrente log (x+1) 10.585 0.173 Altitude log (x+1) -1.332 0.003

70
Máxima intervenção 10m 1.061 0.027 Mínima intervenção 10m -0.546 0.234 Classes de impacto 10m 0.264 0.033 Distâncias dos charcos às ribeiras -0.003 0.015
Presença Ambos anos 2003/09 B. Sig. Ribª (1) / Charco (0) 1.008 0.018 Permanente (1) / Temporário (0) 2009 2.217 0.000 Gravilha margem -0.027 0.219 Cascalho fundo arc 0.215 0.057 Vegetação aquática flutuante log (x+1) 0.716 0.075 Vegetação aquática submersa log (x+1) 0.690 0.017 Vegetação aquática emergente log (x+1) 1.098 0.001 Total vegetação aquática log (x+1) 1.319 0.000 Sombra log (x+1) 0.941 0.001 Profundidade/100 log (x+1) 7.648 0,000 Profundidade˂50 (0), ˃ 50cm (1) 2.548 0.000 Percentagem visibilidade arc 0.502 0.000 Largura log (x+1) 1,342 0.000 Comprimento 0.008 0.018 Área log (x+1) 0.224 0.061 Altitude -0.013 0.001 Mínima intervenção 100m 0.838 0.089 Máxima intervenção 10m 0.901 0.060 Classes de impacto 10m 0.190 0.120 Distâncias dos charcos às ribeiras -0.003 0.019 Areia margem arc 0.121 0.186
Ganhou lagostim 2009 B. Sig. Manta morta margem 0.027 0.172 Cascalho fundo 0.025 0.221 Areia fundo 0.018 0.268 Vegetação herbácea log (x+1) -1.873 0.022 Solo nu 0.033 0.116 Área 0.000 0.250 Corrente 7.867 0.084 Média intervenção 100m -1.749 0.171
Perdeu lagostim 2009 B. Sig. Pedra 30cm margem 0.073 0.247 Gravilha margem 0.034 0.032 Gravilha fundo arc 0.213 0.200 Vegetação herbácea -0.017 0.100 Solo nu 0.031 0.023 Vegetação aquática flutuante 0.021 0.160 Profundidade/100 log (x+1) 3.502 0.108 Mínima intervenção 100m -0.749 0.248 Intervenção comparativa 100m 1.241 0.104 Classes de impacto 10m 0.474 0.027 Cascalho fundo arc 0.237 0.153

71
Areia fundo arc 0.192 0.221 Vegetação aquática flutuante log (x+1) 0.704 0.162
Sem lagostim em 2003/09 B. Sig. Ribª (1) / Charco (0) -0.760 0.062 Permanente (1) / Temporário (0) 2009 -1.992 0.000 Manta morta margem -0.014 0.106 Pedra fundo 0.060 0.127 Gravilha fundo 0.017 0.134 Vegetação aquática flutuante log (x+1) -0.986 0.042 Vegetação aquática submersa log (x+1) -0.603 0.032 Vegetação aquática emergente log (x+1) -0.994 0.001 Total vegetação aquática log (x+1) -1.025 0.000 Sombra log (x+1) -0.729 0.008 Profundidade/100 log (x+1) -6.619 0.000 Profundidade ˂50 (0), ˃ 50cm (1) -1.821 0.000 Percentagem visibilidade arc -0.330 0.000 Largura log (x+1) -1.101 0.001 Comprimento log (x+1) -0.523 0.023 Área log (x+1) -0.254 0.040 Corrente -3.784 0.239 Altitude log (x+1) 2.648 0.000 Máxima intervenção 10m -1.471 0.002 Mínima intervenção 10m 0.668 0.156 Classes de impacto 10m -0.336 0.008 Distâncias dos charcos às ribeiras log (x+1) 0.549 0.001 Vegetação herbácea log (x+1) 0.002 0.216 Cascalho fundo arc -0.159 0.154 Vegetação musgo log (x+1) 0.454 0.214
Presença Ambos Outonos 2003/09 B. Sig. Ribª (1) / Charco (0) 1.003 0.046 Permanente (1) / Temporário (0) 2009 4.130 0.000 Areia margem arc 0.198 0.068 Manta morta fundo 0.012 0.198 Vegetação aquática emergente log (x+1) 0.974 0.013 Total vegetação aquática log (x+1) 1.130 0.006 Sombra log (x+1) 1.033 0.004 Profundidade/100 log (x+1) 7.142 0.000 Profundidade ˂50 (0), ˃ 50cm (1) 2.796 0.000 Percentagem visibilidade arc 0.421 0.007 Largura log (x+1) 1.295 0.000 Comprimento 0.010 0.005 Área 0.000 0.057 Altitude -0.018 0.001 Mínima intervenção 100m 2.446 0.020 Classes de impactos 100m -0.209 0.175 Vegetação aquática Submersa log (x+1) 0.500 0.131
Presença Ambos Invernos 2003/09 B. Sig.

72
Ribª (1) / Charco (0) 0.822 0.057 Permanente (1) / Temporário (0) 2009 1.861 0.000 Gravilha margem arc -0.223 0.135 Vegetação aquática flutuante log (x+1) 0.539 0.177 Vegetação aquática submersa log (x+1) 0.514 0.078 Vegetação aquática emergente log (x+1) 1.067 0.002 Total vegetação aquática log (x+1) 1.209 0.000 Profundidade/100 log (x+1) 6.446 0.000 Profundidade ˂50 (0), ˃ 50cm (1) 2.204 0.000 Percentagem visibilidade arc 0.447 0.001 Largura log (x+1) 1.295 0.000 Comprimento 0.009 0.012 Altitude -0.017 0.000 Mínima intervenção 100m 1.138 0.037 Máxima intervenção 10m 0.811 0.102 Classes de impacto 10m 0.162 0.194 Distâncias dos charcos às ribeiras -0.003 0.030 Sombra log (x+1) 0.828 0.005 Área log (x+1) 0.233 0.056 Cascalho fundo arc 0.171 0.137
Presença Ambas Primaveras 2003/09 B. Sig. Ribª (1) / Charco (0) 1.132 0.011 Permanente (1) / Temporário (0) 2009 1.918 0.000 Vegetação aquática flutuante log (x+1) 0.795 0.054 Vegetação aquática submersa log (x+1) 0.657 0.028 Vegetação aquática emergente log (x+1) 1.020 0.003 Total vegetação aquática log (x+1) 1.272 0.000 Profundidade/100 log (x+1) 8.018 0.000 Profundidade ˂50 (0), ˃ 50cm (1) 2.694 0.000 Percentagem visibilidade arc 0.450 0.001 Largura log (x+1) 1.102 0.001 Comprimento 0.002 0.113 Altitude -0.011 0.005 Mínima intervenção 100m 1.061 0.053 Máxima intervenção 10m 1.324 0.013 Média iIntervenção 10m -0.716 0.176 Classes de impacto 10m 0.241 0.062 Distâncias dos charcos às ribeiras -0.003 0.032 Cascalho margem arc -0.372 0.234 Cascalho fundo arc 0.194 0.095 Vegetação musgo log (x+1) -0.580 0.203 Sombra log (x+1) 0.646 0.028

73
Tabela 11: Coeficiente (B) com erro padrão (S.E.) e P-value do Wald teste x2 para as variáveis obtidas na regressão logística multivariada. A variância explicada para cada modelo pelo teste Nagelkerke R2(NR2).
Variáveis Presença em 2009 B. S.E. Wald Sig. NR2 Gravilha margem -0.366 0.221 2.734 0.098 0.897 Veg. aquática emergente log (x+1) 4.691 2.224 4.447 0.035 Altitude log (x+1) -13.903 2.703 3.767 0.05 Distâncias dos charcos às ribeiras log (x+1) -4.029 7.164 4.584 0.032 Constante 27.506 3.501 0.061 Colonização Outono 2009 B. S.E. Wald Sig. NR2 Total Veg. aquática log (x+1) 2.726 1.543 3.121 0.077 0.548 Altitude -0.029 0.011 6.869 0.009 Constante -1.7148 22.4278 2.18 Colonizado Inverno 2009 B. S.E. Wald Sig. NR2 Gravilha margem -0.491 0.25 3.787 0.05 0.907 Veg. aquática emergente log (x+1) 4.621 2.671 2.993 0.084 Profundidade/100 5.47 3.203 2.916 0.088 Altitude log (x+1) -18.37 9.122 4.055 0.044 Distâncias dos charcos às ribeiras log (x+1) -4.26 2.169 3.856 0.05 Constante -0.60077 3.962702 0.046519 Colonizado Primavera 2009 B. S.E. Wald Sig. NR2 Ribª (1) / Charco (0) 9.633 4.782 4.058 0.044 0.871 Gravilha margem -0.353 0.15 5.531 0.019 Profundidade/100 log (x+1) 27.481 12.045 5.205 0.023 Altitude log (x+1) -12.977 5.577 5.414 0.02 Constante 17.604 4.938 0.026 Presença ambos anos 2003 e 2009 B. S.E. Wald Sig. NR2 Ribª (1) / Charco (0) 7.248 2.522 8.258 0.004 0.72 Veg. Aquática emergente log (x+1) 3.875 1.603 5.843 0.016 Altitude -0.037 0.013 8.05 0.005 Constante -4.52 4.096 0.043 Perdeu em 2009 B. S.E. Wald Sig. NR2 Solo nu 0.039 0.018 4.754 0.029 0.294 Classes de impacto 10m 0.525 0.277 3.593 0.05 Areia fundo arc 0.349 0.208 2.83 0.093 Constante -2.079 34.593 0 Sem lagostim em 2003 e 2009 B. S.E. Wald Sig. NR2 Ribª (1) / Charco (0) 14.582 6.043 5.823 0.016 0.869 Pedra fundo 0.476 0.193 6.083 0.014 Gravilha fundo 0.137 0.058 5.58 0.018 Total Veg. aquática log (x+1) -2.401 1.292 3.453 0.063 Largura log (x+1) -11.334 5.06 5.018 0.025 Comprimento log (x+1) -17.01 8.625 3.889 0.049 Área log (x+1) 14.494 6.361 5.191 0.023 Corrente 29.216 13.098 4.976 0.026 Altitude log (x+1) 11.952 4.541 6.929 0.008 Distâncias dos charcos às ribeiras log (x+1) 4.16 1.737 5.737 0.017 Cascalho Fundo arc -2.044 0.822 6.181 0.013 Constante -0.223 0.996 0.318

74
Presença ambos Invernos 2003 e 2009 B. S.E. Wald Sig. NR2 Permanente (1) / Temporário (0) 2009 -5.764 2.958 3.798 0.05 0.795 Profundidade/100 log (x+1) 16.766 8.783 3.644 0.05 Altitude -0.082 0.033 6.026 0.014 Cascalho fundo arc 1.805 0.859 4.415 0.036 Gravilha margem arc -1.76 0.746 5.566 0.018 Sombra log (x+1) 2.787 1.544 3.26 0.071 Constante -0.887 7.807 0.005 Presença ambas Primaveras 2003 e 2009 B. S.E. Wald Sig. NR2 Veg. aquática emergente log (x+1) 2.944 1.311 5.041 0.025 0.702 Altitude -0.035 0.013 7.406 0.007 Cascalho fundo arc 1.172 0.477 6.025 0.014 Constante -1.099 10.863 0.001

75
ANEXO XIX
Tabela 12: Valores de abundância relativa obtidos através dos métodos de armadilha e passagem de
camaroeiro nos habitats amostrados por épocas do ano.
Ribeira Permanente Outono Inverno Primavera
Inds/armadilha 0.2 0.22 0.7
Inds/passagem camaroeiro 1.2 1.85 0.08
Ribeira Temporária Outono Inverno Primavera
Inds/armadilha 0 0.31 0.87
Inds/passagem camaroeiro 0 0.4 0.04
Charco Permanente Outono Inverno Primavera
Inds/armadilha 0 0 2.1
Inds/passagem camaroeiro 0 1.66 0
Charco Temporário Outono Inverno Primavera
Inds/armadilha 0 1.6 2.4
Inds/passagem camaroeiro 0 0.85 0
Tabela 13: Teste de comparação Mann Whitney U para valores de abundância entre habitats
permanentes e temporários no Inverno e Primavera
Inverno Abundância
U test P (sig.)
Ribeiras permanentes vs temporárias 37,000 0,750
Charcos permanentes vs temporários 4,500 0,462
Primavera Abundância
U test P (sig)
Ribeiras permanentes vs temporárias 35,500 0,948
Charcos permanentes vs temporários 5,000 0,724
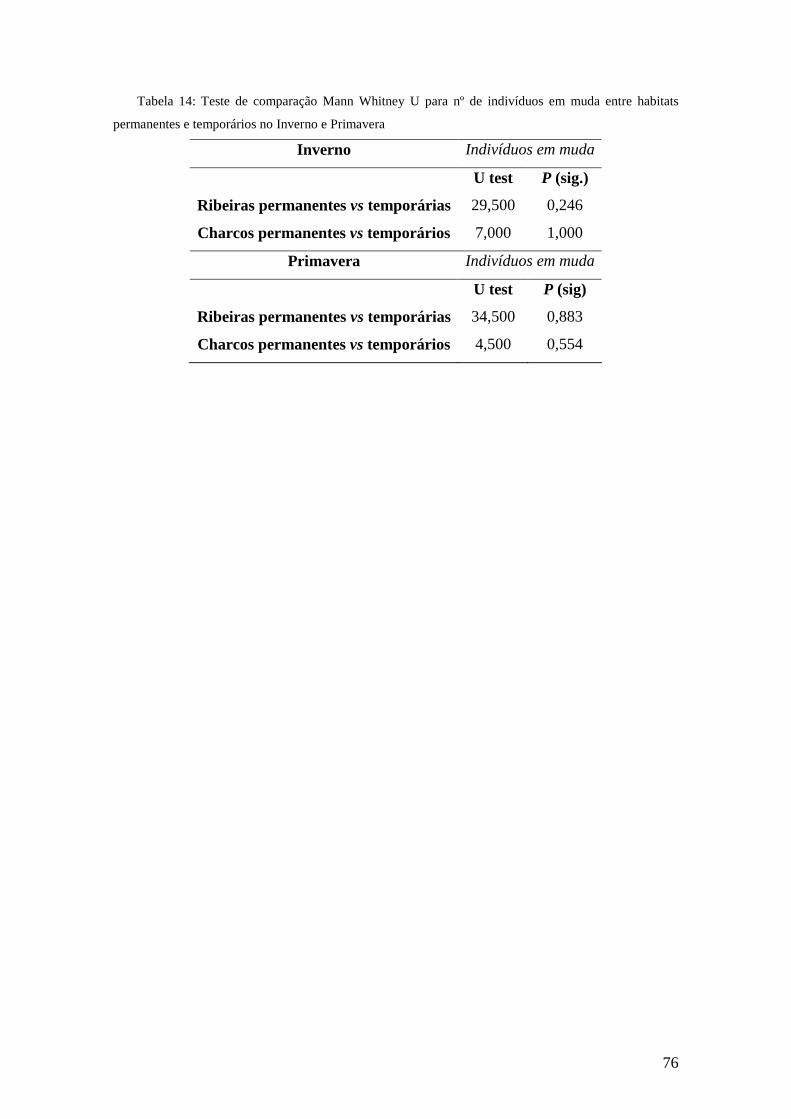
76
Tabela 14: Teste de comparação Mann Whitney U para nº de indivíduos em muda entre habitats
permanentes e temporários no Inverno e Primavera
Inverno Indivíduos em muda
U test P (sig.)
Ribeiras permanentes vs temporárias 29,500 0,246
Charcos permanentes vs temporários 7,000 1,000
Primavera Indivíduos em muda
U test P (sig)
Ribeiras permanentes vs temporárias 34,500 0,883
Charcos permanentes vs temporários 4,500 0,554

77
ANEXO X
Plano de gestão para contenção da espécie invasora de
lagostim, Procambarus clarkii, na Herdade da Ribeira
Abaixo (Grândola - Baixo Alentejo)
“Juntos conseguiremos diminuir sua distribuição e seus impactos”
Autor: Pedro Henrique Sabino de Pereira Leitão

78
O lagostim vermelho Americano (Procambarus clarkii) é uma espécie invasora em
Portugal e tem causado diversos tipos de impacto, tanto para a biodiversidade como
para determinadas culturas agrícolas. A ausência de animais nativos com as mesmas
características ecológicas nos rios e corpos de água, levou a que este lagostim ocupasse
um nicho nunca antes ocupado, não tendo ainda havido tempo para as outras espécies se
adaptarem à sua presença.
O lagostim vermelho adapta-se muito bem a uma variedade de habitats, habitando
tanto grandes rios, ribeiras permanentes e temporárias, como barragens, albufeiras,
açudes, e charcos pequenos, quer permanentes, quer temporários.
Em meados da década de 1980, esta espécie nativa da América do Norte chegou a
Portugal. Na Europa foi inicialmente introduzida em Espanha, em 1973, com vista à
produção em aquacultura. A partir deste país expandiu-se facilmente para Portugal
através da bacia do rio Guadiana e devido à introdução humana. Esta espécie foi
referenciada pela primeira vez na bacia do rio Sado no começo da década de 1990, e
está hoje expandida numa vasta gama de habitats.
Diversos estudos comprovam que a presença de P. clarkii em campos de produção
de arroz pode causar diversos prejuízos aos agricultores, primeiro por impedir a
germinação das sementes, e depois porque a actividade de escavação de buracos nos
canteiros pode vazar a água. O reconhecimento dos seus impactos levou os agricultores
e a comunidade científica a tentar diversas formas de erradicação, mas até agora
nenhuma teve grande sucesso.
A escolha da Herdade Ribeira Abaixo (HRA) (38º 8´N – 8º 33´W, UTM
29SNC3717) como área de aplicação deste plano deve-se a várias razões: é a Estação de
Campo do Centro de Biologia Ambiental (CBA) e da Faculdade de Ciências da
Universidade de Lisboa, e possui um Centro Temático de Educação Ambiental; possui
uma localização geográfica privilegiada - está situada na falda nascente da Serra de
Grândola, a cerca de 7 km a Sul de Grândola, estando limitada a nascente pela Ribeira
dos Castelhanos que desagua na Ribeira de Grândola, sendo esta um afluente do Rio
Sado (importante fonte de lagostim); é bastante rica em espécies de diferentes grupos
faunísticos e florísticos; alberga uma diversidade de habitats e corpos de água estudados
em 2003/04 e 2008/09. O trabalho não se limitará à HRA abrangendo também alguns
pontos importantes na região envolvente, local onde verificou maior alteração nas
colonizações entre os anos de 2003/04 e 2008/09.

79
O plano de contenção apresentado não visa erradicar os lagostins da HRA e
arredores; está sim direccionado para evitar a colonização de charcos temporários
(devido à sua importância ecológica) e de charcos permanentes, pretendendo também
diminuir a densidade de indivíduos nas ribeiras e diminuir assim a sua distribuição e
abrangência dentro da pequena bacia da Ribeira dos Castelhanos. Visa também
trabalhar em conjunto com as escolas e a comunidade local para consciencializar as
pessoas sobre a problemática da introdução irresponsável e não controlada de espécies
exóticas.
Para a elaboração do plano de contenção do lagostim, foram levados em
consideração trabalhos sobre o tema na literatura, o anexo 1 da Directiva Habitats, os
resultados obtidos por Cruz & Rebelo em 2003/04 e o estudo actual em 2008/09 na
bacia do rio Sado, visando a integração entre os trabalhos científicos e as problemáticas
reais, e tendo como resultado uma aplicação prática do conhecimento gerado na
Universidade no campo da Gestão Ambiental e Conservação da Natureza.
Segundo especialistas, as espécies exóticas passam por 3 etapas até se tornarem
invasoras e causarem algum dano ecológico ou económico: 1-) Transporte de
indivíduos; 2-) Estabelecimento da população (aumento em abundância); 3-) Dispersão.
P. clarkii já passou por essas etapas na bacia do rio Sado sendo considerada uma
espécie invasora. O Plano de contenção proposto vai actuar nos pontos 1 e 3 descritos
acima. A consciencialização dos alunos e da comunidade local para a problemática da
introdução de espécies exóticas poderá ajudar a evitar futuros acidentes (etapa nº1); o
plano ir-se-á concentrar na etapa nº 3, para evitar e dispersão dos indivíduos nas ribeiras
e impedir a colonização de charcos temporários. É muito difícil de actuar sobre a etapa
nº 2, visto que a população de P. clarkii já está estabelecida na área.
O plano de contenção está centrado em duas vertentes principais:
1º A diminuição da distribuição do lagostim na área através da contenção nas
ribeiras, evitando a colonização dos charcos temporários;
2º Uma componente educacional, com a consciencialização dos jovens e da
comunidade a respeito da problemática actual da introdução de espécies exóticas.
A inclusão das escolas visa a interacção dos alunos com o seu ambiente próximo, a
interacção com a comunidade, uma melhor divulgação da causa e apoio na realização do
trabalho de campo, além de proporcionar aos alunos o contacto com o dia-a-dia da
profissão de biólogo, suas abrangências e potencialidades.

80
A escolha dos melhores locais para a realização do trabalho terá de cumprir
determinados critérios, sendo que serão escolhidos locais não colonizados por lagostim
e locais colonizados para a realização da acção de contenção.
ACÇÕES NO TERRENO
Escolha da época do ano
O melhor momento para a realização da intervenção é no final do Verão/ começo do
Outono (antes das primeiras chuvas), pois é quando existe menor quantidade de água
nas ribeiras. No final do Outono a intervenção será mais complicada pois já começa a
registar-se maior actividade nas ribeiras e indícios de reprodução, com maior presença
de indivíduos juvenis. No trabalho realizado em 2008/09 na bacia do rio Sado, apenas
23 dos 104 pontos amostrados continham lagostins no Outono, já no Inverno passaram a
41.
O clima pode contribuir para uma maior eficácia da intervenção e consequente
diminuição da área de distribuição. O ano hidrológico de 2008/09 foi mais seco que o
de 2003/04, o que contribuiu para que muitos charcos estivessem secos e muitas ribeiras
secassem, perdendo assim lagostim, o que pode facilitar o trabalho de colheita e
controlo.
O início do Outono tem também a vantagem de coincidir com o início do ano
escolar, permitindo visitas integradas em actividades escolares.
Métodos de Intervenção
Prevenção da colonização nos pontos isolados:
Construção de barreiras físicas para impedir a chegada dos indivíduos aos charcos
com instalação de tela de vedação e armadilhas “pitfall”. Armadilhas de queda
(“pitfall”) são baldes enterrados no chão (para os lagostins o tamanho dos baldes
caseiros já é suficiente) nos quais os indivíduos caiem, não conseguindo sair. A
colocação das telas deverá ser feita nas margens dos pontos que fiquem voltadas para os
rios. As armadilhas de queda deverão estar colocadas nas pontas das telas e algumas no
meio da vedação, sendo que o nº de armadilhas varia consoante a área do ponto. As
armadilhas devem ser verificadas diariamente para a retirada dos lagostins e de outros
animais que eventualmente lá tenham caído. Em alguns casos, barreiras físicas naturais

81
podem ser incrementadas com pedras, cascalhos e material vegetal (galhos e troncos),
pois os lagostins têm dificuldade em atravessar esse tipo de obstáculo.
Colheita intensiva de indivíduos nas ribeiras:
A melhor forma de capturar indivíduos é com a conjugação de diferentes métodos,
cada um mais eficaz para uma determinada ocasião e classe etária. Serão conjugados
três métodos de captura:
Armadilha de funil iscada, mais eficaz para indivíduos adultos;
Passagens de camaroeiro, mais eficaz para juvenis;
Colecta com identificação visual.
As armadilhas podem ser construídas artesanalmente (muito eficaz), com a
utilização de garrafas de plástico de 5 e 1,5 L de água, onde a tampa é cortada e
invertida de forma a transformar-se em um funil. Para o isco pode ser utilizado comida
de gato com sabor peixe. Em cada ponto deverão ser colocadas 50% de armadilhas de 5
L e 50% de 1.5 L. O número de armadilhas por ponto é determinado pelo tamanho do
ponto, não esquecendo que é um trabalho de erradicação e quanto maior o número de
indivíduos capturados melhor. As armadilhas terão de ser colocadas no início da noite e
retiradas durante o dia seguinte, sendo deixadas nos pontos durante 12 h.
A passagem de camaroeiro e a colecta com identificação visual deverão ser
realizados durante o dia e de forma intensiva a fim de capturar o máximo de indivíduos
possível e nos variados microhabitats.
Os lagostins colectados serão mortos de modo a ser o menos doloroso possível para
os animais e o mais rápido quanto possível, usando-se geralmente a decapitação. Os
animais poderão ser doado aos proprietários das herdades vizinhas, visto que é
comestível (razão de sua introdução na Europa) e pode servir de alimentação aos
animais domésticos. É importante garantir que todos os indivíduos devem ser
transportados mortos.
Escolha dos Pontos
Este plano tem um carácter demonstrativo e visa concentrar a contenção numa área
restrita para torná-lo mais facilmente aplicável, devido aos menores custos e maiores
facilidades. Está direccionado para uma acção de educação ambiental e para o maior
controlo da população de lagostim na pequena bacia da ribeira de Castelhanos, HRA e
alguns pontos próximos.

82
Prevenção da colonização nos pontos isolados:
Estes pontos foram escolhidos de acordo com os seguintes critérios:
-Charcos temporários localizados a uma distância menor que 300 metros de um rio
colonizado;
-Charcos temporários que perderam lagostim entre 2003 e 2009;
-Charcos temporários que foram em algum dos anos colonizados por P. clarkii;
-Charcos permanentes que perderam lagostim e que estão a uma distância menor
que 300 metros de um rio colonizado.
Colheita intensiva de indivíduos nas ribeiras:
As ribeiras serão escolhidas consoante a proximidade a algum ponto isolado que
cumpra os critérios acima explicitados. Nas ribeiras que secam parcialmente durante o
verão serão escolhidos os locais que mantêm água durante o período de colheita (pontos
identificados como “fonte” de indivíduos colonizadores de lagostim). No caso das
ribeiras manterem água corrente todo o ano, serão escolhidos os locais mais próximo
aos pontos isolados.
Com o conhecimento do terreno e da tabela das distâncias dos pontos isolados
(Anexo VII), foram seleccionados os locais que estão sob maior risco e que serão
intervencionadas.
Para a HRA foram seleccionados 5 charcos isolados (C03, C04, C05, C06 e C07a)
onde serão construídas as barreiras; toda a colheita intensiva de indivíduos será feita na
ribeira de Castelhanos em diferentes pontos, consoante a proximidade aos charcos.
Para a região envolvente detectámos uma série de pontos isolados que devem ser
intervencionados, são eles: C25 (Fonte dos Narizes); E02, E04 (Barranco de Palhotas) e
C09; C10 (Ribeira de Grândola).
Todos os pontos acima referidos já foram alguma vez colonizados por P. clarkii e
estão a uma distância que lhe confere risco de recolonização. Os pontos C06, E04, C09
e C10 foram colonizados em 2008/09, os restantes estiveram colonizados em 2003/04 e
estão em risco.
O tempo de duração dos trabalhos de captura nas ribeiras será desde o final do
Verão/começo do Outono até ao período no qual os pontos temporários já estão todos
cheios. No entanto a verificação das armadilhas de “pitfall” posicionadas nos pontos
isolados devem ser diárias até ao final do Inverno de acordo com o cronograma.

83
Componente educacional:
O trabalho de contenção será muito mais eficaz se realizado em associação com as
escolas locais. Deverão ser realizadas palestras para chamar a atenção para a
problemática das espécies invasoras, introdução de animais exóticos e os seus impactos
e para evitar futuras introduções. Os alunos acompanharão algumas das saídas de campo
organizadas para as colheitas, onde terão contacto com o campo e com habitats naturais.
Para além dos benefícios óbvios de uma saída de campo os alunos acompanharão o dia
a dia de um profissional da área da biologia, aprenderão sobre as abrangências do
trabalho do biólogo o que poderá ajudá-los a decidir as suas futuras profissões.
Deve-se ter algumas precauções ao incentivar ou estimular a colheita de uma
determinada espécie em determinado habitat, pois temos de garantir que apenas aquela
espécie será capturada e não as outras. É preciso um trabalho de educação cauteloso
antes do trabalho prático, onde será ensinado a identificar o lagostim e outras espécies
que se encontram na área. Como não há espécies de lagostim nativo na região e nem
outra espécie de lagostim exótica, é baixa a possibilidade de confusão.
O professor de Biologia da escola deve aproveitar esse momento para enfatizar os
conceitos leccionados anteriormente ou que ainda serão leccionados. Poderá aproveitar-
se principalmente na área de Ciências da Natureza e Biologia, dependendo do ano
escolar. Há ainda a possibilidade de saídas de campo integradas com outras áreas
disciplinares, como por exemplo a Geografia.
No momento do trabalho de campo poderão ser formadas duas equipas
supervisionadas de alunos, uma com o trabalho de colheita dos indivíduos nas ribeiras e
outra na verificação das armadilhas nos charcos.
O trabalho pode vir a ser uma actividade anual da escola, direccionada por exemplo
a um ano escolar, visto que as colonizações e recolonizações acontecem todos os anos.
A ideia principal deste plano é proporcionar meios para o controlo imediato da
população de lagostim em alguns locais indicados com elevado valor ecológico. No
entanto, sabendo da impossibilidade de erradicação dessa espécie invasora em locais já
colonizados, uma acção possível a longo prazo é proporcionar os meios ideais para a
incorporação desta espécie na alimentação de espécies autóctones, de modo a que haja
um controle populacional e diminuição dos seus impactos. Uma forma adequada para
integrá-lo na cadeia alimentar dos animais autóctones seria a preservação e restauro das
galerias ripícolas e mantendo uma boa qualidade da água dos rios, condições

84
importantes para a manutenção e aumento das populações de carnívoros médios, como a
lontra e a geneta, predadores comprovados de lagostim na HRA.
Para a realização desse plano de contenção indicamos como necessários a
permanência de 2 biólogos. Serão eles os responsáveis por entrar em contacto com as
escolas, pela educação ambiental nas escolas e no campo, principalmente, sobre o tema
da introdução das espécies exóticas. Serão também os encarregados do trabalho prático,
incluindo a colocação e verificação das armadilhas e da captura dos indivíduos nas
ribeiras. No momento da educação ambiental no campo será necessário a presença de no
mínimo 1 biólogo para, com o apoio dos professores envolvidos, realizarem a aula e a
colheita de lagostim, enquanto o outro biólogo verifica os outros pontos.
Cronograma de tarefas:
Tarefas Setembro Outubro Novembro Dezembro Janeiro Fevereiro Março Abril Maio
Verificação das
armadilhas nos
charcos
(Diariamente)
Colheita intensiva de
lagostim nas ribeiras
(Diariamente)
Monitorização dos
pontos
(Uma vez ao mês)

85
Protocolo prático
Passo a passo
-Visitar os locais para verificar acessibilidade e determinar o número de armadilhas
por ponto;
- Construir as armadilhas;
- Armar as barreiras e armadilhas “pitfall” nos pontos isolados;
- Capturas nas ribeiras e verificações diárias das armadilhas “pitfall”.
Construção das armadilhas
Para a construção das armadilhas serão necessários os seguintes materiais:
- Garrafas plásticas de água de 1,5 e 5 Litros;
- Tesoura e Faca;
- Corda.
As garrafas devem ser cortadas na parte superior de modo que, quando invertidas
formem um funil, fique bem presa e com espaço no interior da garrafa (figura 1).
Devem ser feitos furos na parte inferior da armadilha para manter a circulação da água
(muito importante para oxigenação). A corda serve para amarrar a armadilha a locais
seguros nas margens.
Notificações importantes:
- Aprender a identificar correctamente P. clarkii;
- Construir correctamente as armadilhas, não esquecendo dos furos para a circulação
da água;
- Armar correctamente as barreiras e armadilhas de queda “pitfall” nos pontos
isolados;
- Após a abertura das armadilhas de queda “pitfall” é necessário verificá-las todos os
dias até ao seu fechamento ou retirada;
- As armadilhas nas ribeiras devem ficar no máximo 12h e incluir uma noite;
- Preservar as outras espécies;
- As armadilhas devem estar bem presas à margem;
- Não deixar nenhum lixo nos locais de trabalho.

86
Figura 1: Construção de armadilhas
Esquema do local de amostragem de vedação com armadilhas “pitfall”:
Tabela de coordenadas geográficas dos pontos escolhido para a intervenção Ponto Latitude (N) Longitude (W) ( Y_PROJ) (X_PROJ) C03 38.10580434 8.56886236 4217642.08169776 537798.36971623 C04 38.10583401 8.56790791 4217645.76273796 537882.03258783 C05 38.10012686 8.58953951 4217003.92112770 535988.35558486 C06 38.10813275 8.57435360 4217898.22642900 537315.75923732 C07a 38.11345719 8.56997908 4219578.07833659 537410.20818394 C09 38.16173444 8.53884945 4203459.03559976 539518.37255351 C10 38.16352389 8.54273060 4217935.75868760 537680.34911647 C25 38.11746282 8.55997553 4218939.32118281 538571.36104581 E02 38.07144986 8.47977773 4213870.29987718 545629.92712650 E04 38.06173204 8.45927375 4212802.33289858 547434.66679015

87
Exemplo de panfleto de divulgação nas escolas: ----------------------------------------------------------------------------------------------------------
----------------------------------------------------------------------------------------------------------
ESPÉCIE INVASORA Vamos acabar com ela… Procambarus clarkii, o lagostim vermelho americano é a causa de diversos prejuízos aos agricultores, tem impactos na biodiversidade e esta bem ao nosso lado. Consegue espalhar-se rapidamente por todos os corpo de água, incluindo rios e represas.
JUNTE-SE NESSA CAMPANHA
Não introduza essa espécie nos rios e lagos de sua cidade ou herdade… VAMOS JUNTOS AJUDAR A COMBATER ESSA PRAGA !!!