transmissão sináptica - azevedolab.netazevedolab.net/resources/biofisica6.pdf · na figura ao...
TRANSCRIPT

Biofísica
Transmissão SinápticaProf. Dr. Walter F. de Azevedo Jr.
©2
01
5 D
r. W
alte
r F.
de
Aze
ve
do
Jr.
1

O aumento de pesquisas focadas no
funcionamento do cérebro, criou uma
quantidade enorme de informações
científicas relevantes dispersas em
milhares de artigos científicos. A proposta
de um grupo de pesquisadores
americanos da Carnegie Mellon
University, é disponibilizar a informação
sobre dados fisiológicos de neurônios de
forma organizada e integrada. A partir de
uma ferramenta computacional, foi
organizada uma base de dados com
esses resultados e disponibilizada no site:
http://www.neuroelectro.org/. Este site
funciona como uma base de dados sobre
a fisiologia dos neurônios.
2
Notícia Relacionada
Base de dados http://www.neuroelectro.org/ .
Disponível em: < http://www.kurzweilai.net/a-wikipedia-for-
neurons >.
Acesso em: 10 de setembro de 2015.

O que é a singularidade tecnológica ?
O que é o modelo de Hogkin-Huxley?
O que é sinapse?
Como funcionam os fármacos anti-Alzheimer?
Fonte: http://www.kurzweilai.net/
3

Uma grande parte de cientistas da área
de inteligência artificial, acredita que
vivemos um momento especial da história
do desenvolvimento científico. Devido à
importância deste momento, destaco nos
meus modestos cursos alguns aspectos
relevantes do processo da singularidade
tecnológica. Uma das características
desta última é o aumento expressivo da
expectativa de vida. Se compararmos a
expectativa de vida hoje, com a de um
brasileiro do início do século XX, vemos
que mais que dobramos nossa
expectativa. No gráfico ao lado, vemos
que a expectativa de vida do brasileiro em
1910 era de 34 anos, e hoje está acima
de 70 anos. O aumento deve-se a
diversos fatores, tais como o
desenvolvimento no saneamento básico e
as conquistas científicas da medicina
moderna.4
Ano
Expectativa de vida do Brasileiro entre 1910 e 2009. Fonte
dos dados: Informe da Previdência Social. Disponível em: <
http://www.previdencia.gov.br/arquivos/office/4_110525-
171625-908.pdf >.
Acesso em: 10 de setembro de 2015.
0
10
20
30
40
50
60
70
80
1900 1920 1940 1960 1980 2000 2020
Singularidade Tecnológica
Ex
pe
cta
tiva
de
vid
a

Dados para expectativa de vida, antes de
1910, indicam números ainda piores.
Segundo algumas fontes, a expectativa
de vida no Brasil em 1900 era inferior a 30
anos. Fonte: Laboratório de Demografia e
Estudo Populacionais. Disponível em: <
http://www.ufjf.br/ladem/2012/02/28/aume
nto-da-longevidade-e-estancamento-da-
esperanca-de-vida-artigo-de-jose-
eustaquio-diniz-alves/ >. Acesso em: 10
de setembro de 2015.
Um gráfico da evolução da expectativa de
vida ano a ano (2000-2012), mostra
aspectos curiosos do aumento. Vemos um
avanço considerável entre 2002 e 2004.
Este pulo na melhora da expectativa de
vida, é, também, uma consequência direta
de políticas públicas de redução da
pobreza.
5
Expectativa de vida do Brasileiro entre 2000 e 2012. Fonte
dos dados: Index Mundi. Disponível em:
< http://www.indexmundi.com/g/g.aspx?c=br&v=30&l=pt >.
Acesso em: 10 de setembro de 2015.
60
62
64
66
68
70
72
74
76
78
80
1998 2000 2002 2004 2006 2008 2010 2012 2014
Ex
pe
cta
tiva
de
vid
a
Ano
Singularidade Tecnológica

Olhando para o futuro, a expectativa de
vida traz grandes promessas. Um
geneticista da Cambridge University -
Reino Unido, prevê que a primeira pessoa
a viver mais de 1000 anos já está entre
nós (Site da BBC. Disponível em: <
http://news.bbc.co.uk/2/hi/uk_news/40030
63.stm >. Acesso em: 10 de setembro de
2015.
Isto mesmo, mil anos! Não é erro de
digitação. Eu sou cético com relação a
este número, mas acredito, baseado na
aceleração do desenvolvimento científico,
que ultrapassaremos o limite de 120 anos
nas próximas décadas.
6
Página de entrada do site da Strategies for Engineered
Negligible Senescence (SENS) Foundation.
Disponível em: < http://sens.org/>.
Acesso em: 10 de setembro de 2015.
Singularidade Tecnológica

A evolução da ciência médica, nos deu
nas últimas décadas desenvolvimentos
como transplantes, vacinas, novos
fármacos etc. Além disso, temos a
expectativa da substituição de órgãos,
como o rim crescido artificialmente
mostrado ao lado (Song et al., 2013).
Baseado neste cenário, podemos ser
otimistas quanto à expectativa de vida do
ser humano. Esperamos que, nas
próximas décadas, teremos a
possibilidade de substituição de nossos
órgãos conforme envelhecemos. A
substituição do rim por um crescido
artificialmente tem uma perspectiva de ser
possível numa década. Outros órgãos
apresentam equivalente biomecânico,
como o coração.
7
Rim artificial testado em ratos.
Disponível em: < http://www.bbc.co.uk/news/science-
environment-22149844>.
Acesso em: 10 de setembro de 2015.
Referência:
Song JJ, Guyette JP, Gilpin SE, Gonzalez G, Vacanti JP, Ott
HC.
Regeneration and experimental orthotopic transplantation of
a bioengineered kidney. Nat Med. 2013 Apr 14. doi:
10.1038/nm.3154
Singularidade Tecnológica

Além do aumento expressivo do número
de anos vividos, a humanidade usufruirá
de facilidades tecnológicas, cada vez
mais baratas.
A evolução da medicina e da cibernética,
permitirá o desenvolvimento de um
equivalente computacional ao cérebro
humano. Como o desenvolvimento
concomitante da neurociência, espera-se
que tenhamos a capacidade tecnológica
de transferirmos o conjunto de nossas
sinapses para um cérebro eletrônico, ou
seja, a substituição do cérebro humano,
por um equivalente computacional. Nessa
fase a humanidade atingirá virtualmente a
imortalidade. A situação onde esta
transição ocorrerá, é chamada de
singularidade tecnológica.Visão artística da modelagem matemática do cérebro.
Disponível em: <http://www.kurzweilai.net/mind-uploading-
featured-in-academic-journal-for-first-time>.
Acesso em: 10 de setembro de 2015.
8
Singularidade Tecnológica

O gráfico ao lado ilustra a lei de Moore,
que estabelece que aproximadamente
entre 18 e 24 meses o número de
transistores por chip dobra. Esta lei foi
proposta por Gordon Moore cofundador
da Intel. Ou seja, considerando-se os
processadores hoje, esperamos que em
aproximadamente entre 18 e 24 meses
teremos disponíveis, pelo mesmo preço,
computadores com o dobro da
capacidade de processamento. Uma
extrapolação da lei de Moore para 2030,
ou um pouco depois, indica que teremos
computadores com a complexidade do
cérebro humano.
Disponível em: <
http://www.kurzweilai.net/the-law-of-
accelerating-returns >. Acesso em: 10 de
setembro de 2015.
Evolução do número de transistores por chip em função do
ano.
Disponível em:
http://library.thinkquest.org/4116/Science/moore%27s.htm.
Acesso em: 10 de setembro de 2015.
9
Singularidade Tecnológica

A pesquisa em singularidade tecnológica
é uma atividade multidisciplinar, cujo o
foco é o entendimento dos sistemas
biológicos e computacionais,
especificamente a interface do ser
humano com máquinas. A partir deste
conhecimento, teremos condições de
prolongar nossa expectativa de vida, até
termos condições tecnológicas de
transferirmos nossa consciência para um
sistema computacional, o que abre a
possibilidade da imortalidade, bem como
uma nova fase da evolução humana. Tal
fase da evolução permitirá a integração
das consciências computacionais, o que
abre um amplo espectro de
possibilidades. Tais tecnologias ainda não
existem, mas se consideramos a lei de
Moore, vemos que o rápido
desenvolvimento tecnológico nos levará
até este estágio.
Visão artística do cérebro digital.
Disponível em: < http://www.kurzweilai.net/critique-of-
against-naive-uploadism#!prettyPhoto>.
Acesso em: 10 de setembro de 2015.
10
Singularidade Tecnológica

Muitos autores destacam que, as
pesquisas mais importantes e
desafiadoras nos dias de hoje, estão
relacionadas com a singularidade
tecnológica. A biofísica pode contribuir
nesta área em duas frentes de atuação.
Uma frente para entendermos as bases
moleculares do funcionamento do
cérebro, que permitirá seu entendimento e
então sua modelagem computacional.
Noutra frente, ao vivermos mais (aumento
da expectativa de vida), nos tornamos
sujeitos a novas enfermidades, que
podem ser combatidas com abordagens
do desenho de fármacos baseado em
computadores.
11
Página de entrada do site Kurzweil Accelerating Intelligence.
Disponível em:<http://www.kurzweilai.net/ >.
Acesso em: 10 de setembro de 2015.
Singularidade Tecnológica

12
Ao atingirmos a singularidade
tecnológica, abandonaremos as
limitações biológicas do nosso ser e
atingiremos um universo de novas
possibilidades que tal fase nos
trará.
Singularidade Tecnológica

Maiores informações sobre a singularidade tecnológica podem ser encontradas nos
artigos de Ray Kurzweil disponíveis on-line no site Kurzweil Accelerating Intelligence.
► Kurzweil responds: Don’t underestimate the Singularity. Disponível em: <
http://www.kurzweilai.net/kurzweil-responds-dont-underestimate-the-singularity>.
Acesso em: 10 de setembro de 2015.
► The new era of health and medicine as an information technology is broader than
individual genes. Disponível em: < http://www.kurzweilai.net/the-new-era-of-health-
and-medicine >. Acesso em: 10 de setembro de 2015.
► How my predictions are faring — an update by Ray Kurzweil. Disponível em: <
http://www.kurzweilai.net/how-my-predictions-are-faring-an-update-by-ray-kurzweil >.
Acesso em: 10 de setembro de 2015.
► The Law of Accelerating Returns. Disponível em: < http://www.kurzweilai.net/the-
law-of-accelerating-returns >. Acesso em: 10 de setembro de 2015.
13
Singularidade Tecnológica

14
Singularity University

15
Singularity University

O modelo de Hodgkin-Huxley foi proposto em 1952 para modelar o potencial de ação
do axônio de sépia. Os dados sobre a corrente iônica foram obtidos a partir do uso da
técnica de “voltage clamp”. Nesta técnica, o potencial elétrico da célula é mantido
constante, independente das concentrações iônicas. Na situação abaixo as correntes
iônicas são medidas. O diagrama esquemático abaixo ilustra a técnica. Um sistema de
retroalimentação permite que o potencial seja mantido num valor de referência, onde
toda vez que o potencial de membrana desvia-se um pouco do valor ajustado, uma
tensão adicional, fornecida pela fonte de tensão, leva o potencial de membrana de
volta ao valor ajustado.
Axônio
Amperímetro
Eletrodo
extracelular
Eletrodo
intracelular
Amplificador do potencial
de membrana
Fonte de tensão
Amplificador de
retroalimentação
16
Modelo de Hodgkin-Huxley
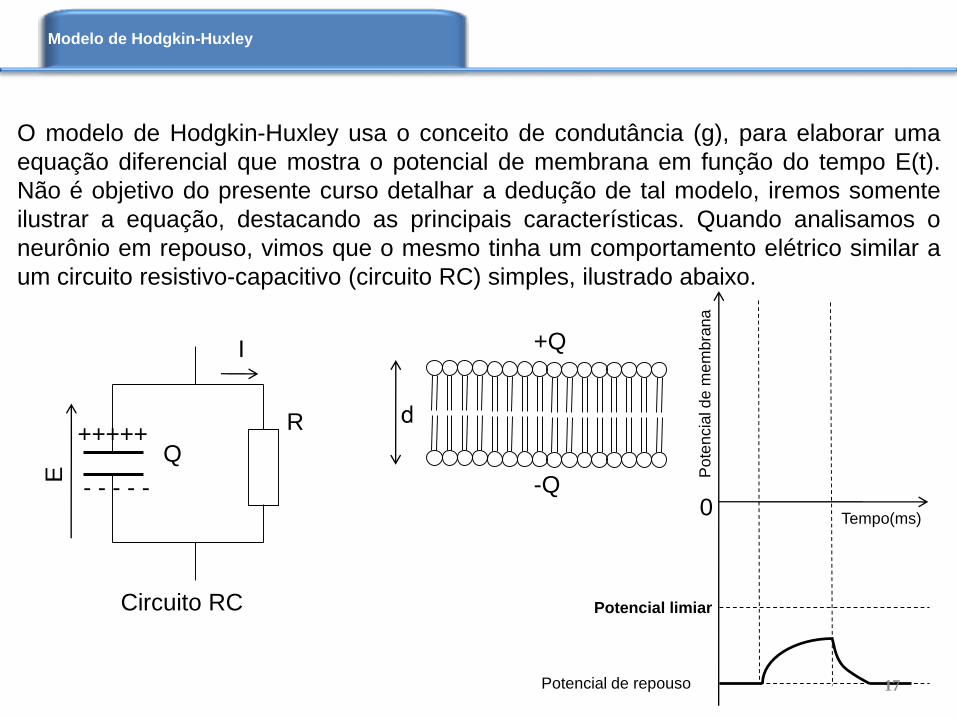
O modelo de Hodgkin-Huxley usa o conceito de condutância (g), para elaborar uma
equação diferencial que mostra o potencial de membrana em função do tempo E(t).
Não é objetivo do presente curso detalhar a dedução de tal modelo, iremos somente
ilustrar a equação, destacando as principais características. Quando analisamos o
neurônio em repouso, vimos que o mesmo tinha um comportamento elétrico similar a
um circuito resistivo-capacitivo (circuito RC) simples, ilustrado abaixo.
R
Circuito RC
+++++
- - - - -
E
I
Q
+Q
-Q
Po
ten
cia
l d
e m
em
bra
na
Tempo(ms)
Potencial de repouso
Potencial limiar
0
17
Modelo de Hodgkin-Huxley

Na figura ao lado, temos o circuito elétrico
equivalente da membrana celular durante
o potencial de ação, chamado de modelo
de Hodgkin-Huxley. No circuito temos 3
tipos de correntes iônicas, a corrente do
Sódio (INa), a corrente do Potássio (IK) e
uma terceira corrente chamada em inglês
de “leak current” (IL), que é composta
principalmente de íons de cloro. A
principal contribuição do modelo Hodgkin-
Huxley, foi a introdução das condutâncias
na análise do comportamento elétrico do
axônio. A condutância (g) é o inverso
da resistência elétrica (R), conforme a
equação abaixo:
A unidade de condutância é o Siemens (1
S = 1/Ohm).
gNa gK gL
ENa EK EL
Cm
IC
INa IK IL
I
IÍon
R
1 g
Circuito elétrico equivalente a membrana celular, segundo
o modelo de Hodgkin-Huxley.
Meio extracelular
Meio intracelular
18
Modelo de Hodgkin-Huxley

Ao incluirmos as condutâncias (g), temos
a possibilidade de modelar o potencial de
ação, considerando o circuito equivalente
ao lado. Temos as condutâncias para
Sódio (gNa), Potássio (gK) e uma terceira
para o termo “leak” (gL), ou seja,
vazamento de íons. A corrente total (I) é a
soma da corrente iônica (Iíon) e a corrente
capacitiva (IC), como segue:
I = IC + Iíon .
A corrente iônica total (Iion) é dada pela
soma de todas as correntes devido a cada
canal (INa e IK) e a terceira corrente (IL) é
constante e minoritária. Assim temos,
Iion = INa + IK + IL
gNa gK gL
ENa EK EL
Cm
IC
INa IK IL
I
IÍon
Circuito elétrico equivalente a membrana celular, segundo
o modelo de Hodgkin-Huxley.
Meio extracelular
Meio intracelular
19
Modelo de Hodgkin-Huxley

Vemos no circuito equivalente a presença
de 3 fontes de potencial, nominalmente
Ena , EK e EL, relativas ao Sódio, Potássio
e vazamento (leak). Tais potenciais
podem ser determinados a partir da
equação de Nernst, vista anteriormente.
Assim, temos as seguintes equações para
as correntes que formam a corrente
iônica,
INa = gNa (V - ENa ) ,
Ik = gk (V - Ek )
e
Il = gL (V - EL )
onde V é o potencial da membrana.
Veja no circuito, que as condutâncias do
Sódio (gNa) e do Potássio (gK) são
variáveis e a condutância de vazamento é
constante.
gNa gK gL
ENa EK EL
Cm
IC
INa IK IL
I
IÍon
Circuito elétrico equivalente a membrana celular, segundo
o modelo de Hodgkin-Huxley.
Meio extracelular
Meio intracelular
20
Modelo de Hodgkin-Huxley
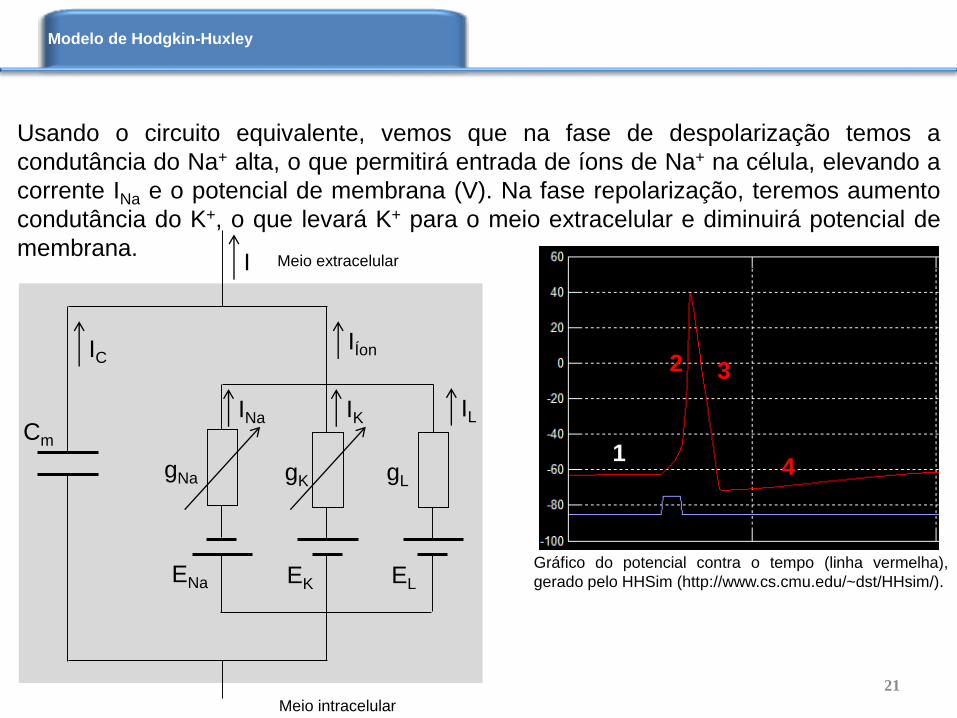
Usando o circuito equivalente, vemos que na fase de despolarização temos a
condutância do Na+ alta, o que permitirá entrada de íons de Na+ na célula, elevando a
corrente INa e o potencial de membrana (V). Na fase repolarização, teremos aumento
condutância do K+, o que levará K+ para o meio extracelular e diminuirá potencial de
membrana.
1
2 3
4
Gráfico do potencial contra o tempo (linha vermelha),
gerado pelo HHSim (http://www.cs.cmu.edu/~dst/HHsim/).
gNa gK gL
ENa EK EL
Cm
IC
INa IK IL
I
IÍon
Meio extracelular
Meio intracelular
21
Modelo de Hodgkin-Huxley

O modelo de Hodgkin-Huxley é um
modelo computacional, sendo
considerado o primeiro modelo da
abordagem de biologia de sistemas. Tal
modelo descreve a resposta do axônio de
sépia a diferentes estímulos elétricos.
Temos a implementação do modelo
computacional de Hodgkin-Huxley
(modelo HH) em diversos programas.
Apresentaremos aqui um que foi
implementado na linguagem MatLab,
chamado HHSim que está disponível no
site http://www.cs.cmu.edu/~dst/HHsim/ .
Esse simulador do potencial de ação
possibilita testarmos diferentes tipos de
estímulos elétricos aplicados ao axônio,
bem como o efeito de moléculas que
interagem com os canais iônicos. Axônio pré-sináptico da sépia, colorido em rosa para
destaque.
Disponível em: http://dels-old.nas.edu/USNC-IBRO-
USCRC/resources_methods_squid.shtml
Acesso em: 10 de setembro de 2015.
Diagrama esquemático de uma seção do axônio de sépia.
22
Modelo de Hodgkin-Huxley

O diagrama esquemático abaixo ilustra o arranjo experimental, simulado no HHSim.
Temos o cilindro representando uma seção do axônio da sépia, onde foram inseridos 2
eletrodos. Temos o eletrodo 1 responsável pelo estímulo, que será medido em
unidades de corrente elétrica, nA (nanoAmpére, 10-9 A).
Eletrodo 1
Gerador de corrente elétrica (estímulo)
Seção do axônio de sépia.23
Modelo de Hodgkin-Huxley

Temos um segundo eletrodo (eletrodo 2), inserido após o eletrodo 1. O
posicionamento de eletrodo 2 indica que ele está mais próximo do terminal axonal que
o eletrodo 1. Assim, o estímulo gerado no eletrodo 1 pode propagar-se ao longo do
axônio e ser registrado no eletrodo 2. O eletrodo 2 está ligado a um voltímetro, que
registra o potencial de membrana em mV em função do tempo (eixo horizontal).
Eletrodo 1
Eletrodo 2
Gerador de corrente elétrica (estímulo) Voltímetro (eixo vertical em mV)
Seção do axônio de sépia.24
Modelo de Hodgkin-Huxley

Na situação ilustrada abaixo, temos que o voltímetro mostra a evolução temporal do
potencial de membrana, num período de 20 ms, suficiente para vermos todas as fases
do potencial de ação (despolarização, repolarização e hiperpolarização).
Eletrodo 1
Eletrodo 2
Gerador de corrente elétrica (estímulo) Voltímetro (eixo vertical em mV)
Seção do axônio de sépia.25
Modelo de Hodgkin-Huxley

Vamos usar o HHSim para destacar as características do potencial de ação. Na figura
abaixo temos a situação de potencial de repouso. A linha vermelha indica o potencial
da membrana (em repouso), a linha roxa indica o estímulo aplicado, a linha amarela a
condutância do Na+ e a verde a condutância do K+ .
Gráfico do potencial contra o tempo
(linha vermelha), gerado pelo HHSim
(http://www.cs.cmu.edu/~dst/HHsim/).
Acesso em: 10 de setembro de 2015.
26
Modelo de Hodgkin-Huxley

Aplicamos um estímulo, linha roxa, temos que o potencial de membrana atinge uma
valor acima do potencial limiar (linha vermelha). Em tal situação, abrem-se os canais
de Na+ dependentes de voltagem. Cofirmarmos a situação, verificando a condutância
do Na+ (linha amarela), que começa a subir, indicado o influxo de Na+. O eixo
horizontal é o do tempo. Todo evento está registrado em pouco mais de 20 ms.
27
Modelo de Hodgkin-Huxley
Gráfico do potencial contra o tempo
(linha vermelha), gerado pelo HHSim
(http://www.cs.cmu.edu/~dst/HHsim/).
Acesso em: 10 de setembro de 2015.

Comparando-se as condutâncias, vemos que a condutância do Na+ (linha amarela)
atinge o valor máximo, antes da a condutância do K+ (linha amarela). Isto deve-se ao
fato do canal de Na+ dependente de voltagem abrir-se antes do canal de K+
dependente de voltagem.
28
Modelo de Hodgkin-Huxley
Gráfico do potencial contra o tempo
(linha vermelha), gerado pelo HHSim
(http://www.cs.cmu.edu/~dst/HHsim/).
Acesso em: 10 de setembro de 2015.

Depois de poucos milisegundos, ambos canais estão fechados, como vemos com as
condutâncias retornando para o valor zero. Depois de mais alguns milisegundos, o
potencial de membrana (linha vermelha) retorna ao valor de repouso.
29
Modelo de Hodgkin-Huxley
Gráfico do potencial contra o tempo
(linha vermelha), gerado pelo HHSim
(http://www.cs.cmu.edu/~dst/HHsim/).
Acesso em: 10 de setembro de 2015.

O cérebro humano é considerado por
muitos como o mais capaz entre os
animais do planeta Terra. Considerando-
se que, um maior número de neurônios
significa maior poder cognitivo, espera-se
que o cérebro humano seja o campeão
entre os animais em número de
neurônios. Na verdade, apesar de muitos
livros textos estabelecerem o número
redondo de 100 bilhões de neurônios no
cérebro (1011 ) ( Williams & Herrup, 1988),
tal número ainda é motivo de grande
debate.
Fonte: Williams RW, Herrup K. The control of neuron number.
Annu Rev Neurosci. 1988;11:423-53.
Imagem de CAT scan do cérebro.
Disponível em : <
http://netanimations.net/Moving_Animated_Heart_Beating_L
ungs_Breathing_Organ_Animations.htm#.UXKY4rXvuSp >.
Acesso em: 10 de setembro de 2015. 30
Cérebro Humano

Se considerarmos que o cérebro humano
é formado por aproximadamente 1011
neurônios, sendo que cada uma dessas
células pode formar até 10.000 conexões,
temos que o cérebro humano pode
apresentar até 1014 sinapses. Tomamos
um valor médio de 103 sinapses por
neurônio. Um estudo sobre o assunto
(Azevedo et al., 2009), estimou o número
em 86,1 ± 8,1 bilhões de neurônios (
8,61.1010) num adulto do sexo masculino.
Revisões posteriores, sobre o número de
neurônios no cérebro humano, ficam entre
75 e 124 bilhões (Lent, 2012), assim, o
número de 100 bilhões, é um valor médio
das estimativas.
Fonte: Azevedo FA, Carvalho LR, Grinberg LT, Farfel JM,
Ferretti RE, Leite RE, Jacob Filho W, Lent R, Herculano-Houzel
S. Equal numbers of neuronal and nonneuronal cells make the
human brain an isometrically scaled-up primate brain
J Comp Neurol. 2009;513(5):532-41.
Concepção artística do cérebro humano.
Disponível em : < http://www.whydomath.org/ >
Acesso em: 10 de setembro de 2015.
31
Cérebro Humano

A complexidade dos pensamentos e do
tráfego de sinais no organismo humano
são resultados da interação entre
neurônios conectados. O impressionante
número de conexões entre os neurônios
cria um sistema altamente complexo
envolvendo 1014 sinapses. Os resultados
da ação desse sistema vemos a cada
segundo de nossas vidas, pensando,
criando e aprendendo... As interações,
que geram padrões complexos, são
resultados das sinapses entre as células.
Resumindo, tudo que pensamos e
lembramos é resultado das interações
dessa rede complexa de sinapses. Iremos
ver as principais características das
sinapses.
Segundo alguns autores, o cérebro humano tem
aproximadamente 1014 sinapses, uns apresentam um
número menor...
Fonte da imagem:
http://images.fanpop.com/images/image_uploads/Homer-
Brain-X-Ray-the-simpsons-60337_1024_768.jpg
Acesso em: 10 de setembro de 2015.
32
Cérebro Humano

As sinapses são junções estruturalmente especializadas, em que uma célula pode
influenciar uma outra célula diretamente por meio do envio de sinal químico ou
elétrico. A forma mais comum de sinapse é a sinapse química. Na sinapse temos a
participação das células pré-sináptica e pós-sináptica.
Célula pré-sináptica: É a célula que envia o sinal nervoso.
Célula pós-sináptica: É a célula que recebe o sinal da célula pré-sináptica.
Neurotransmissor
Ca+2
Receptor
Canal de Ca+2
Vesícula
Membrana pós-sináptica
Membrana pré-sináptica
Fenda sináptica
33
Sinapses

Na sinapse química, a comunicação entre a célula pré-sináptica e pós-sináptica dá-se
por meio de neurotransmissores, que passam da célula pré-sináptica, ligando-se a
receptores específicos na célula pós-sináptica. Os neurotransmissores ficam
armazenados em vesículas e, uma vez que um potencial de ação chega ao terminal
axonal, esses são liberados na fenda sináptica. Os neurotransmissores ligam-se a
uma classe especial de proteínas transmembranares, chamadas receptores.
Neurotransmissor
Ca+2
Receptor
Canal de Ca+2
Vesícula
Membrana pós-sináptica
Membrana pré-sináptica
Fenda sináptica
34
Sinapse Química

De uma forma geral, as células nervosas
comunicam-se através de
neurotransmissores, que são pequenas
moléculas que se difundem facilmente
pela fenda sináptica. Os
neurotransmissores ligam-se às proteínas
transmembranares. Tal ligação promove
uma mudança estrutural, permitindo a
abertura dos receptores. A abertura
permite um influxo de íons. Todo o
processo demora milisegundos e há
diversos tipos de neurotransmissores,
como a acetilcolina e serotonina. O
fechamento dos receptores também
ocorre rapidamente, uma vez fechados, a
entrada de íons para célula pós-sináptica
é interrompida.
Fonte
: http://w
ww
.rcsb.o
rg/p
db/s
tatic.d
o?p=
education_dis
cussio
n/m
ole
cule
_of_
the_m
onth
/pdb71_1.h
tml
Membrana
Receptor
(vista lateral)
Receptor
(vista superior)
35
Sinapse Química

Um tipo especial de receptor é o receptor
de acetilcolina, encontrado nas células do
músculo esquelético e em neurônios do
sistema nervoso central. A acetilcolina,
indicada em vermelho na figura ao lado,
liga-se nas cadeias alfa do pentâmero,
indicadas em laranja, que formam o
receptor de acetilcolina. A faixa cinza na
figura indica a posição da membrana
celular, vemos claramente que o receptor
de acetilcolina atravessa a membrana
celular. Como sempre, a parte de cima da
figura indica o meio extracelular e a parte
inferior o meio intracelular. A faixa cinza é
a bicamada fosfolipídica.
Membrana
Receptor
(vista lateral)
Receptor
(vista superior)
Fonte
: http://w
ww
.rcsb.o
rg/p
db/s
tatic.d
o?p=
education_dis
cussio
n/m
ole
cule
_of_
the_m
onth
/pdb71_1.h
tml
36
Sinapse Química

A acetilcolina (ACh) é liberada por
exocitose da célula pré-sináptica, ligando-
se ao seu receptor na célula pós-
sináptica. O receptor de acetilcolina é
uma proteína transmembranar composta
por 5 cadeias polipeptídicas (pentâmero),
sendo duas cadeias alfa, uma beta, gama
e delta. A ligação da acetilcolina promove
uma mudança conformacional na
estrutura do pentâmero, abrindo um poro
no centro da estrutura. Tal poro permite a
passagem de íons do meio exterior para o
citoplasma, o influxo de íons de sódio
eleva o potencial de membrana na região
próxima à fenda sináptica. Na figura ao
lado vemos o receptor de ACh visto de
cima, do meio extracelular e de perfil,
numa imagem deslocada 90º com relação
à figura de cima.Código PDB: 2BG9
37
Sinapse Química

Observando-se o pentâmero que forma o
receptor de ACh por cima, vê-se
claramente o poro, em formato de estrela
no centro da estrutura. Cada cadeia
polipeptídica está colorida de forma
distinta.
Na figura ao lado, temos uma visão de
perfil do receptor de acetilcolina, o trecho
transmembranar tem uma predominância
de hélices alfa, um padrão comum em
proteínas transmembranares.
Código pdb: 2BG9
38
Sinapse Química

A estrutura mostrada no slide anterior é o
receptor de acetilcolina da raia elétrica
(Torpedo marmorata), similar ao
encontrado na junção neuromuscular de
mamíferos. As arraias e enguias elétricas
apresentam órgãos especializados
capazes de gerar pulsos de eletricidade.
Tais pulsos são capazes de paralisar suas
presas. Os órgãos elétricos são células
musculares modificadas de forma plana,
que encontram-se empilhadas. O
pequeno potencial gerado através de
cada membrana celular, controlada pela
grande densidade de receptores de
acetilcolina, somam-se, gerando choques
elétricos capazes de paralisar uma presa.
Cada célula funciona como um gerador,
que são colocados em série, o que tem
como resultado, a soma dos potenciais
individuais.
Torpedo marmorata (Risso, 1810) fotografia de ©Bernard
Picton.
Disponível em: <
http://www.habitas.org.uk/marinelife/photo.asp?item=tormar
>
Acesso em: 10 de setembro de 2015.
39
Sinapse Química

Fo
nte
: P
urv
es e
t a
l.,
Vid
a. A
ciê
ncia
da
Bio
logia
. 6
a. E
d.
Art
me
d e
dito
ra, 2
00
2 (
pg
. 7
87
).
40
Sinapse Química

A presença de ACh na fenda sináptica, deixaria os receptores de ACh abertos por um
tempo maior que o necessário após a repolarização da célula pré-sináptica. Para
evitar tal situação, entra em ação a enzima acetilcolinaesterase (EC 3.1.1.7), que
catalisa a clivagem da molécula de ACh. A clivagem ocorre rapidamente, para garantir
que a célula pós-sináptica retorne ao repouso, uma vez cessado o potencial de ação
na célula pré-sináptica. A figura abaixo mostra a estrutura da enzima
acetilcolinaesterase (AChE), onde vemos a tríade catalítica no sítio ativo. Em média a
acetilcolinaesterase catalisa a clivagem de uma molécula de ACh em 80
microssegundos.
Tríade catalíca formada pelo resíduos: Ser 200,
Glu 327 e His 440.
Código de acesso PDB: 1ACJ
H440
E327
S200
41
Acetilcolinaesterase
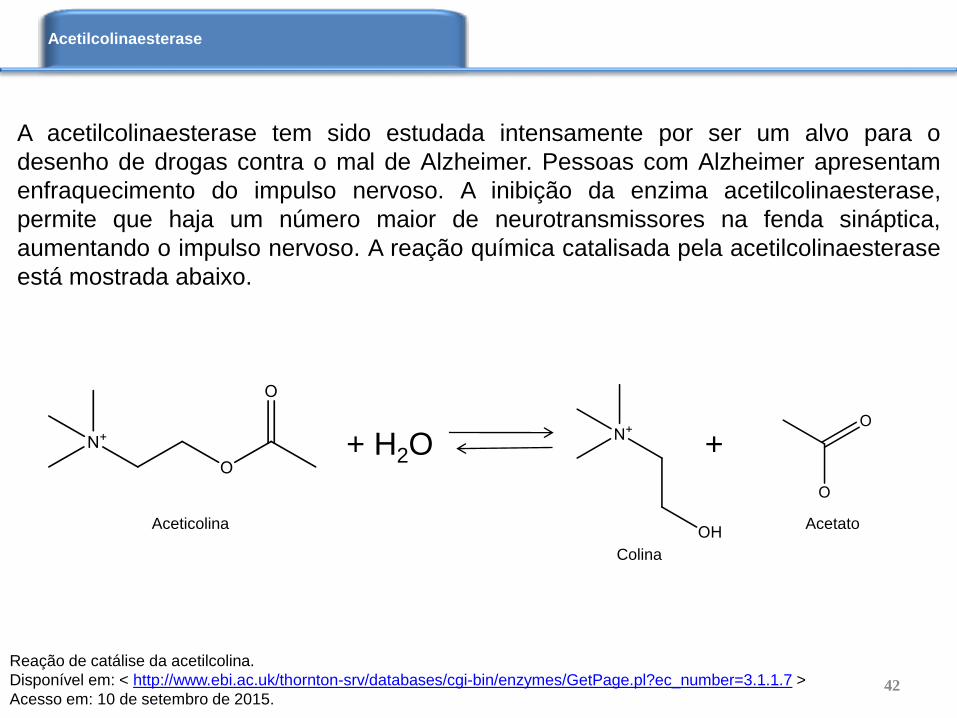
A acetilcolinaesterase tem sido estudada intensamente por ser um alvo para o
desenho de drogas contra o mal de Alzheimer. Pessoas com Alzheimer apresentam
enfraquecimento do impulso nervoso. A inibição da enzima acetilcolinaesterase,
permite que haja um número maior de neurotransmissores na fenda sináptica,
aumentando o impulso nervoso. A reação química catalisada pela acetilcolinaesterase
está mostrada abaixo.
+ H2O +
Reação de catálise da acetilcolina.
Disponível em: < http://www.ebi.ac.uk/thornton-srv/databases/cgi-bin/enzymes/GetPage.pl?ec_number=3.1.1.7 >
Acesso em: 10 de setembro de 2015.
Aceticolina
Colina
Acetato
42
Acetilcolinaesterase

A estrutura abaixo mostra o complexo da aceticolinaesterase com a droga aricept,
resolvido a partir da técnica de cristalografia por difração de raios X. Na estrutura
temos a droga (aricept) bloqueando o sítio ativo da enzima, o que impossibilita a
ligação da ACh, inibindo a reação de catálise da ACh. O inibidor aricept é um inibidor
competitivo, pois compete com a acetilcolina, impedindo sua ligação. Usando o
modelo chave-fechadura, o sítio ativo da enzima é a fechadura e o inibidor a chave.
Droga aricept bloqueando o sítio ativo da
enzima e prevenindo a clivagem de ACh.
Código de acesso PDB: 1EVE43
Acetilcolinaesterase

Sinapses excitatórias levam a célula pós-sináptica a aumentar a probabilidade de
disparo de potencial de ação, ou seja, há entrada de íons positivos na célula pós-
sináptica. É o caso da junção neuromuscular no músculo esquelético, onde a célula
pós-sináptica (fibra muscular) sofre despolarização devido à liberação do
neurotransmissor acetilcolina da célula pré-sináptica.
Acetilcolina
Ca+2
Receptor de acetilcolina
(canal de sódio)
Canal de Ca+2
Vesícula
Membrana pós-sináptica
Membrana pré-sináptica
Fenda sináptica
44
Sinapses Excitatórias
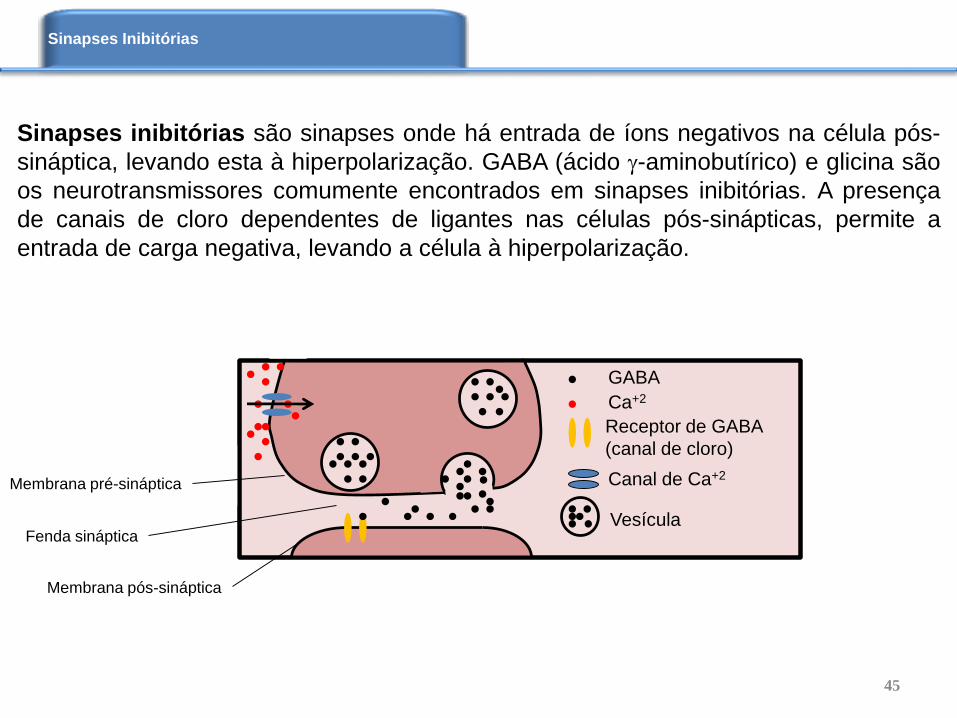
Sinapses inibitórias são sinapses onde há entrada de íons negativos na célula pós-
sináptica, levando esta à hiperpolarização. GABA (ácido -aminobutírico) e glicina são
os neurotransmissores comumente encontrados em sinapses inibitórias. A presença
de canais de cloro dependentes de ligantes nas células pós-sinápticas, permite a
entrada de carga negativa, levando a célula à hiperpolarização.
GABA
Ca+2
Receptor de GABA
(canal de cloro)
Canal de Ca+2
Vesícula
Membrana pós-sináptica
Membrana pré-sináptica
Fenda sináptica
45
Sinapses Inibitórias

As sinapses elétricas têm participação minoritária no sistema nervoso, contudo estão
presentes inclusive no cérebro de mamíferos. São diferentes das sinapses químicas,
porque acoplam neurônios eletricamente. Nesse tipo de sinapse, as membranas das
células pré-sináptica e pós-sináptica estão separadas por uma distância entre 20 e 30
Å.
46
Sinapses Elétricas
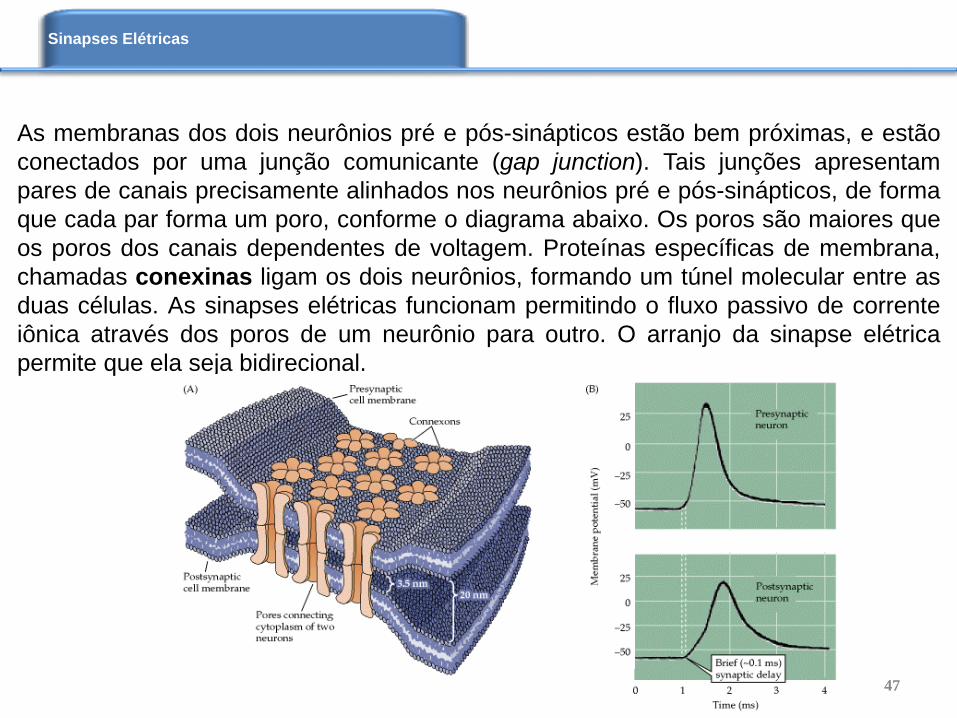
As membranas dos dois neurônios pré e pós-sinápticos estão bem próximas, e estão
conectados por uma junção comunicante (gap junction). Tais junções apresentam
pares de canais precisamente alinhados nos neurônios pré e pós-sinápticos, de forma
que cada par forma um poro, conforme o diagrama abaixo. Os poros são maiores que
os poros dos canais dependentes de voltagem. Proteínas específicas de membrana,
chamadas conexinas ligam os dois neurônios, formando um túnel molecular entre as
duas células. As sinapses elétricas funcionam permitindo o fluxo passivo de corrente
iônica através dos poros de um neurônio para outro. O arranjo da sinapse elétrica
permite que ela seja bidirecional.
47
Sinapses Elétricas

O estudo da sinapse elétrica em crayfish determinou a rapidez da sinapse elétrica,
quando comparada com a sinapse química (Furshpan & Potter, 1959). Um sinal pós-
sináptico é observado em uma fração de milisegundo, após a geração do potencial de
ação pré-sináptico. No caso do crayfish, as inteconecções das sinpases elétricas,
permitem uma rápida resposta ao ataque de um predador. Sinapses elétricas também
são usadas para sincronizar a atividade de populações de neurônios, como em
neurônios de secreção de hormônio localizados no hipotálamo de mamíferos.
Referência: Furshpan, E. J. & Potter, D. D. (1959). J. Physiol. 145:289-325.
48
Sinapses Elétricas

Na aula de hoje vimos detalhes sobre a
sinapse e detalhes sobre a estrutura do
neurônio, um tópico relacionado com a
disciplina Biologia Celular e Tecidual.
Vimos como os fármacos contra
Alzheimer funcionam, um assunto que
está relacionado com a disciplina
Farmacologia, do sétimo semestre do
curso de Biologia. Ao estudarmos as
bases moleculares do sinapses química e
elétrica, vimos um tópico de estudo da
Química e Bioquímica Estrutural. O
estudo do comportamento da raia elétrica
está relacionada com as disciplinas
Zoologia I e II.
Aula de
hoje
QuímicaBioquímica
Estrutural
Biologia
Celular e
Tecidual
Zoologia I e
II
Farmacologia
49
Relação com Outras Disciplinas

Selecionei 5 artigos, 3 trazem resultados interessantes sobre aplicações
farmacológicas da inibição da acetilcolinaesterase, um quarto descreve a polêmica
sobre o número de neurônios. O último artigo descreve a mudança do canal de Na+
dependente de voltagem da serpente que preda a salamandra.
1) Discovery of a novel acetylcholinesterase inhibitor by structure-based virtual screening techniques. Chen
Y, Fang L, Peng S, Liao H, Lehmann J, Zhang Y. Bioorg Med Chem Lett. 2012 May 1;22(9):3181-7.
2) Synthesis, characterization, X-ray crystallography, acetyl cholinesterase inhibition and antioxidant activities
of some novel ketone derivatives of gallic hydrazide-derived Schiff bases. Gwaram NS, Ali HM, Abdulla
MA, Buckle MJ, Sukumaran SD, Chung LY, Othman R, Alhadi AA, Yehye WA, Hadi AH, Hassandarvish P,
Khaledi H, Abdelwahab SI. Molecules. 2012 Feb 28;17(3):2408-27.
3) In silico methods to assist drug developers in acetylcholinesterase inhibitor design. Bermúdez-Lugo JA,
Rosales-Hernández MC, Deeb O, Trujillo-Ferrara J, Correa-Basurto J. Curr Med Chem. 2011;18(8):1122-
36. Review.
4) How many neurons do you have? Some dogmas of quantitative neuroscience under revision. Lent R,
Azevedo FA, Andrade-Moraes CH, Pinto AV. Eur J Neurosci. 2012;35(1):1-9.
5) Mechanisms of adaptation in a predator-prey arms race: TTX-resistant sodium channels. Geffeney S,
Brodie ED, Ruben PC, Brodie ED. Science 2002; 297 (5585): 1336–9.
50
Material Adicional (Artigos Indicados)

A sugestão de filme relacionado com a
aula de hoje é a excelente comédia de
Todd Phillips, Parto de Viagem. No elenco
temos Robert Downey Jr e Zach
Galifianakis. Durante o filme vemos os
efeitos do THC no cérebro humano, de
forma não científica, mas bem divertida.
Cartaz do filme Parto de Viagem (Due Date) de 2010.
Fonte da imagem:
http://www.imdb.com/title/tt1231583/
Acesso em: 10 de setembro de 2015. 51
Material Adicional (Filme Indicado)

Segue uma breve descrição de um site
relacionado à aula de hoje. Se você tiver
alguma sugestão envie-me
http://www.whydomath.org/node/HHneuro/
index.html. O site www.whydomath.org
apresenta modelos computacionais
para simulação da dinâmica do
cérebro. O site está em inglês.
52
Animação com o disparo do potencial de ação.
Disponível em: <
http://www.whydomath.org/node/HHneuro/index.html >.
Acesso em: 10 de setembro de 2015.
Material Adicional (Site Indicado 2)

Explique a sinapse química.
53
Questão
Acetilcolina
Ca+2
Receptor de acetilcolina
(canal de sódio)
Canal de Ca+2
Vesícula

HODGKIN, ALAN L; HUXLEY, ANDREW F. "A quantitative description of membrane
current and its application to conduction and excitation in nerve". Journal of Physiology,
1952; 117 (4): 500-544.
OLIVEIRA, Jarbas Rodrigues de; WACHTER, Paulo Harald; AZAMBUJA, Alan Arrieira.
Biofísica para ciências biomédicas. Porto Alegre: EDIPUCRS, 2002. 313 p.
OKUNO, Emiko; CALDAS, Iberê Luiz; CHOW, Cecil. Física para ciências biológicas e
biomédicas. São Paulo: Harper & Row do Brasil, 1982. 490 p.
PURVES, W. K., SADAVA, D., ORIANS, G. H., HELLER, H. G. Vida. A Ciência da
Biologia. 6a ed. Artmed editora. 2002.
VOET, Donald; VOET, Judith G. Bioquímica. 3ª edição. Porto Alegre: Artmed, 2006.
1596 p.
54
Referências