transmissão dos genes entre geraçõesw3.ufsm.br/geneticavegetal/images/anexos/textosgenetica/texto...
TRANSCRIPT

CENTRO DE CIÊNCIAS NATURAIS E EXATAS – CCNE
DEPARTAMENTO DE BIOLOGIA
DISCIPLINA DE GENÉTICA AGRONOMIA
14
Unidade 2 – Transmissão dos Genes Entre as Gerações
1. Introdução
A Biologia Molecular estuda atualmente genes que pertencem aos organismos com técnicas que permitem
o sequenciamento das bases nucleotídicas desse gene. É possível se conhecer a sequência de bases
nucleotídicas do gene das plantas de arroz, aveia, soja, dos bovinos e até mesmo de humanos. O projeto
Genoma Humano abriu campo para o conhecimento intimo dos genes de todas as espécies. Isso possibilita
conhecer também o que esses genes produzem nas plantas. Os genes ditos “estruturais” produzem proteínas
que atuam no desenvolvimento de todas as etapas por que passam os vegetais desde a germinação das
sementes até a produção de novas sementes.
Para que a planta cresça e se desenvolva proteínas do tipo hormônios, como a citocinina, são produzidas,
permitindo, entre outras ações, as divisões celulares. Essas divisões celulares promovem a multiplicação das
células formando os tecidos da planta. Essas divisões são a mitose e a meiose.
A mitose é a divisão celular que permite alongamento da planta e ocorre nos tecidos meristemáticos que
estão nas extremidades das plantas. Enquanto que a meiose está restrita ao tecido reprodutivo e forma gameta
que formarão novas sementes através da fecundação, levando os genes de uma geração para outra.
Ambos os processos de divisão celular possuem características próprias com referência à movimentação
cromossômica no interior do núcleo e com o tipo de célula formada.
O presente texto fará uma abordagem das estruturas cromossômicas, das divisões celulares e de suas
consequências, evidenciando, por fim, a formação de gametas e a fecundação por ser esse o objetivo de se
entender como os genes passam de uma geração para outra.
2. A Célula, Um breve histórico
Em 1664, o inglês Robert Hooke descreveu a estrutura microscopia dos tecidos vegetais analisando
cortiça, medula velha de cenoura e a essa estrutura deu nome de célula (do latim Cellula) relacionando o
espaço por ele visto aos pequenos quartos dos monges no mosteiro. Célula sob esse ponto de vista é o espaço
delimitado, em forma de caixa observado por Hooke (HARRINSON, 1975). Ainda no século XVII outros
investigadores como Van Leeuwenhoek, na Holanda, Malpighi, na Itália e Grew, na Inglaterra contribuíram
para com os estudos de Hooke fazendo descrições das células em outros organismos.
Somente no século XIX é que o estudo da célula por ter avanços. Isso devido aos fabricantes de
microscópios Carl Zeiss e Ernest Leitz, de Westzlar, na Alemanha, que produziram equipamento com alto
poder de resolução.
Johannes Muller que viveu entre 1801 e 1858, trabalhando na Universidade de Berlim, fundou a disciplina
de fisiologia comparada baseado nos estudos celulares de várias espécies e, em 1838, um dos seus alunos,
Matthias Jakob Schleiden (1804 – 1881), publicou um tratado denominado “Contribuição para a Fitogênese”,
demonstrando que a célula era um elemento comum em todos os tecidos das plantas. Além disso, reconheceu
a importância do núcleo celular descoberto em 1831, pelo investigador britânico escocês Robert Brown.
Theodor Schwan (1810 – 1882) estendeu o trabalho de Schleiden para todo o reino animal e em 1839
publicou seu livro intitulado “Pesquisas Microscópicas em Conformidade com a Estrutura e Crescimento de
Plantas e Animais” onde descreve os itens dos escritos de Schleiden e seus, relativo ao conceito de células
como descrito a seguir (HARRINSON, 1975):

Unidade 2
Transmissão dos Genes entre Gerações 15
1) Os organismos são constituídos por células microscópicas que são unidades organizadas
distintamente;
2) Dentro de um organismo as células se diferenciam por formarem tipos distintos, que têm
propriedades características próprias de determinados tecidos;
3) O núcleo é uma característica comum a todas as células, embora algumas, tal como o floema e os
glóbulos vermelhos dos mamíferos, o núcleo possa desaparecer durante a diferenciação;
4) O conteúdo vivo da célula ou protoplasma determina a atividade da célula e assim, coletivamente,
de todo o organismo;
5) O crescimento é atingido pelo aumento do número de células; só a divisão das células existentes
pode dar origem a novas células.
Nesse breve histórico da descoberta ao aperfeiçoamento do conceito de célula e o resumo da Teoria
Celular de Schleiden e Schwan é possível se perceber pontos de importância capital. Segundo Virchow, em
1885 (citado por HARRINSON, 1975) as células derivam sempre de células já existentes, ocorre
diferenciação entre as células para formarem tecidos também diferentes entre si dentro de um mesmo
organismo. O núcleo é uma característica comum em todas as células, portanto se torna o constituinte
principal.
Quase no final do século XIX, precisamente em 1865, Gregor Mendel elaborou as leis do
comportamento gênico em vegetais sem entender como ocorria a divisão celular, principalmente a meiose que
possibilita a formação de gametas. Daí o termo que usou em seus estudos “fatores”.
Para entender a movimentação cromossômica dentro da célula que vai se dividir é necessário se
estudar como a célula e, principalmente o núcleo, se prepara para os processos de divisão nuclear e celular.
3. O núcleo interfásico
O núcleo interfásico é o núcleo celular entre os períodos G1 e G2 do ciclo celular (Ver item 4 da Unidade
1). O seu conteúdo principal é a molécula de DNA que está na forma desespiralizada com partes ligadas à
carioteca. A esse estado do DNA dá-se o nome de cromatina e está presente entre as divisões celulares no
tecido meristemático que possui continua divisão. Com corantes específicos, que coram somente o DNA
como o corante de Feulgen, é possível se analisar duas regiões diferenciadas da cromatina, a heterocromatina
que é densamente corada e a eucromatina que é pouco corada. A posição de grande parte da heterocromatina
nos cromossomos é constante, sendo então uma característica hereditária. A eucromatina é a região onde se
encontra a maioria dos genes. Por ser menos helicoidizada é alta a atividade gênica nessa região, devido à
transcrição dos genes.
Os três períodos citados, G1, S e G2 são os que caracterizam o núcleo interfásico. Porém, para que a
célula tenha material genético (DNA) para dividir entre as células filhas resultantes da divisão nuclear, é
necessário que haja a duplicação, ficando, portanto com o conteúdo celular dobrado. Guerra (1988) relata que
o núcleo G1 possui 1C de DNA e em G2 2C de DNA, como consequência da duplicação da molécula.
Todo esse DNA está desespiralizado no núcleo da célula, entretanto é necessário que haja uma
organização de toda essa cromatina para que ocorra a divisão igualitária para as células filhas. Para isso a
cromatina se transforma em cromossomos, pelo processo de condensação.
4. Os cromossomos e suas estruturas
O cromossomo sendo uma molécula de DNA enrolada contendo todos os genes do indivíduo é o
instrumento que leva os genes através das gerações. Pode-se considerar geração celular, como no caso dos
tecidos meristemáticos, como geração entre indivíduos.

Unidade 2
Transmissão dos Genes entre Gerações 16
Devido à compactação da molécula de DNA transformando-se em cromossomos, estes, por sua vez,
possuem estruturas que o definem e caracterizam cada espécie. Cada cromossomo terá sempre o centrômero.
Também chamado de constrição primária é a região aonde os fios do fuso de divisão irão se ligar para haver a
segregação. O centrômero é um marco cromossômico que divide o cromossomo em braços. Esses braços
poderão ser do mesmo tamanho ou de tamanhos distintos. A análise da proporção entre os braços dos
cromossomos é uma característica de cada espécie.
Pela posição do centrômero os cromossomos podem ser assim classificados (Figura 2.1). Dentro da
classificação exposta pode haver cromossomos longos e curtos.
Nas extremidades de cada cromossomo está a região
do telômero. Essa região possui a capacidade de impedir
que os cromossomos se agrupem uns aos outros durante
o processo de divisão. Entretanto, quebras
cromossômicas são possíveis e caso isso venha a acontecer à parte que quebrou pode-se ligar a outro
cromossomo, ficando esse com dois centrômeros. Em Cyparaceae há cromossomos policêntricos, com vários
centrômeros, cujo fenômeno de quebra e reunião deva ter sido um dos motivos da evolução dessa espécie. A
parte do cromossomo que quebrou e ficou sem centrômero se torna um micronúcleo que acaba por se perder
nas divisões celulares seguintes, alterando, dessa forma, o cariótipo da planta.
Observa-se também nas células que vão entrar em divisão e estão “empacotando” o DNA na forma de
cromossomos, a Região Organizadora do Nucléolo (RON). Os nucléolos são considerados organelas do
núcleo que possuem RNA que originarão os ribossomos. Essa RON está associada aos cromossomos e em
qual cromossomo está associado é uma característica da espécie, por exemplo, em tomate a RON está no
cromossomo 2.
Voltando ao centrômero dos cromossomos, pode-se afirmar que é a estrutura mais importante sob aspecto
da divisão das células, assim como para o estabelecimento e estudo do cariótipo.
5. O Cariótipo
Cariótipo de uma espécie significa a representação clara e específica do seu conjunto cromossômico. A
representação do cariótipo pode ser feita na forma de cariograma (imagem dos cromossomos) ou idiograma
(esquema dos cromossomos). O cariograma é construído após a preparação citogenética de uma lâmina de
microscópio com tecido vegetal. Ponta de raiz é o mais utilizado. No microscópio procura-se a fase de
metáfase, onde os cromossomos estão com sua máxima condensação e com as estruturas definidas.
Fotografa-se-os, revela-se a foto e procede-se o corte dos cromossomos de forma manual colando-os aos
pares numa folha de papel branca e em ordem de tamanho e forma. A Figura 2.2 demonstra o cariótipo de
Crotalaria sp e de Passiflora sp.
Figura 2.1 – Tipos de cromossomos de acordo
com a posição do centrômero (Fonte:
http://www.virtual.epm.br/cursos/genetica/htm/
base.htm ).

Unidade 2
Transmissão dos Genes entre Gerações 17
5.1. A importância do estudo de Cariótipos
A ciência aplicada não pode se dissociar da ciência básica e o estudo de cromossomos evidencia tal fato.
Na ciência aplicada, no caso do Melhoramento de Plantas, por exemplo, o cruzamento entre espécies com a
finalidade de se obter genótipos desejáveis é um dos instrumentos de seu estudo. Supõe-se nesse caso a
necessidade da realização de hibridação interespecífica entre duas espécies de mesmo gênero. Uma delas pode
ser chamada de X0 e a outra X1. X0 é de porte ereto, mas com baixa produção de sementes, enquanto que X1 é
de boa produtividade, porém com hábito de crescimento decumbente.
Ambas as espécies são cruzadas. Fazem-se os cruzamentos recíprocos (X0 >> X1) e (X1 >> X0) e se obtém
plantas entre tantas com características desejadas. O pesquisador verifica que poucas sementes se formam,
algumas ficam chochas e algumas flores abortam. É um sinal de esterilidade.
As sementes então, das duas espécies são levadas ao citogeneticista com a finalidade de estudar o
cariótipo. A análise do cariótipo mostra que as duas espécies possuem 2n = 14 cromossomos, por exemplo,
porém ocorre variação entre os tipos de cromossomos. A diferença entre os tipos de cromossomos dificulta
ou, até mesmo, impede a formação de híbridos férteis.
No caso presente, pode-se supor que a fórmula cromossômica da espécie X0 é constituída dos seguintes
tipos de cromossomos: F = 2 ml + 2 mc + 1 al + 2 ac, enquanto que a da espécie X1 é: F = 2 ml + 2 smc +3 al.
Ambas as fórmulas cromossômicas são diferentes, porém alguns pares de cromossomos possuem a mesma
estrutura. Se nesses cromossomos semelhantes estiverem os genes homólogos é possível um pareamento entre
eles, todavia nem todos os cromossomos vão parear na meiose, dificultando a segregação e a formação de
tétrades e posteriormente de gametas.
A dificuldade no pareamento e na segregação irregular dos cromossomos leva a infertilidade, nesse caso,
parcial, pois há cromossomos iguais em sua estrutura. O entendimento das fases iniciais da meiose, onde
inicia o engrossamento dos cromossomos, o pareamento de homólogos e a troca de partes entre as cromátides
homólogas, resulta na compreensão de que pode ocorrer infertilidade total ou parcial entre as espécies
cruzadas pelas diferenças estruturais dos cromossomos. Além disso, o acompanhamento das fases seguintes
permite a compreensão das consequências dos fenômenos que ocorrem na prófase I, embora todas elas
estejam sujeitas as variações que podem ocorrer de forma aleatória.
(a)
(b)

Unidade 2
Transmissão dos Genes entre Gerações 18
Vários híbridos interespecíficos que são produzidos apresentam variados graus de esterilidade. Oriza
sativa L., arroz de origem asiática mostrou alguma fertilidade quando cruzada com Oriza glaberrima Steud,
de origem africana. Ambas as espécies apresentam 2n = 24 cromossomos (Ver Unidade 3).
6. A divisão nuclear. Considerações iniciais
Ao se pensar sobre as divisões celulares normais, mitose e meiose, percebe-se a necessidade da formação
do fuso de divisão e a ligação de seus filamentos ao centrômero dos cromossomos.
Na metáfase da mitose os cromossomos deverão se posicionar no plano equatorial da célula e já estarem
ligados aos fios do fuso ou concluindo essa ligação. O posicionamento dos cromossomos no meio da célula é
para que haja divisão equitativa, enquanto que a ligação aos fios do fuso é para que ocorra migração correta
para um dos polos, na mesma velocidade, evitando atraso de algum dos cromossomos.
Tendo em vista que a fase de anáfase é a divisão propriamente dita à velocidade de chegada das
cromátides irmãs deve ser a mesma, possibilitando que a formação da nova parede celular divida a célula em
duas metades.
Quando se refere à divisão meiótica o fenômeno é o mesmo, com exceção do complexo metáfase –
anáfase na meiose I, devido ocorrer o pareamento de homólogos, entre outros fenômenos como preparatório
da divisão celular. É imprescindível que cada cromossomo homólogo esteja ligado a um dos fios do fuso.
Desta forma a separação de homólogos é correta e o encaminhamento para a meiose II estará pronto. A
ausência da prófase II é devido a não haver novas sínteses de DNA entre os estágios e o desenrolamento total
de todos os cromossomos. Esses agora se encaminham para o complexo metáfase II – anáfase II e os
fenômenos da mitose se repetem nessa fase da meiose.
A conclusão final da meiose é o aparecimento das tétrades e cada uma com o número cromossômico da
espécie reduzido, devido à etapa reducional da meiose, que é a meiose I.
É na forma de cromossomos que o material genético e os genes estão “arrumados” para se dividirem
igualitariamente para as células filhas.
Os fenômenos sequenciais da mitose e da meiose basicamente são os mesmos em todos os organismos,
mesmo que se estude um porífero ou um vertebrado, um musgo ou uma angiosperma. Guerra (1988) sugere
por isso que esses processos teriam se originado antes dos organismos pluricelulares e que esses teriam tido
origem comum.

Unidade 2
Transmissão dos Genes entre Gerações 19
6.1. Mitose
A mitose (do grego mitos = fios, filamentos) se caracteriza por ser uma divisão equacional servindo para a
multiplicação das células, permitindo o crescimento vegetal através do tecido meristemático. As fases da
mitose e seus fenômenos são:
Prófase: (pró = antes da divisão) ocorre a condensação dos cromossomos;
Metáfase: (meta = centro ou metade) os centrômeros se localizam no centro das células e estão ligados
aos fios do fuso de divisão;
Anáfase: (ana = divisão; fase de divisão) é o período de divisão propriamente dito. As cromátides irmãs
migram para polos opostos da célula;
Telófase: (telo = final; término) as cromátides irmãs chegam aos polos da célula, reorganiza-se a
carioteca, forma-se a parede celular e os plasmodesmos e a célula volta ao período G1.
6.1.1. As Consequências da Mitose
A consequência principal é o crescimento em número de células permitindo o alongamento do vegetal
através do meristema primário, na parte aérea, e nas extremidades das raízes. As duas células derivadas de
uma só, com 2n cromossomos, podem seguir dois caminhos após a divisão, ou duplica novamente o DNA no
período S e retorna a divisão ou para no período G1 da interfase e inicia o processo de diferenciação.
O termo merista deriva do grego Meristo, que significa divisível. Esse tipo de tecido está em todos os
ápices de raízes e brotos, principais ou laterais.
A mitose também é responsável pelos apêndices como tricoma, gavinhas, que se formam a partir da
epiderme (SACCO, 1970). Outros tecidos que se mantêm vivos, mas não mais se dividem por mitose são os
parênquimas, colênquimas e esclerênquimas. Esses tecidos, por não se dividirem mais, podem ser
considerados no período G0 (BURNS e BOTTINO, 1991; SUZUKI et al., 1992).
O parênquima do corpo primário da planta, parênquima do córtex, medula, do mesófilo foliar e floral
se diferenciam a partir do meristema fundamental. O parênquima fotossintético tem elevado número de
cloroplastos, enquanto que os do córtex e da medula são ricos em amido (SACCO, 1970). Nesses tecidos
diferenciados a expressão gênica tem a sua manifestação.
Os genes específicos para a produção de proteínas, enzimáticas ou de estruturação, são transcritos e
traduzidos produzidos fenótipos referentes à função de cada um desses tecidos (Ver Unidade 1).
Além disso, as proteínas WRKY são as responsáveis pela formação das sépalas, pétalas, estames,
androceu e gineceu.
Em um mesmo corte de raiz para se
observar e estudar a mitose pode aparecer todas
ou algumas fases da mitose. A figura 2.5
demonstra que as células estão em todas as
fases de divisão num único corte. Além disso, a
Figura 2.4 – Fases da mitose. Da esquerda
para direita (em cima) prófase, anáfase,
(embaixo) metáfase, telófase (Fonte:
http://med.mui.ac.ir/slide/genetic/mitosis7.
gif ).

Unidade 2
Transmissão dos Genes entre Gerações 20
anáfase, principalmente, os cromossomos poderão ser fixados em vários estágios.
6.2. Meiose. Considerações iniciais
A mitose por ser uma divisão mais simples parece que antecedeu, no processo evolutivo das espécies, a
meiose, que é um processo mais longo e complexo. Apesar da sua complexidade ocorre da mesma maneira em
todos os eucariontes.
Diferentemente da mitose, onde os cromossomos se contraem e se movimentam independentemente, a
meiose, por sua vez, se caracteriza pela reunião dos cromossomos levando em conta sua homologia, portanto,
comportam-se como um conjunto. Essa reunião de homólogos é o que permite a separação igualitária dos
cromossomos para as células resultantes.
O processo de meiose ocorre nos organismos de reprodução sexuada levando a produção de gametas.
Esses gametas possuem o conteúdo cromossômico dividido pela metade devido ao fato de que a meiose é uma
divisão reducional. Sua primeira etapa inclui-se nessa definição, pois há a separação dos cromossomos
homólogos na anáfase I, dividindo o número somático da espécie pela metade. A segunda etapa da meiose, a
meiose II, possui a movimentação cromossômica igual à mitose, portanto essa etapa se caracteriza por ser
equacional.
Como essa divisão nuclear leva a formação dos gametas com “n“ número de cromossomos, o
restabelecimento do conteúdo “2n” cromossomos será restabelecido após a fertilização, com a formação do
embrião e do endosperma.
6.2.1. Caracterização das fases da meiose
A meiose se caracteriza por ter uma prófase longa no início da primeira etapa, meiose I, onde vários
fenômenos de importância citológica acontecem. Todos eles são do interesse do pesquisador da área e
citogenética, entretanto para cumprir os objetivos desse trabalho somente dois fenômenos serão aqui citados e
comentados sua importância. A figura 2.6 caracteriza o pareamento dos cromossomos homólogos e a troca de
partes entre as cromátides homólogas.
a) Prófase
O pareamento dos cromossomos homólogos – Cada cromossomo possui duas cromátides derivadas
da duplicação do DNA na fase S da interfase. Na célula mãe do grão de pólen (CMP) existe os cromossomos

Unidade 2
Transmissão dos Genes entre Gerações 21
homólogos que são entidades individuais com correlações genéticas. Esses cromossomos, um de origem
paterna e outro de origem materna se aproximam e pareiam entre si, gene a gene, locus a locus.
Esse pareamento ocorre no zigóteno possibilitando que os genes fiquem lado a lado. Esses genes
trazem informações alélicas iguais ou diferentes. Suponho que a característica “cor da flor” seja determinada
pelo gene A; e o seu alelo “A” diz que a cor será violeta, porém seu alelo complementar “a” diz que será
branca. Esses dois alelos se encontram durante o pareamento. O mesmo raciocínio se pode fazer para o gene
B, que determina, por exemplo, posição da inflorescência. O alelo “B” diz que será terminal, enquanto que o
alelo “b” diz que será axilar. Ambos os alelos também se encontrarão no pareamento dos cromossomos
homólogos.
Seguindo esse raciocínio se pode concluir que todos os cromossomos homólogos pareiam entre si,
formando o que se denomina de bivalentes (JOHN e LEWIS, 1979). O pareamento dos homólogos se dá
semelhante a um fecho “éclair”, iniciando pela região telomérica.
A Troca de Partes – No diplóteno são bem visíveis os cromossomos com as duas cromátides e,
portanto, o bivalente. Entre as cromátides homólogas ocorre a formação de uma placa de natureza proteica
que une os dois homólogos e possibilita a quebra e reunião invertida das partes das cromátides homólogas.
Esse conjunto de cromossomos homólogos e placa proteica recebe o nome de complexo sinaptonêmico e a
permuta das partes dos cromossomos de quiasmas.
A troca de partes se dá entre os genes e como resultado disso se obtém variabilidade na espécie. Esse
fenômeno recebe o nome de permuta (= “crossing-over”) e é de grande importância, pois seu estudo levou a
conclusão de que dois genes podem estar ligados no mesmo cromossomo (Ver Unidade 6).
A figura 2.6 demonstra o processo de pareamento e troca de partes entre os bivalentes. Salienta-se que
quaisquer cromátides homólogas podem trocar material genético entre si. Além disso, ocorrem vários
quiasmas ao longo do mesmo cromossomo dependendo da distância entre os genes. O impedimento para a
ocorrência de uma permuta é a realização, antes, de outra permuta nas proximidades. Da mesma forma,
próximo à região centromérica a incidência de permuta é reduzida, todavia na região telomérica a ocorrência é
bastante alta.
b) Metáfase I
Essa metáfase é designada de metáfase I para diferenciá-la da metáfase da mitose. O fenômeno que
ocorre na metáfase I é a movimentação dos bivalentes para o plano equatorial da célula. Ainda esses
bivalentes permanecem unidos pelos pontos de permuta e também ligados aos fios do fuso de divisão para
posterior segregação. A medição quantitativa do DNA nessa fase resultará em dobro em relação ao período G1
da interfase.

Unidade 2
Transmissão dos Genes entre Gerações 22
c) Anáfase I
Nessa fase ocorre a movimentação dos homólogos para os polos opostos da célula, acontecendo à
redução. É importante salientar que é nessa fase que se dá o início da redução do número somático de
cromossomos da espécie para o número haploide de cromossomos. Todas as células ficarão com “n”
cromossomos.
d) Telófase I
As células telofásicas apresentam-se com “n” cromossomos completando a etapa de segregação
iniciada na fase anterior. Os cromossomos nessa fase estão com duas cromátides e se nesse ponto for também
medida a quantidade de DNA as células terão 1C de DNA, comprovando a existência do número reduzido de
cromossomos.
É nessa fase que se encerra a etapa reducional que caracteriza a meiose. A partir de então as fases
subsequentes possuem movimentação cromossômica semelhante à mitose.
e) Metáfase II
Os cromossomos voltam ao plano equatorial da célula. Essa fase é diferente da metáfase mitótica
porque cada uma das células possui “n” cromossomos e as cromátides irmãs poderão ser diferentes entre si
devido à permuta ocorrida na prófase I.
f) Anáfase II
Ocorre a segregação as cromátides irmãs para os polos das células pelo encurtamento dos fios de
divisão celular.
g) Telófase II
O processo de citocinese acontece. Reaparece a carioteca e se formam as tétrades. Cada cromátide
irmã pode ser considerada nessa fase como um cromossomo unifilamentar. Inicia-se a descondensação da
cromatina.
Os processos de divisão nuclear são os instrumentos que levam os vegetais a crescerem e
diferenciarem seus tecidos, como demonstrado na mitose. A meiose tem o objetivo de reprodução e
manutenção da espécie. Mas para que essa manutenção ocorra é necessário que as células da telófase II
sofram modificações morfológicas e que, pela fecundação, se encontrem formando o embrião e restaurando o
número somático da espécie. Essas transformações recebem o nome de microsporogênese e
macrosporogênese (= megasporogênese).
7. A reprodução sexuada das plantas
Embora existam pontos de vista diferentes sobre a origem evolutiva da flor e de suas partes, há consenso
geral de que é um ramo condensado que contém estruturas altamente condensadas e especializadas para a
produção de sementes (GEMMELL, 1981). É nessa estrutura especializada na reprodução sexuada das plantas
que ocorre a formação de grão de pólen (microsporogênese) e do óvulo (macrosporogênese).
7.1. Microsporogênese e Macrosporogênese
Os processos de microsporogênese e de macrosporogênese produzem o grão de pólen e a oosfera,
respectivamente. Quando ambos se unem pelo processo de fertilização formam o embrião e o endosperma
que, pela diferenciação, formarão a semente. Em todas as etapas ambos os tipos de divisão celular, mitose e
meiose estão participando conjuntamente.
a) Microsporogênese
Unidade 2
Transmissão dos Genes entre Gerações 23
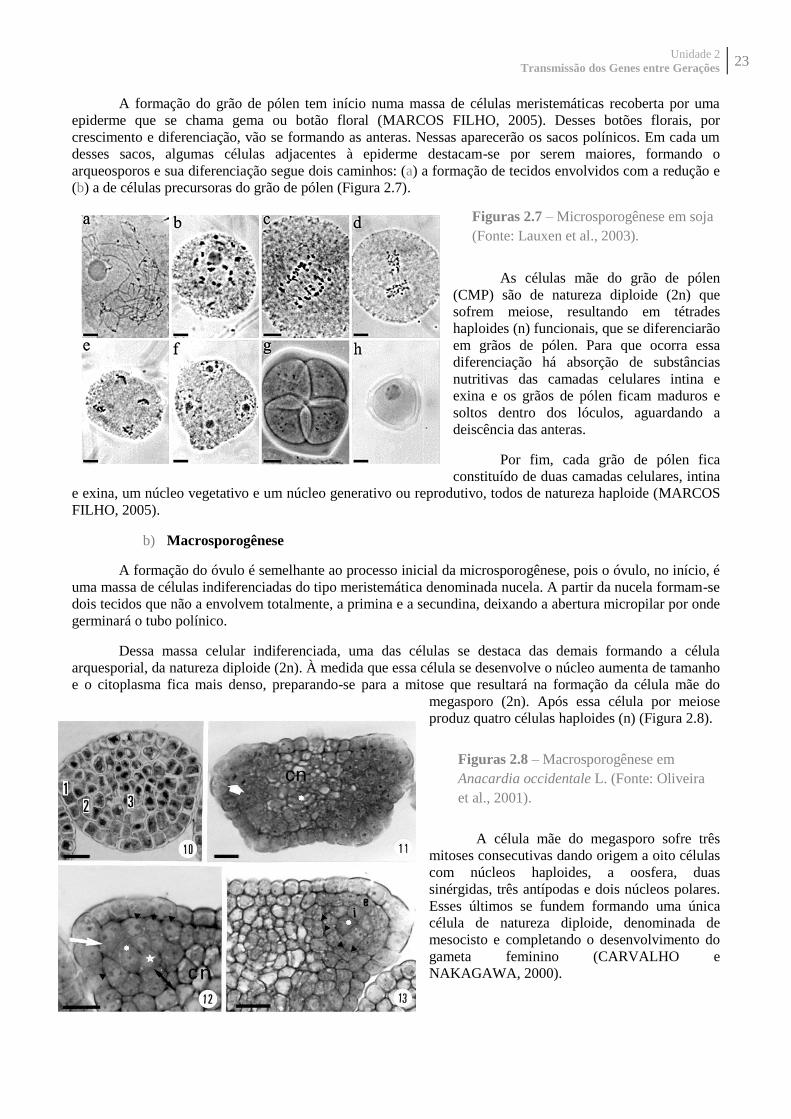
A formação do grão de pólen tem início numa massa de células meristemáticas recoberta por uma
epiderme que se chama gema ou botão floral (MARCOS FILHO, 2005). Desses botões florais, por
crescimento e diferenciação, vão se formando as anteras. Nessas aparecerão os sacos polínicos. Em cada um
desses sacos, algumas células adjacentes à epiderme destacam-se por serem maiores, formando o
arqueosporos e sua diferenciação segue dois caminhos: (a) a formação de tecidos envolvidos com a redução e
(b) a de células precursoras do grão de pólen (Figura 2.7).
As células mãe do grão de pólen
(CMP) são de natureza diploide (2n) que
sofrem meiose, resultando em tétrades
haploides (n) funcionais, que se diferenciarão
em grãos de pólen. Para que ocorra essa
diferenciação há absorção de substâncias
nutritivas das camadas celulares intina e
exina e os grãos de pólen ficam maduros e
soltos dentro dos lóculos, aguardando a
deiscência das anteras.
Por fim, cada grão de pólen fica
constituído de duas camadas celulares, intina
e exina, um núcleo vegetativo e um núcleo generativo ou reprodutivo, todos de natureza haploide (MARCOS
FILHO, 2005).
b) Macrosporogênese
A formação do óvulo é semelhante ao processo inicial da microsporogênese, pois o óvulo, no início, é
uma massa de células indiferenciadas do tipo meristemática denominada nucela. A partir da nucela formam-se
dois tecidos que não a envolvem totalmente, a primina e a secundina, deixando a abertura micropilar por onde
germinará o tubo polínico.
Dessa massa celular indiferenciada, uma das células se destaca das demais formando a célula
arquesporial, da natureza diploide (2n). À medida que essa célula se desenvolve o núcleo aumenta de tamanho
e o citoplasma fica mais denso, preparando-se para a mitose que resultará na formação da célula mãe do
megasporo (2n). Após essa célula por meiose
produz quatro células haploides (n) (Figura 2.8).
A célula mãe do megasporo sofre três
mitoses consecutivas dando origem a oito células
com núcleos haploides, a oosfera, duas
sinérgidas, três antípodas e dois núcleos polares.
Esses últimos se fundem formando uma única
célula de natureza diploide, denominada de
mesocisto e completando o desenvolvimento do
gameta feminino (CARVALHO e
NAKAGAWA, 2000).
Figuras 2.7 – Microsporogênese em soja
(Fonte: Lauxen et al., 2003).
Figuras 2.8 – Macrosporogênese em
Anacardia occidentale L. (Fonte: Oliveira
et al., 2001).

Unidade 2
Transmissão dos Genes entre Gerações 24
Pelas descrições de ambos o processo pode-se verificar a existência de semelhança entre eles. O que
os difere são os genes que têm diferentes regulações em função do tecido que estão e o destino deles. A união
dos dois gametas formará a semente que originando outra planta concluirá a transferência de genes de uma
geração para outra.
8. Referências Bibliográficas
BURNS, G.W; BOTTINO, P.J. Genética. Rio de Janeiro: Guanabara-Koogan. 6.ed. 1991. p.35-38.
CARVALHO, N.M.; NAKAGAWA, J. Sementes: ciência, tecnologia e produção. Jaboticabal: Funep. 4.ed.
2000. p.588.
CUCO, S.M.; MONDIN, M.; VIEIRA, M.L.C. et al. Técnicas para a obtenção de preparações citológicas com
alta frequência de metáfases mitóticas em plantas: Passiflora (Passifloraceae) e Crotalaria (Leguminosae).
Acta Botânica Brasílica, v.17, n.3, p.363-370, 2003.
GEMMELL, A.R. Anatomia do vegetal em desenvolvimento. São Paulo: EPU/EDUSP. Coleção Temas de
Biologia. v.12. 1981. p.58-61.
GUERRA, M.S. Introdução à citogenética vegetal. Rio de Janeiro: Guanabara-Koogan. 1988. p.137.
HARRINSON, D. Biologia. Lisboa: Editora Presença. 1975. p.15-46.
JOHN, B.; LEWIS, K.R. Hierarquia cromossômica: Introdução à biologia dos cromossomos. São Paulo:
LTD/EDUSP. 1979. p.1-58.
LAUXEN, M.S.; KALTCHUK-SANTOS, E.; CHYNG-YEH HU et al. Association between floral bud size
and developmental stage in soybean microspores. Brazilian Archives of Biology and Technology, v.46, n.4,
p.515-520, 2003.
MARCOS FILHO, J. Fisiologia de sementes de plantas cultivadas. Piracicaba: FEALQ. 2005. p.27-40.
OLIVEIRA, J.M.S.; MARIATH, A.J.E.; BUENO, D.M. Desenvolvimento floral e estaminal no clone CP76
de Anacardium occidentale L. cajueiro-anão precoce (Anacardiaceae). Revista Brasileira de Botânica, v.24,
n.4, p.377-388, 2001.
SACCO, J.C. Anatomia vegetal. Caderno Didático do Departamento de Biologia. UFPEL, v.1. 1970. p.16.
SUZUKI, D.T.; GRIFFITHS, A.J.F.; MULLER, J.H.; LEWONTIN, R.C. Introdução à genética. Rio de
Janeiro: Guanabara-Koogan, 4.ed. 1992. p.638.
WEBERLING, R.; SCHWANTES, H. M O. Taxonomia vegetal. São Paulo: EPU. 1986, p.28-35 e 284.
Exercícios
1. A microsporogênese é dependente da perpendicularidade do complexo metáfase I – anáfase I em relação
ao complexo metáfase II – anáfase II. Explique por motivo isso ocorre?
2. Qual a importância do estudo do cariótipo das espécies vegetais? Quando é mais necessária a sua análise?
3. Qual a origem do endosperma?

Unidade 2
Transmissão dos Genes entre Gerações 25
4. Explique a razão pela qual se pode afirmar que a divisão celular tem início no período “S” da interfase.
5. Qual a função do centrômero com relação à fórmula cariotípica?
6. O embrião e o endosperma da semente são formados pela dupla fertilização após processos de
microsporogênese e megasporogênese. Considerando as características, forma da semente e teor de amido
no endosperma, verifique a proporção, em F2, do cruzamento das seguintes gerações paternais: CC x cc,
sendo que C determina sementes grandes e é dominante sobre c que condiciona sementes pequenas. Para
o endosperma o alelo C adiciona 1,56 µg de amido à semente à quantidade básica de 0,35µg. R: 1/4 CC –
3,47 µg; 2/4 Cc – 1,91 µg e 1/4 cc – 0,35 µg.
7. Considerando que uma planta possua 2n = 32 cromossomos, diga:
a. Quantos cromossomos terão no final da mitose e da meiose? R: Mitose – 32 e meiose 16
b. Quantas cromátides tem cada célula desse vegetal que sofre divisão? R: 64 cromátides
c. Quantos centrômeros uma célula dessa planta possui quando está em metáfase? R: 32 centrômeros
d. Quantos cromossomos terão as células mãe do grão de pólen e quantos no micrósporo? R: CMP – 32 e
micrósporo – 16
8. Em Coleus blumei as células somáticas são diploides e possuem 24 cromossomos. Quantos, de cada um
dos seguintes, estarão presentes em cada célula no estágio de meiose ou mitose indicados:
a. Centrômeros na anáfase? R: 48
b. Centrômero na anáfase I? R: 24
c. Cromátides na metáfase I? R: 48
d. Cromátides na anáfase? R: 48
e. Cromossomos na anáfase? R: 48
f. Cromossomos na metáfase? R: 24
g. Cromossomos no final da telófase I e telófase
II? R: telófase I – 12 e telófase II - 12
(Fonte: BUNS, G.W., 1984, p.94).
9. Certa planta tem oito cromossomos nas células de suas raízes, um par metacêntrico comprido, um par
metacêntrico curto, um par acrocêntrico longo e um par acrocêntrico curto. Em sua meiose quantos destes
cromossomos aparecerão nas células resultantes. Demonstre o processo por desenho. R: Um de cada tipo
10. Estabeleça uma relação entre a duplicação do DNA e as cromátides irmãs.
11. Quais os pontos semelhantes entre a microsporogênese e a megasporogênese?
12. O que são cromossomos homólogos e como se comportam na meiose para a formação das tétrades.
13. Na embriogênese vegetal a disposição dos cromossomos nas fases de divisão resulta na formação das
tétrades. Relacione a divisão de cromossomos homólogos e a divisão de cromátides irmãs com as tétrades
para originar os grãos de pólen.