resultados - biblioteca digital de teses e … · me60 110µg 0,40,0 ma80 150µg ac16 123µg 3,5...
TRANSCRIPT

RESULTADOS

______________________________________________Resultados e Discussão 58
4. RESULTADOS E DISCUSSÃO
4.1 Seleção da espécie de Penicillium frente a conidiogênese
Para selecionar, através da produção de conídios, uma espécie para
estudo, cinco espécies de Penicillium foram inoculadas em vários meios de
cultura semi-sólidos e incubadas por três tempos distintos em estufa a 30°C.
A suspensão conidial obtida de cada cultura foi avaliada por contagem
em Câmara de Neübauer. De acordo com os resultados apresentados nas
tabelas 8, 9 e 10 (p. 5 e 56) foi possível estabelecer as melhores condições
de cultivo para cada espécie e selecionar uma para ser quimicamente
estudada.
Tabela 8. Média da contagem de conídios dos fungos incubados por 7 dias a 30°C (conid/mL X 107)
Tabela 9. Média da contagem de conídios dos fungos incubados por 10 dias
a 30°C (conid/mL X 107)
YES CYA MEA AVEIA PDA P. verrucosum 0,15 0,15 5,04 2,61 5,18 P. ochrochlorum
0,04 0,01 0,92 0,56 2,15
P. waksmanii 3,10 0,27 3,24 2,48 3,43 P. simplicissimum
0,02 0,01 0,72 0,34 0,98
P. viridicatum 0,42 0,88 4,37 9,78 4,70

______________________________________________Resultados e Discussão 59
Tabela 10. Média da contagem de conídios dos fungos incubados por 16
dias a 30°C (conid/mL X 107)
A conidiogênese de um fungo é ideal quando este apresenta maior
concentração de conídios em menor tempo, otimizando a fase inicial do
cultivo e conseqüentemente a produção de metabólitos secundários. Nesta
etapa de desenvolvimento, os fungos P. verrucosum e P. viridicatum são os
mais promissores. A figura 21 (p. 56) resume as melhores condições para
cada espécie de Penicillium.
YES CYA MEA AVEIA PDA P. verrucosum 0,98 0,24 3,06 2,59 3,54 P. ochrochlorum
0,10 0,02 1,49 0,23 3,53
P. waksmanii 1,19 0,49 10,26 4,89 5,91 P. simplicissimum
0,07 0,01 0,96 0,37 0,25
P. viridicatum 0,32 0,81 2,02 8,51 3,11
YES CYA MEA AVEIA PDA P. verrucosum 2,03 0,16 2,11 3,10 4,65 P. ochrochlorum
0,01 0,02 1,84 0,16 1,15
P. waksmanii 0,98 0,04 4,25 6,62 6,95 P. simplicissimum
0,07 0,01 0,92 0,61 0,72
P. viridicatum 1,04 0,02 1,66 2,71 7,24
0
2
4
6
8
10
12
0 7 10 16
Tempo (dias)
Nú
mer
o d
e co
níd
ios
(x 1
07 co
nid
/ml)
P. verrucosum (PDA)
P. ochrochlorum (PDA)
P. waksmanii (MEA)
P. simplicissimum (PDA)
P. viridicatum (AVEIA)

______________________________________________Resultados e Discussão 60
Figura 21. Resumo das melhores condições de conidiogênese das 5
espécies de Penicillium
A obtenção de massa micelial, no meio pré-fermentativo, capaz de
produzir quantidades detectáveis de substâncias, em meio fermentativo, é
dependente da quantidade de conídios inoculada (Freitas, 2002). Portanto,
P. verrucosum cultivado em meio PDA por 7dias a 30°C foi selecionado para
cultivo e estudo químico neste trabalho devido à rapidez na produção de
conídios e facilidade no preparo do meio de cultura selecionado.
Os fungos P. viridicatum e P. waksmanii foram cultivados e estão
sendo estudados quimicamente por outros alunos do Laboratório de Química
Farmacêutica da FCFRP-USP sob orientação da Profa. Dra. Mônica Tallarico
Pupo.
O fungo P. simplicissimum apresentou coloração branca com mínima
produção de conídios. Não houve alteração neste quadro quando deixado
por alguns dias em contato direto com a luz e nem por permanecer até 20
dias em estufa a 30°C. Este fungo foi descartado para estudos posteriores
devido à instabilidade apresentada e ao baixo número de conídios.

______________________________________________Resultados e Discussão 61
4.2 Triagem dos extratos de P. verrucosum pelos ensaios, in vitro,
sobre a forma tripomastigota de T. cruzi, bioquímicos sobre as
enzimas GAPDH de T. cruzi e APRT de L. tarentolae.
4.2.1 Ensaios sobre a forma tripomastigota de T. cruzi
Os 115 extratos brutos obtidos dos cultivos, em pequena escala, de P.
verrucosum foram testados na forma tripomastigota do parasita T. cruzi, com
objetivo de selecionar o(s) mais promissor(es) para isolamento e
identificação estrutural da(s) substância(s) (tabela 11, p. 58).
Os extratos que promoveram uma porcentagem de lise das formas
tripomastigotas de T. cruzi acima de 50% foram considerados ativos.
Tabela 11. Extratos testados no ensaio com a forma tripomastigota de T.
cruzi.
N° (%) Lise 1mg/mL N°
(%) Lise 1mg/mL N°
(%) Lise 1mg/mL
Ac1 41,7 Aq39 41,5 Ac77 7,5 Ac2 37,1 Dc40 17,1 Ac78 11,6 Ac3 55,4 Dc41 50,6 Ma79 17,1 Ma4 50,8 Dc42 31,7 Aq82 20,5
Ma5 48,3 Me43 67,4 Aq83 6,8 Ma6 51,7 Me44 30,3 Me85 23,3 Aq7 52,9 Me45 41,6 Me86 8,2 Aq8 47,9 Ac46 9,0 Dc88 13,0 Aq9 56,7 Ac47 77,5 Dc89 5,5 Dc10 53,3 Ac48 46,1 Ac91 2,7 Dc11 35,8 Ma49 14,6 Ac92 30,1 Dc12 50,0 Ma50 13,5 Ac93 27,4 Me13 43,3 Ma51 1,7 Ma94 21,4 Me14 45,8 Aq52 39,0 Ma95 28,6 Me15 35,0 Aq53 39,0 Ma96 13,1

______________________________________________Resultados e Discussão 62
Ac16 1,7 Aq54 45,7 Aq97 51,1 Ac17 41,6 Dc55 38,4 Aq98 28,6 Ac18 31,5 Dc56 43,9 Aq99 14,9 Ma19 19,1 Dc57 22,6 Me100 4,1 Ma20 4,5 Me58 27,0 Me101 6,5 Ma21 6,6 Me59 61,2 Me102 53,6 Aq22 17,1 Me60 61,8 Dc103 39,0 Aq23 46,3 Ac61 27,4 Dc104 9,2 Aq24 43,3 Ac62 6,6 Dc105 14,0 Dc25 46,3 Ac63 41,5 Ac106 16,1 Dc26 45,1 Ma64 61,0 Ac107 25,3 Dc27 50,0 Ma65 27,4 Ac108 8,9 Me28 30,3 Ma66 1,2 Ma109 0,7 Me29 1,1 Aq67 51,2 Ma110 19,2 Me30 0,0 Aq68 29,3 Ma111 26,7 Ac31 8,4 Aq69 58,5 Aq112 8,6 Ac32 15,2 Me70 43,9 Aq113 10,6 Ac33 24,2 Me71 12,3 Aq114 0,0 Ma34 7,9 Me72 31,5 Me115 30,1 Ma35 3,9 Dc73 17,8 Me116 17,8 Ma36 1,7 Dc74 2,1 Me117 29,1 Aq37 14,6 Dc75 31,5 Dc118 21,2 Aq38 21,9 Ac76 17,1 Dc119 27,4 Dc120 10,96
Os extratos Ac47, Me59, Me60, Ma64 e Me43 apresentaram as
atividades antiparasitárias mais promissoras (figura 22, p. 60). Os três
primeiros são provenientes do cultivo de P. verrucosum em meio
fermentativo Takeuchi, o penúltimo em meio Czapeck e o último em meio
Jackson. É importante destacar que os extratos Me43, Me59 e Me60 foram
obtidos da massa micelial e apresentam o mesmo perfil químico,
evidenciado através de análise em CCDA. Estes resultados indicam que em
meio Takeuchi, tanto em 72 horas quanto em 144 horas, os metabólitos
secundários produzidos são muito semelhantes. Além disso, quando a
massa micelial de P. verrucosum foi cultivada por 48 horas em meio Jackson

______________________________________________Resultados e Discussão 63
e por 72 e 144 horas em meio Takeuchi, os metabólitos produzidos nestas
diferentes condições apresentaram-se em misturas análogas.
Figura 22. Extratos com atividade antiparasitária significativa no ensaio
biológico, in vitro, realizado com a cepa Y da forma
tripomastigota do Trypanosoma cruzi
Alguns extratos foram testados em menores concentrações e tiveram
seus respectivos IC50 calculados. Estes extratos apresentaram-se ativos
contra a forma tripomastigota de T. cruzi, confirmando a presença de
substâncias ativas (tabela 12, p. 61).
5551 52 53
5753
50 50 51
68
77
61 62 61
5158
51 54
0102030405060708090
% d
e lis
e do
par
asita
Ac3
Ma4
Ma6
Aq7
Aq9
Dc10
Dc12
Dc27
Dc41
Me43
Ac47
Me59
Me60
Ma64
Aq67
Aq69
Aq97
Me102

______________________________________________Resultados e Discussão 64
Tabela 12. Comparação da atividade tripanocida em diferentes
concentrações e IC 50 dos extratos brutos.
4.2.2 Ensaios bioquímicos com a enzima GAPDH de T. cruzi e enzima
APRT de L. tarentolae
Os 115 extratos brutos obtidos em pequena escala foram avaliados
frente a inibição das enzimas GAPDH de T. cruzi (tabela 13, p. 61) e APRT
de L. tarentolae (tabela 15, p. 65).
Tabela 13. Resultados obtidos no ensaio da atividade enzimática da GAPDH de T. cruzi frente aos extratos brutos obtidos de culturas de P. verrucosum
N°
[ ] % de inibição
N°
[ ] % de inibição
Ac1 100µg 0,0 Dc55 180µg 0,0 Ac2 80µg 0,0 Dc56 140µg 0,0 Ac3 75µg 0,0 Dc57 100µg 0,0 Ma4 110µg 0,0 Ac61 100µg 5,8
Ma5 120µg 0,0 Ac62 70µg 21,1 Ma6 140µg 0,0 Ac63 90µg 2,5 Aq7 150µg 0,0 Ma64 120µg 0,0 Aq8 135µg 0,0 Ma65 150µg 0,0 Aq9 90µg 0,0 Ma66 140µg 0,0 Dc10 80µg 0,0 Aq67 150µg 0,0 Dc11 135µg 0,0 Aq68 150µg 0,0 Dc12 75µg 0,0 Aq69 160µg 2,9 Me13 90µg 0,0 Me70 150µg 0,0
N°
(%) 1mg/mL
(%) 0,5mg/mL
(%) 0,25mg/mL
(%) 0,1mg/mL
IC50
(µg/ml)
Ac3 55,4 47,9 23,1 0,7 513,3 Dc10 53,3 63,3 23,4 4,9 401,5 Me43 67,4 51,7 43,3 39,9 463,7 Ac47 77,5 56,6 37,8 12,3 393,2 Ac48 46,0 64,3 59,4 10,5 263,9 Me59 61,2 42,7 37,1 11,2 597,6 Me60 61,8 59,4 27,3 6,6 411,7

______________________________________________Resultados e Discussão 65
Me14 100µg 0,0 Me71 140µg 0,0 Me15 105µg 0,0 Me72 110µg 13,6 Me43 100µg 13,1 Ac76 90µg 2,3 Ac47 85µg 2,0 Ac77 90µg 11,6 Me59 110µg 0,0 Ma79 130µg 0,0 Me60 110µg 0,0 Ma80 150µg 0,4 Ac16 123µg 3,5 Aq82 110µg 3,8 Ac17 225µg 4,0 Aq83 180µg 11,4 Ac18 225µg 0,0 Me85 130µg 14,5 Ma19 125µg 0,0 Me86 150µg 11,1 Ma20 110µg 0,0 Ac91 130µg 10,2 Ma21 225µg 0,0 Ac92 90µg 11,1 Me28 140µg 0,0 Ac93 120µg 14,1 Me29 120µg 0,0 Ma94 160µg 22,1 Me30 120µg 0,0 Ma95 110µg 15,3 Ac31 105µg 6,4 Ma96 110µg 16,6 Ac32 85µg 3,5 Aq97 100µg 20,0 Ac33 250µg 6,4 Aq98 130µg 15,5 Ma34 170µg 1,1 Aq99 110µg 23,7 Ma35 210µg 0,0 Me100 110µg 11,2 Ma36 220µg 0,0 Me101 150µg 11,2 Me44 200µg 0,0 Me102 120µg 16,4 Me45 150µg 0,0 Ac106 90µg 16,8 Ac46 100µg 0,0 Ac107 80µg 0,0 Ac48 250µg 0,0 Ac108 90µg 0,0 Ma49 245µg 0,0 Ma109 130µg 0,0 Ma50 160µg 0,0 Ma110 130µg 0,0 Ma51 187µg 0,0 Ma111 110µg 0,0 Me58 140µg 0,0 Aq112 110µg 0,0 Aq22 150µg 0,0 Aq113 140µg 0,0 Aq23 130µg 0,0 Aq114 150µg 0,0 Aq24 140µg 0,0 Me115 120µg 0,0 Dc25 110µg 0,0 Me116 120µg 0,0 Dc26 100µg 0,0 Me117 130µg 0,0 Dc27 100µg 0,0 Dc118 120µg 0,0 Aq37 170µg 0,0 Dc119 160µg 0,0 Aq38 140µg 0,0 Dc120 100µg 0,0 Aq39 120µg 0,0 Dc73 190µg 0,0 Dc40 300µg 0,0 Dc74 140µg 0,0 Dc41 150µg 0,0 Dc75 80µg 0,0 Dc42 130µg 0,0 Dc88 100µg 0,0 Aq52 140µg 0,0 Dc89 100µg 0,0 Aq53 160µg 3,9 Dc103 130µg 0,0 Aq54 160µg 1,2 Dc104 130µg 0,0 Dc105 130µg 0,0
Controle-DMSO

______________________________________________Resultados e Discussão 66
No ensaio com a enzima GAPDH observou-se que os extratos mais
ativos (tabela 13, p. 61) foram provenientes do cultivo do fungo em meio pré-
fermentativo 48 horas. Além disso, o meio fermentativo Jackson foi o mais
promissor para a produção de substâncias potencialmente ativas frente à
enzima GAPDH de T. cruzi.
O ensaio foi realizado com um alvo específico em baixa concentração,
por isso para extratos brutos foi difícil estimar a porcentagem de inibição da
atividade enzimática, uma vez que um extrato pode conter substâncias
ativas em concentrações muito baixas.
Um exemplo disto pôde ser ilustrado pela inibição de 3,9% da
atividade enzimática apresentada pelo extrato hexânico dos galhos de
Pilocarpus spicatus (Vieira et al., 2001). Deste extrato foi isolada uma
cumarina, chalepina, que apresentou IC50 = 64µM (Vieira et al., 2001)
tornando-se um promissor protótipo para o desenho de inibidores mais
ativos. A obtenção da estrutura do complexo cristalino chalepina - GAPDH
(figura 23, p. 64) indicou as interações do produto natural com aminoácidos
do sítio catalítico da enzima (Pavão et al., 2002) e possibilitou o
entendimento dos mecanismos moleculares envolvidos na inibição, bem
como a proposição de novas estruturas cumarínicas potencialmente ativas,
que vêm sendo sintetizadas em projeto paralelo coordenado pela Profa. Dra.
Mônica Tallarico Pupo.
Comparando-se os resultados de inibição da enzima GAPDH com os
obtidos na lise das formas tripomastigotas de T. cruzi, observou-se que os
extratos Aq97 e Me102, oriundos do meio fermentativo Jackson, mostraram-

______________________________________________Resultados e Discussão 67
se promissores, o que sugere uma futura investigação com isolamento e
identificação de substâncias bioativas com mecanismo de ação
possivelmente via inibição da enzima GAPDH (tabela 14, p. 65).
Figura 23. Comparação da estrutura monomérica nativa da GAPDH de
T.cruzi (rosa) e complexada (azul) com o inibidor chalepina
Dos 115 extratos brutos ensaiados no teste bioquímico para inibição
da enzima APRT de L tarentolae, os extratos Dc73, Dc75, Dc88, Dc103 e
Dc104 apresentaram as atividades inibitórias mais promissoras (tabela 15, p.
65). Os dois primeiros foram provenientes do cultivo de P. verrucosum em
meio fermentativo Czapeck, Dc88 em meio Vogel e os dois últimos em meio
Jackson. É importante destacar que estes extratos foram cultivados em meio
OO
HO
Chalepina

______________________________________________Resultados e Discussão 68
pré-fermentativo por 48 horas e que foram obtidos da massa micelial do
fungo extraída com CH2Cl2. Estes dados ilustram o potencial de culturas de
P. verrucosum na produção de metabólitos ativos inibidores da enzima
APRT de L. tarentolae.
Tabela 14. Comparação dos extratos com atividade superior a 15% de inibição da enzima GAPDH com a % de lise da forma tripomastigota de T. cruzi
Tabela 15. Ensaio da atividade enzimática da APRT de L. tarentolae frente aos extratos brutos obtidos de culturas de P. verrucosum
N°
[ ] % de inibição
N°
[ ] % de inibição
Ac1 50µg 1,5 Me58 70µg 3,9 Ac2 40µg 0,0 Me59 55µg 0,0 Ac3 38µg 12,7 Me60 55µg 1,2 Ma4 50µg 0,0 Ac61 50µg 0,0
Ma5 65µg 0,0 Ac62 80µg 0,0 Ma6 70µg 0,0 Ac63 75µg 0,0 Aq7 75µg 3,4 Ma64 50µg 8,7 Aq8 65µg 0,0 Ma65 75µg 3,6 Aq9 45µg 3,4 Ma66 60µg 5,8 Dc10 48µg 0,0 Aq67 60µg 4,1 Dc11 65µg 0,0 Aq68 80µg 4,1 Dc12 38µg 0,0 Aq69 85µg 0,4 Me13 45µg 3,9 Me70 75µg 0,0
N°
% de lise da forma tripomastigota
(1mg/mL)
% de inibição da GAPDH (≈100 µg/mL)
Ac62 6,6 21,1 Ma94 21,4 22,1 Ma95 28,6 15,3 Ma96 13,1 16,6 Aq97 51,1 20,0 Aq98 28,6 15,5 Aq99 14,9 23,7
Me102 53,6 16,4 Ac106 16,1 16,8
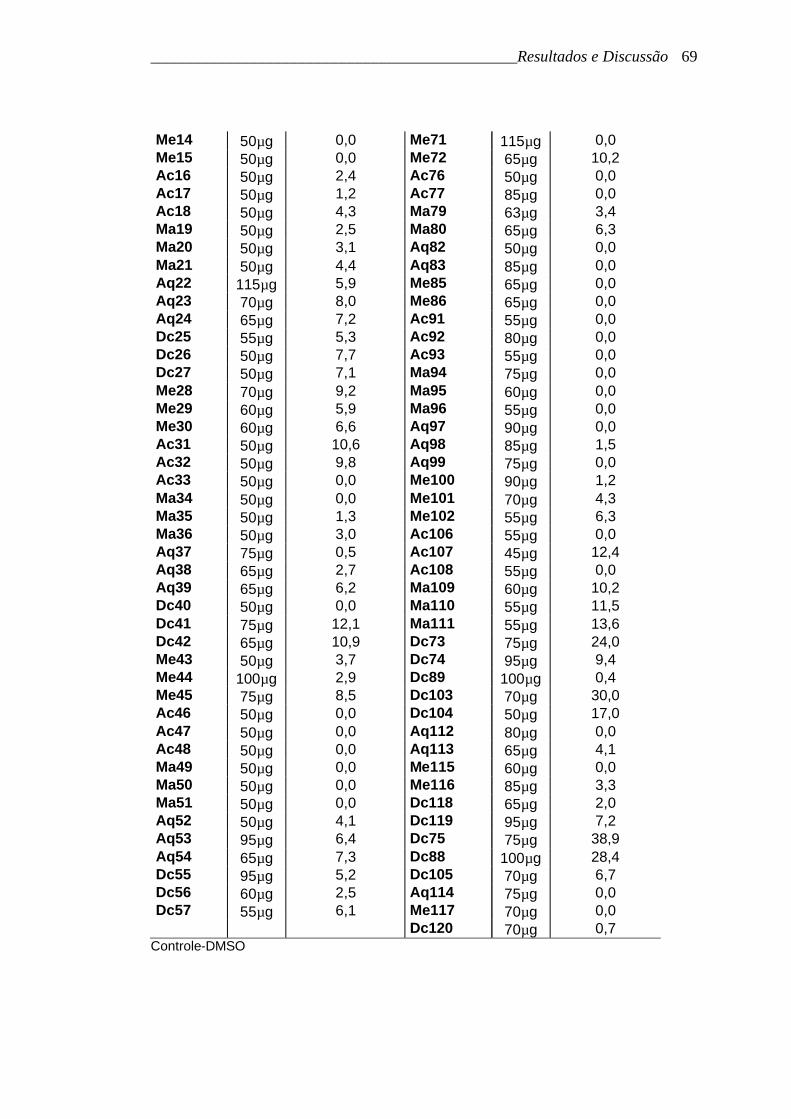
______________________________________________Resultados e Discussão 69
Me14 50µg 0,0 Me71 115µg 0,0 Me15 50µg 0,0 Me72 65µg 10,2 Ac16 50µg 2,4 Ac76 50µg 0,0 Ac17 50µg 1,2 Ac77 85µg 0,0 Ac18 50µg 4,3 Ma79 63µg 3,4 Ma19 50µg 2,5 Ma80 65µg 6,3 Ma20 50µg 3,1 Aq82 50µg 0,0 Ma21 50µg 4,4 Aq83 85µg 0,0 Aq22 115µg 5,9 Me85 65µg 0,0 Aq23 70µg 8,0 Me86 65µg 0,0 Aq24 65µg 7,2 Ac91 55µg 0,0 Dc25 55µg 5,3 Ac92 80µg 0,0 Dc26 50µg 7,7 Ac93 55µg 0,0 Dc27 50µg 7,1 Ma94 75µg 0,0 Me28 70µg 9,2 Ma95 60µg 0,0 Me29 60µg 5,9 Ma96 55µg 0,0 Me30 60µg 6,6 Aq97 90µg 0,0 Ac31 50µg 10,6 Aq98 85µg 1,5 Ac32 50µg 9,8 Aq99 75µg 0,0 Ac33 50µg 0,0 Me100 90µg 1,2 Ma34 50µg 0,0 Me101 70µg 4,3 Ma35 50µg 1,3 Me102 55µg 6,3 Ma36 50µg 3,0 Ac106 55µg 0,0 Aq37 75µg 0,5 Ac107 45µg 12,4 Aq38 65µg 2,7 Ac108 55µg 0,0 Aq39 65µg 6,2 Ma109 60µg 10,2 Dc40 50µg 0,0 Ma110 55µg 11,5 Dc41 75µg 12,1 Ma111 55µg 13,6 Dc42 65µg 10,9 Dc73 75µg 24,0 Me43 50µg 3,7 Dc74 95µg 9,4 Me44 100µg 2,9 Dc89 100µg 0,4 Me45 75µg 8,5 Dc103 70µg 30,0 Ac46 50µg 0,0 Dc104 50µg 17,0 Ac47 50µg 0,0 Aq112 80µg 0,0 Ac48 50µg 0,0 Aq113 65µg 4,1 Ma49 50µg 0,0 Me115 60µg 0,0 Ma50 50µg 0,0 Me116 85µg 3,3 Ma51 50µg 0,0 Dc118 65µg 2,0 Aq52 50µg 4,1 Dc119 95µg 7,2 Aq53 95µg 6,4 Dc75 75µg 38,9 Aq54 65µg 7,3 Dc88 100µg 28,4 Dc55 95µg 5,2 Dc105 70µg 6,7 Dc56 60µg 2,5 Aq114 75µg 0,0 Dc57 55µg 6,1 Me117 70µg 0,0 Dc120 70µg 0,7
Controle-DMSO

______________________________________________Resultados e Discussão 70
Entre os 18 extratos brutos tripanocidas encontrados (figura 22, p.
60), os mais promissores foram oriundos do meio pré-fermentativo incubado
por 24 horas, meio fermentativo Takeuchi e provenientes da massa micelial
extraída com metanol e fluido da cultura extraído com AcOEt. Já os extratos
ativos nas enzimas GAPDH e APRT, todos foram oriundos do meio pré-
fermentativo incubado por 48 horas, mas para GAPDH houve o predomínio
de extratos cultivados em meio fermentativo Jackson e extraídos do fluido da
cultura, para a APRT, houve predomínio dos extratos obtidos da massa
micelial do fungo cultivado em meios Jackson, Czapeck e Vogel.
Estes resultados demonstraram que a produção de diferentes
metabólitos secundários bioativos varia de acordo com as condições de
cultivo do fungo. Além disso, diferentes melhorias no cultivo poderiam ser
futuramente introduzidas na tentativa de aumentar a produção dos
metabólitos ativos.
4.3 Considerações gerais sobre o isolamento das substâncias
Os extratos Ac47 do fluido da cultura e Me59 da massa micelial foram
selecionados para ter sua escala ampliada por apresentarem lise
significativa das formas tripomastigotas de T. cruzi (tabela 22, p. 60). Além
disso, são oriundos da mesma condição de cultivo de P. verrucosum.
O fracionamento do extrato Ac47, produzido em escala intermediária
resultou em muitas frações com baixo grau de pureza que não foram
ensaiadas e purificadas devido às pequenas massas obtidas. Os espectros
de RMN 1H das frações finais, obtidas em escala intermediária, mostraram-

______________________________________________Resultados e Discussão 71
se em misturas complexas contendo sinais na região entre δ 0,8 e 1,4
indicando a possibilidade da presença de unidades de leucina, isoleucina,
valina, alanina ou treonina nas estruturas. Além disso, observaram-se vários
sinais entre δ 2,0 e 4,5 característicos de hidrogênio α-carbonílicos e ligados
a carbono ligado a heteroátomo.
Visto que o estudo químico e o biomonitoramento estavam limitados devido
às pequenas quantidades de massas e complexidade das misturas obtidas,
o fungo P. verrucosum foi cultivado em escala ampliada para a obtenção de
frações com massa suficiente para o isolamento. Novamente, os espectros
de RMN 1H das frações obtidas dos processos cromatográficos de Ac47
mostraram que as mesmas eram compostas por misturas de substâncias,
constituídas principalmente por aminoácidos. Também observou-se em
algumas frações sinais característicos de sistema aromático para
dissubstituído (AA’XX’), possíveis para uma unidade de tirosina.
A complexidade das misturas sugeriu que as técnicas cromatográficas
usuais não estariam sendo eficazes, devido a possível semelhança nas
estruturas químicas dos aminoácidos precursores e conseqüentemente nas
características das moléculas produzidas no metabolismo secundário de P.
verrucosum.
Cromatografia líquida de alta eficiência reciclante (CLAE - R), foi uma
alternativa usada neste trabalho. Este cromatógrafo foi utilizado no
laboratório de Produtos Naturais do Departamento de Química da
Universidade Federal de São Carlos-UFSCar, disponibilizado pelo Prof. Dr.
Paulo Cezar Vieira. CLAE - R distingue-se das CLAE usuais principalmente

______________________________________________Resultados e Discussão 72
pelo fato de retornar a amostra a ser separada para a coluna. O número de
vezes (ciclos) que a amostra volta para a coluna é ilimitado. Isto possibilita
maior eficiência na separação.
Assim, optou-se por adotar esta técnica para algumas frações. A escolha foi
feita em função das quantidades das frações, da análise prévia dos
espectros de RMN 1H e atividades de inibição da enzima GAPDH de T.
cruzi.
As frações Ac479.4.4 e Ac479.4.5 (escala ampliada) foram separadas por
CLAE – R (figuras 24, p. 70; 25a, p. 71 e 25b, p. 72) das quais foi isolada a
substância F1 (p. 73).
As frações obtidas por CLAE - R que se constituíam por mistura aparente de
2 ou 3 substâncias foram analisadas via ESI-EM/EM, na tentativa de se
identificar as substâncias em mistura, mas a ionização por electrospray
destas pequenas moléculas não ocorreu, limitando a utilização desta
técnica. Este experimento foi realizado no pelo Prof. Dr. Norberto Peporine
Lopes no laboratório de Química Orgânica do Departamento de Ciências
Farmacêuticas da FCFRP-USP.
O extrato Me59 obtido da massa micelial de P. verrucosum em escala
ampliada foi fracionado por técnicas usuais, levando ao isolamento das
substâncias M2, M3, M4, M5 e M6 (p. 73). A mistura de substâncias M2 foi
obtida por esterificação com diazometano, as substâncias M3 e M4 foram
acetiladas e a mistura de substâncias M6 foi transesterificada.

Figura 24. Cromatograma da fração AC47.9.4.4 em CLAE - R no tempo de retenção 0 a 150 minutos. Condições experimentais: coluna polimérica Shodex, φ =21, 5 mm e h = 500 mm, fluxo de 5 mL/min, MeOH:CH2Cl2 1:1, 225 nm, 6 ciclos.
Minutes
0 20 40 60 80 100 120 140
Vo
lts0,
00,
10,
2
0,0
0,1
0,2
ciclos 1 2 3 4 5 6
Substância F1

Figura 25a. Cromatograma da fração AC47.9.4.5 em CLAE - R no tempo de retenção 0 a 450 minutos. Condições experimentais: coluna polimérica Shodex, φ =21,5 mm e h = 500 mm, fluxo de 3 mL/min, MeOH:CH2Cl2 1:1, 225 nm, 22 ciclos. (continuação na próxima página como figura 25b).
Minutes
0 100 200 300 400
Vol
ts0,
00
,10,
2
0,0
0,1
0,2
ciclos 1 2 3 4 5 6 7 8 9 10 11

Minutes
450 500 550 600 650 700 750 800 850
Vol
ts0,
00,
10,
2
0,0
0,1
0,2 Figura 25b: (continuação da figura 25a) Cromatograma da fração Ac47.9.4.5 em CLAE – R no tempo de retenção 450
a 860 minutos. Condições experimentais: coluna polimérica Shodex, φ =21,5 mm e h = 500 mm, fluxo de 3 mL/min, MeOH:CH2Cl2 1:1, 225 nm, 22 ciclos.
12 13 14 15 16 17 18 19 20 21 22
Substância F1

______________________________________________Resultados e Discussão
73
4.4 Substâncias Isoladas
HO
OH
OH
OH
OH
OH
(F1) Alcalóide dicetopiperazínico
(M3) Manitol
(M2) Ácidos graxos
(M6) Triglicerídeos
(M5) Glicerilfosfocolina
O
O
O
OO
On
n
n
O
OH
n
O
OH
n
O
H
HO
H
HO
H
O
OHHH
OH
O
H
OH
H
OH
H
HO HH
OH
(M4) α, α - Trealose
NH
HN O
HO O
O
POO
-O
N+
OH
OH

______________________________________________Resultados e Discussão
74
4.4.1 Alcalóide dicetopiperazínico (F1)
A substância F1 foi isolada do fluido da cultura de P. verrucosum
extraído com AcOEt. O sólido branco apresentou-se insolúvel em água.
O espectro de RMN 1H apresentou sinais referentes a 20 hidrogênios
(tabela 16, p. 78 e figura 26, p. 82). A presença de dois dubletos em δ 7,38
(J= 8,6Hz; 2H) e δ 7,08 (J= 8,6Hz; 2H), caracterizando um sistema de spins
AA’XX’, sugeriu a presença de uma unidade de tirosina na molécula, o que
já era esperado através da análise prévia dos espectros das frações que
originaram a substância F1 e do conhecimento da composição do meio de
cultura rico em aminoácidos.
O espectro de massas indicou a presença de número par de átomos
de nitrogênio na molécula, apresentando íon em m/z 277 ([M+H]+).
Os sinais em δ 3,56 (dd; J= 13,6; 5,1Hz; 1H) e δ 3,24 (dd; J= 13,6;
4,5Hz; 1H) foram atribuídos aos hidrogênios benzílicos da unidade de
tirosina. O acoplamento entre estes hidrogênios foi confirmado no mapa de
contornos COSY 1H - 1H (figura 27, p. 83), que mostrou também o
acoplamento destes hidrogênios com um hidrogênio na região em δ 4,64 (m;
1H), atribuído ao hidrogênio metínico α ao grupo amino e carbonílico da
unidade de tirosina. Também neste mapa evidenciou o acoplamento do
Tirosina
COOH
NH2
HO

______________________________________________Resultados e Discussão
75
hidrogênio em δ 4,64 com um hidrogênio em δ 9,26 (d; J= 2,0Hz; 1H),
atribuído a um hidrogênio ligado a nitrogênio. A presença de um hidrogênio
ligado a nitrogênio indicou que o nitrogênio deveria estar dissubstituído.
O espectro de RMN 13C (tabela 16, p. 77 e figura 28, p. 84)
apresentou 15 sinais, dos quais 2 apresentaram valores de deslocamento
químico característicos de carbonos carbonílicos de amidas (δ 167,7 e
168,9), o que é esperado em substâncias originadas de condensação entre
aminoácidos.
Os demais sinais no espectro de RMN 1H sugeriram a presença de
uma unidade de leucina: 2 dubletos em δ 0,84 (J= 6,3Hz; 3H) e δ 0,86 (J=
6,3Hz; 3H); um multipleto entre δ 1,86-1,96 (1H), 3 duplo duplo dubletos em
δ 1,62 (J= 4,6; 9,6; 13,6Hz; 1H), δ 0,93 (J= 4,8; 9,8; 13,6Hz; 1H) e δ 4,14 (J=
3,3; 3,8; 9,6Hz; 1H). Os acoplamentos entre estes hidrogênios foram
evidenciados no mapa de contornos COSY 1H -1H. Novamente observou-se
o acoplamento entre o sinal referente ao hidrogênio α carbonílico e α amino
(δ 4,14) da unidade de leucina e o sinal em δ 9,16 (d; J= 2,0Hz; 1H) atribuído
ao hidrogênio ligado ao nitrogênio da unidade de leucina.
HO O R'
R
HN
Tirosina com N dissubstituído

______________________________________________Resultados e Discussão
76
Leucina com nitrogênio dissubstituído.
Além destes sinais foi observado um singleto largo em δ 11,40
atribuído ao hidrogênio fenólico da unidade de tirosina.
Com base nos dados discutidos foi proposta a estrutura de um
alcalóide dicetopiperazínico (F1) formado pela condensação entre os
aminoácidos tirosina e leucina.
Proposta para substância F1- C15H20N2O3
O mapa de contornos HMQC (figura 29, p. 85) possibilitou as
atribuições dos carbonos da molécula. O mapa HMBC (figura 30, p. 86;
figura 31, p. 87 e figura 32, p. 88) permitiu a confirmação da proposta
estrutural, além de confirmar as atribuições dos deslocamentos químicos de
carbono e hidrogênio.
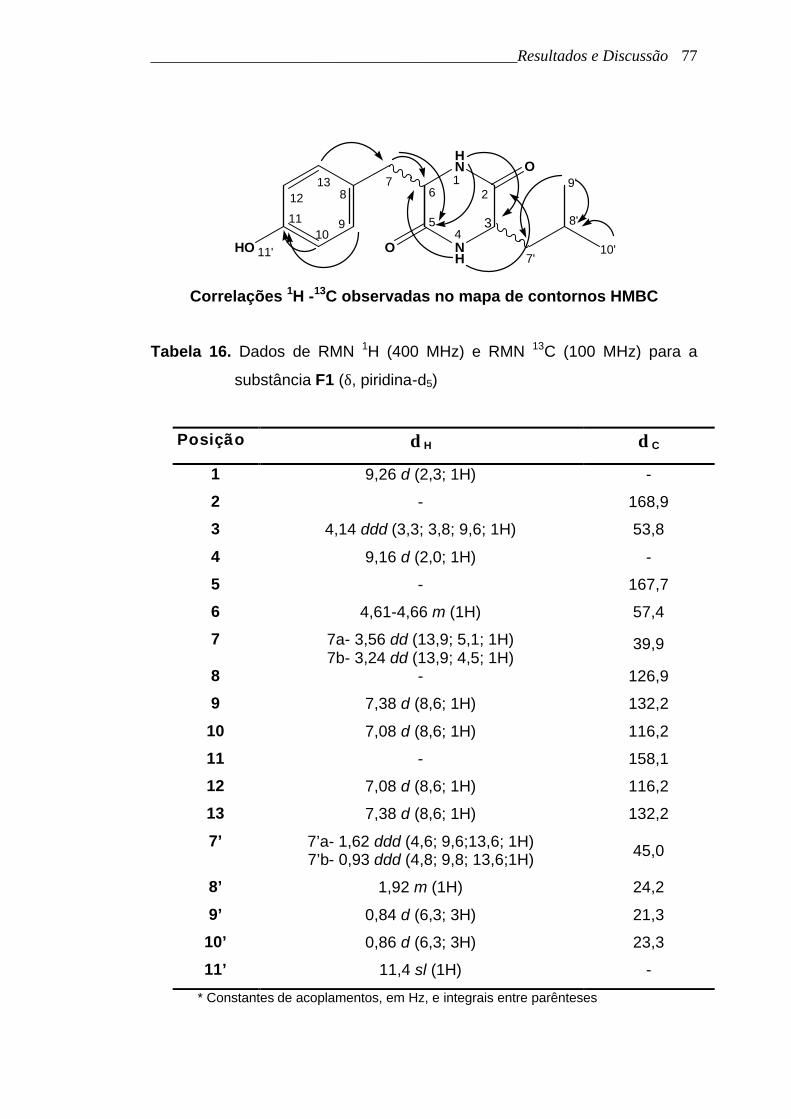
As principais correlações observadas no mapa de contornos HMBC
para a comprovação do anel dicetopiperazínico foram as correlações do H-1
com C-3 e H-4 com C-6. As correlações destes hidrogênios ligados aos
nitrogênios com os carbonos metínicos só seriam possíveis numa estrutura
contendo o anel dicetopiperazínico.
R'
NH
R O
NH
HN O
HO O
______________________________________________Resultados e Discussão
77
Correlações 1H -13C observadas no mapa de contornos HMBC
Tabela 16. Dados de RMN 1H (400 MHz) e RMN 13C (100 MHz) para a
substância F1 (δ, piridina-d5)
Posição δ H δ C
1 9,26 d (2,3; 1H) -
2 - 168,9
3 4,14 ddd (3,3; 3,8; 9,6; 1H) 53,8
4 9,16 d (2,0; 1H) -
5 - 167,7
6 4,61-4,66 m (1H) 57,4
7 7a- 3,56 dd (13,9; 5,1; 1H) 7b- 3,24 dd (13,9; 4,5; 1H)
39,9
8 - 126,9
9 7,38 d (8,6; 1H) 132,2
10 7,08 d (8,6; 1H) 116,2
11 - 158,1
12 7,08 d (8,6; 1H) 116,2
13 7,38 d (8,6; 1H) 132,2
7’ 7’a- 1,62 ddd (4,6; 9,6;13,6; 1H) 7’b- 0,93 ddd (4,8; 9,8; 13,6;1H) 45,0
8’ 1,92 m (1H) 24,2
9’ 0,84 d (6,3; 3H) 21,3
10’ 0,86 d (6,3; 3H) 23,3
11’ 11,4 sl (1H) -
* Constantes de acoplamentos, em Hz, e integrais entre parênteses
10'
9'
8'
7'
34NH
HN O
O
16
5
7
HO
2813
12
1110
9
11'

______________________________________________Resultados e Discussão
78
Na tentativa de se determinar a configuração relativa do alcalóide
dicetopiperazínico foi realizado o experimento de NOE-diff. Neste
experimento, irradiou-se H-6, H-3 e H-7b, porém os NOE observados não
permitiram a proposição da configuração relativa da molécula.
Alcalóides dicetopiperazínicos constituem uma classe de metabólitos
secundários amplamente distribuída em fungos, principalmente em espécies
de Penicillium (Laws & Mantle, 1985). Mais de 40 substâncias
dicetopiperazínicas estão listadas como metabólitos de fungos e a mais
freqüente biossíntese envolve a condensação de dois ou três aminoácidos
como precursores (Laws & Mantle, 1985; Kozlovsky et al., 2000).
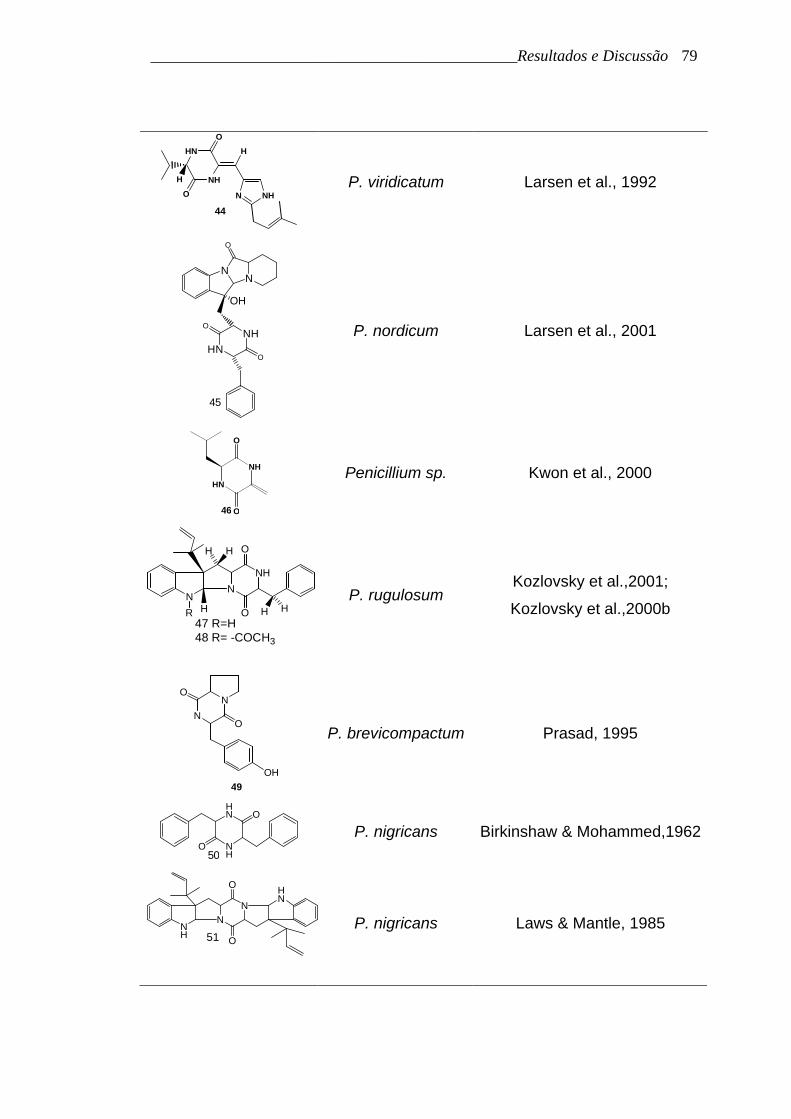
A tabela 17 (p. 78) apresenta as estruturas dos alcalóides
dicetopiperazínicos isolados de espécies de Penicillium encontrados na
literatura até Maio de 2003, incluindo as substâncias 2, 3, 33, 34, 37, 38 e 39
apresentadas anteriormente
Tabela 17. Alcalóides dicetopiperazínicos isolados de espécies de
Penicillium
Estrutura Espécie Referência
HN
NHO
O
42
P. megasporum Nozawa et al., 1989
HN
NH
H
O
O N NH
H
43
P. aurantiogriseum Larsen et al., 1992
______________________________________________Resultados e Discussão
79
HN
NH
H
O
H
O N NH
44
P. viridicatum Larsen et al., 1992
NN
HNNH
OH
45
O
O
O
P. nordicum Larsen et al., 2001
HN
NH
O
O46
Penicillium sp. Kwon et al., 2000
NN
NH
H
H
H
O
O
H
R47 R=H48 R= -COCH3
H
P. rugulosum Kozlovsky et al.,2001;
Kozlovsky et al.,2000b
N
NO
O
49OH
P. brevicompactum Prasad, 1995
O NH
HN O
50 P. nigricans Birkinshaw & Mohammed,1962
NH
NN
HN
O
O51
P. nigricans Laws & Mantle, 1985
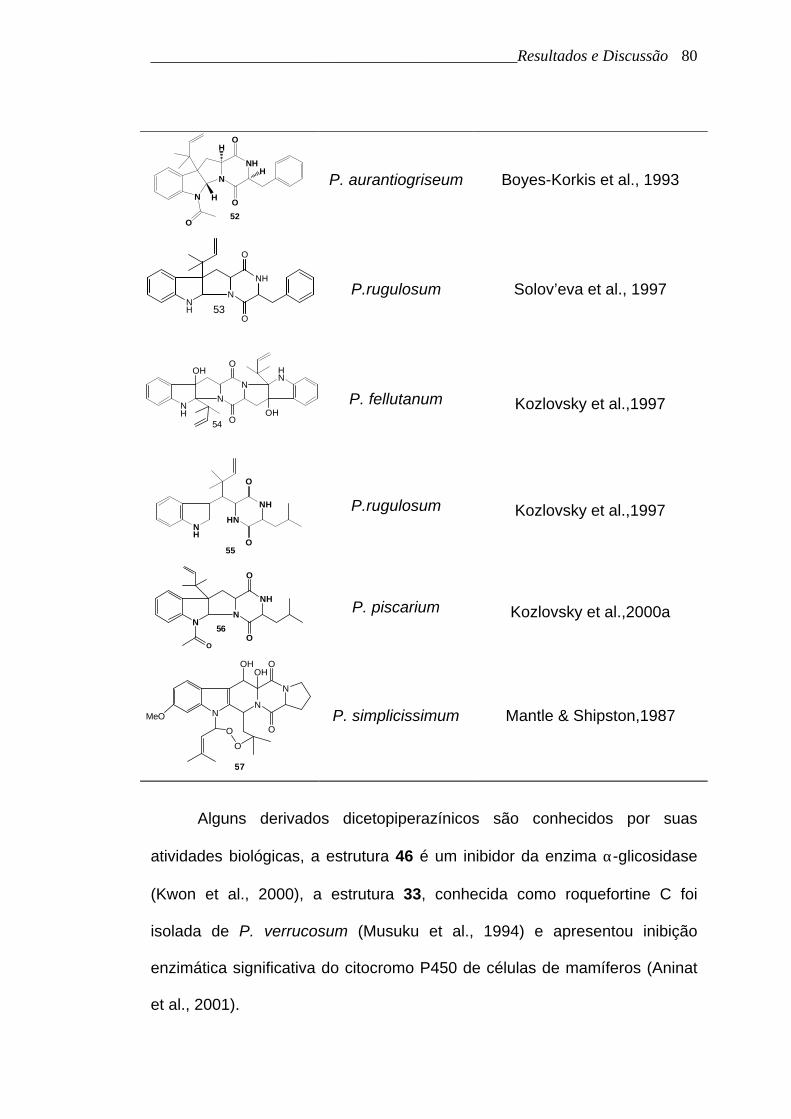
______________________________________________Resultados e Discussão
80
52
N
N
NH
O
O
O
H
H
H
P. aurantiogriseum Boyes-Korkis et al., 1993
NH
N
NH
O
O53
P.rugulosum Solov’eva et al., 1997
NH
N
N
HN
OHO
OOH
54
P. fellutanum Kozlovsky et al.,1997
NH
HN
NH
O
O55
P.rugulosum Kozlovsky et al.,1997
NN
NH
O
O56
O
P. piscarium Kozlovsky et al.,2000a
NN
N
MeO
OHOH
O
O
57
O
O
P. simplicissimum Mantle & Shipston,1987
Alguns derivados dicetopiperazínicos são conhecidos por suas
atividades biológicas, a estrutura 46 é um inibidor da enzima α-glicosidase
(Kwon et al., 2000), a estrutura 33, conhecida como roquefortine C foi
isolada de P. verrucosum (Musuku et al., 1994) e apresentou inibição
enzimática significativa do citocromo P450 de células de mamíferos (Aninat
et al., 2001).

______________________________________________Resultados e Discussão
81
Dicetopiperazinas isoladas de outros microrganismos apresentaram
atividade antifúngica (Yang, et al., 2002) e antagonista da substância P
(Barrow & Sedlock, 1994). Algumas dicetopiperazinas isoladas de plantas
são potentes agentes sedativos (Lee & Liu, 2001), imunosupressores
(Ravikanth, et al., 2001) e antitumorais (Pearce, 1994). Além disso, muitas
dicetopiperazinas têm sido sintetizadas baseadas em protótipos naturais
apresentando inibição de proteína tirosina quinase (Li & Peng, 1998) e
inibição de HIV protease (Falorni et al, 1996).
Até o momento, não se encontraram relatos na literatura sobre a
presença da substância F1 em espécies de Penicillium ou qualquer outra
fonte natural de metabólitos secundários.

Figura 26. Espectro de RMN 1H da substância F1 (Piridina-d5, 400 MHz)
1234567891011
7.007.107.207.307.40
9.159.209.259.30
4.104.204.304.404.504.604.70
3.103.203.303.403.503.603.70
1.601.701.801.902.00
0.850.900.95
1 (1H) 4 (1H)
9 e 13 (2H) 10 e 12 (2H)
7b’ (1H)
6 (1H)
3 (1H)
7a (1H) 7b (1H)
8’ (1H)
7a’ (1H)
10’ e 9’ (6H)
11’ (1H)
1
9'
8'
7'
34
2
NH
HN O
OHO
16
5
78
910
11
1213
11'

Figura 27. Mapa de contornos COSY 1H -1H da substância F1 (Piridina-d5, 400 MHz)
H1 H4 H6 H3
10'
9'
8'
7'
34
2
NH
HN O
OHO
16
5
78
910
11
1213
11'

Figura 28. Espectro de RMN 13C da substância F1 (Piridina-d5, 100 MHz)
20203030404050506060707080809090100100110110120120130130140140150150160160170170
2
5
11 9 e 13
10 e 12
8 6
3 7’
7
8’ 9’
10’
10'
9'
8'
7'
34
2
NH
HN O
OHO
16
5
78
910
11
1213
11'

Figura 29. Mapa de contornos HMQC da substância F1 (Piridina-d5)
10'
9'
8'
7'
34
2
NH
HN O
OHO
16
5
78
910
11
1213
11'
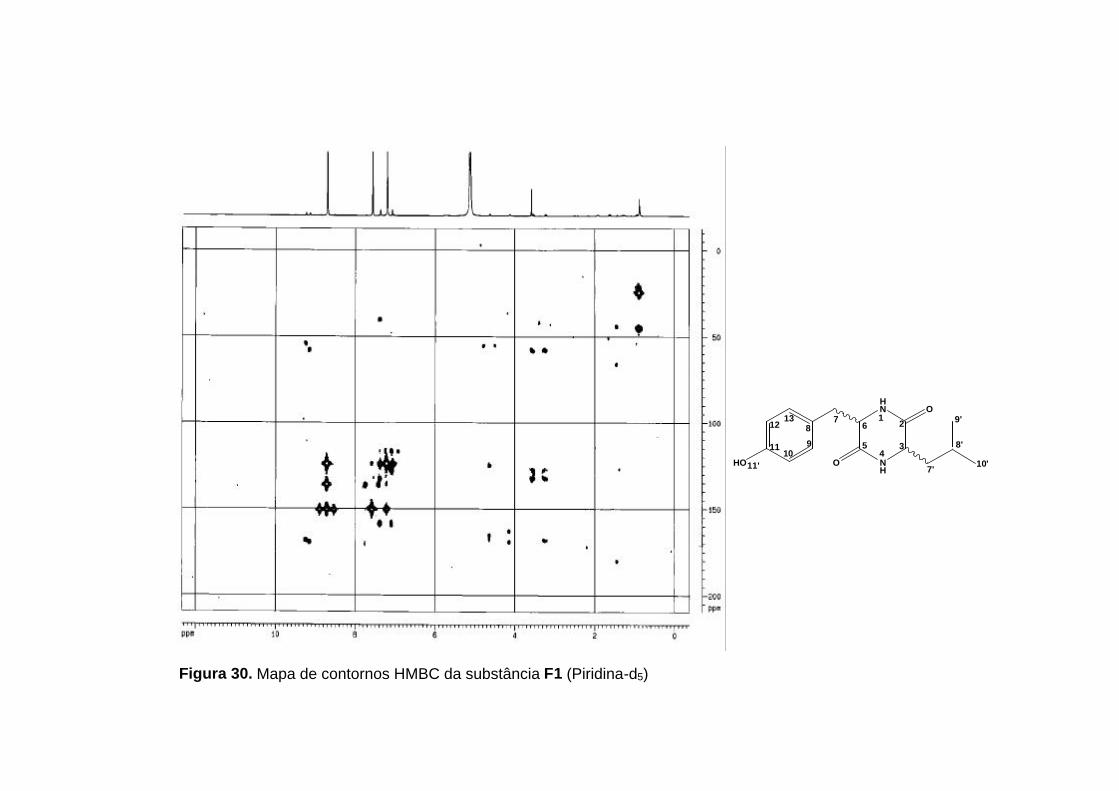
Figura 30. Mapa de contornos HMBC da substância F1 (Piridina-d5)
10'
9'
8'
7'
34
2
NH
HN O
OHO
16
5
78
910
11
1213
11'

Figura 31. Ampliação do mapa de contornos HMBC da substância F1 (Piridina-d5)
9'
8'
7'
34
2
NH
HN O
OHO
16
5
78
910
11
1213
11'
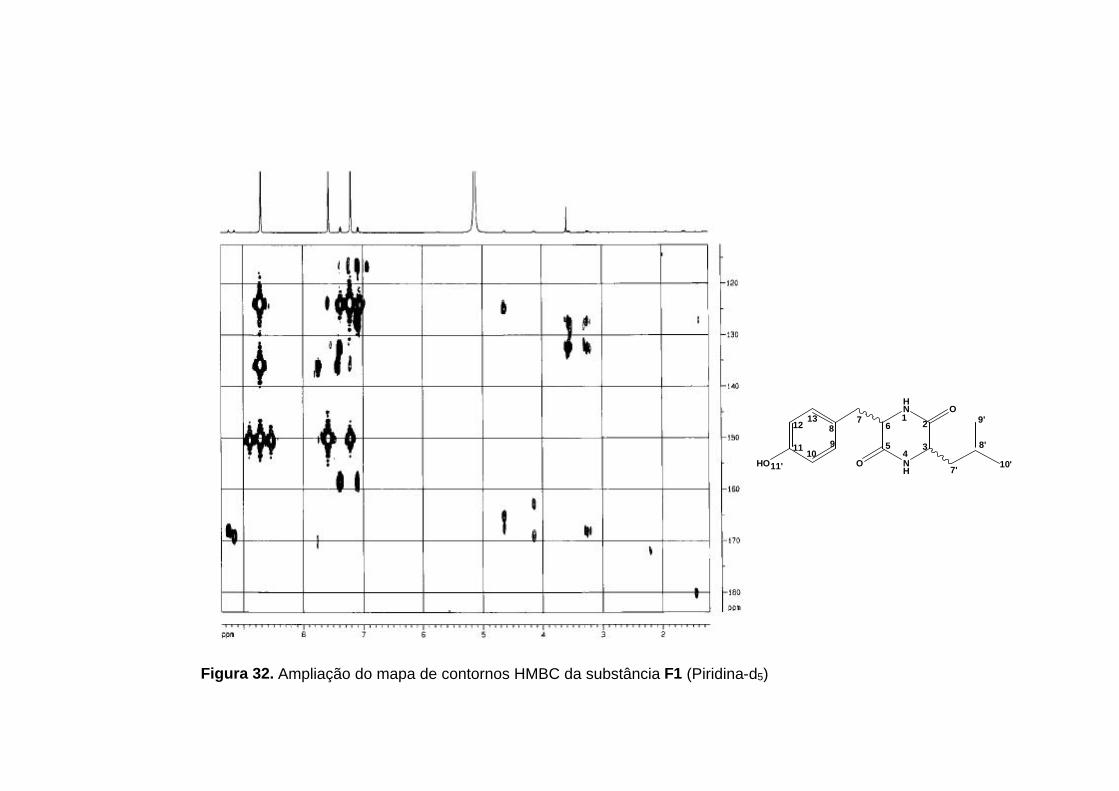
Figura 32. Ampliação do mapa de contornos HMBC da substância F1 (Piridina-d5)
10'
9'
8'
7'
34
2
NH
HN O
OHO
16
5
78
910
11
1213
11'

______________________________________________Resultados e Discussão
89
4.4.2 Ácidos graxos (M2)
A mistura dos ácidos graxos foi isolada da fração n-BuOH do extrato
metanólico obtido do micélio de P. verrucosum.
No espectro no infravermelho da fração Me59.2.8 (figura 33, p. 91)
foram observadas absorções de alguns grupos funcionais, listados na tabela
18 (p. 89).
Tabela 18. Principais absorções observadas no espectro no infravermelho da fração Me59.2.8
3283,54 cm-1 Deformação axial de O-H
2923,79 cm-1 Deformação axial de C-H
1736,27 cm-1 Deformação axial de C=O
1083,88 cm-1 Deformação axial de C-O
1458,85 cm-1 Deformação angular de C-O-H
As substâncias foram identificadas em mistura, após esterificação
com diazometano. A determinação estrutural baseou-se em RMN 1H e
principalmente CG-EM dos ésteres metílicos (M2b) obtidos.
Na análise do espectro de RMN 1H (figura 34, p. 92) dos ésteres
metílicos alguns sinais característicos foram observados, como, tripleto em δ
2,30 (J= 7,8 Hz; 2H), referente a hidrogênios α às carbonilas, um singleto em
δ 3,65 (3H) referentes aos hidrogênios metílicos do grupo carbometoxila,
O
OH
n
O
OH
n

______________________________________________Resultados e Discussão
90
vários sinais entre δ 1,0 - 2,0, caracterizando hidrogênios da cadeia
metilênica e um tripleto em δ 0,9 (3H) referente as metilas terminais.
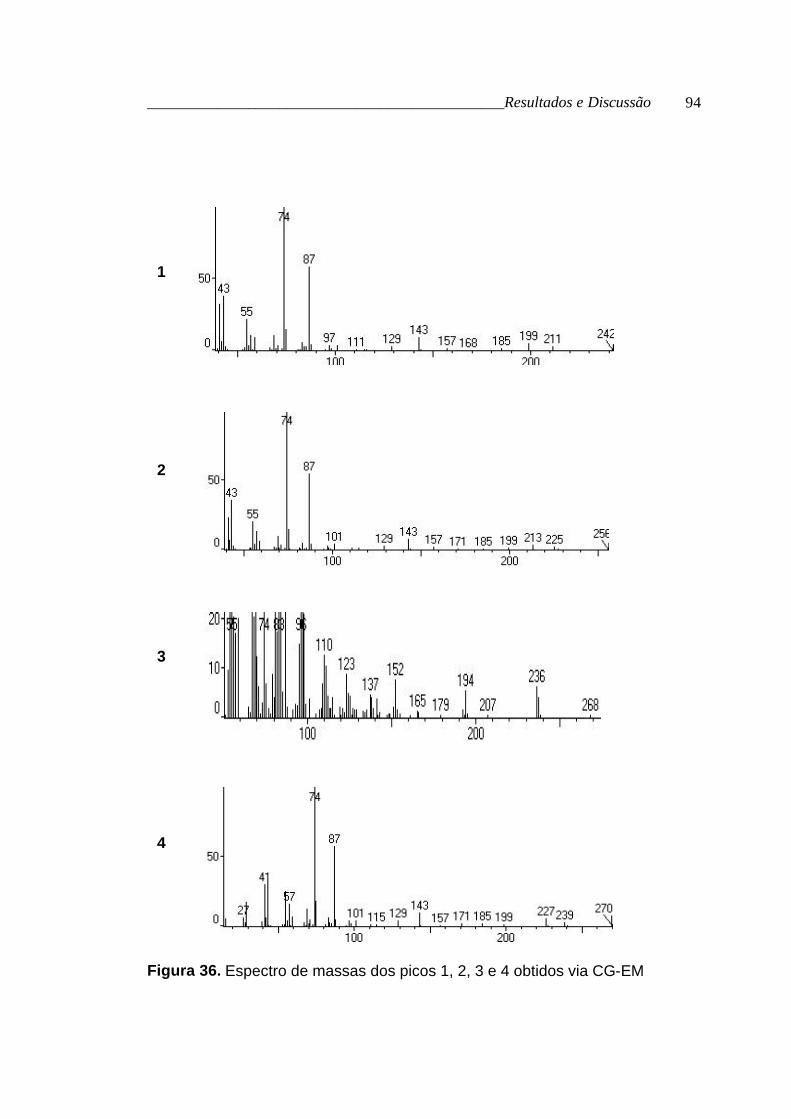
A análise da mistura via CG-EM (figuras 35, p. 93; figura 36, p. 94 e
figura 37, p. 95) possibilitou a determinação do tamanho das cadeias
metilênicas. Os espectros de massas obtidos para cada substância do
cromatograma foram comparados com os espectros da biblioteca do
aparelho. Aqueles com índice de correlação superior a 90% foram
indiretamente caracterizados (tabela 19, p. 90).
Tabela 19. Dados obtidos via CG-EM para os ésteres metílicos dos ácidos
graxos
Picos Tr (min.) [M+] Fórmula
molecular nome
1 16,76 242 C15H30O2 tetradecanoato de metila
2 18,20 256 C16H32O2 pentadecanoato de metila
3 19,67 268 C17H32O2 9-hexadecenoato de metila
4 20,16 270 C17H34O2 hexadecanoato de metila
5 22,73 284 C18H36O2 heptadecanoato de metila
6 25,11 294 C19H34O2 9,12-octadecadienoato de metila
7 25,30 296 C19H36O2 9-octadecenoato de metila
8 26,29 298 C19H38O2 octadecanoato de metila
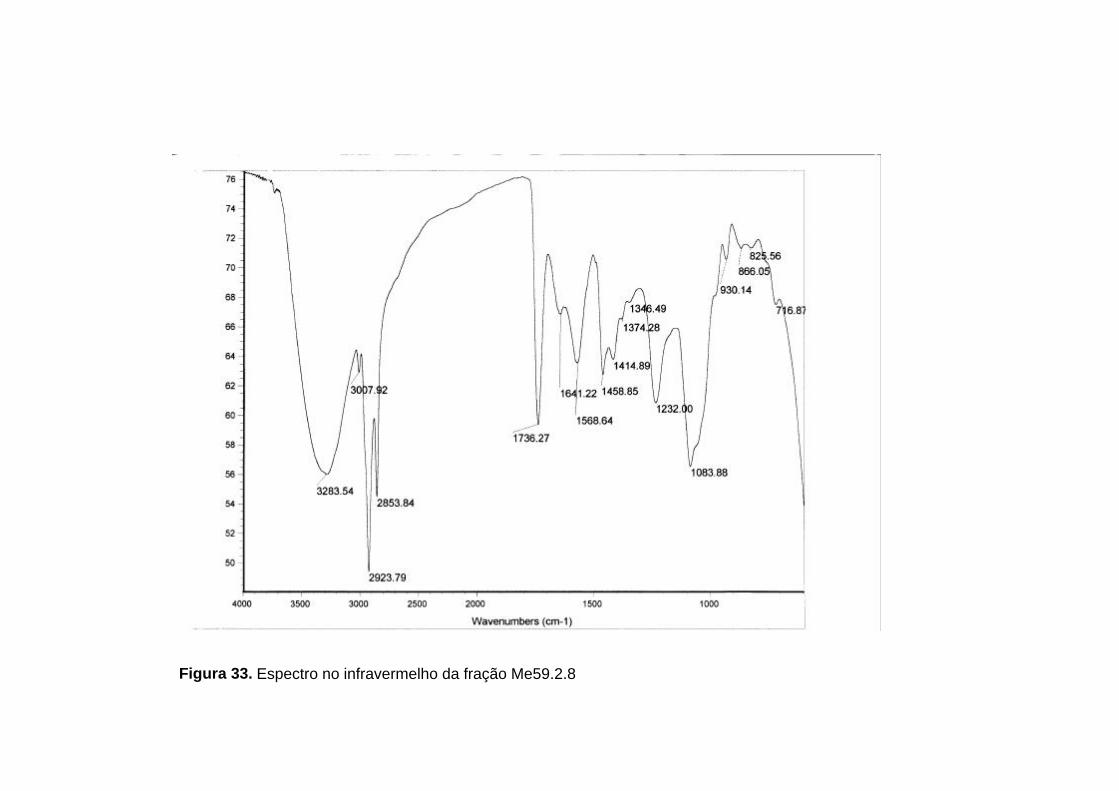
Figura 33. Espectro no infravermelho da fração Me59.2.8

Figura 34. Espectro de RMN 1H dos ésteres metílicos (M2b) obtidos

Figura 35. Cromatograma, obtido via CG, dos ésteres metílicos da mistura M2b (coluna DB - 5; 25 m, 0,25 cm, 0,25 µm, detector de massas)
1
2
4
5
6 7
8
3
______________________________________________Resultados e Discussão
94
Figura 36. Espectro de massas dos picos 1, 2, 3 e 4 obtidos via CG-EM
1
2
3
4

______________________________________________Resultados e Discussão
95
Figura 37. Espectro de massas dos picos 5, 6, 7 e 8 obtidos via CG-EM
5
6
7
8

______________________________________________Resultados e Discussão
96
4.4.3 Triglicerídeos (M6)
Os triglicerídeos foram isolados da fração AcOEt do extrato metanólico
de P. verrucosum e identificados através de RMN 1H.
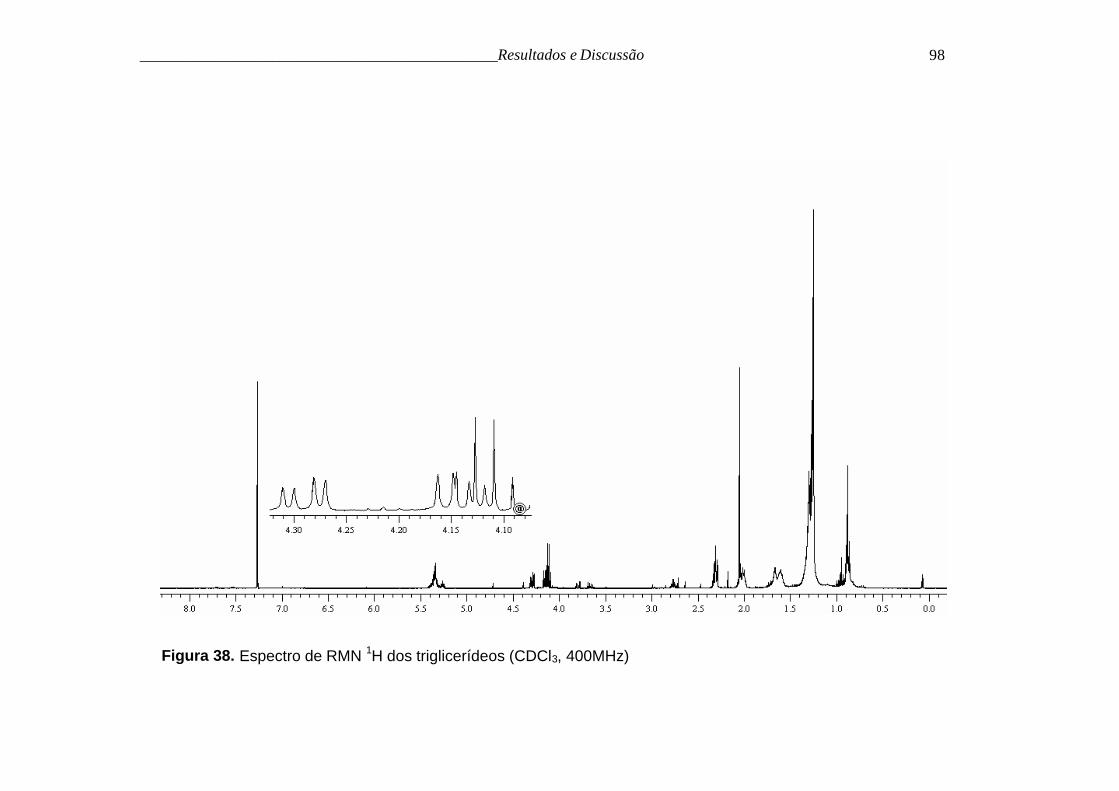
O espectro de RMN 1H (figura 38, p. 98), apresentou alguns sinais
característicos de triglicerídeos, como tripleto em δ 0,88 (J= 7,0 Hz), referente
as metilas terminais, sinais intensos na região de δ 1,25 referentes aos
metilenos da cadeia alquílica, multipleto em δ 2,13 referente aos metilenos α às
carbonilas, dois duplos dubletos em δ 4,14 (J= 2,3; 7,4 Hz) e δ 4,29 (J= 4,3; 7,6
Hz), relativos aos metilenos acilcarbinólicos.
Os ácidos graxos que esterificavam o glicerol, após reação de
transesterificação (M6b), com MeOH e H2SO4, foram identificados indiretamente
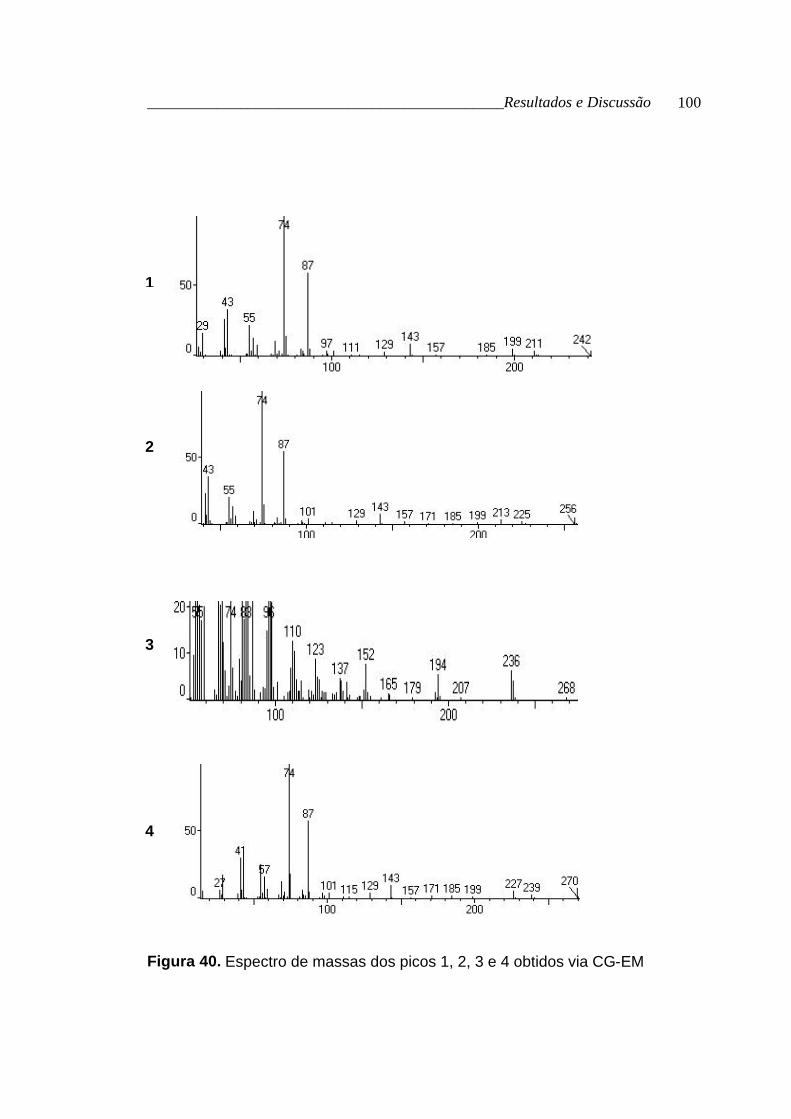
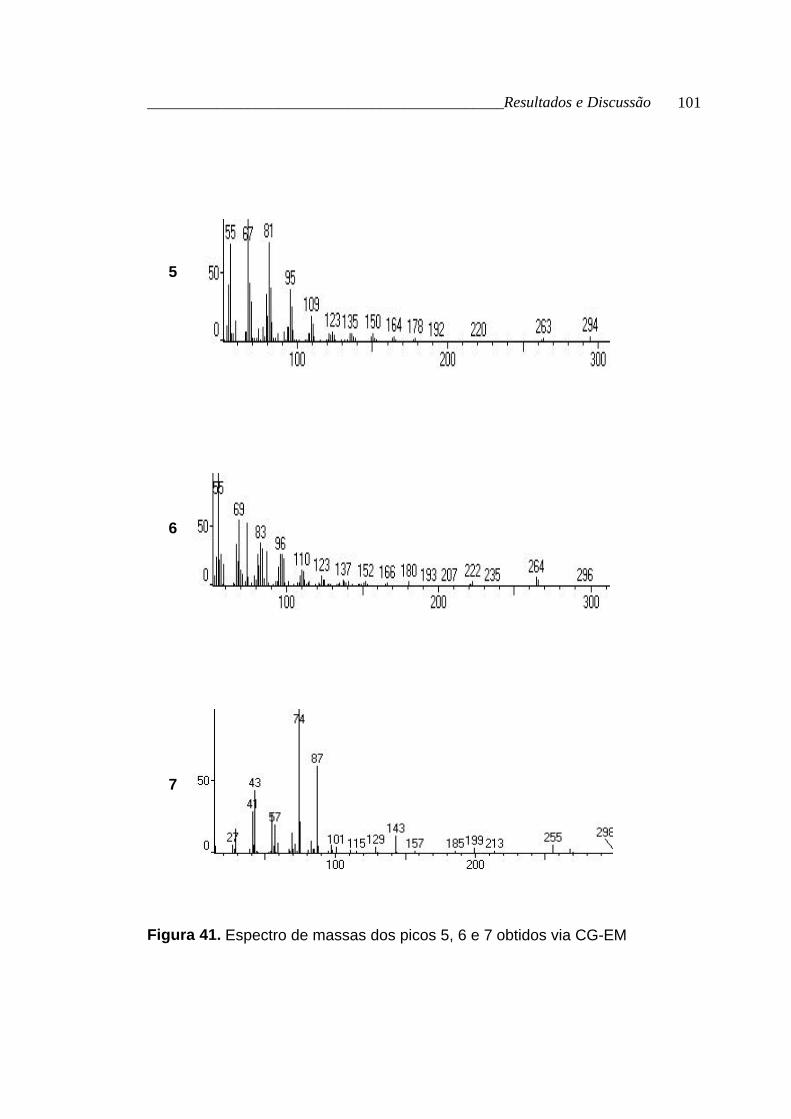
por CG-EM (tabela 20, p. 97; figura 39, p. 99; figura 40, p. 100 e figura 41, p.
101). Além disso, os ésteres metílicos foram confirmados e quantificados
usando padrões, via CG (tabela 20, p. 97; figura 42, p. 102 e figura 43, p. 103).
O
O
O
OO
On
n
n

______________________________________________Resultados e Discussão
97
Tabela 20. Dados obtidos via CG e CG-EM para os ésteres metílicos dos
ácidos graxos que esterificam o glicerol
Picos Tr (min.) [M+] FM nome %
1 16,68 242 C15H30O2 tetradecanoato de metila *
2 18,28 256 C16H32O2 pentadecanoato de metila 1,06
3 19,76 268 C17H32O2 9-hexadecenoato de metila 2,62
4 20,07 270 C17H34O2 hexadecanoato de metila 20,52
5 25,06 294 C19H34O2 9,12-octadecadienoato de metila 14,55
6 25,30 296 C19H36O2 9-octadecenoato de metila 35,84
7 26,27 298 C19H38O2 octadecanoato de metila 11,56
* não encontrado nos padrões do CG
______________________________________________Resultados e Discussão
98
Figura 38. Espectro de RMN 1H dos triglicerídeos (CDCl3, 400MHz)

______________________________________________Resultados e Discussão
99
Figura 39. Cromatograma, obtido via CG, dos ésteres metílicos da mistura M6b (coluna DB - 5; 25 m, 0,25 cm, 0,25 µm, detector de massas)
1 23
4
5
6
7
______________________________________________Resultados e Discussão
100
Figura 40. Espectro de massas dos picos 1, 2, 3 e 4 obtidos via CG-EM
1
2
3
4
______________________________________________Resultados e Discussão
101
Figura 41. Espectro de massas dos picos 5, 6 e 7 obtidos via CG-EM
5
6
7

Figura 42. Cromatograma dos ésteres metílicos da mistura M6, via CG (coluna HP - 1; 25 m, 0,25 cm, 0,25 µm, detector FID 300°C)
2
3
4 5
6 7
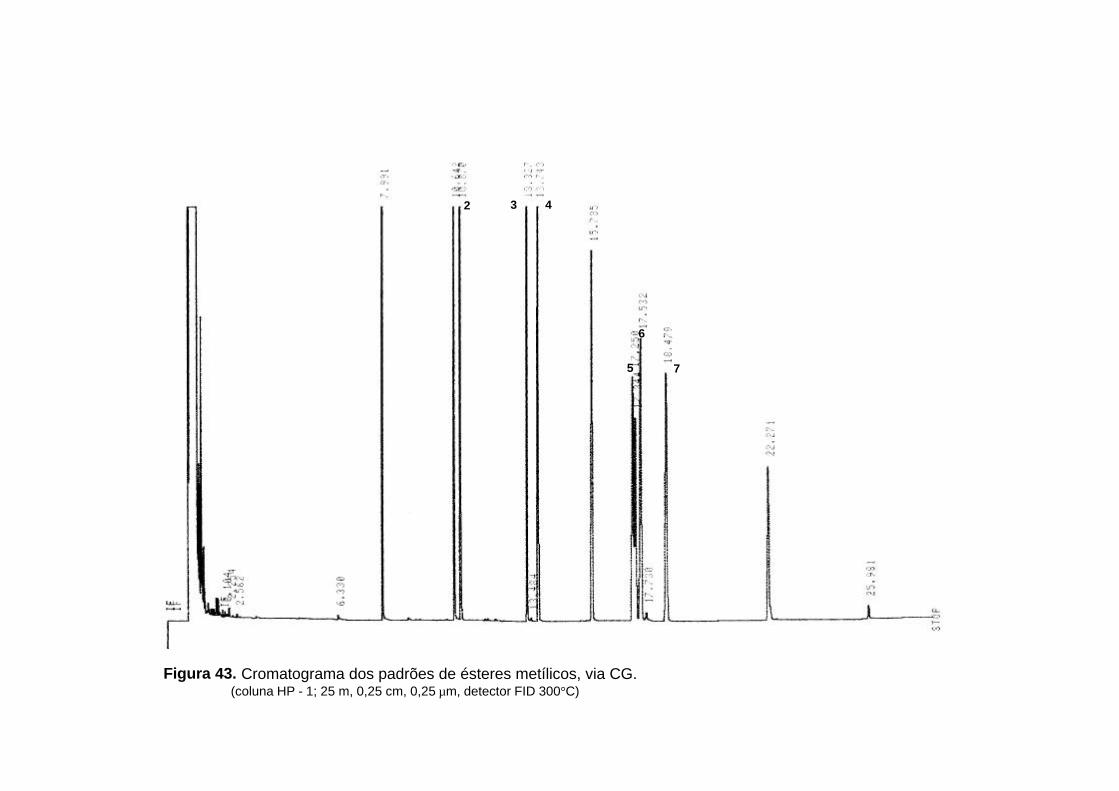
Figura 43. Cromatograma dos padrões de ésteres metílicos, via CG. (coluna HP - 1; 25 m, 0,25 cm, 0,25 µm, detector FID 300°C)
2 3 4
5
6
7

104
4.4.4 Manitol (M3) O manitol foi isolado da fração aquosa do extrato metanólico do
micélio do fungo P. verrucosum cultivado em meio Takeuchi.
No espectro no infravermelho (figura 44, p. 107) foram observadas
absorções de alguns grupos funcionais listados na tabela 21 (p. 104).
Tabela 21. Principais absorções observadas no espectro no infravermelho da subtância M3
3288,65 cm-1 Deformação axial de O-H
2948,29 cm-1 Deformação axial de C-H
1082,11 cm-1 Deformação axial de C-O
1421,75 cm-1 Deformação angular de C-H
O espectro de RMN 1H (figura 45, p. 108) apresentou quatro sinais
referentes a 8 hidrogênios carbinólicos, dois duplos dubletos em δ 3,67 (J=
11,6; 5,8 Hz) e δ 3,86 (J= 11,6; 2,5 Hz), um duplo duplo dubleto em δ 3,76
(J= 8,6; 5,8; 2,5 Hz) e um dubleto em δ 3,79 (J= 8,6 Hz). No espectro de
RMN 13C (figura 46, p. 109 e figura 47, p. 110) observou-se três carbonos
carbinólicos em δ 71,0 (CH), δ 69,5 (CH) e δ 63,5 (CH2), referentes aos seis
carbonos presentes na molécula (tabela 22, p. 105). A existência de apenas
HOOH
OH
OH
OH
OH
1 2 3 3' 2' 1'

105
três sinais de carbonos é explicada pela simetria observada na molécula
(Collins & Ferrier, 1996).
Os valores encontrados referentes aos dados de RMN 1H e 13C do
manitol foram confrontados com modelos da literatura, tabela 22 (p. 105)
(www.aist.go.jp e Bock & Pedersen, 1983).
Tabela 22. Dados de RMN 1H e RMN 13C para a substância M3
δ C, Manitol
(Bock &
Pedersen, 1983)
δ C,
M3
δ H, Manitol
(www.aist.go.jp)
δ H,
M3
1 – 72,2 71,0 1a- 3,38 dd (10,7; 5,6; 2H) 1b- 3,61 dd (10,7; 3,3; 2H)
1a- 3,67 dd (11,6; 5,8; 2H) 1b- 3,86 dd (11,6; 2,5; 2H)
2 – 70,7 69,5 3,45 ddd (5,6; 3,3 2H) 3,76 ddd (8,6; 5,8; 2,5 2H)
3 – 64,6 63,5 3,54 d (2H) 3,79 d (8,6 2H)
* Constantes de acoplamentos, em Hz, e integrais entre parênteses.Solvente da literatura DMSO. Solvente da substância M3, D2O, 400 MHz e 100 MHz.
Para confirmar a estrutura da molécula, a substância M3 foi acetilada
utilizando-se anidrido acético e piridina em quantidades catalíticas e os
espectros do respectivo acetato (M3a) foram obtidos.
Na análise do espectro de RMN 1H da substância M3a (figura 48, p.
111), foi observada a presença de dois duplos dubletos em δ 4,07 (J= 12,4;
5,3 Hz) e δ 4,22 (J= 12,6; 3,0 Hz), referente a H-1a e H-1b, um multipleto em
δ 5,07 referente a H-2, um dubleto em δ 5,45 (J= 8,6 Hz) referente a H-3 e a
presença de três singletos em δ 1,98, δ 2,01 e δ 2,07, referentes às metilas
dos grupos acetato. Os acoplamentos entre estes hidrogênios foram
evidenciados no mapa de contornos COSY 1H -1H (figura 50, p. 113).

106
O espectro de RMN 13C (figura 49, p. 112) apresentou sinais de três
carbonos carbonílicos de éster em δ 170,3, três sinais de carbonos
carbinólicos em δ 68,7, δ 68,3 e δ 62,5 e três sinais de carbonos metílicos
dos grupos acetato em δ 20,5, δ 20,6 e δ 20,8. Estes dados foram
confrontados com Bock & Pedersen, 1983 na tabela 23 (p. 106).
Tabela 23. Dados de RMN 13C dos carbonos carbinólicos da substância M3a
δ C, Acetato de manitol
(Bock & Pedersen, 1983)
δ C, substância M3a
1 – 62,0 62,5
2 – 68,1 68,3
3 – 67,7 68,7
Solvente da literatura DMSO. Solvente da substância M3a, Me2CO, 100 MHz
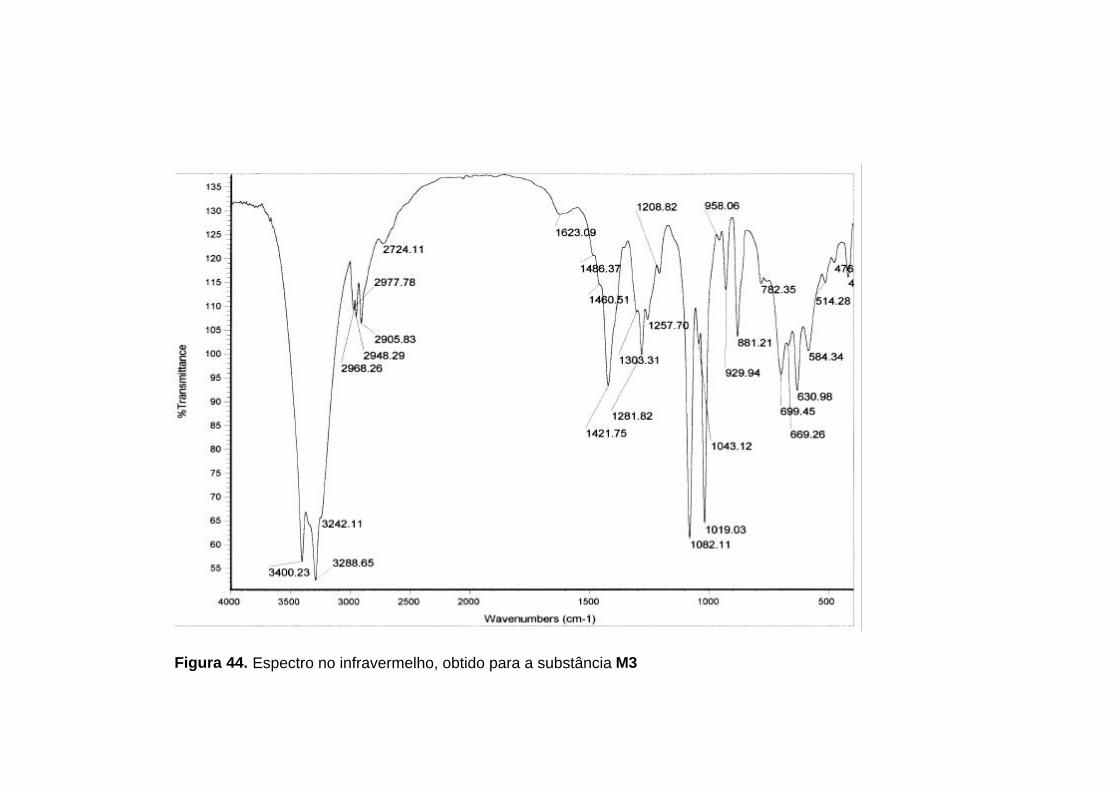
Figura 44. Espectro no infravermelho, obtido para a substância M3

Figura 45. Espectro de RMN 1H obtido da substância M3 (D2O, 400MHz)
HOOH
OH
OH
OH
OH
1 2 3 3' 2' 1'
1a (2H) 1b (2H)
3 (2H) 2 (2H)

Figura 46. Espectro de RMN 13C obtido da substância M3 (D2O, 100MHz)
HOOH
OH
OH
OH
OH
1 2 3 3' 2' 1'1
3
2
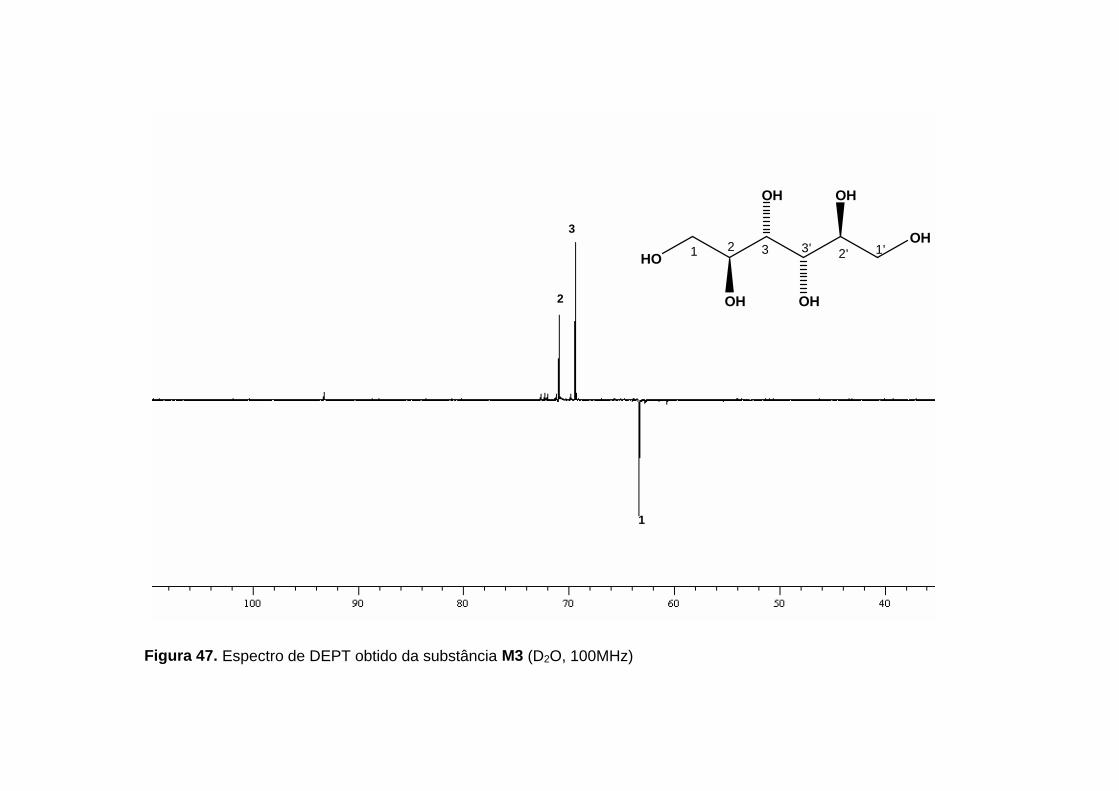
Figura 47. Espectro de DEPT obtido da substância M3 (D2O, 100MHz)
HOOH
OH
OH
OH
OH
1 2 3 3' 2' 1'
1
3
2

Figura 48. Espectro de RMN 1H obtido da substância M3a (Me2CO, 400MHz)
AcOOAc
OAc
OAc
OAc
OAc
1 2 3 3' 2' 1'
1a (2H) 1b (2H) 2 (2H)
3 (2H)
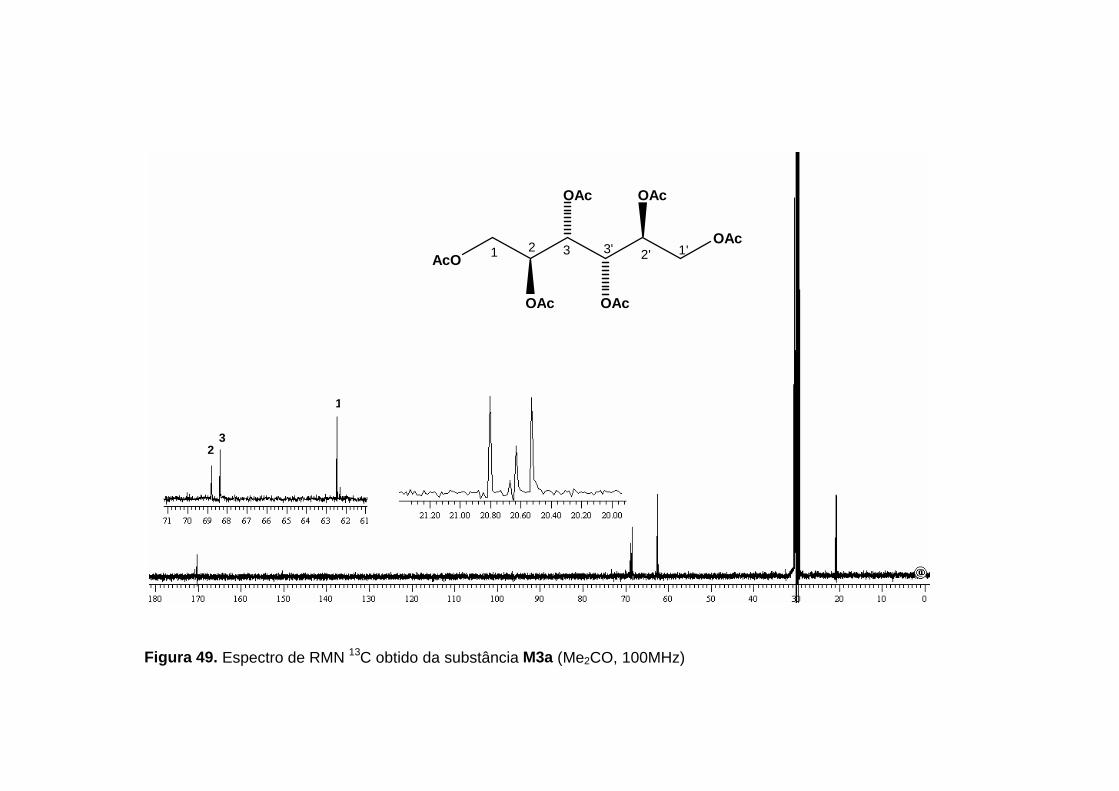
Figura 49. Espectro de RMN 13C obtido da substância M3a (Me2CO, 100MHz)
AcOOAc
OAc
OAc
OAc
OAc
1 2 3 3' 2' 1'
1
3 2

AcO
OAc
OAc
OAc
OAc
OAc
1 2 3 3' 2' 1'

Figura 50. Mapa de contornos COSY 1H -1H da substância M3a (Me2CO, 400MHz)

______________________________________________Resultados e Discussão
114
4.4.5 α,α-Trealose (M4)
A substância M4 foi isolada do micélio do fungo P. verrucosum,
apresentando alta polaridade sendo solúvel apenas em água.
Na análise do espectro de RMN 1H (figura 51, p. 119) foi observada
a presença de vários sinais na região de hidrogênios de açúcares e um
dubleto em δ 5,20 (J= 3,8 Hz) característico de hidrogênio anomérico.
No experimento de RMN 1H, os sinais desdobraram-se em um
tripleto em δ 3,45 (J= 9,3 Hz) correspondente ao H-4 e H-4’, um duplo
dubleto (H-2 e H-2’) em δ 3,65 (J= 9,9 e 3,8 Hz) e um multipleto entre δ 3,74
e 3,90 (H-3 e H-3’, H-5 e H-5’, H-6a e H-6a’ e H-6b e H-6b’).
No espectro de RMN 13C (figura 52, p.120), foram observados os
sinais de 6 átomos de carbonos, que somados aos de hidrogênios sugeriram
que a molécula fosse a glicose.
Após análise destes espectros a substância M4 foi então acetilada,
utilizando anidrido acético e piridina em quantidades catalíticas, para a
confirmação da estrutura. Novos espectros foram obtidos para o produto
M4a.
Na análise do espectro no infravermelho (figuras 53, p. 121), foram
observadas absorções de alguns grupos funcionais (tabela 24, p. 115) e
ausência de deformação axial de O-H, confirmando a acetilação.
No espectro de RMN 1H de M4a (figuras 54, p. 122), pode–se
observar na região de δ 3,99 a 4,08 a sobreposição de um duplo dubleto (H-
6a) com um duplo duplo dubleto (H-5) referentes a hidrogênios que acoplam
entre si (J= 2,3 Hz). Além disso, um duplo dubleto (H-2) em δ 5,03 (J= 3,8;

______________________________________________Resultados e Discussão
115
10,2 Hz) apresentou-se coalescente com um tripleto (H-4) em δ 5,05 (J= 9,8
Hz). Observou-se ainda, um duplo dubleto em δ 4,23 (J= 5,8; 12,1 Hz)
referente a H-6b e um tripleto em δ 5,49 (J= 9,4 Hz) referente a H-3.
Todos os sinais corresponderam ao da glicose acetilada, com
exceção de um dubleto em δ 5,29 (J= 4,0 Hz) referente ao hidrogênio
anomérico. Este fato, aliado à presença de apenas quatro metilas referentes
a grupos acetato, sugeriu que a molécula não fosse a glicose, e sim um
dissacarídeo dímero da glicose, conhecido como trealose. Analisando a
constante de acoplamento de H-1 e H-2 (J= 4,0 Hz), conclui-se que estes
hidrogênios estão em relação cis, sendo que H-1 está em posição equatorial,
o que só é possível para a α,α-trealose.
Tabela 24. Principais absorções observadas no espectro no infravermelho da substância M4a
3026,19 cm-1 Deformação axial de C-H
2960,15 cm-1 Deformação axial assimétrica de CH3
2852,78 cm-1 Deformação axial simétrica de CH2
1753,46 cm-1 Deformação axial de C=O
1433,60 cm-1 Deformação angular simétrica de CH2
1370,22 cm-1 Deformação angular simétrica de CH3
1222,44 cm-1 Deformação axial de C-C(=O)-O acetato
1039,76 cm-1 Deformação axial assimétrica de O-C-C
O espectro de RMN 13C de M4a (figura 55, p. 123), apresentou
quatro sinais de carbonos carbonílicos de éster na região de δ 169,7 a 170,8,
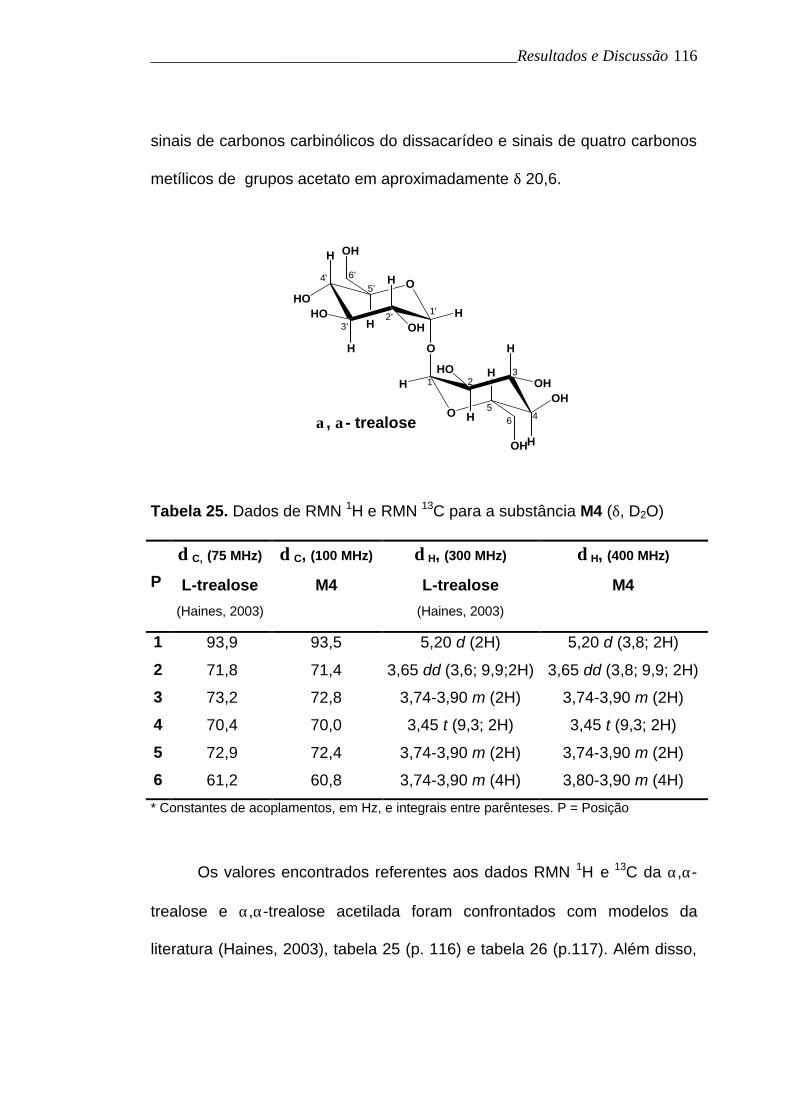
______________________________________________Resultados e Discussão
116
sinais de carbonos carbinólicos do dissacarídeo e sinais de quatro carbonos
metílicos de grupos acetato em aproximadamente δ 20,6.
Tabela 25. Dados de RMN 1H e RMN 13C para a substância M4 (δ, D2O)
P
δ C, (75 MHz)
L-trealose
(Haines, 2003)
δ C, (100 MHz)
M4
δ H, (300 MHz)
L-trealose
(Haines, 2003)
δ H, (400 MHz)
M4
1 93,9 93,5 5,20 d (2H) 5,20 d (3,8; 2H)
2 71,8 71,4 3,65 dd (3,6; 9,9;2H) 3,65 dd (3,8; 9,9; 2H)
3 73,2 72,8 3,74-3,90 m (2H) 3,74-3,90 m (2H)
4 70,4 70,0 3,45 t (9,3; 2H) 3,45 t (9,3; 2H)
5 72,9 72,4 3,74-3,90 m (2H) 3,74-3,90 m (2H)
6 61,2 60,8 3,74-3,90 m (4H) 3,80-3,90 m (4H)
* Constantes de acoplamentos, em Hz, e integrais entre parênteses. P = Posição Os valores encontrados referentes aos dados RMN 1H e 13C da α,α-
trealose e α,α-trealose acetilada foram confrontados com modelos da
literatura (Haines, 2003), tabela 25 (p. 116) e tabela 26 (p.117). Além disso,
α, α- trealose
O
H
HO
H
HO
H
O
OHHH
OH
O
H
OH
H
OH
H
HO HH
OH
1
4
32
4'
3'2' 1'
65
6'5'

______________________________________________Resultados e Discussão
117
os dados RMN 13C da α, α - trealose tiveram concordância satisfatória com
Breitmaier & Voelter,1989.
Tabela 26. Dados de RMN 1H e RMN 13C para a substância M4a (δ, CDCl3)
Constantes de acoplamentos, em Hz, e integrais entre parênteses. δ C (100 MHz).P=Posição
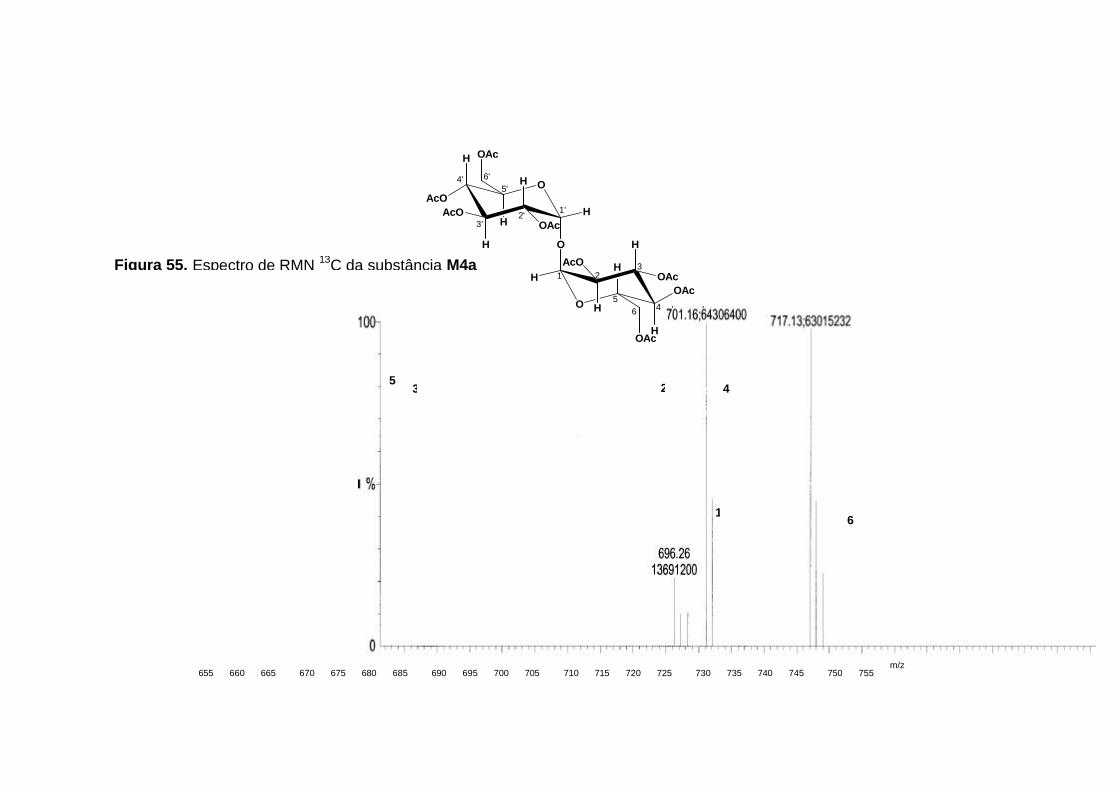
No espectro de massas pode-se confirmar a formação de M4a, esta
foi analisada por ESI-EM, nos modos positivo e negativo. A presença dos
íons em m/z 701 ([M+Na]+) e m/z 717 ([M+K]+) no modo positivo, e do íon
em m/z 677([M-H]-) no modo negativo, estão coerentes e corroboram com a
estrutura proposta (figura 56, p. 124 e figura 57, p. 125).
O dissacarídeo comumente chamado de α,α-trealose é composto da
ligação 1-1’ de duas α-glicopiranoses, sendo um de uma família de três
P δ C,
L- trealose
(Haines, 2003)
δ C,
M4a
δ H, (300 MHz)
L- trealose,
(Haines, 2003)
δ H, (400 MHz)
M4a
1 92,4 92,2 (2C) 5,22 d (3,8; 2H) 5,29 d (4,0; 2H)
2 68,7 68,6 (2C) 4,97 dd (3,8; 9,3; 2H) 5,03 dd (3,8; 10,4; 2H)
3 70,6 70,0 (2C) 5,43 t (9,4; 2H) 5,49 t (9,4; 2H)
4 68,4 68,3 (2C) 4,98 t (9,4; 2H) 5,05 t (9,8; 2H)
5 70,0 69,9 (2C) 3,98 ddd (2,0;5,7;10,3; 2H) 4,05 ddd (2,3;5,6;10,1;2)
6a 61,9 61,8 (1C) 3,94 dd (2,0; 12,1; 2H) 4,01 dd (2,3; 12,0; 2H)
6b 61,9 61,8 (1C) 4,18 dd (2,0; 12,1; 2H) 4,23 dd (5,8; 12,1; 2H)
OCOMe 170,8 170,6 (2C) - -
OCOMe 170,2 169,9 (2C) - -
OCOMe 169,8 169,6 (2C) - -
OCOMe 169,7 169,5 (2C) - -
OCOMe 20,9 20,7 (2C) 1,97 s (6H) 2,03 s (6H)
OCOMe 20,8 20,6 (2C) 1,99 s (6H) 2,05 s (6H)
OCOMe 20,8 20,6 (2C) 2,01 s (6H) 2,07 s (6H)
OCOMe 20,7 20,5 (2C) 2,02 s (6H) 2,08 s (6H)

______________________________________________Resultados e Discussão
118
trealoses: α-α, α-β e β-β. Encontra-se difuso na natureza principalmente em
leveduras, fungos, insetos, bactérias e em algumas plantas e algas (Gil et
al.,1996).
A α,α-trealose é comumente encontrada em organismos que
suportam calor excessivo e desidratação, como certas plantas desérticas,
camarões, nematóides e leveduras desidratadas. Este dissacarídeo constitui
mais de 20% do peso seco destes organismos, permitindo que mesmo após
anos desidratados tenham a ativação da sua via metabólica apenas com a
hidratação (Haines, 2003).

Figura 51. Espectro de RMN 1H da substância M4 (D2O, 400MHz)
1 (2H)
3, 5, 6a e 6b (8H)
4 (2H) 2 (2H)
O
H
HO
H
HO
H
O
OHHH
OH
O
H
O
H
OH
H
HO HH
OH
1
4
32
4'
3'2' 1'
65
6'5'
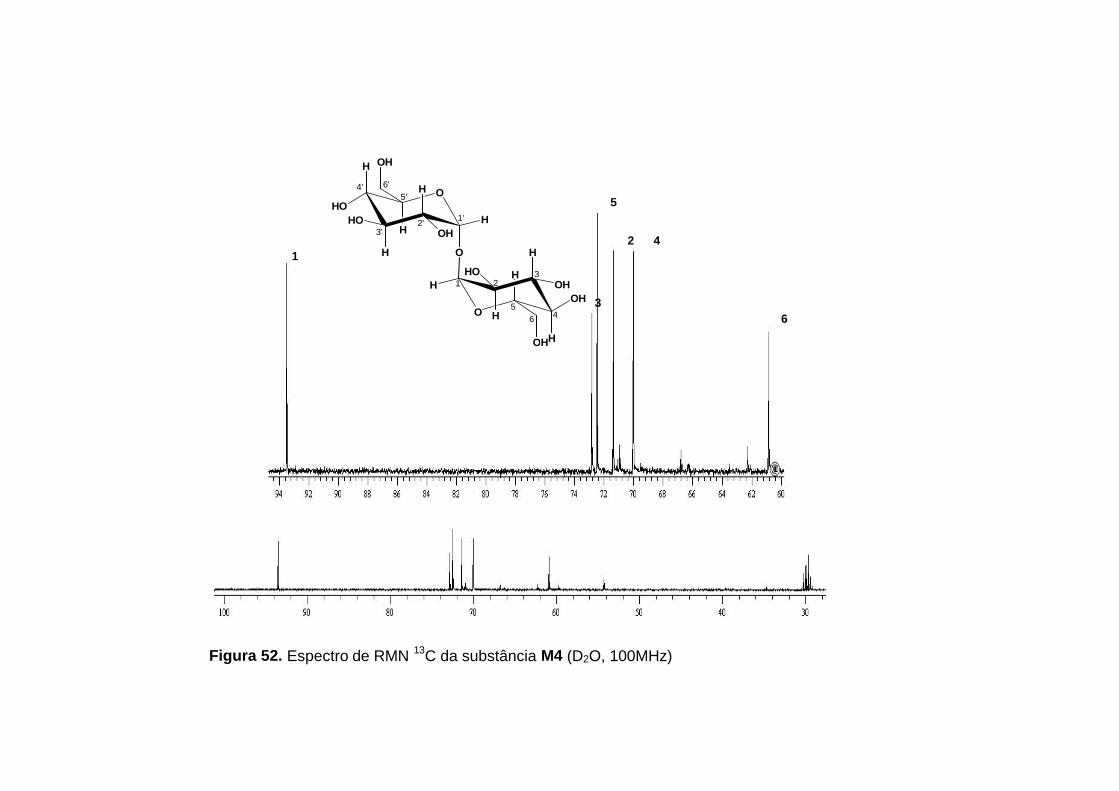
Figura 52. Espectro de RMN 13C da substância M4 (D2O, 100MHz)
1 2 4
3
5
6
O
H
HO
H
HO
H
O
OHHH
OH
O
H
OH
H
OH
H
HO HH
OH
1
4
32
4'
3'2' 1'
65
6'5'

Figura 53. Espectro no infravermelho, obtido para a substância M4a

Figura 54. Espectro de RMN 1H da substância M4a (CDCl3, 400MHz)
6b (2H)
5 (2H) e 6a (2H)
3 (2H)
1 (2H)
2 (2H) e 4 (2H)
O
H
AcO
H
AcO
H
O
OAcHH
OAc
O
H
OAc
H
OAc
H
AcO HH
OAc
1
4
32
4'
3'2' 1'
65
6'5'
655 660 665 670 675 680 685 690 695 700 705 710 715 720 725 730 735 740 745 750 755 m/z
6
5 3 2 4
1
O
H
AcO
H
AcO
H
O
OAcHH
OAc
O
H
OAc
H
OAc
H
AcO HH
OAc
1
4
32
4'
3'2'
1'
65
6'5'
Figura 55. Espectro de RMN 13C da substância M4a

Figura 56. Espectro de massas, no modo positivo, da substância M4a, obtido via ESI-EM.

Figura 57. Espectro de massas, no modo negativo, da substância M4a, obtido via ESI-EM.

______________________________________________Resultados e Discussão
126
4.4.6 Glicerilfosfocolina (M5) A substância M5 foi isolada da fração aquosa do extrato metanólico
obtido do micélio de P. verrucosum cultivado em meio Takeuchi.
O espectro de RMN 1H (figura 59, p. 132, tabela 28, p. 129)
apresentou sinais entre δ 3,20 e 4,30, referentes a 18 hidrogênios, indicando
que todos hidrogênios da molécula estariam desblindados pela presença de
heteroátomos. Dois duplos dubletos que consistem em um sistema de
acoplamento de spins ABX foram observados em δ 3,52 (J= 11,3; 5,6 Hz;
1H) e δ 3,56 (J= 11,3; 5,4 Hz; 1H), referentes a H-8a e H-8b. Estes dois
hidrogênios acoplam entre si e com H-7 em δ 3,73 (qui; J= 5,4 Hz; 1H), que
por sua vez acopla com outros dois hidrogênios em δ 3,83 (ddd; J= 10,8, 6,5,
5,7 Hz; 1H) e δ 3,89 (ddd, J= 10,8, 6,4, 4,5 Hz; 1H), referentes aos H-6a e H-
6b. Estes acoplamentos foram confirmados no mapa de contornos COSY 1H
-1H (figura 63, p. 136) e permitiram a proposição do fragmento I para a
molécula:
O infravermelho (figura 58, p. 131, tabela 27, p. 128), apresentou
absorção típicas de hidroxilas em 3379,06 cm-1 referente à deformação axial
de O-H.
Inicialmente, o fragmento proposto não satisfez a multiplicidade dos
sinais observados para os dois hidrogênios da posição 6, duplos duplos
RO OH
OH
87
6
I

______________________________________________Resultados e Discussão
127
dubletos, sugerindo um acoplamento adicional que não foi observado no
mapa de contornos COSY 1H -1H.
Os outros dois sinais observados no espectro de RMN 1H, referem-
se a H-4 e H-5, multipletos em δ 4,25 (2H) e δ 3,60 (2H) correspondentes a
hidrogênios metilênicos que acoplam entre si, conforme evidenciado no
COSY 1H -1H. O singleto em δ 3,20 (9H) indicou a presença de três metilas
que poderiam estar ligadas a um nitrogênio quaternário.
No espectro de RMN 13C (figura 60, p. 133; 61,p. 134; tabela 28, p.
129) foi observado um sinal em δ 54,7 que no mapa de contornos HMQC
(figura 64, p. 137) mostrou correlação com os nove hidrogênios em δ 3,20.
Estas informações levaram a proposição do fragmento II para a molécula:
Um fato que chamou atenção no espectro de RMN 13C,
desacoplado de hidrogênio, foi o desdobramento das linhas dos sinais em δ
54,7; δ 72,5; δ 67,9; δ 67,6 e δ 60,5, o que sugeriu a existência de
acoplamento dos carbonos com outro núcleo, possivelmente 31P.
A presença de fósforo na molécula foi comprovada pelo espectro de
RMN 31P (figura 62, p. 135), com sinal em δ 0,70, apresentando-se como um
tripleto alargado. A presença de fósforo, na forma de grupo fosfato, foi ainda
confirmada pelo espectro no infravermelho com absorções em 1225,32 cm-1,
referente a P=0 e absorções em 1086-1054 cm-1, referente a deformação
axial de P-O-C alifático.
RO
N+4 5
II

______________________________________________Resultados e Discussão
128
Com isso, propôs-se que os fragmentos I e II estivessem unidos
por um grupo fosfato:
Proposta para substância M5- C8H20NO6P
Para confirmar a estrutura da substância M5, a mesma foi analisada
por ESI-EM, no modo positivo, e a presença do íon em m/z 280 ([M+Na]+)
está coerente com a estrutura proposta (figura 66, p. 139).
Tabela 27. Principais absorções observadas para a substância M5 no
espectro no infravermelho
3379,06 cm-1 Deformação axial de O-H
2956,59 cm-1 Deformação axial de C-H
1225,32cm-1 Grupo (RO)3P=O
A presença de fósforo na molécula justifica os acoplamentos
observados no espectro de RMN 13C para os carbonos distantes até três
ligações do átomo de fósforo. Além disso, a multiplicidade de H-6a e H-6b
sendo um duplo duplo dubleto é explicada pelo acoplamento adicional com
31P. Vale ressaltar, que as multiplicidades observadas para C-4 e para os
carbonos metílicos indicam um acoplamento destes carbonos com o
nitrogênio quaternário. Portanto, o desdobramento do sinal atribuído ao C-4
pode ser explicado pelo seu acoplamento com o fósforo e com o nitrogênio
quaternário.
O
POO
-O
N+
OH
OH
1
765
43
2
8

______________________________________________Resultados e Discussão
129
O mapa de contornos HMQC possibilitou a atribuição correta de
todos os carbonos da molécula. No mapa de contornos HMBC (figura 65, p.
138), observou-se a correlação dos carbonos metílicos com os hidrogênios
em δ 3,60 (m; 2H), que confirmou que estes hidrogênios estavam ligados a
C-4, α grupo amônio quaternário, enquanto que os H-5 encontram-se mais
desblindados em δ 4,25.
Tabela 28. Dados de RMN 1H (500 MHz) e RMN 13C (100 MHz) para a
substância M5 (δ, Metanol-d4).
Posição δ H δ C
1,2 e 3 3,20 s (9H) 54,7
4 3,60 m (2H) 67,6
5 4,25 m (2H) 60,5
6 6a- 3,83 ddd (10,8; 6,5; 5,7; 1H) 6b- 3,89 ddd (10,8; 6,4; 4,5; 1H)
67,9
7 3,73 qui (5,4; 1H) 72,5
8 8a- 3,52 dd (11,3; 5,6; 1H) 8b- 3,56 dd (11,3; 5,4; 1H)
63,8
* Constantes de acoplamentos, em Hz, e integrais entre parênteses A estrutura proposta corresponde a glicerilfosfocolina, cujo
precursor é a fosfocolina que também é precursor de moléculas biológicas,
principalmente fosfolipídeos complexos de membrana. Recentemente, duas
classes de substâncias análogas a estas moléculas foram identificadas
como substâncias bioativas; as alquilfosfocolinas, onde a fosfocolina é
diretamente esterificada com longas cadeias alquílicas e as
alquilglicerofosfocolinas, que possuem substituições alquílicas no glicerol

______________________________________________Resultados e Discussão
130
(Croft et al., 2003). A classe das alquilglicerofosfocolinas apresentou
substâncias com atividades anticancer, tripanocidas e leishmanicidas (Croft
et al., 2003). A classe alquilfosfocolina é composta por substâncias que
agem na membrana e apresentam atividades anticâncer, antiviral,
tripanocida (Konstantinov et al., 1997) e leishmanicida (Escobar, et al.,
2002).
Apesar da miltefosina, uma alquilfosfocolina, ter sido aprovada na
fase III dos testes clínicos para tratamento da leishmaniose visceral, não há
evidências de que esta seja a substância perfeita, pois poucos compostos
desta classe foram estudados até o momento (Croft et al., 2003), o que
mostra o grande potencial biológico de estruturas como a substância M5 que
ainda não foi avaliada em ensaios biológicos.

Figura 58. Espectro no infravermelho da substância M5
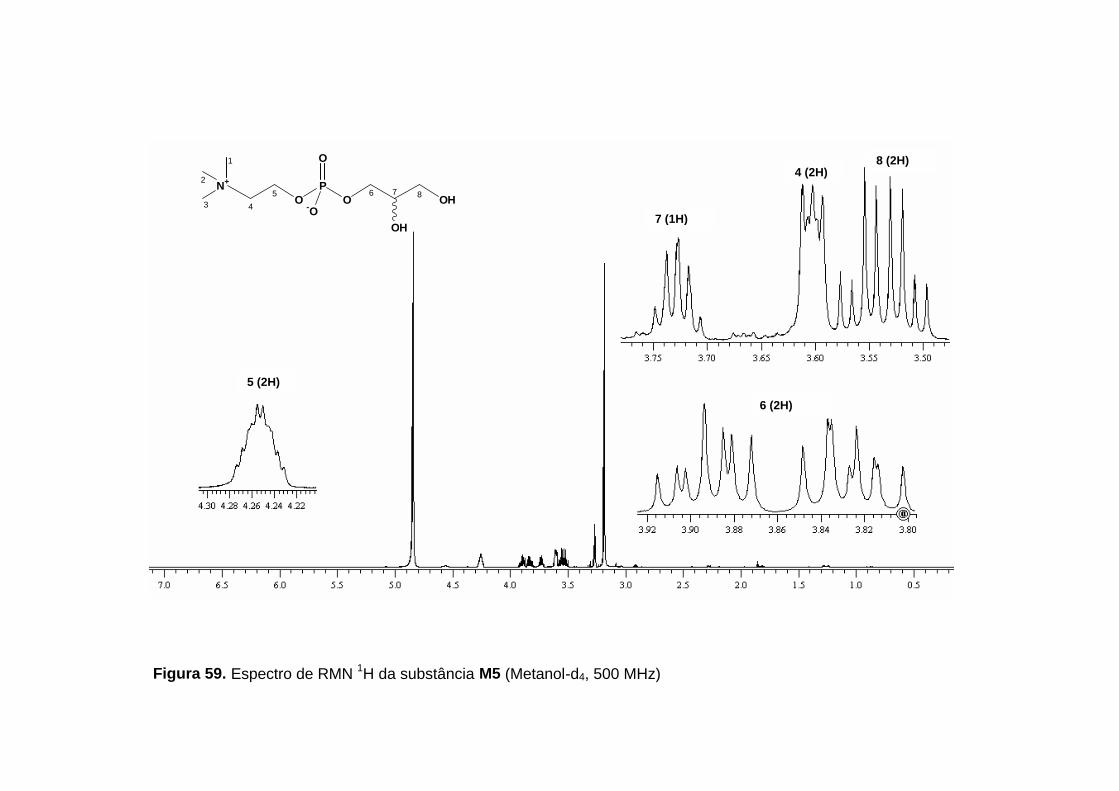
Figura 59. Espectro de RMN 1H da substância M5 (Metanol-d4, 500 MHz)
5 (2H)
7 (1H)
4 (2H) 8 (2H)
6 (2H)
O
POO
-O
N+
OH
OH
1
765
43
2
8
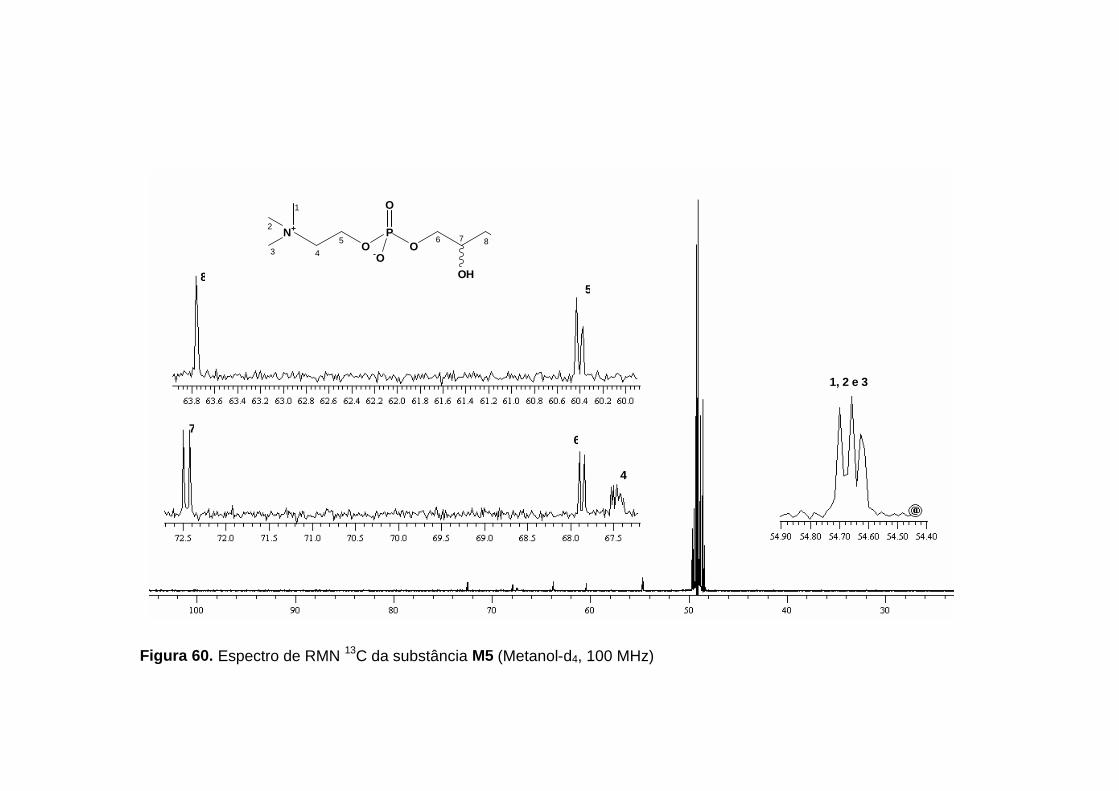
Figura 60. Espectro de RMN 13C da substância M5 (Metanol-d4, 100 MHz)
76
4
85
1, 2 e 3
O
POO
-O
N+
OH
1
765
43
2
8

Figura 61. Espectro de DEPT da substância M5 (Metanol-d4, 100 MHz)
7
6
4
8
5
1, 2 e 3
O
POO
-O
N+
OH
OH
1
765
43
2
8
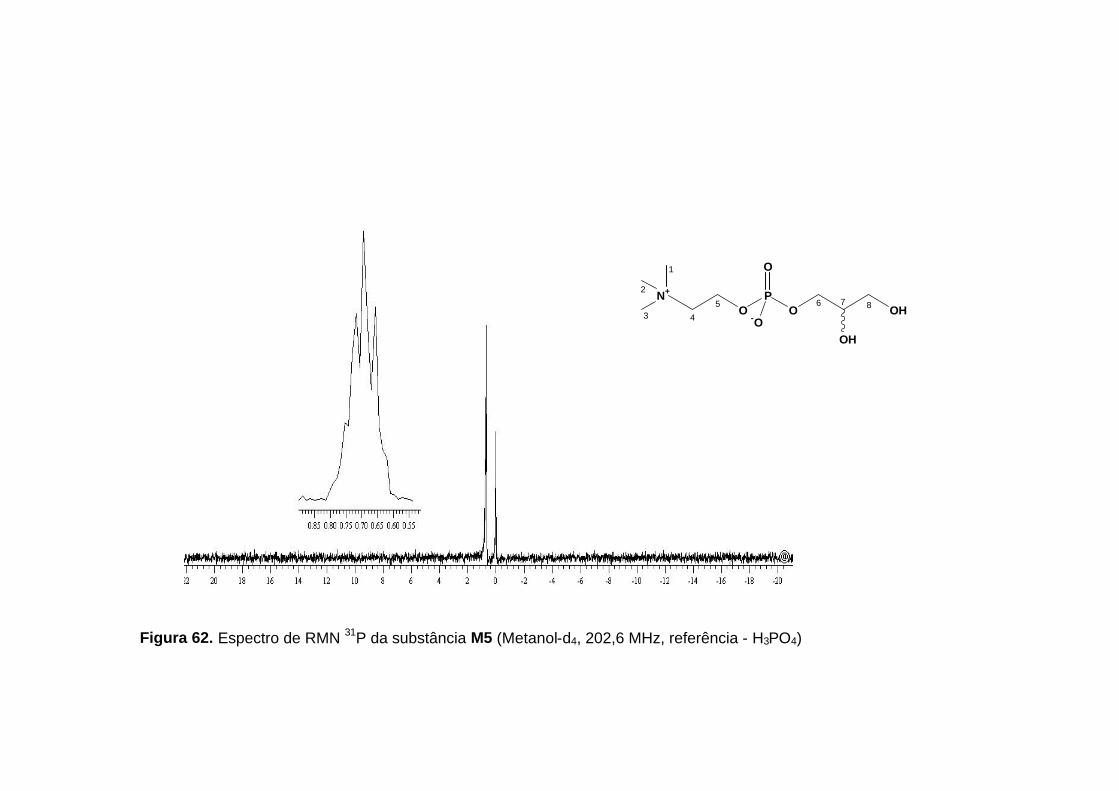
Figura 62. Espectro de RMN 31P da substância M5 (Metanol-d4, 202,6 MHz, referência - H3PO4)
O
POO
-O
N+
OH
OH
1
765
43
2
8

Figura 63. Mapa de contornos COSY 1H -1H da substância M5 (Metanol-d4)
O
POO
-O
N+
OH
OH
1
765
43
2
8

Figura 64. Mapa de contornos HMQC da substância M5 (Metanol-d4)
O
POO
-O
N+
OH
OH
1
765
43
2
8

Figura 65. Mapa de contornos HMBC da substância M5 (Metanol-d4)
O
POO
-O
N+
OH
OH
1
765
43
2
8

Figura 66. Espectro de massas da substância M5, via ESI-EM

_______________________________________Resultados e Discussão
140
4.5 Atividade tripanocida das substâncias isoladas
As substâncias puras foram testadas frente às formas tripomastigotas de
T. cruzi apresentando baixa atividade tripanocida.
Tabela 29. Atividade tripanocida das substâncias isoladas
Substância Concentração (µM) x % de lise IC50 (µM)
8,0 32,0 128,0
F1 4,8 11,7 26,3 547,8
M3 17,0 36,8 38,6 343,0
M4 11,7 16,4 38,6 217,3
M5 8,2 33,3 39,2 231,0
Controle positivo - violeta genciana a 250µg/mL; negativo - sangue infectado + 5% de DMSO
4.6 Análise da influência das condições de cultivo de P. verrucosum na
produção de metabólitos secundários, via perfis cromatográficos
O principal objetivo deste experimento foi investigar se as diferentes
condições de cultivo de P. verrucosum interferiam nos perfis químicos dos
extratos brutos obtidos. Dessa forma, não foram utilizados parâmetros como
padrões internos ou externos, comprimento de onda máximo no UV e relação
entre as intensidades de absorção nos comprimentos de onda, pois o objetivo
não foi identificar substâncias indiretamente.

_______________________________________Resultados e Discussão
141
Apesar dos cromatogramas terem sido analisados por 60min, 30 min
foi tempo suficiente para a análise completa dos perfis químicos. Além disso,
apesar de três comprimentos de onda terem sido analisados, 220nm foi o
mais apropriado para estas amostras.
Dos 115 extratos brutos de P. verrucosum obtidos neste trabalho 30
foram selecionados para serem analisados via CLAE, entre eles Ac47, Me59
que foram produzidos em escala ampliada e seus similares. Um método
geral padronizado para detecção de metabólitos secundários de fungos foi
previamente apresentado (Frisvad & Thrane, 1987; Frisvad, 1987). O
método é baseado na análise, via CLAE, da produção dos metabólitos
secundários de extratos obtidos de culturas cultivadas em diferentes
substratos.
A nutrição de fungos filamentosos é feita por absorção, através de
enzimas que hidrolisam grande variedade de substratos tornando-os
assimiláveis. Para o desenvolvimento e manutenção, os fungos necessitam
de fontes de carbono e nitrogênio. O carbono tem a função de produzir
energia, sendo os carboidratos mais utilizados glicose, sacarose, maltose,
amido e celulose. Como fonte de nitrogênio algumas espécies utilizam sais
de amônio ou nitratos (nitrogênio inorgânico) e outras exigem substratos
nitrogenados como as peptonas. Além disso, elementos como ferro, zinco,
manganês, cobre, molibdênio e cálcio são necessários em pequenas
quantidades (Pearce, 1997). Os meios de cultura utilizados neste trabalho
foram descritos anteriormente e apresentam-se bem distintos quanto às
fontes de nutrientes (tabela 5, p. 33).

_______________________________________Resultados e Discussão
142
A água é fundamental para o desenvolvimento fúngico, sendo um
fator determinante no ciclo de vida destes microrganismos, por isso o fungo
foi cultivado em meio de cultura líquido. Outro fator que altera a velocidade
de crescimento do fungo é a oxigenação. Já que P. verrucosum é um fungo
aeróbio, as culturas foram cultivadas sob agitação constante de 120 rpm,
aumentando a aeração. Espécies de Phoma, fungo fitopatogênico, também
foram cultivados em cultura líquida sob agitação constante de 130 rpm e a
produção dos metabólitos foi analisada, via CLAE, com 48, 72, 96 e 168
horas de incubação. A produção máxima das substâncias padrões
detectadas nos extratos variou de dois a cinco dias de incubação (Pedras &
Biesenthal, 2000).
A produção de metabólitos secundários de P. verrucosum foi
influenciada pelo tempo de incubação na pré-cultura (MPF), 24 ou 48 horas,
e durante o cultivo em meio fermentativo, 48, 72 e 144 horas. Os meios de
cultivo fermentativos Czapeck, Takeuchi, Jackson e Vogel interferiram na
produção qualitativa destes metabólitos. Estas condições de cultivo foram
utilizadas na produção de substâncias tripanocidas e antimicrobianas por
Talaromyces flavus (Freitas et al., 2002).
Os fungos podem acumular substâncias de reserva, como
carboidratos e lipídeos em seu micélio (Pearce, 1997). P. verrucosum,
apresentou diferenças marcantes na produção de metabólitos intra e
extracelulares presentes nos extratos MeOH e extratos AcOEt,
respectivamente.
_______________________________________Resultados e Discussão
143
Os diferentes extratos extracelulares obtidos de um mesmo meio de
cultura, sob as mesmas condições, apresentaram perfis cromatográficos
com semelhanças qualitativas, não importando o tempo de incubação no
meio fermentativo (figuras 67-81, p. 143).
Takeuchi 72 horas
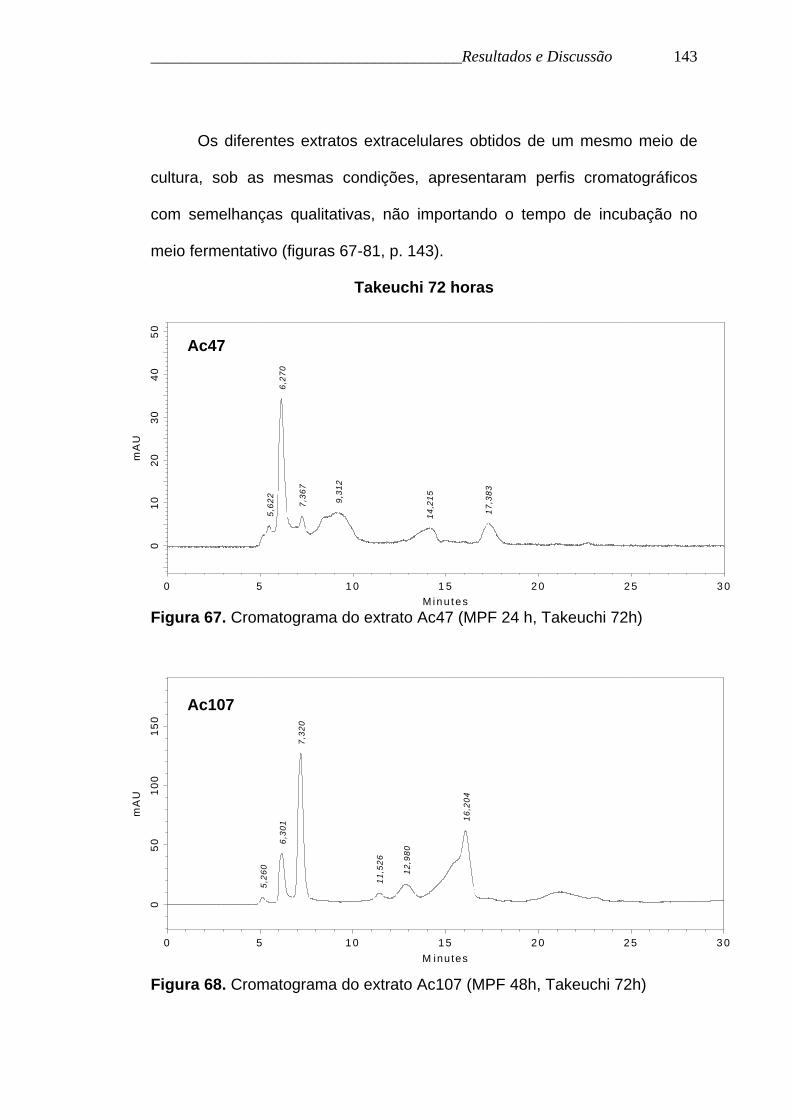
Figura 67. Cromatograma do extrato Ac47 (MPF 24 h, Takeuchi 72h)
Figura 68. Cromatograma do extrato Ac107 (MPF 48h, Takeuchi 72h)
Minu tes0 5 1 0 1 5 2 0 2 5 3 0
mA
U
01
02
03
04
05
0
5,6
22
6,2
70
7,3
67
9,3
12
14
,21
5
17
,38
3
D e t e c t o r A - 2 2 0 n mB a r b a r aV E 4 7 - 1
R e t e n t i o n T i m e
M inutes0 5 1 0 1 5 2 0 2 5 3 0
mA
U
05
01
00
15
0
5,2
60
6,3
01
7,3
20
11
,52
6
12
,98
0
16
,20
4
Detec tor A -220 nmB a r b a r aVE 107 -1
R e t e n t i o n T i m eAc107
Ac47

_______________________________________Resultados e Discussão
144
Takeuchi 144 horas
Figura 69. Cromatograma do extrato Ac48 (MPF 24h, Takeuchi 144h)
Figura 70. Cromatograma do extrato Ac108 (MPF 48h, Takeuchi 144h)
M inutes0 5 1 0 1 5 2 0 2 5 3 0
mA
U
02
04
06
08
0
5,4
26
6,2
64
7,4
28
8,4
41
11
,29
4
15
,34
5
16
,89
1
18
,64
2
20
,70
6
23
,23
3
29
,78
5
Detec to r A -220 nmB a r b a r aVE 108 -1
Reten t i on T ime
Ac48
Minu tes0 5 1 0 1 5 2 0 2 5 3 0
mA
U
01
02
03
04
05
0
5,6
22
6,2
70
7,3
67
9,3
12
14
,21
5
17
,38
3
D e t e c t o r A - 2 2 0 n mB a r b a r aV E 4 7 - 1
R e t e n t i o n T i m e
Ac48
Ac108
_______________________________________Resultados e Discussão
145
Czapeck 72 horas
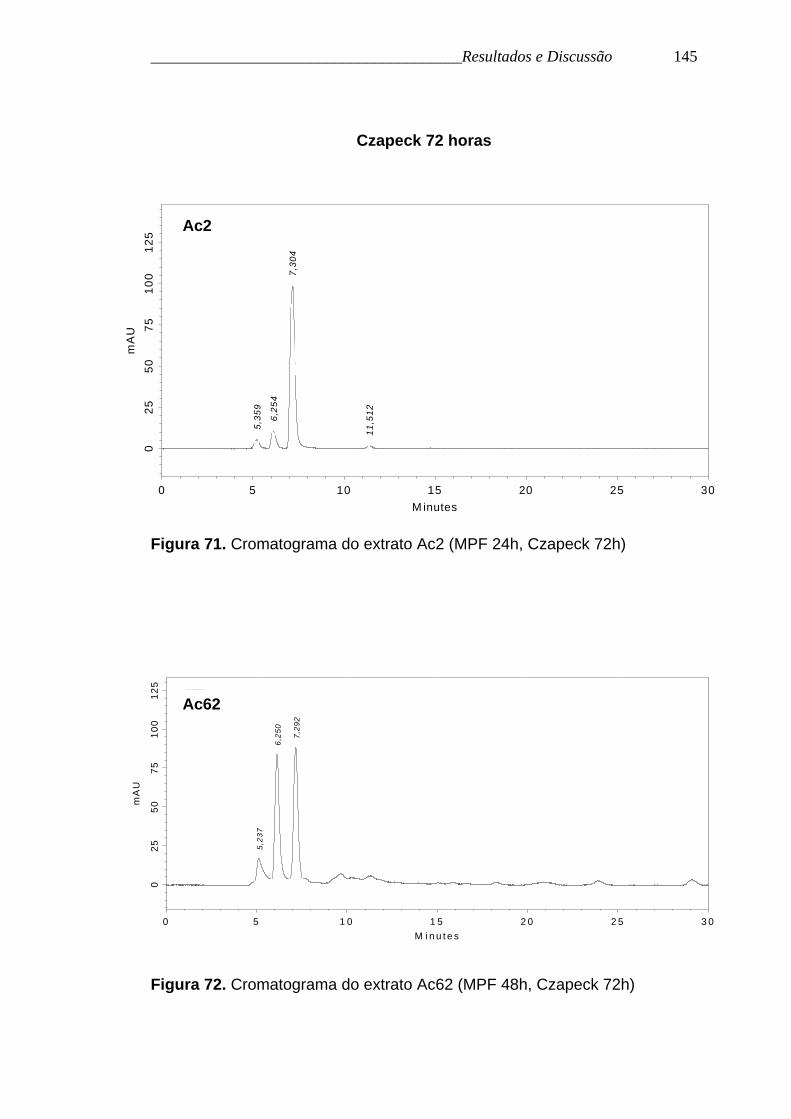
Figura 71. Cromatograma do extrato Ac2 (MPF 24h, Czapeck 72h)
Figura 72. Cromatograma do extrato Ac62 (MPF 48h, Czapeck 72h)
M inutes0 5 10 15 20 25 30
mA
U
02
55
07
51
00
12
5
5,3
59
6,2
54
7,3
04
11
,51
2
Detector A-220 nmBarbaraVE2 -1
Retention Time
M i n u t e s0 5 1 0 1 5 2 0 2 5 3 0
mA
U
02
55
07
51
00
12
5
5,2
37
6,2
50
7,2
92
D e t e c t o r A - 2 2 0 n mB a r b a r aV E 6 2 - 1
R e t e n t i o n T i m e
Ac2
Ac62

_______________________________________Resultados e Discussão
146
Czapeck 144 horas
Figura 73. Cromatograma do extrato Ac3 (MPF 24h, Czapeck 144h)
Figura 74. Cromatograma do extrato Ac63 (MPF 48h, Czapeck 144h)
M inutes0 5 1 0 1 5 20 2 5 3 0
mA
U
010
2030
5,2
16
5,3
77
6,2
73
7,3
98
7,8
02
16
,50
1
18
,12
5
23
,43
0
28
,81
2Detector A-220 nmBarbaraVE 3 - 2
Reten t i on T imeAc63
M in u t e s0 5 1 0 1 5 2 0 2 5 3 0
mA
U
01
02
03
0
5,21
6
6,2
73
7,8
02
8,5
38
17
,89
7
23
,43
0
28
,67
0
D e t e c t o r A - 2 2 0 n mB a r b a r aV E 3 - 2
R e t e n t i o n T i m eAc3

_______________________________________Resultados e Discussão
147
Jackson 72 horas
Figura 75. Cromatograma do extrato Ac32 (MPF 24h, Jackson 72h)
Figura x. Cromatograma do extrato Ac32 (MPF 24h)
Figura 76. Cromatograma do extrato Ac92 (MPF 48h, Jackson 72h)
M inu tes0 5 1 0 1 5 2 0 2 5 3 0
mA
U
01
02
03
0
6,3
57
6,9
22
7,4
02
12
,69
0
15
,31
3 16
,75
0
17
,74
2 25
,58
3
D e t e c t o r A - 2 2 0 n mB a r b a r aV E 3 2 - 1
R e t e n t i o n T i m e
M inutes0 5 1 0 1 5 2 0 2 5 3 0
mA
U
02
04
06
08
0
6,3
15 6
,94
8 12
,66
8
15
,32
4
16
,62
3
17
,73
8
25
,58
2
Detec to r A -220 nmB a r b a r aV E 9 2 - 1
R e t e n t i o n T i m e
Ac32
Ac92

_______________________________________Resultados e Discussão
148
Jackson 144 horas
Figura 77. Cromatograma do extrato Ac33 (MPF 24h, Jackson 144h)
Figura 78. Cromatograma do extrato Ac93 (MPF 48h,Jackson 144h)
M in u t e s0 5 1 0 1 5 2 0 2 5 3 0
mA
U
050
10
01
50
6,9
06
12
,60
5
15,1
44
16
,41
9
17
,63
5
25
,38
2
D e t e c t o r A - 2 2 0 n mB a r b a r aV E 9 3 - 1
R e t e n t i o n T i m eAc93
M in u t e s0 5 1 0 1 5 2 0 2 5 3 0
mA
U
02
46
81
0
5,6
49
6,3
77
6,9
20
8,5
61
12
,80
8
17
,92
0
26
,18
0
D e t e c t o r A - 2 2 0 n mB a r b a r aV E 3 3 - 1 1 1
R e t e n t i o n T i m eAc33

_______________________________________Resultados e Discussão
149
Vogel 72 horas
Figura 79. Cromatograma do extrato Ac17 (MPF 24h, Vogel 72h)
Figura 80. Cromatograma do extrato Ac77 (MPF 48h, Vogel 72h)
M inu tes0 5 1 0 1 5 20 2 5 3 0
mA
U
01
2
5,3
71
D e t e c t o r A - 2 2 0 n mBarbaraVE 17 -2
R e t e n t i o n T i m e
M inu tes0 5 1 0 1 5 20 2 5 3 0
mA
U
02
46
81
0
5,3
16
6,2
67
7,3
88
23
,62
0
29
,05
1D e t e c t o r A - 2 2 0 n mBarbaraVE 77 -1
R e t e n t i o n T i m e
Ac17
Ac77

_______________________________________Resultados e Discussão
150
Vogel 144 horas
Figura 81. Cromatograma do extrato Ac18 (MPF 24h, Vogel 144h)
No que se refere à intensidade de absorção dos picos relacionada ao
tempo de incubação na pré-cultura (24 e 48 horas), foi observada uma
relativa diferença quantitativa. Nos cromatogramas dos extratos obtidos do
fungo pré-cultivado por 48 horas há uma tendência ao aumento da
intensidade de picos já existentes e ao surgimento de novos picos,
sugerindo que o aumento do tempo de incubação na pré-cultura possa estar
relacionado à produção de maior quantidade de metabólitos na cultura
fermentativa.
A análise detalhada dos dados cromatográficos dos 15 extratos
extracelulares demonstrou uma nítida diferença qualitativa no perfil químico
obtido dos quatro diferentes meios fermentativos. Esta diferença está bem
M inutes0 5 10 1 5 2 0 2 5 30
mA
U
01
02
03
04
0
5,4
21
6,2
51
7,4
26
8,0
16
12
,04
3
D e t e c t o r A - 2 2 0 n mB a r b a r aV E 1 8 - 1
R e t e n t i o n T i m eAc18

_______________________________________Resultados e Discussão
151
representada pelos picos intensos com tempo de retenção distintos nos
cromatogramas já apresentados. Isto indica que mudanças nos substratos
presentes nos meios fermentativos estão diretamente relacionadas à
diversidade na produção de metabólitos secundários por P. verrucosum.
Observou-se pequena diferença qualitativa e grande similaridade
entre os perfis cromatográficos dos 15 extratos intracelulares avaliados
(anexos 1-15, p. 177).
Espectros de RMN 1H dos extratos MeOH mostraram que a maioria
dos metabólitos encontrados dentro da célula de P. verrucosum, cultivados
nas condições apresentadas neste trabalho, são primários (triglicerídeos,
ácidos graxos e açúcares).
Analisando os perfis químicos dos extratos intra e extracelulares
observou-se que P. verrucosum tende a excretar seus metabólitos
secundários. Todos os cromatogramas dos extratos intracelulares,
independente da condição de cultivo, apresentaram uma grande semelhança
enquanto os extratos extracelulares apresentaram-se distintos. Além disso,
as quantidades de extratos intracelulares foram superiores às dos
extracelulares.
Através das análises dos espectros de RMN 1H do extrato Ac47 e de
frações extracelulares, obtidas das culturas cultivadas por 72 horas em meio
Takeuchi, detectou-se ainda grandes quantidades dos constituintes do meio
de cultivo no fluido da cultura. Isto indica que estas substâncias não foram
totalmente metabolizadas durante o cultivo e encontram-se em mistura com
os metabólitos secundários produzidos pelo fungo. O meio de cultivo

_______________________________________Resultados e Discussão
152
Takeuchi é composto por uma mistura de peptídeos de tamanhos variados.
Dessa forma, o isolamento e identificação dos metabólitos biossintetizados a
partir de aminoácidos foi dificultada pela semelhança no comportamento
cromatográfico e espectroscópico. Isto pode ser exemplificado pelos
inúmeros ciclos necessários para o isolamento da substância F1 via CLAE-R
(p. 70-72).
Os cromatogramas dos extratos do fungo cultivado por 48 horas na
pré-cultura e 72 e 144 horas em meio Takeuchi mostraram perfis químicos
diferentes. O extrato Ac108, cultivado por 144 horas em meio Takeuchi,
apresentou grande variedade na produção de metabólitos quando
comparado com o extrato Ac107 cultivado por 72 horas. Por outro lado,
quando a pré-cultura é de 24 horas, não se observa diferença qualitativa
entre o cultivo do fungo no meio Takeuchi por 72 (Ac47) ou 144 horas
(Ac48).
A grande quantidade de metabólitos primários intracelulares sugere
que após 72 horas de cultivo, em meio Takeuchi, P. verrucosum ainda
estivesse no início da idiofase, ou seja, na fase de produção de metabólitos
secundários. Isto permite inferir que um eventual acréscimo no tempo de
incubação do fungo na pré-cultura aumentaria a quantidade de massa
micelial inoculada no meio fermentativo, podendo levar a uma elevação na
produção de metabólitos secundários por aumentar o tempo de permanência
do fungo na idiofase. A biossíntese de substâncias em meio fermentativo
está diretamente relacionada à massa micelial (Gaden & Elmer, 2000). P.
verrucosum apresentou uma razoável variação na quantidade de massa

_______________________________________Resultados e Discussão
153
micelial produzida na pré-cultura, quando incubada por 24 ou 48 horas.
Estas diferentes massas foram inoculadas na mesma quantidade de meio
fermentativo, o que pode ter influenciado a velocidade de produção dos
metabólitos secundários. Provavelmente, a massa cultivada por 48 horas na
pré-cultura (tabela 30, p. 153) atingiu mais rapidamente a idiofase, quando
inoculada em meio fermentativo.
Tabela 30. Massas miceliais úmidas obtidas na pré-cultura de P. verrucosum
TEMPO DE INCUBAÇÃO
(horas)
PESO DA MASSA ÚMIDA (g)
24 4,33 48 6,62
Como já mencionado, o meio Takeuchi é um meio muito rico em
nutrientes o poderia levar o fungo a permanecer mais tempo na tropofase,
ou seja, na fase exponencial do crescimento. Isto pode ser demonstrado
pela quantidade de massa micelial seca produzida pelo fungo em meio
Takeuchi comparada com os outros meios de cultura sob as mesmas
condições de cultivo (tabela 31, p. 154).
Os meios Takeuchi e Czapeck foram os mais promissores para a
produção de metabólitos secundários tripanocidas extracelulares por P.
verrucosum, indicados pelo extrato Ac47 com 77% e pelo extrato Ac3 com
55% de lise do T. cruzi (figura 67, p. 143 e figura 73, p. 146
respectivamente). Além disso, os dois extratos apresentam o pico majoritário
no mesmo tempo de retenção (tR - 6,27 min), o que merece futura
investigação.
_______________________________________Resultados e Discussão
154
Tabela 31. Efeito das condições de cultivo sobre a produção de massa micelial seca por P. verrucosum
MEIO FERMENTATIVO TEMPO DE INCUBAÇÃO
(horas) PESO DA MASSA
SECA (g) MEIO PRÉ-FERMENTATIVO INCUBADO POR 24 HORAS
Czapeck 48 1,27 Czapeck 72 1,60 Czapeck 144 1,51 Vogel 48 1,80 Vogel 72 1,55 Vogel 144 0,70 Jackson 48 1,45 Jackson 72 1,82 Jackson 144 2,62 Takeuchi 48 4,82 Takeuchi 72 5,31 Takeuchi 144 3,66
MEIO PRÉ-FERMENTATIVO INCUBADO POR 48 HORAS Czapeck 48 3,51 Czapeck 72 4,86 Czapeck 144 3,58 Vogel 48 3,08 Vogel 72 5,32 Vogel 144 * Jackson 48 2,91 Jackson 72 1,83 Jackson 144 2,57 Takeuchi 48 4,64 Takeuchi 72 5,74 Takeuchi 144 6,58
* Perda durante o processo. Massas de P. verrucosum produzido em pequena escala.
Os quatro extratos que apresentaram picos com tempo de retenção
maior que 30 minutos são extracelulares e foram obtidos de P. verrucosum
cultivado em meio Jackson. Estes extratos apresentam-se singulares na
produção de metabólitos em relação a todos os outros extratos analisados, o
que também merece futura investigação (anexos 16-19, p. 184).