oxidativo causado por cádmio e a resposta adaptativa a...
TRANSCRIPT

“Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este estresse em Saccharomyces cerevisiae”
Luciana Mara Costa Moreira
Dissertação apresentada como parte dos requisitos para obtenção do grau de Mestre em Ciência e Tecnologia das Radiações, Minerais e Materiais
2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
Comissão Nacional de Energia Nuclear
Centro de Desenvolvimento da Tecnologia Nuclear
Mestrado em Ciência e Tecnologia das Radiações, Minerais e Materiais
Papel da trealose na proteção durante o estresse oxidativo causado por cádmio
e a resposta adaptativa a este estresse em Saccharomyces cerevisiae
Luciana Mara Costa Moreira
Dissertação apresentada ao Curso de Pós-graduação em Ciência e Tecnologia das Radiações, Minerais e Materiais, com requisito parcial à obtenção do Grau de Mestre em
Ciência e Tecnologia das Radiações, Minerais e Materiais
Área de concentração: Ciências e Tecnologia das Radiações
Orientadora: Dra. Maria José Neves
Belo Horizonte 2007

iii
“Um sonho que se sonha só, é
um sonho que se sonha só, mas
um sonho que se sonha junto, é
REALIDADE”.
Raul Seixas

iv
“Aos meus avós, Maria e João, por criarem o caminho,
Aos meus pais, Clarice e João, por me ensinarem a caminhar,
Ao Túlio por caminhar ao meu lado”.
Dedico-lhes

v
À minha orientadora, Dra Maria José
Neves, que durante toda orientação
deste trabalho me fez sentir no
caminho certo.
Você fez a diferença!

vi
Agradecimentos
A Deus, por cada um dos dias vividos e por viver, pelas experiências boas e também pelas ruins que nos permitem crescer durante o curso da existência, e principalmente por colocar em minha vida, pessoas especiais que me acompanharam em cada momento dessa dissertação! À minha orientadora, Dra Maria José Neves que de chefe passou a fazer parte da minha vida, que me apresentou o lindo mundo do laboratório, me acompanhou em todo o momento me apoiando e não me deixando desistir, me dando força e principalmente me ensinando a amar o que jamais vou esquecer: a pesquisa. Muito obrigada por acreditar em mim!!!!! Aos meus pais, base da existência, luz do meu caminho, vocês não me deram apenas a vida, vocês são a minha vida!!! Obrigada por tudo, e essa vitória mais uma vez é nossa. Ao Túlio, obrigada por ser meu mundo, meu tudo e nada mais! Obrigada por me acompanhar e me apoiar em mais um momento de minha vida! Te Amo Pê! “Ainda bem que você vive comigo, por que senão como seria essa vida... sei lá! Sei lá!” Cris, irmão que eu amo, obrigada pelo silêncio que diz muito... A tia Fátima, carinho constante, apoio essencial, força nas horas tristes, sorriso certo em momentos felizes... A tia Vilma, por acreditar em mim, e ser um exemplo no mundo da pesquisa; a Dindinha Dora, apoio essencial sempre! Amo vocês! Aos tios, tias, primos e primas... Obrigada por fazerem parte de minha vida em cada momento dessa dissertação, obrigada por compreender o mau humor e sorrir em cada vitória. Família Costa eu amo todos vocês! A toda família Bacelette pala alegria contagiante; A Carol, fiel amiga nas horas alegres e difíceis, apoio essencial na hora da escrita... valeu de coração o companheirismo e a motivação!!! “E viva as leveduras” A Dani Foschetti, que aparece sempre na minha vida, seguindo os mesmos caminhos e passando pelos mesmos sufocos, dando aquela força especial... Ao Flaviano e Ariane pelo carinho, ensinamentos, e ajuda constante em todos os momentos... Adoro vocês! As três pessoas queridas da Radiobiologia, que caminharam junto comigo, compartilhando ensinamentos e principalmente sentimentos: A Estefânia, amiga fiel e alegre, que me fez sorrir, acreditar na alegria de viver, que me acompanhou em cada vitória e cada tristeza. Tetê, você mora no meu coração! À Marina por sempre ter a calma que tanto preciso, por

vii
me ouvir, me emprestar o ombro nas horas difíceis e ter alegria sempre. A Marcella pela amizade que significa muito e por conselhos financeiros sempre complexos... Ao pessoal do laboratório de Radiobilogia, que foram essenciais na realização deste trabalho, que além de amigos viraram irmãos, me acompanharam em cada vitória e em cada frustação... A Karine por me apoiar em todos os momentos lúcidos e não lúcidos, ao Fred por sobreviver a minha chatice e ser meu melhor EX estagiário, ao Edinho meu irmão de coração que me alegrou por vários momentos, ao Nino que me ensinou com seu “vasto” conhecimento, a Priscilla por entender a minha grande necessidade de falar, ao Paulo por seus diagnósticos precisos, a Ju Soprani pelo carinho, a Thaíssa por me ajudar quando o mundo está rodando, a Ju Lage pela simpatia, ao Rômulo por me alegrar sempre, a Pryscila
Rodrigues e Bárbara... “Amigos são anjos que nos levantam, quando nossas asas se esquecem como voar”
Aos sobreviventes do mestrado, a todas as pessoas especiais que estiveram ao meu lado, me apoiando, me dando força, e alegrando minha vida: Jaqueline, Nelson, Carlos Renato, Janúbia, Carol Bracinni, Karynne, Leo, Janúbia, Vanessa, Val... Ao Antero Silva Ribeiro e Raquel Gouveia dos Santos que acompanharam meu trabalho contribuindo com informações técnicas valiosas. A todos os professores da pós graduação do CDTN/CNEN A Suely Gomes Figueiredo do laboratório de Química de Proteínas da UFV por me receber de forma carinhosa. E a todos do laboratório: Manu, Ju Cassoli, Ju Carnielli, Gigi, Rafael... muito obrigada pela convivência e carinho; Ao Willian da Microanálise do ICEX/UFMG, por toda paciência e pelo excelente trabalho de analise da superfície das minhas células; A professora Jaqueline Leite, do laboratório de Bioquímica Nutricional ICB/UFMG e a Érica pela ajuda no espectrofluorímetro; Ao Zacarias, Geraldinho, Oliene e Dovenir por todo o carinho e ajuda. A equipe do reator (Fausto Moretti Junior, Amir Zacarias Mesquita, Paulo Fernandes de Oliveira, Luiz Otávio I. Sette Câmara, Wagner Souza) pela irradiação das amostras. A Maria Ângela de Barros Correia Menezes, da Radioquímica pelas determinações das concentrações de cádmio incorporadas pelas células; Ângela Maria Amaral pelas análises e também pela simpatia constante e pelos longos papos durante o almoço. Ao Adair e Noil pela grande ajuda com nossos rejeitos de cádmio; Ao Wagner pela simpatia constante e pela paciência com as fotografias das minhas placas;

viii
As bibliotecárias Lenira, Nívea e Virgínia por terem me recebido sempre com muita simpatia e atenção. Aos secretários da pós-graduação: Andréia, Roseli e Fulgêncio; A todas as pessoas do CDTN que direta ou indiretamente contribuíram para a realização deste trabalho.

ix
Resumo
A poluição por metais pesados é um dos mais sérios problemas ambientais da atualidade devido tanto ao aumento de sua aplicação, como de sua natureza imutável. A importância em se estudar os metais, deve-se aos seus intensos efeitos tóxicos para o homem e todos os outros seres vivos. O cádmio é um metal não essencial e representa um grande potencial de danos a seres humanos e ao meio ambiente. A trealose é um dissacarídeo que participa como protetor em microrganismos em diferentes condições de estresses. Neste trabalho foi analisada a influência da trealose em resposta ao estresse oxidativo induzido pelo cádmio em linhagens de Saccharomyces cerevisiae. As linhagens utilizadas neste estudo são deficientes em genes importantes no metabolismo de trealose, na síntese (S. cerevisaie tps1) e na degradação (S. cerevisaie nth1) do dissacarídeo, desta forma foi possível analisar o papel da trealose frente ao estresse induzido pelo cádmio. Inicialmente, analisou-se o crescimento das linhagens em meio líquido com diferentes concentrações de cádmio, onde observou-se inibição no crescimento. A tolerância ao CdCl2 foi realizada através do plaqueamento, verificou-se que não existe formação de colônias em concentração superior a 25ppm. A incorporação de cádmio é maior quando foram usadas células viáveis coletadas na fase logarítmica. A linhagem tps1 possui uma incorporação de cádmio 3 vezes menor quando comparada às demais linhagens. A adição de glutationa reduzida ao meio extracelular não provocou alteração de incorporação de cádmio, porém ao adicionarmos aminoácidos (que formam a glutationa) ocorreu diminuição da incorporação em todas as linhagens. Quanto a formação de radicais livres intracelulares obtivemos que a linhagem tps1, possui um alto nível de oxidação celular mesmo na condição sem cádmio e quando exposta ao mesmo, apresentou um aumento na oxidação apesar de uma incorporação menor do metal que as demais linhagens. Análise da peroxidação lipídica realizada antes e após exposição das células ao cádmio, mostraram danos aos lipídeos. Observamos que células da linhagem nth1 (excesso de trealose) também sofrem esses danos, ou seja, a presença de um excesso de trealose não impediu a presença de danos aos lipídeos. Células da linhagem tps1 apresentaram maior dano lipídico que as demais linhagens evidenciando que, a ausência da trealose deixa os lipídeos mais susceptíveis a alterações deletérias. Procedeu-se também a análise da carbonilação de proteína onde observamos que na presença de cádmio não ocorreu carbonilação de proteínas, apenas a linhagem nth1 na concentração de 800ppm apresentou proteínas carboniladas, fato este relacionado a grande quantidade de radicais livres formados. As três linhagens após exposição à concentração de 50ppm de cádmio, apresentaram um aumento nos níveis de resíduos sulfidrílicos enquanto que na concentração de 800ppm apresentaram queda dos resíduos sulfidrílicos. Os resultados apresentados neste trabalho ajudaram a compreender o papel da trealose em células de S.
cerevisiae frente ao estresse causado pelo cloreto de cádmio. Palavras-chave: Trealose; Estresse oxidativo; Saccharomyces cerevisiae; Cádmio.

x
Abstract
Pollution with heavy metals is one of the most serious environmental problems nowadays, which had in such a way to the increase of its application as, of its imutable nature. The importance of studying metals is directly related to intensive toxic effects to all living beings in our world. Cadmium is a non essential metal and represents a great danger to the human beings and also to the environmental. Trehalose is a disaccharide that participates as a protector in different conditions of stress. In this work the influence of trehalose in oxidative stress induced by cadmium was analyzed in different strains of Saccharomyces
cerevisiae. The strains used in this study were not effective in important genes of the metabolism of trehalose, in the synthesis (S. cerevisaie tps1) and also in the degradation (S.
cerevisaie nth1), so it was possible to analyze the performance of the trehalose in relation to the stress induced for cadmium. Initially the growth with different cadmium concentrations in medium liquid was analyzed, and the growth inhibition was observed. The tolerance to the CdCl2 was carried out by the growth on plates and one could observe that the formation was not present in concentrations up to 25ppm. Cells collected in the logarithmic phase had bigger cadmium incorporation. The strains tps1 presented a lower cadmium incorporation (3 times) when compared with other strains. The addition of reduced glutathione to the extracellular did not cause any alteration of in the cadmium incorporation, however, by adding amino acids, the ones which form glutathione, a reduction of the cadmium incorporation in all the strains occurred. The tps1 strains showed high level of cellular oxidation in control condition (without cadmium), and the cadmium incorporation showed an increase in the oxidation.Analyses of the lipid peroxidation carried out before and after exposition of the cells to cadmium showed damages to the lipids. We observe that cells of the strain nth1 (excess of trehalose) also suffer these damages. The presence of an excess of trehalose protected the lipids. Cells of the strain tps1 presented greater lipidic damage than the other strains, making clear that the absence of trehalose make the lipids more probable to have deleterious alterations. It was also performed a protein carbonilation analysis at cadmium presence and it did not induce a carbonilation of proteins. The stains nth1, whose concentration was 800ppm, presented carbonilated protein due to the high levels of ROS. The three strains after the exposition of 50ppm of cadmium showed an increase while on the concentration of 800ppm, it presented a drop on the levels of sulphydryl residues. The results presented in this work helped to understand the role of trehalose on Saccharomyces
cerevisiae cells during cadmium stress. Key-word: Trehalose; Oxidative stress; Saccharomyces cerevisiae; cadmium.

xi
Lista de Figuras
FIGURA 01 – Formação das espécies reativas de oxigênio. ................................................ 3
FIGURA 02 – Resultado do estresse oxidativo pelo desbalanço entre antioxidantes e ROS.
................................................................................................................................................ 4
FIGURA 03 – Ativação de vias de sinalização em resposta ao estresse oxidativo. .............. 5
FIGURA 04 – Respostas das leveduras às condições de estresses........................................ 7
FIGURA 05 – Possíveis mecanismos de indução de estresse oxidativo por metais
pesados. ................................................................................................................................ 15
FIGURA 06 – Danos aos lipídeos após contato com espécies reativas de oxigênio........... 18
FIGURA 07 – Estruturas dos derivados carbonilados produzidos através da oxidação de
resíduos de aminoácidos das proteínas. ................................................................................ 19
FIGURA 08 – Geração de proteínas carboniladas. ............................................................. 21
FIGURA 09 – Estrutura da trealose, α,α1-1-trealose. ........................................................ 23
FIGURA 10 – Papel da trealose no choque térmico............................................................ 25
FIGURA 11 – Ciclo da Trealose em Saccharomyces cerevisiae. ....................................... 28
FIGURA 12 – Células de Saccharomyces cerevisiae.......................................................... 30
FIGURA 13 – Câmara de Neubeuer.................................................................................... 41
FIGURA 14 – Sonda fluorescente 2,7-diclorofluoresceína................................................. 46
FIGURA 15 – Reagente Ellman.......................................................................................... 49
FIGURA 16 – Esquema “scavenging” do radical DPPH• pelo antioxidante. ..................... 49
FIGURA 17 – Influência de diferentes concentrações de cloreto de cádmio no crescimento
das linhagens WT (A), tps1 (B), e nth1 (C). ......................................................................... 54
FIGURA 18 – Determinação da sobrevivência das células em meio sólido em ausência de
cádmio (A) e presença de 5 ppm (B) e 10 ppm (C) e 25ppm (D) de cloreto de cádmio...... 55

xii
FIGURA 19 – Determinação do cádmio incorporado pelas células das linhagens WT, tps1 e
nth1 coletadas na fase LOGARÍTMICA (A) de crescimento celular, e na fase
ESTACIONÁRIA (B). ......................................................................................................... 58
FIGURA 20 – Determinação de cádmio incorporado pelas células viáveis e não viáveis no
período de 16 horas. Linhagens WT (A), tps1 (B) e nth1 (C)............................................... 62
FIGURA 21 – Incorporação de cádmio em tempos variáveis por células viáveis de
Saccharomyces cerevisiae, WT, tps1 e nth1. ........................................................................ 64
FIGURA 22 – Complexo formado entre cádmio e a glutationa, Bis(glutationato) de cádmio
– Cd(GH)2............................................................................................................................. 65
FIGURA 23 – Incorporação de cádmio durante 16 horas em células não tratadas, tratadas
aminoácidos (aa) e tratadas com glutationa reduzida (GSH) nas linhagens S. cerevisiae WT
(A), S. cerevisiae tps1 (B) e S. cerevisiae nth1 (C). ............................................................. 68
FIGURA 24 – Determinação de cádmio na superfície celular de Saccharomyces cerevisiae
WT em células mortas – Biosorção (A), e em células viáveis – Bioacumulação (B)........... 71
FIGURA 25 – Determinação de cádmio na superfície celular de Saccharomyces cerevisiae
tps1 em células mortas – Biosorção (A), e em células viáveis – Bioacumulação (B).......... 72
FIGURA 26 – Determinação de cádmio na superfície celular de Saccharomyces cerevisiae
nth1 em células mortas – Biosorção (A), e em células viáveis – Bioacumulação (B). ........ 73
FIGURA 27 – Diferença nos níveis de incorporação de cádmio em 16 horas, nos meios
YPGalactose e YPGlicose. Linhagem WT (A) e nth1 (B).................................................... 75
FIGURA 28 – Influência das concentrações de cloreto de cádmio sobre os níveis de
trealose da linhagem S. cerevisiae WT (A) e S. cerevisiae nth1 (B). ................................... 79
FIGURA 29 – Mecanismo de ação das esterases na 2,7-diclorodihidrofluoresceína
diacetato (DCFH-DA) formando 2,7- diclorohidrofluoresceína, e a futura oxidação pela
ROS formando assim a 2,7-diclorofluoresceína (DCF). ...................................................... 80
FIGURA 30 – Determinação do status antioxidante total das diferentes linhagens
Saccharomyces cerevisiae utilizando o radical DPPH•........................................................ 94

xiii
Lista de Tabelas
TABELA 01 – Linhagens de Saccharomyces cerevisiae utilizadas. ...................................38
TABELA 02 – Incorporação de cádmio durante 16 horas por células viáveis de
Saccharomyces cerevisiae (µg Cd incorporado/g peso seco)...............................................63
TABELA 03 – Determinação dos níveis de oxidação intracelular em linhagens de
Saccharomyces cerevisiae ....................................................................................................83
TABELA 04 - Determinação dos níveis de peroxidação de lipídeos (picomoles MDA/mg
proteína/mL) em linhagens Saccharomyces cerevisiae........................................................86
TABELA 05 – Determinação dos níveis de carbonilação de proteínas (nanomoles/mg
proteína/mL) em linhagens de Saccharomyces cerevisiae. ..................................................89
TABELA 06 – Determinação dos níveis resíduos sulfidrílicos totais (µmoles/mg
proteína/mL) em linhagens de Saccharomyces cerevisiae. ..................................................92

xiv
Lista de Abreviaturas
µµµµg micrograma µµµµL microlitro mL mililitro mg miligrama nm Nanômetro aa Aminoácido Abs. Absorbância Al Alumínio Arg Arginina BER Base Excision repair BSA Soro albumina bovina CaCl2 Cloreto de cálcio Cd Cádmio CdCl2 Cloreto de cádmio Cd(GS)2 Complexo bis (glutationato) de cádmio CdSO4 Sulfato de cádmio CDTN Centro de Desenvolvimento da Tecnologia Nuclear CNEN Comissão Nacional de Energia Nuclear Cu Cobre Cys Cisteína Da Dalton DCH diclorofluoresceína DCFH 2,7 – diclorofluoresceína DCFH-DA 2,7 – diclorohidrofluoresceína diacetato DNA Ácido desoxiribonucléico DNP 2,4 Dinitrofenilhidrazona DNPH 2,4 Dinitrofenilhidrazina D.O. Densidade óptica DPPH α,α-difenil-β-picrylladrazil DTNB 5’5-ditiobis-2-ácido nitrobenzóico ERK Kinase regulada por sinais oxidativos extracelulares EDS Espectometria por dispersão de energia (Energy Dispersive Espectrometer) EDTA Ácido etilenodiaminotetraacético Fe Ferro Glu Ácido glutâmico GME glutationa monoetil éster GSH Glutationa reduzida GSH1 α-glutamilcisteína sintase GSH2 Cistenilglicina dipeptidase GSSG Glutationa oxidada HCl Ácido Clorídrico Hg Mercúrio HNE Hidroxinoneal His Histidina

xv
H2O2 Peróxido de hidrogênio HSP Proteínas de choque térmico (heat shock proteins) IPEN Instituto de Pesquisas Energéticas e Nucleares JNK Kinase c-jun amino-terminal kJ Kilo Joule Lys Lisina MAPK Proteína kinase ativada por mitógeno MDA Malondialdeído MMR Mistmach repair
Mn Manganês MT Metalotioneína NADPH Nicotinamida Adenina Nucleotídeo Fosfato (forma reduzida) Na2CO3 Carbonato de sódio NER Nucleotide excision repair Ni Níquel nth1∆∆∆∆ Células deficientes de trealase neutra Pb Chumbo PMSF Phenylmethysulphonylfluoride
ppm Partes por milhão Pro Prolina PUFA Ácidos graxos polinsaturados O2 Molécula de oxigênio O2
- Ânion superóxido O3 Ozônio OH•••• Radical Hidroxil RNA Ácido ribonucleico ROS Espécies reativas de oxigênio (Reactive oxygen species) r.p.m Rotação por minuto Se Selênio TBA Ácido Tiobarbitúrico TBARS Espécies que reagem com ácido tiobarbitúrico (thiobarbituric acid reagent
species) TCA Ácido Tricloracético Thr Treonina tps1∆∆∆∆ células deficientes de trealose-6-fosfato sintase Zn Zinco YPGalactose Meio contendo peptona, estrato de levedura e galactose YPGlicose Meio contendo peptona, estrato de levedura e glicose

xvi
Sumário
Agradecimentos ...................................................................................................................vi
Resumo .................................................................................................................................ix
Abstract .................................................................................................................................x
Lista de Figuras ...................................................................................................................xi
Lista de Tabelas .................................................................................................................xiii
Lista de Abreviaturas........................................................................................................xiv
Sumário ..............................................................................................................................xvi
1 - Introdução........................................................................................................................ 1
1.1 – Espécies reativas de oxigênio e estresse oxidativo.................................................... 2
1.2 – Defesas Antioxidantes ............................................................................................... 8
1.3 – Bioremediação ........................................................................................................... 8
1.4 – Cádmio .................................................................................................................... 11
1.5 – Cádmio como causador de estresse oxidativo ......................................................... 13
1.6 – Danos causados pelas ROS ..................................................................................... 16
1.6.1 – Peroxidação de Lipídeos................................................................................... 16
1.6.2 – Carbonilação de proteínas ................................................................................ 19
1.6.3 – Danos ao DNA ................................................................................................. 21
1.7 – Trealose ................................................................................................................... 22
1.8 – Saccharomyces cerevisiae como modelo de estudo................................................ 28
2 - Justificativa .................................................................................................................... 31
3 - Objetivos ........................................................................................................................ 33
3.1 - Objetivo geral ........................................................................................................... 34
3.2 - Objetivos específicos ............................................................................................... 34

xvii
4 - Estratégia Experimental ............................................................................................... 35
5 - Materiais e Métodos ...................................................................................................... 37
5.1 - Microrganismo Utilizado ......................................................................................... 38
5.2 - Meios de cultura ....................................................................................................... 38
5.3 - Manutenção das linhagens em laboratório ............................................................... 38
5.4 – Estresse com cloreto de cádmio .............................................................................. 39
5.5 – Obtenção de extratos celulares ................................................................................ 40
5.6 - Dosagem de proteínas .............................................................................................. 40
5.7 - Curva de crescimento ............................................................................................... 40
5.8 - Tolerância ao cádmio ............................................................................................... 41
5.9 - Dosagem de Trealose ............................................................................................... 42
5.10 - Extração da trealase do Humicola grisea var. thermoidea .................................... 42
5.11 - Determinação de cádmio incorporado pelas células .............................................. 43
5.12 - Preparação das amostras para determinação do cádmio incorporado .................... 44
5.13 - Incorporação de CdCl2 na presença de Glutationa reduzida adicionada ao meio.. 44
5.14 - Determinação de cádmio na superfície celular ...................................................... 45
5.15 - Determinação da oxidação intracelular .................................................................. 45
5.16 - Carbonilação de Proteínas...................................................................................... 46
5.17 - Peroxidação de lipídeos.......................................................................................... 47
5.18 - Resíduos Sulfidrílicos Totais ................................................................................. 48
5.19 - Determinação do status antioxidante total pelo radical DPPH .............................. 49
5.20 – Descarte de Rejeitos .............................................................................................. 50
5.21 - Análise Estatística .................................................................................................. 50
6 - Resultados e Discussão.................................................................................................. 51

xviii
6.1 – Curva de crescimento e tolerância ao cádmio ......................................................... 52
6.2 - Determinação da incorporação de cádmio nas diferentes fases de crescimento
celular ............................................................................................................................... 56
6.3 - Determinação de cádmio incorporado pelas células viáveis e não viáveis .............. 59
6.4 – Determinação de incorporação de cádmio em células suplementadas com glutationa
reduzida e aminoácidos .................................................................................................... 65
6.5 – Determinação de cádmio na superfície celular........................................................ 69
6.6 - Determinação da incorporação de cádmio em meio suplementado com galactose e
glicose............................................................................................................................... 74
6.7 – Síntese de trealose em resposta à presença de cádmio ............................................ 76
6.8 – Determinação do nível da oxidação intracelular ..................................................... 80
6.9 – Determinação dos níveis de peroxidação lipídica ................................................... 84
6.10 - Carbonilação de Proteína ....................................................................................... 87
6.11 – Resíduos Sulfidrílicos Totais................................................................................. 90
6.12 - Determinação do status antioxidante total das linhagens pelo radical DPPH........ 93
7 – Conclusão ...................................................................................................................... 95
8 - Referências Bibliográficas ............................................................................................ 97

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Sacchamoryces cerevisiae
1
1 - Introdução

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
2
1.1 – Espécies reativas de oxigênio e estresse oxidativo
As moléculas de oxigênio diatômico na atmosfera terrestre são as maiores
promotoras de reações redox nas células vivas. A molécula de O2 é a principal receptora
biológica de elétrons e desempenha importante papel nas funções celulares, incluindo o
processo de respiração aeróbica. Exceto para aqueles organismos que são especialmente
adaptados a viverem sob condições anaeróbicas todos os seres vivos, como os animais,
plantas e microrganismos requerem oxigênio para uma eficiente produção de energia. O
surgimento do oxigênio deve ter sido acompanhado pelo aparecimento da camada de
ozônio (O3) na alta atmosfera, com a conseqüente absorção da radiação ultravioleta (UV)
pela camada de ozônio, evitando assim, os efeitos nocivos da radiação UV. Tal situação
provavelmente permitiu a evolução dos organismos terrestres mais complexos.
(HALLIEWELL, 1989).
Normalmente, em torno de 98% do oxigênio absorvido pelos organismos aeróbicos
é reduzido através da cadeia de transporte de elétrons na mitocôndria, no processo
conhecido por fosforilação oxidativa, onde ocorre à redução do O2 pelo sistema citocromo
oxidase, fornecendo simultaneamente quatro elétrons para o oxigênio, que se reduz
diretamente à água (HALLIEWELL, 1989). Entretanto, juntamente com as propriedades
benéficas do O2, tem-se à formação de espécies reativas de oxigênio (do inglês Reactive
Oxygen Species, ou seja, ROS), que incluem o ânion superóxido (O2-), peróxido de
hidrogênio (H2O2) e radical hidroxil (OH•). A diferença das ROS, para a molécula de
oxigênio é que elas possuem um ou mais elétrons, não pareados. (Figura 01) (KILEY et al.,
2004).

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
3
FIGURA 01 – Formação das espécies reativas de oxigênio. Adaptado de: KILEY et al., 2004.
Os radicais livres tais como O2•-, e OH• e o não radical H2O2, são produzidos em
todos os organismos aeróbicos, e normalmente existem na célula em um balanço com
moléculas antioxidantes. De acordo com esta visão, o estresse oxidativo ocorre quando este
balanço é interrompido devido à depleção de antioxidantes ou acúmulo de ROS (Figura
02). A formação de ROS pode ser acelerada como conseqüência de várias condições de
estresse do meio, incluindo radiação UV, alta intensidade de luz, exposição a herbicidas,
temperaturas extremas, toxinas, poluentes e metais (SCANDALIOS, 2005).
Avanços nos estudos da ação das ROS levaram a uma nova definição do estresse
oxidativo que considera o papel vital das ROS como moléculas sinalizadoras, responsáveis
pela modulação de vias de transdução de sinais, do balanço redox e de seus controles
(JONES, 2006). Alterações no controle redox e na sinalização podem ocorrer com ou sem
mudança no balanço total dos oxidantes e antioxidantes. Por exemplo: agentes redox como
metais podem romper seletivamente vias de sinalização, e a adição de antioxidantes não

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
4
mudará este panorama, inclusive dependendo da concentração, alguns antioxidantes podem
se tornar um pro-oxidante em total antagonismo à definição antiga de desbalanço entre
oxidantes e antioxidantes (HANSEN et al., 2006)
FIGURA 02 – Resultado do estresse oxidativo pelo desbalanço entre antioxidantes e ROS. Fonte:
SCANDALIOS, 2005.
As espécies reativas de oxigênio participam também da comunicação bioquímica
das células e organismos (GHEZZI et al., 2005). As ROS atuam em vias de sinalização
especificas, localizadas tanto na resposta ao estresse, como em processos fisiológicos
normais (INOUNE et al., 2004; WU et al., 2005). Entre as principais vias de sinalização ao
estresse oxidativo estão as ERK (kinase regulada por sinais oxidativos extracelular), JNK
(kinase c-jun amino-terminal) e p38 ativada por MAPK (proteína kinase ativada por
mitógeno) e a resposta ao choque térmico. A ativação destas vias não é exclusiva do
estresse oxidativo, elas também participam na regulação das respostas celulares a outros

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
5
estresses, bem como da regulação do metabolismo e do crescimento normal. Em muitas
situações a resposta aos oxidantes pode envolver uma super estimulação das vias de
sinalização normais regulada pelas ROS (Figura 03) (FINKEL et al., 2000).
FIGURA 03 – Ativação de vias de sinalização em resposta ao estresse oxidativo. Fonte: FINKEL et
al., 2000.
A geração de ROS pode ocorrer através de um grande número de processos
fisiológicos e não fisiológicos, sendo um produto normal do metabolismo celular. Estudos
comprovam que a maior fonte intracelular de ROS é a mitocôndria. A produção de radicais
ocorre principalmente em dois pontos da cadeia de transporte de elétrons, chamadas de
complexo I (NADH desidrogenase) e o complexo III (ubiquitina – citocromo c redutase)
(TURRENTS, 1997). As ROS podem causar danos a vários tipos de moléculas biológicas,
são os chamados danos oxidativos às proteínas, lipídios ou DNA, que podem ser seriamente
deletérios. (DALLE-DONNE et al., 2003). Os danos causados a estes componentes
celulares causam a perda de suas funções, podendo assim levar a morte celular (PEREIRA

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
6
et al., 2003). Recentemente, o estresse oxidativo tem sido um grande foco de estudos
devido a sua correlação com várias doenças letais. Dentre estas encontra-se o câncer,
desordens neurodegenerativas e também o envelhecimento, considerado como um processo
normal e inevitável de acúmulo de ROS nos seres vivos (FINKEL et al., 2000).
Vários estudos demonstram que metais como ferro, cobre, cádmio, cromo,
mercúrio, níquel e vanádio possuem habilidades em produzir radicais livres de forma direta
ou indireta, resultando assim em danos celulares (VALKO et al., 2005). Brennan e
colaboradores (1996) demonstraram que a exposição de células da levedura Saccharomyces
cerevisiae ao cádmio provocava estresse oxidativo. Entretanto o mecanismo pelo qual isto
ocorria, ainda não era bem conhecido uma vez que o cádmio não gera diretamente o
estresse oxidativo. Aparentemente o metal monopoliza as defesas celulares e interfere com
os mecanismos de reparo da levedura.
Células de leveduras submetidas a diferentes condições de estresse, tais como:
choque térmico, exposição ao etanol, metais pesados, alta osmolaridade e oxidantes
apresentam um mecanismo comum de defesa (Figura 04). Atualmente, sabe-se que o
aumento da produção de ROS, devido à exposição das células a diferentes formas de
estresses, estaria envolvido com ativação da resposta celular e aquisição de tolerância
contra um eventual estresse oxidativo severo (COSTA et al., 2001). Normalmente, a
levedura Saccharomyces cerevisiae responde a diferentes fontes de estresse oxidativo com
alterações nas defesas antioxidantes, sejam enzimáticas e/ou não enzimáticas. Assim, na
presença de cádmio, ocorre um aumento de atividade das enzimas glutationa peroxidase e
superóxido dismutase, enzimas responsáveis pela detoxificação de radicais livres gerados
(LIU et al., 2005). Células expostas à menadiona, gerador de radical livre intracelular,
apresentam aumento nos níveis de superóxido dismutase e glutationa redutase (CYRNE et
al., 2003).

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
7
FIGURA 04 – Respostas das leveduras às condições de estresses. Fonte: COSTA et al. (2001).

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
8
1.2 – Defesas Antioxidantes
As células possuem sistemas de defesa antioxidantes conhecidos como: enzimáticos
e não enzimáticos que, são necessários para proteger seus constituintes celulares, e manter
o estado redox da célula. O sistema de defesa não enzimático consiste tipicamente de
pequenas moléculas que são solúveis em qualquer meio aquoso ou, em alguns casos, em
meios lipídicos. Eles agem em geral, como “scavenging” de radicais e removem os
oxidantes da solução (HALLIWELL et al., 1993).
As defesas antioxidantes enzimáticas incluem a atividade da superóxido dismutase
(SOD), catalase (CAT), glutationa peroxidase (GPx), glutationa redutase (GR) e glutationa
S-tranferase (GTS). Para minimizar os efeitos tóxicos dos radicais livres formados, é
necessário um adequado equilíbrio das enzimas antioxidantes citadas acima
(SCANDALIOS, 2005).
Dentre as defesas não enzimáticas, o mais conhecido exemplo é a glutationa, um
tripeptídeo (L-γ-glutamil -L- cistinilglicina). A glutationa reduzida (GSH) apresenta um
grupo sulfidril com ação redox, reagindo com oxidantes para produção de glutationa
oxidada (GSSG). A glutationa é possivelmente a mais abundante molécula “scavenger” -
redox nas células (MARCHLER et al., 1993) e conseqüentemente é a substância mais
importante na manutenção do estado redox celular. Os genes envolvidos na biossíntese da
GSH já foram identificados em S. cerevisiae são: GSH1 e GSH2, que codificam a γ-
glutamilcisteína sintetase e glutationa sintetase, respectivamente (OHTAKE et al.,1991)
1.3 – Bioremediação
A poluição por metais pesados representa um importante problema ambiental
devido aos efeitos tóxicos dos metais. O acúmulo de metais nos vários elos da cadeia
alimentar pode acarretar sérios problemas ecológicos e de saúde (MALIK et al., 2004).
Existe um crescente entendimento que a contaminação por metais no meio ambiente
funcione como um agente seletivo na proliferação da resistência à antibióticos. Associação
entre tipos e níveis de contaminação metálica e vias específicas de resistência a antibióticos

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
9
sugerem que muitos mecanismos sofreram um processo de co-seleção. Assim, a
contaminação por metais representa a persistência por longo tempo de uma pressão seletiva
que potencialmente contribui para a manutenção e a propagação da resistência a
antibióticos (BAKER-AUSTIN et al., 2006).
Altos níveis de metais pesados no meio ambiente estão associados com atividades
antropogênicas. Este tipo de poluição geralmente diminui a biodiversidade nos
ecossistemas terrestres e aquáticos. Para sobreviver à exposição ao metal, a bactéria ou
fungo desenvolveram numerosos mecanismos de resistência. Fungos podem sequestrar,
mobilizar ou transformar vários íons. Assim, esses microrganismos podem ser usados para
reduzir o impacto ambiental causado pelos metais. Aplicações potenciais da bioremediação
tem renovado o interesse do efeito dos metais em fungos (BRAHA et al., 2007).
Para remoção de metais pesados de efluentes industriais são usados métodos
convencionais tais como precipitação, separação por membranas, filtração e extração de
íons. Esses métodos convencionais possuem significativas desvantagens, incluindo a
remoção incompleta, equipamentos de alto custo, sistemas de monitoramento, gasto de
energia, geração de lodo tóxico, entre outros. Estas desvantagens dos sistemas
convencionais, junto com a necessidade de economia e eficiência dos métodos de remoção
de metais, têm resultado no desenvolvimento de novas tecnologias de tratamento de
efluentes (GÖKSUNGUR et al., 2005). Decorrente destes problemas tornou-se necessário o
desenvolvimento de alternativas tecnológicas eficientes na remoção destes poluentes do
meio ambiente.
A bioremediação tem recebido uma grande atenção recentemente, não apenas como
novidade cientifica, mas principalmente como uma eficiente tecnologia de tratamento de
aplicação industrial. Microrganismos, incluído algas (ROMERA, et al., 2006;
KADUKOVÁ, et al., 2005), bactérias, fungos filamentosos e leveduras (KAPOOR, et al.,
1995; GÖKSUNGUR, et al., 2005) são eficientes bioremediadores, removendo metais
através de mecanismos ativos e passivos denominados respectivamente, bioacumulação e
biosorção.
A biosorção e a bioacumulação são técnicas que usam organismos biológicos
adequados para a remoção de metais de efluentes. As grandes vantagens destes métodos são

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
10
o baixo custo de operação, diminuição do volume de lodo biológico gerado e
principalmente a alta eficiência na detoxificação de efluentes muito diluídos, abaixo de
100ppm (KADUKOVÁ et al., 2005)
A biosorção ou bioadsorção é uma forma passiva de imobilização de metais pela
biomassa. O mecanismo de sorção na superfície celular é independente do metabolismo da
célula; eles são baseados em interações entre o metal e grupos funcionais presentes na
parede celular. A parede celular dos microrganismos consiste principalmente de
polissacarídeos, lipídeos e proteínas que são os principais sítios de ligação dos metais.
(KADUKOVÁ et al, 2005)
A bioacumulação é o acumulo intracelular dos metais. Este processo envolve a
ligação dos metais em componentes superficiais, transporte para dentro da célula e, fixação
em componentes citoplasmáticos ou, vacuolares (KADUKOVÁ et al., 2005). No interior da
célula os metais podem se ligar a várias espécies moleculares ou componentes celulares. As
ligações mais comuns são as realizadas com as metalotioneínas (proteínas com vários
resíduos SH disponíveis para a ligação com o metal) e a glutationa (tripeptídeo formado por
cisteína, ácido glutâmico e glicina). O metal também pode ser transportado e ficar
sequestrado em vacúolos.
Na bioacumulação, o acúmulo de metais pela célula depende do metabolismo desta,
portanto a célula deve estar viva. Entretanto, para a biosorção a célula pode estar morta
pois, a ligação será feita em grupamentos químicos da parede celular. Outra grande
diferença entre os processos de bioacumulação e biosorção são, as cinéticas e os valores de
energia de ativação. A energia de ativação necessária para a biosorção é cerca de 21kJ/mol,
de acordo com a natureza física do processo. A energia de ativação para bioacumulação é
cerca de 63kJ/mol, que corresponde aos processos bioquímicos (KADUKOVÁ et al, 2005).
A biosorção é um processo mais rápido, independente de nutrientes específicos, não é
necessária a regulação de pH e temperatura, enquanto a bioacumulação é lenta e
dependente de nutrientes específicos (KAPOOR et al, 1995).
Fungos e leveduras podem acumular micronutrientes, tais como Cu, Zn e Mn, e
metais não nutrientes, tais como U, Ni, Cd, Sn, Hg (KAPOOR et al, 1995). O potencial da

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
11
biomassa de fungos como adsorventes para a remoção de metais de efluentes poluídos tem
sido analisadas em vários estudos (GÖKSUNGUR et al., 2005; KAPOOR et al., 1995)
1.4 – Cádmio
O termo metal pesado vem sendo aplicado a um grupo heterogêneo de elementos
incluindo metais, semi-metais e não metais que possuem numero atômico maior que 20 ou,
peso específico maior que 5 g cm3. Alguns deles são nutrientes essenciais aos organismos,
como o Cu, Fe, Mn, Ni e Zn, Se, são também conhecidos por elementos traços ou, metais
traços por serem encontrados em concentrações de poucas partes por milhão. Outros metais
são considerados não essenciais ou, não apresentam funções, tais como Cd, Al, Hg e Pb
(MELO et al., 1997).
A importância de se estudar estes metais, deve-se aos seus intensos efeitos tóxicos
para o homem e, todos os outros seres vivos, associados a sua ampla liberação no ambiente.
Com o aumento das atividades industriais, a poluição do ambiente com metais pesados
tóxicos têm ocorrido em todo o mundo, levando a deterioração de vários ecossistemas
(DÖNMEZ et al., 1999). De todos os metais poluentes, o cádmio é um dos mais tóxicos
para os seres humanos, animais, plantas e tem apresentado elevadas taxas de emissão no
ambiente nas últimas décadas (KEFALA et al., 1998). O cádmio é um metal do grupo IIB
na tabela periódica e não tendo qualquer função fisiológica no organismo biológico, com a
única exceção conhecida, da enzima anidrase carbônica da alga Thalassiosira weissflogi
que é dependente do metal (LANE et al., 2005).
Em 2005, o cádmio foi o 8º classificado na lista de “Substâncias Perigosas” da
ATSDR (Agency for Toxic Susbstances and Disease Registry), do CERCLA
(Comprehensive Ennvioremental Response, Coompensation and Liability Act) juntamente
com a classificação na EPA (Ennviromental Protection Agency). As substâncias são
classificadas de acordo com sua toxidade, potencial de risco à saúde e exposição destes aos
organismos vivos (ATSDR, 2007).
Geralmente, as contaminações ambientais pelo cádmio são resultantes de atividades
como mineração, metalurgia e uso de manufaturados industriais como, baterias e pilhas de

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
12
níquel-cádmio, pigmentos, estabilizantes plásticos e de produtos anticorrosivos. A
intoxicação humana ocorre principalmente por fumantes, devido a altas concentrações de
cádmio em cigarros, mas também através da água, alimentos e contaminação do ar
(BERTIN et al., 2006).
Em geral, o cloreto de cádmio e o acetato de cádmio são os mais absorvidos e mais
tóxico que outros compostos (ATSDR, 2007). Como medida de segurança, nos EUA o
nível seguro de cádmio na água para consumo humano é de 10ppb (parte por bilhão). No
Brasil a concentração permitida pelo Conselho Nacional de Meio Ambiente (CONAMA)
varia de 40ppb à 1 ppb, dependendo da classe de água. Por exemplo: para a água classe 1
(água destinada ao abastecimento doméstico após tratamento simplificado, recriação de
contato, irrigação de hortaliças, criação natural e/ou intensiva de espécies destinadas à
alimentação humana) o limite permitido de cádmio é de 1ppb enquanto para água classe 3
(água destinada ao abastecimento doméstico após tratamento convencional, irrigação de
culturas arbóreas e dessendentação de animais), o limite permitido é de 10 ppb – Resolução
nº 357 – CONAMA (CONAMA, 2007)
O cádmio é um potente carcinogênico humano, sua exposição ocupacional tem sido
associada com câncer de pulmão, próstatas, pâncreas e rins. Devido a sua característica
como um carcinogênico de pulmão (WAISBERG et al., 2005), é classificado na categoria
carcinogênico 1 (carcinogenese humana) pelo International Agency for Research on Cancer
(INARC) (American Cancer Society, 2007).
O mecanismo ou mecanismos de carcinogênese do cádmio não são bem
conhecidos. Vários modelos celulares têm sido desenvolvidos para definir o potencial
carcinogênico deste metal. O cádmio não é um metal redox ativo, produzindo estresse
oxidativo de forma indireta, que pode resultar indiretamente no ataque ao DNA. Os fatores
que contribuem para oncogenicidade do cádmio incluem: ativação de genes aberrantes,
repressão da apoptose e/ou alteração dos reparos de DNA (WAALKES, 2003).
Estudos recentes comprovaram as mudanças que o cádmio causam no metabolismo
de plantas (DECKERT, 2005) e, em microrganismos como leveduras e bactérias. Estas
mudanças levam a inibição do crescimento e a síntese de vários compostos com a função de

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
13
combater a toxidade, tais como metalotioneinas (LIU et al., 2005), glutationa, além de
várias enzimas como superóxido dismutase e glutationa peroxidase (LIU et al., 2005).
1.5 – Cádmio como causador de estresse oxidativo
Muitos estudos têm apresentado os efeitos tóxicos e carcinogênicos induzidos em
humanos e animais expostos a certos metais (Figura 05). É bem conhecido, que vários
metais de transição tais como: zinco, ferro, cobre, cobalto e manganês participam do
controle de várias vias metabólicas e de sinalização. De acordo com suas coordenações
químicas e propriedades redox, os metais não essenciais são capazes de mimetizar metais
essenciais sendo transportados, compartimentalizados em tecidos e constituintes celulares,
promovendo alterações na homeostase celular e nos mecanismos de controle. A ruptura
desses mecanismos de controle pode levar os metais não essenciais a se ligarem à diversas
proteínas e ao DNA. O acúmulo de informação com metais não essenciais na literatura
evidencia o fato que metais tóxicos e carcinogênicos são capazes de interagir com proteínas
nucleares e DNA, causando deterioração oxidativa das macromoléculas biológicas
(VALKO et al., 2005).
De acordo com Ercal e colaboradores (2001), metais pesados causam estresse
oxidativo; metais como ferro e cobre, participam de reações redox, doando ou recebendo
elétrons e são diretamente ligados a formação de radicais livres pela reação de Harber
Weiss e Fenton. Metais como cádmio, chumbo, arsênico e mercúrio são metais não redox e
causam um aumento do estresse oxidativo de forma indireta, através dos danos causados no
sistema de defesa antioxidante.
O cádmio tem múltiplos efeitos na célula (WAISBERG et al., 2003). Os
mecanismos propostos para a indução de estresse oxidativo pelo cádmio são classificados
em quatro grupos: 1) efeitos adversos do cádmio no sistema de defesa celular e status tiol;
2) peroxidação de lipídeos; 3) efeitos deletérios do cádmio nas enzimas celulares (ERCAL
et al., 2001) e 4) alterações no DNA (BERTIN et al., 2006)
De acordo com Bertin e colaboradores (2006), a intoxicação por cádmio causa
modificação nos níveis de expressão de muitas proteínas que respondem ao estresse, tais

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
14
como as proteínas de choque térmico (do inglês Heat Shock Proteins, ou HSPs). As HSPs
são proteínas celulares que podem ser induzidas por alterações na temperatura e também
como resposta à várias situações de estresse, incluindo a exposição ao cádmio. A indução
da HSPs é geralmente considerada como uma resposta adaptativa das células ao estresse,
assim como a sobrevivência celular, ou seja, a indução de HSPs após exposição ao cádmio
está ligado ao aumento do estresse oxidativo.
Estresse oxidativo em sistemas biológicos exclui a formação de diferentes espécies
reativas do oxigênio. Devidos às suas propriedades redox alguns metais como Fe+2 e Cu+2
catalisam a formação de ROS. Em contraste, metais sem esta capacidade redox como o
Cd+2 agem principalmente influenciando o status redox da célula reduzindo os níveis de
glutationa. Tióis são essenciais para a sinalização redox das células e são submetidos a
grandes controles em animais, plantas e fungos (POCSI et al., 2004).
De acordo com Baudouin-Cornu e colaboradores (2006), outra hipótese para a
toxidade do cádmio é a depleção de glutationa do citosol. A via de detoxificação do cádmio
envolve a quelação de glutationa e subseqüentemente, o transporte do complexo Cd(GS)2
para o vacúolo, assim contribuindo para a diminuição da glutationa livre no citosol. Como
conseqüência pode haver redução da atividade de enzimas dependentes de glutationa, tais
como glutationa peroxidase, glutationa S-tranferase e glutaredoxinas, que são envolvidas na
defesa do estresse oxidativo e outras funções essenciais das células.
A peroxidação de lipídeos tem sido bastante proeminente na exposição ao cádmio.
Reações de peroxidação de lipídeos após a exposição ao Cd não são totalmente conhecidas,
mas acredita-se que distúrbios nas concentrações de GSH (glutationa) e MT
(metalotioneina) permitam que radicais livres, tais como radical HO• e O2•, ataquem a
“dupla ligação” dos lipídeos de membrana e resultem assim em um aumento na
peroxidação de lipídeos. Esses trabalhos concluíram que o cádmio causa estresse oxidativo
de forma indireta, por meio da diminuição das defesas antioxidantes enzimáticas e não
enzimáticas.

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
15
FIGURA 05 – Possíveis mecanismos de indução de estresse oxidativo por metais pesados. Fonte:
ERCAL, et al., 2001.
Metal Tóxico
Danos as defesas
antioxidantes
Depleção dos grupamentos
tiois
ROS
Peroxidação de
Lipídeos
Oxidação de Ácidos
Nucleicos
Oxidação de proteínas
Danos na
membrana
Perda da função das proteínas
MORTE
CELULAR
CARCINOGENESE
MUTAGENICIDADE
LIPÍDEOS DNA PROTEÍNAS

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
16
1.6 – Danos causados pelas ROS
A exposição de células a fontes geradoras de radicais livres, provoca danos muitas
vezes irreparáveis ao DNA e às membranas, neste caso tornando-se letal para as células.
Estes radicais também pode atacar moléculas de DNA, RNA, lipídeos e proteínas (FARR et
al., 1991). Os danos causados a estes componentes celulares causam a perda de suas
funções, podendo assim levar a morte celular (PEREIRA et al., 2003). Abaixo são descritos
de maneira breve a peroxidação de lipídeos, carbonilação de proteínas e alterações no
DNA.
1.6.1 – Peroxidação de Lipídeos
A peroxidação de lipídeos ocorre quando um componente pró-oxidante reage com
um ácido graxo insaturado da membrana biológica; sua modificação causa mudança nas
propriedades físicas e químicas das membranas, alterando assim a fluidez e a
permeabilidade com conseqüente aumento no risco de ruptura da membrana. Desta forma, a
determinação dos níveis de lipídeos peroxidados das membranas celulares também é
considerada uma forma de monitoramento do estresse oxidativo sofrido pela célula. As
modificações no estado redox do lipídeo podem também afetar propriedades específicas da
membrana, tais como transdução de sinais e alterações na permeabilidade íonica (Figura
06) (VENDEMIALE et al, 1999).
O processo de peroxidação de lipídeos é uma reação em cadeia dirigida por um
radical livre que pode induzir a oxidação de um grande número de moléculas lipídicas
(LH), principalmente fosfolipídeos contendo ácidos graxos polinsaturados (PUFA). Os
ácidos graxos monoinsaturados e saturados são menos reativos, mas também podem sofrer
oxidação.
O processo de peroxidação lipídica é iniciado por um radical (R•), com a abstração
de um elétron do átomo de hidrogênio de um PUFA (reação a).

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
17
LH + R• → L•+ RH (Reação a)
Rapidamente uma molécula de oxigênio adiciona-se a este radical lipídico formando um
radical livre centrado no carbono (L•) levando posteriormente, a formação de um radical
peróxi-lipídico (reação b).
L• + O2 → LOO• (Reação b)
Este por sua vez, pode abstrair um hidrogênio de outra PUFA, similar a equação (a)
LH + LOO• → L• + LOOH (Reação c)
A reação c, chamada de propagação, resulta na conversão de vários moléculas de
PUFA à lipídeos hidroperóxidos. O lipídeo hidroperóxido (LOOH) é o primeiro produto da
reação de peroxidação de lipídeo. Sob condições onde a peroxidação de lipídeo é
continuamente iniciada, a terminação da reação leva a formação de produtos não radicais
(PNR) e destruição de dois radicais ao mesmo tempo. Estes produtos (PNR), conhecidos
como produtos secundários, não são inócuos, pelo contrário, são bem reativos e podem
atacar outras proteínas, lipídeos e o DNA.
LOO• + LOO• → PNR (Reação d)
Na presença de metais de transição, o LOOH pode aumentar a geração de radicais
capazes de reiniciar a peroxidação de lipídeos pelo ciclo redox de alguns metais. Os
lipídeos hidroperóxidos, na ausência ou presença de metais também podem gerar uma
grande variedade de produtos, incluindo aldeídos de cadeias curtas e longas (CATALÁ,
2006).
Uma vez formado, o radical peroxil (ROO•) pode também se rearranjar via reação
de ciclização com endoperóxidos (precursores do malondialdeído) com o produto final do

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
18
processo de peroxidação, formando o malondialdeído (MDA), que pode causar danos ao
reagir com as bases de DNA (MARNETT, 1999). Outro aldeído produzido através do
processo de peroxidação de lipídeos é o 4 hidroxi-2-noneal (HNE) que é um mutagênico
fraco mas, aparece como o principal produto tóxico da peroxidação de lipídeos (VALKO et
al., 2006).
Os produtos formados pela peroxidação de lipídeos são altamente reativos, e
disseminam e intensificam os efeitos dos radicais livres por meio de danos ao DNA e
proteínas (COSTA et al., 2001).
FIGURA 06 – Danos aos lipídeos após contato com espécies reativas de oxigênio. Fonte: VALKO
(2006).

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
19
1.6.2 – Carbonilação de proteínas
As proteínas podem ser modificadas por um grande número de reações envolvendo
espécies reativas de oxigênio (ROS) que estão presentes em poluentes na atmosfera, tais
como metais ou, geradas como um produto de um metabolismo celular normal formados
durante a exposição a agentes como radiação ultravioleta (STADTMAN et al., 2003). As
ROS podem causar graves e irreversíveis danos às biomoléculas, sendo as proteínas os
principais alvos dos danos oxidativos (DALLE-DONNE et al., 2003).
Grupos carbonilas (aldeídos e cetonas) são gerados nas proteínas pela oxidação dos
grupamentos laterais, principalmente os resíduos prolina, arginina, lisina e treonina. Os
derivados gerados pela oxidação destes aminoácidos são quimicamente estáveis, são de
fácil detecção e possuem um fácil armazenamento para as amostras (Figura 07) (DALLE-
DONNE et al., 2003).
FIGURA 07 – Estruturas dos derivados carbonilados produzidos através da oxidação de resíduos
de aminoácidos das proteínas. Adaptado: DALLE-DONE et al., 2003
A oxidação por radicais livres de vários aminoácidos resulta da formação de grupos
carbonil nas proteínas. Devido a extensão desta reação, os grupos carbonilas são
comumente usados como indicadores para avaliação da intensidade do ataque de ROS nas
proteínas (STADTMAN et al., 2000)
O aumento de proteínas carboniladas em resposta ao estresse oxidativo não ocorre
ao acaso, muitas proteínas são mais susceptíveis a carbonilação do que outras (DALLE-

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
20
DONNE et al., 2006). Segundo Standman (2000, 2003), a presença de metais de transição
nestas proteínas são a chave para uma maior susceptibilidade e portanto, podem sofrer
maior carbonilação via oxidação catalisada por metais. Os metais de transição em proteínas,
são fontes de radicais livres que iniciam a cascata de reações resultando na adição de
grupos carbonil aos resíduos de aminoácidos da cadeia proteica. Acredita-se ainda que
muitas proteínas, como por exemplo, enzimas que atuam no ciclo de Krebs e na cadeia de
transporte de elétrons, podem também ser susceptíveis à oxidação principalmente por
estarem localizadas na mitocôndria, um dos principais sítios geradores de espécies reativas
de oxigênio (NYSTRÖM, 2005).
Além da reação das ROS especificamente com resíduos dos aminoácidos das
proteínas, os grupos carbonil (CO) podem também ser introduzidos nas proteínas através de
reações secundárias dos sítios de cadeias nucleofílicas de resíduos Cys, His e Lys, com
produtos da peroxidação de lipídeos (4-hidroxi-2-noneal, malondialdeído), com a reação de
redução de açúcares ou ainda, os produtos da redução de açúcares podem se ligar aos
resíduos de lisina das proteínas (Figura 08) (DALLE-DONNE et al., 2003).
Os grupos carbonilas são utilizados como um indicador da intensidade dos ataques
dos radicais livres nas proteínas. Existem vários métodos de dosagens dos níveis de
carbonilação de proteína, todos são baseados na interação da dinitrofenilhidrazina (DNPH)
com as proteínas carboniladas, resultando na formação da dinitrofenilhidrazona (DNP) que
é dosado espectofotometricamente (LUSHCHAK, 2006).

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
21
FIGURA 08 – Geração de proteínas carboniladas. Fonte: SINGER, 1998.
1.6.3 – Danos ao DNA
A exposição das células às espécies reativas de oxigênio e à radiação ionizante,
resulta em diversas lesões na molécula de DNA, tanto nas bases nitrogenadas como nas
moléculas de açúcar constituintes do DNA (COSTA et al., 2001).
A integridade do genoma é essencial para a manutenção das funções celulares
corretas, principalmente para replicação, expressão gênica e síntese protéica. A indução de
danos ao DNA pode gerar uma parada no ciclo celular, mutagênese, instabilidade
genômica, câncer e também morte celular programada (apoptose).
Dentre os danos oxidativos mais perigosos, as alterações nas bases nitrogenadas
possuem uma posição de destaque pois são freqüentemente associados à formação de
mutações e a perda da integridade do DNA. Observa-se ainda que durante os danos
oxidativos ao DNA, ocorre formação de sítios abásicos (sítios com perda de bases
nitrogenadas) e também quebra de fitas simples. Recentes estudos relacionam a perda da

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
22
integridade genômica de células humanas com o processo de envelhecimento, doenças
degenerativas e câncer (HALLIELL et al., 1993).
Em S. cerevisiae, o peróxido de hidrogênio e superóxidos geram compostos que
induzem a oxidação das bases, gerando quebra das fitas simples e o aumento da frequência
de recombinação intracromossomal (FRANKENBERG et al., 1993; LEE et al., 1998). As
células possuem mecanismos capazes de remover e substituir as bases oxidadas através de
um sistema secundário de defesa antioxidante (COSTA, et al., 2001).
Os efeitos do cádmio na instabilidade genômica surgem de forma indireta
principalmente via o aumento do estresse oxidativo, podendo assim levar à danos ao DNA.
Vários estudos indicam que o cádmio inibe o reparo do DNA. Assim, tem sido
demonstrado que o cádmio causa alterações em diferentes sistemas de reparo do DNA,
sistemas de reparo envolvidos com danos em quebra de fita simples (HOEIJMAKERS,
2001), como por exemplo: no sistema de reparo Mistmatch repair (MMR), que se trata do
reparo do pareamento incorreto das bases nas fitas de DNA, quando o MMR é inibido não
pode haver a correção ou troca da base adicionada errada na fita que está sendo sintetizada
e assim, não pode haver a interação com a fita molde de DNA através das ligações
hidrogênio; NER - Nucleotide excision repair, que consiste na retirada de alguns
nucleotideos erroneamente adicionados ao DNA que causam uma distorção no mesmo;
BER - Base excision repair, ou seja, é o reparo onde a base errada é retirada através da
quebra da ligação glicosídica entre a base e a ribose formando um sítio abásico ocorrendo a
substituição da base (GIAGINIS et al., 2006; BERTIN et al., 2006). Se esses sistemas de
reparos são inibidos ou impedidos de funcionar, dado a presença do cádmio, diversas
alterações no DNA ocorreram e assim o cádmio pode exercer seu papel mutagênico e
genotóxico
1.7 – Trealose
A trealose (α-D-glicopiranosil-1,1-α-glicopiranosídio) é um dissacarídeo não
redutor formado por dois resíduos de D-glicose que são ligadas por átomos do carbono alfa
reduzidos (Figura 09). É encontrado em bactérias, fungos, plantas e invertebrados, e

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
23
ausente em mamíferos (BENARDOUJ et al., 2001). As leveduras acumulam grande
quantidade de trealose, tornando-se assim, excelentes modelos para o estudo da resposta
das células eucarióticas a diversos estresses (PANEK et al., 1995).
FIGURA 09 – Estrutura da trealose, α,α1-1-trealose. Fonte: ELBEIN et al., 2003.
Elbein (1974) descreveu o papel da trealose como fonte de armazenamento de
glicose para energia e/ou para síntese de componentes celulares. Porém, o papel da trealose
não se limita apenas a reserva de carboidratos, está relacionado principalmente, à proteção
contra estresse como: desidratação (FRANÇA et al., 2007), dessecação (LIU, et al., 2005),
congelamento, estresse térmico (ATTFIELD et al., 1987), estresse osmótico (HOUNSA, et
al., 1998), produtos químicos como metanol e peróxido de hidrogênio (BENAROUDJ et al,
2001), metais pesados. Estudos in vitro e in vivo demonstram a extraordinária propriedade
desta molécula na proteção biológica de membranas e proteínas desta maneira, a trealose
apresenta um duplo papel: como reserva de carbono e energia mas, principalmente, como
um protetor a diferentes estresses em células da Saccharomyces cerevisiae (SINGER et
al.,1998).
O aumento nos níveis de tolerância às várias formas de estresse, conferido pelo
aumento da concentração da trealose, pode ser explicado não somente pela estabilização de
estruturas proteicas e manutenção das atividades enzimáticas, como também pela
estabilização das biomembranas (CROWE et al., 1984). Existem outros fatores
responsáveis pela resistência ao estresse, entre eles, as proteínas de choque térmico e,

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
24
outros mais específicos a determinados estresses tais como níveis glutationa, tioredoxina,
glutaredoxina, metalotioneinas, glicerol dentre outros.
Estudos sobre o mecanismo protetor da trealose durante o choque térmico
mostraram que a presença de trealose é fundamental na proteção a este estresse, entretanto,
quando é feita a recuperação do choque térmico (volta das células à temperatura normal) a
trealose deve ser rapidamente degradada para permitir o acesso das proteínas de choque
térmico (HSPs) às proteína e assim, as HSP contribuem para a manutenção da estrutura
proteica normal, impedindo que as proteínas sofram desnaturação pelo calor (Figura 10).
Estudos com mutantes tps1, (mutantes que possuem deleção no gene que codifica para a
enzima de síntese de trealose, a trealose-6-fostato sintase) evidenciam o grande papel da
trealose no choque térmico, estas linhagens não sintetizam trealose, comprometendo assim
sua termotolerância e prejudicando também a síntese de proteínas de choque térmico (DE
VIRGILIO et al., 1994).

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
25
FIGURA 10 – Papel da trealose no choque térmico - (A) altas temperaturas causam desnaturação
de proteínas, que podem se associar e para formar agregados. (B) S. cerevisiae sintetizam grandes
quantidades de trealose durante o choque térmico. Este dissacarídeo estabiliza proteínas em seu
estado nativo sobre estas condições, e também suprime a agregação de proteínas que estão
desnaturadas. (C) Células de leveduras normalmente degradam trealose rapidamente após choque
térmico, prevenindo a agregação de proteínas. (D) A persistência de altos níveis de trealose,

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
26
interferem com a reativação de substratos desnaturados, explicando assim a necessidade de
degradar a trealose após o retorno da temperatura (SINGER et al., 1998).

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
27
Embora o possível envolvimento da trealose na proteção contra radicais livres não
tenha sido inteiramente demonstrado, vários trabalhos sugerem a hipótese de seu papel na
defesa aos ataques das ROS (ELBEIN et al., 2003). O único trabalho que claramente
demonstra o importante papel que a trealose desempenha na defesa contra radicais livres foi
o de BENAROUDJ (2001). O autor mostrou que a presença de trealose protegeu as
proteínas contra os radicais livres gerados após tratamento com peróxido de hidrogênio.
Na levedura S. cerevisiae, a trealose é formada no citosol por uma reação em duas
etapas (CABIB & LELOIR., 1958). Essa reação é conduzida por um complexo de três
subunidades: (1) uma subunidade catalítica de 56 kDa que é a trealose-6-fosfato sintase
codificada por TPS1 (ou C1F1) (BELL et al., 1992) (EC2.4.1.15), (2) uma subunidade de
102,8 kDa que é a trealose-6-fosfato fosfatase codificada por TPS2 (DE VIRGILIO et al.,
1993) (EC.3.1.3.12), (3) a maior subunidade de 123-kDa codificada redundantemente por 2
genes, TSL1 (trealose sintase de cadeia longa) e TPS3 (homologo a TSL1) com atividade
regulatória (VUORIO et al., 1993). Os mutantes com dupla deleção, TSL1 e TPS3, exibem
uma grande desestabilização no complexo de síntese da trealose (BELL et al., 1998).
Na primeira etapa de formação da trealose ocorre a condensação da uridina 5’-α-D-
glicopiranosil pirofosfato (UDP glicose) e glicose-6-fosfato pela ação da trealose-6-fosfato
sintase (tps1). Essa enzima transfere o resíduo glicosil do UDGP para a glicose-6-fosfato
formando a trealose-6-fosfato. Na segunda etapa, a trealose-6-fosfato é defosforilada por
uma fosfatase específica (trealose-6-fosfato-fosfatase/tps2) para produzir a trealose (Figura
11). A quebra da trealose gera duas moléculas de glicose sendo realizada pelas enzimas
trealases neutra ou ácida. Em S. cerevisiae a síntese da trealose é feita no citoplasma pelo
complexo trealose-6-fosfato sintase/fosfatase (TPS) que contem trealose-6-fosfato sintase
(Tps1p), trealose-6-fosfato fosfatase (Tps2p) e várias subunidades, tais como: Tps3p e
Tsl1p, que aparecem tanto para estabilizar quanto para regular o complexo TPS
(SAMPEDRO & URIBE, 2004).
Três enzimas são capazes de degradar trealose: duas trealases neutra ou intracelular,
e a trealase ácida, ou extracelular. As enzimas neutra e ácida diferem em função da sua
localização celular, propriedades catalíticas e, regulação. (PARROU et al., 2005;
SAMPEDRO & URIBE 2004). A trealase neutra, sintetizada pelo gene NTH1 é uma

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
28
enzima citoplasmática com atividade ótima em pH 7,0, o gene NTH2 sintetiza a outra
trealase neutra que não se sabe na realidade qual a sua função. A trealase ácida, parece ser
uma enzima encontrada na superfície celular, possui atividade ótima em pH 4,5, é
responsável pela utilização da trealose externa como fonte de carbono (PARROU, et al.,
2005; TERESHINA, 2005).
FIGURA 11 – Ciclo da Trealose em Saccharomyces cerevisiae. Os sinais representam (-) inibição
e (+) ativação. Fonte: VOIT (2003).
1.8 – Saccharomyces cerevisiae como modelo de estudo
As células de S. cerevisiae (Figura 12) têm diversas similaridades com as células de
mamíferos nas suas macromoléculas e organelas e, diversas proteínas destas leveduras, têm
sido funcionalmente capaz de substituir proteínas homólogas humanas. Outros fatores que
colaboraram para que as leveduras se tornassem um modelo experimental interessante
foram: a facilidade de obtenção de mutantes por meio de técnicas de genética e biologia
molecular, o fácil cultivo em um meio de cultura de composição simples, o pequeno tempo

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
29
de geração, quando comparada com outras células, o completo seqüenciamento do seu
genoma e, ao fato desta levedura não estar associada a características patogênicas como as
bactérias. Assim, a levedura Saccharomyces cerevisiae é um modelo relevante que, tem
contribuído para o entendimento de mecanismos bioquímicos e moleculares de diversos
problemas desafiadores da ciência atual, entre eles, os ligados ao estresse oxidativo
(COSTA & MORADAS-FERREIRA, 2001).
As concentrações de Fe, Mg e Zn, Cu e outros metais de importância fisiológica é
finamente regulada em células vivas. O maior determinante da homeostase dos metais é a
atividade dos transportadores de membrana. Metais tóxicos como Cd, Pb, Hg e Ni, não
possuem qualquer função benéfica conhecida mas, competem com os transportadores dos
metais biológicos essenciais. Assim, a atividade e, especificidade dos transportadores
fisiologicamente importantes de metais também controlam a letalidade dos metais tóxicos.
Em todas as espécies vivas, numerosas proteínas transportam metais através de todas as
membranas celulares. A levedura S. cerevisiae tem sido o organismo de escolha para
investigar o papel dos transportadores de metais na homeostase em eucariotos.
Além de todos os fatores relacionados acima, outra grande vantagem que a levedura
S. cerevisiae possui é o seu uso em vários processos de fermentação industrial sendo capaz
de gerar abundante matéria prima. Esta matéria prima está sendo submetida à várias
investigações para ser usada como material absorvente para metais e, tem sido estudada em
diferentes propostas de pesquisa, como por exemplo, células vivas/ células mortas
(KAPOOR & VIRARAGHAVAN, 1995), células imobilizadas e células livres (VEGLIO
et al., 1997); células pré tratadas por processos físico-químicos e não tratadas (MARQUES
et al., 1999), células laboratoriais e, células obtidas de diferentes processos industriais
fermentativos (PARK et al., 2003).

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
30
FIGURA 12 – Células de Saccharomyces cerevisiae. Fonte: www.bath.ac.uk/bio-sci/wheals2.htm

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
31
2 - Justificativa

Justificativa
32
As atividades industriais, a agropecuária e a mineração contaminam o solo, ar e a
água devido à liberação de metais pesados no ambiente, este fato tem intensificado os
problemas da poluição ambiental com vários reflexos na saúde humana. A remoção de
metais pesados dos efluentes industriais normalmente realizada por métodos químicos é
eficiente, porém, no caso de efluentes diluídos, tem custo elevado, é ineficaz e, leva a
geração de lodo tóxico. Novas alternativas têm sido estudadas, entre elas, a bioremediação.
Bioremediação é um processo que utiliza microrganismos na remoção de metais pesados do
ambiente.
Entre os metais pesados poluentes, o cádmio é um dos mais tóxicos para seres
humanos, animais, plantas sendo seus efeitos visíveis mesmo à baixas concentrações. Em
geral, no interior dos organismos, os metais induzem um forte estresse oxidativo. O estresse
oxidativo é causado pela excessiva produção de radicais livres. Os radicais livres estão
relacionados ao processo fisiológico de envelhecimento e, a doenças tais como: Alzheimer,
câncer, arteriosclerose, doença de Parkinson, doenças cardiovasculares, doenças
autoimunes e diabetes.
O estudo dessas adaptações bioquímicas é de grande utilidade para um possível uso
dessas células na incorporação de metais de efluentes líquidos e, para o conhecimento dos
mecanismos de proteção intracelulares uma vez que, várias doenças que afetam a
humanidade, são causadas ou, são agravadas pelo estresse oxidativo. Além dos efeitos
deletérios na saúde humana, um dos grandes efeitos biológicos da radiação ionizante é o
efeito indireto das radiações, ou seja, a geração de radicais livres que leva ao estresse
oxidativo fato de grande interesse no âmbito da instituição em que foi realizada esta
dissertação.

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
33
3 - Objetivos

Objetivos
34
3.1 - Objetivo geral
Avaliar a influência da trealose na resposta ao estresse oxidativo induzido pelo cádmio na
levedura Saccharomyces cerevisiae.
3.2 - Objetivos específicos
Usar linhagens de Saccharomyces cerevisiae com diferentes níveis de trealose para
verificar:
• a influência da exposição à concentrações crescentes de cloreto de cádmio sobre o
crescimento;
• a tolerância das linhagens às concentrações crescentes de cádmio;
• a influência da exposição à concentrações crescentes de cloreto de cádmio nos níveis de
trealose;
• a influência da fonte de carbono na incorporação de cádmio;
• a influência das diferentes fases de crescimento celular na incorporação de cádmio;
• a incorporação de cádmio em células viáveis e células mortas;
• a influência da glutationa reduzida e de aminoácidos formadores de glutationa na
incorporação ao cádmio;
• a influência do cádmio oxidação intracelular;
• a influência do cádmio na peroxidação de lipídeos;
• a influência do cádmio na carbonilação de proteína;
• a influência do cádmio no conteúdo de resíduos sulfidrílicos totais;
• o status antioxidante total de cada linhagem.

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
35
4 - Estratégia Experimental

Estratégia Experimental
36
Foram utilizadas três linhagens para observação da influência da trealose no estresse
oxidativo induzido pelo cádmio. A linhagem selvagem possui todas as enzimas necessárias
para a síntese e degradação da trealose. A linhagem tps1 não possui o gene que codifica a
enzima trealose-6-fosfato sintase, portanto, não pode, sob qualquer circunstância, sintetizar
o carboidrato. A linhagem nth1 não possui o gene que codifica a enzima trealase neutra 1
portanto, acumula a trealose sintetizada.
As células das três linhagens foram incubadas na presença ou, ausência (controle)
de cloreto de cádmio. Foi determinada a influência que a incorporação de cádmio teve no
crescimento celular, na tolerância das células ao metal e, a quantidade de cádmio
incorporado pelas diferentes linhagens. Foi verificado se a presença de trealose tem
influência na incorporação de cádmio e, na resposta ao estresse oxidativo. A indução do
estresse oxidativo devido à geração de radicais livres formados após a exposição ao metal
foi avaliada pela incorporação de uma sonda fluorescente e, analisado os efeitos deletérios
dos radicais livres em lipídeos, proteínas e conteúdo de resíduos sulfidrílicos totais.

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a
este estresse em Saccharomyces cerevisiae
37
5 - Materiais e Métodos

Materiais e Métodos
38
5.1 - Microrganismo Utilizado
As linhagens de Saccharomyces cerevisiae utilizadas e os genótipos em estudo estão
apresentados na tabela abaixo:
Tabela 01 – Linhagens de Saccharomyces cerevisiae utilizadas.
LINHAGENS GENÓTIPO CARACTERÍSTICAS W303-1A (WT) Can1-100 leu2-3trp1-1ura3-
1ade2-1his3-11,15 Linhagem laboratorial
W303-829 (tps1∆) W303-1A tps1�::TRP1 Linhagem com deleção no gene da trealase neutra
W303-648 (nth1∆) W303-1A nth1�::URA3 Linhagem com deleção no gene da trealose-6-fosfato sintase
Fonte: Stefan Hohmann
5.2 - Meios de cultura
O crescimento das células foi realizado nos seguintes meios: YPGlicose (2%
glicose, 2% peptona e 1% extrato de levedura) para as linhagens WT e nth1∆, e
YPGalactose (2% galactose, 2% peptona, 1% extrato) para a linhagem tps1. A linhagem
mutante tps1 não pode crescer na presença de glicose por isso, para o crescimento desta
linhagem, foi usada a galactose como fonte de carbono.
5.3 - Manutenção das linhagens em laboratório
A manutenção das linhagens foi realizada utilizando três técnicas diferentes para
armazenagem de leveduras em laboratório:
• Armazenamento das células em óleo mineral na geladeira:
Uma alça contendo células foi inoculada em tubos com meio YPGlicose ou, YPGalactose
sólido, previamente inclinado. Após 48 horas a 30ºC, acrescenta-se à superfície das
leveduras, óleo mineral devidamente esterilizado. Os tubos foram estocados a 4ºC.

Materiais e Métodos
39
• Manutenção em placa de petri:
As leveduras foram mantidas em placa de petri contendo meio sólido YPGlicose ou,
YPGalactose e, repicadas periodicamente, a partir do original mantido em tubo inclinado
com óleo mineral. As placas também foram mantidas a 4ºC.
• Manutenção em meio glicerol:
As linhagens foram pré-crescidas em meio YPGlicose ou, YPGalactose sob agitação a
30ºC, coletadas em fase estacionária, por centrifugação e, ressuspensas em meio contendo
1% extrato de leveduras, 2% peptona, 25% glicerol. 1000 µL deste meio contendo as
células foi transferido para tubos criogênicos, e estocados em freezer a –70ºC.
5.4 – Estresse com cloreto de cádmio
As células foram pré incubadas a uma temperatura de 30ºC, agitadas a 160 rpm
(rotação por minuto) por um período de aproximadamente 16 horas, em meio líquido
YPGlicose ou YPGalactose, dependendo da linhagem utilizada. Após este período, o meio
foi centrifugado por 5 minutos em 3000 rpm, o sobrenadante descartado e, as células eram
transferidas para um novo meio líquido YPG, por um período de 4 horas, para que assim as
células entrassem em fase exponencial de crescimento. A fase exponencial do crescimento
foi verificada pela presença de glicose no meio, utilizando fitas indicadoras de glicose na
urina (UROFITA G – Biobrás Diagnóstico). De acordo com a necessidade experimental, as
células foram incubadas em meio YPG acrescido de concentrações pré-estabelecidas de
cloreto de cádmio (50ppm e 800ppm) e também na condição controle (ausência de cádmio).
O cloreto de cádmio foi autoclavado isoladamente, em solução aquosa concentrada e,
adicionado ao meio YPG. Posteriormente, as células foram incubadas em meio YPG a
30ºC, sob agitação a 160 rpm, durante períodos de tempo preestabelecidos. O tempo de
incubação e a concentração de cloreto de cádmio foram variáveis, de acordo com a
necessidade de cada experimento. Após o tempo estabelecido, as células foram coletadas
por filtração a vácuo usando filtros de nitrocelulose de 0,45 µm de porosidade, 47 mm de
diâmetro. As células foram lavadas três vezes com água destilada fria. A massa celular foi
removida do filtro com espátula e, transferida para papel alumínio, congeladas em

Materiais e Métodos
40
nitrogênio líquido e, estocadas em freezer – 20ºC. Posteriormente, esta massa de células
congelada foi utilizada para as determinações experimentais desejadas.
5.5 – Obtenção de extratos celulares
Após a realização da incubação descrita no item 5.4, onde as células foram
coletadas em fase logarítmica e, incubadas em meio YPGlicose novo, na ausência
(controle) e presença de 50 e 800 ppm de cloreto de cádmio, as células foram ressuspensas
em tampão de lise (Tampão de Fosfato de Sódio 50mM, pH 7,0; EDTA 1 mM e PMSF 1
mM), adicionou-se, a seguir, pérolas de vidro. O rompimento das células foi realizado por
seis ciclos de 30 segundos de agitação vigorosa em vórtex, intercalados com períodos de
repouso em gelo. Após a lise celular, a suspensão foi centrifugada a 3000 rpm por 5
minutos. Utilizou-se este sobrenadante (extrato bruto) para a realização das dosagens
necessárias.
5.6 - Dosagem de proteínas
O conteúdo protéico dos extratos brutos de cada amostra foi determinado de acordo
com o método descrito por Lowry e colaboradores (1951), utilizando uma solução de
soroalbumina bovina (BSA) como padrão.
5.7 - Curva de crescimento
Para curva de crescimento das linhagens WT e nth1, um pré inoculo foi crescido por
cerca de 16 horas a 30ºC, a 160 rpm, em meio YPGlicose. Para a linhagem tps1, um pré
inoculo foi crescido nas mesma condições acima citadas em meio YPGalactose. Após este
período, foi verificada a densidade óptica da cultura em espectrofotômetro, a 600 nm. Os
experimentos foram iniciados com uma D.O. de 0,15. Nos tempos pré estabelecidos, uma
alíquota, usualmente 100µl, foi retirada e, colocada em 900µl de água. O crescimento foi

Materiais e Métodos
41
seguido pela medida da turbidez da suspensão a 600 nm, em espectrofotômetro. Caso
necessário, foram feitas diluições em água para registro da turbidez.
5.8 - Tolerância ao cádmio
A análise de sobrevivência das células ao cloreto de cádmio foi feita por
plaqueamento em meio YPGalactose acrescido de ágar. Foram preparados pré inóculos das
linhagens S. cerevisiae WT, S. cerevisiae nth1 e S. cerevisiae tps1 em meio YPGalactose e,
após 48 horas de crescimento foi feita contagem do número de células, com uso da câmara
de Neubeuer (Figura 13). Foram coletados 100µL de cada pré-inóculo e, realizadas
diluições sucessivas em água destilada estéril, obtendo assim as concentrações de 10-3, 10-4,
10-5, 10-6, 10-7, 10-8 células/ml. Em seguida, foram inoculados 5µL de cada suspensão em
placas contendo YPGalactose + ágar com concentrações de cádmio de 5, 10 e 25ppm e, na
ausência do metal, como controle. Após 72 horas de incubação a 300C as placas foram
fotografadas.
FIGURA 13 – Câmara de Neubeuer (Fonte: MANNHEIM, 1998)

Materiais e Métodos
42
5.9 - Dosagem de Trealose
A quantificação de trealose acumulada pelas células foi realizada utilizando a
metodologia descrita por NEVES e colaboradores (1994).
As células foram coletadas em fase logarítmica e, incubadas em meio YPGlicose
novo, na ausência (controle) e presença de 50 e 800 ppm de cloreto de cádmio.
Posteriormente, a massa celular congelada foi transferida para tubos cônicos, pesados e
mantidos em gelo. Em seguida, acrescentou-se 1000µL de 0,25 M Na2CO3. Imediatamente,
a suspensão foi fervida durante 20 minutos, centrifugada 5 minutos a 3000 rpm. Após esta
etapa, 200µL do sobrenadante foi acidificado com ácido acético 1,2 N (Concentração final
de 0,30 N) até que o pH 5,0 – 5,5 fosse atingido (verificado com fita indicadora de pH) e,
tamponado com 75 mM de acetato de sódio contendo 7,5 mM de CaCl2. A solução foi
misturada em vórtex e, 100µL da mistura foi usada para determinação da trealose. A
trealose foi determinada enzimaticamente com uma preparação de trealase extraída do
fungo Humicola grisea var. thermoidea. A reação foi processada incubando o extrato
tamponado e a preparação de enzima por 2 horas a 40ºC. Após este período, usou-se 50µL
desta solução para a dosagem de glicose formada. A glicose formada foi determinada com
auxílio de um kit comercial da Bioclin (Glicose GOD- Clin). A reação desenvolveu-se a
37ºC, durante 15 minutos. O produto colorido formado foi determinado
espectofometricamente a 505 nm. Para cada amostra foram realizadas incubações paralelas,
na presença de trealase desnaturada, para verificação de possível contaminação por glicose
do meio.
Uma unidade de trealase foi definida como a quantidade de enzima que liberou um
µmole de glicose por minuto nessas condições. O acúmulo de trealose foi expresso em
µmoles de glicose liberada por grama de peso úmido (µmoles glicose liberada / g peso
úmido).
5.10 - Extração da trealase do Humicola grisea var. thermoidea

Materiais e Métodos
43
A trealase foi extraída do fungo Humicola grisea var. thermoidea, cultivado em
ágar aveia (4% farinha de aveia, 1,8% ágar), durante 20 dias a 40ºC. Após esse período, os
frascos foram deixados durante 5 dias a temperatura de 4ºC, procedimento que aumenta a
quantidade de trealase a ser extraída. Adicionou-se aos erlemeyers, 10 ml de água destilada
e, raspou-se a superfície do ágar com bastão de vidro. A suspensão formada foi filtrada em
funil contendo gaze em seu interior. Posteriormente, a suspensão foi centrifugada a 3000
rpm por 40 minutos. Adicionou-se ao sobrenadanete 1% de Kaolin (silicato de alumínio
hidratado/Sigma) e a mistura foi deixada a temperatura ambiente durante 20 minutos com
agitação esporádica. A solução foi centrifugada por 20 minutos a 3000 rpm e, o
sobrenadante foi dialisado toda noite contra H2O à 4ºC. A suspensão foi distribuída em
tubos e, congelada. Para cada lote de enzima preparada, foram realizados testes de
atividade, utilizando como padrões a trealose comercial (Sigma) nas concentrações de 2, 5
e 10 mM.
5.11 - Determinação de cádmio incorporado pelas células
Foi utilizada a análise por ativação neutrônica para determinação do cádmio
incorporado pelas células. A análise por ativação neutrônica tem sido reconhecida como
uma das mais importantes ferramentas analíticas na determinação da composição química
elementar em teores traços. A técnica tem como princípio à indução de radioatividade
artificial, em uma amostra, através da irradiação com nêutrons e, posterior medida da
atividade induzida mediante a detecção da radiação gama. Os fenômenos físicos, na qual
está fundamentada esta análise são, as propriedades do núcleo, radioatividade e a interação
da radiação com a matéria, via reação de neutrons-gama (n, γ). Os raios gama emitidos,
denominados de raios gama de decaimento, possuem energias que são características de
cada radionuclídeo. Assim, quando detectados por espectroscopia gama, podem ser
utilizados para identificar e, quantificar os elementos químicos presentes em uma amostra.
A técnica tem capacidade de análise de matrizes no estado sólido, líquido e gasoso:
a reação nuclear (n, γ) independe do estado físico da matriz. O método é não destrutivo, a
amostra não é visivelmente ou, quimicamente alterada.

Materiais e Métodos
44
A ativação neutrônica é realizada em reatores de pesquisa. No Brasil, apenas dois
Centros de Pesquisa utilizam a ativação neutrônica. O Centro de Desenvolvimento de
Tecnologia Nuclear (CDTN) em Belo Horizonte e, o Instituto de Pesquisas Energéticas e
Nucleares (IPEN), em São Paulo, ambos pertencentes à Comissão Nacional de Energia
Nuclear (CNEN).
O Centro de Desenvolvimento da Tecnologia Nuclear possui o reator TRIGA IPR-
1, dotado de mesa giratória e dispositivo de irradiação especialmente apropriado para esta
técnica analítica. Durante a irradiação, a mesa gira em torno do núcleo do reator a uma
velocidade constante, de modo a garantir um fluxo de nêutrons uniforme nas amostras.
Atualmente, a potência do reator é de 110 kW, nesta potência o fluxo de nêutrons atinge um
valor em torno 1,65 x 1012n/cm2 s. a meia-vida do 115Cd é de 12,2 dias (MENEZES et al.,
2003)
5.12 - Preparação das amostras para determinação do cádmio incorporado
Após realização do item 5.4, a massa celular foi coletada e transferida para tubos
apropriados para a Ativação Neutrônica. Os tubos permaneceram por três dias a 70ºC. Após
este período, toda água presente no material evaporou-se e assim foram pesadas para
determinação do peso seco, e enviadas ao Reator Triga (CDTN/CNEN) para irradiação. A
irradiação foi realizada durante 8 horas, nas condições do reator acima especificadas.
Posteriormente, as amostras foram submetidas à espectrometria gama e, os picos
característicos do cádmio foram determinados e, usados para a determinação da quantidade
do metal presente na célula. Os valores são expressos em µgrama de Cd incorporado/grama
de peso seco.
5.13 - Incorporação de CdCl2 na presença de Glutationa reduzida adicionada
ao meio
Para determinar a influência da glutationa na incorporação do cádmio nas linhagens
estudadas, foram realizados dois experimentos independentes: primeiramente, foi

Materiais e Métodos
45
observado se a adição de glutationa reduzida (concentração final de 80µM) influenciaria a
incorporação de cádmio. O segundo experimento foi realizado da mesma forma, apenas
que, no lugar da glutationa, foi adicionado aminoácidos (10Mm ácido glutâmico, 18mM
glicina e 3,35mM cisteína). Estes aminoácidos (aa) são os formadores da glutationa.
O experimento foi realizado na situação controle (ausência de cádmio) e exposição
ao cádmio (50ppm e 800ppm). As células primeiramente cresceram em meio
YPGlicose/YPGalactose suplementado com GSH e com aminoácidos, por cinco horas.
Após este tempo, foi ressuspendido em novo meio e realizado o estresse com cádmio
(procedimento descrito no item 5.4). As amostras foram preparadas de acordo com o item
5.12.
5.14 - Determinação de cádmio na superfície celular
A presença de metal na superfície celular foi determinada utilizando a técnica de
espectrometria por dispersão de energia (do inglês, Energy Dispersive Spectrometer –
EDS), o equipamento consiste de uma detetor, uma fonte polarizadora de alta-voltagem
para o detetor, um amplificador, um analisador multicanal, um reservatório de nitrogênio e
uma fonte de força. Este sistema proporciona análises rápidas, precisas e não destrutivas de
elementos químicos presentes nas amostras que estão sendo analisadas.
Após a realização do procedimento descrito no item 5.4, as células foram incubadas
por três dias a 70ºC para obtenção do peso seco e, enviadas ao Laboratório de Microanálise
do Instituto de Ciências Exatas (ICEX/UFMG), para determinação do cádmio presente na
superfície celular.
5.15 - Determinação da oxidação intracelular
A sonda fluorescente 2,7-diclorofluoresceína (Figura 14) é uma molécula que
penetra na membrana celular por difusão passiva. Em seu interior, ela sofre sucessivos
ataques pelos radicais livres, produzindo assim um composto fluorescente. Os níveis de
fluorescência podem ser monitorados e relacionados com os níveis de oxidação intracelular
(GOMES, et al., 2002).

Materiais e Métodos
46
A fluorescência foi determinada utilizando espectrofluorímetro, calibrado com os
comprimentos de onda para excitação de 504 nm e, comprimento de onda para emissão de
524 nm. As células foram pré incubadas durante 48 horas na temperatura de 30ºC, agitadas
a 160 rpm em meio líquido YPGlicose/ YPGalactose. As células foram transferidas para
um novo meio YPGlicose ou YPGalactose e, após 4 horas nestas condições, adicionou-se a
sonda fluorescente ao meio (concentração final 10µM de solução preparada em etanol
absoluto). As amostras foram incubadas com a sonda fluorescente durante 20 minutos para
permitir sua absorção por difusão passiva. Em seguida, adicionou-se cádmio e, a incubação
prosseguiu por 16 horas, tanto na situação controle, como nas que tiveram adição do metal
(50ppm e 800ppm).
Após as 16 horas de incubação, as células foram centrifugadas, lavadas três vezes
com água estéril gelada e, ressuspensas em 1000µL de água estéril gelada. Foram então
lisadas com pérolas de vidro (10 ciclos de 30 segundos em vórtex seguidos por
resfriamento em gelo). Após o rompimento das células, a amostra foi centrifugada (13000
rpm x 5 min) e transferido para tubo eppendorf. O pellet contendo as pérolas de vidro foi
lavado em 400µL de água estéril gelada e esta suspensão foi adicionada ao sobrenadante
reservado e usado para determinar a fluorescência após diluição em água. Os resultados
foram expressos como uma relação entre a fluorescência de células tratadas com cádmio
(50ppm e 800 ppm) e, a fluorescência das células não submetidas a situação de estresse
(controle).
FIGURA 14 – Sonda fluorescente 2,7-diclorofluoresceína
5.16 - Carbonilação de Proteínas

Materiais e Métodos
47
A determinação da carbonilação de proteína foi realizada utilizando o método
descrito em Luschchak e colaboradores (2005). A carbonilação das proteínas foi
determinada utilizando dinitrofenilhidrazina (DNPH). A dinitrofenilhidrazina reage com os
grupos carbonil das proteínas para formar dinitrofenilhidrazonas (DNP). Este composto
formado possui cor característica que pode ser determinada espectrofotometricamente a 370
nm.
Os extratos foram preparados de acordo com o item 5.5. Após centrifugação (13000
rpm por 5 minutos) utilizou-se o pellet para análise de carbonilação de proteínas. O pellet
foi ressuspenso em solução 10mM de DNPH preparada em 2M HCl e deixado em
temperatura ambiente durante 1 hora. Logo após foi adicionado TCA (concentração final de
10%) e deixado em gelo durante 15 minutos. Após este período, a suspensão foi
centrifugada à 10.000 rpm por 10 minutos, o sobrenadante foi descartado e, adicionado ao
pellet Etanol/Butila Acetato (1:1) esse procedimento foi repetido três vezes, com
centrifugações nas mesmas condições acima descritas. O pellet foi então ressuspenso em
6M Guanidina-HCl e centrifugado (3 minutos a 10.000 rpm) para remoção das partículas
insolúveis. O sobrenadante foi usado para determinação da dinitrofenilhidrazona formada,
usando um comprimento de onde de 370 nm. Os cálculos de carbonilação de proteínas
foram realizados usando um coeficiente de extinção molar de 22 x 103 M-1 cm-1 para o
DNPH (LUSCHCHAK, et al., 2005) e os resultados foram expressos como nanomoles/mg
proteína/ mL.
5.17 - Peroxidação de lipídeos
A determinação da peroxidação lipídica foi feita pelo método TBARS (do inglês
Thiobarbituric Acid Reactive Species) ou, substâncias reativas com o ácido tiobarbiturico.
Os níveis de peroxidação lipídica foram determinados de acordo com as substâncias que
reagiram com o ácido tiobarbitúrico. As substâncias reativas ao ácido tiobarbitúrico
(TBARS) são compostas de diversos produtos finais de baixo peso molecular formados via
decomposição de certos produtos primários e secundários da peroxidação lipídica. A
análise da quantidade de TBARS é utilizada como um índice de estresse oxidativo. O

Materiais e Métodos
48
cálculo foi realizado usando-se uma curva padrão de malondialdeído (MDA) um dos
produtos da peroxidação lipídica.
Os extratos celulares foram preparados de acordo com item 5.5. Ao sobrenadante
previamente separado adicionou-se ácido tricloracético (TCA concentração final de 10%) e,
a suspensão foi deixada em gelo durante 30 minutos. Após este período, foi centrifugada
(3000 rpm por 5 minutos) e, retirou-se 600µL do sobrenadante ao qual se adicionou a
solução de TBA (0,5% de ácido tiobarbitúrico preparado em TCA 20%), aqueceu-se por 60
minutos a 100ºC. Em seguida, a amostra foi deixada a temperatura ambiente para
resfriamento. Após esta etapa, a absorbância foi determinada espectofotometricamente a
532nm As medidas foram corrigidas para turbidez inespecífica pela subtração da
absorbância a 600nm. O índice de peroxidação lipídica foi expresso em picomoles de
malondialdeído (produto da reação) por miligrama de proteína por mL (picomoles MDA/
mg proteína/ mL).
5.18 - Resíduos Sulfidrílicos Totais
Os resíduos sulfidrílicos totais foram determinados pelos métodos de ELLMAN
(1959) e DAVIDSON e colaboradores (2001), usando 5-5´dithio-bis 2-ácido-nitrobenzoico
- DTNB. Na presença de grupamento tiol a solução de DTNB forma 5-Mercapto-2-
Nitrobenzoico, produzindo uma coloração amarela característica que pode ser determinada
espectrofotometricamente a 412 nm. (Figura 15).
Os extratos celulares foram preparados de acordo com o item 5.5. Utiliza-se 0,5 mg
de proteína diluída em tampão 0,1M de fosfato de sódio com pH 7,0. Retirou-se 200µL de
cada preparação e, adicionou-se tampão 0,01M fosfato de sódio, p.H. 8,0 e 7µL de reagente
Ellman (0,01M DTNB em 10mL de tampão 0,1M de fosfato de sódio pH 7,0). A
absorbância foi determinada a 412nm após 20 minutos de incubação à temperatura
ambiente. O coeficiente de extinção molar para o DTNB é 13.600 M-1 cm-1, e foi usado
para determinar a concentração de resíduos sulfidrílicos totais.
Os resultados foram expressos em micromoles de resíduos sulfidrílicos por
miligrama de proteína por mL (µmoles/ mg proteína/ mL)
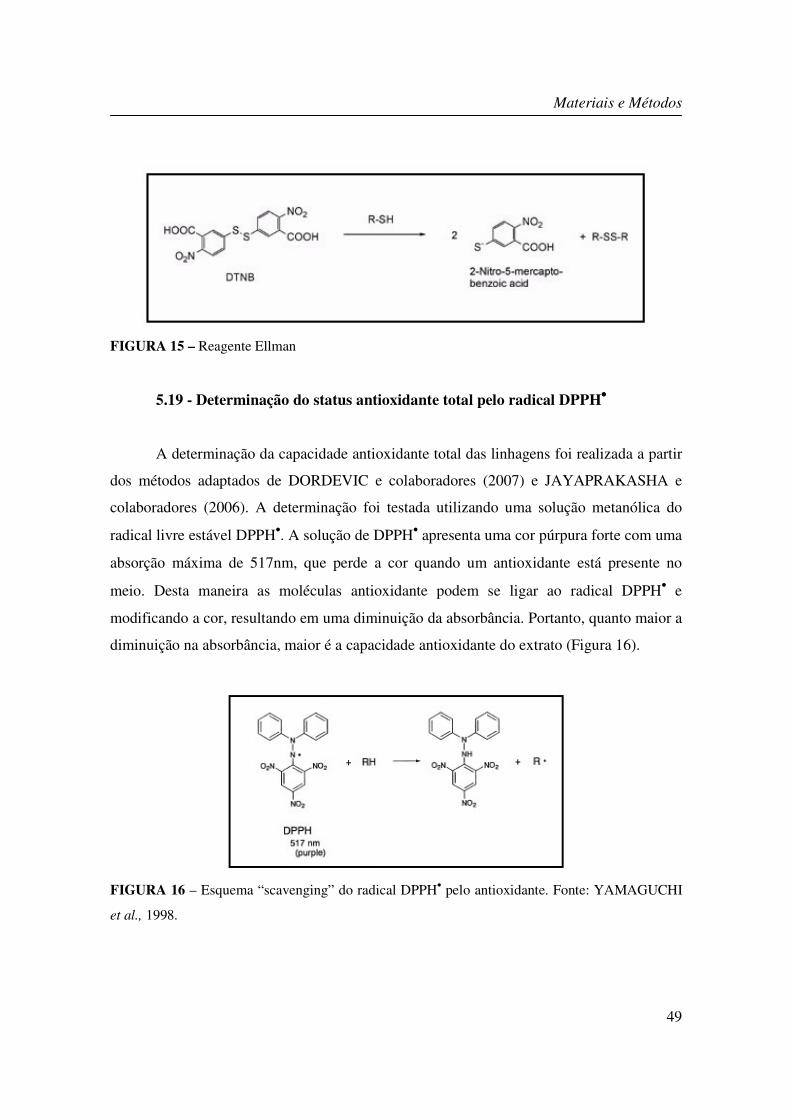
Materiais e Métodos
49
FIGURA 15 – Reagente Ellman
5.19 - Determinação do status antioxidante total pelo radical DPPH••••
A determinação da capacidade antioxidante total das linhagens foi realizada a partir
dos métodos adaptados de DORDEVIC e colaboradores (2007) e JAYAPRAKASHA e
colaboradores (2006). A determinação foi testada utilizando uma solução metanólica do
radical livre estável DPPH•. A solução de DPPH• apresenta uma cor púrpura forte com uma
absorção máxima de 517nm, que perde a cor quando um antioxidante está presente no
meio. Desta maneira as moléculas antioxidante podem se ligar ao radical DPPH• e
modificando a cor, resultando em uma diminuição da absorbância. Portanto, quanto maior a
diminuição na absorbância, maior é a capacidade antioxidante do extrato (Figura 16).
FIGURA 16 – Esquema “scavenging” do radical DPPH• pelo antioxidante. Fonte: YAMAGUCHI
et al., 1998.

Materiais e Métodos
50
Os extratos celulares foram preparados de acordo com o item 5.5. Utilizou-se 2 mg
de proteína diluída em metanol absoluto e 0,1mM de DPPH• dissolvido em solução
metanólica preparada na hora. A preparação foi agitada vigorosamente e, mantida no escuro
por 30 minutos a temperatura ambiente. A leitura foi feita usando 1000µL do sobrenadante.
A absorbância controle foi determinada usando DPPH• 0,1 mM em solução metanólica
diluído em 4000µL de metanol.
A inibição do radical DPPH foi calculada utilizando a equação:
controleaAbsorbânci
padrãoaAbsorbâncicontroleaAbsorbânciInibição
)(100
−×=
5.20 – Descarte de Rejeitos
Todos meios de cultivo foram estocados no laboratório, em bombonas plásticas
colocadas dentro de caixas plásticas, com etiquetas identificando rejeito químico contendo
cádmio e, quando repletas, foram enviadas ao Serviço de Gerência de Rejeitos do CDTN
que o manterá sob sua guarda até a destinação final do rejeito.
5.21 - Análise Estatística
Os resultados foram expressos como média e ± desvio padrão de pelo menos três
experimentos diferentes. A diferença estatística dos resultados foi realizada utilizando o test
T (T Student) com níveis de significância de < 0,05.

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a
este estresse em Saccharomyces cerevisiae
51
6 - Resultados e Discussão

Resultados e Discussão
52
6.1 – Curva de crescimento e tolerância ao cádmio
A presença de metais no interior das células, sua incorporação e seu acúmulo, são
fenômenos submetidos à intensa regulação, mesmo para os metais essenciais como Zinco e
Ferro. Em concentrações que ultrapassem aqueles limites considerados normais, os metais
essenciais são nocivos para as células, que precisam se adaptar à sua presença, ativando
mecanismos de defesa para se tornarem mais resistentes e viáveis. Se tal fato ocorre para
metais essenciais, para os metais não essenciais como cádmio por exemplo, esta regulação
também deve ser altamente controlada, assim metais não essenciais são considerados
tóxicos para as células, mesmo em baixas concentrações.
A análise do crescimento das células das diferentes linhagens na presença de
concentrações crescentes de cloreto de cádmio (ppm) foi comparada com a situação
controle (ausência de cloreto de cádmio) em cada linhagem. De acordo com os resultados
apresentados na Figura 17, as linhagens WT, tps1 e nth1, na situação controle, apresentam
perfil de crescimento esperado, ou seja, aumento do número de células à medida que o
tempo progrida enquanto que todas as linhagens suplementadas com aumentos gradativos
da concentração de cádmio (10, 25, 50, 75 e 100ppm), apresentaram um crescimento
reduzido ou inibição do crescimento, quando comparadas à situação controle no período de
tempo estudado (8 horas).
Após a realização das curvas de crescimento em meio líquido foi analisada a
tolerância das linhagens WT, tps1 e nth1 ao CdCl2 através do plaqueamento das células em
meio sólido (YPGalactose) suplementado com 0, 5, 10 e 25 ppm de cloreto de cádmio e a
posterior observação das colônias formadas.
Analisando o crescimento das colônias em placas com concentrações crescentes de
cloreto de cádmio (Figura 18), observa-se que as células das três linhagens apresentam o
mesmo perfil de crescimento. Na situação controle e nas concentrações de 5ppm e 10ppm,
não foi observado qualquer influência no crescimento, enquanto na concentração de 25ppm
a célula apresentou uma inibição do crescimento.
Gharieb (2001) trabalhando com o fungo Fusarium sp mostrou que a inibição do
crescimento aumenta com a elevação da concentração de cádmio no meio (0, 0, 25, 0,5 e
1,0 mmol l-1) (0,005 – 0,020ppm). Todas as concentrações testadas no presente trabalho

Resultados e Discussão
53
levaram ao bloqueio do crescimento, durante o período estudado, entretanto, as
concentrações usadas neste trabalho foram superiores aos citados. De acordo com Momose
e colaboradores (2001), o efeito inibitório do cádmio sobre o crescimento celular, se deve a
vários fatores, todos eles ligados ao bloqueio de grupos funcionais de moléculas
importantes tais como: enzimas, polinucleotídeos, sistema de transporte de íons ou
nutrientes e, substituição de íons essenciais em sítios ativos, os autores observaram em
estudos com S. cerevisiae submetido a 0,3 mM de cloreto de cádmio (0,06ppm) durante 2
horas, uma diminuição na taxa de crescimento. Romandini e colaboradores (1992) usando
três linhagens de S. cerevisiae observaram queda no crescimento das células em meio
líquido na presença de 40µM de CdSO4 (0,01ppm), mostrando que mesmo em baixas
concentrações o cádmio pode causar inibição do crescimento celular.
No presente trabalho foi observado que o cádmio pode prejudicar o crescimento das
células de S. cerevisiae em meio líquido, a sua tolerância ao metal em meio sólido e, que o
metal pode diminuir a sobrevivência celular. Interessou-nos também saber se as células de
S. cerevisiae W303-WT (linhagem laboratorial) tinham capacidade de incorporar cádmio e
qual seria a influência das fases de crescimento na incorporação do cádmio pelas diferentes
linhagens. É conhecido que os níveis de trealose intracelulares sofrem forte influência da
fase de crescimento, são baixos na fase logarítmica e altos na fase estacionária
(THEVELEIN, 1984). Optamos por usar as concentrações de 50ppm e 800ppm de cádmio
como forma de submeter a levedura a um estresse sob concentração moderada de cádmio
(50 ppm) e estresse em alta concentração de cádmio (800 ppm). Utilizando duas
concentrações de cádmio (50 e 800 ppm) nosso objetivo foi detectar e acompanhar
mudanças bioquímicas e fisiológicas importantes que pudessem ocorrer dependendo da
concentração usada e da linhagem estudada.

Resultados e Discussão
54
A
0
0,5
1
1,5
2
2,5
0 2 4 6 8
Tempo (horas)
D.O
. 600
nm
Controle 10ppm 25ppm
50ppm 75ppm 100ppm
0
0,5
1
1,5
2
2,5
0 2 4 6 8Tempo (horas)
D.O
. 600
nm
Controle 10ppm 25ppm
50ppm 75ppm 100ppm
0
0,5
1
1,5
2
2,5
0 2 4 6 8Tempo (horas)
D.O
. 600
nm
Controle 10ppm 25ppm
50ppm 75ppm 100ppm
FIGURA 17 – Influência de diferentes concentrações de cloreto de cádmio no crescimento das
linhagens WT (A), tps1 (B), e nth1 (C).
B
C

Resultados e Discussão
55
FIGURA 18 – Determinação da sobrevivência das células em meio sólido em ausência de cádmio
(A) e presença de 5 ppm (B) e 10 ppm (C) e 25ppm (D) de cloreto de cádmio.

Resultados e Discussão
56
6.2 - Determinação da incorporação de cádmio nas diferentes fases de
crescimento celular
As células de leveduras possuem duas fases de crescimento: logarítmica (ou
exponencial) e estacionária quando crescem em glicose, como fonte de carbono. Na
primeira fase, a fase exponencial do crescimento, as células usam glicose para produção de
energia através da via glicolítica. Na fase diauxica (ou pré-estacionária) há o início da
carência de alguns nutrientes e a célula se prepara para respirar. Nesta etapa, as enzimas da
cadeia respiratória encontram-se sintetizadas, a taxa de crescimento diminui e a célula
passa a utilizar o etanol ou outra fonte de carbono disponível para obtenção de energia.
Durante esta fase, glicogênio e trealose são sintetizados e estocados, assim as células se
tornam muito mais resistentes a qualquer forma de estresse. Na total carência de nutrientes,
as células de leveduras entram na verdadeira fase estacionária. Não há mais divisão celular
e a taxa metabólica diminui. As células podem sobreviver nesta situação de semanas a
meses (LONGO et al., 1996).
Inicialmente, foi determinada a influência da fase de crescimento na incorporação
de cádmio. Para tanto realizamos o experimento onde as células da linhagem WT, tps1 e
nth1 foram coletadas em fase logarítmica e estacionária, incubadas com diferentes
concentrações de cádmio (50 e 800ppm). Em todas as linhagens estudadas observa-se que
com o aumento da concentração de cádmio (50ppm para 800ppm), ocorre um aumento da
incorporação, tanto na fase logarítmica quando na fase estacionária.
Na linhagem WT observa-se que não há diferença na incorporação de cádmio na
concentração de 50ppm entre as fases logarítmica e estacionária. Porém, quando se
compara a incorporação na concentração de 800ppm tanto no tempo de duas horas quanto
no tempo de 4 horas, observa-se que na fase logarítmica a incorporação é 51% e 70% maior
do que na fase estacionária. O mesmo ocorre na linhagem com excesso de trealose (nth1),
onde se observa que apenas na concentração de 800ppm ocorre um aumento significativo
de 27% e 20% (duas e 4 horas, respectivamente) de incorporação do metal entre a fase
logarítmica e estacionária (Figura 19).
A linhagem tps1 não apresentou diferença significativa de incorporação de cádmio
entre as fases de crescimento, em nenhuma das concentrações estudadas (50 e 800ppm).

Resultados e Discussão
57
Em comparação com as outras duas linhagens, incorpora menos cádmio, conforme pode ser
observado nos resultados apresentados na figura 19.
De maneira geral, as células coletadas em fase logarítmica incorporam maior
quantidade de cádmio do que células coletadas em fase estacionária, este resultado não foi
observado na linhagem tps1. Observa-se pela análise da figura 19, que existe uma
correlação entre o aumento da quantidade de cádmio incorporado pelas células e o aumento
da concentração de cádmio na qual as células foram expostas. Este resultado indicaria que
em efluentes muito diluídos, a incorporação do metal pela levedura seria menor. Outro
resultado que se deve ressaltar é que o mutante tps1 incorpora pequena quantidade de
cádmio quando comparado às outras duas linhagens. Nossa hipótese para esta diferença de
incorporação entre as linhagens WT, tps1 e nth1 é que a trealose exerceria alguma função
na captação e/ou transporte do cádmio para o interior da célula. É bem conhecido que
mutantes com deleção do gene TPS1 apresentam diversas alterações fisiológicas e
bioquímicas entre elas, uma série de alterações na regulação do transporte da glicose e na
metabolização desta molécula (VAN AELST et al., 1993). Assim, não é de todo
improvável que a ausência de trealose tenha reflexos na captação e/ou transporte do
cádmio. Entretanto, para confirmação desta suposição, outros experimentos deverão ser
realizados.
Os resultados apresentados na figura 19 indicaram que a incorporação de metais
depende do microrganismo utilizado (ou seja, da linhagem utilizada), e da fase do
crescimento celular em que as células se apresentam. Condições como fase de coleta das
células (logarítmica ou, estacionária), e tempo de exposição ao metal influenciam na
capacidade das células em incorporar os metais. Assim, em todos os experimentos
subsequentes foram utilizadas células coletadas na fase logarítmica do crescimento, uma
vez que, estas apresentaram uma maior capacidade de incorporação de cádmio do meio
extracelular.

Resultados e Discussão
58
0
5000
10000
15000
20000
50ppm (2hrs) 50ppm (4hrs) 800ppm (2 hrs) 800ppm (4hrs)
µµ µµg
Cd
inco
rpor
ado/
g pe
so s
eco
WT tps1 nth1
0
5000
10000
15000
20000
50ppm (2hrs) 50ppm (4hrs) 800ppm (2 hrs) 800ppm (4hrs)
g C
d in
corp
orad
o/ g
pes
o se
co
WT tps1 nth1
FIGURA 19 – Determinação do cádmio incorporado pelas células das linhagens WT, tps1 e nth1
coletadas na fase LOGARÍTMICA (A) de crescimento celular, e na fase ESTACIONÁRIA (B).
A
B

Resultados e Discussão
59
6.3 - Determinação de cádmio incorporado pelas células viáveis e mortas
De acordo com os dados apresentados na figura 20, usando-se a concentração de
50ppm não foram observadas diferença na incorporação de cádmio. Na concentração de
800ppm observou-se que existe uma maior incorporação de cádmio utilizando células
vivas. Houve um aumento de 45%, 17% e 52% na incorporação de cádmio usando células
vivas das linhagens WT, tps1 e nth1 respectivamente, quando comparado com a
incorporação pelas células mortas por autoclavação. Muito provavelmente, a diminuição na
incorporação do cádmio em células mortas ocorre devido aos seguintes fatores: em células
mortas a incorporação ocorre somente na superfície celular, devido a dificuldade de
penetração do metal, causada pelos danos aos complexos celulares responsáveis pelo
transporte; os sítios de adsorção podem ter sido desnaturados devido ao choque da alta
temperatura durante o processo de autoclavação.
Em estudos realizados por Suh e colaboradores (1998), células de S. cerevisiae
vivas incorporam 10 vezes mais Pb2+, do que células mortas por autoclavação. Utilizando a
técnica de microscopia eletrônica os autores observaram que células vivas incorporam Pb2+
tanto na superfície celular quanto nos componentes intracelulares. Enquanto que, em
células mortas observa-se a presença do metal apenas na membrana da célula. Desta forma
os autores concluíram que o acúmulo de Pb2+ em células mortas de S. cerevisiae é menor
devido à diminuição do número de sítios ligantes do metal, causados pela autoclavação.
Adamis e colaboradores (2003), trabalhando com a levedura S. cerevisiae,
observaram que células mortas não incorporam cádmio. Esses resultados são contrários aos
encontrados neste trabalho. De fato, foi observado que as células mortas têm uma menor
incorporação de cádmio quando comparadas as células viáveis, porém o fato das células
estarem mortas não implica na incapacidade de incorporação do cádmio (Figura 20).
A figura 19, mostra que havia uma dependência do tempo de incubação das células
com o metal. Para certificar que o fato ocorria, aumentou-se o tempo de exposição das
células ao metal para 8 e 16 horas. Os resultados, apresentados na figura 21, mostraram que
o aumento do tempo de exposição ao metal aumenta a quantidade de cádmio incorporado.
Após a verificação que células viáveis apresentam uma maior incorporação de
cádmio que células mortas por autoclavação, todos os experimentos subseqüentes foram

Resultados e Discussão
60
realizados com células viáveis. O protocolo usado na obtenção das células se manteve
constante: células viáveis coletadas em fase logarítmica, expostas por 16 horas ao cádmio,
adicionado no meio extracelular. A opção por 16 horas se deveu ao fato que, quanto maior
o tempo de exposição ao metal, maior é a incorporação de cádmio pelas células e o
protocolo de estresse com cloreto de cádmio neste tempo era exeqüível, permitindo que as
células fossem obtidas após entrarem em fase logarítmica do crescimento.
De acordo com os resultados apresentados na tabela 02, a incorporação de Cd
durante 16 horas por células viáveis, incubadas na concentração de 50ppm, não apresenta
diferença significativa entre as três linhagens estudadas. Na concentração de 800 ppm
observou-se que a célula com excesso de trealose (nth1) não apresentou diferença na
incorporação de cádmio, quanto comparada com a linhagem WT. Porém, observa-se que
quando não existe a presença do dissacarídeo (linhagem tps1), as células têm uma queda na
incorporação de cádmio de 59%, comparada à incorporação obtida nas células da linhagem
WT na presença de 800 ppm. Este resultado mostra que a falta de trealose tem uma
importante participação na incorporação do metal sob altas concentrações.
Martínez-Esparza e colaboradores (2007), usando mutante que apresenta deleção do
gene que codifica a enzima trealose-6-fosfato sintase (tps1/tps1) na levedura Candida
albicans, observaram a existência de modificações estruturais e/ou químicas na parede
celular destas células. Da mesma forma, é possível considerar que tais alterações também
ocorram na linhagem tps1 de S. cerevisiae, de modo a dificultar a incorporação de cádmio,
ou ainda que exista uma diminuição do transporte deste metal para o interior das células
(hipótese já considerada anteriormente). Situações estas que poderiam justificar a
diminuição da incorporação do cádmio pela mutante tps1. Além do mais, a presença de um
excesso de trealose intracelular (linhagem nth1) não promove uma maior incorporação de
cádmio, quando comparados com os resultados obtidos na linhagem selvagem. Em
conjunto, pode-se considerar que na ausência de trealose, as células incorporam menos
cádmio, e que a presença de um excesso de trealose não tem qualquer efeito na
incorporação do metal. Esse resultado parece estar em acordo com resultados bem
conhecidos sobre o papel desempenhado pela trealose na proteção ao choque térmico: a
trealose parece ser importante durante o choque térmico, mas, sua permanência na
recuperação do estresse térmico atrapalha a ação das proteínas do choque térmico (SINGER

Resultados e Discussão
61
et al.,1998). Nos resultados aqui apresentados não foi observado que o excesso de trealose
prejudica a incorporação do metal, entretanto, não foi observado aumento na incorporação
de cádmio proporcionado pelo excesso do carboidrato.

Resultados e Discussão
62
0
5000
10000
15000
20000
25000
30000
35000
50ppm 800ppm
µµ µµg
Cd
inco
rpor
ado/
g pe
so s
eco
BIOSORÇÃO BIOACUMULAÇÃO
0
5000
10000
15000
20000
25000
30000
35000
50ppm 800ppmµµ µµg
Cd
inco
rpor
ado/
g p
eso
seco
BIOSORÇÃO BIOACUMULAÇÃO
0
5000
10000
15000
20000
25000
30000
35000
50ppm 800ppmµµ µµg
Cd
inco
rpor
ado/
g p
eso
seco
BIOSORÇÃO BIOACUMULAÇÃO
FIGURA 20 – Determinação de cádmio incorporado pelas células viáveis e não viáveis no período
de 16 horas. Linhagens WT (A), tps1 (B) e nth1 (C).
A
C
B

Resultados e Discussão
63
TABELA 02 – Incorporação de cádmio durante 16 horas por células viáveis de Saccharomyces
cerevisiae (µg Cd incorporado/g peso seco)
WT tps1 nth1
50ppm CdCl2 1100(a) ± 173(b) 1100 ± 173 1210 ± 420
800ppm CdCl2 24666 ± 2041 10233 ± 680 25500 ± 2121
a – média de experimento realizado em triplicata
b – desvio padrão

Resultados e Discussão
64
0
5000
10000
15000
20000
25000
30000
35000
50ppm (2hrs)
50ppm (4hrs)
50ppm (8hrs)
50ppm(16 hrs)
800ppm(2 hrs)
800ppm(4 hrs)
800ppm(8 hrs)
800ppm(16 hrs)
µµ µµg
Cd
inco
rpor
ado/
g p
eso
seco
WT tps1 nth1
FIGURA 21 – Incorporação de cádmio em tempos variáveis por células viáveis de Saccharomyces
cerevisiae, WT, tps1 e nth1.

Resultados e Discussão
65
6.4 – Determinação de incorporação de cádmio em células suplementadas com
glutationa reduzida e aminoácidos
Para verificar se, além da presença da trealose, outro composto conhecido como
importante na defesa ao papel nocivo desencadeado por metais em altas concentrações teria
influência na incorporação do cádmio, foi realizado a incorporação do metal na presença de
glutationa, e dos aminoácidos formadores de glutationa, adicionados previamente ao meio
de incubação.
De acordo com a literatura, a glutationa (GSH) é a principal molécula implicada no
seqüestro de metais, dentre eles o cádmio. Glutationa é um tripeptídeo formado pelos
aminoácidos: L-ácido glutâmico, L-cisteina e glicina, é provavelmente o mais importante
grupo tiol de baixo peso molecular encontrado em sistemas biológicos. Está envolvido em
numerosas reações bioredutoras, processos de transporte, proteção contra os radicais livres
prejudiciais e, detoxificação de diferentes xenobióticos (PENNINCKX et al., 1993).
A via para a detoxificação do cádmio pela levedura S. cerevisiae envolve sua
quelação pelas moléculas de glutationa e a formação de complexos chamados bis-
glutationato de cádmio - Cd(GS)2 e seu transporte para o vacúolo (Figura 22)
(BAUDOUIN-CORNU et al., 2006). Algumas leveduras têm a habilidade de acumular
glutationa intracelular e certos aminoácidos têm um importante papel no efeito da
acumulação de glutationa pelas leveduras (WEN, 2005).
FIGURA 22 – Complexo formado entre cádmio e a glutationa, Bis(glutationato) de cádmio –
Cd(GH)2. Fonte: LI et al. (1997).

Resultados e Discussão
66
Com o intuito de verificar a influência da glutationa e, dos aminoácidos que
constituem a mesma, na incorporação de cádmio, nas linhagens de S. cerevisiae,
adicionado, em uma série, glutationa reduzida e, em outra, os aminoácidos (cisteína, ácido
glutâmico e glicina) ao meio externo. Após cinco horas de incubação as células foram
transferidas para um novo meio e expostas ao cádmio durante 16 horas.
Analisando os resultados apresentados na figura 23, foi observado que as células
tratadas com glutationa reduzida suplementada ao meio externo, não apresentaram
alteração na incorporação de cádmio quando comparada com as células não suplementadas.
De acordo com a literatura, a glutationa adicionada ao meio não é capaz de ser transportada
pelas células efetivamente (ANDERSON et al., 1989). Em estudo realizado por Meister e
colaboradores (1983) foi observado que a administração de glutationa exógena leva a um
pequeno aumento nos níveis intracelulares de glutationa, porém esse aumento não é
resultado de um transporte da glutationa, mas sim, de sua degradação extracelular, seguida
pelo transporte dos produtos e, posterior síntese intracelular. Para uma melhor incorporação
de glutationa, a literatura recomenda o uso de um derivado da glutationa, a glutationa
monoetil éster (GME), que atravessa a membrana celular por difusão passiva, dentro da
célula sofre uma hidrólise por esterases e, assim, aumenta os níveis de glutationa
intracelular (ANDERSON et al., 1989).
Após constatar que a presença de glutationa reduzida ao meio externo não alterou a
incorporação de cádmio, optou-se por verificar se, os aminoácidos que formam a
glutationa, (10mM ácido glutâmico, 18 mM glicina e 3,35 mM cisteína) seriam capazes de
influenciar na incorporação do metal pelas células. Em estudo realizado por Wen e
colaboradores (2005), após a incubação de Saccharomyces cerevisiae com as concentrações
acima citadas de aminoácidos, o nível de glutationa intracelular aumentou em 72,2% em
relação ao controle.
De acordo com a figura 23 observa-se que nas três linhagens estudadas, as células
tratadas com aminoácidos, tiveram diminuição significativa na incorporação de cádmio,
tanto na concentração de 50ppm quanto, na concentração de 800ppm. Gomes e
colaboradores (2002) formularam a hipótese que a concentração do complexo cádmio-
glutationa pode controlar a incorporação do metal pela célula. O estudo concluiu que,

Resultados e Discussão
67
quanto maior for a concentração do complexo Cd(GS)2 no citoplasma menor é a
incorporação de cádmio, por outro lado, quando a concentração do complexo for baixa a
célula incorpora uma maior quantidade do metal. Está hipótese foi confirmada pelos
autores utilizando uma linhagem de S. cerevisiae que apresenta deleção no gene GSH1
(linhagem incapaz de sintetizar glutationa, portanto, não pode formar o complexo cádmio-
glutationa). Esta linhagem incorporou duas vezes mais Cd que a linhagem controle, porém
ao tratar essas células com GME (glutationa monoetil éster), o nível de glutationa no
interior da célula foi restaurado e, observou-se que a capacidade de incorporação do metal
diminuía aos mesmos níveis da linhagem controle. Estes dados confirmam os resultados
obtidos na figura 23, após a incorporação dos aminoácidos responsáveis pela formação da
glutationa, o nível de cádmio absorvido pelas linhagens diminuiu significativamente,
provavelmente, ocorreu um aumento nos níveis de glutationa e assim, um aumento na
formação do complexo Cd(GS)2 que resultaram no controle da entrada do metal pela célula.
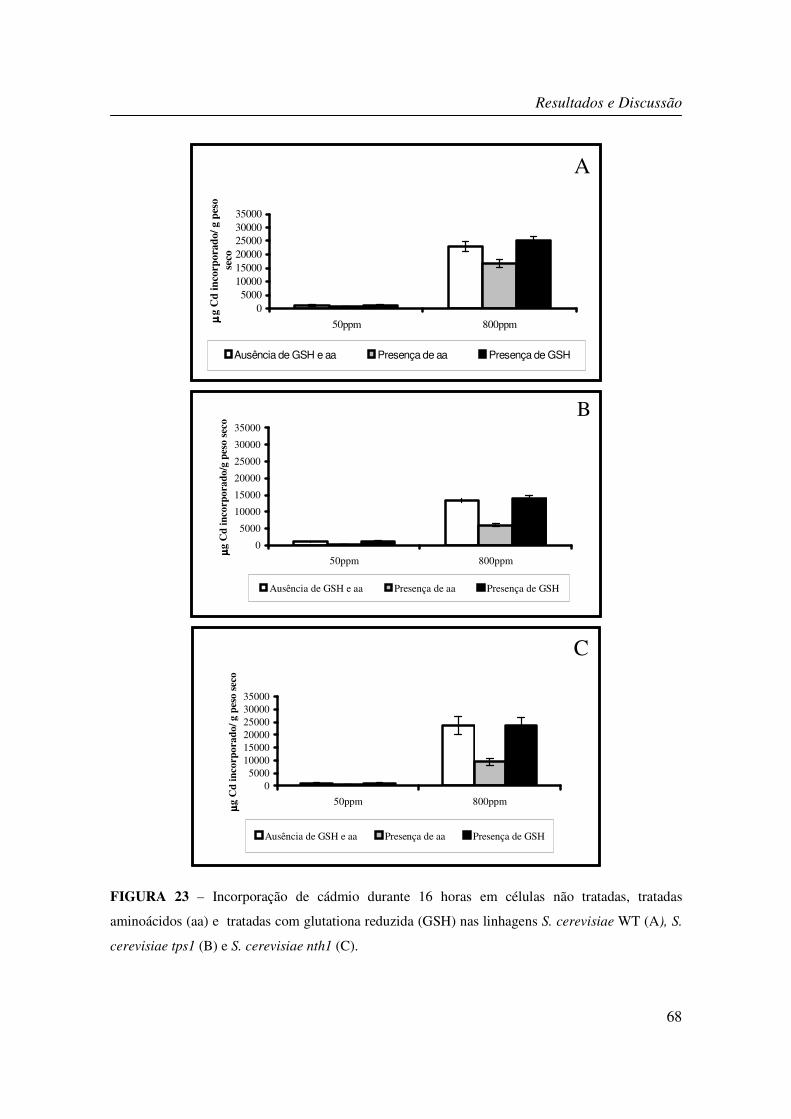
Resultados e Discussão
68
A
05000
100001500020000250003000035000
50ppm 800ppmµµ µµg
Cd
inco
rpor
ado/
g p
eso
seco
Ausência de GSH e aa Presença de aa Presença de GSH
B
0
5000
10000
15000
20000
25000
30000
35000
50ppm 800ppm
µµ µµg
Cd
inco
rpor
ado/
g pe
so s
eco
Ausência de GSH e aa Presença de aa Presença de GSH
C
05000
100001500020000250003000035000
50ppm 800ppm
µµ µµg
Cd
inco
rpor
ado/
g p
eso
seco
Ausência de GSH e aa Presença de aa Presença de GSH
FIGURA 23 – Incorporação de cádmio durante 16 horas em células não tratadas, tratadas
aminoácidos (aa) e tratadas com glutationa reduzida (GSH) nas linhagens S. cerevisiae WT (A), S.
cerevisiae tps1 (B) e S. cerevisiae nth1 (C).

Resultados e Discussão
69
6.5 – Determinação de cádmio na superfície celular
O papel da parede dos microrganismos no processo de biosorção é adsorver os
metais em sua parede ou transportar os metais através de sua membrana celular. Volesky e
colaboradores (1993) observaram que o cádmio não pode ser adsorvido na superfície da
parede celular de S. cerevisiae, mas sim acumulado em vacúolos no interior das células. No
entanto, Suh e colaboradores (1998) mostraram que o cádmio foi adsorvido na parede
celular de S. cerevisiae e também acumulado dentro da célula, e que o chumbo foi
acumulado apenas na superfície da célula de Aureobasidium pullulans devido a influencia
de polímeros extracelulares.
A determinação de cádmio na superfície celular foi realizada utilizando do uso da
técnica de espectometria por dispersão de energia (EDS) que permite análise na superfície
da amostra e a determinação da presença ou não de cádmio na parede celular das células
vivas (bioacumulação) e das células mortas (biosorção). A técnica de espectometria é capaz
de determinar a composição de materiais em volumes de até no mínimo 1µm cúbico
aproximadamente.
Os resultados apresentados nas figuras 24, 25 e 26, mostram que nas linhagens WT,
tps1 e nth1, na concentração de 50ppm de cádmio, o metal é encontrado apenas na
superfície de células mortas. Enquanto que na concentração de 800ppm, o metal é
encontrado tanto na superfície das células mortas quanto em células viáveis. Esta mesma
técnica foi utilizada no trabalho de PARK (2003), onde duas linhagens de S. cerevisiae
foram analisadas por EDS e, o autor verificou que o cádmio tem a capacidade de se ligar na
superfície celular.
Em células mortas o metal se liga a componentes presentes na parede celular,
enquanto em células vivas o metal é incorporado pelas células por processos ativos. Foi
observado que na concentração de 50ppm, as células viáveis não apresentam o pico
característico de cádmio, mas nas células não viáveis este pico foi detectado. Na
concentração de 800ppm, os picos de cádmio foram identificados nas células viáveis e nas
células mortas. Quando foi empregada a técnica da ativação neutrônica foi determinado
cádmio nas células vivas, na concentração de 50 ppm (vide figura 16). Usando a técnica
EDS o cádmio não pode ser determinado (Ver figura 24B). Como o EDS é uma técnica que

Resultados e Discussão
70
permite somente detectar o que está ligado na superfície, pode-se considerar que em células
viáveis, na concentração de 50 ppm, não existe fixação de cádmio na parede celular. Desta
forma, conclui-se que, nesta concentração, as células são capazes de incorporar todo o
metal e depositá-lo no interior.
O mesmo perfil dos resultados descritos para a linhagem WT (Figura 24) foi
observado nas linhagens tps1 e nth1 (vide Figura 25 e 26)

Resultados e Discussão
71
FIGURA 24 – Determinação de cádmio na superfície celular de Saccharomyces cerevisiae WT em
células mortas – Biosorção (A), e em células viáveis – Bioacumulação (B).

Resultados e Discussão
72
FIGURA 25 – Determinação de cádmio na superfície celular de Saccharomyces cerevisiae tps1 em
células mortas – Biosorção (A), e em células viáveis – Bioacumulação (B).

Resultados e Discussão
73
FIGURA 26 – Determinação de cádmio na superfície celular de Saccharomyces cerevisiae nth1 em
células mortas – Biosorção (A), e em células viáveis – Bioacumulação (B).

Resultados e Discussão
74
6.6 - Determinação da incorporação de cádmio em meio suplementado com
galactose e glicose
O acúmulo intracelular de trealose além de ser muito importante na proteção às
diferentes condições de estresse, sua presença está relacionada com a capacidade da célula
em crescer em meios contendo açúcares rapidamente fermentáveis (THEVELEIN et al.,
1995). A linhagem mutante tps1 de S. cerevisiae (linhagem que não sintetiza trealose) é
incapaz de crescer em fontes de carbono fermentáveis como glicose e frutose. Sabe-se que
os níveis mínimos de trealose-6-fosfato sintase são necessários para o controle do influxo
de glicose.
Estudos comprovam que a linhagem tps1 não é capaz de crescer em glicose devido
a uma desregulação no influxo do açúcar, culminando com um grande acúmulo de glicose
livre intracelular (glicose na forma não fosforilada), uma rápida depleção no ATP e em
particular, uma diminuição acentuada na concentração dos fosfatos livres (HOHMANN et
al., 1993).
A linhagem tps1 utilizada neste estudo teve como fonte de carbono para seu
crescimento galactose, as demais linhagens (WT e nth1) tiveram como fonte de carbono a
glicose. Para avaliar se a fonte de carbono influenciaria na incorporação do metal pelas
leveduras, as linhagens WT e nth1 foram crescidas em meio com galactose e incubadas com
cádmio durante 16 horas.
Como pode ser observado nas figuras 27A e B, o crescimento das linhagens usando
como fonte de carbono galactose ou glicose, não influenciou na incorporação do metal.
Portanto, prosseguiram-se os experimentos utilizando a glicose como fonte de carbono para
o crescimento das linhagens WT e nth1, e galactose para a linhagem tps1.

Resultados e Discussão
75
0
5000
10000
15000
20000
25000
30000
35000
50ppm 800ppm
µµ µµg
Cd
inco
rpor
ado/
g pe
so s
eco
Galactose Glicose
0
5000
10000
15000
20000
25000
30000
35000
50ppm 800ppm
µµ µµg
Cd
inco
rpor
ado/
g pe
so s
eco
Galactose Glicose
FIGURA 27 – Diferença nos níveis de incorporação de cádmio em 16 horas, nos meios
YPGalactose e YPGlicose. Linhagem WT (A) e nth1 (B).
A
B

Resultados e Discussão
76
6.7 – Síntese de trealose em resposta à presença de cádmio
Em várias espécies de leveduras as células respondem à presença de estresses no
meio com um grande acúmulo de trealose. Este acúmulo de trealose intracelular parece ser
uma forma importante para a proteção da integridade da célula e se correlaciona com uma
maior possibilidade de sobrevivência das mesmas (ARGÜELES, 2000; SINGER et al.,
1998). No escopo deste trabalho, era de grande importância conhecer se a presença de
cádmio no meio extracelular induziria o acúmulo de trealose pelas células. Para tanto
realizou-se o experimento apresentado na figura 28-A. Deve-se ressaltar que as células
foram coletadas em fase logarítmica, fase em que os níveis de trealose são muito baixos,
isto pode ser observado pelos níveis de trealose determinado nas células na situação
controle (ausência de cádmio). Observa-se na linhagem WT, que com o aumento gradativo
das concentrações de cádmio (50, 100, 350 e 800ppm) ocorre uma elevação significativa
nos níveis de trealose quando comparados aos níveis apresentados pelas células na situação
controle. Este acúmulo de trealose foi obtido na fase logarítmica, significando que as
células responderam a presença de cádmio com o acúmulo de trealose. Observa-se ainda
que quando as células foram incubadas por 2 horas na presença de 800ppm de cádmio,
houve um aumento na síntese de trealose maior que o determinado para as outras
concentrações. O aumento do tempo de incubação de 2 para 4 horas faz com que os níveis
de trealose fossem altos, semelhantes aos valores obtidos com 800 ppm em 2 horas.
Aparentemente, este valor máximo de trealose intracelular acumulada, obtido à 800ppm,
parece ser a capacidade máxima da célula em acumular trealose intracelular na presença de
CdCl2 nas condições deste estudo.
Os resultados obtidos usando-se a linhagem nth1 foram bastante diferentes daqueles
observados para a linhagem WT. Inicialmente, notou-se que os níveis de trealose na
ausência de cádmio (situação controle) são em média 5 vezes maiores do que os
apresentados pela linhagem selvagem na ausência de estresse. Este alto valor obtido é
facilmente compreendido pelo fato da linhagem nth1 não apresentar o gene que codifica a
enzima trealase neutra (NTH1) responsável pela quebra da trealose. Quando as células
foram expostas por 2 horas às concentrações aumentadas de cloreto de cádmio, não
apresentaram alterações na síntese de trealose quando comparado aos seus controles. Este

Resultados e Discussão
77
resultado pode indicar que o tempo de 2 horas foi curto para esta linhagem identificar a
presença de cádmio no meio como uma situação nociva, que deveria ser combatida com o
aumento na síntese de moléculas que possam ajudar a sua proteção. Este resultado foi o
mesmo obtido na linhagem WT, excetuando o resultado obtido a 800 ppm, e ao fato dos
níveis de trealose sintetizados serem menores. Com o aumento do tempo de exposição das
células nth1 ao metal para 4 horas, observa-se que houve um aumento significativo de
síntese do dissacarídeo em todas as concentrações de cádmio quando comparados aos seus
controles (ausência de cádmio). Este resultado indica que o tempo de 4 horas de exposição
ao metal, foi suficiente para a célula conseguir detectá-lo e responder com um pequeno
aumento na síntese de trealose (pequeno se comparado com o aumento verificado na
linhagem WT na mesma situação).
Outro fato que chama a atenção nos resultados obtidos com a linhagem nth1 é que
na situação controle (ausência de cádmio), verifica-se uma diminuição nos níveis de
trealose tanto no tempo de 2 horas como no tempo de 4 horas. Deve-se ressaltar que apesar
desta diminuição, os níveis de trealose acumulados são 4 vezes maiores que os obtidos na
situação controle para a linhagem WT. Acredita-se que isto se deva ao fato da linhagem
nth1 apresentar apenas deleção no gene NTH1, e conservar intacto o gene NTH2. Dados na
literatura mostram que S. cerevisiae possui 2 genes que codificam para a trealase neutra
NTH1 e NTH2. Estes genes codificam duas proteínas isoformas com 77% de identidade.
Tem sido notado que a deleção no gene NTH1 em leveduras é associado com a ausência da
dosagem “in vitro” da atividade trealásica (NWAKA et al., 1995). Recentemente, foi
detectada atividade residual trealásica com degradação de trealose “in vivo”, na fase
estacionária de crescimento, usando glicose ou trealose como fonte de carbono (PARROU
et al., 2005). Esta atividade residual foi atribuída a expressão do gene NTH2 uma vez que,
os autores usaram a mesma linhagem utilizada em nosso estudo. Assim, acredita-se que a
diminuição dos níveis de trealose obtidos na situação controle, nesta linhagem, se deva à
presença do gene NTH2.
O acúmulo de trealose intracelular aparece na literatura como sendo importante na
defesa de células de S. cerevisiae submetidas a diferentes condições de estresse
(ATTFIELD, 1987; VAN LAERE, 1989). Entretanto, existem poucos trabalhos que
demonstram o papel da trealose na proteção contra o estresse oxidativo. Uma exceção é o

Resultados e Discussão
78
trabalho de Benaroudj e colaboradores (2001). Estes autores demonstraram a importância
deste carboidrato em células de S. cerevisiae expostas ao peróxido de hidrogênio. Neste
trabalho os autores também trabalharam com mutante tps1, entretanto, devemos ressaltar
que são bem conhecidos os fatores de transcrição ativados pelo peróxido de hidrogênio
(YAP1 e SKN7) enquanto para o cádmio apenas um fator de transcrição é ativado (YAP1)
(LEE et al., 1999; VIDO et al., 2001). Outros trabalhos sugerem a importância deste
carboidrato no estresse oxidativo, mas não demonstram o papel da trealose neste estresse,
por exemplo, Elbein e colaboradores (2003). Já tinha sido demonstrado por Brennan (1996)
que o cádmio é um indutor de estresse oxidativo na levedura S. cerevisiae, assim, decidiu-
se verificar o papel da trealose no estresse oxidativo induzido pelo cádmio.
Para melhor entender o papel da trealose no quadro do estresse oxidativo induzido
pelo cádmio, foi determinado os níveis de oxidação intracelular gerados pelo metal com o
auxílio de uma sonda fluorescente. Foram usadas células em situação controle (ausência de
cádmio), células expostas a 50ppm e 800ppm de cádmio e, linhagens que apresentam níveis
diferenciados de trealose.
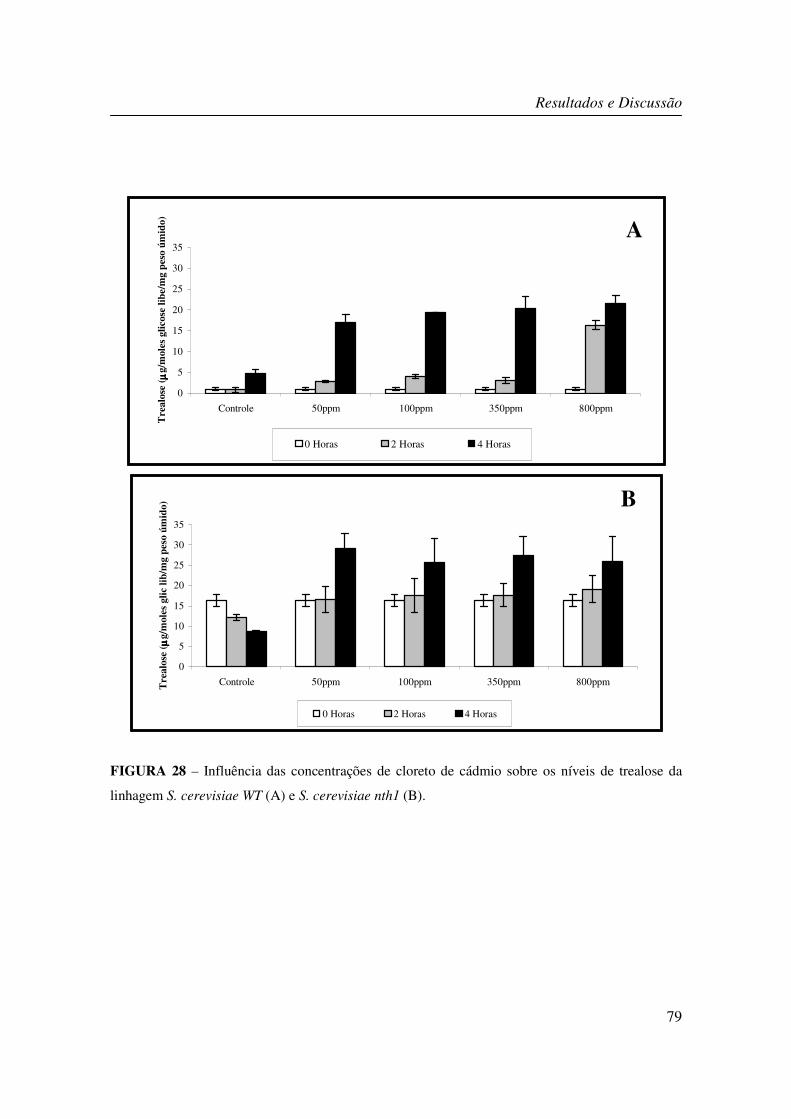
Resultados e Discussão
79
0
5
10
15
20
25
30
35
Controle 50ppm 100ppm 350ppm 800ppm
Tre
alos
e ( µµ µµ
g/m
oles
glic
ose
libe/
mg
peso
úm
ido)
0 Horas 2 Horas 4 Horas
0
5
10
15
20
25
30
35
Controle 50ppm 100ppm 350ppm 800ppmTre
alos
e ( µµ µµ
g/m
oles
glic
lib/
mg
peso
úm
ido)
0 Horas 2 Horas 4 Horas
FIGURA 28 – Influência das concentrações de cloreto de cádmio sobre os níveis de trealose da
linhagem S. cerevisiae WT (A) e S. cerevisiae nth1 (B).
A
B

Resultados e Discussão
80
6.8 – Determinação do nível da oxidação intracelular
Para determinar o aumento da quantidade de ROS gerados nas células, após a
incorporação de cádmio, foi utilizada a sonda fluorescente 2´,7`- diclorodihidrofluoresceína
diacetato (DCFH-DA). Esta sonda é absorvida e transformada no interior das células após
clivagem dos diacetatos por esterases intracelulares, perdendo assim a capacidade de ser
exteriorizada da célula. No interior das células o produto resultante, a 2´,7´-
diclorofluoresceína, se torna mais fluorescente quanto mais oxidante for o ambiente
intracelular, assim o aumento na fluorescência se correlaciona com o aumento da oxidação
intracelular (FIGURA 29) (DAVIDSON et al., 1996). A sonda fluorescente adicionada ao
meio é capaz de detectar qualquer tipo de oxidação presente na solução. O excesso da
sonda fluorescente, que não foi absorvida pelas células, foi eliminado pela lavagem do
extrato, antes da ruptura da membrana celular, realizado com o auxílio de pérolas de vidro.
FIGURA 29 – Mecanismo de ação das esterases na 2,7-diclorodihidrofluoresceína diacetato
(DCFH-DA) formando 2,7- diclorohidrofluoresceína, e a futura oxidação pela ROS formando assim
a 2,7-diclorofluoresceína (DCF). Fonte: GOMES et al. (2005).

Resultados e Discussão
81
O monitoramento da fluorescência obtido nas linhagens WT, tps1 e nth1 foi usado
para estimar o nível de oxidação intracelular das células e está apresentado na tabela 3.
Observa-se na tabela 03 que as linhagens WT e nth1 não apresentam diferença nos níveis de
oxidação intracelular na concentração de 50ppm, quando comparado às suas situações
controle (ausência de cloreto de cádmio). Apenas na concentração de 800ppm ocorreu um
aumento de 2,9 (WT) e 6,6 (nth1), nos níveis da oxidação intracelular, quando comparados
à condição controle de cada linhagem. Estes dados são parcialmente coerentes com o
resultado de incorporação de cádmio, apresentado na tabela 02, (página 62), onde
observou-se que as linhagens WT e nht1 apresentam a mesma incorporação de cádmio.
Porém, obtivemos níveis de oxidação intracelular, em 800 ppm, muito maior na nth1. No
presente caso, os níveis de trealose são altos devido a uma mutação e não devido à presença
de estresse, assim, a resposta nestas duas situações seria diferente. Na célula nth1 o nível
endógeno natural de trealose é muito alto, sendo pouco aumentados pela presença de 800
ppm de cádmio (Figura 28), sendo assim, não pode contrabalançar a grande indução de
radicais livres muito alta nestas condições.
Observa-se ainda na tabela 03 que a linhagem tps1 possui um alto nível de oxidação
na condição controle, quando comparados com os controles das linhagens WT e nth1.
Verifica-se um aumento de 61% e 74% em relação às linhagens WT e nth1,
respectivamente. Este resultado mostra que, na ausência de qualquer estresse, existe um
aumento na produção e/ou geração de radicais livres determinados na linhagem tps1.
Acredita-se que isto se deva ao fato das células serem incubadas em presença de oxigênio e
os radicais livres gerados pela redução ineficiente desta molécula. O fato desta linhagem
não apresentar trealose parece proporcionar aumento na geração de radicais livres em
ausência de estresse. Observa-se ainda que na linhagem tps1 houve um aumento
significativo de 47% e 83% nos níveis de oxidação intracelular, tanto a 50ppm quanto a
800ppm, comparados com o resultado obtido na ausência de cádmio. Correlacionando este
resultado com a incorporação do cádmio (Tabela 02, página 62), observa-se que a linhagem
tps1 incorpora menos cádmio que as linhagens WT e nth1, porém, a geração de radicais
livres é maior.
Concluiu-se que na linhagem que não apresenta trealose ocorreu uma diminuição da
incorporação de cádmio e um aumento nos níveis oxidativos intracelulares. Na linhagem

Resultados e Discussão
82
com excesso de trealose (nth1), a incorporação de cádmio é a mesma verificada na
linhagem selvagem (Tabela 02), porém, a geração de radicais livre é muito maior (tabela
03).
Em estudo realizado por ADAMIS e colaboradores (2003) observou-se que o
aumento dos níveis de oxidação intracelular é diretamente relacionado com a quantidade de
cádmio incorporado pelas células de S. cerevisiae. Neste estudo não foi obtido esta
correlação, o fato da tps1 incorporar menos cádmio não implicou em menores níveis de
oxidação, e o fato da nth1 incorporar a mesma quantidade de cádmio que a WT não
implicou na mesma determinação de radicais livres gerados.
Após verificar os níveis oxidativos das linhagens estudadas, foram avaliados os
danos causados pelos radicais livres usando alguns biomarcadores do estresse oxidativo. A
peroxidação de lipídeos, a carbonilação de proteínas e o conteúdo de resíduos sulfidrílicos
totais.

Resultados e Discussão
83
TABELA 03 – Determinação dos níveis de oxidação intracelular em linhagens de Saccharomyces
cerevisiae
WT tps1 nth1
Controle 84,9(a) ± 10,06(b) 137,0 ± 9,83 78,7 ± 5,51
50ppm CdCl2 98,6 ± 15,66 202,2 ± 13,65 89,2 ± 4,97
800ppm CdCl2 247,5 ± 60,68 251,9 ± 33,30 518 ± 31,71
a – média de experimento realizado em triplicata
b – desvio padrão

Resultados e Discussão
84
6.9 – Determinação dos níveis de peroxidação lipídica
Um dos principais alvos dos radicais livres formados durante o estresse oxidativo
são os ácidos graxos poli insaturados (PUFA – Poli Unsaturated Fat Acid) que compõem a
bicamada lipídica das membranas celulares (VALKO et al., 2005). Apesar das células de S.
cerevisiae possuírem pouco PUFA, é possível determinar os danos lipídicos, conforme
trabalho realizado por Howllet e colaboradores (1997), que demonstraram que a
peroxidação de lipídeos foi induzida por diferentes concentrações dos metais cádmio e
cobre em linhagem de S. cerevisiae.
A formação de peróxidos lipídicos causam grandes prejuízos ao organismo, pois
promove uma alteração na capacidade seletiva das membranas, favorecendo a entrada e a
saída indiscriminada de metabólitos e detritos da célula, fato este freqüentemente associado
à morte celular (PEREIRA et al., 2003). É importante mencionar que a manutenção da
integridade da membrana citoplasmática ou das organelas, é vital para o funcionamento
celular
Os níveis de peroxidação lipídica foram determinados de acordo com a quantidade
de malondialdeído (MDA), produto final do aquecimento de peróxidos lipídicos. Cada
molécula de MDA reage com duas moléculas de ácido tiobarbitúico, formando uma
substância cromófora que é passível de ser determinada espectrofotometricamente
(ESTEUBAUER et al., 1991). Os resultados obtidos foram expressos na forma de relação
entre os níveis de MDA de células expostas a 50ppm e 800ppm de cádmio e células
controles (ausência de cádmio).
A análise dos resultados da tabela 04 mostra que na linhagem WT, na concentração
de 50ppm não ocorre aumento dos danos lipídicos quando comparado a situação controle,
porém na concentração de 800ppm foi determinado um aumento de dano lipídico em 1,6
vezes em relação ao seu controle. Assim, pode-se considerar que altas concentrações do
metal causam danos aos lipídeos.
A linhagem nth1 na situação controle apresenta um nível de peroxidação de lipídica
maior que a situação controle da linhagem WT, ou seja, as células da linhagem com excesso
de trealose, mesmo sem estresse tem um alto nível de danos lipídicos. Porém, comparando
as concentrações de 50ppm e 800ppm com seu controle (ausência de cádmio), não

Resultados e Discussão
85
apresentam diferença significativa entre si. Aparentemente, um excesso de trealose não
permite um aumento nos danos lipídicos quando a célula está exposta ao cádmio.
A linhagem que não apresenta síntese de trealose (tps1) mostra um alto nível de
danos aos lipídeos já na situação controle, ou seja, ausência de qualquer estresse quando
comparado com a linhagem WT também na situação controle. A presença do cádmio
promove um aumento de 1,2 e 1,4 vezes nos danos lipídicos, nas concentrações de 50ppm e
800ppm quando comparada a sua situação controle. Observa-se ainda que a linhagem (tps1)
incorpora menos Cd (Tabela 02) que as demais linhagens, porém mesmo com uma menor
incorporação de Cd, os danos aos lipídeos são maiores (Tabela 04). Observa-se assim que a
ausência desse dissacarídeo causa um aumento nos danos lipídicos e demonstra o
importante papel protetor da trealose aos lipídeos.
Comparando-se as linhagens tps1 e nth1 em ausência de cádmio, verificou-se que as
duas linhagens possuem duas vezes mais lipídeos peroxidados que os determinados na
linhagem WT, na mesma situação (controle). Possivelmente, as células na ausência do
cádmio (situação controle) necessitam de um nível normal de trealose, níveis esse
encontrados em situações normais de células sem deleções, para modularem os ataques de
radicais livres provenientes da respiração celular e, conseguirem proteger seus lipídeos. A
total ausência de trealose (tps1) ou, o excesso (nth1), deixam a célula exposta as ROS
verificando-se assim, um maior dano aos lipideos.
De acordo com dados da literatura, a trealose tem o papel de estabilizar membranas
durante exposição ao estresse oxidativo (HERDEIRO et al., 2006). Estudos realizados por
Pereira e colaboradores (2003) mostraram que sob situação de desidratação, a tolerância
das leveduras é altamente influenciada pela presença de trealose, que em muitos casos
protege a membrana plasmática contra os danos. Oku e colaboradores (2003) demonstraram
em estudos in vitro que parte da molécula de trealose, interage especificamente com uma
dupla ligação cis de um ácido graxo insaturado, por meio de ligações de ponte de
hidrogênio, que envolve grupos hidroxilas OH-6 e OH-3 ou OH-2 com a trealose.

Resultados e Discussão
86
TABELA 04 - Determinação dos níveis de peroxidação de lipídeos (picomoles MDA/mg
proteína/mL) em linhagens Saccharomyces cerevisiae.
WT tps1 nth1
Controle 28,1(a) ± 5,93(b) 51,4 ± 1,34 49,03 ± 1,46
50ppm CdCl2 23,8 ± 1,65 63,3 ± 3,28 50,70 ± 1,14
800ppm CdCl2 45,6 ± 3,35 77,8 ± 1,31 53,10 ± 1,59
a – média de experimento realizado em triplicata
b – desvio padrão

Resultados e Discussão
87
6.10 - Carbonilação de Proteína
As proteínas são as maiores indicadoras de danos causados pelas ROS e dos
processos de estresse oxidativos que ocorrem in vivo, seja nos meios intracelulares ou
extracelulares. De acordo com Davis e colaboradores (1999), durante o estresse oxidativo
cerca 50-75% das proteínas podem ser danificadas. Como as proteínas são os maiores
componentes dos sistemas biológicos, elas são as macromoléculas que mais sofrem com o
ataque das espécies reativas de oxigênio.
Os danos causados pelos radicais livres às proteínas levam a formação de
grupamentos carbonilas. A carbonilação de proteína é produzida por meio da oxidação de
grupos funcionais presentes em cadeias laterais. Os grupos carbonilados podem ser
formados em proteínas por uma reação secundária entre grupamentos da cadeia lateral
(cisteína, lisina e histidina) com aldeídos (malondialdeído e hidroxinoneal) produzidos
durante a peroxidação lipídica ou derivados carbonilados reativos, produzidos pela redução
de açúcares (BERLLET et al., 1997).
Os resultados apresentados na tabela 05 mostraram que a linhagem selvagem na
condição controle, apresentou maior quantidade de proteínas carboniladas que o observado
na mesma situação para as outras linhagens. As linhagem tps1 e nth1, na situação controle,
apresentaram menor dano às proteínas (1,6 e 2,2 vezes respectivamente) quando,
comparadas a situação controle da linhagem WT. A presença de cádmio (nas concentrações
de 50 e 800 ppm) não alterou significativamente os danos às proteínas. Na concentração de
800 ppm, na linhagem nth1, houve um aumento das proteínas carboniladas quando
comparadas ao seu controle. Estes resultados são compatíveis com os resultados obtidos
com a técnica que usa diclorofluoresceína (níveis de oxidação intracelular), vide tabela 03.
Acredita-se que os altos níveis de ROS gerados com 800 ppm na linhagem nth1 foram
responsáveis pelos danos verificados às proteínas.
As proteínas carboniladas determinadas nas três linhagens em ausência de estresse
(condição controle) mostraram que a linhagem com excesso de trealose (nth1) apresentou
uma menor carbonilação de proteínas quando comparada a linhagem WT. Verificou-se
assim, o papel protetor da trealose contra os radicais livres normalmente formados na

Resultados e Discussão
88
célula, porém na ausência de trealose não foi verificado aumento na carbonilação de
proteínas.
Singer e colaborador (1998) demonstraram o importante papel da trealose no
estresse térmico. Com o aumento da temperatura, foi observado um aumento nos níveis de
trealose e seu papel fundamental na estabilização das proteínas no seu estado natural,
reduzindo a agregação e desnaturação proteica. No caso do estresse induzido pelo cádmio,
as proteínas não parecem ser o alvo principal. Apesar da literatura (Singer e colaborador,
1998) considerar que as proteínas são os alvos mais vulneráveis aos ataques pelas ROS,
neste trabalho pode-se observar, que os danos a proteínas são causados apenas quando, os
níveis de ROS são excessivamente altos (vide tabela 03, linhagem nth1, 800ppm).

Resultados e Discussão
89
TABELA 05 – Determinação dos níveis de carbonilação de proteínas (nanomoles/mg proteína/mL)
em linhagens de Saccharomyces cerevisiae.
WT tps1 nth1
Controle 2,67(a) ± 0,39(b) 1,68 ± 0,39 1,19 ± 0,040
50ppm CdCl2 3,17 ± 0,46 1,76 ± 0,62 0,99 ± 0,36
800ppm CdCl2 3,18 ± 0,021 2,24 ± 0,34 1,35 ± 0,21
a – média de experimento realizado em triplicata
b – desvio padrão

Resultados e Discussão
90
6.11 – Resíduos Sulfidrílicos Totais
Os resíduos sulfidrilicos representados principalmente pela glutationa são
abundantes em células e podem ser oxidados pelas espécies reativas de oxigênio, podendo
ser facilmente utilizado como indicador de estresse oxidativo (DEMASI et al., 2006). Um
dos mecanismos da toxicidade ao cádmio em leveduras é sua ligação às proteínas,
resultando na inibição de enzimas essenciais. O cádmio tem afinidade pelo grupo tiol, o que
resulta em sua fácil ligação com resíduos cisteínas em proteínas. Grande parte dos
grupamentos sulfidrílicos da célula se refere à presença de glutationa, que pode ser
encontrada em concentrações de até 10mM em leveduras. A glutationa oxidada (GSSG)
não apresenta grupamentos SH livres, somente quando está na forma reduzida (GSH) que
apresenta os grupamentos SH livres que podem ser usados para se ligar em algumas
substâncias, como por exemplo o cádmio, fazendo o completo cádmio-glutationa –
Cd(GS)2 (BAUDUIN-CORNU et al., 2006). Assim, na presença de algum ligante neste
grupo SH, os resíduos sulfidrílicos determinados serão menores. O reagente Ellman pode
reagir com todos os grupos tiois livres presentes no extrato celular, sendo o mais abundante
a forma reduzida de glutationa (DAVIDSON et al., 2001).
De acordo com os resultados apresentados na tabela 06, pode-se observar que a
linhagem WT possui um aumento de 12% nos níveis de resíduos sulfidrílicos totais, após
tratamento com 50ppm de cádmio, quando comparada com a situação controle. Após
exposição a 800ppm foi determinado uma diminuição de 39% nos resíduos sulfidrílicos. A
linhagem nth1 apresenta o mesmo comportamento observado na linhagem WT, após o
tratamento com 50ppm de cádmio obteve um aumento de 35% nos resíduos sulfidrilicos e,
uma queda de 36% na concentração de 800 ppm
A linhagem tps1 apresentou um aumento de 41% nos níveis de resíduos sulfidrílicos
quando tratada com 50ppm de cádmio comparado com o seu controle. E uma diminuição
de 25% em relação ao seu controle, quando tratada com 800ppm de cádmio. Observa-se
ainda que a linhagem com deleção no gene responsável pela síntese de trealose (tps1), na
situação controle, apresenta um baixo nível de resíduos sulfidrílicos quando comparada
com as linhagens WT e nth1 entretanto, os níveis de resíduos após incorporação de cádmio
se tornam semelhantes às demais linhagens. Pode-se ainda observar que esta linhagem

Resultados e Discussão
91
incorpora menos cádmio quando comparada as linhagens WT e nth1 (Tabela 02), porém
seus níveis de resíduos sulfidrílicos não apresentam diferença nas células tratadas com
cádmio (50 e 800ppm) (Tabela 06).
As células quando tratadas com concentração de 50ppm, apresentaram um aumento
nos níveis de resíduos sulfidrílicos nas três linhagens enquanto, na concentração severa de
cloreto de cádmio (800ppm) provocou diminuição significativa dos resíduos sulfidrílicos
totais, quando comparadas a situação controle (ausência de CdCl2). Observou-se que altas
concentrações de trealose (nth1) ou, ausência de trealose (tps1) não interfere no conteúdo
de resíduos sulfidrílicos em presença de cloreto de cádmio. Na situação controle verifica-se
uma alteração quando as três linhagens são comparadas: os níveis de resíduos sulfidrílicos
totais é bem menor na linhagem tps1, provavelmente, isto se deva ao fato de que na
ausência de trealose a célula deva usar muito do seu estoque de glutationa reduzida na
defesa contra os radicais livres, normalmente formados na célula. Desta forma quando esta
determinação é realizada na linhagem tps1, obteve-se níveis bem menores que nas
linhagens que não têm a mutação apresentada pela linhagem tps1.
Segundo Brennan (1996), baixas concentrações de cádmio podem aumentar a
resistência, e consequentemente, induzir os mecanismos de defesa. Assim, quando as
células são incubadas com 50 ppm de cádmio, o metal deve induzir um aumento nas
defesas antioxidantes não enzimáticas, determinando assim, uma maior concentração de
grupos tiois livres. A incubação das células com 800 ppm de cádmio, considerada
excessivamente tóxica implica nas células não conseguirem manter uma resposta
antioxidante (não enzimática) suficiente e continuada para combater esta concentração.
Assim, não foi observada a indução das defesas antioxidantes sendo portanto determinado
uma menor concentração de resíduos sulfidrílicos totais. O mesmo efeito foi observado por
Davidson e colaboradores (2001) em células de S. cerevisiae exposta ao estresse térmico,
onde observaram queda de 50% nos níveis de resíduos sulfidrílicos totais, mostrando assim
os danos provocados pelo choque térmico nas defesas antioxiadantes não enzimáticas.

Resultados e Discussão
92
TABELA 06 – Determinação dos níveis resíduos sulfidrílicos totais (µmoles/mg proteína/mL) em
linhagens de Saccharomyces cerevisiae.
WT tps1 nth1
Controle 9,16(a) ± 0,41(b) 6,54 ± 1,04 9,46 ± 1,15
50ppm CdCl2 10,33 ± 0,62 9,25 ± 1,32 12,86 ± 2,72
800ppm CdCl2 5,63 ± 0,32 4,94 ± 0,61 5,96 ± 1,15
a – média de experimento realizado em triplicata
b – desvio padrão

Resultados e Discussão
93
6.12 - Determinação do status antioxidante total das linhagens pelo radical
DPPH
Recentemente, vários trabalhos sugerem que alguns biopolimeros como glucanos
possuem atividade antioxidante (Jaehrig et al., 2007). A parede celular de S. cerevisiae
consiste de aproximadamente 29-64% -glucanos, 31% manose, 13% proteínas, 9% lipideos,
e 1-2% quitina. Se de fato os glucanos têm atividade antioxidantes, este fato poderia indicar
que estes compostos poderiam proteger os organismos vivos dos ataques das ROS.
Saccharomyces cerevisiae possuem várias substâncias que agem como antioxidantes
enzimáticos (SOD, catalase, glutationa redutase) ou, não enzimáticos (glutationa
ubiquinona, aminoácidos com resíduos sulfirilicos e selênio). Em adição aos antioxidantes
que estão dentro das células, potencialmente, a parede da levedura poderia também exercer
atividade antioxidante. O interesse ao realizar este experimento foi determinar a capacidade
antioxidante das linhagens, WT, tps1 e nth1 usando uma solução metanólica do radical
livre estável DPPH•. O objetivo de usar este método foi verificar se, a ausência ou, excesso
de trealose, promoveria alteração no status antioxidante total das linhagens.
A essência do método DPPH• consiste na reação dos antioxidantes presentes na
amostra com o radical livre estável α,α-difenil-β-picrylladrazil - DPPH• (com forte cor
violeta) e se converte a α,α-difenil-β-picriladrazina com descoloração, o grau de
descoloração indica o potencial “scavenging” da amostra. Os antioxidantes presentes nas
amostras reduzem e descolorem o DPPH através da doação de elétrons. A solução
metanólica de DPPH foi preparada na hora e apresenta uma cor púrpura com uma absorção
máxima de 517 nm (JAYAPRAKASHA et al., 2006).
Segundos os resultados obtidos na figura 30, observa-se que não há diferença
significativa entre as três linhagens quanto aos níveis de defesas antioxidantes. A linhagem
nth1 apresenta um pequeno aumento em relação à linhagem controle, porém não é
significativo. Pode-se observar assim que a diferenças obtidas neste trabalho não parecem
ser devidas às diferenças nos níveis de defesas antioxidantes totais como determinado pelo
radical DPPH•, e sim pelo excesso ou ausência da trealose avaliado pelas linhagens nth1 e
tps1, respectivamente.

Resultados e Discussão
94
0
5
10
15
20
25
30
35
40
45
50
WT tps1 nth1
% In
ibiç
ão
do
rad
ical D
PP
H
FIGURA 30 – Determinação do status antioxidante total das diferentes linhagens Saccharomyces
cerevisiae utilizando o radical DPPH•.

Papel da trealose na proteção durante o estresse oxidativo causado por cádmio e a resposta adaptativa a este
estresse em Saccharomyces cerevisiae
95
7 – Conclusão

Conclusão
96
• A presença de cádmio (10, 25, 50, 75 e 100ppm) inibe o crescimento das células de S.
cerevisiae;
• Em meio sólido observou-se a formação de colônias até a concentração de 25ppm;
• Células coletadas em fase logarítmica incorporaram maior quantidade de cádmio;
• Células viáveis incorporaram mais cádmio que células mortas;
• A presença de glutationa reduzida, no meio extracelular, não interferiu na incorporação
de cádmio
• A presença de aminoácidos formadores da glutationa, no meio extracelular,
proporcionaram diminuição da incorporação de cádmio;
• As células respondem à presença de cádmio no meio extracelular com o acúmulo de
trealose intracelular;
• A linhagem com ausência de trealose (tps1) possui uma maior oxidação intracelular; as
linhagens WT e nth1 apresentaram níveis de oxidação intracelular aumentados apenas na
concentração de 800ppm;
• A presença de cádmio induz a peroxidação de lipídeos;
• Na presença de cádmio não foi verificado o aumento de proteínas carboniladas;
• Altas concentrações de cádmio causaram diminuição dos resíduos sulfidrílicos
enquanto que baixas concentrações causaram aumento;
• Nas três linhagens não foi verificado alteração no status antioxidante total determinado
pelo método DPPH•.

Papel da trealose na proteção durante estresse oxidativo causado por cádmio e a resposta adaptativa ao
estresse em Saccharomyces cerevisiae
97
8 - Referências Bibliográficas

Referências Bibliográficas
98
ADAMIS, P. D. B.; PANEK, A. D.; LEITE, SELMA, G. F.; ELEUTHERIO, ELIS, C. A.
Factores involved with cadmium absorption by a wild-type strain of Saccharomyces
cerevisiae. Brazilian Journal of Microbiology, v. 34, p. 55-60, 2003.
American Cancer Society. Lista dos prováveis carcinorgênicos e suas categorias.
Disponível em:http://www.cancer.org> acesso em 14 de Maio de 2007
ANDERSON, M. E.; MEISTER, A. Glutathione Monoester. Analytical Biochemistry, v.
183, p. 16-20, 1989.
ARGÜELLES, J. C. Physiological roles of trehalose in bacteria and yeast: a comparative
analysis. Archives of Microbiology, v. 174, p. 217-224, 2000.
ATTFIELD, P. V. Trehalose accumulates in Saccharomyces cerevisiae during exposure to
agents that induce heat shock response. FEBS Letters, v. 225, n. 1, p. 259-263, 1987.
ATSDR. Agency for Toxic Substances and Disease Registry. Lista de substâncias
perigosas classificadas de acordo com sua toxidade, potencial de risco a saúde e exposição
aos organismos vivos. Disponível em: <http://www.atsdr.cdc.gov/cercla/05list.html>
Acesso em 14 de maio de 2007.
BAKER-AUSTIN, C.; WRIGHT, M. S.; STEPANAUSKAS, R. McARTHUR, J. V. Co-
selection of antibiotic and metal resistance. TRENDS in Microbiology, v. 14, n. 4, p.176-
182, 2006.
BAUDOUIN-CORNU, P.; LABARRE, J. Regulation of the cadmium stress response
through SCF-like ubiquitin ligases: comparison between Saccharomyces cerevisiae,
Schizosaccharomyces pombe and mammalian cells. Biochimie, v. 88, n. 11, p. 1673-1685,
2006.

Referências Bibliográficas
99
BELL, W.; SUN, W.; HOHMANN, S.; WERA, S.; REINDERS, A.; DE VIRGILIO, C.;
WIEMKEN, A.; THEVELEIN, J. M. Composition and functional analysis of
Saccharomyces cerevisiae trehalose synthase complex. The Journal of Biological
Chemistry, v. 273, n. 50, p. 33311-33319, 1998.
BELL, W.; KLAASSEN, P.; OHNACKER, M.; BOLLER, T.; HERWEIJER, M.;
SCHOPPINK, P.; VAN DER ZEE, P.; WIEMKEN, A. Characterization of 56-kDa subunit
of yeast trehalose-6-phosphatate synthase and clonig of its gene reveal its identity with the
product of CIF1, a regulator of carbon catabolite inactivation. Europen Journal of
Biochemistry, v. 209, p. 951-959, 1992.
BENARDOUJ, N.; LEE, D. H.; GOLDBERG, A. L. Trehalose accumulation during
cellular stress protects cells and cellular proteins from damage by oxygen radicals. The
Journal of Biological Chemistry, v. 276, n. 26, p. 24261-24267, 2001.
BERLETT, B. S.; STADTMANT, E. R. Protein Oxidation in Aging, Disease, and
Oxidative Stress. The Journal of Biological Chemistry, v. 272, n. 33, p. 20313-20316,
1997.
BERTIN, G.; AVERBECK, D. Cadmium: cellular effects, modifications of biomolecules,
modulation of DNA repair and genotoxic consequences (a review). Biochimie, v. 88, p.
1549-1559, 2006.
BRAHA, B.; TINTEMANN, H.; KRAUSS, G.; EHRMAN, J.; BÄRLOCHER, F.;
KRAUSS, G-J. Stress response in two strains of the aquatic hyphomycete Heliscus
lugdunensis after exposure to cadmium and copper ions. BioMetals, v. 20, p. 93-105, 2007.
BRENNAN, R. J.; SCHIESTL, R.H. Cadmium is an induce of oxidative stress in yeast.
Mutation Research. v. 356, p. 171-178, 1996.

Referências Bibliográficas
100
CABIB, E.; LELOIR, L. F. The biosynthesis of trehalose phosphate. The Journal of
Biological Chemistry, v. 231, p. 259-275, 1958.
CATALÁ, A. An overview of lipid peroxidation with emphasis in outer segments of
photoreceptors and the chemiluminescence assay. The International Journal of
Biochemistry & Cell Biology, v. 38, p. 1482-1495, 2006.
CONAMA. Conselho Nacional do Meio Ambiente. Resolução CONAMA nº357:
estabelece a classificação das águas e os níveis de qualidade exigidos. Disponível em:
<http://www.mma.gov.br/port/conama/res/res86/res2086.html.> Acesso em 14 de maio de
2007.
COSTA, V.; MORADA-FERREIRA, P. Oxidative stress and signal transduction in
Saccharomyces cerevisiae: insights into ageing, apoptosis and disease. Molecular Aspects
of Medicine, v. 22, p. 217-246, 2001.
CROWE, J. H.; CROWE, L. M.; CHAPMAN, D. Preservation of membranes in
anhydrobiotic organisms: the role of trehalose. Science, v. 223, p. 701-703, 1984.
CYRNE, L. et al. Regulation of antioxidant enzymes gene expression in the yeast
Saccharomyces cerevisiae during stationary phase. Free Radical Biology & Medicine, v.
34, n. 3, p. 385-393, 2003.
DALLE-DONNE, I.; ROSSI, R.; GIUSTARINI, D.; MILZANI, A.; COLOMBO, R.
Protein carbonyl groups as biomarkers of oxidative stress. Clinica Chimica Acta;
International Journal of Clinical Chemistry, v. 329, p. 23-38, 2003.
DALLE-DONNE, I.; ALDINI, G.; CARINI, M.; COLOMBO, R.; ROSSI, R.; MILZANI,
A. Protein carbonylation, cellular dysfunction, and disease progression. Journal of
Cellular and Molecular Medicine, v. 10, n. 2, p. 389-406, 2006.

Referências Bibliográficas
101
DAVIDSON, J. F.; WHITE, B.; BISSINGER, P. H.; SCHIESTL, R. H. Oxidative stress is
involved in heat-induced cell death in Saccharomyces cerevisiae. Proceedings of the
National Academy of Sciences of the United States of America, v. 93, p. 5116-5121,
1996.
DAVIDSON, J. F.; SCHIESTL, R. H. Cytotoxic and Genotoxic consequences of heat stress
are dependent on the presence of oxygen in Saccharomyces cerevisiae. Journal of
Bacteriology, p. 4580 – 4587, 2001.
DAVIS, M. J.; FU, S.; WANG, H.; DEAN, R. T. Stable markers of oxidant damage to
proteins and their application in study of human disease. Free Radical Biology &
Medicine, v, 27, p. 1151-1161, 1999.
DECKERT, J. Cadmium toxicity in plants: Is there any analogy to its carcinogenic effect in
mammalian cells? BioMetals, v. 18, p. 475-481, 2005.
DEMASI, A. P. D.; PEREIRA, G. A. G.; NETTO, A. E. S. Yeast oxidative stress response
– Influences of cytosolic thioredoxin peroxidase I and of the mitochondrial functional state.
FEBS Journal, v. 273, p. 805-816, 2006.
DE VIRGILIO, C.; BÜRCKERT, N.; BELL, W.; JENO, P.; BOLLER, T.; WIEMKEN, A.
Disruption of TPS2, the gene encoding the 100-kDa subunit of the trehalose-6-phosphate
synthase/phosphatase complex in Saccharomyces cerevisiae, causes accumulation of
trehalose-6-phosphate and loss of trehalose-6-phosphate phosphatase activity. European
Journal of Biochemistry, v. 212, p. 315-323, 1993.
DE VIRGILIO, C.; HOTTIGER, T.; DOMINGUEZ, J.; BOLLER, T.; WIEMKEN, A. The
role of trehalose synthesis for the acquisition of thermotolerance in yeast. I. Genetic
evidence that trehalose is a thermoprotectant. European Journal of Biochemistry, v. 219,
p. 179-186, 1994.

Referências Bibliográficas
102
DÖNMEZ, G.; AKSU, Z. The effects of copper (II) ions on the growth and
bioaccumulation properties of some yeast. Process Biochemistry, v. 35, p. 135-142, 1999.
DORDEVIC, S.; PETROVIC, S.; DOBRIC, S.; MILENKOVIC, M.; VUCICEVIC, D.;
ZIZIC, S.; KUKIC, J. Antimicrobial, anti-inflammatory, anti-ulcer and antioxidant
activities of Carlina acanthifolia root essential oil. Journal of Ethnopharmacology, v.
109, n. 3, p. 458-463, 2007.
ELBEIN, A. D. The metabolism of α, α Trehalose. Advances in Carbohydrate
Chemistry and Biochemistry, v. 30, p. 227,256, 1974.
ELBEIN, A.D.; PAN, Y.T.; PASTUSZAK,I.; CARROLL, D. New insights on trehalose: a
multifunctional molecule. Glycobiology, v. 13, n.4, p. 17-27, 2003.
ELLMAN, G. L. Tissue Sulfhydryl groups. Archives of Biochemistry and Biophycisc, v.
82, p. 70-77, 1959.
ERCAL, N.; GURER-ORHAN, H.; AYKIN-BURNS, N. Toxic Metals and oxidative stress
Part I: Mechanisms Involved in Metal induced Oxidative Damage. Current Topics in
Medicinal Chemistry, p. 529-539, 2001.
ESTERBAUER, H.; SCHAUR, R. J.; ZOLLNER, H. Chemistry and biochemistry of 4-
hydroxynonenal, malonaldehyde and related aldehydes. Free Radical Biology &
Medicine, v.11, n.1, p. 81-128, 1991.
FARR, S.; KOGOMA, T. Oxidative stress response in Escherichia coli and Salmonella
typhimurium. Microbiology and Molecular Biology Review, v. 55, p. 561-585, 1991.
FINKEL, T.; HOLBROOK, N. J. Oxidants, oxidative stress and the biology of ageing.
Nature, v. 408, p. 239-241, 2000.

Referências Bibliográficas
103
FRANÇA, M. B. M; PANEK, A. D.; ELEUTHERIO, E. C. A. Oxidative stress and its
effects during dehydration. Comparative Biochemistry and Physiology, v. 146, p. 621-
631, 2007.
FRANKENBERG, D.; FRANKENBERG-SCHWAGNER, M.; HARBICH, R.
Mechanisms of oxygen radiosensitization in irradiated yeast. I. DNA double-strand
breakage. International Journal of Radiation Biology, v. 64, p. 511-521,1993.
GHARIEB, M. M.; Pattern of cadmium accumulation and essential cations during growth
of cadmium-tolerant fungi. Biometals, v. 14, p. 143-151, 2001.
GHEZZI, P.; BONETTO, V.; FRATELLI, M. Thiol-disulfite balance: from the concept of
oxidative stress to that of redox regulations. Antioxidants and Redox Signaling, v.7, p.
964-972, 2005.
GIAGINIS, C.; GATZIDOU, E.; THEOCHARIS, S. DNA repair systems as targets of
cadmium toxicity. Toxicology and Applied Pharmacology, V. 213, n. 3, p. 282-290,
2006.
GÖKSUNGUR, Y.; ÜREN, S.; GÜVENÇ, U. Biosorption of cadmium and lead ions by
ethanol treated waste baker’s yeast biomass. Bioresource Technology, v. 96, p. 103-109,
2005.
GOMES, A.; FERNANDES, E.; LIMA, J. L. F. C. Fluorescence probes used for detection
of reactive oxygen species. Journal of Biochemical and Biophysical Methods, v. 65,
p.45-80, 2005.
GOMES, D.S.; FRAGOSO, L.C.; RIGER, C.J.; RIGER, A.; ELEUTHERIO, E.C.A.
Regulation of cadmium uptake by Saccharomyces cerevisae. Biochimica et Biophysica
Acta, v. 1573, p. 21-25, 2002.

Referências Bibliográficas
104
HALLIWELL, B.; GUTTERIDGE, J. M. C. Free radicals in Biology and Medicine.
Oxford: Clarendon Press, p. 543, 1989.
HALLIWELL, B.; ARUOMA, O. I. DNA and Free Radicals. Ellins Horwood, London
(1993)
HERDEIRO, R.S.; PEREIRA, M. D.; PANEK, M.A. D.; ELEUTHERIO, E.C.A. Trehalose
protects Saccharomyces cerevisiae from lipid peroxidation during oxidative stress.
Biochimica et Biophysica Acta, v. 1760, p. 340-346, 2006.
HOEIJMAKERS, J. H. Genome maintenance mechanisms for preventing cancer. Nature,
v. 411, p. 366-347, 2001.
HOHMANN, S.; NEVES, M. J.; KONING, W.; ALIJO, R.; RAMOS, J.; THEVELEIN, J.
M. The growth and signalling defects of the ggs1 (fdp1/byp1) deletion mutant on glucose
are suppressed by a deletion of the gene encoding hexokinase PII. Current Genetics, v. 23,
p. 281-289, 1993.
HOUNSA, C. G.; BRANDT, E. V.; THEVELEIN, J.; HOHMANN, S.; PRIOR, B. A. Role
of trehalose in survival of Saccharomyces cerevisiae under osmotic stress. Microbiology,
v. 144, p. 671-680, 1998.
HOWLETT, N. G.; AVERY, S. V. Induction of Lipid peroxidation during heavy metal
stress in Saccharomyces cerevisiae and influence of plasma membrane fatty acid
unsaturation. Applied and Environment Microbiology, p. 2971-2976, 1997.
INOUE, M.; SATO, E. F.; NICHIKAWA, M.; HIRAMOTO, K. KASHIWAGI, A.
UTSUMI, K. Free radicals theory of apoptose and metamorphosis. Redox Report, v. 9, p.
238-248, 2004.

Referências Bibliográficas
105
JAYAPRAKASHA, G. K.; NEGI, P. S.; JENA, B. S. Antioxidative and antimutagenic
actives of the extracts from the rinds of Garcinia pedunculata. Innovative Food Science
and Emerging Technologies, v. 7, p. 246 – 250, 2006.
JAEHRIG, S. C.; ROHN, S.; KROH, L. W.; FLEISCHER, L.; KURZ, T. In Vitro Potential
Antioxidant Activity of (13),(16)- D-Glucan and Protein Fractions from Saccharomyces
cerevisiae cell walls. Journal of Agricultural and Food Chemistry, v.55, n. 12, p. 4710-
4716, 2007.
JONES, D. P. Redefining Oxidative Stress. Antioxidantes & Redox signaling, v. 8, n. 9,
p. 1865-1879, 2006
KADUKOVÁ, J.; VIRCIKOVÁ, E. Comparison of differences between copper
bioaccumation and biosorption. Environment International, v. 31, p. 227-232, 2005.
KAPOOR, A.; VIRARAGHAVAN T. Fungal Biosorption – an alternative treatment option
for heavy metal bearing wasters: a Rewiew. Bioresource Technology, v. 55, p. 195-206,
1995.
KEFALA, M. I.; ZOURBOULIS, A. I.; MATIS, K. A. Biosorption of cadmium ions by
Actinomycetes and separation by flotation. Environmental Pollution, v. 104, p. 283-293,
1998.
KILEY, P. J; STORZ, G. Exploiting thiol modifications. Plos Biology, v. 2; p. 1714-1717,
2004.
KOPP, M.; MÜLLER, H.; HOLZER, H. Molecular analysis of the neutral Trehalases gene
from Saccaromyces cerevisiae. The Jounal of Biological Chemistry, v. 268, n. 7, p. 4766-
4774, 1993.

Referências Bibliográficas
106
LANE, W. T.; SAITO, A. M.; GEORGE, N, G.; PICKERING, J. I.; PRINCE, C. R.;
MOREL, M. M. F. Biochemistry: A cadmium enzyme from a marine diatom. Nature, v.
435, p.42-42, 2005.
LEE, J.; GODON, C.; LAGNIEL, G.; SPECTOR, D.; GARDIN, J.; LABARRE, J.;
TOLEDANO, M. B. Yap1 and Skn7 Control Two Specialized Oxidative Stress Response
Regulators in Yeast. The Journal of Biological Chemistry, v. 274, p. 16040-16046, 1999.
LEE, S. M.; PARK, J. W. A yeast mutant lacking thiol-dependent protector protein in
hypersensitive to menadione. Biochimica Biophysica Acta, v. 1382, p. 167-175, 1998.
LI, Z. S.; Lu Y.P.; Zhen R.G.; Szczypka M.; Thiele D.J.; Rea P.A.; A new pathway for
vacuolar cadmium sequestration in Saccharomyces cerevisiae: YCF1 – catalyzed transport
of bis (glutathinato) cadmium. Proceedings of the National Academy of Sciences of the
United States of America., v. 94, p. 42-47, 1997
LIU, H-J.; LIU, D-H.; ZHONG, J-J. Interesting physiological response of the osmophilic
yeast Candida krusei to heat shock. Enzyme and Microbiol Technology, v. 36, p. 409-
416, 2005.
LIU, JIANGHAI; ZHANG, YINGMEI; HUANG, DEJUN; SONG, GANG. Cadmium
induced MTs synthesis via oxidative stress in yeast Saccharomyces cerevisiae. Molecular
and Cellular Biochemistry, v. 280, p. 139-145, 2005.
LONGO, V. D.; GRALLA, E. B.; VALENTINE, J. S. Superoxide dismutase activity is
essential for stationary phase survival in Saccharomyces cerevisiae. The Journal of
Biological Chemistry, v. 271, n. 21, p. 12275-12280, 1996.
LOWRY, O. H.; ROSEBROUGH, N. J.; FARR, A. L.; RANDALL, R. J. Protein
measurement with the Folin reagent. The Journal of Biological Chemistry, 193, p. 265-
275, 1951.

Referências Bibliográficas
107
LUSHCHAK, V. I. Budding yeast Saccharomyces cerevisiae as a model to study oxidative
modification of proteins in eukaryotes. Acta Biochimica Polonica, v. 53, n. 4, p. 679-684,
2006.
LUSHCHAK, V.; GOSPODARYOV, D. V. Catalases protect cellular proteins from
oxidative modification in Saccharomyces cerevisiae. Cell Biology International, v. 29, p.
187-192, 2005.
MALIK, A. Metal Bioremediation through growing cells. Environment International, v.
30, p. 261-278, 2004.
MANNHEIM B. Cell death – Cell proliferation and viability. In: Apoptosis and cell
proliferation. Biochemica, 1998.
MARCHLER, G.; SCHÜLLER, C.; ADAM, G.; RUIS, H. A. Saccharomyces cerevisiae
UAS element controlled by protein kinase A activates transcription in response to a variety
of stress conditions. The EMBO Journal, v. 12, p. 1997-2003, 1993.
MARNETT, L. J. Chemistry and biology of DNA damage by malondialdehyde. IARC
Scientific Publications, v. 150, p. 17-27, 1999.
MARQUES, P. A.; PINHEIRO, H. M. TEIXEIRA, J. A.; ROSA, M. F. Removal efficiency
of Cu2+, Cd2+ and Pb2+ by waste brewery biomass: pH and cation association effects.
Desalination, v. 124, p. 137-144, 1999.
MARTÍNEZ-ESPARZA, M.; AGUINAGA, A.; GONZÁLES-PÁRRAGA, P.; GARCÍA-
PEÑARRUBIA, P.; JOAULT, T.; ARGÜELLES, J. C. Role of trehalose in resistance to
macrophage killing: study with a tps1/tps1 trehalose-deficient mutant of Candida albicans.
Clinical Microbiology and Infection, v. 13, p. 384-394, 2007.

Referências Bibliográficas
108
MEISTER, A.; ANDERSON, M. E. Glutathione. Annual Review of Biochemistry, v. 52,
p. 711-760, 1983.
MENEZES, M. A. B. C.; SABINO, C. V. S.; FRANCO, M. B.; KASTNER, G. F.;
MONTOYA, E. H. R. K0-intrumental neutron activation establishment at CDTN, Brazil: a
successfum story. Journal of Radioanalytical and Nuclear Chemistry, v. 257, n. 3, p.
627-632, 2003. EU NÃO TENHO ESSE
MELO, L. C.; DOS SANTOS, J. B.; RAMALHO, M. A. P. Choice of parents to obtain
common bean (Phasealus vulgaris) cultivars tolerance to low temperatures at the adult
stage. Brazilian Journal of Genetics, v. 20, p. 283-293, 1997.
MOMOSE, Y.; IWAHASHI, H. Biossay of cadmium using a DNA microarray: genome-
wide expression patterns of Saccharomyces cerevisiae response to cadmium.
Environmental Toxicology and Chemistry, v. 20, n. 10, p. 2353-2360, 2001.
NEVES, M. J.; TERENZI, H. F.; LEONE, F. A; JORGE, J. A. Quantification of trehalose
in biological samples with conidial trehalase from thermophilic fungus Humicola grisea
var. thermoidea. World Journal of Microbiology & Biotechnology, v. 10, p. 17-19, 1994.
NWAKA, S.; MECHLER, B.; DESTRUELLE, M.; HOLZER, H. Phenotypic features of
trehalase mutants in Saccharomyces cerevisiae. FEBS Letters, v. 360, p. 286-290, 1995.
NYSTRÖM, T. Role of oxidative carbonylation in protein quality control and senescence.
The EMBO Journal, v. 24, p. 1311-1317, 2005.
OHTAKE, Y.; YABUUCHI, S. Molecular cloning of the gamma-glutamylcysteine
synthetase gene of Saccharomyces cerevisiae. Yeast, v. 7, p. 953-961, 1991.
OKU, K.; WATANABE, H.; KUBOTA, M.; FUKUDA, S.; KURIMOTO, M.;
TSUJISAKA, Y. KOMORI, M.; INOUE, Y. SAKURAI, M. NMR e quantum chemical

Referências Bibliográficas
109
study on the OH…pi and CH…O interaction between trehalose and unsaturated fatty acids:
implication for the mechanism of antioxidant function of trehalose. Journal of the
American Chemical Society, v. 125, p. 12739-12748, 2003.
PANEK, A. D. Trehalose metabolism – new is in technological applications. Brazilian
Journal of Medical and Biological Research, v. 28, p. 169-181, 1995.
PARK, J. K.; LEE, J. W.; JUNG, J. Y. Cadmium uptake capacity of two strains of
Saccharomyces cerevisiae cells. Enzyme and Microbial Technology, v. 33, p. 371-378,
2003.
PARROU, J. L.; JULES, M.; BELTRAN, G.; FRANÇOIS, J. Acid trehalase in yeast and
filamentous fungi: Localization, regulation and physiological function. FEMS Yeast
Research, v.5, p.503-511, 2005.
PENNINCKX, M. J.; ELSKENS, M. T. Metabolism and functions of glutathione in
microorganisms. Advances in Microbiology and Physiology, v. 34, p. 239-301, 1993.
PEREIRA, E. J.; PANEK, A. D.; ELEUTHERIO, E. C. A. Protection against oxidation
during dehydrationof yeast. Cell Stress & Chaperones, v. 8, p. 120-124, 2003.
PEREIRA, M. D.; HERDEIRO, R. S.; FERNANDES, P. N.; ELEUTHERIO, E. C. A.;
PANEK, A. D. Targets of oxidative stress in yeast sod mutants. Biochimica et Biophysica
Acta, v. 1620, p. 245-251, 2003.
POCSI, I.; PRADE, R.A.; PENNINCHKX, M. J. Glutathione, altruistic metabolite in fungi.
Advances in Microbial Physiology, v. 49, p. 1-76, 2004.
ROMANDINI, P. , TALLANDINI, L.; BELTRAMINI, M.; SALVATO, B.; MANZANO,
M.; BERTOLDI, M.; ROCCO, G. P. Effects of copper and cadmium on growth, superoxide

Referências Bibliográficas
110
dismutase and catalase activities in different yeast strains. Comparative Biochemistry and
Physiology, v. 103, n. 2, p. 255-262, 1992.
ROMERA, E.; GONZALEZ, F.; BALLESTER, A.; BLAZQUEZ, M. L.; MUNOZ, J. A.
Biosorption with algae: a statistical review. Critical Reviews in Biotechnology, v. 26, n. 4,
p. 223- 235, 2006.
SAMPEDRO, J. G.; URIBE, S. Trehalose-enzyme interactions result in structure
stabilization and activity inhibition. The role of viscosity. Molecular and Cellular
Biochemistry, v. 256/257, p. 319-327, 2004.
SCANDALIOS, J. G. Oxidative stress: molecular perception and transduction of signals
triggering antioxidant gene defense. Brazilian Journal of Medical and Biological
Research, v. 38, p. 995-1014, 2005.
SINGER, M.A.; LINDQUIST, S. Multiple of trehalose on protein folding in vitro and in
vivo. Molecular Cell, v. 1, p. 639-648, 1998.
SINGER, M.A.; LINDQUIST, S. Thermotolerance in Saccharomyces cerevisiae: the yin
and yang of trehalose. Trends in Biotechnology, v. 16, p. 460-468, 1998.
STANDMAN, E. R.; LEVINE, R. L. Protein Oxidation. Annals New York Academy of
Sciences, v. 899, p. 191-208, 2000.
STANDMAN, E. R.; LEVINE, R. L. Free radical-mediated oxidation of free amino acids
and amino acid residues in proteins. Amino Acids, v. 25, p. 207-218, 2003.
SUH, J. H.; YUN, J. W.; KIM, D. S. Comparison of Pb2+ characteristics between live and
dead cells of Saccharomyces cerevisiae and Aureobsidium pullulans. Biotechnology
Letters, v. 20, p. 247-251, 1998.

Referências Bibliográficas
111
SUH, J.H.; YUN, J.W.; KIM, D.S. Cátion (K+, MG2+, Ca2+) exchange in Pb2+ accumulation
by Saccharomyces cerevisiae. Bioprocess Engineering, v. 21, p. 383-387, 1999.
THEVELEIN, J. M. Regulation of trehalose mobilization in fungi. Microbiological
Review, v.48, p. 42-59, 1984.
THEVELEIN, J. M.; HOHMANN, S. Trehalose synthase: guard to the gate of glycolysis in
yeast? Trends in Biochemical Sciences, v. 20, p. 3-10, 1995.
TERESHINA, V. M. Thermotolerance in Fungi: The role of heat proteins and Trehalose.
Microbiology, v. 74, p. 247-257, 2005.
TURRENS, J. F. Superoxide production by the mitochondrial respiratory chain. Bioscience
Reports., v. 17, p. 3-8, 1997.
VALKO, M.; RHODES, C. J.; MONCOL, J.; IZAKOVIC, M.; MAZUR, M. Free radicals,
metals and antioxidant in oxidative stress-induced cancer. Chemical Biological
Interactions, v. 160, p. 1-40, 2006.
VALKO, M.; MORRIS, H.; CRONIN, M. T. D. Metals, toxicity and oxidative stress.
Current Medicinal Chemistry, v. 12, p. 1161-1208, 2005.
VAN AEIST, L.; HOHMANN, S.; BULAYA, B.; KONIN W.; SIERKSTRA, L.; NEVES,
M. J.; LUYTEN, K.; ALIJO, R.; RAMOS, J.; COCCETTI, P. Molecular cloning of a gene
involved in glucose sensing in the yeast Saccharomyces cerevisiae. Molecular
Microbiology, v. 8, p. 927-943, 1993.
VAN LAERE, A. Trehalose, reserve and/or stress metabolite? FEMS Microbiological
Review, v. 63, p. 201-210, 1989.

Referências Bibliográficas
112
VEGLIO, F.; BEOLCHINI, F. Removal of metals by biosorption: a review.
Hidrometallurgy, v. 44, p. 301-316, 1997.
VENDEMIALE, G.; GRATTAGLIANO, I.; ALTOMARE, E. An uptake in the role of free
radicals and antioxidant defense in human disease. International Journal of Clinical &
Laboratory Research, v. 29, p. 49-55, 1999.
VIDO, K.; SPECTOR, D.; LAGNIEL, G.; LOPEZ, S.; TOLEDANO, M. B.; LABARRE, J.
A Proteome Analysis of the Cadmium Response in Saccharomyces cerevisiae. The
Journal of Biological Chemistry, v. 276, n. 11, p. 8469-8474, 2001.
VOIT, E. O. Biochemical and genomic regulation of the trehalose cycle in yeast: review of
observations and canonical model analysis. Journal of Theoretical Biology, v. 223, p. 55-
78, 2003.
VOLESKY, B.; MAY, H.; HOLAN, G. R. Cadmium biosorption by Saccharomyces
cerevisiae. Biotechnology and Bioengineering, v. 41, p. 826-829, 1993.
VUORIO, O. E.; KALKKINEN, N.; LONDESBOROUGH, J. Cloning of two related genes
enconding the 56-kDa and 123-kDa subunits of trehalose synthase from the yeast
Saccharomyces cerevisiae. European Journal of Biochemistry, v. 216, p. 849-861, 1993.
WAALKES, M. P. Cadmium carcinogenesis. Mutation Research, v. 533, p. 107-120,
2003.
WAISBERG, M.; JOSEPH, P.; HALE, B.; BEYERSANN, D. Molecular and cellular
mechanisms of cadmium carcinogenesis. Toxicology, v. 192, p. 95-117, 2003.
WANG, J.; CHEN, C. Biosorption of heavy metals by Saccharomyces cerevisiae: A
review. Biotechnology Advances, v. 24, p. 427-451, 2006.

Referências Bibliográficas
113
WEN, S.; ZHANG, T.; TAN, T. Optimization of the amino acid composition in glutathione
fermentation. Process Biochemistry, v. 40, p. 3474-3479, 2005.
WU, R. F.; XU, Y. C.; MA, Z.; NWARIAKU, F. E.; SAROSI, J. G. A.; TERADA, L. S.
Subcellular targeting of oxidants during endothelial cell migration. Journal of Cell
Biology, v. 171, p. 893-904, 2005.
YAMAGUCHI, T.; TAKAMURA, H.; MATOBA, T.; TERAO, J. HPLC Method for
evaluation of the free radical-scavenging Activity of foods by using 1,1-Diphenyl-2-
picrylhydrazyl. Bioscience, Biotechnology and Biochemistry, v. 62, n.6, p. 1201-1204,
1998.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo