obtenção de um antígeno para ensaio imunoenzimático (elisa
TRANSCRIPT

Universidade Federal de Minas Gerais
Instituto de Ciências Biológicas
Departamento de Parasitologia
Obtenção de um antígeno para Ensaio
Imunoenzimático (ELISA) a ser utilizado no
diagnóstico da Leishmaniose Visceral Canina pelos
órgãos de Saúde Pública no Brasil
Eloísa de Freitas
Belo Horizonte
2010

II
Eloísa de Freitas
Obtenção de um antígeno para Ensaio Imunoenzimático
(ELISA) a ser utilizado no diagnóstico da Leishmaniose
Visceral Canina pelos órgãos de Saúde Pública no Brasil
Tese apresentada ao Programa de Pós
Graduação em Parasitologia do Instituto de
Ciências Biológicas da Universidade Federal
de Minas Gerais, como requisito parcial para
a obtenção do título de Doutor em Ciências
Área de Concentração: Protozoologia
Orientadora: Profª. Drª. Maria Norma Melo
Co- orientadora: Profª. Drª. Marilene Suzan Marques Michalick
Belo Horizonte
Minas Gerais - Brasil
2010

III
043 Freitas, Eloísa de.
Obtenção de um antígeno para ensaio imunoenzimático (ELISA) a ser utilizado no
diagnóstico da Leishmaniose visceral canina pelos órgãos de Saúde Pública no Brasil.
[manuscrito] / Eloísa de Freitas. – 2010.
127 f. : il. ; 29,5 cm
Orientadora: Maria Norma Melo. Co-orientadora: Marilene Suzan Marques
Michalick.
Tese (doutorado) – Universidade Federal de Minas Gerais, Departamento de
Parasitologia.
1. Leishmaniose visceral canina. 2. Leishmania chagasi. 3. Calazar - Teses. 4.
Teste imunoenzimático - Teses. 5. Antígenos – Teses. 6. Diagnóstico humoral. 7.
Parasitologia – Teses. I. Melo, Maria Norma. II. Michalick, Marilene Suzan Marques.
III. Universidade Federal de Minas Gerais, Departamento de Parasitologia. III. Título.
CDU: 576.88/.89

IV
Trabalho desenvolvido nos Laboratórios de Biologia de
Leishmania e de Sorologia do Departamento de
Parasitologia do Instituto de Ciências Biológicas da
UFMG, Financiado em parte pelo CNPq, e em parceria
com o Setor de Pesquisas da Fundação Ezequiel Dias-
FUNED

V
A todos os cães que foram submetidos à
eutanásia vítimas da Leishmaniose Visceral,
Com todo o meu amor dedico este trabalho.

VI
Agradecimentos
A Deus, pela cumplicidade em cada momento da minha vida.
A minha família pelo apoio e amor incondicional.
Às minhas orientadoras, Profas. Dra. Maria Norma Melo e Dra Marilene Suzan
Marques Michalick, pela oportunidade oferecida na orientação deste trabalho, por
acompanharem passo a passo cada etapa, sempre otimistas e confiantes, contagiando a
todos ao seu redor e, principalmente, pela compreensão e apoio oferecidos durante meus
momentos de dificuldades, demonstrando serem, além de excelentes orientadoras,
grandes amigas e seres humanos exemplares. A vocês, minha eterna admiração.
A Profa. Hélida Monteiro de Andrade por me acolher em seu laboratório e aceitar
comigo o desafio, revelando-se uma ótima e competente orientadora, assim como uma
grande amiga. Agradeço também a Simone, pela prestatividade na transmissão de
conhecimentos, paciência e apoio técnico. Angélica, Junior e Felipe, obrigada demais!
Talita, Aliani e Jordanna, obrigada pela amizade.
Ao professor Dr.George Luiz Lins Machado Coelho pela ajuda na análise estatística dos
resultados.
As amigas de laboratório Elza Moreira da Silva, Rosálida Estevam Nazar Lopes,
Ludmila Gomes Muniz e Miriam Costa pela enorme colaboração e apoio técnico,
ofertados sempre com muita prestatividade. No início, colegas de trabalho, mas agora,
amigas para sempre!
A Rosangela Gomes e Soraia de Oliveira Silva, técnicas e amigas, sempre com um
sorriso e respostas certas e naturalmente competentes. Sem a ajuda de vocês nunca
teriam sido possível a realização deste trabalho.
A Sumara, amiga de longa data e companheira desta e outras jornadas que já se
completaram. Não tenho palavras para expressar sua bondade de coração, o apoio e
carinho a mim dedicados, que revelam sua nobreza de caráter. Obrigada por tudo!
A Elizabeth Castro Moreno, pela ajuda no momento importante, pela paciência e
transmissão de conhecimentos.
A Leonardo T. Ituassu pela gentileza na disponibilização dos animais de seu
experimento para coleta de material.
Aos professores e funcionários do Departamento de Parasitologia, pelos conhecimentos
ofertados e momentos de descontração ao longo deste período. Afonso, Sr Neri,
Joãozinho, Edna, vou sentir saudades.
A Dra Adriane Pimenta da Costa Val, companheira de jornada, amiga, mestre, irmã.
A Dra Junia Maria Cordeiro de Menezes, exemplo de pessoa e de profissional, sempre
acessível e nos ensinando com muita humildade e boa vontade. Aprendi e tenho muito a
aprender com você.

VII
A toda turma do Hospital Veterinário, companheiros nesta caminhada e com os quais
escrevemos uma estória... Elizete, Eliana, Ronaldo, Joaquim, Rubens, Gleidice, Luiz
Carlos, Creide, Bretas, Erlaine, Fábio. Vocês são a minha melhor parte nesta escola.
Aos professores Fabíola Paes Leme e Paulo Ricardo, assim como a todos os residentes
que passaram pelo LAC, todos os funcionários, estagiários e bolsistas, em especial a
querida Renata, no apoio pela causa animal. Serei sempre grata a vocês.
A Roxane Rafaela Macedo dos Santos e Evair Nunes da Costa. Obrigada pela ajuda
incessante e pelo exemplo de desprendimento, a amizade de vocês sempre levarei
comigo. Mais um ciclo se fecha, outros se iniciam. E se hoje sou mais forte, vocês me
ajudaram a crescer. Obrigada!
A Juliana Cabral Gomes, Vanessa Jareno Arão e Wylliana Gonçalves Fontes, obrigada
pelos serviços oferecidos, pelo carinho e paciência, pela amizade que se iniciou e por
acreditarem em meu potencial e na minha recuperação.
Ao Sr. Edson, D. Jane, Juliana, Eduardo e Fernanda por me acolherem de forma tão
especial e me fazerem sentir em família e entre amigos.
A Eliane Gonçalves Paiva Lopes, eterna amiga e irmã em Cristo. Sempre pronta com
um conselho e uma palavra de sabedoria. Amiga, te adoro! Obrigada!
A Daniella Naves Paglioni e Sandra Naves Paglionni. Mesmo distantes, sempre estamos
juntas, pois um sentimento muito forte nos une. Benção de Deus em minha vida.
A Sra. Terezinha Antunes Abbas e Salim Abbas, pelo carinho dedicado e a acolhida em
sua casa.
Ao Dr. Silvio Leite, parceiro pra todos os momentos, excelente profissional e grande
amigo.
A Dra Gabrielle Márcia Cury, pela confiança no empréstimo de seu patrimônio que
tanto me ajudou, pela amizade e apoio técnico constantes.
Aos amigos do bairro Jaqueline, Sandro, Leo, Kenia, Sonia, Miriam, Sidnei, Ana
Cristina, Paula, pela convivência e pelos momentos de descontração e amizade.
Aos amigos Cláudia Capistrano, Ana Paula, Angélica, Renata, Tae, Lucia (in
memoriam), Moema, Gregório, Iara, Nino, Sara, Luciana, Márcia, Aparecida, Ivonete,
Vânia, Cida, Ariane, Leoneide, que sempre me apoiaram, acreditaram e torceram por
mim.
A todos que não estão aqui citados, e que são os muitos outros mestres, com ou sem
título, que me ensinaram as diversas facetas da vida. A eles, serei sempre grata.
A Fundação Ezequiel Dias, pela parceria no desenvolvimento deste estudo
A FAPEMIG e ao CNPq, pelo suporte financeiro e disponibilização da bolsa de estudos,
sem os quais não teria sido possível a conclusão deste trabalho.

VIII
Resumo
No Brasil, dentre as medidas de controle preconizadas pelo Ministério da Saúde para
controle da Leishmaniose visceral, a identificação de cães positivos para leishmaniose
visceral canina (LVC), é recomendada, integradas com outras, visto ser este animal um
dos principais elos da cadeia epidemiológica. Os métodos sorológicos utilizados para
identificação dos animais, não são 100% eficazes que pode comprometer esta medida de
controle. Visando à padronização de um antígeno que permita separar de maneira
adequada animais acometidos ou não pela LVC, cento e três animais oriundos do
Centro de Controle de Zoonoses da cidade de Betim foram categorizados como
positivos ou negativos para LVC através de testes parasitológicos e sorológicos,
resultando em cinqüenta e quatro positivos e quarenta e nove negativos para esta
patologia. Estes animais foram doadores de soro para os ensaios de padronização de um
antígeno para o teste de ELISA. Duas cepas de Leishmania (L.) chagasi, BH46 e
BH400, isoladas de paciente humano e canino respectivamente, e uma cepa de
Leishmania (L.) amazonensis, BH6, isolada de paciente humano foram cultivadas em
meio LIT e Alfa-MEM e as massas antigênicas utilizadas para preparar antígenos para
sensibilização de placas de ELISA e lâminas de RIFI, assim como para obtenção de um
extrato antigênico a ser utilizado na eletroforese em gel bidimensional seguida de
Immunobloting. As placas de ELISA foram testadas com 24 horas, 3 meses e 6 meses
de armazenamento em freezer a -20ºC. As lâminas de RIFI foram armazenadas da
mesma forma e utilizadas após 30 dias. Em todas as fases foram determinados para
ELISA e para RIFI, os valores de sensibilidade, especificidade, valor preditivo positivo,
valor preditivo negativo e índice de concordância de Youden. Os resultados foram
comparados com aqueles obtidos com os kits de ELISA e RIFI de Biomanguinhos. A
cepa BH400, independente do meio de crescimento, foi a que apresentou melhor
desempenho em caracterizar corretamente animais positivos e negativos. Esta cepa
mostrou, durante o experimento, valores de sensibilidade e especificidade entre 98-
100%, valor preditivo positivo de 100%, valor preditivo negativo entre 98-100% e
índice de concordância de Youden de 98-100%. Houve reação cruzada com soros de
cães com Trypanosoma cruzi e com Leishmaniose tegumentar, mas o antígeno não
apresentou reação cruzada com Babesia canis e Ehrlichia canis. O meio Alfa-MEM foi
escolhido para cultivo desta cepa e caracterização do perfil proteômico-imunogênico,
devido à facilidade de obtenção comercial e preparo do mesmo. O Immunobloting foi

IX
realizado com pool de soros de alto e baixo títulos de anticorpos e pool de soros
negativos, e revelou mais de cinqüenta “spots” reativos sem reação com soros
negativos, mostrando alta imunogenicidade do antígeno. No gel bidimensional foram
localizados 37 “spots” que poderão futuramente ser identificados por espectrometria de
massa, visando obtenção de proteínas que possam ser utilizadas como antígenos para
ELISA e conferir aumento da sensibilidade e especificidade deste teste.

X
Abstract
In Brazil, control measures of visceral leishmaniasis (VL) recommended by the
Ministry of Health include identification of positive dogs, integrated with others
procedures, since this animal is one of epidemiological chain main links. Serological
methods used for animal identification, are not 100% effective and this fact may
compromise this control measure. Aiming at obtaining an antigen that allows separate
adequately infected animals one hundred and seven animals were categorized as
positive or negative for LV by parasitological and serological tests resulting in fifty-four
positive and forty-nine negative. These animals were serum donors for ELISA and
IFAT serological tests. Two strains of Leishmania (L.) chagasi, BH46 and BH400,
isolated from human and canine patients respectively, and one strain of Leishmania (L.)
amazonensis, BH6, isolated from human patients were grown in LIT and alpha-MEM
mediuns and used to prepare antigens for sensitization of ELISA plates and glass slides
of IFAT, as well as for obtaining an antigenic extract to be used in two-dimensional gel
electrophoresis followed by immunoblotting. ELISA plates were tested at 24 hours, 3
months and 6 months of storage at -20 ° C. IFAT slides were stored in the same way
and used after 30 days. At all stages were determined for ELISA and IFAT, sensitivity,
specificity, positive predictive value, negative predictive value and Youden index of
agreement. The results were compared with those obtained with IFAT and ELISA
Biomanguinhos commercial kits. Cultivation of BH400 strain, regardless of medium,
showed the best performance in correctly characterizes positive and negative animals.
This strain showed during the experiment, 98-100% sensitivity and specificity, 100%
positive predictive value, 98-100% negative predictive value and Youden index of
agreement of 98-100%. There was ELISA cross-reaction with sera from dogs with
Trypanosoma cruzi and cutaneous leishmaniasis, but the antigen showed no cross
reactivity with Babesia canis and Ehrlichia canis. The Alpha-MEM medium was
chosen for cultivation and proteomic -immunogenic profile characterization of this
strain due to the ease of obtaining and preparing this medium. The immunoblotting was
performed with pooled sera from high and low antibody titers and pool of negative sera
and revealed more than fifty “spots” reactive with no reaction with negative sera,
showing high antigen immunogenicity. The two-dimensional gel revealed 37 “spots”
that may be identified by mass spectrometry in order to obtain proteins that can be used
as antigens for ELISA and provide increased sensitivity and specificity of this test.

XI
Lista de Figuras
Figura 1 Curva de crescimento da cepa BH400 de Leismania (Leishmania) chagasi, (A) cultivada em meio LIT (Liver Infusion Tryptose) e (B) em meio α- MEM (Minimum
Essential Medium) acrescido de 10% de SFB .................................................................. 54
Figura 2 Teste de ELISA realizado em soros de animais positivos para leishmaniose visceral
canina, com os antígenos A, B, C, D, E e F, testados 24 horas após sensibilização das placas. ............................................................................................................................. 56
Figura 3 Teste de ELISA realizado em soros de animais negativos para leishmaniose visceral
canina, com os antígenos A, B, C, D, E e F, testados 24 horas após sensibilização das placas .............................................................................................................................. 56
Figura 4 Teste de ELISA realizado em soros de animais positivos para leishmaniose visceral
canina, com os antígenos A, B, C, D, E e F, testados três meses após sensibilização das placas. ............................................................................................................................. 56
Figura 5 Teste de ELISA realizado em soros de animais negativos para leishmaniose visceral
canina, com os antígenos A, B, C, D, E e F, testados três meses após sensibilização das
placas .............................................................................................................................. 56
Figura 6 Teste de ELISA realizado em soros de animais positivos para leishmaniose visceral
canina, antígenos A, B, C, D, E e F, ................................................................................. 56
Figura 7 Teste de ELISA realizados em soros de animais negativos para leishmaniose visceral canina, com o antígenos A, B, C, D, E e F, testados seis meses após sensibilização das
placas .............................................................................................................................. 56
Figura 8 Comparação da sensibilidade, especificidade, valor preditivo positivo e valor preditivo negativo dos antígenos utilizados no teste ELISA 24 horas após a sensibilização das placas.
(A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400 (Leishmania (L) chagasi),
cultivadas em meio LIT; B, BH6 (Leishmania (L.) amazonensis); D e F, BH46 e BH400
Leishmania (L) chagasi, cultivadas em meio α-MEM SFB 10%) ..................................... 60
Figura 9 Comparação da sensibilidade, especificidade, valor preditivo positivo e, valor preditivo
negativo dos antígenos utilizados no teste ELISA três meses após a sensibilização das
placas. (A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400 (Leishmania (L) chagasi), cultivadas em meio LIT; B, BH6 (Leishmania (L.) amazonensis); D e F, BH46 e
BH400 Leishmania (L) chagasi, cultivadas em meio α-MEM SFB 10%) ......................... 61
Figura 10 Comparação da sensibilidade, especificidade, valor preditivo positivo e, valor
preditivo negativo dos antígenos utilizados no teste ELISA seis meses após a sensibilização das placas. A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400
(Leishmania (L) chagasi), cultivadas em meio LIT; B, BH6 (Leishmania (L.)
amazonensis); D e F, BH46 e BH400 Leishmania (L) chagasi, cultivadas em meio α-MEM SFB 10%) ....................................................................................................................... 61
Figura 11 Teste da RIFI realizados em soros de cães portadores de leishmaniose visceral com
diferentes antígenos. (A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400 (Leishmania (L) chagasi), cultivadas em meio LIT; B, BH6 (Leishmania (L.)
amazonensis); D e F, BH46 e BH400 Leishmania (L) chagasi, cultivadas em meio α-MEM
SFB 10%) ....................................................................................................................... 63
Figura 12 Teste da RIFI realizados em soros de cães negativos para leishmaniose visceral com diferentes antígenos. (A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400
(Leishmania (L) chagasi), cultivadas em em meio LIT; B, BH6 (Leishmania (L.)
amazonensis); D e F, BH46 e BH400 Leishmania (L) chagasi, cultivadas em em meio α-MEM SFB 10%) ............................................................................................................. 63

XII
Figura 13 Comparação da sensibilidade, especificidade, valor preditivo positivo e, valor
preditivo negativo dos antígenos utilizados na Reação de Imunofluorescência Indireta –
RIFI. ............................................................................................................................... 64
Figura 14 Teste de ELISA realizado em soros verdadeiramente positivos e negativos para
leishmaniose visceral canina com o kit Biomanguinhos, LOTE 086EL007Z .................... 65
Figura 15 Teste de RIFI realizado em amostras de soros positivos e negativos para leishmaniose visceral canina como kit Biomanguinhos, LOTE 086LC004Z. ......................................... 65
Figura 16 Comparação da sensibilidade, especificidade, valor preditivo positivo e, valor
preditivo negativo dos antígenos dos kit-ELISA Biomanguinhos LOTE 086EL007Z e kit-
RIFI Biomanguinhos LOTE 086LC004Z ......................................................................... 66
Figura 17 Relação entre os valores de absorbância do ELISA, após 24 horas de sensibilização
das placas, e os títulos das diluições da RIFI e a tendência das linhas de regressão para os
antígenos A, B, C, D, E e F.............................................................................................. 67
Figura 18 Relação entre os valores de absorbância do ELISA, após três meses de sensibilização
das placas, e os títulos das diluições da RIFI e a tendência das linhas de regressão para os
antígenos A, B, C, D, E e F.............................................................................................. 68
Figura 19 Relação entre os valores de absorbância do ELISA, após seis meses de sensibilização
das placas, e os títulos das diluições da RIFI e a tendência das linhas de regressão para os
antígenos A, B, C, D, E e F ............................................................................................. 69
Figura 20 Comparação das linhas de tendência dos antígenos E e F, mostrando a relação entre os valores de absorbância do ELISA, 24 horas após a sensibilização das placas, e os títulos
das diluições da RIFI ....................................................................................................... 70
Figura 21 Comparação das linhas de tendência dos antígenos E e F, mostrando a relação entre os valores de absorbância do ELISA, três meses após a sensibilização das placas, e os
títulos das diluições da RIFI ............................................................................................ 71
Figura 22 Comparação das linhas de tendência dos antígenos E e F, mostrando a relação entre
os valores de absorbância do ELISA, seis meses após a sensibilização das placas, e os títulos das diluições da RIFI ............................................................................................ 72
Figura 23 Gel 2-DE contendo 600g de proteínas de Leishmania chagasi (cepa BH400 crescida em meio α-MEM) fracionadas em tiras de 13 cm pH 4-7 (direita para esquerda) e SDS-
PAGE a 12%. O gel foi corado por comassie (A) ou transferido para membrana de nitrocelulose e incubadas com soros de animais não infectados (B) e infectados que
apresentavam baixos títulos de anticorpos (C) e altos títulos de anticorpos (D). O
conjugado utilizado foi anti-IgG total de cão. Em cada membrana estão destacados os
“spots” detectados por cada grupo de soros. ..................................................................... 74
Figura 24 Gel 2-DE contendo 600g de proteínas de Leishmania chagasi (cepa BH400 crescida em meio α MEM) fracionadas em tiras de 13 cm pH 4-7 (direita para esquerda) e SDS-
PAGE a 12% corado por comassie. Em destaque em vermelho, os “spots” reativos frente a
soros de cães não infectados, em azul “spots” reativos frente a cães que apresentam baixos
títulos de anticorpos e em preto os “spots” reativos frente a soros de cães que apresentam altos títulos de anticorpos. ............................................................................................... 76

XIII
Lista de Tabelas
Tabela 1: Categorização dos animais pré-selecionados como portadores ou não de
leishmaniose visceral, após avaliação parasitológica e sorológica. ........................ 53
Tabela 2 Codificação dos antígenos produzidos com as diferentes amostras de
Leishmania crescidas nos meios de cultura LIT (Camargo, 1966) e Alfa-MEM
acrescido de 20% de soro fetal bovino.................................................................. 55
Tabela 3 Teste de ELISA nos tempos de 24 horas, três e seis meses após a
sensibilização das placas, com os antígenos A, B, C, D, E e F, em soros caninos
padrões positivo e negativo para leishmaniose visceral. ........................................ 58
Tabela 4 Valores percentuais e absolutos de número de amostras classificadas como
Zona Cinza, acima e abaixo do ponto de corte do ELISA, durante todas as fases do
experimento. ........................................................................................................ 59
Tabela 5 Teste de ELISA em soro de cães portadores de Leishmaniose Tegumentar,
Babesiose, Erlichiose e Doença de Chagas com o antígeno F ............................... 73
Tabela 6 “spots” observados na análise proteômica do antígeno da cepa BH400 crescida
em meio Alfa-MEM. ............................................................................................ 75

XIV
LISTAS DE ABREVIATURAS E SIGLAS
2-DE – eletroforese bidimensional em gel 2-D
- alfa
X – vezes
® - Registrado
nm – nanômetro
µl – microlitro
µm – micrometro
mm – milímetro
n – Número de animais por grupo
g – micrograma
mg – miligrama
g – grama
g/dl – grama por decilitro
g/l – grama por litro
kg – kilograma
ml – mililitro
rpm – rotação por minuto
-MEM - -Minimum Essential Medium
CCZ – Centro de Controle de Zoonoses
CD4 - Marcador de superfície celular da subpopulação de linfócitos T auxiliares
CD8 - Marcador de superfície celular da subpopulação de linfócitos T citotóxicos
CO2 – Dióxido de carbono
DNA – Ácido desoxirribonucléico

XV
EM- espectrometria de Massa
ELISA – Enzyme Linked Immunosorbent Assay (Ensaio Imunoenzimático)
EDTA – Ácido etilenodiaminotetracético (anticoagulante)
IC – intervalo de confiança
ICB – Instituto de Ciências Biológicas
IFN-γ- Interferon-gama
IgA – Imunoglobulina da classe A
IgE – Imunoglobulina da classe E
IgG - Imunoglobulinas G
IgG1 – Imunoglobulinas da subclasse G1
IgG2 – Imunoglobulinas da subclasse G2
IgM – Imunoglobulina da classe M
IL-2 – Interleucina 2
IM – Intramuscular
IV – Intravenosa
K39 – Antígeno recombinante K39
LIT - Liver Infusion Tryptose
LV - Leishmaniose Visceral
LVC - Leishmaniose Visceral Canina
MG – Minas Gerais
MS – Ministério da Saúde
NNN - NICOLE, NOVY e NEAL
OMS – Organização Mundial de Saúde
PBS – Phosphate Buffer Saline
PCR - Polymerase Chain Reaction (Reação em Cadeia da Polimerase)
pH – Potencial hidrogeniônico

XVI
PVC - Cloreto de polivinil
PVPI - Polivinil pirrolidona iodo 10%
RIFI - Reação de Imunofluorescência Indireta
RNA – Acido Ribonucléico
ROC - Receiver operating characteristic
SC – Subcutânea
SFB- soro fetal bovino
SFM - Sistema fagocítico mononuclear
SRD - Sem Raça Definida
Th – Célula T helper
TNF - Fator de Necrose Tumoral
UFMG – Universidade Federal de Minas Gerais
UI/L - Unidade Internacional por litro
WHO – World Health Organization (Organização Mundial de Saúde)

XVII
Sumário
1. INTRODUÇÃO ..................................................................................................... 15
1.1 Leishmanioses ....................................................................................................... 15
1.2 Ciclo de vida da Leishmania sp. ............................................................................ 16
1.3. Leishmanioses no Brasil ....................................................................................... 18
1.3.1 Leishmaniose Tegumentar (LT).......................................................................... 18
1.3.2. Leishmaniose Visceral....................................................................................... 20
1.3.2.1 Leishmaniose Visceral Canina .................................................................. 22
1.3.2.2 Manifestações Clínicas ............................................................................. 22
1.4 Diagnóstico da Leishmaniose Visceral Canina ...................................................... 25
1.4.1 Diagnóstico Clínico ............................................................................................ 25
1.4.2 Diagnóstico Laboratorial .................................................................................... 26
1.4.2.1 Diagnóstico Parasitológico ....................................................................... 26
1.4.2.2 Diagnóstico Imunológico ......................................................................... 27
1.5 Controle da Leishmaniose Visceral no Brasil ........................................................ 33
2. JUSTIFICATIVA.................................................................................................. 35
3. OBJETIVOS ......................................................................................................... 38
3.1.Objetivo Geral ....................................................................................................... 38
3.2. Objetivos Específicos ........................................................................................... 38
4. MATERIAIS E MÉTODOS ................................................................................. 39
4.1 Delineamento Experimental .................................................................................. 39
4.1.2 Seleção dos Animais .......................................................................................... 39
4.1.3 Obtenção dos parasitos ....................................................................................... 39
4.2 Seleção dos Animais e Coleta de Material ............................................................. 40
4.3 Avaliação Parasitológica dos Animais ................................................................... 41
4.3.1 Esfregaço por extensão ....................................................................................... 41
4.3.2 Cultura de medula óssea ..................................................................................... 41
4.3.3 Reação em cadeia da polimerase - PCR .............................................................. 41
4.4 Preparo dos Antígenos........................................................................................... 44
4.4.1 Curva de Crescimento ........................................................................................ 44
4.4.2 Obtenção de parasitos para o Preparo dos Antígenos .......................................... 45
4.4.2.1 Preparação do Antígeno para Sensibilização das Placas de ELISA ........... 45
4.4.2.2 Obtenção de Parasitos e Preparação do antígeno para RIFI ....................... 46
4.4 2.3 Preparação do Antígeno para Eletroforese-Extração de Proteínas ............. 46
4.5 Testes dos Antígenos ............................................................................................. 47
4.5.1 Reação de Imunofluorescência Indireta- RIFI ..................................................... 47
4.5.2 Enzime Linked Immunosorbent Assay - ELISA ................................................. 48
4.5.2.1 Curva ROC - Receiver Operating Characteristic ....................................... 49
4.5.3 Eletroforese Bidimensional (2-DE) ..................................................................... 50
4.5.3.1 Focalização Isoelétrica (IEF) – ................................................................. 50

XVIII
4.5.3.2 SDS-PAGE .............................................................................................. 50
4.5.3.3 Western blot - 2D gel ............................................................................... 50
4.5.3.4 Análise dos Géis- ..................................................................................... 51
4.6 Análise Estatística ................................................................................................. 51
5. RESULTADOS ..................................................................................................... 53
5.1 Seleção dos animais .............................................................................................. 53
5.2 Curva de Crescimento e Produção dos Antígenos .................................................. 53
5.3 Teste dos Antígenos .............................................................................................. 55
5.3.1 Reação de ELISA ............................................................................................... 55
5.3.2 Reação de Imunofluorescência Indireta .............................................................. 61
5.4 Reações de RIFI e ELISA com os kits de Biomanguinhos ..................................... 64
5.5 Relação entre os títulos da RIFI e os valores de absorbância do ELISA ................. 66
5.5.1 Avaliação dos antígenos com melhor desempenho nos testes de ELISA com 24
horas, três meses e seis meses após a sensibilização das placas. ............................ 70
5.6 Teste de Reações Cruzadas .................................................................................... 73
5.7 Escolha do melhor antígeno................................................................................... 73
5.8 Western blot - 2D gel ............................................................................................ 73
6. DISCUSSÃO ......................................................................................................... 77
7. PERSPECTIVAS FUTURAS ............................................................................... 94
8. CONCLUSÕES ..................................................................................................... 95
9. REFERENCIAS BIBLIOGRAFICAS ................................................................. 96
10 . ANEXOS........................................................................................................... 111
10.1 Depósito da Patente do Antígeno ....................................................................... 111
10.2 Certificado do Comitê de Ética em Experimentação Animal – CETEA ............. 114
10.3 Artigo Publicado ............................................................................................... 115

1. Introdução
1.1 Leishmanioses
Leishmanioses são doenças parasitárias geralmente, de caráter zoonótico,
causadas por protozoários do gênero Leishmania, Ross, 1903, pertencentes à Família
Trypanosomatidae (Woodcock, 1906), Ordem Kinetoplastida (Honigberg, 1963;
Vickerman, 1976), Filo Sarcomastigophora (Halstead, 1965) e Subreino Protozoa. Estes
parasitos digenéticos são transmitidos ao homem pela picada da fêmea infectada do
inseto vetor, pertencente à Ordem Diptera, Família Psychodidae e Subfamília
Phlebotominae e aos gêneros Lutzomyia em países do Novo Mundo, e Phlebotomus, em
países do Velho Mundo (Lainson & Rangel, 2005).
Com uma estimativa de prevalência global de 14 milhões de casos, e largamente
distribuídas em cinco continentes, as leishmanioses são endêmicas em 90 países.
Destes, 72 estão em desenvolvimento e neste grupo, estão incluídos 13 com a menor
taxa de desenvolvimento mundial (Desjeux, 2004). Cerca de 90% dos casos de
leishmaniose visceral em todo mundo são registrados na Índia, Nepal, Sudão
Bangladesh e com menos evidência no Brasil, sendo as áreas suburbanas e rurais destes
países as mais afetadas.
A leishmaniose tegumentar, por sua vez, afeta principalmente sete países: Brasil,
Afeganistão, Iran, Peru, Arábia Saudita, Síria e Argélia, sendo os mesmos responsáveis
por cerca de 90% dos casos desta forma da doença (Monteiro et al., 1994; WHO, 2000;
Desjeux, 2004). A estimativa é que 12 milhões de pessoas estejam acometidas pela
doença e que 350 milhões estejam expostas ao risco de infecção. Acredita-se que, a cada
ano, surjam 1,5 milhões de casos de leishmaniose tegumentar e 500 mil novos casos de
leishmaniose visceral (WHO, 2000; Alvar et al., 2004; Desjeux, 2004).
Considerada pela Organização Mundial de Saúde como uma das principais
doenças negligenciadas, as leishmanioses são endêmicas em regiões tropicais,
subtropicais e temperadas do Velho e do Novo Mundo. O Programa Especial para
Pesquisa e Treinamento em Doenças Tropicais (TDR), que tem como foco as doenças
negligenciadas, que afetam populações pobres e marginalizadas, coloca as
leishmanioses como afecções prioritárias entre as endemias abordadas pelo programa,

16
sendo considerada a segunda doença, em importância médica, entre as doenças causadas
por protozoários, vindo logo depois da malária (WHO, 2007).
Aproximadamente 21 espécies de Leishmania, pertencentes aos subgêneros
Leishmania e Viannia (esse último presente apenas no Novo Mundo), podem infectar o
homem, dando lugar a uma ampla variedade de quadros clínicos. As manifestações
clínicas são dependentes de vários fatores, como a espécie de Leishmania envolvida, sua
virulência, vetores e aspectos ligados ao hospedeiro, como sua resposta imune e seu
estado nutricional, entre outros (Reithinger et al., 2007; Piscopo et al., 2006).
A infecção caracteriza-se pelo parasitismo das células do sistema fagocítico
mononuclear (SFM) da derme, das mucosas ou de órgãos como o fígado, baço, medula
óssea e outros órgãos do hospedeiro vertebrado, estando associada à alta morbidade na
forma tegumentar e a alta mortalidade na forma visceral. As manifestações clínicas da
infecção variam desde infecções inaparentes ou subclínicas, a um amplo espectro de
manifestações, envolvendo o tegumento (pele e mucosas) com lesões de diferentes
graus de gravidade, podendo se apresentar sob forma de lesão cutânea localizada, com
cura espontânea, até lesões mucosas desfigurantes, graves e ainda, apresentar-se sob a
forma de lesões nos órgãos internos, sobretudo as vísceras, com quadros graves muitas
vezes fatais, quando não tratados (Alvar et al., 2004).
1.2 Ciclo de vida da Leishmania sp.
O gênero é constituído por protozoários que possuem um ciclo de vida
heteroxeno, sendo encontrados no trato digestório do hospedeiro invertebrado (insetos
hematófagos da família Psychodidae da subfamília Phlebotominae), nas formas
flageladas denominadas promastigotas e paramastigotas, reproduzindo-se por divisão
binária. Estas formas passam por mudanças morfológicas, fisiológicas e bioquímicas,
diferenciando-se no final do ciclo, pelo menos em parte, em promastigotas metacíclicas
infectantes que invadem o esôfago e faringe do inseto, mas não se multiplicam e não se
ligam ao epitélio intestinal do tubo digestório dos flebotomíneos (Killick-Kendrick,
1979; Bates & Rogers, 2004; Glossage et al., 2003; Bates, 2007). A infecção da fêmea
de L. longipalpis, por exemplo, ocorre quando esta, ao exercer o repasto sanguíneo em
um hospedeiro vertebrado infectado, ingere juntamente com componentes tissulares,
formas amastigotas livres ou presentes no interior de macrófagos da derme. (Lainson &
Rangel, 2005). As fêmeas tornam-se infectantes três a quatro dias após o repasto
sanguíneo (Lainson & Rangel, 2005).

17
Os vetores são específicos, bem definidos e variações nas moléculas de
superfície dos parasitos facilitam o sucesso da infecção no vetor apropriado (Sacks,
1992). Nos hospedeiros mamíferos os parasitos são encontrados na forma aflagelada
denominada amastigota, no interior de células fagocíticas mononucleares,
principalmente macrófagos. Como hospedeiros mamíferos estão incluídos grande
variedade de animais: roedores, edentados (tatu, tamanduá, preguiça), canídeos,
marsupiais e primatas, incluindo o homem. Quando o flebotomíneo infectado com
Leishmania sp. alimenta-se por telmatofagia (sangue, líquido intersticial e restos
celulares) em um mamífero susceptível, inocula junto com sua saliva as formas
infectantes promastigotas metacíclicas presentes na probóscide. A infecção dependerá
do número de formas metacíclicas presentes nas peças bucais neste momento e da
imunocompetência do hospedeiro. A saliva de L. longipalpis, por exemplo, apresenta
importância fundamental no processo de infecção, pois contém maxidilam, que entre
outras atividades apresenta capacidade vasodilatadora e antiplaquetária, mantendo o
fluxo sanguíneo e dificultando a coagulação, facilitando desta forma a inoculação dos
parasitos (Killick-Kendrick, 2002).
Os parasitos que são então fagocitados, primeiramente, por neutrófilos, perdem o
flagelo transformando-se em amastigotas, e horas depois são fagocitados por
macrófagos, processo mediado por receptores (Cunningham, 2002; de Almeida, 2003).
Os parasitos são internalizados no vacúolo parasitóforo, que logo se fusiona com os
lisossomos, dando origem ao vacúolo fagolissosomal, onde o parasito permanece em
um ambiente hostil, contendo enzimas lisossomais e metabólitos reativos do oxigênio,
mecanismos de sinalização celular, produção de óxido nítrico e citocinas (Cunningham,
2002).
No entanto, Leishmania sp. está adaptada para sobreviver neste ambiente, pois é
capaz de inibir diversos mecanismos de defesa celular que deveriam causar sua lise, e
então se multiplicar por divisão binária, até levar a ruptura da célula infectada e liberar
formas amastigotas que são fagocitadas por outros macrófagos (Peters, 2008). Ocorre
então a disseminação linfática ou sanguínea para outros tecidos ricos em células do
sistema mononuclear fagocitário (Sacks & Noben-Trauth, 2002; Lainson & Rangel,
2005; Little, 2006). A infecção, nas formas viscerais, dissemina-se para linfonodos,
baço e medula óssea dentro das primeiras horas (Kontos, 1993; Baneth, 2006).

18
Em camundongos demonstrou-se que os macrófagos, as células dendríticas e as
células de Langerhans, infectadas por parasitas, atuam como células apresentadoras de
antígenos face às células imunocompetentes, particularmente aos linfócitos T-auxiliares
ativando linfócitos (CD4+) Th1 ou Th2. Se forem ativadas preferencialmente as células
Th1 ocorre a produção de citocinas pró-inflamatórias, as quais aumentam a eficiência
das células fagocíticas e de linfócitos citotóxicos, podendo haver controle do
parasitismo, com a eliminação da infecção. Em contraste, quando a infecção está
associada à indução de linfócitos Th2, ocorre a produção de citocinas antiinflamatórias,
proliferação de células B com subseqüente produção de imunoglobulinas, sobretudo
IgG e IgM e desenvolvimento da infecção (Wilson, 1993; Pinelli et al., 1994; Feitosa et
al., 2000; Ferrer, 2002a; Ciaramella, 2003). Potencialmente, quando há ativação de
linfócitos Th2 há uma atividade exuberante de linfócitos B com a geração de uma
grande quantidade de imunocomplexos circulantes, causando danos em vários órgãos
(Kontos, 1993; Ferrer et al., 1995; Noli, 1999; Feitosa et al., 2000; Ciaramella, 2003;
Baneth, 2006). Na enfermidade natural, estão ativados tanto os subtipos Th1 quanto
Th2, sendo que a variedade dos sintomas e a gravidade da doença dependem do
equilíbrio entre esses subtipos de linfócitos T(Pinelli et al., 1994; Feitosa et al., 2000;
Ezquerra, 2001).
1.3. Leishmanioses no Brasil
No Brasil as leishmanioses podem provocar quadros de lesões cutâneas,
mucosas, ou viscerais, e pode ser classificada em grupos, de acordo com suas
manifestações clínicas (Silveira et al., 2004).
1.3.1 Leishmaniose Tegumentar (LT)
A Leishmaniose Tegumentar (LT) é uma doença infecciosa, não contagiosa,
causada por diferentes espécies de Leishmania sp. que acometem pele e mucosas. O
número de casos no mundo tem aumentado durante as últimas décadas, e este aumento
pode ser explicado, em parte, pelos diagnósticos não confirmados, pela ausência de
notificações de casos, mas é também resultado do controle inadequado de vetores ou
reservatórios, o que aumenta a detecção da LT associada a infecções oportunistas.
Contudo, pelo fato de algumas infecções serem assintomáticas ou não diagnosticadas, o
número global de casos de LT pode estar sendo subestimado.

19
Os ciclos de transmissão têm se adaptado ao peridomicílio e expandido para
áreas não endêmicas, como resultado da urbanização e desmatamento, sendo os animais
domésticos potenciais reservatórios. Adicionalmente, problemas econômicos, desastres
naturais, conflitos entre povos e o turismo, causam migrações de populações
susceptíveis para áreas endêmicas de LT. Estas populações tornam-se expostas ao risco
de infecção, o que facilmente resulta em epidemias (Reithinger et al., 2007).
Nas Américas, a LT, globalmente conhecida como Leishmaniose Tegumentar
Americana (LTA) é primariamente, uma infecção zoonótica, afetando outros animais
que não o ser humano, o qual pode ser envolvido secundariamente. Apresenta ampla
distribuição, com registro de casos em todas as regiões brasileiras (Brasil – Ministério
da Saúde, 2007).
A LTA clinicamente, se apresenta sob diferentes formas que podem ser
subdivididas em leishmaniose cutânea localizada (LCL), leishmaniose cutânea difusa
(LCD), leishmaniose cutânea disseminada (LCDs) e leishmaniose mucocutânea (LMC)
(Silveira et al., 2004).
Dentre os aspectos clínicos da LTA destacam-se lesões de pele com úlceras
localizadas ou múltiplas, o que representa a forma mais freqüente da doença. A LTA
pode ter como agente etiológico qualquer espécie dos subgêneros que podem causar
formas tegumentares no homem, no Novo Mundo. No entanto, a Leishmania (Viannia)
braziliensis é considerada como a mais prevalente, associada a esta forma de doença.
Apresenta acometimento primário da pele, com lesão única ou lesões múltiplas,
geralmente do tipo ulcerado, que algumas vezes tem tendência à cura espontânea ou
apresenta boa resposta ao tratamento.
A LCD é caracterizada por lesões nodulares disseminadas por todo o corpo, ricas
em amastigotas (Silveira et al., 2004). Constitui uma forma clínica rara, porém grave,
que ocorre em pacientes com anergia e deficiência específica na resposta imune celular
a antígenos de Leishmania (Brasil, Ministério da Saúde - 2007).
A LCDs, também é uma manifestação relativamente rara, caracterizada pelo
aparecimento de múltiplas e até centenas de lesões papulares e de aparência acneiforme,
que acometem vários segmentos corporais, envolvendo com freqüência a face e o
tronco.

20
A LMC manifesta-se por lesões destrutivas localizadas nas mucosas das vias
aéreas superiores (Silveira et al., 2004). Geralmente surge após a cura clínica da LTA e
na maioria dos casos, resulta de uma LTA de evolução crônica e clinicamente resolvida
(lesões cicatrizadas) com ou sem tratamento ou com tratamento inadequado, mas pode
aparecer concomitantemente à lesão cutânea. (Brasil, Ministério da Saúde - 2007).
Aproximadamente 30 espécies ou subespécies de flebotomíneos são vetores
comprovados de leishmanioses, e mais de 40 espécies estão provavelmente envolvidas
na transmissão (Lainson & Rangel, 2005). No Brasil, as principais espécies de
flebotomíneos envolvidas na transmissão da LTA são: Lutzomyia flaviscutellata, L.
whitmani, L. umbratilis, L. intermedia, L. wellcomei e L. migonei (Shaw, 2003).
1.3.2. Leishmaniose Visceral
A Leishmaniose Visceral (LV) ou Calazar é considerada a forma mais grave da
doença, estando associada à alta mortalidade quando não tratada. É causada por espécies
de Leishmania, pertencentes ao complexo L. (Leishmania) donovani: L. (L.) donovani,
L. (L.) infantum e L.(L.) chagasi, sendo que estas duas últimas foram consideradas por
alguns autores como sendo a mesma espécie, pois a comparação destes parasitos
utilizando-se de métodos bioquímicos e moleculares não demonstrou diferenças entre os
mesmos (Maurício et al., 2000; 2001).
No Brasil o agente etiológico da LV é a L. (L) chagasi (=L. (L.)
infantum). Os insetos vetores vivem em ambientes variados, mas suas formas imaturas
se desenvolvem em ambientes terrestres úmidos, ricos em matéria orgânica e de baixa
incidência luminosa (Rey, 2001b; Neves, 2005). Têm atividade no crepúsculo matutino
e vespertino para picar os seus hospedeiros (Little, 2006). Duas espécies estão
relacionadas com a transmissão de L. chagasi: Lutzomyia longipalpis, que é considerada
a principal transmissora e L. cruzi, descrita como vetora no Estado de Mato Grosso do
Sul (Elkhoury, 2005).
Os reservatórios estão restritos a um grupo específico de mamíferos,
particularmente os pertencentes à família Canidae, podendo ainda ter envolvimento de
outros animais reservatórios secundários. As raposas Dusicyon vetulus (=Lycalopex
vetulus), no Nordeste (Deane, 1956) e Cerdocyon thous, na Amazônia (Curi et al., 2006;
Lainson & Shaw, 1987) são os principais reservatórios silvestres. Como reservatório
doméstico, o cão (Canis familiaris) é a principal fonte de infecção para os
flebotomíneos, sendo considerado o responsável pela persistência da LV nas regiões

21
neotropicais (Melo, 2004), atribuindo-se aos demais canídeos a responsabilidade pela
transmissão e manutenção do ciclo silvestre da doença (Melo, 2004). L. chagasi já foi
descrita também em outros animais como os marsupiais (Didelphis albiventris e D.
marsuialis), roedores (Rattus rattus, Nectomys squamipes, Proechimys canicollis) e
gato doméstico (Felis catus) (Dantas-Torres & Brandão-Filho, 2006; Silva et al., 2010).
Entretanto, o papel destes animais em relação à epidemiologia da LV carece de mais
estudos, para determinar sua relevância no contexto da transmissão ao homem (Dantas-
Torres & Brandão-Filho, 2006; Silva et al., 2010).
Ao contrário do que ocorre com L. donovani na Índia, na transmissão de L.
chagasi, o homem serve apenas como fraca fonte de infecção para os flebotomíneos. Já
o cão e as raposas por apresentarem intenso parasitismo cutâneo, serem altamente
susceptíveis à infecção são os principais reservatórios, soma-se a isto, no caso dos cães
que sua convivência e proximidade ao homem, preenchem as condições necessárias
para serem considerados reservatórios da doença no ambiente peridomiciliar e urbano
(Deane & Deane, 1954; Melo, 2004; Dantas-Torres & Brandão-Filho, 2006). Outros
fatores a serem considerados, de igual importância no favorecimento da infecção dos
vetores, baseiam-se nas evidências que a LV é mais prevalente na população canina que
na humana, e também a constatação de que os casos humanos normalmente são
precedidos por casos caninos (Castro, 1996; Santa Rosa, 1997; Scott et al, 2001;
Baneth, 2006).
No passado a LV era descrita como uma doença de ambientes silvestres ou
rurais, entretanto, as transformações ambientais provocadas pelo intenso processo
migratório e o processo de urbanização crescente levaram a uma expansão das áreas
endêmicas, com o aparecimento de novos focos, apontando a enfermidade como uma
doença reemergente (Castro, 1996; Alves, 2004; Melo, 2004).
Cerca de 90% dos casos humanos de LV das Américas ocorrem no Brasil, sendo
que a doença já foi identificada em 25 unidades da Federação, em todas as regiões
geográficas. Mostra ainda, nítida expansão com crescente número de casos, resultado de
fatores como urbanização acelerada e alterações do meio ambiente. A doença
concentrou no Nordeste, na década de 1990-2000, onde ocorreram quase 90% de todos
os casos registrados no país, especialmente nos estados do Piauí, Maranhão, Bahia e
Ceará, devido a fatores climáticos favoráveis, temperatura e vegetação (Bavia et al.,
2005). Entre 2000 e 2009 foram registrados 44.440 casos em todo o país (Brasil-

22
Ministério da Saúde, 2010). Entretanto, a região Nordeste representou 47,5% dos casos,
seguida pelas regiões Norte (19,2%), Sudeste (17,4%), Centro-Oeste (7,4%) e Sul
(0,2%).
Após o registro da primeira epidemia em Terezina, no Piauí, na década de 1980,
o processo de urbanização se intensificou com a ocorrência de importantes epidemias
em várias cidades da região Nordeste, como São Luís, Natal, e Aracaju; Norte (Boa
Vista e Santarém); Sudeste (Belo Horizonte, Montes Claros e Araçatuba) e Centro
Oeste (Cuiabá e Campo Grande). Atualmente são 1235 municípios brasileiros atingidos
pela doença e em 130 a condição de transmissão é considerada intensa pela
classificação do Ministério da Saúde (Brasil-Ministério da Saúde, 2010).
Em Minas Gerais a doença ocorre desde a década de 1940, quando foram
registrados os primeiros casos humanos na região norte do estado. Nessa época a doença
era tipicamente rural. Atualmente, 75% dos casos notificados em Minas Gerais
correspondem à pacientes residentes em zonas urbanas. Em 1989 a doença foi
notificada na região metropolitana da capital mineira (Melo, 2004). Belo Horizonte
experimenta hoje duas situações epidêmicas que caracterizam a LV, a epidemia humana
e a epidemia canina. De 1994 a 2009 foram notificados 1.215 casos humanos de
leishmaniose visceral, sendo que de 2005 a 2009 ocorreram 658 casos com 85 óbitos.
Embora os dados possam não representar a realidade do município de Belo Horizonte,
dadas às dificuldades de coleta e diagnóstico, no mesmo período foram coletadas
705.592 amostras de material canino e destas, 57.570 (8,1%) estavam positivas para
leishmaniose (PBH, 2010 http//portalpbh.pbh.gov.br)
No Estado de São Paulo, a doença canina foi identificada pela primeira vez no
município de Araçatuba no ano de 1998 e, desde então, já foi descrita em 41
municípios, difundindo-se para outras regiões do estado, inclusive com casos autóctones
identificados na região metropolitana da cidade de São Paulo a partir do ano de 2005
(São Paulo, 2003; Reichman, 2006).
1.3.2.1 Leishmaniose Visceral Canina
1.3.2.2 Manifestações Clínicas
Admite-se, para a Leishmaniose Visceral Canina (LVC), um período de
incubação de três meses a vários anos e uma apresentação clínica nos animais

23
acometidos, partindo de total ausência de sinais até graves sintomas clínicos (Alvar et
al., 2004; Grima, 2005). Os animais podem ser classificados em: assintomáticos,
oligossintomáticos e sintomáticos (Mancianti et al.,1988). Uma importante
característica da doença canina é a habilidade do cão permanecer clinicamente sem
sinais da doença por longos períodos (Marzochi et al.,1986).
Cerca de 50% a 60% de todos os cães portadores de formas amastigotas não
exibem qualquer sinal clínico da doença, e 20% destes animais apresentam parasitos na
pele (Alvar et al., 2004; Baneth, 2006). Com o passar do tempo, pode haver progressão
para doença sintomática, ao menos que a replicação das amastigotas seja cessada por
mecanismos imunológicos. Aproximadamente 15% dos animais infectados são
clinicamente resistentes, ou seja, capazes de recuperar dos sintomas clínicos e eliminar
os parasitos espontaneamente (Alvar et al., 2004), enquanto outros, considerados
suscetíveis, são predispostos à desenvolverem uma infecção e doença sintomática
(Little, 2006).
As manifestações clínicas da LVC geralmente são classificadas como sendo do
tipo visceral ou cutânea, sendo que em alguns casos há sobreposição destas
manifestações (Amusategui et al., 2003; Baneth, 2006).Os sintomas variam em
decorrência dos mecanismos imunológicos ativados pelo hospedeiro, e também em
relação aos órgãos acometidos. O período inicial da infecção, geralmente cursa como
uma síndrome geral inespecífica, que pode passar despercebida (Grima, 2005). Os sinais
clínicos no cão e no homem são muito parecidos e, freqüentemente, incluem alterações
inespecíficas tais como, apatia, emagrecimento progressivo, hiporexia ou anorexia,
linfoadenomegalia, hepatoesplenomegalia e hipertermia (Feitosa et al, 2000).
Linfoadenomegalia generalizada e esplenomegalia são alterações bastante comuns
causadas por uma proliferação intensa de linfócitos B, histiócitos e macrófagos em
resposta à presença do parasito (Kontos, 1993; Strauss-Ayali et al., 2000; Ciaramella,
2003). O aumento da produção de imunoglobulinas não é protetor e potencialmente
pode ser prejudicial para o animal, sendo um dos resultados da hipergamaglobulinemia,
a formação de grande quantidade de imunocomplexos circulantes que, uma vez
depositados na parede dos vasos sanguíneos, resulta em vasculites e predispõe a eventos
hemorrágicos e alterações da função do órgão (Lopez et al., 1996). Estas manifestações
são acompanhadas por alterações hematológicas e bioquímicas como anemia
normocítica e normocrômica com caráter regenerativo ou arregenerativo,

24
trombocitopenia, leucopenia. Uma leucocitose por neutrofilia, quando presente está
normalmente associada a um quadro de infecção bacteriana secundária. As alterações
em linfócitos são mais raras, porém a presença de monocitose é comum, muitas vezes
acompanhada de grande ativação de monócitos. As alterações da urinálise ou de
avaliações bioquímicas refletem, normalmente, um comprometimento renal ou hepático
em decorrência da doença. O aumento das proteínas totais do soro com inversão da taxa
albumina/globulina é um achado freqüente. Eletroforese de proteínas revela um
decréscimo significante na concentração de albumina e um aumento combinado de e
globulinas (Burillo et al., 1994; Campino, 2003; Baneth, 2006).
A perda de peso está quase sempre relacionada ao envolvimento visceral,
podendo ocorrer mesmo em cães com normorexia (Feitosa et al, 2000). Os cães podem
ainda apresentar uma atrofia muscular proeminente, particularmente em musculatura
facial e temporal (Kontos, 1993). É comum a ocorrência de hipertermia intermitente,
com valores de temperatura variando entre 40,5ºC e 41ºC (Feitosa et al, 2000).
As alterações dermatológicas são de variável extensão e caracterização. A
maioria dos autores afirma que em entre 60% e 90% dos cães infectados apresentam
algum tipo de lesão dermatológica (Amusategui et al., 2003; Alvar et al., 2004; Baneth,
2006). O cão pode apresentar tanto um pêlo seco como uma seborréia oleosa,
acompanhada ou não de prurido e, muitas vezes, com contaminação bacteriana
secundária quanto excessiva descamação cutânea que pode, eventualmente, tornar-se
disseminada por todo o corpo. É comum a observação de áreas de rarefação pilosa e
alopecia, associadas ou não à ocorrência de úlceras cutâneas (Slappendel, 1988; Feitosa
et al, 2000; Scott et al., 2001). Em locais correspondentes a saliências ósseas pode-se
observar tanto áreas de hiperqueratose e lignificação como áreas ulceradas, as quais
estão, freqüentemente, relacionadas a uma ação direta do parasita ou a uma vasculite
necrosante causada pela deposição de imunocomplexos (Ciaramella et al., 1997).
Alguns cães apresentam despigmentação cutânea, principalmente no plano nasal. A
onicogrifose é um achado relativamente comum e é causada pela presença do parasita
estimulando a matriz ungueal (Slappendel, 1988; Feitosa et al., 2000; Scott et al., 2001).
As alterações oftálmicas acometem preferencialmente o segmento anterior do
globo ocular, sendo mais comum a blefarite associada à dermatite facial, embora não
seja raro observar ceratoconjuntivite seca (Ciaramella et al., 1997; Noli, 1999). A
uveíte, geralmente bilateral, pode ser observada em associação com edema de córnea e

25
formação de sinéquias devido à presença de Leishmania ou como conseqüência da
deposição de imunocomplexos na íris e corpo ciliar (Feitosa, 2006). Em Belo Horizonte,
Fulgêncio (2006) encontrou uma prevalência de manifestações oculares e perioculares
em 48% de 100 cães recolhidos pelo Centro de controle de Zoonoses da Prefeitura
Municipal de Belo Horizonte, com diagnóstico de LV.
Muitos cães com LV apresentam epistaxe, normalmente unilateral, moderada e
intermitente. A provável causa é uma combinação de lesões ulcerativas e inflamatórias
na mucosa nasal (Kontos, 1993; Ciaramella & Corona, 2003). É possível ainda,
observar animais com quadro de diarréia crônica devido à presença de ulcerações de
mucosa gástrica e intestinal, resultando em fezes aquosas acompanhadas de
hematoquezia ou melena (Ciaramella & Corona, 2003).
A deposição de imunocomplexos nos rins eventualmete resulta em
glomerulonefrite membranoproliferativa e nefrite tubulointersticial, podendo evoluir
para uma insuficiência renal crônica (Kontos, 1993; Ferrer, 2002b).
Dentre os sintomas neurológicos já observados em cães naturalmente
acometidos por leishmaniose visceral destacam-se as convulsões, as alterações em pares
de nervos cranianos (estrabismo, ptose facial e disfagia), alterações de locomoção tais
como andar compulsivo e em círculos, lesões vestibulares evidenciadas por ataxia e
nistagmo e, alterações cerebelares com ocorrência de tremor de intenção. Tais sintomas
podem ser decorrentes da deposição de imunocomplexos ou por infecções oportunistas
no sistema nervoso central (Ciaramella et al., 1997; Feitosa et al., 2005).
1.4 Diagnóstico da Leishmaniose Visceral Canina
1.4.1 Diagnóstico Clínico
O diagnóstico da LVC só deve ser estabelecido após uma combinação da
sintomatologia clínica apresentada pelo animal com o resultado de exames
parasitológicos, sorológicos e moleculares (Ferrer, 1999; Alvar et al., 2004). O
diagnóstico puramente clínico da LVC torna-se impossível devido ao fato de que os
sinais clínicos, quando presentes, podem ser confundidos com os de outras doenças
(Feitosa et al., 2000; Gradoni, 2002; Singh et al., 2003; Feitosa, 2006). Embora a
prevalência da infecção em áreas endêmicas possa chegar a mais de 50%, a prevalência
da doença varia entre 3 e 10%, demonstrando que a maioria dos cães infectados não

26
desenvolve sintomas, dificultando mais ainda o diagnóstico (Ferrer, 2002a). Esses
animais podem permanecer assintomáticos por toda a vida ou desenvolver sintomas
após períodos que variam de três meses a alguns anos (Ferrer et al., 1995). Assim o
diagnóstico laboratorial torna-se necessário para confirmar a suspeita clínica, sendo que
fatores como a agilidade e confiabilidade dos resultados são importantes para as
medidas de controle, por ser o cão um elo importante da cadeia epidemiológica,
sobretudo, em áreas urbanas de LV.
1.4.2 Diagnóstico Laboratorial
1.4.2.1 Diagnóstico Parasitológico
Embora haja discordância entre alguns autores, o exame parasitológico é ainda
considerado, o padrão ouro no diagnóstico da doença (Dye et al., 1993; Leontides et al.,
2002; Singh et al., 2003). Podem ser observadas formas amastigotas do parasita em
esfregaços de linfonodos, medula óssea, aspirado esplênico, biópsia hepática e
esfregaços sangüíneos corados com corantes de rotina derivados de Romanowsky,
Giemsa ou Leishman, como o Wright e Panótico. Na dependência do tempo despendido
procurando o parasita, a sensibilidade passa a ser de, no máximo, 80% em cães
sintomáticos e menor em cães assintomáticos. A sensibilidade depende do grau de
parasitismo e seu valor relativo dificilmente pode ser analisado devido à falta de
uniformidade no critério de escolha do tipo de material biológico coletado e do tempo
de leitura da lâmina (Wolschrijn et al., 1996). Em alguns pacientes a visualização de
parasitas é muito laboriosa e os resultados negativos não são incomuns, especialmente
nos casos crônicos (Slappendel, 1988; Ikeda-Garcia et al, 2006). A semeadura, em meio
de cultura apropriado para o crescimento de Leishmania, é capaz de aumentar a
sensibilidade destes testes em até 20%. Estes exames permitem, diante da positividade,
o diagnóstico de certeza da infecção (Alvar et al., 2004). A inoculação de amostras de
tecidos em animais de laboratório, sobretudo em hamsters (Mesocricetus auratus),
necessita de longo tempo para a positividade (na nossa experiência 1 a 4 meses).
O exame histopatológico de fragmentos da pele e linfonodos pode ser usado
como método de diagnóstico, mas deve-se considerar que o infiltrado inflamatório
encontrado não é específico de LVC e que apenas o encontro das formas amastigotas dá
o diagnóstico de positividade (Costa Val, 2004). As técnicas de imunohistoquímica
(IHQ) podem melhorar os problemas relacionados à evidenciação do parasito, pois

27
anticorpos específicos marcados detectam com maior sensibilidade e especificidade as
formas amastigotas nos cortes de tecidos (Tafuri et al., 2004).
O xenodiagnóstico é uma técnica aplicada para detecção e isolamento do
parasito utilizando-se o vetor artrópode natural (Guarga et al., 2000). Embora não possa
ser considerada como uma técnica de rotina no diagnóstico da Leishmania ela
representa grande papel na resolução de questões epidemiológicas relacionadas ao
quadro clínico e evolução no tratamento de cães acometidos por LV (Gradoni, 2002;
Alvar et al., 2004).
Uma vez que a sensibilidade das técnicas sorológicas e de demonstração do
parasita, geralmente empregadas no diagnóstico da LV canina em áreas endêmicas é
geralmente insuficiente (Fisa et al., 2001), e que os métodos sorológicos necessitam de
níveis elevados de anticorpos e não fazem distinção entre as fases da doença quando os
níveis de imunoglobulinas estão próximo ou no ponto de corte (Aisa et al., 1998), os
testes que identificam o antígeno por métodos que empregam a biologia molecular
assumem grande importância (Tavares et al, 2003). Dentre as técnicas existentes, a
Reação em Cadeia da Polimerase (PCR) tem se mostrado muito útil, tanto no
diagnóstico quanto no acompanhamento de cães com LV (Ferrer, 1997), pelo aumento
da sensibilidade associado á grande especificidade (Fisa et al., 2001).
A PCR que se baseia na amplificação de uma seqüência conhecida de
oligonucleotídeos específicos do patógeno (DNA genômico ou do cinetoplasto), mostra
sensibilidade e especificidade que podem variar dependendo de fatores como tipo de
iniciadores, número de cópias do alvo dos iniciadores, método de extração do DNA,
natureza do material biopsiado, densidade parasitária da amostra e protocolo escolhido
para a reação. Apresenta como vantagem sua realização em grande variedade de
materiais clínicos, como sangue, aspirados de medula ou linfonodos, biópsias de pele,
urina, material colhido na conjuntiva do animal com swab (swab conjuntival), dentre
outros (Alvar et al., 2002; Tavares et al, 2003; Alvar et al, 2004).
1.4.2.2 Diagnóstico Imunológico
A hipergamaglobulinemia, principalmente de IgG, apresentada pelos animais
permite que técnicas menos invasivas identifiquem sorologicamente o animal acometido
por LV através da detecção de anticorpos anti-Leishmania (Bray, 1980; Tavares et al.,

28
2003). A soroconversão ocorre aproximadamente três meses após a infecção e os títulos
permanecem elevados por, pelo menos, dois anos. Entretanto, os testes sorológicos
devem ser interpretados com cautela, uma vez que não são 100% sensíveis e falham em
detectar cães infectados no período pré-patente e, portanto, antes da soroconversão, cães
que nunca farão soroconversão e cães soropositivos que se convertem em soronegativos,
mas ainda permanecem infectados (Ferrer et al., 1995; Ferrer, 2002a; Leontides et al.,
2002; Ikeda-Garcia et al, 2006;). Animais com menos de três meses de idade não devem
ser avaliados através de métodos sorológicos, pois podem apresentar resultados
positivos pela presença de anticorpos maternos (Braga et al., 1998).
Várias técnicas podem ser utilizadas para o diagnóstico do calazar, tais como, a
Reação de Fixação do Complemento (Hockmeyer et al., 1984), Hemaglutinação Direta
(Harith et al., 1988 Harith et al., 1996), Hemaglutinação Indireta (Bray & Lainson,
1967), Reação de Imunofluorescência Indireta (RIFI) (Badaró et al.,1983 e 1986),
Enzyme Linked Imunosorbent Assay (ELISA) (Ho et al., 1983; Badaró et al.,
1986).Com o uso de diferentes antígenos nas mais variadas técnicas sorológicas, o que
se observa em vários trabalhos é uma grande variação de resultados. A técnica
sorológica mais utilizada é o teste de ELISA, que é altamente sensível e a especificidade
depende do antígeno utilizado, o antígeno mais comumente utilizado é o bruto, derivado
de um lisado de promastigotas de Leishmania spp. Este teste apresenta, no entanto
reatividade cruzada com outras patologias causadas por tripanosomatídeos, como
leishmaniose tegumentar e Doença de Chagas e mesmo com outras espécies
geneticamente distintas de tripanosomatídeos.
Nos últimos anos têm surgido vários "kits" comerciais para diagnóstico rápido
da LVC, sobretudo no mercado veterinário europeu. A maioria desses "kits" consiste de
ensaios imunocromatográficos que empregam anti imunoglobulinas monoclonais (IgG)
caninas marcadas com ouro coloidal e antígeno de diferentes fontes. Eles são atrativos
porque são testes simples de serem realizados, fáceis de utilizar e a resposta é obtida
rapidamente, em aproximadamente 10 minutos. Entretanto, há informações de vários
autores que estes "kits" são usualmente limitados, mostrando resultados que carecem de
maiores evidências científicas para sua utilização (Bern et al., 2000; Grandoni,2002;
Reithinger et al., 2002; Sundar et al., 2002; Carvalho et al., 2003). Por exemplo, a
sensibilidade e a especificidade destes testes, utilizando o antígeno recombinante rK39
para o diagnóstico da LV humana variam, de acordo com a literatura, de 90 a 100% e de

29
93 a 100%, respectivamente (Bern et al., 2000; Sundar et al., 2002; Carvalho et al.,
2003).
Na avaliação do teste de imunocromatografia para detecção de LVC, enquanto
alguns autores descreveram valores de sensibilidade entre 84 e 92,1% e de
especificidade entre 99 e 100% (Melo, 2004; Laurenti et al., 2005), outros observaram
uma sensibilidade de 100% e especificidade de 75% (Reithinger et al., 2002). Embora
os relatos de que os testes de imunocromatografia possam levar a uma grande proporção
de diagnósticos falso positivos, existem discordâncias quanto à ocorrência de reação
cruzada do antígeno rK39 com Leishmania braziliensis ou Trypanosoma cruzi
(Reithinger et al., 2002).
A RIFI e o ELISA são as técnicas mais utilizadas para avaliação da
soropositividade canina, além de serem também recomendadas pelo Ministério da
Saúde do Brasil em inquéritos caninos amostrais e censitários, devido à alta
sensibilidade e especificidade das mesmas, em comparação às demais (Badaró, 1983;
Guimarães et al., 1990; Brasil-Ministério da Saúde, 2003).
A RIFI utiliza como antígeno promastigotas de Leishmania spp., podendo ser
inclusive de espécies tais como L. mexicana e L. braziliensis (Costa et al., 1991). A
preparação do antígeno exige pessoal qualificado (Tavares et al., 2003) e, além disto,
quando da análise de um grande número de amostras, o tempo despendido pelo
profissional para a leitura das lâminas é muito grande (Rachamin et al., 1991). Embora
existam “kits” comerciais, as lâminas preparadas nos próprios laboratórios onde os
diagnósticos são realizados, produzem resultados melhores e mais repetitivos (Ferrer,
1997). Como teste diagnóstico a RIFI demonstra sensibilidade variando de 83% a
100%, e especificidade, dependendo da região geográfica em que o teste é aplicado,
entre 74% a 100% (Almeida et al., 2005; Da Silva et al., 2006). A especificidade desta
prova é prejudicada devido à ocorrência de reações cruzadas com doenças causadas por
outros tripanosomatídeos, tais como o agente etiológico da Doença de Chagas. Portanto,
seus resultados não devem ser utilizados como indicadores de infecção leishmaniótica
específica, particularmente em áreas onde a Doença de Chagas é endêmica (Costa et al.,
1991; Alves et al, 2004;). Existem inúmeros relatos da ocorrência de reação sorológica
cruzada entre os membros da família Trypanosomatidae, envolvendo principalmente, os
protozoários L. chagasi, L. braziliensis e Trypanosoma. cruzi, os quais possuem uma
relação filogenética muito estreita. No entanto, a ocorrência de reação cruzada entre

30
LVC e outros agentes etiológicos, quando se utilizam métodos sorológicos, ainda é
motivo de discordâncias na medicina veterinária. Antígenos de superfície e antígenos
dos microtúbulos do citoesqueleto do protozoário são comuns a todos os
tripanosomatídeos o que justifica a ocorrência de reação cruzada entre eles (Badaró et
al., 1986).
Já, com relação à ocorrência de reação cruzada com outras enfermidades,
Vercammen et al. (1997), utilizando amostras de soro de três cães infectados por
Babesia sp. e de um cão infectado por Ehrlichia sp., não observaram resultados
positivos quando da realização da RIFI para LV.
A técnica de RIFI é adotada como padrão ouro pelo Ministério da Saúde,
portaria nº 1943, que determina que a Leishmaniose como uma das doenças de
notificação obrigatória em todo território nacional. O kit diagnóstico oficial, produzido
por Biomanguinhos, para a reação de RIFI recomenda como resultado positivo o título
igual ou superior a 1:40 (Instituto Adolfo Lutz, 2003).
Os antígenos brutos utilizados no ELISA são derivados de promastigotas
cultivadas in vitro, geralmente rompidas por ultrasom, expondo cerca de 30 antígenos
somáticos e inúmeros componentes de superfície (Tavares et al., 2003). O teste, possui
sensibilidade de 97% a 100% (Laurenti et al., 2005) e especificidade de 88% a 100%,
quando são usados antígenos brutos (Rosário et al., 2005). Há, contudo, discordâncias
entre alguns autores, os quais descreveram valores de sensibilidade e especificidade de
80 e 81%, respectivamente (Ashford et al., 1995) e outros que observaram valores de
100% para ambos os índices (Vercammen et al., 1997) .Quando se utilizam antígenos
totais as técnicas tornam-se limitadas em termos de especificidade, apresentando
reações cruzadas não somente com outras espécies da família Trypanosomatidae, mas
também com organismos filogeneticamente distantes (Rachamin et al., 1991; Tavares et
al., 2003; Melo, 2004). Enquanto alguns trabalhos apontam para a ocorrência de reação
cruzada entre LV e babesiose canina ou Doença de Chagas por meio da técnica de
ELISA (Rosário et al., 2005), outros afirmam não existir reação cruzada com erlichiose
e babesiose (Lima et al., 2005; Vercammen et al.,1997), toxoplasmose (Lima et al.,
2005) e Doença de Chagas (Vercammen et al.,1997).
A sensibilidade e especificidade do ELISA podem também variar dependendo
do tipo de antígeno empregado (espécie ou forma evolutiva do parasita) e de mudanças

31
no protocolo experimental padrão (tempo de incubação ou tipo de microplacas
utilizadas) (Reinthinger et al., 2002).
O teste de ELISA é recomendado para a triagem de cães sorologicamente
negativos, e a RIFI para a confirmação dos cães sororeagentes, ou como uma técnica
diagnóstica de rotina (Brasil-Ministério da Saúde, 2003). Para melhorar a especificidade
do método tem-se buscado a caracterização molecular dos componentes do antígeno de
Leishmania (Gradoni, 2002).
O Immunoblotting ou Western blotting é uma técnica mais sensível que a RIFI e
o ELISA, porém como o processo de extração de antígenos ainda não é padronizado e a
necessidade laboratórios especializados para a sua realização fazem com que não seja
usado como ferramenta primária de diagnóstico (Alvar, 2004).
A busca para superar as limitações destas técnicas tem como principal desafio a
preparação do antígeno. Como fonte de produção de antígenos tem sido usado o parasita
completo ou extratos solúveis dos mesmos. Com isso, observa-se nos diversos
trabalhos, uma grande variedade de resultados e uma dificuldade de padronização dos
métodos. Geralmente, métodos que usam os parasitas íntegros oferecem maior
sensibilidade, reprodutibilidade e resultados mais confiáveis em relação aqueles que
usam antígenos solúveis (Ferrer, 1997; Alvar, 2004). A tendência é a utilização de
antígenos purificados e/ou recombinantes para assegurar uma maior sensibilidade e
especificidade dos testes (Kar, 1995, Ferreira et al., 2003, Tavares et al., 2003).
As espécies de Leishmania apresentam homologia gênica que varia de 69 a 90%
(Maurício et al., 2001). Os parasitos possuem na sua membrana mais de quarenta
moléculas distintas, com pesos variando de 12 a 220 kDa, das quais aproximadamente
30 são glicoproteínas (Dwyer, 1974; Mukherjee et al., 2002). A preparação de antígenos
de glicoproteínas purificadas de membrana como gp63, gp72, gp70 específicas do
gênero Leishmania têm sido utilizada na tentativa de elevar a sensibilidade e a
especificidade das provas sorológicas. Entretanto, ainda assim podem ocorrer reações
cruzadas com outros tripanosomatídeos (Badaró et al., 1986; Vexenat et al., 1996;
Scalone et al., 2002; Alves et al, 2004).
Alguns antígenos recombinantes, como o rK39, o rK9 e o rK26, são antígenos
recombinantes desenvolvidos para conferir grande sensibilidade quando da realização
da sorologia (Bhatia et al., 1999; Rosati et al., 2003).O antígeno ligante fucose manose

32
(FML) é uma glicoproteina de superfície, isolada de promastigotas de L. donovani,
necessária para interações com macrófagos in vitro. Quando este antígeno foi utilizado
no diagnóstico sorológico de cães de área endêmica do Rio Grande do Norte, Cabrera et
al. (1999) demonstraram taxas de sensibilidade e de especificidade de 100% para o
ELISA, enquanto que a sensibilidade da RIFI foi de apenas 2,9%. O FML também
demonstrou valor preditivo de 100%, pois dos 27 animais assintomáticos que foram
positivos ao ELISA FML, 21 morreram de leishmaniose em um período de seis meses.
O antígeno recombinante K39 (rK39), derivado purificado de epítopo
imumodominante repetitivo de uma região altamente conservada entre as espécies
viscerotropicas de Leishmania, foi validado em estudo multicêntrico na Itália. Cerca de
5.500 soros de cães foram utilizados por Scalone et al. (2002) neste estudo, que
demonstrou maior especificidade e maior sensibilidade do ELISA rK39, quando
comparado ao RIFI e ao ELISA de antígeno convencional. Os autores concluem que o
ELISA rK39 pode ser usado como alternativa ao RIFI, especialmente em estudos com
grande quantidades de amostras.
Outro antígeno recombinante a proteína A2 , expressa na forma amastigota de L.
donovani, composta predominantemente por um elemento conservado altamente
repetitivo, foi investigada por Carvalho et al. (2002). O autor pesquisou anticorpos
antiproteína A2 em soros de cães positivos para LV, previamente diagnosticados por
RIFI ou por diagnóstico parasitológico, usando este antígeno recombinante em reações
de ELISA. O método forneceu 87% de concordância com RIFI, porém não foram
observadas reações cruzadas com doenças comuns dos cães, bem como com cães
portadores de Doença de Chagas. Os autores concluíram que a presença de anticorpos
anti A2 pode ser considerada como marcador de doença ativa nos cães e que o método
per si será de grande aplicabilidade no Brasil, onde localidades distantes ou rurais não
dispõem de laboratórios ou equipamentos próprios para o diagnóstico preciso da
doença.
Os antígenos recombinantes até agora desenvolvidos, pelo menos no Brasil,
ainda não são utilizados na rotina do diagnóstico da LVC em inquéritos sorológicos
censitários, que têm como objetivo ações de controle.
A proteômica representa o desafio do futuro para aumentar nosso conhecimento
da composição molecular da célula e, a eletroforese bi-dimensional (2-DE), é uma das
ferramentas para estudar a expressão de proteínas em larga escala (Haseloff et al.,

33
2003). A análise das proteínas utilizando 2-DE envolve a separação sistemática,
identificação e quantificação de muitas proteínas simultaneamente a partir de uma única
amostra. Essa técnica também é capaz de detectar modificações traducionais e pós-
traducionais, as quais não podem ser observadas por seqüenciamento do genoma. A 2-
DE é uma ferramenta poderosa e amplamente utilizada para análise de misturas
complexas de proteínas extraídas de células, tecidos ou outras amostras biológicas e é
complementada pela espectrometria de massa (EM) para identificação das proteínas
(Haseloff et al. 2003). As proteínas mais imunogênicas, responsáveis por maior
antigenicidade, podem ser seqüenciadas e empregadas, por exemplo, na sensibilização
de placas de ELISA visando uma maior sensibilidade e especificidade deste ensaio
Estudos do proteoma de Leishmania foram realizados principalmente para
comparação dos diferentes estágios do ciclo de vida de L. donovani (Thiel & Bruchhaus
2001; Avila et al. 2003), L. infantum (El Fakhry et al., 2002), L. (Viannia) guyanensis e
L. (V.) panamensis (Gongora et al., 2003). Recentemente, um estudo utilizando 2-DE de
extrato de promastigotas de L. infantum, seguido de Western Blot com soro de coelhos
imunizados e análise por espectrometria de massa, permitiu a identificação de várias
proteínas antigênicas relevantes (Dea-Ayuela et al., 2006). Numa abordagem
semelhante, foram identificados vários antígenos de L. donovani utilizando soros e
parasitos isolados de pacientes indianos permitindo concluir que a abordagem
proteoma-sorologia produz uma compreensiva e altamente resolvida representação da
antigenicidade de L. donovani e da especificidade da resposta imune anti-Leishmania
em pacientes com LV (Forgber et al. 2006).
Em síntese, estudos proteômicos com Leishmania já demonstraram a diferença
de expressão protéica entre as formas amastigotas e promastigotas e a possibilidade de
utilização da sorologia proteômica como uma abordagem adequada para mapeamento
da antigenicidade na leishmaniose. Entretanto, os trabalhos utilizando soros de cães,
principais reservatórios, são ainda inéditos, apesar da potencial aplicação.
1.5 Controle da Leishmaniose Visceral no Brasil
Em virtude das características epidemiológicas e do conhecimento, ainda
insuficiente, sobre os vários elementos que compõem a cadeia de transmissão da LV no
Brasil, sobretudo nas áreas urbanas, as estratégias de controle desta endemia têm sido
pouco efetivas. Estão centradas no diagnóstico e tratamento precoces dos casos

34
humanos, redução da população de flebotomíneos vetores, eliminação dos reservatórios
domésticos positivos para leishmaniose e atividades de educação em saúde (Brasil-
Ministério da Saúde, 2003).
Embora o programa brasileiro de controle de Leishmanioses exista desde a
década de 1960, a incidência da LVH permanece alta. A ausência de impacto na
aplicação destas medidas tem sido atribuída principalmente, a baixa sensibilidade dos
testes diagnósticos, a demora na retirada dos animais soropositivos e na desconfiança no
diagnóstico, e conseqüente resistência dos proprietários em entregar os cães
sororeagentes nas últimas décadas (Romero & Boelaert, 2010)
O diagnóstico dos animais sororeagentes tem sido realizado pelos serviços de
controle utilizando “kits” de diagnóstico para as técnicas de RIFI e de ELISA
produzidos por Biomanginhos/Fiocruz/ Ministério de Saúde.
As divergências entre os exames sorológicos realizados pelos laboratórios
públicos, privados e naqueles existentes em universidades consistem em grande
problema para a execução das ações de controle da LV. Esta dificuldade em relação ao
diagnóstico da LVC está diretamente relacionada, entre outros fatores, a diferentes
técnicas de produção dos antígenos pertencentes aos kits diagnósticos comerciais.

35
2. Justificativa
A leishmaniose visceral é uma protozoose que determina uma alta taxa de
mortalidade, quando não tratada, e está em expansão geográfica, principalmente no
Brasil, invadindo áreas antes livres da doença. Um conjunto de fatores contribuiu para o
aumento da ocorrência de leishmaniose visceral nos centros urbanos, já assinalada por
Deane em 1956, dentre eles, as mudanças que ocorreram nos últimos anos na estrutura
agrária do país, o aumento das migrações internas da área rural para grandes centros, as
pressões econômicas ou sociais, a adaptação do vetor ao ambiente modificado pelo
homem, as alterações climáticas, investimentos precários na saúde e as ações de
controle pouco eficientes (Gontijo e Melo, 2004).
A urbanização da leishmaniose visceral tem decorrido tanto devido às condições
precárias de vida existentes nas periferias das cidades, para onde vieram grandes
deslocamentos de populações de áreas rurais. As cidades tiveram suas periferias
expandidas, sem que este crescimento tenha sido acompanhado da consolidação de sua
estrutura de serviços urbanos. Estas periferias mantiveram algumas características de
áreas rurais de origem dos migrantes, como a presença de mosaicos de vegetação em
áreas ainda não completamente ocupadas, a criação de animais domésticos nos
peridomicílios e as condições precárias das habitações, permitindo, em certas áreas do
país, a presença da leishmaniose no espaço urbano.
O cão é o principal reservatório urbano da doença. Normalmente verifica-se que
o maior número de casos humanos ocorre naqueles municípios ou áreas com as maiores
prevalências de LVC. Há uma relação espaço temporal na qual a doença canina, na
grande maioria das vezes, precede a detecção de casos humanos.
O processo de expansão geográfica e urbanização da LV levaram à necessidade
de se estabelecer medidas eficazes de controle, hoje focadas no diagnóstico e tratamento
dos casos humanos, diagnóstico e eliminação dos cães positivos e controle dos
flebotomíneos vetores, baseado no uso de inseticida de ação residual, direcionado para
as formas adultas, uma vez que os criadouros da espécie são pouco conhecidos. Neste
contexto, a correta identificação dos cães infectados constitui uma medida
estrategicamente importante num programa de saúde pública.

36
O diagnóstico sorológico da LV, tanto humano quanto canino tem sido realizado
no Brasil de acordo com orientações do Ministério da Saúde, através dos testes de RIFI
e ELISA. O teste de ELISA é usado para uma triagem inicial, principalmente nos
inquéritos epidemiológicos caninos, devido a sua maior especificidade e sensibilidade, e
a confirmação é feita pela RIFI. Por utilizarem antígenos brutos, podem apresentar
reações cruzadas com outras espécies da família Trypanosomatidae e mesmo com
organismos filogeneticamente distantes.
As divergências existentes entre os exames realizados pelos laboratórios
públicos e privados e a falta de integração entre os diversos órgãos envolvidos direta ou
indiretamente com o controle desta zoonose, fez com que em julho de 2005 se
reunissem representantes do Ministério da Agricultura Pecuária e Abastecimento,
Ministério da Saúde e Conselho Regional de Medicina Veterinária, com o propósito de
ajustar condutas que pudessem otimizar a detecção da doença canina e
conseqüentemente interferir na ocorrência da doença humana. Foi determinada uma
uniformização de condutas, padronizando entre outras medidas, o diagnóstico
sorológico da LVC realizado pela rede pública e privada. Esta padronização define que
somente “kits” diagnósticos devidamente registrados pelo Ministério da Agricultura
Pecuária e Abastecimento poderão ser utilizados no diagnóstico da doença canina e
propõem que o diagnóstico sorológico para LVC fosse acompanhado pela FUNED,
respaldado na Portaria 2.031 de 23 de setembro de 2004/Ministério da Saúde.
Os serviços de diagnóstico laboratorial da Leishmaniose Visceral realizados pela
rede pública são possíveis, através de uma ação integrada com a Fundação Ezequiel
Dias (FUNED), referência nacional para o diagnóstico da leishmaniose canina e
humana. Esta instituição é quem monitora e controla a qualidade tanto dos “kits”
produzidos para o diagnóstico, quanto dos exames realizados, por meio de amostragem
previamente estabelecida. Os “kits” diagnósticos utilizados pela rede pública são
produzidos pela Biomanguinhos/Ministério da Saúde e registrados na Agência Nacional
de Vigilância Sanitária (ANVISA) e Mistério da Agricultura Pecuária e Abastecimento.
Este órgão não tem até o presente momento capacidade para atender a demanda oficial
de produção de “kits” diagnósticos exigida tanto pelas secretarias estaduais e municipais
de saúde, quanto pela rede de laboratórios particulares.
Tendo em vista esta dificuldade, a FUNED estabeleceu uma parceria com o
Departamento de Parasitologia do Instituto de Ciências Biológicas da Universidade

37
Federal de Minas Gerais, para um programa de cooperação técnico-científica na
realização de estudos, testes e ensaios direcionados na produção de um antígeno que
permitisse diagnóstico precoce da doença, fosse estável, apresentasse reprodução de
resultados em qualquer local que fosse utilizado e que pudesse ser produzido em escala
industrial para atender a demanda nacional. Assim, espera-se eliminar a divergência de
resultados sorológicos decorrentes da utilização de “kits” diagnósticos confeccionados
com diferentes cepas de Leishmania e por diferentes métodos, e contribuir para atender
a demanda nacional de diagnóstico. A produção deste antígeno seria feita segundo as
normas internacionais de Good Manufacturation Practice (GMP), pela FUNED.
A Universidade Federal de Minas Gerais através do Departamento de
Parasitologia do ICB tem, desde o início do ano 2000, buscado a obtenção do um
antígeno que apresentasse estabilidade, reprodutibilidade de resultados, além de
sensibilidade e especificidade, que pudesse ser incorporado a um “kit” diagnóstico para
suprir a demanda de Minas Gerais e talvez, implementar a dinâmica do diagnóstico da
LVC no país. A FUNED através de suas atividades de pesquisa e desenvolvimento,
produção e de vigilância sanitária, epidemiológica e ambiental e principalmente, por sua
tradição na produção de soros hiperimunes, tem uma estrutura de produção industrial de
medicamentos e soros que lhe permite a produção, em escala do antígeno, o
desenvolvimento, teste e produção de um “kit” de diagnóstico de LVC. Como o
Laboratório Central de Saúde Pública (LACEN/FUNED) é referência nacional para
muitas doenças negligenciadas e de grande impacto econômico, incluindo a
leishmaniose, esse apresenta a capacitação para certificação e validação dos métodos.
Os testes sorológicos realizados com os diferentes antígenos produzidos e
descritos neste projeto foram realizados no Departamento de Parasitologia do
ICB/UFMG. A seguir o antígeno escolhido neste trabalho será encaminhado à FUNED
para testes multicêntrico. A meta a ser atingida é a produção do antígeno em escala
industrial pela FUNED, para fornecimento ao Ministério da Saúde, de “kit” diagnóstico
para LVC, validado pela LACEN.

38
3. Objetivos
3.1.Objetivo Geral
Obter um antígeno bruto, a partir de promastigotas de Leishmania spp.de cultura
"in vitro", para composição de um "kit" diagnóstico para Leishmaniose Visceral
Canina a ser utilizado na técnica de ELISA
3.2. Objetivos Específicos
Testar as cepas (MHOM/BR/1972/BH46) e (MHOM/BR/2004/BH400) de
Leishmania (Leishmania) chagasi e (MHOM/BR/1960/BH6) de L. (L.)
amazonensis cultivadas em meio LIT e em meio α-MEM, na produção de
antígenos para o teste de ELISA;
Determinar a reatividade de soros de animais comprovadamente positivos e
negativos para LV frente aos antígenos produzidos;
Testar a estabilidade da preparação antigênica nos tempos de 24 horas, 3 e 6
meses após a sensibilização das placas de ELISA com os antígenos;
Testar os antígenos produzidos em RIFI para diagnóstico de LVC;
Avaliar ocorrências de reações cruzadas pelo teste de ELISA com outras
patologias, empregando o antígeno considerado de melhor desempenho nas
reações;
Determinar o perfil proteômico imunogênico do melhor antígeno.
.

39
4. Materiais e Métodos
4.1 Delineamento Experimental
4.1.2 Seleção dos Animais
4.1.3 Obtenção dos parasitos
110 animais provenientes CCZ Prefeitura Municipal de Betim:
20 mL de sangue
RIFI e ELISA
Coleta de medula óssea sob anestesia
Esfregaço em lâmina de vidro
Semeadura em meio de cultura
PCR
Cepas
BH 46 BH 400 BH 6
Meios LIT e -MEM
Curva de Crescimento
Massa de parasitos para a obtenção de ANTÍGENOS
Ultrassonicação e Dosagem de Proteínas
ELISA
Dosagem de Proteína e Eetroforese 2D
Imunoblotting
RIFI

40
4.2 Seleção dos Animais e Coleta de Material
Os cães doadores do soro para os testes de avaliação dos antígenos produzidos
para este trabalho foram oriundos do projeto “Estudo clínico para avaliação de vacinas
contra as leishmanioses em cães usando antígenos recombinantes de Leishmania
definidos pela resposta imune humana”, que teve como responsável o professor Evaldo
Nascimento na orientação do aluno de mestrado Leonardo T. Ituassu do Departamento
de Parasitologia do Instituto de Ciências Biológicas da UFMG. Os animais eram
trazidos do Centro de Controle de Zoonoses da Prefeitura Municipal de Betim e
alojados no canil do ICB/UFMG. No momento da coleta de material destes cães,
amostras foram cedidas para nosso projeto. O projeto foi certificado pelo Comitê de
Ética em Experimentação Animal – CETEA- sob protocolo 140/2006, em anexo.
Como critério de inclusão do soro animal para os testes, estabeleceu-se positivo
o animal que se apresentasse positividade em um dos testes parasitológicos associado ao
resultado, também positivo, de pelo menos um dos testes sorológicos. O critério de
amostra negativa implicava na negatividade de todos os testes sorológicos e
parasitológicos realizados.
Os animais foram sedados e pré-anestesiados com Xilazina®
– 2,2 mg/kg,
Ketamina® - 11mg/kg e Diazepam® – 1mL/10kg, por via intramuscular. Quando o cão
apresentou discreta sedação, foram coletados de cada animal 20mL de sangue total
através de punção da veia jugular. Ainda com o animal sob anestesia foi realizada a
depilação da região esternal, numa área suficiente para garantir assepsia da coleta e
após, uma agulha de 40 gauges, acoplada a uma seringa de 10mL foi introduzida na
região do manúbrio, seguindo-se a aspiração da medula óssea.
A amostra de sangue foi centrifugada a 1500g durante dez minutos para
obtenção de soro. Os soros foram divididos em alíquotas de 100mL, codificadas e
armazenadas a -20ºC. O aspirado de medula óssea foi utilizado para a avaliação
parasitológica, sendo colocado em meio de cultura NNN/Alfa-MEM (GIBCO-BRL,
USA).
Os soros foram avaliados em relação à positividade ou negatividade para LVC
pelos testes de Enzime Linked Immunosorbent Assay-ELISA (Voller et al, 1979) e

41
Reação de Imunofluorescência Indireta-RIFI (Camargo, 1966), segundo rotina
diagnóstica já estabelecida no Laboratório de Sorologia de Leishmania, do
Departamento de Parasitologia do Instituto de Ciências Biológicas da UFMG.
4.3 Avaliação Parasitológica dos Animais
Para a comprovação parasitológica da infecção animal, o material puncionado da
medula óssea, foi submetido a esfregaço por extensão em lâmina de vidro, semeadura
em meio de cultura e aplicado em papel de filtro próprio para extração de DNA.
4.3.1 Esfregaço por extensão
Uma gota de material puncionado de medula óssea foi colocada em cada uma de
duas lâminas de vidro limpas e desengorduradas (Invicta®, Brasil). A extensão do
material foi realizada com outra lâmina de vidro, e as mesmas deixadas para secar em
temperatura ambiente, sendo posteriormente armazenadas, para coloração através do
método Panótico Rápido (Bioclin, Brasil). Cada amostra foi imersa por 20 segundos em
cada solução (fixador, corante básico e corante ácido, respectivamente), sob discreta
agitação, seguindo-se posterior lavagem em água corrente. A secagem foi realizada em
temperatura ambiente. As lâminas foram examinadas em microscópio ótico no aumento
ocular de 10X e objetiva de 100X, em imersão.
4.3.2 Cultura de medula óssea
Para o isolamento do parasito, parte da amostra da medula óssea foi semeada em
2 tubos contendo meio de cultura bifásico NNN (Novy, McNeal e Nicolle) enriquecido
com meio Alfa-MEM (Minimum Essential Medium, Gibco, USA) acrescido de 10% de
Soro Fetal Bovino, que foram mantidos a 24+1oC em estufa biológica (FANEM,
Brasil). A cada sete dias todas as culturas eram examinadas em câmara asséptica e
repicadas, num total de três repiques, para a avaliação da possível presença e
crescimento de parasitos. Ao final de trinta dias, o diagnóstico foi concluído e, todos os
tubos, foram examinados e descartados.
4.3.3 Reação em cadeia da polimerase - PCR

42
As amostras de medula óssea depositadas em papel de filtro próprio para
extração de DNA (FTA®
Classic Card) e deixadas para secar a temperatura ambiente,
foram armazenadas a 4ºC e utilizadas como recurso para confirmação da
positividade/negatividade sorológica de cada animal avaliado.
A extração do DNA foi realizada pela técnica descrita por da Silva et al. (2001)
com modificações. Cinco fragmentos do papel de filtro embebido com material de
medula óssea foram obtidos, de forma aleatória, com auxílio de um “punch” de dois
milímetros de diâmetro (Punch para Biópsia®, Kolplastci LTDA, Brasil). As amostras
foram transferidas para tubos de microcentrífuga de 1,5L de capacidade contendo
200L de solução tampão de lise (FTA® Purification Reagent, Whatman, EUA) e foram
submetidas à agitação com auxílio de um aparelho vórtex por 20 segundos, sendo
homogeneizadas por cinco minutos à temperatura ambiente, em movimentos constantes
com auxílio de um roteador (Lab Rotator®), sendo posteriormente descartado o
sobrenadante. Este procedimento foi repetido por mais duas vezes. Após a terceira
lavagem, 200L de solução tampão TE (10 mM Tris-HCL, 1mM EDTA, pH 7.5) foi
adicionada ao tubo de microcentrífuga contendo os discos de papel, seguido por nova
agitação em vórtex por vinte segundos e incubação à temperatura ambiente por mais
cinco minutos com posterior descarte do sobrenadante. Este procedimento foi repetido
duas vezes. Os discos de papel de filtro contendo DNA foram estocados a -4oC para
serem utilizados na realização da técnica de PCR.
A amplificação do K-DNA através da PCR específica, o desenho dos iniciadores
da reação bem como o protocolo da PCR foram realizados segundo a técnica descrita
por Piarroux et al. (1993), com modificações. A amplificação do DNA foi realizada em
termociclador (PTC-100®, M.J. Research Inc., EUA), utilizando-se o par de iniciadores
LV1 e LV2 (Quadro 1) específicos para o complexo Leishmania donovani (Invitrogen,
Brasil).
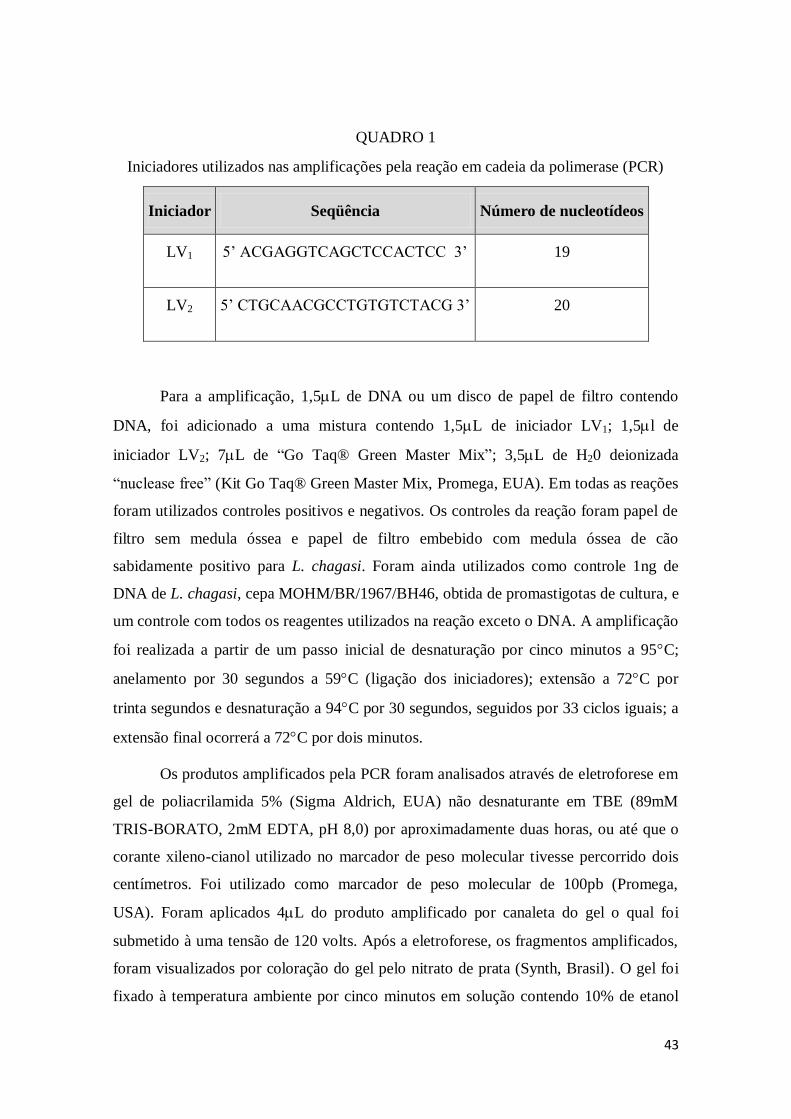
43
QUADRO 1
Iniciadores utilizados nas amplificações pela reação em cadeia da polimerase (PCR)
Iniciador Seqüência Número de nucleotídeos
LV1 5’ ACGAGGTCAGCTCCACTCC 3’ 19
LV2 5’ CTGCAACGCCTGTGTCTACG 3’ 20
Para a amplificação, 1,5L de DNA ou um disco de papel de filtro contendo
DNA, foi adicionado a uma mistura contendo 1,5L de iniciador LV1; 1,5l de
iniciador LV2; 7L de “Go Taq® Green Master Mix”; 3,5L de H20 deionizada
“nuclease free” (Kit Go Taq® Green Master Mix, Promega, EUA). Em todas as reações
foram utilizados controles positivos e negativos. Os controles da reação foram papel de
filtro sem medula óssea e papel de filtro embebido com medula óssea de cão
sabidamente positivo para L. chagasi. Foram ainda utilizados como controle 1ng de
DNA de L. chagasi, cepa MOHM/BR/1967/BH46, obtida de promastigotas de cultura, e
um controle com todos os reagentes utilizados na reação exceto o DNA. A amplificação
foi realizada a partir de um passo inicial de desnaturação por cinco minutos a 95C;
anelamento por 30 segundos a 59C (ligação dos iniciadores); extensão a 72C por
trinta segundos e desnaturação a 94C por 30 segundos, seguidos por 33 ciclos iguais; a
extensão final ocorrerá a 72C por dois minutos.
Os produtos amplificados pela PCR foram analisados através de eletroforese em
gel de poliacrilamida 5% (Sigma Aldrich, EUA) não desnaturante em TBE (89mM
TRIS-BORATO, 2mM EDTA, pH 8,0) por aproximadamente duas horas, ou até que o
corante xileno-cianol utilizado no marcador de peso molecular tivesse percorrido dois
centímetros. Foi utilizado como marcador de peso molecular de 100pb (Promega,
USA). Foram aplicados 4L do produto amplificado por canaleta do gel o qual foi
submetido à uma tensão de 120 volts. Após a eletroforese, os fragmentos amplificados,
foram visualizados por coloração do gel pelo nitrato de prata (Synth, Brasil). O gel foi
fixado à temperatura ambiente por cinco minutos em solução contendo 10% de etanol

44
absoluto (v/v) (Merck, Alemanha) e 0,5% de ácido acético (v/v) (Merck, Alemanha),
seguida de incubação por 5 minutos em 150mL de solução contendo 0,7% de nitrato de
prata (p/v) (Synth, Brasil), e de nova incubação com uma solução reveladora contento
3% de NaOH (p/v) (Merck, Alemanha) e 0,1% de formaldeído 37% (v/v) (Merck,
Alemanha), por 10 minutos ou até o aparecimento das bandas, sempre sob agitação
suave (Orbit Shaker
, Lab-Line). Posteriormente o gel foi transferido para a solução
fixadora e documentado por fotografia para análise. A contaminação da reação por
amplicons foi evitada pelo uso de diferentes ambientes para o processamento das
amostras, além dos processos rotineiros de descontaminação das áreas de trabalho.
4.4 Preparo dos Antígenos
Os antígenos foram preparados a partir das promastigotes das cepas de
Leishmania, que se encontravam criopreservadas em nitrogênio líquido no Laboratório
de Biologia de Leishmania do Departamento de Parasitologia do Instituto de Ciências
Biológicas da Universidade Federal de Minas Gerais, descritas a seguir.
Cepa BH46: Leishmania (Leishmania) chagasi (MHOM/BR/1972/BH46) isolada
de um paciente com leishmaniose visceral, natural de Mantena, Minas Gerais,
Brasil, em 07/06/1972;
Cepa BH6: Leishmania (L.) amazonensis (MHOM/BR/1960/BH6) isolada de um
paciente com leishmaniose cutânea, no Estado do Ceará, Brasil, 1960.
Cepa BH400: Leishmania (L.) chagasi (MCAN/BR/2002/BH400): isolada de um
cão com LVC, na metropolitana de Belo Horizonte, Brasil, em 2002.
4.4.1 Curva de Crescimento
Cada cepa foi cultivada em 10 mL de cada um dos meios de cultivo Liver
Infusion Triptose (LIT) (Camargo, 1966) e Minimum Essential Medium (Alfa-MEM)
acrescido de 10% de Soro Fetal Bovino, a partir de um inóculo de 106 promastigotas,
em fase log de crescimento, provenientes do mesmo meio.
Quando adicionadas ao meio de cultivo, as Leishmania inicialmente se adaptam,
fase lag, período onde, embora os parasitos estejam metabolicamente ativos, supõe-se
não ocorrer divisão binária. A seguir, os parasitos iniciam uma fase de crescimento
logarítmico ou exponencial, na qual se dividem por fissão binária e ocorre o pico de

45
suas atividades metabólicas e fisiológicas, chamada fase log. Entretanto, quando os
nutrientes são utilizados e/ou quando os produtos de excreção metabólica se acumulam
em quantidade no ambiente, isto parece intervir no crescimento estabelecendo-se uma
fase estacionária. Esta fase é seguida de morte parasitária, diminuição dos parasitos
viáveis de forma exponencial e caracteriza a fase de declínio ou de morte. As cepas
foram caracterizadas quanto ao crescimento nos diferentes meios através da contagem
diária de promastigotes em câmara de Neubauer e foi delineada uma curva gráfica para
visualização das diferentes fases de crescimento.
4.4.2 Obtenção de parasitos para o Preparo dos Antígenos
Para obtenção de uma massa de promastigotas, visando o preparo dos antígenos,
foram distribuídos em garrafas Roux de 1000 mL, contendo 50mL de meio LIT ou
50mL do meio Alfa-MEM, acrescido de 10% soro fetal bovino, 10mL de cultura de
cada cepa na fase logarítmica de crescimento. As culturas foram mantidas em estufa
biológica (FANEM –Brasil) a 24º±1ºC. Após 4 dias, foi avaliada a viabilidade dos
parasitos, a ausência de contaminantes e 10mL de cada cultura foram repassados para
mais quatro garrafas de Roux contendo 50mL de meio LIT ou Alfa-MEM. As culturas
foram mantidas nas mesmas condições descritas. Decorridos quatro dias as
promastigotas foram examinadas, lavadas por centrifugação a 1000g por 10 minutos a
4ºC, o sedimento foi ressuspenso em solução salina 0,85% e foram realizadas mais três
centrifugações nas mesmas condições. A massa de parasitos foi então dividida em três
alíquotas em tubos Falcon com capacidade para 50mL, uma destinada à produção de
antígenos para sensibilização de placas de ELISA, outra para realização de eletroforese
bidimensional em gel 2-D (2-DE) e a terceira para o preparo do antígeno de RIFI. Até o
momento do processamento as alíquotas foram mantidas em freezer a -80 ºC, exceto
aquelas destinadas ao preparo de lâminas para a reação de RIFI, que foram utilizadas no
preparo do antígeno imediatamente, como descrito no item 4.4.2.2.
4.4.2.1 Preparação do Antígeno para Sensibilização das Placas de ELISA
Considerando uma relação de 100mg de massa úmida, para 10mg de massa seca,
foram utilizados 10mL de PBS 15M, pH 7,2, com mertiolate na concentração final de
1:10.000, contendo ainda 130 mg de iodoacetamida por 5mL de antígeno. O antígeno
foi ultrassonicado 3 vezes por 1 minuto a 40 Hertz com intervalo de 1 minuto entre as

46
ultrassonicações,em banho de gelo, em tubo Falcon. Em seguida foi agitado lentamente
durante 12 horas a 4ºC. Seguiu-se centrifugação a 5000g a 4
ºC. A concentração de
proteínas foi dosada pelo método de Lowry (Lowry et al., 1951) e ajustada para 2mg
por mL. O antígeno foi adicionado de Tioglicolato de Sódio na concentração final de
0,01% e aliquotado em tubos Eppendorf em volumes de um 1mL e armazenado a -20ºC
até o momento do uso. As placas de ELISA planas com fundo plano (Inlab®) foram
sensibilizadas diluindo o antígeno em tampão carbonato, segundo metodologia já
descrita.
4.4.2.2 Obtenção de Parasitos e Preparação do antígeno para RIFI
A preparação do antígeno para a Reação de Imunofluorescência Indireta seguiu
o Protocolo do Laboratório de Sorologia do Departamento de Parasitologia do Instituto
de Ciências Biológicas da UFMG, resumido a seguir:
Os parasitos foram cultivados em meio de cultura LIT ou Alfa-MEM. Quando as
Leishmania atingiram a fase logarítmica de crescimento, colocou-se diretamente no
meio de cultura, solução de formaldeído em PBS a 10% na razão de 1:1. Após
centrifugação a 400g durante 10 minutos, o sobrenadante foi desprezado e adicionado
de 10mL de tampão fosfato pH 7,4 , homogeinizado e centrifugado a 400g para a
lavagem dos parasitos. Este procedimento foi executado por três vezes. Após o último
procedimento, a suspensão foi homogeneizada com pipeta de Pasteur estéril, e uma gota
(0,25 mL) foi colocada em lâmina de vidro para observação ao microscópio óptico em
objetiva de 40X. A suspensão foi ajustada para se obter em um campo microscópico o
número de 30 a 50 formas promastigotas. Estando em número satisfatório, a solução foi
homogeneizada e uma gota (0,25mL) foi colocada em cada campo previamente
marcado nas lâminas de vidro especiais para a RIFI. As lâminas foram deixadas para
secar à temperatura ambiente, sendo posteriormente enroladas em papel absorvente e a
seguir em papel alumínio, para estocagem a -20ºC até o momento do uso.
4.4 2.3 Preparação do Antígeno para Eletroforese-Extração de Proteínas
A massa de parasitas foi homogeneizada em tampão de lise (8M uréia, 2M
tiouréia, 4% CHAPS, 65 mM dithiothreitol - DTT, 40mM Tris-base e coquetel de
inibidor de protease da GE Healhtcere) na proporção de 500 μL de tampão de lise para
109 parasitas. A amostra foi incubada por 2 horas a temperatura ambiente sob agitação

47
lenta, e posteriormente passada em seringa de insulina por 20 vezes. Seguiu-se
centrifugação a 20.000g por 30 minutos a temperatura ambiente. O sobrenadante
(extrato protéico) foi aliquotado em volume de 100µL e mantido a -80°C. A proteína
foi quantificada por 2D-Quant kit (GE Healthcare, USA) de acordo com as instruções
do fabricante.
4.5 Testes dos Antígenos
Foram realizados pelas reações de ELISA e Imunofluorescência Indireta. Os
antígenos foram testados em três momentos: 24 horas, 3 meses e 6 meses após o
armazenamento das placas de ELISA em freezer a -20C. Na reação de ELISA, para
cada momento foram sensibilizadas 16 placas com cada antígeno. Na RIFI foram
sensibilizadas 50 lâminas de cada antígeno, cada uma com capacidade de teste de
dezesseis soros, e dois controles, positivo e negativo. Da mesma forma as lâminas
foram armazenadas em freezer a -20ºC.
Os soros dos animais foram testados na diluição de 1:400, sempre em duplicata
nas placas de ELISA. Na RIFI o título inicial foi de 1:40, executando-se diluições
sucessivas até a obtenção da negatividade (diluição total). Os testes foram realizados
com as amostras de soro e dos antígenos codificados de forma que apenas um indivíduo
que não fazia parte do trabalho tinha a codificação.
Finalizando o período de teste, foram realizadas as análises estatísticas para
escolha do melhor antígeno. A seguir, todos foram decodificados e, apenas com o
antígeno que demonstrou melhor desempenho, foram realizados testes de reação
cruzada com soros de animais sabidamente positivos para Doença de Chagas (fase
crônica), Leishmaniose Tegumentar, Babesiose e Erliquiose caninas. Os soros positivos
para realização dos testes de reação cruzada foram gentilmente cedidos pelos
professores Dr. George Luiz Lins Machado Coelho – UFOP (Doença de Chagas), Dr.
Múcio Flávio Barbosa Ribeiro – UFMG (Babesiose e Erliquiose) e Prof. Dr. Ricardo
Fujiwara – UFMG (Leishmaniose Tegumentar).
4.5.1 Reação de Imunofluorescência Indireta- RIFI
Os soros testados foram diluídos na razão dois, a partir de 1:40 e até a diluição
final, em solução tampão fosfato (PBS, pH 7,4) . Foram colocados 25μL da diluição

48
obtida sobre cada região demarcada de uma lâmina onde o antígeno, constituído por
formas promastigotas íntegras de Leishmania, havia sido previamente fixado. Após a
incubação das lâminas em câmara úmida por 30 minutos em estufa a 37oC, estas foram
lavadas com PBS, cobertas com o tampão fosfato por 5 minutos, lavadas em água
destilada e secadas sob ventilação artificial (Ventilador Britânia B20, Brasil). A seguir,
em cada região demarcada da lâmina foram acrescentados 25μL do conjugado diluído a
seu título em PBS com Tween 2% (Tween®
20, Merck, Alemanha). O conjugado
utilizado foi anti IgG de cão, marcado com Isotiocianato de Fluoresceína (Bethyl Lab.
Inc., EUA) na proporção de 1:1500, acrescido de Azul de Evans 1% (EVANS BLUE®,
Sigma Aldrich, EUA). Após nova incubação, lavagem e secagem, a lâmina foi então
coberta com glicerina (Merck PA, Alemanha) tamponada, pH 7,4 e sobreposta por uma
lamínula. A leitura foi procedida em microscópio de luz ultravioleta (Olympus BX 41®,
Japão). Soros conhecidamente positivos e negativos foram usados em cada lâmina,
como controle positivo e negativo da reação.
4.5.2 Enzime Linked Immunosorbent Assay - ELISA
Foram utilizadas microplacas para ELISA de polietileno (Inlab®) de 96 orifícios
e fundo plano. Cada orifício foi sensibilizado com 2μg de antígeno, diluídos em 100μL
em tampão carbonato, durante um período mínimo de 24 horas. Para a realização do
teste, o excesso de antígeno foi retirado da placa pela lavagem de cada orifício por duas
vezes com solução de lavagem contendo 0,9% (p/v), de cloreto de sódio e 0,05% (v/v),
de Tween 20®. A seguir, para bloqueio dos sítios inespecíficos, foram adicionados
150μL solução de PBS-Caseína 2% contendo 2% (p/v) de caseína (Sigma Aldrich,
USA) em PBS, por orifício. A placa foi incubada por 60 minutos em estufa a 37°C. O
excesso de solução de bloqueio foi então retirado por duas lavagens sucessivas. Em
seguida as placas foram colocadas para secar por inversão à temperatura ambiente e
posteriormente congelada em freezer a -20ºC. No momento do uso, após a retirada do
freezer, 100μL dos soros, diluídos em tampão de incubação e na razão de 1:400, foram
aplicados em cada orifício e a placa novamente colocada em estufa a 37ºC por 60
minutos. O excesso do soro diluído foi retirado logo após por uma série de cinco
lavagens. Foi utilizado como conjugado, anti-imunoglobulina de cão IgG, marcada com
Peroxidase VI (Bethyl Lab. Inc., USA). O conjugado foi previamente titulado e após ser
diluído ao seu título, 100μL da diluição foi acrescentado a cada orifício, e a placa

49
deixada em incubação por mais 60 minutos a 37°C. Posteriormente, esta solução foi
retirada por nova série de cinco lavagens e em seguida, foram adicionados a cada
orifício 100μL de solução de OPD - Orto-Fenilenediamino dihidrocloridro (Sigma
Aldrich, USA). A reação ocorreu em estufa à 37ºC durante 8-10 minutos, e
interrompida por adição de 25μL de H2SO4 4N (Merck,PA,Alemanha). Em qualquer
etapa, após as lavagens, as placas foram secadas por inversão sobre papel absorvente. A
leitura foi efetuada em leitor de ELISA (Bio-Rad 2550®
, USA), a 495nm. O ponto de
corte ou “cut off” foi determinado pela curva ROC (Receiver Operating Characteristic)
para cada placa.
4.5.2.1 Curva ROC - Receiver Operating Characteristic
Pela possibilidade de definir um ponto de corte, ou um limiar de decisão para se
classificar e contabilizar o número de predições positivas e negativas na comparação
relativa da fração de verdadeiros positivos (FVP), fração de falsos positivos (FFP),
fração de verdadeiros negativos (FVN) e fração de falsos negativos (FFN) foi escolhida
a curva ROC na obtenção do ponto de corte do teste de ELISA.
A curva ROC facilita o entendimento entre a relação de sensibilidade e
especificidade de um teste diagnóstico quantitativo ao longo de valores contínuos de um
ponto de corte (Martinez, et al., 2003).
Esta situação pode ser descrita graficamente pela curva ROC apresentada a
seguir.

50
4.5.3 Eletroforese Bidimensional (2-DE)
Para realizar a caracterização do perfil proteômico Imunogênico do antígeno de
melhor desempenho foram utilizadas as técnicas de Eletroforese Bidimensional (2-DE)
e Western Blot. A 2-DE foi feita em triplicata experimental, sendo que, em cada
experimento foram produzidos 4 géis simultaneamente, um deles era destinado a
coloração e os demais à realização de western blot.
4.5.3.1 Focalização Isoelétrica (IEF) –
As tiras IPG (gradiente de pH imobilizado) de 13 cm, pH 4-7 (GE Healthcare)
foram hidratadas com 600µg de amostra protéica em tampão de re-hidratação IEF
(Uréia 8M, Tiuréia 2M, CHAPS 4%, azul de bromofenol 0,0025%) e anfólitos IPG
Buffer pH 4-7, 1% v/v em um volume final de 250L. Em seguida as tiras foram
focalizadas usando o aparelho IEF Cell System (GE Healthcare), com os seguintes
passos: 250V por 15 minutos; 300V por 4 horas; 1000V até acumular 300Vh; 5000V
até acumular 4000Vh; 5000V até 1250Vh e 500V por 1 hora. Finalizada a focalização
isoelétrica, as strips foram submetidas à redução e alquilação por incubação
consecutiva, por 15 minutos, em 5 mL de DTT 125 mM primeiro e Iodacetamida 125
mM depois, em tampão de equilíbrio (50mM Tris-HCl pH 8.8, 6M uréia, 30% glicerol,
2% SDS e 0.002% bromophenol blue).
4.5.3.2 SDS-PAGE
Para separação na segunda dimensão foi realizada eletroforese desnaturante em
gel de poliacrilamida a 12% usando o sistema Ettan DALT 6 unit (GE Healthcare)
conectado a um sistema de resfriamento. As strips foram posicionadas horizontalmente
no topo dos géis e foi adicionado 1mL de agarose 0,5% contendo azul de bromofenol.
Após a corrida, os géis eram corados por comassie colloidal - CBB G-250 (Neuhoff, et
al., 1988) ou transferidos para membrana de nitrocelulose.
4.5.3.3 Western blot - 2D gel
As proteínas de géis 2-DE foram transferidas para a membrana de nitrocelulose
(Amersham Biosciences) utilizando o “semidry transfer Unit” (GE Healthcare) e
incubadas com uma solução de bloqueio (5% de leite desnatado Molico em 20mM Tris,

51
500mM NaCl, pH 7.4) a 4C por 16 a 18h. Cada experimento foi feito com três
membranas simultaneamente, sendo uma incubada com “pool” (n=10) de soros de cães
naturalmente infectados com altos títulos de anticorpos (RIFI > 1:1.280), outra com
“pool” (n=10) de soro de cães naturalmente infectados com baixos títulos de anticorpos
(RIFI 1:160), e a terceira com “pool” (n=10) de soros de cães não infectados por L.
chagasi. O anticorpo primário foi utilizado em diluição 1:500 em solução de bloqueio,
por 2 horas, à temperatura ambiente. Após 3 lavagens com 0.05% PBS–Tween por 15
minutos, as membranas foram incubadas com anti-IgG total, conjugados com
peroxidase (Sigma) na diluição de 1:10.000. Para revelação da reação foi utilizada a
mistura de duas soluções: 6 mg de 3,3'-diaminobenzidina DAB (D-5637, Sigma) em 12
ml do tampão PBS e 12mL da solução contendo 10mL de PBS, 2mL de metanol e 10 μl
de peróxido de hidrogênio (Towbin, H. & Gordon, J., 1984).
4.5.3.4 Análise dos Géis-
Os géis corados e membranas provenientes do WB foram digitalizados utilizando o
equipamento ImageScanner (GE Healthcare) e as imagens foram analisadas com o
programa Image Master 2D Platinum 6 (GE Healthcare).
As imagens das membranas foram analisadas para seleção dos “spots” que são
reconhecidos por soros com baixos e altos títulos de anticorpos individualmente e
simultaneamente, descartando-se aqueles que são reconhecidos por soros de animais
não infectados (reações inespecíficas).
Dessa forma foi possível localizar nos géis 2-DE as proteínas do antígeno que
apresentou melhor performance e que são imunogênicas para soros caninos com baixos
e altos títulos de anticorpos individualmente e simultaneamente. Os “spots”
selecionados como imunogênicos e sem reações inespecíficas serão posteriormente
retirados dos géis para identificação por EM.
4.6 Análise Estatística
Os valores de sensibilidade, especificidade, valor preditivo positivo e valor
preditivo negativo foram obtidos através do programa Open Epi, com intervalo de
confiança de 95%. A concordância entre os resultados das reações de cada um dos

52
antígenos com o verdadeiro estatus (positivo ou negativo) da amostra foi feita através
do índice de Youden (J).
Para as reações de RIFI e ELISA/RIFI de Biomanguinhos, os valores de
sensibilidade, especificidade, valor preditivo positivo e valor preditivo negativo foram
obtidos também através do programa Open Epi, com intervalo de confiança de 95%. A
concordância entre os resultados das reações de cada um dos antígenos com o
verdadeiro estatus (positivo ou negativo) da amostra foi feito pelo Índice Kappa de
Cohen.
As curvas de variação das absorbâncias obtidas na ELISA em função dos títulos
de anticorpos detectados na RIFI foram analisadas individualmente pela regressão linear
simples, assumindo como significativas aquelas com p<0,05.
Em cada um dos gráficos foi realizada a comparação das linhas de tendência da
variação da absorbância da ELISA em função do título obtido na RIFI de cada antígeno,
utilizando a análise de variância (ANOVA), assumindo como diferenças significativas
aquelas com valor p<0,05.
Foi calculado também o Coeficiente de Determinação (R2) para evidenciar o
quanto da variação total é comum aos elementos que constituem os pares analisados.

53
5. Resultados
5.1 Seleção dos animais
Cento e dez cães com testes sorológicos convencionais (ELISA e RIFI) positivos
e negativos para LV foram pré-selecionados como doadores de soro para o experimento.
Após esta avaliação sorológica, foi realizada a parasitológica, e considerando os
critérios estabelecidos para a inclusão da amostra como verdadeiramente positiva ou
verdadeiramente negativa, foram descartados os soros de sete animais, por apresentarem
resultado parasitológico positivo no PCR e negativo nos testes de ELISA e RIFI. Assim,
foram incluídos 54 doadores de soro positivo e 49 doadores de soro negativo. A
caracterização final dos animais está descrita na Tabela 1.
Tabela 1: Categorização dos animais pré-selecionados como portadores ou não de
leishmaniose visceral, após avaliação parasitológica e sorológica.
Testes Parasitológicos Testes
Sorológicos
Número
de
amostras
Categorização
Crescimento em
meio NNN/Alfa
MEM
PCR
Exame do
Esfregaço de
Medula Óssea
ELISA RIFI
Neg Pos Neg Neg Neg 07 Excluído
Pos Pos Pos Pos Pos 54 Positivo
Neg Neg Neg Neg Neg 49 Negativo
Pos – Positivo Neg – Negativo
5.2 Curva de Crescimento e Produção dos Antígenos
Os gráficos obtidos com os dados das contagens reproduzem o já descrito na literatura e
apontam padrões diferentes de crescimento para a mesma amostra em cada um dos dois
meios de cultura. A Figura 1 mostra, como representativo, as curvas de crescimento
obtidas nos dois meios de cultura com a amostra BH400. As características do
crescimento foram observadas para a obtenção dos parasitos em fase exponencial e
início da fase estacionária, segundo rotina já pré-estabelecida na preparação de
antígenos.

54
Figura 1 Curva de crescimento da cepa BH400 de Leishmania (Leishmania) chagasi, (A)
cultivada em meio LIT (Liver Infusion Tryptose) e (B) em meio α- MEM
(Minimum Essential Medium) acrescido de 10% de SFB

55
Os antígenos produzidos para os testes de ELISA e RIFI foram codificados e
testados como tal, conforme o apresentado na Tabela 2.
Tabela 2 Codificação dos antígenos produzidos com as diferentes amostras de
Leishmania crescidas nos meios de cultura LIT (Camargo, 1966) e Alfa-
MEM acrescido de 20% de soro fetal bovino.
5.3 Teste dos Antígenos
5.3.1 Reação de ELISA
Para a realização da reação de ELISA, os soros e os antígenos foram testados
codificados, sem o conhecimento de quais eram os negativos e positivos. Para cada
placa testada foram utilizados soros, como controle positivo e negativo, e a
determinação do ponto de corte positivo/negativo de leitura foi realizado através da
Curva ROC. A zona cinza foi estabelecida para todas as placas como as leituras
compreendidas entre 10% acima e abaixo do ponto de corte. Os resultados são
mostrados por teste dos soros positivos e negativos, nos tempos 24 horas, três e seis
meses após a sensibilização das placas. As Figuras, 2 e 3, 4 e 5, 6 e 7 mostram
respectivamente, os resultados obtidos para os soros positivos e negativos testados 24
horas, três e seis meses após a sensibilização das placas.
ESPÉCIE DE Leishmania CÓDIGO DO
ANTÍGENO Meio de Cultura
Leishmania ( Leishmania)
amazonensis
(MHOM/BR/1960/BH6)
A
MEIO LIT Leishmania (Leishmania) chagasi
(MHOM/BR/1972/BH46) C
Leishmania (Leishmania) chagasi
(MCAN/BR/2002/BH400 E
Leishmania (Leishmania)
amazonensis
(MHOM/BR/1960/BH6)
B
MEIO ALFA MEM Leishmania (Leishmania) chagasi
(MHOM/BR/1972/BH46) D
Leishmania (Leishmania) chagasi
(MCAN/BR/2002/BH400) F

56
Figura 2 Teste de ELISA realizado em soros
de animais positivos para
leishmaniose visceral canina, com
os antígenos A, B, C, D, E e F, testados 24 horas após
sensibilização das placas.
Figura 4 Teste de ELISA realizado em soros
de animais positivos para
leishmaniose visceral canina, com os antígenos A, B, C, D, E e F,
testados três meses após
sensibilização das placas.
Figura 6 Teste de ELISA realizado em soros
de animais positivos para
leishmaniose visceral canina,
antígenos A, B, C, D, E e F, testados seis meses após
sensibilização das placas
Figura 3 Teste de ELISA realizado em soros
de animais negativos para
leishmaniose visceral canina, com
os antígenos A, B, C, D, E e F, testados 24 horas após
sensibilização das placas
Figura 5 Teste de ELISA realizado em soros
de animais negativos para
leishmaniose visceral canina, com
os antígenos A, B, C, D, E e F, testados três meses após
sensibilização das placas
Figura 7 Teste de ELISA realizados em
soros de animais negativos para leishmaniose visceral canina, com
o antígenos A, B, C, D, E e F,
testados seis meses após sensibilização das placas
0 20 40 60 80 100
A
B
C
D
E
F
%
An
tíge
no
s
POSITIVO
NEGATIVO
ZONA CINZA
0 20 40 60 80 100
A
B
C
D
E
F
%
An
tíge
no
s
POSITIVO
NEGATIVO
ZONA CINZA
0 20 40 60 80 100
A
B
C
D
E
F
%
An
tíge
no
s
POSITIVO
NEGATIVO
ZONA CINZA
0 20 40 60 80 100
A
B
C
D
E
F
%
An
tíge
no
s
POSITIVO
NEGATIVO
ZONA CINZA
0 20 40 60 80 100
A
B
C
D
E
F
%
An
tíge
no
sPOSITIVO
NEGATIVO
ZONA CINZA
0 20 40 60 80 100
A
B
C
D
E
F
%
An
tíge
no
s POSITIVO
NEGATIVO
ZONA CINZA
A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400 (Leishmania (L) chagasi), cultivadas em em meio LIT; B, BH6 (Leishmania (L.)
amazonensis); D e F, BH46 e BH400 Leishmania (L) chagasi, cultivadas em em meio Alfa-MEM SFB 10%)

57
Os resultados mostraram que os todos os antígenos apresentam alto indice de
concordância com a categoria verdadeiro positivo e negativo para LVC dos soros
testados, quando o teste de ELISA foi realizado 24 horas após sensibilização das placas.
O antígeno E foi capaz de identificar 100% dos soros positivos (54/54). Já os
antígenos A, C, D e F identificaram 98% (52/54) das amostras positivas, e o antígeno B
96% (52/54). Estes valores foram observados em decorrencia de resultados falso
negativos. Com os soros negativos o diagnóstico foi confirmado em todos os antígenos
testados, sem resultados falsos.
Após três meses de conservação das placas, os resultados mostraram que os
antígenos E e F identificaram 100% (54/54) dos soros verdadeiramente positivos e
nenhum deles apresentou resultados falso negativos. Os demais antígenos A 92,6%
(50/54), B 87% (47/54), C 96,2% (52/54), e D 89% (48/54) identificaram dezenove
soros como falso negativos (4; 7; 2 e 6 respectivamente). Exceto o antígeno B, todos os
outros mostraram leituras na zona cinza. Nos testes com os soros negativos, os
antígenos A, C e D identificaram falso positivos (4, 2 e 1 amostras, respectivamente).
Os antígenos A, C, D, E e F mostraram leituras na zona cinza.
Após seis meses, os testes se mostraram muito semelhantes aqueles apresentados
pelos antígenos após os 3 meses de conservação das placas para os soros positivos.
Dentre os soros positivos, os antígenos A e F identificaram 100% (54/54) das amostras
corretamente e nenhum deles presentou resultados falso negativos. Os antígenos D e E
identificaram 98% (53/54) com 1 resultado falso negativo cada. O antígeno C
identificou 96,2% (52/54) e o antígeno B 91% (49/54). Todos apresentaram resultado
em zona cinza (Tabela 4).
Em relação aos soros negativos D, E e F apresentaram concordancia total de
100% (49/49). Os demais apresentaram A 96% (47/49), B 92% (45/49) e C 98%
(48/49). Houve 7 resultados falso positivos detectados pelos antígenos A, B e C. Todos
apresentaram leituras em zona cinza.
A Tabela 3 mostra um consolidado dos resultados dos testes realizados nos três
tempos.

58
Tabela 3 Teste de ELISA nos tempos de 24 horas, três e seis meses após a
sensibilização das placas, com os antígenos A, B, C, D, E e F, em soros
caninos padrões positivo e negativo para leishmaniose visceral.
Antígeno A
24 horas 3 meses 6 meses
Positivo Negativo Total de
Soros Positivo Negativo
Total de
Soros Positivo Negativo
Total de
Soros
Positivo 53 0 53 50 4 54 54 2 56
Negativo 1 49 50 4 45 49 0 47 47
Total de Testes 54 49 103 54 49 103 54 49 103
Antígeno B
24 horas 3 meses 6 meses
Positivo Negativo Total de
Soros Positivo Negativo
Total de
Soros Positivo Negativo
Total de
Soros
Positivo 52 0 52 47 0 47 49 4 53
Negativo 2 49 51 7 49 56 5 45 50
Total de Testes 54 50 103 54 49 103 54 49 103
Antígeno C
24 horas 3 meses 6 meses
Positivo Negativo Total de
Soros Positivo Negativo
Total de
Soros Positivo Negativo
Total de
Soros
Positivo 53 0 53 52 2 54 52 1 53
Negativo 1 49 50 2 47 49 2 48 50
Total de Testes 54 49 103 54 49 103 54 49 103
Antígeno D
24 horas 3 meses 6 meses
Positivo Negativo Total de
Soros Positivo Negativo
Total de
Soros Positivo Negativo
Total de
Soros
Positivo 53 0 53 48 1 49 53 0 53
Negativo 1 49 50 6 48 54 1 49 50
Total de Testes 54 49 103 54 49 103 54 49 103
Antígeno E
24 horas 3 meses 6 meses
Positivo Negativo Total de
Soros Positivo Negativo
Total de
Soros Positivo Negativo
Total de
Soros
Positivo 54 0 54 54 0 54 53 0 53
Negativo 0 49 49 0 49 49 1 49 50
Total de Testes 54 49 103 54 49 103 54 49 103
Antígeno F
24 horas 3 meses 6 meses
Positivo Negativo Total de
Soros Positivo Negativo
Total de
Soros Positivo Negativo
Total de
Soros
Positivo 53 0 53 54 0 54 54 0 54
Negativo 1 49 50 0 49 49 0 49 49
Total de Testes 54 49 103 54 49 103 54 49 103

59
Os resultados apresentados na Tabela 3 mostram que o antígeno E e F
mostraram resultados mais coerentes com a condição positivo ou negativo do soro
testado, uma vez que apresentaram apenas um resultado falso, entretanto 17 soros
mostraram resultados de leitura na zona cinza para cada um. Os antígenos B, e D
apresentaram menos resultados de leitura na zona cinza, porém, mostraram resultados
falsos, tanto negativos quanto positivos. Os antígenos A e C apresentaram 32 e 20
resultados em zona cinza respectivamente e da mesma forma que os anteriores,
mostraram testes falsos positivos e negativos.
Tabela 4 Valores percentuais e absolutos de número de amostras classificadas como
Zona Cinza, acima e abaixo do ponto de corte do ELISA, durante todas as
fases do experimento.
Antígenos A (%) B (%) C (%) D (%) E (%) F (%)
Tempo
24 horas
ZC+ 9 (5/54) 7 (4/54) 4 (2/54) 2 (1/54) 6 (3/54) 7 (4/54)
ZC- 2 (1/49) 2 (1/49) 0 (0/49) 2 (1/49) 0 (0/49) 0 (0/49)
Total 6 (6/103) 5 (5/103) 2 (2/103) 2 (2/103) 3 (3/103) 4 (4/103)
3 meses
ZC+ 15 (8/54) 0 (0/54) 9 (5/54) 6 (3/54) 6 (3/54) 7 (4/54)
ZC- 12 (6/49) 0 (0/49) 4 (2/49) 0 (0/49) 6 (3/49) 4 (2/49)
Total 14 (14/103) 0 (0/103) 7 (7/103) 3 (3/103) 6 (6/103) 6 (6/103)
6 meses
ZC+ 11 (6/54) 7 (4/54) 13 (7/54) 9 (5/54) 7 (4/54) 7 (4/54)
ZC- 12 (6/49) 10 (5/49) 8 (4/49) 4 (2/49) 8 (4/49) 6 (3/49)
Total 12 (12/103) 9 (9/103) 11 (11/103) 7 (7/103) 8 (8/103) 7 (7/103)
Total
Geral 10 (32/309) 5 (14/309) 6 (20/309) 4 (12/309) 6 (17/309)
6
(17/309)
ZC+ amostras situadas acima do ponto de corte ZC- amostras situadas abaixo do ponto de corte
A maior porcentagem dos animais em ZC ficou representada em todos os tempos pelo
antígeno A (10%). Valores menores foram encontrados, nas 24 horas após a
sensibilização das placas pelos antígenos C e D, nos três meses de sensibilização com o
antígeno B e aos seis meses pelos antígenos D e F.

60
Nas reações realizadas com 24 horas, nas amostras em zona cinza não houve soros
falsos positivos ou negativos. Já nos três meses ocorreram resultados falsos positivos
nas zonas cinzas dos antígenos A (uma amostra) e C (duas amostras). Nas placas
conservadas por seis meses, todos os antígenos com exceção do F apresentaram
resultados falsos, sendo dois falso positivos para o antígeno A e um falso negativo para
cada um dos antígenos B, C, D e E.
Os valores da sensibilidade, especificidade, valor preditivo positivo e valor
preditivo negativo do teste ELISA nos três tempos foram avaliados com intervalo de
confiança de 95% e os resultados estão mostrados nas figuras 8, 9 e 10, a seguir:
Figura 8 Comparação da sensibilidade, especificidade, valor preditivo positivo e valor
preditivo negativo dos antígenos utilizados no teste ELISA 24 horas após a
sensibilização das placas. (A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400
(Leishmania (L) chagasi), cultivadas em meio LIT; B, BH6 (Leishmania (L.) amazonensis); D e
F, BH46 e BH400 Leishmania (L) chagasi, cultivadas em meio α-MEM SFB 10%)
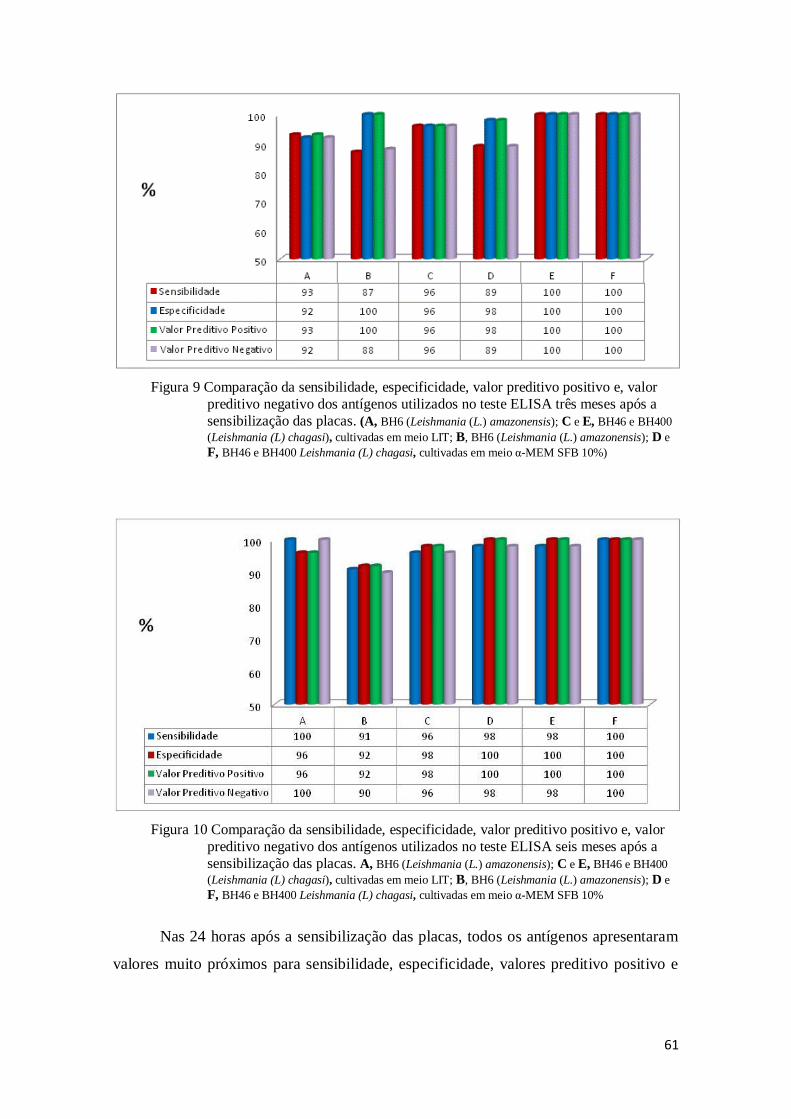
61
Figura 9 Comparação da sensibilidade, especificidade, valor preditivo positivo e, valor
preditivo negativo dos antígenos utilizados no teste ELISA três meses após a
sensibilização das placas. (A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400
(Leishmania (L) chagasi), cultivadas em meio LIT; B, BH6 (Leishmania (L.) amazonensis); D e
F, BH46 e BH400 Leishmania (L) chagasi, cultivadas em meio α-MEM SFB 10%)
Figura 10 Comparação da sensibilidade, especificidade, valor preditivo positivo e, valor
preditivo negativo dos antígenos utilizados no teste ELISA seis meses após a
sensibilização das placas. A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400
(Leishmania (L) chagasi), cultivadas em meio LIT; B, BH6 (Leishmania (L.) amazonensis); D e
F, BH46 e BH400 Leishmania (L) chagasi, cultivadas em meio α-MEM SFB 10%
Nas 24 horas após a sensibilização das placas, todos os antígenos apresentaram
valores muito próximos para sensibilidade, especificidade, valores preditivo positivo e

62
preditivo negativo, variando de 96-100% (Figura 8). O índice de concordância de
Youden foi de 0,96 (antígeno B); 0,98 (antígenos A, C, D e F) e 1,00 (antígeno E).
Após três meses os antígenos E e F apresentaram valores de 100% para todos os
quesitos avaliados. Para os outros antígenos a sensibilidade variou de 87 a 96%, a
especificidade de 92 a 100%, o valor preditivo positivo de 93 a 100% e o valor preditivo
negativo de 88 a 96% (Figura 9). O índice de concordância de Youden variou de 0,85
(antígeno A); 0,87 (antígenos B e D); 0,98 (antígeno C) e 1,00 (antígeno E e F).
Aos seis meses o antígeno F foi o único que mostrou valor de 100% para todas
as variáveis avaliadas. Para os outros antígenos a sensibilidade variou de 91 a 100%, a
especificidade de 92 a 100%, o valor preditivo positivo de 92 a 100% e o valor preditivo
negativo de 96 a 96% (Figura 10). O índice de concordância de Youden foi de 0,81
(antígeno B); 0,96 (antígeno C) e 1,00 (antígeno A, D, E e F).
5.3.2 Reação de Imunofluorescência Indireta
A Reação de Imunofluorescência Indireta foi realizada com cada um dos
antígenos em apenas um momento do experimento, 30 dias, após a preparação dos
antígenos. Quando reagente, o soro foi diluído até o limite para a negatividade Os
resultados estão expressos nas figuras 11 e 12.

63
Figura 11 Teste da RIFI realizados em soros de cães portadores de leishmaniose visceral com diferentes antígenos. (A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400
(Leishmania (L) chagasi), cultivadas em meio LIT; B, BH6 (Leishmania (L.) amazonensis); D e
F, BH46 e BH400 Leishmania (L) chagasi, cultivadas em meio α-MEM SFB 10%)
Figura 12 Teste da RIFI realizados em soros de cães negativos para leishmaniose visceral
com diferentes antígenos. (A, BH6 (Leishmania (L.) amazonensis); C e E, BH46 e BH400
(Leishmania (L) chagasi), cultivadas em em meio LIT; B, BH6 (Leishmania (L.) amazonensis);
D e F, BH46 e BH400 Leishmania (L) chagasi, cultivadas em em meio α-MEM SFB 10%)
Nas reações realizadas com os soros verdadeiramente positivos foram
observados sete soros falso negativo quando o teste foi realizado com os antígenos C e
D. Para aos soros verdadeiramente negativos os resultado do teste foi coincidente para
100% dos soros com todos os antígenos.
0 20 40 60 80 100
A
B
C
D
E
F
%
An
tíge
no
s
POSITIVO
NEGATIVO
0 20 40 60 80 100
A
B
C
D
E
F
%
An
tíge
no
s
POSITIVO
NEGATIVO

64
A comparação entre a sensibilidade, especificidade, valores preditivo negativo e
positivo está apresentada na figura 13.
Figura 13 Comparação da sensibilidade, especificidade, valor preditivo positivo e, valor preditivo negativo dos antígenos utilizados na Reação de Imunofluorescência
Indireta –RIFI.
Os valores de sensibilidade, especificidade, valor preditivo positivo e valor
preditivo negativo, que foram de 100% para os antígenos A, B, E, F. Os antígenos C e
D, por apresentarem sete resultados falsos negativos, apresentaram valores de
sensibilidade e valor preditivo negativo de 87%. Para especificidade e valor preditivo
positivo estes valores foram de 100%. O índice de concordância Kappa foi de 0,87 para
os antígenos C e D e de 1,00 para os demais.
5.4 Reações de RIFI e ELISA com os kits de Biomanguinhos
Os soros selecionados para o experimento foram também testados pelo kit
ELISA de Biomanguinhos (LOTE 086EL007Z). Este kit é atualmente utilizado por
laboratórios da rede pública para o diagnóstico da LVC. Os resultados são mostrados na
Figura14
80%
85%
90%
95%
100%
A B C D E F
Sensibilidade 100% 100% 87% 87% 100% 100%
Especificidade 100% 100% 100% 100% 100% 100%
Valor Preditivo Positivo 100% 100% 100% 100% 100% 100%
Valor Preditivo Negativo 100% 100% 87% 87% 100% 100%
%
Antígenos
Sensibilidade Especificidade Valor Preditivo Positivo Valor Preditivo Negativo

65
Figura 14 Teste de ELISA realizado em soros verdadeiramente positivos e negativos para
leishmaniose visceral canina com o kit Biomanguinhos, LOTE 086EL007Z
Não houve resultados falso negativos entretanto, entre os soros negativos 10
amostras foram identificadas como falso positivos e 5 mostraram valores de leitura
dentro da zona cinza.
Estes soros foram testados também com o kit de RIFI de Biomanguinhos (LOTE
086LC004Z), que utiliza como antígeno a cepa BH6 - Leishmania (L.) amazonensis
(MHOM/BR/1960/BH6), crescida em meio LIT. Os resultados são mostrados na
Figuras 15.
Figura 15 Teste de RIFI realizado em amostras de soros positivos e negativos para
leishmaniose visceral canina como kit Biomanguinhos, LOTE 086LC004Z.
Os resultados mostraram um soro falso negativo entre aqueles comprovadamente
positivos para LVC. Os soros comprovadamente negativos mostraram doze resultados
falso positivos com o kit RIFI de Biomanguinhos.
0
20
40
60
80
100
Soros positivos Soros Negativos
%
Zona Cinza
Negativo
Positivo
0
20
40
60
80
100
Soros Positivos Soros Negativos
% Negativo
Positivo

66
Os valores da sensibilidade, especificidade, valor preditivo positivo, valor preditivo
negativo das reações sorológicas realizadas com os kits ELISA e RIFI Biomanguinhos
são mostrados na Figura 16 O índice de concordância Kappa foi de 0,80 para o ELISA e
de 0,90 para a RIFI.
Figura 16 Comparação da sensibilidade, especificidade, valor preditivo positivo e, valor preditivo negativo dos antígenos dos kit-ELISA Biomanguinhos LOTE
086EL007Z e kit- RIFI Biomanguinhos LOTE 086LC004Z
5.5 Relação entre os títulos da RIFI e os valores de absorbância do
ELISA
Nos gráficos a seguir (Figuras 17, 18 e 19) são mostradas as linhas de regressão
representando as médias de absorbância do ELISA em relação aos títulos da RIFI. Foi
realizada para cada tempo a Análise de Variância (ANOVA) para evidenciar possíveis
diferenças significativas entre as médias de absorbância do ELISA, mostradas com cada
um dos antígenos, e calculado o Coeficiente de Determinação (R2) para evidenciar o
quanto da variação total é comum aos elementos que constituem os pares analisados, ou
seja, se quando um dos pares varia o outro também sofre variação. Quando o valor de
R2 é maior do que 0,60 indica uma boa correlação.
0
20
40
60
80
100
ELISA RIFI
Sensibilidade 100 98
Especificidade 80 76
Valor Predetivo Positivo 84 82
Valor Preditivo negativo 100 97
Sensibilidade Especificidade Valor Predetivo Positivo Valor Preditivo negativo

67
Figura 17 Relação entre os valores de absorbância do ELISA, após 24 horas de
sensibilização das placas, e os títulos das diluições da RIFI e a tendência das
linhas de regressão para os antígenos A, B, C, D, E e F.
1-Negativo 2-1:40 a 1:80 3-1:160 a 1:320
4-1:640 a 1:1280 5-1:2560 a 1:5120 6-1:10240 a 1:20480
Considerando-se um valor de diluição da RIFI e correlacionando-se com o valor
correspondente do ELISA, percebe-se que as absorbâncias são bem maiores para os
antígenos E e F. Isto é particularmente interessante na RIFI para os soros negativos e
para os primeiros títulos positivos (códigos 1 e 2 respectivamente), o que significa que
estes dois antígenos são capazes de apresentar valores baixos de absorbância para
amostras negativas e valores altos para positivas e desta forma, diferenciam melhor os
soros. O gráfico mostra também que as absorbâncias do ELISA estão positivamente
correlacionadas com os títulos da RIFI, com exceção dos antígenos B e F para títulos de
RIFI maiores que 1:2560 (p=0,000 e R2= 0,685).

68
Figura 18 Relação entre os valores de absorbância do ELISA, após três meses de sensibilização das placas, e os títulos das diluições da RIFI e a tendência das
linhas de regressão para os antígenos A, B, C, D, E e F.
1-Negativo 2-1:40 a 1:80 3-1:160 a 1:320
4-1:640 a 1:1280 5-1:2560 a 1:5120 6-1:10240 a 1:20480
Nos três meses, a ANOVA também mostrou que as diferenças das médias de
absorbância do ELISA são significativas no nível de 5% (p=0,000 e R2=0,692). Os
antígenos que mostraram melhor desempenho em diferenciar resultados positivos dos
negativos foram C e E, apresentando para leituras de RIFI entre 1:40-1:80 valores de
ELISA acima de 0,300. As absorbâncias de ELISA estão diretamente correlacionadas
com os títulos da RIFI, com exceção do antígeno F, para títulos acima de 1:2560.

69
Figura 19 Relação entre os valores de absorbância do ELISA, após seis meses de
sensibilização das placas, e os títulos das diluições da RIFI e a tendência das
linhas de regressão para os antígenos A, B, C, D, E e F.
1-Negativo 2-1:40 a 1:80 3-1:160 a 1:320
4-1:640 a 1:1280 5-1:2560 a 1:5120 6-1:10240 a 1:20480
Após os seis meses a ANOVA mostrou ainda significância entre médias de
absorbância do ELISA obtidas com os antígenos C, D, E e F (p=0,000 e R2=0,697). Já
para os antígenos A e B não foram encontradas diferenças (p=0,266 e R2=0,692).
Os antígenos C e E apresentaram melhor capacidade para distinguir soros
positivos e negativos, porém, considerando-se o título dos negativos, o antígeno C
apresenta maiores valores de absorbância, tornando-se inferior ao antígeno E.
Independente do antígeno, as absorbâncias aumentam significativamente com o
aumento dos títulos da RIFI.
Considerando-se que os antígenos E e F apresentaram uma melhor performance
em relação aos demais, demonstrada por seus valores de sensibilidade, especificidade,
valores preditivos positivos e negativos e índice de concordância de Youden, foi
realizada uma análise isolada dos mesmos em relação aos títulos de RIFI, mostradas nas
Figuras 20 e 21 e 22.

70
5.5.1 Avaliação dos antígenos com melhor desempenho nos testes de
ELISA com 24 horas, três meses e seis meses após a sensibilização das
placas.
Considerando-se que os antígenos E e F apresentaram uma melhor performance
em relação aos demais, demonstrada por seus valores de sensibilidade, especificidade,
valores preditivos positivos e negativos e índice de concordância de Youden, foi
realizada uma análise isolada dos mesmos em relação aos títulos de RIFI., mostradas
nas Figuras 20 e 21 e 22.
Figura 20 Comparação das linhas de tendência dos antígenos E e F, mostrando a relação entre os valores de absorbância do ELISA, 24 horas após a sensibilização das
placas, e os títulos das diluições da RIFI
Nas reações do ELISA, realizadas 24 horas após a sensibilização das placas,
quando comparadas com os resultados da RIFI observou-se que as médias das
absorbâncias são significativamente diferentes (ANOVA p=0,000; R2
2=0,671) para
ambos os antígenos, com valores maiores para o antígeno E, que apenas nos títulos mais
altos da RIFI discrimina melhor os positivos dos negativos (Códigos 4 e 6). A tendência
do aumento da absorbância em função dos títulos da RIFI é semelhante para os dois
antígenos. Portanto, baseados neste resultado, poderia ser escolhido o antígeno E ou F.
1-Negativo 2-1:40 a 1:80 3-1:160 a 1:320
4-1:640 a 1:1280 5-1:2560 a 1:5120 6-1:10240 a 1:20480

71
Figura 21 Comparação das linhas de tendência dos antígenos E e F, mostrando a relação entre os valores de absorbância do ELISA, três meses após a sensibilização das
placas, e os títulos das diluições da RIFI
1-Negativo 2-1:40 a 1:80 3-1:160 a 1:320
4-1:640 a 1:1280 5-1:2560 a 1:5120 6-1:10240 a 1:20480
A Figura 21 mostra resultados semelhantes aos da Figura 20 para a
significância da diferença das médias de absorbâncias do ELISA, realizado três meses
após a sensibilização das placas, (ANOVA p=0,000; R2
2=676). O antígeno F discrimina
menos os negativos dos positivos nos códigos 2 e 6 da RIFI, e nestes pontos observa-se
também que a tendência do aumento da absorbância em função dos títulos da RIFI é
diferente. Em vista deste resultado, a escolha deve ser do antígeno E. Entretanto, não
deve ser descartada a possibilidade de erro ou interferência de um fator externo, uma
vez que esta etapa apresenta perfil diferente do mostrado para a reação nas 24 horas.

72
Figura 22 Comparação das linhas de tendência dos antígenos E e F, mostrando a relação
entre os valores de absorbância do ELISA, seis meses após a sensibilização das placas, e os títulos das diluições da RIFI
1-Negativo 2-1:40 a 1:80 3-1:160 a 1:320
4-1:640 a 1:1280 5-1:2560 a 1:5120 6-1:10240 a 1:20480
Seis meses depois de sensibilizadas as placas de ELISA, na comparação das
médias de absorbância do ELISA entre os dois antígenos os resultados mostraram
significância (ANOVA p=0,000; R2=710), e tendências semelhantes do aumento da
absorbância em função dos títulos da RIFI. A capacidade de diferenciação de positivos e
negativos também foi igual e, portanto, pode-se escolher entre os antígenos E ou F.
Como a literatura descreve que não existe regra de similaridade para aumento da
absorbância do ELISA em função dos títulos da RIFI, este fator não foi considerado
como desclassificatório para o antígeno F em relação ao antígeno E, uma vez que
obteve excelente desempenho quando calculados os valores de sensibilidade,
especificidade, valores preditivos positivos e negativos e índice de concordância de
Youden nas técnicas de ELISA e de RIFI.

73
5.6 Teste de Reações Cruzadas
As reações com soros de cães portadores de outras patologias, para testar a
presença de reações cruzadas foram realizadas com o antígeno F e os resultados estão
expressos na Tabela 5.
Tabela 5 Teste de ELISA em soro de cães portadores de Leishmaniose Tegumentar,
Babesiose, Erlichiose e Doença de Chagas com o antígeno F
Soro de Cães/ Patologia
Número de Amostras
Resultados Positivos
% Resultados Negativos
%
Leishmaniose Tegumentar 18 18/18 100 0 0
Babesiose 7 0/7 0 7 100
Erlichiose 8 0/8 0 8 100
Doença de Chagas 18 15/18 83 3 17
Ocorreram reações cruzadas com os soros de cães portadores de Doença de
Chagas (15/18) e Leishmaniose Tegumentar (18/18). Os soros de animais portadores de
Erlichiose e Babesiose não mostraram reações positivas nos teste frente ao antígeno F.
5.7 Escolha do melhor antígeno
Tendo em vista que a cepa BH400 foi considerada como a de melhor
desempenho em todas as análises realizadas, seja crescida no meio LIT ou no alfa -
MEM, a mesma foi escolhida como candidata a ser utilizada em kit diagnóstico
utilizando de ELISA para LVC. O perfil proteômico imunogênico da mesma foi feito na
cepa crescida em meio de cultura Alfa-MEM, que foi escolhido pela maior facilidade de
preparo e praticidade na obtenção os componentes. A análise proteômica-imunogênica é
de fundamental importância na identificação da proteína majoritária responsável por
maior antigenicidade, sendo que em estudos futuros a mesma pode ser seqüenciada e
empregada na sensibilização de placas de ELISA visando uma maior sensibilidade e
especificidade deste ensaio.
5.8 Western blot - 2D gel
As proteínas do antígeno obtido do crescimento da cepa BH400 em meio alfa -
MEM foram fracionadas em gel bidimensional que após corado por comassie, foi

74
transferido para membrana de nitrocelulose e posteriormente, incubado com um“pool”
de soros de cães com baixos títulos de anticorpos (RIFI 1:160) e outro “pool” de soros
com altos títulos de anticorpos (RIFI > 1:1280), assim como, um “pool” de soros de
animais não infectados. Após a incubação com conjugado anti IgG total de cão,
marcado com peroxidase (Sigma®), foi realizada uma sobreposição das membranas e
dos géis e feitas as comparações mostradas na Figura 23.
Figura 23 Gel 2-DE contendo 600g de proteínas de Leishmania chagasi (cepa BH400 crescida em meio α-MEM) fracionadas em tiras de 13 cm pH 4-7 (direita para esquerda) e SDS-PAGE a 12%. O gel foi corado por comassie (A) ou
transferido para membrana de nitrocelulose e incubadas com soros de animais
não infectados (B) e infectados que apresentavam baixos títulos de anticorpos
(C) e altos títulos de anticorpos (D). O conjugado utilizado foi anti-IgG total de cão. Em cada membrana estão destacados os “spots” detectados por cada grupo
de soros.
Observou-se que cerca de 350 “spots” foram visualizados nos géis (Figura 23A),
32 foram detectados nas membranas incubadas com soros negativos (Figura 23B) e nas
membranas incubadas com soros positivos, após exclusão das reações comuns ao
controle negativo, 41 “spots” foram reconhecidos por soros com baixos títulos de
A B
C D
MM
(kDa)
216
132
78
65
45
32
18
7.6
4 7 4 7
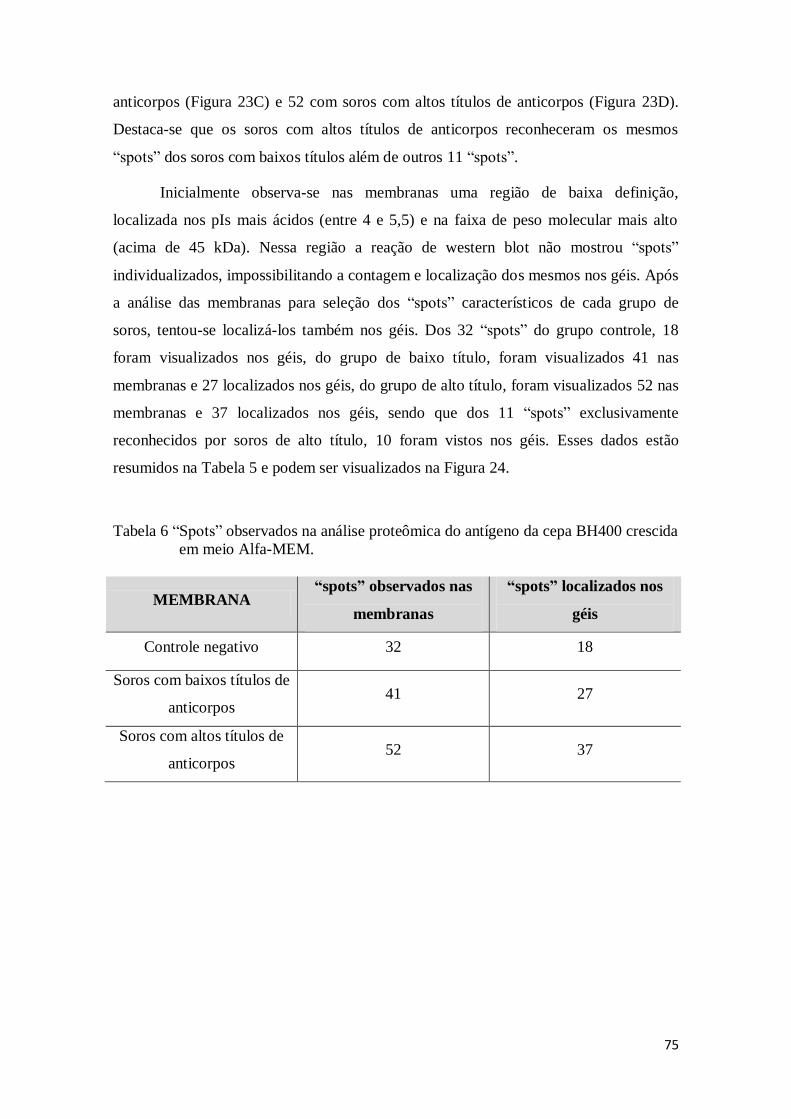
75
anticorpos (Figura 23C) e 52 com soros com altos títulos de anticorpos (Figura 23D).
Destaca-se que os soros com altos títulos de anticorpos reconheceram os mesmos
“spots” dos soros com baixos títulos além de outros 11 “spots”.
Inicialmente observa-se nas membranas uma região de baixa definição,
localizada nos pIs mais ácidos (entre 4 e 5,5) e na faixa de peso molecular mais alto
(acima de 45 kDa). Nessa região a reação de western blot não mostrou “spots”
individualizados, impossibilitando a contagem e localização dos mesmos nos géis. Após
a análise das membranas para seleção dos “spots” característicos de cada grupo de
soros, tentou-se localizá-los também nos géis. Dos 32 “spots” do grupo controle, 18
foram visualizados nos géis, do grupo de baixo título, foram visualizados 41 nas
membranas e 27 localizados nos géis, do grupo de alto título, foram visualizados 52 nas
membranas e 37 localizados nos géis, sendo que dos 11 “spots” exclusivamente
reconhecidos por soros de alto título, 10 foram vistos nos géis. Esses dados estão
resumidos na Tabela 5 e podem ser visualizados na Figura 24.
Tabela 6 “Spots” observados na análise proteômica do antígeno da cepa BH400 crescida
em meio Alfa-MEM.
MEMBRANA “spots” observados nas
membranas
“spots” localizados nos
géis
Controle negativo 32 18
Soros com baixos títulos de
anticorpos 41 27
Soros com altos títulos de
anticorpos 52 37

76
Figura 24 Gel 2-DE contendo 600g de proteínas de Leishmania chagasi (cepa BH400
crescida em meio α MEM) fracionadas em tiras de 13 cm pH 4-7 (esquerda para
direita) e SDS-PAGE a 12% corado por comassie. Em destaque em vermelho,
os “spots” reativos frente a soros de cães não infectados, em azul “spots” reativos frente a cães que apresentam baixos títulos de anticorpos e em preto os
“spots” reativos frente a soros de cães que apresentam altos títulos de
anticorpos.
MM
(kDa)216132
78
65
45
32
18
7.6
4 7

77
6. Discussão
Levando em consideração as características epidemiológicas da leishmaniose
visceral, onde faltam conhecimentos fundamentais sobre os fatores de risco, sobretudo
relacionados às mudanças no meio ambiente naturais ou introduzidas pelo homem, a
resposta imune protetora e ao aumento de casos resistentes ao tratamento de primeira
escolha (antimoniais pentavalentes), a LV é um problema preocupante no que diz
respeito à Saúde Pública, principalmente quando se trata das formas zoonóticas, como
as que ocorrem na América
Um dos principais fatores de risco da LV são os fenômenos da urbanização da
doença, sobretudo na América. No Brasil as estratégias de controle da doença estão
focadas no diagnóstico e tratamento precoce dos casos humanos, redução da população
de flebotomíneos vetores, eliminação do reservatório canino e atividades de educação
em saúde, lembrando sempre que o diagnóstico e tratamento dos doentes é prioritário,
ao lado das atividades de educação em saúde, mas estas medidas devem estar sempre
integradas para que possam ser efetivas.
O reservatório nos ambientes doméstico e peridoméstico é o cão (Canis
familiaris) que é um dos elos mais importantes da cadeia epidemiológica, para o ciclo
de manutenção da doença. Uma das ferramentas recomendadas oficialmente pelo
Ministério da Saúde nos inquéritos epidemiológicos para diagnóstico dos animais
positivos são os testes de ELISA e RIFI. Portanto, a precocidade do diagnóstico tem
grande importância no sucesso das medidas de controle, que devem ser aplicadas
integradas.
O cão, como o homem, pode apresentar um espectro de manifestações clínicas
variadas, condicionadas à imunocompetência individual. Muitos animais podem
permanecer sem sinais clínicos por longo período de tempo (Deane 1956; Marzochi,
1994, Almeida et al., 2005). O diagnóstico da LVC, em especial, se apresenta como um
problema para os serviços de saúde pública, principalmente devido à inexistência de um
teste diagnóstico 100% específico e sensível. O diagnóstico clínico é difícil, os médicos
veterinários têm muitas vezes problemas, pois há um amplo espectro de sinais clínicos e
muitos sinais são comuns a várias doenças prevalentes nos cães. Atualmente, testes
sorológicos, exames parasitológicos e técnicas moleculares que possuem sensibilidade e
especificidade distintas, ajudam na elucidação da positividade para LVC (Feitosa et al.,

78
2000; Singh et al., 2003; Feitosa, 2006). Avanços têm ocorrido nos últimos anos, e há
uma série de métodos sorológicos para avaliação “in vitro” de anticorpos presentes em
humanos e cães, mas estes ainda requerem melhoramentos, sobretudo quanto à
reatividade cruzada com outros tripanossomatídeos e no caso do homem, sorologia
residual pós-terapêutica.
A utilização de testes sorológicos para detecção de anticorpos circulantes para
Leishmania spp. vem sendo utilizada principalmente em estudo soro-epidemiológicos,
evitando assim os métodos parasitológicos, que são invasivos e de pouca praticidade
(Gontijo e Melo, 2004).
No Brasil, o diagnóstico sorológico canino era realizado com a técnica de RIFI,
considerando-se reagentes todos os soros com positividade a partir da diluição 1:40. Em
2002, o Ministério da Saúde recomendou a introdução do teste de ELISA para a triagem
inicial e da RIFI para a confirmação dos resultados positivos (Ministério da Saúde,
2003). Estes métodos nem sempre alcançam sensibilidades e especificidades
satisfatórias, já que muitos animais infectados apresentam testes sorológicos negativos.
Além disso, esses dois testes podem apresentar, em alguns casos, resultados divergentes
no diagnostico da LVC (Zanette, 2006). Sabe-se que os resultados de qualquer prova
sorológica são influenciados por inúmeros fatores como o tipos de antígeno utilizado, a
coexistência de infecções que propiciam reações cruzadas e a magnitude da resposta
imune individual (Lainson & Rangel 2005). De maneira geral, o maior obstáculo para a
interpretação de dados sorológicos é a ausência de informações parasitológicas
(Madeira et al., 2006).
A RIFI apresenta como principais limitações diagnósticas a reatividade cruzada
com outras doenças, como leishmaniose tegumentar, doença de Chagas, malária,
esquistossomose e tuberculose pulmonar, além de exigir muito tempo para a leitura das
lâminas quando da análise de um grande número de amostras, além da subjetividade dos
examinadores na leitura de reações fracamente positivas. Possui também sensibilidade
reduzida para detecção de cães assintomáticos (Braga et al., 1998; Sundar et al., 2002;
Almeida et al., 2004)
A técnica de ELISA pode apresentar em alguns casos reações cruzadas e
inespecíficas, mas possui certas vantagens como leitura automatizada não dependente
do observador, necessita de apenas uma diluição do soro, é de fácil execução e permite
que vários testes possam ser processados rapidamente ao mesmo tempo, possui baixo

79
custo e sensibilidade e especificidade adequadas quando comparada a outros métodos
(Dantas-Torres et al., 2006; Ikeda-Garcia e Marcondes, 2007). Estes facilitadores
contribuem para que esta técnica venha sendo utilizada em levantamentos
epidemiológicos (Sundar et al., 2002).
Métodos de ELISA utilizando o antígeno total do parasito são descritos para o
diagnóstico da doença canina e humana (Mohammed et al., 1985; Martin et al., 1998;
Lima et al., 2003). A preparação de antígeno total é simples, grandes quantidades são
produzidas sob condições padronizadas e apresenta determinados epítopos antigênicos
que não estão presentes no antígeno solúvel. (Evans et al., 1990). Recentemente, vários
antígenos recombinantes de Leishmania têm sido testados como ferramentas para
melhorar o diagnóstico da LV em cães. Rosário (2005) recomenda a associação de
antígenos brutos e recombinantes como uma estratégia interessante a ser utilizada em
programas de controle da doença. Gontijo e Melo (2004) afirmam que deve ser dada
ênfase à pesquisa de componentes antigênicos purificados que possam ser utilizados
como ferramenta de diagnóstico específico, assegurando maior sensibilidade e
especificidade aos testes sorológicos. Os antígenos purificados têm mostrado maior
sensibilidade que os antígenos brutos, mas sua preparação muitas vezes demorada pode
requerer métodos sofisticados de purificação, além de mostrar muitas vezes problema na
reprodução dos resultados. Antígenos recombinantes são mais fáceis de obter e alguns
já se encontram disponíveis para o diagnóstico da LV humana e canina (Gontijo e Melo,
2004).
Em nosso estudo, para a padronização de um antígeno a ser utilizado no
diagnóstico da LVC pelos órgãos de saúde pública, decidiu-se trabalhar com ELISA e
RIFI por serem os métodos recomendados pelo Ministério da Saúde em inquéritos soro-
epidemiológicos (Brasil-Ministério da Saúde, 2003).
Na seleção inicial dos animais doadores de soro, para melhor categorizar as
amostras, nós utilizamos a própria sorologia RIFI e ELISA, além da pesquisa direta do
parasito em esfregaços por extensão de material de medula óssea, semeadura da medula
óssea em meio alfa - MEM acrescido de 10% de soro fetal bovino, e ainda a PCR. Só
consideramos positivas ou negativas as amostras com concordância entre os testes
sorológicos e parasitológicos. Sete cães apresentaram-se positivos no exame
parasitológico pela técnica de PCR, mas negativos sorologicamente nas reações de
ELISA e RIFI e tiveram que ser excluídos do experimento, pois a padronização de um

80
antígeno só permite trabalhar com animais devidamente categorizados como positivos
ou negativos. Estes dados corroboram com os observados por Moreira et al. (2002),
Brasil (2003) e Barrouin-Mello et al.(2006), que evidenciam a existência de animais
portadores do parasito mas ainda negativos sorologicamente. De acordo com estes
autores tratam-se de indivíduos na fase inicial da doença e, portanto, com títulos de
anticorpos baixos que não podem ser detectados sorologicamente. Com o passar do
tempo e de acordo com a magnitude da resposta imune individual, estes animais
poderão soroconverter-se ou progredir pra autocura, ou ainda permanecerem neste
estado indefinidamente (Neves, 2005).
Desde o início dos anos 1990, o Departamento de Parasitologia, através do setor
de Leishmanioses, que abriga vários laboratórios, iniciou um estudo para obtenção de
um antígeno bruto, que pudesse ser utilizado nas reações de ELISA e que mostrasse ser
mais específico e sensível que o então empregado no laboratório de Sorologia de
Leishmania. O antígeno inicialmente empregado era obtido a partir da cepa BH6,
mesma que foi depois empregada pelo laboratório de Biomanguinhos na confecção dos
kits diagnósticos de ELISA e RIFI para os inquéritos epidemiológicos. Mesmo sendo
uma cepa de L. amazonensis ela foi utilizada, pois há uma reação cruzada das
Leishmaniae entre si nestas duas técnicas utilizando antígeno bruto. Já as cepas BH46 e
BH400 não são utilizadas na confecção dos kits diagnósticos de saúde pública, mas seu
uso neste estudo se explica pelo fato de se tratarem de Leishmania chagasi (sinômino de
L. infantum), agente específico da LV humana e canina, respectivamente.
Nesses estudos ao longo do tempo as cepas BH6 e BH46 começaram a ser
testadas no preparo de antígenos, com diferentes concentrações dos mesmos, diferentes
concentrações de conjugados e diferentes conservantes, mertiolate 1:10.000,
iodacetamida 1:10.000 e o acréscimo de tioglicolato de sódio em pequenas
concentrações, abaixo de 0,5%, como adjunto para a conservação. Foram ainda testadas
diferentes concentrações de soros, 1:80, 1:100, 1:200 e 1:400 (dados não mostrados).
Desses estudos não resultaram publicações, porque o setor de leishmanioses não tinha
interesse de publicar resultados incompletos, além de tratar-se de um trabalho técnico na
busca de um antígeno para ELISA, que não mostrasse reações cruzadas com outras
patologias caninas e que fosse capaz de indicar com segurança os cães realmente
positivos e negativos para LVC. Esses trabalhos contribuíram para mostrar, por
exemplo, que a cepa BH46, que também é utilizada nesta tese, na diluição do soro

81
1:400, com o antígeno na concentração de 2µg/ml mostrava melhores resultados em
discriminar cães com LVC positivos e negativos na reação de ELISA (dados não
mostrados). Utilizando este antígeno publicamos recentemente um trabalho, anexado a
esta tese: “Humoral Immunological profile and Parasitological Statuses of Leishmune®
Vaccinated and Visceral Leishmaniasis Infected Dogs from an Endemic Area” onde
foram utilizados antígenos de BH46 nos testes comparativos entre cães vacinados com
Leishmune®
e cães naturalmente infectados.
Este antígeno passou a ser utilizado nos trabalhos de Sorologia de Leishmania
no setor de Leishmanioses do Departamento de Parasitologia. A literatura já havia
mostrado que testes de ELISA com antígenos homólogos apontavam, usualmente, uma
sensibilidade mais alta (revistos por Romero & Boleart, 2010).
No presente trabalho nós incorporamos a cepa BH400 também de L. chagasi,
isolada de um cão com LVC no município de Belo Horizonte, área endêmica para LV
humana e canina. Esta cepa foi testada porque na nossa rotina de manutenção, já
havíamos observado que ela cresce bem em meios de cultura e mostrava maior
infecciosidade para hamsters, em relação a cepa BH46.
Nós testamos os antígenos com promastigotas das cepas escolhidas em dois
diferentes meios de cultura “Liver Infusion Tryptose”-LIT e meio Alfa-MEM acrescido
de 10% de Soro Fetal Bovino. Nos últimos anos diferentes meios de cultura têm sido
desenvolvidos para cultivo de Leishmania sp. e podem ser classificados em duas
grandes categorias: bifásicos semi-sólidos e monofásicos líquidos. Assim, o meio Novy-
MacNeal-Nicolle (NNN) (Nicolle, 1908) e outros do tipo bifásico são habitualmente
usados para manutenção inicial de formas promastigotas de Leishmania (Schuster e
Sullivan, 2002; Chouihi et al., 2009; Rodrigues 2010). A relativa facilidade com que
formas promastigotas de Leishmania sp. podem ser cultivadas axenicamente in vitro,
em uma série de meios de cultura bifásicos ou monofásicos, levou ao conhecimento
aprofundado sobre os mecanismos metabólicos e moleculares de promastigotas de
Leishmania sp.(Somanna et al. 2002). Diversos autores descrevem o comportamento de
cepas de Leishmania sp. em diferentes meios de cultura e mostram que pode ocorrer
expressão de diferentes epítopos antigênicos quando os parasitos são cultivados em
meios com formulações diferentes, e que isto se dá devido à composição química do
meio que vai favorecer ou não esta expressão (Schuster e Sullivan, 2002; Somanna et al.
2002; Muniaraj et al., 2007; Cuervo et al., 2009; Rodrigues, 2010).

82
Decidimos trabalhar com os meios de cultura LIT e Alfa-MEM, que já são
utilizados nas rotinas laboratoriais de isolamento e crescimento de Leishmania sp., no
Setor de Leishmanioses do Departamento de Parasitologia do Instituto de Ciências
Biológicas da UFMG, e com efetividade comprovada em diversos trabalhos de pesquisa
(comunicação pessoal, M.N.Melo).
O meio LIT é tradicionalmente usado para identificação, isolamento e
crescimento de Leishmania sp. por diversas instituições de pesquisa inclusive pelo
laboratório de Biomanguinhos na confecção de kits diagnósticos de ELISA e RIFI. O
preparo do meio envolve uma série de etapas, como a utilização de componentes do
sangue bovino, ajuste de pH para a faixa situada entre 7,2 e 7,4 e inserção de
antibióticos. É composto de triptose, infuso de fígado, soro e hemoglobina bovina
(Camargo 1964), substâncias de origem biológica que nem sempre são passíveis de
padronização bioquímica, uma vez que são provenientes de diferentes animais que
podem ser de raças diferentes, diferentes idades e sexo, embora da mesma espécie. É
um meio artesanal, que consome muito tempo na sua preparação, e não estando
disponível comercialmente.
O meio α-MEM é um meio disponível no comércio na forma de uma mistura
seca, contém ribonucleosídeo, deoxiribonucleosídeo, sais minerais, aminoácidos,
vitaminas e bicarbonato de sódio, e para sua confecção basta ser dissolvido e adicionado
de soro fetal bovino, também disponível comercialmente. Este meio é muito mais
simples e fácil de ser preparado quando se quer crescer Leishmania sp. em larga escala.
Da mesma forma que o meio de LIT, é também largamente utilizado por diversas
instituições de pesquisa.
A sensibilidade e a especificidade do teste de ELISA são diretamente
influenciadas pela qualidade do antígeno empregado na sensibilização das placas
(Liberal et al.,1992; Tully, 1996). Placas sensibilizadas com um maior número de
antígenos, como os presentes em antígenos brutos, proporcionam bons resultados,
especialmente em testes de triagem, dando oportunidade de identificação de diferentes
indivíduos que respondem de maneira individual a um mesmo agente, ou seja, as
variedades antigênicas determinam as diferentes avidezes dos anticorpos. Porém, a
baixa estabilidade do tipo de antígeno pode comprometer seu uso (Brown et al., 1996).
Em nosso trabalho, as placas de ELISA foram sensibilizadas com antígenos alcalinos de
promastigotas de diferentes cepas adicionados de um inibidor de proteases, sendo

83
bloqueadas para posterior estocagem em freezer a -20ºC. Esta estocagem foi feita
segundo o protocolo desenvolvido neste projeto para produção de antígenos para ELISA
e os diferentes tempos de uso das placas permitiram realizar considerações sobre a
reprodutibilidade dos ensaios.
Não foram feitos neste projeto avaliações periódicas do antígeno ao longo do
tempo de estocagem para verificar se houve mudanças na estrutura física do mesmo.
Testes bioquímicos e moleculares poderão futuramente ser conduzidos como
continuidade do estudo destas cepas para melhor compreensão deste processo.
Existem inúmeros trabalhos na literatura comparando a sensibilidade e
especificidade das reações de ELISA utilizando diferentes antígenos. Além do tipo de
antígeno empregado (espécie ou forma evolutiva do parasita) estes índices podem variar
também com mudanças no protocolo experimental padrão (tempo de incubação ou tipo
de microplacas utilizadas) (Reinthinger et al., 2002).
Como a maioria das preparações antigênicas para o teste de ELISA perde na
especificidade, em inquéritos sorológicos costuma-se usar este teste como triagem e a
RIFI como confirmatório. Entretanto, os testes sorológicos devem ser interpretados com
cautela, uma vez que não são 100% sensíveis e falham ao detectar cães infectados no
período pré-patente e, portanto, antes da soroconversão, cães que nunca farão
soroconversão e cães soropositivos que se convertem em soronegativos, mas ainda
permanecem infectados (Ferrer et al., 1995; Ferrer, 2002a; Leontides et al., 2002; Ikeda-
Garcia, 2006).
O teste de ELISA para pesquisa de anticorpos anti-Leishmania sp. apresenta,
dependendo do antígeno empregado, uma sensibilidade que varia entre 95% e 99,5% e
uma especificidade entre 97,1% a 100% (Laurenti et al., 2005; Mancianti et al., 1995).
Há, contudo, discordâncias entre alguns autores, os quais descreveram valores de
sensibilidade e especificidade de 80 e 81%, respectivamente (Ashford et al., 1995) e
outros que observaram valores de 100% para ambos os índices (Vercammen et al.,
1997).
Barbosa-de-Deus et al.(2002) utilizou antígeno bruto de L. major-like para o
teste de ELISA tendo a RIFI como padrão ouro. A análise de 188 soros caninos
positivos e 1582 negativos para LVC, além de 55 soros de animais portadores de
doenças que podem potencialmente acusar reação cruzada com LV apresentou uma

84
sensibilidade de 98%, especificidade de 95%, valor preditivo positivo de 66%, valor
preditivo negativo de 100% e uma taxa de concordância de 93%. Não houve número
significante de reações falso positivas e falso negativas nem reações cruzadas com
tumor venéreo transmissível, erliquiose, riquetsiose e Doença de Chagas. Concluiu-se
que o ELISA é um pouco mais sensível do que a RIFI, mas esta parece ser mais
específica.
Machado et al. (2006) trabalhando com antígeno bruto de Leishmania major e
utilizando imprint de baço dos animais soropositivos como padrão ouro, testou 86 soros
positivos e 57 negativos, encontrando 98,84% de sensibilidade e 100% de
especificidade para o ELISA. A concordância do ELISA com o padrão ouro foi ótima,
de acordo com os critérios do teste de Kappa. Os resultados demonstraram que a técnica
de ELISA, utilizando a L. major, apresentou valores elevados de sensibilidade e
especificidade, constituindo uma ferramenta importante para o diagnóstico e controle da
doença. Estes valores concordam com Oliveira et al. (2005) que encontraram
sensibilidade e a especificidade de 90% e 100% respectivamente utilizando antígeno
bruto de L. chagasi. Valores menores foram encontrados por Zanette (2006) usando
antígeno bruto de L. chagasi em área endêmica para LVC. O método de ELISA
apresentou 94% de sensibilidade e 84,4% de especificidade, além de boa concordância
com o método parasitológico (Kappa = 0,788).
Nossos resultados mostraram sensibilidade de 96 a 100% para todos os
antígenos testados na primeira fase do experimento (24 horas após sensibilização das
placas). Na segunda fase (três meses após sensibilização das placas), este parâmetro
variou de 87 a 100% e na terceira fase (seis meses após sensibilização das placas)
variou de 91 a 100%. Todos os dados foram calculados com 95% de intervalo de
confiança. De maneira geral, todos os antígenos testados revelaram uma boa
sensibilidade nos tempos testados, sendo que estes valores caíram de uma forma mais
significativa ao longo do tempo para os antígenos da cepa BH6. Os antígenos das cepas
BH400 crescidas nos meios LIT e α-MEM respectivamente mostraram sensibilidade de
98 a 100% em todas as fases do experimento evidenciando boa estabilidade ao tempo de
estocagem. Esta é a nosso ver uma característica que é desejável, quando se quer
produzir um kit para ser usado em inquéritos epidemiocensitários e epidemiológicos
pelos órgãos de Saúde Pública, quando os antígenos devem ter estabilidade para serem

85
transportados e usados por um tempo maior, sobretudo num país como o Brasil que tem
dimensões continentais.
A especificidade, calculada com 95% de intervalo de confiança, variou de 98 a
100% nos testes realizados 24 horas depois de sensibilizadas as placas e de 92 a 100%
após três e seis meses de conservação a -20°C. O antígeno da cepa BH400 crescida em
meio LIT e alfa - MEM mostrou especificidade de 100% em todos os momentos do
experimento.
Assim, a cepa BH400, Leishmania chagasi isolada de cão, independente do
meio de cultivo, foi considerada como a de melhor desempenho e escolhida como
candidata a ser utilizada em kit diagnóstico utilizando a técnica de ELISA para LVC.
Resolvemos trabalhar esta cepa crescida em meio alfa - MEM para a caracterização do
perfil proteômico-imunogênico, devido à praticidade de preparo da solução e da
facilidade de obtenção comercial da mistura solúvel. A análise proteômico-imunogênica
desta cepa crescida em meio LIT será uma das perspectivas de continuidade deste
trabalho.
A RIFI demonstra sensibilidade que varia entre 90 e 100% e especificidade entre
80 e 100% (Mancianti et al., 1996; Alves et al., 2004; Mettler et al., 2005). Mancianti et
al. (1995), trabalhando com L. infantum, encontrou valores de sensibilidade e
especificidade de 98,4% e 100%, respectivamente e afirma que a variação destes valores
depende do antígeno empregado e das condições da RIFI. A especificidade dessa prova
apresentou reações cruzadas com outras doenças, principalmente aquelas causadas por
tripanosomatídeos, como o agente causador da doença de Chagas. Portanto, seus
resultados não devem ser utilizados como indicadores de infecção leishmaniótica
específica, particularmente em áreas onde a Doença de Chagas é endêmica ao lado das
leishmanioses (Mancianti et al.,1995; Mancianti et al.,1996).
Zanette (2006), trabalhando com 50 cães parasitologicamente positivos, mostrou
sensibilidade de 98% e especificidade de 91% para a RIFI utilizando como antígeno
promastigotas de L. chagasi, além de ótima concordância com o diagnóstico
parasitológico (Kappa=0,893). Oliveira et al. (2005) encontrou sensibilidade e
especificidade de 40% e 98,6 respectivamente e valores preditivos positivos e negativos
de 92,3% e 79,5% pra este teste. Esta autora relata que os valores encontrados ficaram
muito aquém do que normalmente é citado na literatura e discorda de alguns autores que

86
afirmam a semelhança de desempenho entre os testes de ELISA e RIFI (Mancianti et
al., 1995; Scalone et al., 2002; Vercammen et al., 2002).
Figueiredo et al. (2010) sugere que a maior precisão do ELISA faz com que este
método de diagnóstico seja o mais adequado para as campanhas de controle da LV,
reduzindo dois grandes problemas que aparecem quando se usa apenas o teste de RIFI:
o número de cães infectados que permanecem após a identificação dos soropositivos e a
eutanásia desnecessária de cães identificados erroneamente como positivos, devido ao
grande número de reações falsos-positivos neste teste.
Na RIFI os antígenos da cepa BH6 crescida em meio LIT e α-MEM, assim como
da cepa BH400 crescida nos mesmos meios, apresentaram 100% de concordância com a
classificação dos animais para LVC. Os valores de sensibilidade, especificidade, valor
preditivo positivo e valor preditivo negativo para as reações de imunofluorescência
indireta foram de 100% com 95% de intervalo de confiança (kappa=1). Os antígenos da
cepa BH46 crescida em meio LIT e α-MEM por apresentarem sete resultados falsos
negativos, mostraram 87% de sensibilidade, 100% de especificidade, valor preditivo
positivo de 100% e valor preditivo negativo de 87% (IC=95%, Kappa=0,87).
As reações de RIFI foram realizadas em apenas um momento do experimento,
30 dias após a confecção das lâminas, pois nossa intenção foi apresentar um antígeno
alternativo para ser utilizado na produção dos kits de ELISA pela rede pública. A
realização da RIFI com os diferentes antígenos estudados reforçou a caracterização das
cepas e escolha de uma delas como uma excelente opção na melhoria da qualidade do
kit diagnóstico.
Os dados apresentados neste trabalho sobre o desempenho dos testes de ELISA e
RIFI foram obtidos em laboratórios de pesquisa e referem-se a um número reduzido de
amostras de soro canino examinadas. Essas condições ideais de execução da técnica
nem sempre podem ser reproduzidas na rotina dos laboratórios da rede pública de saúde,
em virtude do fornecimento irregular de materiais de consumo, da dificuldade na
realização da técnica em laboratórios, muitas vezes inadequados, e da subjetividade na
interpretação dos resultados, freqüentemente agravada pela urgência no processamento
de um número elevado de amostras coletadas durante os inquéritos sorológicos caninos
nas áreas endêmicas.

87
Tendo em vista a possibilidade de utilização do antígeno da cepa BH400 como
componente de kit diagnóstico para ELISA, outro fator que deve ser considerado, como
perspectiva de continuidade de nosso estudo, é a realização de testes para se observar o
comportamento do antígeno, quando o sangue for colhido em papel filtro. A literatura
relata a diminuição dos índices de sensibilidade e especificidade quando as amostras são
coletadas em papel de filtro, o que é a prática mais comum nos inquéritos soro-
epidemiológicos. Esta observação foi confirmada em um estudo realizado por
Figueiredo et al. (2009) na região de Carapiá no Rio de Janeiro, na qual 34,9% de cães
positivos foram identificados pela RIFI utilizando-se soro, entre 146 amostras
soronegativas previamente testadas utilizando-se amostras de sangue coletadas em papel
de filtro. Aspectos relacionados à coleta, transporte e armazenamento das amostras,
podem explicar, em parte, o baixo desempenho dos eluatos quando comparado a
amostras de soro, mas outros problemas não estudados podem estar influindo nos
resultados.
De acordo com a literatura, os testes sorológicos de RIFI e ELISA realizados
com os kits produzidos por Biomanguinhos apresentam menor sensibilidade e
especificidade quando comparados com testes in house. Em nosso experimento as
reações com estes kits foram realizadas em um só momento, respeitando a data de
validade dos mesmos, pois nossa intenção era apenas verificar como nossos soros
seriam classificados nestes kits.
Quaresma (2007) mostrou que houve concordância ruim entre o PCR e a
sorologia (ELISA e RIFI de Biomanguinhos), utilizando este método parasitológico
como padrão ouro. O animal era considerando sorologicamente positivo se apresentasse
reatividade em pelo menos uma das técnicas sorológicas. A autora trabalhou com 147
(68%) cães positivos e 70 (32%) cães negativos pelas técnicas sorológicas, mas quando
estes resultados foram comparados com o PCR, evidenciou-se índices de 65,4% de
verdadeiros positivos, 0,92% de verdadeiros negativos, 2,3% falso positivos e 31,34%
falso negativos.
Em outro trabalho, Ferreira et al., (2007) trabalhando com 112 cães positivos
verificou para o ELISA e RIFI de Biomanguinhos valores de sensibilidade de 95 e 72%,
e especificidade de 64 e 52% respectivamente. Os autores relatam ainda que ambos os
testes apresentaram reação cruzada com soros de cães infectados por T. cruzi, L.
braziliensis e E. canis.

88
Silva (2005) trabalhou com amostras de soro e papel de filtro coletadas de cães
de área endêmica para LVC, com diagnóstico clínico e parasitológico positivos. A RIFI
foi avaliada utilizando-se o kit comercial produzindo por Biomanguinhos e um ensaio in
house utilizando promastigotas de L. (L.) chagasi. O ensaio in house com os soros
mostrou-se capaz de separar todos os verdadeiros negativos dos verdadeiros positivos e
apontou para uma eficiência de 60% a 76% para o kit comercial. Quando comparados
os resultados da RIFI do kit comercial empregada em soros e papel de filtro, observou-
se que o melhor ponto de corte para o papel de filtro seria a diluição de 1/80, o que
seguramente diminuiria o número de falsos positivos.
Távora et al. (2007) usando os kits RIFI e ELISA de Biomanguinhos mostrou
que em 370 amostras de soros coletadas de cães errantes, 6 (1,6%) foram positivas e
355 (95,9%) negativas para ambos os testes, 2 (0,5%) obtiveram positividade para RIFI
e negatividade para ELISA e 7 (1,9%) negatividade para RIFI e positividade para
ELISA. A relação de co-positividade ELISA/RIFI foi de 46,2%, e a de co-negatividade
ELISA/RIFI foi de 99,4%; a co-positividade RIFI/ELISA foi de 75% e a co-
negatividade RIFI/ELISA foi de 98,1%. Através destes resultados pode-se realizar o
teste de Kappa, para avaliar a concordância entre as duas técnicas diagnósticas, onde se
verificou uma concordância observada de 97,6% e esperada 94,5%, altamente
significativa (p<0,01) entre os resultados positivos e negativos, sendo esta concordância
classificada como forte (0,8 ≤ kappa < 1).
Queiroz et al. (2010) avaliou pelos kits de Biomanguinhos 34 cães naturalmente
infectados e dentre estes o ELISA detectou 64,7% e a RIFI 55,9%. Figueiredo et al.
(2010) encontraram valores mais altos de sensibilidade e especificidade com os kits de
Biomanguinhos, pois o ELISA mostrou sensibilidade de 100% e especificidade de
96,6%. Já para a RIFI utilizando-se como ponto de corte a diluição de 1:40, estes
valores foram de 100% e 65,5%, e com ponto de corte 1:80, 100% e 83,4%,
respectivamente.
Em nosso estudo, nas reações sorológicas realizadas com o kit de
Biomanguinhos, quando comparados aos nossos resultados, o ELISA não mostrou
resultados falso negativos, porém o kit detectou dez resultados falso positivos. Cinco
soros permaneceram na zona cinza, todos negativos para LVC. Já na RIFI foram
detectados doze soros falsos positivos e um falso negativo..

89
Para o kit de Biomanguinhos foi encontrado valor de sensibilidade de 100% para
ELISA e 98% para RIFI. A especificidade foi de 80% para o ELISA e 76% para a RIFI.
O valor preditivo positivo foi de 84% para ELISA e 82% para RIFI. O valor preditivo
negativo foi de 100% para o ELISA e de 97% para RIFI. O teste de concordância Kappa
foi de 0,8 para o ELISA e 0,9 para a RIFI. Todos os índices foram calculados com 95%
de intervalo de confiança.
Em nosso experimento, houve uma boa correlação entre tendências do aumento
da absorbância do ELISA em função do aumento dos títulos da RIFI, com exceção de
alguns casos onde esta correlação foi contrária. O fato de uma amostra possuir altos
valores de absorbância no ELISA e títulos baixos ou negativos na RIFI vem sendo
demonstrado na literatura e se justifica pela alta sensibilidade do teste ELISA, que pode
detectar precocemente animais infectados (Barbosa-de-Deus et al.,2002; Laurenti et al.,
2005; Oliveira et al., 2005; Machado et al., 2006; Zanette, 2006; Figueiredo et al.,
2010). Aliado a este fato está a individualidade da resposta imune de cada animal e a
antigenicidade dos diferentes antígenos testados.
No início da infecção natural pode ocorrer produção de altas taxas de anticorpos
contra antígenos somáticos, em detrimento de baixas taxas contra antígenos de
superfície, e desta forma, o ELISA é o primeiro teste a sinalizar a doença. A avidez dos
anticorpos também se explica, considerando a expressão antigênica de cada cepa
utilizada nos diferentes meios de cultura, favorecendo ou não o reconhecimento das
proteínas.
A Erliquiose e Babesiose são doenças transmitidas aos cães por carrapatos e que
são muito comuns no Brasil e segundo alguns autores podem dar reações cruzadas na
ELISA e RIFI com LVC (Mancianti et al, 1996;. Gomes, 2004). Babesiose é uma
doença hemolítica causadas por Babesia canis, parasitas intraeritrocitários (Lobetti,
1998) e Ehrlichia canis é uma riquétsia intracitoplasmática obrigatória que se replica
nos linfonodos, no interior dos macrófagos mononucleares, como a Leishmania (Guillén
Llera et al., 2002). Sinais clínicos e hematológicos destas três doenças são semelhantes,
e a sorologia pode ser essencial para se realizar o diagnóstico diferencial. Algumas
pesquisas têm relatado que os anticorpos contra E. canis e B. canis apresentam reação
cruzada com Leishmania sp. nos testes sorológicos para detecção de cães infectados por
Leishmania sp. (Mancianti et al, 1996;. Gomes, 2004).

90
A reação cruzada de Leishmania sp. com Doença de Chagas e leishmaniose
tegumentar já tem sido descritas na literatura e se explicam pela relação filogenética
entre estes parasitos. Já o cruzamento com Babesiose, Erliquiose e outras doenças como
Toxoplasmose e Neosporose tem sido objeto de pesquisa e ainda apresentam resultados
divergentes quando se utilizam diferentes técnicas sorológicas e diferentes antígenos
(Vercammen et al., 1997; Rosário et al., 2005; Lima et al., 2005).
Barbosa de Deus (2002) testou pelo método de ELISA soros de cães com
diagnóstico clínico e sorológico e também animais com diagnóstico clínico e
parasitológico de LVC. O antígeno bruto utilizado foi L major e não houve reatividade
cruzada com soros de cães sadios e de cães com tumor venéreo transmissível, doença de
Chagas, erliquiose, ou rickettsioses.
Zanette (2006) testou amostras de cães portadores de doenças que
potencialmente possam acusar reação cruzada com leishmaniose visceral. Foram
testados 14 animais com Doença de Chagas e destes, 9 (64,3%) apresentram reações
positivas no teste ELISA e 6 (42,9%) na RIFI. Para erliquiose canina, de 13 animais
avaliados, apenas 1 (7,7%) foi reativo pelo teste ELISA. O mesmo não ocorreu para
babesiose, que não acusou reatividade em nenhuma das técnicas. Já para 6 animais com
infecção concomitante por Ehrliquia sp e Babesia sp, foi encontrada reatividade cruzada
apenas pelo teste de ELISA em 5 (83,3%) animais.
O mesmo autor testou com kit próprio para diagnóstico sorológico de doença de
Chagas, erliquiose e babesiose, 50 animais positivos para LVC e encontrou positividade
em 28 animais (56%) no teste sorológico para erliquiose e em 42 animais (85,7%) no
teste sorológico para babesiose.
Ferreira et al., (2007) testou pelos métodos de ELISA e RIFI soros de cães com
doenças parasitárias incluindo os de 7 cães com T. cruzi, , 5 cães com L. braziliensis e 3
cães com E. canis. No teste de ELISA os valores de reatividade foram de 57,14% (4
amostras) para T. cruzi, 60% (3 amostras) para L. braziliensis e 33,33% (1 amostra)
para E. canis. Já no teste de RIFI estes valores foram de 100% (7 amostras), 40% (2
amostras) e 66,67% (2 amostras) respectivamente. Os kits diagnósticos utilizados
foram produzidos pelo laboratório de Biomanguinhos, sendo o antígeno de ELISA
parasitas pertencentes ao complexo L. donovani e da RIFI formas promastigotas de L.
infantum.

91
Oliveira et al.( 2008) demonstraram a não reatividade cruzada pelos métodos de
ELISA e RIFI de soros de cães positivos para L. (L.) chagasi utilizando como antígeno
E. canis e B. canis. Porém, testando para as três patologias 62 soros de cães com LV
naturalmente infectados encontrou positividade na RIFI de 51% para Babesiose e 43%
para Erliquiose, evidenciando a presença de anticorpos contra as três doenças nos cães
e não reatividade cruzada, e desta forma chama a atenção para a co-infecção entre B.
canis, E. canis e L. chagasi nas áreas endêmicas.
No presente trabalho, nossos resultados mostraram que em dezoito cães com
Leishmaniose Tegumentar experimentalmente inoculados, dezoito com doença de
Chagas, sete com babesiose e oito com erliquiose, a reatividade cruzada ocorreu apenas
para os 18 animais com leishmaniose tegumentar e 15 animais com Doença de Chagas.
Nossos dados concordam com Vercammen et al. (1997), Barbosa de Deus (2002) que
afirmam não existir reação cruzada entre leishmaniose visceral e erliquiose, mas estes
autores avaliaram o soro de um único cão, o que certamente prejudica a confiabilidade
dos resultados. Por outro lado, Lima et al. (2005) testaram amostras de soro de 15 cães
com erliquiose, um número um pouco maior que o testado no presente experimento, e
não obtiveram reações cruzadas.
A ausência de reação cruzada com soros de animais infectados por Babesia sp.
também não foi observada em nosso estudo, discordando de Vercammen et al.(1997),
Lima et al. (2005) e Zanette (2006), Ferreira et al., (2007) e concordando com Barbosa
de Deus (2002), Rosário et al (2005) e Oliveira et al., 2008.
É importante ressaltar, no entanto, que nos nossos resultados, a maioria das
reações cruzadas era fracamente positivas, com valores de absorbância de ELISA
próximos ao ponto de corte estabelecido e muito inferiores aos demonstrados pelos
controles positivos. Algumas amostras de soro positivas para doença de Chagas
atingiram valores elevados de absorbância pelo método de ELISA, fato este justificado
pela relação filogenética entre L. chagasi e T. cruzi, confirmando os relatos de Badaró et
al. (1986). Segundo Melo (2004), o emprego de antígenos totais nos métodos de ELISA
e RIFI, aparentemente, prejudica a especificidade e predispõe à ocorrência de reação
cruzada com o T. cruzi. Uma desvantagem do uso de antígeno total é que ele pode
compartilhar epítopos comuns com outros antígenos microbianos e pode reagir com
soros positivos para outras doenças infecciosas.

92
A ocorrência de reações cruzadas é um fato que gera preocupação, pois estas
duas enfermidades apresentam sintomas que, freqüentemente, assemelham-se aos
observados na leishmaniose visceral canina, além de coexistirem em grande parte do
país. Por este motivo, devemos atentar aos resultados positivos para o calazar canino e
quando houver suspeita de co-infecções com B. canis e E. canis estas devem ser
descartadas por meio da pesquisa de mórulas de Ehrlichia sp. ou pela presença de
Babesia sp. em esfregaços de sangue periférico antes de se firmar o diagnóstico
sorológico.
Uma vez que não existe um método para o diagnóstico da LVC, que ofereça
100% de especificidade e sensibilidade, a escolha da técnica empregada deve depender
do objetivo da investigação (Ferreira et al., 2007). Gomes et al. (2008) afirmam que as
dificuldades em obter um teste altamente confiável ilustra a necessidade de uma melhor
compreensão e padronização de procedimentos e reagentes moleculares, para garantir
níveis adequados de sensibilidade e especificidade que são particularmente desejáveis.
O acúmulo de informação de genomas de patógenos, acompanhado de avanços
na área de química de proteínas e tecnologia recombinante, certamente revolucionaram
o campo de desenvolvimento de testes diagnóstico. O uso dessas novas tecnologias
facilita a identificação e a caracterização estrutural e ainda, permite a produção de
grandes quantidades e vários testes destes antígenos no laboratório. No entanto, extrato
protéico bruto ainda consiste no antígeno mais utilizado no diagnóstico sorológico da
LVC. No presente trabalho, caracterizamos antígenos imunodominantes de Leishmania
chagasi utilizando técnicas proteômicas e de ensaios imunológicos, que poderão
contribuir com a melhoria do antígeno.
Semelhante a outros autores (Dea-Ayuela et al., 2006) observamos baixa
definição dos “spots” no western blot na faixa de pI 4 a 5,4 e acima de 45 kDa, o que
impossibilitou a análise detalhada dessa região, e esse tipo de reação ocorreu também
com soros negativos. Para nós isso pode indicar a provável localização de proteínas que
diminuem a especificidade do antígeno. Dessa forma, futuramente, uma pré-purificação
do antígeno bruto excluindo proteínas de massa molecular acima de 45 kDa pode levar a
obtenção de um antígeno de alta especificidade. Esse tipo de purificação é simples, de
baixo custo e mantém uma grande quantidade de proteínas imunogênicas, direcionando
também a obtenção de um teste sensível.

93
Outro resultado interessante da análise proteômica, foi a observação que os soros
de cães com baixo títulos de anticorpos reconhecem os mesmo antígenos dos soros de
cães com altos títulos, e estes por sua vez, reconhecem outros 11“spots a mais. Os
“spots” mais interessantes para estudos posteriores são aqueles comuns a ambos os
grupos. A identificação desses “spots” indicará proteínas que deverão ser produzidas de
forma recombinante e testadas juntas ou separadamente para o desenvolvimento de
novos ensaios imunológicos.
Dos 52 “spots” selecionados como reconhecidos somente por soros de cães
infectados, 37 deles foram localizados no gel e as proteínas correspondentes poderão ser
identificadas por espectrometria de massa. O fato de terem sido observados no western
blot “spots” que não foram detectados no gel, pode ser explicada porque a detecção de
antígeno no Western blotting é mais sensível que a detecção da proteína pela coloração
por comassie. Esse mesmo fato foi também relatado por Forgber et al, (2006) que
utilizaram coloração pela prata nos géis.
Estudos do proteoma de Leishmania foram realizados principalmente para
comparação dos diferentes estágios do ciclo de vida de L. donovani (Thiel et al., 2001;
Avila et al., 2003), L. infantum (El Fakhry et al, 2002), L. (Viannia) guyanensis e L. (V.)
panamensis (Gongora et al, 2003). Recentemente, um estudo utilizando 2-DE de extrato
de promastigotas de L. infantum, seguido de Western Blot com soro de coelhos
imunizados e análise por EM, permitiu a identificação de várias proteínas antigênicas
relevantes (Dea-Ayuela et al, 2006). Gupta et al., (2007), utilizaram abordagem
proteômica para identificação e caracterização de novas proteínas imunoestimuladoras
de antígenos solúveis de promastigotas de Leishmania donovani. Numa abordagem
semelhante a nossa, foram identificados vários antígenos de L. donovani utilizando
soros e parasitos isolados de pacientes indianos permitindo concluir que a abordagem
proteoma-sorologia produz uma compreensiva e altamente resolvida representação da
antigenicidade de L. donovani e da especificidade da resposta imune anti-leishmania em
pacientes com LV (Forgber et al, 2006).

94
7. Perspectivas Futuras
O antígeno escolhido para a produção do kit será testado em experimentos
multicêntricos em diferentes instituições, antes de sua produção em larga escala.
Também serão completados os estudos com a proteína predominante na identificação
através do Immunoblotting, utilizando maior número de soros de cães com LVC,
sintomáticos e assintomáticos, na identificação da proteína majoritária que será
seqüenciada e empregada como antígeno para sensibilizar as placas de ELISA.

95
8. Conclusões
Dentre as cepas utilizadas, a BH400 (Leishmania (L.) chagasi
MCAN/BR/2002/BH400) isolada de cão, apresento um melhor desempenho em
todas as etapas do estudo e foi a cepa escolhida para o preparo de antígeno bruto
para o teste de ELISA, a ser utilizado no diagnóstico da LVC;
Os antígenos produzidos para o teste de ELISA com a cepa BH400 não mostraram
diferenças significativas de sensibilidade, especificidade, valor preditivo positivo e
valor preditivo negativo independente do meio de cultura utilizado;
As reações de ELISA realizadas com os antígenos produzidos com a cepa BH400,
em qualquer meio de cultura, identificou de maneira mais eficiente os animais
verdadeiramente positivo e negativo, quando comparada as reações realizadas com
o kit de ELISA de Biomanguinhos;
O antígeno produzido com a cepa BH400 não apresentou reações cruzadas com
Babesia canis e Ehrlichia canis nas reações de ELISA;
O antígeno produzido com a cepa BH400 apresentou reações cruzadas com
leishmaniose tegumentar canina e doença de Chagas canina experimental;
No extrato de antígeno total da cepa BH400 cultivada em meio Alfa-MEM foram
detectados mais de 50 “spots” reativos e sem reação com soros negativos,
mostrando alta imunogenicidade do antígeno;
Foi possível na análise proteômica da cepa BH400 cultivada em meio Alfa-MEM
observar que antígenos de alto peso molecular (70 Kd) e pH entre 4 e 5,5 parecem
ser o de menor especificidade;
No gel bidimensional do extrato de antígeno da cepa BH400 cultivada em meio
Alfa-MEM foi possível localizar 37 “spots” que deverão ser identificados por
espectrometria de Massa para melhor caracterização do antígeno;
Os “spots” reconhecidos ao mesmo tempo pelos soros de alto e baixo título são os
mais interessantes para estudos de futura produção de antígenos.

96
9. Referencias Bibliográficas
Abranches, P., Santos-Gomes, G.; Rachamim, N.; Campino, L.;Schnur, L.F.; Jaffe, C.L.
An experimental model for canine leishmaniasis. Parasite Immunology, v.13, n.5,
p.537-550, 1991.
Aisa, M.J.; Castillejo, S.; Gallego, M.; Fisa, R.; Riera, M.C.; De Colmenares, M.;
Torras, S.; Roura, X.; Sentis, J.; Portus, M. Diagnostic potential of Western Blot
analysis of sera from dogs with leishmaniasis in endemic areas and significance of
pattern. American Journal of Tropical Medicine and Hygiene, v.58, n.2, p. 154-159,
1998
Almeida, M. A.; Jesus, E. E.; Souza-Atta, M. L.; Alves, L. C.; Berne, M. E.; Atta, A. M.
Clinical and serological aspects of visceral leishmaniasis in northeast Brazilian dogs
naturally infected with Leishmania chagasi. Vet. Parasitol., v. 127 (3-4), p. 227-232,
2005.
Alvar, J., Cañavate, C., Molina, R., Moreno, J., Nieto, J. Canine leishmaniasis. Adv.
Parasitol., v. 57, p. 1-87, 2004.
Alvar, J.; Cruz, I.; Morales, M. A.; Cañavate, C. Molecular biology tools in
leishmaniasis diagnosis and epidemiology. In: Canine leishmaniasis: moving towards a
solution. Proceedings… Sevilha, Espanha, 2002. p. 25-30, 2002.
Alves, W.A.; Bevilacqua, P.D. Reflexões sobre a qualidade do diagnóstico da
leishmaniose visceral canina em inquéritos epidemiológicos: o caso da epidemia de
Belo Horizonte, Minas Gerais, Brasil, 1993-1997. Caderno de Saúde Pública, v.20,
n.1, p.259-265, 2004.
Amusategui, I.; Sainz, A.; Rodríguez, F.; Tesouro, M.A. Distribution and relationships
between clinical and pathological parameters in canine leishmaniasis. European
Journal of Epidemiology, v.18, [s.p.], p.147-156, 2003.
Ashford, D.A.; Bozza, M.; Freire, M.; Miranda, J.C.; Sherlock, I.; Eulálio, C.; Lopes,
U.; Fernandes, O.; Degrave, W.; Barker Jr, R.H.; Badaró, R.; David, J.R. Comparison of
the polymerase chain reaction and serology for 55 the detection of canine visceral
leishmaniasis. American Journal of Tropical Medicine and Hygiene, v.53, n.3,
p.251-255, 1995.
Ashford, R.W. The leishmaniasis as emerging and reemerging zoonoses. Int J
Parasitol, v.30, p.1269-1281, 2000.
Avila HA., Pereira JB, Thiemann O, De Paiva E, DeGrave W, Bente CMM, Harder S,
Wiesgigl M, Heukeshoven J, Gelhaus C, Krause Clos J, Bruchhaus I. Developmentally
induced changes of the proteome in the protozoan parasite Leishmania donovani.
Proteomics 3(9): 1811-29, 2003.
Badaró R, Jones T.C., Lorenco R, Cerf BJ, Sampaio D, Carvalho EM, Rocha H,
Teixeira R, Jonhson-Jr WD. A prospective study of visceral leishmaniasis in an
endemic area of Brazil. J Infec Dis 154: 639-649, 1986.

97
Badaró R, Reed S.G., Barral A, Orge G., Jones T.C. Evaluation of the micro enzyme-
linked immunosorvent assay (ELISA) for antibodies in american visceral leishmaniasis.
Antigen selection for detection of infection specific responses. Am J Trop Med; 35:72-
8, 1986.
Badaró, R.; Lourenço, R.; Carvalho E.M.; Teixeira, R.; Cerf, B.; Jones, T.C. Estudo
longitudinal soro-epidemiológico de leishmaniose visceral americana (LVA) em área
endêmica de Jacobina-BA. In: Reunião Anual sobre Doença de Chagas, Caxambu,
1983.
Baneth, G. Leishmaniases. In: Greene, C.E. Infectious diseases of the dog and cat.
3.ed. Philadelphia: Elsevier, 2006. p.685-698.
Barbosa-de-Deus, R.; Mares-Guia, M.L.; Nunes, A.Z.; Costa, K.M.; Junqueira, R.G.;
Mayrink, W.; Genaro, O.; Tavares, C.A.P. Leishmania major-Like Antigen for specific
and sensitive serodiagnosis of human and canine visceral leishmaniasis. Clin. Diagn.
Lab. Immunol. v. 9, n. 6, p. 1361-1366, 2002.
Barrouin-Melo, S.M.; Laranjeira, D.F.; Andrade Filho, F.A.; Trigo, J.; Julião, F.S.;
Franke, C.R.; Aguiar, P.H.P.; Dos Santos, W.L.C.; Pontes-de-Carvalho, L. Can spleen
aspirations be safely used for the parasitological diagnosis of canine visceral
leishmaniosis? A study on assymptomatic and polysymptomatic animals. Vet. J. v.
171, n. 2, p. 331-339, 2006.
Bastien, P.; Blaineau, C.; Pages, M. Leishmania: sex, lies and karyotype. Parasitol
Today, May;8(5):174–177, 1992.
Bates, P.A, Rogers, M.E. New insights into the developmental biology and transmission
mechanisms of Leishmania Curr. Mol. Med. 4, 601–609, 2004.
Bates, P.A. Transmission of Leishmania metacyclic promastigotes by flebotomine sand
flies. Int J Parasitol. 37 : 1097-106, 2007.
Bavia, M. E. ; Carneiro, D.D.M.T ; Gurgel, H. C.; Madureira Filho, C. ; Barbosa, M.G.
R. Remote Sensing and Geographic Information Systems and Risk of American
Visceral Leishmaniasis in Bahia, Brazil. Parassitologia, Roma, Itália, v. 47, p. 165-169,
2005.
Brasil. Ministério da Saúde. Secretaria de Vigilância em Saúde. Departamento de
Vigilância Epidemiológica. Manual de Vigilância da Leishmaniose Tegumentar
Americana. Brasília. Editora do Ministério da Saúde. Brasília, Brasil, 182 p., 2007
Brasil. Ministério da Saúde. Manual de Vigilância e Controle da Leishmaniose Visceral,
Ministério da Saúde, Brasília, 122 pp., 2008
Brasil. Ministério da Saúde. Secretaria de Vigilância em Saúde. Manual de Vigilância e
Controle da Leishmaniose Visceral, Brasília, Brasil, 122 p., 2003.
Brasil. Ministério da Saúde. Manual de Vigilância e Controle da Leishmaniose Visceral,
Ministério da Saúde, Brasília, 2010

98
Bern, C., S. N. Jha, A. B. Joshi, G. D. Takur, and M. B. Bista. Use of the recombinant
K39 dipstick test and the direct agglutination test in a setting endemic for visceral
leishmaniasis in Nepal. Am. J. Trop. Med. Hyg. 63:153-157, 2000.
Bhatia, A.; Daifalla, N. S.; Jen. S.; Badaró, R.; Reed, S. G.; Sheiky, Y. A. W. Cloning
characterization and serological evaluation of K9 and K26: two related hydrophilic
antigens of Leishmania chagasi. Molecular and Biochemical Parasitology, v.102, n.2,
p.249-261, 1999.
Braga, M. D. M. et al. Controle do calazar canino: comparação dos resultados de um
programa de eliminação rápida de cães sororreagentes por ensaio imuno-enzimático
com outro de eliminação tardia de cães sororreagentes por teste de imunofluorescência
indireta de eluato de papel de filtro. Revista Sociedade Brasileira de Medicina
Tropical, v. 31, n. 5, p. 419-424, 1998.
Bray, R. S. Leishmaniasis. Immunological Investigations of Tropical Parasitic Diseases,
(Houba V ed) Churchil Livingstone, Edimburgh, p. 65-83, 1980.
Bray, R.S.; Lainson, R. Studies on the immunology and serology of leishmaniasis. V.
The use of particles as vehicles in passive agglutination tests. Trans R Soc Trop Med
Hyg.;61(4):490–505, 1967.
Brown, M.B.; Bradbury, M.J.; Davis, J.K. ELISA in small animall hosts, rodents and
birds. In: Tully, J.G.; RAZIN, S. Molecular and diagnostic procedures in
mycoplasmology. London: Academic, 1996, p.93-103.
Burillo, F.L.; Perez, F.M.G.; Liesa, J.P.; Fabian, M.C.A. Iron status and anemia in
canine leishmaniasis. Revue Med Vet, v. 145, n.3, p.171-176, 1994.
Cabrera, M.A.; Paula, A.A.; Camacho, L.A.; Marzochi, M.C.; Xavier, S.C.; Da Silva,
A.V.; Jansen, A.M. Canine Visceral leishmaniasis in Barra de Guaratiba, Rio de
Janeiro, Brazil: Assesement of risck factors. Revista do Instituto de Medicina
Tropical de São Paulo, São Paulo, v.45, n.2, p.79-83, 1999.
Camargo, E. P. Growth and differentiation in Trypanosoma cruzi. I. Origin of
metacyclic trypanosoma in liquid media. Rev. Inst. Med. trop. S. Paulo, 6:93-100,
1964.
Camargo, M.E. Fluorescent antibody test for the sorodiagnosis of American
Tripanosomiasis. Technical modificationemploying preserved culture forms of
Trypanosoma cruzi in a slide test. Revista do Instituto de Medicina Tropical de São
Paulo, São Paulo, v.8, n.5, p 227-234, 1966.
Campino, L.M. In: Farrel J., ed., World Class Parasites: Leishmania, v.4, Kluwer
Academic Publishers. Boston, Dordrecht, London, 2003.
Carvalho, F.A.; Charest, H.; Tavares, C.A.; Matlashewski, G.; Valente, E.P.; Rabello,
A.; Gazzinelli, R.T.; Fernandes, A.P. Diagnosis of American Visceral leishmaniasis in
humans and dogs using the recombinant Leishmania donovani A2 antigen. Diagnostic
Microbiology and Infectious Diseases, v.43, [s.n.], p.289-295, 2002.

99
Carvalho, S.F.G.; Lemos, E.M.; Corey, R.; Dietze, R. Performance of recombinat k39
antigen in the diagnosis of Brazilian visceral leishmaniasis. American Journal of
Tropical Medicine and Hygiene, v.68, n.3, p.321-324, 2003.
Castro, A.G. Controle, diagnóstico e tratamento da leishmaniose visceral. Brasília:
Fundação Nacional de Saúde, 1996. 86p
Chouihi E, Amri F, Bouslimi N, Siala E, Selmi K, Zallagua N, Ben Abdallah R,
Bouratbine A, Aoun K. Cultures on NNN medium for the diagnosis of leishmaniasis
Pathol Biol (Paris). May;57(3):219-24, 2009.
Ciaramella, P. et al. A retrospective clinical study of canine leishmaniasis in 150 dogs
naturally infected with L. infantum. Veterinary Record, v. 141, n. 21, p. 539-543,
1997.
Ciaramella, P.; Corona, M. Canine Leishmaniasis: clinical and diagnostic aspects.
Compendium on Continuing Education for the Practicing Veterinarian, v.25, n.5,
p.358-368, 2003.
Costa Val, A.P. Tratamento da Leishmaniose Visceral canina com antimonial
pentavalente encapsulado em lipossomas. 2004. 125p. Tese (Doutorado em Ciência
Animal) - Escola de Veterinária, Universidade Federal de Minas Gerais, Belo
Horizonte, 2004.
Costa, C.A.; Genaro, O.; De Lana, M.; Magalhães, P.A.; Dias, M.; Michalick, M.S.;
Melo, M.N.; Da Costa, R.T.; Magalhães-Rocha, N.M.; Mayrink, W. Leishmaniose
visceral canina: avaliação da metodologia sorológica utilizada em inquéritos
epidemiológicos. Revista da Sociedade Brasileira de Medicina Tropical, v. 24, n.1,
p.21-25, 1991.
Cunningham, A.C. Parasitic adaptative mechanisms in infection by Leishmania. Exp
Mol Pathol, v.72, n.2, p.132-41, 2002.
Curi, N. H.; Miranda, I.; Talamoni, S. A. Serologic evidence of Leishmania infection in
free-ranging wild and domestic canids around a Brazilian National Park. Mem. Inst.
Oswaldo Cruz, v. 101, n.1, p. 99-101, 2006.
da Silva, E. S.; Van Der Meide, W. F.; Schoone, G. J.; Gontijo, C. M.; Schallig, H. D.;
Brazil, R. P. Diagnosis of canine leishmaniasis in the endemic area of Belo Horizonte,
Minas Gerais, Brazil by parasite, antibody and DNA detection assays. Vet. Res.
Commun, v. 30, n. 6, p. 637-643, 2006.
Dantas-Torres, F.; Brandão-Filho, S.P. Visceral leishmaniasis in Brazil: revisiting
paradigms of epidemiology and control. Rev. Inst. Med. Trop. S. Paulo, n. 48, v. 3, p.
151-156, 2006.
de Almeida, M.C. et al. Leishmanial Infection: Analysis of its First Steps. A Review
Mem Inst Oswaldo Cruz, Rio de Janeiro, v. 98(7): 861-870, 2003.
Dea-Ayuela MA, Rama-Iñiguez S and Bolás-Fernández F. Proteomic analysis of
antigens from Leishmania infantum promastigotes. Proteomics 6, 4187–4194, 2006.

100
Deane, L. M.; Deane, M. P. Encontro de cães naturalmente infectados por Leishmania
donovani, no Ceará. O Hospital, v.55, n.6, p.43-47, 1954.
Deane, L., M. Leishmaniose visceral no Brasil – estudo sobre reservatórios e
transmissores realizados no Estado do Ceará. 1a ed. Rio de Janeiro: Serviço Nacional de
Educação Sanitária, 1956, 161 p
Desjeux P. Leishmaniasis: current situation and new perspectives. Comp Immunol
Microbiol Infect Dis, 27(5): 305-18, 2004.
Dea-Ayuela MA, Rama-Iñiguez S and Bolás-Fernández F. Proteomic analysis of
antigens from Leishmania infantum promastigotes. Proteomics 6, 4187–4194, 2006.
Dwyer. D.M. Lectin binding saccharides on a parasite protozoan. Science, 184: 471-3,
1974.
Dye C, Williams BG Malnutrition, age and the risk of parasitic disease: visceral
leishmaniasis revisited. Proc R Soc Lond 254: 33-9, 1993
El Fakhry Y, Ouellette M, Papadopoulou B. A proteomic approach to identify
developmentally regulated proteins in Leishmania infantum. Proteomics, 2(8): 1007-17,
2002
Elkhoury, A. N. S. M. Vigilância e controle da leishmaniose visceral no Brasil. In:
Consulta de Expertos OPS/OMS Sobre Leishmaniasis Visceral em Las Americas, 23 a
25 de novembro de 2005, Brasília, 2005.
Evans, D.A. Leishmania. In vitro methods for parasite cultivation (ed. Taylor, A.E.R. &
Baker, J.R.), London: Academic Press., p.52-75, 1987.
vans, T.G.; Vasconcelos, I.A.B.; Lima, J.W.; Teixeira, J.M.; Macaullife, I.T.; Lopes,
U.G.; Pearson, R.D.; Vasconcelos, A.W. Canine visceral leishmaniasis in Northast
Brazil: Assesment of serodiagnostic methods. Am. J. Trop. Med. Hyg. v. 42, n. 2, p.
118-123, 1990
Ezquerra, J. P. A. Lãs leishmaniasis: de la biologia al control. Philadelphia: W.B.
Saunders. (2a ed) 2001. Cap. 6, p. 157-159.
Feitosa, M. M.; Ikeda, F. A.; Bonello, F. L.; Ciarlini, P. C.; Gonçalves, M. E.; Lima, V.
M. F.; Perri, S. H. V. Avaliação liquórica de cães, com e sem sintomatologia
neurológica, naturalmente acometidos por leishmaniose visceral. Veterinária Notícias,
Uberlândia, v. 11, n. 2, p. 61-69, 2005.
Feitosa, M.M. Leishmaniose visceral: facetas da doença. In: 6º Congresso Paulista de
Clínicos Veterinários de Pequenos Animais, 2006, São Paulo, p 57-58.
Feitosa, M.M.; Ikeda, F.A.; Luvizotto, M.C.R.; Perri, S.H.V. Aspectos clínicos de cães
com leishmaniose visceral no município de Araçatuba – São Paulo. Clin Vet, v.5, n.28,
p.36-44, 2000.
Ferreira, S. A. Avaliação do swab conjuntival para o diagnóstico da leishmaniose
visceral canina por PCR-hibridização. Dissertação (Mestrado) - Centro de

101
Desenvolvimento da Tecnologia Nuclear. Comissão Nacional de Energia Nuclear, Belo
Horizonte, 2008
Ferreira, E. C.; Lana, M.;Carneiro, M.; Reis, A. B.; Paes Dv ; Silva, E. S.; Schallig, H. ;
Gontijo CMF. Comparison of serological assays for the diagnosis of canine visceral
leishmaniasis in animals presenting diffrent clinical manifestations. Veterinary
Pathology, v. 146, p. 235-241, 2007
Ferreira, W.A.; Mayrink, W.; Mares-Guia, M.L.; Tavares, C.A. Detection and
characterization of leishmania antigens from an American cutaneous leishmaniasis
vaccine for diagnosis of visceral leishmaniasis. Diagn Microbiol Infect Dis.
Jan;45(1):35-43, 2003.
Ferrer, L. Clinical aspects of canine leishmaniasis. In: Canine Leishmaniasis: an update,
1, 1999, Barcelona. Proceedings of the International Canine Leishmaniasis Forum.
Sumène,: [s.ed.], agosto, 1999. p.6-10.
Ferrer, L. Leishmaniasis update in diagnosis and treatment of canine leishmaniasis.
Veterinary Record, London, v. 136, n.3, p. 514-516, 1997.
Ferrer, L. M. The pathology of canine leishmaniasis. In: Canine Leishmaniasis: Moving
Towards a Solution. Proceeding… Sevilha, Espanha,. p. 21-24, 2002.
Ferrer, L.; Aisa, M. J.; Roura, X.; Portús, M. Serological diagnosis and treatment of
canine leishmaniasis. Veterinary Record, v.136, p.514-516, 1995.
Figueiredo, F.B. et al. Efficacy of an indirect immunofluorescence test in the diagnosis
of canine leishmaniosis. Rev. Inst. Med. Trop. S. Paulo vol.52 no.4 São Paulo
July/Aug. 2010
Figueiredo, F.B.; Madeira, M.F.; Menezes, R.C.; Pacheco, R.S.; Pires, M.Q.; Furtado,
M.C.; et al. Efficacy of an indirect immunofluorescence test in the diagnosis of canine
leishmaniosis. Vet J. 2009; In press. (Epub ahead of print).
Fisa, R.; Riera, C.; Gállego, M.; Manubens, J.; Portús, M. Nested PCR for diagnosis of
canine leishmaniosis in peripheral blood, lymph node and bone marrow aspirates.
Veterinary Parasitology, v. 99, [s.n.], p.105-111, 2001.
Forgber M, Basu R, Roychoudhury K, Theinert S, Roy S, Sundar S, Walden P.
Mapping the Antigenicity of the Parasites in Leishmania donovani Infection by
Proteome Serology. PLoS ONE | www.plosone.org 1 December 2006 | Issue 1 | e40,
2006.
Fulgêncio, G.O. Prevalência de Oftalmopatias em cães naturalmente infectados com
Leishmania (Leishmania) chagasi no município de Belo Horizonte - Estudo clínico e
histopatológico. Dissertação (Mestrado em Medicina e Cirurgia Veterinárias) -
Universidade Federal de Minas Gerais,2006
Gomes, A.P.S.; Cordeiro, R.L.R. Reação cruzada no diagnóstico sorológico de
leishmaniose canina. Revista Brasileira de Parasitologia Veterinária, v. 23, Supl. 1, p.
238, 2004

102
Gongora R, Acestor N, Quadroni M, Fasel N, Saraiva NG, Walker J. Mapping the
proteome of Leishmania Viannia parasites using two-dimensional polyacrylamide gel
electrophoresis and associated technologies. Biomedica, 23(2): 153-60, 2003.
Gontijo, C.M.F.; Melo, M.N. Visceral leishmaniasis in Brazil: current status, challenges
and prospects. Rev. Bras. Epidemiol., v. 7, n. 3, p. 338-349, 2004.
Gossage, S.M.; Rogers, M.E.; Bates, P.A. Two separate growth phases during the
development of Leishmania in sand flies: implications for understanding the cycle. Int J
Parasitol 33: 1027-1034, 2003.
Gradoni, L. The diagnosis of canine leishmaniasis. In: Canine leishmaniasis: moving
towards a solution. Proceeding… Sevilha, Espanha,2002. p. 7-14, 2002.
Grima, M. Z. La Leishmaniosis canina. 1a Parte. Panorâmica general de la enfermedad.
Inf. Vet. Revista oficial del consejo general de colégios veterinários de Españal,
junho, p.14-18, 2005.
Guarga, J.L. et al. Experimental infection of Plebothomus perniciosus and
determination of the natural infection rates of Leishmania infantum in dogs. Acta
Tropica, v.77, [s.n.], p.203-207, 2000.
Guillén Llera, J.L.; García, M.L.L.; Martin Reinoso, E.; De Vivar González, R.
Differential serological testing by simultaneus indirect immunofluorescent antibody test
in canine leishmaniosis and erlichiosis. Veterinary Parasitology, v. 109, n. 3, p.185-
190, 2002.
Guimarães, M.C.S.; Celeste, B.J. & Franco, E.L. - Diagnostic performance indices for
immunolluorescent tests and enzyme immunoassays of leishmaniasis sera from
Northern and Northeastern Brazil. Bull. Wld. Htlh. Org., 68: 39-43, 1990.
Gupta SK, Sisodia BS, Sinha S, Hajela K, Naik S, Shasany AK, Dube A. Proteomic
approach for identification and characterization of novel immunostimulatory proteins
from soluble antigens of Leishmania donovani promastigotes. Proteomics;7(5):816-23,
2007
Halstead, B. W. Poisonous and Venomous Marine Animais of the World. 2.Vertebrates.
US Govt Printing Office, Washington, De., 1967.
Harith, A E.; Chowdhury, S.; Al-Masum, A.; Semião-Santos, S.; Das, P.K.; Akhter, S.;
Vetter, J.C.; HAQ, I. Reactivity of various leishmanial antigens in a direct agglutination
test and their value in differentiating postkala-azar dermal leishmaniasis from leprosy
and other skin conditions. J Med Microbiol. Feb; 44(2):141-6, 1996.
Harith, A.E.; Kolk, A.H.J.; Minter Goedbloed, E.; Kager, P.A. & Kolk, A.H.J. -
Improvement of a direct agglutination test for field studies of visceral leishmaniasis. J.
Clin. Microbiol., 26: 1321-1325, 1988.
Haseloff R, Krause E, Blasig I 2003. Proteomics of brain endotelium: Separation of
protein by two-dimensional gel electrophoresis and identification by mass spectrometry.
Methods Mol Med 89: 465-77, 2003.

103
Ho M, Leeuwenburg J, Mbungua G, Wamachi A, Voller A. An enzyme-linked
immunosorbent assay (ELISA) for field diagnosis of visceral leishmaniasis. Am J Trop
Med Hyg 32 : 943-946, 1983.
Hockmeyer, W.T.; Wellde, B.T.; Sabwa, C.L.A. Complement fixation test for visceral
leishmaniasis using homologous parasite antigen I. Ann Trop Med Parasitol; 78:489-
93 1984.
Honigberg, B.M. Evolutionary and systematic relationships in the flagellate Order
Trichomonadida Kirby, J. Protozool. 10 , pp. 20–63, 1963.
Ikeda-Garcia, F. A.; Feitosa, M. M. Métodos de diagnóstico da leishmaniose visceral
canina. Clínica Veterinária (São Paulo), v. 62, p. 32-38, 2006.
Instituto Adolfo Lutz, II Informe Técnico: Leishmaniose visceral americana. Secretaria
de Estado da Saúde do Estado de São Paulo, ed. MS, p.13-14, 2003.
Kar, K. Serodiagnosis of Leishmaniasis. Critical Reviews In Microbiology, 21 (2):
123-152, 1995.
Killick-Kendrick, R. Biology of Leishmania in phlebotomine sandflies. In: Biology of
Kinetoplastida (W. H. R. Lumsden & D. A. Evans, eds.), v. 2, pp. 395-460, London:
Academic Press. 1979
Killicck-Kendrick, R. The life cycles of Leishmania in the sand fly and transmission of
leishmaniasis by bite. In: Canine Leishmaniasis: moving towards a solution.
Proceeding… Sevilha, Espanha, 2002. p. 57-69, 2002.
Kontos, V. J.; Koutinas, A. F. Old world canine leishmaniasis. Continuing Education
Articles, n.15, p.949-959, 1993.
Lainson, R.; Shaw, J. J. Evolution, classification and geographical distribution. In:
Peters, W.; Killick-Kendrick, R. The leishmaniasis in biology and medicine, v. 1. 1a ed.
Londres: Academic Press, 1987. cap. 7, p. 291-364.
Laison, R.; Rangel, E. F. Lutzomyia longipalpis and the eco-epidemiology of American
visceral leishmaniasis, with particular reference to Brazil - A Review. Mem. Inst.
Oswaldo Cruz, v. 100, n. 8, p. 811-827, 2005.
Laurenti, M.D.; Lemos, E.M.; Reis, A.B.; Moreira, M.A.B.; Luvizotto, M.C.R.; Corbett,
C.E.P.; Dietze, R. Evaluation of Kalazar detect rapid test for serodiagnosis of canine
visceral leishmaniasis in Brazil. In: World Congress on Leishmaniasis, 3., 2005. Italy.
Abstract book...Italy, 2005. p.160.
Leontides L.S., Saridomichelakis M.N., Billinis C., Kontos V., Koutinas A.F., Galatos
A.D. & Mylonakis M.E. A cross-sectional study of Leishmania spp. infection in
clinically healthy dogs with polymerase chain reaction and serology in Greece. Vet.
Parasitol. 109:19-27, 2002.
Liberal, M.H.; Boughton, E. Standardization of an enzyme-linked immunossorbent
assay (ELISA) for the serodiagnosis of Mycoplasma bovis. Braz. J Microbiol, v.23,
p.146-150, 1992.

104
Lima, V.M.F.; gonçalves, M.E.; Ikeda, F.A.; Luvizotto, M.C.R.; Feitosa, m.m. Anti-
leishmania antibodies in cerebrospinal fluid from dogs witn visceral leishmaniasis.
Braz. J. Med. Biol. Res. v. 36, n.4, p. 485-489, 2003.
Lima, V.M.F.; Biazzono, L.; Silva, A.C.; Correa, A.P.F.L.; Luvizotto, M.C.R.
Serological diagnosis of visceral leishmaniasis by an enzyme immunoassay using
protein A in naturally infected dogs. Pesquisa Veterinária Brasileira, v.25, n.4, p.215-
218, 2005.
Little, S. E. Protozoal Diseases. Leishmaniases. In Greene, C. E. Infectious diseases of
the dog and cat. 3th ed. St. Louis: Saunders, 2006. p. 685-698.
Lobetti, R. G. Canine babesiosis. The Compendium on Continuing Education for the
Practicing Veterinarian, Princeton, v. 20, n. 4, p. 418-431,1998
Lopez, R.; Lucena, R.; Novalez, M. circulating immune complexes and renal function
in canine leishmaniasis. Zent Vet, v. 43, p. 460-474, 1996.
Lowry, O. H.; Rosebrough, N. J.; Lewis Farr, A.; Randall, R. J. Protein measurement
with the folin phenol reagent. J. Biol. Chem., v. 193, p. 265-275, 1951.
Machado, J. G.; Troncarelli, M. Z. ; Hoffmann, J. L. ; Camargo, L. B. et al. Comparação
do Ensaio Imunoenzimático com a Imunofluorescência Indireta no Diagnóstico da
Leishmaniose Visceral Canina. Veterinária e Zootecnia, v. 15, p. 85-90, 2008.
Madeira, M. F. et al. Post mortem parasitological evaluation of dogs seroreactive for
Leishmania from Rio de Janeiro, Brazil. Veterinary Parasitology, v. 138, n. 3-4, p.
366-370, 2006.
Mancianti F, Pedonese F, Poli A. Evaluation of dot enzyme-linked immunosorbent
assay (dot-ELISA) for the serodiagnosis of canine leishmaniosis as compared with
indirect immunofluorescence assay. Veterinary Parasitology. 1996;65:1-9, 1996.
Mancianti F, Falcone ML, Giannelli C, Poli A. Comparison between an enzyme-linked
immunosorbent assay using a detergentsoluble Leishmania infantum antigen and
indirect immunofluorescence for the diagnosis of canine leishmaniosis. Veterinary
Parasitology. 1995;59:13-21, 1995.
Mancianti, F.; Gramiccia, M.; Gradoni, L.; Pieri, S. Studies on canine leishmaniasis
control. I. Evolution of infection of different clinical forms of canine leishmaniasis
following antimonial treatment. Trans. R, Soc. Trop. Med. Hyg., 82: 566-567, 1988.
Martinez E.Z., Louzada Neto F., Pereira B.B. A Curva ROC para Testes Diagnósticos.
Cadernos de Saúde Coletiva, v. XI, n. 1, p. 7-31, 2003
Marzochi, M.C.A.; Coutinho, S.G.; Toledo, L.M.; et al. Leishmaniose visceral canina
no município do Rio de Janeiro. Fundação Oswaldo Cruz, Rio de Janeiro, 12-22,
1986.
Marzochi, M.C.A., Marzochi, K.F.B. Tegumentary and visceral leishmaniasis in Brazil:
emerging anthropozoonosis and possibilities for their control. Cad Saúde Publ 10:
359-375, 1994.

105
Martin, S.K.; Thuita-Harun, L.; Adoyo-Adoyo, M.; Wasunna, K.M. A diagnostic
ELISA for visceral leishmaniasis, based on antigen from media conditioned by
Leishmania donovani promastigotes. Ann Trop Med Parasitol; 92:571-7, 1998
Mauricio, I. L.; Gaunt, M. W.; Stodard, J. R.; Miles, M. A. Genetic typing and
phylogeny of the Leishmania donovani complex by restriction analysis of PCR
amplified gp63 intergenic regions. Parasitology, v. 122, p. 393-403, 2001.
Mauricio, I. L.; Stothard, J. R.; Miles, M. A. The strange case of Leishmania chagasi.
Parasitol. Today, v. 16, n. 5, p. 189, 2000.
Melo, M.N. Leishmaniose visceral no Brasil: desafios e perspectivas. In: Congresso de
Parasitologia Veterinária & I Simpósio Latino-Americano de Ricketisioses, Ouro
Preto, MG, 2004.
Mettler, M.; Grimm, F.; Capelli, G.; Heinrich, C.; Deplazes, P. Evaluation of Enzyme-
Linked Immunosorbent Assay, an Immunofluorescent-Antibody Test, and two rapid
test (Immunochromatographic-Dipstick and Gel Test) for serological diagnosis of
symptomatic and asymptomatic Leishmania Infections in dogs. J. Clinic. Microbiol.
v. 43, n. 11, p. 5515-5519, 2005.
Metz, C. E. Statistical analysis of ROC data in evaluating diagnostic performance.
Multiple Regression Analysis: Applications in the Health Sciencs, n.13, p.365-384,
1986.
Mohammed, A.R.; Wright, E.P.; Abdel Rahman, A.M.; Kolk, A.; Laarman, J.J.;
Pondmsn, K.W... Seroadiagnosis of Sudanese visceral and mucosal leishmaniasis:
comparación of Elisa-imunofluorescence and indirect haemagglutination. Trans R Soc
Trop Med Hyg; 80:271-4, 1986.
Moreira, M.A.B.; Luvizotto, M.C.R.; Nunes, C.M.; Silva, T.C.C.; Laurenti, M.D.;
Corbett, C.E.P. Aplicação da técnica de imunofluorescência direta para o diagnóstico
da leishmaniose visceral canina em aspirado de linfonodo. Braz. J. Vet. Res. Anim.
Sci. v. 39, n. 2, p. 103-106, 2002.
Monteiro, P. S.; Lacerda, M. M.; Arias, J. R. Controle da Leishmaniose no Brasil.
Revista da Sociedade Brasileira de Medicina Tropical, Rio de Janeiro, v. 27, p. 67-
72, 1994.
Mukherjee, M.; Bhattacharyya, A.; Duttagupta, S. Serodiagnostic and
immunoprophylactic potencial of a 78kDa protein of Leishmania donovani of Indian
origin. Med. Sci. Monit., v8, n.4, p.117-122, 2002.
Muniaraj, M.; Lal, C.S.; Kumar, S.; Sinha, P.K.; Das, P. Milk of cow (Bos taurus),
buffalo (Bubalus bubalis), and goat (Capra hircus): a better alternative than fetal bovine
serum in media for primary isolation, in vitro cultivation, and maintenance of
Leishmania donovani promastigotes. J Clin Microbiol 45:1353–1356, 2007.
Neuhoff V, Arold N, Taube D, Ehrhardt W. Improved staining of proteins in
polyacrylamide gels including isoelectric focusing gels with clear background at

106
nanogram sensitivity using Coomassie Brilliant Blue G-250 and R-250.
Electrophoresis. Jun;9(6):255-62, 1988
Nicolle C (1908) Culture du parasite du bouton d'Orient. C R Acad Sci 146:842–843
Noli, C. Leishmaniosis canina. Waltham Focus. v.9, n.2, p.16-24, 1999.
Oliveira, L.S. et al. A utilização da imunofluorescência indireta no diagnóstico de rotina
da leishmaniose visceral canina e suas implicações no controle da doença. Ciência
Animal Brasileira, Goiânia, v.6, n.1, p.41-47, 2005
Oskam, L., Slappendel, R.J., Beijer, E.G., Kroon, N.C.M., van Ingen, C.W., Ozensoy,
S.. O¨ zbel, Y. and Terpstra, W.J. Dog-DAT: a direct agglutination test using stabilized,
freeze-dried antigen for the serodiagnosis of canine visceral leishmaniasis. FEMS
Immunology and Medical Microbiology, 16, 235–239, 1996.
Oskam, L.; Nieuwenhuijs, L. & Hailu, A. Evaluation of the direct agglutination test
(DAT) using freeze-dried antigen for the detection of anti-Leishmania antibodies in
stored sera from various patient groups in Ethiopia. Transactions of the Royal Society
of Tropical Medicine and Hygiene 93, 275–277, 1999
Peters NC et al. In Vivo Imaging Reveals an Essential Role for Neutrophils in
Leishmaniasis Transmitted by Sand Flies Science Vol. 321. no. 5891, pp. 970 – 974,
2008.
Piarroux, R., Azaiez, R., Lossi, A.M., Reynier, P., Muscatelli, F., Gambarelli, F.,
Fontes, M., Dumon, H., Quilici, M. Isolation and characterization of a repetitive DNA
sequence from Leishmania infantum: development of a visceral leishmaniasis
polymerase chain reaction. Am. J. Trop. Med. Hyg. 49, 364–369, 1993
Pinelli, E.; Killick-KENDRICK, R.; Wagenaar, J.; Bernadina, W.; Del Real, G.;
Ruitenberg, J. Cellular and humoral immune responses in dogs experimentally and
naturally infected with Leishmania infantum. Infec Imm, v.62, n.1, p.229-335, 1994.
Piscopo T.V.; Azzopardi C.M. Leishmaniasis. In.: Postgrad Med J. v. 82, 2006. p.
649–657
Prefeitura Municipal de Belo Horizonte. Disponível em http//portalpbh.pbh.gov.br.
Acesso em novembro de 2010.
Quaresma, P.F, Diagnóstico molecular da leishmaniose visceral canina e quantificação
da carga parasitária através da Reação em Cadeia da Polimerase. 2007. 121p. Tese
(Mestrado em Ciências da Saúde) Fundação Oswaldo Cruz, Centro de Pesquisas René
Rachou, 2007
Queiroz, N. M. G. P.; Assis, J. ; Oliveira, T. M. F. De S.; Machado, R. Z. ; Nunes, C.
M. ; Starke-Buzetti, W. A. . Diagnóstico da Leishmaniose Visceral Canina pelas
técnicas de imunoistoquímica e PCR em tecidos cutâneos em associação com a RIFI e
ELISA-teste. Revista Brasileira de Parasitologia Veterinária, v. 19, p. 34-40, 2010.

107
Rachamin, N.; Jaffes, C. L.; Abranches, P.; Silva-Pereira, M.C.D.; Schnur, L.F.;
Jacobson, R.L. Serodiagnosis of canine visceral leishmaniasis in Portugal: comparison
of three methods. Annal of Tropical and Parasitology, v.85, n.5, p.503-508, 1991.
Reichmann, M.L.A.B. Leishmaniose visceral canina – uma zoonose reemergente. In: 1º
Fórum sobre Leishmaniose Visceral Canina, Jaboticabal, SP, 2006.
Reithinger R.; Dujardin J.; Louzir H.; Pirmez C.; Alexander B.; Brooker S. Cutaneous
leishmaniasis. Lancet Infectious Disease, v.7 n.9 p.581-596, 2007.
Reithinger, R.; Quinnell, R.J.; Alexander, B.; Davies, C.R. Rapid detection of
Leishmania infantum infection in dogs: comparative study using na
immunochromatografic dipstick test, enzyme-linked immunosorbent assay, and PCR.
Journal of Clinical Microbiology, v.40, n.7, p.2352-2356, 2002
Rodrigues Ide A., da Silva, B.A., dos Santos, A.L., Vermelho, A.B., Alviano, C.S.,
Dutra, P.M., Rosa, M.do S. A new experimental culture medium for cultivation of
Leishmania amazonensis: its efficacy for the continuous in vitro growth and
differentiation of infective promastigote forms. Parasitol Res. Apr;106(5):1249-52,
2010.
Romero, G.A.; Boelaert, M Control of visceral leishmaniasis in latin america-a
systematic review. Plos Negl Trop Dis. Jan 19;4(1): e584, 2010.
Rosario, E. Y., Genaro O.; França-Silva, J. C.;Da Costa, R. T.;Mayrink, W.; Reis, A.
B.; Carneiro, M. Evaluation of enzyme-linked immunosorbent assay using crude
Leishmania and recombinant antigens as a diagnostic marker for canine visceral
leishmaniasis. Mem. Inst. Oswaldo Cruz, v. 100, n. 2, p. 197-203, 2005.
Rosati, S.; Ortoffi, M.; Profiti, M.; Manelli, A.; Mignoni, W.; Bollo, E.; Gradoni, L.
Prokaryotic expression and antigenic characterization of three recombinant Leishmania
antigens for serological diagnosis of canine leishmaniasis. Clinical and Diagnostic
Laboratory Immunology, v.10, n.6, p.1153-1156, 2003.
Ross, R. (1) Note on the bodies recently described by Leishman and Donovan and (2)
further notes no Leishmn’s bodies. Brit Med J, v.2, p.1261-1401, 1903.
Sacks D.L. The structure and function of the surface lipophosphoglycan on different
developmental stages of Leishmania promastigotes. Infect Agents Dis. Aug;1(4):200–
206, 1992.
Sacks, D., Kamhawi, S. Molecular aspects of Parasite-Vector and Vector Host
Interactions in Leishmaniasis. Annual Reviews in Microbiology, v. 55, p. 453-483,
2001.
Sacks, D.; Noben-Trauth, N. The immunology of susceptibility and resistance to
Leishmania major in mice. Nat Rev Immunol 2(11), 845-858, 2002.
Santa Rosa, I.C.A.; Oliveira, I.C.S. Leishmaniose visceral: breve revisão sobre uma
zoonose reemergente. Clinica Veterinária, v.2, n.11, p.24-28, 1997.

108
Santos, F. R.; Pena, S. D. J.; Epplen, J. T. Genetic and population study of a Y-linked
tetranucleotide repeat DNA polymorphism with a simple non-isotopic technique. Hum.
Genet., v. 90, p. 655-656, 1993.
São Paulo (Estado). Secretaria de Estado da Saúde. Leishmaniose Visceral Americana:
II Informe Técnico. São Paulo: Secretaria de Estado da Saúde, 2003. 48p. Disponível
em: www.sucen.sp.gov.br/doencas/leish_visc/LVA24ago03.pdf Acesso em 13 de
ago.de 2006.
Scalone, A.; De Luna, R.; Oliva, G.; Baldi, L.; Satta, G.; Vesco, G.; Mignone, W.;
Turilli, C.; Mondesire, R.R.; Simpson, D.; Donoghue, A.R.; Frank, G.R.; Gradoni, L.
Evaluation of the Leishmania recombinant k39 antigen as a diagnostic marker for
canine leishmaniasis and validation of a standardized enzyme-linked immunosorbent
assay. Veterinary Parasitology, v.104, n.4, p.275-285, 2002.
Scott, D.W.; William, H.M.; Griffin, G.E. Viral, rickettisial and protozoal skin disease.
In: Small Animal Dermatology, 6 ed, Philadelphia:Saunders Company, 2001, p.517-
542
Schoone, G.J.; Hailu, A.; Kroon, C.C.M.; Nieuwenhuys, J.L.; Schallig, H.D.F.H.;
Oskam, L. A fast agglutination-screening test (FAST) for the detection of anti-
Leishmania antibodies. Trans R Soc Trop Med Hyg 95:400-401, 2001.
Shaw, J.J. New World Leishmaniasis: the ecology of leishmaniasis and the diversity of
leishmanial species in Central and South America. In J Farrell, World Class Parasites:
Leishmania, Kluwer Academic Publishers, Boston, Vol. 4, p. 11-31, 2003
Schuster F.L., Sullivan J.J. Cultivation of clinically significant hemoflagellates. Clin
Microbiol Rev 15:374–389, 2002.
Silva, M.R. Estudo comparativo entre os métodos de Elisa e IFI na análise de amostras
de sangue de cães provenientes de municípios endêmicos e enzoóticos para LVC [tese
de doutorado]. São Paulo: Faculdade de Saúde Pública da USP; 2005
Silva, E. S.; Gontijo, C. M. F.; Pirmez, C.; Fernandes, O.; Brazil, R. P. Short report:
detection of Leishmania DNA by polymerase chain reaction on blood samples from
dogs with visceral leishmaniasis. Am. J. Trop. Med. Hyg., v. 65, n. 6, p. 896-898,
2001.
Silva, S.M.; Rabelo, P.F.B.; Gontijo, N.F.; Ribeiro, R.R.; Melo, M.N.; Ribeiro, V.M.;
Michalick, M.S.M. First report of infection of Lutzomyia longipalpis by Leishmania
(Leishmania) infantum from a naturally infected cat of Brazil Vet Parasitol. Nov
24;174 (1-2):150-4, 2010.
Silveira, F.T.; Laison, R.; Corbett, C.E.P. Further observations on clinical,
histopathological, and immunological features of borderline disseminated cutaneous
leishmaniasis caused by Leishmania (Leishmania) amazonensis. Mem. Inst. Oswaldo
Cruz, v. 100, n. 5, p. 525-534, 2005.
Singh S, Sivakumar R. Recent advances in the diagnosis of leishmaniasis. J Postgrad
Med, 49:55-60, 2003

109
Slappendel, R. J. Canine leishmaniasis. Vet. Q., v. 10, n. 1, p. 1-16, jan.1988.
Somanna A.; Mundodi, V.; Gedamu, L. In vitro cultivation and characterization of
Leishmania chagasi amastigote-like forms. Acta Trop 83:37–42, 2002
Strauss-Ayali, D.; Baneth, G. Canine Visceral Leishmaniasis. In: Recent Advances in
Canine Infections Diseases. International Veterinary Information Service, 2000.
Sundar S, Pai K, Sahu M, Kumar V & Murray W. Immunochromatographic strip-test
detection of anti-K39 antibody in Indian visceral leismaniasis. Ann Trop Med
Parasitol 96: 19-23, 2002.
Swets, J. Measuring the accuracy of diagnostic systems. Science, 240:1285-1293, 1988
Tafuri, W. L.; Santos, R. L.; Arantes, R. M.; Gonçalves, R.; Melo, M.N.; Michalick, M.
S. M.; Tafuri, W. L. An alternative immunohistochemical method for detecting
Leishmania amastigotes in paraffin-embedded canine tissues. J. Immunol. Methods, v.
292, n. 1-2, p. 17-23, 2004.
Tavares, C. A; Fernandes, A.P. e Melo, M.N. Molecular diagnosis of leishmaniasis.
Expert Review Molecular Diagnosis, v.3, n5, p.657-667, 2003.
Tavora, M.P.F. et al Estudo de validação comparativo entre as técnicas ELISA e RIFI
para diagnosticar Leishmania sp em cães errantes apreendidos no município de Campos
dos Goytacazes, Estado do Rio de Janeiro. Revista da Sociedade Brasileira de Medicina
Tropical 40(4):482-483, jul-ago, 2007
Thiel M, Bruchhaus I. 2001. Comparative proteome analysis of Leishmania donovani at
different stages of transformation from promastigotes to amastigotes. Med Microbiol
Immunol (Berl). 190 (1-2): 33-6. 2001
Towbin, H and Gordon, J. Immunoblotting and dot immunobinding-current status and
outlook. J. Immunol. Methods 72: 313-340, 1984
Tully, J.G. Immunological tools – Introductory remarks. In: Tully, J.G.; RAZIN, S.
Molecular and diagnostic procedures in mycoplasmology. London: Academic, 1996,
p.93-103.
Vercammen, F.; Berkvens, D.; Leray, D.; Jacquet, D.; Vervoort, T. Development of a
slide ELISA for canine leishmaniasis and comparison with four serological tests.
Veterinary Record, v.141, n.13, p.328-330, 1997.
Vexenat, A. C.; Santana, J. M.; Teixeira, A. R. L. Cross-reactivity in human infections
by the kinetoplastid protozoa Trypanosoma cruzi, Leishmania chagasi and Leishmania
(viannia) braziliensis. Revista do Instituto de Medicina Tropical, v.38, n.3, p.177-
185, 1996.
Vickerman, K. The diversity of the kinetoplastid flagellates. In: LUMSDEN, W.H.R.;
EVANS, D.A. Biology of the kinetoplastida. London/New York/San Francisco:
Academic, p.1-34, 1976.

110
Voller, A., Bidwell, D. E. & Barlett, A. The enzyme linked immunossorbent assay
(ELISA): A guide with abstracts of microplate applications, Guernsey: Dynatech
Europe, 124p., 1979.
Wilson, M.E. Leishmaniasis. Current Opinion In Infectious Diseases. 6: 331-341,
1993.
Wolschrijn, C.F.; Meyer, H.P.; Hazewinkel, H.A.W.; Wolvekamp, W.T. Destructive
polyarthritis in a dog with leishmaniasis. Journal Small Animal Practice, v.37, n.12,
p.601-603, 1996.
Woodcock, H.M. The life-cycle of Cystobia irregularis (Minch.) together with
observations on other ´neogamous`gregarines. J.Microsc. Sci. 50:1-100, 1906.
WHO World Health Organization 2000 Leishmaniasis and Leishmania/HIV co-
infection. WHO report on global surveillance of epidemic-prone infectious diseases
Geneva:World Health Organization . (document WHO/CDS/CSR/ISR/2000.
WHO - World Health Organization 2007. Leishmaniasis [accession 2007 Apr 11].
Available from: http//www.who.int.
Zanette MF. Comparação entre os métodos de ELISA, imunofluorescência indireta e
imunocromatografia para o diagnóstico da leishmaniose visceral canina [dissertação de
mestrado]. Araçatuba (SP): Faculdade de Odontologia, Curso de Medicina
Veterináriada Unesp; 2006.

111
10 . Anexos
10.1 Depósito da Patente do Antígeno

112

113

114
10.2 Certificado do Comitê de Ética em Experimentação Animal –
CETEA

115
10.3 Artigo Publicado

116

117
118

119

120

121

122

123