observações sobre a dispersão de sementes por primatas no ... · com instituto de biologia da...
TRANSCRIPT

UNIVERSIDADE VEIGA DE ALMEIDA COORDENAÇÃO DE BIOLOGIA
JEHNNIFER RIBEIRO DE MENDONÇA
Observações sobre a dispersão de sementes por primatas no Jardim Botânico do Rio de Janeiro
Rio de Janeiro – RJ
2016

II
JEHNNIFER RIBEIRO DE MENDONÇA
Observações sobre a dispersão de sementes por primatas no Jardim Botânico do Rio de Janeiro
Monografia apresentada à Coordenação de Ciências Biológicas da Universidade Veiga de Almeida como parte dos requisitos para conclusão do curso de Bacharelado em Ciências Biológicas, ênfase em Ecologia. Orientadoras: Profa. Dra. Uiara Gomes Drª Tânia Sampaio Pereira Msª Cristiane Rangel
Rio de Janeiro – RJ
2016

III
Dedico este trabalho ao meu pai,
minha mãe e meu irmão, pois sem
vocês nada disso seria possível.
Obrigada pelo apoio, carinho e
compreensão. Essa vitória não é só
minha, é nossa!

IV
AGRADECIMENTOS
Agradeço primeiramente a Deus, por ser essencial na minha vida, autor de
meu destino, meu guia, socorro presente na hora da angústia, pois sem ele não teria
forças para concluir essa jornada.
Agradeço especialmente aos meus pais, Marcílio F. de Mendonça e Josefa R.
de Mendonça, e meu irmão, Johnatan R. de Mendonça, que sempre me
incentivaram e proporcionaram as melhores condições para que eu pudesse vencer
cada etapa desta e de todas as jornadas que percorri até hoje. Este trabalho é
dedicado a vocês, que são as pessoas mais importantes da minha vida.
As minhas orientadoras, Uiara Gomes Cabral e Tânia Sampaio Pereira, que
foram ótimas orientadoras, com ótimas criticas construtivas e por ter me ajudando
sempre que precisei. Muito obrigada por terem me aceito, foi muito construtivo e
prazeroso poder trabalhar com vocês.
As minhas orientadoras no estágio Projeto Fauna JBRJ, Gabriela Heliodoro e
Cristiane Rangel, muito obrigada por vocês me ensinarem importância e
responsabilidade que nós temos em estudar e proteger a fauna do JBRJ. Por todos
os conhecimentos passados, de biologia e de vida.
Ao Gustavo e Caco que trabalham na sala com o pessoal do Projeto Fauna,
tornando o ambiente de trabalho o melhor possível. Sempre dispostos a ajudar e de
bom humor, obrigada por serem sempre tão acolhedores e amigos.
A minha amiga e irmã, Tatiane Brito, que sempre esteve comigo nos bons e
maus momentos, me apoiando em tudo que fosse preciso.
Aos meus amigos do estágio, companheiros de trabalho e amigos de
verdade, Marina Bordin, Guilherme Nunan, Mônica Brito, Simone Nogueira e Yasmin
Balzi. Obrigada por sempre estarem comigo e me apoiarem em tudo. Obrigada por
terem se tornado uma nova família para mim.
Aos meus amigos, (não dá pra escrever o nome de todos), pelas alegrias,
tristezas e dores compartilhadas. Com vocês, as pausas entre um parágrafo e outro
de produção melhora tudo o que tenho produzido na vida.
Aos meus professores, que dividiram seu conhecimento, e me deram a

V
oportunidade de ser uma profissional melhor.
Aos meus amigos e colegas de faculdade, Christiane Silveira, Monira Bicalho,
Flavia Tebaldi, Tiago Sousa e Paulo Vitor, por terem me ajudado e terem passado
comigo todos os problemas que enfrentamos desde o início da graduação, e terem
tido força para continuar até aqui.
Ao Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, por ter permitido
realizar meu trabalho e todo o meu estágio.

VI
Deus não escolhe os capacitados, capacita os escolhidos. Fazer ou não fazer algo só depende de nossa vontade e perseverança!
Albert Einstein

VII
RESUMO
Observações sobre a dispersão de sementes por primatas no Jardim Botânico
do Rio de Janeiro
O macaco-prego (Sapajus nigritus) - nativo- e o sagüi (Callithrix sp.) – exótico-, são primatas residentes e frequentadores do Jardim Botânico do Rio de Janeiro (JBRJ) e da área de Mata Atlântica local. Sua presença pode trazer danos à coleção científica de plantas vivas do JBRJ ao dispersarem sementes e/ou propágulos no Arboreto e ao consumirem espécimes exóticos da coleção viva do JBRJ e os introduzindo na Mata Atlântica local. O objetivo deste trabalho foi verificar e analisar a possibilidade de dispersão de sementes através das fezes dos primatas Sapajus nigritus, Callithrix jacchus, Callithrix penicillata e híbridos. As sementes foram obtidas entre dezembro de 2014 e setembro de 2016 (estações secas e úmidas), diretamente das fezes dos animais coletadas em campo. As fezes foram filtradas em gaze com soro fisiológico e triadas na lupa. As sementes e/ou propágulos encontrados foram parcialmente identificados e a germinação das sementes foi realizada no Laboratório de Sementes do JBRJ. Foram coletadas 36 de amostras de sagui com os seguintes resultados: 14 amostras possuíam sementes ou propágulos de 8 famílias diferentes. Macaco-prego foram 25 amostras com 30 tipos de sementes e/ou propágulos distribuídos em 14 famílias. As amostras de sementes de espécies/famílias nativas e exóticas germinaram bem atestando a viabilidade das mesmas. Os resultados obtidos com este trabalho indicam que S. nigritus é dispersor mais eficiente que o Callithrix sp., diante da diversidade de vegetais com sementes pequenas e maiores nas amostras de fezes ao longo de todas as estações (seca e úmida).
Palavras-chaves: Callitrirx sp., Sapajus nigritus e Mata Atlântica.

VIII
ABSTRACT
Observations on the dispersal of seeds by primates in the Botanic Garden of
Rio de Janeiro.
The tufted capuchin (Sapajus nigritus) - native - and the marmoset (Callithrix sp.) - exotic - are residents primates and regulars Rio de Janeiro Botanical Garden (JBRJ) and the local Atlantic Forest area. Their presence may damage the JBRJ's scientific collection of living plants by dispersing seeds and / or propagules in the Arboretum and by consuming exotic specimens from the JBRJ living collection by introducing them into the local Atlantic Forest. The objective of this work was to verify and analyze the possibility of seed dispersal through primate feces (Sapajus nigritus, Callithrix jacchus, Callithrix penicillata and hybrids). The seeds were obtained between December 2014 and September 2016 (dry and humid seasons), directly from the feces of the animals collected in the field. The feces were gauze filtered with physiological saline and screened in the magnifying glass. The seeds and / or propagules were partially identified and seed germination was performed at the JBRJ Seed Laboratory. Thirty - six samples were collected with the following results: 14 samples had seeds or seedlings from 8 different families. Monkey-nail were 25 samples with 30 types of seeds and / or propagules distributed in 14 families. Seed samples from native and exotic species / families germinated in well attesting to their viability. The results obtained with this work indicate that S. nigritus is a more efficient disperser than Callithrix sp., Due to the diversity of plants with small and larger seeds in stool samples throughout all seasons (dry and humid).
Keywords: Callithrix sp., Sapajus nigritus, Atlantic forest.

IX
LISTA DE FIGURAS
FIGURA 1: Vista aérea do JBRJ................................................................................19
FIGURA 2: (A) Sapajus nigritus (macaco-prego) e (B) Callithrix jacchus, Callithrix
penicillata e híbridos (sagui).........................................................................................22
FIGURA 3: Mapa das plataformas de ceva no JBRJ.................................................23
FIGURA 4: (A) Preparação das amostras (B) Filtragem das amostras de fezes......24
FIGURA 5: (A) Plataforma de ceva. (B) Estufa B.O.D. (C) Separação de sementes na lupa. Imagem (D) Laboratório de sementes do JBRJ...........................................25
FIGURA 6: Gráfico de frequência de estruturas presentes em amostras de fezes de Sagui..........................................................................................................................27
FIGURA 7: Gráfico de sementes e/ou propágulos encontrados em amostras de fezes de Sagui.....................................................................................................................28
FIGURA 8: Gráfico de quantidade de sementes e/ou propágulos encontrados nas amostras de fezes do sagui nas estações secas e úmida.........................................29
FIGURA 9: (A) Amostra S004 (B) Amostra S005.......................................................29
FIGURA 10: Gráfico de frequência de estruturas presentes em amostras de fezes de macaco-prego.............................................................................................................32
FIGURA 11: Gráfico das sementes e/ou propágulos encontrados em amostras de fezes de macaco-prego..............................................................................................33
FIGURA 12: Gráfico da quantidade de sementes e/ou propágulos encontrados nas amostras de fezes do Macaco-prego nas estações secas e úmidas.........................34
FIGURA 13: (A) Amostra P001 (B) Amostra P014.....................................................34
FIGURA 14: Gráfico das sementes e/ou propágulos encontrados em amostras dos primatas......................................................................................................................35
FIGURA 15: Gráfico representativo das sementes e/ou propágulos semeados e germinados em amostras de fezes dos primatas.......................................................36
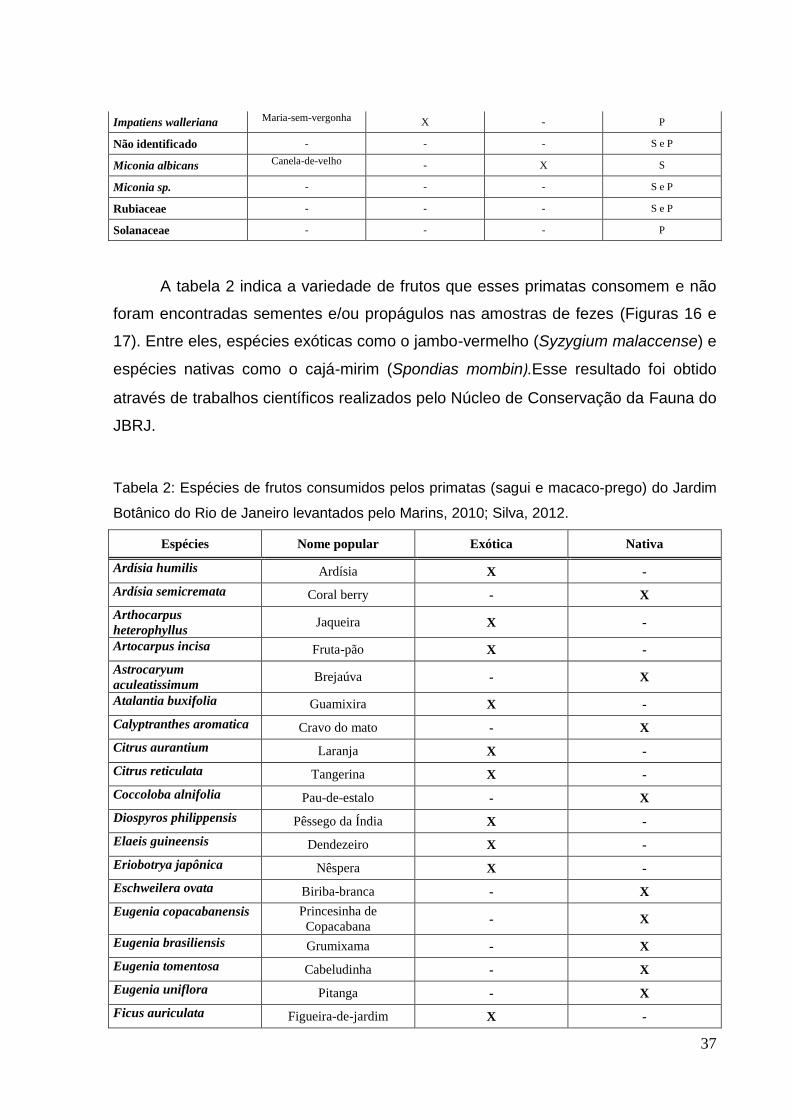
FIGURA 16: (A) macaco-prego (Sapajus nigritus) alimentando-se do fruto Arthocarpus heterophyllus. (B) sagui (Callithrix jacchus, Callithrix penicillata e híbridos) alimentando-se do fruto Eugenia uniflora (pitanga).....................................................................................................................38 FIGURA 17: (A) macaco-prego (Sapajus nigritus) alimentando-se do fruto Psidium guajava (Goiaba) no Jardim Botânico do Rio de Janeiro - JBRJ. (B) sagui (Callithrix jacchus, Callithrix penicillata e híbridos) alimentando-se do fruto Arthocarpus heterophyllus (Jaca) no Jardim Botânico do Rio de Janeiro – JBRJ...........................................................................................................................39

X
LISTA DE TABELAS
TABELA 1: Lista das sementes e/ou propágulos encontradas nas fezes dos primatas......................................................................................................................36
TABELA 2: Espécies de frutos consumidos pelos primatas.......................................37
TABELA 3: Relação das estruturas encontradas nas amostras de fezes de Sagui no período de estação seca¹ de abril a set/2015............................................................56
TABELA 4: Relação das estruturas encontradas nas amostras de fezes de Sagui no período de estação úmida de out/2015 a mar/2016..................................................57
TABELA 5: Relação das estruturas encontradas nas amostras de fezes de Sagui no período de estação seca² de abril a set/2016............................................................58
TABELA 6: Relação das estruturas encontradas nas amostras de fezes de Macaco-prego no período de estação úmida¹ de dez/2014 a mar/2015.................................59
TABELA 7: Relação das estruturas encontradas nas amostras de fezes de Macaco-prego no período de estação seca¹ abril a set/2015..................................................59
TABELA 8: Relação das estruturas encontradas nas amostras de fezes de Macaco-prego no período de estação úmida² de out/2015 a mar/2016..................................60
TABELA 9: Relação das estruturas encontradas nas amostras de fezes de macaco-prego no período de estação seca² de abr a set/2016..............................................61
TABELA 10: Relação das sementes e/ou propágulos encontrados nas amostras de Sagui quanto à germinação no período de estação seca e úmida............................62
TABELA 11: Relação das sementes e/ou propágulos encontrados nas amostras de Macaco-prego quanto à germinação no período de estação seca e úmida..............63

XI
SUMÁRIO
1. INTRODUÇÃO..................................................................................................12
1.1 ZOOCORIA POR PRIMATAS.........................................................................12
1.2 MATA ATLÂNTICA E PRIMATAS DO JBRJ...................................................14
2. HIPÓTESE........................................................................................................17
3. OBJETIVO GERAL..........................................................................................18
3.1 OBJETIVOS ESPECÍFICOS...........................................................................18
4. METODOLOGIA...............................................................................................19
4.1 ÁREA DE ESTUDO........................................................................................19
4.2 DELINEAMENTO AMOSTRAL.......................................................................21
5. RESULTADOS.................................................................................................26
5.1 AMOSTRAS DE FEZES DE SAGUI...............................................................26
5.2 AMOSTRAS DE FEZES DE MACACO-PREGO............................................30
6. DISCUSSÃO.....................................................................................................40
7. CONCLUSÃO...................................................................................................47
8. REFERÊNCIAS BIBLIOGRÁFICAS................................................................48
9. ANEXOS...........................................................................................................56

12
1. INTRODUÇÃO
1.1 ZOOCORIA POR PRIMATAS
O desenvolvimento das plantas inicia-se com a germinação das sementes,
que contêm, no seu interior, o embrião (Guerreiro & Glória, 2003). A dispersão de
sementes é importante para regeneração das florestas tropicais e colonização de
habitats (Van Der Pijl, 1982). Ela também é importante para a recuperação de áreas
degradadas por ações antrópicas e pode beneficiar o estabelecimento,
desenvolvimento e evolução das espécies florestais (Rondon-Neto et al., 2001). De
acordo com Figliolia (1993), o processo de dispersão é complexo e envolve as
relações entre planta e agente dispersores. Acredita-se que entre 50% a 90%, de
todas as árvores das florestas tropicais foram dispersas por animais (Fleming, 1987).
Zoocoria é o termo utilizado para dispersão de sementes por animais e de acordo
com Instituto de Biologia da Universidade de Uberlândia – UFU (2016) a dispersão
por animais (zoocoria) pode ser dividida em três tipos: Endozoocoria: quando a
dispersão é feita através da ingestão e depois liberação da semente nas fezes ou
por regurgito; Sinzoocoria: quando as sementes são deliberadamente carregadas,
principalmente na boca; Epizoocoria: quando as sementes são carregadas
acidentalmente. A variabilidade de padrões de dispersão de sementes fica mais
evidente à medida que os estudos tornam-se mais numerosos e detalhados (Garber
& Lambert, 1998).
As sementes que possuem grandes características de sobrevivência deram
às plantas que as produzem a mais importante vantagem seletiva (Raven, 2001). A
dispersão e a predação de sementes são fortemente relacionados aos fatores
bióticos e aos eventos reprodutivos fenológicos (Neves & Viana, 2008). A dispersão
de sementes é um processo pelo qual as sementes são removidas de perto da
planta mãe para distâncias maiores, em locais seguros, onde a predação e
competição não são tão eficientes, para que a semente ou propágulo tenha maiores
chances de sobrevivência e de gerar descendentes férteis, portanto sendo mais
propício à germinação. É um processo importante dentro do ciclo de vida da maioria
das plantas, especialmente em ambientes tropicais (Howe & Miriti, 2004). Se as
sementes forem dispersas em uma área mais ampla, aumentará a chance de

13
algumas sementes caírem em local favorável facilitando o estabelecimento do
propágulo. A dispersão é feita por diásporos que são as sementes, os frutos, a
planta inteira ou parte dela, ou a junção desses diásporos (Instituto de Biologia da
Universidade de Uberlândia – UFU, 2016).
Os diásporos dispersos por animais são muito importantes para a
manutenção da tentativa de obter recursos para a fauna ao longo do ano (Figliolia &
Kageyama 1995), principalmente em locais que predominam clima sazonal, onde
ocorrem períodos de escassez de frutos como alimentos (Galetti & Pedroni 1994;
Develey & Peres 2000).
A dispersão das sementes é um processo demográfico importante na vida
das plantas por representar a união da polinização com o recrutamento que levará
ao estabelecimento de novas plantas adultas (Harper, 1977). Em união com a
polinização, a dispersão das sementes é um ponto chave para o movimento dos
genes das plantas. Sementes novas apresentadas em uma população não
representam somente novos indivíduos, mas também genótipos distintos. Em geral
sementes dispersas pelo vento prevalecem em florestas secas e a dispersão por
zoocoria ganha mais importância em florestas úmidas (Howe & Smallwood, 1982).
A dispersão das sementes por zoocoria tem grande potencial para influenciar
os padrões de fluxo gênico e a estrutura genética garantindo a viabilidade genética
das populações (Sork et al., 1999; Jordano & Godoy, 2002). Segundo Parron e
colaboradores, (2015) a dispersão de sementes por animais (zoocoria) está entre os
processos naturais mais importantes para que outros processos ambientais existam,
sendo considerado um serviço de suporte.
Os primatas são importantes dispersores de sementes de muitas espécies
frutíferas (Rímoli et al, 2008). A qualidade da dispersão depende do local e do
padrão de deposição das sementes (Howe et al., 1985). Os frutos possuem
características próprias como variação de tamanho, variações de formas e cores que
podem servir para atrair o predador. Assim as sementes menores possuem
vantagens na dispersão quando comparada com sementes maiores devido às
sementes menores possuírem maiores chances de serem engolidas e defecadas
intactas do que as maiores onde o animal retira um pedaço da semente que pode
danificar o embrião e assim não ocorrer à germinação (Lucas & Corlett, 1998;

14
Norconk et al., 1998). Primatas de uma mesma espécie podem dispersar ou predar
sementes de uma mesma espécie de planta em épocas diferentes, dependendo da
disponibilidade de recursos alimentares na época (Gautier Hion et al., 1993; Kaplin
et al., 1998).
1.2 MATA ATLÂNTICA E PRIMATAS DO JBRJ
Da Mata Atlântica nos dias atuais restam apenas 8,5% de remanescentes
florestais acima de 100 hectares do que existia originalmente (SOS Mata Atlântica,
2015). No estado do Rio de Janeiro, a Mata Atlântica equivale a 17% de sua
cobertura original (Rocha et al. 2003). De acordo com a Secretaria Municipal de
Meio Ambiente – SMAC (2016), o município do Rio de Janeiro está todo inserido no
bioma Mata Atlântica, com paisagem de elevações montanhosa cobertas de
vegetação florestal.
Hoje, a Mata Atlântica é considerada um hot spot mundial, devido à sua
grande biodiversidade e por ser considerada ameaçada. Sua composição é um
mosaico de vegetações definidas como florestas ombrófilas densas, abertas e
mistas; florestas estacionais deciduais e semideciduais; campos de altitude,
mangues e restingas (SOS Mata Atlântica, 2015). A Mata Atlântica ocupa o território
do município do Rio de Janeiro o valor de 35.290 ha, ou seja, 28,9% são formados
por Floresta Ombrófila Densa, Restinga, Manguezal (SMAC, 2016).
Na Mata Atlântica, cerca de 90% das árvores produzem frutos carnosos
capazes de atrair animais dispersores. Essas árvores dependem de animais para a
dispersão de suas sementes (Howe & Smallwood, 1982; Jordano & Schupp, 2000).
De acordo com SOS Mata Atlântica (2015), na Mata Atlântica vivem mais de 20 mil
espécies de plantas, sendo 8 mil endêmicas, 270 espécies conhecidas de
mamíferos, 992 espécies de pássaros, 197 répteis, 372 anfíbios e 350 peixes.
O Jardim Botânico do Rio de Janeiro (JBRJ) possui uma área de vegetação
natural situada nas encostas da Mata Atlântica adjacente ao arboreto e pertencente
ao complexo montanhoso que compõe o Maciço da Tijuca (Reis, 2008). Assim, o
JBRJ tem importância significativa, por ser um local que abriga um remanescente
florestal de Mata Atlântica (Kato, 2013). Das espécies nativas, em se tratando de
primatas, ocorre a espécie Sapajus nigritus (Goldfuss, 1809), conhecido

15
popularmente como macaco-prego (Silva Junior, 2001).
O macaco-prego (S. nigritus) é endêmico da Mata Atlântica e ocorre no
Sudeste e Sul do Brasil, desde a margem direita do rio Doce (MG e ES) até o Rio
Grande do Sul, e na Argentina na região nordeste (Silva Junior, 2005; Vilanova et
al., 2005). São animais de porte médio, possuindo peso médio para adultos de 3,2
Kg (machos) e 2,3 Kg (fêmeas) (Reis et al, 2008), tem hábitos diurnos, vivem em
grupos sociais, podendo conter vários indivíduos, entre machos e fêmeas jovens,
adultos e infantes e um macho alfa (Freese & Oppenheimer, 1981; Izawa, 1990).
Locomovem-se com grande facilidade em troncos e galhos de árvores,
principalmente para seu forrageio (Silva Junior, 2001).
A espécie alimenta-se de frutos, insetos, flores, pequenos vertebrados,
néctar, brotos e ovos de pássaros. Possui a capacidade de manipulação e destreza
manual utilizando ferramentas para acessar objetos indisponíveis (Reis et al, 2008).
O macaco-prego está diretamente relacionado a dois importantes processos
ecológicos, a dispersão de sementes e o controle populacional de insetos porque
que consomem em grande quantidade desses itens (Parron et al, 2015). Entre os
mamíferos de médio porte os macacos-prego são capazes de sobreviver adaptando-
se em pequenos fragmentos de floresta (Chiarello, 1999), porque possuem uma
grande capacidade de aprendizado e plasticidade comportamental, que permitem
sua adaptação a ambientes antropizados, com alguma cobertura florestal (Ludwig et
al., 2005; Rocha, 2000; Vidolin & Mikich, 2004). Segundo Ludwig e colaboradores
(2005), esse primata pode ser um dos poucos mamíferos provedores de alguns
serviços ambientais de suporte e de provisão em áreas degradadas ou produtivas.
Outros primatas que compõe a fauna da Mata Atlântica são o Callithrix
jacchus (Linnaeus, 1758) e o Callithrix penicillata (E. Greoffroy 1812), conhecidos
popularmente como “sagui” que são consideradas espécies invasoras, por terem
sido introduzidas nas regiões sudeste e sul através do tráfico de animais (Rylands &
Faria, 1993). Segundo a Convenção Internacional sobre Diversidade Biológica –
CDB (1992) na 6ª Conferência das Partes (CDB COP-6, Decisão VI/23, 2002),
Espécie Exótica Invasora é toda espécie que se desenvolve fora da sua área de
origem (distribuição natural) e que pode afetar de forma negativa ecossistemas,
habitats ou espécies. Espécies exóticas representam um risco para espécies

16
nativas, devido à predação, competição, hibridação e transmissão de agentes
patogênicos (Rangel, 2010). As duas espécies invasoras de primatas, o C. jacchus e
o C. penicillata e os híbridos, que são o resultado do cruzamento das duas espécies
exóticas (Rangel, 2010), são encontradas no Jardim Botânico do Rio de Janeiro
(JBRJ).
Callithrix jacchus, também conhecido como “sagui-do-nordeste”, tem sua
distribuição original pelos estados do Rio Grande do Norte, Alagoas, Pernambuco,
Paraíba, Ceará e Piauí, com uma variedade de tipos de florestas, ocupando os
biomas de Mata Atlântica e Caatinga. (Rylands, 1996; Reis et al, 2008). Callithrix
penicillata, também conhecido como “sagui-de-tufos-pretos” ou “mico-estrela” tem
sua distribuição desde os estados do Maranhão e sudeste do Piauí ate o norte de
São Paulo, incluindo a maior parte do estado da Bahia, Minas Gerais e Goiás,
ocupando o bioma do cerrado (Rylands, 1996). As duas espécies podem conviver,
formando grupos híbridos que podem gerar descendentes férteis (Rangel, 2010). De
acordo com Coimbra-Filho e colaboradores (1993) tal fato foi observado em cativeiro
e em vida livre. Callithrix sp. são animais diurnos e vivem em grupos, usam galhos
de árvores para se protegerem contra intempéries e predadores. São onívoros por
consumirem em grande quantidade uma variedade de itens alimentares e gomívoros
por consumirem exsudatos das árvores (Rangel, 2010). Alimentam-se também de
frutos, flores, néctar, insetos, aranhas, caramujos, lagartos, anuros, ovos e filhotes
de aves (Digby & Barreto, 1998; Maier et al, 1982; Rizzini & Coimbra-Filho, 1981;
Rothe, 1999; Stevenson & Rylands, 1988).
Callithrix jacchus e C. penicillata são espécies bem adaptadas a florestas
secundarias por terem uma grande versatilidade na dieta e sistema de acasalamento
eficaz possuindo uma taxa alta de reprodução e poucos predadores nas áreas que
foram introduzidas. Por isso os grupos das duas espécies e híbridos são bem
distribuídas no Estado do Rio de Janeiro. De acordo com Rangel (2010), a
população de saguis que vive no JBRJ é numerosa e em crescimento. Danificam a
coleção do JBRJ, interagem com funcionários e visitantes e competem por alimento
com outras espécies nativas.
Primatas macaco-prego (S nigritus) e os saguis (C. jacchus, C. penicillata e
híbridos) são residentes e frequentadores do JBRJ e da área de Mata Atlântica local

17
remanescente do Jardim Botânico, também conhecido como Serra da Carioca (Reis,
2008). Por consumirem alimentos de origem vegetal (Reis, et al 2008) podem
prejudicar a coleção científica de plantas vivas do JBRJ e a área de mata
remanescente da instituição. Danos podem acontecer de várias formas: ao
predarem e introduzirem por dispersão de sementes espécies da Mata adjacente na
coleção viva do JBRJ como também podem dispersarem espécies exóticas da
coleção do JBRJ na Mata Atlântica contígua. Diante do exposto algumas questões
podem ser levantadas: esses primatas, macaco-prego (S. nigritus) e o sagui (C.
jacchus, C. penicillata e híbridos) são dispersores viáveis? Quais seriam as espécies
vegetais exóticas ou nativas dispersadas? Essa dispersão tem potencial de impacto
no JBRJ e na Mata Atlântica remanescente?
A importância ecológica da pesquisa reside no fato de serem escassas as
informações sobre a dispersão de sementes por primatas residentes e
frequentadores do JBRJ e da Mata Atlântica contígua. Com esse estudo será
verificado e analisado se os primatas macacos-prego e sagui são reais dispersores
de sementes e quais espécies eles estariam dispersando. Além disso, pretende-se
avaliar se a dispersão pode ser prejudicial ao JBRJ ou a Serra da Carioca, causando
danos à coleção ou à mata contígua ao dispersarem espécies exóticas à Flora local.
2. HIPÓTESE
O macaco-prego (Sapajus nigritus) e os saguis (Callithrix jacchus, Callithrix
penicillata e híbridos) são reais dispersores de sementes exóticas e nativas no JBRJ
e na Mata Atlântica. Assim, alteram a coleção viva do Jardim Botânico dispersando
espécies que não fazem parte da coleção botânica e promovem a dispersão de
plantas exóticas, presentes na referida coleção, na Serra da Carioca, introduzindo
espécies exóticas e com possível potencial invasor na mata.

18
3. OBJETIVO GERAL
O objetivo do presente trabalho é verificar e analisar a possibilidade de
dispersão de sementes por primatas (Sapajus nigritus, Callithrix jacchus, Callithrix
penicillata e híbridos) residentes ou visitantes no Instituto de Pesquisas Jardim
Botânico do Rio de Janeiro (JBRJ), através das fezes dos animais, tanto no JBRJ
quanto em mata contígua.
3.1 OBJETIVOS ESPECÍFICOS:
Para alcançar o objetivo geral pretende-se:
- Verificar a presença de sementes nas fezes dos primatas;
- Testar a germinação e viabilidade das sementes encontradas, quando em número
suficiente nas amostras;
- Identificar os propágulos e/ou sementes das espécies germinadas e não
germinadas;
- Listar espécies nativas e exóticas encontradas nas amostras de fezes.
- Listar espécies de frutos que foram observados por trabalhos científicos do Núcleo
da Fauna do JBRJ e não foram encontradas sementes e/ou propágulos nas
amostras de fezes.

19
4. METODOLOGIA
4.1 ÁREA DE ESTUDO
O estudo foi realizado no arboreto do Instituto de Pesquisas Jardim Botânico
do Rio de Janeiro (JBRJ), localizado na Rua Jardim Botânico, número 1008, Rio de
Janeiro, RJ (22º57’a 22º59’S e 43º13’a 43º14’W) (Figura 1). Sua área abrange 137
hectares, sendo 54 hectares de arboreto plantado com espécies do mundo todo e 83
hectares de remanescentes da Floresta Atlântica (Mendonça et al., 2004).
Figura 1: Vista aérea do Jardim Botânico do Rio de Janeiro - JBRJ. Crédito Google Earth, acesso em 19 de novembro de 2016.
O Instituto de Pesquisa do Jardim Botânico foi fundado em 13 de junho de
1808 pelo príncipe regente D. João que decidiu instalar no local uma fábrica de
pólvora e um jardim para aclimatização de espécies vegetais originárias de outras
partes do mundo. De acordo com Bediaga e colaboradores (2008), o desafio foi
aclimatar as espécies botânicas que eram consideradas especiarias do Oriente:
baunilha, canela, pimenta e outras. Assim, o JBRJ foi um local de experiências com

20
vegetais enviados de províncias portuguesas. Em geral, aclimatizar uma espécie de
planta significava que o Jardim teria que aperfeiçoar o transporte das sementes e
mudas, construir viveiros para semeá-las e finalmente transplantar os vegetais para
o solo e observar a necessidade de sol, sombra, água e etc. (Bediaga et al. 2008).
Em 21 de abril de 1821, D. João VI retornou para Portugal e seu filho D.
Pedro I, ao assumir o trono, deu prosseguimento às obras iniciadas pelo pai. Em
1824, nomeou seu primeiro diretor botânico, Frei Leandro do Sacramento que era
carmelita e professor de botânica na Academia de Medicina e Cirurgia (Lavôr, 1983).
Frei Leandro do Sacramento aumentou a área cultivada, aterrou alguns locais,
traçou uma cascata, construiu um lago, projetou aléias, inaugurou um relógio do sol,
plantou mangueiras, nogueiras, longanas, pitombas e etc. (Lavôr, 1983).
Atualmente, o Jardim Botânico do Rio de Janeiro (JBRJ), fora das estufas,
possui uma área cultivada cerca de 9 mil exemplares botânico pertencentes a cerca
de 1500 espécies, sendo que apenas 30% representa a flora nacional (Reis, 2008).
Nos canteiros predominam espécies de porte arbóreo, sendo Leguminosae,
Myrtaceae, Palmae (Arecaceae) e Bignoniaceae as famílias botânicas mais bem
representadas. Já nas estufas e nas coleções temáticas a realidade se inverte. Por
exemplo, as coleções de orquídeas e de bromélias juntas somam mais de 8 mil
vasos e mais de 1000 espécies, a maioria absoluta brasileira e obtida em
expedições botânicas realizadas por pesquisadores da Casa nos últimos 40 anos
(Reis, 2008). Bignoniaceae, Passifloraceae e Convolvulaceae são as famílias de
trepadeiras mais encontradas no JBRJ.
A fauna dentro do JBRJ é bastante diversificada. Existem aqueles animais
que são vistos com mais facilidade, que é o caso dos macacos-prego (Sapajus
nigritus), caxinguelês (Guerlinguetus ingrami) e dos saguis (Callithrix jacchus,
Callithrix penicillata e híbridos), 200 espécies aves (dentre as migratórias, residentes
e aquelas que nunca pousam, mas passam pelo alto), alguns répteis como o
calango (Tropidurus torquatus), teiú (Salvator merianae), jiboias (Boa constrictor) e
cobras verdes (Liophis sp), espécies de anfíbios, peixes e muitas espécies de
artrópodes (Nunan, 2015). Apesar da maioria dos animais não serem vistos com
tanta facilidade, existem registros de mamíferos de médio e pequeno porte que
habitam o Jardim, como cuícas (Caluromys philander e Monodelphis americana),

21
gambás (Didelphis aurita), ouriços-caxeiros (Coendou villosus), mãos-peladas
(Procyon cancrivorus), preguiças (Bradypus variegatus), tamandua-mirím
(Tamandua tetradactyla), pacas (Cuniculus paca), quatis (Nasua nasua), lontra
(Lutra sp.), diversas espécies de morcegos e capivara (Hydrochaeris hydrochaeris)
(Rangel, 2005).
O JBRJ abriga também o Núcleo de Conservação da Fauna, que tem como
objetivos o atendimento de socorro à fauna de vertebrados presentes no território do
JBRJ, a identificação taxonômica, observação de animais silvestres em vida livre
(comportamento e/ou dieta e/ou ecologia) para desenvolvimento de pesquisas
científicas e ainda colabora para a orientação ao público e funcionários em relação
ao correto comportamento com a fauna (Núcleo de Conservação da Fauna, 2016).
Este núcleo é lotado regimentalmente na Coordenação de Conservação de Áreas
Verdes / Diretoria de Ambiente e Tecnologia.
4.2 DELINEAMENTO AMOSTRAL
A pesquisa foi realizada com grupos de primatas macacos-prego (S. nigritus)
e de saguis (C. jacchus, C. penicillata e hibridos), residentes e frequentadores do
JBRJ (Figura 2). De acordo com núcleo de Conservação Fauna, (2016) estima-se
que o grupo de macaco-prego possui em torno de 30 indivíduos entre machos e
fêmeas, adultos, jovens e filhotes. Nesse caso optou-se por usa o grupo todo. A
coleta de fezes dos macacos-prego foi feita através de busca ativa e
acompanhamento do grupo. Para a coleta de fezes de sagui foram utilizadas três
plataformas feitas de bambu suspensas nas árvores. Essas plataformas são
cevadas com bananas e armadilhas modelo tomahawk foram colocadas para
captura dos indivíduos e, assim, a coleta das fezes foi realizada nos animais que
frequentam as plataformas e/ou nos que são capturados.

22
Figura 2: (A) indivíduo de Sapajus nigritus (macaco-prego), (B) indivíduo de Callithrix
sp. (sagui) no Jardim Botânico do Rio de Janeiro – JBRJ (Fotos de Alexandre Machado).
Existem cerca de 12 grupos mistos de Callithrix spp. no arboreto do Jardim
Botânico do Rio de Janeiro (Rangel et al., 2009). Para a coleta de fezes de saguis
foram selecionados três grupos de áreas diferentes (Figura 3). Os três grupos
possuem entre 5 a 7 indivíduos Essas plataformas estão localizadas nos canteiros
32C, orquidário e Região Amazônica (o arboreto do Jardim Botânico é dividido em
sessões e canteiros), dentro do JBRJ (Núcleo de Conservação da Fauna, 2016). As
áreas selecionadas para colocação das plataformas foram locais onde já existia
acompanhamento dos grupos para outras pesquisas. A maioria dos indivíduos de
saguis é híbrida, não sendo possível a separação entre espécies, portanto, no atual
estudo adotaremos a identificação como Callithrix sp.

23
Figura 3: Mapa do Jardim Botânico do Rio de Janeiro – JBRJ com a localização das plataformas de capturas de fezes de sagui (Canteiro 32C, Orquidário e Região Amazônica).
As amostras coletadas foram armazenadas em potes de vidros e colocadas
na geladeira para conservação das mesmas. As amostras de fezes desses primatas
foram filtradas através de gaze dobrada em cinco partes, 100 ml de soro e um funil
pequeno de plástico (Figura 4). A filtragem realiza separação das partículas com
maior granulometria das fezes e permite a separação dos propágulos e/ou
sementes, que foram analisados na lupa. Por fim as amostras foram triadas na lupa
(modelo MoticMicroscops smz168), separando possíveis propágulos/sementes para
identificação e germinação (Figura 5).
A coleta e verificação da presença dos propágulos e/ou sementes nas
amostras de sagui e macaco-prego foram divididas entre estações secas (outono e
inverno) e úmidas (primavera e verão). As amostras de sagui foram divididas entre:
Estação seca¹: abril a setembro de 2015 e estação seca²: abril a setembro de 2016;
Estação úmida: entre outubro de 2015 a março de 2016. As amostras de sagui
foram dividas dessa forma, para que facilitasse a coleta, a identificação das
amostras e por terem uma quantidade maior de amostras de saguis do que de
macaco-prego. As amostras de macaco-prego foram divididas entre: Estação
úmida¹: dezembro de 2014 a março de 2015; Estação seca¹: abril a setembro de

24
2015; Estação úmida²: outubro de 2015 a março de 2016 e Estação seca²: abril a
setembro de 2016. O grupo de macaco-prego obteve-se mais dificuldade para a
coleta de fezes, devido os mesmos não possuir ponto fixo e se locomovem por todo
o Jardim Botânico e mata contigua.
As amostras de sementes foram distribuídas em uma caixa plástica
transparente do tipo Gerbox, com papel filtro umedecido com água destilada para
que germinasse. As caixas foram tampadas com a identificação na tampa ou na
parte de baixo do recipiente. As sementes foram expostas à luz artificial em
períodos de 8-16 horas luz em uma estufa de B.O.D a 25 °C para geminação. As
sementes colocadas para germinar foram umedecidas com água destilada três
vezes por semana. A identificação das sementes e/ou propágulos foi realizada com
auxilio de um especialista em sementes e pesquisas bibliográficas. As amostras que
possuíam frutos, como aquênios, e/ou tinham pouca quantidade de sementes e/ou
propágulos (1 a 2) não foram semeadas, a fim de não comprometer uma possível
identificação no futuro, com a ajuda de especialista.
Figuras 4: (A) Becker com a medida do soro e funil pequeno com gaze. (B) Filtragem das amostras de fezes. Núcleo de Conservação da Fauna do Jardim Botânico do Rio de Janeiro – JBRJ. (Fotos: Equipe Fauna – JBRJ, 2016).

25
Figura 5: (A) Plataforma de ceva e acompanhamentos com armadilhas. (B) Estufas B.O.D. (C) Separação de sementes na lupa. (D) Laboratório de sementes do JBRJ. (Fotos: Jehnnifer Mendonça e Simone Nogueira, 2015-2016).
Para cada semente e/ou propágulo identificado foi feita uma pesquisa
bibliográfica (Flora do Brasil) para o reconhecimento se planta nativa ou exótica.
Foram consultados trabalhos científicos desenvolvidos por voluntários do Núcleo de
Conservação da Fauna no JBRJ para listar outros frutos que os primatas
consomem, mas que no presente trabalho não foram encontrados sementes ou
propágulos nas fezes desses animais.

26
5. RESULTADOS
5.1 AMOSTRAS DE FEZES DE SAGUI
Foram coletadas um total de 36 amostras de fezes de saguis durante o
período de abril de 2015 a setembro de 2016 e numeradas S001 a S036. Nas
amostras foram encontradas várias estruturas, entre elas, propágulos (frutos com
sementes), sementes e fragmentos de insetos (Tabelas 3; 4 e 5 – anexo).
No período de estação seca1 foram analisadas 10 amostras (Tabela 3 –
Anexo). Na amostra S001 registrou-se um (01) aquênio de Cyperaceae. Nas
amostras S002, S003, S006 e S007 não foram encontradas sementes, somente
outras estruturas. Nas amostras S004 e S005 foram encontradas sementes
pertencentes à família Rubiaceae (Figura 9). Das 56 sementes encontradas na
amostra S004 40 sementes foram semeadas, germinando 39 em 8-10 dias após a
semeadura. Na amostra S005 foram encontradas 34 sementes, destas 20 foram
semeadas. Quinze (15) plântulas foram obtidas após 8-10 dias. Na amostra S008
foram encontradas seis sementes, que foram semeadas, contudo não houve
germinação, dificultando a identificação. Já na amostra S009 foram encontrados 10
sementes, semeadas e identificadas como Rubiaceae. Somente quatro propágulos
germinaram nesta amostra após 8-10 dias. Na S010 não foram encontrados
sementes e/ou propágulos.
Já na estação úmida foram coletadas e analisadas 10 amostras (Tabela 4 e
10 – em anexos). Na amostra S011 foram encontrados 12 propágulos identificados
como Miconia sp. (Melastomataceae), seis foram semeados e após 13 dias obteve-
se duas plântulas. Foi encontrado somente um (1) propágulo (aquênio) de
Cyperaceae na amostra S012. Foram encontradas dois tipos de sementes na
amostra S013. Dez (10) sementes de Clidemia sp. (Melastomataceae) foram
encontradas na amostra S013A, semeadas e germinadas três sementes entre 8-10
dias. Já na amostra S013B foram encontradas 83 sementes de Miconia albicans.
Quarenta (40) sementes foram semeadas desta amostra e germinaram 23 sementes
após 8-10 dias. Nas amostras S014, S015, S017, S018, S019 e S020 não foram
encontradas sementes ou propágulos apenas outras estruturas, entre elas,
fragmentos de inseto e goma Enquanto na amostra S016 foram encontradas 33
sementes de uma espécie de Rubiaceae. Vinte sete (27) foram semeadas, mas não
27
houve germinação.
Na estação seca2 foram coletadas e analisadas 16 amostras (Tabela: 5 –
Anexos). Nas amostras S021, S022, S023, S024, S028, S029, S030, S031, S032,
S033, S035 e S036 não foram encontradas sementes ou propágulos. Foi encontrada
na amostra S025 uma (1) semente, porém não se obteve identificação tampouco
germinação. Um aquênio de Cyperaceae foi encontrado na amostra S026. Na
amostra seguinte (S027) foram encontradas 35 sementes de Cecropia sp. Após
semear 27 sementes não houve germinação. Uma semente de Fabaceae foi
encontra na amostra S034, semeada, porém não obteve germinação.
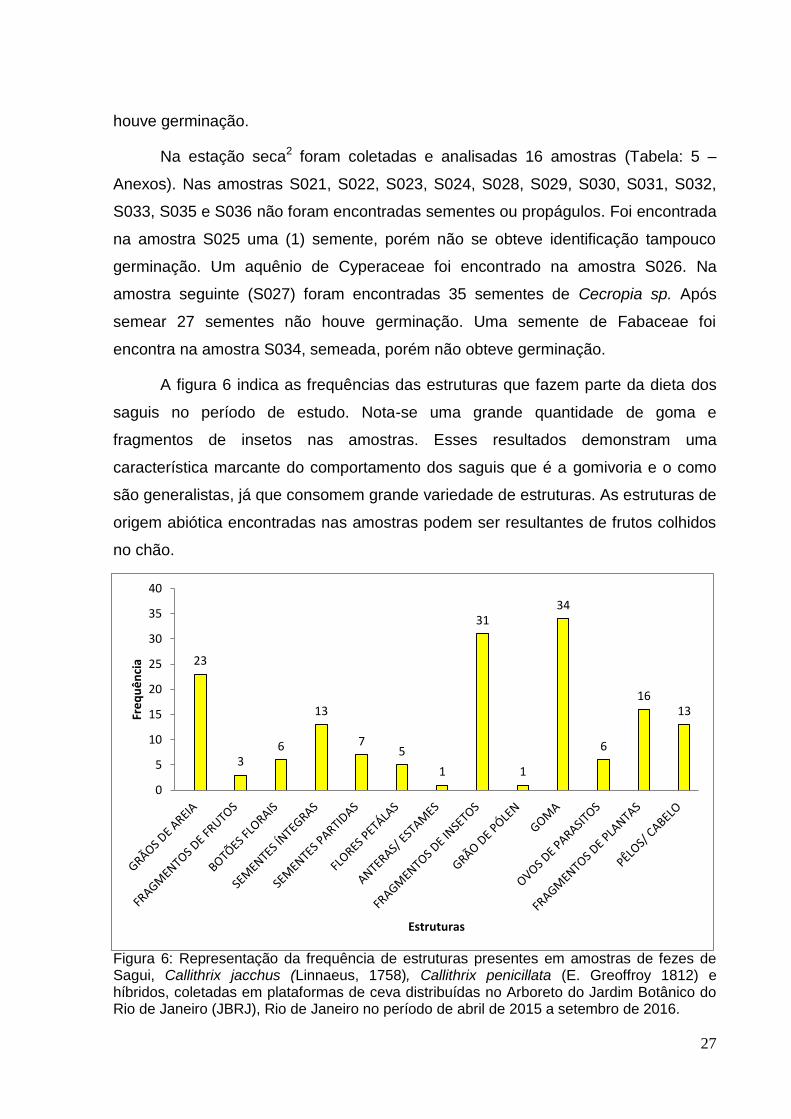
A figura 6 indica as frequências das estruturas que fazem parte da dieta dos
saguis no período de estudo. Nota-se uma grande quantidade de goma e
fragmentos de insetos nas amostras. Esses resultados demonstram uma
característica marcante do comportamento dos saguis que é a gomivoria e o como
são generalistas, já que consomem grande variedade de estruturas. As estruturas de
origem abiótica encontradas nas amostras podem ser resultantes de frutos colhidos
no chão.
Figura 6: Representação da frequência de estruturas presentes em amostras de fezes de Sagui, Callithrix jacchus (Linnaeus, 1758), Callithrix penicillata (E. Greoffroy 1812) e híbridos, coletadas em plataformas de ceva distribuídas no Arboreto do Jardim Botânico do Rio de Janeiro (JBRJ), Rio de Janeiro no período de abril de 2015 a setembro de 2016.
23
3 6
13
7 5
1
31
1
34
6
16 13
0
5
10
15
20
25
30
35
40
Fre
qu
ên
cia
Estruturas
28
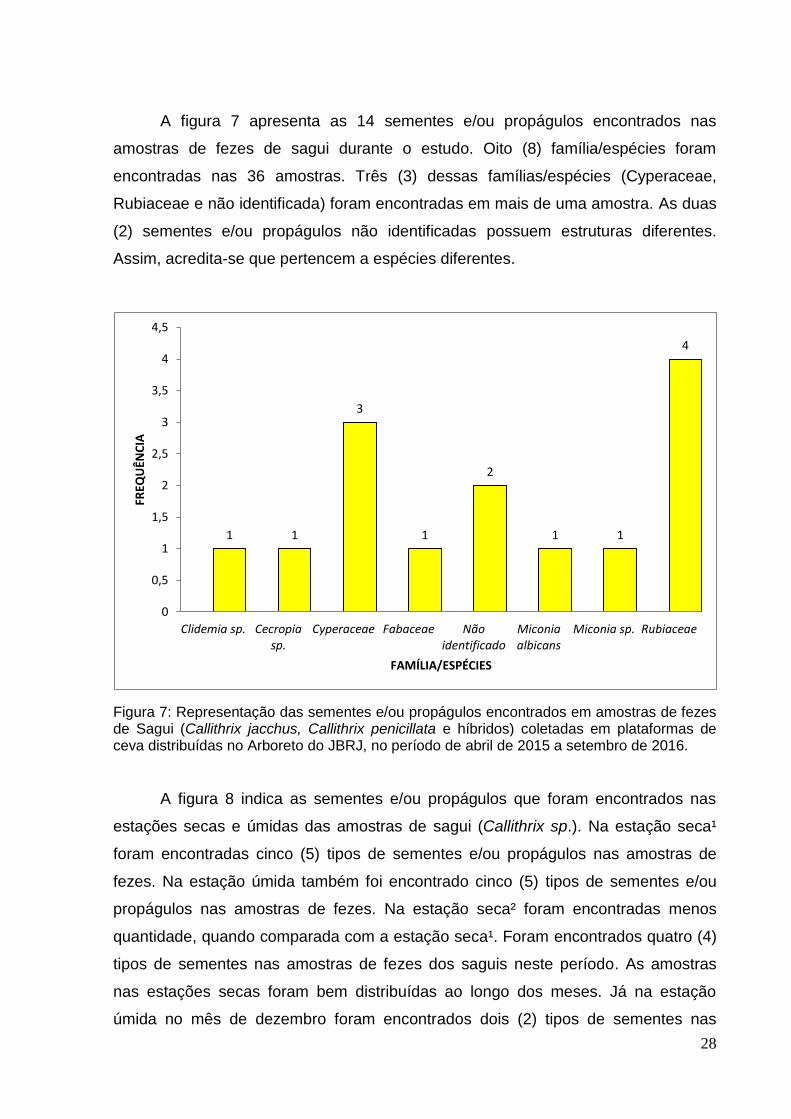
A figura 7 apresenta as 14 sementes e/ou propágulos encontrados nas
amostras de fezes de sagui durante o estudo. Oito (8) família/espécies foram
encontradas nas 36 amostras. Três (3) dessas famílias/espécies (Cyperaceae,
Rubiaceae e não identificada) foram encontradas em mais de uma amostra. As duas
(2) sementes e/ou propágulos não identificadas possuem estruturas diferentes.
Assim, acredita-se que pertencem a espécies diferentes.
Figura 7: Representação das sementes e/ou propágulos encontrados em amostras de fezes de Sagui (Callithrix jacchus, Callithrix penicillata e híbridos) coletadas em plataformas de ceva distribuídas no Arboreto do JBRJ, no período de abril de 2015 a setembro de 2016.
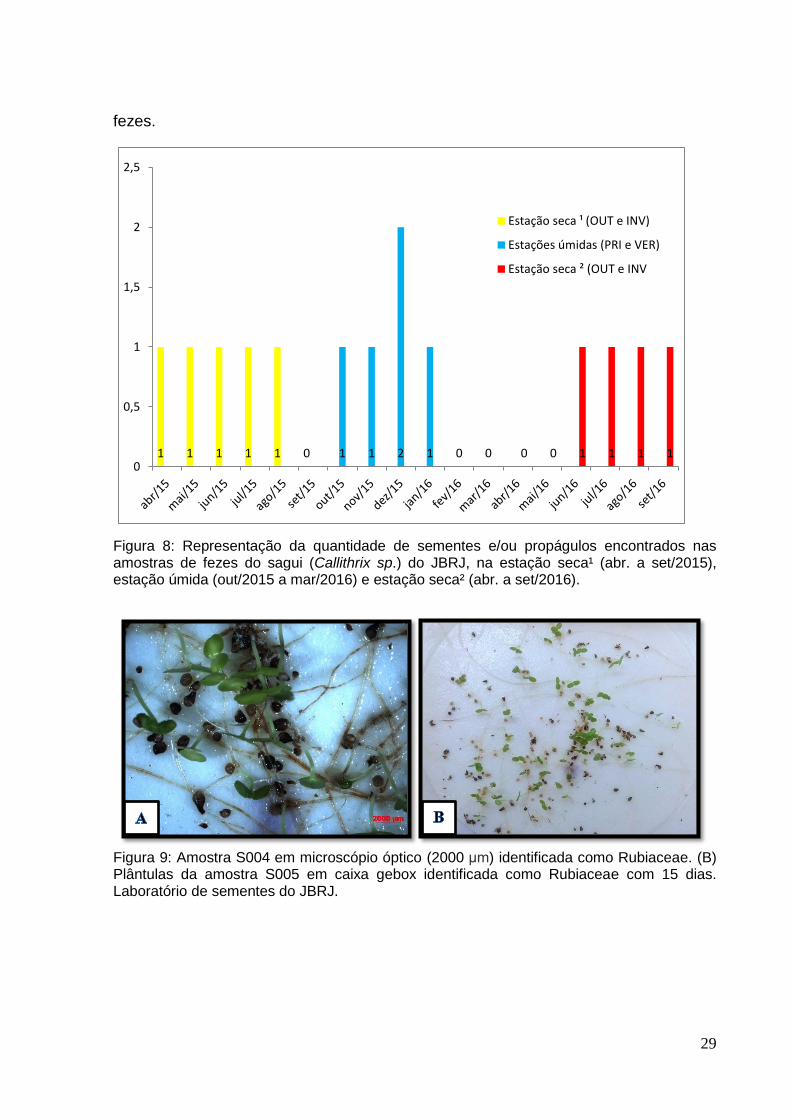
A figura 8 indica as sementes e/ou propágulos que foram encontrados nas
estações secas e úmidas das amostras de sagui (Callithrix sp.). Na estação seca¹
foram encontradas cinco (5) tipos de sementes e/ou propágulos nas amostras de
fezes. Na estação úmida também foi encontrado cinco (5) tipos de sementes e/ou
propágulos nas amostras de fezes. Na estação seca² foram encontradas menos
quantidade, quando comparada com a estação seca¹. Foram encontrados quatro (4)
tipos de sementes nas amostras de fezes dos saguis neste período. As amostras
nas estações secas foram bem distribuídas ao longo dos meses. Já na estação
úmida no mês de dezembro foram encontrados dois (2) tipos de sementes nas
1 1
3
1
2
1 1
4
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
Clidemia sp. Cecropiasp.
Cyperaceae Fabaceae Nãoidentificado
Miconiaalbicans
Miconia sp. Rubiaceae
FREQ
UÊN
CIA
FAMÍLIA/ESPÉCIES
29
fezes.
Figura 8: Representação da quantidade de sementes e/ou propágulos encontrados nas amostras de fezes do sagui (Callithrix sp.) do JBRJ, na estação seca¹ (abr. a set/2015), estação úmida (out/2015 a mar/2016) e estação seca² (abr. a set/2016).
Figura 9: Amostra S004 em microscópio óptico (2000 μm) identificada como Rubiaceae. (B) Plântulas da amostra S005 em caixa gebox identificada como Rubiaceae com 15 dias. Laboratório de sementes do JBRJ.
1 1 1 1 1 0 1 1 2 1 0 0 0 0 1 1 1 1 0
0,5
1
1,5
2
2,5
Estação seca ¹ (OUT e INV)
Estações úmidas (PRI e VER)
Estação seca ² (OUT e INV

30
5.2 AMOSTRAS DE FEZES DE MACACO-PREGO
Foram coletadas um total 25 amostras de fezes de macacos-prego durante o
período de dezembro de 2014 a setembro de 2016, numeradas de P001 a P025.
Nas amostras foram encontradas várias estruturas, entre elas propágulos (frutos
com sementes), sementes e fragmentos de insetos. (Tabela 6; 7; 8 e 9 – em anexo).
Na estação úmida foram coletadas e analisadas cinco (5) amostras de fezes
de macaco-prego. Na primeira amostra de macaco-prego (P001) foram encontradas
16 sementes de Impatiens walleriana. Foram semeadas e germinaram 11 sementes
em 8-10 dias. Nas amostras P002 e P003 não foram encontrados propágulos e/ou
sementes, no entanto, outras estruturas florais foram encontradas. Analisando a
amostra P004 foram encontradas quatro tipos de sementes. Quatro (4) sementes de
Erythoxylum pulchrum foram encontradas na amostra P004A. Desta amostra foi
semeada uma (1) semente, que germinou em 15 dias. Na amostra P004B uma (1)
semente de Rubiaceae. Já na amostra P004C foram encontradas duas sementes de
Cecropia sp. que semeadas não germinaram. Apenas uma (1) semente de
Arecaceae foi encontrada na amostra P004D. Foi encontrada uma (1) semente na
amostra P005 não identificada, que semeada não germinou.
Na estação seca¹ foram coletadas e analisadas cinco (5) amostras de fezes
de macaco-prego. Na P006 não foram encontradas sementes e/ou propágulos.
Enquanto nas amostras P007 e P008 foram encontradas dois tipos de sementes.
Uma (1) semente de Erythoxylum pulchrum na amostra P007A, que foi semeada e
não germinou. Já na amostra P007B foram encontradas 12 sementes de Cecropia
hololeuca, semeadas e germinadas 12 sementes em 15 dias. Um (1) aquênio de
Cyperaceae foi encontrado na amostra P008A. Contudo, na amostra P008B, 56
sementes de Miconia sp foram encontradas. Desta amostra, 44 sementes foram
semeadas e germinam quatro (4) em 8-10 dias. Uma (1) semente de espécie de
Rubiaceae foi encontrada na amostra P009, semeada, mas não houve germinação.
Na amostra seguinte (P010) foram encontrados dois tipos de sementes: Cecropia
sp. na P010A e Rubiaceae na P010B. Seis (6) sementes de Cecropia sp. foram
encontradas, semeadas duas (2), porém não obteve-se germinação. Já na P010B
foram encontradas 68 sementes. Desta amostra 40 sementes de Rubiaceae foram
semeadas, contudo, apenas 23 sementes germinaram em 8-10 dias.
Já na estação úmida² foram coletadas e analisadas sete (7) amostras de

31
fezes de macaco-prego. Nas amostras P011 e P0012 foram encontradas sementes
de Rubiaceae. Entretanto na amostra P011 em menos quantidades (quatro
sementes). Elas foram semeadas e germinaram duas (2) em 8-10 dias. Ao contrário
da amostra P012 que foram encontradas 114 sementes. Destas 114 foram
semeadas 28, germinando 16 sementes em 8-10 dias. Sementes de Myrtaceae e
Solanaceae foram encontradas na amostra P013. Duas (2) Eugenia sp. na amostra
P013A e uma (1) semente de Solanaceae na P013B. Amostras P014 e P015A foram
encontradas sementes não identificadas. Na amostra P014 foram encontradas
sementes em uma boa quantidade (pelo menos 247 sementes). Destas 43 foram
semeadas e germinando 23 sementes. Na amostra P015A foram menos
quantidades (20). Desta amostra 11 sementes foram semeadas, germinando seis (6)
sementes. Já na amostra P015B foram encontradas duas sementes de
Cucurbitaceae. Uma (1) semente foi semeada e germinada em 52 dias. Na amostra
seguinte (P016), não foi encontrada sementes e/ou propágulos, apenas estruturas
florais. Duzentas (200) sementes foram encontradas na amostra P017. Desta
amostra volumosa, 30 sementes foram semeadas e germinaram 26 em 8-10 dias.
Na estação seca² foram coletadas e analisadas oito (8) amostras de fezes de
macaco-prego. Na primeira amostra (P018) de desse período foram encontrados
dois tipos de sementes e/ou propágulos. Três (3) Gramíneae foram encontradas na
amostra P018A. Já na amostra P018B foram encontradas 16 sementes não
identificadas. Desta amostra, 10 foram semeadas não havendo germinação. Duas
(2) sementes de Fabaceae foram encontradas na amostra P019. Na amostra
seguinte (P020) foram achadas 36 sementes não identificadas. Desta amostra,
foram semeadas 27 e gerou três (3) plântulas em 12 dias. Já na amostra P021 foram
encontradas sementes de três espécies (Cecropia sp, Miconia sp. e não
identificada). Na amostra P021A foram encontradas duas (2) sementes de Cecropia
sp. Na P021B foram encontradas duas (2) Miconia sp. Na P021C foram encontradas
quatro (4) sementes não identificadas. Nas amostras P022 e P024 não foram
encontradas sementes e/ou propágulos. Quando analisada a amostra P023 foram
identificadas três sementes de Cecropia sp. Desta amostra duas (2) sementes foram
semeadas e germinadas em 45 dias. Foram encontradas 89 sementes de
Rubiaceae na amostra P025. Desta amostra 36 foram semeadas e germinaram 19
sementes em 12 dias.
32
As amostras P014, P015A, P017, P018B, P020 e P021C acredita-se que as
sementes encontradas pertençam à mesma espécie, devido à semelhança do
material analisado, porém não foi possível a sua identificação. A amostra P005 (não
identificada) possuía estruturas diferentes das demais não identificadas, portanto,
pertence à outra espécie.
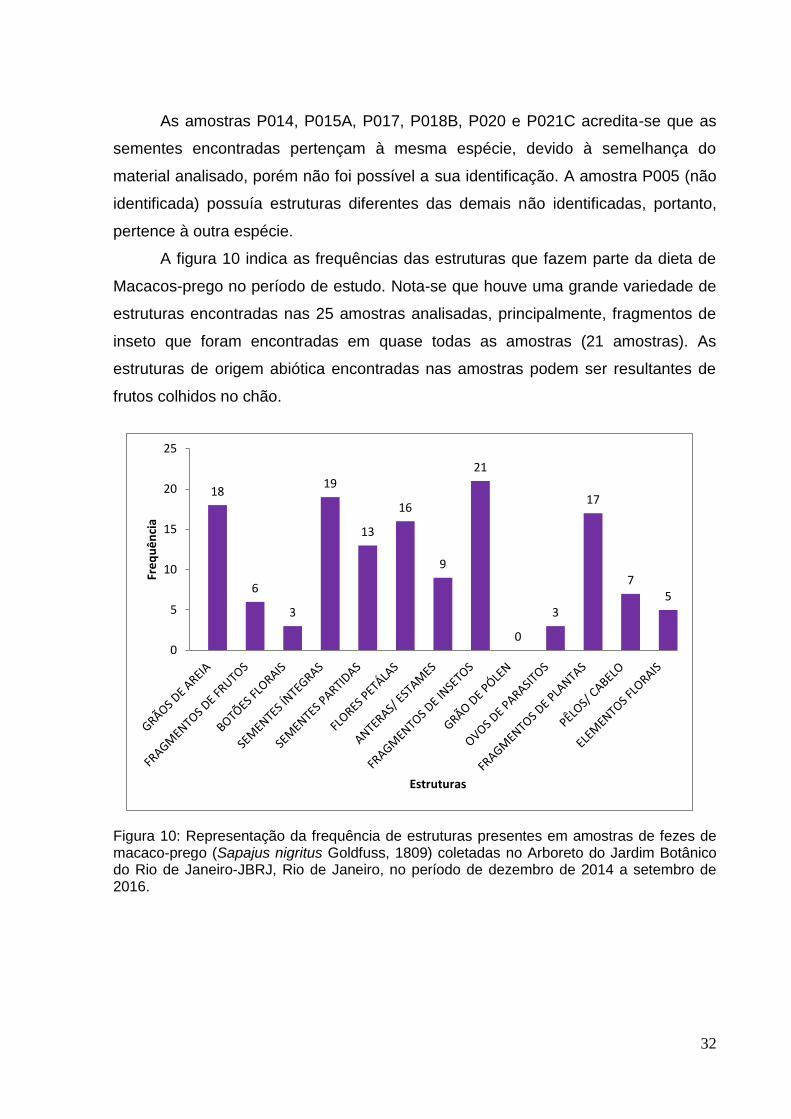
A figura 10 indica as frequências das estruturas que fazem parte da dieta de
Macacos-prego no período de estudo. Nota-se que houve uma grande variedade de
estruturas encontradas nas 25 amostras analisadas, principalmente, fragmentos de
inseto que foram encontradas em quase todas as amostras (21 amostras). As
estruturas de origem abiótica encontradas nas amostras podem ser resultantes de
frutos colhidos no chão.
Figura 10: Representação da frequência de estruturas presentes em amostras de fezes de macaco-prego (Sapajus nigritus Goldfuss, 1809) coletadas no Arboreto do Jardim Botânico do Rio de Janeiro-JBRJ, Rio de Janeiro, no período de dezembro de 2014 a setembro de 2016.
18
6
3
19
13
16
9
21
0
3
17
7 5
0
5
10
15
20
25
Fre
qu
ên
cia
Estruturas
33
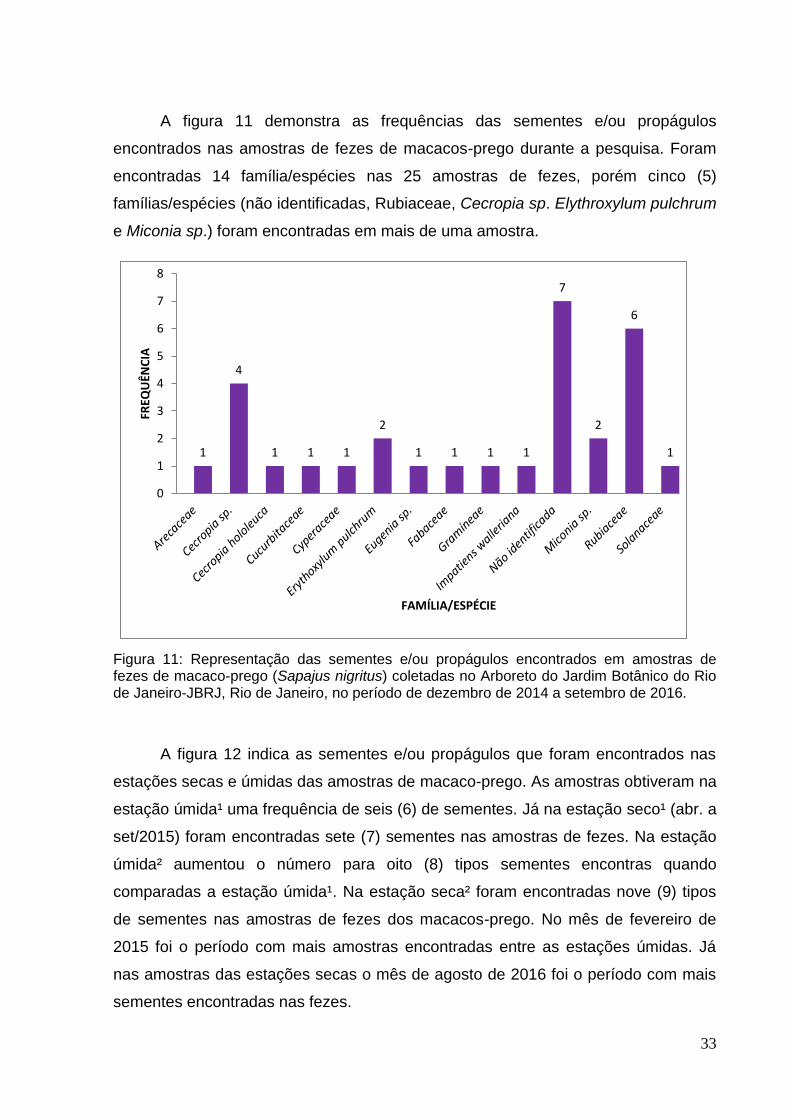
A figura 11 demonstra as frequências das sementes e/ou propágulos
encontrados nas amostras de fezes de macacos-prego durante a pesquisa. Foram
encontradas 14 família/espécies nas 25 amostras de fezes, porém cinco (5)
famílias/espécies (não identificadas, Rubiaceae, Cecropia sp. Elythroxylum pulchrum
e Miconia sp.) foram encontradas em mais de uma amostra.
Figura 11: Representação das sementes e/ou propágulos encontrados em amostras de fezes de macaco-prego (Sapajus nigritus) coletadas no Arboreto do Jardim Botânico do Rio de Janeiro-JBRJ, Rio de Janeiro, no período de dezembro de 2014 a setembro de 2016.
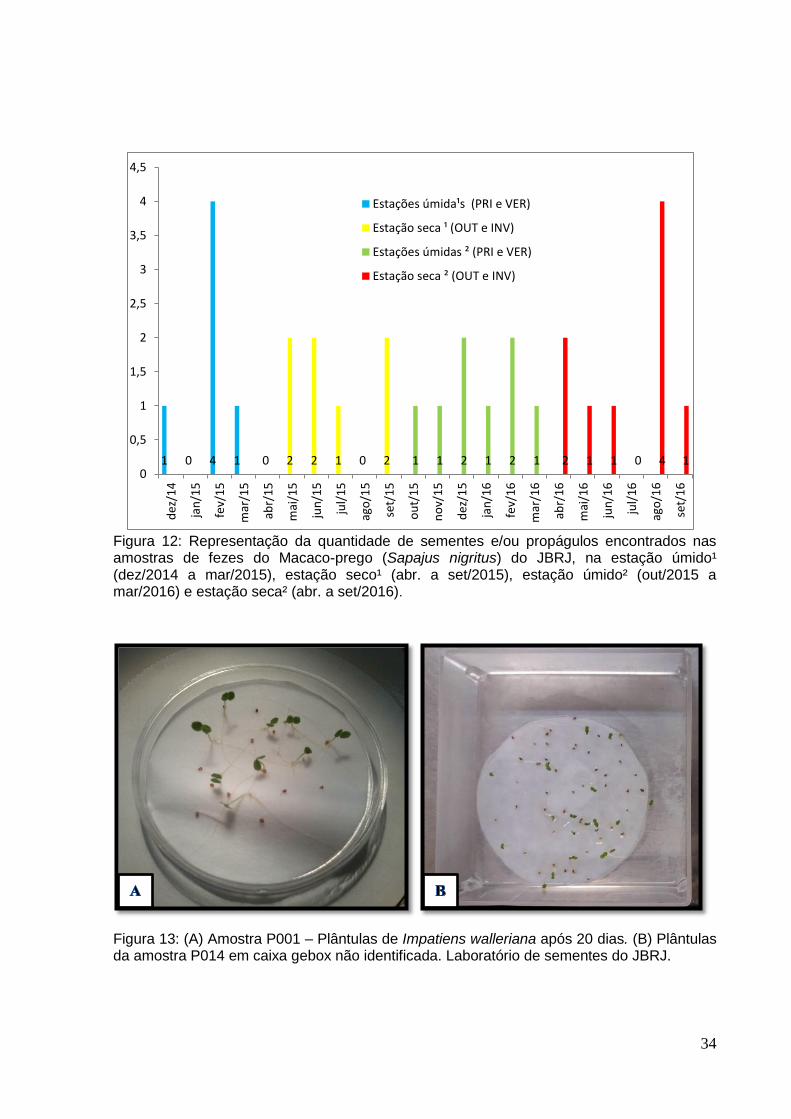
A figura 12 indica as sementes e/ou propágulos que foram encontrados nas
estações secas e úmidas das amostras de macaco-prego. As amostras obtiveram na
estação úmida¹ uma frequência de seis (6) de sementes. Já na estação seco¹ (abr. a
set/2015) foram encontradas sete (7) sementes nas amostras de fezes. Na estação
úmida² aumentou o número para oito (8) tipos sementes encontras quando
comparadas a estação úmida¹. Na estação seca² foram encontradas nove (9) tipos
de sementes nas amostras de fezes dos macacos-prego. No mês de fevereiro de
2015 foi o período com mais amostras encontradas entre as estações úmidas. Já
nas amostras das estações secas o mês de agosto de 2016 foi o período com mais
sementes encontradas nas fezes.
1
4
1 1 1
2
1 1 1 1
7
2
6
1
0
1
2
3
4
5
6
7
8
FREQ
UÊN
CIA
FAMÍLIA/ESPÉCIE
34
Figura 12: Representação da quantidade de sementes e/ou propágulos encontrados nas amostras de fezes do Macaco-prego (Sapajus nigritus) do JBRJ, na estação úmido¹ (dez/2014 a mar/2015), estação seco¹ (abr. a set/2015), estação úmido² (out/2015 a mar/2016) e estação seca² (abr. a set/2016).
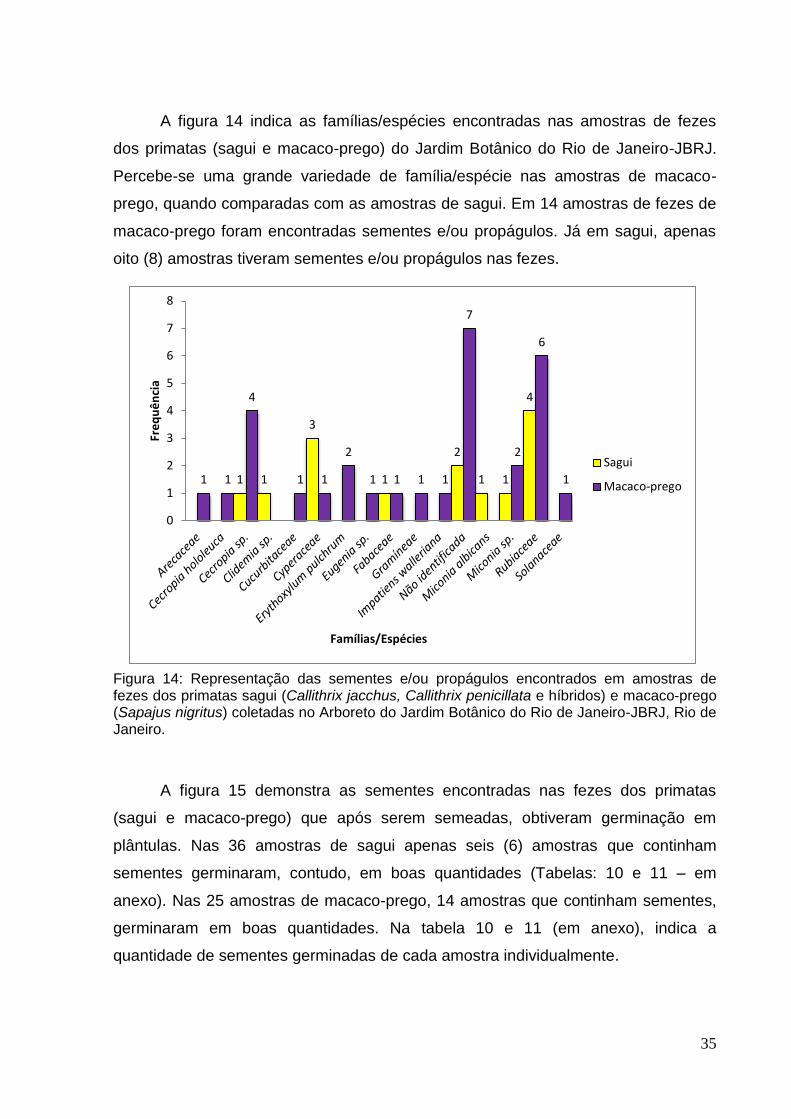
Figura 13: (A) Amostra P001 – Plântulas de Impatiens walleriana após 20 dias. (B) Plântulas da amostra P014 em caixa gebox não identificada. Laboratório de sementes do JBRJ.
1 0 4 1 0 2 2 1 0 2 1 1 2 1 2 1 2 1 1 0 4 1 0
0,5
1
1,5
2
2,5
3
3,5
4
4,5d
ez/1
4
jan
/15
fev/
15
mar
/15
abr/
15
mai
/15
jun
/15
jul/
15
ago
/15
set/
15
ou
t/1
5
no
v/1
5
dez
/15
jan
/16
fev/
16
mar
/16
abr/
16
mai
/16
jun
/16
jul/
16
ago
/16
set/
16
Estações úmida¹s (PRI e VER)
Estação seca ¹ (OUT e INV)
Estações úmidas ² (PRI e VER)
Estação seca ² (OUT e INV)
35
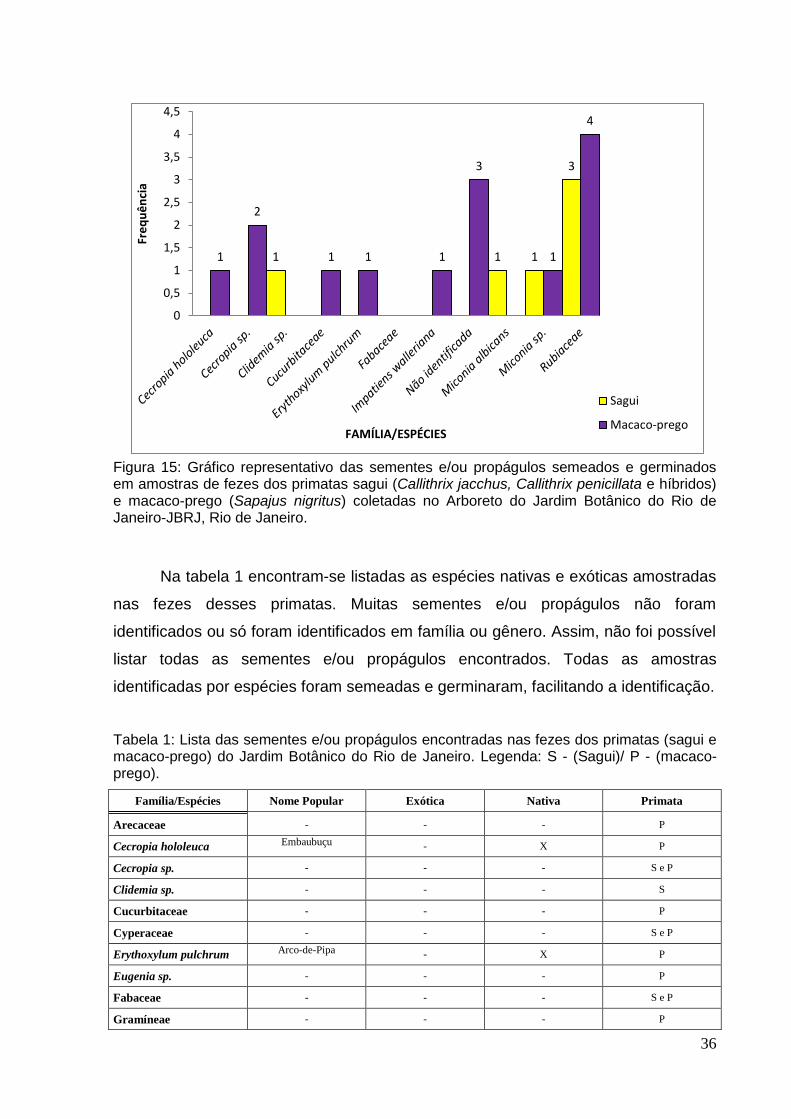
A figura 14 indica as famílias/espécies encontradas nas amostras de fezes
dos primatas (sagui e macaco-prego) do Jardim Botânico do Rio de Janeiro-JBRJ.
Percebe-se uma grande variedade de família/espécie nas amostras de macaco-
prego, quando comparadas com as amostras de sagui. Em 14 amostras de fezes de
macaco-prego foram encontradas sementes e/ou propágulos. Já em sagui, apenas
oito (8) amostras tiveram sementes e/ou propágulos nas fezes.
Figura 14: Representação das sementes e/ou propágulos encontrados em amostras de fezes dos primatas sagui (Callithrix jacchus, Callithrix penicillata e híbridos) e macaco-prego (Sapajus nigritus) coletadas no Arboreto do Jardim Botânico do Rio de Janeiro-JBRJ, Rio de Janeiro.
A figura 15 demonstra as sementes encontradas nas fezes dos primatas
(sagui e macaco-prego) que após serem semeadas, obtiveram germinação em
plântulas. Nas 36 amostras de sagui apenas seis (6) amostras que continham
sementes germinaram, contudo, em boas quantidades (Tabelas: 10 e 11 – em
anexo). Nas 25 amostras de macaco-prego, 14 amostras que continham sementes,
germinaram em boas quantidades. Na tabela 10 e 11 (em anexo), indica a
quantidade de sementes germinadas de cada amostra individualmente.
1 1
3
1
2
1 1
4
1 1
4
1 1
2
1 1 1 1
7
2
6
1
0
1
2
3
4
5
6
7
8
Fre
qu
ên
cia
Famílias/Espécies
Sagui
Macaco-prego
36
Figura 15: Gráfico representativo das sementes e/ou propágulos semeados e germinados em amostras de fezes dos primatas sagui (Callithrix jacchus, Callithrix penicillata e híbridos) e macaco-prego (Sapajus nigritus) coletadas no Arboreto do Jardim Botânico do Rio de Janeiro-JBRJ, Rio de Janeiro.
Na tabela 1 encontram-se listadas as espécies nativas e exóticas amostradas
nas fezes desses primatas. Muitas sementes e/ou propágulos não foram
identificados ou só foram identificados em família ou gênero. Assim, não foi possível
listar todas as sementes e/ou propágulos encontrados. Todas as amostras
identificadas por espécies foram semeadas e germinaram, facilitando a identificação.
Tabela 1: Lista das sementes e/ou propágulos encontradas nas fezes dos primatas (sagui e macaco-prego) do Jardim Botânico do Rio de Janeiro. Legenda: S - (Sagui)/ P - (macaco-prego).
Família/Espécies Nome Popular Exótica Nativa Primata
Arecaceae - - - P
Cecropia hololeuca Embaubuçu - X P
Cecropia sp. - - - S e P
Clidemia sp. - - - S
Cucurbitaceae - - - P
Cyperaceae - - - S e P
Erythoxylum pulchrum Arco-de-Pipa - X P
Eugenia sp. - - - P
Fabaceae - - - S e P
Gramíneae - - - P
1 1 1
3
1
2
1 1 1
3
1
4
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
Fre
qu
ên
cia
FAMÍLIA/ESPÉCIES
Sagui
Macaco-prego
37
Impatiens walleriana Maria-sem-vergonha X - P
Não identificado - - - S e P
Miconia albicans Canela-de-velho - X S
Miconia sp. - - - S e P
Rubiaceae - - - S e P
Solanaceae - - - P
A tabela 2 indica a variedade de frutos que esses primatas consomem e não
foram encontradas sementes e/ou propágulos nas amostras de fezes (Figuras 16 e
17). Entre eles, espécies exóticas como o jambo-vermelho (Syzygium malaccense) e
espécies nativas como o cajá-mirim (Spondias mombin).Esse resultado foi obtido
através de trabalhos científicos realizados pelo Núcleo de Conservação da Fauna do
JBRJ.
Tabela 2: Espécies de frutos consumidos pelos primatas (sagui e macaco-prego) do Jardim
Botânico do Rio de Janeiro levantados pelo Marins, 2010; Silva, 2012.
Espécies Nome popular Exótica Nativa
Ardísia humilis Ardísia X -
Ardísia semicremata Coral berry - X
Arthocarpus
heterophyllus Jaqueira X -
Artocarpus incisa Fruta-pão X -
Astrocaryum
aculeatissimum Brejaúva - X
Atalantia buxifolia Guamixira X -
Calyptranthes aromatica Cravo do mato - X
Citrus aurantium Laranja X -
Citrus reticulata Tangerina X -
Coccoloba alnifolia Pau-de-estalo - X
Diospyros philippensis Pêssego da Índia X -
Elaeis guineensis Dendezeiro X -
Eriobotrya japônica Nêspera X -
Eschweilera ovata Biriba-branca - X
Eugenia copacabanensis Princesinha de
Copacabana - X
Eugenia brasiliensis Grumixama - X
Eugenia tomentosa Cabeludinha - X
Eugenia uniflora Pitanga - X
Ficus auriculata Figueira-de-jardim X -
38
Genipa americana Genipapo X -
Gmelina asiática Guimelina X -
Guarea guidonia Carrapeta-verdadeira - X
Hymenaea courbaril Jatobá - X
Lucuma caimito Abiú - X
Mammea americana Abricó do Pará X -
Manilkara zapota Sapoteiro X -
Mangifera indica Mangueira X -
Myrciaria cauliflora Jabuticabeira - X
Psidium guajava Goiabeira X -
Rhipsalis baccifera Cacto-macarrão - X
Rhipsalis sp. Ripsalis - X
Spondias mombin Cajá-mirim - X
Syzygium cumini Jamelão X -
Syzygium malaccense Jambeiro-vermelho X -
Talisia esculenta Pitomba - X
Tapirira guianensis Pau-bombo - X
Theobroma cação Cacau X -
Thevetia peruviana Chapéu-de-napoleão - X
Trema micrantha Crindiúva X -
Triphasia trifólia Limeberry X -
Vitex flavens Tarumã X -
FIGURA 16: (A) macaco-prego (Sapajus nigritus) alimentando-se do fruto Arthocarpus heterophyllus (Jaca) no Jardim Botânico do Rio de Janeiro - JBRJ. (B) sagui (Callithrix sp.) alimentando-se do fruto Eugenia uniflora (pitanga) no Jardim Botânico do Rio de Janeiro – JBRJ. (Fotos: Fernando Pereira e Luciana Silva).

39
FIGURA 17: (A) macaco-prego (Sapajus nigritus) alimentando-se do fruto Psidium guajava
(Goiaba) no Jardim Botânico do Rio de Janeiro - JBRJ. (B) sagui (Callithrix sp.) alimentando-se do fruto Arthocarpus heterophyllus (Jaca) no Jardim Botânico do Rio de Janeiro – JBRJ. (Fotos: Alexandre Machado e Cristiane Rangel).

40
6. DISCUSSÃO
Os resultados indicam que os primatas sagui (Callithrix jacchus, Callithrix
penicillata e híbridos) e macaco-prego (Sapajus nigritus) consomem uma variedade
de estruturas, principalmente frutos com sementes pequenas e medianas. Os
primatas são dispersores importantes por disseminarem várias sementes de
espécies frutíferas (Poulsen et al., 2001; Stevenson et al., 2002; Wehncke et al.,
2003). Realizam um serviço de suporte para os processos naturais quando
dispersam sementes e ao consumirem insetos que atuam como pragas (Guedes &
Seehusen, 2011). As informações descritas na literatura são corroboradas pelos
resultados deste estudo, onde foram encontrados vários fragmentos de insetos nas
estações secas e úmidas (Figuras: 6 e 10).
Observou-se que muitas espécies de frutos possuem sementes grandes
impossibilitando a ingestão (Cáceres & Monteiro-Filho 2007; Lessa . Alguns autores
& Costa 2010) consideram a relação entre tamanho de sementes e tamanho do
dispersor importante, pois podem influenciar no tamanho das sementes ingeridas.
Sendo assim, observou-se que o tamanho do dispersor pode influenciar na
quantidade de vegetais (família/espécies) encontrados nas amostras de fezes. Os
primatas (saguis e macacos-prego) do Jardim Botânico do Rio de Janeiro
compartilham vários tipos de alimentos. Os macacos-prego por possuírem um porte
maior, acabam “ajudando” os saguis indiretamente quando abrem frutos muito
grandes. Segundo Vilela (2008), os primatas mordem o alimento/fruto, consomem a
parte da poupa e/ou parte comestível do fruto e liberam a semente no solo, por isso
são considerados dispersores dessas sementes. Os Callithrix sp., apesar de serem
animais de pequeno porte, consomem frutos de diferentes tamanhos, porém, quanto
à ingestão de sementes, conseguem apenas consumir sementes pequenas
compatíveis com seu tamanho. Isto pode explicar a pouca variedade de vegetais
(famílias/espécies) encontrados nas amostras de fezes. Observou-se também uma
grande frequência de goma nas amostras de fezes de sagui que está relacionado
com o comportamento de gomivoria. De acordo com Rangel (2010) tal
comportamento é realizado especialmente no período de escassez de alimento,
onde o animal alimenta-se dos exsudatos das árvores e por isso, este
comportamento pode influenciar na quantidade de sementes encontradas nas
amostras de fezes. Alguns autores (Vilela, 2007; Marins, 2010) acreditam que a

41
principal fonte alimentar dos Callithrix é o exsudato, devido ser fonte carboidratos.
Outros estudos sobre comportamento e dieta de sagui do Jardim Botânico (Santos,
2009; Marins, 2010; Silva, 2012) indicam que os animais teriam tendência a
deslocar-se menos para economizar energia ao invés de procurar os recursos
alimentares que fornecessem melhor retorno energético.
De acordo com Izar, (2008) os primatas dispersam sementes relativamente
pequenas, o que pode ser em parte reflexo da metodologia de análise de fezes, e
não de alimentação, pois os animais podem descartar sementes relativamente
grandes, ao invés de ingeri-las. Para Vilela (2007), ocorre uma baixa competição por
alimento entre espécies de S. nigritus e Callithrix sp., ocorrendo assim utilização da
mesma área de uso sem maiores conflitos entre as espécies de primatas.
Os macacos-prego (S. nigritus) consomem frutos maiores ou menores em
várias quantidades (Parron et al., 2015). Por isso, observou-se nas suas amostras
de fezes uma variedade de famílias/espécies. Autores como Janson et al. (1986) e
Izar, 2008, determinaram que o macaco-prego prefere espécies de frutos com alto
valor energético e espécies com alta densidade. Segundo Cunha et al. (2006), no
Parque Nacional da Tijuca, Artocarpus heterophyllus (jaqueira) é o item alimentar
mais importante da dieta do macaco-prego. No Jardim Botânico não é diferente, no
período de frutificação (estação úmida) eles são vistos frequentemente consumindo
esses frutos que possuem sementes grandes (Marins, 2010). Entretanto nem
fragmentos destes aparecem nas amostras de fezes desse primata, pois as
sementes são descartadas sem ingestão. Segundo Vilela, (2007) macacos-prego
possuem uma grande capacidade de manejar e explorar recursos diferentes, sendo
considerados eficientes oportunistas e generalistas por consumir uma variedade de
itens. Por apresentarem essa característica de comportamento podem explorar itens
mais pesados e/ou protegidos por uma casca dura, tais como sementes, meristemas
de palmeiras, nozes e etc. (Visalberghi, 1988). Em vista disso, pode ser este o
motivo de ter se encontrado uma maior diversidade de vegetais (sementes e/ou
propágulos) em suas amostras de fezes do que nas amostras de sagui, que não
utilizam esse recurso de manejar e explorar itens alimentares.
As estruturas abióticas encontradas nas amostras de fezes de macaco-prego
podem ser em razão de coletas obtidas do chão. Macacos-prego são animais bem

42
criteriosos quando se trata de alimento (Núcleo de Conservação da Fauna, 2016).
Já as amostras de fezes de sagui foram coletadas nas armadinhas em contato com
o chão de terra do arboreto do JBRJ.
A observação da germinação das sementes viáveis é importante para a
obtenção de plântulas, que além de auxiliar na identificação das sementes e/ou
propágulos, revelam que a dispersão foi efetivamente realizada (Schupp, 1993).
Mesmo em pequenas quantidades as sementes de famílias/espécies obtidas em
todo período, germinaram bem e produziram plântulas saudáveis. Amostras com
sementes encontradas em grandes quantidades, também estavam viáveis e
revelaram uma boa germinação.
Alguns autores (Janzen 1983; Schupp, 1993) acreditam que a qualidade da
dispersão depende do local e do padrão de deposição das sementes, ainda que a
quantidade seja boa, as sementes podem perder a viabilidade após a passagem
pelo trato digestivo dos vertebrados (Traveset & Verdú 2002; Rodrigues-Pérez et
al., 2005), mas não foi o caso da maioria das espécies vegetais encontradas nas
amostras de fezes neste estudo. Outros fatores que podem influenciar na
germinação das sementes são as características das plantas e o potencial dos
dispersores quando as sementes passam pelo trato digestivo (Lessa, 2013). Jordano
(1992) avaliou que sementes menores podem permanecer presas no sistema
digestivo por um período de tempo maior do que as sementes maiores quando
ingeridas, influenciando na germinação das sementes. Quando a semente fica retida
em um período de tempo maior no trato digestivo, pode reduzir capacidade de
germinação (Murray 1994) ou o tempo de germinação. Algumas sementes podem
ter diversos tipos de bloqueio da germinação e só evoluíram de acordo com a
diversidade de climas e hábitats, germinando quando estiverem em um local
apropriado (Finch-Savage & Leubner-Metzger, 2006).
As características próprias dos frutos e sementes podem afetar a dispersão
de sementes menores, por exemplo, os quais têm maior chance de ser engolidas e
defecadas intactas do que as maiores (Lucas & Corlett, 1998; Norconk et al., 1998),
e probabilidade maior de escapar da predação (Izar, 2008). Todavia, sementes
pequenas têm poucas chances de se estabelecer como plântulas, devido às baixas
reservas de nutrientes e probabilidade menor de suportar as condições

43
desfavoráveis do ambiente (Schupp 1995). Isso pode explicar as diferenças na
capacidade e no tempo de germinação de sementes muito pequenas que foram
encontradas nas amostras de fezes (Figura: 15).
A variação ambiental pode influenciar na germinação de sementes, pois
favorece a sobrevivência das plantas em determinados locais mais do que em outros
(Schupp 1995), portanto, pode ser um fator limitante para a germinação das
sementes. Os experimentos de germinação revelaram que, embora as sementes
permanecessem firmes (sem danos aparentes), após passagem pelo trato digestivo,
elas podem não germinar (Figura: 15), devido às condições ambientais e por ter tido
contato com a acidez do trato digestivo dos primatas.
As sementes da família Rubiaceae foram encontradas com maior frequência
nas fezes desses primatas (Figura: 14). Muitos frutos de Rubiaceae são tipo baga,
globoso a subgloboso, carnoso, epicarpo amarelo a vermelho, com duas a várias
sementes por fruto, envolvidas em polpa doce (Zappi et al., 2007), por isso, são
frutos muito dispersos por aves e mamíferos (Lima et al, 2010). Plantas com frutos
que são dispersos por animais apresentam características, tais como, presença de
uma porção comestível, envolvendo a semente e cores atrativas, que estimulam e
facilitam seu consumo e, consequentemente, a dispersão de suas sementes (Howe
& Smallwood, 1982). Várias famílias /espécies que foram encontradas nas amostras
de fezes desses primatas possuem essa características marcantes em seus frutos.
Também foram encontras muitas sementes não identificadas nas amostras, que
germinaram bem (Tabela 10 e 11- em anexo).
Esses dados indicam que os animais estudados são dispersores de sementes
o ano todo (Figura 8 e 12). Todas as espécies de sementes que foram encontradas
nas amostras fazem parte da vegetação, sejam da coleção viva ou plantas de
ocorrência espontânea presentes no Jardim Botânico do Rio de Janeiro (JBRJ,
2016).
As famílias Melastomataceae e Urticaceae também apresentaram boa
frequência nas amostras de fezes desses primatas. Espécies da família
Melastomataceae possuem frutos tipo cápsula loculicida ou baga, com sementes
numerosas e de várias formas e raro alados (Martins, 2009) e seus frutos escuros,
podem servir de atrativos para aves e mamíferos. Miconia sp., Miconia albincans e

44
Clidemia sp. (Melastomataceae) foram gêneros encontrados nas amostras de fezes
de Sagui e Macaco-prego, estes possuem frutos ricos em carboidratos e sementes
pequenas, facilitando assim a dispersão por vários animais (Snow, 1981). As plantas
da família Urticacea possuem os frutos finos, alongados ou tipo aquênio e quando
maduros a polpa fica estufada e macia com gosto doce (Sato et al., 2008). Cecropia
sp. (Urticaceae) é o gênero mais encontrado nas amostras de fezes dos primatas
analisados. Esse gênero é característico da região Neotropical (Berg, 1978) que
inclui árvores de crescimento rápido e são dispersas por várias aves e mamíferos
(Sato et al, 2008).
Aquênios de Cyperaceae foram encontrados nas amostras de sagui e
macaco-prego. A família Cyperaceae é constituída de plantas herbáceas, de
pequeno a grande porte, frequentemente associadas a áreas alagadas e seu fruto, é
tipo aquênio (seco e simples) (Prata et al., 2008). Por serem frutos secos e simples,
os primatas podem ter se alimentado por estar no período seco e/ou escassez de
alimento, já que todas as sementes encontradas foram nas estações secas tanto de
macaco-prego como de sagui.
As sementes não identificadas de macaco-prego possuíam os mesmos tipos
de estruturas, por isso, acredita-se pertencer à mesma planta que frutifica em todas
as estações (secas e úmidas). Pelas características trata se de um fruto carnoso,
atrativo e com muitas sementes. As plântulas desses frutos não identificados não
obtiveram tamanho suficiente para serem identificadas.
Nas estações seca¹ e seca² as sementes e/ou propágulos foram contínuos,
cinco e quatro tipos respectivamente. Já na estação úmida foram encontrados cinco
tipos de sementes e/ou propágulos nas amostras de fezes. De acordo com Reys et
al., (2005) as comunidades vegetais apresentam período de floração e frutificação
maior na estação úmida (outubro a março), devido ao início das chuvas. Isto não foi
visto no estudo, pois a quantidade de amostras encontradas nas estações seca¹ e
seca² foram relativamente iguais da estação úmida. As sementes encontradas nas
amostras de Macaco-prego foram relativamente bem distribuídas entre estações
úmidas e secas. Assim, constata-se que pode haver uma diversidade vegetal no
Jardim Botânico no período seco e úmido, por se tratar de uma coleção de plantas
nativas e exóticas.

45
As espécies encontradas nas amostras de fezes dos primatas do Jardim
Botânico do Rio de Janeiro – JBRJ que foram identificadas são na maioria nativas,
contudo, foi encontrada uma espécie exótica (Impatiens walleriana). As espécies
nativas podem ter sido consumidas da Mata Atlântica local, sendo dispersas no
JBRJ. Os levantamentos feitos por Marins, 2010; Silva, 2012 do Núcleo de
Conservação da Fauna mostram vários frutos que os primatas consomem e não foi
encontrada nenhuma estrutura nas fezes (Tabela: 2). Nesta tabela constam
espécies nativas e exóticas. Isso pode indicar que os primatas Callithrix sp. e S.
nigritus estão consumindo espécies do Jardim Botânico e da Mata Atlântica
adjacente. O predomínio de algumas espécies vegetais na dieta dos animais pode
ter sido reflexo do tipo do ambiente utilizado (Galetti e Pedroni, 1994),
provavelmente porque as espécies mais utilizadas são muito abundantes no JBRJ
(Figura 16).
O processo mutualístico entre o agente dispersor e a planta, pode beneficiar
espécies de plantas exóticas que foram introduzidas em novos hábitats, ajudando
estas plantas a superar os obstáculos ao estabelecimento para se tornarem
naturalizadas e, em alguns casos, invasoras (Richardson et al., 2000). Plantas
exóticas introduzidas em alguns habitats podem causar grandes impactos e se
tornam invasoras (Levine et al., 2003). As plantas exóticas produzem descendentes
em número muito elevado, assim, conseguem se dispersar a grandes distâncias e
prontamente competem com espécies nativas (Schneider, 2007). De acordo com
Rejmánek & Richardson (1996) plantas com frutos carnosos são geralmente
consideradas com alta capacidade de invasão. De acordo com por Marins, 2010;
Silva, 2012 muitas espécies de plantas exóticas são consumidas pelos primatas em
grande quantidade.
As interações estabelecidas entre a planta invasora e a fauna nativa também
podem ser negativas para a planta invasora, considerando a herbivoria e a predação
de sementes (Aslan e Rejmánek, 2010). Portanto, apesar da presença de potenciais
dispersores da vegetação exótica, esta última pode não se tornar invasora por haver
uma grande competição com as nativas que também produzem frutos carnosos
(Richardson et al., 2000). Assim, torna-se necessária a realização de um estudo
específico para verificar essa competição na Mata Atlântica contigua no JBRJ.
Segundo Richardson et al., (2000); Aslan & Rejmánek, (2010) a introdução de uma

46
espécie de planta exótica com frutos carnosos pode fornecer novos recursos
alimentares para espécies nativas da fauna em épocas de escassez de alimentos
nativos. Isso pode ser uma vantagem para os primatas que vivem na Mata Atlântica
contígua e não utilizam o Jardim Botânico como local de alimentação e forrageio.

47
7. CONCLUSÃO
Este trabalho se mostra pioneiro no estudo de dispersão de sementes. Os
resultados obtidos com este trabalho indicam que Sapajus nigritus é dispersor mais
eficiente que o Callithrix sp., diante da diversidade de vegetais com sementes
pequenas e maiores nas amostras de fezes ao longo de todas as estações (seca e
úmida). Portanto os primatas podem estar não só colaborando com a dispersão de
espécies nativas na área do Arboreto do Jardim Botânico do Rio de Janeiro - JBRJ,
como também ampliando a distribuição de espécies exóticas (possivelmente
invasoras) à Flora da Serra da Carioca nas imediações do Jardim Botânico do Rio
de Janeiro.
O JBRJ oferece grande diversidade de alimentos tanto de origem vegetal,
quanto de origem animal para os grupos de S. nigritus e Callithrix sp. não sendo
necessário que funcionários e visitantes os alimentem.
Os primatas (S. nigritus e Callthrix sp.) são dispersores de forma direta,
quando dispersam sementes e/ou propágulos pelas fezes e indiretamente, quando
consomem os frutos e liberam as sementes pelo chão. Os primatas são bons
dispersores de sementes, já que são generalistas em sua alimentação.

48
8. REFERÊNCIAS BIBLIOGRÁFICAS
ASLAN, C. E. & REJMÁNEK, M. Avian use of introduced plants: ornithologist records illuminate interspecific associations and research needs. Ecological Applications, 20(4): 1005-1020. 2010. APPEZZATO-DA-GLÓRIA, B. & CARMELLO-GUERREIRO, S. M. Anatomia Vegetal. Viçosa: Editora UFV. 438p. 2003 BEDIAGA, B et al. 200 anos Jardim Botânico do Rio de Janeiro. 1. ed. Rio de Janeiro. 2008. BERG, C.C. Espécies de Cecropia da Amazônia Brasileira. Acta Amazônica, 8(2):149-182. 1978. CÁCERES, NC & MONTEIRO-FILHO, ELA. Germinação em Espécies de Sementes
Ingeridas por Opossums: Implicações para Dispersão de Sementes e Conservação
Florestal. Braz. Arco. Biol. Techol. 50 (6): 921-928. 2007.
CDB. Conservação da diversidade biológica, 1992. <www.mma.gov.br/destaques/item/7513-conven%C3%A7%C3%A3o-sobre-diversidade-biol%C3%B3gica-cdb> Acessado em 10/10/2016 CHIARELLO, A.G. Effects of fragmentation of the Atlantic forest on mammal communities in southeastern Brazil. Biological Conservation, 89: 71-82. 1999. COIMBRA-FILHO, A. F., A. PISSINATTI AND A. B. RYLANDS. Experimental multiple hybridism among Callithrix species from eastern Brazil, pp. 95–120 in Marmosets and Tamarins: Systematic, Ecology and Behaviour, edited by A. B. Rylands. Oxford University Press, Oxford, 1993. CUNHA, A. A.; VIEIRA, M.V.; GRELLE, C.E.V. Preliminary observations on habitat, support use and diet in two non-native primates in an Urban Atlantic Forest Fragment: the capuchin monkey (Cebus sp.) and the common marmoset (Callithrix jacchus) in Tijucas Forest, Rio de Janeiro. Urban Ecosyst, 9 : 351-359. 2006. DEVELEY, P.F. & PERES, C.A. Resource seasonality and the structure of mixed species bird flocks in a coastal Atlantic forest of southeastern Brazil. J. Trop. Ecol. 16: 33-53. 2000 DIGBY, L. e BARRETO, C. E. Vertebrate predation in common marmosts. Neotropical Primates, v.6, n.4, p.124-126. 1998. FIGLIOOLIA, M. B. Maturação de sementes de Inga uruguensis Hook et Arn. Associada à fenologia reprodutiva e a dispersão de sementes em floresta ripária do rio Mogi Guaçu – município de Mogi Guaçu. 1993. 150 f. Dissertação de Mestrado

49
em Engenharia Florestal – Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo, 1993. FIGLIOLIA, M.B. & KAGEYAMA, P.Y. Dispersão de sementes de Inga uruguensis Hook. Et Arn. em floresta ripária do rio Mogi Guaçu, município de Mogi Guaçu – SP. Rev. Inst. Florestal 7: 65-80. 1995. FINCH-SAVAGE, WILLIAM E., and GERHARD LEUBNER-METZGER. "Seed dormancy and the control of germination." New Phytologist 171.3 (2006): 501-523. FLEMING, T. H. 1987. Patterns of tropical vertebrate frugivore diversity. Annual Review of Ecology and Systematics 18: 91-109. 1987. FREESE, C. H., & OPPENHEIMER, J. R. The capuchin monkey, genus Cebus. Em A. F. Coimbra-Filho & R. A. Mittermeier (Eds.), Ecology and Behaviour of Neotropical Primates 1 (pp. 331-390). Rio de Janeiro: Academia Brasileira de Ciências. 1981. GALETTI, M. & PEDRONI, F. Seasonal diet of capuchin monkeys (Cebus apella) in a semideciduous Forest in south-east Brazil. J. Trop. Ecol. 10: 27-39. 1994. GARBER, P. A. & LAMBERT, J.E. Primates as seed dispersers: ecological processes and directions for future research. American Journal of Primatology 45: 3-8. 1998. GAUTIER-HION, A., GAUTIER, J.P. & MAISELS F. Seed dispersal versus seed predation: an inter-site comparison of two related African monkeys. Vegetatio 107/108: 237-244. 1993. GUEDES, F. B. & SEEHUSEN, S. E. Pagamento por Serviços Ambientais na Mata Atlântica: lições aprendidas e desafios. Brasília: MMA, p. 272, 2011. Guerreiro, S.M.C. & Gloria, B.A. Anatomia Vegetal, 3 ed., Viçosa. 2003. HARPER, J. L. The population biology of plants. London: Academic Press. 1977. HOWE, H. F. & SMALLWOOD, J.Ecology of seed dispersal. Annual Review of Ecology and Systematics 13: 201-228. 1982. HOWE, H.F.; MIRITI, M.N. When seed dispersal matters. BioScience, 651-660, 2004. HOWE, H.F., SCHUPP, E.W.; WESTLEY, L.C. Early consequences of seed dispersal for Neotropical tree (Virola surinamensis). Ecology, v. 66, p. 781-791, 1985. Instituto de Biologia-UFU, Anatomia Vegetal. Disponível em: < www.anatomiavegetal.ib.ufu.br/pdf-recursos-didaticos/morfvegetalorg DISPERSAO.pdf>. Acessado em: 24/08/2016.

50
IZAR, P. Dispersão de sementes por Cebus nigritus e Brachyteles arachnoides em área de Mata Atlântica, Parque Estadual Intervales, SP pp. 8-24. In: A Primatologia no Brasil - 9 (S.F. Ferrari & J. Rímoli, Eds.) Aracaju, Sociedade Brasileira de Primatologia, Biologia Geral e Experimental – UFS. 2008. IZAWA, K. Foods and feeding behavior of wild back-capped capuchin (Cebus apella). Primates, 20(1), 57-76. 1990. JANSON, C.H., E.W. STILES & D.W. WHITE. Selection on plant fruiting traits by brown capuchin monkeys: a multivariate approach pp. 83-92. In: Frugivores and Seed Dispersal (A. Estrada & T.H. Fleming, Eds.) Dr W. Junk Publishers, Dordrecht. 1986. JANZEN, D.H. Dispersal of seed by vertebrate guts pp. 232-262. In: Coevolution (D.J. Futuyma & M. Slatkin, Eds.) Sinauer Associates Inc., Sunderland. 1983. JBRJ, Jardim Botânico do Rio de Janeiro. Disponível em: <www.jbrj.gov.br/historia> Acessado em: 10/06/2016 JORDANO, P. Frutos e frutos. Pp. 105-156. In:. Fenner M. (Ed.) Sementes: a ecologia da regeneração de comunidades vegetais. Nova Iorque, CABI Publishing. 1992. JORDANO, P. & GODOY, J. A. Frugivore-generated seed shadows: a landscape view of demographic and genetic effects, p. 305-321. In Levey, D. J., Silva, W. & Galetti, M. (eds.). Frugivores and seed dispersal: ecological, evolutionary, and conservation. CAB International, Wallingford, UK. 2002. JORDANO, P. & SCHUPP, E. W. Determinants of seed disperser effectiveness: the quantity component and patterns of seed rain for Prunus mahaleb. Ecological Monographs 70. 2000. KAPLIN, B.A.,MUNYALIGOGA, V. & MOERMOND, T.C. The influence of temporal changes in fruit availability on diet composition and seed handling in blue monkeys (Cercopithecus mitis doggetti). Biotropica 30: 56-71. 1998. KATO, M. M. Composição faunística de Anfíbios no Jardim Botânico do Rio de Janeiro. Rio de Janeiro. Monografia (ciências biológicas). Universidade Estácio de Sá. 2013. LAVÔR, João Conrado Niemeyer. Histórico do Jardim Botânico do Rio de Janeiro. Rio de Janeiro: IBDF/JBRJ. Disponível em <www.jbrj.gov.br>, acessado em 15 de maio de 2015. 1983. LESSA, L. G & COSTA, F. N. Dieta e dispersão de sementes por cinco marsupiais (Didelphimorphia: Didelphidae). Em uma reserva de cerrado brasileiro Mammalian Biology 75: 10-16. 2010. LESSA, L. G. ; GEISE, L.; COSTA, F. N. Effects of gut passage on the germination of

51
seeds ingested by didelphid marsupials in a neotropical savanna. Acta Botanica Brasilica 27(3): 519-525. 2013. LEVINE, J.M., VILA, M., D’ANTONIO, C., DUKES, J.S., GRIGULIS, K. & LAVOREL, S. Mechanisms underlying the impact of exotic plant invasions. Phil. Trans. Roy. Soc., 270, 775–781. 2003. LIMA, L.F., LIMA, P.B., ALMEIDA Jr., E.B. & ZICKEL, C.S. Morphology of fruits, seeds and seedlings Guettarda platypoda DC. (Rubiaceae). Biota Neotrop. 10(1) Listas de espécies da flora do Brasil. Disponível em: <www.floradobrasil.jbrj.gov.br/> Acessado em: 07/11/2016. LUCAS, P.W. & CORLETT, R. T. Seed dispersal by long-tailed macaques. American Journal of Primatology 45: 29-44. 1998. LUDWIG, G.L., AGUIAR, M. and ROCHA, V. J. Uma Avaliação da Dieta, da Área de Vida e das Estimativas Populacionais de Cebus nigritus (Goldfuss, 1809) em um Fragmento Florestal no Norte do Estado do Paraná. Neotropical Primates 13(3):12-18. 2005. MAIER, W., ALONSO, C., and LANGGUTH, A. Field observations on Callithrix jacchus jaccus L.Z. SÄugetierkunde 47: 334–346. 1982. MARINS, J. R. G. A. Ecologia alimentar de macaco-prego (Cebus nigritus nigritus - Goldfuss, 1809) (Cebidae) no Jardim Botânico do Rio de Janeiro. Monografia (ciências biológicas). Universidade Santa Úrsula. 46p. 2010. MARTINS, A.B. (coord.). Melastomataceae In: Martins, S.E., Wanderley, M.G.L., Shepherd, G.J., Giulietti, A.M., Melhem, T.S. (eds.) Flora Fanerogâmica do Estado de São Paulo. Instituto de Botânica, São Paulo, vol. 6, pp: 1-168. 2009 MENDONÇA, D. T. N. F. et al. Ministério do Meio Ambiente. 2004. 7p. MURRAY, KG. Laxantes frutas e taxas de passagem de sementes em frugívoros:. Consequências para o sucesso reprodutivo da planta. Ecology 75: 989-994. 1994. NEVES, E.L. & VIANA B.F. Dispersão e predação de sementes de três espécies de Jatropha L. (Euphorbiaceae) da Caatinga, semi-árido do Brasil. revista virtual Candombá. 146-157, 2008. NORCONK, M.A., B.W. GRAFTON & CONKLIN-BRITTAIN, N. L. Seed dispersal by Neotropical seed predators. American Journal of Primatology 45: 103-126. 1998. Núcleo de Conservação da Fauna, 2016.
NUNAN, G. W. Dieta e comportamento alimentar dos Psitacídeos Brotogeris tirica e Pyrrhura frontalis no Jardim Botânico do Rio de Janeiro. Rio de Janeiro. Monografia

52
(ciências biológicas). Universidade Veiga de Almeida. 38p. 2015. PARRON, L. M.; GARCIA, J. R.; OLIVEIRA, E. B. de; BROWN, G. G.; PRADO, R. B. (Ed.). Serviços ambientais em sistemas agrícolas e florestais do Bioma Mata Atlântica. Brasília, DF: Embrapa, 2015. POULSEN, C.J., PIERREHUMBERT, R.T. and JACOB, R.L. Impact of ocean dynamics on the simulation of the neoproterozoic “snowball Earth”. Geophysical Research Letters 28: 2001. PRATA, A. P. N.; THOMAS, W. W.; WANDERLEY, G. L. Micromorphology of achene surface in Bulbostylis Kunth (Cyperaceae). Rev. bras. Bot. 10p. 2008. RANGEL, C. H. Ecologia e comportamento de Callitrichidae (Primates) no Jardim Botânico do Rio de Janeiro. Dissertação de Mestrado, Universidade Estadual do Rio de Janeiro, Rio de Janeiro, BR. 2010. RANGEL, C. H. Levantamento preliminar da Fauna do Jardim Botânico do Rio de Janeiro. 4p. 2005. RANGEL, C. H.; SOUSA, F. S. F.; GRELLE, C. E. V. Dieta de Callitrichidae no Jardim Botânico do Rio de Janeiro. In: XIII Congresso de Brasileiro de Primatologia, Blumenau. Resumos de trabalhos apresentados no XIII Congresso de Brasileiro de Primatologia. 1 CD-ROM. 2009. RAVEN, P.H; EVERT, R.F; EICHHORN, S.E. Biologia Vegetal. 6 ed. Rio de Janeiro: Guanabara Koogan. 906 p. 2001. REIS, N. R., PERACCHI, A. L., ANDRADE, F. R. Primatas Brasileiros. 1. ed. Londrina: Technical Books, 260 p. 2008. REIS, R. C. C. 200 anos Jardim Botânico do Rio de Janeiro. 1. ed. Rio de Janeiro. 2008. REJMÁNEK M. & RICHARDSON D.M. "What attributes make some plant species more invasive?" Ecology, 77: 1655–1661. 1996. REYS, P.; GALETTI, L. M.; MORELLATO, P.C.; SABINO, J. fenologia reprodutiva e disponibilidade de frutos de espécies arbóreas em mata ciliar no rio formoso, mato grosso do sul. Biota neotropica. 10p. 2005. RICHARDSON, D. M.; ALLSOPP, N.; D’ANTONIO, C. M.; MILTON, S. J. & REJMÁNEK, M. Plant invasions — the role of mutualisms. Biological Reviews, 75, 65–93. 2000. RÍMOLI, J., STRIER, K. B. e FERRARI, S. F. Seasonal and longitudinal variation in the behavior of free-ranging black tufted capuchins Cebus nigritus (Goldfuss, 1809) in a fragment of Atlantic Forest in Southeastern Brazil pp. 130-146. In: A Primatologia no Brasil - 9 (S.F. Ferrari & J. Rímoli, Eds.) Aracaju, Sociedade Brasileira de Primatologia, Biologia Geral e Experimental – UFS. 2008.

53
RIZZINI, C.T., COIMBRA-FILHO, A.F. Lesões produzidas pelos saguis, Callithrix p. penicillata (E. Geoffroy, 1982), em árvores do cerrado (Callitrichadae, Primates). Revista Brasileira de Biologia 41: 579- 583. 1981. ROCHA, C. F. D, BERGALHO, H.G., ALVES, M. A. S. & SLUYS, M.V. A biodiversidade nos grandes remanescentes florestais do estado do Rio de Janeiro e nas restingas da Mata Atlântica. São Carlos. RiMA, 160p. 2003. ROCHA, V. J. Macaco-prego, como controlar esta nova praga florestal. Floresta 30:1–295–99. 2000. RODRÍGUEZ-PÉREZ, J.; RIERA, N. & TRAVESET, A. Efeito da passagem de sementes através de aves e lagartos sobre a taxa de surgimento de espécies mediterrânicas: Diferenças entre as condições naturais e controlados Ecologia Funcional 19: 699-706. 2005. RONDON-NETO, R. M.; WATZLAWICK, L. F.; CALDEIRA, M. V. W. Diversidade florística e síndromes de dispersão de diásporos das espécies arbóreas de um fragmento de floresta ombrófila mista. Revista Ciências Exatas e Naturais, Guarapuava, 209-216, 2001. ROTHE, H. Adaptation to natural food resources by semi-free common marmosets (Callithrix jacchus): preliminary results. Neotropical Primates, 54-7. 1999. RYLANDS, A. B. Habitat and the evolution of social and reproductive behavior in Callitrichidae. American Journal of Primatology, New York, v. 38, p. 5-18, 1996. RYLANDS, A.B. e FARIA, D. S. D. Habitats, feeding ecology, and home range size in the genus Callithrix. In: RYLANDS, A. B. (Ed.) Marmosets and Tamarins: systematics. Behaviour, and ecology. Oxford University Press, p262-272. 1993. SANTOS, D. C. Ecologia e alimentação de sagui-de-tufo-branco (callithrix sp.) selvagem no jardim botânico do rio de janeiro, brasil. Monografia (ciências biológicas). Universidade Estácio de Sá. 30p. 2009. SATO, T.M.; PASSOS, F. C.; NOGUEIRA, A. C. Frugivoria de morcegos (Mammalia, Chiroptera) em CeCropia paChystaChya (Urticaceae) e seus efeitos na germinação das sementes. Museu de Zoologia da USP. 48(3):19 26, 9p. 2008. SCHNEIDER, A.A. A flora naturalizada no estado do Rio Grande do Sul, Brasil: herbáceas subespontâneas. Biociências 15(2): 257-268. 2007. SCHUPP, E.W. Quantity, quality and the effectiveness of seed dispersal by animals. Vegetatio 107/108: 15-29. 1993. SCHUPP, E.W., Seed seedling conflicts, habitat choice and patterns of plant recruitment. Am. J. Bot. 82: 399–409. 1995. SCHUPP, E.W., FUENTES, M. Spatial patterns of seed dispersal and the unification

54
of plant population ecology. E´ coscience 2, 267–275. 1995. SILVA JÚNIOR, J. S. Especiação nos macacos-prego e caiararas, gênero Cebus Erxleben, 1777 (Primates, Cebidae). Boletim da Sociedade Brasileira de Mastozoologia. 42: 11-12, 2005. SILVA JÚNIOR, J. S. Especiação nos macacos-prego e caiararas, gênero Cebus Erxleben, 1777 (Primates, Cebidae). Tese de Doutoramento, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2001. SILVA, L. A. Fenologia e variações na dieta e área de vida de saguis (callithrix sp.) no jardim botânico do rio de janeiro. Monografia (ciências biológicas). Universidade São José. 57p. 2012. SMAC, Secretaria Municipal do Meio Ambiente. Disponível em:< www.rio.rj.gov.br/web/smac> Acessado em: 10/06/2016 SNOW, DW., Tropical frugivorous birds and their food plants: a world survey. Biotropica, vol. 13, no. 1, p. 1-14. 1981. SORK, V. L., NASON, J., CAMPBELL, D. R. & FERNÁNDEZ, J. F. Landscape approaches to historical and contemporary gene flow in plants. Trends in Ecology and Evolution 14: 224. 1999 SOS Mata Atlântica, Nossa causa, a mata atlântica. Disponível em: <www.sosma.org.br/nossa-causa/a-mata-atlantica/> Acessado em: 10/06/ 2015. STEVENSON, M. F. & RYLANDS, A. B.,The marmosets, genus Callithrix. In: R. A. Mittermeier, A. B. Rylands, A. F. Coimbra-Filho & G. A. B. Fonseca (eds.), Ecology and behavior of Neotropical primates, 2nd vol. W.W.F., Washington, pp. 131-222.1988. STEVENSON, C. S., WHITMONT S., BORNHOLT L., et al. A cognitive remediation programme for adults with attention deficit hyperactivity disorder. Austral NZ Psychiatry. 36:610–16. 2002. TRAVESET, A. & VERDÚ, M. Uma meta-análise do efeito do tratamento intestinal na germinação de sementes. Pp. 339-350. Em: Levey, DJ; Silva, WR & Galetti, M. (Eds.) A dispersão de sementes e frugivoria:. Ecologia, evolução e conservação. Wallingford, CABI Publishing. 2002. VAN DER PIJL, L. Principles of dispersal in higher plants. 2. ed. Berlim: Springer-Verlag, 1972. ______. Principles of dispersal in higher plants. 3. ed. New York: Springer-Verlag, 1982. VIDOLIN, G. P.; MIKICH, S. B. Cebus nigritus (Primates: Cebidae) no P. E. Vila Rica doEspírito Santo, Fênix – PR: estimativa populacional e área de vida, composição e dinâmica dos grupos. Anais do IV Congresso Brasileiro de Unidades de Conservação. Curitiba: Fundação O Boticário de Proteção à Natureza e

55
Rede Pró-Unidades de Conservação, 2004. VILANOVA, R.; SILVA JUNIOR, J. S.; GRELLE, C. E. V.; CERQUEIRA, G. M. R. Limites climáticos e vegetacionais das distribuições de Cebus nigritus e Cebus robustus (Cebinae, Platyrrhini). Neotropical Primates, 2005. VILELA, S.L. Simpatia e dieta de Callithrix penicillata (Hershkovitz) (Callitrichidae) e Cebus libidinosus (Spix) (Cebidae) em matas de galeria do Distrito Federal, Brasil. Revista Brasileira Zoologia 24:601-607. 2007. VISALBERGHI, E. "Responsiveness to objects in two social groups of tufted capuchin monkeys (Cebus apella)." American Journal of Primatology 15.4: 349-360. 1988. WEHNCKE, E. V.; HUBELL, S. P.; FOSTER, R. B.; DALLING, J. W. Seed dispersal patterns proudced by whitefaced monkeys: implications for the dispersal limitation of neotropical trees species. Journal of Ecology, 677, 2003.
ZAPPI D. et al. Flora Fanerogâmica do Estado de São Paulo, vol. 5, pp: 259-460. 2007.
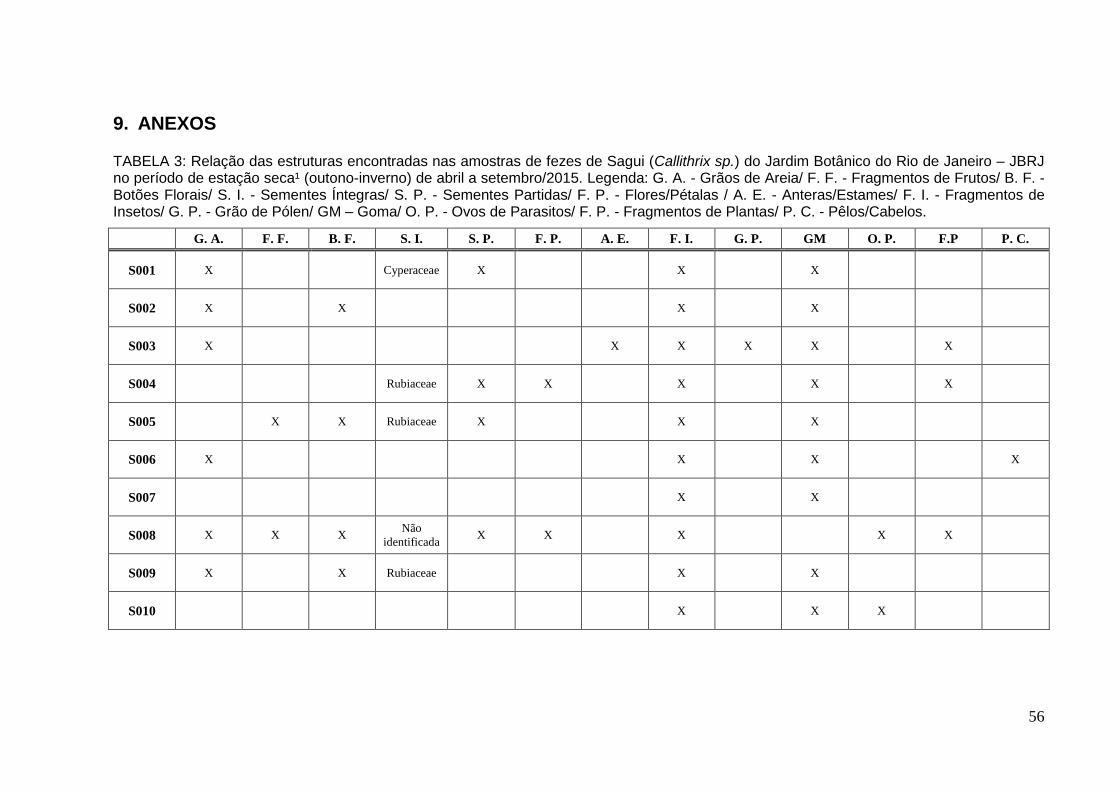
56
9. ANEXOS
TABELA 3: Relação das estruturas encontradas nas amostras de fezes de Sagui (Callithrix sp.) do Jardim Botânico do Rio de Janeiro – JBRJ no período de estação seca¹ (outono-inverno) de abril a setembro/2015. Legenda: G. A. - Grãos de Areia/ F. F. - Fragmentos de Frutos/ B. F. - Botões Florais/ S. I. - Sementes Íntegras/ S. P. - Sementes Partidas/ F. P. - Flores/Pétalas / A. E. - Anteras/Estames/ F. I. - Fragmentos de Insetos/ G. P. - Grão de Pólen/ GM – Goma/ O. P. - Ovos de Parasitos/ F. P. - Fragmentos de Plantas/ P. C. - Pêlos/Cabelos.
G. A. F. F. B. F. S. I. S. P. F. P. A. E. F. I. G. P. GM O. P. F.P P. C.
S001 X Cyperaceae X X X
S002 X X X X
S003 X X X X X X
S004 Rubiaceae X X X X X
S005 X X Rubiaceae X X X
S006 X X X X
S007 X X
S008 X X X Não
identificada X X X X X
S009 X X Rubiaceae X X
S010 X X X
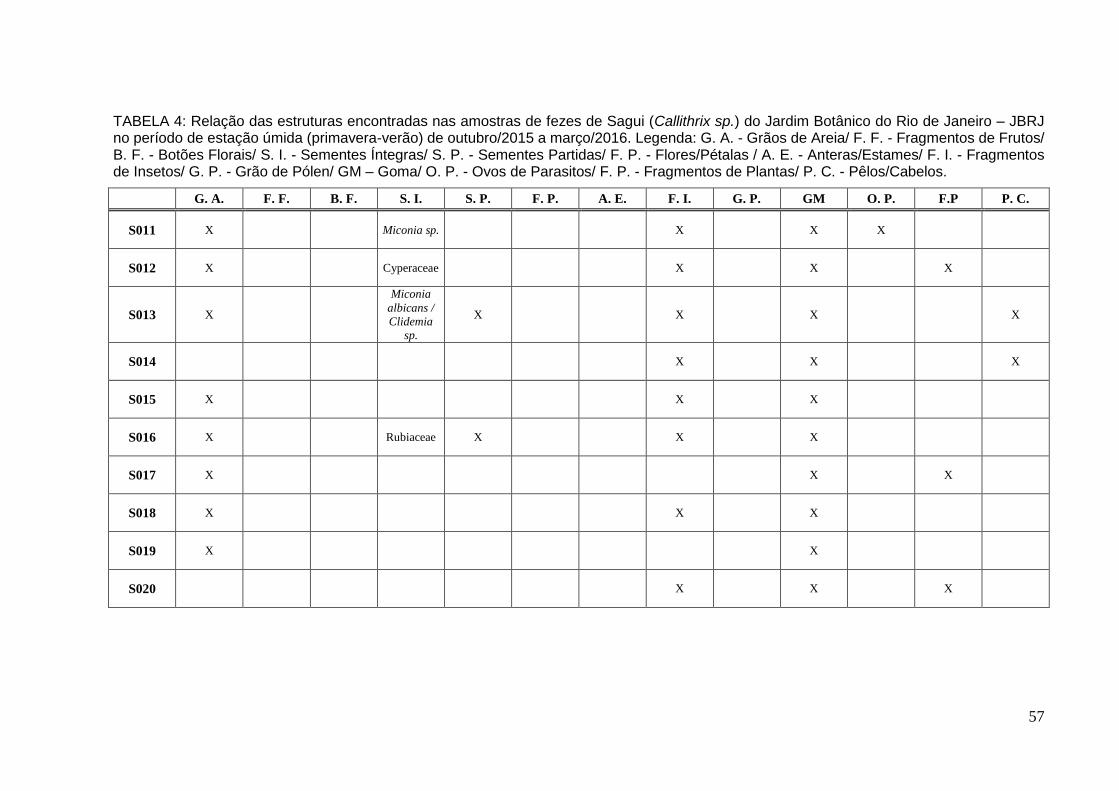
57
TABELA 4: Relação das estruturas encontradas nas amostras de fezes de Sagui (Callithrix sp.) do Jardim Botânico do Rio de Janeiro – JBRJ no período de estação úmida (primavera-verão) de outubro/2015 a março/2016. Legenda: G. A. - Grãos de Areia/ F. F. - Fragmentos de Frutos/ B. F. - Botões Florais/ S. I. - Sementes Íntegras/ S. P. - Sementes Partidas/ F. P. - Flores/Pétalas / A. E. - Anteras/Estames/ F. I. - Fragmentos de Insetos/ G. P. - Grão de Pólen/ GM – Goma/ O. P. - Ovos de Parasitos/ F. P. - Fragmentos de Plantas/ P. C. - Pêlos/Cabelos.
G. A. F. F. B. F. S. I. S. P. F. P. A. E. F. I. G. P. GM O. P. F.P P. C.
S011 X Miconia sp. X X X
S012 X Cyperaceae X X X
S013 X
Miconia
albicans /
Clidemia
sp.
X X X X
S014 X X X
S015 X X X
S016 X Rubiaceae X X X
S017 X X X
S018 X X X
S019 X X
S020 X X X
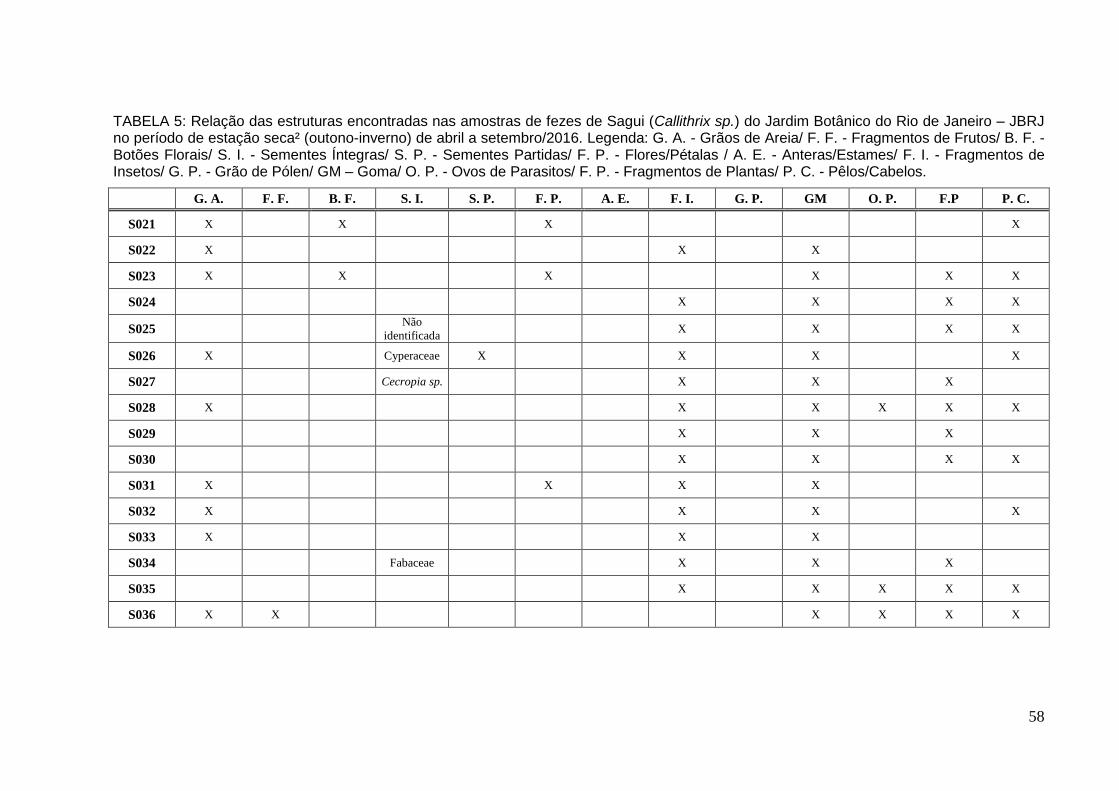
58
TABELA 5: Relação das estruturas encontradas nas amostras de fezes de Sagui (Callithrix sp.) do Jardim Botânico do Rio de Janeiro – JBRJ no período de estação seca² (outono-inverno) de abril a setembro/2016. Legenda: G. A. - Grãos de Areia/ F. F. - Fragmentos de Frutos/ B. F. - Botões Florais/ S. I. - Sementes Íntegras/ S. P. - Sementes Partidas/ F. P. - Flores/Pétalas / A. E. - Anteras/Estames/ F. I. - Fragmentos de Insetos/ G. P. - Grão de Pólen/ GM – Goma/ O. P. - Ovos de Parasitos/ F. P. - Fragmentos de Plantas/ P. C. - Pêlos/Cabelos.
G. A. F. F. B. F. S. I. S. P. F. P. A. E. F. I. G. P. GM O. P. F.P P. C.
S021 X X X X
S022 X X X
S023 X X X X X X
S024 X X X X
S025 Não
identificada X X X X
S026 X Cyperaceae X X X X
S027 Cecropia sp. X X X
S028 X X X X X X
S029 X X X
S030 X X X X
S031 X X X X
S032 X X X X
S033 X X X
S034 Fabaceae X X X
S035 X X X X X
S036 X X X X X X
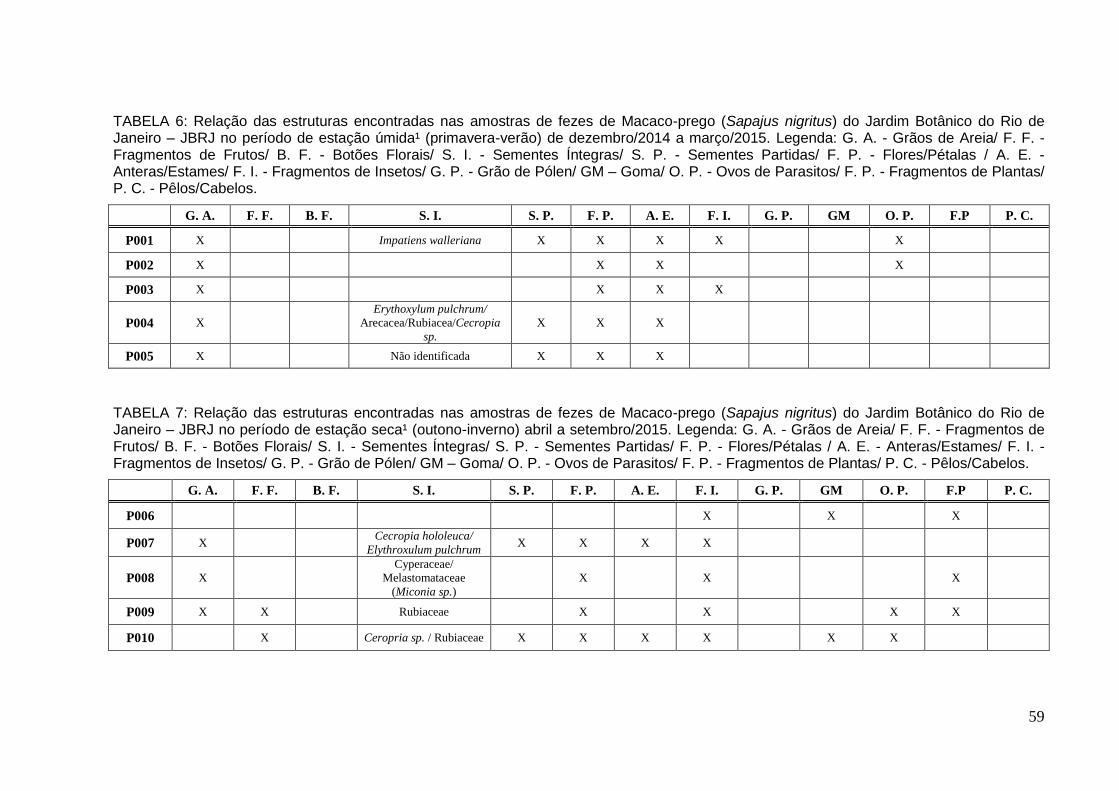
59
TABELA 6: Relação das estruturas encontradas nas amostras de fezes de Macaco-prego (Sapajus nigritus) do Jardim Botânico do Rio de Janeiro – JBRJ no período de estação úmida¹ (primavera-verão) de dezembro/2014 a março/2015. Legenda: G. A. - Grãos de Areia/ F. F. - Fragmentos de Frutos/ B. F. - Botões Florais/ S. I. - Sementes Íntegras/ S. P. - Sementes Partidas/ F. P. - Flores/Pétalas / A. E. - Anteras/Estames/ F. I. - Fragmentos de Insetos/ G. P. - Grão de Pólen/ GM – Goma/ O. P. - Ovos de Parasitos/ F. P. - Fragmentos de Plantas/ P. C. - Pêlos/Cabelos.
G. A. F. F. B. F. S. I. S. P. F. P. A. E. F. I. G. P. GM O. P. F.P P. C.
P001 X Impatiens walleriana X X X X X
P002 X X X X
P003 X X X X
P004 X
Erythoxylum pulchrum/
Arecacea/Rubiacea/Cecropia
sp.
X X X
P005 X Não identificada X X X
TABELA 7: Relação das estruturas encontradas nas amostras de fezes de Macaco-prego (Sapajus nigritus) do Jardim Botânico do Rio de Janeiro – JBRJ no período de estação seca¹ (outono-inverno) abril a setembro/2015. Legenda: G. A. - Grãos de Areia/ F. F. - Fragmentos de Frutos/ B. F. - Botões Florais/ S. I. - Sementes Íntegras/ S. P. - Sementes Partidas/ F. P. - Flores/Pétalas / A. E. - Anteras/Estames/ F. I. - Fragmentos de Insetos/ G. P. - Grão de Pólen/ GM – Goma/ O. P. - Ovos de Parasitos/ F. P. - Fragmentos de Plantas/ P. C. - Pêlos/Cabelos.
G. A. F. F. B. F. S. I. S. P. F. P. A. E. F. I. G. P. GM O. P. F.P P. C.
P006 X X X
P007 X Cecropia hololeuca/
Elythroxulum pulchrum X X X X
P008 X
Cyperaceae/
Melastomataceae
(Miconia sp.)
X X X
P009 X X Rubiaceae X X X X
P010 X Ceropria sp. / Rubiaceae X X X X X X
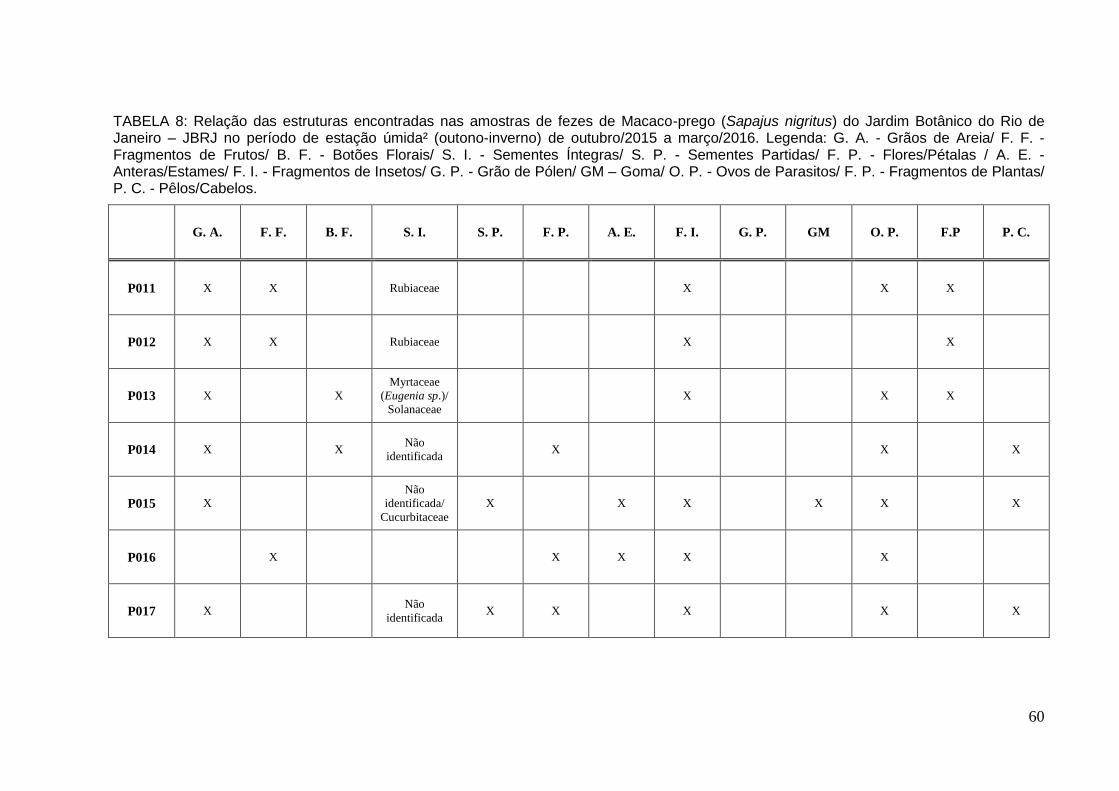
60
TABELA 8: Relação das estruturas encontradas nas amostras de fezes de Macaco-prego (Sapajus nigritus) do Jardim Botânico do Rio de Janeiro – JBRJ no período de estação úmida² (outono-inverno) de outubro/2015 a março/2016. Legenda: G. A. - Grãos de Areia/ F. F. - Fragmentos de Frutos/ B. F. - Botões Florais/ S. I. - Sementes Íntegras/ S. P. - Sementes Partidas/ F. P. - Flores/Pétalas / A. E. - Anteras/Estames/ F. I. - Fragmentos de Insetos/ G. P. - Grão de Pólen/ GM – Goma/ O. P. - Ovos de Parasitos/ F. P. - Fragmentos de Plantas/ P. C. - Pêlos/Cabelos.
G. A. F. F. B. F. S. I. S. P. F. P. A. E. F. I. G. P. GM O. P. F.P P. C.
P011 X X Rubiaceae X X X
P012 X X Rubiaceae X X
P013 X X
Myrtaceae
(Eugenia sp.)/
Solanaceae
X X X
P014 X X Não
identificada X X X
P015 X
Não
identificada/
Cucurbitaceae
X X X X X X
P016 X X X X X
P017 X Não
identificada X X X X X
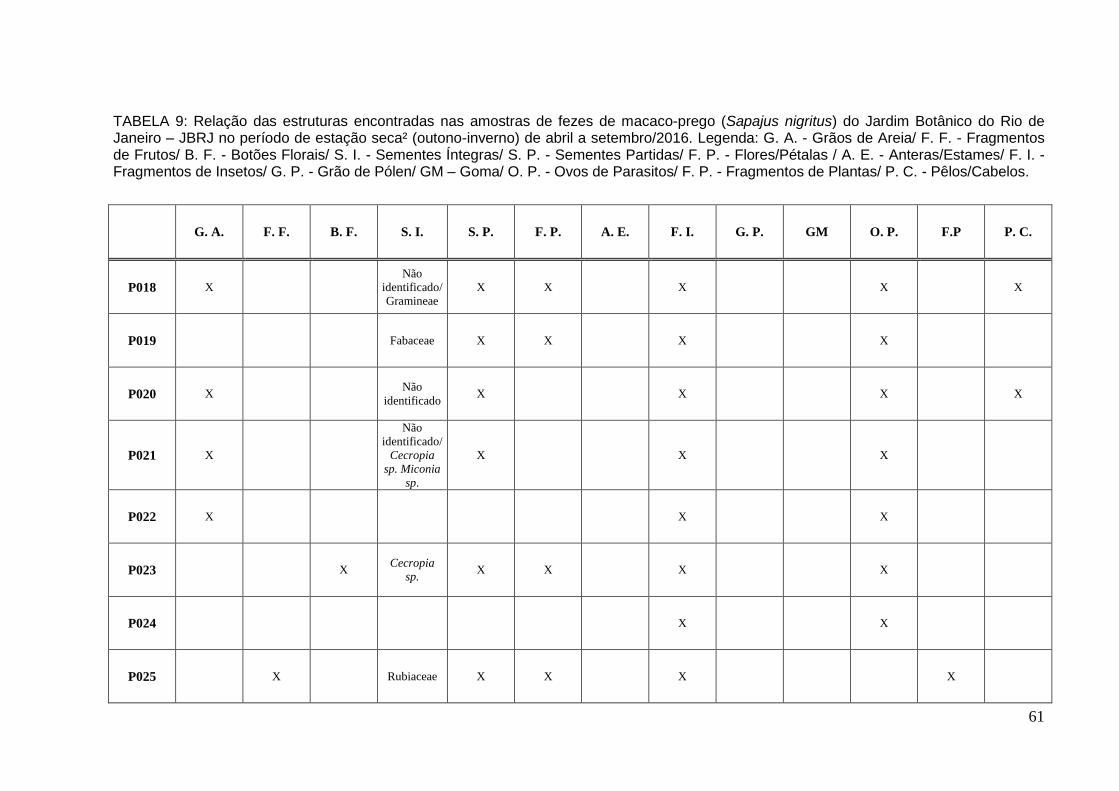
61
TABELA 9: Relação das estruturas encontradas nas amostras de fezes de macaco-prego (Sapajus nigritus) do Jardim Botânico do Rio de Janeiro – JBRJ no período de estação seca² (outono-inverno) de abril a setembro/2016. Legenda: G. A. - Grãos de Areia/ F. F. - Fragmentos de Frutos/ B. F. - Botões Florais/ S. I. - Sementes Íntegras/ S. P. - Sementes Partidas/ F. P. - Flores/Pétalas / A. E. - Anteras/Estames/ F. I. - Fragmentos de Insetos/ G. P. - Grão de Pólen/ GM – Goma/ O. P. - Ovos de Parasitos/ F. P. - Fragmentos de Plantas/ P. C. - Pêlos/Cabelos.
G. A. F. F. B. F. S. I. S. P. F. P. A. E. F. I. G. P. GM O. P. F.P P. C.
P018 X
Não
identificado/
Gramineae
X X X X X
P019 Fabaceae X X X X
P020 X Não
identificado X X X X
P021 X
Não
identificado/
Cecropia
sp. Miconia
sp.
X X X
P022 X X X
P023 X Cecropia
sp. X X X X
P024 X X
P025 X Rubiaceae X X X X
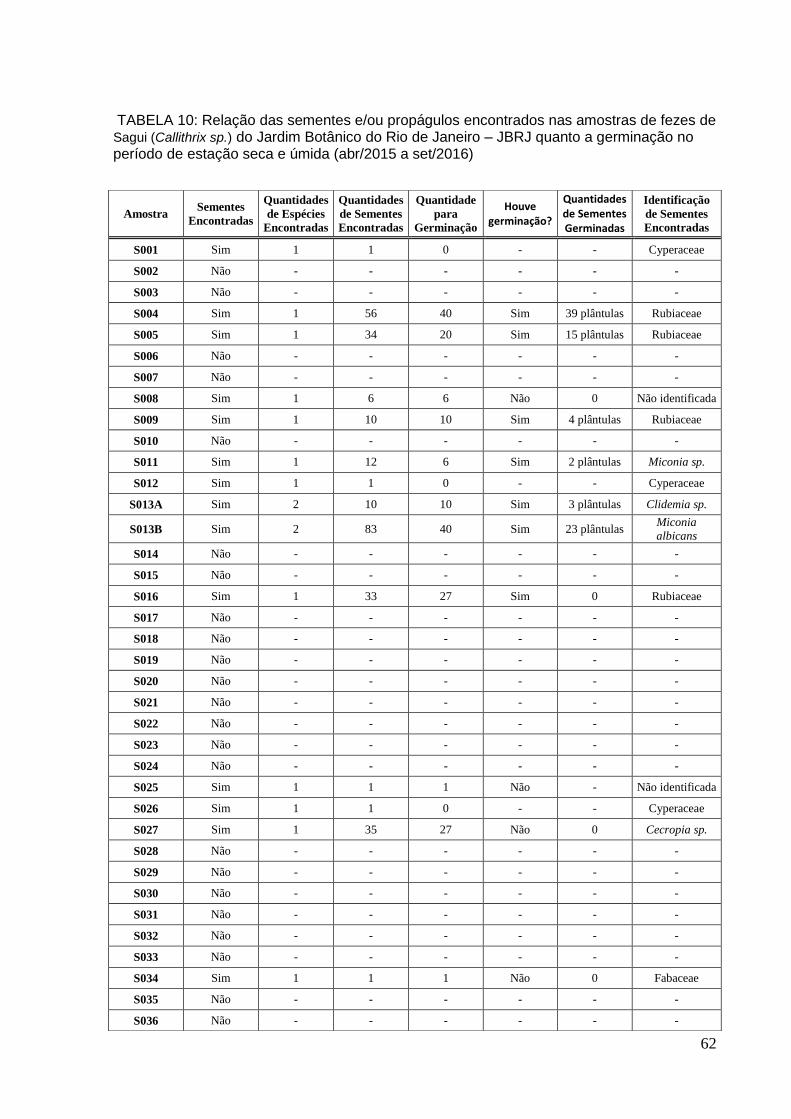
62
TABELA 10: Relação das sementes e/ou propágulos encontrados nas amostras de fezes de Sagui (Callithrix sp.) do Jardim Botânico do Rio de Janeiro – JBRJ quanto a germinação no período de estação seca e úmida (abr/2015 a set/2016)
Amostra Sementes
Encontradas
Quantidades
de Espécies
Encontradas
Quantidades
de Sementes
Encontradas
Quantidade
para
Germinação
Houve germinação?
Quantidades de Sementes Germinadas
Identificação
de Sementes
Encontradas
S001 Sim 1 1 0 - - Cyperaceae
S002 Não - - - - - -
S003 Não - - - - - -
S004 Sim 1 56 40 Sim 39 plântulas Rubiaceae
S005 Sim 1 34 20 Sim 15 plântulas Rubiaceae
S006 Não - - - - - -
S007 Não - - - - - -
S008 Sim 1 6 6 Não 0 Não identificada
S009 Sim 1 10 10 Sim 4 plântulas Rubiaceae
S010 Não - - - - - -
S011 Sim 1 12 6 Sim 2 plântulas Miconia sp.
S012 Sim 1 1 0 - - Cyperaceae
S013A Sim 2 10 10 Sim 3 plântulas Clidemia sp.
S013B Sim 2 83 40 Sim 23 plântulas Miconia
albicans
S014 Não - - - - - -
S015 Não - - - - - -
S016 Sim 1 33 27 Sim 0 Rubiaceae
S017 Não - - - - - -
S018 Não - - - - - -
S019 Não - - - - - -
S020 Não - - - - - -
S021 Não - - - - - -
S022 Não - - - - - -
S023 Não - - - - - -
S024 Não - - - - - -
S025 Sim 1 1 1 Não - Não identificada
S026 Sim 1 1 0 - - Cyperaceae
S027 Sim 1 35 27 Não 0 Cecropia sp.
S028 Não - - - - - -
S029 Não - - - - - -
S030 Não - - - - - -
S031 Não - - - - - -
S032 Não - - - - - -
S033 Não - - - - - -
S034 Sim 1 1 1 Não 0 Fabaceae
S035 Não - - - - - -
S036 Não - - - - - -
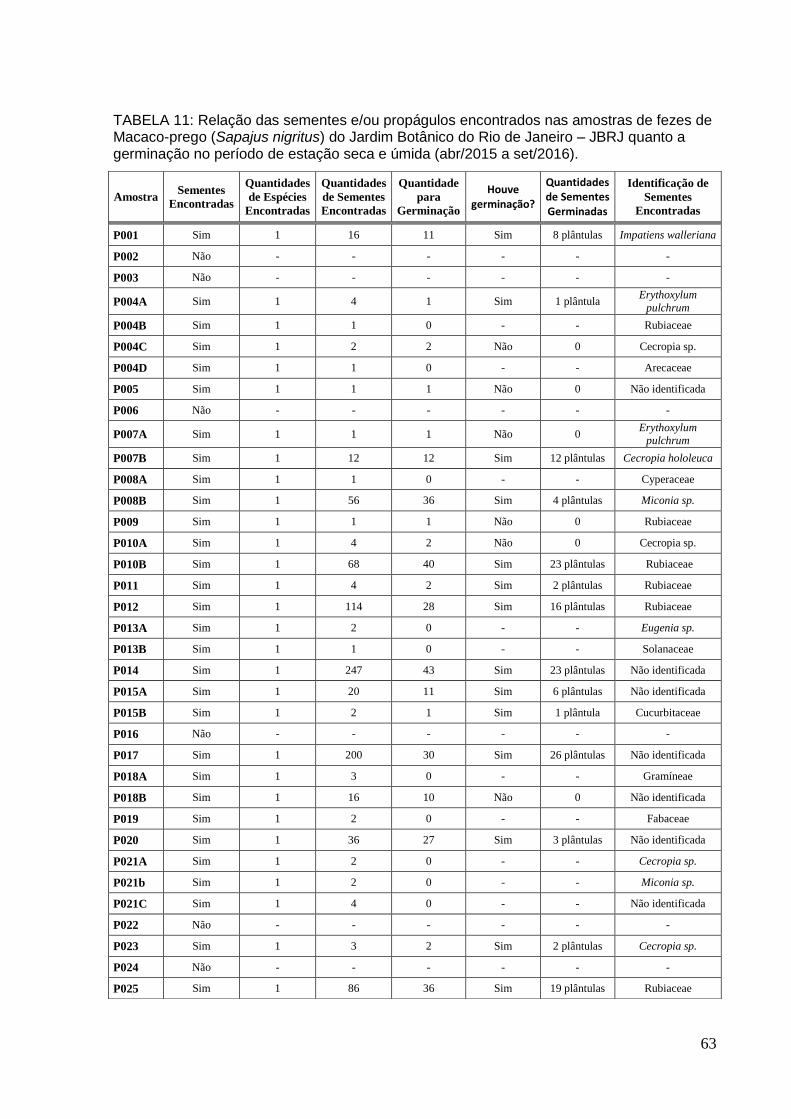
63
TABELA 11: Relação das sementes e/ou propágulos encontrados nas amostras de fezes de Macaco-prego (Sapajus nigritus) do Jardim Botânico do Rio de Janeiro – JBRJ quanto a germinação no período de estação seca e úmida (abr/2015 a set/2016).
Amostra Sementes
Encontradas
Quantidades
de Espécies
Encontradas
Quantidades
de Sementes
Encontradas
Quantidade
para
Germinação
Houve germinação?
Quantidades de Sementes Germinadas
Identificação de
Sementes
Encontradas
P001 Sim 1 16 11 Sim 8 plântulas Impatiens walleriana
P002 Não - - - - - -
P003 Não - - - - - -
P004A Sim 1 4 1 Sim 1 plântula Erythoxylum
pulchrum
P004B Sim 1 1 0 - - Rubiaceae
P004C Sim 1 2 2 Não 0 Cecropia sp.
P004D Sim 1 1 0 - - Arecaceae
P005 Sim 1 1 1 Não 0 Não identificada
P006 Não - - - - - -
P007A Sim 1 1 1 Não 0 Erythoxylum
pulchrum
P007B Sim 1 12 12 Sim 12 plântulas Cecropia hololeuca
P008A Sim 1 1 0 - - Cyperaceae
P008B Sim 1 56 36 Sim 4 plântulas Miconia sp.
P009 Sim 1 1 1 Não 0 Rubiaceae
P010A Sim 1 4 2 Não 0 Cecropia sp.
P010B Sim 1 68 40 Sim 23 plântulas Rubiaceae
P011 Sim 1 4 2 Sim 2 plântulas Rubiaceae
P012 Sim 1 114 28 Sim 16 plântulas Rubiaceae
P013A Sim 1 2 0 - - Eugenia sp.
P013B Sim 1 1 0 - - Solanaceae
P014 Sim 1 247 43 Sim 23 plântulas Não identificada
P015A Sim 1 20 11 Sim 6 plântulas Não identificada
P015B Sim 1 2 1 Sim 1 plântula Cucurbitaceae
P016 Não - - - - - -
P017 Sim 1 200 30 Sim 26 plântulas Não identificada
P018A Sim 1 3 0 - - Gramíneae
P018B Sim 1 16 10 Não 0 Não identificada
P019 Sim 1 2 0 - - Fabaceae
P020 Sim 1 36 27 Sim 3 plântulas Não identificada
P021A Sim 1 2 0 - - Cecropia sp.
P021b Sim 1 2 0 - - Miconia sp.
P021C Sim 1 4 0 - - Não identificada
P022 Não - - - - - -
P023 Sim 1 3 2 Sim 2 plântulas Cecropia sp.
P024 Não - - - - - -
P025 Sim 1 86 36 Sim 19 plântulas Rubiaceae

64