o cerradÃo e o cerrado sentido …repositorio.unb.br/bitstream/10482/14998/1/2013...iii o cerradÃo...
TRANSCRIPT

UNIVERSIDADE DE BRASÍLIA
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE BOTÂNICA
PROGRAMA DE PÓS-GRADUAÇÃO EM BOTÂNICA
O CERRADÃO E O CERRADO SENTIDO RESTRITO
NO JARDIM BOTÂNICO DE BRASÍLIA
RENATA ALVES CASEIRO
ORIENTADOR: MANOEL CLÁUDIO DA SILVA JÚNIOR
BRASÍLIA, ABRIL/2013

UNIVERSIDADE DE BRASÍLIA
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE BOTÂNICA
PROGRAMA DE PÓS-GRADUAÇÃO EM BOTÂNICA
RENATA ALVES CASEIRO
ORIENTADOR: MANOEL CLÁUDIO DA SILVA JÚNIOR
O CERRADÃO E O CERRADO SENTIDO RESTRITONO JARDIM BOTÂNICO DE BRASÍLIA
Dissertação submetida ao
Departamento de Botânica, do
Instituto de Ciências Biológicas da
Universidade de Brasília, como
requisito parcial para a obtenção do
título de Mestre em Botânica.
BRASÍLIAABRIL/2013

iii
O CERRADÃO E O CERRADO SENTIDO RESTRITONO JARDIM BOTÂNICO DE BRASÍLIA
Aprovado por:
______________________________________Dr. Manoel Cláudio da Silva Júnior
Departamento de Engenharia FlorestalUniversidade de Brasília – UnB
Presidente da Banda Examinadora
______________________________________Dr. José Roberto Rodrigues Pinto
Departamento de Engenharia FlorestalUniversidade de Brasília – UnB
Membro Titular da Banca
______________________________________Dr. Sabrina do Couto de Miranda
Departamento de BiologiaUniversidade Estadual de Goiás – Unidade de Itapuranga
Membro Titular
______________________________________Dra. Cássia Beatriz Rodrigues Munhoz
Departamento de BotânicaUniversidade de Brasília – UnB
Membro Suplente

iv
Ao meu pai Moacir(in memoriam)

v
Agradecimentos
Ao Professor Manoel Cláudio pela orientação, pela amizade, pelas agradáveis saídas de
campo, pelos conselhos sábios e por tudo o que você me ensinou sobre o Cerrado!
À Universidade de Brasília por oferecer cursos de pós-graduação e a CAPES por meconceder a bolsa de estudos!
Aos Professores Zé Roberto e Cássia Munhoz por terem participado de etapas importantes
do mestrado. Obrigada por todas as sugestões e correções!
À Kadja Milena, do herbário, por ter sido muito simpática em nos ajudar a identificar asLauraceae.
Ao Matheus, do curso da Engenharia Florestal, bolsista PIC da Engenharia Florestal emembro integrante do projeto, sempre presente nas saídas de campo e nas identificações dasplantas no herbário. Garoto GPS, nosso guia! Muito Obrigada!
Ao Felipe, aluno de mestrado também “desorientando” do Manoel Claudio, por nosacompanhar em algumas saídas de campo e por nos ajudar com as questões estatísticas do trabalho.
Ao meu colega do mestrado Thiago DeRoure, sempre muito gentil, me ajudou em váriasetapas do mestrado, a começar pela companhia nas saídas de campo, nas análises estatísticas, nasminhas escritas, na busca de artigos, e principalmente pelas palavras amigas durante os meusmomentos de desespero. Obrigada pela paciência!
À Milena e outros alunos da graduação da Engenharia Florestal que participaram de algumasdas saídas de campo e colaboraram com as coletas e identificação das árvores.
À minha querida amiga Thaís Costa, que nos acompanhou numa saída de campo quando
ninguém mais podia ir. Obrigada pela companhia em vários almoços pela UnB e pela bela amizadeque temos!
Aos meus colegas de curso pelas trocas de conhecimento e pelas prosas presenciais e viainternet!
Às minhas amigas Flávia e Irene, simplesmente por serem minhas amigas e fiéis companheirasde tantos anos. Amo vocês!
Agradeço com muito carinho a minha mãe, que sempre me ajudou, direta ou indiretamente!Obrigada por ser essa mãe tranquila, amiga e inteligente, que me ensinou a viver a vida comdignidade e respeito pelo próximo.

vi
Aos meus irmãos: Gabriela, Claudia e Roberto pela amizade fraterna e por vários quebra
galhos. Nossa amizade não tem preço, e sempre estaremos unidos! Agradeço também aos meussobrinhos Juju e Henrique, crianças lindas, e responsáveis por muitos momentos de felicidade naminha vida.
À minha tia Sandra por me ajudar na finalização desse trabalho! Obrigada pelo tempodisponibilizado.
Ao meu companheiro Luiz Felipe, pela parceria, carinho, amizade, paciência e, principalmente,te agradeço por me ajudar a produzir o presente mais lindo do mundo, nosso filho!
À minha querida Zefinha, que me ajudou e cuidou do meu filho durante os seis últimosmeses que eu me dediquei à escrita da dissertação! Sem ela tudo seria muito mais difícil. Obrigadademais!
À Deus por nos dar a vida e fornecer todos os recursos naturais que nos permitem viver.Agradeço a Ele pela criação dessa natureza exuberante que me inspira e me consola em meio amuitos problemas da sociedade humana!
E, por fim, agradeço a todos que fazem parte do meu dia-a-dia! Incluindo os gatos, o cachorro
e os pássaros do meu jardim!

vii
"O mundo não foi feito em alfabeto. Senão queprimeiro em água e luz. Depois árvore (...) "
Manoel de Barros

viii
Resumo Geral
O Cerrado no Brasil Central se destaca pela elevada diversidade em espécies e figura entre as
savanas mais ricas no mundo. A ampla variedade de fitofisionomias coexiste e se mistura mesmo
em condições edáficas e posições topográficas semelhantes. O Jardim Botânico de Brasília (JBB)
é considerado área importante de conservação da natureza, pois abriga as mais diversas
fitofisionomias do Cerrado. O objetivo deste estudo foi comparar a florística, a estrutura da
vegetação lenhosa e as propriedades químicas e físicas dos solos do cerradão (Cr) e do cerrado
sentido restrito denso (Cd), localizados no Centro de Visitantes do Jardim Botânico de Brasília
(CVJBB), protegidas do fogo há mais de 20 anos. Para isso, foram instaladas em cada área dez
parcelas permanentes de 20x50 m (1.000 m²). Foram registrados, identificados e medidos, quanto
a altura e diâmetro, todos os indivíduos com diâmetro da base, medido a 30 cm do solo, (Db30
cm) e” 5 cm, inclusive os indivíduos mortos em pé. Foram coletadas amostras compostas da
camada superficial do solo (10-20 cm de profundidade) de cada parcela nas duas áreas. Nestas
amostras foram realizadas análises químicas e físicas. Foram realizadas análises multivariadas a
fim de relacionar as propriedades químicas e físicas dos solos com a composição florística de
cada área. Na comparação florística Cr/Cd foram representados por 40/32 famílias, 14/3 famílias
exclusivas, 71/48 gêneros, 99/68 espécies. De um total de 122 espécies amostradas, 43 foram
compartilhadas pelas comunidades, entre elas apenas Emmotum nitens e Copaifera langsdorffii
são consideradas espécies florestais, o restante é comumente encontrado em áreas de cerrado
sentido restrito. Este resultado definiu os baixos valores encontrados para os índices de similaridade
de Sørensen e Chao-Sørensen com variação entre 0,06/0,47 e 0,03/0,50, respectivamente. As
famílias com maior representatividade foram Fabaceae, Vochysiaceae e Myrtaceae em ambas as
áreas. As espécies com maior IVI em Cr/Cd foram Emmotum nitens/Dalbergia miscolobium,
Siphoneugena densiflora/Miconia burchellii, Nectandra reticulata Poliouratea hexasperma,
Miconia cuspidata/Miconia ferruginata e Xylopia sericea/Qualea parviflora, que somaram 29,9%
no Cr e 28,3% no Cd do IVI total. A diversidade (H’) e a equabilidade (J’) foram, respectivamente,
para Cr/Cd de 3,55/3,43 e 0,76/0,81. Na estrutura, as diferenças Cr/Cd foram anotadas com
densidade total estimada em 2.412/2.193 ind.ha-1, área basal de 27,92/15,98 m².ha-1, altura média
(m) 6,6±2,10/3,9±2,7, altura máxima (m) 18/12, número de árvores mortas 272 (11%)/152 (7%)
e área basal das árvores mortas (m².ha-1) 3,14 (11%)/1,61 (10%). Dentre os parâmetros estruturais

ix
citados acima, calculados para cada área, apenas a densidade total não foi estatisticamente distinta
nas comparações Cr/Cd. A distribuição dos diâmetros nas comunidades apresentaram curvas
com aspecto J-reverso, indicando balanço positivo entre recrutamento e mortalidade. Ambas as
comunidades possuem solos argilosos, com baixos teores de bases trocáveis e alta saturação de
Al. Os solos no Cr apresentaram teores estatisticamente maiores de Al, Zn, Fe, K, Na, silte, CTC
e saturação por bases (m%), enquanto os níveis de P e saturação por bases (V%) foram
estatisticamente maiores nos solos do Cd. As comunidades Cr/Cd se apresentaram floristicamente
e estruturalmente diferenciadas apesar da proximidade geográfica. A classificação pelo TWINSPAN
separou o Cr do Cd e apontou homogeneidade florística entre as parcelas em cada área. Os
resultados obtidos pela PCA explicaram 78% da variância total e distinguiram em parte dos
resultados do TWINSPAN. A PCA não apontou relação das variáveis do solo com a distribuição
das espécies em cada área. Os maiores níveis de cátions no Cr não esclarecem a maior área basal
e altura da vegetação que teoricamente necessitaria de maior aporte de nutrientes para seu
crescimento, como maiores níveis de macronutrientes e, consequentemente, maior saturação por
bases. Os níveis mais elevados de silte, matéria orgânica e argila nas parcelas do Cr podem
contribuir para a conservação da água e dos nutrientes nas camadas superiores no perfil dos solos
e estes seriam, então, melhor utilizados na vegetação do Cr.
Palavras-chave – Bioma Cerrado, fitofisionomias, índice de valor de importância (IVI), diversidade, similaridadeflorística e solos.

x
Abstract General
The Cerrado in central Brazil stands out for its high diversity in species and ranks among the
richest savanna in the world. Wide variety of vegetation coexists and mixes, even in similar edaphic
conditions. Brasilia’s Botanic Garden (JBB) is a unit of nature conservation which houses the
most diverse vegetation types, being considered an important area for research on the biodiversity
of the Cerrado. The purpose of this work was to compare the floristic, the structure of woody
vegetation and the chemical and physical properties of the soil of cerradão (Cr) and of dense
cerrado strict sensu (Cd), located at the Brasilia Botanic Garden’s visitor center (DF) protected
from fire for over twenty years. Ten permanent plots were installed at the Cr and Cd of 20x50 m
(1.000 m²). All individuals with base diameter were registered, identified and measured, as for
height and diameter, measured at 30 cm of soil, (Db30 cm) e” 5 cm, including standing dead
individuals. Composed samples were collected from the surface soil layer (0-20 cm deep) of each
plot. Chemical and physical analyzes were performed. PCA and TWISNPAN classification related
analysis were carried out in order to establish the relationships among tree species sampled with
5 or more individuals and statistically distinct soil properties. In multivariate analyzes included all
species shown. Cr/Cd floristic comparison showed 40/32 families, 14/3 exclusive families, 71/48
genera, 99/68 species. A total of 43 species were shared by the communities, among them only
Emmotum nitens and Copaifera langsdorffii are considered forest species, the rest is commonly
found in areas of cerrado strictu sensu. This result set the low values found for the similarity
indices of Sorensen and Chao-Sorensen respectively varying between 0,06/0,47 and 0,03/0,50.
The most representative families were Fabaceae, Vochysiaceae and Myrtaceae in both areas. The
species with the highest IVI in Cr/Cd were Emmotum nitens/Dalbergia miscolobium,
Siphoneugena densiflora/Miconia burchellii, Nectandra reticulate/Poliouratea hexasperma,
Miconia cuspidata/Miconia ferruginata e Xylopia sericea/Qualea parviflora, IVI totaling in
29,9 Cr and 28,3% Cd. The (H’) diversity and (J’) equability were, respectively, for Cr/Cd of
3,55/3,43 and 0,76/0,81. In structure, the Cr/Cd differences were noted with a total estimated
density in 2.412/2.193 ind.ha--1, basal area of 27,92/15,98 m².ha-1, 6,6±2,10/3,9±2,7 (m) medium
height, 18/12 (m) maximum height, number of dead trees 272 (11%)/152 (7%) and basal number
of dead trees (m².ha-1) 3,14 (11%)/1,61 (10%). Among the structural parameters mentioned above,
calculated for each area, only the total density was not statistically distinct in Cr/Cd comparisons.
Both Cr and Cd community diameter distribution showed an inverted J curve which revealed a

xi
positive balance among recruitment and mortality. Cr and Cd were associated to clayey soils,
showing low exchangeable bases levels and high A1 saturation. The cerradão soils showed
significantly higher A1, Zn, Fe, K, Na, silte, CTC and (m%) base saturation, while the levels of P
and (V%) base saturation were statically larger on the soil of Cd. The Ca and Mg showed exactly
the same levels in all plots of both areas. The communities Cd/Cr showed floristically and
structurally differentiated despite the geographical proximity. The TWINSPAN classification
separated both communities and pointed floristic homogeneity among Cr and Cd plots. The results
by APC explained 78% of the total variance and distinguished in part of the TWINSPAN results.
The PCA showed no relationship of soil variables with the distribution of species in each area.
The highest levels of cations in the Cr did not explain the higher basal area and height of vegetation
which theoretically would require higher nutrient for growth, as increased levels of macronutrients
and saturation. The higher levels of silt organic matter and clay on portions of Cr may contribute
to the conservation of water and nutrients in the upper layers of the soil profile which would be,
then, best used in the vegetation Cr.
Key words – Cerrado biome, phytofisiognomies, importance value index (IVI), diversity, floristic similarityand soil.

Sumário
Resumo Geral ..........................................................................................................................viii
Abstract General ........................................................................................................................ x
Introdução Geral ........................................................................................................................ 3
Revisão Bibliográfica ................................................................................................................. 6
As savanas e o bioma Cerrado ................................................................................................. 6
Principais fatores que influenciam as paisagens do bioma Cerrado ......................................... 7
O Cerradão e o cerrado sentido restrito: semelhanças florísticase diferenças estruturais da vegetação ...................................................................................... 11
Capítulo 1................................................................................................................................. 16Comparação da vegetação lenhosa do cerradãoe do cerrado denso no Jardim Botânico de Brasília ............................................................ 16
Resumo ................................................................................................................................... 16
Introdução............................................................................................................................... 17
Material e Métodos ................................................................................................................. 18
Resultados .............................................................................................................................. 25
Discussão................................................................................................................................ 37
Conclusões .............................................................................................................................. 41
Capítulo 2................................................................................................................................. 42Relações solo e vegetação no cerradão eno cerrado denso no Jardim Botânico de Brasília ............................................................... 42
Resumo ................................................................................................................................... 42
Introdução............................................................................................................................... 43
Material e Métodos ................................................................................................................. 44
Resultados .............................................................................................................................. 46
Discussão................................................................................................................................ 50
Conclusões .............................................................................................................................. 54
Conclusões Gerais .................................................................................................................... 55
Referências Bibliográficas ........................................................................................................ 56

Índice de figuras e tabelas
Capítulo 1Figura 1. Carta imagem da Localização do Centro de Visitantes do Jardim Botânico de Brasília(CVJBB-JBB) (área delimitada em vermelho) e locais de edificações existentes (área delimitadaem branco e pontos amarelos). Fonte: Geo Lógica - [email protected]. ..................................................... 20Figura 2. Vista da estrutura interna do cerradão (Cr), localizado no Jardim Botânico de Brasília (DF).Em detalhe o adensamento da cobertura arbórea, galhos emaranhados e a camada de serapilheirasobre o solo. Foto: Manoel Cláudio. ............................................................................................................................. 21Figura 3. Imagem da cobertura arbórea do cerradão (Cr), localizado no Jardim Botânicode Brasília (DF). Foto: Manoel Cláudio. ...................................................................................................................... 21Figura 4. Vista da estrutura externa do cerrado denso (Cd), localizado no Jardim Botânicode Brasília (DF). Em detalhe a baixa estatura das árvores, galhos baixos e densos e o solode cor avermelhada e pouca camada de serapilheira. Foto: Manoel Cláudio. ............................................................. 22Figura 5. Imagem da cobertura arbórea do cerrado denso (Cd), localizado no Jardim Botânicode Brasília (DF). Foto: Manoel Cláudio. ...................................................................................................................... 22Figura 6. Localização das áreas amostradas no Centro de Visitantes do Jardim Botânicode Brasília (CVJBB-DF). Cerradão (Cr) e cerrado denso (Cd) distam 2 km de distânciaem linha reta (laranja). Fonte: Google Earth. ............................................................................................................... 23Figura 7. Curvas de rarefação baseadas no número de indivíduos amostrados e nonúmero de parcelas (10) de cada área, localizados no Jardim Botânico de Brasília (DF).Cr= cerradão e Cd= cerrado denso. ............................................................................................................................... 25Figura 8. Distribuição dos indivíduos por classes de diâmetro no cerradão (Cr) eno cerrado denso (Cd) do Jardim Botânico de Brasília (DF). ...................................................................................... 29Figura 9. Distribuição dos indivíduos por classes de altura no cerradão (Cr) eno cerrado denso (Cd) do Jardim Botânico de Brasília, DF. ........................................................................................ 30Figura 10. Distribuição dos indivíduos mortos em pé por classes de diâmetrono cerradão (Cr) e cerrado denso (Cd) no Jardim Botânico de Brasília (DF). ............................................................ 30
Tabela 1. Comparação das características fitossociológicas do cerradão (Cr) ecerrado denso (Cd), localizados no Jardim Botânico de Brasília (DF). ....................................................................... 26Tabela 2. Índices de similaridade de Chao-Sørensen (abaixo) e Sørensen (acima)entre as 20 parcelas amostradas nas áreas de cerradão (Cr) e cerrado denso (Cd),localizados no Jardim Botânico de Brasília (DF). .................................................................................................... 27Tabela 3. Fitossociologia das espécies lenhosas, com suas respectivas famílias, do cerradão(Cr) e do cerrado denso (Cd), localizados no Jardim Botânico de Brasília (DF).DA= densidade absoluta; DR= densidade relativa; FA: frequência absoluta; FR= frequência relativa;DoA: dominância absoluta; DOR= dominância relativa, e IVI= índice do valor de importância. ........................ 31
Capítulo2Figura 1. Análise de Componentes Principais de oito variáveis do solo Al, H+Al, K, Fe, Zn. Na,P e silte, que foram estatisticamente distintas entre o cerradão (Cr) e o cerrado denso (Cd),localizados no Jardim Botânico de Brasília (DF). ........................................................................................................ 49Figura 2. Classificação pelo método TWINSPAN das 20 parcelas amostradas, dez no cerradão(Cr) e dez no cerrado denso (Cd), localizados no Jardim Botânico de Brasília (DF). Os valoresentre parênteses referem-se aos autovalores. ................................................................................................................ 50
Tabela 1. Propriedades químicas e físicas dos solos (10-20 cm de profundidade) amostradasno cerradão (Cr) e no cerrado denso (Cd), localizados no Jardim Botânico de Brasília (DF). MO:matéria orgânica; CTA: capacidade de troca catiônica; V: saturação por bases; m: saturação poralumínio; DP: desvio padrão; p: nível de significância (p < 0,05) pelo teste t de Mann-Whitney;***: significativo e ns: não significativo. ...................................................................................................................... 47

3
Introdução Geral
Uma associação de fatores ambientais, tais como clima estacional, geomorfologia, classes
de solos, disponibilidade de água para as plantas, fogo, herbivoria e ação antrópica figuram entre
os principais determinantes da paisagem no Cerrado (Eiten, 1972; Lopes & Cox, 1977; Coutinho,
1982; Cole, 1986; Furley, 1999; Coutinho, 2002; Ludwing, 2004; Ribeiro & Walter, 1998; 2008).
Segundo Eiten (1972) e Cole (1986) o clima e o solo exerceriam os efeitos mais significativos na
fisionomia e na distribuição das savanas.
O clima sazonal, com invernos secos e verões chuvosos, tem influência temporal na origem
do Cerrado, pois as chuvas ao longo do tempo geológico intemperizaram os solos deixando-os
pobres em nutrientes essenciais (Walter, 2006). Os períodos secos permitem que as constantes
queimadas causem impactos importantes na estrutura e na composição florística da vegetação
(Coutinho, 1982). Contudo, regiões do Cerrado no limite com a Caatinga e na interface com a
Amazônia têm características climáticas próprias, com precipitações que variam entre 600 a
2.200mm por ano e duração do período seco que varia de três a oito meses (Reatto & Martins,
2005).
Diante das particularidades climáticas, existe também grande variedade de classes de solos
que suportam as diferentes paisagens do Cerrado (Reatto & Martins, 2005). No geral, as
fitofisionomias apresentam solos em sua maioria ácidos, com baixa capacidade de troca catiônica
e alto nível de saturação por alumínio, que por sua vez, afetam os sistemas radiculares e inibem a
absorção de cálcio e fósforo (Furley & Ratter, 1988).
Aproximadamente 46% do território do Cerrado (Reatto et al., 2008) é ocupado por
Latossolos, a mais extensa classe de solos na região (Reatto et al., 2008). Estes são, em geral,
profundos e bem drenados (Haridasan, 2000), frequentemente distróficos com níveis muito baixos
de N, P e outros cátions e níveis extremamente altos de Al (Goodland, 1971; Haridasan, 1982;
Sarmiento, 1984). As paisagens mais comuns que ocupam esses solos são o cerrado sentido
restrito e o cerradão (Reatto & Martins, 2005). Estas fitofisionomias podem ocorrer em curtas
distâncias geográficas, ao longo de gradientes de vegetação ou em áreas contíguas (Costa &
Araújo 2001; Marimom-Júnior & Haridasan, 2005; Campos et al., 2006). O cerradão é considerado
fisionomia florestal distinta, apesar de não apresentar identidade florística (Felfili et al., 1994;
Ribeiro & Walter, 2008; Mendonça et al., 2008). A composição florística varia de acordo com a

4
fertilidade do solo, que podem ser considerados distróficos ou mesotróficos (Ratter et al., 1977,
Araújo & Haridasan, 1988). Dados na literatura apontam algumas espécies que são consideradas
indicadoras para cada ambiente (Ratter et al., 1971; 1977; 1978; Araújo & Haridasan, 1988;
Felfili et al. 1994).
A fitofisionomia do cerrado sentido restrito apresenta subtipos: denso, típico, ralo e rupestre,
onde a cobertura de árvores e arbustos pode variar de 5% a 70% (Eiten, 1972; Ribeiro & Walter,
2008). Os subtipos denso, típico e ralo se subdividem de acordo com a densidade arbórea (Ribeiro
& Walter, 2008) e, o rupestre se diferencia por habitar solos rochosos. No geral, o cerrado
sentido restrito ocorre em solos distróficos e, raramente, em solos mesotróficos, mas quando
ocorre em solos mais férteis tem sua composição florística variada (Haridasan, 2000).
A comparação entre cerradão e o cerrado sentido restrito em áreas geograficamente
próximas, apontam similaridade florísticas e semelhanças nas propriedades dos solos que chamam
a atenção dos pesquisadores (Solorzano et al., 2012). No cerradão ocorrem espécies florestais,
savânicas e generalistas (Solorzano et al., 2012), mas caracteriza-se preferencialmente pela presença
de espécies que ocorrem no cerrado sentido restrito (Walter & Ribeiro, 2010). A similaridade
florística entre estas fitofisionomias variam e não seguem um padrão fitogeográfico (Solorzano,
2011; Solorzano et al., 2012). Costa & Araújo (2000) indicaram que no cerradão distrófico,
amostrado por eles em Minas Gerais, a vegetação foi grandemente influenciada pelas comunidades
vizinhas, o que resultou na elevada similaridade florística com o cerrado sentido restrito adjacente.
De acordo com Marimon-Junior & Haridasan (2005) as diferenças florísticas e estruturais entre
o cerradão e o cerrado sentido restrito, podem ocorrer apesar da proximidade geográfica e das
semelhanças na composição química dos solos.
No Centro de Visitantes do Jardim Botânico de Brasília ocorre o subtipo cerrado denso,
bem como a fitofisionomia de cerradão, em áreas geograficamente próximas, protegidas do fogo
há mais de vinte anos e com características climáticas iguais. Dentro deste contexto, os fatores
que parecem exercer influências na distribuição do cerradão e do cerrado denso são o solo e a
disponibilidade de água para as plantas, bem como as oscilações do lençol freático. A proximidade
entre essas comunidades se apresenta como excelente experimento natural para a investigação
sobre as relações ecológicas determinantes destas paisagens e, dessa forma, possibilitou a realização
deste estudo comparativo e também levantar as seguintes questões:
1. Qual é a composição florística e a estrutura da vegetação lenhosa no cerrado sentido
restrito denso (Cd) e cerradão (Cr) no Centro de Visitantes do Jardim Botânico de Brasília
(CVJBB)?

5
2. Qual é a similaridade florística entre as duas comunidades vizinhas?
3. Existem diferenças significativas entre os parâmetros estruturais (densidade, área basal e
altura) nessas comunidades?
4. Como os diâmetros e alturas dos indivíduos lenhosos se distribuem nessas comunidades?
5. Existem diferenças significativas entre as propriedades químicas e físicas dos solos nessas
comunidades?
6. Existem associações entre a vegetação e as propriedades dos solos nessas comunidades?
Para tanto, foram alocadas dez parcelas de 20x50 (1.000 m²) em cada área, e analisados os
parâmetros fitossociológicos convencionais da vegetação lenhosa (Müller-Dombois & Ellenberg,
1974). Foram calculados também a riqueza florística pelos estimadores Chao 1 e Chao 2, a
diversidade alfa por meio do índice de Shannon e da equabilidade de Pielou (Magurran, 2004) e
a similaridade florística pelos índices de Sørensen e Chao-Sørensen (Kent & Coker, 1992; Chao
et al., 2005). A estrutura de tamanhos (diâmetro e altura) também foi verificada a partir de intervalos
de classes fixos.
A vegetação lenhosa das áreas comparadas foi classificada pelo método TWINSPAN (Hill,
1979) (Two-Way Indicator Species Analysis). Também foram coletadas em cada parcela, amostras
compostas da camada superficial do solo (10-20 cm), e assim, analisadas quanto às propriedades
químicas e físicas. As variáveis do solo, estatisticamente distintas entre as áreas, foram ordenadas
pela PCA e, posteriormente, relacionadas com os resultados obtidos pelo TWINSPAN entre as
áreas e para cada área.
Os resultados serão apresentados e discutidos da forma que se segue:
Capítulo 1. Comparação da vegetação lenhosa do cerradão e do cerrado denso no Jardim
Botânico de Brasília.
Capítulo 2. Relações solo e vegetação no cerradão e no cerrado denso no Jardim Botânico
de Brasília.

6
Revisão Bibliográfica
As savanas e o bioma Cerrado
As savanas são caracterizadas por paisagem com predominância do estrato rasteiro e com
densidades variáveis dos componentes arbóreo e arbustivo, ocorrem comumente nos trópicos e
subtrópicos em áreas com forte sazonalidade, onde o verão é chuvoso e o inverno é seco (Cole,
1986). São ecossistemas considerados dinâmicos, determinados por fatores físicos, químicos e
biológicos em diferentes escalas espaciais e temporais. Além disso, o uso das savanas pelo homem
também é considerado fator determinante de suas diferentes paisagens (Mistry, 2000). Os longos
períodos de seca propiciam a propagação do fogo, que frequentemente ocorre pela ação antrópica.
O fogo não só influencia a biomassa total nas savanas, como também o microclima e a distribuição
de recursos em escala local, como a umidade do solo e os nutrientes (Cole, 1986; Mistry, 2000;
Ludwing, 2004).
O Cerrado é a savana brasileira que ocorre principalmente na região central do Brasil. Segundo
as observações de Cole (1986), Eiten (1972), Lopes & Cox (1979) e Ribeiro & Walter (2008), o
Cerrado exibe diversas formas fisionômicas que variam desde campos graminosos (campo limpo)
até formações florestais com dossel parcialmente fechado (cerradão). Coutinho (1978) sugere
que o Cerrado apresenta dois extremos: o cerradão, com vegetação arbóreo-arbustiva, e o campo
limpo com componentes herbáceos e subarbustivos. Entre estes extremos figuram as formações
campestres e savânicas que se incluem no conceito de Cerrado sentido amplo ou sensu lato
(Eiten, 1972).
Segundo Coutinho (1978), alguns autores defendem que o termo savana não está de acordo
com a enorme variedade de fitofisionomias que compõem o bioma Cerrado, em especial as
fisionomias florestais. Para Walter (2006), o bioma como um todo não é savana, uma vez que
nele ocorrem florestas (as matas de galeria, matas secas e cerradão) e campos puros, mas é
caracterizado primordialmente por típica vegetação de savana, com solo coberto por vegetação
graminosa onde se distribuem arbustos e árvores em diferentes densidades. De acordo com Mistry
(2000), as savanas são heterogêneas entremeadas com outros tipos de vegetações, como por
exemplo, as matas de galeria e ciliares. No geral, o conceito de Cerrado designa a vegetação
xeromorfa arbórea, arbustiva, savânica e campestre (Eiten, 1978). Dentre as fitofisionomias

7
existentes, a mais característica e de maior ocupação territorial é denominada cerrado sentido
restrito (Ribeiro & Walter, 2008) ou strictu sensu (Eiten, 1972).
Recentemente, Ribeiro & Walter (2008) definiram o Cerrado como enorme mosaico
vegetacional composto por formações savânicas (cerrado sentido restrito, parque cerrado, palmeiral
e vereda), campestres (campo rupestre, campo sujo e campo limpo) e florestais (mata seca, mata
de galeria, mata ciliar e cerradão) que coexistem e se misturam na paisagem do Brasil Central.
Principais fatores que influenciam as paisagens do bioma Cerrado
Entre os fatores que influenciam o mosaico de fitofisionomias que representam o Cerrado
destacam-se o clima estacional, a disponibilidade de água para as plantas, o histórico de fogo, as
condições edáficas e a geomorfologia das regiões (Lopes & Cox, 1977; Coutinho, 1982; Furley,
1999; Ribeiro & Walter, 2008). Segundo Eiten (1972) e Cole (1986) o clima e o solo exerceriam
os efeitos mais significativos na fisionomia e na distribuição das savanas.
O clima do Cerrado é estacional, com período chuvoso de outubro a março e período seco de
abril a setembro. A precipitação média anual é de 1.500 mm e as temperaturas variam ao longo do
ano entre 18ºC e 28ºC em média (Dias, 1992; Klink & Machado, 2005). Assim como em todas as
savanas, o clima sazonal com invernos secos e verões chuvosos permite que as constantes queimadas
causem impactos importantes na estrutura e na composição florística da vegetação (Coutinho,
1982). O clima também tem influência temporal na origem do Cerrado, pois as chuvas, ao longo do
tempo geológico intemperizaram os solos, deixando-os pobres em nutrientes essenciais (Walter,
2006). Contudo, regiões do Cerrado no limite com a Caatinga e na interface com a Amazônia têm
características climáticas próprias, com precipitações que variam entre 600 a 2.200mm por ano e
duração do período seco que varia de três a oito meses (Reatto & Martins, 2005).
Os períodos secos na região do Cerrado permitem que as constantes queimadas causem
impactos importantes na estrutura e na composição florística das formações vegetais (Coutinho,
1982). O fogo é coadjuvante antigo no Cerrado e nas savanas no mundo. Na América do Sul é
datada a ocorrência de queimadas muito antes da chegada dos humanos (Miranda et al., 2010),
de forma que a maioria dos organismos já estava adaptada aos incêndios, não como evento
casual, mas como um regime de queima definido pela época e frequência de ocorrência (Eiten,
1972; Miranda et al., 2010). O registro mais antigo de fogo na flora do Cerrado foi encontrado
no Planalto Central brasileiro e remonta aproximadamente 32.000 anos AP (Ledru, 2002).

8
A velocidade e a combustão do fogo dependem da composição e da dinâmica do combustível,
como serapilheira, galhos de árvores e arbustos, que estão relacionados com a fisionomia da
vegetação. Dependendo da época de queima, do período sem chuva e da velocidade dos ventos,
os incêndios podem atingir altas velocidades e destruir grandes áreas das vegetações no Cerrado
(Miranda et al., 2010).
De acordo com as diferentes formações no bioma, as fitofisionomias florestais estão menos
sujeitas aos incêndios naturais do que as savânicas e campestres. Entre as formações florestais, as
matas de galerias representam a vegetação menos adaptada ao fogo, enquanto a flora das savanas
e dos campos possuem inúmeras espécies adaptadas com características morfológicas, anatômicas
e fisiológicas ao fogo (Walter & Ribeiro, 2010). O fogo influencia a distribuição e a composição
florística das fitofisionomias por afetar a estrutura dos trechos de vegetação. Tal fator é
determinante para os ritmos fenológicos na comunidade rasteira, além de alterar a dinâmica de
comunidades vegetais, afetar a frequência de espécies raras e alterar ou não a dominância de
espécies comuns (Munhoz & Amaral, 2010; Walter, 2006).
O fogo também exerce influência positiva e negativa ao solo (Coutinho, 1990), pois, após a
sua passagem as concentrações de certos nutrientes minerais nas camadas superficiais do solo
podem aumentar, assim como alguns nutrientes são perdidos por volatização (Cavalcanti, 1978;
Brinkley et al., 1994; Pomianoski, 2005). Com a queimada da biomassa vegetal e da matéria
orgânica do solo, as cinzas com alta concentração de P, K e Ca (Owensby & Wyrill, 1973; Coutinho,
1990) são arrastadas pelas águas ou lixiviados em profundidades que prejudicam o processo de
ciclagem biogeoquímica que diretamente afetam a produtividade vegetal (Jordan, 1987; Oliveira,
1994).
Durante muito tempo pesquisadores consideraram a água como fator limitante para as
plantas nativas do Cerrado devido aos longos períodos de seca e pelas características xéricas da
vegetação (Goodland & Ferri, 1979). Tais características nas plantas do Cerrado podem estar
associadas à origem evolutiva da vegetação do tipo savana, quando a disponibilidade de água,
que se tornou muito baixa em algum ponto durante as glaciações no período Quaternário (Ferri,
1977), teria selecionado as espécies mais adaptadas às condições secas.
Estudos recentes sobre a variação da umidade nos solos indicaram que nas camadas mais
profundas, o mesmo permanecia úmido mesmo na estação seca (Quesada et. al, 2004). Os níveis
do lençol freático são determinados pela quantidade de chuva na última estação chuvosa e a água
armazenada se torna disponível para as plantas durante a estação seca (Schubart &Rawitscher,

9
1950 apud Goodland & Ferri, 1979). Dessa forma, a estrutura da vegetação também estaria
relacionada com a disponibilidade de água e com as oscilações no nível do lençol freático e não
com a escassez de água (Haridasan, 1991).
Várias espécies lenhosas do Cerrado têm raízes profundas (Rawitscher, 1948 apud Franco,
2004; Jackson et al., 1999), mas o simples acesso às reservas de água no subsolo não garante
água suficiente para fazer frente à demanda evaporativa da atmosfera, mesmo durante a estação
chuvosa (Franco, 2004). Portanto, o sucesso de plantas no bioma depende da sua capacidade de
manter um balanço hídrico adequado durante o período seco e de minimizar os efeitos do déficit
hídrico no seu balanço anual de carbono. Os ritmos fenológicos, especialmente a produção e
perda de folhas, e o potencial de aclimatação do seu aparato fotossintético às condições
contrastantes de luminosidade, que podem ocorrer ao longo do seu ciclo de vida, são estratégias
utilizadas pelas plantas para minimizar ou escapar dos efeitos deletérios da alternância do regime
hídrico (Franco, 2004).
As características e as propriedades dos solos variam bastante entre os trópicos e entre
regiões nos trópicos. Os solos do Cerrado, conforme critérios utilizados na agronomia apresentam
baixa fertilidade e, em geral, são profundos e bem drenados (Haridasan, 2000). O nitrogênio, o
fósforo e vários cátions ocorrem em níveis muito baixos e os teores de alumínio no solo podem
ser extremamente altos (Haridasan, 1982; Sarmiento, 1984).
O Latossolo é a classe de solo mais extensa na região do Cerrado e cobre cerca de 50% da
região (Reatto et al., 2008). Os Latossolos são solos muito intemperizados, em geral, profundos,
bem drenados, fortemente ácidos, com baixa saturação por bases, distróficos ou alumínicos
(EMBRAPA, 2006). No entanto, ocorrem Latossolos com média e até mesmo alta saturação por
bases, encontrados geralmente em zonas com seca pronunciada, semiáridas ou não, ou ainda por
influência de rochas básicas ou calcárias (EMBRAPA, 2006). Apresentam pH baixo e elevados
teores de sesquióxidos de ferro e alumínio, características que aumentam a capacidade de retenção
do fósforo em formas pouco solúveis, sendo este nutriente de baixa disponibilidade para as plantas
(Goodland, 1971). Essa baixa fertilidade dos solos pode ser observada nas pequenas concentrações
de nutrientes nas folhas das espécies nativas em comunidades associadas aos solos distróficos
(Haridasan, 1982; Haridasan & Araújo, 1988).
Os Latossolos são representados por: Latossolo Vermelho, no qual ocupa 22,1% da área do
Cerrado; Latossolo Vermelho-Amarelo, com 21,6% da área; e Latossolo Amarelo que ocupa 2%
da área do Cerrado. Os Latossolos Vermelhos nas áreas mais altas do Planalto Central do Brasil

10
têm boa drenagem interna, como mostrado por sua cor vermelha, que indica um ambiente oxidante
devido à hematite, um mineral de óxido de ferro à fracção de argila (Cornell & Schwertmann,
1996). Nas áreas mais baixas o Latossolo Vermelho-Amarelo têm as cores vermelho-amarelo do
mineral óxido de ferro, goetita, o que indica condições de solo mais úmido do que em solos
avermelhados (Oliveira-Filho & Ratter, 2002).
Além dos Latossolos, as outras classes de solo – Neossolos, Argissolos, Cambissolos,
Plintossolos, Litólicos, Planossolos e Gleissolos – estão associadas às condições de clima e
estabelecem grande diversidade de espécies vegetais, seja do estrato graminoso, arbustivo ou
arbóreo (Oliveira-Filho & Ratter, 2002; Reatto et al., 2008). Segundo Haridasan (2000), os
fatores edáficos como profundidade do solo, presença de concreções no perfil, proximidade à
superfície do lençol freático, drenagem e fertilidade influenciam as diferenças entre fitofisionomias
no bioma.
O Cerrado ocorre ao longo de inúmeras formações geológicas, tipos de topografias e em
altitudes que variam de aproximadamente 300m, na Baixada Cuiabana (MT), a mais de 1.600 m
na Chapada dos Veadeiros (GO) (Cole, 1986; Ribeiro & Walter, 2008). A topografia da maior
parte na área central do Brasil consiste de plano a suavemente ondulado além dos interflúvios,
mais acidentados, onde ocorrem campos, matas de galeria e ciliares ao longo dos córregos (Eiten,
1972). A associação geral das formações savânicas, florestais e campestres, com superfícies de
aplainamento e com terrenos inclinados sugerem que suas distribuições podem ser influenciadas
por condições de solo e de águas subterrâneas.

11
O Cerradão e o cerrado sentido restrito:
semelhanças florísticas e diferenças estruturais da vegetação
Diante dos fatores ambientais que influenciam a distribuição das fitofisionomias do Cerrado,
o cerradão e o cerrado sentido restrito ocorrem, na maioria das vezes, em áreas adjacentes seguindo
gradientes vegetativos (Marimon-Junior e Haridasan, 2005). Muitos cerradões associados a solos
distróficos possuem nível de fertilidade, pH e teores de alumínio semelhantes aos dos solos
distróficos colonizados por cerrado sentido restrito. São fitofisionomias estruturalmente diferentes
onde composição e quantidade das espécies comuns variam de local para outro (Costa & Araújo
2000; Marimom-Junior & Haridasan, 2005).
Na descrição de Ribeiro & Walter (2008) o cerradão é formação florestal que possui
cobertura arbórea que varia de 50% a 90%, semelhante à de mata de galeria. Geralmente apresenta
três estratos distinguíveis: arbóreo, arbustivo e herbáceo (Ribeiro& Walter, 2008). O estrato
arbóreo pode variar de 6 a 7 m de altura até apresentar árvores com 15 a 20 m (Ratter et al.,
1977). O segundo estrato composto por arbustos e arvoretas apresenta altura entre 2 e 5 m, já o
estrato herbáceo geralmente é pouco desenvolvido, e está quase ausente nas porções com maior
cobertura do dossel e, consequentemente, com maior sombreamento (Oliveira-Filho & Ratter,
2002). O cerradão coloniza locais bem drenados, em solos profundos (Ribeiro & Walter, 2008),
com características de posicionamento topográfico semelhante ao do cerrado sentido restrito.
A composição florística dos cerradões varia conforme a fertilidade do solo. Podem ocorrer
tanto em solos distróficos quanto mesotróficos (Ratter et al., 1977; Haridasan & Araújo, 1988).
Quando mesotrófico é caracterizado por ocorrer em solos com níveis médios de nutrientes: teor
de cálcio maior que 2 meq/100 g de solo e pH entre 5,5 e 7,0. Já em solo distrófico é caracterizado
por apresentar pH entre 4,0 e 4,8 e teores de cálcio menores que 0,5 meq/100 g de solo (Ratter
et al., 1978). Em relação às espécies, os cerradões foram caracterizados como cerradão mesotrófico
ou cerradão de Callisthene fasciculata e Magonia pubescens (Ratter et al., 1971; 1977) e cerradão
distrófico ou cerradão de Hirtella gandulosa (Ratter et al., 1971). Em cerradões distrófico e
mesotrófico de Goiás, Haridasan & Araújo (1988) registraram alta dissimilaridade florística entre
os dois cerradões, com maior área basal por hectare das espécies arbóreas em solo mesotrófico,
fato este associado pelos autores à maior fertilidade do solo e maior número de árvores por
hectare no cerradão distrófico associado à cobertura menos compacta de copas e presença de
espécies de menor porte.

12
O cerradão apresenta árvores comuns às formações savânicas e florestais, com maiores
valores de similaridade florística com o cerrado sentido restrito do que com as matas (Mendonça
et al., 2008). Segundo Ribeiro & Walter (2008), o cerradão é floresta, mas floristicamente é mais
similar ao cerrado sentido restrito, justamente por conter elevado número de espécies típicas do
cerrado sentido restrito. Solorzano et al., (2012) defendeu que o padrão co-ocorrente para o
cerradão é a presença de espécies das categorias florestais, savânicas e generalistas, de acordo
com sua preferência por habitat.
O conjunto de dados analisados por Solorzano et al., (2012) em seis áreas distribuídas no
MT, MS, MG, TO, PI e DF, destacou algumas espécies que quando ocorrem em conjunto podem
ser usadas para tipificar o cerradão: Emmotum nitens, Qualea grandiflora, Tachigali subvelutina,
Copaifera langsdorffii, Vatairea macrocarpa e Bowdichia virgilioides. Outras espécies como
Blepharocalyx salicifolius, Simarouba versicolor e Ocotea spixiana foram apontadas por Felfili
& Silva Júnior (1992) como características desta fitofisionomia. Além das espécies já citadas,
Furley & Ratter (1988) também destacaram um grupo de espécies como indicadoras do cerradão
distrófico, as quais são: Callisthene major, Cardiopetalum calophyllum, Cinnamomum erythropus,
Hirtella glandulosa, Tachigali subvelutina, Siparuna guianensis, Siphoneugena densiflora,
Vochysia haenkeana e Virola sebifera. Destas, Hirtella glandulosa, Emmotum nitens, Virola
sebifera, Cinnamomum erythropus, Vochysia haenkeana e Callisthene major ocorrem
preferencialmente em formações florestais, inclusive no cerradão; Blepharocalyx salicifolius,
Siphoneugena densiflora, Siparuna guianensis e Tachigali subvelutina ocorrem com mais
frequência em matas e cerradão, mas também aparecem no cerrado sentido restrito; e Ocotea
spixiana e Cardiopetalum calophyllum são comuns tanto em matas e cerradões quanto em
formações savânicas (Walter, 2006; Mendonça et al., 2008).
De acordo com Solorzano et al., (2012), a vegetação do cerradão varia conforme a
região e são poucas as espécies que ocorrem com a mesma importância nos diferentes locais
estudados. Castro & Martins (1999 apud Solorzano, 2011) chamaram atenção para dois tipos
de cerradões fitogeograficamente distintos: (1) meridionais, que são mais florestais e têm poucas
conexões florísticas com as formações savânicas; e (2) setentrionais (do Nordeste), que seriam
mais savânicos e floristicamente mais similares ao cerrado lato sensu, que também foi chamado
de “cerradão de cerrado”.
Diferente do cerradão que ocupava menos de 1% do Cerrado (Haridasan, 2000), o cerrado
sentido restrito é a fitofisionomia mais extensa e ocupava originalmente cerca de 70% da área

13
geográfica do Cerrado na porção central do Brasil (Eiten, 1972, Ribeiro e Walter 1998; 2008). É
formação savânica caracterizada pela presença de árvores baixas, com altura entre 3 e 8 m,
algumas espécies podem chegar até 10 m (Ribeiro & Walter, 2008; Marimon-Junior & Haridasan,
2005). As árvores geralmente são retorcidas, com ramificações irregulares, cascas de cortiça gros-
sa, fendida ou sulcada, onde os arbustos e subarbustos encontram-se espalhados pela área (Ribeiro
& Walter, 2008). A fitofisionomia do cerrado sentido restrito apresenta subtipos: denso, típico, ralo e
rupestre, onde a cobertura de árvores e arbustos pode variar de 5% a 70% (Eiten, 1972; Ribeiro &
Walter, 2008). De acordo com esses autores, os três primeiros subtipos refletem variações na
forma dos agrupamentos e espaçamentos entre os indivíduos lenhosos, seguindo gradientes de
densidade decrescente do cerrado denso ao cerrado ralo. O cerrado denso tem cobertura arbórea
que varia de 50% a 70% e altura média de 5 a 8 m, o cerrado típico atinge cobertura arbórea de
50% e altura média que varia de 3 a 6 m e, o cerrado ralo com cobertura arbórea que varia de
5% a 20% tem altura média variada entre 2 e 3 m. O cerrado rupestre diferencia-se dos demais
pelo substrato, tipicamente de solos rasos com afloramentos de rocha e pelas espécies típicas e
adaptadas a este ambiente (Ribeiro & Walter, 2008; Walter & Ribeiro, 2010).
O cerrado sentido restrito abriga elevado número de espécies, entre as quais pode-se destacar
Qualea parviflora, Qualea grandiflora, Curatella americana, Davilla elliptica, Ouratea
hexasperma, Caryocar brasiliensis, Hymenaea stignocarpa e Dalbergia miscolobium (Mendonça
et al., 1998, 2008; Felfili et al., 2000; Andrade et al., 2002; Assunção & Felfili, 2004; Felfili et al.,
2007; Costa et al., 2010; Mews, 2011). Assim como em outros ambientes tropicais, no cerrado
sentido restrito o número de espécies é elevado e a distribuição é desigual, poucas espécies ocorrem
com elevado número de indivíduos (Felfili & Felfili, 2001). Essa tendência, onde um grupo pequeno
de espécies prevalece sobre as demais, foi verificada em diversos trabalhos no cerrado sentido
restrito (Felfili et al., 2002). A elevada riqueza encontrada nesta fitofisionomia está em torno de 70
espécies por hectare (Haridasan, 2005) e varia dependendo do tamanho da área amostrada.
As principais classes de solos que suportam o cerrado sentido restrito são os Latossolos
Vermelhos e Neossolos Quartzarênicos (Haridasan, 2005). Em geral esses solos são profundos e
bem drenados e não apresentam restrições ao crescimento radicular (Haridasan, 2005).
Comunidades de cerrado sentido restrito também são encontradas sobre as classes de solos
Cambissolos, Chernossolos, Argissolos, entre outras (Reatto & Martins, 2005).
As diferenças químicas e físicas das propriedades dos solos e a similaridade florística estão
entre os assuntos mais tratados entre as fitofisionomias de cerradão e cerrado sentido restrito.

14
Estudos desta natureza visam entender a relação ecológica e os fatores que influenciam as diferenças
estruturais e florísticas entre essas comunidades geograficamente próximas.
Marimon-Junior & Haridasan (2005) compararam a fitossociologia entre o cerradão e
cerrado sentido restrito em solos distróficos, em Nova Xavantina (MT), a fim de determinar se a
ocorrência do cerradão estaria associada à maior fertilidade nos solos. Entretanto, não foi
constatada qualquer diferença na composição química, mas o cerradão apresentou solo mais
argiloso ao longo de todo o perfil. Os autores concluíram que o solo mais argiloso no cerradão
apresentava maior capacidade de retenção de água disponível e esta disponibilidade de água
regulava a dinâmica dos nutrientes no solo e sua absorção pelas plantas.
Costa & Araújo (2000), na Reserva do Panga, em Uberlândia (MG), compararam a
fitossociologia e os solos de cerradão e cerrado sentido restrito. As análises químicas dos solos
apontaram que ambas as fitofisionomias ocorrem em solos distróficos, onde nenhuma diferença
química foi constatada entre as áreas. Em relação à fitossociologia, a espécie Qualea grandiflora
apresentou maior valor de importância tanto no cerradão quanto no cerrado. As espécies Matayba
guianensis, Coussarea hydrangeaefolia, Virola sebifera e Cardiopetallum callophylum
apresentaram altos índices de valor de importância no cerradão, mas não foram exclusivas desse
ambiente, ocorreram também no cerrado sentido restrito, porém em menor densidade e frequência.
Os autores concluíram que, além da similaridade com o cerrado sentido restrito, as espécies de
matas que foram encontradas no cerradão parecem ocorrer no limite de distribuição da área
amostrada, fato que caracteriza o cerradão como possível área de transição entre as matas e o
cerrado sentido restrito.
Mesmo em solos distróficos, com propriedades químicas semelhantes e em áreas contíguas
ou relativamente próximas, ambas fitofisionomias podem apresentar estruturas fitossociológicas
diferentes. Com o intuito de entender as relações ecológicas e os fatores que influenciam nas
diferenças estruturais e florísticas entre essas comunidades de solos semelhantes, Haridasan (1992)
investigou as diferenças nas concentrações de nutrientes foliares entre cerrado sentido restrito e
cerradões em solos distróficos e mesotróficos, e constatou que o cerrado sentido restrito e o
cerradão de mesma fertilidade não apresentaram diferenças significativas. A partir deste resultado,
o autor sugeriu que cerradões em solos distróficos se estabeleceram quando o solo ainda poderia
fornecer nutrientes em quantidades maiores para o estabelecimento de maior biomassa, ou seja,
o cerradão e o cerrado sentido restrito se estabeleceram em períodos diferentes.
Ao comparar a área original do cerradão que é menor que 1% com o cerrado sentido
restrito de aproximadamente 70%, Marimon Junior & Haridasan (2005) indicaram a urgência de
sua conservação. Devido ao aspecto florestal, o cerradão é considerado por agricultores como

15
vegetação que ocorre em solos férteis e, por isso, sofre intensa especulação antrópica (Goodland,
1971; Sano et al., 2010). De modo geral, o conhecimento da flora do Cerrado como a riqueza,
distribuição das espécies raras, endêmicas e ameaçadas e da diversidade de habitats, faz parte dos
critérios de prioridade para o estabelecimento de ações de conservação do bioma (Klink &
Machado, 2005).

16
Capítulo 1
Comparação da vegetação lenhosa do cerradãoe do cerrado denso no Jardim Botânico de Brasília
Resumo – O objetivo deste capítulo foi comparar a florística e a estrutura do cerradão (Cr) e do
cerrado sentido restrito denso (Cd), ambos sobre solos distróficos e geograficamente próximos
na área de visitação no Jardim Botânico de Brasília (DF). Em cada área foram estabelecidas dez
parcelas permanentes de 20x50 m cada (1.000 m²). Foram registrados as alturas e os diâmetros
da base (Db30 cm) e” 5 cm de todos os indivíduos inclusive os mortos em pé. Foram calculados
os parâmetros estruturais (densidade, frequência, área basal e IVI), o índice de diversidade de
Shannon e os índices de similaridade de Chao-Sørensen e Sørensen. A estrutura de tamanhos
também foi verificada por intervalos fixos de altura e diâmetro. Na comparação florística Cr/Cd,
as áreas foram representadas por 40/32 famílias, 14/3 famílias exclusivas, 71/48 gêneros, 99/68
espécies. Das 43 espécies compartilhadas apenas duas são consideradas tipicamente florestais,
Emmotum nitens e Copaifera langsdorffii, o restante é comumente encontrado em áreas de
cerrado sentido restrito. Além destas, 56/26 espécies foram exclusivas. Estes resultados definiram
os baixos índices de similaridade que variaram de 0,06 a 0,47 (Sørensen) e 0,03 a 0,50 (Chao-
Sørensen). As famílias com maior representatividade nas duas áreas foram Fabaceae, Vochysiaceae
e Myrtaceae. As espécies de maior IVI: Emmotum nitens / Dalbergia miscolobium, Siphoneugena
densiflora / Miconia burchellii, Nectandra reticulata / Poliouratea hexasperma, Miconia
cuspidata / Miconia ferruginata e Xylopia sericea / Qualea parviflora somaram 29,9/28,3% do
IVI total. A diversidade (H’) e a equabilidade (J’) foram, respectivamente, de 3,55 / 3,43 e 0,76
/ 0,81. Na estrutura as diferenças Cr/Cd foram anotadas para densidade total 2.412 / 2.193 ind.ha-
1, área basal de 27,92 / 15,98 m².ha-1, altura média (m) 6,6±2,10 / 3,9±2,7, altura máxima (m) 18
/ 12, número de árvores mortas 272 (11%) / 152 (7%) e área basal das árvores mortas (m².ha-1)
3,14 (11%) / 1,61 (10%). Dentre os parâmetros estruturais citados acima apenas a densidade
total não foi estatisticamente diferente na comparação entre as áreas. A distribuição dos diâmetros
nas comunidades apresentou curvas de aspecto J-reverso com balanço positivo entre recrutamento
e mortalidade. As comunidades Cd/Cr se apresentaram floristicamente e estruturalmente
diferenciadas apesar de sua localização aproximada.
Palavras-chave – Bioma Cerrado, fitofisionomias, índice de valor de importância (IVI), diversidade,distribuição diamétrica.

17
Introdução
As savanas ocorrem comumente nos trópicos e subtrópicos e são ecossistemas dinâmicos
determinados por fatores físicos, químicos e biológicos em diferentes escalas espaciais e temporais
(Cole, 1986; Mistry, 2000). O termo savana é aplicado a distintas formas fisionômicas, dado a
sua grande variedade florística e aspectos estruturais (Cole, 1986; Walter, 2006). Em geral, o
estrato rasteiro domina a paisagem e há variações nos componentes arbóreos e arbustivos (Cole,
1986; Ribeiro & Walter 2008;Walter, 2006).
No Brasil, Cole (1986) utiliza o termo savana para classificar a vegetação do bioma Cerrado,
em que considera os campos limpos, campos sujos, campos cerrados, cerradão assim como descrito
por Coutinho (1978). Para Ribeiro & Walter (1998; 2008) o bioma Cerrado é um complexo
vegetacional que possui relações ecológicas e fisiológicas com as savanas da América Tropical,
da África, do Sudeste Asiático e da Austrália. No entanto, o bioma como um todo não pode ser
considerado savana, uma vez que o termo Cerrado inclui todas as formações vegetais nele
encontradas (Ribeiro & Walter, 2010). O cerrado sentido amplo, que não inclui as matas de
galeria, ciliares e secas, é caracterizado por vegetação típica de savana e ocupa a maior porção
do mosaico fitofisionômico que compõe o bioma Cerrado (Walter, 2006; Ribeiro & Walter, 2010).
Diante da enorme variedade de fitofisionomias do bioma Cerrado, Ribeiro & Walter (2008)
criaram um sistema formal e definiram três grandes formações vegetacionais: florestais, savânicas
e campestres. Para essas formações os autores listaram onze tipos fitofisionômicos, onde o cerrado
sentido restrito e o cerradão fazem parte da composição fitofisionômica das formações savânicas
e florestais, respectivamente. O cerradão e o cerrado sentido restrito podem ocorrer em curtas
distâncias geográficas, ao longo de gradientes de vegetação ou em áreas contiguas (Costa &
Araújo 2001; Marimom-Júnior & Haridasan, 2005; Campos et al., 2006).
O cerradão apresenta dossel contínuo e cobertura arbórea que pode oscilar de 50% a 90%. A
altura média do estrato arbóreo varia de 8 a 15 m, o que proporciona condições de luminosidade
que permitem a formação de estratos arbustivos e herbáceos pouco desenvolvidos (Ribeiro & Walter,
2008; 2010). Ocorre com composição florística distinta em solos distróficos e mesotróficos (Ratter,
1971; Ratter et al., 1973; Araújo & Haridasan, 1988). Já o cerrado sentido restrito ocorre princi-
palmente associado aos solos distróficos (Haridasan, 1992). Esta fitofisionomia é composta por
estrato arbóreo-arbustivo e herbáceo-graminoso e de acordo com a variação da densidade arbórea
pode ser classificado como ralo, típico e denso (Eiten, 1994; Ribeiro & Walter, 2008).

18
Estudos realizados por Felfili (2001) e Ribeiro & Walter (2008) afirmaram que cerradão e
cerrado sentido restrito de fato abrigam espécies comuns, mas caracterizam-se preferencialmente
por espécies savânicas. Com base nos levantamentos de Mendonça et al., (2008), o cerradão
compartilha espécies principalmente com o cerrado sentido restrito, seguido pelas matas de galeria
e em menor número pelas matas secas. Embora o cerradão apresente vegetação mista (Felfili,
2001; Ribeiro & Walter, 2008), Felfili & Silva-Júnior (1992) alegaram que algumas espécies são
características, associadas aos solos distróficos, como Emmotum nitens, Blepharocalyx salicifolius,
Simaurouba versicolor e Ocotea spixiana. Ribeiro & Haridasan (1984) apontaram que há variação
florística entre estas duas fitofisionomias devido às diferenças nas estruturas da vegetação.
Marimom-Junior & Haridasan (2005) compararam trechos adjacentes de cerradão e cerrado
sentido restrito, no leste mato-grossense, ambos em solos distróficos. As duas fitofisionomias fo-
ram distintas quanto à florística, estrutura e fitossociologia. Dentre as espécies mais importantes
nenhuma foi comum às duas áreas, o que evidenciou a baixa similaridade florística. Ribeiro &
Haridasan (1984), em Planaltina (DF), registraram baixo índice de similaridade (0,36) entre cerradão
e cerrado denso e nenhuma das espécies encontradas foram simultaneamente importantes nas duas
áreas, assim como registrado por Marimon-Junior & Haridasan (2005). Já Costa & Araújo (2001),
na Reserva do Panga, em Uberlândia (MG), Qualea grandiflora foi a espécie mais importante nas
duas fitofisionomias. O índice de similaridade florística de Sørensen foi alto (0,73). Segundo os
autores esta elevada similaridade resultou da contiguidade das áreas o que permitiu que espécies
com alto potencial de dispersão ocupassem fisionomias distintas. Embora a proximidade entre
áreas possa permitir a ampla colonização por diferentes espécies, o mesmo resultado não foi
encontrado por Marimon-Junior & Haridasan (2005) e Ribeiro & Haridasan (1984).
Neste sentido, o presente capítulo objetivou comparar a florística e a estrutura fitossociológica
da vegetação lenhosa em áreas adjacentes de cerradão (Cr) e de cerrado denso (Cd), amostrados
no Jardim Botânico de Brasília (JBB) (DF).
Material e Métodos
O cerradão e o cerrado denso estão localizados no Centro de Visitantes do Jardim Botânico
de Brasília (CVJBB), situado nas coordenadas 15°50’-15°55’S, 47°49’-47°55’O, com altitude
média de 1.056 m (Figura 1). O Jardim Botânico de Brasília (JBB) possui aproximadamente 526
ha para visitação (CVJBB) e, junto com a Estação Ecológica (EEJBB) compreendem

19
aproximadamente 5.000 ha. Parte da composição florística do CVJBB já foi estudada por Azevedo
et. al. (1990). O JBB apresenta variações fitofisionômicas que incluem representantes das três
formações vegetacionais descritas por Ribeiro e Walter (1998): florestais, savânicas e campestres
(Heringer Salles et al., 2007, Proença et al., 2001).
O cerradão (Figuras 2 e 3) ocorre em fragmento localizado no limite da cerca com a Escola
de Administração Fazendária (ESAF) e com as estradas DF-001 e DF-035, situado entre as
coordenadas 15º51’-15º52’S e 47º49’O. O cerrado sentido restrito ocorre próximo a cerca que
divide o CVJBB e a EEJBB, onde ocorrem os subtipos do cerrado sentido restrito, entre eles o
cerrado denso situado entre as coordenadas 15º52’-15º53’S e 47º49’-47º50’O (Figuras 4 e 5).
No total, o cerrado sentido restrito abrange área de 2.970 hectares e ocorre de forma descontínua
(Heringer Salles et al., 2007). O cerradão e o cerrado denso distam, em linha reta, aproximadamente
2 km entre si (Figura 6). As suas áreas ocorrem onde a declividade do terreno não ultrapassa os
5% (Azevedo et al. 1990).
O clima do Distrito Federal é do tipo Aw ou tropical chuvoso, de acordo com a classificação
de Köppen, com invernos secos e verões chuvosos (CODEPLAN, 1984). A partir dos dados
meteorológicos entre 2007 a 2012, fornecidos pelo banco de dados do INMET
(www.inmet.gov.br), a estação seca que ocorre no período de maio a setembro obteve temperatura
média mínima anual de 15°C e média máxima de 27°C, a umidade relativa média anual neste
período foi de 50%. Na estação chuvosa, período que se estende de outubro a abril, a umidade
relativa média anual foi de 72%, com temperatura mínima média anual de 18°C e média máxima
de 27°C. Julho é o mês mais frio, com mínimas que variaram de 13°C a 15°C. A precipitação
média anual foi de 1.503 mm.ano-1 (www.inmet.gov.br).
Cavedon & Sommer (1990) estudaram os diferentes tipos de solo do JBB e concluíram que
a vegetação traduz o tipo de solo, especialmente quanto à textura. Os autores classificaram o tipo
de vegetação que ocorre sobre os Latossolos de acordo com a granulometria do solo, a saber:
textura muito argilosa ocorrem sob matas, textura argilosa sob cerrado com árvores de maior
porte e os de textura média sob cerrado do tipo arbustivo. Os solos predominantes nas duas áreas
estudadas são do tipo Latossolo Vermelho e Amarelo, ambos considerados distróficos (Cavedon
& Sommer, 1990). Além desses solos também são encontrados no JBB Cambissolos (campo
limpo, campo cerrado e floresta) e Gleissolos (campo limpo) (Cavedon & Sommer, 1990). O
JBB apresenta rochas pré-cambrianas superior da formação Paranoá (Cavedon & Sommer, 1990).
O Plano de Prevenção e Combate aos Incêndios Florestais do Distrito Federal, criado em
junho de 1996 pelo Decreto Federal nº 17.431, incluiu o JBB entre as áreas de proteção (SEMARH,
2004). O plano contempla a prevenção, vigilância da área, pré-supressão e medidas estratégicas

20
para o combate a eventuais incêndios. Na área de visitação do JBB foram construídos talhões eestradas que funcionam como aceiros que impedem a propagação do fogo (observação pessoal).Diante de tais ações contra incêndios, o Cr e o Cd estão protegidos do fogo há mais de 20 anos(Informação pessoal do professor Manoel Cláudio da Silva Júnior).
Para o estudo da vegetação foram instaladas dez parcelas permanentes de 20x50 m cada(1.000 m²) de forma sistemática no cerradão e dez parcelas aleatórias no cerrado denso, deacordo com Felfili et. al. (2005) (Figura 6). Em cada parcela foram registrados, identificados emedidos todos os indivíduos com diâmetro igual ou superior a 5 cm, medido a 30 cm do solo,(Db30 cm) e” 5 cm, inclusive os indivíduos mortos em pé, com exceção das lianas e palmeiras.Indivíduos bifurcados desde a base ou em touceiras foram incluídos desde que pelo menos umdos indivíduos apresentasse diâmetro mínimo de inclusão. As alturas foram medidas com varagraduada em metros. Considerou-se a projeção vertical da copa até o solo. Os dados da vegetaçãolenhosa do cerrado denso foram cedidos pela professora Sabrina de Couto Miranda e parte destesdados já foram publicados na forma de resumo expandido no 60º Congresso Nacional de Botânica(Miranda et al., 2009).
As espécies amostradas foram nomeadas a nível específico de acordo com o sistema denomenclatura do Angiosperm Phylogeny Group III (APG III, 2009). Os nomes foram consultadosnos sites W3 Tropicos (http://www.tropicos.org/) e Lista da Flora do Brasil (http://
floradobrasil.jbrj.gov.br).
Figura 1. Carta imagem da Localização do Centro de Visitantes do Jardim Botânico de Brasília (CVJBB-JBB)(área delimitada em vermelho) e locais de edificações existentes (área delimitada em branco e pontos amarelos).Fonte: Geo Lógica - [email protected].

21
Figura 2. Vista da estrutura interna do cerradão (Cr), localizado no Jardim Botânico de Brasília (DF). Emdetalhe o adensamento da cobertura arbórea, galhos emaranhados e a camada de serapilheira sobre o solo. Foto:Manoel Cláudio.
Figura 3. Imagem da cobertura arbórea do cerradão (Cr), localizado no Jardim Botânico de Brasília (DF). Foto:Manoel Cláudio.

22
Figura 4. Vista da estrutura externa do cerrado denso (Cd), localizado no Jardim Botânico de Brasília (DF). Emdetalhe a baixa estatura das árvores, galhos baixos e densos e o solo de cor avermelhada e pouca camada deserapilheira. Foto: Manoel Cláudio.
Figura 5. Imagem da cobertura arbórea do cerrado denso (Cd), localizado no Jardim Botânico de Brasília (DF).Foto: Manoel Cláudio.

23
Figura 6. Localização das áreas amostradas no Centro de Visitantes do Jardim Botânico de Brasília (CVJBB-DF). Cerradão (Cr) e cerrado denso (Cd) distam 2 km de distância em linha reta (laranja). Fonte: Google Earth.
Os parâmetros estruturais (densidade, frequência, dominância e IVI) foram calculados de
acordo com Mueller-Dombois e Ellemberg (1974), com o auxílio do programa Excell 2007. Os
dados de densidade, riqueza, área basal, altura (média, máxima e mínima) foram verificados
quanto à normalidade e a homocedasticidade pelos testes de Shapiro-Wilk (Shapiro & Wilk,
1965) e Levenne (Hartley, 1950), respectivamente. Os dados que não apresentaram normalidade
foram transformados na base logarítmica ou raiz quadrada, a fim de normalizar os dados (Kent &
Coker, 1992; Palmer, 2005). Uma vez que as variáveis não apresentaram normalidade, aplicou-se
o teste não paramétrico de Mann-Whitney, utilizado para comparação entre as médias obtidas.
Tanto a normalidade e a homocedasticidade dos dados quanto a aplicação do teste de Mann-
Whitney foram executados no programa PAST versão 2.17.
Para avaliar a abrangência florística das amostragens foi elaborada a curva de rarefação
construída com base na média de “n” repetições calculadas, com intervalo de 95% de confiança
(Felfili et. al. 2011). Para a elaboração da curva de rarefação utilizou-se o programa PAST versão
2.17. Também foi calculado o erro padrão, coeficiente de variação e intervalo de confiança a

24
95% de probabilidade da média para densidade e área basal por parcela em cada área, conforme
sugerido por Felfili & Imaña-Encinas (2001).
A riqueza florística foi avaliada com o uso dos estimadores não paramétricos Chao1 e Chao
2, onde o primeiro se baseia no número de espécies raras em uma amostra e o segundo é uma
variação do primeiro e baseia-se na frequência de espécies únicas na amostra (Magurran, 2004).
Os estimadores foram calculados no programa Estimate versão 8.0.
A diversidade alfa e a equabilidade foram calculadas, respectivamente, pelos índices de
Shannon (H’) (Magurran, 1988) e de uniformidade de Pielou (J’) (Kent & Coker, 1992) por meio
do programa Estimate versão 8.0. A diversidade alfa está relacionada com a riqueza ou número
de espécies por unidade de área (Felfili & Felfili, 2001). O índice de diversidade varia de 0 a
valores positivos e, situa-se, usualmente, entre 1,5 e 3,5 (Felfili & Rezende, 2003). A equabilidade
representa a percentagem máxima de diversidade que uma amostra pode atingir, e está relacionada
com a distribuição do número de indivíduos por espécie (Felfili & Felfili, 2001). Os valores
variam de 0 a 1, quanto mais próximo de 1, mais homogenia é a distribuição dos indivíduos das
espécies na área analisada (Felfili & Rezende, 2003).
A similaridade florística entre as áreas foi verificada pelo índice quantitativo “Chao-Sørensen-
Raw Abundance-based” (Chao et al., 2005), para isso foi utilizado os dados de densidade. Esse
índice é baseado na probabilidade de dois indivíduos escolhidos aleatoriamente, um de cada uma
das duas amostras, pertencerem à lista de espécies compartilhadas pelas mesmas (Chao et al.,
2005). Para fins de comparação com outros estudos, a similaridade florística também foi verificada
pelo tradicional índice de Sørensen com base nos dados de presença e ausência (Mueller-Dombois
& Ellemberg, 1974). Ambos os índices de similaridade foram calculados no programa Estimate
versão 8.0.
Para verificar e comparar a estrutura de tamanhos nas áreas estudadas foram adotados
intervalos fixos para as classes de diâmetro e altura. Estes intervalos foram de 5 cm e 1 m, para
diâmetro e altura, respectivamente.

25
Resultados
A curva de rarefação de cada área tendeu a assíntota. No cerradão (Cr), a curva mostrou quea amostra de 50% das parcelas resultou em mais de 80% das espécies levantadas (Figura 7),resultado semelhante foi encontrado no cerrado denso (Cd). A inclusão de outras cinco parcelasculminou no acréscimo de outras dezesseis espécies no Cr e oito no Cd.
A amostragem nas duas áreas foi considerada suficiente para estimar a densidade e a áreabasal, visto que os erros-padrões foram 7,4% (IC= ±19,0) e de 6,2% (IC= ±0,1), respectivamente,para a densidade e área basal no cerradão. E de 4,5% (IC= ±9,9) para a densidade e 7% para aárea-basal (IC= ±0,1) no cerrado denso. A riqueza estimada por Chao 1 e Chao 2 foi 137/78 parao cerradão e 137/76 para o cerrado denso. Assim, conseguiu-se amostrar mais de 80% da riqueza
florística estimada para as áreas, o que corrobora que a metodologia utilizada foi satisfatória.
Figura 7. Curvas de rarefação baseadas no número de indivíduos amostrados e no número de parcelas (10) decada área, localizados no Jardim Botânico de Brasília (DF). Cr= cerradão e Cd= cerrado denso.
Na comparação florística entre Cr/Cd foram encontrados 99/68 espécies, 40/32 famílias e71/48 gêneros (Tabela 1). As espécies raras, representadas por apenas um ou dois indivíduos nascomunidades, somaram 31/25% do total de espécies no Cr/Cd. As famílias mais importantes emnúmero de espécies (em parênteses) no Cr/ Cd foram: Fabaceae (17/12), Melastomataceae (8/5),Lauraceae (6/0), Malpighiaceae (3/4), Myrtaceae (6/3), Vochysiaceae (5/5) e Calophyllaceae (0/3). Estas representaram 39% do total de famílias amostradas no Cr e 43% no Cd e estão entre asdez mais ricas em ambas as áreas. Fabaceae esteve representada em Cr/Cd por 14/10 gêneros dototal de 71/48 gêneros presentes. Miconia (Melastomataceae) foi o gênero mais rico nas duasáreas. Embora Fabaceae seja a mais rica em espécies, Myrtaceae, Melastomataceae e Emmotaceaeforam as mais densas no cerradão, por somar 41% (887) dos indivíduos. Já no cerrado densoFabaceae, Melastomataceae e Ochnaceae contribuíram com 43% (877) dos indivíduos.

26
Das 29 famílias de ocorrência comum, as dez mais ricas representaram 65/54% do total de
espécies em Cr/Cd. No Cr 30% das famílias foram exclusivas: Aquifoliaceae, Clusiaceae,
Cunoniaceae, Dichapetalaceae, Euphorbiaceae, Lauraceae, Myristicaceae, Primulaceae, Rubiaceae,
Sapindaceae e Simaroubaceae. Já no Cd menos de 10% foram exclusivas, a saber: Calophyllaceae,
Connaraceae e Ebenaceae. As famílias representadas por apenas uma espécie somaram 26,7/62%
na comparação Cr/Cd, o que reflete grande diversidade de espécies nas duas comunidades.
Os valores de H’ e J’ calculados foram 3,55 e 0,76 para o Cr e 3,43 e 0,81 para o Cd (Tabela
1). Os índices de similaridade de Chao-Sørensen entre as comunidades foram inferiores a 0,50,
onde o menor valor foi encontrado entre as parcelas Cr1 e Cd8 (Chao-Sørensen=0,03) (Tabela
2). O baixo índice de similaridade florística está relacionado à presença de espécies exclusivas,
que somaram 61% no cerradão e 38% no cerrado denso (Tabela 1). No índice de similaridade
tradicional de Sørensen, os valores também foram inferiores a 0,50, o menor valor foi entre as
parcelas Cr1-Cr3 e Cd8 e entre as parcelas Cr8 e Cd3 (Sørensen=0,06).
Tabela 1. Comparação das características fitossociológicas do cerradão (Cr) e cerrado denso (Cd), localizadosno Jardim Botânico de Brasília (DF).
Características
Área amostral (hectare)
Número de espécies
Espécies exclusivas
Espécies comuns
Número de famílias
Famílias exclusivas
Área amostral (hectare)
Densidade (árvores/ha)
Área basal (m².ha-1)
Altura média (m)
Altura máxima (m)
Número de árvores mortas
Área basal das árvores mortas (m².ha-1)
Cerradão (Cr)
1
99
56
43
40
14
1
2.412a
27,92a
6,6±2,10*a
18a
272a
3,14a
Cerrado denso (Cd)
1
68
26
42
32
3
1
2.193a
15,98b
3,9±2,7*b
12b
152b
1,61b
*Desvio padrão. As médias seguidas pela mesma letra, nas colunas, não diferem entre si, a 5% de probabilidade,pelo teste de Mann-Whitney (p < 0,05).

27
Tabela 2. Índices de similaridade de Chao-Sørensen (abaixo) e Sørensen (acima) entre as 20 parcelas amostradasnas áreas de cerradão (Cr) e cerrado denso (Cd), localizados no Jardim Botânico de Brasília (DF).
0,11
0,47
0,19
0,29
0,4
0,33
0,28
0,17
0,33
0,34
Cd6
0,11
0,38
0,16
0,29
0,37
0,35
0,31
0,14
0,33
0,32
Cd1
0,11
0,45
0,18
0,27
0,44
0,38
0,32
0,25
0,36
0,35
Cd2
0,06
0,26
0,12
0,24
0,3
0,23
0,14
0,06
0,28
0,22
Cd3
0,11
0,36
0,16
0,24
0,35
0,31
0,23
0,14
0,29
0,32
Cd4
0,16
0,38
0,17
0,31
0,27
0,33
0,25
0,22
0,33
0,27
Cd7
0,12
0,41
0,16
0,3
0,33
0,36
0,24
0,21
0,27
0,3
Cd5
0,06
0,36
0,17
0,26
0,37
0,33
0,3
0,16
0,38
0,27
Cd8
0,11
0,38
0,19
0,27
0,45
0,31
0,23
0,14
0,34
0,37
Cd9
0,11
0,41
0,21
0,3
0,36
0,29
0,25
0,14
0,34
0,31
Cd10
Cr1
Cr2
Cr3
Cr4
Cr5
Cr6
Cr7
Cr8
Cr9
Cr10
Sørensen
Cd1
Cd2
Cd3
Cd4
Cd5
Cd6
Cd7
Cd8
Cd9
Cd10
0,05
0,11
0,03
0,05
0,5
0,05
0,1
0,03
0,05
0,05
Cr1
0,37
0,42
0,25
0,33
0,37
0,43
0,36
0,36
0,38
0,36
Cr2
0,1
0,14
0,04
0,1
0,09
0,09
0,14
0,09
0,1
0,11
Cr3
0,19
0,24
0,1
0,17
0,19
0,19
0,2
0,16
0,18
0,2
Cr4
0,28
0,35
0,2
0,26
0,25
0,25
0,23
0,25
0,29
0,25
Cr5
0,31
0,41
0,16
0,26
0,27
0,29
0,27
0,36
0,28
0,26
Cr6
0,21
0,2
0,07
0,15
0,13
0,16
0,16
0,17
0,15
0,13
Cr7
0,05
0,16
0,01
0,08
0,07
0,07
0,07
0,07
0,07
0,07
Cr8
0,34
0,34
0,13
0,25
0,24
0,26
0,32
0,37
0,34
0,28
Cr9
0,23
0,26
0,16
0,26
0,25
0,27
0,26
0,19
0,26
0,26
Cr10
Chao-Sørensen
*Valores mínimos em sublinhado e máximo em negrito para os índices de Chao-Sørensen e Sørensen.

28
Foram encontrados para Cr e Cd, respectivamente, 2.412/2.193 ind.ha-1. A área basal foi
calculada em 27,92 m².ha-¹ no Cr e 15,98 m².ha-¹ no Cd (Tabela 1). Os indivíduos mortos em pé
representaram 11% (272 ind.ha-1) e 7% (152 ind.ha-1) da densidade relativa total e 11% (25 m².ha-
¹) e 10% (20 m².ha-¹) da área basal total ocupada em Cr e Cd, respectivamente. As espécies raras
(em parênteses o número de indivíduos) que ocorreram com um ou dois indivíduos na amostra
representaram 31% (32) e 25% (16) do total de espécies e 2% (26) e 1% (22) da densidade total
e 2,3% (0,65 m².ha-¹) e 2,6 % (0,10 m².ha-¹) da área basal total em Cr e Cd, respectivamente.
As espécies mais importantes (IVI) em Cr foram: Emmotum nitens (27,48), Siphoneugena
densiflora (22,6), Nectandra reticulata (17,3), Miconia cuspidata (12,5) e Xilopia sericea (11,1)
que somaram 30,3% do IVI total (Tabela 3). S. densiflora, E. nitens, N. reticulata, M. cuspidata
e X. sericea foram também as espécies mais densas e representaram 36,5% da densidade total e
E. nitens e N. reticulata juntas representaram 26% da dominância total. Em Cd aquelas de maior
importância foram: Dalbergia miscolobium (23,1), Miconia burchellii (19,7), Ouratea hexasperma
(17,8), Miconia ferruginata (13,4) e Qualea parviflora (10,8), as quais representaram 28,3% do
IVI total (Tabela 4). As espécies mais densas D. miscolobium, P. hexasperma, M. burchellii, M.
ferruginata e S. macrocarpa representaram 37% da densidade total e D. miscolobium e M.
burchellii, com maior área basal, representaram 20% da dominância total.
Do total de espécies amostradas nas duas áreas (122), 43 (34%) foram comuns em Cr e Cd
(Tabela 3). Dessas espécies, 22 (69%) foram mais abundantes em Cd e sete (24%) mais abundantes
em Cr. Entre as espécies compartilhadas apenas duas, Emmotum nitens e Copaifera langsdorffii,
são consideradas tipicamente florestais (Mendonça et al., 2008). Das 122 espécies, 79 (60%) são
comuns em cerrado sentido restrito e 46 (35%) são comuns às matas de galeria e ciliares e ao
cerradão (Mendonça et al., 2008). Entre as espécies exclusivas do Cr, 58% (33) são mais frequentes
em matas de galeria e ciliares do que em áreas de cerrado sentido restrito (Mendonça et al.,
2008). No Cd apenas a espécie Symplocos rhamnifolia ocorre tipicamente em matas de galeria,
segundo Mendonça et al., (2008).
A distribuição de diâmetros nas duas comunidades apresentou aspecto “J-reverso” ou
exponencial negativo (Figura 6). Em Cr foram registradas árvores em 10 classes diamétricas com
amplitude de 50 cm, as maiores árvores foram Nectandra reticulata (53,8 cm) e Emmotum nitens
(53,3 cm). No Cd os indivíduos foram distribuídos em sete classes diamétricas com amplitude de
35 cm, Vochysia thyrsoidea com 40,4 cm e Qualea grandiflora com 34 cm foram os maiores
indivíduos.

29
As duas áreas apresentaram principalmente indivíduos jovens, pois 66% do total amostrado
no Cr e 75% em Cd apresentaram diâmetros de até 10 cm. Nas duas áreas mais de 90% dos
indivíduos apresentaram diâmetro até 25 cm (Figura 8). As maiores classes foram representadas
por poucos indivíduos.
No Cr os indivíduos lenhosos apresentaram altura média de 6,6 m com 83% destes nas
classes 2 e 3, alturas entre 3,6 e 9,5 m (Figura 9). As árvores mais altas foram Bowdichia virgilioides
e Pterodon emarginatus, ambas com 18 metros, seguidas por Virola sebifera que atingiu 16
metros. Os menores indivíduos foram Plenckia populnea e Qualea parviflora com 0,5 m e
Schefflera macrocarpa com 1 m.
Na comunidade Cd, 76% dos indivíduos ocuparam as classes 1 e 2 até 3,5 m de altura. A
maioria das árvores, 54,4%, ocorreu na classe 2 com 2,7 a 3,5 m de altura (Figura 9). Os indivíduos
mais altos foram Tachigali subvelutina com 12 metros, seguida por Pouteria torta, Schefflera
macrocarpa e Vochysia thyrsoidea todas com 10 m. A menor árvore registrada foi de um indivíduo
de Poliouratea hexasperma com 0,8 m. Foram registrados 67 indivíduos de diferentes espécies
com altura de 1 m na comunidade Cd.
Figura 8. Distribuição dos indivíduos por classes de diâmetro no cerradão (Cr) e no cerrado denso (Cd) doJardim Botânico de Brasília (DF).

30
Figura 9. Distribuição dos indivíduos por classes de altura no cerradão (Cr) e no cerrado denso (Cd) doJardim Botânico de Brasília, DF.
A distribuição de diâmetros para os indivíduos mortos em pé mostrou representantes em
sete classes no Cr e cinco no Cd (Figura 10). A maior mortalidade ocorreu nas menores classes de
diâmetro, até 15 cm, com 85,2% em Cr e 80,2% em Cd. A mortalidade foi de 100% nas parcelas
amostradas do Cr e do Cd.
Figura 10. Distribuição dos indivíduos mortos em pé por classes de diâmetro no cerradão (Cr) e cerrado denso(Cd) no Jardim Botânico de Brasília (DF).

31
Tabe
la 3
. Fito
ssoc
iolo
gia
das
espé
cies
lenh
osas
, com
sua
s re
spec
tivas
fam
ílias
, do
cerr
adão
(Cr)
e d
o ce
rrad
o de
nso
(Cd)
, loc
aliz
ados
no
Jard
im B
otân
ico
de B
rasí
lia (D
F).
DA
= de
nsid
ade
abso
luta
; DR
= de
nsid
ade
rela
tiva;
FA
: fre
quên
cia
abso
luta
; FR
= fr
equê
ncia
rela
tiva;
DoA
: dom
inân
cia
abso
luta
; DO
R= d
omin
ânci
a re
lativ
a, e
IVI=
índi
cedo
val
or d
e im
portâ
ncia
.
Emm
otac
eae
***
Myr
tace
aeLa
urac
eae
Mel
asto
mat
acea
eA
nnon
acea
eVo
chys
iace
aeSi
mar
ouba
ceae
Myr
istic
acea
eFa
bace
aeM
yrta
ceae
Voch
ysia
ceae
Mel
asto
mat
acea
eFa
bace
aeM
alpi
ghia
ceae
Euph
orbi
acea
eLa
urac
eae
Rubi
acea
eM
yrta
ceae
Com
bret
acea
eFa
bace
ae
Emm
otum
nite
ns (B
enth
.) M
iers
Mor
tas
Siph
oneu
gena
den
siflo
ra O
.Ber
gNe
ctan
dra
retic
ulat
a (R
uiz
& P
av.)
Mez
Mic
onia
cus
pida
ta N
audi
nXy
lopi
a se
ricea
A.S
t.-H
il.Q
uale
a gr
andi
flora
Mar
t.Si
mar
ouba
ver
sicol
or A
.St.-
Hil.
Viro
la se
bife
ra A
ubl.
Copa
ifera
lang
sdor
ffii (
Hay
ne)
Kunt
zeM
yrci
a sp
lend
ens (
Sw.)
DC.
Calli
sthen
e m
ajor
Mar
t. &
Zuc
c.M
icon
ia p
eper
icar
pa D
C.D
albe
rgia
misc
olob
ium
Ben
thBy
rson
ima
laxi
flora
Gris
eb.
Map
roun
ea g
uian
ensis
Aub
l.Ne
ctan
dra
turb
acen
sis (K
unth
) Ne
esCo
rdie
ra m
acro
phyl
la (K
.Sch
um.)
Kunt
zeBl
epha
roca
lyx
salic
ifoliu
s (Ku
nth)
O.B
erg
Term
inal
ia fa
gifo
lia M
art.
Hym
enae
a sti
gono
carp
a M
art.
ex H
ayne
0.4
19.6 * * * * 9.3 * * 0.3 * * * 23.1 * * * * 3.8
0.7
3.4
27.5
24.9
22.7
17.0
12.5
11.2
11.2
9.5
8.4
7.8
7.8
7.7
7.2
5.5
5.4
5.2
5.0
4.4
4.3
3.9
3.7
0.1
10.1 * * * * 5.1 * * 0.0 * * * 10.1 * * * * 0.7
0.0
0.9
16.7
11.2
8.9
10.0
3.8
2.6
4.6
4.0
2.8
3.6
1.5
3.4
1.2
2.7
1.0
1.6
2.5
0.7
1.0
1.4
0.9
0.0
1.6 * * * * 0.8 * * 0.0 * * * 1.6 * * * * 0.1
0.0
0.1
4.7
3.1
2.5
2.8
1.1
0.7
1.3
1.1
0.8
1.0
0.4
1.0
0.3
0.8
0.3
0.4
0.7
0.2
0.3
0.4
0.3
0.3
2.6 * * * * 2.3 * * 0.3 * * * 2.6 * * * * 1.8
0.5
1.8
2.4
2.4
2.4
2.4
2.4
2.2
2.4
2.2
2.4
2.4
2.4
1.9
2.2
1.9
2.4
2.2
1.2
2.2
1.9
1.2
1.9
1 10 * * * * 9 * * 1 * * * 10 * * * * 7 2 7
10 10 10 10 10 9 10 9 10 10 10 8 9 8 10 9 5 9 8 5 8
0.0
6.9 * * * * 1.9 * * 0.0 * * * 10.5 * * * * 1.3
0.1
0.6
8.4
11.3
11.4
4.6
6.3
6.4
4.1
0.3
3.2
1.8
3.8
2.4
3.8
0.9
2.0
1.5
1.3
1.5
1.3
1.3
0.8
1 152 * * * * 42 * * 1 * * * 230 * * * * 29 2 13
202
272
274
112
152
155
100
80 76 43 92 58 92 21 49 35 31 35 32 31 20
Fam
ília
Esp
écie
s C
rC
dC
rC
dC
rC
dC
rC
dC
rC
rC
rC
dC
dC
d
DA
DR
FAFR
DoA
DoR
IVI

32
Tabe
la 3
. (C
ontin
uaçã
o)
Ara
liace
aeA
quifo
liace
aeM
elas
tom
atac
eae
Mal
vace
aePe
race
aePr
otea
ceae
Voch
ysia
ceae
Prim
ulac
eae
Voch
ysia
ceae
Faba
ceae
Nyc
tagi
nace
aeC
alop
hylla
ceae
Laur
acea
eC
elas
trace
aeN
ycta
gina
ceae
Mel
asto
mat
acea
eC
alop
hylla
ceae
Styr
acac
eae
Salic
acea
eVo
chys
iace
aeFa
bace
aeM
yrta
ceae
Apo
cyna
ceae
Mel
asto
mat
acea
e
Fam
ília
Esp
écie
s C
rC
dC
rC
dC
rC
dC
rC
dC
rC
rC
rC
dC
dC
d
DA
DR
FAFR
DoA
DoR
IVI 10
.3 * 2.0
3.9 * 7.8
10.9 * 5.0 * 1.7
3.8 * * 6.5
19.7
1.6
10.3 * 2.9
5.0 * 3.5 *
3.4
3.0
2.5
2.5
2.4
2.4
2.4
2.4
2.1
2.0
2.0
1.9
1.9
1.9
1.8
1.7
1.7
1.7
2.0
1.7
2.2
1.6
1.6
1.5
3.7 * 0.3
1.3 * 1.8
4.9 * 2.7 * 0.3
0.5 * * 1.9
9.1
0.2
4.1 * 0.4
2.8 * 0.5 *
0.6
0.5
0.3
0.5
0.2
0.3
0.5
0.3
0.2
0.5
0.2
0.2
0.4
0.2
0.2
0.4
0.3
0.3
0.1
0.1
0.9
0.2
0.1
0.1
0.6 * 0.0
0.2 * 0.3
0.8 * 0.4 * 0.0
0.1 * * 0.3
1.4
0.0
0.7 * 0.1
0.5 * 0.1 *
0.2
0.1
0.1
0.1
0.1
0.1
0.1
0.1
0.0
0.1
0.1
0.1
0.1
0.1
0.0
0.1
0.1
0.1
0.0
0.0
0.2
0.1
0.0
0.0
2.6 * 1.3
1.6 * 2.3
2.1 * 1.6 * 1.0
2.3 * * 2.3
2.6
1.0
2.3 * 1.8
1.3 * 2.1 *
1.7
1.7
1.4
1.7
1.7
1.2
1.2
1.4
1.4
1.2
1.2
1.2
1.2
1.2
1.2
0.7
1.0
1.0
1.4
1.2
1.0
1.0
1.2
1.2
10 * 5 6 * 9 8 * 6 * 4 9 * * 9 10 4 9 * 7 5 * 8 *
7 7 6 7 7 5 5 6 6 5 5 5 5 5 5 3 4 4 6 5 4 4 5 5
4.0 * 0.5
1.0 * 3.6
3.9 * 0.7 * 0.4
0.9 * * 2.2
8.0
0.3
3.9 * 0.6
0.9 * 0.9 *
1.1
0.7
0.8
0.4
0.5
0.9
0.7
0.6
0.5
0.3
0.6
0.5
0.3
0.4
0.4
0.6
0.5
0.5
0.4
0.3
0.3
0.4
0.2
0.2
88 * 10 22 * 79 85 * 16 * 8 20 * * 49 176 7 86 * 14 19 * 20 *
26 18 20 9 13 22 17 14 11 7 14 13 8 10 10 15 12 11 10 8 8 10 6 6
Sche
ffler
a m
acro
carp
a (C
ham
. & S
chltd
l.) F
rodi
nIle
x af
finis
Gar
dner
Mic
onia
alb
ican
s (Sw
.) Tr
iana
Erio
thec
a pu
besc
ens (
Mar
t. &
Zuc
c.) S
chot
t & E
ndl.
Pera
gla
brat
a (S
chot
t) Po
epp.
ex
Baill
.Ro
upal
a m
onta
na A
ubl.
Qua
lea
parv
iflor
a M
art.
Myr
sine
cor
iace
a (S
w.)
R.Br
. ex
Roem
. & S
chul
t.Vo
chys
ia th
yrso
idea
Poh
lBo
wdi
chia
virg
ilioi
des K
unth
Gua
pira
gra
cilif
lora
(Mar
t. ex
Sch
mid
t) Lu
ndel
lK
ielm
eyer
a co
riac
ea M
art.
& Z
ucc
Oco
tea
pom
ader
roid
es (M
eisn
.) M
ezC
heilo
clin
ium
cog
natu
m (M
iers
) A.
C.S
m.
Gua
pira
nox
ia (N
etto
) Lu
ndel
lM
icon
ia b
urch
ellii
Tri
ana
Kie
lmey
era
spec
iosa
A.S
t.-H
il.St
yrax
ferr
ugin
eus N
ees
& M
art.
Cas
eari
a gr
andi
flora
Cam
bess
.Q
uale
a m
ultif
lora
Mar
t.Ta
chig
ali
subv
elut
ina
(Ben
th.)
Oliv
eira
-Filh
oPi
men
ta p
seud
ocar
yoph
yllu
s (G
omes
) La
ndru
mAs
pido
sper
ma
tom
ento
sum
Mar
t.M
icon
ia se
llow
iana
Nau
din

33
Tabe
la 3
. (C
ontin
uaçã
o)
Ann
onac
eae
Laur
acea
eD
icha
peta
lace
aeFa
bace
aeSa
pota
ceae
Apo
cyna
ceae
Big
noni
acea
eA
naca
rdia
ceae
Car
yoca
race
aeA
nnon
acea
eC
hrys
obal
anac
eae
Faba
ceae
Laur
acea
ePr
imul
acea
eC
unon
iace
aeO
chin
acea
eLo
gani
acea
eN
ycta
gina
ceae
Sapi
ndac
eae
Dic
hape
tala
ceae
Sym
ploc
acea
eSy
mpl
ocac
eae
Rub
iace
aeFa
bace
ae
Fam
ília
Esp
écie
s C
rC
dC
rC
dC
rC
dC
rC
dC
rC
rC
rC
dC
dC
d
DA
DR
FAFR
DoA
DoR
IVI * * * 2.
3 * 3.0
0.9 * 9.1
3.0
0.7
0.6 * * * 17.8
2.1 * * * * * * 2.0
1.5
1.4
1.3
1.3
1.2
1.2
1.1
1.1
1.1
1.0
1.0
1.0
0.9
0.9
0.9
0.9
0.9
0.9
0.9
0.9
0.8
0.7
0.7
0.7
* * * 0.2 * 0.6
0.1 * 4.3
0.8
0.1
0.0 * * * 6.3
0.5 * * * * * * 0.4
0.1
0.2
0.1
0.2
0.4
0.2
0.1
0.2
0.4
0.1
0.1
0.7
0.1
0.7
0.0
0.1
0.1
0.1
0.5
0.1
0.2
0.1
0.0
0.1
* * * 0.0 * 0.1
0.0 * 0.7
0.1
0.0
0.0 * * * 1.0
0.1 * * * * * * 0.1
0.0
0.1
0.0
0.0
0.1
0.1
0.0
0.0
0.1
0.0
0.0
0.2
0.0
0.2
0.0
0.0
0.0
0.0
0.1
0.0
0.1
0.0
0.0
0.0
* * * 1.6 * 1.6
0.5 * 2.6
1.6
0.5
0.5 * * * 2.6
1.3 * * * * * * 1.3
1.2
1.0
1.0
1.0
0.7
0.7
0.7
0.7
0.5
0.7
0.7
0.2
0.7
0.2
0.7
0.5
0.7
0.7
0.2
0.5
0.5
0.5
0.5
0.5
* * * 6 * 6 2 * 10 6 2 2 * * * 10 5 * * * * * * 5
5 4 4 4 3 3 3 3 2 3 3 1 3 1 3 2 3 3 1 2 2 2 2 2
* * * 0.5 * 0.9
0.3 * 2.1
0.6
0.1
0.1 * * * 8.9
0.4 * * * * * * 0.3
0.2
0.2
0.2
0.2
0.2
0.2
0.3
0.2
0.1
0.2
0.2
0.0
0.2
0.0
0.2
0.3
0.1
0.1
0.2
0.2
0.1
0.1
0.2
0.1
* * * 10 * 19 6 * 47 14 2 2 * * * 195 8 * * * * * * 6
5 5 6 4 4 6 7 4 3 5 5 1 4 1 4 8 3 3 5 6 3 3 4 3
Gua
tteri
a se
llow
iana
Sch
ltdl.
Oco
tea
spix
iana
(Nee
s) M
ezTa
pura
am
azon
ica
Poep
p. &
End
l.D
imor
phan
dra
mol
lis B
enth
.C
hrys
ophy
llum
mar
gina
tum
(Hoo
k. &
Arn
.) Ra
dlk.
Aspi
dosp
erm
a m
acro
carp
on M
art.
Han
droa
nthu
s oc
hrac
eus
(Cha
m.)
Mat
tos
Tapi
rira
obt
usa
(Ben
th.)
J.D
.Mitc
h.C
aryo
car b
rasi
liens
e C
ambe
ss.
Anno
na c
rass
iflor
a M
art.
Cou
epia
gra
ndifl
ora
(Mar
t. &
Zuc
c.) B
enth
Pter
odon
em
argi
natu
s Vog
elN
ecta
ndra
mol
lis (K
unth
) N
ees
Cyb
iant
hus g
ardn
eri (
A.D
C.)
G.Ag
ostin
iLa
man
onia
tern
ata
Vell.
Our
atea
hex
aspe
rma
(A.S
t.-H
il.)
Baill
.St
rych
nos
pseu
doqu
ina
A.St
.-Hil
Gua
pira
are
olat
a (H
eim
erl)
Lund
ell
Mat
ayba
gui
anen
sis
Aubl
.Ta
pura
gui
anen
sis A
ubl.
Sym
ploc
os m
osen
ii Br
and
Sym
ploc
os n
itens
(Poh
l) Be
nth.
Amai
oua
guia
nens
is A
ubl.
Ente
rolo
bium
gum
mife
rum
(Mar
t.) J
.F.M
acbr
.

34
Tabe
la 3
. (C
ontin
uaçã
o)
Loga
niac
eae
Faba
ceae
Lyth
race
aeM
alpi
ghia
ceae
Mel
asto
mat
acea
eFa
bace
aeD
illen
iace
aeSt
yrac
acea
eFa
bace
aeFa
bace
aeA
ralia
ceae
Faba
ceae
Mel
asto
mat
acea
eC
elas
trace
aeFa
bace
aeFa
bace
aeFa
bace
aeB
igno
niac
eae
Cel
astra
ceae
Mel
asto
mat
acea
eR
osac
eae
Cel
astra
ceae
Laur
acea
eM
alpi
ghia
ceae
Fam
ília
Esp
écie
s C
rC
dC
rC
dC
rC
dC
rC
dC
rC
rC
rC
dC
dC
d
DA
DR
FAFR
DoA
DoR
IVI * * 0.
81.
013
.40.
46.
9 * * * * 4.9 * 0.7 * * 4.7 * * 1.1 * * * *
0.6
0.6
0.6
0.6
0.6
0.6
0.6
0.4
0.4
0.4
0.4
0.4
0.4
0.8
0.3
0.3
0.3
0.3
0.3
0.3
0.3
0.3
0.3
0.3
* * 0.2
0.1
5.5
0.1
1.5 * * * * 1.6 * 0.1 * * 1.5 * * 0.1 * * * *
0.1
0.1
0.0
0.0
0.0
0.0
0.0
0.0
0.1
0.1
0.0
0.1
0.1
0.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
* * 0.0
0.0
0.9
0.0
0.2 * * * * 0.3 * 0.0 * * 0.2 * * 0.0 * * * *
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
* * 0.5
0.8
2.6
0.3
2.6 * * * * 2.3 * 0.5 * * 2.1 * * 0.8 * * * *
0.2
0.5
0.5
0.5
0.5
0.5
0.5
0.2
0.2
0.2
0.2
0.2
0.2
0.5
0.2
0.2
0.2
0.2
0.2
0.2
0.2
0.2
0.2
0.2
* * 2 3 10 1 10 * * * * 9 * 2 * * 8 * * 3 * * * *
1 2 2 2 2 2 2 1 1 1 1 1 1 2 1 1 1 1 1 1 1 1 1 1
* * 0.1
0.1
5.3
0.1
2.7 * * * * 1.0 * 0.1 * * 1.1 * * 0.2 * * * *
0.2
0.1
0.1
0.1
0.1
0.1
0.1
0.2
0.0
0.0
0.1
0.0
0.0
0.2
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
* * 3 3 116 2 60 * * * * 22 * 2 * * 25 * * 4 * * * *
6 2 2 2 2 2 2 4 1 1 2 1 1 5 1 1 1 1 1 1 1 1 1 1
Anto
nia
ovat
a Po
hlPi
ptoc
arph
a m
acro
poda
(DC
.) Ba
ker
Lafo
ensi
a pa
cari
A.S
t.-H
il.By
rson
ima
cocc
olob
ifolia
Kun
thM
icon
ia fe
rrug
inat
a D
C.
Mac
haer
ium
acu
tifol
ium
Vog
elD
avill
a el
liptic
a A.
St.-H
il.St
yrax
cam
poru
m P
ohl
Hym
enae
a co
urba
ril L
.Pl
atyp
odiu
m e
lega
ns V
ogel
Sche
ffler
a mo
roto
toni
(Aub
l.) M
agui
re, S
teyer
m. &
Fro
din
Pter
odon
pub
esce
ns (B
enth
.) Be
nth.
Tibo
uchi
na c
ando
llean
a (M
art.
ex D
C) C
ogn.
Sala
cia
cras
sifo
lia (M
art.
ex S
chul
t.) G
. Don
Plat
hym
enia
ret
icul
ata
Bent
h.Va
taire
a m
acro
carp
a (B
enth
.) D
ucke
Mac
haer
ium
opa
cum
Vog
elJa
cara
nda
pube
rula
Cha
m.
Plen
ckia
pop
ulne
a Re
isse
kM
icon
ia r
ubig
inos
a (B
onpl
.) D
C.
Prun
us s
ello
wii
Koe
hne
Sala
cia
ellip
tica
(Mar
t. ex
Sch
ult.)
G. D
onCr
ypto
cary
a as
cher
soni
ana
Mez
Byrs
onim
a pa
chyp
hylla
A. J
uss.
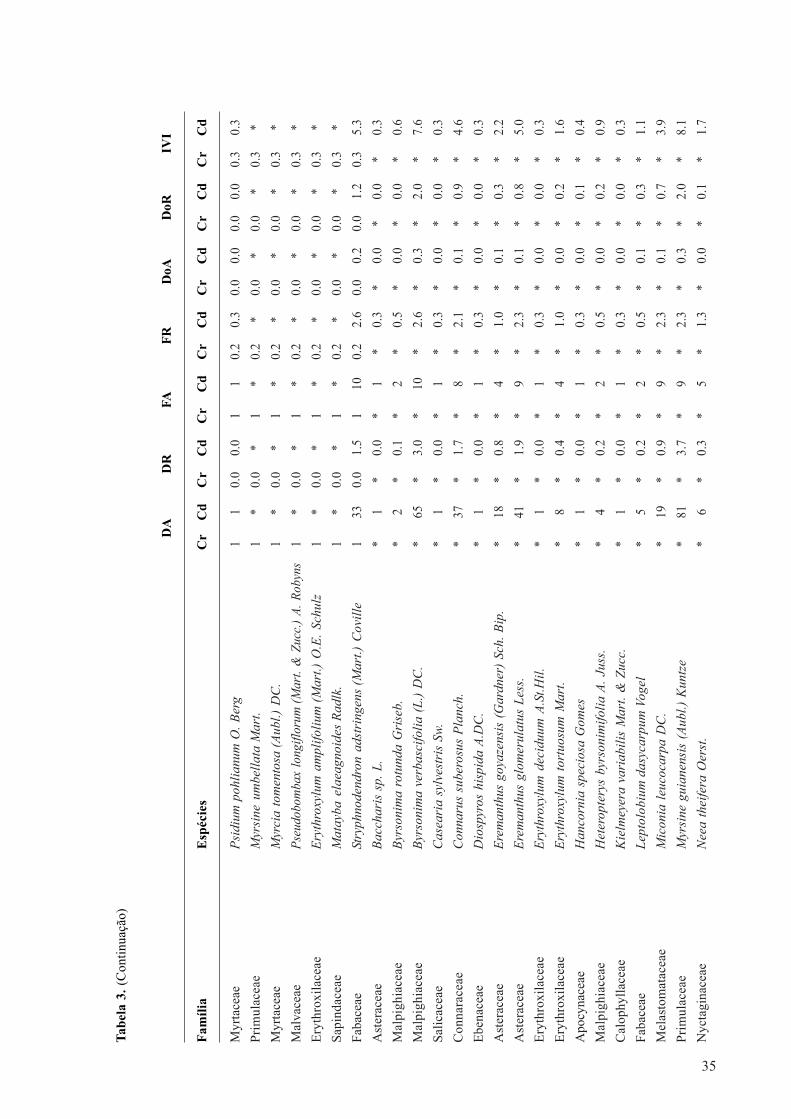
35
Tabe
la 3
. (C
ontin
uaçã
o)
Myr
tace
aePr
imul
acea
eM
yrta
ceae
Mal
vace
aeEr
ythr
oxila
ceae
Sapi
ndac
eae
Faba
ceae
Ast
erac
eae
Mal
pigh
iace
aeM
alpi
ghia
ceae
Salic
acea
eC
onna
race
aeEb
enac
eae
Ast
erac
eae
Ast
erac
eae
Eryt
hrox
ilace
aeEr
ythr
oxila
ceae
Apo
cyna
ceae
Mal
pigh
iace
aeC
alop
hylla
ceae
Faba
ceae
Mel
asto
mat
acea
ePr
imul
acea
eN
ycta
gina
ceae
Fam
ília
Esp
écie
s C
rC
dC
rC
dC
rC
dC
rC
dC
rC
rC
rC
dC
dC
d
DA
DR
FAFR
DoA
DoR
IVI 0.
3 * * * * * 5.3
0.3
0.6
7.6
0.3
4.6
0.3
2.2
5.0
0.3
1.6
0.4
0.9
0.3
1.1
3.9
8.1
1.7
0.3
0.3
0.3
0.3
0.3
0.3
0.3 * * * * * * * * * * * * * * * * *
0.0 * * * * * 1.2
0.0
0.0
2.0
0.0
0.9
0.0
0.3
0.8
0.0
0.2
0.1
0.2
0.0
0.3
0.7
2.0
0.1
0.0
0.0
0.0
0.0
0.0
0.0
0.0 * * * * * * * * * * * * * * * * *
0.0 * * * * * 0.2
0.0
0.0
0.3
0.0
0.1
0.0
0.1
0.1
0.0
0.0
0.0
0.0
0.0
0.1
0.1
0.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0 * * * * * * * * * * * * * * * * *
0.3 * * * * * 2.6
0.3
0.5
2.6
0.3
2.1
0.3
1.0
2.3
0.3
1.0
0.3
0.5
0.3
0.5
2.3
2.3
1.3
0.2
0.2
0.2
0.2
0.2
0.2
0.2 * * * * * * * * * * * * * * * * *
1 * * * * * 10 1 2 10 1 8 1 4 9 1 4 1 2 1 2 9 9 5
1 1 1 1 1 1 1 * * * * * * * * * * * * * * * * *
0.0 * * * * * 1.5
0.0
0.1
3.0
0.0
1.7
0.0
0.8
1.9
0.0
0.4
0.0
0.2
0.0
0.2
0.9
3.7
0.3
0.0
0.0
0.0
0.0
0.0
0.0
0.0 * * * * * * * * * * * * * * * * *
1 * * * * * 33 1 2 65 1 37 1 18 41 1 8 1 4 1 5 19 81 6
1 1 1 1 1 1 1 * * * * * * * * * * * * * * * * *
Psid
ium
poh
lianu
m O
. Ber
gM
yrsi
ne u
mbe
llata
Mar
t.M
yrci
a to
men
tosa
(Aub
l.) D
C.
Pseu
dobo
mba
x lo
ngifl
orum
(Mar
t. &
Zuc
c.) A
. Rob
yns
Eryt
hrox
ylum
am
plifo
lium
(Mar
t.) O
.E. S
chul
zM
atay
ba e
laea
gnoi
des R
adlk
.St
ryph
node
ndro
n ad
stri
ngen
s (M
art.)
Cov
ille
Bacc
hari
s sp
. L.
Byrs
onim
a ro
tund
a G
rise
b.By
rson
ima
verb
asci
folia
(L.)
DC
.C
asea
ria
sylv
estr
is S
w.C
onna
rus
sube
rosu
s Pl
anch
.D
iosp
yros
his
pida
A.D
C.
Erem
anth
us g
oyaz
ensi
s (G
ardn
er) S
ch. B
ip.
Erem
anth
us g
lom
erul
atus
Les
s.Er
ythr
oxyl
um d
ecid
uum
A.S
t.Hil.
Eryt
hrox
ylum
tort
uosu
m M
art.
Han
corn
ia s
peci
osa
Gom
esH
eter
opte
rys
byrs
onim
ifolia
A. J
uss.
Kie
lmey
era
vari
abili
s M
art.
& Z
ucc.
Lept
olob
ium
das
ycar
pum
Vog
elM
icon
ia le
ucoc
arpa
DC
.M
yrsi
ne g
uian
ensi
s (A
ubl.)
Kun
tze
Nee
a th
eife
ra O
erst
.

36
Tabe
la 3
. (C
ontin
uaçã
o)
Rub
iace
aeA
ster
acea
eSa
pota
ceae
Myr
tace
aeSy
mpl
ocac
eae
Voch
ysia
ceae
Fam
ília
Esp
écie
s C
rC
dC
rC
dC
rC
dC
rC
dC
rC
rC
rC
dC
dC
d
DA
DR
FAFR
DoA
DoR
IVI 3.
46.
23.
90.
32.
12.
1
* * * * * *
0.3
1.5
1.4
0.0
0.3
0.4
* * * * * *
0.1
0.2
0.2
0.0
0.1
0.1
* * * * * *
2.3
2.3
1.6
0.3
1.3
1.3
* * * * * *
9 9 6 1 5 5
* * * * * *
0.7
2.3
1.0
0.0
0.5
0.5
* * * * * *
15 51 21 1 11 10
* * * * * *
Palic
oure
a ri
gida
Kun
thPi
ptoc
arph
a ro
tund
ifolia
(Les
s.) B
aker
Pout
eria
ram
iflor
a (M
art.)
Rad
lk.
Psid
ium
myr
sini
tes
DC
.Sy
mpl
ocos
rha
mni
folia
A. D
C.
Voch
ysia
elli
ptic
a M
art.

37
Discussão
Nas duas áreas o aumento no esforço amostral a partir da 5ª parcela resultou em pequena
adição no número de espécies. O padrão encontrado para as curvas de rarefação corroboram que
para a vegetação do Cerrado há acréscimos mais acentuados de riqueza até a primeira metade em
amostragens de um hectare, quando então, a inclinação da curva diminui até a estabilização
(Assunção & Felfili, 2004; Fonseca & Silva Júnior, 2004). Felfili & Imaña-Encinas (2001) afirmam
que uma amostragem é considerada suficiente se a curva espécie-área, no caso do presente estudo
a curva de rarefação, mostrar sinais de estabilização e se os erros padrões forem inferiores a 10%
das médias estimadas. Portanto, a amostragem nas duas áreas foi considerada suficiente.
As famílias Fabaceae e Vochysiaceae foram relatadas com destaque na maioria dos
levantamentos realizados em cerradão (Costa & Araújo, 2001; Ribeiro & Haridasan, 1984; Pereira-
Silva et al., 2004) e em cerrado sentido restrito (Teixeira et.al., 2004; Silva et al., 2002; Costa et
al., 2010; Assunção & Felfili, 2004; e Guarin-Neto et al., 1994). A região central do Brasil foi
considerada o principal centro de diversificação da família Fabaceae (Polhill et al., 1981). A
enorme representatividade desta família no Cerrado (Mendonça et al., 2008) pode ser atribuída à
associação de algumas das suas espécies com bactérias fixadoras de nitrogênio, fato que lhe
proporciona vantagem competitiva nos solos distróficos do Cerrado (Goodland, 1979). Já para a
Vochysiaceae muitas de suas espécies são acumuladoras de alumínio (Haridasan & Araújo, 1988)
o que pode configurar também vantagem competitiva para crescer, com sucesso, em solos
distróficos, ácidos e ricos em alumínio predominantes no Cerrado (Felfili & Silva Júnior, 1993).
Melastomataceae é a sexta maior família de Angiospermas no Brasil, com 68 gêneros e mais
de 1.500 espécies, que se distribuem desde a Amazônia até o Rio Grande do Sul, estando presente
em praticamente todas as formações vegetacionais com número variável de espécies (Romero &
Martins, 2002). Muitas de suas espécies também são acumuladoras de alumínio (Haridasan &
Araújo, 1987), componente comum que geralmente ocorre em elevadas concentrações nos solos
da região (Eiten, 1972; Goodland 1979; Haridasan, 1982), isto, em parte, poderia justificar o
sucesso no estabelecimento de espécies da família no Brasil Central (Felfili &Silva-Júnior, 1993).
Entretanto, algumas sementes e mudas de espécies desta família são sensíveis ao fogo, e a
intensificação da frequência com que ocorrem os incêndios aumenta significativamente a
mortalidade de suas plântulas (Hoffmann, 1998), tendo em vista que o fogo é agente comum no
Cerrado. Estudos apontam que a espécie Miconia albicans tem dificuldades de floração, produção
de frutos e estabelecimento de mudas após a passagem do fogo (Hoffmann, 1998; Sato, 2003).

38
Hoffmann (1998), ao estudar os efeitos do fogo no ciclo reprodutivo de espécies lenhosas do
Cerrado, na reserva do IBGE (Instituto de Brasileiro de Geografia de Estatística), em Brasília
(DF), observou mortalidade de 100% para mudas de M. albicans mesmo depois de dois anos de
idade das plântulas. Segundo Silveira (2011), um ambiente livre de fogo deve aumentar o
recrutamento de sementes e mudas sensíveis ao fogo. A riqueza dessa família nas áreas do presente
estudo levanta a hipótese de que a proteção de mais de vinte anos nas áreas pode ter favorecido
o enriquecimento. Estudos específicos de comparação podem demonstrar relação entre proteção
ao fogo e enriquecimento de espécies ou de densidade de indivíduos da família.
As comunidades apresentaram alta diversidade de espécies e distribuição equitativa dos
indivíduos entre as espécies amostradas (Margalef, 1972; Margurran, 1988). Os índices de
diversidade e equabilidade equivalem aos encontrados para outras áreas de cerradão (Ribeiro &
Haridasan, 1984; Costa & Araújo, 2001; Marimon-Junior& Haridasan, 2005; Felfili et al., 2007;
Silva, 2009) e cerrado sentido restrito (Felfili & Silva-Júnior, 1993; Costa & Araújo, 2001; Andrade
et al., 2002; Fonseca & Silva-Júnior, 2004; Marimon-Junior& Haridasan, 2005) estudados no
Brasil Central.
Os valores dos índices de similaridade de Chao-Sørensen e Sørensen entre as comunidades
indicaram baixa similaridade florística. Os trabalhos tomados como referência para o presente estu-
do utilizaram o índice de similaridade de Sørensen tradicional. No estudo comparativo entre cerradão
e cerrado denso conduzido por Ribeiro & Haridasan (1984), em Planaltina (DF), foi encontrado
baixo valor de similaridade florística (0,36) e foi apontada como causas a variação florística e a
participação das espécies acumuladoras de alumínio que foi sete vezes maior no somatório do IVI
no cerrado denso que no cerradão. Para o cerradão e o cerrado sentido restrito estudados por
Marimon-Junior & Haridasan (2005), em Nova Xavantina (MT), foi obtido baixo valor de similari-
dade florística (0,58) que, de acordo com os autores, o resultado reforçou as diferenças entre a
composição de espécies entre as áreas. Na Reserva do Panga, em Uberlândia (MG), este índice foi
de 0,73 (Costa & Araújo, 2001), a contiguidade entre estas áreas que facilitaria a dispersão das
espécies foi considerada fator determinante da elevada similaridade. No presente estudo a proximi-
dade entre áreas parece não ter afetado a composição em espécies nas duas áreas.
A soma da densidade e da área basal dos indivíduos encontrados no Cr é comum em
comunidades florestais (Marimon-Júnior & Haridasan, 2005) e figuram entre os mais altos
encontrados para o cerradão que até então variaram entre 716 e 2.178 ind.ha-¹ (Felfili et al., 2007;
Franczak, 2006 apud Solorzano, 2011) e 17,47 e 24 m².ha-¹ (Felfili et. al. 1994; Franczak, 2006

39
apud Solorzano, 2011; Felfili et al., 2007), respectivamente. Esses trabalhos utilizaram a mesma
metodologia (Db30 cm) e” 5 cm de inclusão dos diâmetros.
Apesar dos altos valores de densidade nas duas áreas, a área basal em Cr foi o dobro daquela
em Cd. De fato o Cr apresenta árvores com diâmetros e alturas maiores que o Cd e, portanto,
figura no grupo das fitofisionomias florestais de Ribeiro & Walter (2008). No Cd a densidade e a
área basal estão entre as maiores já registradas para outras áreas de cerrado sentido restrito no
Brasil Central (Azevedo et al., 1990; Felfili & Silva-Júnior, 1993; Andrade et al., 2002; Assunção
& Felfili, 2004). No cerrado sentido restrito na Floresta Nacional de Paraopeba (MG), Balduino
et al., (2005) registraram área basal de 18,13 m².ha-1 resultante da elevada densidade de 1.990
ind.ha-1, sendo a maior área basal já registrada para esta fitofisionomia.
Foram observadas diferenças significativas na comparação Cr/Cd para os parâmetros: área
basal (m².ha-1), altura média (m), altura máxima (m), número de árvores mortas e área basal das
árvores mortas (m².ha-1). Estes resultados indicam para a comunidade Cr níveis mais elevados de
competição por luz e por recursos nos solos.
O Cr inclui a maioria das espécies descritas por Furley e Ratter (1988) como indicadoras da
condição distrófica do cerradão, as quais são: Blepharocalyx salicifolius, Callisthene major,
Emmotum nitens, Ocotea spixiana, Tachigali subvelutina, Siphoneugena densiflora e Virola
sebifera. A composição de espécies no Cr é semelhante aos cerradões estudados por Ribeiro &
Haridasan (1984), Costa & Araújo (2001), Marimon Júnior & Haridasan (2005) e Silva et al.,
(2008). As principais espécies amostradas em Cd evidenciam o padrão florístico típico observado
para cerrado sentido restrito em outras áreas no Distrito Federal (Ribeiro et al., 1983; Ribeiro &
Haridasan, 1984; Felfili, 2001; Andrade et al., 2002; Assunção & Felfili, 2004; Silva Júnior &
Sarmento, 2009). A comunidade arbórea registrada por Azevedo et al. (1990) para o cerrado
denso é mais parecida com a composição florística do Cr do que com a do Cd.
O número de espécies compartilhadas entre as áreas e o total de espécies exclusivas do Cr
que ocorre frequentemente em comunidades de cerrado sentido restrito e de matas de galeria e
ciliares corroboram com as argumentações de Felfili et al., (1992) e Ribeiro & Walter (2008).
Felfili et al., (1992) indicaram que o cerradão apresenta identidade fisionômica, mas não florística
por não terem encontrado qualquer espécie lenhosa exclusiva ao cerradão na Chapada Pratinha
(MG, DF e GO). Concluíram, então, que a composição florística nesta comunidade resulta da
combinação de espécies lenhosas do cerrado sentido restrito e das formações florestais na
vizinhança. De acordo com Ribeiro & Walter (2008), do ponto de vista fisionômico o cerradão é

40
floresta, mas floristicamente é similar aos cerrados sentido restrito. Apesar dessas semelhanças,
algumas espécies como Emmotum nitens, Blepharocalyx salicifolius, Simaurouba versicolor e
Ocotea spixiana, entre outras, ocorrem com maior importância no cerradão (Felfili & Silva Júnior,
1992), já Virola sebifera e Copaifera langsdorffii que figuraram entre as dez mais importantes
em Cr são algumas vezes encontradas com maior importância nas matas de galeria. Costa &
Araújo (2001) encontraram no cerradão espécies de cerrado sentido restrito e de matas mesófilas
semidecíduas, que parecem ocorrer no limite de distribuição da área estudada. Dessa forma os
autores caracterizaram o cerradão como possível área de transição entre as matas e cerrado. A
composição florística encontrada no Cr indica que este habitat satisfaz os requisitos básicos para
o estabelecimento conjunto de espécies típicas de fitofisionomias diferentes.
O número elevado de indivíduos com menor diâmetro encontrado nas duas áreas indica
balanço positivo entre recrutamento e mortalidade (Felfili, 1997; Silva Júnior, 2004; 2005a) e
sugere que as comunidades são autorregenerativas (Assunção & Felfili, 2004). Resultados
semelhantes foram obtidos em outras áreas do cerrado sentido restrito (Felfili et al., 2000, Pereira-
Silva et al., 2004; Silva Júnior, 2004, 2005; Marimon-Júnior & Haridasan, 2005) e cerradão
(Marimon-Júnior & Haridasan, 2005, Silva, 2009). A alta densidade de indivíduos menores
caracteriza comunidades com grande potencial para a regeneração natural (Pires & Prance, 1977).
A análise da distribuição das alturas reflete a estrutura vertical da comunidade e suas
características na competição por luz (Miranda, 2008). Portanto, fica claro a maior competitividade
por luz no Cr, devido as elevadas alturas dos indivíduos em relação as alturas encontradas no Cd.
A comunidade Cd por ser mais aberta, ou seja, menor cobertura de dossel sugere que as árvores
nesta área não necessitam competir por luz. Além disso, muitos indivíduos no Cd foram registrados
com 1 m de altura e o mesmo não foi encontrado no Cr.
De acordo com Felfili & Silva Júnior (1992) alta frequência de árvores mortas pode indicar
perturbações recentes na área. Porém, as comunidades estão em áreas protegidas de distúrbios
como o fogo e degradações antrópicas há mais de 20 anos, o que sugere que a competição intra
e interespecífica pode estar regulando as populações nas duas áreas. A competição por luz e por
espaço, devido à alta densidade de árvores, pode ser considerada fator limitante nas duas áreas,
provocando alta mortalidade do componente arbóreo.

41
Conclusões
O cerradão (Cr) e o cerrado denso (Cd) amostrados no Jardim Botânico de Brasília (DF)
são comunidades floristicamente diferentes apesar de compartilharem 43 (34%) espécies, visto
que as similaridades máximas de Chao-Sørensen e Sørensen entre áreas foram de 0,47 e 0,50,
respectivamente. A estrutura das comunidades também diferiu entre as áreas. Dentre os parâmetros
estruturais analisados, apenas a densidade foi estatisticamente semelhante. O Cr, por apresentar
comunidade mais fechada onde a competição por luz é maior, teve a área basal e as alturas médias
e totais das árvores estatisticamente maiores que no Cd.
A florística e a estrutura registradas no Cr e no Cd são compatíveis com a classificação de
cerradão e cerrado sentido restrito denso de Ribeiro & Walter (2008). A composição florística do
Cr foi considerada mista, composta por espécies florestais e savânicas. O Cr está entre os cerradões
mais ricos já amostrados, tendo como base os estudos com a mesma metodologia de mensuração.
O Cd é composto basicamente por espécies comuns a esse habitat. Esse resultado acentua a
importância do JBB para a conservação das espécies do bioma. Um quarto das espécies, em
ambas as áreas, são localmente raras, o que indica elevada diversidade de espécies.
As áreas estudadas são representadas principalmente por indivíduos jovens, o que caracteriza
balanço positivo entre recrutamento e mortalidade nas duas áreas. Os níveis de distúrbio são
permanentemente administrados pelo JBB em sua área de visitação, justamente por se tratar de
área de conservação. A alta densidade de árvores mortas sugere alta competição inter e
intraespecífica nas duas áreas.
Diferenças nos níveis de fertilidade nos solos podem determinar diferenças florísticas,
fitossociológicas e estruturais na vegetação. Esta hipótese foi discutida no capítulo 2, onde as
propriedades químicas e físicas dos solos analisados foram relacionadas com as características
estruturais e florísticas encontradas em cada fitofisionomia.

42
Capítulo 2
Relações solo e vegetação no cerradão e no cerrado denso no JardimBotânico de Brasília
Resumo – As comunidades de cerradão (Cr) e cerrado denso (Cd) localizadas no Jardim Botânico
de Brasília (DF) estão sobre solos distróficos com predomínio de Latossolos Vermelho e Vermelho
Amarelo. O presente capítulo teve como objetivos relacionar a estrutura das comunidades arbóreas
do Cr e do Cd com as características edáficas das parcelas amostradas. Foram alocadas dez
parcelas de 20 x 50 (1.000 m²) em cada área para o registro de indivíduos com diâmetro igual ou
acima de 5 cm, medido a 30 cm do solo, (Db30 cm) e” 5 cm, inclusive os indivíduos mortos em
pé. Nas duas áreas foram coletadas amostras compostas dos solos, 10-20 cm de profundidade,
que foram analisadas quanto as suas propriedades químicas e físicas. As análises multivariadas
TWINSPAN e PCA foram conduzidas buscando relacionar a vegetação com as variáveis dos
solos. Ambas as comunidades possuem solos argilosos, com baixos teores de bases trocáveis e
alta saturação por Al. O Cr apresentou teores estatisticamente maiores de Al, H+Al, Zn, Fe, K,
Na, silte, CTC e saturação por Al (m%). Já o solo do Cd foi considerado estatisticamente mais
fértil devido a maior concentração de saturação por bases (V%). O TWINSPAN separou as duas
comunidades e agrupou as parcelas floristicamente semelhantes. Os resultados obtidos pela PCA
explicaram 78% da variância total e distinguiram em parte dos resultados gerados pelo
TWINSPAN. A distribuição das espécies dentro de cada área não mostrou relação direta com as
variáveis do solo. As parcelas do Cr devido aos maiores níveis de silte, argila e MO apresentaram-
se mais úmidas em relação às parcelas do Cd, e foi associado a este fato, o maior adensamento do
componente arbóreo.
Palavras-chave – Solos distróficos, variáveis dos solos, TWINSPAN e PCA.

43
Introdução
O Cerrado apresenta grande diversidade edáfica com variações nas características físicas e
químicas. Os Latossolos são predominantes nesses ambientes por cobrir aproximadamente 50% de
sua área geográfica (Reatto et al., 2008; Haridasan, 2007). Essa classe de solos é caracterizada por
solos profundos, bem drenados, ricos em argila e pobres em nutriente (Furley & Ratter, 1988),
devido à baixa capacidade de troca catiônica, elevada acidez e os altos níveis de saturação por Al
(Furley & Ratter, 1988; EMBRAPA, 2006; Reatto et al., 2008). Os Latossolos podem ser distróficos
ou mesotróficos, existindo sítios de média a alta saturação por bases (EMBRAPA, 2006).
A fertilidade nos solos distróficos e mesotróficos foram verificadas nas baixas concentrações
de nutrientes foliares em espécies nativas do Cerrado (Haridasan, 1982; Haridasan &Araújo,
1988; Haridasan et al., 1997). O cerradão associado a solos distróficos ou mesotróficos (Ratter,
1971; Ratter et al., 1977; 1978; Araújo & Haridasan, 1988), tem a sua estrutura e composição
florística variada conforme tal fertilidade. Diante das diferenças na composição florística, Furley
& Ratter (1988) destacaram grupos de espécies indicadoras da condição distrófica e mesotrófica
no cerradão.
Alguns cerradões com níveis médios de fertilidade podem estar associados a áreas de
transição de floresta decíduas e cerrado sentido restrito (Ratter et al., 1977, 1978). Também
foram relacionados níveis de fertilidade com a posição topográfica inferior a outras áreas de
vegetação (Moreno & Schiavini, 2001). Nessas condições topográficas os cátions de áreas mais
elevadas são lixiviados e transportados para as áreas mais baixas.
Ao contrário das variações na fertilidade dos solos encontradas para o cerradão, a maioria
dos cerrados sentido restrito estudados foi associada aos solos distróficos (Silva Júnior &
Sarmiento, 2009; Assunção & Felfili, 2004; Felfili et al., 2000; Felfili, et al., 2000), frequentemente
em Latossolos, mas também podem ocorrer em Cambissolos, Neossolos e Chernossolos, entre
outros (Reatto & Martins, 2005). Segundo Haridasan (2000), o cerrado sentido restrito raramente
ocorre em solos mesotróficos, mas tem sua composição florística variada quando ocorre em
solos mais férteis (Haridasan, 2000).
As divisões dos subtipos do cerrado sentido restrito em denso, típico e ralo, com exceção do
rupestre, indicam gradiente decrescente de densidade arbóreo-arbustiva. Este gradiente é
influenciado por vários fatores ambientais, entre eles estão as condições edáficas (fertilidade, pH,
saturação de alumínio) e hídricas, a profundidade do solo e a frequência de queimadas (Coutinho,

44
1992, Meirelles et al., 1997; Ribeiro & Walter, 2010; Eiten, 1978). Meirelles et al., (1997) sugeriram
que o cerrado sentido restrito, quando protegido do fogo, teria aumentada a densidade arbórea e
culminaria na forma de cerradão. Já Haridasan (2000) alegou que isto não seria provável, pois a
reserva de nutrientes na biomassa e no solo de um cerrado sentido restrito não seria suficiente
para o desenvolvimento da biomassa maior de árvores. Além do mais, as espécies exclusivas do
cerradão precisariam de maior disponibilidade de nutrientes para o seu estabelecimento. Lopes &
Cox (1977) sugeriram que o cerradão poderia ocorrer em sítios mais férteis que os solos do
cerrado sentido restrito sem, no entanto, serem considerados mesotróficos.
No Jardim Botânico de Brasília, o solo e a disponibilidade de água para as plantas são alguns
dos fatores que possivelmente poderiam influenciar na distribuição florística e nos aspectos
estruturais do cerradão e do cerrado denso, tendo em vista que o clima e a geomorfologia são os
mesmos para ambos, e o fogo não ocorre há mais de vinte anos nessas áreas. De acordo com as
considerações para ambas as fitofisionomias o objetivo deste capítulo foi comparar as propriedades
químicas e físicas dos solos entre cerradão e cerrado denso, sobre solos distróficos e
predominantemente classificados como Latossolos Vermelhos e Vermelhos Amarelos (Cavedon
& Sommer, 1990). Pretendeu-se também apontar as variáveis dos solos com maior influencia nas
diferenças estruturais e florísticas anotadas neste capítulo e no capítulo 2.
Material e Métodos
As áreas estudadas estão localizadas no Centro de Visitantes do Jardim Botânico de Brasília
(CVJBB), Distrito Federal, entre as coordenadas 15°52’21"S e 47°50’50"W. Ali foram instaladas
dez parcelas permanentes de 20 x 50 m (1.000 m²) de forma sistemática no cerradão (Cr) e dez
parcelas aleatórias no cerrado denso (Cd), conforme recomendado por Felfili et al., (2005). Foram
registrados, identificados e medidos todos os indivíduos arbóreos quanto a sua altura total e
diâmetro igual ou superior a 5 cm, medido a 30 cm do solo, (Db30 cm) e” 5 cm, inclusive os
indivíduos mortos em pé. A distância entre Cr e Cd é de aproximadamente 2 km em linha reta.
Para mais detalhes da descrição das áreas de estudos ver o capítulo 1.
Os solos predominantes no Cr e no Cd são do tipo Latossolo Vermelho e Vermelho-Amarelo,
ambos considerados distróficos (Cavedon & Sommer, 1990). Para amostragem da fertilidade do
solo foram coletadas em cada parcela três amostras da camada superficial do solo (10-20 cm de
profundidade) no centro e nos vértices das parcelas que, misturadas, compuseram uma amostra

45
composta. As análises químicas e físicas das amostras foram feitas na “Solocria Laboratório
Agropecuário Ltda”, em Goiânia (GO) de acordo com a metodologia proposta pela Embrapa
(1997). Foram realizadas as análises de pH em CaCl², teor percentual de matéria orgânica (MO),
acidez potencial (H+Al), saturação por alumínio (m) e de bases (V), capacidade de troca catiônica
(CTC) e concentrações disponíveis dos elementos: Al, Ca, Mg, P, K, Mn, Cu, Zn, Fe e Na para as
20 parcelas estudadas. As estruturas granulométricas areia, silte e argila também foram analisadas.
Os testes de Shapiro-Wilk e Levenne foram utilizados para a verificação da existência de
normalidade (Shapiro & Wilk, 1965) e homocedasticidade (Hartley, 1950), respectivamente, para
os dados de todas as variáveis dos solos analisadas. Os dados que não apresentaram normalidade
foram transformados na base logarítmica ou raiz quadrada, a fim de normalizar os dados (Kent &
Coker, 1992; Palmer, 2005). Uma vez que as variáveis não apresentaram normalidade, utilizou-
se o teste não paramétrico de Mann-Whitney, utilizado para comparação entre as médias obtidas.
Tanto a normalidade e a homocedasticidade dos dados, quanto a aplicação do teste de Mann-
Whitney foram executados no programa PAST versão 2.17.
Para correlacionar e obter a percentagem explicativa das variáveis dos solos com as áreas
amostradas foi conduzida a técnica de ordenação indireta PCA (Principal Component Analysis).
A PCA condensa informações contidas em grandes grupos de variáveis a um pequeno grupo de
novas composições dimensionais (Kent & Coker, 1992; McGarigal et al., 2000). Para essa análise
foi construída a matriz com as vinte parcelas amostradas, dez em cada área, e as propriedades
químicas e físicas, estatisticamente diferentes entre as parcelas com base no teste t de Mann-
Whitney (p < 0,05) com exceção dos dados complementares CTC, V e m .
A vegetação lenhosa das duas áreas amostradas foi classificada por meio do método
TWINSPAN (Two Way Indicator Species Analysis) (Hill, 1979) com o objetivo de identificar as
espécies que caracterizam as comunidades. O método TWINSPAN parte do princípio de que,
para um grupo de amostras que constituem um tipo de comunidade, haverá um grupo
correspondente de espécies que caracterizam aquele tipo, denominadas espécies indicadoras (Kent
& Coker, 1992). Neste estudo a 1ª divisão separou Cr do Cd. As 2ª e 3ª divisões incluíram Cr e
Cd em separado. Para cada divisão é calculado um autovalor, que varia de 0 a 1, no qual representa
a contribuição relativa de cada componente para a explicação do total da variação nos dados
(Felfili et al., 2007). Quando o autovalor é maior que 0,3 o componente é considerado relevante
na determinação da variação dos dados, quando for abaixo de 0,3 os dados são considerados
homogêneos (Gauch, 1982; Felfili & Venturoli, 2000).

46
Para a análise de classificação TWINSPAN foi construída uma matriz com as 20 parcelas
amostradas (dez do Cr e dez do Cd) e, utilizou-se como variável a densidade de cada espécie com
densidade igual ou superior a cinco indivíduos, esta estratégia foi adotada com o intuito de
diminuir o peso das espécies raras (Kent & Coker, 1992). As análises multivariadas foram realizadas
no programa PC-ORD versão 5.12 (McCune & Mefford, 1997). Os resultados obtidos pela PCA
foram relacionados com a classificação gerada pelo TWINSPAN.
Resultados
As áreas de Cr e Cd apresentaram solos ácidos e distróficos, com saturação por bases <
50%, saturação por alumínio > 50% e baixa capacidade de troca catiônica (CTC) < 27 cmolc/Kg-
1, de acordo com o Sistema Brasileiro de Classificação dos Solos (EMBRAPA, 2006) (Tabela 1).
Ambos os solos apresentaram textura argilosa, presente na composição granulométrica mais de
50% de argila (IBGE, 2007). Os níveis de areia variaram de 23 a 62% no Cr e de 19 a 82% em
Cd, ambos com coeficiente de variação relativamente altos (Tabela 1). A parcela 1 do Cd apresentou
82% de areia e 14% de argila, considerada, portanto, de textura média a arenosa (IBGE, 2007).
Os valores de Ca e Mg, 0,2 e 0,1 cmolc/dm³, respectivamente, foram os mesmos para as
duas áreas. Os níveis de MO, Cu, Mn e argila foram maiores nos solos do Cr e os valores de pH
e a percentagem de areia foram mais elevados nos solos do Cd (Tabela 1). A saturação por bases
(V) e o teor de P foram estatisticamente distintos entre os solos das áreas estudadas, com níveis
mais elevados no Cd (Tabela 1). A saturação por Al, os níveis de Al, H+Al, Fe, Zn, K, Na, silte e
a capacidade de troca catiônica (CTC) também diferiram estatisticamente, sendo maiores nos
solos do Cr (Tabela 1).

47
Tabe
la 1
. Pro
prie
dade
s qu
ímic
as e
físi
cas
dos
solo
s (1
0-20
cm
de
prof
undi
dade
) am
ostra
das
no c
erra
dão
(Cr)
e n
o ce
rrad
o de
nso
(Cd)
, loc
aliz
ados
no
Jard
im B
otân
ico
deB
rasí
lia (D
F). M
O: m
atér
ia o
rgân
ica;
CTA
: cap
acid
ade
de tr
oca
catiô
nica
; V: s
atur
ação
por
bas
es; m
: sat
uraç
ão p
or a
lum
ínio
; DP:
des
vio
padr
ão; p
: nív
el d
e si
gnifi
cânc
ia (p
< 0,
05) p
elo
test
e t d
e M
ann-
Whi
tney
; ***
: sig
nific
ativ
o e
ns: n
ão s
igni
ficat
ivo.
p ***
***
***
***
ns ***
***
ns ***
ns ns ns ***
ns ***
***
***
CV
%
10,3
5
10,6
7
15,0
7 1
10,3
8
16,7
4
38,4
9
52,0
1
68,7
7
17,4
2
42,2
2
3,2
2
24,2
1
15,8
3
37,3
6
10,3
4
9,6
5
2,9
7
DP
0
,14
1
,13
7
,22
0
,85
6
,28
1
,15
0
,50
0
,43
2
3,52
2
,08
0
,12
1
3,07
1
,65
1
3,30
1
,14
0
,38
2
,26
méd
ia
1
,37
10
,57
47
,90
0
,77
37
,50
3
,00
0
,96
0
,62
134
,99
4
,92
3
,82
54
,00
10
,40
35
,60
10
,99
3
,97
76
,22
Cr1
0 1,4
11,9 59 0,5 39 3 0,8
0,3
153,
65,
63,
9 68 9 23 12,4
3,7
75,6
Cr9 1,2
9,5 47 0,8 27 5 0,8
0,2
137,
75,
93,
9 47 10 43 9,9
4,4
74,0
Cr8 1,4
12,4 57 0,8 48 3 1,1
0,2
165,
54,
93,
8 65 10 25 12,9
3,6
75,6
Cr7 1,2
10,6 54 0,5 43 2 1,3
0,2
166,
37,
3 4 67 9 24 11,0
4,0
73,1
Cr6 1,2
9,7 51 1,8 39 3 1,7
0,2
139,
49 4 65 10 25 10,1
4,3
73,6
Cr5 1,5
9,8 39 0,1 31 2 0,5
1,1
133
2,7
3,7 57 13 30 10,2
4,0
78,9
Cr4 1,3
11,6 41 0,1 37 2 0,4 1
100,
63 3,7 57 13 30 12,0
3,4
76,4
Cr3 1,5 10 39 2,7 33 3 0,5
0,9
109,
42,
73,
7 42 11 47 10,4
4,0
78,9
Cr2 1,4
9,1 47 0,3 35 5 0,7 1 106
4,2
3,8 30 8 62 9,5
4,6
76,9
Cr1 1,6
11,1 45 0,1 43 2 1,8
1,1
138,
43,
93,
7 42 11 47 11,5
3,7
79,2
Vari
ávei
s/Pa
rcel
as A
l cm
olc/
dm³
H+A
L cm
olc/
dm³
K m
g/dm
³P
mg/
dm³
MO
g/d
m³
NA
g/d
m³
Zn m
g/dm
³C
u m
g/dm
³Fe
mg/
dm³
Mn
mg/
dm³
pH(c
acl2
)A
rg %
silte
%A
reia
%C
TCV
%m
%

48
Tabe
la 1
. (C
ontin
uaçã
o)
Vari
ávei
s/Pa
rcel
as
Al c
mol
c/dm
³H
+AL
cmol
c/dm
³K
mg/
dm³
P m
g/dm
³M
O g
/dm
³N
A g
/dm
³Zn
mg/
dm³
Cu
mg/
dm³
Fe m
g/dm
³M
n m
g/dm
³pH
(cac
l2)
Arg
%si
lte %
Are
ia %
CTC
V%
m%
Cd1 0,8
5,3 21 0,8 26 2 0,2
0,2 75 2,3
3,8 14 3,3 82 5,6
6,34
69,7
5
Cd2 0,8
5,7 27 1,2 33 2,2
0,5
0,4
108,
25,
83,
8 59 5 36 6 6,24
68,5
7
Cd3 0,9
6,8 27 0,8 35 1,8
0,2
0,4
87,5
2,4
3,8 74 7 19 7,1
5,26
70,8
Cd4 0,8
7,5 26 1,2 39 2 0,2
0,4
107,
54,
83,
8 64 6,7 29 7,8
4,8
68,3
Cd5 1 7,2 33 1,2 39 1,6
0,2
0,6
82,4
4,4
3,8 67 6,7 26 7,6
5,1
72,4
Cd6 0,8
7,8 37 1,5 35 2 0,2
0,5
88,7
2,4
3,8 69 8,3 22 8,2
4,8
67,2
Cd7 0,8
6,7 47 1,5 35 2 0,3
0,4
85,7
2,1
3,8 69 6,7 24 7,1 6 65,5
Cd8 0,9
4,9 39 1,2 27 1,8
0,3
0,2
80,8
2,7
3,9 37 3,3 59 5,3
7,6
69,2
Cd9 0,7
4,1 31 1,2 25 2 0,2
0,4 83 3,8
3,9 37 3,3 59 4,5
8,6
64,8
Cd1
0 0,6
5,2 37 1,2 39 1,6
0,5
0,5
97,9
6,2
3,9 49 3,3 47 5,6 7 60,6
méd
ia
0,81
6,12
32,5
01,
1833
,30
1,90
0,28
0,40
89,6
73,
693,
8353
,90
5,36
40,3
06,
48 6
,17
67,7
1
DP
0,11
1,25
7,68
0,23
5,46
0,19
0,12
0,12
11,2
6 1
,54
0,05
19,2
81,
9420
,78
1,24
1,27
3,39
CV
% 13
,59
20,3
523
,62
19,8
916
,39
10,2
343
,90
31,1
812
,56
41,6
61,
2635
,78
36,1
751
,58
19,
1220
,51
5,0
0
p ***
***
***
***
ns ***
***
ns ***
ns ns ns ***
ns ***
***
***

49
A ordenação das análises dos solos pela PCA explicou 78% da variância total e separou as
parcelas de Cr das parcelas de Cd. Apenas o eixo I incluiu 64% da variância total, negativamente
correlacionado com as variáveis Al, H+Al, Na, Fe, Zn, K e silte e, positivamente correlacionado
com P. O eixo II incluiu 14% da variância total e correlacionou-se negativamente com K, Na, Zn
e Fe e, positivamente com as variáveis Al, H+Al e silte (Figura 1).
Figura 1. Análise de Componentes Principais de oito variáveis do solo Al, H+Al, K, Fe, Zn. Na, P e silte, queforam estatisticamente distintas entre o cerradão (Cr) e o cerrado denso (Cd), localizados no Jardim Botânico deBrasília (DF).
A ordenação das parcelas mostrou maior homogeneidade na comunidade Cd, onde todas as
parcelas estão positivamente correlacionadas com o eixo I. As parcelas 1, 3, 4 e 5 estão
positivamente correlacionadas com o eixo II, enquanto as parcelas 2, 6, 7, 8, 9 e 10 estão
negativamente correlacionadas com este eixo e influenciadas pelo nutriente P.
A comunidade Cr, negativamente correlacionada com o eixo I, é influenciada por todas as
outras variáveis incluídas na PCA. As parcelas 1, 4 e 5 estão positivamente correlacionadas com
o eixo II e influenciadas por A, H+Al e silte. Já as parcelas 2, 3, 6, 7, 8, 9 e 10 são influenciadas
por Fe, Zn, K e Na e negativamente correlacionadas com o eixo II.
A classificação pelo TWISPAN separou as duas áreas com autovalor de 0,709 (Figura 2). A
espécie indicadora desta divisão foi Byrsonima laxiflora que ocorreu apenas no Cr. Entre as

50
espécies preferências do Cr estão Siphoneugena densiflora, Nectandra reticulata e Emmotum
nitens com densidades acima de 20 ind.ha-1 e, abaixo de 10 ind.ha-1. estão Tapirira guianensis e
Lamanonia ternata. Em Cd, as espécies preferenciais com densidades acima de 20 ind.ha-1. foram
Dalbergia miscolobium, Poliouratea hexasperma e Davilla elíptica, entre outras, e apenas a
espécie Leptolobium dasycarpum obteve densidade abaixo de 10 ind.ha-1.
Das 81 espécies analisadas 19 (23%) não foram preferenciais a qualquer uma das duas
comunidades, a saber: Annona crassiflora, Aspidosperma tomentosum, Blepharocalyx salicifolius,
Dalbergia miscolobium, Eriotheca pubescens, Guapira graciliflora, Guapira noxia,
Handroanthus ochraceus, Hymenaea stigonocarpa, Kielmeyera coriacea, Kielmeyera speciosa,
Miconia albicans, Qualea grandiflora, Qualea multiflora, Qualea parviflora, Roupala montana,
Schefflera macrocarpa, Tachigali subvelutina e Vochysia thyrsoidea. As 2ª e 3ª divisões não
tiveram autovalores significativos o que indicou homogeneidade dentro das áreas (Figura 2).
Figura 2. Classificação pelo método TWINSPAN das 20 parcelas amostradas, dez no cerradão (Cr) e dez nocerrado denso (Cd), localizados no Jardim Botânico de Brasília (DF). Os valores entre parênteses referem-se aosautovalores.
Discussão
Os Latossolos, em geral, são solos fortemente ácidos, distróficos, alumínicos e com baixa
saturação por bases (EMBRAPA, 2006). Diante da condição distrófica encontrada nos solos
estudados e das diferenças nas propriedades químicas e físicas (Tabela 1), o Cr exibiu solos mais

51
ácidos, devido aos elevados valores de saturação por alumínio, alumínio e acidez potencial, mais
ricos em cátions, com maiores proporções de silte e maior capacidade de troca catiônica. Enquanto
os solos do Cd apresentaram-se estatisticamente mais férteis e com maiores níveis de fósforo.
Os valores de matéria orgânica, pH, zinco e potássio encontrados no Cr e no Cd estão
dentro da variação conhecida para o Cerrado (Furley & Ratter, 1988). Os baixos valores de
cálcio e magnésio encontram-se abaixo de 2 cmolc/Kg-1, corroborando a condição distrófica dos
solos estudados, pois esse valor é usado como critério para separar solos mesotróficos de solos
distróficos (Haridasan & Araújo, 1988; Ratter et al., 1977).
A elevada saturação por alumínio registrada tanto no Cr quanto no Cd está dentro da variação
encontrada por Ratter e Furley (1988) para o Cerrado, que foi de 1,1 a 89,4%. Maior teor de
alumino no cerradão também foi encontrado nos estudados de Moreno & Schiavini (2001), Costa
& Araújo (2001), Ruggiero et al., (2002) e Marimom Junior & Haridasan (2005). Altos teores
desse elemento no solo determinam menor disponibilidade de nutrientes para a vegetação (Brady
& Weil, 2013), uma vez que, a precipitação lixívia o cálcio e outras bases e, o alumínio em
contato com os coloides do solo se adere rapidamente à superfície destes e não sofre lixiviação,
ou seja, o alumínio compete com as bases trocáveis pelos mesmos sítios de troca no complexo
coloidal. O empobrecimento do solo diretamente relacionado com o alumínio ficou conhecido
como hipótese do oligotrofismo aluminotóxico (Goodland, 1971b). Este fato foi observado por
Goodland & Pollard (1973), em gradiente fisionômico de cerrado sentido amplo, no Triângulo
Mineiro (MG), onde os autores encontraram relação negativa entre o alumínio e os teores de
cálcio, magnésio e potássio.
O clima tropical úmido e os solos bem drenados favorecem a abundância de Al nos solos do
Cerrado. Assim, altas quantidades desse metal nos solos dos cerradões contribuem para a menor
fertilidade dos mesmos. Dessa forma, os solos do Cd são relativamente mais férteis por terem
menos alumínio trocável, uma vez que, os valores de cálcio e magnésio não diferiram entre as
áreas, e os elementos potássio e sódio foram maiores no Cr. Estes elementos fazem parte da
somatória necessária para obter o valor de saturação por bases (IBGE, 2007).
A maior quantidade de matéria orgânica, argila e silte indica que os solos do Cr são mais
úmidos, pois são variáveis capazes de reter água e permitir que o solo fique úmido por mais
tempo. O silte é descrito como uma micropartícula de areia que possui certa viscosidade e capa-
cidade de adsorção de água devido a película adesiva de argila que a envolve. Quando somado ao
poder coloidal da argila, lhe confere textura fina e movimentação lenta de ar e água (Brady &

52
Weil, 2013). A argila, que existe em estado coloidal, é caracterizada por tamanhos extremamente
pequenos, com grande área externa por unidade de peso e presença de cargas de superfície que
atraem íons e água (Brady & Weil, 2013). As frações de argila, mediante a atração de íons para
suas superfícies, protegem temporariamente as substâncias nutritivas essenciais contra a lixiviação
e as liberam então lentamente para a utilização pelas plantas (Brady & Weil, 2013). A matéria
orgânica aumenta o volume de água que um solo pode absorver e a proporção assimilável desta
água para o crescimento vegetal. Além disso, é a principal fonte de energia para os micro-orga-
nismos que são os principais agentes da ciclagem de nutrientes no solo (Brady & Weil, 2013).
Dessa forma, a característica úmida e as maiores concentrações de cátions presentes nos
solos do Cr, podem ter favorecido o adensamento do componente arbóreo, assim como o
estabelecimento de espécies florestais, como anotado no capítulo 1, tendo em vista que as espécies
florestais de matas de galeria e ciliares possuem estreita relação com o gradiente de umidade e de
fertilidade do solo (Felfili, 1998; Silva Júnior, 2001).
Resultados semelhantes quanto à granulometria do solo, foram observados por Marimom
Junior & Haridasan (2005) no cerradão em Nova Xavantina, MT. Segundo os autores, solos com
maior capacidade de retenção de água são capazes de suprir mais adequadamente as plantas nos
processos de síntese de biomassa, bem como possuir maior fertilidade ativa, pois a disponibilidade
de água regula a dinâmica dos nutrientes no solo e sua absorção pelas plantas. Ruggiero &
Pivello (2005) ao estudarem os fatores que poderiam influenciar as diferenças estruturais e
florísticas entre o cerrado sentido amplo e a floresta estacional semi-decídua, na Gleba Pé-de-
Gigante (MG), encontraram maior percentual de argila no solo florestal. Os autores concluíram
que a umidade do solo proporcionada pela capacidade que a argila tem de reter água seria um dos
fatores de influencia na distinção das fitofisionomias. A saturação por bases também foi fortemente
apontada como fator importante nessa distinção.
Mudanças no regime de água e fertilidade no solo já foram apontadas por outros pesquisadores
como principais fatores na distinção entre savanas e florestas (Lopes & Cox, 1977; Furley, 1992).
De acordo com Ruggiero & Pivello (2005) a disponibilidade de água para as plantas deve ter
influência direta na estrutura e na fisionomia da vegetação e na disponibilização dos nutrientes no
solo. Estudos mais específicos neste âmbito podem complementar os resultados do presente estudo.
De acordo com Haridasan (1987; 2000), a maior biomassa presente nos cerradões em relação
à biomassa dos cerrados sentido restrito, sugere que no passado os solos dos cerradões eram
mais férteis, mantendo-se até o presente por meio da ciclagem de nutrientes. Os resultados

53
encontrados no presente estudo concordam em parte, com as observações de Lopes & Cox
(1977) e Goodland (1971), nas quais, o cerradão estaria relacionado ao gradiente de fertilidade
dos solos, de forma a ocupar sítios mais férteis. De fato os solos do Cr têm maiores níveis de
nutrientes, cátions importantes para o crescimento vegetal. No entanto, a maior saturação por
bases encontrada no Cd impede que os resultados encontrados para o Cr, sejam esclarecedores
quanto a maior área basal e altura da vegetação que teoricamente necessitariam de maior aporte
de nutrientes para o seu crescimento (Haridasan, 2000; Brady & Weil, 2013).
A ordenação da PCA corroborou com os resultados do TWINSPAN quando separou o Cr
do Cd e indicou possível influência das variáveis ambientais na separação da composição florística
entre as áreas. O agrupamento das parcelas para cada área no TWINSPAN não foi distinguido na
PCA, o que sugeriu que as variáveis dos solos estatisticamente distintas parece não influenciar na
distribuição florística dentro de cada comunidade. As espécies consideradas preferenciais do Cr
são tipicamente florestais e ocorrem em abundância no cerradão, com exceção de Tapirira
guianensis que ocorre também em cerrado sentido restrito. As espécies preferenciais do Cd são
frequentemente encontradas nas formações savânicas (Mendonça et al., 2008; Ribeiro & Walter,
2008). Dalbergia miscolobium e Davilla elíptica são consideradas típicas de cerrado sentido
restrito (Mendonça et al., 2008). As espécies não preferenciais são consideradas de ampla
distribuição entre as duas áreas.
Segundo Haridasan (2000; 1982) e Haridasan & Araújo (1988) a fertilidade do solo também
está representada nas diferentes concentrações de nutrientes foliares, assim, estudos dessa
magnitude podem indicar exigências nutricionais das espécies que representaram cada comunidade
amostrada e sua estreita relação com os fatores nutricionais. Outros fatores ambientais podem
estar atuando na distribuição florística de cada área. Corroborando com Ruggiero & Pivello
(2005) no qual acreditam que em cada local um fator ambiental específico pode mostrar-se mais
relevante do que outro, sugerimos que novos estudos nas parcelas marcadas de forma permanente
são necessários para entender melhor as diferenças estruturais e a distribuição da composição
florística em cada área estudada.

54
Conclusões
As comunidades de cerradão (Cr) e cerrado denso (Cd) no Jardim Botânico de Brasília
(JBB) apresentam solos ácidos, com baixa saturação por bases e altos níveis de alumínio no solo.
A comunidade Cr apresentou maiores níveis de cátions, maior acidez potencial e solos com maior
saturação por alumínio. A análise PCA apontou forte influência dessas variáveis na distribuição
das parcelas, onde explicaram 78% da variação total das amostras. Os maiores teores dessas
variáveis nos solos apontam o Cr como sítios de maior capacidade de troca catiônica em relação
ao Cd. Por sua vez, o Cd apresentou maiores valores de saturação por bases e teores mais
elevados de fósforo.
A maior concentração de silte no Cr, somado aos teores elevados de argila e matéria orgânica,
indica maior umidade nos solos. Essa condição mais úmida cria um microclima favorável à
conservação dos nutrientes para as plantas e aparentemente favoreceu o adensamento do
componente arbóreo culminando no aspecto florestal do Cr.
A técnica TWINSPAN separou claramente as duas áreas, mas indicou homogeneidade
entre as parcelas de cada comunidade. As variáveis dos solos ordenadas pela PCA parece não
influenciar na distribuição das espécies dentro das comunidades, mas enfatizou as diferenças
químicas e físicas entre as áreas.
Apesar da proximidade entre áreas o Cr difere florística e estruturalmente do Cd fato que
pode ser relacionado com as propriedades edáficas. Sugerimos, portanto, que a vegetação Cr no
JBB, que ocorre nos solos com maiores níveis de alumínio, acidez potencial, ferro, zinco, potássio
e sódio, pode ser favorecida por esses elementos que estariam conservados pela maior umidade
conferida pelos distintos níveis de silte encontrados nas camadas superiores dos seus solos.

55
Conclusões Gerais
O cerradão (Cr) e o cerrado denso (Cd) amostrados no Jardim Botânico de Brasília (JBB -
DF) são comunidades floristicamente e estruturalmente diferentes. Dentre os parâmetros estruturais
analisados, apenas a densidade foi estatisticamente semelhante entre as áreas. O Cr, por apresentar
comunidade mais fechada onde a competição por luz é maior, teve a área basal e as alturas médias
e totais das árvores estatisticamente maiores que no Cd.
A composição florística do Cd é composta basicamente por espécies comuns a esse habitat,
enquanto a flora do Cr possui composição mista de espécies savânicas e florestais, assim como
no descrito na literatura. Ambas as comunidades são ricas em espécies e abrigam elevado número
de espécies raras, o que acentua a importância do JBB para a conservação da biodiversidade do
Cerrado. Além disso, são comunidades auto regenerativas com balanço positivo entre recrutamento
e mortalidade.
Os solos do Cr e Cd apresentam-se ácidos, com baixa saturação por bases e altos níveis de
alumínio no solo. A comunidade Cr apresentou maiores níveis de cátions, elevada acidez potencial
e solos com alta saturação por alumínio. Os maiores teores dessas variáveis nos solos apontam o
Cr como sítios de maior capacidade de troca catiônica em relação ao Cd. Por sua vez, o Cd
apresentou solos mais férteis e teores mais elevados de fósforo.
A análise multivariada TWINSPAN reforçou as diferenças florísticas e apontou
heterogeneidade entre as áreas e homogeneidade dentro das comunidades. A PCA reforçou a
separação das áreas com base nas variáveis dos solos estatisticamente distintas. Entretanto, essas
variáveis parecem não influenciar na distribuição das espécies dentro das comunidades.
Sendo assim, apesar da proximidade entre áreas o Cr difere florística e estruturalmente do
Cd fato que pode ser relacionado com as propriedades edáficas. Sugerimos que a vegetação do
Cr, que ocorre nos solos com maiores níveis de cátions pode ser favorecida por esses elementos
que estariam conservados pela maior umidade conferida pelos distintos níveis de silte encontrados
nas camadas superiores dos seus solos.

56
Referências Bibliográficas
Adámoli, J.; Macedo, J.; Azevedo, L. G. & Madeira Neto, J. 1985. Pp 33-73. Caracterização da
região dos Cerrados. In: Goedert, W. J. Solos dos cerrados. EMBRAPA/ Nobel, Brasília.
Alencar, A. L.; Silva, M. A. P. & Barros, L. M. 2007. Florística e Fitossociologia de uma Área de
cerradão na Chapada do Araripe, Crato - CE. Revista Brasileira de Biociências 5 (2): 18-20.
Almeida, S. P.; Proença, C. E. B; Sano, S. M. & Ribeiro, J. F. 1998. Cerrado: espécies vegetais
úteis. EMBRAPA-CPAC. Planaltina, DF. 464p.
Andrade, L. A. Z.; Felfili, J. M. & Violatti, L. 2002. Fitossociologia de uma área de cerrado
denso na RECOR-IBGE, Brasília-DF. Acta Botanica Brasilica 16 (2): 225-240.
Angiosperm Phylogeny Group (APG III). 2009. An update of the Angiosperm Phylogeny Group
classification for the orders and families of flowering plants: APG III. Botanical Journal of
the Linnean Society 161: 105-121.
Araújo, G. M.; Nunes, J. J.; Rosa, A. G. & Resende, E. J. 1997a. Estrutura comunitária de vinte
áreas de cerrados residuais no município de Uberlândia, MG. Daphne 7: 7-14.
Assunção, S. L. & Felfili1, J. M. 2004. Fitossociologia de um fragmento de cerrado sensu stricto
na APA do Paranoá, DF, Brasil. Acta Botanica Brasilica18 (4): 903-909.
Azevedo, L. G.; Ribeiro, J. R.; Schiavini, I. & Oliveira, P. E. A. M. 1990. Levantamento da
Vegetação do Jardim Botânico de Brasília-DF. Fundação Zoobotânica. Brasília, DF. 93p.
Azevedo, L. G.; Ribeiro, J. R.; Schiavini, I. & Oliveira, P. E. A. M. 1990. Levantamento da
vegetação do Jardim Botânico de Brasília, DF. Fundação Zoobotânica do Distrito Federal,
Brasília.
Balduino, A. P. D. C.. Souza, A. L. D.; Neto, M.; Alves, J. A.; Silva, A. F. D. & Silva Júnior, M.
C. D. 2005. Fitossociologia e análise comparativa da composição florística do cerrado da
flora de Paraopeba-MG. Revista Árvore 29(1): 25-34.
Brady, N. C.; Weil, R. R. 2012. Elementos da Natureza e Propriedades dos Solos. Ed. Bookman
Companhia Editora Ltda. 716 p.

57
Brinkley, D.; Richter, D.; David, M. B.; Caldwell, B. 1994. Soil chemistry in a loblolly/longleaf
pine forest with interval burning. Ecological Applications 2: 157-164.
Bucci, F. F. B. 1997. Floração de algumas espécies de Melastomataceae do Distrito Federal:
uso de dados de herbário para obter padrões sazonais. Brasília, Departamento de Ecologia,
Universidade de Brasília. Dissertação de Mestrado.
Campos, E. P.; Duarte, T. G.; Neri, A. V.; Silva, A. F.; Meira-Neto, J. A. A. & Valente, G. E. 2006.
Composição florística de um trecho de cerrado sensu stricto e cerradão e sua relação com o
solo na Floresta Nacional (FlONA) de Paraopeba, MG, Brasil. Revista Árvore 30 (3): 471-
479.
Castro, A. A. J. F. & Martins, F. R. 1999. Cerrados do Brasil e do Nordeste: caracterização, área
de ocupação e considerações sobre a sua fitodiversidade. Pesquisa Foco 7(9): 147-17. In:
Solorzano, A. 2011. Análise fitogeográfica do cerradão: conexões florísticas, padrões
estruturais, relações ecológicas e modelagem de sua distribuição potencial. Brasília:
Instituto de Ciências Biológicas, Universidade de Brasília. Tese de Doutorado.
Cavalcanti, L. M. 1978. Efeito das cinzas resultantes da queimada sobre a produtividade do
estrato herbáceo-subarbustivo do Cerrado de Emas. Tese de doutorado, São Paulo. Instituto
de Biociências, Universidade de São Paulo. 219 p.
Cavedon, D. S. & Sommer, S. 1990. Levantamento semidetalhado dos solos do Jardim
Botânico de Brasília. Fundação Zoobotânica do Distrito Federal, Brasília. 95 p.
Chao, A.; Chazdon, R. L.; Colwell, R. K. & Shein, T. J. 2005. A new statistical approach for
assessing similarity of species composition with incidence and abundance data. EcologyLetters
8: 148-159.
Clarke, K. R. 1993. Non-parametric multivariate analyses of changes in community
structure. Australian Journal of Ecology 18: 117-143.
Codeplan. 1984. Atlas do Distrito Federal, GDF, Brasília.
Cole, M. M. 1986. The savannas biogeography and geobatany. Academic Press. 438p.
Cornell, R. M. & Schwertmann, U. 1996. The iron oxides. Structure, properties, reactions,
occurrence and uses. Weinheim, VHC. 573 p. [acesso em 02/2013]. Disponível em: http://
books.google.com.br.

58
Costa, A. A. & Araújo, G. M. 2001. Comparação da vegetação arbórea de cerradão e de cerrado
na Reserva do Panga, Uberlândia, Minas Gerais. Acta Botanica Brasilica 15 (1): 63-72.
Costa, F. V.; Oliveira, K. N.; Nunes, Y. R. F.; Menino, G. C. O; Brandão, D. O.; Araújo, L. S.;
Miranda, W. O. & Neto, S. D. 2010. Florística e estrutura da comunidade arbórea de duas
áreas de cerrado sentido restrito no norte de Minas Gerais. Revista Cerne 16(3): 267-281.
Coutinho L. M. 1982.Ecologicaleffectsofthefire in Brazilian cerrado. In: Huntley B. J & Walker
B. H. Ecologyof tropical savannas. Springer Verlag273–291.
Coutinho, L. M. 1990. O cerrado - a ecologia do fogo. Ciência Hoje 12 (68): 23-30.
Coutinho, L. M. 1992. O cerrado e a ecologia do fogo. Ciência Hoje, vol. Especial Eco-Brasil.
Pp. 130-138.
Coutinho, L.M. 1978. O conceito de cerrado. Revista Brasileira de Botânica 1: 17-23.
Dias, B.F.S. 1992. Alternativas de desenvolvimento dos Cerrados: manejo e conservação
dos recursos naturais renováveis. Instituto Brasileiro do Meio Ambiente e dos Recursos
Naturais Renováveis (IBAMA), Fundação Pró-Natureza (FUNATURA), Brasília.
Dietzsch, L.; Rezende, A. V.; Pinto, J. R. R & Pereira, B. A. S. 2006. Caracterização da flora
arbórea de dois fragmentos de mata de galeria do Parque Canjerana, DF. Cerne, Lavras 12
(3):201-210.
Eiten, G. 1978. Delimitation of the Cerrado Concept. Vegetation 36(3): 169-178.
Eiten, G. 1994. Vegetação do Cerrado. Pp. 17-74. In: Pinto. M. N. Cerrado: caracterização,
ocupação e perspectivas. Editora Universidade de Brasília.
Eiten, G., 1972. The Cerrado Vegetation of Brazil. Botanical Review 38: 201–341
Empresa Brasileira de PesquisaAgropecuária (EMBRAPA). 2006. Sistema brasileiro de
classificação de solos. 2ª ed. Centro Nacional de Pesquisa de Solos, Rio de Janeiro. 306p.
Empresa Brasileira de PesquisaAgropecuária (EMBRAPA). 1997. Manual de Método de Análise
de Solo. Rio de Janeiro: EMBRAPA Solos. 212p.
Felfili, J. M. & Silva Júnior, M. C. 1988. Distribuição dos diâmetros numa faixa de cerrado na
Fazenda Água Limpa (FAL) em Brasília-DF. Acta Botanica Brasilica 2 (1-2): 85-104.

59
Felfili, J. M. & Silva Júnior, M. C. 1992. Floristic composition, phytosociology and comparison
of cerrado and gallery forests at Fazenda Agua Limpa, Federal District, Brazil. Pp. 393–416.
In: Proctor, J. A.; Ratter, J. A.& Furley, P. A. Nature and dynamics of forest-savanna
boundaries. London, Chapman & Hall.
Felfili, J. M. & Silva Júnior, M. C. 1993. A comparative study of cerrado (sensu stricto) vegetation
in Central Brazil. Journal of Tropical Ecology 9: 277-289.
Felfili, J. M. &Imaña-Encinas, J. 2001. Suficiência da amostragem no cerradão sensu stricto das
quarto areas estudadas na Chapada do Espigão Mestre do São Francisco. Pp. 31-35. In: Felfili,
J. M. & Silva Júnior, M. C. Biogeografia do bioma Cerrado: estudo fitofisionômico da
Chapada do EspigãoMestre do São Francisco. Universidade de Brasília, Faculdade de
Tecnologia, Departamento de EngenhariaFlorestal.
Felfili, J. M. 1997. Diameter and height distributions of a gallery forest community and some of
its main species in central Brazil over a six-year period (1985-1991). Revista Brasileira de
Botânica 20: 155-162.
Felfili, J. M. 2001. Principais fisionomias do Espigão Mestre do São Francisco. Pp. 18-30. In: J.
M. Felfili & M.C. Silva Júnior. Biogeografia do Bioma Cerrado: Estudo Fitofisionômicona
Chapada do Espigão Mestre do São Francisco. Departamento de Engenharia Florestal.
Universidade de Brasília, Brasília, DF.
Felfili, J. M.; Carvalho, F. A., Haidar, R. F. 2005. Manual para o monitoramento de Parcelas
permanentes nos biomas Cerrado e Pantanal. Brasília: Universidade de Brasília,
Departamento de Engenharia Florestal. Pp. 20-24.
Felfili, J. M.; Filgueiras, T. S.; Haridasan, M.; Silva Júnior, M. C.; Mendonça, R. & Rezende, A.
V. 1994. Projeto biogeografia do bioma cerrado: Vegetação e solos. Caderno de Geociências
12: 75-166.
Felfili, J. M.; Filgueiras, T. S.; Haridasan, M.; Silva Júnior, M. C.; Mendonça, R. & Rezende, A.
V. 1994. Projeto Biogeografia do Bioma Cerrado: vegetação e solos. Caderno de Geociências
do IBGE (12): 75-166.
Felfili, J. M.; Nogueira, P. E.; Silva Júnior, M. C.; Marimon, B. S. &Delitti, W. B. C. 2002.
Composição florística e fitossociologia do cerrado sentido restrito no município de Água Boa
– MT. Acta Botanica Brasilica 16(1): 103-112.

60
Felfili, J. M.; Rezende, A. V. & Silva-Júnior, M. C. 2007. Biogeografia do bioma Cerrado:
vegetação e solos da Chapada dos Veadeiros. Ed. Universidade de Brasília, FINATEC –
Brasília.
Felfili, J. M.; Rezende, A. V.; Silva Júnior, M. C. & Silva, M. A. 2000. Changes in the Floristic
Composition of Cerrado sensustricto in Brazil over a nine-year period. Journal of Tropical
Ecology 16(4): 579-590.
Felfili, J.M. 1998. Determinação de padrões de distribuição de espécies em uma mata de galeria
no Brasil Central com a utilização de técnicas de análise multivariada. Boletim do Herbário
Ezechias Paulo Heringer2: 35-48.
Felfili, J.M.; Mendonça, R.C.; Walter, B.M.T.; Silva Júnior, M.C.; Nóbrega, M.G.G.; Fagg, C.W.;
Sevilha, A.C. & Silva, M.A. 2001. Flora fanerógama das matas de galeria e ciliares do Brasil
Central. Pp. 195-206.In: Ribeiro, J. F.; Fonseca, A. E. L. & Sousa-Silva. Cerrado:
caracterização e recuperação de Matas de Galeria. Planaltina, Emprapa Cerrados.
Felfili, M. C. &Felfili, M. J. 2001. Diversidade alfa e beta no cerradão sensu stricto da Chapada
Pratinha, Brasil. Acta botanica Brasilica 15 (2): 243-254.
Ferri, M. G. 1977. Ecologia dos cerrados. Pp. 15-33. In: IV Simpósio sobre o cerrado.
Universidade de São Paulo, Editora Itatiaia.
Fonseca, M. S. & Silva Júnior, M. C. 2004. Fitossociologia e similaridade florística entre trechos
de cerrado sentido restrito em interflúvio e em vale no Jardim Botânico de Brasília, DF. Acta
botânica brasílica 18 (1): 19-29.
Franco, A. C. 2004. Estratégias funcionais de plantas lenhosas das savanas do Brasil Central:
relação ao déficit hídrico e ao regime luminoso. Pp. 173-178. In: Cabrera, H. M. Fisiología
ecológica en plantas: Mecanismos y Respuestas a Estrésenlos Ecosistemas. EUV,
Valparaíso, Chile.
Franczak, D. 2006. Dinâmica da comunidade arbórea em um cerradão do Parque do Bacaba,
Nova Xavantina-MT. Monografia de Graduação, Universidade do Estado Mato Grosso (MT).
52 p.
Furley, P. A. & Ratter, J. A. 1988. Soil resources and plant communities of the central Brazilian
cerrado and their development. Journal of Biogeography 15: 97-108.

61
Furley, P. A. 1992. Edaphic changes at the forest-savanna bondary with particular reference to
the neotropics. Pp. 91-115. In: Furley, P. A.; Proctor, J. & Ratter, J. A. Nature and Dynamics
of Forest–Savanna Boundaries.
Furley, P. A. 1999. The nature and diversity of neotropical savanna vegetation with particular
reference to the Brazilian cerrados. Global Ecology and Biogeography 8: 223-241.
Gomes, B. Z.; Martins, F. R. & Tamashiro, J. Y. 2004. Estrutura do cerradão e da transição entre
cerradão e floresta paludícola num fragmento da International Paper do Brasil Ltda., em Brotas,
SP. Revista Brasileira de Botânica 27 (2): 249-262.
Goodland, R. & Ferri, M. G. 1979. Ecologia do Cerrado. Ed. Itatiaia, Belo Horizonte, MG.
193p.
Goodland, R. & Pollard, R. 1973. The Brazilian cerrado vegetation: a fertility gradient. Journal
of Ecology 61: 219-224.
Goodland, R. 1971. Oligotrofismo e alumínio no cerrado. Pp. 44-60. In: Ferri, M. G. III Simpósio
sobre o cerrado. EdUSP, São Paulo.
Goodland, R. 1971a. A physionomic analyses of the “cerrado” vegetation of the central Brazil.
Journal of Ecology 59: 411-419. [acesso em 03/2013]. Disponível em: http://www.jstor.org.
Goodland, R. 1971b. A physiognomic analysis of the cerrado vegetation of central Brazil. Journal
of Ecology 59: 411-419.
Goodland, R. 1979. Análise ecológica da vegetação do cerrado. Pp.61-160. In: Goodland, R. &
Ferri, M. G. Ecologia do Cerrado. Ed. Itatiaia, Belo Horizonte, MG.
Guarim-Neto, G.; Guarim, V. L. M. S. & Prance, G. T. 1994. Structure and floristic composition
of the trees of an area of cerrado near Cuiabá, Mato Grosso, Brazil. Kew Bulletin 49(3): 499-
509.
Haridasan, M, 1992. Observation on soils, foliar nutrientes concentrations and floristic composi-
tion of cerrado and cerradão communities in Central Brazil. Pp. 174-184. In: Proctor, J.;
Ratter, J. A. & Furley, P. A. The nature and dynamics forest-savanna boundaries. Chapman
& Hall, London.
Haridasan, M. &Araújo,G.M. 1988. Aluminium-acummulating species in two forest communities
in the cerrado region of central Brazil. Forestry Ecology Management 24:15-26.

62
Haridasan, M. 1982. Aluminiu-accumulationby some cerrado native species of central Brazil.
Plantand Soil 65: 265-273.
Haridasan, M. 2000. Nutrição mineral das plantas nativas do Cerrado. Revista Brasileira de
Fisiologia Vegetal 12(1): 54-64.
Haridasan, M. 2007. Solos. Pp. 27-43. In: Felfili, J. M.; Rezende, A. V. & Silva Júnior, M. C.
Biogeografia do bioma cerrado: Chapada dos Veadeiros. Ed. UnB, Brasília.
Haridasan, M.; Pinheiro, A. A. M. C. & Torres, F. R. R. 1997. Resposta de algumas espécies do
estrato rasteiro de um cerrado à calagem e à adubação. Pp. 87-91. In: Leite, L. L. & Saito,
C. H.Contribuição ao conhecimento ecológico do cerrado. Universidade de Brasília,
Brasília, Brasil.
Hartley, H. O. 1950. The maximum F-ratio as a short cut test for heterogeneity of variances.
Biometrika 37: 308-312.
Heringer Salles, A. 2007. Jardim Botânico de Brasília: diversidade e conservação. Sociedade
Amigos do Jardim Botânico de Brasília. Dupligráfica Editora LTDA, Brasília. 355p.
Hill, M. O. 1979. TWINSPAN- a FORTRAN program for arranging multivariate data in an
ordered two-way table by classification of the individuals and attributes. Cornell University.
Ithaca, New York.
Hoffmann, W.A. 1998. Post-burn reproduction of woody plants in a neotropical savanna: the
relative importance of sexual and vegetative reproduction. Journal of Applied Ecology 35:
422-433.
Instituto Brasileiro de Geografia e Estatística. 2007. Manual técnico de pedologia. IBGE, Rio
de Janeiro. 316p.
Jackson, P. C.; Meinzer, F. C.; Bustamante, M.; Goldstein, G.; Franco, A.; Rundel, P. W.; Caldas,
L.; Igler, E. &Causin, F. 1999. Partitioning of soil water among tree species in a Brazilian
Cerrado ecosystem. Tree Physiology 19: 717-724. In: Franco, A. C. 2004. Estratégias
funcionais de plantas lenhosas das savanas do Brasil Central: relação ao déficit hídrico e ao
regime luminoso. Pp. 173-178. In: Cabrera, H. M. Fisiología ecológica en plantas:
Mecanismos y Respuestas a Estrésenlos Ecosistemas. EUV, Valparaíso, Chile.
Jordan, C. F. 1987. Amazonian rainforests: ecosystem disturbance and recovery. New York:
Springer-Verlag. 133 p.

63
Kent, M. & Coker, P. 1992. Vegetation description and analysis: a pratical Approach. Belhaven
Press, London. 363 p.
Klink, C. A. & Machado, R. 2005. Conservation of the Brazilian Cerrado. Conservation Biology
19 (3): 707-713.
Klink, C. A.; Machado, R. B. 2005. Conservation of the Brazilian Cerrado. Conservation Biology
19: 707-713.
Ledru, M. P. 2002. Late quaternary history and evolution of the cerrados as revealed by
palinological records. Pp. 33-50. In: Oliveira, P. S. & Marquis, R. J. The cerrados of Brazil.
Ecologyand natural historyof a neotropical savanna. Columbia University Press, New York.
Lopes, A. S. & Cox, F. R. 1977. Cerrado vegetation in Brazil: na edaphic gradient. Agronomy
Journal 69: 828-831.
Ludwig, F., H. de Kroon, F. Berendse, and H. H. T. Prins. 2004. The influence of savanna trees on
nutrient, water and light availability and theun derstorey vegetation. Plant Ecology 170:93-
105.
Magalhães, G. M. Contribuição ao estudo fitogeográfico do médio São Francisco. Boletim de
Agricultura 1 (5): 26-39, 1952.
Magurran, A. E. 1988. Ecological diversity and its measurement. Chapman and Hall, London.
179p.
Magurran, A. E. 2004. Measuring biological diversity. Blackwell publishing. Pp. 100-130.
Malavolta, E. 1979. Nutrição mineral. Pp. 97-116. In: Ferri, M. G. Fisiologia vegetal 1. EDUSP,
São Paulo.
Marimon-Junior, B. H. & Haridasan, M. 2005. Comparação da vegetação arbórea e características
edáficas de um cerradão e um cerrado sensu stricto em áreas adjacentes sobre solo distrófico
no leste de Mato Grosso, Brasil. Acta Botanica Brasilica 19 (4): 913-926.
McCune, B. &Mefford, M. J. 2006. PC-ORD: multivariate analysis of ecological data. MjM
Software Design, Oregon.
Meirelles, M. L.; Klink, C. A. & Sousa-Silva, J. C. 1997. Um modelo de estados y transiciones
para el cerrado brasileño. Ecotropicos10: 45-50.

64
Mendonça, R. C., Felfili, J. M., Walter, B. M. T., Silva Júnior, M. C., Rezende, A. V., Filgueiras,
T. S. & Nogueira, P. E. 1998. Pp. 289-556. Flora vascular do Cerrado. In: Sano, S. M. &
Almeida, S. P. Cerrado, Ambiente e Flora. EMBRAPA-CPAC, Planaltina.
Mendonça, R. C.; Felfili, J. M.; Walter, B. M. T.; Silva Junior, M. C.; Rezende, A. B.; Filgueiras,
T. S.; Nogueira, P. E. & Fagg, C. W. 2008. Flora vascular do bioma Cerrado: checklist com
12.356 espécies. 1279p.
Mews, H. A.; Marimon, B. S.; Maracahipes, L.; Franczak. D. D. & Marimon-Junior, B. H.2011.
Dinâmica da comunidade lenhosa de um Cerrado Típico na região Nordeste do Estado de
Mato Grosso, Brasil. Biota Neotropica 11(1) 73-82.
Miranda, H. S. Efeitos do regime de fogo sobre a estrutura de comunidades de Cerrado:
Projeto Fogo. Brasília: IBAMA.
Miranda, H. S.; Neto, W. N. & Neves, B. M. C. 2010. Pp. 23-33. Caracterização das queimadas
de Cerrado. In: Miranda, H. S. Efeitos do regime de fogo sobre a estrutura de comunidades
de cerrado: Projeto Fogo. Brasília-IBAMA.
Miranda, I. S.; Almeida, S. S.; Dantas, P. J. 2006. Florística e estrutura de comunidades arbóreas
em cerrados de Rondônia, Brasil. Acta Amazonica 36 (4): 419 – 430.
Miranda, S. C. 2008. Comunidades lenhosas de cerrado sentido restrito na Serra Dourada
em dois substratos. Brasília: Instituto de Ciências Biológicas, Universidade de Brasília.
Dissertação de Mestrado. 104 p.
Miranda, S. C.; Silva Júnior, M. C.; Carvalho, P. S.; Salles, L. A.; Oliveira, A. M. & Corezzi, A.
C. 2009. O cerrado sentido restrito no Jardim Botânico de Brasília 24 anos sem incêndios. In:
Anais do 60º Congresso Nacional de Botânica, Bahia.
Mistry, J. 2000. World Savannas: ecology and human use. London, Pearson Education Ltd.
344p.
Moreno, M. I. C. & Schiavini, I. 2001. Relação entre vegetação e solo em um gradiente florestal
na Estação Ecológica do Panga, Uberlândia (MG). Revista brasileira de Botânica 24 (4):
537-544.
Müller-Dombois, H. & Ellenberg, D. 1974. Aims and methods of vegetation ecology. Wiley,
New York.

65
Munhoz, C. B. R. & Amaral, A. G. 2010 – Efeito do fogo no estrato herbáceo-subarbustivo do
Cerrado. Pp. 93-102. In: Miranda, H. S. Efeitos do regime de fogo sobre a estrutura de
comunidades de Cerrado: Projeto Fogo. Brasília: IBAMA.
Oliveira, E. C. L. & Felfili1, J. M. 2005 Estrutura e dinâmica da regeneração natural de uma mata
de galeria no Distrito Federal, Brasil. Acta Botanica Brasilica 19 (4): 801-811.
Oliveira-Filho, A. T. & Martins, F. R. 1986. Distribuição, caracterização e composição florística
das formações vegetais da região da Salgadeira, na Chapada dos Guimarães (MT). Revista
brasileira de Botânica 9: 207-223
Oliveira-Filho, A. T. &Ratter, J. A. 2002. Vegetation physiognomies and wood flora of the Cerrado
biome. Pp. 91-120. In: Oliveira, O. S. & Marquis, R. J. The cerrados of Brazil: ecologyand
natural historyof a neotropical savana. Columbia University Press, New York.
Owensby, C. & Wyrill, J. 1973. Effects of range burning on Kansas Flint Hills soil. Journal of
Range Management 26 (3): 185-188.
Palmer, M. W. 2005. Ordination methods for ecologists. Oklahoma State University, USA. [Acesso
em 12/2012]. Disponível em http://ordination.okstate.edu.
Peck J. E. 2010. Multivariate analysis for community ecologists: step-by-stepusing PCord.
MjM Software, Gleneden Beach, Oregon.
Pennington, R. T.; Prado, D. E. & Colin, A. 2000. Neotropical Seasonally Dry Forests and
Quaternary Vegetation Changes. Journal of Biogeography 27 (2): 261-273.
Pereira-Silva, E. F. L.; Santos, J. E.; Kageyama, P. Y. & Hardt, E. 2004. Florística e ûtossociologia
dos estratos arbustivo e arbóreo de um remanescente de cerradão em uma unidade de
conservação do Estado de São Paulo. Revista Brasileira de Botânica 27: 533-544.
Pereira-Silva, E. F. L.; Santos, J. E.; Kageyama, P. Y. &Hardt, E. 2004. Florística e fitossociologia
dos estratos arbustivo e arbóreo de um remanescente de cerradão em uma Unidade de
Conservação do Estado de São Paulo. Revista Brasileira de Botânica 27(3): 533-544.
Pereira-Silva, E.F.L.; Santos, J. E.; Kageyama, P. Y. &Hardt, E. 2004. Florística e fitossociologia
dos estratos arbustivo e arbóreo de um remanescente de cerradão em uma unidade de
conservação do Estado de São Paulo. Revista Brasileira de Botânica 27 (3): 533-544.

66
Pires, J. M. & Prance, G. T. 1977. The Amazon Forest: a natural heritage to be preserved. Pp.158-194.
In: Prance, G.T. & Elias, T. S. Extinction is forever. New York Botanical Garden, New York.
Polhill, R. M.; Raven, P. H. & Stirton, C. H. 1981. Evolution and systematics of the Leguminosae.
Pp. 1-26. In: Polhill, R. M. & Raven, P. H. Advances in legume systematics. Kew:Royal
Botanical Gardens, part1.
Pomianoski, D. J. W. 2005. Perdas de solo e água no sistema agroflorestal da bracatinga
(Mimosa scabrella Bertham) em diferentes declividades e manejos. Dissertação de Mestrado
em Ciências do solo – Universidade Federal do Paraná. 92 p.
Proença, C. E. B.; Munhoz, C. B. R.; Jorge, C. L.; Nóbrega, M. G. G. 2001. Listagem e nível de
proteção das espécies de fanerógamas do Distrito Federal, Brasil. Pp. 89-359. In: Cavalcanti,
T. B.; Ramos, A. E. Flora do Distrito Federal, Brasil. Brasília (DF) Embrapa Recursos
Genéticos e Biotecnologia.
Quesada, C.A., Miranda, A.C., Hodnett, M.G. et al. 2004. Seasonal and depth variation of soil
moisture in a burned open savanna (campo sujo) in central Brazil. Ecological Applications 14
(suppl.): 33–41
Ratter, J. A. 1971. Some notes on two types of cerradão occurring in northeastern MatoGrosso.
Pp.110-112. In: M.G. Ferri. III Simpósio Sobre o Cerrado. São Paulo, EDUSP/ Edgard
Blücher.
Ratter, J. A.; Askew, G. P.; Montgomery, R. F. & Gifford, D. R. 1977. Observações adicionais
sobre o cerradão de solos mesotróficos no Brasil central. Pp. 303-316. In: Ferri, M. G. IV
Simpósio sobre o Cerrado: Bases para a Utilização Agropecuária. São Paulo, EDUSP.
Ratter, J. A.; Askew, G. P.; Montgomery, R. F. & Gifford, D. R. 1978. Observations on forests of
some mesotrophic soils in central Brazil. Revista Brasileira de Botânica 1:47-58.
Ratter, J. A.; Bridgewater, S.; Atkinson, R. & Ribeiro, J. F. 1996. Analysis of the floristic
composition of the Brazilian cerrado vegetation II: comparision of the woody vegetation of
98 areas. Edinburgh Journal of Botanic 53: 153-180.
Ratter, J. A.; Richards, P. W.; Argent, G. & Gifford D. R. 1973. Observations on the vegetation of
north eastern MatoGrosso I: The woody vegetation types of the Xavantina-Cachimbo Expedition
Area. Philosophical Transactions of the Royal Society of London B 226: 449-492.

67
Rawitscher, F. 1948. The water economy of the vegetation of the “campos cerrados” in southern
Brazil. Journal of Ecology 36: 237-268. In: Franco, A. C. 2004. Estratégias funcionais de
plantas lenhosas das savanas do Brasil Central: relação ao déficit hídrico e ao regime luminoso.
Pp. 173-178. In: Cabrera, H. M. (ed.) Fisiología ecológica en plantas: Mecanismos y
Respuestas a Estrésenlos Ecosistemas. EUV, Valparaíso, Chile.
Reatto, A. & Martins, E. S. 2005. Classes de solo em relação aos controles de paisagem do bioma
Cerrado. Pp. 49-59. In: Scariot, A.; Sousa-Silva, J. C. &Felfili, J. M. Cerrado: Ecologia,
Biodiversidade e Conservação. Brasília: Ministério do Meio Ambiente.
Reatto, A., Correia, J. R., Spera, S. T. & Martins, E. S. 2008. Solos do bioma Cerrado: aspectos
pedológicos. Pp.107-150. In: Sano, S. M., Almeida, S. P. & Ribeiro, J. F. Cerrado: ecologia
e flora. EMBRAPA-CPAC, Planaltina.
Ribeiro, J. F. & Haridasan, M. 1984. Comparação fitossociológica de um cerrado denso e um
cerradão em solos distróficos no Distrito Federal. In: Anais do 35º Congresso Nacional
Botânica, Manaus.
Ribeiro, J. F. & Walter, B. M. T. 2008. As principais fitofisionomias do bioma Cerrado. Pp.151-
212. In: S. M. Sano; S. P. Almeida & J. F. Ribeiro. Cerrado: Ecologia e Flora (2). Embrapa
cerrados/Embrapa Informação Tecnológica. Brasília, DF.
Ribeiro, J. F. & Walter, B. M. T. Fitofisionomias do bioma Cerrado. In: Sano, S. M. & Almeida,
S. P. Cerrado: ambiente e flora. Planaltina: EMBRAPA-CPAC, 1998. Pp. 89-152.
Ribeiro, J. F.; Sano, S. M.; Macedo, J.; Silva, J. A. 1983. Os principais tipos fisionômicos da
região dos cerrados. Boletim de Pesquisas 21. EMBRAPA-CPAC, Planaltina-DF.
Ribeiro, J. F; Bridgewater, s.; Ratter, J. A. & Souza-Silva, J. C. 2005. Ocupação do bioma Cerrado
e conservação da sua diversidade vegetal. Pp. 385-399. In: Scariot, A.; Souza-Silva, J. C. &
Felfili, J. M. Cerrado: Ecologia, Biodiversidade e Conservação. Brasília: Ministério do
Meio Ambiente.
Rizzini, C. T. 1979. Tratado de fitogeografia do Brasil - aspectos sociológicos e florísticos
(2). Editora HUCITEC / EDUSP. São Paulo, SP.
Romero, R. & Martins, A. B. 2002. Melastomataceae do Parque Nacional da Serra da Canastra,
Minas Gerais, Brasil. Revista Brasileira de Botânica 25: 19-24.

68
Ruggiero, P. G. C & Pivello, V. R. 2005. Pp. 173-188. CAP 13 - O Solo e a Comunidade Vegetal.
In: Pivello, V. R. & Varanda, E. M. O Cerrado Pé-de-Gigante (Parque Estadual de Vassununga,
SP) - Ecologia e Conservação. São Paulo, Secretaria de Estado do Meio Ambiente.
Ruggiero, P. G. C.; Batalha, M.A.; Pivelo, V. R. & Meireles, S. T. 2002. Soil-vegetation relationships
in cerrado (Brazilian savanna) and semideciduous forest, Southeastern Brazil. Plant Ecology
160: 1-16.
Sano, E. E.; Rosa, R.; Brito, J. L. S. & Ferreira, L. G. 2010. Land cover mapping of the tropical
savanna region in Brazil. Environmental Monitoring and Assessment 166:113-124.
Sarmento, T. R. & Silva Júnior, M. C. 2009. Comunidades lenhosas no cerrado sentido restrito
em duas posições topográficas na estação ecológica do Jardim Botânico de Brasília, DF, Brasil.
Rodriguésia 60 (2): 277-294.
Sarmiento, G. 1984. The Ecology of Neotropical Savannas. Harvard University Press, Cambridge.
Sassaki, R. M.; Rodoni, J. N.; Zaidan,L. B. P. & Felippe, G. M. 1999. Number of buried seeds and
seedlings emergence in cerradão, cerrado and gallery Forest soil sat Pedregulho, Itirapina
(SP), Brazil. Revista brasileira de Botânica 22 (2): 147-152.
Sato, M. N. 2003. Efeito a longo prazo de queimadas na estrutura da comunidade de lenhosas da
vegetação do cerrado sensu stricto. Tese de Doutorado, Universidade de Brasília, Brasília,
Brasil.
Schubart, O. & Rawitscher, F. K. 1950. Notas sobre o movimento de água subterrânea de Emas.
Bol. Fac. Filos. Cienc. Univ. São Paulo, 109, Botânica 8: 69-73. In: Goodland, R. &Ferri, M.
G. 1979. Ecologia do Cerrado. Ed. Itatiaia, Belo Horizonte, MG.
SEMARH. 2004. Prevenção e combate aos incêndios florestais em Unidades de Conservação.
Caderno Técnico. Brasília. Pp. 96. In: Heringer Salles, A. Sociedade Amigos do Jardim
Botânico de Brasília. 2007. Jardim Botânico de Brasília: diversidade e conservação.
Dupligráfica Editora LTDA, Brasília, pp. 355.
Shapiro, S. S. & Wilk, M. B. 1965. An analysis of variance test for normality (complete samples).
Biometrika 52: 591-611.
Silva Júnior, M. C. 2004. Fitossociologia e estrutura diamétrica da mata de galeria do Taquara,
na Reserva Ecológica do IBGE, DF. Revista Árvore 28 (3): 419-428.

69
Silva Júnior, M. C. 2005. Fitossociologia e estrutura diamétrica na mata de galeria do córrego
Pitoco, na Reserva Ecológica do IBGE, DF. Revista Cerne 11 (2): 147-15.
Silva, J. S. & Felfili, J. M. 2008. Floristic composition of the conservation unit “ARIE do cerradão”
in the Federal District, Brazil. IX Simpósio Nacional Cerrado.
Silva, J. S. 2009. Diversidade alfa, florística e fitossociológica na ARIE do cerradão na APA
Gama e Cabeça de Veado. DF. Dissertação de Mestrado. Instituto de Ciências Biológicas,
Universidade de Brasília.
Silva, L. O.; Costa, D. A.; Espírito Santo Filho, K. D.; Ferreira, H. D. & Brandão, D. 2002.
Levantamento florístico e fitossociológico em duas áreas de cerrado sensu stricto no Parque
Estadual da Serra de Caldas Novas, Goiás. Acta Botanica Brasilica 16(1): 43-53.
Silveira, F. A. L. 2011. Ecofisiologia evolutiva de sementes de Melastomataceae de campos
rupestres. Tese de Doutorado. Universidade Federal de Minas Gerais Instituto de Ciências
Biológicas Departamento de Botânica.
Solorzano, A. 2011. Análise fitogeográfica do cerradão: conexões florísticas, padrões
estruturais, relações ecológicas e modelagem de sua distribuição potencial. Brasília:
Instituto de Ciências Biológicas, Universidade de Brasília. Tese de Doutorado.
Solorzano, A.; Pinto, J. R. R.; Felfili, J. M. & Hay, J. D. V. 2012. Perûl florístico e estrutural do
componente lenhoso em seis áreas de cerradão ao longo do bioma Cerrado. Acta Botanica
Brasilica 26 (2): 328-341.
Teixeira, M. I. J. G.; Araujo, A. R. B.; Valeri, S. V. & Rodrigues, R. R. 2004. Florística e
fitossociologia de área de cerrado s.s. no município de Patrocínio Paulista, nordeste do estado
de São Paulo. Bragantia 63 (1): 1-11.
Walter, B. M. T. & Ribeiro, J. F. 2010. Diversidade fisionômica e o papel do fogo no bioma
Cerrado. Pp. 59-76. In: Miranda, H. S. Efeitos do regime de fogo sobre a estrutura de
comunidades de Cerrado: Projeto Fogo. Brasília: IBAMA.
Walter, B. M. T. 2006. Fitofisionomias do bioma Cerrado: síntese terminológica e relações
florísticas. Brasília: Instituto de Ciências Biológicas, Universidade de Brasília. Tese de
Doutorado.