nayra da silva resende...6 agradecimentos a deus, antes de tudo, por ter me permitido vir a esse...
TRANSCRIPT

1
NAYRA DA SILVA RESENDE
DISTRIBUIÇÃO DOS GRUPAMENTOS NEURONAIS COLINÉRGICOS NO
ENCÉFALO DO MOCÓ (Kerodon rupestris)
Dissertação apresentada a Universidade
Federal do Rio Grande do Norte, para a
obtenção do título de Mestre em Biologia
Estrutural e Funcional.
Natal-RN
2017

1
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
DEPARTAMENTO DE MORFOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA ESTRUTURAL E FUNCIONAL
DISTRIBUIÇÃO DOS GRUPAMENTOS NEURONAIS COLINÉRGICOS NO
ENCÉFALO DO MOCÓ (Kerodon rupestris)
ORIENTADORA:
Profa Dra Miriam Stela Maris de Oliveira Costa
Natal-RN
2017

1
Universidade Federal do Rio Grande do Norte - UFRN
Sistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Setorial Prof. Leopoldo Nelson - Centro de
Biociências - CB
Resende, Nayra da Silva.
Distribuição dos grupamentos neuronais colinérgicos no
encéfalo do mocó (Kerodon rupestris) / Nayra da Silva Resende. - Natal, 2017.
96 f.: il.
Dissertação (Mestrado) - Universidade Federal do Rio Grande do
Norte. Centro de Biociências. Programa de Pós-Graduação em Biologia Estrutural e Funcional.
Orientadora: Profa. Dra. Miriam Stela Maris de Oliveira
Costa.
1. Kerodon rupestris - Dissertação. 2. Acetilcolina -
Dissertação. 3. Sistema colinérgico - Dissertação. I. Costa, Miriam Stela Maris de Oliveira. II. Universidade Federal do Rio
Grande do Norte. III. Título.
RN/UF/BSE-CB CDU 599.322/.324
Elaborado por KATIA REJANE DA SILVA - CRB-15/351

1
RESENDE, N. S. Distribuição dos grupamentos neuronais colinérgicos no encéfalo do mocó
(Kerodon rupestris). Dissertação apresentada à Universidade Federal do Rio Grande do Norte
para a obtenção do título de Mestre em Biologia Estrutural e Funcional.
DATA DA DEFESA: 14 de dezembro de 2017.
BANCA EXAMINADORA
Prof. Dr. Francisco Gilberto Oliveira Universidade Regional do Cariri
Prof. Dr. Fernando Vagner Lobo Ladd Universidade Federal do Rio Grande do Norte
Prof. Dra. Miriam Stela Maris de Oliveira Costa - Orientadora Universidade Federal do Rio Grande do Norte

4
Dedico a todos que já tiveram um
momento de fraqueza.
Não vai doer ‘pra’ sempre, então
não deixe isso afetar o que há de
melhor em você.

5
“O cientista não é o homem que fornece
as verdadeiras respostas; é quem faz as
verdadeiras perguntas”.
(Claude Lévi-Strauss)

6
AGRADECIMENTOS
A Deus, antes de tudo, por ter me permitido vir a esse mundo, crescer e aprender a cada
dia mais e mais.
A minha mãe Vera Lúcia, quem me ensinou desde pequena, a não desistir nos primeiros
obstáculos, a sempre seguir com garra, a ela que me ensinou a ter coragem, mesmo defronte à
um abismo. Todo e qualquer agradecimento seria insuficiente, Te amo!
Ao meu pai Wilson, por me ensinar que sou capaz de superar todas os medos e desafios,
dos mais simples aos mais complicados.
Aos meus irmãos Hudson Resende, Wertson Resende e Kelyson Resende, que me
dedicaram horas e horas de conselhos, que se despuseram a me ajudar nos momentos em que eu
acreditei não haver mais ajudas, as broncas, as discussões, os abraços e os sorrisos que me
deram, minha vida sem vocês três, não teria o menor sentido.
Aos meus sobrinhos Guilherme Resende, Lucas Resende e Arthur Resende, que me
proporcionam um retorno a infância a cada momento em que passamos juntos, agradeço por me
alegrarem sempre com seus sorrisos de criança, e com a sinceridade no olhar, obrigada por
fazerem as nossas vidas serem mais leves. Titia ama vocês demais, mais que tudo!
A minha madrinha Hilma Bico – “in memoriam”, por ter me proporcionado excelentes
momentos em vida, por ter me ensinado a “nunca descer do salto”, e principalmente por ter me
ensinado a generosidade para com o próximo. Uma pena a senhora não ter me acompanhado
nessa etapa, sei que onde quer que esteja sua energia boa está me acolhendo sempre. Minha
saudade e meu amor eterno.
Ao meu avô Manoel Marques – “in memoriam”, que me ensinou, no fim da sua vida a
lutar até o último instante, sua força , garra, e vontade de viver me mostraram o quanto somos
pequenos diante dos problemas dos outros, e que nessa vida não somos absolutamente nada além
do que pequenos grãos de areia, prestes a se dissipar na imensidão do vento, me inspiro a cada
dia na sua força e fé na vida.
As minhas amigas: Rafaella e Luanna, que mesmo distante estiveram presentes em cada
momento que passei, fosse de tristeza ou alegria, estiveram me apoiando, obrigada pelas palavras
de apoio e pela torcida.
As minhas amigas: Amanda, Bárbara, Simone e Thaísa, que me acompanham desde a
graduação, e mesmo com a minha ausência em vários momentos, não me deixaram de lado, não
me abandonaram, e estão sempre torcendo pelo meu sucesso.
A minha amiga Janine, quem pude contar a todo momento durante a jornada da pós, e até
mesmo antes dela, agradeço imensamente toda torcida e apoio, conversas, incentivos, enfim,
TUDO.

7
As amigas que o LabNeuro me deu e que levarei para a vida, Karen e Luísa, não há
palavras para descrever nossa amizade, é aquela coisa, estamos longe, porém juntas ao mesmo
tempo, não importa a maneira, nem a circunstância, sabemos que estaremos lá.
Agradeço aos amigos do “TinderCapes#NE” que me proporcionaram momentos de
divertimento e risadas, nos momentos de angústia, vocês foram de grande valia para que que me
espelhasse em seus exemplos, e pudesse levantar a cabeça e seguir em frente.
Ao Profº Jeferson Cavalcante, por ter me ensinado o amor pela “neuro”, por ter me
inserido no LabNeuro, e por ter me proporcionado vários momentos de aprendizagem, não sou
capaz de descrever meus agradecimentos, apenas quero que saibas que sou muito grata.
Aos amigos do Lenq ou “LAPJE” (Rovena, Kayo, André, Janaína e Felipe). Que sempre
se dispuseram a me ajudar, fosse tirando uma dúvida, me auxiliando a manusear algum
equipamento tecnológico, ou até mesmo nas conversas informais.
Aos Professores do LabNeuro, pelo tempo dedicado, pelo conhecimento e sabedoria
compartilhados, meu imenso, obrigada!
A Regina, por nunca nos deixar sem as soluções, por sempre tentar pôr ordem no
ambiente, embora algumas vezes não conseguisse, pela amizade dedicada, o meu agradecimento.
Aos colegas do LabNeuro, por tornarem o ambiente do laboratório, um pouco mais leve.
Em especial a Alane, Daniel, Naryllene, Paloma, Brenna, Mayara, Melquisedec, Nayana e
Joacil, pela parceria e por me auxiliarem nos experimentos e a Nelyane, por sempre acreditar no
meu “potencial” e mostrar que sou capaz de alcançar meus objetivos, meu muito obrigada!
Aos colegas de turma (Dáfiny, Edson, Epifânio, Jonas, Kadigna, Mardem, Marília,
Natan e Rodrigo), pela companhia, pelas conversas e momentos de descontração. Em especial,
agradeço a Mario, Manu e Wylqui, que no meio do caminho passaram a me acompanhar e a
ajudar nessa jornada que foi o mestrado, transformando o trabalho com os nossos “pequenos
serumaninhos” (os mocós), muito mais leve e divertido, vocês são excelentes pessoa, agradeço
por tudo, principalmente pelos risos.
Agradeço a banca examinadora, pela disponibilidade de tempo e enriquecimento de
conhecimento ao meu trabalho.
Por fim, em especial, agradeço a minha orientadora, Profª Miriam Stela, por ter me
acolhido desde a iniciação cientifica até o mestrado, muito obrigada pela paciência, pelos
ensinamentos, por não ter me deixado ficar sem esperanças, pelo auxílio nos momentos de
dificuldade, por ter me mostrando alternativas, me encorajado e principalmente pela confiança a
cada dia e a cada experimento. Tudo isso foi imprescindível para que pudéssemos estar aqui
hoje.
A todos vocês meu singelo e de coração, OBRIGADA!

8
SUMÁRIO
Página
LISTA DE ABREVIAÇÕES ......................................................................................... 10
RESUMO ....................................................................................................................... 13
ABSTRACT .................................................................................................................. 14
1. INTRODUÇÃO .......................................................................................................... 15
1.1. A Acetilcolina...................................................................................................... 15
1.2. Modelo animal..................................................................................................... 18
2. OBJETIVOS ............................................................................................................... 22
3. METODOLOGIA ....................................................................................................... 23
3.1. Coleta dos animais ............................................................................................. 23
3.2. Anestesia ............................................................................................................. 23
3.3. Perfusão .............................................................................................................. 23
3.4. Emblocamento estereotaxico............................................................................... 24
3.5. Microtomia.......................................................................................................... 24
3.6. Coloração de citoarquitetônica ......................................................................... 25
3.7. Imunoistoquimica ............................................................................................... 25
3.8 Obtenção de Imagens ........................................................................................... 26
4. REFERÊNCIAS BIBLIOGRÁFICAS........................................................................ 27
5. RESULTADOS (Artigo) ........................................................................................... 32
Abstract ..................................................................................................................... 34
1. Introduction .......................................................................................................... 35
2. Materials and Methods.......................................................................................... 38
3. Results ....................... .......................................................................................... 39
4. Discussion ............................................................................................................ 42
Acknowlegments ...................................................................................................... 48
References ................................................................................................................ 49
Figure captions ......................................................................................................... 60
Abbreviations ........................................................................................................... 61
Figure 1 .................................................................................................................... 64
Figure 2 ..................................................................................................................... 70
Figure 3 ..................................................................................................................... 71
Figure 4 ..................................................................................................................... 72
Figure 5 ..................................................................................................................... 73
Figure 6 ..................................................................................................................... 74
6. CONSIDERAÇÕES FINAIS .................................................................................... 75

9
ANEXO A – Protocolo do Comitê de Ética para uso de Animais (CEUA-UFRN)
ANEXO B – Autorização SISBIO/IBAMA para coleta de animais
ANEXO C – Instruções para os autores Journal of Chemical Neuroanatomy

10
LISTA DE ABREVIAÇÕES
3N Núcleo oculomotor
3V Terceiro ventrículo
A Área amigdaloide
ac Comissura anterior
aca Comissura anterior, anterior
Acb Núcleo accumbens
AcetilCoA Acetil-coenzima A
Ach Acetilcolina
AchE Acetilcolinesterase
acp Comissura anterior, posterior
AD Núcleo talâmico anterodorsal
AHA Área hipotalâmica anterior
APT Núcleo pré-tectal anterior
Aq Aqueduto
Arc Núcleo arqueado hipotalâmico
B Núcleo basal
BIC Núcleo do braço do colículo inferior
bic Braço do colículo inferior
cc Corpo caloso
ChAT Colina-acetiltransferase
Cl Claustrum
CLi Núcleo linear caudal da rafe
Co Núcleo amigdalóide cortical
cp Pedúnculo cerebral
CPu Caudado-putamen
csc Comissura do colículo superior
CxA Córtex de transição da amígdala
DLG Núcleo geniculado leteral dorsal
DM Núcleo hipotalâmico dorsomedial
DR Núcleo dorsal da rafe
ec Cápsula externa
En Núcleo endopiriforme
EP Núcleo entopeduncular
f Fórnix
fr Fascículo retroflexo

11
GP Globo pálido
hbc Comissura habenular
HDB Núcleo da banda diagnonal de Broca, braço horizontal
IC Colículo inferior
ic Cápsula interna
ICj Ilhas de Calleja
IGL Folheto intergeniculado
IP Núcleo interpeduncular
LD Núcleo talâmico laterodorsal
LDTg Núcleo tegmental laterodorsal
lfp Fasciculo longitudinal da ponte
LH Área hipotalâmica lateral
LHb Núcleo habenular lateral
lo Trato olfatório lateral
LP Núcleo talâmico lateral posterior
LPO Área pré-optica lateral
LV Ventrículo lateral
M Núcleo mammilar
Me5 Núcleo trigeminal mesencefálico
MG Núcleo geniculado medial
MHb Núcleo habenular medial
ml Lemnisco medial
mlf Fascículo longitudinal medial
MnPO Núcleo pré-óptico mediano
MnR Núcleo mediano da rafe
MPA Área pre-óptica medial
MS Núcleo medial septal
mt Trato mamilotalâmico
och Quiasma óptico
opt Trato óptico
Pa Núcleo paraventricular hipotalâmico
PAG Substância cinzenta periaquedutal
PBG Núcleo parabigeminal
PF Núcleo parafascicular talâmico
Pir Córtex piriforme
PMnR Núcleo paramediano da rafe

12
Pn Núcleos pontinos
PT Núcleo paratenial talâmico
PTg Núcleo tegmental pedunculopontino
PV Núcleo talâmico paraventricular
PVP Núcleo paraventricular talâmico posterior
RCh Área retroquiasmática
Rt Núcleo reticular talâmico
SC Colículo superior
SCh Núcleo supraquiasmático
sm Estria medular talâmica
SNC Substância negra compacta
SNR Substância negra reticular
SO Núcleo supraóptico
ST Núcleo intersticial da estria terminal
st Estria terminal
STh Núcleo subtalâmico
tfp Fibras transversais da ponte
TT Tenia tecta
Tu Tubérculo olfatório
VDB Núcleo diagonal de Broca, banda vertical
VLG Núcleo geniculado ventral lateral
VLH Núcleo hipotalâmico ventrolateral
VLPO Núcleo pré-optico ventrolateral
VMH Núcleo hiptalâmico ventromedial
VP Pálido ventral
xscp Decussação do pedúnculo cerebelar superior
ZI Zona incerta

13
RESUMO
A acetilcolina (Ach) foi o primeiro neurotransmissor descoberto, nos neurônios
motores somáticos e neurônios autonômicos, e depois observado em vários aglomerados
neuronais no sistema nervoso central, sob a forma de interneurônios e grandes neurônios de
projeção. No sistema nervoso central, Ach está envolvido no controle de certas atividades
motoras e processos de aprendizado e memória. O objetivo deste estudo foi
citoarquitetonicamente e por imunoistoquimica de colina-acetiltransferase (ChAT) delimitar
os grupos colinérgicos no encéfalo do mocó (Kerodon rupestris), um roedor Caviidae
crepuscular do Nordeste brasileiro. Para isso, três animais adultos foram anestesiados e
transcardiacamente perfundidos. Os encéfalos foram congelados, seccionados no plano
coronal, obtendo-se 6 séries de seções de 30 μm. As seções de uma série foram submetidas
à coloração com Nissl. Outra série foi submetida a imunoistoquímica para revelar a
acetilcolina presente em diversos centros neurais do mocó, utilizando a enzima sintetizadora
ChAT como marcador. As lâminas foram analisadas sob microscópio óptico e os resultados
documentados por descrição e fotomicrografias digitais. Os neurônios imunorreativos a
ChAT foram identificados no telencefalo (núcleo accumbens, caudado-putamen, globo
pálido, núcleo entopeduncular, globo pálido ventral, tubérculo olfatório e Ilhas de Calleja,
núcleo da banda diagonal de Broca, núcleo basal e núcleo septal medial), diencéfalo
(núcleos pré-optico ventrolateral, hipotalâmico ventrolateral e habenular medial) e tronco
encefálico (núcleos parabigeminal, tegmentais laterodorsal e pedunculopontino). Esses
achados são discutidos em uma perspectiva funcional e filogenética, pois embora haja
semelhanças entre as espécies anteriormente estudadas, no nosso trabalho foi possível
observar diferenças e novos resultados antes não descritos em outras espécies de mamíferos.
PALAVRAS CHAVE: acetilcolina, colina acetiltransferase, Kerodon rupestris,
neurotransmissores, sistema colinérgico.

14
ABSTRACT
The acetylcholine (Ach) was the first discovered neurotransmitter, in the somatic
motor neurons and autonomic neurons, and then observed in several neuronal clusters in the
central nervous system, in the form of interneurons and large projection neurons. In the
central nervous system, Ach is involved in the control of certain motor activities and
processes to learning and memory. The aim of this study was to cytoarchitectonicly and by
cholineacetyltransferase (ChAT)-immunohistochemistry delimit the cholinergic groups in
the encephalon of the rock cavy (Kerodon rupestris), a crepuscular Caviidae rodent from
Brazilian Northeast. For this, threeyoung adult animals were anesthetized and transcardially
perfused. The encephala were frozen-cut in the coronal plane, obtaining 6 series of 30 µm
sections. The sections from one series were subjected to Nissl staining. Another series was
subjected to immunohistochemistry to develop the acetylcholine putatively present in
diverse neural centers of the rock cavy, using the synthesizing enzyme ChAT as marker.
The slides were analyzed under light microscope and the results documented by description
and digital photomicrographs. ChAT-immunoreactive neurons were identified in the
telencephalon (nucleus accumbens, caudate-putamen, globus pallidus, entopeduncular
nucleus and ventral globus pallidus, olfatory tubercle and Islands of Calleja, diagonal band
of Broca nucleus, nucleus basalisand medial septal nucleus), diencephalon (ventrolateral
preoptic, hypothalamic ventrolateralandmedial habenularnuclei) and brainstem
(parabigeminal, laterodorsal tegmental and pedunculopontine tegmental nuclei). These
findings are discussed in a functional and phylogenetic perspective, because although there
are similarities between the species previously studied, in our work it was possible to
observe differences and new results previously not described in other species of mammals.
KEY WORDS: acetylcholine choline acetyltransferase, Kerodon rupestris, Cholinergic
Sistem.

15
1. INTRODUÇÃO
A Acetilcolina
A acetilcolina (Ach), cuja a estrutura encontra-se representada na figura 1, foi o primeiro
neurotransmissor a ser identificado, inaugurando o conceito moderno de transmissão química no
sistema nervoso (MCGEER et al., 1985). Inicialmente identificada como o neurotransmissor
utilizado por neurônios motores somáticos e autônomos, foi posteriormente encontrada em
vários grupamentos neuronais no sistema nervoso central, na forma de interneurônios e grandes
neurônios de projeção (VON BOHLEN UND HALBACH e DERMIETZEL, 2006). Os
neurônios capazes de sintetizar, armazenar e liberar a Ach para a neurotransmissão, são
denominados neurônios colinérgicos (ODA e NAKANISHI, 2000)
Figura 1. Estrutura da acetilcolina.
A síntese de Ach ocorre a partir dos substratos colina e acetil-coenzima A (acetilCoA),
sendo catalisada pela enzima colina acetil-transferase (ChAT), a qual é sintetizada no corpo
celular e transportada até o terminal axônico, onde ocorre a síntese. A ChAT sintetiza a Ach no
citosol do terminal axônico e o neurotransmissor é, a seguir, concentrado dentro de vesículas
sinápticas, mediante um transportador de Ach (Fig. 2). Somente os neurônios colinérgicos
possuem ChAT, de forma que esta enzima é um bom marcador para células que usam Ach como
neurotransmissor. A acetilcoA é derivada, em sua maior parte, do metabolismo da glicose e, em
menor quantidade, dos fosfolipídios. A colina provém principalmente da degradação do
fosfolipídio fosfatidilcolina da dieta. Como a disponibilidade de colina limita a quantidade de
Ach que pode ser sintetizada no terminal axônico, o transporte de colina para dentro do neurônio
é a etapa limitante da velocidade da síntese de Ach (VON BOHLEN UND HALBACH e
DERMIETZEL, 2006).

16
Figura 2. Síntese e degradação da acetilcolina. Fonte: http://arquivobioqui.blodspot.com.br
As sinapses colinérgicas são muito conservadoras. Após hidrólise da Ach pela
acetilcolinesterase (AchE), do que resulta colina e acetato livre, a colina é recaptada pela
terminação pré-sináptica, sendo mais de 50% reutilizada na síntese de Ach (Fig. 2). A AchE é
secretada na fenda sináptica e está associada às membranas dos terminais axônicos colinérgicos.
A AchE também é produzida por alguns neurônios não colinérgicos, portanto esta enzima não é
marcador confiável para o sistema colinérgico (VON BOHLEN UND HALBACH e
DERMIETZEL, 2006).
A Ach exerce efeitos a partir de interações com dois tipos de receptores encontrados nas
membranas, pré e pós-sinápticas, sendo estes, os receptores múscarínicos metabotrópicos e
nicotínicos ionotrópicos, os múscarínicos encontrados acoplados a proteína Gq, as quais ativam
as fosfolipases ou acoplads a proteína Gi, as quais se acoplam negativamente ao adenilato ciclase
(Figura 3), ligando a Ach à uma cascata de sinalização bioquímica. Já os receptores nicotínicos
funcionam de forma não seletiva (PICCIOTTO et al., 2012). Os efeitos da Ach são
preferencialmente de natureza excitatória, mas também pode ser inibitória em alguns casos,
como por exemplo, no núcleo reticular do tálamo e no núcleo parabraquial (VON BOHLEN
UND HALBACH e DERMIETZEL, 2006).
Figura 3. Sítio de ação dos receptores nicotínicos e muscarínicos. (Adaptado de PICCIOTTO et al. 2012)

17
Ach é um neurotransmissor de ação rápida na junção neuromuscular e nos gânglios
autonômicos. Porém, a Ach parece atuar como um neuromodulador, no encéfalo, apesar de seu
papel como neurotransmissor excitatório primário, na periferia. No cérebro, Ach modifica a
excitabilidade neuronal, alterando a liberação pré-sináptica de outros neurotransmissores e
coordenando o disparo de outros grupamentos neuronais (PICCIOTTO et al., 2012). Deste modo
em conjunto com a sua atividade de regulação motora (CALABRESI et al., 2000), a modulação
colinérgica intergere em funções como a plasticidade sináptica (GIOCOMO e HANSSELMO,
2007; McKAY et al., 2007; DREVER et al., 2011), desenvolvimento neuronal (ROLE e BERG,
1996), Ach, interfere também na modulação de sistemas cerebrais mesolímbicos como o
dopaminérgico, associados ao vício e recompensa ( OMEICHENKI e SESACK, 2006),
regulação da atividade ccortical, (HANSSELMO e SARTER, 2011). O sistema colinérgico pode
alterar também funções hipotalâmicas, como a termorregulação (MYERS e WALLER, 1973),
regulação do sono (STERIADE, 2004), ingestão de alimentos (MINEUR et al., 2011), liberação
de insulina e glucagon (ISHIKAWA et al., 1982). Crescentes evidências mostram a sinalização
de Ach em várias áreas cerebrais, envolvidas na resposta ao estresse (MARK et al., 1996), e em
processos ligados a aprendizagem e memória (HASSELMO, 1999; 2006; GOLD, 2003; GAIS e
BORN, 2004).
No sistema nervoso central, a Ach é o neurotransmissor encontrado em algumas
populações de interneurônios colinérgicos que podem ser encontrados no prosencéfalo e no
tronco encefálico, como também os considerados grandes neurônios colinérgicos no
prosencéfalo basal e no tegmento mesopontino, que originam longas projeções ascendentes.
Entre os interneurônios, os neurônios mais representativos são aqueles no striatum. Estes
interneurônios interagem com os terminais dopaminérgicos de neurônios que para aí se projetam
a partir da substância negra. Há também interneurônios colinérgicos esparsamente distribuídos
no córtex cerebral, no hipocampo e no bulbo olfatório. Entre os neurônios de projeção, os grupos
colinérgicos do prosencéfalo basal incluem o núcleo septal medial (Ch1), as porções horizontal e
vertical do núcleo da banda diagonal (Ch2-Ch3) e o núcleo basal de Meynert (Ch4), que
originam longas projeções ascendentes, inervando topograficamente todo o córtex cerebral,
incluindo o hipocampo e a amígdala. Os neurônios colinérgicos mesopontinos são divididos
entre uma coluna ventrolateral (grupo celular Ch6, ou o núcleo pedúnculo-pontino), próximo à
margem lateral do pedúnculo cerebelar superior, e uma coluna dorsomedial (grupo celular Ch5,
ou o núcleo tegmental látero-dorsal), um componente da substância cinzenta periaquedutal,
posicionado justamente rostral ao locus coeruleus. Esses dois grupos celulares enviam uma
importante projeção descendente para a formação reticular ponto-bulbar, os núcleos vestibulares,

18
o locus coeruleus e vários núcleos da rafe, além de fornecerem uma extensa inervação
colinérgica ascendente para o tálamo e hipotálamo. Acredita-se que essas projeções têm um
papel de destaque na regulação do ciclo sono-vigília. Os neurônios do grupo Ch7 ocorrem na
habênula e se projetam para o núcleo interpeduncular. Finalmente, neurônios do grupo Ch8 estão
localizados no núcleo parabigeminal e enviam projeções para o colículo superior (SAPER, 2000;
VON BOHLEN UND HALBACH e DERMIETZEL, 2006).
Os núcleos colinérgicos foram delimitados imunoistoquimicamente em várias espécies de
mamíferos, como rato (ARMSTRONG et al., 1983; ICHIKAWA et al., 1997; ROGHANI et al.,
1998; SCHÄFER et al., 1998), monotremados (MANGER et al., 2002), macaco Rhesus (Macaca
mulatta) (KUS et al., 2003), toupeira (DA SILVA et al., 2006), morcegos (MASEKO e
MANGER, 2007; MASEKO et al., 2007; DELL et al., 2010; KRUGER et al., 2010), porco-
espinho (LIMACHER et al., 2008), damão do cabo (GRAVETT et al., 2009), girafa (BUX et al.,
2010), musaranho-elefante (PIETERS et al., 2010), demônio da tasmânia (PATZKE et al., 2014)
.
Modelo animal
O mocó (Kerodon rupestris) (Fig. 3) é classificado taxonomicamente como representante
do filo Chordata, classe Mammalia, superordem Glires, ordem Rodentia, subordem
Hystricomorpha, família Caviidae e subfamília Caviinae (SILVA NETO, 2000). A subordem
Hystricomorpha compreende várias famílias com um número de espécies encontradas no Brasil,
que inclui, além do mocó, a cutia (família Dasyproctidae), a paca (família Cuniculidae), a
capivara (Hydrochaeridae), todas elas utilizadas como modelos experimentais por pesquisadores
brasileiros (ver por exemplo, SILVEIRA, 1985; SILVEIRA et al., 1989; ROCHA et al., 2009;
PICANÇO-DINIZ et al, 1991; 2011). Estudos filogenéticos usando abordagem molecular
relacionam o gênero Kerodon com o gênero Hydrochaeris, o qual inclui a capivara (família
Hydrochaeridae), e é estreitamente alinhado com o gênero Dolicotis da subfamília Dolicothinae,
cujo principal representante da América do Sul é a lebre da Patagônia (Dolichotis patagonum)
(ROWE e HONEYCUTT, 2002). O mocó é uma espécie nativa da região Nordeste, sendo
encontrada desde o Piauí até o norte de Minas Gerais (CABRERA, 1961).

19
Figura 4. O mocó (Kerodon rupestris). Fonte:http://otempo.com.br
Os mocós são animais endêmicos da caatinga, habitando a região do semi-árido
nordestino, onde estão localizadas as rochas graníticas, que lhes servem como refúgio e abrigo
contra predadores, os quais são principalmente os gatos macambira e vermelho, as raposas, o
gavião pé de serra e o jacurutu (CARVALHO, 1969; LACHER Jr, 1981). Eles são altamente
adaptados às condições ecológicas regionais, caracterizadas pelo calor, a escassez de água e
alimentos, principalmente nos períodos de grandes secas nas regiões do semi-árido nordestino.
São herbívoros e costumam se alimentar de cascas de árvores, brotos, pequenos arbustos e ramos
de algumas espécies de plantas trepadeiras (MENDES, 1985; 1987). Em cativeiro aceitam bem
frutas (banana, mamão, melancia, manga) e raízes. Atingem a fase adulta aos 200 dias de vida,
podendo atingir 50 cm de comprimento e 1 quilo de peso corporal (ROBERTS et al., 1984).
Os mocós apresentam coloração cinzenta, com pelos pretos e amarelos ou esbranquiçados
na parte superior, marrons na região posterior e um pouco acastanhada nas pernas, e brancos na
região do pescoço (CARVALHO, 1969). Animais do gênero Kerodon possuem o mesmo corpo
básico dos outros cavíneos, embora apresentem algumas características morfológicas corporais e
craniais únicas para a subfamília. As mãos e os pés são acolchoados com epiderme semelhante a
pele-couro e não possuem garras. O pé tem unhas subcutâneas sobre todos os dígitos, exceto um
no interior dos pés onde a unha foi modificada como uma pequena garra usada para pentear o
pelo. O crânio, especialmente o focinho, é maior e mais estreito do que nos outros cavíneos,
assim como a distância entre os incisivos e os pré-molares é proporcionalmente maior (LACHER
Jr, 1981).
O mocó é um animal trepador extremamente ágil, apesar de não possuir garra e nem
cauda, duas adaptações normalmente associadas com arborealidade. Tem o olfato bem
desenvolvido, principalmente nos machos, que podem detectar a presença do homem e de outros

20
animais a longas distâncias. A audição é também bastante aguçada, de modo que qualquer ruído
curto, de pequena intensidade, como o de um assobio, um pequeno galho quebrado, ou o barulho
de folhas roçando no solo, costuma atrair o mocó (CARVALHO, 1969). Quanto à reprodução,
ocorrem nascimentos ao longo do ano, exceto no período de abril a junho. As fêmeas apresentam
estro pós-parto, podendo acasalar-se poucas horas após o nascimento dos filhotes. Embora as
proles sejam pequenas a cada parto (cerca de 1 a 2 filhotes), o curto período gestacional (75 dias)
garante uma elevada produção de crias a cada ano. Os animais atingem o tamanho adulto aos 200
dias, embora a primeira concepção das fêmeas possa ocorrer aos 115 dias de vida (LACHER Jr,
1981; ROBERTS et al., 1984).
A partir de observações próprias no campo e de informações de moradores da região,
CARVALHO (1969) relatou que o mocó sai para alimentar-se de manhã e à tarde nos dias mais
escuros e à noite nos dias claros. Nos dias nublados, depois de uma chuva, costumam sair a
qualquer hora do dia, permanecendo fora da toca o tempo necessário para a secagem das rochas,
o que algumas vezes pode durar o dia inteiro. LACHER Jr (1981) verificou que, no campo, esse
roedor sai para forragear ao longo do dia e da noite, porém a maior parte da atividade ocorre
durante o dia, com picos de atividade no crepúsculo. De acordo com MENDES (1987), os mocós
são animais gregários, de hábitos crepusculares, que durante o dia ficam abrigados em tocas
rochosas, saindo à tardinha e ao amanhecer para alimentar-se. Em linha com essas observações,
SOUSA e MENEZES (2006), estudando o mocó sob condições controladas de laboratório,
registraram para essa espécie a expressão de atividade locomotora ao longo de 24 horas do dia,
com picos nas fases de transição de luminosidade, caracterizando um comportamento
predominantemente crepuscular.
Estudos neuroanatômicos no mocó foram iniciados em nosso laboratório, quando esta
espécie foi adotada como um modelo regional de roedor em estudos de ritmos circadianos. Além
da caracterização do ritmo de atividade e outras respostas circadianas (SOUSA e MENEZES,
2006), as estruturas controladoras da ritmicidade circadiana – o núcleo supraquiasmático e o
folheto intergeniculado – foram caracterizadas com relação ao seu conteúdo neuroquímico e
projeções retinianas (CAVALCANTE et al., 2008; NASCIMENTO Jr, et al., 2010b). Do mesmo
modo, projeções retinianas diretas para os núcleos talâmicos paraventricular (NASCIMENTO Jr,
et al., 2008) e médio-dorsal (NASCIMENTO Jr, et al., 2010a) e para a zona incerta caudal
(MORAIS et al., 2014) foram demonstradas. O estudo do sistema serotoninérgico permitiu a
identificação dos núcleos da rafe e outros núcleos extra-rafe (SOARES et al., 2012). Além disso,
a distribuíção dos terminais serotoninérgicos para núcleos talâmicos da linha média e
intralaminares foi descrita (SILVA et al., 2014), os grupos dopaminérgicos mesencefálicos
(CAVALCANTI et al., 2014) e os grupamentos nitrérgicos do diencéfalo (REIS et al., 2018)

21
foram delineados e a anatomia do olho e estrutura retiniana foram descritas (OLIVEIRA et al.,
2014). Um estudo sobre os alvos das projeções retinianas constituintes dos sistemas visual
primário e óptico acessório, além de outros alvos hipotalâmicos, está em andamento.
Sendo o mocó (Kerodon rupestris), um modelo experimental regional, já consolidado em
estudos neuroanatômicos no Laboratório de Neuroanatomia-UFRN, este trabalho vem agregar
com a proposta de estudar o sistema colinérgico nesta espécie. Sabe-se que este sistema de
neurotransmissor tem sido estudado em várias espécies de mamíferos, mostrando certa
conservação filogenética. Considerando a importância de estudar os sistemas neurais do ponto de
vista comparativo-evolutivo, aliado à importância de aprofundar o conhecimento acerca do papel
da Ach como neurotransmissor no sistema nervoso central, é imperativo estender o estudo ao
maior número possível de espécies. Por estas razões é plenamente justificado este estudo no
mocó.

22
2. OBJETIVOS
Objetivo Geral
Este trabalho tem como objetivo caracterizar e delimitar os núcleos colinérgicos no
encéfalo do mocó (Kerodon rupestris).
Objetivos Específicos
Caracterizar citoarquitetonicamente os núcleos colinérgicos no encéfalo do mocó;
Identificar imunoistoquimicamente os núcleos colinérgicos no encéfalo do mocó;
Descrever a morfologia neuronal em cada núcleo.

23
3. MATERIAIS E MÉTODOS
Coletas dos animais
Foram utilizados 3 mocós adultos jovens, capturados nos municípios da região do Seridó
do estado do Rio Grande do Norte, mediante a autorização do Instituto Brasileiro do Meio
Ambiente e Recursos Naturais Renováveis – IBAMA, licença SISBIO no. 42960-1 de
10/03/2014 e aprovado pelo Comitê de Ética para Uso de Animais da UFRN (CEUA-UFRN,
protocolo no 004/2014). Após capturados, os animais foram acomodados, por um curto período
para aclimatação, na fazenda SAMISA de propriedade da UFRN, no município de Extremoz-
RN, em uma sala ampla de quatro paredes de alvenaria com porta e janela gradeadas, teto de
telhas de cerâmica e piso de solo natural com vegetação rasteira e pedras, simulando o habitat
natural do animal. Neste ambiente os animais ficaram expostos às condições ambientais naturais
de temperatura, umidade do ar e luminosidade, com alimentação e água ad libitum.
Todos os cuidados foram tomados no sentido de evitar dor e sofrimento aos animais
durante os procedimentos experimentais, seguindo estritamente as normas estabelecidas pela
National Research Council of the National Academy, disponível no formato pdf, no site da
Sociedade Brasileira de Neurociências e Comportamento – SBNeC
(http://www.sbnec.gov.br/links).
Anestesia
Os animais foram anestesiados por via intramuscular, com cloridrato de ketamina 10%
(1ml) e xilazina 2% (0,1ml) por kg de peso do animal.
Perfusão
Após atingirem o plano anestésico, os animais foram submetidos a perfusão
transcardíaca, seguindo estes passos:
1. O animal foi posicionado em decúbito dorsal, sobre uma tela de arame e sob ponto de água;
2. Toracotomia, com incisão de pele, músculos e gradil costal, removidos para a exposição do
coração;
3. Cardiopunção no ventrículo esquerdo, utilizando uma agulha com dimensões 17 mm x 1,5
mm, direcionada para o vestíbulo da artéria aorta ascedente, seguindo-se de uma incisão no átrio
direito. A agulha foi conectada a uma bomba peristáltica (Cole-Parmer) (Inserir Modelo),
passando-se 300 ml da solução salina a 0,9% em tampão fosfato 0,1M, pH 7,4 com heparina
(Parinex, Hipolabor, 2 ml/1000 ml de solução salina) durante um tempo estimado de seis
minutos. Em seguida foram infundidos 700 ml de solução fixadora, consistindo de

24
paraformaldeído 4% em tampão fosfato 0,1M, pH 7.4, sendo que metade desta solução foi
bombeada num fluxo de 70 ml/min e a outra metade num fluxo de 17,5 ml/min, com tempo de
duração aproximado de 30 minutos.
Emblocamento estereotáxico
Finalizado o processo de perfusão, os animais foram posicionados no aparelho
estereotáxico específico para roedores (INSIGHT MME-930). Após fazer uma incisão
longitudinal na pele e rebatê-la, foi realizada a limpeza da superfície óssea, facilitando a
visualização do bregma e do lambda, então nivelados na mesma altura dorsoventral, ajustando-se
as barras dos incisivos, padronizando desta maneira o plano de corte coronal para todos os
animais. Anotadas as coordenadas do bregma e do lambda, o osso da calota craniana foi
removido com o uso de broca e trocater, expondo o encéfalo. Ainda no aparelho estereotáxico, o
encéfalo foi seccionado em três blocos, através de duas secções coronais, sendo uma a nível do
bregma e outra no nível do lambda.
Os encéfalos foram retirados delicadamente para evitar danos, preservando bem sua
estrutura, em seguida foram medidos (da extremidade anterior do bulbo olfatório ao limite bulbo-
espinal) e fotografados. Logo em seguida, os três blocos foram armazenados em solução
contendo sacarose a 30% em tampão fosfato 0,1M, pH 7,4, a 4°C, até serem submetidos a
microtomia.
Microtomia
Os encéfalos mantidos na solução com sacarose, foram submetidos a microtomia,
obtendo secções coronais com espessura de 30 µm.
Os encéfalos foram congelados pelo dióxido de carbono solidificado (gelo seco), e
seccionados em micrótomo horizontal de deslizamento (Leica SM2000R). As secções foram
coletadas em um meio liquido de solução anti-congelante, distribuídas sequencialmente em seis
compartimentos, de maneira cíclica, de modo a manter a distância entre uma secção e a outra
imediatamente seguinte de um mesmo compartimento de aproximadamente 180 µm.
Os cortes do primeiro compartimento foram imediatamente montados em lâminas de
vidro silanizadas e submetidas a coloração pelo método de Nissl para o estudo citoarquitetônico
dos grupamentos neuronais colinérgicos. Os cortes dos demais compartimentos foram
conservados a -20°C para a realização de procedimentos posteriores de imunoistoquímica.

25
Coloração citoarquitetônica (Método de Nissl)
Uma série foi submetida à coloração pelo método de Nissl, utilizando o acetato de tionina
como corante, para a análise citoarquitetônica. Neste procedimento, os cortes previamente
montados em lâminas silanizadas, passaram por um período de secagem, para posteriormente
serem submetidas ao processo de coloração, onde inicialmente passaram por uma bateria de
álcoois etílicos em concentrações crescentes, a saber: primeiramente foram mergulhados em
álcool a 70% por um período de no mínimo 2 horas, posteriormente foi seguido de 2 banhos em
álcool 95%, de 3 minutos cada, 2 banhos em álcool absoluto, durante 3 minutos cada. Ao final da
bateria, as lâminas foram mergulhadas em xilol, num primeiro banho de 3 minutos e
sequencialmente imersos num segundo banho de 30 minutos. Na sequência, os cortes foram
reidratados em um banho de xilol durante 2 minutos, seguindo de banhos em álcoois etílicos em
concentrações decrescentes (a mesma utilizada na primeira bateria, porém em sentido inverso, e
no tempo de 2 minutos cada), acrescido do álcool a 50% e água destilada. Posteriormente foram
imersos em Acetato de Tionina por 40 segundos, para novamente serem desidratados e
diafanizados como descrito anteriormente. Ao final do processo, os cortes foram cobertos com
lamínula, utilizando o meio de montagem ERV-Mount (mistura sintética, a base de polímeros
plásticos).
Imunoistoquímica
Os compartimentos restantes foram submetidos ao processo de imunoistoquímica, o qual
ocorreu em sua totalidade em agitador orbital. O material utilizado foi submetido a um pré-
tratamento para a eliminação das peroxidases endógenas. Este processo dá-se através de 4
lavagens em tampão fosfato 0,1M, pH 7,4, de 10 minutos cada, posteriormente uma imersão em
peróxido de hidrogênio (H2O2) 0,3% e tampão fosfato 0,1M, pH 7,4, por 20 minutos. Após esse
tratamento os cortes passaram por 4 lavagens em tampão fosfato 0,1M, pH 7,4, de 10 minutos
cada.
No próximo passo os cortes foram incubados no anticorpo primário, uma solução
formada pelo anticorpo anti-ChAT obtido em cabra (Millipore), à concentração de 1:1000,
acrescido de soro normal de asno (Sigma) a 2% em Triton X-100 a 0,4% permanecendo
incubado em torno de 18 horas, no agitador orbital, a 40 (quarenta) rpm (rotações por minuto).
Ao fim desse período, os cortes passaram por quatro lavagens em tampão fosfato 0,1M,
pH 7,4, por 10 minutos, em agitador orbital, em seguida os cortes foram incubados em anticorpo
secundário anti-cabra obtido em asno, à concentração de 1:1000, em Triton X-100 a 0,4%,
permanecendo incubado por 90 minutos, no agitador orbital, a 40 rpm.

26
Em seguida os cortes foram lavados quatro vezes em tampão fosfato 0,1M, pH 7,4, por
10 minutos, e depois colocados na solução do complexo avidina-biotina-HRP (Protocolo ABC,
Kit elite da Vector), numa diluição de 1:100 em Triton X-100 a 0,4%, contendo NaCl, por 90
minutos à temperatura ambiente, sob agitação lenta, no agitador orbital. Terminada esta fase as
secções foram novamente submetidas a quatro lavagens em tampão fosfato 0,1M, pH 7,4, por 10
minutos, em agitador orbital.
Para a visualização da reação, as secções foram postas em meio contendo H2O2 como
substrato e a 3,3‟4,4‟ tetrahidrocloreto-diaminobenzidina (DAB), utilizada como cromógeno.
Esta reação ocorre em torno de 15 minutos. Ao final deste processo, os cortes foram lavados
mais quatro vezes em tampão fosfato 0,1M, pH 7,4, por 10 minutos cada, em agitador orbital.
Os cortes foram montados em lâminas silanizadas, que após secas foram imersas em
solução de tetróxido de ósmio a 0,05% com o intuito de intensificar a reação. Esta etapa foi
seguida por uma bateria de desidratação em álcoois de concentrações crescentes até o álcool
absoluto, e diafanização em xilol. As lamínulas foram montadas sobre os cortes com o uso do
meio de montagem ERV-Mount.
Obtenção de imagens e análise microscópica
As secções, coradas pelo método de Nissl, e as secções submetidas ao processo de
imunoistoquímica para ChAT, foram examinadas ao microscópio óptico (Nikon Eclipse Ni) em
campo claro. Imagens digitais foram obtidas de secções representativas usando uma videocâmera
digital (Nikon DSRe1), acoplada ao microscópio óptico, usando o software NIS – Elements AR.
As imagens foram analisadas, corrigidas minimamente para brilho e contraste, usando os
programas Canvas 12 (ACD System of America, inc.) e Adobe Photoshop (PS CC 2015).
Diagramas foram obtidos de imagens de secções coronais coradas pela técnica de Nissl
usando o programa Canvas 12 (ACD System of America, inc.). As secções imunocoradas
correspondentes foram superpostas aos gráficos e os neurônios ChAT-IR foram plotados.
O formato dos neurônios marcados nos diversos grupamentos neuronais foi delineado
com o auxílio de câmara clara (Alltech, Leica) acoplada a um microscópio Olympus BX41.

27
4. REFERÊNCIAS BIBLIOGRÁFICAS
ARMSTRONG, D. M.; SAPER, C. B.; LEVEY, A. I.; WAINER, B. H.; TERRY, R. D.
Distribution of cholinergic neurons in rat brain: demonstrated by the immunocytochemical
localization of cholineacetyltransferase. J. Comp. Neurol., v. 216, p. 53-68, 1983.
BHAGWANDIN, A.; FUXE, K.; BENNETT, N. G., MANGER, P. R. Nuclear organization and
morphology of cholinergic, putative catecholaminergic and serotonergic neurons in the brains of
two species of African mole-rat. J. Chem. Neuroanat., v. 35, p. 371-387, 2008.
BUX, F.; BHAGWANDIN, A.; FUXE, K.; MANGER, P. R. Organization of cholinergic,
putative catecholaminergic and serotonergic nuclei in the diencephalon, midbrain and pons of
sub-adult male giraffes. J. Chem. Neuroanat., v. 39, p. 189-203, 2010.
CABRERA, A. Catálogo de los mamíferos de America del Sur. Rev. Mus. Argentino Cien.
Nat. “Bernardo Rivadavia”, v.4, p.1-732, 1961.
CALABRESI, P., CENTONZE, D., GUBELLINI, P., PISANI, A., BERNARDI, G.
Acetylcholine-mediated modulation of striatal function. Trends Neurosci. 23, 120-126, 2000.
CARVALHO, J. C. M. Notas de viagem de um zoólogo à região das caatingas e áreas limítrofes.
Fortaleza, Imprensa universitária do Ceará, 1969.
CAVALCANTE, J. S.; BRITTO, L. R. G.; TOLEDO, C. A. B.; NASCIMENTO Jr, E. S.;
LIMA, R. R. M.; PONTES, A. L. B.; COSTA, M. S. M. O. Calcium-binding proteins in the
circadian centers of the common marmoset (Callithrix jacchus) and the rock cavy (Kerodon
rupestris) brains. Brain Res. Bull., v. 76, p. 354-360, 2008.
CAVALCANTI, J. R. L. P.; SOARES, J. G.; OLIVEIRA, F. G.; GUZEN, F. P.; PONTES, A. L.
B.; SOUSA, T. B.; CAVALCANTE, J. S.; NASCIMENTO Jr, E. S. A cytoarchitectonic and TH-
immunohistochemistry characterization of the dopamine cell groups in the substantia nigra,
ventral tegmental area and retrorubral field in the rock cavy (Kerodon rupestris). J. Chem.
Neuroanat., v. 55, p. 58-66, 2014.
CUI, H.; MALPELI, J.G.; Activity in the parabigeminal nucleus during eye movements directed
at moving and stationary target., J. Neurophysiol., v. 89, p. 3128-3142, 2003.
DA SILVA, J. N.; FUXE, K.; MANGER, P. R. Nuclear parcellation of certain
immunohistochemically identifiable neuronal systems in the midbrain and pons of the Highveld
molerat (Cryptomys hottentotus). J. Chem. Neuroanat., v. 31, p. 37-50, 2006.
DELL, L. –A.; KRUGER, J. –L.; BHAGWANDIN, A.; JILLANI, N. E.; PETTIGREW, J. D.;
MANGER, P. R. Nuclear organization of cholinergic, putative catecholaminergic and
serotonergic systems in the brains of two megachiropteran species. J. Chem. Neuroanat., v. 40,
p. 177-195, 2010.
DREVER, B.D., RIEDEL, G., PLATT, B. The cholinergic system and hippocampal plasticity.
Behav. Brain Res. 221, 505-514, 2011.
GAIS, S.; BORN, J. Low acetylcholine during slow-wave sleep is critical for declarative
memory consolidation. PNAS, v. 101, p. 2140-2144, 2004.
GIOCOMO, L.M., HASSELMO, M.E. Neuromodulation by glutamate and acetylcholine can
change circuit dynamics by regulating the relative influence of afferent input and excitatory
feedback. Mol. Neurobiol. 36, 184-200, 2007.
GODDARD, C. A.; KNUDSEN, E. I.; HUGUENARD, J. R. Intrinsic excitability of cholinergic
neurons in the rat parabigeminal nucleus. J. Neurophysiol., v.98, p. 3486-3493, 2007.

28
GOLD, P. E. Acetylcholine modulation of neural systems involved in learning and memory.
Neurobiol. Learn. Mem., v. 80, p. 194-210, 2003.
GRAVETT, N.; BHAGWANDIN, A.; FUXE, K.; MANGER, P. R. Nuclear organization and
morphology of cholinergic, putative catecholaminergic and serotonergic neurons in the brain of
the rock hyrax, Procavia capensis. J. Chem. Neuroanat., v. 38, p. 57-74, 2009.
HASSELMO, M. E. Neuromodulation: acetylcholine and memory consolidation. Trends Cogn
Sci., v. 3, p. 351-359, 1999.
HASSELMO, M. E. The role of acetylcholine in learning and memory. Curr. Opin. Neurobiol.,
v. 16, p. 710-715, 2006.
HASSELMO, M.E., SARTER, M. Modes and models of forebrain cholinergic neuromodulation of
cognition. Neuropsychopharmacology 36, 52-73, 2011.
ICHIKAWA, T.; AJIKI, K.; MATSURA, J.; MISAWA, H. Localization of two cholinergic
markers, choline acetyltransferase transporters in the central nervous system of the rat: in situ
hybridization histochemistry and immunohistochemistry. J. Chem. Neuroanat., v. 13, p. 23-29,
1997.
KRUGER, J. L.; DELL, L. A.; BHAGWANDIN, A.; JILLANI, N. E.; PETTIGREW, J. D.;
MANGER, P. R. Nuclear organization of cholinergic, putative catecholaminergic and
serotonergic systems in the brains of five microchiropteran species. J. Chem. Neuroanat., v. 40,
p. 210-222, 2010.
KUS, L.; CHU, Y. P.; FERGUSON, S. M.; BLAKELY, K. D.; EMBORGE, M. E.;
KORODOWER, J. H.; LEVEY, A. I.; MUFSON, E. J. Distribution of hight affinity choline
transporter immunoreactivity the primate central nervous system. J. Comp. Neurol., v. 463, p.
341-357, 2003.
LACHER Jr, T. E. The comparative social behavior of Kerodon rupestris and Galea spixii and
the evolution of behavior in the cavidiae. Bulletin of Carnegie Museum of Natural History
Pittsburg, v.17, p.5-71, 1981.
LIMACHER, A. „M.; BHAGWANDIN, A.; FUXE, K.; MANGER, P. R. Nuclear organization
and morphology of cholinergic, putative catecholaminergic and serotonergic neurons in the brain
of the Cape porcupine (Hystrix africae australis): Increased brain size does not lead to increased
organizational complexity. J. Chem. Neuroanat., v. 36, p. 33-52, 2008.
MAGALHÃES, M. A. F. O trato retino-hipotalâmico no mocó (Kerodon rupestris): um
estudo de traçado anterógrado com a subunidade B da toxina colérica. Programa de pós-
graduação em Psicobiologia, UFRN, Natal, RN. Dissertação de Mestrado. 2008. 93 pp.
MANGER, P. R.; FAHRINGER, H. M.; PETTIGREW, J. D.; SIEGEL, J. M., The distribution
and morphological characteristics of cholinergic cells the brain of monotremes as revealed by
ChAT immunohistochemistry. Brain Behav. Evol., v. 60, p. 275-297, 2002.
MASEKO, B. C.; MANGER, P. R. Distribution and morphology of cholinergic,
catecholaminergic and serotonergic neurons in the brain of Schreiber‟s long-fingered bat,
Miniopterus schreibersii. J. Chem. Neuroanat., v. 34, p. 80-94, 2007.
MASEKO, B. C.; BOURNE, J. A.; MANGER, P. R. Distribution and morphology of
cholinergic, putative catecholaminergic and serotonergic neurons in the brain of the Egyptian
rousette flying fox, Rousettus aegyptiacus. J. Chem. Neuroanat., v. 34, p. 108-127, 2007.
MENDES, B. V. Alternativas tecnológicas para a agropecuária do semi-árido. São Paulo:
Nobel, 171 pp., 1985.
MENDES, B. V. Plantas e animais para o nordeste. Rio de Janeiro: Globo, 167 pp., 1987.

29
McGEER, P. L.; McGEER, E. G.; MIZUKAWA, K.; TAGO, H.; PENG, J.H. Distribution of
cholinergic neurons in human brain. In: DUN, N. J. and PERLMAN R. L. Neurobiology of
acetylcholine. Maywood, Illinois: [s.n], p. 13-16, 1985.
MCKAY, B.E., PLACZEK, A.N., DANI, J.A. Regulation of synaptic transmission and plasticity
by neuronal nicotinic acetylcholine receptors. Biochem. Pharmacol. 74, 1120-1133, 2007.
MINEUR, Y.S., ABIZAID, A., RAO, Y., SALAS, R., DILEONE, R.J., GÜNDISCH, D.,
DIANO, S., DE BIASI, M., HORVATH, T.L., GAO, X.-B., PICCIOTTO, M.R. Nicotine
decreases food intake through ativation of POMC neurons. Science 332, 1330-1332, 2011.
MORAIS, P. L. A. G.; SANTANA, M. A. D.; CAVALCANTE, J. C.; COSTA, M. S. M. O.;
CAVALCANTE, J. S.; NASCIMENTO Jr, E. S. Retinal projections into the Zona Incerta of the
rock cavy (Kerodon rupestris): A CTb study. Neurosci. Res., v. 89, p. 75-80, 2014.
MOTTS, S.D.; SLUSARCZYK, A.S.; SOWICK, C.S.; SCHOFIELD, B. R.; Distribution of
cholinergic cells in guinea pig brainstem. Neuroscience. v. 154(1): p.186-195, 2008.
MYERS, R.D., WALLER, M.B. Differential release of acetylcholine from the hypothalamus and
mesencephalon of the monkey during thermoregulation. J. Physiol. 230, 273-293, 1973.
NASCIMENTO Jr, E.S.; DUARTE, R. B.; SILVA, S. F.; ENGELBERTH, R. C. G. J.;
TOLEDO, C. A. B.; CAVALCANTE, J. S.; COSTA, M. S. M. O. Retinal projections to the
thalamic paraventricular nucleus in the rock cavy (Kerodon rupestris). Brain Res., v. 1241, p.
56-61, 2008.
NASCIMENTO Jr, E. S.; CAVALCANTE, J. S.; CAVALCANTE, J. C.; COSTA, M. S. M. O.
Retinal afferents to the thalamic mediodorsal nucleus in the rock cavy (Kerodon rupestris).
Neurosci. Lett., v.475, p. 38-43, 2010a.
NASCIMENTO Jr, E. S.; SOUZA, A. P. M.; DUARTE, R. B.; MAGALHÃES, M. A. F.;
SILVA, S. F.; CAVALCANTE, J. C.; CAVALCANTE, J. S.; Costa, M. S. M. O. The
suprachiasmatic nucleus and the intergeniculate leaflet in the rock cavy (Kerodon rupestris):
Retinal projections and immunohistochemical characterization. Brain Res., v.1320, p. 34-46,
2010b.
ODA, Y. and NAKANISHI, I., The distribution of cholinergic neurons in the human central
nervous system. Histol. Histopathol., v. 15, p. 825-834, 2000.
OMEICHENKO, N., SESACK, S.R. Cholinergic axons in the rat ventral tegmental area synapse
preferentially onto mesoaccumbens dopamine neurons. J. Comp. Neurol. 494, 863-875, , 2006.
OLIVEIRA, F. G.; ANDRADE-DA-COSTA, B. L. S.; CAVALCANTE, J. S.; SILVA, S. F.;
SOARES, J. G.; LIMA, R. R. M.; NASCIMENTO Jr, E. S.; CAVALCANTE, J. C.; RESENDE,
N. S.; COSTA, M. S. M. O. The eye of the crepuscular rodent rock cavy (Kerodon rupestris)
(Wied, 1820). J. Morphol. Sci., v. 31, p. 89-97, 2014.
PANG, X., LIU, L., NGOLAB, J., ZHAO-SHEA, R., MCINTOSH, J.M., GARDNER, P.D.,
TAPPER, A.R., Habenula cholinergic neurons regulate anxiety during nicotine withdrawal via
nicotinic acetylcholine receptors, Neuropharmacology, v. 107, p. 294-304, 2016.
PATZKE, N.; BERTELSEN, M. F.; FUXE, K.; MANGER, P. R., Nuclear organization of
cholinergic, catecolaminergic, serotonergic and orexinergic systems in the brain of Tasmanian
devil (Sarcophilus harrisii). J. Chem. Neuroanat., v. 61-62, p. 94-106, 2014.
PICANÇO-DINIZ, C. W.; SILVEIRA, L. C. L.; CARVALHO, M. S. P.; OSWALDO-CRUZ, E.
Contralateral visual field representation in area 17 of the cerebral cortex of the agouti: A

30
comparison between the cortical magnification factor and retinal ganglion cell distribution.
Neuroscience, v. 44, p. 325-333, 1991.
PICANÇO-DINIZ, C. W.; ROCHA, E. G.; SILVEIRA, L. C. L.; ELSTON, G.; OSWALDO-
CRUZ, E. Cortical representation of the horizon in V1 and peripheral scaling in mammals with
lateral eyes. Psychol. Neurosci., v. 4, p. 19-27, 2011.
PICCIOTTO, M.R., HIGLEY, M.J., MINEUR, Y.S. Acetylcholine as a neuromodulator:
cholinergic signaling shapes nervous system function and behavior. Neuron 76, 116-129, 2012.
PIETERS, R. P.; GRAVETT, N.; FUXE, K.; MANGER, P. R. Nuclear organization of
cholinergic, putative catecholaminergic and serotonergic nuclei in the brain of the eastern rock
elephant shrew, Elephantulus myurus. J. Chem. Neuroanat., v. 39, p. 175-188, 2010.
PRATT W. E.; KELLEY, A. E., Nucleus accumbens acetylcholine regulates appetitive learning and
motivation for food via activation of muscarinic receptors. Behav. Neurosci., v. 118, n. 4, p. 730-739,
2004.
ROBERTS, M. E.; MALINIAK, E.; DEAL, M. The reproductive biology of the rock cavy,
Kerodon rupestris in captivity: a study of reproductive adaptation in a trophic specialist.
Mammalia, v. 48, p. 253-266, 1984.
ROCHA, F. A. F.; AHNELT, P. K.; PEICHL, L.; SAITO, C. A.; SILVEIRA, L. C. L.; DE
LIMA, S. M. A. The topography of cone photoreceptors in the retina of a diurnal rodent, the
agouti (Dasyprocta aguti). Vis. Neurosci., v. 26, p. 167-175, 2009.
ROGHANI, A.; SHIRZADI, A.; BUTCHER, L. L.; EDWARDS, R. H. Distribution of the
vesicular transporter for acetylcholine in the rat central nervous system. Neuroscience, v. 82, p.
1195-1212, 1998.
ROLE, L.W., BERG, D.K., 1996. Nicotinic receptors in the development and modulation of
CNS synapses. Neuron 16, 1077-1085.
ROWE, D. L.; HONEYCUTT, R. L. Phylogenetic relationships, ecological correlates, and
molecular evolution within the cavioidea (Mammalia, Rodentia). Mol. Biol. Evol., v. 19, p. 263-
277, 2002.
SAPER, C. A modulação da sensação, movimento e consciência pelo tronco encefálico. In
KANDEL, E. R., SCHWARTZ, J. H., JESSEL, T. M. (eds.) Principles of Neural Science., P.
889-909, 2000.
SATOH, K.; FIBIGER, H. C., Distribution of central cholinergic neurons in the baboon (Papio
papio). II. A topographic atlas correlated with catecholamine neurons. J. Comp. Neurol., v. 236,
p. 215-233, 1985.
SCHÄFER, M. K. –H.; EIDEN, L. E.; WEIHE, E. Cholinergic neurons and terminal fields
revealed by immunohistochemistry for the vesicular acetylcholine transporter. I. Central nervous
system. Neuroscience, v. 84, p. 331-359, 1998.
SILVA, A. M. Padrão de distribuição e caracterização morfológica de fibras serotonérgicas
nos núcleos da linha média/intralaminares do tálamo do mocó (Kerodon rupestris).
Programa de pós-graduação em Psicobiologia, UFRN, Natal, RN. Dissertação de mestrado. 73
pp., 2013.
SILVA, S. F. Núcleos dos sistemas visual primário e óptico acessório no mocó (Kerodon
rupestris): Caracterização pela projeção retiniana e citoarquitetura. Programa de pós-
graduação em Psicobiologia, UFRN, Natal, RN. Tese de doutorado, 87 pp., . 2009.
SILVA NETO, E. J. Morphology of the regions ethmoidalis and orbitotemporalis in Galea
musteloides Meyen 1832 and Kerodon rupestris (Wied-Neuwied 1820) (Rodentia: Caviidae)

31
with comments on the phylogenetic systematic of the Caviidae. J. Zool. Syst. Evol. Res., v.38,
p. 219-229, 2000.
SILVEIRA, L. C. L. Organização do Sistema Visual de Roedores da Amazônia: Óptica
Ocular e Distribuição das Células Ganglionares Retinianas. Instituto de Biofísica,
Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil. Tese de doutorado. 1985.
SILVEIRA, L. C.; YAMADA, E. S.; PICANÇO-DINIZ, C. W. Displaced horizontal cells and
biplexiform horizontal cells in the mammalian retina. Visual Neurosci., v. 3, p. 483-488, 1989.
SOARES, J. G.; CAVALCANTI, J. R. L. P.; OLIVEIRA, F. G.; PONTES, A. L. B.; SOUSA, T.
B.; FREITAS, L. M.; CAVALCANTE, J. S.; NASCIMENTO Jr, E. S.; CAVALCANTE, J. C.;
COSTA, M. S. M. O. Nuclear organization of the serotonergic system in the brain of the rock
cavy (Kerodon rupestris). J. Chem. Neuroanat., v. 43, p. 112-119, 2012.
SOUSA, R. A.; MENEZES, A. A. L. Circadian rhythm of motor activity of the Brazilian rock
cavy (Kerodon rupestris) under artificial photoperiod. Biol. Rhythm Res., v. 37, p. 443-450,
2006.
STERIADE, M. Acetylcholine systems and rhythmic activities during the waking–sleep cycle.
Prog. Brain Res. v. 145, p. 179-196, 2004.
VON BOHLEN UND HALBACH, O.; DERMIETZEL, R. Neurotransmitters and
Neuromodulators. Wiley-VCH, Germany, Pp. 46-59, 2006.
WARNER-SCHMIDT, J. L.; SCHMIDT, E. F.; MARSHALL, J. J.; RUBIN, A. J.; ARANGO-
LIEVANO, M.; KAPLITT, M. G.; IBAÑEZ-TALLON, I.; HEINTZ, N.; GREENGARD, P.
Cholinergic interneurons in the nucleus accumbens regulate depression-like behavior.
Neuroscience, v. 109 (28), p. 11360-11365, 2012.
Síntese e degradação da acetilcolina: Figura retirada de: http://arquivobioqui.blodspot.com.br/,
Acessado em: Novembro de 2017.
O mocó (Kerodon rupestris). Figura retirada de: http://otempo.com.br. Acessado em Novembro
de 2017.
*De acordo com:
ASSOCIAÇÃO BRASILEIRA DE NORMAS TÉCNICAS. Nbr 6023: Informação e
Documentação – Referências – Elaboração. Rio de Janeiro: ABNT, 2000. 22 p.

32
5. RESULTADOS
Artigo: NUCLEAR ORGANIZATION AND MORPHOLOGY OF CHOLINERGIC
NEURONS IN THE BRAIN OF THE ROCK CAVY (Kerodon rupestris) (Wied, 1820)
Periódico: Jornal of Chemical Neuroanatomy

33
NUCLEAR ORGANIZATION AND MORPHOLOGY OF CHOLINERGIC NEURONS
IN THE BRAIN OF THE ROCK CAVY (Kerodon rupestris) (Wied, 1820)
Nayra da Silva Resende1, Péricles Lasfir Soares Filho1, Paloma Pinheiro de Aquino Peixoto1,
Alane de Medeiros Silva1, Sebastião Franco da Silva1, Joacil Germano Soares1, Expedito Silva
do Nascimento Jr1, Judney Cley Cavalcante1, Jeferson de Souza Cavalcante2, Miriam Stela Maris
de Oliveira Costa1*.
Department of Morphology, Laboratory of Neuroanatomy, Biosciences Center, Federal
University of Rio Grande do Norte, Natal, RN, Brazil.
Running head: The rock cavy cholinergic neurons
Associate editor:
Keywords: acetylcholine, choline acetyltransferase, cholinergic system, Kerodon rupestris,
neurotransmitters.
*Corresponding author:
Miriam Stela Maris de Oliveira Costa
Department of Morphology, Laboratory of Neuroanatomy, Biosciences Center, Federal
University of Rio Grande do Norte, 59072-970, Natal, RN, Brazil.
Telephone number: 55 84 32153431
Fax number: 55 84 32119207
E-mail addresses: [email protected]; [email protected]
Financial support: CNPq, CAPES, FAPERN

34
ABSTRACT
The aim of this study was to delimit cytoarchitectonicly and by
cholineacetyltransferase (ChAT)immunohistochemistry the cholinergic groups in the
encephalon of the rock cavy (Kerodon rupestris), a crepuscular Caviidae rodent from the
Brazilian Northeast. For this, three young adult animals were anesthetized and transcardially
perfused. The encephala were frozen-cut in the coronal plane, obtaining 6 series of 30 µm
sections. The sections from one series were subjected to Nissl staining. Another series was
subjected to immunohistochemistry to develop the acetylcholine putatively present in
diverse neural centers of the rock cavy, using the synthesizing enzyme ChAT as marker.
The slides were analyzed under light microscope and the results documented by description
and digital photomicrographs. ChAT-immunoreactive neurons were identified in the
telencephalon (nucleus accumbens, caudate-putamen, globus pallidus, entopeduncular
nucleus and ventral globus pallidus, olfatory tubercle and Islands of Calleja, diagonal band
of Broca nucleus, nucleus basalis and medial septal nucleus), diencephalon (ventrolateral
preoptic, hypothalamic ventrolateral and medial habenular nuclei) and brainstem
(parabigeminal, laterodorsal tegmental and pedunculopontine tegmental nuclei). These
findings are discussed through both a functional and phylogenetic perspective.

35
1. Introduction
Acetylcholine (Ach) was the first neurotransmitter to be found as the neurotransmitter
used by somatic motoneurons and autonomic neurons. It was then identified in several neuronal
clusters in the central nervous system, such as interneurons and large projection neurons (Von
Bohlen und Halbach and Dermietzel, 2006). Neurons that synthesize and release acetylcholine
for neurotransmission are referred to as cholinergic neurons (Oda and Nakanishi, 2000).
Ach is a fast-acting, point-to-point neurotransmitter at the neuromuscular junction and in
the autonomic ganglia. Ach, however, also appears to act as a neuromodulator in the brain,
despite its role as the primary excitatory neurotransmitter at the periphery. In the brain, Ach
modifies neuronal excitability, alters the presynaptic release of other neurotransmitters, and
coordinates the firing of neuronal groups (Picciotto et al., 2012). In this sense, alongside its role
in known motor regulation (Calabresi et al., 2000), the central cholinergic modulation interferes
in functions such as synaptic plasticity (Drever et al., 2011; Giocomo and Hasselmo, 2007;
McKay et al., 2007) and neuronal development (Role and Berg, 1996). Ach also interferes in the
modulation of brain systems such as the mesolimbic dopaminergic system, associated with
addiction and reward (Omeichenko and Sesack, 2006) and cortical activity regulation (Hasselmo
and Sarter, 2011). Cholinergic signaling may also alter hypothalamic functions, such as
thermoregulation (Myers and Waller, 1973), sleep patterns (Steriade, 2004), food intake (Mineur
et al., 2011), pancreatic insulin and glucagon release (Ishikawa et al., 1982). Increasing evidence
suggests that Ach signaling in a number of brain cells is important for stress response (Mark et
al., 1996), and in learning and memory processes (Gais and Born, 2004; Gold, 2003; Hasselmo,
1999; 2006).
In the central nervous system, the Ach, besides its classical presence in the cholinergic
somatic and autonomic motoneurons of the spinal cord and brainstem, is the neurotransmitter
found in some populations of cholinergic interneurons, which may be found in the
prosencephalon and the brainstem, as well as the so-called large cholinergic neurons in the basal
prosencephalon and in the mesopontine tegment, which give rise to large ascending projections.
Among the interneurons, the most representative are those in the striatum. These interneurons
interact with the dopaminergic terminals of neurons that project to the striatum from the
substantia nigra. There also are cholinergic interneurons sparsely distributed in the cerebral
cortex, hippocampus and olfactory bulb. Among projection neurons, the cholinergic groups of
the basal prosencephalon include the medial septal nucleus (Ch1), the horizontal and vertical
limb of the diagonal band nucleus (Ch2-Ch3) and the basal nucleus of Meynert (Ch4), which are

36
responsible for large ascending projections, innervating topographically throughout the cerebral
cortex, including the hippocampus and the amygdala. The mesopontine cholinergic neurons are
divided between a ventrolateral column (cell group Ch6, or the pedunculopontine nucleus), close
to the lateral border of the superior cerebellar peduncle, and a dorsomedial column (cell group
Ch5, or the laterodorsal tegmental nucleus), a component of the periaqueductal gray, located just
rostral to the locus coeruleus. Both these nuclei send an important descending projection to the
pontobulbar reticular formation, vestibular nuclei, locus coeruleus and several raphe nuclei,
besides providing an extensive ascending cholinergic innervation to the thalamus and
hypothalamus. It is believed that these projections have a prominent role in regulating the sleep-
wake cycle. The neurons of the Ch7 group are present in the habenula and project to the
interpeduncular nucleus. Finally, neurons of the Ch8 group are located in the parabigeminal
nucleus and send projections to the superior colliculus (Mesulam et al., 1983a; Von Bohlen und
Halbach & Dermietzel, 2006).
The cholinergic nuclei are delimited by the ChAT or Ach vesicular transporter
immunohistochemistry in the brain or part of it in several species of mammals, such as rat
(Armstrong et al., 1983; Ichikawa et al., 1997; Roghani et al., 1998; Schäfer et al., 1998);
monotremes (Manger et al., 2002), mole rat (Bhagwandin et al., 2008; Da Silva et al., 2006), bats
(Dell et al., 2010; Kruger et al., 2010; Maseko and Manger, 2007; Maseko et al., 2007),
porcupine (Limacher et al., 2008), guinea pig (Motts et al., 2008), rock hyrax (Gravett et al.,
2009), giraffe (Bux et al., 2010), elephant shrew (Pieters et al., 2010), three species of
Afrotherian species (Calvey et al., 2013), Tasmanian devil (Patzke et al., 2014), two species of
Euarchontoglires (Calvey et al., 2015a), five species of insectivore (Calvey et al., 2016), the
Goettingen miniature pig (Mahady et al., 2017), and non-human (Calvey et al., 2015b; Kus et al.,
2003; Satoh and Fibiger, 1985a; 1985b), and human (Oda and Nakanishi, 2000) primates.
Considering the importance of studying the neural systems from a comparative-evolutionary
point of view, it is imperative to extend the study to the greatest number of species, that is why
we chose to undertake this study in the rock cavy.
The rock cavy (Kerodon rupestris) is classified taxonomically as representative of the
phylum Chordata, class Mammalia, superorder Glires, order Rodentia, suborder Hystricomorpha,
family Caviidae and subfamily Caviinae (Silva Neto, 2000). The suborder Hystricomorpha
includes several families with a number of species found in Brazil. These include, besides the
rock cavy, the agouti (family Dasyproctidae), the paca (family Cuniculidae), the capybara
(Hydrochaeridae), all of them used as experimental models by Brazilian researchers (see for
example, Freire et al., 2010; PicançoDiniz et al., 1991; 2011; Rocha FAF et al., 2009; Rocha EG
et al., 2012; Silveira, 1985; Silveira et al., 1989). Phylogenetic studies using a molecular

37
approach have connected the genus Kerodon with the genus Hydrochaeris, which includes the
capybara (family Hydrochaeridae), and is closely related to the genus Dolicotis of the subfamily
Dolicothinae, whose representative in South America is the Patagonian hare (Dolichotis
patagonum) (Rowe and Honeycutt, 2002). The rock cavy inhabits the semiarid Caatinga of the
Brazilian Northeast, although it can be found in the Southeast region as far as the state of Minas
Gerais. Colonies of rock cavies usually live in cracks and crevices of granitic rocks, which serve
as refuge and shelter from predators (Lacher Jr, 1981). This species reaches adulthood at 200
days and can reach up to 50 cm in length and 1 kilogram in body weight (Roberts et al., 1984).
Behavioral studies carried out in the field have reported that this rodent species emerges
to forage throughout both day and night, but most of the activity occurs during the day, with
peaks of activity at dawn and dusk (Carvalho, 1969; Lacher Jr, 1981). In line with these
observations, an investigation performed under controlled laboratory conditions showed that the
rock cavy was active throughout the 24-h day, with peaks during sunrise and sunset, featuring a
predominantly crepuscular behavior (Sousa and Menezes, 2006).
Neuroanatomical studies in the rock cavy were started in our laboratory, when this
species was adopted as a regional rodent model in circadian rhythms studies. Besides the
characterization of the activity rhythm and other circadian responses (Sousa and Menezes, 2006),
the circadian rhythmicity controlling structures – the suprachiasmatic nucleus and the
intergeniculate leaflet – have been identified for their neurochemical content, as well as the
retinal projection (Cavalcante et al., 2008; Nascimento Jr et al., 2010b). Direct retinal projections
to paraventricular (Nascimento Jr et al., 2008) and mediodorsal (Nascimento Jr et al., 2010a)
thalamic nuclei, and caudal zona incerta (Morais et al., 2014) were identified. The study of the
serotonergic system enabled the identification of the raphe nuclei and other extra-raphe nuclei
(Soares et al., 2012) and the distribution of serotonergic terminals on the thalamic midline and
intralaminar nuclei (Silva et al., 2014). The midbrain dopaminergic (Cavalcanti et al., 2014) and
the diencephalon nitrergic (Reis et al., 2018) groups were also outlined. As well, the eye
anatomy and retinal structure (Oliveira et al., 2014) and other retinal specializations (Oliveira et
al., 2018) were described. Studies on targets of retinal projections constituting the primary visual
and accessory optic systems, as well as projections to other hypothalamic targets have been
ongoing. The present study aimed to outline the cholinergic cell groups in the brain of the rock
cavy by ChAT-immunohistochemistry, as well as to describe the neuronal morphology in each
nucleus.

38
2. Materials and methods
Three young adult rock cavies from the countryside municipalities in the state of Rio
Grande do Norte, Brazil, were used. The animals were captured after permission from the
Brazilian Environmental Agency (IBAMA, license SISBIO 42960-1 de 10/03/2014) and housed
in a large room with ceramic tile ceiling and natural soil floor with creeping vegetation and
rocks, simulating the animal´s natural habitat. The animals were exposed to environmental
temperature, air humidity and light, with unlimited access to food and water. All efforts were
made to minimize the number of animals and their suffering, the procedures following strictly
the norms established by the National Research Council of the National Academy published in
the “Guidelines for the Care and Use of Mammals in Neuroscience and Behavioral Research”
and recommended by the Brazilian Society of Neuroscience and Behavior (SBNeC),
http://www.sbnec.gov.br/links. The project was approved by the local Ethics Committee (CEUA-
UFRN, protocol 004/2014).
All experimental procedures were made in the Neuroanatomy Laboratory, Department of
Morphology, UFRN.
Each animal was anesthetized with an intramuscular injection of ketamine hydrochloride
10% (Agener União, Brazil, 1 ml) and xylazine hydrochloride 2% (Agener União, Brazil, 0.1
ml), both per kilogram of animal weight, and perfused using a cannula positioned in the aorta
and connected to a peristaltic pump (Cole-Parmer) with 300 ml of 0.9% saline solution in 0.1M
phosphate buffer, pH 7.4, containing heparin (Parinex, Hipolabor, Sabará, MG, Brazil, 2
ml/1000 of saline solution) for around 5 minutes. Next, 700 ml of a 4% paraformaldehyde in
0.1M phosphate buffer, pH 7.4 fixative solution was pumped, half of the solution with a 70
ml/minute flow rate and the other half 17,5 ml/minute, the entire procedure lasting 30 minutes,
After perfusion, the animals were placed in a stereotaxic frame and the incisor bar was
adjusted until the lambda and bregma were at the same height. The skull bones were removed to
expose the dorsal surface of the encephalon, which was sectioned into 3 blocks by means of two
coronal sections: one at the bregma level and the other at the lambda level. Finally, the
encephalon was removed from the skull, stored in 30% sucrose solution in 0.1M phosphate
buffer, pH 7.4, for 24 to 48 hours, and then sectioned by dry ice freezing in sliding microtome,
obtaining coronal sections of 30 µm. The sections were grouped sequentially into 6
compartments, each containing one of every 6 sections, thereby representing a serial sequence
with an interval of 180 µm between one section and the next one. Sections from one series were
immediately mounted on silanized glass slides and Nissl stained with thionin, to visualize the
cytoarchitectonic delimitation of neuronal groups. Sections from another series were submitted

39
to immunohistochemistry to reveal Ach, using the antibody against ChAT, the synthesizing
enzyme. The sections, previously submitted to the pretreatment with hydrogen peroxide (H2O2),
were placed in contact with goat anti-ChAT polyclonal antibody (Millipore, cat #AB144P, Lot
#NG1780580) at 1:1000 dilution and 2% normal rabbit serum in 0.4% Triton X-100 for around
18 h, in a rotator. The sections were then immersed in rabbit biotinylated anti-goat secondary
antibody (Jackson Immunoresearch Labs, West Grove, PA), diluted at 1:1000 in the same
vehicle as above, in a rotator, for 90 minutes. To visualize the reaction, the sections were put in
contact with the avidin-biotin-HRP complex (Vector Elite ABC kit). This was followed by the
final reaction in a medium containing H2O2 as substrate and tetrahydrochloride
diaminobenzidine (DAB) as chromogen. At the beginning, between one step and another and at
the end, the sections were thoroughly washed with 0.1M phosphate buffer, pH 7.4. The sections
were mounted on silanized glass slides, which, after drying at room temperature, were
submersed rapidly in a 0.05% osmium tetroxide solution to intensify the reaction, then
dehydrated in an increasing concentrations ethanol series, cleared with xylene and cover slipped
with ERV-mount®. All the immunohistochemical procedures were performed at room
temperature. As control for staining specificity, some sections were submitted to
immunohistochemical reactions omitting the primary antibody. In these cases, no ChAT-
immunoreactivity was obtained. The sections of the remaining compartments were stored in an
antifreeze solution for eventual repetitions of immunohistochemical reaction to ChAT or other
substances.
Nissl- and ChAT-immuno-stained sections were examined under a light microscope
(Nikon Eclipse NI) on a bright field. Digital images were obtained from representative sections
using a digital videocamera (Nikon DS Ri1), attached to a light microscope, using the software
NISElements AR to capture the images. The digitized images were converted to gray scale,
corrected minimally for brightness and contrast, and mounted using Adobe Photoshop CC2017
software (Adobe Systems, Mountain View, CA, USA). Diagrams were obtained from images of
Nissl-stained coronal sections using Canvas version 12 software (ACD Systems of America,
Inc.). The corresponding immunostained sections were then matched to the drawings and the
immunopositive neurons marked.
3. Results
ChAT-immunoreactive (ChAT-IR) neurons forming groups were found in the
telencephalon, diencephalon and brainstem (midbrain-pons). This analysis did not deal with
those cholinergic neurons found in cranial nerve nuclei present in the brainstem, whose

40
axons exit from the peripheral nervous system, entering the composition of the respective
cranial nerves.
INSERT FIGURE 1 HERE
Telencephalic groups
Nucleus accumbens. The nucleus accumbens (Acb) locates ventromedially to the
caudate-putamen nucleus (CPu), extended from the anterior end of the anterior horn of the
lateral ventricle, rostrally, to the anterior commissure. ChAT-IR neurons were found
sparsely but evenly distributed throughout the Acb in both portions, core and shell, without
distinction (Figs. 1A-C and 2A) and exhibited predominantly a fusiform shape with the
presence of some triangular shaped neurons (Fig. 2B).
Caudate-putamen nucleus. The CPu is located laterally to the lateral ventricle,
extended approximately from the corpus callosum to the middle level of the habenular
nuclei. ChAT-IR neurons were found sparsely distributed, albeit evenly, throughout this
nucleus (Figs. 1A-I and 2C) and exhibited predominantly triangular and rounded shape with
the presence of some fusiform shaped neurons (Fig. 2D).
Olfactory tubercle and Islands of Calleja. These nuclei are located ventrally to the
Acb, extending from the level of the anterior horn of the lateral ventricle to the level of the
anterior commissure. ChAT-IR neurons were seen forming clusters in the regions of the
Islands of Calleja (ICj) and granule cell layer of the olfactory tubercle (Tu) (Figs. 1A-D and
2E) and exhibited predominantly triangular shaped neurons (Fig. 2F).
INSERT FIGURE 2 HERE
Diagonal band nucleus. The diagonal band nucleus (DB) resides at the medial and
ventral contour of the telencephalon at rostral levels. ChAT-IR neurons were seen in both
the horizontal (HDB) (Figs. 1C and 3A) and the vertical (VDB) (Fig. 1C and 3C) portions
of that nucleus, forming a continuum. These neurons exhibited triangular and rounded shape
in the HDB (Fig. 3B) and fusiform in the VDB (Fig. 3D).
Medial septal nucleus. The medial septal nucleus (MS) is the medialest nucleus of
the septal complex, located in the medial wall of the cerebral hemisphere. ChAT-IR neurons
were seen filling that nucleus almost completely (Figs. 1C-F and 3E). These were fusiform
or triangular shaped (Fig. 3F).

41
INSERT FIGURE 3 HERE
Ventral globus pallidus. The ventral globus pallidus (VP) is a neuronal cluster
located ventrally to Acb and anterior comissure, which contains widely distributed ChAT-
IR neurons (Fig. 1A-F).
Globus pallidus. The globus pallidus (GP) is located ventromedially to the CPu. It
starts rostrally together with the posterior part of the anterior comissure and extends to the
end of the habenular nuclei and is characterized by poor dye afinity. ChAT-IR neurons were
seen in this nucleus, mainly concentrated in its lateral contour, bordering the CPu (Figs. 1E-
H and 4A). These neurons were characterized by their triangular, fusiform or rounded shape
(Fig. 4B).
Entopeduncular nucleus. The entopeduncular nucleus (EP) is a cluster of neurons
included in the ventromedial end of the internal capsule and is considered equivalent to the
inner segment of the globus pallidus of primates in carnivores and rodents. This nucleus was
seen to contain sparse ChAT-IR neurons (Figs 1H-I and 4C), which exhibited elongated
format (Fig. 4D).
Basal nucleus. The basal nucleus (B) is found in the vicinity of the VP. This nucleus
is strongly labeled by the presence of ChAT-IR neurons (Figs. 1E-F and 4E), exhibiting
triangular, rounded or fusiform shape (Fig. 4F).
INSERT FIGURE 4 HERE
Diencephalic groups
Ventrolateral preoptic nucleus. The ventrolateral preoptic nucleus (VLPO) is a small
cell cluster in the region of the lateral preoptic nucleus, located dorsally to the supraoptic
nucleus. ChAT-IR neurons were seen in this locus (Figs. 1F and 5A), which exhibited
triangular or rounded format (Fig. 5B).
Ventrolateral hypothalamic nucleus. The ventrolateral hypothalamic nucleus (VLH)
is a small cell cluster in the region of the lateral hypothalamus, located dorsally to the
supraoptic nucleus, caudally to the VLPO, where ChAT-IR neurons were detected (Figs. 1G
and 5C). These neurons were triangular or rounded shaped (Fig. 5D).
Medial habenular nucleus. The medial habenular nucleus (MHb) is the medial
subdivision of the habenula, which is part of the epithalamus. It is found in the dorsal part of
the diencephalon, on the floor of the third ventricle. The MHb is strongly labeled by the

42
massive presence of the ChAT-IR neurons, mainly in its ventral part (Figs. 1I-J and 5E).
These neurons were characterized by their fusiform shape (Fig. 5F).
INSERT FIGURE 5 HERE
Pontomesencephalic nuclei
Parabigeminal nucleus. The parabigeminal nucleus (PBG) is a small oval-shaped
neuronal collection located in the pontomesencephalic region, in the lateral aspect of the
tegmentum, ventrally to the inferior colliculus. This nucleus is strongly labeled by the
massive presence of ChAT-IR neurons (Figs. 1K-M and 6A). These neurons exhibited
predominantly fusiform shape, whith dendritic expansions along the largest axis of the
nucleus, although some rounded neurons were also present (Fig. 6B).
Laterodorsal tegmental nucleus. The laterodorsal tegmental nucleus (LDTg) is
incrusted in the dorsal portion of the periaqueductal gray, laterally to the dorsal nucleus of
raphe. This nucleus is characterized by a considerable presence of ChAT-IR neurons (Figs.
1L-M and 6C), which exhibited triangular or rounded shape (Fig. 6D).
Pedunculopontine tegmental nucleus. The pedunculopontine tegmental nucleus
(PTg) is found in the pontomesencephalic tegmentum, laterally to the superior cerebellar
peduncle. A large number of ChAT-IR neurons is present in this nucleus (Figs. 1L-M and
6C), exhibiting fusiform or triangular shape (Fig. 6E).
INSERT FIGURE 6 HERE
4. Discussion
According to our results, ChAT-IR neurons forming groups were found in the
telencephalon, diencephalon and brainstem (midbrain-pons) of the rock cavy brain. The
results revealed that the distribution of the cholinergic neurons is similar to what has
previously been described in other mammals, with some minor differences.
Telencephalic groups
As in our study, the presence of cholinergic neurons in the striatal complex
comprising nucleus accumbens, ventral pallidum, caudate-putamen and globus pallidus, as
well as the olfactory tubercle and Islands of Calleja, the diagonal band nucleus, the medial
septal nucleus and the basal nucleus was reported in all studied species, such as rat
(Armstrong et al., 1983; Ichikawa et al., 1997; Roghani et al., 1998), monotremes (Manger

43
et al., 2002), mole rat (Bhagwandin et al., 2008), bats (Dell et al., 2010; Kruger et al., 2010;
Maseko and Manger, 2007; Maseko et al., 2007), Cape porcupine (Limacher et al., 2008),
rock hyrax (Gravett et al., 2009), elephant shrew (Pieters et al., 2010), Afrotherian species-
Potomogale velox, Amblysomus hottetotus and Petrodromus tetradactylus (Calvey et al.,
2013), Tasmanian devil (Patzke et al., 2014), a lagomorph Lepus capensis and a Scadentia
Tupaia belangeri (Calvey et al., 2015a), insectivore Crocidura cyanea, Crocidura olivieri,
Sylvisorex ollula, Paraechinus aethiopicus and Atelerix frontalis (Calvey et al., 2016),
Goetting miniature pig (Mahady et al., 2017), nonhuman (Calvey et al., 2015b; Kus et al.,
2003; Satoh and Fibiger, 1985a; 1985b) and human (Oda and Nakanishi, 2000) primates.
We also detected a hitherto not reported presence of cholinergic neurons in the
entopeduncular nucleus
In a classical conception, in primates, the principal components of the basal ganglia
are the striatum, represented by the caudate and putamen nuclei, the pallidum, that includes
the outer and inner segments. In all primates and many non-primate mammals the striatum
is subdivided by a plate-like internal capsule into two districts, the dorsomedial caudate
nucleus and the ventrolateral putamen. In other mammalian species, including the rat, the
anterior part of the internal capsule passes to the striatum in the form of a brush rather than a
plate; in such species the striatum lacks clear subdivision and is therefore referred to as
caudate-putamen. Furthermore, in the most of non-primate species, the entopeduncular
nucleus, a cell cluster incrusted in the ventral end of the internal capsule, is the equivalent to
the primate inner segment of the globus pallidus (Parent and Hazrati, 1995). Hodological
findings in the rat suggested that the striatum is subdivided into a ventromedial, limbic
system-connected region, and a dorsolateral, “nonlimbic” region largely connected with
motor cortical areas. It is suggested that the limbicconnected striatal sector acts as an
interface between the motivational and the more strictly motor aspects of movement (Nauta
and Domesick, 1984). Acb and VP compose the ventral striatum (Nauta and Domesick,
1984), which has also been implicated as the critical neuroanatomic substrate for the
anticipation of rewards in mammals. Acb is a crucial component of the so-called reward
circuit, receiving dopaminergic inputs from the ventral tegmental area, signaling predictive
reward (Newman and Winans, 1980a; Rodriguez-López et al., 2017; Schultz, 1998), and
projecting through medium spiny neurons to regions that modulate motor output, such as the
VP and the substantia nigra (Heimer et al., 1991; Nauta and Domesick, 1984; Newman and
Winans, 1980a). Acb also receives glutamatergic inputs from cortical areas related to
learning and memory processes (Mc George and Faull, 1989). Several studies have
confirmed the participation of Acb cholinergic interneurons, and consequently of the Ach

44
and its receptors in several of these processes, such as modulation of appetitive learning,
performance, and motivation for food (Pratt and Blackstone, 2009; Pratt and Kelley 2004).
One study showed that Acb cholinergic neuronal activity regulates depressive behavior,
reinforcing that the activity of Acb cholinergic interneurons is fundamental for the
regulation of mood and motivation (WarnerSchmidt et al., 2012).
CP and GP compose the dorsal striatum (Nauta and Domesick, 1984). ChAT
neurons in the dorsal striatum are proven interneurons, receiving cortical glutamatergic and
substantia nigra dopaminergic inputs, connecting to their own projecting neurons (Woolf,
1991). These ChAT interneurons represent the main source of Ach in the striatum. Ach-
mediated neurotransmission plays a crucial role in the control of voluntary movements
exerted by the striatum, having been implicated in the pathogenesis of the Parkinson
disease. Thus, it has been proposed that an underlying defect in this disorder would be a
functional imbalance between dopaminergic and cholinergic systems subsequent to the loss
of dopaminergic neurons (Barbeau, 1962; Calabresi et al., 2000).
Tu has been considered part of the olfactory cortex, receiving fibers of the olfactory
bulb by the lateral olfactory tract (Newman and Winans, 1980b; Scott et al., 1980).
However, it has been suggested that the most of the Tu of the rat is an extension of the
ventral striatum, based on cytological, histochemical, immunohistochemical and
hodological evidence, considering similarities with Acb (Newman and Winans, 1980b;
Switzer et al., 1982). Furthermore, experimental research has implicated Tu in reward
actions of cocaine, morphine and the rewarding effects of brain stimulation (Kornetsky et
al., 1991). ICj are clusters of granule cells located relative to Tu and neighborhoods of
septum and Acb. Connectional studies have shown that these cells are connected with
olfactory and non-olfactory components of the basal forebrain and are afferented by
dopaminergic neurons of the ventral tegmental area and medial substantia nigra (Fallon,
1983; Fallon et al., 1978). Cytoarchitectonic, hodological, histochemical and
immunohistochemical studies also situate ICj into the striatopallidal system (Fallon et al.,
1983; Meyer et al., 1989; Millhouse, 1987).
The cell bodies of the basal forebrain cholinergic neurons are interspersed with
noncholinergic neurons and distributed in a series of nuclei, including the DB, with vertical
and horizontal portions, the MS and the B (Woolf, 1991), which collectively serve as the
major sources of cholinergic projection neurons to neocortex, hippocampus, and amygdala
(Mesulam et al., 1983b). Cholinergic signaling from these neurons appears to provide
important control over circuit dynamics underlying cognitive processing, with their deficit

45
linked to cognitive decline, and subsequent neurodegenerative diseases, such as Alzheimer
and Parkinson diseases (Ballinger et al., 2016).
Diencephalic groups
Cholinergic neurons have been detected forming dorsal, lateral and ventral
hypothalamic clusters in several species (Bhagwandin et al., 2008; Calvey et al., 2013;
2015a; 2015b; 2016; Dell et al., 2010; Gravett et al., 2009; Kruger et al., 2010; Limacher et
al., 2008; Maseko and Manger, 2007; Maseko et al., 2007; Patzke et al., 2014; Pieters et al.,
2010; Satoh and Fibiger, 1985a). Cholinergic neurons were also reported in the arcuate
nucleus in other species (Jeong et al. 2016; Mahady et al., 2017). In the rock cavy (present
study) cholinergic neurons were not visualized in these locations, but we have detected a not
hitherto reported presence of cholinergic neuronal soma in the VLPO and VLH. The VLPO,
identified in many species, is considered a critical component of the sleep circuitry (Saper et
al., 2005; Szymusiak et al., 1998; 2007). However, the functional significance of these
cholinergic neurons is unknown.
The MHb have been described as marked by the presence of cholinergic neurons in
most of the mammalian species studied (Bhagwandin et al., 2008; Calvey et al., 2013;
2015a; 2015b; 2016; Dell et al., 2010; Gravett et al., 2009; Kruger et al., 2010; Kus et al.,
2003; Limacher et al., 2008; Mahady et al., 2017; Manger et al., 2002; Maseko and Manger,
2007; Maseko et al., 2007; Patzke et al., 2014; Pieters et al., 2010; Roghani et al., 1998). In
mammals the habenular complex is composed of two separate nuclei, one on each side, a
medial nucleus, and a lateral one, in turn subdivided into a lateral and a medial portion
(Andres, 1999; Paxinos and Watson, 2007). Each of these subdivisions has different
anatomic connections, serving different functions (Herkenham and Nauta, 1977; 1979).
Thus, the cholinergic MHb receives input from the septum via stria medularis and its
efferents reach the single midline interpeduncular nucleus via the habenulo-interpeduncular
tract, or fasciculus retroflexus. The MHb is a key link between the limbic forebrain and the
midbrain, composing the so-called dorsal diencephalic conduction system (Sutherland,
1982). Experimental evidence suggests that, besides Ach, this pathway also utilizes
substance P and glutamate as neurotransmitter (Qin and Luo, 2009). Functionally, this
system has been implicated in the control of emotional behaviors including anxiety and fear
responses (Yamaguchi et al., 2013). More recent studies have evidenced a link between the
habenulo-interpeduncular pathway in the context of nicotine aversion and withdrawal
(AntolinFontes et al., 2015; Pang et al., 2016) and psychiatric disorders, such as mood

46
disorders, schizophrenia, and substance use disorder (Fakhoury, 2017), as well as cocaine,
methamphetamine and alcohol addiction (Viswanath et al., 2014).
Pontomesencephalic nuclei
In line with several studies in many mammalian species (Bhagwandin et al., 2008;
Bux et al., 2010; Calvey et al., 2013; 2015a; 2015b; 2016; Da Silva et al., 2006; Dell et al.,
2010; Gravett et al., 2009; Kruger et al., 2010; Kus et al., 2003; Limacher et al., 2008;
Mahady et al., 2017; Manger et al., 2002; Maseko and Manger, 2007; Maseko et al., 2007;
Motts et al., 2008; Oda and Nakanishi, 2000; Patzke et al., 2014; Pieters et al., 2010;
Roghani et al., 1998), the rock cavy PBG is heavily marked by the presence of Ach-
producing neurons. The PBG is strongly interconnected with the superior colliculus, of
which it has been characterized as a satellite (Graybiel, 1978; Henderson, 1989; Mufson et
al., 1986). In addition, the PBG also projects to other brain structures such as the dorsal
lateral geniculate nucleus (Harting et al., 1986; Hashikawa et al., 1986) and the amygdala
(Usunoff et al., 2007). Recording of the activity of PBG neurons shows that they fire during
saccadic movements directed at moving targets but not at spontaneous saccadic movements,
suggesting that PBG is an integral part of the mesencephalic circuit that generates target
location information (Cui and Malpeli, 2003). It has also been shown that PBG neurons in
vitro trigger spontaneous action potentials, even in the presence of synaptic blockers,
indicating the existence of spontaneous activity (Godard et al., 2007).
We detected Ach-producing neurons in two cell clusters in the rock cavy pontine
tegmentum, the LDTg and PTg, corroborating findings in several mammalian species
studied (Armstrong et al., 1983; Bhagwandin et al., 2008; Bux et al., 2010; Calvey et al.,
2013; 2015a; 2015b; 2016; Da Silva et al., 2006; Dell et al., 2010; Gravett et al., 2009;
Kruger et al., 2010; Kus et al., 2003; Limacher et al., 2008; Mahady et al., 2017; Manger et
al., 2002; Maseko and Manger, 2007; Maseko et al., 2007; Motts et al., 2008; Oda and
Nakanishi, 2000; Patzke et al., 2014; Pieters et al., 2010; Roghani et al., 1998; Satoh and
Fibiger, 1985a; 1985b).
Based on connectional data, the LDTg appears to be a highly differentiated part of
the ascending reticular activating system, concerned not only with specific cortical and
thalamic regions, especially those associated with the limbic system, but also with the basal
ganglia, and oculomotor visual mechanisms (Cornwall et al., 1990). According to the
pattern of their firing rates, differentiated neurons in the LDTg are related to diverse phases
of the sleep-wake cycle (Kayama et al., 1992). Part of the hypothalamus-LDTg projections
was shown to be from hypocretin containing-neurons (Peyron et al., 1998), which plays a

47
role in both the promotion of wakefulness and the suppression of active sleep (Xi et al.,
2001). Furthermore, a lesion study showed that the LDTg cholinergic cell loss does not
significantly alter the rewarding effects of cocaine, but LDTg lesion can reduce the rat´s
responsiveness to cocaine-predictive stimuli (Steidl et al., 2015).
Classically, the PTg has been identified by its population of cholinergic neurons, the
labeling of which is still the best way to define it. This neuronal population is constituted
predominantly of projection neurons with ascending and descending trajectories. The
ascending projections follow both a dorsal stream and a ventral stream. The ascending
dorsal stream innervates the superior and inferior colliculi and parafascicular, reticular and
anterior thalamic nuclei, also reaching all major motor, sensory, associative and limbic
thalamic nuclei. The ascending ventral stream innervates some parts of the basal ganglia in
the midbrain, substantia nigra pars compacta and pars reticulata, ventral tegmental area,
subthlamic nucleus, lateral hypothalamus, ventral pallidal area, amygdala and medial and
lateral septum (Garcia-Rill, 1991; Inglis and Winn, 1995; Mena-Segovia and Bolam, 2017).
The PTg as well as the LDTg also contain other cell groups, including glutamatergic and
GABAergic neurons, but their role remains to be better clarified (Wang and Morales, 2009).
Like the LDTg, the PTg has been traditionally associated with the functions of the reticular
activating system, because of its ascending cholinergic projections to the thalamus that
modulate cortical activation (Garcia-Rill et al., 2013; Steriade, 2004) and with the
functions of the mesencephalic locomotor region through the modulation of goal-directed
locomotion (Mena Segovia, 2016). Furthermore, regarding connections with the basal
ganglia, the PTg is in a position to influence activity in the whole of the basal ganglia,
considering that their interconnections form complex feedback and feedforward circuits,
allowing opposing effects on basal ganglia output nuclei, suggesting that the PTg should be
considered as part of the basal ganglia (Mena-Segovia et al., 2004). Thus, through its
connections with basal ganglia structures and the spinal cord, the PTg has been conceived as
a locomotor center that also processes sensory and behavioral information. Thus, its
dysfunction may be associated with movement disorders, such as those found in Parkinson
disease (Hamani et al., 2007; Lee et al., 2000; Pahapill and Lozano, 2000; Wen et al., 2015;
Winn, 2008). It has also been reported that PTg lesions blocked the locomotor excitation
and the conditioned hyperactivity produced by amphetamine, as well as the conditioned
increase in locomotion, but not the catalepsy, produced by morphine. These data suggest
that the neural circuits mediating the acute rewarding effects of drug stimuli acting at
forebrain sites exit the limbic system in the PTg region of the brainstem, where motivation
gain access to motor systems that initiate approach and exploration (Bechara and Van Der

48
Kooy, 1992). Also, bilateral lesions of the PTg double dissociate the rewarding and aversive
motivational effects of nicotine and blocked a nicotine reward signal and reveal the aversive
motivational properties of nicotine injected into the ventral tegmental area (Laviolette et al.,
2002). However, PTg lesions do not block cocaine reward (Parker and Van Der Kooy,
1995).
In summary, the present findings indicate that, although there are some differences,
the overall distribution and morphological characteristics of cholinergic neurons as
identified in the rock cavy are similar to those previously found in other species. It is worth
mentioning that, at the telencephalic level, the presence of ChAT-immunopositive neurons
in the nucleus accumbens, caudate-putamen, ventral pallidum, globus pallidum, diagonal
band nucleus, medial septal nucleus, olfactory tubercle/Islands of Calleja and basal nucleus
appears to be a constant in all species. Exception is made to our recording in the rock cavy
of the presence of ChATimmunopositive neurons in the entopeduncular nucleus, not
mentioned in any other report. In the diencephalon, the medial habenular nucleus with a
significant population of cholinergic neurons also appears to be a constant. However, the
descriptions are discrepant, as for the cholinergic neurons in the hypothalamus, with a rather
different location in the rock cavy, in relation to other descriptions in other species, and
even to their absence, as is the case of the monotremes. Several authors have also referred
to the presence of cholinergic neurons in the thalamic nuclei of some species, which we did
not find in our material. At the midbrain-pons level, for the rock cavy, we found cholinergic
neurons in the parabigeminal, laterodorsal tegmental and pedunculopontine tegmental
nuclei. The presence of these neurons in the laterodorsal tegmental and pedunculopontine
tegmental nuclei also appears to be constant; however, in some species studied, its absence
is related in the parabigeminal nucleus, as previously described. Thus, it can be concluded
that, for the most part, the cholinergic system is largely maintained in the mammal species.
Acknowledgments
We would like to thank Miriam Regina Celi de Escala Oliveira for expert technical assistance.
The English text of this paper has been revised by Sidney Pratt, Canadian, MAT (The Johns
Hopkins University), RSAdip - TESL (Cambridge University). This study was supported by the
National Council for Scientific and Technological Development (CNPq), Coordination for High
Level Staff Improvement (CAPES), Rio Grande do Norte Research Support Foundation
(FAPERN) and Research and Projects Financing (FINEP), Brazil grants.

49
References
Andres, K.H., Von During, M., Veh, R.W., 1999. Subnuclear organization of the rat habenular
complexes. J. Comp. Neurol. 407, 130-150.
Antolin-Fontes, B., Ables, J., Görlich, A., Ibañes-Tallon, I., 2015. The habenulo-interpeduncular
pathway in nicotine aversion and withdrawal. Neuropharmacology 96, 213-222.
Armstrong, D.M., Saper, C.B., Levey, A.I., Wainer, B.H., Terry, R.D., 1983. Distribution of
cholinergic neurons in rat brain: demonstrated by the immunocytochemical localization of
cholineacetyltransferase. J. Comp. Neurol. 216, 53-68.
Ballinger, E.C., Ananth, M., Talmage, D.A., Role, L.W., 2016. Basal forebrain cholinergic
circuits and signaling in cognition and cognitive decline. Neuron 91, 1199-1218.
Barbeau, A., 1962. The pathogenesis of Parkinson´s disease: a new hypothesis. Can. Med.
Assoc. J. 87, 802-807.
Bechara, A., Van Der Kooy, D., 1992. Lesions of the tegmental pedunculopontine nucleus:
effects on the locomotor activity induced by morphine and amphetamine. Pharmacol. Biochem.
Behav. 42, 9-18.
Bhagwandin, A., Fuxe, K., Bennet, N.C., Manger, P.R., 2008. Nuclear organization and
morphology of cholinergic, putative catecholaminergic and serotonergic neurons in the brains of
two species of African mole-rat. J. Chem. Neuroanat. 35, 371-387.
Bux, F., Bhagwandin, A., Fuxe, K., Manger, P.R., 2010. Organization of cholinergic, putative
catecholaminergic and serotonergic nuclei in the diencephalon, midbrain and pons of sub-adult
male giraffes. J. Chem. Neuroanat. 39, 189-203.
Calabresi, P., Centonze, D., Gubellini, P., Pisani, A., Bernardi, G., 2000. Acetylcholine-mediated
modulation of striatal function. Trends Neurosci. 23, 120-126.
Calvey, T., Patzke, N., Kaswera, C., Gilissen, E., Bennet, N.C., Manger, P.R., 2013. Nuclear
organisation of some immunohistochemically identifiable neural systems in three Afroterian
speciesPotomagale velox, Amblysomus hottentotus and Petrodomus tetradactylus. J. Chem.
Neuroanat. 5051, 48-65.
Calvey, T., Alagaili, A.N., Bertelsen, M.F., Bhagwandin, A., Pettigrew, J.D., Manger, P.R.,
2015a. Nuclear organization of some immunohistochemically identifiable neural systems in two
species of the Euarchontoglires: A Lagomorph, Lepus capensis, and a Scandentia, Tupaia
belangeri. J. Chem. Neuroanat. 70, 1-19.
Calvey, T., Patzke, N., Kaswera-Kyamakya, C., Gilissen, E., Bertelsen, M.F., Pettigrew, J.D.,
Manger, P.R., 2015b. Organization of cholinergic, catecholaminergic, serotonergic and

50
orexinergic nuclei in three strepsirrhine primates: Galago demidoff, Perodicticus potto and
Lemur catta. J. Chem. Neuroanat. 70, 42-57.
Calvey, T., Patzke, N., Bennett, N.C., Consolate, K-K., Gilissen, E., Alagaili, A.N., Mohammed,
O.B., Pettigrew, J.D., Manger, P.R., 2016. Nuclear organisation of some immunohistochemically
identifiable neural systems in five species of insectivore-Crocidura cyanea, Crocidura olivieri,
Sylvisorex ollula, Paraechinus aethiopicus and Atelerix frontalis. J. Chem. Neuroanat. 72, 34-52.
Carvalho, J.C.M. 1969. Notas de viagem de um zoólogo à região das caatingas e áreas limítrofes.
Fortaleza, Imprensa universitária do Ceará.
Cavalcante, J.S., Britto, L.R.G., Toledo, C.A.B., Nascimento Jr, E.S., Lima, R.R.M., Pontes,
A.L.B., Costa, M.S.M.O., 2008. Calcium-binding proteins in the circadian centers of the
common marmoset (Callithrix jacchus) and the rock cavy (Kerodon rupestris) brains. Brain Res.
Bull. 76, 354-360.
Cavalcanti, J.R.L.P., Soares, J.G., Oliveira, F.G., Guzen, F.P., Pontes, A.L.B., Sousa, T.B.,
Cavalcante, J.S., Nascimento Jr, E.S., Cavalcante, J.C., Costa, M.S.M.O., 2014. A
cytoarchitectonic and TH-immunohistochemistry characterization of the dopamine cell groups in
the substantia nigra, ventral tegmental area and retrorubral field in the rock cavy (Kerodon
rupestris). J. Chem. Neuroanat. 55, 58-66.
Cornwall, J., Cooper, J.D., Phillipson, O.T., 1990. Afferent and efferent connections of the
laterodorsal tegmental nucleus in the rat. Brain Res. Bull. 25, 271-284. Cui, H., Malpeli, J.G.,
2003. Activity in the parabigeminal nucleus during eye movements directed at moving and
stationary targets. J. Neurophysiol. 89, 3128-3142.
Da Silva, J.N., Fuxe, K., Manger, P.R., 2006. Nuclear parcellation of certain
immunohistochemically identifiable neuronal systems in the midbrain and pons of the Highveld
mole rat (Cryptomys hottentotus). J. Chem. Neuroanat. 31, 37-50.
Dell, L.A., Kruger, J.L., Bhagwandin, A., Jillani, N.E., Pettigrew, J.D., Manger, P.R., 2010.
Nuclear organization of cholinergic, putative catecholaminergic and serotonergic systems in the
brains of two megachiropteran species. J. Chem. Neuroanat. 40, 177-195.
Drever, B.D., Riedel, G., Platt, B., 2011. The cholinergic system and hippocampal plasticity.
Behav. Brain Res. 221, 505-514. Fakhoury, M., 2017. The habenula in psychiatric disorders:
More than three decades of translational investigation. Neurosci. Biobehav. Rev. In press.
Fallon, J.H., 1983. The Islands of Calleja complex of rat basal forebrain II: Connections of
médium and large sized cells. Brain Res. Bull. 10, 775-793.
Fallon, J.H., Loughlin, S.E., Ribak, C.E., 1983. The Islands of Calleja complex of rat basal
forebrain. III. Histochemical evidence for a striatopallidal system. J. Comp. Neurol. 218, 91-120.

51
Fallon, J.H., Riley, J.N., Sipe, J.C., Moore, R.Y., 1978. The Islands of Calleja: Organization and
connections. J. Comp. Neurol. 181, 375-396.
Freire, M.A.M., Rocha, E.G., Oliveira, J.L.F., Guimarães, J.S., Silveira, L.C.L., Elston, G.N.,
Pereira, A., Picanço-Diniz, C.W., 2010. Morphological variability of NADPH diaphorase
neurons across areas V1, V2, and V3 of the common agouti. Brain Res. 1318, 52-63. Gais, S.,
Born, J., 2004. Low acetylcholine during slow-wave sleep is critical for declarative memory
consolidation. PNAS, 101, 2140-2144.
Garcia-Rill, E., 1991. The pedunculopontine nucleus. Prog. Neurobiol. 36, 363-389.
Garcia-Rill, E., Kezunovic, N., Hyde, J., Simon, C., Beck, P., Urbano, F.J., 2013. Coherence and
frequency in the reticular activating system (RAS). Sleep Med. Rev. 17, 227-238.
Giocomo, L.M., Hasselmo, M.E., 2007. Neuromodulation by glutamate and acetylcholine can
change circuit dynamics by regulating the relative influence of afferent input and excitatory
feedback. Mol. Neurobiol. 36, 184-200.
Goddard, C.A., Knudsen, E.I., Huguenard, J.R., 2007. Intrinsic excitability of cholinergic
neurons in the rat parabigeminal nucleus. J. Neurophysiol. 98, 3486-3493. Gold, P.E., 2003.
Acetylcholine modulation of neural systems involved in learning and memory. Neurobiol. Learn.
Mem. 80, 194-210.
Gravett, N., Bhagwandin, A., Fuxe, K., Manger, P.R., 2009. Nuclear organization and
morphology of cholinergic, putative catecholaminergic and serotonergic neurons in the brain of
the rock hyrax, Procavia capensis. J. Chem. Neuroanat. 38, 57-74.
Graybiel, A.M., 1978. A satellite system of the superior colliculus: the parabigeminal nucleus
and its projections to the superficial collicular layers. Brain Res. 145, 365-374.
Hamani, C., Stone, S., Laxton, A., Lozano, A.M., 2007. The pedunculopontine nucleus and
movement disorders: Anatomy and the role for deep brain stimulation. Parkinsonism and Related
Disorders 13, S276-S280.
Harting, J.K., Hashikawa, T., Van Lieshout, D., 1986. Laminar distribution of tectal,
parabigeminal and pretectal inputs to the primate dorsal lateral geniculate nucleus: connectional
studies in Galago crassicaudatus. Brain Res. 366, 358-363.
Hashikawa, T., Van Lieshout, D., Harting, J.K., 1986. Projections from the parabigeminal
nucleus to the dorsal lateral geniculate nucleus in the tree shrew Tupaia glis. J. Comp. Neurol.
246, 382-394.
Hasselmo, M.E., 1999. Neuromodulation: acetylcholine and memory consolidation. Trends Cogn
Sci. 3, 351-359. Hasselmo, M.E., 2006. The role of acetylcholine in learning and memory. Curr.
Opin. Neurobiol. 16, 710-715.

52
Hasselmo, M.E., Sarter, M. 2011. Modes and models of forebrain cholinergic neuromodulation
of cognition. Neuropsychopharmacology 36, 52-73.
Heimer, L., Zahm, D.S., Churchill, L., Kalivas, P.W., Wohltmann, C., 1991. Specificity in the
projection patterns of accumbal core and shell in the rat. Neuroscience 41, 89-125.
Henderson, Z., 1989. The cholinergic input to the superficial layers of the superior colliculus: an
ultrastructural immunocytochemical study in the ferret. Brain Res. 476, 149-153.
Herkenham, M., Nauta, W.J.H., 1977. Afferent connections of the habenular nuclei in the rat. A
horseradish peroxidase study, with a note on the fiber-of-passage problem. J. Comp. Neurol. 173,
123146.
Herkenham, M., Nauta, W.J.H., 1979. Efferent connections of the habenular nuclei in the rat. J.
Comp. Neurol. 187, 19-48.
Ichikawa, T., Ajiki, K., Matsura, J., Misawa, H., 1997. Localization of two cholinergic markers,
choline acetyltransferase transporters in the central nervous system of the rat: in situ
hybridization histochemistry and immunohistochemistry. J. Chem. Neuroanat. 13, 23-29. Inglis,
W.L., Winn, P., 1995. The pedunculopontine tegmental nucleus: where the striatum meets the
reticular formation. Prog. Neurobiol. 47, 1-29.
Ishikawa, K., Suzuki, M., Shimazu, T., 1982. Effects of acetylcholine injection into the
hypothalamus on the insulin and glucagon release. Neuroendocrinology 34, 310-314.
Jeong, J.H., Woo, Y.J., Chua Jr, S.C., Jo, Y. H., 2016. Single-cell gene expression analysis of
cholinergic neurons in the arcuate nucleus of the hypothalamus. PlosOne DOI:
10.1371/journal.pone.0162839, 1-14.
Kayama, Y., Ohta, M., Jodo, E., 1992. Firing of “possibly” cholinergic neurons in the rat
laterodorsal tegmental nucleus during sleep and wakefulness. Brain Res, 569, 210-220.
Kornetsky, C., Huston-Lyons, D., Porrino, L.J., 1991. The role of the olfactory tubercle in the
effects of cocaine, morphine and brain-stimulation reward. Brain Res. 541, 75-81.
Kruger, J. L., Dell, L. A., Bhagwandin, A., Jillani, N.E., Pettigrew, J.D., Manger, P.R., 2010.
Nuclear organization of cholinergic, putative catecholaminergic and serotonergic systems in the
brains of five microchiropteran species. J. Chem. Neuroanat. 40, 210-222.
Kus, L., Borys, E., Chu, Y.P., Ferguson, S.M., Blakely, R.D., Emborg, M.E., Kordower, J.H.,
Levey, A.I., Mufson, E.J., 2003. Distribution of high affinity choline transporter
immunoreactivity in the primate central nervous system. J. Comp. Neurol. 463, 341-357.
Lacher Jr, T.E., 1981. The comparative social behavior of Kerodon rupestris and Galea spixii
and the evolution of behavior in the cavidiae. Bulletin of Carnegie Museum of Natural History
Pittsburg 17, 5-71.

53
Laviolette, S.R., Alexson, T.O., Van der Kooy, D., 2002. Lesions of the tegmental
pedunculopontine nucleus block the rewarding effects and reveal the aversive effects of nicotine
in the ventral tegmental area. J. Neurosci. 22, 8653-8660.
Lee, M.S., Rinne, J.O., Marsden, D., 2000. The pedunculopontine nucleus: its role in the genesis
of movement disorders. Yonsei Med. J. 41, 167-184.
Limacher, A.„M., Bhagwandin, A., Fuxe, K., Manger, P.R., 2008. Nuclear organization and
morphology of cholinergic, putative catecholaminergic and serotonergic neurons in the brain of
the Cape porcupine (Hystrix africae australis): Increased brain size does not lead to increased
organizational complexity. J. Chem. Neuroanat. 36, 33-52.
Mahady, L.J., Perez, S.E., Emerich, D.F., Wahlberg, L.U., Mufson, E.J., 2017. Cholinergic
profiles in the Goettingen miniature pig (Sus scrofa domesticus) brain. J. Comp. Neurol. 525,
553-573.
Manger, P.R., Fahringer, H.M., Pettigrew, J.D., Siegel, J.M., 2002. The distribution and
morphological characteristics of cholinergic cells in the brain of monotremes as revealed by
ChAT immunohistochemistry. Brain Behav. Evol. 60, 275-297.
Mark, G.P., Rada, P.V., Shors, T.J., 1996. Inescapable stress enhances extracelular acetylcholine
in the rat hippocampus and prefrontal cortex but not the nucleus accumbens or amygdala.
Neuroscience 74, 767-774.
Maseko, B.C., Manger, P.R., 2007. Distribution and morphology of cholinergic,
catecholaminergic and serotonergic neurons in the brain of Schreiber‟s long-fingered bat,
Miniopterus schreibersii. J. Chem. Neuroanat. 34, 80-94.
Maseko, B.C., Bourne, J.A., Manger, P.R., 2007. Distribution and morphology of cholinergic,
putative catecholaminergic and serotonergic neurons in the brain of the Egyptian rousette flying
fox, Rousettus aegyptiacus. J. Chem. Neuroanat. 34, 108-127.
McGeorge, A.J., Faull, R.L., 1989. The organization of the projection from the cerebral cortex to
the striatum in the rat. Neuroscience 29, 503-537.
McKay, B.E., Placzek, A.N., Dani, J.A., 2007. Regulation of synaptic transmission and plasticity
by neuronal nicotinic acetylcholine receptors. Biochem. Pharmacol. 74, 1120-1133.
Mena-Segovia, J., 2016. Structural and functional considerations of the cholinergic brainstem. J.
Neural Transm. 123, 731-736.
Mena-Segovia, J., Bolam, J.P., 2017. Rethinking the pedunculopontine nucleus: from cellular
organization to function. Neuron 94, 7-18.
Mena-Segovia, J., Bolam, J.P., Magill, P.J., 2004. Pedunculopontine nucleus and basal ganglia:
distant relatives or part of the same family? Trends Neurosci. 27, 585-588.

54
Mesulam, M.-M., Mufson, E.J., Wainer, B.H., Levey, A.I., 1983a. Central cholinergic pathways
in the rat: An overview based on an alternative nomenclature (Ch1-Ch6). Neuroscience 10,
1185-1201.
Mesulam, M.-M., Mufso, E.J., Levey, A.I., Wainer, B.H., 1983b. Cholinergic innervation of
cortx by the basal forebrain: Cytochemistry and cortical connections of the septal area, diagonal
band nuclei, nucleus basalis (substantia innominata), and hypothalamus in the Rhesus monkey. J.
Comp. Neurol. 214, 170-197
Meyer, G., Gonzalez-Hernandez, T., Carrillo-Padilla, F., Ferres-Torres, R., 1989. Aggregations
of granule cells in the basal forebrain (Islands of Calleja): Golgi and cytoarchitectonic study in
different mammals, including man. J. Comp. Neurol. 284, 405-428.
Millhouse, O.E., 1987. Granule cells of the olfactory tubercle and the question of the Islands of
Calleja. J. Comp. Neurol. 265, 1-24.
Mineur, Y.S., Abizaid, A., Rao, Y., Salas, R., DiLeone, R.J., Gündisch, D., Diano, S., De Biasi,
M., Horvath, T.L., Gao, X.-B., Picciotto, M.R., 2011. Nicotine decreases food intake through
ativation of POMC neurons. Science 332, 1330-1332.
Morais, P.L.A.G., Santana, M.A.D., Cavalcante, J.C., Costa, M.S.M.O., Cavalcante, J.S.,
Nascimento Jr, E.S., 2014. Retinal projections into the Zona incerta of the rock cavy (Kerodon
rupestris): a CTb study. Neurosci. Res. 89, 75-80.
Motts, S.D., Slusarczyk, A.S., Sowick, C.S., Schofield, B.R., 2008. Distribution of cholinergic
cells in guinea pig brainstem. Neuroscience 154, 186-195.
Mufson, E.J., Martin, T.L., Mash, D.C., Wainer, B.H., Mesulam, M.-M., 1986. Cholinergic
projections from the parabigeminal nucleus (Ch8) to the superior colliculus in the mouse: a
combined analysis of horseradish peroxidase transport and choline acetyltransferase
immunohistochemistry. Brain Res. 370, 144-148.
Myers, R.D., Waller, M.B., 1973. Differential release of acetylcholine from the hypothalamus
and mesencephalon of the monkey during thermoregulation. J. Physiol. 230, 273-293.
Nascimento Jr, E.S., Duarte, R.B., Silva, S.F., Engelberth, R.C.G.J., Toledo, C.A.B., Cavalcante,
J.S., Costa, M.S.M.O., 2008. Retinal projections to the thalamic paraventricular nucleus in the
rock cavy (Kerodon rupestris). Brain Res. 1241, 56-61.
Nascimento Jr, E.S., Cavalcante, J.S., Cavalcante, J.C., Costa, M.S.M.O., 2010a. Retinal
afferents to the thalamic mediodorsal nucleus in the rock cavy (Kerodon rupestris). Neurosci.
Lett. 475, 3843.
Nascimento Jr, E.S., Souza, A.P.M., Duarte, R.B., Magalhães, M.A.F., Silva, S.F., Cavalcante,
J.C., Cavalcante, J.S., Costa, M. S. M. O., 2010b. The suprachiasmatic nucleus and the
intergeniculate leaflet in the rock cavy (Kerodon rupestris): Retinal projections and

55
immunohistochemical characterization. Brain Res. 1320, 34-46. Nauta, W.J., Domesick, V.B.,
1984. Afferent and efferent relationships of the basal ganglia. CIBA Foundation Symposium
107, 3-29.
Newman, R., Winans, S.S., 1980a. An experimental study of the ventral striatum of the Golden
hamster. I. Neuronal connections of the nucleus accumbens. J. Comp. Neurol. 191, 167-192.
Newman, R., Winans, S.S., 1980b. An experimental study of the ventral striatum of the Golden
hamster. I. Neuronal connections of the olfactory tubercle. J. Comp. Neurol. 191, 193-212.
Oda, Y., Nakanishi, I., 2000. The distribution of cholinergic neurons in the human central
nervous system. Histol. Histopathol. 15, 825-834.
Oliveira, F.G., Andrade-da-Costa, B.L.S., Cavalcante, J.S., Silva, S.F., Soares, J.G., Lima,
R.R.M., Nascimento Jr, E.S., Cavalcante, J.C., Resende, N.S., Costa, M.S.M.O., 2014. The eye
of the crepuscular rodent rock cavy (Kerodon rupestris) (Wied, 1820). J. Morphol. Sci. 31, 89-
97.
Oliveira, F.G., Nascimento Jr, E.S., Cavalcante, J.C., Guzen, F.P., Cavalcante, J.S., Soares, J.G.,
Cavalcanti, J.R.L.P., Freitas, L.M., Costa, M.S.M.O., Andrade-da-Costa, B.L.S., 2018.
Topographic specializations of catecholaminergic cells and ganglion cells and distribution of
calcium binding proteins in the crepuscular rock cavy (Kerodon rupestris) retina. J. Chem.
Neuroanat. 90, 57-69.
Omeichenko, N., Sesack, S.R., 2006. Cholinergic axons in the rat ventral tegmental area synapse
preferentially onto mesoaccumbens dopamine neurons. J. Comp. Neurol. 494, 863-875.
Pahapill, P.A., Lozano, A.M., 2000. The pedunculopontine nucleus and Parkinson´s disease.
Brain 123, 1767-1783. Pang, X., Liu, L., Ngolab, J., Zhao-Shea, R., McIntosh, J.M., Gardner,
P.D., Tapper, A.R., 2016. Habenula cholinergic neurons regulate anxiety during nicotine
withdrawal via nicotinic acetylcholine receptors. Neuropharmacology 107, 294-304.
Parent, A., Hazrati, L.-N., 1995.Functional anatomy of the basal ganglia. I. The cortico-basal
gangliathalamo-cortical loop. Brain Res. Rev. 20, 91-127.
Park, J.L., Kooy, Van Der Kooy, D., 1995. Tegmental pedunculopontine nucleus lesions do not
block cocaine reward. Pharmacol. Biochem. Behav. 52, 77-83.
Patzke, N., Bertelsen, M.F., Fuxe, K., Manger, P.R., 2014. Nuclear organization of cholinergic,
catecholaminergic, serotonergic and orexinergic systems in the brain of the Tasmanian devil. J.
Chem. Neuroanat. 61-62, 94-106.
Paxinos, G., Watson, C., 2007. The rat brain in stereotaxic coordinates. Elsevier, Amsterdam.
Peyron, C., Tighe, D.K., van den Pol, A.N., de Lecea, L., Heller, H.C., Sutcliffe, J.G., Kilduff,
T.S., 1998. Neurons containing hypocretin (orexin) project to multiple neuronal systems. J.
Neurosci. 18, 9996-10015.

56
Picanço-Diniz, C.W., Silveira, L.C.L., Carvalho, M.S.P., Oswaldo-Cruz, E., 1991. Contralateral
visual field representation in area 17 of the cerebral cortex of the agouti: A comparison between
the cortical magnification factor and retinal ganglion cell distribution. Neuroscience 44, 325-333.
Picanço-Diniz, C.W., Rocha, E.G., Silveira, L.C.L., Elston, G., Oswaldo-Cruz, E., 2011. Cortical
representation of the horizon in V1 and peripheral scaling in mammals with lateral eyes.
Psychol. Neurosci. 4, 19-27.
Picciotto, M.R., Higley, M.J., Mineur, Y.S., 2012. Acetylcholine as a neuromodulator:
cholinergic signaling shapes nervous system function and behavior. Neuron 76, 116-129.
Pieters, R.P., Gravett, N., Fuxe, K., Manger, P.R., 2010. Nuclear organization of cholinergic,
putative catecholaminergic and serotonergic nuclei in the brain of the eastern rock elephant
shrew, Elephantulus myurus. J. Chem. Neuroanat. 39, 175-188.
Pratt, W.E., Blackstone, K., 2009. Nucleus accumbens acetylcholine and food intake: Decreased
muscarinic tone reduces feeding but not food-seeking. Behav. Brain Res. 198, 252-257.
Pratt, W.E., Kelley, A.E., 2004. Nucleus accumbens acetylcholine regulates appetitive learning
and motivation for food via activation of muscarinic receptors. Behav. Neurosci. 118, 730-739.
Qin, C., Luo, M., 2009. Neurochemical phenotypes of the afferent and efferent projections of the
mouse medial habenula. Neuroscience 161, 827-837.
Reis, M.E.M., Araújo, L.T.F., Andrade, W.M.G., Resende, N.S., Lima, R.R.M., Nascimento Jr,
E.S., Costa, M.S.M.O., Cavalcante, J.C., 2018. Distribution of nitric oxide synthase in the rock
cavy (Kerodon rupestris) brain I: The diencephalon. Brain Res. 1685, 60-78.
Roberts, M.E., Maliniak, E., Deal, M., 1984. The reproductive biology of the rock cavy,
Kerodon rupestris in captivity: a study of reproductive adaptation in a trophic specialist.
Mammalia 48, 253266.
Rocha, F.A.F., Ahnelt, P.K., Peichl, L., Saito, C.A., Silveira, L.C.L., De Lima, S.M.A., 2009.
The topography of cone photoreceptors in the retina of a diurnal rodent, the agouti (Dasyprocta
aguti). Vis. Neurosci. 26, 167-175.
Rocha, E.G., Freire, M.A.M., Bahia, C.P., Pereira, A., Sosthenes, M.C.K., Silveira, L.C.L.,
Elston, G.N., Picanço-Diniz, C.W., 2012. Dendritic structure varies as a function of eccentricity
in V1: A quantitative study of NADPH diaphorase neurons in the diurnal South American rodent
agouti, Dasyprocta prymnolopha. Neuroscience 216, 94-102.
Rodriguez-López, C., Clascá, F., Prensa, L., 2017. The mesoaccumbens pathway: A retrograde
labeling and single cell axon tracing analysis in the mouse. Frontiers in Neuroanatomy 11, 1-15.
Roghani, A., Shirzadi, A., Butcher, L.L., Edwards, R.H., 1998. Distribution of the vesicular
transporter for acetylcholine in the rat central nervous system. Neuroscience 82, 1195-1212.

57
Role, L.W., Berg, D.K., 1996. Nicotinic receptors in the development and modulation of CNS
synapses. Neuron 16, 1077-1085.
Rowe, D.L., Honeycutt, R.L., 2002. Phylogenetic relationships, ecological correlates, and
molecular evolution within the cavioidea (Mammalia, Rodentia). Mol. Biol. Evol. 19, 263-277.
Saper, C.B., Cano, G., Scammel, T.E., 2005. Homeostatic, circadian, and emotional regulation of
sleep. J. Comp. Neurol. 493, 92-98.
Satoh, K., Fibiger, H.C., 1985a. Distribution of central cholinergic neurons in the baboon (Papio
papio). I. General morphology. J. Comp. Neurol. 236, 197-214. Satoh, K., Fibiger, H.C., 1985b.
Distribution of central cholinergic neurons in the baboon (Papio papio). II. A topographic atlas
correlated with catecholamine neurons. J. Comp. Neurol. 236, 215233.
Schäfer, M.K.–H., Eiden, L.E., Weihe, E., 1998. Cholinergic neurons and terminal fields
revealed by immunohistochemistry for the vesicular acetylcholine transporter. I. Central nervous
system. Neuroscience 84, 331-359.
Schultz, W., 1998. Predictive reward signal of dopamine neurons. J. Neurophysiol. 80, 1-27.
Scott, J.W., McBride, R.L., Schneider, S.P., 1980. The organization of projections from the
olfactory bulb to the piriform cortex and olfactory tubercle in the rat. J. Comp. Neurol. 194, 519-
534.
Silva, A.M., Santana, M.A.D., Morais, P.L.A.G., Sousa, T.B., Engelberth, R.C.G.J., Lucena,
E.E.S., Campêlo. C.L.C., Cavalcante, J.S., Cavalcante, J.C., Costa, M.S.M.O., Nascimento Jr,
E.S., 2014. Serotonergic fibers distribution in the midline and intralaminar thalamic nuclei in the
rock cavy (Kerodon rupestris). Brain Res. 1586, 99-108.
Silva Neto, E.J., 2000. Morphology of the regions ethmoidalis and orbitotemporalis in Galea
musteloides Meyen 1832 and Kerodon rupestris (Wied-Neuwied 1820) (Rodentia: Caviidae)
with comments on the phylogenetic systematic of the Caviidae. J. Zool. Syst. Evol. Res. 38, 219-
229.
Silveira, L.C.L., 1985. Organização do Sistema Visual de Roedores da Amazônia: Óptica Ocular
e Distribuição das Células Ganglionares Retinianas. Instituto de Biofísica, Universidade Federal
do Rio de Janeiro, Rio de Janeiro, Brazil. Tese de doutorado.
Silveira, L.C., Yamada, E.S., Picanço-Diniz, C.W., 1989. Displaced horizontal cells and
biplexiform horizontal cells in the mammalian retina. Visual Neurosci. 3, 483-488.
Soares, J.G., Cavalcanti, J.R.L.P., Oliveira, F.G., Pontes, A.L.B., Sousa, T.B., Freitas, L.M.,
Cavalcante, J.S., Nascimento Jr, E.S., Cavalcante, J.C., Costa, M.S.M.O., 2012. Nuclear
organization of the serotonergic system in the brain of the rock cavy (Kerodon rupestris). J.
Chem. Neuroanat. 43, 112-119.

58
Sousa, R.A., Menezes, A.A.L., 2006. Circadian rhythm of motor activity of the Brazilian rock
cavy (Kerodon rupestris) under artificial photoperiod. Biol. Rhythm Res. 37, 443-450.
Steidl, S., Cardiff, K.M., Wise, R.A., 2015. Increased latencies to initiate cocaine self-
administration following laterodorsal tegmental nucleus lesions. Behav. Brain Res. 287, 82-88.
Steriade, M., 2004. Acetylcholine systems and rhythmic activities during the waking–sleep
cycle. Prog. Brain Res. 145, 179-196.
Sutherland, R.J., 1982. The dorsal diencephalic conduction system: A review of the anatomy and
functions of the habenular complex. Neurosci. Biovehav. Rev. 6, 1-13.
Switzer, R.C., Hill, J.M., Heimer, L., 1982. The globus pallidus and its rostroventral extension
into the olfactory tubercle of the rat: A cyto- and chemoarchitectural study. Neuroscience 7,
1891-1904.
Szymusiak, R., Alam, N., Steininger, T.L., McGinty, D., 1998. Sleep-waking discharge patterns
of ventrolateral preoptic/anterior hypothalamic neurons in rats. Brain Res. 803, 178-188.
Szymusiak, R., Gvilia, I., McGinty, D., 2007. Hypothalamic control of sleep. Sleep Med. 8, 291-
301.
Usunoff, K.G., Schmitt, O., Itzeu, D.E., Rolfs, A., Wree, A., 2007. Efferent connections of the
parabigeminal nucleus to the amygdala and the superior colliculus in the rat: A double-labeling
fluorescent retrograde tracing study. Brain Res. 1133, 87-91.
Viswanath, H., Carter, A.Q., Baldwin, P.R., Molfese, D.L., Salas, R., 2014. The medial
habenula: still neglected. Front. Hum. Neurosci. 7, 1-6.
Von Bohlen und Halbach, O., Dermietzel, R., 2006. Neurotransmitters and Neuromodulators.
WileyVCH, Germany, Pp. 46-59.
Wang, H.-L., Morales, M., 2009. Pedunculopontine and laterodorsal tegmental nuclei contain
distinct populations of cholinergic, glutamatergic and GABAergic neuron in the rat. Eur. J.
Neurosci. 29-340358.
Warner-Schmidt, J.L., Schimidt, E.F., Marshall, J.J., Rubin, A.J., Arango-Lievano, M., Kaplitt,
M.G., Ibañez-Tallon, I., Heintz, N., Greengard, P., 2012. Neuroscience 109, 11360-11365.
Wen, P., Li, M., Xiao, H., Ding, R., Chen, H., Chang, J., Zhou, M., Yang, Y., Wang, J., Zheng,
W., Zhang, W., 2015. Low-frequency stimulation of the pedunculopontine nucleus affects gait
and the neurotransmitter level in the ventrolateral thalamic nucleus in 6-OHDA Parkinsonian
rats. Neurosci. Let. 600, 62-68.
Winn, P., 2008. Experimental studies of pedunculopontine functions: are they motor, sensory or
integrative? Parkinsonism and Related Disorders 14, S194-S198.
Woolf, N.J., 1991. Cholinergic systems in mammalian brain and spinal cord. Prog. Neurobiol.
37, 475-524.

59
Xi, M.-C., Morales, F.R., Chase, M.H., 2001. Effects on sleep and wakefulness of the injection
of hypocretin-1 (orexin-A) into the laterodorsal tegmental nucleus of the cat. Brain Res. 901,
259-264.
Yamaguchi, T., Danjo, T., Pastan, I., Hikida, T., Nakanishi, S., 2013. Distinct roles of segregated
neurotransmission of septo-habenular pathway in anxiety and fear. Neuron 78, 573-544.

60
FIGURE CAPTIONS
Figure 1. Canvas serial drawings of Nissl stained coronal sections through the rock cavy brain.
ChATimmunoreactive neurons (dots) were superimposed on the right half and the acronyms on
the left half of the figure. Numbers at right indicate the distance of the section in relation to the
previous section. For the meaning of each acronym, see the list of abbreviations.
Figure 2. Photomicrographs of coronal ChAT-immunostained sections showing (A) ChAT-IR
neurons in the nucleus accumbens (Acb), (B) Magnification of the square in A, (C) ChAT-IR
neurons in the caudate-putamen nucleus (CPu), (D) Magnification of the square in C, (E) ChAT-
IR neurons in the olfactory tubercle/Islands of Calleja (Tu/ICj), (F) Magnification of the square
in E. Bar: 500 µm in A, C, E and 100 µm in B, D, F.
Figure 3. Photomicrographs of coronal ChAT-immunostained sections showing (A) ChAT-IR
neurons in horizontal portion of the diagonal band nucleus (HDB), (B) Magnification of the
square in A, (C) ChAT-IR neurons in vertical portion of the diagonal band nucleus (VDB), (D)
Magnification of the square in C, (E) ChAT-IR neurons in medial septal nucleus (MS), (F)
Magnification of the square in E. Bar: 500 µm in A, C, E and 100 µm in B, D, F.
Figure 4. Photomicrographs of coronal ChAT-immunostained sections showing (A) ChAT-IR
neurons in the globus pallidus (GP), (B) Magnification of the square in A, (C) ChAT-IR neurons
in entopeduncular nucleus (EP), (D) Magnification of the box in C, (E) ChAT-IR neurons in
basal nucleus (B), (F) Magnification of the box in E. Bar: 500 µm in A, C, E and 100 µm in B,
D, F.
Figure 5. Photomicrographs of coronal ChAT-immunostained sections showing (A) ChAT-IR
neurons in the ventrolateral preoptic nucleus (VLPO), (B) Magnification of the square in A, (C)
ChAT-IR neurons in ventrolateral hypothalamic nucleus (VLH), (D) Magnification of the square
in C, (E) ChAT-IR neurons in medial habenular nucleus (MHb), (F) Magnification of the square
in E. Bar: 500 µm in A, C, E and 100 µm in B, D, F.
Figure 6. Photomicrographs of coronal ChAT-immunostained sections showing (A) ChAT-IR
neurons in the parabigeminal nucleus (PBG), (B) Magnification of the square in A, (C) ChAT-IR
neurons in the laterodorsal tegmental nucleus (LDTg) and the pedunculopontine tegmental
nucleus (PTg), (D) Magnification of square “d” in C, (E) Magnification of square “e” in C. Bar:
500 µm in A, C, and 100 µm in B, D, E.

61
ABBREVIATIONS
3N Oculomotor nucleus
3V Third ventricle
A Amygdaloid área
ac Anterior comissure
aca Anterior comissure, ant
Acb Accumbens nucleus
Ach Acetylcholine
acp Anterior comissure, post
AD Anterodorsal thalamic nucleus
AHA Anterior hypothalamic área
APT Anterior pretectal nucleus
Aq Aqueduct
Arc Arcuate hypothalamic nucleus
B Basal nucleus
BIC Nucleus brachium inferior colliculus
bic Brachium inferior colliculus
cc Corpus callosum
ChAT Choline acetyltransferase
Cl Claustrum
CLi Caudal linear nucleus of the raphe
Co Cortical amygdaloid nucleus
cp Cerebral peduncle
CPu Caudate putamen
csc Comissure superior collicullus
CxA Cortex-amygdala transition
DLG Dorsal lateral geniculate nucleus
DM Dorsomedial hypothalamic nucleus
DR Dorsal raphe nucleus
ec External capsule
En Endopiriform nucleus
EP Entopeduncular nucleus
f Fornix
fr Fasciculus retroflexus
GP Globus pallidus

62
hbc Habenular comissure
HDB Diagonal nucleus of Broca, horizontal limb
IC Inferior colliculus
ic Internal capsule
ICj Islands of Calleja
IGL Intergeniculate leaflet
IP Interpeduncular nucleus
LD Laterodorsal thalamic nucleus
LDTg Laterodorsal tegmental nucleus
lfp Longitudinal fasciculus pons
LH Lateral hypothalamic área
LHb Lateral habenular nucleus
lo Lateral olfactory tract
LP Lateral posterior thalamic nucleus
LPO Lateral preoptic área
LV Lateral ventricle
M Mammillary nucleus
Me5 Mesencephalic trigeminal nucleus
MG Medial geniculate nucleus
MHb Medial habenular nucleus
ml Medial lemniscos
mlf Medial longitudinal fasciculus
MnPO Median preoptic nucleus
MnR Median raphe nucleus
MPA Medial preoptic área
MS Medial septal nucleus
mt Mammillothalamic tract
och Optic chiasm
opt Optic tract
Pa Paraventricular hypothalamic nucleus
PAG Periaqueductal gray
PBG Parabigeminal nucleus
PF Parafascicular thalamic nucleus
Pir Piriform córtex
PMnR Paramedian raphe nucleus
Pn Pontine nuclei

63
PT Paratenial thalamic nucleus
PTg Pedunculopontine tegmental nucleus
PV Paraventricular thalamic nucleus
PVP Paraventricular thalamic nucleus, posterior
RCh Retrochiasmatic área
Rt Reticular thalamic nucleus
SC Superior collicullus
SCh Suprachiasmatic nucleus
sm Stria medullaris thalamus
SNC Substantia nigra, compacta
SNR Substantia nigra, reticular
SO Supraoptic nucleus
ST Bed nucleus stria terminalis
st Stria terminalis
STh Subthalamic nucleus
tfp Transverse fibers pons
TT Tenia tecta
Tu Olfactory tubercle
VDB Diagonal nucleus of Broca, vertical limb
VLG Ventral lateral geniculate nucleus
VLH Ventrolateral hypothalamic nucleus
VLPO Ventrolateral preoptic nucleus
VMH Ventromedial hypothalamic nucleus
VP Ventral pallidum
xscp Decussation superior cerebellar peduncle
ZI Zona incerta
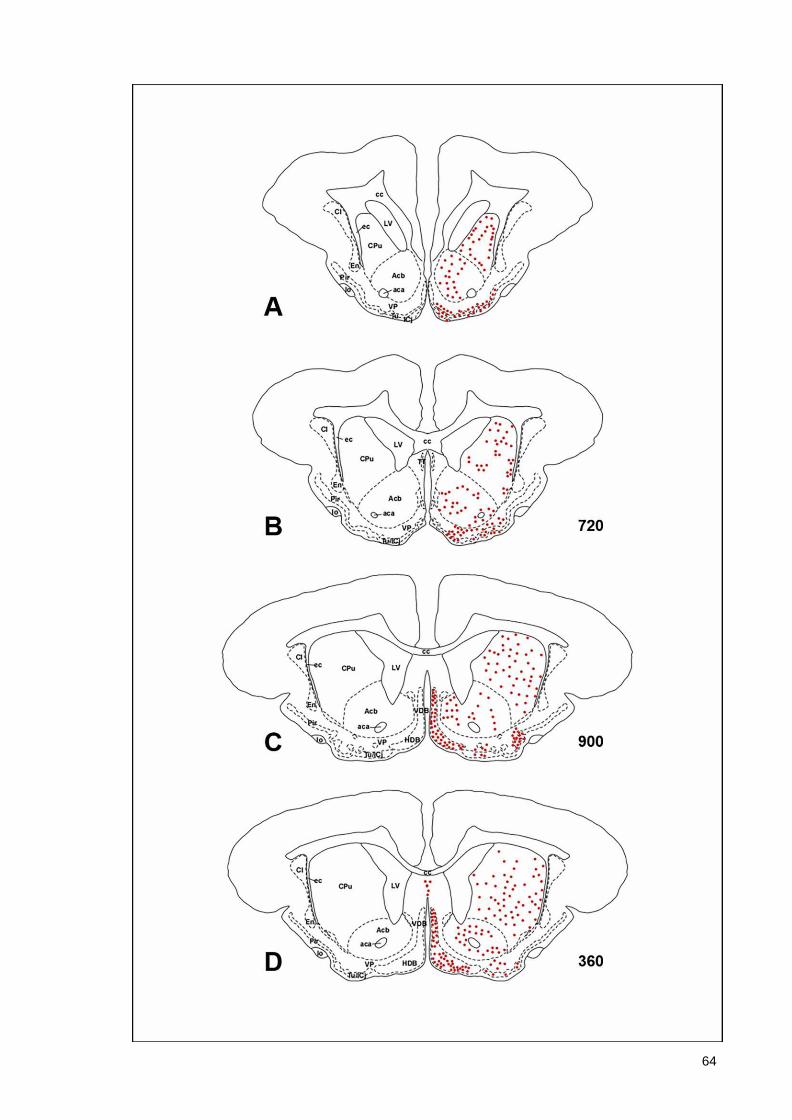
64
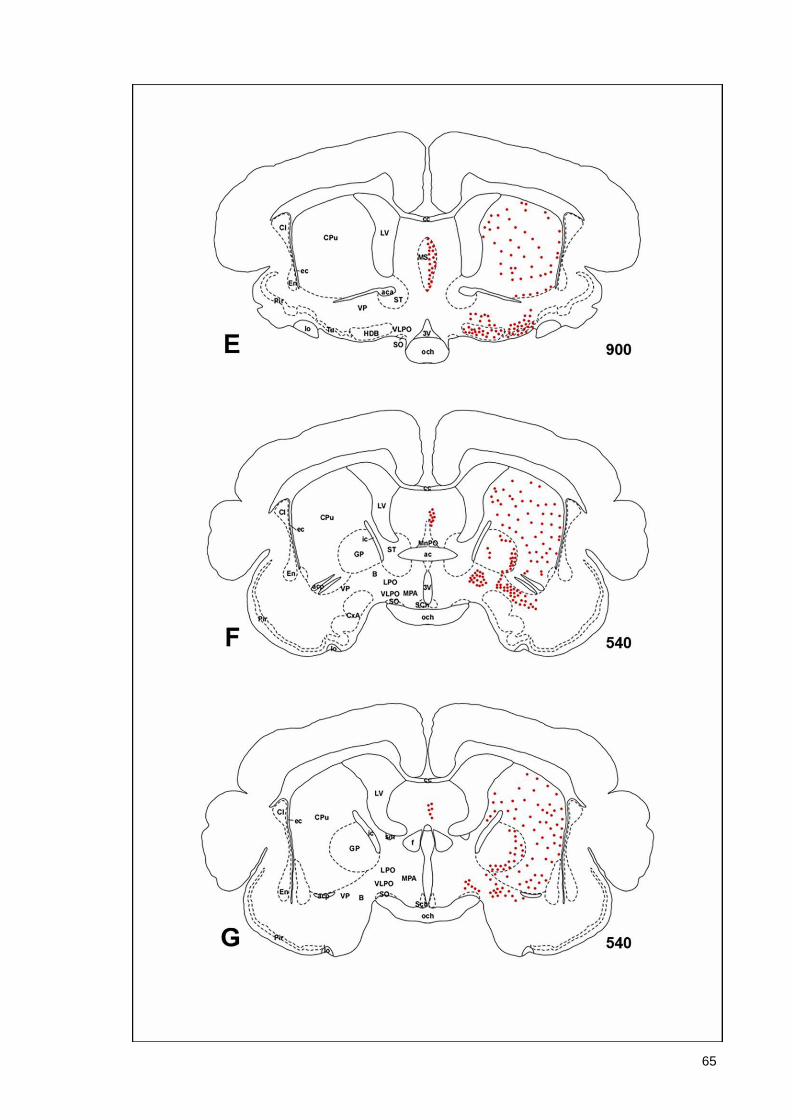
65
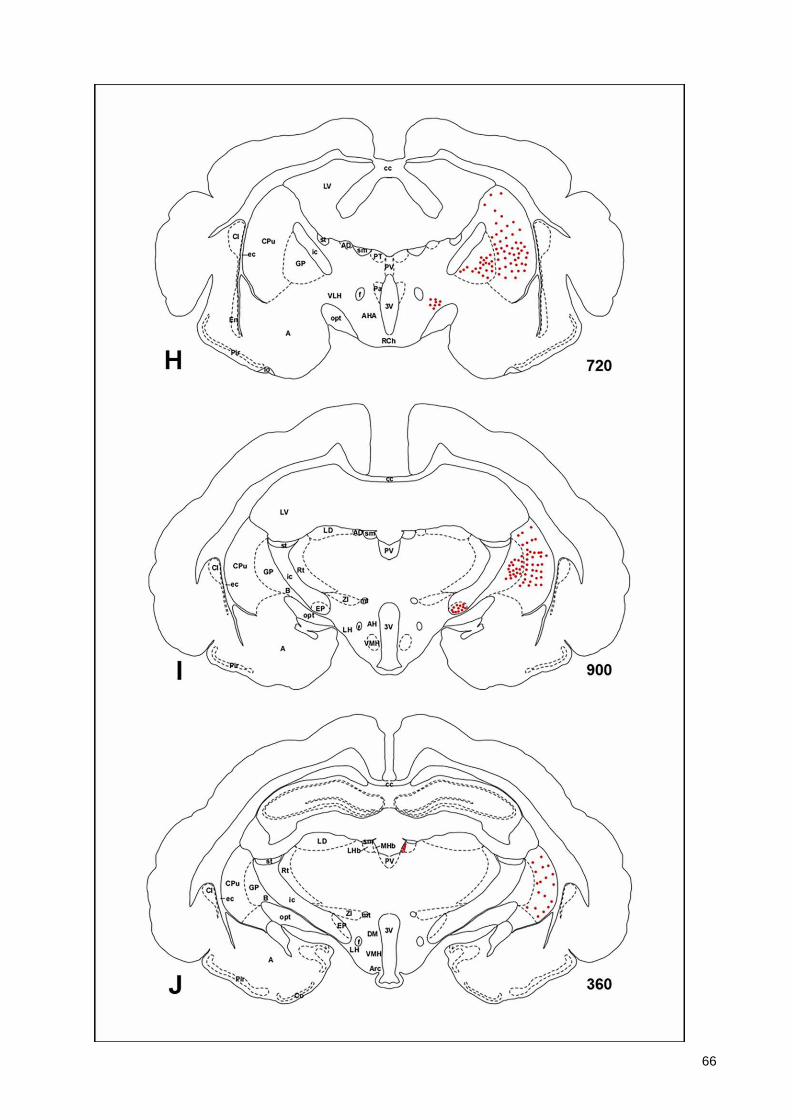
66
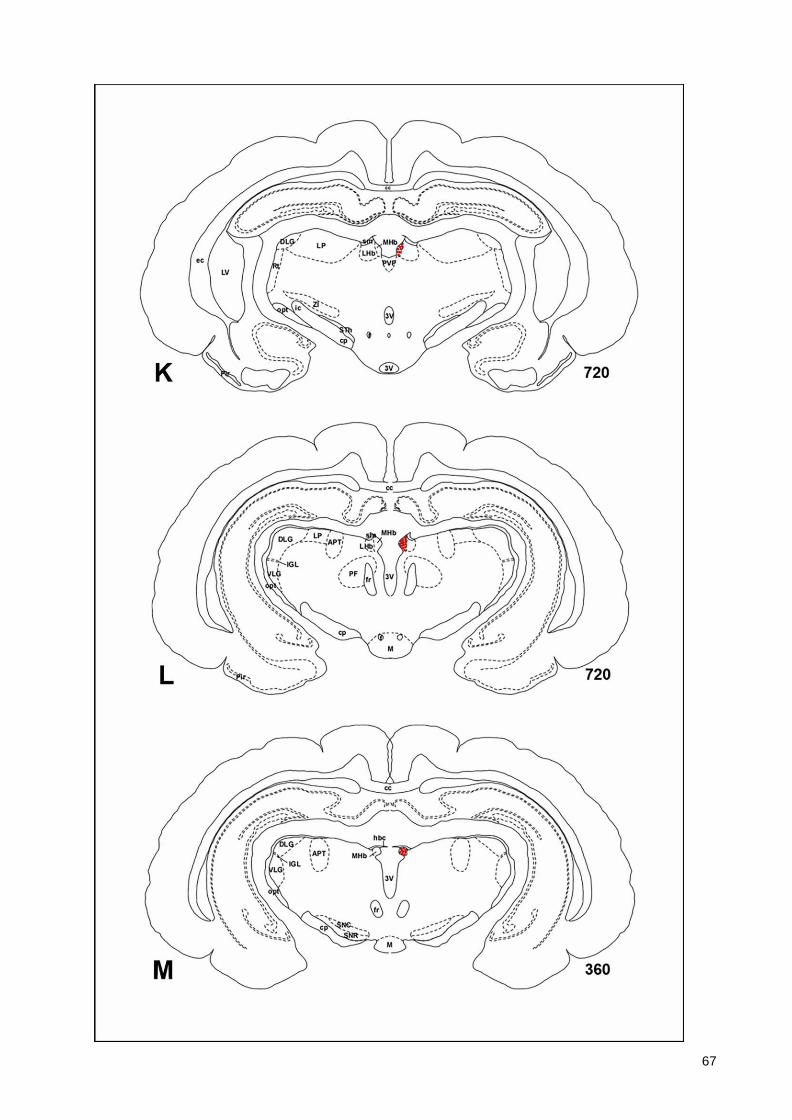
67
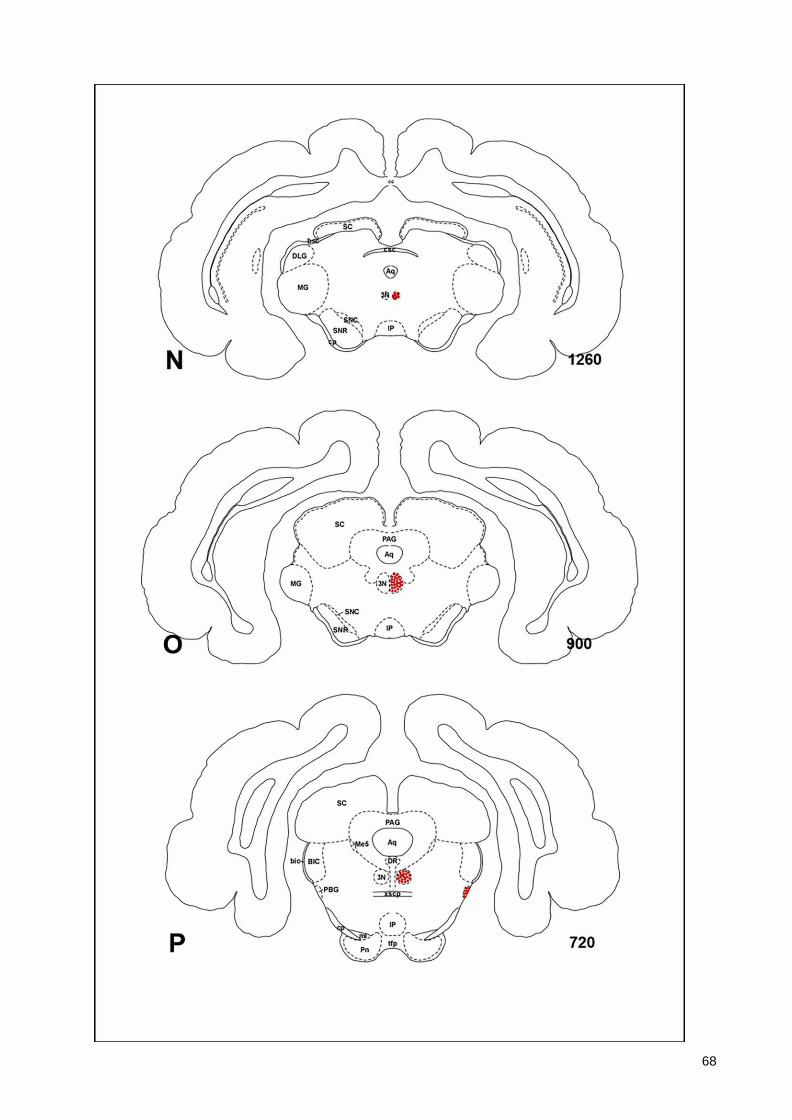
68
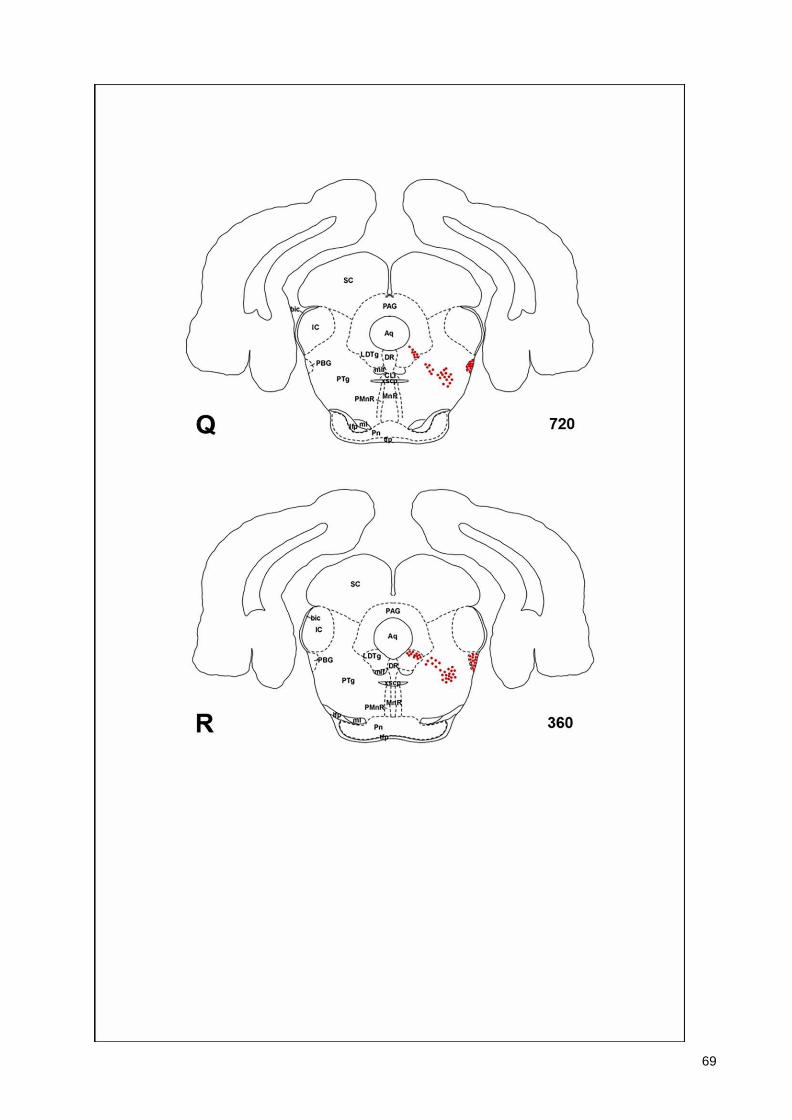
69
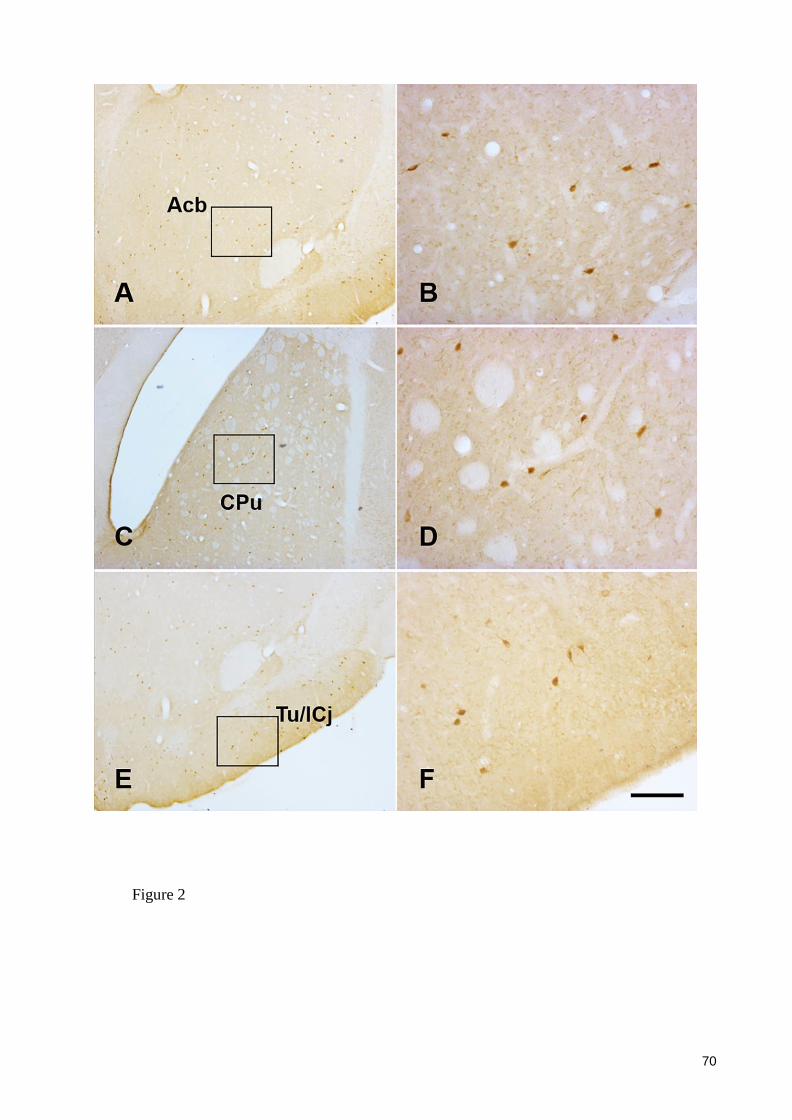
70
Figure 2

71
Figure 3
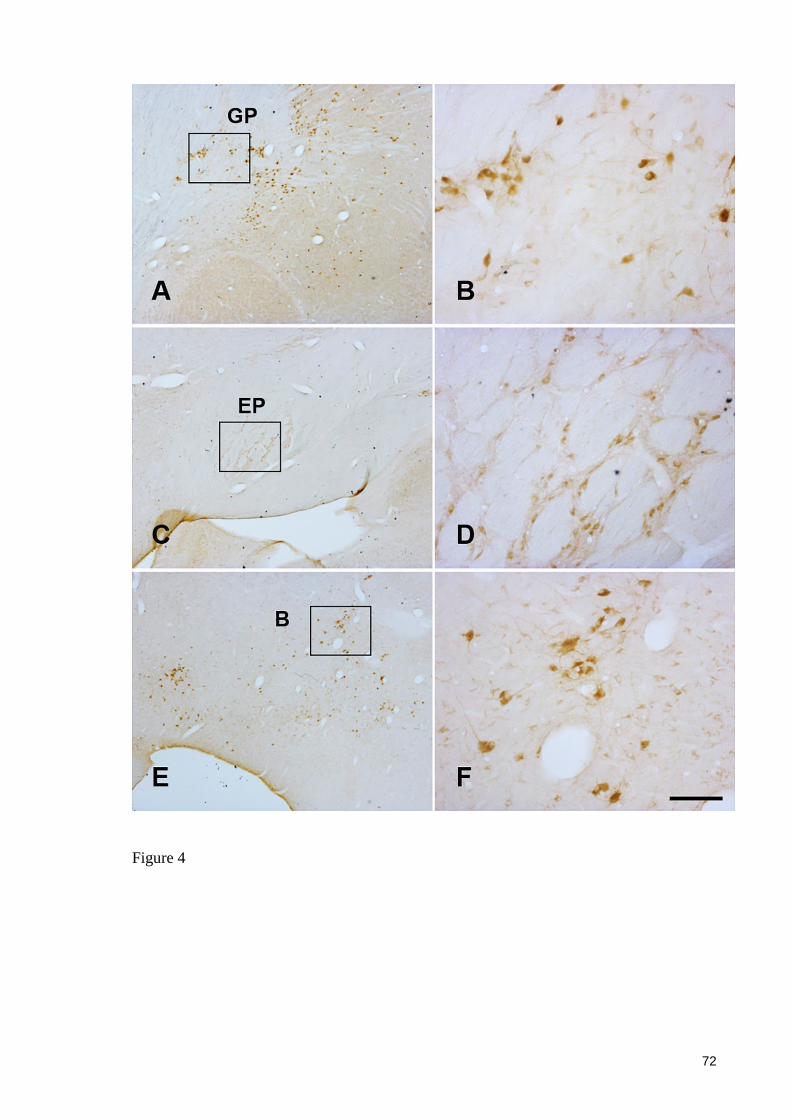
72
Figure 4
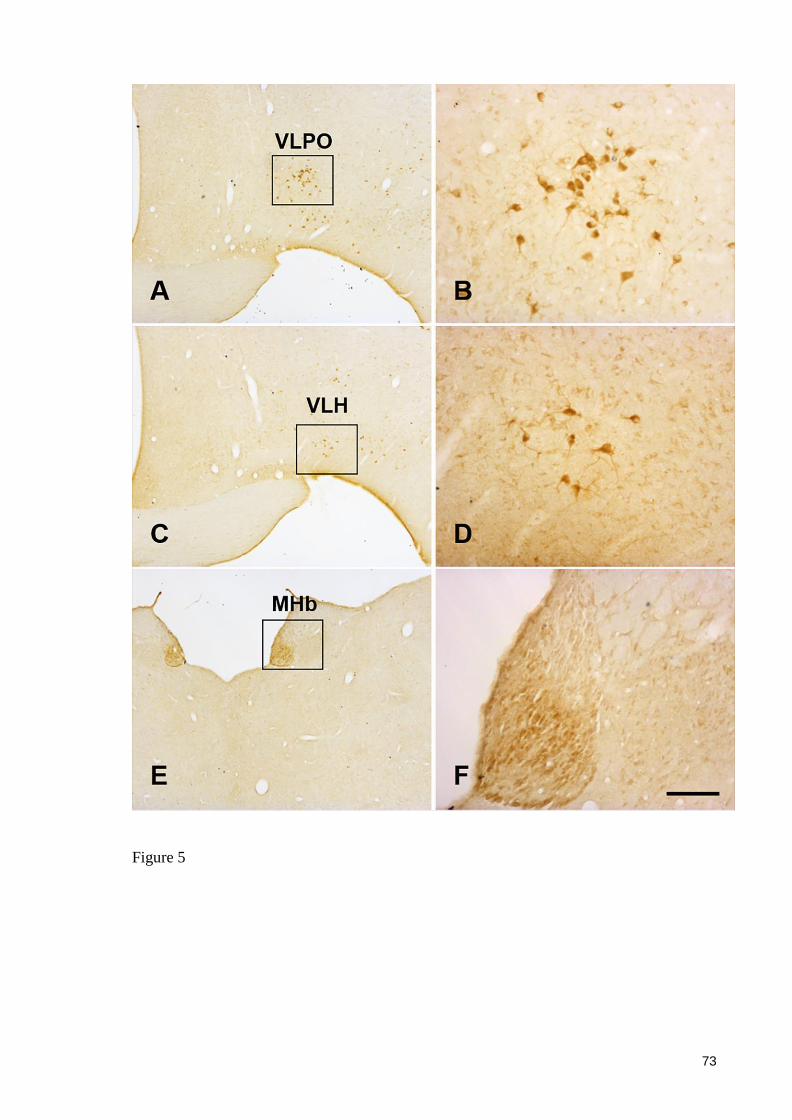
73
Figure 5
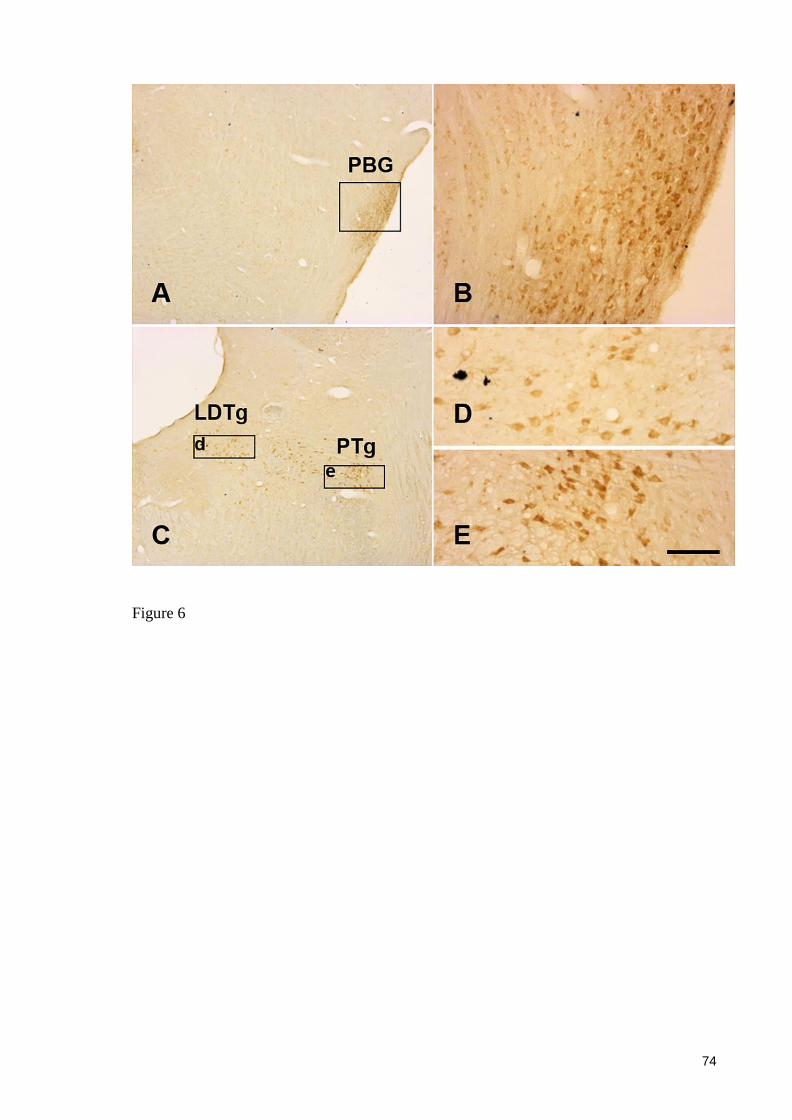
74
Figure 6

75
6. CONSIDERAÇÕES FINAIS
A partir dos nossos resultados podemos inferir que, apesar de constantes semelhanças, com
relação a distribuição global e as características morfológicas de neurônios colinérgcos, nós
identificamos estruturas e neurônios colinérgicos no mocó, não identificados em espécies
anteriormentes estudadas. No nível telencefálico, a presença de neurônios imunorreativos a
ChAT, nos núcleos accumbens, caudado-putamen globo pálido ventral, globo pálido, núcleo da
banda diagonal, septal medial, tubérculo olfativo/ Ilhas de Calleja e núcleo basal, estes
encontrados constantemente em outras espécies estudadas. Em nosso trabalho a exceção foi a
presença de neurônios imunorreativos a ChAT no núcleo entopeduncular, não mencionado em
nenhum estudo anterior. No diencéfalo, o núcleo habenular medial, apresentou uma população
significativa de neurônios ChAT imunorreativos, descrito de forma constante em outros estudos
já realizados. Porém, as descrições são discrepantes, para neurônios colinérgicos no hipotálamo,
bastante diferente no mocó, em relação a descrições em outras espécies, ou até mesmo ausência,
como nos monotremados. Estudos anteriores mostraram presença de neurônios colinérgicos em
núcleos talâmicos em algumas espécies, porém não encontradas em nosso estudo. Na região
mesesncéfalo-ponte, encontramos no mocó, neurônios colinérgicos nos núcleos parabigeminal,
tegmental laterodorsal e tegmental pedunculopontino, estes descritos em outras espécies, porém
em algumas espécies o núcleo parabigeminal não contém neurônios colinérgicos. Concluímos
então que os grupamentos neuronais colinérgicos, mesmo em sua maioria permanecendo
amplamente conservados dentre as espécies de mamíferos, no mocó há diferenças bem como
presença de núcleos anteriormente não encontrados em estudos realizados anteriormente.

76
ANEXO A

77

78

79
ANEXO B

80

81

82
ANEXO C

83

84

85

86

87

88

89

90

91

92

93

94

95

96