nÁdia das dores moreira - repositorio.ufop.br · À kátia, a alegria de viver desta garota não...
TRANSCRIPT

NÁDIA DAS DORES MOREIRA
HISTÓRIA NATURAL DA LEISHMANIOSE VISCERAL EM HAMSTER “Mesocricetus auratus”
EXPERIMENTALMENTE INFECTADOS POR DUAS CEPAS DE Leishmania infantum COM
PERFIS DISTINTOS DE VIRULÊNCIA E PATOGENICIDADE
OURO PRETO, MG
2012

NÁDIA DAS DORES MOREIRA
HISTÓRIA NATURAL DA LEISHMANIOSE VISCERAL EM HAMSTER “Mesocricetus auratus”
EXPERIMENTALMENTE INFECTADOS POR DUAS CEPAS DE Leishmania infantum COM
PERFIS DISTINTOS DE VIRULÊNCIA E PATOGENICIDADE
Tese de doutorado apresentada ao Programa de Pós-Graduação em Ciências Biológicas do Núcleo de Pesquisas em Ciências Biológicas como parte das exigências para obtenção do grau de Doutor em Ciências Biológicas pela Universidade Federal de Ouro Preto na área de concentração Imunobiologia de Protozoários
Orientador: Alexandre Barbosa Reis
Co-Orientadores: Cláudia Martins Carneiro
Rodolfo Cordeiro Giunchetti
OURO PRETO, MG 2012

Catalogação: [email protected]
M838h Moreira, Nádia das Dores.
História natural da leishmaniose visceral em hamster “Mesocricetus
auratus” experimentalmente infectados por duas cepas de Leishmania
infantum com perfis distintos de virulência e patogenicidade [manuscrito] / Nádia das Dores Moreira. – 2012.
145 f.: il., color; grafs.; tabs. Orientador: Prof. Dr. Alexandre Barbosa Reis.
Tese (Doutorado) - Universidade Federal de Ouro Preto. Instituto de Ciências Exatas e Biológicas. Núcleo de Pesquisas em Ciências Biológicas. Programa de Pós-Graduação em Ciências Biológicas.
Área de concentração: Imunobiologia de protozoários.
1. Leishmaniose visceral - Teses. 2. Leishmania - Estudos experimentais - Teses. 3. Hamster como animal de laboratório - Teses. 4. Baço - Teses. 5. Fígado - Teses. I. Universidade Federal de Ouro Preto. II. Título.
CDU: 616.993.161


i
“Nenhum de nós chegou onde está exclusivamente através do impulso de nossos próprios pés. Chegamos aqui porque alguém se inclinou e nos
alavancou. ’’ Thurgood Marshal

Moreira, N. D. Dedicatória
ii
Dedico,
A Deus, amor infinito
Aos meus pais, Manoel e Maria, pelo apoio irrestrito, amor imensurável e exemplos de honestidade e caráter, dos quais sempre procurei
espelhar-me.
À minha Irmã e segunda mãe, Meire, pelo amor incondicional, compreensão, bondade e apoio nas horas difíceis.
Ao meu sobrinho Gustavo, luz da minha vida.
À minha cunhada Elaine e meu irmão Marcone por estarem sempre perto da minha mãe, fazendo por ela o que eu não pude fazer, por estar tão distante.
Ao meu noivo Danilo, foi tudo mais fácil com você ao meu lado.
A Paula, Carol e em especial ao Bruno e Juliana vocês são fundamentais em minha vida. Ao Alexandre, por me conceder um trabalho tão importante, objeto de minha tese. Palavras não exprimem o que você fez por mim. Eterna gratidão A Jamile, minha cópia... sem palavras!

Moreira, N. D. Agradecimentos
iii
Agradecimentos
A Deus, em primeiro lugar, pelo dom da vida. Hoje devo esta vitória a ti e tenho a certeza
de que, sem tua presença, teria sido impossível continuar e chegar aonde cheguei.
Agradeço a ti pelas provocações, pois através delas pude amadurecer; pelas lágrimas
derramadas, pois elas me tornaram mais fortes; pela ausência das pessoas que amo, pois
através dela pude ver o quanto são importantes na minha vida. Obrigada Senhor!
Ao Papai e Mamãe, amores da minha vida, e que fazem de tudo pela minha felicidade,
realizando todos os meus sonhos. Só tenho a agradecer a Deus todos os dias por tudo que
fazem por mim. Obrigada pela compreensão em suportar pacientemente uma filha distante
da vida familiar durante anos. No entanto, são vocês a razão de tudo isso, e é a vocês que
ofereço esta conquista. Papai e Mamãe, amo vocês.
À Elaine minha cunhada, amiga, irmã. Ela que me deu forças para continuar e não desistir
no primeiro ano de doutorado, momento mais difícil que enfrentei até hoje. O meu sonho e
vontade de fazer o doutorado não eram suficientes para que eu prosseguisse se não fosse o
apoio dela.
Ao Danilo, pelas palavras de incentivo, seu gesto de compreensão, sua atitude de
segurança, mesmo nos períodos de desânimo. Obrigada por caminhar ao meu lado.
Obrigada pelo amor, pela paciência e pelo companheirismo.
Ao Dr. Alexandre, exemplo de dedicação e profissionalismo, de quem tive a honra de ser
orientada. Aliado aos seus profundos conhecimentos científico você tem contribuído
sobremaneira em minha formação profissional e humana.
À Drª. Cláudia Martins Carneiro, a ausência do professor Alexandre foi menos dolorida
com você por perto. As inúmeras tarefas não a impediram de desejar sempre boa sorte a
cada experimento, pra mim isso foi fundamental. Obrigada por todos os ensinamentos
desde o mestrado. Muita coisa do que sou hoje devo a você.
Ao Dr. Rodolfo Cordeiro Giunchetti pelo convívio e ensinamentos que foram (e são) muito
importantes para meu crescimento profissional e pessoal. Obrigada por tudo.

Moreira, N. D. Agradecimentos
iv
À Drª. Marta de Lana pela colaboração e imenso carinho.
À Maria Chaves, o que seria de mim sem ela no LIMP, é minha mãe... Amo muito.
À Juliana, pela transparência, sinceridade e por ter me ensinado o valor de uma amizade
verdadeira, onde sempre existiu o respeito e a confiança mútua e também por ter estado
presente nos momentos mais críticos e mais felizes destes últimos anos... as pessoas
entram na nossa vida por acaso, mas não é por acaso que elas permanecem... Você foi e é
um presente de Deus em minha vida. Muito obrigada por tudo, minha amiga.
Ao Bruno, o que dizer de uma pessoa como o Bruno, sempre disposto a ajudar todo
mundo, uma pessoa com enorme motivação pela ciência, me ajudou desde o primeiro
experimento até o ultimo, abraçou este projeto como se fosse seu. Não existem palavras
para expressar tudo o que você fez por mim durante estes nove anos de convivência, mas
se pudesse caracterizaria como um respeito mútuo, admiração e confiança. A você Bruno
minha eterna gratidão.
À Paula, querida amiga, que fiz durante essa longa caminhada e não me esquecerei nunca,
muito obrigada, por todo o apoio é pouco. Estou certa de que, sem você, a minha
caminhada seria mais difícil. Você é um exemplo a ser seguido, tem um coração do
tamanho do mundo... Alegria imensa por tê-la como minha amiga.
À Jamille, impossível descrever tudo que Jam fez e faz por mim. Além do que, ela é minha
cópia. Jam é a única pessoa que consegue ler meus pensamentos e entender meu jeito
introspectivo e volúvel de ser. É amiga, irmã. Amo demais minha filha!
À Carolzinha, uma doçura de pessoa, obrigada pelo carinho, ensinamentos, convivência,
ajuda, paciência e pelas tardes de descontração em que demos boas risadas. É um
privilégio ser sua amiga. Obrigada por tudo. Você mora em meu coração. Te Adooooooro!
À Kátia, a alegria de viver desta garota não existe, é muito cativante. É bom demais ter
pessoas como você ao nosso lado principalmente em um ambiente de trabalho. Tudo fica
mais fácil quando você está por perto. Muito obrigada pelo auxílio nos experimentos.

Moreira, N. D. Agradecimentos
v
Ao Henrique, Rodrigo, Levi, Flávia e Fernando pelo imensurável apoio durante o
doutorado. A vocês minha eterna gratidão.
À Larissa, no momento em que mais precisei lá estava ela, não mediu esforços em me
ajudar... Quanta dedicação, quanta responsabilidade, a você minha eterna gratidão.
À Amanda, Luiza, Lucilene, Ana, Nathália, Aline, Renata, Leoneide, Sheller, e Wendel
pela agradávél convivência e colaboração em todos os momentos.
À Tânia, pela presteza e precisão nos serviços burocráticos do LIMP.
À Renata Guerra por permitir que eu realizasse os experimentos de qRT-PCR em seu
laboratório.
À todos do Laboratório de Enzimologia e Proteômica, em especial à Karina pela presteza.
À todos do Laboratório de Doença de Chagas em especial a Lívia e Ivo Caldas.
Aos funcionários do biotério, pelo carinho que sempre tiveram comigo nos momentos em
que precisei. Luciana muito obrigada, eu te adoro.
À Érika, menina de Ouro, orgulho de ser sua amiga. Palavras não exprimem tudo que você
fez por mim. Não foi nada fácil montar a colônia de hamsters se não fosse sua dedicação e
calma, não sei o que seria de mim. Muito Obrigada.
Aos docentes e colegas do Curso de Pós-Graduação do Núcleo de Pesquisas em Ciências
Biológicas, pelos ensinamentos e convivência.
A CAPES pela concessão da bolsa. A FAPEMIG e CNPq pelo financiamento do projeto.

Moreira, N. D. Agradecimentos
vi
A todos aqueles que por um momento de distração deixei de mencionar os nomes, mas de
uma maneira ou de outra, colaboraram para que este trabalho se concretizasse,
contribuindo de modo efetivo para a sua realização.
A todos,
minha sincera gratidão.

Moreira, N. D. Sumário
vii
1. INTRODUÇÃO ................................................................................................................. 1
2. REVISÃO DE LITERATURA ......................................................................................... 4
2.1 Aspectos gerais da leishmaniose ................................................................................. 5
2.2 Aspectos imunopatológicos e curso da infecção em modelos experimentais na LV .. 8
2.3 O hamster como modelo experimental na leishmaniose ........................................... 14
3. JUSTIFICATIVA ............................................................................................................ 22
4. OBJETIVOS .................................................................................................................... 24
4.1 Objetivo geral ............................................................................................................ 25
4.2 Objetivos específicos ................................................................................................. 25
5. MATERIAL E MÉTODOS ............................................................................................. 26
5.1 Animais ...................................................................................................................... 27
5.2 Delineamento e protocolo experimental .................................................................... 27
5.3 Crescimento in vitro de formas Promastigotas .......................................................... 29
5.4 Parasitos e inóculos ................................................................................................... 29
5.5 Taxa de sobrevida ...................................................................................................... 30
5.6 Procedimentos de coleta de amostras biológicas ....................................................... 30
5.7 Avaliação sorológica ................................................................................................. 31
5.7.1 Avaliação de IgG-total por ELISA ..................................................................... 31
5.8 Avaliação do quadro hematológico e bioquímico ..................................................... 32
5.9 Necropsia dos animais e obtenção do material biológico.......................................... 32
5.10 Processamento e avaliação histológica do fígado e baço ........................................ 33
5.11 Avaliação da densidade parasitária – LDU (Leishman Donovan Units) ................. 34
5.12 Análise Molecular para quantificação da carga parasitária no fígado e baço pela técnica de qPCR .............................................................................................................. 34
5.12.1 Extração de DNA das amostras de Fígado ....................................................... 34
5.12.2 Extração de DNA das amostras de baço ........................................................... 35
5.12.3 Clonagem dos produtos da amplificação do gene de L. infantum .................... 36
5.12.4 Reação em cadeia da Polimerase - PCR ........................................................... 36
5.12.5 Clonagem .......................................................................................................... 36
5.12.6 Preparo de bactérias competentes e transformação bacteriana ......................... 37
5.12.7 Purificação dos vetores por lise alcalina .......................................................... 38
5.12.8 Construção de curvas-padrão para a qPCR em tempo real .............................. 39
5.13 Avaliação da expressão gênica por qRT-PCR ......................................................... 41
5.13.1 Desenho dos iniciadores (primers) ................................................................... 41
5.13.2 Extração e purificação do RNA total das amostras de baço ............................. 42
5.13.3 Quantificação e avaliação do grau de pureza do RNA total extraído ............... 43
5.13.4 Integridade do RNA extraído ........................................................................... 44
5.13.5 Transcrição reversa (síntese da 1ª fita de DNA complementar- cDNA) .......... 44
5.13.6 Avaliação da curva de eficiência dos primers .................................................. 45
5.13.7 Expressão de citocinas por qRT-PCR .............................................................. 46
5.14 Análises estatísticas ................................................................................................. 47
6. RESULTADOS ............................................................................................................... 48
6.1 Avaliação do perfil de crescimento in vitro das formas promastigotas de L. infantum das cepas PP75 e selvagem (C46) ................................................................................... 49
6.2 Avaliação da taxa de sobrevida nos diferentes grupos experimentais....................... 50
6.3 Avaliação clinicopatológica ...................................................................................... 50
6.4 Avaliação da resposta humoral .................................................................................. 53
6.5 Avaliação de parâmetros clínicos laboratoriais ......................................................... 54
6.5.1 Hemograma: série branca e vermelha ........................................................................ 54

Moreira, N. D. Sumário
viii
6.6 Avaliação dos parâmetros Bioquímicos .................................................................... 60
6.6.1 Provas de função renal ........................................................................................ 60
6.6.2 Prova de função hepática .................................................................................... 62
6.6.3 Proteinograma ..................................................................................................... 63
6.6.4 Proteína total ....................................................................................................... 64
6.6.5 Albumina ............................................................................................................ 64
6.6.6 Globulina ............................................................................................................ 66
6.6.7 Relação Albumina Globulina: A/G .................................................................... 67
6.8 Avaliação anatomopatológica do fígado ................................................................... 69
6.8.1 Alterações macroscópicas e microscópicas ........................................................ 69
6.9 Avaliação anatomopatológica do baço ...................................................................... 76
6.9.1 Alterações macroscópicas e microscópicas ........................................................ 76
6.10 Avaliação da carga parasitária no fígado e baço: LDU (Leishmam donovam Units) e PCR em tempo real .......................................................................................................... 82
6.10.1 LDU (Leishmam Donovam Units) .................................................................... 82
6.11 Análise de correlação: parasitismo no baço X IgG total ......................................... 85
6.12 PCR em tempo real .................................................................................................. 86
6.13 Avaliação da expressão relativa de mRNA das citocinas proinflamatórias TNF-α e IFN-γ e anti-inflamatórias (IL-10 e TGF-β) no baço ...................................................... 92
6.13.1 Citocinas proinflamatórias TNF-α e IFN-γ ...................................................... 92
6.13.2 Citocinas anti-inflamatórias IL-10 e TGF-β ..................................................... 94
6.14 Razão de TNF-α/TGF-β e TNF-α/IL-10 no baço de hamsters infectados com promastigotas de L. infantum .......................................................................................... 96
6.15 Razão de IFN-γ/TGF-β e IFN-γ/IL-10 no baço de hamsters infectados com promastigotas de L. infantum .......................................................................................... 98
7. DISCUSSÃO ................................................................................................................. 101
8. CONCLUSÃO ............................................................................................................... 124
9. REFERÊNCIAS BIBLIOGRÁFICAS .......................................................................... 128
10. ANEXO ....................................................................................................................... 144

Moreira, N. D. Lista de Abreviaturas
ix
ALT Alaninaminotransferase APCs Células apresentadoras de antígeno AST Aspartatoaminotransferase BOD Biochemical oxygen demand
CaCl Cloreto de cálcio CCA Centro de Ciência Animal CCL2 Chemokine (C-C motif) ligand 2
CCL3 Chemokine (C-C motif) ligand 3
CCZ Centro de Controle de Zoonoses Ct Ciclo Threshold CTAB Brometo de Cetiltrimetilamônio CXCL10 C-X-C motif chemokine 10
DEPC Dietilpirocarbonato DNA Ácido desoxirribonucléico dNTP Desoxirribonucleotídeos EDTA Ácido etilenodiamino tetra-acético ELISA Enzyme linked immunosorbent assay g Gramas GAPDH Gliceraldeído 3-fosfato desidrogenase HCl Ácido Clorídrico HE Hematoxilina eosina IC Intracardíaca ID Intradérmica IFN-γ Interferon gama IgG Imunoglobulina da classe G IL Interleucina IL-10 Interleucina 10 IL-12 Interleucina 12 IL-2 Interleucina 2 IL-4 Interleucina 4 iNOS Óxido nítrico sintase induzida IP Intraperitoneal IPTG Isopropyl β-D-1-thiogalactopyranoside
LB Luria Bertani LDU Leishman Donovan Units
LIT Liver infusion tryptose medium LT Linfócitos T LV Leishmaniose Visceral LVC Leishmaniose Visceral Canina LVH Leishmaniose Visceral Humana MCP-1 Monocyte chemotactic protein-1 mg Miligrama MgCl Cloreto de maganésio MIP-1α Macrophage Inflammatory Proteins alfa
mL Mililitro MnCl Cloreto de manganês MOPS 3-(N-morpholino) propanesulfonic acid
mRNA RNA mensageiro mRNA RNA Mensageiro

Moreira, N. D. Lista de Abreviaturas
x
NaCl Cloreto de sódio NCB National Center for Biotechnology Information
ng Nanograma NK Natural Killer
NNN Nicole, Novy & Neal (Meio de cultivo bifásico) NO Óxido Nítrico NUPEB Núcleo de Pesquisas em Ciências Biológicas OMS Organização Mundial de Saúde OPD Orthophenylenediamine
PBS Phosphate buffer saline (tampão fosfato salina) PCLV Programa de Controle da Leishmaniose Visceral PCR Reação em Cadeia da Polimerase pmol Picomol qRT-PCR Real Time quantitative Reverse Transcription PCR
RFLP Restriction Fragment Length Polymorphism
RIFI Reação de Imunofluorescência Indireta RNA Ácido ribonucléico RPM Rotação por minuto SFB Soro fetal bovino TGF-β Fator de transformação do crescimento beta Th1 Células T CD4+ secretoras do padrão 1 de citocinas Th2 Células T CD4+ secretoras do padrão 2 de citocinas Tm Temperatura de fusão TNF-α, Fator de necrose tumoral-α Tris Tris (Hidroximetil aminometano) UFOP Universidade Federal de Ouro Preto WHO World Health Organization (OMS) X-Gal 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside
µm Micrômetro µg Micrograma µL Microlitro

Moreira, N. D. Lista de Figuras
xi
Figura 1: Mapa do vetor p-GEM®-T Vector.. ................................................................... 37 Figura 2: Exemplo da curva padrão referente ao gene da DNA polimerase de L. infantum.. ............................................................................................................................................. 41 Figura 3: Curva padrão referente ao gene GAPDH utilizando diluição seriada de 5x de cDNA. .................................................................................................................................. 45 Figura 4: Plot de amplificação referente à curva de eficiência do GAPDH utilizando diluição seriada de 5x de cDNA. ......................................................................................... 46 Figura 5: Curva de crescimento das formas promastigotas de L. infantum das cepas PP75 e selvagem acompanhadas por 20 dias, com contagens diárias.. ........................................... 49 Figura 6: Taxa de sobrevida de hamsters pertencentes ao grupo controle e experimentalmente infectados com as cepas de L. infantum. .............................................. 50 Figura 7: Alterações macroscópicas observadas em hamsters após a infecção com a cepa selvagem em 9 meses após a infecção. ................................................................................ 51 Figura 8: Reatividade de IgG total no soro de hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum. ................................................................ 54 Figura 9: Níveis séricos de uréia e creatinina de hamsters dos grupos controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ......................................................................... 61 Figura 10: Níveis séricos de AST e ALT de hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum. ........................................................................ 63 Figura 11: Níveis séricos de proteínas totais de hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ......................................................................... 64 Figura 12: Níveis séricos de albumina de hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ...................................................................................................... 66 Figura 13: Níveis séricos globulina de hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ...................................................................................................... 67 Figura 14: Relação de albumina globulina de hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ................................................................. 68

Moreira, N. D. Lista de Figuras
xii
Figura 15: Peso dos hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L.
infantum ............................................................................................................................... 69 Figura 16: Alterações macroscópicas do fígado de hamsters controle e experimentalmente inoculados pela via intracardíaca com as cepas PP75 e selvagem de L. infantum, avaliados no tempo de 9 meses após infecção ..................................................................................... 70 Figura 17: Peso do fígado e peso relativo (peso fígado/peso corporal) de hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ............................................ 71 Figura 18: Avaliação do processo inflamatório no fígado de hamsters experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ...................................................................................................... 73 Figura 19: Fotomicrografias de secções histológicas do fígado de hamsters Mesocricetus
auratus inoculados com as cepas PP75 e C46 (selvagem) de L. infantum pelas vias intradérmica, intraperitoneal ou intracardíaca avaliados aos 6 ou 9 meses após inóculo ... 74 Figura 20: Fotomicrografias de secções histológicas do fígado de hamsters Mesocricetus
auratus inoculados com as cepas PP75 e selvagem de L. infantum pelas vias intraperitoneal ou intracardíaca avaliados aos 6 ou 9 meses após inóculo .................................................. 75 Figura 21: Alterações macroscópicas do baço de hamsters controle e experimentalmente inoculados pela via intracardíaca com as cepas PP75 e selvagem de L. infantum, avaliados no tempo de 9 meses após infecção ..................................................................................... 76 Figura 22: Peso absoluto e relativo do baço (peso baço/peso corporal) de hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ........................................... 77 Figura 23: Percentual de animais apresentando capsula normal e periesplenite de hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ........................................ 79 Figura 24: Percentual de animais apresentando aspecto histológico normal; hiperplasia da polpa vermelha variando de discreta, moderada e intensa de hamsters do grupo controle e experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. ............................................................................................................................ 79 Figura 25: Fotomicrografias de secções histológicas do baço de hamsters Mesocricetus
auratus inoculados com a cepa PP75 de L. infantum pelas vias intradérmica, intraperitoneal ou intracardíaca avaliados aos 6 ou 9 meses após a infecção ............................................. 80 Figura 26: Fotomicrografias de secções histológicas do baço de hamsters Mesocricetus
auratus infectados com a cepa selvagem de L. infantum pelas vias intradérmica, intraperitoneal ou intracardíaca avaliados aos 6 ou 9 meses após a infecção. .................. 81

Moreira, N. D. Lista de Figuras
xiii
Figura 27: Densidade parasitária no fígado de hamsters experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ........................................................................................................................... 83 Figura 28: Densidade parasitária no baço de hamsters experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L.
infantum ............................................................................................................................... 84 Figura 29: Densidade parasitária no fígado e baço de hamsters experimentalmente infectados com as cepas PP75 ou selvagem de L. infantum inoculados pelas vias IP e IC, avaliados aos 9 meses após a infecção ................................................................................ 85 Figura 30: Análise de correlação entre o número de amastigotas no baço de hamsters experimentalmente inoculados pela via intracardíaca com as cepas PP75 e selvagem de L.
infantum. Os índices de correlação de Pearson (r) e os valores p estão representados nos gráficos. ............................................................................................................................... 86 Figura 31: Número de amastigotas no fígado de hamsters experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca ...................................................... 88 Figura 32: Número de amastigotas no baço de hamsters experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca....................................................... 90 Figura 33: Expressão relativa de mRNA das citocinas proinflamatórias TNF-α e IFN-γ no baço de hamsters experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ............................................ 93 Figura 34: Expressão relativa de mRNA das citocinas anti-inflamatórias IL-10 e TGF-β no baço de hamsters experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ............................................ 96 Figura 35: Razão TNF-α/TGF-β e TNF-α/IL-10 no baço de hamsters experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum ...................................................................................................... 98 Figura 36: Razão IFN-γ/TGF-β e IFN-γ/IL-10 no baço de hamsters experimentalmente inoculados pelas vias intradérmica, intraperitoneal e intracardíaca com as cepas PP75 ou selvagem de L. infantum .................................................................................................... 100

Moreira, N. D. Lista de Tabelas
xiv
Tabela 1: Sequências de primers forward e reverse referente aos genes avaliados, função, tamanho do amplicon gerado e número de acesso das sequências no GeneBank. .............. 42 Tabela 2: Análise anatomoclínica de hamsters experimentalmente infectados com L.
infantum. .............................................................................................................................. 52 Tabela 3: Leucograma de hamsters controle e experimentalmente infectados com as cepas PP75 e selvagem de L. infantum. ......................................................................................... 56 Tabela 4: Eritrograma de hamsters controle e experimentalmente infectados com as cepas PP75 e selvagem de L. infantum. ......................................................................................... 59 Tabela 5: Número de amastigotas a partir de 20ng de DNA em amostras de fígado e baço de hamsters experimentalmente infectados com promastigotas de L. infantum.................. 91

Moreira, N. D. Resumo
xv
A leishmaniose visceral humana (LVH) é a mais temida e devastadora dentre as várias formas clínicas das leishmanioses. Esta doença negligenciada, tem ganhando grande importância devido as altas taxas de mortalidade. A espécie do parasito, imunidade, características genéticas e estado nutricional do hospedeiro desempenham papel crucial no desenvolvimento espectral das formas clínicas, podendo evoluir de uma infecção assintomática inaparente para uma forma clínica letal, em casos extremos. O tratamento quimioterápico, não é suficiente para controlar e deter a LVH em todo mundo. Um agravante importante, é a ausência de uma vacina que possa ser empregada em programas de controle da LV. Portanto, o desenvolvimento de novas estratégias profiláticas e terapêuticas contra a LV se faz necessário e urgente. Muitos modelos animais experimentais, como roedores e primatas não humanos ja foram avaliados, cada um apresentando características específicas, sendo que nenhum destes, reproduz com precisão o que acontece na LV humana e canina. Neste contexto, no presente estudo, o modelo hamster (Mesocricetus auratus) foi usado, e os animais foram divididos em dois grandes grupos experimentais: um grupo infectado com a cepa padrão da OMS (MHOM/BR/74/PP75; n=120) e outro infectado com a cepa selvagem (n=120) de L.
infantum, além de um grupo controle não infectado (C; n=80). Estes dois grandes grupos foram subdivididos em três subgrupos referentes às vias de inóculo utilizada: intradérmica (ID), intracardíaca (IC) e intraperitoneal (IP). Os animais foram avaliados durante nove meses após a infecção para a análise da sobrevida. Para cada tempo, dez animais/grupo foram avaliados. Amostras de sangue, soro, fragmentos de fígado e baço foram colhidos para realização de diferentes análises laboratoriais. As características clinicopatológicas, hematológicas, o perfil bioquímico da função renal e hepática além do quadro histopatológico foram alvos deste trabalho. Além disso, foram utilizadas abordagens moleculares empregando a PCR em tempo real quantitativa (qPCR e qRT-PCR) para determinar a carga parasitária no fígado e baço, bem como, para avaliar a expressão de citocinas no baço, respectivamente. Nossos principais resultados revelaram o aparecimento de sinais clínicos e a presença de biomarcadores laboratoriais típicos em ambas as cepas empregadas. No entanto, de um modo em geral, a cepa selvagem foi capaz de induzir doença grave, com evolução fatal principalmente nos hamsters inoculados pela via IC. As principais alterações hematológicas encontradas em hamsters que evoluíram para infecção ativa da LV foram: anemia, leucopenia acompanhada de neutropenia, linfopenia, e em alguns casos monocitopenia e eosinopenia. Os parâmetros bioquímicos relacionados à avaliação da função renal mostraram elevação de uréia e creatinina em ambas às cepas utilizadas. Já em relação à função hepática observou-se elevação nos níveis de ALT e AST nos diferentes grupos e tempos avaliados. No compartimento hepático, observou-se a instalação de processo inflamatório nos espaços-porta com a evolução da infecção, acompanhada pela formação de granulomas, com maior frequência e intensidade no grupo IC. A hipoplasia dos folículos linfóides, associada à proliferação de macrófagos da polpa vermelha, destruição da arquitetura esplênica foram mais evidentes no grupo IC, por apresentar maior gravidade da doença. A qPCR mostrou-se sensível na detecção do DNA de L. infantum tanto no fígado como no baço, em todos os grupos experimentais independente da cepa utilizada e do período de avaliação. Como observado na análise do índice de LDU o qPCR também evidenciou maior parasitismo no fígado e baço principalmente nos animais infectados com a cepa selvagem, pela via IC. Além disso foi verificado forte correlação entre parasitismo tecidual e produção de anticopos anti-IgG. A predominância da expressão de mRNA para IFN-γ, nos animais infectados com a cepa PP75 no fim do período de avaliação no grupo IC, sugere maior expressão desta citocina em resposta à replicação parasitária, induzindo um perfil inflamatório concomitante e

Moreira, N. D. Resumo
xvi
intenso. A resposta imune observada em hamsters infectados com a cepa selvagem pela via IC durante o período inicial (3 meses) foi relacionada com a expressão aumentada de IL-10 e TGF-β. Este fenômeno parece suprimir estes animais permitindo o estabelecimento da infecção ativa bem como a proliferação do parasito no baço. Neste sentido, a inoculação por via IC é rapidamente seguida pela proliferação de parasitos, exibindo sinais clínicos típicos da LV semelhantes aos observados em seres humanos e/ou cães naturalmente infectados. Os resultados obtidos no presente estudo, sugerem o uso de hamsters como modelo experimental para avaliar os aspectos imunopatológicos durante o curso da LV ativa. Além disto, considerando que os hamsters experimentalmente infectados apresentaram muitas das características clínicas e imunopatológicas como observado na LVH e LVC, este modelo experimental é altamente relevante em avaliações pré-clínicas de potenciais drogas leishmanicidas e vacinas.

Moreira, N. D. Abstract
xvii
Human Visceral Leishmaniasis (HVL) is the most dreaded and devastating parasitic disease amongst the various forms of leishmaniasis. This neglected disease, has gained immense importance because of its high mortality rate. The parasite acts on the hosts immune status and plays a crucial role in the manifestation of a spectrum of clinical syndromes, which can, in most extreme cases, go from an asymptomatic infection to a fatal form of HVL. Chemotherapy alone is not sufficient to control and contain VL spreading around of the world. The lack of vaccines to prevent and/or treat this infection, as well as the emergence of drug resistant parasites, is a serious impediment to controlling VL. Therefore, developing new prophylactic and therapeutic strategies against VL is urgently required. Many experimental animal models like rodents and monkeys have been developed, each with specific features, but none accurately reproduce what happens in humans and canine VL. The prerequisite of a reliable animal model is that it should have a considerably good correlation with the clinical situation. Moreover, it also should mimic the pathological features and immunological responses observed in humans and dogs when exposed to a strain variety of L. infantum with different virulent and pathogenicity characteristics. In this respect, herein we use hamsters (Mesocricetus auratus) subdivided into two major experimental groups according to the strain applied: 120 animals were infected with the reference L. infantum strain (WHO/MHOM/BR/74/PP75) and 120 animals were infected with the wild strain. For each strain, three different routes of inoculation were used: intradermal (ID, n=40), intraperitoneal (IP, n=40) and intracardiac (IC, n=40). Animals not infected were used as a control group (C, n=80). The animals were followed for a nine month post-infection evaluation for survival analysis. For each time point of the follow-up, ten animals per group were evaluated. Samples of blood, serum, liver and spleen were collected for the different laboratorial analysis. Clinicalpathological, hematological, the seric biochemical profile (renal and hepatic) and histopathological picture were also evaluated. In addiction we quantified the parasite burden in the liver and spleen and the cytokines immunological profiles in the spleen were also investigated throughout the study. Furthermore, molecular approaches using quantitative Real Time PCR for DNA and RNA, respectively were carried out. Ours major results revealed the appearance of the many typical clinical laboratorial biomarkers in both strains. However, in general the wild strain was more able to induce a severe disease with fatal evolution mainly occurring in hamsters inoculated by the IC route. Anemia, leukopenia accompanied by neutropenia, lymphopenia, and in some cases monocytopenia and eosinopenia were the main hematological alteration found in hamsters that evolved into an active VL infection. Considering the biochemical parameters related to the renal function an increase in urea and creatinine in both strains, were detected. Regarding liver function, it was observed an increase in the levels of ALT and AST in different groups and times points evaluated. The installation of inflammatory process in the liver compartment occurred in the portal spaces with the evolution of infection, accompanied by the formation of granulomas, with higher frequency and intensity in the IC group. Hypoplasia of lymphoid follicles was associated with the macrophages proliferation in red pulp and the destruction of tissue architecture was more evident in the IC group. The qPCR showed a higher sensitive enough to detect the presence of DNA of L. infantum in both, spleen and liver, in all experimental groups (ID, IP and IC) independent of the strain used and the period evaluated. As observed in LDU index analysis, the qPCR also demonstrated that the higher parasitism in liver and spleen when the animals were infected with the wild strain by IC route. Moreover it was found a strong correlation between tissue parasitism and production of anti-IgG total L.
infantum. The predominant expression of mRNA for IFN-γ in hamster infected with PP75 strain were detected at the end of evaluation in IC group, suggesting higher expression of

Moreira, N. D. Abstract
xviii
this cytokine in response to the parasite replication inducing an intense concomitant inflammatory profile. The cytokine immune response observed in hamsters infected with the wild strain in the IC group during the initial period (3 months) was related to highly increased expression of IL-10 and TGF-β. Τhese finding appear to suggest suppression of these animals and the establishment of the active infection and parasitic tissue proliferation. In this sense, the inoculation by IC route is rapidly followed by a fulminating parasite proliferation and displaying a typical VL clinical signs similar of the pattern observed in humans or dogs. Taken together the results obtained in the present study, it is possible suggest the use of hamsters as experimental model to evaluate the immunopathological findings during ongoing active VL. Considering that the infected hamster presented many the clinicoimmunopathological features as observed in HVL and CVL we recommend this experimental model for preclinical evaluation of potential antileishmanial drugs and vaccine.

1. INTRODUÇÃO

Moreira, N. D. Introdução
2
Um dos grandes desafios atuais dos pesquisadores para vencer a crescente
urbanização da leishmaniose visceral (LV), em diversas áreas do mundo, é a busca de
estratégias imunoprofiláticas empregando vacinas imunoprotetoras e/ou esquemas
terapêuticos capazes de promover a cura parasitológica que possam ser empregados como
alternativas aos tratamentos de primeira escolha. Entretanto, poucas são as opções para o
tratamento da LV, considerando que os atuais fármacos têm perdido a capacidade de cura
devido ao surgimento de cepas resistentes e, necessariamente, precisam compor um
tratamento de baixo custo, fundamental aos países pobres, onde a doença é mais
prevalente. No que diz respeito à imunoprofilaxia, as vacinas existentes para LV ainda não
conquistaram confiança da comunidade científica nem dos órgãos de saúde pública para
serem utilizadas em cães e humanos, em campanhas de controle governamentais (WHO,
2010; Reis et al., 2010).
Neste cenário, os investimentos realizados nos últimos anos em pesquisas pré-
clínicas e clínicas para testes com novas drogas e vacinas ainda se mostram insuficientes.
Neste sentido, torna-se evidente a necessidade da caracterização de modelos experimentais
que reflitam a história natural da LV favorecendo o desenvolvimento de ensaios pré-
clínicos e clínicos, visando avaliar novos fármacos e vacinas contra o agente etiológico da
LV e/ou LVC (Leishmaniose Visceral Canina). Neste contexto, o hamster (Mesocricetus
auratus) também conhecido como hamster sírio dourado tem sido resgatado como um dos
mais promissores modelos experimentais para o estudo da LV. No entanto, até algumas
décadas atrás este modelo só havia sido explorado no estudo de sítios imunológicos
privilegiados, em meados dos anos 40. As décadas de 60, 70 até meados da década de 80
representaram o apogeu das pesquisas utilizando a bolsa alimentar de hamster,
posteriormente substituída pela utilização de camundongos isogênicos e knokout.
Assim, é de fundamental importância ampliar os esforços com ênfase no
estabelecimento de testes de drogas e vacinas oferecendo a comunidade científica um
modelo experimental que antecedesse os testes em cães e em humanos e que permitisse
avaliar a infecção em todo seu curso crônico. Neste sentido, o presente estudo buscou
caracterizar a infecção experimental por L. infantum empregando o hamster (Mesocricetus
auratus) como modelo e duas cepas com perfis distintos de virulência e patogenicidade.
Sendo assim, buscou avaliar o perfil clínico-patológico, bioquímico-hematológico,
sorológico, parasitológico e imunológico, além da taxa de sobrevida, durante nove meses
de infecção experimental. Este trabalho buscou contribuir para a ampliação do

Moreira, N. D. Introdução
3
conhecimento da história natural da infecção experimental em hamster em seus diversos
aspectos evidenciando novos biomarcadores de monitoração laboratorial que permitirão
uma aplicação mais racional do modelo hamster em testes de drogas e vacinas contra LV.

2. REVISÃO DE LITERATURA

Moreira, N. D. Revisão de Literatura
5
2.1 Aspectos gerais da leishmaniose
As leishmanioses são um complexo de doenças causadas por protozoários da ordem
Kinetoplastida, família Trypanosomatidae e do gênero Leishmania (Ross, 1903; Laison &
Shaw, 1987). Este complexo de doenças apresenta amplo espectro de formas clínicas
dependente de vários aspectos: espécie do parasito, fatores eco-epidemiológicos,
imunidade, características genéticas e estado nutricional do hospedeiro (McMahon-Pratt &
Alexander, 2004).
A Organização Mundial de Saúde (OMS/WHO) classifica as leishmanioses em
quatro formas clínicas: cutânea, mucosa, difusa e visceral. Entretanto, a LV é a forma mais
grave, e é caracterizada em sua forma típica (Calazar) por febre alta, perda de peso,
hepatoesplenomegalia, linfadenopatia. A esse quadro, associam-se alterações laboratoriais
caracterizadas por pancitopenia, leucopenia hipergamaglobulinemia e hipoalbuminemia
(Desjeux, 2004; Piscopo & Mallia Chappuis, 2007). Além disso, a presença de anticorpos
formadores de complexos imunes depositados nos rins pode levar ao desenvolvimento de
glomerulonefrites graves (Sartori et al., 1987). A LV por sua vez pode ser subdividida em
quatro formas clínicas distintas: assintomática ou inaparente, subclínica ou
oligossintomática, LV aguda e LV crônica ou Calazar propriamente dito (Badaró et
al.,1986). No Sudão, na Etiópia e principalmente na Índia encontra-se, ainda, a
leishmaniose dérmica pós-calazar, que se manifesta após a cura da doença com o
tratamento e é caracterizada por lesões cutâneas com presença do parasito (Berman, 2006;
Piscopo & Mallia, 2006).
A leishmaniose é uma das doenças mais negligenciadas do mundo, afetando
principalmente regiões pobres e países em desenvolvimento. Atualmente as leishmanioses
afetam cerca de 12 milhões de pessoas em 88 países, estimando-se que 350-400 milhões
encontra-se em áreas de risco, sendo a incidência anual de 500.000 novos casos para LV. A
incidência da leishmaniose visceral humana é de 1,6 milhões ao ano e apesar deste enorme
número de casos, apenas 500 são notificados (Desjeux, 2004; WHO, 2010). Neste sentido,
vale ressaltar que a LV é uma doença de notificação compulsória na maioria dos países
onde ocorre, entretanto estima-se que mesmo assim, ainda ocorram muitas subnotificações
(WHO, 2010).
Assim, como em outras doenças tropicais, as leishmanioses estão relacionadas às
mudanças ambientais de caráter antroponótico que promovem o contato entre hospedeiros

Moreira, N. D. Revisão de Literatura
6
vertebrados e invertebrados. Embora as leishmanioses sejam consideradas doenças
tipicamente rurais, a recente emergência e reemergência em várias cidades do mundo as
posicionam como um grave problema de saúde pública (Ashford, 2000; WHO, 2010).
A LV possui ampla distribuição geográfica ocorrendo na Índia, Bangladesh, Nepal,
Brasil e Sudão, sendo que o Brasil contribui com 90% dos registros que ocorrem no
continente americano (Monteiro et al., 1994; Desjeux, 1996). No Brasil, em função da sua
ampla distribuição geográfica, a LV apresenta aspectos climáticos, geográficos e sociais
diferenciados, envolvendo as regiões Norte, Nordeste, Centro-oeste, Sudeste e Sul. Na
década de 90, aproximadamente 90% dos casos notificados de LV ocorreram na região
Nordeste. Na medida em que a doença expandiu para as outras regiões, essa situação vem
se modificando e, recentemente a região Nordeste representa 48% dos casos do país
(Brasil, 2009). Estes dados mostram uma forte mudança no perfil epidemilógico da LV em
nosso país que se expande e urbaniza nos últimos anos, passando a ocorrer em diversos
estados da federação que eram livres da doença.
O agente etiológico da LV no velho mundo (Índia e Leste da África) é a
Leishmania (Leishmania) donovani; na China, Ásia Central, Sudeste da Europa,
Mediterrâneo é a Leishmania (L.) infantum e América Latina a Leishmania (L.) chagasi
(Lainson & Shaw, 1987; Lukes et al., 2007). Embora diferentes no nome e localização
geográfica, técnicas moleculares e bioquímicas confirmaram que a L. infantum e L.
chagasi tratam-se da mesma espécie (Maurício et al., 2000; Lukes et al., 2007).
Atualmente a OMS tem empregado a denominação de L. infantum para o agente etiológico
da LV nas Américas, passando então a considerar apenas estas duas espécies (L. donovani
e L. infantum) como agente etiológico da LV no mundo (WHO, 2010). Diante do exposto,
neste estudo optou-se pela utilização de L. infantum.
A principal forma de transmissão do parasito ocorre por meio da picada de fêmeas
de dípteros da família Psychodidae, subfamília Phebotominae, sendo a Lutzomyia
longipalpis a principal espécie transmissora da L. infantum no Novo Mundo, embora já
tenha sido demonstrado que outras espécies de Lutzomyia, como L. evansi e L. cruzi
estejam atuando como vetores (Travi et al., 1996; dos Santos et al., 1998). No ciclo
silvestre da L. infantum as raposas são conhecidas até o momento, como supostos
reservatórios silvestres do parasito no Brasil. Duas espécies de raposas foram encontradas
naturalmente infectadas: Lycalopex vetulus no Ceará; e Cerdocyun thous no Pará e em
Minas Gerais (Shaw, 2003).

Moreira, N. D. Revisão de Literatura
7
É importante resaltar que os cães (Canis familiaris) são considerados o principal
reservatório doméstico do parasito, ocupando uma posição de destaque na cadeia
epidemiológica devido à sua proximidade com o homem no ambiente doméstico
principalmente em áreas rurais e urbanas, apresentando elevado parasitismo cutâneo
observado até mesmo nos animais assintomáticos (Tesh, 1995; Giunchetti et al, 2006; de
Queiroz et. al, 2011). Este hospedeiro apresenta uma série de manifestações da doença,
variando desde animais aparentemente sadios (assintomáticos), passando por uma fase de
sinais moderados como pequena perda de peso, adenopatia linfóide e pêlo opaco
(oligosintomáticos), podendo atingir estágios graves da doença, com intenso parasitismo
cutâneo, diferentes alterações clínicas na pele (alopecia, dermatite furfurácia, úlceras e
hiperqueratose), ceratoconjuntivite, onicogrifose, paresia dos membros posteriores,
emagrecimento e caquexia (polissintomáticos) (Abranches et al., 1991; Costa et al., 1999;
Reis, 2001; Reis et al., 2006a, 2006b, 2006c ; Giunchetti et al., 2006; Reis et al., 2009).
Além disso, podem apresentar alterações hematológicas e bioquímicas com aumento sérico
das atividades das enzimas hepáticas e elevação da uréia e creatinina (Ciaramella et al.,
1997). Além disto, tem sido observado que o agravamento dos sinais clínicos no cão
apresenta relação direta com a carga parasitária em diferentes tecidos (Reis, 2001; Reis et
al., 2006a, 2006b, 2006c; Giunchetti et al., 2006, 2008a, 2008b, 2008c).
Sendo assim, o cão representa uma abundante fonte de infecção para os
hospedeiros invertebrados, confirmando a sua importância na manutenção da transmissão
da infecção para o homem, sendo encontrado em todos os focos da doença humana,
caracterizando o principal elo na cadeia de transmissão da LV (Deplazes et al., 1995; Maia
et al., 2010).
O processo de expansão geográfica e urbanização da LV têm levado à necessidade
de se estabelecer medidas mais eficazes de controle. Hoje as estratégias de controle são
centradas no diagnóstico e tratamento precoce dos casos humanos, redução da população
de flebotomíneos no peri e intradomicilio, eliminação dos reservatórios e atividades de
educação em saúde. Além disso, o Programa de Controle da Leishmaniose Visceral
(PCLV) busca uma melhor definição das áreas de transmissão ou de risco e propõe ações
de vigilância para os municípios considerados silenciosos (Tesh, 1995).
O tratamento da LVC vem sendo estudado por vários pesquisadores, desde a
descoberta de que LV humana e canina são causadas pelo mesmo agente (Oliva et al.,
2004). Entretanto, as drogas leishmanicidas usadas com sucesso na terapia da LV humana,

Moreira, N. D. Revisão de Literatura
8
tem se mostrado de eficácia controversa no tratamento da LVC. Este fato sustenta a
posição da OMS que não recomenda a terapia antimonial convencional em cães
naturalmente infectados por Leishmania infantum (WHO, 2010). Além disto, tem sido
descrito que o uso de fármacos anti-Leishmania em cães, induz a remissão temporária dos
sinais clínicos, mas não previne a ocorrência de recidivas, por não levar a cura
parasitológica, o que pode acarretar séria consequência no âmbito epidemiológico, como
desenvolvimento de cepas resistentes a estes fármacos (Baneth et al., 2002; Moreno et al.,
2002; Oliva et al., 2004).
Diante do exposto, fica clara a necessidade do estabelecimento de um modelo
experimental que possa contribuir em estudos de terapêutica e imunoprofilaxia contra a LV
humana e canina (Nieto et al., 2011; Reis et al., 2010). Isto poderia subsidiar estudos que
possam contribuir diretamente com as estratégias de controle da LV em nosso país e no
mundo. Neste sentido, destaca-se em importância o modelo experimental hamster, que é o
foco central deste estudo.
2.2 Aspectos imunopatológicos e curso da infecção em modelos experimentais na LV
Na leishmaniose a resposta imune protetora é mediada pela imunidade celular e
resulta na morte do parasito por macrófagos ativados Scott (2003). A utilização de modelos
experimentais em estudos da leishmaniose tem apontado questões importantes relacionadas
à evolução da doença e desenvolvimento da resposta imune de resistência e
susceptibilidade. Assim, estes modelos têm admitido a avaliação de componentes do
sistema imune envolvidos na resolução da infecção e permitido obter e ampliar as
perspectivas e propor novas estratégias terapêuticas e vacinais (Hommel et al., 1995;
Carríon et al., 2006). Entretanto, os mecanismos que influenciam o direcionamento da
resposta imune ainda precisam ser esclarecidos no modelo hamster. Evidências sugerem
que, fatores como as citocinas presentes no meio durante os eventos iniciais da
diferenciação celular, a interação com células apresentadoras de antígeno (que podem
preferencialmente apresentar diferentes classes de antígenos) podem comprometer esse
processo durante a infecção por Leishmania.
Modelos experimentais como o cão e o hamster geralmente exibem diversos sinais
clínicos tipicos de LV durante a infecção experimental, à semelhança da doença humana;
em contrapartida, camundongos apresentam poucos ou nenhum sinal (Melby et al., 2001;
Requena et al., 2000; Garg & Dub, 2006; Carríon et al., 2006; Nieto et al., 2011). A

Moreira, N. D. Revisão de Literatura
9
condição de susceptibilidade ou de resistência em cada animal é controlada por fatores
genéticos próprios, mas também é característica de cada espécie de Leishmania (Hommel
et al., 1995; Nieto et al., 2011). Sabe-se que a evolução da doença visceral experimental é
dependente de uma série de fatores. Dados indicam que a capacidade de infectividade do
parasito, as vias de inóculo e a dose utilizada, bem como a forma biológica do parasito
(amastigota e promastigota) podem influenciar no curso da história natural da LV
experimental. Estes aspectos dificultam a seleção de um modelo experimental ideal, que
seja capaz de reproduzir a doença na sua forma visceral exibindo diferentes manifestações
clínicas e evoluindo da forma aguda a crônica (Melby et al., 1998; Kaye et al., 2004;
Carríon et al., 2006). Entretanto, a influência do inóculo é particularmente importante em
modelos experimentais e uma variedade de vias tem sido utilizada. Embora as vias
subcutânea e intradérmica sejam as mais próximas do que ocorre naturalmente, são poucos
os estudos com a sua utilização (Miralles et al., 1994; Handman, 2001; Rolão et al., 2007).
Nas infecções experimentais da LV, as vias intracardíaca e intraperitoneal, são as mais
empregadas para inoculação dos parasitos. A maioria dos estudos que empregam o modelo
hamster e mesmo camundongos utiliza preferencialmente as formas promastigotas do
parasito para compor a infecção experimental.
Considerando os diferentes modelos existentes nos estudos da LV experimental, o
camundongo Balb/c e o hamster (Mesocricetus auratus) tem sido os mais utilizados (Garg
& Dube, 2006). Em camundongos Balb/c a infecção por Leishmania pode variar
consideraravelmente em diferentes órgãos (baço e fígado) no mesmo animal (Engwerda &
Kaye, 2000).
Neste sentido, camundongos Balb/c são geralmente utilizados para estudar a doença
visceral devido à sua susceptibilidade. Entretanto, em camundongos infectados com L.
donovani ou L. infantum a LV crônica é estabelecida com sucesso após a infecção
endovenosa ou intradérmica, mas estes não reproduzem as características da LV humana
ou canina ativa. Neste sentido, embora a infecção progrida clinicamente durante as duas
primeiras semanas após o desafio, o parasito é controlado pela resposta imune do
hospedeiro, possivelmente devido a uma resposta do tipo 1 inerente a estes camundongos
que é montada após o inóculo experimental (Nieto et al., 2011). Desta forma, acredita-se
que seja determinante para o padrão auto-resolutivo do curso da infecção por Leishmania
no modelo murino a produção de IFN-γ pelas células T (Murray et al., 1982; Engwerda &
Kaye, 2000; Zanin et al.,2007; Nieto et al., 2011) estimuladas por uma produção elevada

Moreira, N. D. Revisão de Literatura
10
de IL-12 (Murray, 1997). Além disto, a geração de óxido nítrico sintase induzida (iNOS)
por IFN-γ seria um dos mecanismos efetores crítico envolvido no controle da replicação do
parasito (Stenger et al.,1994).
Este curso auto-limitante da LV no modelo murino pela infecção por L. donovani
ou L. infantum tem sido considerado um bom modelo de replicação inicial do parasito,
seguido por controle imunológico e infecção subclínica, mas não é um bom modelo para a
progressão da doença capaz de reproduzir a história natural da LV humana e/ou canina
ativa (Melby et al., 2001). As diferenças entre a carga parasitária no fígado, baço e medula
óssea após a infecção tem se mostrado como importante marcador de susceptibilidade e
resistência no modelo murino (Kaye et al., 2004). Na maioria das linhagens de
camundongos, os parasitos são eliminados do fígado, e a resistência à infecção hepática
resulta de uma resposta coordenada do hospedeiro envolvendo uma variedade de vias
efetoras e reguladoras específicas dentro das estruturas definidas, os granulomas.
Entretanto, os parasitos persistem no baço e medula óssea por mecanismos que ainda não
são totalmente compreendidos. Sugere-se que a persistência do parasito no baço esteja
realacionado a um defeito na formação de granulomas, acompanhada por uma variedade de
alterações patológicas, incluindo esplenomegalia, destruição da microarquitetura do tecido
linfóide e atividade hematopoiética aumentada (Kaye et al., 2004). Estudos experimentais
em camundongos utilizando a via intracardíaca de infecção, tem mostrado que o controle
da infecção no fígado provavelmente esteja relacionado a síntese de IFN- γ e TNF-α,
enquanto que a progressão da infecção no baço, a síntese inadequada destas citocinas
principalmente IFN- γ durante o estágio inicial da infecção (Mukherjee et al., 2003).
Buscando avaliar biomarcadores de susceptibilidade e resistência na LV no modelo
BALB/c foi possível determinar que tanto IFN-γ como IL-2 são fatores chave para o
desenvolvimento de uma resposta imune protetora. Contrariamente as descobertas em
modelos de leishmaniose cutânea, a IL-4 não parece ser importante fator em mediar à
susceptibilidade na LV. Outra citocina, a IL-10 com perfil anti-inflamatório e
imunomodulador, encontrada na infecção ativa também pode estar relacionada à
susceptibilidade no BALB/c assim como no homem (Goto et al., 2004). Modelos
experimentais tem claramente demonstrado o papel central desempenhado pela IL-10 na
imunopatologia e persistência de parasitos durante a história natural da LV (Kane et al.,
2001; Murphy et al., 2001). Esta citocina suprime muitas funções de células NK e de

Moreira, N. D. Revisão de Literatura
11
células T por inibir a apresentação de antígenos e a produção de citocinas proinflamatórias
como IFN-γ e IL-12 (Trinchieri et al., 2007; Nylén & Sacks et al., 2007).
A citocina TGF-β também tem revelado importante papel na modulação de
moléculas co-estimulatórias na LV murina (Gomes et al., 2000). Estudos mostraram que
TGF-β inibe a produção de óxido nítrico (NO) em macrófagos de camundongos infectados
por via intracardíaca, sugerindo que sobrevivência de parasitos no baço destes animais
esteja relacionada à elevada síntese desta citocina (Mukherjee et al., 2003).
Entretanto, a infecção experimental do hamster dourado reproduz vários aspectos
da doença humana e canina, tais como: pancitopenia marcada por leucopenia e anemia,
caquexia, hipergamaglobulinemia e ausência de resposta de células T Leishmania-
específicas após a infecção experimental (Bunn-Moreno et al., 1985; Gifawesen et al.,
1989; Mathias et al.,2001). Sendo assim, esta última disfunção tem sido atribuída à
incapacidade de células apresentadoras de antigéno (APCs) em estimular células T
específicas (Fruth et al., 1993). Isto tem sido considerado por diversos pesquisadores como
elemento chave na progressão da LV no modelo hamster, o que poderia reproduzir a forma
grave da doença humana bem como a LVC (Rodrigues et al., 1998, Vasconcellos et al.,
1996; Nieto et al., 2011). Portanto, apesar de uma forte produção de citocinas Tipo 1 (IL-2,
IFN-γ, e TNF-α) no fígado, baço e medula óssea destes animais, há replicação
descontrolada do parasito, conduzindo a um padrão de doença progressiva neste modelo
(Melby et al., 2001). Por outro lado, a produção de citocinas proinflamatórias durante a LV
ativa, não é um achado exclusivo de hamster, sendo também já demonstrado por alguns
pesquisadores em esplenócitos de cães com elevação de INF-γ simultaneamente a IL-10
(Lage et al., 2007) e em células mononucleares do sangue periférico de pacientes com LV
(Peruhipe-Magalhães et al., 2006).
Neste sentido, a hipoplasia da polpa branca do baço, granulomas hepáticos e a
deposição de substância amilóide secundária tanto no baço como no fígado tem sido
relatada como as principais características histopatológicas detectadas durante a LV
experimental em hamster (Wilson et al., 1987; Riça-Capela et al., 2003). Também foi
demonstrado que a infecção visceral em hamsters induz alterações nos hepatócitos,
principalmente no sistema endomembrana e compartimento peroxissomal, levando a
distúrbios no metabolismo do fígado (Vianna et al., 2002). Estudos recentes realizados por
Nieto et al. (2011) mostraram que após a infecção por L. infantum por via intracardíaca,
hamsters exibiram alterações histopatológicas graves tanto no baço quanto no fígado com o

Moreira, N. D. Revisão de Literatura
12
aumento da carga parasitária. Entre essas alterações, detectou-se a presença de granulomas
hepáticos em diferentes estágios de maturação, granulomas com células gigantes
parasitadas por amastigotas, assim como destruição da arquitetura esplênica a qual foi
acompanhada por uma depleção linfóide. Além disso, Mathias et al. (2001) demonstraram
que o aumento na carga parasitária no baço e fígado e dos níveis de anticorpos em
hamsters infectados com amastigotas de L. infantum aumentou progressivamente com a
evolução da infecção.
A LV experimental é marcada pelo desenvolvimento de uma resposta imune órgão-
específica, sendo o fígado e baço os principais órgãos alvos atingidos (Rousseau et al.,
2001). O fígado é resistente a reinfecção, apresenta poucos danos teciduais, sendo um local
onde a resolução da infecção aguda está associada ao desenvolvimento de granulomas
inflamatórios em torno das células de Kupffer (Stanley & Engweda, 2007). Neste sentido,
uma resposta imune eficiente em infecções por Leishmania no fígado depende da formação
destes granulomas, processo dependente da produção de quimiocinas, subsequente
recrutamento de monócitos e neutrófilos, além de linfócitos T CD4+e T CD8+ e produção
de citocinas inflamatórias gerando ativação das células infectadas (Murray et al., 2000;
Stanley & Engweda, 2007). Vale ressaltar que as células de Kupffer, macrófagos
encontrados no fígado são os principais alvos para infecção por Leishmania, possivelmente
secretam quimiocinas incluindo CCL3 (MIP-1α), CCL2 (MCP-1) e CXCL10 (IP-10) após
a infecção experimental, resultando no recrutamento inicial de monócitos e neutrófilos,
críticos no controle efetivo do crescimento do parasito (Smelt et al., 2000).
Segundo Giunchetti et al. (2008c) as principais alterações histopatológicas
observadas no compartimento hepático de cães com LV, são inflamação da cápsula,
inflamação portal, hipertrofia e hiperplasia das células de Kupffer e presença de granuloma
intralobular hepático. Em um estudo realizado com cães com classificação clinica definida
de regiões endêmicas para LV no Brasil, foram observados granulomas de tamanhos
variáveis e estes eram constituídos por macrófagos, parasitados ou não, com formas
amastigotas de L. infantum, algumas células epitelióides, pequeno número de linfócitos e
plasmócitos, e raros neutrófilos (Sant’Ana et al., 2007). Este estudo demonstrou que, cães
assintomáticos apresentaram maior número de granulomas do que os animais
oligossintomáticos e sintomáticos (Sant’Ana et al., 2007). Estudos experimentais em
modelos murinos têm mostrado que, embora o fígado seja resistente à reinfecção após a
cura, o baço mostra nenhuma proteção contra reinfecção (Murray et al., 1992; Engwerda &

Moreira, N. D. Revisão de Literatura
13
Kaye, 2000; Rousseau et al., 2001). Os parasitos infectam o baço em período mais tardio,
proliferando mais lentamente neste e na medula óssea, onde podem persistir por toda a
vida do animal (Engwerda & Kaye, 2000).
O baço desempenha um papel central na LV, pois é um sítio inicial para a geração
de resposta imune específica contra o parasito mediada por células, podendo se tornar
também um local de persistência do parasito apresentando inúmeras alterações
imunopatológicas. O baço é um órgão linfóide muito parasitado, representando um local de
interação entre a resposta do parasito e o sistema imune do hospedeiro. Esse cenário torna
o baço um importante órgão para analisar as alterações imunológicas, que levam à
disfunção da imunidade celular e à ativação de uma resposta humoral ineficaz, que são de
extrema importância na patogênese da LV (Riça-Capela et al., 2003; Santana et al., 2008).
A persistência do parasito é acompanhada por um defeito na formação do granuloma
esplênico, esplenomegalia, alteração da microarquitetura do tecido linfóide incluindo
folículos e zona marginal. O desenvolvimento das alterações histopatológicas do baço vem
sendo associado aos altos níveis de TNF-α e IL-10, simultaneamente. Neste contexto,
Stanley & Engwerda (2007) relataram que parece existir certo equilíbrio imunológico
determinando respostas que efetivamente promoverão a eliminação do parasito no fígado e
a persistência da infecção no baço.
Na leishmaniose visceral canina (LVC) os parasitos induzem uma desorganização
na estrutura celular do órgão, com a hiperplasia da polpa branca e polpa vermelha,
formação de granulomas e atrofia de folículos linfóides, que determinam a esplenomegalia
em diversos graus. A esplenomegalia parece estar associada, também, ao aumento de
células do órgão, em especial no número de monócitos e macrófagos, juntamente com
alterações na estrutura da microvasculatura e aumento do número das fibras reticulares
(Santana et al., 2008).
Diante do exposto, faz- se necessário um estudo mais detalhado da LV, com ênfase
especial nas alterações da arquitetura esplênica e hepática, bem como a investigação dos
perfis de citocinas no modelo hamster buscando evidenciar semelhanças e diferenças entre
estas alterações e biomarcadores com a doença humana e canina.

Moreira, N. D. Revisão de Literatura
14
2.3 O hamster como modelo experimental na leishmaniose
O hamster (Mesocricetus auratus) é uma espécie também conhecida como hamster
sírio dourado ou esquilo libanês porque sua origem é de uma região no norte da Síria perto
do rio Eufrates, chamado Aleppo. O hamster sírio dourado foi descrito como uma nova
espécie e originalmente nomeado “Cricetus auratus” por Walterhouse em 1839
(Walterhouse, 1840) e por razões desconhecidas no início da década de 20 ele foi
considerado extinto em seu ambiente natural. A sua utilização como modelo experimental
para estudos na LV advém da descoberta da susceptibilidade à infecção por L. donovani,
fato observado inicialmente por Yong et al. (1919) ao utilizar o hamster chinês (Cricetulus
griseus) em estudos do Kala-azar no norte da China, momento em que demonstrou vários
aspectos importantes da patologia. No final de 1920 Dr. Saul Adler famoso médico
tropicalista da Universidade Hebraica de Jerusalém foi estudar o calazar indiano. O único
modelo experimental adequado para tal naquela época era o hamster chinês. Entretanto,
este modelo tinha que ser importado do oriente e não reproduzia em cativeiro. Devido a
necessidade de um modelo para uso experimental surgiu a idéia de obter o “Cricetulus
phaeus”, um parente próximo do hamster chinês.
Em 1930, o Professor Aharoni, catedrático do Departamento de Zoologia da
Universidade Hebraica de Jerusalém, foi em uma expedição à Síria e concordou em coletar
e fornecer espécimes de “Cricetulus Phaeus” para a pesquisa do Dr. Saul Adler. Neste
mesmo ano ele coletou uma fêmea de hamster dourado com sua ninhada e apresentou a
Benachen na Universidade Hebraica. Um mês após, Benachen conseguiu produzir as
primeiras ninhadas, nascidas em cativeiro. Depois disso, uma colônia foi estabelecida no
laboratório. Dr. Saul Adler, rapidamente percebeu a importância do estabelecimento e
manutenção do hamster sírio em condições de laboratório. Alguns dos exemplares desta
linhagem foram enviados para diversos países (Inglaterra, França, Reino Unido e Estados
Unidos), afim de evitar problemas com a perda da colônia. As espécies facilmente
adaptáveis a países ocidentais tornaram-se mais comuns entre todos os hamsters (Adler,
1948). Hoje em dia, todos os animais existentes das espécies de hamsters são descendentes
dessa linhagem criada em Jerusalém (Adler, 1989).
Desde a sua descoberta, o animal tem sido largamente empregado na manutenção
de parasitos, nos estudos de seus ciclos biológicos e da relação parasito-hospedeiro
(Pearson et al., 1990). Em 1931, após um ano de criação, Adler e Theodor introduziram o
hamster “Mesocricetus auratus” na comunidade científica. Dentre os inúmeros

Moreira, N. D. Revisão de Literatura
15
protozoários, Leishmania infantum foi o primeiro parasito a ser experimentalmente
inoculado neste animal e, com êxito obtiveram a infecção do hamster para seus estudos do
calazar indiano. Ele foi rapidamente aceito e posteriormente utilizado como animal
experimental em diversos trabalhos, como estudos em febre amarela (Findlay, 1934), sífilis
(Bessemans & De Wilde, 1935), brucelose (Hoffman et al., 1968), hanseníase (Adler,
1937), pneumonia virótica (Pearson & Eaton, 1940) e tuberculose (Tchernomoretz et
al.,1940).
Desde então, o hamster “Mesocricetus auratus” se estabeleceu junto a diversos
modelos (gato, camundongo, rato, coelho e cão) como um trunfo importante entre os
animais comumente usados em trabalhos experimentais. Bruce e Hindle (1934) elegeram o
hamster como um excelente modelo para estudos em leishmaniose. Antes de 1954, foram
estudados praticamente todos aspectos da relação parasito-hospedeiro infectados por cepas
do complexo donovani, o que proporcionou uma vasta bibliografia, cobrindo aspectos
relacionados com a clínica, biologia, fisiologia, imunologia, patologia, diagnóstico e
terapêutica (Magalhaes, 1954). Aproximadamente em 1963 a utilização do hamster em
estudos da área microbiológica, odontológica e parasitológica começou a declinar, mas
estavam sendo ainda ativamente usados na pesquisa de câncer e fisiologia reprodutiva
(Magalhaes, 1968). Um estudo sistematizado do hamster em relação a outros animais foi
publicado em 1958 por Stauber. Neste trabalho, o hamster foi considerado, ao lado do
chinchila (Chinchila laniger), como extremamente susceptível à infecção por Leishmania,
ao contrário de ratos brancos e coelhos, que são resistentes a infecção (Stauber et al.,
1958).
A bolsa alimentar do hamster foi utilizada em um trabalho pioneiro para estudos da
relação parasito-hospedeiro e foi consagrado bom modelo para estudos experimentais com
Leishmania (Michalick, 1996). Neste sentido, Michalick (1996) avaliou duas espécies de
Leishmania pertencentes aos complexos braziliensis e mexicana. Neste estudo, foi
inoculado promastigotas de L. braziliensis e L. amazonensis na porção terminal da bolsa
alimentar e na pele do focinho do hamster. Resultados demonstraram que L. braziliensis,
além de determinar a infecção de ambos os locais foi capaz de disseminar para os
linfonodos, baço, fígado e pele com aumento dos níveis de IgG independente do local de
ínóculo. Em contrapartida, L. amazonensis não foi infectante para este sítio embora na pele
do focinho do animal tenha mostrado evolução clínica.

Moreira, N. D. Revisão de Literatura
16
O hamster apresenta características clínico-patológicas da LV que se assemelha a
doença humana ativa. A infecção sistêmica do hamster com amastigotas de L. donovani
resulta em aumento na carga parasitária tecidual (baço e fígado), caquexia,
hepatoesplenomegalia, pancitopenia, hipergamaglobulinemia e consequente morte
(Pearson et al., 1990). Estudos realizados por Melby et al. (2001) mostraram aumento da
carga parasitária em órgãos viscerais (baço, fígado e medula óssea) de hamsters após a
infecção com promastigotas de L. donovani. Embora detectada uma forte expressão de
citocinas Th1 (IL-2 e IFN-γ) no fígado, baço e medula óssea, a replicação de parasitos
nestes sítios levaram a uma doença progressiva, onde não foi detectada expressão de
mRNA para a enzima iNOS. Além disto, foi observada produção de TGF-β e IL-10, que
possivelmente estariam agindo na inibição dos macrófagos na geração de NO,
possibilitando a evolução da doença neste modelo experimental (Melby et al., 2001).
Além disto, estudos avaliando a cinética histopatológica da celularidade na pele
de hamsters infectados por L. donovani foram importantes para ampliar a compreensão dos
aspectos imunopatológicos envolvidos nos eventos iniciais do estabelecimento da infecção.
Neste sentido, Wilson et al. (1987) observaram que o inóculo de promastigotas de L.
donovani em hamster por via intradérmica, proporcionou a ocorrência de um infiltrado
inflamatório caracterizado por presença de fagócitos mononucleares e polimorfonucleares
após 24 horas do inóculo, com uma modificação 48 horas após o inóculo, passando a
observar então predomínio de células mononucleares (Wilson et al., 1987). Neste estudo, a
formação do granuloma foi observada entre a quarta e sexta semana após a infecção, com
predomínio de células gigantes, células epitelióides, eosinófilos e parasitos no interior de
macrófagos levando a ocorrência de necrose (Wilson et al., 1987). Este perfil do infiltrado
inflamatório dérmico seria compatível com a multiplicação de L. donovani no local do
inóculo, com subsequente visceralização do parasito e evolução da doença no modelo
hamster.
A formação do granuloma foi à principal alteração histopatológica observada no
pulmão, intestino e em especial no fígado de hamsters experimentalmente infectados com
promastigotas de L. infantum por via intradérmica (Mangoud et al., 1997). Neste estudo
observou-se que o infiltrado inflamatório no espaço porta teve início quando surgiram os
primeiros granulomas, aleatoriamente distribuídos no parênquima. Com a evolução da
infecção os granulomas mostraram-se aumentados em número e tamanho constituídos
principalmente por macrófagos com áreas de necrose e poucos linfócitos. Embora não

Moreira, N. D. Revisão de Literatura
17
tenha sido observado aumento significativo de granulomas no pulmão e intestino nas duas
últimas semanas de infecção, a lâmina própria do intestino e placas de Peyer apresentaram-
se infiltradas por macrófagos parasitados causando a ulceração da mucosa (Mangoud et al.,
1997).
A fim de avaliar a sequência da formação de granulomas hepáticos após a
infecção com amastigotas de L. infantum por via subcutânea, Laurenti et al. (1990)
demonstraram a presença de infiltrado inflamatório com muitos parasitos 7 e 15 dias após a
infecção. Neste estudo, foi observada formação inicial do granuloma após 30 dias do
inóculo com a presença de granulomas epitelióides bem definidos contendo poucos
neutrófilos entre os macrófagos e poucos parasitos após 45 dias da infecção. Neste mesmo
período foram observadas células gigantes com citoplasma contendo corpos de
Schaumann. Estes resultados apontam o importante papel do granuloma na tentativa de
controlar a infecção no sítio do inóculo, indicando que a disseminação do parasito deve
ocorrer nos primeiros 45 dias, antes do estabelecimento dos granulomas.
Em outro estudo, foi avaliada a migração de parasitos para órgãos viscerais de
hamsters após a infecção subcutânea de 107 promastigotas de L. infantum (Corbett &
Laurenti, 1998). Estes autores mostraram macrófagos parasitados no linfonodo poplíteo
logo nas primeiras 2-24 horas após a infecção, sugerindo que existe uma migração de
parasitos da pele para o linfonodo com persistência e replicação inicial nos mesmos, antes
da disseminação para outros órgãos.
Vale ressaltar que a susceptibilidade do hamster à infecção por Leishmania
depende de alguns fatores do hospedeiro (sexo e idade), os quais desempenham papel
importante na modulação do sistema imune (Anuradha, et al., 1990; Singh, et al., 2007).
Deste modo, os hormônios sexuais, tais como andrógenos, estrógenos e progesterona
podem interagir diretamente com as células do sistema imunológico, afetando assim o
desenvolvimento da resposta imune. Neste contexto, macrófagos e linfócitos, os dois
principais tipos celulares envolvidos na resposta imune frente à infecção por Leishmania,
possuem receptores para os hormônios sexuais. Assim, macrófagos e células T possuem
receptores de andrógeno (Cutolo et al., 1996; Benten et al., 1999; Wunderlich et al., 2002),
receptores de estrógeno (Cutolo et al., 1996) e progesterona (Piccinni et al., 2000; Jones et
al., 2008). Com base nestes dados, não é surpreendente inferir que o sexo pode de certo
modo afetar o curso da infecção por Leishmania (Snider et al., 2009). Corroborando com
esta hipótese, o estudo realizado por Travi et al. (2002) mostrou que hamsters machos

Moreira, N. D. Revisão de Literatura
18
infectados com L. (V.) panamensis eram mais susceptíveis à infecção, apresentavam lesões
maiores e mais graves e maior carga parasitária em linfonodos, comparados as fêmeas.
Neste sentido, a doença mais grave em hamsters machos foi associada ao aumento da
produção das citocinas IL-4, IL-10 e TGF-β, que favorecem um microambiente favorável a
manutenção e replicação do parasito no hospedeiro. De forma semelhante, Anuradha et al.
(1990) verificaram que hamsters machos infectados com L. donovani são mais susceptíveis
à doença e apresentam maior carga parasitária em relação as fêmeas. No entanto, o papel
dos hormônios sexuais específicos na exacerbação da doença/resistência é um fator que
precisa ser mais estudado.
Poucos estudos da infecção experimental utilizando L. infantum no modelo hamster
tem sido realizados e geralmente a infecção tem sido avaliada usando tanto formas
amastigotas quanto promastigotas destes parasitos (Bories et al., 1998; Rica-Capela et al.,
2003). Sabe-se que a via de inóculo é um fator determinante no curso da infecção e de uma
maneira geral, o desafio de hamsters com L. infantum e L. donovani é realizado pela via
intraperitoneal ou intracardíaca geralmente com 107-108 parasitos, um número
relativamente elevado. A infecção é geralmente estabelecida em todos os animais, mas a
disseminação do parasito e o desenvolvimento de sinais clínicos da doença dependem da
concentração de parasitos no inóculo e da via utilizada (Binhazim et al., 1993; Requena et
al., 2000; Rica-Capela et al., 2003, Kaye et al., 2004). A infecção de hamsters utilizando a
via intracardíaca, com L. donovani, foi demonstrada por Stauber (1955, 1958), em um
estudo pioneiro. Neste estudo foi demonstrado que o inóculo utilizando a via intracardíaca
geralmente induz infecções reproduzíveis enquanto a intraperitoneal promove resultados
mais variáveis (Stauber, 1955, 1958). Entretanto, a preparação e administração de inóculo
pela via intracardíaca é um procedimento que requer treinamento para ser realizado
(Wyllie et al., 2006; Dea Ayuela et al., 2007).
Estudo recente foi realizado em hamster utilizando a veia safena para inóculo de
promastigotas de L. chagasi, com o objetivo de determinar a aplicabilidade desta via de
infecção experimental em relação à via intracardíaca. Resultados mostraram êxito na
infecção em quase 100% dos hamsters avaliados (Lei et al., 2010). Por outro lado, a
facilidade na realização da infecção utilizando a via intraperitoneal, tem sido pré-requisito
para sua preferencial utilização (Ott et al., 1967). Em um estudo comparativo realizado por
Wyllie et al. (2006), utilizando as vias intraperitoneal e intracardíaca na infecção de
hamsters com amastigotas de L. donovani, consideraram a via intraperitoneal como a via

Moreira, N. D. Revisão de Literatura
19
de escolha por ser mais simples de ser utilizada, uma vez que ambas as vias foram capazes
de promover sinais clínicos nos animais. Assim, embora a infecção pela via intracardíaca
tenha induzido uma infecção mais rápida, hamsters infectados pela via intraperitoneal
demonstraram maior número de parasitos no baço.
O curso da infecção experimental utilizando diferentes doses (103-107) de L.
donovani em hamster inoculados por via intraperitoneal foi demonstrado em um trabalho
publicado por Ott et al. (1967). Neste estudo, o tempo médio de morte foi comparado a
outro estudo utilizando a via intracardíaca de infecção, Stauber (1955, 1962). Os resultados
demonstraram que em ambas as vias, o tamanho da dose inoculada é inversamente
proporcional ao tempo médio de morte. Além disso, para qualquer inóculo o tempo médio
de morte foi maior quando os parasitos foram inoculados pela via intraperitoneal a via
intracardíaca. Em outro estudo empregando-se a mesma via de inóculo (intracardíaca) e
um grande número de formas amastigotas de L. infantum (107) de origem canina,
resultaram na reprodução da LV no hamster, com elevada carga parasitária no fígado no
início da infecção em relação ao baço, hepatoesplenomegalia, caquexia e reações
inflamatórias granulomatosas (Benhazim et al., 1993).
Outro estudo com hamsters infectados utilizando um número variável (103-105) de
promastigotas de L. infantum, realizado por Requena et al. (2000) verificaram a existência
de uma associação entre a quantidade de parasitas inoculados e a sintomatologia clínica
bem como os parâmetros imunológicos. Sendo assim, alguns dos animais apresentaram
infecções subclínicas, mantendo-se assintomáticos, enquanto outros inoculados com igual
número de parasitos desenvolveram infecções sintomáticas, com sinais imunológicos e
clínicos evidentes. Dea Ayuela et al. (2007) afim de estabelecer parâmetros
imunobiológicos para teste in vivo de novos compostos antileishmanicidas, infectaram
hamsters utilizando alta (107) e baixa (105) dose de formas promastigotas de L. infantum
pela via intracardíaca. Os resultados deste estudo mostraram o estabelecimento da infecção
independente da dose administrada, porém com evolução mais rápida da doença nos
animais inoculados com alta dose (107), com sinais clínicos evidentes três meses após a
infecção e elevada carga parasitária esplênica. Estes resultados demonstram que altas doses
utilizando formas promastigotas metacíclicas por via intracardíaca, induzem infecção
sintomática em todos os animais, permitindo o estabelecimento de grupos uniformes para
tratamentos, de acordo com a condição clínica e seus constituintes.

Moreira, N. D. Revisão de Literatura
20
Rica-Capela et al. (2003) realizaram um estudo comparativo empregando formas
amastigotas e promastigotas de L. infantum inoculadas por via intraperitoneal em hamsters,
a fim de avaliar a infectividade e virulência. Estes autores observaram que em comparação
a infecção com promastigotas, o desafio com amastigotas leva a um aumento mais rápido
da carga parasitária tanto no baço quanto no fígado, sendo o baço o primeiro órgão a
revelar o parasito, sugerindo uma invasão precoce deste na infecção visceral. Além disto,
foi relatado que a presença de alguns sinais clínicos como ascite, anemia e caquexia era
independente da forma de infecção utilizada.
A distribuição do parasito em diferentes órgãos de hamsters inoculados com
amostras de medula óssea de pacientes com LV por via intraperitoneal, foi mostrada
recentemente por Oliveira et al. (2011). Os resultados obtidos por estes autores
confirmaram a indução de esplenomegalia, hepatomegalia, perda de peso e ascite, com a
presença do parasito no baço e fígado de todos os animais, além de 83,3% apresentarem
formas amastigotas no encefálo. Poot et al. (2006) infectaram hamsters com promastigotas
de L. infantum e testaram diferentes doses (108, 106, 104 e 102) e vias de inóculo
(intradérmica, intraperitoneal e subcutânea) a fim de determinar a melhor estratégia para
estudos de eficácia vacinal. Os resultados demonstraram que o desafio com alta dose de L.
infantum resultaram na disseminação do parasito independente da via de infecção. Além
disto, sinais clínicos não foram observados, mas os parasitos foram detectados facilmente
em culturas de baço, fígado, linfonodo e medula óssea.
Estes diferentes estudos que procuraram definir aspectos relacionados à infecção
experimental em hamster podem contribuir com a realização de ensaios pré-clínicos para
teste de estratégias vacinais na LV, empregando-se este modelo experimental. Neste
sentido, estudos conduzidos por Gomes et al. (2008), demonstraram que a imunização de
hamsters com plasmídeo que codifica uma proteína da saliva do vetor Lutzomyia
longipalpis, LMJ19, foi capaz de induzir proteção contra a infecção por L.infantum.
Kumari et al. (2008) avaliaram o desempenho da imunogenicidade de frações solúveis de
promastigotas de L. donovani como vacina de subunidades em hamster. Dentre as sete
frações testadas de 97.4-68 kDa, quatro administradas individualmente ou em subgrupos,
estimularam intensa resposta Th1, óxido nítrico, IFN-γ e IL-12 (Kumari et al., 2008).
Neste sentido, considerando a comprovada susceptibilidade do hamster
(Mesocricetus auratus) a diferentes espécies de Leishmania e por desenvolver sinais
clínicos e alterações histopatológicas semelhantes aos observados na LV canina e humana,

Moreira, N. D. Revisão de Literatura
21
o presente estudo propõe a avaliação da infecção experimental neste modelo utilizando
diferentes vias de inóculo (intradérmica, intraperitoneal e intracardíaca) e cepas de L.
infantum (PP75 e selvagem) apresentando diferentes graus de virulência e patogenicidade,
empregando análises histopatológicas, bioquímica/hematológicas, imunológicas e
parasitológicas.

3. JUSTIFICATIVA

Moreira, N. D. Justificativa
23
A LV é considerada um grave problema de saúde pública, pois o tratamento de
escolha desta doença emprega a mesma droga utilizada desde o início do século passado,
extremamente tóxica, e que já vem apresentando diferentes níveis de eficácia em várias
regiões do mundo. Um agravante importante, consiste no fato de não haver ainda uma
vacina que possa ser empregada em programas de controle da LV. Desse modo, a busca
por novas estratégias terapêuticas e/ou imunoprofiláticas que favoreçam o controle do
parasito torna-se de grande relevância em doenças negligenciadas como a LV. No entanto,
uma das grandes limitações para os ensaios pré-clínicos destinados ao teste de esquemas
terapêuticos ou vacinais contra LV consiste na avaliação empregando-se modelos
experimentais que reproduzam o curso progressivo da história natural da infecção. Neste
sentido, destaca-se o hamster como modelo experimental que mimetiza tanto a evolução da
doença humana como canina, podendo desta forma ser empregado em ensaios pré-clínicos
para triagem de novos fármacos ou candidatos vacinais contra LV. Neste contexto, o
presente estudo buscou estabelecer e ampliar o conhecimento de biomarcadores de
prognóstico e monitoração da infecção experimental no modelo hamster. Esta estratégia
permitirá agregar a este modelo novas abordagens investigativas no âmbito das alterações:
anatomoclínicas, bioquímico-hematológicas, histopatológicas, parasitológicas e
imunológicas (resposta celular e humoral), aumentando a amplitude laboratorial nas
avaliações terapêuticas ou vacinais contra LV em ensaios pré-clínicos. Desta forma, o
presente estudo foi determinante para obter uma caracterização detalhada da infecção
experimental do hamster empregando-se num âmbito global análises imunopatológicas que
pretendem subsidiar estudos pré-clínicos com vistas a futuras estratégias terapêuticas e
vacinais contra LV humana e canina.

4. OBJETIVOS

Moreira, N. D. Objetivos
25
4.1 Objetivo geral
• Estudar a história natural da leishmaniose visceral em hamsters (Mesocricetus
auratus) experimentalmente infectados por duas cepas de Leishmania infantum
com perfis distintos de virulência e patogenicidade.
4.2 Objetivos específicos
Nos tempos de 1, 3, 6 e 9 meses após infecção com promastigotas de L.infantum,
selecionar o melhor protocolo de infecção experimental em hamster (Mesocricetus
auratus) empregando diferentes vias de inóculo: intradérmica, intraperitoneal e
intracardíaca e duas cepas de L.infantum: MHOM/BR/74/PP75 e Selvagem com a
finalidade de:
• Avaliar a taxa de sobrevida nos diferentes modelos experimentais;
• Avaliar a presença de sinais clínicos;
• Avaliar os níveis de IgG-total;
• Avaliar os parâmetros hematológicos e bioquímicos;
• Determinar as alterações histopatológicas no fígado e no baço;
• Determinar o parasitismo no fígado e baço;
• Determinar a expressão de mRNA das citocinas IL-10 TGF-β, IFN-γ e TNF-α no
baço.

5. MATERIAL E MÉTODOS

Moreira, N. D. Material e Métodos
27
5.1 Animais
Para realização deste estudo, foram utilizados 320 hamsters (Mesocricetus auratus)
machos, com idade variando de 4-6 semanas e peso corporal de entre 50-80g. Os animais
foram nascidos e criados em racks climatizadas no Setor de Criação e após um mês foram
transferidos para o Setor de Experimentação em hamster do Centro de Ciência Animal
(CCA) da Universidade Federal de Ouro Preto (UFOP).
Os animais foram mantidos em temperatura ambiente e alojados em grupos de oito
animais em gaiolas de plástico com dimensões de 40x30x10cm, forradas com maravalha
autoclavada, sendo oferecidos água e ração comercial ad libitum, diariamente.
Este trabalho foi aprovado pela Comissão de Ética no uso de Animais da
Universidade Federal de Ouro Preto/UFOP, registrado sob o parecer Nº. 009/2009 (Anexo
1).
5.2 Delineamento e protocolo experimental
Para a realização da infecção experimental foram utilizadas duas cepas de L.
infantum, sendo uma de referência da Organização Mundial de Saúde (OMS) e outra
selvagem (C46).
A cepa selvagem foi isolada por pesquisadores de nosso grupo de pesquisa de um
cão assintomático e naturalmente infectado procedente do Centro de Controle de Zoonoses
(CCZ) do Município de Governador Valadares, Minas Gerais. Os parasitos foram isolados
por punção de medula óssea e para o isolamento e crescimento foi utilizado meio
NNN/LIT. Posteriormente os parasitos foram expandidos em meio LIT para
criopreservação e posterior caracterização e estudo de infecção experimental, segundo o
procedimento operacional padrão de cultivo de promastigotas de Leishmania do
Laboratorio de Imunopatologia, NUPEB/UFOP. O diagnóstico sorológico deste animal foi
confirmado por três técnicas: RIFI, ELISA e na imunocromatografia em papel com
antígeno rK39. Além disto, foi realizada análise molecular por meio de PCR-RFLP (de
Andrade et al., 2006; Coura-Vital et al., 2011) dos isolados do parasito e os resultados
desta reação confirmaram infecção por L. infantum em função do perfil de digestão das
bandas originadas a partir do produto obtido na PCR convencional.

Moreira, N. D. Material e Métodos
28
Desta forma, estudos prévios realizados em nosso laboratório caracterizaram a cepa
PP75 como de baixa virulência e patogenicidade e a cepa Selvagem (C46) com alta
virulência e patogenicidade.
No Fluxograma 01 encontra-se o delineamento experimental deste estudo. Os
animais foram divididos em dois grandes grupos experimentais: um grupo infectado com a
cepa (MHOM/BR/74/PP75; n=120) padrão de L. infantum da OMS e o segundo grupo
infectado com a cepa selvagem (n=120), além de um grupo controle não infectado (C;
n=80). Os dois grandes grupos foram subdivididos em três grupos referentes às vias de
inóculo utilizada: intradérmica (ID), intracardíaca (IC) e intraperitoneal (IP). A dose
empregada foi de 1 x 107 promastigotas de L. infantum em fase estacionária de crescimento
Os hamsters inoculados com ambas as cepas de L. infantum e controles foram necropsiados
(mínimo de n=10 por grupo em cada período de tempo) e avaliados 1, 3, 6 e 9 meses após
a infecção.
Fluxograma 1: Delineamento experimental relacionado ao estabelecimento do modelo de infecção para L.
infantum em hamster, utilizando 3 vias de inóculo: intradérmica (ID); intracardíaca (IC) e intraperitoneal (IP).
1x107 promastigotas de L. infantum
(fase estacionária de crescimento)
320 Hamsters (Mesocricetus auratus)
ID (n
=40)
IC (n
=40)
IP (n
=40)
C (n
=40)
MHOM/BR/74/PP75 n=160
Selvagem (C46) n=160
ID (n
=40)
C (n
=40)
Necropsia
Alterações macroscópicas/ anatomoclínicas
1 3 6 9 meses após a infecção
IP (n
=40)
IC (n
=40)
Hemograma e bioquímica
sérica
Alterações laboratoriais
Avaliação parasitológica (baço e fígado)
Alterações microscópicas
histopatológicas
RespostaImune
Histologia: baço e fígado
Microscopia ótica - LDU
qPCR
IgG-ELISAqRT/PCR- baço (TNF-α, IFN-γ, TGF-β e
IL-10)
Sinais e sintomas

Moreira, N. D. Material e Métodos
29
5.3 Crescimento in vitro de formas Promastigotas
O crescimento das formas promastigotas de L.infantum das cepas
MHOM/BR/1974/PP75 e selvagem foi realizado a partir de um inóculo inicial padrão de
107 promastigotas/mL em meio LIT acrescido de 10% de soro fetal bovino. Foi realizada
contagem diária dos parasitos por 20 dias em câmara de Neubawer a fim de analisar a
curva de crescimento das respectivas cepas definindo qual o período (intervalo de dias) em
que os parasitos se encontrassem em fase estacionária de crescimento para posterior
retirada e inóculo nos hamsters.
5.4 Parasitos e inóculos
Neste estudo foram utilizadas formas promastigotas de L. infantum em fase
estacionária de crescimento em meio LIT (Liver Infusion tryptose) e NNN (McNeal, Novy
& Nicolle). Para obtenção das formas promastigotas em fase estacionária de crescimento a
cultura foi expandida a partir de um inóculo inicial de 10 mL de meio LIT contendo entre
107 a 108 promastigotas/mL de L. infantum em crescimento logarítmico. As promastigotas
foram adicionadas a 40mL de meio de cultura NNN/LIT, distribuídas em condições
estéreis em capela de fluxo laminar (Veco, Campinas, São Paulo, Brasil), sendo
acondicionada em seguida a temperatura de 23°C ± 1°C. Após sete dias do inóculo inicial,
foi realizado o primeiro repique, depois de constatadas a viabilidade e ausência de agentes
contaminantes com auxílio do microscópio óptico, e distribuídos os 50mL de cultura em 5
novos Erlenmeyers nas mesmas condições do inóculo inicial (10mL de cultura em 40mL
de meio LIT). Ao término dos sete dias, foram obtidos 250mL da cultura, na concentração
de 107 a 108 promastigotas/mL. A cultura foi removida em capela de fluxo laminar e
transferida para tubos estéreis de polipropileno de 50mL (Falcon, Becton Dickinson,
EUA), que foram submetidos à centrifugação a 900 x g durante 15 minutos em temperatura
ambiente. Após descartar o sobrenadante, o sedimento de promastigotas foi
homogeneizado em solução salina estéril (NaCl 0,85%), e em seguida foi realizada a
centrifugação na mesma condição. Este procedimento de lavagem foi repetido por mais 1
vez. Após as etapas de lavagem, realizou-se a contagem do número de promastigotas, as
quais encontravam em quinta (cepa PP75) e sexta (Cepa selvagem) passagem.
Para os inóculos com as cepas (MHOM/BR/74/PP75) e selvagem, foi utilizado à
concentração de 1x107 promastigotas de L. infantum em fase estacionária de crescimento

Moreira, N. D. Material e Métodos
30
240 Hamsters Mesocricetus auratus
Cepa PP75 ou Selvagem (C46)
1x107 promastigotas de L. infantum
(fase estacionária de crescimento)
Intradérmica IntracardíacaIntraperitoneal
20µL 500µL 200µL
de cultura que foram concentradas em um volume final de acordo com cada uma das três
vias utilizadas como demonstrado no fluxograma a seguir:
Fluxograma 2: Vias de infecção experimental, ilustrando os diferentes inóculos empregados neste estudo: intradérmica (ID), intraperitoneal (IP) e intracardíaca (IC).
5.5 Taxa de sobrevida
Para a determinação da taxa de sobrevida os animais foram acompanhados por um
período de 9 meses, através de visitas diárias onde os animais eram vistoriados, contados e
a taxa de sobrevida determinada.
5.6 Procedimentos de coleta de amostras biológicas
A-Obtenção de amostras de sangue Antes do procedimento de necropsia dos animais foi coletado sangue em seringas
descartáveis estéreis de 1mL, através de punção intracardíaca. A coleta foi realizada após
administração de anestésico (tiopental sódico 2,5%) na dose de 30mg/kg de peso por via
intraperitoneal. Do volume total de sangue coletado, 50µL de sangue foram transferidos
para um tubo eppendorf (Hamburgo, Alemanha) destinado à realização do hemograma

Moreira, N. D. Material e Métodos
31
completo (leucograma, eritrograma e contagem de plaquetas), em contador automático de
células Auto Hematology Analyser (BC-2800 Vet, Bio-Medical Eletronics Co., Ltd.,
Mindray, China). Aproximadamente 10µL de sangue foram destinados a confecção dos
esfregaços sanguíneos em lâminas que depois de coradas pelo Panótico Rápido InstantProv
(Newprov®) serviram para contagem diferencial de células à microscopia óptica. O
restante do volume de sangue coletado foi imediatamente centrifugado a 10000rpm por 10
minutos para obtenção das amostras de soro, que foram acondicionadas em freezer -80oC
até o momento do uso para a realização dos testes sorológicos e dosagens bioquímicas.
5.7 Avaliação sorológica
5.7.1 Avaliação da IgG-total por ELISA
Foram realizadas reações imunoenzimáticas de ELISA para avaliação do perfil de
imunoglobulinas da classe IgG (Goat Anti-Hmster - Caltag Laboratories). A diluição ótima
dos conjugados foi determinada por titulação, empregando-se soros padrões positivos e
negativos, obtendo-se as diluição padrão de 1:3000 do conjugado anti -IgG.
A sensibilização das placas de poliestireno de fundo chato (MaxiSorpTM Surface,
Nunc-Immuno Plate, USA) foi procedida com 2µg/orifício de antígeno solúvel de L.
infantum (MHOM/BR/1972/BH46) em tampão carbonato e em seguida incubadas
overnight (18 horas) à temperatura de 4°C. O excesso de antígeno foi removido das placas
através de lavagens sucessivas com solução de lavagem (solução salina contendo Tween -
20 a 0,05%). Posteriormente, foi realizado o bloqueio de ligações inespecíficas aplicando
100µL/orifício de solução de PBS com 5% SFB. Em seguida, as placas foram incubadas a
37°C por 1 hora e 30 minutos, e após o término seguiu-se nova lavagem. Os soros, em
duplicata e na diluição de 1:80, foram aplicados às placas e incubados overnight (18 horas)
à temperatura de 4°C. No dia seguinte, procedeu-se a lavagem e subsequentemente o
conjugado foi aplicado. As placas foram incubadas por 2 horas a 21-25ºC seguida de nova
lavagem. Posteriormente, foram adicionados 100µL/orifício da solução de substrato
[tampão citrato fosfato 0,1M + 1µg de O-fenilenodiamino (Tablete – OPD, SIGMA, USA)
e 2µL de H2O2 a 30%], mantendo-as a 37°C, por 10 minutos. Após este período foi
adicionado 32µL/orifício de solução de ácido sulfúrico (H2SO4) a 2,5M para interromper a
reação. As leituras foram realizadas no leitor de ELISA (ELX800 Biotek Instruments VT,
USA) a 490nm para IgG e os resultados foram expressos em valores de densidade óptica

Moreira, N. D. Material e Métodos
32
(absorbância). Em todas as placas foram utilizados soros controle positivos e negativos de
hamsters. Em todos os ensaios controles (negativo e positivo) foram utilizados. Para cada
placa, o ponto de corte ou “cut off” foi estabelecido a partir da média das leituras de
absorbância de oito soros de hamsters não infectados.
5.8 Avaliação do quadro hematológico e bioquímico
Para a avaliação do perfil hematológico, foi utilizado o aparelho Auto Hematology
Analyzer (Mindray BC-2800 Vet, Hamburgo, Alemanha) para a análise global de
leucócitos, eritrócitos, hematócrito, hemoglobina e plaquetas. O leucograma foi
determinado pelo aparelho hematológico em número de leucócitos/mm3. A contagem
diferencial de células foi realizada em esfregaços sanguíneos corados com Panótico Rápido
InstantProv (Newprov®) e avaliada por microscopia óptica em objetiva de imersão. Desta
forma, foi determinado o número absoluto de neutrófilos, eosinófilos, linfócitos e
monócitos, por meio da contagem de 100 leucócitos/lâmina.
As avaliações bioquímicas consistiram das seguintes análises: dosagem de proteína
total; dosagem de albumina; globulina; relação albumina/globulina; prova de função renal
a partir da dosagem de uréia e creatinina; provas de função hepática, compreendendo a
dosagem das enzimas alanina amino transferase (ALT), aspartato amino transferase (AST).
Para a avaliação dos parâmetros foi utilizado o Sistema Bioquímico Automático (CELM
SBA-200, Barueri, SP, Brasil) e empregado Kits comerciais do Labtest (Labtest
Diagnóstica S.A., Lagoa Santa, MG, Brasil), seguindo o método descrito pelo fabricante.
5.9 Necropsia dos animais e obtenção do material biológico
Para os procedimentos de eutanásia e necropsia dos hamsters foi realizada a
administração de anestésico (tiopental sódico 2,5%) na dose de 30 mg/kg via
intraperitoneal. Atingido o plano anestésico adequado foi então administrado a dose letal
(120 mg/kg). Imediatamente após a comprovação da morte do animal, os mesmos foram
pesados e em seguida iniciados os procedimentos de necropsia e obtenção do material
biológico. Os hamsters foram necropsiados nos tempos de 1, 3, 6 e 9 meses após infecção.
Os animais foram pesados individualmente e identificados por grupo. Os hamsters foram
colocados em decúbito dorsal e a cavidade abdominal aberta com auxílio de uma tesoura
cirúrgica. O fígado e baço foram removidos de forma asséptica, analisados

Moreira, N. D. Material e Métodos
33
macroscopicamente e pesados. Baço e fígado foram subdivididos em fragmentos de
aproximadamente 20mg, sendo estes destinados a: confecção de esfregaços por aposição
para avaliação da densidade parasitária por microscopia óptica em lâminas coradas pelo
Panótico Rápido InstantProv (Newprov®), preservação em formol tamponado a 10% pH
7,2 para avaliação histopatológica e ainda fragmentos para avaliação da expressão gênica
por qRT-PCR e qPCR. Os fragmentos para qRT-PCR e qPCR foram retirados e
previamente lavados com água tratada com Dietilpirocarbonato (DEPC) 1%, envoltos em
papel alumínio autoclavado, identificados e imediatamente imersos em nitrogênio líquido
onde permaneceram até o fim da necropsia. Após conclusão do procedimento da necropsia
os fragmentos foram retirados do nitrogênio e armazenados em freezer a temperatura de -
80ºC até o momento da extração de RNA e DNA total utilizado para avaliação da
expressão gênica por qRT-PCR e carga parasitária por qPCR, respectivamente.
5.10 Processamento e avaliação histológica do fígado e baço
Fragmentos de fígado e baço de hamsters fixados em formol 10% tamponado pH
7,2 foram processados e embebidos em parafina. Sobre lâminas de vidro, previamente
desengorduradas com solução álcool-éter, foram colocados cortes histológicos com
espessura de cinco µm. Estes procedimentos operacionais são padrões do laboratório de
Imunopatologia do Núcleo de Pesquisas em Ciências Biológicas (NUPEB/UFOP),
seguindo as normas de controle de qualidade do laboratório. As lâminas obtidas foram
coradas pelo método de Hematoxilina e Eosina (HE) para análise rotineira das alterações
histopatológicas. As imagens de fígado visualizadas pela objetiva de 40x foram
digitalizadas através da microcâmera Leica DFC340FX associada ao microscópio Leica
DM5000B e todas as imagens foram analisadas pelo software de análise e processamento
de imagem Leica Qwin no Laboratório Multiusuários do Núcleo de Pesquisas em Ciências
Biológicas/NUPEB da UFOP.
O processo inflamatório no fígado foi quantificado em 20 imagens aleatórias (área
total percorrida igual a 1,5 x 106 µm2). A escolha pela análise em 20 imagens foi devido a
um estudo piloto no laboratório, onde foi demonstrado que esse número de campos era
suficiente para a obtenção de uma leitura representativa da lâmina inteira.
As alterações histopatológicas do baço e fígado avaliadas de forma qualitativa e
realizadas levando-se em consideração a extensão das alterações observadas em todo o

Moreira, N. D. Material e Métodos
34
corte histológico. As alterações histopatológicas avaliadas foram: Baço: hiperplasia
nodular, depleção de polpa branca, hiperplasia de polpa vermelha. Fígado: presença de
granulomas, inflamação periportal, fenômenos degenerativos, hiperemia.
5.11 Avaliação da densidade parasitária – LDU (Leishman Donovan Units)
Para a avaliação da carga parasitária por microscopia optica, no momento da
necropsia fragmentos de fígado e baço foram impressos em lâminas previamente limpas e
desengorduradas por método de esfregaços por aposição em lâmina. Em seguida as
lâminas foram coradas pelo Panótico Rápido InstantProv (Newprov®) e após secagem foi
determinado em cada lâmina, por microscopia óptica em objetiva de imersão, o número de
amastigotas em 1000 núcleos de células. O índice de LDU foi determinado conforme
Stauber et al. (1955), e consiste na razão do número de amastigotas/1000 células
nucleadas, multiplicando-se pelo peso do órgão (g).
5.12 Análise Molecular para quantificação da carga parasitária no fígado e baço pela
técnica de qPCR
5.12.1 Extração de DNA das amostras de Fígado
A extração do DNA foi realizada a partir de amostras de tecido do fígado. Para
obtenção do DNA das amostras foi utilizado o método de CTAB (brometo de
Cetiltrimetilamônio). Foram utilizadas quantidades de tecidos com peso de 20mg. Aos
tubos contendo os fragmentos de fígado foram adicionados 250µL de tampão de lise (Tris
5mM pH 7,5; EDTA 1mM; n-lauril sarcosina 1%). A mistura foi posteriormente incubada
em banho seco a 37°C por uma hora, sendo a suspensão homogeneizada em vórtex em
intervalos de 20 minutos. Terminado o tempo, foi adicionado 20µL de Proteinase K
(20mg/mL), homogeneizadas novamente em vórtex e incubadas overnight (18 horas) a
55ºC. Posteriormente foi adicionado 100µL de NaCl 5M e mantido por 10 minutos à 65ºC
e em seguida adicionou-se 50µL de solução CTAB 10% (v/v). As amostras foram
mantidas por 20 minutos a 65ºC e em seguida foi adicionado 400µL de Clorofórmio
(Sigma, St. Louis, MO, USA) sob agitação em vortex e, seguida as amostras foram
centrifugadas a 12000 x g em microcentrífuga (Eppendorf- Modelo 5418, NY, USA) por
10 minutos. A fase aquosa foi transferida para novo tubo e, adicionados 400µL de

Moreira, N. D. Material e Métodos
35
isopropanol (Merck, Darmstad, Alemanha), sendo precipitado por cerca de uma hora no
freezer. Após precipitação do DNA centrifugou-se novamente por 10 minutos a 12000 x g
descartando o sobrenadante. Em seguida, foi efetuada a lavagem do pellet com etanol 70%
(Merck, Darmstad, Alemanha) gelado. Subsequentemente as amostras foram
homogeneizadas suavemente por inversão do tubo durante um minuto. Em seguida as
amostras foram centrifugadas por cinco minutos a 12000 x g e o sobrenadante descartado.
Posteriormente, o precipitado foi homogeneizado em 50µL de água livre de nucleases. As
amostras foram armazenadas em geladeira a 4°C até o momento da análise da qualidade do
DNA extraído e o início da reação de qPCR.
5.12.2 Extração de DNA das amostras de baço
A extração do DNA foi realizada a partir de fragmentos de tecido do baço. Para
obtenção do DNA das amostras um kit de extração WizardTM Genomic DNA Purification
Kit (Promega, Madison, WI, USA) foi utilizado seguindo recomendações do fabricante
com modificações, conforme brevemente descrito. Foram utilizadas quantidades de tecidos
com peso de 20mg. Aos tubos contendo os fragmentos de baço foram adicionadas
proteinase K (20mg/mL) e a solução de lise nuclear. Em seguida as amostras foram
incubadas overnight (18 horas) em banho seco a 55°C. No dia seguinte foi adicionado 3µL
da solução de RNase ao lisado nuclear, seguida de homogeneização e incubação em banho
seco a 37ºC por 30 minutos. As amostras foram resfriadas por cinco minutos a 25ºC e em
seguida foi adicionado 200µL da solução de precipitação protéica. Com auxílio de um
homogeneizador tipo vórtex, as amostras foram homogeneizadas por 20 segundos em baixa
velocidade e incubadas no gelo por cinco minutos. Em seguida foram centrifugadas a
16000 x g em microcentrífuga (Eppendorf- Modelo 5418, NY, USA) durante cinco
minutos. O sobrenadante foi transferido para novo tubo, onde se adicionou 600µL de
isopropanol (Merck, Darmstad, Alemanha) e as amostras foram homogeneizadas por
inversão dos tubos. Após este procedimento, foi realizada centrifugação a 16000 x g
durante dois minutos e o sobrenadante descartado. Posteriormente foi acrescentado 600µL
de etanol 70% (Merck, Darmstad, Alemanha), a amostra homogeneizada e centrifugada a
16000 g por dois minutos, sendo o sobrenadante desprezado. As amostras foram deixadas
por 1 hora a 21-25ºC para total evaporação do etanol. Posteriormente, foi adicionado
100µL de solução de hidratação e as amostras foram mantidas em banho seco a 37º C por

Moreira, N. D. Material e Métodos
36
uma hora. Após este tempo as amostras foram mantidas a 21-25ºC por duas horas e em
seguida foram armazenadas em geladeira a 4°C até o momento da análise da qualidade do
DNA extraído e o início da reação de PCR.
5.12.3 Clonagem dos produtos da amplificação do gene de L. infantum Para a construção da curva padrão foram obtidos controles que possuíam um
número conhecido de cópias do DNA de L. infantum. O produto de amplificação
específico obtido foi clonado. A amplificação do DNA pela PCR convencional foi
realizada de uma amostra de DNA de L. infantum, utilizando o par de iniciadores (direto:
5’ TGT CGC TTG CAG ACC AGA TG 3’, e reverso: 5’GCA TCG CAG GTG TGA GCA
C 3’) que foram descritos por Bretagne et al., 2001 e tem como alvo o gene da DNA
polimerase de L. infantum (acesso no GenBank: AF009147) que é um gene de cópia única
e amplificam um fragmento de 90pb.
5.12.4 Reação em cadeia da Polimerase - PCR
Para a reação, foi utilizado DNA de L. infantum (1,0ng/µL). A reação constituiu de:
tampão 1X, 1,5mM de MgCl2, 2,0µM de dNTP, primer (5pmol), 0,76µL U Taq DNA
polimerase (Fermentas -Sinapse®), 2,5µL de DNA e H2O Milli Q totalizando um volume
de 12,5µL por poço da placa (MicroAmp® Fast Optical 96-Well, Applied Biosystems). As
condições utilizadas na reação de PCR foram: desnaturação inicial a 94°C por um minuto,
seguida por 40 ciclos a 93ºC por 30 segundos, 64°C por 1 minuto, 72°C por 1 minuto e
uma extensão final a 72°C por sete minutos. O equipamento utilizado para a realização de
PCR foi o Verit Termal Cycler 96 well (Applied Biosystems®, Califórnia, USA). Parte do
produto final da reação foi aplicada em um gel de agarose 1,5% (Invitrogen by Life
Technologies, EUA), para comprovar o resultado da amplificação pela visualização da
banda de interesse (90 pb).
5.12.5 Clonagem
Após confirmação da presença e integridade da banda de interesse, e a ausência de
outros amplicons, o produto da PCR foi inserido em um vetor plasmidial com objetivo de

Moreira, N. D. Material e Métodos
37
realizar a clonagem gênica usando o kit pGEM® T Vector Systems (Promega, EUA). O
Mapa do vetor utilizado encontra-se na Figura 1.
Para cada reação de ligação, foi preparado um mix contendo 5µL de “2X Rapid
Ligation Buffer”, 1µL de “p-GEM-T Easy Vector” e 1µL de “T4 DNA Ligase” e a razão
molar foi 8 de inserto:1 de vetor. Após ser homogeneizado, o tubo foi mantido overnight
4ºC por 16 horas para ocorrer à reação de ligação do inserto no vetor.
Figura 1: Mapa do vetor p-GEM®-T Vector. A sequência representada mostra o sitio de clonagem, região codificadora para o gene de resistência a ampicilina e β galactosidase.
5.12.6 Preparo de bactérias competentes e transformação bacteriana
A- Preparo de bactérias competentes As bactérias DH-5α foram inoculadas em 5mL de meio Luria Bertani (LB)1X
(Bacto triptona 1%p/v; extrato de levedura 0,5%p/v; NaCl 171mM) e incubadas sob
agitação 180rpm (Shaker, Innova 44), a 37ºC por 16 horas. A cultura bacteriana foi
transferida para 500mL de meio LB e novamente incubada a 37ºC a 180rpm. O
crescimento foi acompanhado até a cultura atingir uma A600 nm igual a 0,6,
correspondendo ao crescimento equivalente à metade da fase logarítmica. As bactérias
foram centrifugadas a 5000rpm durante 10 minutos a 4ºC e o sedimento homogeneizado
em 150mL de 75mM de CaCl2,10mM de Tris-HCl pH 8,0. Subsequentemente, a suspensão
bacteriana foi incubada em gelo por 20 minutos e centrifugada, como descrito previamente.
O sedimento celular foi ressuspenso em 30mL da mesma solução, acrescida de 15% de
glicerol e congelado rapidamente em mistura, gelo seco álcool e mantidas a -80ºC, até o
momento do uso.

Moreira, N. D. Material e Métodos
38
B-Transformação
A transformação bacteriana foi realizada utilizando 5µL da reação de ligação,
adicionadas a 50µL de bactéria Escherichia coli DH5-α quimiocompetentes,
descongeladas em gelo. Esta mistura foi homogeneizada levemente e mantida em gelo por
30 minutos, subsequentemente foi transferida para um banho aquecido a 42ºC por 2
minutos, e em seguida, colocada em banho de gelo por 2 minutos, sendo imediatamente
adicionado a ela 1µL de meio LB 1X sem antibióticos a cada tubo, os quais foram
incubados por 1 hora no Shaker a 200-300rpm a 37ºC. A cultura bacteriana foi
sedimentada em microcentifuga, a 1000rpm por 5 minutos. Em seguida, foi
homogeneizada em 100µL de meio LB e um volume de 50µL foi semeada em placas de
Petri contendo LB-ágar (LB acrescido de 1% de ágar) acrescido de ampicilina
(100mg/mL), 0,4µL de Isopropyl β-D-1-thiogalactopyranoside (IPTG) e 4µL de 5-bromo-
4-chloro-3-indolyl-β-D-galactopyranoside (X-Gal) 4%. Após este procedimento foi
incubada em estufa B.O.D. (Estufa D.B.O., EletroLab, Brasil) a 37ºC overnight (16 horas).
No dia seguinte, as colônias brancas, consideradas positivas, foram submetidas à
triagem por PCR conforme especificado no item 5.7.4, para a confirmação do inserto. As
colônias positivas para o inserto foram selecionadas para purificação do DNA plasmidial.
5.12.7 Purificação dos vetores por lise alcalina
Para obtenção de DNA plasmidial em pequena escala foi utilizado o kit Wizard
Miniprep (Promega,USA). O crescimento bacteriano foi feito em 10mL de meio LB com
ampicilina a 37ºC por 16 horas. Posteriormente, a cultura bacteriana foi centrifugada por 5
minutos a 5000rpm, a 4ºC. O sobrenadante foi descartado e o sedimento foi ressuspendido
em 250µL de solução de ressuspensão. Em seguida, foram adicionados 250µL de solução
de lise e a mistura foi homogeneizada por inversão 4 vezes. Subsequentemente foram
adicionados 10µL de solução de protease alcalina, a mistura foi invertida quatro vezes e
incubada por cinco minutos a temperatura ambiente. Posteriormente, foram adicionados
350µL de solução de neutralização e a mistura foi homogeneizada por inversão,
centrifugada a 13000rpm por 10 minutos a temperatura ambiente. Em seguida, o
sobrenadante foi removido e transferido para uma coluna contida em tubo coletor.
Posteriormente, a coluna foi acoplada a um tubo de microcentrífuga e centrifugada a
13000rpm por 1 minuto, a temperatura ambiente. Foram adicionados 750µL de solução de

Moreira, N. D. Material e Métodos
39
pmol/µL = µg/µL x 106pg x 1pmol x 11 µg 660pg N
lavagem a coluna, centrifugada a 13000rpm por 1 minuto. Este procedimento de lavagem
foi repetido por mais uma vez. O DNA plasmidial foi eluído em 50µL de água livre de
nucleases e mantido a - 20ºC, para posterior análise por qPCR em tempo real.
5.12.8 Construção de curvas-padrão para a qPCR em tempo real
O produto da clonagem do gene de L. infantum, foi utilizado para a construção da
curva-padrão. Os vetores contendo os insertos foram inicialmente dosados utilizando o
equipamento Nanovue (GE Healthcare Bio Sciences AB Sweden) a 260nm e 280nm. A
partir da dosagem obtida, calculou-se a concentração de DNA em µg/µL, de acordo com a
fórmula:
Onde;
� N é o número de nucleotídeos (plasmídeo e inserto);
� 660pg é a massa molecular média de um ácido nucléico.
Levando-se em consideração o número de Avogadro: 1mol = 6,02x1023 moléculas, e
sabendo-se que a concentração de cada solução de plasmídeo+inserto de interesse em
pmol/µL, foi possível determinar o número de moléculas presentes por microlitro de cada
amostra.
Depois de realizados estes cálculos, as amostras foram diluídas sucessivamente,
sendo que aquelas contendo de 107 até 100 moléculas foram utilizadas para a construção
das curvas-padrão. As reações de qPCR foram realizadas em placas de 96 poços -
MicroAmp®Optical 96 - Well Reaction Plate with Barcode (Applied Biosystems by Life
Technologies, EUA), cobertas com adesivos ópticos- Optical Adhesive Covers (Applied
Biosystems, EUA) e processadas em termociclador ABI Prism 7500 Sequence Detection
System (Applied Biosystems, EUA) do Laboratório de Equipamentos Multiusuários do
NUPEB/UFOP.
Os controles positivos de cada placa foram às amostras diluídas de plasmídeo
contendo o inserto utilizado na curva padrão; e como controle negativo foi usado água livre
de nucleases, ao invés de DNA. A reação de cada amostra foi realizada em duplicata, com
as seguintes condições: uma desnaturação inicial a 95ºC por 10min, seguida por 40 ciclos

Moreira, N. D. Material e Métodos
40
de 95ºC por 15segundos e 60ºC por 1minuto. Ao final das reações a temperatura da
máquina é elevada gradualmente até que todas as fitas duplas de material amplificado se
dissociem, para a verificação de possível contaminação dos DNA’s das amostras em
estudo com DNA genômico ou dímeros dos iniciadores. As reações foram realizadas
utilizando-se SYBR Green PCR Master Mix (Applied Biosystems, EUA); DNA (5ng/µL);
iniciadores (5pmol/µL) e água livre de nucleases em quantidade suficiente para um volume
final de 10µL por poço. Para a quantificação do número de moléculas de DNA de
Leishmania nas amostras, inicialmente foi determinado para cada poço o número de ciclos
em que a fluorescência cruzou uma linha limiar arbitrária, denominada threshold (Ct),
calculada pelo programa 7500 Software v.2.0.1 for 7500 and 7500 Fast Real-Time PCR
Systems (Applied Biosystems, EUA). A quantidade de cópias de DNA em cada amostra foi
determinada a partir de uma regressão linear usando os valores do Ct das amostras
utilizadas na curva padrão, gerada com quantidades conhecidas dos plasmídeos com os
insertos de interesse.
O procedimento descrito acima foi também realizado para a amplificação e
quantificação do gene de TNF-α, que é expresso de forma constitutiva e por este motivo
foi utilizado para verificar a integridade dos DNA’s analisados. Para a amplificação do
gene TNF-α, foram utilizados os iniciadores direto: 5’ GAG TGG CTG AGC CAT CGT
3’ e reverso: 5’ TGG TTG TCT TTG AGA GAC ATG 3’, que amplificam um fragmento
de 86pb (acesso no GenBank: AF046215). As reações foram realizadas utilizando-se SYBR
Green PCR Master Mix (Applied Biosystems, EUA); DNA (5ng/µL); iniciador (2,5
pmol/µL) e água livre de nucleases em quantidade suficiente para um volume final de
10µL por poço. Para a análise dos resultados foram consideradas as reações com eficiência
entre 98-100% e curva padrão com valores satisfatórios de coeficiente de linearidade (r2 =
0,99). Segue abaixo como exemplo, a curva padrão referente ao gene da DNA polimerase
utilizada neste estudo (Figura 2).
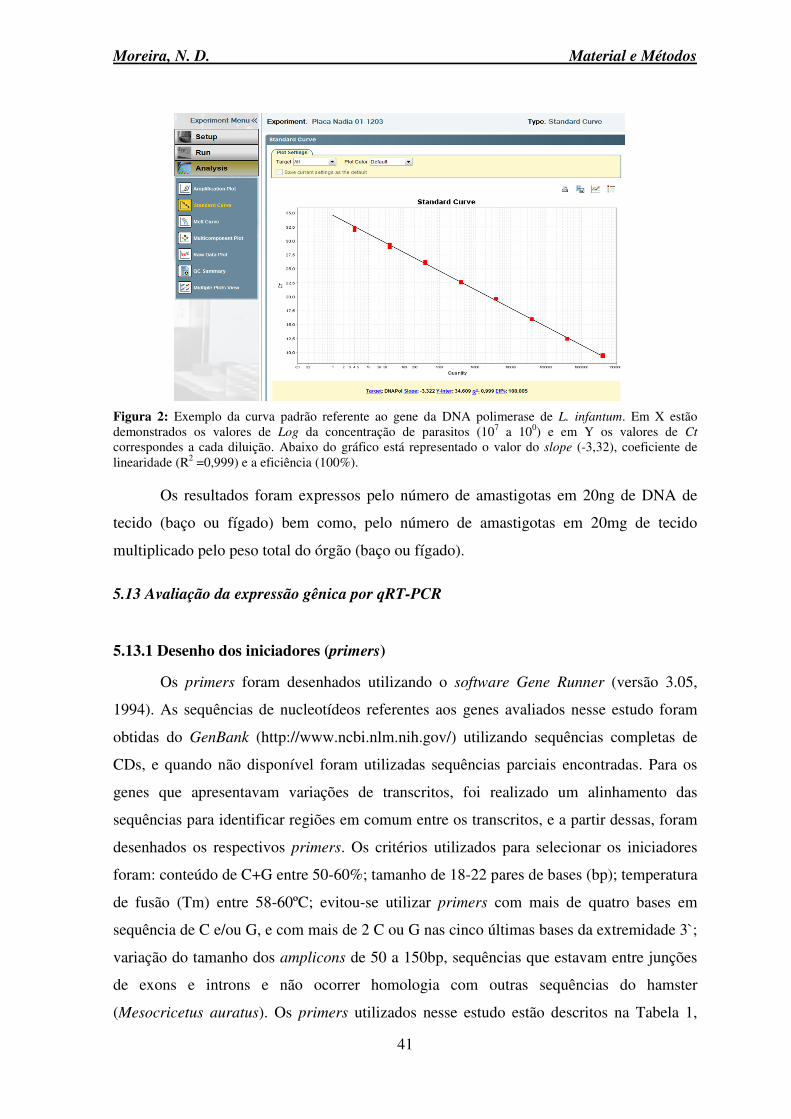
Moreira, N. D. Material e Métodos
41
Figura 2: Exemplo da curva padrão referente ao gene da DNA polimerase de L. infantum. Em X estão demonstrados os valores de Log da concentração de parasitos (107 a 100) e em Y os valores de Ct
correspondes a cada diluição. Abaixo do gráfico está representado o valor do slope (-3,32), coeficiente de linearidade (R2 =0,999) e a eficiência (100%).
Os resultados foram expressos pelo número de amastigotas em 20ng de DNA de
tecido (baço ou fígado) bem como, pelo número de amastigotas em 20mg de tecido
multiplicado pelo peso total do órgão (baço ou fígado).
5.13 Avaliação da expressão gênica por qRT-PCR
5.13.1 Desenho dos iniciadores (primers)
Os primers foram desenhados utilizando o software Gene Runner (versão 3.05,
1994). As sequências de nucleotídeos referentes aos genes avaliados nesse estudo foram
obtidas do GenBank (http://www.ncbi.nlm.nih.gov/) utilizando sequências completas de
CDs, e quando não disponível foram utilizadas sequências parciais encontradas. Para os
genes que apresentavam variações de transcritos, foi realizado um alinhamento das
sequências para identificar regiões em comum entre os transcritos, e a partir dessas, foram
desenhados os respectivos primers. Os critérios utilizados para selecionar os iniciadores
foram: conteúdo de C+G entre 50-60%; tamanho de 18-22 pares de bases (bp); temperatura
de fusão (Tm) entre 58-60ºC; evitou-se utilizar primers com mais de quatro bases em
sequência de C e/ou G, e com mais de 2 C ou G nas cinco últimas bases da extremidade 3`;
variação do tamanho dos amplicons de 50 a 150bp, sequências que estavam entre junções
de exons e introns e não ocorrer homologia com outras sequências do hamster
(Mesocricetus auratus). Os primers utilizados nesse estudo estão descritos na Tabela 1,

Moreira, N. D. Material e Métodos
42
assim como a sequência de nucleotídeos, a função, o número de acesso ao GeneBank
(NCBI) e o tamanho dos amplicons.
Tabela 1: Sequências de primers forward e reverse referente aos genes avaliados, função, tamanho do amplicon gerado e número de acesso das sequências no GeneBank.
5.13.2 Extração e purificação do RNA total das amostras de baço
Na etapa de extração do RNA total, foram utilizadas quantidades de tecidos com o
peso de 20mg. Aos tubos contendo os fragmentos de baço foram adicionados 1mL de
TRIzol® Reagent (Invitrogen São Paulo, SP, Brasil). As amostras foram maceradas
utilizando um homogeneizador de tecidos e todos os instrumentos utilizados no processo
de maceração, foram lavados com água tratada com Dietilpirocarbonato (DEPC) e
posteriormente limpos com gaze embebida com um removedor de nucleases (RNAses)
RNAseAway (Invitrogen São Paulo). Posteriormente, o homogeneizado de cada amostra
foi transferido para um tubo “eppendorf” de 1,5mL e mantidos no gelo até que todas
fossem maceradas. Após o término as amostras foram retiradas do gelo e incubadas à
temperatura de 21-23ºC para permitir a completa dissociação dos complexos de
nucleoproteínas. Foram então adicionados 0,2mL de clorofórmio (Sigma, St. Louis, MO,
USA) para cada 1mL de TRIzol® aos tubos contendo o homogeneizado e em seguida
Gene Sequência de Nucleotídeos (5`-3`) Função Tamanho do
Amplicon (pb)
Número de acesso
GeneBank
GAPDH Constitutivo 80 U10983 F TGGAGTCTACTGGCGTCTTC R GGAGATGATGACCCTCTTG
IFN-γγγγ Citocina 82 AF034482 F CAGATCGTCTCCTTCTACTTC R CCTTGATGGTGTCTATGC
TNF- αααα Citocina 86 AF046215 F GAGTGGCTGAGCCATCGT R TGGTTGTCTTTGAGAGACATG
IL-10 Citocina 150 AF046210 F CTTGAAGACGCCTTTCTC R GAGCATTTGAACTCCCTAG
TGF-ββββ Citocina 125 AF046214 F GGCTGGAAGTGGATTCAC R GGGTTGTGTTGGTTGTAG

Moreira, N. D. Material e Métodos
43
foram fechados e agitados vigorosamente por 15 segundos e incubado por cinco minutos a
temperatura ambiente. Posteriormente, as amostras foram centrifugadas por 10 minutos a
12000 x g em centrífuga (Eppendorf centrifuge 5810 R, NY, USA). A fase aquosa foi
transferida para um novo tubo de 1,5mL, seguido da adição de volume equivalente de
etanol 95% (v/v) (preparado com água tratada com DEPC) e homogeneizado suavemente
invertendo o tubo por três vezes para precipitação do RNA total. Para purificação do RNA
total contido na amostra, utilizou-se o kit de extração de RNA total (SV total RNA
Isolation System - Promega, Madison, WI, USA) com algumas adaptações descritas a
seguir. O RNA obtido como descrito acima, foi transferido para um “spin basket” acoplado
a um tubo coletor de 2mL e centrifugado a 12000 x g por dois minutos em centrífuga. O
líquido residual no tubo coletor foi descartado após as centrifugações. Foram então
adicionados 600µL de solução de lavagem (SV RNA Wash Solution) e em seguida
centrifugado a 12000 x g por dois minutos. As amostras de RNA foram tratadas com
DNase para assegurar a ausência de contaminação por DNA genômico. Para isso, foi
preparado um mix contendo 40µL de “yellow core buffer”, 5,0µL MnCl2 0,09M e 5µL de
DNAse e aplicado sobre a membrana do spin e incubado por 25 minutos a temperatura de
21-25ºC. Posteriormente, foi adicionado 200µL de “SV DNAse Stop Solution” para inibição
da atividade enzimática da DNAse. Realizou-se então a centrifugação a 12000 x g por dois
minutos e o líquido contido no tubo coletor foi descartado. Foram adicionados 600µL de
solução de lavagem (SV RNA Wash Solution) e centrifugado por 12000 x g por dois
minutos. Adicionou-se então, 250µL de solução de lavagem (SV RNA Wash Solution),
seguido de centrifugação a 14000 x g por três minutos. Posteriormente, o spin foi
transferido para um tubo de eluição e adicionou-se 100µL de água livre de nucleases
seguido de centrifugação a 12000 x g por 2 minutos. As amostras foram armazenadas em -
80ºC até o momento do uso.
5.13.3 Quantificação e avaliação do grau de pureza do RNA total extraído
Para a quantificação e avaliação do grau de pureza do RNA extraído as amostras
foram dosadas utilizando o equipamento Nanovue (GE Healthcare Bio Sciences AB
Sweden). Foi utilizado 2µL de RNA total e o grau de pureza em relação à presença de
proteínas, foi avaliado utilizando a relação dos valores de absorbância obtidos
A260/280nm. O grau de contaminação por outros compostos (como sais, polissacarídeos e

Moreira, N. D. Material e Métodos
44
compostos orgânicos como fenol) foi avaliado pela absorbância A260/230nm. As amostras
que apresentaram absorbância A260/280nm maior ou igual a 2 foram consideradas com
um grau de pureza satisfatório.
5.13.4 Integridade do RNA extraído
A separação eletroforética das amostras de RNA total foi realizada em gel de
agarose em condições desnaturantes e específicas para RNA. Sendo assim, 0,6g de agarose
foram dissolvidas em 40mL de água tratada com DEPC. A mistura foi aquecida com
auxílio de um microondas convencional. Após a agarose atingir uma temperatura de 45º foi
adicionado na mesma uma segunda solução composta por 12,8mL de água tratada com
DEPC, 6mL de tampão MOPS 10X e 1,2mL de formaldeído (Sigma). Após
homogeneização, a solução foi vertida sobre o aparato de eletroforese com pente
apropriado e aguardado a completa solidificação. Paralelamente, foi preparado o tampão de
amostra contendo 93,75µL de formamida (Sigma), 13,75µL de formaldeído, 18,75µL de
MOPS 10X, 21µL de água (tratada com DEPC), 0,25µL de azul de bromofenol (100
mg/mL) e 0,25µL de brometo de etídio (Sigma). Foi utilizado 5µg das amostras de RNA
para 10µL de tampão de amostra e incubado por cinco minutos a 65ºC (desnaturação do
RNA) seguido de banho de gelo por três minutos. Posteriormente, as amostras foram
aplicadas ao gel e corridas a 90V em tampão MOPS 1X. As amostras consideradas
adequadas para avaliação dos níveis de transcritos de mRNA foram as que apresentavam
bandas íntegras referentes as subunidades do RNA ribossomal.
5.13.5 Transcrição reversa (síntese da 1ª fita de DNA complementar- cDNA)
A primeira fita de cDNA, foi sintetizada utilizando o kit “High Capacity cDNA
Reverse Transcription kit” (PE Applied Biosystems, Foster City, CA, USA) de acordo com
as recomendações do fabricante. Para isso, adicionou-se em um tubo 1µg de RNA total
presente em um volume final máximo de 10µL. Posteriormente, foi adicionado 10µL do
mix de transcrição reversa (RT Buffer 10X, dNTP Mix (100Mm) 25X, RT Random
primers 10X, enzima Multiscribe, Rnase inhibitor e água livre de nucleases) em cada tudo
de 0,2mL contendo o RNA. Além disso, foi adicionado um controle negativo contendo
todos os reagentes menos a amostra, com o objetivo de se verificar possíveis
contaminações dos reagentes. Subsequentemente, os tubos contendo as amostras foram
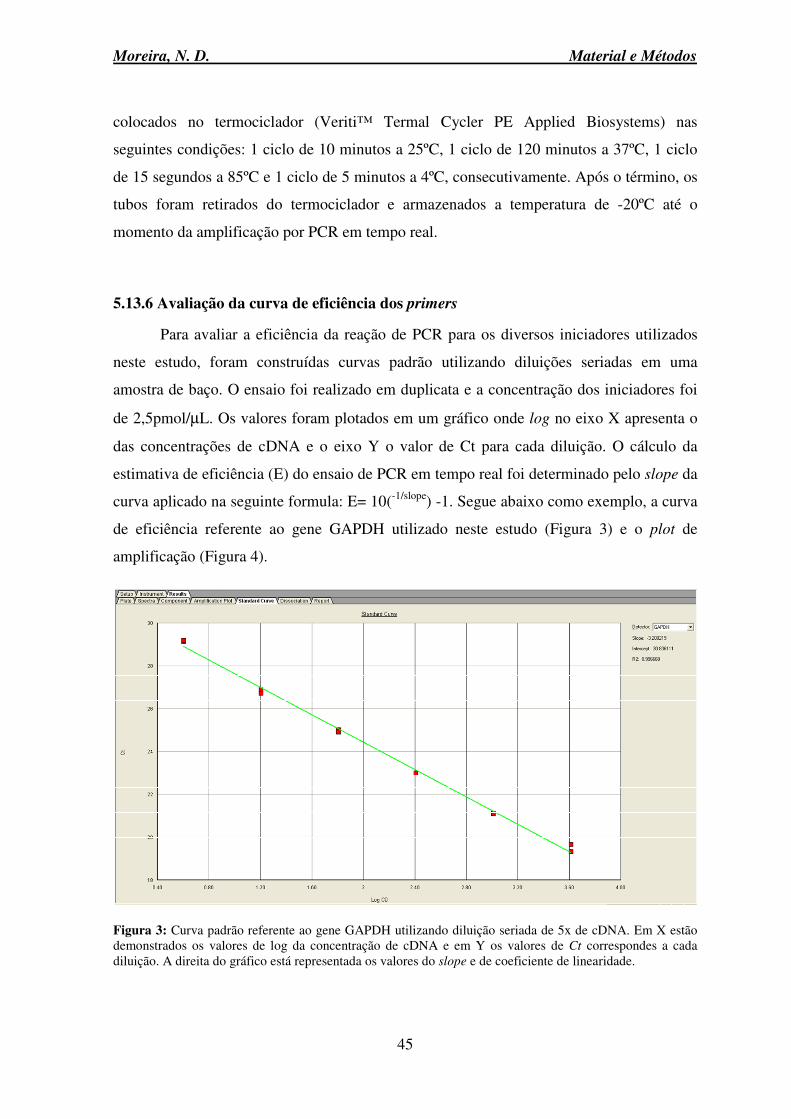
Moreira, N. D. Material e Métodos
45
colocados no termociclador (Veriti™ Termal Cycler PE Applied Biosystems) nas
seguintes condições: 1 ciclo de 10 minutos a 25ºC, 1 ciclo de 120 minutos a 37ºC, 1 ciclo
de 15 segundos a 85ºC e 1 ciclo de 5 minutos a 4ºC, consecutivamente. Após o término, os
tubos foram retirados do termociclador e armazenados a temperatura de -20ºC até o
momento da amplificação por PCR em tempo real.
5.13.6 Avaliação da curva de eficiência dos primers
Para avaliar a eficiência da reação de PCR para os diversos iniciadores utilizados
neste estudo, foram construídas curvas padrão utilizando diluições seriadas em uma
amostra de baço. O ensaio foi realizado em duplicata e a concentração dos iniciadores foi
de 2,5pmol/µL. Os valores foram plotados em um gráfico onde log no eixo X apresenta o
das concentrações de cDNA e o eixo Y o valor de Ct para cada diluição. O cálculo da
estimativa de eficiência (E) do ensaio de PCR em tempo real foi determinado pelo slope da
curva aplicado na seguinte formula: E= 10(-1/slope) -1. Segue abaixo como exemplo, a curva
de eficiência referente ao gene GAPDH utilizado neste estudo (Figura 3) e o plot de
amplificação (Figura 4).
Figura 3: Curva padrão referente ao gene GAPDH utilizando diluição seriada de 5x de cDNA. Em X estão demonstrados os valores de log da concentração de cDNA e em Y os valores de Ct correspondes a cada diluição. A direita do gráfico está representada os valores do slope e de coeficiente de linearidade.

Moreira, N. D. Material e Métodos
46
Figura 4: Plot de amplificação referente à curva de eficiência do GAPDH utilizando diluição seriada de 5x de cDNA.
5.13.7 Expressão de citocinas por qRT-PCR
O ensaio para avaliar a expressão gênica das citocinas (TNF-α, IFN-γ, TGF-β, IL-
10) nas amostras de baço, foi realizado utilizando os iniciadores (primers) na concentração
de 2,5p/mol, SYBR™ Green (Applied Biosystems, EUA) e cDNA diluído 5x em água
livre de nucleases, perfazendo um volume final de 10µL de reação. O ensaio foi realizado
em duplicata para todos os genes e o gene normalizador presente na mesma placa que os
genes avaliados. Os resultados foram expressos pelo método CT comparativo (2-∆∆Ct) que
consiste de uma quantificação relativa que utiliza fórmulas aritméticas para determinar as
diferenças na expressão de um alvo de uma amostra comparando com a sua expressão em
uma amostra controle. Sendo assim, neste tipo de análise é realizada uma normalização de
expressão deste alvo para cada amplificação, ou seja, sua expressão é subtraída pela
expressão de um gene constitutivo que é expresso de maneira semelhante em diferentes
tecidos e em diferentes condições. Assim, para cada amostra alvo foram amplificados os
genes das citocinas (TNF-α, IFN-γ, IL-10 e TGF-β) e o gene constitutivo (GAPDH), de
forma a se obter os valores dos Threshold Cycles (Ct) correspondentes e a realização dos
cálculos para obtenção dos resultados. Para se realizar uma análise com base nos valores
do ∆∆Ct faz-se necessário que as eficiências entre alvos (citocinas) e o gene endógeno
sejam semelhantes. Assim, os cálculos para a determinação do ∆∆Ct foram realizados a
partir do método, onde as diferenças na expressão gênica são calculadas baseando-se

Moreira, N. D. Material e Métodos
47
apenas nas eficiências das amplificações e as diferenças de Cts do alvo na amostra a no
controle.
5.14 Análises estatísticas
As análises estatísticas foram realizadas usando o GraphPad Prism 5.0 (Prism
Software, Irvine, CA, USA). Depois de demonstrada a normalidade dos dados usando o
teste Kolmogorov-Smirnoff, foi realizada análise de variância (ANOVA one-way) seguida
do teste de Tukey para determinar as diferenças específicas entre os grupos. Além disto,
foram realizadas análises de correlações entre as variáveis pelo método de correlação de
Pearson. Em todas as análises estatísticas as diferenças foram consideradas significativas
quando o valor de P foi menor que 0,05.

6. RESULTADOS
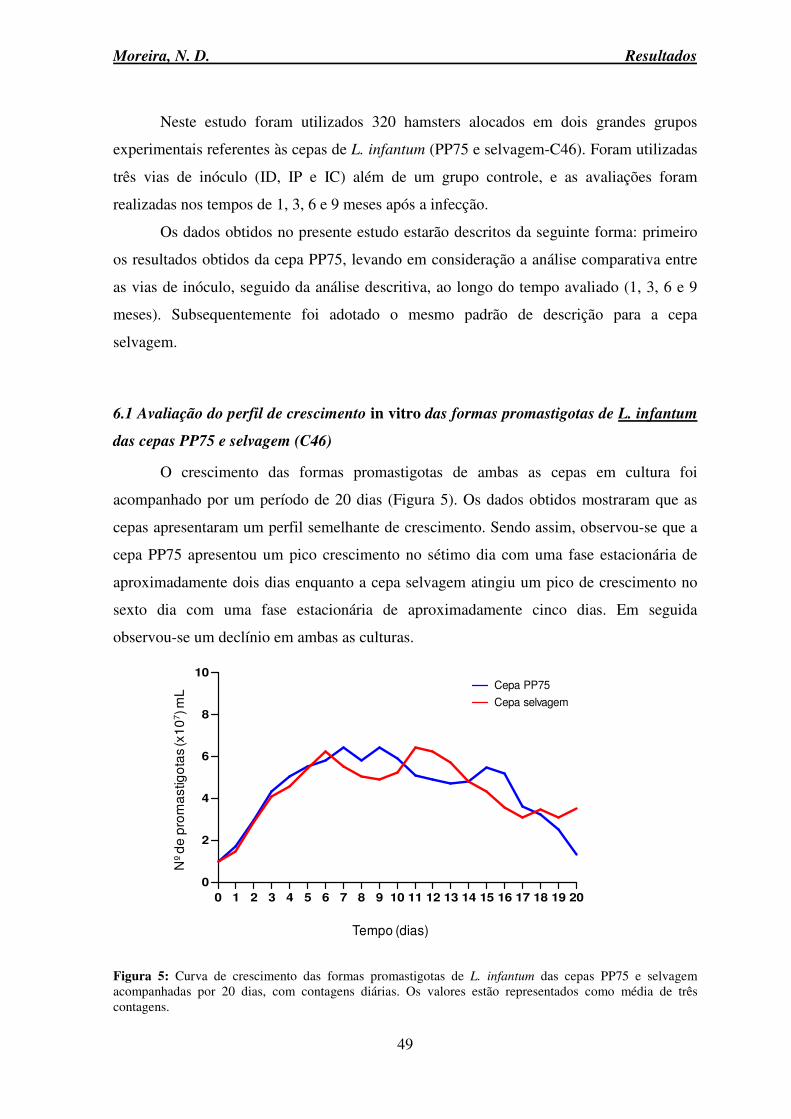
Moreira, N. D. Resultados
49
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
0
2
4
6
8
10
Nº d
e p
rom
ast
igo
tas
(x1
07) m
L
Tempo (dias)
Cepa PP75
Cepa selvagem
Neste estudo foram utilizados 320 hamsters alocados em dois grandes grupos
experimentais referentes às cepas de L. infantum (PP75 e selvagem-C46). Foram utilizadas
três vias de inóculo (ID, IP e IC) além de um grupo controle, e as avaliações foram
realizadas nos tempos de 1, 3, 6 e 9 meses após a infecção.
Os dados obtidos no presente estudo estarão descritos da seguinte forma: primeiro
os resultados obtidos da cepa PP75, levando em consideração a análise comparativa entre
as vias de inóculo, seguido da análise descritiva, ao longo do tempo avaliado (1, 3, 6 e 9
meses). Subsequentemente foi adotado o mesmo padrão de descrição para a cepa
selvagem.
6.1 Avaliação do perfil de crescimento in vitro das formas promastigotas de L. infantum
das cepas PP75 e selvagem (C46)
O crescimento das formas promastigotas de ambas as cepas em cultura foi
acompanhado por um período de 20 dias (Figura 5). Os dados obtidos mostraram que as
cepas apresentaram um perfil semelhante de crescimento. Sendo assim, observou-se que a
cepa PP75 apresentou um pico crescimento no sétimo dia com uma fase estacionária de
aproximadamente dois dias enquanto a cepa selvagem atingiu um pico de crescimento no
sexto dia com uma fase estacionária de aproximadamente cinco dias. Em seguida
observou-se um declínio em ambas as culturas.
Figura 5: Curva de crescimento das formas promastigotas de L. infantum das cepas PP75 e selvagem acompanhadas por 20 dias, com contagens diárias. Os valores estão representados como média de três contagens.
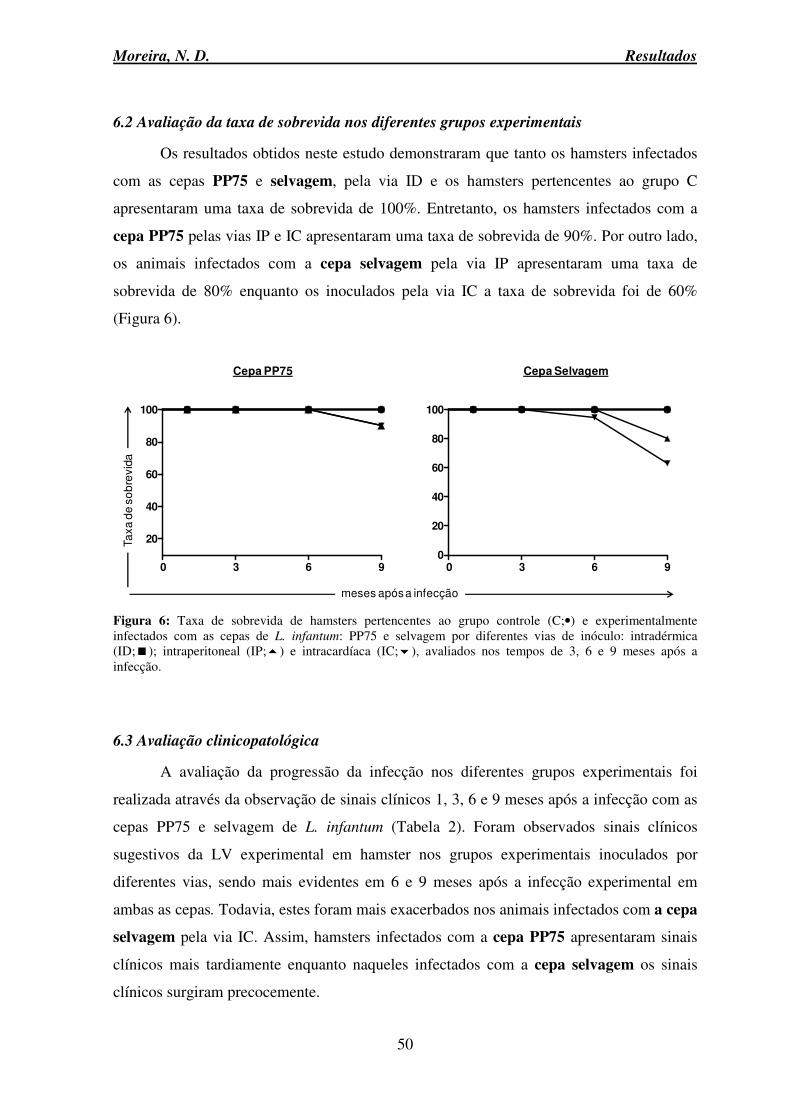
Moreira, N. D. Resultados
50
0 3 6 9
20
40
60
80
100
0 3 6 90
20
40
60
80
100
Taxa
de
so
bre
vida
meses após a infecção
Cepa SelvagemCepa PP75
6.2 Avaliação da taxa de sobrevida nos diferentes grupos experimentais
Os resultados obtidos neste estudo demonstraram que tanto os hamsters infectados
com as cepas PP75 e selvagem, pela via ID e os hamsters pertencentes ao grupo C
apresentaram uma taxa de sobrevida de 100%. Entretanto, os hamsters infectados com a
cepa PP75 pelas vias IP e IC apresentaram uma taxa de sobrevida de 90%. Por outro lado,
os animais infectados com a cepa selvagem pela via IP apresentaram uma taxa de
sobrevida de 80% enquanto os inoculados pela via IC a taxa de sobrevida foi de 60%
(Figura 6).
Figura 6: Taxa de sobrevida de hamsters pertencentes ao grupo controle (C;•) e experimentalmente infectados com as cepas de L. infantum: PP75 e selvagem por diferentes vias de inóculo: intradérmica (ID;�); intraperitoneal (IP;�) e intracardíaca (IC;�), avaliados nos tempos de 3, 6 e 9 meses após a infecção.
6.3 Avaliação clinicopatológica
A avaliação da progressão da infecção nos diferentes grupos experimentais foi
realizada através da observação de sinais clínicos 1, 3, 6 e 9 meses após a infecção com as
cepas PP75 e selvagem de L. infantum (Tabela 2). Foram observados sinais clínicos
sugestivos da LV experimental em hamster nos grupos experimentais inoculados por
diferentes vias, sendo mais evidentes em 6 e 9 meses após a infecção experimental em
ambas as cepas. Todavia, estes foram mais exacerbados nos animais infectados com a cepa
selvagem pela via IC. Assim, hamsters infectados com a cepa PP75 apresentaram sinais
clínicos mais tardiamente enquanto naqueles infectados com a cepa selvagem os sinais
clínicos surgiram precocemente.
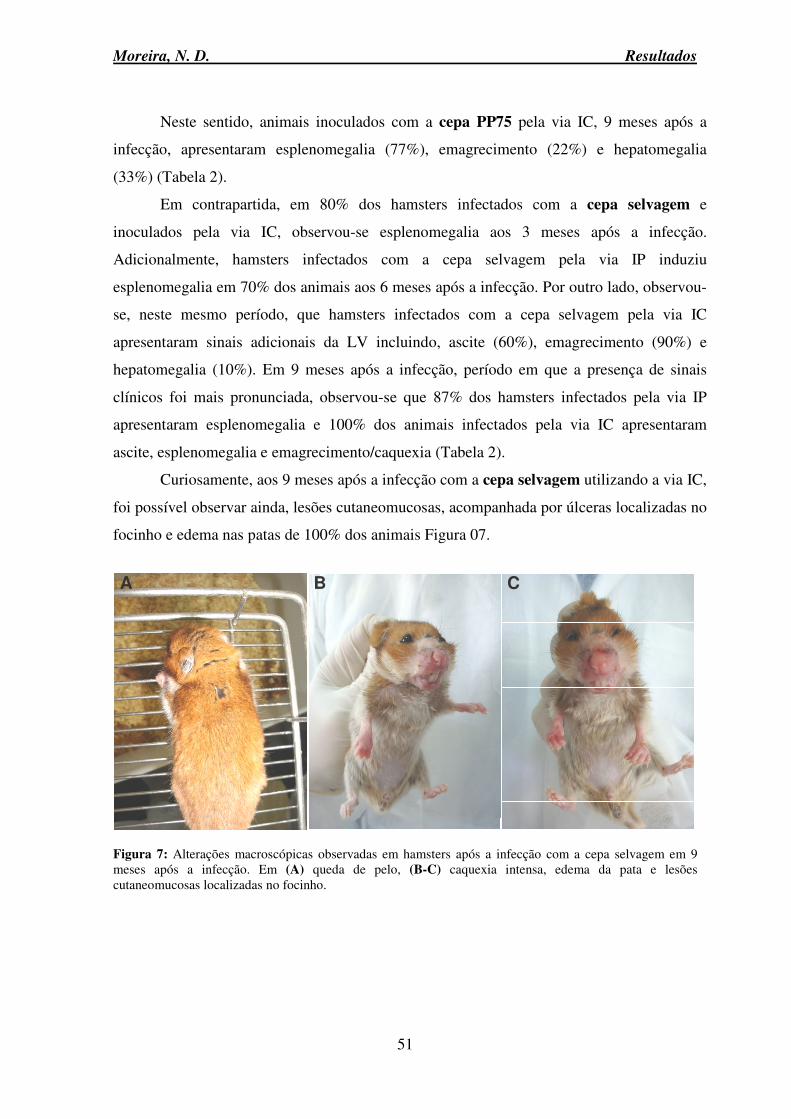
Moreira, N. D. Resultados
51
A B C
Neste sentido, animais inoculados com a cepa PP75 pela via IC, 9 meses após a
infecção, apresentaram esplenomegalia (77%), emagrecimento (22%) e hepatomegalia
(33%) (Tabela 2).
Em contrapartida, em 80% dos hamsters infectados com a cepa selvagem e
inoculados pela via IC, observou-se esplenomegalia aos 3 meses após a infecção.
Adicionalmente, hamsters infectados com a cepa selvagem pela via IP induziu
esplenomegalia em 70% dos animais aos 6 meses após a infecção. Por outro lado, observou-
se, neste mesmo período, que hamsters infectados com a cepa selvagem pela via IC
apresentaram sinais adicionais da LV incluindo, ascite (60%), emagrecimento (90%) e
hepatomegalia (10%). Em 9 meses após a infecção, período em que a presença de sinais
clínicos foi mais pronunciada, observou-se que 87% dos hamsters infectados pela via IP
apresentaram esplenomegalia e 100% dos animais infectados pela via IC apresentaram
ascite, esplenomegalia e emagrecimento/caquexia (Tabela 2).
Curiosamente, aos 9 meses após a infecção com a cepa selvagem utilizando a via IC,
foi possível observar ainda, lesões cutaneomucosas, acompanhada por úlceras localizadas no
focinho e edema nas patas de 100% dos animais Figura 07.
Figura 7: Alterações macroscópicas observadas em hamsters após a infecção com a cepa selvagem em 9 meses após a infecção. Em (A) queda de pelo, (B-C) caquexia intensa, edema da pata e lesões cutaneomucosas localizadas no focinho.
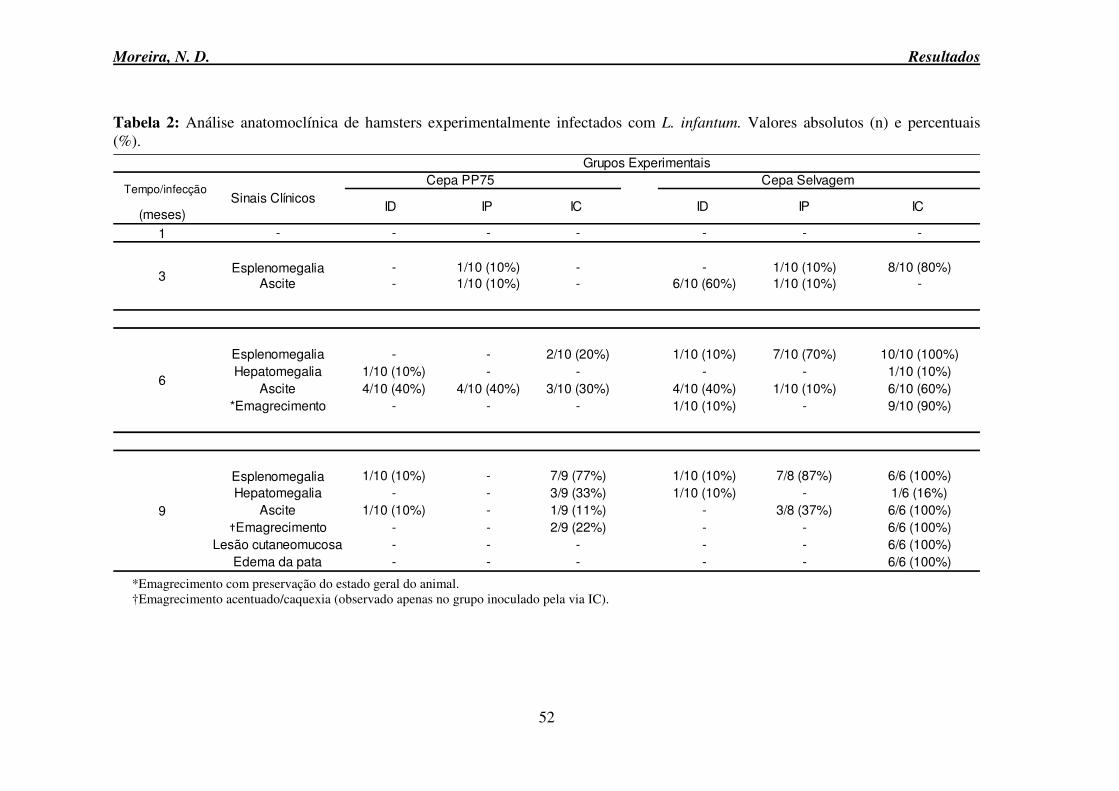
Moreira, N. D. Resultados
52
(meses)1 - - - - - - -
Esplenomegalia - 1/10 (10%) - - 1/10 (10%) 8/10 (80%)Ascite - 1/10 (10%) - 6/10 (60%) 1/10 (10%) -
Esplenomegalia - - 2/10 (20%) 1/10 (10%) 7/10 (70%) 10/10 (100%)Hepatomegalia 1/10 (10%) - - - - 1/10 (10%)
Ascite 4/10 (40%) 4/10 (40%) 3/10 (30%) 4/10 (40%) 1/10 (10%) 6/10 (60%)*Emagrecimento - - - 1/10 (10%) - 9/10 (90%)
Esplenomegalia 1/10 (10%) - 7/9 (77%) 1/10 (10%) 7/8 (87%) 6/6 (100%)Hepatomegalia - - 3/9 (33%) 1/10 (10%) - 1/6 (16%)
Ascite 1/10 (10%) - 1/9 (11%) - 3/8 (37%) 6/6 (100%)†Emagrecimento - - 2/9 (22%) - - 6/6 (100%)
Lesão cutaneomucosa - - - - - 6/6 (100%)Edema da pata - - - - - 6/6 (100%)
Grupos Experimentais
Tempo/infecçãoSinais Clínicos
Cepa PP75 Cepa Selvagem
ID IP IC ID IP IC
3
6
9
Tabela 2: Análise anatomoclínica de hamsters experimentalmente infectados com L. infantum. Valores absolutos (n) e percentuais (%).
*Emagrecimento com preservação do estado geral do animal. †Emagrecimento acentuado/caquexia (observado apenas no grupo inoculado pela via IC).

Moreira, N. D. Resultados
53
6.4 Avaliação da resposta humoral
A resposta imune humoral foi avaliada no soro dos animais dos diferentes grupos
experimentais (C, ID, IP e IC). A Figura 8 mostra que os hamsters infectados com a cepa
PP75 pelas vias IP e IC apresentaram aumento nos níveis de IgG-total quando comparados
tanto aos hamsters do grupo C quanto aqueles inoculados pela via ID, 1 mês após a
infecção. Em 3 meses após a infecção este aumento se manteve somente no grupo
infectado pela via IC quando comparado ao grupo C. Adicionalmente, 6 e 9 meses após a
infecção foram observados elevados níveis de IgG nos hamsters infectados pela via IC
quando comparados aos demais grupos.
Na avaliação dos tempos observou-se aumento nos níveis de IgG nos animais
inoculados com a cepa PP75 pela via IC nos períodos de 6 e 9 meses em relação aos
períodos de 1 e 3 meses após a infecção.
Em contrapartida, hamsters infectados com a cepa selvagem apresentaram aumento
significativo nos níveis de IgG nos três grupos experimentais (ID, IP e IC) em relação ao
grupo C durante todo período avaliado (1, 3, 6 e 9 meses após a infecção). Nestes animais,
foi ainda evidenciado, aumento da reatividade de IgG nos grupos infectados pelas vias IP e
IC quando comparados ao grupo infectado pela via ID nos tempos de 3, 6 e 9 meses após a
infecção (Figura 8).
Na avaliação dos tempos observou-se que, hamsters infectados com a cepa
selvagem apresentaram aumento significativo nos níveis de IgG nos grupos inoculados
pelas vias IC e IP nos tempos de 3, 6 e 9 meses quando comparados ao período de 1 mês
após a infecção.
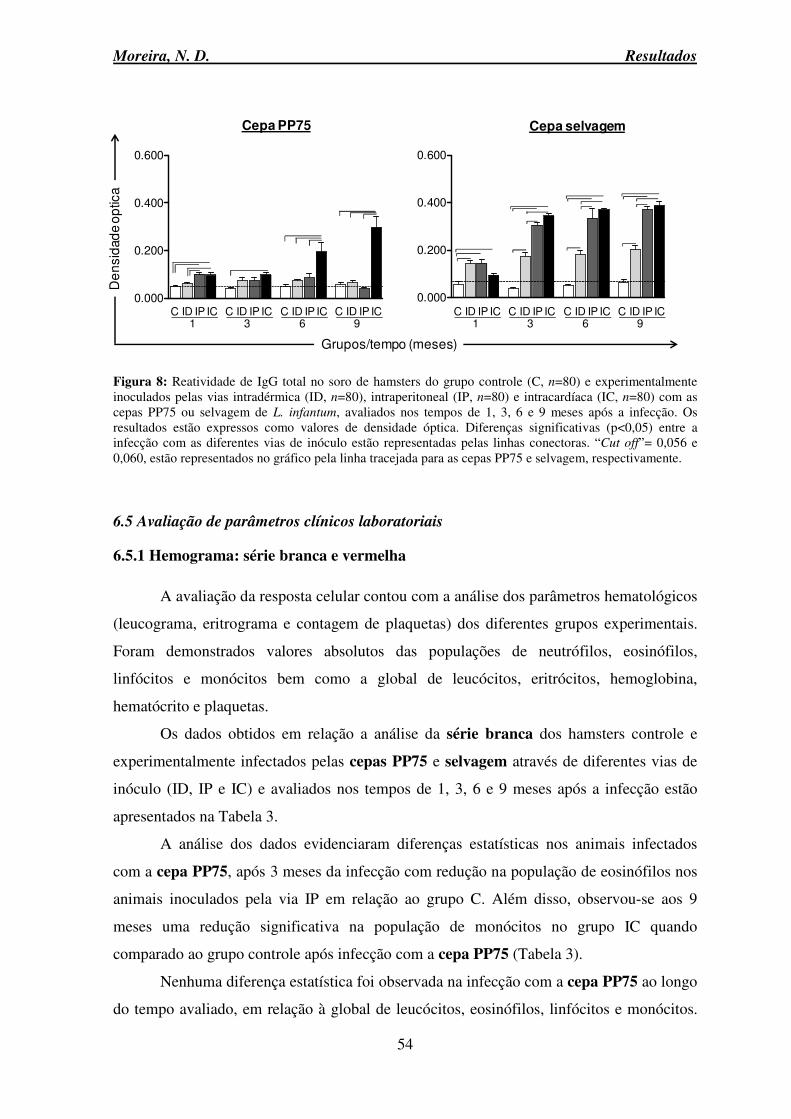
Moreira, N. D. Resultados
54
Figura 8: Reatividade de IgG total no soro de hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como valores de densidade óptica. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras. “Cut off”= 0,056 e 0,060, estão representados no gráfico pela linha tracejada para as cepas PP75 e selvagem, respectivamente.
6.5 Avaliação de parâmetros clínicos laboratoriais
6.5.1 Hemograma: série branca e vermelha
A avaliação da resposta celular contou com a análise dos parâmetros hematológicos
(leucograma, eritrograma e contagem de plaquetas) dos diferentes grupos experimentais.
Foram demonstrados valores absolutos das populações de neutrófilos, eosinófilos,
linfócitos e monócitos bem como a global de leucócitos, eritrócitos, hemoglobina,
hematócrito e plaquetas.
Os dados obtidos em relação a análise da série branca dos hamsters controle e
experimentalmente infectados pelas cepas PP75 e selvagem através de diferentes vias de
inóculo (ID, IP e IC) e avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção estão
apresentados na Tabela 3.
A análise dos dados evidenciaram diferenças estatísticas nos animais infectados
com a cepa PP75, após 3 meses da infecção com redução na população de eosinófilos nos
animais inoculados pela via IP em relação ao grupo C. Além disso, observou-se aos 9
meses uma redução significativa na população de monócitos no grupo IC quando
comparado ao grupo controle após infecção com a cepa PP75 (Tabela 3).
Nenhuma diferença estatística foi observada na infecção com a cepa PP75 ao longo
do tempo avaliado, em relação à global de leucócitos, eosinófilos, linfócitos e monócitos.
Grupos/tempo (meses)
De
nsid
ade
optic
a
ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C 0.000
0.200
0.400
0.600
0.000
0.200
0.400
0.600
Cepa PP75 Cepa selvagem

Moreira, N. D. Resultados
55
Observou-se apenas aumento na população de neutrófilos no grupo IP no período de 3
meses quando comparado ao período de 1 mês após a infecção.
Por outro lado, após um mês de infecção com a cepa selvagem pelas vias ID e IP
foi observado aumento no número de monócitos em relação ao grupo C. Após 3 meses da
infecção observou-se aumento na população de linfócitos do grupo inoculado pela via IP
quando comparado ao grupo C. Neste mesmo tempo, animais infectados pela via IC
apresentaram leucopenia em relação aos grupos C, ID e IP. Esta leucopenia apresentou
uma marcada redução da população de monócitos, neste mesmo período (após 3 meses da
infecção), para o grupo IC quando comparado ao grupo C. Neste sentido, este grupo
apresentou após 6 meses de infecção redução do número de neutrófilos totais em relação
ao grupo ID; redução do número de eosinófilos em relação aos grupos C e ID, redução do
número de linfócitos em relação aos grupos C, ID e IP; além de redução do número de
monócitos em relação aos grupos C e IP. De forma similar 9 meses após a infecção com a
cepa selvagem, hamsters inoculados pelas vias IP e IC apresentaram leucopenia em
relação aos grupos C e ID, que foi caracterizada pela diminuição de neutrófilos totais,
eosinófilos e monócitos em relação aos grupos C e ID (Tabela 3).
Contudo, na avaliação ao longo do tempo, hamsters infectados com a cepa
selvagem pela via IC, apresentaram redução significativa na global de leucócitos
(leucopenia) no período de 9 meses após infecção quando comparados ao período de 1
mês.
Em relação à população de linfócitos observou-se redução no período de 9 meses
após a infecção nos animais infectados pela via IP quando comparados ao período de 3
meses após a infecção. Ainda em relação a esta população celular, observou-se redução
significativa nos animais infectados pela via IC no período de 3 meses após a infecção
quando comparados ao período de 1 mês.
Já em relação aos monócitos os dados obtidos mostraram redução significativa
desta população nos animais infectados pelas vias ID, IP e IC no período de 9 meses após a
infecção quando comparado ao período de 1 mês após a infecção. No período de 3 e 6
meses em animais inoculados pela via IC observou-se redução significativa desta
população em relação ao periodo de 1 mês após a infecção.
Nenhuma diferença estatística foi observada ao longo do tempo avaliado, em
relação a população de neutrófilos e eosinófilos nos hamsters infectados com a cepa
selvagem.

Moreira, N. D. Resultados
56
1 3 6 9 1 3 6 9
Global de Leucócitos 3640±521 4810±1638 2700±614 3950±994 3120±1029 3590±1199 4240±1102 3610±1005
Neutrófilos Totais 1597±442 2490±1258 1106±448 1783±803 1467±570 2291±1240 1473±511 1848±774Eosinófilos C 53±16 96±50 25±8 43±12 41±22 53,40±29 47±18 56±32Linfócitos 1943±484 2173±932 1470±470 2066±621 1577±617 1183±495 2655±755 1848±774
Monócitos 47±19 52±18 27±25 52±16 35±16 63±28 65±28 41±13
1 3 6 9 1 3 6 9
Global de Leucócitos 4250±846 4990±1049 3930±1421 3440±1561 4225±504 3678±1266 4910±1171 3980±691
Neutrófilos Totais 1296±776 2349±695 1963±1126 1695±882 1837±363 2487±1315 2022±962 1784±546Eosinófilos ID 42±8 68±24 61±44 30±10 57±18 51±29 61±18 57±26Linfócitos 2859±731 2485±606 1802±803 1665±741 1837±719 1589±812 2274±618 2090±405
Monócitos 53±29 68±21 50±28 33±15 94±31 a 53±32 58±31 49±22
Global de Leucócitos 3890±1366 5590±1664 3690±1002 3867±1280 3889±1554 3867±1057 3210±769 b
2200±1035 a, b
Neutrófilos Totais 1267±685 2722±1060 1394±661 1777±627 1537±454 1879±742 852±422 b
798±337 a, b
Eosinófilos IP 38±15 55±17 a 52±23 38±13 62±43 55±25 39±17
b22±10
a, b
Linfócitos 2516±879 2751±947 2185±880 1995±796 2320±957 2376±1261 a 2259±472 1130±622
Monócitos 43±18 56±17 40±9 41±9 88±41 a 50±23 61±30 18±8
a, b
Global de Leucócitos 3580±1604 4480±1383 3000±1159 3056±2496 3770±1078 2220±797 a, b, c
2480±843 a, b
1617±643 a, b
Neutrófilos Totais 1196±660 2156±932 1305±499 1168±1391 1605±490 1185±696 1082±435 b
565±322 a, b
Eosinófilos IC 40±16 68±32 44±31 36±27 54±33 26±11 25±8 a, b
16±6 a, b
Linfócitos 2303±1058 2203±733 1612±859 1373±677 2168±563 981±450 1346±831 a, b, c 1020±379
Monócitos 41±18 53±20 31±11 26±27 a 74±43 28±13 a
27±11 a, c
16±6 a, b
Selvagem
Parâmetros
hematológicosGrupo
Tempo/meses
Parâmetros
hematológicosGrupos
Cepa/meses após a infecção
PP75
Tabela 3: Leucograma de hamsters do grupo controle e experimentalmente infectados com as cepas PP75 e selvagem de L. infantum.
Valores absolutos (média±desvio padrão) do leucograma de hamsters controle (C) e infectados com as cepas PP75 e selvagem de L. infantum por diferentes vias de inóculo: intradérmica (ID); intraperitoneal (IP); intracardíaca (IC). As diferenças significativas (P<0,05) estão representadas pelas letras a, b, c relacionadas ao grupo C, ID, IP, respectivamente.

Moreira, N. D. Resultados
57
As alterações relacionadas à série vermelha estão apresentadas na Tabela 4. Os
valores do número de eritrócitos, níveis de hemoglobina e hematócrito foram os
parâmetros analisados no âmbito do eritrograma a partir do hemograma completo.
Os resultados obtidos neste estudo demonstraram no primeiro mês de infecção com
a cepa PP75, redução significativa dos níveis de hematócrito considerando as três vias de
inóculo ID, IP e IC, em relação aos animais do grupo C. Além disto, 6 meses após a
infecção foi observado um quadro compatível com anemia, nos animais inoculados pelas
diferentes vias de inóculo (ID, IP e IC), que apresentaram redução significativa do número
de eritrócitos, níveis de hemoglobina e hematócrito em relação ao grupo C. Por outro lado,
9 meses após a infecção, os hamsters infectados pela via IP apresentaram aumento do
número de eritrócitos em relação aos grupos C e ID (Tabela 4).
Outro parâmetro analisado neste estudo focou na contagem de plaquetas sendo
possível identificar trombocitose após 3 meses da infecção com a cepa PP75 no grupo IP
em relação ao grupo C, e trombocitopenia no grupo IC em relação ao grupo IP (Tabela 4).
A avaliação ao longo do tempo demonstrou que animais infectados com a cepa
PP75 apresentaram aumento do número de eritrócitos e nos níveis de hemoglobina nos 3
grupos experimentais (ID, IP e IC) no período de 9 meses quando comparado aos períodos
de 1, 3 e 6 meses após a infecção. Já em relação aos níveis de hematócrito, observou-se
aumento nos períodos de 3 e 9 meses após a infecção no grupo inoculado pela via IP
quando comparado ao período de 1 mês após a infecção. Consequente redução nos níveis
de hematócrito foi observada no período de 6 meses em relação aos períodos de 3 e 9
meses após a infecção. No período de 9 meses após a infecção, o grupo IC apresentou
aumento nos níveis de hematócrito quando comparado aos períodos de 1, 3 e 6 meses.
Em relação à contagem de plaquetas, foi observado aumento no período de 3 meses
após a infecção no grupo inoculado pela via IP quando comparado aos períodos de 1, 6 e 9
meses após a infecção.
As principais alterações da série vermelha nos hamsters infectados com a cepa
selvagem foram observadas 6 e 9 meses após a infecção no grupo IC. Neste sentido, um
quadro de anemia grave foi observado após 6 meses de infecção no grupo IC, com redução
significativa do número de eritrócitos, dos níveis de hemoglobina e hematócrito em relação
aos grupos C, ID e IP. De forma semelhante, após 9 meses da infecção, o grupo IC
apresentou um quadro de anemia, caracterizado pela redução do número de eritrócitos em
relação aos grupos C, ID e IP; redução dos níveis de hemoglobina em relação aos grupos C

Moreira, N. D. Resultados
58
e ID; bem como redução do hematócrito em relação aos grupos C e ID. Adicionalmente,
quando se avaliou a contagem de plaquetas, foi observada trombocitose no grupo IC, após
6 e 9 meses, em relação aos grupos C, ID e IP (Tabela 4).
Na avaliação dos tempos, hamsters infectados com a cepa selvagem apresentaram
redução no número de eritrócitos no período de 9 meses no grupo inoculado pela via IP
quando comparado ao período de 1 mês após a infecção. Ainda observou-se redução no
período de 9 meses após a infecção no grupo inoculado pela via IC em relação aos
períodos de 1, 3 e 6 meses após a infecção.
Em relação aos níveis de hemoglobina, houve redução no período de 6 meses após
a infecção no grupo inoculado pela via ID quando comparado ao período de 1 mês. Ainda
em relação aos níveis de hemoglobina, observou-se redução nos períodos de 3, 6 e 9 meses
após a infecção no grupo inoculado pela via IC quando comparado ao período de 1 mês
após a infecção. Além disso, observou-se redução no período de 6 meses após a infecção
quando comparado aos períodos de 3 e 9 meses após a infecção.
Por outro lado, em relação aos níveis de hematócrito, observou-se redução no
período de 6 meses no grupo infectado pela via IC quando comparado aos tempos de 1 e 3
meses após a infecção. Ainda mostrou redução destes níveis no período de 9 meses em
relação ao período de 1 mês após a infecção.
Em relação a contagem de plaquetas observou-se aumento nos períodos de 6 e 9
meses após a infecção no grupo inoculado pela via IC quando comparado aos períodos de
1e 3 meses após a infecção.

Moreira, N. D. Resultados
59
1 3 6 9 1 3 6 9
Eritrócitos (106/mm
3) 7,2±0,3 7,2±1,1 8,5±1,2 8,4±1,0 7,4±0,4 7,0±0,3 7,1±0,2 6,7±0,3
Hemoglobina (g/dL) 13,9±0,6 13,5±0,8 17,6±2,1 17,5±2,3 15,0±1,0 13,3±0,9 13,3±0,5 13,2±0,7Hematócrito (%) 35,5±3,6 34,6±7,2 39,5±5,5 38,4±6,7 32,6±1,5 30,5±2,3 32,5±2,3 33,2±2,0Plaquetas (10
6/mm
3) 165,8±37,4 142,6±33,8 154,9±143 127,4±12,5 139,9±20,8 136,4±16,9 164,7±15,5 174,0±20,5
1 3 6 9 1 3 6 9
Eritrócitos (106/mm
3) 6,5±0,4 7,2±0,2 6,5±0,4
a 8,5±0,7 7,4±0,3 6,8±0,3 6,7±0,8 6,9±0,3
Hemoglobina (g/dL) 13,7±1,1 13,6±0,5 12,7±1,0a 19,7±4,5 15,2±0,6 13,3±0,7 12,6±1,6 13,7±0,3
Hematócrito (%) 29,3±2,1a 36,3±3,0 31,1±2,7
a 38,3±3,9 33,9±1,5 31,2±3,0 33,6±3,6 34,0±2,0Plaquetas (10
6/mm
3) 123,8±16,2 168,70±25,2
6138,3±19,2 108,1±18,0 156,8±27,5 141,1±24,4 185,5±24,4 189,4±27,3
Eritrócitos (106/mm
3) 6,1±0,1 6,8±0,7 6,7±0,2
a9,8±0,5
a,b 7,5±1,0 7,1±0,4 7,0±0,5 6,2±1,0
Hemoglobina (g/dL) 13,4±0,5 13,1±1,2 13,4±0,4a 21,2±1,3 14,4±0,6 13,6±1,2 12,8±1,1 12,0±2,0
Hematócrito (%) 28,9±1,5a 38,7±3,2 31,2±2,7
a 43,9±2,2 32,0±2,6 31,4±3,2 36,1±2,6 31,0±4,2Plaquetas (10
6/mm
3) 134,1±45,3 221,6±75,0
a 132,2±23,4 126,3±14,5 169,3±35,4 145,6±32,1 184,4±32,7 174,6±87,4
Eritrócitos (106/mm
3) 6,6±0,4 7,2±0,7 6,8±0,3
a 9,2±1,0 7,1±0,4 6,7±0,3 4,3±1,4a,b,c
4,7±1,9a,b, c
Hemoglobina (g/dL) 13,7±0,8 13,8±1,2 13,6±0,7a 20,0±2,4 14,5±0,9 12,3±1,6 7,0±1,8
a,b,c10,1±4,2
a,b
Hematócrito (%) 30,1±1,8a 35,0±3,6 31,5±2,4
a 41,5±4,3 31,6±2,1 29,0±4,4 21,0±6,9a,b,c
25,0±9,4a,b
Plaquetas (106/mm
3) 134,1±21,5 144,6±22,3
c 135,4±13,5 108,5±17,5 156,5±17,0 110,9±29,8 329,4±153,5a,b,c
291,3±136,4a,b,c
Parâmetros
hematológicosGrupo
Tempo/meses
C
ID
IP
IC
Parâmetros
hematológicosGrupos
Cepa/meses após a infecção
PP75 Selvagem
Tabela 4: Eritrograma de hamsters controle e experimentalmente infectados com as cepas PP75 e selvagem de L. infantum.
Valores (média±desvio padrão) do eritrograma de hamsters controle (C) e infectados com as cepas PP75 e selvagem de L. infantum por diferentes vias de inóculo: intradérmica (ID); intraperitoneal (IP); intracardíaca (IC). As diferenças significativas (P<0,05) estão representadas pelas letras a, b, c relacionadas ao grupo C, ID, IP, respectivamente.

Moreira, N. D. Resultados
60
6.6 Avaliação dos parâmetros Bioquímicos
Os dados referentes ao perfil bioquímico dos hamsters do grupo controle e
submetidos à inóculos com as cepas PP75 e selvagem estão demonstrados nos respectivos
gráficos abaixo. Neste estudo, os valores obtidos nos diferentes grupos experimentais (ID,
IC e IP) acima dos valores obtidos no grupo controle C, foram considerados alterados.
6.6.1 Provas de função renal
Para a avaliação da função renal foram realizadas dosagens de uréia e creatinina no
soro de hamsters do grupo C e infectados com as diferentes cepas de L.infantum. Os
resultados obtidos estão demonstrados na Figura 9.
Os resultados demonstraram no sexto mês de infecção com a cepa PP75, aumento
significativo dos níveis de uréia no grupo IC, em relação aos animais do grupo C e IP. Aos 9
meses após a infecção, foi observado aumento dos níveis de uréia nos 3 grupos
experimentais (ID, IP e IC) quando comparado ao grupo C. Além disso, foi detectado neste
mesmo período níveis aumentados de uréia nos grupos IP e IC em relação ao grupo ID
(Figura 9).
A avaliação ao longo do tempo revelou maiores níveis de uréia no grupo IC nos
períodos de 6 e 9 meses quando comparado ao período de 1 mês após a infecção com a cepa
PP75.
As principais alterações nos níveis de uréia nos hamsters infectados com a cepa
selvagem foram observadas em 3 e 6 meses após a infecção no grupo IC, onde foi
observado aumento significativo quando comparado ao grupo C e aos grupos inoculados
pelas vias ID e IP (Figura 9).
Nenhuma diferença significativa foi observada quando se avaliou cada grupo ao
longo dos tempos, na infecçao experimental pela cepa selvagem.
Os resultados obtidos a partir da dosagem dos níveis séricos de creatinina, revelaram
aumento dos níveis em 9 meses após a infeção com a cepa PP75 nos três grupos
experimentais (ID, IP e IC) quando comparados ao grupo C (Figura 9).

Moreira, N. D. Resultados
61
A avaliação ao longo do tempo demonstrou que animais infectados com a cepa PP75
apresentaram aumento dos níveis de creatinina nos 3 grupos experimentais (ID, IP e IC) no
período de 9 meses quando comparado aos períodos de 1, 3 e 6 meses após a infecção.
Por outro lado, após 3 e 9 meses de infecção com a cepa selvagem pela via IC foi
observado aumento nos níveis de creatinina em relação aos grupos C, ID e IP (Figura 9).
Na avaliação dos tempos, observou-se redução nos níveis de creatinina em hamsters
inoculados pelas vias ID e IP no período de 3 meses quando comparado ao período de 1 mês
após a infecção, sendo que em 6 e 9 meses observou-se aumento destes níveis quando
comparado ao período de 3 meses após a infecção. De forma semelhante, no período de 9
meses observou-se aumento nos níveis de creatina no grupo IC quando comparado aos
períodos de 1 e 3 meses após a infecção.
Figura 9: Níveis séricos de uréia e creatinina de hamsters dos grupos controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
0
1
2
3
Cepa PP75 Cepa selvagem
0
20
40
60
80
0
1
2
3
0
20
40
60
80
Grupos/tempo (meses)
Cre
atin
ina
) (m
g/d
L)
Uré
ia (m
g/d
L)
ID IPIC1
C ID IPIC3
C ID IPIC6
C ID IPIC9
C ID IPIC1
C ID IPIC3
C ID IPIC6
C ID IPIC9
C

Moreira, N. D. Resultados
62
6.6.2 Prova de função hepática
A avaliação da função hepática foi realizada a fim de verificar possível
comprometimento hepático nos hamsters infectados com as diferentes cepas de L. infantum
Dentre as principais enzimas hepáticas, foram avaliadas as transaminases: aspartato
aminotranferase (AST) e alanina aminotransferase (ALT). Os resultados estão demonstrados
na Figura 10.
A partir dos valores obtidos da dosagem de AST, observou-se que em animais
infectados com a cepa PP75, houve aumento nos níveis de AST no grupo IC em 1 mês após
a infecção em relação aos grupos C e ID. De forma semelhante, foi demonstrado em 9 meses
após a infecção aumento significativo nos níveis de AST no grupo IC quando comparado
aos grupos C, ID e IP (Figura 10).
Não foram observadas diferenças quando se avaliou cada grupo ao longo dos
tempos.
Por outro lado, em animais inoculados com a cepa selvagem pela via IC observou-se
aumento nos níveis de AST em 3 meses após a infecção quando comparado aos grupos C,
ID e IP. De forma diferente, em 6 meses após a infecção foi observado aumento
significativo nos níveis de AST no grupo IP quando comparado aos grupos C, ID e IP. Aos 9
meses após a infecção, aumento destes níveis foi observado nos grupos IP e IC, quando
comparado tanto ao grupo C quanto ao grupo ID (Figura 10).
Em relação à avaliação ao longo do tempo, observou-se que, no período de 3 meses
após a infecção com a cepa selvagem pela via IC houve aumento nos níveis de AST, quando
comparado ao período de 1 mês após a infecção.
A avaliação dos níveis de ALT revelou que animais inoculados com a cepa PP75
pelas vias IP e IC 1 mês após a infecção, apresentaram aumento nos níveis de ALT, quando
comparado ao grupo C. Aos 9 meses após a infecção foi observado aumento significativo de
ALT no grupo IC quando comparado aos grupos C, ID e IP (Figura 10).
Em relação à avaliação ao longo do tempo, observou-se que, no período de 1 mês
após a infecção com a cepa PP75 pela via IP houve aumento significativo nos níveis de
ALT, quando comparado aos período de 3, 6 e 9 meses após a infecção. Ainda, observou-se
aumento dos níveis de ALT no grupo IC no período de 1 mês quando comparado ao período
de 3 meses após a infecção.
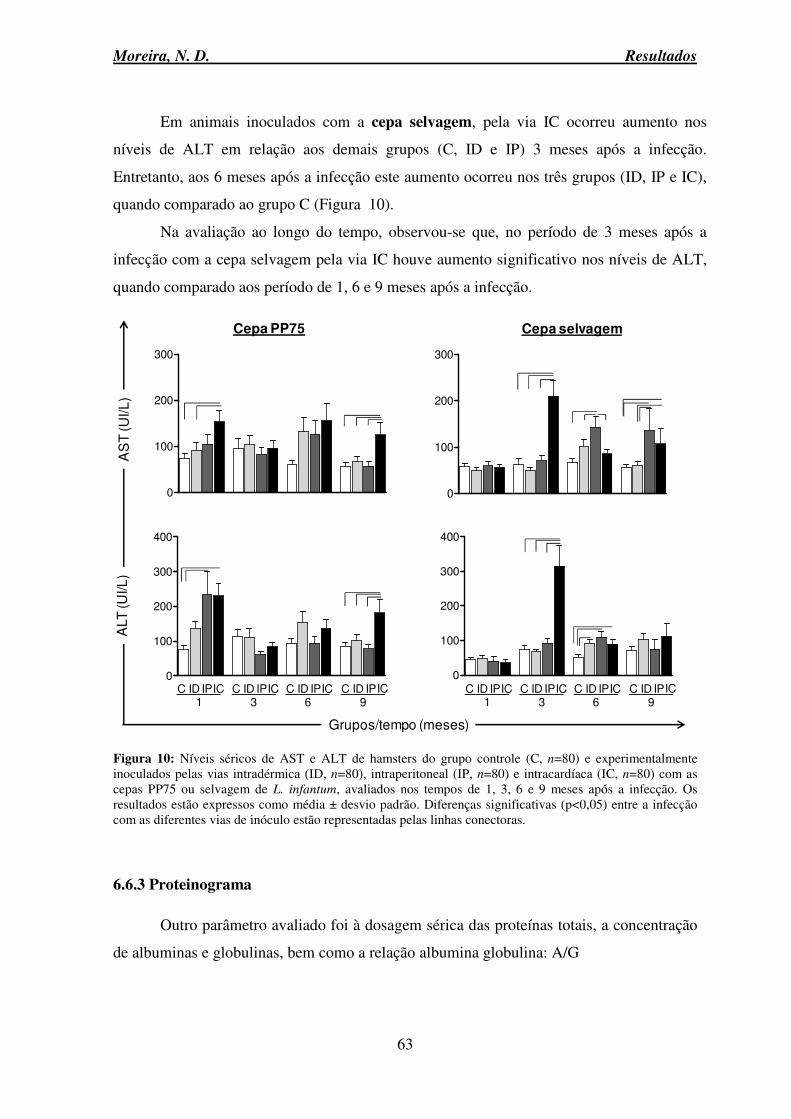
Moreira, N. D. Resultados
63
Em animais inoculados com a cepa selvagem, pela via IC ocorreu aumento nos
níveis de ALT em relação aos demais grupos (C, ID e IP) 3 meses após a infecção.
Entretanto, aos 6 meses após a infecção este aumento ocorreu nos três grupos (ID, IP e IC),
quando comparado ao grupo C (Figura 10).
Na avaliação ao longo do tempo, observou-se que, no período de 3 meses após a
infecção com a cepa selvagem pela via IC houve aumento significativo nos níveis de ALT,
quando comparado aos período de 1, 6 e 9 meses após a infecção.
Figura 10: Níveis séricos de AST e ALT de hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
6.6.3 Proteinograma
Outro parâmetro avaliado foi à dosagem sérica das proteínas totais, a concentração
de albuminas e globulinas, bem como a relação albumina globulina: A/G
Cepa PP75 Cepa selvagem
0
100
200
300
400
0
100
200
300
0
100
200
300
400
0
100
200
300
Grupos/tempo (meses)
ID IPIC1
C ID IPIC3
C ID IPIC6
C ID IPIC9
C
ALT
(UI/L
)A
ST
(UI/L
)
ID IPIC1
C ID IPIC3
C ID IPIC6
C ID IPIC9
C
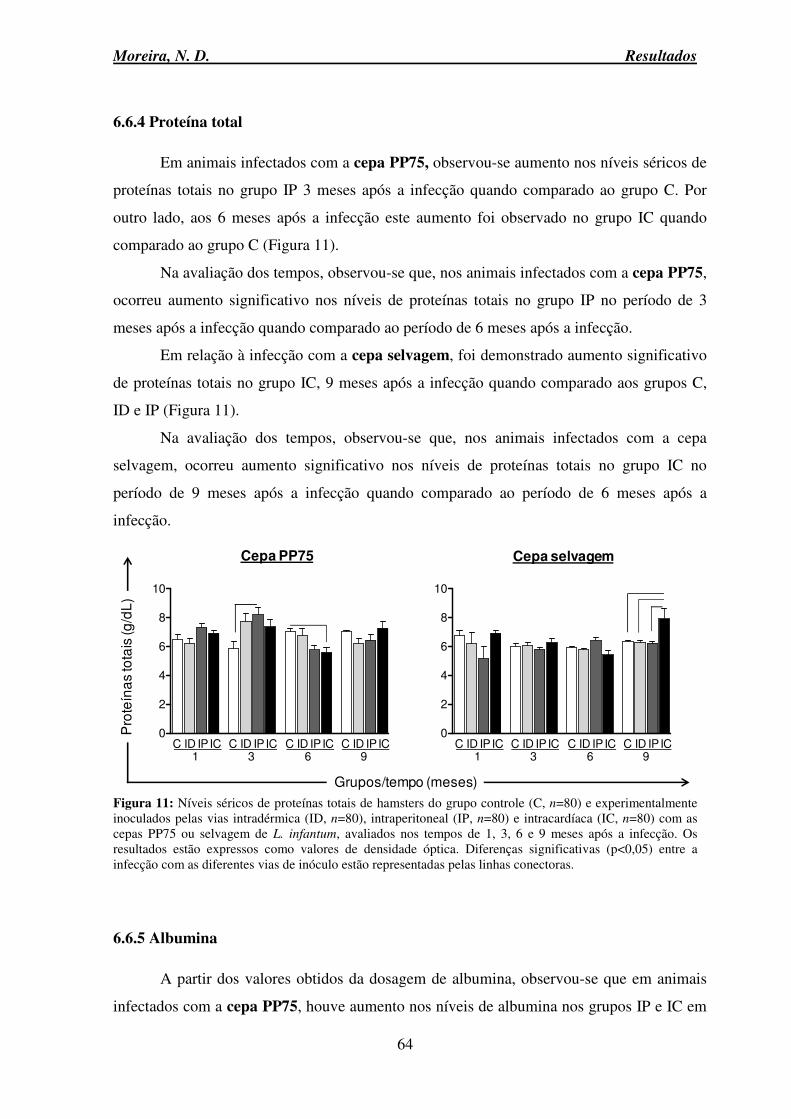
Moreira, N. D. Resultados
64
Cepa PP75 Cepa selvagem
Grupos/tempo (meses)
Pro
teín
as
tota
is (g
/dL
)
0
2
4
6
8
10
0
2
4
6
8
10
ID IPIC1
C ID IPIC3
C ID IPIC6
C ID IPIC9
C ID IPIC1
C ID IPIC3
C ID IPIC6
C ID IPIC9
C
6.6.4 Proteína total
Em animais infectados com a cepa PP75, observou-se aumento nos níveis séricos de
proteínas totais no grupo IP 3 meses após a infecção quando comparado ao grupo C. Por
outro lado, aos 6 meses após a infecção este aumento foi observado no grupo IC quando
comparado ao grupo C (Figura 11).
Na avaliação dos tempos, observou-se que, nos animais infectados com a cepa PP75,
ocorreu aumento significativo nos níveis de proteínas totais no grupo IP no período de 3
meses após a infecção quando comparado ao período de 6 meses após a infecção.
Em relação à infecção com a cepa selvagem, foi demonstrado aumento significativo
de proteínas totais no grupo IC, 9 meses após a infecção quando comparado aos grupos C,
ID e IP (Figura 11).
Na avaliação dos tempos, observou-se que, nos animais infectados com a cepa
selvagem, ocorreu aumento significativo nos níveis de proteínas totais no grupo IC no
período de 9 meses após a infecção quando comparado ao período de 6 meses após a
infecção.
Figura 11: Níveis séricos de proteínas totais de hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como valores de densidade óptica. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
6.6.5 Albumina
A partir dos valores obtidos da dosagem de albumina, observou-se que em animais
infectados com a cepa PP75, houve aumento nos níveis de albumina nos grupos IP e IC em

Moreira, N. D. Resultados
65
1 e 9 meses após a infecção em relação aos grupos C e ID. Entretanto aos 6 meses após a
infecção, houve uma inversão com redução nos grupos IP e IC, quando comparado aos
grupos C e ID. Aos 3 meses após a infecção, observou-se aumento nos níveis de albumina
nos três grupos experimentais (ID, IP e IC), quando comparado ao grupo C (Figura 12).
Na avaliação dos tempos, observou-se que, nos animais infectados com a cepa PP75,
ocorreu aumento significativo nos níveis de albumina no grupo ID nos períodos de 3, 6 e 9
meses após a infecção quando comparado ao período de 1 mês após a infecção. Ainda
mostrou aumento no período de 3 meses quando comparado aos períodos de 6 e 9 meses
após a infecção. Além disso, foi demonstrado aumento nos níveis de albumina no período de
3 meses no grupo IP em relação aos períodos de 1 e 6 meses após a infecção; no período de
1 mês em relação a 6 meses; e no período de 9 meses quando comparado ao período de 6
meses após a infecção. Logo, no período de 3 meses após a infecção observou-se aumento
nos níveis de albumina no grupo IC quando comparado aos períodos de 1, 6 e 9 após a
infecção. Este aumento foi observado também no período de 1 e 9 meses, porém, quando
comparado aos períodos de 6 meses após a infecção.
Em relação à infecção com a cepa selvagem, foi demonstrado aumento significativo
de albumina nos grupos ID e IP, 3 meses após a infecção quando comparado aos grupos C e
IC. Logo, aos 9 meses após a infecção, aumento foi observado apenas no grupo IC em
relação ao grupo IP (Figura 12).
Na avaliação ao longo do tempo, observou-se que, no período de 3 meses após a
infecção com a cepa selvagem pela via ID houve aumento significativo nos níveis de
albumina, quando comparado ao período de 9 meses após a infecção. Além disso, foi
demonstrado aumento nos níveis de albumina no período de 3 meses no grupo IP em relação
aos períodos de 1 e 9 meses após a infecção.
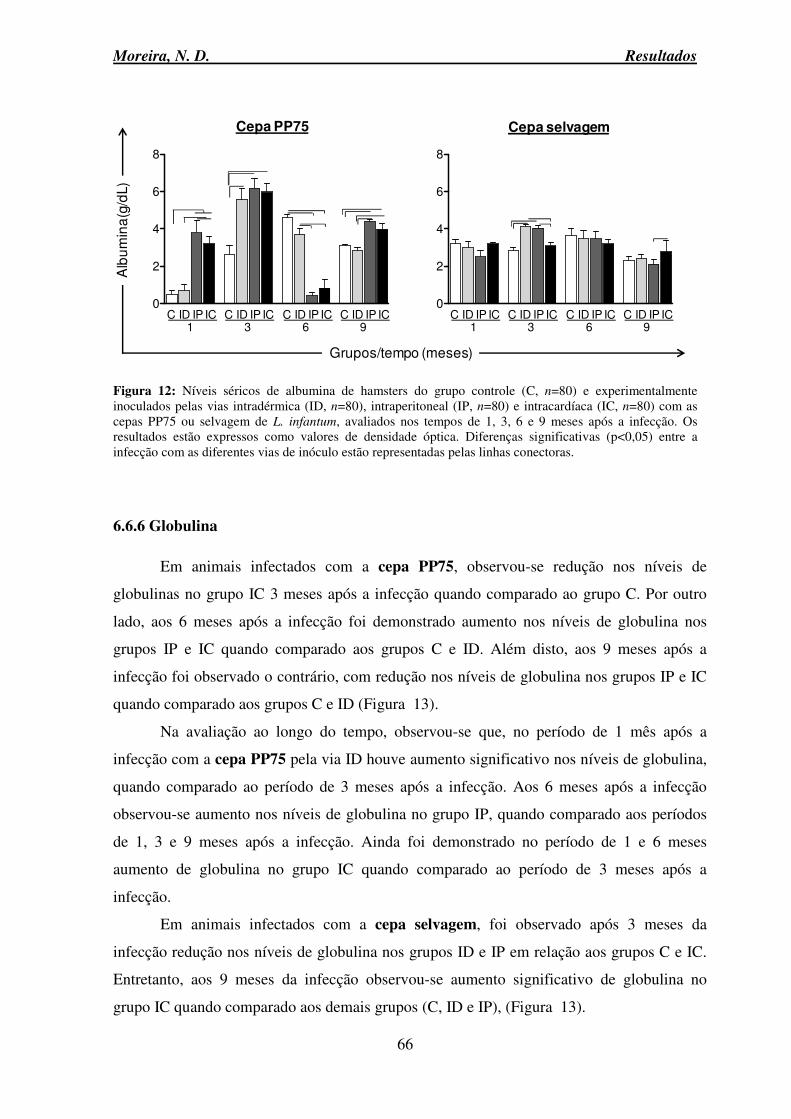
Moreira, N. D. Resultados
66
Figura 12: Níveis séricos de albumina de hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como valores de densidade óptica. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
6.6.6 Globulina
Em animais infectados com a cepa PP75, observou-se redução nos níveis de
globulinas no grupo IC 3 meses após a infecção quando comparado ao grupo C. Por outro
lado, aos 6 meses após a infecção foi demonstrado aumento nos níveis de globulina nos
grupos IP e IC quando comparado aos grupos C e ID. Além disto, aos 9 meses após a
infecção foi observado o contrário, com redução nos níveis de globulina nos grupos IP e IC
quando comparado aos grupos C e ID (Figura 13).
Na avaliação ao longo do tempo, observou-se que, no período de 1 mês após a
infecção com a cepa PP75 pela via ID houve aumento significativo nos níveis de globulina,
quando comparado ao período de 3 meses após a infecção. Aos 6 meses após a infecção
observou-se aumento nos níveis de globulina no grupo IP, quando comparado aos períodos
de 1, 3 e 9 meses após a infecção. Ainda foi demonstrado no período de 1 e 6 meses
aumento de globulina no grupo IC quando comparado ao período de 3 meses após a
infecção.
Em animais infectados com a cepa selvagem, foi observado após 3 meses da
infecção redução nos níveis de globulina nos grupos ID e IP em relação aos grupos C e IC.
Entretanto, aos 9 meses da infecção observou-se aumento significativo de globulina no
grupo IC quando comparado aos demais grupos (C, ID e IP), (Figura 13).
Cepa PP75 Cepa selvagem
Grupos/tempo (meses)
Alb
umin
a(g/
dL)
ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C 0
2
4
6
8
0
2
4
6
8
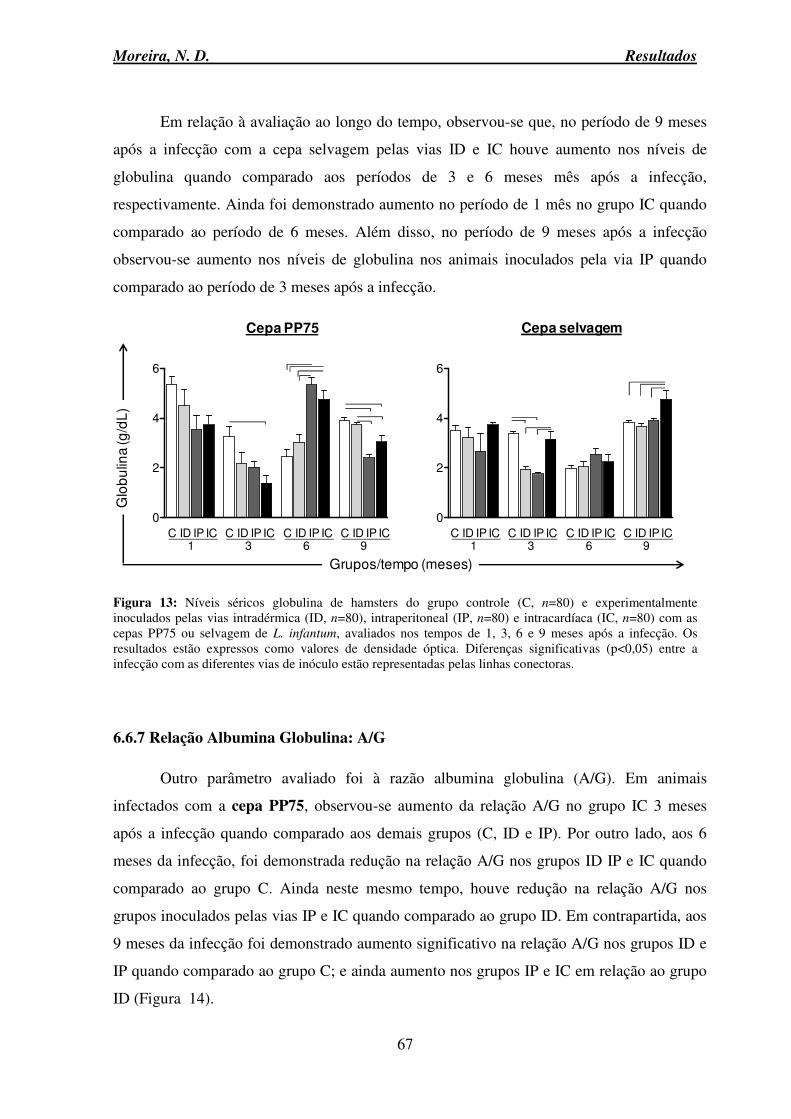
Moreira, N. D. Resultados
67
Em relação à avaliação ao longo do tempo, observou-se que, no período de 9 meses
após a infecção com a cepa selvagem pelas vias ID e IC houve aumento nos níveis de
globulina quando comparado aos períodos de 3 e 6 meses mês após a infecção,
respectivamente. Ainda foi demonstrado aumento no período de 1 mês no grupo IC quando
comparado ao período de 6 meses. Além disso, no período de 9 meses após a infecção
observou-se aumento nos níveis de globulina nos animais inoculados pela via IP quando
comparado ao período de 3 meses após a infecção.
Figura 13: Níveis séricos globulina de hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como valores de densidade óptica. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
6.6.7 Relação Albumina Globulina: A/G Outro parâmetro avaliado foi à razão albumina globulina (A/G). Em animais
infectados com a cepa PP75, observou-se aumento da relação A/G no grupo IC 3 meses
após a infecção quando comparado aos demais grupos (C, ID e IP). Por outro lado, aos 6
meses da infecção, foi demonstrada redução na relação A/G nos grupos ID IP e IC quando
comparado ao grupo C. Ainda neste mesmo tempo, houve redução na relação A/G nos
grupos inoculados pelas vias IP e IC quando comparado ao grupo ID. Em contrapartida, aos
9 meses da infecção foi demonstrado aumento significativo na relação A/G nos grupos ID e
IP quando comparado ao grupo C; e ainda aumento nos grupos IP e IC em relação ao grupo
ID (Figura 14).
Cepa PP75 Cepa selvagem
Grupos/tempo (meses)
0
2
4
6
0
2
4
6
Glo
bu
lina
(g/d
L)
ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C
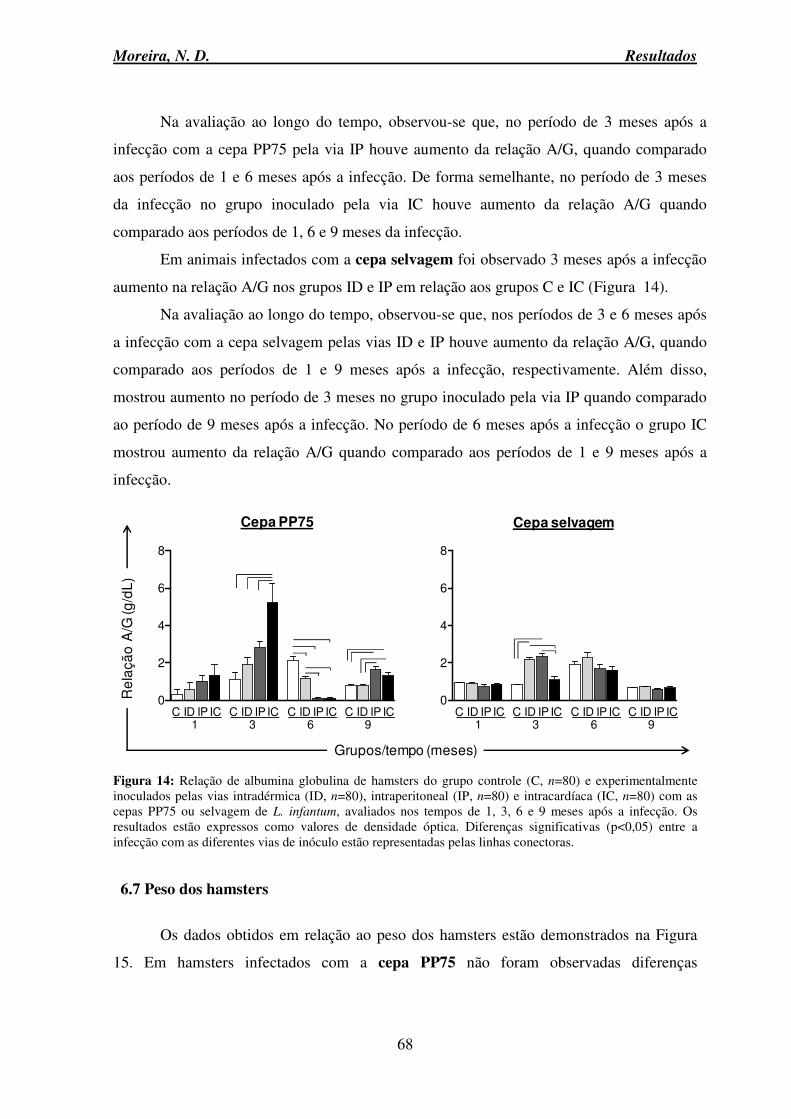
Moreira, N. D. Resultados
68
Cepa PP75 Cepa selvagem
Grupos/tempo (meses)
Rel
ação
A/G
(g/d
L)
ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C 0
2
4
6
8
0
2
4
6
8
Na avaliação ao longo do tempo, observou-se que, no período de 3 meses após a
infecção com a cepa PP75 pela via IP houve aumento da relação A/G, quando comparado
aos períodos de 1 e 6 meses após a infecção. De forma semelhante, no período de 3 meses
da infecção no grupo inoculado pela via IC houve aumento da relação A/G quando
comparado aos períodos de 1, 6 e 9 meses da infecção.
Em animais infectados com a cepa selvagem foi observado 3 meses após a infecção
aumento na relação A/G nos grupos ID e IP em relação aos grupos C e IC (Figura 14).
Na avaliação ao longo do tempo, observou-se que, nos períodos de 3 e 6 meses após
a infecção com a cepa selvagem pelas vias ID e IP houve aumento da relação A/G, quando
comparado aos períodos de 1 e 9 meses após a infecção, respectivamente. Além disso,
mostrou aumento no período de 3 meses no grupo inoculado pela via IP quando comparado
ao período de 9 meses após a infecção. No período de 6 meses após a infecção o grupo IC
mostrou aumento da relação A/G quando comparado aos períodos de 1 e 9 meses após a
infecção.
Figura 14: Relação de albumina globulina de hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como valores de densidade óptica. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
6.7 Peso dos hamsters
Os dados obtidos em relação ao peso dos hamsters estão demonstrados na Figura
15. Em hamsters infectados com a cepa PP75 não foram observadas diferenças
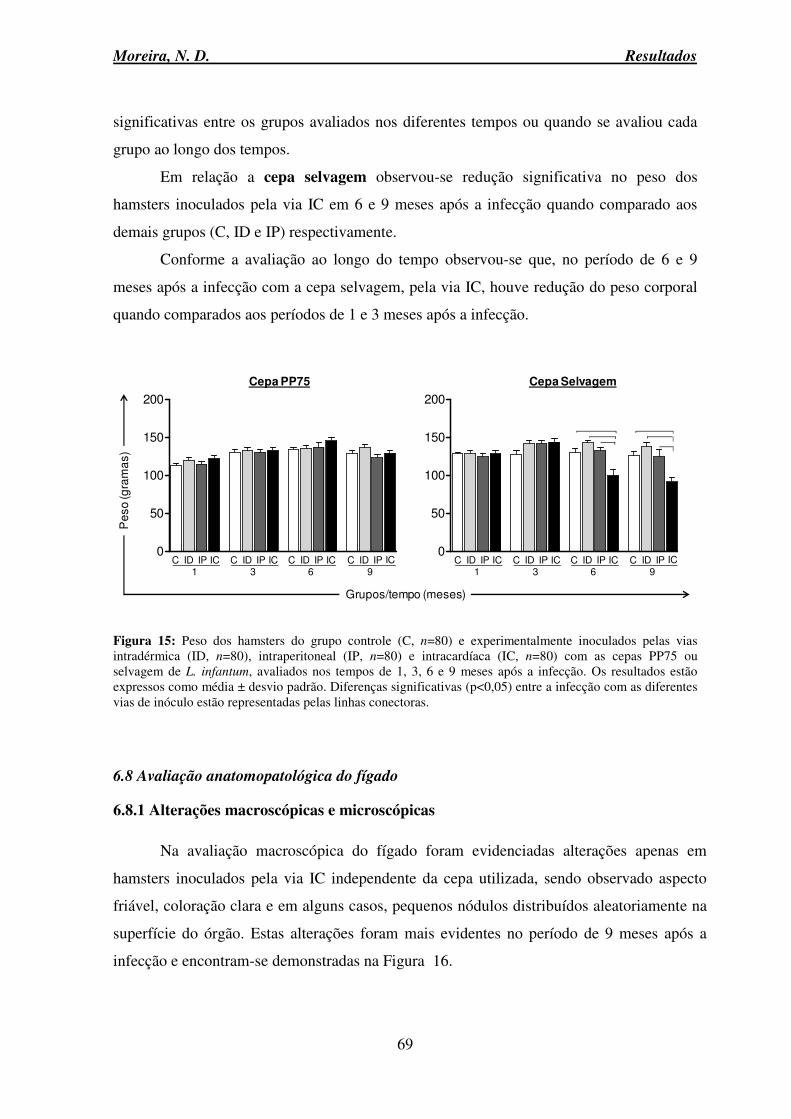
Moreira, N. D. Resultados
69
significativas entre os grupos avaliados nos diferentes tempos ou quando se avaliou cada
grupo ao longo dos tempos.
Em relação a cepa selvagem observou-se redução significativa no peso dos
hamsters inoculados pela via IC em 6 e 9 meses após a infecção quando comparado aos
demais grupos (C, ID e IP) respectivamente.
Conforme a avaliação ao longo do tempo observou-se que, no período de 6 e 9
meses após a infecção com a cepa selvagem, pela via IC, houve redução do peso corporal
quando comparados aos períodos de 1 e 3 meses após a infecção.
Figura 15: Peso dos hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
6.8 Avaliação anatomopatológica do fígado
6.8.1 Alterações macroscópicas e microscópicas
Na avaliação macroscópica do fígado foram evidenciadas alterações apenas em
hamsters inoculados pela via IC independente da cepa utilizada, sendo observado aspecto
friável, coloração clara e em alguns casos, pequenos nódulos distribuídos aleatoriamente na
superfície do órgão. Estas alterações foram mais evidentes no período de 9 meses após a
infecção e encontram-se demonstradas na Figura 16.
Pes
o (g
ram
as)
Cepa SelvagemCepa PP75
Grupos/tempo (meses)
ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C0
50
100
150
200
0
50
100
150
200
ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C
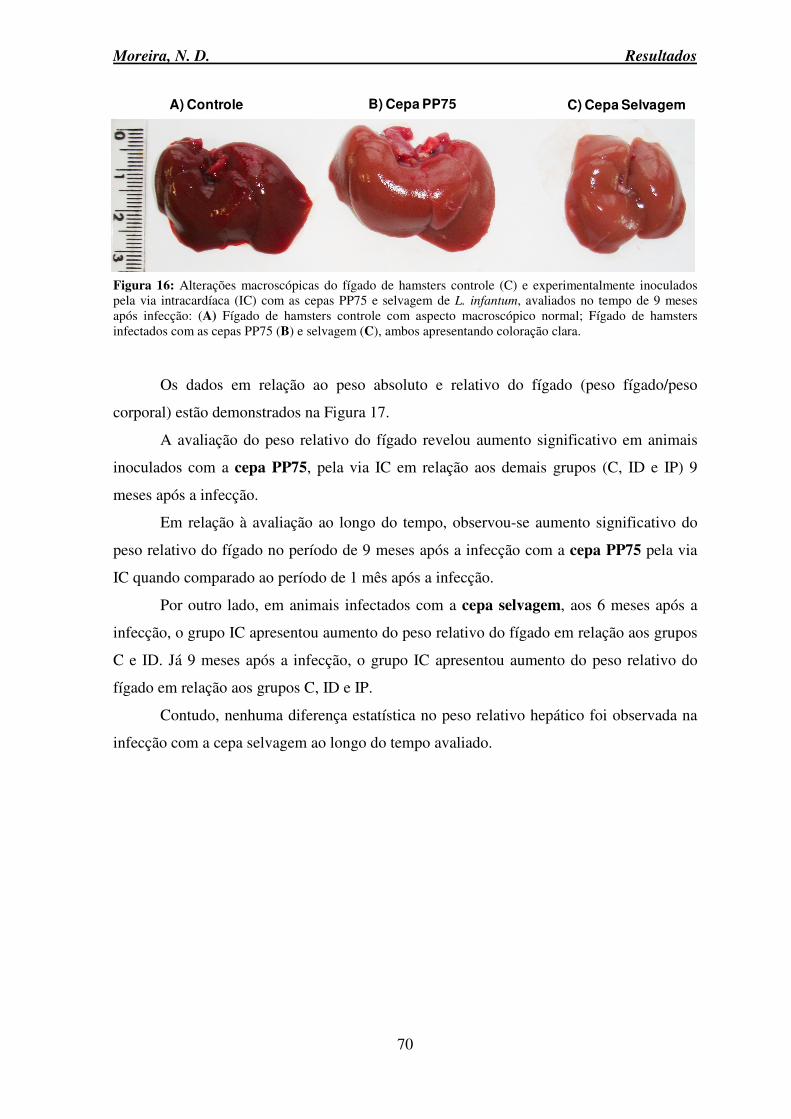
Moreira, N. D. Resultados
70
A) Controle B) Cepa PP75 C) Cepa Selvagem
Figura 16: Alterações macroscópicas do fígado de hamsters controle (C) e experimentalmente inoculados pela via intracardíaca (IC) com as cepas PP75 e selvagem de L. infantum, avaliados no tempo de 9 meses após infecção: (A) Fígado de hamsters controle com aspecto macroscópico normal; Fígado de hamsters infectados com as cepas PP75 (B) e selvagem (C), ambos apresentando coloração clara.
Os dados em relação ao peso absoluto e relativo do fígado (peso fígado/peso
corporal) estão demonstrados na Figura 17.
A avaliação do peso relativo do fígado revelou aumento significativo em animais
inoculados com a cepa PP75, pela via IC em relação aos demais grupos (C, ID e IP) 9
meses após a infecção.
Em relação à avaliação ao longo do tempo, observou-se aumento significativo do
peso relativo do fígado no período de 9 meses após a infecção com a cepa PP75 pela via
IC quando comparado ao período de 1 mês após a infecção.
Por outro lado, em animais infectados com a cepa selvagem, aos 6 meses após a
infecção, o grupo IC apresentou aumento do peso relativo do fígado em relação aos grupos
C e ID. Já 9 meses após a infecção, o grupo IC apresentou aumento do peso relativo do
fígado em relação aos grupos C, ID e IP.
Contudo, nenhuma diferença estatística no peso relativo hepático foi observada na
infecção com a cepa selvagem ao longo do tempo avaliado.

Moreira, N. D. Resultados
71
Cepa SelvagemCepa PP75
Grupos/tempo (meses)
ID IPIC1
C ID IPIC3
C ID IPIC6
C ID IPIC9
C ID IPIC1
C ID IPIC3
C ID IPIC6
C ID IPIC9
C
0
2
4
6
0
2
4
6
0.00
0.01
0.02
0.03
0.04
0.05
0.00
0.01
0.02
0.03
0.04
0.05
Pes
o re
lativ
o (%
)P
eso
fíg
ado
(g)
Figura 17: Peso do fígado e peso relativo (peso fígado/peso corporal) de hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
Ao exame microscópico, os principais achados histopatológicos foram: processo
inflamatório localizado no espaço-porta hepático, sendo a inflamação caracterizada por um
exsudato celular composto por macrófagos, plasmócitos e linfócitos e raros neutrófilos;
granulomas, sendo estes constituídos, predominantemente, por macrófagos, algumas
células epitelióides, linfócitos e raros granulócitos neutrófilos. Além disto, outras
alterações foram observadas, porém com menor freqüência como a presença de fenômenos
degenerativos, dilatação dos sinusóides e hiperemia em alguns casos.
O fígado dos animais do grupo C de ambas as cepas (PP75 e selvagem)
apresentou-se de um modo geral, ao longo de todo o tempo avaliado (1, 3, 6 e 9 meses),
aspecto histológico normal.
Em relação aos animais infectados com a cepa PP75, foi observado aspecto
histológico normal, em 1 e 3 meses após a infecção nos três grupos experimentais (ID, IC e
IP). Contudo no sexto e no nono mês após a infecção utilizando as vias ID, IP e IC, foi

Moreira, N. D. Resultados
72
possível observar a presença de alguns fenômenos degenerativos, sendo estes em grau
variado, ora apresentando um perfil discreto ora um perfil intenso com dilatação dos
capilares sinusóides. Os grupos ID e IP apresentaram um infiltrado inflamatório discreto
(Figura 18), (Figura 19 A e B) e (Figura 19 E e F). Além disso, foi observado ainda em 6 e
9 meses no grupo IC um infiltrado inflamatório que variou de discreto a moderado (Figura
19 I e J). A partir do sexto mês após a infecção utilizando a via IC, observou-se a presença
de granulomas intralobulares em alguns casos (Figura 20 A e B).
Neste sentido, quando a cepa selvagem foi inoculada utilizando a via ID, em 6 e 9
meses após a infecção, foi observado o infiltrado inflamatório discreto (Figura 19 C e D).
Por outro lado, em animais infectados com via IP, foi demonstrado infiltrado inflamatório
que variou de discreto a moderado (Figura 18) e (Figura 19 G e H), presença de alguns
granulomas intralobulares durante todo o período de avaliação (1, 3, 6 e 9 meses após a
infecção) sendo mais evidentes no sexto e nono mês (Figura 20 C e D), e ainda, fenômenos
degenerativos e dilatação dos capilares sinusóides em alguns casos. Contudo, em animais
infectados com a cepa selvagem pela via IC, foram observadas as maiores alterações
hepáticas, sendo estas, mais pronunciadas no sexto e nono mês após a infecção. Dentre as
principais alterações histológicas observados neste grupo (IC), destacam-se, presença de
granulomas intralobulares observada logo no primeiro e terceiro mês da infecção,
totalizando 40% do grupo. Em 6 e 9 meses após a infecção a presença de granulomas
abrangeu 90% do grupo (Figura 20 E e F). A avaliação histopatológica evidenciou também
infiltrado inflamatório, o qual variou de moderado a intenso (Figura 18), conforme a
evolução da infecção, além disso, observou-se áreas necrose (Figura 20 E e F), dilatação
dos capilares sinusóides e hiperemia. Ainda, foi possível observar neste grupo a presença
de formas amastigotas.
As células inflamatórias no fígado foram quantificadas por morfometria e a Figura
18 ilustra os resultados da quantificação do processo inflamatório hepático, sendo
representado cada um dos grupos (ID, IP e IC) infectados com as diferentes cepas de L.
infantum (PP75 e Selvagem) e avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção.
Em relação aos animais infectados com a cepa PP75, foi observado inflamação,
somente no grupo inoculado pela via IC no sexto mês de infecção, em relação aos grupos
ID e IP. Além disto, o grupo IC apresentou processo inflamatório moderado aos 9 meses
após a infecção quando comparado aos grupos ID e IC.
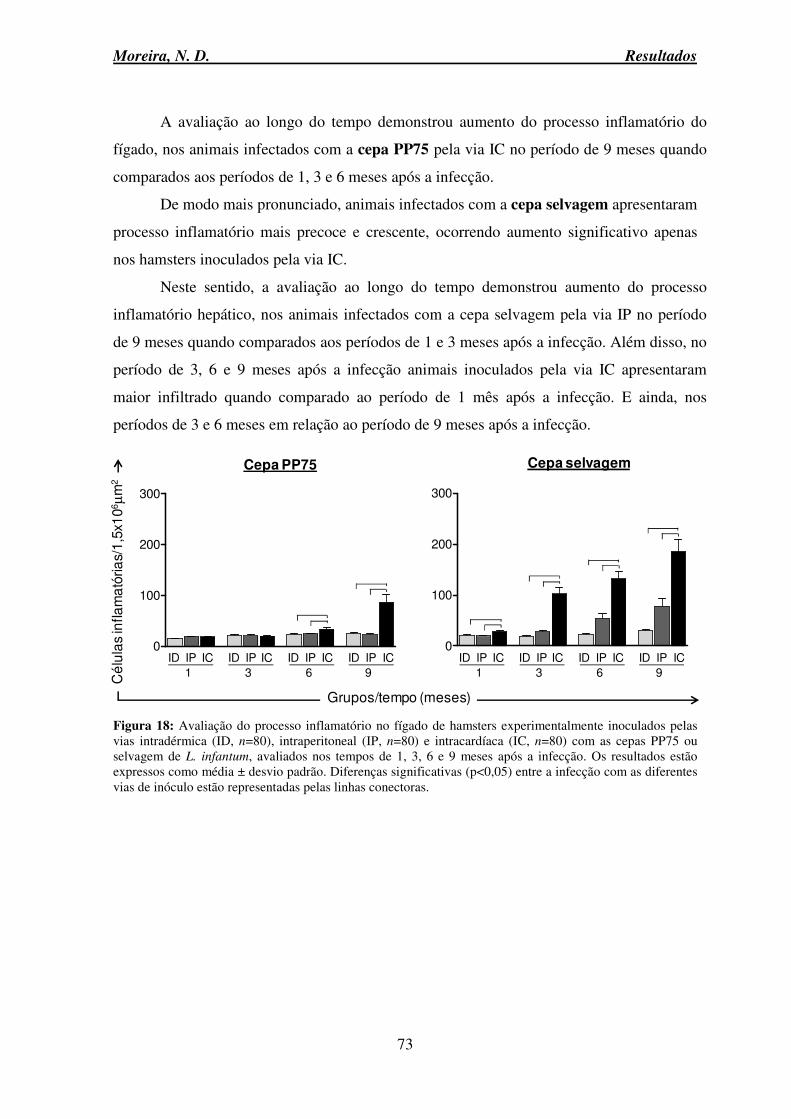
Moreira, N. D. Resultados
73
Cepa PP75 Cepa selvagem
Grupos/tempo (meses)
ID IP IC1
ID IP IC3
ID IP IC6
ID IP IC9C
élul
asin
flam
atór
ias/
1,5x
106 µ
m2
0
100
200
300
0
100
200
300
ID IP IC1
ID IP IC3
ID IP IC6
ID IP IC9
A avaliação ao longo do tempo demonstrou aumento do processo inflamatório do
fígado, nos animais infectados com a cepa PP75 pela via IC no período de 9 meses quando
comparados aos períodos de 1, 3 e 6 meses após a infecção.
De modo mais pronunciado, animais infectados com a cepa selvagem apresentaram
processo inflamatório mais precoce e crescente, ocorrendo aumento significativo apenas
nos hamsters inoculados pela via IC.
Neste sentido, a avaliação ao longo do tempo demonstrou aumento do processo
inflamatório hepático, nos animais infectados com a cepa selvagem pela via IP no período
de 9 meses quando comparados aos períodos de 1 e 3 meses após a infecção. Além disso, no
período de 3, 6 e 9 meses após a infecção animais inoculados pela via IC apresentaram
maior infiltrado quando comparado ao período de 1 mês após a infecção. E ainda, nos
períodos de 3 e 6 meses em relação ao período de 9 meses após a infecção.
Figura 18: Avaliação do processo inflamatório no fígado de hamsters experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
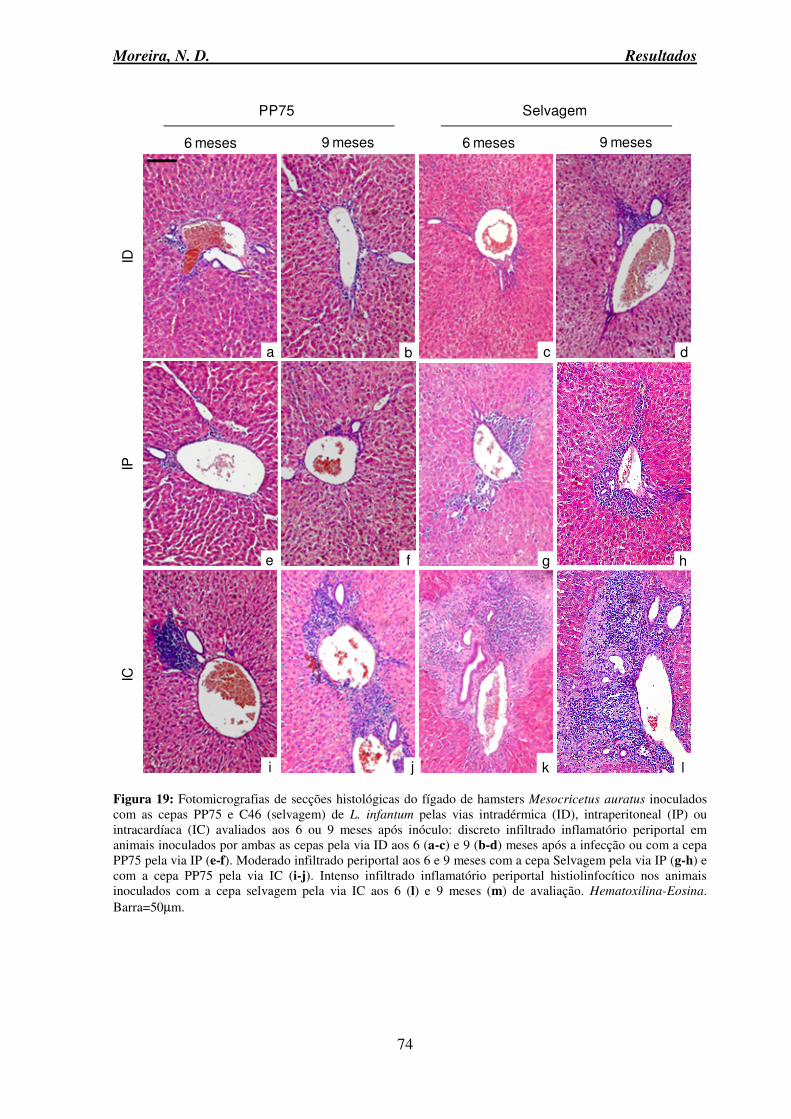
Moreira, N. D. Resultados
74
PP75ID
IPIC
6 meses 9 meses
Selvagem
6 meses 9 meses
a b c d
hgfe
i j k l
Figura 19: Fotomicrografias de secções histológicas do fígado de hamsters Mesocricetus auratus inoculados com as cepas PP75 e C46 (selvagem) de L. infantum pelas vias intradérmica (ID), intraperitoneal (IP) ou intracardíaca (IC) avaliados aos 6 ou 9 meses após inóculo: discreto infiltrado inflamatório periportal em animais inoculados por ambas as cepas pela via ID aos 6 (a-c) e 9 (b-d) meses após a infecção ou com a cepa PP75 pela via IP (e-f). Moderado infiltrado periportal aos 6 e 9 meses com a cepa Selvagem pela via IP (g-h) e com a cepa PP75 pela via IC (i-j). Intenso infiltrado inflamatório periportal histiolinfocítico nos animais inoculados com a cepa selvagem pela via IC aos 6 (l) e 9 meses (m) de avaliação. Hematoxilina-Eosina. Barra=50µm.
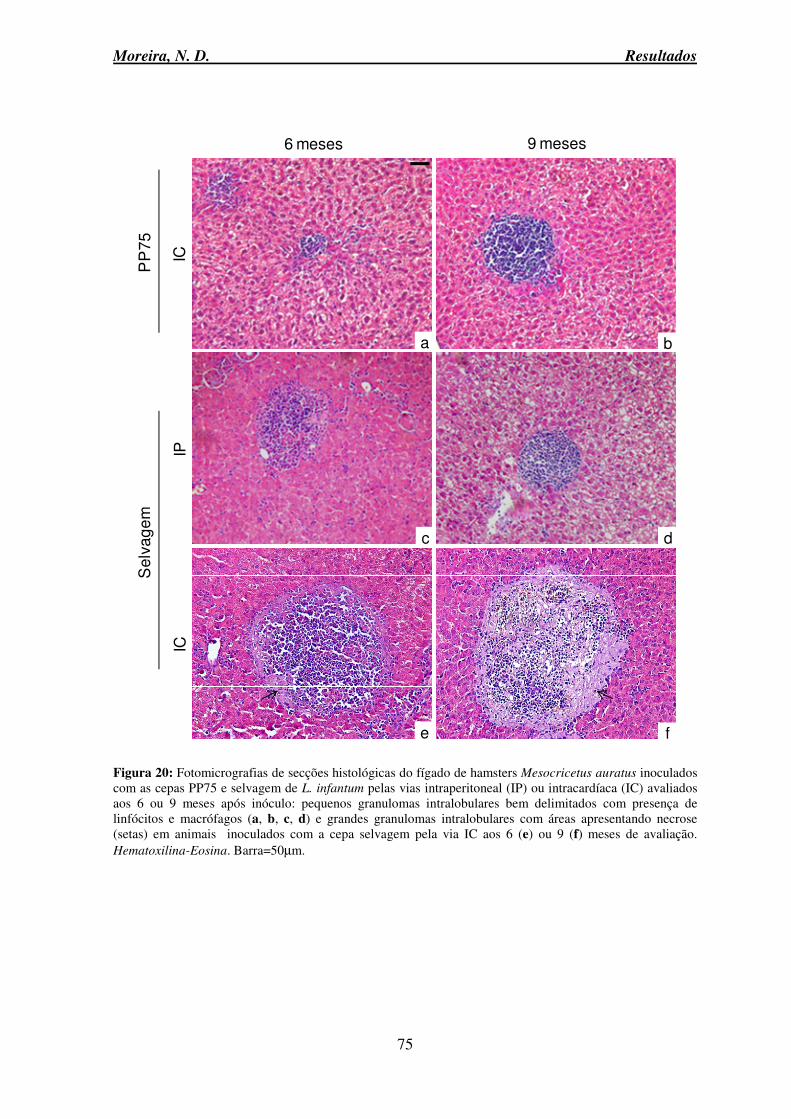
Moreira, N. D. Resultados
75
a b
c
e f
d
6 meses 9 mesesP
P75
Sel
vage
m
ICIP
IC
Figura 20: Fotomicrografias de secções histológicas do fígado de hamsters Mesocricetus auratus inoculados com as cepas PP75 e selvagem de L. infantum pelas vias intraperitoneal (IP) ou intracardíaca (IC) avaliados aos 6 ou 9 meses após inóculo: pequenos granulomas intralobulares bem delimitados com presença de linfócitos e macrófagos (a, b, c, d) e grandes granulomas intralobulares com áreas apresentando necrose (setas) em animais inoculados com a cepa selvagem pela via IC aos 6 (e) ou 9 (f) meses de avaliação. Hematoxilina-Eosina. Barra=50µm.

Moreira, N. D. Resultados
76
Controle Cepa PP75 Cepa selvagem
6.9 Avaliação anatomopatológica do baço
6.9.1 Alterações macroscópicas e microscópicas
As alterações esplênicas macroscópicas observadas com maior frequência incluíram:
aumento de volume do órgão (esplenomegalia), espessamento das bordas, aspecto rugoso e
irregular da superfície e coloração escura, sugerindo um quadro de hiperemia (Figura21).
Figura 21: Alterações macroscópicas do baço de hamsters controle (C) e experimentalmente inoculados pela via intracardíaca (IC) com as cepas PP75 e selvagem de L. infantum, avaliados no tempo de 9 meses após infecção: Controle: baço com aspecto macroscópico normal; Cepas PP75 e selvagem: baço apresentando aspecto rugoso, coloração escura e diferentes níveis de esplenomegalia.
Os dados obtidos em relação ao peso do baço e, ao peso relativo (peso do baço/peso
corporal) entre os grupos analisados estão demonstrados na Figura 22.
Aos 3 meses de infecção, observou-se um discreto aumento no peso relativo do
baço nos animais infectados com a cepa PP75 pela via IP quando comparado aos grupos
C, ID e IC, normalizando já aos 6 meses de infecção. Em contrapartida, aos 9 meses após a
infecção foi observado aumento expressivo no peso relativo do baço no grupo IC em
relação aos demais grupos.
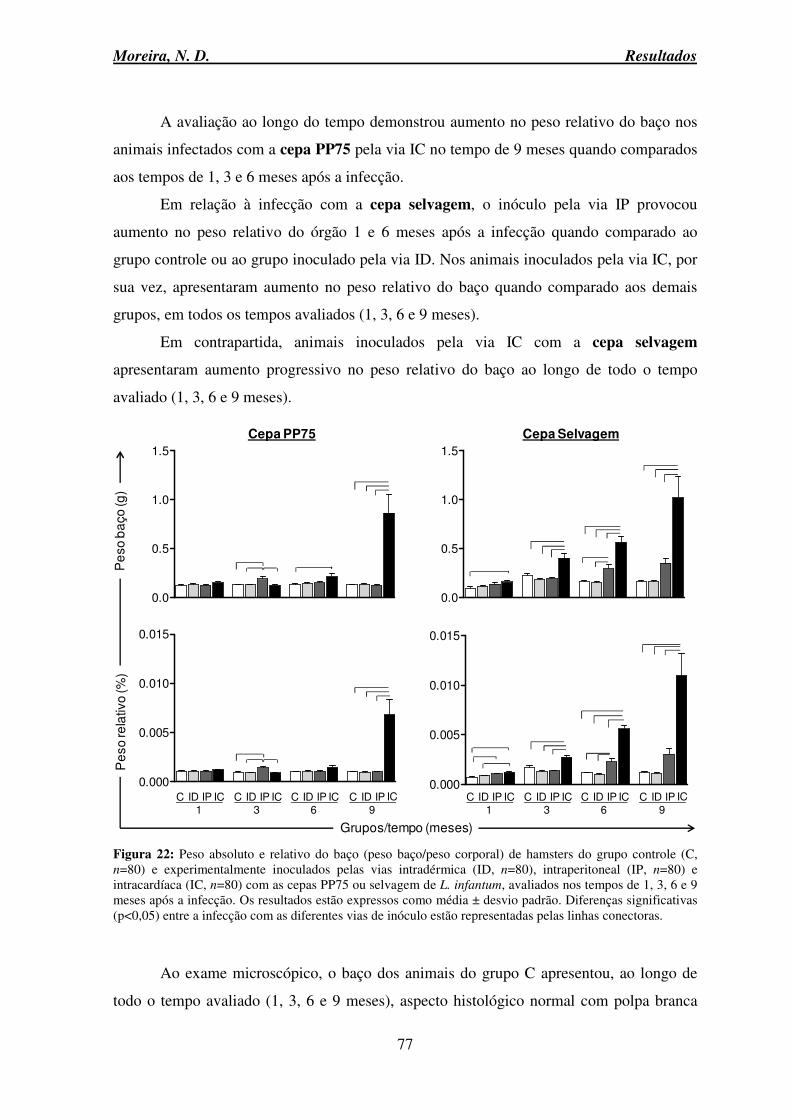
Moreira, N. D. Resultados
77
Grupos/tempo (meses)
Cepa SelvagemCepa PP75
Pes
o re
lativ
o (%
)P
eso
baç
o (g
)
0.0
0.5
1.0
1.5
0.000
0.005
0.010
0.015
0.0
0.5
1.0
1.5
0.000
0.005
0.010
0.015
ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C ID IP IC1
C ID IP IC3
C ID IP IC6
C ID IP IC9
C
A avaliação ao longo do tempo demonstrou aumento no peso relativo do baço nos
animais infectados com a cepa PP75 pela via IC no tempo de 9 meses quando comparados
aos tempos de 1, 3 e 6 meses após a infecção.
Em relação à infecção com a cepa selvagem, o inóculo pela via IP provocou
aumento no peso relativo do órgão 1 e 6 meses após a infecção quando comparado ao
grupo controle ou ao grupo inoculado pela via ID. Nos animais inoculados pela via IC, por
sua vez, apresentaram aumento no peso relativo do baço quando comparado aos demais
grupos, em todos os tempos avaliados (1, 3, 6 e 9 meses).
Em contrapartida, animais inoculados pela via IC com a cepa selvagem
apresentaram aumento progressivo no peso relativo do baço ao longo de todo o tempo
avaliado (1, 3, 6 e 9 meses).
Figura 22: Peso absoluto e relativo do baço (peso baço/peso corporal) de hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
Ao exame microscópico, o baço dos animais do grupo C apresentou, ao longo de
todo o tempo avaliado (1, 3, 6 e 9 meses), aspecto histológico normal com polpa branca

Moreira, N. D. Resultados
78
(PB) organizada em folículos linfóides, bem delimitados apresentando, na maioria das
vezes, centro germinativo e polpa vermelha (PV) constituída principalmente por linfócitos
(Figura 25 e 26 A e E).
De um modo geral, quando a cepa PP75 foi inoculada utilizando as via ID e IP, a
arquitetura histológica do baço foi mantida, ao longo dos nove meses avaliados, com polpa
branca restrita a folículos linfóides primários e secundários. No entanto, aos 3 meses após a
infecção, a polpa vermelha sofreu modificações em decorrência do aumento considerável
de macrófagos. Além disso, na polpa branca, folículos hipertrofiados apresentaram centro
germinativo com depleção de linfócitos e intensa substituição por macrófagos. A partir do
sexto mês após a infecção, apesar de a reatividade estar mantida, a arquitetura apresentou-
se em franca recuperação (Figura 25 B e F) e (Figuras 25 C e G). A infecção com a cepa
PP75 pela via IC provocou acentuada reatividade com aumento do volume e da densidade
linfocitária ao 1, 3 e 6 meses após a infecção. Alterações na polpa vermelha, como
proliferação de macrófagos (Figura 24), variando de discreta a moderada foram
identificadas a partir terceiro mês após a infecção. Estas alterações foram mantidas até o
nono mês, sendo mais frequente neste último mês (Figuras 24) e (Figuras 25 D e H), onde
macrófagos densamente parasitados por formas amastigotas de L. infantum e periesplenite
foram constantemente observados (Figuras 23) e (Figuras 25 D e H).
Na infecção com a cepa selvagem observa-se reatividade e hipertrofia folicular no
primeiro e terceiro mês após o inóculo pelas três vias avaliadas (ID, IP e IC), sendo
mantidas até o nono mês apenas nos grupos onde foram utilizadas as vias ID e IP. Nestes
dois grupos houve proliferação de macrófagos da polpa vermelha, variando de leve a
moderada, a partir do terceiro mês após o inóculo, sendo mais frequente no sexto mês
(Figura 26 B e C). No nono mês, animais inoculados pela via ID apresentaram melhores
níveis de organização tecidual (Figura 26 F) enquanto animais inoculados pela via IP,
mantiveram hiperplasia da polpa vermelha em 60% dos animais (Figura 26 G).
Adicionalmente, aos 6 e 9 meses após inóculo da cepa selvagem pela via IP foi observado
periesplenite (Figura 23) e parasitismo tecidual em 10% e 75% dos animais,
respectivamente. No grupo IC observa-se, nestes dois tempos de avaliação (3 e 6 mês),
atrofia linfóide, relacionada à desorganização histológica onde ocorre perda da definição
ou hipoplasia das regiões da polpa branca, frequentemente associada à proliferação de
macrófagos da PV (Figura 24). Esta associação foi evidenciada principalmente no sexto
mês após o inóculo, onde todos os animais avaliados apresentaram perda da arquitetura
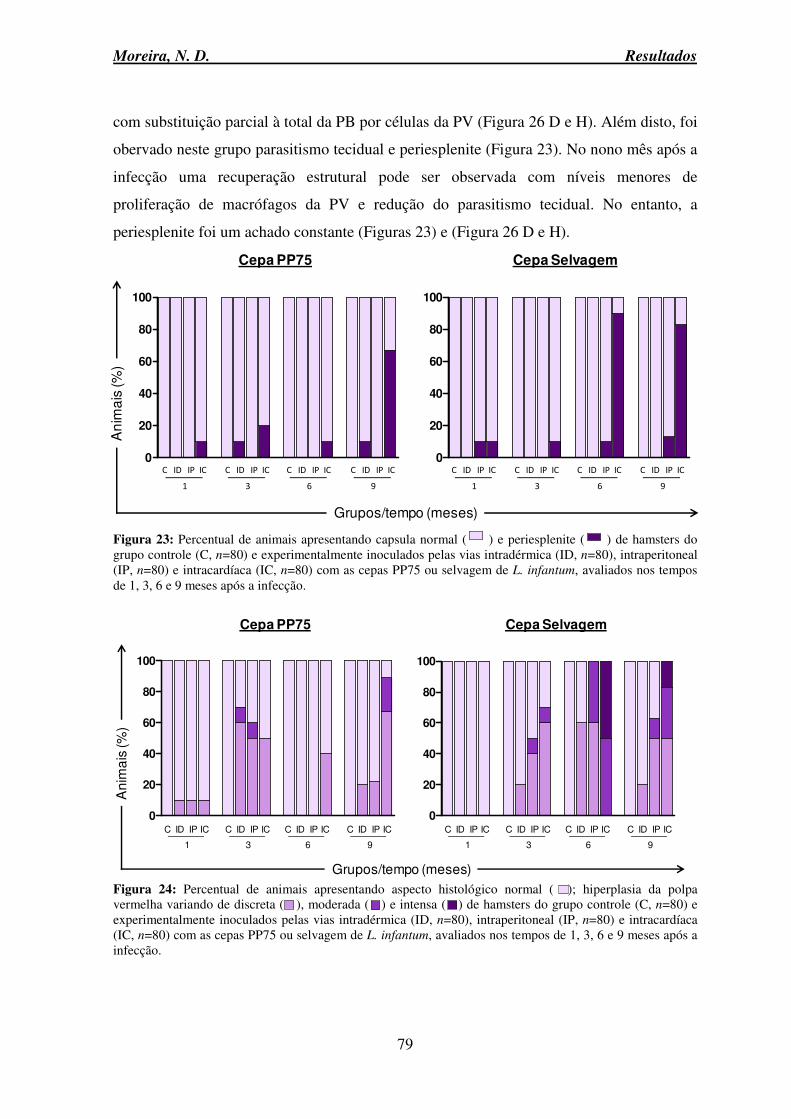
Moreira, N. D. Resultados
79
0
20
40
60
80
100
0
20
40
60
80
100
ID ICC IP
1
ID ICC IP
3
ID ICC IP
6
ID ICC IP
9
ID ICC IP
1
ID ICC IP
3
ID ICC IP
6
ID ICC IP
9
Cepa PP75 Cepa Selvagem
Grupos/tempo (meses)
An
imai
s (%
)com substituição parcial à total da PB por células da PV (Figura 26 D e H). Além disto, foi
obervado neste grupo parasitismo tecidual e periesplenite (Figura 23). No nono mês após a
infecção uma recuperação estrutural pode ser observada com níveis menores de
proliferação de macrófagos da PV e redução do parasitismo tecidual. No entanto, a
periesplenite foi um achado constante (Figuras 23) e (Figura 26 D e H).
Figura 23: Percentual de animais apresentando capsula normal ( ) e periesplenite ( ) de hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção.
Figura 24: Percentual de animais apresentando aspecto histológico normal ( ); hiperplasia da polpa vermelha variando de discreta ( ), moderada ( ) e intensa ( ) de hamsters do grupo controle (C, n=80) e experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção.
0
20
40
60
80
100
0
20
40
60
80
100
Cepa PP75 Cepa Selvagem
ID ICC IP
1
ID ICC IP
3
ID ICC IP
6
ID ICC IP
9
ID ICC IP
1
ID ICC IP
3
ID ICC IP
6
ID ICC IP
9
Grupos/tempo (meses)
Ani
ma
is (%
)
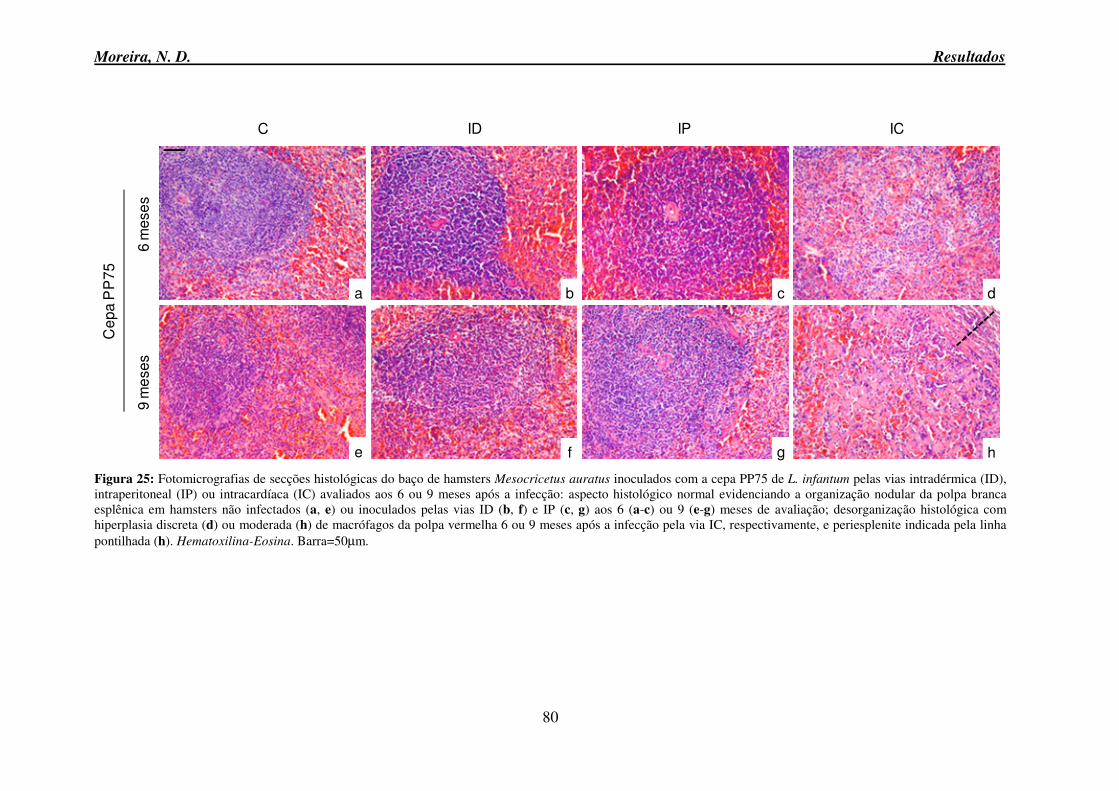
Moreira, N. D. Resultados
80
6 m
eses
9 m
eses
ICIPIDCC
epa
PP
75
a b c d
e f g h
Figura 25: Fotomicrografias de secções histológicas do baço de hamsters Mesocricetus auratus inoculados com a cepa PP75 de L. infantum pelas vias intradérmica (ID), intraperitoneal (IP) ou intracardíaca (IC) avaliados aos 6 ou 9 meses após a infecção: aspecto histológico normal evidenciando a organização nodular da polpa branca esplênica em hamsters não infectados (a, e) ou inoculados pelas vias ID (b, f) e IP (c, g) aos 6 (a-c) ou 9 (e-g) meses de avaliação; desorganização histológica com hiperplasia discreta (d) ou moderada (h) de macrófagos da polpa vermelha 6 ou 9 meses após a infecção pela via IC, respectivamente, e periesplenite indicada pela linha pontilhada (h). Hematoxilina-Eosina. Barra=50µm.
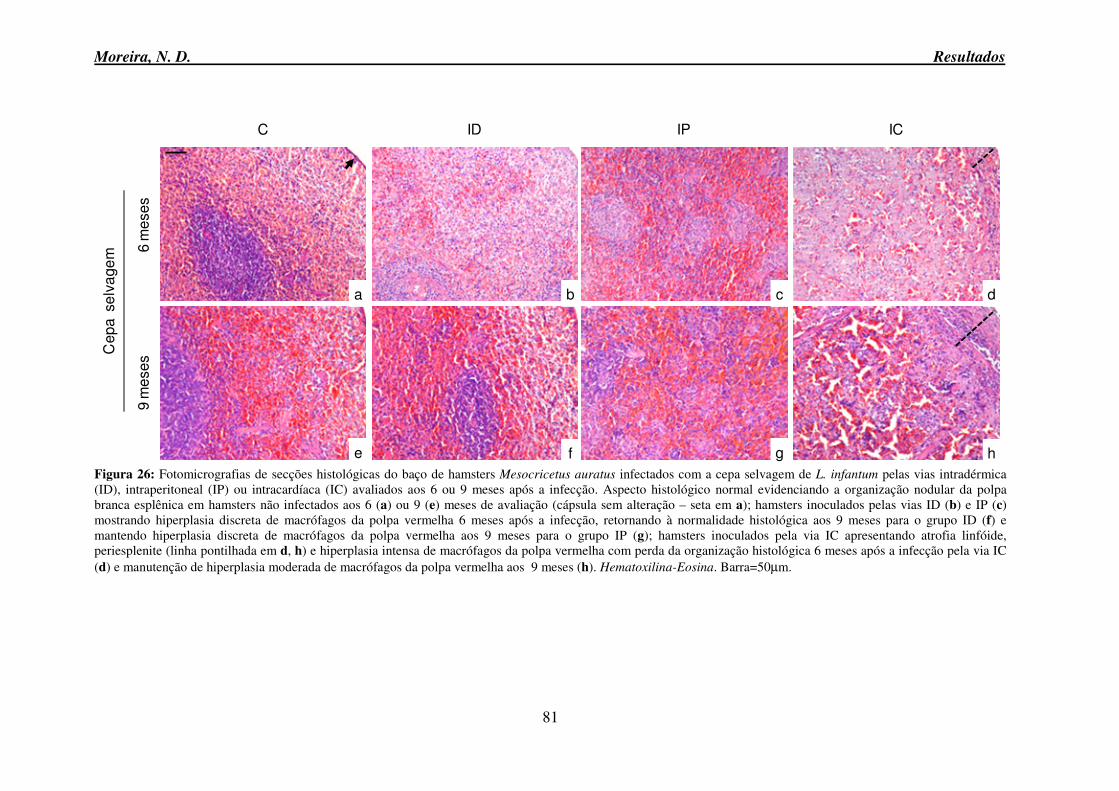
Moreira, N. D. Resultados
81
6 m
eses
9 m
eses
ICIPIDCC
epa
sel
vage
m
a b c d
e f g h
Figura 26: Fotomicrografias de secções histológicas do baço de hamsters Mesocricetus auratus infectados com a cepa selvagem de L. infantum pelas vias intradérmica (ID), intraperitoneal (IP) ou intracardíaca (IC) avaliados aos 6 ou 9 meses após a infecção. Aspecto histológico normal evidenciando a organização nodular da polpa branca esplênica em hamsters não infectados aos 6 (a) ou 9 (e) meses de avaliação (cápsula sem alteração – seta em a); hamsters inoculados pelas vias ID (b) e IP (c) mostrando hiperplasia discreta de macrófagos da polpa vermelha 6 meses após a infecção, retornando à normalidade histológica aos 9 meses para o grupo ID (f) e mantendo hiperplasia discreta de macrófagos da polpa vermelha aos 9 meses para o grupo IP (g); hamsters inoculados pela via IC apresentando atrofia linfóide, periesplenite (linha pontilhada em d, h) e hiperplasia intensa de macrófagos da polpa vermelha com perda da organização histológica 6 meses após a infecção pela via IC (d) e manutenção de hiperplasia moderada de macrófagos da polpa vermelha aos 9 meses (h). Hematoxilina-Eosina. Barra=50µm.

Moreira, N. D. Resultados
82
6.10 Avaliação da carga parasitária no fígado e baço: LDU (Leishmam donovam Units)
e PCR em tempo real
Com a finalidade de avaliar a evolução da infecção em hamsters experimentalmente
infectados com as diferentes cepas (PP75 e selvagem) de L. infantum por diferentes vias
de inóculo (ID, IP e IC), a quantificação da carga parasitária foi realizada em amostras de
fígado e baço, sendo determinada pelas técnicas de LDU e qPCR em tempo real e
avaliados nos tempos de 1,3, 6 e 9 meses após a infecção.
6.10.1 LDU (Leishmam Donovam Units)
Os dados obtidos através do índice de LDU no fígado dos animais infectados com a
cepa PP75 mostraram maior parasitismo em 6 e 9 meses (com 40 e 44% dos animais
apresentado o parasito, respectivamente) após a infecção, nos animais inoculados pela via
IC, quando comparados aos grupos ID e IP. Não foi observado amastigota no fígado dos
hamsters inoculados pelas vias ID e IP após a infecção com a cepa PP75 (Figura 27) e
(Figura 29).
Por outro lado, em animais infectados com a cepa selvagem do grupo IC o
parasitismo do fígado, analisado pelo índice LDU, apresentou aumento em relação aos
grupos ID e IP, em todos os tempos avaliados (1, 3, 6 e 9 meses após o inóculo), com
100% de positividade no sexto e nono mês após a infecção. Entretanto, em hamsters
inoculados pela via IP o parasitismo no fígado iniciou apenas a partir do sexto mês,
seguindo até o nono mês após a infecção, sendo superior ao do grupo ID. Sendo que em 6
meses 60% dos animais apresentaram o parasito e em 9 meses, 87% (Figura 27) e (Figura
29).
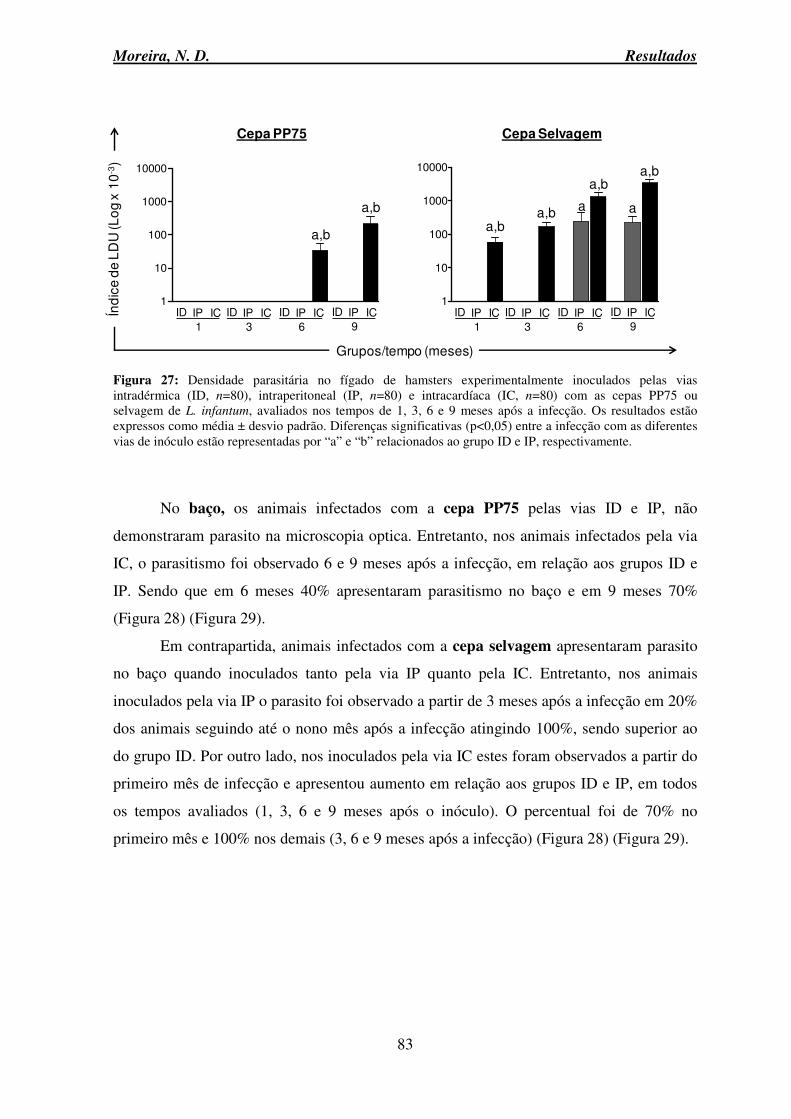
Moreira, N. D. Resultados
83
Cepa SelvagemCepa PP75
Grupos/tempo (meses)
Índ
ice
de
LD
U (L
og
x 1
0-3
)
a,b
a,ba,b
a,b a
a,ba,b
a
ID IP IC9
ID IP IC6
ID IP IC3
ID IP IC1
ID IP IC9
ID IP IC6
ID IP IC3
ID IP IC1
1
10
100
1000
10000
1
10
100
1000
10000
Figura 27: Densidade parasitária no fígado de hamsters experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas por “a” e “b” relacionados ao grupo ID e IP, respectivamente.
No baço, os animais infectados com a cepa PP75 pelas vias ID e IP, não
demonstraram parasito na microscopia optica. Entretanto, nos animais infectados pela via
IC, o parasitismo foi observado 6 e 9 meses após a infecção, em relação aos grupos ID e
IP. Sendo que em 6 meses 40% apresentaram parasitismo no baço e em 9 meses 70%
(Figura 28) (Figura 29).
Em contrapartida, animais infectados com a cepa selvagem apresentaram parasito
no baço quando inoculados tanto pela via IP quanto pela IC. Entretanto, nos animais
inoculados pela via IP o parasito foi observado a partir de 3 meses após a infecção em 20%
dos animais seguindo até o nono mês após a infecção atingindo 100%, sendo superior ao
do grupo ID. Por outro lado, nos inoculados pela via IC estes foram observados a partir do
primeiro mês de infecção e apresentou aumento em relação aos grupos ID e IP, em todos
os tempos avaliados (1, 3, 6 e 9 meses após o inóculo). O percentual foi de 70% no
primeiro mês e 100% nos demais (3, 6 e 9 meses após a infecção) (Figura 28) (Figura 29).
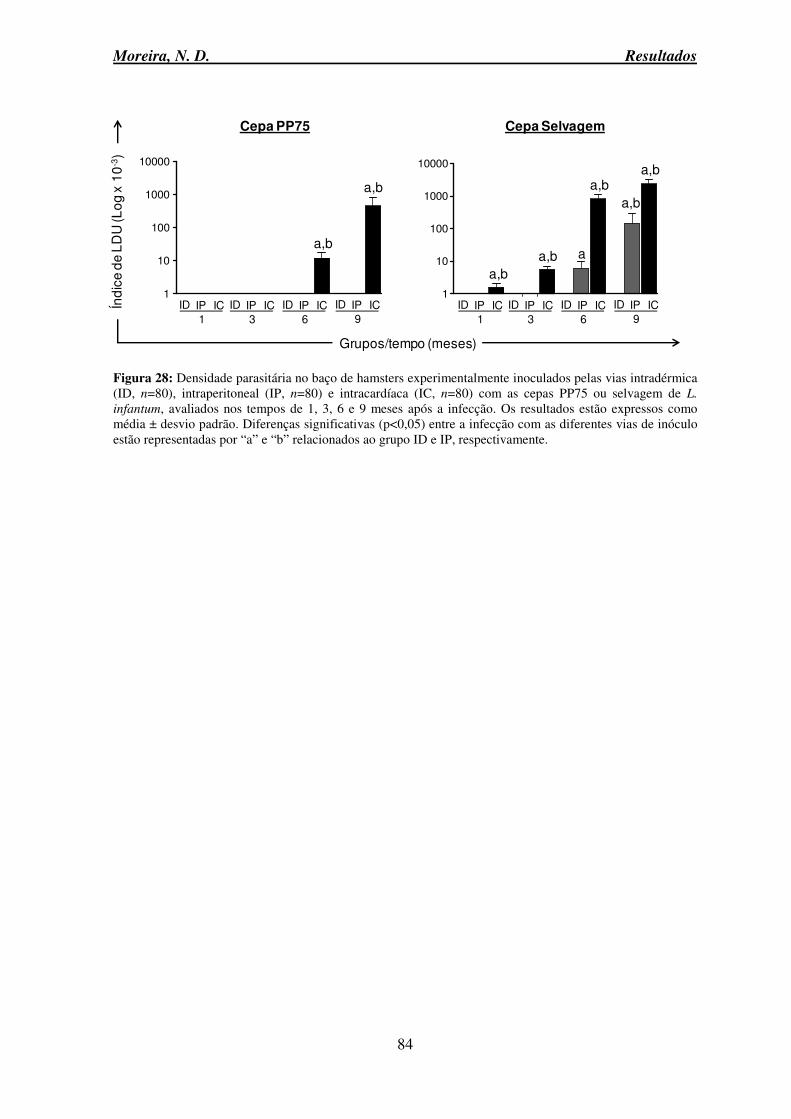
Moreira, N. D. Resultados
84
Cepa SelvagemCepa PP75
Grupos/tempo (meses)
Índ
ice
de
LD
U (L
og
x 1
0-3
)
1
10
100
1000
10000
*
1
10
100
1000
10000
a,b
a,b
a,ba,b a
a,ba,b
a,b
ID IP IC9
ID IP IC6
ID IP IC3
ID IP IC1
ID IP IC9
ID IP IC6
ID IP IC3
ID IP IC1
Figura 28: Densidade parasitária no baço de hamsters experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80) com as cepas PP75 ou selvagem de L.
infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas por “a” e “b” relacionados ao grupo ID e IP, respectivamente.

Moreira, N. D. Resultados
85
Fíg
ado
Baç
o
Cepa PP75 Cepa selvagem
a b c
d e f
Figura 29: Densidade parasitária no fígado e baço de hamsters experimentalmente infectados com as cepas PP75 ou selvagem de L. infantum inoculados pelas vias IP e IC, avaliados aos 9 meses após a infecção. Presença de poucas amastigotas em animais infectados com a cepa PP75 pela via IC (A) e infectados com a cepa selvagem pela via IP (B-E) e IC (C). Intenso parasitismo esplênico em animais infectados com a cepa PP75 (D) ou selvagem (F) pela via IC. Giemsa. Barra = 10µm.
6.11 Análise de correlação: parasitismo no baço X IgG total
Visando identificar associação entre parasitismo tecidual no baço e fígado medido
pela LDU versus os níveis de IgG, foi realizada uma análise de correlação entre estes
parâmetros.
A Figura 30 representa a correlação entre IgG total e o parasitismo no baço de
hamsters experimentalmente infectados com a cepa selvagem utilizando a via IC. Foi
observada uma associação positiva forte (P= 0,001; r=0,7882); (P= 0,001; 0,7559) entre
estes dois parâmetros no baço, tanto para cepa PP75, quanto para a cepa selvagem,

Moreira, N. D. Resultados
86
IgG
tota
l
amastigotas no baço
r=0,7882p=0,0001
r=0,7559p=0,0001
0 3000 60000.00
0.25
0.50
0 1500 30000.00
0.25
0.50
Cepa SelvagemCepa PP75
respectivamente. Entretanto não foi evidenciada correlação entre IgG total e o parasitismo
determinado pelo índice de LDU no fígado, com as diferentes vias de inóculo avaliadas.
Figura 30: Análise de correlação entre o número de amastigotas no baço de hamsters experimentalmente inoculados pela via intracardíaca (IC;▼) com as cepas PP75 e selvagem de L. infantum. Os índices de correlação de Pearson (r) e os valores p estão representados nos gráficos.
6.12 PCR em tempo real
Os resultados obtidos pela qPCR em tempo real encontram-se apresentados
utilizando dois métodos de análise. No primeiro, foi determinado o número de amastigotas
obtidas a partir de 20ng de DNA e no segundo, os resultados obtidos foram demonstrados a
partir do cálculo do número de amastigotas em 20mg de tecido multiplicado pelo peso do
órgão.
Os dados obtidos do fígado após a infecção com as cepas PP75 e selvagem estão
representados na Figura 31, sendo que no painel superior, referente às cepas (PP75 e
selvagem), encontram-se os resultados obtidos a partir do número de amastigotas em 20ng
de DNA no fígado e, no painel inferior, o número de amastigotas em 20mg de tecido x
peso do fígado.
Conforme demonstrado na Figura 31A, animais infectados com a cepa PP75,
mostraram redução na carga parasitária no grupo IC em 3 meses após a infecção quando
comparados ao grupo IP. Portanto, em uma segunda análise, onde foi obtido o número de
amastigotas pelo peso do órgão, observou-se ainda neste mesmo período diferença
significativa na carga parasitária no grupo IP em relação ao grupo ID. Além disso, no sexto
mês foi possível observar redução na carga parasitária no grupo IP quando comparado ao

Moreira, N. D. Resultados
87
grupo ID, além de aumento da carga parasitária no grupo IC em relação aos grupos ID e
IP.
Por outro lado, em animais infectados com a cepa selvagem, no primeiro mês de
infecção foi observado aumento do número de amastigotas no IC quando comparado ao
grupo IP. Além disso, em 3, 6 e 9 meses observou-se aumento significativo no número de
amastigotas no grupo IC quando comparado aos grupos ID e IP. Entretanto quando o
número de amastigotas foi calculado multiplicando pelo peso do fígado, o aumento no
número de amastigotas foi significativo apenas em 6 e 9 meses no grupo inoculado pela via
IC quando comparado aos grupos ID e IP (Figura 31 B).
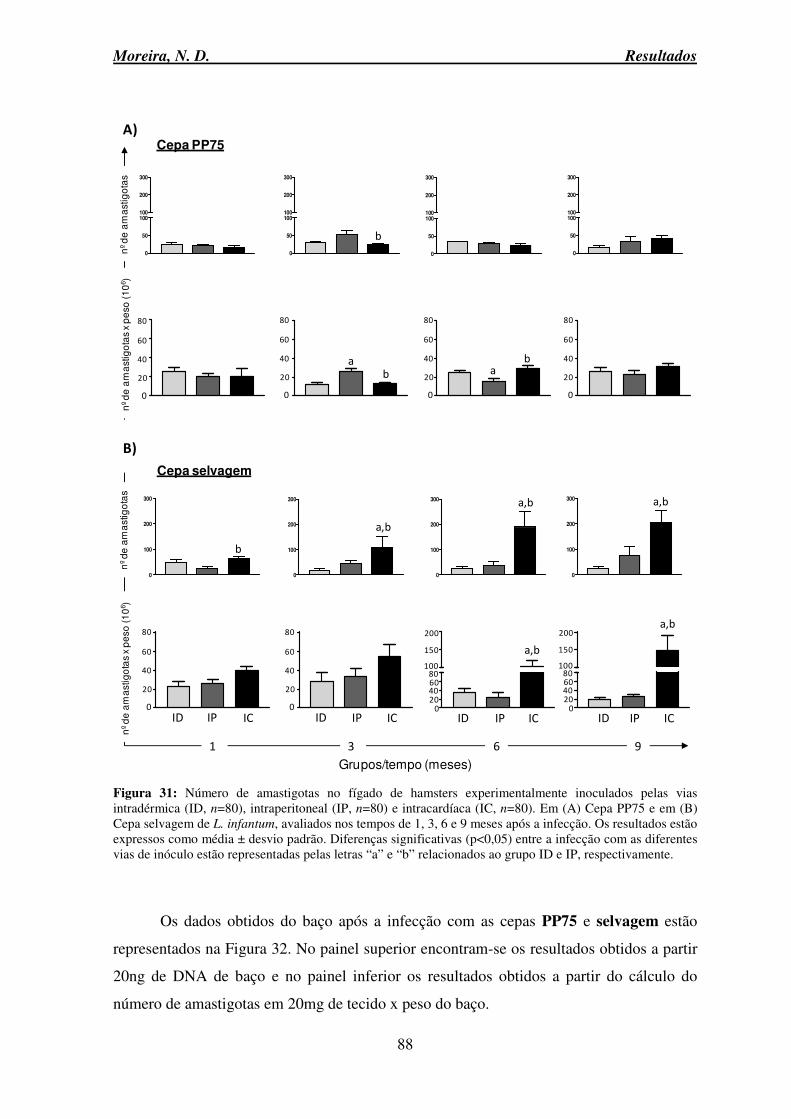
Moreira, N. D. Resultados
88
A)
B)
Cepa PP75
Cepa selvagem
nº d
e a
ma
stig
otas
0
20
40
60
80
0
20
40
60
80
0
20
40
60
80
0
20
40
60
80
a
b ab
0
50
100100
200
300
0
50
100100
200
300
0
50
100100
200
300
0
50
100100
200
300
nº d
e a
ma
stig
otas
x p
eso
(10
6 )
0
20
40
60
80
0
100
150
200
40
80
20
60
0
20
40
60
80
0
100
150
200
40
80
20
60
a,b
a,b
0
100
200
300
0
100
200
300
0
100
200
300
0
100
200
300
nº d
e a
ma
stig
ota
sn
º de
ama
stig
ota
s x
pes
o (1
06 )
1 3 6 9
ID IP IC ID IP IC ID IP IC ID IP IC
Grupos/tempo (meses)
b
b
a,b a,b
a,b
Figura 31: Número de amastigotas no fígado de hamsters experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80). Em (A) Cepa PP75 e em (B) Cepa selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas letras “a” e “b” relacionados ao grupo ID e IP, respectivamente.
Os dados obtidos do baço após a infecção com as cepas PP75 e selvagem estão
representados na Figura 32. No painel superior encontram-se os resultados obtidos a partir
20ng de DNA de baço e no painel inferior os resultados obtidos a partir do cálculo do
número de amastigotas em 20mg de tecido x peso do baço.

Moreira, N. D. Resultados
89
Conforme demonstrado na Figura 32 A, animais infectados com a cepa PP75
apresentaram aumento na carga parasitária no grupo IC em 3 e 6 meses após a infecção
quando comparado aos grupos ID e IP. Por outro lado, em uma segunda análise onde foi
determinado o número de amastigotas em 20mg de tecido x peso do baço, observou-se no
primeiro mês aumento da carga parasitária no baço dos animais inoculados pela via ID em
relação ao grupo IP.
Por outro lado, em animais infectados com a cepa selvagem, pela via IC apresentou
aumento na carga parasitária em relação aos grupos ID e IP, em todos os tempos avaliados
(1, 3, 6 e 9 meses após o inóculo). Entretanto em uma segunda análise quando o número de
amastigotas foi calculado multiplicando pelo peso do baço, o aumento no número de
amastigotas foi significativo apenas em 6 e 9 meses no grupo inoculado pela via IC quando
comparado aos grupos ID e IP (Figura 32 B).

Moreira, N. D. Resultados
90
0
0.5
1
1.5
2
0
0.5
1
1.5
2
0
0.5
1
1.5
2
0
0.5
10
20
2
30
1.5
1.0
0
1
2
3
5
4
00.5
11.5
2
200
100
300
400
00.5
11.5
2
200
100
300
400
0
1
2
3
5
4
ID IP IC ID IP IC ID IP IC ID IP IC
a,bb
a
a,ba,b
a,b
0
100
200
2000
4000
0
100
200
2000
4000
0
100
200
2000
4000
0
100
200
2000
4000
0
100
200
2000
4000
0
100
200
2000
4000
0
100
2002000
3000
4000
0
100
200
2000
4000
A)
B)
Cepa PP75
Cepa selvagem
nº d
e a
ma
stig
ota
snº
de
ama
stig
otas
nº d
e am
ast
igot
as x
pe
so (
106 )
1 3 6 9
Grupos/tempo (meses)
nº d
e a
mas
tigo
tas
x p
eso
(106 )
a,b
a,b
a,ba,b
a,b a,b
a
Figura 32: Número de amastigotas no baço de hamsters experimentalmente inoculados pelas vias intradérmica (ID, n=80), intraperitoneal (IP, n=80) e intracardíaca (IC, n=80). Em (A) Cepa PP75 e em (B) Cepa selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas letras “a” e “b” relacionados ao grupo ID e IP, respectivamente
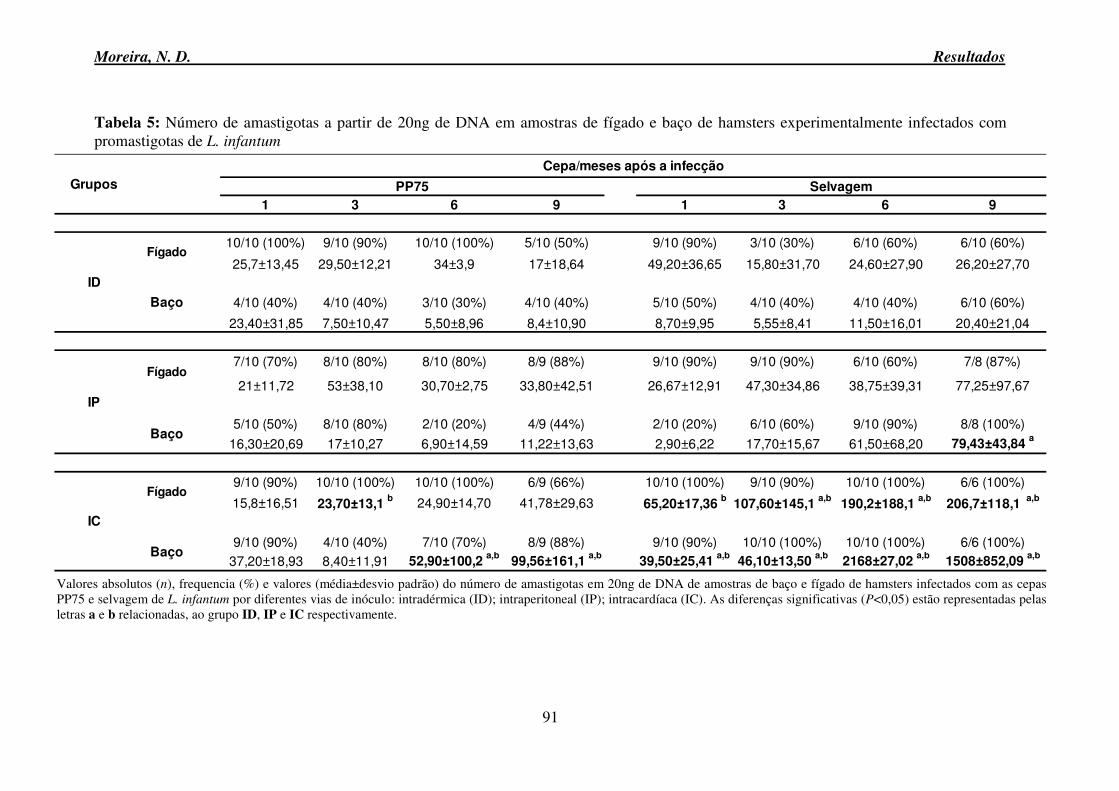
Moreira, N. D. Resultados
91
1 3 6 9 1 3 6 9
10/10 (100%) 9/10 (90%) 10/10 (100%) 5/10 (50%) 9/10 (90%) 3/10 (30%) 6/10 (60%) 6/10 (60%)
25,7±13,45 29,50±12,21 34±3,9 17±18,64 49,20±36,65 15,80±31,70 24,60±27,90 26,20±27,70
4/10 (40%) 4/10 (40%) 3/10 (30%) 4/10 (40%) 5/10 (50%) 4/10 (40%) 4/10 (40%) 6/10 (60%)
23,40±31,85 7,50±10,47 5,50±8,96 8,4±10,90 8,70±9,95 5,55±8,41 11,50±16,01 20,40±21,04
7/10 (70%) 8/10 (80%) 8/10 (80%) 8/9 (88%) 9/10 (90%) 9/10 (90%) 6/10 (60%) 7/8 (87%)
21±11,72 53±38,10 30,70±2,75 33,80±42,51 26,67±12,91 47,30±34,86 38,75±39,31 77,25±97,67
5/10 (50%) 8/10 (80%) 2/10 (20%) 4/9 (44%) 2/10 (20%) 6/10 (60%) 9/10 (90%) 8/8 (100%)
16,30±20,69 17±10,27 6,90±14,59 11,22±13,63 2,90±6,22 17,70±15,67 61,50±68,20 79,43±43,84 a
9/10 (90%) 10/10 (100%) 10/10 (100%) 6/9 (66%) 10/10 (100%) 9/10 (90%) 10/10 (100%) 6/6 (100%)
15,8±16,51 23,70±13,1 b 24,90±14,70 41,78±29,63 65,20±17,36
b107,60±145,1
a,b190,2±188,1
a,b206,7±118,1
a,b
9/10 (90%) 4/10 (40%) 7/10 (70%) 8/9 (88%) 9/10 (90%) 10/10 (100%) 10/10 (100%) 6/6 (100%)37,20±18,93 8,40±11,91 52,90±100,2
a,b99,56±161,1
a,b39,50±25,41
a,b46,10±13,50
a,b2168±27,02
a,b1508±852,09
a,b
Grupos
Cepa/meses após a infecção
PP75 Selvagem
ID
Fígado
Baço
IC
Baço
IP
Baço
Fígado
Fígado
Tabela 5: Número de amastigotas a partir de 20ng de DNA em amostras de fígado e baço de hamsters experimentalmente infectados com promastigotas de L. infantum
Valores absolutos (n), frequencia (%) e valores (média±desvio padrão) do número de amastigotas em 20ng de DNA de amostras de baço e fígado de hamsters infectados com as cepas PP75 e selvagem de L. infantum por diferentes vias de inóculo: intradérmica (ID); intraperitoneal (IP); intracardíaca (IC). As diferenças significativas (P<0,05) estão representadas pelas letras a e b relacionadas, ao grupo ID, IP e IC respectivamente.

Moreira, N. D. Resultados
92
6.13 Avaliação da expressão relativa de mRNA das citocinas proinflamatórias TNF-αααα e
IFN-γγγγ e anti-inflamatórias (IL-10 e TGF-ββββ) no baço
6.13.1 Citocinas proinflamatórias TNF-αααα e IFN-γγγγ
A expressão relativa de mRNA das citocinas TNF-α e IFN-γ foi avaliada
quantitativamente pela técnica de qRT-PCR no baço de hamsters experimentalmente
infectados com L. infantum e os resultados estão representados em expressao relativa de
mRNA/citocinas na Figura 33.
Após 1 mês do inóculo, nos animais infectados com a cepa PP75 observou-se que a
expressão relativa de mRNA da citocina TNF-α α α α nos animais inoculados pela via IC foi
significativamente superior que no grupo ID (Figura 33). Não foi observado diferença
significativa entre os grupos experimentais nos tempos de 3, 6 e 9 meses após a infecção
com esta cepa.
De forma diferente, em hamsters infectados com a cepa selvagem a expressão
relativa de mRNA da citocina TNF-α nos animais inoculados pela via IC foi maior que no
grupo ID, apenas no terceiro mês de infecção(Figura 33).
Não foram observadas diferenças significativas quando se avaliou cada grupo ao
longo dos tempos.
Em relação à citocina IFN-γγγγ, observou-se maior expressão de mRNA nos animais
infectados com a cepa PP75 pela via IC em relação aos demais grupos (ID e IP) nos
tempos de 6 e 9 meses após a infecção (Figura 33).
A avaliação ao longo do tempo mostrou que animais infectados com a cepa PP75
pela via IP, no período de 9 meses após a infecção, apresentam aumento na expressão de
mRNA de IFN-γ quando comparado aos períodos de 1, 3 e 6 meses. Desta maneira, no
período de 6 meses observou-se aumento significativo na expressão de mRNA de IFN-γ
nos animais inoculados pela via IC em relação aos períodos de 1 e 3 meses após a infecção.
Da mesma forma, foi observado maior expressão de mRNA de IFN-γ no grupo IC no
período de 9 meses em relação aos períodos de 1, 3 e 6 meses após a infecção.
Por outro lado, os resultados obtidos em relação à cepa selvagem monstraram
aumento na expressão do mRNA de IFN-γ, nos animais inoculados pela via IP em relação
a via ID, 3 meses após a infecção. Além disto, neste mesmo tempo, observou-se maior
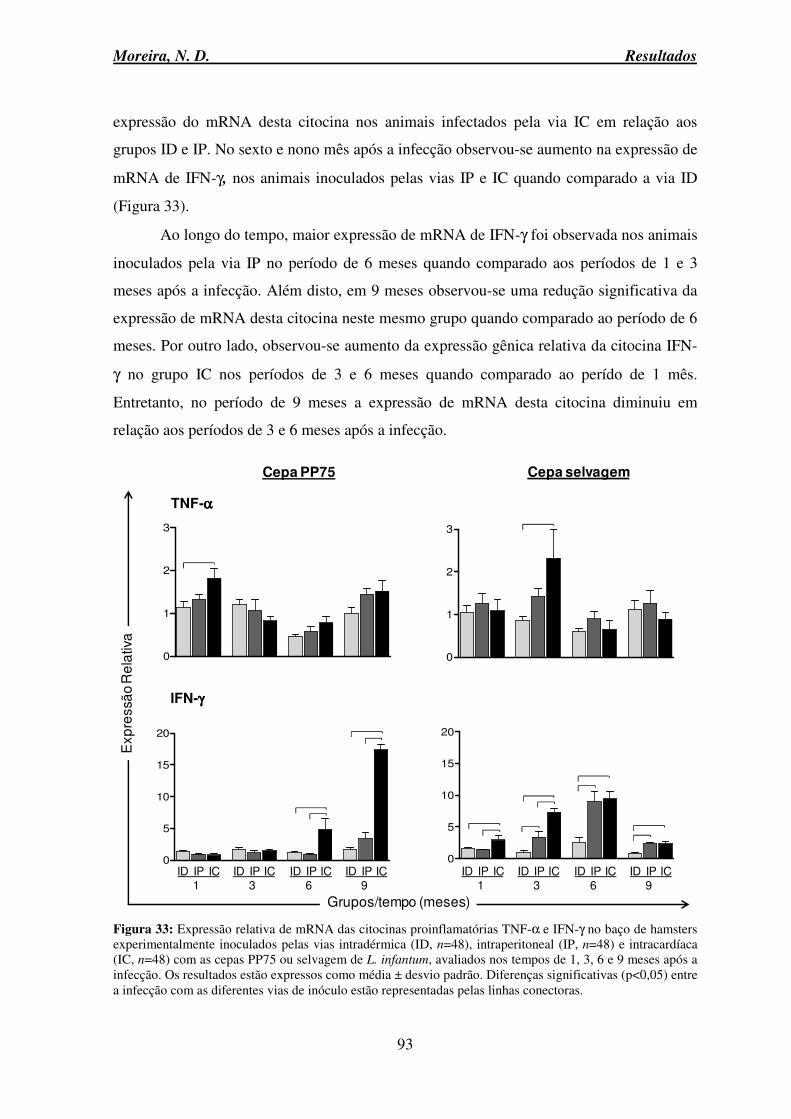
Moreira, N. D. Resultados
93
Cepa PP75 Cepa selvagem
Grupos/tempo (meses)
0
1
2
3
0
1
2
3
0
5
10
15
20
0
5
10
15
20
Exp
ress
ão
Re
lativ
a
TNF-αααα
IFN-γγγγ
ID IP IC1
ID IP IC3
ID IP IC6
ID IP IC9
ID IP IC1
ID IP IC3
ID IP IC6
ID IP IC9
expressão do mRNA desta citocina nos animais infectados pela via IC em relação aos
grupos ID e IP. No sexto e nono mês após a infecção observou-se aumento na expressão de
mRNA de IFN-γ, nos animais inoculados pelas vias IP e IC quando comparado a via ID
(Figura 33).
Ao longo do tempo, maior expressão de mRNA de IFN-γ foi observada nos animais
inoculados pela via IP no período de 6 meses quando comparado aos períodos de 1 e 3
meses após a infecção. Além disto, em 9 meses observou-se uma redução significativa da
expressão de mRNA desta citocina neste mesmo grupo quando comparado ao período de 6
meses. Por outro lado, observou-se aumento da expressão gênica relativa da citocina IFN-
γ no grupo IC nos períodos de 3 e 6 meses quando comparado ao perído de 1 mês.
Entretanto, no período de 9 meses a expressão de mRNA desta citocina diminuiu em
relação aos períodos de 3 e 6 meses após a infecção.
Figura 33: Expressão relativa de mRNA das citocinas proinflamatórias TNF-α e IFN-γ no baço de hamsters experimentalmente inoculados pelas vias intradérmica (ID, n=48), intraperitoneal (IP, n=48) e intracardíaca (IC, n=48) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.

Moreira, N. D. Resultados
94
6.13.2 Citocinas anti-inflamatórias IL-10 e TGF-ββββ
A expressão relativa de mRNA das citocinas anti-inflamatórias IL-10 e TGF-β foi
avaliada quantitativamente pela técnica de qRT-PCR no baço de hamsters
experimentalmente infectados com L. infantum e os resultados estão representados na
Figura 34.
Nos animais inoculados com a cepa PP75 pela via IC foi observado aumento
significativo na expressão relativa da citocina IL-10 quando comparado ao grupo ID no
primeiro mês após a infeção e ID e IP no sexto mês após a infecção. Ao contrário, aumento
de mRNA desta citocina foi observado nos animais inoculados pela via IP quando
comparado aos demais grupos 9 meses após a infecção (Figura 34).
Ao longo do tempo, observou-se que animais inoculados com a cepa PP75
apresentaram redução significativa da expressão de mRNA para de IL-10 no período de 6
meses após a infecção nos animais inoculados pelas vias ID e IP quando comparados aos
períodos de 1 e 3 meses. Por outro lado, no período de 9 meses foi observado aumento
significativo na expressão de mRNA para IL-10 no grupo inoculado pela via IP quando
comparado ao período de 6 meses após a infecção. Além disso, nos períodos de 6 e 9
meses após a infecção o grupo inoculado pela via IC apresentou redução significativa na
expressão de mRNA quando comparado aos perídos de 1 e 3 meses.
Por outro lado, animais infectados com a cepa selvagem apresentaram aumento
significativo na expressão mRNA para IL-10 nos animais inoculados pela via IC, quando
comparado aos demais grupos 3 meses após a infeção. Entretanto, no nono mês após a
infecção, foi observado uma diminuição na expressão de mRNA para essa citocina nos
animais inoculados via IC em relação aos grupos ID e IP (Figura 34).
Animais infectados com a cepa selvagem, apresentaram aumento de mRNA para
IL-10 nos grupos inoculados pelas vias ID e IP no período de 9 meses após a infecção
quando comparados aos tempos de 1, 3 e 6 meses após a infecção. Por outro lado,
observou-se maior expressão de mRNA para IL-10 no grupo IC no período de 3 meses
quando comparado aos períodos de 6 e 9 meses após a infecção.
Animais infectados com a cepa PP75, apresentaram aumento significativo na
expressão de mRNA para a citocina TGF-β, no sexto mês após a infecção, no grupo
inoculado pela via IC em relação aos grupos ID e IP (Figura 34).

Moreira, N. D. Resultados
95
A análise ao longo do tempo mostrou que em hamsters infectados com a cepa PP75
pelas 3 vias de inóculo nos períodos de 3, 6, 9 apresentaram redução significativa na
expressão relativa de mRNA para a citocina TGF-β quando comparado ao período de 1
mês após a infecção.
De forma diferente, no terceiro mês após a infecção, os animais infectados com a
cepa selvagem pela via IC apresentaram aumento significativo de mRNA para TGF-β
quando comparados aos grupos ID e IP. Entretanto, no nono mês após a infecção foi
observado uma diminuição na expressão de mRNA para essa citocina nos animais
inoculados via IC em relação aos grupos ID e IP (Figura 34).
Ao longo do tempo, observou-se que animais infectados com a cepa selvagem
pelas vias ID e IP apresentaram aumento na expressão de mRNA para a citocina TGF-β no
período de 9 meses quando comparado ao período de 1, 3 e 6 meses. Por outro lado, o
grupo inoculado pela via IC no período de 3 meses apresentou maior expressão mRNA
para TGF-β quando comparado aos períodos de 1 e 3 meses após a infecção.
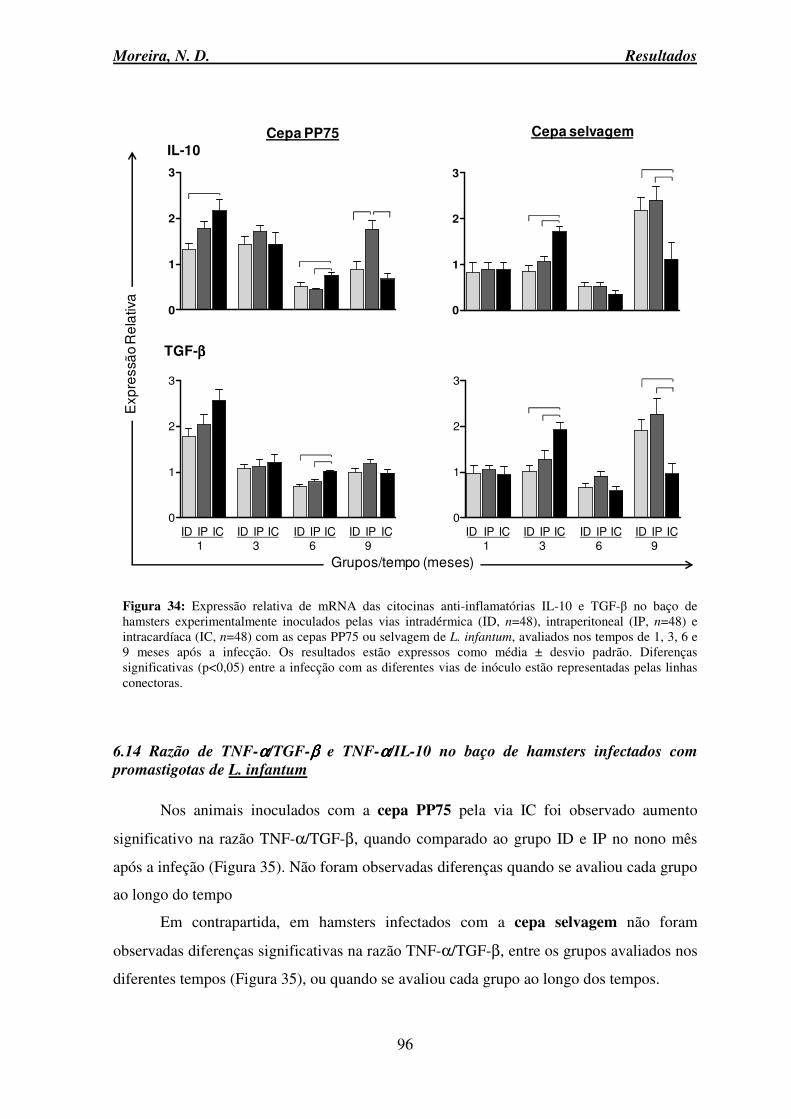
Moreira, N. D. Resultados
96
Cepa PP75 Cepa selvagem
Grupos/tempo (meses)
Exp
ress
ão
Re
lativ
a
TGF-ββββ
IL-10
ID IP IC1
ID IP IC3
ID IP IC6
ID IP IC9
ID IP IC1
ID IP IC3
ID IP IC6
ID IP IC9
0
1
2
3
0
1
2
3
0
1
2
3
0
1
2
3
Figura 34: Expressão relativa de mRNA das citocinas anti-inflamatórias IL-10 e TGF-β no baço de hamsters experimentalmente inoculados pelas vias intradérmica (ID, n=48), intraperitoneal (IP, n=48) e intracardíaca (IC, n=48) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
6.14 Razão de TNF-αααα/TGF-ββββ e TNF-αααα/IL-10 no baço de hamsters infectados com
promastigotas de L. infantum
Nos animais inoculados com a cepa PP75 pela via IC foi observado aumento
significativo na razão TNF-α/TGF-β, quando comparado ao grupo ID e IP no nono mês
após a infeção (Figura 35). Não foram observadas diferenças quando se avaliou cada grupo
ao longo do tempo
Em contrapartida, em hamsters infectados com a cepa selvagem não foram
observadas diferenças significativas na razão TNF-α/TGF-β, entre os grupos avaliados nos
diferentes tempos (Figura 35), ou quando se avaliou cada grupo ao longo dos tempos.

Moreira, N. D. Resultados
97
Em relação à razão TNF-α/IL-10 foi observado redução na razão, nos animais
infectados com a cepa PP75 pela via IP quando comparado aos grupos ID, em 3 meses
após a infecção. Além disso, no nono mês após a infecção foi observado aumento
significativo na razão TNF-α/IL-10 no grupo IC quando comparado aos grupos ID e IP
(Figura 35).
A avaliação ao longo do tempo mostrou que animais infectados com a cepa PP75
pela via IC, no período de 9 meses após a infecção, apresentam aumento na razão TNF-
α/IL-10 quando comparado aos períodos de 1, 3 e 6 meses infecção.
Contudo, em hamsters infectados com a cepa selvagem não foram observadas
diferenças na razão TNF-α/IL-10 entre os grupos avaliados nos diferentes tempos (Figura
35).
A avaliação ao longo do tempo mostrou que animais infectados com a cepa
selvagem pela via IP, no período de 9 meses após a infecção, apresentam redução na razão
TNF-α/IL-10 quando comparado aos períodos de 1 e 6 meses após a infecção.
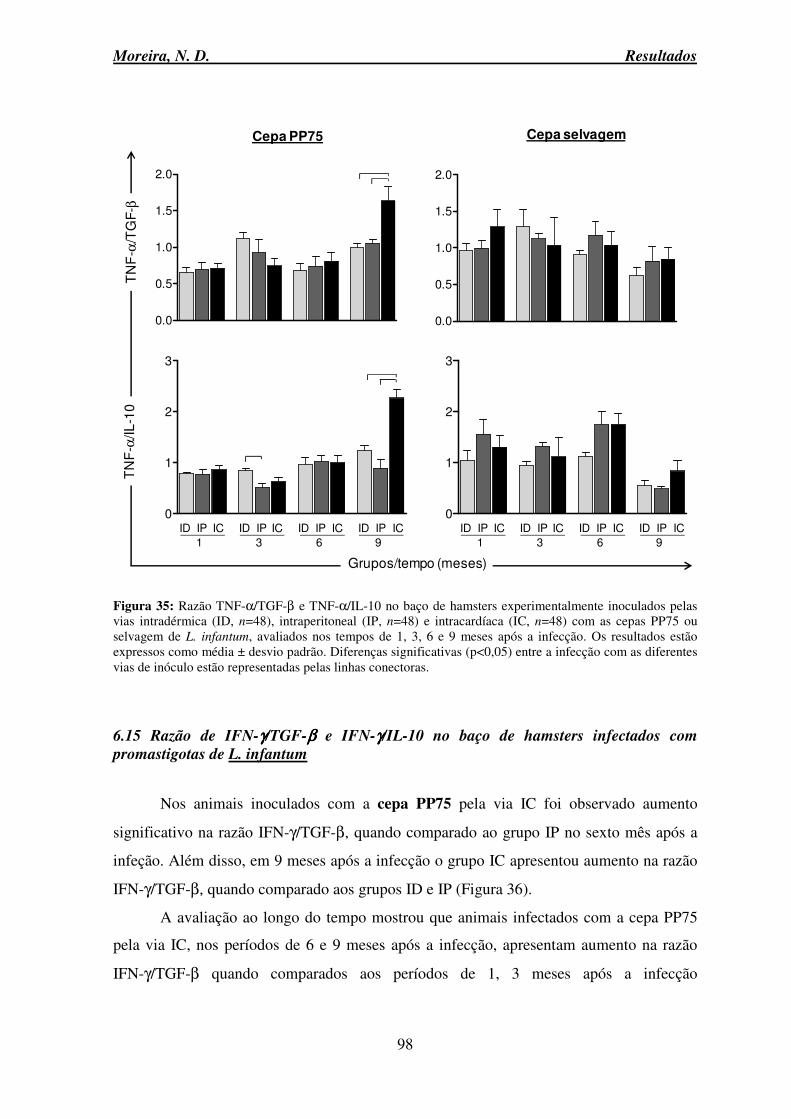
Moreira, N. D. Resultados
98
Cepa PP75 Cepa selvagem
Grupos/tempo (meses)
ID IP IC1
ID IP IC3
ID IP IC6
ID IP IC9
TN
F-α
/IL-1
0
ID IP IC1
ID IP IC3
ID IP IC6
ID IP IC9
TN
F-α
/TG
F-β
0
1
2
3
0
1
2
3
0.0
0.5
1.0
1.5
2.0
0.0
0.5
1.0
1.5
2.0
Figura 35: Razão TNF-α/TGF-β e TNF-α/IL-10 no baço de hamsters experimentalmente inoculados pelas vias intradérmica (ID, n=48), intraperitoneal (IP, n=48) e intracardíaca (IC, n=48) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.
6.15 Razão de IFN-γγγγ/TGF-ββββ e IFN-γγγγ/IL-10 no baço de hamsters infectados com
promastigotas de L. infantum
Nos animais inoculados com a cepa PP75 pela via IC foi observado aumento
significativo na razão IFN-γ/TGF-β, quando comparado ao grupo IP no sexto mês após a
infeção. Além disso, em 9 meses após a infecção o grupo IC apresentou aumento na razão
IFN-γ/TGF-β, quando comparado aos grupos ID e IP (Figura 36).
A avaliação ao longo do tempo mostrou que animais infectados com a cepa PP75
pela via IC, nos períodos de 6 e 9 meses após a infecção, apresentam aumento na razão
IFN-γ/TGF-β quando comparados aos períodos de 1, 3 meses após a infecção

Moreira, N. D. Resultados
99
respectivamente. Ainda mostrou aumento no período de 6 meses quando comparado ao
período de 9 meses após a infecção.
Por outro lado, em animais infectados com a cepa selvagem foi observado aumento
da razão IFN-γ/TGF-β no grupo inoculado pela via IC no terceiro mês após a infecção,
quando comparado as vias ID e IP, além disso, no sexto e nono mês após a infecção este
aumento foi observado no grupo IC apenas em relação ao grupo ID (Figura 36).
A avaliação ao longo do tempo mostrou que animais infectados com a cepa
selvagem pelas vias IP e IC, no período de 6 meses após a infecção, apresentam aumento
na razão IFN-γ/TGF-β quando comparado aos períodos de 1, 3 e 9 meses após a infecção.
Em relação à razão IFN-γ/IL-10 foi observado aumento significativo nos animais
infectados com a cepa PP75 pela via IC quando comparado aos grupos ID e IP, em 9
meses após a infecção (Figura 36).
A avaliação ao longo do tempo mostrou que animais infectados com a cepa PP75
pela via IC, no período de 9 meses após a infecção, apresentam aumento na razão IFN-
γ/IL-10 quando comparado aos períodos de 1, 3 e 6 meses após a infecção.
Os animais infectados com a cepa selvagem demonstraram aumento da IFN-γγγγ/IL-
10 no terceiro mês da infecção no grupo inoculado pela via IC em relação aos demais
grupos (ID e IP). Ainda, no sexto e nono mês este aumento foi observado no grupo IC,
porém em relação apenas ao grupo ID (Figura 36).
A avaliação ao longo do tempo mostrou que animais infectados com a cepa
selvagem pelas vias IP e IC, no período de 6 meses após a infecção, apresentam aumento
na razão IFN-γ/IL-10 quando comparado aos períodos de 1, 3 e 9 meses após a infecção.

Moreira, N. D. Resultados
100
Cepa PP75 Cepa selvagem
Grupos/tempo (meses)
ID IP IC1
ID IP IC3
ID IP IC6
ID IP IC9
IFN
-γ/IL
-10
ID IP IC1
ID IP IC3
ID IP IC6
ID IP IC9
0
10
20
30
40
50
0
10
20
30
40
50
0
5
10
15
20
25
0
5
10
15
20
25
IFN
-γ/T
GF
-β Figura 36: Razão IFN-γ/TGF-β e IFN-γ/IL-10 no baço de hamsters experimentalmente inoculados pelas vias intradérmica (ID, n=48), intraperitoneal (IP, n=48) e intracardíaca (IC, n=48) com as cepas PP75 ou selvagem de L. infantum, avaliados nos tempos de 1, 3, 6 e 9 meses após a infecção. Os resultados estão expressos como média ± desvio padrão. Diferenças significativas (p<0,05) entre a infecção com as diferentes vias de inóculo estão representadas pelas linhas conectoras.

7. DISCUSSÃO

Moreira, N. D. Discussão
102
A elevada prevalência, as dificuldades na intervenção em nível de controle e
profilaxia no ciclo de transmissão e a repercussão sócio econômica tornam as
leishmanioses, em especial a LV, um grande problema de saúde pública no Brasil e no
mundo. Um dos atuais e grandes desafios dos pesquisadores para vencer a crescente
urbanização da LV, em diversas áreas do mundo, é a busca de estratégias imunoprofiláticas
e terapêuticas capazes de promover a melhora clínica, a cura parasitológica e reverter
quadros graves da doença. No entanto, uma das grandes limitações para atingir êxito
dessas metas são os ensaios pré-clínicos destinados a testes de esquemas terapêuticos ou
vacinais para LV. Estes testes consistem basicamente na avaliação em modelos
experimentais que reproduzam os principais eventos clínicopatológicos decorrentes do
curso natural da infecção. A necessidade de caracterizar modelos, que reproduzam a LV
humana e canina, na sua forma ativa, é um dos fatores limitantes e extremamente
importante na sua escolha, seja para testes de drogas e ensaios vacinais.
O modelo experimental hamster desempenha importante papel no estudo das
leishmanioses uma vez que diversos autores tem demonstrado em trabalhos anteriores uma
elevada susceptibilidade deste modelo à infecção por espécies de Leishmania, responsáveis
pelas formas clínicas, seja por L. donovani (Duarte et al., 1988; Gifawesen & Farrell,
1989; Ghose et al., 1999), responsável pela LV no Velho Mundo ou por L. infantum
(Bories et al., 1998; Requena et al., 2000) agente etiológico da LV no Novo Mundo. Neste
contexto, o hamster se torna extremamente importante uma vez que, observam-se
caracteristicamente, uma associação entre parasitismo, títulos elevados de anticorpos e
ausência de respostas linfoproliferativas (Evans et al., 1990; Requena et., 2000; Melby et
al., 2001; Riça-Capela et al., 2003). Tais biomarcadores de diagnóstico, prognóstico e
monitoração apresentam-se com perfis muito semelhantes aos observados na LV humana e
canina. Portanto, um melhor entendimento do desempenho do hamster como modelo
experimental para LV, torna-se fundamental, uma vez que reproduz diversos aspectos
clínico-patológicos da doença humana e canina.
Assim, no presente estudo avaliou-se o estabelecimento da infecção experimental
no modelo hamster, utilizando três diferentes vias de inóculo (intradérmica, intraperitoneal
e intracardíaca) e duas cepas de L. infantum (PP75 e selvagem) com perfis distintos de
virulência e patogenicidade buscando assim, ampliar as abordagens investigativas na busca
de marcadores de prognóstico e monitoração, através da avaliação das alterações
histopatológicas no baço e fígado, bem como análises imunológicas e parasitológicas.

Moreira, N. D. Discussão
103
Em uma primeira abordagem foi avaliado o perfil de crescimento in vitro das
formas promastigotas pertencentes às cepas PP75 e selvagem de L.infantum. Os dados
obtidos demonstraram que as formas promastigotas de ambas as cepas avaliadas
apresentaram perfis de crescimento semelhante e com um perfil clássico de crescimento e
variando de uma fase lag para logarítmica, passando para estácionária e por fim declínio.
Vale ressaltar que as condições de cultivo de ambas as cepas foram às mesmas durante
todo o experimento. Embora tenha sido demonstrado no trabalho publicado por Schuster et
al. (2002) que cepas de mesma espécie de Leishmania sp., mantidos sob as mesmas
condições de cultivo podem apresentar diferenças em seus perfis de crescimento, em nosso
trabalho não foi observado nenhuma diferença neste perfil. Esta análise inicial do
crescimento das formas promastigotas de ambas as cepas foi extremamente importante
para certificarmos de que as cepas utilizadas para o inóculo no modelo hamster fossem
capazes de induzir uma infecção eficaz, importante para a realização das análises
subsequentes.
Posteriormente ao estabelecimento da infecção foi avaliada a presença de sinais
clínicos em hamsters infectados com as cepas PP75 e selvagem de L. infantum, por meio
de uma cinética de infecção experimental, nos tempos de 1, 3, 6 e 9 meses após a infecção.
Os hamsters apresentaram sinais clínicos clássicos similares aos observados na LV
sintomática humana, tais como: emagrecimento acentuado e esplenomegalia. Estes
achados foram mais evidentes no nono mês após a infecção com a cepa PP75, no grupo IC,
porém com baixa frequência. Logo, a cepa selvagem foi mais infectiva, os sinais clínicos
foram mais frequentes e com maior exacerbação no grupo IC, com 80% (6 meses) e 100%
(9 meses) dos animais apresentando esplenomegalia, sendo este o achado mais prevalente
do grupo, seguido de emagrecimento com consequente caquexia no ultimo mês da
infecção. No entanto, animais infectados pela via IP a presença de sinais foi mais evidente
a partir do sexto mês e com uma frequência inferior ao grupo IC. O achado mais prevalente
no grupo IP foi à esplenomegalia, acometendo 70% do grupo no sexto mês e 87% no nono
mês da infecção. A frequência de sinais clínicos no grupo ID foi relativamente baixa
durante todo período avaliado quando ambas as cepas foram utilizadas.
Sinais clínicos semelhantes aos observados neste estudo, têm sido também
demonstrados por diversos outros pesquisadores (Melby et al., 2001; Requena et al., 2000;
Poot et al., 2006; Riça-Capela et al., 2003; Dea-Ayuela et al., 2007, Nieto et al., 2011),
utilizando este modelo. Embora a frequência destes tenha variado nos trabalhos citados

Moreira, N. D. Discussão
104
acima, porém a ocorrência dos mesmos está de acordo com nossos resultados, uma vez que
os sinais mais prevalentes encontrados no presente estudo foram: ascite, esplenomegalia e
emagrecimento. Nos animais inoculados pela via ID a presença de sinais clínicos foi
menos frequente e apareceram geralmente no período mais tardio da infecção,
principalmente quando a cepa PP75 foi utilizada. Estudos conduzidos por (Wilson et al.
(1987) ao utilizarem a mesma via de inóculo na infecção experimental em hamster
demonstram presença de sinais clínicos da LV somente a partir de 10 meses após infecção
experimental.
Em ambas as cepas empregadas no presente estudo, a hepatomegalia foi um achado
pouco frequente. Assim, foi relatado aumento no peso relativo do fígado dos animais
infectados com a cepa selvagem pela via IC, no sexto mês após a infecção. Todavia ao
contrário do esperado, ou seja, um quadro clássico de hepatomegalia propriamente dito, os
resultados de nosso estudo apontam sim para uma hepatomegalia em reflexo ao
emagrecimento dos animais, pois não houve diferença em relação ao peso do órgão entre
os hamsters infectados ao grupo controle não infectado neste período. No entanto, a
hepatomegalia não é necessariamente sempre observada durante a doença (Melo et al.,
2008). Estas diferenças, bem como acontece com outras alterações, provavelmente sejam
inerentes da complexidade das alterações geradas pelo parasito e resposta imune do
hospedeiro. Macroscopicamente, o fígado dos animais inoculados pela via IC apresentou
aspecto friável com vários pontos brancos distribuídos em toda a superfície do órgão e ao
longo do parênquima hepático, independente da cepa utilizada no nono mês da infecção.
Estas mesmas alterações macroscópicas foram também observadas por Vianna et al.
(2002), na infecção experimental em hamsters por L.donovani utilizando a via
intracardíaca de inóculo.
Aos 9 meses após a infecção com a cepa selvagem utilizando a via IC, foi possível
observar, lesões cutaneomucosas, acompanhada por úlceras localizadas no focinho bem
como, edema nas patas em 100% dos animais. Estes achados também foram observados
nos estudos conduzidos por Nieto et al. (2011). Estes autores descreveram as mesmas
lesões observadas em nosso estudo, aos 7 meses após a infecção experimental intracardíaca
com 107 promastigotas de L. infantum. Estes dados reforçam a capacidade da via de
inóculo intracardíaca em causar lesões graves em hamsters incluindo tegumentização das
lesões e edemas de membros inferiores. A tegumentização de lesões e edemas de membros
inferiores é um achado frequente na leishmaniose visceral canina (Reis et al., 2009) e

Moreira, N. D. Discussão
105
também pode aparecer em casos graves da LV humana principalmente em pacientes co-
infectados pelo HIV (Catorze et al., 2006).
Diante do exposto, podemos concluir que sinais clínicos clássicos já observados e
relatados por diversos autores na LV humana (Badaró et al., 1986; Pastorino et al., 2002,
Pedrosa, 2004; Oliveira et al., 2010) e canina (Santos-Gomes et al., 2002; Paranhos-Silva,
et al., 2003; Reis et al., 2006; da Costa-Val et al., 2007; Ikeda-Garcia et al.,2007; Baneth
et al., 2008; Alves et al., 2009; Solano-Gallego et al., 2009; Solano-Gallego et al., 2011)
também ocorrem na LV experimental em hamster. Desta forma, podemos inferir que a
cepa utilizada, a quantidade de parasitos e a via de inóculo podem favorecer na
mimetização da infecção experimental e permitir uma evolução para formas crônicas
graves da doença permitindo assim empregar o modelo para testes de drogas
leishmanicidas, de modo que se possa avaliar a remissão destes sinais clínicos após a
terapêutica específica contra o agente etiológico da LV. Além disso, a avaliação das duas
cepas foi extremamente importante para o entendimento dos distintos padrões de virulência
e patogenicidade apresentados por cada uma.
Um marcador fundamental na LV são os anticorpos anti-Leishmania, de um modo
geral, estes quando produzidos em grande quantidade, não são associados à proteção,
apresentando neste sentido, grande importância como biomarcador de progressão da
doença. Altos títulos de anticorpos são observados em pacientes com LV (Nylén &
Gautam, 2010). Na LVC, os cães sintomáticos geralmente apresentam altos títulos de
anticorpos anti-Leishmania e os cães resistentes à infecção exibem baixos níveis ou
ausência destes como recentemente demonstrados em cães assintomáticos soronegativos e
PCR+ (Carrera et al., 1996; Reis et al., 2006; Solano-Gallego et al., 2009; Reis et al., 2009;
Coura-Vital et al., 2011). Muitos aspectos da patogênese da LVC são atribuídos aos
anticorpos produzidos durante a história natural da doença, os quais formam
imunocomplexos que se depositam em diversos tecidos, principalmente nos glomérulos
renais, gerando lesões inflamatórias graves (Miles et al., 2005). A elevada produção de
anticorpos na LV está intimamente relacionada à expansão policlonal de linfócitos B,
conduzindo um quadro típico de hipergamaglobulinemia e gerando quadros graves de
disfunção renal devido à grande deposição de imunocomplexos, ocasionando óbito de
indivíduos e cães com LV grave (Galvão-Castro et al., 1984; Dutra et al., 1985; Prasad et
al 1992; Alvar et al., 2004; Plevraki et al., 2006; Lima-Verde et al., 2007; Costa-Val et al.,
2007; Reis et al., 2009; Agenor et al., 2009; Clementi et al., 2011).

Moreira, N. D. Discussão
106
No presente estudo, foi demonstrado que, os elevados níveis de IgG detectados nos
animais inoculados com a cepa PP75 foram observados mais tardiamente, com maiores
picos em 6 e 9 meses após a infecção experimental no grupo IC. Entretanto, em animais
infectados com a cepa selvagem os níveis de IgG foram elevados em todos os grupos
infectados comparado ao grupo controle, apresentando maiores picos nos grupos IP e IC a
partir do terceiro mês. Estes dados demonstraram que a cepa selvagem foi capaz de induzir
níveis mais elevados de IgG e em tempo mais precoce que a cepa PP75, com maior
exacerbação nos grupos IP e IC, o que foi associado a maior densidade parasitária
esplênica observada nestes grupos, principalmente em estágios mais avançados da doença,
além da presença de sinais clínicos evidentes. Estes dados reforçam a idéia de que, no caso
de cepas menos virulentas, como a cepa PP75, a escolha da via de inóculo é extremamente
importante para o estabelecimento da infecção, de forma a reproduzir quadros de LV ativa
neste modelo. Assim, tanto a cepa PP75 quanto a selvagem foram capazes de induzir
aumento nos níveis de IgG, principalmente no grupo IC, demonstrando que
semelhantemente a LV humana e canina, a avaliação de anticorpos anti-Leishmania é uma
ferramenta importante no monitoramento do estabelecimento da infecção no modelo
hamster.
Nossos resultados, estão de acordo com Requena et al (2000) que associaram a
forte resposta humoral em hamsters experimentalmente infectados por L. infantum a
presença de sinais clínicos evidentes e a elevada densidade parasitária tecidual. Além
destes autores, outros estudos tem demonstrado uma relação entre a carga parasitária e
altos títulos de anticorpos em hamster experimentalmente infectados com diferentes formas
e espécies de Leishmania (Evans et al., 1990; Melby et al., 2001; Riça-Capela et al., 2003,
Dea-Ayuela et al., 2007). Achados semelhantes também foram observados em cães
experimentalmente e naturalmente infectados e na LV humana (Genaro, 1993; Reis et al.,
2006; Silveira et al., 2009).
Na literatura, as informações que dizem respeito à mensuração de parâmetros
bioquímicos e hematológicos em hamster são extremamente limitadas. A avaliação
hematológica e bioquímica representa uma importante área de estudo da leishmaniose
visceral seja humana ou canina como fonte de monitoração do diagnóstico e prognóstico
da evolução do quadro clínico bem como na identificação de formas graves da doença.
Embora, a avaliação do quadro hematológico forneça pouco valor para o diagnóstico na
LV, estes parâmetros podem apresentar relevância no acompanhamento do estado clínico,

Moreira, N. D. Discussão
107
com um forte valor prognóstico, sendo importantes parâmetros de definição de condutas
terapêuticas em protocolos de consenso médico (Nasir et al., 1995; Feitosa et al., 2003;
Reis et al., 2006a; Costa-Val et al., 2007; Reis et al., 2009; Solano-Gallego et al. 2009;
Brasil, 2009; Oliveira et al., 2010).
Vale ressaltar que na LV humana e canina, observa-se a formação de um quadro
hematológico grave, marcado por uma (pancitopenia) que na série vermelha é
caracterizada por anemia normocítica, nomocrômica, e ainda com valores diminuídos de
hemoglobina e hematócrito. Com relação à série branca o quadro se agrava ainda mais,
pois geralmente na LV ativa humana e canina a doença evolui para um nítido quadro de
imunossupressão marcada por uma forte queda nos valores globais de leucócitos
(leucopenia) caracterizados por diminuição de monócitos (monocitopenia), linfócitos
(linfopenia), eosinófilos (eosinopenia) e por vezes ocorre aumento de neutrófilo
(neutrofilia) ou mesmo um esquerda à direita, ou seja, queda de neutrófilo (neutropenia),
como relatado por diversos autores (Pearson, 1996; Bourdoiseau et al.,1997; Ciaramella et
al., 1997; Pastorino et al., 2002; Feitosa et al., 2003; Pedrosa, 2004; Reis et al., 2006a;
Costa-Val et al., 2007; Maia et al., 2010; Oliveira et al., 2010).
Diferentes padrões nos parâmetros hematológicos e bioquímicos podem ser
observados como resultado de lesão celular ou disfunção orgânica. Estes padrões refletem
tanto o extravasamento de constituintes celulares para o soro quanto à produção e excreção
de vários componentes séricos. Grandes variações nos valores hematológicos e
bioquímicos podem alterar em função da idade, sexo, criação, dieta, temperatura, além da
excitação proveniente da contenção do animal no momento da coleta de sangue (Thall,
2007). Além disso, os procedimentos laboratoriais não estão padronizados o que pode
acarretar resultados divergentes. Devido à falta de padronização nos laboratórios e as
diferentes intercorrências para emissão de resultados hematológicos e bioquímicos com
elevada acurácia e qualidade, é recomendável que cada laboratório desenvolva seus
próprios intervalos de referência a partir de amostras de animais normais, levando em
consideração todos estes fatores.
Neste estudo, as alterações nos parâmetros hematológicos dos hamsters infectados
com a cepa PP75 foram pouco expressivas, com redução apenas na população de
monócitos (monocitopenia) no último mês da infecção no grupo IC. Entretanto, em
hamsters infectados com a cepa selvagem as alterações foram evidentes principalmente nos
hamsters infectados pela via IC e IP. O grupo IC, com aparecimento de leucopenia já partir

Moreira, N. D. Discussão
108
do terceiro mês de infecção acompanhada de redução de monócitos. No sexto mês de
infecção experimental, foi evidenciada uma típica imunossupressão da LV nestes animais,
com uma leucopenia intensa e marcada pela redução de neutrófilos, eosinófilos, linfócitos
e monócitos. Aos 9 meses após a infecção experimental os hamsters inoculados tanto pela
via IP quanto pela via IC a leucopenia foi caracterizada pela diminuição de neutrófilos,
eosinófilos e monócitos, em relação ao grupo C e ID. Neste sentido, as alterações
hematológicas clássicas observadas na LV humana e canina foram aqui reproduzidas nos
animais infectados pela cepa selvagem. Estes dados sugerem que as alterações observadas
nos grupos IP e IC acompanhadas pela presença de sinais clínicos sugestivos da LV ativa
(em especial a esplenomegalia) e ao elevado parasitismo tecidual (baço e fígado)
observado nestes grupos deve-se a um complexo de interações relacionadas ao potencial de
virulência e patogenicidade de cada uma das duas cepas utilizadas.
Na LV humana, caracteristicamente a leucopenia, neutropenia e eosiponenia são
encontradas com grande frequência em estudos clínicos e laboratoriais envolvendo
pacientes com LV (Campos, 1995; Pearson, 1996; Berman, 1997). Estudos conduzidos por
Queiroz et al. (2004) relataram que 85% das crianças evoluíram com leucopenia e 74%
com neutropenia, provavelmente devido a esplenomegalia, além de hipoplasia ou
depressão medular e hemofagocitose. Da mesma forma a leucopenia esteve presente em
85,5% dos pacientes com LV, no momento da internação, e assim como no trabalho acima
citado, é possível que a esplenomegalia registrada em 71% dos pacientes tenha contribuído
para esta alteração (Maltezou et al., 2000; Cascio et al., 2002; Collin et al., 2006).
Segundo Reis et al. (2006a) o comprometimento do quadro hematológico na LVC,
está associado com manifestações clínicas graves, evidenciadas por leucopenia marcada
por linfopenia, eosinopenia, monocitopenia e intensa anemia. Alguns autores tem
demonstrado ausência de variações no leucograma na maioria dos cães com LVC,
enquanto outros demonstram a ocorrência de leucopenia moderada ou grave (Costa-Val et
al., 2007; Reis et al., 2006a). Sendo assim, Alvar et al. (2004) demonstraram que durante a
fase sintomática da LVC, há uma diminuição na contagem de leucócitos devido
principalmente à queda nos valores relativos e absolutos de monócitos, eosinófilos e
principalmente de linfócitos, sendo este último um dos achados mais relevantes.
Recentemente Coura-Vital et al., (2011) demonstraram que cães assintomáticos
soronegativos e PCR+ apresentam valores normais na contagem diferencial de monócitos e
que esta população declina no sangue circulante logo após a soroconversão caindo

Moreira, N. D. Discussão
109
dramaticamente no cães sintomáticos. Por outro lado, Bourdoiseau et al. (1997) revelaram
que na fase inicial da doença ocorre leucocitose associada à neutrofilia, enquanto que nos
estágios mais avançados, há leucopenia associada à linfopenia. De fato, é possível obervar
leucocitose na LVC assintomática como a observda por Reis et al. (2006a) que também
evidenciaram linfocitose nesta mesma forma clínica.
No eritrograma, os animais infectados com a cepa PP75 pelas 3 vias de inóculo
apresentaram diminuição de hematócrito em 1 mês após a infecção, provavelmente em
função de alguma alteração que tenha conduzido à disfunção medular. Além disso, foi
evidenciado um quadro de anemia nos grupos ID, IP e IC no sexto mês da infecção. Por
outro lado, nos animais infectados com a cepa selvagem pela via IC foi observado um
quadro de anemia grave, marcada pela redução de eritrócitos, hematócrito e hemoglobina a
partir do sexto mês da infecção. Estes dados mostram que as principais alterações no
eritrograma dos hamsters infectados com a cepa PP75 ocorreram no primeiro e no sexto
mês da infecção nos três grupos experimentais (ID, IP e IC), enquanto nos animais
infectados com a cepa selvagem as alterações foram observadas no sexto e nono mês da
infecção, restritas apenas ao grupo IC. A cepa PP75 por induzir uma infecção mais branda,
quando o parâmetro de avaliação é o eritrograma, não existe diferença nas alterações
apresentada em relação às três vias de inóculo utilizada. Entretanto, quando se usa uma
cepa virulenta e patogênica a resposta a estas alterações tem influência sob a via de inóculo
que é utilizada.
A anemia foi também demonstrada por Riça-Capela et al. (2003) em 70-80% dos
hamsters experimentalmente infectados com diferentes formas parasitárias de L. infamtum
(amastigotas e promastigotas). A anemia ocorre devido à perda de sangue, lise de hemácias
e diminuição da eritropoiese, devido à hipoplasia e aplasia medular (Feitosa et al., 2003).
Na LVH a anemia normocítica e normocrômica é um achado frequente nos pacientes que
apresentam forma crônica da doença (Pearson, 1996). Já na LVC, Costa-Val et al.(2007),
relataram em seu estudo que 42% dos cães avaliados apresentaram anemia normocítica
normocrômica e não regenerativa, estes dados coincidem com os descritos em outros
trabalhos que apontam a anemia normocítica e normocrômica como um achado comum na
LVC ativa (Ciaramella et al.,1997; Feitosa et al., 2003; Reis et al., 2006a).
Nos hamsters infectados com a cepa PP75, no terceiro mês da infecção foi
observada trombocitose no grupo IP. De forma similar, nos animais infectados com a cepa
selvagem o grupo IC, em 6 e 9 meses após a infecção apresentou trombocitose. A

Moreira, N. D. Discussão
110
trombocitose nos grupos IP e IC podem ser conseqüência do quadro de anemia grave
apresentado ou pode ser também que não tenha nenhuma relação com este quadro clínico.
Os parâmetros bioquímicos relacionados à avaliação da função renal consistem
principalmente na dosagem dos níveis séricos de uréia e creatinina. As análises
bioquímicas do presente estudo revelaram alteração renal uma vez que houve aumento nos
níveis de uréia e creatina em alguns grupos avaliados. Um dos achados observados está
relacionado aos níveis de uréia nos animais infectados com a cepa PP75 que demonstraram
aumentados no sexto mês após a infecção apenas no grupo IC e nono mês nos três grupos
(ID, IP e IC), além de apresentar aumento nos grupos IP e IC em relação ao ID. Além
disso, no nono mês ainda foi demonstrado aumento nos níveis de creatinina nos três grupos
(ID, IP e IC). Por outro lado, nos infectados com a cepa selvagem o aumento tanto de uréia
quanto de creatinina foram observados nos períodos de 3 e em 9 meses após a infecção
apenas no grupo IC. Estes dados sugerem injúrias nos rins destes animais, devido talvez à
deposição de imunocomplexos. A insuficiência renal aguda ou crônica pode estar
relacionada com a deposição dos imunocomplexos em tecidos renais, constituindo uma
importante manifestação patológica na LV (Zatelli et al. 2003).
Em estudo com 50 pacientes humanos portadores da LV sintomática foi
demonstrado que 8,5% destes apresentaram dosagem de uréia elevada e 16,3% com
dosagem de creatinina aumentada (Dutra et al.,1985). Segundo, Nieto et al. (1992), em
estudo com 10 cães infectados, sete apresentaram dosagem sérica de uréia aumentada e três
com dosagem de creatinina elevada. Devido a elevada prevalência de patologia renal, a
azotemia na LVC, típica de insuficiência renal é um achado laboratorial comum, e é
evidente somente quando a maioria dos néfrons torna-se disfuncionais com a progressão da
doença. As glomerulonefrites são alterações mais comuns, enquanto a amiloidose é menos
frequente (Zatelli et al., 2003). Estudos conduzidos por Riça-Capela et al. (2003)
mostraram grande quantidade de amilóide no fígado e baço de hamster experimentalmente
infectados com formas amastigotas e promastigotas de L. infantum.
As alterações na concentração das enzimas aminotransferases séricas (ALT e AST)
são considerados importantes indicadores de lesões nas células hepáticas. A dosagem da
concentração das enzimas ALT, AST permitem averiguar a presença de alterações da
permeabilidade dos hepatócitos quando se observa elevação de níveis séricos de ALT e
AST (Kaneko, 1997).

Moreira, N. D. Discussão
111
No presente estudo, observou-se que a infecção com a cepa PP75 induziu aumento
nos níveis de AST no grupo IC, no início da infecção (1 mês) e no final (9 meses).
Entretanto nos animais infectados com a cepa selvagem este aumento foi demonstrado no
terceiro e nono mês no grupo IC e a partir do sexto mês no grupo IP. Quanto aos níveis de
ALT, os animais infectados com a cepa PP75 induziu aumento no inicio da infecção (1
mês) nos grupos IC e IP e aos 9 meses apenas no grupo IC. Entretanto, a cepa selvagem
induziu aumento de ALT aos 3 meses após a infecção no grupo IC e em 6 meses em todos
os grupos (ID, IP e IC). Estes dados fornecem subsídios de alterações na função hepática e
que a elevação da atividade de AST e ALT nos grupos IP e IC pode ser indicativa de
alguma lesão, uma vez que a AST e ALT são enzimas que estão presentes em altas
concentrações no compartimento hepático (hepatócitos). O envolvimento hepático tem sido
relatado com maior frequência quando o diagnóstico da LV humana é mais tardio e indica
uma maior gravidade (Queiroz et al., 2004).
A ALT é encontrada principalmente no citoplasma do hepatócito e é considerada
hepatoespecífica, sendo o aumento sérico um indicador sensível e específico de dano
hepatocelular, enquanto que 80% da AST encontram-se presente na mitocôndria dos
hepatócitos. Consequentemente, a ALT sérica eleva-se mais rapidamente e usualmente está
em níveis mais altos que a AST. Em danos hepatocelulares leves a forma predominante no
soro é a citoplasmática, enquanto que em lesões graves há liberação da enzima
mitocondrial. O aumento destas enzimas hepáticas implica na tentativa de manutenção da
viabilidade e regeneração dos hepatócitos, podendo ser usada na interpretação do avanço
da doença (Kaneko, 1997).
Apesar de escassos os estudos que buscaram avaliar alterações nos níveis de AST e
ALT no modelo hamster para LV, diversos autores relatam aumento nos níveis séricos
destas enzimas na LV humana (Queiroz et al., 2004; Oliveira et al., 2010) bem como na
LV canina (Nieto et al., 1992; Solano-Gallego et al., 2009; Solano-Gallego et al., 2011).
Assim, acreditamos que o monitoramento laboratorial do perfil bioquímico enzimático de
AST e ALT deva ser considerado quando se emprega este modelo principalmente para
estudos de testes de drogas para LV. Embora tenham sido demonstradas algumas
diferenças em relação aos níveis de proteínas, albumina, globulina bem como na razão
albumina/globulina não foi possível estabelecer um padrão destas alterações em relação à
sintomatologia observada e as vias de inóculo utilizada neste estudo, onde foi possível
detectar alterações em graus variáveis.

Moreira, N. D. Discussão
112
A LV experimental é marcada pelo desenvolvimento de uma resposta imune órgão-
específica, sendo o fígado e o baço os principais órgãos alvos atingidos (Rousseau et al.,
2001). O fígado é resistente a reinfecção, poucos danos teciduais, é o local onde a
resolução da infecção aguda esta associada ao desenvolvimento de granulomas hepáticos
intralobulares tipicamente inflamatórios em torno das células de Kupffer (Stanley &
Engweda, 2007). A resposta imune eficiente em infecções por Leishmania no fígado é
intimamente dependente da formação de granulomas hepáticos. Em hamsters com LV, as
alterações hepáticas em geral são decorrentes não somente pela presença de um infiltrado
inflamatório como também pela formação destes granulomas (Wilson et al., 1987; Vianna
et al., 2002). A formação do granuloma foi à principal alteração histopatológica observada
no fígado de hamsters experimentalmente infectados com promastigotas de L. infantum por
via intradérmica (Mangoud et al., 1997).
No presente estudo, o infiltrado inflamatório no espaço porta e parênquima hepático
e formação de granulomas foram às principais alterações observadas. Nos animais
infectados com a cepa PP75 as alterações histopatológicas hepáticas ocorreram a partir do
sexto mês de infecção com maior frequência no grupo IC. Nestes animais foi observado o
desenvolvimento de um processo inflamatório caracterizado pela presença de infiltrado de
células mononucleares nos espaço-porta que variou de discreto a moderado e alguns
granulomas distribuídos aleatoriamente pelo parênquima. Estudos realizados por Riça-
Capela et al. (2003), demonstraram discreto infiltrado inflamatório no espaço porta,
desorganização de trabéculas hepáticas com formação de pequenos granulomas hepáticos
em hamsters infectados com promastigotas de L.infantum.
No entanto, nos animais infectados com a cepa selvagem a presença de granulomas
ocorreu nos grupos IP e IC. No grupo IP os granulomas estavam presentes durante todo o
período da avaliação, porém com frequência menor comparado ao grupo IC. No grupo IC
foi observado granulomas durante todo o período de avaliação e em maior número no sexto
e nono mês da infecção. Além disso, foi possível observar presença de amastigotas, áreas
intensas de necrose, destruição na morfologia dos hepatócitos e infiltrado inflamatório que
variou de discreto a intenso de acordo com a evolução da infecção. Juntos estes dados
sugerem que as intensidades das alterações histopatológicas observadas no parênquima
hepático neste estudo estão diretamente relacionadas ao parasitismo apresentado por cada
um dos grupos avaliados (ID, IC e IP). O aumento do processo inflamatório e a presença
de maior número de granulomas apresentado no grupo IC podem estar relacionados ao

Moreira, N. D. Discussão
113
intenso parasitismo que foi observado neste grupo. A infecção visceral em hamsters induz
alterações em hepatócitos, principalmente no sistema endomembrana e no compartimento
peroxissomal levando a distúrbios no metabolismo do fígado (Vianna et al., 2002).
Mangoud et al. (1997) observaram em hamsters experimentalmente infectados por
L.infantum que, o infiltrado inflamatório no espaço porta teve início quando surgiram os
primeiros granulomas, os quais apresentaram distribuídos aleatoriamente pelo parênquima
hepático. Com a evolução da infecção os granulomas mostraram aumentados em número e
tamanho e eram formados principalmente de macrófagos com áreas de necrose e poucos
linfócitos de forma semelhante ao observado por nós. A marcada necrose nos granulomas
hepáticos pode ser devido a efeitos citotóxicos de linfócitos T nos estágios avançados da
infecção (Murray et al., 1982). Nieto et al. (2011) relataram que as alterações, como
formação de granulomas em diferentes estágios de maturação e granulomas com
amastigotas no fígado de hamsters infectados por via IC, foram mais frequentes com o
pico da carga parasitária. Na LVH a presença de granulomas no fígado parece estar
associada ao controle espontâneo e manutenção da infecção em um estado subclínico
(Murray et al., 2001). Dentre as principais alterações histopatológicas observadas no
fígado de cães infectados com LV, tem sido destacada a reação inflamatória exsudativa
com infiltrado linfoplasmocitário nos espaços-porta e interlobulares, inflamação da
capsula, além da presença de granulomas intralobulares, com células epitelióides,
parasitados ou não (Genaro, 1993; Tafuri et al., 1996; Tafuri et al., 2001; Giunchetti et al.,
2008c).
O baço desempenha um papel central na LV, pois é um sítio inicial para a geração
de resposta imune específica contra o parasito mediada por células, podendo se tornar
também um local de persistência do mesmo apresentando inúmeras alterações
imunopatológicas (Kaye et al., 2004).
No presente estudo a avaliação histopatológica esplênica revelou de um modo
geral, que em animais inoculados com cepa PP75 pelas vias ID e IP, a arquitetura
histológica do baço foi mantida ao longo dos 9 meses avaliados. Modificações na polpa
vermelha no terceiro mês da infecção nestes mesmos grupos ocorreram em decorrência do
aumento considerável de macrófagos. Os folículos linfóides do baço de hamsters
inoculados pela via IC mostraram acentuada reatividade folicular com aumento do volume
e da densidade linfocitária e moderada proliferação de macrófagos na polpa vermelha aos 9
meses após a infecção. Além disso, macrófagos densamente parasitados por formas

Moreira, N. D. Discussão
114
amastigotas de L. infantum e periesplenite foram observados. Estudos conduzidos por
Binhazim et al., (1993), mostraram macrófagos densamente parasitados formando
agregados na polpa vermelha esplênica evidentes após a infecção experimental em
hamsters submetidos a inoculação por via intracardíaca. Apesar de não se conhecerem os
mecanismos que levam a periesplenite, sugere-se que a esplenomegalia pode predispor ao
estresse e trauma mecânico da capsula, mas a gênese real deste achado na LV permanece
desconhecida. A periesplenite é um achado comum na LVC, associada com intenso
parasitismo na região subcapsular. Estes achados tem sido observados em alguns estudos
(Tafuri et al., 1996; Xavier et al., 2006; Santana et al., 2008) e podem indicar o papel da
inflamação na disseminação de parasitos durante o curso clínico da LVC (dos-Santos et al.,
2008) ou que o parasito provoca uma reação inflamatória. De acordo com Santana et al.
(2008), periesplenite esteve presente em 28% dos casos avaliados, sendo mais frequente no
grupo dos animais sintomático.
Na cepa selvagem a proliferação de macrófagos da polpa vermelha, nos grupos IP e
ID foi mais frequente no sexto mês da infecção. Além disso, periesplenite no grupo IP foi
observada a partir do sexto mês da infecção. O grupo IC apresentou atrofia linfóide no
terceiro e sexto mês, relacionada à desorganização histológica com perda da definição ou
hipoplasia das regiões da polpa branca, frequentemente associada à proliferação de
macrófagos da polpa vermelha. Estes achados foram mais evidentes no sexto mês, onde
todos os animais avaliados apresentaram perda da arquitetura com substituição parcial à
total da polpa branca por células da polpa vermelha. Além disso, a maioria dos animais do
grupo IC apresentou intenso parasitismo tecidual e periesplenite.
A atrofia linfóide e a desorganização da arquitetura esplênica é muito frequente
durante a infecção ativa por Leishmania (Veress et al., 1983; Engwerda et al., 2002). As
alterações esplênicas nos animais infectados com a cepa PP75 foram mais evidentes no
grupo IC e embora tenham ocorrido durante todo período de avaliação estas se mostraram
com maior intensidade nos períodos finais da infecção (6 e 9 meses). Entretanto nos
animais infectados com a cepa selvagem, as alterações no grupo IC ocorreram em todo
período avaliado, porém com uma exacerbação ainda maior no sexto e nono mês. Estes
dados sugerem que a hipoplasia da polpa branca, hiperplasia da polpa vermelha, perda da
arquitetura e periesplenite demonstradas nos períodos de 6 e 9 meses no grupo IC estejam
associadas ao intenso parasitismo e sinais mais severos apresentados pelo grupo tais como:
emagrecimento e esplenomegalia.

Moreira, N. D. Discussão
115
Existem associações entre alterações morfológicas no baço e emagrecimento,
inicialmente inerente à desorganização da estrutura da polpa branca. Estudos conduzidos
por Santana et al. (2008) demonstraram que 70% dos cães com caquexia apresentaram
desorganização da polpa branca. A perda de definição dos folículos linfóides, periesplenite
e alta densidade parasitária foram mais frequentes nos animais que estavam abaixo do
peso. O baço desempenha um importante papel na LVC, sendo acometido em todos os
casos da doença. Em contraste a outros órgãos como o fígado, o baço mantém a infecção
durante todo o curso da doença canina. Entretanto a resposta imunológica no baço dos
animais infectados não está muito bem definida (Sanchez et al., 2004).
Estudos recentes conduzidos por Silva et al. (2012), ao avaliar dois grupos de cães
naturalmente infectados por Leishmania infantum com distinta organização histológica
esplênica, mostraram que cães que apresentavam uma desorganização estrutural do baço
eram os que apresentavam doença mais grave que aqueles que mativeram certa
organização. Neste estudo observou-se que a polpa branca foi reduzida em tamanho, os
folículos e a zona marginal foram os compartimentos da polpa branca mais severamente
afetados. Estas alterações foram associados com diminuição do número de células T e
células dendríticas nos folículos, e as células B nos folículos e zonas marginais.
NA LVH o baço é marcado por esplenomegalia resultante da reatividade do sistema
fagocítico monuclear e congestão sinusoidal esplênica. A hipertrofia e hiperplasia da polpa
branca é uma alteração esplênica frequente em indivíduos portadores da LV. É observado
diminuição significativa nos linfócitos dos folículos linfóides, infiltração de células
plasmáticas e macrófagos densamente parasitados por amastigotas (Drumond & Costa,
2011). De fato um índice de parasitismo esplênico é usado como critério clínico da
resposta terapêutica na LVH.
A hipoplasia dos folículos linfóides, associada à proliferação de macrófagos da
polpa vermelha, destruição da arquitetura esplênica, tem sido demonstrada por alguns
autores na infecção experimental em hamsters (Veress et al., 1983; Mangoud et al., 1997;
Riça-Capela et al, 2002, Nieto et al., 2011). A atrofia e desaparecimento de áreas de células
B esplênicas, foram evidentes com a evolução e progressão da doença (Veress et al., 1997).
Com base no exposto acima podemos afirmar que de fato o modelo experimental
hamster é altamente atrativo para os estudos de imunopatologia experimental, testes de
drogas e vacinas anti-LV uma vez que parece reproduzir diversos achados frequentes na

Moreira, N. D. Discussão
116
LV humana e canina mesmo no que diz respeito a imunopatologia e histopatologia no
compartimento hepático e esplênico.
No presente estudo foram utilizadas duas técnicas com abordagens investigativas
extremamente distintas para quantificar o parasitismo nos diferentes órgãos alvo (baço e
fígado): LDU (Leishman Donovan Units) e qPCR. O desenvolvimento de uma técnica que
permita quantificar parasitos e que possibilite a comparação dos valores obtidos no baço e
fígado de hamsters experimentalmente infectados por diferentes vias de inóculo (ID, IP e
IC) é de fundamental importância nas investigações científicas que empregam este modelo
experimental. Certamente, a quantificação da carga parasitária é de fundamental
importância para um melhor entendimento das manifestações clínico-patológicas durante a
história natural na LV experimental. Adicionalmente, o desenvolvimento de uma
abordagem molecular para quantificar a carga parasitária no fígado e baço de hamsters
experimentalmente infectados é extremamente relevante, pois, este modelo tem sido
utilizado em estudos de testes de drogas e formulações vacinais contra a leishmaniose
visceral (Carrión et al., 2011; Reimão et al., 2012; Gupta et al., 2012). Neste contexto,
avaliamos o impacto da carga parasitária no baço e fígado após a infecção com diferentes
cepas de L. infantum exibindo perfis distintos de virulência e patogenicidade.
O índice de LDU conforme descrito por Stauber em 1955, apesar de ser uma
técnica que identifica e quantifica a real presença de amastigotas em tecidos tem a
desvantagem por apresentar baixa sensibilidade (Stauber, 1955). Além disso, um agravante
neste método é que a detecção das formas amastigotas requer profissional extremamente
treinado e paciente, o que pode acarretar resultados divergentes quando não realizados pelo
mesmo microscopista. Contudo, possui ainda a desvantagem de depender da dispersão dos
parasitos nos esfregaços por aposição corados pelo Giemsa já que estes se encontram
distribuídos no tecido de uma forma não uniformizada (Bretagne et al., 2001).
Entretanto a técnica de qPCR em tempo real vem sendo utilizada por vários
pesquisadores e tem se tornado nos últimos anos uma ferramenta de fundamental
importância na aplicação de técnicas de biologia molecular para diagnosticar/monitorar a
evolução da LV através da quantificação da carga parasitária tecidual, em substituição a
outras técnicas, por apresentar alta sensibilidade, acurácia e reprodutibilidade (Manna et
al., 2008; Bretagne et al., 2001).
Conforme esperado, em nosso estudo, o LDU apresentou baixa sensibilidade
detectando parasitos apenas nos grupos IP e IC em alguns tempos avaliados. Sendo assim,

Moreira, N. D. Discussão
117
em animais infectados com a cepa PP75 as amastigotas foram detectadas no sexto e nono
mês de infecção nas amostras de fígado e de baço, apenas no grupo IC. Por outro lado, em
animais infectados com a cepa selvagem, foi possível detectar o parasito a partir do
terceiro mês de infecção no grupo IP e no grupo IC a partir do primeiro mês de infecção.
Assim, foi possível verificar que tanto em animais infectados com a cepa PP75 quanto pela
selvagem, à carga parasitária evoluiu com a progressão da infecção. Além disso, o
parasitismo no baço e fígado com a cepa selvagem foi muito maior comparado aos da cepa
PP75. De acordo com os resultados obtidos pela enumeração de amastigotas por 1000
núcleos de células sem considerar o peso do órgão, o baço foi o órgão mais parasitaddo
que o fígado.
Segundo Ott et al. (1967), o índice de parasitismo no baço é mais consistente em
relação ao fígado uma vez que existe grande diferença de tamanho e peso entre os mesmos.
Estudos conduzidos por Binhazim et al. (1993) utilizando a via IC de infecção,
demonstraram maior parasitismo no fígado de hamster. Por outro lado, maior parasitismo
no baço tem sido demonstrado por diversos pesquisadores em estudos de infecção
experimental em hamsters por diferentes espécies de Leishmania, formas (amastigotas e
promastigotas) e vias de inóculo (Stauber, 1955; Ott et al., 1967; Melby et al., 2001;
Requena et al., 2000; Riça-Capela et al., 2003; Dea-Ayuela et al., 2007). A variabilidade
destes resultados podem ser explicada porque muitos desses autores empregam outra
técnica, a diluição limitante, utilizada para quantificar a carga parasitária no fígado e no
baço, e que não inclui o peso do órgão, dificultado muitas vezes, a comparação entre os
resultados divulgados na literatura.
As diferenças no parasitismo observado neste estudo variaram não apenas em
função da técnica empregada para enumerá-los (LDU ou qPCR), como também da cepa
infectante e as vias de inóculo utilizadas. Isto reforça mais uma vez que nos casos em que
se utilizam técnicas de baixa sensibilidade como a LDU, a via de inóculo e a cepa
infectante que é empregada refletem muito no resultado obtido. No caso dos animais do
grupo IP, foi necessário um maior tempo de infecção para que fosse possível a detecção do
parasito pela microscopia óptica. Embora os parasitos quando inoculados no peritoneo
tenha um período de pré-patência maior, um grande número deles não sobrevive para se
estabelecer no baço ou no fígado (Ott et al., 1967), dificultando assim a sua visualização.
Neste sentido, o tempo após a infecção com a via IP, foi determinante para que parasitos
fossem visualizados e quantificados por microscopia óptica no baço e fígado. Os resultados

Moreira, N. D. Discussão
118
obtidos através da densidade parasitária demonstraram que a via IC foi capaz de induzir
uma infecção progressiva nos animais, com intensa carga parasitária tecidual. Desde os
trabalhos pioneiros conduzidos por Stauber, tem sido demonstrado em hamsters que a via
intracardíaca, geralmente, induz infecções reproduzíveis com valores mais elevados na
carga parasitária (Stauber, 1955). Estes achados corroboram com os obtidos neste trabalho,
uma vez que em animais infectados com a cepa selvagem, foram demonstradas diferenças
significativas em relação ao número de parasitos obtidos, entre os dois grupos (IP e IC) em
amostras tanto de baço quanto de fígado.
A literatura ainda é escassa em trabalhos que avaliam o uso da PCR quantitativa em
Tempo Real (qPCR) para a quantificação da densidade parasitária em diferentes órgãos na
LV experimental em hamster. Para a quantificação da carga parasitária no baço e fígado de
hamsters experimentalmente infectados, utilizamos curvas-padrão construídas a partir de
concentrações conhecidas do DNA de L. infantum ligado a um plasmídeo, que é uma das
formas mais eficientes na determinação dos resultados obtidos pela qPCR (Cikos et al.
2007). Neste estudo optamos por mostrar os resultados de qPCR empregando duas
abordagens de análise: pelo número de amastigotas em 20ng de DNA e pelo número de
amastigotas multiplicado pelo peso do órgão em cada tempo da cinética avaliada. Estas
abordagens tiveram como finalidade evidenciar melhor o resultado que foi obtido com os
achados clínico-patológicos observados nestes órgãos. Ao comparar estes resultados, o que
se observa é uma maior evidência no número de amastigotas em cada grupo com o peso do
órgão durante a evolução da história natural da infecção experimental em hamster.
No fígado de animais infectados com a cepa PP75, embora tenha sido observado
um percentual alto de animais que efetivamente se infectaram ao longo do período
avaliado a carga parasitária foi baixa, e a infecção ocorreu de forma mais lenta nos três
grupos ID, IP e IC. Por outro lado, em animais infectados com a cepa selvagem pela via
ID, observou-se um pico no período inicial da infecção e à medida que a doença evoluiu
apresentou-se oscilantes no parasitismo durante o período da cinética avaliada, até mesmo
com redução da carga parasitária. O grupo IP atingiu seu pico máximo aos 9 meses após a
infecção. No grupo IC, o aumento do parasitismo foi progressivo e significativo em relação
aos grupos ID e IP a partir do terceiro mês, atingindo pico máximo no sexto e nono mês
após a infecção experimental com L. infantum.
Em relação ao parasitismo no baço mensurados pela qPCR dos animais infectados
com a cepa PP75, observou-se que os grupos ID e IP o mesmo oscilou bastante durante

Moreira, N. D. Discussão
119
todo o período avaliado, se estabelecendo de forma mais lenta. Entretanto, o grupo IC o
parasitismo progrediu conforme a doença evoluiu atingindo seu pico máximo em 6 e 9
meses após a infecção. Em animais infectados com a cepa selvagem o parasitismo evolui
nos grupos ID e IP, com pico da carga parasitária no sexto e nono mês da infecção,
enquanto o grupo IC atinge um pico máximo em 6 meses com pequena redução no nono
mês após a infecção.
Estes dados mostram mais uma vez que a evolução da infecção experimental
depende de alguns fatores, como a virulência do parasito, a via de inoculação, o órgão
avaliado e até mesmo o tempo após a infecção (Ott et al., 1967; Hommel et al., 1995;
Melby et al., 1998; Kaye et al., 2004; Carríon et al., 2006, Garg &Dube, 2006). No caso de
uma cepa mais virulenta e patogência, como a cepa selvagem a infecção evolui de forma
mais agressiva de acordo com as particularidades de cada uma das vias de inóculo e órgãos
avaliados. A variação encontrada no percentual e número de parasitos nas amostras de
baço e fígado nos diferentes grupos experimentais provavelmente está relacionada à
resposta imunológica dos hamsters à evolução da doença, o que possivelmente contribui
para uma não distribuição homogênea dos parasitos em todos os órgãos acometidos.
Em nosso estudo, a qPCR mostrou-se superior a LDU e sensível o suficiente para
detectar a presença de parasitos em amostras tanto de baço quanto de fígado em todos os
grupos experimentais (ID, IP e IC) independente da cepa empregada, além de acompanhar
a evolução da carga parasitária nestes órgãos durante todo o período de avaliação. Assim
como no LDU, pela qPCR o maior parasitismo no fígado e no baço foi observado quando
utilizou-se a cepa selvagem e a via intracardiáca. A qPCR também demonstrou que a via
IC induziu maior parasitismo no baço e consequentemente uma infecção mais rápida e
sucedida com sinais clínicos evidentes a partir do terceiro mês de infecção, indicando que a
utilização de tal via de infecção pode-se constituir como a mais apropriada para a obtenção
de uma infecção mais vigorosa e progressiva com a espécie L. infantum e em curto
período, principalmente quando o objetivo é testar drogas numa plataforma in vivo
empregando-se este modelo. (Stauber, 1955; Dea Ayuela et al., 2007).
O grupo IP, embora tenha produzido valores variáveis e baixos na carga parasitária
principalmente quando a cepa PP75 foi utilizada, nos períodos finais da infecção com a
cepa selvagem atingiu alguns picos com valores relativamente maiores e presença de sinal
clínico sugestivo da LV. Estes dados sugerem que um maior tempo no estabelecimento da
infecção experimental é requerido quando esta via é empregada (Ott et al., 1967).

Moreira, N. D. Discussão
120
Embora a via intradérmica, seja considerada a que mais assemelha o que acontece
na infecção natural pela picada de flebotomíneos, observa-se que a doença manifesta de
forma lenta e os animais permanecem sem qualquer sinal clínico por um longo período
(Wilson et al., 1987; Paranhos-Silva et al., 2003; Goto et al., 2004; Gomes et al., 2008).
Sendo assim, apesar de os animais do grupo ID não terem apresentado sinais clínicos
evidentes durante o período avaliado, foi demonstrado parasito no baço e fígado mostrando
que o parasito teve capacidade de se desenvolver e multiplicar, o que comprova a
virulência das cepas utilizadas e sensibilidade da técnica empregada.
As diferenças detectadas entre os resultados do parasitismo obtidos no baço e
fígado de hamsters experimentalmente infectados pelo LDU e os obtidos pela qPCR nos
diferentes grupos experimentais (ID, IC e IP), reforçam o uso da qPCR para quantificar e
determinar a densidade parasitária nestes órgãos.
Finalmente, no presente estudo avaliou-se a expressão de mRNA das citocinas
proinflamatórias (TNF-α e IFN-γ) e anti-inflamatórias (TGF-β e IL-10) exclusivamente no
baço por RT-PCR quantitativa em tempo Real (qRT-PCR). Os fatores que determinam o
controle ou a evolução da infecção na LV não estão totalmente esclarecidos, mas sabe-se
que a resposta imune celular desempenha importante papel na evolução da infecção
(Bittencourt & Barral-Netto, 1995). Diversos trabalhos já demonstraram que o IFN-γ,
desempenha importante papel em limitar a progressão da LV uma vez que é conhecido
como um dos principais ativadores de macrófagos, através da ativação iNOS e
consequentemente produção de óxido nítrico, mecanismos efetores críticos envolvidos no
controle da replicação do parasito (Liew et al., 1989, Melby et al., 1998; Melby et al.,
2001; Basu et al., 2005).
Nos animais infectados com a cepa PP75, observou-se maior expressão de mRNA
para as citocinas TNF-α e IL-10 no grupo IC no primeiro mês após infecção experimental.
Contudo, no sexto mês o grupo IC observou-se tanto a expressão de mRNA para IFN-γ
quanto de IL-10 e TGF-β, no entanto no nono mês da infecção observou-se apenas
expressão de mRNA para IFN-γ. Além disso, neste mesmo grupo foi evidenciado aumento
nas razões TNF-α/TGF-β; TNF-α/IL-10 e IFN-γ/IL-10.
Assim, verificamos que apesar da presença de INF-γ e TNF-α, ocorre um marcante
perfil de resposta tipo 2 gerada pela elevação na expressão de IL-10 e TGF-β conforme
evidenciado nos dados de razões destas citocinas. Sabe-se que a IL-10 desempenha

Moreira, N. D. Discussão
121
importante papel na modulação da resposta inflamatória e preservação da homeostase
imune (Bacellar et al., 2000). Estes resultados sugerem que, a produção das citocinas está
intimamente associada às variações da carga parasitária nos animais do grupo (IC), tendo
sido observados valores mais elevados de citocinas proinflamatórias e anti-inflamatórias
nos mesmos tempos em que foi evidenciado aumento da densidade parasitária esplênica
neste grupo. O fato de ter sido observado valores aumentados na expressão de mRNA para
IFN-γ, IL-10 e TGF-β no período correspondente aos valores máximos de parasitismo (6
meses após a infecção) seguida da diminuição na expressão de mRNA das citocinas IL-10
e TGF-β, sugere maior expressão da citocina IFN-γ em resposta ao parasito, o que pode ter
contribuído para a menor evidência dos sinais clínicos observados nos animais infectados
com a cepa PP75, sugerindo um controle da multiplicação do parasito em resposta a
imunomodulação observada neste tempo.
Estudos na LV experimental canina, conduzidos por Maia & Campino, (2011)
demonstraram, que IFN-γ foi expresso apenas pelos tecidos com alta carga parasitária
(baço, medula óssea, e fígado) sugerindo que a presença desta citocina não significa
remoção do parasito. Uma possível explicação para a associação entre a presença de IFN-γ
concomintante as altas cargas parasitárias pode ser devido ao fato que os parasitos estão
constantemente migrando para outros tecidos infectados estimulando sua expressão
(Strauss-Ayali et al., 2007).
Osório et al., (2003), compararam a expressão de mRNA de citocinas no focinho e
na pata de hamsters experimentalmente infectados com L. panamensis e observaram que a
expressão de mRNA para IFN-γ, IL-10 e TGF-β foi maior nos animais que apresentaram
lesões mais severas no focinho, indicando que o local de inóculo influência na evolução da
infecção na leishmaniose cutânea.
Nos animais infectados com a cepa selvagem pela via IC, verificou-se que a mesma
induziu aumento na expressão de mRNA para a citocina IFN-γ durante todo o período
avaliado (1, 3 e 6 meses após a infecção), nestes mesmos tempos também foi observado
um aumento gradativo do parasitismo esplênico. Entretanto aos 9 meses houve uma
redução na expressão de mRNA para IFN-γ neste grupo, fato que pode estar diretamente
relacionado a redução na carga parasitária esplênica observada no grupo IC neste período,
evidenciado pela qPCR.
No terceiro mês da infecção o grupo IC apresentou aumento na expressão de
mRNA tanto para as citocinas TNF-α, como para TGF-β e IL-10. Além disso, em 3, 6 e 9

Moreira, N. D. Discussão
122
meses, verificou-se aumento nas razões IFN-γ/TGF-β e também IFN-γ/IL-10. Estes dados
sugerem que o perfil de resposta imune observada em hamsters infectados com a cepa
selvagem no grupo IC no período inicial (3 meses), com elevada expressão de IL-10 e
TGF-β indica uma supressão destes animais, permitindo o estabelecimento da infecção
ativa e proliferação parasitária tecidual, onde foi observado elevado parasitismo esplênico
implicando uma relação direta entre a expressão de IFN-γ e o aumento de parasitos no
grupo IC, com um perfil unicamente inflamatório.
Estudos experimentais tem demonstrado a coexistência de valores elevados de IFN-
γ com elevado parasitismo em infecções experimentais com espécies viscerotrópicas de L.
donovani, L. infantum (Wilson et al., 1996; Rousseau et al., 2001; Ahmed et al., 2003).
Sendo assim, a expressão de mRNA das citocinas TGF-β e IL-10 detectados no presente
estudo não foram capaz de inibir a produção de IFN-γ nem mesmo a eliminação dos
parasitos. Estes dados corroboram com os de Melby et al (2001), visto que foi
demonstrado aumento na expressão das citocinas (IFN-γ), bem como de (TGF-β e IL-10)
no baço de hamsters experimentalmente infectados por L.donovani.
O grupo IP apresentou aumento na expressão de mRNA para a citocina IFN-γ a
partir do terceiro mês da infecção, com aumento na expressão de mRNA para as citocinas
TGF-β e IL-10 apenas no nono mês da infecção, momento em que foi constatado maior
parasitismo neste grupo pela qPCR.
Contudo, no grupo ID verificou-se aumento na expressão de mRNA apenas para
TGF-β e IL-10 no nono mês da infecção. Este dado sugere que o aumento na expressão
destas citocinas estaria favorecendo o estabelecimento da infecção com multiplicação do
parasito neste grupo e podem justificar o maior parasitismo neste período no grupo ID
como demonstrados pela qPCR . TGF-β tem mostrado desempenhar importante papel na
indução de imunossupressão em hamster com LV progressiva (Rodrigues et al.,1998).
Em estudos conduzidos por pesquisadores de nosso grupo de pesquisa, mostraram
que durante a evolução da LVC cães sintomáticos com altas cargas parasitárias esplênicas
apresentaram aumento de IFN-γ e que 100% destes animais possuíam expressão
aumentada de IL-10 (Lage et al., 2007). Na LVH também foi demonstrado por Peruhipe-
Magalhães et al. (2006) aumento destas citocinas em células mononucleares do sangue
periférico. Apesar de escassos estudos terem avaliado o perfil de citocinas anti e

Moreira, N. D. Discussão
123
proinflamatória no modelo hamster com LV podemos concluir que muitos dos fenômenos
imunomodulatórios aqui observados também coincidem com os relatados na LVH e LVC.
Neste contexto, buscou avaliar e comparar o comportamento das diferentes vias de
inóculo (intradérmica, intracardíaca e intraperitoneal) em infecções por L. infantum no
modelo hamster. Além disto, o presente estudo foi determinante para obter uma
caracterização detalhada da infecção experimental neste modelo empregando-se num
âmbito global, análises que pretendem subsidiar estudos pré-clínicos com vistas a futuras
estratégias terapêuticas e vacinais contra LV humana e canina. Neste sentido, foi possível
estabelecer um modelo de infecção capaz de reproduzir a fase aguda e crônica da LV com
parâmetros imunopatológicos, hematológico-bioquímicos e carga parasitária similares ao
observado à infecção humana e canina. A qPCR demonstrou-se como uma ferramenta
sensível e importante, pois possibilitou a comparação de resultados de carga parasitária
entre os diferentes grupos experimentais infectados por diferentes cepas de L.infantum,
particularmente aqueles com reduzido número de parasitos, bem como entre os órgãos
(fígado e o baço). Adicionalmente, a ampliação do espectro de análises investigativas por
meio da imunologia molecular empregando a técnica de PCR em tempo Real para avaliar
perfis de citocinas em diferentes tecidos, permitiu um melhor entendimento da história
natural da infecção experimental no modelo hamster para LV. Neste sentido, acreditamos
que o presente estudo irá permitir ampliar o emprego do modelo hamster para testes pré-
clínicos de drogas e vacinas anti-LV, contribuindo para as pesquisas nestas áreas.

8. CONCLUSÃO

Moreira, N. D. Conclusão
125
Neste estudo foi avaliado o estabelecimento da infecção experimental no hamster,
utilizando diferentes vias de inóculo (intradérmica, intraperitoneal e intracardíaca) e duas
cepas de L. infantum (PP75 e selvagem-C46) com perfis distintos de virulência e
patogenicidade. Os resultados obtidos mostraram:
• maior infectividade com a utilização da cepa selvagem e menor taxa de sobrevida
nos animais inoculados pela via intraperitoneal e intracardíaca;
• sinais clínicos similares aos observados na LV sintomática humana e canina
(emagrecimento acentuado e esplenomegalia);
• exacerbação dos sinais clínicos na infecção pela via IC, sendo estes mais
frequentes e intensos em hamsters infectados com a cepa selvagem, reforçando a
influência da via de inóculo na manifestação de sinais clínicos;
• níveis mais elevados e mais precoces de IgG nos animais infectados com a cepa
selvagem e com maiores níveis nos grupos IP e IC;
• alterações hematológicas clássicas da LV humana e canina: anemia e leucopenia,
acentuados em hamsters infectados com a cepa selvagem pelas vias IP e IC;
• alterações renais com elevação nos níveis de uréia e creatina além de alteração
hepática com elevação nos níveis de AST e ALT independente da cepa, via de
inóculo e tempo avaliado;
• intensidade das alterações histopatológicas do parênquima hepático intimamente
relacionadas ao parasitismo apresentado por cada um dos grupos avaliados (ID,
IC e IP). O aumento do processo inflamatório e a presença de maior número de
granulomas apresentado no grupo IC parece estar relacionado ao intenso
parasitismo;
• alterações histopatológicas esplênicas nos animais infectados com a cepa PP75
foram mais evidentes no grupo IC e, embora tenham ocorrido durante todo período
de avaliação, apresentaram ser com maior intensidade nos períodos finais da
infecção (6 e 9 meses). Entretanto nos animais infectados com a cepa selvagem, as
alterações esplênicas no grupo IC ocorreram em todo período avaliado, porém
com maior exuberância a partir do sexto mês.
• a hipoplasia da polpa branca, hiperplasia da polpa vermelha, perda da arquitetura
e periesplenite demonstradas nos períodos de 6 e 9 meses no grupo IC infectado

Moreira, N. D. Conclusão
126
com a cepa selvagem estão associadas ao intenso parasitismo e sinais mais
severos, tais como: emagrecimento e esplenomegalia;
• alta sensibilidade e elevada performance da qPCR em relação a LDU na detecção
e quantificação de DNA de L.infantum em amostras tanto de baço quanto de
fígado, independente da via de inóculo, do tempo e da cepa empregada;
• semelhante ao observado pela LDU, a qPCR também evidenciou maior parasitismo
no fígado e no baço quando utilizou-se a cepa selvagem e a via intracardíaca;
• aumento da expressão de mRNA de citocinas anti e pró-inflamatórias em hamsters
inoculados com a cepa PP75 associada ao aumento do parasitismo esplênico no
sexto e nono mês da infecção;
• predomínio da expressão de mRNA para IFN-γ, nos animais infectados com a cepa
PP75 no fim do período de avaliação no grupo IC, possivelmente em resposta ao
parasitismo;
• O perfil de resposta imune observada em hamsters infectados com a cepa selvagem
no grupo IC no período inicial (3 meses), com elevada expressão de IL-10 e TGF-β
indica uma supressão destes animais, permitindo o estabelecimento da infecção
ativa e proliferação parasitária tecidual;
• durante os períodos em que foram observados picos de alta densidade parasitária
ocorreru elevação de INF-γ, mostrando um perfil tipicamente inflamatório.
Diante do conjunto de evidências acima exposto, pode-se conclui que:
• O modelo hamster reproduz diversas das características clínicas e laboratoriais da
LV humana e canina, principalmente nas alterações clínico e histopatológicas,
bioquímico-hematológicas, parasitológicas e imunopatológicas. A via de inóculo e
a cepa utilizada são elementos fundamentais para o estabelecimento da infecção e
são determinantes importantes na definição de estratégias empregadas em estudos
pré-clinicos seja para testes de drogas e/ou vacinas.
• Nossos resultados permitem também preconizar que para os estudos pré-clínicos de
testes de drogas leishmanicidas é recomendável usar a cepa selvagem (C46) de L.
infantum buscando o estabelecimento da infecção experimental neste modelo,

Moreira, N. D. Conclusão
127
preferencialmente pela via intracardíaca. Por outro lado, em estudos de testes de
vacinas no modelo hamster é recomendável o emprego da via intradérmica
preferencialmente, e ambas as cepas podem ser utilizadas na infecção desafio desde
que se empreguem métodos moleculares quantitativos na avaliação da densidade
parasitária tecidual após o desafio experimental.

9. REFERÊNCIAS BIBLIOGRÁFICAS

Moreira, N. D. Referências Bibliográficas
129
ABRANCHES, P.; SILVA-PEREIRA, M. C. et al. Canine leishmaniasis: pathological and ecological factors influencing transmission of infection. J Parasitol, v.77, n.4, Aug, p.557-6, 1991. ADLER, S. Origin of the golden hamster Cricetus auratus as a laboratory animal. Nature, v.162, p.256, 1948. ADLER, J. H.The origin of the golden hamster as a laboratory animal. Isr J Med Sci, v.25, p.206-9, 1989. ADLER, S. Inoculation of human leprosy into Syrian hamster. Lancet, v.233, p.714-715, 1937. AGENOR, A. L. V.; LIMA-VERDE, F. A. et al. Renal tubular dysfunction in human visceral leishmaniasis (Kala-azar). Clin Nephrol, v.71, n.5, May, p.492-500, 2009. AHMED, S.; COLMENARES, M. et al. Intradermal infection model for pathogenesis and vaccine studies of murine visceral leishmaniasis. Infect Immun, v.71, n.1, Jan, p.401-10, 2003. ALVAR, J.; CANAVATE C. et al. Canine leishmaniasis. Adv Parasitol, v.57, p.1-88, 2004. ALVES, C. F.; DE AMORIM, I. F. et al. Expression of IFN-gamma, TNF-alpha, IL-10 and TGF-beta in lymph nodes associates with parasite load and clinical form of disease in dogs naturally infected with Leishmania (Leishmania) chagasi. Vet Immunol
Immunopathol, v.128, n.4, April, p.349-58, 2009. ANURADHA; PAL, R. et al. Sex-influenced population kinetics of Leishmania donovani in hamsters. Indian J Exp Biol, v.28, n.9, Sep, p.876-9, 1990. ASHFORD, R. W. The leishmaniasis as emerging and reemerging zoonoses. Int J
Parasitol, v.30, p.1269-81, 2000. BACELLAR, O.; D'OLIVEIRA, A, Jr, A. et al. IL-10 and IL-12 are the main regulatory cytokines in visceral leishmaniasis. Cytokine, v.12, n.8, Aug, p.1228-31, 2000. BADARO, R.; JONES, T. C. et al. New perspectives on a subclinical form of visceral leishmaniasis. J Infect Dis, v.154, n.6, Dec, p.1003-11, 1986. BADARO, R.; JONES, T. C. et al. A prospective study of visceral leishmaniasis in an endemic area of Brazil. J Infect. Dis, v.154, n.4, Oct, p.639-49, 1986. BANETH, G., AROCH, I. Canine leishmaniasis: a diagnostic and clinical challenge. Vet J, v.175, n.1, Jan, p.14-5, 2008. BANETH, G.; KOUTINAS, A. F. et al. Canine leishmaniosis - new concepts and insights on an expanding zoonosis: part one. Trends Parasitol, v.24, n.7, Jul, p.324-30, 2008.

Moreira, N. D. Referências Bibliográficas
130
BANETH, G, E; SHAW, S. Chemotherapy of canine leishmaniosis. Vet Parasitol, v.106, n.4, Jul, p.315-24, 2002. BASU, R.; BHAUMIK, S. et al. Kinetoplastid membrane protein-11 DNA vaccination induces complete protection against both pentavalent antimonial-sensitive and -resistant strains of Leishmania donovani that correlates with inducible nitric oxide synthase activity and IL-4 generation: evidence for mixed Th1- and Th2-like responses in visceral leishmaniasis. J Immunol, v.174, n.11, Jun, p.7160-71, 2005. BENTEN, W. P.; LIEBERHERR, M. et al. Testosterone signaling through internalizable surface receptors in androgen receptor-free macrophages. Mol Biol Cell, v.10, n.10, Oct, p.3113-23, 1999. BERMAN, J. D. Human leishmaniasis: clinical, diagnostic and chemotherapeutic developments in the last 10 years. Clin Infect Dis, v.24, p.684-703, 1997. BERMAN, J. Visceral leishmaniasis in the New World & Africa. Indian J Med Res, v.123, n.3, Mar, p.289-94, 2006. BESSEMANS, A.; DE WILDE, H. Receptivite inapparente de I´ hamster ã Treponema pallidum vains essais de syphilisation du chien, du porc et de la grenouile: Inaparent receptivity of the hamster to treponema pallidum tested by syphilizaton of dog, pig and frog. C.R.Soc. Biol, v.119, p.326-330, 1935. BINHAZIM, A. A.; CHAPMAN, W. L. Jr. et al. Determination of virulence and pathogenesis of a canine strain of Leishmania (Leishmania) infantum in hamsters and dogs.
Am J Vet Res, v.54, n.1, Jan, p.113-21, 1993. BITTENCOURT; BARRAL-NETTO, M. Leishmaniasis. Tropical Pathology, p.597-651. 1995. BORIES, C.; COFFIN, C. et al. Lack of a nitric-oxide response during the course of Leishmania infantum infection in the golden hamster (Mesocricetus auratus), with or without treatment with liposomal amphotericin B. Ann Trop Med Parasitol, v.92, n.6, Sep, p.685-92, 1998. BOURDOISEAU, G.; BONNEFONT, C. et al. Lymphocyte subset abnormalities in canine leishmaniasis. Vet Immunol Immunopathol, v.56, n.3-4, May, p.345-51, 1997. BRASIL. Guia de Vigilância Epidemiológica, Secretaria de Vigilância em Saúde, Departamento de Vigilância Epidemiológica, Ministério da Saúde. Brasília, p.816, 2009. BRETAGNE, S.; DURAND, R. et al. Real-time PCR as a new tool for quantifying Leishmania infantum in liver in infected mice. Clin Diagn Lab Immunol, v.8, n.4, Jul, p.828-31, 2001. BRUCE, H. M.; HINDLE, E. The golden hamster, Cricetus (Mesocricetus) auratus Waterhouse: notes on its breeding and gowth. Proc Zool. Soc, v.104, p.361-366, 1934.

Moreira, N. D. Referências Bibliográficas
131
BUNN-MORENO, M. M.; MADEIRA, E. D. et al. Hypergammaglobulinaemia in Leishmania donovani infected hamsters: possible association with a polyclonal activator of B cells and with suppression of T cell function. Clin Exp Immunol, v.59, n.2, Feb, p.427-34, 1985. CAMPOS. Características do calazar na criança. Estudo de 75 casos. Pediatr (Rio J), v.71, p.261-5, 1995. CARRERA, L.; FERMIN, M. L. et al. Antibody response in dogs experimentally infected with Leishmania infantum: infection course antigen markers. Exp Parasitol, v.82, n.2, Mar, p.139-46, 1996. CARRIÓN, J.; NIETO, A. et al. Immunohistological features of visceral leishmaniasis in BALB/c mice. Parasite Immunol, v.28, n.5, May, p.173-83, 2006. CARRIÓN, J.; FOLGUEIRA, C. et al. Leishmania infantum HSP70-II null mutant as candidate vaccine against leishmaniasis: a preliminary evaluation. Parasit Vectors, v.4, p.150, 2011. CASCIO, A.; COLOMBA, C. et al. Pediatric visceral leishmaniasis in Western Sicily, Italy: a retrospective analysis of 111 cases. Eur J Clin Microbiol Infect Dis, v.21, n.4, Apr, p.277-82, 2002. CATORZE, G., ALBERTO, J. et al. Leishmania infantum/HIV co-infection: cutaneous lesions following treatment of visceral leishmaniasis. Ann Dermatol Venereol, v.133, n.1, Jan, p.39-42, 2006. CIARAMELLA, P.; OLIVA, G. et al. A retrospective clinical study of canine leishmaniasis in 150 dogs naturally infected by Leishmania infantum. Vet Rec, v.141, n.21, Nov 22, p.539-43, 1997. CIKOS, S.; BUKOVSKÁ, A. et al. Relative quantification of mRNA: comparison of methods currently used for real-time PCR data analysis. BMC Mol Biol, v.8, p.113, 2007. CLEMENTI, A.; BATTAGLIA, G. et al. Renal involvement in leishmaniasis: a review of the literature. NDT Plus, v.4, p.147-152, 2011. COLLIN, S. M.; COLEMAN, P. G., et al. Unseen Kala-azar deaths in south Sudan (1999-2002). Trop Med Int Health, v.11, n.4, Apr, p.509-12, 2006. CORBETT, C. E. P.; LAURENTI, M. D. Early detection of Leishmania (Leishmania)
chagasi in draining lymph node after subcutaneous inoculation in hamster. Parasitology
International, v.47, p.307-310, 1998. COSTA, C. H; PEREIRA, H. F. et al. Is the household dog a risk factor for American visceral leishmaniasis in Brazil? Transactions of the Royal Society of Tropical Medicine
and Hygiene, v.93, p.464, 1999. COURA-VITAL, W.; MARQUES M. J. et al. Humoral and cellular immune responses in

Moreira, N. D. Referências Bibliográficas
132
dogs with inapparent natural Leishmania infantum infection. Vet J, v.190, n.2, Nov, p.e43-7, 2011. CUTOLO, M.; ACCARDO S. et al. Androgen and estrogen receptors are present in primary cultures of human synovial macrophages. J Clin Endocrinol Metab, v.81, n.2, Feb, p.820-7, 1996. DA COSTA-VAL, A. P.; CAVALCANTI, R. R. et al. Canine visceral leishmaniasis: relationships between clinical status, humoral immune response, haematology and Lutzomyia (Lutzomyia) longipalpis infectivity. Vet J, v.174, n.3, Nov, p.636-43, 2007. DE ANDRADE, H. M.; REIS A. B. et al. Use of PCR-RFLP to identify Leishmania
species in naturally-infected dogs. Vet Parasitol, v.140, n.3-4, Sep 10, p.231-8, 2006. DE QUEIROZ, N. M.; DA SILVEIRA, R. C. et al. Detection of Leishmania (L.) chagasi
in canine skin. Vet Parasitol, v.178, n.1-2, May 31, p.1-8, 2011. DEA-AYUELA, M. A.; RAMA-INIGUEZ, S. et al. Setting new immunobiological parameters in the hamster model of visceral leishmaniasis for in vivo testing of antileishmanial compounds. Vet Res Commun, v.31, n.6, Aug, p.703-17, 2007. DEPLAZES, P.; SMITH, N. C. et al. Specific IgG1 and IgG2 antibody responses of dogs to Leishmania infantum and other parasites. Parasite Immunol, v.17, n.9, Sep, p.451-8, 1995. DESJEUX, P. Leishmaniasis: Public health aspects and control. Clin Dermatol, v.14, n.5, Sep-Oct, p.417-23, 1996. DESJEUX, P. Leishmaniasis: current situation and new perspectives. Comp Immunol Microbiol Infect Dis, v.27, n.5, Sep, p.305-18, 2004. DOS SANTOS, S. O.; ARIAS J. et al. Incrimination of Lutzomyia cruzi as a vector of American visceral leishmaniasis. Med Vet Entomol, v.12, p.315-7, 1998. DOS-SANTOS, W. L.; JESUS, E. E. et al. Associations among immunological, parasitological and clinical parameters in canine visceral leishmaniasis: Emaciation, spleen parasitism, specific antibodies and leishmanin skin test reaction. Vet Immunol
Immunopathol, v.123, n.3-4, Jun 15, p.251-9, 2008. DRUMOND, K. O.; COSTA, F. A. Forty years of visceral leishmaniasis in the State of Piaui: a review. Rev Inst Med Trop Sao Paulo, v.53, n.1, Jan-Feb, p.3-11, 2011. DUARTE, M. I.; LAURENT, M. D. et al. Comparative study of the biological behaviour in hamster of two isolates of Leishmania characterized respectively as L. major-like and L.
donovani. Rev Inst Med Trop Sao Paulo, v.30, n.1, Jan-Feb, p.21-7, 1988. DUTRA, M.; MARTINELLI, R. et al. Renal involvement in visceral leishmaniasis. Am J
Kidney Dis, v.6, n.1, Jul, p.22-7, 1985.

Moreira, N. D. Referências Bibliográficas
133
ENGWERDA, C. R.; KAYE, P. M. Organ-specific immune responses associated with infectious disease. Immunol Today, v.21, n.2, Feb, p.73-8, 2000. ENGWERDA, C. R., ATO, M. et al. A role for tumor necrosis factor-alpha in remodeling the splenic marginal zone during Leishmania donovani infection. Am J Pathol, v.161, n.2, Aug, p.429-37., 2002. EVANS, T. G.; SMITH, D. et al. Humoral factors and nonspecific immune suppression in Syrian hamsters infected with Leishmania donovani. J Parasitol, v.76, n.2, Apr, p.212-7, 1990. FEITOSA, M. M. I.; LUVIZOTTO, F.A. et al. Aspectos clínicos de cães com leishmaniose visceral no município de Araçatuba – São Paulo., v.5, p.36-44, 2000. FINDLAY, G. M. The infectivity of neurotropic yellow fever virus for animals. J. Path.
Bact, v.38, p.1-6, 1934. FRUTH, U.; SOLIOZ, N. et al. Leishmania major interferes with antigen presentation by infected macrophages. J Immunol, v.150, n.5, Mar 1, p.1857-64, 1993. GALVAO-CASTRO, B.; SÁ FERREIRA, J. A. et al. Polyclonal B cell activation, circulating immune complexes and autoimmunity in human american visceral leishmaniasis. Clin Exp Immunol, v.56, n.1, Apr, p.58-66, 1984. GARG, R.; DUBE, A. Animal models for vaccine studies for visceral leishmaniasis. Indian J Med Res, v.123, n.3, Mar, p.439-54, 2006. GENARO, O. Leishmaniose visceral canina experimental. Tese (Doutorado em Ciências), Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, p.202, 1993. GHOSE, A. C.; MOOKERJEE, A. et al. Therapeutic and prophylactic uses of protein A in the control of Leishmania donovani infection in experimental animals. Immunol Lett, v.65, n.3, Feb, p.175-81, 1999. GIFAWESEN, C.; FARRELL, J. P. Comparison of T-cell responses in self-limiting versus progressive visceral Leishmania donovani infections in golden hamsters. Infect Immun, v.57, n.10, Oct, p.3091-6, 1989. GIUNCHETTI, R. C., W. MAYRINK. et al. Relationship between canine visceral leishmaniosis and the Leishmania (Leishmania) chagasi burden in dermal inflammatory foci. J Comp Pathol, v.135, n.2-3, Aug-Oct, p.100-7, 2006. GIUNCHETTI, R. C., R. CORREA-OLIVEIRA. et al. Immunogenicity of a killed Leishmania vaccine with saponin adjuvant in dogs. Vaccine, v.25, n.44, Nov 1, p.7674-86, 2007. GIUNCHETTI, R. C.; CORREA-OLIVEIRA, R. et al. A killed Leishmania vaccine with sand fly saliva extract and saponin adjuvant displays immunogenicity in dogs. Vaccine,

Moreira, N. D. Referências Bibliográficas
134
v.26, n.5, Jan, p.623-38, 2008a. GIUNCHETTI, R. C.; MARTINS-FILHO O. A. et al. Histopathology, parasite density and cell phenotypes of the popliteal lymph node in canine visceral leishmaniasis. Vet Immunol
Immunopathol, v.121, n.1-2, Jan 15, p.23-33, 2008b. GIUNCHETTI, R. C.; MAYRINK W. et al. Histopathological and immunohistochemical investigations of the hepatic compartment associated with parasitism and serum biochemical changes in canine visceral leishmaniasis. Res Vet Sci, v.84, n.2, Apr, p.269-77, 2008c. GOMES, R.; TEIXEIRA C. et al. Immunity to a salivary protein of a sand fly vector protects against the fatal outcome of visceral leishmaniasis in a hamster model. Proc Natl
Acad Sci U S A, v.105, n.22, Jun 3, p.7845-50, 2008. GOTO, H.; LINDOSO. J. A. Immunity and immunosuppression in experimental visceral leishmaniasis. Braz J Med Biol Res, v.37, n.4, Apr, p.615-23, 2004. GUPTA, R.; KUSHAWAHA, P. K. et al. Treatment of Leishmania donovani-infected hamsters with miltefosine: analysis of cytokine mRNA expression by real-time PCR, lymphoproliferation, nitrite production and antibody responses. J Antimicrob Chemother, v.67, n.2, Feb, p.440-3, 2012. HANDMAN, E. Leishmaniasis: current status of vaccine development. Clin Microbiol
Rev, v.14, n.2, Apr, p.229-43, 2001. HOFFMANN, R. A.; ROBINSON, P.F. et al. The golden hamster, its biology and use in Medical Research. Iowa: The Iowa University Press, p.545, 1968. HOMMEL, M.; JAFFE, C. L. et al. Experimental models for leishmaniasis and for testing anti-leishmanial vaccines. Ann Trop Med Parasitol, v.89 Suppl 1, Dec, p.55-73, 1995. IKEDA-GARCIA, F. A.; LOPES, R. S. et al. Clinical and parasitological evaluation of dogs naturally infected by Leishmania (Leishmania) chagasi submitted to treatment with meglumine antimoniate. Vet Parasitol, v.143, n.3-4, Feb 28, p.254-9, 2007. JONES, L. A.; ANTHONY, J. P. et al. Toll-like receptor-4-mediated macrophage activation is differentially regulated by progesterone via the glucocorticoid and progesterone receptors. Immunology, v.125, n.1, Sep, p.59-69, 2008. KANE, M. M.; MOSSER, D. M. The role of IL-10 in promoting disease progression in leishmaniasis. J Immunol, v.166, n.2, Jan 15, p.1141-7, 2001. KANEKO, J. J.; HARVEY, J. W. et al. Clinical Biochemestry of domestic animals. Academic Press Inc, San Diego, p.932, 1997. KAYE, P. M.; SVENSSON, M., et al. The immunopathology of experimental visceral leishmaniasis. Immunol Rev, v.201, Oct, p.239-53, 2004.

Moreira, N. D. Referências Bibliográficas
135
KUMARI, S.; SAMANT, M. et al. Th1-stimulatory polyproteins of soluble Leishmania
donovani promastigotes ranging from 89.9 to 97.1 kDa offers long-lasting protection against experimental visceral leishmaniasis. Vaccine, v.26, n.45, Oct 23, p.5700-11, 2008. LAGE, R. S.; OLIVEIRA, G. C. et al. Analysis of the cytokine profile in spleen cells from dogs naturally infected by Leishmania chagasi. Vet Immunol Immunopathol, v.115, n.1-2, Jan 15, p.135-45, 2007. LAINSON, R.; SHAW, J. J. Evolution, classification and geographical distribution. The Leishmaniasis in Biology and Medicine. Academic Press Inc, London, p.1-120, 1987. LAURENTI, M. D.; SOTTO, M. N. et al. Experimental visceral leishmaniasis: sequential events of granuloma formation at subcutaneous inoculation site. Int J Exp Pathol, v.71, n.6, Dec, p.791-7, 1990. LEI, S. M.; RAMER-TAIT, A. E. et al. Reduced hamster usage and stress in propagating Leishmania chagasi promastigotes using cryopreservation and saphenous vein inoculation. J Parasitol, v.96, n.1, Feb, p.103-8, 2010. LIEW, F. Y.; MILLOTT, S. et al. Macrophage activation by interferon-gamma from host-protective T cells is inhibited by interleukin (IL) 3 and IL4 produced by disease-promoting T cells in leishmaniasis. Eur J Immunol, v.19, n.7, Jul, p.1227-32, 1989. LIMA VERDE, F. A.; LIMA VERDE, F. A. et al. Evaluation of renal function in human visceral leishmaniasis (kala-azar): a prospective study on 50 patients from Brazil. J
Nephrol, v.20, n.4, Jul-Aug, p.430-6, 2007. LUKES, J., MAURICIO I. L. et al. Evolutionary and geographical history of the Leishmania donovani complex with a revision of current taxonomy. Proc Natl Acad Sci U
S A, v.104, n.22, May, p.9375-80, 2007. MAIA, C.; CAMPINO, L. Cytokine and Phenotypic Cell Profiles of Leishmania infantum Infection in the Dog. J Trop Med, v.2012, p.541571, 2012. MAIA, C.; NUNES M. et al. Experimental canine leishmaniasis: clinical, parasitological and serological follow-up. Acta Trop, v.116, n.3, Dec, p.193-9, 2010. MALTEZOU, H. C.; SIAFAS C. et al. Visceral leishmaniasis during childhood in southern Greece. Clin Infect Dis, v.31, n.5, Nov, p.1139-43, 2000. MANGOUD, A. M.; RAMADAN, M. E. et al. Histopathological studies of Syrian golden hamsters experimentally infected with Leishmania infantum. J Egypt Soc Parasitol, v.27, n.3, Dec, p.689-702, 1997. MANNA, L.; REALE S. et al. Leishmania DNA load and cytokine expression levels in asymptomatic naturally infected dogs. Vet Parasitol, v.142, n.3-4, Dec, p.271-80, 2006. MATHIAS, R.; COSTA F. A. et al. Detection of immunoglobulin G in the lung and liver of hamsters with visceral leishmaniasis. Braz J Med Biol Res, v.34, n.4, Apr, p.539-43,

Moreira, N. D. Referências Bibliográficas
136
2001. MAURICIO, I. L.; STOTHARD J. R. et al. The strange case of Leishmania chagasi. Parasitol Today, v.16, n.5, May, p.188-9, 2000. MCMAHON-PRATT, D.; ALEXANDER, J. Does the Leishmania major paradigm of pathogenesis and protection hold for New World cutaneous leishmaniases or the visceral disease? Immunol Rev, v.201, Oct, p.206-24, 2004. MELBY, P. C.; CHANDRASEKAR, B. et al. The hamster as a model of human visceral leishmaniasis: progressive disease and impaired generation of nitric oxide in the face of a prominent Th1-like cytokine response. J Immunol, v.166, n.3, Feb 1, p.1912-20, 2001. MELBY, P. C.; TABARES, A. et al. Leishmania donovani: evolution and architecture of the splenic cellular immune response related to control of infection. Exp Parasitol, v.99, n.1, Sep, p.17-25, 2001. MELBY, P. C.; TRYON, V. V. et al. Cloning of Syrian hamster (Mesocricetus auratus) cytokine cDNAs and analysis of cytokine mRNA expression in experimental visceral leishmaniasis. Infect Immun, v.66, n.5, May, p.2135-42, 1998. MELO, F.; AMARAL, M., et al. Diffuse intralobular liver fibrosis in dogs naturally infected with Leishmania (Leishmania) chagasi. Am J Trop Med Hyg, v.79, n.2, Aug, p.198-204, 2008. MICHALICK, M. S. M. Comportamento Biológico de Amostras de Leishmania dos Complexos braziliensis e mexicana na Bolsa Alimentar do Hamster (Mesocricetus
auratus). Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, Tese de Doutorado, Belo Horizonte, MG, p.145, 1996. MILES, S. A.; CONRAD, S. M. et al. A role for IgG immune complexes during infection with the intracellular pathogen Leishmania. J Exp Med, v.201, n.5, Mar 7, p.747-54, 2005. MIRALLES, G. D.; STOECKLE, M. Y. et al. Th1 and Th2 cell-associated cytokines in experimental visceral leishmaniasis. Infect Immun, v.62, n.3, Mar, p.1058-63, 1994. MONTEIRO, P. S., LACERDA, M. M., et al. Controle da leishmaniose Visceral no Brasil. Rev. Soc. Bras. Med. Trop, v.27, p.67-72, 1994. MORENO, J.; ALVAR, J. Canine leishmaniasis: epidemiological risk and the experimental model. Trends Parasitol, v.18, n.9, Sep, p.399-405, 2002. MUKHERJEE, P.; GHOSH, A. K. et al. Infection pattern and immune response in the spleen and liver of BALB/c mice intracardially infected with Leishmania donovani
amastigotes. Immunol Lett, v.86, n.2, Apr 3, p.131-8, 2003. MURPHY, M. L.; WILLE, U. et al. IL-10 mediates susceptibility to Leishmania donovani infection. Eur J Immunol, v.31, n.10, Oct, p.2848-56, 2001.

Moreira, N. D. Referências Bibliográficas
137
MURRAY, H. W. Endogenous interleukin-12 regulates acquired resistance in experimental visceral leishmaniasis. J Infect Dis, v.175, n.6, Jun, p.1477-9, 1997. MURRAY, H. W. Tissue granuloma structure-function in experimental visceral leishmaniasis. Int J Exp Pathol, v.82, n.5, Oct, p.249-67, 2001. MURRAY, H. W.; LU, C. M. et al. Interleukin-10 (IL-10) in experimental visceral leishmaniasis and IL-10 receptor blockade as immunotherapy. Infect Immun, v.70, n.11, Nov, p.6284-93, 2002. MURRAY, H. W.; MASUR H. et al. Cell-mediated immune response in experimental visceral leishmaniasis. I. Correlation between resistance to Leishmania donovani and lymphokine-generating capacity. J Immunol, v.129, n.1, Jul, p.344-50, 1982. MURRAY, H. W.; MONTELIBANO C. et al. Interleukin-12 regulates the response to chemotherapy in experimental visceral Leishmaniasis. J Infect Dis, v.182, n.5, Nov, p.1497-502, 2000. MURRAY, H. W.; SQUIRES, K. E., et al. Acquired resistance and granuloma formation in experimental visceral leishmaniasis. Differential T cell and lymphokine roles in initial versus established immunity. J Immunol, v.148, n.6, Mar 15, p.1858-63, 1992. NASIR, A.M, AL-NASSER, M.N. et al. The haematological manifestations of visceral leishmaniasis in infancy and childhood. J Trop Pediatr (Rio J), v.41, p.143-8, 1995. NIETO, A., G.; DOMINGUEZ-BERNAL. et al. Mechanisms of resistance and susceptibility to experimental visceral leishmaniosis: BALB/c mouse versus syrian hamster model. Vet Res, v.42, n.1, Jan-Feb, p.39, 2011. NIETO, C. G.; NAVARRETE, I. et al. Pathological changes in kidneys of dogs with natural Leishmania infection. Vet Parasitol, v.45, n.1-2, Dec, p.33-47, 1992. NYLEN, S.; GAUTAM, S. Immunological perspectives of leishmaniasis. J Glob Infect
Dis, v.2, n.2, May, p.135-46, 2010. NYLEN, S.; SACKS, D. Interleukin-10 and the pathogenesis of human visceral leishmaniasis. Trends Immunol, v.28, n.9, Sep, p.378-84, 2007. OLIVA, G.; FOGLIA MANZILLO, V. et al. Canine leishmaniasis: evolution of the chemotherapeutic protocols. Parasitologia, v.46, n.1-2, Jun, p.231-4, 2004. OLIVEIRA, E.; OSHIRO, E. T. et al. Presence of amastigotes in the central nervous system of hamsters infected with Leishmania sp. Rev Bras Parasitol Vet, v.20, n.2, Apr-Jun, p.97-102, 2011. OLIVEIRA, J. M.; FERNANDES, A.N. et al. Mortality due to visceral leishmaniasis: clinical and laboratory characteristics. Revista da Sociedade Brasileira de Medicina
Tropical, v.43, p.188-193, 2010.

Moreira, N. D. Referências Bibliográficas
138
OSORIO, Y.; MELBY, P. C. et al. The site of cutaneous infection influences the immunological response and clinical outcome of hamsters infected with Leishmania
panamensis. Parasite Immunol, v.25, n.3, Mar, p.139-48, 2003.
OTT, K. J.; HANSON, W. L. et al. Course of infection of Leishmania donovani in hamsters inoculated by the intraperitoneal route. J Parasitol, v.53, n.3, Jun, p.641-5, 1967. PARANHOS-SILVA, M.; OLIVEIRA, G. G. et al. A follow-up of Beagle dogs intradermally infected with Leishmania chagasi in the presence or absence of sand fly saliva. Vet Parasitol, v.114, n.2, May 30, p.97-111, 2003. PASTORINO, A. C.; JACOB, C. M. et al. Visceral leishmaniasis: clinical and laboratorial aspects. J Pediatr (Rio J), v.78, n.2, Mar-Apr, p.120-7, 2002. PEARSON, H. E.; EATON, M.D. A virus pneumonia of Syrian hamsters. Proc.Soc. Exp.
Biol. Med, v.45, p.677-679, 1940. PEARSON, R. D.; ROBERTS, D. Host immunoglobulin on spleen-derived Leishmania
donovani amastigotes. Am J Trop Med Hyg, v.43, n.3, Sep, p.263-5, 1990. PEARSON, R. D.; SOUSA, A. Q. Clinical spectrum of Leishmaniasis. Clin Infect Dis,
v.22, n.1, Jan, p.1-13, 1996. PEDROSA, C. M. S. Aspectos clínicos e epidemiológicos da leishmaniose visceral em menores de 15 anos procedentes de Alagoas, Brasil. Rev Soc Bras Med Trop, v.37, p.300-304, 2004. PERUHYPE-MAGALHAES, V.; MARTINS-FILHO, O. A. et al. Mixed inflammatory/regulatory cytokine profile marked by simultaneous raise of interferon-gamma and interleukin-10 and low frequency of tumour necrosis factor-alpha(+) monocytes are hallmarks of active human visceral Leishmaniasis due to Leishmania
chagasi infection. Clin Exp Immunol, v.146, n.1, Oct, p.124-32, 2006. PICCINNI, M. P.; SCALETTI, C. et al. Role of hormone-controlled Th1- and Th2-type cytokines in successful pregnancy. J Neuroimmunol, v.109, n.1, Sep 1, p.30-3, 2000. PISCOPO, T. V.; MALLIA, A. C. Leishmaniasis. Postgrad Med J, v.82, n.972, Oct, p.649-57, 2006. PISCOPO, T. V.; MALLIA A. C. Leishmaniasis. Postgrad Med J, v.83, n.976, Feb, p.649-57, 2007. PLEVRAKI, K.; KOUTINAS, A. F. et al. Effects of allopurinol treatment on the progression of chronic nephritis in Canine leishmaniosis (Leishmania infantum). J Vet
Intern Med, v.20, n.2, Mar-Apr, p.228-33, 2006. POOT, J.; SPREEUWENBERG, K. et al. Optimization of a Leishmania infantum challenge model in hamsters. Intervet International B.V, p.57-70, 2006.

Moreira, N. D. Referências Bibliográficas
139
PRASAD, L. S.; SEN, S. et al. Renal involvement in kala-azar. Indian J Med Res, v.95, Jan, p.43-6, 1992. QUEIROZ, M. J.; ALVES, J. G., et al. Visceral leishmaniasis: clinical and epidemiological features of children in an endemic area. J Pediatr (Rio J), v.80, n.2, Mar-Apr, p.141-6, 2004. REIMAO, J. Q.; COLOMBO, F. A. et al. Effectiveness of liposomal buparvaquone in an experimental hamster model of Leishmania (L.) infantum chagasi. Exp Parasitol, v.130, n.3, Mar, p.195-9, 2012. REIS, A. B. Avaliação de parâmetros laboratoriais e imunológicos de cães naturalmente infectados com Leishmania (Leishmania) chagasi, portadores de diferentes formas clínicas da infecção. Belo Horizonte: Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, Tese de Doutorado, p.180, 2001. REIS, A. B.; MARTINS-FILHO, O. A. et al. Parasite density and impaired biochemical/hematological status are associated with severe clinical aspects of canine visceral leishmaniasis. Res Vet Sci, v.81, n.1, Aug, p.68-75, 2006a. REIS, A. B., TEIXEIRA-CARVALHO, A. et al. Isotype patterns of immunoglobulins: hallmarks for clinical status and tissue parasite density in Brazilian dogs naturally infected by Leishmania (Leishmania) chagasi. Vet Immunol Immunopathol, v.112, n.3-4, Aug 15, p.102-16, 2006b. REIS, A. B.; TEIXEIRA-CARVALHO A. et al. Phenotypic features of circulating leucocytes as immunological markers for clinical status and bone marrow parasite density in dogs naturally infected by Leishmania chagasi. Clin Exp Immunol, v.146, n.2, Nov, p.303-11, 2006c. REIS, A. B.; MARTINS-FILHO O. A. et al. Systemic and compartmentalized immune response in canine visceral leishmaniasis. Vet Immunol Immunopathol, v.128, n.1-3, Mar 15, p.87-95, 2009. REIS, A. B.; GIUNCHETTI, R. C. et al. Immunity to Leishmania and the rational search for vaccines against canine leishmaniasis. Trends Parasitol, v.26, n.7, Jul, p.341-9, 2010. REQUENA, J. M.; SOTO, M. et al. Immune and clinical parameters associated with Leishmania infantum infection in the golden hamster model. Vet Immunol Immunopathol, v.76, n.3-4, Oct 31, p.269-81, 2000. RICA-CAPELA, M. J.; CORTES, S. et al. Immunological and histopathological studies in a rodent model infected with Leishmania infantum promastigotes or amastigotes. Parasitol
Res, v.89, n.3, Feb, p.163-9, 2003. RODRIGUES, V. Jr.; SANTANA DA SILVA, J. et al. Transforming growth factor beta and immunosuppression in experimental visceral leishmaniasis. Infect Immun, v.66, n.3, Mar, p.1233-6, 1998.

Moreira, N. D. Referências Bibliográficas
140
ROLAO, N.; CORTES, S. et al. Leishmania infantum: mixed T-helper-1/T-helper-2 immune response in experimentally infected BALB/c mice. Exp Parasitol, v.115, n.3, Mar, p.270-6, 2007. ROSS, R. Note on the bodies recently described by Leishman and Donovan and further notes no Leishmn’s bodies. Brit Med J, v.2, p.1261-1401, 1903. ROUSSEAU, D.; DEMARTINO, S. et al. Sustained parasite burden in the spleen of Leishmania infantum-infected BALB/c mice is accompanied by expression of MCP-1 transcripts and lack of protection against challenge. Eur Cytokine Netw, v.12, n.2, Apr-Jun, p.340-7, 2001. SANCHEZ, M. A.; DIAZ, N. L. et al. Organ-specific immunity in canine visceral leishmaniasis: analysis of symptomatic and asymptomatic dogs naturally infected with Leishmania chagasi. Am J Trop Med Hyg, v.70, n.6, Jun, p.618-24, 2004. SANT`ANA, J. A. P.; OLIVEIRA W. G. L. et al. Hepatic granulomas in canine visceral leishmaniasis and clinical status. Arq. Bras. Med. Vet. Zootec, v.59, p.1137-1144, 2007. SANTANA, C. C.; VASSALLO, J. et al. Inflammation and structural changes of splenic lymphoid tissue in visceral leishmaniasis: a study on naturally infected dogs. Parasite
Immunol, v.30, n.10, Oct, p.515-24, 2008. SANTOS-GOMES G. M.; ROSA R. et al. Cytokine expression during the outcome of canine experimental infection by Leishmania infantum. Vet Immunol Immunopathol, v.88, n.1-2, Sep 6, p.21-30, 2002. SARTORI, A.; DE OLIVEIRA, A. V. et al. Immune complex glomerulonephritis in experimental kala-azar. Parasite Immunol, v.9, n.1, Jan, p.93-103, 1987. SCHUSTER, F. L. & SULLIVAN, J. J. Cultivation of clinically significant hemoflagellates. Clin Microbiol Rev, v.15, n.3, Jul, p.374-89, 2002. SCOTT, P. Development and regulation of cell-mediated immunity in experimental leishmaniasis. Immunol Res, v.27, n.2-3, p.489-98, 2003. SHAW, S. Leishmaniasis. Tijdschr Diergeneeskd, v.128, n.10, May 15, p.320-1, 2003. SILVA, J. S.; ANDRADE, A. C. et al. Low CXCL13 expression, splenic lymphoid tissue atrophy and germinal center disruption in severe canine visceral leishmaniasis. PLoS One, v.7, n.1, p.e29103, 2012. SILVEIRA, F. T., LAINSON, R. et al. A longitudinal study on the transmission dynamics of human Leishmania (Leishmania) infantum chagasi infection in Amazonian Brazil, with special reference to its prevalence and incidence. Parasitol Res, v.104, n.3, Feb, p.559-67, 2009. SINGH, N.; SAMANT, M. et al. Age-influenced population kinetics and immunological

Moreira, N. D. Referências Bibliográficas
141
responses of Leishmania donovani in hamsters. Parasitol Res, v.101, n.4, Sep, p.919-24, 2007. SMELT, S. C.; COTTERELL, S. E. et al. B cell-deficient mice are highly resistant to Leishmania donovani infection, but develop neutrophil-mediated tissue pathology. J
Immunol, v.164, n.7, Apr, p.3681-8, 2000. SNIDER, H.; LEZAMA-DAVILA, C. et al. Sex hormones and modulation of immunity against leishmaniasis. Neuroimmunomodulation, v.16, n.2, p.106-13, 2009. SOLANO-GALLEGO, L.; KOUTINAS, A. et al. Directions for the diagnosis, clinical staging, treatment and prevention of canine leishmaniosis. Vet Parasitol, v.165, n.1-2, Oct 28, p.1-18, 2009. SOLANO-GALLEGO, L.; MIRO, G. et al. LeishVet guidelines for the practical management of canine leishmaniosis. Parasit Vectors, v.4, p.86, 2011. STANLEY, A. C.; ENGWERDA, C. R. Balancing immunity and pathology in visceral leishmaniasis. Immunol Cell Biol, v.85, n.2, Feb-Mar, p.138-47, 2007. STAUBER, L. A. Leishmaniasis in the hamster. p.76-90, 1955. STAUBER, L. A. Host resistance of the Khartoum strain of Leishmania donovani. Rice
Inst Pamphlet, v.45, p.80-96, 1958. STENGER, S.; THURING, H. et al. Tissue expression of inducible nitric oxide synthase is closely associated with resistance to Leishmania major. J Exp Med, v.180, n.3, Sep, p.783-93, 1994. STRAUSS-AYALI, D.; BANETH G. et al. Splenic immune responses during canine visceral leishmaniasis. Vet Res, v.38, n.4, Jul-Aug, p.547-64, 2007. TAFURI, W. L.; TAFURI, W. L. et al. Histopathology and immunocytochemical study of type 3 and type 4 complement receptors in the liver and spleen of dogs naturally and experimentally infected with Leishmania (Leishmania) chagasi. Rev Inst Med Trop Sao
Paulo, v.38, n.2, Mar-Apr, p.81-9, 1996. TAFURI, W. L., DE OLIVEIRA, M. R. et al. Canine visceral leishmaniosis: a remarkable histopathological picture of one case reported from Brazil. Vet Parasitol, v.96, n.3, Apr 2, p.203-12, 2001. TCHERNOMORETZ, I.; BERGMAN, F. et al. A note on the susceptibility of the hamster Cricetus auratus to Mycobacterium tuberculosis of bovine origin. Refuah Vet, v.2, p.46, 1940. TESH, R. B. Control of zoonotic visceral leishmaniasis: is it time to change strategies? Am
J Trop Med Hyg, v.52, p.287-92, 1995. THALL. Hematologia e Bioquímica Clínica Veterinária. Roca: São Paulo, p.114-117,

Moreira, N. D. Referências Bibliográficas
142
2007. TRAVI, B. L.; MONTOYA J. et al. Bionomics of Lutzomyia evansi (Diptera: Psychodidae) vector of visceral leishmaniasis in northern Columbia. J Med Entomol, v.33, n.3, May, p.278-85, 1996. TRAVI, B. L.; OSORIO, Y. et al. Gender is a major determinant of the clinical evolution and immune response in hamsters infected with Leishmania spp. Infect Immun, v.70, n.5, May, p.2288-96, 2002. TRINCHIERI, G. Interleukin-10 production by effector T cells: Th1 cells show self control. J Exp Med, v.204, n.2, Feb, p.239-43, 2007. VASCONCELLOS, R. C.; URAGO K. P. et al. Suppressor activity in Leishmania
donovani-infected hamster serum: reversion by delipidated bovine serum albumin and role in cell cycle events. Braz J Med Biol Res, v.29, n.5, May, p.615-22, 1996. VERESS, B.; ABDALLA, R. E. et al. Visceral spreading depletion of thymus-dependent regions and amyloidosis in mice and hamsters infected intradermally with Leishmania isolated from Sudanese cutaneous leishmaniasis. Br J Exp Pathol, v.64, n.5, Oct, p.505-14, 1983. VIANNA, V. L.; TAKIYA, C. M. et al. Histopathologic analysis of hamster hepatocytes submitted to experimental infection with Leishmania donovani. Parasitol Res, v.88, n.9, Sep, p.829-36, 2002. WATERHOUSE, G. T. The Hamster: Reprodution and behavior: Proceedings of learned societies. In: Siegel, H.I. New York: Plenum Press, c1985, p.6, 1940. WHO. Working to overcome the global impact of neglected tropical diseases: First WHO reported on neglected tropical diseases. In WHO Geneva, p.184, 2010. WILSON, M. E.; INNES, D. J. et al. Early histopathology of experimental infection with Leishmania donovani in hamsters. J Parasitol, v.73, n.1, Feb, p.55-63, 1987. WILSON, M.; WEINSTOCK, J. V. Hepatic granulomas in murine visceral leishmaniasis caused by Leishmania chagasi. Methods, v.9, n.2, Apr, p.248-54, 1996. WUNDERLICH, F.; BENTEN W. P. et al. Testosterone signaling in T cells and macrophages. Steroids, v.67, n.6, May, p.535-8, 2002. WYLLIE, S.; FAIRLAMB, A. H. Refinement of techniques for the propagation of Leishmania donovani in hamsters. Acta Trop, v.97, n.3, Mar, p.364-9, 2006. XAVIER, S. C.; DE ANDRADE, H. M. et al. Comparison of paraffin-embedded skin biopsies from different anatomical regions as sampling methods for detection of Leishmania infection in dogs using histological, immunohistochemical and PCR methods. BMC Vet Res, v.2, p.17, 2006.

Moreira, N. D. Referências Bibliográficas
143
YOUNG, C. W.; SMILY. H. J.; BROWN, C. Experimental cala-azar a hamster (Cricetus
griseus). Nat. Met. J. China, v.20, p.357-9, 1919. ZANIN, F. H.; COELHO, E. A. et al. Evaluation of immune responses and protection induced by A2 and nucleoside hydrolase (NH) DNA vaccines against Leishmania chagasi and Leishmania amazonensis experimental infections. Microbes Infect, v.9, n.9, Jul, p.1070-7, 2007. ZATELLI, A.; BORGARELLI M. et al. Glomerular lesions in dogs infected with Leishmania organisms. Am J Vet Res, v.64, n.5, May, p.558-61, 2003.

10. ANEXO

Moreira, N. D. Anexo
145