mudanças do uso da terra sobre a comunidade de formigas e a...
TRANSCRIPT

Universidade de Brasília
Instituto de Ciências Biológicas
Programa de Pós-graduação em Ecologia
Mudanças do uso da terra sobre
a comunidade de formigas e a retenção dos
serviços ecossistêmicos no Cerrado
Aluno: Tiago Luiz Massochini Frizzo
Orientador: Edison Ryoiti Sujii
Brasília - 2016

1
Agradecimentos
Agradeço em primeiro lugar a todos educadores que passaram pela minha vida,
conhecimento, compreensão e aprendizado são partes fundamentais para admirar a beleza do
mundo.
Em especial gostaria de agradecer ao orientador desta tese, Edson Ryoitti Sujii, pela
presença, apoio e discussões que levaram ao melhoramento da tese. Ao meu primeiro
orientador durante a graduação, Heraldo L. Vasconcelos, que na prática foi quem me inseriu
na vida acadêmica. Aos meus estagiários, sem os quais não conseguiria ter completado a parte
física do trabalho: Maísa Meneses, Antônio, Karina e Letícia. Aos colegas da Embrapa
Cenargen: Lucas, Érica, Pedro, entre muitos outros, além dos pesquisadores que também
contribuíram muito para o desenvolvimento desta tese. Agradeço também aos meus colegas
do Laboratório de Ecologia em Insetos Sociais da Universidade Federal de Uberlândia,
principalmente a Renata Pacheco, pela ajuda na identificação das espécies de formigas e
conversas a respeito da tese. Aos inúmeros proprietários de terras que permitiram que eu
fizesse amostragens em suas propriedades, incluindo também o Instituto Brasília Ambiental
(IBRAM), Fazenda Água Limpa da UnB, Estação Ecológica do Jardim Botânico de Brasília,
Estação Ecológica de Águas Emendadas, Parque Nacional de Brasília, Embrapa Hortaliças,
Embrapa Cerrados e o Instituto Federal de Brasília. Também agradeço a Capes e CNPq pelo
financiamento do projeto e bolsa de estudo.
Por último, gostaria de ressaltar meus agradecimentos especiais aos meus pais, Luiz
Antônio Frizzo e Leoni Massochini, que me encorajaram e financiaram durante o doutorado,
sem os quais com certeza teria tido uma trajetória muito mais difícil. Além da minha noiva,
Carla Érica Oliveira Ferreira, que me deu e me dá suporte emocional, tornando a vida mais
fácil, diminuindo às angústias e buscando um futuro melhor.

2
Resumo
Muitos estudos têm relatado que vivenciamos um período de extinção em massa,
principalmente devido à expansão dos sistemas agrícolas. Aliado a isso, tem-se demonstrado
que a diminuição na biodiversidade pode causar perda ou redução das funções do
ecossistema, gerando problemas de diversas naturezas. Por isso nós buscamos explorar os
efeitos da conversão de áreas nativas do Cerrado em sistemas agrícolas sobre a fauna de
formigas. Para isso realizamos amostragens e experimentos que contemplaram 74 localidades
diferentes no Distrito Federal. Os locais de coleta foram cultivos convencionais de soja,
pastagens, áreas de agricultura orgânica e áreas com vegetação nativa de campo, cerrado
stricto sensu e mata. Foram feitos 5.298 registros de formigas e encontradas 265 espécies.
Constatamos que as pastagens e áreas de agricultura orgânica abrigam 61% e 56% da
mirmecofauna nativa respectivamente, enquanto que as áreas de cultivo convencional de soja
abrigam apenas 17% das espécies nativas de formigas. Detectamos que as espécies raras (i.e.
com baixa frequência) foram as principais promotoras de diversidade de formigas no Cerrado,
e potencialmente as mais ameaçadas de extinção local, devido à baixa densidade e raridade
dentro dos agrossistemas. Quanto às funções do ecossistema desempenhadas pelas formigas,
utilizamos uma abordagem experimental e encontramos que a remoção de biomassa de
recursos que simulam organismos animais (sardinha) e atrativos vegetais (banana), na
superfície do solo, está correlacionada à riqueza de formigas. Além disso, mostramos que nas
áreas de produção orgânica de hortaliças as formigas predam mais do que o dobro de ovos do
inseto praga Anticarsia gemmatalis (Lepidoptera: Noctuidae) que os outros predadores.
Vimos também que os efeitos das formigas sobre as funções do ecossistema dependem da
função do ecossistema avaliada, do tipo de uso da terra e da riqueza de formigas no local.
Resumidamente encontramos que a expansão da agricultura reduz a diversidade de formigas,
sendo esse efeito particularmente maior na soja. Concluímos que a perda dessa diversidade
pode afetar negativamente funções e serviços do ecossistema e que medidas de conservação
devem focar na proteção de múltiplas fitofisionomias nativas.
Palavras-chave: Hymenoptera, Formicidae, Controle biológico, Intensificação do uso da terra,
perturbações, agroecossistemas.

3
Abstract
Human activities, especially the expansion of agroecosystems, are appointed by
studies as the mainly cause of extinction. The ecological consequences of biodiversity loss
usually are the reduction of ecosystem functions. However, these responses vary according to
the type of land-use change and the ecological setting. This study measured the impact of
land-use type and ecosystem functions on the ant assemblage of Cerrado biome. Ant
composition was assessed in 74 different locations in the Federal District (Brazil). The
collected sites were conventional crops of soybeans, pastures, organic farms and native
vegetation, including grassland, cerrado stricto sensu (savanna) and forests. We record 5,298
ants and 265 species. The pastures and organic farm hold 61% and 56% of the native ant
fauna respectively, while soybean crop have only 17% of the native species of ants. We found
that rare species (i.e. low frequency) were the main promoters of ant diversity in the Cerrado,
and potentially the most threatened by local extinction due to low density and rarity in the
agroecosystems. For measure functions performed by the ants, we use an experimental
approach. We found that removing biomass resource simulating animal organisms (sardines)
and vegetables attractive (banana) on the soil surface, is correlated with ant species richness.
Furthermore, we show that ants prey more than twice folds the eggs of insect pest Anticarsia
gemmatalis (Lepidoptera: Noctuidae) than the other predators in organic farms. Indicating
that the effects on ecosystem functions, provided by ants are dependent to the: ecosystem
function feature, the land-use type and ant richness. Overall, our results found that agricultural
expansion reduces the ant diversity, particularly in soybeans crops, and can affect ecosystem
functions. To mitigate the reduction in the ant assemblage, we recommend the conservation of
multiple natural habitats.
Keywords: Hymenoptera, Formicidae, Biological Control, Land use intensification,
Disturbance, Savanna.

4
Sumário
Lista de Figuras ....................................................................................................................................... 5
Lista de Tabelas ....................................................................................................................................... 6
Introdução Geral ...................................................................................................................................... 7
Capítulo I – Efeitos da mudança no uso da terra sobre a comunidade de formigas do solo ................. 14
Resumo .............................................................................................................................................. 14
Introdução .......................................................................................................................................... 15
Metodologia ...................................................................................................................................... 18
Resultados ......................................................................................................................................... 28
Discussão ........................................................................................................................................... 44
Conclusão .......................................................................................................................................... 51
Capítulo II – Variação nas taxas de remoção de biomassa pela mirmecofauna de solo sobre diferentes
tipos de uso da terra ............................................................................................................................... 53
Resumo .............................................................................................................................................. 53
Introdução .......................................................................................................................................... 54
Metodologia ...................................................................................................................................... 57
Resultados ......................................................................................................................................... 67
Discussão ........................................................................................................................................... 77
Conclusão .......................................................................................................................................... 84
Capítulo III – Contribuição das formigas (Hymenoptera: Formicidae) para a predação de ovos de
lagarta em sistemas orgânicos de produção de hortaliças ..................................................................... 85
Resumo .............................................................................................................................................. 85
Introdução .......................................................................................................................................... 86
Metodologia ...................................................................................................................................... 90
Resultados ......................................................................................................................................... 98
Discussão ......................................................................................................................................... 106
Conclusão ........................................................................................................................................ 111
Considerações finais ............................................................................................................................ 112
Referências Bibliográficas .................................................................................................................. 116
Anexo I. ............................................................................................................................................... 140
Anexo II. .............................................................................................................................................. 143
Anexo III. ............................................................................................................................................ 150
Anexo IV. ............................................................................................................................................ 151

5
Lista de Figuras
Capítulo I
Figura 1.1 Mapa da área de coleta no DF..................................................................... 25
Figura 1.2 Fotografias das áreas de coleta.................................................................... 26
Figura 1.3 Esquema da metodologia de coleta............................................................. 27
Figura 1.4 Curva de acumulo de espécies.................................................................... 32
Figura 1.5 Curva de riqueza estimada.......................................................................... 32
Figura 1.6 Número médio de espécies amostradas....................................................... 33
Figura 1.7 Ocorrência de formigas por tipo de habitat.................................................
Figura 1.8 Distribuição por habitat da frequência de ocorrência..................................
33
34
Figura 1.9 Ocorrência das espécies.............................................................................. 35
Figura 1.10 NMDS.......................................................................................................
Figuira 1.11 Grupos funcionais....................................................................................
36
37
Figura 1.12 Análise da partição aditiva........................................................................ 38
Figura 1.13 Análise da partição multiplicativa............................................................. 39
Figura 1.14 Relação entre a diversidade e distância geográfica................................... 40
Capítulo II
Figura 2.1 Esquema da metodologia............................................................................ 64
Figura 2.2 Fotografias da metodologia......................................................................... 65
Figura 2.3 Proporção de estações de remoção com formigas....................................... 70
Figura 2.4 Biomassa removida..................................................................................... 71
Figura 2.5 Correlação entre biomassa e riqueza de formigas....................................... 72
Figura 2.6 Matriz de correlação.................................................................................... 73
Figura 2.7 Matriz de correlação sem os dados para soja.............................................. 74
Figura 2.8 CCA para todas as espécies......................................................................... 75
Figura 2.9 CCA apenas para as espécies abundantes................................................... 76
Capítulo III
Figura 3.1 Mapa da localização dos experimentos....................................................... 95
Figura 3.2 Fotografia da metodologia.......................................................................... 96
Figura 3.3 Proporção das guildas tróficas..................................................................... 100
Figura 3.4 Proporção dos táxons de predadores........................................................... 101
Figura 3.5 Número de ovos predados........................................................................... 102
Figura 3.6 Regressões e correlação com a abundância................................................. 103

6
Lista de Tabelas
Capítulo I
Tabela 1.1 Localidades amostradas........................................................................ 23-24
Tabela 1.2 Valores indicativos (IndVal)............................................................... 41-43
Capítulo II
Tabela 2.1 Número de estações de biomassa......................................................... 66
Capítulo III
Tabela 3.1 Localização das propriedades............................................................... 97
Tabela 3.2 Porcentagem de ocorrência das paisagens............................................ 97
Tabela 3.3 Lista de espécies................................................................................... 104
Tabela 3.4 Resultados da análise de modelos........................................................ 105

7
Introdução Geral
Atualmente dois dos maiores desafios da ciência são identificar os impactos da atual
taxa de extinção de espécies e garantir a segurança alimentar da população humana. Apesar de
serem dois temas de pesquisa sem aparente conexão, a expansão dos sistemas agrícolas
aliados à fragmentação resultante deste processo é a maior causa de extinção de espécies
conhecida (Pimm et al. 1995; Didham et al. 1996; Sala et al. 2000; Fahrig 2003; Rockströn et
al. 2009; Barnosky et al. 2011; Bregman et al. 2015; Newbold et al. 2015). Ao mesmo tempo,
a extinção também pode ter efeitos negativos sobre a produção agrícola (e.g. Larsen et al.
2005; Brittain & Potts 2011). Isso aponta para a necessidade de se aprimorar ou criar
tecnologias que permitam o aumento da produção agrícola reduzindo a possibilidade de
extinção de espécies nativas, bem como a de gerar dados consistentes para tomada de decisão
visando à sustentabilidade.
Pimm et al. (1995) já alertavam que, para alguns táxons, entre 5 e 20% das espécies já
teriam sido extintas. Mais recentemente, Barnosky et al. (2011) afirmam que, nos próximos
três séculos, podemos atingir o status de extinção em massa com a perda de 75% das espécies
do planeta, sendo que as estimativas mais recentes apontam que conhecemos apenas 13% das
aproximadamente 10.960.000 espécies (eucariotas) existentes no planeta. Portanto estamos
perdendo espécies que ainda sequer foram descritas (Mora et al. 2011).
Hoje, muitos cientistas acreditam que estamos vivenciando o sexto período de
extinção em massa devido à atividade humana principalmente no que concerne à agricultura e
sua expansão (Chapin et al. 2000; Wake & Vredenburg 2008). Em vista disso, é necessário
destacar que essa expansão é e deve continuar sendo um fato, já que há um aumento da
demanda por alimento, devido não só ao crescimento populacional do planeta, mas também
ao aumento no consumo de alimento per capita (FAO 2009; UNPD 2011). Além disso, há

8
outros fatores que contribuem para a expansão da agricultura, tais como o incremento da
produção de biocombustíveis e outros produtos não alimentícios (Groom et al. 2008; Popp et
al. 2013), da degradação de zonas agrícolas existentes, dos imprevisíveis efeitos das
mudanças climáticas sobre a produção de alimento nos próximos anos, das restrições cada vez
maiores sobre o uso de agrotóxicos e o aumento do custo de transporte (Tscharntke et al.
2012). Logo, esse cenário apresenta muitos atributos fortes para agravar as taxas de extinção
de espécies.
Mas quais são as consequências de acelerar essas taxas de extinção de espécies? O
primeiro, e talvez menos considerado pela sociedade, é: qual o direito que nós (humanos)
temos de eliminar uma espécie do planeta? Este é ou pelo menos deveria ser um argumento
poderoso para a preservação de espécies (Ehrlich 2002). O segundo é que os estudos apontam
que a extinção de espécies geralmente causa a perda ou redução de funções do ecossistema,
incluindo serviços ecológicos, no entanto esse não é um padrão universal, o que gera muito
debate (De Fries et al. 2004; Hopper et al. 2005; Larsen et al. 2005; Balvanera et al. 2006;
Loreau et al. 2008; Cardinale et al. 2006; Benayas et al. 2009; Duffy 2009; Allan et al. 2013).
Funções do ecossistema podem ser definidas como a capacidade de processos naturais
e componentes de prover bens e serviços que satisfazem as necessidades humanas, direta ou
indiretamente (De Groot 1992). Primariamente esses processos e bens ocorrem com a
assimilação da energia solar pelos fotossintetizantes e a redistribuição desta energia por toda
cadeia alimentar até chegar aos decompositores. Durante essa passagem de energia há uma
infinidade de processos e trocas físico-químicas (envolvendo também o meio abiótico) que
moldam o nosso mundo da forma como é hoje. São exemplos de funções do ecossistema: a
produção primária, a decomposição, a ciclagem de nutrientes, a filtragem da água, a predação,
refúgio para a vida silvestre, alimento que não é cultivado, recreação entre outros (De Groot
1992; Daily et al. 1997). Logo, quando uma determinada função do ecossistema passa a

9
produzir um benefício capaz de ser valorado para os humanos, passamos a chamá-la de
serviço ou bens do ecossistema (De Groot et al. 2002). Como exemplos destes, podemos
citar: a polinização de culturas, o controle biológico de pragas, a fertilização promovida por
agentes bióticos, a disponibilidade de alimentos nas áreas naturais, a disponibilidade de água
potável entre outros (Daily et al. 1997).
Apesar da extinção de espécies gerar a perda ou redução de serviços ecológicos (e.g.
Larsen et al. 2005; Cadotte et al. 2012; Boyer & Jetz 2014; Birkhofer et al. 2015; Mensens et
al. 2015; Van der Plas et al. 2016), há também estudos que indicam a possibilidade de, por
meio do uso de diferentes tipos de métodos, aumentar a riqueza de espécies incrementando as
funções do ecossistema e serviços ecológicos em ambientes manejados (e.g. Langellotto &
Denno 2004; Tiemann et al. 2015).
No entanto, nem sempre é encontrada uma relação entre o número de espécies e
funções do ecossistema (e.g. Radchuck et al. 2016), pois as extinções podem não ter efeitos
imediatos, porque as espécies mais propensas à extinção já são populações naturalmente
pequenas e de distribuição restrita ou porque outras espécies já desempenham a mesma
função (redundância) no ecossistema (Hooper et al. 2005). Entretanto, parece ser um
consenso que uma alta riqueza de espécies é importante em termos de estabilidade de funções
ou serviços, em caso de distúrbios ou mudanças ambientais (Hooper et al. 2005; Tscharntke et
al. 2005; Loreau et al. 2008).
Na natureza, em contrapartida, as espécies desaparecem de acordo com sua
sensibilidade a estresses ambientais (e.g. Bregman et al. 2015). Mensens et al. (2015), em um
estudo utilizando comunidades de diatomáceas expostas ao herbicida Atrazina, observaram
uma pequena perda de biodiversidade ocasionando uma perda desproporcional das funções no
ecossistema (produção primária, conteúdo energético e estabilização de sedimento). Isso
porque Atrazina afeta principalmente a espécie que apresenta a maior produtividade dentro do

10
sistema, demostrando que a sensibilidade de uma espécie específica deve ser levada em
consideração para se prever efeitos de distúrbios sobre as funções do ecossistema em áreas
sobre estresse antrópico.
Para o Brasil, a expansão da agricultura devido à crescente demanda do mercado de
exportação do agronegócio, que já atingiu 43% do valor total exportado pelo país e 26% do
PIB (MAPA 2016a), acaba se tornando a maior ameaça para o Cerrado (Klink & Machado
2005; Silva et al. 2006; Sano et al. 2010). Essa ameaça, aliada ao alto número de espécies
endêmicas nesse bioma, faz do Cerrado um dos hotspots do mundo, contendo 1,5% das
espécies de plantas vasculares do planeta, o que corresponde a 10.000 espécies, dentre as
quais 4.400 são endêmicas (Myers et al. 2000).
Antes de 1970, o uso da terra no Cerrado era basicamente para a produção de gado
sobre pastagens naturais (Sano et al. 2010). A partir de então, se iniciou uma intensa expansão
da produção agrícola mecanizada para exportação de soja, milho, algodão e café (Jepson
2005; Klink & Machado 2005). Estima-se que, devido à expansão agrícola, entre 40% e 80%
da área natural de Cerrado foi convertida em agrossistemas durante as últimas cinco décadas
(Cavalcanti & Joly 2002; Klink & Moreira 2002; Machado et al. 2004; Brannstrom et al.
2008). No entanto, essas estimativas são muito variáveis devido a dificuldades para se
discriminar nas imagens de satélite o que são áreas de vegetação nativa e áreas de pastagem
(Ferreira et al. 2004).
Segundo Sano et al. (2010), em 2002, pastagens eram o maior tipo de uso da terra do
Cerrado, pois ocupavam uma área de 54.150.000 de ha, representando cerca de 26% da área
desse bioma. Já o segundo maior uso da terra, desse mesmo bioma, é para a agricultura, na
qual a soja ocupa um pouco menos da metade da área (MAPA 2016b), totalizando 21.590.000
de ha (mais de 10% da área do Cerrado). E, apesar de áreas agrícolas de produção orgânica
não serem representativas nas estatísticas, a agricultura orgânica é, seguramente, um dos

11
setores que mais crescem no Brasil e no mundo (Rigby & Cáceres 2001; Blanc 2009;
ABRASCO 2015).
Esses três tipos de agrossistemas se diferenciam pelo tipo de manejo e composição da
vegetação, sendo que as principais características das áreas de plantio de soja são seu manejo
intensivo com agrotóxicos e o plantio de uma monocultura extensiva, que pode ser alternada
principalmente com o plantio de milho, sorgo ou milheto, e que geralmente está restrita às
épocas de chuva. Inversamente, em áreas de agricultura orgânica, geralmente os cultivos
ocupam poucos ha, o uso de agrotóxicos é ausente, há um intenso manejo do solo com capina
e, devido à irrigação constante, há o plantio de uma grande variedade de hortaliças ao longo
de todo ano. Já as pastagens certamente são as áreas com menor manejo ou, na maior parte do
tempo, sem manejo que, apesar de serem originalmente plantadas como uma monocultura de
capim (sendo o Urochloa (Syn. Brachiaria) decumbens, uma espécie exótica e a mais
comum), podem ser colonizadas por algumas ervas, arbustos e/ou árvores nativas em seus
campos. Mesmo sendo a soja a cultura que mais cresce no Cerrado (Fearnside 2001; MAPA
2016a), espera-se que as áreas de agricultura orgânica ocupem cada vez mais espaço na
produção de alimentos (Rigby & Cáceres 2001; Blanc 2009; ABRASCO 2015).
Tendo em vista que as perspectivas para 2050 são de expansão dos agrossistemas
sobre 120 milhões de hectares de áreas de vegetação nativa, nos países em desenvolvimento
(FAO 2009), é imperativo que se conheça os efeitos da conversão de terras em agrossistemas
sobre a biodiversidade e se descubra como reduzir a perda de espécies (Tscharntke et al.
2005; Balmeford et al. 2012). Sendo assim, como apresentado por Tscharntke et al. (2005)
em uma revisão, já sabemos que os efeitos da mudança no uso da terra podem ser percebidos
de formas diferentes, dependendo do grupo taxonômico, da espécie e do tipo de uso da terra
(e.g. Perfecto et al. 2004; Armbrecht et al. 2006; Kessler et al. 2009; Frizzo & Vasconcelos
2013).

12
Dentre os organismos encontrados nos ecossistemas terrestres, as formigas são um
grupo de particular importância ecológica (Folgarait 1998). Na Amazônia a sua biomassa é
maior do que a biomassa somada de todos os vertebrados, incluindo-se pássaros, mamíferos,
anfíbios e répteis (Fittkau & Klinge 1973). Hölldobler e Wilson (2009) chegam a afirmar que
a biomassa de formigas no mundo é tão grande que pode equiparar-se com a dos humanos.
Porém, estes organismos não são apenas abundantes, geralmente eles apresentam hábitos
predatórios e, assim, podem atuar no controle de insetos, incluindo aqui aqueles que são
prejudiciais à agricultura (Risch & Carroll 1982; López & Potter 2000; Philpott & Armbrecht
2006; Armbrecht & Gallego 2007; Howe et al. 2009; Styrsky & Eubanks 2010; Gonthier et
al. 2013; Offenberg 2015). Há também espécies consideradas importantes pragas agrícolas,
como as formigas da tribo Attini, que funcionalmente atuam como herbívoras (Hernandez &
Jaffé 1995; Costa et al. 2008; Zanetti et al. 2014), e várias outras espécies que se associam
mutualisticamente com afídeos (insetos parasitas de plantas) (Delabie 2001; Styrsk &
Eubanks 2007). Por essa importância ecológica, aliada a taxonomia relativamente bem
conhecida e ampla ocorrência, esse grupo foi escolhido como objeto de estudo do presente
trabalho.
Frizzo & Vasconcelos (2013), em um estudo no Cerrado, apontam que a conversão da
vegetação nativa em pastagem não afeta a mirmecofauna (fauna de formigas). Por outro lado,
a conversão dessa vegetação em plantio de soja tem um efeito negativo no número de
espécies, ocasionando ainda mudanças na composição de espécies de formigas (Frizzo &
Vasconcelos 2013; Pacheco et al. 2013). De modo geral, os estudos apontam que mudanças
na estrutura do habitat e o manejo são as principais causas dessas mudanças. Mais
especificamente, a redução de local para nidificação das formigas, arar o solo, uso de
agrotóxicos, além de efeitos indiretos, como a interação interespecífica com formigas
dominantes que limitam a coexistência com outras espécies, têm sido apontados como os

13
principais fatores da redução do número de espécies de formigas nos agrossistemas do
Cerrado (Frizzo & Vasconcelos 2013; Pacheco et al. 2013). Pacheco et al. (2013) também
indicam que as espécies que persistem nos agrossistemas de soja em geral são um
subconjunto das espécies associadas às savanas mais abertas, além de que a maior parte das
formigas que desaparecem são aquelas que apresentam comportamento mais especializado
(sensu Didham et al 1996; Weiner et al. 2014; Bregman et al. 2015). No entanto, ambos os
trabalhos já realizados no Cerrado, de Frizzo & Vasconcelos (2013) e Pacheco et al. (2013),
relatam que o número de espécies perdidas com a mudança no uso da terra podem estar
subestimados devido à coleta próxima de locais potencialmente fontes de espécies de
formigas.
Em vista disso, o objetivo geral desse estudo foi determinar como as mudanças no uso
da terra afetam a comunidade de formigas e suas funções ecológicas. Especificamente
procuramos determinar quantas e quais são as espécies que conseguem persistir em áreas
convertidas em agrossistemas comparativamente às áreas de vegetação nativa e quais os tipos
de agrossistema têm um menor impacto sobre essa fauna. Além disso, queremos determinar
se após a conversão da vegetação nativa em agrossistemas há uma redução nos serviços
ecológicos prestados pela mirmecofauna de remoção de biomassa e predação.

14
Capítulo I – Efeitos da mudança no uso da terra sobre a comunidade de formigas do
solo
Resumo
É um consenso entre os pesquisadores que os sistemas agrícolas reduzem a
biodiversidade. No entanto, a magnitude desse efeito depende das características do sistema
agrícola implantado e do grupo taxonômico sujeito a essa perturbação. Por isso foi analisado
como três diferentes tipos de uso da terra (cultivo de soja, pastagem e produção orgânica de
hortaliças) afetam a comunidade de formigas, amostrando, para isso, 69 localidades, que
incluíram áreas nativas de campo, cerrado stricto sensu e mata. Ao todo, coletamos 264
espécies de formigas, na superfície do solo no Distrito Federal, onde se estima ocorrer em
torno de 342 a 354 espécies. As pastagens e áreas de agricultura orgânica abrigam 61% e 56%
da mirmecofauna nativa, respectivamente, enquanto que as áreas de cultivo convencional de
soja abrigam apenas 17% das espécies nativas de formigas. As áreas de formação florestal
apresentam a comunidade mais singular. Já as áreas de soja apresentam o padrão mais forte de
homogeneização biótica. Detectamos também que são as espécies raras (de baixa frequência)
os principais promotores de riqueza no Cerrado e, potencialmente, as mais ameaçadas de
extinção local, devido a essa baixa densidade e pouca ocorrência nos agrossistemas. Nossos
dados sugerem de modo geral que o cultivo de soja convencional é a principal ameaça à
mirmecofauna no Cerrado e medidas de conservação devem focar na proteção de múltiplas
fitofisionomias nativas.

15
Introdução
Tradicionalmente os esforços de conservação têm sido focados em grupos de plantas e
vertebrados. Myers et al. (2000), por exemplo, usaram apenas plantas vasculares e
vertebrados para determinar os “hotspots” do planeta. No entanto, estes grupos representam
proporcionalmente uma pequena parte da biodiversidade e a mais conhecida em termos de
espécies descritas (Mora et al. 2011). Já os insetos, que representam seguramente a maior
parte da biodiversidade animal, têm sido negligenciados nos estudos de conservação, talvez
devido ao grande número de espécies e/ou à dificuldade para se identificar estas espécies, que
podem, em muitos casos, ainda não terem sido descritas (Mora et al. 2011).
Os insetos são um grupo de grande importância nos ecossistemas, influenciando a
composição e tamanho das populações de plantas, herbívoros, predadores e detritívoros
(Wheeler 1990; Didham et al. 1996). Especificamente no Cerrado, a comunidade de insetos é
influenciada pelo fogo (Frizzo et al. 2011; Maravalhas & Vasconcelos 2014),
heterogeneidade da vegetação (Silva et al. 2006; Pacheco & Vasconcelos 2012), sazonalidade
(Pinheiro et al. 2008), fragmentação (Brandão et al. 2011), introdução de espécies exóticas
(Pivello et al. 1999; Hoffmann et al 2004) e, principalmente nas últimas décadas, pelos
efeitos da mudança no uso da terra (Almeida et al. 2011; Frizzo & Vasconcelos 2013;
Pacheco et al. 2013), entre outros fatores.
Em geral, a conversão de terras em agrossistemas tem sido vista, tradicionalmente,
como antagônica à conservação da biodiversidade (Tscharntke et al. 2005). Em parte, porque
a maioria dos sistemas agrícolas é derivada da conversão de ecossistemas naturais complexos
para os ecossistemas simplificados, com uso intenso de maquinário, insumos químicos e
biológicos, incluindo culturas melhoradas e geneticamente modificadas (e.g. Pimental et al.
1992; Groot & Dicke 2002; Armbrecht et al. 2006; Haddad et al. 2009; Frizzo & Vasconcelos

16
2013). Estes agrossistemas são a maior causa de perda de biodiversidade devido à
fragmentação e destruição da vegetação nativa (Didham et al 1996; Fahrig 2003; Bregman et
al. 2015). No entanto, os efeitos da mudança no uso da terra sobre a fauna nativa dependem
do tamanho da divergência das condições ecológicas pré e pós-conversão do uso da terra
(Kessler et al. 2009). Existindo, assim, sistemas agrícolas que tem um grande impacto sobre a
fauna e flora nativa, e outros tipos de sistemas agrícolas de baixo impacto, que podem ter um
papel fundamental na conservação de espécies (Tscharntke et al. 2005), já que a paisagem
agrícola de baixo impacto pode abrigar grande parte da biodiversidade do mundo (Pimentel et
al. 1992) e a contribuição relativa de cada tipo de agrossistema para a conservação é pouco
conhecida (Tscharntke et al. 2005).
Estudos recentes mostram que, no Cerrado, mudanças no uso da terra para sistemas
simplificados e mais intensamente manejados, como plantações de soja ou milho, reduzem a
riqueza de formigas (Frizzo & Vasconcelos 2013; Pacheco et al. 2013). Todavia,
agrossistemas menos intensivos, como pastagem, podem comportar uma riqueza de espécies
semelhante à de áreas naturais (Frizzo & Vasconcelos 2013), mostrando que áreas de
produção agrícola podem ser utilizadas para a conservação de pelo menos parte da
biodiversidade e/ou manejadas para reduzir a perda de espécies.
Porém o fato de que alguns tipos de agricultura abriguem muitas espécies nativas por
si só não garante a conservação integral da biodiversidade. Introduzida por Whittaker (1960;
1972), a ideia de que o número de espécies de uma região (diversidade gama) é determinado
pelo número de espécies em um local (diversidade alfa), mais a troca de espécies com outros
locais (diversidade beta) é de suma importância para compreender como a diversidade está
distribuída na paisagem e determinar ações conservacionistas.
Mais recentemente, Baselga (2010) criou uma metodologia para decompor a
diversidade beta ou troca de espécies entre locais em dois processos distintos, chamado por

17
ele de substituição de espécies ou perda de espécies. Basicamente isso retrata se a diversidade
beta aumenta devido a uma troca de espécies entre os locais (por espécies diferentes) ou se o
padrão de diversidade beta esta ligado a uma diferença no número de espécies devido ao
aninhamento. Este nada mais é do que um padrão em que a comunidade com menos espécies
é um subconjunto das comunidades com mais espécies (Almeida-Neto et al. 2008; Atmar &
Patterson 1993).
Por exemplo, no estudo de Pacheco e Vasconcelos (2012) no Cerrado, foi observado
um aumento no número de espécies de formigas à medida que a estrutura da vegetação se
tornava mais complexa, partindo de um gradiente de vereda até cerradão. Porém, a
preservação apenas destes habitats complexos (com mais espécies localmente) está longe de
garantir a conservação integral das espécies de formigas, já que 50% da riqueza (diversidade
beta) é explicada pela troca de espécies entre fitofisionomias do Cerrado e 25% por diferentes
áreas de mesma fitofisionomia (Pacheco & Vasconcelos 2012).
Porém, estes estudos são restritos a poucas áreas, e os padrões de diversidade, assim
como a capacidade de manutenção destas espécies em diferentes tipos de ambientes
modificados pelo homem, ainda não foram avaliados. Desse modo, este capítulo busca
determinar como a conversão de áreas de vegetação nativa afeta a riqueza e composição de
espécies de formigas de acordo com o tipo de uso da terra. Nesse caso, foram avaliados
sistemas de plantio de soja, pastagem e agricultura orgânica, levando-se em conta os padrões
de diversidade local, troca de espécies entre locais e diversidade regional.

18
Metodologia
Área de estudo
As coletas foram realizadas de fevereiro a junho de 2013 e de fevereiro a maio de
2014, no Distrito Federal (abrangendo uma área de aproximadamente 5.800 Km², localizada
centralmente a 15°47’S, 47°55’O), em áreas como pastagem, plantio de soja e áreas de
agricultura orgânica, além de áreas de vegetação nativa (Tabela 1.1). Ao todo foram
amostradas 69 localidades, sendo: 14 plantios de soja convencional, 15 pastagens, 16 áreas de
agricultura orgânica (destas, 14 são plantios de hortaliças e dois, AO-15 e AO-16, são plantios
de soja orgânica) e 24 áreas de vegetação nativa. Das 24 áreas de vegetação nativa, optou-se
por coletar nas três fitofisionomias mais comuns, as quais são: sete formações campestres,
oito cerrado stricto sensu e nove formações florestais (Figura 1.1 e 1.2).
Segundo a classificação de Köppen (1948), o clima nessa região é do tipo Aw,
apresentando forte sazonalidade com um verão chuvoso e o inverno seco. As temperaturas
médias mensais podem variar de 22 a 27°C e a precipitação anual média é de 1.500 mm
(Klink & Machado 2005). Todas as localidades estão situadas no bioma Cerrado (para mais
informações sobre a vegetação veja Oliveira-Filho & Ratter 2002).
Protocolo amostral
Em cada uma das localidades foram amostrados três pontos distantes 20 metros entre
si (chamados posteriormente como parcela amostral) (Figura 1.3 - A). Cada parcela amostral
era composta por um grid de 12 armadilhas de queda do tipo pitfall, sendo dispostos em três
fileiras com quatro armadilhas cada, e com espaçamento de dois metros entre si (Figura 1.3 -
B). As armadilhas de queda são copos plásticos de 200 ml, preenchidos até 1/3 com água e

19
detergente e ficaram em atividade no campo por 24 horas (Figura 1.3 - C). Ao todo foram
instalados 12 pitfalls por parcela, 36 por localidade e 2.484 nas 69 localidades.
Após a coleta, o conteúdo de cada pitfall foi peneirado (em uma malha fina que
impossibilita a perda de pequenos invertebrados) e armazenado em um recipiente com álcool
92,8%, até ser finalmente triado. Todos os 12 pitfalls de cada parcela amostral foram reunidos
em uma única amostra composta, para reduzir o tempo de triagem. Portanto, cada localidade
possui ao todo três subamostras, e cada uma destas foi triada, de modo que para Formicidae
foi realizada uma separação até o nível de espécie ou morfoespécie, sendo os gêneros
separados seguindo a chave de Bolton et al. (2007) e Fernández (2003).
Para diminuir a possível influência do clima sobre a amostragem, as coletas foram
realizadas apenas em dias sem a previsão de chuvas. Além disso, de duas a três localidades
eram amostradas ao mesmo tempo (Tabela 1.1), diluindo possíveis efeitos climáticos entre as
diferentes categorias analisadas.
Análise dos dados
A riqueza de espécies de formigas foi comparada entre ambientes de vegetação nativa
e os diferentes sistemas agrícolas visando estimar a perda potencial de espécies devido à
conversão de áreas nativas em sistemas agrícolas. Curvas de acúmulo de espécies e a
estimativa de riqueza utilizando Jack 1 e Chao 2 foram calculadas para saber quanto do total
esperado de espécies de formigas foi amostrado e se a riqueza por tipo de habitat é diferente
entre os sistemas. Para todas essas análises foi utilizado o programa EstimateS 9.1.0 (Colwell
2013). Para saber se mudanças no uso da terra afetou de forma significativa a riqueza de
formigas na localidade (diversidade alfa) foi utilizada uma Anova com teste a posteriore de
Tukey (Gotelli & Ellison 2011), sendo o fator habitat dividido em área de vegetação nativa,
pastagem, soja ou agricultura orgânica.

20
Para todas as análises de composição de espécies entre os habitats, foi utilizada uma
matriz de presença e ausência por localidade (264 espécies x 69 localidades). A primeira
análise a ser utilizada foi uma PERMANOVA, com 999 permutações, usando a função adonis
do pacote vegan (Oksanen et al. 2015) no programa R (R Development Core Team, 2008).
Esta foi utilizada por ser a análise mais robusta para dados não balanceados (Anderson &
Walsh 2013). Porém, como a análise de PERMANOVA não é capaz de definir quais habitats
são diferentes entre si, foi realizada uma ordenação de NMDS (Non-metric multidimensional
scaling) (Anderson 2001), utilizando a função metaMDS do pacote vegan (Oksanen et al.
2015), agrupando os pontos de coleta por suas similaridades. E, por fim, foi utilizada a análise
de Valores Indicativos (IndVal) de Dufrene e Legendre (1997), que mostra quais espécies
estão associadas e são indicativas de um determinado habitat com um valor de significância.
Para esta última análise foi utilizada a função indval do pacote labdsv (Roberts 2016).
Além da diferença na ocorrência de espécies, foi realizada uma análise por grupo
funcional, adaptando a classificação proposta por Frizzo et al. (2012) e Brandão et al. (2012).
As espécies foram classificadas em: (1) Arborícola, espécies que nidificam estritamente na
vegetação arbórea e arbustiva; (2) Attini, tribo que inclui todas as formigas que cultivam
fungo; (3) Camponotini, tribo que inclui todas as espécies do gênero Camponotus; (4)
Correição, grupo que contempla os gêneros Labidus, Neivamyrmex, Eciton e Nomamyrmex;
(5) Especialista, grupo que contempla gêneros e algumas espécies com baixa frequência de
ocorrência, geralmente apresentando aspectos ecológicos e/ou morfológicos incomuns (por
ex: alimentar-se principalmente de sementes, possuir olhos muito grandes ou a ausência dos
mesmos); (6) Generalista, grupo que contempla espécies com grande plasticidade ecológica e
onívoras, sendo composto pela maioria das espécies do gênero Pheidole, Solenopsis,
Brachymyrmex, além de espécies exóticas; (7) Generalista dominante, grupo composto por
três espécies onívoras (Pheidole gertrudae (Forel, 1886), Pheidole sp.01 e Solenopsis

21
saevissima (Smith, F., 1855)), que possuem a característica, devido ao grande recrutamento
de indivíduos, de monopolizar o recurso encontrado frente aos demais competidores; (8)
Oportunista, espécies do gênero Dorymyrmex e Linepthema, que forrageiam nos períodos
mais quentes e que geralemente não monopolizam a fonte de alimento diante de
competidores; (9) Predador grande, grupo que contempla a maioria das Ponerinaes e o gênero
Ectatomma (para mais informações e a lista da classificação das espécies veja o Anexo I).
Após a classificação, foi avaliada qual a frequência de ocorrência de cada um dos
grupos funcionais nos diferentes tipos de habitats. Para isso foi realizado um ánalise de
modelos lineares generalizados, utilizando a função glm do R (R Development Core Team
2008). Nesta análise foi utilizado o número de registros por localidade, ou seja, foi
contabilizado em quantas parcelas cada espécies foi registrada dentro de cada localidade.
Para avaliar se a riqueza de espécies de formigas é mais dependente da riqueza alfa ou
beta, foi empregada uma análise de partição aditiva da diversidade em três níveis
hierárquicos: parcela amostral, localidade e tipo de habitat. A soma da diversidade alfa
(número médio de espécies na parcela amostral) mais cada um dos componentes da
diversidade beta (número de espécies diferentes entre as localidades e tipos de habitats) deve
ser igual à diversidade total de espécies de formigas coletada (ou seja, 264). Com a intenção
avaliar se existe algum fator influenciando a diversidade de formigas, ou seja, alterando os
valores de diversidade alfa e beta de forma significativamente diferente do que seria esperado
ao acaso, foi criado um modelo nulo. Este modelo nulo foi feito atráves de 2.000
aleatorizações dos dados obtidos em campo, e a partir destas aleatorização é gerado um
padrão que definimos como sendo ao acaso. Após esta etapa, comparamos o padrão
encontrado em campo, com o padrão considerado ao acaso, caso a probabilidade do padrão
encontrado em campo seja menor do que 5% de chance de ter ocorrido devido ao acaso,
consideramos este padrão como significativamente diferente, sendo este influenciado por

22
algum fator (Veech et al. 2002). Para esse tipo de análise foi utilizada a função adipart, do
pacote vegan (Oksanen et al. 2015).
Adicionalmente, diversidade beta (β) foi decomposta em dois componentes que são
processos ecológicos distintos: aninhamento (βnes) e troca de espécies (βsim) (Baselga et al.
2010). Simplificadamente, a diversidade β foi calculada par a par entre as localidades com
índice de Sorensen (βsor) e Simpson (βsim). βsor é calculado levando-se em consideração a
identidade das espécies e absorve a informação tanto da troca (turnover) como do
aninhamento das espécies. βsim é calculado a partir da probabilidade de se capturar
determinada espécie em função do total de espécies (independente da identidade delas),
calculando apenas a troca de espécies. Por isso, a diferença entre os índices calculados
fornece o valor de diversidade beta devido ao aninhamento: βnes = βsor - βsim (Baselga et al.
2010).
Para essa análise foi utilizada a função beta.multi do pacote betapart (Baselga et al.
2015). Visando detectar diferenças entre agrossistemas e a vegetação nativa, foram feitas
comparações entre os valores de βsor, depois entre os de βsim e, por fim, entre os de βnes,
através de um teste-t. A mesma comparação foi realizada entre os habitats (soja, pastagem,
agricultura orgânica, campo, cerrado e mata), mas utilizando o teste não paramétrico de
Kruskall-Wallis com um teste a posteriori usando a função posthoc.kruskal.nemenyi.test do
pacote PMCMR (Pohlert 2016). Nesse teste de Kruskall-Wallis, os valores iguais dentro do
banco de dados foram ranqueados aleatoriamente para evitar nós (ties) na análise. Em
seguida, por meio da análise de Mantel (com o pacote vegan), entre a matriz de
dissimilaridade gerada pela função beta.multi e uma matriz de distância entre as localidades,
foi avaliada se a distância entre os pontos afeta a troca de espécies. Nos casos em que o
resultado foi significativo, foi realizado uma correlação de Pearson para determinar a direção
e a força da correlação.

23
Tabela 1.1. Localidades amostradas no Distrito Federal com suas respectivas coordenadas, data de coleta e
altitude em metros. Os termos entre parênteses referem-se à fitofisionomia nas áreas de vegetação nativa, sendo
que “Campo” é referente a formações campestres, “Cerrado” a áreas de cerrado stricto sensu e “Mata” referente
a formações florestais.
Sistema Localidade Latitude Longitude Data Altitude
Agricultura orgânica AO-01 S 15° 58' 10.6'' W 047° 29' 41.8'' 03/04/2013 890
Agricultura orgânica AO-02 S 15° 45' 44.2'' W 047° 38' 30.5'' 03/04/2013 948
Agricultura orgânica AO-03 S 15° 33' 55.6'' W 048° 01' 50.8'' 29/04/2013 1250
Agricultura orgânica AO-04 S 15° 49' 30.1'' W 048° 15' 07.2'' 07/05/2013 1.086
Agricultura orgânica AO-05 S 15° 56' 32.9'' W 048° 08' 24.9'' 07/05/2013 989
Agricultura orgânica AO-06 S 15° 49' 42.7'' W 048° 04' 16.4'' 11/05/2013 1.153
Agricultura orgânica AO-07 S 15° 38' 40.0'' W 048° 09' 58.5'' 11/05/2013 1.134
Agricultura orgânica AO-08 S 15° 56' 06.8'' W 047° 56' 14.8'' 14/05/2013 1.045
Agricultura orgânica AO-09 S 16° 00' 49.7'' W 047° 44' 45.8'' 23/05/2013 1.018
Agricultura orgânica AO-10 S 15° 58' 14.7'' W 047° 53' 27.9'' 23/05/2013 1.125
Agricultura orgânica AO-11 S 15°39'07.1" W 048°06'30.0" 07/04/2014 1.209
Agricultura orgânica AO-12 S 15°55'36.5" W 047°46'02.5" 12/05/2014 923
Agricultura orgânica AO-13 S 15°39'02.0" W 048°12'03.3" 15/05/2014 1.105
Agricultura orgânica AO-14 S 15°36'44.5" W 048°04'42.7" 19/05/2014 1.257
Agricultura orgânica AO-15 S 15° 57' 55.3'' W 047° 30' 08.0'' 14/03/2013 895
Agricultura orgânica AO-16 S 15° 40' 03.8'' W 048° 10' 00.5'' 19/03/2013 1.219
Pastagem P-01 S 15° 42' 00.9'' W 047° 55' 02.9'' 11/03/2013 1.053
Pastagem P-02 S 15° 58' 27.9'' W 047° 29' 50.1'' 03/04/2013 879
Pastagem P-03 S 15° 35' 51.5'' W 047° 43' 20.2'' 17/04/2013 1.002
Pastagem P-04 S 15° 35' 59.3'' W 047° 36' 32.6'' 17/04/2013 1046
Pastagem P-05 S 15° 33' 53.7'' W 047° 31' 16.8'' 17/04/2013 1.102
Pastagem P-06 S 15° 47' 45.6'' W 047° 38' 59.5'' 22/04/2013 1031
Pastagem P-07 S 15° 52' 58.4'' W 047° 34' 55.1'' 22/04/2013 1.001
Pastagem P-08 S 15° 56' 28.0'' W 047° 54' 52.8'' 14/05/2013 1.010
Pastagem P-09 S 15° 57' 05.1'' W 048° 08' 17.1'' 20/05/2013 987
Pastagem P-10 S 15° 49' 37.9'' W 048° 14' 56.6'' 20/05/2013 1.104
Pastagem P-11 S 15° 39' 06.4" W 048 °06' 09.1" 07/04/2014 1.196
Pastagem P-12 S 15° 55' 17.3" W 048° 02' 04.4" 23/04/2014 1.213
Pastagem P-13 S 15° 39' 16.1" W 047° 42' 10.6'' 01/05/2014 967
Pastagem P-14 S 15° 55' 42.0" W 047° 44' 50.1" 12/05/2014 959
Pastagem P-15 S 15° 36' 32.0" W 048° 09' 48.5" 15/05/2014 1.254
Plantação de soja S-01 S 15° 34' 26.0'' W 047° 32' 48.6'' 07/02/2013 1.159
Plantação de soja S-02 S 15° 40' 52.0'' W 047° 20' 46.4'' 10/02/2013 928
Plantação de soja S-03 S 15° 42' 25.8'' W 047° 27' 38.6'' 10/02/2013 998
Plantação de soja S-04 S 15° 43' 06.9'' W 047° 35' 24.8'' 10/02/2013 1.175
Plantação de soja S-05 S 15° 58' 32.8'' W 047° 34' 33.2'' 14/02/2013 1.041
Plantação de soja S-06 S 15° 58' 10.6'' W 047° 51' 39.3'' 22/02/2013 1.098
Plantação de soja S-07 S 15° 47' 18.5'' W 047° 38' 21.6'' 14/02/2013 1.002
Plantação de soja S-08 S 15° 46' 37.8'' W 047° 23' 17.8'' 26/02/2013 946
Plantação de soja S-09 S 15° 55' 13.2'' W 047° 22' 57.9'' 01/03/2013 879
Continua...

24
Sistema Localidade Latitude Longitude Data Altitude
Plantação de soja S-10 S 16° 00' 37.6'' W 047° 27' 36.3'' 01/03/2013 984
Plantação de soja S-11 S 15° 46' 40.2" W 047° 33' 55.9" 03/02/2014 946
Plantação de soja S-12 S 15° 47' 58.1" W 047° 32' 54.5" 03/02/2014 988
Plantação de soja S-13 S 15° 36' 28.3" W 047° 44' 38.7" 14/04/2014 1.121
Plantação de soja S-14 S 15° 54' 48.1" W 048° 02' 20.7" 23/04/2014 1.226
Vegetação nativa (Campo) VN-01 S 15° 58' 35.0'' W 047° 56' 55.9'' 02/05/2013 1.174
Vegetação nativa (Mata) VN-02 S 15° 55' 53.7'' W 047° 54' 09.9'' 02/05/2013 1.062
Vegetação nativa (Cerrado) VN-03 S 15° 57' 55.9'' W 047° 55' 41.7'' 02/05/2013 1.153
Vegetação nativa (Cerrado) VN-04 S 15° 43' 49.4'' W 047° 57' 41.6'' 29/05/2013 1.108
Vegetação nativa (Mata) VN-05 S 15° 43' 59.6'' W 048° 00' 53.8'' 29/05/2013 1.101
Vegetação nativa (Campo) VN-06 S 15° 39' 37.2'' W 047° 58' 33.9'' 05/06/2013 1.112
Vegetação nativa (Cerrado) VN-07 S 15° 38' 24.1'' W 048° 01' 32.5'' 05/06/2013 1.238
Vegetação nativa (Campo) VN-08 S 15° 32' 15.4'' W 047° 36' 45.6'' 10/06/2013 1.038
Vegetação nativa (Cerrado) VN-09 S 15° 35' 01.1'' W 047° 36' 51.2'' 10/06/2013 1.041
Vegetação nativa (Mata) VN-10 S 15° 34' 40.3'' W 047° 41' 55.7'' 10/06/2013 982
Vegetação nativa (Campo) VN-11 S 15° 56' 46.1'' W 048° 09' 11.4'' 13/06/2013 1.070
Vegetação nativa (Mata) VN-12 S 15° 56' 35.1'' W 048° 08' 25.6'' 13/06/2013 984
Vegetação nativa (Mata) VN-13 S 15° 53' 15.2" W 047° 50' 28.7" 12/02/2014 1088
Vegetação nativa (Campo) VN-14 S 15° 52' 37.3" W 047° 49' 31.9" 12/02/2014 1104
Vegetação nativa (Cerrado) VN-15 S 15° 53' 11.1" W 047° 49' 37.4" 12/02/2014 1125
Vegetação nativa (Cerrado) VN-16 S 15° 55' 40.5" W 048° 01' 38.1" 17/03/2014 1185
Vegetação nativa (Mata) VN-17 S 15° 54' 24.1" W 048° 00' 40.3" 17/03/2014 1036
Vegetação nativa (Campo) VN-18 S 15° 36' 18.1" W 047° 44' 58.5" 14/04/2014 1153
Vegetação nativa (Cerrado) VN-19 S 15° 36' 38.4" W 047° 43' 43.4" 17/04/2014 1065
Vegetação nativa (Mata) VN-20 S 15° 36' 28.6" W 047° 42' 49.4" 17/04/2014 1000
Vegetação nativa (Campo) VN-21 S 15° 39' 11.0" W 047° 42' 48.2" 28/04/2014 962
Vegetação nativa (Cerrado) VN-22 S 15° 39' 43.8" W 047° 42' 22.4" 28/04/2014 955
Vegetação nativa (Mata) VN-23 S 15° 39' 33.0" W 047° 41' 18.1" 01/05/2014 929
Vegetação nativa (Mata) VN-24 S 15° 33' 49.7'' W 047° 32' 25.4'' 07/02/2013 1.149
25
Figura 1.1. Mapa das 69 localidades amostradas dentro do Distrito Federal no ano de 2013 e 2014.

26
Figura 1.2. Fotografia das áreas de coleta no Distrito Federal no ano de 2013 e 2014. A) Área de agricultura
orgânica. B) Área de pastagem. C) Área de plantio de soja. Fotografias das áreas de vegetação nativa: D) área de
formação campestre, E) área de cerrado stricto sensu e F) área de formação florestal.

27
Figura 1.3. Metodologia empregada para a coleta de formigas. A) Distribuição das parcelas amostrais em cada
localidade. B) Distribuição dos pitfalls em cada parcela amostral. C) Fotografia de um pitfall no campo.

28
Resultados
Foram coletadas ao todo 264 espécies, o que representa de 74% a 78% das espécies
estimadas na superfície do solo para a região do Distrito Federal, utilizando os estimadores de
diversidade Jack 1 (342 espécies) e Chao 2 (354 espécies). Registrou-se 3.965 ocorrências de
formigas, sendo estas distribuídas em 58 gêneros e oito subfamílias (Anexo II). Myrmicinae
foi a subfamília que apresentou maior riqueza, com 151 espécies de formigas, e o gênero
Pheidole o mais diverso, com 41 espécies e mais três complexos de espécies (ocorrendo em
98% das localidades). A subfamília Formicinae foi a segunda mais diversa, com 48 espécies,
e que apresenta o segundo gênero mais diverso, Camponotus, com 29 espécies. Os gêneros
Solenopsis, Crematogaster, Brachymyrmex e Trachymyrmex também se destacam por ter
entre 17 e 10 espécies, todos pertencentes à subfamília Myrmicinae, com exceção de
Brachymyrmex, da subfamília Formicinae.
Comparando a diversidade rarefeita de formigas, entre a vegetação nativa e sistemas
de produção agropecuários, nota-se que há uma queda na riqueza de formigas de 39% para
pastagens, 44% para áreas de agricultura orgânica e 83% em áreas de monocultura de soja
(Figura 1.4). Esses valores são semelhantes quando utilizado o estimador de riqueza Jack 1,
de 40% para pastagem, 47% para áreas de agricultura orgânica e 82% para o cultivo de soja
(Figura 1.5).
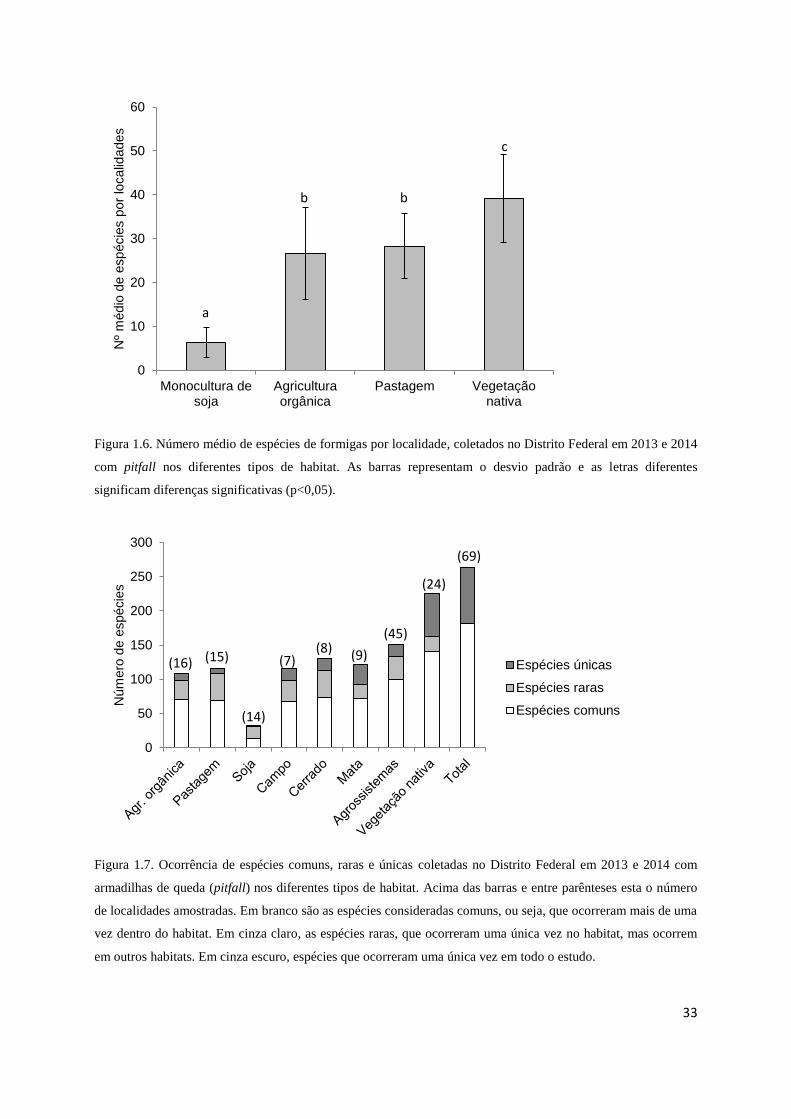
A análise da riqueza local, que corresponde ao número médio de espécies coletadas
em cada uma das 69 localidades, também revela que há uma diferença entre as áreas de
vegetação nativa e os sistemas agrícolas. As áreas de soja são significativamente mais pobres
em espécies do que os sistemas orgânicos e pastagens (F3,65=66,96 p<0,001), que apresentam
o mesmo número de espécies entre si (Figura 1.6). Já as áreas de vegetação nativa são mais
ricas em espécies do que todas as áreas manejadas.

29
Por fim, detectamos que tanto a riqueza como a composição de espécies nas áreas de
vegetação nativa são influenciadas pela ocorrência de espécies únicas e raras (Figura 1.7, 1.8
e 1.9). Consideramos como espécies raras aquelas que ocorreram apenas uma vez em um
determinado tipo de sistema agrícola ou fitofisionomia nativa, e como espécies únicas aquelas
que ocorreram apenas uma vez em todo o estudo. As espécies raras e únicas corresponderam a
aproximadamente 30% das espécies que ocorrem tanto em agrossistemas como em áreas de
vegetação nativa (Figura 1.7). Porém as áreas de vegetação nativa apresentam
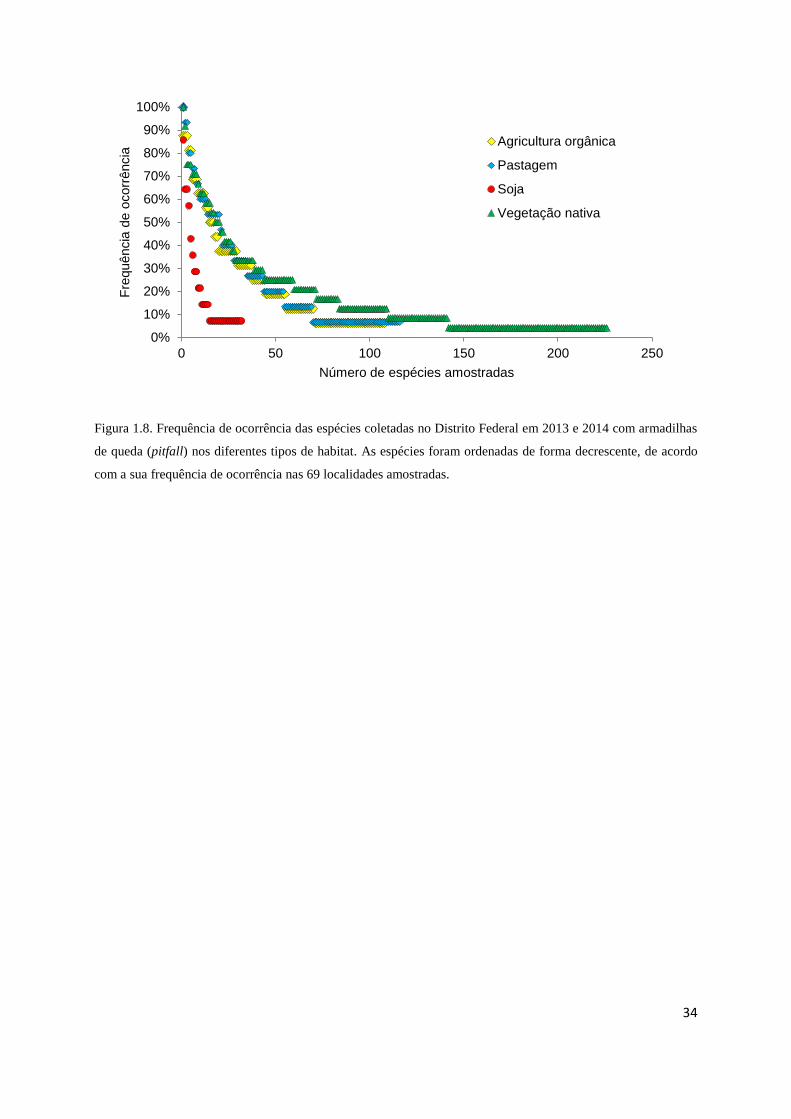
proporcionalmente muito mais espécies raras e únicas que os sistemas agrícolas (Figura 1.8).
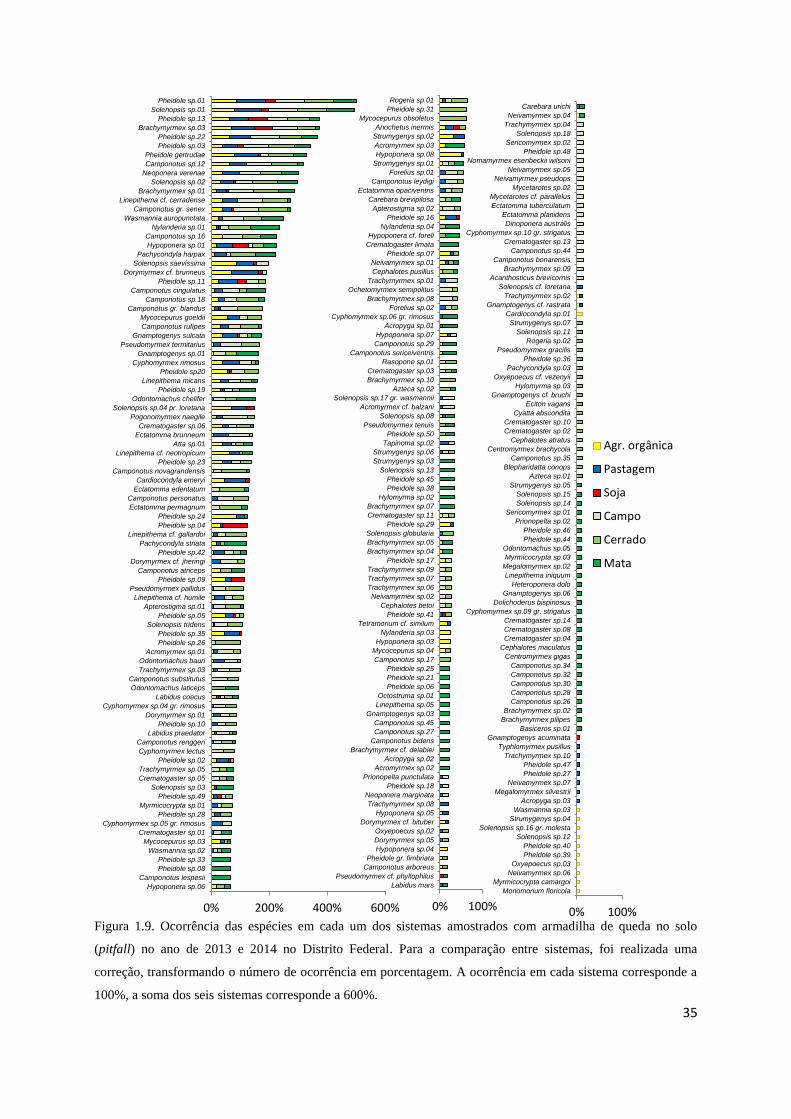
É interessante notar também que, foram amostradas espécies com alta frequência de
ocorrência, capazes de sobreviver tanto em áreas de vegetação nativa, como nos
agrossistemas, mas há medida que as espécies diminuem em frequência elas demonstram uma
preferencia ou exclusividade por determinado tipo de habitat (figura 1.9).
A análise PERMANOVA indica que há uma diferença significativa na composição de
espécies (F3,68=10,697 p<0,001). A ordenação (Figura 1.10) mostra que, além da riqueza, há
também diferenças entre a composição de espécies dos sistemas de vegetação nativa e a dos
sistemas agropecuários. Fica, então, visível um gradiente de mudança, na qual monocultura de
soja e formações florestais são comunidades singulares (diferentes de todas aos outras). Além
destes, mais dois grupos se formam, cerrado e campo, duas fitofisionomias naturais muito
mais similares entre si do que entre as áreas de mata. Pastagem e áreas de agricultura orgânica
também possuem uma comunidade relativamente similar.
Uma segunda evidência da alteração no padrão de distribuição e abundância das
espécies está nos resultados obtidos na análise com o Indval, que detectou 91 espécies com
valores de indicação significativos, variando de 1% até 75% (Tabela 1.2). Por exemplo:
Pheidole sp.04 tem 59% de chance de ocorrer em uma área de monocultura de soja, Pheidole
sp.35 tem 61% de chance de ocorrer em áreas de agricultura orgânica e Dorymyrmex cf.

30
brunneus (Forel, 1908) tem 46% de chance de ocorrer em pastagens. É importante notar
também que o número de espécies indicadoras é quase três vezes e meia maior nas
fitofisionomias nativas comparativamente às áreas agrícolas.
Fica evidente também que os grupos funcionais são afetados pela conversão de áreas
nativas em sistemas agrícolas ou pelo tipo de uso da terra (Figura 1.11). Dos diversos grupos
funcionais, Correição foi o único grupo que não mostrou diferença significativa de ocorrência
entre os diferentes tipos de habitats, porém este é um grupo nômade e relativamente raro. O
grupo das Generalistas Dominantes foi o único grupo que aumentou em número de registros
nas áreas de agricultura orgânica e pastagem, comparativamente às áreas de vegetação nativa.
Para todos os demais grupos, as áreas de vegetação nativa possuem significativamente mais
registros que os sistemas agrícolas, com exceção das Generalistas e Oportunistas.
Realizando a análise de partição aditiva de diversidade (Figura 1.12) com todos os
sistemas e fitofisionomias hierarquizados, observou-se que a riqueza alfa (α1) é maior que o
esperado (5,8%). Já a troca de espécies entre as parcelas de cada localidade é 29% menor do
que o esperado, assim como a troca de espécies entre localidades dentro do mesmo sistema ou
fitofisionomia (14% menor que o esperado). A troca de espécies entre os diferentes sistemas e
fitofisionomias (β3) passa, então, a ter a maior importância para o padrão de diversidade
encontrado, sendo 15,9% maior que o esperado ao acaso.
Análises de partição aditiva da diversidade também foram feitas em separado para
cada sistema ou fitofisionomia (Anexo III). Entretanto, independentemente do sistema ou
fitofisionomia, os padrões de partição aditiva da diversidade são semelhantes entre si. Todos
foram diferentes do esperado ao acaso, de modo que, quando se analisa cada sistema ou
fitofisionomia em separado, a diversidade alfa (riqueza dentro de cada parcela) é em torno de
2,3% mais alta que o esperado. Já a diversidade beta1, ou troca de espécie entre as parcelas de

31
pitfalls dentro de cada localidade, é 4,1% menor que o esperado, mostrando homogeneidade
nesta escala. Enquanto a troca de espécies entre localidades é 4,1% maior que o esperado.
Por outro lado, a diversidade beta (βsor) pode estar agrupando dois processos
distintos: a troca de espécies (turnover) e/ou o aninhamento (perda de espécies). A análise de
diversidade multiplicativa separou estes dois fatores e demostra que a maior parte da
diversidade é devida à troca de espécies (βsim), tendo pouca influência do aninhamento
(βnes) (Figura 1.13). Porém, o aninhamento para os agrossistemas é o triplo do encontrado em
áreas nativas, o que demonstra maior homogeneização nas áreas agrícolas. Além disso, a
análise de Mantel entre o nível de dissimilaridade e a distância entre os pontos, demonstra que
apenas para os agrossistemas quanto maior a distância maior a troca de espécies (Figura 1.14).

32
Figura 1.4. Número acumulado de espécies de formigas por localidades amostradas com o respectivo desvio
padrão baseadas em coletadas realizadas em 2013 e 2014 no Distrito Federal com pitfall.
Figura 1.5. Número estimado de espécies de formigas por número de localidades amostradas utilizando o
estimador Jack 1 com o respectivo desvio padrão baseado em coletas realizadas em 2013 e 2014 no Distrito
Federal com pitfall.
0
50
100
150
200
250
0 5 10 15 20 25
Núm
ero
de e
spécie
s o
bserv
ado
Número de localidades amostradas
Vegetação nativa
Monocultura de soja
Agricultura orgânica
Pastagem
0
50
100
150
200
250
300
350
0 5 10 15 20 25
Núm
ero
de e
spécie
s e
stim
ado
Número de localidades amostradas
Vegetação nativa
Monocultura de soja
Agricultura orgânica
Pastagem
33
Figura 1.6. Número médio de espécies de formigas por localidade, coletados no Distrito Federal em 2013 e 2014
com pitfall nos diferentes tipos de habitat. As barras representam o desvio padrão e as letras diferentes
significam diferenças significativas (p<0,05).
Figura 1.7. Ocorrência de espécies comuns, raras e únicas coletadas no Distrito Federal em 2013 e 2014 com
armadilhas de queda (pitfall) nos diferentes tipos de habitat. Acima das barras e entre parênteses esta o número
de localidades amostradas. Em branco são as espécies consideradas comuns, ou seja, que ocorreram mais de uma
vez dentro do habitat. Em cinza claro, as espécies raras, que ocorreram uma única vez no habitat, mas ocorrem
em outros habitats. Em cinza escuro, espécies que ocorreram uma única vez em todo o estudo.
0
10
20
30
40
50
60
Monocultura desoja
Agriculturaorgânica
Pastagem Vegetaçãonativa
Nº
médio
de e
spécie
s p
or
localid
ades
0
50
100
150
200
250
300
Núm
ero
de e
spécie
s
Espécies únicas
Espécies raras
Espécies comuns
(16) (15)
(14)
(7) (8) (9)
(45)
(24)
(69)
a
b b
c
c
c
34
Figura 1.8. Frequência de ocorrência das espécies coletadas no Distrito Federal em 2013 e 2014 com armadilhas
de queda (pitfall) nos diferentes tipos de habitat. As espécies foram ordenadas de forma decrescente, de acordo
com a sua frequência de ocorrência nas 69 localidades amostradas.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 50 100 150 200 250
Fre
quência
de o
corr
ência
Número de espécies amostradas
Agricultura orgânica
Pastagem
Soja
Vegetação nativa
35
Figura 1.9. Ocorrência das espécies em cada um dos sistemas amostrados com armadilha de queda no solo
(pitfall) no ano de 2013 e 2014 no Distrito Federal. Para a comparação entre sistemas, foi realizada uma
correção, transformando o número de ocorrência em porcentagem. A ocorrência em cada sistema corresponde a
100%, a soma dos seis sistemas corresponde a 600%.
0% 100%
Monomorium floricola
Myrmicocrypta camargoi
Neivamyrmex sp.06
Oxyepoecus sp.03
Pheidole sp.39
Pheidole sp.40
Solenopsis sp.12
Solenopsis sp.16 gr. molesta
Strumygenys sp.04
Wasmannia sp.03
Acropyga sp.03
Megalomyrmex silvestrii
Neivamyrmex sp.07
Pheidole sp.27
Pheidole sp.47
Trachymyrmex sp.10
Typhlomyrmex pusillus
Gnamptogenys acuminata
Basiceros sp.01
Brachymyrmex pilipes
Brachymyrmex sp.02
Camponotus sp.26
Camponotus sp.28
Camponotus sp.30
Camponotus sp.32
Camponotus sp.34
Centromyrmex gigas
Cephalotes maculatus
Crematogaster sp.04
Crematogaster sp.08
Crematogaster sp.14
Cyphomyrmex sp.09 gr. strigatus
Dolichoderus bispinosus
Gnamptogenys sp.06
Heteroponera dolo
Linepithema iniquum
Megalomyrmex sp.02
Myrmicocrypta sp.03
Odontomachus sp.05
Pheidole sp.44
Pheidole sp.46
Prionopelta sp.02
Sericomyrmex sp.01
Solenopsis sp.14
Solenopsis sp.15
Strumygenys sp.05
Azteca sp.01
Blepharidatta conops
Camponotus sp.35
Centromyrmex brachycola
Cephalotes atratus
Crematogaster sp.02
Crematogaster sp.10
Cyatta abscondita
Eciton vagans
Gnamptogenys cf. bruchi
Hylomyrma sp.03
Oxyepoecus cf. vezenyii
Pachycondyla sp.03
Pheidole sp.36
Pseudomyrmex gracilis
Rogeria sp.02
Solenopsis sp.11
Strumygenys sp.07
Cardiocondyla sp.01
Gnamptogenys cf. rastrata
Trachymyrmex sp.02
Solenopsis cf. loretana
Acanthosticus brevicornis
Brachymyrmex sp.09
Camponotus bonarensis
Camponotus sp.44
Crematogaster sp.13
Cyphomyrmex sp.10 gr. strigatus
Dinoponera australis
Ectatomma planidens
Ectatomma tuberculatum
Mycetarotes cf. parallelus
Mycetarotes sp.02
Neivamyrmex pseudops
Neivamyrmex sp.05
Nomamyrmex esenbeckii wilsoni
Pheidole sp.48
Sericomyrmex sp.02
Solenopsis sp.18
Trachymyrmex sp.04
Neivamyrmex sp.04
Carebara urichi
Agr. orgânica
Pastagem
Soja
Campo
Cerrado
Mata
0% 100%
Labidus mars
Pseudomyrmex cf. phyllophilus
Camponotus arboreus
Pheidole gr. fimbriata
Hypoponera sp.04
Dorymyrmex sp.05
Oxyepoecus sp.02
Dorymyrmex cf. bituber
Hypoponera sp.05
Trachymyrmex sp.08
Neoponera marginata
Pheidole sp.18
Prionopelta punctulata
Acromyrmex sp.02
Acropyga sp.02
Brachymyrmex cf. delabiei
Camponotus bidens
Camponotus sp.27
Camponotus sp.45
Gnamptogenys sp.03
Linepithema sp.05
Octostruma sp.01
Pheidole sp.06
Pheidole sp.21
Pheidole sp.25
Camponotus sp.17
Mycocepurus sp.04
Hypoponera sp.03
Nylanderia sp.03
Tetramorium cf. similum
Pheidole sp.41
Cephalotes betoi
Neivamyrmex sp.02
Trachymyrmex sp.06
Trachymyrmex sp.07
Trachymyrmex sp.09
Pheidole sp.17
Brachymyrmex sp.04
Brachymyrmex sp.05
Solenopsis globularia
Pheidole sp.29
Crematogaster sp.11
Brachymyrmex sp.07
Hylomyrma sp.02
Pheidole sp.38
Pheidole sp.45
Solenopsis sp.13
Strumygenys sp.03
Strumygenys sp.06
Tapinoma sp.02
Pheidole sp.50
Pseudomyrmex tenuis
Solenopsis sp.08
Acromyrmex cf. balzani
Solenopsis sp.17 gr. wasmannii
Azteca sp.02
Brachymyrmex sp.10
Crematogaster sp.03
Rasopone sp.01
Camponotus sericeiventris
Camponotus sp.29
Hypoponera sp.07
Acropyga sp.01
Cyphomyrmex sp.06 gr. rimosus
Forelius sp.02
Brachymyrmex sp.08
Ochetomyrmex semipolitus
Trachymyrmex sp.01
Cephalotes pusillus
Neivamyrmex sp.01
Pheidole sp.07
Crematogaster limata
Hypoponera cf. foreli
Nylanderia sp.04
Pheidole sp.16
Apterostigma sp.02
Carebara brevipilosa
Ectatomma opaciventris
Camponotus leydigi
Forelius sp.01
Strumygenys sp.01
Hypoponera sp.08
Acromyrmex sp.03
Strumygenys sp.02
Anochetus inermis
Mycocepurus obsoletus
Pheidole sp.31
Rogeria sp.01
0% 200% 400% 600%
Hypoponera sp.06
Camponotus lespesii
Pheidole sp.08
Pheidole sp.33
Wasmannia sp.02
Mycocepurus sp.03
Crematogaster sp.01
Cyphomyrmex sp.05 gr. rimosus
Pheidole sp.28
Myrmicocrypta sp.01
Pheidole sp.49
Solenopsis sp.03
Crematogaster sp.05
Trachymyrmex sp.05
Pheidole sp.02
Cyphomyrmex lectus
Camponotus renggeri
Labidus praedator
Pheidole sp.10
Dorymyrmex sp.01
Cyphomyrmex sp.04 gr. rimosus
Labidus coecus
Odontomachus laticeps
Camponotus substitutus
Trachymyrmex sp.03
Odontomachus bauri
Acromyrmex sp.01
Pheidole sp.26
Pheidole sp.35
Solenopsis tridens
Pheidole sp.05
Apterostigma sp.01
Linepithema cf. humile
Pseudomyrmex pallidus
Pheidole sp.09
Camponotus atriceps
Dorymyrmex cf. jheringi
Pheidole sp.42
Pachycondyla striata
Linepithema cf. gallardoi
Pheidole sp.04
Pheidole sp.24
Ectatomma permagnum
Camponotus personatus
Ectatomma edentatum
Cardiocondyla emeryi
Camponotus novagrandensis
Pheidole sp.23
Linepithema cf. neotropicum
Atta sp.01
Ectatomma brunneum
Crematogaster sp.06
Pogonomyrmex naegile
Solenopsis sp.04 pr. loretana
Odontomachus chelifer
Pheidole sp.19
Linepithema micans
Pheidole sp20
Cyphomyrmex rimosus
Gnamptogenys sp.01
Pseudomyrmex termitarius
Gnamptogenys sulcata
Camponotus rufipes
Mycocepurus goeldii
Camponotus gr. blandus
Camponotus sp.18
Camponotus cingulatus
Pheidole sp.11
Dorymyrmex cf. brunneus
Solenopsis saevissima
Pachycondyla harpax
Hypoponera sp.01
Camponotus sp.16
Nylanderia sp.01
Wasmannia auropunctata
Camponotus gr. senex
Linepithema cf. cerradense
Brachymyrmex sp.01
Solenopsis sp.02
Neoponera verenae
Camponotus sp.12
Pheidole gertrudae
Pheidole sp.03
Pheidole sp.22
Brachymyrmex sp.03
Pheidole sp.13
Solenopsis sp.01
Pheidole sp.01

36
Figura 1.10. Escalonamento multidimensional não métrico (NMDS) baseado na matriz de presença e ausência
que aproxima as localidades com a composição de formigas mais similares entre si. Baseado em coletas
realizadas em 2013 e 2014 no Distrito Federal com armadilhas de queda (pitfall).

37
Figura 1.11. Número médio de registro por localidade de formigas pertencentes a diferentes grupos funcionais,
coletadas em 2013 e 2014 no Distrito Federal com armadilhas de queda (pitfall). Para mais informação sobre as
espécies e características dos grupos funcionais veja o Anexo I. Letras diferentes indicam diferenças
significativas entre os habitats e barras o desvio padrão.
0
5
10
15
20
25
30
35N
úm
ero
médio
de r
egis
tro p
or
localid
ade
Soja
Agri. org.
Pastagem
Veg. nativa
a a a
b
a
b b c
a
b b
c
a a a a a
b b
c
a
b b b
a
b b
c
a
b b b
a a
b
c

38
Figura 1.12. Análise de partição aditiva de diversidade de formigas no Distrito Federal. α1 é o número médio de
espécies nas parcelas. β1 é a troca de espécies entre parcelas dentro da mesma localidade. β2 é a troca de
espécies entre localidades pertencentes ao mesmo tipo de agrossistema ou fitofisionomia. β3 é a troca de
espécies entre os diferentes tipos de agrossistemas e fitofisionomias. Para todos os valores observados a
diferença foi significativa (p<0,001) em comparação ao modelo esperado.
0
50
100
150
200
250
300
Observado Esperado
Núm
ero
de e
spécie
s
β3
β2
β1
α1

39
Figura 1.13. Valores da diversidade β baseados em dados de presença e ausência de espécies de formigas
coletadas por armadilha de queda (pitfall) em 2013 e 2014 no Distrito Federal. βsor representa a soma de βsim
(troca de espécies, ou seja, turnover) e βnes (aninhamento). Letras representam diferenças significativas. Para as
categorias Agrossistemas e Vegetação nativa foi feito um teste t com os valores das comparações par a par
obtida nas diferentes matrizes. Para as demais categorias foi realizado o teste não paramétrico de Kruskal-Wallis.

40
Figura 1.14. Relação entre a dissimilaridade da fauna de formiga (βsor, βsim e βnes) e a relação com a distância
entre as localidades da Vegetação nativa e dos Agrossistemas. A Correlação de Pearson (r) e a significância
computada usando análise de Mantel estão dispostas dentro dos gráficos. Os pontos correspondem aos valores de
comparação par a par entre as localidades amostradas em 2013 e 2014 com armadilhas de queda (pitfall) no
Distrito Federal.
Dis
sim
ilari
da
de (
βsor)
Dis
sim
ilari
da
de (
βsor)
Dis
sim
ilari
da
de (
βsim
)
Dis
sim
ilari
da
de (
βsim
)
Dis
sim
ilari
da
de (
βn
es)
Dis
sim
ilari
da
de (
βn
es)

41
Tabela 1.2. Espécies de formigas e seus respectivos valores de indicação para diferentes tipos de habitats. Os
dados foram obtidos a partir de armadilhas de queda (pitfall) nos anos de 2013 e 2014 no Distrito Federal. Todas
as espécies e índices abaixo foram obtidos de resultados significativos. Habitats: AO = agricultura orgânica e
Past= pastagem. Cores brancas, amarelas e verdes indicam um gradiente de baixo a alto valor de indicação.
Espécie Soja AO Past Campo Cerrado Mata Valor p
Pheidole sp.04 0,59 0,08 0,00 0,00 0,00 0,00 0,00
Pheidole sp.09 0,16 0,22 0,04 0,00 0,00 0,00 0,03
Pheidole sp.35 0,00 0,61 0,06 0,00 0,00 0,01 0,01
Camponotus gr. blandus 0,00 0,44 0,01 0,00 0,00 0,00 0,00
Solenopsis saevissima 0,00 0,39 0,18 0,09 0,00 0,00 0,00
Solenopsis sp.04 pr. loretana 0,05 0,31 0,19 0,00 0,00 0,00 0,00
Nylanderia sp.01 0,00 0,28 0,08 0,00 0,01 0,08 0,00
Camponotus gr. senex 0,00 0,25 0,00 0,00 0,00 0,00 0,00
Brachymyrmex sp.01 0,00 0,25 0,00 0,00 0,00 0,00 0,02
Dorymyrmex cf. brunneus 0,01 0,25 0,46 0,01 0,00 0,00 0,00
Solenopsis sp.02 0,00 0,20 0,19 0,02 0,23 0,06 0,04
Pheidole gertrudae 0,00 0,20 0,01 0,00 0,00 0,00 0,04
Pheidole sp.24 0,00 0,18 0,27 0,00 0,00 0,00 0,00
Hypoponera sp.08 0,00 0,11 0,15 0,27 0,15 0,08 0,00
Cardiocondyla emeryi 0,02 0,15 0,41 0,00 0,00 0,00 0,00
Pheidole sp.11 0,04 0,03 0,24 0,10 0,03 0,00 0,04
Pheidole sp.02 0,01 0,04 0,20 0,00 0,02 0,00 0,04
Acromyrmex sp.01 0,00 0,00 0,23 0,12 0,00 0,00 0,01
Solenopsis tridens 0,00 0,00 0,20 0,00 0,00 0,00 0,01
Linepithema cf. gallardoi 0,00 0,00 0,20 0,36 0,01 0,00 0,00
Camponotus novagrandensis 0,00 0,00 0,10 0,28 0,05 0,00 0,00
Pachycondyla striata 0,00 0,02 0,03 0,49 0,04 0,00 0,00
Linepithema cf. neotropicum 0,00 0,06 0,04 0,11 0,22 0,01 0,02
Nylanderia sp.03 0,00 0,05 0,10 0,27 0,28 0,00 0,02
Hypoponera sp.03 0,00 0,05 0,04 0,27 0,36 0,00 0,02
Pheidole sp.29 0,00 0,03 0,07 0,11 0,26 0,15 0,03
Pheidole sp.22 0,00 0,03 0,02 0,10 0,30 0,03 0,03
Cyphomyrmex sp06 gr. rimosus 0,00 0,00 0,00 0,35 0,15 0,00 0,01
Trachymyrmex sp.08 0,00 0,00 0,02 0,32 0,06 0,00 0,05
Pseudomyrmex termitarius 0,00 0,00 0,04 0,31 0,23 0,00 0,01
Solenopsis sp.03 0,00 0,01 0,04 0,26 0,27 0,11 0,00
Cyphomyrmex sp05 gr. rimosus 0,00 0,00 0,03 0,26 0,20 0,00 0,02
Dorymyrmex cf. jheringi 0,00 0,00 0,01 0,23 0,00 0,00 0,01
Myrmicocrypta sp.01 0,00 0,00 0,01 0,23 0,00 0,00 0,05
Acropyga sp.01 0,00 0,00 0,00 0,23 0,18 0,00 0,01
Acromyrmex sp.03 0,00 0,02 0,01 0,21 0,02 0,01 0,00
Trachymyrmex sp.01 0,00 0,00 0,01 0,21 0,16 0,00 0,03
Wasmannia auropunctata 0,00 0,02 0,01 0,20 0,16 0,24 0,04
Gnamptogenys sp.01 0,00 0,00 0,04 0,19 0,00 0,00 0,00
Camponotus sericeiventris 0,00 0,01 0,01 0,18 0,43 0,00 0,01
Continua...

42
Espécie Soja AO Past Campo Cerrado Mata p value
Labidus praedator 0,00 0,01 0,04 0,17 0,03 0,24 0,03
Cyphomyrmex sp04 gr. rimosus 0,00 0,00 0,00 0,17 0,24 0,00 0,04
Brachymyrmex sp.10 0,00 0,00 0,00 0,17 0,23 0,01 0,00
Pseudomyrmex pallidus 0,00 0,00 0,00 0,16 0,35 0,00 0,00
Camponotus personatus 0,00 0,00 0,01 0,07 0,46 0,00 0,03
Camponotus sp.17 0,00 0,00 0,00 0,02 0,75 0,00 0,02
Camponotus substitutus 0,00 0,00 0,00 0,00 0,63 0,00 0,00
Cyphomyrmex lectus 0,00 0,00 0,00 0,00 0,63 0,00 0,03
Pheidole sp.31 0,00 0,00 0,00 0,06 0,60 0,01 0,00
Mycocepurus obsoletus 0,00 0,00 0,00 0,06 0,57 0,01 0,00
Pheidole sp.26 0,00 0,00 0,00 0,06 0,45 0,04 0,00
Odontomachus chelifer 0,00 0,00 0,00 0,00 0,38 0,00 0,01
Pheidole sp.28 0,00 0,01 0,07 0,01 0,35 0,20 0,05
Ectatomma edentatum 0,00 0,00 0,00 0,00 0,26 0,21 0,00
Apterostigma sp.01 0,00 0,00 0,00 0,00 0,25 0,00 0,03
Rogeria sp.01 0,00 0,01 0,00 0,03 0,24 0,42 0,03
Camponotus sp.29 0,00 0,01 0,01 0,03 0,22 0,00 0,01
Ectatomma brunneum 0,00 0,00 0,06 0,03 0,20 0,00 0,00
Camponotus sp.18 0,00 0,02 0,06 0,00 0,20 0,00 0,01
Camponotus rufipes 0,00 0,03 0,01 0,00 0,01 0,49 0,04
Linepithema cf. cerradense 0,00 0,03 0,00 0,00 0,00 0,35 0,02
Pogonomyrmex naegile 0,00 0,02 0,01 0,00 0,00 0,41 0,00
Camponotus cingulatus 0,00 0,01 0,00 0,00 0,00 0,28 0,04
Pachycondyla harpax 0,00 0,01 0,00 0,00 0,00 0,28 0,00
Acromyrmex cf. balzani 0,00 0,00 0,01 0,00 0,00 0,28 0,02
Solenopsis sp17 gr. wasmannii 0,00 0,00 0,01 0,00 0,00 0,28 0,02
Ectatomma permagnum 0,00 0,00 0,00 0,00 0,00 0,22 0,00
Odontomachus laticeps 0,00 0,00 0,00 0,00 0,00 0,22 0,02
Camponotus sp.45 0,00 0,00 0,00 0,00 0,00 0,22 0,03
Pheidole sp.25 0,00 0,00 0,00 0,00 0,00 0,22 0,03
Camponotus bidens 0,00 0,00 0,00 0,00 0,00 0,22 0,03
Linepithema sp.05 0,00 0,00 0,00 0,00 0,00 0,22 0,04
Acropyga sp.02 0,00 0,00 0,00 0,00 0,00 0,22 0,04
Gnamptogenys sp.03 0,00 0,00 0,00 0,00 0,00 0,22 0,04
Pheidole sp.21 0,00 0,00 0,00 0,00 0,00 0,22 0,04
Brachymyrmex cf. delabiei 0,00 0,00 0,00 0,00 0,00 0,22 0,04
Pheidole sp.06 0,00 0,00 0,00 0,00 0,00 0,22 0,04
Camponotus sp.27 0,00 0,00 0,00 0,00 0,03 0,24 0,04
Octostruma sp.01 0,00 0,00 0,00 0,00 0,03 0,24 0,05
Hypoponera cf. foreli 0,00 0,00 0,00 0,12 0,09 0,29 0,02
Nylanderia sp.04 0,00 0,00 0,00 0,00 0,00 0,33 0,02
Brachymyrmex sp.07 0,00 0,00 0,00 0,00 0,00 0,33 0,00
Hylomyrma sp.02 0,00 0,00 0,00 0,00 0,00 0,33 0,00
Solenopsis sp.13 0,00 0,00 0,00 0,00 0,00 0,33 0,00
Continua...
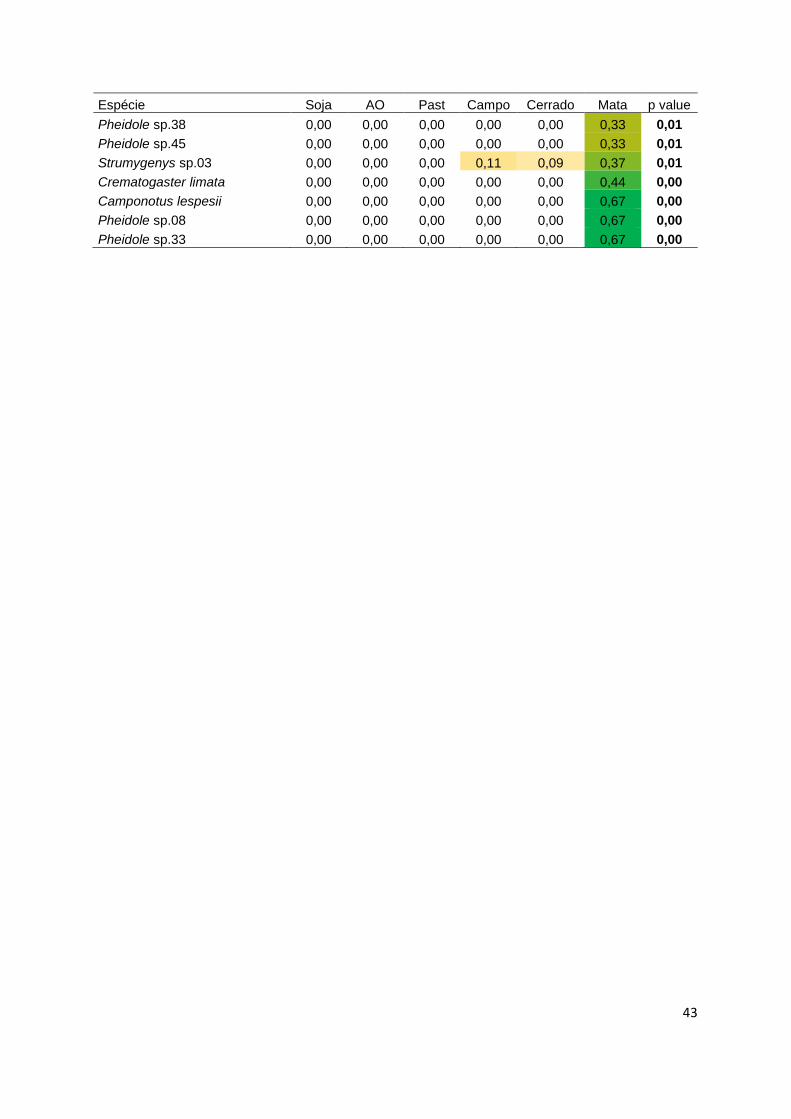
43
Espécie Soja AO Past Campo Cerrado Mata p value
Pheidole sp.38 0,00 0,00 0,00 0,00 0,00 0,33 0,01
Pheidole sp.45 0,00 0,00 0,00 0,00 0,00 0,33 0,01
Strumygenys sp.03 0,00 0,00 0,00 0,11 0,09 0,37 0,01
Crematogaster limata 0,00 0,00 0,00 0,00 0,00 0,44 0,00
Camponotus lespesii 0,00 0,00 0,00 0,00 0,00 0,67 0,00
Pheidole sp.08 0,00 0,00 0,00 0,00 0,00 0,67 0,00
Pheidole sp.33 0,00 0,00 0,00 0,00 0,00 0,67 0,00

44
Discussão
Efeitos sobre a riqueza
Os resultados mostram que as conversões de áreas nativas do Cerrado em
agrossistemas causam a perda da diversidade local. Porém, a magnitude desta perda está
condicionada ao tipo de agrossistema implantado, pois monoculturas de soja causam maior
queda na diversidade do que pastagens ou agricultura orgânica. Kessler et al. (2009) utiliza o
termo “divergência das condições ecológicas” para explicar porque alguns tipos de uso da
terra têm maior impacto sobre a biodiversidade. Basicamente esse termo tenta retratar que
quanto mais diferente é a condição pós-conversão em relação à vegetação original, geralmente
simplificando ou aumentando a intensidade do manejo do sistema, menor é a diversidade
esperada. Estes são resultados semelhantes aos encontrados na literatura científica, que
apresenta diversos estudos, com diferentes táxons, mostrando que a conversão de habitats
naturais e intensificação dos agrossistemas acarretam em redução e mudanças na composição
das espécies, além de mudanças na estrutura do funcionamento do ecossistema (e.g. Jones et
al. 2003; Hole et al. 2005; Tylianakis et al. 2005; Flynn et al. 2009; Marichal 2014; Uchida &
Ushimaru 2014; Van Meerbeek et al. 2014; Hausberger & Korb 2016; Lu et al. 2016;
Ribeiro-Neto et al. 2016).
Dos diversos exemplos na literatura de como a conversão de áreas nativas para a
produção de alimentos impactaram a fauna local, talvez o mais bem estudado seja a produção
de café na América Latina, segundo o qual em uma revisão feita por Philpott et al. (2008) foi
notada uma redução na diversidade de formigas. Porém, a magnitude dessa redução depende
do tipo de manejo da terra (monocultivo de café ou café à sombra de árvores nativas), além de
refletir nos serviços ecológicos (Perfecto & Vandermeer 2001; Armbrecht & Perfecto 2003;
Philpott & Armbrecht 2006; Armbrecht & Gallego 2007). Padrões similares de redução na

45
diversidade ou riqueza de formigas em sistemas de plantio são encontrados também na
Amazônia (Solaris et al. 2015), Argentina (Bestelmeyer & Wiens 1996), Austrália (Hoffmann
& Andersen 2003), África (Lemessa et al. 2015), China (Lu et al. 2016) e México (Cuatle et
al. 2016).
Então, apesar da perda de espécies devido à conversão da vegetação nativa em
agrossistemas ser um padrão global bem conhecido, a magnitude desse efeito e sua interação
com os diversos tipos de agrossistemas ainda são, em sua maioria, desconhecidos. A redução
na diversidade, encontrada ao longo do presente trabalho, é maior que a descrita por Pacheco
et al. (2013) e Frizzo & Vasconcelos (2013).
De certo modo, este trabalho retrata como monoculturas de soja com aplicação de
agrotóxicos, aração e gradagem do solo, períodos de entresafra sem cobertura viva do solo e a
dessecação pré-plantio são formas de manejo que tornam esses campos um ambiente de difícil
sobrevivência, tanto para insetos praga (sendo sua existência a principal causa para realização
de parte destes manejos, como aplicação de agrotóxico e dessecação pré-plantio) como para
todos os outros seres vivos. Para esse processo muitos autores tem utilizado a expressão
intensificação do uso da terra, uma forma de, independentemente do tipo de cultura plantada,
caracterizar o manejo destas áreas como causa para as diferentes riquezas encontradas nestes
ambientes (Tscharntke et al. 2005).
Em contrapartida, as pastagens praticamente não são manejadas, possuindo uma
riqueza de espécies de formigas maior que as de áreas de soja (como já descrito por Frizzo e
Vasconcelos 2013), já que aparentemente o pastejo em si pouco afeta a diversidade de
formigas (Bestelmeyer & Wiens 2001). Nessas áreas, o efeito mais importante é o efeito
indireto da ocupação de espécies exóticas de gramíneas (Barton et al. 2016), que são
competitivamente superiores às gramíneas nativas e reduzem a diversidade da flora e fauna
(Pivello et al. 1999; Almeida et al. 2011). A expansão das gramíneas exóticas de origem

46
africana é considerada uma das grandes ameaças ao Cerrado (Hoffmann et al. 2004; Klink &
Machado 2005).
Já as áreas de agricultura orgânica surpreendentemente são tão diversas como as de
pastagem, sugerindo que os agrotóxicos, assim como as técnicas de manejo, são os grandes
responsáveis pela redução da diversidade em áreas de agricultura convencional (Hole et al.
2005; Winqvist et al. 2011). Além da restrição no uso de agrotóxicos em geral, as áreas de
orgânicos possuem uma grande heterogeneidade local, o plantio de flores para fornecer
recurso alternativo para os predadores e irrigação constante ao longo do ano, de modo que
sempre há matéria verde e, consequentemente, herbívoros e outros organismos que podem
servir de fonte de alimentação para as formigas (Frizzo, observação pessoal). Além disso,
diferente do que encontrado na soja, as áreas de agricultura orgânica possuem faixas de terra
com plantas perenes e árvores, onde o solo não é revolvido nessa região, potencialmente
agindo como refúgio para formigas que nidificam no solo ou nas próprias árvores (Frizzo &
Vasconcelos 2013).
Efeito sobre a composição
Quanto à composição, as espécies que conseguem persistir nos sistemas agrícolas
geralmente são caracterizadas por serem de hábito generalista, recrutarem para monopolizar
os recursos disponíveis em campo e/ou apresentarem comportamento dominante, como os
gêneros Pheidole e Solenopsis (Pacheco et al. 2013). Outro exemplo de espécies comuns nos
agrossistemas analisados são as formigas oportunistas com tolerância a temperaturas altas,
como as do gênero Dorymyrmex e Linepithema (Cuezzo 2011; Farji-Brener et al. 2002).
Além destas há as formigas generalistas de tamanho muito reduzido (~ 1,5 mm), como
Brachymyrmex e as chamadas “thief ants”, também do gênero Solenopsis, que ocorrem tanto
em ambientes naturais como em perturbados, incluindo residências em áreas urbanas.

47
Já as formigas arborícolas, os predadores grandes, as especialistas e as tribos Attini e
Camponotini, com a conversão de áreas nativas em sistemas agrícolas, sofrem uma redução
na sua ocorrência (Figura 1.11). Isso ocorre tanto devido as mudanças ambientais, como ao
combate direto (manejo para reduzir a população), no caso das formigas pertencentes a tribo
Attini. É evidente também que o tipo de uso da terra afeta a ocorrência de determinados
grupos funcionais de formigas em diferentes graus de magnitude. Em geral, o plantio de soja
é o tipo de uso da terra que tem mais efeitos negativos para quase todos os grupos funcionais,
enquanto agricultura orgânica e pastagem apresentam valores intermediários entre soja e
vegetação nativa.
Mas a grande diferença na composição das formigas que ocorre entre os agrossistemas
e a vegetação nativa está associada às espécies raras ou às que ocorreram apenas uma vez.
Como pode se observar na figura 1.7, as áreas de vegetação nativa têm 3,5 vezes mais
espécies únicas (amostradas uma única vez) do que os agrossistemas, apesar do esforço
amostral nestas áreas ser quase a metade do ocorrido nos agrossistemas. Estas espécies raras
geralmente apresentam algum aspecto da morfologia, ecologia ou comportamento
especializado (Didham et al. 1996; Gibb et al. 2015). Campos et al. (2011) demonstraram que
a fauna de formigas do Cerrado é caracterizada por uma grande proporção de formigas raras
(45% das espécies ocorrem em menos do que 5% das amostras), comparativamente à savana
Australiana (onde 27% das espécies são raras). Resultados semelhantes também foram
encontrados para Lepidópteros (Price et al. 1995). Esse fato torna ainda mais preocupante o
avanço dos agrossistemas sobre as áreas nativas, visto que as espécies raras são as mais
suscetíveis à extinção devido a baixa densidade e pouca flexibilidade que possuem para
persistir em ambientes perturbados (Didham et al. 1996).
Muitos trabalhos discutem quais são os fatores que podem ser limitantes para a
colonização de uma área agrícola, e a maioria parece supor o local para nidificação como

48
fator limitante, que pode ser, além do solo, serapilheira, galhos caídos sobre o solo e o estrato
arbóreo arbustivo (Philpott & Foster 2005; Armbrecht et al. 2006). Porém, Frizzo et al.
(2011) relataram que a passagem do fogo no cerrado stricto sensu, não altera a comunidade de
formigas no solo, mesmo esse consumindo toda a serapilheira, galhos caídos e o estrato
arbustivo, o que indica que para o Cerrado, talvez esse não seja o efeito primordial. Tem-se
também, como outro provável fator limitante, o favorecimento das espécies generalistas e
dominantes que podem ter um efeito supressivo no estabelecimento e ação de diversas
espécies, gerando exclusão competitiva devido às interações interespecíficas (Pacheco et al.
2013). Também foi encontrado na literatura que, arar o solo pode reduzir em até cinco vezes a
ocorrência de formigas na folhagem de culturas (Pereira et al. 2010). No entanto, com
exceção do último, estudos que isolem cada um destes possíveis fatores que afetam as
formigas nos sistemas agrícolas são inexistentes ou não foram encontrados.
Do ponto de vista conservacionista, também é importante ressaltar a mudança na
composição em função da fitofisionomia nativa coletada (Pacheco & Vasconcelos 2012), o
que aumenta ainda mais a importância de conservação de múltiplas áreas nativas. De certo
modo, com exceção das espécies invasoras, em tese, todas as espécies encontradas nos
sistemas agrícolas são capazes de sobreviver nos sistemas nativos, no entanto, o inverso não é
verdadeiro. Visto que ambientes convertidos em sistemas agrícolas não contêm espécies, que
são relativamente comuns em ambientes naturais. Isso fica claro quando analisamos a
composição de espécies da formação florestal, a qual apresenta o maior número de espécies
indicadoras singulares (dados do IndVal, Tabela 1.2). Isso significa que nas áreas de mata
existem muitas espécies únicas que não são capazes de persistir em sistemas agrícolas ou nas
áreas de vegetação nativas mais abertas. Resultados semelhantes no Cerrado também foram
encontrados para Drosophilideos, no qual Mata & Tidon (2013) discutem que, apesar das
matas de galeria ocuparem menos de 10% do Cerrado, uma grande parte da diversidade

49
demonstra uma preferência ou associação a esse ambiente. Isso também foi observado para
outros táxons, incluindo árvores (Hoffman et al. 2003), mamíferos (Redford & Fonseca,
1986), formigas (Pacheco & Vasconcelos 2012) besouros (Durães et al. 2005), algumas
famílias de lepidópteros (Brown & Gifford 2002), entre outros.
Áreas de vegetação nativa de cerrado e campo também apresentam algumas espécies
únicas, que provavelmente seriam extintas localmente se houvesse a conversão destas em
sistemas agrícolas. Mesmo pastagem e agricultura orgânica sendo sistemas menos
impactantes sobre a biodiversidade de formigas, estas apresentam uma composição distinta
em relação às áreas nativas, as quais contêm um número maior de espécies arborícolas,
crípticas, predadores especialistas e espécies de tamanho relativo, pequeno e grande. Essas
mudanças no número de espécies com diferentes tamanhos, também é relatado por Senior et
al. (2013) para aves, besouros e formigas.
Partição aditiva e multiplicativa da diversidade
A comunidade de formigas do Cerrado é caracterizada pela maior presença de espécies
raras (Campos et al. 2011), o que influencia o não aninhamento da comunidade como um
todo (Almeida-Neto et al. 2008). De certo modo, as espécies mais abundantes persistem na
maioria dos habitats e, à medida que se tornam raras, elas demostram um padrão de
preferência de habitat (Figura 1.13). O que a análise de partição aditiva de diversidade revela
é que o fator mais importante para a diversidade de formigas no Distrito Federal é a existência
de uma maior diversidade de habitats (sensu Pacheco e Vasconcelos 2012).
Separando a diversidade β em dois fatores, que afetam a troca de espécies entre as
diferentes localidades, verificou-se que a magnitude da diferença de βsor entre sistemas
agrícolas e as fitofisionomias nativas é pequena, apesar de significativa (figura 1.13). Pode-se
notar que há uma troca maior de espécies (dissimilaridade βsor) nos sistemas agrícolas em

50
comparação às fitofisionomias nativas, e que essa dissimilaridade é mais em função do
aninhamento (βnes) do que da troca de espécies propriamente dita (βsim). Resultados
semelhantes foram encontrados por Solar et al. (2015) na Amazônia, porém com o dobro dos
valores de aninhamento encontrados no Cerrado. Isso demonstra que no Cerrado as áreas de
produção agrícola, com o aumento da intensificação do uso da terra, têm causado a
homogeneização biótica (McKinney & Lockwood, 1999), porém não tão drástica como as
encontradas na Amazônia.
Além disso, um segundo padrão na diversidade βsor foi encontrado, mas este
associado apenas à troca de espécies (βsim) e aos agrossistemas. Na figura 1.14, observou-se
que a dissimilaridade entre as localidades agrícolas aumenta à medida que estas localidades
ficam mais distantes entre si. Isso significa que a composição de espécies nos agrossistemas é
influenciada em função do local em que se encontra. A mudança da composição em função da
posição geográfica é um fato esperado, porém não a distâncias tão pequenas quanto 100 km.
Baselga (2010) também encontrou influência da distância sobre as variáveis de
dissimilaridade (βsor, βsim e βnes), mas em um gradiente com quase 3.000 km de distância.

51
Conclusão
A conversão da vegetação nativa do Cerrado em sistemas de produção agricola causa
uma perda local na diversidade e mudança na composição da mirmecofauna. Esta perda está
associada ao tipo de sistema agrícola, sendo a monocultura de soja, em comparação a áreas de
agricultura orgânica e pastagens, o que causa maior decréscimo na riqueza local.
De modo geral, os grupos de formigas que apresentam maior especificidade, como
nidificar em árvores ou ter uma especialização alimentar, são os que têm a maior redução em
relação a sua frequência de ocorrência. Estes grupos que apresentam especializações, na
maioria dos casos já são espécies naturalmente raras (de baixa frequência), sendo estes os
principais promotores de biodiversidade e, potencialmente, as espécies mais ameaçadas de
extinção local no Cerrado.
Outro fator importante em termos de conservação é que diferentes fitofisionomias
nativas do Cerrado possuem uma composição de espécies distintas. As áres de formação
florestal apresentam a fauna mais exclusiva, com muitas espécies únicas que não ocorrem nas
áreas nativas de cerrado stricto sensu, campo e principalmente nos agrossistemas. Já as
formações savanicas (cerrado stricto sensu e campo) apesar de também apresentam espécies
exclusivas, possuem um subconjunto de espécies que são capazes de persistir ou colonizar os
sistemas agrícolas.
Duas implicações práticas deste trabalho são: (1) reavaliar como está sendo realizado o
manejo das áreas agrícolas, principalmente monocultura de soja, para, assim, diminuir seu
impacto; (2) considerar que, em futuras ações conservacionistas, áreas de pastagem e
agricultura orgânica possam ser sistemas de produção preferenciais ou incentivados no
entorno de áreas de conservação ou nas de relevante biodiversidade, como zonas de

52
amortização de impacto ou até mesmo corredores entre fragmentos de cerrado stricto sensu e
campo.

53
Capítulo II – Variação nas taxas de remoção de biomassa pela mirmecofauna de solo
sobre diferentes tipos de uso da terra
Resumo
Um dos principais efeitos da redução na biodiversidade sugerido pelos pesquisadores
tem sido a redução ou perda de funções do ecossistema, mas esse não é um efeito universal.
No Cerrado, já observamos que sistemas agrícolas causam uma redução na biodiversidade de
formigas, mas não se sabe como as funções do ecossistema desempenhadas por estas são
afetadas. O principal objetivo deste estudo foi avaliar se a conversão de áreas nativas em
sistemas agrícolas, afeta a remoção de biomassa realizada por formigas, uma importante
função do ecossistema ligada à decomposição e predação. Para isso, foram utilizadas iscas de
sardinha (simulando organismos animais), banana (simulando recurso atrativo vegetal) e
sementes de gramíneas, em áreas nativas (campo, cerrado stricto sensu e mata) e áreas
agrícolas (plantio de soja, pastagens e agricultura orgânica). Exceto para soja, os dados
mostram que formigas removem mais sardinha, do que banana, e removem muito pouca
semente. Porém, existe um efeito relacionado ao tipo de uso da terra, já que apenas as áreas de
soja tiveram níveis significativamente baixos de remoção de sardinha. Para a isca de sementes
não foi detectado nenhum efeito porque pouca biomassa é removida, tanto nas áreas nativas
como nos sistemas agrícolas. Já para a isca de banana, os dados sugerem existir um efeito de
redundância com outro grupo da macrofauna. No entanto, a biomassa da macrofauna
(excluindo-se formigas) não explica essa remoção de biomassa. Também não foi detectado
efeito da composição de espécies de formigas sobre as biomassas removidas. A remoção de
biomassa de sardinha e banana está correlacionada à riqueza de formigas, indicando que os
efeitos sobre a função do ecossistema dependem das particularidades de cada função avaliada
(como tipo de recurso), do tipo de uso da terra e da riqueza de formigas no local.

54
Introdução
A expansão dos sistemas agrícolas tem efeitos sobre a biodiversidade local (Pacheco et
al. 2013) e global (Newbold et al. 2015), sendo a magnitude destes efeitos dependente das
características locais, táxon e tipo de sistema agrícola implantado (Flynn et al. 2009; Kessler
et al. 2009; Bommarco et al. 2014). Porém, tão ou mais importante quanto o efeito sobre a
biodiversidade é responder qual é o efeito da expansão dos sistemas agrícolas sobre o
funcionamento do ecossistema.
A hipótese da biodiversidade-função do ecossistema, uma teoria que começou a ser
amplamente debatida no começo da década de 1990, prediz que uma redução na
biodiversidade (número de espécies, variabilidade genética e etc...) causa decréscimo nos
processos do ecossistema (Srivastava & Vellend 2005). Porém, este não é um efeito universal
(e.g. Birkhofen et al. 2015), revisões e metanálises a respeito deste assunto têm chegado a
uma mesma conclusão: a redução da biodiversidade, reduz a funcionalidade do ecossistema
dependendo das características locais, da função do ecossistema analisada e do tipo de
perturbação (Hopper et al. 2005; Cardinale et al. 2006; Allan et al. 2013).
Essa grande quantidade de resultados divergentes, encontrados nos estudos sobre
serviços do ecossistema devido aos efeitos da antropização, torna a previsibilidade destes
impactos complexas e dependentes de estudos que foquem em diferentes ambientes,
processos e tipos de perturbação. No entanto, além de existirem relativamente poucos estudos,
boa parte destes tem analisado comunidades de plantas ou comunidades aquáticas em
experimentos manipulativos (Srivastava & Vellend 2005; Allan et al. 2013; Cardinale et al.
2006), dando pouca importância aos processos ecológicos que dependem da fauna terrestre.
Dentre esses processos ecológicos, pode-se citar a predação e a decomposição,
exercidos pela fauna terrestre, de fundamental importância para o funcionamento e regulação

55
do ecossistema (De Groot et al. 2002). Isso porque a predação possibilita o controle das
diversas populações, incluindo consumidores e plantas. Já a fauna detritívora, atua no
processo de decomposição e ciclagem da matéria orgânica contribuindo para a fertilidade do
solo e nutrição das plantas.
Existem poucos estudos que avaliam como a expansão da agricultura afeta a
biodiversidade e como isso reflete nos processos ecológicos do Cerrado, cujo alto grau de
endemismo e de degradação é um dos maiores no mundo (Myers et al. 2000). A expansão da
fronteira agrícola no Cerrado é uma realidade, com áreas densamente ocupadas,
principalmente ao sul deste bioma (Sano et al. 2010). Os impactos desta ocupação têm sido
avaliados utilizando diversos táxons, sendo as formigas um dos principais objetos de estudo
(revisado por Folgarait 1998; Ribas et al. 2012), isso porque é um dos táxons mais
abundantes e de maior biomassa em ambientes tropicais (Fittkau & Klinge 1973). Só no
Cerrado, estima-se que elas consomem de 13 a 17% da biomassa foliar de plantas lenhosas
por ano (Costa et al. 2008). As formigas também podem atuar como predadores, podendo
controlar a população de pragas agrícolas (Risch & Carroll 1982; López & Potter 2000;
Philpott & Armbrecht 2006; Armbrecht & Gallego 2007; Howe et al. 2009; Styrsky &
Eubanks 2010; Gonthier et al. 2013; Offenberg 2015) ou como predadores e/ou dispersores de
sementes (Wall et al. 2005), principalmente das que contenham elaiosoma (um apêndice
atrativo da semente) (Ferreira et al. 2011). Outro importante serviço ecológico é a remoção de
biomassa orgânica da superfície para o subsolo, melhorando as qualidades nutricionais do
solo (Folgarait 1998).
As formigas possuem diferentes hábitos alimentares, sendo que a maioria das espécies
é onívora, existindo algumas poucas espécies estritamente predadoras, granívoras ou que
atuam como herbívoros (Fernández 2003). Além disso, de toda a gama de recursos utilizados
pelas formigas, sempre há algum elemento nutricional limitante para seu desenvolvimento e o

56
da colônia e, por isso, estudos apontam que as formigas têm demonstrado preferência
alimentar justamente por aqueles recursos que contenham uma proporção maior desses
elementos limitantes (Kaspariz & Yanovlak 2001; Bihn et al. 2008; Kaspari et al. 2008;
Vieira & Vasconcelos 2014).
Em vista disso, este estudo tem o objetivo de avaliar as taxas de remoção de biomassa
por formigas de diferentes tipos de recursos (sensu Vieira & Vasconelos 2014) entre
ecossistemas nativos e agrícolas. Para tanto, foram selecionadas áreas de agricultura orgânica,
plantações de soja, pastagens, além de áreas nativas de campo, cerrado stricto sensu e
formações florestais. Especificamente procuramos responder às seguintes perguntas: (1)
existe diferença na biomassa removida entre recursos que simulam organismos animais,
apêndices atrativos de sementes e sementes de gramíneas? (2) Mudanças no uso da terra
afetam as taxas de remoção de biomassa? (3) Quais variáveis locais explicam melhor a
quantidade de biomassa removida, riqueza de formigas, composição da assembleia de
formigas ou biomassa do restante da macrofauna?

57
Metodologia
Área de estudo
O estudo foi realizado no Distrito Federal que abrange uma área de mais de 5.800
Km², localizado centralmente a 15°47’S e 47°55’O. As localidades amostradas foram as
mesmas descritas no Capítulo I (Tabela 1.1 e Figura 1.1). O experimento foi instalado em 69
localidades, porém o ponto P-03 foi pisoteado por gado e os pontos VN-18 e VN-21 foram
atacados de forma massiva por formigas de correição, por isso estes foram excluídos das
análises estatísticas. Assim, ao todo, foram analisados: 14 plantios de soja convencional, 14
pastagens, 16 áreas de agricultura orgânica (sendo duas destas, AO-15 e AO-16, plantio de
soja orgânica) e 22 áreas de vegetação nativa. A escolha dos sistemas agrícolas, pastagem e
plantio de soja, se devem a esses serem os tipos de uso da terra mais comuns no Cerrado,
representando 27,6% e 11,7% da área do bioma respectivamente (Sano et al. 2010). Já as
áreas de agricultura orgânica representam uma forma alternativa de produção de alimento
vegetal sem o uso de agrotóxicos e cujo mercado esta em rápida expansão (Rigby & Cáceres
2001; Blanc 2009; ABRASCO 2015). Das 22 áreas de vegetação nativa, optou-se por coletar
nas três fitofisionomias também mais comuns da região, as quais são: cinco localidades de
formações campestres, oito de cerrado stricto sensu e nove de formações florestais (Figuras
1.1 e 1.2).
Segundo a classificação de Köppen (1948), o clima nessa região é do tipo Aw,
apresentando forte sazonalidade, com um verão chuvoso e o inverno seco. As temperaturas
médias mensais podem variar de 22 a 27°C e a precipitação anual média é de 1.500 mm
(Klink & Machado 2005). Todas as localidades estão situadas dentro do bioma Cerrado (para
mais informações sobre a vegetação nativa veja Oliveira-Filho & Ratter 2002).

58
Protocolo amostral
Nesse experimento, foram desenvolvidas “estações de remoção de biomassa” para
medir as taxas de remoção de recurso por formigas de forma concomitante com a amostragem
da fauna descrita no Capítulo I (Figura 2.1). Essas estações consistiam em potes plásticos de
um litro de volume, com tampa e com vários furos laterais, em forma de semicírculo com 1
cm de altura e 2 cm de largura, para a macrofauna ter acesso ao interior do pote (Figura 2.2 -
A). Dentro de cada estação foi disponibilizada uma quantidade de recurso (também
denominado de isca) de massa conhecida e, após 24 horas, essas estações foram recolhidas e a
biomassa do recurso foi novamente mensurada, a fim de se determinar a quantidade de
biomassa removida pela macrofauna. Apenas para o tratamento contendo semente estas foram
secas em estufa a 50 Cº, por 24 horas, antes e depois do experimento em campo, devido à
grande absorção de água pelas sementes no campo. Para as iscas de sardinha e banana, estas
eram preparadas em laboratório, individualizadas e congeladas, em porções de massa
variando entre 20 a 30 gramas por estação de remoção de biomassa, até serem utilizadas no
experimento. As sementes de gramíneas também foram individualizadas em porções de 15
gramas por estação de remoção de biomassa.
Em campo, este experimento foi montado ao mesmo tempo, que a instalação das
armadilhas de queda do Capítulo I. Para cada parcela amostral citadas no Capítulo I, também
foram instaladas 12 estações de remoção de biomassa que ficara em campo por 24 horas
(Figura 2.2 - B). Para cada localidade as parcelas amostrais foram istaladas no mesmo
momento, sendo a escolha do tipo de isca em cada parcela escolhida de forma aleatória. Ao
final do experimento, as estações foram removidas e as formigas presentes dentro das
estações de remoção de biomassa foram coletadas e identificadas em laboratório.
Nos extremos da parcela foram colocadas duas estações de remoção de biomassa,
denominadas “controle”. Isso porque os recursos poderiam perder massa por dessecação,

59
então foi criado um controle para abater estes valores dos tratamentos. Essas estações controle
possuiam o mesmo número e tamanho de abertura que as demais, porém estas eram
posicionadas na lateral próximas à tampa do pote (parte superior). Isto foi feito para diminuir
o acesso da fauna em geral ao recurso. Além disso, foi instalada uma tira com três centímetros
de altura de armadilha adesiva amarela no lado interno do pote, esta circundava o pote
impedindo o acesso de animais à isca (Figura 2.2 - C). Então a massa perdida após 24 horas
foi considerada uma perda de umidade do recurso para o ambiente e, por isso, foi
proporcionalmente descontada da massa inicial das outras estações de remoção de biomassa.
Adicionalmente, as tampas de todas as estações de remoção de biomassa e controle foram
cobertas por uma camada de aproximadamente 3 mm de impermeabilizante emborrachado
branco, a fim de diminuir o aumento interno de temperatura.
Dentro de cada localidade, as três parcelas amostrais foram diferenciadas pelo tipo de
recurso contido nas estações de remoção de biomassa. Foram utilizados três tipos de recurso
que simulam os alimentos mais comuns disponíveis para as formigas e que poderiam ser
comprados em grande quantidade e baixo custo. Esses recursos são sardinha enlatada
(simulando organismos animais), banana da variedade prata (simulando alimentos como o
arilo de sementes ou frutos) e um composto de sementes não beneficiadas, em proporção de
massa iguais, de arroz, alpiste e painço (simulando sementes de gramíneas nativas do
cerrado). Em termos nutricionais, sardinha é muito rica em sódio, proteína e gorduras totais,
banana é rica em açúcar e as sementes em carboidratos (USDA 2016). Para a preparação da
sardinha, o óleo foi drenado e depois foi esmagada com um garfo, já a banana foi descascada
e esmagada.
Considerou-se que a utilização de iscas exógenas ao ambiente não inviabilizaria os
resultados porque todas as localidades são réplicas idênticas umas das outras. Portanto,
comparação entre localidades diferentes refletem diferenças reais. Além disso,

60
independentemente da diferença de intensidade do odor entre os tipos de isca, observações de
campo indicam que todas as iscas são rapidamente encontradas e monopolizadas, apenas para
sementes, não foi observado monopolização (ou defesa do recurso). Outros estudos similares
usaram atrativos exógenos para formigas, como: atum e biscoito (Fayle et al. 2011) e isca
para pesca feitas de pelotas de minhoca (Gray et al. 2015).
Junto com o experimento também foram instalados, nas parcelas, um sistema de
choque elétrico para evitar o acesso de animais (vertebrados) atraídos pelas iscas. Esse
sistema foi montado a partir de pedaços de tela de galinheiro presos à parte superior das
estações de remoção de biomassa (Figura 2.2 - D) e ligados, por fios elétricos encapados, a
um aparelho de cerca elétrica para gado. Este aparelho era alimentado por uma bateria de 12
volts e estava conectado por meio de fios a todas as estações de remoção de biomassa. Após a
instalação desse sistema, nenhuma estação de controle ou de tratamento foi destruída, ao
contrário de como ocorreu no experimento piloto.
Cálculo da biomassa
As taxas de remoção de biomassa por estação foram calculadas, descartando a massa
dos potes, de acordo com a função:
Br = (PI x Fc)-(PF)
Onde:
Br é biomassa removida;
PI é a massa inicial de alimento posto dentro da estação;
Fc é o fator de correção de biomassa perdida por dessecação (fórmula do cálculo
abaixo);
PF é a massa do conteúdo restante do alimento após 24 horas;

61
Para corrigir a massa de alimento perdida devido à dessecação nas estações de
remoção de biomassa foi calculado um fator de correção (Fc), com as estações de biomassa
controle, a partir da seguinte fórmula:
Fc = PFc / PI
Onde:
Fc é o fator de correção de biomassa perdida por dessecação;
PFc é a massa do conteúdo restante do alimento após 24 horas;
PI é a massa inicial de alimento posto dentro da estação controle.
Ainda após esses cálculos foi realizada a soma dos valores de alimento removidos por
parcela amostral. Como algumas poucas estações de remoção de biomassa foram
acidentalmente perdidas (Tabela 2.1), foi realizada uma estimativa do valor total de biomassa
removida por parcela amostral. Esse valor foi obtido através da fórmula:
Brestimada= (ΣBr por ponto amostral / Nt) x 12
Onde:
ΣBr por ponto amostral é o somatório de toda biomassa removida em uma parcela amostral;
Nt é o número de estações de remoção de biomassa resgatado por parcela amostral e;
12 seria o número de estações que deveria existir em cada parcela amostral.
Além dos dados de riqueza já obtidos no capítulo I, para essas mesmas áreas também
foram utilizados dados de biomassa das formigas amostradas nos pitfalls e do restante da
macrofauna amostrada. Para isso, todos os invertebrados capturados nos pitfalls, por parcela
amostral, foram separados entre formigas e outros invertebrados. Logo após isso, as amostras
foram secas a 60Cº por 48 horas e depois pesadas em uma balança analítica com precisão de
0,001 g.

62
Análise dos dados
Os dados foram separados em quatro categorias de habitats: plantio de soja,
agricultura orgânica, pastagem e vegetação nativa. Para a vegetação nativa foi realizado o
agrupamento dos pontos que contemplam diferentes fitofisionomias por que: (1) entre eles
não há diferenças significavas nos padrões de remoção de biomassa e; (2) a separação por
fitofisionomia formar grupos com número de amostras muito pequenos para a realização das
análises estatísticas subsequentes.
Todas as análises foram realizadas utilizando o programa R (R Development Core
Team 2008). A primeira análise visou verificar se o número de estações de remoção de
biomassa com formigas, ao final das 24 horas em campo, diferiria entre os distintos habitats.
Para esta análise, o número de estações de remoção de biomassa com formigas, em cada
parcela amostral foi contabilizado. Em seguida foi realizada uma análise de modelos lineares
generalizados, utilizando a função glm do R.
Para determinar qual é o tipo de isca (banana, sardinha ou semente) mais removido no
sistema, foi realizado o teste não paramétrico de Friedman em blocos (devido à
heterogeneidade de variâncias), com um teste a posteriori (Hollander & Wolfe 1973)
utilizando a função posthoc.friedman.nemenyi.test do pacote PMCMR (Pohlert 2016). Devido
às áreas de soja apresentarem um padrão visualmente diferente dos demais, quanto à
preferência por recurso (Figura 2.4), para estas foi realizada a mesma análise em separado.
Em seguida, para determinar se a biomassa removida de cada tipo de isca, também varia em
função dos diferentes tipos de habitats avaliados foi realizada uma análise de modelos lineares
generalizados, utilizando a função glm do R.
Foi realizada também uma correlação Pearson entre a riqueza e massa de formigas
amostradas por localidade. Devido à não normalidade de parte dos dados e problemas de
heterocedasticidade e presença constante de pontos tendenciando os resíduos, não foi possível

63
realizar nenhuma análise englobando múltiplos fatores e suas interações para explicar quais as
variáveis influenciam a remoção de biomassa de sardinha, banana e semente. Por isso, foram
feitas análises de correlação de Pearson entre as variáveis: riqueza de formigas, biomassa da
macrofauna, biomassa removida de sardinha, biomassa removida de banana e biomassa
removida de semente. Novamente, como as áreas de soja apresentam padrões diferentes na
avaliação visual dos gráficos, outra matriz de correlação de Pearson foi construída sem a
presença das áreas de soja.
Para determinar se a composição de espécies de formigas influencia nas taxas de
remoção de biomassa, foi realizada uma análise de correspondência canônica (CCA),
utilizando a função cca do pacote vegan (Oksanen et al. 2016). Essa análise foi feita baseada
em uma matriz de espécies por locais, sendo contabilizada a frequência de ocorrência por
parcela (com três parcelas por localidade). As variáveis avaliadas na CCA foram: tipo de
habitat, número de espécies de formigas por localidade, biomassa da macrofauna e biomassa
removida de sardinha, banana e semente.
Para validar a CCA foi realizado um teste para detectar multicolinearidade entre as
variáveis analisadas usando a função vif.cca, do pacote vegan (Oksanen et al. 2016) – neste
teste, se os valores de VIF (fator de inflação da variância) forem maiores que 10 indicam
multicolinearidade entre duas variáveis (Belsley et al. 1980). Em seguida, foi utilizada a
função anova.cca (pacote vegan), com 999 permutações, para determinar se o modelo, as
variáveis e os eixos de explicação são significativos. Estas análises (CCA) foram repetidas
mais uma vez, com uma matriz utilizando apenas as espécies consideradas abundantes (que
ocorreram mais de 30 vezes).

64
Figura 2.1. Esquema da metodologia de amostragem realizada em 2013 e 2014 no Distrito Federal. A) Como as
três parcelas amostrais estavam distribuídas ao longo de cada localidade, sendo que em cada uma das parcelas
foi utilizado um tipo de isca escolhido de forma aleatória (sardinha, banana e semente). B) Esquema da parcela
amostral, tendo 12 estações de remoção de biomassa, 12 pitfalls e duas estações de biomassa controle nas
extremidades.

65
Figura 2.2. Fotografias da metodologia de estudo realizada em 2013 e 2014 no Distrito Federal. A) Estação de
remoção de biomassa com isca de sardinha. B) Fotografia da parcela amostral em uma área de agricultura
orgânica. C) Estação de biomassa controle. D) Estação de remoção de biomassa com uma tela de arame ligada a
um aparelho de choque elétrico.
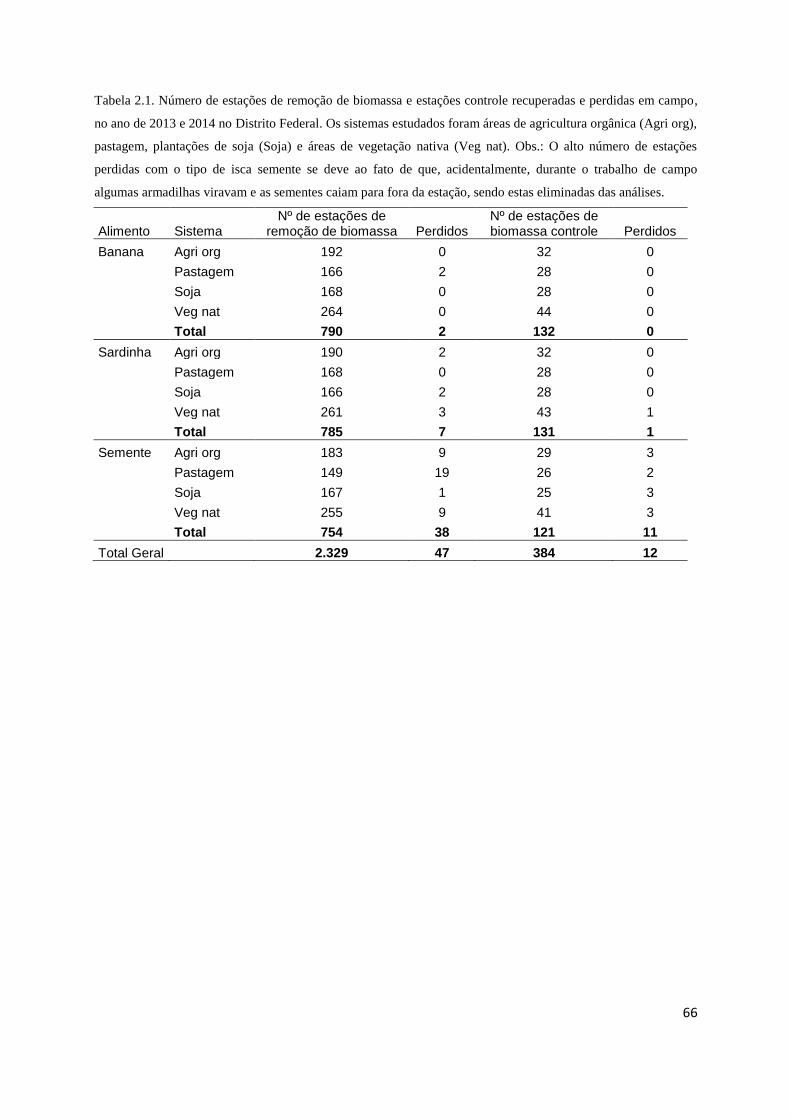
66
Tabela 2.1. Número de estações de remoção de biomassa e estações controle recuperadas e perdidas em campo,
no ano de 2013 e 2014 no Distrito Federal. Os sistemas estudados foram áreas de agricultura orgânica (Agri org),
pastagem, plantações de soja (Soja) e áreas de vegetação nativa (Veg nat). Obs.: O alto número de estações
perdidas com o tipo de isca semente se deve ao fato de que, acidentalmente, durante o trabalho de campo
algumas armadilhas viravam e as sementes caiam para fora da estação, sendo estas eliminadas das análises.
Alimento Sistema Nº de estações de
remoção de biomassa Perdidos Nº de estações de biomassa controle Perdidos
Banana Agri org 192 0 32 0
Pastagem 166 2 28 0
Soja 168 0 28 0
Veg nat 264 0 44 0
Total 790 2 132 0
Sardinha Agri org 190 2 32 0
Pastagem 168 0 28 0
Soja 166 2 28 0
Veg nat 261 3 43 1
Total 785 7 131 1
Semente Agri org 183 9 29 3
Pastagem 149 19 26 2
Soja 167 1 25 3
Veg nat 255 9 41 3
Total 754 38 121 11
Total Geral 2.329 47 384 12

67
Resultados
Ocorrência de formigas nas estações de remoção de biomassa
As formigas foram o principal grupo de insetos observado dentro das estações de
remoção de biomassa após 24 horas de coleta (Anexo IV). Apenas em 55 estações de remoção
de biomassa, das mais de 2.300 em campo, haviam outros insetos, como moscas, mariposas e
gafanhotos. Isso indica que a maior parte da biomassa removida é devido à ação das formigas,
principalmente porque em observações de campo anteriores notou-se que formigas
monopolizam iscas atrativas atacando qualquer outro organismo que se aproxime. Porém, é
importante destacar que, no término do experimento, a porcentagem de estações de remoção
de biomassa ocupadas por formigas variaram significativamente entre os habitats, sendo
menores nas áreas de monocultura de soja (variação na ocupação de 0 a 57%, com média de
13%, Figura 2.3).
Taxas de remoção de biomassa entre os tipos de recursos
As formigas foram capazes de remover relativamente grandes quantidades de
biomassa por parcela (máximo removido de 237g, toda sardinha disponível na localidade VN-
18), mas isso depende do tipo de alimento (de acordo com o teste em blocos de Fridman,
X²=85,534, df=2, p<0,001) (Figura 2.4-A). Assim, detectou-se que sardinha foi o recurso
mais removido, com uma média de 87,2g (σ ±22,8), seguido de banana (média de 70,1g, σ
±30,5) e, por último, de semente (média de 7,6g, σ ±7,1) (maior p=0,017).
Porém apenas para soja o padrão é diferente dos demais (Figura 2.4-B), pois há uma
diferença estatística entre a biomassa removida para os diferentes tipos de recurso (teste em
blocos de Fridman, X²=26,143, df=2, p<0,001), sendo a banana o mais removido (média de
74,8g, σ ±13,6). Já a diferença entre a biomassa removida de banana e sardinha é

68
marginalmente significativa (p=0,06), pois em média foi removido 44,5g (σ ±12) de sardinha.
Enquanto que, para semente, continuou havendo uma remoção significativamente (p<0,01)
inferior aos demais tipos de recursos (média de 5,3, σ ±7,1).
Taxas de remoção de biomassa entre os habitats
Em relação à biomassa removida por tipo de habitat (soja, pastagem, agricultura
orgânica ou vegetação nativa) não foram encontradas diferenças significativas para banana
(GLM, F3,62=0,4565, p=0,7136) e para semente (GLM, F3,62=1,2561, p=0,2972) (Figura 2.4-
C). Já a remoção de sardinha está significativamente ligada ao tipo de habitat (GLM,
F3,62=9,1307, p<0,001), afinal, apenas na soja há uma remoção menor de biomassa de
sardinha em comparação a todos os outros sistemas (p<0,001). Entre agricultura orgânica,
pastagem e vegetação nativa os valores de biomassa removida de sardinha não apresentaram
diferenças (p>0,05).
Influência da riqueza de formigas e biomassa da fauna
A correlação mais forte encontrada neste estudo é entre a riqueza de formiga e sua
biomassa (r=0,68, p<0,001) (Figura 2.5). Já para as biomassas removidas de recursos,
encontramos que: (a) para remoção de semente não existe correlação com nenhuma das
variáveis apresentadas (p>0,05); (b) as correlações detectadas para os recursos de sardinha e
banana foram relativamente fracas; e (c) para remoção de biomassa de sardinha foi encontrada
uma correlação com a riqueza de formigas (r=0,54, p<0,001) (Figura 2.6). Entretanto,
excluindo os dados da soja (Figura 2.7), para a remoção de biomassa de banana surge uma
correlação desta com a riqueza de formigas (r=0,32, p=0,02).

69
Influência da composição de espécies
Quanto à composição de espécies de formigas analisadas nas CCAs, os modelos foram
significativos (ANOVA.CCA completa F8,57=1,7478, p<0,001 e ANOVA.CCA apenas para espécies
abundantes F8,57= 3,5721, p<0,001), assim como os dois primeiros eixos que formam as figuras
2.8 e 2.9. Também não foi detectado multicolinearidade entre as variáveis dos modelos (maior
VIF foi igual a 2,25). Para os dois modelos, o fator tipo de uso da terra afeta a composição de
espécies (para a variável uso da terra ANOVA.CCA completa F3,57=2,7151, p<0,001 e
ANOVA.CCA apenas para espécies abundantes F3,57= 7,7473, p<0,001). Mas apenas para a análise das
espécies abundates é que a riqueza influência significativamente na composição das espécies
(ANOVA.CCA para espécies abundantes F8,57= 1,5914, p=0,044).

70
Figura 2.3. Proporção de estações de remoção de biomassa ocupadas por formigas nas coletas de 2013 e 2014 no
Distrito Federal. Letras diferentes significam diferenças significativas (p<0,05) entre as proporções de ocupação
dentro de cada tipo de recurso, barras representam o desvio padrão.
a a
a
b b
c
b
b,c
c
b
b
b
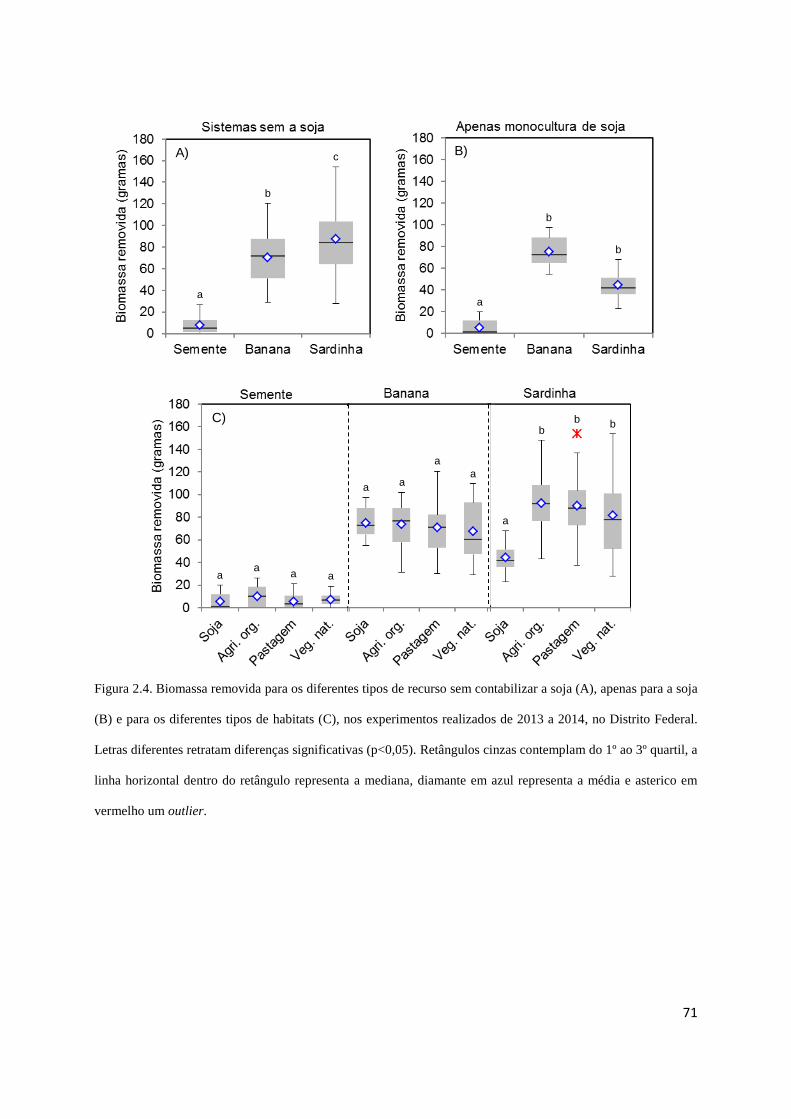
71
Figura 2.4. Biomassa removida para os diferentes tipos de recurso sem contabilizar a soja (A), apenas para a soja
(B) e para os diferentes tipos de habitats (C), nos experimentos realizados de 2013 a 2014, no Distrito Federal.
Letras diferentes retratam diferenças significativas (p<0,05). Retângulos cinzas contemplam do 1º ao 3º quartil, a
linha horizontal dentro do retângulo representa a mediana, diamante em azul representa a média e asterico em
vermelho um outlier.
A) B)
C)
a a
b
c
b
b
a a
a a
a a
a a
a
b b b
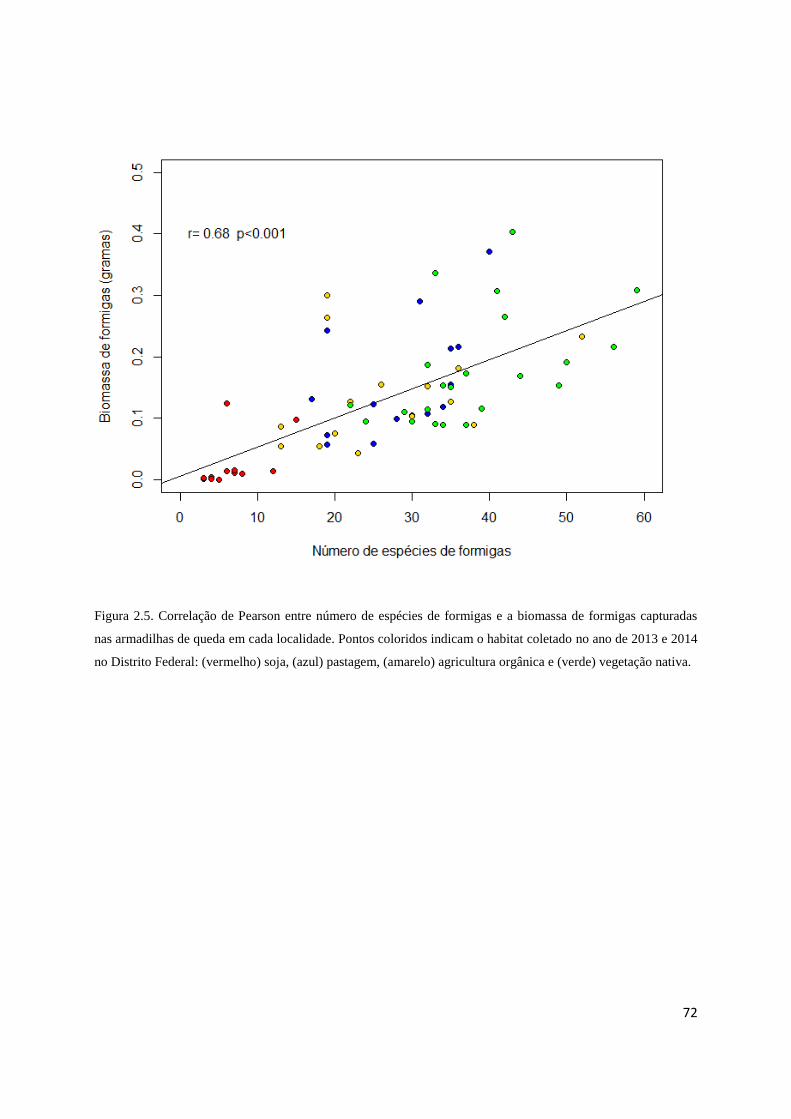
72
Figura 2.5. Correlação de Pearson entre número de espécies de formigas e a biomassa de formigas capturadas
nas armadilhas de queda em cada localidade. Pontos coloridos indicam o habitat coletado no ano de 2013 e 2014
no Distrito Federal: (vermelho) soja, (azul) pastagem, (amarelo) agricultura orgânica e (verde) vegetação nativa.

73
Figura 2.6. Matriz de correlação de Pearson entre número de espécies de formigas, massa da macrofauna (exceto
formigas) e biomassa removida de banana, sardinha e semente. Os valores de correlação significativos (p<0,05)
são indicados em negrito. Linha em vermelho representa a tendência não linear de dispersão dos dados. Pontos
coloridos indicam o habitat coletado no ano de 2013 e 2014 no Distrito Federal: (vermelho) soja, (azul)
pastagem, (amarelo) agricultura orgânica e (verde) vegetação nativa.
Riqueza
0.0 0.6 1.2
r= 0.11
p= 0.364
r= 0.15
p= 0.245
20 80 140
r= 0.54
p<0.001
10
30
50
r= 0.23
p= 0.064
0.0
0.6
1.2
Massa_Fauna
r= 0.13
p= 0.313
r= 0.12
p= 0.333
r= 0.15
p= 0.245
Banana
r= 0.29
p= 0.02
40
80
12
0
r= 0.20
p= 0.103
20
80
14
0
Sardinha
r= 0.21
p= 0.088
10 30 50 40 80 120 0 10 200
10
20Semente
Massa da
fauna
Riqueza de
formigas
Banana
Sardinha
Semente
p<0.001
p=0.002

74
Figura 2.7. Matriz de correlação Pearson (neste caso, sem os dados da soja) entre número de espécies de
formigas, massa da macrofauna (exceto formigas) e biomassa removida de banana, sardinha e semente. Os
valores de correlação significativos (p<0,05) são indicados em negrito. Linha em vermelho representa a
tendência não linear de dispersão dos dados. Pontos coloridos indicam o habitat coletado no ano de 2013 e 2014
no Distrito Federal: (azul) pastagem, (amarelo) agricultura orgânica e (verde) vegetação nativa.
Riqueza
0.0 0.6 1.2
r= 0.016
p= 0.911
r= 0.32
p= 0.02
40 100
r= 0.25
p= 0.076
20
40
60
r= 0.18
p= 0.206
0.0
0.6
1.2
Massa_Fauna
r= 0.20
p= 0.159
r= 0.31
p= 0.023
r= 0.22
p= 0.114
Banana
r= 0.42
p= 0.002
40
80
12
0
r= 0.20
p= 0.164
40
10
0
Sardinha
r= 0.19
p= 0.167
20 40 60 40 80 120 0 10 200
10
20Semente
Massa da
fauna
Riqueza de
formigas
Banana
Sardinha
Semente
p=0.02
p=0.002
p=0.023

75
Figura 2.8. Análise de correspondência canônica que verifica a influência de variáveis sobre a composição de
espécies de formigas, coletadas no ano de 2013 e 2014 no Distrito Federal. A única variável significativa nesta
análise foi o sistema em que as formigas foram coletadas (F3,57=2,7151, p<0,001). A posição das espécies nesta
imagem foi suprimida para facilitar a visualização dos dados.
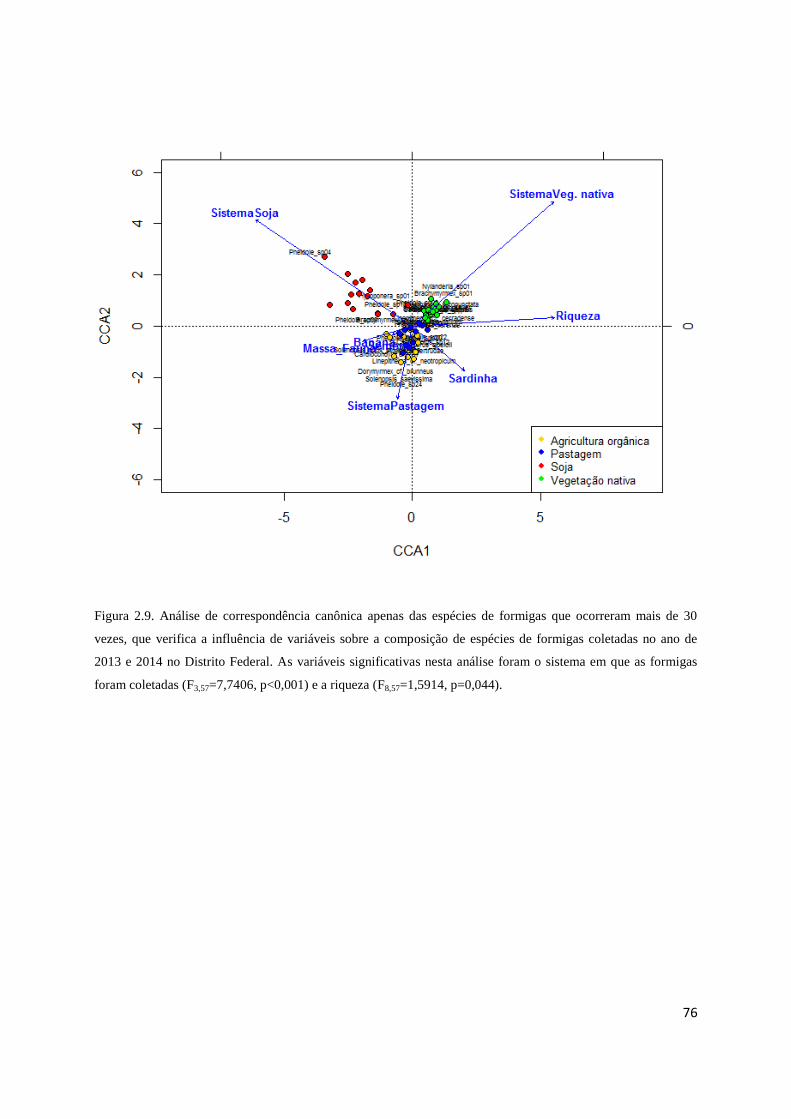
76
Figura 2.9. Análise de correspondência canônica apenas das espécies de formigas que ocorreram mais de 30
vezes, que verifica a influência de variáveis sobre a composição de espécies de formigas coletadas no ano de
2013 e 2014 no Distrito Federal. As variáveis significativas nesta análise foram o sistema em que as formigas
foram coletadas (F3,57=7,7406, p<0,001) e a riqueza (F8,57=1,5914, p=0,044).

77
Discussão
Ocorrência de formigas nas estações de remoção de biomassa
No Cerrado, as formigas são o grupo dominante sobre a superfície do solo,
monopolizando os recursos por elas utilizados. Prova disso é o número de estações de
remoção de biomassa, ocupadas por formigas após 24 horas (Figura 2.3), que foram
encontradas e monopolizadas por elas, logo nos primeiros minutos. No entanto, detectou-se
um efeito do tipo de uso da terra, pois os agrossistemas e, em especial, os plantios de
monocultura de soja apresentaram uma menor proporção de estações ocupadas após 24 horas
em comparação com as áreas de vegetação nativa.
A existência de estações de remoção de biomassa com alimento, mas sem a presença
de formigas, está ligada a três fatores: a saciedade, o territorialismo e/ou a ausência de
formigas. De modo geral, após as 24 horas de experimento em campo, é provável que para
algumas colônias de formigas tenha ocorrido a saciedade do consumo de alimento (sensu
Galen 2005). Além disso, devido ao comportamento territorialista das formigas, é pouco
provável que formigas venham de longe remover estas iscas (Hölldobler & Lumsden 1980).
Sendo assim, a ausência de formigas dentro das estações de remoção de biomassa, refletem a
quantidade de formigas no local.
Taxas de remoção de biomassa entre os tipos de recursos
Vários estudos apontam que formigas apresentam preferência alimentar de acordo com
o nutriente mais limitante naquele ambiente (Kaspariz & Yanovlak 2001; Bihn et al. 2008;
Kaspari et al. 2008; Vieira & Vasconcelos 2014). Neste estudo, nota-se que a sardinha foi a
isca com maior consumo por formigas, sendo esse tipo de isca, amplamente utilizado como
atrativo para formigas no Cerrado (e.g. Lopes & Vasconcelos 2008) e em outros biomas,

78
como Mata Atlântica (e.g. Diehl et al. 2004), Caatinga (e.g. Ulysséa & Brandão 2013) e
Amazônia (e.g. Vasconcelos et al. 2008). Logo, nossos resultados contrariam o estudo de
Vieira & Vasconcelos (2014), também realizado no Cerrado (mas utilizando iscas diferentes),
no qual formigas de solo demostram preferência por alimentos que contenham sacarose,
enquanto são as formigas arborícolas as que utilizam mais recurso rico em sódio (abundante
no recurso sardinha), resultados similares ao encontrado por Kaspariz & Yanovlak (2001),
utilizando carne e açucar. Segundo esses estudos, isso ocorre aparentemente devido ao hábito
alimentar das formigas, já que espécies no solo raramente teriam contato com alimentos
açucarados, mais comuns na vegetação arbórea (Vieira & Vasconcelos 2014).
Os alimentos simulando organismos animais (sardinha), em geral, foram os que as
formigas mais removeram ao final das 24 horas (com exceção dos plantios de soja). Segundo
Fisher et al. (2005), o recurso alimentar consumido pelos adultos e larvas de formigas são
diferentes. Os recursos proteicos e com lipídios capturados pelos forrageadores são
preferencialmente utilizados para alimentar as larvas no ninho e garantir o crescimento da
colônia. Já os alimentos ricos em açúcares e carboidratos são geralmente utilizados pelas
operárias para os gastos diários de energia (Fisher et al. 2005). Esse pode ser o fator
determinante para uma maior remoção de sardinha e não das outras iscas.
Os valores removidos de biomassa de sardinha e de banana foram próximos entre si, o
que demonstra que esta pode ser um importante complemento alimentar para a mirmecofauna
do solo, como propôs Vieira & Vasconcelos (2014). Logo, essa proximidade também é um
claro indicativo de que, assim como relatado por Hunter (2009), a “promiscuidade trófica”
nos artrópodes parece ser muito mais ampla do que imaginado a priori pelos pesquisadores.
Além de ser uma possível explicação para o paradoxo formiga-biomassa de Tobin (1994), no
qual ele discute que a biomassa de formigas em alguns ecossistemas é tão grande que apenas

79
a alimentação via predação de outros animais, não é capaz de explicar a quantidade de
formigas observada.
No entanto, verifica-se também que, apesar de estudos indicarem que formigas têm um
importante papel na predação e dispersão de sementes (Ferreira et al. 2011), estas parecem ser
um alimento, relativamente, de baixa importância. Isso se confirma mesmo também tendo
capturado espécies que consomem predominantemente sementes, como Pogonomyrmex
naegelii (Belchior et al. 2012).
Um último fato que também pode influênciar a taxa de remoção de biomassa dos
diferentes tipos de recurso é o comportamento durante a remoção. Nas observações de campo,
notou-se que a isca de sardinha era removida carregando-se os pedaços individualmente ou
em grupo, quando estes eram muito grandes. Já para banana, assim como sardinha, ocorreu o
recrutamento, geralmente com a monopolização do recurso, mas a remoção deste era sempre
feita de forma individual e as formigas aparentemente sugavam a banana ao invés de carregar
pedaços entre as mandíbulas. Já para sementes, não há recrutamento e nem monopolização,
isso provavelmente devido ao fato de que esse tipo de recurso na natureza não tem ocorrência
agregada (por isso, não requer o recrutamento) e também não é um recurso raro.
Taxas de remoção de biomassa entre os habitats
Tanto para iscas de banana como para de semente não foram encontradas diferenças
significativas nos valores removidos de biomassa entre os distintos tipos de habitats. Para
semente isso provavelmente se deve ao fato de que o consumo desse alimento é muito
reduzido, se comparado a outros tipos de alimento. Nesses casos em que as diferenças podem
ter magnitudes pequenas, uma metodologia com maior precisão, como contar o número de
sementes, pode encontrar tais diferenças (como a realizada por Ferreira et al. 2011). Mas com

80
nossos dados não se constata nenhum fator que explique a remoção de semente, pois, para 36
das 67 localidades analisadas, a remoção de sementes foi menor do que cinco gramas.
Já para banana, os valores removidos foram independentes do tipo de habitat. Porém,
levando em consideração que os monocultivos de soja praticamente não têm formigas (em
número de espécies e massa), comparativamente aos outros habitats, é ilógico admitir que esta
remoção no habitat soja tenha sido realizada por formigas. Além disso, sabe-se que muitas das
pragas de soja, em especial lagartas da família Noctuidae, alimentam-se à noite (Lingren et al.
1977; Gregory Jr 1989), por isso supomos a existência de um efeito de substituição por uma
espécie ou grupo de espécies que desempenhe função similar às formigas de remoção de
biomassa de banana. Existe a suposição também de que baratas (Ordem: Blattodea) podem ser
um importante consumidor das iscas de banana nas áreas de soja. Mas, como não se capturou
nenhum invertebrado que justifique essa afirmação, já que poucas moscas, mariposas e
gafanhotos foram encontrados no final do experimento, dentro das estações de remoção de
biomassa na soja, essa suposição é apenas uma hipótese. Isso porque esta se assemelha ao
resultado descrito por Hooper et al. (2005), o qual afirma que, na ausência da espécie ou
grupo de espécies que desempenha determinada função do ecossistema, uma segunda espécie
ou grupo de espécies pode assumir a função destas.
Influência da riqueza de formigas e biomassa da fauna
Como discutido, a soja possivelmente é um habitat no qual a remoção de biomassa de
banana está sendo realizada por outro grupo de invertebrados. Por isso, excluindo a soja da
matriz de correlação (Figura 2.7), observa-se que existe uma correlação entre a riqueza de
formigas (r=0,32) com a biomassa removida de banana. O mesmo padrão, só que mais forte,
também foi encontrado para a biomassa removida de sardinha (Figura 2.6). Os dados também
demonstram que essa riqueza de formigas esta correlacionada positivamente com a biomassa

81
de formigas (Figura 2.5). Três hipóteses explicam o porquê destes fatores afetarem a função
do ecossistema de remoção de biomassa. Primeiramente, em relação à riqueza de formigas,
sugere-se a hipótese do efeito amostral, que prediz que, quanto maior o número de espécies,
mais provável é que alguma destas desempenhe um papel importante nessa função (Hopper et
al. 2005; Balvanera et al. 2006). Existe também a hipótese da complementariedade, a qual
intui que a soma quantitativa do papel de cada nova espécie encontrada aumenta na mesma
proporção a função a elas correlacionadas (Tscharntke et al. 2005). Contudo, Loreau et al.
(2008) discutem que tanto o efeito amostral como a complementariedade não são mutuamente
exclusivos, portanto, essas duas hipóteses provavelmente afetam a remoção de biomassa ao
mesmo tempo. Por último, a respeito da biomassa de formigas, há a hipótese de mais
indivíduos (Srivastava & Lawton 1998), segundo a qual quanto mais recurso, maior a
quantidade de consumidores (neste caso, influenciando a biomassa removida). Em estudos
similares, Fayle et al. (2011) também encontraram uma relação positiva entre riqueza de
formiga e taxa de remoção de biomassa, porém, Gray et al. (2015) não encontrou influência
da riqueza e nem do recrutamento de formigas sobre as taxas de remoção de isca. Neste
estudo de Gray et al. (2015) o que influenciou os valores de remoção de biomassa, foram a
quantidade de isca que as formigas conseguiam transportar.
Influência da composição de espécies
As correlações encontradas foram de moderado a fracas, com r < 0,7, muitas vezes
valores característicos de experimentos realizados em condições reais, mas que também pode
indicar a potencial existência de outras variáveis explicativas. Uma dessas potenciais
variáveis, que aumentam o erro, provavelmente, é a identidade das espécies de formigas que
realizam esta função de remoção de biomassa. Essa questão da identidade tabém é abordada
por Del-Claro & Marquis (2015), Armbrecht & Gallego (2007) e Gonthier et al. (2013), de

82
acordo com os quais a identidade da espécie tem grande relevância para determinar os níveis
de predação.
De modo similar, apesar da análise de correspondência canônica indicar que somente
o tipo de habitat ou a riqueza de espécies têm relação com a composição da assembleia de
formigas, é inegável que algumas espécies podem desempenhar o papel de remoção de
biomassa melhor que outras. Como exemplo disso pode-se citar as espécies do gênero
Brachymyrmex, todas de tamanho diminuto (em média 1,25 mm) que, independentemente do
tamanho da colônia, dificilmente conseguiriam remover quantidades significativas das iscas
disponibilizadas. No entanto, formigas do gênero Ectatomma (em média 130 mm) conseguem
remover pedaços relativamente grandes das iscas de uma única vez e ter potencialmente
grande influência sobre o total de biomassa removida. Assim, dentre essas formigas citadas,
também é importante destacar o papel das de tamanho intermediário, que têm o
comportamento de recrutar, podendo remover grandes quantidades das iscas devido ao
número de indivíduos que trabalham em conjunto, como algumas espécies do gênero
Pheidole, Solenopsis e as formigas de correição.
As três espécies de formigas mais encontradas dentro das estações de remoção de
biomassa após as 24 horas em campo, foram Pheidole sp. 01, Pheidole gertrudae (Forel,
1886) e Solenopsis saevissima (Smith, F., 1855) (Anexo IV), todas pertencentes ao grupo
funcional descrito anteriormente no Capítulo I como generalistas dominates. Além destas, em
eventos esporádicos, estas iscas foram encontradas por formigas de correição, um grupo que
abrange os gêneros Eciton, Labidus, Neivamyrmex e Nomamyrmex, que são espécies nômades
(sem colônia fixa), as quais podem ter colônias relativamente grandes e são reconhecidamente
vorazes (Palacio 2003). Em decorrência de eventos como esses, duas localidades tiveram de
ser excluídas das análises estatísticas – o ponto VN-18 e o VN-21 –, pois a ocorrência de
correição da espécie Labidus praedator (Smith, F., 1858) foi tão grande que a massa

83
desidratada de formigas capturadas em cada um desses pontos foi mais de 7,5 vezes maior
que o terceiro ponto com mais formigas. Isso levou a ocorrer, nesses pontos específicos,
praticamente a remoção total de biomassa de sardinha disponível, cerca de 237g e 224g.
Em vista disso, sugere-se que futuros estudos usem uma abordagem que leve em
consideração a abundância espécie/específica. Assim como já indicado, a presença abundante
de L. praedator tem grandes efeitos sobre a biomassa removida, mas essa espécie também foi
capturada em outras localidades e, provavelmente devido a uma abundância menor de
indivíduos dela nessas localidades, não ocorreu uma remoção tão intensa como a da
amostrada nas localidades VN-18 e VN-21.

84
Conclusão
As formigas têm preferência por remover iscas de sardinha (que simula uma dieta
baseada em organismos animais), em média, 20% a mais do que de banana, recurso que
simula apêndices atrativos de sementes e frutas do Cerrado. Já as sementes, relativamente às
outras duas iscas, praticamente não foram removidas. Isso evidencia que formigas selecionam
os recursos preferenciais e, ao mesmo tempo, possuem uma grande “promiscuidade trófica”
(Hunter 2009).
Foi detectado também uma capacidade de resistência das funções do ecossistema,
frente as mudanças no uso da terra. Isso, porque apenas nas áreas de soja detectamos uma
redução significativa na remoção de biomassa de sardinha. Mais especificamente foi
encontrado que a riqueza de formigas está correlacionada à função do ecossistema de remoção
de biomassa orgânica (sensu Fayle et al. 2011). E, assim como descritos nas revisões feitas
por Hopper et al. (2005), Cardinale et al. (2006) e Allan et al. (2013), também encontrou-se
que a intensificação da agricultura pode ou não levar à redução de determinada função do
ecossistema, dependendo das características da função citada e do tipo de uso da terra.

85
Capítulo III – Contribuição das formigas (Hymenoptera: Formicidae) para a predação
de ovos de lagarta em sistemas orgânicos de produção de hortaliças
Resumo
Devido à crescente demanda por alimentos ecológicos e sem agrotóxicos, tem ocorrido
um aumento no número de agricultores orgânicos. No entanto, para viabilizar ou aumentar a
eficiência da produção de orgâncicos, se faz necessário uma série de conhecimentos. Dentre
estes se incluem a identificação das espécies que realizam o controle biológico das pragas
agrícolas e qual a influência da paisagem. Já é consenso que Coleoptera, Araneae, Diptera e
Hemiptera são os principais predadores nos sistemas orgânicos, mas, no que diz respeito ao
Cerrado, há poucos estudos sobre a taxa de predação exercida por Formicidae (formigas). Por
isso, este estudo avaliou as taxas de predação de ovos de lagarta e como essas taxas são
influenciadas pelo tipo de uso da terra no entorno das propriedades. Para isso, foi realizado
um experimento em 18 áreas de agricultura orgânica no Distrito Federal envolvendo testes de
predação de ovos de Anticarsia gemmatalis e amostragem da fauna. Este estudo mostrou que
formigas são capazes de reduzir a população de ovos em 26,8% ao dia, enquanto outros
táxons de predadores, representados principalmente por Coleoptera (das famílias
Coccinellidae e Staphylinidae), removeram 13% dos ovos. Verificou-se também que as
formigas do gênero Linepithema, Dorymyrmex e Camponotus foram as mais frequentes,
porém os gêneros Pheidole e Solenopis são as com maior poder de recrutamento. A proporção
de área plantada num raio de 500 metros, além da interação das outras categorias da paisagem
entre si, tem uma pequena influência para a predação, apenas quando os predadores não são
formigas. Para a predação realizada por predadores em geral, incluindo formigas, ou apenas
por formigas, a paisagem não tem influência.

86
Introdução
Um dos oito objetivos do milênio lançados pela ONU é garantir o desenvolvimento
sustentável (UNPD 2015). Isso implica em mudar a forma como se produz alimento
atualmente, pois o que ococorre hoje é, em sua maioria, o processo de intensificação do uso
da terra para aumentar a produtividade, o que consequentemente amplifica os efeitos
negativos sobre a biodiversidade (e.g. Flynn et al. 2010; Jones et al. 2003). Por outro lado,
aumentar a intensificação do uso da terra não é a única maneira de aumentar a produtividade.
Em uma revisão, Tscharntke et al. (2005) apontaram que a intensificação do uso da
terra significa a conversão de sistemas naturais complexos em direção a ecossistemas
manejados e simplificados, dependentes de insumos artificiais, da mecanização e da frequente
intervenção humana. Aliado à consequente perda de espécies pela intensificação, encontra-se
a redução nos serviços ecológicos, incluindo a polinização (Brittain & Potts 2011), controle
biológico (Langellotto & Denno 2004), entre outros, o que aumenta ainda mais a dependência
destes sistemas em relação à intervenção humana. Porém, existem alternativas que reduzem
esse impacto, como a produção orgânica (Winqvist et al. 2011), apesar de exitirem algumas
incertezas quanto a seus reais benefícios ambientais (Hole et al. 2005). Nesse tipo de
produção agrícola, destaca-se a produção de hortaliças orgânicas, que tem uma “pegada
ecológica” menor do que a produção de grãos e carne (Wackernagel et al. 1999) e ainda com
o benefício de estar associado a alimentos mais saudáveis, devido a não utilização de
agrotóxicos (e.g. Chrisman et al. 2009).
No Brasil 85% do alimento consumido internamente é produzido por agricultores
familiares, em áreas de, no máximo, quatro módulos fiscais e nas quais se cultiva mais do que
uma cultura comercial (IPD 2010). Com o intuito de agregar ainda mais valor ao seu produto,
muito desses agricultores têm migrado para o sistema orgânico, cujo mercado, só em 2015,

87
cresceu 35%, atingindo R$ 2,5 bilhões em produtos comercializados (BioFach 2016; MAPA
2016c).
Mas essa transição, da agricultura convencional para a orgânica, exige mudanças de
manejo que envolvem certos riscos, pois a produção orgânica depende da restauração e
conservação de diferentes serviços ecológicos, os quais viabilizam uma produção agrícola
sustentável (Sujii et al. 2010). Em vista disso, uma das principais demandas dos agricultores
está relacionada com os problemas fitossanitários com insetos praga, que não podem ser
controlados com agrotóxicos nos sistemas orgânicos, por isso, recorre-se ao controle
biológico (Landis et al. 2000). Por exemplo, o uso de insetos parasitoides de pragas agrícolas
já é uma ferramenta utilizada comercialmente, na qual o parasitoide efetivamente regula o
tamanho populacional de seus hospedeiros (Parra & Zucchi 2004).
Para incrementar o controle biológico e para não favorecer insetos praga, a técnica
mais amplamente utilizada e pesquisada nas últimas décadas tem sido o aumento da
diversidade vegetal em escalas, as quais variam desde o talhão de cultivo até a diversidade da
paisagem agrícola (Gurr et al. 2003; Zhender et al. 2004; Sujii et al. 2010; Letourneau et al.
2011). O aumento da diversidade vegetal no primeiro nível trófico, em geral, tende a ter um
efeito positivo nos inimigos naturais e um efeito negativo nos herbívoros (Haddad et al. 2009;
Letourneau et al. 2011). Em contrapartida, o aumento da diversidade vegetal gera um
aumento da complexidade do habitat, o que diminui a chance de exclusão dos insetos praga
deste ambiente, mas, ao mesmo tempo, diminui a probabilidade de explosão populacional
destes insetos (Karban et al. 2013).
No que diz respeito à paisagem de zonas agrícolas, estas têm sido estudadas para
explicar o porquê de algumas áreas abrigarem mais predadores e/ou possuírem níveis maiores
de predação sobre insetos praga. Até agora, nesses estudos tem sido percebido que paisagens
agrícolas, onde há um aumento da complexidade ou da proporção de áreas nativas, têm mais

88
espécies (revisado por Tscharntke et al. 2005) e/ou maiores taxas de predação e produtividade
(Leiere et al. 2015; Balzan et al. 2016), apesar desse efeito estar ligado também ao táxon
(Lemessa et al. 2015). Porém, há alguns estudos que não detectaram efeito da paisagem (e.g.
Silva & Marco Jr 2014).
Comumente os organismos benéficos mais estudados dentro das áreas orgânicas são os
coleópteros da Família Coccinellidae, popularmente conhecidos como joaninhas (e.g.
Harterreiten-Souza et al. 2012; Togni et al. 2016), e os micro-hymenópteras (e.g. Vieira et al.
2013). Porém as formigas, além de comportamento predatório e organização eusocial,
intituladas até de superorganismo (Hölldobler & Wilson 2009), possuem todas as
prerrogativas para serem consideradas agentes de controle biológico. Offenberg et al. (2015),
em uma revisão, afirmaram que formigas do gênero Oecophilla reduzem a abundância das
pragas e de seus danos e aumentam os ganhos financeiros em diversos cultivos. A eficiência
delas foi considerada até maior do que a do uso de pesticidas químicos e com a vantagem de
ter baixo custo.
Antagonicamente, algumas espécies de formigas podem causar prejuízo aos
agricultores. Um exemplo comum disso, é a associação mutualística de várias espécies de
formigas com afídeos, na qual a planta hospedeira pode ou não ser prejudicada (revisado por
Styrsk & Eubanks 2007). Além disso, formigas do gênero Atta e Acromyrmex, que cortam
folhas para cultivar fungos dos quais se alimentam (Fernández 2003), são classificadas como
importantes pragas agrícolas. E por serem pragas que demandam constante monitoramento e
com grande potencial de dano econômico (Hernández & Jaffé 1995; Zanetti et al. 2014)
acabam sendo associadas pelos agricultores a todas as outras espécies de formigas, incluindo
as outras 2.000 espécies que se estima existir no Brasil (Baccaro et al. 2015).
Com base nestas informações, buscou-se avaliar qual a contribuição das formigas para
o controle biológico natural de insetos herbívoros em agrossistemas e determinar quais são os

89
fatores mais importantes que influenciam as taxas de predação nas áreas orgânicas.
Especificamente, procurou-se determinar: (1) se formigas são um grupo que efetivamente
reduz a população de herbívoros através da predação de ovos de lagarta; e (2) se estas taxas de
predação são explicadas pela abundância de predadores (incluindo táxons diferentes das
formigas), ocorrência de formigas e/ou uso da terra no entorno e dentro das propriedades.

90
Metodologia
Áreas de estudo
As coletas foram realizadas dentro do Distrito Federal nos meses de junho e agosto de
2015, em 18 áreas de produção orgânica de hortaliças, chamadas de localidades (Figura 3.1 e
Tabela 3.1). A distância mínima entre as localidades foi de 1,5 km, sendo que todas as áreas
pertencem a agricultores diferentes, com exceção das localidades amostradas Malunga 1 e
Malunga 2. As culturas plantadas em cada localidade são muito variadas, mas incluem
principalmente: alface, couve, cebolinha, salsinha, berinjela, quiabo, jiló, cenoura, cará,
inhame, milho, abobrinha, tomate e pimentão. O método de cultivo também é muito variado,
há localidades em que o talhão todo é de um único tipo de cultivo, outras em que cada linha
de cultivo é de uma espécie diferente de hortaliça e, por último, algumas propriedades
misturam, na mesma linha de cultivo, mais de um tipo de hortaliça. Há ainda o tipo de manejo
agronômico que varia de uma propriedade para outra – como o tipo de adubação ou o uso de
produtos naturais para combate às pragas –, sendo inviável avaliar como isso afeta o controle
biológico pela falta de repetição ou gradiente de mudança entre as propriedades.
Protocolo amostral
Para medir as taxas de predação, foi elaborado um experimento com 24 horas de
duração, para o qual foi desenvolvido um dispositivo (chamado de estaca de predação) que
simula uma planta (Figura 3.2-A), neste eram disponibilizados ovos de Anticarsia gemmatalis
(Hübner, 1818)(Lepidoptera: Noctuidae), um inseto praga de várias culturas (especialmente a
soja), criado em laboratório, dentro da Embrapa Cenargen, para ser utilizado em pesquisas.
Esse dispositivo, com altura de 25 a 30 cm, possui no topo seis “ramos”, os quais seguram,
nas pontas, retângulos de cartolina verde com aproximadamente 5 x 3 cm, simulando folhas.

91
Em cada um desses dispositivos existia: a) um ramo com uma folha com ovos, estes poderiam
ser acessados por todos os possíveis predadores – esta folha foi chamada de tratamento; e b)
um segundo ramo coberto por graxa de lítio (também conhecida como graxa branca), uma
substância inodora, insolúvel e que impede o trânsito de animais não alados, principalmente
formigas, e, na ponta deste ramo, uma folha, chamada de controle. Nesta segunda folha,
também foram disponibilizados ovos, com o cuidado de, em cada estaca de predação, se
colocar um número de ovos igual ou aproximado ao da cartela tratamento.
Devido a variação na densidade de ovos vindo da criação de A. gemmatalis, de acordo
com a idade da colônia, o número de ovos disponibilizado em cada cartela variou de um a 90,
para evitar que isso influencie os resultados, as estacas com baixa densidade (≤20 ovos) foram
alternadas com as de alta densidade (>20 ovos). Além disso, para simular as condições
naturais de postura da maioria dos insetos praga, todos os ovos foram disponibilizados na
página inferior da folha (voltados para baixo). Por fim, todas as cartelas, antes de irem a
campo, tiveram o número de ovos contados e escrito na própria cartela (Figura 3.2-B).
Portanto, pode-se determinar o número de ovos predados logo após a exposição da cartela em
campo.
Além desses dispositivos para medir a predação, foram criadas estacas de amostragem,
com a finalidade de amostrar a fauna que acessa as estacas de predação (Figura 3.2-C). Para
isso, no topo de uma estaca, também com altura de 25 a 30 cm, foi instalado um coletor
universal de 100 ml, que em campo era preenchido até a metade de seu volume por água com
detergente, na borda deste coletor foi derramada uma pequena quantidade de mel como
atrativo. Além deste coletor, havia um ramo isolado por graxa de lítio, com um pedaço de
cartolina verde, de 5 x 5 cm, na sua ponta, o qual continha ao menos 20 ovos de A.
gemmatalis e um pedaço de 5 x 3 cm de armadilha adesiva amarela (Figura 3.2-D). Portanto,
foi possível amostrar a fauna que sobe nas estacas, separando-se a que potencialmente acessa

92
os ovos do tratamento (capturadas no coletor universal) ou do controle (capturadas na
armadilha adesiva amarela).
Desenho amostral
Em cada localidade foram instaladas 60 estacas, sendo, 40 estacas de predação e 20
estacas de amostragem, totalizando 1.080 estacas nas 18 localidades. As estacas foram
distribuídas seguindo as linhas de cultivo, a uma distância de sete metros uma da outra, e
sendo sempre instaladas dentro do canteiro na seguinte ordem: estaca de amostragem, estaca
de predação com cartelas contendo de 1 a 10 ovos, estaca de predação com cartelas contendo
de 11 a 90 ovos e, assim, repetidamente. Portanto, foram instaladas, em cada localidade, 20
estacas de amostragem e 40 de predação, o que em linha reta teria um comprimento de 413
metros. No entanto, como os talhões geralmente são menores do que 40 metros de
comprimento, uma nova linha era instalada paralelamente a esta, a sete metros de distância
também. Por fim, tomou-se o cuidado de alternar as estacas de predação com baixas e com
altas densidades de ovos, para evitar a concentração de ovos em determinada região da
localidade e enviesar os resultados.
Análise da paisagem
Para realizar a categorização da paisagem e testar se esta influencia nas taxas de
predação de ovos, foram utilizadas imagens disponibilizadas pela Codeplan (Companhia de
Planejamento do Distrito Federal) no site ortofoto.mapa.codeplan.df.gov.br/demo/tms. Essas
imagens foram capturadas por um VANT (veículo aéreo não tripulado), na mesma época da
coleta (junho a agosto de 2015), e com uma resolução de 24 cm. Em seguida, utilizando-se o
programa ArcGIS 10.2.2 e ArcGIS Pro, foram mapeadas as áreas de: cobertura arbórea,
construção, com plantio e sem plantio (Tabela 3.2).

93
A cobertura arbórea foi definida como toda árvore ou conjunto de árvores, com
diâmetro da copa superior a aproximadamente 4 metros nas imagens aéreas. As áreas de
construção foram definidas como toda e qualquer área com edificações cobertas, cimentada
ou asfaltada, incluindo açudes e estradas de terra bem definidas. Plantio foi determinado
como a área plantada, a qual era identificada pela presença de linhas de plantio, geralmente
apresentando formas geométricas que facilitaram sua identificação. No entanto, para essa
classificação não foi possível determinar se esta área corresponde ao plantio com manejo
convencional (usando agrotóxicos) ou ao orgânico. O restante, que foi chamado de sem
plantio, basicamente, trata-se de pastagens, terrenos abandonados, potencialmente abrigando
também áreas de vegetação nativa de campo ou de cerrado stricto sensu, com diversos graus
de perturbação. Estas áreas não puderam ser classificadas de forma mais precisa porque estão
distribuídas em um gradiente imperceptível pelas imagens aéreas.
Posteriormente a essa classificação, foi contabilizada a área em m² de cada uma dessas
categorias em “buffers” (ou seja, uma área) com raios de 1.000, 500, 250 e 125 metros, a
partir do ponto central do conjunto de estacas instalado em cada localidade.
Análise dos dados
Todas as análises foram realizadas no programa R (R Development Core Team 2008).
Inicialmente foi aplicado um teste-t pareado, a fim de testar se as formigas efetivamente
contribuem para o aumento das taxas de predação nas cartelas tratamento em relação ao
controle, sendo a estaca o fator de pareamento. Logo após isso foram realizadas seleções de
modelos para verificar se a paisagem influencia as taxas de predação de ovos. Nesta análise
foram avaliadas três variáveis respostas: (1) o número de ovos predados no tratamento
(predação realizada por formigas e outros predadores), (2) o número de ovos predados no
controle (predação causada por artrópodes alados, excluindo-se formigas) e (3) o número de

94
ovos predados no tratamento menos o controle (predação atribuída às formigas). As variáveis
explanatórias foram as áreas das categorias da paisagem (cobertura arbórea, plantio, sem
plantio e construção) e todas as possíveis interações entre duas destas variáveis. Para cada
uma destas variáveis respostas foram construídos quatro modelos, um para cada buffer (1.000
m, 500 m, 250 m e 125 m).
O procedimento de seleção de modelos foi realizado pelo processo “backward”,
utilizando a função lm do R (R Development Core Team 2008). Nesse processo começou-se
com um modelo contendo todas as variáveis, e então a variável com maior p foi removida, e
novamente o modelo foi testado, até que restassem somente variáveis significativas. Além
desse processo, a cada novo modelo com uma variável removida, este foi comparado ao
anterior para verificar se ele sofre alterações significativas, utilizando-se, para isso, a função
anova do R (R Development Core Team 2008). Por fim, para determinar o melhor modelo, foi
aplicado o critério de Akaike, com a função AIC (Sakamoto et al. 1986) do R (R
Development Core Team 2008).
Para determinar se o número de ovos predados está relacionado ao número de
predadores, foram realizadas três regressões simples: (a) número de ovos predados no
tratamento em função do número total de predadores; (b) número de ovos predados no
controle em função dos predadores excluindo-se as formigas; e (c) número de ovos predados
no tratamento menos o controle em função do número de ocorrência de formigas. É
necessário esclarecer que, como Formicidae é um táxon social, o que leva a superestimava da
abundância quando ocorre o recrutamento, apenas para esse grupo foi considerado como
abundância o número de ocorrências registradas nos diferentes pontos (e.g. Vasconcelos et al.
2014). Além desses testes, como Formicidae foi o único táxon identificado até gênero,
seguindo a chave de Bolton et al. (2007) e Fernández (2003), e posteriormente até morfotipo
ou espécie, apenas para esse táxon foi realizada uma correlação entre ocorrência e riqueza.

95
Figura 3.1. Mapa das 18 localidades de agricultura orgânica amostradas no Distrito Federal no ano de 2015 e
dois exemplos de buffers categorizados em cobertura arbórea, plantio e construção.

96
Figura 3.2. Imagens da metodologia empregada nas 18 localidades de produção de hortaliças orgânicas do
Distrito Federal no ano de 2015. (A) Fotografia de uma das 720 estacas de predação em campo, onde eram
disponibilizadas duas cartelas com ovos de A. gemmatalis. (B) Fotografia das cartelas de ovos após 24 horas em
campo, com o número de ovos iniciais escritos a lápis, sendo que na cartela à esquerda ocorreu a predação de
todos os ovos. (C) Fotografia de uma das 320 estacas de amostragem, contendo um coletor universal e uma
cartela com armadilha amarela adesiva. (D) Fotografia de uma das cartelas de amostragem que ficaram em
campo, com uma tira contendo ovos como atrativo na parte superior e coberto por plástico transparente para
armazenagem. Setas indicam os locais em (A) e (C) que contêm graxa branca impedindo a passagem de
formigas.

97
Tabela 3.1. Localização e data de coleta das propriedades amostradas no Distrito Federal no ano de 2015.
Localidade Latitude Longitude Data de coleta
Juã 15°33'53.99"S 48° 1'53.87"O 11-06-2015 Corujinha 15°37'14.28"S 48° 3'52.44"O 06-08-2015 Cleide e Marlene 15°36'43.64"S 48° 4'42.64"O 11-06-2015 Chácara Guaruja 15°40'19.19"S 48° 9'52.90"O 06-08-2015 Valdir 15°39'2.36"S 48°12'2.68"O 29-06-2015 Valdimar 15°49'31.68"S 48°15'7.85"O 29-06-2015 Massae 15°49'49.08"S 48° 4'13.74"O 18-06-2015 Geranium 15°50'44.80"S 48° 4'20.94"O 18-06-2015 Heleno 15°53'9.91"S 48° 7'57.98"O 17-08-2015 FAL (Faz. Água Limpa) 15°56'59.04"S 47°56'0.81"O 19-08-2015 Buriti Alegre 15°58'15.59"S 47°53'38.28"O 22-06-2015 Idalercio 15°58'4.08"S 47°50'27.10"O 22-06-2015 Valter 15°55'36.51"S 47°46'1.40"O 25-06-2015 Alaercio 16° 0'50.76"S 47°44'47.17"O 25-06-2015 Zé Preto 15°44'57.67"S 47°38'54.86"O 15-06-2015 José Balde 15°45'44.00"S 47°38'30.58"O 15-06-2015 Malunga 1 15°58'40.96"S 47°29'46.19"O 03-08-2015 Malunga 2 15°57'59.41"S 47°30'17.73"O 03-08-2015
Tabela 3.2. Porcentagem mínima e máxima da ocorrência de diferentes categorias da paisagem no entorno das
18 localidades de produção de hortaliças orgânicas amostradas no Distrito Federal, no ano de 2015.
Buffer 1000m Buffer 500m Buffer 250m Buffer 125m
Mínimo Máximo Mínimo Máximo Mínimo Máximo Mínimo Máximo
Construção 2,8% 37,8% 2,2% 43,9% 2,8% 40,4% 1,8% 21,7%
Cobertura arbórea 2,7% 38,7% 3,4% 46,9% 3,2% 35,9% 2,8% 41,4%
Plantio 0,2% 31,2% 0,5% 45,3% 2,0% 57,0% 7,9% 79,7%
Sem plantio 33,1% 84,5% 24,3% 86,7% 18,3% 81,7% 9% 78,3%

98
Resultados
Desconsiderando as formigas, foram coletados 3.631 espécimes de artrópodes (Figura
3.3), entre estes, 406 predadores (Figura 3.4). Os predadores mais comuns foram Coleoptera
(178 indivíduos), destacando-se a família Staphylinidae (105) e Coccinellidae (71). A ordem
Diptera foi a segunda mais comum entre os predadores, com a família Dolichopodidae (128),
porém estes são predadores preferenciais de animais em voo e de pequeno porte (Harterreiten-
Souza, comunicação oral), não sendo observadas em campo a predação de ovos de A.
gemmatalis, por isso não foram incorporados às análises dos predadores. Os outros
predadores encontrados foram Hemiptera-Heteroptera (35), Araneae (27), Hymenoptera-
Vespoidea (25), Neuroptera (6), Psocoptera (2), Pseudoscorpionida (1), Coleoptera-Melyridae
(1) e Coleoptera-Carabidae (1).
A utilização de graxa branca para evitar a passagem de formigas demonstrou ser
eficiente, apesar de não ser infalível (Figura 3.4). Ao todo, foram capturados 1.831 indivíduos
de formigas, em 244 ocorrências (Tabela 3.3), e distribuídos em 35 espécies com potencial
para atuar como predadoras dos ovos. Além destas, foi realizado um registro de Atta laevigata
(Smith, 1858), uma espécie de formiga que atua funcionalmente como herbívora. As espécies
de formigas mais frequentes foram Linepithema micans (Forel, 1908), Dorymyrmex cf.
brunneus (Forel, 1908) e Linepithema cf. cerradense (Wild, 2007), todas da subfamília
Dolichoderinae. Porém, a espécie com maior número de indivíduos foi Pheidole gertrudae
(Forel, 1886), da subfamília Myrmicinae. De modo geral, as dez espécies mais comuns de
formigas representaram mais de 80% da ocorrência de formigas.
O número de ovos predados entre controle e tratamento (Figura 3.5) demostra que
formigas são seus principais predadores (teste-t pareado, t = 12.445, df = 717, p < 0,0001).
Em média a porcentagem estimada de ovos predados é de aproximadamente 39,8% por dia,

99
dos quais 26,8% são por formigas e 13% por outros predadores. Há, porém, muita variação
entre as propriedades, tanto na predação total, variando de 9,4% até 63,2% dos ovos, como na
predação só por formigas (de 5,3% até 52,4%) ou só por outros predadores (4,2% até 21%).
Análises de modelos lineares mostram que o número total de ovos predados e o de
ovos predados por formigas em cada localidade orgânica não sofre influência da paisagem
(Tabela 3.4). No entanto, área plantada no raio de 500 metros influencia positivamente os
ovos predados no controle, além da influência de interações com o resto da paisagem.
Em contrapartida, as regressões entre o número de ovos predados e o número de
predadores foram todas significativas (Figura 3.6). O número de ovos predados no tratamento
é positivo e significativamente relacionado com o número total de predadores (Nº de ovos
predados = 129,4 + 4,878 x Nº de predadores, R²=0,41, p=0,004). Assim como o controle (Nº
de ovos predados = 49,8 + 2,6 x Nº de predadores não formigas, R² = 0,27, p = 0,025) e o
tratamento menos o controle (Nº de ovos predados = 110,5 + 5,185 x Nº de ocorrência de
formigas, R² = 0,30, p = 0,017). Por fim, também se encontrou uma correlação para formigas
entre sua ocorrência e a riqueza de espécies (t = 7,5254, R=0,88 , p<0,001).
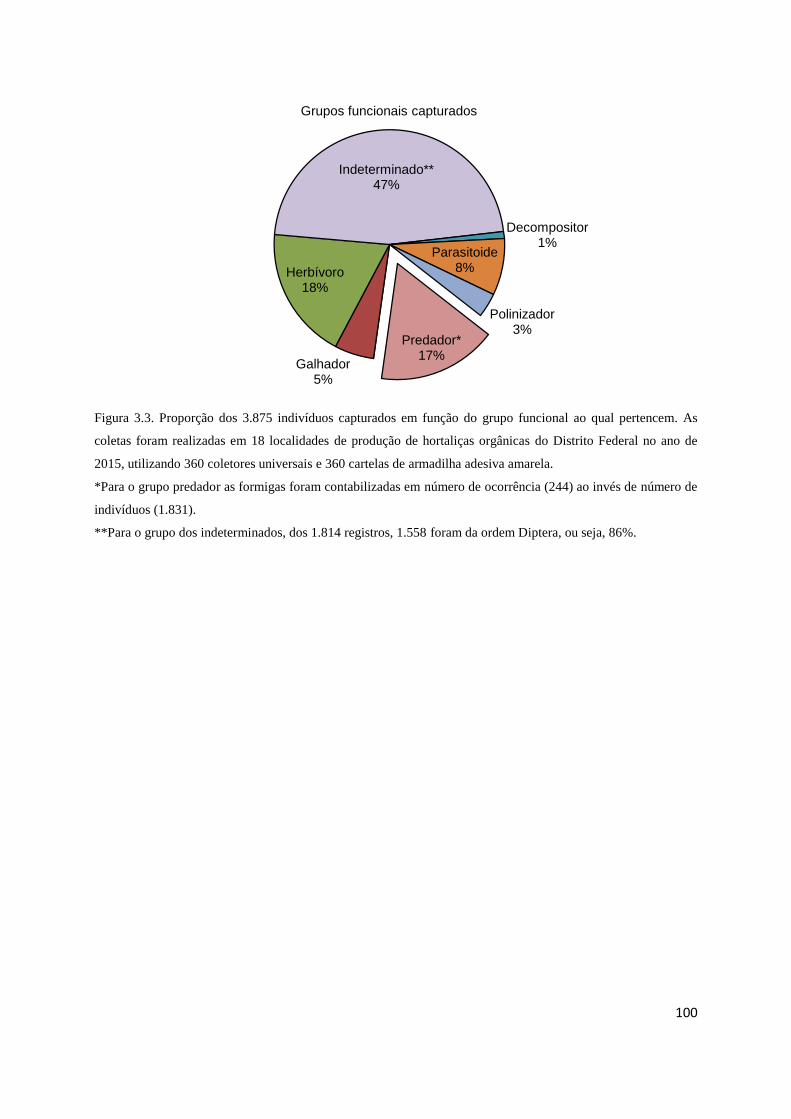
100
Figura 3.3. Proporção dos 3.875 indivíduos capturados em função do grupo funcional ao qual pertencem. As
coletas foram realizadas em 18 localidades de produção de hortaliças orgânicas do Distrito Federal no ano de
2015, utilizando 360 coletores universais e 360 cartelas de armadilha adesiva amarela.
*Para o grupo predador as formigas foram contabilizadas em número de ocorrência (244) ao invés de número de
indivíduos (1.831).
**Para o grupo dos indeterminados, dos 1.814 registros, 1.558 foram da ordem Diptera, ou seja, 86%.
Galhador 5%
Herbívoro 18%
Indeterminado** 47%
Decompositor 1%
Parasitoide 8%
Polinizador 3%
Predador* 17%
Grupos funcionais capturados

101
Figura 3.4. Proporção do táxon de predadores capturados na armadilha adesiva amarela (170 registros) e no
coletor universal (352 registros). As coletas foram realizadas em 18 localidades de produção de hortaliças
orgânicas do Distrito Federal no ano de 2015, utilizando 360 coletores universais e 360 cartelas de armadilha
adesiva amarela. Nessa figura, a família Dolichopodidae, com 128 indivíduos, foi suprimida.
*Para a Família Formicidae as formigas foram contabilizadas em número de ocorrência (244) ao invés de
número de indivíduos (1.831).
Araneae 4%
Vespoidea 6%
Coccinellidae 35%
Heteroptera 15%
Staphylinidae 29%
Formicidae* 7%
Outros 4%
Predadores amostrados na armadilha adesiva amarela
Araneae 6%
Vespoidea 4%
Coccinellidae 3%
Heteroptera 3%
Staphylinidae 16%
Formicidae* 66%
Outros 2%
Predadores amostrados no coletor universal

102
Figura 3.5. Número de ovos de A. gemmatalis disponibilizados e predados nas cartelas de controle (sem acesso
às formigas) e tratamento (com acesso às formigas). O experimento foi realizado em 18 localidades, dentro do
Distrito Federal no ano de 2015. Barras representam desvio padrão e letras diferentes indicam diferenças
significativas (p<0,05).
0
2000
4000
6000
8000
10000
12000
14000
Inicial Predado Inicial Predado
Controle Tratamento
Núm
ero
de o
vos
a a
b
c

103
Figura 3.6. Relações lineares entre abundância, predação ou riqueza, de modo que: (a) ovos predados nas
cartelas tratamento em relação a todos os predadores; (b) ovos predados nas cartelas controle em relação aos
predadores exceto formigas; (c) ovos predados no tratamento menos o controle (predação atribuída às formigas)
em relação à ocorrência de formigas; e (d) correlação entre a riqueza de formigas e sua ocorrência. Para todas as
regressões e a correlação, os resultados foram significativos (p<0,05). Esses dados são baseados no experimento
realizado em 18 localidades, dentro do Distrito Federal no ano de 2015.
0
40
80
120
160
200
0 20 40
Núm
ero
de o
vos p
redados
Número de predadores (exceto formigas)
b) Controle
0
2
4
6
8
10
12
14
16
0 10 20 30 40
Riq
ueza d
e f
orm
igas
Ocorrência de formigas
d) Correlação
0
100
200
300
400
500
600
0 20 40 60
Núm
ero
de o
vos p
redados
Número de predadores
a) Tratamento
0
50
100
150
200
250
300
350
0 10 20 30 40
Núm
ero
de o
vos p
redados
Ocorrência de formigas
c) T - C

104
Tabela 3.3. Lista de espécies de formigas potencialmente predadoras de ovos de A. gemmatalis capturadas nas
360 estacas de amostragem. As coletas foram realizadas em 18 localidades de produção de hortaliças orgânicas
do Distrito Federal no ano de 2015.
Espécies
Nº de localidades
com ocorrência
Nº de indivíduos capturados
Ocorrência Porcentagem
acumulativa de ocorrência
Linepithema micans 11 371 44 18%
Dorymyrmex cf. brunneus 10 245 33 32%
Linepithema cf. cerradense 7 154 22 41%
Camponotus sp.12 9 26 22 50%
Camponotus sp.16 8 57 20 58%
Pheidole gertrudae 7 493 14 64%
Dorymyrmex cf. jheringi 5 82 14 69%
Pheidole sp.01 7 104 12 74%
Pheidole sp.24 5 102 8 77%
Solenopsis saevissima 4 44 8 81%
Camponotus gr. senex 6 13 6 83%
Brachymyrmex sp03 4 47 5 85%
Pheidole sp.23 3 14 4 87%
Camponotus gr. blandus 3 7 4 89%
Pheidole sp.13 1 9 2 89%
Nylanderia sp.03 1 8 2 90%
Pheidole sp.20 2 7 2 91%
Pheidole sp.22 2 7 2 92%
Brachymyrmex sp.01 2 6 2 93%
Labidus praedator 2 6 2 93%
Brachymyrmex sp.02 2 2 2 94%
Pseudomyrmex termitarius 1 6 1 95%
Camponotus sp.17 1 4 1 95%
Crematogaster sp.10 1 3 1 95%
Pheidole sp.03 1 3 1 96%
Camponotus arboreus 1 2 1 96%
Camponotus atriceps 1 1 1 97%
Camponotus sp.18 1 1 1 97%
Camponotus sp.46 1 1 1 98%
Cardiocondyla sp.01 1 1 1 98%
Pheidole sp.02 1 1 1 98%
Pheidole sp.04 1 1 1 99%
Pheidole sp.37 1 1 1 99%
Pseudomyrmex rufiventris 1 1 1 99%
Solenopsis sp.02 1 1 1 100%
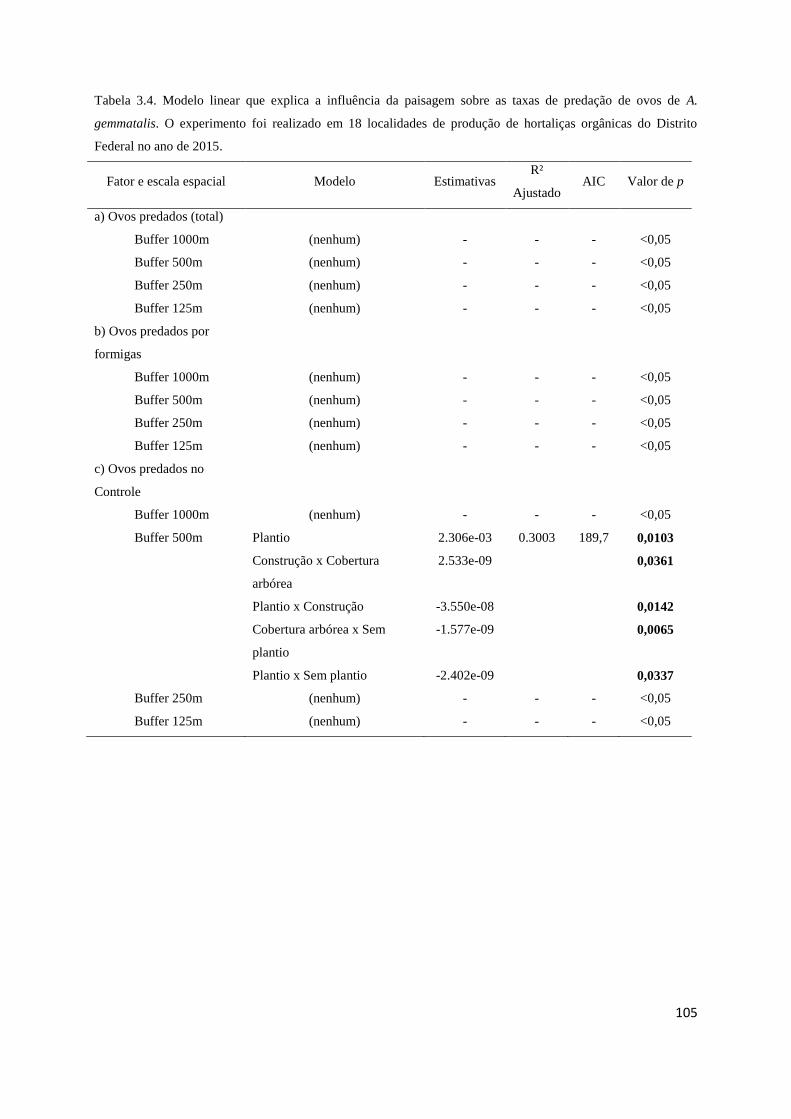
105
Tabela 3.4. Modelo linear que explica a influência da paisagem sobre as taxas de predação de ovos de A.
gemmatalis. O experimento foi realizado em 18 localidades de produção de hortaliças orgânicas do Distrito
Federal no ano de 2015.
Fator e escala espacial Modelo Estimativas R²
Ajustado AIC Valor de p
a) Ovos predados (total)
Buffer 1000m
(nenhum)
-
-
-
<0,05
Buffer 500m (nenhum) - - - <0,05
Buffer 250m (nenhum) - - - <0,05
Buffer 125m (nenhum) - - - <0,05
b) Ovos predados por
formigas
Buffer 1000m
(nenhum)
-
-
-
<0,05
Buffer 500m (nenhum) - - - <0,05
Buffer 250m (nenhum) - - - <0,05
Buffer 125m (nenhum) - - - <0,05
c) Ovos predados no
Controle
Buffer 1000m
(nenhum)
-
-
-
<0,05
Buffer 500m Plantio
Construção x Cobertura
arbórea
Plantio x Construção
Cobertura arbórea x Sem
plantio
Plantio x Sem plantio
2.306e-03
2.533e-09
-3.550e-08
-1.577e-09
-2.402e-09
0.3003 189,7 0,0103
0,0361
0,0142
0,0065
0,0337
Buffer 250m (nenhum) - - - <0,05
Buffer 125m (nenhum) - - - <0,05

106
Discussão
Conseguiu-se, neste estudo, isolar os efeitos de predadores alados em relação aos não
alados, atráves do uso de graxa branca, assim como também realizado por Rabelo & Francini
(2014) (Figuras 3.4 e 3.5). Apesar deste isolamento não ser completo – visto que 12 formigas
também foram encontradas nas armadilhas adesivas amarelas –, podemos considera-lo
eficiente. Evidentemente formigas não foram os únicos predadores isolados durante o
experimento, além destas, em tese também podem ter sido isoladas aranhas, ninfas e larvas
predadoras, as quais também podem ter contribuído para as taxas de predação no tratamento.
No entanto, a ocorrência destes predadores foi similar à ocorrida no controle, e desprezível se
comparada à de formigas.
Sendo assim, os dados sugerem fortemente que as formigas são os principais
predadores de ovoposições de A. gemmatalis em hortaliças ou arbustos nas áreas de
agricultura orgânica (Figura 3.5). O mesmo também foi observado em outros estudos em
diferentes países: na Colômbia, Armbrecht & Gallego (2007) mediram uma taxa de predação
por formigas de 30,5% sobre a broca-do-café (Hypothenemus hampei) em cinco dias; nos
EUA, Karban et al. (2013) encontraram de 20% a 40% de predação sobre a lagarta
Platyprepia virginalis em oito dias; e López & Potter (2000) encontraram uma predação de
65,5% a 71,8% sobre ovos da lagarta (Agrotis ipsilon), durante 22 horas em campos de golfe.
Offberg (2015) faz uma revisão, com estudos da Oceania e sudeste Asiático, demonstrando
que, de 17 publicações diferentes, formigas do gênero Oecophilla só não reduziram o
tamanho populacional de pragas em um dos estudos.
Dentre as espécies de formiga amostradas, fica evidente que poucas são responsáveis
pela maior parte da predação (Tabela 3.3). Surpreendentemente as três espécies mais
frequentes capturadas nas estacas foram da subfamília Dolichoderinae, pertencentes aos

107
gêneros Linepthema e Dorymyrmex, reconhecidamente um grupo que apresenta tolerância
térmica a altas temperaturas e preferência por locais abertos (Farji-Brener et al. 2002; Cuezzo
2011), comuns nos agrossistemas do Cerrado. Geralmente, os estudos têm apontado os
gêneros Pheidole e Solenopsis como os principais predadores nas áreas agrícolas neotropicais
(Risch & Carroll 1982: Armbrecht et al. 2007), principalmente por terem características de
predadoras generalistas, grande capacidade de recrutamento e subjugarem presas muitas vezes
maiores que elas próprias. Mas o que se encontrou é que, apesar destes gêneros dominarem de
fato a superfície do solo (Frizzo, dados do Capítulo I e II), a habilidade das espécies dos
gêneros Dorymyrmex e Linepithema, assim como secundariamente o gênero Camponotus, de
subir nas plantas arbustivas, confere a esses o papel de principais predadores de ovos de A.
gemmatalis sobre as hortaliças. Porém, as formigas da subfamília Myrmicinae, que somam
24% das ocorrências e são representadas principalmente pelo gênero Pheidole, tiveram as
maiores médias de indivíduos recrutados por ocorrência (média de 13,4 indivíduos) em
comparação à subfamília Dolichoderinae (7,5) e Formicinae (2,6).
Complementando o papel desempenhado pelas formigas, uma metanálise com estudos
em agrossistemas encontrou que predadores generalistas controlam a abundância de
herbívoros em 79% das publicações analisadas, e em 65% dos estudos reduzem o dano na
planta ou aumentam o rendimento da produção (Symondson et al. 2002). De fato, a existência
de cultivos orgânicos comerciais só é possível devido à presença dos predadores que
controlam a ocorrência de pragas (e.g. Birkhofer et al. 2016). Mas além das formigas, outros
predadores das ordens Coleoptera, Dermaptera, Neuroptera e Diptera também contribuem
substancialmente para esse serviço ecológico (Harterreiten-Souza et al. 2014; Togni et al.
2016).
Quando se analisa o controle de pragas em sistemas agrícolas, é importante ressaltar
que a identidade da praga afeta a capacidade desta de ser controlada por um determinado

108
táxon de predador. Por exemplo, formigas evidentemente são ótimos predadores de ovos,
como visto aqui, porém pragas aladas, como adultos da mosca branca (Bemisia tabaci,
Gennadius 1889), em tese, dificilmente seriam predadas por formigas, devido ao seu tamanho
reduzido e à capacidade de voo, sendo este alvo preferencial de alguns predadores das ordens
Hemiptera, Neuroptera e Coleoptera (Naranjo 2001). Outro exemplo disso é presença de
afídeos, um fitófago que pode se associar mutualisticamente às formigas sendo protegido por
estas (Styrsk & Eubanks 2007). Assim, joaninhas (Coccinellidae) atuariam como predadores
complementares para afídeos e redundantes para ovos (Togni et al. 2016).
Para melhor demonstrar o quanto a predação pode ser benéfica aos produtores,
podemos simular os cálculos de herbívoria foliar. Segundo Bueno et al. (2011), em toda a
faze larval da A. gemmatalis, ela consome em média 89 cm² de área folhar de soja (variando
entre tipos de cultivar). Supondo que todos os ovos predados deste experimento gerassem
larvas, que tivessem consumido 89 cm²/indivíduo de área foliar, em média, cada localidade
sofreria um dano de 2,4m²/dia de perda de área foliar. Sendo, um terço desta perda foliar
evitado por predadores não alados e o restante pela predação exercida por formigas.
Muitos estudos têm sugerido que a paisagem tem importância, sendo ela provedora de
biodiversidade e podendo incrementar a presença de predadores de múltiplos táxons e
consequentemente aumentar o potencial do serviço ecológico de controle biológico (Rusch et
al. 2013). No entanto, não foi isso que se encontrou neste estudo, já que se observa que a
paisagem não tem influência para o total de ovos predados e aqueles predados apenas por
formigas (Tabela 3.5). Assim como visto em outros estudos, a influência e importância
relativa de cada fator da paisagem variam de acordo com o táxon estudado (Winqvist et al.
2011; Lemessa et al. 2015). Segundo Bikhofer et al. (2016), em sistemas orgânicos é possível
manter o controle biológico de pragas, independente da paisagem, através do manejo delas.

109
Mas esse manejo depende do tipo de praga, do predador e da interação que existe entre esses
dois.
Ao contrário das nossas expectativas, a cobertura arbórea na paisagem, que está ligada
principalmente a ocorrência de formações florestais, não teve efeito isolado do resto da
paisagem sobre a predação de ovos exercida por nenhum predador. Para formigas, isso reflete
a especialização de habitats, assim como visto no Capítulo I e por Pacheco & Vasconcelos
(2012), pois a comunidade de formigas das áreas florestais é um grupo de espécies diferente
das que ocorrem nos agrossistemas. No entanto, outro estudo no Cerrado demonstrou que a
presença de árvores espalhadas nos sistemas agrícolas aumenta a riqueza local de espécies de
formigas (Frizzo & Vasconcelos 2013), o que está correlacionado com a ocorrência de
formigas (Figura 3.6) e possivelmente pode gerar um incremento nas taxas de predação, mas
ainda sem comprovação científica.
Para os predadores, excluindo-se formigas, a cobertura arbórea tem efeito sobre as
taxas de predação interagindo com outras categorias da paisagem (Tabela 3.4). Assim como
visto por Harterreiten-Souza et al. (2014), nessa região, a formação florestal aumenta a
diversidade tanto de herbívoros como de predadores nos cultivos do entorno, servindo de
fonte de inimigos naturais. A presença de formações florestais na paisagem está relacionada
ao aumento de recursos alternativos e à disponibilidade de local de refúgio para períodos de
baixa disponibilidade de recurso, tanto para predadores como para pragas (revisado por
Landis et al. 2000; Langellotto & Denno 2004).
Já a presença de plantio na paisagem, significativamente aumenta as taxas de predação
por predadores alados (e.g. Liere et al. 2015), provavelmente, porque está ligada a hipótese de
concentração de recurso (Root 1973) e por manter um conjunto de espécies regional já
adaptado aos agrossistemas. Porém é importante resaltar que proporções muito altas de
plantio convencional na paisagem – acima dos 31,2% da paisagem, como analisado aqui –,

110
têm demonstrado efeito negativo sobre a biodiversidade e seus serviços (Langellotto & Denno
2004; Tscharntke et al. 2005). Além disso, como visto na Tabela 3.4, existem várias
interações entre as categorias da paisagem, indicando que o resto da paisagem também
influencia a predação de ovos por predadores alados, mesmo que essa influência para as
interações sejam padrões complexos e com um modelo que se ajuste relativamente mal aos
dados (r² ajustado=0,30).
Entretanto, para explicar as taxas de predação foram encontradas relações
significativas com a abundância de todos os predadores, além dos predadores alados e
formigas (Figura 3.6). Isso provavelmente está ligado à hipótese de mais indivíduos
(Srivastava & Lawton 1998), segundo a qual, quanto mais recurso, maior é quantidade de
consumidores. Neste caso, uma quantidade maior de recurso nas propriedades implica em um
número maior de predadores e, consequentemente, em maiores taxas de predação. Isso sugere
um sucesso na capacidade das propriedades com altas taxas de predação em conseguir
aumentar o número de predadores, porém não se foi capazes de determinar o que causa essa
maior abundância. Na literatura, já é sabido que a disponibilidade de recursos florais é um
alimento alternativo para predadores como joaninhas (Lundgren 2009). Além disso, as
fazendas podem ser manejadas para criar uma “infraestrutura ecológica” adequada para
aumentar o controle biológico (revisado por Landis et al. 2000). Também se observou que,
aparentemente, o uso de matéria orgânica (folhas, serragem, casca de arroz entre outros) para
adubação pode estar ligado a um aumento na disponibilidade de presas alternativas
(detritívoros) para as formigas. Entretanto, isso é baseado em observações de campo,
realizadas principalmente na propriedade Juã. Além disso, dados na literatura sugerem que o
aumento da complexidade do ambiente com o incremento de serrapilheira teria o efeito
inverso ao sugerido aqui, reduzindo a probabilidade de predação (Gibb & Parr 2010; Karban
et al. 2013).

111
Conclusão
As formigas, apesar do desconhecimento por parte dos produtores, é um importante
grupo de insetos predadores, capazes de reduzir a população de ovos de uma lagarta praga nas
plantas de fazendas orgânicas numa média de 26,8% ao dia. Destaca-se também que os outros
predadores amostrados consistem principalmente de duas famílias da ordem Coleoptera
(Coccinellidae e Staphylinidae) que, juntamente com outros táxons de predadores, removeram
13% dos ovos disponíveis.
A paisagem apresenta um efeito limitado apenas sobre a predação realizada pelos
predadores alados, sendo influenciados positivamente por áreas de plantio e por interações
entre o resto da paisagem, em um raio de 500 metros. Foi encontrado por fim, uma relação
positiva entre a abundância dos predadores e suas respectivas taxas de predação. O que indica
que o sucesso da conservação do controle biológico, dentro de áreas orgânicas, está associado
a capacidade desses sistemas de conseguir manter uma grande quantidade de predadores.

112
Considerações finais
Padrões encontrados
A redução da biodiversidade local, regional e global, devido à atividade agrícola, é
aparentemente um padrão universal, mas a intensidade dessa redução é dependente da
proporção e tipo de uso da terra (e.g. Sala et al. 2000; Rockströn et al. 2009; Newbold et al.
2015). Isso porque tornar sistemas complexos com milhares a milhões de anos de evolução
em sistemas simplificados causa a exclusão de espécies (Tscharntke et al. 2005). Assim como
confirmado por este estudo, no Cerrado não é diferente, mas tão importante quanto detectar o
efeito – redução da biodiversidade – é determinar a magnitude deste efeito, já que ele varia
em função do táxon, tipo de uso da terra (Kessler et al. 2009; Bommarco et al. 2014) e da
região geográfica (Newbold et al. 2015). Isso se evidencia porque se mostrou que o cultivo de
soja convencional é o mais restritivo para a mirmecofauna nativa do Cerrado, contendo
apenas 17% das espécies da região, e mesmo áreas de pastagens e agricultura orgânica sendo
menos impactantes comportam aproximadamente metade da fauna nativa.
A diversidade de espécies de formigas no Cerrado é em grande parte dependente de
espécies raras, comparativamente, por exemplo, à savana Australiana (Campos et al. 2011).
Essas espécies raras são as mais suscetíveis à extinção (Didham et al. 1996; Weiner et al.
2014). E, assim como se mostrou, a riqueza de formigas está correlacionada com: a biomassa
de formigas; a ocorrência de formigas; e determinadas funções ou serviços do ecossistema
como remoção de biomassa e predação. Portanto, ressalta-se que a perda dessas espécies raras
pode ter implicações negativas na biomassa de artrópodes e funções do ecossistema.
As matas comportaram o grupo de espécies de formigas mais singular do estudo. Essas
espécies florestais geralmente não ocorrem nas paisagens agrícolas e, por isso, para as
formigas do Cerrado, áreas de mata na paisagem não aumentam o controle biológico em áreas

113
agrícolas adjacentes. No entanto, destaca-se que para outros táxons matas podem ser fonte de
agentes de controle biológico (Harterreiten-Souza et al. 2014).
Detectaram-se também mecanismos de redundância entre formigas e outros
predadores (sendo os principais pertencentes à Ordem Coleoptera). Mecanismos de
redundância também existem nos cultivos de soja, no qual, na ausência de formigas a
remoção de biomassa de banana (recurso que simula apêndices atrativos de semente e frutas)
é realizada por outros táxons.
Implicações
Na prática, porém, a expansão da agricultura sobre áreas de vegetação nativa no
Cerrado ainda é uma realidade, com apenas 2,2% da área sobre proteção legal e com taxas de
desmatamento superior às da Amazônia (Klink & Machado 2005). Por isso é imperativo que
as ações conservacionistas passem a incluir os sistemas agrícolas (Tscharntke et al. 2015).
Sugere-se também que determinados tipos de uso da terra (como pastagem ou agricultura
orgânica) sejam preferencialmente usados no entornos de áreas de proteção ambiental ou de
relevante biodiversidade como forma de amortização dos efeitos de sistemas intensificados,
como a produção de soja.
Os motivos pelos quais, nos cultivos de soja, o número de espécies e biomassa de
formigas é menor comparativamente aos outros sistemas ainda não foi conclusivamente
estudado. Mas se aponta que arar o solo (Pereira et al. 2010; Köhl et al. 2014) e uso de
agrotóxicos (revisado por Aktar et al. 2009) afetam vários organismos. Além destes, a
redução de local para nidificação das formigas e efeitos indiretos, como competição
interespecífica, também são sugeridos (Philpott & Foster 2005; Armbrecht et al. 2006; Frizzo
& Vasconcelos, 2013; Pacheco et al. 2013).

114
Comparações entre os cultivos orgânicos e de soja são um indicativo de que o fator
mais importante para redução da biodiversidade na soja talvez seja o uso de agrotóxico.
Entretanto isso não é conclusivo, visto que as propriedades de produção orgânica são
produtivas durante todo ano e entremeadas por plantas perenes, o que pode influenciar
positivamente na capacidade das formigas de persistirem nestes ambientes.
Todavia, é um fato, segundo dados da ABRASCO (2015), que nos últimos anos há um
aumento desproporcional no uso de inseticidas, que de 2000 a 2012 teve um crescimento
acumulado de 162,3%, sendo hoje, o Brasil o maior consumidor de agrotóxicos do mundo
(MMA 2016). Em uma revisão, Popp et al. (2013) indicam algumas formas para se reduzir os
possíveis impactos de sistemas altamente manejados como a soja. A primeira, seria aumentar
o uso de cultivos geneticamente modificados, que são resistentes a determinados tipos de
praga, e, consequentemente, demandam um menor consumo de agrotóxicos. A segunda é,
assim como ocorre na Noruega, os agrotóxicos poderiam ser taxados de acordo com sua
toxicidade, elevando o custo daqueles que são potencialmente mais perigosos. Por último,
pode-se utilizar o MIP (manejo integrado de pragas), uma técnica com mais de três décadas
no Brasil. Essa técnica consiste no combate às pragas utilizando várias metodologias, e
recorrendo aos agrotóxicos somente quando a população de pragas atinge um determinado
tamanho (chamado de nível de controle). Assim como discutido no XXVI Congresso de
Entomologia, os agricultores brasileiros abandonaram o uso do MIP e têm realizado
pulverizações preventivas, com a mistura de diferentes produtos e em quantidades
desnecessárias, aumentando inclusive os casos de resistência de pragas.
Em relação às pastagens, elas apresentam desvantagem em comparação com as áreas
de agricultura orgânica. Primeiro que para se produzir 1 kg de carne é necessário utilizar 303
m² de pastagem, o que equivale a 83 vezes mais área do que para produzir 1 kg de cereal, ou
382 vezes mais área do que para produzir 1 kg de raiz ou tubérculo, ou, ainda, 545 vezes mais

115
área do que para produzir 1 kg de hortaliça (Wackernagel et al. 1999). Outro fator de
preocupação é a utilização do capim exótico Melinis minutiflora como forrageira, que é
considerado mais danoso ao meio ambiente que o Urochloa (Syn. Brachiaria) decumbens,
principalmente pela dominância competitiva em relação às espécies nativas e pelos efeitos
indiretos em caso de queimada (Pivello et al. 1999; Hoffmann et al. 2004).
Em contrapartida, a agricultura orgânica contempla muitos pequenos produtores, o que
traz benefícios sociais, ambientais e aumenta a segurança alimentar (Tscharntke et al. 2012).
No entanto, mesmo áreas de agricultura orgânica têm sido submetidas ao que Hall &
Mogyorody (2001) chamam de “convencionalização”, que é um processo de aumento da
produção, respeitando as normas dos produtos orgânicos, mas não os princípios (Blanc 2009).
Isso demonstra que a evolução do agronegócio e a expansão de suas atividades são a principal
ameaça à mirmecofauna do Cerrado, o que provavelmente ocorre com os outros táxons (Silva
et al. 2006).
Por fim, em uma revisão, De Fries et al. (2004) aponta que há um balanço entre as
necessidades humanas imediatas e a manutenção das funções do ecossistema, sendo que esse
equilíbrio é afetado pelos valores da sociedade. Ou seja, o uso exacerbado dos recursos
naturais pode causar prejuízos à sociedade, assim como trazer benefícios imediatos (como
conforto ou lucro financeiro). A medida desse equilíbrio é dada pela informação recebida pela
sociedade, importância que ela dá a estas informações e como esta é interpretada e influencia
suas decisões.

116
Referências Bibliográficas
ABRASCO (2015) Dossiê ABRASCO um alerta sobre os impactos dos agrotóxicos na saúde/
Organização Carneiro FF, Rigotto RM, Augusto LGS, Freidrich K, Búrgico AC.
Associação brasileira de saúde coletiva – ABRASCO. EPSJV Rio de Janeiro.
Aktar MW, Sengupta D, Chowdhury A (2009) Impact of pesticides use in agriculture: their
benefits and hazards. Interdisciplinary Toxicology. 2:1-13.
Allan E, Weisser WW, Fischer M, Schulze ED, Weigelt A, Roscher C, Baade J, Barnard RL,
Beßler H (2013) A comparison of the strength of biodiversity effects across multiple
functions. Oecologia, 173: 223-237.
Almeida S, Louzada J, Sperber C, Barlow J (2011) Subtle land-use change and tropical
biodiversity: dung beetle communities in Cerrado grasslands and exotic pastures.
Biotropica. 43:704–710.
Almeida-Neto M, Guimarães P, Guimarães Jr PR, Loyola RD, Ulrich W (2008) A consistente
metric for nestedness analysis in ecological systems: reconciling concept and
measurement. Oikos. 117:1227-1239.
Andersen MJ, Walsch DCI (2013) PERMANOVA, ANOSIM, and the Mantel test in the face
of heterogeneous dispersions: What null hypothesis are you testing? Ecological
Monographs. 83:557-574.
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance.
Austral Ecology. 26:32-46.
Armbrecht I, Gallego MC (2007) Testing ant predation on the coffee berry borer in shaded
and sun coffee plantations in Colombia. Entomologia Experimentalis et Applicata.
124:261–267

117
Armbrecht I, Perfecto I (2003) Litter-twig dwelling ant species richness and predation
potential within a forest fragment and neighboring coffee plantations of contrasting
habitat quality in Mexico. Agriculture Ecosystems & Environment. 97:107-115.
Armbrecht I, Perfecto I, Silverman E (2006) Limitation of nesting resources for ants in
Colombian forests and coffee plantations. Ecological Entomology. 31:403-410.
Atmar W, Patterson BD (1993) The measure of order and disorder in the distribution of
species in fragmented habitat. Oecologia. 96: 373-382.
Baccaro FB, Feitosa RM, Fernandez F, Fernandes IO, Izzo TJ, de Souza JLP, Solar R (2015)
Guia para os gêneros de formigas do Brasil. INPA, Manaus.
Balmford A, Green R, Phalan B (2012) What conservationists need to know about farming.
Proc Biol Sci. 279:2714–2724.
Balvanera P, Pfisterer AB, Buchmann N, He JS, Nakashizuka T, Raffaelli D, Schmid B
(2006) Quantifying the evidence for biodiversity effects on ecosystem functioning and
services. Ecology letters. 9:1146-1156.
Balzan MV, Bocci G, Moonen AC (2016) Landscape complexity and field margin vegetation
diversity enhance natural enemies and reduce herbivory by Lepidoptera pests on
tomato crop. BioControl. 61:141-154.
Barnosky AD, Matzke N, Tomiya S, Wogan GOU, Swartz B, Quental TB, Marshall C,
McGuire JL, Lindsey EL, Maguire KC, Mersey B, Ferrer EA (2011) Has the Earth’s
sixth mass extinction already arrived? Nature. 471:51-57.
Barton PS, Sato CF, Kay GM, Florance D, Lindenmayer DB (2016) Effects of environmental
variation and livestock grazing on ant community structure in temperate eucalypt
woodlands. Insect Conservation and Diversity. 9:124-134.
Baselga A (2010) Partitioning the turnover and nestedness components of beta diversity.
Global Ecology and Biogeography. 19:134-143.

118
Baselga A, Leprieur F (2015) Comparing methods to separate components of beta diversity.
Methods in Ecology and Evolution. 6:1069-1079.
Belchior C, Del-Claro K, Oliveira PS (2012) Seasonal patterns in the foraging ecology of the
harvester ant Pogonomyrmex naegelii (Formicidae, Myrmicinae) in a Neotropical
savanna: daily rhythms, shifts in granivory and carnivory, and home range. Arthropod-
Plant Interactions. 6:571-582.
Belsley, D. A., Kuh, E. & Welsch, R. E. (1980). Regression Diagnostics: Identifying
Influential Data and Sources of Collinearity. New York: John Wiley.
Benayas LMR, Newton AC, Diaz A, Bullock JM (2009) Enhancement of Biodiversity and
Ecosystem Services by Ecological Restoration: A Meta-Analysis. Science. 325:1221-
1224.
Bestelmeyer BT, Wiens JA (1996) The effects of land use on the structure of ground-foraging
ant communities in the Argentine chaco. Ecological Applications. 6:1225-1240.
Bestelmeyer BT, Wiens JA (2001) Ant Biodivesity in Semiarid Landscape Mosaics: The
consequences of grazing vs. natural heterogeneity. Ecological Applications. 11:123-
1140.
Bihn JH, Gebauer G, Brandl R (2010) Loss of functional diversity of ant assemblages in
secondary tropical forests. Ecology. 91:782-792.
Biofach (2016) Brazilian organic market trends and news. Organizador Ming Liu.
IPD/Organics Brasil.
Birkhofer K, Arvidsson F, Ehlers D, Mader VL, Bengtsson J, Smith HG (2016) Organic
farming affects the biological control of hemipteran pests and yields in spring barley
independent of landscape complexity. Landscape Ecology. 31:567-579.

119
Blanc J (2009) Family farmers and major retail chains in the Brazilian organic sector:
Assessing new development pathways. A case study in a peri-urban district of São
Paulo. Journal of Rural Studies. 25:322-332.
Bolton B, Alpert G, Ward PS, Naskrecki P (2007) Bolton’s catalogue of ants of the world:
1758-2005. Harvard University Press, MA Cambridge.
Bommarco R, Lindborg R, Marini L, Öckinger E (2014) Extinction debt for plants and
flower-visiting insects in landscapes with contrasting land use history. Diversity and
Distributions. 20:591-599.
Boyer AG, Jetz W (2014) Extinctions and the loss of ecological function in island bird
communities. Global Ecology and Biogeography. 23:679–688.
Brandão CRF, Silva RR, Feitosa RM (2011) Cerrado ground-dwelling ants (Hymenoptera:
Formicidae) as indicators of edge effects. Zoologia. 28:379–387.
Brandão CRF, Siva RR, Delabie JHC (2012) Neotropical Ants (Hymenoptera) Functional
Groups: Nutritional and Applied Implications. Editores: Panizzi AR, Parra JRP. Insect
Bioecology and Nutrition for Integrated Pest Manegement. CRC Press. Boca Roton
NW.
Brannstrom C, Jepson W, Filippi AM, Redo D, Xu ZW, Ganesh S (2008) Land change in the
Brazilian Savanna (Cerrado), 1986-2002: Comparative analysis and implications for
land-use policy. Land Use Policy. 25:579-595.
Brittain C, Potts SG (2011) The potential impacts of insecticides on the life-history traits of
bees and the consequences for pollination. Basic and Applied Ecology. 12:321–331.
Brown-Jr KS, Gifford DR (2002) Lepidoptera in the Cerrado Landscape and the Conservation
of Vegetation, Soil, and Topographical Mosaics. Editores: Oliveira PS, Marquis RJ.
The Cerrados of Brazil: Ecology and Natural History of a Neotropical Savanna.
Columbia University Press, New York.

120
Bueno RCOF, Bueno AF, Moscardi F, Parra JRP, Hoffmann-Campo CB (2011) Lepidopteran
larva consumption of soybean foliage: basis for developingmultiple-species economic
thresholds for pestmanagement decisions. Pest Management Science. 67: 170-174.
Cadotte MW, Dinnage R, Tilman D (2012) Phylogenetic diversity promotes ecosystem
stability. Ecology. 93: 223-233.
Campos RI, Vasconcelos HL, Andersen AN, Frizzo TLM, Spena KC (2011) Multi-scale ant
diversity in savanna woodlands: an intercontinental comparison. Austral Ecology.
36:983-992.
Cardinale BJ, Srivastava DS, Duffy JE, Wright JP, Downing AL, Sankaran M, Jouseau C
(2006) Effects of biodiversity on the functioning of trophic groups and
ecosystems. Nature. 443:989-992.
Cavalcanti RB, Joly CA (2002) Biodiversity and conservation priorities in the Cerrado region.
Pp 351–367 in P. S. Oliveira, and R. J. Marquis, editors. The Cerrados of Brazil:
Ecology and Natural History of a Neotropical Savanna. Columbia University Press,
New York.
Chapin FS, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM, Reynolds HL, Hooper DU,
Lavorel S, Sala OE, Hobbie SE, Mack MC, Diaz S (2000) Consequences of changing
biodiversity. Nature. 405:234-242.
Chrisman JR, Koifman S, Sarcinelli PN, Moreira JC,Koifman RJ, Meyer A (2009) Pesticide
sales and adult male cancer mortality in Brazil. Int J Hyg Environ Health. 212:310-
321.
Colwell RK (2013) EstimateS: Statistical estimation of species richness and shared species
from samples. Version 9. Persistent URL <purl.oclc.org/estimates>.

121
Costa AN, Vasconcelos HL, Vieira-Neto EHM, Bruna EM (2008) Do herbivores exert top-
down effects in Neotropical savannas? Estimates of biomass consumption by leaf-
cutter ants. Journal of Vegetation Science. 19: 849–854.
Cuautle M, Vergara CH, Badano EI (2016) Comparison of Ant Community Diversity and
Functional Group Composition Associated to Land Use Change in a Seasonally Dry
Oak Forest. Neotropical Entomology. 45:170-179.
Cuezzo F (2003) Subfamilia Dolichoderinae. In: (Ed) Fernández F. Idroducción a las
Hormigas de la Región Neotropical. Instituto de Investigación de Recursos Biológicos
Alexander von Humboldt, Bogotá.
Daily GC, Alexander A, Ehrlich PR, Goulder L, Lubchenco J, Matson PA, Mooney HA,
Postel S, Schnneider SH, Tilman D, Woodwell GM (1997) Ecosystem Services:
Benefits Supplied to Human Societies by Natural Ecosystems. Issues in Ecology. 2:1-
16.
De-Fries RS, Foley JA, Asner GP (2004) Land-use choices: balancing human needs and
ecosystem function. Frontiers in Ecology and Envirnment. 2:249-257.
De-Groot RS (1992) Functions of Nature: Evaluation of Nature in Environmental Planning,
Management and Decision Making. Wolters-Noordhoff, Groningen.
De-Groot RS, Wilson MA, Boumans RMJ (2002) A typology for the classification,
description and valuation of ecosystem functions, goods and services. Ecological
Economics. 41:393-408.
Delabie, J. H. 2001. Trophobiosis between Formicidae and Hemiptera (Sternorrhyncha and
Auchenorrhyncha): an overview. Neotropical Entomology. 30:501-516.
Del-Claro K, Marquis RJ (2015) Ant Species Identity has a Greater Effect than Fire on the
Outcome of an Ant Protection System in Brazilian Cerrado. Biotropica. 47:459-467.

122
Didham RK, Ghazoul J, Stork NE, Davis AJ (1996) Insects in fragmented forests: a
functional approach. Trends in Ecology & Evolution. 11:255-260.
Diehl E, Sanhudo CED, Diehl-Fleig ED (2004) Ground-dwelling ant fauna of sites with high
levels of copper. Brazilian Journal of Biology. 64:33-39.
Duffy JE (2009) Why biodiversity is important to the functioning of real-world ecosystems.
Frontiers in Ecology and the Environment. 7:437–444.
Dufrêne M, Legendre P (1997) Species assemblages and indicator species: the need for a
flexible asymmetrical approach. Ecological Monographs. 67:345-366.
Durães R, Martins WP, Vaz-De-Mello FZ (Dung Beetle (Coleoptera: Scarabaeidae)
Assemblages across a Natural Forest-Cerrado Ecotone in Minas Gerais, Brazil.
Ecology, Behavior and Bionomics. 34:721-731.
Ehrlich PR (2002) Human Natures, Nature Conservation, and Environmental Ethics.
BioScience. 52: 31-43.
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annual Review of Ecology
Evolution and Systematics. 34:487-515.
FAO (2009) State of Food Insecurity in the World. Food and Agriculture Organization of the
United Nations. Rome, Italy.
Farji-Brener AG, Corley JC (2002) The effects of fire on ant communities in northwestern
Patagonia: The importance of habitat structure and regional context. Diversity and
Distribution. 8:235-243.
Fayle TM, Bakker L, Cheah C, Ching TM, Davey A, Dem F, Earl A, Huaimei Y, Hyland S,
Johansson B, Ligtermoet E, Lim R, Lin LK, Luangyotha P, Martins BH, Palmeirim
AF, Paninhuan S, Rojas SK, Sam L, Sam PTT, Susanto D, Wahyudi A, Walsh J,
Weigl S, Craze PG, Jehle R, Metcalfe D, Trevelyan R (2011) A positive relationship
between ant biodiversity (Hymenoptera: Formicidae) and rate of scavenger-mediated

123
nutrient redistribution along a disturbance gradient in a southeast Asian rain forest.
Myrmecological News. 14:5-12.
Fearnside PM (2001) Soybean cultivation as a threat to the environment in Brazil.
Environmental Conservation. 28:23-38.
Fernández F (2003) Introducción a las hormigas de la región Neotropical. Instituto de
Investigación de Recursos Biológicos Alexander von Humboldt. Bogota
Ferreira AV, Bruna EM, Vasconcelos HL (2011) Seed predators limit plant recruitment in
Neotropical savannas. Oikos. 120:1013-1022.
Ferreira LG, Yoshioka H, Huete Y, Sano EE (2004) Optical characterization of the Brazilian
savanna physiognomies for improved land cover monitoring of the cerrado biome:
preliminary assessments from an airborne campaign over an LBA core site. Journal of
Arid Environments. 56:425-447.
Fittkau EJ, Klinge H (1973) On biomass and trophic structure of the central Amazonian rain
forest ecosystem. Biotropica. 5:2-14.
Flynn DFB, Gogol-Prokurat M, Nogeire T, Molinari N, Richers BT, Lin BB, Simpson N,
Mayfield MM, DeClerck F (2010) Loss of functional diversity under land use
intensification across multiple taxa. Ecology Letters. 12:22-33.
Folgarait PJ (1998) Ant biodiversity and its relationship to ecosystem functioning: a review.
Biodiversity and Conservation. 7: 1221–1244.
Frizzo TLM, Campos RI, Vasconcelos HL (2012) Contrasting Effects of Fire on Arboreal and
Ground‐Dwelling Ant Communities of a Neotropical Savanna. Biotropica. 44:254-
261.
Frizzo TLM, Vasconcelos HL (2013) The Potential Role of Scattered Trees for Ant
Conservation in an Agriculturally Dominated Neotropical Landscape. Biotropica. 45:
644–651.

124
Fusaro S, Gavinelli F, Sommaggio D, Paoletti MG (2016) Higher efficiency in organic than in
conventional management of biological control in horticultural crops in north-eastern
Italy. Biological Control. 97:89-101.
Galen C (2005) Catching ants with honey: an experimental test of distraction and satiation as
alternative modes of escape from flower-damaging ants. Oecologia. 144:80-7.
Gibb H, Parr CL (2010) How does habitat complexity aff ect ant foraging success? A test
using functional measures on three continents. Oecologia. 164:1061-1073.
Gibb H, Stoklosa J, Warton DI, Brown AM, Andrew NR, Cunningham SA (2015) Does
morphology predict trophic position and habitat use of ant species and assemblages?
Oecologia. 177:519-531.
Gonthier DJ, Ennis KK, Philpott SM, Vandermeer J, Perfecto I (2013) Ants defend coffee
from berry borer colonization. BioControl. 58:815-820.
Gotelli NJ, Ellison AM (2011) Princípios de estatística em ecologia. Tradução: Fabrício
Beggiato Baccaro. Artmed. Porto Alegre.
Gray CL, Lewis OT, Chung AYC, Fayle TM (2015) Riparian reserves within oil palm
plantations conserve logged forest leaf litter ant communities and maintain associated
scavenging rates. Journal of Applied Ecology. 52:31-40.
Gregory Jr B (1989) Field observations of mating, oviposition, and feeding behavior of
Anticarsia gemmatalis (Lepidoptera: Noctuidae) in a soybean field. Journal of Insect
Behavior. 2:761-774.
Groom MJ, Gray EM, Townsend AA (2008) Biofuels and Biodiversity: Principles for
Creating Better Policies for Biofuel Production. Conservation Biology. 22:602-609.
Groot AT Dicke M (2002) Insect-resistant transgenic plants in a multi-trophic context. Plant
Journal. 31: 387–406.

125
Gurr GM, Wratten SD, Luna JM (2003) Multi-function agricultural biodiversity: pest
management and other benefits. Basic and Applied Ecology. 4:107-116.
Haddad NM, Crutsinger GM, Gross K, Haarstad J, Knops JM, Tilman D (2009). Plant species
loss decreases arthropod diversity and shifts trophic structure. Ecology Letters. 12:
1029-1039.
Hall A, Mogyorody V (2001) Organic farmers in Ontario: an examination of the
conventionalisation argument. Soc Ruralis. 41:399–422.
Harterreiten-Souza ES, Togni PHB, Milane PVGN, Cavalcante KR, Medeiros MA, Pires
CSS, Sujii ER (2012) Seasonal fluctuation in the population of Harmonia axyridis
(Pallas, 1773) (Coleoptera: Coccinellidae) and co-occurrence with other Coccinellidae
in the Federal District of Brazil. Papéis Avulsos de Zoologia. 52:133-139.
Harterreiten-Souza ES, Togni PHB, Pires CSS, Sujii ER (2014) The role of integrating
agroforestry and vegetable planting in structuring communities of herbivorous insects
and their natural enemies in the Neotropical region. Agroforest Systems. 88:205-2019.
Hausberger B, Korb J (2016) The Impact of Anthropogenic Disturbance on Assembly
Patterns of Termite Communities. Biotropica. 48:356-364.
Hernandez JV, Jaffé K (1995) Dano econômico causado por populações de formigas Atta
laevigata (F. Smith) em plantações de Pinus caribaea Mor. e elementos para o manejo
de praga. An Soc Entomol Brasil. 24: 287-298.
Hoffmann BD, Andersen AN (2003) Responses of ants to disturbance in Australia, with
particular reference to functional groups. Austral Ecology. 28, 444–464.
Hoffmann WA, Lucatelli VMPC, Silva FJ, Azeuedo INC, Marinho MS, Albuquerque ANS,
Lopes AO, Moreira SP (2004) Impact of the invasive alien grass Melinis minutiflora at
the savanna-forest ecotone in the Brazilian Cerrado. Diversity and Distributions.
10:99-103.

126
Hoffmann WA, Orthen B, Nascimento PKV (2003) Comparative fire ecology of tropical
savanna and forest trees. Functional Ecology. 17:720-726.
Hole DG, Perkins AJ, Wilson JD, Alexander IH, Grice PV, Evans AD (2005) Does organic
farming benefit biodiversity? Biological Conservation. 122:113-130.
Hollander M, Wolfe DA (1973) Nonparametric Statistical Methods. John Wiley & Sons. New
York.
Hölldobler B, Lumsden DJ (1980) Territorial Strategies in Ants. Science. 210:732-739.
Hölldobler B, Wilson EO (2009) The Superorganism: The Beauty, Elegance, and Strangeness
of Insect Societies. W. W. Norton & Company Inc, New York.
Hooper DU, Chapin III FS, EwelJJ, Hector A, Inchausti P, Lavorel S, Lawton JH, Lodge DM,
Loreau M, Naeem S, Schmid B, Setala H, Symstad AJ, Vandermeer J, Wardle DA
(2005) Effects of biodiversity on ecosystem functioning: a consensus of current
knowledge. Ecological Monographs. 75:3-35.
Howe A, Lövei GL, Nachman G (2009) Dummy caterpillars as a simple method to assess
predation rates on invertebrates in a tropical agroecosystem. Entomologia
Experimentalis et Applicata. 131:325–329.
Hunter MD (2009) Trophic promiscuity, intraguild predation and the problemof omnivores.
Agricultural and Forest Entomology. 11:125–131.
IPD (2010) Perfil do mercado orgânico brasileiro como processo de inclusão social. Instituto
de Promoção do Desenvolvimento. IPD orgânicos, Curitiba.
IWMI (2007) Water for food, water for life: a comprehensive assessment of water
management in agriculture. Earthscan and Colombo. International Water Management
Institute, London
Jepson W (2005) A disappearing biome? Reconsidering land-cover change in the Brazilian
savanna. Geographical Journal. 171:99-111.

127
Jones DT, Susilo FX, Bignell DE, Hardiwinoto S, Gillison AN, Eggleton P (2003) Termite
assemblage collapse along a land-use intensification gradient in lowland central
Sumatra, Indonesia. Journal of Applied Ecology. 40:380-391.
Karban R, Mata TM, Grof‐Tisza P, Crutsinger G, Holyoak MA (2013) Non‐trophic effects of
litter reduce ant predation and determine caterpillar survival and distribution. Oikos.
122:1362–1370.
Kaspari M. Yanoviak SP, Dudley R (2008) On the biogeography of salt limitation: a study of
ant communities. Proceedings of the National Academy of Scienes of the United States
of America. 105:17848-17851.
Kaspariz M, Yanovlak SP (2001) Bait Use in Tropical Litter and Canopy Ants‐Evidence of
Differences in Nutrient Limitation. Biotropica. 33:207-211.
Kessler M, Abrahamczyk S, Bos M, Buchori D, Putra DD, Gradstein SR, Hohn P, Kluge J,
Orend F, Pltopang R, Saleh S, Schulze CH, Sporn SG, Steffan-Dewenter I,
Tjitrosoedirdjo SS, Tscharntke T (2009) Alpha and beta diversity of plants and
animals along a tropical land-use gradient. Ecological Applications. 19:2142-2156.
Klink CA, Machado RB (2005) Conservation of the Brazilian Cerrado. Conservation Biology.
19:707-713.
Klink CA, Moreira AG (2002) Past and current human occupation, and land use. Pp 69–88 in
P. S. Oliveira, & R. J. Marquis, editors. The Cerrados of Brazil: Ecology and Natural
History of a Neotropical Savanna. Columbia University Press, New York.
Köppen W (1948) Climatologia: con un estudio de los climas de la tierra. Fondo de Cultura
Econômica, México.
Kremen C (2005) Managing ecosystem services: what do we need to know about their
ecology? Ecology Letters. 8:468–879.

128
Landis DA, Wratten SD, Gurr GM (2000) Habitat management to conserve natural enemies
of arthropod pests in agriculture. Annual Review of Entomology. 45:175-201.
Langellotto GA, Denno RF (2004) Responses of invertebrate natural enemies to complex-
structured habitats: a meta-analytical synthesis. Oecologia. 139:1-10.
Larsen TH, Williams NM, Kremen C (2005) Extinction order and altered community
structure rapidly disrupt ecosystem functioning. Ecology Letters. 8:538–547.
Layman CA, Araujo MS, Boucek R, Hammerschlag-Peyer CM, Harrison E, Jud ZR, Matich
P, Rosenblatt A, Vaudo JJ, Yeager L, Post DM Bearhop S (2011) Applying stable
isotopes to examine food web structure: an overview of analytical tools. Biological
Reviews. 87: 545–562.
Lemessa D, Hambäck PA, Hylander K (2015) The effect of local and landscape level land-use
composition on predatory arthropods in a tropical agricultural landscape. Landscape
Ecology. 30:167-180.
Letourneau DK, Armbrecht I, Rivera BS, Lerma JM, Carmona EJ, Daza MC, Escobar S,
Galindo V, Gutiérrez C, López SD, Mejía JL, Rangel AMA, Rangel J H, Rivera L,
Saavedra CA, Torres AM, Trujillo AR (2010) Does plant diversity benefit
agroecosystems? A synthetic review. Ecological Applications. 21:9-21.
Lieri H, Kim TN, Werling BP, Meehan TD, Landis DA, Gratton C (2015) Trophic cascades in
agricultural landscapes: indirect effects of landscape composition on crop yield.
Ecological Applications. 25:652-661.
Lingren PD, Greene GL, Davis DR, Baumhover AH, Henneberry TJ (1977) Nocturnal
Behavior of Four Lepidopteran Pests That Attack Tobacco and Other Crops. Annals of
the Entomological Society of America. 161-167.

129
Lopes CT, Vasconcelos HL (2008) Evaluation of Three Methods for Sampling Ground-
Dwelling Ants in the Brazilian Cerrado. Ecology, Behavior and Bionomics. 37:399-
405.
López R, Potter DA (2000) Ant Predation on Eggs and Larvae of the Black Cutworm
(Lepidoptera: Noctuidae) and Japanese Beetle (Coleoptera: Scarabaeidae) in
Turfgrass. Environmental Entomology. 29:116-125.
Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime JP, Hector A, Hooper DU, Huston MA,
Raffaelli S, Schmid B, Tilman D, Wardle DA (2008) Biodiversity and Ecosystem
Functioning: Current Knowledge and Future Challenges. Science. 294:804-808.
Lu Z, Hoffmann BD, Chen Y (2016) Can reforested and plantation habitats effectively
conserve SW China’s ant biodiversity? Biodiversyt and Conservation. 25:753-770.
Lumley T (2009) Package leaps: regression subset selection. R package version 2.9.
http://CRAN.R-project.org/package=leaps
Lundgren JG (2009) Nutritional aspects of non-prey foods in the life histories of predaceous
Coccinellidae. Biological Control. 51:294-305.
Machado RB, Neto MBR, Pereira PGP, Caldas EF, Gonçalves DA, Santos NS, Tabor K,
Steininger M (2004) Estimativas de perda da área do Cerrado brasileiro. Relatório
técnico não publicado. Conservação Internacional, Brasília - DF.
MAPA (2016a) Ministério da Agricultura Pecuária e Abastecimento. (documento on line)
http://www.agricultura.gov.br/arq_editor/Pasta%20de%20Maio%20-%202016.pdf
/Acessado em: 24/06/2016
MAPA (2016b) Ministério da Agricultura Pecuária e Abastecimento. (documento on line)
http://www.agricultura.gov.br/vegetal/culturas/soja /Acessado em: 24/06/2016

130
MAPA (2016c) Ministério da Agricultura Pecuária e Abastecimento. (documento on
line)http://www.agricultura.gov.br/comunicacao/noticias/2015/09/mercado-brasileiro-
de-organicos-deve-movimentar-rs-2-bi-em-2016 /Acessado em: 24/06/2016
Maravalhas J, Vasconcelos HL (2014) Revisiting the pyrodiversity–biodiversity hypothesis:
long-term fire regimes and the structure of ant communities in a Neotropical savanna
hotspot. Journal of Applied Ecology. 51:1661-1668.
Marichal R, Grimaldi M, Feijo AM, Oszwald J, Praxedes C, Cobo MHR, Hurtado MP,
Desjardins T, Silva-Junior ML, Costa LGS, Miranda IS, Oliveira MND, Brownh J,
Tsélouikoa S, Martins MB, Decaëns T, Velasquez E, Lavelle P (2014) Soil
macroinvertebrate communities and ecosystem services indeforested landscapes of
Amazonia. Applied Soil Ecology. 83:177-185.
Mata RA. Tidon R (2013) The relative roles of habitat heterogeneity and disturbance in
drosophilid assemblage (Diptera, Drosophilidae) in the Cerrado. Insect Conservation
and Diversity. 6:663-670.
McKinney ML, Lockwood JL (1999) Biotic homogenization: a few winners replacing many
losers in the next mass extinction. Trends in Ecology & Evolution. 14:450–453.
Mensens C, Frederik De Laender, Colin R. Janssen, Koen Sabbe and Marleen De Troch
Stressor-induced biodiversity gradients: revisiting biodiversity–ecosystem functioning
relationships. Oikos. 124:677–684.
MMA (2016) Ministério do Meio Ambiente. (documento on line)
http://www.mma.gov.br/seguranca-quimica/agrotoxicos /Acessado em: 24/06/2016
Mora C, Tittensor DP, Adl S, Simpson AGB, Worm B. 2011. How Many Species Are There
on Earth and in the Ocean? PLoS Biology. 9:e1001127.
Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GA, Kent J (2000) Biodiversity
hotspots for conservation priorities. Nature. 403: 853-858.

131
Naranjo SE (2001) Conservation and evaluation of natural enemies in IPM systems for
Bemisia tabaci. Crop Protection. 20:835-852.
Newbold T, Hudson LH, Hill SLL, Contu S, Lysenko I, Senior RA, Börger L, Bennett DJ,
Choimes A, Collen B, Day J, De Palma A, Díaz S (2015) Global effects of land use on
local terrestrial biodiversity. Nature. 520: 45-50.
Offenberg J (2015) Ants as tools in sustainable agriculture. Journal of Applied Ecology. 52:
1197–1205.
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RR, Simpson GL,
Solymos P, Henry M, Stevens H, Wagner H (2015) vegan: Community Ecology
Package. R package version 2.3-1. http://CRAN.R-project.org/package=vegan
Oliveira-Filho T, Ratter JA (2002) Vegetation physiognomies and woody flora of the Cerrado
Biome. Pp 91-120 in P. S. Oliveira, & R. J. Marquis, editors. The Cerrados of Brazil -
Ecology and Natural History of a Neotropical Savanna. Columbia University Press,
New York.
Pacheco R, Vasconcelos HL (2012) Habitat diversity enhances ant diversity in a naturally
heterogeneous Brazilian landscape. Biodiversity and Conservation. 21:797–809.
Pacheco R, Vasconcelos HL, Groc S, Camacho GP, Frizzo TLM (2013) The importance of
remnants of natural vegetation for maintaining ant diversity in Brazilian agricultural
landscapes. Conservation Biology. 22: 983-997.
Parra JR, Zucchi RA (2004) Trichogramma in Brazil: feasibility of use after twenty years of
research. Neotropical Entomology. 33:271-281.
Perfecto I, Vandermeer J (2002) Quality of Agroecological Matrix in a Tropical Montane
Landscape: Ants in Coffee Plantations in Southern Mexico. Conservatio Biology.
16:174-182.

132
Perfecto I, Vandermeer JH, Bautista GL, Nunez GI, Greenberg R, Bichier P, Langridges S
(2004) Greater predation in shaded coffee farms: the role of resident neotropical.
Ecology. 85:2677–2681.
Pereira JL, Picanc MC¸ Pereira EJG, Silva AA, Jakelaitis A, Pereira RR, Xavier VM (2010)
Influence of crop management practices on bean foliage arthropods. Bulletin of
Entomological Research. 100:679-688.
Philpott SM, Arendt WJ, Armbrecht I, Bichier P, Diestch TV, Gordon C, Greenberg R,
Perfecto I, Reynoso-Santos R, Soto-Pinto L, Tejeda-Cruz C, Williams-Linera G,
Valenzuela J, Zolotoff JM (2008) Biodiversity Loss in Latin American Coffee
Landscapes: Review of the Evidence on Ants, Birds, and Trees. Conservation Biology.
22:1093-1105.
Philpott SM, Armbrecht I (2006) Biodiversity in tropical agroforests and the ecological role
of ants and ant diversity in predatory function. Ecological Entomology. 31:369–377.
Philpott SM, Foster PF (2005) Nest-site limitation in coffe agroecosystems: Artificial nests
maintain diversity of arboreal ants. Ecologial Aplication. 15:1478-1485.
Pimental D, Stachow U, Takacs A, Brubaker W, Dumas AR, Meaney JS et al. (1992)
Conserving biological diversity in agricultural/forestry systems. Bioscience. 432:354–
362.
Pimm SL, Russell GJ, Gittleman JL, Brooks TM (1995) The future of biodiversity. Science.
269:347-350.
Pinheiro F, Diniz IR, Coelho D, Bandeira MPS (2008) Seasonal pattern of insect abundance
in the Brazilian Cerrado. Austral Ecology. 27:132–136.
Pivello VR, Shida CN, Meirelles ST (1999) Alien grasses in Brazilian savannas: a threat to
the biodiversity. Biodiversity and Conservation. 8:1281-1294.

133
Pohlert T (2014). The Pairwise Multiple Comparison of Mean Ranks Package (PMCMR). R
package, <URL:http://CRAN.R-project.org/package=PMCMR>.
Popp J, Pető K, Nagy J (2013) Pesticide productivity and food security. A review. Agron.
Sustain. Dev. 33:243-255.
Price PW, Diniz IR, Morais HC, Marques ESA (1995) The abundance of insect herbivore
species in the tropics: the hight local richness of rare species. Biotropica. 27: 468-478.
R Development Core Team (2008). R: A language and environment for statistical computing.
R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL
http://www.R-project.org.
Redford KH, Fonseca GAB (1986) The role of gallery forests in the zoogeography of the
Cerrado’s non-volant mammalian fauna. Biotropica. 18:126–135.
Ribas CR, Campos RBF, Schmidt FA, Solar RRC (2012) Ants as indicators in Brazil: A
review with suggestions to improve the use of ants in envirnmental monitoring
programs. Psyche. 2012:1-23.
Ribeiro-Neto JD, Arnan X, Tabarelli M, Leal IR (2016) Chronic anthropogenic disturbance
causes homogenization of plant and ant communities in the Brazilian Caatinga.
Biodiversyt and Conservation. 25:943-956.
Rigby D, Cáceres D (2001) Organic farming and the sustainability of agricultural
systems. Agricultural systems. 68: 21-40.
Risch SJ, Carroll CR (1982) Effect of a keystone predaceous ant, solenopsis-geminata, on
arthropods in a tropical agroecosystem. Ecology. 63:1979-1983.
Roberts DW (2016). labdsv: Ordination and Multivariate Analysis for Ecology. R package
version 1.8-0. http://CRAN.R-project.org/package=labdsv
Rockström J, Steffen W, Noone K, Persson A, Chapin FS, Lambin EF, Lenton TM, Scheffer
M, Folke C, Schellnhuber HJ, Nykvist B, de Wit CA, Hughes T, Van der Leeuw S,

134
Rodhe H, Sörlin S, Snyder PK, Costanza R, Svedin U, Falkenmark M, Karlberg L,
Corell RW, Fabry VJ, Hansen J, Walker B, Liverman D, Richardson K, Crutzen P,
Foley JA (2009) A safe operating space for humanity. Nature. 46:472-475.
Root RB (1973) Organization of a plant-arthropod association in simple and diverse habitats:
the fauna of collards (Brassica oleraceae). Ecological Monography. 43:95–124.
Sakamoto Y, Ishiguro M, Kitagawa G (1986) Akaike Information Criterion Statistics. D.
Reidel Publishing Company. Tokyo.
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald R,
Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney AH,
Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH (2000)
Biodiversity - Global biodiversity scenarios for the year 2100. Science. 287:1770-
1774.
Sanders D, Platner C (2007) Intraguild Interactions between Spiders and Ants and Top-Down
Control in a Grassland Food Web. Oecologia. 150:611-624.
Sano EE, Rosa R, Brito JLS, Ferreira LG (2010) Land cover mapping of the tropical savanna
region in Brazil. Environmental Monitoring and Assessment. 166:113-124.
Senior MJM, Hamer KC, Bottrell S, Edwards DP, Fayle TM, Lucey JM, Mayhew PJ, Newton
R, Peh KSH, Sheldon FH, Stewart C, Styring AR, Thom MDF, Woodcock P, Hill JK
(2013) Trait-dependent declines of species following conversion of rain forest to oil
palm plantations. Biodiversity and Conservation. 22:253-268.
Silberman RE, Gordon D, Ingram KK (2016) Nutrient stores predict task behaviors in diverse
ant species. Insectes Sociaux. 63:299-307.
Silva DP, Marco Jr P (2014) No evidence of habitat loss Affecting the Orchid Bees
Eulaema nigrita Lepeletier and Eufriesea auriceps Friese (Apidae: Euglossini) in the
Brazilian Cerrado Savanna. Ecology, Begavior and Bionomics. 43:509-518.

135
Silva JF, Farinãs MR, Felfili JM, Klink CA (2006) Spatial heterogeneity, land use and
conservation in the cerrado region of Brazil. Journal of Biogeography. 33:536-548.
Solar RRC, Barow J, Ferreira J, Berenguer E, Lees AC, Thomson JR, Louzada J, Maués M,
Moura NG, Oliveira, VHF, Chaul JCM, Schoereder JH, Vieira ICG, Nally RM,
Gardner TA (2015) How pervasive is biotic homogenization in human-modified
tropical forest landscape? Ecology Letters. 18:1108-1118.
Srivastava D, Lawton J (1998) Why more productive sites have more species: an
experimental test of theory using tree-hole communities. American Naturalist. 152:
510–529.
Srivastava DS, Vellend M (2005) Biodiversity-Ecosystem Function Research: Is It Relevant
to Conservation? Annual Review Ecology and Systematic. 36:267-294.
Styrsky JD, Eubanks MD (2007) Ecological consequences of interactions between ants and
honeydew-producing insects. Proc R Soc B. 274:151-164.
Styrsky JD, Eubanks MD (2010) A facultative mutualism between aphids and an invasive ant
increases plant reproduction. Ecological Entomology. 35:190-199.
Sujii ER, Venzon M, Medeiros MA, Pires CSS, Togni PHB (2010) Práticas culturais no
manejo de pragas na agricultura orgânica. In: Venzon M, Júnior TJP, Pallini A (eds.)
Controle alternativo de pragas e doenças na agricultura orgânica. EPAMIG, Viçosa
Suzuki R, Shimodaira H (2015). pvclust: Hierarchical Clustering with P-Values via
Multiscale Bootstrap Resampling. R package version 2.0-0. http://CRAN.R-
project.org/package=pvclust
Symondson WOC, Sunderland KD, Greenstone MH (2002) Can generalist predators be
effective biocontrol agents? Annual Review Entomology. 47:561-594.

136
Tiemann LK, Grandy AS, Atkinson EE, Marin-Spiotta E, McDaniel MD (2015) Crop
rotational diversity enhances belowground communities and functions in an
agroecosystem. Ecology Letters. 18:761-771.
Tobin JE (1994) Ants as primary consumers: diet and abundance in the Formicidae. In: Hunt
JH & Nalepa CA (eds.) Nourishment and Evolution in Insect Societies. Westview
Press, Oxford.
Togni PHB, Venzon M, Muniz CA, Martins EF, Pallini A, Sujii ER (2016) Mechanisms
underlying the innate attraction of an aphidophagous coccinellid to coriander plants:
Implications for conservation biological control. Biological Control. 92:77-84.
Tscharntke T, Clough Y, Wanger TC, Jackson L, Motzke I, Perfecto I, et al. (2012) Global
food security, biodiversity conservation and the future of agricultural intensification.
Biological Conservation. 151:53–59.
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C (2005) Landscape
perspectives on agricultural intensification and biodiversity – ecosystem service
management. Ecology Letters. 8:857-874.
Uchida K, Ushimaru A (2014) Biodiversity declines due to abandonment and intensification
of agricultural lands: patterns and mechanisms. Ecological Monographs. 84:637-658.
Ulysséa MA, Brandão CRF (2013) Ant species (Hymenoptera, Formicidae) from the
seasonally dry tropical forest of northeastern Brazil: a compilation from field surveys
in Bahia and literature records. Revista Brasileira de Entomologia. 57:217-224.
UNPD (2011) World population prospects: the 2010 revision. United Nations Population
Division. New York: United Nations. Website https://ndb.nal.usda.gov/ndb/search.
Acessado em: 20 maio de 2016.
USDA (2016) United States Department of Agriculture. Agriculture Reserch Service. USDA
Food Composition Databases. Acessado em: 20 maio de 2016.

137
Van der Plas F, Manning P, Soliveres S, Allana E, Scherer-Lorenzen M, Verheyen K, Wirth
F, Zavalag MA, Ampoorterd E, Baeten L, Barbaro L, Bauhus J, Benavides R,
Benneter A, Bonal D, Bouriaud O, Bruelheide H, Bussotti F, Carnol M, Castagneyrol
B, Charbonnier Y, Coomes DA, Coppi A, Bestias CC, Dawud SM, Wandeler H,
Domisch T, Finér L, Gessler A, Granier A, Grossiord C, Guyot V, Hättenschwiler S,
Jactel H, Jaroszewicz B, Joly FX, Jucker T, Korichevaa J, Milligana H, Mueller S,
Muyst B, Nguyenb D, Pollastrinio M, Ratcliffee S, Raulund-Rasmussens K, Selvi F,
Stenlidb J, Valladares F, Vesterdal L, Zielínski D, Fische M (2016). Biotic
homogenization can decrease landscape-scale forest multifunctionality. Proceedings of
the National Academy of Sciences. 113:3557-3562.
Van Meerbeek K, Helsen K, Hermy M (2014) Impact of land-use intensity on the
conservation of functional and phylogenetic diversity in temperatesemi-natural plant
communities. Biodiversyt and Conservation. 23:2259-2272.
Vasconcelos HL, Frizzo TLM, Pacheco R, Maravalhas JB, Camacho GP, Carvalho KS, Koch
EBA, Pujol-Luz JR (2014) Evaluating sampling sufficiency and the use of surrogates
forassessing ant diversity in a Neotropical biodiversity hotspot. Ecological Indicators.
46:286-292.
Vasconcelos HL, Leite MF, Vilhena JMS, Lima AP, Magnusson WE. 2008. Ant diversity in
an Amazonian savanna: Relationship with vegetation structure, disturbance by fire,
and dominant ants. Austral Ecology. 33:221-231.
Veech JA, Summerville KS, Crist TO, Gering JC (2002) The additive partitioning of species
diversity: recent revival of an old idea. Oikos. 99:3-9.
Vieira CR, Moraes MCB, Borges M, Sujii ER, Laumann RA (2013) cis-Jasmone indirect
action on egg parasitoids (Hymenoptera: Scelionidae) and its application in biological

138
control of soybean stink bugs (Hemiptera: Pentatomidae). Biological Control. 64:75-
82.
Vieira J, Vasconcelos HL (2014) Limitação por sódio em assembleia de formigas no Cerrado.
Dissertação de mestrado. Programa de Pós-graduação em ecologia e conservação de
recursos naturais. Instituto de Biologia. Universidade Federal de Uberlândia.
Wackernagel M, Onisto L, Bello P, Linares AC, Falfán ISL, García JM, Guerrero AIS,
Guerrero MGS (1999) National natural capital accounting with the ecological footprint
concept. Ecological Economics. 29:375-390.
Wake DB, Vredenburg VT (2008) Are we in the midst of the sixth mass extinction? A view
from the world of amphibians. Proceedings of the National Academy of Sciences. 105:
11466-11473.
Wall SBV, Kuhn KM, Beck MJ (2005) Seed Removal, Seed Predation, And Secondary
Dispersal. Ecology. 86:801-806.
Weiner CN, Werner M, Linsenmair KE, Blüthgen N (2014) Land-use impacts on plant–
pollinator networks: interaction strength and specialization predict pollinator declines.
Ecology. 95:466-474.
Whittaker RH (1960) Vegetation of the Siskiyou Mountains, Oregon and California.
Ecological Monographs. 30:279-338.
Whittaker RH (1972) Evolution and measurement of species diversity. Taxon. 21:213-251.
Winqvist C, Bengtsson J, Aavik T, Berendse F, Clement LW, Eggers S, Fischer C, Flohre A,
Geiger F, Liira J, Pärt T, Thies C, Tscharntke T, Weisser WW, Bommarco R (2011)
Mixed effects of organic farming and landscape complexity on farmland biodiversity
and biological control potential across Europe. Journal of Applied Ecology. 48:570-
579.

139
Zanetti R, Zanuncio JC, Santos JC, Silva WLP, Ribeiro GT, Lemes PG (2014) An Overview
of Integrated Management of Leaf-Cutting Ants (Hymenoptera: Formicidae) in
Brazilian Forest Plantations. Forests. 5:439-454.
Zehnder G, Gurr GM, Kühne S, Wade MR, Wratten SD, Wyss E (2007) Arthropod pest
management in organic crops. Annual Reviews Entomology. 52:57-80.

140
Anexo I. Definição dos grupos funcionais e lista das respectivas espécies. Grupo funcional / Descrição Espécies
Arborícola
Espécies que nidificam
obrigatoriamente em árvores e
arbustos.
Azteca sp.01, Azteca sp.02, Cephalotes atratus, Cephalotes betoi,
Cephalotes maculatus, Cephalotes pusillus Crematogaster limata,
Crematogaster sp.01, Crematogaster sp.02, Crematogaster sp.03,
Crematogaster sp.04, Crematogaster sp.05, Crematogaster sp.06,
Crematogaster sp.08, Crematogaster sp.10, Crematogaster sp.11,
Crematogaster sp.13, Crematogaster sp.14, Pseudomyrmex cf.
phyllophilus, Pseudomyrmex gracilis, Pseudomyrmex pallidus e
Pseudomyrmex tenuis.
Attini
Tribo da subfamília Myrmicinae,
cuja as espécies cultivam fungo
em matéria vegetal recolhida
pelas operárias. Algumas destas
são consideradas pragas
agrícolas com as formigas do
gênero Atta e Acromyrmex.
Acromyrmex cf. balzani, Acromyrmex sp.01, Acromyrmex sp.02,
Acromyrmex sp.03, Apterostigma sp.01, Apterostigma sp.02, Atta
sp.01, Cyatta abscondita, Cyphomyrmex lectus, Cyphomyrmex
rimosus, Cyphomyrmex sp.04 gr. rimosus, Cyphomyrmex sp.05 gr.
rimosus, Cyphomyrmex sp.06 gr. rimosus, Cyphomyrmex sp.09 gr.
strigatus, Cyphomyrmex sp.10 gr. strigatus, Mycetarotes cf.
parallelus, Mycetarotes sp.02, Mycocepurus goeldii, Mycocepurus
obsoletus, Mycocepurus sp.03, Mycocepurus sp.04, Myrmicocrypta
camargoi, Myrmicocrypta sp.01, Myrmicocrypta sp.03, Sericomyrmex
sp.01, Sericomyrmex sp.02, Trachymyrmex sp.01, Trachymyrmex
sp.02, Trachymyrmex sp.03, Trachymyrmex sp.04, Trachymyrmex
sp.05, Trachymyrmex sp.06, Trachymyrmex sp.07, Trachymyrmex
sp.08, Trachymyrmex sp.09 e Trachymyrmex sp.10.
Camponotini
Tribo da subfamília Formicinae,
representada aqui por um único
gênero, Camponotus. Esse grupo
apresenta espécies capazes de
nidificar no solo e/ou na
vegetação arbórea. Apresentam
um comportamento oportunista,
sendo facilmente encontradas em
interações mutualísticas com
afídeos. Porém, diferente do
grupo funcional oportunista
(com espécies de pequeno
porte), as espécies apresentam
um tamanho corporal de médio a
grande.
Camponotus arboreus, Camponotus atriceps, Camponotus bidens,
Camponotus bonarensis, Camponotus cingulatus, Camponotus gr.
blandus, Camponotus gr. senex, Camponotus lespesii, Camponotus
leydigi, Camponotus novagrandensis, Camponotus personatus,
Camponotus renggeri, Camponotus rufipes, Camponotus
sericeiventris, Camponotus sp.12, Camponotus sp.17, Camponotus
sp.18, Camponotus sp.26, Camponotus sp.27, Camponotus sp.28,
Camponotus sp.29, Camponotus sp.30, Camponotus sp.32,
Camponotus sp.34, Camponotus sp.35, Camponotus sp.44,
Camponotus sp.45, Camponotus sp.16 e Camponotus substitutus.
Correição
Grupo de espécies nômades, que
forrageiam em grandes colunas
(muitos indivíduos, um ao lado
do outro) e são extremamente
vorazes, sendo encontrado na
literatura registros de predação
de até pequenos roedores.
Eciton vagans, Labidus coecus, Labidus mars, Labidus praedator,
Neivamyrmex pseudops, Neivamyrmex sp.01, Neivamyrmex sp.02,
Neivamyrmex sp.04, Neivamyrmex sp.05, Neivamyrmex sp.06,
Neivamyrmex sp.07 e Nomamyrmex esenbeckii wilsoni.
Especialista
Grupo de espécies que
apresentam características
Acanthosticus brevicornis, Acropyga sp.01, Acropyga sp.02,
Acropyga sp.03, Basiceros sp.01, Blepharidatta conops, Carebara
brevipilosa, Carebara urichi, Centromyrmex brachycola,

141
morfológicas ou ecológicas
muito específicas. Em geral, sua
frequência de registro é baixa,
sendo a maioria de predadores
especialistas de um determinado
táxon.
Centromyrmex gigas, Dolichoderus bispinosus, Gnamptogenys
acuminata, Gnamptogenys cf. bruchi, Gnamptogenys cf. rastrata,
Gnamptogenys sp.01, Gnamptogenys sp.03, Gnamptogenys sp.06,
Gnamptogenys sulcata, Heteroponera dolo, Hylomyrma sp.02,
Hylomyrma sp.03, Hypoponera cf. foreli, Hypoponera sp.01,
Hypoponera sp.03, Hypoponera sp.04, Hypoponera sp.05,
Hypoponera sp.06, Hypoponera sp.07, Hypoponera sp.08,
Megalomyrmex silvestrii, Megalomyrmex sp.02, Octostruma sp.01,
Oxyepoecus cf. vezenyii, Oxyepoecus sp.02, Oxyepoecus sp.03,
Pogonomyrmex naegile, Prionopelta punctulata, Prionopelta sp.02,
Pseudomyrmex termitarius, Rasopone sp.01, Rogeria sp.01, Rogeria
sp.02, Strumygenys sp.01, Strumygenys sp.02, Strumygenys sp.03,
Strumygenys sp.04, Strumygenys sp.05, Strumygenys sp.06,
Strumygenys sp.07 e Typhlomyrmex pusillus.
Generalista
Grupo que contempla espécies
onívoras e com grande
plasticidade ecológica. A
maioria das espécies nidificam
no solo (ex: Pheidole spp.),
porém algumas também
conseguem nidificar na
serapilheira, em galhos caídos e
em árvores e arbustos.
Apresentam o comportamento de
recrutar outros indivíduos da
colônia quando encontram
alimento. Participam deste grupo
algumas espécies exóticas como:
Cardiocondyla emeryi,
Monomorium florícola e
Tetramorium cf. similum.
Brachymyrmex cf. delabiei, Brachymyrmex pilipes, Brachymyrmex
sp.01, Brachymyrmex sp.02, Brachymyrmex sp.03, Brachymyrmex
sp.04, Brachymyrmex sp.05, Brachymyrmex sp.07, Brachymyrmex
sp.08, Brachymyrmex sp.09, Brachymyrmex sp.10, Cardiocondyla
emeryi, Cardiocondyla sp.01, Forelius sp.01, Forelius sp.02,
Monomorium florícola, Nylanderia sp.01, Nylanderia sp.03,
Nylanderia sp.04, Ochetomyrmex semipolitus, Pheidole gr. fimbriata,
Pheidole sp.02, Pheidole sp.03, Pheidole sp.04, Pheidole sp.05,
Pheidole sp.06, Pheidole sp.07, Pheidole sp.08, Pheidole sp.09,
Pheidole sp.10, Pheidole sp.11, Pheidole sp.13, Pheidole sp.16,
Pheidole sp.17, Pheidole sp.18, Pheidole sp.19, Pheidole sp.21,
Pheidole sp.23, Pheidole sp.24, Pheidole sp.25, Pheidole sp.26,
Pheidole sp.27, Pheidole sp.28, Pheidole sp.29, Pheidole sp.31,
Pheidole sp.33, Pheidole sp.35, Pheidole sp.36, Pheidole sp.38,
Pheidole sp.39, Pheidole sp.40, Pheidole sp.41, Pheidole sp.42,
Pheidole sp.44, Pheidole sp.45, Pheidole sp.46, Pheidole sp.47,
Pheidole sp.48, Pheidole sp.49, Pheidole sp.50, Pheidole sp.20,
Pheidole sp.22, Solenopsis cf. loretana, Solenopsis globularia,
Solenopsis sp.03, Solenopsis sp.04 pr. loretana, Solenopsis sp.08,
Solenopsis sp.11, Solenopsis sp.12, Solenopsis sp.13, Solenopsis
sp.14, Solenopsis sp.15, Solenopsis sp.16 gr. molesta, Solenopsis
sp.17 gr. wasmannii, Solenopsis sp.18, Solenopsis sp.01, Solenopsis
sp.02, Solenopsis tridens, Tapinoma sp.02, Tetramorium cf. similum,
Wasmannia auropunctata, Wasmannia sp.02 e Wasmannia sp.03.
Generalista dominante
Grupo composto por três
espécies onívoras, que possuem
a característica, devido ao
grande recrutamento de
indivíduos, de monopolizar o
recurso encontrado frente aos
demais competidores.
Pheidole gertrudae, Pheidole sp.01 e Solenopsis saevissima.
Oportunista
Grupo capaz de forragear nos
períodos mais quentes do dia, e
que geralemente não
monopolizam a fonte de
Dorymyrmex cf. bituber, Dorymyrmex cf. brunneus, Dorymyrmex cf.
jheringi, Dorymyrmex sp01, Dorymyrmex sp05, Linepithema cf.
cerradense, Linepithema cf. gallardoi, Linepithema cf. humile,
Linepithema cf. neotropicum, Linepithema iniquum, Linepithema
micans e Linepithema sp05.

142
alimento diante de competidores,
apresentando um
comportamento conhecido como
insinuators.
Predador grande
Espécies de grande porte, sendo
algumas predadoras
especialistas.
Anochetus inermis, Dinoponera australis, Ectatomma brunneum,
Ectatomma edentatum, Ectatomma opaciventris, Ectatomma
permagnum, Ectatomma planidens, Ectatomma tuberculatum,
Neoponera marginata, Neoponera verenae, Odontomachus bauri,
Odontomachus chelifer, Odontomachus laticeps, Odontomachus
sp.05, Pachycondyla harpax, Pachycondyla sp.03 e Pachycondyla
striata.

143
Anexo II. Lista de espécies amostradas no Capítulo I.
Agrossistemas Vegetação nativa
Subfamílias/Espécies Ocorrências
Nº médio de
registros por
localidade Ocorrências
Nº médio de
registros por
localidade
Dolichoderinae 254 5,64 143 5,96
Azteca sp.01 0 0 4 0,17
Azteca sp.02 0 0 6 0,25
Dolichoderus bispinosus 0 0 3 0,13
Dorymyrmex cf. bituber 9 0,2 0 0
Dorymyrmex cf. brunneus 112 2,49 3 0,13
Dorymyrmex cf. jheringi 11 0,24 11 0,46
Dorymyrmex sp.01 20 0,44 7 0,29
Dorymyrmex sp.05 1 0,02 1 0,04
Forelius sp.01 3 0,07 3 0,13
Forelius sp.02 2 0,04 2 0,08
Linepithema cf. cerradense 19 0,42 38 1,58
Linepithema cf. gallardoi 3 0,07 15 0,63
Linepithema cf. humile 9 0,2 13 0,54
Linepithema cf. neotropicum 43 0,96 14 0,58
Linepithema iniquum 0 0 1 0,04
Linepithema micans 14 0,31 10 0,42
Linepithema sp.05 0 0 11 0,46
Tapinoma sp.02 8 0,18 1 0,04
Dorylinae 29 0,64 35 1,46
Acanthosticus brevicornis 0 0 1 0,04
Eciton vagans 0 0 1 0,04
Labidus coecus 16 0,36 7 0,29
Labidus mars 1 0,02 1 0,04
Labidus praedator 6 0,13 14 0,58
Neivamyrmex pseudops 0 0 2 0,08
Neivamyrmex sp.01 3 0,07 3 0,13
Neivamyrmex sp.02 0 0 3 0,13
Neivamyrmex sp.04 1 0,02 1 0,04
Neivamyrmex sp.05 0 0 1 0,04
Neivamyrmex sp.06 1 0,02 0 0
Neivamyrmex sp.07 1 0,02 0 0
Nomamyrmex esenbeckii
wilsoni 0 0 1 0,04
Ectatomminae 26 0,58 65 2,71
Ectatomma brunneum 24 0,53 13 0,54
Ectatomma edentatum 0 0 29 1,21
Ectatomma opaciventris 2 0,04 6 0,25
Ectatomma permagnum 0 0 14 0,58
Ectatomma planidens 0 0 1 0,04
Ectatomma tuberculatum 0 0 2 0,08
continua...

144
Agrossistemas Vegetação nativa
Subfamílias/Espécies Ocorrências
Nº médio de
registros por
localidade Ocorrências
Nº médio de
registros por
localidade
Formicinae 357 7,93 636 26,5
Acropyga sp.01 1 0,02 4 0,17
Acropyga sp.02 0 0 3 0,13
Acropyga sp.03 1 0,02 0 0
Brachymyrmex cf. delabiei 0 0 7 0,29
Brachymyrmex pilipes 0 0 1 0,04
Brachymyrmex sp.01 12 0,27 45 1,88
Brachymyrmex sp.02 0 0 1 0,04
Brachymyrmex sp.03 116 2,58 45 1,88
Brachymyrmex sp.04 2 0,04 5 0,21
Brachymyrmex sp.05 1 0,02 8 0,33
Brachymyrmex sp.07 0 0 4 0,17
Brachymyrmex sp.08 0 0 3 0,13
Brachymyrmex sp.09 0 0 1 0,04
Brachymyrmex sp.10 0 0 4 0,17
Camponotus arboreus 1 0,02 2 0,08
Camponotus atriceps 8 0,18 12 0,5
Camponotus bidens 0 0 2 0,08
Camponotus bonarensis 0 0 2 0,08
Camponotus cingulatus 18 0,4 33 1,38
Camponotus gr. blandus 11 0,24 48 2
Camponotus gr. senex 30 0,67 57 2,38
Camponotus lespesii 0 0 11 0,46
Camponotus leydigi 4 0,09 3 0,13
Camponotus novagrandensis 2 0,04 45 1,88
Camponotus personatus 5 0,11 26 1,08
Camponotus renggeri 1 0,02 14 0,58
Camponotus rufipes 23 0,51 40 1,67
Camponotus sericeiventris 2 0,04 5 0,21
Camponotus sp.12 65 1,44 55 2,29
Camponotus sp.17 0 0 2 0,08
Camponotus sp.18 8 0,18 15 0,63
Camponotus sp.26 0 0 1 0,04
Camponotus sp.27 0 0 3 0,13
Camponotus sp.28 0 0 1 0,04
Camponotus sp.29 1 0,02 4 0,17
Camponotus sp.30 0 0 1 0,04
Camponotus sp.32 0 0 1 0,04
Camponotus sp.34 0 0 1 0,04
Camponotus sp.35 0 0 1 0,04
Camponotus sp.44 0 0 1 0,04
Camponotus sp.45 0 0 2 0,08
Camponotus sp.16 16 0,36 53 2,21
continua...

145
Agrossistemas Vegetação nativa
Subfamílias/Espécies Ocorrências
Nº médio de
registros por
localidade Ocorrências
Nº médio de
registros por
localidade
Camponotus substitutus 0 0 10 0,42
Nylanderia sp.01 6 0,13 42 1,75
Nylanderia sp.03 22 0,49 0 0
Nylanderia sp.04 0 0 10 0,42
Prionopelta punctulata 1 0,02 1 0,04
Prionopelta sp.02 0 0 1 0,04
Heteroponerinae 0 0 3 0,13
Heteroponera dolo 0 0 3 0,13
Myrmicinae 1693 37,62 1381 57,54
Acromyrmex cf. balzani 1 0,02 3 0,13
Acromyrmex sp.01 5 0,11 22 0,92
Acromyrmex sp.02 0 0 2 0,08
Acromyrmex sp.03 4 0,09 16 0,67
Apterostigma sp.01 1 0,02 11 0,46
Apterostigma sp.02 1 0,02 3 0,13
Atta sp.01 20 0,44 14 0,58
Basiceros sp.01 0 0 1 0,04
Blepharidatta conops 0 0 3 0,13
Cardiocondyla emeryi 40 0,89 0 0
Cardiocondyla sp.01 3 0,07 0 0
Carebara brevipilosa 0 0 7 0,29
Carebara urichi 1 0,02 1 0,04
Cephalotes atratus 0 0 1 0,04
Cephalotes betoi 0 0 2 0,08
Cephalotes maculatus 0 0 1 0,04
Cephalotes pusillus 4 0,09 3 0,13
Crematogaster limata 0 0 30 1,25
Crematogaster sp.01 2 0,04 10 0,42
Crematogaster sp.02 0 0 3 0,13
Crematogaster sp.03 0 0 8 0,33
Crematogaster sp.04 0 0 2 0,08
Crematogaster sp.05 0 0 24 1
Crematogaster sp.06 22 0,49 20 0,83
Crematogaster sp.08 0 0 2 0,08
Crematogaster sp.10 0 0 7 0,29
Crematogaster sp.11 1 0,02 4 0,17
Crematogaster sp.13 0 0 1 0,04
Crematogaster sp.14 0 0 1 0,04
Cyatta abscondita 0 0 1 0,04
Cyphomyrmex lectus 0 0 7 0,29
Cyphomyrmex rimosus 28 0,62 8 0,33
Cyphomyrmex sp04 gr. rimosus 1 0,02 12 0,5
Cyphomyrmex sp05 gr. rimosus 8 0,18 4 0,17
continua...

146
Agrossistemas Vegetação nativa
Subfamílias/Espécies Ocorrências
Nº médio de
registros por
localidade Ocorrências
Nº médio de
registros por
localidade
Cyphomyrmex sp06 gr. rimosus 1 0,02 5 0,21
Cyphomyrmex sp.09 gr.
strigatus 0 0 1 0,04
Cyphomyrmex sp.10 gr.
strigatus 0 0 1 0,04
Gnamptogenys acuminata 1 0,02 0 0
Gnamptogenys cf. bruchi 0 0 1 0,04
Gnamptogenys cf. rastrata 2 0,04 0 0
Gnamptogenys sp.01 2 0,04 28 1,17
Gnamptogenys sp.03 0 0 3 0,13
Gnamptogenys sp.06 0 0 1 0,04
Gnamptogenys sulcata 23 0,51 6 0,25
Hylomyrma sp.02 0 0 5 0,21
Hylomyrma sp.03 0 0 1 0,04
Megalomyrmex silvestrii 2 0,04 0 0
Megalomyrmex sp.02 0 0 1 0,04
Monomorium floricola 1 0,02 0 0
Mycetarotes cf. parallelus 0 0 1 0,04
Mycetarotes sp.02 0 0 1 0,04
Mycocepurus goeldii 29 0,64 12 0,5
Mycocepurus obsoletus 0 0 11 0,46
Mycocepurus sp.03 14 0,31 3 0,13
Mycocepurus sp.04 2 0,04 1 0,04
Myrmicocrypta camargoi 1 0,02 0 0
Myrmicocrypta sp.01 4 0,09 7 0,29
Myrmicocrypta sp.03 0 0 1 0,04
Ochetomyrmex semipolitus 0 0 6 0,25
Octostruma sp.01 0 0 2 0,08
Oxyepoecus cf. vezenyii 0 0 1 0,04
Oxyepoecus sp.02 1 0,02 1 0,04
Oxyepoecus sp.03 1 0,02 0 0
Pheidole gertrudae 155 3,44 70 2,92
Pheidole gr. fimbriata 1 0,02 1 0,04
Pheidole sp.02 38 0,84 3 0,13
Pheidole sp.03 63 1,4 85 3,54
Pheidole sp.04 68 1,51 0 0
Pheidole sp.05 49 1,09 3 0,13
Pheidole sp.06 0 0 8 0,33
Pheidole sp.07 10 0,22 1 0,04
Pheidole sp.08 0 0 16 0,67
Pheidole sp.09 51 1,13 0 0
Pheidole sp.10 3 0,07 13 0,54
Pheidole sp.11 43 0,96 13 0,54
continua...

147
Agrossistemas Vegetação nativa
Subfamílias/Espécies Ocorrências
Nº médio de
registros por
localidade Ocorrências
Nº médio de
registros por
localidade
Pheidole sp.13 97 2,16 59 2,46
Pheidole sp.16 19 0,42 0 0
Pheidole sp.17 2 0,04 3 0,13
Pheidole sp.18 1 0,02 1 0,04
Pheidole sp.19 7 0,16 62 2,58
Pheidole sp.21 0 0 4 0,17
Pheidole sp.23 15 0,33 8 0,33
Pheidole sp.24 70 1,56 1 0,04
Pheidole sp.25 0 0 7 0,29
Pheidole sp.26 0 0 33 1,38
Pheidole sp.27 3 0,07 0 0
Pheidole sp.28 7 0,16 4 0,17
Pheidole sp.29 8 0,18 0 0
Pheidole sp.31 0 0 7 0,29
Pheidole sp.33 0 0 10 0,42
Pheidole sp.35 24 0,53 0 0
Pheidole sp.36 0 0 1 0,04
Pheidole sp.38 0 0 4 0,17
Pheidole sp.39 1 0,02 0 0
Pheidole sp.40 2 0,04 0 0
Pheidole sp.41 3 0,07 1 0,04
Pheidole sp.42 10 0,22 9 0,38
Pheidole sp.44 0 0 1 0,04
Pheidole sp.45 0 0 3 0,13
Pheidole sp46 0 0 1 0,04
Pheidole sp.47 1 0,02 0 0
Pheidole sp.48 0 0 1 0,04
Pheidole sp.49 8 0,18 5 0,21
Pheidole sp.50 0 0 3 0,13
Pheidole sp.01 184 4,09 187 7,79
Pheidole sp.20 63 1,4 33 1,38
Pheidole sp.22 58 1,29 51 2,13
Pogonomyrmex naegile 11 0,24 14 0,58
Rogeria sp.01 2 0,04 6 0,25
Rogeria sp.02 0 0 1 0,04
Sericomyrmex sp.01 0 0 6 0,25
Sericomyrmex sp.02 0 0 1 0,04
Solenopsis cf. loretana 2 0,04 0 0
Solenopsis globularia 1 0,02 2 0,08
Solenopsis saevissima 141 3,13 13 0,54
Solenopsis sp.03 4 0,09 26 1,08
Solenopsis sp.04 pr. loretana 67 1,49 0 0
Solenopsis sp.08 6 0,13 4 0,17
continua...

148
Agrossistemas Vegetação nativa
Subfamílias/Espécies Ocorrências
Nº médio de
registros por
localidade Ocorrências
Nº médio de
registros por
localidade
Solenopsis sp.11 0 0 1 0,04
Solenopsis sp.12 1 0,02 0 0
Solenopsis sp.13 0 0 4 0,17
Solenopsis sp.14 0 0 1 0,04
Solenopsis sp.15 0 0 1 0,04
Solenopsis sp.16 gr. molesta 1 0,02 0 0
Solenopsis sp.17 gr. wasmannii 1 0,02 2 0,08
Solenopsis sp.18 0 0 1 0,04
Solenopsis sp.01 82 1,82 115 4,79
Solenopsis sp.02 20 0,44 28 1,17
Solenopsis tridens 6 0,13 14 0,58
Strumygenys sp.01 2 0,04 5 0,21
Strumygenys sp.02 10 0,22 0 0
Strumygenys sp.03 0 0 3 0,13
Strumygenys sp.04 1 0,02 0 0
Strumygenys sp.05 0 0 1 0,04
Strumygenys sp.06 1 0,02 2 0,08
Strumygenys sp.07 0 0 1 0,04
Tetramorium cf. similum 10 0,22 0 0
Trachymyrmex sp.01 4 0,09 2 0,08
Trachymyrmex sp.02 4 0,09 0 0
Trachymyrmex sp.03 4 0,09 9 0,38
Trachymyrmex sp.04 0 0 1 0,04
Trachymyrmex sp.05 0 0 7 0,29
Trachymyrmex sp.06 0 0 2 0,08
Trachymyrmex sp.07 0 0 3 0,13
Trachymyrmex sp.08 5 0,11 0 0
Trachymyrmex sp.09 0 0 2 0,08
Trachymyrmex sp.10 1 0,02 0 0
Typhlomyrmex pusillus 1 0,02 0 0
Wasmannia auropunctata 12 0,27 52 2,17
Wasmannia sp.02 2 0,04 5 0,21
Wasmannia sp.03 1 0,02 0 0
continua...

149
Agrossistemas Vegetação nativa
Subfamílias/Espécies Ocorrências
Nº médio de
registros por
localidade Ocorrências
Nº médio de
registros por
localidade
Ponerinae 158 3,51 178 7,42
Anochetus inermis 9 0,2 1 0,04
Centromyrmex brachycola 0 0 1 0,04
Centromyrmex gigas 0 0 1 0,04
Dinoponera australis 0 0 2 0,08
Hypoponera cf. foreli 0 0 4 0,17
Hypoponera sp.01 36 0,8 14 0,58
Hypoponera sp.03 7 0,16 0 0
Hypoponera sp.04 3 0,07 0 0
Hypoponera sp.05 3 0,07 0 0
Hypoponera sp.06 3 0,07 4 0,17
Hypoponera sp.07 4 0,09 2 0,08
Hypoponera sp.08 17 0,38 0 0
Neoponera marginata 4 0,09 1 0,04
Neoponera verenae 37 0,82 43 1,79
Odontomachus bauri 13 0,29 9 0,38
Odontomachus chelifer 1 0,02 30 1,25
Odontomachus laticeps 0 0 11 0,46
Odontomachus sp.05 0 0 3 0,13
Pachycondyla harpax 14 0,31 29 1,21
Pachycondyla sp.03 0 0 1 0,04
Pachycondyla striata 7 0,16 17 0,71
Rasopone sp.01 0 0 5 0,21
Pseudomyrmecinae 12 0,27 39 1,63
Pseudomyrmex cf. phyllophilus 1 0,02 1 0,04
Pseudomyrmex gracilis 0 0 1 0,04
Pseudomyrmex pallidus 1 0,02 11 0,46
Pseudomyrmex tenuis 0 0 5 0,21
Pseudomyrmex termitarius 10 0,22 21 0,88
Total Geral 2529 56,2 2480 103,33

150
Anexo III.
Figuras das análises de partição aditiva de diversidade de formigas no Distrito Federal separada por tipo de
sistema agrícola ou fitofisionomia da vegetação nativa. α1 é quantidade de espécies por parcela. β1 é a troca de
espécies entre parcelas dentro da mesma localidade. β2 é a troca de espécies entre localidades pertencentes ao
mesmo tipo de agrossistema ou fitofisionomia. β3 é a troca de espécies entre os diferentes tipos de agrossistemas
e fitofisionomias.
0%
20%
40%
60%
80%
100%
Observado Esperado
Soja
0%
20%
40%
60%
80%
100%
Observado Esperado
Campo
0%
20%
40%
60%
80%
100%
Observado Esperado
Agricultura orgânica
0%
20%
40%
60%
80%
100%
Observado Esperado
Cerrado
0%
20%
40%
60%
80%
100%
Observado Esperado
Pastagem
0%
20%
40%
60%
80%
100%
Observado Esperado
Mata
0%
20%
40%
60%
80%
100%
Observado Esperado
Agrossistemas
0%
20%
40%
60%
80%
100%
Observado Esperado
Vegetação nativa
β3
β2
β1
α1

151
Anexo IV. Lista de espécies capturadas dentro das estações de remoção de biomassa e ordenadas por sua ocorrência.
Espécies Banana Sardinha Semente
Frequequência acumulada
de ocorrência
Pheidole sp. 01 74 94 31 10,7%
Pheidole gertrudae 48 78 13 18,1%
Solenopsis saevissima 33 44 18 23,2%
Solenopsis sp. 01 15 54 15 27,8%
Pheidole sp. 13 29 32 8 31,5%
Pheidole sp. 03 17 41 10 35,1%
Pheidole sp. 20 20 33 13 38,6%
Brachymyrmex sp. 03 19 40 3 42,0%
Camponotus sp. 12 14 24 10 44,6%
Dorymyrmex cf. brunneus 18 23 5 47,0%
Pheidole sp. 19 12 10 18 49,2%
Camponotus gr. blandus 4 27 7 51,2%
Camponotus sp. 16 5 25 8 53,2%
Pheidole sp. 05 11 16 11 55,3%
Camponotus gr. senex 12 20 3 57,2%
Pheidole sp. 24 21 5 7 58,9%
Pheidole sp. 04 12 17 3 60,7%
Pheidole sp. 22 16 13 3 62,4%
Camponotus rufipes 3 19 8 64,0%
Solenopsis sp. 03 11 11 8 65,6%
Camponotus novagrandensis 3 21 5 67,1%
Linepithema cf. neotropicum 13 11 5 68,7%
Crematogaster limata 9 15 2 70,1%
Pheidole sp. 02 5 16 2 71,3%
Camponotus cingulatus 5 15 2 72,5%
Crematogaster sp. 05 5 12 5 73,7%
Crematogaster sp. 06 3 15 3 74,8%
Wasmannia auropunctata 8 6 5 75,8%
Brachymyrmex sp. 01 5 10 2 76,8%
Camponotus personatus 6 3 8 77,7%
Linepithema cf. cerradense 9 7 1 78,6%
Acromyrmex sp. 01 10
5 79,4%
Pheidole sp. 11 6 7 2 80,2%
Nylanderia sp. 01 5 7
80,8%
Nylanderia sp. 03 3 6 3 81,5%
Pheidole sp. 26 6 4 2 82,1%
Solenopsis sp. 04 pr. loretana 4 6 2 82,8%
Pheidole sp. 09 1 9 1 83,4%
Camponotus sp. 18 2 5 3 83,9%
Dorymyrmex sp. 01 4 5 1 84,4%
Pheidole sp. 23 1 6 2 84,9%
Pseudomyrmex termitarius 1 7 1 85,4%
Acromyrmex sp. 03 6 1 1 85,8%
Continua...
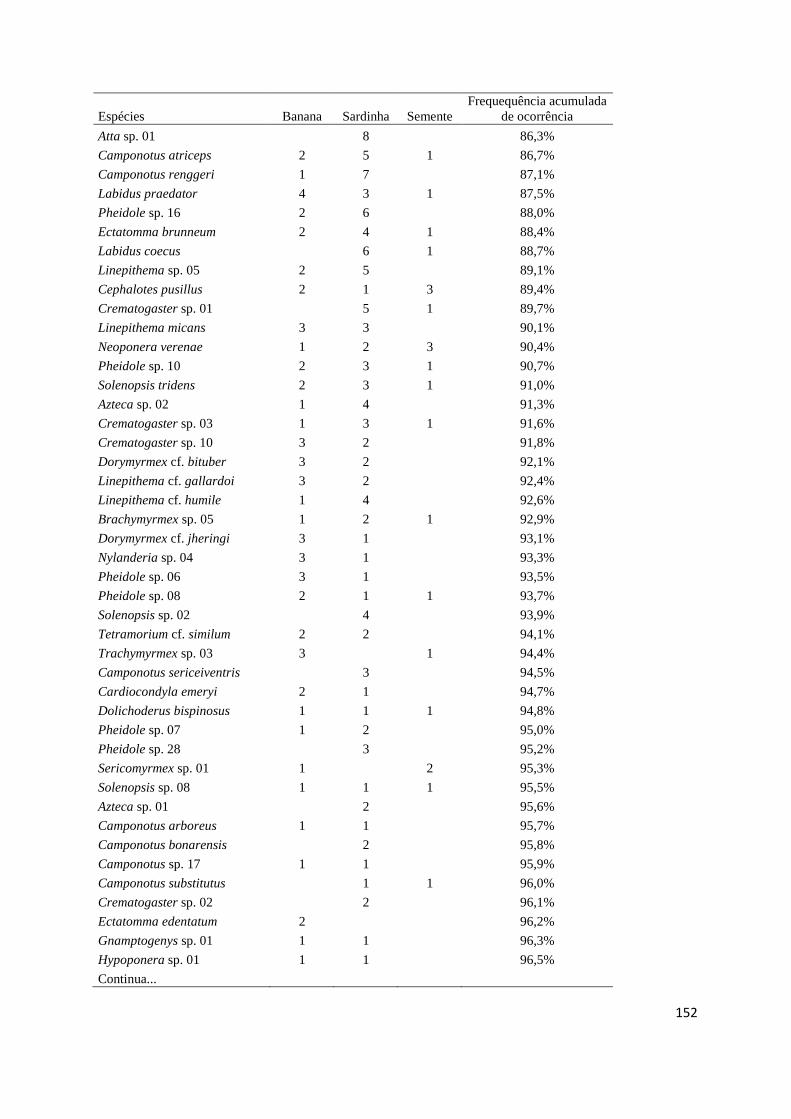
152
Espécies Banana Sardinha Semente
Frequequência acumulada
de ocorrência
Atta sp. 01
8
86,3%
Camponotus atriceps 2 5 1 86,7%
Camponotus renggeri 1 7
87,1%
Labidus praedator 4 3 1 87,5%
Pheidole sp. 16 2 6
88,0%
Ectatomma brunneum 2 4 1 88,4%
Labidus coecus
6 1 88,7%
Linepithema sp. 05 2 5
89,1%
Cephalotes pusillus 2 1 3 89,4%
Crematogaster sp. 01
5 1 89,7%
Linepithema micans 3 3
90,1%
Neoponera verenae 1 2 3 90,4%
Pheidole sp. 10 2 3 1 90,7%
Solenopsis tridens 2 3 1 91,0%
Azteca sp. 02 1 4
91,3%
Crematogaster sp. 03 1 3 1 91,6%
Crematogaster sp. 10 3 2
91,8%
Dorymyrmex cf. bituber 3 2
92,1%
Linepithema cf. gallardoi 3 2
92,4%
Linepithema cf. humile 1 4
92,6%
Brachymyrmex sp. 05 1 2 1 92,9%
Dorymyrmex cf. jheringi 3 1
93,1%
Nylanderia sp. 04 3 1
93,3%
Pheidole sp. 06 3 1
93,5%
Pheidole sp. 08 2 1 1 93,7%
Solenopsis sp. 02
4
93,9%
Tetramorium cf. similum 2 2
94,1%
Trachymyrmex sp. 03 3
1 94,4%
Camponotus sericeiventris
3
94,5%
Cardiocondyla emeryi 2 1
94,7%
Dolichoderus bispinosus 1 1 1 94,8%
Pheidole sp. 07 1 2
95,0%
Pheidole sp. 28
3
95,2%
Sericomyrmex sp. 01 1
2 95,3%
Solenopsis sp. 08 1 1 1 95,5%
Azteca sp. 01
2
95,6%
Camponotus arboreus 1 1
95,7%
Camponotus bonarensis
2
95,8%
Camponotus sp. 17 1 1
95,9%
Camponotus substitutus
1 1 96,0%
Crematogaster sp. 02
2
96,1%
Ectatomma edentatum 2
96,2%
Gnamptogenys sp. 01 1 1
96,3%
Hypoponera sp. 01 1 1
96,5%
Continua...

153
Espécies Banana Sardinha Semente
Frequequência acumulada
de ocorrência
Odontomachus bauri 2
96,6%
Odontomachus chelifer
1 1 96,7%
Pachycondyla harpax
2
96,8%
Pheidole sp. 17
2
96,9%
Pheidole sp. 21
2
97,0%
Pheidole sp. 29
2 97,1%
Pheidole sp. 42 1 1
97,2%
Pogonomyrmex naegile 1 1
97,3%
Pseudomyrmex cf. phyllophilus
2 97,4%
Pseudomyrmex pallidus
2
97,5%
Pseudomyrmex tenuis 2
97,6%
Solenopsis cf. loretana 1 1
97,7%
Trachymyrmex sp. 02 2
97,9%
Acromyrmex sp. 02
1 97,9%
Apterostigma sp. 01
1
98,0%
Blepharidatta conops
1
98,0%
Brachymyrmex cf. delabiei
1
98,1%
Brachymyrmex sp. 02 1
98,1%
Brachymyrmex sp. 04
1
98,2%
Brachymyrmex sp. 07 1
98,2%
Camponotus lespesii
1
98,3%
Camponotus leydigi
1
98,3%
Camponotus sp. 26
1 98,4%
Camponotus sp. 27 1
98,4%
Camponotus sp. 29
1
98,5%
Camponotus sp. 45
1
98,6%
Carebara brevipilosa
1
98,6%
Cephalotes betoi
1
98,7%
Crematogaster sp. 04
1 98,7%
Crematogaster sp. 08
1 98,8%
Crematogaster sp. 11 1
98,8%
Cyphomyrmex rimosus 1
98,9%
Ectatomma opaciventris
1
98,9%
Ectatomma tuberculatum 1
99,0%
Forelius sp. 01 1
99,0%
Heteroponera dolo
1
99,1%
Hypoponera sp. 06 1
99,1%
Linepithema iniquum
1
99,2%
Mycocepurus goeldii
1 99,2%
Neivamyrmex sp. 01
1
99,3%
Ochetomyrmex semipolitus
1
99,4%
Odontomachus laticeps 1
99,4%
Pheidole sp. 18 1
99,5%
Pheidole sp. 25
1
99,5%
Continua...
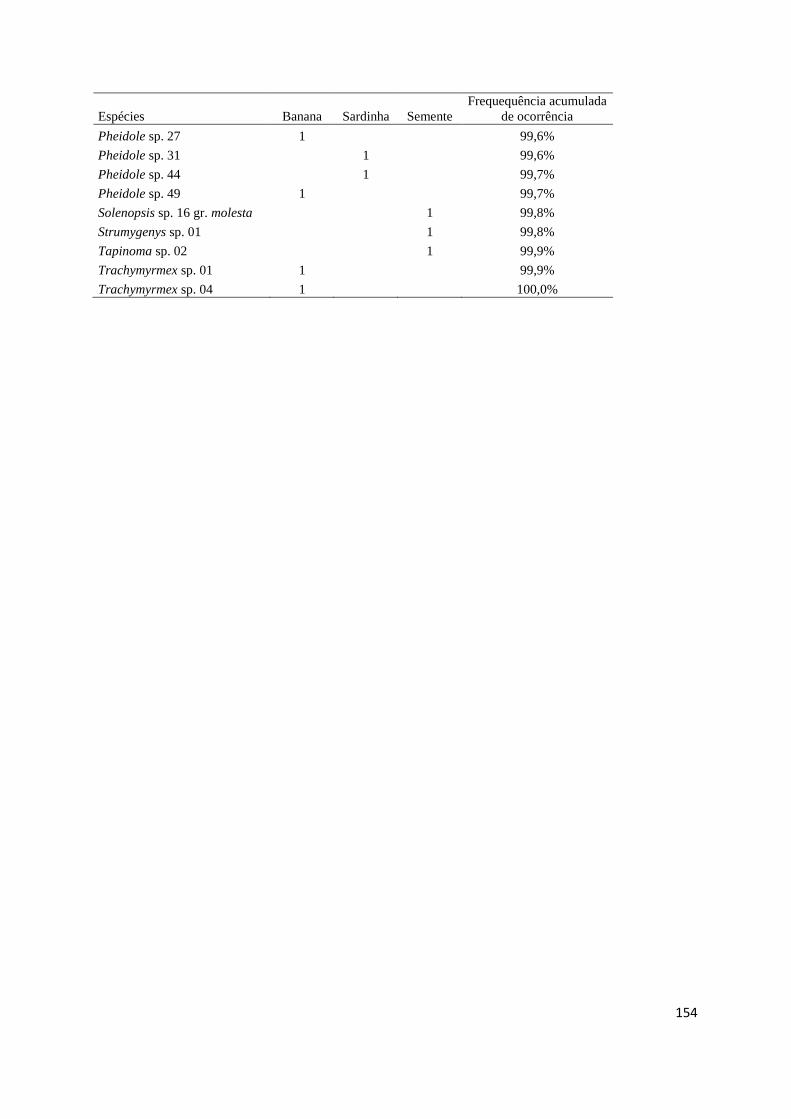
154
Espécies Banana Sardinha Semente
Frequequência acumulada
de ocorrência
Pheidole sp. 27 1
99,6%
Pheidole sp. 31
1
99,6%
Pheidole sp. 44
1
99,7%
Pheidole sp. 49 1
99,7%
Solenopsis sp. 16 gr. molesta
1 99,8%
Strumygenys sp. 01
1 99,8%
Tapinoma sp. 02
1 99,9%
Trachymyrmex sp. 01 1
99,9%
Trachymyrmex sp. 04 1
100,0%