modelo de dissertaÇÃo de mestrado no ppga · 2019. 5. 23. · rc 2: de tomateiro para...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DO CENTRO-OESTE, UNICENTRO-PR
DESENVOLVIMENTO DE GENÓTIPOS F2RC2 DE
TOMATEIRO PARA PROCESSAMENTO COM ALTOS
TEORES DE ACIL-AÇÚCARES E RESISTENTES A
ARTRÓPODES-PRAGA
TESE DE DOUTORADO
DIEGO MUNHOZ DIAS
GUARAPUAVA-PR
2019

DIEGO MUNHOZ DIAS
DESENVOLVIMENTO DE GENÓTIPOS F2RC2 DE TOMATEIRO PARA
PROCESSAMENTO COM ALTOS TEORES DE ACIL-AÇÚCARES E
RESISTENTES A ARTRÓPODES-PRAGA
Tese apresentada à Universidade Estadual do
Centro-Oeste, como parte das exigências do
Programa de Pós-Graduação em Agronomia -
Doutorado, área de concentração em Produção
Vegetal, para obtenção do título de Doutor.
Prof. Dr. Juliano Tadeu Vilela de Resende
Orientador
GUARAPUAVA-PR
2019

Catalogação na Publicação Biblioteca Central da Unicentro, Campus Santa Cruz
Dias, Diego Munhoz
D541d Desenvolvimento de genótipos F2RC2: de tomateiro para processamento com altos teores de acil-açúcares e resistentes a artrópodes-praga / Diego Munhoz Dias. – – Guarapuava, 2019.
x, 82 f. : il. ; 28 cm
Tese (doutorado) - Universidade Estadual do Centro-Oeste, Programa
de Pós-Graduação em Agronomia, área de concentração em Produção Vegetal, 2019
Orientador: Juliano Tadeu Vilela de Resende
Banca examinadora: Vanderlei Aparecido de Lima, Leandro Simões Azeredo Gonçalves, Paulo Roberto da Silva, Cristiano Kopanski Camargo
Bibliografia
1. Agronomia. 2. Produção vegetal. 3. Solanum lycopersicum. 4.
Solanum pennellii. 5. Aleloquímico. 6. Resistência às pragas. I. Título. II. Programa de Pós-Graduação em Agronomia.
CDD 630


AGRADECIMENTOS
Agradecer primeiramente a Deus pelo dom da vida.
A minha família, Geni Munhoz Dias, José Aparecido Veiga Dias, Jéssica Hellmann,
Diniara Munhoz Dias e Giovana Munhoz Dias, pela confiança e incentivo aos estudos.
A minha mãe, Geni, agradeço em especial, por todo investimento e carinho que
somente uma mãe pode oferecer.
Ao meu pai, José, que sempre evidenciou a importância da agricultura e as primeiras
aulas práticas da vida no campo.
Ao meu amor, Jéssica Hellmann, agradeço pelo companheirismo, confiança, amor,
paciência e por aceitar estar ao meu lado.
As minhas irmãs, Diniara e Giovana, pela parceria, amizade e confiança.
A Universidade Estadual do Centro-Oeste e ao Programa de Pós-Graduação em
Agronomia pela estrutura, professores, amigos, funcionários e serviços prestados que
possibilitaram a minha formação de Engenheiro Agrônomo, título de mestre em Produção
Vegetal e agora o título de doutor em Produção Vegetal.
A Coordenação de Aperfeiçoamento de Pessoal de Nível Superior-CAPES pela
concessão da bolsa de estudos, pois sem o auxílio, dificilmente conseguiria concluir o
mestrado.
Ao meu orientador Juliano Tadeu Vilela de Resende, pelo incentivo, orientação,
paciência, confiança, transferência de conhecimento, exemplo e amizade.
Ao Núcleo de Pesquisa em Hortaliças pelo incentivo da pesquisa científica e a todos
os integrantes que direta ou indiretamente contribuíram para a pesquisa e proporcionaram
novas amizades.
Aos integrantes do Laboratório de Genética e Biologia Molecular Vegetal da
Universidade Estadual do Centro-Oeste (UNICENTRO), e especial ao professor Paulo
Roberto da Silva e a Claudia Gobatto, pela paciência, auxilio e ensinamentos.
Aos colegas de graduação, mestrado e doutorado pelas histórias, convívio, amizade,
festas e parceria.

SUMÁRIO
1. INTRODUÇÃO ................................................................................................................ 8
2. OBJETIVOS ................................................................................................................... 10 2.1 Geral ......................................................................................................................................................... 10
2.2 Específicos ................................................................................................................................................ 10
3. REFERENCIAL TEÓRICO .......................................................................................... 11 3.1 Origens e histórico do tomateiro ................................................................................................................ 11
3.2 Taxonomia, classificação e botânica do tomateiro ...................................................................................... 12
3.3 Aspectos gerais de produtividade do tomateiro ........................................................................................... 13
3.4 Aspectos gerais de produtividade do tomateiro para processamento ............................................................ 14
3.5 Artrópodes-praga do tomateiro................................................................................................................... 15
3.5.1 Traça-do-tomateiro - Tuta absoluta (Meyrick, 1917) ............................................................................... 15
3.5.2 Mosca-branca- Bemisia tabaci (Genn.) biótipo B e/ou Bemisia argentifolii (Bellows e Perring, 1994) ...... 17
3.5.3 Ácaro rajado - Tetranychus urticae (Koch, 1836) .................................................................................... 19
3.6 Manejo integrado de pragas ....................................................................................................................... 20
3.7 Melhoramento genético do tomateiro visando resistência a artrópodes-praga .............................................. 21
3.8 Seleção assistida por marcadores moleculares. ........................................................................................... 23
3.9 Solanum pennellii LA 716 como fonte de resistência a artrópodes-praga..................................................... 24
4. REFERÊNCIAS BIBLIOGRÁFICAS .......................................................................... 28
ACIL-AÇÚCARES MEDIADORES DE RESISTÊNCIA À MOSCA BRANCA E À
FUMAGINA EM POPULAÇÕES SEGREGANTES DE TOMATEIRO ....................... 36 1. INTRODUÇÃO .............................................................................................................. 38
2. MATERIAL E MÉTODOS ........................................................................................... 40 2.1 Local do experimento ................................................................................................................................ 40
2.2 Obtenção das populações segregantes de tomateiro F2RC2 .......................................................................... 40
2.3 Medida indireta dos teores de acil-açúcares e seleção dos genótipos de tomateiro F2RC2 ............................. 41
2.4 Teste de resistência à mosca-branca, distribuição da mosca-branca na planta e severidade da fumagina em
genótipos de tomateiro com teores contrastantes de acil-açúcares ..................................................................... 41
2.5 Análises estatísticas ................................................................................................................................... 43
3. RESULTADOS E DISCUSSÕES .................................................................................. 44 4. CONCLUSÕES .............................................................................................................. 52
5. REFERÊNCIAS ............................................................................................................. 53
ACIL-AÇÚCARES MEDIADORES DE RESISTÊNCIA À TRAÇA DO TOMATEIRO
EM POPULAÇÕES SEGREGANTES DE TOMATEIRO.............................................. 57
1. INTRODUÇÃO .............................................................................................................. 59 2. MATERIAL E MÉTODOS ........................................................................................... 61 2.1 Local do experimento ................................................................................................................................ 61
2.2 Obtenção das populações segregantes de tomateiro F2RC2 .......................................................................... 61
2.3 Quantificação dos teores de acil-açúcares e seleção dos genótipos de tomateiro F2RC2 ................................ 62

2.5 Teste de resistência à traça do tomateiro ..................................................................................................... 62
2.5.1 Análises estatísticas ................................................................................................................................ 64
2.4 Avaliação da recuperação do genoma recorrente ........................................................................................ 64
2.4.1 Marcadores moleculares (ISSR e SSR) .................................................................................................... 65
3. RESULTADOS E DISCUSSÕES .................................................................................. 68 3.3 Teste de resistência a traça do tomateiro ..................................................................................................... 68
3.1 Uso de marcadores moleculares ISSR e SSR na seleção de genótipos com altos teores de acil-açúcares ...... 74
4. CONCLUSÕES .............................................................................................................. 76
5. CONSIDERAÇÕES FINAIS. ........................................................................................ 77 5. REFERÊNCIAS ............................................................................................................. 78

LISTA DE FIGURAS
FIGURA 1- Círculo de correlação das variáveis teor de acil-açúcares (Teor de acil-açúcares, número de ovos
(Nº ovos), número de ninfas (Nº ninfas), número de adultos (Nº adultos) e números de exúvias (Nº exúvias) e
distribuição dos genótipos em plano RVTA-2010-31-177pl#28 (G28), RVTA-2010-31-177pl#39 (G39), RVTA-
2010-83-347pl#61 (G61), RVTA-2010-83-347pl#77 (G77), RVTA-2010-31-177pl#113 (G113), RVTA-2010-
31-177pl#177 (G177), RVTA-2010-31-177pl#180 (G180), RVTA-2010-31-310 pl#205 (G205), RVTA-2010-31-319pl#214 (G214), RVTA-2010-83-347pl#257 (G257), RVTA-2010-83-347pl#301 (G301) e RVTA-2010-
83-347pl#359 (G359). Guarapuava-PR –UNICENTRO, 2019. ......................................................................... 49
FIGURA 2- Círculo de correlação das variáveis teor de acil-açúcares (Teor de acil-açúcares (TAA), número de
ovos (TTO), número de larvas (TL), Danos na planta (DFM) e Lesões nos folíolos (LFM) e Porcentagem de
folíolos atacados (PFM) e distribuição dos genótipos em plano Solanum pennellii (G1), RVTA-2010pl#28
(G28), RVTA-2010pl#39 (G39), RVTA-2010pl#113 (G113), RVTA-2010pl#177 (G177), RVTA-2010pl#180
(G180), RVTA-2010pl#232 (G232), RVTA-2010pl#257 (G257), RVTA-2010pl#359 (G359), RVTA-
2010pl#205 (G205), RVTA-2010pl#227 (G227), RVTA-2010pl#378 (G378), RVTA-2010pl#385 (G385) e
cultivar Redenção (G1). Guarapuava-PR –UNICENTRO, 2019. ...................................................................... 73

LISTA DE TABELAS
TABELA 1- Dados sobre os genótipos, classificação do teor de acil-açúcar e genótipo
utilizados no experimento. Guarapuava-PR –UNICENTRO, 2019. ...................................... 44
TABELA 2- Teor de acil-açúcares, número médio de ovos, número médio de ninfas, número
médio de exúvias, número médio de adultos, correlações e contraste em genótipos F2RC2 de
tomateiro com diferentes teores de acil-açúcares, Solanum pennellii (acesso LA716) e
Solanum lycopersicum (Cultivar Redenção) submetidos à infestação da mosca-branca.
Guarapuava-PR –UNICENTRO, 2019. ................................................................................ 45
TABELA 3- Porcentagem de cobertura fumagina presente nas folhas, correlações lineares e
contraste, em genótipos F2RC2 de tomateiro, Solanum pennellii e cultivar Redenção.
Guarapuava-PR –UNICENTRO, 2019. ................................................................................ 51
TABELA 4- Primers ISSR utilizados para avaliação da similaridade genética entre os
parentais e as linhagens de tomateiro com alto de teor de acil-açúcares. Temperatura de
anelamento (TºCa). Guarapuava-PR –UNICENTRO, 2019. ................................................. 66
TABELA 5- Primers SSR indicando cromossomo onde foi mapeado, sequência e a indicação
se o primer é polimórfico. Guarapuava-PR –UNICENTRO, 2019. ....................................... 67
TABELA 6- Dados sobre os genótipos, classificação do teor de acil-açúcar e genótipo
utilizados no experimento. Guarapuava-PR –UNICENTRO, 2019. ...................................... 68
TABELA 7- Teores de acil-açúcares, número médio de ovos, número médio de larvas e
contraste, durante 28 dias de avaliações, em genótipos F2RC2 de tomateiro com diferentes
teores de acil-açúcares, Solanum pennellii (acesso LA716) e Solanum lycopersicum (Cultivar
Redenção) submetidos à infestação da traça do tomateiro. Guarapuava-PR –UNICENTRO,
2019. .................................................................................................................................... 69
TABELA 8- Teores de acil-açúcares, danos nas plantas (DP) lesões nos folíolos (LF),
porcentagem de folíolos atacados (PF) e contraste, durante 28 dias de avaliações, em
genótipos F2RC2 de tomateiro com diferentes teores de acil-açúcares, Solanum pennellii
(acesso LA716) e Solanum lycopersicum (Cultivar Redenção) submetidos à infestação da
traça do tomateiro. Guarapuava-PR –UNICENTRO, 2019. .................................................. 71
TABELA 9- Similaridade genética entre linhagens F2RC2 de tomateiro oriundas do
cruzamento entre o genótipo selvagem Solanum pennellii e a cultivar Redenção (Solanum
lycopersicum). Guarapuava-PR-UNICENTRO, 2019. .......................................................... 75

i
Resumo
Dias, Diego Munhoz. Desenvolvimento de genótipos F2RC2 de tomateiro para
processamento com altos teores de acil-açúcares e resistentes a artrópodes-praga.
Guarapuava: UNICENTRO, 2019. 88p. (Tese - Doutorado em Produção Vegetal).
A cultura do tomateiro apresenta alta suscetibilidade a vários artrópodes-praga, que causam
uma série de prejuízos. Estudos com a incorporação de genes advindos de espécies silvestres
são importantes, dentro do manejo integrado de pragas (MIP), como forma de resistência as
principais pragas. Objetivou-se neste estudo comparar os teores de acil-açúcares (AAs)
presentes nos folíolos quanto a resistência de genótipos F2RC2 de tomateiro para a mosca
branca, fumagina e traça do tomateiro, além do uso de marcadores moleculares ISSR e SSR
para seleção de genótipos com maior recuperação do parental recorrente. Nos experimentos
incluíram genótipos F2RC2 com altos teores de AAs, genótipos F2RC2 com baixos teores de
AAs, além dos parentais Solanum pennellii (padrão de resistência, altos teores de AAs) e
cultivar Redenção (linhagem comercial, padrão suscetibilidade). Os genótipos F2RC2 foram
clonados via enraizamento de brotações axilares e os parentais cultivados a partir de
sementes. Os testes de resistência à mosca branca, traça do tomateiro e para a fumagina, os
experimentos foram instalados em casa de vegetação em delineamento em blocos ao acaso
com (DBC), compreendendo os tratamentos (genótipos) com 4 repetições. Foram feitas
criações programada da mosca-branca e traça do tomateiro em gaiolas com tela antiafídica em
plantas hospedeiras de cada inseto para testes isolados para cada praga. No estágio
reprodutivo os genótipos F2RC2 foram infestados com as pragas e avaliados fatores
reprodutivos dos insetos e os danos causados pelas pragas nos diferentes experimentos.
Simultaneamente, os genótipos F2RC2 foram analisados com o uso de marcadores ISSR e
SSR. Foram utilizados seis primers ISSR e seis primers SSR. Geralmente, os genótipos com
altos teores de AAs apresentaram maiores níveis de resistência à mosca-branca, traça do
tomateiro, quando comparados aos com as testemunhas sem o aleloquímico. Foram obtidos
correlações e contrastes significativos entre os teores de AAs e os parâmetros de resistência às
pragas utilizadas no experimento, confirmando a associação entre os altos teores de AAs com
a resistência as principais pragas. A seleção direta em laboratório foi eficiente na
quantificação dos teores de AAs com a diferenciação dos genótipos contrastantes, além de
facilitar a seleção de genótipos F2RC2 de tomateiro. A utilização de marcadores moleculares
ISSR e SSR possibilitou a seleção de plantas com maior proporção genética do parental

ii
recorrente. Os genótipos F2RC2 de tomateiro RVTA-2010pl# 39, RVTA-2010pl#232 e
RVTA-2010pl#257 com características de resistência similar ao parental silvestre e maiores
proporção do parental Redenção serão utilizadas para prosseguir o programa de
melhoramento genético.
Palavras-chave: Solanum lycopersicum, Solanum pennellii, aleloquímico, resistência às
pragas.

iii
Abstract
Dias, Diego Munhoz. Development of tomato F2RC2 genotypes for processing with high
levels of acyl-sugars and resistant to pest arthropods. Guarapuava: UNICENTRO, 2019.
92p. (Thesis - PhD in Plant Production).
Tomato cultivation presents high susceptibility to several pest arthropods, which is a series of
losses. Studies with the incorporation of genes from wild species are important, as part of
integrated pest management (IPM), as a form of resistance as the main pests. The objective of
this study was to compare the levels of acyl-sugars (AAs) present in leaflets on the resistance
of tomato F2RC2 genotypes to whitefly, fumagine and tomato moth, as well as the use of
molecular markers ISSR and SSR for the selection of genotypes with higher recurrent parental
recovery. The experiments included F2RC2 genotypes with high levels of AA, F2RC2
genotypes with low levels of AAs, besides the Solanum pennellii genotypes (resistance
pattern, high levels of AAs) and cultivar Redenção (commercial strain, data exploitability).
F2RC2 genotypes were cloned via axillary sprouting and parental sprouting from seeds. The
experiments were carried out in the greenhouse in a randomized complete block design
(DBC), comprising the treatments (genotypes) with four replicates. Programmed creations of
the whitefly and the tomato moth were carried out in cages with antiphlogistic screens on host
plants of each insect for testicles, for each pest. In the reproductive stage, the F2RC2
genotypes were infested with pests and the pests in the different experiments evaluated the
insect factors and the negative effects. Simultaneously, F2RC2 genotypes were used with the
use of ISSR and SSR. Six ISSR primers and six SSR primers were used. Generally, genotypes
with high acid content decrease levels of resistance to whitefly, the tomato moth, when
compared to non-allelochemical controls. Correlated testimonies and contrasts between AA
theorists and the main situational indicators in experiments, confirming an association
between the high levels of AAs with war as major pests. The direct selection in the laboratory
was efficient in quantifying the AA contents with a differentiation of the contrasting
genotypes, besides facilitating the selection of tomato F2RC2 genotypes. The use of molecular
markers ISSR and SSR allowed the selection of plants with a higher genetic proportion of the
recurrent parent. The F2RC2 genotypes of RVTA-2010pl # 39, RVTA-2010pl # 232 and
RVTA-2010pl # 257 with parental resistance characteristics and a higher proportion of
Redenção parents are used for the progress of the breeding program.
Keywords: Solanum lycopersicum, Solanum pennellii, allelochemical, resistance to pests.

8
1. INTRODUÇÃO
O tomateiro cultivado (Solanum lycopersicum) é classificado como uma hortaliça fruto
de grande importância na agricultura nacional. No Brasil, especialmente no ano de 2017,
foram cultivados aproximadamente 64,6 mil hectares com produção 4,37 milhões de
toneladas, sendo cultivado em praticamente todas as regiões brasileiras (IBGE, 2018).
O mercado consumidor do tomate é amplamente versátil, apresentando grande
aceitação por parte dos consumidores, tanto para consumo tanto in natura, bem como o uso
de seus derivados por meio da industrialização (ABCSEM, 2010; CNA, 2017). Além de fazer
parte dos hábitos alimentares dos brasileiros o tomate possui importante papel econômico e
social para o país (ABCSEM, 2010). Na questão econômica, somente o tomate representa
16% do PIB produzido pelas hortaliças. Na questão social, a cultura possibilita a fixação do
homem no campo com a geração de cerca de 300 mil empregos na área agrícola, sem
considerar as atividades industriais que possuem elevada demanda de mão de obra
(GERALDINI et al., 2011).
O tomateiro cultivado para fins industriais no Brasil representa cerca de um terço da
produção com 1,45 milhões de toneladas no ano de 2017 (WPTC, 2017). O consumo per
capita de tomate processado é de cerca 0,665 kg de massa de tomate e 0,634 kg de molho de
tomate por ano. Porém, estima-se para os próximos anos, crescimento no consumo de
derivados do tomate. Os maiores centros consumidores nas regiões Sul e Sudeste,
possivelmente, devido ao maior poder de compra da população (CLEMENTE e BOITEUX,
2012).
Todavia, a cultura do tomateiro apresenta uma série de fatores que desfavorecem o
aumento do seu cultivo, dentre eles, a grande suscetibilidade ao ataque de vários artrópodes
que causam danos a cultura. O manejo destes é complexo e oneroso, resultando em aumento
do custo de produção sem, muitas vezes, apresentam resultados satisfatórios. Dentre as
principais pragas que acometem danos a cultura, a mosca-branca (Bemisia tabaci biótipo B) e
a traça-do-tomateiro (Tuta absoluta) são consideradas pragas chave, muitas vezes,
impossibilitando o cultivo do tomate em algumas regiões. Porém, em ambiente protegido
outras pragas de importância secundária podem ser o principal problema, como é o caso dos
ácaros do gênero Tetranychus.
A principal forma de manejo utilizada pelos agricultores para contornar os entraves
causados pelas principais pragas é o uso agroquímicos que quando pulverizados de forma
indiscriminada, favorecem a seleção de populações resistentes dificultando ainda mais o seu

9
manejo. Neste contexto, devido as pulverizações excessivas e o uso de produtos não
registrados para a cultura, o tomateiro é considerado como uma hortaliça que apresenta níveis
consideráveis de resíduos de agroquímicos que podem acarretar severos danos à saúde do
trabalhador rural, ao consumidor e ao meio ambiente (ANVISA, 2015).
Pesquisas que busquem reduzir o uso de agroquímicos com métodos alternativos é de
extrema importância. A seleção e uso de genótipos de tomateiros com um nível adequado de
resistência a artrópodes-praga (TOSCANO et al., 2002; FANCELLI et al., 2003) tem
demonstrado ser promissor. Dentro do manejo integrado de pragas, a resistência varietal surge
como uma alternativa viável para reduzir os resíduos de agroquímicos e os custos de
produção.
As espécies silvestres de tomateiro podem ser usadas como fontes de genes de
resistência às principais doenças e pragas. A resistência as principais pragas apresentadas pelo
genótipo silvestre Solanum pennellii acesso LA 716 tem sido associada a presença de um
fitoquímico chamado acil-açúcares (AAs). Este aleloquímico possibilita efeitos deletérios no
desenvolvimento de um artrópode-praga, podendo impedir ou dificultar a alimentação ou, até
mesmo, a ovoposição por meio da liberação de aleloquímicos (RESENDE et al., 2002a;
LUCINI et al., 2015). Em programas de melhoramento visando a resistência às pragas o uso
do S. pennellii na introgressão de genes de resistência em cultivares comerciais é promissor,
principalmente por meio de retrocruzamentos para obter linhagens avançadas para a formação
de novos híbridos (RESENDE et al., 2009). Neste sentido, o uso simultâneo do
retrocruzamento assistido por marcadores moleculares apresentam-se como alternativa viável
para aumentar a eficiência do processo de retrocruzamento e identificação precisa e precoce
dos indivíduos que apresentam maior recuperação do genoma do genitor recorrente
(OPENSHAW et al., 1994).
Por fim, devido a necessidade de cultivares de tomate para processamento com um
nível de resistência satisfatório as principais pragas, pesquisas com a espécie silvestre S.
pennellii são de extrema importância na introgressão de novos genes em tomates, visando o
manejo ecologicamente sustentável, economicamente viável e socialmente justo.

10
2. OBJETIVOS
2.1 Geral
Selecionar genótipos F2RC2 de tomateiro para processamento com altos teores de acil-
açúcares, provenientes do cruzamento entre S. lycopersicum L. (cultivar Redenção) x S.
pennellii acesso LA-716, resistentes a artrópodes-praga.
2.2 Específicos
Realizar o avanço da geração F2RC1 em genótipos de tomateiro por meio do uso de
retrocruzamentos para a geração F2RC2;
Quantificar o teor de acil-açúcares em genótipos F2RC2 de tomateiro pertencentes do
cruzamento interespecífico entre Solanum lycopersicum L. “Redenção” x Solanum pennellii
acesso LA-716.
Selecionar os genótipos F2RC2 de tomateiro com altos teores de acil-açúcares
resistentes à traça-do-tomateiro (Tuta absoluta) e à mosca-branca (Bemisia tabaci biótipo B).
Selecionar genótipos F2RC2 de tomateiro com altos teores de acil-açúcares que
apresentam maior proporção do genoma do pai recorrente com base nos marcadores SSR e
ISSR.

11
3. REFERENCIAL TEÓRICO
3.1 Origens e histórico do tomateiro
O tomateiro (S. lycopersicum) é cultivado na maior parte do Mundo, sendo
descendente da espécie silvestre Solanum lycopersicum var. cerasiforme que produz frutos
menores, do tipo cereja. A região andina da Colômbia, Bolívia, Equador, Peru e parte do
Chile são considerados como o possível centro de origem do tomateiro. Contudo, a
domesticação das espécies silvestres ocorrida no México antes da colonização espanhola fez
com que a região fosse considerada o centro secundário da cultura do tomateiro (NAIKA et
al., 2006; SANTOS, 2009; ALVARENGA, 2013).
Em meados do século XVI ocorreu a introdução do tomateiro na Europa pelos
espanhóis e a partir de então, a cultura foi difundida para os demais países do mundo. Nestas
regiões, inicialmente, as plantas de tomate eram utilizadas apenas para ornamentação de
jardins, não sendo utilizado para alimentação. Os frutos eram considerados tóxicos por
pertencer à mesma família de algumas plantas tóxicas já conhecidas pelos Europeus e também
devido a coloração vermelha de seus frutos. A toxidade destas plantas estava ligada a
presença de um alcaloide chamado tomatina, presente em maiores níveis nas folhas e frutos
imaturos. Porém, em frutos maduros é possível o consumo devido a tomatina ser degradada
em outros compostos inertes (ALVARENGA, 2013; BERGOUGNOX, 2014).
Os primeiros relatos de cultivos do tomate foram feitos pelos italianos devido ao
potencial ornamental e a beleza dos frutos do tomateiro. Na Itália em 1554, Matthiolus
realizou a primeira referência do uso do tomate para consumo, no qual a primeira espécie
introduzida apresentava frutos amarelos conhecidos popularmente como maça dourada ou
Pomi d’oro. Na América latina, especialmente no Brasil, o tomateiro foi introduzido por
imigrantes europeus no século XIX. Entretanto, o surgimento do tomate Santa Cruz no estado
do Rio de Janeiro foi considerado o marco inicial da trajetória do cultivo de tomate no Brasil,
em 1940. O tomate é considerado como uma das hortaliças mais prestigiadas e cultivadas no
Mundo (FILGUEIRA, 2008; ALVARENGA, 2013).

12
3.2 Taxonomia, classificação e botânica do tomateiro
O tomateiro é uma espécie dicotiledônea da ordem Tubiflorae, família Solanacea e
atualmente ao gênero Solanum e subgênero Eulycopersicon e Euriopersicon (PERALTA e
SPOONER, 2000; PERALTA et al., 2006; FILGUEIRA, 2008; ALVARENGA, 2013). O
primeiro descritor do gênero Solanum foi o pesquisador Carl Van Linnaeus no livro “Species
Plantarum”, porém, ocorreu a reclassicação do tomate como sendo do gênero Lycopersicum
por Miller e mais tarde descreveu a espécie Lycopersicon esculentum como sendo o tomateiro
cultivado. Posteriormente, diversas pesquisas evidenciaram alta correlação genética entre as
espécies do gênero Solanum com o tomate cultivado (L. esculentum). Devido a semelhança
encontrada com o outro gênero o gênero Lycopersicon caiu em desuso e, novamente, foi
reconhecido a espécie cultivada como do gênero Solanum e reclassificada como Solanum
esculentum (PERALTA e SPOONER, 2000; PERALTA et al., 2006). Por fim, após uma série
de estudos filogenéticos com a utilização de sequências de DNA, distribuição e detalhadas
pesquisas de morfologia, a maioria dos pesquisadores entraram em consenso sobre a espécie
cultivada de tomateiro como sendo Solanum, anteriormente proposta por Linnaeus,
apresentando mudança apenas no final do nome, sendo classificado pelos pesquisadores como
Solanum lycopersicum (SPOONER et al., 2005; PERALTA et al., 2006).
O tomateiro mesmo sendo uma planta herbácea perene, possui comportamento similar
a plantas anuais desde a semeadura até a produção de sementes (NAIKA et al., 2006;
FILGUEIRA, 2008). Possuem inúmeras ramificações laterais com arquitetura que lembra
uma moita em seu desenvolvimento natural. Porém, fazendo o uso do manejo cultural da
desbrota a característica pode mudar drasticamente. O crescimento em altura e o número de
ramificações estão intimamente ligados a arquitetura da planta, que possui dois tipos de
crescimento como o tipo determinado e indeterminado (ALVARENGA, 2013).
A parte comercializável da planta são os frutos que quando maduros apresentam
coloração vermelha, devido ao acúmulo de um pigmento carotenoide chamado licopeno. O
licopeno por atuar como antioxidante natural vem sendo ligado à prevenção do câncer de
próstata entre outras funções (FILGUEIRA, 2008).
Atualmente existem cerca de 13 espécies de tomateiro catalogadas e identificadas.
Dentre as espécies de tomateiro, apenas a espécie S. lycopersicum é cultivada, as demais são
consideradas silvestre, sendo elas, S. cheesmaniae, S. chilense, S. chmielewskii, S.
habrochaites, S. neorickii, S. pennellii, S. peruvianum, S. pimpinellifolium (ALVAREGA,
2013). Por fim, as últimas espécies identificadas foram S. huaylasense, S. galapagense, S.

13
corneliomuelleri e S. arcanum (PERALTA et al. 2006).
As espécies silvestres não são comercializáveis devido uma série de características
como baixa produção, frutos extremamente pequenos e, às vezes, pubescentes. Entretanto,
apresentam várias características de interesse no melhoramento genético como resistência à
artrópodes-praga, resistência a doenças, estresse abióticos e melhoria da qualidade nutricional.
No melhoramento genético do tomateiro estas espécies silvestres vem sendo utilizadas como
fonte de genes visando introgressão destes genes benéficos incorporados na cultivar comercial
por meio de cruzamentos interespecíficos (GONÇALVES et al., 1998; ZORZOLI et al., 2000;
MALUF et al., 2001; ARAGÃO et al., 2002; RESENDE et al, 2002b; AZEVEDO et al.,
2003; GONÇALVES et al., 2006; RESENDE et al., 2008; RESENDE et al., 2009;
GONÇALVES NETO et al., 2010; MALUF et al., 2010; DIAS et al., 2013; BAIER et al.,
2015).
3.3 Aspectos gerais de produtividade do tomateiro
Na safra de 2016, a produção mundial de tomate somou 177 milhões de toneladas em
uma área cultivada de 4,78 milhões de hectares com rendimento médio de 37 toneladas por
hectare. Dentre 173 países produtores de tomate no Mundo, merecem destaque os maiores
produtores mundiais, sendo em ordem decrescente a China com 56,3 milhões de toneladas,
Índia com 18,4 milhões de toneladas, Estados Unidos com 13,0 milhões de toneladas, Turquia
com 12,6 milhões de toneladas, Egito com 7,9 milhões de toneladas, Itália com 6,5 milhões de
toneladas, Iran com 6,4 milhões de toneladas, Espanha com 4,7 milhões de toneladas e Brasil
com 4,17 milhões de toneladas (FAOSTAT, 2016).
Com relação à área cultivada mundial, a China é o país que possui a maior área
cultivada de tomate com cerca de 999 mil hectares, seguida pela Índia com 760 mil hectares,
Nigéria com 574 mil hectares, Egito com 199 mil hectares e Turquia com 188 mil hectares.
No ranking mundial o Brasil é o 9° colocado em produção, 14° em área de cultivo e 40° no
rendimento (65,15 toneladas por hectare) (FAOSTAT, 2016).
Na safra 2017, o Brasil obteve produção aproximada de 4,37 milhões de toneladas em
uma área de cultivo 64,6 mil hectares e rendimento médio de 67,6 toneladas por hectare. As
regiões de maior destaque em produção de tomate do país são o Sudeste (44,8 %), Centro-
Oeste (29,7 %), Sul (13,1 %) e Nordeste (12,0 %). Em relação aos estados com maior
destaque na produção de tomate são Goiás (28.9 %), São Paulo (21,5 %), Minas Gerais (15,5
%), Bahia (7,0 %) e Paraná (6,0 %) (IBGE, 2017). No Brasil a cultura possui importante

14
papel, sendo amplamente utilizada pelo brasileiro no consumo in natura, assim como em
produtos industrializados, com consumo per capita de 39 kg ano-1 (CNA, 2017).
3.4 Aspectos gerais de produtividade do tomateiro para processamento
Segundo Word Production Estimate of Tomatoes for Processing- WPTC (2017), na
safra de 2017 houve redução na produção mundial de 1,9 %, obtendo assim o valor de 37,4
milhões de toneladas de tomate para processamento. O maior crescimento na produção foi
constatado na Rússia, com aumento de 176 % em comparação com 2016, porém, com
produção de apenas 400 mil toneladas. As maiores reduções nas produções ocorreram em
países como Austrália (- 32,7 %), Estados Unidos (- 17 %) e Polônia (- 35 %).
Contudo, o maior produtor de tomate para processamento continua sendo a Estados
Unidos, com previsão de produção de 9,52 milhões de toneladas, cerca de 32 % da produção
mundial, seguido pela China (5,6 milhões de toneladas), Itália (5,2 milhões de toneladas),
Espanha (3,3 milhões de toneladas), Turquia (1,9 milhões de toneladas), Portugal (1,55
milhões de toneladas) e Brasil (1,45 milhões de toneladas). Atualmente, o Brasil ocupa a
sétima posição na produção de tomate para processamento (WPTC, 2017).
No Brasil o mercado nacional de atomatados está em constante expansão,
impulsionado pela melhoria da renda per capita, crescimentos das redes “fast food”, inserção
de outras empresas de processamento, mudanças nos hábitos alimentares da população e,
principalmente, aumento da entrada da mulher no mercado de trabalho onde procura soluções
práticas de alimentação (CLEMENTE e BOITEUX, 2012).
O fruto do tomateiro possui de 90 a 95 % de água sendo considerado como um fruto
altamente perecível, tolerando assim poucos transtornos desde sua retirada do campo até o
consumidor. Uma alternativa para agregar valor e reduzir as perdas é a industrialização do
tomate com posterior comercialização de seus derivados, possibilitando que o produto
permaneça disponível por um maior período de tempo, aumentando a vida (GAMEIRO et al.,
2007). Além do fruto altamente perecível a alta suscetibilidade a diversas pragas torna o
manejo do tomateiro amplamente complexo.

15
3.5 Artrópodes-praga do tomateiro
3.5.1 Traça-do-tomateiro - Tuta absoluta (Meyrick, 1917)
Nativa da América Central a traça-do-tomateiro (Tuta absoluta) pertence à família
Gelechiidae, sendo considerada como um inseto holometabólico, por apresentar metamorfose
completa, com ciclo de vida composto pelas fases de ovo, larva, pupa e adulto (INCAPER,
2010; CLEMENTE e BOITEUX, 2012).
Os ovos apresentam formatos elípticos, minúsculos e de coloração branca que com o
passar do tempo tornam-se amarelados. Embora ocorra divergências entre a preferência do
inseto para depositar seus ovos entre a superfície adaxial e abaxial das folhas é sabido que
ocorrem a presença de ovos em ambas as faces da folha (INCAPER, 2010; CLEMENTE e
BOITEUX, 2012). Porém, podem ser encontrados ovos nas flores, frutos, cálice e hastes das
plantas de tomateiro (SILVA e CARVALHO, 2004; CLEMENTE e BOITEUX, 2012). O
período de incubação dos ovos, larvas, pupas e adultos são variáveis e dependentes das
condições climáticas. Em média, a incubação dos ovos varia de 2 a 4 dias e após a eclosão dos
ovos as larvas penetram imediatamente nos frutos, ponteiros ou folíolos, onde permanecem
consumindo o tecido vegetal por 8 a 10 dias (SILVA e CARVALHO, 2004; INCAPER,
2010).
A coloração das larvas são variáveis de verde-claro ao rosado com a presença de uma
placa quitinosa de coloração escura no primeiro segmento torácico. Além de cinco pares de
pernas abdominais e três pares de pernas torácicas. Após a eclosão dos ovos as larvas passam
por 4 instares, variam o seu tamanho com o avanço dos instares, variando de 6 a 9 mm de
comprimento em 13 a 17 dias. Após este período a larva torna-se pupa com fase variando de 7
a 10 dias. As pupas podem ser encontradas nos frutos, gema apical e folhas, sempre
envolvidas por um casulo de seda. No solo, as pupas podem ser encontradas em detritos
vegetais (INCAPER, 2010).
A traça-do-tomateiro na fase adulta é uma pequena mariposa de hábito crepuscular de
cerca de 11 mm de comprimento de coloração cinza-prateada e asas franjadas, logo após a
emergência acasalam-se. Na fase adulta a longevidade do inseto pode chegar até 15 dias
(INCAPER, 2010). Na literatura é verificada divergência entre a quantidade de ovos que uma
fêmea consegue ovopositar, variando de 50 até 260 ovos. O ciclo biológico do inseto é muito
dependente das condições ambientais do local de avaliação e tipo de cultivo, podendo variar
de 30 até 38 dias (INCAPER, 2010; CLEMENTE e BOITEUX, 2012; ALVARENGA, 2013).
A traça-do-tomateiro está presente em qualquer sistema de cultivo e durante todo o

16
ciclo da cultura sendo considerada uma praga-chave. Acredita-se que por meio da importação
do tomate chileno ocorreu a introdução desta praga no Brasil (OLIVEIRA, 2004). No Brasil, a
primeira constatação foi na cidade de Jaboticabal-SP, disseminando, desde então, para as
demais regiões produtoras de tomate (SOUZA e REIS, 2000).
Os níveis de infestação deste artrópode são severamente influenciados pelas condições
climáticas. As condições de altas temperaturas com baixa precipitação pluviométrica e
veranicos em épocas chuvosas favorecem a proliferação desta praga (INCAPER, 2010). Em
contrapartida, os danos são reduzidos em cultivos sob precipitação pluvial e podem
desaparecer em períodos chuvosos. Os sistemas de irrigação podem influenciar fortemente na
população da traça-do-tomateiro. Normalmente, sistemas de irrigação por aspersão e por pivô
central ocorrem menores infestações quando comparados aos sistemas de irrigação localizada
por gotejamento e sulcos. Possivelmente, a redução na taxa de infestação ocorre devido a
água proveniente da irrigação derrubar os ovos, lagartas e pupas presentes na planta,
dificultando o ciclo e reduzindo o potencial reprodutivo do inseto (SILVA e GIORDANO,
2000; CLEMENTE e BOITEUX, 2012).
Os danos causados pela traça-do-tomateiro ocorrem no seu período larval, logo após a
eclosão dos ovos. Nas folhas, as lagartas penetram na mesma e consomem o parênquima
foliar, deixando apenas a epiderme superior e inferior da folha. Os sintomas nas folhas são
caracterizados por minas irregulares nos folíolos. Os principais danos causados pela praga
ocorrem no terço médio da planta, mas os primeiros danos ocorrem no terço superior. Nos
ponteiros as lagartas consumem a gema apical e penetram no caule no sentido descendente,
impedindo o fluxo de seiva no ponteiro, causando o travamento do crescimento da gema
apical ou até a sua morte. A constatação dos danos nos ponteiros pode ser feita pela
constatação de fezes escuras da traça do tomateiro próximas aos pequenos folíolos da gema
apical.
Nos frutos, o inseto faz galerias que tendem a ficar escuras devido a deposição de
fezes, provenientes da alimentação da polpa do fruto, onde os sintomas são notados próximos
das sépalas (INCAPER, 2010). Vale ressaltar que todos os frutos que apresentam perfurações
e galerias junto do cálice devido ao ataque de lagartas são descartados pelos produtores
devido a perda do valor comercial. Além da redução da produção devido ao superbrotamento
causado pelos danos no ponteiro e alimentação de botões florais que reduzem
consideravelmente a formação de frutos comercializáveis (SOUZA e REIS, 2003; SILVA e
CARVALHO, 2004).

17
3.5.2 Mosca-branca- Bemisia tabaci (Genn.) biótipo B e/ou Bemisia argentifolii
(Bellows e Perring, 1994)
A mosca-branca é uma praga chave da cultura do tomateiro pertencente à família
Aleyrodidae e a ordem hemíptera. Apresenta desenvolvimento do tipo hemimetabolia com
ciclo biológico dependente das condições ambientais que dura em média 15 dias
compreendendo as fases ovo, ninfa e adulto (INCAPER, 2010).
Os adultos são pequenos insetos de 1 mm de comprimento com dois pares de asas
membranosas com uma pulverulência branca. A pulverulência modifica o aspecto do inseto e
causa o nome falho apresentado pela mesma, mosca-branca. Os adultos apresentam
preferência pela ovoposição pela superfície abaxial dos folíolos do tomateiro, fixando por
meio de um curto pedúnculo os ovos na folha com duração de 3 a 6 dias até o inicio do
estágio ninfa (ALVARENGA, 2013).
Após a eclosão dos ovos surgem as ninfas com corpo de formato ovalado e
translúcidas. A coloração das ninfas é variável podendo apresentar cores do amarelo ao
amarelo pálido, onde possuem pernas que possibilitam sair do local da eclosão dos ovos para
outro local da folha que facilite a introdução do seu estilete para o início do processo de
sucção de seiva (INCAPER, 2010).
Na tomaticultura ocorre, principalmente, a presença de duas espécies de mosca-
branca. A Bemisia tabaci é considerada a espécie mais comum com ovoposição média de 110
ovos por fêmea. No entanto, outro biótipo conhecido como Bemisia tabaci biótipo ou B.
argentifolii tem causado maiores problemas para o cultivo do tomateiro, principalmente,
devido a maior capacidade de ovoposição da fêmea com cerca de 300 ovos (ALVARENGA,
2013).
A mosca-branca possui elevada capacidade de redução da produtividade e elevado
potencial de danos nas lavouras. É considerada como uma das pragas-chave da cultura do
tomateiro (FRANÇA et al., 2000). Na região de Goiás, devido aos prejuízos que este sugador
vem ocasionando, desde 2002, esta praga é considerada como o principal inseto na cultura
(MARTINS, 2013). Em testes de preferências com diversas chances de escolha entre as
principais culturas, demonstraram, entre as culturas estudadas o tomateiro e a abobrinha são
as preferidas para ovoposição pela mosca branca (VILLAS BÔAS et al., 2002).
A coloração branca é predominante nos adultos, principalmente pelas asas que cobrem

18
a maior parte do corpo inseto. Entretanto, o dorso do inseto apresenta a coloração amarelo-
claro. Em relação ao tamanho, os adultos apresentam cerca de 1 mm de comprimento, sendo
considerados insetos pequenos. Nos adultos ocorre dimorfismo sexual, do qual os machos são
menores em relação as fêmeas. O aparelho bucal da mosca-branca é do tipo “picador-
sugador” com elevada capacidade de sucção de seiva, principalmente, pelo número de insetos
presentes na folha (INCAPER, 2010).
A mosca-branca é capaz de causar diversos danos ao tomateiro, sendo estes danos
divididos entre danos diretos e indiretos. Os danos indiretos causados pela B. tabaci ocorrem
no momento em que é inserido o aparelho bucal no interior do tecido vegetal para alimentação
e, concomitantemente, pode ocorrer a transmissão de partículas virais, em especial, ao grupo
geminivírus que a mosca-branca é especializada na sua transmissão. Independente do período
de desenvolvimento da planta a transmissão pode ocorrer, sendo mais drásticas nos estágios
iniciais da cultura. Os danos causados pela geminivirose variam de 40 a 70 %, principalmente,
devido a paralização do crescimento normal da planta e redução de sua produção (FRANÇA
et al., 2000). Nas plantas infectadas ocorrem mosaicos amarelados nas folhas com posterior
enrijecimento, além de apresentar o fenômeno de curvatura da folha para baixo conhecido
como epinastia. Nos frutos o vírus pode causar perda da atratividade no consumidor final
devido ao amadurecimento desuniforme reduzindo seu valor comercial. Na alimentação deste
artrópode-praga ocorre a liberação de uma secreção açucarada no limbo foliar que serve de
substrato para o crescimento de um fungo responsável pelo escurecimento dos órgãos
atacados (folhas e frutos) e redução da capacidade fotossintética da planta (HAJI et al., 2005).
Os danos diretos causados pela mosca-branca são os danos causados apenas pela
alimentação do inseto, do qual, o mesmo insere o aparelho bucal e suga a seiva. Em altas
infestações, a retirada considerável de seiva causa uma série de modificações fisiológicas na
planta com redução de até 50 % da produtividade da cultura. Em várias regiões a mosca-
branca é considerada como uma ameaça para a produção de tomate por todos os prejuízos
associados a esse inseto, além a alta capacidade de adaptação a diversos ambientes e ataque a
diversas culturas, condicionam uma situação ideal para o seu desenvolvimento (FRANÇA et
al.,2000).

19
3.5.3 Ácaro rajado - Tetranychus urticae (Koch, 1836)
O ciclo de vida do ácaro rajado compreende as fases de ovo, larva, protoninfa,
deutoninfa e adulto. A característica diferenciadora da espécie em contraste as demais são a
presença nas fêmeas da coloração esverdeada na lateral de seu dorso com um par de manchas
escuras. O dimorfismo sexual apresentado por esta espécie é considerado alto, no qual as
fêmeas apresentam corpo ovalado e maior tamanho quando comparado ao macho. A espécie é
um haplo-diplóide, em que as fêmeas são advindas da reprodução sexuada e os machos de
partogênese arrenótoca (SILVA e CARVALHO, 2004).
A superfície abaxial das folhas é preferida para a ovoposição pelas fêmeas do ácaro
rajado, que ocorrem próximos as nervuras ou sobre as teias. Os ovos são de aproximadamente
0,14 mm de diâmetro com formato esférico, primeiramente de coloração quase transparente
passando para a coloração opaca e quando estão próximos da eclosão das larvas apresentam
coloração amarelada (GALLO et al., 2002). As larvas possuem coloração incolor ou
translúcidas com tamanho semelhante ao dos ovos, mas com três pares de pernas e
apresentando duas manchas laterais com coloração avermelhada. A coloração da larva vai
sendo modificada de acordo com a sua alimentação. Nas fases de protoninfa e deutoninfa são
diferenciadas somente pelo tamanho, cuja deutoninfa é pouco maior que a protoninfa
(MORAES e FLECHTMANN, 2008)
Os ácaros podem apresentar elevada capacidade de explosão populacional, devido a
sua alta capacidade reprodutiva de cerca de 20 a 25 gerações em um mesmo ano. Portanto, o
manejo do ácaro rajado deve apresentar grande eficiência, sempre mantendo a população de
ácaros presentes na cultura abaixo do nível de dano econômico (MARUYAMA et al, 2002;
MARUYAMA e TOSCANO, 2003).
No mundo, o ácaro rajado é considerado como o único representante das seis espécies
de Tetraniquídeos de importância econômica e o principal ácaro fitófago, devido ao grande
número de hospedeiros e os sérios danos causados às plantas (BOOM et al., 2003; SILVA e
CARVALHO, 2004; SATO et al., 2007). Os ácaros pertencentes ao gênero Tetranychus spp.
por tecerem uma determinada quantidade de teia são conhecidos como ‘spider mites’. As teias
possuem diversas funções, sendo utilizadas principalmente para dispersão, marcar território e
proteção contra predadores (MORAES e FLECHTMANN, 2008). Em condições ambientais
favoráveis como baixa umidade e alta temperatura os ácaros podem se tornar a principal praga
na cultura do tomateiro (MALUF et al., 2007). Em ambiente protegido ocorrem os principais
danos causados pelos ácaros na cultura do tomateiro. Neste tipo de sistema os ácaros

20
encontram os fatores ideais ao pleno desenvolvimento e podem ser uma das principais pragas
presentes (SILVA e CARVALHO, 2004).
Na alimentação o ácaro insere o seu estilete no limbo foliar, alimentando-se de todo o
conteúdo celular e, posteriormente, formando bolsas. No decorrer da alimentação ocorre a
formação de manchas branca-prateadas que com a oxidação adquirem coloração escura
(bronzeadas). Em ataque severo é comum a ocorrência de manchas necróticas com posterior
secamento e queda prematura da folha. Nos frutos o ataque severo nas folhas leva a
diminuição do número e tamanho dos frutos, além do amadurecimento prematuro dos frutos
remanescentes (MARUYAMA e TOSCANO, 2003; SILVA e CARVALHO, 2004;
MORAES e FLECHTMANN, 2008).
Neste sentido, o desenvolvimento de cultivares com nível satisfatório de resistência ao
ácaro é de grande valia para redução do número de pulverizações com produtos químicos. As
pesquisas com espécies silvestres do gênero Solanum tem procurado novos mecanismos e
fontes de resistência aos principais artrópodes-praga entre eles, os ácaros (KENNEDY, 2002).
3.6 Manejo integrado de pragas
O manejo integrado de pragas (MIP) no tomateiro é de extrema importância na cultura
do tomateiro, visto que, a cultura é danificada por uma infinidade de artrópodes-pragas, que
podem causar severos danos e, por vezes, impossibilitar a comercialização do tomate pelo
agricultor. Visando garantir um produto de qualidade ao comércio os produtores utilizam
produtos fitossanitários como principal forma de manejo. Contudo, o uso indiscriminado pode
elevar os custos de produção sem um manejo eficiente da praga. O controle químico na
cultura do tomateiro é oneroso e representa cerca de 8,1 % do custo total da lavoura (FAEG,
2013).
O MIP na cultura do tomateiro surgiu com o intuito de buscar uma série de medidas
eficientes para o manejo de pragas. O MIP é reconhecido como o uso simultâneo de medidas
diferentes de manejo de artrópodes-praga, visando reduzir os danos até um nível que não
cause prejuízo econômico, procurando conciliar que o inseto sobreviva na cultura sem
prejuízos significativos. Dentre as diversas medidas utilizadas, merece destaque o uso de
cultivares resistentes, medidas preventivas, medidas culturais, medidas de manejo biológico e
uso do manejo químico racional (FRANÇA et al., 2000; GALLO et. al., 2002).
A resistência genética é considerada por vários autores como uma medida importante

21
no controle de pragas por apresentar custo relativamente baixo e possibilitar a manutenção
dos insetos abaixo do nível de dano econômico. O uso de cultivares resistentes possibilitaria a
manutenção do equilíbrio ecológico entre os inimigos naturais e as pragas, além de não
oferecer risco à saúde humana e ao meio ambiente (VENDRAMIM e NISHIKAWA, 2001;
GALLO et al., 2002). Por não influenciar nos tratos culturais do tomateiro e ainda facilitar o
manejo, as cultivares resistentes de tomateiro a artrópodes-pragas podem ser inseridas em
novas áreas sem resultar em dificuldades de manejo para o agricultor, sendo uma alternativa
economicamente viável, não resultando em resíduos e podendo ser usada em conjunto com
outros métodos do manejo integrado de pragas (VENDRAMIM, 1990).
3.7 Melhoramento genético do tomateiro visando resistência a artrópodes-
praga
A resistência de cultivares de tomateiro aos danos provocados por artrópodes-praga
ocorrem devido ao somatório relativo das características hereditárias presentes na planta, que
influenciam na redução dos danos provocados pela praga (PAINTER, 1951). Neste sentido,
quando uma população, com diferentes indivíduos da mesma espécie, é submetida ao ataque
de uma determinada praga que causa uma série de danos, podemos considerar plantas
resistentes aquelas que possuem a capacidade de manter ou aumentar as características
agronômicas desejáveis em relação a maioria das plantas (GALLO et al., 2002). Diversos
programas de melhoramento genético vegetal exploram três tipos de resistências de plantas às
pragas que são conhecidos como antibiose, não preferência e tolerância (OLIVEIRA, 2013;
LUCINI et al., 2015).
A resistência do tipo antibiose é presente quando ocorre a alimentação dos tecidos da
planta hospedeira pelo artrópode-praga. No entanto, quando em contato ou ingeridos os
tecidos da planta com a praga ocorrem uma cadeia de efeitos deletérios sobre a praga. Os
efeitos podem alterar alguns dos seus padrões biológicos, como fecundidade, ovoposição,
duração das fases imaturas e mortalidade. Neste tipo de resistência a ingestão dos metabólitos
deletérios produzidos na planta pela praga são a causa de resistência. A antixenose ou não
preferência é uma modalidade de resistência quando a planta possui alguma característica que
cause uma resposta negativa pela praga, reduzindo a atratividade da planta para determinadas
funções básicas do artrópode-praga como abrigo, alimentação e ovoposição. Possivelmente,
os fatores característicos nesta resistência sejam aspectos químicos, físicos e morfológicos

22
apresentados pela planta. Por fim, a tolerância é um tipo de resistência que ocorre quando a
planta não modifica a biologia ou o comportamento dos insetos, porém consegue manter as
principais características produtivas quando comparadas a outras plantas sobre o mesmo
ataque de pragas. As plantas consideradas tolerantes compensam os danos causados pelas
pragas com o aumento da área foliar, brotações ou espessura da cutícula (GALLO et al.,
2002).
As resistências de plantas às pragas podem ter origem de diferentes causas podendo
ser agrupadas em fatores químicos, físicos e morfológicos. Os fatores químicos podem ser
representados por enzimas, antimetabólitos e substâncias tóxicas que agem no metabolismo
e/ou comportamento da praga. Diferente dos fatores morfológicos e físicos que são associados
aos aspectos estruturais presentes na epiderme da planta como cerosidade, textura, espessura
e, principalmente, a presença de tricomas grandulares (NORRIS e KOGAN, 1980).
Na natureza é possível observar diferentes níveis de resistência entre as plantas
podendo ser diferenciadas por possuírem alta suscetibilidade, suscetibilidade, resistência
moderada, alta resistência e imunidade. As plantas que apresentam dano médio superior ao da
média da população em que foi comparada são chamadas de plantas altamente suscetíveis.
Posteriormente, quando o dano presente em uma planta é similar ao ocorrido pela média geral
a mesma é reconhecida como plantas suscetíveis. Na seleção de plantas resistentes é
interessante plantas com danos menores ao dano médio ocorrido pelas demais, sendo
chamadas de plantas resistência moderada. As plantas classificadas com alta resistência são as
que apresentam poucos danos em relação à média geral. O ideal, porém, difícil, seria a seleção
de plantas imunes que são as plantas que em qualquer condição não sofrem nenhum dano
(Lara, 1991).
A introgressão de novos genes que promovam a resistência às principais pragas é
importante. Normalmente, os programas de melhoramento genético do tomateiro utilizam
cruzamentos interespecíficos entre algumas espécies silvestres que apresentam características
de resistência químicas ou morfológicas, para o cruzamento com o tomateiro cultivado,
visando a incorporação desta característica de resistência na espécie de uso comercial
(RESENDE et al., 2008; RESENDE et al.,2009; SILVA et al., 2009; OLIVEIRA et al., 2012;
DIAS et al., 2013; NEIVA et al., 2013; BAIER et al., 2015).

23
3.8 Seleção assistida por marcadores moleculares.
Dentro dos diferentes métodos de melhoramento, virou rotina nos programas de
melhoramento a utilização da seleção assistida por marcadores moleculares (SAM). O SAM é
definido como sendo a seleção de uma determinada característica com base no genótipo
marcador correlacionado com a característica de interesse (FOOLAD; PANTHEE, 2012).
A utilização da SAM de forma eficiente pode aumentar a precisão de seleção, diminuir
os custos e ainda aumentar a eficiência de seleção, quando comparada aos métodos que levam
em consideração as características fenotípicas. Entre as diversas vantagens na utilização de
marcadores moleculares como critério de seleção vale destacar o fato de que eles são
independentes das perturbações do ambiente, sendo assim mais precisos. A maior eficácia tem
sido documentada em programas de melhoramento no qual as características de interesse são
de controle monogênico, sendo a maior eficiência da SAM em estudos com herdabilidade
moderada ou baixa em relação a seleção fenotípica (FOOLAD, 2007).
O uso de marcadores moleculares em programas de melhoramento com o uso de
retrocruzamento tem sido bem trabalhada, pois além de monitorar a introgressão do gene de
interesse, a utilização da genotipagem molecular dos indivíduos permite a seleção dos
genótipos mais semelhantes ao pai recorrente e com melhor conversão na região próxima ao
gene recentemente incorporado (DUARTE et al., 2003). Portanto, o número de ciclos de
retrocruzamentos necessários para a recuperação do pai recorrente é reduzido, resultando na
maximização do desenvolvimento de variedades melhoradas (OPENSHAW et al., 1994).
Atualmente, vários marcadores moleculares têm estreita ligação ou mesmo fazendo parte de
alguns destes genes, chamados de marcadores funcionais, estes já estão disponíveis para
serem utilizados na seleção assistida (TANKSLEY, 2004).
Nos diferentes processos de transferência de alelos de resistência, os marcadores
moleculares do DNA podem servir como uma ferramenta de grande valia. Esses marcadores
quando estão ligados aos alelos de resistência podem ser usados no processo de SAM,
principalmente nas etapas iniciais e intermediárias do programa de melhoramento. A
identificação de marcadores moleculares ligados aos alelos de resistência a doenças são um
dos principais objetivos dos atuais programas de melhoramento genético da cultura do
tomateiro. Podemos citar como exemplo o marcador molecular microssatélite SSR-47 que é
mapeado no cromossomo 6 do tomateiro a uma distância menor que 1 cM do loco Mi (SOL
GENOMICS NETWORK, 2008). Este marcador molecular também está intimamente ligado

24
ao loco Ty-1, possuindo grande importância na caracterização de genótipos de tomateiro
quanto a presença do alelo Ty-1 de resistência a begomovírus (ZAMIR et al., 1994).
Em relação a virose conhecida como tospovírus (TSWV), uma importante ferramenta
para a resistência a esta virose conferida pelo alelo Sw-5 e a sua detecção feita através do uso
dos marcadores moleculares funcionais. O marcador SCAR codominante correspondente a
uma certa região dentro do lócus Sw-5, sua utilização possibilitou identificar tomateiros
resistentes e suscetíveis de maneira inequívoca ao tospovírus. O uso destes marcadores é um
facilitador do programa de melhoramento, pois não é necessário o emprego de técnicas
biológicas de inoculação de doenças, que são demoradas, além de consumir recursos e espaço
com casas de vegetação (DIANESE et al., 2010). O gene Sw-7 supera a resistência conferida
do Sw-5, sendo que o lócus Sw-7 foi localizado próximo ao marcador SSR20 no cromossomo
12 (DOCKTER et al., 2009), também facilitando a detecção.
Nas pesquisas com a cultura do tomateiro tem sido desenvolvido mapas genéticos
extremamente densos, possuindo um número elevados de marcadores, estes tem permitido a
localização genômica/isolamento dos genes. Além de proporcionar o uso de diversos
marcadores moleculares em programas de seleção assistia em genótipos superiores
(BARONE et al., 2009). Vários genes do tomateiro relacionados a resistência a doenças têm
sido isolados via mapeamento genético de lata resolução (BARONE & FRUSCIANTE,
2007).
As técnicas genético-moleculares têm contribuído em diagnose, pesquisas com estudos
de herança e mecanismos de resistência contra pragas e patógenos. As informações genômicas
tem oferecido maior segurança no processo de seleção de plantas superiores nos programas de
melhoramento. Nas sementes, a detecção mais precisa de patógenos usando técnicas
moleculares tem permitido aumento na qualidade sanitária de sementes comerciais evitando
danos aos produtores por sementes contaminadas (CARVALHO et al., 2014).
3.9 Solanum pennellii LA 716 como fonte de resistência a artrópodes-praga
As espécies silvestres são as principais fontes de genes de resistência às principais
limitações causadas por pragas na cultura do tomateiro. As principais espécies utilizadas são a
S. pennellii, S. habrochaites, S. peruvianum e S. pimpinellifolium (MARUYAMA et al., 2002;
FANCELLI e VENDRAMIM, 2002; SUINAGA et al., 2004; ESCOBAR et al., 2010;
GONÇALVES-NETO et al., 2010; ORIANI et al., 2011). No entanto, as espécies silvestres S.

25
habrochaites e S. pennellii merecem destaque entre as demais quando se objetiva a resistência
pragas.
A espécie S. pennellii acesso LA 716 possui diversas pesquisas que demonstraram a
sua importância na cultura do tomateiro, principalmente, devido aos altos níveis de resistência
aos principais artrópodes-praga, dentre eles pragas-chave da cultura como a mosca-branca e
traça-do-tomateiro e outras secundárias de maior importância em cultivo protegido como os
ácaros tetranichídeos (PAMPLOMA, 2001; NEVES et al., 2003; GONÇALVES et al., 2006;
RESENDE et al., 2006; MACIEL et al., 2009; PEREIRA et al., 2008; RESENDE et al., 2008;
RESENDE et al., 2009; SILVA et al., 2009; GONÇALVES- NETO et al., 2010; MALUF et
al., 2010; DIAS et al., 2013; BAIER et al. 2015).
Em relação a mosca-branca (B. tabaci e B. tabaci biótipo B) diversas pesquisas com
acessos de S. pennellii LA-716 demonstraram a menor preferência para ovoposição da praga
por este acesso silvestre. Demonstrando a presença de resistência do tipo antixenose que
possivelmente, é promovida pela presença de tricomas na superfície dos folíolos desta espécie
(PAMPLOMA, 2001; TOSCANO et al., 2002; FANCELLI e VENDRAMIM, 2002). Nas
ninfas os efeitos deletérios ocorrem no período do seu desenvolvimento, não sendo descartado
a atuação da resistência do tipo antibiose (FANCELLI e VENDRAMIM, 2002). Em adultos,
foi observado maior mortalidade de adultos submetidos a espécie silvestre e os genótipos F2
selecionados para a característica de resistência a insetos (PAMPLOMA, 2001). A maioria
das características de resistência da espécie silvestre S. pennellii está envolvida aos tricomas
foliares e a concentração de acil-açúcares presentes nos folíolos.
A espécie S. pennellii em pesquisa com a traça-do-tomateiro possibilitou maior
rejeição para a ovoposição, sendo que a menor quantidade de ovos presentes na folha favorece
a diminuição dos danos causamos pela praga quando comparada a uma cultivar comercial,
sendo considerada uma espécie importante para o melhoramento (DIAS et al., 2013). A
resistência do tipo não preferência da espécie S. pennellii também foi constatada em pesquisas
com o ácaro T. evansi, provavelmente, pela presença de fitoquímicos conhecidos como acil-
açúcares presentes na espécie silvestre em contraste a espécie comercial (RESENDE et al.,
2008).
A presença de tricomas glandulares do tipo IV na espécie S. pennellii têm sido
associados a resistência do tipo antixenose e antibiose. Os tricomas glandulares presentes nos
folíolos da espécie silvestre produzem um fitoquímico que atua exercendo um efeito deletério
sobre a alimentação, ovoposição e no desenvolvimento da praga (GIANFAGNA et al., 1992;
ECOLE et al., 1999; MALUF et al., 2007; GONÇALVES-NETO et al., 2010; LUCINI et al.,

26
2015). Este aleloquímico pode ser encontrado em outros tecidos da planta como por exemplo
a lamela média das folhas e nos frutos (e ISMAN, 1981).
Os acil-açúcares (AAs) são um dos composto fitoquímicos mais estudados na
resistência as principais pragas do tomateiro, sendo considerado um éster de ácido graxo
(PEREIRA et al., 2008; RESENDE et al., 2009; GONÇALVES-NETO et al., 2010; MACIEL
et al., 2010, 2011; DIAS et al., 2013; BAIER et al., 2015). Na espécie S. pennellii os AAs é o
principal fitoquímico (aleloquímico) presente (RESENDE et al., 2006; GONÇALVES-NETO
et al., 2010; MALUF et al., 2010; MACIEL et al., 2011). Estudos utilizando plantas com
teores contrastantes do aleloquímico demonstraram correlação positiva dos altos teores do
aleloquímico com a maior resistência às pragas (PEREIRA et al., 2008; DIAS et., 2013).
A utilização de cruzamentos interespecífico entre a espécie silvestre S. pennellii com
altos teores de AAs com uma cultivar comercial que naturalmente possuem baixos teores de
AAs é de grande valia no melhoramento genético do tomateiro, visto que na geração F2 do
cruzamento alguns descendentes podem ser selecionados para teores altos e médios de acil-
açúcares visando a resistência às pragas (RESENDE et al., 2006).
Vale destacar que, geralmente, os genótipos F2 selecionados para altos teores de AAs
provenientes do cruzamento interespecífico entre a espécie silvestre e uma cultivar comercial
foram mais eficientes em reduzir os danos causados pela traça-do-tomateiro, semelhante ao
parental S. pennellii, sendo, portanto, promissores para a continuação do programa de
melhoramento genético por meio de retrocruzamentos em tomateiro (DIAS et al., 2013).
Em relação a mosca-branca, quando avaliados diferentes genótipos F2 quanto a
resistência a ovoposição e mortalidade de adultos, verificaram que os genótipos com altos
teores de AAs foram mais efetivos nos fatores relativos a maior resistência quando
comparados com os genótipos com menores teores de AAs. As principais diferenças presentes
entre os genótipos estavam ligadas, principalmente, aos teores de AAs presentes nos folíolos e
a densidade de tricomas foliares (PAMPLOMA, 2001). Em ácaros foi verificada uma
correlação negativa e significativa entre os teores de AAs e as médias de distâncias
percorridas por ácaros, sendo a menor distância percorrida aos genótipos com maiores
concentrações de AAs em seus folíolos. A seleção indireta realizada para quantificar altos
teores de AAs nos diferentes genótipos de tomateiro foi efetiva no aumento da resistência
(PEREIRA et al., 2008).
Há estudos genéticos com o aleloquímico acil-açúcar que demonstraram herança
monogênica, facilitando a seleção de plantas com os alelos de interesses nas diferentes etapas
de seleção (RESENDE et al., 2002b; BAIER et al., 2015). A seleção de plantas com médios e

27
altos teores de AAs provenientes do cruzamento interespecífico pode ser realizada de maneira
eficiente e com níveis satisfatórios de resistência as principais pragas. Visto que os teores do
aleloquímico AAs se mantém com o avanço das gerações de retrocruzamentos, possibilitando
ao melhoristas a seleção de novos indivíduos com elevados teores de AAs e com bons níveis
de resistência a pragas (GONÇALVES et al., 2006).

28
4. REFERÊNCIAS BIBLIOGRÁFICAS
ABCSEM - Associação Brasileira do Comércio de Sementes e Mudas. Tomate lidera
crescimento e lucratividade no setor de hortaliças. A Lavoura. p. 29-31, 2010.
ALVARENGA, M.A.R. Tomate: produção em campo, em casa de vegetação e em
hidroponia. 2. ed. Lavras: UFLA, 2013, p. 457.
ANVISA, 2016. Programa de análise de resíduos de agrotóxicos em alimentos. Disponível
em http://www.anvisa.gov.br. Acessado em 10 fev. 2017.
ARAGÃO, F.A.S.; RIBEIRO, C.S.C.; CASALI, V.W.D.; GIORDANO, L.B. Cultivo de
embriões de tomate in vitro visando a introgressão de genes de Lycopersicon peruvianum
em L. esculentum. Horticultura Brasileira, Brasília, v. 20, n. 4, p. 605-610, 2002.
BAIER, J.E.; RESENDE, J. T. V.; FARIA, M. V.; SCHWARZ, K.; MEERT, L. Indirect
selection of industrial tomato genotypes resistant to spider mite Tetranychus urticae.
Genetics and Molecular Research. V. 14. p. 244-252. 2015.
BARONE, A. & FRUSCIANTE, L. Molecular marker-assisted selection for resistance to
pathogens in tomato. Marker-Assisted Selection, Current status and future
perspectives in crops, livestock, forestry and fish, 153- 164, 2007.
BARONE, A., DI MATTEO, A., CARPUTO, D. & FRUSCIANTE, L. High-throughput
genomics enhances tomato breeding efficiency. Current genomics, 10, 1, 2009.
BOOM, C. E. M.; BEEK, T. A.; DICKE, M. Differences among plant species in acceptance
by the spider mite Tetranychus urticae Koch. Journal of Applied Entomology,
Hamburg, v. 127, n. 3, p. 177-183, 2003.
CARVALHO, R.C.P.; TOBAR, L.L.M.; DIANESE, E.C.; FONSECA, M.E.N.; BOITEUX,
L.S. Melhoramento genético do tomateiro para resistência a doenças de etiologia viral:
avanços e perspectivas. RAPP, v 22, 2014.
CLEMENTE, F.M.V.T.; BOITEUX, L.S. Produção de tomate para processamento
industrial. Embrapa. p. 344. 2012.
CNA-Confederação de Agricultura e Pecuária do Brasil. Mapeamento e qualificação da
cadeia produtiva das hortaliças do Brasil. 79p. 2017. Disponível em:
http://www.cnabrasil.org.br/sites/default/files/sites/default/files/uploads/mapeamento_e_q
uantificacao_da_cadeia_de_hortalicas.pdf. Acesssado 07 fev. 2017.
DIANESE, É. C., DE FONSECA, M. E. N., GOLDBACH, R., KORMELINK, R., INOUE-
NAGATA, A. K., RESENDE, R. O. & BOITEUX, L. S. Development of a locus-
specific, co-dominant SCAR marker for assisted-selection of the Sw-5 (Tospovirus

29
resistance) gene cluster in a wide range of tomato accessions. Molecular Breeding, 25,
133-142, 2010.
DIAS, D.M.; RESENDE, J.T.V.; FARIA, M.V.; CAMARGO, L.K.P.; LIMA, I.P. Selection
of processing tomato genotypes with high acyl sugar content that are resistant to the
tomato pinworm. Genetics and Molecular Research. v.12. p. 381-389. 2013.
DOCKTER, K. G., O’NEIL, D. S., PRICE, D. L., SCOTT, J. & STEVENS, M. R. Molecular
mapping of the Tomato spotted wilt virus resistance gene Sw-7 in tomato. Hort Science
44, 1123, 2009.
DUARTE, J.M; GUIMARÃES, C.T.; LANA, U.G.P.; PACHECO, C.A.P.; GUIMARÃES,
P.E.; PAIVA. Retrocruzamento assistido por marcadores moleculares para conversão de
linhagem de milho normal em QPM. In: 2o Congresso Brasileiro de Melhoramento de
Plantas, Porto Seguro, BA. 2003.
DUFFEY, S.S.; ISMAN, M.B. Inhibition of insect larval growth by phenolics in glandular
trichomes of tomato leaves. Experientia, Berlin, v.37, p. 574-576, 1981.
ECOLE, C.C.; PICANÇO, M.; JHAN, G.M.; GUEDES, R.N.C. Variability of Lycopersicum
hirsutum f. typicum and possible compounds involved in its resistance to Tuta absoluta.
Agricultural and Forest Entomology, Londres, v.1, p. 249-254, 1999.
ESCOBAR, R.; LÓPEZ, M. J.R.; CANO, J.M.A.; FERNÁNDEZ-MUÑOZ, R.; FONSECA,
M.E.N. Resistencia a Tuta absoluta en una entrada de la especie selvagem de tomate
Solanum pimpinellifolium. Phytoma España, La Rioja, v. 217, p. 126-127, 2010.
FAEG. Custos de produção tomate industrial. http://sistemafaeg.com.br/mercados-e-
cotacoes/outros/custo-de-producao?acao=custoProducao .Acesso em 02 set. 2014.
FANCELLI, M., J.D. VENDRAMIM, A.L. LOURENÇÃO e C.T.S. Dias. Atratividade e
preferência para ovoposição de Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae)
biótipo B em genótipos de tomateiro. Neotropical Entomology v.32, p. 319-328. 2003.
FANCELLI, M.; VENDRAMIM, J.D. Development of Bemisia tabaci (GENNADIUS, 1889)
biotype B on Lycopersicon spp. genotypes. Scientia Agricola, Piracicaba, v.59, n.4
p.665-669, 2002.
FAO -FAOSTAT - FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED
NATIONS. Database Results. Disponível em: <
http://www.fao.org/faostat/en/#data/QC>. Acesso em: 08 fev. 2018.
FILGUEIRA, F.A.R. Novo manual de olericultura: Agrotecnologia moderna na produção e
comercialização de hortaliças. 2 ed. Viçosa, MG: UFV, 412p. 2003.
FOOLAD, M.R. Genome mapping and molecular breeding of tomato. 2007.

30
FOOLAD, M.R.; PANTHEE, D.R. Marker-assisted selection in tomato breeding. Critical
rewiews in plant scienses, v. 31, p. 93-123, 2012.
FRANÇA, F. H.; VILLAS BÔAS, G. L.; CASTELO BRANCO, M.; MEDEIROS, M. A. de.
Manejo integrado de pragas. In: SILVA, J. B. C. da; GIORDANO, L. B. Tomate para
processamento industrial. Brasília: EMBRAPA, p. 37-59, 2000.
GALLO, D.; NAKANO, O.; SILVEIRA NETO, S.; CARVALHO, R. P. L.; BAPTISTA, G.
C. de; BERTI FILHO, E.; PARRA, J. R. P.; ZUCCHI, R. A.; ALVES, S. B.;
VENDRAMIM, J. D.; MARCHINI, L. C.; LOPES, J. R. S.; OMOTO, C. Entomologia
agrícola. Piracicaba: FEALQ, 2002, p. 920 (FEALQ. Biblioteca de Ciências Agrárias
Luiz de Queiroz, 10).
GAMEIRO, A. H.; CAIXETA FILHO, J. V.; ROCCO, C. D.; RANGEL, R. Estimativa de
perdas no suprimento de tomates para processamento industrial no estado de Goiás.
Informações Econômicas, São Paulo, v. 37, n. 7, p. 07-16, 2007.
GERALDINI, F.; JULIÃO, L.; BORGATO, E. Procuram-se agroindústrias. Hortifruti
Brasil. n.104, p. 8-23, 2011.
GIANFAGNA, T.J.; CARTER, C.D.; SACALIS, J.N. Temperature ans photoperiod influence
on trichome density and sesquiterpeno content of Lycopersicum hirsutum f. hirsutum.
Plant Phisiology, Rockville, v.100, n.4, p. 1403-1405, 1992.
GONÇALVES, L.D.; MALUF, W.R.; CARDOSO, M. das G.; RESENDE, J.T.V. de;
CASTRO, E.M. de.; SANTOS, N.M.; NASCIMENTO, I.R do.; FARIA, M.V. Relação
entre zingibereno, tricomas foliares e repelência de tomateiros a Tetranychus evansi.
Pesquisa Agropecuária Brasileira, Brasília, v.41, n.2, p.267-273, 2006.
GONÇALVES, M.I.F.; MALUF, W.R.; GOMES, L.A.A.; BARBOSA, L.V. Variation of 2-
tridecanone level in tomato plant leaflets and resistance to two mite species (Tetranychus
sp.). Euphytica, Wageningen, v.104, p.33-38, 1998.
GONÇALVES-NETO, A.C.; SILVA, V. de F.; MALUF, W.R.; MACIEL, G.M.; NÍZIO,
D.A.C.; GOMES, L.A.A.; AZEVEDO, S.M. de. Resistência à traça-do-tomateiro em
plantas com altos teores de acil-açúcares nas folhas. Horticultura brasileira, Brasília,
v.28, n.2, p.203-208, 2010.
HAJI, F. N. P.; MATTOS, M. M. de A.; ALENCAR, J. A. de; BARBOZA, F. R.;
PARANHOS, B. J. Manejo da mosca branca na cultura do tomate. Brasília: EMBRAPA,
p. 16, 2005. (Circular técnica, 81).
IBGE- INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Levantamento
sistemático da produção agrícola. Rio de Janeiro, 2018.

31
INCAPER- INSTITUTO CAPIXABA DE PESQUISA, ASSISTÊNCIA TÉCNICA E
EXTENSÃO RURAL. Tomate. Vitória: DCM/Incaper, 2010, 430 p. ISBN 978-85-
89724-17-3.
KENNEDY, G.G. Tomato, pests, parasitoids, and predators: tritrophic interactions involving
the genus Lycopersicon. Annual Review Entomology, Palo Alto, v. 48, p. 51-72, 2002.
LARA, F.M. Princípios de resistência de plantas. São Paulo: Ícone. 336p. 1991.
LUCINI, T.; FARIA, M. V.; ROHDE, C.; RESENDE, J. T. V.; OLIVEIRA, J. R. F.
Acylsugar and the role of tricomes in tomato genotypes resistence to Tetranychus urticae.
Arthopod-Plant Interactions. v. 9, p. 45-53, jan., 2015.
MACIEL G.M; MALUF, W.R; SILVA, V.F; GONÇALVES NETO, A.C; GOMES, L.A.A.
Híbridos pré-comerciais resistentes a Tuta absoluta obtidos de linhagem de tomateiro rica
em acilaçúcares. Horticultura Brasileira, 29: 151-156. 2011.
MALUF, W. R.; CAMPOS, G. A.; CARDOSO, M. G. Relationships between trichome types
and spider mite (Tetranychus evansi) repellence in tomatoes with respect to foliar
zingiberene contents. Euphytica, Wageningen, v. 121, n. 1, p.73-80, Out. 2001.
MALUF, W.R.; INOUE, I.F.; FERREIRA, R. de P.D.; GOMES, L.A.A.; CASTRO, E.M. de.;
CARDOSO, M. das G. Higher glandular trichome density in tomato leaflets and
repellence to spider mites. Pesquisa agropecuária brasileira, Brasília, v.42, n.9, p.1227-
1235, set., 2007.
MALUF, W.R.; MACIEL, G.M.; GOMES, L.A.A.; CARDOSO, M.G.; GONÇALVES, L.D.;
SILVA, E.C.; KNAPP, M. Broad-spectrum arthropod resistance in hybrids between high-
and low-acylsugar tomato lines. Crop Science, Madison, v.50, p.439-450, 2010.
MARTINS, A. M. B. Programa de prevenção e controle de pragas em tomate. Disponível em:
<http://www.agrodefesa.go.gov.br/index.php?option=com_content&view=article&id=77
>. Acesso em: 06 abr. 2013.
MARUYAMA, W.I.; TOSCANO, L.C. Tomate: reação aos ácaros. In: Revista Cultivar
Hortaliças e Frutas. Cultivar Hortaliças e Frutas. v. 19, p. 15-18, 2003.
MARUYAMA, W.I.; TOSCANO, L.C.; BOIÇA JÚNIOR, A.L.; BARBOSA, J.C. Resistência
de genótipos de tomateiro ao ácaro rajado. Horticultura Brasileira, Brasília, v.20, n.3,
p.480-484, set., 2002.
MORAES, G.J. de.; FLECHTMANN, C.H.W. Manual de Acarologia: acarologia básica e
ácaros de plantas cultivadas no Brasil. Ribeirão Preto: Editora Holos, 2008. 308p.
NAIKA, S.; JEUDE, J. V. L. de.; GOFFAU, M. de.; HILMI, M.; DAM, B. V. A cultura do
tomate: produção, processamento e comercialização. Agrodok 17, 2006, 104 p.

32
NEIVA, I.P.; ANDRADE JÚNIOR, V.C.; MALUF, W.R.; OLIVEIRA, C.M.; MACIEL,
G.M. Role of allelochemicals and trichome density in the Resistance of tomato to
whitefly. Ciência e Agrotecnologia, Lavras, v. 37, n. 1, p. 61-67, jan./fev., 2013
NEVES, L.G.; LEAL, N.R.; RODRIGUES, R.; PEREIRA, N.E. Estimativa de parâmetros
genéticos e correlação entre componentes de resistência à traça-do-tomateiro em
progênies de Lycopersicon esculentum x L. hirsutum f. glabratum. Horticultura
Brasileira, Brasília, v. 21, n. 3, p. 458-461, 2003.
NORRIS, D.M.; KOGAN, M. Biochemical and morphological bases of resistance. In:
MAxWELL, F.G.; JENNINGS, P.R. (Ed.). Breeding plants resistance to insects. New
York: J. Wiley. p. 23-61. 1980.
OLIVEIRA, C. M.; ANDRADE JUNIOR, V. C.; MALUF, W. R.; NEIVA, I. P.; MACIEL,
G.M. Resistance of tomato strains to the moth tuta absoluta imparted by allelochemicals
and trichome density. Ciência e Agrotecnologia, Lavras, v. 36, n. 1, p. 45-52, jan./fev.,
2012.
OLIVEIRA, F. A Antixenose em acessos de tomateiro do banco de germoplasma de
hortaliças da UFV a Tuta absoluta e suas possíveis causas químicas. Dissertação de
mestrado em genética e melhoramento na UFV. Viçosa: MG. p. 49, 2004.
OLIVEIRA, J. R. F de. Parâmetros associados à resistência ao ácaro rajado de genótipos
de tomateiro com altos teores de zingibereno. 2013. 50 p. Dissertação (Mestrado em
Agronomia) -Universidade Estadual do Centro-Oeste, Guarapuava-PR.
OPENSHAW, S.J.; JARBOE, S.G. e BEAVIS, W.D. Marker-assisted selection in backcross
breeding. In: ASHS/CSSA Joint Plant Breeding Symposium, 2, Corvallis Proceedings...
Corvallis: Oregon State University. 1994.
ORIANI, M.A. de G.; VENDRAMIM, J.D.; VASCONCELOS, C.J. Biology of Bemisia
tabaci (Genn.) B biotype (Hemiptera, Aleyrodidae) on tomato genotypes. Scientia
Agrícola, Piracicaba, v.68, n.1, p.37-41, 2011.
PAINTER, R.H. Insect resistance in croup plants. New York: The Macmillan. 520p. 1951.
PAMPLOMA, A. M. S. R. Avaliação de genótipos de tomate Lycopersicon ssp. com
diferentes concentrações de acil-açúcares, quanto a resistência a Bemisia tabaci
(Gennadius, 1889) (Hemitera: Aleyrodidae). 2001. 70 p. Dissertação (Mestrado em
Entomologia) - Universidade Federal de Lavras, Lavras, MG.
PERALTA, I.E. KNAPP, S. SPOONER, D.M. Nomenclature for wild and cultivated
tomatoes. Tomato Genetics Cooperative Report, Flórida, v.56, p. 6-12, 2006.
PERALTA, I.E.; SPOONER, D.M. Classification of wild tomatoes: a review. Kurtziana,

33
Córdoba, v.28, n.1, p.45-54, 2000.
PEREIRA, G.V.N.; MALUF, W.R.; GONÇALVES, L.D.; NASCIMENTO, I.R. do.;
GOMES, L.A.A.; LICURSI, V. Seleção para alto teor de acil-açúcares em genótipos de
tomateiro e sua relação com a resistência ao ácaro vermelho (Tetranychus evansi) e à
traça (Tuta absoluta). Ciência e Agrotecnologia, Lavras, v.32, n.3, p.996-1004, 2008.
RESENDE, J,T.V. de.; MALUF. W.R.; CARDOSO, M. G.; FARIA, M.V.; GONÇALVES,
L.D.; NASCIMENTO, I. R. do. Resistance of tomato genotypes with high level of
acylsugars to Tetranychus evansi Baker & Pritchard. Scientia Agrícola, Piracicaba, v.65,
n.1, p.31-35, 2008.
RESENDE, J,T.V. de.; MALUF. W.R.; CARDOSO, M. G.; GONÇALVES, L.D.; FARIA,
M.V.; NASCIMENTO, I. R. do. Resistance of tomato genotypes to the silverleaf whitefly
mediated by acylsugars. Horticultura brasileira, Brasília, v.27, n.3, p. 345-348, 2009.
RESENDE, J.T.V. de.; MALUF, W.R.; CARDOSO, M. G.; NELSON, D.L.; FARIA, M.V.
Inheritance of acylsugar contents in tomatoes derived from an interspecific cross with the
wild tomato Lycopersicon pennellii and their effect on spider mite repellence. Genetics
and Molecular Research, Ribeirão Preto, v.1, p.106-116, 2002b.
RESENDE, J.T.V.; CARDOSO, M.G.; MALUF, W.R.; SANTOS, C.D.; GONÇALVES,
L.D.; RESENDE, L.V.; NAVES, F.O. Método colorimétrico para quantificação de acil-
açúcar em genótipos de tomateiro. Ciência e Agrotecnologia, Lavras, v. 26, n. 6, p.
1204-1208, nov./dez. 2002a.
RESENDE, T.V. de R.; MALUF, W.R.; FARIA, M.V.; PFANN, A.Z.; NASCIMENTO, I.R.
do. Acylsugars in Tomato Leaflets Confer Resistance to the South American Tomato
Pinworm, Tuta absoluta Meyr. Scientia Agricola, Piracicaba, v.63, n.1, p.20-25, 2006.
SANTOS, F.F.B. dos. Obtenção e seleção de híbridos de tomate visando à resistência ao
Tomato yellow vein streak virus (ToYVSV). 2009. 86 p. (Mestrado em Agricultura
Tropical e Subtropical) – Instituto Agronômico de Campinas, São Paulo.
SATO, M.E.; SILVA, M.Z. da.; CANGANI, K.G.; RAGA, A. Seleções para resistência e
suscetibilidade, detecção e monitoramento da resistência de Tetranychus urticae ao
acaricida clorfenapir. Bragantia, Campinas, v.66, n.1, p.89-95, 2007.
SILVA, A. C.; CARVALHO, G. A. Manejo integrado de pragas. In: ALVARENGA, A. R.
Tomate: produção em campo, em casa-de-vegetação e em hidroponia. Lavras:
Editora UFLA. p. 309-366, 2004.
SILVA, F. DE A. S. E. e AZEVEDO, C. A. V. de. Principal Components Analysis in the
Software Assistat-Statistical Attendance. In:WORLD CONGRESS ON COMPUTERS

34
IN AGRICULTURE, 7, Reno-NV-USA: American Society of Agricultural and
Biological Engineers, 2009.
SILVA, J. B. C.; GIORDANO, L. B. Tomate para processamento industrial. Brasília:
Embrapa Comunicação para Transferência de Tecnologia/Embrapa Hortaliças, 2000, 167
p., ISBN 85-7358-078-6.
SILVA, V.F.; MALUF, W.R.; CARDOSO, M.G.; GONÇALVES NETO, A.C.; MACIEL,
G.M.; NIZIO, D.A.C.; SILVA, V.A. Resistência mediada por aleloquímicos de genótipos
de tomateiro à mosca-branca e ao ácaro-rajado. Pesquisa Agropecuária Brasileira,
Brasília, v.44, n.9, p.1262-1269, 2009.
SOL GENOMICS NETWORK. International Solanaceae Genome Project (SOL): systems
approach to diversity and adaptation. Available at: <http://solgenomics.net/solanaceae-
project/index.pl >. Accessed on: 20 abril. 2018.
SOUZA, J. C.; REIS, P. R. Traça-do-tomateiro: histórico, reconhecimento, biologia, prejuízos
e controle. 3ed. ver. aum. Belo Horizonte: EPAMIG, Boletim técnico 57, p. 32. 2000.
SUINAGA, F.A.; PICANÇO, M.C.; MOREIRA, M.D.; SEMEÃO, A.A.; MAGALHÃES,
S.T.V. Resistência por antibiose de Lycopersicon peruvianum a traça do tomateiro.
Horticultura Brasileira. Brasília, v. 22, n. 2, p. 281-285, 2004.
TANKSLEY, S.D. The genetic, developmental, and molecular bases of fruit size and shape
variation in tomato. The plant cell, v.16, p. S181-S189, 2004.
TOSCANO, L.C.; BOIÇA-JÚNIOR, A.L.; MARUYAMA, W.I. Non-preference of whitefly
for oviposition in tomato genotypes. Scientia Agrícola, Piracicaba, v.59, n.4, p.677-681,
2002.
VENDRAMIM, J. D.; NISHIKAWA, M. A. N. Melhoramento para resistência a insetos.
In: NASS, L. L.; VALOIS, A. C. C.; MELO, I. S. de, VALADARES-INGLIS, M. C.
Recursos genéticos e melhoramento de plantas. Rondonópolis: Fundação MT. p. 1183.
2001.
VENDRAMIM, J.D.A. Resistência de plantas e o manejo de pragas, In: CROCOMO, W.B.
(Ed.). Manejo integrado de pragas. São Paulo: Unesp. p. 177-197. 1990.
VILLAS BÔAS, G. L.; FRANÇA, F. H.; MACEDO, N.; MOITA, A. W. Avaliação da
preferência de Bemisia argentifolii por diferentes espécies de plantas. Horticultura
Brasileira, Brasília, v. 19, n. 2, p. 130-134, 2002.
WPTC – WORLD PROCESSING TOMATO COUNCIL. World production estimate as of
June 2014. Disponível em:< http://www.wptc.to/index.php>. Acesso em: 27 ago. 2014.
ZALOM, F.G. Pests endangered pesticides and processing tomatoes. Acta horticulturae, v.

35
613, p. 223-233. 2003.
ZAMIR, D., EKSTEIN-MICHELSON, I., ZAKAY, Y., NAVOT, N., ZEIDAN, M.,
SARFATTI, M., ESHED, Y., HAREL, E., PLEBAN, T. & VAN-OSS, H. 1994. Mapping
and introgression of a Tomato yellow leaf curl virus tolerance gene, Ty-1. TAG
Theoretical and Applied Genetics, 88, 141-146.
ZORZOLI, R.; PRATTA, G. R.; PICARDI, L. A. Variabilidad genética para la vida
postcosecha y el peso de los frutos en tomate para familias F3 de un híbrido
interespecífico. Pesquisa agropecuária Brasileira, Brasília, v. 35, n.12, p. 2423-2427,
2000.

36
CAPÍTULO I
ACIL-AÇÚCARES MEDIADORES DE RESISTÊNCIA À MOSCA BRANCA E À
FUMAGINA EM POPULAÇÕES SEGREGANTES DE TOMATEIRO
Resumo- A mosca branca é considerada como uma das principais pragas capazes de causar
danos na cultura do tomateiro. Os prejuízos provocados pela mosca branca apresentam-se
desde distúrbios fisiológicos no momento da alimentação, como também danos indiretos via
transmissão de viroses para as plantas e exsudação de substâncias açucaradas que servem de
substrato para o crescimento da fumagina. A fumagina causa na planta escurecimento dos
tecidos e pode interferir nas trocas gasosas, sendo mais um fator para análise. Objetivou-se
nesta pesquisa selecionar genótipos F2RC2 de tomateiro com altos teores de acil-açúcares
(AAs) e com resistência à mosca-branca e à fumagina. Para tanto, foram utilizados genótipos
F2RC1 de tomateiro com altos teores AAs, provenientes do cruzamento interespecífico entre a
cultivar Redenção e a espécie silvestre Solanum pennellii. Os genótipos foram retrocruzados
com a cultivar Redenção e autofecundados até a geração F2RC2, que incluía um total de 500
genótipos com teores contrastantes do aleloquímico. Posteriormente, os genótipos foram
submetidos a quantificação dos teores de AAs mediante análises de laboratório, sendo
selecionados 14 genótipos para o teste de resistência para a mosca branca e a severidade da
fumagina. No experimento foi utilizado o delineamento experimental em blocos ao acaso com
14 tratamentos e 4 repetições. Dentre os tratamentos foram utilizados nove genótipos com
altos teores de AAs, três genótipos com baixos teores de AAs, e os parentais como padrão de
resistência (Solanum pennellii) e suscetibilidade (cultivar Redenção), sendo cultivados em
casa de vegetação. A infestação com a praga ocorreu 45 dias após o transplante. Nas
avaliações foram contabilizados o número médio de ovos, número médio de ninfas, número
médio de exúvias e número médio de adultos presentes nos folíolos do terço superior,
mediano e inferior em um período de 28 dias de análises. Os adultos foram contabilizados no
folíolo e os ovos, ninfas e exúvias foram contabilizados em uma superfície de 2 cm2 do
folíolo. A avaliação referente a porcentagem da folha com a fumagina foi realizada com a
retirada de uma folha do terço superior, mediano e inferior, escaneadas e analisadas no
programa QUANT. Ocorreram diferenças significativas na redução do número de ovos,
ninfas, exúvias e adultos da mosca-branca em relação ao parental comercial Redenção. Em
relação a fumagina, os genótipos com altos teores de AAs obtiveram a menor severidade de
cobertura foliar com a presença da fumagina quando comparados com a cultivar Redenção.

37
Houveram correlações negativas e significativas entre os teores de AAs e os ovos, ninfas,
exúvias, adultos e severidade de cobertura da fumagina na folha. O programa QUANT
possibilitou quantificar a porcentagem de fumagina presente nas folhas. Os genótipos RVTA-
2010pl# 28, RVTA-2010pl# 39 e RVTA-2010pl# 113 serão utilizados na continuidade do
programa de melhoramento genético visando obtenção de genótipos com altos teores de AAs.
Palavras-chave: Solanum pennellii, Solanum lycopersicum, Bemisia tabaci, Capnodium sp.

38
1. INTRODUÇÃO
O tomateiro (Solanum lycopersicum) é uma cultura suscetível a artrópodes-praga, que
causam uma série de limitações que dificultam o sistema de cultivo. O manejo das pragas é
complexo, sendo necessário o uso do manejo integrado de pragas para obter um resultado
satisfatório. Os danos causados pelas pragas elevam os custos de produção por causar
distúrbios que diretamente ou indiretamente afetam o produto comercial, ou seja, os frutos
(ALVARENGA, 2013).
Dentre as principais pragas que causam danos significativos na cultura do tomateiro
destaque-se a mosca-branca (Bemisia tabaci e Bemisia tabaci biótipo B) pela sua expressiva
capacidade de provocar danos e redução do potencial produtivo do tomateiro (SRINIVASAN
et al., 2012). Os diversos prejuízos causados pela mosca-branca podem ser agrupados em
danos de origem direta e indireta (VILLAS BÔAS et al., 2002). Os danos de origem direta
ocorrem no momento da sucção da seiva presente no floema, e, em alta população da praga,
provocam uma série de distúrbios fisiológicos na planta, como a maturação irregular dos
frutos (FIRDAUS et al.,2012). Por sua vez, os danos de origem indireta são aqueles
decorrentes dos danos diretos. Na alimentação dos adultos da mosca-branca ocorre o
extravasamento da seiva para fora da epiderme da planta, servindo de substrato para o
aparecimento do patógeno fumagina (Capnodium sp.) que provoca o escurecimento dos
tecidos da planta, principalmente folhas e frutos, interferindo as trocas gasosas da planta e,
consequentemente, na produtividade (FIRDAUS et al., 2012). Porém, a maior preocupação é
com a geminivirose, uma virose que é transmitida pela mosca-branca no momento da sucção
de seiva causando uma série de distúrbios fisiológicos irreversíveis, sendo considerado outro
dano indireto causado por esta praga (TAVARES, 2002).
A principal forma de manejo utilizado pelos produtores para contornar os danos
causados pelos artrópodes-praga é o uso de produtos fitossanitários. No entanto, quando
usados de forma indiscriminada e sem o auxílio de outros métodos de manejo, provocam a
seleção de indivíduos com resistência as principais moléculas químicas, perigo de
contaminação ambiental e o risco à saúde humana (FORGET et al., 1993; ISMAN, 2006). A
utilização desenfreada de produtos não seletivos pode causar desequilíbrios biológicos pela
morte de polinizadores e insetos predadores de outras pragas (SILVA et al., 2009). Neste
sentido, o uso de métodos alternativos de manejo da mosca-branca que visem reduzir a
quantidade de produtos químicos é de grande importância (SAYEDA et al., 2009). Dentre os

39
diferentes métodos, o melhoramento visando a seleção de genótipos de tomateiro resistentes
aos principais artrópodes-praga tem apresentado resultados promissores (TOSCANO et al.,
2002; FANCELLI et al., 2003; MALUF et al, 2007; DIAS et al., 2013; LUCINI et al., 2015).
Vale destacar que dentro do manejo integrado de pragas a resistência genética surge como
uma forma importante de auxílio no manejo da mosca-branca, uma vez que os níveis de
resistência presentes nas cultivares comerciais são insuficientes (DIAS et al.,2016).
Na domesticação do tomateiro e seleção de espécies mais produtivas sem levar em
conta outros fatores de seleção resulta na perda de alelos importantes, especialmente os
responsáveis pela resistência a pragas e doenças. O estreitamento da base genética é um dos
fatores prováveis da suscetibilidade das cultivares atuais a inúmeras pragas e doenças
(GONÇALVES NETO et al., 2010; MALUF et al., 2010).
Diversas espécies de tomateiro silvestre são utilizadas como fonte de resistência a
doenças e pragas, como o Solanum pennellii. A principal característica de resistência para as
principais pragas presentes no S. pennellii vem sendo associada a fitoquímico chamado acil-
açúcares (AAs), principalmente 2,3,4-tri-O-éster de glicose (BURKE et al., 1987). Estes
fitoquímicos que atuam causando algum efeito deletério no ciclo de vida de determinadas
fases dos artrópodes-praga e contribuem na redução da preferência para ovoposição e a
alimentação da praga (MUIGAI et al., 2002; RESENDE et al., 2002a; SILVA et al., 2009;
LUCINI et al., 2015).
A utilização de genótipos selecionados para altos teores de AAs dentro de um
programa de melhoramento para pragas tem apresentado resultados satisfatórios, que por
meio de retrocruzamentos podem-se obter linhagens avançadas resistentes para o
desenvolvimento de novos híbridos de tomateiro de mesa (RESENDE et al., 2009).
A seleção de genótipos de tomateiro com altos teores de AAs presentes em seus
tecidos e com nível satisfatório de resistência as principais pragas são de grande valia para
redução do uso de produtos fitossanitários, aumento de renda do agricultor e favorecer o
manejo consciente. Atualmente ocorre a necessidade de novas cultivares de tomateiros
resistentes as principais pragas, devido não ter referências sobre o uso destes materiais para
uso comercial (DIAS et al., 2016). Diante do exposto, objetivou-se avaliar a resistência dos
genótipos F2RC2 de tomateiro para processamento com teores contrastantes de AAs quanto a
resistência a mosca-branca e a severidade da fumagina.

40
2. MATERIAL E MÉTODOS
2.1 Local do experimento
A obtenção das populações segregantes da geração F2RC2 e os testes de resistência à
mosca-branca e à fumagina foram conduzidos em casa de vegetação pertencentes ao Setor de
Olericultura e a quantificação dos diferentes teores de acil-açúcares foram realizadas no
Laboratório de Fisiologia Vegetal/Núcleo de Pesquisa em Hortaliças, ambos presentes ao
Departamento de Agronomia da Universidade Estadual do Centro-Oeste, UNICENTRO,
Guarapuava-PR. A região possui altitude média de 1.100 m, 25°23’01’’ de latitude Sul e
51°29’46’’ de longitude Oeste.
2.2 Obtenção das populações segregantes de tomateiro F2RC2
Foram utilizados no experimento os genótipos de tomateiro F2RC1 com altos teores de
acil-açúcares (AAs) 177, 310, 319 e 345 anteriormente selecionados por Dias et al. (2016)
para resistência à mosca-branca. Os genótipos utilizados no experimento foram provenientes
do cruzamento interespecífico entre a cultivar Redenção (Solanum lycopersicum, linhagem
que apresenta naturalmente baixos teores de AAs e com características favoráveis para o
processamento) e a espécie silvestre Solanum pennellii acesso LA-716 (acesso silvestre que
apresenta altos teores de AAs e fonte de resistência para artrópodes-praga) submetidos a
vários testes de resistência a pragas nas populações segregantes e retrocruzadas na cultivar
Redenção.
Os genótipos de tomateiro F2RC1 com altos teores de acil-açúcares (AAs) RVTA-
2010-31-pl#177, RVTA-2010-31-pl#310, RVTA-2010-31-pl#319 e RVTA-2010-83-pl#345
foram clonadas e retrocruzadas com o pai recorrente, cultivar Redenção. A clonagem foi
realizada com a intenção de aumentar os indivíduos de cada genótipo sem a perda da
característica de resistência. Na clonagem foram utilizados brotos axilares retirados de cada
genótipo e transplantados para bandejas de poliestireno de 60 células preenchidas com
substrato comercial, separadamente e devidamente identificados. Simultaneamente a
clonagem foi realizada a semeadura da cultivar Redenção em bandejas de 128 células com
substrato comercial. Após 30 dias da semeadura os clones e a cultivar Redenção foram
transplantados para vasos com capacidade de 7 dm3.
Os retrocruzamentos foram realizados no estágio de pleno florescimento dos genótipos

41
selecionados e da cultivar Redenção. Os frutos provenientes de cada retrocruzamento foram
identificados e colhidos separadamente. As sementes F1RC2 foram retiradas dos frutos,
secadas e semeadas em bandeja de 128 células. As plantas F1RC2 foram cultivadas em vasos
de 7 dm3 para a coleta dos frutos e obtenção da semente F2RC2.
2.3 Medida indireta dos teores de acil-açúcares e seleção dos genótipos de
tomateiro F2RC2
Na quantificação dos teores de acil-açúcares (AAs), os genótipos foram cultivados em
casa de vegetação em vasos de 2 dm3, utilizou-se 420 genótipos da geração F2RC2, sendo 105
plantas de cada parental, 40 plantas da cultivar Redenção (S. lycopersicum, testemunha para
baixo teor AAs) e 40 plantas de Solanum pennellii (testemunha para altos teores de AAs). Os
genótipos foram distribuídos em bancada elevada sequencialmente e identificadas no croqui.
No início do florescimento dos genótipos foram retirados seis discos foliares do terço
superior com auxílio de um vazador 3/8 de diâmetro de cada planta com posterior
acondicionamento em tubos de ensaio com identificação. Em seguida, foram extraídos os
AAs com a adição de diclorometano e reação colorimétrica pelo teste de Somogy-Nelson. As
amostras foram submetidas a leitura no espectrofotômetro e a absorbância obtida no
comprimento de onda de 540 nm (NELSON, 1944) com 3 repetições, segundo a metodologia
proposta por Resende et al. (2002a). Considera-se que as concentrações de AAs presentes nos
folíolos são diretamente proporcionais as leituras de absorbância, com maiores teores de AAs
refletindo em maiores valores de absorbância. Por meio da concentração, foram selecionadas
as plantas com teores contrastantes de AAs da população F2RC2.
2.4 Teste de resistência à mosca-branca, distribuição da mosca-branca na planta
e severidade da fumagina em genótipos de tomateiro com teores contrastantes de
acil-açúcares
O bioensaio foi conduzido em casa de vegetação com delineamento experimental em
blocos ao acaso com 14 tratamentos e 4 repetições. Os tratamentos que fizeram parte do
experimento foram 9 genótipos de tomateiro F2RC2 selecionados para altos teores de acil-
açúcares (AAs) (RVTA-2010-31-177pl#28, RVTA-2010-31-177pl#39, RVTA-2010-83-
347pl#61, RVTA-2010-31-177pl#113, RVTA-2010-31-177pl#177, RVTA-2010-31-

42
177pl#180, RVTA-2010-31-319pl#214, RVTA-2010-83-347pl#257 e RVTA-2010-83-
347pl#359), 3 genótipos de tomateiro F2RC2 selecionados para baixos teores de AAs (RVTA-
2010-83-347pl#77, RVTA-2010-31-310pl#205 e RVTA-2010-83-347pl#301) e como
testemunhas os parentais Redenção (S. lycopersicum) e Solanum pennellii acesso LA-716.
Anteriormente ao bioensaio foi realizado a criação programada da mosca-branca em
gaiola de madeira com tela antiafídica, onde a população de mosca branca foi criada em
plantas de feijão comum (Phaseolus vulgaris). Os genótipos de tomateiro F2RC2 utilizados no
experimento foram clonados com a utilização de brotos axilares, em bandeja de polipropileno
de 45 células com uso de substrato comercial. Os parentais utilizados como testemunhas de
resistência foram semeados em bandejas de poliestireno de 128 células. Após os clones
apresentarem adequado desenvolvimento radicular e os parentais com 3 a 5 folhas definitivas,
foram realizados os transplantes para vasos de polipropileno com capacidade de 7 dm3,
preenchidos com solo de subsuperfície corrigido mediante análise do solo e substrato
comercial, na proporção 1:1.
Na implantação do experimento foi realizada o sorteio dos vasos de cada genótipo
dentro de cada bloco, considerando-se cada tratamento um vaso com um genótipo diferente
presente em cada bloco. A infestação com a mosca-branca nos tratamentos foi realizado com a
retirada de todos os vasos de feijão cobertos com tela antiafídica que estavam na criação
programada e levados para a casa de vegetação e distribuídos nas entrelinhas dos blocos dos
tratamentos.
Os tratamentos foram submetidos à contagem do número de ovos, ninfas, exúvias e
adultos presentes em seus folíolos em intervalos correspondentes aos 7, 14, 21, e 28 dias após
a infestação. Na avaliação, primeiramente, foi contado o número de adultos presentes em cada
folíolo identificado em cada terço da planta e após foram retirados os mesmos e
acondicionados em placas de gerbox para o transporte até o Laboratório de Fisiologia
Vegetal/Núcleo de Pesquisa em Hortaliças para posterior contagem do número médio de
ovos, ninfas e exúvias. A contagem foi realizada na superfície abaxial do folíolo e no mesmo
lugar de cada tratamento em uma área correspondente a 2 cm2, com auxílio de um
microscópio estereoscópico. O número de ovos e adultos são correspondentes a média das 4
avaliações, o número de ninfas correspondente a média das 2 últimas avaliações e as exúvias
correspondentes da última avaliação.
Foram realizadas análises da distribuição de ovos, ninfas, exúvias e adultos na planta
utilizando cada terço da planta como um tratamento e utilizado a média das 14 plantas em
cada terço da planta para o estudo da preferência de cada estágio da praga pelo terço superior,

43
terço mediano e inferior da planta. O número de ovos e adultos são correspondentes a média
das 4 avaliações, o número de ninfas correspondente a média das 2 últimas avaliações e as
exúvias correspondentes a média da última avaliação.
A avaliação da fumagina foi realizada na última avaliação dos testes de resistência
para a mosca-branca com a retirada de uma folha proveniente do terço superior, terço
mediano e terço inferior de cada tratamento. As folhas foram devidamente identificadas e
levadas para o Laboratório de Fisiologia Vegetal/Núcleo de Pesquisa em Hortaliças.
Posteriormente, foi feito o escâner destas folhas em 300 dpi e salva em computador. A análise
da porcentagem de fumagina na superfície adaxial foi realizada pelo programa QUANT
v.1.0.1 licenciado pela Universidade Federal de Lavras (UFLA). A avaliação foi realizada
com a utilização de uma foto da folha correspondente a um dos tratamentos que era aberta no
programa. Em seguida, no programa a foto era selecionada para 300 dpi. Após era reduzido as
cores da imagem para apenas 3 cores, sendo uma do fundo da imagem, outra da folha normal
e outra da fumagina presente na superfície adaxial. O programa QUANT v.1.0.1 após a
redução para 3 cores, era excluído o fundo da imagem e o programa calculava a porcentagem
da folha com a presença da fumagina. Foram consideradas as médias de porcentagem de
fumagina em cada um dos genótipos de tomateiro F2RC2 para avaliar a severidade do fungo
na planta.
Por fim, a análise da distribuição da fumagina na planta foi realizada utilizando 3
folhas de cada terço da planta como um tratamento e utilizado a média das 14 plantas em cada
terço da planta para o estudo da distribuição da fumagina pelo terço superior, terço mediano e
inferior da planta. A porcentagem de fumagina é correspondente a média das 14 folhas
presentes em cada bloco.
2.5 Análises estatísticas
Mediante distribuição normal e a homogeneidade dos dados, estes foram
submetidos à análise de variância e posteriormente foi realizado análise de
agrupamento pelo teste de Scott-Knott, usando o programa estatístico SISVAR (FERREIRA,
2008).
A associação dos teores de acil-açúcar e a resistência dos genótipos foram estimadas
por meio da correlação de Pearson e a significância das correlações foi calculada por meio do
teste t, utilizando o programa ASSISTAT (SILVA e AZEVEDO, 2009).
No bioensaio, foram estimados contrastes entre os grupos de genótipos com teores

44
contrastantes de AAs, com o intuito de caracterizar diferenças nos níveis de resistência a
pragas em função dos teores de acil-açúcar, por meio do programa estatístico SISVAR, em
ambas as gerações (FERREIRA, 2008). A análise de componentes principais foi processada
no software STATISTICA versão 7.0 (STATSOFT, 2004).
3. RESULTADOS E DISCUSSÕES
A partir da concentração foliar do aleloquímico, foram selecionadas as plantas com
teores contrastantes de acil-açúcares (AAs) da população F2RC2. Para altos teores de AAs
foram selecionados os genótipos RVTA-2010-31-177pl#28 (28), RVTA-2010-31-177pl#39
(39), RVTA-2010-83-347pl#61(61), RVTA-2010-31-177pl#113 (113), RVTA-2010-31-
177pl#177 (177), RVTA-2010-31-177pl#180 (180), RVTA-2010-31-319pl#214 (214),
RVTA-2010-83-347pl#257 (257) e RVTA-2010-83-347pl#359 (359). Enquanto que para
baixos teores de AAs foram selecionados os genótipos RVTA-2010-83-347pl#77 (77),
RVTA-2010-31-310 pl#205 (205) e RVTA-2010-83-347pl#301 (301) (Tabela 1).
TABELA 1- Dados sobre os genótipos, classificação do teor de acil-açúcar e genótipo
utilizados no experimento. Guarapuava-PR –UNICENTRO, 2019.
Dados sobre os genótipos Teor de acil-açúcar Genótipo
Solanum pennellii acesso LA-716 Alto Solanum pennellii
RVTA-2010-31-177pl#28 Alto RVTA-2010pl#28
RVTA-2010-31-177pl#39 Alto RVTA-2010pl#39
RVTA-2010-83-347pl#61 Alto RVTA-2010pl#61
RVTA-2010-31-177pl#113 Alto RVTA-2010pl#113
RVTA-2010-31-177pl#177 Alto RVTA-2010pl#177
RVTA-2010-31-177pl#180 Alto RVTA-2010pl#180
RVTA-2010-31-319pl#214 Alto RVTA-2010pl#214
RVTA-2010-83-347pl#257 Alto RVTA-2010pl#257
RVTA-2010-83-347pl#359 Alto RVTA-2010pl#359
RVTA-2010-83-347pl#77 Baixo RVTA-2010pl#77
RVTA-2010-31-310 pl#205 Baixo RVTA-2010pl#205
RVTA-2010-83-347pl#301 Baixo RVTA-2010pl#301
Linhagem Redenção (Solanum lycopersicum) Baixo Cultivar Redenção
A preferência da mosca-branca foi significantemente influenciada pelos teores de AAs
presentes nos tecidos foliares dos genótipos F2RC2, provenientes do cruzamento entre o
acesso silvestre LA-716 (Solanum pennellii) e a linhagem comercial Redenção (Solanum
lycopersicum). Nas avaliações foram observadas diferenças significativas (p<0,05) entre os
genótipos do presente estudo em todas as características avaliadas (Tabela 2).

45
Em relação ao número de ovos, observou-se a ocorrência de 5 grupos distintos, com o
número médio de ovos variando de 0,00 (Solanum pennellii) até 25,69 (cultivar Redenção)
(Tabela 2). Dentre os diferentes grupos, merece destaque o acesso silvestre S. pennellii e o
genótipo com altos teores de AAs RVTA-2010pl#39 que apresentaram a menor preferência
pela mosca-branca para ovoposição em relação aos demais. Porém, vale destacar, que todos
os genótipos com alto teores de AAs obtiveram, estatisticamente, menor preferência de
ovoposição quando comparados a cultivar comercial Redenção e os genótipos com baixos
teores de AAs RVTA-2010pl#205 e RVTA-2010pl#301.
TABELA 2- Teor de acil-açúcares, número médio de ovos, número médio de ninfas, número médio de exúvias,
número médio de adultos, correlações e contraste em genótipos F2RC2 de tomateiro com diferentes teores de
acil-açúcares, Solanum pennellii (acesso LA716) e Solanum lycopersicum (Cultivar Redenção) submetidos à
infestação da mosca-branca. Guarapuava-PR –UNICENTRO, 2019.
Genótipo Valores de
absorbância
Ovos
(2 cm2)
Ninfas
(2 cm2)
Exúvias
(2 cm2)
Adultos
(por
folíolo)
Solanum pennellii (alto) 0,4833 0,00 e 0,00 f 0,00 e 0,31 f
RVTA-2010pl#28 (alto) 0,3614 10,00 d 44,87 d 4,00 d 6,38 c
RVTA-2010pl#39 (alto) 0,4564 3,13 e 27,38 e 3,75 d 4,81 d
RVTA-2010pl#61 (alto) 0,2930 11,50 d 66,38 c 10,25 c 8,50 b
RVTA-2010pl#113 (alto) 0,3119 9,94 d 48,00 d 1,50 e 9,13 b
RVTA-2010pl#177 (alto) 0,2251 16,56 c 11,75 f 0,25 e 5,63 c
RVTA-2010pl#180 (alto) 0,3478 11,63 d 58,38 c 5,75 d 2,69 e
RVTA-2010pl#214 (alto) 0,4212 9,56 d 43,00 d 0,50 e 2,94 e
RVTA-2010pl#257 (alto) 0,3112 15,75 c 63,13 c 4,00 d 8,81 b
RVTA-2010pl#359 (alto) 0,2107 11,06 d 74,38 b 2,75 d 8,50 b
RVTA-2010pl#77 (baixo) 0,1094 11,75 d 99,75 a 23,50 b 7,19 c
RVTA-2010pl#205 (baixo) 0,0643 22,38 b 76,25 b 10,50 c 7,63 b
RVTA-2010pl#301 (baixo) 0,1017 13,79 c 89,50 a 36,75 a 17,81 a
Cultivar Redenção (baixo) 0,1104 25,69 a 102,38 a 10,75 c 8,06 b
CV (%) 22,38 16,82 22,72 14,96
C1- Genótipos (altos) vs. Genótipos (baixos) - 8,34** - 48,24 ** - 17,10 ** - 4,40 **
Correlação de Pearson - 0,78* -0,77* -0,65* -0,64*
Médias seguidas pela mesma letra na coluna pertencem ao mesmo grupo pelo teste de Scott-Knott a 5 % de
probabilidade. 1Valores de absorbância a 540 nm.
* Significativo ao nível de 5% de probabilidade.
** Significativo ao nível de 1% de probabilidade.
Possivelmente, os teores de AAs presentes nos folíolos dos genótipos testados foram
os fatores determinantes na menor preferência da mosca-branca pela ovoposição. Tal fato
possibilita demonstrar os AAs como uma importante forma de manejo da mosca-branca,
sendo os altos teores de AAs presentes nos genótipos F2RC2 responsáveis pela redução da
ovoposição, fator este relacionado a resistência do tipo antixenose ou não preferência (Tabela
2). Os acil-açúcares são ésteres de sacarose e glicose liberados pelos tricomas glandulares do
tipo IV, presentes naturalmente nos folíolos da espécie silvestre Solanum pimpinellifolium e

46
Solanum pennellii que possibilitam uma série de efeitos deletérios no ciclo de vida dos
artrópodes-praga (SLOCOMBE et al., 2008; SCHILMILLER et al., 2009; RODRIGUEZ-
LOPEZ et al., 2011). Os aleloquímicos AAs quando presentes nos folíolos tornam a planta
menos atrativa à praga para as funções básicas do seu ciclo de vida como alimentação, abrigo,
ovoposição e entre outras características essenciais (LUCINI et al. 2015).
A redução da taxa de ovoposição da mosca-branca em genótipos com altos teores de
AAs é de grande valia dentro do manejo integrado de pragas, pois contribui com o manejo,
reduzindo o uso de produtos químicos e, concomitantemente, favorecendo a redução do custo
de produção (DIAS et al., 2016). Lucini et al., (2015) realizando estudos com genótipos F2
com teores contrastantes de AAs quanto a resistência ao ácaro, provenientes do mesmo
cruzamento entre a cultivar Redenção e a espécie silvestre S. pennellii. Os principais fatores
envolvidos com a resistência destes genótipos foram a alta concentração de AAs nos folíolos
e a densidade de tricomas do tipo IV.
Diversos estudos genéticos indicaram altas correlações entre os tricomas do tipo IV
com a maior resistência à mosca-branca (FANCELLI e VENDRAMIM 2002; MUIGAI et al.,
2003; ORIANI e VENDRAMIM 2011; RODRIGUEZ-LOPEZ et al., 2011). Os trabalhos de
pesquisa de Maciel et al. (2009), Resende et al. (2009) e Silva et al. (2009), Dias et al. (2016)
relatam os menores níveis de ovoposição da mosca-branca em genótipos de tomateiro com
altos teores de AAs, corroborando, portanto, com o presente estudo.
Na avaliação relativa ao número médio de ninfas presentes em 2 cm2 na superfície
abaxial dos folíolos ocorreu a presença de 6 diferentes grupos pelo teste de Scott Knott, com
variação de 0,00 (S. pennellii) a 102,38 ninfas (cultivar Redenção) (Tabela 2). O grupo com a
menor média de ninfas era composto pelo acesso silvestre S. pennellii e o genótipo com alto
teor de AAs RVTA-2010pl#177. Nos genótipos com baixos teores de AAs (RVTA-
2010pl#77, RVTA-2010pl#205 e RVTA-2010pl#301) e na cultivar Redenção ocorreram o
maior número médio de ninfas diferindo, estatisticamente, dos genótipos com altos teores de
AAs RVTA-2010pl#28, RVTA-2010pl#39, RVTA-2010pl#61, RVTA-2010pl#113, RVTA-
2010pl#177, RVTA-2010pl#180, RVTA-2010pl#214, RVTA-2010pl#257 e do acesso
silvestre Solanum pennellii com a menor presença da praga.
Com exceção do genótipo RVTA-2010pl#359, as maiores médias de ninfas presentes
nos folíolos foram encontradas nos genótipos que apresentavam baixos teores de AAs nos
folíolos (Tabela 2). Fato este que pode estar ligado a menor taxa de ovoposição da mosca-
branca ou a efeitos deletérios que o aleloquímico acil-açúcar pode causar no desenvolvimento
das ninfas. Resultados semelhantes também foram encontrados por Freitas et al. (2000) e

47
Fancellii et al. (2005) que comprovaram a importância dos AAs. Dias et al. (2016) relatam o
maior número de ninfas na maioria dos genótipos com baixos teores de AAs e,
principalmente, na cultivar Redenção, quando comparados com os genótipos contrastantes.
Em relação ao número médio de exúvias foi observado a ocorrência de 4 grupos
distintos de médias com valores entre 0,00 (S. pennellii) a 36,75 exúvias (RVTA-2010pl#301,
genótipo com baixos teores de AAs) (Tabela 2). Dentre os diversos grupos, podemos destacar
o grupo composto por S. pennellii, RVTA-2010pl#113, RVTA-2010pl#177 e RVTA-
2010pl#214 que apresentaram, o menor número médio de exúvias diferindo dos demais
genótipos.
Entretanto, quando comparamos os grupos de médias referentes aos genótipos com
altos teores de AAs pode ser observado diferença significativa entre os genótipos com baixos
teores de AAs (RVTA-2010pl#77, RVTA-2010pl#205 e RVTA-2010pl#301). Sendo os
genótipos com altos teores de AAs apresentaram menor média de exúvias quando comparado
com os demais. O número de exúvias é um importante fator de resistência para a mosca-
branca, pois demonstra o número de ninfas que efetivamente passaram para o estágio adulto.
Na avaliação referente ao número médio de adultos presente nos folíolos na superfície
abaxial constatou-se a ocorrência de 6 grupos de médias com valores entre 0,31 (S. pennellii)
até 17,81 adultos (RVTA-2010pl#301, genótipo com baixos teores de AAs) (Tabela 2). O
grupo com menor média de adultos foi compostos pelo acesso silvestre S. pennellii. Porém,
vale destacar os genótipos com altos teores de AAs RVTA-2010pl#180, RVTA-2010pl#214 e
RVTA-2010pl#39 que apresentaram menor média de adultos na folha quando comparados
com os genótipos com baixos teores de AAs (RVTA-2010pl#77, RVTA-2010pl#205 e
RVTA-2010pl#301) e a cultivar Redenção.
O contraste C1 demonstrou a relação inversamente proporcional e significativa (p
<0,01) entre os genótipos com altos teores de AAs quando comparados aos genótipos de
baixos teores com resultados significativos para todas as variáveis analisadas (Tabela 2).
Neste sentido o contraste C1 comprova a importância dos AAs presentes nos folíolos no
aumento de resistência a mosca-branca.
Pela análise de correlação de Person foi observado que o teor de AAs é inversamente
proporcional ao número médio de ovos (-0,78), número médio de ninfas (- 0,77), número
médio de exúvias (-0,65) e número médio de adultos (- 0,64), corroborando com a análise de
agrupamento de Scott-Knott e indicando a importância dos aleloquímicos AAs na redução das
taxas destas variáveis nos genótipos analisados (Tabela 2).
Resultados semelhantes foram encontrados por Dias et al. (2013) que obtiveram

48
correlações negativas e significativas ao comparar os teores de AAs presentes nos genótipos
de tomateiro com o número de ovos e lagartas da traça-do-tomateiro. Porém, Resende et al.
(2009) em trabalhos com mosca-branca obtiveram correlações negativas e significativas em
relação aos teores de AAs e o número médio de ninfas, não ocorrendo diferença significativa
em relação ao número de ovos.
O acesso silvestre foi utilizado como doador de genes de resistência para a mosca-
branca e apresentou comportamento similar aos genótipos com altos teores de AAs em
relação aos fatores de resistência avaliados. Resultado contrário ao ocorrido pelos genótipos
com baixos teores de AAs, com comportamento de suscetibilidade semelhante a cultivar
Redenção. Dias et al. (2016) evidenciam a importância dos altos teores de AAs como sendo o
principal fator de resistência apresentado pelo acesso S. pennellii para a mosca-branca, onde o
comportamento similar dos genótipos com altos teores de AAs são a garantia que o programa
de melhoramento foi eficiente na manutenção do padrão de resistência no avanço das
gerações.
Na pesquisa podemos observar que mesmo com uma maior quantidade de adultos
presente no folíolo como o ocorrido pelo genótipo RVTA-2010pl#61, RVTA-2010pl#113,
RVTA-2010pl#257 e RVTA-2010pl#359, não resultaram em maior número de ovos.
Possivelmente, algum fator desfavorável reduziu a preferência da mosca-branca pela
ovoposição nestes genótipos. Dentre outros fatores, os teores de AAs presentes nos folíolos
podem tem causado este fator desfavorável ao adulto, resultando na repulsa pela maior taxa de
ovoposição. Fato diferente ao ocorrido na cultivar Redenção com baixos teores de acil-
açúcares, pois poucos adultos resultaram em maior número de ovos.
A ACP foi realizada sobre o conjunto de dados e indica que os dois primeiros
componentes explicam 87,46% da variabilidade dos dados. A análise de componentes
principais (ACP) mostrou a formação de quatro grupos. O grupo 2 com os genótipos G1,
G205, G257 e G359 apresentam elevados números de ovos e de ninfas (Figura 1). O grupo 3
(Figura 1) com os genótipos G61, G77 e G301 apresentam alto números de adultos e de
exúvias. O grupo 4 com os genótipos G2, G28, G39 e G113 (Figuras 1) apresentam altos
teores de acil-açúcares. O grupo 1 (Figuras 1) formado pelos genótipos G177, G180 e G214
apresentam os medianos teores de acil-açúcares e os menores valores para os números de
ovos, de ninfas, de exúvias e números de adultos entre todos os genótipos selecionados.

49
FIGURA 1- Círculo de correlação das variáveis teor de acil-açúcares (Teor de acil-açúcares,
número de ovos (Nº ovos), número de ninfas (Nº ninfas), número de adultos (Nº adultos) e
números de exúvias (Nº exúvias) e distribuição dos genótipos em plano RVTA-2010-31-
177pl#28 (G28), RVTA-2010-31-177pl#39 (G39), RVTA-2010-83-347pl#61 (G61), RVTA-
2010-83-347pl#77 (G77), RVTA-2010-31-177pl#113 (G113), RVTA-2010-31-177pl#177
(G177), RVTA-2010-31-177pl#180 (G180), RVTA-2010-31-310 pl#205 (G205), RVTA-
2010-31-319pl#214 (G214), RVTA-2010-83-347pl#257 (G257), RVTA-2010-83-347pl#301
(G301) e RVTA-2010-83-347pl#359 (G359). Guarapuava-PR –UNICENTRO, 2019.

50
Em relação a porcentagem de fumagina na folha, foram identificados 4 grupos
distintos com valores variando de 2,96 (RVTA-2010pl#177) até 70,29 % (cultivar Redenção)
(Tabela 3). Dentre os genótipos merecem destaque o grupo composto pelo acesso silvestre S.
pennellii, RVTA-2010pl#39 e RVTA-2010pl#177 que diferiram significativamente dos
demais genótipos com a menor porcentagem de área foliar atacada pela fumagina. Vale
destacar que todos os genótipos com altos teores de AAs obtiveram menor área foliar com a
presença da fumagina quando comparados com o cultivar comercial. Quando comparados
com os genótipos com baixos teores de AAs, os genótipos com altos teores de AAs RVTA-
2010pl#28, RVTA-2010pl#39, RVTA-2010pl#61, RVTA-2010pl#113, RVTA-2010pl#177,
RVTA-2010pl#214, RVTA-2010pl#257 e RVTA-2010pl#359 obtiveram menor porcentagem
da folha com a presença da fumagina.
No tomateiro o desenvolvimento da fumagina nas folhas é causado, principalmente,
pela liberação de excreções açucaradas (honeydrew) que servem como substrato para o
desenvolvimento da fumagina na folha, que por sua vez, reduz o aproveitamento
fotossintético, prejudicando a qualidade dos frutos e a produção (FIRDAUS et al., 2012).
Na pesquisa, observamos a liberação de excreções açucaradas pela mosca branca
(Bemisia tabaci) nos estádios de ninfa e adulto. A presença do inseto é predominante na
superfície abaxial da folha, enquanto que a maior severidade de fumagina é encontrada no
terço inferior da planta e na superfície adaxial. Possivelmente, a mosca em diferentes estádios
(adulto e ninfa) quando realiza a excreção de substâncias açucaradas na superfície abaxial das
folhas do terço superior e mediano estes excrementos por gravidade se depositam na
superfície adaxial das folhas do terço inferior favorecendo o aumento da severidade do fungo.
A presença da fumagina (Capnodium sp.) sobre as folhas ocasiona o aumento da
temperatura e o acúmulo de radiação solar pode acarretar queima das folhas. Além de
bloquear boa parte da radiação solar, prejudicando a fotossíntese (OLIVEIRA et al., 2001). O
bloqueio da radiação fotossinteticamente ativa (PAR) nos comprimentos de onda entre 400 e
700 nm é significativo e com potencial de suprimir em até 40% a fotossíntese líquida (WOOD
et al., 1988).
Pela análise de correlação de Person foi observado correlações negativas (- 0,64) e
significativas (p<0,01) entre os teores de AAs e a área foliar com a presença da fumagina
(Tabela 3). Possibilitando inferir que os maiores teores de AAs presentes nos folíolos
possibilitaram menores áreas foliares com a presença da fumagina. Fato este, devido à
redução no número de ninfas e adultos presentes no folíolo.
O contraste C2 evidenciou, significativamente, a redução de área foliar afetada pela

51
fumagina na folha dos genótipos com altos teores de AAs, quando comparados aos baixos
teores de AAs presentes nos demais genótipos (Tabela 3). Dias et al. (2016) obtiveram
resultados significativos e negativos quando contrastaram plantas com altos teores de AAs e
plantas com baixos teores de AAs, sendo menores as médias de ovos e ninfas nos genótipos
selecionados para maiores teores do aleloquímico.
TABELA 3- Porcentagem de cobertura fumagina presente nas folhas, correlações lineares e contraste, em
genótipos F2RC2 de tomateiro, Solanum pennellii e cultivar Redenção. Guarapuava-PR –UNICENTRO, 2019.
Genótipo Valores de absorbância Fumagina
Solanum pennellii (alto) 0,4557 16,72 e
RVTA-2010pl#28 (alto) 0,3614 14,40 d
RVTA-2010pl#39 (alto) 0,4564 4,58 e
RVTA-2010pl#61 (alto) 0,2930 32,55 c
RVTA-2010pl#113 (alto) 0,3119 26,29 c
RVTA-2010pl#177 (alto) 0,2251 2,96 e
RVTA-2010pl#180 (alto) 0,3478 40,61 b
RVTA-2010pl#214 (alto) 0,4212 24,68 c
RVTA-2010pl#257 (alto) 0,3112 28,09 c
RVTA-2010pl#359 (alto) 0,2107 29,49 c
RVTA-2010pl#77 (baixo) 0,1094 42,10 b
RVTA-2010pl#205 (baixo) 0,0643 40,14 b
RVTA-2010pl#301 (baixo) 0,1017 51,21 b
Cultivar Redenção (baixo) 0,1017 70,29 a
CV (%) 23,49
Correlação linear -0,64 **
C2- Genótipos (alto) vs. Genótipos (baixo) -28,90**
Médias seguidas pela mesma letra na coluna pertencem ao mesmo grupo pelo teste de Scott-Knott a 5 % de
probabilidade. 1Valores de absorbância a 540 nm.
** Significativo ao nível de 1% de probabilidade (p<0,01).
A redução da área foliar atacada com a fumagina é uma característica interessante no
manejo da mosca-branca, visto que, o aparecimento deste fungo está relacionado com a
alimentação deste inseto. Firdaus et al. (2012) relatam que com o avançar do escurecimento
dos tecidos vão afetar ativamente na taxa fotossintética da planta que vai refletir na
diminuição da produtividade.
Silva et al. (2009) conduziram pesquisas com híbridos de tomateiro, provenientes de
cruzamentos de linhagens com altos teores de aleloquímicos AAs e zingibereno para
observarem fatores envolvidos na resistência para a mosca-branca. Os autores concluíram não
ocorrer efeito sinérgico entre híbridos heterozigotos para AAs ou zingibereno quando a
resistência à mosca-branca. Neste sentido, apenas os usos de genótipos com altos teores de
AAs seriam eficientes no manejo da mosca-branca.
Neste estudo a seleção direta em laboratório, realizada anteriormente aos testes de

52
resistência visando a seleção de plantas com teores contrastantes de AAs, demonstrou
eficiente na seleção de plantas com diferentes níveis de resistência. Principalmente na
facilidade de descarte de genótipos poucos promissores dentro do programa de melhoramento
genético, possibilitando acelerar os trabalhos de pesquisa com o avanço dos genótipos.
Entretanto são necessárias pesquisas com exposição dos genótipos selecionados para altos
teores de AAs com as principais pragas para confirmar a resistência e selecionar genótipos
com outros fatores relacionados ao aumento de resistência. Resende et al. (2002b) destacam
que plantas com altos teores de AAs nas folhas de tomateiro possuem padrão de herança
simples com valores moderadamente altos de herdabilidade, possibilitando em populações
segregantes aumentar a eficiência de seleção de genótipos com altos teores do aleloquímico.
4. CONCLUSÕES
Os genótipos F2RC2 com altos teores de AAs RVTA-2010pl# 28, RVTA-2010pl# 39 e
RVTA-2010pl# 113 apresentaram maiores características de resistência devem ser usamos
como fonte de genes para continuar o programa de melhoramento.

53
5. REFERÊNCIAS
ALVARENGA, M.A.R. Tomate: produção em campo, em casa de vegetação e em
hidroponia. 2. ed. Lavras: UFLA, 2013, p. 457.
BURKE, A.B.; GOLDSBY, G.; MUDD, J.B. Polar epicuticular lipds of Lycopersicon
pennellii. Phytochemistry. v. 26, n.9, p. 2567-2571, 1987.
DIAS, D.M.; RESENDE, J.T.V.; FARIA, M.V.; CAMARGO, L.K.P.; LIMA, I.P. Selection
of processing tomato genotypes with high acyl sugar content that are resistant to the
tomato pinworm. Genetics and Molecular Research. v.12. p. 381-389. 2013.
DIAS, D.M.; RESENDE, J.T.V.; MARODIN, J.C.; MATOS, R.; LUSTOSA, I.F.;
RESENDE, N.C.V. Acyl sugars and whitefly (Bemisia tabaci) resistance in segregating
populations of tomato genotypes. Genetics and Molecular Research. v. 15. 2016.
FANCELLI, M.; VENDRAMIM, J.D.; FRIGHETTO, R.T.S.; LOURENCÃO, A.L
.Glandular exudate of tomato genotypes and development of Bemisia tabaci (Genn.)
(Sternorryncha: Aleyrodidae) biotype B. Neotrop. Entomol. 34: 659-665, 2005.
FANCELLI, M.; VENDRAMIM, J.D.; LOURENÇÃO, A.L.; DIAS, C.T.S. Atraividade e
preferência para ovoposição de Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae)
biótipo B em genótipos de tomateiro. Neot. Ento. 32: 319-328, 2003.
FERREIRA, D. SISVAR: um programa para análises e ensino de estatística. Revista
Symposium. v. 6, p. 36-41, 2008.
FIRDAUS, S.; HEUSDENAW, VAN.; HIDAYATI, N.; SUPENA EDJ, et al.. Resistance to
Bemisia tabaci in tomato wild relatives. Euphytica 187: 31-45, 2012.
FORGET, G.; GOODMAN, T.; VILLIERS, A. Impact of pesticide use on health in
developing countries. Ottawa: Int. Dev. Res. Centre. 335 pp, 1993.
GONÇALVES-NETO, A.C.; SILVA, V. de F.; MALUF, W.R.; MACIEL, G.M.; NÍZIO,
D.A.C.; GOMES, L.A.A.; AZEVEDO, S.M. de. Resistência à traça-do-tomateiro em
plantas com altos teores de acil-açúcares nas folhas. Horticultura brasileira, Brasília,
v.28, n.2, p.203-208, 2010.
ISMAN, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an
increasingly regulated world. Annu. Rer. Entomol. 51: 45-66. 2006.
LUCINI, T.; FARIA, M. V.; ROHDE, C.; RESENDE, J. T. V.; OLIVEIRA, J. R. F.
Acylsugar and the role of tricomes in tomato genotypes resistence to Tetranychus urticae.
Arthopod-Plant Interactions. v. 9, p. 45-53, jan., 2015.

54
MACIEL, G.M. NÍZIO, D.A.C. SILVA, V.A. Resistência mediada por aleloquímicos de
genótipos de tomateiro à mosca-branca e ao ácaro-rajado. Pesquisa agropecuária
brasileira, Brasília, v.44, n.9, p.1262-1269, 2009.
MALUF, W.R.; INOUE, I.F.; FERREIRA, R. de P.D.; GOMES, L.A.A.; CASTRO, E.M. de.;
CARDOSO, M. das G. Higher glandular trichome density in tomato leaflets and
repellence to spider mites. Pesquisa agropecuária brasileira, Brasília, v.42, n.9, p.1227-
1235, set., 2007.
MALUF, W.R.; MACIEL, G.M.; GOMES, L.A.A.; CARDOSO, M.G.; GONÇALVES, L.D.;
SILVA, E.C.; KNAPP, M. Broad-spectrum arthropod resistance in hybrids between high-
and low-acylsugar tomato lines. Crop Science, Madison, v.50, p.439-450, 2010.
MUIGAI, S.G.; BASSETT, M.J.; SCHUSTER, D.J.; SCOTT ,J.W. Greenhouse and field
screening of wild Lycopersicon germplasm for resistance to the whitefly Bemisia
argentifolii. Phytoparasitica 31: 27-38, 2003.
NELSON, N. A photometric adaptation of the Sommogy method for the determination of
glucose. Journal of Biological Chemistry, v.153, p.375-380, 1944.
OLIVEIRA, M.R.V. et al. History, current status, and collaborative research projects for
Bemisia tabacii. Crop Protection, v.20, p.709-723, 2001.
ORIANI, M.A. de G.; VENDRAMIM, J.D.; VASCONCELOS, C.J. Biology of Bemisia
tabaci (Genn.) B biotype (Hemiptera, Aleyrodidae) on tomato genotypes. Scientia
Agrícola, Piracicaba, v.68, n.1, p.37-41, 2011.
RESENDE, J,T.V. de.; MALUF. W.R.; CARDOSO, M. G.; GONÇALVES, L.D.; FARIA,
M.V.; NASCIMENTO, I. R. do. Resistance of tomato genotypes to the silverleaf whitefly
mediated by acylsugars. Horticultura brasileira, Brasília, v.27, n.3, p. 345-348, 2009.
RESENDE, J.T.V. de.; MALUF, W.R.; CARDOSO, M. G.; NELSON, D.L.; FARIA, M.V.
Inheritance of acylsugar contents in tomatoes derived from an interspecific cross with the
wild tomato Lycopersicon pennellii and their effect on spider mite repellence. Genetics
and Molecular Research, Ribeirão Preto, v.1, p.106-116, 2002b.
RESENDE, J.T.V.; CARDOSO, M.G.; MALUF, W.R.; SANTOS, C.D.; GONÇALVES,
L.D.; RESENDE, L.V.; NAVES, F.O. Método colorimétrico para quantificação de acil-
açúcar em genótipos de tomateiro. Ciência e Agrotecnologia, Lavras, v. 26, n. 6, p.
1204-1208, nov./dez. 2002a.
RODRIGUEZ-LOPEZ, M.J.; GARZO, E.; BONANI, J.P.; FERERES, A. Whitefly resistance
traits derived from the wild tomato Solanum pimpinellifolium affect the preference and

55
feeding behavior of Bemisia tabaci and reduce the spread of tomato yellow leaf curl
virus. Phytopathology 101: 1191-1201, 2011.
SAYEDA, F.F.; TORKEY, H.M.; HALA, M. Natural extracts and their chemical constituents
in relation to toxicity against whitefly (Bemisia tabaci) and aphid (Aphis craccivora).
Aust. J. Basic Appl. Sci. 3: 3217-3223, 2009.
SCHILMILLER, A.L.; SCHAUVINHOLD, I.; LARSON, M.; XU, R. Monoterpenes in the
glandular trichomes of tomato are synthesized from a neryl diphosphate precursor rather
than geranyl diphosphate. Proc. Natl. Acad. Sci. U. S. A. 106: 10865-10870, 2009.
SILVA, F. DE A. S. E. e AZEVEDO, C. A. V. de. Principal Components Analysis in the
Software Assistat-Statistical Attendance. In:WORLD CONGRESS ON COMPUTERS
IN AGRICULTURE, 7, Reno-NV-USA: American Society of Agricultural and
Biological Engineers, 2009.
SILVA, V.F.; MALUF, W.R.; CARDOSO, M.G.; GONÇALVES NETO, A.C.; MACIEL,
G.M.; NIZIO, D.A.C.; SILVA, V.A. Resistência mediada por aleloquímicos de genótipos
de tomateiro à mosca-branca e ao ácaro-rajado. Pesquisa Agropecuária Brasileira,
Brasília, v.44, n.9, p.1262-1269, 2009.
SLOCOMBE, S.P.; SCHAUVINHOLD, I.; MCQUINN, R.P.; BESSER, K. Transcriptomic
and reverse genetic analyses of branched-chain fatty acid and acyl sugar production in
Solanum pennellii and Nicotiana benthamiana. Plan. Physiol. 148: 1830-1846, 2008.
SRINIVASAN, R.; RILEY, D.; DIFFIE, S.; SPARKS, A.; e ADKINS, S. Whitefly population
dynamics and evaluation of whitefly transmitted Tomato yellow leaf curl virus (TYLCV)
- resistant tomato genotypes as whitefly and TYLCV reservoirs. J. Econ. Entomol. 105:
1447-1456. 2012.
STATSOFT, Inc. (2004). STATISTICA (data analysis software system), version 7.
Disponível em:<www.statsoft.com> .
TAVARES, C. A. M. Perspectivas econômicas da tomaticultura frente aos problemas
causados pelo Geminivirus. Biológico, São Paulo, v. 64, n. 2, p. 157-158, 2002.
TOSCANO, L.C.; BOIÇA-JÚNIOR, A.L.; MARUYAMA, W.I. Non-preference of whitefly
for oviposition in tomato genotypes. Scientia Agrícola, Piracicaba, v.59, n.4, p.677-681,
2002.
VILLAS BÔAS, G. L.; FRANÇA, F. H.; MACEDO, N.; MOITA, A. W. Avaliação da
preferência de Bemisia argentifolii por diferentes espécies de plantas. Horticultura
Brasileira, Brasília, v. 19, n. 2, p. 130-134, 2002.
WOOD, B.W.; TEDDERS, W.L.; REILLY, C. C. Sooty mold fungus on pecan foliage

56
suppresses light penetration and net photosynthesis. HortScience, Alexandria, v.23,
p.851-853, 1988.

57
CAPÍTULO II
ACIL-AÇÚCARES MEDIADORES DE RESISTÊNCIA À TRAÇA DO TOMATEIRO
EM POPULAÇÕES SEGREGANTES DE TOMATEIRO
Resumo- A traça do tomateiro (Tuta absoluta) é considerada praga-chave na cultura do
tomateiro, causando uma série de prejuízos na produção desta hortaliça-fruto. O
desenvolvimento de cultivares resistentes é uma alternativa importante para reduzir os danos
causados por esta praga. Objetivou-se com este trabalho selecionar genótipos de tomateiro
com elevados teores de acil-açúcares (AAs), resistentes a traça do tomateiro e com maior
proporção do genoma do parental recorrente via utilização de marcadores moleculares. Foram
utilizados genótipos da segunda geração de retrocruzamentos (F2RC2) interespecíficos entre a
espécie silvestre Solanum pennellii acesso LA 716 com altos teores de AAs, e a cultivar
Redenção com características comerciais de tomate para processamento. Foram realizados
dois experimentos distintos visando a seleção de plantas. O primeiro experimento foi
realizado contendo 14 tratamentos, contendo genótipos com teores contrastantes de AAs,
além dos parentais. Os tratamentos foram submetidos a um ensaio de resistência a traça do
tomateiro visando avaliar as características de resistência presentes em cada genótipo.
Houveram correlações e contrastes significativos e inversamente proporcionais entre os teores
de AAs e as características de resistência avaliadas. A análise dos componentes principais
(ACP) possibilitou a formação de dois grupos distintos, sendo cada grupo representante de
cada parental. Os genótipos RVTA-2010pl#39, RVTA-2010pl#359, RVTA-2010pl#232 e
RVTA-2010pl#257 comportou-se de maneira similar ao parental silvestre reduzindo os danos
causados pela traça do tomateiro. O segundo experimento possuiu 15 tratamentos, sendo
utilizado os genótipos F2RC2 de tomateiro com altos teores de AAs presentes nos folíolos,
juntamente com seus parentais. Os genótipos foram analisados em laboratório com a
utilização de 6 primers ISSR e 6 primers SSR. Os fragmentos polimórficos amplificados
foram utilizados para a montagem da tabela de similaridade. A maior similaridade ocorreu
entre os genótipos RVTA-2010pl#257, RVTA-2010pl#152 e RVTA-2010pl#28 e os com
menores, RVTA-2010pl#177 e RVTA-2010pl#180. A análise conjunta dos dados de
resistência e molecular evidência que os genótipos RVTA-2010pl#232 e RVTA-2010pl#257
são os com maior potencial para utilização para avanço do programa de melhoramento para
obtenção de linhagens com boas características comerciais e com resistência a traça do
tomateiro.

58
Palavras-chave: Solanum pennellii, Solanum lycopersicum, Tuta absoluta, Marcadores
moleculares.

59
1. INTRODUÇÃO
No mundo, o tomateiro (Solanum lycopersicum) é considerado a principal hortaliça-
fruto, sendo cultivado nas regiões tropicais e subtropicais (EL BALLA et al. 2013). No Brasil,
a produção de tomate é distribuída em diversas classes de produtores rurais. Geralmente, nas
grandes áreas de cultivo ocorre o predomínio empresarial com preferência de produção para o
processamento industrial, diferindo de pequenas e médias propriedades rurais onde a
produção é voltada para o consumo in natura (GAMEIRO et al., 2007).
A suscetibilidade a diversas pragas que a cultura do tomateiro possui dificultam a
logística produtiva, influenciando na redução da produção e aumento de frutos fora do padrão
comercial. Na tentativa de reduzir os prejuízos ocasionados pelas pragas o uso de produtos
fitossanitários que aumentam os custos de produção, podendo ocasionar desequilíbrios
ecológicos e problemas de saúde do consumidor e do produtor rural (SILVA et al., 2009;
MAMTA et al., 2016).
Dentre as diferentes pragas, a traça do tomateiro (Tuta absoluta) é considerada como o
inseto com maior capacidade de dano na tomaticultura brasileira, sendo considerada como
praga-chave da cultura. O manejo traça do tomateiro é complexo, sendo necessários diversos
métodos de controle para obter êxito. Dentro do manejo integrado de pragas (MIP) a
utilização de métodos alternativos como a utilização de genótipos resistentes seriam uma
opção viável e de fácil adaptação para o produtor rural (MOREIRA et al., 2013; SILVA et al.,
2016).
Em diversas pesquisas científicas as espécies silvestres de tomateiro são utilizadas
como doadoras de genes de resistências em programas de melhoramento para as principais
pragas e doenças (LUCINI et al., 2015; LIMA et al., 2016; DIAS et al., 2016). A espécie
Solanum pennellii, acesso LA-716, apresenta efetiva resistência a pragas (MOREIRA et al.,
2013; DIAS et al., 2016). Os aleloquímicos presentes neste material tem sido associado aos
mecanismos de resistência do tipo antixenose e antibiose (MALUF et al., 2010; TIAN et al.,
2012; DIAS et al., 2016).
Um dos fitoquímicos presente no acesso LA-716 é os acil-açúcares (AAs), que são
exsudados por tricomas glandulares do tipo IV (ANDRADE et al., 2017), quando os insetos
entram em contato com os tricomas. Os AAs agem ocasionando uma série de efeitos
deletérios no ciclo de vida do inseto, assim como, reduzindo a alimentação e/ou a ovoposição
(MOREIRA et al., 2013; LUCINI et al., 2015). Estudos com a herança dos AAs

60
demonstraram que a obtenção de maiores teores do aleloquímico devem-se, principalmente, a
ação de um gene maior associado a outros de efeito menor e com ação aditiva. Em razão da
herança da característica envolvida, é possível a obtenção de híbridos com nível satisfatório
de resistência a pragas pela sequência de cruzamentos e seleção de linhagens com altos teores
de AA foliares (RESENDE et al., 2009).
A utilização de retrocruzamentos para seleção de genótipos com altos teores de AAs
são importantes nos programas de melhoramento visando resistência a pragas. Em cada ciclo
de retrocruzamento é possível obter linhagens resistentes e com maiores níveis de
características comerciais, que futuramente, poderão ser utilizadas para o desenvolvimento de
híbridos de tomateiro (RESENDE et al., 2009). Porém, somente o uso do retrocruzamento
apresenta algumas limitações, como longo período de tempo e custo relativamente alto para
terminar o processo e obter maior recuperação do genitor recorrente (MESQUITA et al,
2005).
Em comparação aos métodos convencionais, a seleção assistida por marcadores
moleculares apresenta melhor custo benefício (DREHER et al., 2003). Os marcadores
moleculares não são influenciados pelo ambiente, neste sentido, a introgressão de genes com
o uso de retrocruzamentos combinado ao uso de marcadores moleculares é uma forma viável
de incorporação de um alelo desejável de uma linhagem doadora e também para selecionar
maior número de alelos do genoma do recorrente, visto que o genoma doador apresenta
características não comerciais (ALZATE-MARIN et al., 2001).
Os marcadores moleculares ISSR (inter simple sequence repeat) são amplamente
utilizados em estudos de polimorfismo por ser uma técnica eficiente, relativamente simples e
com alta reprodutibilidade, além dos altos índices de polimorfismo (REDDY et al., 2002).
Porém, dentre os marcadores moleculares, os microssatélites ou SSR (simple sequence
repeats) além de utilizar a agilidade da técnica de PCR, apresentam-se codominantes com
uma frequência alta e distribuição uniforme ao longo de todo o genoma da planta, sendo
preferidos em estudos (AKKAYA et al., 1992).
A maior parte das pesquisas realizadas para a obtenção de linhagens com altos teores
de aleloquímicos nos folíolos com resistência a pragas são realizados com linhagens com
características para consumo in natura (SILVA et al., 2009; GONÇALVES NETO et al.,
2010; MALUF et al., 2010). No tomateiro para processamento industrial faltam trabalhos para
a incorporação de genes de resistência a pragas com utilização de marcadores moleculares e
que exploram a variabilidade interespecífica com aptidão para indústria aliado aos altos teores
de AAs. Portanto, objetivou-se selecionar genótipos de segunda geração do segundo

61
retrocruzamento (F2RC2) com elevados teores de AAs presentes nos folíolos e resistentes à
traça do tomateiro com auxílio de marcadores moleculares.
2. MATERIAL E MÉTODOS
2.1 Local do experimento
A obtenção das populações segregantes da geração F2RC2 e os testes de resistência à
traça do tomateiro foram conduzidos em casa de vegetação pertencentes ao Setor de
Olericultura e a quantificação dos diferentes teores de acil-açúcares foram realizadas no
Laboratório de Fisiologia Vegetal/Núcleo de Pesquisa em Hortaliças, ambos presentes ao
Departamento de Agronomia da Universidade Estadual do Centro-Oeste, UNICENTRO,
Guarapuava-PR. A região possui altitude média de 1.100 m, 25°23’01’’ de latitude Sul e
51°29’46’’ de longitude Oeste.
2.2 Obtenção das populações segregantes de tomateiro F2RC2
Foram utilizados no experimento os genótipos de tomateiro F2RC1 com altos teores de
acil-açúcares (AAs) RVTA-2010-31-pl#177, RVTA-2010-31-pl#310, RVTA-2010-31-pl#319
e RVTA-2010pl#345 anteriormente selecionados por Dias et al. (2016) para resistência a
mosca-branca. Os genótipos utilizados no experimento foram provenientes do cruzamento
interespecífico entre a cultivar Redenção (Solanum lycopersicum, linhagem que apresenta
naturalmente baixos teores de AAs e com características favoráveis para o processamento) e a
espécie silvestre Solanum pennellii acesso LA-716 (acesso silvestre que apresenta altos teores
de AAs e fonte de resistência para artrópodes-praga) submetidos a várias testes de resistência
a pragas nas populações segregantes e retrocruzadas na cultivar Redenção.
Os genótipos de tomateiro F2RC1 com altos teores de acil-açúcares (AAs) RVTA-
2010-31-pl#177, RVTA-2010-31-pl#310, RVTA-2010-31-pl#319 e RVTA-2010pl#345
foram clonadas e retrocruzadas com o pai recorrente, cultivar Redenção. A clonagem foi
realizada com a intenção de aumentar os indivíduos de cada genótipo sem a perda da
característica de resistência. Na clonagem foram utilizados brotos axilares retirados de cada
genótipo e transplantados para bandejas de poliestireno de 60 células preenchidas com
substrato comercial, separadamente e devidamente identificados. Simultaneamente a

62
clonagem foi realizada a semeadura da cultivar Redenção em bandejas de 128 células com
substrato comercial. Após 30 dias da semeadura os clones e a cultivar Redenção foram
transplantados para vasos com capacidade de 7 dm3.
Os retrocruzamentos foram realizados no estágio de pleno florescimento dos genótipos
selecionados e da cultivar Redenção. Os frutos provenientes de cada retrocruzamento foram
identificados e colhidos separadamente. As sementes F1RC2 foram retiradas dos frutos,
secadas e semeadas em bandeja de 128 células. As plantas F1RC2 foram cultivadas em vasos
de 7 dm3 para a coleta dos frutos e obtenção da semente F2RC2.
2.3 Quantificação dos teores de acil-açúcares e seleção dos genótipos de
tomateiro F2RC2
Na quantificação dos teores de acil-açúcares (AAs), os genótipos foram cultivados
dentro na casa de vegetação em vasos de 2 dm3, utilizando 420 genótipos da geração F2RC2,
sendo 105 genótipos de cada genótipo selecionado para altos teores de AAs, 40 plantas da
cultivar Redenção (S. lycopersicum, testemunha para baixo teor AAs) e 40 plantas de
Solanum pennellii (testemunha para altos teores de AAs). Os genótipos foram distribuídos em
bancada elevada sequencialmente e identificadas no croqui.
No início do florescimento dos genótipos foram retirados seis discos foliares do terço
superior com auxílio de um vazador 3/8 de diâmetro de cada planta com posterior
acondicionamento em tubos de ensaio com identificação. Em seguida, foram extraídos os
AAs com a adição de diclorometano e reação da reação colorimétrica pelo teste de Somogy-
Nelson. As amostras foram submetidas a leitura no espectrofotômetro e a absorbância obtida
no comprimento de onda de 540 nm (NELSON, 1944) com 3 repetições, segundo a
metodologia proposta por Resende et al. (2002a). Considera-se que as concentrações de AAs
presentes nos folíolos são diretamente proporcionais as leituras de absorbância, com maiores
teores de AAs refletindo em maiores valores de absorbância. Por meio da concentração, foram
selecionadas as plantas com teores contrastantes de AAs da população F2RC2.
2.5 Teste de resistência à traça do tomateiro
O bioensaio foi conduzido em casa de vegetação com delineamento experimental em

63
blocos ao acaso com 14 tratamentos e 4 repetições. Os tratamentos que fizeram parte do
experimento foram 8 genótipos de tomateiro F2RC2 selecionados para altos teores de acil-
açúcares (AAs), 4 genótipos de tomateiro F2RC2 selecionados para baixos teores de AAs e
como testemunhas os parentais Redenção (S. lycopersicum) e Solanum pennellii acesso LA-
716. Em relação ao teste de resistência a mosca branca, para os genótipos de altos teores de
AAs foram descartados os genótipos RVTA-2010-83-347pl#61 e RVTA-2010-31-
319pl#2014 e adicionados os genótipos RVTA-2010-31-177pl# 232. Para baixo teor AAs
foram descartados os genótipos RVTA-2010-83-347pl#77 e RVTA-2010-83-347pl#301 e
adicionado o genótipo RVTA-2010-31-177pl#227, RVTA-2010-31-177pl#376 e RVTA-
2010-31-310pl#385. Esta troca de genótipos foi em função da perda das plantas quando estas
estavam sendo clonadas para a realização do experimento de resistência a traça do tomateiro.
Anteriormente ao bioensaio foi realizado a criação programada da traça do tomateiro
em gaiola de madeira com tela antiafídica, onde a população foi criada em plantas de tomate
(Solanum lycopersicum). Os genótipos de tomateiro F2RC2 utilizados no experimento foram
clonados com a utilização de brotos axilares, em bandeja de polipropileno de 45 células com
uso de substrato comercial. Os parentais utilizados como testemunhas de resistência foram
semeados em bandejas poliestireno de 128 células. Após os clones apresentarem adequado
desenvolvimento radicular e os parentais com 3 a 5 folhas definitivas, foram realizados os
transplantes para vasos de polipropileno com capacidade de 7 dm3, preenchidos com solo de
subsuperfície corrigido mediante análise do solo e substrato comercial, na proporção 1:1.
Na implantação do experimento foi realizada com sorteio dos vasos de cada genótipo
dentro de cada bloco, considerando-se cada tratamento um vaso com um genótipo diferente
presente em cada bloco. A infestação com a traça do tomateiro nos tratamentos foi feita com a
retirada de todos os vasos de tomate cobertos com tela antiafídica que estavam na criação
programada e levados para a casa de vegetação e distribuídos nas entrelinhas dos blocos dos
tratamentos.
Os tratamentos foram submetidos à contagem do número de ovos e lagartas presentes
em seus folíolos na superfície adaxial e abaxial em intervalos correspondentes aos 7, 14, 21, e
28 dias após a infestação. Na avaliação, primeiramente, foi contado o número de adultos
presentes em cada folíolo identificado em cada terço da planta e após foram retirados os
mesmos e acondicionados em placas de gerbox para o transporte até o Laboratório de
Fisiologia Vegetal/Núcleo de Pesquisa em Hortaliças para posterior contagem do número de
ovos e lagartas. A contagem foi realizada na superfície abaxial e adaxial do folíolo com
auxílio de um microscópio estereoscópico. O número de ovos e adultos são correspondentes a

64
média das 4 avaliações, o número de lagartas correspondente a média das 3 últimas
avaliações.
A severidade dos danos ocasionados pela traça-do-tomateiro foi avaliada por meio de
escala de notas (Labory et al. 1999), conforme segue:
A) intensidade de danos nas plantas (IDP): 0- nenhum dano; 1- lesões pequenas (0,1 a 5% de
dano); 2- lesões pequenas e não coalescentes (5,1 a 20% de dano); 3- lesões médias a grandes
(20,1 a 50% de dano); 4- lesões grandes e coalescentes (50,1 a 80% de danos); e 5- plantas
totalmente destruídas (acima de 80% de danos).
B) Tipos de lesões nos folíolos (TLF): 0- sem lesões; 1- poucas e pequenas lesões; 2- lesões
pequenas e médias; 3- lesões de tamanho médio, sem coalescência; 4- lesões grandes e
coalescentes, folíolos deformados; e 5- folíolo totalmente destruído.
C) Porcentagem de folíolos atacados (PFA): 0- sem folíolos atacados; 1- 0,1 a 5% de folíolos
atacados; 2- 5,1 a 20% de folíolos atacados; 3- 20,1 a 50% de folíolos atacados; 4- 50,1 a 80%
de folíolos atacados; e 5- mais de 80% dos folíolos atacados. As avaliações ocorreram aos 14
e 21 dias após a infestação, por cinco avaliadores previamente treinados.
2.5.1 Análises estatísticas
Mediante distribuição normal e a homogeneidade dos dados, os dados foram
submetidos à análise de variância e as variáveis ovos e larvas transformados (raiz de X+1) e,
posteriormente, foi realizado análise de agrupamento pelo teste de Scott-Knott, usando o
programa estatístico SISVAR (FERREIRA, 2008).
A associação dos teores de acil-açúcar e a resistência dos genótipos foram estimadas
por meio da correlação de Pearson e a significância das correlações foi calculada por meio do
teste t, utilizando o programa ASSISTAT (SILVA e AZEVEDO, 2009).
No bioensaio, foram estimados contrastes entre os grupos de genótipos com teores
contrastantes de AAs, com o intuito de caracterizar diferenças nos níveis de resistência a
pragas em função dos teores de acil-açúcar, por meio do programa estatístico SISVAR, em
ambas as gerações (FERREIRA, 2008). A análise de componentes principais foi processada
no software STATISTICA versão 7.0 (STATSOFT, 2004).
2.4 Avaliação da recuperação do genoma recorrente
O experimento foi conduzido no Laboratório de genética e Biologia Molecular

65
Vegetal da Universidade Estadual do Centro-Oeste (UNICENTRO). No trabalho foram
utilizados 13 genótipos F2RC2 selecionados para altos teores de acil-açúcares (AAs) (todos os
genótipos utilizados para o teste de resistência a mosca branca e traça do tomateiro) e os
parentais da linhagem comercial Redenção (Solanum lycopersicum) e o acesso silvestre
Solanum pennellii LA 716. Nesta etapa não foram utilizados os genótipos com baixo teor de
AAs uma vez que estes são somente controles para os testes de teores de AAs e resistência a
praga. Para as análises moleculares foram coletadas folhas de cada genótipo e armazenadas
em nitrogênio líquido para posterior extração do DNA. A extração do DNA foi realizada
seguindo o método descrito por Sharma et al. (2008). Primeiramente foi realizada o
esmagamento do material vegetal coletado em nitrogênio líquido e separado em tubo 100 mg
de tecido. A este tecido foi adicionado 1 mL de tampão de extração (100 mM Tris-HCl pH
8.0, 200 mM EDTA, 2 M NaCl, 2%PVP, 2% CTAB, 2% β-mercaptoethanol), posteriormente
os tubos foram colocados em banho-maria por 30 min a 65°C. Após, com a utilização de
fenol:clorofórmio:alcoolisoamílico (proporção:25:24:1, respectivamente) o DNA foi separado
da solução pelo processo de centrifugação. Em seguida, o DNA foi precipitado com PEG
(Polyethylene Glycol) e para obter um alto grau de pureza foram realizadas sucessivas
lavagens com etanol. No final da extração o DNA foi ressuspendido em TE (10mM Tris-HCl;
pH 8,0; 1mM EDTA) com a RNAse, incubado a 37 °C por 30 min e armazenado em freezer a
–20 °C até o uso. O DNA foi quantificado por eletroforese em gel de agarose 0,8%, corado
com brometo de etídio (0,5 μg mL-1). A determinação da quantidade de DNA foi feita pela
comparação com quantidades conhecidas do DNA do fago Lambda (50, 100 e 200 ng).
2.4.1 Marcadores moleculares (ISSR e SSR)
No estudo foram utilizados seis primers ISSR (Tabela 4) e seis pares de primers SSR.
Estes primers foram anteriormente selecionados por Secco (2013) como os mais informativos
considerando os parentais utilizados neste estudo. Para os marcadores ISSR, nas reações de
amplificação do DNA de cada genótipo via PCR foram conduzidas em volume final de 12,5
μL que continham: 20 ng de DNA, 0,2 μM de primer, 200 μM de cada dNTP, 1,5 mM de
MgCl2 e 1U de Taq DNA polimerase e tampão para PCR 1X. A programação do
termociclador para amplificação seguiu os seguintes passos, para desnaturação inicial a 94 °C
por 5 minutos, em seguida com 35 ciclos de 94 °C por 45s, a temperatura de anelamento dos
primers por 45s e 72°C por 90s, e um passo a 72 °C por 5 minutos para extensão final dos
fragmentos. Os produtos de amplificação foram resolvidos por eletroforese em gel de agarose
1,8% corado com brometo etídio (0,5 μg mL-1). A determinação do tamanho dos fragmentos

66
amplificados fui por comparação com o marcador de peso molecular DNA Ladder 100 pb. O
resultante da eletroforese foi visualizado com o auxílio de luz UV e fotodocumentador com
sistema digital.
Tabela 4- Primers ISSR utilizados para avaliação da similaridade genética entre os parentais
e as linhagens de tomateiro com alto de teor de acil-açúcares. Temperatura de anelamento
(TºCa). Guarapuava-PR –UNICENTRO, 2019.
Primer Sequência (5’-3’) TºCa
807 (AG)8T 52º
809 (AG)8G 55º
835 (AG)8YC 54º
864 (ATG)6 50º
873 (GACA)4 50º
878 (GGAT)4 54º
Os pares de primers SSR utilizados estão descritos na Tabela 5. Para cada genótipo as
reações de amplificação do DNA via PCR foram conduzidas em volume final de 10 μL
contendo: 0,5 ng de DNA, 0,8 μM de cada primer, 0,2 mM de cada dNTP, 3 mM de MgCl2 e
0,4U de Taq DNA polimerase, tampão para PCR 1X e água ultrapura para completar o
volume. Para amplificação o termociclador foi programado para uma desnaturação inicial a
94ºC por 3 minutos, seguida de 9 ciclos de 94ºC por 30s, 68ºC por 30s e 72ºC por 30s,
seguido de mais 21 ciclos de 94ºC por 30s, 62ºC por 30s e 72ºC por 30s, para o anelamento
dos primers, e por foi feito um passo a 72°C por 10 minutos para extensão final dos
fragmentos.
Os produtos da amplificação foram separados em gel de poliacrilamida a 12%. Após a
aplicação das amostras no gel, estas migraram por 40 min a 2000 V 55 MA 5 W para
empilhamento das amostras de DNA. Em seguida os parâmetros foram ajustados para 2000 V
55 MA 55W e corridos por 4 horas. O resultado da eletroforese foi visualizado por coloração
com nitrato de prata, utilizando o seguinte protocolo: Após desprendimento das placas, o gel
foi mantido em solução de fixação contendo 1 L de água ultrapura, 200 mL de etanol e 10 mL
de ácido acético durante 5 min. Em seguida, transferido para solução de coloração com 1 L de
água ultrapura, 1,5 g de nitrato de prata e 5 mL de formaldeído por 7 min, após isso, uma
lavagem em 1 L de água ultrapura por 15 s. Por fim, o gel foi revelado em solução contendo 1
L de água ultrapura, 15 g de hidróxido de sódio e 10 mL de formaldeído até o surgimento das
bandas.
As análises das fotografias dos géis foram realizadas a olho nu cuidadosamente,

67
seguida pela genotipagem dos indivíduos de acordo com a presença (1) ou ausência (0) de
bandas. A partir desses dados foi gerado uma matriz binária para análise de similaridade
genética entre os genótipos, utilizou-se o coeficiente de Jaccard, com utilização do programa
NTSYS 2.2. A análise foi feita em conjunto com os dados dos marcadores ISSR e SSR.
TABELA 5- Primers SSR indicando cromossomo onde foi mapeado, sequência e a indicação
se o primer é polimórfico. Guarapuava-PR –UNICENTRO, 2019.
Cromossomo Primer Forwad/Reverse Sequência (5’-3’)
1 TES1444 TES1444F GATTTTTGTTTTGTCGCGTGC
TES1444R AAAGCTAACCGTGCAAAGGA
1 TES1242 TES1242F CCACCGCAACAAACCTTATT
TES1242R GGGTGGTGAGAAGGATCTGA
2 TES1925 TES1925F GAGCTTTTAACATGGCGGATG
TES1935R CCACAGCTGAATCTCCAACA
3 TES1935 TES1935F GTTCAAAGGAGTCCTCATGGC
TES1935R CCACAGCTGAATCTCCAACA
3 TES1203 TES1203F GAAGACTGCAGGCGATCCTTA
TES1203R CCTCACAAACAGGTTTCGGT
8 TES1884 TES1884F GCCTTCCTTACTTCGCCACTG
TES1884R TTCCGTGAAATACTTTGCCC

68
3. RESULTADOS E DISCUSSÕES
3.3 Teste de resistência a traça do tomateiro
A partir da concentração foliar do aleloquímico, foram selecionadas as plantas com
teores contrastantes de acil-açúcares (AAs) da população F2RC2. Para altos teores de AAs
foram selecionados os genótipos RVTA-2010-31-177pl#28, RVTA-2010-31-177pl#39,
RVTA-2010-31-177pl#113, RVTA-2010-31-177pl#177, RVTA-2010-31-177pl#180, RVTA-
2010-31-177pl#232, RVTA-2010-83-347pl#257 e RVTA-2010-83-347pl#359. Enquanto que
para baixos teores de AAs foram selecionados os genótipos RVTA-2010-31-310 pl#205,
RVTA-2010-31-177pl#227, RVTA-2010-31-177pl#276 e RVTA-2010-31-177pl#385 (Tabela
6).
TABELA 6- Dados sobre os genótipos, classificação do teor de acil-açúcar e genótipo
utilizados no experimento. Guarapuava-PR –UNICENTRO, 2019.
Dados sobre os genótipos Teor de acil-
açúcar Genótipo
Solanum pennellii acesso LA-716 Alto Solanum pennellii
RVTA-2010-31-177pl#28 Alto RVTA-2010pl#28
RVTA-2010-31-177pl#39 Alto RVTA-2010pl#39
RVTA-2010-31-177pl#113 Alto RVTA-2010pl#113
RVTA-2010-31-177pl#177 Alto RVTA-2010pl#177
RVTA-2010-31-177pl#180 Alto RVTA-2010pl#180
RVTA-2010-31-177pl# 232 Alto RVTA-2010pl#232
RVTA-2010-83-347pl#257 Alto RVTA-2010pl#257
RVTA-2010-83-347pl#359 Alto RVTA-2010pl#359
RVTA-2010-31-310 pl#205 Baixo RVTA-2010pl#205
RVTA-2010-31-177pl#227 Baixo RVTA-2010pl#227
RVTA-2010-31-177pl#376 Baixo RVTA-2010pl#376
RVTA-2010-31-310pl#385 Baixo RVTA-2010pl#385
Linhagem Redenção (Solanum lycopersicum) Baixo Cultivar Redenção
Na pesquisa a preferência da traça do tomateiro foi significativamente influenciada
pelos teores de AAs presentes nos genótipos F2RC2, provenientes do cruzamento entre a
linhagem comercial Redenção (Solanum lycopersicum) e o acesso silvestre LA-716 (Solanum
pennellii). Nas avaliações foram observadas diferenças significativas (p<0,05) entre os
genótipos em estudo em todas as características avaliadas (Tabela 7).
Em relação a quantidade média de ovos presentes nos folíolos foram identificados 6
grupos distintos, com valores médios variando de 1,00 (Solanum pennellii) até 2,85 (cultivar
Redenção) (Tabela 7). Dentre os diferentes grupos, o genótipo com menor preferência pela

69
praga foi o Solanum pennellii. Os genótipos provenientes do cruzamento interespecífico e
com altos teores de AAs RVTA-2010pl#39, RVTA-2010pl#177, RVTA-2010pl#232, RVTA-
2010pl#257 e RVTA-2010pl#359 obtiveram menores valores de ovoposição quando
comparados com os genótipos selecionados para baixos teores de AAs (RVTA-2010pl#205,
RVTA-2010pl#205, RVTA-2010pl#376 e RVTA-2010pl#385) e a cultivar Redenção.
TABELA 7- Teores de acil-açúcares, número médio de ovos, número médio de larvas e
contraste, durante 28 dias de avaliações, em genótipos F2RC2 de tomateiro com diferentes
teores de acil-açúcares, Solanum pennellii (acesso LA716) e Solanum lycopersicum (Cultivar
Redenção) submetidos à infestação da traça do tomateiro. Guarapuava-PR –UNICENTRO,
2019.
Genótipo Valores de absorbância Ovos Larvas
Solanum pennellii (alto) 0,4557 1,00 f 1,00 d
RVTA-2010pl#28 (alto) 0,3614 2,38 b 1,55 c
RVTA-2010pl#39 (alto) 0,4564 1,74 e 1,60 c
RVTA-2010pl#113 (alto) 0,3119 2,34 b 1,48 c
RVTA-2010pl#177 (alto) 0,2251 2,02 c 1,82 c
RVTA-2010pl#180 (alto) 0,3478 2,43 b 1,52 c
RVTA-2010pl#232 (alto) 0,3969 1,76 e 1,25 d
RVTA-2010pl#257 (alto) 0,3112 1,68 e 1,32 d
RVTA-2010pl#359 (alto) 0,2107 1,80 d 1,20 d
RVTA-2010pl#205 (baixo) 0,0643 2,43 b 2,35 b
RVTA-2010pl#227 (baixo) 0,1080 2,55 b 2,56 a
RVTA-2010pl#376 (baixo) 0,1052 2,34 b 2,21 b
RVTA-2010pl#385 (baixo) 0,0955 2,38 b 2,51 a
Cultivar Redenção (baixo) 0,1104 2,85 a 2,57 a
CV (%) 6,14 10,09
C1- Genótipos (altos) vs. Genótipos (baixos) -0,40** -0,90**
Correlação de Pearson -0,65** -0,82** Médias seguidas pela mesma letra na coluna pertencem ao mesmo grupo pelo teste de Scott-Knott a 5 % de probabilidade. 1Valores de absorbância a 540 nm.
** Significativo ao nível de 1% de probabilidade.
O número médio de larvas presentes nos folíolos formou 4 grupos distintos, com
valores médios variando de 1 (Solanum pennellii) até 2,57 (cultivar Redenção) (Tabela 7). O
grupo que apresentou menor quantidade de ovos foi formado pelo Solanum pennellii e os
genótipos com altos teores de AAs RVTA-2010pl#232, RVTA-2010pl#257 e RVTA-
2010pl#359. Porém, os genótipos que apresentaram maiores quantidade de larvas presentes
nos folíolos foram os genótipos com baixos teores AAs (RVTA-2010pl#205, RVTA-
2010pl#227, RVTA-2010pl#376 e RVTA-2010pl#385) e a cultivar Redenção quando
comparados com os genótipos que apresentavam maiores teores de AAs (Solanum pennellii,
RVTA-2010pl#28, RVTA-2010pl#39, RVTA-2010pl#113, RVTA-2010pl#177, RVTA-

70
2010pl#180, RVTA-2010pl#232, RVTA-2010pl#257 e RVTA-2010pl#359).
Os genótipos F2RC2 selecionados para altos teores de AAs apresentaram, geralmente,
os menores valores de ovos e larvas presentes nos folíolos. No entanto, alguns genótipos com
altos teores do aleloquímico foram similares estatisticamente aos genótipos contrastantes em
relação ao número de ovos. Possivelmente, estes resultados têm relação com a perda de alelos
ligados a resistência da planta, possuindo ou não relação com a presença dos AAs, fatores
estes que com o avançar das gerações podem ser perdidos. Neste sentido, a exposição dos
genótipos ao inseto-praga favorece a seleção dos genótipos com maior quantidade de genes
responsáveis pela resistência.
Resultados semelhantes foram observados por Dias et al. (2013) em relação ao número
de ovos e larvas em que alguns genótipos com altos teores de AAs que não diferiram dos
genótipos selecionados para baixos teores. Maluf et al. (2010) relatam que em programas de
melhoramento genético visando a resistência a pragas que a divergência de resultados é
comumente observada nos genótipos retrocruzados obtidos a partir do cruzamento entre uma
espécie silvestre e uma linhagem de S. lycopersicum.
Lucini et al. (2015) utilizaram genótipos F2 com teores contrastantes de AAs e
provenientes do cruzamento interespecífico entre S. lycopersicum e S. pennellii acesso LA
716 para estudos relativos com o mecanismo de resistência. Foram observados os
mecanismos de resistência do tipo antibiose e antixenose nos genótipos F2 selecionados para
altos teores de AAs em testes com e sem chances de escolha.
O contraste C1 demonstrou relação significativa (p<0,05) e inversamente proporcional
entre os genótipos com altos teores de AAs quando comparados com os genótipos que
apresentavam baixos teores do aleloquímico para a quantidade de ovos (-0,40) e larvas (-0,90)
presentes nos folíolos (Tabela 7). Neste sentido o contraste C1 evidencia os teores de AAs
presentes nos folíolos como um fator de resistência para a traça do tomateiro.
Os prejuízos causados pela larva da traça do tomateiro foram estatisticamente
influenciados pelos teores de AAs presentes nos folíolos dos genótipos quando comparados os
danos na planta (DP), lesões nos folíolos (LF) e porcentagem de folíolos atacados (PF)
(Tabela 9). Em relação aos DP, LF e PF, foram observados a presença de 3 grupos distintos, o
grupo com a menor quantidade de danos causado pela traça do tomateiro que foi composto
exclusivamente pelo acesso silvestre Solanum pennellii LA- 716. O segundo grupo composto
pelos genótipos com altos teores de AAs (RVTA-2010-31-177pl#28, RVTA-2010-31-
177pl#39, RVTA-2010-31-177pl#113, RVTA-2010-31-177pl#177, RVTA-2010-31-
177pl#232, RVTA-2010-83-347pl#257 e RVTA-2010-83-347pl#359), exceto o genótipo
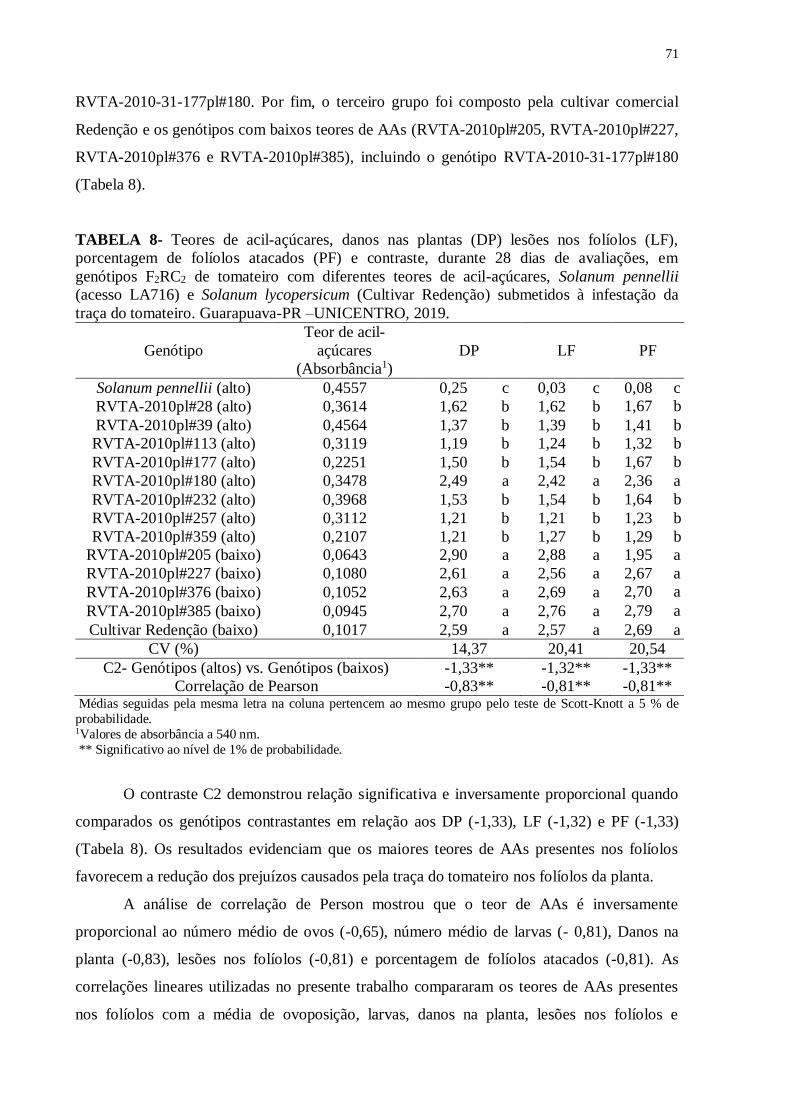
71
RVTA-2010-31-177pl#180. Por fim, o terceiro grupo foi composto pela cultivar comercial
Redenção e os genótipos com baixos teores de AAs (RVTA-2010pl#205, RVTA-2010pl#227,
RVTA-2010pl#376 e RVTA-2010pl#385), incluindo o genótipo RVTA-2010-31-177pl#180
(Tabela 8).
TABELA 8- Teores de acil-açúcares, danos nas plantas (DP) lesões nos folíolos (LF),
porcentagem de folíolos atacados (PF) e contraste, durante 28 dias de avaliações, em
genótipos F2RC2 de tomateiro com diferentes teores de acil-açúcares, Solanum pennellii
(acesso LA716) e Solanum lycopersicum (Cultivar Redenção) submetidos à infestação da
traça do tomateiro. Guarapuava-PR –UNICENTRO, 2019.
Genótipo
Teor de acil-
açúcares
(Absorbância1)
DP LF PF
Solanum pennellii (alto) 0,4557 0,25 c 0,03 c 0,08 c
RVTA-2010pl#28 (alto) 0,3614 1,62 b 1,62 b 1,67 b
RVTA-2010pl#39 (alto) 0,4564 1,37 b 1,39 b 1,41 b
RVTA-2010pl#113 (alto) 0,3119 1,19 b 1,24 b 1,32 b
RVTA-2010pl#177 (alto) 0,2251 1,50 b 1,54 b 1,67 b
RVTA-2010pl#180 (alto) 0,3478 2,49 a 2,42 a 2,36 a
RVTA-2010pl#232 (alto) 0,3968 1,53 b 1,54 b 1,64 b
RVTA-2010pl#257 (alto) 0,3112 1,21 b 1,21 b 1,23 b
RVTA-2010pl#359 (alto) 0,2107 1,21 b 1,27 b 1,29 b
RVTA-2010pl#205 (baixo) 0,0643 2,90 a 2,88 a 1,95 a
RVTA-2010pl#227 (baixo) 0,1080 2,61 a 2,56 a 2,67 a
RVTA-2010pl#376 (baixo) 0,1052 2,63 a 2,69 a 2,70 a
RVTA-2010pl#385 (baixo) 0,0945 2,70 a 2,76 a 2,79 a
Cultivar Redenção (baixo) 0,1017 2,59 a 2,57 a 2,69 a
CV (%) 14,37 20,41 20,54
C2- Genótipos (altos) vs. Genótipos (baixos) -1,33** -1,32** -1,33**
Correlação de Pearson -0,83** -0,81** -0,81**
Médias seguidas pela mesma letra na coluna pertencem ao mesmo grupo pelo teste de Scott-Knott a 5 % de
probabilidade. 1Valores de absorbância a 540 nm.
** Significativo ao nível de 1% de probabilidade.
O contraste C2 demonstrou relação significativa e inversamente proporcional quando
comparados os genótipos contrastantes em relação aos DP (-1,33), LF (-1,32) e PF (-1,33)
(Tabela 8). Os resultados evidenciam que os maiores teores de AAs presentes nos folíolos
favorecem a redução dos prejuízos causados pela traça do tomateiro nos folíolos da planta.
A análise de correlação de Person mostrou que o teor de AAs é inversamente
proporcional ao número médio de ovos (-0,65), número médio de larvas (- 0,81), Danos na
planta (-0,83), lesões nos folíolos (-0,81) e porcentagem de folíolos atacados (-0,81). As
correlações lineares utilizadas no presente trabalho compararam os teores de AAs presentes
nos folíolos com a média de ovoposição, larvas, danos na planta, lesões nos folíolos e

72
porcentagem de folíolos atacados. Possibilitando a averiguação do importante papel dos
aleloquímicos AAs na redução das taxas destas variáveis nos genótipos analisados (Tabelas
8).
Foram observados resultados nesta pesquisa semelhantes a inferências de outros
autores (RESENDE et al., 2006; SAEIDI et al., 2007; RESENDE et al., 2008, PEREIRA et
al., 2008; GONÇALVES NETO et al., 2008, 2010, MALUF et al., 2010; MACIEL et al.,
2011; DIAS et al., 2013; DIAS et al., 2016; LUCINI et al., 2015). O principal provedor de
resistência a pragas foram a presença de altos teores de AAs nos folíolos. A seleção em
laboratórios dos teores de AAs para a obtenção de materiais com bons níveis de resistência é
satisfatória, principalmente, pelo tipo de herança simples responsável pela expressão da
resistência. A herança do tipo monogênica com a presença de outros genes modificadores de
efeito aditivo favorece a introdução dos AAs em linhagens elite de tomateiro (RESENDE et
al., 2002; GONÇALVES et al., 2007).
A ACP foi realizada sobre o conjunto de dados e mostrou que as duas primeiras
componentes explicam 94,77% da variabilidade dos dados. A análise de componentes
principais (ACP) mostrou a formação de dois grupos (Figura 2). O grupo 1 formado pelos
genótipos RVTA-2010pl#205 (G205), RVTA-2010pl#227 (G227), RVTA-2010pl#378
(G378), RVTA-2010pl#385 (G385) e a cultivar Redenção com baixos teores de AAs e
maiores quantidades da presença de ovos, larvas, danos na planta, lesões nos folíolos e
porcentagem de folíolos atacados (Figura 2). Todavia, o Grupo 2 formado pelos genótipos
Solanum pennellii (G1), RVTA-2010pl#28 (G28), RVTA-2010pl#39 (G39), RVTA-
2010pl#113 (G113), RVTA-2010pl#177 (G177), RVTA-2010pl#180 (G180), RVTA-
2010pl#232 (G232), RVTA-2010pl#257 (G257), RVTA-2010pl#359 (G359) com maiores
teores de AAs e menores números de ovos, larvas e danos provocados pela praga (Figuras 2).

73
-1.0 -0.5 0.0 0.5 1.0
-1.0
-0.5
0.0
0.5
1.0
Variables factor map (PCA)
Dim 1 (88.40%)
Dim
2 (6
.37%
)
TAA
TTO
TL
DFMLFM
PFM
-4 -2 0 2 4
-10
12
Factor map
Dim 1 (88.40%)
Dim
2 (
6.3
7%
)
G2 G257
G39
G232G359
G113
G180
G28
G177
G376
G227
G385 G205
G1
cluster 1
cluster 2
FIGURA 2- Círculo de correlação das variáveis teor de acil-açúcares (Teor de acil-açúcares
(TAA), número de ovos (TTO), número de larvas (TL), Danos na planta (DFM) e Lesões nos
folíolos (LFM) e Porcentagem de folíolos atacados (PFM) e distribuição dos genótipos em
plano Solanum pennellii (G1), RVTA-2010pl#28 (G28), RVTA-2010pl#39 (G39), RVTA-
2010pl#113 (G113), RVTA-2010pl#177 (G177), RVTA-2010pl#180 (G180), RVTA-
2010pl#232 (G232), RVTA-2010pl#257 (G257), RVTA-2010pl#359 (G359), RVTA-
2010pl#205 (G205), RVTA-2010pl#227 (G227), RVTA-2010pl#378 (G378), RVTA-
2010pl#385 (G385) e cultivar Redenção (G1). Guarapuava-PR –UNICENTRO, 2019.

74
Maluf et al. (2010), Maciel et al. (2011), Dias et al. (2013) e Baier et al. (2015),
relatam a importância da presença de altos teores de AAs nos folíolos como provedor de
resistência a pragas em genótipos obtidos a partir do cruzamento interespecífico entre o
acesso silvestre LA 716 e S. lycopersicum e posteriores retrocruzamentos com a linhagem
comercial. Dentre os genótipos segregantes F2RC2 destacou-se os genótipos RVTA-
2010pl#39, RVTA-2010pl#232, RVTA-2010pl#257 e RVTA-2010pl#359 com a presença de
altos teores de AAs foliares e com maiores padrões de resistência à traça do tomateiro, visto
que apresentou menor preferência para ovoposição, larvas e reduzida severidade dos danos
ocasionados pela praga similares ao Solanum pennellii.
3.1 Uso de marcadores moleculares ISSR e SSR na seleção de genótipos com
altos teores de acil-açúcares
Os seis primers ISSR amplificaram 40 fragmentos polimórficos entre a cultivar
Redenção (genoma recorrente) e o acesso silvestre Solanum pennellii LA 716 (genoma
doador). Em relação aos seis primers SSR testados foram obtidas 10 bandas polimórficas. A
avaliação conjunta dos 50 fragmentos polimórficos permitiu estabelecer a similaridade
genética entre todos os genótipos avaliados (Tabela 9). Houve diferença na similaridade com
a cultivar Redenção quando comparado as diferentes linhagens (Tabela 9). A maior
similaridade ocorreu entre os genótipos RVTA-2010pl#257 (12), RVTA-2010pl#152 (5) e
RVTA-2010pl#28 (1) e os com menores, RVTA-2010pl#177 (8) e RVTA-2010pl#180 (9)
(Tabela 9).
A avaliação da similaridade entre as linhagens e os parentais evidenciou que todas as
linhagens apresentaram maior similaridade com a cultivar redenção do que com o parental
selvagem (Tabela 9). Esta condição provavelmente é devido a seleção de plantas com
características mais semelhantes com a cultivar redenção. Embora ainda as plantas não
estejam sendo selecionadas de forma criteriosa para as características da cultivar redenção, no
momento de escolher as plantas a serem avaliadas, é dado preferência para as com o fenótipo
mais semelhante ao tomate cultivado.

75
TABELA 9- Similaridade genética entre linhagens F2RC2 de tomateiro oriundas do
cruzamento entre o genótipo selvagem Solanum pennellii e a cultivar Redenção (Solanum
lycopersicum). Guarapuava-PR-UNICENTRO, 2019.
Solanum
Pennellii Redenção 1 2 3 4 5 7 8 9 10 11 12 13
Redenção (RE) 0,00
RVTA-2010pl#28 (1) 0,23 0,62
RVTA-2010pl#39 (2) 0,33 0,54 0,65
RVTA-2010pl#61 (3) 0,24 0,57 0,61 0,73
RVTA-2010pl#113 (4) 0,32 0,54 0,66 0,69 0,65
RVTA-2010pl#152 (5) 0,23 0,62 0,66 0,78 0,83 0,62
RVTA-2010pl#155 (7) 0,29 0,54 0,58 0,65 0,69 0,74 0,70
RVTA-2010pl#177 (8) 0,40 0,42 0,60 0,60 0,56 0,58 0,57 0,60
RVTA-2010pl#180 (9) 0,32 0,50 0,62 0,58 0,57 0,72 0,58 0,70 0,64
RVTA-2010pl#214 (10) 0,14 0,56 0,45 0,53 0,60 0,53 0,53 0,57 0,51 0,53
RVTA-2010pl#232 (11) 0,22 0,60 0,56 0,63 0,68 0,64 0,73 0,68 0,48 0,52 0,46
RVTA-2010pl#257 (12) 0,21 0,64 0,68 0,75 0,81 0,63 0,86 0,72 0,62 0,56 0,58 0,70
RVTA-2010pl#359 (13) 0,12 0,55 0,44 0,40 0,42 0,45 0,51 0,56 0,50 0,60 0,53 0,49 0,57
RVTA-2010pl#160 (17) 0,18 0,59 0,55 0,55 0,58 0,55 0,63 0,59 0,50 0,48 0,44 0,57 0,69 0,52
O uso dos marcadores ISSR e SSR foram eficientes na identificação genótipos com maior
recuperação do genoma recorrentes neste estudo. Na literatura o uso de marcadores
moleculares para seleção assistida tem sido uma das aplicações mais eficientes da
biotecnologia na agricultura. Goswami et al. (2018), em estudos para seleção de linhagens de
milho com maior teor de provitamina A (proA), a introgressão assistida por meio de
marcadores moleculares foram eficientes no desenvolvimento acelerado de progênies elite
com utilização de 100 SSRs e recuperação média de 91,07 % do genoma recorrente.
Pukalentthy et al. (2019), em pesquisas com linhagens de milho para incorporar o gene
recessivo opaco-2 (o2) responsável pelo maior conteúdo de lisina e triptofano em suas
proteínas, relatam a importância da seleção assistida com 250 SSR aliado a seleção fenotípica
possibilitou a seleção de 3 linhas avançadas de milho com 97% de recuperação do genoma
recorrente e com a presença do gene recessivo o2. Babu et al. (2005), relatam o uso de
marcadores moleculares SSR foram eficientes na seleção de 3 plantas com 93 a 96 % de
conteúdo genômico recorrente auxiliando na redução do tempo envolvido para obtenção de
uma linhagem com alta concentração de triptofano em proteína (0,85 %) em suas sementes.
Iftekharuddaula et al. (2011), em estudos para incorporação da tolerância a submersão em
arroz irrigado por meio de cruzamentos assistidos por marcadores moleculares, entre eles os
marcadores SSR, possibilitaram em apenas dois retrocruzamentos e uma geração de
autofecundação a obtenção de uma linhagem com 99,8% do genitor recorrente, produtiva e

76
tolerante a submersão. Rai et al. (2017) relatam o uso combinado de três locos de
características quantitativas (QTLs), diferentes marcadores SSR e seleção fenotípica que
resultaram na seleção de 5 variedades de trigo com potencial uniformidade, estabilidade e
tolerância ao estresse hídrico e com 89,2 % a 95, 4 % de recuperação do parental recorrente.
Neste sentido, o uso simultâneo do retrocruzamento assistido por marcadores moleculares
apresentam-se como alternativa viável para aumentar a eficiência do processo de
retrocruzamento e identificação precisa e precoce dos indivíduos que apresentam maior
recuperação do genoma do genitor recorrente (OPENSHAW et al., 1994).
4. CONCLUSÕES
Nos testes de resistência a traça do tomateiro os genótipos com maiores níveis de
resistência foram RVTA-2010pl#39, RVTA-2010pl#359, RVTA-2010pl#232 e RVTA-
2010pl#257.
A utilização de marcadores moleculares possibilitou a identificação dos genótipos
RVTA-2010pl#257, RVTA-2010pl#152, RVTA-2010pl#28 e RVTA-2010pl#113 como os
com maior recuperação do genoma recorrente.
A análise conjunta dos dados de resistência e molecular evidência que os genótipos
RVTA-2010pl#232 (11) e RVTA-2010pl#257 (12) são os com maior potencial para utilização
para avanço do programa de melhoramento para obtenção de linhagens com boas
características morfoagronômicas e com resistência a traça do tomateiro.

77
5. CONSIDERAÇÕES FINAIS.
O teste de resistência para a mosca branca, análise dos componentes principais e teste
de avaliação da severidade de fumagina presente nos folíolos possibilitaram a seleção dos
genótipos F2RC2 de tomateiro RVTA-2010pl# 28, RVTA-2010pl# 39 e RVTA-2010pl# 113
que devem ser usados como fonte de genes para continuar o programa de melhoramento para
resistência a mosca branca.
O teste de resistência para a traça do tomateiro, análise dos componentes principais
(ACP) e a seleção assistida por marcadores moleculares evidenciaram que os genótipos F2RC2
de tomateiro com maiores níveis de resistência a traça do tomateiro foram os genótipos
RVTA-2010pl#232 e RVTA-2010pl#257 que devem ser usados como fonte de genes para
continuar o programa de melhoramento para resistência a traça do tomateiro.
O programa de melhoramento genético do tomateiro visando resistência as principais
pragas presentes no Núcleo de Pesquisa em Hortaliças na Universidade Estadual do Centro-
Oeste devem continuar os retrocruzamentos e a seleção assistida visando a seleção de
genótipos com maiores características do parental recorrente e resistência para artrópodes-
pragas com os genótipos F2RC2 de tomateiro RVTA-2010pl# 39, RVTA-2010pl#232 e
RVTA-2010pl#257.

78
5. REFERÊNCIAS
AGUILERA, J.G.; PESSONI, L.A.; RODRIGUES, G. B.; ELSAYED, A. Y.; SILVA, D.J.H.
DA; BARROS, E.G. DE. Genetic variability by ISSR markers in tomato (Solanum
lycopersicon Mill.) Rev. Bras. Ciênc. Agrár., Recife, v.6, n.2, p.243-252, 2011.
AKKAYA, M.S., BHAGWAT, A.A.; CREGAN, P.B. Length polymorphisms of simple
sequence repeat DNA in soybean. Genetics, v.132, p.1131-1139, 1992.
ALZATE-MARIN, A.L.; COSTA, M.R.; SARTORATO, A.; RAVA, C.A.; BARROS, E.G.;
MOREIRA, M.A. Use of markers as a tool to investigate the presence of disease
resistance genes in common bean cultivars. Crop Breeding and Applied
Biotechnology, Londrina, v.1, n.2, p.125-133, 2001.
ANDRADE, M.C.; SILVA, A.A. da.; NEIVA, I.P.; OLIVEIRA, I.R.; CASTRO, E.M.;
FRANCIS, D.M.; MALUF, W.R. Inheritance of type IV glandular trichome density and
its association with whitefly resistance from Solanum galapagense accession LA140.
Euphytica 213: 52- 64. 2017.
ANTONIO, R.P.; SANTOS, J.B. dos; ALVES, F.C.; GONÇALVES, P.R.C.; LARA, L.A.C.
Seleção assistida por marcadores de dna em retrocruzamento visando resistência ao
mofo branco em feijoeiro. Revista Caatinga, Mossoró, v. 25, n. 4, p. 61-67, out-dez.,
2012.
BABU, R.; NAIR, S. K.; KUMAR, A.;VENKATESH, S.; SEKHAR, J. C.; SINGH, N. N.;
SRINIVASAN, G.; GUPTA, H. S. Two-generation marker-aided backcrossing for rapid
conversion of normal maize lines to quality protein maize (QPM). Theor Appl Genet
111: 888–897. 2005.
DREHER, K.; KHAIRALLAH, M.; RIBAUT, J. M.; MORRIS, M. Money matters (I): costs
of field and laboratory procedures associated with conventional and marker-assisted
maize breeding at CIMMYT. Molecular Breeding, Dordrecht, v. 11, p. 221-234, 2003.
DIAS, D.M.; RESENDE, J.T.V.; FARIA, M.V.; CAMARGO, L.K.P.; LIMA, I.P. Selection
of processing tomato genotypes with high acyl sugar content that are resistant to the
tomato pinworm. Genetics and Molecular Research. v.12. p. 381-389. 2013.
DIAS, D.M.; RESENDE, J.T.V.; MARODIN, J.C.; MATOS, R.; LUSTOSA, I.F.;
RESENDE, N.C.V. Acyl sugars and whitefly (Bemisia tabaci) resistance in segregating
populations of tomato genotypes. Genetics and Molecular Research 15: 1-11. 2016.

79
EL BALLA, M.M.A.; HAMID, A.A.; ABDELMAGEED, A.H.A. Effects of time of water
stress on flowering, seed yield and seed quality of common onion (Allium cepa L.)
under the arid tropical conditions of Sudan. Agricultural Water Management, 121:
49-157. 2013.
FERREIRA, D. SISVAR: um programa para análises e ensino de estatística. Revista
Symposium. v. 6, p. 36-41, 2008.
GAMEIRO, A. H.; CAIXETA FILHO, J. V.; ROCCO, C. D.; RANGEL, R. Estimativa de
perdas no suprimento de tomates para processamento industrial no estado de Goiás.
Informações Econômicas, São Paulo, v. 37, n. 7, p. 07-16, 2007.
GONÇALVES-NETO, A.C.; SILVA, V. de F.; MALUF, W.R.; MACIEL, G.M.; NÍZIO,
D.A.C.; GOMES, L.A.A.; AZEVEDO, S.M. de. Resistência à traça-do-tomateiro em
plantas com altos teores de acil-açúcares nas folhas. Horticultura brasileira, Brasília,
v.28, n.2, p.203-208, 2010.
GOSWAMI, R.; UTTAMRAO, R. Z.; KHAN, S.; BAVEJA, A.; MUTHUSAMY, V.;
HOSSAIN, F. Marker‐assisted introgression of rare allele of β‐carotene hydroxylase
(crtRB1) gene into elite quality protein maize inbred for combining high lysine,
tryptophan and provitamin A in maize. Plant Breeding. 1–10. 2019.
IFTEKHARUDDAULA, K. M.; NEWAZ, M. A.; SALAM, M. A.; AHMED, H. U.;
MAHBUB, M. A. A.; SEPTININGSIH, E. M.; COLLARD, B. C. Y.; SANCHEZ, D.
L.; PAMPLONA, A. M.; MACKILL, D. J. Rapid and high-precision marker assisted
backcrossing to introgress the SUB1 QTL into BR11, the rainfed lowland rice mega
variety of Bangladesh. Euphytica, 178:83–97. 2017.
LABORY, C.R.G.; SANTA-CECÍLIA, L.V.C.; MALUF, W.R.; CARDOSO, C.G.;
BEARZOTTI, E.; SOUZA, J.C. de. Seleção indireta para teor de 2-tridecanona em
tomateiros segregantes e sua relação com a resistência à traça-do-tomateiro. Pesquisa
Agropecuária Brasileira 34: 733-740. 1999.
LIMA, I.P.; RESENDE, J.T.V.; OLIVEIRA, J.R.F.; FARIA, M.V.; DIAS, D.M.; RESENDE,
N.C.V. Selection of tomato genotypes for processing with high zingiberene content,
resistant to pests. Horticultura Brasileira, 34: 387-391. 2016.
LUCINI, T.; FARIA, M. V.; ROHDE, C.; RESENDE, J. T. V.; OLIVEIRA, J. R. F.
Acylsugar and the role of tricomes in tomato genotypes resistence to Tetranychus
urticae. Arthopod-Plant Interactions. v. 9, p. 45-53, jan., 2015.
MACIEL G.M; MALUF, W.R; SILVA, V.F; GONÇALVES NETO, A.C; GOMES, L.A.A.
Híbridos pré-comerciais resistentes a Tuta absoluta obtidos de linhagem de tomateiro

80
rica em acilaçúcares. Horticultura Brasileira, 29: 151-156. 2011.
MALUF, W.R.; MACIEL, G.M.; GOMES, L.A.A.; CARDOSO, M.G.; GONÇALVES, L.D.;
SILVA, E.C.; KNAPP, M. Broad-spectrum arthropod resistance in hybrids between
high- and low-acylsugar tomato lines. Crop Science, Madison, v.50, p.439-450, 2010.
MAMTA, K.; REDDY, R.K.; RAJAM, M.V. Targeting chitinase gene of Helicoverpa
armigera by host-induced RNA interference confers insect resistance in tobacco and
tomato. Plant Molecular Biology, 90: 281-292. 2016.
MESQUITA, A. G. G.; GUIMARÃES,C. T. PARENTONI, S. N.; PAIVA, E. Recuperação
do genitor recorrente em milho utilizando retrocruzamento assistido por marcadores
microssatélites. Revista brasileira de milho e sorgo, v.4, n.3, p.275-285 , 2005.
MOREIRA, G.R.; SILVA, D.J.H. da.; CARNEIRO, P.C.S.; PICANÇO, M.C.;
VASCONCELOS, A. A.; PINTO, C.M.F. Herança de caracteres de resistência por
antixenose de Solanum pennellii à traça-do-tomateiro em cruzamento com ‘Santa
Clara’. Horticultura Brasileira, 31: 574-581. 2013.
NELSON, N. A photometric adaptation of the Sommogy method for the determination of
glucose. Journal of Biological Chemistry, v.153, p.375-380, 1944.
OPENSHAW, S.J.; JARBOE, S.G.; BEAVIS, W.D. Marker assisted selection in backcross
Corvallis, Oregon. Proceedings...Corvallis: American Society for Horticultural
Science/Crop Science of America, 1994. p.41-43.
PEREIRA, G.V.N.; MALUF, W.R.; GONÇALVES, L.D.; NASCIMENTO, I.R. do.;
GOMES, L.A.A.; LICURSI, V. Seleção para alto teor de acil-açúcares em genótipos de
tomateiro e sua relação com a resistência ao ácaro vermelho (Tetranychus evansi) e à
traça (Tuta absoluta). Ciência e Agrotecnologia, Lavras, v.32, n.3, p.996-1004, 2008.
PUKALENTHY, B.; MANICKAM, D.; CHANDRAN, S.; ADHIMOOLAM, K.;
SAMPATHRAJAN, V.; RAJASEKARAN, R.; ARUNACHALAM, K.;
GANAPATHYSWAMY, H.; CHOCKLINGAM, V.; MUTHUSAMY, V. Incorporation
of opaque-2 into ‘UMI 1200’, an elite maize inbred line, through marker-assisted
backcross breeding, Biotechnology & Biotechnological Equipment. 2019.
RAI,N.; BELLUNDAGI, A.; KUMAR, P.K.C.;THIMMAPPA, R.K.; RANI,S.; SINHA,
N.;HARIKRISHNA.; JAIN, N.; SINGH, G.P.; SINGH, P.K.; CHAND, S.; PRABHU,
K.V. Marker-assisted backcross breeding for improvement of drought tolerance in
bread wheat (Triticum aestivum L. em Thell). Plant Breeding. 1–13. 2018.
REDDY, M. P.; SARLA, N.; SIDDIQ, E. A. Inter simple sequence repeat (ISSR)
polymorphism and its application in plant breeding. Euphytica, Dordrecht, v. 128, p. 9-

81
17, 2002.
RESENDE, J.T.V.; CARDOSO, M.G.; MALUF, W.R.; SANTOS, C.D.; GONÇALVES,
L.D.; RESENDE, L.V.; NAVES, F.O. Método colorimétrico para quantificação de acil-
açúcar em genótipos de tomateiro. Ciência e Agrotecnologia, Lavras, v. 26, n. 6, p.
1204-1208, nov./dez. 2002a.
RESENDE, J,T.V. de.; MALUF. W.R.; CARDOSO, M. G.; FARIA, M.V.; GONÇALVES,
L.D.; NASCIMENTO, I. R. do. Resistance of tomato genotypes with high level of
acylsugars to Tetranychus evansi Baker & Pritchard. Scientia Agrícola, Piracicaba,
v.65, n.1, p.31-35, 2008.
RESENDE, J,T.V. de.; MALUF. W.R.; CARDOSO, M. G.; GONÇALVES, L.D.; FARIA,
M.V.; NASCIMENTO, I. R. do. Resistance of tomato genotypes to the silverleaf
whitefly mediated by acylsugars. Horticultura brasileira, Brasília, v.27, n.3, p. 345-
348, 2009.
RESENDE, T.V. de R.; MALUF, W.R.; FARIA, M.V.; PFANN, A.Z.; NASCIMENTO, I.R.
do. Acylsugars in Tomato Leaflets Confer Resistance to the South American Tomato
Pinworm, Tuta absoluta Meyr. Scientia Agricola, Piracicaba, v.63, n.1, p.20-25, 2006.
SAEIDI, Z.; MALLIK, B.; KULKARNI, R.S. Inheritance of glandular trichomes and two-
spotted spider mite resistance in cross Lycopersicon esculentum ‘Nandi’ and L. pennellii
‘LA2963’. Euphytica, 154:231–238. 2007.
SECCO, D. Aplicação de uma metodologia molecular para aceleração na obtenção de
tomate resistente a pragas. 2013. 30p. Monografia (licenciatura em Ciências
Biológicas)-Universidade Estadual do Centro-Oeste, Guarapuava, PR.
SILVA, A.A.; ANDRADE, M.C.; CARVALHO, R.C.; NEIVA, I.P.; SANTOS, D.C.;
MALUF, W.R. Resistência à Helicoverpa armigera em genótipos de tomateiro obtidos
do cruzamento de Solanum lycopersicum com Solanum galapagense. Pesquisa
Agropecuária Brasileira, 51: 801-808. 2016.
SILVA, F. DE A. S. E. e AZEVEDO, C. A. V. de. Principal Components Analysis in the
Software Assistat-Statistical Attendance. In:WORLD CONGRESS ON COMPUTERS
IN AGRICULTURE, 7, Reno-NV-USA: American Society of Agricultural and
Biological Engineers, 2009.
SILVA, V.F.; MALUF, W.R.; CARDOSO, M.G.; GONÇALVES NETO, A.C.; MACIEL,
G.M.; NIZIO, D.A.C.; SILVA, V.A. Resistência mediada por aleloquímicos de
genótipos de tomateiro à mosca-branca e ao ácaro-rajado. Pesquisa Agropecuária
Brasileira, Brasília, v.44, n.9, p.1262-1269, 2009.

82
STATSOFT, Inc. (2004). STATISTICA (data analysis software system), version 7.
Disponível em:<www.statsoft.com> .
TIAN, D.; TOOKER. J,; PEIFFER, M.; CHUNG, S.H.; FELTON, G.W. Role of trichomes in
defense against herbivores: comparison of herbivore response to woolly and hairless
trichome mutants in tomato (Solanum lycopersicum). Planta, 236: 1053-1066. 2012.