ministÉrio da educaÇÃo universidade federal …livros01.livrosgratis.com.br/cp155681.pdf ·...
TRANSCRIPT

MINISTÉRIO DA EDUCAÇÃO
UNIVERSIDADE FEDERAL RURAL DA AMAZÔNIA
JOSEMAR MOREIRA VASCONCELOS
INDICADORES QUÍMICOS E BIOLÓGICOS DE LATOSSOLO AMARELO
SUBMETIDO AOS SISTEMA DE PREPARO DE ÁREA USANDO A QUEIMA E A
TRITURAÇÃO DA VEGETAÇÃO NO NORDESTE PARAENSE
BELÉM
2010

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

MINISTÉRIO DA EDUCAÇÃO
UNIVERSIDADE FEDERAL RURAL DA AMAZÔNIA
JOSEMAR MOREIRA VASCONCELOS
INDICADORES QUÍMICOS E BIOLÓGICOS DE LATOSSOLO AMARELO
SUBMETIDOS AO SISTEMA DE PREPARO DE ÁREA USANDO A QUEIMA E A
TRITURAÇÃO DA VEGETAÇÃO NO NORDESTE PARAENSE
BELÉM
2010
Dissertação apresentada à Universidade Federal
Rural da Amazônia, como parte das exigências
do programa de pós-graduação em Agronomia,
área de concentração em Solos e Nutrição de
Plantas, para obtenção do título de “Mestre em
Agronomia”.
Orientador: Prof. Dr. Mário Lopes da Silva
Júnior

Vasconcelos, Josemar Moreira
Indicadores químicos e biológicos de latossolo amarelo
submetido ao sistema de preparo de área usando a queima e a
trituração da vegetação no Nordeste Paraense./ Josemar Moreira
Vasconcelos. – Belém, 2010.
105 f.:il.
Dissertação (Mestrado em Agronomia) – Universidade Federal
Rural da Amazônia, 2010.
1. Capoeira 2. Agricultura itinerante 3. Atributos químicos 4.
Atributos biológicos I. Título.
CDD – 631.41

MINISTÉRIO DA EDUCAÇÃO
UNIVERSIDADE FEDERAL RURAL DA AMAZÔNIA
JOSEMAR MOREIRA VASCONCELOS
INDICADORES QUÍMICOS E BIOLÓGICOS DE LATOSSOLO AMARELO
SUBMETIDOS AO SISTEMA DE PREPARO DE ÁREA USANDO A QUEIMA E A
TRITURAÇÃO DA VEGETAÇÃO NO NORDESTE PARAENSE
Dissertação apresentada à Universidade Federal Rural da Amazônia, como parte das
exigências do programa de pós-graduação em Agronomia, área de concentração em Solos e
Nutrição de Plantas, para obtenção do título de “Mestre em Agronomia”.
Aprovado em 24 de Setembro de 2010
Banca Examinadora
_________________________________________________________________
Engenheiro Agrônomo, Professor Dr. Mário Lopes da Silva Júnior
(Orientador)
Universidade Federal Rural da Amazônia – UFRA
________________________________________________________________
Geóloga, Pesquisadora Dra. Maria de Lourdes Pinheiro Ruivo
Museu Paraense Emílio Goeldi
________________________________________________________________
Engenheira Agrônoma, Professora Dra. Rosana Cardoso Rodrigues
Instituto Federal do Pará - IFPA
______________________________________________________________
Engenheiro Agrônomo, Professor Dr. George Rodrigues da Silva
Universidade Federal Rural da Amazônia – UFRA

Ao meu pai José Marçal de Vasconcelos e à minha mãe, Iolanda Moreira
Vasconcelos (in memorian), cujo exemplo dedicação e amor estimularam cada passo da
minha vida, e à minha filha, Joyce Alexandria Vasconcelos.

AGRADECIMENTOS
Primeiramente, agradeço a Deus, que me guiou durante essa etapa de minha vida, por que sem
ele nada é possível e nenhum obstáculo é vencido;
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq, pelo apoio
financeiro ao Projeto Biodiversidade e Funcionamento do Solo em áreas submetidas ao
preparo do solo usando a queima e a não-queima da vegetação na Amazônia Oriental,
Proc. Nº 483696/2009-9, Edital MCT/CNPq 14/2009, o qual deu suporte para o
desenvolvimento desta pesquisa.
Ao meu orientador, Dr. Mário Lopes da Silva Júnior, pela orientação, confiança, e valiosos
conselhos e ensinamentos;
Ao Profº Paulo Cézar Rodrigues Gurjão e família, pelo incentivo no recomeço na caminhada
do ensino;
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), pelo apoio
financeiro nesses dois anos de estudo e pesquisa.
À mãe de minha filha, Lena Cláudia Lobato de Alexandria, pelo incentivo no recomeço.
À avó e avô de minha filha, Elizete Lobato de Alexandria e Paulo de Tarso de Alexandria
pelo incentivo e conselhos;
À minha filhona Joyce Alexandria, pela compreensão dos momentos ausentes;
A Profª. Drª Vânia Silva de Melo, também pela orientação e contribuição na execução desse
trabalho.
Aos meus irmãos, Izabel, Antônia, Cláudio, Maria do Carmo, José Otávio e Márcio, também,
pelo apoio e força nesta caminhada;
À pesquisadora da Embrapa Amazônia oriental, Sônia Maria Botelho, pela oportunidade em
adentrar para o universo da ciência;
Á Raphaël Marichal pela sua contribuição nas análises multivariadas o que enriqueceu este
trabalho.
Aos professores Norberto Noronha, Marcos André Piedade Gama, Gisele Barata, Herdjânia
Veras Lima, Waldiney Travassos, Francisco de Assis Oliveira, Ana Regina Araújo, pelos
ensinamentos repassados durante as disciplinas ministradas na Pós-graduação;
Aos professores e funcionários do Sistema Teorema de Ensino pelo apoio e incentivo;
Aos meus colegas e amigos da pós-graduação, Elaine Santos, Sannah Birani, Josiane Cardoso,
Josiane Castro, Augusto Pedroso, Gilson de Matos, Hérica Santos, Bruno Brabo, Erika
Freires, Alessandra Moraes, Adriane Rocha, Katiane Barros, Kaliene Carvalho, Marcos
Albuquerque, Mariana Oliveira, Andréia Saldanha, Ângela Braga, Márcia Rocha, Rosemary
da Costa, Helen Monique, Francisco.

Aos amigos Bolsistas de iniciação científica, Saime Joaquina, Possidônio Rodrigues, Gerson
barros, Samuel Cohen, Jaílson pereira, José Ailton melo Jr e Naísa Leal e Gaby, que
contribuíram diretamente para que esse trabalho fosse realizado.
À Universidade Federal Rural da Amazônia-UFRA e ao Programa de Pós-graduação em
Agronomia, com todo seu corpo docente e funcionários;
Ao Coordenador, Secretária e técnicos do Laboratório de Química do Solo da Embrapa
Amazônia Oriental, Orivan Teixeira, Maria José de Oliveira, Francisco Gomes da Silva,
Antônio Hidalgo Agostinho Monteiro, João Sérgio B. A. Silva, José Renato Figueiredo, Izaias
Chaves da Silva, Héder Pereira Ferro e Anderson Cleyton pelo apoio e incentivo.
Aos técnicos do Laboratório de Química do solo, da Universidade Federal Rural da
Amazônia-UFRA, Fábio Gomes Moura, Júlio Cezar, Amauri Guilherme, Demócrito salgado,
Samuel Moreira, pelo apoio.
Ao meu compadre e “irmão”, Márcio Roberto, que sempre esteve me incentivando e orando
para que tudo desse certo na minha caminhada.
À secretaria do curso de Mestrado em Agronomia, Gracy Kelly Monteiro, pela atenção e
dedicação em sua função, sempre ajudando quando solicitada.
E a todas as pessoas que participaram ou contribuíram para esse trabalho, mas que não foram
citados, meu muito obrigado, e que Deus abençoe a todos.
E a todos aqueles que acreditaram em mim, meu muito obrigado!!!
TUDO valeu a pena!!

Aos meus pais (in memorian),
a Joyce Alexandria, minha filha,
ao Papai do Céu,
e a todas pessoas que torceram por mim.

AGRADEÇO
À Deus
Por me dá a vida, força e esperança na caminhada neste mundo, e por me fazer chegar
até esse momento que é muito importante para minha vida.

É justamente a possibilidade de realizar um
sonho que torna a vida interessante
(De o Alquimista )

BIOGRAFIA
JOSEMAR MOREIRA VASCONCELOS, filho de José Marçal de Vasconcelos e
Yolanda Moreira Vasconcelos, nasceu no dia 05 de Novembro de 1966, na cidade de Cametá,
Estado do Pará.
Em abril de 2003, iniciou o curso de Agronomia na Universidade Federal Rural da
Amazônia, - graduando-se em novembro de 2007.
Em março de 2008, iniciou estudo ao nível de mestrado no Programa de Pós-
Graduação em Agronomia, área de concentração em Solos e Nutrição de Plantas, na
Universidade Federal Rural da Amazônia - UFRA, em Belém/PA, submetendo-se à defesa da
dissertação em agosto de 2010.

RESUMO: A avaliação da qualidade do solo é uma ferramenta importante para monitorar a
sua degradação. O objetivo deste estudo foi avaliar as alterações ocorridas nas propriedades
químicas e biológicas do solo, em diferentes profundidades, em áreas submetidas ao manejo
utilizando a queima e a não queima da vegetação, sob a influência de variação sazonal. O
estudo foi realizado no município de Igarapé-Açu (PA), nas localidades de Cumaru, Fazenda
Experimental, da Universidade Federal Rural da Amazônia, (FEIGA/UFRA) e comunidade de
Nova Olinda (SAF). O delineamento experimental foi inteiramente casualizado com quatro
repetições dos tratamentos. Amostras de um Latossolo Amarelo foram coletadas nas
profundidades de 0-5 e 5-10 cm, nos meses de janeiro/2009 (época seca), junho/2009 (época
chuvosa), e dezembro/2009 (época seca), sob diferentes tipos de cobertura vegetal: capoeira;
plantio com trituração da vegetação e plantio com queima da vegetação. Os dados foram
submetidos à análise de variância com médias comparadas pelo teste de Duncan a 5% de
probabilidade, considerando três fatores: sistemas de uso do solo, época de amostragem e
profundidade do solo. Também, foi feita a análise dos dados por meio de estatística
multivariada de componentes principais, em que foram consideradas as profundidades de
amostragem para variáveis químicas e microbiológicas e identificação dos atributos que mais
sofreram interferência do sistema de uso do solo, avaliados na amostragem de Jan/2009.
Maiores valores de pH ocorreu no SAF para as duas profundidades; o alumínio apresentou
teor mais elevado para capoeira, enquanto que o P apresentou maior valor no SAF,
apresentando diferença estatística apenas para área preparada com queima FEIGA/UFRA, na
profundidade de 0-5cm. Foram observados maiores valores de Ca, Mg, K, SB e V% na área
preparada com queima FEIGA/UFRA. Maiores teores de carbono orgânico e N total foram
encontrados na capoeira de 15 anos. Dentre as variáveis microbiológicas estudadas, o carbono
da biomassa microbiana foi encontrado em maiores valores na estação chuvosa; a maior
liberação de CO2 ocorreu no período chuvoso (capoeira), sendo que o qCO2 foi maior na
estação chuvosa, nos tratamentos com queima(T2) e trituração(T6). Dentre as variáveis
químicas e microbiológicas estudadas, CTCefetiva, SB, Ca, Mg, K, Mg e CBM:C foram as
mais sensíveis às alterações do uso do solo,
Palavras-chave: Capoeira, agricultura itinerante, atributos químicos, atributos biológicos

ABSTRACT: The evaluation of the soil quality is an important tool to monitory its
degradation. The objective of this study was to evaluate the alterations occurred in the
chemical and biological proprieties of the soil, in different depths, in areas under management
using burn and non burn of vegetation, under influence of seasonal variation. The study has
been done in Igarapé-Açú (PA), in the location called Cumaru, an experimental farm of
FEIGA/UFRA and a community called Nova Olinda. The experimental design was integrally
randomized with four treatment repetitions. Samples from oxisol have been collected in
depths at 0-5 cm and 5-10 cm, in January/2009 (dry season), June/2009 (rainy season), and
December 2009 (dry season), under different kinds of cover: post burning vegetation,
plantation with plant crunching and vegetation burning. The data has been submitted to and
variance analysis with average compared to Duncan test at 5% probability, considering three
factors: soil usage systems, the time of sampling and soil depth. The analysis have also been
made through multivariate statistics with main components, in which the sampling depths
were considered for chemical and biological variables and an identification of the attributes
which have suffered interference from the soil usage more, evaluated ate the January/2009
samplings. The highest pH values were reached at “SAF” for both depths; Aluminum has
been presented in a higher content for post burning vegetation, while phosphor presented a
higher value at “SAF”, also presenting statistic difference only in areas prepared by burning
FEIGA/UFRA, at 0-5cm. Higher values of Ca, Mg, K, SB and V% in the area prepared
through burning FEIGA/UFRA. Higher organic carbon contents and Total nitrogen have been
found at fifteen years “post burning vegetation”. Among the microbiological variables, the
microbial biomass carbon has been found in higher values during rainy seasons; the highest
CO2 release has occurred in the rainy period (post burning vegetation), as long as CO2 was
bigger in within the rainy season, in treatments involving burning (T2) and crunching (T6).
Among the chemical variables studied, effective CTC, SB, Ca, Mg, K, Mg and CBM:C have
been the most sensible in relation to the alterations in soil usage.
Keywords: post burning vegetation, itinerant agriculture, chemical attributes, biological
attributes.

SUMÁRIO
RESUMO..................................................................................................... 12
ABSTRACT................................................................................................. 13
1 INTRODUÇÃO........................................................................................... 20
1.1 Objetivo Geral............................................................................................. 23
2 REVISÃO DA LITERATURA.................................................................. 24
2.1 VEGETAÇÃO SECUNDÁRIA (CAPOEIRA) E O SEU
APROVEITAMENTO .................................................................................
24
2.2 SISTEMAS AGROFLORESTAIS .............................................................. 25
2.3 AGRICULTURA ITINERANTE OU MIGRATÓRIA................................ 28
2.4 INDICADORES DE SUSTENTABILIDADE AMBIENTAL.................... 29
2.5 INDICADORES MICROBIOLÓGICOS..................................................... 31
2.6 VARIABILIDADE SAZONAL NOS ATRIBUTOS QUÍMICOS E
BIOLÓGICOS DO SOLO............................................................................
33
3 MATERIAL E MÉTODOS ....................................................................... 35
3.1 CARACTERIZAÇÃO DA ÁREA DE ESTUDO........................................ 35
3.1.1 Localização da área experimental............................................................. 35
3.1.2 Clima ........................................................................................................... 36
3.1.3 Solo ............................................................................................................... 38
3.1.4 Vegetação .................................................................................................... 38
3.2 DELINEAMENTO EXP, TRAT. E VARIÁVEIS ESTUDADOS............ 39
3.3 HISTÓRICO DE USO DAS ÁREAS EXPERIMENTAIS ........................ 42
3.4 AMOSTRAGEM......................................................................................... 45
3.5 Análises Químicas....................................................................................... 46
3.6 Análises biológicas...................................................................................... 46
3.7 Análises Estatística..................................................................................... 49
4 RESULTADO E DISCUSSÃO ................................................................. 51
4.1 ATRIBUTOS QUÍMICOS DOS SOLOS SOB DIFERENTES
SISTEMAS DE MANEJO............................................................................
51
4.2 ATRIBUTOS BIOLÓGICOS DOS SOLOS SOB DIFERENTES SISTEMAS
DE MANEJO.....................................................................................................
61
4.2.1 Carbono da biomassa microbiana (CBM).................................................... 61

4.2.1.1 Interação sistema de manejo versus profundidade do solo.............................. 63
4.2.1.2 Interação época de coleta versus profundidade do solo.................................... 65
4.2.2. Relação Carbono da biomassa microbiana/carbono orgânico (CBM:C)
Jan/2009............................................................................................................
66
4.2.3 Respiração Basal.............................................................................................. 68
4.2.3.1 Interação sistema de manejo versus profundidade do solo................................ 69
4.2.3.2 Interação época de coleta versus profundidade do solo..................................... 70
4.2.4 Quociente metabólico (qCO2)......................................................................... 72
4.2.4.1 Interação sistema de manejo versus profundidade do solo ............................... 73
4.2.4.2 Interação época de coleta versus profundidade do solo..................................... 75
4.3 ANÁLISE MULTIVARIADA PARA OS ATRIBUTOS QUÍMICOS E
BIOLÓGICOS ...................................................................................................
75
5 CONCLUSÕES ............................................................................................... 83
6 REFERÊNCIAS BIBBLIOGRÁFICAS ....................................................... 84
ANEXOS........................................................................................................... 103

LISTA DE TABELAS
Tabela 1 Atributos químicos do solo sob diferentes sistemas de manejo em localidades
de Igarapé–Açú (PA). Jan/2009. Os valores são médias de quatro repetições
por sistema de manejo .......................................................................................... 54
Tabela 2 Carbono orgânico (C), nitrogênio total (N) e relação carbono:nitrogênio (C/N)
de Latossolo Amarelo, sob diferentes sistemas de manejo e uso do solo em
Igarapé-Açú (PA). Janeiro de 2009; média de quatro repetições.........................
59
Tabela 3 Carbono da biomassa microbiana (CBM) de Latossolo Amarelo, sob diferentes
sistemas de manejo e uso, de Igarapé-Açu (PA), coletado em diferentes épocas.
Média de quatro repetições ................................................................................... 62
Tabela 4 Carbono da biomassa microbiana (CBM) de Latossolo Amarelo, sob diferentes
sistemas de manejo e uso, de Igarapé-Açu (PA), coletado em diferentes
profundidades. Média de quatro repetições ..........................................................
64
Tabela 5 Carbono da biomassa microbiana (CBM) de Latossolo Amarelo de Igarapé-
Açu (PA), coletado em diferentes épocas e profundidades. Média de quatro
repetições .............................................................................................................. 65
Tabela 6 Respiração basal (CO2) da biomassa microbiana de Latossolo Amarelo de
Igarapé-Açu (PA), sob diferentes sistemas de manejo e uso da terra, coletado
em diferentes épocas. Média de quatro repetições ............................................... 68
Tabela 7 Respiração basal (CO2) da biomassa microbiana de Latossolo Amarelo de
Igarapé-Açu (PA), interação sistemas de manejo vs. profundidade do solo.
Média de quatro repetições ................................................................................... 70
Tabela 8 Respiração basal (CO2) da biomassa microbiana de Latossolo Amarelo de
Igarapé-Açu (PA), coletado em diferentes épocas e profundidades. Média de
quatro repetições ................................................................................................... 71
Tabela 9 Quociente metabólico (qCO2) da biomassa microbiana de Latossolo Amarelo
de Igarapé-Açu (PA), coletado sob diferentes sistemas de manejo e uso do solo
e em diferentes épocas. Média de quatro repetições ............................................ 72
Tabela 10 Quociente metabólico (qCO2) da biomassa microbiana de Latossolo Amarelo
de Igarapé-Açu (PA), coletado sob diferentes sistemas de manejo e uso do solo
e diferentes profundidades. Média de quatro repetições 74
Tabela 11 Quociente metabólico (qCO2) da biomassa microbiana de Latossolo Amarelo
de Igarapé-Açu (PA), coletado sob diferentes sistemas de manejo e uso do solo
e diferentes profundidades. Média de quatro repetições ...................................... 75

LISTA DE FIGURAS
Figura 1 Localização da área de estudo no município de Igarapé-Açú,
mesorregião do nordeste paraense........................................................ 35
Figura 2 Distribuição da precipitação pluviométrica média mensal acumulada,
medida no município de Igarapé-Açú, Pará, Jan/dez (2008) e Jan/Dez
(2009)............................................................................................... 37
Figura 3 Distribuição da precipitação pluviométrica média mensal em Igarapé-
Açú, Pará. Período: 1994-2009................................................................. 37
Figura 4 Trituração da capoeira utilizador o trator (Tritucap) nas áreas
experimentais do projeto Tipitamba, município de Igarapé-Açú (PA)...... 40
Figura 5 Fotos das áreas experimentais com diferentes tipos de cobertura:
capoeira, triturada, queimada e sob sistema agroflorestal, município de
Igarapé-Açú (PA) ..............................................................................
41
Figura 6 Localização da área de estudo na comunidade de Cumaru, município de
Igarapé-Açú (PA). .................................................................................... 43
Figura 7 Localização da área de estudo na UFRA/FEIGA, município de Igarapé-
Açú (PA) .................................................................................................. 44
Figura 8 Foto da trincheira utilizada para coletar as amostras nas camadas de 0-5,
5-10 cm, município de Igarapé-Açu (PA) ................................................ 45
Figura 9 Detalhe das amostras fumigadas em dessecador para determinação do
carbono da biomassa microbiana (CBM) .................................................. 47
Figura 10 Detalhe das amostras incubadas para determinação da respiração basal.... 48
Figura 11
Efeito dos tratamentos na relação carbono da biomassa microbiana e carbono
orgânico de solo de Latossolo Amarelo Município de Igarapé-Açú (PA)....... 66
Figura12 Efeito dos tratamentos na relação carbono da biomassa microbiana e carbono
orgânico de Latossolo Amarelo Município de Igarapé-Açú (PA)................. 67
Figura 13 Análise de Componentes Principais (PCA) realizada para as
propriedades químicas de um Latossolo Amarelo de Igarapé-Açu (PA),
sob diferentes sistemas manejo e uso do solo, coletado em janeiro de
2009, na profundidade de 0-5 cm. Média de quatro repetições para cada
atributo............................................................................................... 76

Figura 14 Análise de Componentes Principais (PCA) realizada para as
propriedades biológicas de um Latossolo Amarelo de Igarapé-Açu (PA),
sob diferentes sistemas de manejo e uso do solo, coletado em janeiro de
2009, na profundidade de 0-5 cm. Média de quatro repetições para cada
atributo...................................................................................... 78
Figura 15 Análise de Componentes Principais (PCA) realizada para as
propriedades químicas de um Latossolo Amarelo de Igarapé-Açu (PA),
sob diferentes sistemas manejo e uso do solo, coletado em janeiro de
2009, na profundidade de 5-10 cm. Média de quatro repetições para cada
atributo.............................................................................................. 79
Figura 16 Análise de Componentes Principais (PCA) realizada para as
propriedades biológicas de um Latossolo Amarelo de Igarapé-Açu (PA),
sob diferentes sistemas de manejo e uso do solo, coletado em janeiro de
2009, na profundidade de 5-10 cm. Média de quatro repetições para cada
atributo...................................................................................... 80
Figura 17 Análise de Componentes Principais (PCA) realizada para as
propriedades químicas e biológicas de um Latossolo Amarelo de
Igarapé-Açu (PA), sob diferentes sistemas manejo e uso do solo,
coletado em janeiro de 2009, na profundidade de 0-5
cm..................................................................................................... 81

LISTA DE QUADROS
Quadro 1 Especificidades das áreas em estudos................................................... 42
Quadro 2 Especificidades das áreas em estudos................................................. 44
Quadro 3 Especificidades das áreas em estudos....................................................... 45

20
1 INTRODUÇÃO
O Brasil é o país que possui a maior área de floreta tropical do mundo, segundo
informativo da FAO (2005). Esse ambiente abriga uma infinidade de funções e serviços
necessários e úteis à vida no planeta. Por conseguinte, a sua degradação provoca danos
incalculáveis, tais como perda de biodiversidade, alteração nos ciclos hidrológicos do
solo, da atmosfera e alterações ecológicas, além de atingir milhões de pessoas no
mundo, que têm suas vidas diretamente ligadas aos recursos florestais, onde vivem e de
onde retiram o necessário para sobreviverem (COLFER, et al., 2001; FAO, 2005).
Na Amazônia brasileira estima-se que mais de 50 milhões de hectares do que era
floresta tropical está convertida em outros usos ou são vegetações secundárias em vários
estágios de desenvolvimento, incluindo florestas secundárias e florestas degradadas
(OIMT, 2002; FEARNSIDE, 2005).
A Microrregião Homogênea Bragantina é uma das mais antigas a sofrer a ação
antrópica na Amazônia Oriental, sendo a primeira área do estado do Pará a ser
colonizada maciçamente. Sua ocupação iniciou-se por volta de 1891 com a abertura da
ferrovia Belém-Bragança, cuja finalidade era promover a colonização da área e
assegurar o transporte de produtos agrícolas da região para Belém (DENICH, 1991),
destinados a abastecer de alimentos os grandes contingentes populacionais atraídos para
a produção de borracha, sendo as cidades de Belém e Manaus os principais centros
comerciais e de serviços da região, as quais passavam por grande desenvolvimento em
função desse produto.
Atualmente, a região é quase totalmente destituída de floresta primitiva,
observando-se vegetação secundária, conhecida também como capoeira, em vários
estágios de desenvolvimento e algumas áreas degradadas. Em estudos de paisagem
realizados nos municípios de Nova Timboteua, Peixe-Boi e Capanema, do Nordeste
Paraense, foram demonstrados que havia apenas 15% da cobertura original da floresta
(VIEIRA; SALOMÃO; ROSA, 1996), enquanto que no município de Igarapé-Açu foi
identificado apenas 5,3% da floresta primitiva remanescente (WATRIN;
VENTURIERI; SAMPAIO, 1998).
No município de Igarapé-Açu, a agricultura familiar, como em toda região
amazônica, baseia-se no sistema de derruba e queima da capoeira (vegetação

21
secundária) para o preparo das áreas a serem cultivadas. Segundo Kato et al, (1999) os
produtores cultivam a área durante dois anos em média, então a área entra novamente
em pousio para posterior crescimento e regeneração da capoeira.
Com o uso deste sistema de preparo da área ocorre à perda da biomassa e
alterações no ciclo do carbono no ecossistema como um todo, haja vista a diminuição da
produtividade ao longo do tempo.
A queima contribui para emissão de dióxido de carbono (CO2) para atmosfera,
elevando assim o efeito estufa e, conseqüentemente, o aquecimento do planeta, Pode,
também, acarretar a perda de nutrientes do solo na forma de gases para atmosfera, ou a
remoção física pelo vento e enxurradas (HÖLSCHER et al., 1997), levando ao
empobrecimento do solo, podendo este ser degradado ainda, pela sua exposição a raios
solares e as chuvas intensas.
Na perspectiva de melhorar o uso da terra na região, a Embrapa Amazônia
Oriental desenvolve e repassa a agricultores diversas tecnologias que contribuem com o
seqüestro de carbono e o aumento da fertilidade desses solos degradados pela
agricultura itinerante. Entre as tecnologias está o plantio direto na capoeira (Projeto
Tipitamba), que propõe a substituição do fogo pela trituração da capoeira – vegetação
secundária ou de pousio, usada no processo tradicional de derruba e queima no preparo
de áreas para a agricultura; evitando assim a emissão de carbono para a atmosfera. Esse
sistema completa-se com o enriquecimento da capoeira com espécies leguminosas de
rápido crescimento que depositam nutrientes no solo, aumentam a biomassa da
vegetação e retiram o carbono presente na atmosfera.
Com a trituração da capoeira, em substituição ao uso do fogo, os resíduos
orgânicos de origem vegetal, tanto protegem o solo, como são fonte de materiais
orgânicos que permanecem no sistema, contribuindo assim para um melhor equilíbrio
nutricional e, também, aumentando a diversidade biológica desses solos. Nesse sistema
toda a biomassa aérea da vegetação secundária retorna como fonte de matéria orgânica
para o solo. A quantidade dessa biomassa produzida varia depende de sua idade, o
sistema de uso da terra e da intensificação do uso da terra, podendo variar, segundo
Kato et al. (2006), de 8 ton/ha-1
(capoeira de um ano) a 90 ton/ha-1
(capoeira de 10
anos).
A conservação da biodiversidade, a dinâmica de água e nutrientes (manutenção
das taxas de recarga dos estoques de água subterrânea, manutenção das vazões de

22
igarapés e rios e redução de impactos nos sistemas aquáticos ocasionados pela
lixiviação de nutrientes) são contribuições já comprovadas pela prática da trituração de
corte sem queima, preconizada pelo projeto Tipitamba.
Neste contexto, o tipo de vegetação e as condições ambientais são fatores que
determinam a quantidade e a qualidade do material que será depositado no solo. As
decomposições destes materias dependem dos processos de transformação da matéria
orgânica pelos microrganismos do solo, por meio dos quais se podem avaliar qualidade
do solo (MIELNICZUK et al., 2003). Assim, a matéria orgânica do solo é o mais
importante reservatório de energia e fonte de carbono para os microrganismos e também
de nutrientes para os vegetais.
A matéria orgânica do solo contribui de forma benéfica para o estabelecimento
vegetal afetando propriedades físicas, químicas e biológicas do solo (STEVENSON,
1982). O aporte de matéria orgânica no solo contribui para o aumento de estabilidade de
agregados, melhora a aeração, a drenagem e a capacidade de retenção da solução do
solo. Entre os atributos químicos, a matéria orgânica é um estoque de nutrientes
disponível para a mineralização, além de aumentar a capacidade de troca de catiônica do
solo. Com isso a matéria orgânica serve para avaliar a preservação dos ecossistemas
naturais a e os desequilíbrios dos agroecossistemas. Portanto, a sustentabilidade dos
agroecossistemas dessa região depende, principalmente, da ciclagem de nutrientes
contidos na matéria orgânica do solo.
A biomassa microbiana, não fornece indicação sobre os níveis de atividade das
populações microbianas do solo, contudo, é importante para avaliar outros parâmetros, a
como a respiração basal e a atividade enzimática, que servem para estimar o estado
metabólico das comunidades de microrganismos do solo (NANNIPIERI; MUCCINI;
CIARDI, 1983). A respiração basal, é um indicador ecológico de ciclagem de
nutrientes, visto que quanto mais elevada for à atividade dos microorganismos, maior
será a liberação de nutrientes e a mineralização da matéria orgânica (ANDERSON,
1982; PARKINSON; COLEMAN, 1991) o quociente metabólico (qCO2) é outro
parâmetro que pode ser utilizado para melhor entendimento da atividade da biomassa
microbiana do solo, pois uma biomassa microbiana eficiente perde menos carbono
como CO2 pela respiração e incorpora maior proporção de carbono ao tecido
(ANDERSON; DOMSCH, 1985; SANTRUCKOVA; STRASKRABA, 1991).

23
Portanto a biomassa microbiana, a respiração basal e o quociente metabólico são
indicadores biológicos da qualidade do solo. E são parâmetros que avaliam as mudanças
que ocorrem em um sistema alterado por ações antrópicas (sistema de uso do solo), e
variações sazonais de umidade e temperatura.
Este trabalho tem como objetivo avaliar os atributos químicos (pH, C orgânico, N
total, P disponível, K, Ca, Mg, Na e Al trocáveis, H titulável, CTC, soma de bases,
porcentagem de bases e porcentagem de alumínio) e biológicas (C da biomassa
microbiana, respiração basal, quociente metabólico e relação C biomassa microbiana:C
orgânico) de um Latossolo Amarelo do município de Igarapé-Açu, sob sistemas de
manejo envolvendo plantios com queima da vegetação, sem queima e com trituração da
vegetação, e sistema agroflorestal (SAF), em duas profundidades (0-5 e 5-10cm) e três
períodos de amostragem (seco-chuvoso-seco), e compará-los entre si e com as mesmas
variáveis em condições de vegetação secundária (capoeira), a fim de determinar se o
sistema de preparo do solo sem queima e com trituração da vegetação, se configura
como uma alternativa viável de manejo sustentado, capaz de promover a regeneração de
áreas exauridas pelo uso inadequado.
1.1 OBJETIVO GERAL
Avaliar o efeito do sistema de preparo do solo usando a não queima da
vegetação sobre as propriedades químicas e biológicas de solos submetidos a
diferentes tipos de sistemas de manejos no nordeste paraense.

24
2 REVISÃO DE LITERATURA
2.1 VEGETAÇÃO SECUNDÁRIA (CAPOEIRA) E O SEU APROVEITAMENTO
A floresta secundária (FS) é um tipo de vegetação que aparece após a retirada ou
modificação drástica na composição e estrutura da vegetação primária, seja pela ação
humana ou por fenômenos naturais (COMISION NACIONAL DE CERCITICACIÓN
FORESTAL, 1999; CHOKKALINGAM; DE JONG, 2001). Porém, existem outras
definições nas quais se utilizam diferentes critérios de análise (CARAZO et al, 1997).
Para efeito deste estudo, FS foi considerada aquela vegetação surgida após corte raso da
floresta primária, num sistema de uso da terra predominante na Amazônia brasileira
(agricultura de corte-e-queima).
No pousio, a vegetação secundária (capoeira) se refaz por meio de rebrotas de
tocos, raízes e sementes, principalmente aquelas que sobrevivem ao corte e à queimada.
As taxas de rotação exigem períodos de pousio longos de modo que a nova vegetação
recomposta possa contar, pelo menos parcialmente, com a diversidade florística, a
ciclagem de água e nutrientes (HÖLSCHER et al., 1997 a, b; SOMMER et al., 2004),
proporcionando, ao mesmo tempo, o acúmulo de carbono e nutrientes na sua biomassa
(DENICH, 1991; DENICH et al., 1999; TIPPMANN; DENICH; VIELHAUER, 2000),
reciclagem e recuperação de nutrientes de camadas profundas do solo (SOMMER,
2000), controle da erosão (HOANG FAGERSTROM et al., 2002; MACDONALD et
al., 2002), supressão de plantas invasoras (ROUW, 1995; GALLAGHER et al., 1999),
suprimento de madeira e lenha (SANCHEZ, 1995) e manutenção da biodiversidade
(BAAR, 1997).
Nesses solos a eficiência da reciclagem de nutrientes aumenta em relação aos
que sofrem corte-e-queima. Nesses ecossistemas tanto o dossel das árvores quanto a
espessa liteira protegem o solo da erosividade das chuvas, possibilitando fluxo de
nutriente mais rápido e ciclagem mais eficiente. Portanto, esse sistema funciona como
importante reservatório de nutrientes para as plantas, regulando as transformações da
matéria orgânica, além de atuar na manutenção da estrutura do solo, dando
sustentabilidade para os ecossistemas.

25
Para a agricultura, os principais efeitos negativos da queima da vegetação
durante a fase de preparo de área para o plantio nos sistemas de produção são as perdas
de nutrientes retidos na biomassa da vegetação que atingem valores de 96% do
nitrogênio, 47% do potássio, 35% do cálcio, 40% do magnésio e 76% do enxofre,
comprometendo a sustentabilidade do sistema de produção da agricultura familiar
(EMBRAPA, 2001).
Há, portanto, a possibilidade do manejo de vegetação secundária com o objetivo
de torná-las mais produtivas, o que é possível com a adaptação dos princípios técnicos
para esse tipo de vegetação. Com a existência de florestas secundárias produtivas,
possivelmente haverá uma menor pressão sobre as florestas primárias e poderá
contribuir com a permanência de famílias de agricultores no campo (DENICH, 1991;
FINEGAN, 1992).
2.2 SISTEMAS AGROFLORESTAIS
São sistemas que já foram praticados há milênios, tanto na Ásia, como na
América Latina, mas essa ciência desenvolveu-se mais intensamente nas décadas de 80
e 90. Sua abrangência é muito grande, tem sido adotado com sucesso em diversos
ambientes biofísicos e sócioecônomicos, desde regiões de clima úmido, semi-árido ou
temperado e sistemas de baixo nível tecnológico e uso de insumos à alta tecnologia,
tanto em pequenas como em grandes áreas de produção, áreas degradadas ou de alto
potencial produtivo (Nair, 1989).
Em 1977 foi criado o Centro Internacional de Pesquisa Agroflorestal (ICRAF),
em Nairobi, Kenya. Por volta de 1983, o agricultor e pesquisador suíço Ernst Gotsch
iniciou seus trabalhos no Brasil. A REBRAF, Instituto Rede Brasileira Agroflorestal
que promove a adoção no Brasil de alternativas agroflorestais, foi criada em 1990.
Existe vários conceito de sistemas agroflorestais como: plantar árvores em
fazendas para melhorar a qualidade de vida dos agricultores pobres e proteger os
recursos naturais, transmitida ao público pelo ICRAF (2000); bem como outras mais

26
ricas e complexas percepções do sistema de vida; consórcios de culturas agrícolas com
espécies arbóreas que recuperam aspectos dos ecossistemas florestais, como a estrutura
da cobertura vegetal e a biodiversidade, restabelecendo funções ecológicas como a
ciclagem de nutrientes e a proteção do solo (MACDICKEN; VERGARA, 1990);
formas de uso e manejo dos recursos naturais nas quais espécies lenhosas (árvores,
arbustos, palmeiras) são utilizadas em associação deliberada com cultivos agrícolas ou
com animais no mesmo terreno, de maneira simultânea ou em seqüência temporal (
OTS; CATIE, 1986).
Outros autores concordam com esta prática, citando que os benefícios dos
sistemas agroflorestais são evidenciados pela diversidade de formas de uso da terra,
onde árvores e arbustos são cultivados de forma interativa com cultivos agrícolas,
pastagens e/ou animais, com múltiplas finalidades, constituindo-se numa opção viável
de manejo sustentável da terra (RIBASKI; MONTOYA; RODIGHERI, 2002).
Os sistemas agroflorestais classificam-se em sistemas silvipastoris (animais e
árvores ou arbustos); agrossilviculturais (plantas anuais e árvores ou arbustos) e
agrossilvipastoris (animais, plantas anuais e árvores ou arbustos). Podem ainda ser
classificados de acordo com sua estrutura no espaço, seu desempenho ao longo do
tempo, a importância relativa, a função dos diferentes componentes, seus objetivos de
produção e com as características sociais e econômicas que prevalecem.
Os modelos de SAF‟s mais importantes, segundo (LEITE; VIRGENS,
2002) são: consórcio com o cultivo perene que é o resultado de arranjo espacial de
espécies em caráter permanente, com benefícios mútuos e ainda com melhor proteção
dos solos, ciclagem de nutrientes, aumento da receita, melhor aproveitamento da área,
de insumos e da mão-de-obra; sistema em aléia ou alley cropping caracterizados pelo
plantio de espécies alimentares de ciclo curto entre ruas ou fileiras de árvores, arbustos
ou palmeiras em caráter permanente; cultivo de encosta ou taungya que corresponde ao
cultivo de espécies alimentares e anuais, conjuntamente com espécies florestais ou
arbóreas, durante os primeiros anos de estabelecimento destas, visando à recomposição
de encostas degradadas ou em vias de degradação; sistema silvopastoril consistindo no
consórcio de espécies lenhosas ou palmáceas com pastagem e animais, com finalidade
de produzir sombra, alimento para os animais, banco de proteínas, melhorar a fertilidade
do solo e madeira; sistemas agrossilvipastoris que além das espécies e finalidades

27
contidas no sistema silvopastoril, emprega arbustos e plantas herbáceas, também, com a
finalidade de produção.
Os componentes arbóreos adicionados nesse sistema incrementam a entrada
de matéria orgânica no solo que beneficia as características físicas, químicas e
biológicas do solo, conduzindo o sistema para uma situação mais sustentável. O
aumento da diversidade vegetal contribui para uma maior diversidade comunidade
microbiológica e da fauna do solo que atuam como agentes de controle biológico e
condicionadores de solo (YOUNG, 1994).
Além do caráter conservacionista, esses sistemas visam uma produção
contínua e diversificada, aumentando a produção e a renda do agricultor, além de
reduzir o risco de prejuízo na perda de uma cultura (MACDICKEN; VERGARA,
1990). Os sistemas agroflorestais são uma prática indicada para a agricultura familiar
devido à grande demanda de mão de obra. A diversificação da produção, junto com o
cuidado de manter o manejo orgânico, é um importante passo para a segurança e a
soberania alimentar no meio rural. Devido aos benefícios promovidos por sistemas
agroflorestais no solo, essa prática é indicada para a recuperação de áreas degradadas e
para a preservação de áreas de entorno de reservas ambientais (REINERT, 1998;
MENDONÇA; LEITE; FERREIRA NETO, 2001).
Para a Amazônia, alguns modelos de SAF podem ser considerados
promissores, dentre os quais se destacam: o sistema taungay modificado, que poderia
ser utilizado no reflorestamento de áreas de terras firmes degradadas (EMBRAPA,
2005); o pousio melhorado, com adoção de árvores leguminosas fixadoras de nitrogênio
ou espécies perenes regionais de valor comercial, que dessem rápido retorno
econômico, como as fruteiras; o cultivo perenes em faixas, que poderiam ser
empregados em áreas declivosas, objetivando minimizar a erosão e controlar as
invasoras através do fornecimento de "mulch" (cobertura morta); os sistemas
multiestratos, que seriam a associação de fruteiras e espécies florestais, com a
introdução de cobertura e, ou componente animal dentro do sistema; os sistemas
silvipastoris, através do estabelecimento de espécies madeiráveis em pastos, que
aumentaria o retorno econômico em médio prazo e justificaria incentivos a curto prazo
para o melhoramento de pastagens e; cercas vivas, que poderiam ser utilizadas em
substituição às cercas tradicionais, provavelmente, com impacto positivo sobre o

28
desmatamento. Acrescentando que a pesquisa nessa área é fundamental para que se
possa acelerar o desenvolvimento e extensão de tecnologias agroflorestais apropriadas.
Portanto com incremento da diversidade vegetal espera-se que disponibilize substratos
com características bastante variadas, diversificadas também ao longo do tempo, o que
estimularia a maior diversidade de espécies na microbiota do solo, aumentando a
magnitude dos serviços ambientais disponíveis nesses sistemas (BALIEIRO et al.,
2004; NICODEMO; PRIMAVESI, 2007; DUBOC, 2009).
2.3 AGRICULTURA ITINERANTE OU MIGRATÓRIA
A agricultura itinerante de derruba-e-queima é a forma de uso de terra mais
importante nos trópicos em geral e mais ainda nos trópicos úmidos. A sua importância
socioeconômica é central, pois ela sustenta uns 300-500 milhões de pessoas e representa
cerca de 30% da área agrícola do mundo ou 40% da área agrícola dos trópicos (BRADY
1996). Somente na Amazônia Brasileira, esta forma de uso da terra afeta mais de 10
milhões de hectares, sustenta meio milhão de famílias e providencia cerca de 80% da
produção alimentar da região (SERRÃO, 1995).
Na Amazônia, é um dos principais sistemas de uso da terra, tanto pelo ponto de
vista econômico como também pela quantidade de pessoas envolvidas nesse processo
de produção agrícola. Segundo Costa (2003) esse é um sistema tradicional de
agricultura desenvolvido e praticado em quase toda região por, pelo menos, 600 mil
pequenos agricultores, produzindo principalmente, feijão, mandioca, arroz, milho,
malva, juta, frutas e algodão, entre outros produtos.
Vielhauer e Sá (2000) citam que devido a este tipo de agricultura na região, foi
estabelecendo-se um equilíbrio entre o uso agrícola e a vegetação secundária, chamada
de capoeira, que se manteve por algumas dezenas de anos, até a aparência óbvia da
degradação dos solos inadequadamente manejados. Todavia, a região nordeste paraense
vem sofrendo essa prática de derruba-e-queima, a mais antiga na área de exploração
agrícola, desde 1875, devendo-se ao processo de colonização, que objetivou aumentar e

29
abastecer de produtos agrícolas a cidade de Belém, capital do estado do Pará e região,
incentivado pelo governo da época (CARDOSO JÚNIOR, 2001).
Para o agricultor familiar, com baixo grau de capitalização e pouco acesso a
outra alternativa, o preparo de área para plantio com queima ainda se constitui no
instrumento mais eficaz ao seu alcance por ser um processo menos oneroso, que
promove a fertilização imediata e gratuita do solo e obtém produções para sua
subsistência (KATO; KATO, 2000).
O lado negativo e degradante da agricultura itinerante é intimamente associado
aos efeitos deletérios da queima (KLEINMAN; PIMENTEL; BRYANT, 1995).
Benéfica no curto prazo como acima delineado, a queima causa perdas pesadas de
matéria orgânica e de nutrientes, notavelmente dos nutrientes mais voláteis como N e S
(MACKENSEN et al., 1996; HÖLSCHER et al., 1997). As emissões de CO2 e de N2O
contribuem significantemente para o „efeito estufa‟ (HOUGHTON et al., 2000;
GRACE, 2004). No Brasil, as mudanças de uso dos solos e as queimadas são
responsáveis por 70% das emissões de CO2 para a atmosfera (BORN; PICCHIONI;
PIVA, 2007), contribuindo mais decisivamente para a produção de gases de efeito
estufa do que a queima de combustíveis fósseis. O fogo também reduz a biodiversidade,
elimina muitas espécies florestais não adaptadas ao fogo, e causa a dominância e
persistência de espécies ruderais agressivas e bem adaptadas a estas condições, como
Imperata brasiliensis e Paspalum melanospermum (gramíneas), Pteridium aquilinum
(samambaia) ou Orbygnia phalerata (palmeira babaçu).
2.4 INDICADORES DE SUSTENTABILIDADE AMBIENTAL
O solo em seu estado natural encontra-se coberto pela vegetação, que o protege
da erosão e contribui para manter o equilíbrio entre os fatores de sua formação e aqueles
que provocam sua degradação. O rompimento dessa relação provoca alterações físicas,
químicas e biológicas, as quais se não forem adequadamente monitoradas e controladas,
levam à queda de produtividade e à degradação do ecossistema (SIQUEIRA et al.,
1994).

30
Atualmente, são desenvolvidas práticas agrícolas que protegem o ecossistema
da degradação, melhorando sua sustentabilidade e, por conseguinte, a qualidade do solo,
configurados por sistemas de manejo caracterizados pela considerável quantidade de
resíduos vegetais sobre ou próximo à superfície, protegendo contra a erosão e
minimizando as necessidades do solo para o plantio (BRADY; WEIL, 2002).
O manejo conhecido como plantio direto é caracterizado pelo não-
revolvimento do solo e pela manutenção de restos culturais sobre a superfície. Ele é
considerado um sistema de manejo mais adequado às condições tropicais e subtropicais,
com reflexos diretos sobre a redução de custos de produção e aumento da rentabilidade,
além de promover a manutenção ou, até mesmo, o aumento da matéria orgânica.
Roldán et al. (2007) verificaram que o sistema plantio direto pode ser
considerado eficiente para alcançar a agricultura sustentável em condições subtropicais,
devido ao aumento da qualidade física e bioquímica do solo e ao seqüestro de carbono.
Estudos têm mostrado que a adoção do sistema de plantio direto afeta as propriedades
físicas (KARLEN et al., 1994; KUSHWAHA; TRIPATHI;SINGH, 2001; PEIXOTO et
al., 2006), químicas (DORAN, 1980; ANGERS et al., 1993; EKENLER; TABATABAI,
2003) e biológicas (ALVAREZ et al., 1998; MATSUOKA; MENDES; LOUREIRO,
2003; FRANCHINI et al., 2007) do solo, sendo que, na maioria das vezes, as
modificações são favoráveis (BRADY; WEIL, 2002).
Com a trituração da vegetação e, com a decomposição da mesma, o solo recebe
todos esses benefícios através de resíduos orgânicos incorporados; impedindo também a
emissão de dióxido de carbono e aumentando a produtividade. Na fase inicial há a
imobilização de nutrientes pelos microorganismos durante a decomposição da camada
de cobertura morta provenientes da trituração da capoeira (Kato et al., 1999b). Portanto,
a ciclagem de nutrientes ocorre como conseqüência da atividade microbiana e é,
especialmente, importante em ecossistemas de baixa fertilidade. Segundo Sanchez
(1976) cerca de 80% dos solos dos trópicos úmidos são considerados de baixa
fertilidade.

31
2.5 INDICADORES MICROBIOLÓGICOS
Nos solos tropicais, a matéria orgânica tem papel central na manutenção de sua
fertilidade. Com isso a ciclagem de nutrientes nesses solos ocorre como conseqüência
da atividade microbiana e é especialmente importante em ecossistemas de baixa
fertilidade. A biomassa microbiana do solo é responsável pela decomposição da matéria
orgânica e pela ciclagem de nutrientes. É fonte de nutrientes e ao mesmo tempo os
imobiliza, tornando-os menos imediatamente acessíveis para as plantas e impedindo sua
lixiviação do solo (CARDOSO, 2004).
Além de atuar na ciclagem de nutrientes a microbiota do solo atua, também, na
formação e estabilização de agregados, sendo considerada bioindicadora da qualidade
do solo (SILVEIRA et al., 2006; VARGAS; SCHOLLES, 2000). As comunidades
biológicas do solo, compostas de microrganismos e de fauna, ocorrem em ordens de
grandeza várias vezes maiores que as comunidades encontradas sobre sua superfície
(BARDGETT, 2002).
As bactérias e fungos são os integrantes predominantes da biota do solo
(JASTROW; AMONETTE; BAILEY, 2007). A capacidade de responder rapidamente a
mudanças no ambiente do solo derivadas das alterações no manejo justifica o uso de
microrganismos e processos microbiológicos como indicadores de qualidade do solo
(KENNEDY; PAPENDICK, 1995).
A qualidade dos resíduos influencia a taxa de decomposição e a composição da
comunidade microbiana (GUPTA; GERMIDA, 1988; CATTELAN; VIDOR, 1990;
ANGERS et al., 1993). Portanto, a decomposição da manta orgânica é uma das etapas
mais importantes na ciclagem de nutrientes e é um processo essencialmente biológico
(CORREA; ANDRADE, 1999). Dentre os organismos responsáveis por esse processo
destacam-se os microrganismos que compõem a biomassa microbiana.
A biomassa microbiana é definida como a parte viva da matéria orgânica do
solo, incluindo bactérias, actinomicetos, fungos, protozoários, algas e macrofauna.
Excluindo-se raízes de plantas e animais do solo maiores do que 5,10 μm, a biomassa
microbiana corresponde em média, de 2 a 5% do C orgânico do solo (JENKINSON;
LADD, 1981) e de 1 a 5% do N total do solo (SMITH; PAUL, 1990). Portanto, pode ser
um parâmetro fundamental para o estudo de ciclagem de nutrientes em diferentes

32
ecossistemas. Assim sendo, estimativa da biomassa microbiana pode fornecer
informações úteis sobre a disponibilidade de nutrientes imobilizados, assim como, no
entendimento do funcionamento desses ecossistemas e na qualidade do solo, pois ela
reage rapidamente às perturbações sofridas decorrentes do manejo e do uso do solo.
A biomassa microbiana é estreitamente influenciada pelo tipo de utilização do
solo, e a relevância de sua quantificação se deve, portanto, à conveniência de usá-la
como índice mais sensível às mudanças ocorridas no sistema, comparado aos conteúdos
de C orgânico e N total (GAMA-RODRIGUES et al., 2005). A biomassa microbiana
nunca deve ser analisada isoladamente como uma única maneira de se estimar a
situação das populações de microrganismos, devendo ser analisada juntamente com a
atividade dos microrganismos (respiração basal), nas mesmas amostras de solo, face à
extrema heterogeneidade do ambiente natural da microbiota e da sua biodiversidade
(GRISI, 1996).
Os microrganismos do solo são os principais componentes do sistema de
decomposição de matéria orgânica, sendo os principais contribuintes para a respiração
basal do solo, atuando como reguladores do ciclo de nutrientes e, conseqüentemente, da
produção primária e do fluxo de energia. A respiração basal é definida como a
respiração sem adição de substratos orgânicos ao solo, e pode ser avaliada através da
produção de CO2 ou consumo de O2 (ALEF, 1995).
A respiração é o parâmetro mais antigo utilizado na quantificação da atividade
metabólica nos solos. Assim como outras atividades metabólicas, a respiração é
dependente do estado fisiológico da célula e influenciada por diversos fatores no solo,
tais como, umidade, temperatura, estrutura e disponibilidade de nutrientes (ALLEN;
SCHLESINGER, 2004; GONÇALVES et al, 2002).
A interpretação dos resultados da atividade biológica deve ser feita com
critério, uma vez que elevados valores de respiração nem sempre indicam condições
desejáveis: uma alta taxa de respiração pode significar, em curto prazo, liberação de
nutrientes para as plantas e, em longo prazo, perda de carbono orgânico do solo para a
atmosfera (PARKIN; DORAN; FRANCO-VISCAINO, 1996; DALAL, 1998).
A biomassa microbiana é determinada através da quantificação do carbono
orgânico solúvel em amostras fumigadas com clorofórmio, onde o material celular está
disponível para extração, subtraído de amostras não fumigadas (VANCE; BROOKES;
JENKINSON, 1987). Esses organismos atuam como agentes finais da decomposição da

33
matéria orgânica. A estimativa da BMS, ao contrário do carbono orgânico total do solo
(COT), foi sugerida como um parâmetro sensível às mudanças iniciais no processo de
transformação de matéria orgânica do solo, sendo um indicador útil para avaliar
alterações em ecossistema com interferência antrópica (JENKINSON; RAINER, 1977;
POWLSON; BROOKES; CHRISTENSEN, 1987). Insam e Domsch (1988) sugerem
que a razão CBMS / COT é um índice apropriado para a comparação de diferentes
ambientes.
Para compreensão da atividade dos microrganismos do solo na mineralização
da matéria orgânica, Anderson e Domsch (1985) propuseram a determinação do
quociente metabólico microbiano (qCO2), que é a razão entre a respiração basal do solo
(CO2), obtida a partir da incubação do solo em laboratório, e o C-BMS por unidade de
tempo, como um indicador das mudanças da atividade microbiana após um distúrbio
ambiental sobre a atividade microbiana do solo. O qCO2 pode ser usado como
parâmetro de avaliação do empobrecimento dos solos em termos de matéria orgânica.
Assim, quanto menor o qCO2 mais eficiente se torna a biomassa microbiana do solo, ou
ainda menos C é perdido pela respiração e uma fração significativa de carbono é
incorporado no tecido microbiano.
De acordo com a teoria da sucessão ecológica de Odum (1969; 1985), o qCO2
decai durante a sucessão ou durante a regeneração de um distúrbio, visto que no estado
de equilíbrio dinâmico do clímax sucessional, a comunidade microbiana do solo se
tornaria mais eficiente em conservar carbono.
2.6 VARIABILIDADE SAZONAL NOS ATRIBUTOS QUÍMICOS E BIOLÓGICOS
DO SOLO
Após a retirada da vegetação natural para uso na agricultura o solo sofre
alterações em suas propriedades químicas e biológicas. Essas propriedades são
dependentes das características do solo, do clima e do tipo de vegetação. A interação
destes fatores estabelece uma nova condição de equilíbrio no sistema solo.
A precipitação é a característica climática mais importante para o crescimento
das plantas na região tropical. Ao contrário da temperatura que é relativamente

34
constante, a precipitação pluviométrica é muito variável, constituindo-se no principal
fator a determinar a produtividade biológica (BARROS; COMERFORD, 2002), pois é
uma das variáveis mais importantes na produção de biomassa e, conseqüentemente, de
matéria orgânica (VASCONCELOS, 2002).
Nas regiões de clima tropical úmido, tanto a temperatura como a precipitação
pluviométrica, tendem a estimular a proliferação e a atividade da população microbiana
do solo. Esses dois fatores climáticos favorecem o desenvolvimento dos
microrganismos que atuam diretamente na decomposição da matéria orgânica. De
acordo com Keeney e Sahrawat (1986), a alternância de ciclos com maior e menor
umidade no solo, é responsável pelo aumento na taxa de mineralização de nutrientes.
Muitas vezes os teores de C orgânico e N total ao longo das diferentes estações
passam a impressão de que a matéria orgânica do solo manteve-se estável ao longo do
ano. No entanto, De-Polli e Guerra (1996) destaca que nem sempre esses parâmetros se
mostram índices sensíveis a mudanças sofridas no conteúdo da matéria orgânica do solo
quando comparados à biomassa microbiana, pois a interpretação dos resultados
observados em relação à variação sazonal da biomassa microbiana pode ser entendida a
partir das condições climáticas dominantes no local e seus efeitos sobre a decomposição
dos resíduos.
O aumento do CBM e do NBM pode ser entendido pela elevação da
precipitação pluviométrica, o que acarreta condições favoráveis ao aumento da
biomassa microbiana do solo, que segundo Gama-rodrigues (1999), tais elevações
conferem função de reserva à biomassa microbiana, acumulando grandes quantidades
de nutrientes. A respiração e o quociente metabólico do solo, também, costumam sofrer
flutuações sazonais, mostrando-se intimamente associados com o teor de umidade do
solo. Maiores níveis de atividade microbiana têm sido relatados com o aumento da
umidade do solo (SANTRUCKOVA ; STRASKRAVA, 1991; ALVAREZ et al., 1995).

35
3 MATERIAL E MÉTODOS
3.1 CARACTERIZAÇÃO DA ÁREA DE ESTUDO
3.1.1 Localização da área experimental
As áreas experimentais situam-se nas comunidades de Cumaru (Travessa
Cumaru) e Nova Olinda (em propriedades de pequenos produtores), e Fazenda
experimental FEIGA/UFRA, com as seguintes coordenadas geográficas: 1º 11‟ 23,5” S
e 47º 31‟ 59,5” W; 1° 02‟ 33,8” S e 47° 29‟ 47” W, mais 1° 07‟ 15,5” S e 47° 36‟ 12,7”
W, respectivamente, (Figuras 1) no município de Igarapé-Açú, localizado na
microrregião Bragantina, pertencente à mesorregião do nordeste paraense. A sede
municipal tem as seguintes coordenadas geográficas: 01°07‟33‟‟ de latitude sul e
47°37‟27‟‟ de longitude a oeste de Greenwich. O município possui uma área de 797
km² e tem uma população de 32.361 habitantes, com uma densidade demográfica de
40,6 habitantes/km² (IBGE, 2009).
Figura 1 - Localização da área de estudo no município de Igarapé-Açú, mesorregião do nordeste
paraense. (Foto: Google Earth, acesso: 09 ago 2010).

36
3.1.2 Clima
O clima está incluído na categoria do equatorial megatérmico úmido,
correspondente ao tipo Ami, da classificação de Köppen. A temperatura media anual
está entre 25 ºC e 27 ºC, com máximas que podem chegar a 40 ºC, e pequena amplitude
térmica. Apresenta precipitação pluviométrica abundante, com média anual em torno de
2.500 mm e umidade relativa do ar média em torno de 85%, com estação chuvosa entre
os meses de dezembro a maio e, a menos chuvosa, de junho a novembro (BASTOS;
PACHECO, 2000; SEPOF, 2008). A Figura 2 mostra dados da precipitação
pluviométrica média mensal do período de janeiro de 2008 à dezembro de 2009, no
município de Igarapé-Açú (PA). Na Figura 3 encontram-se os dados da média mensal
dos últimos quinze anos. No período de janeiro de 2009, primeira coleta, verificou-se
que houve um prolongamento da estiagem, o que pode ser explicado pela influência dos
eventos El Niño – Oscilação Sul (ENSO).
Ropelewski e Halpert (1987) e Kiladis e Diaz (1989) estudaram a variabilidade
interanual das precipitações associada ao ENSO, mostrando que a Amazônia é uma das
regiões do globo significativamente influenciada pelos eventos El Niño e La Niña. A
fase quente do ENSO (El Niño) está associada a déficit e a fria (La Niña) a excesso de
precipitação na região. Marengo e Hastenrath (1993), usando resultados de modelos,
mostraram que em anos de grande aquecimento das águas equatoriais no oceano
Pacífico central, há uma redução de umidade que entra na Amazônia. O El Niño
influencia negativamente a precipitação no norte do Nordeste brasileiro e leste da
Amazônia devido à inibição da formação de nuvens convectivas.

37
Figura 2 - Distribuição da precipitação pluviométrica média mensal acumulada, medida no município de
Igarapé-Açú, Pará, Jan/dez (2008) e Jan/Dez (2009). (Fonte: Laboratório de Climatologia/Embrapa
Amazônia Oriental).
Figura 3 - Distribuição da precipitação pluviométrica média mensal acumulada em Igarapé-Açú, Pará.
Período: 1994-2009. (Fonte: Laboratório de Climatologia/Embrapa Amazônia Oriental).

38
3.1.3 Solo
Os solos da Região Bragatina apresentam características pedológicas
relativamente uniformes em grande parte da região. Devido às características do
material parental e em virtude de processos de lixiviação, os sedimentos são
extremamente oligotróficos, dando origem a solos com baixa oferta de nutrientes
(DENICH, 1989; DENICH, 1991; NUNEZ, 1995; KATO, 1998).
No município de Igarapé-Açu, o solo dominante é o Latossolo Amarelo com
horizonte A moderado, textura média, sendo que algumas porções apresentam o caráter
concrecionário em terra firme, além da presença de solos hidromórficos encontrados nas
várzeas dos rios que servem o município. De acordo com Gama (2002), com o uso
intensivo da agricultura itinerante e com a erosão laminar do horizonte superficial, parte
dos solos da Região Bragantina começaram a apresentar uma relação textural do
horizonte B sob o A bastante elevada (abrúptica). Em estudo realizado por Kanashiro e
Denich (1998), o solo foi classificado como Podzólico Amarelo álico, que pela atual
classificação brasileira é denominado de Argissolo Amarelo distrófico, com textura
variando de arenosa a média (EMBRAPA, 1999; EMBRAPA, 2006).
3.1.4 Vegetação
A vegetação predominante em Igarapé-Açu é representada pela floresta
secundária, proveniente da remoção da floresta primária, para a implantação de cultivos
de subsistência. Na região há uma tendência quanto à conversão das florestas primárias
e secundárias em extensas pastagens. Nas margens dos rios e riachos encontra-se a
vegetação de galeria. Antes da intervenção humana, predominava na região a vegetação
primária de Floresta Equatorial Perenifólia, as várzeas e igapós e campos inundados
que, atualmente, estão restritos a algumas localidades. Hoje, a região tem uma paisagem
agrícola bastante expressiva e a cobertura vegetal é dominada por lavouras que se
alternam com a capoeira, devido ao corte-e-queima a agricultura familiar.

39
Nas propriedades rurais de Igarapé-Açu, mais da metade da área é ocupada por
capoeira em diferentes estágios de desenvolvimento, provenientes da remoção da
floresta primária para a implantação de cultivos de subsistência (16% até quatro anos de
idade e 50% mais de quatro anos), enquanto as florestas (naturais e plantadas) ocupam
apenas 7,5% da área útil (DENICH; KANASHIRO, 1993; RODRIGUES, 1996; KATO,
1998).
3.2 DELINEAMENTO EXPERIMENTAL, TRATAMENTOS E VARIÁVEIS
ESTUDADAS
O experimento foi conduzido em delineamento inteiramente casualizado.
Foram avaliados os atributos químicos pH, carbono orgânico (C), nitrogênio total
(N), fósforo disponível (P), potássio (K), cálcio (Ca), magnésio (MG e alumínio (Al)
trocáveis, CTC pH 7,0, soma de bases (SB), porcentagem de bases (%V) e
porcentagem de alumínio (%m), em amostras coletadas em janeiro/2009 (período
seco atípico), nas camadas de 0-5 e 5-10cm de um Latossolo Amarelo, sob seis
diferentes sistemas de manejo e uso, descritos a seguir:
T1 – plantio sem queima, com trituração da vegetação – Cumaru;
T2 – Plantio com queima da vegetação – Cumaru;
T3 – Capoeira (vegetação secundária);
T4 – Plantio sem queima, com trituração da vegetação – FEIGA/UFRA;
T5 – Plantio com queima da vegetação – FEIGA / UFRA;
T6 – SAF sem queima, com trituração da vegetação – Nova Olinda.
Os atributos biológicos carbono da biomassa microbiana (CBM), respiração
basal (CO2) e quociente metabólico (qCO2) foram avaliados por meio de tratamentos,
em quatro repetições, distribuídos em sistema fatorial 6x3x2, compreendendo os seis
sistemas acima citados, três épocas de coleta do solo em 2009 (janeiro e dezembro -
período seco; junho – período chuvoso) e duas profundidade do solo (0-5 cm e 0-1-
cm). Na avaliação da relação carbono da biomassa microbiana:carbono orgânico
(CBM:C), foi considerada somente a amostragem realizada em janeiro/2009 (período

40
seco atípico), nas duas camadas do solo, já descritas, sob os seis sistemas de manejo
e uso estudados.
Figura 4 – Trituração da capoeira utilizando o trator (Tritucap), nas áreas experimentais do projeto
Tipitamba, município de Igarapé-Açú (PA).
Foi realizada análise multivariada (Análise de Componente Principais) com a
finalidade de determinar as variáveis químicas e biológicas do solo mais sensíveis às
modificações na cobertura vegetal, proporcionadas pelos diferentes sistemas de
manejo e uso do solo, coletado em janeiro/2009, nas profundidades de 0-5 cm e 5-
10cm.
Na Figura 4, observa-se detalhe do preparo da área de Capoeira derrubada, sem
queima e com trituração da vegetação.

41
A Figura 5 mostra as áreas experimentais com as diferentes coberturas vegetais
estudadas.
Figura 5 – Visão das áreas experimentais com diferentes tipos de cobertura: capoeira, triturada,
queimada e sob sistema agroflorestal, município de Igarapé-Açú (PA).

42
3.3 HISTÓRICO DE USO DAS ÁREAS EXPERIMENTAIS
Na área referente à localidade de Cumaru (Quadro 1), após os preparos com
derruba e queima da vegetação e sem queima e com trituração da vegetação, foram
cultivados arroz (Oriza sativa), caupi (Vigna unguiculata) e mandioca (Manihot
esculeta Crantz), nos períodos de 1995 a 1998. A área de cultivo entrou em pousio
de 1999 a 2001, sendo que no período de 2002 a 2005 voltou a receber as mesmas
culturas. Entrou em pousio novamente no período de 2006 a 2009, recebendo o
cultivo da mandioca em 2010/2011.
Quadro 01- Especificidades das áreas em estudos.
Experimento Cumaru
Sem
Queima(triturado) Com Queima
1995 e 1996 1995 e 1996
arroz, caupi e
mandioca
arroz, caupi e
mandioca
1997 e 1998 1997 e 1998
arroz, caupi e
mandioca
arroz, caupi e
mandioca
1999 a 2001 1999 a 2001
pousio pousio
2002 e 2003 2002 e 2003
arroz, caupi e
mandioca
arroz, caupi e
mandioca
2004 e 2005 2004 e 2005
arroz, caupi e
mandioca
arroz, caupi e
mandioca
2006, 2007, 2008 e
2009
2006, 2007, 2008 e
2009
pousio pousio
2010 e 2011 2010 e 2011
mandioca mandioca Fonte. Pesquisa de campo, município de Igarapé-açu – PA, 2009

43
A Figura 6 mostra a localização da área experimental em Cumaru, Igarapé-
Açu.
Figura 6 - Localização da área de estudo na comunidade de Cumaru, município de Igarapé-Açú (PA).
(Foto: Google Earth, acesso: 09/agosto/2010).
O experimento na FEIGA/UFRA contemplou área com vegetação de capoeira
com 15 anos de idade, além de áreas preparadas com queima da vegetação, e sem
queima e com trituração da vegetação (Quadro 2). Em 2002/2003, na área sem
queima, foram plantados milho (Zea mays) e mandioca, e promovido o
enriquecimento com Acacia mangium Willd; em 2003/2004, pousio e
enriquecimento com Acacia mangium Willd; de 2005 a 2006, cultivo de milho e
mandioca; 2006 a 2007, plantio de caupi e mandioca e enriquecimento com Inga
edulis e Acacia mangium Willd; pousio e enriquecimento com Acacia mangium
Willd em 2008 e 2009; em 2010 a 2011, cultivo de milho e mandioca.
Na área queimada, plantio de milho e mandioca, de 2002 a 2003; pousio
natural em 2005 e 2006; cultivo de milho e mandioca, de 2007 a 2008; pousio de
2008 a 2011.

44
Quadro 02 - Especificidades das áreas em estudos.
Fonte. Pesquisa de campo, município de Igarapé-açu – PA, 2009
A localização da área estudada na FEIGA/UFRA encontra-se na Figura 7.
Figura 7 - Localização da área de estudo na UFRA/FEIGA, município de Igarapé-Açú (PA). (Foto:
Google Earth, acesso: 09/agosto/2010).
Experimento FEIGA/UFRA
Capoeira Sem queima(triturado) Com Queima
15 anos
2002 e 2003 2002 e 2003
milho e mandioca + enriquecimento com Acacia mangium Willd milho e
mandioca
2003 e 2004 2003 e 2004
pousio e enriquecimento pousio natural
2005 e 2006 2005 e 2006
milho e mandioca pousio natural
2006 e 2007 2007 e 2008
caupi e mandioca + enriquecimento com Inga edulis e a Acacia mangium Willd milho e
mandioca
2008 e 2009 2008 e 2009
pousio e enriquecimento pousio natural
2010 e 2011 2010 e 2011
milho emandioca pousio natural

45
Na área referente à localidade de Nova Olinda (Quadro 3), após os preparos com
derruba e trituração da vegetação de quatro anos, foi implantado o sistema agroflestal
com Cupuaçu (Theobroma grandiflorum), Açaí (Euterpe oleracea), Gliricídia
(Gliricídia sepium) e Paricá (Schizolobium amazonicum), em 2007.
Quadro 03 - Especificidades das áreas em estudos.
SAF – Nova Olinda
2007
Cupuaçu, Açaí, Gliricídia e Paricá Fonte. Pesquisa de campo, município de Igarapé-açu – PA, 2009
3.4 AMOSTRAGEM
As amostras de solo foram coletadas em janeiro de 2009, junho de 2009 e
Dezembro de 2009, em áreas pré-selecionadas em propriedades de agricultores
familiares, no âmbito do Projeto Tipitamba. Nessas áreas foram abertos miniperfis com
25 cm de profundidade e coletadas fatias de solo nas profundidades de 0 a 5 cm, e 5 a
10 cm (Figura 8), com quatro repetições formando-se cinco amostras simples para
formar uma amostra composta.
Figura 8 – Foto da trincheira utilizada para coletar as amostras nas camadas de 0-5, 5-10 cm, município
de Igarapé-Açu (PA).

46
As amostras foram acondicionadas sob baixa temperatura e submetidas à
análises químicas no Laboratório de Química do Solo da Embrapa Amazônia Oriental
conforme metodologia preconizada por Embrapa (1997). A análise de respiração basal e
carbono da biomassa microbiana foram efetuados no Laboratório de Microbiologia da
Universidade Federal Rural da Amazônia.
3.5 ANÁLISES QUÍMICAS
O pH foi medido com potenciômetro na relação solo:água 1:2,5. O fósforo
disponível, o potássio e o sódio trocáveis foram extraídos com uma solução Mehlich-1;
sendo que o fósforo foi determinado por colorimetria (comprimento de onda 660µm); o
potássio e o sódio foram determinados por fotometria de chama. O cálcio, o magnésio e
o alumínio trocáveis foram extraídos por uma solução de KCl 1 M; o cálcio e o
magnésio foram determinados por meio de titulação com EDTA 0,025 M e o alumínio
por titulação com NaOH 0,025 M. O hidrogênio + alumínio foram extraídos com
CH3COO2Ca 1M pH 7,0 e determinado por titulação com NaOH 0,025 M.
O carbono e nitrogênio totais do solo foram determinados por combustão via
seca com um autoanalisador, de alíquotas moídas e peneiradas a 0,2 mm. A partir dos
dados obtidos foram calculados a soma de bases, CTC a pH 7,0, CTCefetiva, porcentagem
de bases (%V) e porcentagem de alumínio (% m) e, relação carbono da
biomassa/carbono orgânico (CBM/C).
3.6 ANÁLISES BIOLÓGICAS
Utilizou-se o método da fumigação-extração para estimar o carbono da
biomassa microbiana (CBM) conforme Vance, Brookes e Jenkinson (1987) e Tate, Ross
e Feltham (1988). Amostras de aproximadamente 12,5 g (peso fresco) foram
acondicionadas em dessecador e submetidas à fumigação com clorofórmio livre de
álcool por 24 horas, agitadas por 30 minutos em extrato de K2SO4 0,5M e,

47
posteriormente, filtradas (Figura 9). Amostras não-fumigadas foram pesadas ao mesmo
tempo das amostras fumigadas e, posteriormente, conservadas em geladeira até a
retirada das amostras fumigadas do dessecador, para a extração simultânea dessas
amostras. Em seguidas, os extratos foram armazenados em frascos plásticos sob
congelamento até o início das análises químicas.
Figura 9 - Detalhe das amostras fumigadas em dessecador para determinação do carbono da biomassa
microbiana (CBM).
O CBM foi oxidado em meio ácido, com dicromato de potássio (0,4 N), sendo
a determinação realizada por titulação de oxi-redução, com sulfato de ferro ferroso
amoniacal (0,4 N).
Para o cálculo do CBM, foi utilizado o coeficiente Kc de 0,26 determinado
especificamente para solos da Amazônia, por Feigl et al. (1995), E expresso pela
quantidade de 14
C adicionado (como glicose), menos o respirado, menos o extraído
antes da fumigação; a partir destes dados foi calculado a diferença entre 14
C extraído
após e antes da fumigação, para finalmente ser obtido o Kc por meio da relação entre o
fluxo de 14
C extraível e 14
C microbiano.

48
Para o cálculo do CBM, utilizou-se a seguinte equação:
CBM = (CF – CNF)/ KC
Onde:
CBM = C da biomassa microbiana em µg de C por g de solo seco;
CF = quantidade de C extraído na amostra fumigada em µg de C/g de solo
seco;
CNF = quantidade de C extraído na amostra não fumigada em µg de C/g de
solo seco; e
KC = eficiência de extração de carbono;
A respiração basal do solo foi determinada fazendo-se a incubação do solo
durante 10 dias (Figura 10), sendo quantificada a partir do CO2 produzido pela
respiração dos microrganismos, seguindo a metodologia proposta por Gregorich et al.
(1990).
Figura 10 – Detalhe das amostras incubadas para determinação da respiração basal.
O ensaio constou de um kit formado por um frasco plástico com capacidade de
2 L, fechado hermeticamente, contendo um copo com 30 g de solo na umidade de
campo, juntamente com um béquer contendo 10 mL de NaOH 0,5 mol/L, sem entrar em
contato direto com a amostra de solo. Fez-se necessária também uma unidade deste kit
sem a adição de solo, para ser utilizada como prova em branco. O CO2 capturado pelo
hidróxido de sódio foi determinado por titulação, com solução de HCl 0,5 mol/L. Para
isto, colocou-se uma alíquota de 4 mL de solução de BaCl2 1 mol/L e 5 gotas de
fenolftaleína, como indicador. Todas as determinações foram feitas em duplicatas e os
resultados expressos com base no solo seco.

49
A quantidade de CO2 liberada foi calculada pela fórmula:
AT
xNxfxVT)-(Bsolo de /gCO g 2
em que:
B = mL da titulação da prova em branco;
T = mL da titulação da amostra;
N = normalidade do ácido (eqg/L);
f = fator de correção do HCl 0,5 M;
V = mL de NaOH (0,5M) usado na captura do CO2;
AT = mL da aliquota a ser titulada
O quociente metabólico (qCO2), que é a quantidade de CO2 liberada por
unidade de biomassa microbiana pelo tempo, foi determinado como proposto por
Anderson e Domsch (1993).
Os resultados foram expressos em µg CO2 µg CBM-1
h-1
3.7 ANÁLISE ESTATÍSTICA
As médias obtidas nas análises dos atributos químicos e biológicos foram
submetidas à análise de variância (ANAVA), com a finalidade de avaliar os efeitos da
mudança da cobertura vegetal, épocas de amostragem e da profundidade do solo nas
variáveis estudadas. A comparação das médias obtidas das variáveis estudadas foi feita
pelo teste de Duncan, com 5% de probabilidade de erro, utilizando o programa
estatístico SAEG 8.1.
Os dados da primeira amostragem foram submetidos a uma análise
multivariada (análise de componentes principais), sendo utilizado o software R-
Development-Core-Team 2009 (IHAKA; GENTLEMAN, 1996) . A Análise dos
Componentes Principais, PCA (do inglês “Principal Component Analysis”), tem por
finalidade básica, a redução de dados a partir de combinações lineares das variáveis

50
originais. PCA é um método exploratório porque ajuda na elaboração de hipóteses
gerais a partir dos dados coletados, contrastando com estudos direcionados nos quais
hipóteses prévias são testadas. Ela é capaz de separar a informação importante da
redundante. Na análise de componentes principais (ACP), o primeiro componente
principal explica o maior grau de variabilidade, que vai diminuindo para cada
componente à medida que sua ordem aumenta (CP 2, CP 3...). A significância estatística
dos agrupamentos (tratamentos) foi avaliada pelo test de permutação de Monte Carlo.
Os resultados encontrados utilizando-se a análise multivariada (ACP) foram
confrontados com os resultados obtidos pela análise de variância para a primeira coleta,
buscando encontrar as variáveis químicas e microbiológicas que se mostraram boas
indicadoras das mudanças de uso do solo para ambas as análises estatísticas.

51
4 RESULTADOS E DISCUSSÃO
4.1 ATRIBUTOS QUIMICOS DOS SOLOS SOB DIFERENTES SISTEMAS DE
MANEJO
As médias dos atributos químicos dos solos analisados nas áreas com diferentes
sistemas de uso e manejo encontram-se na Tabela 1. O resumo da análise de variância
destas variáveis encontra-se nas tabelas correspondentes ao Anexo 1 e 2.
O valor do pH (H2O) dos solos variou de 4,8 a 5,28 na profundidade de 0-5 cm e
de 4,55 a 5, 32 na profundidade de 5-10 cm. Esses resultados são típicos de Latossolos
da Amazônia, que são fortemente ácidos. Ressalta-se que na região de estudo os solos
apresentam acidez natural elevada, em decorrência do material de origem (VIEIRA;
SANTOS, 1987) e da intensa precipitação pluviométrica. Na área sob SAF, em que o
preparo do solo foi feito com trituração da vegetação (T6), e na área de plantio com
corte e queima da vegetação, da FEIGA/ UFRA (T5), com pousio de três anos, os
valores de pH foram respectivamente, de 5,28 e 5,05, sendo classificados como de
acidez média, enquanto que nas mesmas áreas, os valores de pH ficaram abaixo de 5,0,
considerados como acidez elevada (TOMÉ JR., 1997).
O menor valor de pH na camada 0-5 cm do solo (4,80) foi encontrado na
capoeira T3, resultado esse que se aproxima dos encontrados por Kato et al. (2005), em
solos da mesma área de estudo, em que observaram valores mais elevados de pH, de 5,1
e 5,2 em capoeiras de 4 e 10 anos, respectivamente, na camada de 0-30 cm, porém sobre
um Argissolo Amarelo distrófico.
Na camada superficial, o T6 (SAF-trituração) apresentou o maior valor de pH
(5,28), com maior valor apresentam diferença significativa para os demais tratamentos,
tanto para capoeira como para os sistemas que foram derrubados e queimados. Neste
estudo, verificou-se também acidez média para o tratamento T5 (UFRA-queima) que
sofreu corte e queima, o que pode ser explicado pelo aumento significativo do pH logo
após a queima da biomassa, no entanto, outros tratamentos não tiveram os mesmos
efeitos.

52
Soares (2007), estudando solos em ecossistemas de terra firme de duas
comunidades tradicionais do município de Benjamin Constant (AM), observou que a
queima não afetou o pH dos sistemas estudados em ambas as comunidades, exceto no
solo da capoeira de 20 anos. O autor observou que esses resultados podem estar
relacionados a vários fatores, como a intensidade da queimada, pois em geral os
agricultores da região do Alto Solimões realizam a queima de suas áreas com a
vegetação derrubada ainda não totalmente seca, com isso as áreas ficam “mal
queimadas” restando muitos troncos e galhos pouco queimados. Silva (1998) também
não observou, nas condições edafoclimáticas da mata Atlântica, um aumento de pH no
solo após a queima de uma capoeira de 5 anos em Ilha Grande, Rio de janeiro.
Todavia, outros autores observaram aumento significativo no pH do solo logo
após a queima da vegetação. Holscher et al., (1997) observaram aumento de 11% com a
queima de duas capoeiras de 7 anos para a implantação de roças de mandioca e milho,
sob um Argissolo em Igarapé-Açu, Pará. Sánchez et al. (1993) e Sampaio (1997)
observaram aumentos acima de 15% após a queima de florestas sobre Latossolo em São
Carlos do Rio Negro, na Venezuela.
Aumentos nos valores de pH na profundidade de 5-10 cm, também foram
encontrados em alguns tratamentos, o que é corroborado pelos resultados encontrados
por Falcão e Silva (2000), onde observaram ligeiras diferenças no aumento do pH com a
profundidade do solo, em estudo realizado com Latossolo Amarelo em Manacapuru-
AM.
Na profundidade avaliada 0-5 cm as concentrações de K variaram de 0,04 a 0,1
cmolc.dm-3
sendo os maiores valores encontrados nas áreas T4 (FEIGA/UFRA-
trituração), T5 (FEIGA/UFRA-queima), T6 (SAF-trituração) e T3 (capoeira),
respectivamente, sem diferença significativa entre si, com tendência de superioridade no
T4 sem queima, com 0,1 cmolc.dm-3
. Na profundidade de 5-10 cm as concentrações de
K variaram de 0,04 a 0,07 cmolc.dm-3
, sendo que nos tratamentos T4 (FEIGA/UFRA-
trituração) e T6 (SAF-trituração) os valores obtidos de 0,07 cmolc.dm-3
superaram
significativamente os encontrados nos demais tratamentos.
Foi observado tendência de decréscimo nos valores de K com o aumento da
profundidade. Resultado semelhante ao encontrado por Sena (2006) estudando
diferentes sistemas agroflorestais sobre Latossolo Amarelo em Marituba-PA, verificou
também decréscimo do conteúdo de K com o aumento da profundidade. Em todos os

53
casos, as concentrações de K são consideradas de níveis baixos, conforme Tomé jr.
(1997). No entanto, Kato et al. (2005) estudando Argissolo do Oeste do Pará,
observaram valores mais elevados de K (0,30 a 0,80 cmolc.kg-1
) em capoeiras de 4 e 10
anos. O maior conteúdo de K de 0,1 cmolc.dm-3
encontrado no tratamento T4
(FEIGA/UFRA-trituração) pode ser explicado pelo maior acúmulo de matéria orgânica
na camada superficial do solo, promovendo aumento da disponibilidade dos nutrientes
essenciais às plantas. Além de possuir K+
trocável, preso aos grupos carboxílicos e
fenólicos, a matéria orgânica apresenta potássio no seu interior o qual é liberado com a
mineralização de seus constituintes, processo que é mais intenso com o aumento do teor
de umidade do solo, (MALAVOLTA, 1980), o que foi observado por Sena (2006).
As concentrações de Ca variaram de 0,92 a 2,92 cmolc.dm-3
e de 0,68 a 2,48
cmolc.dm-3
nas profundidades de 0-5 cm e 5-10 cm, respectivamente. Os maiores
valores de Ca a 0-5 cm, ocorreram nos tratamentos T5 (FEIGA/UFRA-queima), T6
(SAF-trituração) e T4 (UFRA-trituração), respectivamente, 2,92 cmolc.dm-3
, 2,60
cmolc.dm-3
e 2,58 cmolc.dm-3
, sem diferenças significativas. Na profundidade de 5-10
cm, foram encontrados nos tratamentos T6 (SAF-trituração) e T5 (FEIGA/UFRA-
queima), os maiores valores de 2,48 cmolc.dm-3
e 2,25 cmolc.dm-3
, respectivamente,
sem diferenças significativas. Esses resultados estão de acordo com muitos estudos na
Amazônia que tem mostrado que a derruba e queima ocasiona um incremento deste
nutriente no solo, pois a maior quantidade de cálcio no sistema encontra-se na biomassa
vegetal e é liberada junto às cinzas, na ocasião da queima (SMYTH, 1996; HOLSCHER
et al. 1997).
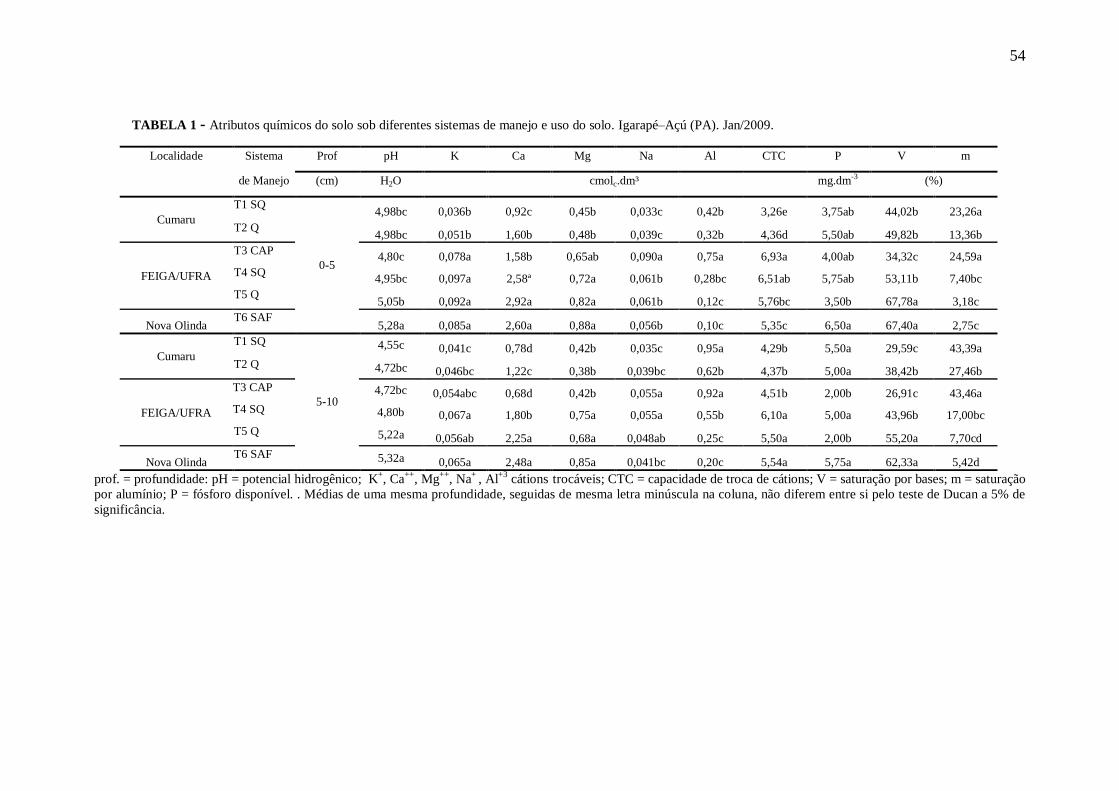
54
TABELA 1 - Atributos químicos do solo sob diferentes sistemas de manejo e uso do solo. Igarapé–Açú (PA). Jan/2009.
prof. = profundidade: pH = potencial hidrogênico; K+, Ca
++, Mg
++, Na
+ , Al
+3 cátions
trocáveis; CTC = capacidade de troca de cátions; V = saturação por bases; m = saturação
por alumínio; P = fósforo disponível. . Médias de uma mesma profundidade, seguidas de mesma letra minúscula na coluna, não diferem entre si pelo teste de Ducan a 5% de
significância.
Localidade Sistema Prof pH K Ca Mg Na Al CTC P V m
de Manejo (cm) H2O cmolc.dm³ mg.dm-3 (%)
Cumaru
T1 SQ
0-5
4,98bc 0,036b 0,92c 0,45b 0,033c 0,42b 3,26e 3,75ab 44,02b 23,26a
T2 Q 4,98bc 0,051b 1,60b 0,48b 0,039c 0,32b 4,36d 5,50ab 49,82b 13,36b
FEIGA/UFRA
T3 CAP 4,80c 0,078a 1,58b 0,65ab 0,090a 0,75a 6,93a 4,00ab 34,32c 24,59a
T4 SQ 4,95bc 0,097a 2,58ª 0,72a 0,061b 0,28bc 6,51ab 5,75ab 53,11b 7,40bc
T5 Q 5,05b 0,092a 2,92a 0,82a 0,061b 0,12c 5,76bc 3,50b 67,78a 3,18c
Nova Olinda T6 SAF
5,28a 0,085a 2,60a 0,88a 0,056b 0,10c 5,35c 6,50a 67,40a 2,75c
Cumaru
T1 SQ
5-10
4,55c 0,041c 0,78d 0,42b 0,035c 0,95a 4,29b 5,50a 29,59c 43,39a
T2 Q 4,72bc 0,046bc 1,22c 0,38b 0,039bc 0,62b 4,37b 5,00a 38,42b 27,46b
FEIGA/UFRA
T3 CAP 4,72bc 0,054abc 0,68d 0,42b 0,055a 0,92a 4,51b 2,00b 26,91c 43,46a
T4 SQ 4,80b 0,067a 1,80b 0,75a 0,055a 0,55b 6,10a 5,00a 43,96b 17,00bc
T5 Q 5,22a 0,056ab 2,25a 0,68a 0,048ab 0,25c 5,50a 2,00b 55,20a 7,70cd
Nova Olinda T6 SAF 5,32a 0,065a 2,48a 0,85a 0,041bc 0,20c 5,54a 5,75a 62,33a 5,42d

55
Nos tratamentos que sofreram trituração T4 (UFRA/FEIGA-trituração) e T6 (SAF-
trituração), os valores elevados de Ca, apresentados podem ser explicados pela maior
concentração de matéria orgânica e, como conseqüência, ocorrerá menor eficiência na
ciclagem desse nutriente. Apesar da acidez natural dos solos das áreas tropicais, a matéria
orgânica bem manejada é um dos principais recursos dos sistemas agrícolas, sendo
responsáveis por alterações químicas na camada superficial do solo, como a elevação do pH e
do teor de Ca, em detrimento do Al (SANCHES; VILLACHICA; BANDY, 1983; AMIEN,
1989; BESSHO; BELL, 1992). Também, o tipo de vegetação e as condições ambientais são
fatores que determinam a quantidade e a qualidade do material que se deposita no solo,
influenciando a heterogeneidade e a taxa de decomposição do material depositado à superfície
(MOREIRA; SIQUEIRA, 2002).
Todavia, o acúmulo de nutrientes na vegetação em pousio está associado com a
diversidade de espécies, o tamanho da vegetação e a fertilidade do solo (SZOTT; PALM;
BURESH, 1999). Soares (2007), estudando áreas com diferentes sistemas de uso da terra, na
região do Alto Solimões (AM), verificou que o tempo de pousio das capoeiras contribuiu para
um incremento significativo nos teores de Ca no solo. Os maiores aumentos foram verificados
nas capoeiras com maior tempo de pousio e, os menores, nas florestas e capoeiras de três
anos.
Os teores de Mg variaram de 0,45 a 0,88 cmolc.dm-3
e de 0,38 a 0,85 cmolc.dm-3
nas
profundidades de 0-5 cm e 5-10 cm, respectivamente. Os maiores valores para as duas
profundidades foram encontradas no T6 (SAF-trituração), porém, sem diferença estatística
significativa com os tratamentos T3 (capoeira), T4 (FEIGA/UFRA-trituração) e T5
(UFRA/FEIGA-queima). Em solos ácidos, como os solos da Amazônia, normalmente o Mg
encontra-se em baixos teores na solução do solo, seja devido as perdas de bases ao longo do
perfil, à pobreza do material de origem ou ao baixo pH do solo (LUCHESE; FAVERO,
2001).
Os teores de magnésio encontrados no presente estudo são considerados de baixo a alto
(TOMÉ JR., 1997; RAIJ et al, 1996). Os teores de Mg decresceram com maior profundidade
estudada, semelhantes aos resultados encontrados por Araújo et al. (2004), Ferreira (2006), e
Sena (2006), em solos da Amazônia. Assim como observado para o Ca, os teores de Mg
também foram maiores nos tratamentos T3 (capoeira), T4 (FEIGA/UFRA-trituração) e T5
(UFRA/FEIGA-queima), o que pode ter as mesmas explicações anteriores.

56
Kato et al. (2005) observaram teores médios de magnésio para um Argissolo Amarelo
distrófico, na Amazônia Oriental, que variaram de 0,2 a 0,4 cmolc. dm-3
, em capoeiras de
quatro e dez anos de pousio, respectivamente.
Os teores de alumínio dos solos estudados variaram de 0,10 a 0,75 cmolc.dm-3
e de 0,20
a 0,95 cmolc.dm-3
. Os maiores valores foram encontrados nas profundidades de 5-10 cm nos
tratamentos T1 (Cumaru-trituração) e T3 (capoeira), valores estes considerados de nível
médio, conforme Tomé Junior (1997) e Raij et al. (1996). Foi observado, também, que os
valores são menores na camada superficial e aumentam com a profundidade. Isto se deve a
maior presença de matéria orgânica em superfície, que possuem radicais carboxílicos e
hidroxílicos, que formam complexos com o alumínio e impedem que este possa atingir teores
trocáveis elevados e tóxicos aos vegetais (LUCHESE; FAVERO, 2001).
Os menores valores de alumínio trocável foram encontrados nos tratamentos T6 (SAF-
trituração) e T5 (FEIGA/UFRA-queima) nas profundidades estudadas, respectivamente, 0,10
e 0,12 cmolc.dm-3
e 0,20 e 0,25 cmolc.dm-3
. Tais resultados podem ser explicados pela maior
deposição e qualidade de resíduos orgânicos no solo do SAF com trituração, e na deposição
de cinza no solo do T5 com queima, esta disponibilizando bases trocáveis no primeiro
momento para o solo. Portanto, é de se esperar que ocorra redução dos teores de Al trocável
após a queima da vegetação, pois as cinzas resultantes da combustão da biomassa agem como
corretivo da acidez do solo e precipitam (insolubilizam) os íons de Al do solo, na forma de
Al(OH)3 não absorvível pelas plantas, em um claro efeito benéfico para o crescimento e
desenvolvimento das culturas (JUO; MANU, 1996; SMYTH, 1996). Por sua vez, matéria
orgânica presente nos solos forma complexos com o Al trocável, diminuindo desta forma o
seu teor na solução do solo, aumentando à disponibilidade de nutrientes as plantas (SZOTT;
PALM, 1999).
Foi observado, também, efeito, da queima e trituração da biomassa vegetal, nos
tratamentos T1 (Cumaru-trituração), T2 (Cumaru-queima) e T4 (FEIGA/UFRA-trituração),
que apresentaram teores de alumínio classificados como baixo, mas que apresentaram
diferenças estatísticas significativas para os tratamentos T5 (UFRA/FEIGA-queima) e T6
(SAF-trituração). Holscher et al. (1997), observaram o efeito da queima da biomassa vegetal
nas propriedades químicas de solo do Nordeste Paraense, com redução nos teores de Al
trocável após a queima de uma capoeira de sete anos, para a implantação de roças de
mandioca e milho.

57
Os teores de fósforo no solo variaram de 3,50 a 6,50 mg.dm-3
na profundidade de 0-5cm
e de 2,0 a 5,75 mg.dm-3
a 5-10 cm de profundidade, sendo esses valores considerados baixos
(TOMÉ JUNIOR, 1997), evidenciando a escassez de fósforo nos solos intemperizados da
Amazônia. O tratamento T6 (SAF-trituração) apresentou valor mais elevado (6,50 mg.dm-3
),
porém não havendo diferença estatística entre os demais tratamentos. A trituração da
vegetação produziu incremento significativo na disponibilidade de fósforo no solo nas duas
camadas estudadas. Provavelmente, esse acréscimo de P nos tratamentos com trituração seja
devido à presença de P orgânico, o qual é mineralizado e adicionado ao solo. Esses valores
evidenciam a escassez de P nos solos de terra firme da Amazônia, apesar dos teores obtidos
situarem-se um pouco acima dos Latossolos da região, que são da ordem de 1 a 3 mg.kg-1
(SMYTH, 1996; ALFAIA; SOUZA, 2002).
A CTC a pH 7,0 variou de 3,26 a 6,93 cmolc.dm-3
e de 4,29 a 6,10 cmolc.dm-3
, nas
camadas de 0-5 cm e 5-10 cm, respectivamente, valores considerados de baixo a médio,
conforme Mello et al. (1985). O maior valor foi encontrado na profundidade superficial, no
tratamento T3 (capoeira) igual a 6,93 cmolc.dm-3
, mostrando que em alguns tratamentos
houve decréscimo da CTC com o aumento da profundidade de amostragem, porém, em outros
tratamentos não foi observado uma redução na CTC.
Sena (2006) estudando diferentes sistemas agroflorestais sobre um Latossolo Amarelo
em Marituba-PA, em época seca, verificou valores que corroboram com os encontrados nesse
trabalho, sendo de 5,02 cmolc.dm-3
a 0-5 cm e 5,42 cmolc.dm-3
a 5-10 cm. O autor verificou
resultado semelhante quando comparou os diferentes sistemas de uso; tendo a floresta
secundária nos dois períodos de amostragem (estação seca e chuvosa) superado os demais no
parâmetro para CTC. A maioria dos valores encontrados ficaram entre 3,0 e 5,0 cmolc.dm-3
,
segundo Lopes e Guilherme (1992), os solos com CTC entre 1 a 5 cmolc.dm-3
são compostos
por alta percentagem de areia e, ou baixo teor de matéria orgânica, e apresentam alta
lixiviação de N e P e menor capacidade de retenção de umidade.
A saturação por bases (V%) mostrou valores entre 34,3% a 67,8% e de 26,9% a 62,3%,
nas profundidades estudadas. Os valores mais baixos de V% foram encontrados em T3
(capoeira), independentes da profundidade estudada. A explicação para isto é que esse
ecossistema dependem da ciclagem interna de nutrientes, e este solo encontra-se com pH
baixo. Os valores mais elevados foram observados nos tratamentos T5 (FEIGA/UFRA-
queima) e T6 (SAF-trituração), porém, sem diferença estatística entre os mesmos; para ambas
as profundidades. A saturação por bases é um excelente indicativo das condições gerais de

58
fertilidade do solo e, quando se apresenta com valores menores que 50%, indica que o solo
está com baixa fertilidade; acima de 50% está com boa fertilidade.
A saturação por alumínio (m%) variou de 2,75% a 24,59% na profundidade de 0-5 cm e
5,42% a 43,46% na profundidade de 5-10 cm, nos sistemas de manejo estudados. Os maiores
valores foram encontrados no tratamento T3(capoeira), para as duas profundidades estudadas,
porém, sem diferenças significativas para o sistema T1(Cumaru-trituração), onde ocorreu o
corte e trituração da capoeira. Os menores valores observados na camada de 0-5 cm foram
encontrados nos sistemas T6 (SAF-trituração)), T5 (FEIGA/UFRA-queima) e T4
(FEIGA/UFRA-trituração, respectivamente 2,75, 3,18 e 7,40 m%, considerados valores
baixos, ou não-prejudicias (TOMÉ JUNIOR, 1997). Para a profundidade 5-10 cm, apenas o
sistema T6 (SAF-trituração) se manteve dentro do nível que vai de 0-15 %V. Valores entre
16-35 % são considerados de nível médio (levemente prejudicial), segundo o autor. Gama
(2002), encontrou valores para m% de 11 na camada de 0-10 em área de vegetação secundária
de 4 anos, e de 30,1 em vegetação secundária de 10 anos
Os teores de carbono orgânico (C) encontrados nas duas profundidades na primeira
coleta (período seco) variaram de 5,33 a 13,03 g.kg-1
, (Tabela 2), considerados de baixo a
médio (TOMÉ JUNIOR, 1997). O teor mais elevado foi observado no T3 (capoeira), camada
de 0-5 cm, o qual difere estatisticamente dos demais tratamentos. À exceção dos sistemas T1
(Cumaru-trituração) e T2 (cumaru-queima), nos demais os teores de C decresceram com a
profundidade. Maiores teores de C nas camadas mais superficiais são comumente observados
em diversos tipos de solos sob diferentes tipos de vegetação, e estão relacionados com
maiores concentrações de matéria orgânica, que contém, em média, cerca de 58% de C
(RECCO et al., 2000; RESENDE, 1995). Rodrigues (2006) estudando solos em diferentes
tipos de cobertura vegetal, SAFs e capoeira no município de Aurora do Pará (PA), verificou
redução significativa do C em função da época de coleta e profundidade de amostragem, com
as maiores quantidades de C encontradas na estação seca (8,7 g.kg-1
) e na profundidade de 0-5
cm (8,0 g.kg-1
), corroborando com os resultados encontrados neste estudo.
Estudando a dinâmica da matéria em diferentes sistemas de manejo na Amazônia
ocidental, Moreira e Malavolta (2004), também verificaram que os maiores valores foram
encontrados na camada superficial, semelhantes aos resultados encontrados no presente
trabalho.
O teor de nitrogênio total (N) variou de 0,9 a 2,2 g.k-1
na profundidade de 0-5 cm, e de
1,2 a 1,5 g.k-1
a 5-10 cm, observando-se, também, os maiores valores no T3 (capoeira). Como
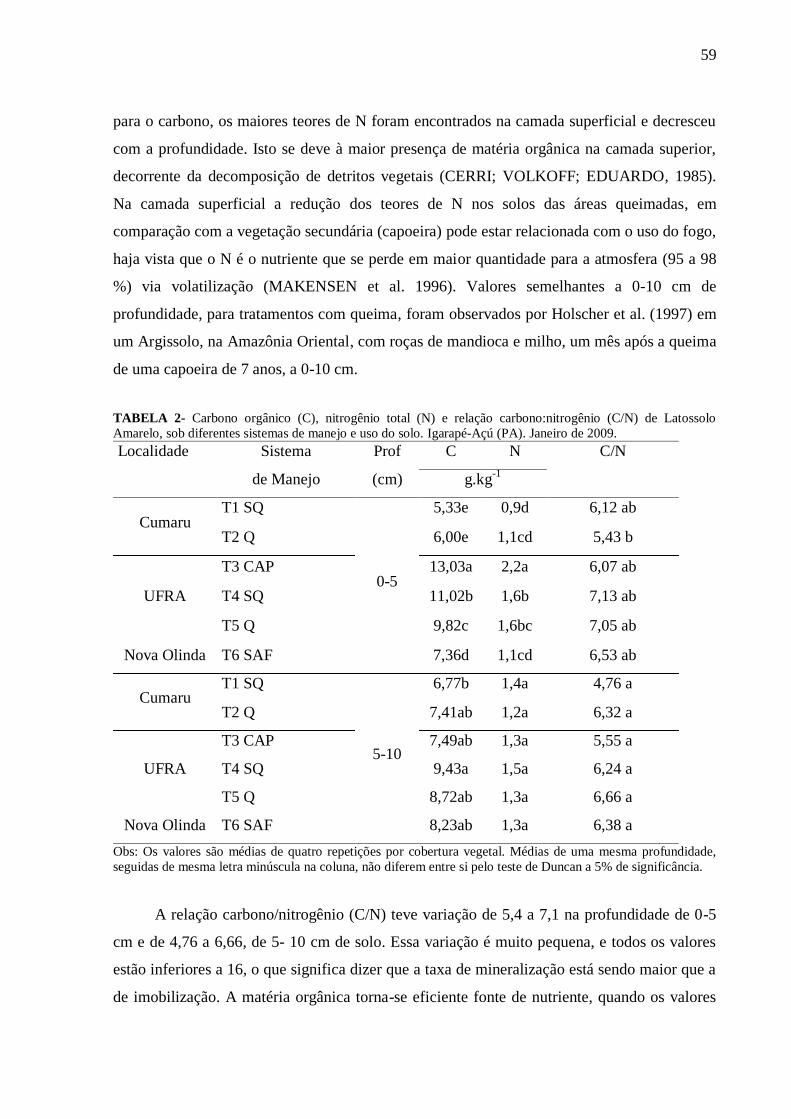
59
para o carbono, os maiores teores de N foram encontrados na camada superficial e decresceu
com a profundidade. Isto se deve à maior presença de matéria orgânica na camada superior,
decorrente da decomposição de detritos vegetais (CERRI; VOLKOFF; EDUARDO, 1985).
Na camada superficial a redução dos teores de N nos solos das áreas queimadas, em
comparação com a vegetação secundária (capoeira) pode estar relacionada com o uso do fogo,
haja vista que o N é o nutriente que se perde em maior quantidade para a atmosfera (95 a 98
%) via volatilização (MAKENSEN et al. 1996). Valores semelhantes a 0-10 cm de
profundidade, para tratamentos com queima, foram observados por Holscher et al. (1997) em
um Argissolo, na Amazônia Oriental, com roças de mandioca e milho, um mês após a queima
de uma capoeira de 7 anos, a 0-10 cm.
TABELA 2- Carbono orgânico (C), nitrogênio total (N) e relação carbono:nitrogênio (C/N) de Latossolo
Amarelo, sob diferentes sistemas de manejo e uso do solo. Igarapé-Açú (PA). Janeiro de 2009.
Localidade Sistema Prof C N C/N
de Manejo (cm) g.kg-1
Cumaru T1 SQ
0-5
5,33e 0,9d 6,12 ab
T2 Q 6,00e 1,1cd 5,43 b
UFRA
T3 CAP 13,03a 2,2a 6,07 ab
T4 SQ 11,02b 1,6b 7,13 ab
T5 Q 9,82c 1,6bc 7,05 ab
Nova Olinda T6 SAF 7,36d 1,1cd 6,53 ab
Cumaru T1 SQ
5-10
6,77b 1,4a 4,76 a
T2 Q 7,41ab 1,2a 6,32 a
UFRA
T3 CAP 7,49ab 1,3a 5,55 a
T4 SQ 9,43a 1,5a 6,24 a
T5 Q 8,72ab 1,3a 6,66 a
Nova Olinda T6 SAF 8,23ab 1,3a 6,38 a
Obs: Os valores são médias de quatro repetições por cobertura vegetal. Médias de uma mesma profundidade,
seguidas de mesma letra minúscula na coluna, não diferem entre si pelo teste de Duncan a 5% de significância.
A relação carbono/nitrogênio (C/N) teve variação de 5,4 a 7,1 na profundidade de 0-5
cm e de 4,76 a 6,66, de 5- 10 cm de solo. Essa variação é muito pequena, e todos os valores
estão inferiores a 16, o que significa dizer que a taxa de mineralização está sendo maior que a
de imobilização. A matéria orgânica torna-se eficiente fonte de nutriente, quando os valores

60
da relação C/N estão entre 11 e 16, indicando que o processo de mineralização está
predominando (COLOZZI-FILHO; ANDRADE; BALOTA, 2001). O maior valor foi
encontrado na camada superficial, no sistema T4 (FEIGA/UFRA-trituração) correspondente a
área que sofreu trituração e enriquecimento com leguminosas, com diferença significativa
apenas para o tratamento T2 (Cumaru-queima), onde ocorreu a queima da biomassa vegetal.
Este resultado não é comum, porém, essa área encontrava-se em pousio na época da
amostragem, podendo-se inferir sinais de recuperação do solo. Kristensen (2000), encontrou
uma relação C/N mais elevada na matéria orgânica total do solo, sob plantio direto, atribuindo
o fato à presença de compostos em estado menos avançado de humificação, o que não ocorreu
neste estudo. Calderón (2001) justifica que compostos com baixa relação C/N podem estar
protegidos em agregados no sistema de plantio direto, sendo, eventualmente mineralizado.
Para as amostras subsuperficial, os valores da relação C/N tiveram um pequeno
decréscimo, porém não se observou diferença estatística entre os sistemas de uso estudados.
Segundo Colozzi-Filho, Andrade e Balota (2001) os valores encontrados para a relação C/N
(< 20) são considerados baixos, indicando, possivelmente, que os resíduos culturais destes
sistemas em estudos são ricos em N, o que resultou em maior taxa de mineralização do
nutriente do que sua imobilização pela microbiota do solo. Valores próximos a estes foi
encontrado por Silva (2006) em sistemas agroflorestais em Paraty, (RJ), que variou entre 8,9 e
7,4, respectivamente em SAF e mata nativa. Em Argissolo de Igarapé-Açú (PA), em
diferentes sistemas agroflorestais, Duarte (2003) encontrou uma relação C/N na estação seca,
de 11,44 na profundidade 0-5 cm, e de 10,65 para a profundidade de 5-10 cm, valores estes
mais elevados que os encontrados neste estudo, mas que ficam dentro da faixa considerado
baixo, onde está ocorrendo uma maior taxa de mineralização da matéria orgânica. Os valores
encontrados nesta pesquisa, considerados baixos, leva a crer que os organismos
decompositores não são limitados pelo N, o que resulta em uma liberação líquida de N
inorgânico para a solução do solo.

61
4.2 ATRIBUTOS BIOLÓGICOS DOS SOLOS SOB DIFERENTES SISTEMAS DE
MANEJO
4.2.1 Carbono da biomassa microbiana (CBM)
Nas tabelas constantes dos anexos 3 e 4, encontram-se as análises de variância dos
atributos biológicos estudados.
Na Tabela 3 encontram-se as médias de carbono da biomassa microbiana (CBM) de
Latossolo Amarelo submetido a diferentes sistemas de uso e coletado em diferentes épocas.
Houve efeito marcante de sazonalidade nos sistemas de manejo, com os maiores
valores de CBM observados no período chuvoso (Jun/2009), sendo que os sistemas T5
(FEIGA/UFRA-queima), T4 (FEIGA/UFRA-sem queima) e embora sem diferirem entre si,
T3 (capoeira), foram superiores aos demais.
Os valores de CBM encontrados no período chuvoso, nos sistemas T2 (Cumaru-
queima) e T6 (Nova Olinda-SAF), mostraram tendência de superioridade sobre aqueles
obtidos nos períodos secos, sem que essas diferenças apresentassem significância estatística.
No período seco (Dez/2009) os valores de CBM foram reduzidos, em relação à
amostragem anterior,mantendo-se nos sistemas de manejo T3, T5 e T4,os maiores valores,
sem diferenças significativas.
Os maiores valores de CBM encontrados no período chuvoso, são respaldados na
literatura sobre o assunto. Conforme Luizão (1989), provavelmente, o aumento da
precipitação pluvial provocou aumento do carbono microbiano, tendo em vista que este
mantém uma correlação positiva com o aumento da disponibilidade de água do solo, que
favorece a atividade da população microbana.

62
TABELA 3 - Carbono da biomassa microbiana (CBM) de Latossolo Amarelo, em função de diferentes sistemas
de manejo e uso do solo e épocas de amostragem. Igarapé-Açú (PA).
Época T1 T2 T3 T4 T5 T6 Média
CBM (µg.g-1
de solo)
Jan/2009
Jun/2009
Dez/2009
41,65 aA
37,05 aB
44,62aBC
17,06 aA
33,64aB
27,73 aC
43,00 bA
134,03aA
92,77 aA
27,21 bA
146,51aA
73,82bBC
36,43 bA
172,83aA
75,76bB
20,38aA
42,38 aB
33,03 aC
30,95 b
78,23 a
74,13 a
Média 41,11 B 26,14 B 89,93 A 82,51 A 95,01 A 31,93 B
Obs: Médias seguidas da mesma letra minúscula dentro da coluna e maiúscula na linha não diferem entre si pelo
Teste de Duncan a 5% de probabilidade.
Facci (2008) estudando vários sistemas de uso do solo em São Paulo, tais como mata
nativa, plantio direto e convencional, também encontrou os maiores valores de CBM nas
amostragens ocorridas no verão (período chuvoso), fazendo supor que algumas características
associadas a essa estação, tais como maiores temperaturas e umidade, e a cultura, com distinto
padrão de exsudação radicular, influenciaram a microbiota, resultando em maiores teores
desta variável.
Os valores mais elevados dos teores de carbono microbiano implicam maior
imobilização temporária de nutrientes e, conseqüentemente, em menores perdas de nutrientes
no sistema solo-planta (MERCANTE, 2004). O CBM é um dos indicadores mais promissores
da qualidade do solo, pois ele responde prontamente às mudanças ambientais, muitas vezes,
muito mais cedo do que os parâmetros físicos e químicos, incluindo o carbono orgânico total
/COT, e até mesmo a produtividade das culturas, de forma consistente ao longo flutuações
sazonais devido às condições climáticas (SPARLING, 1997; BALOTA et al., 1998;
SEYBOLD; HERRICK; BREJDA, 1999; NOGUEIRA et al., 2006; ROSCOE et al., 2006;
FRANCHINI et al., 2007; HUNGRIA et al., 2009).
Uma provável explicação para o maior valor ocorrido no período seco (Dez/2009), no
tratamento T5 (FEIGAUFRA-queima), é que nesse sistema encontra-se, também, uma
vegetação de gramíneas invasoras, densa, porte médio, o que pode estar relacionado com
maiores teores de matéria orgânica obtidos na estação seca, pois é amplamente conhecido que
o CBM aumenta com aumento da matéria orgânica no solo. Oliveira et al. (2001) justificam
que os maiores valores de CBM encontrados em pastagem, é devido à maior densidade de
raízes e, conseqüentemente, maior efeito rizosférico, aumentando a disponibilidade de

63
substratos orgânicos para a comunidade microbiana do solo. Rodrigues (2006) estudando
SAFs na Amazônia Oriental, encontrou valores de CBM na época seca que corroboram com
os resultados desta pesquisa. O autor ressalta, ainda, que outro fato que também contribuiu
para o aumento da matéria orgânica, foi a manutenção nos SAF‟s dos restos culturais do caupi
cultivado por dois anos consecutivos.
O SAF estudado neste trabalho apresentou valores de CBM que não diferiram
significativamente entre as épocas de amostragem. Vasconcelos (2002) obteve respostas
similares, quantificando maiores teores de CBM na estação seca em vegetação secundária, o
que também foi observado neste estudo. Na média geral de CBM, entre os tratamentos,
verificou-se a superioridade do sistema T5 (FEIGA/UFRA-queima), sobre os demais, porém,
não sendo verificada diferença significativa em relação aos sistemas T3 (capoeira) e T4
(FEIGA/UFRA-trituração). A maior média geral de CBM entre as três épocas de amostragem,
ocorreu no período chuvoso (Jun/2009), não apresentando diferença significativa para o
período seco (Dez/2009).
Kaschuk, Alberton e Hungria (2009), em estudo sobre biomassa microbiana do solo,
em experimento de três décadas em ecossistemas brasileiros, verificaram que o aumento do
CBM em plantio direto foi de 58% no período de 10-15 anos, permanecendo estável até aos
25 anos. Os autores ressaltam, também, as tendências para o aumento da CBM, em média, em
uma série de estudos em todo o Brasil. No entanto, os valores obtidos por Kaschuk, Alberton
e Hungria (2009) foram inferiores aos determinados por Balota, Collozi-Filho e Andrade
(1998), iguais a 118%, Pereira et al. (2007), de 114%, e Franchini et al. (2007),
correspondente a 80%, em plantio direto em Latossolos no Sul do Brasil, mas foram
altamente significativos e positivos, e de forma consistente ao longo dos anos.
Neste estudo foi observado que o tratamento com corte e trituração-FEIGA/UFRA
(T4) foi tão eficiente quanto a vegetação secundária (T3), no tocante ao CBM, o que pode ser
explicado e entendido pelos resultados dos autores citados acima, indicando que ocorre um
aumento consistente de CBM ao longo dos anos nos sistemas de manejo mais sustentávies.
4.2.1.1 Interação sistema de manejo versus profundidade do solo
Na Tabela 4 estão apresentadas às médias de carbono da biomassa microbiana, sob os
efeitos da interação entre os sistemas de manejo e a profundidade do solo.

64
Nos sistemas de manejo T3, T4, e T5, todos na FEIGA/UFRA, ocorreram os maiores
valores de CBM, nas duas camadas do solo, amostrados, com decréscimo significativo dos
valores, com a profundidade. Dentro de cada camada, os maiores valores de T3, T4 e T5, não
diferiram entre si.
Os sistemas T1, T2 e T6 apresentaram os menores valores de CBM, sem diferenças
significativas, independente da profundidade do solo.
O maior aporte de resíduos orgânicos na capoeira de 15 anos, como foi verificado por
Vasconcelos (2002), em Castanhal (PA), evidencia o aumento do conteúdo de CBM de
acordo com o aumento da idade da capoeira. A redução do CBM, com o aumento da
profundidade de amostragem, é decorrente, provavelmente, da diminuição da população
microbiana nas camadas inferiores, devido às condições mais desfavoráveis ao seu
desenvolvimento, como a diminuição da aeração do solo e a menor disponibilidade de matéria
orgânica de mais fácil decomposição.
TABELA 4 - Carbono da biomassa microbiana (CBM) de Latossolo Amarelo, em função de diferentes sistemas
de manejo e uso do solo e profundidade de amostragem. Igarapé-Açu (PA).
Sistema de Manejo Prof 0-5 cm Prof 5-10 cm Média
CBM (µg.g-1
de solo)
T1
T2
T3
T4
T5
T6
35,51 bA
23,47 bA
122,73 aA
103,90 aA
92,59 aA
38,48 bA
46,71 bA
28,82 bA
57,13 abB
61,12 abB
97,42 aA
25,38 bA
41,11
26,14
89,93
82,51
95,00
31,93
Média 69,45 A 52,76 B
Obs: Médias seguidas da mesma letra minúscula dentro da coluna e maiúscula na linha não diferem entre si pelo
Teste de Duncan a 5% de probabilidade.
Observou-se interação significativa apenas nos tratamentos T3 (capoeira) e T4
(UFRA/FEIGA-trituração), nas duas profundidades estudadas, o que pode ser explicado pela
maior atividade microbiana nesses primeiros centímetros do solo, como já foi discutido
anteriormente. Considera-se que a biomassa microbiana concentra-se nos cinco centímetros
superficiais, onde há substrato mais abundante (PICCOLO et al., 2009, citados por
NICODEMO, 2009). Por outro lado, Rodrigues (2006), não verificou efeito da interação entre

65
a cobertura vegetal e as profundidades estudadas no conteúdo de CBM de um Latossolo
Amarelo, no Município de Aurora do Pará-PA.
4.2.1.2 Interação época de coleta versus profundidade do solo
Os teores de CBM em relação à interação épocas de amostragem e profundidades,
encontram-se na Tabela 5. Verifica-se que o maior valor de CBM ocorreu no período de
maior umidade do solo, na profundidade de 0-5 cm (104,03 µg.g-1
solo), apresentando
diferença significativa para a camada de 5-10 cm (52,42 µg.g-1
).
Na profundidade de 0-5 cm, o maior valor de CMB ocorrido no período chuvoso,
diferente significativamente das demais épocas de amostragem, evidencia a importância da
disponibilidade de água no solo para a atividade e crescimento da população microbiana. Na
profundidade de 5-10 cm foi observou maior valor no período seco (Dez/2009), sem diferença
significativa para o valor obtido no período chuvoso.
TABELA 5 - Carbono da biomassa microbiana (CBM) de Latossolo Amarelo coletado em diferentes épocas e
profundidades. Igarapé-Açu (PA)
Época Prof 0-5 cm Prof 5-10 cm Média
CBM (µg.g-1
de solo)
Janeiro/2009
Junho/2009
Dezembro/2009
30,87 cA
104,03 aA
73,44 bA
31,04 bA
52,42 abB
74,83 aA
30,95 b
78,23 a
74,13 a
Média 69,45 A 52,76 B
Obs: Médias seguidas da mesma letra minúscula dentro da coluna e maiúscula na linha não diferem entre si pelo
Teste de Duncan a 5% de probabilidade.
Dentro da mesma profundidade de 0-5 cm, o maior valor de CMB ocorreu no período
chuvoso (Jun/2009) com diferença estatística significativa para as demais épocas de
amostragem. Esses resultados evidenciam a importância da disponibilidade de água no solo
para a atividade e crescimento da população microbiana. Na profundidade de 5-10 cm foi
observou maior valor no período seco (Dez/2009), no entanto não foi observou diferença
significativa para o período chuvoso.

66
Os resultados de CBM mostrando que a variação sazonal e a profundidade interferiram
na imobilização de C pela comunidade microbiana. Rodrigues (2006) não verificou efeito da
interação entre a época da coleta e as profundidades estudadas (0-5 e 5-20 cm) de um
Latossolo Amarelo, no Município de Aurora do Pará-Pa, sobre o CBM do solo, diferente do
que foi verificado neste estudo.
4.2.2 Relação carbono da biomassa microbiana/carbono orgânico (CBM:C)
Os valores para a relação carbono da biomassa microbiana / carbono orgânico obtidos
em amostragem realizada em Janeiro/2009, encontram-se na Figura 11 e 12.
Os valores de CBM/C para a camada de 0-5 cm do solo variaram de 2,29 a 6,28%,
respectivamente, nos tratamentos T2 (Cumaru-queima) e T1 (Cumaru-trituração), conforme
mostra a Figura 11. Para a camada de 5-10 cm a variação foi 2,31 a 6,97%, respectivamente,
em T6 (SAF-trituração) e T1 (Cumaru-trituração). Não foi verificada diferença estatística
significativa na camada superior entre os tratamentos T1 (Cumaru-trituração) e T3 (capoeira)
com os maiores valores, e na camada inferior entre os tratamentos T1 (Cumaru-trituração), T3
(capoeira), T5 (UFRA/FEIGA-queima) e T2 (Cumaru-queima). Os valores encontrados no
presente estudo estão de acordo com os propostos por Anderson e Domsch (1989), que
consideram normal os valores desta relação variar entre 0,3 a 7%.
Figura 11 – Efeito de sistemas de manejo e uso do solo na relação carbono da biomassa microbiana / carbono
orgânico de Latossolo Amarelo, 0-5 cm de profundidade. Igarapé-Açú (PA).

67
Figura 12 – Efeito de sistemas de manejo e uso do solo na relação carbono da biomassa microbiana / carbono
orgânico de Latossolo Amarelo, 5-10 cm de profundidade. Igarapé-Açú (PA).
A relação CBM/C pode ser usada como indicador preliminar das alterações do solo,
uma vez que a biomassa microbiana é um constituinte da matéria orgânica do solo. A
biomassa microbiana representa até 5% do C orgânico total do solo (POWLSON;
JENKINSON, 1981). Nos tratamentos T1 (Cumaru-trituração) e T3 (capoeira 15 anos) na
profundidade de 0-5 cm os maiores índices CBM/C, indicam que houve maior acúmulo de
biomassa microbiana nesses sistemas. Para a profundidade 5-10 estes sistemas (T1eT3)
apresentam, também, os maiores índices da relação CBM/C.
Nunes et al. (2009) encontraram resultados parecidos em Sobral (CE), em
profundidade de 0-10 cm, onde verificaram maior relação CBM/C nessa ordem: em vegetação
primária, área de pousio de 5 anos e áreas com enleiramento dos resíduos de corte de pousio
de 5 anos. Os outros tratamentos que sofreram corte e trituração não diferiram
significativamente da capoeira. Em circunstâncias de desequilíbrio ambiental, com matéria
orgânica de baixa qualidade ou em situação em que a biomassa experimenta algum fator de
estresse (deficiência de nutrientes, acidez, déficit hídrico, etc), a capacidade de utilização de C
é diminuída e, neste caso a relação CBM/C diminui (WARDLE, 1994).

68
4.2.3 Respiração Basal
Os valores da respiração basal do solo coletado em diferentes épocas, em função do
sistema de manejo e uso do solo, são apresentados na Tabela 6.
Verificou-se que as maiores perdas de C-CO2 por atividade microbiana ocorreram nos
tratamentos T3 (capoeira) igual a 186,26 ug C-CO2.g-1
solo em época de amostragem mais
chuvosa (jun/2009), porém não houve diferença significativa para os tratamentos T4
(UFRA/FEIGA-trituração) e T2 (Cumaru-queima), respectivamente, 160,99 e 156,46 ug C-
CO2 g-1
solo. No entanto, verificaram-se para todos os sistemas em estudo, maiores valores
nesse período de amostragem.
TABELA 6 - Respiração basal (CO2) da biomassa microbiana de Latossolo Amarelo sob diferentes sistemas de
manejo e uso do solo, coletado em diferentes épocas. Igarapé-Açu (PA).
Época T1 T2 T3 T4 T5 T6 Média
CO2 (ug C-CO2 g-1
solo)
Jan/2009
Jun/2009
Dez/2009
40,07 bA
126,51aB
58,18 bA
27,04 bA
156,46aA
43,06 bA
39,47 bA
186,26aA
60,23 bA
35,31 bA
160,99aA
43,32 bA
33,24 aA
41,49 aD
52,22 aA
59,40 aA
81,22 aC
62,54 aA
39,09 c
125,49 a
53,24 b
Média 74,92 B 75,52 B 95,32 A 79,87 AB 42,32 C 67,69 B
Obs: Médias seguidas da mesma letra minúscula dentro da coluna e maiúscula na linha, não diferem entre si pelo
teste de Duncan a 5% de probabilidade.
Os resultados sugerem que a maior atividade microbiana nestes sistemas ocorreu no
período de maior umidade do solo. Teores mais elevados de perdas de CO2 do solo via
respiração microbiana, na estação chuvosa, foi observado por Rodrigues (2006), em sistemas
de uso do solo na Amazônia Oriental, sendo justificados pelo fato de ser na camada
superficial do solo que se concentra a maior quantidade de matéria orgânica e nutrientes, os
quais são imprescindíveis para o desenvolvimento da macrofauna do solo. Resultados que
corroboram essas justificativas de Rodrigues (2006), foram encontrados por Balota et al.
(1998) e Luizão, Costa e Luizão (1999). Além disso, Nogueira et al. (2006) relataram que o
solo que apresentou maior atividade microbiana tinha, também, o maior teor de água.
Observou-se, também, que ocorreu menor atividade microbiana em todos os
tratamentos no período da amostragem da época seca (Dezembro/2009). Em Igarapé-Açú,

69
Nordeste Paraense, Rangel et al. (2005) estudando vegetação secundária de diversas idades,
verificaram também, menor atividade microbiana no período seco, em ecossistemas florestais
no litoral paranaense.
A menor atividade microbiana nos outros tratamentos (T1, T5 e T6), pode ser devido a
menor quantidade de substrato disponível para ação dos microrganismos, como foi observado,
também, por Monteiro e Gama-Rodrigues (2000), estudando diferentes ecossistemas florestais
sob solo de eucalipto. Vasconcelos (2002) observou que a menor atividade microbiana e
elevada BMS nos períodos secos indicam que, neste período, a BMS imobiliza mais nutriente
em seu tecido, do que mineraliza.
Para a média geral entre os tratamentos, observou-se maior atividade microbiana no
tratamento T3 (capoeira), sem diferença significativa para o tratamento T4 (UFRA/FEIGA-
trituração), o que está de acordo com os resultados obtidos por Rodrigues (2006), o qual
verificou que a maiores perdas de CO2 foram observados na capoeira, seguido da pastagem.
Peña et al. (2005) conclui em seu trabalho que a maior produção de CO2 ocorre na fase
avançada da sucessão. Entre as épocas de amostragem, a maior média foi observada no
período chuvoso, que diferiu significativamente das demais amostragens, o que já foi
discutido anteriormente.
4.2.3.1 Interação sistema de manejo versus profundidade do solo
Com base nos resultados da Tabela 7, verificou-se que as maiores perdas de C-CO2
por atividade microbiana ocorreram nos tratamento T3 (capoeira) na camada de 0-5 cm, sem
diferenças significativas para os sistemas T1 (Cumaru-trituração), T2 (Cumaru-queima) e T4
(UFRA/FEIGA-trituração), respectivamente.
Houve interação significativa entre as duas profundidades estudadas, dentro do
tratamento T3 (capoeira), onde se observou que ocorreu maior perda de CO2, na camada de 0-
5 cm, o que pode ser explicado pelo maior acúmulo de material orgânico que é depositado
pelas plantas e animais neste sistema. Para Vargas e Scholles (2000), as variações da
atividade respiratória ao longo do perfil estão associadas a variações na disponibilidade de
carbono orgânico, e a maior deposição de resíduos na superfície, que leva a uma maior
concentração de substratos orgânicos nos primeiros centímetros do solo. A produção de CO2
nas camadas superficiais do solo sofre influência direta da quantidade e da qualidade da

70
matéria orgânica presente (PEÑA et al., 2005). Rodrigues (2006) verificou que as maiores
perdas de CO2 ocorreram em capoeira, porém, independente das profundidades estudadas (0-5
e 5-20 cm) de um Latossolo Amarelo, no Município de Aurora do Pará(PA).
TABELA 7 - Respiração basal (CO2) da biomassa microbiana de Latossolo Amarelo, em função de diferentes
sistemas e uso do solo e profundidade de amostragem. Igarapé-Açú-(PA).
Sistema de Manejo Prof 0-5 cm Prof 5-10 cm Média
CO2 (ug C-CO2 g-1
solo)
T1
T2
T3
T4
T5
T6
83,99 abA
86,59 abA
109,51 aA
86,79 abA
40,16 cA
74,48 bA
65,85 abA
64,44 abA
81,13 aB
72,97 aA
44,47 bA
60,90 abA
74,92
75,52
95,32
79,88
42,46
67,69
Média 80,25 A 64,96 B
Obs: Médias seguidas da mesma letra minúscula dentro da coluna e maiúscula na linha não diferem entre si pelo
Teste de Duncan a 5% de probabilidade.
Quanto à menor atividade nos demais sistemas de manejo da terra, pode ser devido à
menor quantidade de substrato disponível para ação dos microorganismos, como foi
observado também por Monteiro e Gama-Rodrigues (2000), estudando diferentes
ecossistemas florestais sob solo de eucalipto. Vargas e Scholles (2000) em estudo em sistemas
de plantio direto verificaram aumento na respiração do solo, justificada pelo acúmulo de
matéria orgânica rica em frações lábeis na superfície.
4.2.3.2 Interação época da coleta versus profundidade do solo
Foi verificado o efeito da interação entre época e profundidade de amostragem sobre a
respiração basal do solo Tabela 8. Os maiores valores de CO2 desprendidos do solo ocorreram
em Jun/2009, para ambas as profundidade estudadas, sendo diferentes estatisticamente das
demais épocas de amostragem, resultados estes que têm relação direta com a umidade do solo
e com o carbono da biomassa microbiana.

71
Entre as duas profundidades observa-se diferença estatística para as épocas de coleta
de junho e dezembro de 2009, em que os maiores valores ocorreram na camada de 0-5 cm.
Analisando as médias gerais tanto para profundidade como para época de amostragem obtida
para o efeito da interação, observa-se maior CO2 na camada de 0-5 cm , na estação chuvosa
(80,25 e 125,49 μg de CO2/g de solo).
TABELA 8 - Respiração basal (CO2) da biomassa microbiana de Latossolo Amarelo coletado em diferentes
épocas e profundidades. Igarapé-Açu (PA).
Época Prof 0-5 cm Prof 5-10 cm Média
CO2 (ug C-CO2 g-1
solo)
Janeiro/2009
Junho/2009
Dezembro/2009
44,29 cA
134,01 aA
62,45 bA
33,88 bA
116,97 aB
44,04 bB
39,09 c
125,49 a
53,24 b
Média 80,25 A 64,96 B
Obs: Médias seguidas da mesma letra minúscula dentro da coluna e maiúscula na linha não diferem entre si pelo
Teste de Duncan a 5% de probabilidade.
Para Vargas e Scholles (2000) os microrganismos são muito sensíveis e podem ser
influenciados pelos fatores bióticos e abióticos, dentre estes está a umidade do solo. Segundo
Santos e Camargo (1999), dentre as variáveis climáticas, a precipitação pluvial e a
temperatura são as que exercem maior influência na atividade microbiana do solo. Ainda para
Vargas e Scholles (2000), as diferenças nas atividades do perfil do solo refletem a distribuição
dos resíduos vegetais Nogueira et al. (2006) relataram que o solo que apresentou maior
atividade microbiana tinha, também, o maior teor de água, o que foi observado também neste
trabalho.
Os resultados encontrados neste estudo, juntamente com os de outros autores citados
acima, mostram que, a despeito da disponibilidade de nutrientes e matéria orgânica no
ambiente solo, aqui na região tropical como em outras regiões, o fator que mais influencia e
limita o desenvolvimento da população microbiana do solo, é a água. Depreende-se, portanto,
que não só a matéria orgânica do solo deva ser manejada de forma eficiente, mas a água
também, pois a mesma passa a ser fator que deve ser considerado para o bom equilíbrio do
ecossistema. Então, em sistemas de manejo que conservem a matéria orgânica do solo e o
protejam da ação direta do sol e da chuva, como o sistema de corte e trituração, deve ser
recomendado para o aumento da sustentabilidade dos solos tropicais.

72
4.2.4 Quociente metabólico (qCO2)
Os valores do quociente metabólico (qCO2) de amostras de solo coletadas em
diferentes épocas e em áreas sob diferentes sistemas de manejo e uso da terra são
apresentados na Tabela 9.
TABELA 9 - Quociente metabólico (qCO2) da biomassa microbiana de Latossolo Amarelo sob diferentes
sistemas de manejo e uso do solo, coletado em diferentes épocas. Igarapé-Açu (PA).
Data T1 T2 T3 T4 T5 T6 Média
qCO2 (x 10-5
C-BM-1
h-1
)
Jan/2009
Jun/2009
Dez/2009
5,62 bAB
17,09aAB
5,91 bA
11,59bAB
24,49 aA
8,30 bA
4,84 aB
7,78aCD
2,96 aA
8,07abAB
11,60aBC
2,93 bA
4,78 aB
2,61 aD
1,31 aA
14,07 aA
13,23aBC
9,30 Aa
8,16 b
12,80 a
5,12 b
Média 9,54 BC 14,79 A 5,19 CD 7,53 BCD 2,90 D 12,20 AB
Obs: Médias seguidas da mesma letra minúscula dentro da coluna e maiúscula na linha não diferem entre si pelo
Teste de Duncan a 5% de probabilidade.
Os maiores valores de qCO2, sem diferenças significativas, foram observados nos
tratamentos T2 (Cumaru-queima) (24,49 x 10-5
C-BM-1
h-1
), T1 (cumaru-trituração) (17,09 x
10-5
C-BM-1
h-1
), os quais ocorreram no período chuvoso. De um modo geral, em todos os
sistemas os valores de qCO2 foram maiores no período chuvoso. Rodrigues (2006)
trabalhando em sistemas de SAF‟s e capoeira encontrou, também, valores mais elevados na
estação chuvosa, o que corrobora com os resultados encontrados nesta pesquisa, sugerindo
que há maior eficiência da biomassa microbiana em imobilizar o carbono durante a estação
seca, ou ocorre uma redução da atividade da população microbiana do solo, provocando a
redução do quociente metabólico, porque, como explica De Polli et al. (2000), à medida que a
biomassa microbiana se torna mais eficiente, menos carbono é perdido na forma de CO2 pela
respiração e como conseqüência, uma fração do C é incorporada ao tecido microbiano.
Uma possível explicação para as maiores perdas de CO2 nos sistemas T2 e T3 é que a
melhoria temporária de alguns atributos químicos proporcionados pela queima, como, por
exemplo, aumento de K+, Ca
2+, Mg
2+ e aumento de pH e redução nos teores de Al
3+
(RHEINHEIMER et al., 2003), permite que uma população de oportunistas se multiplique
rapidamente, gerando desequilíbrio e, conseqüentemente, maiores perdas de C no sistema na
forma de CO2.

73
Para os sistemas de corte e trituração o que também pode ser inferido para capoeira de
15 anos, a possível explicação para os altos valores de qCO2 é que aumento no teor de matéria
orgânica na superfície do solo, de certo modo, juntamente com a maior disponibilidade de
água nesse ecossistema, contribuiu significativamente para o aumento da população
microbiana e sua atividade, conseqüentemente, aumentando a atividade respiratória dos
microrganismos do solo. De acordo com Insam e Haselwandter (1989), citados por Kaschuk,
Alberton e Hungria (2009), sucessões de vegetação secundária acumulam mais matéria
orgânica, porque há uma melhor eficiência metabólica microbiana (medida pelo qCO2) na
utilização de resíduos de C. Da mesma forma, em longo prazo o plantio direto acumularia
mais carbono orgânico devido ao menor qCO2 da CBM. A autora relata que qCO2 foi
reduzida em plantio direto ao longo do tempo. Ademais, tem sido demonstrado que o plantio
direto favorece uma maior proporção de fungos procariontes porque não quebra suas redes de
hifas (BEARE, 1997; FREY; ELLIOTT; PAUSTIAN, 1999; BAILEY; SMITH; BOLTON
JR., 2002). Os fungos têm uma exigência menor de energia para a sua manutenção, do que
procariontes, e assim, transformam o substrato em C-microbiano-C de forma mais eficiente
(ALVAREZ et al., 1995; HAYNES, 1999).
O T5 (FEIGA/UFRA-queima) foi o que apresentou menor qCO2 em todas as épocas de
amostragem, sem diferenças significativas. O qCO2 do T5 (FEIGA/UFRA-queima) no
período chuvoso (Jun/2009), não diferiu significativamente do valor obtido no sistema T3
(capoeira), se configurando como um sistema mais conservacionista, com menores perdas de
C e de maior sustentabilidade, no que se refere ao fluxo de C. Assim, o quociente metabólico
permitiu discriminar os sistemas quanto à sustentabilidade de C no solo. Os valores
relativamente mais elevados de qCO2, na estação chuvosa, sugerem, que a biomassa
microbiana está mineralizando mais C, do que imobilizando.
4.2.4.1 Interação sistema de manejo versus profundidade do solo
Os maiores valores de quociente metabólico (qCO2) para todos os períodos de
amostragem, ocorreram nos tratamentos T2 (Cumaru-queima) e T6 (SAF-trituração),
enquanto os menores valores foram observados nos tratamentos T5 (UFRA/FEIGA-queima) e
T3 (capoeira), nas duas profundidades (Tabela 10).

74
TABELA 10 - Quociente metabólico (qCO2) da biomassa microbiana de Latossolo Amarelo sob diferentes
sistemas de manejo e uso do solo coletado em diferentes profundidades. Igarapé-Açu (PA).
Sistema de Manejo Prof 0-5 cm Prof 5-10 cm Média
qCO2 (x 10-5
C-BM-1
h-1
)
T1
T2
T3
T4
T5
T6
10,31 abA
15,58 aA
3,57 cA
7,74 bcA
2,95 cA
10,60 abA
8,77 abA
14,01 aA
6,82 abA
7,32 abA
2,85 bA
13,80 abA
9,54
14,80
5,20
7,53
2,90
12,20
Média 8,45 A 8,93 A
Obs: Médias seguidas da mesma letra minúscula dentro da coluna e maiúscula na linha não diferem entre si pelo
Teste de Duncan a 5% de probabilidade.
Verifica-se que o sistema de uso do solo influenciou na atividade da biomassa
microbiana do solo, conforme mostram os valores de quociente metabólico (qCO2) para as
profundidades estudadas. Os maiores valores de qCO2 indicam maiores perdas de C em um
sistema, na forma de respiração, o que foi verificado no T2 (Cumaru-queima) e no T6 (SAF-
trituração). Valores mais altos de qCO2 indicam maior necessidade de energia de mantença
para os microrganismos. Quando o qCO2 é baixo, mais carbono estará disponível para a
produção de biomassa, o que deve se refletir em maior porcentagem de C microbiano em
relação ao C orgânico total (ANDERSON, 2003; KASCHU; ALBERTON; HUNGRIA,
2009).
Nicodemo (2009) verificou que na camada superficial o solo sob mata nativa
apresentou menor quociente metabólico (mas semelhante ao sistema agroflorestal), o que
indicava maior atividade microbiana e maior fixação de C, o que foi, também, verificado
neste trabalho apenas na capoeira de 15 anos e ao sistema T5 (FEIGA/UFRA-queima). No
entanto, os tratamentos T1 (Cumaru-triturado), T2 (Cumaru-Queima) e T6 (SAF-triturado)
apresentaram valores elevado de qCO2, o que pode ser explicado pelo fato desses sistemas,
que sofreram queima e trituração, ainda não se encontrarem em equilíbrio, conforme já
explicado acima. Quanto à interação entre as duas profundidades em estudo, não houve
significância em todos os tratamentos observados neste trabalho, verificando-se, que na
maioria dos sistemas ocorreu um decréscimo no qCO2, porém, nos tratamentos T3 e T6 houve
um pequeno aumento que não foi significativo.

75
4.2.4.2 Interação época da coleta versus profundidade
O efeito interação época da coleta versus profundidade, para qCO2, não apresentou
diferença significativa para a profundidade 0-5 cm, porém, o menor valor foi verificado em
Dez/2009 (Tabela 11). Na profundidade de 5-10 cm, também na estação seca (Dez/2009),
ocorreu o menor valor de qCO2, o qual diferiu somente do período de amostragem de
Jun/2009, época de maior umidade no solo.
Verificou-se que não houve efeito significativo da profundidade de coleta dentro de
cada período de amostragem avaliado. Mas entre os períodos de amostragem as médias gerais
apresentaram diferença significativa, observando-se novamente, menores valores na estação
seca. Rodrigues (2006) verificou interação entre a época de coleta e profundidade de
amostragem, com maior qCO2 verificado na estação chuvosa e na profundidade de 5-20cm, o
que corrobora com os resultado deste trabalho.
TABELA 11 - Quociente metabólico (qCO2) da biomassa microbiana de Latossolo Amarelo sob diferentes
sistemas de manejo e uso do solo coletado em diferentes profundidades. Igarapé-Açu (PA).
Data Prof 0-5 cm Prof 5-10 cm Média
qCO2 (x 10-5
C-BM-1
h-1
)
Janeiro/2009
Junho/2009
Dezembro/2009
8,20 aA
10,91 aA
6,27 aA
8,12 bA
14,69 aA
3,97 bA
8,16 b
12,80 a
5,12 b
Média 8,45 A 8,93 A
Obs: Médias seguidas da mesma letra minúscula dentro da coluna e maiúscula na linha não diferem entre si pelo
Teste de Duncan a 5% de probabilidade.
4.3 ANÁLISE MULTIVARIADA PARA OS ATRIBUTOS QUÍMICOS E BIOLÓGICOS
Figura 13, que mostra os resultados da análise de componentes principais (ACP) para
os atributos químicos do solo na profundidade de 0-5 cm mostra que a distribuição das
variáveis selecionadas apresentou variação acumulada de 84,2% para os eixos F1 e F2, sendo
que o eixo F1 foi capaz de explicar 49,3% e o F2, 34,9% dessa variância. Os atributos
químicos do solo mais fortemente correlacionados com o primeiro eixo de ordenação foram

76
CTCefetiva, SB, Ca, Mg, K e Mg. Os demais atributos apresentaram menor correlação com esse
eixo, tendo, portanto, menor importância para explicar esse componente principal.
O segundo eixo teve como atributos mais relevantes o C, N, H e Al. As demais
variáveis apresentaram baixa correlação com este eixo. Os atributos CTCefetiva, SB, Ca, Mg, K
e Mg, foram os mais discriminantes na avaliação da variação total entre os sistema de uso do
solo avaliados. A dispersão gráfica dos tratamentos em relação aos dois eixos demonstra que
houve dissimilaridade entre os sistemas de manejo adotados em relação aos atributos
químicos na profundidade de 0-5 cm do solo. Os tratamentos T1 (Cumaru-trituração) e T2
(Cumaru-queima) localizam-se à direita do diagrama, próximos a CP1, enquanto os demais
tratamentos agruparam-se à esquerda. Estes resultados são decorrentes dos menores valores
de CTCefetiva, SB, Ca, Mg, K e Mg, observados nestes tratamentos, tendo em vista que estas
variáveis foram as mais importantes para explicação da variação total dos resultados.
Figura 13 - Análise de Componentes Principais (ACP) realizada para as propriedades químicas de um Latossolo
Amarelo de Igarapé-Açu (PA), sob diferentes sistemas manejo e uso do solo, coletado em janeiro de 2009, na
profundidade de 0-5 cm. Média de quatro repetições para cada atributo químico. (a) Circulo de correlações entre
as variáveis químicas do solo. (b) Diagrama de ordenação das amostras em função dos tratamentos.
Significiância estatisticas dos agrupamentsos, p = 0,001, observação = 0,78

77
O posicionamento do tratamento T3 (capoeira) à direita do gráfico, próximos da CP2,
demonstra que C, N, H, e Al, atributos mais correlacionados com o segundo eixo, foram
características de maior influência para diferenciar este tratamento dos demais, conforme
pode ser observado nas Tabelas 1 e 2. Observa-se comportamento antagônico entre as
variáveis V, pH, P e Al, m%, o que evidencia a relação indireta entre elas, em que os maiores
valores de V, pH, P correspondem aos menores valores percentuais de Al, m%, estando de
acordo com o preconiza a literatura (MELLO et al., 1989; RAIJ, 1981).
Por meio da Figura 14, verifica-se que a análise de componentes principais (ACP) para
os atributos microbianos (CBM, CBM:C, CO2 e qCO2) do solo, profundidade de 0-5 cm,
apresentou autovalores de 60,2 % para o primeiro eixo (horizontal CP1) e 30,3 % para o
segundo eixo (vertical, CP2 ), o que representa 90,5% da variação acumulada nos dois
primeiros eixos. Os atributos do solo mais fortemente correlacionados com o primeiro eixo de
ordenação foram CBM, CBM:C e qCO2. O segundo eixo teve como atributo mais relevante o
CO2. De maneira geral, pode-se visualizar tendência para o tratamento T6 (SAF-trituração)
ficar do lado direito do gráfico, em correlação mais com CO2 e qCO2, o que indica maiores
valores no SAF. Observou-se no T6 (SAF-trituração) menores relações CBM:C e o maior
qCO2 (Tabela 9), sugerindo menor eficiência da biomassa microbiana na imobilização de C.
O posicionamento dos tratamentos T1 (Cumaru-trituração) e T4 (FEIGA/FEIGA-
trituração) próximos do CP2 indicam, também, que estes tratamentos apresentam elevada
atividade microbiana. O posicionamento dos tratamentos T3 (capoeira), T1 (Cumaru-
trituração), T5 FEIGA/UFRA-queima) e T4 (FEIGA/UFRA-trituração) à esquerda do gráfico,
próxima da CP2, demonstra que CBM e CBM:C, foram as características de maior influência
para diferenciação destes tratamentos. A dispersão gráfica dos tratamentos em relação os dois
eixos demonstra que houve dissimilaridade entre os sistemas de uso do solo em relação aos
atributos avaliados.
Quanto à análise de componentes principais (ACP) para os atributos químicos na
profundidade de 5-10 cm, verificou-se que a distribuição das variáveis selecionadas
apresentou variação acumulada de 73,3% para os eixos F1 e F2, sendo que o eixo F1 foi capaz
de explicar 51,1% e o F2, 22,2% dessa variância (Figura 15). Os atributos químicos do solo
mais fortemente correlacionado com o primeiro eixo de ordenação à esquerda do gráfico
foram CTCefetiva, SB, Ca, Mg, K e Mg, e à direita do gráfico foram m% e Al, mostrando
claramente a relação inversa que existe entre esses atributos. Os demais atributos

78
apresentaram menor correlação com esse eixo, tendo, portanto, menor importância para
explicar esse componente principal. O segundo eixo teve como atributos mais relevantes o N,
H, Na e C. As demais variáveis apresentaram baixa correlação com este eixo. Os teores de
CTCefetiva, SB, Ca, Mg, K e Mg, foram novamente as variáveis mais discriminantes na
avaliação da variação total entre os sistema de uso do solo avaliados. Observam-se para o T3
(capoeira) valores maiores de m% e Al seguido dos tratamentos T1 (Cumaru-trituração). A
dispersão gráfica dos tratamentos entre os dois eixos demonstra que houve dissimilaridade
entre as variáveis estudadas nesta profundidade. O tratamento que mais se correlacionou com
a CP2 foi o T4 (FEIGA/UFRA-trituração), o que evidencia maiores teores dessas variáveis
nesse tratamento e na profundidade estudada, conforme pode ser melhor observados por meio
dos dados apresentados na Tabela 1.
Figura 14 - Análise de Componentes Principais (PCA) realizada para as propriedades biológicas de um
Latossolo Amarelo de Igarapé-Açu (PA), sob diferentes sistemas de manejo e uso do solo, coletado em janeiro
de 2009, na profundidade de 0-5 cm. Média de quatro repetições para cada atributo. (a) Circulo de correlações
entre as variáveis biológicas do solo. (b) Diagrama de ordenação das amostras em função dos tratamentos.
Significiância estatisticas dos agrupamentsos, p = 0,019, a observação = 0,41

79
A análise de componentes principais (ACP) para os atributos microbianos (CBM,
CBM:C, CO2 e qCO2) do solo, profundidade de 5-10 cm, apresentou autovalores de 64,6%
para o primeiro eixo (horizontal CP1) e 26,3% para o segundo eixo (vertical, CP2 ), o que
representa 90,9% da variação acumulada nos dois primeiros eixos (Figura 16). Os atributos do
solo mais fortemente correlacionados com o primeiro eixo de ordenação foram CBM, CBM:C
e qCO2. O segundo eixo teve como atributo mais relevante o CO2. De maneira geral, pode-se
visualizar tendência para agrupamento de todos os tratamentos o que caracteriza que não
ocorreu dissimilaridade entre os mesmos.
Quanto à análise de componentes principais (ACP) para os atributos químicos e
biológicos na profundidade de 0-5 cm, verificou-se que a distribuição das variáveis
selecionadas apresentou variação acumulada de 71,5% para os eixos F1e F2, sendo que o eixo
F1 foi capaz de explicar 40,0% e o F2, 31,5% dessa variância (Figura 17).
Figura 15 - Análise de Componentes Principais (PCA) realizada para as propriedades químicas de um Latossolo
Amarelo de Igarapé-Açu (PA), sob diferentes sistemas manejo e uso do solo, coletado em Janeiro de 2009, na
profundidade de 5-10 cm. Média de quatro repetições para cada atributo(a) Circulo de correlações entre as
variáveis químicas do solo. (b) Diagrama de ordenação das amostras em função dos tratamentos. Significiância
estatisticas dos agrupamentsos, p = 0,001, a observação = 0,72

80
Os atributos químicos e biológicos do solo que mais correlacionaram com o primeiro
eixo de ordenação foram CTCefetiva, SB, Ca, Mg, K, Mg e CBM:C. Os demais atributos
apresentaram menor correlação com esse eixo, tendo, portanto, menor importância para
explicar esse componente principal.
O segundo eixo teve como atributos mais relevantes para o quadrante superior o qCO2,
CO2, P e o pH; porém estes dois últimos atributos com menor relevância. Para o segundo
quadrante ainda na CP2 verificaram-se as variáveis CBM, H, Al, N e C. As demais variáveis
apresentaram baixa correlação com este eixo. Os teores de CTCefetiva, SB, Ca, Mg, K e Mg e
CBM:C foram as variáveis mais discriminantes na avaliação da variação total entre os sistema
de uso do solo avaliados.
Figura 16 - Análise de Componentes Principais (PCA) realizada para as propriedades biológicas de um
Latossolo Amarelo de Igarapé-Açu (PA), sob diferentes sistemas de manejo e uso do solo, coletado em janeiro
de 2009, na profundidade de 5-10 cm. Média de quatro repetições para cada atributo. (a) Circulo de correlações
entre as variáveis biológicas do solo. (b) Diagrama de ordenação das amostras em função dos tratamentos.
Significiância estatisticas dos agrupamentsos, p = 0,145, a observação = 0,31

81
A dispersão gráfica dos tratamentos em relação aos dois eixos demonstra que houve
dissimilaridade entre os sistemas de manejo adotados em relação aos atributos químicos e
biológicos na profundidade de 0-5 cm do solo. Os tratamentos T1 (Cumaru-trituração) e T2
(Cumaru-queima) localizam-se a esquerda do diagrama, próximo da CP1, demonstrando que
CTCefetiva, SB, Ca, Mg, K, Mg, foram os atributos que menos se correlacionaram com estes
tratamentos, tendo em vista que estas variáveis foram as mais importantes para explicação da
variação total dos resultados.
No entanto, o T1 (Cumaru-trituração) foi o que apresentou maior similaridade com
quociente microbiano (CBM;C), conforme se observa na Figura 17, o que é evidenciado
também na análise de variância (Anexo 4). Para a CP2 o tratamento que mais se
correlacionou foi T3 (capoeira de 15 anos), observando-se maior correlação com os atributos
Al, CBM, H, N e C.
Figura 17 - Análise de Componentes Principais (PCA) realizada para as propriedades químicas e biológicas de
um Latossolo Amarelo de Igarapé-Açu (PA), sob diferentes sistemas manejo e uso do solo, coletado em janeiro
de 2009, na profundidade de 0-5 cm. Média de doze repetições para cada atributo. (a) Circulo de correlações
entre as variáveis químicas e biológicas do solo. (b) Diagrama de ordenação das amostras em função dos
tratamentos. Significiância estatisticas dos agrupamentsos, p = 0,001, a observação = 0,70

82
Os tratamentos que mais apresentaram similaridade com as variáveis mais
discriminantes, na avaliação da variação total, entre os diferentes usos do solo foram
tratamento T5, T4 e T6, respectivamente (FEIGA/UFRA-queima), (FEIGA/UFRA-
trituração) e (SAF). Ao contrário dos resultados encontrados neste estudo DINESH et al.
(2003), também utilizando a análise de componentes principais para interpretar a relação entre
atributos químicos e microbianos de solos sob diferentes coberturas florestais, verificaram no
primeiro eixo maiores correlações entre CBM, C e N, e sugeriram que a associação entre
essas variáveis caracterizaria a atividade microbiana e a dependência dos microrganismos do
solo pelos teores de matéria orgânica e N total. Barreto et al., (2008) verificaram que os teores
de C orgânico e N total foram as variáveis mais discriminantes na avaliação da variação total
entre os solos sob eucalipto de diferentes idades. Segundo os autores esses resultados
contrariam comprovações de que a biomassa microbiana é um indicador mais sensível de
mudanças nos teores de MOS do que o teor de C orgânico do solo (POWLSON; BROOKES;
CHRISTENSEN, 1987; ANDERSON; DOMSCH, 1989; SPARLING, 1992, GAMA-
RODRIGUES, 1999). No entanto, Zatorre et al., (2009), também utilizando a análise de
componentes principais para interpretar a relação entre atributos químicos, físicos e
microbianos de solos sob diferentes coberturas florestais: mata, pastagem e capoeira no Acre,
verificaram que as variáveis biológicas mostraram-se bem mais sensíveis às mudanças na
cobertura do solo.
. Os teores de CTCefetiva, SB, Ca, Mg, K, Mg e CBM:C foram as variáveis mais
discriminantes na avaliação de variação total entre os diferentes usos do solo nesta
profundidade e na época de amostragem (período seco-Jan2009).

83
5 CONCLUSÕES
Os dados obtidos no presente estudo permitem as seguintes conclusões:
a) Soma de bases e teores de Ca, Mg e K trocáveis foram os atributos químicos mais
influenciados pelos sistemas que sofreram corte (com queima e com trituração da
vegetação);
b) CTC efetiva, soma de bases, Ca, Mg, K trocáveis e razão CBM:C, foram as variáveis mais
sensíveis às mudanças dos sistemas de uso do solo, quando a amostragem foi realizada em
janeiro/2009 (período seco);
c) A variação sazonal limitou a disponibilidade de nutrientes e matéria orgânica no solo das
áreas estudadas;
d) A variação sazonal e a profundidade de amostragem interferiram na imobilização de C
pela comunidade microbiana do solo;
e) A umidade foi o fator que mais influenciou e limitou o desenvolvimento das
populações microbianas no solo;
f) Os valores de CBM foram superiores na época mais chuvosa, independente do sistema
e da profundidade de amostragem;
g) Os altos valores para a razão CBM:C encontrados nos sistemas utilizando corte e
trituração da vegetação, e na vegetação secundária, indicam que está havendo acúmulo de C
nesses sistemas;
h) O sistema de manejo que utilizou corte e trituração da vegetação, no preparo do solo, se
assemelhou à floresta secundária e ao sistema com queima da vegetação, quanto à eficiência
na ciclagem de nutrientes e aporte de matéria orgânica, configurando-se como alternativa
viável de manejo sustentável.

84
REFERÊNCIAS
ALEF, K Nitrogen mineralization in soils. In: ALEF, K.; NANNIPIERI, P. (Ed). Methods in
applied soil microbiology and biochemistry. London: Academic Press, 1995. p.234-245.
ALFAIA, S. S.; SOUZA, L. A. G. Perspectivas de uso e manejo dos solos na Amazônia. In:
ARAÚJO, Q. R. (Ed). 500 Anos de uso do solo no Brasil. Ilhéus: Editus, 2002. p.311-327.
ALLEN, A. S.; SCHLESINGER, H. Nutrient limitations to soil microbial biomass and
activity in loblolly pine forests. Soil Biology and Biochemistry, v.36, p.581-589, 2004.
ALVAREZ, C. R. ; ALVAREZ, R. ; GRIGERA, M. S. ; LAVADO, R. S. Associations
between organic matter frations and the active soil microbial biomass. Soil Biology &
Biochemistry, v.30, n.6, p.767-773, 1998.
ALVAREZ, R.; DÍAZ, R.A.; BARBERO, N.; SANTANATOGLIA, O.J.; BLOTTA, L. Soil
organic carbon, microbial biomass and CO2–C production from three tillage systems. Soil &
Tillage Research, v.33, p.17–28, 1995.
ALVAREZ, R.; SANTANATOGLIA, O. J.; P. E.; GARCIA, R. Respiration and specific
activity of soil microbial biomass under conventional and reduced tillage. Pesquisa
Agropecuária Brasileira, v.30, n.5, p.701-709, 1995.
ANDERSON, J. P. Soil respiration. In: PAGE, A. L. ; MILLER, R. H.; KEENEY, D. R. (Ed)
Methods of soil analysis. American Society of Agronomy. p.831-871, 1982.
ANDERSON, T. H.; DOMSCH, K. H. Ratios of microbial biomass carbon to total organic
carbon in arable soils. Soil Biology Biochemistry, v.21, p.471-479, 1989.
ANDERSON, T. H.; DOMSCH, K.H. Determination of eco-physiological maintenance
requirements of soil microorganisms in a dormant stage. Biology and Fertility of Soils, v.11,
p.81-89, 1985.
ANDERSON, T. H.; DOMSH, K. H. The metabolic quotient for CO2 (qCO2) as a specific
activity parameter to assess the effects of environmental conditions, such as pH, on the
microbial biomass of forest soils. Soil Biology and Biochemistry, v.25, p.393-395, 1993.

85
ANDERSON, T. H. Microbial eco-physiological indicators to assess soil quality.
Agriculture, Ecosystems and Environment, v.98, p.285-293, 2003.
ANGERS, D. A.; BISSONNETTE, N.; LÉGÈRE, A.; SAMSON, N. Microbial and
biochemical changes induced by rotation and tillage in a soil under barley production.
Canadian Journal of Soil Science, v.73, p.39-50, 1993.
ARAÚJO, E. A.; LANI, J. L.; AMARAL, E. F.; GUERRA, A. Uso da terra e propriedades
físicas e químicas de Argissolo Amarelo distrófico na Amazônia Ocidental. Revista
Brasileira de Ciência do Solo, Viçosa, v.28, n.2, Mar./Apr. 2004.
BAILEY, V. L.; SMITH, J. L.; BOLTON- JR., H. Fungal-to-bacterial ratios in soils
investigated for enhanced C sequestration. Soil Biology & Biochemistry, v.34, p.997–1007,
2002.
BALIEIRO, F. C.; FRANCO, A. A.; DIAS, P. F.; SOUTO, S. M.; CAMPELLO, E. F. C.
Sistemas agrossilvipastoris: a importância das leguminosas arbóreas para as pastagens na
região centro-sul. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE
ZOOTECNIA, 41., 2004, Campo Grande. Anais... Campo Grande: SBZ, 2004.
BALOTA, E. L.; COLOZZI-FILHO, A.; ANDRADE, D. S.; HUNGRIA, M. Biomassa
microbiana e sua atividade em solos sob diferentes sistemas de preparo e sucessão de culturas.
Revista Brasileira Ciência Solo, v.22, p.641-649, 1998.
BALOTA, E. L.; COLOZZI-FILHO, A.; ANDRADE, D. S.; HUNGRIA, M. Biomassa
microbiana e sua atividade em solos sob diferentes sistemas de preparo e sucessão de culturas.
Revista Brasileira de Ciência do Solo, v.22, p.641–649, 1998.
BARDGETT, R. D. Causes and consequences of biological diversity in soil. Zoology, v.105,
p.367-374, 2002.
BARRETO, P. A. B.; GAMA-RODRIGUES, E. F.; GAMA-RODRIGUES, A. C.; BARROS,
N. F.; FONSECA, S.; Atividade microbiana, carbono e nitrogênio da biomassa microbiana
em plantações de eucalipto, em seqüência de idades. Revista Brasileira de Ciência do Solo,
v.32, p.611-619, 2008.
BARROS, N. F.; COMERFORD, N. B. Sustentabilidade da produção de florestas plantadas
na região tropical. In: ALVAREZ V. V. H.; SCHAEFER, C. E. G. R.; BARROS, N. F.;
MELLO, J. W. V.; COSTA, L. M. (Ed). Tópicos em ciência do solo. Sociedade Brasileira de
Ciência do Solo, Viçosa, 2002. v.2. p.487-592.

86
BEARE, M. H. Fungal and bacterial pathways of organic matter decomposition and nitrogen
mineralization in arable soils. In: BRUSSAARD, L.; FERRERA-CERRATO, R. (Ed), Soil
Ecology in Sustainable Agricultural Systems. CRC/Lewis Publishers, Boca Raton, FL,
1997. p.37–70.
BESSHO, T.; BELL, L. C. Soil solid and solution phase changes and mung bean response
during amelioration of aluminium toxicity with organic matter. Plant and soil, v.140, n.2.
p.183-196, 1992.
BORN, R. H.; PICCHIONI, S.; PIVA, L. Mudanças climáticas e o Brasil. Contribuições e
diretrizes para incorporar questões de mudanças de clima em políticas públicas, 2007.
Disponível em: <http://www.vitaecivilis.org.br/anexos/
mudancas%20climaticas%5B1%5D.pdf>. Acesso em:10 mar 2010.
BRADY, N. C.; WEIL, R. R. The Nature and properties of soils. Thirteenth ed. Prentice
Hall, New Jersey, 2002, 960p.
BRADY, N. C. Alternatives to slash-and-burn: a global imperative. Agriculture,Ecosystems
and Environment, v.58, p. 3-11, 1996.
BROKES, P. C.; LANDMAN, A.; PRUDEN, G.; JENKINSON, D.S. Chloroform fumigation
and the release of soil nitrogen; a rapid direct extraction method to measure microbial
biomass nitrogen in soil. Soil Biology and Biochemistry, v.17, p.837-842, 1985.
CALDERÓN, F. J. Short-term dynamics of nitrogen, microbial activity, and phosphoolipid
fatty acids after tillage. Soil Science Society of America Journal, Madison, v.65, n.1, p.118-
126, 2001.
CARAZO, V. R. et al (Ed). Taller internacional sobre el estado actual y potencial de manejo
ydesarrollo del bosque secundario tropical em america latina. 1997. Pucallpa. Memórias...
Pucallpa: Secretaria Pro Tempore Venezuela. 1997.
CARDOSO JÚNIOR, E. Q. Alterações físicas e químicas de solos sob efeito de manejos
para recuperação de pastagem (Brachiaria humidicola, Rendle), no município de
castanhal, Pará. 2001. 105f. Dissertação (Mestrado em Agronomia) - Faculdade de Ciências
Agrárias do Pará, Belém, 2001.
CARDOSO, M. O. Métodos para quantificação da biomassa microbiana no solo.
Agropecuária Técnica, v.25, n.1, p.1-12, 2004.

87
CATTELLAN, A. J.; VIDOR, C. Sistemas de culturas e a população microbiana do solo.
Revista Brasileira de Ciência do Solo, v.14, n.2, p.125-132, 1990.
CERRI, C. C.; VOLKOFF, B.; EDUARDO, B. P. Efeito do desmatamento sobre a biomassa
microbiana em Latossolo amarelo da Amazônia. Revista Brasileira de Ciência do Solo, v.9,
p.1-14, 1985.
CHOKKALINGAM, U.; DE JONG, W. Secondary forest: a working definition and typology.
International Forestry Review, v.3, n.1, p.19-26, 2001.
COLFER, C. J. et al. Introduction: History and conceptual framework. In: COLFER, C. J. ;
BYRON, Y. (Ed). People managing forests. Resources for the future, Washington, DC:
Center for International Forestry Research, 2001, p 1-49.
COLOZZI-FILHO, A.; ANDRADE, D. V.; BALOTA, E. L. Atividade microbiana em solos
cultivados em sistemas de plantio direto. Informe Agropecuário, Belo Horizonte, v.22. n.
208, p.84-91, 2001.
COMISION NACIONAL DE CERTIFICACION FORESTAL - CNCF. Estándares y
procedimientos para el manejo sostenible y la certificación forestal en Costa Rica. San
José, 1999. 54p.
CORREA, M. E. F.; ANDRADE, A. G. Formação de serrapilheira e ciclagem de nutrientes.
In: SANTOS, G. A.; CAMARGO, F. A. O. Fundamentos da matéria orgânica do solo:
ecossistemas tropicais e subtropicais. Porto Alegre: Genesis, 1999. 508p.
COSTA, F. S. et al. Propriedades físicas de um Latossolo Bruno afetadas pelos sistemas
plantio direto e preparo convencional. Revista Brasileira de Ciência do solo, v.27, n.3,
p.527-535, 2003.
DALAL, R. C. Soil microbial biomass – what do the numbers really mean? Australian
journal of Experimental Agriculture, v.38, p.649-665, 1998.
DENICH, M. Estudo da importância de uma vegetação secundária nova para o
incremento da produtividade do sistema de produção na Amazônia oriental brasileira.
EMBRAPA/ CPATU; GTZ: Eschborn, 1991. 284p.
DE-POLLI, H.; GUERRA, J. G. M. Biomassa: Perspectivas para o uso e manejo do solo. In:
ALVAREZ V.; V. H.; FONTES, L. E. F.; FONTES, M. P. F. (Ed). O solo nos grandes

88
domínios morfoclimáticos do Brasil e o desenvolvimento sustentado. Viçosa: Sociedade
Brasileira de Ciência do Solo/UFV, 1996, p.551-564.
DE-POLLI, H.; GAMA-RODRIGUES, E.F da; GUERRA, J.G.M. Determinação da
Biomassa Microbiana do Solo: Avanços e Limitações. XXIV Reunião Brasileira de
Fertilidade do Solo e nutrição de Plantas, Santa Maria-RS. Simpósio, 2000.
DENICH, M. Untersuchungen zur Bedeutung junger Sekundärvegetation für die
Nutzungssystemproduktivität im östlichen Amazonasgebiet,Brasilien. Göttingen:
Universität Göttingen, 1989 (Göttinger Beiträge zur Land- und Forstwirtschaft in den Tropen
und Subtropen 46).
DINESH, R.; GHOSHAL CHAUDHURI, S.; GANESHAMURTHY, A. N.; CHANCHAL
DEY. Changes in soil microbial indices and their relationships following deforestation and
cultivation in wet tropical forests. Soil Biology Biochemistry, v.24, p.17-26, 2003.
DENICH, M.; KANASHIRO, M. Secondary Vegetation in the Agricultural Landscape of
Northeast Pará, Brazil. In: PARROTTA, J. A.; KANASHIRO, M. (Ed.). Proceedings of an
International Symposium/Workshop Management and Rehabilitation of Degraded
Lands and Secondary Forests in Amazonia“, 18-22 April 1993, Santarém. Rio Piedras:
EMBRAPA-CPATU, International Institute of Tropical Forestry, USDA Forest Service,
1995.
DENICH, M; KANASHIRO, M.; VLEK, P. L. G. The potential and dynamics of carbon
sequestration in traditional and modified fallow systems of the Eastern Amazon region,
Brazil. In: Lal, R.; Kimble, M. J.; Stewart, B. A. (Editors). Global climate change and
tropical ecosystems. CRC Press, Boca Raton, p.213-229. 1999.
DORAN, J. W. Soil microbial and biochemical changes associated with reduced tillage. Soil
Science Society American Journal, v.44, p.765-771, 1980.
DUARTE, A. N. Avaliação da biomassa microbiana de um Argissolo sob diferentes
sistemas agroflorestais instalados no município de Igarapé-Açú - Pará. 2003. 84f.
Dissertação (Mestrado em Agronomia) - Universidade Federal Rural da Amazônia, Belém.
2003.
DUBOC, E. Sistemas agroflorestais e o cerrado, 2009. Disponível em:
<http://simposio.cpac.embrapa.br/simposio/projeto/palestras/ capitulo_31.pdf>. Acesso em:
09 fev. 2010.

89
EKENLER, M.; TABATABAI, M. A. Tillage and residue management effects on
ßglucosaminidase activity in soils. Soil Biology & Biochemistry, v.35, p.871-874, 2003.
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA /CPATU. Folder Produzir
sem queimar – Tipitamba. Belém-Pa, 2001.
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Sistema Brasileiro de
Classificação de Solos. 421p. Brasília, DF: Embrapa Produção de Informação; Rio de
Janeiro: EMBRAPA – SNPS, 2006.
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Sistemas agroflorestais na
Amazônia brasileira, 2003. Disponível em: http:// www.cpafro.embrapa.br/
Artigos/sist_agro_amaz.html. Acesso em: 09 mai. 2009.
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Sistema Brasileiro de
Classificação de Solos. 412p. Brasília: Serviço de Produção de Informação/Embrapa –
Centro Nacional de Pesquisa de Solos, 1999.
FACCI, L. D. 2008. 101f. Variáveis Microbiológicas como Indicadoras da Qualidade do
Solo SOB Diferentes Usos. Dissertação (Mestre em Agricultura Tropical e Subtropical) -
Instituto Agronômico de Campinas. Campinas, SP. 2008.
FAO. Situación de los bosques del mundo.: Situación y acontecimientos recientes em el
sector forestal. 2005, part. I, 20p.
FEARNSIDE, P. M. Desmatamento na Amazônia brasileira: história, índices e
conseqüências. Megadiversidade, v.1, n.1, p.114-123, 2005.
FEIGL, B. J.; SPARLING, G. P.; ROSS, D. J.; CERRI, C. C. Soil microbial biomass in
Amazon soils: evaluation of methods andestimates of pool sizes. Soil Biology and
Biochemistry, v.27, n.11, p.1467-1472, 1995.
FERREIRA, C. P.; KATO, O. R.; COSTA, C. A. C. Carbono orgânico, nitrogênio e a razão
C/N em solo sob sistemas agroflorestais no nordeste paraense. In: Congresso Brasileiro de
Sistemas Agroflorestais, 5, 2004, Curitiba. CD-Rom.Curitiba: EMBRAPA, 2004
FERREIRA, C. P. Atributos físicos-hídricos e químicos do solo em sistemas agrícolas na
microrregião de Castanhal, Pará. 2006. 145f. Tese (Doutorado em Ciências Agrárias) -
Universidade Federal Rural da Amazônia. Belém, PA. 2006.

90
FINEGAN, B. The managememt potemtial of neotropical secondary lowand rain forest.
Forest Ecology and Managememt, v.47, p.295-321, 1992.
FRANCHINI, J. C.; CRISPINO, C. C.; SOUZA, R. A.; TORRES, E.; HUNGRIA, M.
Microbiological parameters as indicators of soil quality under various soil management and
crop rotation systems in southern Brazil. Soil & Tillage Research, v.92, p.18–29, 2007.
FREY, S. D.; ELLIOTT, E. T.; PAUSTIAN, K. Bacterial and fungal abundance and biomass
in conventional and no-tillage agroecosystems along two climatic gradients. Soil Biology &
Biochemistry, v.31, p.573–585, 1999.
GALLARDO, A.; SCHLESINGER, W.H. Estimating microbial biomass nitrogen using the
fumigation-incubation and fumigation-extration method in awarm-temperate forest soil. Soil
Biology and Biochemistry, v.22, p.927-932, 1990.
GAMA, M. A. P. Dinâmica do fósforo em solo submetido a sistemas de preparo
alternativos ao de corte e queima no nordeste paraense. 2002. 110 f. Tese Doutorado.
ESALQ, Piracicaba, 2002.
GAMA-RODRIGUES, A. C.; BARROS, N. F.; GAMA-RODRIGUES, E. F.; FREITAS, M.
S. M.; VIANA, A. P.; JASMIN, J. M.; MARCIANO, C. R.; CARNEIRO, J. G. A. (Org.).
Sistemas Agroflorestais: Bases Cientificas para o Desenvolvimento Sustentável. Campos
dos Goytacazes, RJ: Universidade Estadual do Norte Fluminense Darcy Ribeiro, 2006. 365 p.
GAMA-RODRIGUES, E. F.; BARROS, N. F.; RODRIGUES, A. C. G.; SANTOS, G. A.
Nitrogênio, carbono, e atividade da biomassa microbiana do solo em plantações de eucalipto.
Revista Brasileira de Ciência do Solo, v.29, n.6, p.823-902, 2005.
GAMA-RODRIGUES, E. F. Biomassa microbiana e ciclagem de nutrientes. In: SANTOS, G.
A.; CAMARGO, F. A. O.(Ed). Fundamentos da matéria orgânica do solo: Ecossistemas
tropicais e subtropicais. Porto Alegre : Gênesis, 1999. p.227-243.
GAMA-RODRIGUES, E. F. Biomassa microbiana e ciclagem de nutrients, In: SANTOS, G.
A.; CAMARGO, F. A. O. Fundamentos da matéria orgânica do solo: ecossistema tropicais
e subtropicais. Porto Alegre : Gênesis, 1999. p.227-243.
GONÇALVES, A. S.; MONTEIRO, M. T.; GUERRA, J. G. M.; DE-POLLI, H. Biomassa
microbiana em amostras de solos secos secadas ao ar e reumedecidas. Pesquisas
Agropecuária Brasileira. v.37, n.5, p.651-658, 2002.

91
GRACE, J. Understanding and managing the global carbon cycle. Journal of Ecology, v.92,
p.189-202, 2004.
GREGORICH, E. G.; WEN, G.; VORONEY, R. P.; KACHANOSKI, R. G. Calibration of
rapid direct chloroform extraction method for measuring soil microbial biomass C. Soil
Biology and Biochemistry, v.22, p.1009-1011, 1990.
GRISSI, B. M.; GRAY, T. R. G. Comparação dos métodos de fumigação, taxa de respiração
em resposta à glicose e conteúdo de ATP para estimar a biomassa microbiana do solo.
Revista Brasileira de Ciência do Solo, v.10, p.109-115, 1996.
GUPTA, V. V. S. R. ; GERMIDA, J. J. Distribuition of microbial biomass and its activity in
different soil aggregate size classes as affected by cultivation. Soil Biology and
Biochemistry, v.20, p.777-786, 1988.
HAYNES, R. J. Size and activity of the soil microbial biomass under grass and arable
management. Biology and Fertility of Soils, v.30, p.210–216, 1999.
HÖLSCHER , D. ; LUDWIG, B. ; MOLLER, M. R. F. ; FOLSTER, H.H Dynamic of soil
chemical parameters in shifting agriculture in the Eastern Amazon Agric Ecosyst. Environ,
v.66, p.153-163, 1997.
HOLSCHER, D.; LUWING, B.; MOLLER, R. F.; FOLSTER, H. Dynamic of soil chemical
parameters in shifting agriculture in the Eastern Amazon. Agriculture, Ecosystems and
Environment, v.66, p.153-163, 1997.
HÖLSCHER, D.; MOELLER, R. F.; DENICH, M.; FÖLSTER, H. Nutrient input-output
budget of shifting agriculture in eastern Amazonia. Nutrient Cycling in Agroecosystems
v.47, p.49-57, 1997.
HOUGHTON, R. A.; SKOLE, D. L.; NOBRE, C. A.; HACKLER, J. L.; LAWRENCE, K. T.
CHOMETOWSKI, W. H. Annual fluxes of carbon from deforestation and regrowth in the
Brazilian Amazon. Nature, v.403, p.301-304, 2000.
HUNGRIA, M.; FRANCHINI, J. C.; BRANDÃO-JUNIOR, O.; KASCHUK, G.; SOUZA, R.
A. Soil microbial activity and crop sustainability in a long-term experiment with three soil-
tillage and two crop-rotation systems. Applied Soil Ecology, v.42, p.288–296, 2009.
ICRAF. Trees of change – Corporate Report 2000. 73p. Nairobi: ICRAF, 2000.

92
IHAKA, R.; GENTLEMAN, R. A language for data analysis and graphics. Journal of
Computational Graphical Statistics. v.5, p.299-314, 1996.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Estimativas da
População. Disponível em:.< http://www.ibge.gov.br>. Acesso em: 20 nov. 2009.
INSAM, H.; DOMSCH, K. H. Relationship between soil organic C and microbial biomass on
chronosequeces of reclamation sites. Microbial Ecology v.15, p.177-188, 1988.
INSAM, H. ; HASELWANDTER, K. Metabolic quotient of the soil microflora in relation to
plant succession. Oecologia, v.79, p.174–178, 1989.
JASTROW, J. D.; AMONETTE, J. E.; BAILEY, V. L. Mechanisms controlling soil carbon
turnover and their potential application for enhancing carbon sequestration. Climatic
Change, v.80, p.5-23, 2007.
JENKINSON, D. S. ; LADD, J. N. Microbial biomass in soil: measurement and turnover. In:
PAUL, E.A.; LADD, J.N. (Ed). Soil biochemistry. New York: Marcel Dekker, 1981. v.5,
p.415-471.
JENKINSON, D. S. ; RAYNER, J. H. The turnover of organic matter in some of the
Rothamsted Classical Experiments. Soil Science, v.123, p.298-305, 1977.
JOHNSON, C. M. ; VIEIRA, I. C. G. ; ZARIN, J. F. ; JOHNSON, A. H. Carbon and nutrient
storage in primary and secondary forests in eastern Amazonia. Forest Ecology and
Management. v.147, p.245-252, 2001.
JUO, A. S. R.; MANU, A. Chemical dynamics in slash and burn agriculture. Agriculture,
Ecosystems and Environmentn, v.58, p.49-60, 1996.
KANASHIRO, M.; DENICH, M. Possibilidades de utilização e manejo adequado de áreas
alteradas e abandonadas na Amazônia brasileira. Brasília: MCT/CNPq, 1998. 157p.
(Estudos dos impactos humanos nas florestas e áreas inundadas nos trópicos).
KARLEN, D. L. ; WOLLENHAUPT, N. C. ; ERBACH, D. C. ; BERRY, E. C. ; SWAN, J.
B.; EASH, N. S. ; JORDAHL, J. L. Long-tem tillage effects on soil quality. Soil & Tillage
Research, v.32, p.313-327, 1994.

93
KASCHUK, G.; ALBERTON, O.; HUNGRIA, M. Three decades of soil microbial biomass
studies in Brazilian ecosystems: lessons learned about soil quality and indications for
improving sustainability. Soil Biology & Biochemistry, v.42, n.1, p.1-13, January, 2009.
KATO, M. S. A. ; KATO, O. R. ; DENICH, M. ; VLEK, P.L.G. Fire-free alternatives to
slash-and-burn for shifting cultivation in the eastern Amazon region: the role of fertilizers.
Field Crops Research. v.62, p.225-237, 1999.
KATO, O. R.; KATO, M. S. A.; CARVALHO, C. R. Figueiredo. Manejo de vegetação
secundaria na Amazônia visando ao aumento da sustentabilidade do uso agrícola do solo. In:
CONGRESSO BRASILEIRO DE CIÊNCIAS DO SOLO. 30., 2005. Recife. Anais... Recife:
Sociedade Brasileira d Ciência do Solo,2005.
KATO, O. R.; KATO, M. S. A.; SÁ, T. D. A.; FIGUEIREDO, R. Plantio direto na capoeira.
Ciência e ambiente, v.29, p.99-111, 2000.
KATO, O. R. et al. Método de preparo de área sem queima: uma alternativa para
agricultura tradicional da Amazônia Oriental. Belém: EMBRAPA/CPATU, 1999(b). 3p.(
Comunicado Técnico, 13).
KATO, O. R. Preparo de área sem queima, uma alternativa para a agricultura de
derruba e queima da Amazônia Oriental: Aspectos agroecológicos. In: SEMINÁRIO
SOBRE MANEJO DA VEGETAÇÃO SECUNDÁRIA PARA A SUSTENTABILIDADE
DA AGRICULTURA FAMILIAR DA AMAZÔNIA ORIENTAL, 1999, Belém, PA. Anais...
Belém: Embrapa Amazônia Oriental/CNPq, 2000. p.35-37. (Embrapa Amazônia Oriental.
Documentos, 69). 2000.
KATO, M.S.A. Fire-free land preparation as an alternative to slash-and-burn
agriculture in the bragantina region, eastern Amazon: Crop performance and
phosphorus dynamics. Doctoral dissertation. University Göttingen, 144 p. 1988.
Kato, O. R.; Kato, M. S. A.; Carvalho, C. R. Figueiredo. Manejo de vegetação secundaria
na Amazônia visando ao aumento da sustentabilidade do uso agrícola do solo. In: XXX
Congresso Brasileiro de Ciências do Solo. 2005.
KEENEY, D. R.; SAHRAWAT, K.L. Nitrogen transformations in floodedrice soils.
Fertilizer Research, v.9, p.15-38, 1986.

94
KENNEDY, A. C.; PAPENDICK, R.I. Microbial characteristics of soil quality. Journal of
soil and water conservation, v.50, n.3, p.243-248, 1995.
Kiladis, G.; Diaz, H. F. Globall climatic anomalies associated with extremes in the Southern
Oscillation. Journal of Climate, v. 2, n. 9, p. 1069-1089, Sept. 1989.
KLEINMAN, P. J. A.; PIMENTEl, D.; BRYANT, R.B. The ecological sustainability of
slashand-burn agriculture. Agriculture, Ecosystems and Environment, v.52, p.235-249,
1995.
KRISTENSEN, H. L. Effects of soil disturbance on mineralization of organic soil nitogen..
Soil Science Society of America Journal, Madison, v,64, n.1, p.371-378, 2000.
KUSHWAHA, C. P.; TRIPATHI, S. K.; SINGH, K.P. Soil organic matter and waterstabe
aggregates under different tillage and residue conditions in a tropical dryland agroecosystem.
Applied Soil Ecology, v.16, p.229-241, 2001.
LEITE, J. B. V.; VIRGENS, A. C. Sistemas agroflorestais no Sudeste da Bahia: uma revisão
conceitual, In: CONGRESSO BRASILEIRO DE SISTEMAS AGROFLORESTAIS, 4, 2002,
Ilhéus. Anais... Ilhéus: EMBRAPA, 2002. CD-Rom.
LOPES. A. S.; GUILHERME, L. R. G. Interpretação de análise do solo: conceitos e
aplicações. São Paulo: ANDA, 1992. (Boletim Técnico, 2).
LUCHESE, E. B.; FAVERO, L. O. B.; LENZI, E. A matéria organica nos solos. In:
Fundamentos da química do solo. Rio de Janeiro, 2001. p. 36-45.
LUIZÃO, R. C. C.; COSTA, E. S.; LUIZÃO, F. J. Mudanças na biomassa microbiana e nas
transformações de nitrogênio do solo em uma sequência de idades de pastagem após a derruba
e queima da floresta na Amazônia Central. Acta Amazônica, v.29, n.1, p.43-56. 1999.
LUIZÃO, R. C. C. Variações temporais da biomassa microbiana e aspectos da ciclagem
do nitrogênio em solos de floresta natural e de sistemas manejados na Amazônia central.
1989. 67f. Dissertação (Mestrado) - PPG INPA/FUA. 1989.
MACDICKEN, K. G.; VERGARA, N.T. Introduction to agroforestry. In: MACDICKEN,
K.G.; VERGARA, N. T. (Ed). Agroforestry: classification and management. New York:
John Wiley & Sons, 1990. p.1-30.

95
MACKENSEN, J.; HÖLSCHER, D.; KLINGE, R.; FÖLSTER, H. Nutrient transfer to the
atmosphere by burning of debris in eastern Amazonia. Forest Ecology and Management.
v.86, p.121-128, 1996.
MCDONALD M.A., HEALEY J.R.; STEVENS P.A. The effects of secondary forest
clearance and subsequent land-use on erosion losses and soil properties in the Blue Mountains
of Jamaica. Agric Ecosyst Environ, v92, p.1–19. 2002.
MALAVOLTA, E. Elementos de nutrição mineral de planta. São Paulo: Agronômica
Ceres. 251pp. 1980.
MARUMOTO, T.; ANDERSON, J. P. E. ; DOMSCH, K.H. Mineralization of nutrients from
soil microbial biomass. Soil Biology and Biochemistry, v.14, p.469-475, 1982.
MATSUOKA, M.; MENDES, I. C.; LOUREIRO, M. F. Biomassa microbiana e atividade
enzimática em solos sob vegetação nativa e sistemas agrícolas anuais e perenes na região de
Primavera do Leste (MT). Revista Brasileira de Ciência do Solo, v. 27, p.425-433, 2003.
MELLO, F. A. F.; SOBRINHO, M. O. C. B.; ARZOLLAS, S.; SILVEIRA, R. I. ; NETTO, A.
C.; KIEHL, J. C. Fertilidade do Solo. 3. ed. Piracicaba, 1985. 400p.
MENDONÇA, E. S.; LEITE, L. F. C.; FERREIRA NETO, P. S. Cultivo de café em sistema
agroflorestal: uma opção para a recuperação de solos degradados. Revista Árvore, v. 25,
p.375-383. 2001.
MERCANTE, F. M. Parâmetros microbiológicos como indicadores da qualidade do solo
sob sistemas integrados de produção agropecuária. Dourados: Embrapa Agropecuária
Oeste, 2004. (Boletim de Pesquisa e Desenvolvimento, 20).
MIELNICZUK, J.; BAYER, C.; BESAN, F. M.; LOVATO, T.; FERNÁNDEZ, F. F.;
DEBARBA, L. Manejo do solo e culturas e sua relação com os estoques de carbono e
nitrogênio do solo. In: CURI, N.; MARQUES, J. J.; GUILHERME, L. R. G.; LIMA, J. M.;
LOPES, A. S.; ALVAREZ, V. H. (Ed). Tópicos em ciência do solo. Viçosa, MG, Sociedade
Brasileira de Ciência do Solo, 2003, v. 3, p. 209-248.
MONTEIRO, M. T.; GAMA-RODRIGUES, E. F. Carbono, nitrogênio e atividade da
biomassa microbiana do solo sob diferentes ecossistemas florestais da região norte
fluminense, 2000, e-mail: [email protected].

96
MOREIRA, A., MALAVOLTA, E. Dinâmica da matéria orgânica e da biomassa microbiana
em solo submetido a diferentes sistemas de manejo na Amazônia Ocidental. Pesquisa
Agropecuária. Brasileira, Brasília, v. 39, n. 11, nov, 2004.
MOREIRA, F. M. S.; SIQUEIRA, J. O. Microbiologia e bioquímica do solo. Lavras: Ufla,
2002. 625p.
NAIR, P. K. R. Agroforestry systems in the tropics. London: Kluwer, 1989. 664p.
NANNIPIERI, P.; MUCCINI, L.; CIARDI, C. Microbial biomass and enzyme activities:
production and persistence. Soil Biology and Biochemistry, v.15, p. 679-685, 1983.
NICODEMO, M. L. F.; PRIMAVESI, O. Sistemas agroflorestais – em busca de sistemas de
produção sustentáveis. In: SEMANA DO ESTUDANTE, 18., 2007, São Carlos. Anais... São
Carlos: Embrapa Pecuária Sudeste, 2007. p.11.
NICODEMO, M. L., Uso de biomassa microbiana para avaliação de qualidade do solo
em sistemas silvipastoris. São Carlos, SP: Embrapa Pecuária Sudeste, 2009. (Embrapa
Pecuária Sudeste. Documentos, 93).
NOGUEIRA, M. A.; ALBINO, U. B.; BRANDÃO-JÚNIOR, O.; BRAUN, G.;CRUZ, M. F.;
DIAS, B. A.; DUARTE, R. T. D.; GIOPPO, N. M. R.; MENNA, P.; ORLANDI, J. M.;
RAIMAN, M. P.; RAMPAZO, L. G. L.; SANTOS, M. A.; SILVA, M. E. Z.; VIEIRA, F. P.;
TOREZAN, J. M. D.; HUGRIA, M.; ANDRADE, G. Promising indicators for assessment of
agroecosystems alteration among natural, reforested and agricultural land use in southern
Brazil. Agriculture, Ecosystems and Environment, v. 115, p. 237-247, 2006.
NUNEZ, J. B. H. Fitomassa e estoque de bioelementos das diversas fases da vegetação
secundária proveniente de diferentes sistemas de uso da terra no nordeste
paraense.1995.183 p. Dissertação (Mestrado em Ciências Biológicas). UFPA. 1995.
ODUM, E. P. The strategy of ecosystem development. Science, v.164, p. 262-270, 1969.
ODUM, E. P. Trends expected in stressed ecosystems. Bioscience, v. 35, p. 419-422, 1985.
OIMT. Diretrices de la OIMT para la restauración, ordenación y rehabilitación de los
bosques tropicales secondários y degradados. 2002. 89p. (Série de Pliticas Forestales, n.
13).

97
OLIVEIRA, J.R.A.; MENDES, I.C. & VIVALDI, L. Carbono da biomassa microbiana em
solos de cerrado sob vegetação nativa e sob cultivo: avaliação dos métodos
fumigaçãoincubação e fumigação-extração. R. Bras. Ci. Solo, v. 25, p. 863- 871, 2001.
OTS; CATIE. Sistemas Agroflorestales: Princípios e aplicaciones en los trópicos. San José-
Costa Rica: CATIE, 1986.
PARKIN, T. B.; DORAN, J.W.; FRANCO-VIZCAÍNO, E. Field and laboratory tests of soil
respiration. In: DORAN, J.W.; JONES, A. (Ed). Methods for assessing soil quality.
Madison: Soil Science Society of América, 1996. p. 231-245.
PARKINSONS, D.; COLEMAN, D.C. Methods for assessing soil microbial populations,
activity and biomass-microbial communities, activity and biomass. Agriculture, Ecosystems
and Environment, v. 34, p. 3-33, 1991.
PEIXOTO, R. S.; COUTINHO, H. L. C.; MADARI, B.; MACHADO, P. L. O. A.;
RUMJANEK, N. G.; VAN ELSAS, J. D. ; SELDIN,L.; ROSADO, A. S. Soil aggregation and
bacterial community structure as affected by tillage and cover cropping in the Brazilian
Cerrados. Soil & Tillage Research, v.90, n.1-2, p.16-28, 2006.
PEÑA, M. L. P.; MARQUES, R.; JAHNEL, M. C.; ANJOS, A. Respiração microbiana como
indicador da qualidade do solo em ecossistema florestal. Floresta, Curitiba, PR, v. 35, n.1, p.
117-127, 2005.
PEREIRA, A. A. ; HUNGRIA, M. ; FRANCHINI, J. C. ; KASCHUK, G. ; CHUEIRE, L. M.
O. ; CAMPO, R. J. ; TORRES, E. Variações qualitativas e quantitativas na microbiota do solo
e na fixação biológica do nitrogênio sob diferentes manejos com soja. Revista Brasileira de
Ciência do Solo, v. 31, p. 1397–1412, 2007.
PICCOLO, M. C.; FEIGL, B. J. ; CERRI, C. C. Avaliação da biomassa microbiana do solo:
revisão. Disponível em: <http:// sbcs.solos.ufv.br/solos/artigos/971_06/971_06.doc>. Acesso
em: 10 jan 2010.
POWLSON, D.S. & JENKINSON, D.S. A comparison of the organic matter, biomass,
adenosine triphosphate and mineralizable nitrogen contents of ploughed and directdrilled
soils. J.Agric. Sci., v. 97, p. 713-721, 1981.
POWLSON, D. S.; BROOKES, P. C.; CHRISTENSEN, B.T. Measurement of soil microbial
biomass provides an early indication of changes in the total soil organic matter due to straw
incorporation. Soil Biology Biochemistry, v.19, p.159-164, 1987.

98
RAIJ, B. V. Avaliação da fertilidade do solo. Campinas, SP: Instituto Agronômico do estado
de São Paulo, 1981.
RAIJ, B. V; CANTARELLA, H.; QUAGGIO, J. A.; FURLANI, A. M. C. Recomendações
de adubação e calagem para o Estado de São Paulo. 2.ed. 285p. Campinas: Instituto
Agronômico: Fundação IAC, 1996.
RANGEL VASCONCELOS, L. G. T.; ZARIN, D. J. ; CARVALHO, C. J. R.; S. SANTOS,
M. M. L.; VASCONCELOS, S. S.; OLIVEIRA, F. A. Carbono, nitrogênio e atividade da
biomassa microbiana de um solo sob vegetação secundária de diferentes idades na Amazônia
oriental. Revista de Ciências Agrárias, n. 44. p.49-63, 2005.
RECCO, R. D. ; AMARAL, E. F.; PINTO, E. M.; MELO, A. W. F. Avaliação do nível de
carbono orgânico em solos tropicais submetidos a plantio de sistemas agroflorestais em
diferentes idades na Amazônia Ocidental. In: CONGRESSO BRASILEIRO DE SISTEMAS
AGROFLORESTAIS. 3., 2000, Manaus. Anais... Manaus, AM. 2000.
REINERT, D. J. Recuperação de solos em sistemas agropastoris. In: DIAS, L. E.; MELLO, J.
W. V. (Ed). Recuperação de áreas degradadas. Viçosa: UFV/SOBRADE, 1998. p.163-176.
RESENDE, M. Matéria orgânica e seus componentes. In: PEDOLOGIA: base para distinção
de ambientes. Viçosa : Neput, 1995. p.79 – 85.
RHEINHEIMER, D.S.; SANTOS, J.C.P.; FERNANDES, V.B.B.; MAFRA, A.L. &
ALMEIDA, J.A. Modificações nos atributos químicos de solo sob campo nativo submetido à
queima. Ci. Rural, v. 33, p. 49-55, 2003.
RIBASKI, J.; MONTOYA, L. J.; RODIGHERI, H. R. Sistemas Agroflorestais: aspectos
ambientais e sócio-econômicos. In: CONGRESSO BRASILEIRO DE SISTEMAS
AGROFLORESTAIS, 4, 2002, Ilhéus. Anais... Ilhéus: EMBRAPA, 2002. CD-Rom.
Rodrigues, T. E. Solos da Amazônia. In: O solo nos grandes domínios morfoclimáticos
doBrasil e o desenvolvimento sustentado. Viçosa. MG. 19-60. 1996.
RODRIGUES, R, C. Biomassa microbiana e acúmulo de liteira em sistemas
agroflorestais composto por Meliaceas utilizadas como indicadores biológicos de
qualidade do solo. 2006. 101f. Tese (Doutorado em Ciências Agrárias) - Universidade
Federal Rural da Amazônia. Belém, PA. 2006.
ROLDÁN, A.; SALINASGARCÍA, J. R.; ALGUACIL, M. M. ; CARAVACA, F. Soil
sustainability indicators following conservation tillage practices under subtropical maize and
bean crops. Soil & Tillage Research, v. 93, p.273-282, 2007.

99
ROSCOE, R.; MERCANTE, F. M.; MENDES, I. C.; REIS-JÚNIOR, F. B.; SANTOS, J. C.
F.; HUNGRIA, M. Biomassa microbiana do solo: fração mais ativa da matéria orgânica. In:
ROSCOE, R.; MERCANTE, F. M.; SALTON, J. C. (Ed). Dinâmica da Matéria Orgânica
do Solo em Sistemas Conservacionistas – Modelagem Matemática e Métodos Auxiliares.
Embrapa Agropecuária Oeste, Dourados, MS, 2006. p. 163–200.
SAMPAIO, F. A. R. Balanço de nutrientes em um sistema de agricultura migratória no
município de Ji-Paraná – RO. 1997. Dissertação (Mestrado em Agronomia, solos e nutrição
de plantas) – Universidade Federal de Viçosa, MG, 1997.
ROUW A. The fallow period as a weed-break in shifting cultivation (tropical wet forests).
Agric Ecosyst Environ, v54, p.31– 43. 1995.
SANCHEZ, P. A.. Properties and management of soils in the tropic. New York, Wiley.
1976.
SÁNCHEZ, P. A.; GARRITY, D. P.; BRANDY, D. E.; TORRES, F. ; SWIFT, M. J.
Alternativas sustentáveis a agricultura migratória e a recuperação de áreas degradadas nos
trópicos úmidos. In: SIMPÓSIO DE ÁREAS DEGRADADAS E FLORESTAS
SECUNDÁRIAS NA AMAZÔNIA. 1993, Santarém. Anais… Santarém, USDA, 1993. p.1-
13.
SANCHEZ P. A. Science in agroforestry. Agroforest Syst, v 30, p 5–55. 1995.
SÁNCHEZ, P. A.; VILLACHICA, J. H. ; BRANDY, D. E.; Soil fertility dynamics after
clearing a tropical rainforest in Peru. Soil Science Society of America Journal, v.47. p.1171-
1178, 1983.
SANTOS, G. A.; CAMARGO, F.A.O. (Ed). Fundamentos da matéria orgânica do solo:
ecossistemas tropicais e subtropicais. Porto Alegre: Gênesis, 1999. 491p.
SANTRUCKOVA, H. ; STRASKRABA, M. On the relationship between specific respiration
activity and microbial biomass in soils. Soil Biology and Biochemistry, v.23, p.525-532,
1991.
SENA, W. L. Avaliação dos atributos químicos e carbono microbiano de um Latossolo
Amarelo sob sistemas agroflorestais em comparação com a floresta secundária,
Marituba, Pará. 2006. 101f. Tese (Doutorado em Ciências Agrárias) - Universidade Federal
Rural da Amazônia. Belém, PA. 2006.

100
SERRÃO, E. A. S. Possibilities for sustainable agricultural development in the Brazilian
Amazon: an EMBRAPA proposal. pp 250-273 in: CLUESENER-GODT, M.; SACHS, I.
(Ed). Brazilian perspectives on sustainable development of the Amazon region. London:
UNESCO, 1995.
SEYBOLD, C. A.; HERRICK, J. E.; BREJDA, J. J. Soil resilience: a fundamental component
of soil quality. Soil Science, v. 164, p. 224–234. 1999.
SIQUEIRA, J.O. et al. Microrganismos e processos biológicos do solo: perspectiva
ambiental. EMBRAPA-SPI, Brasília DF, 1994. 142 p.
SILVA, M. S. C. Indicadores de qualidade do solo em sistemas agroflorestais em Paraty,
RJ. 2006. 54f. Dissertação (Mestrado em Ciência em agronomia) – Universidade Federal
Rural do Rio de Janeiro. Seropédica, RJ, 2006.
SILVA, R. F. Roça caiçara: dinâmica de nutrientes, propriedades físicas e fauna do solo em
um ciclo de cultura.1998. 192f. Dissertação (Mestrado em agronomia ciências do solo) -
Universidade Federal Rural do Rio de Janeiro. Seropédica, RJ, 1998.
SMITH, J. L.; PAUL, E. A. The significance of soil microbial biomass estimations. In:
BOLLAG, J.M.; STOTZKY, G. (Ed.). Soil Biochemistry, New York, v.6, p.357-396, 1990.
SMYTH, T. J. Manejo da fertilidade do solo para a produção sustentada de cultivos na
Amazônia. In: O solo nos grandes domínios morfoclimáticos do Brasil e o
desenvolvimento sustentado. Viçosa. MG, 1996. p.71-94.
SOARES, A. E. S. Analises químicas de nutrientes em áreas com diferentes sistemas de
uso da terra no município de Benjamin Constant, Amazonas. 2007. Dissertação
(Mestrado) – Instituto Nacional de Pesquisa da Amazônia/Universidade Federal do
Amazônas, 2007.
SOMMER, R. No fires! Slash-and-mulch- a promising alternative. ZEFnews, n. 6, p. 8v,
2004.
SOMMER, R. Water and nutrient balance in deep soils under shifting cultivation with
and without burning in the Eastern Amazon. PhD (thesis) - Göttingen: Universität,
Cuvillier, p.240, 2004.
SPARLING, G. P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive
indicator of changes in soil organic matter. Australian Journal of Soil Researsh, v. 30,
p.195-207, 1992.

101
SPARLING, G. P. Soil microbial biomass, activity and nutrient cycling as indicators of soil
health. In: PANKHURST, C. E. ; DOUBE, B. M. ; GUPTA, V. V. S. R. (Ed), Biological
Indicators of Soil Health. CAB International : Wallingford, UK, 1997. p. 9-119.
STEVENSON, F. J. Humus chemistry: genesis, composition, reactions. New York: John
Wiley, 1982, 443p.
SWIFT, M. J.; HEAL, O. W.; ANDERSON, J. M. Decomposition in terrestrial ecosystems.
Berkeley: University of California Press, 1979. 372p.
SZOTT, L. T.; PALM, C. A. Dinamica del suelo y la vegetacion alternando cultivos y
periodos de descanso. Contribution of the tropical soil program in collaboration with the
Instituto Nacional de Investigacion y Promocion Agropecuaria of Peru. 1999.
SZOTT, L. T.; PALM, C. A.; BURESH, R. J. Ecosystem fertility and fallow function in the
humid and subhumid tropics. Agroforestry Systems, Netherlands, v. 47. p.163-196, 1999.
TATE, K. R.; ROSS, D. J.; FELTHAM, C. W. A direct extraction method to estimate soil
microbial C: Effects of experimental variables and some different calibration procedures. Soil
Biology and Biochemistry, v. 20, p. 329-335, 1988.
TIPPMANN, R. ; DENICH, M. ;VIELHAUER, K. Integration of geo- and remote sensing
data for the assessment and monitoring of changes in smallholder land-use systems at farer
level. In: GERMAN-BRAZILIAN WORKSHOP ON NEOTROPICAL ECOSYSTEMS-
ACHIEVEMENTS AND PROSPECTS OF COOPERATIVE RESEARCH. 2000, Hamburg.
Abstracts,... Hamburg, 2000. p.297.
TOMÉ JÚNIOR, J. B. Manual para interpretação de análise de solo. Guaíba:
Agropecuária, 1997. 247 p.
VANCE, E. D. ; BROOKES, P. C. ; JENKINSON, D. S. An extraction method for measuring
soil microbial biomass C. Soil Biology and Biochemistry, v. 19, p. 703-707, 1987.
VARGAS, L. K.; SCHOLLES,D. Biomassa microbiana e produção de C-CO2 e N mineral de
um podzólico vermelho-escuro submetido a diferentes sistemas de manejo. Revista
Brasileira de Ciência do Solo, v.24, n.1, p. 35-42, 2000.

102
VASCONCELOS, L. G. T. R. Biomassa microbiana de um solo sob vegetação secundária
na Amazônia Oriental. 2002. 80f. Dissertação (Mestrado) – Faculdade de Ciências Agrárias
do Pará, Belém, 2002.
VIEIRA, L.S.; SANTOS, P.C.T. Amazônia: seus solos e outros recursos naturais. São Paulo:
Agronômica Ceres, 1987. 416p.
VIEIRA, I. C.; SALOMÃO, R. P.; ROSA, N. A. O renascimento da floresta no rastro da
agricultura. Como a floresta Amazônica sobrevive ao desmatamento e à queimadas. Ciencia
Hoje, v. 20, n. 119, p. 1996.
VIELHAUER, K.; SÁ, T. D. de A. Efeito do enriquecimento de capoeiras com árvores
leguminosas de rápido crescimento para a produção agrícola no Nordeste Paraense. In:
SEMINÁRIO SOBRE MANEJO DA VEGETAÇÃO SECUNDÁRIA PARA A
SUSTENTABILIDADE DA AGRICULTURA FAMILIAR DA AMAZÔNIA ORIENTAL,
1., 1999, Belém. Anais..., Belém: Embrapa Amazônia Oriental, 2000. p.27-34. Embrapa
Amazônia oriental. Documentos, 69.
WATRIN, O. S.; VENTURIERI, A.; SAMPAIO, S. M. N. Analise multitemporal do uso da
terra e suas interrelações com a cobertura vegetal em comunidades rurais do Nordeste
Paraense. In: SIMPÓSIO BRASILEIRO DE SENSORIAMENTO REMOTO, 9., Santos.
Anais... Santos: INPE, 1998. p.1573-1583.
YOUNG, A. Agroforestry for soil conservation. Wallingford: CAB International, 1994,
276p.
ZATORRE, N. P.; TEIXEIRA, R. O. ; FELÍCIO, N. C.; FRANÇA, A. F.; WADT, P. G. S.;
BERBARA, R. ; SANTOS, G. Influência das Modificações da Cobertura em Algumas
Propriedades de Solos do Acre, Revista Brasileira de Agroecologia, v. 4, n. 2, p. 2545-2549,
2009.
WARDLE, D.A. Metodologia para quantificação de biomassa microbiana do solo. In:
HUNGRIA, M. & ARAÚJO, R.S. (Eds.) Manual de métodos empregados em estudos de
microbiologia agrícola. Brasília, EMBRAPA, 1994, P. 419-436.

103
ANEXOS
104
ANEXO 1 – Valores de F e nível de significância da análise de variância para atributos
químicos do solo de áreas submetidas a diferentes sistemas de manejo da 1ª coleta de solo
(janeiro/ 2009) na profundidade de 0-5 cm, município de Igarapé-Açú (PA).
Atributos
químicos Fonte de Variação
Tratamento
(0-5 cm) CV (%)
pH 5,43* 2,67
C 96,02** 7,05
N 21,35** 13,87
P 2,11 35,33
H 19,98** 18,55
Al 17,0** 34,65
K 14,51** 17,49
Ca 24,42** 15,50
Mg 6,41** 20,91
Na 20,33** 15,59
CTC 20,99** 11,14
CTCefetiva 19,09** 12,12
SB 19,28 15,64
V 19,94** 11,17
m 14,46** 44,23
ns – não significativo; * e ** - significativo a (P< 0,05) e (P< 0,01)
ANEXO 2 – Valores de F e nível de significância da análise de variância para atributos
químicos do solo de áreas submetidas a diferentes sistemas de manejo da 1ª coleta de solo
(janeiro/ 2009) na profundidade de 5-10 cm, município de Igarapé-Açú (PA).
Atributos
químicos Fonte de Variação
Tratamento
(5-10 cm) CV (%)
pH 19,18** 2,89
C 2,36 15,79
N 1,20 16,00
P 10,97** 24,89
H 4,79** 14,00
Al 14,60** 28,71
K 4,73** 17,32
Ca 42,29** 15,21
Mg 7,05** 25,87
Na 5,99** 15,49
CTC 10,98** 9,05
CTCefetiva 46,97** 6,77
SB 56,61** 11,40
V 26,56** 12,73
m 21,69** 30,13
ns – não significativo; * e ** - significativo a (P< 0,05) e (P< 0,01)
105
ANEXO 3 – Valores de F e nível de significância da análise de variância para relação
carbono da biomassa microbiana (CBM): carbono total (C) do solo de áreas submetidas a
diferentes sistemas de manejo da 1ª coleta de solo (janeiro/ 2009), município de Igarapé-Açú
(PA).
Fonte de Variação Profundidade (cm)
0-5 5-10
Tratamento 2,70* 2,15*
CV% 47,3 59,4
ns – não significativo; * e ** - significativo a (P < 0,05) e (P < 0,01)
ANEXO 4 – Valores de F e nível de significância da análise de variância para atributos
biológicos do solo de áreas submetidas a diferentes sistemas de manejo, coletado em
diferentes épocas de coleta de solo, município de Igarapé-Açú (PA).
Atributos
Fonte de Variação
Época
(Ep)
Tratamento
(Trat)
Profundidade
(Prof)
Ep x
Trat
Ep x
Prof
Trat x
Prof CV%
CO2 124,42** 8,83* 10,16* 10,94** 0,27ns
0,87ns
39,6%
CBM 14,59** 10,44** 4,44* 4,76** 4,86** 2,56* 77,7%
qCO2 11,79** 7,68** 0,13ns
1,59* 1,86* 0,48ns
89,7%
ns – não significativo; * e ** - significativo a (P < 0,05) e (P < 0,01)

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo