jessika brenda pinheiro fungos produtores de ... 2014.pdf · fungos produtores de...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE SÃO JOÃO DEL-REI
CAMPOS SETE LAGOAS – CSL BACHARELADO INTERDICIPLINAR EM BIOSSISTEMAS
JESSIKA BRENDA PINHEIRO
FUNGOS PRODUTORES DE EXOPOLISSACARÍDEOS E SUAS APLICAÇÕES FUNCIONAIS E INDUSTRIAIS
Sete Lagoas - MG 2014

UNIVERSIDADE FEDERAL DE SÃO JOÃO DEL-REI
CAMPOS SETE LAGOAS – CSL BACHARELADO INTERDICIPLINAR EM BIOSSISTEMAS
JESSIKA BRENDA PINHEIRO
FUNGOS PRODUTORES DE EXOPOLISSACARÍDEOS E SUAS APLICAÇÕES FUNCIONAIS E INDUSTRIAIS
Trabalho de Conclusão de Curso apresentado ao Curso de Bacharelado Interdisciplinar em Biossistemas da Universidade Federal de São João del-Rei como requisito parcial para a obtenção do título de Bacharel em Biossistemas. Orientador: Prof. Dr.º Juliano de Carvalho Cury
Sete Lagoas – MG 2014

1
Monografia de Bacharelado Interdisciplinar em Biossistemas do Campus Sete Lagoas
da Universidade Federal de São João Del-Rei/UFSJ defendida em 16 de Julho de 2014
pela seguinte banca examinadora:
Prof. Dr. Juliano de Carvalho Cury Orientador Departamento de Ciências Exatas e Biológicas - DECEB Campus Sete Lagoas Universidade Federal de São João Del-Rei _______________________________________ Leonardo Henrique França Lima Campus Sete Lagoas Universidade Federal de São João Del-Rei _______________________________________ Amanda Miranda de Souza Campus Sete Lagoas Universidade Federal de São João Del-Rei

2
Dedico esta monografia a Deus, minha família e a todos que acreditaram e que de
alguma forma foram instrumentos da minha conquista.
Aos meus pais, Isabel e Adenaudo que sempre estiveram presentes me auxiliando e
dando forças quando tudo parecia não caminhar, me instruindo no amor, na paciência
e na confiança de que tudo acontece não na hora exata do nosso desejo, mas no tempo
de Deus!
Á minha tia Maria José, que confiou e acreditou no meu sonho e desde então se fez
presente, investindo no meu futuro profissional possibilitando cada passo sonhado
nesta caminhada do saber.
À minha irmã Gabriella, avós, tios, primos e amigos que sempre estiveram comigo,
apoiando e incentivando cada passo rumo à concretização desse sonho.
Jordan, e sua família que se fizerem presentes desde o início, me apoiando e
amparando nas horas mais difíceis, atuando como uma verdadeira família.

3
Agradecimentos
Agradeço a Deus, que me guiou e me deu forças para que eu não desviasse dos
meus sonhos, e pudesse sempre ao final de uma longa jornada de aprendizado
alcançar meus objetivos.
Agradeço a minha Mãe, que sempre em momentos insertos, de angustia e
dificuldades me mostrou que obstáculos são feitos para serem superados e que em
cada um deles aprendemos a ter mais confiança em nós mesmos, a ter fé e esperar em
Deus; a ser humilde para encarar os erros e humilde também para lidar com vitórias.
Ao meu Pai, que sempre se fez presente acreditando e sonhando comigo com
cada novo objetivo, compartilhando sempre com a máxima atenção e entusiasmo cada
novo passo que eu dava, cada nova conquista, cada experiência nova, me
impulsionando sempre a ir além, a sempre buscar por novos conhecimentos.
A minha tia Maria José, que sempre presente, me deu suporte nestes anos
todos de estudos, possibilitando a realização de um sonho em busca de um futuro
profissional, sempre acreditando e apoiando cada passo rumo a esta conquista.
Ao meu Orientador Juliano, que me deu a oportunidade de enriquecer meus
conhecimentos, me ensinando como dar cada passo em busca da concretização de um
trabalho. E aos demais professores por todo o conhecimento transmitindo sempre
aptos a ajudar no que fosse necessário. E a todos que de certa forma me auxiliaram
nesta caminhada.
“Tudo posso naquele
que me fortalece”
Filipenses 4:13

4
Resumo
Exopolissacarídeos (EPSs) são polissacarídeos extracelulares, podendo ser produzidos por alguns fungos e bactérias. São produzidos durante a fase estacionária do crescimento microbiano, sendo considerados metabólitos secundários. Podem ser encontrados na forma de cápsulas ao redor da célula, ou na forma de limos (mucos) que são excretados para o meio extracelular provocando alteração de viscosidade no meio de cultivo. Esta revisão teve como objetivo abordar a produção de EPSs por fungos à partir de cultivo submerso, avaliando diferentes condições de cultivo e suas potencialidades industriais e funcionais. Foi verificado nesta revisão que os EPSs produzidos por microrganismos são amplamente utilizados em atividades industriais dos ramos alimentício, médico e de cosméticos, sendo aplicados para a formação de géis, emulsões e biofilmes, além de terem funções de absorção e coesão. No processamento de alimentos os EPSs são utilizados para conferir características de textura tais como corpo, viscosidade e consistência, possuindo ainda diversas atividades biológicas como ação anti-tumoral, antiviral, antiinflamatória, hipoglicêmica e imuno-estimulante. Sua produção por fungos tem a vantagem de ser independente de condições climáticas, sendo realizada sob condições controladas, resultando em EPSs de alta regularidade estrutural, ao contrário de sua obtenção a partir de algas e plantas. A utilização do cultivo submerso proporciona um aumento potencial na produção de biomassa micelial ocupando um pequeno espaço e menor tempo de fermentação, diminuindo as chances de contaminação, possibilitando a utilização de matérias-primas regionais e de subprodutos industriais. Não só a fonte de carbono utilizada, mas também sua concentração são fatores interferentes na produção de EPSs, sendo as fontes de carbono e nitrogênio os principais componentes do meio nutriente. Visto a vantagem da produção de EPSs por fungos sob cultivo submerso, as indústrias de alimentos e farmacêutica têm demonstrado grande interesse no desenvolvimento de pesquisas nessa área. Dessa forma, o estudo de novos organismos vivos que possam produzir moléculas com propriedades tecnológicas relevantes é bastante interessante e estratégico para o Brasil, visto que é um país rico em biodiversidade.
Palavras-chave: Exopolissacarídeos, Fungos, Biotecnologia, Condições de Cultivo, Meios de Cultura Alternativos

5
Abstract
Exopolysaccharides (EPSs) are extracellular polysaccharides may be produced by some fungi and bacteria. They are produced during the stationary phase of microbial growth, and are considered secondary metabolites. They can be found in capsule form around the cell, or in the form of slime (mucus) that are excreted into the extracellular space causing change in viscosity in the culture medium. This review aimed to address the production of EPSs by fungi from the submerged cultivation, evaluating different growing conditions and its industry and functional capabilities. It was found in this review that the EPSs produced by microorganisms are widely used in industrial activities in the food, cosmetic and medical branches being applied to the formation of gels, emulsions and biofilms, and have functions of absorption and cohesion. In food processing the EPSs are used to impart texture features such as the body, viscosity and consistency, yet having various biological activities such as antitumor activity, antiviral, anti-inflammatory, immunostimulant and hypoglycemic. Its production by fungi has the advantage of being independent of weather conditions, being carried out under controlled conditions, resulting in high structural regularity of EPSs, unlike obtaining from algae and plants. The use of submerged cultivation provides a potential increase in the production of mycelial biomass occupying a small space and shorter fermentation, reducing the chances of contamination, enabling the use of regional raw materials and industrial byproducts. Not only the carbon source used, but its concentration is interfering factors in the production of EPSs, and the carbon and nitrogen sources of the main components of the nutrient medium. Wear advantage of producing EPSs yeast under submerged cultivation, the food and pharmaceutical industries have shown great interest in the development of research in this area. Thus, the study of new living organisms that can produce molecules with relevant technological properties is quite interesting and strategic for Brazil, since it is a country rich in biodiversity.
Keywords: Exopolysaccharides, Fungi, Biotechnology, Growing Conditions, Means of Alternative Culture

6
Sumário
1- INTRODUÇÃO ........................................................................................................................ 7
2- OBJETIVO .............................................................................................................................. 8
3- METODOLOGIA ..................................................................................................................... 9
4- REVISÃO BIBLIOGRÁFICA ...................................................................................................... 9
4.1- DESCRIÇÃO BIOLÓGICA DOS FUNGOS .............................................................................. 9
4.2- POLISSACARÍDEOS E EXOPOLISSACARIDEOS .................................................................. 12
4.3- HISTÓRICO ........................................................................................................................ 13
4.4 - EFEITO DO MEIO E CONDIÇÕES DE CULTURA ................................................................. 16
4.4.1- FONTE DE CARBONO ................................................................................................ 16
4.4.2- FONTES DE NITROGENIO ........................................................................................... 18
4.4.3- FERMENTAÇÃO SUBMERSA ...................................................................................... 20
4.5- RECUPERAÇÃO E QUANTIFICAÇÃO DE EXOPOLISSACARÍDEOS ....................................... 21
4.6- BIOATIVIDADE E ESTRUTURA DOS EXOPOLISSACARÍDEOS .............................................. 22
4.7- PROCESSOS BIOTECNOLÓGICOS NA PRODUÇÃO DE EXOPOLISSACARÍDEOS E SUAS
APLICAÇÕES ............................................................................................................................. 24
4.8- UTILIZAÇÃO DE REJEITOS INDUSTRIAIS NA PRODUÇÃO DE EPSs .................................... 25
4.8.1- RESÍDUO DE CERVEJARIA .......................................................................................... 26
4.8.2- BAGAÇO DE MAÇÃ .................................................................................................... 26
4.8.3- LICOR CITRICO ........................................................................................................... 27
4.8.4- MELAÇO CÍTRICO....................................................................................................... 27
5- CONSIDERAÇÕES FINAIS ..................................................................................................... 28
6- REFERÊNCIAS BIBLIOGRÁFICAS .......................................................................................... 28

7
1- INTRODUÇÃO
Polissacarídeos (PSs) são macromoléculas constituídas por unidades
monoméricas unidas por ligações glicosídicas. Os PSs se diferem em função de suas
características estruturais como unidade e o grau de ramificação, dos tipos de ligações
e do comprimento de suas cadeias (Silva et al., 2006). Podem ser classificados como
homopolissacarídeos ou heteropolissacarídeos, sendo os homopolissacarídeos
constituídos de um único tipo de unidade monomérica, como é o caso do amido e do
glicogênio, e os heteropolissacarídeos constituídos de duas ou mais unidades
monoméricas diferentes, como os peptideoglicanos (Silva et al., 2006).
Exopolissacarídeos (EPSs) são polissacarídeos extracelulares, podendo ser
produzidos por alguns fungos e bactérias. São encontrados na forma de cápsulas ao
redor da célula, tornando-se parte da parede celular, ou na forma de limos (mucos)
que são excretados para o meio extracelular, difundindo-se na fase líquida durante o
processo de fermentação provocando um aumento significativo na viscosidade do
meio (Silva et al., 2006).
Os EPSs produzidos por microrganismos são utilizados em atividades
industriais dos ramos alimentício, médico e de cosméticos, dentre outros (Lee et al.
2003; Liu et al. 1997; Mansell, 1994; Ricciardi et al. 2002; Sugihara et al. 2001),
aplicados para a formação de géis, emulsões e biofilmes, além de terem funções de
absorção e coesão (Paul et al. 1986; Sutherland, 1999; Sutherland, 2001). No
processamento de alimentos os EPSs são utilizados para conferir características de
textura tais como corpo, viscosidade e consistência aos mesmos (Souza et al., 2004).
Um objetivo comum da aplicação dos EPSs em alimentos é o de facilitar a dissolução
dos ingredientes, aumentando a viscosidade e evitando assim a formação de cristais, o
que poderia comprometer a textura e homogeneidade do alimento.
Alguns destes EPSs são modificados quimicamente. Como exemplo, podemos
citar a carboximetilcelulose, que assim como o monoesterato glicerol, é amplamente
utilizada como emulsificante artificial na indústria de alimentos. Porém, tem sido cada
vez maior o interesse pelos EPSs biogênicos que possam substituir os aditivos
sintéticos (Borges et al., 2004). Este é o caso dos EPSs produzidos por fungos, que têm

8
sido também bastante estudados devido suas diversas atividades biológicas como ação
anti-tumoral, antiviral, antiinflamatória, hipoglicêmica e imuno-estimulante (Franz,
1989; Borchers et al., 1999; Babitskatya et al., 2000; Wasser, 2002; Yang et al., 2002).
Os EPSs constituem uma importante porcentagem da biomassa dos fungos,
podendo chegar a 75% dos PSs constituintes da parede das hifas, além de poderem
constituir capa extracelular na forma de muco ao redor das hifas (Silva et al., 2006).
A produção de EPSs por fungos tem a vantagem de ser independente de
condições climáticas, visto que sua produção é controlada laboratorialemte, ao
contrário de algas e plantas, além de possibilitar a utilização de matérias-primas
regionais e mesmo de subprodutos industriais, apresentar grande agilidade de
obtenção do produto acabado e necessitar de um espaço relativamente pequeno
(Souza et al., 2004). Além disso, por serem produzidos sob condições controladas, com
espécies selecionadas, os problemas com variação estrutural dos PSs podem ser
evitados, obtendo-se PSs com alta regularidade estrutural, o que não é comum em PSs
vindos de outras fontes (Souza et al., 2004). Para poderem ser utilizados
industrialmente é preciso que os EPSs sejam produzidos em grandes quantidades pelos
microrganismos, estando estes adaptados a variações de pH e temperatura
relativamente amplas, além de apresentarem composição química compatível com o
metabolismo humano (Borges et al., 2004). Entretanto, segundo Silva et al. (2005),
muitos EPSs fúngicos ainda não foram adequadamente explorados, sendo poucos
ainda os que são produzidos em escala industrial.
2- OBJETIVO
Esta monografia tem por objetivo realizar uma revisão de literatura
abordando o potencial de produção de exopolissacarídeos a partir do cultivo submerso
de fungos em diferentes condições e suas potencialidades funcionais e tecnológicas.

9
3- METODOLOGIA
Foi realizada uma pesquisa bibliográfica abordando os principais meios e
técnicas de produção de exopolissacarídeos e suas capacidades funcionais e
tecnológicas descritas nos últimos 25 anos, utilizando as seguintes bases de dados:
ScientificElectronic Library Online-SCIELO e Google acadêmico. As palavras-chave
utilizadas foram: exopolissacarídeos, fungos, fermentação, extração, atividade
funcional, aplicações, biotecnologia e resíduos industriais.
4- REVISÃO BIBLIOGRÁFICA
4.1- DESCRIÇÃO BIOLÓGICA DOS FUNGOS
Os fungos fazem parte de um grande e diverso grupo de microrganismos,
incluindo os bolores, cogumelos e leveduras. Atualmente já foram descritas
aproximadamente 100.000 espécies de fungos, estimando-se a possibilidade de
existência de mais de 1,5 milhão de organismos (Brock, 2010). Podem possuir
diferentes habitats, estando presentes no meio aquático (água doce e marinha),
terrestre (solo e matéria orgânica em decomposição), além de também poderem atuar
como parasitas de diversos organismos.
Por serem quimiorganotróficos, geralmente apresentam nutrição simples,
alimentando-se por meio de secreções enzimáticas que digerem macromoléculas
(principalmente carboidratos e proteínas) em monômeros aptos para serem
absorvidos (Brock, 2010).
A obtenção de nutrientes efetua-se por absorção através das paredes das
células, sendo que os elementos devem estar na forma de solução. As células
segregam enzimas que atuam sobre as substâncias, degradando-as. No caso dos
fungos filamentosos, a partir do micélio são emitidas estruturas chamadas haustórios,
que penetram no tecido dos organismos hospedeiros, absorvendo os nutrientes (Silva
et al., 2006).
Apesar de poderem ser unicelulares, no caso das leveduras, a maioria dos
fungos são multicelulares, formando uma rede chamada micélio, composta pelo

10
conjunto de filamentos, que são as hifas. As hifas são paredes celulares tubulares que
envolvem a membrana citoplasmática, podendo ser septadas, possuindo paredes
transversais dividindo cada hifa em células separadas, ou cenocíticas, sem as divisões
como mostrado na Figura 1 (Brock, 2010). Em uma das fases do ciclo de vida do fungo
filamentoso, as hifas podem se ramificar atingindo o ar, nessas ramificações aéreas é
que são formados os esporos, que são as estruturas de dispersão e resistência dos
fungos (Brock, 2010).
Figura 1: Diferenciação de hifas, septadas e cenocíticas
Com base em critérios quimiotaxonômicos, estes microrganismos são
classificados como oomicetos, zigomicetos, basidiomicetos, ascomicetos e
deuteromicetos, incluindo espécies parasitas, saprófitas e simbiontes (Silva et al.,
2006).
A estrutura das paredes celulares fúngicas assemelham-se às paredes celulares
vegetais, mas quando observadas quimicamente são diferentes. Ao contrário das
plantas, a parede celular fúngica é composta de quitina, um polímero derivado da N-
acetilglicosamina, sendo disposta como feixes microfibrilares de acordo com a Figura
2. Alguns polissacarídeos como mananas (polímeros de manose), galactosanas
(polímeros de galactose) e glucanas (polímeros de glicose) podem substituir a quitina
nas paredes celulares destes microrganismos, podendo ser responsáveis por 80-90%
da sua composição (Brock, 2010).

11
Figura 2: Composição e estrutura da parede celular fúngica
Durante o crescimento microbiano são observadas três fases: lag (adaptação),
log ou exponencial (crescimento ou multiplicação) e estacionária, como demonstrado
na Figura 3. A formação de compostos extracelulares pode ocorrer tanto durante a
fase de crescimento exponencial como no final dela, na fase estacionária. Neste
contexto, existem dois tipos de metabólitos microbianos, os primários e os
secundários. Um metabólito primário é sintetizando durante a fase exponencial de
crescimento e o secundário é sintetizado próximo ao final da fase exponencial ou até
mesmo na fase estacionária de crescimento (Brock, 2010). Os metabólitos secundários
têm como características não serem essenciais ao crescimento e reprodução do
microrganismo, sendo sua síntese dependente das condições de cultivo,
especialmente no que diz respeita à composição do meio de cultura. Já os metabólitos
primários estão estritamente associados ao crescimento e ao metabolismo energético
do microrganismo (Brock, 2010).
Figura 3: Fases do crescimento microbiano

12
4.2- POLISSACARÍDEOS E EXOPOLISSACARIDEOS
Polissacarídeos (PSs) são macromoléculas constituídas por unidades
monoméricas unidas por ligações glicosídicas. São encontrados em todos os
organismos vivos, estando presentes nas plantas principalmente na forma de celulose
e amido e nos animais na forma de glicogênio, constituindo uma das principais
reservas de energia. Os PSs diferem entre si em função da unidade e o grau de
ramificação, dos tipos de ligações que as unem e do comprimento de suas cadeias
(Silva et al., 2006). Os PSs podem ser classificados como homopolissacarídeos ou
heteropolissacarídeos. Os homopolissacarídeos possuem um único tipo de unidade
monomérica, como é o caso do amido e do glicogênio. Já os heteropolissacarídeos
contêm duas ou mais unidades monoméricas diferentes, como os peptideoglicanos,
que fazem parte da parede celular das células bacterianas (Silva et al., 2006).
Exopolissacarídeos (EPSs) são polissacarídeos extracelulares, podendo ser
produzidos por alguns fungos e bactérias. Por serem produzidos durante a fase
estacionária do crescimento microbiano, estes EPSs são considerados metabólitos
secundários. Dependendo do tipo de microrganismo, os EPSs podem ser encontrados
na forma de cápsulas ao redor da célula, tornando-se parte da parede celular, ou na
forma de limos (mucos) que são excretados para o meio extracelular, sendo capazes
de se difundirem na fase líquida durante o processo de fermentação, havendo
alteração no meio de cultivo, com formação inicial de um fluido Newtoniano de baixa
viscosidade até que se atinja, ao final, um fluido não-Newtoniano de alta densidade
(Silva et al., 2006).
Os polissacarídeos constituem uma importante percentagem da biomassa
fúngica. A parede da hifa contém mais de 75% deste tipo de biomolécula. Além de
atuarem como elemento de suporte para as hifas, alguns polissacarídeos constituem
uma capa extracelular ao redor do micélio, podem ter a função de manter o pH ótimo
para as enzimas ligninolíticas, além de impedirem a desidratação das hifas e de
regularem a concentração de glucose extracelular (Silva et al., 2006).

13
4.3- HISTÓRICO
As primeiras observações sobre casos de polimerização com a formação de
polissacarídeos foram feitas em caldo de cana e descritas por Vauquelin, em 1822
(Barbosa, et al., 2004). Somente 40 anos depois, Pasteur descreveu observações
semelhantes, demonstrando que essa polimerização era provocada por
microrganismos (Barbosa et al., 2004). O polissacarídeo mais importante, obtido por
fermentação, foi a dextrana (C6H10O5)n, descrita por Scheibler em 1874. Produzida pela
bactéria Leuconostoc mesenteroides, tendo sido isolada e nominada pela primeira vez
por Van Tieghen em 1880 (Barbosa et al., 2004), sua estrutura molecular pode ser
observada na Figura 4. Foi a partir da dextrana que se iniciou e se desenvolveu o
estudo dos exopolissacarídeos microbianos, havendo inicialmente um
desenvolvimento mais acentuado sobre a produção de EPSs produzidos por bactérias
e, posteriormente, por fungos (Barbosa et al., 2004).
Figura 4: Estrutura molecular da dextrana
A produção de EPSs por fungos tem a vantagem de ser independente de
condições climáticas, visto que sua produção pode ser controlada em laboratório, ao
contrário de algas e plantas, além de possibilitar a utilização de matérias-primas
regionais e mesmo de subprodutos industriais, apresentar grande agilidade de
obtenção do produto acabado e necessitar de um espaço relativamente pequeno
(Souza et al., 2004). Além disso, por serem produzidos sob condições controladas de
pH, temperatura, taxa de aeração, concentração de nutrientes e utilizando espécies
selecionadas, os problemas com variação estrutural dos PSs podem ser evitados,
obtendo-se PSs com alta regularidade estrutural, o que não é comum em PSs vindos de
14
outras fontes (Souza et al., 2004). Para poderem ser utilizados industrialmente é
preciso que os EPSs sejam produzidos em grandes quantidades pelos microrganismos,
estando estes adaptados a variações de pH e temperatura relativamente amplas, além
de apresentarem composição química compatível com o metabolismo humano (Borges
et al., 2004).
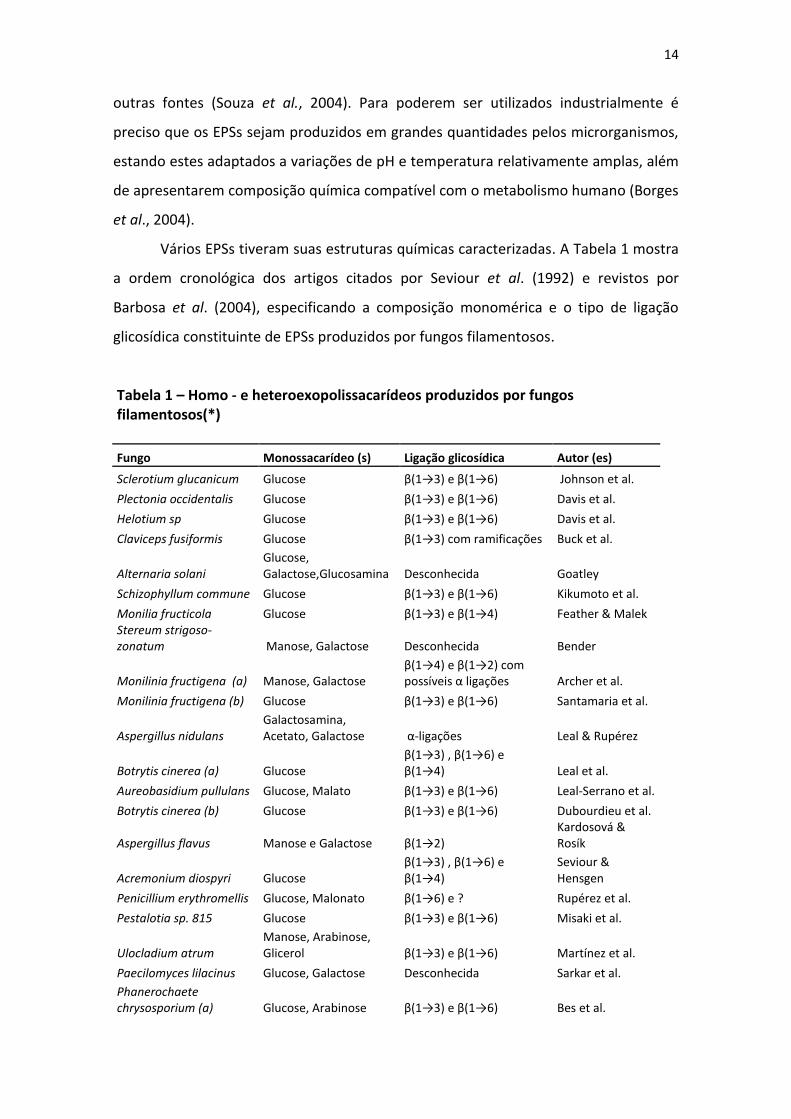
Vários EPSs tiveram suas estruturas químicas caracterizadas. A Tabela 1 mostra
a ordem cronológica dos artigos citados por Seviour et al. (1992) e revistos por
Barbosa et al. (2004), especificando a composição monomérica e o tipo de ligação
glicosídica constituinte de EPSs produzidos por fungos filamentosos.
Tabela 1 – Homo - e heteroexopolissacarídeos produzidos por fungos filamentosos(*)
Fungo Monossacarídeo (s) Ligação glicosídica Autor (es)
Sclerotium glucanicum Glucose β(1→3) e β(1→6) Johnson et al.
Plectonia occidentalis Glucose β(1→3) e β(1→6) Davis et al.
Helotium sp Glucose β(1→3) e β(1→6) Davis et al.
Claviceps fusiformis Glucose β(1→3) com ramificações Buck et al.
Alternaria solani
Glucose, Galactose,Glucosamina Desconhecida Goatley
Schizophyllum commune Glucose β(1→3) e β(1→6) Kikumoto et al.
Monilia fructicola Glucose β(1→3) e β(1→4) Feather & Malek
Stereum strigoso-zonatum Manose, Galactose Desconhecida Bender
Monilinia fructigena (a) Manose, Galactose
β(1→4) e β(1→2) com possíveis α ligações Archer et al.
Monilinia fructigena (b) Glucose β(1→3) e β(1→6) Santamaria et al.
Aspergillus nidulans
Galactosamina, Acetato, Galactose α-ligações Leal & Rupérez
Botrytis cinerea (a) Glucose
β(1→3) , β(1→6) e β(1→4) Leal et al.
Aureobasidium pullulans Glucose, Malato β(1→3) e β(1→6) Leal-Serrano et al.
Botrytis cinerea (b) Glucose β(1→3) e β(1→6) Dubourdieu et al.
Aspergillus flavus Manose e Galactose β(1→2)
Kardosová & Rosík
Acremonium diospyri Glucose
β(1→3) , β(1→6) e β(1→4)
Seviour & Hensgen
Penicillium erythromellis Glucose, Malonato β(1→6) e ? Rupérez et al.
Pestalotia sp. 815 Glucose β(1→3) e β(1→6) Misaki et al.
Ulocladium atrum
Manose, Arabinose, Glicerol β(1→3) e β(1→6) Martínez et al.
Paecilomyces lilacinus Glucose, Galactose Desconhecida Sarkar et al.
Phanerochaete chrysosporium (a) Glucose, Arabinose β(1→3) e β(1→6) Bes et al.

15
Phanerochaete chrysosporium (b) Glucose β(1→3) e β(1→6) Buchala & Leisola
Epicoccum purpurascens Glucose β-ligações Michel et al.
Phytophthora parasitica Glucose β(1→3) e β(1→6) Bruneteau et al.
Acremonium persicinum Glucose β-ligações
Stasinopolous & Seviour
Glomerella cingulata Glucose β(1→3) e β(1→6) Gomaa et al.
Laetisaria arvalis Glucose β(1→3) e β(1→6) Aouadi et al.
Aureobasidium pullulans p56 Glucose α(1→6) Schuster et al.
Aureobasidium pullulans ATCC 9348 Glucose α(1→6) Gibbs & Seviour
Pleurotus ostreatus
Glucose, Manose, Galactose
Gutiérrez et al.
Fusarium solani
Manose, Galactose, Glucose Pentose desconhecida Polycarpo et al.
Aureobasidium pullulans aubasidani Glucose
β(1→3) , β(1→6) e α(1→4) Yurlova & Hoog
Aureobasidium pullulans ATCC 9348 Glucose α(1→6) Gibbs & Seviour
Sclerotium rolfsii ATCC 201126 Glucose β(1→3) , β(1→6) Fariña et al.
Aureobasidium pullulans NRRLY- 6220 Glucose α(1→6) Barnett et al.
Schizophyllum commune Glucose β(1→3) , β(1→6) Maziero et al.
Segundo Silva et al. (2006), muitos EPSs fúngicos ainda não foram
adequadamente explorados, sendo poucos ainda os que são produzidos em escala
industrial. Muitos destes PSs produzidos por bactérias e fungos possuem propriedades
semelhantes às do ágar, enquanto outros possuem propriedades reológicas que são
valiosas para emprego industrial (Barbosa et al., 2004).
Os EPSs microbianos são mais conhecidos por suas propriedades espessantes,
geleificantes e emulsificantes, porém outra possibilidade de aplicação destes
biopolímeros é na saúde humana (Barbosa et al., 2004), devido suas atividades
biológicas com ação anti-tumoral, antiviral, antiinflamatória, hipoglicêmica e imuno-
estimulante (Franz, 1989; Borchers et al., 1999; Babitskatya et al., 2000; Wasser, 2002;
Yang et al., 2002).

16
4.4 - EFEITO DO MEIO E CONDIÇÕES DE CULTURA
Os microrganismos são adaptados ao meio ambiente natural onde vivem, e
nele existem fatores limitantes ao seu crescimento como as fontes de carbono e
nitrogênio, aeração, movimentação, microelementos entre outros, e isso confere a
eles um estado de homeostase. Na produção de EPSs por fungos, deve ser considerada
a cepa fúngica escolhida, como também o seu meio ambiente natural para adequar,
em laboratório, o meio de cultivo em termos de características nutricionais, fatores de
aeração, temperatura, pH, dentre outros fatores que possam ser relevantes para a
produção de EPSs (Barbosa et al, 2004).
As fontes de carbono e nitrogênio constituem os principais componentes do
meio nutriente em um processo de cultivo, as quais precisam assegurar bom
crescimento do microrganismo e maior rendimento do produto desejado, devendo-se
ressaltar que a fonte de carbono que é metabolizada pelo fungo está diretamente
relacionada com a composição monomérica do polissacarídeo produzido, embora nem
sempre na mesma proporção (Cunha et al., 2008).
Para alguns fungos, a aeração pode não afetar a produção, enquanto que para
outros, pode aumentar ou mesmo diminuí-la, já a agitação tem função de melhorar a
distribuição de oxigênio e nutrientes para as células fúngicas no meio de cultivo.
Independentemente da taxa de agitação, a produção de EPS é mais afetada pelo nível
da pressão de oxigênio no fermentador do que pela agitação propriamente dita
(Barbosa et al., 2004).
4.4.1- FONTE DE CARBONO
A produção de EPSs por fungos parece depender do tipo de fonte de carbono e
da sua concentração no meio de cultura, existindo assim um tipo e uma concentração
ótima de fonte de carbono para a produção de EPSs para cada microrganismo (Barbosa
et al., 2004).
A glicose e a sacarose têm sido as fontes de carbono mais utilizadas para a
produção de EPSs fúngicos. As concentrações destes açúcares utilizadas para a
produção de EPSs variam bastante em relação ao microrganismo empregado,

17
indicando a importância da adequação das concentrações iniciais do substrato para
cada cepa estudada (Cunha et al., 2008).
Em estudos realizados por Sudhakaran et al. (1998), a fim de avaliar diferentes
fontes de carbono para a produção de EPSs, verificou-se que a glicose foi a melhor
fonte de carbono utilizada para a produção de EPSs, seguida pela sacarose, maltose,
sorbitol, manitol, rafinose, acetato, manose, glicerol, xilose, arabinose, ribose, lactose
e galactose, onde diferentes microrganismos utilizam da melhor forma diferentes
fontes de carbono. Por exemplo, Aspergillus nidulans prefere frutose e glicose, já
Eusino eleucospila prefere frutose e galactose ao invés de glicose, o que sugere uma
especificidade de cada microrganismo para uma determinada fonte de carbono.
É possível ainda que diferentes fontes de carbono possam ter efeitos diferentes
no metabolismo secundário celular. Kim et al. (2005) verificaram que entre as fontes
de carbono testadas, glicose rendeu o maior crescimento micelial, porém, a máxima
produção de EPS foi alcançada quando maltose foi utilizada.
Quando relacionada à forma de assimilação da fonte de carbono, percebe-se
um consumo gradual durante o crescimento do microrganismo. Em estudos realizados
por Cunha et al. (2008), verificou-se que o substrato glicose foi consumido ao longo do
processo de fermentação, sendo praticamente esgotado ao final de 72 horas.
Em trabalhos semelhantes, Xu et al. (2004) também verificaram que a
concentração de açúcar residual foi decaindo durante a fermentação, concomitante
com o aumento da biomassa e produção de EPSs. Também foi verificado que maiores
concentrações iniciais de glicose contribuem para maior produção de EPSs pelo
microrganismo concomitantemente com a produção de biomassa, sendo verificada
produção de 2,62 g.L-1 de EPS nos cultivos onde foram utilizadas as menores
concentrações iniciais do substrato (20 g.L-1) e 4,32 g.L-1 nos cultivos onde foram
empregadas as maiores concentrações (50 g.L-1) (Xu et al., 2004). Este perfil de
consumo já era esperado, visto que a glicose é a fonte de carbono e energia mais
facilmente assimilada pelos microrganismos, uma vez que a maioria dos
microrganismos possui as enzimas envolvidas no processo de catabolismo dessa
hexose.
Cho et al. (2006), realizaram estudos utilizando Tremella fuciformis e
verificaram que as maiores produções de biomassa e de EPS foram obtidas quando

18
utilizada glicose, frutose e sacarose, sendo que o maior crescimento micelial e
produção de EPS foram obtidos quando glicose foi utilizada como fonte de carbono.
Já Lee et al. (2004), verificaram que tanto a biomassa quanto a produção de EPS
foram maiores quando a glicose foi avaliada, onde a concentração máxima para a
produção de ambos foi de 30 g.L-1.
Porém, em estudos realizados por Kim et al. (2002), onde foram testadas
diferentes fontes de carbono (glicose, frutose, maltose, manitol e sacarose) utilizando
Paecilomyces sinclairii a fim de avaliar a maior produção de biomassa e EPSs, verificou-
se que as maiores produções de biomassa (16,3 g.L-1) e EPS (1,3 g.L-1) foram obtidas
quando a sacarose foi usada como fonte de carbono.
Assim como o tipo de fonte de carbono, as concentrações ideais também
variam bastante em relação ao microrganismo empregado, indicando a importância da
adequação das concentrações iniciais do substrato para cada cepa estudada (Cunha et
al., 2008).
Experimentos realizados com o fungo Paecilomyces japonica, utilizando-se 10,
20, 30, 40 e 50 g.L-1 de maltose demonstraram que o maior crescimento micelial foi
obtido com 30 g.L-1 concomitantemente com a maior produção de EPS (Barbosa et al.,
2004). Isso indica que cada microrganismo possui especificidade tanto para a fonte de
carbono como para a concentração ótima desta a ser utilizada, descartando a hipótese
de que quanto maior a concentração da fonte de carbono maior a produção micelial e
de EPSs até um certo limite.
4.4.2- FONTES DE NITROGENIO
Um outro fator nutricional importante para a produção efetiva de EPSs é a
fonte de nitrogênio. As fontes orgânicas mais utilizadas são: peptona, glutamato, L-
asparagina, extrato de levedura, succinato de amônio, dentre outras. Entre as
inorgânicas, podem ser citadas: sulfato de amônio, nitrato de sódio, nitrato de
potássio, nitrato de amônio, dentre outras (Seviour et al. 1992; Krcmar et al. 1999; Bae
et al. 2000; Barbosa et al. 2004).
Tem sido descrito em vários trabalhos que os PSs são produzidos somente sob
condições de nitrogênio limitantes e que altos níveis de nitrogênio no meio de cultivo

19
reprimem a formação de EPSs. Este efeito tem sido observado em vários fungos
produtores de β-glucanas. Entretanto, acredita-se que este tipo de regulação depende
da fonte de nitrogênio utilizada (Barbosa et al., 2004).
Em estudos realizados por Sudhakaran et al. (1998) foram testadas diferentes
fontes de nitrogênio e verificou-se que houve uma maior produção de EPS (4,8 g.L-1)
quando o extrato de levedura foi utilizando e que a concentração utilizada influenciou
na quantidade de EPS produzido, sendo a concentração de 0,1% de extrato de
levedura a que proporcionou o maior rendimento, sendo que em sua ausência
pequenas quantidades de EPS (0,84 g.L-1) eram obtidas.
Assim como Sudhakaran et al. (1998), Lee et al. (2004) também observaram
que a máxima produção de EPS foi obtida quando utilizado extrato de levedura como
fonte de nitrogênio.
Já nos estudos realizados por Kim et al. (2002) foram testadas 10 fontes de
nitrogênio, algumas orgânicas como extrato de milho fermentado, peptona de carne,
extrato de levedura, peptona de soja e polipeptona, e algumas inorgânicas como
nitrato de amônio, cloreto de amônio, fosfato de amônio, sulfato de amônio e citrato
de amônio. Dentre estas, extrato de milho fermentado, peptona de carne, extrato de
levedura e peptona de soja mostraram-se relativamente favoráveis ao crescimento
micelial de Paecilomyces sinclairii, sendo a produção máxima de EPSs verificada
quando o extrato de milho fermentado foi utilizado como fonte de nitrogênio.
Por outro lado, ao contrário dos resultados obtidos em diversos trabalhos
utilizando fontes orgânicas de nitrogênio, alguns autores verificaram que quando
utilizado nitrogênio inorgânico obtiveram-se menor crescimento micelial e produção
de EPSs (Sudhakaran et al. 1998; Kim et al. 2002; Lee et al. 2004; Kim et al. 2005; Cho
et al. 2006).
Pelos resultados descritos na literatura, acredita-se que existem preferências
específicas quanto à fonte de nitrogênio, as quais estão diretamente vinculadas ao
metabolismo de cada microrganismo (Barbosa et al., 2004).

20
4.4.3- FERMENTAÇÃO SUBMERSA
O cultivo submerso proporciona um aumento potencial na produção de
biomassa micelial, ocupando um pequeno espaço e menor tempo de fermentação,
além de diminuir significativamente as chances de contaminação. Além disso, os EPSs
produzidos desta maneira possuem efeito sinérgico com o micélio, podendo ser
simultaneamente produzidos (Kim et al., 2002).
A agitação/aeração durante o crescimento resulta em uma melhor mistura dos
componentes do meio, ajudando a manter uma concentração gradiente entre o
exterior e o interior da célula, permitindo uma maior difusão dos nutrientes.
Aparentemente uma alta taxa de agitação/aeração é requerida para aumentar a
produção de biomassa micelial de culturas submersas, porém a ótima condição de
agitação/aeração para a produção de EPSs ou outros metabólitos depende de outros
fatores, inclusive dos substratos utilizados, da morfologia e da cepa utilizada (Xu et al.,
2004).
Estudos realizados por Xu et al. (2004) durante a otimização de produção de
EPSs em cultura líquida de Paecilomyces tenuipes indicam que as propriedades
moleculares dos EPSs foram influenciadas pela taxa de aeração. Além disso, também
verificaram que houve um aumento na produção de biomassa (21,56 g.L-1) e EPS (2,36
g.L-1) quando a taxa de agitação/aeração foi aumentada.
Também foi verificado por Xu et al. (2004) que diferentes condições de
agitação/aeração afetaram significativamente o peso molecular e as características
químicas dos EPSs produzidos pelo Paecilomyces tenuipes em cultura submersa.
Xu et al. (2004) verificaram que ao aumentar a taxa de agitação/aeração, a
viscosidade aparente do caldo de fermentação aumentou rapidamente com o
microrganismo ,entrando em sua fase exponencial de crescimento, sendo que a maior
viscosidade (490 cP) foi obtida em uma taxa de aeração de 3,5 vvm depois de 10 dias,
resultando principalmente em um maior acumulo de micélio.
Durante o crescimento, as diversas condições a que o microrganismo é exposto
podem gerar grandes variações no grau de ramificação de alguns dos polímeros de
escleroglucana, uma β-glucana, o que pode afetar muito sua solubilidade. Quando a
escleroglucana é mais ramificada, sua solução é pseudoplástica e sua viscosidade

21
parece não ser afetada na faixa de 20 a 90°C. Em contrapartida, a viscosidade da
solução de pululana produzida por Aureobasidium pullulans ATCC9348 foi avaliada sob
diferentes condições de cultivo em biorreator e observou-se que os valores foram
constantes. Este resultado sugeriu que a massa molecular de pululana e seu grau de
polimerização não são afetados pelo meio físico (Barbosa, et al., 2004).
As características morfológicas do microrganismo também influenciam no
processo de crescimento e produção de EPSs. Em estudos realizados por Barbosa et
al., (2002), verificou-se que as características morfológicas de Paecilomyces japonica
exerceram uma influência significativa sobre a reologia do caldo contendo EPSs, já que
a forma de micélio esférico promoveu menor viscosidade do que a forma de micélio
filamentoso.
4.5- RECUPERAÇÃO E QUANTIFICAÇÃO DE EXOPOLISSACARÍDEOS
Os exopolissacarídeos fúngicos são recuperados do meio de cultivo de forma
semelhante aos de bactérias, ou seja, através de centrifugação, filtração, agentes
precipitantes, diálise e liofilização. As centrifugações são utilizadas para separar as
células, sedimentando-as e separando-as do EPS solúvel presente no meio
extracelular. Também podem ser utilizadas para auxiliar na sedimentação posterior
dos EPSs presentes no sobrenadante, após tratamento com agentes precipitantes
(Barbosa et al., 2004).
O etanol é o agente precipitante mais comum, embora isopropanol ou cetonas
possam também ser utilizados (Barbosa et al., 2004).
A metodologia comumente utilizada para a quantificação de EPSs e biomassa
micelial é através do método gravimétrico, após separação da biomassa e extração dos
EPSs, obtidos após o período de crescimento. Após a centrifugação do meio de cultura
o sobrenadante é transferido para outro frasco e o pelete formado, após lavado com
água destilada, é seco a aproximadamente 70°C até peso constante. Para a
precipitação dos EPSs normalmente são adicionados ao sobrenadante quatro volumes
de etanol, seguido de centrifugação. Após a secagem os EPSs extraídos são pesados
(Kim et al. 2002; Lee et al. 2003; Kim et al. 2004; Xu et al. 2004; Kim et al. 2005; Cho et
al. 2006; Fernandes et al. 2011).

22
Já Selbmann et al. (2002) utilizaram dois volumes de etanol para a precipitação
de EPSs em seus trabalhos utilizando Phoma herbarum CCFEE 5080, Sclerotium
glucanicum NRRL 3006 e Botryosphaeria rhodina DABAC-P82.
A importância de um bom método para a recuperação e quantificação dos EPSs
fúngicos do meio de cultivo reside no fato de possibilitar a comparação da produção
de EPSs por uma ou outra cepa fúngica.
4.6- BIOATIVIDADE E ESTRUTURA DOS EXOPOLISSACARÍDEOS
As propriedades dos PSs são determinadas por características como tipo de
ligação glicosídica e grau de ramificação e composição monossacarídica (Silva et al.,
2006). Estas propriedades podem ser afetadas durante o crescimento do fungo em
função da composição do meio de cultura utilizado, assim como as condições de
cultivo a que esse microrganismo será exposto (Barbosa et al., 2004).
Para avançar na investigação da correlação existente entre a estrutura química
e a bioatividade dos EPSs, frações com alta pureza e diferentes propriedades
moleculares são frequentemente necessárias. Mesmo após vários estudos tendo
examinado os efeitos das condições de cultura na produção microbiana de EPSs, pouco
é sabido sobre a influencia na qualidade do produto, particularmente as características
moleculares (Xu et al., 2004).
Antes que a caracterização química seja iniciada, é necessária a purificação do
EPS, geralmente a parte mais laboriosa, pois, para se analisar a estrutura de um
polissacarídeo é necessário, primeiro, determinar os tipos de resíduos
monossacarídicos que constituem o composto biológico. Métodos espectroscópicos
como ressonância magnética nuclear e infravermelho acoplado ao transformador
Fourier são utilizados para identificar a configuração anomérica das ligações
glicosídicas do EPS (Barbosa, et al., 2004).
Diversos estudo têm demonstrado que as atividades biológicas dos EPSs estão
intimamente ligadas às suas estruturas químicas, como demonstrado nos estudos de
Barbosa et al. (2004), onde verificou-se que as propriedades reológicas e atividade
antitumor apresentadas pelas glucanas extracelulares do tipo β, especialmente
glucanas β (1->3), como escleroglucana (Sclerotium glucanicum), esquizofilana

23
(Schizophyllum commune), cinereana (Botrytis cinerea) e pestalotana (Pestalotia sp),
não eram verificadas nas glucanas do tipo α, β (1->4) ou β (1->6).
Nos estudos realizados por Fernandes et al. (2011), foi verificada a capacidade
antitumoral de EPSs secos por freeze dryer produzidos por Agaricus blazei LPB 03
contra células de Ehrlich in vitro, obtendo-se 28% de inibição utilizando-se 5 mg de
EPSs, sendo que para se obter inibição de 50% das células foram necessários 34 mg de
EPSs, uma pequena dose quando comparado com outros extratos de inibição de
células tumorais. Porém, estes EPSs também mostraram características citotóxicas,
não permitindo seu uso devido a possibilidade de danificar macrófagos.
Tais características também foram verificadas nos trabalhos realizados por Lima
et al. (2009), onde os EPSs obtidos de seis espécies de basidiomicetos apresentaram
forte atividade antitumoral, inibindo em mais de 70% o crescimento das células
tumorais na maior concentração testada (200 μg.mL-1), o que sugere que eles
apresentam potencial para investigação de novos agentes quimioterápicos contra este
tipo de tumor.
Lentinana e esquizofilana, duas glucanas fúngicas β-(1->3; 1->6), tornaram-se
clinicamente relevantes como imunoadjuvantes na terapia contra o câncer no Japão.
Estas glucanas, não citotóxicas, supostamente expressam seu efeito antitumor por
estimulação do sistema imunológico (Silva et al., 2006).
Além das características antitumorais, são observadas também funções
antioxidantes desempenhadas pelos EPSs. Em estudos realizados por Lee et al. (2003),
pode ser verificada a ação antioxidante de quatro PSs obtidos por fermentação de
Grifola frondosa.
Estudos realizados por Cheung (1996) indicaram a capacidade
hipocolesterolemiante de EPSs produzidos por Volvuriella volvacea, sendo que esta
capacidade estaria principalmente ligada à presença de β-glucanas, que seriam as
responsáveis pela diminuição da absorção do colesterol devido a formação de micelas
entre as moléculas de colesterol e os PSs.

24
4.7- PROCESSOS BIOTECNOLÓGICOS NA PRODUÇÃO DE
EXOPOLISSACARÍDEOS E SUAS APLICAÇÕES
A biotecnologia é um dos ramos mais antigos da ciência e pode ser entendida
como a tecnologia que objetiva o desenvolvimento de produtos e serviços por meio de
processos biológicos. O homem já utilizava organismos vivos, ainda que ingenuamente,
para a produção de bebidas e alimentos nos primórdios da civilização. Atualmente a
biotecnologia desperta grande interesse nos pesquisadores e na indústria, e muitos
produtos de interesse para o homem são obtidos por processos biotecnológicos
(Brock, 2010).
As indústrias de alimentos e farmacêutica têm grande interesse no
desenvolvimento de pesquisas nessa área. Dessa forma, o estudo de novos organismos
vivos que possam produzir moléculas com propriedades tecnológicas relevantes é
bastante interessante e estratégico para o Brasil, visto que é um país rico em
biodiversidade (Cunha et al., 2008).
Entretanto, para serem empregados em processos tecnológicos industriais, os
microrganismos utilizados nestes processos devem possuir outras características além
da capacidade de produzir a substância de interesse em altos níveis. O microrganismo
deve ser capaz de crescer e sintetizar o produto em larga escala, produzir esporos ou
algum tipo de estrutura reprodutiva que facilite a inoculação, além de possuir
crescimento relativamente rápido e sintetizar o produto desejado em um espaço curto
de tempo (Brock, 2010).
Os microrganismos (algas, bactérias e fungos), assim como as plantas,
produzem diferentes polissacarídeos com diversas e importantes aplicações
biotecnológicas (Silva et al., 2006). Os EPSs produzidos por microrganismos são
exemplos de produtos de interesse que podem ser obtidos a partir da manipulação de
organismos vivos. A célula microbiana pode ser uma fonte rica de moléculas de
carboidratos, sendo alguns destes componentes da parede celular, enquanto outros
podem ser encontrados completamente dissociados da célula e são conhecidos como
exopolissacarídeos (Cunha et al., 2008).
As aplicações biotecnológicas dos polissacarídeos são limitadas devido aos
problemas relacionados com as modificações físicas, químicas e enzimáticas, que são

25
necessárias para o desenvolvimento de novos produtos. Técnicas de fermentação e
biotecnologia têm sido desenvolvidas com o objetivo de sintetizar e modificar
carboidratos, sendo baseadas essencialmente no uso de polissacarídeos (França et al.,
2007).
Agentes químicos como o monoesterato glicerol e a carboximetilcelulose são
emulsificantes amplamente utilizados na indústria de alimentos. Embora muito
efetivos nas suas funções, estes componentes estão perdendo gradualmente seu
campo devido a maior conscientização dos consumidores em reduzir o uso de aditivos
artificiais ou quimicamente sintetizados em alimentos (Borges, 2004). Por isso
atualmente há considerável interesse em polissacarídeos obtidos pela ação de
microrganismos. Estes polissacarídeos, conhecidos como biopolímeros, são obtidos
por processos fermentativos. Possuem capacidade de formar soluções viscosas e géis
em meio aquoso, mesmo quando aplicados em baixas concentrações.
Um exemplo de utilização biotecnológica é a pululana, uma α-glucana
produzida pelo fungo Aureobasidium pullulans, que é resistente ao óleo e por isso
aplicada em poços de petróleo. Além disso, este EPS forma filmes solúveis em água
com baixa permeabilidade ao oxigênio e pode revestir alimentos, conservando o seu
sabor e a sua aparência (Silva et al., 2006). A pululana é também um bom adesivo e
pode ser usado na preparação de algumas fibras, como um componente de sistemas
aquosos bifásicos. Este EPS também tem sido usado para preparar padrões de massa
molecular de baixa dispersão para calibrar HPLC (Barbosa et al., 2004).
4.8- UTILIZAÇÃO DE REJEITOS INDUSTRIAIS NA PRODUÇÃO DE EPSs
A disposição de resíduos gerados pelo setor agroindustrial no meio ambiente
tem resultado em frequentes relatos de problemas de poluição. Tais problemas
levaram ao desenvolvimento de pesquisas que gerassem medidas efetivas na
minimização dos níveis de poluição. Entre essas medidas, podem ser citadas as
seguintes: minimização da quantidade de resíduo gerada; tratamento adequado dos
resíduos antes da disposição no ambiente; aproveitamento dos resíduos em outras
atividades (Canuto et al., 2006).

26
Muitos processos microbiológicos industriais utilizam rejeitos de carbono
oriundos de outras indústrias como ingredientes principais ou suplementares de meios
de cultura de larga escala, dentre eles incluem-se agua da moagem de milho, que é
rico em nitrogênio e fatores de crescimento, e o leitelho, que é rejeito da indústria de
laticínios contendo alto teor de lactose e minerais.
Analogicamente à produção de EPS, Coelho et al. (2001) utilizaram resíduo do
processamento de coco verde para a produção de enzimas mediante fermentação
semi-sólida utilizando diferentes granulometrias em pó, e verificaram produção de
diferente complexos enzimáticos ricos em diferentes atividades, validando assim a
possibilidade do uso deste resíduo na produção de enzimas, o que possibilita testes de
produção de EPS utilizando tal resíduo agroindustrial.
Selbmann et al. (2002) realizaram estudos comparando diferentes fontes de
carbono alternativas (amido de milho, farinha de trigo mole, farinha de batata, farinha
de mandioca e batata doce e farinha de batata industrial) quanto ao crescimento
micelial e produção de EPS de dois fungos, Sclerotium glucanicum NRRL 3006 e
Botryosphaeria rhodina DABAC-P82, e verificaram que ambos os fungos tiveram bom
crescimento micelial e produção de EPS em todos os meios testados na concentração
de 30 g.L-1.
4.8.1- RESÍDUO DE CERVEJARIA
Segundo Camelini (2012), o resíduo das dornas de produção de cerveja possui
aproximadamente 48,6% de carbono disponível para utilização no processo
fermentativo, indicando serem fontes ricas em açúcares, principalmente na forma de
polissacarídeos e 5% de Nitrogênio, indicando não serem fontes eficientes de
proteínas e aminoácidos, caracterizando este rejeito como sendo fonte alternativa de
carbono.
4.8.2- BAGAÇO DE MAÇÃ
O bagaço de maçã é obtido a partir do processamento da fruta para a obtenção
do suco, podendo apresentar teor de água variável (75-87%), possuindo todos os

27
componentes encontrados na fruta e no suco, sendo concentrados os valores de
açucares por se ligarem às fibras solúveis e insolúveis (Venturini, 2010).
Para ser utilizado em processos fermentativos, o bagaço de maçã deve conter
uma agregação mínima de água, obtendo-se um extrato com o maior teor de sólidos
solúveis possível (Venturini, 2010).
De acordo com Venturini (2010), o bagaço de maçã desidratado apresenta um
elevado teor de açúcares e fibras, o que torna possível sua utilização como fonte de
carbono alternativa na fermentação de fungos visando a produção de EPSs.
4.8.3- LICOR CITRICO
O número de subprodutos da industrialização do suco de laranja cresceu
significativamente nos últimos anos, podendo inclusive superar o valor comercial do
suco de laranja. Dentre os subprodutos primários gerados durante o processamento
da laranja estão cascas (interna e externa), polpa e sementes, que são ricos em
açucares e fibras, sendo passível de utilização em processos fermentativos (Venturini,
2010).
4.8.4- MELAÇO CÍTRICO
O melaço cítrico aparece como subproduto do processamento de tangerina
para a produção de sucos, sendo obtido através da prensagem do bagaço resultante
da extração do suco. O melaço compreende cerca de 8 a 15% de sólidos dissolvidos,
sendo constituído principalmente de 60-70% de carboidratos (glicose, frutose,
sacarose e pentoses) e cerca de 0,1 a 0,5% de óleos ricos em terpenos e outros sólidos
(Venturini, 2010). Devido sua alta concentração de carboidratos, o melaço cítrico pode
ser utilizado em substituição às fontes de carbono convencionais.

28
5- CONSIDERAÇÕES FINAIS
A biotecnologia tem despertado grande interesse nos pesquisadores e na
indústria, devido à obtenção de muitos produtos de interesse para o homem através
dos processos biotecnológicos.
É percebido um crescente aumento no uso de polissacarídeos microbianos nos
ramos alimentício, médio e de cosméticos devido à sua potencialidade na formação de
géis, emulsões e biofilmes, suas características funcionais, trazendo grandes benefícios
tanto ao processo tecnológico quando à saúde dos consumidores, visto que suas
características tecnológicas são semelhantes aos aditivos químicos comumente
utilizados e por possuírem composição química compatível ao metabolismo humano,
possibilitando assim sua aplicação no ramo alimentício.
Devido as propriedades dos EPSs serem afetadas durante o crescimento do
fungo em função da composição do meio de cultura e das condições de cultivo a que
esse microrganismo será exposto, torna-se indispensável a avaliação destas condições
a fim de se obter condições ótimas de crescimento micelial e produção de EPS, de
forma que as características dos EPSs produzidos não sejam afetadas diante das
condições a que o microrganismo será exposto.
Dessa forma, o estudo de novos organismos vivos que possam produzir
moléculas com propriedades tecnológicas relevantes tanto por métodos convencionais
quanto a partir de subprodutos industriais é bastante interessante e estratégico para o
Brasil, visto que é um país rico em biodiversidade.
6- REFERÊNCIAS BIBLIOGRÁFICAS
BABITSKATYA, U.G.; SHERBA, V.V.; MITROPOLSKAYA, N.Y.; BISKO, N.A. 2000. Exopolysaccharides of some medicinal mushroom: production and composition. Int J Med Mushroom. 2:51-54.
BAE, J. T. et al. 2000. Optimization of submerged culture conditions for exo-biopolymer production by Paecilomyces japonica. JournalofMicrobiology and. Biotechnology, Kangnam-Ku, v.10, p.482-487

29
BARBOSA, A.M.; CUNHA, P.D.T.; Pigatto, M.M.; SILVA, M.L.C.2004.Produção e Aplicações de ExopolissacarídeosFúngicos.CiênciasExatas e Tecnológicas, Londrina, v. 25, n. 1, p. 29-42
BORCHERS, A.T.; STERM, J.S.; HACKMAN, R.M.; KEEN, C.L.; GERSHWIN, H.E. Mushrooms, tumors and immunity. 1999. Pro SocExpBiol Med. 221:281-293.
BORGES, C. D.; MOREIRA, A. N.; MOREIRA, A. S.; DEL PINO, F. A. B.; VENDRUSCULO, C. T. 2004. Caracterização de biopolímeros produzidos por Beijerinckiasp. 7070 em diferentes tempos de cultivo. Ciênc. Tecnol. Aliment., Campinas, 24(3): 327-332.
CAMELINI, C.M. 2010. PRODUÇÃO DE BIOMASSA DO FUNGO Agaricussubrufescens POR PROCESSOS FERMENTATIVOS SÓLIDO E SUBMERSO PARA OBTENÇÃO DE POLISSACARÍDEOS BIOATIVOS. Tese, Biotecnologia da Universidade Federal de Santa Catarina CANUTO, A.P.;2006.DESENVOLVIMENTO DE BIOPROCESSO PARA PRODUÇÃO DE GOMA XANTANA POR FERMENTAÇÃO NO ESTADO SÓLIDO A PARTIR DE RESÍDUOS E SUBPRODUTOS AGROINDUSTRIAIS. Dissertação, ProcessosBiotecnológicos da Universidade Federal do Paraná. CHO, E. J.; OH, J. Y.;CHANG, H. Y.; YUN, J. W.2006.Production ofexopolysaccharidesbysubmergedmyceliacultureof a mushroomTremellafuciformis.JournalofBiotechnology 127 (2006) 129–140. CHEUNG, P.C.K. Ph.D. 1996.THE HYPOCHOLESTEROLEMIC EFFECT OF EXTRACELLULAR POLYSACCHARIDE FROM THE SUBMERGED FERMENTATION OF MUSHROOM.Nueition Research, Vol. 16. Nos. 11112, pp. 1953-1957.1 COELHO, M.A.Z.; LEITE, S.G.F.; ROSA, M.F.; FURTADO, A.A.L. 2001. APROVEITAMENTO DE RESÍDUOS AGROINDUSTRIAIS: PRODUÇÃO DE ENZIMAS A PARTIR DA CASCA DE COCO VERDE. B.CEPPA, Curitiba, v. 19, n. 1, p. 3342 CUNHA, M.A.A.; PAWALAK, M.P.; FABBRIS, E.Z.; BARBOSA, A.M.; DEKKER, R.F.H. 2008.PRODUÇÃO DE EXOPOLISSACARÍDEO PELO FUNGO ASCOMICETO BotryosphaeriarhodinaMMPI. ISSN: 1981-366X / v. 02, n. 20 FERNANDES, M. B. A.; HABU, S.; LIMA, M.A.; THOMAZ-SOCCOL, V.; SOCCOL, C.R. 2011. Influence of drying methods over in vitro antitumoral effects of exopolysaccharides produced by Agaricusblazei LPB 03 on submerged fermentation. BioprocessBiosystEng34:253–261 FRANÇA, E.;DALL´ANTONIA, L.H. 2007.Novos compósitos de polianilina e exopolissacarídeos com aplicações em biomateriais: uma revisão. Ciências Exatas e Tecnológicas, Londrina, v. 28, n. 1, p. 37-46

30
FRANZ, G. 1989. Polysaccharides in pharmacy: current applications and future concepts. Planta Med. 55:493-497.
KIM, S.W.; HWANG, H.J.; PARK, J.P.; CHO, Y.J.; SONG, C.H.; YUN, J.W. Mycelial growth and exo-biopolymer production by submerged culture of various edible mushrooms under different media. 2002. Letters in Applied Microbiology, 34, 56±61 KIM, H.O.; LIM, J.M.; JOO, J.H.; KIM, S.W.; HWANG, H.J.; CHOI, J.W.; YUN, J.W. 2004. Optimization of submerged culture condition for the production of mycelial biomass and exopolysaccharides by Agrocybecylindracea.Bioresource Technology 96 1175–1182 Kim, H.O.; Yun, J.W. 2005.A comparative study on the production of exopolysaccharides between two entomopathogenic fungi Cordycepsmilitaris and Cordycepssinensis in submerged mycelial cultures. Journal of Applied Microbiology, 99, 728–738 KRCMAR, P. et al. 1999. Structure of extracellular polysaccharide produced by lignin-degrading fungus Phlebiaradiatain liquid culture. International Journal of Biological Macromolecules, Amsterdam, v.24, p.61- 64 LEE, B.C.; BAE, J.T.; PYO, H.B.; CHOE, T.B.; KIM, S.W.; HWANG, H.J.; Yun, J.W. 2003. Biological activities of the polysaccharides produced from submerged culture of the edible BasidiomyceteGrifolafrondosa. Enzyme Microb Technol. 32:574-581.
LEE, B.C.; BAE, J.T.; PYO, H.B.; CHOE, T.B.; KIM, S.W.; HWANG, H.J.; YUN, J.W. 2004. Submerged culture conditions for the production of mycelial biomass and exopolysaccharides by the edible BasidiomyceteGrifolafrondosa. Enzymeand Microbial Technology 35- 369–376 LIMA, M.P. POTENCIAL BIOTECNOLÓGICO DE BASIDIOMICETOS ISOLADOS NO ESTADO DO PARANÁ. 2009. Dissertação, Setor de Tecnologia da Universidade Federal do Paraná. LIU, F.; OOI, V.E.C.; CHANG. S.T. 1997. Free radical scavenging activities of mushroom polysaccharide extracts. Life Sci. 60:763-771.
MANSELL, P.W.A. 1994. Polysaccharides in skin care. CosmetToiletries. 109:67-72.
RICCIARDI, A.; PARENTE, E.; CRUDELE, M.A.; ZANETTI, F.; SCOLARI, G.; MANNAZZU, I. 2002. Exopolysaccharide production by Streptococcus thermophilusSY: production and preliminary characterization of the polymer. J ApplMicrobiol. 92:297-306.
SELBMANN, L.; CROGNALE, S.; PETRUCCIOLI, M. 2002.Exopolysaccharide production from SclerotiumglucanicumNRRL 3006 and Botryosphaeriarhodina DABAC-P82on raw and hydrolysed starchy materials.Letters in AppliedMicrobiology, 34, 51±55 SELBMANN, L.; ONOFRE, S.; FENICE, M.; FEDERICI, F.; PETRUCCIOLI, M. 2002.Production and structural characterization of the exopolysaccharide of the

31
Antarctic fungus PhomaherbarumCCFEE 5080. Research in Microbiology 153 (2002) 585–592. SEVIOUR, R. J. et al. A. Production of pullulan and other exopolysaccharides by filamentous fungi.CriticalReviews in Biotechnology, Boca Raton, v.12, p.279-298, 1992. SILVA, M. L. C.; MARTINEZ, P. F.; IZELI, N. L.; SILVA, I. R.; VASCONCELOS, A. F. D.; CARDOSO, M. S.; STELUTTI, R. M.; GEISE, E. C.; BARBOSA, A. M. 2005. Caracterização química de glucanasfúngicas e suas aplicações biotecnológicas. Quim. Nova, Vol. 29, No. 1, 85-92, 2006.
SUGIHARA, R.; OISO, Y.; MATSUMOTO, Y.; OHMORI, H. Production of an immunosuppressive polysaccharide, A29, in the culture of Klebsiellaoxytocastrain TNM3.2001. J. BiosciBioeng. 92:485-487.
PAUL, F.; MORIN, A.; MONSAN, P. 1986. Microbial polysaccharides with actual potential industrial applications.Biotechnol Adv. 4:245-259.
SUDHAKARAN, V. K.; SHEWALE, J. G. 1998.Exopolysaccharide production by Nigrosporaoryzae var. glucanicum.EnzymeMicrob.Technol., 1988, vol. 10. SUTHERLAND, I.W. 1999. Polysaccharides for microbial exopolysaccharides.CarbohydrPolym. 38:319-328. SUTHERLAND, I.W. Microbial polysaccharides from gram-negative bacteria.2001.Int Dairy J. 11:663-674.
SOUZA, D. M.; GARCIA-CRUZ, C. H. 2004. Produção fermentativa de polissacarídeos extracelulares por bactérias. Semina: Ciências Agrárias, Londrina, v. 25, n. 4, p. 331-340.
WASSER, S.P. 2002. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. ApplMicrobiolBiotechnol. 60:258-274.
YANG, B.K.; HA, H.Y.; JEONG, S.C.; JEON, Y.J.; RA, K.S.; DAS, S.; Yun, J.W.; Song, C.H. Hypolipidemic effect of an exo-biopolymer produced from submerged mycelial culture of Auriculariapolytrichain rats.2002.BiotechnolLett. 24:319-325.
XU, C.P.; YUN, J.W. Influence of aeration on the production and the quality of the exopolysaccharides from PaecilomycestenuipesC240 in a stirred-tank fermenter. 2004. Enzymeand Microbial Technology 35 -33–39.