instituto nacional de pesquisas da amazÔnia – inpa … · 2015. 5. 5. · instituto nacional de...
TRANSCRIPT

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA UNIVERSIDADE FEDERAL DO AMAZONAS – UFAM
ESTIMATIVA DA VARIABILIDADE GENÉTICA DA REGIÃO CONTROLE DO DNA MITOCONDRIAL DE Nannostomus eques (CHARACIFORMES,
LEBIASINIDAE) DA BACIA DO RIO NEGRO MARIA LEANDRA TERENCIO
MANAUS - AM FEVEREIRO - 2009

MARIA LEANDRA TERENCIO
ESTIMATIVA DA VARIABILIDADE GENÉTICA DA REGIÃO CONTROLE DO DNA MITOCONDRIAL DE Nannostomus eques (CHARACIFORMES,
LEBIASINIDAE) DA BACIA DO RIO NEGRO
Orientador: Jorge Ivan Rebelo Porto, Dr.
Dissertação apresentada ao Programa
Integrado de Pós-Graduação em Biologia
Tropical e Recursos Naturais do convênio
INPA/UFAM, como parte dos requisitos
para obtenção do título de Mestre em
Genética, Conservação e Biologia Evolutiva.
MANAUS - AM FEVEREIRO 2009

ii
Ficha Catalográfica
Sinopse:
Estudou-se a diversidade genética de oito populações naturais de Nannostomus eques do alto, médio e
baixo rio Negro, fundamentada na análise da região controle (D-loop) do DNA mitocondrial. As
análises filogenéticas e populacionais sugerem a existência de pelo menos duas unidades evolutivas
dentro de N. eques sendo que das oito populações analisadas somente a população do rio Demeni,
pertencente a unidade evolutiva 2, encontrou-se estruturada geneticamente.
T316 Terencio, Maria Leandra Estimativa da variabilidade genética da região controle do DNA mitocondrial de Nannostomus eques (Characiformes, Lesbiasinidae) da bacia do rio Negro/ Maria Leandra Terencio .--- Manaus : [s.n.], 2009. xvi, 102 f. : il. color. Dissertação (mestrado)-- INPA/UFAM, Manaus, 2009 Orientador : Jorge Ivan Rebelo Porto Área de concentração : Genética, Conservação e Biologia Evolutiva 1. Peixe ornamental – Negro, Rio (AM) . 2. Genética de populações. 3. DNA mitocondrial. 4. Nannostomus eques. I. Título. CDD 19. ed. 597.50415

iii
Aos meus pais, Flávio e Maria Inês,
minha avó Teresa e ao meu esposo
Igor.

iv
“O que faz da evolução uma teoria tão impecável é o fato
de ela explicar como a complexidade organizada pode
surgir da simplicidade primitiva”
Richard Dawkins

v
Agradecimentos
Ao Programa Integrado de Pós-Graduação em Biologia Tropical e Recursos Naturais
do Instituto Nacional de Pesquisas da Amazônia (INPA) e Universidade Federal do
Amazonas (UFAM), curso de Genética, Conservação e Biologia Evolutiva.
Ao Laboratório Temático de Biologia Molecular (LTBM) do INPA, Coordenação de
Pesquisas (COPE), onde foi desenvolvida a maior parte deste trabalho.
Ao CNPq e FINEP pelo financiamento dos Projetos de pesquisas “Biotecnologia
aplicada ao estudo das populações de peixes de importância econômica para a Amazônia”
(CNPq/CT/Amazônia), Projeto “Navegabilidade, hidrologia, qualidade de água e
biodiversidade do rio Negro (FINEP/CNPq 3724/04) - Projeto Fronteira.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pela
bolsa de estudo durante a realização deste trabalho.
A todas essas pessoas fica a minha eterna gratidão, em especial:
Ao meu orientador, Jorge Porto, que até nas corriqueiras ocasiões do “assina isto pra
mim...” aproveitava o momento para me questionar, plantar dúvidas em minha cabeça.
Foram raras as ocasiões em que não saí de sua sala sem um enorme ponto de interrogação
pairando no ar. No início não entendia muito bem, mas hoje sei que estes momentos foram
fundamentais para meu amadurecimento. Sou extremamente grata por este tempo de
convivência, onde mesmo sabendo que eu nunca “havia pisado” em um laboratório de
biologia molecular, confiou em mim! Obrigada.
À Dra. Eliana Feldberg, pelo apoio, incentivo e “socorro” nas horas difíceis. Por me
incluir muitas vezes nos “momentos família” em sua casa e no sítio, aliviando a saudade da
família.
Ao Dr. Jansen Zuanon pela disponibilidade na identificação dos espécimes e pelo
“presente” coletado no Demeni.
À Maria Claudia Gross, “Claudinha cabeça” por existir! Já que não consigo expressar
em palavras quão importante você foi e é em minha vida acadêmica, desde os tempos que
fazíamos parte do seleto grupo “bóias-frias da ciência”, com o agravante de que a bolsa de
Pibic era dividida para três estudantes, Você, Carlos e Eu. Não é que valeu a pena?
Profissional exemplar e acima de tudo humilde em todos os seus atos. Obrigada por me
deixar fazer parte disto. Não adianta, aceite que você é especial!
Ao grande “tudo” Carlos Henrique Schneider, com você aprendi tudo que sei do
mundo da biologia molecular. Obrigada pela paciência ao ensinar “a cabeçuda”, desde o

vi
simples calibrar de uma pipeta até o não usar luva durante uma reação de seqüenciamento
(sabe que muito de suas neuras tem lógica? não se anime que não são todas). Obrigada, mil
vezes!
Aos técnicos e colegas de excursão, Agenor, Arnóbio e em especial ao Roberval pelos
momentos incríveis que foram as coletas e por me ensinarem o vocabulário do pescador, o
que se torna fundamental quando lhe dizem “Neste não! Vamos pegar a lharga dele!!!!”
Aos amigos de mestrado da “grande” turma GCBEV-2007, Adam, Isaura, Letícia e
Marcos.
A toda turma do projeto Pirada (Jacque, Kyara, Jane, Naiara, Iamíli, Rosa, Luci,
Larissa) pelo auxílio no laboratório e momentos de descontração. Em especial à Giselle,
companheira “do turno das sete da matina” por me ensinar muito e confiar seu filho “o
seqüenciador de DNA” em minhas mãos, e à Fabíola, que de “coleguinha de pós”, tornou-se
grande amiga. Obrigada por ouvir e compreender “meus momentos de ira”.
Ao pessoal do “Peixe-elétrico” pela ajuda nas coletas no alto rio Negro e em especial
à Renata Schmitt pela sua atenção e disponibilidade no campo e laboratório.
Aos amigos do Laboratório de Genética Animal do INPA: Claudia, Carlos, Leila,
Eduardo, Rodrigo, Brenda, Érica, Cacá, Marco, Alexandra e Cebolinha. Obrigado pelas
dicas, sugestões, discussões e festas!
Ao Robertão, Mara, Marcelo, e Miguel e demais colegas do Laboratório de
Citogenética Animal da Universidade Estadual de Ponta Grossa, por me iniciarem na vida
científica.
Aos compadres Carlos e Claudia. Se não fosse pela indicação de vocês, com certeza
eu não estaria em Manaus. Obrigada pela guarida quando Igor e eu ficamos “sem teto”,
pelos momentos de descontração, piscina, churrasco, piscina e mais churrasco! Polacos,
vocês são verdadeiros irmãos!
Ao casal de amigos Waleska e Daniel pelos ótimos momentos de diversão, bem como
pelo apoio nas horas que mais precisei.
Ao meu esposo Igor José Theodorovitz, por fazer parte da minha vida, apoiando,
rindo, chorando, brigando, torcendo. Sempre ao meu lado, sem você eu não seria quem sou e
nem faria o que eu faço. Obrigada!
Aos meus pais Flávio e Maria Inês, onde apesar de maneiras diferentes, e mesmo sem
entender completamente o que eu faço sempre me estimularam e me apoiaram a seguir em
frente. Mãe você sempre foi meu exemplo em tudo que faço!

vii
Este mestrado em Manaus me furtou grandes momentos em família e agradeço muito
a tolerância dos meus parentes mais próximos, principalmente de minha “vozinha Teresa”
em compreender que esta ausência foi muito maior do que eu gostaria.
Obrigada!

viii
Resumo
A bacia do médio rio Negro é a área de maior importância para pesca de peixes
ornamentais no estado do Amazonas e o município de Barcelos é o principal entreposto
comercial. Nannostomus eques (Lebiasinidae: Characiformes), conhecida popularmente
como peixe-lápis, é uma dentre as diversas espécies de peixes liberadas para
comercialização como ornamentais. Este trabalho apresenta um estudo sobre a variabilidade
genética em Nannostomus eques fundamentada na análise de seqüências da região controle
do DNA mitocondrial de 125 indivíduos coletados em oito tributários ao longo do alto
(Açaituba, Miuá, Jaradi e Arixanã), médio (Demeni) e baixo (Jacundá, Maguari e Catalão)
rio Negro. As inferências filogenéticas (máxima parcimônia, máxima verossimilhança e
distância genética) dos dados mitocondriais evidenciaram duas unidades evolutivas dentro
de N. eques, as quais estão separadas por 60 passos mutacionais. O limite geográfico entre as
duas unidades evolutivas parece ser as adjacências do principal tributário do rio Negro, o rio
Branco. A distância genética entre as duas unidades evolutivas variou de 5,50% a 8,30%.
Ambas as unidades diferiram de 8,50% a 11,80% em relação à espécie irmã, N. unifasciatus.
As análises populacionais (polimorfismo de DNA, AMOVA, Teste de Mantel) permitiram a
identificação de uma alta diversidade haplotípica (h > 0,90) em cada uma das unidades
evolutivas, uma forte estruturação genética populacional no rio Demeni e uma correlação
entre divergência genética e distância geográfica em apenas uma das unidades. Com base
nos dados moleculares as corredeiras e cachoeiras nas proximidades de São Gabriel da
Cachoeira (alto rio Negro) são a principal barreira ao fluxo gênico entre as populações da
unidade evolutiva 1. Aparentemente os surgimentos do rio Branco e do Arquipélago das
Anavilhanas foram responsáveis pela distribuição descontínua das duas unidades evolutivas,
exceto no igarapé Jacundá onde houve simpatria entre as unidades evolutivas. Tendo em
vista o grau de diferença entre as unidades evolutivas, N. eques da bacia do rio Negro não
pode ser tratada como um único estoque. Estes resultados podem ser de grande importância
para o manejo da pesca deste peixe ornamental.

ix
Abstract
The middle Negro River Basin is the most important area for ornamental fisheries in
the state of Amazonas being that the city of Barcelos is the main trading post. Nannostomus
eques (Lebiasinidae: Characiformes), commonly known as pencilfish, is one among the
many fish species allowed to be exported as ornamental. This paper presents a study on the
genetic variability of Nannostomus eques based on analysis of sequences of the control
region of mitochondrial DNA of 125 individuals collected in eight tributaries along the
upper (Açaituba, Miuá, Jaradi and Arixanã), middle (Demeni) and lower (Jacundá, Maguari
and Catalão) Negro River. The phylogenetic inferences from mitochondrial DNA data of N.
eques (parsimony, likelihood and genetic distance methods) showed two evolutionary units
separated by 60 mutation steps. The rough geographic boundary between the two units
seems to be the vicinity of the Branco River, the main tributary of the Negro River. The
genetic distance between the two evolutionary units ranged from 5.50% to 8.30%. Both
units differ from 8.50% to 11.80% from sister species, N. unifasciatus. The population
analysis (DNA polymorphism, AMOVA, Mantel test) resulted in the identification of high
haplotype diversity (h > 0.90) in each evolutionary unit, a strong population genetic
structure in the Demeni River and a correlation between genetic divergence and geographic
distance in only one of these units. Based on molecular data the rapids and waterfalls near
São Gabriel da Cachoeira (upper Negro River) are the main barriers to the gene flow among
individuals of the evolutionary unit 1. The emergence of Branco River and the Anavilhanas
Archipelago apparently were responsible for the disjoint distribution of the two evolutionary
units, except at Igarapé Jacundá where the evolutionary units were sympatric. In view of
extent differences between the evolutionary units, N. eques can not be treated as a single
stock in the Negro River basin. These results may have important implications for the
fishery management of this ornamental fish.

x
Sumário
Ficha Catalográfica ......................................................................................................... ii
Sinopse: ............................................................................................................................. ii
Agradecimentos ............................................................................................................... v
Resumo .......................................................................................................................... viii
Abstract ........................................................................................................................... ix
1. Introdução .................................................................................................................... 1 1.1 Bacia amazônica e sub-bacia do rio Negro .............................................................. 2
1.2 A ictiofauna do rio Negro ......................................................................................... 4
1.3 Comércio de peixes ornamentais .............................................................................. 5
1.4 Família Lebiasinidae e o gênero Nannostomus ........................................................ 6
1.5 DNA mitocondrial e variabilidade genética ........................................................... 10
2. Objetivos ..................................................................................................................... 13 2.1 Objetivo geral ......................................................................................................... 13
2.1.2 Objetivos específicos .......................................................................................... 13
2.2 Hipóteses ................................................................................................................ 13
3. Material e métodos .................................................................................................... 14
3.1 Área de estudo e procedimentos de campo ............................................................ 14
3.2 Análises moleculares .............................................................................................. 15
3.2.1 Extração de DNA ............................................................................................... 15
3.2.2 Amplificação e purificação dos fragmentos ....................................................... 16
3.2.3 Seqüenciamento do DNA ................................................................................... 17
3.3 Análise dos dados ................................................................................................... 18
3.3.1 Edição e alinhamento das seqüências ................................................................. 18
3.3.2 Árvore de haplótipos .......................................................................................... 18
3.3.3 Inferências Evolutivas ........................................................................................ 18
3.3.4 Análises populacionais ....................................................................................... 20
4. Resultados .................................................................................................................. 22
4.1 A região controle de Nannostomus eques .............................................................. 22
4.2 Análise e distribuição dos haplótipos ..................................................................... 24
4.3 Árvore de haplótipos .............................................................................................. 24

xi
4.4 Matriz de distância genética ................................................................................... 27
4.5 Análises populacionais ........................................................................................... 31
4.5.1 Análises de polimorfismo de DNA .................................................................... 31
4.5.2 Distribuição da variabilidade genética e diferenciação populacional ................ 32
4.5.3 Teste de Mantel .................................................................................................. 34
5. Discussão .................................................................................................................... 35 5.1 Caracterização da região controle de N. eques ...................................................... 35
5.2 Inferências evolutivas ............................................................................................. 37
5.3 Genética de populações .......................................................................................... 41
7. Referências Bibliográficas ........................................................................................ 46
8. Anexos ......................................................................................................................... 57 Anexo 01 ...................................................................................................................... 57
Anexo 02 ...................................................................................................................... 62
Anexo 03 ...................................................................................................................... 64

xii
Lista de Figuras
Figura 01- Peixe-lápis - Nannostomus eques (barra: 1cm)....................................................... 9
Figura 02 – Distribuição geográfica de N. eques. Os pontos vermelhos indicam os locais de
ocorrência de N. eques obtidos no banco de dados NeoDat II e NeoDat III. ........................... 9
Figura 03 – Genoma mitocondrial (Fonte: modificado de Pereira, 2000).............................. 11
Figura 04- Bacia do rio Negro. Os círculos correspondem à localização dos locais
amostrados. Açaituba (AÇ); Miuá (MI); Jaradi (JA); Arixanã (AR); Demeni (DE); Maguari
(MA); Jacundá (JC); Catalão (CT). (ARN) alto rio Negro; (MRN) médio rio Negro; (BRN)
baixo rio Negro. ...................................................................................................................... 14
Figura 05 - Esquema do local de anelamento dos primers (setas vermelhas) que foram
utilizados para a amplificação do fragmento de interesse da região controle do DNAmt. .... 17
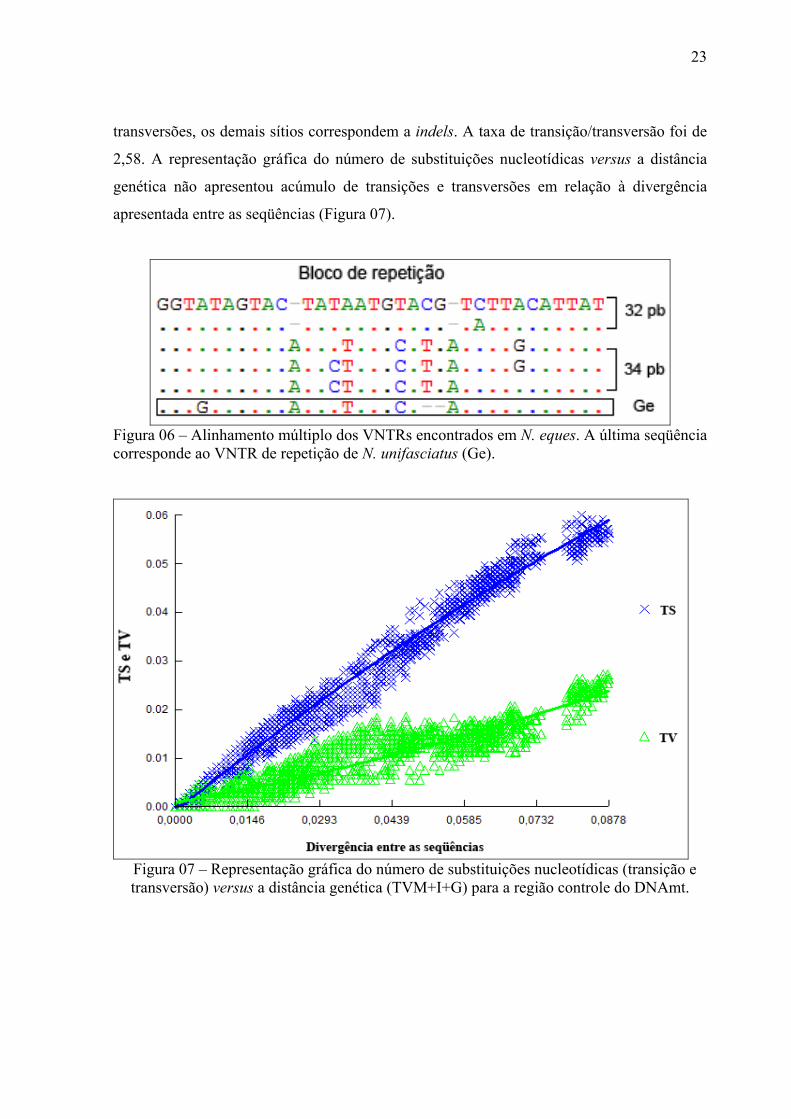
Figura 06 – Alinhamento múltiplo dos VNTRs encontrados em N. eques. A última seqüência
corresponde ao VNTR de repetição de N. unifasciatus (Ge). ................................................ 23
Figura 07 – Representação gráfica do número de substituições nucleotídicas (transição e
transversão) versus a distância genética (TVM+I+G) para a região controle do DNAmt. .... 23
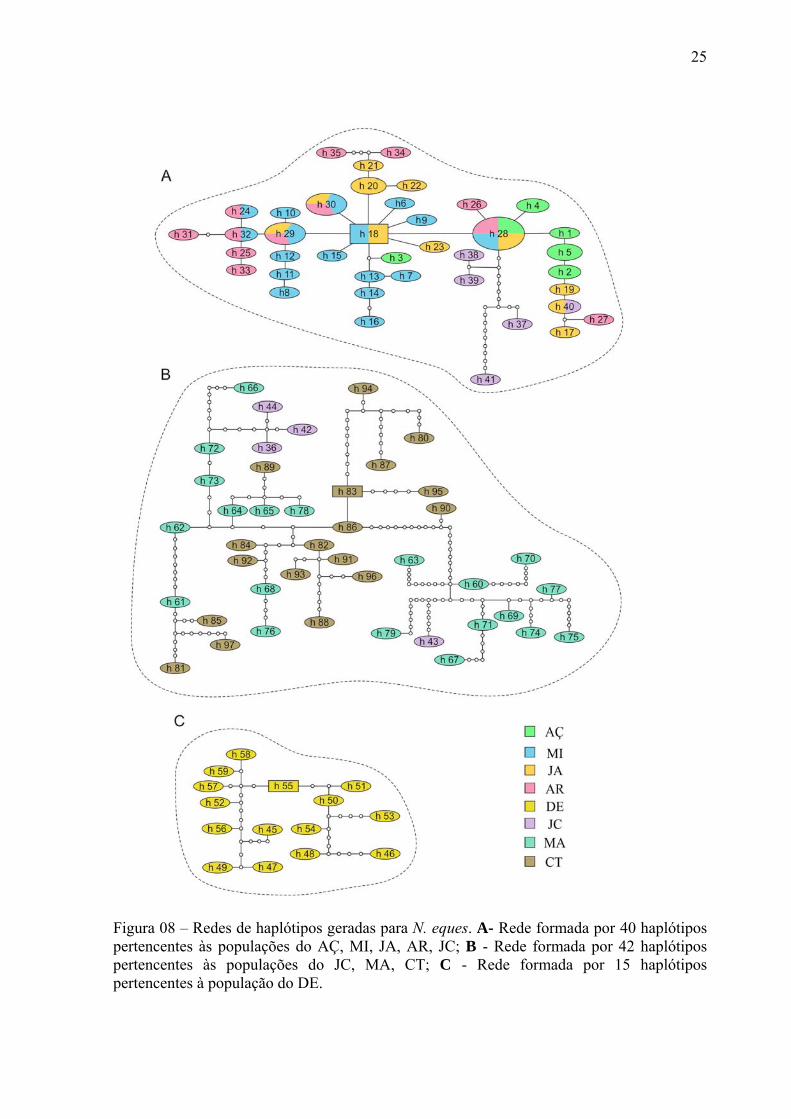
Figura 08 – Redes de haplótipos geradas para N. eques. A- Rede formada por 40 haplótipos
pertencentes às populações do AÇ, MI, JA, AR, JC; B - Rede formada por 42 haplótipos
pertencentes às populações do JC, MA, CT; C - Rede formada por 15 haplótipos
pertencentes à população do DE. ............................................................................................ 25
Figura 09 – Árvore gerada pela MP. Valores de bootstrap estão acima dos ramos. Grupo 1
(Açaituba, Miuá, Jaradi, Arixanã e Jacundá); Grupo 2 (Demeni, Jacundá, Maguari e
Catalão). Ge = N. unifasciatus. Os valores em porcentagem refere-se à distância genética
entre os ramos. ........................................................................................................................ 28
Figura 10 – Árvore gerada pela MV (Ln=3304.7620). Valores de bootstrap estão acima dos
ramos. Em destaque o grupo 1 (Açaituba, Miuá, Jaradi, Arixanã e Jacundá) e a população do
Demeni.. Ge = N. unifasciatus. Os valores em porcentagem refere-se à distância genética
entre os ramos. ........................................................................................................................ 29

xiii
Lista de Tabelas
Tabela 01 – Identificação dos locais de coleta de N. eques. ................................................... 15
Tabela 02 – Distância genética média entre e dentro das populações (em porcentagem)
obtida pela distância p não corrigida. Destacado em azul a distancia média entre as
populações do grupo 1; destacado em rosa a distancia média entre as populações do grupo
dois; em vermelho a elevada distância genética nas comparações envolvendo o Demeni. ... 27
Tabela 03 – Índices de polimorfismo de DNA para os espécimes de N. eques das oito
localidades amostradas. N=número amostral; H= número de haplótipos; HU=haplótipos
únicos; S=sítios polimórficos; ETA=número de mutações; HD=diversidade haplotípica; PI=
diversidade nucleotídica; K=média das diferenças nucleotídicas par a par. .......................... 32
Tabela 04 – AMOVA para o grupo 1 de N. eques (Açaituba, Miuá, Jaradi, Arixanã e
Jacundá 1). .............................................................................................................................. 33
Tabela 05 – Diferenciação genética (Φst) acima da diagonal e estimativa indireta de fluxo
gênico (Nm) abaixo da diagonal para o grupo 1. Os valores sublinhados correspondem a
ausência de fluxo gênico efetivo. ........................................................................................... 33
Tabela 06 – AMOVA para o grupo 2 de N. eques (Demini, Jacundá, Maguari e Catalão). .. 33
Tabela 07- Diferenciação genética (Φst) acima da diagonal estimativa indireta de fluxo
gênico (Nm) abaixo da diagonal para o grupo 2. ns = não significativo. Os valores
sublinhados correspondem a ausência de fluxo gênico efetivo. ............................................. 33
Tabela 08 – Matriz de distância geográfica (km) entre as localidades amostradas. ............... 34
Tabela 09 – Valores de correlação obtidos no teste de Mantel para os grupos 1 e 2. ............ 34
Tabela 10 - Comparação da seqüência tipo TAS encontrada em N. eques com seqüência de
outros peixes. .......................................................................................................................... 36

1
1. Introdução
Com grande potencial para servir como modelo de manejo sustentável, os peixes
ornamentais têm sido considerados recursos aquáticos renováveis na Amazônia,
especialmente nos tributários do rio Negro. A grande riqueza de espécies transformou
algumas regiões desta bacia em áreas de comércio de peixes ornamentais, que atualmente
tem um papel importante na região, particularmente para comunidades ribeirinhas, que
através da comercialização no mercado internacional, utilizam desse recurso como fonte de
renda (Chao, 2001). Entretanto, muito pouco é conhecido sobre a história natural e a
estrutura genética das populações, dificultando o entendimento do modo de dispersão de
muitos organismos e a formulação de hipóteses que reflitam a filogenia das espécies. Esta
carência de estudos compromete as estratégias de preservação e manejo racional dos
estoques (Chao, 1993; Harris & Petry, 2001; Porto et al., 2001).
Um problema biológico comum entre os recursos aquáticos é que neles pode conter
uma biodiversidade oculta, representada por complexos de espécies. A exploração de
estoques, sem o conhecimento prévio de sua variabilidade genética e do número real de
espécies existentes pode levar a perda de linhagens isoladas reduzindo a variabilidade
genética total das mesmas (Frankham, 2005). O entendimento da estrutura genética
populacional de uma dada espécie é um passo importante, pois permite conhecer as
diferenças genéticas geradas por processos vicariantes e suas relações com as diferenças
adaptativas das populações (Avise, 1994). Recentemente, dois estudos genético-
populacionais em espécies distintas de peixes do rio Negro claramente demonstram que há
uma biodiversidade oculta (espécies crípticas) em Hipopygus lepturus (Schmitt, 2005) e
Carnegiella strigata (Schneider, 2007).
Dessa forma, através do seqüenciamento de DNA, este trabalho objetivou determinar
se a captura do ornamental Nannostomus eques, conhecido vulgarmente como peixe-lápis,
ao longo do rio Negro é feita sobre um estoque único, geneticamente homogêneo, ou se a
espécie em questão é constituída de várias populações geneticamente diferenciadas. Os
resultados apresentados nesta dissertação trazem informações importantes sobre a estrutura
genética desta espécie muito explorada comercialmente que poderá auxiliar diretamente nas
estratégias de conservação e manejo.

2
1.1 Bacia amazônica e sub-bacia do rio Negro
No começo do Cretáceo, o deslocamento para oeste da placa Sulamericana sob a Placa
de Nazca propiciou o nascimento da futura Cordilheira dos Andes que alcançou seu ápice no
fim do Mioceno. Esta barreira impediu o fluxo livre das águas que circulavam entre os atuais
Oceanos Atlântico e Pacífico represando as águas em um sistema de rios e lagos. Por conta
disto, a bacia amazônica tornou-se o maior ecossistema lacustre pantanoso que a terra já
conheceu. Os sedimentos provenientes dos Andes acumulados ao longo da bacia e as
alterações do nível do mar durante as glaciações do Quaternário (1,6 milhões de anos atrás
até o momento) definiram o episódio que contribuiu para o delineamento do sistema de
escoamento atual (Goulding et al., 1988, Lundberg et al., 1998, Rossetti et al., 2005).
A bacia amazônica está localizada em uma região de planície e possui cerca de 23.000
km de rios navegáveis, sendo o maior sistema de água doce do mundo, com
aproximadamente 7.000.000 km² de área de drenagem. De sua área total, 58% localiza-se no
Brasil, 16% no Peru, 10% na Bolívia e o restante na Colômbia, Equador e Venezuela. É
formada por várias sub-bacias fortemente influenciadas pela sazonalidade das chuvas que
contribuem para a formação dos mais variados habitats. Na época das cheias, os rios
amazônicos transbordam de seus leitos e avançam sobre a vegetação que cresce em volta de
suas margens, alagando uma área de 300.000 km2 (várzea e igapó), formando o mais extenso
ecossistema de áreas alagadas do planeta (Junk & Furch, 1993).
O Solimões-Amazonas é o coletor final deste complexo sistema de drenagem formado
por cerca de sete mil tributários, sendo os principais afluentes de sua margem esquerda o
Japurá, o Negro e o Trombetas, e da direita os rios Juruá, Madeira, Xingu e Tapajós (Santos
& Ferreira, 1999; Goulding et al., 2003).
Os rios amazônicos são diferentes não apenas na morfologia de seus cursos, mas
também nas propriedades físicas e químicas de suas águas. Com base nestas características,
Sioli (1950) classificou as águas da bacia amazônica como claras, brancas e pretas.
As águas pretas recebem este nome devido à coloração escura que suas águas possuem
quando em grande volume. A cor escura é devida à grande concentração de ácidos húmicos
e fúlvicos que se originam da decomposição, em condições de acidez, da matéria orgânica
das florestas que circundam os rios e dos processos edáficos existentes nos solos
amazônicos. Ainda são desconhecidos os fatores que causam a produção destes ácidos. A
acidez (pH entre 3,0 e 5,0) se deve à ausência de cálcio/magnésio e à pobreza de sais
minerais (Sioli, 1990).

3
Dentre as águas pretas destaca-se a bacia do rio Negro, formada por este rio e seus
afluentes. Este sistema de drenagem, com aproximadamente 700.000 km2, é responsável por
14% do volume de água da bacia amazônica (Goulding et al., 2003).
O Negro é um rio relativamente canalizado, com poucos meandros, mas
freqüentemente separado em braços pelas numerosas ilhas que norteiam o rio. Nasce na
região pré-andina da Colômbia, entra em território brasileiro e encontra-se com o rio
Solimões para formar o Amazonas, totalizando 1.700 km de extensão (Goulding et al.,
1988).
De acordo com a geomorfologia da área, o curso do rio Negro foi dividido em três
porções: alto, médio e baixo rio Negro (Franzinelli et al., 2002). O alto rio Negro refere-se à
região de sua nascente até a localidade de Santa Isabel do Rio Negro, onde o rio deixa as
rochas cristalinas do escudo das Guianas revelado pelas cataratas onde os canais atravessam
afloramentos de rochas antigas e flui através de depósitos sedimentares da bacia amazônica.
Nesta região o canal possui seções onde as águas fluem lentamente alternadas por seções de
correnteza entremeada de corredeiras e cachoeiras. A profundidade máxima é de
aproximadamente 12 metros no período da seca.
O médio rio Negro flui nos depósitos sedimentares do Terciário. Nesta região o canal
apresenta numerosos bancos de areia, especialmente perto de sua extremidade ao sul da
confluência com o rio Branco. Alcança uma largura de 20 quilômetros e profundidade
máxima de 18 metros na estação seca, assemelhando-se a uma bacia de sedimentação, com
rochas cristalinas até a boca do rio Branco onde se forma uma barreira causada pelo
acúmulo de sedimentos rio acima.
O baixo rio Negro tem início na região de confluência com o rio Branco e termina no
encontro de suas águas com o rio Solimões. Nesta região o rio Branco é o principal
fornecedor dos sedimentos, os quais não são suficientes para afetar a coloração da água
preta. O neotectonismo nesta área é responsável pela profundidade do rio e pela ocorrência
das íngremes falésias ao longo de suas margens Além disso, o neotectonismo parece ter
influenciado na origem de Anavilhanas, arquipélago formado por cerca de 400 ilhas, que
pelo grande fluxo das águas abriga uma ictiofauna bem diversificada (Franzinelli et al.,
2002).
Posteriormente, Latrubesse & Franzinelli (2005) subdividiram o rio Negro em seis
subunidades geomorfológicas, sendo que a primeira, subdividida em duas porções

4
correspondentes ao alto rio Negro; a segunda e a terceira correspondentes ao médio rio
Negro; e as demais subunidades correspondentes ao baixo rio Negro.
O rio Negro possui flutuações anuais do nível de água, o que determina períodos de
seca e inundação, que controla a biota do canal do rio e de áreas alagáveis (Junk et al.,
1989). Devido a este pulso de inundação, no período da cheia, muitos dos afluentes da
região mediana de seu curso podem se conectar por meio de uma planície, também chamada
zona de interflúvio (Goulding et al., 2003). Ainda, imagens de satélite no período da cheia
revelam que o rio Negro se conecta com o rio Uatumã ao leste e com o rio Japurá a oeste
(Goulding et al., 1988).
1.2 A ictiofauna do rio Negro
A formação da bacia amazônica foi um dos fatores responsáveis pela diversidade da
fauna de peixes de água doce Neotropical (Vari & Malabarba, 1998). Estima-se que existe
mais de 6.000 espécies habitando as águas continentais da região Neotropical (Reis et al.,
2003). A maior parte desta diversidade concentra-se no norte da América do Sul, que
abrange a região ao leste dos Andes compreendida pela bacia amazônica e bacia do Orinoco
(Buckup et al., 2007; Vari, 1997).
A ictiofauna amazônica está representada principalmente pela superordem
Ostariophysi, que agrupa cerca de 85% das espécies amazônicas, em média 43% estão
incluídas na ordem Characiformes, 39% na ordem Siluriformes e 3% na ordem
Gymnotiformes. As demais espécies pertencem a outras 14 famílias de diferentes ordens
(Santos & Ferreira, 1999).
O catálogo das espécies de peixes de água doce do Brasil, recentemente elaborado,
registra a ocorrência de 2.587 espécies pertencentes a famílias de peixes que ocorrem
exclusivamente em ambientes de água doce, onde 2.481 espécies estão formalmente
descritas e 106 em fase de descrição (Buckup et al., 2007). É bem provável que 50% de
todas as espécies de peixes do Brasil ocorram na bacia amazônica, já que apenas para o rio
Negro são creditadas de 700 (Goulding et al., 1998) a 946 espécies (Chao, 2001).
A comunidade de peixes do rio Negro é composta em sua maioria por peixes
pequenos. Goulding et al. (1988) relacionaram este fator à menor quantidade de nutrientes
das águas pretas. Cerca de 40 espécies alcançam sua maturidade sexual com tamanhos
menores que 20 mm, cujos adultos medem no máximo 40 mm de comprimento o que
caracteriza o fenômeno conhecido como miniaturização (Santos et al., 2006).

5
Dentre as inúmeras espécies encontradas na bacia do rio Negro, várias são liberadas
para comercialização como peixes ornamentais: cardinal (Paracheirodon axelrodi), acará-
disco (Shymphysodon spp.), acará-bandeira (Pterophyllum spp.), peixe-borboleta
(Carnegiella spp.), peixe-lápis (Nannostomus spp.), entre outras (Chao, 2001).
1.3 Comércio de peixes ornamentais
O comércio de peixes ornamentais se mantém entre os principais produtos da pauta
de exportações no Estado do Amazonas, o que gera uma renda anual de cerca de 3 milhões
de dólares. Entre 2005 a 2007, aproximadamente 30 milhões de peixes ornamentais foram
exportados do Estado do Amazonas. O volume exportado aumentou a uma taxa de
crescimento média de aproximadamente 28,8% ao ano, passando de 17 milhões em 2002,
para 36,2 milhões em 2005 (Anjos et al.,2007, MMA 2008).
Os peixes ornamentais exportados do Estado do Amazonas correspondem de 130 a
140 espécies, distribuídas em 25 famílias das quais a Characidae, com aproximadamente 26
espécies, representam 79% das exportações. O cardinal é a espécie mais popular e representa
em média 60% do volume exportado. Calcula-se que cerca de 70% dos peixes exportados do
Estado do Amazonas provem da região do rio Negro próximo ao município de Barcelos,
onde grande parte das espécies são capturadas em pequenos riachos de águas pretas e claras
(Anjos et al., 2007).
A bacia do médio rio Negro é a maior área de pesca de peixe ornamental do Estado
do Amazonas e o município de Barcelos é o principal entreposto comercial. Estima-se que
80% da população ribeirinha da região tenha alguma relação econômica com este comércio
sendo a principal fonte de emprego e rendimento para a população dos municípios de
Barcelos e Santa Isabel do Rio Negro (Prang, 2001; 2007).
Apesar da grande importância do comércio de peixes ornamentais para o
desenvolvimento da região amazônica, existem falhas graves nas atividades ligadas à
comercialização. Anjos et al. (2007), analisando declarações de exportações fornecidas pelo
IBAMA, detectaram muitas informações errôneas sobre as espécies comercializadas nos
portos da região amazônica. Freqüentemente os autores encontraram grupos de espécies
sendo exportadas sob uma mesma denominação, isso ocorre porque muitos exportadores
classificam as espécies de acordo com catálogos de peixes ornamentais destinados aos
aquaristas, com base em fotografias. Fica clara a falta de controle que se tem sobre o número
real de espécies que são exploradas pelo comércio de ornamentais. Esta situação não se

6
alterou com a recente publicação da IN 203/2008 pelo Ministério do Meio Ambiente que
normatiza as espécies de peixes ornamentais passíveis de serem comercializadas, pois
muitas delas ainda permanecem indeterminadas (Ancistrus spp; Baryancistrus spp;
Engemannia spp; Farlowella spp; Hiphessobrycon spp; Hypostomus spp; Scobiancistrus
spp).
1.4 Família Lebiasinidae e o gênero Nannostomus
Pertencente à ordem Characiformes, a família Lebiasinidae consiste de 61 espécies
consideradas válidas e se encontram distribuídas em sete gêneros: Lebiasina, Piabucina
(subfamília Lebiasininae), Nannostomus, Copeina, Copella, Pyrrhulina (subfamília
Pyrrhulininae) e o gênero Derhamia, recentemente descrito, mas não incluído em nenhuma
subfamília (Netto-Ferreira, 2005). Desse total de espécies, 16 lebiasinídeos estão
oficialmente liberados pelo IBAMA para serem comercializadas para fins ornamentais:
Copeina (C. guttata; C. arnoldi; C. metae); Copella (C. nattereri; C. nigrofasciata);
Nannostomus (N. beckfordi; N. digrammus; N. eques; N. espei; N. marginatus; N.
trifasciatus; N. unifasciatus); Pyrrhulina (P. brevis; P. laeta; P. rachoviana; P. vittata)
(IBAMA IN 203/2008).
Os lebiasinideos são endêmicos da região Neotropical, sendo encontrados na América
Central e em todos os países da América do Sul, exceto o Chile. São peixes que vivem
próximos às margens e podem ser encontrados em rios de água preta, branca e clara
(Weitzman & Weitzman, 1982).
Os representantes desta família possuem o corpo cilíndrico e alongado, sendo
considerados de pequeno a médio porte com comprimento variando de 1,6 cm
(Nannostomus anduzei) a 20 cm (algumas espécies de Pyrrhulina) (Weitzman, 1964).
As espécies dos gêneros Copeina, Copella e Pyrrhulina são invertívoras, já as espécies
do gênero Nannostomus alimentam-se de organismos bentônicos associados à vegetação
aquática. Os representantes dos gêneros Lebiasina e Piabucina, por sua vez, possuem
hábitos alimentares mais generalistas, alimentando-se de insetos, crustáceos e peixes
(Román -Valencia, 2004).
Os lebiasinídeos apresentam acentuado dimorfismo sexual relacionado principalmente
à coloração e à morfologia da nadadeira anal. Nos machos ela é mais protuberante e
colorida. Tubérculos nupciais aparecem em algumas espécies de Nannostomus e Lebiasina.
A fecundação é externa e as espécies apresentam cuidado parental (Netto-Ferreira, 2005),

7
que é realizado principalmente pelos machos que permanecem aerando os ovos até a eclosão
dos mesmos (Blumer, 1979). Produzem ovos em pequenas quantidades, possivelmente como
resposta às condições oligotróficas dos igarapés amazônicos de terra-firme (Araújo, 2003).
A taxonomia da família Lebiasinidae é complexa devido às inúmeras propostas de
relacionamento entre os gêneros que a compõe e os demais Characiformes. As hipóteses
filogenéticas com base em caracteres morfológicos incluem os lebiasinideos no clado
monofilético denominado Erythrinoidea, juntamente com as famílias Erythrinidae,
Ctenoluciidae e Hepsetidae (Buckup, 1998; Oyokawa, 1998). Netto-Ferreira (2005), com
base em 25 sinapomorfias, considerou a família Lebiasinidae monofilética. Contrariamente,
Moreira (2007) propôs que a família Lebiasinidae é parafilética onde Pyrrhulininae aparece
como grupo-irmão de Erythrinidae. Os estudos relevantes (Oyokawa, 1998; Buckup, 1998;
Netto-Ferreira, 2005; Moreira 2007), concordam com a existência das duas subfamílias,
Pyrrhulininae e Lebiasininae.
Alguns estudos genéticos conduzidos nos Characiformes permitiram inferir sobre o
posicionamento de representantes da família Lebiasinidae (gênero Nannostomus) na
filogenia da ordem. Orti & Meyer (1996) a partir da análise de seqüências de DNA nuclear,
posicionaram o gênero Nannostomus na base do clado que inclui todos os Characiformes.
Novamente, Orti & Meyer (1997) utilizando seqüências de genes mitocondriais do DNA
ribossomal (12S e 16S) apresentaram três árvores de relacionamento de Nannostomus com
os demais Characiformes, sendo que devido a saturação dos genes a árvore mais plausível
agrupou Nannostomus com Pyrrhulina e este clado como sendo grupo irmão dos clados
Hoplias + Hepsetus, Boulengerela + Ctenolucius, Phenacogrammus + Hydrocynus + Alestes
+ Acestrorhynchus. Calcagnotto et al. (2005) em uma análise fundamentada em seqüências
de quatro genes nucleares (RAG2; sai; fkh; trop) e dois genes mitocondriais (12S e Cyt b),
incluíram a família Lebiasinidae na superfamília Erythrinoidea e propõem uma relação mais
estreita com Hepsetidae (Africano) e Ctenoluciidae (Neotropical) e estes formam um grupo
irmão de Crenuchidae e Erythrinidae.
Com relação às espécies do gênero Nannostomus, estas distribuem-se pelas bacias do
Amazonas, Orinoco e bacias costeiras entre a foz destes dois rios, ocorrendo em águas
abertas dos grandes rios, áreas alagadas e pequenos igarapés (Weitzman & Weitzman,
2003). Três gêneros nominais são atualmente considerados sinônimos de Nannostomus:
Archicheir, Poecilobrycon e Nannobrycon (Netto-Ferreira, 2005).

8
Até o momento existem 16 espécies válidas que integram o gênero Nannostomus. São elas:
N. anduzei, N. beckfordi, N. bifasciatus, N. britskii, N. digrammus, N. eques, N. espei, N.
harrisoni, N. limatus, N. marginatus, N. marilynae, N. minimus, N. mortenthaleri, N. nitidus,
N. trifasciatus e N. unifasciatus (Weitzman & Weitzman, 2003).
As relações filogenéticas dentro do gênero Nannostomus ainda permanecem obscuras,
apesar das revisões já efetuadas. Segundo Weitzman (1966) e Weitzman & Cobb (1975), as
espécies N. harrisoni, N. bifasciatus, N. unifasciatus e N. eques formam um grupo mais
relacionado dentro do gênero, com base em caracteres morfológicos.
Os representantes de Nannostomus são pequenos, coloridos e apresentam de uma a
cinco listras escuras na região longitudinal do corpo, e por conta disto, são conhecidas
popularmente por peixes-lápis. Este padrão de coloração é observado somente durante o dia,
pois durante a noite as listras desaparecem e três listras verticais ou perpendiculares são
evidenciadas. Este padrão noturno de coloração é observado em todos os indivíduos juvenis
de muitas espécies, sendo substituído pelo padrão de listras horizontais somente quando os
indivíduos alcançam a idade adulta. Além disso, o padrão de coloração noturna é exibido por
fêmeas de algumas espécies durante a corte reprodutiva (Weitzman & Cobb, 1975;
Weitzman, 1978). Segundo Fujii & Masagaki (1999) a alternância dos padrões de coloração
se deve à mudanças circadianas que atuam nos melanóforos tegumentares destas espécies.
Nannostomus eques (Figura 01), escolhida como objeto de estudo, é uma das mais
exploradas no ramo da aquariofilia, por apresentar uma posição de natação quase vertical e
um padrão de colorido aparentemente especializado. Além disso, é a única espécie do
gênero a apresentar cinco listras horizontais ao longo do corpo (Weitzman & Weitzman,
2003). De acordo com uma compilação no banco de dados NeoDat II e NeoDat III, a
distribuição geográfica desta espécie inclui tributários das bacias amazônica, Orinoco e
Essequibo (Figura 02).
Apesar da exploração das espécies de Nannostomus, as análises genéticas feitas no gênero ainda são
poucas. Há algumas informações que indicam que o número cromossômico haplóide encontrado nas espécies
deste gênero varia de n=11 a n=23 (Scheel, 1973) e o número diplóide de N. beckfordi e N. eques é 2n=42 e
2n=34, respectivamente (Arefjev, 1990). Há também o desenvolvimento de marcadores microssatélites para o
estudo da variabilidade genética de N. unifasciatus de populações do médio rio Negro (Beheregaray et al.,
2004). Estes últimos autores encontraram uma alta variabilidade genética entre as localidades amostradas e
constataram que existe possibilidade de utilização destes marcadores microssatélites em N. eques.

9
Figura 01- Peixe-lápis - Nannostomus eques (barra: 1cm).
Figura 02 – Distribuição geográfica de N. eques. Os pontos vermelhos indicam os locais de ocorrência de N. eques obtidos no banco de dados NeoDat II e NeoDat III.

10
1.5 DNA mitocondrial e variabilidade genética Porto et al. (2001) destacaram que para explorar o recurso pesqueiro, particularmente
os peixes ornamentais do rio Negro, é de fundamental importância conhecer não apenas o
ciclo de vida das espécies mas também a estrutura genética das populações.
As técnicas moleculares, dentre elas a análise de seqüências de DNA, têm sido
eficientes para fornecer informações que visam avaliar e diagnosticar geneticamente grande
parte da biodiversidade amazônica. De acordo com Avise (1994) este nível de detalhamento
tem sido relevante para discutir questões importantes sobre a conservação e o manejo
genético de diversas espécies de peixes.
O DNA mitocondrial (Figura 03) dos animais vem sendo utilizado para aferir o grau de
diversidade genética das espécies e estimar a variabilidade genética entre populações. Isto se
deve a algumas vantagens: à facilidade de ser isolado, pelo seu tamanho reduzido, por
apresentar herança uniparental, ausência de recombinação e elevada taxa de mutação (Avise,
1994). Estas propriedades do DNA mitocondrial facilitam a reconstrução das relações
evolutivas entre haplótipos que muitas vezes pode ajudar a entender o modo de dispersão de
muitos organismos, podendo gerar hipóteses que reflitam a filogenia da espécie (Avise,
1994; 1998).
O genoma mitocondrial é uma molécula circular de fita dupla contida em múltiplas
cópias nas mitocôndrias. São descritos 37 genes, dos quais 13 codificam proteínas, 22
codificam RNAs transportadores (RNAt) e dois codificam RNAs ribossomais (Meyer,
1993). Além disso, o DNA mitocondrial (DNAmt) possui uma região não codificadora
chamada região controle (alça-D ou D-loop), que apresenta seqüências específicas que
determinam o início da replicação da fita pesada (H- Heavy) e transcrição de ambas as fitas
de DNA. A região controle se encontra situada entre o RNAt da prolina e o RNAt da
fenilalanina no DNAmt de vertebrados, variando em algumas bases de tamanho entre as
espécies (Nahum, 2001). De todo o genoma mitocondrial a região controle é a que mais
acumula mutações podendo ser de duas a cinco vezes maior que dos genes que codificam
proteínas (Meyer, 1993).
Em peixes, a região controle apresenta por volta de 1.100 pares de bases, um domínio
central conservado e duas porções ricas em A/T (extremamente variáveis) nas extremidades
5` e 3` (Meyer, 1993). A região controle mitocondrial é dividida em três domínios:
ETAS/TAS (domínio das seqüências estendidas associadas com a terminação da síntese da
fita pesada durante a replicação), CCD (domínio central conservado) que se mostra pouco

11
variável por conter blocos de sequências conservadas, cuja função ainda é desconhecida;
CSB (blocos de seqüências conservados) que contém a origem de replicação da fita pesada e
os promotores da transcrição deste genoma. O domínio I (ETAS/TAS) têm se mostrado
altamente variável e freqüentemente contém seqüências repetidas em tandem (VNTR –
Variable Number of Tandem Repeats) que podem ou não variar no número de repetições,
sua localização corresponde a porção 5’ logo após o tRNA da prolina (Lee et al., 1995; Ray
& Densmore, 2003).
Figura 03 – Genoma mitocondrial (Fonte: modificado de Pereira, 2000).
O DNA mitocondrial de algumas espécies de peixes da bacia amazônica tem sido
utilizado para estudos de relações inter e intra-específicas e para verificar se existe relação
entre a estrutura genética das populações e os locais de amostragem (Alves-Gomes, 1995;
Porto, 1999; Lovejoy & Araújo, 2000; Sivasundar et al., 2001; Batista, 2001; Formiga-
Aquino, 2004; Schmitt, 2005; Hrbek, 2005; Frederico, 2006; Ribeiro, 2006; Santos, 2007;
Schneider, 2007, Souza, 2008).
De maneira geral, os trabalhos acima citados revelam o grau de variabilidade
genética presente nas espécies estudadas e mostram que os fatores que determinam o
isolamento genético de uma determinada população dependem de forças evolutivas como
seleção natural, fluxo gênico, deriva genética, mutação e o tempo viável para que estes
processos ocorram. Ainda, no caso dos peixes estudados, a estrutura genética dentro e entre

12
populações tem sido influenciada principalmente pelo fluxo gênico e o tamanho
populacional efetivo.
Sabe-se que a depauperação da variabilidade genética, dentro e entre populações,
pode ser um componente decisivo para a sobrevivência de uma espécie a médio e longo
prazos, já que a habilidade de se adaptar às constantes modificações do meio ambiente
depende em grande parte do nível de diversidade genética encontrada na mesma (Solé-Cava,
2001).
A variabilidade genética de uma população pode ser estimada por meio de vários
parâmetros: Número médio ou total de alelos/loco; diversidade genética, heterozigosidade
esperada; conteúdo de informação de polimorfismo; índice de fixação de Wright, entre
outros (Ridley, 2004).
O grau de diferenciação genética entre os estoques pesqueiros depende também de
vários fatores que levam as populações à homogeneidade ou à heterogeneidade. A avaliação
da estrutura genética de uma espécie determina a intensidade com que o fluxo gênico entre
suas populações vem ocorrendo. Se a troca de genes entre estas tem sido limitada a uma
determinada população que esteja sob pesca predatória, a possibilidade de sua recuperação
genética pela migração é pequena (Hilsdorf et al., 2006).
Todavia, o isolamento geográfico entre populações pode não significar uma total
heterogeneidade entre elas, pois as freqüências gênicas observadas podem estar sendo
representadas por episódios de fluxo gênico ocorridos em um passado recente. Assim,
diferentes populações podem não apresentar divergências genéticas estatisticamente
significativas (Frankhan, 2005).

13
2. Objetivos
2.1 Objetivo geral
Caracterizar a região controle do DNA mitocondrial das populações do peixe
ornamental Nannostomus eques coletados em tributários ao longo do rio Negro.
2.1.2 Objetivos específicos
a) Estimar e comparar o grau de variabilidade genética de N. eques.
b) Verificar a presença de fluxo gênico entre as populações de N. eques.
c) Relacionar os agrupamentos encontrados com a distribuição geográfica e propor
hipóteses de diferenciação populacional.
2.2 Hipóteses
H01 – N. eques consiste em uma única linhagem genética dentro da área estudada, sendo
formada por uma população panmítica.
H02 - O fluxo gênico entre as populações não é influenciado pela distância ou barreira
geográfica.

14
3. Material e métodos
3.1 Área de estudo e procedimentos de campo
Os locais de amostragens referem-se a tributários das margens direita e esquerda,
localizados no alto, médio e baixo rio Negro. Ao todo, oito localidades foram amostradas:
Açaituba (AÇ), Miuá (MI), Jaradi (JA), Arixanã (AR), Demeni (DE), Jacundá (JC), Maguari
(MA) e Catalão (CT) (Figura 04).
Os 125 espécimes de Nannostomus eques analisados neste trabalho foram coletados
em fevereiro (no período da seca) e outubro (período de cheia) no ano de 2007, com o
auxílio de puçás e rapichés. Um espécime de N. unifasciatus, coletado no Catalão, foi usado
como grupo externo. Os espécimes foram identificados pelo Dr. Jansen Sampaio Zuanon
(INPA) e encontram-se depositados no Laboratório de Genética da Coordenação de
Pesquisas em Biologia Aquática do INPA. Cada ponto de coleta teve a sua localização
georeferenciada (Tabela 01). As coletas foram realizadas com a autorização do IBAMA
(Licença nº 1960108).
Figura 04- Bacia do rio Negro. Os círculos correspondem à localização dos locais amostrados. Açaituba (AÇ); Miuá (MI); Jaradi (JA); Arixanã (AR); Demeni (DE); Maguari (MA); Jacundá (JC); Catalão (CT). (ARN) alto rio Negro; (MRN) médio rio Negro; (BRN) baixo rio Negro.

15
Tabela 01 – Identificação dos locais de coleta de N. eques. rio Negro Localidade N. amostral Coord. S-N Coord. W-E
alto rio Negro Açaituba (AÇ) 15 00º 54' 28,9''N 66º 50' 6,5''W
Ig. Miuá (MI) 20 00º 08' 41''S 66º 52' 23''W Ig. Jaradi (JA) 14 00º 20' 10''S 65º 18' 32''W Ig. Arixanã (AR) 15 00º 16’15,8''S 62º 47'12,9''W
médio rio Negro Rio Demeni (DE) 15 00º 16’15,8''S 62º 47'12,9''W
baixo rio Negro Ig. Jacundá (JC) 09 01º 26'02''S 61º48' 24''W Ig. Maguari (MA) 20 01º 09' 45''S 61º 50' 53''W Lago Catalão (CT) 18 03º 09' 57''S 59º 54' 44''W
3.2 Análises moleculares
3.2.1 Extração de DNA
A extração de DNA foi realizada à partir de tecido muscular preservado em etanol
100%. O tecido foi retirado da região látero-dorsal de cada espécime. O protocolo de
extração de DNA utilizado foi o descrito por Sambrook & Russell (2001), com algumas
modificações.
Seguindo o protocolo, o tecido muscular foi retirado, colocado em tubo eppendorf e a ele
foi adicionado o tampão de lise (Tris-HCl pH 8,0 em 10 mM, NaCl 0,3 M, EDTA 10 mM,
Urea 4 M, SDS 1%) de Estoup et al. (1993) e Asahida et al. (1996), 15 μL de proteinase K
(10 mg/mL) e 6 μL de RNAse (10 mg/mL). As amostras foram levadas à estufa a 60 ºC para
digestão do tecido. Posteriormente, foram feitas três lavagens sucessivas, mediante
centrifugação a 14000 rpm, com 600 μL de cada um dos seguintes reagentes: fenol, fenol-
clorofórmio e álcool isoamílico.
Após a lise, o DNA foi separado das proteínas por precipitação salina (acetato de amônio
0,3M) por meio de centrifugação à 14000 rpm. O sobrenadante de cada amostra foi
precipitado com 400 μL de isopropanol gelado (100%), por meio de centrifugação. Ao final,
o DNA foi hidratado com aproximadamente 100 μL de água milli-Q, dependendo do
precipitado formado.
A análise da quantidade e integridade do DNA foi feita por eletroforese padrão (com
tampão Tris-Borato-EDTA 1X e corrida a 70 V por 40 minutos) em gel de agarose 0,8%
usando-se 1 μL DNA. A visualização e o registro fotográfico do DNA no gel, corado com

16
brometo de etídio (EtBr – 0,5 g/mL), foram feitos no fotodocumentador Eagle Eye
(Stratagene).
3.2.2 Amplificação e purificação dos fragmentos
A região controle do DNAmt foi amplificada por Reação em Cadeia da Polimerase
(PCR - Saiki et al., 1988), utilizando os seguintes oligonucleotídeos (primers): FTTF2 5’ –
CTAACTCCCAAAGCTAGTATT– 3’ (Ortí et al., 2008), HN20 5’ –
GTGTTATGCTTTAGTTAAGC– 3’ (Bernatchez & Danzmann, 1993) e DL-R1 (desenhado
por Jacqueline Batista, dados não publicados). Um esquema do local do anelamento dos
primers é apresentado na Figura 05.
As reações de amplificação foram feitas em termociclador Eppendorf– Mastercycler
Gradient, para um volume final de 25 μL (~100 ng de DNA genômico; Tampão 1X; 0,5
unidades de Taq DNA Polimerase; 0,2 mM de cada dNTP; 0,2 μM de cada primer; 2,0 mM
de cloreto de magnésio e água mili-Q para completar o volume). O perfil de reação utilizado
foi: 35 ciclos de desnaturação a 92°C por 1 minuto, anelamento a 54°C por 1 minuto e
extensão a 72°C por 1 minuto e 30 segundos. Após todos os ciclos, uma extensão final a
72°C por 5 minutos.
Após a amplificação, os produtos da PCR foram submetidos a corrida eletroforética,
em gel de agarose 1,0%, para verificar se houve ou não a amplificação da região controle.
Para visualização das bandas o gel foi corado com EtBr (0,5 μg/mL) e em seguida
fotodocumentada no Eagle Eye.
A purificação dos produtos da PCR foi realizada com uso do kit GFX PCR DNA Kit
(GE HealthCare), seguindo o protocolo sugerido pelo fabricante.

17
Figura 05 - Esquema do local de anelamento dos primers (setas vermelhas) que foram utilizados para a amplificação do fragmento de interesse da região controle do DNAmt.
3.2.3 Seqüenciamento do DNA
O seqüenciamento do DNA foi realizado pelo método de Sanger et al. (1977),
utilizando terminadores marcados com fluorescência. Para as reações de seqüenciamento foi
utilizado o kit DYEnamic ET Terminator Cycle Sequencing (GE HealthCare), onde cada
dideoxinucleotídeo é marcado com fluoresceína e rodamina, cuja fluorescência é captada
pelo seqüenciador automático de DNA MegaBACE 1000 (GE HealthCare).
As reações de seqüenciamento foram feitas em placas com 96 poços para um volume
final de 10 μL (~ 200 ng do produto purificado da PCR; 0,2 mM de cada primer em reações
separadas; 4 μL do premix e água mili-Q para completar o volume). As reações foram feitas
no termociclador Eppendorf – Mastercycler Gradient, em 30 ciclos de temperatura. O perfil
de reação utilizado foi: 30 ciclos de desnaturação a 95°C por 20 segundos, anelamento a
50°C por 15 segundos e extensão a 72°C por 1 minuto e 30 segundos. Os primers utilizados
no seqüenciamento foram os mesmos utilizados para a amplificação.
Após a reação de seqüenciamento as amostras foram submetidas a um tratamento de
precipitação para eliminar resíduos de baixo peso molecular tais como sais, iniciadores e
dNTPs não incorporados durante a reação. Neste processo adiciona-se à placa 1 μL de
acetato de amônio 7,5M (GE HealthCare) e 25 μL de etanol 100%. Após 15 minutos em
temperatura ambiente foram feitas duas lavagens sucessivas, mediante centrifugação a
14000 rpm com etanol 70% (GE HealthCare).
Depois deste processo, as amostras foram injetadas no seqüenciador automático de
DNA MegaBACE 1000 (GE HealthCare), com o perfil de injeção (2 kv /100 s) seguido de

18
corrida eletroforética (6 kv /300 min.). As seqüências de DNA geradas foram salvas na
extensão abd para posterior análise.
3.3 Análise dos dados
3.3.1 Edição e alinhamento das seqüências
As seqüencias salvas no formato abd foram transferidas do seqüenciador para outro
computador. e comparadas com seqüências depositadas no banco de dados público do
GenBank (http://www.ncbi.nlm.nih.gov) com o auxílio do programa BLAST. Este
procedimento foi realizado para determinar se o fragmento seqüenciado realmente
correspondia à região controle do DNAmt. Uma vez confirmada a homologia das
seqüências, as mesmas foram manualmente conferidas com os programas Chromas Lite 2.01
e o BioEdit 7.0.9 (Hall, 1999). Finalmente, foi gerado um alinhamento múltiplo com auxílio
do programa Clustal W (Thompson et al., 1994), incluído no BioEdit.
3.3.2 Árvore de haplótipos
A genealogia entre os haplótipos foi inferida por meio da construção de uma rede de
haplótipos, utilizando o limite de confiança de 95%, elaborada pelo método de parcimônia
estatística com o auxílio do programa TCS 1.21 (Clement et al., 2000). Este programa
estima a relação entre haplótipos, agrupando-os a partir das mutações obtidas em suas
seqüências. Tal genealogia é baseada no cálculo da freqüência dos haplótipos nas
populações amostradas, usando um algoritmo descrito por Templeton et al. (1992). Na rede
gerada o tamanho dos círculos é proporcional ao número de indivíduos que compartilham o
mesmo haplótipo. Os círculos menores, que ligam os haplótipos identificados correspondem
à haplótipos perdidos ou não amostrados (missing haplotypes). O haplótipo representado por
um retângulo refere-se ao haplótipo ancestral entre os indivíduos analisados.
3.3.3 Inferências Evolutivas
Para estimar as relações genéticas entre as populações foram utilizados os métodos
filogenéticos de máxima parcimônia (Henning, 1966), máxima verossimilhança
(Felsenstein, 1981) e agrupamento de vizinhos (Saitou & Nei, 1987). As análises de

19
parcimônia e agrupamento de vizinhos foram realizadas no programa PAUP* 4.0 (Swofford,
1999), já a análise de máxima verossimilhança foi feita no programa Treefinder (Jobb et al.,
2004). O modelo evolutivo adequado para as análises foi gerado pelo programa ModelTest
3.0 (Posada & Crandall, 1998). Para o enraizamento das árvores, foi utilizado como grupo
externo um indivíduo de Nannostomus unifasciatus.
O método da máxima parcimônia (MP) é empregado em análises de seqüências
moleculares com o propósito de reconstrução de árvores filogenéticas. A filogenia preferida
será aquela que envolver o menor número de mudanças, neste caso, substituições
nucleotídicas que seja compatível com as seqüências observadas. Foram estimados o
comprimento em número de passos (L), o índice de consistência (CI), o índice de retenção
(RI) e o índice de retenção reescalonado (RC). A consistência de cada nó interno foi avaliada
através da análise de bootstrap, realizada com 10.000 replicatas. Nesta análise os gaps foram
tratados como um quinto estado.
O princípio da máxima verossimilhança (MV) avalia a maior probabilidade de serem
encontrados grupos monofiléticos, tendo em vista o modelo de evolução molecular que
melhor explica a variância dos dados apresentados. Para a construção da árvore o modelo
escolhido com o auxílio do programa ModelTest 3.7. (Posada & Crandall, 1998) foi o
TVM+I+G (modelo transversional + sítios invariáveis (I) + média de variação entre os sítios
(G)). A procura pela árvore mais verossímil foi realizada pela busca heurística com adição
aleatória de táxons com 1.000 replicatas.
Os métodos de distância, entre eles o agrupamento de vizinhos (neighbour joining =
NJ) convertem as seqüências alinhadas em uma matriz de distância que representa uma
estimativa da distância evolutiva entre duas seqüências enquanto métodos probabilísticos
analisam cada sítio. Utilizando este método foi gerada uma árvore de distância genética
(distância p) com adição randômica de táxons com 10000 réplicas considerando todos os 97
haplótipos de N. eques e o grupo externo N. unifasciatus.
A diferenciação intraespecífica (distância-p) foi estimada considerando os 97
haplótipos de N. eques utilizando-se o programa MEGA 4.0 (Kumar et al., 2004). Para isto,
foi criada uma matriz de distância “p” (número de diferenças par a par dividido pelo número
total de bases) da qual foram extraídos os valores de divergência entre as seqüências.

20
3.3.4 Análises populacionais
Para as análises populacionais cada ponto de coleta (igarapé) foi considerado a priori
como uma população: Açaituba (n = 15), Miuá (n = 20), Jaradi (n = 14), Arixanã (n = 14),
Demeni (n = 15), Maguari (n = 20), Jacundá (n = 9) e Catalão (n = 18).
As estimativas de variabilidade e diferenciação genética calculadas nos programas
ARLEQUIN 3.1 (Excoffier et al., 2006) e DnaSP 4.0 (Rozas et al., 2003) foram:
H – número de haplótipos;
HU – haplótipos únicos;
ETA – número de mutações;
K – média das diferenças par a par;
HD – diversidade haplotípica
PI – diversidade nucleotídica;
S – número de sítios polimórficos;
Nm – número de migrantes por geração entre populações.
Φst – índice de fixação (Weir & Cockerham, 1984)
O gráfico de saturação gerado a partir do cálculo do número de substituições
nucleotídicas (TS/TV) versus a distância genética foi obtido no programa DAMBE 4.5.56.
A Análise da Variância Molecular – AMOVA (Excoffier et al., 1992) foi utilizada para
estimar a significância da variabilidade encontrada dentro e entre as populações amostradas
à partir de informações de uma matriz de distância entre as seqüências para medir o número
de mutações entre as diferentes populações e o número de mutações entre os diferentes
haplótipos de cada população. Posteriormente, estes valores foram utilizados para o cálculo
dos índices de fixação. Por se tratar de uma análise hierárquica foi possível agrupar os
indivíduos dentro de populações (localidades amostradas) e estas dentro de grupos
(Excoffier et al., 2006). Foram testadas as diferenças dentro e entre cada população.
O isolamento genético por distância foi testado mediante metodologia estatística
desenvolvida por Mantel (1967), denominado teste de Mantel, que averigua a significância
da correlação entre uma matriz de distância genética (índice Φst ) e uma matriz de distância
geográfica entre as localidades (seguindo o curso dos rios). Esta análise foi realizada no
programa ARLEQUIN 3.1 (Excoffier et al., 2006) .

21
Para aferir a neutralidade seletiva nas populações foram utilizados dois testes: D de
Tajima (Tajima, 1989) e o Fs de Fu (Fu, 1997), calculados no programa ARLEQUIN 3.1
(Excoffier et al., 2006). O teste D se baseia no modelo dos sítios infinitos sem recombinação
(Kimura, 1969) e leva em consideração a razão do número de sítios segregantes e o número
médio de diferenças nucleotídicas, estimado pela comparação em pares de bases (Tajima, 1989).
O teste Fs baseia-se na probabilidade de observar em uma amostragem ao acaso um número de
alelos similar ou menor do que o número observado de diferenças par-a-par, tirados da
estimativa de theta. É o mais sensível para detectar expansão populacional (Fu, 1997), sua
significância foi testada por comparações da estatística de Fs contra uma distribuição gerada a
partir de 10000 amostras aleatórias sobre as hipóteses de neutralidade seletiva e equilíbrio
populacional (Hartl & Clark, 1989).

22
4. Resultados
4.1 A região controle de Nannostomus eques
A região controle dos 125 espécimes de N. eques gerou uma matriz totalizando 1087
sítios, dos quais 944 foram constantes, 113 variáveis e 30 indels (inserção/deleção). Os sítios
polimórficos são mostrados no anexo 1. Duas regiões correspondentes ao início e ao término
das seqüências foram excluídas das análises porque não foi possível determiná-las para
todos os indivíduos seqüenciados.
Considerando-se os três domínios que caracterizam a região controle do genoma
mitocondrial, o primeiro deles (TAS), mostrou-se altamente variável em N. eques com 101
sítios polimórficos. Além disso, foi encontrado em todos os indivíduos analisados uma
seqüência de 32 pb e 34 pb repetidas em tandem (VNTR) por nove vezes, totalizando um
bloco entre 288 e 306 pb. Os haplótipos das populações de Açaituba, Miuá, Jaradi, Arixanã
e Jacundá (cinco indivíduos), apresentaram blocos de 32 pb enquanto os haplótipos do
Demeni, Jacundá, Maguari e Catalão apresentaram o bloco de 34 pb. A espécie N.
unifasciatus, utilizado como grupo externo, também apresentou a seqüencia repetitiva com
32 pb, entretanto o padrão de variação encontrado foi exclusiva para a espécie. A figura 06
mostra os dois padrões de VNTRs observados em N. eques e N. unifasciatus.
O segundo domínio (CCD) correspondente a região central conservada foi localizado
mediante a comparação com seqüências de outros peixes teleósteos disponíveis no banco de
dados público do GenBank. No presente estudo esta região mostrou-se pouco variável nos
grandes blocos conservados como visto para a maioria dos vertebrados, entretanto, entre os
blocos conservados foram constatados 18 sítios polimórficos.
O 3º domínio (CSB) também apresentou-se variável em N. eques com exceção da
região correspondente aos blocos conservados (CSBs) associados com a origem de
replicação (OH) do genoma mitocondrial. Neste domínio foi possível constatar 24 sítios
polimórficos.
A média da composição nucleotídica calculada foi de 35% para adenina, 15,7% para
citosina, 12,9% para guanina e 36,4% para timina. Nos 143 sítios polimórficos foram
encontradas 122 substituições nucleotídicas, onde 88 foram transições e 34 foram
23
transversões, os demais sítios correspondem a indels. A taxa de transição/transversão foi de
2,58. A representação gráfica do número de substituições nucleotídicas versus a distância
genética não apresentou acúmulo de transições e transversões em relação à divergência
apresentada entre as seqüências (Figura 07).
Figura 06 – Alinhamento múltiplo dos VNTRs encontrados em N. eques. A última seqüência corresponde ao VNTR de repetição de N. unifasciatus (Ge).
Figura 07 – Representação gráfica do número de substituições nucleotídicas (transição e transversão) versus a distância genética (TVM+I+G) para a região controle do DNAmt.

24
4.2 Análise e distribuição dos haplótipos
Um total de 97 haplótipos foram encontrados nas oito populações analisadas (Anexo
2) O haplótipo h28 foi o mais freqüente, compartilhado por nove indivíduos seguidos pelos
haplótipos h29 e h30 compartilhados por cinco indivíduos e os haplótipos h2, h4, h5,, h18,
h20, h24, h32 e h40 foram compartilhados por dois a quatro indivíduos, os demais
haplótipos (86) foram únicos.
4.3 Árvore de haplótipos
O resultado desta análise gerou três redes de haplótipos (Figura 08). A primeira (A)
foi formada por 68 indivíduos representados por 40 haplótipos pertencentes às populações
de Açaituba, Miuá, Jaradi, Arixanã e Jacundá. A segunda (B) foi formada por 42 indivíduos
(haplótipos únicos) das populações do Jacundá, Maguari e Catalão. A terceira (C) foi
formada apenas pelos indivíduos da população do Demeni. As conexões ambíguas foram
resolvidas utilizando as informações das árvores geradas pelo métodos filogenéticos.
Utilizando o limite de confiança de 95% não houve conexão destas três sub-árvores, a qual
só foi possível quando foi estabelecido 60 passos mutacionais. Excluindo-se a rede A, as
redes B e C se conectam com 20 passos. Somente a população de Jacundá apresentou
haplótipos nas rede A (h37-41) e B (h36, h42-44).
25
Figura 08 – Redes de haplótipos geradas para N. eques. A- Rede formada por 40 haplótipos pertencentes às populações do AÇ, MI, JA, AR, JC; B - Rede formada por 42 haplótipos pertencentes às populações do JC, MA, CT; C - Rede formada por 15 haplótipos pertencentes à população do DE.

26
A análise da máxima parcimônia resultou em uma árvore com 703 passos, índice de
consistência de 0,34 (CI), índice de retenção 0,88 (RI) e índice de retenção reescalonado
0,30 (RC) (Figura 09). As transições e transversões (TS/TV) foram consideradas com pesos
iguais. Do número total de caracteres (1.087), 139 foram informativos para parcimônia. O
resultado desta análise separou N. eques em dois clados. O primeiro, suportado por um
bootstrap de 100, englobou haplótipos do alto (Açaituba, Miuá, Jaradi e Arixanã) e baixo rio
Negro (cinco indivíduos da população do Jacundá). O haplótipo mais representativo deste
clado foi o h28, compartilhado por nove indivíduos das populações de Açaituba, Miuá,
Jaradi e Arixanã. O haplótipo basal foi o h41, referente a um indivíduo da população do
Jacundá. O segundo clado (bootstrap de 74,33) englobou todos os indivíduos das
populações do Demeni (médio rio Negro), Maguari, Catalão e quatro indivíduos da
população do Jacundá (baixo rio Negro). Dentro deste clado, ficou evidente o monofiletismo
da população do Demeni (bootstrap de 99,97).
A árvore de máxima verossimilhança mostrou a existência uma grande politomia, e
dentro desta se destacou dois grupos bem distintos, um deles, formado pelos mesmos
haplótipos do alto (Açaituba, Miuá, Jaradi e Arixanã) e baixo rio Negro (cinco indivíduos da
população do Jacundá) e o outro, formado pelos haplótipos do Demeni, enquanto os demais
haplótipos não foram resolvidos em grupos específicos. (Figura 10). O modelo escolhido foi
o TVM+I+G. e os parâmetros indicados pelo modelo foram: Gamma = 0, 3637 e freqüência
de bases nucleotídicas A = 0,3428; C =0,1525; G =0,1287 e T =0,3760. A análise resultou
em uma árvore com Ln = 3304.7620.
A árvore de distância genética, gerada pelo método de agrupamento de vizinhos (NJ)
separou as populações analisadas de N. eques em dois clados. O primeiro, suportado por um
bootstrap de 93,3 englobou todos os indivíduos das populações do Açaituba, Miuá, Jaradi e
Arixanã (alto rio Negro) e cinco indivíduos da população do Jacundá (baixo rio Negro). O
segundo clado (bootstrap 99,5) englobou todos os indivíduos das populações do Demeni,
Maguari, Catalão e quatro indivíduos da população do Jacundá. Com base nesta árvore há
uma indicação de subdivisão neste grupo. Assim, como na árvore de máxima parcimônia, o
monofiletismo da população do Demeni dentro deste clado também é suportado (bootstrap
100) (Figura 11).

27
4.4 Matriz de distância genética Os resultados da matriz de distância genética par a par, determinados pela distância p
não corrigida entre todos os haplótipos, encontram-se no anexo 03. Entre N. eques e N.
unifasciatus (grupo externo) a distância p variou de 8,50% a 11,80%. Considerando os
haplótipos de todas as populações de N. eques a variação foi de 0,00% a 8,30%. Ao tratar N.
eques como dois grupos distintos (representados pelos clados da árvore de NJ), a distância
genética encontrada dentro do grupo 1 variou de 0,00% a 1,00% e dentro do grupo 2 de
0,10% a 5,50%, essa maior variação deve-se ao sub-clado do Demeni que aparece como um
grupo monofilético, excluindo a população do Demeni, a distância p dentro do grupo 2
variou de 1,20% a 1,70%. Porém, ao determinar a distância p entre os dois grupos foi
encontrada uma variação de 5,50% a 8,30%. Dentro de cada população a distância genética
entre os haplótipos foi baixa. Pelo fato da população do Jacundá conter haplótipos em ambos
os grupos formados, a mesma foi dividida em JC 1 (haplotipos do grupo 1) e JC 2
(haplotipos do grupo 2).
Tabela 02 – Distância genética média entre e dentro das populações (em porcentagem) obtida pela distância p não corrigida. Destacado em cinza claro a distancia média entre as populações do grupo 1; destacado em cinza escuro a distancia média entre as populações do grupo dois; sublinhado a elevada distância genética nas comparações envolvendo o Demeni.
Açaituba Miuá Jaradi Arixanã Jacundá 1 Jacundá 2 Demeni Maguari Catalão Dentro das populações
Açaituba 0,1% Miuá 0,21% 0,2% Jaradi 0,16% 0,15% 0,1%
Arixanã 0,27% 0,21% 0,21% 0,2% Jacundá 1 0,62% 0,75% 0,70% 0,08% 0,9% Jacundá 2 5,66% 5,54% 5,59% 5,49% 5,24% 1,0% Demeni 7,98% 7,91% 7,92% 7,91% 7,94% 4,05% 0,6% Maguari 6,19% 6,09% 6,14% 6,06% 5,87% 1,47% 3,59% 1,5% Catalão 6,10% 6,06% 6,09% 6,02% 5,65% 1,39% 4,50% 1,61% 1,1%
N. unifasciatus 10,82% 10,67% 10,72% 10,59% 10,49% 8,98% 11,40% 9,13% 9,07%

28
Figura 09 – Árvore gerada pela MP. Valores de bootstrap estão acima dos ramos. Grupo 1 (Açaituba, Miuá, Jaradi, Arixanã e Jacundá); Grupo 2 (Demeni, Jacundá, Maguari e Catalão). Ge = N. unifasciatus. Os valores em porcentagem refere-se à distância genética entre os ramos.

29
Figura 10 – Árvore gerada pela MV (Ln=3304.7620). Valores de bootstrap estão acima dos ramos. Em destaque o grupo 1 (Açaituba, Miuá, Jaradi, Arixanã e Jacundá) e a população do Demeni.. Ge = N. unifasciatus. Os valores em porcentagem refere-se à distância genética entre os ramos.

30
Figura 11 – Árvore gerada pelo método NJ mostrando a relação entre os haplótipos. Valores de bootstrap estão acima dos ramos. Grupo 1 (Açaituba, Miuá, Jaradi, Arixanã e Jacundá); Grupo 2 (Demeni, Jacundá, Maguari e Catalão). Ge = N. unifasciatus. Os valores em porcentagem refere-se à distância genética entre os ramos.

31
4.5 Análises populacionais
4.5.1 Análises de polimorfismo de DNA
Considerando os resultados das inferências evolutivas (MP, MV, NJ), onde
ficou evidente a presença de pelo menos dois grupos, optou-se por fazer as análises
populacionais considerando cada um dos grupos (Tabela 03).
As estimativas de polimorfismo genético indicaram diferentes níveis de
variabilidade genética ao longo da bacia do rio Negro.
Para o grupo 1 (Açaituba, Miuá, Jaradi, Arixanã e Jacundá 1) a maior
diversidade haplotípica (HD), nucleotídica (PI) e média das diferenças par a par (K)
foi evidenciada na população do Jacundá 1 (baixo rio Negro). O menor valor para
estes índices (HD; PI; K) foram evidenciados na população do Açaituba (alto rio
Negro).
Para o grupo 2 (Demeni, Jacundá 2, Maguari e Catalão) a maior diversidade
haplótípica foi evidenciada na população do Jacundá 2, a maior diversidade
nucleotídica e o maior valor da média das diferenças par a par foi encontrado na
população do Maguari. O menor valor de HD foi evidenciado na população do
Catalão, os menores valores de PI e K foram encontrados na população do Demeni.
Quando analisados os índices de polimorfismo de DNA totais, comparando os
dois grupos, os maiores valores de HD, PI e K foram evidenciados no grupo 2.
Os testes de neutralidade foram estimados para cada população de N. eques.
Nenhuma população do grupo 1 apresentou valores significativos para ambos os testes
de neutralidade (D de Tajima e Fs de Fu) indicando que o polimorfismo genético está
de acordo com o modelo neutro de mutações. Para as populações do grupo 2 o teste D
de Tajima também não apresentou valores significativos, enquanto que o Fs de Fu
mostrou desvio significativo da expectativa neutra das mutações em três populações:
Demeni, Maguari e Catalão (Tabela 03). Quando as localidades de cada grupo foram
consideradas conjuntamente (grupo 1 e grupo 2) o teste D de Tajima não apresentou
valores significativos. No grupo 2 o Fs de Fu indicou um significante desequilíbrio
genético. Como valores não significativos, mas negativos, podem ser um indício de
expansão populacional foi realizado uma análise de mismatch distribution na qual foi
evidenciada um padrão multimodal indicando que não houve expansão súbita.

32
Tabela 03 – Índices de polimorfismo de DNA para os espécimes de N. eques das oito localidades amostradas. N=número amostral; H= número de haplótipos; HU=haplótipos únicos; S=sítios polimórficos; ETA=número de mutações; HD=diversidade haplotípica; PI= diversidade nucleotídica; K=média das diferenças nucleotídicas par a par.
Pop. N H HU S ETA HD PI K D de Tajima Fs de Fu
AÇ 15 6 2 5 5 0,695 +/- 0,006 0,00093 +/- 0,00003 1,4666+/- 0,9404 -0,62465 -0,358
MI 20 17 11 9 9 0,868 +/- 0,004 0,00161 +/- 0,00001 2,8263 +/- 1,5552 0,64246 -7,062
JA 14 11 5 7 7 0,857 +/- 0,005 0,00124 +/- 0,0000 1,8461 +/- 1,1255 -1,08076 -3,466
AR 14 12 7 10 10 0,934+/- 0,003 0,00232 +/- 0,00033 3,2417 +/- 1,7772 0,45490 -7,456
JC 1 5 5 4 22 22 1,000 +/- 0,016 0,00846 +/- 0,00242 10,3333 +/- 5,9933 -0,46204 -0,480
Grupo 1 68 51 29 34 34 0,913 +/- 0.002 0,00266 +/- 0,00049 3,7919 +/- 1,9334 -1,57707 -12,589
DE 15 15 15 22 22 1,000 +/- 0,005 0,00593 +/- 0,00057 6,9904 +/- 3,4804 0,19126 -10,178*
JC 2 5 5 5 20 20 1,000 +/- 0,031 0,00906 +/- 0,00368 10,3333 +/- 5,9933 -0,52388 0,314
MA 20 20 20 55 58 1,000 +/- 0,003 0,01452 +/- 0,00104 16,8736 +/- 7,8388 0,29254 -6,331*
CT 18 18 18 47 48 1,000 +/- 0,000 0,01103 +/- 0,00146 13,4313 +/- 6,3359 -0,09688 -8,294*
Grupo 2 57 58 58 92 99 0,999 +/- 0.000 0,02390 +/- 0,00130 27,4317 +/- 12,1897 1,17069 -34,378
*Resultados significativos (P < 0.05).
4.5.2 Distribuição da variabilidade genética e diferenciação populacional
Por meio da Análise de Variância Molecular verificou-se que para o grupo 1
(Açaituba, Miuá, Jaradi, Arixanã e Jacunda 1) a maior parte da variação foi observada
dentro do grupo (71,17%) do que entre as localidades do grupo (28,83%) (Φst =
0,28835 P < 0,001) (Tabela 04). A comparação entre as localidades do grupo 1
apresentou valores significativos de Φst. O fluxo gênico estimado a partir do número
de migrantes por geração (Nm) foi baixo em todas as combinações envolvendo a
população do Jacundá 1, e a maioria das combinações envolvendo Açaituba. Entre as
demais populações (Miuá, Jaradi, e Arixanã) observou-se um elevado número de
migrantes por geração entre as populações (2,36 e 6,18), indicando assim altos níveis
de fluxo gênico (Tabela 05).
A AMOVA realizada para o grupo 2 (Demeni, Maguari, Jacundá 2 e Catalão), revelou
uma característica distinta do grupo 1, pois a maior variação deveu-se à diferenças encontradas
entre as populações (61,54%), do que dentro das mesmas (38,46%) (Φst = 0,61541 P < 0,001)
(Tabela 06). Os valores de Φst foram significativos para a maioria das combinações, contudo
foram muito altos nas comparações envolvendo a população do Demeni. Entre as populações do
Jacundá 2, Maguari e Catalão observou-se fluxo gênico efetivo, os valores de Nm encontrados
indicaram 1,73 a 3,61 migrantes por geração entre estas populações (Tabela 07).

33
Tabela 04 – AMOVA para o grupo 1 de N. eques (Açaituba, Miuá, Jaradi, Arixanã e Jacundá 1).
Fonte de variância Componentes da variância
Porcentage de variação
Entre populações 0.58261 28,83% Dentro da população 143.791 71,17%
Total 202.051 100%
Tabela 05 – Diferenciação genética (Φst) acima da diagonal e estimativa indireta de fluxo gênico (Nm) abaixo da diagonal para o grupo 1. Os valores sublinhados correspondem a ausência de fluxo gênico efetivo.
Açaituba Miuá Jaradi Arixana Jacundá Açaituba 0.28542 0.36179 0.36111 0.44462
Miuá 1.25180 0.15090 0.07475 0.47611 Jaradi 0.88202 2.81354 0.17448 0.44529
Arixanã 0.88461 6.18867 2.36569 0.43005 Jacundá 0.62455 0.55019 0.62287 0.66267
Tabela 06 – AMOVA para o grupo 2 de N. eques (Demini, Jacundá, Maguari e Catalão).
Fonte de variância Componentes da
variância Porcentagem de
variação Entre populações 10.23217 61,54%
Dentro da população 6.39434 38,46% Total 16.62651 100%
Tabela 07- Diferenciação genética (Φst) acima da diagonal estimativa indireta de fluxo gênico (Nm) abaixo da diagonal para o grupo 2. ns = não significativo. Os valores sublinhados correspondem a ausência de fluxo gênico efetivo.
Demeni Jacundá Maguari Catalão Demeni 0,82995 0,69269 0,79366 Jacundá 0,10245 0,12161ns 0,22349 Maguari 0,22182 3,61134 0,17983 Catalão 0,12999 1.73725 2,28047

34
4.5.3 Teste de Mantel
A hipótese de isolamento genético por distância foi testada somente entre as
populações dos grupos. A distancia geográfica (km) foi medida tomando como base o leito
fluvial (variação entre 43 e 1.200 km) (Tabela 08) e para a distância genética foram
utilizados os valores obtidos das comparações múltiplas entre os pares de populações
obtidos através do índice Φst.
A hipótese de isolamento por distância testada via teste de Mantel indicou correlação
significativa somente para o grupo 1 (r = 0,80904; P = 0,0244), para o grupo 2 não houve
relação entre a variação genética e a distribuição geográfica entre as populações estudadas (r
= 0,340252; P = 0,2955) (Tabela 09).
Tabela 08 – Matriz de distância geográfica (km) entre as localidades amostradas. Açaituba Miuá Jaradi Arixanã Demeni Jacundá Maguari Catalão
Açaituba 0,00 Miuá 239,36 0,00
Jaradi 419,28 199,92 0,00 Arixanã 435,63 216,27 16,35 0,00 Demeni 817,14 597,68 397,86 381,51 0,00 Jacundá 889,98 670,62 470,70 454,35 262,36 0,00 Maguari 904,25 684,89 484.97 468,62 180,89 43,01 0,00
Catalão 1199,95 980,59 780,67 764,32 472,83 330,50 349,95 0,00
Tabela 09 – Valores de correlação obtidos no teste de Mantel para os grupos 1 e 2.
Grupo 1 Grupo 2
r 0,80904 0,340252 P 0,0244 0,2955

35
5. Discussão
5.1 Caracterização da região controle de N. eques Na Amazônia, a importância sócio-econômica dos peixes é notória e vários estudos
genéticos têm utilizado a região controle como ferramenta na investigação da ictiofauna
amazônica (Porto, 1999; Harris & Petry, 2001; Santos, 2004; Schmitt, 2005; Batista et al.,
2006; Ortí et al., 2008; Souza, 2008), sendo estas informações muito importantes para
definir planos de manejo e conservação (Porto et al., 2001). Por esta razão, a região controle
mitocondrial foi utilizada neste trabalho para verificar os níveis de variabilidade genética nas
populações naturais de N. eques na bacia do rio Negro.
De maneira geral, a região controle de N. eques apresentou-se mais variável no
primeiro e terceiro domínio, TAS e CSB, respectivamente, e pouco variável no domínio
CCD.
O domínio CCD, costuma ser muito conservado para a maioria dos vertebrados,
sugerindo que tenha funções críticas para o metabolismo mitocondrial, enquanto que
seqüências altamente polimórficas são encontradas principalmente na região do primeiro
domínio (TAS) (Ray & Densmore, 2003).
Conforme observado na tabela 11 as seqüências TAS encontradas em N. eques são
similares às de outros Characiformes, tais com Mylesinus parashonburgki (Porto, 1999);
Colossoma macropomum (Santos, 2006); Nannostomus unifasciatus (presente estudo) bem
como a de outros grupos neotropicais .
Em N. eques, um bloco repetitivo (VNTR) foi encontrado em todos os indivíduos
analisados neste trabalho. Apesar do padrão da seqüência repetitiva não estar associado com
estrutura populacional, está relacionado aos diferentes clados filogenéticos (grupo 1 e 2)
encontrados. Considerando estudos envolvendo espécies da região amazônica, o mesmo
resultado foi encontrado em populações de Paracheirodon axelrodi (Harris & Petry, 2001),
de Colossoma macropomum (Santos, 2004) e de Pristobrycon striolatus (Freeman et al.,
2007), dentre estes o VNTR de N. eques e P. striolatus são muito similares. Convém
destacar que ao se comparar a seqüência repetitiva entre os dois grupos de N. eques foram
constatados seis sítios polimórficos (2 indels , 3 transições e 1 transversão).
Seqüências repetitivas caracterizadas como VNTRs, estão normalmente localizadas no
primeiro domínio da região controle o qual flanqueia o tRNA da prolina nos peixes
teleósteos, mas também podem ser encontradas no domínio CSB (Lee et al. 1995, Chen et

36
al., 2004). Em Cyprinella spiloptera Broughton & Dowling (1994; 1997) detectaram
VNTRs e sugeriram que a evolução das repetições deve ter ocorrido de forma independente
do resto do genoma mitocondrial.
Dentre os vários modelos que tentam explicar os mecanismos moleculares
responsáveis pela formação destas duplicações os mais aceitos incluem recombinações
intramoleculares (Rand & Harrinson, 1989) e o pareamento desigual das fitas durante a
replicação (slipped-strand mispairing) (Buroker et al., 1990).
No que diz respeito a composição de bases nucleotídicas, a região controle de N.
eques apresentou maiores quantidades de adenina e timina (71,4%) de que citosina e guanina
(28,6%), tais valores também estão dentro do esperado para a região controle de vertebrados
(Lee et al., 1995) e de peixes neotropicais (Ortí et al., 2008; Souza, 2008).
Com relação aos tipos de substituições nucleotídicas, em N. eques ocorreram mais
transições (72,1%) do que transversões (27,8%). De acordo com Meyer (1994) esta
tendência tem sido observada em todos os genes mitocondriais de peixes e tende a ser maior
na região controle.
Tabela 10 - Comparação da seqüência tipo TAS encontrada em N. eques com seqüência de outros peixes.
ESPÉCIE TAS 1 TAS 2 TAS 3 REF
Cyprinella spiloptera GCATAAACCAAAT - - 1
Mylesinus paraschomburgkii GCATAATATGYAA ACATAAAGCATAM MCATAAACTCCTY 2
Colossoma macropomum ACATTATATGCAT ACATAAACCATAA YYATAC - CTYCAT 3
N. unifasciatus ACATATTATGCA- - ACATTATGGTGTA - 4
N. eques GRUPO 1 AC-TATAATGTACG ACATTATGGTATA - 4
N. eques GRUPO 2 ACATATTATGCATG GCATTATGGTATA - 4
1= Broughton & Dowling, 1997; 2= Porto, 1999; 3= Santos, 2004; 4= presente trabalho.

37
5.2 Inferências evolutivas
As análises evolutivas foram realizadas a partir de três métodos distintos (máxima
parcimônia, máxima verossimilhança e agrupamento de vizinhos) para estimar a topologia
mais provável que possa refletir a história evolutiva de Nannostomus eques do rio Negro.
Tais análises (MP, ML e NJ) indicaram a existência de pelo menos dois grupos em N. eques.
O grupo 1, formado por haplótipos do alto (Açaituba, Miuá, Jaradi, Arixanã) e baixo
(Jacundá 1) rio Negro foi corroborado por um alto valor de bootstrap em todas as árvores.
O grupo 2, formado pelas populações do médio (Demeni) e baixo rio Negro (Jacundá
2, Maguari e Catalão), apresentou-se com uma configuração distinta nas três árvores tendo
ficado evidente o monofiletismo da população do Demeni (bootstrap 100). Com base na
árvore de agrupamento de vizinhos há uma indicação de sub-divisão no grupo 2 (bootstrap
99,5) e que quando comparado com a árvore de máxima parcimônia o suporte de bootstrap
foi baixo (55,04). Na máxima verossimilhança o grupo 2 apresentou-se como uma politomia
e apenas a população do Demeni aparece como um grupo monofilético.
A rede de haplótipos, gerada pelo programa TCS, separou as populações analisadas
em três redes de haplótipos disjuntas (A, B e C) que não podem ser conectados
parcimoniosamente (Figura 08). A rede A, correspondente ao grupo 1, e as redes B e C,
correspondentes ao grupo 2, só se conectam entre si com 60 passos mutacionais. As redes B
e C, que são mais próximas geneticamente, só se conectam com 20 passos mutacionais. A
alta diversidade haplotípica encontrada em ambas unidades sugerem que a maioria das
amostras populacionais apresentam elevada variabilidade genética.
Com relação à rede A, formada por haplótipos do alto e baixo rio Negro, foi possível
constatar a homogeneidade das populações deste grupo. Isto se reflete no compartilhamento
de haplótipos entre a maioria das populações que compõem esta rede, como é o caso do
haplótipo h28, compartilhado por nove indivíduos de quatro populações. Os haplótipos do
Jacundá pertencentes a esta rede (h37, h38, h39, h41) só se conectam entre si e com o h28
por meio de haplótipos perdidos ou não amostrados.
A rede B, formada por haplótipos do baixo rio Negro (Jacundá 2, Maguari e Catalão),
apresentou uma característica diferenciada onde a maioria dos haplótipos amostrados só se
conectam por meio de haplótipos perdidos ou não amostrados (missing haplotype). Além
disso, não houve compartilhamento de haplótipos entre as populações que a compõem

38
(Jacundá 2, Maguari e Catalão) refletindo a grande diversidade genética encontrada neste
grupo.
A rede C, fomada exclusivamente pela população do Demeni, também apresentou
uma diferenciação genética acentuada, corroborando as árvores filogenéticas.
Na prática, uma espécie pode consistir de inúmeras populações separadas,
parcialmente isoladas ou com fluxo gênico restrito, permitindo que as populações evoluam,
com uma certa amplitude, de forma independente (Ridley, 2006). O limite geográfico entre
os grupos 1 e 2 parece ser as adjacências do principal tributário do rio Negro, o rio Branco.
O rio Branco delimita o trecho do baixo e médio rio Negro onde seu delta age como um
obstáculo no vale do rio Negro, estreitando o canal, que nesta localidade tem apenas 2 km de
largura (Latrubesse & Franzinelli, 2005). Assim, o grupo 1 parece ter sua distribuição
preferencialmente associada a fisico-química dos tributários de água preta, enquanto que o
grupo 2 parece ter sua distribuição associada tanto à físico-química de água preta quanto de
água branca.
Com base nas elevações tectônicas reveladas em mapas de radar, acredita-se que
durante boa parte do Quaternário o rio Negro drenava na direção N-NE (região do
Essequibo, Guiana). Estes dados indicam que não houve ligação entre esta drenagem e a
bacia do Solimões até pelo menos antes do Pleistoceno tardio (Rossetti et al., 2005). Durante
o Plio-Pleistoceno mudanças geomorfológicas causaram um afundamento da área a oeste de
Manaus, o que mudou o fluxo do rio para o sentido N-O, alcançando o Oceano Atlântico
através do rio Essequibo na planície do Demerara. Este modelo é corroborado pela
orientação leste-oeste das falhas do Pleistoceno tardio, assim como pela captação
subseqüente da drenagem pelas falhas noroeste/sudeste. Dessa forma, o rio Negro está
instalado em uma grande falha (noroeste-sudeste) formada pela associação de movimentos
no sentido leste-oeste, que culminou em uma bacia rombóide situada entre este rio e o
Solimões (Franzinelli & Igreja, 1990; Franzinelli & Latrubesse, 1993; Bezerra, 2003).
Se assumirmos que desde o Plio-Pleistoceno N. eques já se distribuía em toda a bacia
Amazônica e que o fluxo do rio Negro era diferente do atual, conforme explicado acima,
acreditamos que somente o grupo 1 povoava o rio Negro. A conexão do Negro com o
sistema Solimões/Amazonas permitiu que grupo 2, que até então distribuía-se na região
central do Solimões/Amazonas (águas brancas), começasse a ocupar a região do baixo rio
Negro. A contribuição do rio Branco e o surgimento do arquipélago de Anavilhanas talvez

39
tenha favorecido a colonização por indivíduos do grupo 2 no baixo rio Negro, uma vez que o
aporte de águas ricas em partículas provenientes da bacia do rio Branco torna a margem
esquerda do arquipélago de Anavilhanas mais rica em nutrientes que a margem direita.
Os índices de distância genética nos levam a crer que a diversidade constatada
corrobora a existência de pelo menos duas unidades evolutivas dentro de N. eques, uma
referente aos haplótipos pertencentes ao grupo 1 e a outra representada pelos haplótipos do
grupo 2. Porém, foi possível constatar a existência de uma forte estrutura populacional
dentro do gupo 2 representada pela distância genética entre a população do Demeni e as
demais populações. As distâncias p não corrigidas obtidas nesta dissertação mostram valores
elevados de divergência entre os dois grupos de N. eques (5,50 a 8,30%) sendo muito
próximo da divergência em relação à espécie irmã N. unifasciatus (de 8,50% a 11,80%). Se
desconsiderarmos o Demeni a distância p entre os dois grupos é ainda maior (7,5% a 8,3%).
Além disso, o fato de indivíduos dos dois grupos (Jacundá 1 e Jacundá 2) terem sido
encontrados em simpatria no igarapé Jacundá, reforça ainda mais esta evidência da
existência de pelo menos duas unidades evolutivas dentro de N. eques.
Com freqüência muitos trabalhos envolvendo a região controle teem revelado uma
acentuada divergência intraespecífica em peixes neotropicais. Batista (2001) avaliando a
diversidade genética de Brachyplatystoma rousseauxii por meio de seqüências da região
controle do DNAmt encontrou valores de divergência genética que variaram de 0% a 3,5%.
Formiga-Aquino (2004) utilizando a mesma ferramenta em Brachyplatystoma vaillantii
encontrou valores de divergência intraespecífica que variaram de 0% a 4,7%. Schmitt
(2005), analisando a região controle de Hypopygus lepturus da bacia do médio rio Negro,
encontrou valores muito elevados de divergência entre duas linhagens evolutivas (7,34%).
Em média, a distância genética estimada por meio da região controle nos peixes
Characiformes varia de 0,1% a 4,2% em nível intraespecífico e de 1,2% a 25% em nível
interespecífico (Sivasundar et al., 2001; Hubert et al., 2007; Ortí et al., 2008).
O conceito de unidades evolutivas consiste na definição de uma população ou grupo
de populações que se encontram diferenciados em termos genéticos, morfológicos ou
ecológicos de populações próximas (co-específicas), refletindo uma história de isolamento
geográfico em níveis variáveis (Rider, 1986). Estas populações, ao possuírem características
próprias, mereceriam ser consideradas como unidades independentes para fins de
conservação (Eizirik, 1996). Moritz (1994) sugere o seguinte critério genético para a

40
definição de unidades evolutivas: Estas devem ser reciprocamente monofiléticas para
linhagens do mtDNA e apresentar divergência significativa nas freqüências alélicas em loci
nucleares. A utilização de diferentes marcadores se faz necessário devido a diferenças no
modo em que estes são afetados por processos populacionais históricos e deve ser
interpretada de modo a buscar uma compreensão realista da evolução do organismo em
questão. Segundo Crandall et al., (2000) em táxons com baixo fluxo gênico, as populações
que se diferenciaram por deriva genética podem ser designadas como unidades evolutivas
separadas, mesmo que elas não sejam adaptativamente diferentes. Considerando a
divergência genética intraespecífica encontrada em N. eques, passamos a denominar os
grupos de unidade evolutiva 1 e unidade evolutiva 2.
Pelo menos quatro casos de unidades evolutivas foram identificadas em uma mesma
espécie nominal na bacia do rio Negro: Schmitt (2005) utilizando marcadores genéticos
mitocondriais encontrou que a espécie H. lepturus é um complexo de espécies com pelo
menos três linhagens distintas. Schneider (2007) em uma análise fundamentada nas
subunidades 6 e 8 do gene mitocondrial ATPase concluiu que a espécie ornamental
Carnegiella stigata compreende duas linhagens distintas. Willis et al. (2007) relataram a não
monofilia de populações do tucunaré C. orinocensis onde uma unidade evolutiva ocorre no
baixo e médio rio Negro enquanto a outra unidade ocorre em várias localidades nas bacias
do Orinoco e Cassiquiare. Souza (2008) analisando espécies de Fluviphylax relatou que
dentro de F. obscurus há uma nova espécie, Fluviphylax sp n., mais relacionada à espécie F.
simplex e que distribui-se na área macrogeográfica do alto rio Negro/Orinoco e a espécie F.
obscurus stricto sensu distribuida na área macrogeográfica do médio rio Negro e que foi
grupo irmão do clado (F. zonatus, F. pygmaeus, (Fluviphylax sp., F. simplex)).
No que se refere à quantificação de variação genética necessária para discernir duas
espécies, as investigações feitas não são consensuais, assim como permanece sem respostas
a quantidade de diferenças genéticas necessária para individualizar subespécies. O
polimorfismo acentuado na região controle das duas unidades evolutivas de N. eques não se
mostrou relacionado com as características morfológicas externas (Zuanon, informação
pessoal) mas sim à físico-química da água. Dentre os lebiasinídeos, o gênero Nannostomus é
o único que passou por duas revisões taxonômicas (Weitzman, 1966; Weitzman & Cobb,
1975), mas ainda assim existem prováveis complexos de espécies mal definidos
taxonomicamente, incluindo o “complexo Nannostomus eques” que necessita de revisão

41
(Netto-Ferreira, 2005). Futuras investigações morfológicas e sistemáticas serão
fundamentais para confirmar se essas unidades evolutivas de N. eques correspondem à
espécies cripticas.
5.3 Genética de populações O conhecimento dos efeitos da variabilidade genética em uma população de peixes é
de fundamental importância para o entendimento de como essa diversidade está distribuída
em uma espécie. Se ela apresenta diversidade contínua, qualquer área de sua distribuição é
representativa da espécie, enquanto que, havendo estruturação, a representatividade de cada
subpopulação terá de ser preservada (Frankham, 1995).
As estimativas de polimorfismo genético de N. eques indicaram diferentes níveis de
variabilidade genética ao longo de sua distribuição na bacia do rio Negro.Como visto na
tabela 03, a diversidade haplotípica (HD) foi alta e a diversidade nucleotídica (PI) foi baixa
na maioria das populações analisadas, principalmente nas populações do Demeni, Maguari e
Catalão, onde o teste de neutralidade Fs de Fu detectou valores significativos (P < 0,05). A
associação desses resultados indica que estas populações podem ter passado por um
crescimento populacional recente. O grande número de haplótipos únicos encontrados nestas
populações corroboram esta hipótese, uma vez que um rápido crescimento populacional
aumenta a retenção de novas mutações (Avise et al.,1984). A possível detecção de expansão
observada nas populações do Demeni, Maguari e Catalão não foi confirmada pelo mismatch
distribution, mas poderá ser eventualmente confirmada através de marcadores mais sensíveis
como os microssatélites, já que o DNA mitocondial pode fornecer estimativas distorcidas
em populações onde os eventos como dispersão ou migração ocorrem com taxas diferentes
entre machos e fêmeas e tem quatro vezes mais chance de ser fixado por deriva genética nas
populações do que o DNA nuclear (Causse et al., 1994).
Os índices de diversidade totais revelaram uma grande diversidade genética, porém
não homogênea, já que para a unidade evolutiva 1 os valores foram mais baixos quando
comparados com a unidade evolutiva 2. Dois fatores que poderiam estar influenciando a
elevada diversidade genética dentro da unidade evolutiva 2 seriam: 1) Proteção de estoques,
já que a pesca de N. eques no rio Negro para fins ornamentais só é permitida acima da
confluência do rio Negro com o rio Branco e 2) A grande quantidade de microhabitats e
disponibilidade de alimento proporcionada pelo arquipélago de Anavilhanas.

42
Para a unidade evolutiva 1 os resultados da AMOVA revelaram que grande parte da
variação genética ocorre dentro das populações (71,17% ). Os altos índices de Φst
associados ao limitado fluxo gênico (Nm) demonstraram que algumas populações da
unidade evolutiva 1 são diferenciadas geneticamente, principalmente nas comparações
envolvendo o Açaituba e o Jacundá 1. O número de migrantes por geração foi relativamente
alto (1,25 a 6,18 indivíduos) entre as populações do Miuá, Jaradi e Arixanã indicando fluxo
gênico efetivo entre estas populações que estão localizadas abaixo das corredeiras de São
Gabriel da Cachoeira (alto rio Negro). O alto número de migrantes entre estas populações
pode estar sendo facilitado por influência do pulso de inundação durante o período de cheia,
que apesar de não ser tão marcante no alto rio Negro, pode conectar alguns tributários por
meio de uma planície de inundação (Goulding et al., 2003). Entretanto, a maioria das
combinações par a par envolvendo a população do Açaituba resultou em um baixo número
de migrantes por geração. Considerando que sua localização geográfica é acima do
município de São Gabriel da Cachoeira, as corredeiras e cachoeiras do alto rio Negro podem
estar limitando o fluxo gênico efetivo entre esta população e as demais. Segundo o teste de
Mantel o isolamento por distância é o fator responsável para explicar a divergência genética
encontrada na unidade evolutiva 1, já que a correlação da distância genética e distância
geográfica foi significativa. Desta forma, o baixo fluxo gênico observado entre as
comparações envolvendo Açaituba e Jacundá pode estar relacionado com a ampla
distribuição desta unidade.
Para a unidade evolutiva 2 os resultados da AMOVA foram diferentes da unidade
evolutiva 1 e revelaram que a maior parte da variação genética se deve a diferenças
encontradas entre as populações (61,54%) do que dentro delas (38,46%). As populações do
Jacundá 2, Maguari e Catalão apresentaram de moderada a alta divergência genética nas
comparações par a par. Nestas, o fluxo gênico mostrou-se efetivo para homogeneizar as
populações, já que o cálculo do Nm estimou de 1,73 a 3,61 indivíduos migrantes por
geração. Entretanto, a grande divergência genética encontrada na unidade evolutiva 2 se
deve à forte estrutura populacional no Demeni indicada pela diferenciação genética (Φst)
altamente significativa e ausência de fluxo gênico efetivo (inferior a 1) desta população com
as demais. De princípio pensou-se que a grande distância geográfica separando as
populações fosse o fator determinante das diferenças genéticas observadas, porém, a
hipótese de isolamento por distância foi descartada pelo teste de Mantel.

43
Um dos fatores que poderia estar contribuindo significativamente para a divergência
genética encontrada na unidade evolutiva 2 seria a distribuição geográfica das populações
amostradas em ambientes heterogêneos, tanto do ponto de vista geomorfológico da bacia do
rio Negro, quanto das diferenças físico-químicas das águas amazônicas. Além disso, peixes
com ampla distribuição geográfica, como é o caso de N. eques, podem apresentar fluxo
gênico reduzido quando possuem características biológicas e comportamentais que resultam
em baixa dispersão e conseqüentemente, estrutura populacional. Considerando que N. eques
é uma espécie sedentária que não realiza ciclos de migração e que possui uma preferência
por águas paradas e lagoas isoladas, este fator deve ser considerado. Apesar dos fatores
acima citados, grande parte da variação intrapopulacional na maioria dos casos é devida ao
equilíbrio entre mutação e seleção Mayr (1976).
A pronunciada divergência genética entre a população do Demeni e as demais
populações pode ser devido às características físico-químicas do rio Aracá (de água preta)
que como principal tributário do Demeni (de água branca) pode estar restringindo o fluxo
gênico entre a população amostrada e as populações próximas da calha do rio Negro
(Jacundá, Maguari e Catalão).
Diferenças físico-químicas entre os tipos de água podem ser determinantes para
divergências populacionais ou mesmo especiação. Ribeiro (2006) analisando seqüências do
genoma mitocondrial em espécies do gênero Potamotrygon associou os eventos de
especiação à distribuição geográfica das espécies, onde o fator determinante poderia ser as
diferenças físico-químicas entre os rios Amazonas e Negro. Farias & Hrbek (2008)
estudando os padrões de diversificação de Symphysodon spp. sugerem que as diferenças
entre os discos azuis, marrons e heckel são mantidas por preferências por determinados tipos
de água.
Mudanças ocorridas em nível de genoma nem sempre aparecem na morfologia o que
torna fundamental a associação de estudos genéticos, morfológicos e ecológicos,
principalmente na bacia amazônica que é formada por uma rica variedade de ambientes
associados a diferentes tipos de água.
O reconhecimento de diferenças populacionais é de grande importância para a
determinação de estratégias adequadas para a conservação biológica, pois permite a
identificação de estruturação geográfica e subdivisões populacionais históricas ou atuais nas
espécies. Estudos destes padrões tem sido extremamente úteis na definição de unidades para

44
a conservação e baseiam-se primariamente em filogenias intraespecíficas de linhagens do
mtDNA e de microssatélites (Avise et al., 1987; Baker & Palumbi, 1996).
A diversidade dos recursos genéticos aquáticos é um componente importante da
biodiversidade (Solé-Cava, 2001). Contudo, as preocupações e ações referentes aos
impactos sobre a biodiversidade nos ecossistemas aquáticos estão muito mais relacionadas à
percepção do desaparecimento de uma dada espécie do que à diminuição da diversidade
genética das espécies.
A descoberta de espécies crípticas, antes consideradas como parte de uma espécie
taxonomicamente definida torna-se importante quando são alvos de uso comercial. A bacia
do rio Negro é reconhecida por possuir uma riqueza de espécies de peixes singular
(Goulding et al., 1988). Com o uso de dados moleculares uma biodiversidade oculta no rio
Negro começa a ser revelada.
A questão de como definir espécies e do que este conceito representa em termos reais
tem gerado extensas discussões na literatura, talvez porque seja realmente impossível
chegar-se a uma definição consensual que abranja as infindáveis variações encontradas na
natureza. Ao nível intraespecífico, quaisquer subdivisões tendem a ser vistas como
arbitrárias. No entanto, parece claro que existem subdivisões reais, geradas através de
separações geográficas (históricas) e ou ecológicas entre grupos de populações que podem
ser considerados como pertencentes à mesma espécie por grande parte dos conceitos
vigentes. Estes subgrupos comporiam uma fração significativa da biodiversidade presente
nos ecossistemas e podem representar, em termos evolutivos, unidades tão ou mais reais do
que o conjunto total de populações que compõem a espécie em questão. Isto porque,
dependendo do grau de isolamento histórico ou atual entre estes grupos, pode-se supor que
estes representem unidades com “destino” evolutivo independente (Avise et. al., 1987;
Baum, 1992; Moritz, 1994; Rojas, 1995; Baker & Palumbi, 1996).
Apesar da adoção de medidas preventivas, a importância dos aspectos da genética no
manejo de estoques capturados pela pesca comercial raramente está na pauta de discussões
dos programas de manejo e conservação. Considerando que N. eques pode constituir-se em
um possível complexo de espécies com ampla distribuição na bacia Amazônica, a mesma
não pode ser tratada para fins de manejo como um único estoque.

45
6. Conclusão
Os resultados obtidos para Nannostomus eques, fundamentado na análise de
seqüências da região controle do DNA mitocondrial, permitiram chegar às seguintes
conclusões:
1) Pelo menos duas unidades evolutivas foram evidenciadas na bacia do rio Negro.
Uma, encontrada nas localidades de Açaituba, Miuá, Jaradi, Arixanã e Jacundá e a
outra nas localidades do Demeni, Jacundá, Maguari e Catalão.
2) Mudanças geomorfológicas ocorridas no Quaternário e os ambientes heterogêneos da
bacia do rio Negro parecem ter moldado a distribuição e a estrutura genética dentro e
entre as populações destas unidades. O limite geográfico entre as unidades evolutivas
parece ser as adjacências do principal tributário do rio Negro, o rio Branco.
3) As corredeiras e cachoeiras nas proximidades de São Gabriel da Cachoeira (alto rio
Negro) reduzem o fluxo gênico entre as populações da unidade evolutiva 1.
4) Detectou-se valores significativos do índice de fixação Φst e ausência de fluxo
gênico efetivo entre a população do Demeni e as demais populações que compõem a
unidade evolutiva 2. Outros fatores, excluindo o isolamento por distância podem
estar impedindo o fluxo gênico efetivo entre elas.
5) O teste de neutralidade, Fs de Fu, sugere que as populações do Demeni, Maguari e
Catalão, passaram por um recente crescimento populacional, mas essa hipótese foi
descartada pela análise do mismatch distribution.
6) O polimorfismo acentuado da região controle das duas unidades evolutivas, não se
mostrou correlacionado com as características morfológicas externas.
7) N. eques não pode ser tratada para fins de manejo como um único estoque na bacia
do rio Negro, tendo em vista o grau de diferenças genéticas entre as unidades
evolutivas.

46
7. Referências Bibliográficas Anjos, H. D. B.; Siqueira, J. A.; Amorim, R. M. S. 2007. Comércio de peixes ornamentais
do Estado do Amazonas, Brasil. Boletim Sociedade Brasileira de Ictiologia, 87 4-5 p.
Alves-Gomes, J. A. 1995. The phylogeny and Evolutionary history of the south american
electric fishes (order Gymnotiformes). Tese de Doutorado, Universidade da Califórnia,
São Diego.
Araújo M. S. 2003. Existe gradiente de tamanho, massa corpórea e fecundidade de
Pyrrhulina brevis (Characiformes, Lebiasinidae) em igarapés de terra-firme da
Amazônia Central? In: Ecologia da Floresta Amazônica, 132-135. Manaus, 146p.
Arefjev, V. A. 1990. Karyotypic Diversity of Characid Families (Pisces, Characidae).
Caryologia, 43(4): 291-304.
Asahida, T.; Kobayashi, T.; Saitoh, K.; Nakayama, I. 1996. Tissue preservation and total
DNA extraction from fish stored at ambient temperature using buffers containing high
concentration of urea. Fisheries Science, 62(5):727-730.
Avise, J.C.; Arnold, J.; Ball, R.M.; Bermingham, E.; Lamb, T.; Neigel, J.E.; Reeb, C.A. and
Saunders, N.C. 1987. Intraespecific Phylogeography: the mitochondrial DNA bridge
between population genetics and systematic. Ann. Rev. Ecol. Syst. 18: 489-522.
Avise, J. C. 1994. Molecular markers, natural history and evolution. Chapman & Hall, New
York, 511 pp.
Avise, J. C. 1998. The history and purview of phylogeography: a personal reflection.
Molecular Ecology, 7: 371-379.
Avise, J. C. 2000. Phylogeography: The history and formation of species. Harvard
University Press, London, 447 pp.
Baker, C.S. & Palumbi, S.R. 1996. Population structure, molecular systematic, and forensic
identification of whales and dolphins. In: Conservation Genetics: Case Histories from
Nature (Avise, J.C. and Hamrick, J. L., eds.).
Batista, J. S. 2001. Estimativa da variabilidade genética intra-específica da dourada –
Brachyplatystoma flavicans Castelnau 1855 (Pimelodidae – Siluriformes) no sistema
Estuário-Amazona-Solimões. Dissertação de Mestrado. Instituto Nacional de Pesquisas
da Amazônia / Universidade Federal do Amazonas. Manaus, Amazonas. 113pp.
Baum, D. 1992. Phylogenentic species concepts. Trends Ecol. Evol. 7:1.

47
Beheregaray, L. B.; Schwartz, T. S.; Möller, L. M.;Call, D.; Chao, N. L.; Caccones, A.
2004. A set of microsatellite DNA markers for the one-lined pencilfish Nannostomus
unifasciatus, an Amazonian flooded forest fish. Molecular Ecology Notes, 4 (3): 333-
335.
Bernatchez, L. and Danzmann, R. G. 1993. Congruence in Control-Region Sequence and
Restriction-Site Variation in Mitochondrial DNA of Brook Charr (Salvelinus fontinalis
mitchill) Molecular Biology Evolution, 10: 1002-1014.
Bezerra, P. E. L. 2003. Compartimentação morfotectônica do interflúvio Solimões-
Negro.Tese deDoutorado .Universidade Federal do Pará, Belém 335 p.
Blumer, L. S. 1979. Male parental care in the bony fishes. The Quarterly Review of Biology,
2: 149-161 pp.
Broughton R. E., Dowling, T. E., 1994. Length variation in mitochondrial DNA of the
minnow Cyprinella spiloptera. Genetics, 138:179–190
Dowling T. E.; Broughton. E.; Demarais B. D. 1997. Significant role for historical effects in
the evolution of reproductive isolation: Evidence from patterns of introgression between
the cyprinid fishes, Luxilus cornutus and Luxilus chrysocephalus. Evolution, (51) 1574-
1583.
Buckup, P. A. 1998. Relationships of the Characidinae and Phylogeny of Characiform fishes
(Teleostei: Ostariophysi). In: Malabarba, L. R., Reis, R. E., Vari, R. P., Lucena, Z. M.
S., Lucena, C. A. S. Phylogeny and Classification of Neotropical Fishes, 123-144. Porto
Alegre, Edipucrs, 603p.
Buckup, P. A. 1999. Sistemática e biogeografia de peixes de riachos. In: Caramaschi, E. P.;
Mazzoni, R.; Peres-Neto, P. R.. Ecologia de peixes de riachos. Série Oecologia
Brasiliensis, vol. VI. PPGE-UFRJ. Rio de Janeiro, Brasil.
Buckup, P.A.; Menezes, N.A.; Ghazzi, M.S. (eds.) 2007. Catálogo das espécies de peixes de
água doce do Brasil. Rio de Janeiro, Museu Nacional. 195p. (Série Livros, 23).
Buroker, N. E., Brown, J. R., Gilbert, T. A., O'Hara, P. J., Beckenbach, A. T., Thomas, W.
K., Smith, M. J. 1990. Length Heteroplasmy of Sturgeon Mitochondrial DNA: An
Illegitimate Elongation Model. Genetics,124: 157-163
Calcagnoto, D., Schaefer S. A., Desalle R. 2005. Relationships among characiform fishes
inferred from analysis of nuclear and mitochondrial gene sequences. Molecular
Phylogenetics Evolution, 36: 135-153.

48
Causse, M.A.; Fulton, T.M.; Cho, Y.G.; Ahn, S.N.; Chunwongse, J.; Wu, K.; Xiao, J.; Yu,
Z.; Ronald, P.C.; Harrington, S.E.; Second, G.E.; Mccouch, S.R.; Tanksley, S.D. 1994.
Satured molecular map of rice genome based on an interespecific backcross population.
Genetics, 138: 1251-1274.
Chao, N.L. 1993. Conservations of rio Negro ornamental fishes. Tropical Fish Hobbyist,
41(5): 99-114.
Chao, N. L. 2001. The fishery, diversity and conservation of ornamental fishes in the Rio
Negro Basin, Brazil - A Review of Project Piaba (1989-1999). In: Chao, N. L.; Petry, P.;
Prang, G.; Sonneschien, L. e Tlusty, M. (Eds). Conservation and Management of
ornamental fish Resources of the rio Negro Basin. Amazonia, Brazil - Project Piaba.
Editora da Universidade do Amazonas, Manaus, Amazonas. p. 161-204.
Chen, C. A., Anonuevo, M.C., McManus, J.W., Bell, J.D., Tuan,V. S., Cabanban, A.S.,
Shao1, K.T. 2004. Variable Numbers of Tandem Repeats (VNTRs), Heteroplasmy, and
Sequence Variation of the Mitochondrial Control Region in the Threespot Dascyllus,
Dascyllus trimaculatus (Perciformes: Pomacentridae). Zoological Studies, (4): 803-812
(2004)
Clement, M.; Posada, D.; Crandall, K. A. 2000. TCS: a computer program to estimate gene
genealogies. Molecular Ecology, 10: 1657-1660.
Crandall, K.A.; Bininda-Emonds, O.R.P.; Mace, G.M.; Wayne, R.K. 2000. Considering
evolutionary process in conservation biology. Trends in Ecology & Evolution 15:290
295.
Estoup, A.; Presa, P.; Krieg, F.; Vaiman, D.; Guyomard, R. 1993. (CT)n and (GT)n
microsatellites: a new class of genetic markers for Salmo trutta L. (brown trout).
Heredity, 71: 488-496.
Excoffier, L.; Smouse, P.; Quattro, J. 1992. Analysis of molecular variance inferred from
metric distances among DNA haplotypes: application to human mitochondrial DNA
restriction data. Genetics, 131: 479-491.
Excoffier, L.; Laval, G., Schneider S. 2006. An integrated software package for population
genetics data analysis. Computational and Molecular Population Genetics Lab (CMPG),
Institute of Zoology, University of Berne, Switzerland.
Farias IP.; Hrbek, T. 2008 Patterns of diversification in the discus fishes (Symphysodon spp.
Cichlidae) of the Amazon basin. MolecularPhylogenetics and Evolution, 49: 32-43.

49
Felsenstein, J. F. 1981. Evolutionary tree from DNA sequences: a maximum likelihood
approach. Journal of Molecular Evolution, 17: 368-376.
Ferreira, E. J. G.; Zuanon, J.; Forsberg, B. R.; Goulding, M.; Briglia-Ferreira, S. R. 2007.
Rio Branco - Peixes, Ecologia e Conservação de Roraima. 1. ed. Lima: Wust Ediciones,
v. 1. 201 p.
Formiga-Aquino, K. 2004. Filogeografia da piramutaba (Brachyplatystoma vaillantii) na
Bacia Amazônica. Dissertação de Mestrado. Instituto Nacional de Pesquisas da
Amazônia / Universidade Federal do Amazonas. Manaus, Amazonas. 80pp.
Frankham, R. 2005. Genetics and extinction. Biological Conservation, 126: 131-140.
Franzinelli, E., Igreja, H.L.S., 1990. Utilização de sensoriamento remoto na investigação da
área do Baixo Rio Negro e Grande Manaus. VI Simp. Bras. Sens. Rem. An. 3, 641– 648
Franzinelli E., Latrubesse E., 1993. Neotectonic in the central part of the Amazon basin.
Bulletin INQUA Neotectonic Commission, 16: 10– 13.
Franzinelli E., Igreja, H. 2002. Modern sedimentation in the lower Negro River, Amazonas
state, Brazil. Geomorphology, 44 259–271.
Frederico, R. G. 2006. Filogeografia de Paratrygon aiereba Müller & Henle, 1841
(Chondrichtyes: Potamotrygonidae) na região Amazônica. Dissertação de Mestrado.
Instituto Nacional de Pesquisas da Amazônia / Universidade Federal do Amazonas.
Manaus, Amazonas. 80pp.
Freeman,B.; Nico,L.G.; Osentoski,M.; Jelks,H.L.; Collins,T.M.. 2007. Molecular
systematics of Serrasalmidae: deciphering the identities of piranha species and
unraveling their evolutionary histories. Zootaxa, 1484: 1-38.
Fu, Y-X. 1997. Statistical tests of neutrality of mutations against population growth
hitchhiking and background selection. Genetics, 1447: 915-925.
Fujii, R.; Masagaki, A. 1999. Differential Actions of Melatonin on Melanophores of the
Threeline Pencilfish, Nannostomus trifasciatus. Zoological Science, 16: 35-42.
Goulding, M.; Carvalho, M. L.; Ferreira, E. G. 1988. Rio Negro: rich life in the poor water.
The Hague, The Netherlands: SPB Acadmic Publishing, 200pp.
Goulding, M.; Barthem, R.; Ferreira, E. 2003. The Negro and the Trombetas – Balck and
Clear Waters from Ancient Lands. In: The Smithsonian Atlas of the Amazon. Princeton
Editorial Associates, Inc. Hong Kong.

50
Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis
program for Windows 95/96/NT. Nucleic Acids Symposium Series, 41:95-98.
Harris, P.; Petry, H. 2001. Preliminary report on the genetic population structure and
phylogeography of cardinal tetra (Paracheirodon axelrodi) in the rio Negro basin. In:
Chao, N. L.; Petry, P.; Prang, G.; Sonneschien, L. & Tlusty, M. (Eds). Conservation and
Management of ornamental fish Resources of the rio Negro Basin. Amazonia, Brazil -
Project Piaba. Editora da Universidade do Amazonas, Manaus, Amazonas, p. 205-225.
Hartl, D.L. & Clark, A.G. 1989. Principles of Population Genetics, second edition, Sinauer
Associates Inc, Sunderland MA, 481pp.
Hennig, W 1966. Phylogenetic systematics. University of Illinois Press, Urbana, 263 pp.
Hilsdorf, A. W. S.; Marques D. K. S.; Resende E. K. 2006. Genética e conservação de
estoques pesqueiros de águas continentais no Brasil: Situação atual e perspectivas.
Embrapa, Pantanal. Corumbá, Mato Grosso, 44pp.
Hrbek, T., Farias, I. P.; Crossa, M.; Sampaio, I.; Porto, J. I. R.; Meyer, A. 2005. Population
genetic analysis of Arapaima gigas, one the largest freshwater fishes of the Amazon
basin: implications of this conservation. Animal Conservation, 8) 297-308.
Hubert, N.; Torrico, J.P.; Bonhomme, F.; Renno, J.F. 2007. Species polyphyly and mtDNA
introgression among three Serrasalmus sister-species. Molecular Phylogenetics and
Evolution, 46: 375–381.
Jobb, G.; Haeseler, A.V.; Strimmer, K. 2004. TREEFINDER: a powerful graphical analysis
environment for molecular phylogenetics. BMC Evolution Biology, 4: 1-9.
Junk, W. J.; Bayley, P.B.; Sparks, R.E. 1989. The Flood Pulse Concept in River-Floodplain
Systems. In D.P. Dodge. Proceedings of the International Large River Symposium.
Canadian Special Publications of Fisheries and Aquatic Science, 106: 110-127.
Junk, W. J.; Furch K. 1993. A general review of tropical South American floodplains.
Wetlands Ecology and Management, 4: 231-238.
Latrubesse, E. M. 2008 Patterns of anabranching channels: The ultimate end-member
adjustment of mega rivers. Geomorphology, 101: 130-145.
Latrubesse, E. M.; Franzinelli. E. 2005. The late Quaternary evolution of the Negro River,
Amazon, Brazil: Implications for island and floodplain formation in large anabranching
tropical systems, Geomorphology, 70: 372-397

51
Lee, W.J; Conroy, J.; Howell, W.H.; Kocher, T.D. 1995. Structure and evolution of teleost
mitochondrialcontrol regions. Journal of Molecular Evolution, 41:54-66.
Lovejoy, N. R.; de Araújo, M. L. G. 2000. Molecular systematics, biogeography and
population structure of neotropical freshwater needlefishes of the genus Potamorrhaphis.
Molecular Ecology, 9: 259-268.
Mantel N. 1967. The detection of disease clustering and a generalized regression approach.
Cancer Research, 27: 209-220.
Mayr, E. 1976. Evolution and the Diversity of Life. Cambridge, Mass.: Harvard University
Press.
MMA, Instrução Normativa nº 203, de 22 de outubro de 2008. Diário Oficial da União,
Brasília, DF, 22 de outubro de 2008.
Meyer, A. 1993. Evolution of mitochondrial DNA in fishes. In: Hochachka, P. W. &
Mommsen (Eds). Biochemistry and molecular biology of fishes, Vol 2. Elsevier Science
Publishers. p 1-38.
Meyer, A. 1994. DNA technology and phylogeny of fish: molecular phylogenetic studies of
fish. In: Genetic and evolution of aquatic organisms, Chapman & Hall. Lodon. p. 219-
249.
Moreira, C. R. 2007. Relações filogenéticas na ordem Characiformes (Teleostei:
Ostariophysi. Tese de Doutorado. Instituo de Biociências da Universidade de São Paulo.
São Paulo 468pp.
Moritz, C. 1994. Defining ‘Evolutionarily Significant Units’for conservation. Trends Ecol.
Evol. 9: 373-375.
Nahum, L.A. Evolução dos Genomas. In: Matioli S. R., Biologia Molecular e Evolução.
Ribeirão Preto: Holos Ed., 2001.
Netto-Ferreira A. L. 2005. Relações Filogenéticas dos Gêneros de Lebiasinidae
(Ostariophysi, Characiformes). Dissertação de Mestrado. Universidade Federal do Rio
de Janeiro. Rio de Janeiro. 394pp.
Ortí, G.; Meyer, A. 1996. Molecular evolution of ependymin and the phylogenetic
resolutionof early divergences among euteleost fishes. Molecular Biology and Evolution,
13(4):556-573.
Ortí, G.; Meyer, A. 1997. The radiation of characiform fishes and the limits of resolution of
mitochondrial ribosomal DNA sequences. Systematic Biology, 46(1):75-100.

52
Ortí, G., Sivasundar A.; Dietz K.; Jégu M.; 2008. Phylogeny of the Serrasalmidae
(Characiformes) based on mitochondrial DNA sequence. Genetics and Molecular
Biology. 31 343-351.
Oyakawa, O. T. 1998. Relações filogenéticas das famílias Pyrrhulinidae, Lebiasinidae e
Erythrinidae (Osteichthyes: Characiformes). Tese de Doutorado. Universidade de São
Paulo, São Paulo, 200 pp.
Pereira, S. L., 2000. Mitochondrial genome organization and vertebrate phylogenetics.
Genetics and Molecular Biology. 4 745-752.
Porto, J. I. R., Feldeberg, E.; Nakayama, C. M.; Falcão, J. N. 1992. A checklist of
chromosome numbers and karyotypes of amazonian freshwater fishes. Revue D'
Hydrobiologie Tropicale, 4: 287-299.
Porto, J. I. R. 1999. Análises cariotípicas e sequenciamento de mtDNA de populações de
Mylesinus paraschomburgkii (Characiformes, Serrasalmidae) da bacia Amazônica. Tese
de Doutorado. Instituto Nacional de Pesquisas da Amazônia/Universidade Federal do
Amazonas. Manaus, Amazonas. 120 pp.
Porto, J. I. R.; Alves-Gomes, J. A.; Farias, I. P.; Feldberg, E. 2001. Using molecular biology
techniques to characterize the diversity of amazonian ornamental fishes. In: Chao, N. L.;
Petry, P.; Prang, G.; Sonneschien, L. e Tlusty, M. (Eds). Conservation and Management
of ornamental fish Resources of the rio Negro Basin. Amazonia, Brazil Project Piaba.
Editora da Universidade do Amazonas, Manaus, Amazonas, p. 227-244.
Posada, D.; Crandall, K. A. 1998. MODELTEST: testing the model of DNA substitution.
Bioinformatics, 14: 817-818.
Ray, D. A., Densmore, D. L. 2003. Repetitive Sequences in the Crocodilian Mitochondrial
Control Region: Poly-A sequences and heteroplasmic tandem repeats. Molecular
Biology and Evolution. 6. 1006-1013.
Rand, D. M. Harrison, R. G. 1989. Molecular population genetics of mtDNA size variation
in crickets. Genetics, 121: 551-569.
Reis, R. E.; Kullander, S. O.; Ferraris Jr, C. J. 2003. Check list of the freshwater fishes of
south and central america. Editora da Pontifícia Universidade Católica, Porto Alegre,
Brasil. 729 pp.
Ribeiro, D. T. 2006. História evolutiva de espécies do gênero Potamotrygon Garman, 1877
(Potamotrygonidae) na Bacia Amazônica. Dissertação de Mestrado. Instituto Nacional

53
de Pesquisas da Amazônia / Universidade Federal do Amazonas. Manaus, Amazonas,
114pp.
Ridley, M. 2004. Evolution. Cambrige, Blackwell Science. 719pp.
Ridley, M.; Ferreira, H. B.; Passaglia, L.; Fischer, Araújo, M. A. 2006. Evolução. Ed.
Artmed. 752p.
Rojas, M. 1995. The species problem and conservation: what are we protecting? In: Genes,
Populations, Species: Readings from Conservation Biology. Blackwell Science.
Cambridge, MA, pp. 1-9.
Román-Valencia, C. 2004. Sobre la bioecología de Lebiasina panamensis (Pisces:
Lebiasinidae) de los ríos Atrato y La Vieja, Colombia. Dahlia, 7: 33-35.
Rossetti D. F.; Peter, M. T.; Góes, A. M. 2005. New geological framework for Western
Amazonia (Brazil) and implications for biogeography and evolution. Quaternary
Research, 66: 78-89.
Rozas, J.; Sánchez-DelBarrio, J. C.; Messeguer, X.; Rozas, R. 2003. DnaSP, DNA
polymorphism analyses by the coalescent and other methods. Bioinformatics, 19 : 2496-
2497.
Ruffino, M. 2004. A pesca e os recursos pesqueiros na Amazônia brasileira. Manaus,
IBAMA/PróVárzea. 272pp.
Saiki, R. K.; Gelfand, D. H.; Stoffel, S.; Scharf, S. J.; Higuchi, R.; Horn, G. T.; Mullis, K.
B.; Erlich, H. A. 1988. Primer-directed enzymatic amplification of DNA with a
thermostable DNA polymerase. Science, 239: 487-491.
Saitou, M.; Nei, M. 1987. The Neighbor-joining Method: A New Method for Reconstructing
Phylogenetic Trees. Molecular Biology and Evolution, 4 (4): 406-425.
Sambrook, J.; Russell, D. W. 2001. Molecular cloning: a laboratory manual. Vol. I. Cold
Spring Harbor Press. Cold Spring Harbor. NY.
Sanger, F.; Nicklen, S.; Coulson, A. R. 1977. DNA Sequencing with chain terminating
inhibitors. Proceedings of National Academy of Science. USA, 74: 5463-5467.
Santos, G. M.; Ferreira, E. J. G. 1999. Peixes da Bacia Amazônica. In: Lowe-McConnel,
R.H.(Ed.). Estudos ecológicos de comunidades de peixes tropicais. EDUSP, São Paulo,
Brasil. p. 345-373.

54
Santos S, Schneider H and Sampaio I (2003) Genetic differentiation of Macrodon ancylodon
(Sciaenidae, Perciformes) populations in Atlantic coastal waters of South America as
revealed by mtDNA analysis. Genetics and Molecular Biology, 26 :151-161.
Santos, M. C. F. 2004. Estimativa da variabilidade genética intra e interpopulacional de
Tambaqui (Colossoma macropomum, Cuvier) 1818 na bacia Amazônica. Dissertação de
Mestrado. Universidade Federal de São Carlos. 94pp.
Santos, T.; Vosgueritchian, S.B.; Nazareth, T. M.; Costa, J.B.P. 2006. Tipo de habitat
determina a ocorrência de peixes de tamanhos diferentes? In: Ganade, G. Ecologia da
floresta Amazônica, 2006. Instituto Nacional de Pesquisas da Amazônia (INPA);
Floresta Amazônica; Manaus, AM; BRASIL.
Santos, M. C. F.; Ruffino, M. L.; Farias, I. P. 2007. High levels of genetic variability and
panmixia of the tambaqui Colossoma macropomum (Cuvier, 1816) in the main channel
of the Amazon River. Journal of fish Biology.
Scheel, J. J. 1973. Fish chromosomes and their evolution. Internal Report of Danmarks
Akvarium, Charlottenlund. 22pp.
Schmitt, R. 2005. Filogeografia de Hypopygus lepturus Hoedeman, 1962 (Gymnotiformes:
Rhamphycthyidae) ao longo do médio rio Negro, Amazônia. Dissertação de Mestrado.
Instituto Nacional de Pesquisas da Amazônia /Universidade Federal do Amazonas.
Manaus, Amazonas. 143pp.
Schneider, C.H. 2007. Filogeografia do peixe ornamental Carnegiella strigata
(Characiformes, Gasteropelecidae) de três rios de água preta da Amazônia central.
Dissertação de Mestrado. Instituto Nacional de Pesquisas da Amazônia/ Universidade
Federal do Amazonas. Manaus, Amazonas. 70pp.
Sioli H. 1984. The Amazon: Limnology and Landscape Ecology of a Mighty Tropical River
and its Basin. In: Monographiae Biologicae eds. Dumont HJ, Werger MJA), p. 800.
Springer Verlag, New York, NY.
Sioli, H. 1950. Das Wasser im Amazonasgebiet. Forsch. Fortschr. In: Lowe-McConnel,
R.H.(Ed.). Estudos ecológicos de comunidades de peixes tropicais. EDUSP, São Paulo,
Brasil. p. 345-373.
Sioli, H. 1990. Amazônia: fundamentos da ecologia da maior região de florestas tropicais.
Editora Vozes, Petrópolis, Brasil. 73 pp.

55
Sivasundar, A.; Bermingham, E.; Ortí, G. 2001. Population structure and biogeography of
migratory freshwather fishes (Prochilodus: Characiformes) in major South American
rivers. Molecular Ecology, 10: 407-417.
Solé-Cava, A.M. 2001. Biodiversidade molecular e genética da conservação. In: Matioli,
S.R.(Eds.). Biologia Molecular e Evolução. Holos, Ribeirão Preto, p.172-192.
Souza, E. R. 2008. Filogeografia do gênero neotropical Fluviphylax (Cyprinodontiformes:
Poeciliidae) das bacias do Amazonas e do Orinoco. Dissertação de Mestrado. Instituto
Nacional de Pesquisas da Amazônia/ Universidade Federal do Amazonas. Manaus,
Amazonas. 122pp.
Swofford, D. L. 1999. PAUP: Phylogenetics analysis using parsimony, version 4.0. Sinauer
Associates, Sunderland.
Tajima, F. 1989. Statistical method for testing the neutral mutation hypothesis by DNA
polymorphism. Genetics, 123: 585-595.
Templeton, A.R., Crandall, K. A., Sing, C. F. 1992. A cladistic analysis of phenotypic
associations with haplotypes inferred from restriction endonuclease mapping & DNA
sequence data. III. Cladogram estimation. Genetics, (132): 619–633.
Thompson, J. D.; Higging, D. G.; Gibson, T. J. 1994. ClustalW : Improving the sensitivity
of progressive multiple sequence alignment through sequence weighting, position
specific gap penalties and matrix choice. Nucleic Acids Research, 22: 4673-4680.
Vari, R. P.; Malabarba, L. R. 1998. Neotropical ichthyology: An overview In: Malabarba, L.
R.; Reis, R. E.; Vari, R. P.; Lucena, Z. M. S.; Lucena, C. A. S. (Eds). Phylogeny and
classification of neotropical fishes. Editora da Pontifícia Universidade Católica, Porto
Alegre, Brasil. p.1-11.
Weitzman, S.H. 1964. Osteology and relationships of South American characid fishes of
subfamilies Lebiasinidae and Erythrinidae with special references to subtribe
Nannoatomina. Proceedings of the United States National Museum,116 (3538):1-56.
Weitzman, S. H. 1966. Review of South American characid fishes of the subtribe
Nannostomina. Proceedings of the United States National Museum, 119 (3538): 1-56.
Weitzman, S. H.; COBB, J. S. 1975. A revision of the South American fishes of the genus
Nannostomus Günther (Family Lebiasinidae). Smithsonian Contribution Zoology, 186:
1-36.

56
Weitzman, S. H. 1978. Three new species of fishes of the genus Nannostomus from the
Brazilian states of Pará and Amazonas (Teleostei: Lebiasinidae). Smithsonian
Contribution Zoology, 263: 1-14.
Weitzman, H.; Weitzman, M. 1982. Biogeography and Evolutionary Diversification in
Neotropical Freshwates Fishes, with Comments on the Refuge Theory. In: Prance, G.T.
(ed). Biological Diversification in the Tropics. Columbia University Press. New York. p.
404-422.
Weitzman, S. H.; Weitzman, M. 2003. Family Lebiasinadae. In: Reis, R. E.; Kullander, S.
O.; Ferraris Jr, C. J. (Eds). Check list of the freshwater fishes of south and central
America. Editora da Pontifícia Universidade Católica, Porto Alegre, Brasil. p.101-103.
Willis, S.C.; Nunes, M. S.; Montana, C. G.; Farias, I.P.; Lovejoy, N. R. 2007. Systematics,
biogeography, and evolution of the Neotropical peacock basses Cichla (Perciformes:
Cichlidae). Molecular Phylogenetics and Evolution, 44 (1): 291-307.

57
8. Anexos
Anexo 01 Sítios polimórficos das seqüências da região controle dos 97 haplótipos encontrados neste trabalho. h 1 -TATC-CA-TATC-A-TATC-A-TATC-A-TATC-A-TATC-A-TATC-AA-TATAC-TAAA-TTAATC-A-TATC-AAAAATACACCACCTCAACGGCCATGATTAGA-TTTTTATTGCGCGACGTACTAAGGTT-GCAAGT
h 2 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-..........................G.....G..........................-......
h 3 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.T...-....-......-.-.T..-................................-..........................-......
h 4 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-................................G......................A...-......
h 5 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-..........................G.....-..........................-......
h 6 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T..-..........................G.....G..........................-......
h 7 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T..-................................-........-......-..........-......
h 8 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.T...-....-..T...-.-.T..-..........................G.....G......................A...-......
h 9 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.T...-....-......-.-.T..-................................G..........................-......
h 10 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-..T...-.-.T..-................................-..........................-......
h 11 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.T...-....-..T...-.-.T..-..........................G.....G..........................-......
h 12 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-..T...-.-.T..-..........................G.....G..........................-......
h 13 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T..-................................-........-.................-......
h 14 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T..-................................-........-.-...............-......
h 15 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T..-................................G...............-..........-......
h 16 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T..-................................-..........-....-..........-......
h 17 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T..-.........................-......G......................A...-......
h 18 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T..-................................G..........................-......
h 19 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-.........................-G.....G..........................-......
h 20 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T..-.........................-......G..........................-......
h 21 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-..T...-.-.T..-.........................-......G..........................-......

58
h 22 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.T...-....-......-.-.T..-.........................-......G..........................-......
h 23 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T.T-................................G..........................-......
h 24 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.T...-....-..T...-.-.T..-................................G......................A...-......
h 25 -....-..-....-.-....-.-....-.-....-.-....-.-.T..-..-.T...-....-..T...-.-.T..-................................G..........................-......
h 26 -....-..-....-.-....-.-....-.-....-.-.T..-.-....-..-.....-....-......-.-....-................................G..........................-......
h 27 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-.........................-......G...............-......A...-......
h 28 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-................................G..........................-......
h 29 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-..T...-.-.T..-................................G..........................-......
h 30 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-.T..-................................G......................A...-......
h 31 -....-..-....-.-....-.-....-.-....-.-CT..-.-....-..-.T...-....-..T...-.-.T..-................................G..........................-......
h 32 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.T...-....-..T...-.-.T..-................................G..........................-......
h 33 -....-..-....-.-....-.-....-.-....-.-....-.-.T..-..-.....-....-..T...-.-.T..-................................G..........................-......
h 34 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-..T...-.-.T..-................................G...............-..........-......
h 35 -....-..-....-.-....-.-....-.-....-.-.T..A.-.T..-..-.....-....-..T...-.-.T..-.........................-......G...............-..........-......
h 36 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..T.CTAGA.TCTAG......GGT...T.T...A...CC.GA..A.G.C...T...T..A.T.....T-....AAT..A.
h 37 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-.......GGT...T.T...A...CC.......G....................T.....-....A.
h 38 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-................................G....................T-A...-....A.
h 39 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-.........................-......G....................T-....-....A.
h 40 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-.........................-......G..........................-......
h 41 -....-..-....-.-....-.-....-.-....-.-....-.-....-..-.....-....-......-.-....-.............T.T...A...CC.GA..A.G.C...T...T..A.T.....T-....-A...A.
h 42 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA....A..T.CTAGA.TCTAG......GGT...T.T...A...CC.GA..A.G.C...T......A.T.....T-...CAAT..A.
h 43 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGA.TCTAGA.TCTAG.A.TC.TA....A..T.CTAGA.TCTAG......GGT...T.T...A...CC..AA..GG.A...T...T.......T...-A..CA.T..A.
h 44 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..T.CTAGA.TCTAG......GGT...T.T...A...CC-GA..A.G.C...T...T....T.....T-...CAAT..A.
h 45 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGA.TCTAGA.TCTAG.A.CC.TAAG.TA.C.TCTAGA.TCTAG..GTCGA..TTTTA..GTT.TTCC..AAT.GG.A...T.A.T....T..TT-.-A.A.A.TGCAC
h 46 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.A.TCTTA.GGTA.C.TCTAGA.TCTAG..GTC.A..TTTTA...TTATTCC.GAAT.GG.A...T.A.T.T..T..TT-..A.A.A.TGCAC

59
h 47 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TAAG..A.C.TCTAGA.TCTAG..GTCGA..TTTTA..GTTATTCC..AAT.GG.A...T.A.T....T..TT-..A.A.A.TGCAC
h 48 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.A.TC.TA.G..A.C.TCTAGACTCTAG..GTCGA..TTTTA...TTATTCC.GAAT.GG.A...T.A.T.T..T..TT-..A.A.A.TGCAC
h 49 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A.C.TCTAGA.TCTAG..GTCGA..TTTTA..GTT.TTCC..AAT.GG.A...T.A.T....T..TT-..A.A.A.TGCAC
h 50 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A.C.TCTAGA.TCTAG..GTCGA..TTTTA...TTATTCC.GAAT.GG.A...T.A......T..TT-.-A.A.G.TGCAC
h 51 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A.C.TCTAGA.TCTAG..GTCGA..TTTTA...TT.TTCC.GAAT.GG.A...T.A......T..TT-.-A.A.A.TGCAC
h 52 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTCTTAAG.TA.C.TCTAGA.TCTAG..GTCGA..TTTTA..GTTATTCC..AAT.GG.A...T.A......T..TT-.-.AA.A.TGCAC
h 53 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TAAG.TA.C.TCTAGA.TCTAG..GTCGA..TTTTA...TT.TTCC.GAAT.GG.A...T.A.T.T..T..TT-.-A.A.G.TGCAC
h 54 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A.C.TCTAGA.TCTAG..GTCGA..TTTTA...TTATTCC.GAAT.GG.A...T.A.T....T..T.-.-A.A.A.TGCAC
h 55 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A.C.TCTAGA.TCTAG..GTCGA..TTTTA...TTATTCC.GAAT.GG.A...T.A......T..TT-.-.AA.A.TGCAC
h 56 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TAAG.TA.C.TCTAGA.TCTAG..GTCGA..TTTTA..GTT.TTCC..AAT.GG.A...T.A.T....T..TT..-AAA.A.TGCAC
h 57 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A.C.TCTAGA.TCTAG..GTCGA..TTTTA..GTT.TTCC..AAT.GG.A...T.A......T..TT-T-.AA.A.TGCAC
h 58 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.A.TC.TA.G..A.C.TCTAGA.TCTAG..GTCGA..TTTTA..GTTATTCC..AAT.GG.A...T.A.T....T..TT-.-.AA.A.TGCAC
h 59 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A.C.TCTAGACTCTAG..GTCGA..TTTTA..GTTATTCC..AAT.GG.A...T.A.T....T..TT-.-.AA.A.TGCAC
h 60 ACTCTA.GACTCTAGACTCTAGACTCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..T.CTAGA.TCTAG......G.T...T.T...A...CC.GA...GG.....TA..T....T..T...-A.A.A.T..AC
h 61 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A..C.CTAGA.TCTAG......G.T...T.T...A...CC.GA..A.G.....TA..T..A.T.CT..T-...CAAT..A.
h 62 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..C.CTAGA.TCTAG......G.T...T.T...A...CC.GA..A.G.....TA..T..A.T.CT..T-...CAAT..A.
h 63 ACTCTA.GACTCTAGACTCTAGACTCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..T.CTAGA.TCTAG......G..T..T.T...TATTCC.GAAT..G.A...TA..T....T.CT...-A.A.A.T..AC
h 64 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.ACTC.TA.G..A..C.CTAGA.TCTAG.G....G.T...T.T...A...CC.GA..A.G.C...TA..T..A.T.CT..T-...CAAT..A.
h 65 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.ACTC.TA....A..C.CTAGA.TCTAG.G....G.T...T.T...A...CC-GA..A.G.C.C.T-..T..A.T.CT..T-...CAAT..A.
h 66 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGACTCTAGGACTC.TA.G..A.CT.CTAGA.TCTAG......G.T...T.T...A...CC..A..A.G-.C.CT...T..A.T.CT..T-...CAAT..A.
h 67 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A.CT.CTAGACTCTAG.G....G.T...T.....A.TTCC.GAAT.GG.A...TA..T....T..T...-A.A.A.T..AC
h 68 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA..G.A..C.CTAGA.TCTAG......G.T...T.T...A...CC.GA..A.G.C.C.T...T..A.T.CT..T-...CAAT..A.
h 69 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.A.TC.TA.G..A..T.CTAGA.TCTAG......G.T.....T...A...CC-GA.T.GG.....TA..T.......T...-A.A.A.T..AC
h 70 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.GG.A..C.CTAGA.TCTA.......G.T...T.T...A...CC.GA.T.GG.....TA..T....T..T..T-A.A.A.T..AC
h 71 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A.CT.CTAGA.TCTAG......G.T...T.T...A...CC.GA.T.GG.A...TA..T.......T...-A.A.A.T..AC

60
h 72 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A.CT.CTAGA.TCTAG......G.T...T.T...A...CCAGA..A.G.C...T...T..A.T.CT..T-...CAAT..A.
h 73 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A.CT.CTAGA.TCTAG......G.T...T.T...A...CC-GA..A.G.C...TA..T..A.T.CT..T-...CAAT..A.
h 74 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.A.TC.TA.GG.A..T.CTAGA.TCTAG......G.T...T..T..A...CC-GA.T.GG.....TA..T....T..T...-A...A.T..A.
h 75 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A..T.CTAGA.TCTAG......G.T...T.T...A...CC.GA..A.G.C.C.TA..T....T..T...-....A.T..AC
h 76 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA..G.AC.C.CTAGA.TCTAG......G.T...T.T...A...CC.GA..A.G.CC..TA..T..A.T.CT..T-...CAAT..A.
h 77 ACTCTA..ACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.A.TC.TA.G..A..T.CTAGA.TCTAG......G.T...T.T...A...CC.GA.T.GG.A...TA..T....T..T...-A...A.T..AC
h 78 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGGACTC.TA.G..A..T.CTAGA.TCTAG......G.T...T.T...A...CC-GA..A.G.C.C.T-..T..A.T.CT..T-...CAAT..A.
h 79 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGA.TCTAG.A.TC.TA.G..A..T.CTAGA..CTAG......G.T...TAT..TA...CC.GAA..GG.A...T.A.T....T..TT....AA.A.T..A.
h 80 -....-A.A.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..T.CTAGA.T.TA..G....G.T...T.T...A...CC.GA..A.G.C...T...T.TAGT.CT..T-...CAAT..A.
h 81 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A.CT.CTAGACTCTAG......G.T...T.T...A...CC.GA..A.G.C...T-..T.TAGT.CT..T-...CAAT..A.
h 82 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..C.CTAGA.CCTAGC.....G.T...T.T...A...CC.GA..A.G.C...T...T.TA.T.CT..T-...CAAT..A.
h 83 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..T.CTAGA.CCTAGCG....G.T...T.T...A...CC.GA..A.G.CC..TA..T..A.T.CT..T-...CAAT..A.
h 84 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..T.CTAGA.CCTAGC.....G.T...T.T...A...CC.GA..A.G.C.C.T-..T..A.T.CT..T-...CAAT..A.
h 85 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTC.TA.G..A..T.CTAGA.TCTAG......G.T...T.....A...CC-GA..A.G.CC..TA..T..A.T.CT..T-...CAAT..A.
h 86 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..T.CTAGA.CCTAGCG....G.T...T.T...A...CC.GA..A.G.C...TA..T..A.T.CT..T-...CAAT..A.
h 87 -.T..AA.A.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.GG.A..C..TAGA.CCTAG.G....G.T...T.T...A...CC.GA..A.G.CC..TA..T.TAGT..T..T-...CAAT..A.
h 88 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA..CTAGA..CTAGA..CTAG.A.TC.TA....A..T.CTAGA.CCTAGCG....G.T...T.T...A...CC.GA..A.G.C...T...T.TAGT.CT..T-...CAAT..A.
h 89 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.GG.A..C.CTAGA.CCTAG.G....G.T...T.T...A...CC-GA..A.G.C.C.T-..T..A.T.CT..T-...CAAT..A.
h 90 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGA.TCTAGA.TCTAG.A.TC.TA....A..T.CTAGA.CCTAGCG....G.T...T.T.......CC.GA....G.....TA..T..A.T..T..T-....AAT..AC
h 91 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..-..C.CTAGA.CCTAGC.....G.T...T.T...A...CC.GA..A.G.C...T...T.TAGT.CT..T-...CAAT..A.
h 92 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA..G.A..C.CTA.A.CCTAGC.....G.T...T.T...A...CC.GA..A.G.C.C.T-..T..A.T.CT..T-...CAAT..A.
h 93 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..C.CTAGA.CCTA.C.....G.T...T.T...A...CC.GA..A.G..C..T...T.TAGT.CT..T-...CAAT..A.
h 94 -....AA.A.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.G..A..T.CTAGA.TCTAG.G....G.T...T.T...A...CC..A..A.G.CC..T-..T..A.T..T..T-...CAAT..A.
h 95 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.GG.A..C.CTAGA.CCTAGC.....G.T...T.....A...CC-GA..A.G.CC..TA..T..A.T.CT..T-...CAAT..A.
h 96 A.TCTA.GA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAGA.TCTAG.A.TC.TA.GG.A..C.CTAGA.TCTAGCG....G.T...T.T.......CC.GA..A.G.C...T...T.TAGT.CT..T-...CAAT..A.

61
h 97 ACTCTA.GACTCTAGACTCTAGACTCTAGACTCTAGACTCTAGACTCTAG.ACTCCTA.G..A.CT.CTAGACTCTAGC.....G.T...T.....A...C..GA..A.G.....TA..T..A.T..T..T-....AAT..AC

62
Anexo 02 Freqüência dos haplótipos de N. eques. AÇ= Açaituba; MI= Miuá; JA=Jaradi;
AR=Arixanã; DE=Demeni; MA=Maguari; JC=Jacundá e CT=Catalão.
Alto rio Negro Médio rio Negro Baixo rio Negro Total Haplótipos AÇ MI JA AR DE MA JC CT
h1 1 - - - - - - - 1 h2 2 - - - - - - - 2 h3 1 - - - - - - - 1 h4 2 - - - - - - - 2 h5 3 - - - - - - - 3 h6 - 1 - - - - - - 1 h7 - 1 - - - - - - 1 h8 - 1 - - - - - - 1 h9 - 1 - - - - - - 1 h10 - 1 - - - - - - 1 h11 - 1 - - - - - - 1 h12 - 1 - - - - - - 1 h13 - 1 - - - - - - 1 h14 - 1 - - - - - - 1 h15 - 1 - - - - - - 1 h16 - 1 - - - - - - 1 h17 - - 1 - - - - - 1 h18 - 2 2 - - - - - 4 h19 - - 1 - - - - - 1 h20 - - 3 - - - - - 3 h21 - - 1 - - - - - 1 h22 - - 1 - - - - - 1 h23 - - 1 - - - - - 1 h24 - 1 - 1 - - - - 2 h25 - - - 1 - - - - 1 h26 - - - 1 - - - - 1 h27 - - - 1 - - - - 1 h28 6 1 1 1 - - - - 9 h29 - 1 1 3 - - - - 5 h30 - 3 1 1 - - - - 5 h31 - - - 1 - - - - 1 h32 - 1 - 1 - - - - 2 h33 - - - 1 - - - - 1 h34 - - - 1 - - - - 1 h35 - - - 1 - - - - 1 h36 - - - - - - 1 - 1 h37 - - - - - - 1 - 1 h38 - - - - - - 1 - 1 h39 - - - - - - 1 - 1 h40 - - 1 - - - 1 - 2 h41 - - - - - - 1 - 1 h42 - - - - - - 1 - 1 h43 - - - - - - 1 - 1 h44 - - - - - - 1 - 1 h45 - - - - 1 - - - 1 h46 - - - - 1 - - - 1 h47 - - - - 1 - - - 1 h48 - - - - 1 - - - 1

63
h49 - - - - 1 - - - 1 h50 - - - - 1 - - - 1 h51 - - - - 1 - - - 1 h52 - - - - 1 - - - 1 h53 - - - - 1 - - - 1 h54 - - - - 1 - - - 1 h55 - - - - 1 - - - 1 h56 - - - - 1 - - - 1 h57 - - - - 1 - - - 1 h58 - - - - 1 - - - 1 h59 - - - - 1 - - - 1 h60 - - - - - 1 - - 1 h61 - - - - - 1 - - 1 h62 - - - - - 1 - - 1 h63 - - - - - 1 - - 1 h64 - - - - - 1 - - 1 h65 - - - - - 1 - - 1 h66 - - - - - 1 - - 1 h67 - - - - - 1 - - 1 h68 - - - - - 1 - - 1 h69 - - - - - 1 - - 1 h70 - - - - - 1 - - 1 h71 - - - - - 1 - - 1 h72 - - - - - 1 - - 1 h73 - - - - - 1 - - 1 h74 - - - - - 1 - - 1 h75 - - - - - 1 - - 1 h76 - - - - - 1 - - 1 h77 - - - - - 1 - - 1 h78 - - - - - 1 - - 1 h79 - - - - - 1 - - 1 h80 - - - - - - - 1 1 h81 - - - - - - - 1 1 h82 - - - - - - - 1 1 h83 - - - - - - - 1 1 h84 - - - - - - - 1 1 h85 - - - - - - - 1 1 h86 - - - - - - - 1 1 h87 - - - - - - - 1 1 h88 - - - - - - - 1 1 h89 - - - - - - - 1 1 h90 - - - - - - - 1 1 h91 - - - - - - - 1 1 h92 - - - - - - - 1 1 h93 - - - - - - - 1 1 h94 - - - - - - - 1 1 h95 - - - - - - - 1 1 h96 - - - - - - - 1 1 h97 - - - - - - - 1 1
Total 15 20 14 14 15 20 9 18 125

64
Anexo 03 Distância genética entre os haplótipos (em porcentagem) obtida pela distância p não corrigida. Os haplótipos h 1 até h 41 correspondem à unidade
evolutiva 1 (com exceção do h 36 que pertence à unidade evolutiva 2). Haplótipos h 42 à h 97 pertencem à unidade evolutiva 2. Nu – N. unifasciatus
(Grupo externo).
h1 h2 h3 h4 h5 h6 h7 h8 h9 h10 h11 h12 h13 h14 h15 h16 h17 h18
h1 0,00% h2 0,10% h3 0,20% 0,30% h4 0,10% 0,20% 0,30% h5 0,10% 0,00% 0,30% 0,20% h6 0,40% 0,50% 0,20% 0,30% 0,50% h7 0,20% 0,10% 0,20% 0,30% 0,10% 0,40% h8 0,10% 0,20% 0,10% 0,20% 0,20% 0,30% 0,10% h9 0,50% 0,40% 0,30% 0,40% 0,40% 0,10% 0,30% 0,40% h10 0,20% 0,30% 0,00% 0,30% 0,30% 0,20% 0,20% 0,10% 0,30% h11 0,20% 0,30% 0,20% 0,30% 0,30% 0,20% 0,20% 0,10% 0,30% 0,20% h12 0,40% 0,30% 0,20% 0,50% 0,30% 0,20% 0,20% 0,30% 0,10% 0,20% 0,20% h13 0,30% 0,20% 0,30% 0,40% 0,20% 0,30% 0,10% 0,20% 0,20% 0,30% 0,10% 0,10% h14 0,10% 0,20% 0,10% 0,20% 0,20% 0,30% 0,10% 0,00% 0,40% 0,10% 0,10% 0,30% 0,20% h15 0,10% 0,20% 0,10% 0,20% 0,20% 0,30% 0,10% 0,00% 0,40% 0,10% 0,10% 0,30% 0,20% 0,00% h16 0,10% 0,20% 0,10% 0,20% 0,20% 0,30% 0,10% 0,00% 0,40% 0,10% 0,10% 0,30% 0,20% 0,00% 0,00% h17 0,10% 0,20% 0,10% 0,20% 0,20% 0,30% 0,10% 0,00% 0,40% 0,10% 0,10% 0,30% 0,20% 0,00% 0,00% 0,00% h18 0,20% 0,30% 0,20% 0,10% 0,30% 0,20% 0,20% 0,10% 0,30% 0,20% 0,20% 0,40% 0,30% 0,10% 0,10% 0,10% 0,10% h19 0,10% 0,20% 0,10% 0,20% 0,20% 0,30% 0,10% 0,00% 0,40% 0,10% 0,10% 0,30% 0,20% 0,00% 0,00% 0,00% 0,00% 0,10% h20 0,10% 0,00% 0,30% 0,20% 0,00% 0,50% 0,10% 0,20% 0,40% 0,30% 0,30% 0,30% 0,20% 0,20% 0,20% 0,20% 0,20% 0,30% h21 0,10% 0,20% 0,10% 0,20% 0,20% 0,30% 0,10% 0,00% 0,40% 0,10% 0,10% 0,30% 0,20% 0,00% 0,00% 0,00% 0,00% 0,10% h22 0,20% 0,30% 0,20% 0,30% 0,30% 0,20% 0,20% 0,10% 0,30% 0,20% 0,00% 0,20% 0,10% 0,10% 0,10% 0,10% 0,10% 0,20%

65
h1 h2 h3 h4 h5 h6 h7 h8 h9 h10 h11 h12 h13 h14 h15 h16 h17 h18
h23 0,20% 0,30% 0,00% 0,30% 0,30% 0,20% 0,20% 0,10% 0,30% 0,00% 0,20% 0,20% 0,30% 0,10% 0,10% 0,10% 0,10% 0,20% h24 0,20% 0,30% 0,20% 0,30% 0,30% 0,40% 0,20% 0,10% 0,50% 0,20% 0,20% 0,40% 0,30% 0,10% 0,10% 0,10% 0,10% 0,20% h25 0,20% 0,30% 0,20% 0,30% 0,30% 0,20% 0,20% 0,10% 0,30% 0,20% 0,00% 0,20% 0,10% 0,10% 0,10% 0,10% 0,10% 0,20% h26 0,40% 0,50% 0,20% 0,50% 0,50% 0,20% 0,40% 0,30% 0,30% 0,20% 0,20% 0,20% 0,30% 0,30% 0,30% 0,30% 0,30% 0,40% h27 0,10% 0,20% 0,30% 0,20% 0,20% 0,50% 0,30% 0,20% 0,60% 0,30% 0,30% 0,50% 0,40% 0,20% 0,20% 0,20% 0,20% 0,30% h28 0,10% 0,20% 0,30% 0,00% 0,20% 0,30% 0,30% 0,20% 0,40% 0,30% 0,30% 0,50% 0,40% 0,20% 0,20% 0,20% 0,20% 0,10% h29 0,00% 0,10% 0,20% 0,10% 0,10% 0,40% 0,20% 0,10% 0,50% 0,20% 0,20% 0,40% 0,30% 0,10% 0,10% 0,10% 0,10% 0,20% h30 0,20% 0,30% 0,20% 0,10% 0,30% 0,20% 0,20% 0,10% 0,30% 0,20% 0,20% 0,40% 0,30% 0,10% 0,10% 0,10% 0,10% 0,00% h31 0,50% 0,60% 0,30% 0,60% 0,60% 0,30% 0,50% 0,40% 0,40% 0,30% 0,30% 0,30% 0,40% 0,40% 0,40% 0,40% 0,40% 0,50% h32 0,30% 0,40% 0,10% 0,40% 0,40% 0,10% 0,30% 0,20% 0,20% 0,10% 0,10% 0,10% 0,20% 0,20% 0,20% 0,20% 0,20% 0,30% h33 0,30% 0,40% 0,30% 0,40% 0,40% 0,30% 0,30% 0,20% 0,40% 0,30% 0,10% 0,30% 0,20% 0,20% 0,20% 0,20% 0,20% 0,30% h34 0,20% 0,30% 0,20% 0,30% 0,30% 0,20% 0,20% 0,10% 0,30% 0,20% 0,00% 0,20% 0,10% 0,10% 0,10% 0,10% 0,10% 0,20% h35 0,40% 0,50% 0,40% 0,50% 0,50% 0,40% 0,40% 0,30% 0,50% 0,40% 0,20% 0,40% 0,30% 0,30% 0,30% 0,30% 0,30% 0,40% h36 5,60% 5,50% 5,50% 5,70% 5,50% 5,40% 5,40% 5,60% 5,40% 5,40% 5,50% 5,30% 5,40% 5,60% 5,60% 5,50% 5,60% 5,60% h37 0,90% 1,00% 1,10% 1,00% 1,00% 1,30% 1,10% 1,00% 1,40% 1,10% 1,10% 1,30% 1,20% 1,00% 1,00% 1,00% 1,00% 1,10% h38 0,30% 0,40% 0,50% 0,20% 0,40% 0,50% 0,50% 0,40% 0,60% 0,50% 0,50% 0,70% 0,60% 0,40% 0,40% 0,40% 0,40% 0,30% h39 0,20% 0,30% 0,40% 0,30% 0,30% 0,60% 0,40% 0,30% 0,70% 0,40% 0,40% 0,60% 0,50% 0,30% 0,30% 0,30% 0,30% 0,40% h40 0,00% 0,10% 0,20% 0,10% 0,10% 0,40% 0,20% 0,10% 0,50% 0,20% 0,20% 0,40% 0,30% 0,10% 0,10% 0,10% 0,10% 0,20% h41 1,50% 1,40% 1,70% 1,60% 1,40% 1,90% 1,50% 1,60% 1,80% 1,70% 1,70% 1,70% 1,60% 1,60% 1,60% 1,60% 1,60% 1,70% h42 5,50% 5,40% 5,40% 5,60% 5,50% 5,40% 5,40% 5,50% 5,30% 5,40% 5,40% 5,20% 5,30% 5,50% 5,50% 5,50% 5,50% 5,50% h43 5,90% 6,00% 5,70% 5,80% 6,00% 5,50% 5,90% 5,80% 5,60% 5,70% 5,70% 5,70% 5,80% 5,80% 5,80% 5,80% 5,80% 5,70% h44 5,60% 5,50% 5,50% 5,70% 5,60% 5,50% 5,50% 5,60% 5,40% 5,50% 5,50% 5,30% 5,40% 5,60% 5,60% 5,60% 5,60% 5,60% h45 7,80% 7,90% 7,70% 7,70% 7,90% 7,70% 7,80% 7,70% 7,80% 7,70% 7,80% 7,90% 7,90% 7,70% 7,70% 7,70% 7,70% 7,60% h46 8,20% 8,10% 8,00% 8,10% 8,10% 8,00% 8,00% 8,10% 7,90% 8,00% 8,20% 8,00% 8,10% 8,10% 8,10% 8,10% 8,10% 8,00% h47 8,10% 8,20% 7,90% 8,00% 8,20% 7,90% 8,10% 8,00% 8,00% 7,90% 8,10% 8,10% 8,20% 8,00% 8,00% 8,00% 8,00% 7,90% h48 8,10% 8,00% 7,90% 8,00% 8,00% 7,90% 7,90% 8,00% 7,80% 7,90% 8,10% 7,90% 8,00% 8,00% 8,00% 8,00% 8,00% 7,90% h49 7,90% 8,00% 7,70% 7,80% 8,00% 7,70% 7,90% 7,80% 7,80% 7,70% 7,90% 7,90% 8,00% 7,80% 7,80% 7,80% 7,80% 7,70% h50 7,90% 7,80% 7,70% 7,80% 7,80% 7,70% 7,70% 7,80% 7,60% 7,70% 7,90% 7,70% 7,80% 7,80% 7,80% 7,80% 7,80% 7,70%

66
h1 h2 h3 h4 h5 h6 h7 h8 h9 h10 h11 h12 h13 h14 h15 h16 h17 h18
h51 7,80% 7,70% 7,60% 7,70% 7,70% 7,60% 7,60% 7,70% 7,50% 7,60% 7,80% 7,60% 7,70% 7,70% 7,70% 7,70% 7,70% 7,60% h52 8,20% 8,30% 8,00% 8,30% 8,30% 8,20% 8,20% 8,10% 8,30% 8,00% 8,20% 8,20% 8,30% 8,10% 8,10% 8,10% 8,10% 8,20% h53 8,20% 8,10% 8,00% 8,10% 8,10% 8,00% 8,00% 8,10% 7,90% 8,00% 8,20% 8,00% 8,10% 8,10% 8,10% 8,10% 8,10% 8,00% h54 7,90% 7,80% 7,70% 7,80% 7,80% 7,70% 7,70% 7,80% 7,60% 7,70% 7,90% 7,70% 7,80% 7,80% 7,80% 7,80% 7,80% 7,70% h55 7,90% 7,80% 7,70% 8,00% 7,80% 7,90% 7,70% 7,80% 7,80% 7,70% 7,90% 7,70% 7,80% 7,80% 7,80% 7,80% 7,80% 7,90% h56 8,20% 8,30% 8,00% 8,10% 8,30% 8,00% 8,20% 8,10% 8,10% 8,00% 8,20% 8,20% 8,30% 8,10% 8,10% 8,10% 8,10% 8,00% h57 7,90% 8,00% 7,70% 8,00% 8,00% 7,90% 7,90% 7,80% 8,00% 7,70% 7,90% 7,90% 8,00% 7,80% 7,80% 7,80% 7,80% 7,90% h58 7,90% 8,00% 7,70% 8,00% 8,00% 7,90% 7,90% 7,80% 8,00% 7,70% 7,90% 7,90% 8,00% 7,80% 7,80% 7,80% 7,80% 7,90% h59 8,10% 8,20% 7,90% 8,20% 8,20% 8,10% 8,10% 8,00% 8,20% 7,90% 8,10% 8,10% 8,20% 8,00% 8,00% 8,00% 8,00% 8,10% h60 6,00% 5,90% 5,80% 5,90% 5,90% 5,60% 5,80% 5,90% 5,50% 5,80% 5,80% 5,60% 5,70% 5,90% 5,90% 5,90% 5,90% 5,80% h61 6,60% 6,50% 6,40% 6,70% 6,50% 6,50% 6,40% 6,50% 6,40% 6,40% 6,50% 6,30% 6,40% 6,50% 6,50% 6,50% 6,50% 6,60% h62 5,80% 5,70% 5,60% 5,90% 5,70% 5,70% 5,60% 5,70% 5,60% 5,60% 5,70% 5,50% 5,60% 5,70% 5,70% 5,70% 5,70% 5,80% h63 6,60% 6,50% 6,40% 6,50% 6,50% 6,20% 6,40% 6,50% 6,10% 6,40% 6,40% 6,20% 6,30% 6,50% 6,50% 6,50% 6,50% 6,40% h64 6,10% 6,00% 5,90% 6,20% 6,00% 6,00% 5,90% 6,00% 5,90% 5,90% 6,00% 5,80% 5,90% 6,00% 6,00% 6,00% 6,00% 6,10% h65 6,00% 5,90% 5,80% 6,10% 5,90% 5,90% 5,80% 5,90% 5,80% 5,80% 5,90% 5,70% 5,80% 5,90% 5,90% 5,90% 5,90% 6,00% h66 6,20% 6,30% 6,00% 6,30% 6,30% 6,00% 6,20% 6,10% 6,10% 6,00% 6,00% 6,00% 6,10% 6,10% 6,10% 6,10% 6,10% 6,20% h67 7,00% 6,90% 6,90% 6,90% 7,00% 6,70% 6,90% 7,00% 6,60% 6,90% 6,90% 6,70% 6,80% 7,00% 7,00% 7,00% 7,00% 6,90% h68 5,90% 5,80% 5,70% 6,00% 5,80% 5,80% 5,70% 5,80% 5,70% 5,70% 5,80% 5,60% 5,70% 5,80% 5,80% 5,80% 5,80% 5,90% h69 6,20% 6,10% 6,00% 6,10% 6,10% 5,80% 6,00% 6,10% 5,70% 6,00% 6,00% 5,80% 5,90% 6,10% 6,10% 6,10% 6,10% 6,00% h70 5,80% 5,70% 5,60% 5,70% 5,70% 5,50% 5,60% 5,70% 5,40% 5,60% 5,70% 5,50% 5,60% 5,70% 5,70% 5,70% 5,70% 5,60% h71 6,60% 6,50% 6,40% 6,50% 6,50% 6,20% 6,40% 6,50% 6,10% 6,40% 6,40% 6,20% 6,30% 6,50% 6,50% 6,50% 6,50% 6,40% h72 6,00% 5,90% 5,80% 6,10% 5,90% 5,80% 5,80% 5,90% 5,70% 5,80% 5,80% 5,60% 5,70% 5,90% 5,90% 5,90% 5,90% 5,90% h73 6,00% 5,90% 5,80% 6,10% 5,90% 5,80% 5,80% 5,90% 5,70% 5,80% 5,80% 5,60% 5,70% 5,90% 5,90% 5,90% 5,90% 6,00% h74 6,30% 6,20% 6,10% 6,20% 6,20% 5,90% 6,10% 6,20% 5,80% 6,10% 6,10% 5,90% 6,00% 6,20% 6,20% 6,20% 6,20% 6,10% h75 6,40% 6,30% 6,20% 6,50% 6,30% 6,20% 6,20% 6,30% 6,10% 6,20% 6,20% 6,00% 6,10% 6,30% 6,30% 6,30% 6,30% 6,40% h76 6,10% 6,00% 5,90% 6,20% 6,00% 6,00% 5,90% 6,00% 5,90% 5,90% 6,00% 5,80% 5,90% 6,00% 6,00% 6,00% 6,00% 6,10% h77 6,30% 6,20% 6,10% 6,20% 6,20% 5,90% 6,10% 6,20% 5,80% 6,10% 6,10% 5,90% 6,00% 6,20% 6,20% 6,20% 6,20% 6,10% h78 6,10% 6,00% 5,90% 6,20% 6,00% 5,90% 5,90% 6,00% 5,80% 5,90% 5,90% 5,70% 5,80% 6,00% 6,00% 6,00% 6,00% 6,10%

67
h1 h2 h3 h4 h5 h6 h7 h8 h9 h10 h11 h12 h13 h14 h15 h16 h17 h18
h79 6,50% 6,40% 6,50% 6,60% 6,40% 6,50% 6,50% 6,60% 6,40% 6,50% 6,50% 6,30% 6,40% 6,60% 6,60% 6,60% 6,60% 6,70% h80 5,60% 5,50% 5,50% 5,70% 5,50% 5,40% 5,40% 5,60% 5,40% 5,40% 5,50% 5,30% 5,40% 5,60% 5,60% 5,50% 5,60% 5,60% h81 7,00% 6,90% 6,80% 7,00% 6,90% 6,80% 6,80% 6,90% 6,70% 6,80% 6,80% 6,60% 6,70% 6,90% 6,90% 6,90% 6,90% 7,00% h82 6,00% 5,90% 5,90% 6,10% 5,90% 6,00% 5,90% 6,00% 5,90% 5,90% 6,00% 5,80% 5,90% 6,00% 6,00% 6,00% 6,00% 6,10% h83 6,20% 6,10% 6,10% 6,30% 6,10% 6,10% 6,10% 6,20% 6,00% 6,10% 6,10% 5,90% 6,00% 6,20% 6,20% 6,20% 6,20% 6,30% h84 6,00% 5,90% 5,90% 6,10% 5,90% 5,90% 5,90% 6,00% 5,80% 5,90% 5,90% 5,70% 5,80% 6,00% 6,00% 6,00% 6,00% 6,10% h85 6,70% 6,60% 6,50% 6,80% 6,60% 6,50% 6,50% 6,60% 6,40% 6,50% 6,50% 6,30% 6,40% 6,60% 6,60% 6,60% 6,60% 6,70% h86 6,10% 6,00% 6,00% 6,20% 6,00% 6,00% 6,00% 6,10% 5,90% 6,00% 6,00% 5,80% 5,90% 6,10% 6,10% 6,10% 6,10% 6,20% h87 6,00% 5,90% 5,90% 6,10% 5,90% 6,00% 5,90% 6,00% 5,90% 5,90% 6,00% 5,80% 5,90% 6,00% 6,00% 6,00% 6,00% 6,10% h88 5,80% 5,70% 5,70% 5,90% 5,70% 5,70% 5,70% 5,80% 5,60% 5,70% 5,70% 5,50% 5,60% 5,80% 5,80% 5,80% 5,80% 5,90% h89 6,10% 6,00% 6,00% 6,20% 6,00% 6,10% 6,00% 6,10% 6,00% 6,00% 6,10% 5,90% 6,00% 6,10% 6,10% 6,10% 6,10% 6,20% h90 6,10% 6,00% 6,00% 6,20% 6,00% 6,00% 6,00% 6,10% 5,90% 6,00% 6,00% 5,80% 5,90% 6,10% 6,10% 6,10% 6,10% 6,20% h91 6,10% 6,00% 6,00% 6,20% 6,00% 6,10% 6,00% 6,10% 6,00% 6,00% 6,10% 5,90% 6,00% 6,10% 6,10% 6,10% 6,10% 6,20% h92 5,90% 5,80% 5,80% 6,00% 5,80% 5,90% 5,80% 5,90% 5,80% 5,80% 5,90% 5,70% 5,80% 5,90% 5,90% 5,90% 5,90% 6,00% h93 6,00% 5,90% 5,90% 6,10% 5,90% 6,00% 5,90% 6,00% 5,90% 5,90% 6,00% 5,80% 5,90% 6,00% 6,00% 6,00% 6,00% 6,10% h94 5,60% 5,60% 5,40% 5,60% 5,60% 5,40% 5,50% 5,50% 5,50% 5,40% 5,40% 5,40% 5,50% 5,50% 5,50% 5,50% 5,50% 5,60% h95 6,10% 6,00% 6,00% 6,20% 6,00% 6,10% 6,00% 6,10% 6,00% 6,00% 6,10% 5,90% 6,00% 6,10% 6,10% 6,10% 6,10% 6,20% h96 6,20% 6,10% 6,00% 6,30% 6,10% 6,10% 6,00% 6,10% 6,00% 6,00% 6,10% 5,90% 6,00% 6,10% 6,10% 6,10% 6,10% 6,20% h97 6,70% 6,60% 6,50% 6,80% 6,60% 6,50% 6,50% 6,60% 6,40% 6,50% 6,50% 6,30% 6,40% 6,60% 6,60% 6,60% 6,60% 6,70% Nu 10,80% 10,90% 10,60% 10,70% 10,90% 10,40% 10,80% 10,70% 10,50% 10,60% 10,60% 10,60% 10,70% 10,70% 10,70% 10,70% 10,70% 10,60%

68
h19 h20 h21 h22 h23 h24 h25 h26 h27 h28 h29 h30 h31 h32 h33 h34 h35 h36
h1 h2 h3 h4 h5 h6 h7 h8 h9
h10 h11 h12 h13 h14 h15 h16 h17 h18 h19 h20 h21 h22 0,20% h23 0,00% 0,20% h24 0,10% 0,30% 0,10% h25 0,10% 0,30% 0,10% 0,20% h26 0,10% 0,30% 0,10% 0,20% 0,20% h27 0,10% 0,30% 0,10% 0,00% 0,20% 0,20% h28 0,30% 0,50% 0,30% 0,20% 0,20% 0,40% 0,20%

69
h19 h20 h21 h22 h23 h24 h25 h26 h27 h28 h29 h30 h31 h32 h33 h34 h35 h36 h29 0,20% 0,20% 0,20% 0,30% 0,30% 0,30% 0,30% 0,50% h30 0,20% 0,20% 0,20% 0,30% 0,30% 0,30% 0,30% 0,50% 0,20% h31 0,10% 0,10% 0,10% 0,20% 0,20% 0,20% 0,20% 0,40% 0,10% 0,10% h32 0,10% 0,30% 0,10% 0,20% 0,20% 0,20% 0,20% 0,40% 0,30% 0,10% 0,20% h33 0,40% 0,60% 0,40% 0,30% 0,30% 0,50% 0,30% 0,30% 0,40% 0,60% 0,50% 0,50% h34 0,20% 0,40% 0,20% 0,10% 0,10% 0,30% 0,10% 0,10% 0,40% 0,40% 0,30% 0,30% 0,20% h35 0,20% 0,40% 0,20% 0,10% 0,30% 0,30% 0,10% 0,10% 0,40% 0,40% 0,30% 0,30% 0,40% 0,20% h36 0,10% 0,30% 0,10% 0,00% 0,20% 0,20% 0,00% 0,20% 0,30% 0,30% 0,20% 0,20% 0,30% 0,10% 0,10% h37 0,30% 0,50% 0,30% 0,20% 0,40% 0,40% 0,20% 0,20% 0,30% 0,50% 0,40% 0,40% 0,30% 0,30% 0,10% 0,20% h38 5,50% 5,50% 5,50% 5,50% 5,50% 5,40% 5,40% 5,30% 5,50% 5,70% 5,60% 5,60% 5,40% 5,40% 5,40% 5,50% 5,30% h39 1,00% 1,00% 1,00% 1,10% 1,10% 1,10% 1,10% 1,30% 1,00% 1,00% 0,90% 1,10% 1,40% 1,20% 1,20% 1,10% 1,30% 4,70% h40 0,40% 0,40% 0,40% 0,50% 0,50% 0,50% 0,50% 0,70% 0,40% 0,20% 0,30% 0,30% 0,80% 0,60% 0,60% 0,50% 0,70% 5,50% h41 0,30% 0,30% 0,30% 0,40% 0,40% 0,40% 0,40% 0,60% 0,30% 0,30% 0,20% 0,40% 0,70% 0,50% 0,50% 0,40% 0,60% 5,50% h42 0,10% 0,10% 0,10% 0,20% 0,20% 0,20% 0,20% 0,40% 0,10% 0,10% 0,00% 0,20% 0,50% 0,30% 0,30% 0,20% 0,40% 5,60% h43 1,60% 1,40% 1,60% 1,70% 1,70% 1,70% 1,70% 1,90% 1,60% 1,60% 1,50% 1,70% 2,00% 1,80% 1,80% 1,70% 1,90% 4,10% h44 5,40% 5,50% 5,50% 5,40% 5,40% 5,40% 5,40% 5,20% 5,40% 5,60% 5,50% 5,50% 5,30% 5,30% 5,30% 5,40% 5,20% 0,30% h45 5,80% 6,00% 5,80% 5,70% 5,70% 5,70% 5,70% 5,50% 5,80% 5,80% 5,90% 5,70% 5,60% 5,60% 5,60% 5,70% 5,50% 1,70% h46 5,50% 5,50% 5,50% 5,50% 5,50% 5,50% 5,50% 5,30% 5,50% 5,70% 5,60% 5,60% 5,40% 5,40% 5,40% 5,50% 5,30% 0,20% h47 7,70% 7,90% 7,70% 7,80% 7,70% 7,60% 7,80% 7,70% 7,70% 7,70% 7,80% 7,60% 7,80% 7,80% 7,70% 7,80% 7,60% 4,10% h48 8,10% 8,10% 8,10% 8,20% 8,00% 8,00% 8,20% 8,00% 8,10% 8,10% 8,20% 8,00% 7,90% 8,10% 8,10% 8,20% 8,00% 4,20% h49 8,00% 8,20% 8,00% 8,10% 7,90% 7,90% 8,10% 7,90% 8,00% 8,00% 8,10% 7,90% 7,80% 8,00% 8,00% 8,10% 7,90% 4,30% h50 8,00% 8,00% 8,00% 8,10% 7,90% 7,90% 8,10% 7,90% 8,00% 8,00% 8,10% 7,90% 7,80% 8,00% 8,00% 8,10% 7,90% 4,10% h51 7,80% 8,00% 7,80% 7,90% 7,70% 7,70% 7,90% 7,70% 7,80% 7,80% 7,90% 7,70% 7,60% 7,80% 7,80% 7,90% 7,70% 4,10% h52 7,80% 7,80% 7,80% 7,90% 7,70% 7,70% 7,90% 7,70% 7,80% 7,80% 7,90% 7,70% 7,60% 7,80% 7,80% 7,90% 7,70% 4,20% h53 7,70% 7,70% 7,70% 7,80% 7,60% 7,60% 7,80% 7,60% 7,70% 7,70% 7,80% 7,60% 7,50% 7,70% 7,70% 7,80% 7,60% 4,10% h54 8,10% 8,30% 8,10% 8,20% 8,00% 8,00% 8,20% 8,00% 8,10% 8,30% 8,20% 8,20% 7,90% 8,10% 8,10% 8,20% 8,00% 4,60% h55 8,10% 8,10% 8,10% 8,20% 8,00% 8,00% 8,20% 8,00% 8,10% 8,10% 8,20% 8,00% 7,90% 8,10% 8,10% 8,20% 8,00% 4,30% h56 7,80% 7,80% 7,80% 7,90% 7,70% 7,70% 7,90% 7,70% 7,80% 7,80% 7,90% 7,70% 7,60% 7,80% 7,80% 7,90% 7,70% 4,00%

70
h19 h20 h21 h22 h23 h24 h25 h26 h27 h28 h29 h30 h31 h32 h33 h34 h35 h36 h57 7,80% 8,00% 7,80% 7,90% 7,70% 7,70% 7,90% 7,70% 7,80% 8,00% 7,90% 7,90% 7,60% 7,80% 7,80% 7,90% 7,70% 4,10% h58 7,80% 8,00% 7,80% 7,90% 7,70% 7,70% 7,90% 7,70% 7,80% 8,00% 7,90% 7,90% 7,60% 7,80% 7,80% 7,90% 7,70% 4,10% h59 8,00% 8,20% 8,00% 8,10% 7,90% 7,90% 8,10% 7,90% 8,00% 8,20% 8,10% 8,10% 7,80% 8,00% 8,00% 8,10% 7,90% 4,30% h60 5,90% 5,90% 5,90% 5,80% 5,80% 5,80% 5,80% 5,60% 5,90% 5,90% 6,00% 5,80% 5,70% 5,70% 5,70% 5,80% 5,60% 1,50% h61 6,50% 6,50% 6,50% 6,50% 6,40% 6,40% 6,50% 6,30% 6,50% 6,70% 6,60% 6,60% 6,20% 6,40% 6,40% 6,50% 6,30% 1,40% h62 5,70% 5,70% 5,70% 5,70% 5,60% 5,60% 5,70% 5,50% 5,70% 5,90% 5,80% 5,80% 5,60% 5,60% 5,60% 5,70% 5,50% 0,60% h63 6,50% 6,50% 6,50% 6,40% 6,40% 6,40% 6,40% 6,20% 6,50% 6,50% 6,60% 6,40% 6,30% 6,30% 6,30% 6,40% 6,20% 2,20% h64 6,00% 6,00% 6,00% 6,00% 5,90% 5,90% 6,00% 5,80% 6,00% 6,20% 6,10% 6,10% 5,90% 5,90% 5,90% 6,00% 5,80% 0,70% h65 5,90% 5,90% 5,90% 5,90% 5,80% 5,80% 5,90% 5,70% 5,90% 6,10% 6,00% 6,00% 5,80% 5,80% 5,80% 5,90% 5,70% 0,80% h66 6,10% 6,30% 6,10% 6,00% 6,00% 6,00% 6,00% 5,80% 6,10% 6,30% 6,20% 6,20% 5,90% 5,90% 5,90% 6,00% 5,80% 1,10% h67 6,90% 7,00% 7,00% 6,90% 6,90% 6,90% 6,90% 6,70% 6,90% 7,00% 7,00% 6,90% 6,60% 6,80% 6,80% 6,90% 6,70% 2,60% h68 5,80% 5,80% 5,80% 5,80% 5,70% 5,70% 5,80% 5,60% 5,80% 6,00% 5,90% 5,90% 5,70% 5,70% 5,70% 5,80% 5,60% 0,70% h69 6,10% 6,10% 6,10% 6,00% 6,00% 6,00% 6,00% 5,80% 6,10% 6,10% 6,20% 6,00% 5,70% 5,90% 5,90% 6,00% 5,80% 2,00% h70 5,70% 5,70% 5,70% 5,70% 5,60% 5,60% 5,70% 5,50% 5,70% 5,70% 5,80% 5,60% 5,60% 5,60% 5,60% 5,70% 5,50% 1,40% h71 6,50% 6,50% 6,50% 6,40% 6,40% 6,40% 6,40% 6,20% 6,50% 6,50% 6,60% 6,40% 6,10% 6,30% 6,30% 6,40% 6,20% 2,10% h72 5,90% 5,80% 5,80% 5,70% 5,70% 5,80% 5,80% 5,60% 5,90% 6,00% 6,00% 6,00% 5,70% 5,70% 5,70% 5,80% 5,50% 0,60% h73 5,90% 5,90% 5,90% 5,80% 5,80% 5,80% 5,80% 5,60% 5,90% 6,10% 6,00% 6,00% 5,70% 5,70% 5,70% 5,80% 5,60% 0,60% h74 6,20% 6,20% 6,20% 6,10% 6,10% 6,10% 6,10% 5,90% 6,20% 6,20% 6,30% 6,10% 5,80% 6,00% 6,00% 6,10% 5,90% 1,90% h75 6,30% 6,30% 6,30% 6,20% 6,20% 6,20% 6,20% 6,00% 6,30% 6,50% 6,40% 6,40% 5,90% 6,10% 6,10% 6,20% 6,00% 1,50% h76 6,00% 6,00% 6,00% 6,00% 5,90% 5,90% 6,00% 5,80% 6,00% 6,20% 6,10% 6,10% 5,90% 5,90% 5,90% 6,00% 5,80% 0,90% h77 6,20% 6,20% 6,20% 6,10% 6,10% 6,10% 6,10% 5,90% 6,20% 6,20% 6,30% 6,10% 5,80% 6,00% 6,00% 6,10% 5,90% 1,80% h78 6,00% 6,00% 6,00% 5,90% 5,90% 5,90% 5,90% 5,70% 6,00% 6,20% 6,10% 6,10% 5,80% 5,80% 5,80% 5,90% 5,70% 0,60% h79 6,60% 6,40% 6,60% 6,50% 6,50% 6,50% 6,50% 6,30% 6,40% 6,60% 6,50% 6,70% 6,20% 6,40% 6,40% 6,50% 6,30% 2,00% h80 5,50% 5,50% 5,50% 5,50% 5,50% 5,40% 5,40% 5,30% 5,50% 5,70% 5,60% 5,60% 5,40% 5,40% 5,40% 5,50% 5,30% 1,30% h81 6,90% 6,90% 6,90% 6,80% 6,80% 6,80% 6,80% 6,60% 6,90% 7,10% 7,00% 7,00% 6,50% 6,70% 6,70% 6,80% 6,60% 1,50% h82 6,00% 5,90% 6,00% 6,00% 5,90% 5,90% 6,00% 5,80% 5,90% 6,10% 6,00% 6,10% 5,90% 5,90% 5,90% 6,00% 5,80% 0,70% h83 6,20% 6,10% 6,20% 6,10% 6,10% 6,10% 6,10% 5,90% 6,10% 6,30% 6,20% 6,30% 6,00% 6,00% 6,00% 6,10% 5,90% 0,80%

71
h19 h20 h21 h22 h23 h24 h25 h26 h27 h28 h29 h30 h31 h32 h33 h34 h35 h36 h84 6,00% 5,90% 6,00% 5,90% 5,90% 5,90% 5,90% 5,70% 5,90% 6,10% 6,00% 6,10% 5,80% 5,80% 5,80% 5,90% 5,70% 0,60% h85 6,60% 6,60% 6,60% 6,50% 6,50% 6,50% 6,50% 6,30% 6,60% 6,80% 6,70% 6,70% 6,20% 6,40% 6,40% 6,50% 6,30% 1,40% h86 6,10% 6,00% 6,10% 6,00% 6,00% 6,00% 6,00% 5,80% 6,00% 6,20% 6,10% 6,20% 5,90% 5,90% 5,90% 6,00% 5,80% 0,70% h87 6,00% 5,90% 6,00% 6,00% 5,90% 5,90% 6,00% 5,80% 5,90% 6,10% 6,00% 6,10% 5,90% 5,90% 5,90% 6,00% 5,80% 1,50% h88 5,80% 5,70% 5,80% 5,70% 5,70% 5,70% 5,70% 5,70% 5,90% 5,90% 5,80% 5,90% 5,80% 5,60% 5,80% 5,70% 5,90% 1,20% h89 6,10% 6,00% 6,10% 6,10% 6,00% 6,00% 6,10% 5,90% 6,00% 6,20% 6,10% 6,20% 6,00% 6,00% 6,00% 6,10% 5,90% 0,80% h90 6,10% 6,00% 6,10% 6,00% 6,00% 6,00% 6,00% 5,80% 6,00% 6,20% 6,10% 6,20% 5,90% 5,90% 5,90% 6,00% 5,80% 1,50% h91 6,10% 6,00% 6,10% 6,10% 6,00% 6,00% 6,10% 5,90% 6,00% 6,20% 6,10% 6,20% 6,00% 6,00% 6,00% 6,10% 5,90% 0,80% h92 5,90% 5,80% 5,90% 5,90% 5,80% 5,80% 5,90% 5,70% 5,80% 6,00% 5,90% 6,00% 5,80% 5,80% 5,80% 5,90% 5,70% 1,00% h93 6,00% 5,90% 6,00% 6,00% 5,90% 5,90% 6,00% 5,80% 5,90% 6,10% 6,00% 6,10% 5,90% 5,90% 5,90% 6,00% 5,80% 1,10% h94 5,50% 5,60% 5,50% 5,40% 5,40% 5,40% 5,40% 5,20% 5,50% 5,60% 5,50% 5,50% 5,30% 5,30% 5,30% 5,40% 5,20% 1,00% h95 6,10% 6,00% 6,10% 6,10% 6,00% 6,00% 6,10% 5,90% 6,00% 6,20% 6,10% 6,20% 6,00% 6,00% 6,00% 6,10% 5,90% 1,00% h96 6,10% 6,10% 6,10% 6,10% 6,00% 6,00% 6,10% 5,90% 6,10% 6,30% 6,20% 6,20% 6,00% 6,00% 6,00% 6,10% 5,90% 1,00% h97 6,60% 6,60% 6,60% 6,50% 6,50% 6,50% 6,50% 6,30% 6,60% 6,80% 6,70% 6,70% 6,20% 6,40% 6,40% 6,50% 6,30% 1,70% Nu 10,70% 10,90% 10,70% 10,60% 10,60% 10,70% 10,60% 10,40% 10,70% 10,70% 10,80% 10,60% 10,50% 10,50% 10,50% 10,60% 10,40% 8,80%

72
h37 h38 h39 h40 h41 h42 h43 h44 h45 h46 h47 h48 h49 h50 h51 h52 h53 h54 h1 h2 h3 h4 h5 h6 h7 h8 h9 h10 h11 h12 h13 h14 h15 h16 h17 h18 h19 h20 h21 h22 h23 h24 h25 h26 h27 h28

73
h37 h38 h39 h40 h41 h42 h43 h44 h45 h46 h47 h48 h49 h50 h51 h52 h53 h54 h29 h30 h31 h32 h33 h34 h35 h36 h37 h38 0,80% h39 0,80% 0,10% h40 0,90% 0,30% 0,20% h41 1,10% 1,40% 1,30% 1,50% h42 4,60% 5,40% 5,40% 5,50% 4,20% h43 5,20% 5,80% 5,90% 5,90% 5,60% 1,60% h44 4,70% 5,50% 5,50% 5,60% 4,30% 0,30% 1,50% h45 7,80% 7,70% 7,80% 7,80% 7,60% 4,40% 3,00% 4,20% h46 8,20% 8,10% 8,20% 8,20% 7,80% 4,50% 3,30% 4,20% 1,00% h47 8,10% 8,00% 8,10% 8,10% 7,90% 4,60% 3,20% 4,30% 0,60% 0,80% h48 8,10% 8,00% 8,10% 8,10% 7,70% 4,40% 3,20% 4,20% 0,90% 0,50% 0,60% h49 7,90% 7,80% 7,90% 7,90% 7,70% 4,40% 3,00% 4,20% 0,60% 0,80% 0,20% 0,60% h50 7,90% 7,80% 7,90% 7,90% 7,70% 4,30% 3,30% 4,20% 1,00% 0,70% 0,50% 0,50% 0,50% h51 7,80% 7,70% 7,80% 7,80% 7,60% 4,10% 3,10% 4,10% 0,80% 0,70% 0,50% 0,50% 0,30% 0,20% h52 8,20% 8,30% 8,20% 8,20% 8,20% 4,70% 3,70% 4,60% 0,80% 0,90% 0,50% 1,00% 0,60% 0,70% 0,70% h53 8,20% 8,10% 8,20% 8,20% 7,80% 4,60% 3,40% 4,30% 0,70% 0,60% 0,60% 0,60% 0,60% 0,50% 0,50% 0,80% h54 7,90% 7,80% 7,90% 7,90% 7,50% 4,20% 3,00% 4,00% 0,90% 0,60% 0,40% 0,40% 0,40% 0,30% 0,30% 0,80% 0,60% h55 7,90% 8,00% 7,90% 7,90% 7,70% 4,20% 3,40% 4,20% 1,10% 0,80% 0,60% 0,60% 0,60% 0,30% 0,30% 0,50% 0,70% 0,40% h56 8,20% 8,10% 8,20% 8,20% 8,00% 4,70% 3,30% 4,40% 0,50% 0,90% 0,30% 0,80% 0,30% 0,70% 0,60% 0,40% 0,50% 0,60%

74
h37 h38 h39 h40 h41 h42 h43 h44 h45 h46 h47 h48 h49 h50 h51 h52 h53 h54 h57 7,70% 7,80% 7,70% 7,90% 7,70% 4,20% 3,40% 4,20% 0,90% 1,20% 0,60% 0,90% 0,40% 0,60% 0,50% 0,50% 0,90% 0,70% h58 7,90% 8,00% 7,90% 7,90% 7,70% 4,40% 3,20% 4,20% 0,70% 0,80% 0,40% 0,60% 0,40% 0,60% 0,60% 0,50% 0,90% 0,60% h59 8,10% 8,20% 8,10% 8,10% 7,90% 4,60% 3,40% 4,30% 0,90% 1,00% 0,40% 0,60% 0,40% 0,60% 0,60% 0,50% 0,90% 0,60% h60 5,40% 5,90% 6,00% 6,00% 5,40% 1,70% 1,00% 1,50% 2,90% 3,00% 3,10% 2,90% 2,90% 3,00% 2,90% 3,60% 3,10% 2,80% h61 5,80% 6,50% 6,40% 6,60% 5,30% 1,50% 1,70% 1,40% 4,10% 3,80% 3,70% 3,70% 3,50% 3,60% 3,40% 4,00% 3,70% 3,30% h62 5,10% 5,70% 5,60% 5,80% 4,50% 0,70% 1,80% 0,60% 4,20% 4,30% 4,40% 4,20% 4,20% 4,30% 4,10% 4,70% 4,40% 4,10% h63 6,30% 6,50% 6,60% 6,60% 6,00% 2,50% 1,70% 2,20% 2,50% 2,40% 2,50% 2,30% 2,50% 2,40% 2,40% 2,90% 2,70% 2,10% h64 5,40% 6,00% 5,90% 6,10% 4,60% 0,80% 2,00% 0,70% 4,40% 4,50% 4,40% 4,40% 4,20% 4,30% 4,10% 4,70% 4,40% 4,10% h65 5,30% 5,90% 5,80% 6,00% 4,50% 0,70% 1,90% 0,80% 4,50% 4,60% 4,50% 4,50% 4,30% 4,40% 4,20% 4,80% 4,50% 4,20% h66 5,50% 6,10% 6,00% 6,20% 5,10% 1,20% 2,10% 1,10% 4,40% 4,50% 4,20% 4,40% 4,10% 4,30% 4,20% 4,50% 4,40% 4,10% h67 6,70% 6,90% 7,00% 7,00% 6,60% 2,90% 1,70% 2,60% 2,70% 2,40% 2,30% 2,10% 2,10% 2,20% 2,00% 2,80% 2,30% 1,90% h68 5,20% 5,80% 5,70% 5,90% 4,40% 0,60% 1,80% 0,70% 4,40% 4,30% 4,60% 4,40% 4,40% 4,50% 4,30% 4,90% 4,60% 4,20% h69 5,80% 6,10% 6,20% 6,20% 6,00% 2,30% 1,20% 2,00% 3,10% 2,90% 3,00% 2,80% 2,80% 2,90% 2,70% 3,40% 3,00% 2,60% h70 5,10% 5,50% 5,60% 5,80% 5,10% 1,70% 1,80% 1,40% 3,50% 3,40% 3,70% 3,50% 3,50% 3,60% 3,40% 4,10% 3,70% 3,30% h71 6,00% 6,50% 6,60% 6,60% 6,10% 2,40% 1,20% 2,10% 2,90% 2,70% 2,60% 2,60% 2,40% 2,50% 2,30% 3,00% 2,60% 2,20% h72 5,30% 5,90% 5,70% 5,90% 4,50% 0,60% 1,80% 0,50% 4,10% 4,20% 4,30% 4,10% 4,10% 4,20% 4,10% 4,60% 4,30% 4,00% h73 5,30% 5,90% 5,80% 6,00% 4,50% 0,60% 1,80% 0,60% 4,20% 4,20% 4,30% 4,20% 4,20% 4,20% 4,10% 4,60% 4,30% 4,00% h74 5,90% 6,20% 6,30% 6,30% 5,90% 2,20% 1,30% 1,90% 3,20% 2,80% 3,00% 2,90% 2,90% 3,00% 2,80% 3,50% 3,00% 2,70% h75 5,80% 6,50% 6,40% 6,40% 5,40% 1,70% 1,50% 1,50% 3,60% 3,30% 3,20% 3,20% 3,00% 3,10% 2,90% 3,50% 3,20% 2,90% h76 5,40% 6,00% 5,90% 6,10% 4,60% 0,80% 2,00% 0,90% 4,60% 4,50% 4,80% 4,60% 4,60% 4,70% 4,50% 5,10% 4,80% 4,40% h77 5,70% 6,20% 6,30% 6,30% 5,60% 2,10% 1,10% 1,80% 3,00% 2,80% 2,90% 2,70% 2,70% 2,80% 2,60% 3,30% 2,90% 2,50% h78 5,40% 6,00% 5,90% 6,10% 4,60% 0,70% 1,90% 0,60% 4,40% 4,50% 4,40% 4,40% 4,20% 4,30% 4,20% 4,70% 4,40% 4,10% h79 5,90% 6,60% 6,50% 6,50% 5,80% 2,30% 1,30% 2,00% 2,70% 2,60% 2,70% 2,50% 2,50% 2,60% 2,40% 2,80% 2,70% 2,50% h80 4,90% 5,50% 5,50% 5,60% 4,10% 1,40% 2,60% 1,30% 5,10% 5,00% 5,30% 4,90% 5,10% 5,20% 5,00% 5,50% 5,10% 4,90% h81 6,20% 6,90% 6,80% 7,00% 5,50% 1,60% 1,80% 1,50% 4,20% 3,70% 3,80% 3,40% 3,60% 3,70% 3,50% 4,10% 3,60% 3,40% h82 5,30% 5,90% 5,80% 6,00% 4,50% 0,80% 2,00% 0,70% 4,40% 4,30% 4,60% 4,20% 4,40% 4,50% 4,30% 4,90% 4,40% 4,20% h83 5,40% 6,10% 6,00% 6,20% 4,70% 0,90% 2,10% 0,80% 4,60% 4,70% 4,80% 4,60% 4,60% 4,70% 4,50% 5,10% 4,80% 4,40%

75
h37 h38 h39 h40 h41 h42 h43 h44 h45 h46 h47 h48 h49 h50 h51 h52 h53 h54 h84 5,30% 5,90% 5,80% 6,00% 4,50% 0,70% 1,90% 0,60% 4,40% 4,50% 4,60% 4,40% 4,40% 4,50% 4,30% 4,90% 4,60% 4,20% h85 6,10% 6,60% 6,50% 6,70% 5,40% 1,50% 1,80% 1,40% 4,10% 3,80% 3,70% 3,70% 3,50% 3,60% 3,40% 4,00% 3,70% 3,30% h86 5,40% 6,00% 5,90% 6,10% 4,60% 0,80% 2,00% 0,70% 4,50% 4,60% 4,70% 4,50% 4,50% 4,60% 4,40% 5,00% 4,70% 4,30% h87 5,30% 5,90% 5,80% 6,00% 4,50% 1,60% 2,80% 1,50% 5,20% 4,90% 5,40% 5,00% 5,20% 5,30% 5,10% 5,60% 5,20% 5,00% h88 5,10% 5,70% 5,60% 5,80% 4,30% 1,10% 2,30% 1,20% 5,00% 4,90% 5,20% 4,80% 5,00% 5,10% 4,90% 5,40% 5,00% 4,80% h89 5,40% 6,00% 5,90% 6,10% 4,60% 0,90% 2,10% 0,80% 4,50% 4,40% 4,70% 4,50% 4,50% 4,60% 4,40% 5,00% 4,70% 4,30% h90 5,50% 6,00% 5,90% 6,10% 5,20% 1,60% 1,60% 1,70% 3,80% 3,90% 4,00% 3,80% 3,80% 3,90% 3,70% 4,20% 4,00% 3,60% h91 5,40% 6,00% 5,90% 6,10% 4,60% 0,90% 2,10% 0,80% 4,50% 4,40% 4,70% 4,30% 4,50% 4,60% 4,40% 5,00% 4,50% 4,30% h92 5,20% 5,80% 5,70% 5,90% 4,40% 0,90% 2,10% 1,00% 4,70% 4,60% 4,90% 4,70% 4,70% 4,80% 4,60% 5,20% 4,90% 4,50% h93 5,30% 5,90% 5,80% 6,00% 4,70% 1,20% 2,30% 1,10% 4,70% 4,60% 4,90% 4,50% 4,70% 4,80% 4,60% 5,20% 4,70% 4,50% h94 4,80% 5,50% 5,40% 5,60% 4,20% 1,10% 2,10% 1,00% 4,60% 4,90% 4,80% 4,80% 4,60% 4,90% 4,70% 5,10% 5,00% 4,60% h95 5,50% 6,00% 5,90% 6,10% 4,80% 1,10% 2,30% 1,00% 4,50% 4,40% 4,70% 4,50% 4,50% 4,60% 4,40% 5,00% 4,70% 4,30% h96 5,60% 6,10% 6,00% 6,20% 4,90% 1,10% 2,30% 1,00% 4,60% 4,30% 4,80% 4,40% 4,60% 4,70% 4,50% 5,10% 4,60% 4,40% h97 6,30% 6,60% 6,50% 6,70% 5,70% 2,00% 2,20% 1,90% 4,00% 3,60% 3,60% 3,40% 3,40% 3,50% 3,30% 3,80% 3,60% 3,20% Nu 10,40% 10,50% 10,60% 10,80% 10,00% 8,90% 9,30% 8,90% 11,20% 11,50% 11,40% 11,40% 11,20% 11,50% 11,30% 11,80% 11,50% 11,20%

76
h55 h56 h57 h58 h59 h60 h61 h62 h63 h64 h65 h66 h67 h68 h69 h70 h71 h72 h1 h2 h3 h4 h5 h6 h7 h8 h9
h10 h11 h12 h13 h14 h15 h16 h17 h18 h19 h20 h21 h22 h23 h24 h25 h26 h27 h28

77
h55 h56 h57 h58 h59 h60 h61 h62 h63 h64 h65 h66 h67 h68 h69 h70 h71 h72 h29 h30 h31 h32 h33 h34 h35 h36 h37 h38 h39 h40 h41 h42 h43 h44 h45 h46 h47 h48 h49 h50 h51 h52 h53 h54 h55 h56 0,60%

78
h55 h56 h57 h58 h59 h60 h61 h62 h63 h64 h65 h66 h67 h68 h69 h70 h71 h72 h57 0,40% 0,50% h58 0,40% 0,50% 0,40% h59 0,40% 0,50% 0,40% 0,20% h60 3,10% 3,20% 3,30% 3,10% 3,30% h61 3,50% 3,80% 3,50% 3,70% 3,70% 1,40% h62 4,20% 4,50% 4,20% 4,20% 4,40% 1,40% 0,70% h63 2,50% 2,80% 2,90% 2,50% 2,70% 1,00% 2,00% 2,00% h64 4,20% 4,50% 4,20% 4,40% 4,40% 1,70% 0,80% 0,30% 2,20% h65 4,30% 4,60% 4,30% 4,50% 4,50% 1,80% 1,00% 0,50% 2,40% 0,20% h66 4,20% 4,30% 4,10% 4,20% 4,20% 2,00% 1,20% 0,80% 2,70% 0,90% 1,00% h67 2,30% 2,40% 2,50% 2,50% 2,30% 1,20% 1,80% 2,60% 1,30% 2,40% 2,60% 2,70% h68 4,40% 4,70% 4,40% 4,40% 4,60% 1,80% 1,20% 0,50% 2,40% 0,60% 0,30% 1,10% 2,90% h69 3,00% 3,00% 3,10% 3,00% 3,10% 0,60% 1,40% 1,90% 1,40% 2,20% 2,40% 2,40% 1,00% 2,40% h70 3,70% 3,80% 3,70% 3,70% 3,90% 0,80% 1,80% 1,10% 1,70% 1,40% 1,60% 1,90% 1,80% 1,40% 1,20% h71 2,60% 2,70% 2,80% 2,80% 2,80% 0,70% 1,40% 2,10% 1,40% 2,10% 2,30% 2,20% 0,60% 2,50% 0,40% 1,40% h72 4,10% 4,40% 4,10% 4,10% 4,30% 1,70% 1,20% 0,50% 2,20% 0,60% 0,60% 0,70% 2,60% 0,60% 2,10% 1,60% 2,10% h73 4,20% 4,40% 4,20% 4,20% 4,30% 1,50% 1,00% 0,30% 2,00% 0,40% 0,60% 0,70% 2,40% 0,60% 2,00% 1,40% 1,90% 0,10% h74 3,00% 3,10% 3,20% 3,00% 3,20% 0,80% 1,30% 1,80% 1,70% 2,10% 2,30% 2,30% 1,10% 2,10% 0,60% 1,30% 0,80% 2,00% h75 3,00% 3,30% 3,20% 3,20% 3,20% 0,90% 0,80% 1,60% 1,70% 1,50% 1,50% 1,80% 1,30% 1,70% 0,90% 1,70% 0,80% 1,70% h76 4,60% 4,90% 4,60% 4,60% 4,80% 1,80% 1,20% 0,50% 2,40% 0,60% 0,60% 1,10% 2,90% 0,40% 2,40% 1,40% 2,50% 0,70% h77 2,90% 2,90% 3,00% 2,90% 3,00% 0,60% 1,30% 1,80% 1,30% 2,00% 2,20% 2,30% 0,90% 2,20% 0,50% 1,30% 0,50% 2,00% h78 4,20% 4,50% 4,20% 4,40% 4,40% 1,70% 1,00% 0,50% 2,20% 0,40% 0,40% 0,60% 2,60% 0,50% 2,20% 1,60% 2,10% 0,40% h79 2,30% 2,60% 2,50% 2,30% 2,50% 1,20% 1,80% 2,20% 1,80% 2,40% 2,50% 2,70% 1,70% 2,40% 1,40% 2,00% 1,40% 2,20% h80 5,10% 5,40% 5,10% 5,10% 5,30% 2,40% 1,90% 1,20% 3,00% 1,10% 1,20% 1,70% 3,30% 1,30% 3,00% 2,10% 3,00% 1,10% h81 3,60% 3,90% 3,60% 3,80% 3,60% 1,80% 0,60% 1,30% 2,30% 1,20% 1,40% 1,30% 1,80% 1,50% 1,80% 2,40% 1,50% 1,10% h82 4,40% 4,70% 4,40% 4,40% 4,60% 1,80% 1,20% 0,50% 2,40% 0,60% 0,60% 1,10% 2,90% 0,60% 2,40% 1,60% 2,50% 0,60% h83 4,60% 4,90% 4,60% 4,60% 4,80% 1,70% 1,30% 0,60% 2,30% 0,50% 0,60% 1,00% 2,70% 0,80% 2,30% 1,70% 2,40% 0,60%

79
h55 h56 h57 h58 h59 h60 h61 h62 h63 h64 h65 h66 h67 h68 h69 h70 h71 h72 h84 4,40% 4,70% 4,40% 4,40% 4,60% 1,70% 1,20% 0,50% 2,20% 0,60% 0,60% 1,00% 2,80% 0,50% 2,20% 1,60% 2,30% 0,50% h85 3,50% 3,80% 3,50% 3,70% 3,70% 1,60% 0,40% 1,10% 2,10% 1,00% 1,20% 1,20% 1,80% 1,40% 1,60% 2,20% 1,50% 1,10% h86 4,50% 4,80% 4,50% 4,50% 4,70% 1,70% 1,20% 0,50% 2,20% 0,40% 0,60% 1,10% 2,60% 0,70% 2,20% 1,60% 2,30% 0,60% h87 5,20% 5,40% 5,20% 5,20% 5,40% 2,40% 1,90% 1,20% 3,10% 1,10% 1,30% 1,80% 3,30% 1,30% 3,00% 1,90% 3,00% 1,50% h88 5,00% 5,20% 5,00% 5,00% 5,20% 2,30% 1,80% 1,10% 2,90% 1,00% 0,90% 1,60% 3,20% 1,00% 2,90% 2,20% 2,90% 1,00% h89 4,50% 4,80% 4,50% 4,50% 4,70% 1,80% 1,20% 0,50% 2,40% 0,40% 0,40% 1,20% 2,80% 0,30% 2,40% 1,40% 2,50% 0,60% h90 3,80% 4,10% 3,80% 3,80% 4,00% 1,10% 1,20% 1,40% 1,80% 1,50% 1,50% 2,00% 1,90% 1,70% 1,50% 1,70% 1,70% 1,70% h91 4,50% 4,80% 4,50% 4,50% 4,70% 1,90% 1,30% 0,60% 2,50% 0,60% 0,70% 1,20% 3,00% 0,60% 2,50% 1,70% 2,60% 0,60% h92 4,70% 5,00% 4,70% 4,70% 4,90% 2,00% 1,40% 0,60% 2,60% 0,70% 0,60% 1,40% 3,10% 0,30% 2,60% 1,60% 2,70% 0,80% h93 4,70% 5,00% 4,70% 4,70% 4,90% 2,00% 1,40% 0,60% 2,70% 0,90% 1,00% 1,10% 3,20% 0,90% 2,60% 1,60% 2,80% 0,90% h94 4,80% 4,90% 4,60% 4,60% 4,80% 2,00% 1,80% 1,00% 2,80% 0,90% 1,10% 1,20% 3,00% 1,20% 2,60% 1,90% 2,70% 1,00% h95 4,50% 4,80% 4,50% 4,50% 4,70% 1,90% 1,30% 0,60% 2,50% 0,60% 0,80% 1,20% 2,90% 0,60% 2,50% 1,50% 2,60% 0,70% h96 4,60% 4,90% 4,60% 4,60% 4,80% 2,10% 1,50% 0,70% 2,60% 0,60% 0,70% 1,40% 3,00% 0,60% 2,70% 1,70% 2,80% 0,80% h97 3,40% 3,70% 3,40% 3,60% 3,40% 1,60% 0,90% 1,70% 2,40% 1,70% 1,90% 1,80% 1,50% 2,10% 1,60% 2,20% 1,40% 1,70% Nu 11,50% 11,40% 11,40% 11,40% 11,50% 9,20% 9,50% 8,70% 9,80% 8,70% 8,70% 8,90% 9,80% 8,70% 9,40% 8,50% 9,60% 8,90%

80
h73 h74 h75 h76 h77 h78 h79 h80 h81 h82 h83 h84 h84 h85 h86 h87 h88 h89 h1 h2 h3 h4 h5 h6 h7 h8 h9 h10 h11 h12 h13 h14 h15 h16 h17 h18 h19 h20 h21 h22 h23 h24 h25 h26 h27 h28

81
h73 h74 h75 h76 h77 h78 h79 h80 h81 h82 h83 h84 h84 h85 h86 h87 h88 h89 h29 h30 h31 h32 h33 h34 h35 h36 h37 h38 h39 h40 h41 h42 h43 h44 h45 h46 h47 h48 h49 h50 h51 h52 h53 h54 h55 h56

82
h73 h74 h75 h76 h77 h78 h79 h80 h81 h82 h83 h84 h84 h85 h86 h87 h88 h89 h57 h58 h59 h60 h61 h62 h63 h64 h65 h66 h67 h68 h69 h70 h71 h72 h73 h74 1,90% h75 1,50% 1,00% h76 0,60% 2,10% 1,80% h77 1,80% 0,60% 0,70% 2,20% h78 0,40% 2,10% 1,30% 0,70% 2,00% h79 2,20% 1,50% 1,50% 2,60% 1,30% 2,30% h80 1,10% 2,90% 2,40% 1,50% 2,60% 1,20% 3,00% h81 1,00% 1,70% 1,00% 1,60% 1,60% 1,20% 2,00% 1,70% h82 0,60% 2,30% 1,80% 0,70% 2,20% 0,60% 2,30% 1,10% 1,30% h83 0,50% 2,20% 1,70% 0,60% 2,10% 0,60% 2,40% 1,20% 1,50% 0,50%
83
h73 h74 h75 h76 h77 h78 h79 h80 h81 h82 h83 h84 h84 h85 h86 h87 h88 h89 h84 0,40% 2,10% 1,50% 0,70% 2,00% 0,40% 2,20% 1,20% 1,40% 0,30% 0,30% h85 1,00% 1,30% 0,80% 1,20% 1,40% 1,00% 1,90% 1,90% 0,60% 1,40% 1,10% 1,20% h86 0,40% 2,10% 1,70% 0,70% 2,00% 0,60% 2,30% 1,10% 1,40% 0,40% 0,10% 0,20% 1,20% h87 1,30% 2,70% 2,40% 1,10% 2,60% 1,50% 3,00% 0,90% 1,90% 1,10% 1,00% 1,30% 1,90% 1,10% h88 1,00% 2,80% 2,30% 1,20% 2,70% 1,10% 2,80% 1,20% 1,60% 0,60% 0,70% 0,70% 1,80% 0,60% 1,40% h89 0,60% 2,10% 1,70% 0,60% 2,20% 0,60% 2,40% 1,20% 1,60% 0,50% 0,50% 0,40% 1,40% 0,40% 0,90% 0,90% h90 1,50% 1,60% 1,30% 1,70% 1,40% 1,70% 1,80% 2,20% 1,60% 1,50% 1,20% 1,30% 1,40% 1,10% 2,00% 1,60% 1,50% h91 0,60% 2,40% 1,90% 0,80% 2,30% 0,70% 2,40% 1,00% 1,20% 0,10% 0,60% 0,40% 1,50% 0,50% 1,00% 0,60% 0,60% 0,60% h92 0,70% 2,30% 1,80% 0,60% 2,40% 0,70% 2,60% 1,60% 1,80% 0,50% 0,60% 0,40% 1,60% 0,60% 1,30% 0,90% 0,40% 0,70% h93 0,90% 2,50% 2,20% 0,90% 2,50% 1,00% 2,60% 1,10% 1,50% 0,40% 0,60% 0,60% 1,60% 0,70% 1,10% 0,80% 0,80% 0,90% h94 0,90% 2,50% 2,00% 1,10% 2,20% 1,10% 2,70% 0,60% 1,90% 1,20% 0,80% 1,10% 1,60% 0,90% 0,70% 1,50% 1,10% 0,90% h95 0,60% 2,00% 1,90% 0,50% 2,30% 0,80% 2,60% 1,60% 1,70% 0,50% 0,40% 0,50% 1,10% 0,50% 1,00% 1,10% 0,50% 0,60% h96 0,80% 2,40% 2,10% 0,80% 2,50% 0,90% 2,70% 1,00% 1,40% 0,50% 0,70% 0,70% 1,70% 0,60% 1,00% 0,70% 0,60% 0,80% h97 1,60% 1,50% 1,00% 2,10% 1,50% 1,80% 2,20% 2,70% 0,90% 1,90% 1,80% 1,80% 0,90% 1,70% 2,70% 2,40% 2,10% 1,60% Nu 8,70% 9,20% 9,70% 8,60% 9,10% 8,70% 9,90% 9,00% 9,80% 9,10% 8,70% 8,90% 9,50% 8,70% 9,00% 9,20% 8,70% 8,70%

84
h90 h91 h92 h93 h94 h95 h96 h97 Nu h1 h2 h3 h4 h5 h6 h7 h8 h9 h10 h11 h12 h13 h14 h15 h16 h17 h18 h19 h20 h21 h22 h23 h24 h25 h26 h27 h28

85
h90 h91 h92 h93 h94 h95 h96 h97 Nu h29 h30 h31 h32 h33 h34 h35 h36 h37 h38 h39 h40 h41 h42 h43 h44 h45 h46 h47 h48 h49 h50 h51 h52 h53 h54 h55 h56

86
h90 h91 h92 h93 h94 h95 h96 h97 Nu h57 h58 h59 h60 h61 h62 h63 h64 h65 h66 h67 h68 h69 h70 h71 h72 h73 h74 h75 h76 h77 h78 h79 h80 h81 h82 h83

87
h90 h91 h92 h93 h94 h95 h96 h97 Nuh84 h85 h86 h87 h88 h89 h90 h91 1,60% h92 1,50% 0,60% h93 1,70% 0,30% 0,80% h94 1,80% 1,30% 1,50% 1,40% h95 1,60% 0,60% 0,50% 0,60% 1,20% h96 1,60% 0,40% 0,70% 0,60% 1,30% 0,70% h97 1,20% 2,00% 2,10% 2,10% 2,30% 1,80% 2,20% Nu 9,30% 9,20% 8,70% 9,10% 8,60% 8,90% 9,00% 9,80%