identificação de sorovares de leptospira por por …livros01.livrosgratis.com.br/cp117120.pdf ·...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE MINAS GERAIS
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE BIOLOGIA GERAL
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA
DISSERTAÇÃO DE MESTRADO
Identificação de sorovares de Leptospira Identificação de sorovares de Leptospira Identificação de sorovares de Leptospira Identificação de sorovares de Leptospira
por por por por PCRPCRPCRPCR----RFLP do gene que codifica a RFLP do gene que codifica a RFLP do gene que codifica a RFLP do gene que codifica a
subunidade subunidade subunidade subunidade ββββ da RNA polimerase (rpoB)da RNA polimerase (rpoB)da RNA polimerase (rpoB)da RNA polimerase (rpoB)
ORIENTADO: Lenice Roteia Cardoso Jung
ORIENTADOR: Prof. Dr. Álvaro Cantini Nunes
CO-ORIENTADORA: Dra. Maria Rosa Quaresma Bomfim
BELO HORIZONTE
Outubro - 2009

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

Lenice Roteia Cardoso Jung
Identificação de sorovares de Leptospira por PCR-RFLP do gene que codifica a subunidade ββββ da RNA
polimerase (rpoB)
ORIENTADOR: ÁLVARO CANTINI NUNES
CO-ORIENTADORA: MARIA ROSA QUARESMA BOMFIM
Instituto de Ciências Biológicas
Departamento de Biologia Geral
Belo Horizonte, MG
Outubro/2009
Dissertação apresentada ao Departamento de
Biologia Geral do Instituto de Ciências
Biológicas da Universidade Federal de Minas
Gerais, como requisito para obtenção do grau
de mestre em Genética.

iii
043 Jung, Lenice Roteia Cardoso Identificação de sorovares de Leptospira por PCR-RFLP do gene que codifica a subunidade β da RNA polimerase (rpoB). [manuscrito] / Lenice Roteia Cardoso Jung. – 2009. 57 f. : il. ; 29,5 cm. Orientador: Álvaro Cantini Nunes. Co-orientadora: Maria Rosa Quaresma Bomfim Dissertação (mestrado) – Universidade Federal de Minas Gerais, Departamento de Biologia Geral.
Leptospira – Identificação – Teses. 2. Genética – Teses. 3. Reação em cadeia de polimerase – Teses. 4. Polimorfismo de fragmento de restrição. I. Nunes, lvaro Cantini. II. Bomfim, Maria Rosa Quaresma. III. Universidade Federal de Minas Gerais. Departamento de Biologia Geral. IV. Título. CDU: 575

iv

v
Esse trabalho é dedicado aos meus pais,
“in memoriam”, pois me fizeram compreender
o sentido do trabalho, valorizar o conhecimento
e superar as adversidades, na
busca por meus objetivos.

vi
AgradecimentosAgradecimentosAgradecimentosAgradecimentos
Agradeço ao Departamento de Genética, na figura do meu orientador o
Professor Dr. Álvaro Cantini Nunes, a grande oportunidade, aceitando-
me como mestranda, sua eficiente orientação, a valiosa experiência e a
confiança que me presenteou;
À Secretaria de Educação do Estado de Minas Gerais, representada pela
Dra. Vanessa Guimarães, por autorizar a licença solicitada, para que eu
pudesse freqüentar o mestrado em genética.
À co-orientadora e amiga, Dra Maria Rosa Quaresma Bomfim, agradeço
a disponibilidade, dedicação, altruísmo e sabedoria.
Ao professor Élvio Carlos Moreira e ao técnico Antônio Benjamin de Paula,
do laboratório de Leptospiras do DMP da Escola de Veterinária, sou grata
pela gentileza de fornecer as linhagens de referência, para a pesquisa.
Às professoras Regina Maria Nardi Drummond e Vera Lúcia dos Santos,
agradeço por me permitirem usar o Laboratório de Microbiologia
Aplicada;
Aos colegas: doutorando João Luiz Moreira e mestranda Marlene de
Miranda minha gratidão por guiarem meus “primeiros passos” no
laboratório de genética;
Aos professores, colegas e funcionários, com os quais convivi durante o
Mestrado em Genética e no LGMPP, sou grata pelo aprendizado, a ótima
convivência, a preciosa cooperação e profissionalismo durante os
trabalhos e estudo.
A toda minha família e amigos, agradeço o estímulo, carinho e
paciência, em função da minha constante ausência;
E mais uma vez, agradeço a Deus, por me conceder sempre, esse precioso
tesouro que é ter: saúde, energia e disposição, suficientes para realizar
tudo o que desejo, com muita alegria e paz.
Obrigada!

vii
O conhecimento participa do Infinito;
ele amplia nossas capacidades;
quanto mais alto nele subimos,
mais vastas e magníficas, são as perspectivas,
que se descortinam à nossa frente.
Luiz Roberto Garcia Sierra
Livro: “REFORMA INTERIOR, UM CONVITE À REFLEXÃO”
SUMÁRIO

viii
RESUMO ...................................................................................................................... 1
ABSTRACT .................................................................................................................. 2
1. INTRODUÇÃO ......................................................................................................... 3
1.1. A leptospirose : contextualização histórica............................................................. 3
1.2. Patogênese na leptospirose .................................................................................. 4
1.3. Características do gênero Leptospira..................................................................... 5
1.3.1. As leptospiras: características estruturais e requerimentos nutricionais............. 5
1.3.2. Identificação e taxonomia: a complexidade do gênero Leptospira...................... 7
1.3.3. O genoma de Leptospira spp. ............................................................................ 11
2. OBJETIVO GERAL.................................................................................................... 12
2.1. OBJETIVOS ESPECÍFICOS.................................................................................. 12
3. MATERIAL E MÉTODOS ......................................................................................... 13
3.1. Cultivo dos sorovares de Leptospira de referência ............................................... 13
3.2. Extração do DNA ................................................................................................... 13
3.3. Quantificação do DNA ........................................................................................... 15
3.4. Seleção dos genes para amplificação e estudo de polimorfismo ......................... 15
3.5. Padronização da PCR – específica para o gene rpoB e análise de restrição ...... 16
3.5.1. Análise in silico dos genes ................................................................................. 16
3.5.2. Gene rpoB .......................................................................................................... 17
3.5.3. Digestão com endonucleases de restrição ........................................................ 17
3.5.4. Análise dos produtos da PCR-específica e da digestão enzimática ................. 18
3.5.5. Confecção dos dendogramas ............................................................................ 18
4. RESULTADOS ......................................................................................................... 19
Análise dos dendogramas construídos a partir dos produtos de PCR digeridos com
as enzimas TaqI, Tru1I, Sau3AI, MslI e EcoNI ............................................................ 35
5. DISCUSSÃO ............................................................................................................ 45
6. CONCLUSÃO .......................................................................................................... 51
7. REFERÊNCIAS BIBLIOGRÁFICAS ......................................................................... 52
8. APÊNDICE ............................................................................................................... 56

ix
LISTA DE FIGURAS
FIGURA 1. Ciclo de vida, vias de infecção, o reservatório e a patogenia das leptospiras ..... 4
FIGURA 2. a, b) estrutura das leptospiras; c) posição dos flagelos e eixo de rotação no
espaço periplasmático ............................................................................................................ 6
FIGURA 3. Leptospira interrogans, sorovar Icterohaemorrhagiae, exibindo as estruturas em
gancho. a. eletromicroscopia, b. microscopia em campo escuro ........................................... 6
FIGURA 4. Arquitetura da superfície das leptospiras, mostrando na membrana externa (OM,
outer membrane) a camada de lipopolissacarídeos que define os sorovares ....................... 8
FIGURA 5. Características dos genomas já seqüenciados de leptospiras patogênicas e
saprofíticas ............................................................................................................................ 12
FIGURA 6. Alinhamento das seqüências de nucleotídeos de rpoB depositadas no GenBank
e geradas por seqüenciamento pelo nosso grupo ................................................................ 20
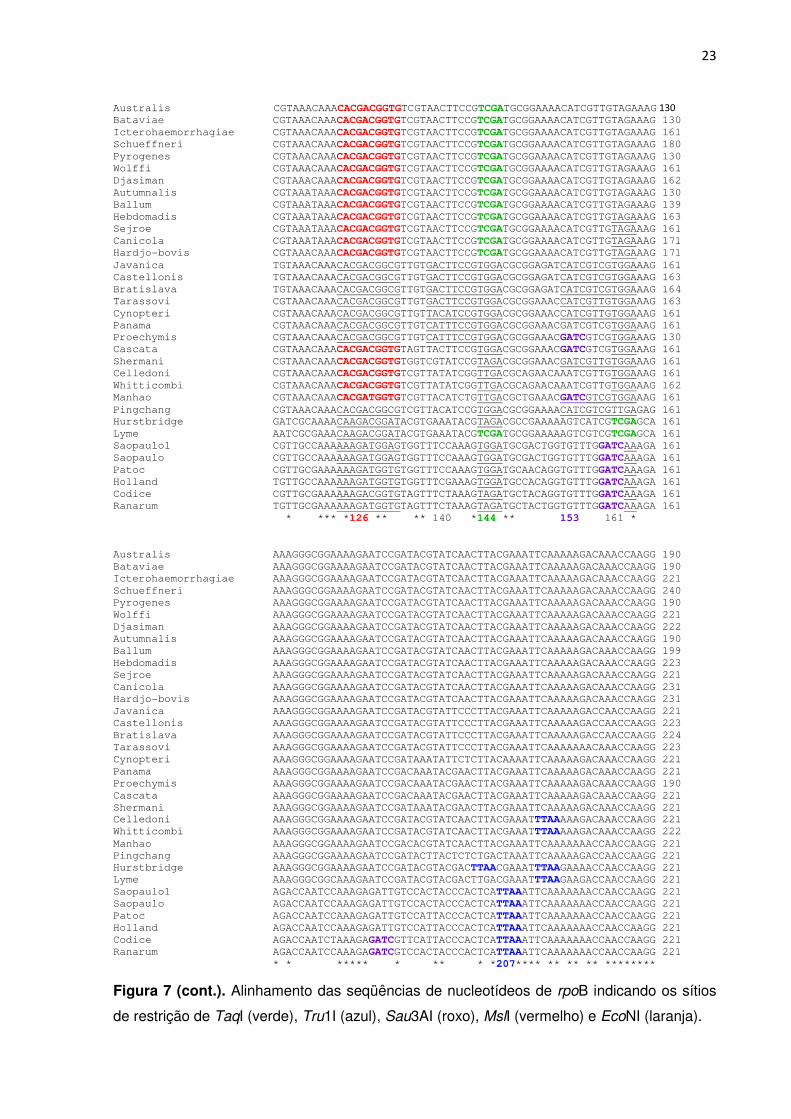
FIGURA 7. Alinhamento das seqüências de nucleotídeos de rpoB indicando os sítios de
restrição de TaqI (verde), Tru1I (azul), Sau3AI (roxo), MslI (vermelho) e EcoNI (laranja)
(continuações) ................................................................................................................. 21-26
FIGURA 8. Eletroforese em gel de poliacrilamida dos produtos de PCR do gene rpoB
digeridos com a endonuclease de restrição TaqI; Mr corresponde ao padrão de massa
molecular 100 pb ladder ....................................................................................................... 30
FIGURA 9. Eletroforese em gel de poliacrilamida dos produtos de PCR do gene rpoB
digeridos com a endonuclease de restrição Tru1I; Mr corresponde ao padrão de massa
molecular 100 pb ladder ....................................................................................................... 31
FIGURA 10. Eletroforese em gel de poliacrilamida dos produtos de PCR do gene rpoB
digeridos com a endonuclease de restrição Sau3AI; Mr corresponde ao padrão de massa
molecular 100 pb ladder ....................................................................................................... 32
FIGURA 11. Eletroforese em gel de poliacrilamida dos produtos de PCR do gene rpoB
digeridos com a endonuclease de restrição MslI; Mr corresponde ao padrão de massa
molecular 100 pb ladder ....................................................................................................... 33
FIGURA 12. Eletroforese em gel de poliacrilamida dos produtos de PCR do gene rpoB
digeridos com a endonuclease de restrição EcoNI; Mr corresponde ao padrão de massa
molecular 100 pb ladder ....................................................................................................... 34
FIGURA 13. Dendograma dos 68 sorovares de Leptospira obtido pela análise das bandas
geradas com a endonuclease de restrição TaqI ................................................................... 39

x
FIGURA 14. Dendograma dos 68 sorovares de Leptospira obtido pela análise das bandas
geradas com a endonuclease de restrição Tru1I ................................................................. 40
FIGURA 15. Dendograma dos 68 sorovares de Leptospira obtido pela análise das bandas
geradas com a endonuclease de restrição Sau3AI .............................................................. 41
FIGURA 16. Dendograma dos 68 sorovares de Leptospira obtido pela análise das bandas
geradas com a endonuclease de restrição MslI ................................................................... 42
FIGURA 17. Dendograma dos 68 sorovares de Leptospira obtido pela análise das bandas
geradas com a endonuclease de restrição EcoNI ................................................................ 43
FIGURA 18. Dendograma dos 68 sorovares de Leptospira obtido pela análise das bandas
geradas com as endonucleases de restrição TaqI, Tru1I, Sau3AI, MslI e EcoNI ................ 44
LISTA DE TABELAS
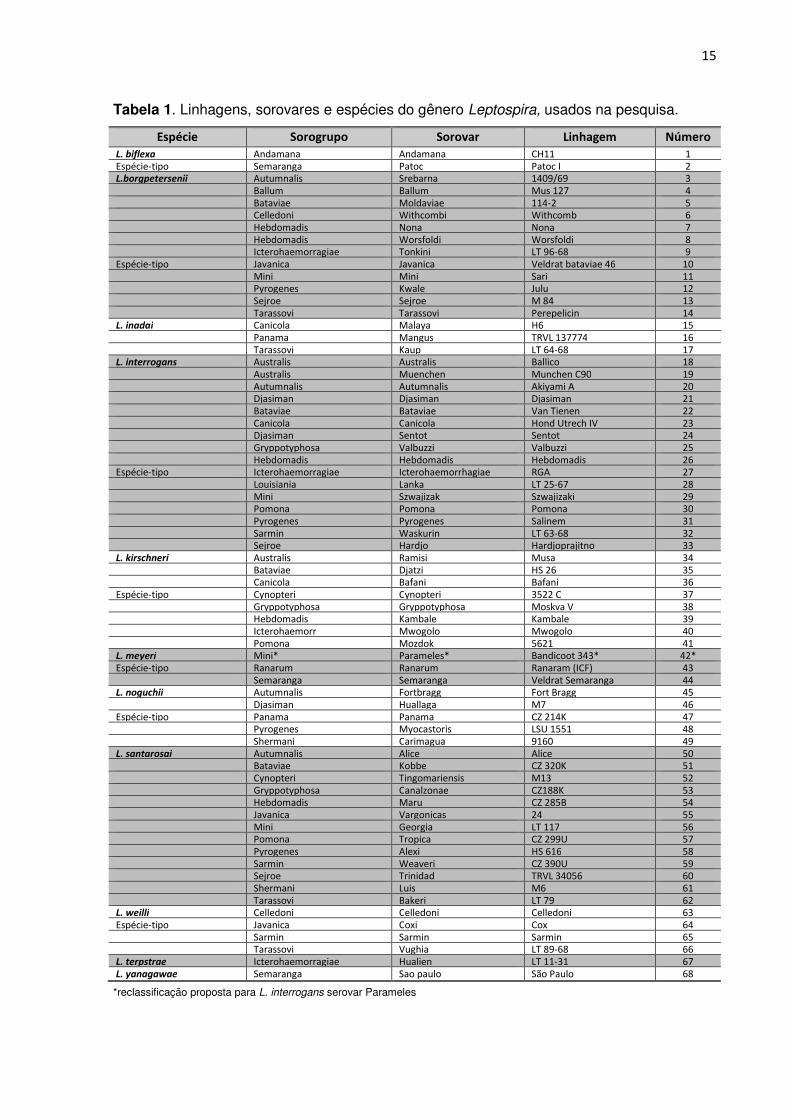
TABELA 1. Linhagens, sorovares, sorogrupos e espécies do gênero Leptospira, usados na
pesquisa ................................................................................................................................ 14
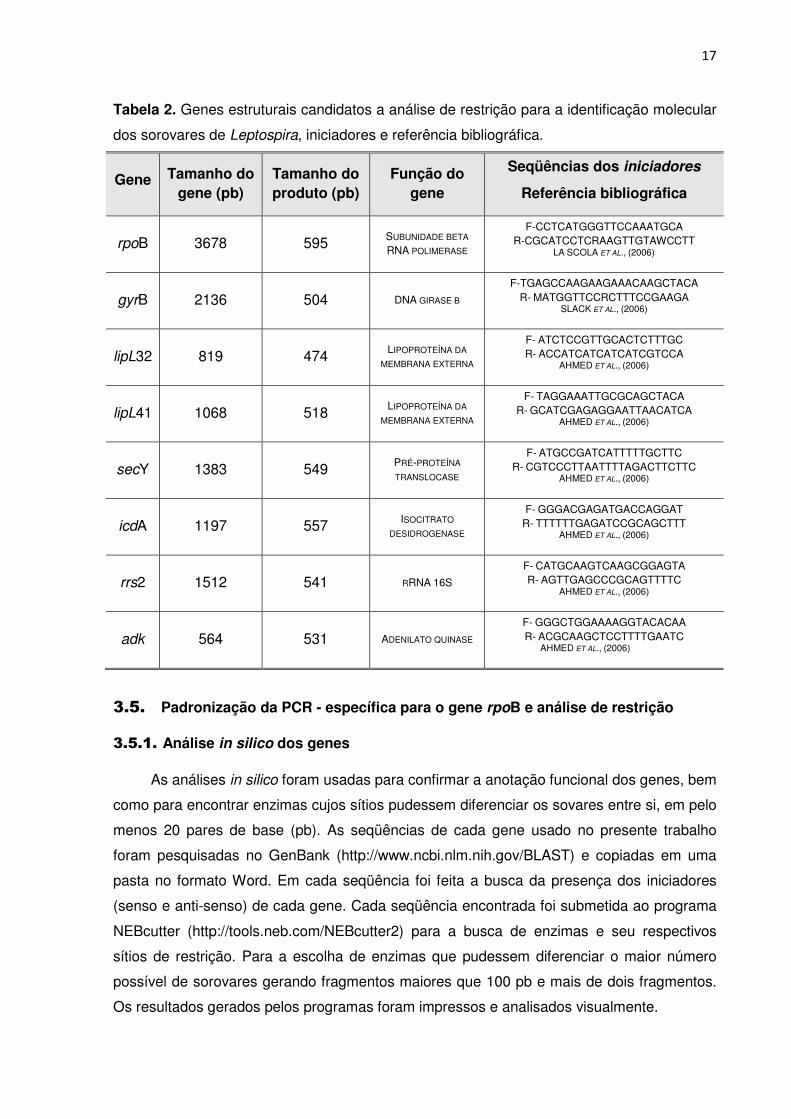
TABELA 2. Genes estruturais candidatos a análise de restrição para a identificação
molecular dos sorovares de Leptospira, iniciadores e referência bibliográfica .................... 16
TABELA 3. Condições da PCR - específica com os iniciadores Lept 1900f e 2500r,
produzidos para amplificação de parte do gene rpoB .......................................................... 17
TABELA 4A. Perfis de restrição dos sorovares de Leptospira com a enzima TaqI ............. 27
TABELA 4B. Perfis de restrição dos sorovares de Leptospira com a enzima Tru1I ............ 27
TABELA 4C. Perfis de restrição dos sorovares de Leptospira com a enzima Sau3AI ........ 28
TABELA 4D. Perfis de restrição dos sorovares de Leptospira com a enzima MslI .............. 28
TABELA 4E. Perfis de restrição dos sorovares de Leptospira com a enzima EcoNI ........... 28
TABELA 5. Agrupamento dos sorovares, sorogrupos e espécies do gênero Leptospira,
usados na pesquisa, pelos perfis de restrição compostos pelas cinco endonucleases ....... 29
TABELA 6. Sorovares de Leptospira identificados pela metodologia de PCR-RFLP do gene
rpoB com as enzimas restrição TaqI, Tru1I, Sau3AI, MslI e EcoNI ...................................... 37

xi
Abreviaturas e Símbolos
% - por cento
µµµµ – micro
BSA – “bovine serum albumin” / albumina de soro bovino
DNA – ácido desoxirribonucléico
dNTP – 2’-desoxirribonucleotídeo 5’-trifosfato
EDTA – ácido etilenodiaminotetracético
ELISA – “enzyme-linked immunosorbent assay” / ensaio de imunoadsorção por ligação
enzimática
EST – “Expressed Sequence Tags” / Marcadores de seqüências expressas
g – grama
h – horas
kDa – quilo Dalton
L – litro
LPS – lipopolissacarídeos
m – mili
M – molar
MLST – “Multi Locus Sequence Typing” / Tipagem por seqüência de múltiplos locos
nm – nanômetro
ºC – graus Célsius
OM – “outer membrane” / membrana externa
OMS – Organização Mundial de Saúde
pb – pares de bases
PCR – “polymerase chain reaction” / reação em cadeia da polimerase
PFGE – “Pulse Field Gel Electrophoresis” / Eletroforese em Gel de Campo Pulsátil
rpm – rotações por minuto
PAGE – eletroforese em gel de poliacrilamida
Tris – tris-(hidroximetil)-aminometano
U – unidade
V – volts
VNTR – “Variable Number of Tandem Repeats” / Número variável de repetições em
tandem
xg – gravidade

1
RESUMO
A leptospirose é uma zoonose de distribuição mundial, com altas taxas de mortalidade e
morbidade em humanos e animais, cujo agente etiológico é classificado em uma variedade
de sorogrupos e sorovares em função de seus determinantes antigênicos. A caracterização
destes sorovares circulantes é essencial para o melhor entendimento da epidemiologia da
doença e, para tanto, até recentemente, a observação da bactéria requeria microscopia de
campo escuro e a principal identificação era baseada em sorotipagem. Atualmente, estudos
envolvendo ferramentas moleculares alternativas para a detecção, identificação e
classificação das linhagens epidemiológicas se fazem necessários, na tentativa de viabilizar
a tipagem de rotina através da aplicação de um método mais rápido, preciso e de fácil
execução. Esta necessidade por rapidez e caracterização específica das leptospiras tem
estimulado o uso de novos testes moleculares tais como amplificação por PCR de regiões
variáveis de genes conservados para a tipagem dos sorovares. O objetivo deste estudo foi
investigar a possível identificação dos sorovares de Leptospira por RFLP do gene rpoB,
codificador da subunidade beta da RNA polimerase. DNA foi extraído de 68 sorovares de
Leptospira, pertencentes a 11 espécies patogênicas e saprofíticas diferentes. Ensaios por
PCR foram realizados usando iniciadores específicos desenhados para amplificar um
fragmento polimórfico do gene rpoB. A digestão enzimática foi realizada usando as
endonucleases de restrição TaqI, Tru1I, Sau3AI, MslI e EcoNI, selecionadas previamente
pelos perfis de digestão in silico de ESTs (Expressed Sequence Tags) e seqüências de
nucleotídeos disponíveis nos bancos de dados. Os fragmentos de restrição foram separados
por eletroforese em géis de poliacrilamida e corados com nitrato de prata. A metodologia
empregada permitiu diferenciar 37% dos sorovares testados. Contudo 43 sorovares de 9
espécies ainda permaneceram agrupados sem um perfil individual. Por outro lado, vale
ressaltar que neste trabalho foram avaliados apenas cerca de 25% das quase 268 sorovares
já descritas na literatura. A tipagem por PCR-RFLP se mostrou viável apesar de manter a
maior parte dos sorovares agrupada em pequenos clusters, podendo ser muito útil no
estudo epidemiológico da leptospirose e para o desenvolvimento de estratégias apropriadas
de prevenção, ou ainda, para a identificação das espécies de Leptospira.

2
ABSTRACT
Leptospirosis is a zoonotic disease with high mortality and morbidity rates in humans and
animals throughout the world, which its causal agent, Leptospira spp., has been classified
into serogroups and serovars as a function of their antigenic determinants. Identification of
Leptospira species is not as important as the determination of the serovar of isolates
because this is essential to an understanding of the epidemiology of leptospirosis. Modern
DNA-based methodologies have greatly improved laboratory procedures for the detection,
identification and classification of epidemiologic strains. To date, observation of these
bacteria requires dark-field microscopy, and their major identification has long relied on
serotyping. The development of alternative tools for identification of Leptospira serovars is
required for routine typing, to circumvent the need of the preparation of rabbit immune sera
against them and become the methodology less time-consuming, more accurate and easy to
be done. This need for rapid and specific characterization of leptospires has stimulated the
use of new molecular tests such as PCR amplification of variable regions of conserved
genes for serovar typing. The aim of this study was to investigate the possible identification
of Leptospira serovars by RFLP typing of rpoB gene, coding the beta subunit of RNA
polymerase. DNA was extracted from 68 Leptospira serovars, belonging to 11 different
pathogenic and saprophytic species. Polymerase chain reaction (PCR) assays were
performed using specific primers designed to amplify a polymorphic fragment of the gene
rpoB. The enzymatic digestion was performed using the restriction endonucleases TaqI,
Tru1I, Sau3AI, MslI and EcoNI, previously selected based on in silico digestion of Expressed
Sequence Tags and nucleotide sequences available in databases. The restriction fragments
were separated in polyacrylamide gels and stained with silver nitrate. We were able to
differentiate 37% of the serovars tested, however 43 serovars from 9 species were not
separated yet, been grouped at small clusters. By use of restriction typing, the phylogenetic
study of extremely fastidious Leptospira species can be carried out without the need to
culture them. In addition, this technique is a promising tool to evaluate genetic diversity of
related strains and may be useful to differentiate Leptospira serovars within the species.

3
INTRODUÇÃO
1.1. A leptospirose: contextualização histórica
Foi em 1886 que WEIL deu seu nome à doença humana que afetava as tropas de
Napoleão Bonaparte, caracterizando-a como uma doença infecciosa que causa
esplenomegalia, icterícia e nefrite (FAINE et al., 1999). O agente causador ainda não era
conhecido, mas estava associado a várias síndromes em animais domésticos e selvagens.
Em 1907, STIMSON usou uma técnica descrita por LEVADITH para corar e observar
espiroquetas nos tecidos de rins de pacientes mortos com sintomas da Febre de Weil. A
coloração foi reproduzida e fotografada por NOGUCHI em 1928, que a denominou de
Spirochaeta ? (interrogans). E foi no Japão, onde a doença de Weil era comum entre os
mineradores, que em 1914 INADA e colaboradores conseguiram infectar, com sucesso,
porquinhos da índia, nos quais observaram uma bactéria espiralada denominando-a
Spirochaeta icterohaemorrhagiae. Estes pesquisadores conseguiram ainda preparar um
soro imune contra a bactéria e demonstraram proteção passiva e bacteriólise intraperitoneal
nas cobaias. Em trabalhos sucessivos foram descritos o modo de infecção, a distribuição
das bactérias nos tecidos, a excreção, multiplicação, filtrabilidade e características
morfológicas, inclusive a degeneração de suas células dentro de formações granulares.
Entre 1914 e 1918, durante a Primeira Guerra Mundial na Europa, houve aumento da
doença, estimulando a pesquisa de campo na Alemanha. Dessa época em diante foram
conhecidos outros agentes da doença, de origem leptospiral, que reagiam com diferentes
soros testados (sorovares) e, ainda, que alguns desses sorovares provocavam icterícia e
outros não.
Nas últimas décadas a leptospirose ficou conhecida como uma importante
antropozoonose mundialmente distribuída causada por espiroquetas patogênicas do gênero
Leptospira (Ordem Spirochaetales, família Leptospiraceae), podendo acometer animais
silvestres, domésticos e o homem. A doença tem distribuição cosmopolita, ocorrendo em
ambientes urbanos de cidades industrializadas e em desenvolvimento, bem como nas áreas
rurais de todo o Planeta (BHARTI et al., 2003). De acordo com LEVETT em 2001 e
MCBRIDE et al. em 2005, trata-se de uma zoonose que tem os roedores como reservatório,
sendo o Rattus norvegicus o principal reservatório urbano (Figura 1).
Em publicação da Organização Mundial de Saúde (OMS) de 1999, a leptospirose foi
considerada uma doença infecciosa reemergente, principalmente de regiões tropicais e
subtropicais, com a ocorrência de extensos surtos na Nicarágua, Índia, Brasil, Sudeste da
Ásia, Estados Unidos da América, China, Malásia, e outros países, com mais de 500.000
casos cada ano e índice de mortalidade superior a 10%.
1.2. Patogênese na leptospirose
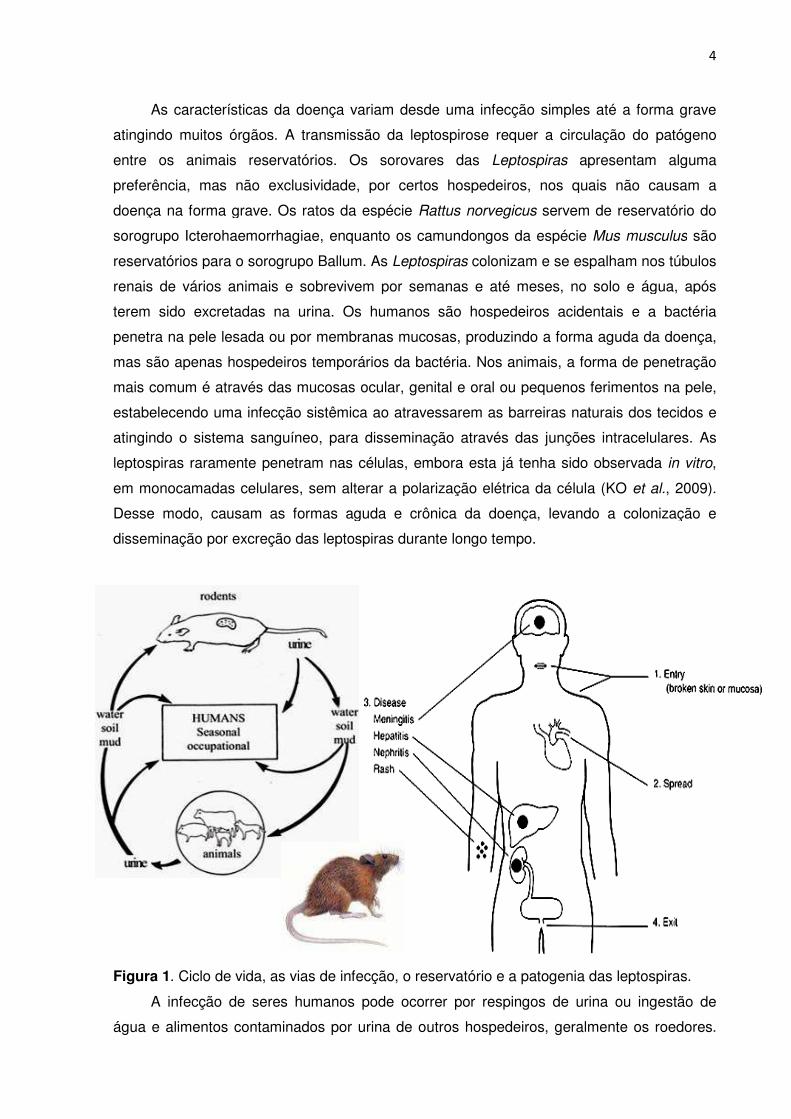
As características da doe
atingindo muitos órgãos. A tran
entre os animais reservatório
preferência, mas não exclusiv
doença na forma grave. Os rato
sorogrupo Icterohaemorrhagiae
reservatórios para o sorogrupo B
renais de vários animais e sob
terem sido excretadas na urin
penetra na pele lesada ou por m
mas são apenas hospedeiros te
mais comum é através das muc
estabelecendo uma infecção sis
atingindo o sistema sanguíneo,
leptospiras raramente penetram
em monocamadas celulares, se
Desse modo, causam as form
disseminação por excreção das
Figura 1. Ciclo de vida, as vias
A infecção de seres hum
água e alimentos contaminados
ença variam desde uma infecção simples até
ansmissão da leptospirose requer a circulação
rios. Os sorovares das Leptospiras aprese
ividade, por certos hospedeiros, nos quais n
atos da espécie Rattus norvegicus servem de r
e, enquanto os camundongos da espécie Mus
Ballum. As Leptospiras colonizam e se espalha
obrevivem por semanas e até meses, no solo
rina. Os humanos são hospedeiros acidentais
r membranas mucosas, produzindo a forma agu
temporários da bactéria. Nos animais, a forma
ucosas ocular, genital e oral ou pequenos ferim
sistêmica ao atravessarem as barreiras naturais
o, para disseminação através das junções intr
m nas células, embora esta já tenha sido obse
sem alterar a polarização elétrica da célula (KO
mas aguda e crônica da doença, levando a
s leptospiras durante longo tempo.
s de infecção, o reservatório e a patogenia das l
manos pode ocorrer por respingos de urina o
os por urina de outros hospedeiros, geralment
4
é a forma grave
ão do patógeno
sentam alguma
não causam a
reservatório do
us musculus são
lham nos túbulos
lo e água, após
ais e a bactéria
guda da doença,
a de penetração
imentos na pele,
is dos tecidos e
ntracelulares. As
servada in vitro,
KO et al., 2009).
a colonização e
leptospiras.
ou ingestão de
nte os roedores.

5
As bactérias após a penetração pela pele lesada ou membranas mucosas alcançam o
sistema circulatório e sendo altamente móveis, migram para o líquido cefalorraquidiano,
produzem hialuronidases, multiplicam-se rápida e intensamente, atingindo vários órgãos,
sendo responsáveis pelo rompimento de pequenos vasos, causando as principais
manifestações da leptospirose. Os fatores de virulência das leptospiras mais conhecidos são
os filamentos axiais (para a penetração em “saca-rolhas”), a capa de proteínas específicas e
lipopolissacarídeos (endotoxinas).
Os sintomas da doença são freqüentemente confundidos com os de uma gripe
comum, indo até a falência renal. Estes sintomas podem ser semelhantes aos da Dengue,
Malária e várias outras doenças, atrasando o diagnóstico que raramente é feito em tempo
de se tratar o paciente com os cuidados necessários e, se a doença não for tratada, o
paciente pode desenvolver meningite, falência dos rins, fígado, respiratória e até morrer.
1.3. Características do gênero Leptospira
1.3.1. As leptospiras: características estruturais e requerimentos nutricionais
As leptospiras formam um grupo de bactérias que divergiu cedo na evolução dos seres
vivos; vivem em vários ambientes como agentes infecciosos de animais ou em vida livre
(Figura 1). As leptospiras são aeróbias obrigatórias, altamente flexíveis e móveis, com forma
alongada, helicoidal e suas células têm diâmetro que varia conforme o grupo de 0,09 µm a
0,75 µm e comprimento de 3 µm a 500 µm. Possuem uma estrutura típica, sendo envoltas
por um envelope de várias camadas (envelope externo), logo abaixo a camada de
peptideoglicanos (parede celular) e a membrana plasmática externa envolvendo o conteúdo
do cilindro protoplasmático da célula (OM) (Figura 2). Entre o envelope externo e a parede
celular estão os flagelos axiais, responsáveis pelo movimento rotatório da bactéria, um em
cada pólo, estendidos para o centro, porém nunca se tocam. As proteínas FlaA e FlaB
formam a bainha e centro dos flagelos, respectivamente. PICARDEAU et al. em 2001,
demonstraram que mutantes para essas proteínas não têm mobilidade. Os
lipopolissacarídeos (LPS) do envelope externo são o alvo principal do sistema anticorpo-
complemento, na reação bactericida do hospedeiro (JOHNSON, 1977). Na maioria das
espécies há uma dobra em forma de gancho em uma das extremidades da célula,
conferindo mais uma característica exclusiva ao grupo (Figura 3).

6
a..
b..
Figura 2. a, b) estrutura das Leptospiras; c) posição dos flagelos e eixo de rotação no
espaço periplasmático.
O seqüenciamento dos genes que codificam a porção 16S do RNA ribossômico indica
que as espiroquetas são o grupos mais antigos entre as bactérias, que evoluíram de um
único ancestral comum, as proto-espiroquetas (MATTHIAS et al., 2008), e contém
ramificações, que refletem a patogenicidade das espécies. Considerando a relação
constante de divergência em 1 a 2% em cada 50 milhões de anos, para o gene rrs, calcula-
se que o tempo de separação entre L. interrogans e L. biflexa esteja entre 590 e 295
milhões de anos (CERQUEIRA E PICARDEAU, 2009).
Figura 3. Leptospira interrogans, sorovar Icterohaemorrhagiae, exibindo as estruturas em
gancho. a. eletromicroscopia, b. microscopia em campo escuro.
Os lipopolissacarídeos (LPS) do envelope externo constituem o principal antígeno de
Leptospira e são estrutural e imunologicamente similares aos LPS de organismos Gram-
negativos. (ADLER E DE LA PEÑA MOCTEZUMA, 2009). Os lipopolissacarídeos são o
primeiro contato da bactéria com os tecidos do hospedeiro e o alvo do sistema anticorpo-
complemento, na reação bactericida do hospedeiro (JOHNSON, 1977).
Os lipídios A das leptospiras possuem características pouco comuns tais como uma
unidade do dissacarídeo glicosamina, fosforilado e metilado. Vários polipeptídios estruturais
e funcionais estão presentes na OM e atuam na interação célula-hospedeiro, juntamente

7
com os LPS. Estes polipeptídios são lipoproteínas, abundantes na superfície celular, LipL32,
LipL21 e LipL41 (NASCIMENTO et al., 2004).
A temperatura ideal para crescimento das leptospiras, em laboratório, é entre 28 e
30ºC, mas já foi comprovado que as bactérias patogênicas se reproduzem melhor em
temperatura corporal, desde que não ultrapasse 42ºC. Estas sobrevivem sob congelamento,
voltando a reproduzirem-se aos 10ºC. Não sobrevivem na ausência de água e são muito
sensíveis a produtos químicos, detergentes, ácidos e metais pesados, mesmo em baixas
concentrações. Utilizam ácidos graxos de cadeia longa como fonte de carbono, sais de
amônio e são exigentes quanto a algumas vitaminas (B1 e B12) e o pH do meio (7,2 a 7,4).
1.3.2. Identificação e taxonomia: a complexidade do gênero Leptospira
O gênero Leptospira pertence ao filo Spirochaetes e inclui três subgrupos, o de
espécies saprófitas com Leptospira biflexa, L. wolbachii, L. kimetyi, L. meyeri, L. vauthielli, L.
terpstrae e L. yanagawae, o grupo de espécies patogênicas com L. interrogans, L.
kirschneri, L. noguchii, L. santarosai, L. borgpetersenii, L. weilii, L. alexanderi e L. alstonii, e
um grupo intermediário onde estão as espécies L. inadai, L. broomi, L. fanei, L. wolffii e L.
licerasiae, cuja patogenicidade ainda não foi definida.
O gênero é dividido em mais de 250 sorovares, pertencentes a duas espécies: L.
interrogans e L. biflexa (classificação sorológica ou sensu lato), contendo as linhagens
patogênicas e saprofíticas, respectivamente. Esta classificação sorológica é baseada em
testes imunológicos como reação de soro-aglutinação microscópica (SAM) e absorção
cruzada de aglutininas. Contudo, o desenvolvimento da biologia molecular tem proposto
mudanças taxonômicas, classificando em três espécies não patogênicas, L. biflexa, L.
meyeri, L. wolbachii, e em sete patogênicas, L. interrogans, L. inadai, L. kirschneri, L.
noguchii, L. santarosai, L. weilii e L. borgpetersenii (OLIVEIRA et al, 2009). A caracterização
das Leptospiras isoladas de pacientes é importante para compreender as propriedades
epidemiológicas da doença e o sorovar é usado para classificar essas bactérias, definindo-
os com base na heterogeneidade estrutural dos carboidratos que compõem a camada de
lipopolissacarídeos (LPS) (CERQUEIRA E PICARDEAU, 2009) presente na membrana
externa (OM), conforme ilustrado na Figura 4.

8
Fig
ura
4.
Arq
uitetura da superfície das leptospiras, mostrando na membrana externa (outer membrane) a
camada de lipopolissacarídeos (LPS) que define os sorovares (ZUERNER et al., 2000).
Com os métodos genéticos, que levam a classificação sensu stricto, ficou claro que o
conceito de “sorovar” já não é satisfatório, pois falha ao definir linhagens
epidemiologicamente importantes. Por exemplo, por tipificação molecular demonstrou-se
que é possível discriminar, com mais eficiência, linhagens do sorogrupo Grippotyphosa, do
que com tipificação sorológica (STEINEN et al., 1992; HARTSKEERT et al., 2004). A
correlação entre classificação sorológica e genotípica é baixa e a identificação é complicada
porque sorovares de um mesmo sorogrupo podem estar distribuídos entre espécies
diferentes. Estudos sobre essa falta de correlação entre espécie e sorovar levam a
suposição que os genes que determinam os sorotipos são transferidos horizontalmente
entre espécies, porém a base da transferência entre diferentes espécies de Leptospira, que
seria responsável pela troca genética de determinantes LPS, ainda é desconhecida
(CERQUEIRA E PICARDEAU, 2009).
A tipificação molecular começou em 1990 com o surgimento dos métodos baseados em
PCR (Reação em Cadeia da DNA Polimerase) e PFGE (Eletroforese em Gel de Campo
Pulsátil). A identificação em nível de espécies auxilia os estudos epidemiológicos apenas no
sentido de diferenciar grupos patogênicos de saprofíticos. Uma vez que cada sorovar está
associado a um tipo de hospedeiro, sua identificação é essencial para o desenvolvimento de
estudos epidemiológicos (CERQUEIRA E PICARDEAU, 2009; WENJUN et al., 2009).
Após o isolamento da bactéria é possível identificar o sorovar por vários métodos
moleculares, como os baseados na análise de REA (Restriction Endonuclease Analysis) e
RFLP (Restriction Fragment Length Polymorphism) de genes específicos (THIERMANN et
al., 1985; PEROLAT et al., 1993), pela comparação dos perfis de bandas da digestão
obtidos após eletroforese em géis de agarose ou poliacrilamida. Estes métodos são
eficientes para diferenciar algumas linhagens de Leptospira, porém são muito trabalhosos e

9
demandam grande volume de cultura dessas bactérias, além de formarem muitas bandas,
dificultando a comparação entre elas (RAMADASS, 2001). A REA pode ser feita em
produtos amplificados por PCR e os genes responsáveis pelas subunidades 16S e 23S do
RNA ribossômico tem sido os favoritos (RALPH et al., 1993; WOO et al., 1997).
A análise por PFGE é considerada um método forte de classificação de linhagens de
Leptospira, pois mostra as diferenças no número de bandas e tamanho dos perfis de
macrorestrição que podem ser originados por rearranjos no genoma, inserção, deleção,
inversão ou substituição de bases, dentro dos sítios de restrição.
Segundo os critérios de classificação genética por hibridização DNA-DNA, duas
linhagens são consideradas pertencentes à mesma espécie genômica se (1) o DNA da
Leptospira isolada for 70% ou mais relacionado com o DNA da amostra de referência, a uma
temperatura ótima de reassociação de 55ºC; (2) os DNAs destas forem 60% ou mais
relacionados entre si, a uma temperatura estringente de reassociação de 70ºC; (3) os DNAs
relacionados entre si apresentarem 5% ou menos de bases não pareadas (RAMADASS et
al., 1992 e WOO et al., 1997).
A metodologia de RAPD (Randomly Amplified Polymorphic DNA), também denominado
“fingerprinting”, usa a amplificação por PCR com iniciadores pequenos de seqüência
conhecida para gerar perfis linhagem- ou espécie-específicos (LEVETT, 2001). RALPH et
al. (1993) associaram a essa técnica o seqüenciamento do gene do RNA ribossômico 16S e
confirmaram a diferenciação entre as espécies de Leptospira.
A metodologia de AP-PCR (Amplified Fragment Length Polymorphism) envolve
procedimentos nos quais o DNA é cortado com duas enzimas de restrição, os fragmentos
ligados a adaptadores que são complementares as extremidades geradas pela restrição, e
amplificados com iniciadores específicos complementares aos adaptadores para gerar
imagens de perfis “footprints”, que são comparadas por programas próprios, em
computador. Atualmente, diversos programas de informática para análise de géis podem ser
usados para observar a relação entre as linhagens, tornando possível a construção de
dendogramas e a comparação com dados de outros laboratórios (GALLOWAY E LEVETT,
2008).
Nos últimos anos as seqüências completas do genoma de várias leptospiras
patogênicas e saprofíticas foram publicadas, bem como seqüências de partes de genes têm
sido depositadas em diversos bancos de dados, para que possam ser acessadas e
comparadas, por pesquisadores. Já são conhecidas diversas seqüências repetitivas e de
inserção no genoma das Leptospiras bem como de estruturas típicas de tandem repeats,
pequenas seqüências de DNA repetitivas; algumas polimórficas, que são denominadas
VNTR (Variable Number of Tandem Repeats). Esse tipo de seqüência é extensivamente
usado para identificar eucariotos e, análises em bancos de dados dessas repetições,

10
mostraram que L. interrogans apresenta muitas delas, com tamanhos menores que 100 pb,
enquanto L. borgpetersenii, possui poucas. A utilização da técnica do MLVA (Multi Locus
VNTR Analysis) requer construção de iniciadores específicos, para cada espécie, pouco
relacionada filogeneticamente. Essa técnica e específica para linhagens patogênicas e pode
distinguir sorovares dos gêneros patogênicos mais pesquisados, entre as leptospiras.
Cada método avaliado tem apresentado sua própria sensibilidade, especificidade e
reprodutibilidade. Provavelmente, alguns importantes fatores tais como: a técnica de
extração do DNA, qualidade do DNA obtido, tipo de amostra clínica, presença de inibidores,
tipo de iniciadores, termocicladores e qualidade dos reagentes, dentre outros, têm
contribuído para variações da taxa de reprodutibilidade de algumas destas técnicas.
Esses fatores, somados à falta de validação internacional e protocolos padronizados
têm dificultado a implementação rotineira desses métodos pelos laboratórios que trabalham
no diagnóstico e identificação das Leptospiras.
Recentemente, a técnica de seqüenciamento de múltiplos locos gênicos tem sido
aplicada na tipagem de bactérias patogênicas gerando resultados com alto grau de precisão
que são adequados para a maioria de estudos epidemiológicos e populacionais. Multi Locus
Sequence Typing (MLST) é um método que mede, diretamente, a variação existente nas
seqüências internas de fragmentos de DNA de um grupo de genes estruturais, geralmente
sete ou oito genes, caracterizando as linhagens pelo perfil obtido. A técnica é baseada na
amplificação por PCR, de partes de seqüências dos genes selecionados, seguida de
seqüenciamento automático de DNA (AHMED et al.,2006).
Com a técnica de MLST pode-se conhecer as linhagens mais relacionadas e definir os
loci, cuja variação na seqüência possibilitará sua utilização na identificação rotineira, de
sorovares patogênicos. A vantagem desse método é a facilidade de acesso às seqüências
dos iniciadores (primers) e a obtenção de diversos protocolos, que estão disponíveis em
bancos de dados, além da deposição das ESTs obtidas de cada sorovar (ADLER & DE LA
PEÑA MOCTEZUMA, 2009).
O MLST também permite que sejam produzidos iniciadores para regiões específicas
dos genes selecionados, que após a amplificação por PCR, são submetidas à digestão com
endonucleases de restrição e analisadas por eletroforese em gel, dispensando o uso do
seqüenciamento de DNA. A visualização dos polimorfismos por esta estratégia sem o
seqüenciador de DNA permite que a técnica seja emprega em mais laboratórios e com
menor custo, por não necessitar de equipamentos especiais.
Vale ressaltar que, a identificação dos sorovares é de suma importância epidemiológica
uma vez as vacinas existentes contra leptospirose animal são preparadas a partir dos
sorovares circulantes em cada região e que o homem é o hospedeiro susceptível, na cadeia
de transmissão.

11
1.3.3. O genoma de Leptospira spp.
Os cromossomas das Leptospiras são caracterizados por seu teor de bases C+G que
fica entre 35 e 41 mol%, dependendo da espécie e o tamanho do genoma está entre 3,0 e
4,6 Mb (Figura 5). As espécies de Leptospira interrogans e L. borgpetersenii (sensu stricto)
possuem um genoma complexo, contendo dois cromossomas circulares de
aproximadamente 4 Mb e 300 Kb, respectivamente. Estudos comparativos do cromossoma
mais longo mostram uma organização genética com grandes rearranjos nas L. interrogans,
que diferencia as duas linhagens relacionadas, demonstrando longo período de mudanças
na escala evolutiva (BORSAUX et al., 2005). NASCIMENTO et al. (2004), descreveram a
presença de seqüências de inserção (elementos IS) no genoma de L. interrogans,
distribuídas ao longo do cromossoma; juntos esses elementos IS e as transposases ocupam
2% do genoma, além disto, descobriram que essas seqüências estão localizadas e podem
ser responsáveis pela variação no grau de patogenicidade e especificidade de alguns
sorovares, para determinados hospedeiros.
Na tentativa de elucidar a diversidade sorológica entre as leptospiras, os genes
envolvidos na biossíntese dos lipopolissacarídeos e lipoproteínas também estão sendo
intensamente estudados, uma vez que Leptospira spp. possui mais genes para lipoproteínas
que os genomas de bactérias não espiraladas. Conhecendo-se a habilidade com a qual as
leptospiras atravessam os tecidos do hospedeiro, deduz-se que seu genoma codifica
diversos tipos de proteases e hemolisinas que facilitam essa penetração. Das proteínas da
parede celular e membrana plasmática, já estão sendo pesquisadas suas funções na
fisiologia da bactéria, tentando desvendar os mecanismos envolvidos nos processos de
especificidade com hospedeiros e patogenia.
Já foram identificados nove genes que codificam hemolisinas, incluindo uma proteína
formadora de poro e uma esfingomielinase, além de uma colagenase microbiana conhecida
por estar envolvida com a destruição de tecidos do hospedeiro em L. interrogans sendo que
esses genes não existem nas L. biflexa.
A origem de replicação do maior cromossoma (replicon) das leptospiras foi localizada
entre os genes dnaA e dnaN, tal como em outros genomas de bactérias. A seqüência total
do genoma de algumas espécies já é conhecida, entretanto, os mecanismos de trocas
genéticas entre essas bactérias ainda é pouco estudado. Já estão sendo desenvolvidas
ferramentas para manipulação genética e estudo dos fatores de virulência e patogenia
(BHARTI et al., 2003). Nas L. biflexa existe um cromossoma adicional de 74 Kb e ainda
podem ter um quarto replicon circular de 74 Kb (bacteriófago LE1).

12

13
Figura 5. Características dos genomas já seqüenciados de leptospiras patogênicas e
saprofíticas (PICARDEAU et al., 2008).
OBJETIVO GERAL
Identificar espécies e sorovares de Leptospira spp. usando a técnica PCR-RFLP do
gene da subunidade β da RNA polimerase (rpoB).
OBJETIVOS ESPECÍFICOS
1. Selecionar na literatura genes com segmentos altamente polimórficos para diferenciação
dos sorovares de Leptospira;
2. Alinhar as seqüências do gene rpoB (codificador da subunidade β da RNA polimerase) de
Leptospira spp. presentes no GenBank e realizar digestão in silico com endonucleases de
restrição;
3. Amplificar por PCR um segmento polimórfico do gene rpoB de 68 sorovares de Leptospira
spp. e realizar digestão com as endonucleases de restrição TaqI, Tru1I, Sau3AI, MslI e
EcoNI.
4. Identificar os diferentes perfis de restrição e construir dendogramas para agrupá-los.

14
MATERIAL E MÉTODOS
Para a execução do presente trabalho, foram selecionados sessenta e oito sorovares
de Leptospira de referência (Tabela 1), distribuídos entre onze espécies, originárias do “Pan
American Institute for Food Protection and Zoonosis” (INNPAZ), gentilmente cedidas pelo
professor Dr. Élvio Carlos Moreira do Departamento de Medicina Preventiva da Escola de
Veterinária da Universidade Federal de Minas Gerais
3.1. Cultivo dos sorovares de Leptospira de referência
Os sorovares foram cultivados em meio de cultivo EMJH (Ellinghausen – McCullogh
– Johnson – Harris, Difco, BD Biosciences), no Laboratório de Microbiologia Aplicada do
Departamento de Microbiologia do Instituto de Ciências Biológicas da UFMG. Os meios
foram preparados e suplementados com Leptospira Enrichment EMJH (Difco, BD
Biosciences). Para o meio semi-sólido foram acrescidos 0,2% de ágar, tornando o meio
adequado para manutenção de culturas de Leptospira. As amostras foram mantidas em
meio semi-sólido EMJH, repicadas de 3 em 3 meses e incubadas a uma temperatura média
de 28-30ºC, até a formação do típico anel de crescimento. Para a obtenção do DNA foram
retiradas alíquotas de 0,5 ml e inoculadas em tubos contendo 6,0 ml do meio EMJH líquido,
no qual o crescimento ocorre de três a cinco dias, para a maioria dos sorovares. O
crescimento no meio líquido foi observado pela turvação delicada e clara e conferido em
microscopia de campo escuro (ELLINGHAUSEN, 1973).
3.2. Extração do DNA
Para a obtenção do DNA dos sorovares de Leptospira a cultura de cada tubo foi
transferida para tubo Falcon de 15 ml e centrifugada por 30 minutos a 3.000 rotações por
minuto (rpm). O sobrenadante foi desprezado e o sedimento suspenso em 1 ml de PBS (140
mM NaCl; 2,7 mM KCl, 10 mM Na2HPO4, 1,8 mM KH2PO4 em 1000ml, pH 7.2 ) e transferido
para tubo Eppendorf de 1,5 ml que foi centrifugado por 5 minutos à 14.000 rpm. O
sobrenadante foi desprezado e o sedimento foi suspenso em 1 ml de PBS e novamente
centrifugado por 2 minutos a 14.000 rpm e o sobrenadante descartado. Esse procedimento
foi repetido até que o sedimento estivesse completamente livre de restos de meio. Em cada
tubo Eppendorf foi adicionado 1 mg de lisozima (10 mg/ml em TE: Tris 50 mM, EDTA 10
mM, pH 8.0) e incubado por 1 hora e trinta minutos à 37°C, depois centrifugado por 2
minutos à 14.000 rpm e o sobrenadante descartado, adicionou-se 400 µl do tampão de lise
do Kit Wizard Genomic DNA Purification System da Promega Co. para rompimento das
células e liberação do DNA.
15
Tabela 1. Linhagens, sorovares e espécies do gênero Leptospira, usados na pesquisa.
Espécie Sorogrupo Sorovar Linhagem Número L. biflexa Andamana Andamana CH11 1 Espécie-tipo Semaranga Patoc Patoc I 2 L.borgpetersenii Autumnalis Srebarna 1409/69 3 Ballum Ballum Mus 127 4 Bataviae Moldaviae 114-2 5 Celledoni Withcombi Withcomb 6 Hebdomadis Nona Nona 7 Hebdomadis Worsfoldi Worsfoldi 8 Icterohaemorragiae Tonkini LT 96-68 9 Espécie-tipo Javanica Javanica Veldrat bataviae 46 10 Mini Mini Sari 11 Pyrogenes Kwale Julu 12 Sejroe Sejroe M 84 13 Tarassovi Tarassovi Perepelicin 14 L. inadai Canicola Malaya H6 15 Panama Mangus TRVL 137774 16 Tarassovi Kaup LT 64-68 17 L. interrogans Australis Australis Ballico 18 Australis Muenchen Munchen C90 19 Autumnalis Autumnalis Akiyami A 20 Djasiman Djasiman Djasiman 21 Bataviae Bataviae Van Tienen 22 Canicola Canicola Hond Utrech IV 23 Djasiman Sentot Sentot 24 Gryppotyphosa Valbuzzi Valbuzzi 25 Hebdomadis Hebdomadis Hebdomadis 26 Espécie-tipo Icterohaemorragiae Icterohaemorrhagiae RGA 27 Louisiania Lanka LT 25-67 28 Mini Szwajizak Szwajizaki 29 Pomona Pomona Pomona 30 Pyrogenes Pyrogenes Salinem 31 Sarmin Waskurin LT 63-68 32 Sejroe Hardjo Hardjoprajitno 33 L. kirschneri Australis Ramisi Musa 34 Bataviae Djatzi HS 26 35 Canicola Bafani Bafani 36 Espécie-tipo Cynopteri Cynopteri 3522 C 37 Gryppotyphosa Gryppotyphosa Moskva V 38 Hebdomadis Kambale Kambale 39 Icterohaemorr Mwogolo Mwogolo 40 Pomona Mozdok 5621 41 L. meyeri Mini* Parameles* Bandicoot 343* 42* Espécie-tipo Ranarum Ranarum Ranaram (ICF) 43 Semaranga Semaranga Veldrat Semaranga 44 L. noguchii Autumnalis Fortbragg Fort Bragg 45 Djasiman Huallaga M7 46 Espécie-tipo Panama Panama CZ 214K 47 Pyrogenes Myocastoris LSU 1551 48 Shermani Carimagua 9160 49 L. santarosai Autumnalis Alice Alice 50 Bataviae Kobbe CZ 320K 51 Cynopteri Tingomariensis M13 52 Gryppotyphosa Canalzonae CZ188K 53 Hebdomadis Maru CZ 285B 54 Javanica Vargonicas 24 55 Mini Georgia LT 117 56 Pomona Tropica CZ 299U 57 Pyrogenes Alexi HS 616 58 Sarmin Weaveri CZ 390U 59 Sejroe Trinidad TRVL 34056 60 Shermani Luis M6 61 Tarassovi Bakeri LT 79 62 L. weilli Celledoni Celledoni Celledoni 63 Espécie-tipo Javanica Coxi Cox 64 Sarmin Sarmin Sarmin 65 Tarassovi Vughia LT 89-68 66 L. terpstrae Icterohaemorragiae Hualien LT 11-31 67 L. yanagawae Semaranga Sao paulo São Paulo 68
*reclassificação proposta para L. interrogans serovar Parameles

16
O material contido em cada tubo foi homogeneizado e transferido para os tubos com
colunas de filtro, devidamente marcadas e seguiu-se o protocolo fornecido com o Kit: os
tubos Eppendorf foram centrifugados por 5 minutos à 14000 rpm e o filtrado foi desprezado.
Foram adicionados 650 µl da solução de lavagem do Kit, previamente acrescida de 95% de
etanol P. A. e centrifugados por 2 minutos à 14000 rpm. Essa lavagem foi repetida por mais
três vezes e cada filtro foi transferido para novo Eppendorf de 1,5 ml e adicionou-se 100 µl
da água do Kit (nuclease free water), em temperatura ambiente, deixando em repouso por 2
minutos, centrifugando-se a seguir por 2 minutos à 14.000 rpm e o DNA obtido foi estocado
à -20°C até o uso.
Para testar a eficiência do método de extração, aplicou-se 5 µl do DNA obtido
acrescido de 1 µl de tampão de amostra 6X (0,25% de azul de bromofenol e 0,25% de
xileno cianol e 4 mL de sacarose a 40%l), em gel de agarose à 0,8% contendo 0,5 µg/ml de
brometo de etídio (10 mg/ml). A eletroforese foi feita a 100 V por 15 minutos o gel foi
exposto à luz Ultra Violeta (UV) e fotografado.
3.3. Quantificação do DNA
O DNA total extraído foi quantificado em aparelho espectrofotômetro Gene Quant RNA/ DNA
Calculator (Pharmacia), usando-se 2 µl de cada amostra diluídos em 498 µl de água Milli-Q
estéril. Para cada amostra de DNA lida foi feito um controle branco (500 µl de água Milli-Q
estéril).
3.4. Seleção dos genes para amplificação e estudo de polimorfismos
Oito genes foram escolhidos por serem conservados (Tabela 2), ou seja, possuírem
seqüências codificadoras de proteínas estruturais, fundamentais à sobrevivência da célula.
Entretanto, esses genes estão sujeitos a mutações, como adição e deleção, transferência e
cópias de seqüências, em regiões do gene que não afetam a sobrevivência da bactéria,
denominadas regiões hipervariáveis. Essas regiões são as candidatas ideais ao estudo
proposto para diferenciação dos sorovares.
O gene rpoB, codificador da subunidade β da enzima RNA polimerase, foi
inicialmente escolhido, por estar sendo bastante estudado em outros organismos e ser
considerado, por vários pesquisadores, um diferenciador entre espécies de bactérias em
substituição ao gene do RNA 16S ribossômico (AHMED et al. 2006) e também por existirem
várias seqüências de Leptospira já depositadas nos bancos de dados, facilitando o acesso e
minimizando o custo do trabalho. Os iniciadores foram confeccionados pela Integrated DNA
Technologies (IDT) por intermédio da Prodimol Biotecnologia S/A.
17
Tabela 2. Genes estruturais candidatos a análise de restrição para a identificação molecular
dos sorovares de Leptospira, iniciadores e referência bibliográfica.
Gene Tamanho do gene (pb)
Tamanho do produto (pb)
Função do gene
Seqüências dos iniciadores
Referência bibliográfica
rpoB 3678 595 SUBUNIDADE BETA
RNA POLIMERASE
F-CCTCATGGGTTCCAAATGCA R-CGCATCCTCRAAGTTGTAWCCTT
LA SCOLA ET AL., (2006)
gyrB 2136 504 DNA GIRASE Β
F-TGAGCCAAGAAGAAACAAGCTACA R- MATGGTTCCRCTTTCCGAAGA
SLACK ET AL., (2006)
lipL32 819 474 LIPOPROTEÍNA DA
MEMBRANA EXTERNA
F- ATCTCCGTTGCACTCTTTGC R- ACCATCATCATCATCGTCCA
AHMED ET AL., (2006)
lipL41 1068 518 LIPOPROTEÍNA DA
MEMBRANA EXTERNA
F- TAGGAAATTGCGCAGCTACA R- GCATCGAGAGGAATTAACATCA
AHMED ET AL., (2006)
secY 1383 549 PRÉ-PROTEÍNA
TRANSLOCASE
F- ATGCCGATCATTTTTGCTTC R- CGTCCCTTAATTTTAGACTTCTTC
AHMED ET AL., (2006)
icdA 1197 557 ISOCITRATO
DESIDROGENASE
F- GGGACGAGATGACCAGGAT R- TTTTTTGAGATCCGCAGCTTT
AHMED ET AL., (2006)
rrs2 1512 541 RRNA 16S
F- CATGCAAGTCAAGCGGAGTA R- AGTTGAGCCCGCAGTTTTC
AHMED ET AL., (2006)
adk 564 531 ADENILATO QUINASE
F- GGGCTGGAAAAGGTACACAA R- ACGCAAGCTCCTTTTGAATC
AHMED ET AL., (2006)
3.5. Padronização da PCR - específica para o gene rpoB e análise de restrição
3.5.1. Análise in silico dos genes
As análises in silico foram usadas para confirmar a anotação funcional dos genes, bem
como para encontrar enzimas cujos sítios pudessem diferenciar os sovares entre si, em pelo
menos 20 pares de base (pb). As seqüências de cada gene usado no presente trabalho
foram pesquisadas no GenBank (http://www.ncbi.nlm.nih.gov/BLAST) e copiadas em uma
pasta no formato Word. Em cada seqüência foi feita a busca da presença dos iniciadores
(senso e anti-senso) de cada gene. Cada seqüência encontrada foi submetida ao programa
NEBcutter (http://tools.neb.com/NEBcutter2) para a busca de enzimas e seu respectivos
sítios de restrição. Para a escolha de enzimas que pudessem diferenciar o maior número
possível de sorovares gerando fragmentos maiores que 100 pb e mais de dois fragmentos.
Os resultados gerados pelos programas foram impressos e analisados visualmente.

18
3.5.2. Gene rpoB
Para a padronização da PCR, inicialmente foi usado o DNA extraído de 23 sorovares
de referência e os iniciadores Lept 1900f (5’ CCT CAT GGG TTC CAA CAT GCA-3’) e 2500r
(5’ CGC ATC CTC RAA GTT GTA WCC TT-3’) descritos por ( LA SCOLA et al., 2006), que
foram gentilmente cedido pela Dra. Maria Rosa Quaresma Bomfim. Nas amplificações por
PCR foram utilizados 1,5 mM de MgCl2, 200µM de dNTPs (MBI Fermentas), 25-50 ng do
DNA extraído, Taq DNA polimerase, 1,5 Unidades; Tampão da Taq DNA polimerase 1x e 50
picomoles de cada iniciador. Os ciclos da amplificação foram efetuados como descrito na
tabela 3.
Tabela 3. Condições da PCR - específica com os iniciadores Lept 1900f e 2500r, produzidos
para amplificação de parte do gene rpoB.
ciclo desnaturação anelamento extensão
Primeiro 95ºC – 2 min 51ºC – 30 seg 72ºC – 2 min
33 subseqüentes 94ºC – 30 seg 51ºC – 30 seg 72ºC – 2 min
Último 94ºC – 30 seg 51ºC – 30 seg 72ºC – 10 min
3.5.3. Digestão com endonucleases de restrição
Os mapas de restrição do gene rpoB apontou sítios de reconhecimento para diferentes
enzimas. As enzimas TaqI, Tru1I, Sau3AI, MslI e EcoNI foram selecionadas por
apresentarem sítios num maior número de sorovares e gerarem fragmentos com tamanhos
adequados à visualização por eletroforese em gel de poliacrilamida. A digestão enzimática
foi feita em volume final de 10 µl, sendo 4 µl do produto amplificado, 1 U da enzima, 1X do
tampão da enzima, acrescido de água Milli-Q estéril até completar o volume final da reação.
Os tubos foram incubados em estufa a 37ºC ou 65ºC, de acordo com a temperatura
sugerida pelo fabricante. O tempo de incubação variou com a enzima usada, sendo de no
mínimo 2 h e máximo 12 h. Após a digestão, o produto digerido foi aplicado em gel de
poliacrilamida 6% e a eletroforese realizada em voltagem constante de 100 volts em tampão
Tris-Borato-EDTA (MANIATIS et al., 1989) por 1,5 h. Para visualisar a migração relativa do
fragmento digerido foi utilizado o padrão molecular 100 pb DNA ladder (fragmentos de 1500,
1000, 900, 800, 700, 600, 500, 400 300, 200 e 100 pares de base). Depois da eletroforese, o
gel foi fixado com etanol 10% e ácido acético 0,5% por 10 minutos e em seguida corado

19
pelo nitrato de prata a 0,2% por 15 minutos. Após coloração, o gel foi lavado com água
bidestilada por 30 segundos. A revelação das bandas ocorreu pelo uso da solução de NaOH
0,75 M e 0,45% de formaldeído em 150 ml de volume final de água bidestilada, até o
aparecimento das bandas. O gel foi transferido para a solução fixador e acondicionado a 4ºC
até ser fotografado (SANGUINETTI et al., 1994).
3.5.4. Análise dos produtos da PCR-específica e da digestão enzimática
Para a análise do perfil de migração dos fragmentos gerados pela PCR - específica,
bem como dos produtos digeridos pelas enzimas de restrição TaqI, Tru1I, Sau3AI, MslI e
EcoNI, em cada 5µl do produto, foram adicionados 5µl do tampão de corrida (xilenocianol e
azul de bromofenol) na concentração 2X. Esta mistura foi aplicada em gel de poliacrilamida
6%, sendo que a eletroforese ocorreu a 100V em tampão TBE 1X (Tris Borato, EDTA 0,5M)
(MANIATIS et al., 1989). Para cálculo da migração relativa do fragmento amplificado, foi
utilizado o padrão molecular 1Kb DNA Ladder. Esse marcador apresenta fragmentos com
tamanhos de 3054, 2036,1636, 1018, 517, 506, 396, 344, 298, 220, 202,154, 134 e 75 pares
de base. Depois da eletroforese, o gel foi fixado com etanol 10% e ácido acético 0,5% por
10 minutos e em seguida corado pelo nitrato de prata a 0,2% por 15 minutos. Após
coloração, o gel foi lavado com água bidestilada por 30 segundos. A revelação das bandas
ocorreu pelo uso da solução de NaOH 0,75 M e 0,45% de formaldeído em 150 ml de volume
final de água bidestilada, por aproximadamente 10 minutos ou até o aparecimento das
bandas. Posteriormente o gel foi transferido novamente para a solução fixadora,
acondicionado a 4ºC até ser fotografado (SANGUINETTI et al., 1994).
3.5.5. Confecção dos Dendogramas
Para a confecção dos dendogramas o tamanho molecular dos fragmentos,
mostrados em gel para cada perfil foram medidos usando-se o programa
LabImage version 2.7.0 (Kapelan GmbH, 1999-2003). Os resultados foram
dispostos em tabelas para a confecção de uma matriz fenotípica baseada na
presença (1) ou na ausência (2) de cada fragmento de DNA obtido das
leptospiras de referência que foram amplificados por PCR-específica, com o
par de iniciadores rpoB. A análise do agrupamento dos fragmentos foi feita
pelo programa TFPGA (Tools for Population Genetic Analyses) software
package (MILLER, 1998), baseada na similaridade como descrito por Nei
(1972). A distância original foi dada pela média aritmética ponderada ou
Unweighted Pair Group Method (UPGMA) algorithm (SNEATH & SOKAL, 1973).

20
4. RESULTADOS
As seqüências de rpoB presentes no GenBank mais aquelas obtidas pela Dra. Maria
Rosa Quaresma Bomfim por seqüenciamento de nucleotídeos (ver Apêndice) – depositadas
no GenBank com os identificadores gi|218750493|gb|EU747302.1| L. borgpetersenii serovar
Ballum, gi|218750509|gb|EU747310.1| L. borgpetersenii serovar Castellonis;
gi|218750499|gb|EU747305.1| L. borgpetersenii serovar Hardjo-bovis;
gi|218750511|gb|EU747311.1| L. borgpetersenii serovar Sejroe; gi|218750503|gb|
EU747307.1| L. borgpetersenii serovar Tarassovi; gi|218750517|gb|EU747314.1| L.
borgpetersenii serovar Whitticombi; gi|218750489|gb|EU747300.1| L. interrogans serovar
Bratislava; gi|218750523|gb|EU747299.1| L. interrogans serovar Canicola; gi|218750513|
gb|EU747312.1| L. interrogans serovar Djasiman; gi|218750497|gb|EU747304.1| L.
interrogans serovar Hebdomadis; gi|218750505|gb|EU747308.1| L. interrogans serovar
Wolffii – foram alinhadas com o programa ClustalW e é mostrado nas figuras 6 e 7.

21
Figura 6. Alinhamento das seqüências de nucleotídeos de rpoB depositadas no GenBank e
geradas por seqüenciamento pelo nosso grupo.

22
Australis --------------------------------------------------TCCGTGAAGA 10
Bataviae --------------------------------------------------TCCGTGAAGA 10
Icterohaemorrhagiae -------------------TCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGA 41
Schueffneri TGTAACCTTCCTAATGGGTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGCGAAGA 60
Pyrogenes --------------------------------------------------TCCGCGAAGA 10
Wolffi -------------------TCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGA 41
Djasiman ------------------TTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGA 42
Autumnalis --------------------------------------------------TCCGTGAAGA 10
Ballum -----------------------------------------TTCCTCTTCTCCGTGAAGA 19
Hebdomadis -----------------GTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGA 43
Sejroe -------------------TCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGA 41
Canicola ---------CCTCATGGGTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGA 51
Hardjo-bovis ---------CCTCATGGGTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGTGAAGA 51
Javanica -------------------TCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGTGAAGA 41
Castellonis -----------------GTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGTGAAGA 43
Bratislava ----------------GGTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGTGAAGA 44
Tarassovi -----------------GTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGA 43
Cynopteri -------------------TCCAACATGCAGCGTCAGGCGGTTCCTCTTCTTCGTGAAGA 41
Panama -------------------TCCAACATGCAGCGTCAGGCGGTTCCTCTTCTCCGTGAAGA 41
Proechymis --------------------------------------------------TCCGTGAAGA 10
Cascata -------------------TCCAACATGCAGCGTCAGGCGGTTCCTCTTCTCCGTGAAGA 41
Shermani -------------------TCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGGGAAGA 41
Celledoni -------------------TCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGCGAAGA 41
Whitticombi ------------------TTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGCGAAGA 42
Manhao -------------------TCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGTGAAGA 41
Pingchang -------------------TCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGGGAAGA 41
Hurstbridge -------------------TCCAACATGCAACGCCAGGCGGTGCCTCTACTCCGCCAAGA 41
Lyme -------------------TCCAACATGCAGCGCCAAGCGGTGCCTTTACTCCGCCAGGA 41
Saopaulo1 -------------------TCCAACATGCAACGCCAAGCAGTGCCTCTTCTTACGGAAGA 41
Saopaulo -------------------TCCAACATGCAACGCCAAGCAGTGCCTCTTCTTACGGAAGA 41
Patoc -------------------TCCAACATGCAACGCCAAGCGGTACCACTCTTAACAGAAGA 41
Holland -------------------TCCAACATGCAACGCCAAGCGGTTCCTCTTTTAACAGAAGA 41
Codice -------------------TCCAACATGCAACGCCAAGCAGTGCCACTACTCACGGAAGA 41
Ranarum -------------------TCCAACATGCAACGCCAAGCAGTTCCTCTTCTCACAGAAGA 41
*41 * **
Australis AGCTCCTTTTGTAGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 70
Bataviae AGCTCCTTTTGTAGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 70
Icterohaemorrhagiae AGCTCCTTTTGTAGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 101
Schueffneri AGCTCCTTTTGTCGGAACCGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 120
Pyrogenes AGCTCCTTTTGTAGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 70
Wolffi AGCTCCTTTTGTAGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 101
Djasiman AGCTCCTTTTGTAGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 102
Autumnalis AGCTCCTTTTGTCGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 70
Ballum AGCTCCTTTTGTCGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 79
Hebdomadis AGCTCCTTTTGTCGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 103
Sejroe AGCTCCTTTTGTCGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 101
Canicola AGCTCCTTTTGTCGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 111
Hardjo-bovis AGCTCCTTTTGTCGGAACCGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 111
Javanica AGCTCCGTTTGTCGGAACAGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 101
Castellonis AGCTCCGTTTGTCGGAACAGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 103
Bratislava AGCTCCGTTTGTCGGAACAGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 104
Tarassovi AGCTCCTTTTGTCGGAACAGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 103
Cynopteri AGCTCCTTTTGTAGGAACGGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 101
Panama AGCTCCTTTTGTCGGAACCGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 101
Proechymis AGCTCCTTTTGTCGGAACAGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 70
Cascata AGCTCCTTTTGTCGGAACCGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 101
Shermani AGCTCCTTTTGTCGGAACAGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 101
Celledoni AGCTCCGTTTGTCGGAACAGGTATGGAAACTAGAGCCGCTTACGATTCCAGAATTTGTAT 101
Whitticombi AGCTCCGTTTGTCGGAACAGGTATGGAAACTAGAGCCGCTTACGATTCCAGAATTTGTAT 102
Manhao AGCTCCTTTTGTCGGAACAGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTAT 101
Pingchang AGCTCCTTTTGTCGGAACAGGTATGGAAACCAGAGCCGCTTACGATTCTAGAATTTGTAT 101
Hurstbridge AGCTCCCTATGTGGGAACGGGAATGGAGAGCCGTGCCGCATACGACTCCAGAATTTGCGT 101
Lyme AGCTCCCTATGTGGGAACCGGAATGGAAAGTCGTGCTGCGTATGATTCCCGAATTTGCGT 101
Saopaulo1 AGCTCCATTTGTGGGAACTGGTATGGAAGCTCGTGCGGCTTATGACGCAGGGGTTTGTAT 101
Saopaulo AGCTCCATTTGTGGGAACTGGTATGGAAGCTCGTGCGGCTTATGACGCAGGGGTTTGTAT 101
Patoc GGCTCCTTTTGTCGGAACTGGTATGGAAGCTCGTGCGGCTTATGACGCAGGGGTTTGTAT 101
Holland AGCTCCTTTTGTGGGAACTGGTATGGAAGCTCGTGCGGCTTATGACGCAGGTGTTTGTAT 101
Codice AGCTCCGTTTGTAGGAACTGGGATGGAAGCTCGTGCGGCTTATGACGCAGGGGTTTGTAT 101
Ranarum AGCGCCGTTTGTAGGAACAGGTATGGAAGCTCGTGCTGCTTATGATGCAGGGGTTTGTAT 101
** ** 60*** ***** ** ***** * ** ** ** ** * * **** *
Figura 7. Alinhamento das seqüências de nucleotídeos de rpoB indicando os sítios de restrição de TaqI (verde), Tru1I (azul), Sau3AI (roxo), MslI (vermelho) e EcoNI (laranja).
23
Australis CGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 130 Bataviae CGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 130
Icterohaemorrhagiae CGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 161
Schueffneri CGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 180
Pyrogenes CGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 130
Wolffi CGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 161
Djasiman CGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 162
Autumnalis CGTAAATAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 130
Ballum CGTAAATAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 139
Hebdomadis CGTAAATAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 163
Sejroe CGTAAATAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 161
Canicola CGTAAATAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 171
Hardjo-bovis CGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAG 171
Javanica TGTAAACAAACACGACGGCGTTGTGACTTCCGTGGACGCGGAGATCATCGTCGTGGAAAG 161
Castellonis TGTAAACAAACACGACGGCGTTGTGACTTCCGTGGACGCGGAGATCATCGTCGTGGAAAG 163
Bratislava TGTAAACAAACACGACGGCGTTGTGACTTCCGTGGACGCGGAGATCATCGTCGTGGAAAG 164
Tarassovi CGTAAACAAACACGACGGCGTTGTGACTTCCGTGGACGCGGAAACCATCGTTGTGGAAAG 163
Cynopteri CGTAAACAAACACGACGGCGTTGTTACATCCGTGGACGCGGAAACCATCGTTGTGGAAAG 161
Panama CGTAAACAAACACGACGGCGTTGTCATTTCCGTGGACGCGGAAACGATCGTCGTGGAAAG 161
Proechymis CGTAAACAAACACGACGGCGTTGTCATTTCCGTGGACGCGGAAACGATCGTCGTGGAAAG 130
Cascata CGTAAACAAACACGACGGTGTAGTTACTTCCGTGGACGCGGAAACGATCGTCGTGGAAAG 161
Shermani CGTAAACAAACACGACGGTGTGGTCGTATCCGTAGACGCGGAAACGATCGTTGTGGAAAG 161
Celledoni CGTAAACAAACACGACGGTGTCGTTATATCGGTTGACGCAGAACAAATCGTTGTGGAAAG 161
Whitticombi CGTAAACAAACACGACGGTGTCGTTATATCGGTTGACGCAGAACAAATCGTTGTGGAAAG 162
Manhao CGTAAACAAACACGATGGTGTCGTTACATCTGTTGACGCTGAAACGATCGTCGTGGAAAG 161
Pingchang CGTAAACAAACACGACGGCGTCGTTACATCCGTGGACGCGGAAAACATCGTCGTTGAGAG 161
Hurstbridge GATCGCAAAACAAGACGGATACGTGAAATACGTAGACGCCGAAAAAGTCATCGTCGAGCA 161
Lyme AATCGCGAAACAAGACGGATACGTGAAATACGTCGATGCGGAAAAAGTCGTCGTCGAGCA 161
Saopaulo1 CGTTGCCAAAAAAGATGGAGTGGTTTCCAAAGTGGATGCGACTGGTGTTTGGATCAAAGA 161
Saopaulo CGTTGCCAAAAAAGATGGAGTGGTTTCCAAAGTGGATGCGACTGGTGTTTGGATCAAAGA 161
Patoc CGTTGCGAAAAAAGATGGTGTGGTTTCCAAAGTGGATGCAACAGGTGTTTGGATCAAAGA 161
Holland TGTTGCCAAAAAAGATGGTGTGGTTTCGAAAGTGGATGCCACAGGTGTTTGGATCAAAGA 161
Codice CGTTGCGAAAAAAGACGGTGTAGTTTCTAAAGTAGATGCTACAGGTGTTTGGATCAAAGA 161
Ranarum TGTTGCGAAAAAAGATGGTGTAGTTTCTAAAGTAGATGCTACTGGTGTTTGGATCAAAGA 161
* *** *126 ** ** 140 *144 ** 153 161 *
Australis AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 190
Bataviae AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 190
Icterohaemorrhagiae AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 221
Schueffneri AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 240
Pyrogenes AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 190
Wolffi AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 221
Djasiman AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 222
Autumnalis AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 190
Ballum AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 199
Hebdomadis AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 223
Sejroe AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 221
Canicola AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 231
Hardjo-bovis AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGG 231
Javanica AAAGGGCGGAAAAGAATCCGATACGTATTCCCTTACGAAATTCAAAAAGACCAACCAAGG 221
Castellonis AAAGGGCGGAAAAGAATCCGATACGTATTCCCTTACGAAATTCAAAAAGACCAACCAAGG 223
Bratislava AAAGGGCGGAAAAGAATCCGATACGTATTCCCTTACGAAATTCAAAAAGACCAACCAAGG 224
Tarassovi AAAGGGCGGAAAAGAATCCGATACGTATTCCCTTACGAAATTCAAAAAAACAAACCAAGG 223
Cynopteri AAAGGGCGGAAAAGAATCCGATAAATATTCTCTTACAAAATTCAAAAAGACAAACCAAGG 221
Panama AAAGGGCGGAAAAGAATCCGACAAATACGAACTTACGAAATTCAAAAAGACAAACCAAGG 221
Proechymis AAAGGGCGGAAAAGAATCCGACAAATACGAACTTACGAAATTCAAAAAGACAAACCAAGG 190
Cascata AAAGGGCGGAAAAGAATCCGACAAATACGAACTTACGAAATTCAAAAAGACAAACCAAGG 221
Shermani AAAGGGCGGAAAAGAATCCGATAAATACGAACTTACGAAATTCAAAAAGACAAACCAAGG 221
Celledoni AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTTAAAAAGACAAACCAAGG 221
Whitticombi AAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTTAAAAAGACAAACCAAGG 222
Manhao AAAGGGCGGAAAAGAATCCGACACGTATCAACTTACGAAATTCAAAAAAACCAACCAAGG 221
Pingchang AAAGGGCGGAAAAGAATCCGATACTTACTCTCTGACTAAATTCAAAAAGACCAACCAAGG 221
Hurstbridge AAAGGGCGGAAAAGAATCCGATACGTACGACTTAACGAAATTTAAGAAAACCAACCAAGG 221
Lyme AAAGGGCGGCAAAGAATCCGATACGTACGACTTGACGAAATTTAAGAAGACCAACCAAGG 221
Saopaulo1 AGACCAATCCAAAGAGATTGTCCACTACCCACTCATTAAATTCAAAAAAACCAACCAAGG 221
Saopaulo AGACCAATCCAAAGAGATTGTCCACTACCCACTCATTAAATTCAAAAAAACCAACCAAGG 221
Patoc AGACCAATCCAAAGAGATTGTCCATTACCCACTCATTAAATTCAAAAAAACCAACCAAGG 221
Holland AGACCAATCCAAAGAGATTGTCCATTACCCACTCATTAAATTCAAAAAAACCAACCAAGG 221
Codice AGACCAATCTAAAGAGATCGTTCATTACCCACTCATTAAATTCAAAAAAACCAACCAAGG 221
Ranarum AGACCAATCCAAAGAGATCGTCCACTACCCACTCATTAAATTCAAAAAAACCAACCAAGG 221
* * ***** * ** * *207**** ** ** ** ********
Figura 7 (cont.). Alinhamento das seqüências de nucleotídeos de rpoB indicando os sítios
de restrição de TaqI (verde), Tru1I (azul), Sau3AI (roxo), MslI (vermelho) e EcoNI (laranja).

24
Australis AGCCTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGT 250
Bataviae AACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGT 250
Icterohaemorrhagiae AACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGT 281
Schueffneri AACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGT 300
Pyrogenes AACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGT 250
Wolffi AACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGT 281
Djasiman AACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGT 282
Autumnalis AACTTGCTTTAATCAGAAACCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGT 250
Ballum AACTTGCTTTAATCAGAAACCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGT 259
Hebdomadis AACTTGCTTTAATCAGAAACCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGT 283
Sejroe AACTTGCTTTAATCAGAAACCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGT 281
Canicola AACTTGCTTTAATCAGAAACCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGT 291
Hardjo-bovis AACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGT 291
Javanica AACCTGCTTCAATCAGAAGCCGATCGTCGGAGTCGTTCACTCAGAGATCAACGGAAAAGT 281
Castellonis AACCTGCTTCAATCAGAAGCCGATCGTCGGAGTCGTTCACTCAGAGATCAACGGAAAAGT 283
Bratislava AACCTGCTTCAATCAGAAGCCGATCGTCGGAGTCGTTCACTCAGAGATCAACGGAAAAGT 284
Tarassovi AACCTGCTTTAATCAGAAGCCGATCGTCGGAGTCGTTCACTCCGAGATCAACGGAAAGGT 283
Cynopteri AACCTGCTTCAACCAAAAGCCGATCGTAGGAGTGGTTCACTCCGAGATCAACGGAAAGGT 281
Panama AACCTGCTTTAACCAAAAGCCGATCGTAGGCGTAGTTCATTCCGAGATCAACGGAAAAGT 281
Proechymis AACCTGCTTTAACCAAAAGCCGATCGTTGGTGTGGTTCATTCCGAGATCAACGGAAAAGT 250
Cascata AACCTGCTTTAACCAAAAGCCGATCGTTGGAGTGGTTCACTCCGAGATCAACGGAAAGGT 281
Shermani AACCTGCTTTAACCAAAAGCCGATCGTGGGAGTGGTTCACTCCGATCGAAACGGTAAAGT 281
Celledoni AACCTGTTTCAATCAGAAGCCGATCGTAGGAGTCGTTCACTCCGAGATCAGCGGAAAGGT 281
Whitticombi AACCTGTTTCAATCAGAAGCCGATCGTAGGAGTCGTTCACTCCGAGATCAGCGGAAAGGT 282
Manhao AACCTGTTTCAATCAGAAACCGATCGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGT 281
Pingchang AACTTGCTTTAACCAAAAGCCAATCGTCGGAGTGGTTCACTCCGATTTTAACGGAAAGGT 281
Hurstbridge CACTTGCTTCAATCAAACTCCCGTCGTCGGAGTGATTCACTCGGAGATCGACGGGAAAGT 281
Lyme CACTTGCTTTAACCAAACTCCCGTAGTCGGAGTGATCCACTCCGAGATAGACGGAAAAGT 281
Saopaulo1 TACTTGTTTTAACCAAAAACCAAACGTATCCATGTTACACACCACAACTGGTGGTAAGGT 281
Saopaulo TACTTGTTTTAACCAAAAACCAAACGTATCCATGTTACACACCACAACTGGTGGTAAGGT 281
Patoc TACTTGTTTTAACCAAAAACCAAACGTATCCATGTTACACACCACAACTGGTGGCAAGGT 281
Holland AACTTGTTTTAACCAAAAACCAAACGTTTCCATGTTGCATACGACGACTGGTGGTAAAGT 281
Codice AACTTGTTTTAACCAAAAACCAAACGTGGCTATGTTACACACGACCACTGGCGGAAAGGT 281
Ranarum TACTTGTTTTAACCAAAAACCGAACGTATCCATGCTTCATACCACAACAGGTGGAAAGGT 281
* ** *240* ** * **252 ** * * ** 276 277 279** ** **
Australis TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 310
Bataviae TTCCAAGGTCTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 310
Icterohaemorrhagiae TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 341
Schueffneri TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 360
Pyrogenes TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 310
Wolffi TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 341
Djasiman TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 342
Autumnalis TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 310
Ballum TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 319
Hebdomadis TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 343
Sejroe TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 341
Canicola TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 351
Hardjo-bovis TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATA 351
Javanica TTCCAAGGTTTCCAAAGAAAAAATCGAAGTGACCGGTGAAAACGGAGAACTGAAAGAATA 341
Castellonis TTCCAAGGTTTCCAAAGAAAAAATCGAAGTGACCGGTGAAAACGGAGAACTGAAAGAATA 343
Bratislava TTCCAAGGTTTCCAAAGAAAAAATCGAAGTGACCGGTGAAAACGGAGAACTGAAAGAATA 344
Tarassovi TTCCAAGGTTTCCAAAGAAAAAATCGAAGTGACCGGTGAAAACGGAGAACTGAAAGAATA 343
Cynopteri TTCCAAGGTTTCTAAAGAAAAAATCGAAGTCACTGGCGAAAACGGAGAAGTAAAAGAATA 341
Panama AAGCAAGGTTTCTAAAGAAAAAATCGAAGTAACCGGCGAAAACGGAGAAGTAAAAGAATA 341
Proechymis AAGCAAGGTTTCTAAAGAAAAAATCGAAGTAACCGGCGAAAACGGAGAAGTAAAAGAATA 310
Cascata TTCCAAGGTTTCTAAAGAAAAAATCGAAGTCACCAGCGAAAACGGAGAAGTAAAAGAATA 341
Shermani TACTAAGGTTTCCAAAGAGAAAATCGAAGTCACCGGCGAAAACGGAGACGTAAAAGAATA 341
Celledoni TACGAAAGTTTCCAAAGAAAAAATCGAAGTAACATCCGAAAACGGAACAGTAAAAGAATA 341
Whitticombi TACGAAAGTTTCCAAAGAAAAAATCGAAGTAACATCCGAAAACGGAACAGTAAAAGAATA 342
Manhao TACGAAAGTTTCCAAAGAAAAAATCGAAGTAACATCCGAAAACGGAGAAGTAAAAGAATA 341
Pingchang TTCCAAGGTTTCCAAAGAAAAAATCGAAGTGACCGGTGAAAACGGACATATCAAAGAATA 341
Hurstbridge CAGTAAGGTGACCAAGGAAAAAATCGAAGTCACTGCGGATAACGGAAGCATAAAAGAATA 341
Lyme CAACAAGGTAACCAGGGAGAAAATTGAAGTCACTGCGGATAACGGAAGCATAAGAGAATA 341
Saopaulo1 GAGTAAGGTTTCCAAAGAACGCATTGAAGTGACTTCTCCCAATGGAGAAAAAGAAATCCA 341
Saopaulo GAGTAAGGTTTCCAAAGAACGCATTGAAGTGACTTCTCCCAATGGAGAAAAAGAAATCCA 341
Patoc AAGTAAGGTTTCGAAAGAACGTGTCGAAGTGACAACTCCTAACGGAGAAAAAGAAACTCA 341
Holland AGGTAAGGTTTCCAAAGAACGTGTGGAGATTACTTCTCCTAACGGAGAAAAAGAAACCCA 341
Codice TAGTAAAGTATCCAAGGAACGAGTGGAACTCACTTCTCCCAACGGGGAGAAAGAAATCCA 341
Ranarum AAGTAAAGTTTCTAAAGAACGGATCGAAGTAACGTCTCCAAACGGAGAAAAAGAAACACA 341
** ** * * ** *315 317** ** ** * *
Figura 7 (cont.). Alinhamento das seqüências de nucleotídeos de rpoB indicando os sítios
de restrição de TaqI (verde), Tru1I (azul), Sau3AI (roxo), MslI (vermelho) e EcoNI (laranja).
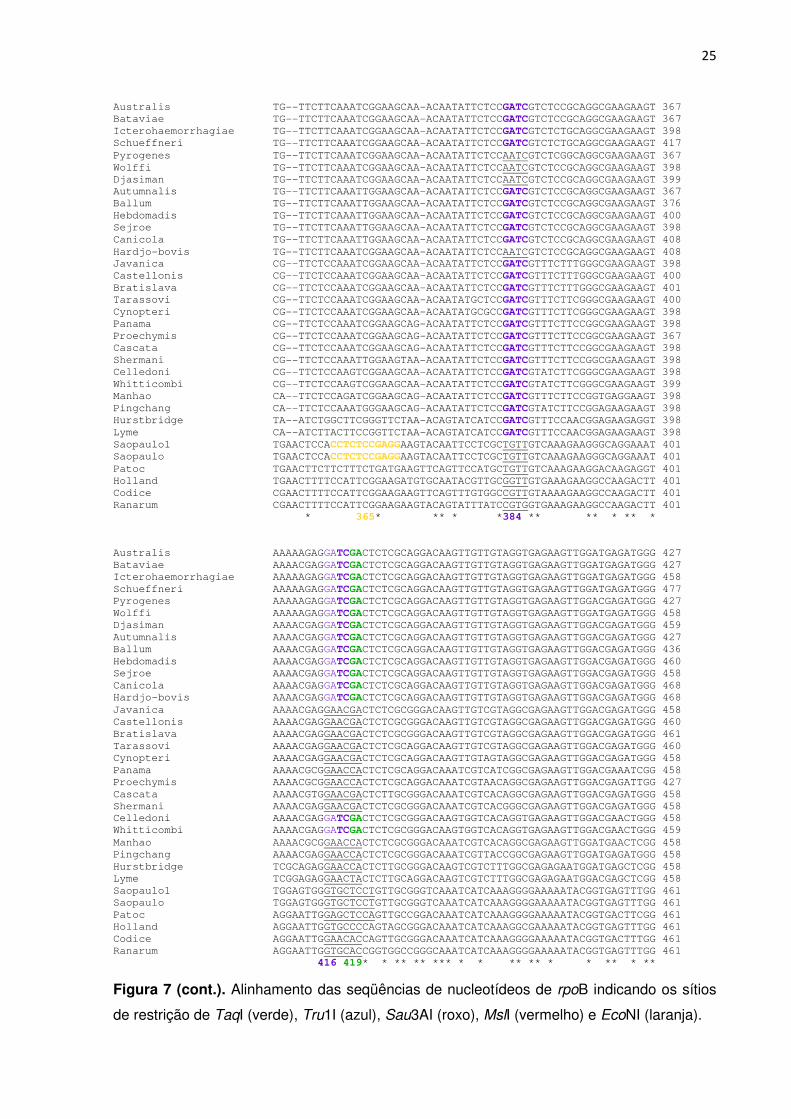
25
Australis TG--TTCTTCAAATCGGAAGCAA-ACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAGT 367
Bataviae TG--TTCTTCAAATCGGAAGCAA-ACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAGT 367
Icterohaemorrhagiae TG--TTCTTCAAATCGGAAGCAA-ACAATATTCTCCGATCGTCTCTGCAGGCGAAGAAGT 398
Schueffneri TG--TTCTTCAAATCGGAAGCAA-ACAATATTCTCCGATCGTCTCTGCAGGCGAAGAAGT 417
Pyrogenes TG--TTCTTCAAATCGGAAGCAA-ACAATATTCTCCAATCGTCTCGGCAGGCGAAGAAGT 367
Wolffi TG--TTCTTCAAATCGGAAGCAA-ACAATATTCTCCAATCGTCTCCGCAGGCGAAGAAGT 398
Djasiman TG--TTCTTCAAATCGGAAGCAA-ACAATATTCTCCAATCGTCTCCGCAGGCGAAGAAGT 399
Autumnalis TG--TTCTTCAAATTGGAAGCAA-ACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAGT 367
Ballum TG--TTCTTCAAATTGGAAGCAA-ACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAGT 376
Hebdomadis TG--TTCTTCAAATTGGAAGCAA-ACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAGT 400
Sejroe TG--TTCTTCAAATTGGAAGCAA-ACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAGT 398
Canicola TG--TTCTTCAAATTGGAAGCAA-ACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAGT 408
Hardjo-bovis TG--TTCTTCAAATCGGAAGCAA-ACAATATTCTCCAATCGTCTCCGCAGGCGAAGAAGT 408
Javanica CG--TTCTCCAAATCGGAAGCAA-ACAATATTCTCCGATCGTTTCTTTGGGCGAAGAAGT 398
Castellonis CG--TTCTCCAAATCGGAAGCAA-ACAATATTCTCCGATCGTTTCTTTGGGCGAAGAAGT 400
Bratislava CG--TTCTCCAAATCGGAAGCAA-ACAATATTCTCCGATCGTTTCTTTGGGCGAAGAAGT 401
Tarassovi CG--TTCTCCAAATCGGAAGCAA-ACAATATGCTCCGATCGTTTCTTCGGGCGAAGAAGT 400
Cynopteri CG--TTCTCCAAATCGGAAGCAA-ACAATATGCGCCGATCGTTTCTTCGGGCGAAGAAGT 398
Panama CG--TTCTCCAAATCGGAAGCAG-ACAATATTCTCCGATCGTTTCTTCCGGCGAAGAAGT 398
Proechymis CG--TTCTCCAAATCGGAAGCAG-ACAATATTCTCCGATCGTTTCTTCCGGCGAAGAAGT 367
Cascata CG--TTCTCCAAATCGGAAGCAG-ACAATATTCTCCGATCGTTTCTTCCGGCGAAGAAGT 398
Shermani CG--TTCTCCAAATTGGAAGTAA-ACAATATTCTCCGATCGTTTCTTCCGGCGAAGAAGT 398
Celledoni CG--TTCTCCAAGTCGGAAGCAA-ACAATATTCTCCGATCGTATCTTCGGGCGAAGAAGT 398
Whitticombi CG--TTCTCCAAGTCGGAAGCAA-ACAATATTCTCCGATCGTATCTTCGGGCGAAGAAGT 399
Manhao CA--TTCTCCAGATCGGAAGCAG-ACAATATTCTCCGATCGTTTCTTCCGGTGAGGAAGT 398
Pingchang CA--TTCTCCAAATGGGAAGCAG-ACAATATTCTCCGATCGTATCTTCCGGAGAAGAAGT 398
Hurstbridge TA--ATCTGGCTTCGGGTTCTAA-ACAGTATCATCCGATCGTTTCCAACGGAGAAGAGGT 398
Lyme CA--ATCTTACTTCCGGTTCTAA-ACAGTATCATCCGATCGTTTCCAACGGAGAAGAAGT 398
Saopaulo1 TGAACTCCACCTCTCCGAGGAAGTACAATTCCTCGCTGTTGTCAAAGAAGGGCAGGAAAT 401
Saopaulo TGAACTCCACCTCTCCGAGGAAGTACAATTCCTCGCTGTTGTCAAAGAAGGGCAGGAAAT 401
Patoc TGAACTTCTTCTTTCTGATGAAGTTCAGTTCCATGCTGTTGTCAAAGAAGGACAAGAGGT 401
Holland TGAACTTTTCCATTCGGAAGATGTGCAATACGTTGCGGTTGTGAAAGAAGGCCAAGACTT 401
Codice CGAACTTTTCCATTCGGAAGAAGTTCAGTTTGTGGCCGTTGTAAAAGAAGGCCAAGACTT 401
Ranarum CGAACTTTTCCATTCGGAAGAAGTACAGTATTTATCCGTGGTGAAAGAAGGCCAAGACTT 401
* 365* ** * *384 ** ** * ** *
Australis AAAAAGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGATGAGATGGG 427
Bataviae AAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGATGAGATGGG 427
Icterohaemorrhagiae AAAAAGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGATGAGATGGG 458
Schueffneri AAAAAGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGATGAGATGGG 477
Pyrogenes AAAAAGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGG 427
Wolffi AAAAAGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGATGAGATGGG 458
Djasiman AAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGG 459
Autumnalis AAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGG 427
Ballum AAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGG 436
Hebdomadis AAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGG 460
Sejroe AAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGG 458
Canicola AAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGG 468
Hardjo-bovis AAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGG 468
Javanica AAAACGAGGAACGACTCTCGCGGGACAAGTTGTCGTAGGCGAGAAGTTGGACGAGATGGG 458
Castellonis AAAACGAGGAACGACTCTCGCGGGACAAGTTGTCGTAGGCGAGAAGTTGGACGAGATGGG 460
Bratislava AAAACGAGGAACGACTCTCGCGGGACAAGTTGTCGTAGGCGAGAAGTTGGACGAGATGGG 461
Tarassovi AAAACGAGGAACGACTCTCGCAGGACAAGTTGTCGTAGGCGAGAAGTTGGACGAGATGGG 460
Cynopteri AAAACGAGGAACGACTCTCGCAGGACAAGTTGTAGTAGGCGAGAAGTTGGACGAGATGGG 458
Panama AAAACGCGGAACCACTCTCGCAGGACAAATCGTCATCGGCGAGAAGTTGGACGAAATCGG 458
Proechymis AAAACGCGGAACCACTCTCGCAGGACAAATCGTAACAGGCGAGAAGTTGGACGAGATTGG 427
Cascata AAAACGTGGAACGACTCTTGCGGGACAAATCGTCACAGGCGAGAAGTTGGACGAGATGGG 458
Shermani AAAACGAGGAACGACTCTCGCGGGACAAATCGTCACGGGCGAGAAGTTGGACGAGATGGG 458
Celledoni AAAACGAGGATCGACTCTCGCGGGACAAGTGGTCACAGGTGAGAAGTTGGACGAACTGGG 458
Whitticombi AAAACGAGGATCGACTCTCGCGGGACAAGTGGTCACAGGTGAGAAGTTGGACGAACTGGG 459
Manhao AAAACGCGGAACCACTCTCGCGGGACAAATCGTCACAGGCGAGAAGTTGGATGAACTCGG 458
Pingchang AAAACGAGGAACCACTCTCGCGGGACAAATCGTTACCGGCGAGAAGTTGGATGAGATGGG 458
Hurstbridge TCGCAGAGGAACCACTCTTGCGGGACAAGTCGTCTTTGGCGAGAGAATGGATGAGCTCGG 458
Lyme TCGGAGAGGAACTACTCTTGCAGGACAAGTCGTCTTTGGCGAGAGAATGGACGAGCTCGG 458
Saopaulo1 TGGAGTGGGTGCTCCTGTTGCGGGTCAAATCATCAAAGGGGAAAAATACGGTGAGTTTGG 461
Saopaulo TGGAGTGGGTGCTCCTGTTGCGGGTCAAATCATCAAAGGGGAAAAATACGGTGAGTTTGG 461
Patoc AGGAATTGGAGCTCCAGTTGCCGGACAAATCATCAAAGGGGAAAAATACGGTGACTTCGG 461
Holland AGGAATTGGTGCCCCAGTAGCGGGACAAATCATCAAAGGCGAAAAATACGGTGAGTTTGG 461
Codice AGGAATTGGAACACCAGTTGCGGGACAAATCATCAAAGGGGAAAAATACGGTGACTTTGG 461
Ranarum AGGAATTGGTGCACCGGTGGCCGGGCAAATCATCAAAGGGGAAAAATACGGTGAGTTTGG 461
416 419* * ** ** *** * * ** ** * * ** * **
Figura 7 (cont.). Alinhamento das seqüências de nucleotídeos de rpoB indicando os sítios
de restrição de TaqI (verde), Tru1I (azul), Sau3AI (roxo), MslI (vermelho) e EcoNI (laranja).

26
Australis AAATATCCTCGTAAAAGGAACCGTTCTTGCTGATGGTCCTGCGGTCGACAACGGAGTTCT 487
Bataviae AAATATCCTCGTAAAAGGAACCGTTCTTGCTGATGGTCCTGCGGTCGACAACGGAGTTCT 487
Icterohaemorrhagiae AAATATCCTCGTAAAAGGAACCGTTCTTGCTGATGGTCCTGCGGTCGACAACGGAGTTCT 518
Schueffneri AAATATCCTCGTAAAAGGAACCGTTCTTGCTGATGGTCCTGCGGTCGACAACGGAGTTCT 537
Pyrogenes AAATATCCTCGTTAAGGGTACGGTTCTTGCCGACGGTCCTGCCGTTGACAATGGAGTTCT 487
Wolffi AAATATCCTCGTAAAAGGTACGGTTCTTGCCGACGGTCCTGCGGTCGACAACGGAGTTCT 518
Djasiman AAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCGGTCGACAACGGAGTTCT 519
Autumnalis AAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGATAATGGAGTTCT 487
Ballum AAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGATAATGGAGTTCT 496
Hebdomadis AAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGATAATGGAGTTCT 520
Sejroe AAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGATAATGGAGTTCT 518
Canicola AAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGATAATGGAGTTCT 528
Hardjo-bovis AAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGACAATGGAGTTCT 528
Javanica AAATATCCTCGTTAAGGGTACGGTTCTTGCGGACGGCCCTGCGGTCGACAACGGGGTTCT 518
Castellonis AAATATCCTCGTTAAGGGTACGGTTCTTGCGGACGGCCCTGCGGTCGACAACGGGGTTCT 520
Bratislava AAATATCCTCGTTAAGGGTACGGTTCTTGCGGACGGCCCTGCGGTCGACAACGGGGTTCT 521
Tarassovi AAATATCCTCGTTAAGGGTACGGTTCTTGCGGACGGCCCTGCGGTCGACAACGGTGTCCT 520
Cynopteri AAATATCCTCGTTAAGGGAACCGTTCTTGCAGACGGTCCCGCGGTTGACAACGGTGTTCT 518
Panama AAATATCCTCGTGAAAGGAACCGTTCTTGCAGACGGTCCTGCGGTTGACAACGGAGTTCT 518
Proechymis AAACATTCTCGTGAAAGGAACCGTTCTTGCGGACGGTCCTGCGGTCGACAACGGAGTTCT 487
Cascata AAATATTCTCGTGAAAGGAACCGTTCTTGCGGACGGTCCCGCCGTTGACAACGGAGTTCT 518
Shermani AAATATCCTCGTTAAGGGAACCGTTCTTGCGGACGGACCTGCGGTCGACAACGGAGTGCT 518
Celledoni AAATATCCTCGTAAAAGGAACGGTCCTTGCGGACGGACCTGCGGTTGACAACGGAGTTCT 518
Whitticombi AAATATCCTCGTAAAAGGAACGGTCCTTGCGGACGGACCTGCGGTTGACAACGGAGTTCT 519
Manhao AAACATTCTCATTAAAGGCACGGTTCTCGCCGACGGTCCTGCGGTGGACAACGGAGTTCT 518
Pingchang AAATATCCTGGTAAAAGGAACCGTTCTTGCGGACGGTCCTGCGGTTGACAACGGAGTACT 518
Hurstbridge AAACATTCTGCAAAAAGGAACCGTTCTTGCGGACGGACCTGCAGTCGATAACGGAACTCT 518
Lyme AAACATTCTGCAAAAAGGAACCGTTCTCGCGGACGGACCTGCCGTCGATAACGGAACTCT 518
Saopaulo1 ACAAATCCTGCAAAAAGGAACTGTCCTTGCAAACGGTCCATCCACTGACGCTGGTTACTT 521
Saopaulo ACAAATCCTGCAAAAAGGAACTGTCCTTGCAAACGGTCCATCCACTGACGCTGGTTACTT 521
Patoc TCAGATCCTTCAAAAAGGAACTGTCCTAGCCAACGGGCCATCCACTGACGCTGGGTATTT 521
Holland ACAAATCCTCCAAAAAGGAACTGTTCTTGCCAATGGACCATCCACTGACGCTGGTTATTT 521
Codice TCAGATCCTACAAAAAGGAACTGTCCTTGCCAACGGTCCATCCACTGACGCTGGTTATTT 521
Ranarum ACAAATCCTCCAAAAAGGAACTGTCCTTGCGAATGGTCCTTCCACTGACGCTGGTTATTT 521
*465 479 480* ** ** ** ** ** * ** ** * 513 ** *
Australis CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGATACAACTT------ 541
Bataviae CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGATACAACTT------ 541
Icterohaemorrhagiae CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGATACAACTT------ 572
Schueffneri CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACTTCGAGGA 597
Pyrogenes CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGATACAACTT------ 541
Wolffi CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACTT------ 572
Djasiman CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGATACA---------- 569
Autumnalis CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGATACAACTT------ 541
Ballum CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACTTCGAGGA 556
Hebdomadis CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACT------- 573
Sejroe CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACT------- 571
Canicola CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACA---------- 578
Hardjo-bovis CGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACTTCGAG-- 586
Javanica CGCTCTCGGTAGAAATGTCCTCGCGGCGTTTATGCCTTGGGAAGGATACAACTT------ 572
Castellonis CGCTCTCGGTAGAAACGTCCTCGCGGCGTTTATGCCTTGGGAAGGATACAACT------- 573
Bratislava CGCTCTCGGTAGAAACGTCCTCGCGGCGTTTATGCCTTGGGAAGGTTACAACTTCGAGGA 581
Tarassovi CGCTCTGGGAAGAAACGTTCTCGCGGCGTTTATGCCTTGGGAAGGTTACAACTTC----- 575
Cynopteri CGCTCTGGGAAGAAACGTTCTCGCGGCGTTTATGCCTTGGGAAGGATACAACTT------ 572
Panama CGCTCTTGGAAGAAACGTTCTCGCGGCTTTCATGCCTTGGGAAGGATACAACTT------ 572
Proechymis CGCTCTTGGAAGAAACGTTCTTGCGGCTTTCATGCCTTGGGAAGGTTACAACTTCGAGGA 547
Cascata CGCTCTGGGAAGAAACGTTCTTGCGGCGTTTATGCCTTGGGAAGGTTACAACTTTGAG-- 576
Shermani CGCACTCGGAAGAAACGTTCTCGCGGCTTTCATGCCTTGGGAAGGATACAACTT------ 572
Celledoni CGCTCTCGGAAGAAACGTTCTCGCTGCGTTCATGCCTTGGGAAGGATACAACTT------ 572
Whitticombi CGCTCTCGGAAGAAACGTTCTCGCTGCGTTCATGCCTTGGGAAGGTTACAA--------- 570
Manhao CGCTCTCGGTAGAAACGTTCTTGCGGCTTTTATGCCTTGGGAAGGATACAACTT------ 572
Pingchang CGCTCTCGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGATACAACTT------ 572
Hurstbridge TGCTTTAGGACGGAACGTGCTCGTAGCGTTTATGCCTTGGGAAGGATACAACTT------ 572
Lyme TGCATTAGGACGAAACGTTCTGGTAGCGTTTATGCCTTGGGAAGGATACAACTT------ 572
Saopaulo1 GGCTCTTGGGAGAAACGTTCTTGTGGCATTTATGCCTTGGGAAGGATACAACTT------ 575
Saopaulo GGCTCTTGGGAGAAACGTTCTTGTGGCATTTATGCCTTGGGAAGGATACAACTT------ 575
Patoc GGCACTTGGACGAAATGTTCTCGTTGCCTTTATGCCTTGGGAAGGATACAACTT------ 575
Holland GGCACTTGGCCGTAACGTTCTTGTGGCATTTATGCCTTGGGAAGGATACAACTT------ 575
Codice GGCTCTAGGCCGAAACGTACTCGTTGCCTTTATGCCATGGGAAGGATACAACTT------ 575
Ranarum GGCACTGGGACGTAATGTACTCGTTGCGTTTATGCCTTGGGAAGGATACAACTT------ 575
** * ** * ** ** ** * ** ** ***** 567***** **** 582
Figura 7 (cont.). Alinhamento das seqüências de nucleotídeos de rpoB indicando os sítios
de restrição de TaqI (verde), Tru1I (azul), Sau3AI (roxo), MslI (vermelho) e EcoNI (laranja).

27
Australis -----------
Bataviae -----------
Icterohaemorrhagiae -----------
Schueffneri TGCGAAGGTTA 608
Pyrogenes -----------
Wolffi -----------
Djasiman -----------
Autumnalis -----------
Ballum -----------
Hebdomadis -----------
Sejroe -----------
Canicola -----------
Hardjo-bovis -----------
Javanica -----------
Castellonis -----------
Bratislava ATGCG------ 586
Tarassovi -----------
Cynopteri -----------
Panama -----------
Proechymis TGC-------- 550
Cascata -----------
Shermani -----------
Celledoni -----------
Whitticombi -----------
Manhao -----------
9Pingchang -----------
Hurstbridge -----------
Lyme -----------
Saopaulo1 -----------
Saopaulo -----------
Patoc -----------
Holland -----------
Codice -----------
Ranarum -----------
Figura 7 (cont.). Alinhamento das seqüências de rpoB indicando os sítios de restrição de
TaqI (verde), Tru1I (azul), Sau3AI (roxo), MslI (vermelho) e EcoNI (laranja).
Dos sessenta e oito sorovares analisados em busca de polimorfismos tipo RFLP em
parte da seqüência codificadora do gene da subunidade β da RNA polimerase, vinte e cinco,
de nove espécies, foram identificados após restrição pelas enzimas TaqI, Tru1I, Sau3AI,
MslI e EcoNI, correspondendo a aproximadamente 37% (Tabela 6).
A enzima TaqI apresentou 10 perfis de restrição distintos esquematizados na Tabela
4A e figura 8, sendo capaz de identificar apenas os sorovares 46 (Huallaga) e 50 (Alice) com
perfis sorovar-específicos. Contudo, o perfil G foi observado em quase todos os sorovares
de Leptospira santarosai, com exceção dos sorovares Alexi, Alice e Tingomariensis, e no
sorovar Muenchen que é da espécie Leptospira interrogans.
A enzima Tru1I também apresentou 10 perfis de restrição distintos, esquematizados na
Tabela 4B e figura 9, identificando os sorovares 46 (Huallaga) e 60 (Trinidad) com perfis
sorovar-específicos. Todavia, a combinação de ambas as enzimas gerou 23 perfis distintos
com algumas observações interessantes: o perfil AA é espécie-específico, pois foi
observado apenas para Leptospira biflexa, o perfil AC é apresentado por todos os sorovares
de Leptospira kirschneri, não sendo espécie-específico devido a presença deste no sorovar
Hualien de L. terpstrae, e os perfis AG, FG, FE, CE, FD, BE e GE são únicos dos sorovares
11 (Mini), 17 (Kaup), 28 (Lanka), 29 (Szwajizak), 32 (Waskurin), 48 (Myocastoris) e 54
(Maru), respectivamente.

28
Tabela 4A. Perfis de restrição dos sorovares de Leptospira com a enzima TaqI.
Perfil Tamanho dos fragmentos (pb)
A 315 277
B 315 198 69 10
C 315 104 173
D 144 171 104 173
E 144 171 104 94 69 10
F 315 104 94 69 10
G 277 38 198 69 10
H 592
I 144 369 69 10
J 277 38 277
Perfil A: sorovares 1, 2, 7, 11, 16, 34, 35, 36, 37, 38, 39, 40, 41, 43, 45, 47, 49, 52 , 65 e 67
Perfil B: sorovares 3, 4, 5, 10, 12, 14 e 48
Perfil C: sorovares 6, 8, 29, 64 e 66
Perfil D: sorovares 9, 20, 23, 25, 26, 30, 31, 33 e 58
Perfil E: sorovares 13, 15, 18, 21, 22, 24, 27 e 42
Perfil F: sorovares 17, 28 e 32
Perfil G: sorovares 19, 51, 53, 54, 55, 56, 57, 59, 60, 61 e 62
Perfil H: sorovares 44, 63 e 68
Perfil I: sorovar 46
Perfil J: sorovar 50
Tabela 4B – Perfis de restrição dos sorovares de Leptospira com a enzima Tru1I.
Perfil Tamanho dos fragmentos (pb)
A 41 166 33 352
B 240 240 112
C 480 112
D 207 33 352
E 240 352
F 240 39 313
G 592
H 279 313
I 41 199 352
J 207 33 240 112
Perfil A: sorovares 1 e 2
Perfil B: sorovares 3, 5, 12, 14, 19, 31, 50, 51, 53, 55, 56, 57, 58, 59, 61 e 62
Perfil C: sorovares 4, 10, 34, 35, 36, 37, 38, 39, 40, 41 e 67
Perfil D: sorovares 6, 8, 32, 44, 63, 64, 66 e 68
Perfil E: sorovares 7, 13, 15, 16, 21, 24, 27, 28, 29, 45, 47, 48, 49, 52 e 54
Perfil F: sorovares 9, 18, 20, 22, 23, 25, 26, 30, 33 e 42
Perfil G: sorovares 11 e 17
Perfil H: sorovares 43 e 65
Perfil I: sorovar 46
Perfil J: sorovar 60

29
Tabela 4C – Perfis de restrição dos sorovares de Leptospira com a enzima Sau3AI.
Perfil Tamanho dos fragmentos (pb)
A 153 312 129
B 252 24 108 208
C 153 99 24 108 208
D 416 176
E 276 108 32 176
F 384 32 176
G 276 140 176
H 153 123 108 32 176
I 252 132 79 129
Perfil A: sorovares 1 e 2 Perfil B: sorovares 3, 5, 12, 14, 35, 36, 37, 38, 39, 40, 41, 46, 48, 55 e 67 Perfil C: sorovares 4, 7, 10, 11, 16, 19, 34, 45, 47, 49, 50, 51, 52, 53, 54, 56, 57, 59, 60, 61 e 62 Perfil D: sorovares 6, 33 e 44 Perfil E: sorovares 8, 27, 28, 64 e 66 Perfil F: sorovares 9, 18, 20, 22, 23, 25, 26, 30, 42, 63 e 68 Perfil G: sorovares 13, 15, 21, 24, 31 e 58 Perfil H: sorovares 17, 29, 32 e 65 Perfil I: sorovar 43
Tabela 4D – Perfis de restrição dos sorovares de Leptospira com a enzima MslI.
Perfil Tamanho dos fragmentos (pb)
A 592
B 161 431
C 126 466
D 140 452
E 317 278
Perfil A: sorovares 1, 2, 34, 44, 46, 54 e 63 Perfil B: sorovares 3, 4, 5, 6, 10, 11, 12, 14, 35, 36, 37, 38, 41 e 67 Perfil C: sorovares 7, 8, 9, 13, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 28, 29, 30, 31, 32, 33, 39, 40, 43, 50, 51, 53, 55, 56, 57, 58, 59, 60, 61, 62, 64, 65 e 66 Perfil D: sorovares 27, 42, 45, 47, 48, 49 e 52 Perfil E: sorovar 68
Tabela 4E – Perfis de restrição dos sorovares de Leptospira com a enzima EcoNI.
Perfil Tamanho dos fragmentos (pb)
A 567 25
B 479 88 25
C 60 507 25
D 60 419 88 25
E 365 205 25
Perfil A: sorovares 1, 2, 6, 7, 8, 9, 11, 16, 17, 20, 23, 26, 30, 32, 33, 43, 44, 45, 46, 47, 48, 49, 52, 54, 57, 63, 64, 65 e 66 Perfil B: sorovares 3, 4, 5, 10, 12, 14, 19, 50, 51, 53, 55, 56, 59, 60, 61 e 62 Perfil C: sorovares 13, 15, 18, 21, 22, 24, 25, 27, 28, 29 e 42 Perfil D: sorovares 31, 34, 35, 36, 37, 38, 39, 40, 41, 58 e 67 Perfil E: sorovar 68
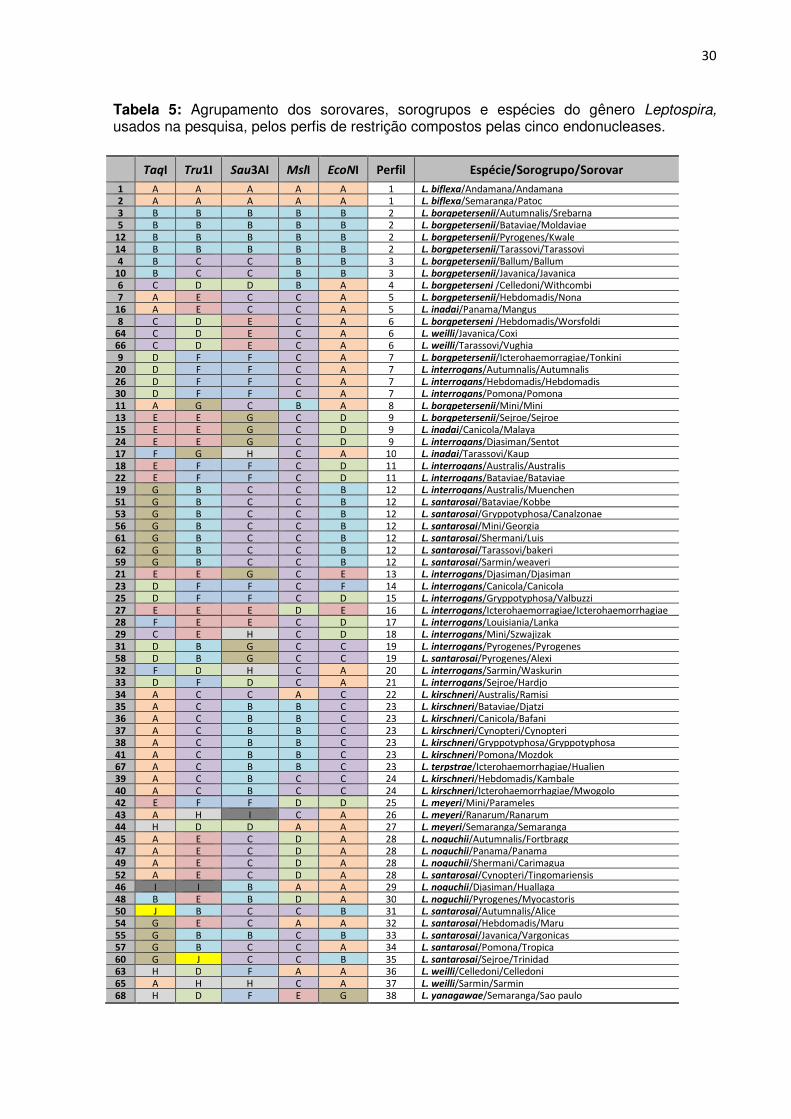
30
Tabela 5: Agrupamento dos sorovares, sorogrupos e espécies do gênero Leptospira, usados na pesquisa, pelos perfis de restrição compostos pelas cinco endonucleases.
TaqI Tru1I Sau3AI MslI EcoNI Perfil Espécie/Sorogrupo/Sorovar
1 A A A A A 1 L. biflexa/Andamana/Andamana 2 A A A A A 1 L. biflexa/Semaranga/Patoc 3 B B B B B 2 L. borgpetersenii/Autumnalis/Srebarna 5 B B B B B 2 L. borgpetersenii/Bataviae/Moldaviae
12 B B B B B 2 L. borgpetersenii/Pyrogenes/Kwale 14 B B B B B 2 L. borgpetersenii/Tarassovi/Tarassovi 4 B C C B B 3 L. borgpetersenii/Ballum/Ballum
10 B C C B B 3 L. borgpetersenii/Javanica/Javanica 6 C D D B A 4 L. borgpeterseni /Celledoni/Withcombi 7 A E C C A 5 L. borgpetersenii/Hebdomadis/Nona
16 A E C C A 5 L. inadai/Panama/Mangus 8 C D E C A 6 L. borgpeterseni /Hebdomadis/Worsfoldi
64 C D E C A 6 L. weilli/Javanica/Coxi 66 C D E C A 6 L. weilli/Tarassovi/Vughia 9 D F F C A 7 L. borgpetersenii/Icterohaemorragiae/Tonkini
20 D F F C A 7 L. interrogans/Autumnalis/Autumnalis 26 D F F C A 7 L. interrogans/Hebdomadis/Hebdomadis 30 D F F C A 7 L. interrogans/Pomona/Pomona 11 A G C B A 8 L. borgpetersenii/Mini/Mini 13 E E G C D 9 L. borgpetersenii/Sejroe/Sejroe 15 E E G C D 9 L. inadai/Canicola/Malaya 24 E E G C D 9 L. interrogans/Djasiman/Sentot 17 F G H C A 10 L. inadai/Tarassovi/Kaup 18 E F F C D 11 L. interrogans/Australis/Australis 22 E F F C D 11 L. interrogans/Bataviae/Bataviae 19 G B C C B 12 L. interrogans/Australis/Muenchen 51 G B C C B 12 L. santarosai/Bataviae/Kobbe 53 G B C C B 12 L. santarosai/Gryppotyphosa/Canalzonae 56 G B C C B 12 L. santarosai/Mini/Georgia 61 G B C C B 12 L. santarosai/Shermani/Luis 62 G B C C B 12 L. santarosai/Tarassovi/bakeri 59 G B C C B 12 L. santarosai/Sarmin/weaveri 21 E E G C E 13 L. interrogans/Djasiman/Djasiman 23 D F F C F 14 L. interrogans/Canicola/Canicola 25 D F F C D 15 L. interrogans/Gryppotyphosa/Valbuzzi 27 E E E D E 16 L. interrogans/Icterohaemorragiae/Icterohaemorrhagiae 28 F E E C D 17 L. interrogans/Louisiania/Lanka 29 C E H C D 18 L. interrogans/Mini/Szwajizak 31 D B G C C 19 L. interrogans/Pyrogenes/Pyrogenes 58 D B G C C 19 L. santarosai/Pyrogenes/Alexi 32 F D H C A 20 L. interrogans/Sarmin/Waskurin 33 D F D C A 21 L. interrogans/Sejroe/Hardjo 34 A C C A C 22 L. kirschneri/Australis/Ramisi 35 A C B B C 23 L. kirschneri/Bataviae/Djatzi 36 A C B B C 23 L. kirschneri/Canicola/Bafani 37 A C B B C 23 L. kirschneri/Cynopteri/Cynopteri 38 A C B B C 23 L. kirschneri/Gryppotyphosa/Gryppotyphosa 41 A C B B C 23 L. kirschneri/Pomona/Mozdok 67 A C B B C 23 L. terpstrae/Icterohaemorrhagiae/Hualien 39 A C B C C 24 L. kirschneri/Hebdomadis/Kambale 40 A C B C C 24 L. kirschneri/Icterohaemorrhagiae/Mwogolo 42 E F F D D 25 L. meyeri/Mini/Parameles 43 A H I C A 26 L. meyeri/Ranarum/Ranarum 44 H D D A A 27 L. meyeri/Semaranga/Semaranga 45 A E C D A 28 L. noguchii/Autumnalis/Fortbragg 47 A E C D A 28 L. noguchii/Panama/Panama 49 A E C D A 28 L. noguchii/Shermani/Carimagua 52 A E C D A 28 L. santarosai/Cynopteri/Tingomariensis 46 I I B A A 29 L. noguchii/Djasiman/Huallaga 48 B E B D A 30 L. noguchii/Pyrogenes/Myocastoris 50 J B C C B 31 L. santarosai/Autumnalis/Alice 54 G E C A A 32 L. santarosai/Hebdomadis/Maru 55 G B B C B 33 L. santarosai/Javanica/Vargonicas 57 G B C C A 34 L. santarosai/Pomona/Tropica 60 G J C C B 35 L. santarosai/Sejroe/Trinidad 63 H D F A A 36 L. weilli/Celledoni/Celledoni 65 A H H C A 37 L. weilli/Sarmin/Sarmin 68 H D F E G 38 L. yanagawae/Semaranga/Sao paulo
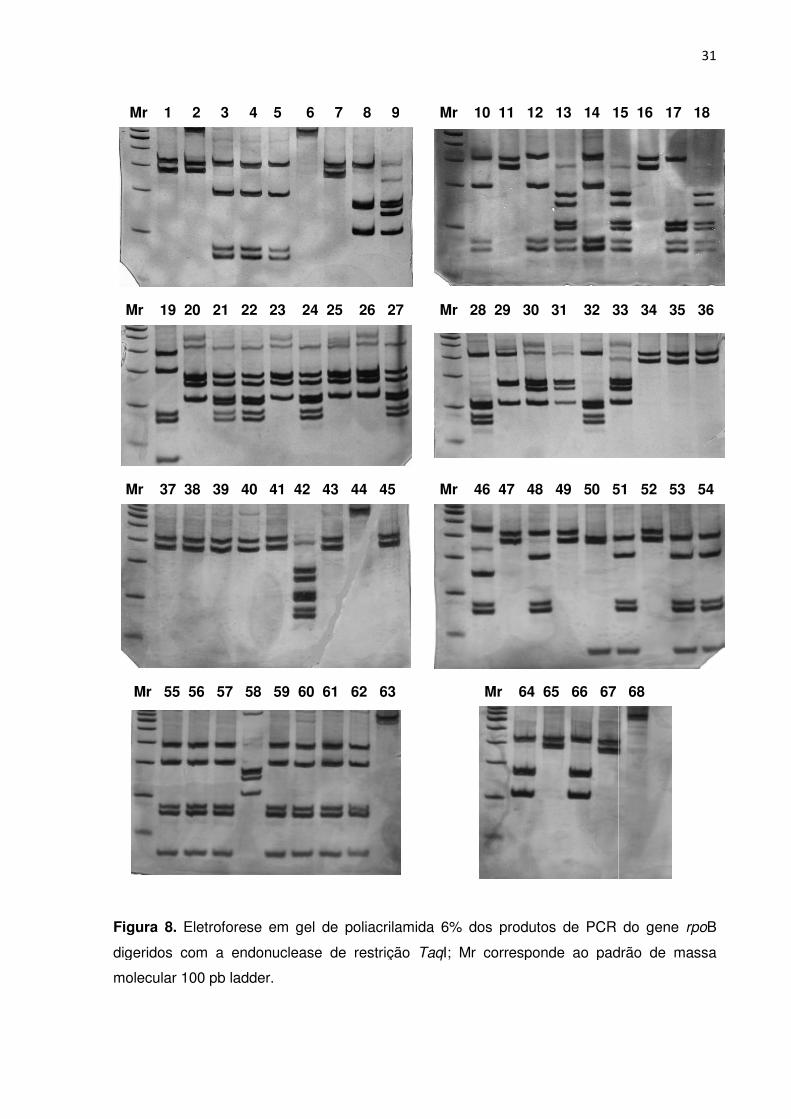
Mr 1 2 3 4 5 6
Mr 19 20 21 22 23 24
Mr 37 38 39 40 41 42
Mr 55 56 57 58 59 60
Figura 8. Eletroforese em gel
digeridos com a endonuclease
molecular 100 pb ladder.
7 8 9 Mr 10 11 12 13 14 1
25 26 27 Mr 28 29 30 31 32 3
43 44 45 Mr 46 47 48 49 50 5
61 62 63 Mr 64 65 66 67
l de poliacrilamida 6% dos produtos de PCR
se de restrição TaqI; Mr corresponde ao pad
31
15 16 17 18
33 34 35 36
51 52 53 54
68
R do gene rpoB
drão de massa
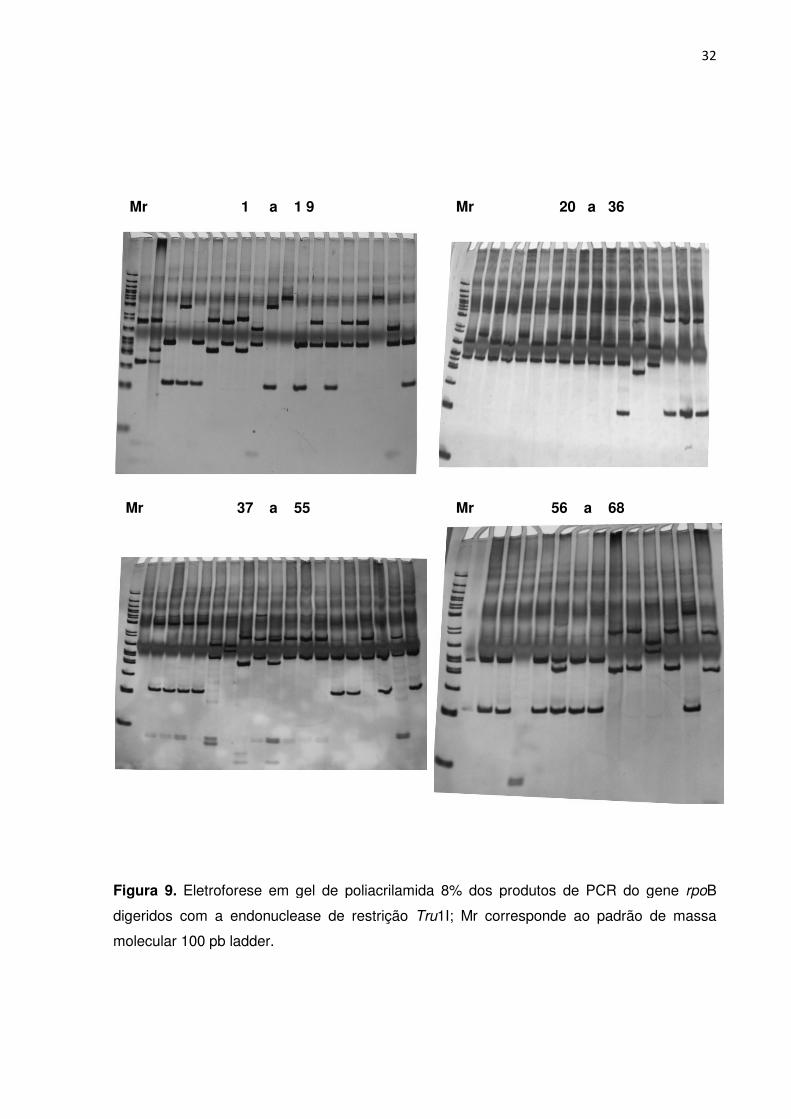
Mr 1 a 1 9
Mr 37 a 55
Figura 9. Eletroforese em gel
digeridos com a endonuclease
molecular 100 pb ladder.
Mr 20 a 36
Mr 56 a 68
l de poliacrilamida 8% dos produtos de PCR
se de restrição Tru1I; Mr corresponde ao pad
32
36
68
R do gene rpoB
adrão de massa
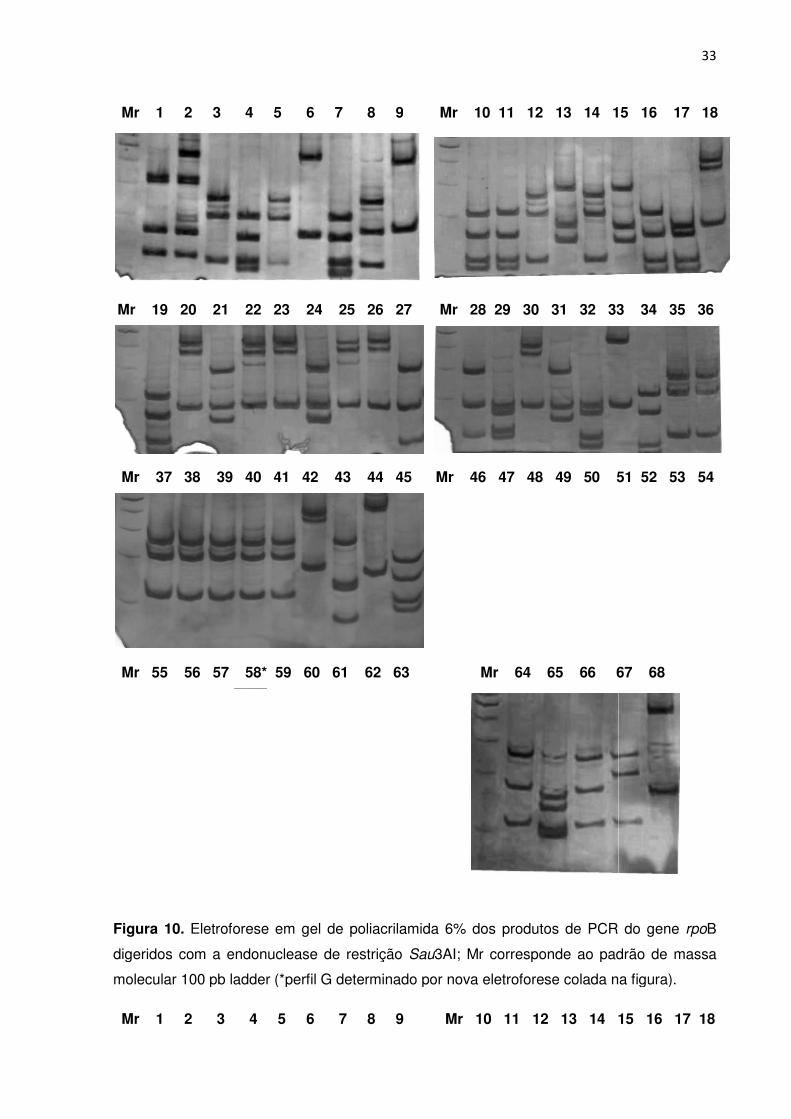
Mr 1 2 3 4 5 6
Mr 19 20 21 22 23 24
Mr 37 38 39 40 41 42
Mr 55 56 57 58* 59 60
Figura 10. Eletroforese em gel
digeridos com a endonuclease
molecular 100 pb ladder (*perfil
Mr 1 2 3 4 5 6
7 8 9 Mr 10 11 12 13 14 1
4 25 26 27 Mr 28 29 30 31 32 33
43 44 45 Mr 46 47 48 49 50 5
0 61 62 63 Mr 64 65 66 6
el de poliacrilamida 6% dos produtos de PCR
e de restrição Sau3AI; Mr corresponde ao pad
fil G determinado por nova eletroforese colada n
7 8 9 Mr 10 11 12 13 14 1
33
15 16 17 18
33 34 35 36
51 52 53 54
67 68
R do gene rpoB
adrão de massa
na figura).
15 16 17 18
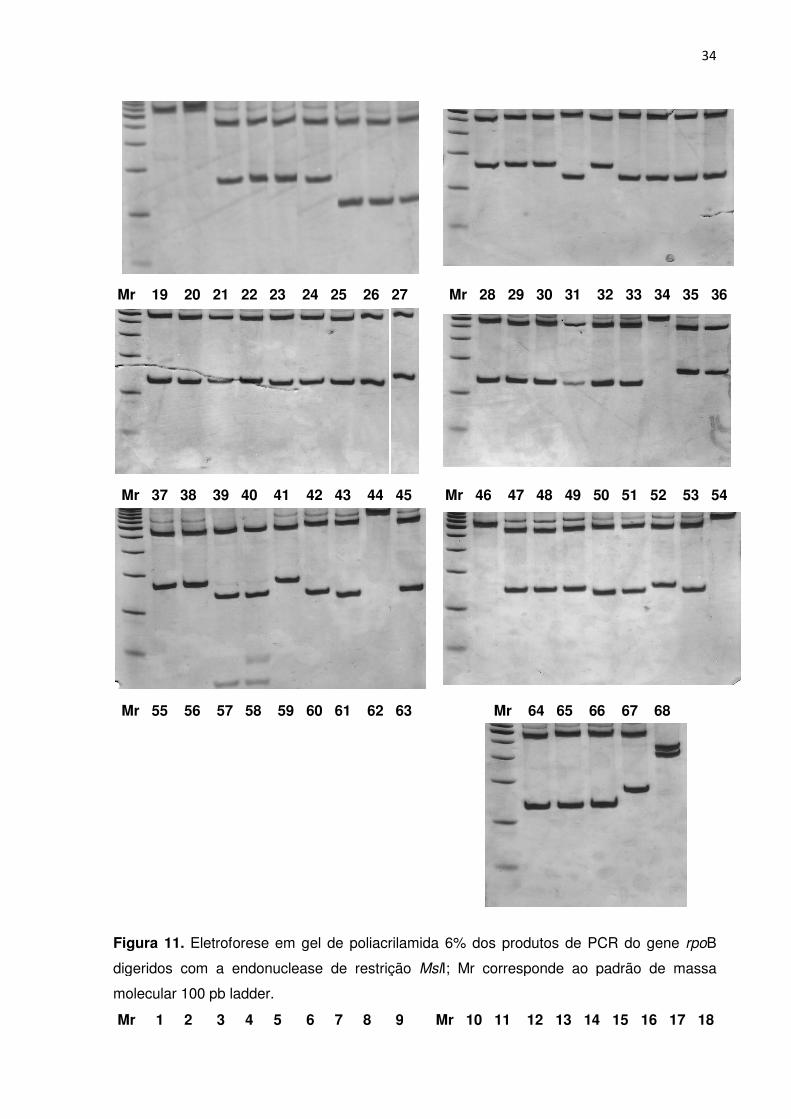
Mr 19 20 21 22 23 24
Mr 37 38 39 40 41 42
Mr 55 56 57 58 59 60
Figura 11. Eletroforese em gel
digeridos com a endonuclease
molecular 100 pb ladder.
Mr 1 2 3 4 5 6
25 26 27 Mr 28 29 30 31 32
2 43 44 45 Mr 46 47 48 49 50
0 61 62 63 Mr 64 65 66
el de poliacrilamida 6% dos produtos de PCR
se de restrição MslI; Mr corresponde ao pad
7 8 9 Mr 10 11 12 13 14 1
34
33 34 35 36
51 52 53 54
67 68
R do gene rpoB
drão de massa
15 16 17 18
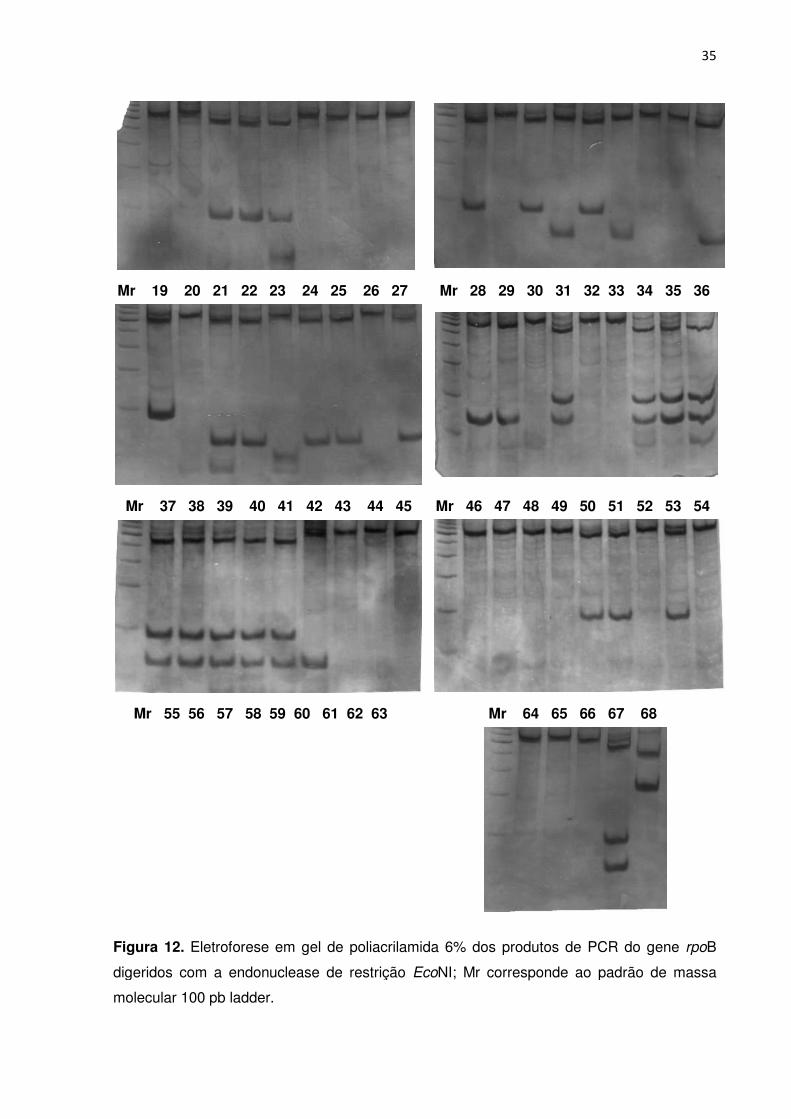
Mr 19 20 21 22 23 24
Mr 37 38 39 40 41 42
Mr 55 56 57 58 59 60
Figura 12. Eletroforese em gel
digeridos com a endonuclease
molecular 100 pb ladder.
25 26 27 Mr 28 29 30 31 32 33
2 43 44 45 Mr 46 47 48 49 50 51
61 62 63 Mr 64 65 66 67
el de poliacrilamida 6% dos produtos de PCR
e de restrição EcoNI; Mr corresponde ao pad
35
33 34 35 36
51 52 53 54
67 68
R do gene rpoB
adrão de massa

36
A enzima Sau3AI apresentou 9 perfis de restrição distintos, esquematizados na Tabela
4C e figura 10, e individualmente foi capaz de identificar apenas o sorovar 43 (Ranarum)
com um perfil sorovar-específico. Todavia, a combinação das três enzimas gerou 30 perfis
distintos com a identificação dos sorovares 6 (Withcombi), 27 (Icterohaemorrhagiae), 33
(Hardjo), 34 (Ramisi), 44 (Semaranga), 55 (Vargonicas) e 65 (Sarmin), que apresentaram os
perfis CDD, EEE, DFD, ACC, HDD, GBB e AHH, respectivamente.
A enzima MslI apresentou apenas 5 perfis de restrição distintos, esquematizados na
Tabela 4D e figura 11, e individualmente foi capaz de identificar apenas o sorovar 68
(Saopaulo) com um perfil sorovar-específico. A combinação das quatro enzimas gerou 34
perfis distintos com a identificação dos sorovares 42 (Parameles) e 63 (Celledoni), que
tiveram os perfis EFFD e HDFA, respectivamente.
Finalmente, a enzima EcoNI apresentou apenas 7 perfis de restrição distintos,
esquematizados na Tabela 4E e figura 12, e individualmente também foi capaz de identificar
apenas o sorovar 68 (Saopaulo) com um perfil sorovar-específico. A combinação das cinco
enzimas gerou 38 perfis distintos com a identificação dos sorovares 21 (Djasiman), 23
(Canicola), 25 (Valbuzzi) e 57 (Tropica), que tiveram os perfis EEGCE, DFFCF, DFFCD e
GBCCA, respectivamente.
Análise dos dendogramas construídos a partir dos produtos de PCR digeridos com as
enzimas TaqI, Tru1I, Sau3AI, MslI e EcoNI
As Figuras 13 a 18 mostram os dendogramas construídos com o auxílio do TFPGA
(Tools for Population Genetic Analyses) software package conforme MILLER (1998). Os
perfis foram agrupados segundo a similaridade usando-se a distância original de NEI (1972)
e segundo a média aritmética ponderada ou agrupamento pelo algoritmo UPGMA. Este
dendograma foi construído baseado nos dados da matriz gerada pelos perfis (presença ou
ausência de fragmentos) da PCR-específica obtidos com os DNAs extraídos de Leptospira
de referência amplificados pelos iniciadores rpoB (Tabelas 4A,4B.4C,4D,4E e Tabela 5).
A análise dos perfis gerados pela digestão com a enzima TaqI (Figura 13) permitiu
agrupar os 68 diferentes sorovares de Leptospira em dois grupos. O grupo 1, formado pelos
agrupamentos A (ramos 1, 2, 3, 4 e 5) e B (ramos 1 e 2). Em todos os ramos do grupo A os
sorovares apresentaram 100% de similaridade, correspondendo aos idênticos perfis de
restrição mostrados no gel. Ramo 1: sorovares das espécies L. santarosai, L. terpstrae
(antiga genomospecie 4), L. noguchii, L. meyeri, L. kirschneri, L. inadai, L. borgpetersenii e
L. biflexa (saprofítica). Ramo 2, sorovares das espécies L. weilli, L. interrogans e L.
borgpetersenii. Ramo 3, sorovares das espécies L. santarosai, L. yanagawae (antiga
genomospecie 5), L. meyeri. Ramo 4, sorovares das espécies L. noguchii e L. borgpetersenii

37
e no ramo 5 do grupo A, sorovares das espécies L. interrogans e L. inadai. No grupo B ramo
1, os sorovares das espécies L. santarosai e L. interrogans apresentaram 100% de
similaridade, e no ramo 2 do grupo B ficou apenas o sorovar Alice pertencente a L.
santarosai. O grupo 2, formado pelos agrupamentos A (ramos 1 e 2) e B mostram a seguinte
distribuição dos sorovares: no ramo 1, espécies L. interrogans, L. santarosai e L.
borgpetersenii. No ramo 2, espécies L. interrogans, L. meyeri, L. inadai e L. borgpetersenii,
apresentaram 100% de similaridade. O grupo 2 B, formado apenas pelo sorovar Huallaga da
espécie L. noguchii.
A Figura 14 mostra a análise fenotípica dos perfis gerados pela digestão com a enzima
Tru1I, permitiu agrupar os 68 sorovares de Leptospira em 3 grupos. O grupo 1, formado
pelos agrupamentos A (ramos 1, 2, 3, 4 e 5), B (ramos 1, 2 e 3) e C (ramos 1 e 2). Nos
ramos (1, 2, 4 e 5) do grupo A os sorovares apresentaram 100% de similaridade,
correspondendo aos idênticos perfis de restrição mostrados no gel. Ramo 1: sorovares das
espécies L. weilli, L. yanagawae, L. meyeri, L. interrogans, L. borgpetersenii. Ramo 2,
sorovares de L. santarosai, L. interrogans e L. borgpetersenii. Ramo 4, sorovares de L.
kirschneri, L. terpstrae e L. borgpetersenii. No ramo 5, sorovares das espécies L.
borgpetersenii e L. inadai. No ramo 3, apenas o sorovar Trinidad de L. santarosai. Nos
ramos (1, 2 e 3) do grupo 2, agrupamento B, todos os sorovares apresentaram 100% de
similaridade entre si. No ramo 1, ficaram agrupados os sorovares das espécies L.
santarosai, L. noguchii, L. interrogans, L. borgpetersenii e L. inadai. No ramo 2, espécies L.
interrogans, L. meyeri e L. borgpetersenii. No ramo 3, espécies L. meyeri e L. weilli. No ramo
1 do grupo 3, do agrupamento C, ficaram os dois sorovares da espécie saprofítica L. biflexa.
No ramo 2 ficou o sorovar Huallaga de L. noguchii.
A Figura 15 mostra a análise fenotípica dos perfis gerados pela digestão com a enzima
Sau3AI que permitiu agrupar os 68 sorovares de Leptospira em 2 grupos. O grupo 1,
formado pelos agrupamentos A (ramos 1, 2, 3, 4 e 5) e B com um agrupamento A contendo
os ramos 1 e 2. Em todos os ramos do grupo A os sorovares apresentaram 100% de
similaridade, no ramo 1 foram agrupados os sorovares das espécies L. weilli, L. interrogans,
L. borgpetersenii. No ramo 2, L. weilli, L. yanagawae e L. meyeri. No ramo 3, L. interrogans,
L. santarosai, L. inadai e L. borgpetersenii. No ramo 4, L. interrogans, L. meyeri e L.
borgpetersenii. No ramo 5, espécies L. interrogans, L. weilli e L. inadai. O agrupamento B,
do grupo 1, no ramo 1 ficou os dois sorovares de L. biflexa e no ramo 2 ficou o sorovar
Ranarum de L. meyeri. No ramo1 do grupo 2, agrupamento A, ficou a maioria dos sorovares
da espécie L. santarosai, L. noguchii, L. kirschneri, L. interrogans e L. borgpetersenii. E no
ramo 2, ficaram os sorovares de L. santarosai, L. weilli, L. noguchii, L. borgpetersenii e L.
kirschneri.

38
Tabela 6: Sorovares de Leptospira identificados pela metodologia de PCR-RFLP do gene
rpoB com as enzimas restrição TaqI, Tru1I, Sau3AI, MslI e EcoNI.
Identificação Espécie Sorovar 06
Leptospira borgpetersenii Withcombi
11 Mini 17 Leptospira inadai Kaup 21
Leptospira interrogans
Djasiman 23 Canicola 25 Valbuzzi 27 Icterohaemorrhagiae 28 Lanka 29 Szwajizak 32 Waskurin 33 Hardjo 34 Leptospira kirschneri Ramisi 42
Leptospira meyeri Parameles
43 Ranarum 44 Semaranga 46
Leptospira noguchii Huallaga
48 Myocastoris 50
Leptospira santarosai
Alice 54 Maru 55 Vargonicas 57 Tropica 60 Trinidad 63
Leptospira weilli Celledoni
65 Sarmin
68 Leptospira yanagawae Saopaulo
A análise dos perfis gerados pela digestão com a enzima MslI (Figura 16) permitiu
agrupar os 68 diferentes sorovares de Leptospira em dois grupos. O grupo 1 formado pelos
agrupamentos A (ramos 1, 2 e 3) e B (ramo 1). Em todos os ramos do grupo A os sorovares
apresentaram 100% de similaridade, correspondendo aos idênticos perfis de restrição
mostrados no gel. Ramo 1 do agrupamento A: espécies L. weilli, a maioria dos sorovares de
L. santarosai e de L. interrogans, L. meyeri, L. kirschneri, L. inadai e L. borgpetersenii. Ramo
2, espécies L. santarosai, L. weilli, L. meyeri, L. kirschneri e os dois sorovares de L. biflexa.
No ramo 3 sorovar Saopaulo da espécie L. yanagawae. Ainda no grupo 1, o agrupamento B
apresentou um ramo contendo os sorovares das espécies L. noguchii, L. santarosai, L.
meyeri e L. interrogans. O grupo 2 apresentou um agrupamento contendo a maioria dos
sorovares da espécie de L. borgpetersenii, L. kirschneri e L. terpstrae.

39
A Figura 17 mostra a análise fenotípica dos perfis gerados pela digestão com a enzima
EcoNI que permitiu agrupar os 68 sorovares de Leptospira em 2 grupos. O grupo 1,
agrupamento A, ramos 1, 2 e 3 e o grupo 2, agrupamento A apresentando os ramos 1 e 2.
Nos dois grupos formados os sorovares apresentaram 100% de similaridade,
correspondendo aos idênticos perfis de restrição mostrados no gel. Grupo 1, ramo 1,
agrupamento A: sorovares das espécies L. weilli, L. santarosai, L. noguchii, L. meyeri, L.
interrogans, L. inadai, L. borgpetersenii e L. biflexa. No ramo 2 ficou o sorovar Sao Paulo de
L. yanagawae. No ramo 3 do grupo 1 ficaram os sorovares de L. inadai, L. meyeri, L.
borgpetersenii e a maioria dos sorovares da espécie L. interrogans. No ramo 1 do grupo 2,
agrupamento A, ficaram os sorovares das espécies L. interrogans e L. borgpetersenii e a
maioria dos sorovares da espécie L. santarosai. No ramo 2 do grupo 2 ficaram os sorovares
de L. santarosai, L. terpstrae, L. interrogans e a maioria dos sorovares de L. kirschneri.
A Figura 18 mostra o dendograma obtido a partir de uma matriz construída usando os
resultados das análises dos fragmentos gerados com as endonucleases de restrição TaqI,
Tru1I, Sau3AI, MslI e EcoNI. Nesta figura, verificou-se que os 68 sorovares de Leptospira
formaram um grande grupo constituído por 3 agrupamentos denominados de A (formado
pelos ramos de 1 a 10); B (ramos de 1 a 10); C (ramos 1 e 2) e D (ramos de 1 a 16). No
ramo 1 do agrupamento A, ficaram os sorovares de L. weilli, L. borgpetersenii. No ramo 2
apenas o sorovar Withcombi de L. borgpetersenii; ramo 3, sorovares de L. inadai e L.
interrogans; ramo 4, L. interrogans; ramo 5, L. weilli, ramo 6, L. meyeri; ramo 7, L. weilli;
ramo 8, L. yanagawae; ramos 9 e 10, L. biflexa. No agrupamento B, ramo 1 ficou 6
sorovares de L. santarosai e 1 da espécie L. interrogans; nos ramos 2, 3 e 4, sorovares de
L. santarosai; no ramo 5, L. noguchii, no ramo 6, L. noguchii e L. santarosai, ramo 7, L.
borgpetersenii e L. inadai; ramo 8, L. kirschneri e L. terpstrae; ramo 9, L. kirschneri; ramo 10,
L. noguchii. No agrupamento C, ramo 1, sorovares de L. interrogans e L. borgpetersenii, no
ramo 2, L. interrogans. No agrupamento D, ramos 1, 2, 3 e 4, sorovares de L. interrogans;
ramo 5, L. meyeri; ramo 6, L. santarosai; ramo 7, L. borgpetersenii; ramo 8, L. noguchii;
ramo 9, L. kirschneri; ramos 10 e 11, L. borgpetersenii, ramo 12, L. interrogans e L.
santarosai; ramo 13, L. interrogans, L. inadai e L. borgpetersenii; ramo 14 a 16, sorovares da
espécie L. interrogans.

Figura 13. Dendograma resulta
Leptospira obtidos pela anális
restrição TaqI. O nível de reprod
repetições.
tante do agrupamento UPGMA dos perfis dos 68
se conjunta das bandas geradas pela end
odutibilidade foi garantido pela análise de “Boot
40
68 sorovares de
ndonuclease de
otstrap” de 1000

Figura 14. Dendograma resulta
Leptospira obtidos pela anális
restrição Tru1I. O nível de repro
repetições.
tante do agrupamento UPGMA dos perfis dos 68
se conjunta das bandas geradas pela end
rodutibilidade foi garantido pela análise de “Boot
41
68 sorovares de
ndonuclease de
otstrap” de 1000

Figura 15. Dendograma resulta
Leptospira obtidos pela anális
restrição Sau3AI. O nível de re
1000 repetições.
tante do agrupamento UPGMA dos perfis dos 68
se conjunta das bandas geradas pela end
reprodutibilidade foi garantido pela análise de
42
68 sorovares de
ndonuclease de
e “Bootstrap” de

Figura 16. Dendograma resulta
Leptospira obtidos pela anális
restrição MslI. O nível de reprod
repetições.
tante do agrupamento UPGMA dos perfis dos 68
se conjunta das bandas geradas pela end
odutibilidade foi garantido pela análise de “Boot
43
68 sorovares de
ndonuclease de
otstrap” de 1000

Figura 17. Dendograma resulta
Leptospira obtidos pela anális
restrição EcoNI. O nível de rep
1000 repetições.
tante do agrupamento UPGMA dos perfis dos 68
se conjunta das bandas geradas pela end
eprodutibilidade foi garantido pela análise de
44
68 sorovares de
ndonuclease de
e “Bootstrap” de

Figura 18. Dendograma resulta
Leptospira obtidos pela anális
restrição Taq1, Tru1I, Sau3AI, M
análise de “Bootstrap” de 1000 r
tante do agrupamento UPGMA dos perfis dos 68
se conjunta das bandas geradas pelas endo
MslI e EcoNI. O nível de reprodutibilidade foi
repetições.
45
68 sorovares de
donucleases de
oi garantido pela

46
5. DISCUSSÃO
De acordo com a Organização Mundial de Saúde, a leptospirose humana tem sido
considerada uma doença reemergente em todo o mundo. Estudos têm demonstrado que o
aumento dos surtos epidêmicos está diretamente relacionado com a explosão demográfica e
com fatores sócio-econômicos e ambientais. Conseqüentemente, as estratégias para o
diagnóstico, identificação e caracterização dos sorovares de Leptospira, envolvidos nas
infecções, precisam ser aprimoradas para o êxito do reconhecimento dos mecanismos de
transmissão, fontes de contaminação e reservatórios, assim como os métodos
epidemiológicos de prevenção e tratamento.
Para esse grupo de bactérias, a tipificação das linhagens isoladas de pacientes é
importantíssima já que existe preferência de determinados sorovares por alguns
hospedeiros, além de diferenças nos sinais clínicos, mecanismos de invasão e evolução da
doença. Portanto é essencial a rápida e eficiente identificação do agente etiológico, para
tratamento adequado e imunização (CERQUEIRA & PICARDEAU 2009; WENJUN et al.,
2009).
As leptospiras têm sido identificadas por métodos que envolvem o estudo das relações
sorológicas: reação de soroaglutinação microscópica (MAT) e testes de absorção cruzada
de aglutininas (CAAT), ambos baseados nos antígenos de superfície da célula. Esta
classificação, denominada sensu lato reconhece duas espécies, a saber: L. interrogans,
patogênica para homens e animais, e L. biflexa, saprofítica (CERQUEIRA & PICARDEAU
2009). As técnicas sorológicas de classificação e identificação apresentam alguns
obstáculos: exigem tempo e paciência na execução e preparação dos soros imunes de
coelhos (TERPSTRA et al., 1985). Poucos laboratórios têm capacidade de realizar CAAT e
a identificação de sorogrupos por MAT não é aceita oficialmente A inabilidade das espécies
patogênicas crescerem em temperaturas de 13 a 30°C durante 3, 7, 10 e 13 dias; e o não
crescimento, em presença de 8-azaguanina (225 µg ml-1) em 30°C, têm sido usado para
diferenciar espécies de Leptospiras patogênicas das não patogênicas (PEROLAT et
al.,1998).
Entre os métodos moleculares, a técnica de hibridização quantitativa de DNA-DNA
para medir a homologia genética tem sido usada como referência, para a distribuição dos
sorovares, dentro das espécies de Leptospira. O método visa comparar os genomas de
linhagens isoladas desconhecidas com vários cuja classificação já tenha sido definida e
publicada (sorovar, sorogrupo ou espécie, de referência). Após cultivo e extração dos DNAs,
eles são marcados in vitro com 32P, desnaturados e depois hibridizados, com sorovares de
referência, à 55°C e em uma temperatura estringente de 70°C, determinando-se também a
composição C+G da linhagem. Para isso (YASUDA, et al., 1987) definiram espécies como

47
um grupo de sorovares cujos DNAs estejam 70% ou mais relacionados na temperatura
ótima de reassociação de 55°C e 55% ou mais relacionados na temperatura estringente de
70°C e ainda aqueles, cujo DNA relacionado tenha menos que 6% bases não pareadas. É
possível também medir o conteúdo C+G das linhagens testadas, de acordo com o grau de
hibridização, (nas temperaturas definidas), com as linhagens de referência. Usando a
técnica de hibridização foi demonstrado que várias linhagens de L. interrogans sensu lato
são na verdade 6 espécies diferentes e não uma como fora classificada (YASUDA et al.,
1987). Atualmente já foram descritas 20 espécies no gênero Leptospira (YASUDA et al.,
1987; LEVETT et al., 2005, 2006; PEROLAT et al., 1998).
A hibridização DNA-DNA foi usada por (YASUDA et al., 1987) para comparar 44
sorovares de 23 sorogrupos conhecidos e outros 3 ainda não tipificados de L. interrogans e
sorovares únicos de L. parva e Leptonema illini. A relação obtida confirmou as novas
espécies L. parva e Leptonema illini e demonstrou que L. interrogans e L. biflexa são muito
heterogêneas. Eles descreveram sete novas espécies: L. borgpetersenii, L. inadai, L.
noguchii, L. santarosai, L. weilii, L. meyeri e L. wolbachii. Analisando, por hibridização em
slot blot, 66 sorovares de espécies de Leptospira potencialmente patogênicas conseguiu-se
classificar 57 desses sorovares, colocando-os em 6 grupos e propor uma nova espécie
Leptospira kirschneri, que agrupa os sorovares Bulgarica, Butembo, Cynopteri, Danin,
Grippotyphosa, Kabura, Kambale, Ramisi e Tsaratsovo. Foram usadas 16 linhagens de
referência como marcadores (RAMADASS et al.,1992).
A hibridização não é empregada rotineiramente, para a identificação das espécies de
Leptospira, por ser de execução tecnicamente complexa, laboriosa e por requerer o uso de
isótopos radioativos, ficando restrita aos laboratórios de referência (MOREY et al., 2006)
além disto, já foi observado que não há muita correlação entre os sistemas de classificação
sorológica e genotípica uma vez que um determinado sorogrupo pode ser freqüentemente
encontrado em várias espécies de Leptospira. Portanto esse método continua sendo
utilizado apenas para determinação de espécies em estudos filogenéticos (YASUDA, et
al.,1987).
A técnica REA (Restriction Endonucleasis Analysis) foi usada por BOLIN & ZUERNER
(1996), no genoma integral de 147 linhagens isoladas de L. interrogans, sorovar Pomona,
linhagem kennewickiem, usando as enzimas EcoRI e HpaII. Com os perfis idênticos obtidos
da digestão com EcoRI foi confirmada a identidade dos sorovares e que L. interrogans,
sorovar Pomona, tipo kennewickiem é linhagem geneticamente diferente; com a enzima
HpaII obteve-se 6 perfis distintos cuja variação foi de acordo com os animais de onde foram
isolados. Várias linhagens de Hardjo e Balcanica, (sorogrupo Hebdomadis de L. interrogans)
foram examinadas por REA e obtidos perfis que possibilitaram sua clara diferenciação
quando restrita pela enzima EcoRI (MARSHALL et al, 1981).

48
A amplificação de material genético por PCR possibilitou o desenvolvimento de uma
gama de protocolos nos quais partes do genoma das Leptospiras podem ser estudados e
comparados. Muitas pesquisas dependentes do PCR, têm sido organizadas com objetivos
de tipificação, estudo do genoma e produtos do metabolismo das Leptospiras.
Utilizando primers pequenos, de seqüências conhecidas RAPD (Polimorfismo do DNA
amplificado ao acaso) foi usado por (RAMADASS et al.,1997) com 14 linhagens de
laboratório dos sorovares (Australis, Autumnalis, Ballum, Bataviae, Canicola,
Grippotyphosa, Hardjoprajitno, Hebdomadis, Icterohaemorrhagiae, Javanica, Pomona,
Pyrogenes, Panama e Tarassovi. As linhagens foram amplificadas com os primers B11
(CCGGAAGAAGGGGCGCCAT) e B12 (CGATTTAGAAGGACTTGCACAC) e o produto
analisado por gel de agarose, resultou em poucas bandas e produziu perfis diferentes para
muitos sorovares. Os resultados mostraram que sorovares Javanica, Hardjoprajitno e
Autumnalis produzem poucas bandas enquanto os outros sorovares, maior número de
bandas porém não os discrimina. Essa mesma técnica foi usada por (ROY et al., 2003),
para comparar isolados clínicos de 1929 até 2002, das Ilhas Andamana, e possibilitou
concluir apenas, que são da mesma linhagem, já identificada por MAT como pertencente ao
sorogrupo Grippotyphosa.
Testando o método de RAPD para tipificação de sorovares, diferentes pesquisadores
não conseguiram grande poder de discriminação, em relação aos métodos sorológicos
(LEVETT, 2001; ROY et al.,2003).
Além da hibridização DNA-DNA, e todos os outros métodos moleculares acima
citados, a amplificação por PCR específica do gene 16s rRNA tem contribuído para a
caracterização molecular de algumas espécies de Leptospira. Neste caso, a vantagem é
que o uso de DNA molde, em especial da região que codifica o gene 16s rRNA, considerada
conservada entre as bactérias, mas ao mesmo tempo, com variabilidade e quantidade de
informações suficientes, pode revelar claramente as relações filogenéticas entre as espécies
(MOREY et al., 2006). Entretanto, a identificação das espécies de Leptospira usando a
amplificação do gene da região 16s rRNA tem mostrado que há significante homologia entre
as seqüências o que torna essencial que a maioria dos genes amplificados, sejam também
seqüenciados, para verificar a diferença entre eles.
A análise filogenética da família Leptospiraceae, realizada por seqüenciamento da
fração 16S do gene do RNA ribossômico, revelou a formação de cinco grupos de espécies:
as leptospiras patogênicas como L. interrogans, L. kirschneri, L. noguchii, L. alexanderi, L.
borgpetersenii e L. santarosai formando um clado diferente das espécies não patogênicas L.
biflexa, L. wolbachii, L. meyeri e genomospecies 3, 4 e 5 e um terceiro clado contendo as
espécies L. fanei, L. broomi e L. inadai, claramente diferenciado das patogênicas e das não
patogênicas; esses três grupos foram ainda diferenciados dos últimos dois clados, que

49
incluem as espécies: Treponema parva e Leptonema illini (PEROLAT et al., 1998, MOREY
et al.,2006, HOOKEY et al., 1993). Foi observado também que alguns sorovares têm
característica de mais de uma espécie e algumas espécies contêm ambos sorovares:
patogênicos e não patogênicos, tal como havia sido observado por (YASUDA, .et al.,1987).
Com a análise por PCR usando a seqüência de 330 pb do gene para o RNA
ribossômico 16S e os primers: (F)fD3 AGAGTTTGATCCTGGCTTAG; (R)UniB
T(AC)AAGGAGGTGATCCAGC; (F)LeptoA* GGCGGCGCGTCTTAAACATG e (R)LeptoB*
TTCCCCCCATTGAGCAAGATT (produzidos a partir do sorovar Semaranga, linhagem
Veldrat Semarang 173), foi testado o poder discriminatório, em linhagens de diferentes
coleções de referência. Essas linhagens foram amplificadas, seqüenciadas, alinhadas e
comparadas entre si. Com os resultados obtidos foram construídas árvores filogenéticas e
os clusters formados, indicaram o posicionamento de linhagens de L alexanderi, separando-
as de linhagens de L. borgpetersenii e L. weilii e de algumas espécies patogênicas que
foram identificadas em nível de sorovar. Foi possível confirmar a similaridade nas
seqüências das linhagens dos sorovares Autumnalis e Icterohaemorrhagiae do Instituto
Pasteur ligadas a L. interrogans, e os sorovares de L. borgpetersenii, Vughia e Celledoni (L.
weilii) e Canalzone (L. santarosai). As linhagens de referência dos sorovares Tarassovi,
Bakeri e Mengdeng, encontrados no banco de dados, mostraram compatibilidade com as
espécies: L. borgpetersenii, L. santarosai e L. weilii, respectivamente. A espécie L. meyeri
de 3 diferentes coleções de referência mostrou distribuição heterogênea. Outras linhagens
como sorovares Parameles e Ranarum diferiram nas coleções de origem, enquanto o
sorovar Sofia de 3 diferentes coleções eram idênticos e pertencentes à L. borgpetersenii
(POSTIC et al., 2000).
A análise por PFGE (Pulse Field Gel Electrophoresis) é considerado um método forte
de classificação de linhagens, já que mostra variação no número e comprimento de bandas
obtidas por macrorestrição, que podem ser originados devido a rearranjos no genoma. A
técnica RAPD, associada à eletroforese em gel de campo pulsátil, foi considerada um ótimo
meio de caracterizar sorovares patogênicos pelo tipo, circular ou linear e tamanho da
molécula de DNA (TAYLOR et al., 1991) e pelo tamanho da molécula, quando usada a
amplificação ex.: L. interrogans possuem apenas uma cópia dos genes rrs. (ZUERNER et
al., 1991; 1993).
A análise por Multilocus VNTR (MLVA) de 99 linhagens de referência e 156 isolados
clínicos foi usada para tentar diferenciar espécies e sorovares de Leptospiras patogênicas
por meio das regiões repetidas em tandem em vários loci do genoma (SALAUN et al. 2006).
Desenharam primers margeando as seqüências VNTL dos dois cromossomos CI e CII das
linhagens Lai e Copenhageni de L. interrogans e Hardjo-bovis de L. borgpetersenii,
encontradas in silico. Em trabalho anterior, (MAJED et al.,2005) mostraram que apenas

50
três marcadores (VNTR-7, VNTR-10 e VNTR-19) seriam necessários para identificar
sorovares de L. interrogans; então redefiniram esses primers e construíram outros para a
análise (VNTR-4, VNTR- Lb4 e VNTR-Lb5). Nas linhagens de referência, verificaram que
alguns dos primers não amplificavam todos os sorovares, que VNTR-10 foi o mais eficiente
e que a espécie L. borgpetersenii foi amplificada quase que exclusivamente quando usaram
VNTR-Lb4 e VNTR-Lb5. Consideraram um quadro de iniciador versus número de sorovares
amplificados em cada espécie, para o resultado. Foi possível diferenciar as espécies (L.
interrogans, L. kirschneri e L. borgpetersenii) nas quais os sorovares foram apenas
relacionados com as respectivas espécies, excetuando-se os sorovares Schueffneri de L.
interrogans e Grippotyphosa, Bogvere, Bim e Valbuzzi de L. kirschneri. L. biflexa não
apresentou tandem repeats, exceto dentro do gene infB1. Conseguiram confirmar 34 das
linhagens isoladas pertencem a sete sorogrupos (Icterohaemorrhagiae, Canicola, Pomona,
Grippotyphosa, Autumnalis, Sejroe e Ballum).
A utilidade da técnica MLVA mostrou limitações, no sentido que possibilita
demonstrar somente que há diversidade genética dentro de alguns sorovares e que as
variações nestas são muito pequenas, nos sorovares das linhagens de coleções, em relação
às variações existentes na população natural impossibilitando a definição de um método
preciso de diferenciação entre eles. Devido aos resultados obtidos do uso de
seqüenciamento e alinhamento de fragmentos de genes estruturais, após amplificação,
surgiu a técnica MLST (Multilocus Sequence Typing). Com MLST (AHMED et al., 2006)
conseguiram diferenciar 120 linhagens de Leptospiras a partir de seqüências obtidas das
regiões hipervariáveis, de 6 genes estruturais. Observaram também a presença de sítios de
polimorfismo nas seqüências e que muitas das linhagens de Leptospira não estão
agrupadas com outras geneticamente similares, quando observadas após serem
organizadas em árvores filogenéticas. Esse pode ser um método adequado para identificar
leptospiras em nível de espécies, com a vantagem de resultar no depósito de seqüências
em bancos de dados, facilitando os trabalhos futuros; com isto a técnica permite também
que após amplificação das seqüências dos genes escolhidos, eles sejam submetidos à
restrição por endonucleases, dispensando o uso de seqüenciamento para análise.
O gene mais usado para identificação baseada em seqüenciamento de DNA é o do
rRNA 16S (rrs). Porém já foi demonstrado (AHMED et al, 2006; LA SCOLA et al., 2006) que
esse gene não possui suficiente polimorfismo e é necessário amplificar um segmento de
1500 pb para se obter uma identificação confiável.
LA SCOLA et al., (2006), alinharam os genomas totais das Leptospiras, que já estão
disponíveis no GenBank (L. biflexa - Patoc AF150880; L. interrogans - Lai, NC-004342; L.
interrogans - Copenhageni, NC-005823) para determinar as regiões mais variáveis, que
margeiam as regiões conservadas do gene rpoB. A partir dessa determinação desenharam

51
o par de iniciadores (Lept 1900f 5’ -CCT-CAT-GGG-TTC-CAA-CAT-GCA-3’ e Lept 2500r 5’-
CGC-ATC-CTC-RAA-GTT-GTA-WCC-TT-3’) que margeiam as posições 1900 e 2500 do
gene e possibilitam a amplificação de um segmento de 600 pares de bases, em 19
linhagens de L. interrogans. As seqüências amplificadas foram alinhadas e analisadas em
suas proporções de similaridades com o gene rpoB e rrs disponíveis no GenBank. A partir
do gene rpoB conseguiu-se distinguir com eficácia 11 sorovares dos 19 sorovares testados,
diferenciando-os dos de outras espécies; e comparando com o gene rrs, observou-se maior
polimorfismo no rpoB o que levou à dedução de que com esse gene é possível distinguir
espécies com um nível de diferença bem maior no número de pares de base.
Em uma alternativa da técnica de MLST, sem usar seqüenciamento, testou-se a
variação de nucleotídeos em parte de um gene estrutural, das espécies e sorovares
estudados e a possibilidade de tipificação de linhagens patogênicas. Nesse estudo foi
utilizada à técnica PCR-RFLP e os iniciadores produzidos por (LA SCOLA et al, 2006), que
amplificam 595 pb do gene rpoB. Foram testados 68 sorovares de referência, obtidos do
INNPAZ. O seqüenciamento foi substituído pela análise dos produtos após restrição com
cinco endonucleases comerciais: TaqI, Tru1I, Sau3AI, MslI e EcoNI, e o resultado
visualizado em gel de poliacrilamida a 6%, corado por prata. Os 38 perfis obtidos da análise
dos fragmentos formados nos géis foram confrontados com os perfis obtidos da restrição in
silico, usando como molde, seqüências do gene rpoB que estão depositadas em bancos de
dados, para confirmar o comprimento das bandas. As enzimas que forneceram maior
número de perfis, no trabalho foram a TaqI e Tru1I, com dez perfis diferentes cada uma,
seguida da enzima Sau3AI que forneceu 9 perfis; EcoNI forneceu 7 perfis e por fim MslI
forneceu 5 perfis diferentes. Com os resultados dos grupos formados e tamanho dos
fragmentos das bandas, nos perfis de cada enzima, foram construídos dendogramas e
observados a formação de clusters, que tal como já tem sido relatado por (PEROLAT et al.,
1998; MOREY et al.,2006; HOOKEY et al., 1993) indicam que sorovares de várias espécies
apresentam perfis idênticos na seqüência analisada, sugerindo que a amplificação desse
método de análise, usando várias seqüências de genes estruturais, poderá mostrar a
precisa classificação sensu stricto desses sorovares. Foi criado um perfil de identificação
para 25 dos sorovares testados, que estão incluídos em 9 espécies de Leptospira, usando
apenas as seqüências que já estão disponíveis nos bancos de dados e conseguiu-se
confirmar que a técnica PCR-RFLP, associada ao gene rpoB, possibilita uma boa
diferenciação no nível de espécies e sorovares.

52
6. CONCLUSÃO
Com esse trabalho comprovou-se que PCR-RFLP é uma metodologia prática e
eficiente, que possibilita a diferenciação de espécies e sorovares com ótimo poder
discriminatório, reprodutibilidade e facilidade de interpretação dos resultados, além de um
custo adequado para a maioria dos laboratórios de pesquisa. A técnica demonstrou, ainda,
ser adequada para estudos filogenéticos e de tipificação de espécies, sorovares e linhagens.
A seleção do gene rpoB e utilizando apenas a seqüência polimórfica contendo 595 pares de
base, do gene, possibilitou a obtenção de perfis de restrição que forneceram a precisa
identificação de 37% dos sorovares testados. Demonstrou-se que a metodologia se adéqua
ao objetivo proposto assim como se demonstrou, mais uma vez, que a tipificação sorológica
fornece uma classificação pouco confiável em se tratando de leptospiras patogênicas. A
adição de novas enzimas, com a produção de diferentes perfis ou a utilização da mesma
técnica, porém com seqüências polimórficas de outros genes estruturais deve, certamente,
aumentar a proporção de sorovares identificados. A análise dos dendogramas feitos com
resultados da restrição de cada enzima e com o resultado de todas as enzimas juntas
indicou a formação de clusters onde sorovares de várias espécies apresentam perfis
idênticos, na seqüência do lócus analisado, sugerindo que à medida que ocorrem estudos
envolvendo métodos genômicos, as linhagens serão corretamente classificadas. Os grupos
formados com os perfis do gene rpoB apresentaram variado grau de similaridade e na
formação dos clados, podem ser observadas etapas nas quais podem ter ocorrido
transferências gênicas entre alguns sorovares, assim como a medida da distância relativa
entre eles, durante a evolução (figuras 13 a 18).

53
7. REFERÊNCIAS BIBLIOGRÁFICAS 1. ADLER B, de la PEÑA MOCTEZUMA A. Leptospira and Leptospirosis. Veterinary
Microbiology 2009; In Press- Elsevier. 2. AHMED N, DEVI SM, VALVERDE, M. DE LOS A., VIJAYACHARI P, MACHANG'U ELLIS
A, HASTSKEERL, R. A. Multilocus sequence typing method for identification and genotypic classification of pathogenic Leptospira species. Annals of Clinical Microbiology and Antimicrobials 2006; 5(28).
3. BHARTI AR, NALLY JE, LOVETT MA. Leptospirosis: a zoonotic disease of global
importance. The Lancet infectious disease 2003; 3. 4. BAROCCHI MA, KO AI, FERRER SR, FARIA MT, REIS MG, RILEY LW. Identification of
new repetitive element in Leptospira interrogans serovar Copenhageni and its application to PCR-based differentiation of Leptospira serogroups. Journal of Clinical Microbiology 2001; 39(1): 191-195.
5. BOMFIM MRQ, KOURY MC. Evaluation of LSSP-PCR for identification of Leptospira spp.
in urine samples of cattle with clinical suspicion of leptospirosis. Veterinary Microbiology 2006; 118(3-4): 278-288.
6. BOURSAUX-EUDE C, SAINT GIRONS I, ZUERNER R. Leptospira genomics.
Electrophoresis. 1998 Apr; 19(4): 589-92. 7. BRENNER, D.J., KAUFMANN, A.F., SULZER, K.R., STEIGERWALT, A.G., ROGERS,
F.C., WEYANT, R.S., 1999. Further determination of DNA relatedness between erogroups and serovars in the family Leptospiraceae with a proposal for Leptospira alexanderi sp. ov. and four new Leptospira genomospecies. Int. J. Syst. Bacteriol. 49, 839–858.
8. CAMERON CE, ZUERNER RL, RAVERTY S, COLEGROVE KA, NORMAN SA,
LAMBOUM DM. Detection of pathogenic Leptospira bacteria in pinniped populations via PCR and identification of a source of transmission for zoonotic leptospirosis in the marine environment. Journal of Clinical Microbiology 2008; 46(5): 1728-1733.
9. CENTER TLI. Overview of the leptospira bacterium itself. In:
http://www.leptospirosis.org/links/,2008 10. CERQUEIRA G, PICARDEAU M. A century of Leptospira strain typing- Review. In:
Infection, Genetics and Evolution 2009. Pelotas, Brazil and Paris, France: Science Direct: 760-768.
11. DEY S, MOHAN CM, RAMADASS P, NACHIMUTHU K. Phylogenetic analysis of LipL32
gene sequence of different pathogenic serovars of Leptospira. Veterinary Research Communications 2007; 31(5): 509-51
12. ELLINGHAUSEN.H.C. Virulence, nutrition and antigenicity of Leptospira interrogans
Serotype Pomona in supplemented and nutrient deleted bovine albumin medium. Annales de Microbiologie 1973; B124(4): 477-493.
13. FONSECA CDA, DE FREITAS VLT, ROMERO EC, SPINOSA C, SANCHES MCA, DA
SILVA MV Polymerase chain reaction in comparison with serological tests for early diagnosis of human leptospirosis. Tropical Medicine & International Health 2006; 11(11): 1699-1707.

54
14. FONSECA CD, TEIXEIRA MMG, ROMERO EC, TENGAN FM, DA SILVA MV, SHIKANAI-YASUDA MA. Leptospira DNA detection for the diagnosis of human leptospirosis. Journal of Infection 2006; 52(1): 15-22.
15. HAAKE DA, SUCHARD MA, KELLEY MM, DUNDOO M, ALT DP, ZUERNER RL.
Molecular evolution and mosaicism of leptospiral outer membrane proteins involves horizontal DNA transfer. Journal of Bacteriology 2004; 186(9): 2818-2828.
16. . HAWLEY JS, REITSTETTER RE, REEB BA, STUART SK, BECKIUS ML, MURRAY
CK. Detection and identification of pathogenic and saprophytic leptospira strains by a PCR matrix approach using multiple primers. American Journal of Tropical Medicine and Hygiene 2005; 73(6): 216-216.
17. HE P, SHENG YY, SHI YZ, JIANG XG, QIN JH, ZHANG ZM. Genetic diversity among
major endemic strains of Leptospira interrogans in China. BMC Genomics 2007; 8. 18. HE P, ZHANG XY, GUO XK, HU BY, HUANG XT, YANG Y. Identification and analysis of
genes present in Leptospira interrogans serovar Lai but absent in L. biflexa serovar Monvalerio (vol 36, pg 832, 2004). Acta Biochimica Et Biophysica Sinica 2005; 37(1): 46-46.
19. HUGH C.Y, HORNBY G, KEY D.W, OSUCH M.R, MAXIE M.G.Preliminary study on
differentiation of Leptospira gryppotyphosa e Leptospira sejroe from other common pathogenic leptospiral serovars incanine urine by Polymerase Chain Reaction Assay. Journal of Veterinary Diagnostic Investigation. 2002; 14: 164-168.
20. JOHSON RC. The Spiroquetes. Annual Reviews Microbiology 1977; 31: 89-106. <www.annualreviews.org/aronline>
21. JOUGLARD SDD, SIMIONATTO S, SEIXAS FK, NASSI FL, DELLAGOSTIN OA. Nested
polymerase chain reaction for detection of pathogenic leptospires. Canadian Journal of Microbiology 2006; 52(8): 747-752.
22. KO A. I., GOARANT C., PICARDEAU M. Leptospira: the dawn of molecular genetics era
for an emerging zoonotic pathogen. Nature reviews/Microbiol. 2009; v.7: 736-747. 23. KOSITANONT U, CHOTINANTAKUL K, PHULSUKSOMBATI D, TRIBUDDHARAT C.
Assessment of Southern blot ribotyping for differentiation of Leptospira strains isolated from field rats. Journal of Microbiological Methods 2007; 69(2): 288-297.
24. KOSITANONT U, RUGSASUK S, LEELAPORN A, PHULSUKSOMBATI D,
TANTITANAWAT S, NAIGOWIT P. Detection and differentiation between pathogenic and saprophytic Leptospira spp. by multiplex polymerase chain reaction. Diagnostic Microbiology and Infectious Disease 2007; 57(2): 117-122.
25. LA SCOLA B, BUI LTM, BARANTON G, KHAMIS A, RAOULT D. Partial rpoB gene
sequencing for identification of Leptospira species. FEMS Microbiology Letters 2006; 263(2): 142-147.
26. LEVETT, P. N. Leptospirosis. Clinical Microbiology Reviews 2001; vol. 14(2): 296-326. 27. LEVETT, P.N., MOREY, R.E., GALLOWAY, R., STEIGERWALT, A.G., ELLIS, W.A.,
Reclassification of Leptospira parva Hovind-Hougen et al., 1982 as Turneriella parva gen. comb. nov. Int. J. Syst. Evol. Microbiol, 2005; 55, 1497–1499.

55
28. INTERNATIONAL COMMITTEE ON SYSTEMATICS OF PROKARYOTES - SUBCOMMITTEE ON THE TAXONOMY OF LEPTOSPIRACEAE; Minutes. Quito-Ecuador. International Journal of Systematic and Evolutionary Microbiology, 2007.
29. LIN M, DAN HH, LI YJ. Identification of a second flagellin gene and functional
characterization of a sigma(70)-like promoter upstream of a Leptospira borgpetersenii flaB gene. Current Microbiology 2004; 48(2): 145-152.
30. LINDSTEDT BA. Multiple-locus variable number tandem repeats analysis for genetic
fingerprinting of pathogenic bacteria. Electrophoresis 2005; 26(13): 2567-2582. 31. LIU DY, LAWRENCE ML, AUSTIN FW, AINSWORTH AJ, PACE LW. PCR detection of
pathogenic Leptospira genomospecies targeting putative transcriptional regulator genes. Canadian Journal of Microbiology 2006; 52(3): 272-277.
32. MAJED Z, BELLENGER E, POSTIC D, POURCEL C, BARANTON G, PICARDEAU M.
Identification of variable-number tandem-repeat loci in Leptospira interrogans sensu stricto. Journal of Clinical Microbiology 2005; 43(2): 539-545.
33. MANIATIS T, FRITCH EEF, SAMBROOK J. Molecular Cloning: a Laboratory Manual.
Cold Spring Harbor, 2nd Ed 1989; 3. 34. MARSHALL R.B, WILTON B.E, ROBINSON A.J. Identification of Leptospira serovars by
restriction endonuclease analysis.Journal of Medical Microbiology. 1981,14:163-166. 35. MERIEN F, PORTNOI D, BOURHY P, CHARAVAY F, BERLIOZ-ARTHAUD A,
BARANTON G. A rapid and quantitative method for the detection of Leptospira species in human leptospirosis. FEMS Microbiology Letters 2005; 249(1): 139-147.
36. MILLER, M. P. TFPGA. Tools for Population Genetic Analyses for Windows. Arizona
State University, 1998, USA. 37. MOREY RE, GALLOWAY RL, BRAGG SL, STEIGERWALT AG, MAYER LW, LEVETT
PN. Species-specific identification of Leptospiraceae by 16S rRNA gene sequencing. Journal of Clinical Microbiology 2006; 44(10): 3510-3516.
38. NASCIMENTO ALTO, KO AI, MARTINS EAL, MONTEIRO-VITORELLO CB, HO PL,
HAAKE DA. Comparative genomics of two Leptospira interrogans serovars reveals novel insights into physiology and pathogenesis. Journal of Bacteriology 2004; 186(7): 2164-2172.
39. NASCIMENTO ALTO, VERJOVSKI-ALMEIDA S, VAN SLUYS MA, MONTEIRO-
VITORELLO CB, CAMARGO LEA, DIGIAMPIETRI LA. Genome features of Leptospira interrogans serovar Copenhageni. Brazilian Journal of Medical and Biological Research 2004; 37(4): 459-478.
40. NEI, M. Genetic distance between populations. American Naturalist, 1972,. 106;, 283-
292. 41. OLIVE MD, P. B. Principles and Applications of Methods for DNA-Based Typing of
Microbial Organisms. Mini review - Journal of Clinical Microbiology 1999; 37(6): 1661-1669.
42. OLIVEIRA, D. S. C., GUIMARÃES, M. J. B., MEDEIROS, Z. Modelo Produtivo para a
Leptospirose. Revista de Patologia Tropical 2009; vol. 38 (1): 17-26.

56
43. PAVAN ME, CAIRO F, BRIHUEGA B, SAMARTINO L. Multiple-locus variable-number
tandem repeat analysis (MLVA) of Leptospira interrogans serovar Pomona from Argentina reveals four new genotypes. Comparative Immunology Microbiology and Infectious Diseases 2008; 31: 37-45.
44. POSTIC D, RIQUELME-SERTOUR N, MERIEN F, PEROLAT P, BARANTON G. Interest
of partial 16S rDNA gene sequences to resolve heterogeneities between Leptospira collections: application to L. meyeri. Res. Microbiol. 2000; 151: 333-341.
45. REITSTETTER RE. Development of species-specific PCR primer sets for the detection of
Leptospira. FEMS Microbiology Letters 2006; 264(1): 31-39. 46. RESCH G, AWAD-MASALMEH M, BAKOSS P, JAREKOVA J. Utility of phylogenetic
studies in the identification of Leptospira strains. Epidemiology and Infection 2007; 135: 1266-1273.
47. ROY, S., BISWAS, D., VIJAYACHARI, P., SUGUNAN, A.P., SEHGAL, S.C., 2004. A 22-
mer primer enhances discriminatory power of AP-PCR fingerprinting technique in characterization of leptospires. Trop. Med. Int. Health 9, 1203–1209..
48. ROMERO EC, YASUDA PH. Molecular characterization of Leptospira sp strains isolated
from human subjects in Sao Paulo, Brazil using a polymerase chain reaction-based assay: a public health tool. Memorias do Instituto Oswaldo Cruz 2006; 101(4): 373-378.
49. SALAUN L, MERIEN F, GURIANOVA S, BARANTON G, PICARDEAU M. Application of
Multilocus Variable-Number Tandem-Repeat Analysis for Molecular Typing of the Agent of Leptospirosis. Journal of Clinical Microbiology 2006; 44(11): 3954-3962.
50. SANGUINETTI CJ, E. DN, SIMPSON AJG. Rapid silver staining and recovery the PCR
products separated on polyacrylamide gels. Biotechnology 1994; 17: 915-918. 51. SLACK AT, SYMONDS ML, DOHNT MF, SMYTHE LD. Identification of pathogenic
Leptospira species by conventional or real-time PCR and sequencing of the DNA gyrase subunit B encoding gene. BMC Microbiology 2006; 6.
52. TAYLOR K.A, BARBOUR A. G, THOMAS D.D. Pulse-Field Gel Electrophoretic Analysis
of Leptospiral DNA. Infection and Immunity,1991; 59(1) 323-329. 53. TERPSTRA, W.J., KORVER, H., VAN LEEUWEN, J., KLATSER, P.R., KOLK, A.H.J.,
The classification of Sejroe group serovars of Leptospira interrogans with monoclonal antibodies. Zentralbl Bakteriol Mikrobiol Hygiene 1985; 259, 498–506
54. WANGROONGSARB P, CHANKET T, GUNLABUN K, LONG DH, SATHEANMETHAKUL
P, JETANADEE S. Molecular typing of Leptospira spp. based on putative O-antigen polymerase gene (wzy), the benefit over 16S rRNA gene sequence. FEMS Microbiology Letters 2007; 271(2): 170-179.
55. WENJUN, L., DIDIER, R., FOURNIER, P-E., Bacterial strain typing in the genomic era.
FEMS microbiology reviews 2009; 33(5): 892-916 56. YANG HL, ZHU YZ, QIN JH, HE P, JIANG XC, ZHAO GP. In silico and microarray-based
genomic approaches to identifying potential vaccine candidates against Leptospira interrogans. BMC Genomics 2006; 7.

57
57. YANG JL, WANG MS, CHENG AC, PAN KC, LI CF, DENG SX. A simple and rapid method for extracting bacterial DNA from intestinal microflora for ERIC-PCR detection. World Journal of Gastroenterology 2008; 14(18): 2872-2876.
58. YASUDA P.H, STEIGERWALT A. G, SULZER, K. R, KAUFMANN A. F, ROGER F,
BRENNER D. J. Desoxiribonucleic acid relatedeness betweenserogrups and serovars in the family Leptospiraceae with proposals for seven new Leptospira species. International Journal of Systematic Bacteriology,1987; 37:407-415.
59. ZUERNER R, HAAKE D, ADLER B, SEGERS R. Technological Advances in the
Molecular Biology of Leptospira. In. JMMB Symposium on Spirochete Molecular Biology: Journal of Microbiology Biotechnology, 2000. p 455-462

58
8. APÊNDICE >gi|83272337|gb|DQ296130.1| Leptospira genomosp. 1 serovar Pingchang RpoB
(rpoB) gene, partial cds
TCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGGGAAGAAGCTCCTTTTGTCGGAACAGGTATGGAAACCAGA
GCCGCTTACGATTCTAGAATTTGTATCGTAAACAAACACGACGGCGTCGTTACATCCGTGGACGCGGAAAACATC
GTCGTTGAGAGAAAGGGCGGAAAAGAATCCGATACTTACTCTCTGACTAAATTCAAAAAGACCAACCAAGGAACT
TGCTTTAACCAAAAGCCAATCGTCGGAGTGGTTCACTCCGATTTTAACGGAAAGGTTTCCAAGGTTTCCAAAGAA
AAAATCGAAGTGACCGGTGAAAACGGACATATCAAAGAATACATTCTCCAAATGGGAAGCAGACAATATTCTCCG
ATCGTATCTTCCGGAGAAGAAGTAAAACGAGGAACCACTCTCGCGGGACAAATCGTTACCGGCGAGAAGTTGGAT
GAGATGGGAAATATCCTGGTAAAAGGAACCGTTCTTGCGGACGGTCCTGCGGTTGACAACGGAGTACTCGCTCTC
GGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGATACAACTT
>gi|83272363|gb|DQ296143.1| Leptospira genomosp. 3 serovar Holland RpoB
(rpoB) gene, partial cds
TCCAACATGCAACGCCAAGCGGTTCCTCTTTTAACAGAAGAAGCTCCTTTTGTGGGAACTGGTATGGAAGCTCGT
GCGGCTTATGACGCAGGTGTTTGTATTGTTGCCAAAAAAGATGGTGTGGTTTCGAAAGTGGATGCCACAGGTGTT
TGGATCAAAGAAGACCAATCCAAAGAGATTGTCCATTACCCACTCATTAAATTCAAAAAAACCAACCAAGGAACT
TGTTTTAACCAAAAACCAAACGTTTCCATGTTGCATACGACGACTGGTGGTAAAGTAGGTAAGGTTTCCAAAGAA
CGTGTGGAGATTACTTCTCCTAACGGAGAAAAAGAAACCCATGAACTTTTCCATTCGGAAGATGTGCAATACGTT
GCGGTTGTGAAAGAAGGCCAAGACTTAGGAATTGGTGCCCCAGTAGCGGGACAAATCATCAAAGGCGAAAAATAC
GGTGAGTTTGGACAAATCCTCCAAAAAGGAACTGTTCTTGCCAATGGACCATCCACTGACGCTGGTTATTTGGCA
CTTGGCCGTAACGTTCTTGTGGCATTTATGCCTTGGGAAGGATACAACTT
>gi|83272349|gb|DQ296136.1| Leptospira genomosp. 5 serovar Saopaulo RpoB
(rpoB) gene, partial cds
TCCAACATGCAACGCCAAGCAGTGCCTCTTCTTACGGAAGAAGCTCCATTTGTGGGAACTGGTATGGAAGCTCGT
GCGGCTTATGACGCAGGGGTTTGTATCGTTGCCAAAAAAGATGGAGTGGTTTCCAAAGTGGATGCGACTGGTGTT
TGGATCAAAGAAGACCAATCCAAAGAGATTGTCCACTACCCACTCATTAAATTCAAAAAAACCAACCAAGGTACT
TGTTTTAACCAAAAACCAAACGTATCCATGTTACACACCACAACTGGTGGTAAGGTGAGTAAGGTTTCCAAAGAA
CGCATTGAAGTGACTTCTCCCAATGGAGAAAAAGAAATCCATGAACTCCACCTCTCCGAGGAAGTACAATTCCTC
GCTGTTGTCAAAGAAGGGCAGGAAATTGGAGTGGGTGCTCCTGTTGCGGGTCAAATCATCAAAGGGGAAAAATAC
GGTGAGTTTGGACAAATCCTGCAAAAAGGAACTGTCCTTGCAAACGGTCCATCCACTGACGCTGGTTACTTGGCT
CTTGGGAGAAACGTTCTTGTGGCATTTATGCCTTGGGAAGGATACAACTT
>gi|83272361|gb|DQ296142.1| Leptospira biflexa serovar Patoc RpoB (rpoB)
gene, partial cds
TCCAACATGCAACGCCAAGCGGTACCACTCTTAACAGAAGAGGCTCCTTTTGTCGGAACTGGTATGGAAGCTCGT
GCGGCTTATGACGCAGGGGTTTGTATCGTTGCGAAAAAAGATGGTGTGGTTTCCAAAGTGGATGCAACAGGTGTT
TGGATCAAAGAAGACCAATCCAAAGAGATTGTCCATTACCCACTCATTAAATTCAAAAAAACCAACCAAGGTACT
TGTTTTAACCAAAAACCAAACGTATCCATGTTACACACCACAACTGGTGGCAAGGTAAGTAAGGTTTCGAAAGAA
CGTGTCGAAGTGACAACTCCTAACGGAGAAAAAGAAACTCATGAACTTCTTCTTTCTGATGAAGTTCAGTTCCAT
GCTGTTGTCAAAGAAGGACAAGAGGTAGGAATTGGAGCTCCAGTTGCCGGACAAATCATCAAAGGGGAAAAATAC
GGTGACTTCGGTCAGATCCTTCAAAAAGGAACTGTCCTAGCCAACGGGCCATCCACTGACGCTGGGTATTTGGCA
CTTGGACGAAATGTTCTCGTTGCCTTTATGCCTTGGGAAGGATACAACTT
>gi|83272339|gb|DQ296131.1| Leptospira santarosai serovar Shermani RpoB
(rpoB) gene, partial cds
TCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGGGAAGAAGCTCCTTTTGTCGGAACAGGTATGGAAACCAGA
GCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGTGTGGTCGTATCCGTAGACGCGGAAACGATC
GTTGTGGAAAGAAAGGGCGGAAAAGAATCCGATAAATACGAACTTACGAAATTCAAAAAGACAAACCAAGGAACC
TGCTTTAACCAAAAGCCGATCGTGGGAGTGGTTCACTCCGATCGAAACGGTAAAGTTACTAAGGTTTCCAAAGAG
AAAATCGAAGTCACCGGCGAAAACGGAGACGTAAAAGAATACGTTCTCCAAATTGGAAGTAAACAATATTCTCCG
ATCGTTTCTTCCGGCGAAGAAGTAAAACGAGGAACGACTCTCGCGGGACAAATCGTCACGGGCGAGAAGTTGGAC
GAGATGGGAAATATCCTCGTTAAGGGAACCGTTCTTGCGGACGGACCTGCGGTCGACAACGGAGTGCTCGCACTC
GGAAGAAACGTTCTCGCGGCTTTCATGCCTTGGGAAGGATACAACTT
>gi|83272355|gb|DQ296139.1| Leptospira kirschneri serovar Cynopteri RpoB
(rpoB) gene, partial cds
TCCAACATGCAGCGTCAGGCGGTTCCTCTTCTTCGTGAAGAAGCTCCTTTTGTAGGAACGGGTATGGAAACCAGA
GCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGCGTTGTTACATCCGTGGACGCGGAAACCATC
GTTGTGGAAAGAAAGGGCGGAAAAGAATCCGATAAATATTCTCTTACAAAATTCAAAAAGACAAACCAAGGAACC
TGCTTCAACCAAAAGCCGATCGTAGGAGTGGTTCACTCCGAGATCAACGGAAAGGTTTCCAAGGTTTCTAAAGAA
AAAATCGAAGTCACTGGCGAAAACGGAGAAGTAAAAGAATACGTTCTCCAAATCGGAAGCAAACAATATGCGCCG
ATCGTTTCTTCGGGCGAAGAAGTAAAACGAGGAACGACTCTCGCAGGACAAGTTGTAGTAGGCGAGAAGTTGGAC
GAGATGGGAAATATCCTCGTTAAGGGAACCGTTCTTGCAGACGGTCCCGCGGTTGACAACGGTGTTCTCGCTCTG
GGAAGAAACGTTCTCGCGGCGTTTATGCCTTGGGAAGGATACAACTT

59
>gi|83272365|gb|DQ296144.1| Leptospira interrogans serovar Australis RpoB
(rpoB) gene, partial cds
TCCGTGAAGAAGCTCCTTTTGTAGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTATCGTAA
ACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAGAAAGGGCGGAAAAGAATCCG
ATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGGAGCCTGCTTTAATCAGAAGCCGATTGTAGGAGTGG
TTCACTCCGAGATTAACGGAAAGGTTTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAAC
TGAAAGAATATGTTCTTCAAATCGGAAGCAAACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAGTAAAAAGAG
GATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGATGAGATGGGAAATATCCTCGTAAAAGGAACCG
TTCTTGCTGATGGTCCTGCGGTCGACAACGGAGTTCTCGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTT
GGGAAGGATACAACTT
>gi|83272367|gb|DQ296145.1| Leptospira interrogans serovar Autumnalis RpoB
(rpoB) gene, partial cds
TCCGTGAAGAAGCTCCTTTTGTCGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTATCGTAA
ATAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAGAAAGGGCGGAAAAGAATCCG
ATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGGAACTTGCTTTAATCAGAAACCGATTGTAGGAGTGG
TTCACTCCGAGATTAACGGAAAGGTTTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAAC
TGAAAGAATATGTTCTTCAAATTGGAAGCAAACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAGTAAAACGAG
GATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGGAAATATCCTCGTAAAAGGAACCG
TTCTTGCCGACGGTCCTGCCGTTGATAATGGAGTTCTCGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTT
GGGAAGGATACAACTT
>gi|83272369|gb|DQ296146.1| Leptospira interrogans serovar Bataviae RpoB
(rpoB) gene, partial cds
TCCGTGAAGAAGCTCCTTTTGTAGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTATCGTAA
ACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAGAAAGGGCGGAAAAGAATCCG
ATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGGAACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGG
TTCACTCCGAGATTAACGGAAAGGTTTCCAAGGTCTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAAC
TGAAAGAATATGTTCTTCAAATCGGAAGCAAACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAGTAAAACGAG
GATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGATGAGATGGGAAATATCCTCGTAAAAGGAACCG
TTCTTGCTGATGGTCCTGCGGTCGACAACGGAGTTCTCGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTT
GGGAAGGATACAACTT
>gi|83272343|gb|DQ296133.1| Leptospira interrogans serovar
Icterohaemorrhagiae RpoB (rpoB) gene, partial cds
TCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGAAGCTCCTTTTGTAGGAACTGGTATGGAAACCAGA
GCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATC
GTTGTAGAAAGAAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGGAACC
TGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGTTTCCAAGGTTTCCAAAGAA
AAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATATGTTCTTCAAATCGGAAGCAAACAATATTCTCCG
ATCGTCTCTGCAGGCGAAGAAGTAAAAAGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGAT
GAGATGGGAAATATCCTCGTAAAAGGAACCGTTCTTGCTGATGGTCCTGCGGTCGACAACGGAGTTCTCGCTCTG
GGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGATACAACTT
>gi|83272371|gb|DQ296147.1| Leptospira interrogans serovar Pyrogenes RpoB
(rpoB) gene, partial cds
TCCGCGAAGAAGCTCCTTTTGTAGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTATCGTAA
ACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAGAAAGGGCGGAAAAGAATCCG
ATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGGAACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGG
TTCACTCCGAGATCAATGGAAAGGTTTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAAC
TGAAAGAATATGTTCTTCAAATCGGAAGCAAACAATATTCTCCAATCGTCTCGGCAGGCGAAGAAGTAAAAAGAG
GATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGGAAATATCCTCGTTAAGGGTACGG
TTCTTGCCGACGGTCCTGCCGTTGACAATGGAGTTCTCGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTT
GGGAAGGATACAACTT
>gi|83272359|gb|DQ296141.1| Leptospira noguchii serovar Panama RpoB (rpoB)
gene, partial cds
TCCAACATGCAGCGTCAGGCGGTTCCTCTTCTCCGTGAAGAAGCTCCTTTTGTCGGAACCGGTATGGAAACCAGA
GCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGCGTTGTCATTTCCGTGGACGCGGAAACGATC
GTCGTGGAAAGAAAGGGCGGAAAAGAATCCGACAAATACGAACTTACGAAATTCAAAAAGACAAACCAAGGAACC
TGCTTTAACCAAAAGCCGATCGTAGGCGTAGTTCATTCCGAGATCAACGGAAAAGTAAGCAAGGTTTCTAAAGAA
AAAATCGAAGTAACCGGCGAAAACGGAGAAGTAAAAGAATACGTTCTCCAAATCGGAAGCAGACAATATTCTCCG
ATCGTTTCTTCCGGCGAAGAAGTAAAACGCGGAACCACTCTCGCAGGACAAATCGTCATCGGCGAGAAGTTGGAC
GAAATCGGAAATATCCTCGTGAAAGGAACCGTTCTTGCAGACGGTCCTGCGGTTGACAACGGAGTTCTCGCTCTT
GGAAGAAACGTTCTCGCGGCTTTCATGCCTTGGGAAGGATACAACTT
>gi|83272345|gb|DQ296134.1| Leptospira borgpetersenii serovar Javanica RpoB
(rpoB) gene, partial cds

60
TCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGTGAAGAAGCTCCGTTTGTCGGAACAGGTATGGAAACCAGA
GCCGCTTACGATTCCAGAATTTGTATTGTAAACAAACACGACGGCGTTGTGACTTCCGTGGACGCGGAGATCATC
GTCGTGGAAAGAAAGGGCGGAAAAGAATCCGATACGTATTCCCTTACGAAATTCAAAAAGACCAACCAAGGAACC
TGCTTCAATCAGAAGCCGATCGTCGGAGTCGTTCACTCAGAGATCAACGGAAAAGTTTCCAAGGTTTCCAAAGAA
AAAATCGAAGTGACCGGTGAAAACGGAGAACTGAAAGAATACGTTCTCCAAATCGGAAGCAAACAATATTCTCCG
ATCGTTTCTTTGGGCGAAGAAGTAAAACGAGGAACGACTCTCGCGGGACAAGTTGTCGTAGGCGAGAAGTTGGAC
GAGATGGGAAATATCCTCGTTAAGGGTACGGTTCTTGCGGACGGCCCTGCGGTCGACAACGGGGTTCTCGCTCTC
GGTAGAAATGTCCTCGCGGCGTTTATGCCTTGGGAAGGATACAACTT
>gi|83272347|gb|DQ296135.1| Leptospira meyeri serovar Ranarum RpoB (rpoB)
gene, partial cds
TCCAACATGCAACGCCAAGCAGTTCCTCTTCTCACAGAAGAAGCGCCGTTTGTAGGAACAGGTATGGAAGCTCGT
GCTGCTTATGATGCAGGGGTTTGTATTGTTGCGAAAAAAGATGGTGTAGTTTCTAAAGTAGATGCTACTGGTGTT
TGGATCAAAGAAGACCAATCCAAAGAGATCGTCCACTACCCACTCATTAAATTCAAAAAAACCAACCAAGGTACT
TGTTTTAACCAAAAACCGAACGTATCCATGCTTCATACCACAACAGGTGGAAAGGTAAGTAAAGTTTCTAAAGAA
CGGATCGAAGTAACGTCTCCAAACGGAGAAAAAGAAACACACGAACTTTTCCATTCGGAAGAAGTACAGTATTTA
TCCGTGGTGAAAGAAGGCCAAGACTTAGGAATTGGTGCACCGGTGGCCGGGCAAATCATCAAAGGGGAAAAATAC
GGTGAGTTTGGACAAATCCTCCAAAAAGGAACTGTCCTTGCGAATGGTCCTTCCACTGACGCTGGTTATTTGGCA
CTGGGACGTAATGTACTCGTTGCGTTTATGCCTTGGGAAGGATACAACTT
>gi|83272341|gb|DQ296132.1| Leptospira weilii serovar Celledoni RpoB (rpoB)
gene, partial cds
TCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGCGAAGAAGCTCCGTTTGTCGGAACAGGTATGGAAACTAGA
GCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGTGTCGTTATATCGGTTGACGCAGAACAAATC
GTTGTGGAAAGAAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTTAAAAAGACAAACCAAGGAACC
TGTTTCAATCAGAAGCCGATCGTAGGAGTCGTTCACTCCGAGATCAGCGGAAAGGTTACGAAAGTTTCCAAAGAA
AAAATCGAAGTAACATCCGAAAACGGAACAGTAAAAGAATACGTTCTCCAAGTCGGAAGCAAACAATATTCTCCG
ATCGTATCTTCGGGCGAAGAAGTAAAACGAGGATCGACTCTCGCGGGACAAGTGGTCACAGGTGAGAAGTTGGAC
GAACTGGGAAATATCCTCGTAAAAGGAACGGTCCTTGCGGACGGACCTGCGGTTGACAACGGAGTTCTCGCTCTC
GGAAGAAACGTTCTCGCTGCGTTCATGCCTTGGGAAGGATACAACTT
>gi|83272353|gb|DQ296138.1| Leptospira wolbachii serovar Codice RpoB (rpoB)
gene, partial cds
TCCAACATGCAACGCCAAGCAGTGCCACTACTCACGGAAGAAGCTCCGTTTGTAGGAACTGGGATGGAAGCTCGT
GCGGCTTATGACGCAGGGGTTTGTATCGTTGCGAAAAAAGACGGTGTAGTTTCTAAAGTAGATGCTACAGGTGTT
TGGATCAAAGAAGACCAATCTAAAGAGATCGTTCATTACCCACTCATTAAATTCAAAAAAACCAACCAAGGAACT
TGTTTTAACCAAAAACCAAACGTGGCTATGTTACACACGACCACTGGCGGAAAGGTTAGTAAAGTATCCAAGGAA
CGAGTGGAACTCACTTCTCCCAACGGGGAGAAAGAAATCCACGAACTTTTCCATTCGGAAGAAGTTCAGTTTGTG
GCCGTTGTAAAAGAAGGCCAAGACTTAGGAATTGGAACACCAGTTGCGGGACAAATCATCAAAGGGGAAAAATAC
GGTGACTTTGGTCAGATCCTACAAAAAGGAACTGTCCTTGCCAACGGTCCATCCACTGACGCTGGTTATTTGGCT
CTAGGCCGAAACGTACTCGTTGCCTTTATGCCATGGGAAGGATACAACTT
>gi|83272351|gb|DQ296137.1| Leptospira fainei serovar Hurstbridge RpoB
(rpoB) gene, partial cds
TCCAACATGCAACGCCAGGCGGTGCCTCTACTCCGCCAAGAAGCTCCCTATGTGGGAACGGGAATGGAGAGCCGT
GCCGCATACGACTCCAGAATTTGCGTGATCGCAAAACAAGACGGATACGTGAAATACGTAGACGCCGAAAAAGTC
ATCGTCGAGCAAAAGGGCGGAAAAGAATCCGATACGTACGACTTAACGAAATTTAAGAAAACCAACCAAGGCACT
TGCTTCAATCAAACTCCCGTCGTCGGAGTGATTCACTCGGAGATCGACGGGAAAGTCAGTAAGGTGACCAAGGAA
AAAATCGAAGTCACTGCGGATAACGGAAGCATAAAAGAATATAATCTGGCTTCGGGTTCTAAACAGTATCATCCG
ATCGTTTCCAACGGAGAAGAGGTTCGCAGAGGAACCACTCTTGCGGGACAAGTCGTCTTTGGCGAGAGAATGGAT
GAGCTCGGAAACATTCTGCAAAAAGGAACCGTTCTTGCGGACGGACCTGCAGTCGATAACGGAACTCTTGCTTTA
GGACGGAACGTGCTCGTAGCGTTTATGCCTTGGGAAGGATACAACTT
>gi|83272357|gb|DQ296140.1| Leptospira inadai serovar Lyme RpoB (rpoB)
gene, partial cds
TCCAACATGCAGCGCCAAGCGGTGCCTTTACTCCGCCAGGAAGCTCCCTATGTGGGAACCGGAATGGAAAGTCGT
GCTGCGTATGATTCCCGAATTTGCGTAATCGCGAAACAAGACGGATACGTGAAATACGTCGATGCGGAAAAAGTC
GTCGTCGAGCAAAAGGGCGGCAAAGAATCCGATACGTACGACTTGACGAAATTTAAGAAGACCAACCAAGGCACT
TGCTTTAACCAAACTCCCGTAGTCGGAGTGATCCACTCCGAGATAGACGGAAAAGTCAACAAGGTAACCAGGGAG
AAAATTGAAGTCACTGCGGATAACGGAAGCATAAGAGAATACAATCTTACTTCCGGTTCTAAACAGTATCATCCG
ATCGTTTCCAACGGAGAAGAAGTTCGGAGAGGAACTACTCTTGCAGGACAAGTCGTCTTTGGCGAGAGAATGGAC
GAGCTCGGAAACATTCTGCAAAAAGGAACCGTTCTCGCGGACGGACCTGCCGTCGATAACGGAACTCTTGCATTA
GGACGAAACGTTCTGGTAGCGTTTATGCCTTGGGAAGGATACAACTT
>gi|83272335|gb|DQ296129.1| Leptospira alexanderi serovar Manhao 3 RpoB
(rpoB) gene, partial cds
TCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGTGAAGAAGCTCCTTTTGTCGGAACAGGTATGGAAACCAGA
GCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGATGGTGTCGTTACATCTGTTGACGCTGAAACGATC

61
GTCGTGGAAAGAAAGGGCGGAAAAGAATCCGACACGTATCAACTTACGAAATTCAAAAAAACCAACCAAGGAACC
TGTTTCAATCAGAAACCGATCGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGTTACGAAAGTTTCCAAAGAA
AAAATCGAAGTAACATCCGAAAACGGAGAAGTAAAAGAATACATTCTCCAGATCGGAAGCAGACAATATTCTCCG
ATCGTTTCTTCCGGTGAGGAAGTAAAACGCGGAACCACTCTCGCGGGACAAATCGTCACAGGCGAGAAGTTGGAT
GAACTCGGAAACATTCTCATTAAAGGCACGGTTCTCGCCGACGGTCCTGCGGTGGACAACGGAGTTCTCGCTCTC
GGTAGAAACGTTCTTGCGGCTTTTATGCCTTGGGAAGGATACAACTT
>gi|167016089|gb|EU349502.1| Leptospira noguchii strain Cascata RpoB (rpoB)
gene, partial cds (serogroup Bataviae)
TCCAACATGCAGCGTCAGGCGGTTCCTCTTCTCCGTGAAGAAGCTCCTTTTGTCGGAACCGGTATGGAAACCAGA
GCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGTGTAGTTACTTCCGTGGACGCGGAAACGATC
GTCGTGGAAAGAAAGGGCGGAAAAGAATCCGACAAATACGAACTTACGAAATTCAAAAAGACAAACCAAGGAACC
TGCTTTAACCAAAAGCCGATCGTTGGAGTGGTTCACTCCGAGATCAACGGAAAGGTTTCCAAGGTTTCTAAAGAA
AAAATCGAAGTCACCAGCGAAAACGGAGAAGTAAAAGAATACGTTCTCCAAATCGGAAGCAGACAATATTCTCCG
ATCGTTTCTTCCGGCGAAGAAGTAAAACGTGGAACGACTCTTGCGGGACAAATCGTCACAGGCGAGAAGTTGGAC
GAGATGGGAAATATTCTCGTGAAAGGAACCGTTCTTGCGGACGGTCCCGCCGTTGACAACGGAGTTCTCGCTCTG
GGAAGAAACGTTCTTGCGGCGTTTATGCCTTGGGAAGGTTACAACTTTGAG
>gi|167016091|gb|EU349503.1| Leptospira noguchii serovar Proechymis strain
1161U RpoB (rpoB) gene, partial cds
TCCGTGAAGAAGCTCCTTTTGTCGGAACAGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTATCGTAA
ACAAACACGACGGCGTTGTCATTTCCGTGGACGCGGAAACGATCGTCGTGGAAAGAAAGGGCGGAAAAGAATCCG
ACAAATACGAACTTACGAAATTCAAAAAGACAAACCAAGGAACCTGCTTTAACCAAAAGCCGATCGTTGGTGTGG
TTCATTCCGAGATCAACGGAAAAGTAAGCAAGGTTTCTAAAGAAAAAATCGAAGTAACCGGCGAAAACGGAGAAG
TAAAAGAATACGTTCTCCAAATCGGAAGCAGACAATATTCTCCGATCGTTTCTTCCGGCGAAGAAGTAAAACGCG
GAACCACTCTCGCAGGACAAATCGTAACAGGCGAGAAGTTGGACGAGATTGGAAACATTCTCGTGAAAGGAACCG
TTCTTGCGGACGGTCCTGCGGTCGACAACGGAGTTCTCGCTCTTGGAAGAAACGTTCTTGCGGCTTTCATGCCTT
GGGAAGGTTACAACTTCGAGGATGC
As seqüências a seguir foram depositadas pelo nosso grupo após seqüenciamento de DNA realizado pela Dra. Maria Rosa Quaresma Bomfim:
>gi|218750493|gb|EU747302.1| Leptospira borgpetersenii serovar Ballum RpoB
(rpoB) gene, partial cds
TTCCTCTTCTCCGTGAAGAAGCTCCTTTTGTCGGAACTGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTT
GTATCGTAAATAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATCGTTGTAGAAAGAAAGGGCGGAA
AAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGGAACTTGCTTTAATCAGAAACCGATTG
TAGGAGTGGTTCACTCCGAGATTAACGGAAAGGTTTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAA
ACGGTGAACTGAAAGAATATGTTCTTCAAATTGGAAGCAAACAATATTCTCCGATCGTCTCCGCAGGCGAAGAAG
TAAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGACGAGATGGGAAATATCCTCGTAA
AAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGATAATGGAGTTCTCGCTCTGGGAAGAAACGTTCTCGCGGCGT
TCATGCCTTGGGAAGGTTACAACTTCGAGGA
>gi|218750509|gb|EU747310.1| Leptospira borgpetersenii serovar Castellonis
RpoB (rpoB) gene, partial cds
GTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGTGAAGAAGCTCCGTTTGTCGGAACAGGTATGGAAACCA
GAGCCGCTTACGATTCCAGAATTTGTATTGTAAACAAACACGACGGCGTTGTGACTTCCGTGGACGCGGAGATCA
TCGTCGTGGAAAGAAAGGGCGGAAAAGAATCCGATACGTATTCCCTTACGAAATTCAAAAAGACCAACCAAGGAA
CCTGCTTCAATCAGAAGCCGATCGTCGGAGTCGTTCACTCAGAGATCAACGGAAAAGTTTCCAAGGTTTCCAAAG
AAAAAATCGAAGTGACCGGTGAAAACGGAGAACTGAAAGAATACGTTCTCCAAATCGGAAGCAAACAATATTCTC
CGATCGTTTCTTTGGGCGAAGAAGTAAAACGAGGAACGACTCTCGCGGGACAAGTTGTCGTAGGCGAGAAGTTGG
ACGAGATGGGAAATATCCTCGTTAAGGGTACGGTTCTTGCGGACGGCCCTGCGGTCGACAACGGGGTTCTCGCTC
TCGGTAGAAACGTCCTCGCGGCGTTTATGCCTTGGGAAGGATACAACT
>gi|218750499|gb|EU747305.1| Leptospira borgpetersenii serovar Hardjo-bovis
RpoB (rpoB) gene, partial cds
CCTCATGGGTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGTGAAGAAGCTCCTTTTGTCGGAACCGGTAT
GGAAACCAGAGCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGC
GGAAAACATCGTTGTAGAAAGAAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAA
CCAAGGAACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGTTTCCAAGGT
TTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATATGTTCTTCAAATCGGAAGCAAACA
ATATTCTCCAATCGTCTCCGCAGGCGAAGAAGTAAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGA
GAAGTTGGACGAGATGGGAAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGACAATGGAGT
TCTCGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACTTCGAG

62
>gi|218750511|gb|EU747311.1| Leptospira borgpetersenii serovar Sejroe RpoB
(rpoB) gene, partial cds
TCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGAAGCTCCTTTTGTCGGAACTGGTATGGAAACCAGA
GCCGCTTACGATTCCAGAATTTGTATCGTAAATAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATC
GTTGTAGAAAGAAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGGAACT
TGCTTTAATCAGAAACCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGTTTCCAAGGTTTCCAAAGAA
AAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATATGTTCTTCAAATTGGAAGCAAACAATATTCTCCG
ATCGTCTCCGCAGGCGAAGAAGTAAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGAC
GAGATGGGAAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGATAATGGAGTTCTCGCTCTG
GGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACT
>gi|218750503|gb|EU747307.1| Leptospira borgpetersenii serovar Tarassovi
RpoB (rpoB) gene, partial cds
GTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGAAGCTCCTTTTGTCGGAACAGGTATGGAAACCA
GAGCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGCGTTGTGACTTCCGTGGACGCGGAAACCA
TCGTTGTGGAAAGAAAGGGCGGAAAAGAATCCGATACGTATTCCCTTACGAAATTCAAAAAAACAAACCAAGGAA
CCTGCTTTAATCAGAAGCCGATCGTCGGAGTCGTTCACTCCGAGATCAACGGAAAGGTTTCCAAGGTTTCCAAAG
AAAAAATCGAAGTGACCGGTGAAAACGGAGAACTGAAAGAATACGTTCTCCAAATCGGAAGCAAACAATATGCTC
CGATCGTTTCTTCGGGCGAAGAAGTAAAACGAGGAACGACTCTCGCAGGACAAGTTGTCGTAGGCGAGAAGTTGG
ACGAGATGGGAAATATCCTCGTTAAGGGTACGGTTCTTGCGGACGGCCCTGCGGTCGACAACGGTGTCCTCGCTC
TGGGAAGAAACGTTCTCGCGGCGTTTATGCCTTGGGAAGGTTACAACTTC
>gi|218750517|gb|EU747314.1| Leptospira borgpetersenii serovar Whitticombi
RpoB (rpoB) gene, partial cds
TTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGCGAAGAAGCTCCGTTTGTCGGAACAGGTATGGAAACTAG
AGCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGTGTCGTTATATCGGTTGACGCAGAACAAAT
CGTTGTGGAAAGAAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTTAAAAAGACAAACCAAGGAAC
CTGTTTCAATCAGAAGCCGATCGTAGGAGTCGTTCACTCCGAGATCAGCGGAAAGGTTACGAAAGTTTCCAAAGA
AAAAATCGAAGTAACATCCGAAAACGGAACAGTAAAAGAATACGTTCTCCAAGTCGGAAGCAAACAATATTCTCC
GATCGTATCTTCGGGCGAAGAAGTAAAACGAGGATCGACTCTCGCGGGACAAGTGGTCACAGGTGAGAAGTTGGA
CGAACTGGGAAATATCCTCGTAAAAGGAACGGTCCTTGCGGACGGACCTGCGGTTGACAACGGAGTTCTCGCTCT
CGGAAGAAACGTTCTCGCTGCGTTCATGCCTTGGGAAGGTTACAA
>gi|218750489|gb|EU747300.1| Leptospira interrogans serovar Bratislava RpoB
(rpoB) gene, partial cds
GGTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTTCGTGAAGAAGCTCCGTTTGTCGGAACAGGTATGGAAACC
AGAGCCGCTTACGATTCCAGAATTTGTATTGTAAACAAACACGACGGCGTTGTGACTTCCGTGGACGCGGAGATC
ATCGTCGTGGAAAGAAAGGGCGGAAAAGAATCCGATACGTATTCCCTTACGAAATTCAAAAAGACCAACCAAGGA
ACCTGCTTCAATCAGAAGCCGATCGTCGGAGTCGTTCACTCAGAGATCAACGGAAAAGTTTCCAAGGTTTCCAAA
GAAAAAATCGAAGTGACCGGTGAAAACGGAGAACTGAAAGAATACGTTCTCCAAATCGGAAGCAAACAATATTCT
CCGATCGTTTCTTTGGGCGAAGAAGTAAAACGAGGAACGACTCTCGCGGGACAAGTTGTCGTAGGCGAGAAGTTG
GACGAGATGGGAAATATCCTCGTTAAGGGTACGGTTCTTGCGGACGGCCCTGCGGTCGACAACGGGGTTCTCGCT
CTCGGTAGAAACGTCCTCGCGGCGTTTATGCCTTGGGAAGGTTACAACTTCGAGGAATGCG
>gi|218750523|gb|EU747299.1| Leptospira interrogans serovar Canicola RpoB
(rpoB) gene, partial cds
CCTCATGGGTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGAAGCTCCTTTTGTCGGAACTGGTAT
GGAAACCAGAGCCGCTTACGATTCCAGAATTTGTATCGTAAATAAACACGACGGTGTCGTAACTTCCGTCGATGC
GGAAAACATCGTTGTAGAAAGAAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAA
CCAAGGAACTTGCTTTAATCAGAAACCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGTTTCCAAGGT
TTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATATGTTCTTCAAATTGGAAGCAAACA
ATATTCTCCGATCGTCTCCGCAGGCGAAGAAGTAAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGA
GAAGTTGGACGAGATGGGAAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGATAATGGAGT
TCTCGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACA
>gi|218750513|gb|EU747312.1| Leptospira interrogans serovar Djasiman RpoB
(rpoB) gene, partial cds
TTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGAAGCTCCTTTTGTAGGAACTGGTATGGAAACCAG
AGCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACAT
CGTTGTAGAAAGAAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGGAAC
CTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGTTTCCAAGGTTTCCAAAGA
AAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATATGTTCTTCAAATCGGAAGCAAACAATATTCTCC
AATCGTCTCCGCAGGCGAAGAAGTAAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGA
CGAGATGGGAAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCGGTCGACAACGGAGTTCTCGCTCT
GGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGATACA

63
>gi|218750497|gb|EU747304.1| Leptospira interrogans serovar Hebdomadis RpoB
(rpoB) gene, partial cds
GTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGAAGCTCCTTTTGTCGGAACTGGTATGGAAACCA
GAGCCGCTTACGATTCCAGAATTTGTATCGTAAATAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACA
TCGTTGTAGAAAGAAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGGAA
CTTGCTTTAATCAGAAACCGATTGTAGGAGTGGTTCACTCCGAGATTAACGGAAAGGTTTCCAAGGTTTCCAAAG
AAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATATGTTCTTCAAATTGGAAGCAAACAATATTCTC
CGATCGTCTCCGCAGGCGAAGAAGTAAAACGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGG
ACGAGATGGGAAATATCCTCGTAAAAGGAACCGTTCTTGCCGACGGTCCTGCCGTTGATAATGGAGTTCTCGCTC
TGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACT
>gi|218750515|gb|EU747313.1| Leptospira interrogans serovar Schueffneri
RpoB (rpoB) gene, partial cds
TGTAACCTTCCTAATGGGTTCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGCGAAGAAGCTCCTTTTGTCGG
AACCGGTATGGAAACCAGAGCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGTGTCGTAACTTC
CGTCGATGCGGAAAACATCGTTGTAGAAAGAAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAA
AAAGACAAACCAAGGAACCTGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGT
TTCCAAGGTTTCCAAAGAAAAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATATGTTCTTCAAATCGG
AAGCAAACAATATTCTCCGATCGTCTCTGCAGGCGAAGAAGTAAAAAGAGGATCGACTCTCGCAGGACAAGTTGT
TGTAGGTGAGAAGTTGGATGAGATGGGAAATATCCTCGTAAAAGGAACCGTTCTTGCTGATGGTCCTGCGGTCGA
CAACGGAGTTCTCGCTCTGGGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACTTCGAGGATGC
GAAGGTTA
>gi|218750505|gb|EU747308.1| Leptospira interrogans serovar Wolffi RpoB
(rpoB) gene, partial cds
TCCAACATGCAACGTCAGGCGGTTCCTCTTCTCCGTGAAGAAGCTCCTTTTGTAGGAACTGGTATGGAAACCAGA
GCCGCTTACGATTCCAGAATTTGTATCGTAAACAAACACGACGGTGTCGTAACTTCCGTCGATGCGGAAAACATC
GTTGTAGAAAGAAAGGGCGGAAAAGAATCCGATACGTATCAACTTACGAAATTCAAAAAGACAAACCAAGGAACC
TGCTTTAATCAGAAGCCGATTGTAGGAGTGGTTCACTCCGAGATCAATGGAAAGGTTTCCAAGGTTTCCAAAGAA
AAAATCGAAGTCACTGGTGAAAACGGTGAACTGAAAGAATATGTTCTTCAAATCGGAAGCAAACAATATTCTCCA
ATCGTCTCCGCAGGCGAAGAAGTAAAAAGAGGATCGACTCTCGCAGGACAAGTTGTTGTAGGTGAGAAGTTGGAT
GAGATGGGAAATATCCTCGTAAAAGGTACGGTTCTTGCCGACGGTCCTGCGGTCGACAACGGAGTTCTCGCTCTG
GGAAGAAACGTTCTCGCGGCGTTCATGCCTTGGGAAGGTTACAACTT
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo