flávia melo rodrigues
TRANSCRIPT

1
Resumo Geral
A genética da conservação é uma nova abordagem no estudo da biodiversidade e utiliza as
análises genéticas moleculares para elucidar aspectos relevantes da biologia das espécies para
fins de manejo e conservação. Neste trabalho, foi avaliada a diversidade genética, utilizando
marcadores RAPD e STR, de duas espécies de canídeos que ocorrem no Parque Nacional das
Emas – GO (PNE), o lobo-guará(Chrysocyon brachyurus) e o cachorro-do-mato(Cerdocyon
thous). O lobo-guará está classificado pela IUCN (2004) como espécie LR/nt, ou seja, com
pequeno risco de extinção, mas próximo a ser considerado vulnerável à extinção, além de estar
incluído na lista de espécies brasileiras ameaçadas de extinção do IBAMA. Enquanto que, o
cachorro-do-mato é o canídeo mais comum e com maior distribuição no Brasil. Os resultados
deste estudo foram divididos em quatro capítulos. O capítulo 1, mostrou um grande potencial de
transferibilidade de marcadores STR entre espécies evolutivamente mais próximas. No capítulo 2,
os resultados obtidos com os marcadores STR e RAPD não indicaram uma significativa redução
da variabilidade genética e nem uma endogamia elevada em ambas espécies. No capítulo 3, a
associação entre a similaridade genética e a similaridade espacial para a população de lobo-guará
mostrou uma estruturação espacial da similaridade genética apenas para os machos, a partir dos
dados RAPD. Enquanto que, os dados gerados pelos marcadores STR mostraram uma
estruturação espacial da similaridade genética de acordo com o padrão de filopatria feminina.
Finalmente, no capítulo 4 a avaliação do tempo de extinção nestas duas espécies, considerando
diferentes cenários sócio-econômico-ambiental da região sudoeste de Goiás, no qual o PNE está
inserido, mostrou que ambas populações podem persistir por um tempo razoável, mesmo quando
considerado o pior cenário. No entanto, sugere-se que estudos posteriores baseados em
parâmetros mais realistas da viabilidade destas populações, bem como do cenário de
desenvolvimento sócio-econômico-ambiental da região, sejam realizados, afim de se obter
avaliações mais precisas da persistência destas populações no PNE e entorno.
Palavras-chave: Genética da Conservação, lobo-guará, Cachorro-do-mato, Parque Nacional das
Emas, Cerrado.

2
Abstract
Conservation Genetics is a new approach concerning biodiversity studies. It uses
molecular genetic analysis to uncover relevant biological aspects of the species for its
management and conservation. This paper evaluated the genetic diversity using RAPD and STR
markers, of two canid species from Ema’s National Park (Parque Nacional das Emas – GO),
Maned wolf (Chrysocyon brachyurus) and Crab-eating fox (Cerdocyon thous). The maned wolf
was classified as an LR/nt species by IUCN (2004); this means that it has a lower probability of
extinction, although susceptible to it. As a matter of fact the Maned wolf is in IBAMA’s list of
Brazilian species that are threatened by extinction. The Crab-eating fox is the most common
canid with the widest distribution in Brazil. The results of this study were divided in 4 chapters.
Chapter one, showed big transferability potential of STR markers among evolutionary related
species. Chapter 2, showed that most results obtained with STR and RAPD markers didn’t
indicate reduction on the genetic variability neither high endogamy in either species. Chapter 3,
the association between genetic and territorial similarities for the Maned wolf population, was
present only among males by the usage of RAPD data. The data generated by STR markers
showed a special structure according to the female philopatry. Chapter 4, analyzed the extinction
time of both species considering different social-economic-environmental sceneries of the
southwest region Goiás where the ENP is located. The results showed that both species could
resist for a reasonable time even if the worst scenery is considered. New studies using more
realistic parameters should be considered to test the viability of these populations. The social
economic environmental development of the region should be studied also, so that a more precise
evaluation of the persistence of these populations at ENP and region are made.
Key words: Conservation Genetics, Crab-eating fox, Maned wolf, Ema’s National Park, Tropical
Savanna (Cerrado).

3
APRESENTAÇÃO
A genética da conservação é uma nova abordagem no estudo da biodiversidade e utiliza as
análises genéticas moleculares para elucidar aspectos relevantes da biologia das espécies para
fins de manejo e conservação (Frankham et al., 2002). Dentre os vários campos da genética da
conservação podemos destacar a detecção dos efeitos deletérios do endocruzamento (depressão
endogâmica), a perda da diversidade genética e conseqüentemente da capacidade de respostas a
mudanças ambientais, a fragmentação das populações e redução do fluxo gênico, processos
estocásticos como a deriva genética, o acumulo de mutações deletérias, problemas enfrentados
pelas populações em cativeiro, sucesso da introdução de novos indivíduos nas populações
naturais, a resolução de incertezas taxonômicas, uso das análises genéticas moleculares na área
forense e definição de unidades de manejo (Frankham et al., 2002; Hedrick, 2001; DeSalle e
Amato, 2004).
Myers et al. (2000) definiu Hotspots como, “ecossistemas que cobrem uma pequena
parcela da superfície da Terra, mas que abrigam uma alta porcentagem da biodiversidade global,
com ênfase às florestas tropicais”. O Cerrado é um dos hotspots e o segundo maior bioma
brasileiro em extensão, ocupando 21% do território brasileiro (Klink e Machado, 2005; Myers et
al., 2000). O Cerrado atualmente é o bioma brasileiro que mais tem sofrido com as pressões
antrópicas impostas pelo avanço das fronteiras agropecuárias (Klink e Moreira, 2002). As
principais ameaças à biodiversidade no Cerrado são fragmentação de habitats, invasões de
espécies exóticas, erosão dos solos, poluição de aqüíferos, alterações nos regimes de queimadas,
desequilíbrios no ciclo do carbono e possivelmente modificações climáticas regionais (Klink e
Machado, 2005).
No Cerrado brasileiro ocorrem 38 espécies de mamíferos de médio e grande porte, sendo
que destes 45% estão classificadas na ordem Carnivora (Marinho-Fillho et al. 2002). A maioria
das espécies da ordem Carnivora possui grandes áreas de vida, de forma que a fragmentação do

4
habitat tem reduzido suas populações a tamanhos que não são viáveis a médio e longo prazo
(Costa et al., 2005). Uma outra ameaça comum aos carnívoros é a mortalidade acidental por
colisão com veículos nas estradas que atravessam o Cerrado do Brasil Central, principalmente em
espécies que percorrem grandes distâncias, como o lobo-guará (Rodrigues et al., 2002). Os
carnívoros representam espécies chaves para os ecossistemas, pois ocupam diversos nichos
disponíveis no ambiente sendo reguladores de populações de presas animais e importantes
dispersores de sementes, e destes uma grande proporção é ameaçada (28%) (IBAMA, 2003;
Costa et al., 2005).
A IUCN (União Internacional para Conservação da Natureza) reconhece a necessidade de
se conservar a diversidade genética devido a dois motivos principais. Primeiro, porque as
mudanças ambientais são processos contínuos e a diversidade genética é importante para que a
população tenha a capacidade de se adaptar a estas mudanças. Segundo, porque a perda da
diversidade genética geralmente está associada com o endocruzamento e com a redução nas taxas
de reprodução e sobrevivência (fitness) (Frankham et al., 2002). No entanto, a falta de
informações genéticas para a maioria das espécies nativas constitui uma das limitações para as
ações conservacionistas e de manejo.
Neste trabalho foi avaliada a diversidade genética de duas espécies de mamíferos
carnívoros (ordem Carnívora) da família Canidae que ocorrem no Parque Nacional das Emas –
GO (PNE), o lobo-guará (Chrysocyon brachyurus) e o cachorro-do-mato(Cerdocyon thous). O
PNE possui uma alta riqueza de espécies e uma boa preservação dos habitats, que pode permitir a
manutenção de populações estáveis de muitas destas espécies. No entanto, a eliminação de
habitats naturais, a interrupção de corredores naturais de dispersão, incêndios, caça ilegal,
atropelamento dos animais e assoreamento de cursos d’água que atingem a sua fauna ameaçam a
sua conservação (Silveira e Jácomo, 1998).

5
O lobo-guará(Chrysocyon brachyurus) é o maior canídeo sul americano, e quando adulto
pode pesar cerca de 25 kg e medir 90 cm de altura nos ombros, com comprimento total entre 145
a 190 cm, sendo a cauda responsável por 45 cm (Eisenberg e Redford, 1999; Rodden et al., 2004).
Vivem em áreas de vegetação aberta, são territoriais, monógamos facultativos e onívoros.
Apresenta um número diplóide de 76 cromossomos e é uma das espécies de carnívoros mais
populares do cerrado brasileiro. Tem orelhas largas e eretas, membros alongados e caminhar
peculiar. Até os três meses apresenta pelo marrom-escuro quase negro, mas após essa idade sua
pelagem muda para castanho-avermelhado, pernas e dorso torna-se pretos, coloração negra
também na extremidade das patas e no focinho e a ponta da cauda permanece esbranquiçada
(Fletchall et al., 2000) (Figura 1).
Figura 1. lobo-guará(Chrysocyon brachyurus) no Parque Nacional das Emas – GO.
A espécie é onívora, abrangendo uma grande variedade de itens na sua dieta, que varia
sazonalmente. Dentre esses itens, a lobeira ou fruta-do-lobo (Solanum lycocarpum) é bastante

6
consumida por este canídeo (Rodden et al., 2004). Além de comer outros tipos de frutos, o
animal se alimenta ainda de pequenos mamíferos (roedores, tatus), ovos de aves, insetos e répteis
(Rodden et al., 2004; Motta-Junior et al., 1996, 2002; Juarez e Marinho-Filho, 2002)
A área de ocorrência original desta espécie compreende a região central do Brasil, o
Paraguai, Bolívia, nordeste da Argentina e Uruguai, sempre em regiões de vegetação aberta,
como campos e Cerrados (Figura 2). São animais territorialistas e ocupam grandes áreas de vida,
variando entre 25 e 115 km² (Rodden et al., 2004).
Figura 2. Área de distribuição do Lobo-gurá. (Fonte: Rodden et al., 2004)
Os Lobos Guará são animais solitários, que formam pares no período de acasalamento. O
número de filhotes varia tipicamente de 1 a 5, sendo cuidados geralmente pela mãe, com um
período de gestação em torno de 65 dias (Rodden et al., 2004; Eisenberg e Redford, 1999). São
animais territorialistas, que ocupam grandes áreas de vida, variando entre 25 a 115 km2 e utilizam
diversos recursos, como fezes, urina e uma vocalização característica para demarcar seu território
diante de outros indivíduos da mesma espécie (Silveira e Jácomo, 1998). Suas populações têm

7
sofrido um considerável declínio, principalmente devido à perda de habitat, como conseqüência
do avanço das fronteiras agrícolas (Rodden et al., 2004; Fonseca et al., 1994; Silveira e Jácomo,
1998). É classificada pela IUCN (2004) como espécie LR/nt, ou seja, com pequeno risco de
extinção, mas próximo a ser considerado vulnerável à extinção. A espécie também está incluída
na lista de espécies brasileiras ameaçadas de extinção do IBAMA - Instituto Brasileiro do Meio
Ambiente e Recursos Naturais Renováveis (IBAMA, 2003).
O cachorro-do-mato(Cerdocyon thous), vulgarmente também conhecido como ‘raposa’ ou
‘graxaim’, é um canídeo de porte médio e seu peso varia entre 5 e 7 kg, com 70 cm de
comprimento na idade adulta e apresenta um número diplóide de 74 cromossomos. Tem a cauda
peluda de cor escura, seu focinho é comprido, apresenta uma cabeça relativamente pequena. Há
uma grande variação de cor da pelagem, de marrom acinzentado ao cinza escuro e alguns a
apresentam áspera e grossa por todo o corpo, as patas podem ser pretas ou cinza escuro, algumas
vezes apresenta manchas amarelo-laranjada (Courtenay e Mafei, 2004) (Figura 3).
Figura 3. cachorro-do-mato(Cerdocyon thous) no Parque Nacional das Emas – GO.

8
Tem hábito preferencialmente noturno e desloca-se solitariamente, aos pares ou mesmo
em grupos maiores (Rocha et al., 2004). É onívoro e é o canídeo mais comum e com maior
distribuição no Brasil, exceto nas áreas baixas da bacia amazônica (Courtenay e Maffei, 2004;
Rocha et al., 2004). Tem uma ampla distribuição geográfica, habita áreas florestais, cerrados,
campos e áreas alteradas e habitadas pelo homem. Ocorre também da Colômbia ao Paraguai e
Uruguai (Figura 4). Está amplamente distribuído no Pantanal, tanto na planície quanto na área de
entorno, na bacia do Alto Paraguai (Courtenay e Maffei, 2004). Podem ser observados nas
margens de estradas, onde procuram restos de animais atropelados e, por isso, são também
vítimas de atropelamentos (Rocha et al., 2004)
Figura 4. Área de distribuição do cachorro-do-mato(Fonte: Courtenay e Maffei, 2004)
Sua dieta inclui insetos, pequenos mamíferos, frutos, plantas, répteis, anfíbios, aves, ovos
e, em regiões litorâneas e no Pantanal, os caranguejos (Juarez and Marinho-Filho, 2002). O
acasalamento ocorre nos meses de Novembro e Dezembro e a gestação é de 52 a 59 dias, onde
nascem de 3 a 6 filhotes, com 120-160 gramas cada (Courtenay e Maffei, 2004, Rocha et al.,
2004). Apesar de ser uma espécie classificada pela IUCN (2004) como espécie LR/lc, ou seja,

9
com pequeno risco de extinção e não dependente de esforços para sua conservação, merece ser
estudada antes que entre na lista de espécies ameaçadas. Os estudos com estas espécies podem
ser importantes, pois elas podem funcionar como espécies-chave no monitoramento do nível de
degradação dos habitats que elas ocupam.
Assim sendo, neste estudo analisamos a diversidade genética, utilizando marcadores
moleculares RAPD (Random Amplified Polymorphic DNA) e Microssatélites, ou STR (Short
Tandem Repeats), nestas duas espécies de canídeos. No Capítulo 1, foi testada a transferibilidade
de 29 marcadores STR, desenvolvidos originalmente para bovinos, búfalos, cachorro doméstico e
gato doméstico para amostras de lobo-guará e Cachorro-do-mato. Essa avaliação inicial permitiu
definir a metodologia básica utilizada nas análises genéticas apresentadas nos Capítulos 2 e 3. Do
total de locos testados, 21% foram transferidos com sucesso e observamos então um grande
potencial de transferibilidade, principalmente entre espécies evolutivamente mais próximas (i.e.
cachorro doméstico).
No Capítulo 2, foi estimada a diversidade genética nestes dois canídeos, utilizando
marcadores RAPD e STR, além de correlacionarmos os dados gerados por estes dois marcadores.
Os resultados obtidos não indicaram uma significativa redução na variabilidade genética e nem
uma endogamia elevada em nenhuma das espécies analisadas. Em relação à associação dos dois
marcadores, as baixas correlações encontradas são corroboradas por outros estudos e devem ser
explicadas pela base genética diferente entre estes, bem como pelo baixo nível da hierarquia
evolutiva analisada (i.e., variabilidade dentro de populações).
No Capítulo 3, foi analisada a associação entre a similaridade genética, baseada nos dados
gerados pelos marcadores RAPD e STR, e a similaridade espacial baseada em dados da área de
vida do lobo-guará no Parque Nacional das Emas e entorno. Os resultados obtidos com os dados
RAPD mostraram uma estruturação espacial da similaridade genética significativa apenas para os
machos. Os resultados obtidos com os marcadores STR mostraram uma estruturação espacial da

10
similaridade genética de acordo com o padrão de filopatria feminina, ou seja, as fêmeas mais
próximas geneticamente tendem a ocupar áreas de vida mais próximas. Como os resultados
obtidos com os marcadores STR são mais esperados, considerando as informações disponíveis
sobre ecologia e história-de-vida da espécie, e os marcadores STR são mais adequados para esse
tipo de análise intra-populacional, acredita-se que os resultados obtidos com os marcadores
RAPD sejam artefatos.
Finalmente, no Capítulo 4 avaliou-se o tempo de extinção nestas duas espécies de
canídeos do PNE, assumindo-se que combinações de efeitos da depressão endogâmica sob
estocasticidade ambiental seriam os possíveis fatores que levariam à extinção organismos em
populações isoladas. Em média, o tempo de extinção foi afetado pelos três parâmetros analisados
e suas interações (taxas de redução da capacidade de suporte, coeficientes de endogamia e
flutuação no tamanho da população). Os cenários foram discutidos em relação à realidade sócio-
econômico-ambiental da região sudoeste do Estado de Goiás, no qual o PNE está inserido. Uma
análise comparativa com base em uma combinação semelhante de parâmetros mostrou que a
população de lobo-guará diminui sempre mais rapidamente do que a população de Cachorro-do-
mato. Porém, os resultados mostraram que as duas populações podem persistir por um tempo
razoável, mesmo quando considerado o pior cenário, ou seja, uma combinação entre elevada
perda de habitat, alta endogamia e grande flutuação populacional. Porém, deve-se levar em
consideração que este estudo é apenas um ponto de partida para estudos posteriores. Esses
estudos deverão ser baseados em parâmetros mais realistas tanto da viabilidade destas populações
quanto do cenário de desenvolvimento sócio-econômico-ambiental da região sudoeste do Estado
de Goiás e, portanto, os resultados podem permitir avaliações mais precisas da persistência destas
populações.

11
REFERÊNCIAS BIBLIOGRÁFICAS
Costa LP, Leite YLR, Mendes SL e Ditchfield AD (2005) Conservação de mamíferos no Brasil.
Megadiversidade 1: 103-112.
Courtenay O e Maffei L (2004) Crab-eating fox Cerdocyon thous (Linnaeus, 1766). In: Sillero-
Zubiri C, Hoffmann M e Macdonald DW (eds) Canids: Foxes, Wolves, Jackals and Dogs:
Status Survey and Conservation Action Plan. IUCN/SSC Canid Specialist Group, IUCN,
Gland, Switzerland and Cambridge, UK, pp 32-38.
DeSalle R e Amato G (2004) The expansion of conservation genetics. Nature 5: 702-712.
Eisenberg JF e Redford KH (1999) Mammals of the Neotropics - The Central Neotropics:
Ecuador, Peru, Bolivia, Brazil. University of Chicago Press, Chicago, 609pp.
Fonseca GAB, Rylands AB, Costa CRM, Machado RB e Leite YLR (1994) Livro Vermelho Dos
Mamíferos Brasileiros Ameaçados de Extinção. Fundação Biodiversitas, Belo Horizonte.
Fletchall NB, Rodden M e Taylor S (2000) Manual de Manejo do lobo-guará(Chrysocyon
brachyurus). CEPREM, São Paulo, 87pp.
Frankham R, Ballou JD e Briscoe DA (2002) Introduction to Conservation Genetics. Cambridge
University Press, 640pp.
Hedrick PW (2001) Conservation genetics: where are we now? Trends in Ecology & Evolution
16: 629-636.
IBAMA (2003) Lista das Espécies da Fauna Brasileira Ameaçadas de Extinção. Anexo à
Instrução Normativa n° 3, de 27 de maio de 2003, do Ministério do Meio Ambiente.
Disponível em: www.ibama.gov.br.
IUCN (2004) 2004 IUCN Red List of Threatened Species. Disponível em: www.redlist.org.

12
Juarez MJ e Marinho-Filho J (2002) Diet, habitat use, and home ranges of sympatric canids in
Central Brazil. Journal of Mammalogy 83 (4): 925-933.
Klink CA e Machado RB (2005) A Conservação do Cerrado Brasileiro. Megadiversidade 1: 143-
155.
Klink CA e Moreira AG (2002) Past and current human occupation and land-use. In: P.S. Oliveira
e R.J. Marquis (eds.). The Cerrado of Brazil. Ecology and natural history of a neotropical
savanna. Columbia University Press, New York, pp 69-88.
Marinho-Filho JS, Rodrigues FHG, Juarez KM. (2002) The Cerrado Mammals: diversity, ecology
and natural history. In: Oliveira PS e Marquis RJ (eds) The Cerrados of Brazil: Ecology and
Natural History of a Neotropical Savanna. Columbia University Press, Irvington, NY, USA,
pp 266-284.
Motta-Junior JC (1996) Diet of the maned wolf, Chrysocyon brachyurus, in central Brazil.
Journal Zoology 240: 277-284.
Motta-Junior JC (2002) Fama injusta: novas informações sobre a dieta do lobo-guarápodem
ajudar a preservá-lo. Ciência Hoje 31: 71-73.
Myers N, Mittermeier RA, Mittermeier CG, Fonseca GAB e Kent J (2000) Biodiversity hotspots
for conservation priorities. Nature 403: 853-858.
Rocha VJ, Reis NR e Sekiama ML (2004) Dieta e dispersão de sementes por Cerdocyon thous
(Linnaeus) (Carnívora, Canidae) em um fragmento florestal no Paraná Brasil. Revista
Brasileira de Zoologia, 21: 871-876.
Rodden M, Rodrigues F e Bestelmeyer S (2004) Maned wolf Chrysocyon brachyurus (Illiger, 1815).
In: Sillero-Zubiri C, Hoffmann M e Macdonald DW (eds) Canids: Foxes, Wolves, Jackals

13
and Dogs: Status Survey and Conservation Action Plan. IUCN/SSC Canid Specialist Group,
IUCN, Gland, Switzerland and Cambridge, UK, pp 38-43.
Rodrigues FHG, Silveira L, Jácomo ATA, Bezerra AP, Coelho AMR, Santos DC, Pagnozzi HG e
Hass A (2002) Composição e caracterização da fauna de mamíferos do Parque Nacional das
Emas, Goiás. Revista Brasileira de Zoologia 19: 589-600.
Silveira L e Jácomo ATA (1998) Ecologia e conservação de mamíferos carnívoros do Parque
Nacional das Emas, GO. Relatório Técnico para o IBAMA/DIREC/DEUC.

14
CAPÍTULO 1

15
Artigo Submetido: Genetics and Molecular Research
Transferibilidade de Marcadores STR para duas Espécies de
Canídeos do Cerrado Brasileiro
FLÁVIA MELO RODRIGUES1,5, MARIANA PIRES DE CAMPOS TELLES2, LUCILEIDE
VILELA RESENDE2, THANNYA NASCIMENTO SOARES2, JOSÉ ALEXANDRE
FELIZOLA DINIZ-FILHO3, 4, 5, ANAH TEREZA DE ALMEIDA JÁCOMO 4 E LEANDRO
SILVEIRA4. 1Universidade Estadual de Goiás, Departamento de Biologia, Anápolis-G0; 2Universidade
Católica de Goiás, Laboratório de Genética e Melhoramento, Departamento de Zootecnia; 3Universidade Federal de Goiás, Departamento de Biologia geral; 4Fundo para a Conservação
da Onça-Pintada (FCOP); 5 Programa de Pós-Graduação em Ciências Ambientais – UFG.
e-mail: [email protected]
Resumo
O lobo-guará (Chrysocyon brachyurus) e cachorro-do-mato (Cerdocyon thous) são duas
espécies de canídeos que ocorrem no cerrado brasileiro. O lobo-guará é uma espécie que está
incluída na lista de espécies brasileiras ameaçadas de extinção, enquanto o cachorro-do-mato é
um canídeo muito comum e abundante nas áreas do cerrado. O objetivo deste trabalho foi testar a
amplificação cruzada de 29 locos STR para amostras de lobo-guará e cachorro-do-mato,
provenientes do Parque Nacional das Emas. Para testar a transferibilidade foram utilizados dois
indivíduos de cada espécie. Em seguida, foram montadas reações de PCR utilizando uma
amplitude de variação na temperatura de anelamento dos primers entre 45 a 65ºC. Os produtos da
amplificação foram separados em gel de poliacrilamida 4% e visualizados por coloração com
nitrato de prata. Os locos transferidos foram utilizados para uma análise com 38 indivíduos de
cada espécie. Do total de locos avaliados, seis foram transferidos com sucesso para as duas
espécies (CSSM-038, PEZ-05, PEZ-20, PEZ-12, PEZ-13 e PEZ-15) . Para o cachorro-do-mato,
foi obtida uma média de 7,8 alelos por locos e para o lobo-guará 4,5 alelos por loco. O loco
CSSM-038 foi monomórfico para as duas espécies e os outros cinco locos apresentaram alto
polimorfismo, permitindo a estimativa de alguns parâmetros genéticos, incluindo a
heterozigosidade média esperada (He = 0.676 para o cachorro-do-mato e He = 0,533 para lobo-

16
guará), e o índice de fixação médio (f = 0,0326 para cachorro-do-mato e f = -0,09078 para lobo-
guará). Com estes resultados, pôde-se confirmar que o potencial de transferibilidade de
marcadores STR é maior entre as espécies evolutivamente mais próximas (i.e. cachorro
doméstico). Essas informações podem ser úteis para outros pesquisadores que trabalham com
estas espécies, pois diminuem os custos e agilizam o processo de obtenção de dados moleculares
para o estudo das populações naturais
Palavras-chave: Marcadores STR, Transferibilidade, Cerdocyon thous, cachorro-do-mato,
Chrysocyon brachyurus, lobo-guará
Enviar correspondência para FM Rodrigues. Universidade Estadual de Goiás, Departamento de
Biologia, Unidade Universitária de Ciências Extas e Tecnológicas, BR 153, Km 3, Campus
Anápolis, CEP: 75110-390, Caixa Postal: 459, Anápolis, GO, Brasil. E-mail: [email protected]

17
Abstract
The Maned-Wolf (Chrysocyon brachyurus) and the Crab-eating fox (Cerdocyon thous)
are two canid species found in Brazilian Cerrado. Maned-Wolf is the largest canid of South
America and is typical of Brazilian Cerrado, whereas Crab-eating fox is a relatively small,
abundant and widely distributed species found in many habitats in Brazil. In this note, we tested
cross-amplification and transferability of 29 STR loci originally developed for cattle and
domestic dogs and cats to these two species, using a total of 38 individuals of each species,
collected in Emas National Park, which is the largest National Park in the Cerrado region. Out of
the 29 STR primers tested, 6 were successfully transferred (CSSM-038, PEZ-05, PEZ-20, PEZ-
12, L-13 and L-15) and 5 showed polymorphism in the population analyzed. The genetic
parameters estimated (number of alleles per loci, expected and observed heterosigosity and
fixation indices) are within the expectation for other canid population wordwide.
Keywords: STR markers, cross-amplification, transferability, canids, Cerdocyon thous,
Chrysocyon brachyurus, Cerrado

18
No cerrado brasileiro ocorrem 38 espécies de mamíferos de médio e maior porte e destas,
45% correspondem a ordem Carnivora (Marinho-Filho et al. 2002). Apesar de sua
representatividade na comunidade faunística do Cerrado, as espécies de Carnivora têm sido
pouco estudadas (Alho, 1990; Garcia-Perea, 1994; Silveira & Jácomo, 1998) e a maioria dos
trabalhos publicados enfocam apenas aspectos ecológicos de uma única espécie (Dietz, 1984;
Dalponte, 1997; Facure, 1996). Dentre os 12 Parques Nacionais no Bioma Cerrado (Brasil), o
Parque Nacional das Emas (PNE) é o que apresenta maior riqueza e abundância faunística de
médios e grandes mamíferos (Erize, 1977; Eisenberg & Redford, 1999). O lobo-guará, ou
simplesmente Guará (Chrysocyon brachyurus), uma das espécies que ocorrem no PNE, é o maior
representante da família dos canídeos da América do Sul. Esta espécie possui uma grande
importância para a manutenção da diversidade do bioma por sua função ecológica de predador de
topo de cadeia e dispersor de sementes, sendo, portanto, importante para a estabilidade deste
ecossistema. Atualmente está classificado pela IUCN (2004) como espécie LR/nt, ou seja, com
pequeno risco de extinção, mas próximo a ser considerado vulnerável à extinção. A espécie
também está incluída na lista de espécies brasileiras ameaçadas de extinção do IBAMA (Instituto
Brasileiro do Meio Ambiente e Recursos Naturais Renováveis) (IBAMA, 2003). Outra espécie de
canídeo encontrada no PNE é o cachorro-do-mato (Cerdocyon thous), que é um mamífero de
porte médio, cuja distribuição estende-se do sul da Amazônia até o Paraguai (EMBRAPA, 2005).
No Brasil, é o canídeo mais comum e com maior distribuição, ocupando uma grande variedade de
habitats. Porém, parece ser mais abundante nas áreas de cerrado do que em áreas de matas
fechadas (Rocha et al. 2004).
Estudos de conservação de espécies nativas freqüentemente requerem o conhecimento da
diversidade genética entre as populações e fluxo gênico. Os marcadores microssatélite ou STR
(Short Tandem Repeats) têm sido freqüentemente escolhidos para quantificar a variabilidade
genética em populações naturais devido ao seu alto índice informativo, quando comparado a

19
outras classes de marcadores moleculares, e pela facilidade com que o polimorfismo pode ser
detectado (Brondani et al. 2003; Wandeler et al. 2003; Li et al. 2002). Os STR’s, estão se
tornando cada vez mais a ferramenta escolhida para estudos de genética de população, fluxo
gênico e análise de paternidade (Iyengar et al. 2005; Klukowska et al. 2003; Wright & Bentzen
1994). Entretanto, avanços no uso dos marcadores STR se tornam difíceis devido ao alto custo e
ao tempo relativamente grande necessários para o desenvolvimento prévio dos marcadores, que
normalmente são desenvolvidos para cada espécie que se pretende estudar. O sucesso na
amplificação cruzada ou transferibilidade de regiões de STR entre espécies, pela Reação em
Cadeia da Polimerase (PCR), tende a estar inversamente relacionada à distância evolutiva entre
as duas espécies consideradas (Navani et al. 2002; Tong et al. 2002; Williamson et al. 2002;
Zucchi et al. 2002). Diversos estudos têm mostrado a possibilidade de se usar alguns marcadores
desenvolvidos para uma outra espécie pertencente ao mesmo gênero (Brondani et al.2003;
Cipriani et al. 1999; Isagi & Suhandono 1997; Zucchi et al.2002), ou até mesmo entre gêneros
diferentes (Roa et al. 2000; Write & Powell 1997).
Neste estudo foi testada a transferibilidade cruzada de marcadores STR desenvolvidos
originalmente para bovinos, búfalos, cachorro doméstico e gato doméstico (Barker et al. 1997,
Cãnón et al. 2001, Denise et al. 2004, Fries et al. 1993, Halverson et al. 1995, Kapes et al. 1997,
Menotti-Raymond et al. 1999, Sunden et al. 1993) (Tabela 1), para duas espécies de canídeos do
cerrado, o cachorro-do-mato (Cerdocyon thous) e o lobo-guará (Chrysocyon brachyurus). O
DNA genômico foi extraído do sangue de 38 indivíduos de cada uma das duas espécies
capturadas no PNE, Goiás, Brasil. No total foram testados 29 primers com dois indivíduos de
cada população e destes, 6 pares de primers (21%) foram transferidos com sucesso e selecionados
para serem utilizados na genotipagem das duas populações (Tabela 2). Para as reações de PCR, o
seguinte protocolo foi utilizado: 5 µl de DNA (~3 ng/µl); 2,2 µl do primers A (~5 ng/µl); 2,2 µl

20
do primers B (~5 ng/µl); 1,3 µl de BSA (10 mg/ml); 1,5 µl tampão da enzima (10 X); 0,5 µl de
MgCl2 (50 mM); 1,3 µl de d’NTP (2,5 mM); 0,2 µl da enzima Taq- Polimerase (5 unidades/µl) e
completando o volume com 0,8 µl de H2O Milli-Q, para um volume final de 15 µl. A
amplificação via PCR foi realizada no termociclador MJ, utilizando o seguinte protocolo: 1º)
desnaturação inicial do DNA a 94ºC por 5 minutos; 2º) 94ºC por 1 minuto; 3º) anelamento dos
“primers” a diferentes temperaturas por 1 minuto; 4º) extensão da molécula pela enzima Taq
polimerase (fornecida pela Amersham Pharmacia Biotech) a 72ºC por 1 minuto; 5º) 30 ciclos
seguindo do 2º ao 4º passos; 6º) etapa final da extensão por 7 minutos a 72ºC para finalizar os
produtos amplificados. Para cada primer foram utilizadas temperaturas de anelamento que
variaram entre 45ºC e 65ºC (Tabela 1). Os fragmentos amplificados foram separados em cuba de
seqüênciamento vertical (BIORAD) em gel de poliacrilamida 4%, utilizando TBE 1X. A
eletroforese teve duração de 2 horas com potência de 90W sob corrente elétrica constante, e
corados com nitrato de prata (Bassan et al. 1991). O padrão de peso molecular, o marcador
Ladder 10bp (Amersham Pharmacia Biotech), foi usado para estimar os tamanhos dos produtos
de PCR
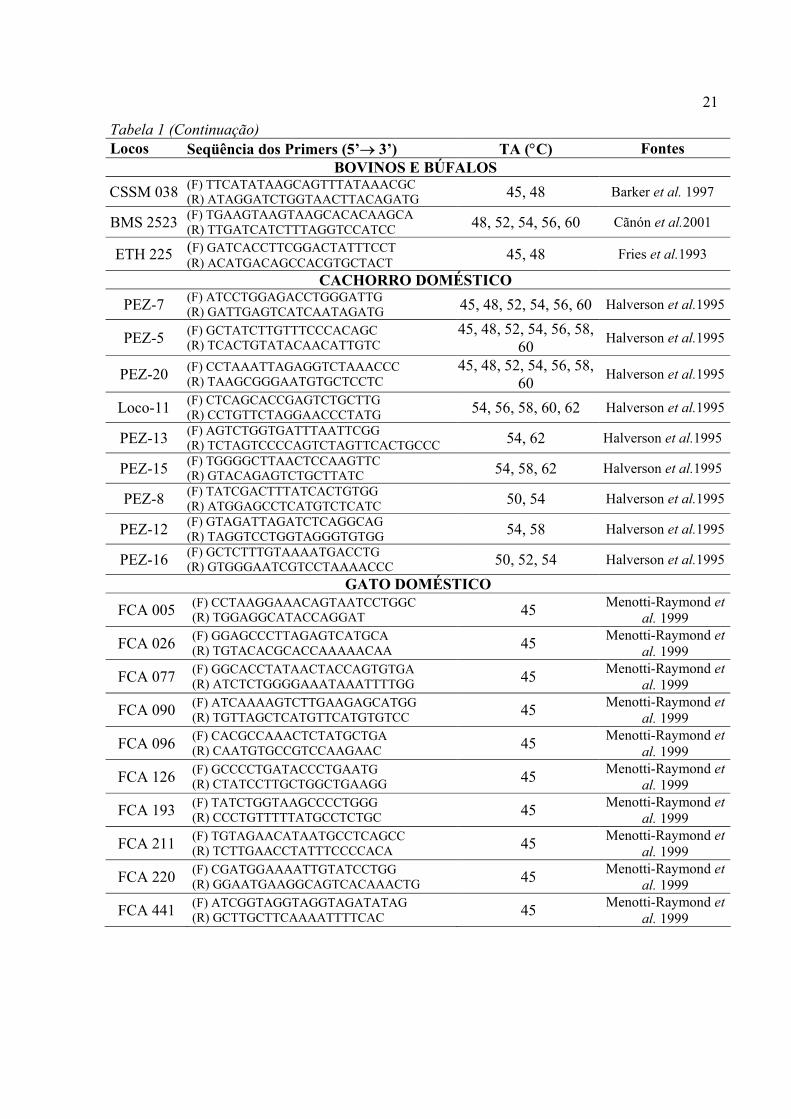
Tabela 1- Identificação dos locos, seqüência dos primers, temperaturas de anelamento (TA) testadas e fontes dos primers utilizados para amplificação cruzada para Cerdocyon thous e Chrysocyon brachyurus. As siglas (F) e (R) significam Forward e Reverse, respectivamente.
Locos Seqüência dos Primers (5’→ 3’) TA (°C) Fontes BOVINOS E BÚFALOS
BM 2113 (F) GCTCCTTCTACCAAATACCC (R) CTTCCTGAGAGAAGCAACACC 45, 48, 52, 54, 56, 60 Sunden et al.1993
MAF (F) ATTGACACTTCAGTAAGTTAACAATGG (R) CAGACACAACTGAGCAACTAGCGC 45, 48, 50 Kappes et al.1997
CSN 032 (F) ATGACACCTTAACCTAATCCC (R) GCACTTTATAAGCACCACAGC 48, 52, 54, 56, 60 Barker et al. 1997
INRA 006 (F) AGGAATATCTGTATCAACCTCAGTC (R) CTGAGCTGGGGTGGGAGCTATAAATA 45, 48, 50 Cãnón et al.2001
INRA 092 (F) GGTTTGACTTCAACTTCAACC (R) GGGAGTATGTGGAGAATGCAAG 45, 48 Cãnón et al.2001
TEXAN 02 (F) ACATTGTCATGTGGTTGCTAAC (R) ACTATGGGTATGTATATGTGCTGCAAG 45, 48 Barker et al. 1997
CSSM 046 (F) TGCACAATCGGAACCTAGAATATT (R) GGCTATTAACTGTTTTCTAGGAAT 45, 48 Barker et al. 1997
21
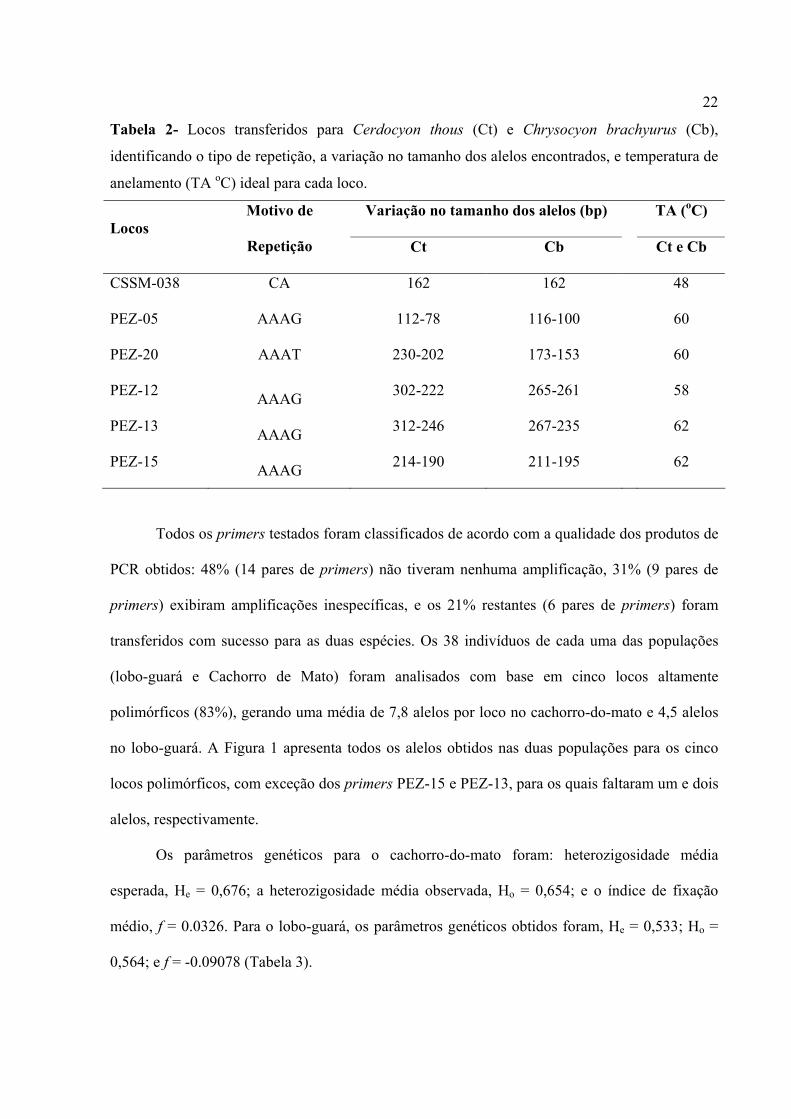
Tabela 1 (Continuação) Locos Seqüência dos Primers (5’→ 3’) TA (°C) Fontes
BOVINOS E BÚFALOS CSSM 038 (F) TTCATATAAGCAGTTTATAAACGC
(R) ATAGGATCTGGTAACTTACAGATG 45, 48 Barker et al. 1997
BMS 2523 (F) TGAAGTAAGTAAGCACACAAGCA (R) TTGATCATCTTTAGGTCCATCC 48, 52, 54, 56, 60 Cãnón et al.2001
ETH 225 (F) GATCACCTTCGGACTATTTCCT (R) ACATGACAGCCACGTGCTACT 45, 48 Fries et al.1993
CACHORRO DOMÉSTICO PEZ-7 (F) ATCCTGGAGACCTGGGATTG
(R) GATTGAGTCATCAATAGATG 45, 48, 52, 54, 56, 60 Halverson et al.1995
PEZ-5 (F) GCTATCTTGTTTCCCACAGC (R) TCACTGTATACAACATTGTC
45, 48, 52, 54, 56, 58, 60 Halverson et al.1995
PEZ-20 (F) CCTAAATTAGAGGTCTAAACCC (R) TAAGCGGGAATGTGCTCCTC
45, 48, 52, 54, 56, 58, 60 Halverson et al.1995
Loco-11 (F) CTCAGCACCGAGTCTGCTTG (R) CCTGTTCTAGGAACCCTATG 54, 56, 58, 60, 62 Halverson et al.1995
PEZ-13 (F) AGTCTGGTGATTTAATTCGG (R) TCTAGTCCCCAGTCTAGTTCACTGCCC 54, 62 Halverson et al.1995
PEZ-15 (F) TGGGGCTTAACTCCAAGTTC (R) GTACAGAGTCTGCTTATC 54, 58, 62 Halverson et al.1995
PEZ-8 (F) TATCGACTTTATCACTGTGG (R) ATGGAGCCTCATGTCTCATC 50, 54 Halverson et al.1995
PEZ-12 (F) GTAGATTAGATCTCAGGCAG (R) TAGGTCCTGGTAGGGTGTGG 54, 58 Halverson et al.1995
PEZ-16 (F) GCTCTTTGTAAAATGACCTG (R) GTGGGAATCGTCCTAAAACCC 50, 52, 54 Halverson et al.1995
GATO DOMÉSTICO
FCA 005 (F) CCTAAGGAAACAGTAATCCTGGC (R) TGGAGGCATACCAGGAT 45 Menotti-Raymond et
al. 1999
FCA 026 (F) GGAGCCCTTAGAGTCATGCA (R) TGTACACGCACCAAAAACAA 45 Menotti-Raymond et
al. 1999
FCA 077 (F) GGCACCTATAACTACCAGTGTGA (R) ATCTCTGGGGAAATAAATTTTGG 45 Menotti-Raymond et
al. 1999
FCA 090 (F) ATCAAAAGTCTTGAAGAGCATGG (R) TGTTAGCTCATGTTCATGTGTCC 45 Menotti-Raymond et
al. 1999
FCA 096 (F) CACGCCAAACTCTATGCTGA (R) CAATGTGCCGTCCAAGAAC 45 Menotti-Raymond et
al. 1999
FCA 126 (F) GCCCCTGATACCCTGAATG (R) CTATCCTTGCTGGCTGAAGG 45 Menotti-Raymond et
al. 1999
FCA 193 (F) TATCTGGTAAGCCCCTGGG (R) CCCTGTTTTTATGCCTCTGC 45 Menotti-Raymond et
al. 1999
FCA 211 (F) TGTAGAACATAATGCCTCAGCC (R) TCTTGAACCTATTTCCCCACA 45 Menotti-Raymond et
al. 1999
FCA 220 (F) CGATGGAAAATTGTATCCTGG (R) GGAATGAAGGCAGTCACAAACTG 45 Menotti-Raymond et
al. 1999
FCA 441 (F) ATCGGTAGGTAGGTAGATATAG (R) GCTTGCTTCAAAATTTTCAC 45 Menotti-Raymond et
al. 1999
22
Tabela 2- Locos transferidos para Cerdocyon thous (Ct) e Chrysocyon brachyurus (Cb),
identificando o tipo de repetição, a variação no tamanho dos alelos encontrados, e temperatura de
anelamento (TA oC) ideal para cada loco.
Variação no tamanho dos alelos (bp) TA (oC) Locos
Motivo de
Repetição Ct Cb Ct e Cb
CSSM-038 CA 162 162 48
PEZ-05 AAAG 112-78 116-100 60
PEZ-20 AAAT 230-202 173-153 60
PEZ-12 AAAG 302-222 265-261 58
PEZ-13 AAAG 312-246 267-235 62
PEZ-15 AAAG 214-190 211-195 62
Todos os primers testados foram classificados de acordo com a qualidade dos produtos de
PCR obtidos: 48% (14 pares de primers) não tiveram nenhuma amplificação, 31% (9 pares de
primers) exibiram amplificações inespecíficas, e os 21% restantes (6 pares de primers) foram
transferidos com sucesso para as duas espécies. Os 38 indivíduos de cada uma das populações
(lobo-guará e Cachorro de Mato) foram analisados com base em cinco locos altamente
polimórficos (83%), gerando uma média de 7,8 alelos por loco no cachorro-do-mato e 4,5 alelos
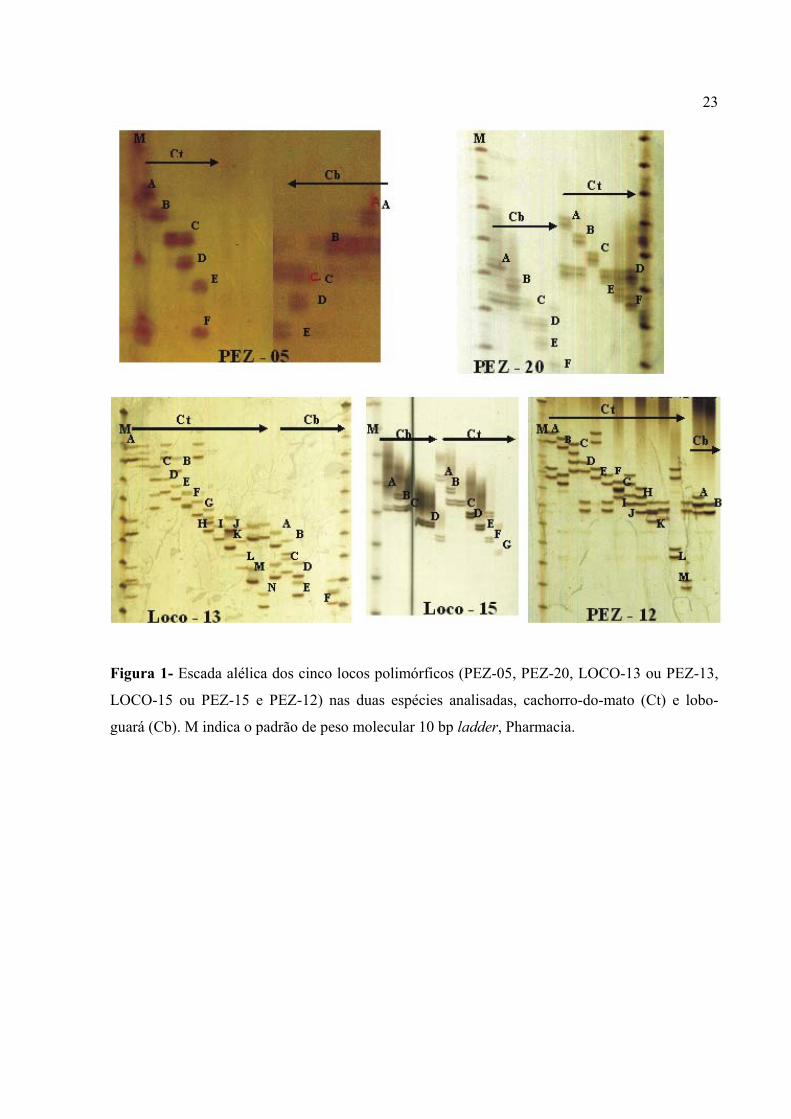
no lobo-guará. A Figura 1 apresenta todos os alelos obtidos nas duas populações para os cinco
locos polimórficos, com exceção dos primers PEZ-15 e PEZ-13, para os quais faltaram um e dois
alelos, respectivamente.
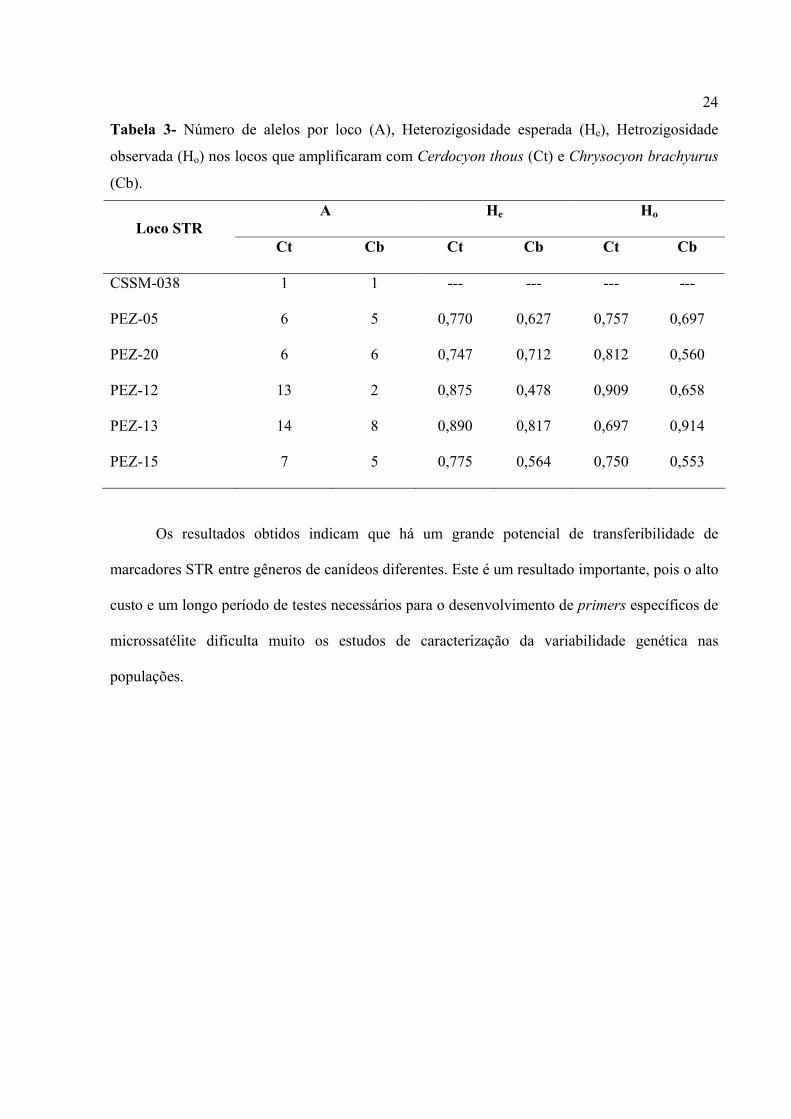
Os parâmetros genéticos para o cachorro-do-mato foram: heterozigosidade média
esperada, He = 0,676; a heterozigosidade média observada, Ho = 0,654; e o índice de fixação
médio, f = 0.0326. Para o lobo-guará, os parâmetros genéticos obtidos foram, He = 0,533; Ho =
0,564; e f = -0.09078 (Tabela 3).
23
Figura 1- Escada alélica dos cinco locos polimórficos (PEZ-05, PEZ-20, LOCO-13 ou PEZ-13,
LOCO-15 ou PEZ-15 e PEZ-12) nas duas espécies analisadas, cachorro-do-mato (Ct) e lobo-
guará (Cb). M indica o padrão de peso molecular 10 bp ladder, Pharmacia.
24
Tabela 3- Número de alelos por loco (A), Heterozigosidade esperada (He), Hetrozigosidade
observada (Ho) nos locos que amplificaram com Cerdocyon thous (Ct) e Chrysocyon brachyurus
(Cb).
A He Ho Loco STR
Ct Cb Ct Cb Ct Cb
CSSM-038 1 1 --- --- --- ---
PEZ-05 6 5 0,770 0,627 0,757 0,697
PEZ-20 6 6 0,747 0,712 0,812 0,560
PEZ-12 13 2 0,875 0,478 0,909 0,658
PEZ-13 14 8 0,890 0,817 0,697 0,914
PEZ-15 7 5 0,775 0,564 0,750 0,553
Os resultados obtidos indicam que há um grande potencial de transferibilidade de
marcadores STR entre gêneros de canídeos diferentes. Este é um resultado importante, pois o alto
custo e um longo período de testes necessários para o desenvolvimento de primers específicos de
microssatélite dificulta muito os estudos de caracterização da variabilidade genética nas
populações.

25
REFERÊNCIAS BIBLIOGRÁFICAS
Alho JRC (1990) Distribuição da fauna num gradiente de recursos em mosaico, Em: Cerrado:
caracterização, ocupação e perspectiva, Maria N. Pinto (Edit.), Editora UnB.
Barker JSF, Moore S, Hetzel S, Evan D, Tan SG, Byrne, K (1997) Genetic diversity of Asian
water buffalo (Bubalus bubalis): microsatellite variation and comparison with protein-coding
loci. Animal Genetics, 28, 103-115.
Bassan BJ, Anollés GCE, Gresshoff PM (1991) Fast and sensitive silver staining of DNA in
polyacrilamide gels. An. Biochem, 196, 80-83.
Brondani C, Rangel PHN, Borba TCO, Brondani RPV (2003) Transferability of microsatellite
and sequence tagged site markers in Orysa species. Hereditas, 138, 187-192.
Cañón, J, Alexandrino P, Bessa I et al. (2001) Genetic diversity measures of local European beef
cattle breeds for conservation purposes. Genetics Selection and Evolution, 33, 311–332.
Cipriani G, Lot G, Huang WG et al. (1999) R. AC/GT and AG/CT microsatellite repeats in peach
[Prunus persica (L.) Batsch]: isolation, characterization and cross-species amplification in
Prunus . Theoretical Applied Genetics, 99, 65-72.
Daponte, JC (1997) Diet horary fox, Lycalopex ventulus, in Mato Grosso, Central Brasil.
Mammalia, 61, 537-546.
Denise S, Johnston E, Halverson J, Marshall K, Rosenfeld D, Mckenna S, Sharp T, Edwards J
(2004) Power of exclusion for parentage verification and probability of match for identity in
American kennel club breeds using 17 canine microsatellite markers. Animal Genetics, 35,
14–17.
Dietz JM (1984) Ecology and social organization of maned wolf Chrysocyon brachyurus. Tese
Doctor of Philosophy, Michigan State University, Washington.

26
Eisenberg JF, Redford KH (1999) Mammals of the Neotropics - The Central Neotropics: Ecuador,
Peru, Bolivia, Brazil. University of Chicago Press, Chicago, 609pp.
EMBRAPA (2005) Monitoramento por Satélite. Campinas, [S.d.]. Cachorro-do-mato. Disponível
em: <http://www.cnpm.embrapa.br/projetos/fauna/mamifero/cac_mato.html
Erize F (1977) Brazil´s finest National Park. Oryx, 13, 457-462.
Facure GF (1996) Ecologia alimentar do cachorro-do-mato Cerdocyon thous (Carnivora –
Canidade), no Parque Florestal do Itapetinga, município de Atibaia, sudeste do Brasil.
Dissertação de Mestrado em Biologia, Universidade Estadual de Campinas.
Fries R, Eggen A, Womack JE (1993) The bovine genome map. Mammals Genome, 4, 405-428.
Garcia-Perea, R (1994) The pampas cat group (genus Lynchailurus Severtzov, 1858) (Carnivora
Felidae): a systematic and biogeographic review. American Museum Novitates.
Halverson J, Dvorak J, Stevenson T (1995) Microsatellite sequences for canine genotyping. US
Patent 05874217.
IBAMA (2003) Lista das Espécies da Fauna Brasileira Ameaçadas de Extinção. Anexo à
Instrução Normativa n° 3, de 27 de maio de 2003, do Ministério do Meio Ambiente.
Disponível em: www.ibama.gov.br.
Isagi Y, Suhandono S (1997) PCR primers amplifying microsatellite loci of Quercus mysinifolia
Blume and their conservation between oak species. Molecular Ecology, 6, 897-899.
IUCN (2004) 2004 Red List of Threatened Species. Disponível em: www.redlist.org.
Iyengar A, Babu VN, Hedges S, Venkataraman AB, Maclean N, Morin PA (2005)
Phylogeography, genetic structure, and diversity in dhole (Cuon alpinus). Molecular Ecology,
14, 2281-2297.

27
Kappes SM, Keele JM, Stone RT, Mc Graw RA, Sonstegard TS, Smith TP, Lopes-Corrales NL,
Beattie CW (1997) Second generation linkage map of the bovine genome. Genome Research,
7 (3), 235-249.
Klukowska BJ, Strabel, T, Mackowski M, Switonski M (2003) Microsatellite polymorphism and
genetic distances betwen the dog, red fox and artic fox. J. Anim. Breed. Genet., 120, 88-94.
Li YC, Korol AB, Fahima T, Beiles A, Nevo E (2002) Microsatellites: genomic distribution,
putative functions and mutational mechanisms: a review. Molecular Ecology, 11, 2453-2465.
Marinho-Filho JS, Rodrigues FHG, Juarez KM. (2002). The Cerrado Mammals: diversity,
ecology and natural history. In: The Cerrados of Brazil: Ecology and Natural History of a
Neotropical Savanna (eds Oliveira PS, Marquis RJ) pp. 266-284. Columbia University Press,
Irvington, NY, USA.
Menotti-Raymond MV, David A, Lyons LA, Schaffer AA, Tomlin JF, Hutton MK, O' Brien SJ
(1999) A genetic linkage map of microsatellites in the domestic cat (Felis catus). Genomics,
57, 9-23.
Navani N, Jain PK, Gupta S, Sisodia BS, Kumar S (2002) A set of cattle microsatellite DNA
markers for genome analysis of riverine buffalo (Bubalus bubalis). Short communication, 33,
149-154.
Roa AC, Chavarriaga-Aguirre P, Duque MC et al. (2000) Cross-species amplification of cassava
(Manhihot esculenta) (Eurphorbiaceae) microsatellites: allelic polymorphism and degree of
realtionship. American Journal of Botany, 87 (11), 1647-1655.
Rocha VJ, Reis NR, Sekiama ML (2004) Dieta e dispersão de sementes por Cerdocyon thous
(Linnaeus) (Carnívora, Canidae) em um fragmento florestal no Paraná Brasil. Revista
Brasileira de Zoologia, 21 (4), 871-876.

28
Silveira L, Jácomo ATA (1998) Ecologia e conservação de mamíferos carnívoros do Parque
Nacional das Emas, GO. Relatório Técnico para o IBAMA/DIREC/DEUC.
Sunden SLF, Stone RT, Bishop MD, Kappes SM, Keele JW, Beattie CW (1993) A highly
polymorphic bovine microsatellite locus: BM2113. Animal Genetics, 24, 69.
Tong J, Wang Z, Yu X, Wu Q, Chu KH (2002) Cross-species amplification in silver carp and
bighead carp with microsatellite primers of common carp. Molecular Ecology Notes, 2, 245-
247.
Wandeler P, Funk SM, Largiader CR, Gloor S, Breitenmoser U (2003) The city-fox phenomenon:
genetic consequences of a recent colonization of urban habitat. Molecular Ecology, 12, 647-
656.
Williamson KS, Cordes JF, May B (2002) Characterization of microsatellite loci in Chinook
salmon (Oncorhynchus tshawytscha) and cross-species amplification in other salmonids.
Molecular Ecology Notes, 2, 17-19.
Wright JH, Bentzen P (1994) Microsatellites: genetic markers of the future. Review. Fish Biology
and Fisheries, 4, 384-388.
Write G, Powell W (1997) Cross-species amplification of SSR loci in the Meliaceae family.
Molecular Ecology, 6, 1195-1197.
Zucchi MI, Brondani RPV, Pinheiro JB, Brondani C, Vencovsky R (2002) Transferability of
microsatellite markers from Eucalyptus spp. to Eugenia dysenterica (Myrtaceae family).
Molecular Ecology Notes, 2, 512–513.

29
CAPÍTULO 2

30
Artigo a ser submetido: Genetics and Molecular Biology
Diversidade Genética do lobo-guará (Chrysocyon brachyurus) e
cachorro-do-mato (Cerdocyon thous) Utilizando Marcadores RAPD
e STR
FLÁVIA MELO RODRIGUES1, 6, MARIANA PIRES DE CAMPOS TELLES4, 5, JOSÉ
ALEXANDRE FELIZOLA DINIZ FILHO2, 3, 5, 6, LUCILEIDE VILELA RESENDE4, ANAH
TEREZA DE ALMEIDA JÁCOMO5, LEANDRO SILVEIRA5 E THANNYA NASCIMENTO
SOARES4 1Departamento de Biologia, Universidade Estadual de Goiás (UEG), Anápolis, GO; 2Departamento de Biologia, Universidade Católica de Goiás (UCG); 3Departamento de Biologia
Geral, ICB, Universidade Federal de Goiás (UFG); 4Laboratório de Genética e Melhoramento,
Departamento de Zootecnia/ITS, Universidade Católica de Goiás (UCG); 5Fundo para a
Conservação da Onça-Pintada (FCOP),6 Programa de Pós-Graduação em Ciências
Ambientais – UFG.
e-mail: [email protected]
Resumo
Quanto maior é a variação genética de uma espécie, maior será a sua chance de sobreviver
a mudanças do ambiente e, por isso, maiores são as suas chances de evitar a extinção. Os
marcadores moleculares, tais como, RAPD e os STR’s, são técnicas da biologia molecular que
têm sido amplamente utilizadas no monitoramento da variabilidade genética das populações. O
objetivo deste trabalho foi estimar a diversidade genética de populações de cachorro-do-mato
(Cerdocyon thous) e de lobo-guará (Chrysocyon brachyurus) do Parque Nacional das Emas
(PNE), Goiás, utilizando os marcadores STR e marcadores RAPD. Foram coletadas amostras de
sangue para as análises genéticas de 38 indivíduos de cada uma das duas espécies estudadas. Para
as análises RAPD foram utilizados nove primers da OPERONTM e em seguida os fragmentos
amplificados foram separados em gel de agarose 1,4%, visualizados por coloração com brometo
de etídio. Para as análises com os marcadores STR, foram utilizados seis primers nas reações de
PCR. Os produtos amplificados foram submetidos a uma eletroforese em gel de acrilamida 4% e

31
este foi corado com nitrato de prata para a obtenção dos genótipos. Em seguida, foram realizadas
as estimativas de diversidade genética. Para estimar a correlação entre os resultados gerados
pelos dois marcadores utilizados, os genótipos obtidos com os dados STR foram transformados
em dados binários. Em média, o número total de locos e número de locos polimórficos por
primer, foi maior no cachorro-do-mato do que no lobo-guará . A similaridade genética obtida
com os marcadores RAPD foi em média maior na população de lobo-guará (0,75) do que na
população de cachorro-do-mato (0,68). Dentre os primers STR, 83% foram altamente
polimórficos. O índice de fixação médio estimado para cada população foi 0,032 e -0,091 para o
cachorro-do-mato e lobo-guará , respectivamente. As correlações matriciais entre as
similaridades genéticas obtidas pelos dois marcadores foram significativas pelo teste de Mantel,
porém com valores de r baixos, 0,242 (P = 0,0056) na população de lobo-guará e 0,2465 (P =
0,0126) na população de cachorro-do-mato. A baixa correlação entre os dois marcadores
moleculares utilizados corrobora outros estudos o que pode ser explicado pela base genética
diferente entre eles. Nossos dados não indicam uma significativa redução na variabilidade
genética e nem endogamia elevada em nenhuma das espécies estudadas. Portanto, deve-se pensar
em propostas adequadas de conservação e manejo destas espécies, principalmente do lobo-guará ,
que é uma espécie vulnerável a extinção, a fim de aumentar a viabilidade das mesmas em longo
prazo.
Palavras-chave: lobo-guará , cachorro-do-mato, Marcadores STR, Marcadores RAPD,
Diversidade Genética.
Enviar correspondência para FM Rodrigues. Universidade Estadual de Goiás, Departamento de
Biologia, Unidade Universitária de Ciências Extas e Tecnológicas, BR 153, Km 3, Campus
Anápolis, CEP: 75110-390, Caixa Postal: 459, Anápolis, GO, Brasil. E-mail: [email protected]

32
Abstract
High genetic variability of a species gives it a higher chance of avoiding extinction.
Molecular markers RAPD and STR are molecular biology techniques that have been widely used
in order to monitor genetic variability of the populations. This research’s goal was to estimate the
genetic diversity of the Crab-eating fox (Cerdocyon thous) population and the Maned wolf
(Chrysocyon brachyurus) population from Ema’s National Park in Goiás. Blood samples of 38
individuals of each species were drawn for genetic analysis. For the RAPD analysis, nine
OPERONTM primers were used and the amplified fragments were separated in Agaroses gel 1,4%,
with bromite etidium coloration. For the STR markers, six primers were used on the PCR
reations, the amplified fragments were submited to eletrphorese in actilamida gel 4%, colored
with silver nitrate, to obtain the genotypes. Then the estimates of the genetic diversity were
calculated. To estimate correlation between the results found with the markers, the genotypes
obtained by STR markers were transformed in binary data. In average, the toral number os locus
and the number of polimorph locus for each primer was higher for the Crab-eating fox. The
genetic similarity obtained with the RAPD market was in average higher for the Maned Wolf
(0,75) and (0,68) for the Crab-eating fox. Comparing primers, 83% of the STR was highly
polimorphic. The average fixation iundex estimated for the population was 0.032 for the Crab-
eating fox and 0,091 for the Maned wolf. The matricial correlations between genetic similarities
were significant with the Mantel test but the values of r were low, r=0,242 (p=0,0056) for Maned
wof population and r=0,2465 (p=0,0126) for Crab-eating fox population. The low correlation
between the two markers agrees with other studies, and can be explained by the different genetic
base between them. The data doesn´t indicate a reduction in the genetic variability, neither high
endogamy in either species. One should think of proper ways to conserve and manage these
species, specially the Maner wolf, which is vulnerable to extinction, so that they become viable in
the long run.
Key words: Maned Wolf, Crab-eating fox, STR marker, RAPD marker, genetic diversity.

33
INTRODUÇÃO
Cada vez mais o conhecimento genético e ecológico das espécies se torna importante para
sua conservação, porque é na variação de seus genes que está a possibilidade de sobrevivência
das espécies. Uma nova abordagem no estudo da biodiversidade, a genética da conservação,
surgiu há alguns anos atrás e se tornou importante em programas de conservação (DeSalle e
Amato, 2004; Manel et al., 2003; Soulé e Wilcox, 1980; Schonewald-Cox et al., 1998). A
genética da conservação prevê o estudo da variabilidade genética das populações naturais e
artificiais com fins de determinar o potencial adaptativo e evolutivo de uma espécie (DeSalle e
Amato, 2004). A genética da conservação é uma área bastante ampla, abrangendo desde estudos
de expressão gênica, adaptações locais, genética quantitativa e de populações, até fatores
ecológicos de seleção. Os aspectos ecológicos da genética da conservação são fundamentais, pois
deles provêm à paisagem seletiva e demográfica que, combinada com a história evolutiva,
produziram os padrões de variação dentro das espécies (Moran, 2002).
Quanto maior é a variação genética de uma espécie, maior será sua chance de sobreviver
se adaptando a um novo ambiente e evitando a extinção (Hedrick, 2001). A genética da
conservação, bem como os outros campos interessados na conservação biológica, está sendo
motivada pela necessidade de se reduzir a atual taxa de extinção e preservar a biodiversidade.
Para atingir tal objetivo, são estudados os fatores genéticos que afetam o risco de extinção e o
controle genético que é necessário para minimizar esse risco (Frankham et al., 2002).
Os efeitos do tamanho das populações são alguns dos grandes problemas enfrentados na
conservação biológica das espécies ameaçadas. Pequenas populações sofrem com o
endocruzamento e a perda da variabilidade genética mais rapidamente, resultando em um elevado
risco de extinção. Conseqüentemente, o principal objetivo do estudo genético é minimizar esse
endocruzamento e a perda da variabilidade genética (DeSalle e Amato, 2004; Frankham et al.,
2002; Hedrick, 2001). O endocruzamento reduz a fecundidade, a sobrevivência e acaba

34
diretamente aumentando o risco de extinção (Frankham et al., 2002). Já a redução da
variabilidade genética compromete as espécies na sua habilidade de persistir frente à mudanças
ambientais, reduzindo suas chances de sobrevivência e persistência em um dado ambiente.
Estudos em genética da conservação tentam identificar as populações onde o endocruzamento e a
variabilidade genética podem interferir nas suas perspectivas de sobrevivência em longo prazo
(DeSalle e Amato, 2004; Frankham et al., 2002; Manel et al., 2003).
As técnicas moleculares têm sido amplamente utilizadas para monitorar a variabilidade
genética das populações (Parker et al., 1998). Os avanços na biologia molecular abriram novas
perspectivas para a pesquisa em conservação de espécies e para estudos de biologia populacional
como um todo. A partir desses avanços as variações encontradas nas espécies naturais podem ser
analisadas em nível de DNA. Vários estudos nesse sentido vem sendo desenvolvidos no Brasil e
no mundo (De Mattos et al., 2004; Farrel et al., 2000; Iyengar et al., 2005; Mengoni et al., 2000;
Ratnayeke et al., 2002; Zucchi et al., 2002).
Os marcadores moleculares compõem essas técnicas da biologia molecular e existe hoje
uma grande variedade de classes de marcadores moleculares (Ferreira e Grattapaglia, 1998). Em
adição as técnicas baseadas em RFLP (Restriction Fragment Length Polymorphism) (Bostein et
al., 1980), muitas tecnologias baseadas na reação em cadeia da polimerase, a PCR (Polymerase
Chain Reaction) (Mullis e Faloona, 1987), que permite a síntese enzimática de milhões de cópias
de um segmento específico de DNA, têm sido desenvolvidas. Essa técnica provocou uma
verdadeira revolução na genética molecular, facilitando bastante os trabalhos desenvolvidos nos
laboratórios. Desde então, várias técnicas baseadas em PCR vêm sendo desenvolvidas e dentre
elas se destacam os Microssatélites (Tautz, 1989) e o polimorfismo de DNA amplificado ao acaso,
o RAPD (Random Amplified Polymorphic DNA) (Williams et al., 1990; Welsh e McClelland,
1990). A preferência por estas duas técnicas em relação ao RFLP é justificada devido a grande

35
quantidade de DNA exigida pela técnica de RFLP, bem como pelo elevado tempo e custo
despendido pelo desenvolvimento do procedimento (Mengoni et al., 2000)
Marcadores Microssatélite ou STR (Short Tandem Repeats), são definidos como
seqüências repetidas em tandem compostas de números variáveis de motivo com 1 a 6 pares de
bases (pb) encontrados no genoma de vertebrados, insetos e outros animais (Charlesworth et al.,
1994; Mengoni et al., 2000, Li et al., 2002). São marcadores genéticos codominantes
multialélicos e evolutivamente neutros. No entanto, alguns estudos discutem o papel dos
marcadores STR em regiões codificantes do genoma tais como, regulação da expressão gênica,
regulação da replicação e recombinação do DNA, organização da cromatina, etc (Li et al., 2002).
As seqüências que flanqueiam os STR são conservadas, o que permite a seleção de um par de
pequenos fragmentos de nucleotídeos iniciadores (Pimers), compostos de 20 a 30 pares de bases
e sua amplificação via PCR.
Os polimorfismos dos marcadores STR são detectados nos comprimentos das seqüências
dos fragmentos amplificados, pois o número de repetições em cada microssatélite é altamente
variável. Essas regiões possuem taxas elevadas de mutações, geralmente do tipo adição e deleção
das repetições, que podem chegar até um a cada 1000 pb (Tautz, 1989). Devido ao alto grau de
polimorfismo, os STR´s tornaram-se marcadores ideais para estudos de mapeamento genético,
discriminação entre acessos e cultivares de bancos de germoplasma, determinação de grau de
parentesco entre indivíduos e estrutura genética entre indivíduos, populações e espécies (Denise
et al., 2004; Francisco et al., 1996; Neff et al., 1999; Yang et al., 1994). Contudo, todas as
facilidades deste marcador dependem da disponibilidade das seqüências de DNA que flanqueiam
os STR, os primers. Por esta razão, o uso destes marcadores é limitado, pois são escassas as
informações sobre seqüências de DNA de espécies naturais e pelo alto custo e trabalho
envolvidos na construção de bibliotecas genômicas, clonagens e sequenciamentos necessários
para o isolamento de STR e desenvolvimento dos primers (Zucchi et al., 2002). No entanto,

36
diversos estudos têm mostrado a possibilidade de se usar alguns marcadores STR (primers)
desenvolvidos para uma outra espécie pertencente ao mesmo gênero (Brondani et al., 2003;
Cipriani et al., 1999; Isagi e Suhandono 1997; Zucchi et al., 2002), ou até mesmo entre gêneros
diferentes (Roa et al., 2000; Write e Powell 1997).
Os marcadores RAPD também têm sido bastante empregados no estudo de populações
naturais, pois são tecnicamente mais simples de se obter do que outros marcadores, tais como o
RFLP e STR, o que facilita o estudo de um grande número de locos, e fornece uma amostragem
aleatória maior do DNA (Ferreira e Grattapaglia, 1998). Estes marcadores são gerados pela
amplificação de segmentos desconhecidos do genoma, utilizando um simples e curto primer de
seqüência arbitrária, contendo 10 pb. São inúmeras as vantagens dos marcadores RAPD tais
como, não utilizar radioatividade, necessita de pequenas quantidades de DNA, não requerem um
conhecimento prévio da seqüência alvo a ser amplificada, e ser aplicável a qualquer espécie, etc
(Ferreira e Grattapaglia, 1998). Entretanto o uso do RAPD também tem limitações, pois são
marcadores de caráter dominante, ou seja, ele não permite a identificação do genótipo
heterozigoto, limitando a estimativa de parâmetros genéticos. Alguns trabalhos discutem alguns
problemas quanto à sua reprodutibilidade (Karp et al., 1996; Lynch e Milligan, 1994). No entanto,
alguns estudos têm demonstrado que os marcadores RAPD tendem a fornecer marcas
relativamente específicas de populações, raças e espécies, o que o torna um importante marcador
para estudos de populações visando à conservação genética (Powell et al., 1996).
O presente trabalho teve por objetivo utilizar os marcadores STR e marcadores RAPD
para estimar a diversidade genética de uma população de cachorro-do-mato (Cerdocyon thous) e
uma população de lobo-guará (Chrysocyon brachyurus) do Parque Nacional das Emas (PNE),
Goiás. O lobo-guará , ou simplesmente guará (Chrysocyon brachyurus), uma das espécies que
ocorrem no PNE, é o maior representante da família dos canídeos da América do Sul. Atualmente
está classificado pela IUCN (2004) como espécie LR/nt, ou seja, com pequeno risco de extinção,

37
mas próximo a ser considerado vulnerável à extinção. A espécie também está incluída na lista de
espécies brasileiras ameaçadas de extinção do IBAMA (Instituo Brasileiro do Meio Ambiente e
Recursos Naturais Renováveis) (IBAMA, 2003). O cachorro-do-mato (Cerdocyon thous), que é
um mamífero de porte médio, cuja distribuição estende-se do sul da Amazônia até o Paraguai
(EMBRAPA, 2005). No Brasil, é o canídeo mais comum e com maior distribuição, ocupando
uma grande variedade de habitats. Porém, parece ser mais abundante nas áreas de cerrado do que
em áreas de matas fechadas (Rocha et al.,2004). O conhecimento dos níveis de variabilidade
genética nessas espécies é importante a fim de estabelecer estratégias de conservação e manejo na
região, que venham a minimizar posteriores riscos de extinção ligado à perda da diversidade em
nível genético.
MATERIAL E MÉTODOS
Coleta do Material Animal e Extração de DNA
Os indivíduos das duas espécies estudadas, lobo-guará e o cachorro-do-mato, foram
capturados no Parque Nacional das Emas (PNE) situado no extremo sudoeste do Estado de Goiás,
próximo ao limite com o Mato Grosso do Sul e Mato Grosso, a 18o19’S e 52o45’E. A maior parte
do PNE é pertencente ao município de Mineiros (GO) e o restante, aos municípios de Chapadão
do Céu (GO), Costa Rica (MS) e Alto Taquari (MT) (Figura 1).
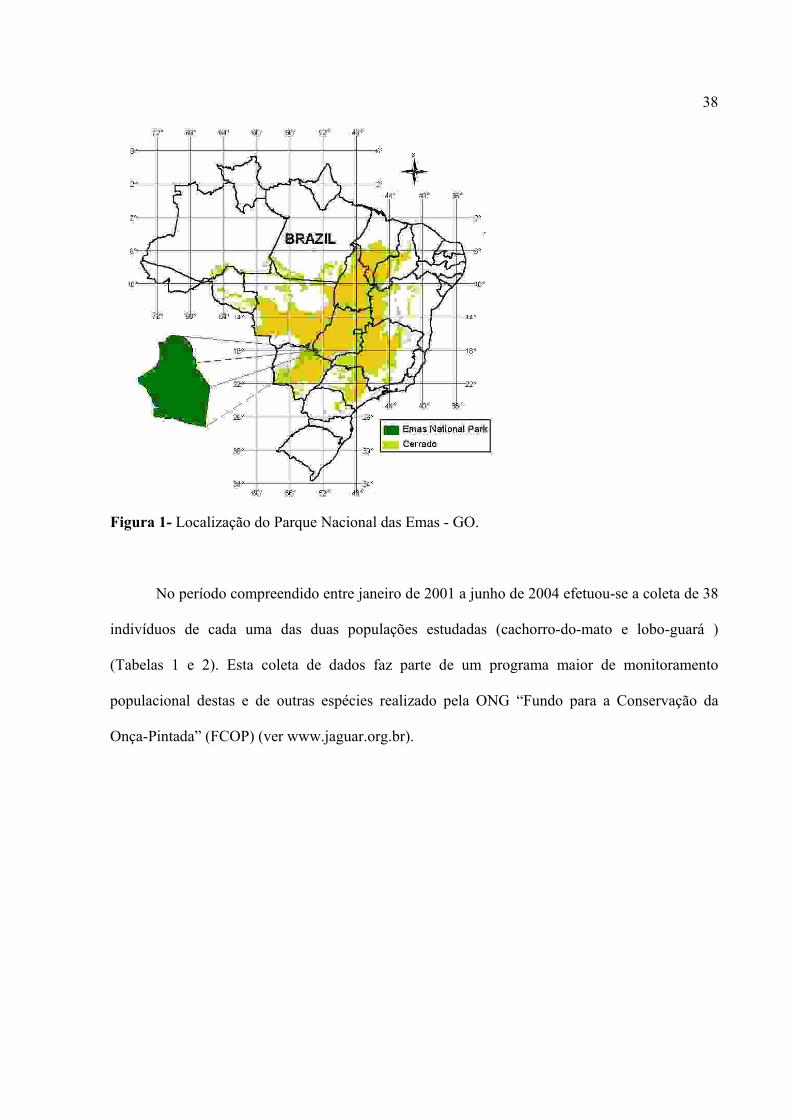
38
Figura 1- Localização do Parque Nacional das Emas - GO.
No período compreendido entre janeiro de 2001 a junho de 2004 efetuou-se a coleta de 38
indivíduos de cada uma das duas populações estudadas (cachorro-do-mato e lobo-guará )
(Tabelas 1 e 2). Esta coleta de dados faz parte de um programa maior de monitoramento
populacional destas e de outras espécies realizado pela ONG “Fundo para a Conservação da
Onça-Pintada” (FCOP) (ver www.jaguar.org.br).
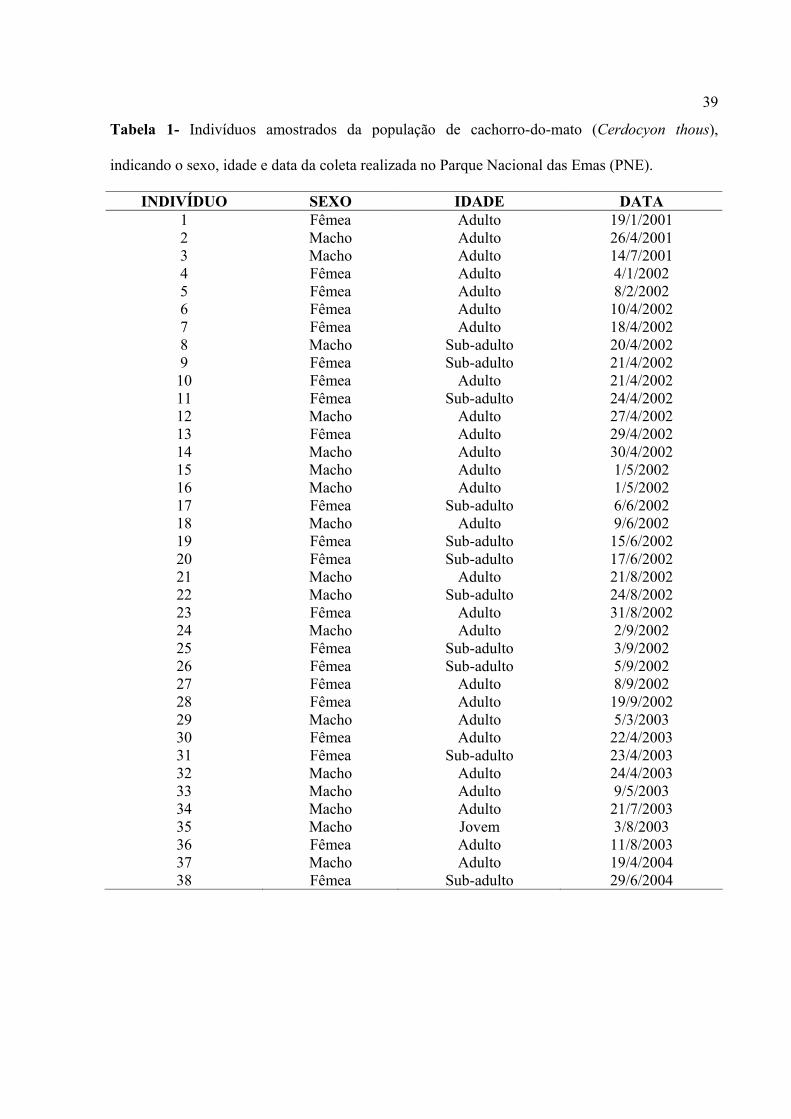
39
Tabela 1- Indivíduos amostrados da população de cachorro-do-mato (Cerdocyon thous),
indicando o sexo, idade e data da coleta realizada no Parque Nacional das Emas (PNE).
INDIVÍDUO SEXO IDADE DATA 1 Fêmea Adulto 19/1/2001 2 Macho Adulto 26/4/2001 3 Macho Adulto 14/7/2001 4 Fêmea Adulto 4/1/2002 5 Fêmea Adulto 8/2/2002 6 Fêmea Adulto 10/4/2002 7 Fêmea Adulto 18/4/2002 8 Macho Sub-adulto 20/4/2002 9 Fêmea Sub-adulto 21/4/2002 10 Fêmea Adulto 21/4/2002 11 Fêmea Sub-adulto 24/4/2002 12 Macho Adulto 27/4/2002 13 Fêmea Adulto 29/4/2002 14 Macho Adulto 30/4/2002 15 Macho Adulto 1/5/2002 16 Macho Adulto 1/5/2002 17 Fêmea Sub-adulto 6/6/2002 18 Macho Adulto 9/6/2002 19 Fêmea Sub-adulto 15/6/2002 20 Fêmea Sub-adulto 17/6/2002 21 Macho Adulto 21/8/2002 22 Macho Sub-adulto 24/8/2002 23 Fêmea Adulto 31/8/2002 24 Macho Adulto 2/9/2002 25 Fêmea Sub-adulto 3/9/2002 26 Fêmea Sub-adulto 5/9/2002 27 Fêmea Adulto 8/9/2002 28 Fêmea Adulto 19/9/2002 29 Macho Adulto 5/3/2003 30 Fêmea Adulto 22/4/2003 31 Fêmea Sub-adulto 23/4/2003 32 Macho Adulto 24/4/2003 33 Macho Adulto 9/5/2003 34 Macho Adulto 21/7/2003 35 Macho Jovem 3/8/2003 36 Fêmea Adulto 11/8/2003 37 Macho Adulto 19/4/2004 38 Fêmea Sub-adulto 29/6/2004
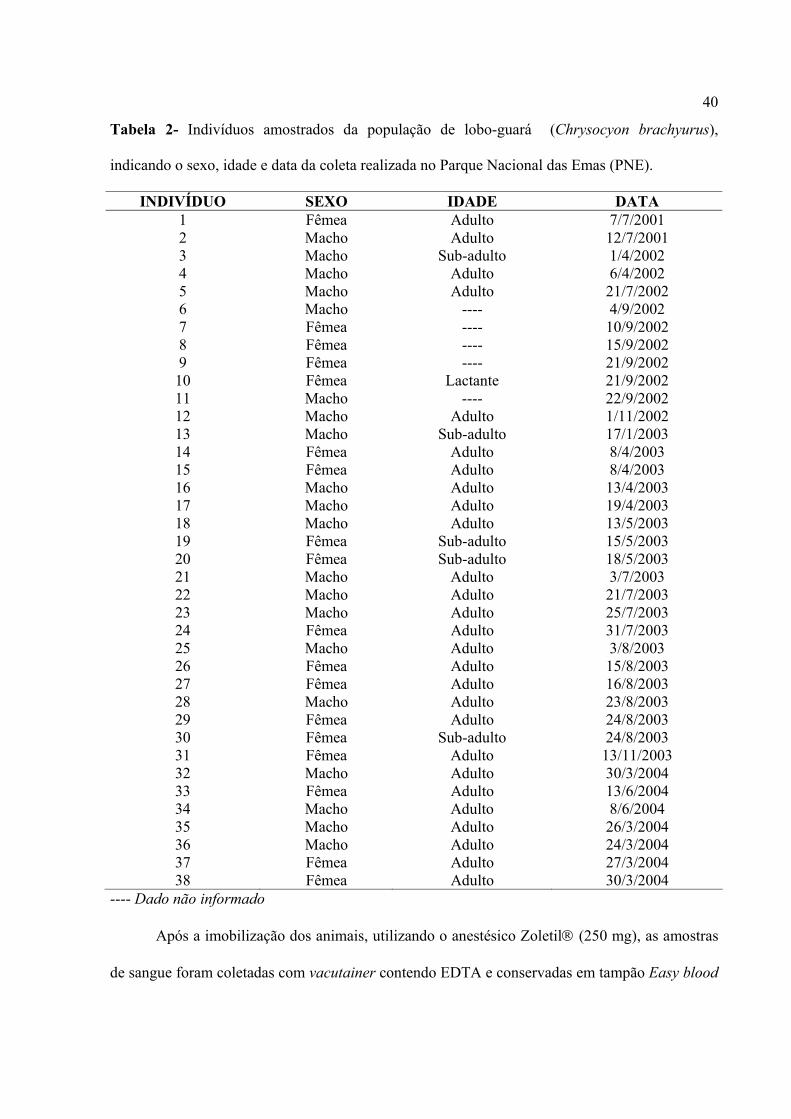
40
Tabela 2- Indivíduos amostrados da população de lobo-guará (Chrysocyon brachyurus),
indicando o sexo, idade e data da coleta realizada no Parque Nacional das Emas (PNE).
INDIVÍDUO SEXO IDADE DATA 1 Fêmea Adulto 7/7/2001 2 Macho Adulto 12/7/2001 3 Macho Sub-adulto 1/4/2002 4 Macho Adulto 6/4/2002 5 Macho Adulto 21/7/2002 6 Macho ---- 4/9/2002 7 Fêmea ---- 10/9/2002 8 Fêmea ---- 15/9/2002 9 Fêmea ---- 21/9/2002
10 Fêmea Lactante 21/9/2002 11 Macho ---- 22/9/2002 12 Macho Adulto 1/11/2002 13 Macho Sub-adulto 17/1/2003 14 Fêmea Adulto 8/4/2003 15 Fêmea Adulto 8/4/2003 16 Macho Adulto 13/4/2003 17 Macho Adulto 19/4/2003 18 Macho Adulto 13/5/2003 19 Fêmea Sub-adulto 15/5/2003 20 Fêmea Sub-adulto 18/5/2003 21 Macho Adulto 3/7/2003 22 Macho Adulto 21/7/2003 23 Macho Adulto 25/7/2003 24 Fêmea Adulto 31/7/2003 25 Macho Adulto 3/8/2003 26 Fêmea Adulto 15/8/2003 27 Fêmea Adulto 16/8/2003 28 Macho Adulto 23/8/2003 29 Fêmea Adulto 24/8/2003 30 Fêmea Sub-adulto 24/8/2003 31 Fêmea Adulto 13/11/2003 32 Macho Adulto 30/3/2004 33 Fêmea Adulto 13/6/2004 34 Macho Adulto 8/6/2004 35 Macho Adulto 26/3/2004 36 Macho Adulto 24/3/2004 37 Fêmea Adulto 27/3/2004 38 Fêmea Adulto 30/3/2004
---- Dado não informado
Após a imobilização dos animais, utilizando o anestésico Zoletil (250 mg), as amostras
de sangue foram coletadas com vacutainer contendo EDTA e conservadas em tampão Easy blood

41
(12,114g de Tris, 37,224g de EDTA e 20g de SDS) a 4°C. Em seguida foi realizada a extração do
DNA através do Kit de purificação de DNA (GFX) da Amersham Pharmacia Biotech. Este
procedimento pode processar mais de 300µl de sangue, com um rendimento de 4,5 a 7,5µg de
DNA, proveniente de 300µl de sangue total (o qual contém aproximadamente 3x106 de células
nucleadas). As etapas do procedimento de extração do DNA são descritas a seguir:
1- Lise das células
- Adicionou-se 3X do volume da amostra de tampão RBC Lysis Solution e misturou-se por
inversão;
- Incubou-se a mistura de lise por 5 minutos em temperatura ambiente e em seguida centrifugou
(entre 1200 e 16000 rpm) em microcentrífuga por 20 segundos;
- Removeu-se o sobrenadante e o resíduo (aproximadamente 20-50µl) utilizou-se para
ressuspender as células antes da extração.
2- Extração
- Ressuspendeu-se as células brancas do sangue, no mesmo tubo de lise e adicionou 500µl de
tampão de extração (Extraction Solution), pré aquecido a 70oC, e misturou com o auxílio de um
vortex;
- Incubou-se a mistura de extração por 5 minutos em temperatura ambiente.
3- Recuperação do DNA
- Transferiu-se toda a mistura da extração para uma coluna (GFX Column) e em seguida
centrifugou-se a 8000rpm por 1 minuto;
- Descartou-se o líquido do tubo coletor e repetiu a etapa de extração.
4- Lavagem
- Descartou-se o líquido do tubo coletor e adicionou-se 500µl de solução de lavagem (Wash
Solution) e em seguida, centrifugou-se a 12000-16000rpm por 3 minutos;

42
- Descartou-se o tubo coletor e transferiu a coluna para outro tubo.
5- Eluição
- Adicionou-se 100 µl de tampão de eluição (TE) pré-aquecido a 70oC e incubou a amostra a
temperatura ambiente por 1 minuto;
- Centrifugou-se a 8000rpm por 1 minuto para recuperar o DNA purificado.
A quantificação do DNA foi realizada com o auxílio do marcador de peso molecular Low
DNA Mass, e diluído para a concentração ideal de uso. O DNA foi utilizado para a realização de
diversas reações de PCR utilizando os marcadores RAPD e STR.
Condições de Amplificação dos Marcadores RAPD
Primeiramente, foi realizada uma seleção dos melhores marcadores utilizando o DNA de
três indivíduos de cada uma das duas espécies. Os testes de amplificação foram realizados com
119 primers das séries OPA, OPB, OPC, OPH, OPM e OPP desenvolvidos pela OPERON.
Destes, um total de nove marcadores apresentaram um melhor padrão de amplificação e foram
selecionados para as reações de PCR com os 38 indivíduos de cada espécie.
As reações de PCR foram montadas da seguinte forma: 3 µl de DNA (~3 ng/µl); 2,0 µl de
primer (~10 ng/µl); 2,6 µl tampão da enzima (10 X); 0,78 µl de MgCl2 (50 mM); 2,08 µl de
d’NTP (2,5 mM); 0,2 µl da enzima Taq-Polimerase (5 unidades/µl) e completando o volume com
9,34 µl de H2O Milli-Q. Foram utilizadas as seguintes condições nas reações de amplificação: (1º)
2’ a 94°C; (2º) 1’30’’ a 92°C, (3º) 1’ a 37°C, (4º) 2’30’’ a 72°C, (5º) 40 ciclos (da 2º a 4º) e (6º)
5’ a 72°C para a extensão final. Os fragmentos de DNA amplificados foram separados através de
eletroforese em gel de agarose 1,5% e em seguida corados com brometo de etídeo. Foi utilizado o
marcador de peso molecular ladder 100 bp (Gibco), para estimar o tamanho dos fragmentos. Os
fragmentos, nos géis, foram visualizados através de um transluminador de luz ultravioleta e

43
fotografados com auxílio do fotodocumentador KODAK EDAS – 120, (Ferreira e Grattapaglia,
1998).
Condições de Amplificação dos Marcadores STR
Após a realização dos testes de transferibilidade dos marcadores STR (ver Capítulo 1), os
locos selecionados foram utilizados nas reações de PCR. Para as reações de PCR o seguinte
protocolo foi utilizado: 5 µl de DNA (~3 ng/µl); 2,2 µl do primers A (forward) (~5 ng/µl); 2,2 µl
do primers B (reverse) (~5 ng/µl); 1,3 µl de BSA (10 mg/ml); 1,5 µl tampão da enzima (10 X);
0,5 µl de MgCl2 (50 mM); 1,3 µl de d’NTP (2,5 mM); 0,2 µl da enzima Taq- Polimerase (5
unidades/µl) e completando o volume com 0,8 µl de H2O Milli-Q, para um volume final de 15 µl.
Para cada reação, realizada no termociclador MJ Research Inc, utlizou-se o seguinte
programa: (1º) desnaturação do DNA a 94ºC por 5 minutos e (2º) a 94ºC por 1 minuto; (3º)
anelamento dos “primers” a XºC por 1 minuto; (4º) extensão da molécula pela enzima Taq
polimerase (fornecida pela Amersham Pharmacia Biotech) a 72ºC por 1 minuto; (5º) 30 ciclos
seguindo do 2º ao 4º passos; (6º) passo final de extensão de 7 minutos a 72ºC para finalizar os
produtos amplificados. Para cada primer foram testadas temperaturas de anelamento que
variaram entre 45ºC e 65ºC, pois este é um dos fatores críticos nas análises com estes marcadores
(Hillis et. al, 1996; Ferreira e Grattapaglia, 1998) e após os resultados dos testes foi utilizado a
temperatura de anelamento específica de cada primer (ver cap. 1).
Em uma primeira etapa dos testes de amplificação realizamos uma eletroforese horizontal,
utilizando o gel de agarose 4% com tampão de corrida TBE 1X, com o intuito de verificar se
houve ou não amplificação de algum fragmento. Os fragmentos foram corados em uma solução
contendo brometo de etídeo visualizados no transiluminador de luz ultravioleta e fotografados
com o fotodocumentador KODAK EDAS 120.

44
As amostras que amplificaram foram, em seguida, submetidas à eletroforese em gel de
poliacrilamida a 4% em TBE 1X durante 2 horas, mantendo a corrente elétrica em 90W constante.
Nesse gel são aplicados 3µl do produto da PCR previamente desnaturado (94°C por 5 min),
contendo o tampão de carregamento. O padrão de peso molecular, o marcador Ladder 10bp
(Amersham Pharmacia Biotech) foi usado para estimar os tamanhos dos produtos de PCR. Após
a eletroforese, o gel de acrilamida foi corado com nitrato de prata (Bassan et al., 1991). Após a
revelação das bandas e a secagem das placas, estas foram colocadas sobre a luz branca para a
obtenção dos genótipos.
Análises dos Dados
Dados RAPD e STR
A partir dos produtos amplificados via RAPD, foi construída uma matriz de dados
binários obtida pela codificação dos géis em genótipos, no qual o valor 1,0 indica presença de
banda no indivíduo e zero a sua ausência. Em seguida, foi realizada uma análise descritiva destes
dados, anotando-se o número total de locos e número de locos polimórficos.
As matrizes binárias contendo dados dos nove primers selecionados foram combinadas a
fim de se obter uma única matriz, na qual foi estimada a similaridade entre os indivíduos para
cada uma das duas populações separadamente e conjuntamente. A similaridade genética entre os
pares de indivíduos foi estimada com base no coeficiente de Jaccard (J) (Sneath e Sokal, 1973)
que é dado por
PMJ =
onde M é o número de bandas em comum e P é o número total de bandas, excluindo-se as bandas
ausentes nos dois indivíduos comparados. Essas análises foram realizadas utilizando a rotina
SIMQUAL (similarity for qualitative data) do programa NTSYS-PC, versão 1.8 (Rohlf, 1991).

45
A partir da interpretação dos genótipos gerados pelos marcadores STR, foram obtidas as
freqüências alélicas (p) e genotípicas em cada loco para cada espécie. Estas freqüências foram
submetidas a estatísticas descritivas, tais como número de alelos observados por loco (na),
número efetivo de alelos (ne), calculados de acordo com Crow e Kimura (1970), e a proporção de
locos polimórficos (lp) para cada espécie analisada.
Para a verificação da hipótese nula de aderência ao Equilíbrio de Hardy-Weinberg (EHW)
foi feito o Teste Exato de Fisher por meio do método de cadeias de Markov, utilizando 10
batches com 1000 permutações por batch, utilizando o programa TFPGA 1.3 (Miller, 1997).
Para estimar a diversidade genética foi calculada a heterozigosidade média observada (Ho),
obtida pela média aritmética das proporções do número de heterozigotos em relação ao número
total de indivíduos entre os locos analisados. A Heterozigosidade esperada por locos (He), foi
calculada por
∑−= 21 ipHe
onde p é a freqüência populacional do i-ésimo alelo de cada loco.
Com base nas estimativas das heterozigosidades esperadas e observadas (He e Ho),
calculou-se o índice de fixação ou coeficiente de endogamia (F) para cada população, como
medida da deficiência ou excesso de heterozigotos, dada pela fórmula ( )He
HoHeF −= . Todas
essas análises foram realizadas no programa GENEPOP 3.3 (Raymond e Rousset, 1995).
Correlação Matricial
A fim de se verificar a associação entre a similaridade genética obtida a partir dos dois
marcadores utilizados para cada espécie, os genótipos gerados pelos marcadores STR foram
transformados em uma matriz binária, onde a presença do alelo no genótipo foi indicada por 1 e a

46
ausência por zero (Kosman e Leonard, 2005). A transformação foi feita de acordo com o exemplo
abaixo:
Genótipos Indivíduos
Primer 1 Primer 2
1 A B A A
2 B B A D
3 A C B C
Alelos / Primer 1 Alelos / Primer 2 Indivíduos
A B C D A B C D
1 1 1 0 0 1 0 0 0
2 0 1 0 0 1 0 0 1
3 1 0 1 0 0 1 1 0
Para avaliar a correlação entre as matrizes de similaridade obtidas a partir dos dados
RAPD e STR das espécies foi utilizado o teste de Mantel (Manly, 1991), que é dado por:
( )∑ ∑ ⋅=i ijijj
MEZ
onde Eij e Mij são o i-ésimo e j-ésimo elementos das matrizes E e M a serem comparadas. A
significância da relação entre as matrizes pode ser testada recalculando o valor de Z diversas
vezes, permutando aleatoriamente, em cada uma delas, a ordem dos elementos de uma das
matrizes. Deste modo, é possível obter uma distribuição de probabilidades à qual o valor de Z
observado pode ser comparado (Manly, 1991). O valor de Z não possui amplitude de variação
definida, mas de fato ele possui uma relação monotônica com o r de Pearson calculado entre as
matrizes. Assim, pode-se utilizar o Z de Mantel para testar a significância da correlação matricial,
Genótipos gerados pelos
locos STR.
Genótipos transformados em
dados binários para cada loco
analisado.

47
que varia entre 1,0 e –1,0. Foram utilizadas 5000 permutações aleatórias para estabelecer a
significância de cada correlação matricial. Essas análises foram realizadas utilizando a rotina
MXCOMP (Matrix Comparison-Graphics) do programa, NTSYS-PC 1.8 (Rohlf, 1991).
RESULTADOS
Dados RAPD
Dentre os 119 primers testados foram selecionados nove para as análises posteriores, por
apresentarem um melhor padrão de amplificação. Os critérios adotados para seleção dos
marcadores foram: 1º) número razoável de locos; 2º) o máximo possível de bandas fortes; 3º)
presença de polimorfismo. Estes critérios facilitam o processo de codificação e maximizam a
chance de obtenção de dados com maior qualidade, repetibilidade e menor custo. Portanto, com
base no padrão de amplificação e da resolução das bandas no gel, foram selecionados os
seguintes primers: OPH-04, OPH-19, OPP-07, OPP-08, OPP-09, OPC-05, OPC-11, OPA-18,
OPP-15 (Figuras 2 e 3). O primer OPH-05 não foi selecionado devido a pouca quantidade.
Figura 2- Perfil eletroforético dos fragmentos RAPD amplificados para ilustrar alguns dos primers selecionados (OPH-04, OPH-05, OPH-19, OPP-07, OPP-08, OPP-09), exceto OPH-05, utilizando três amostras de lobo-guará (Chrysocyon brachyurus). As colunas “M 100bp” indicam o marcador de peso molecular (100 bp ladder, Pharmacia).
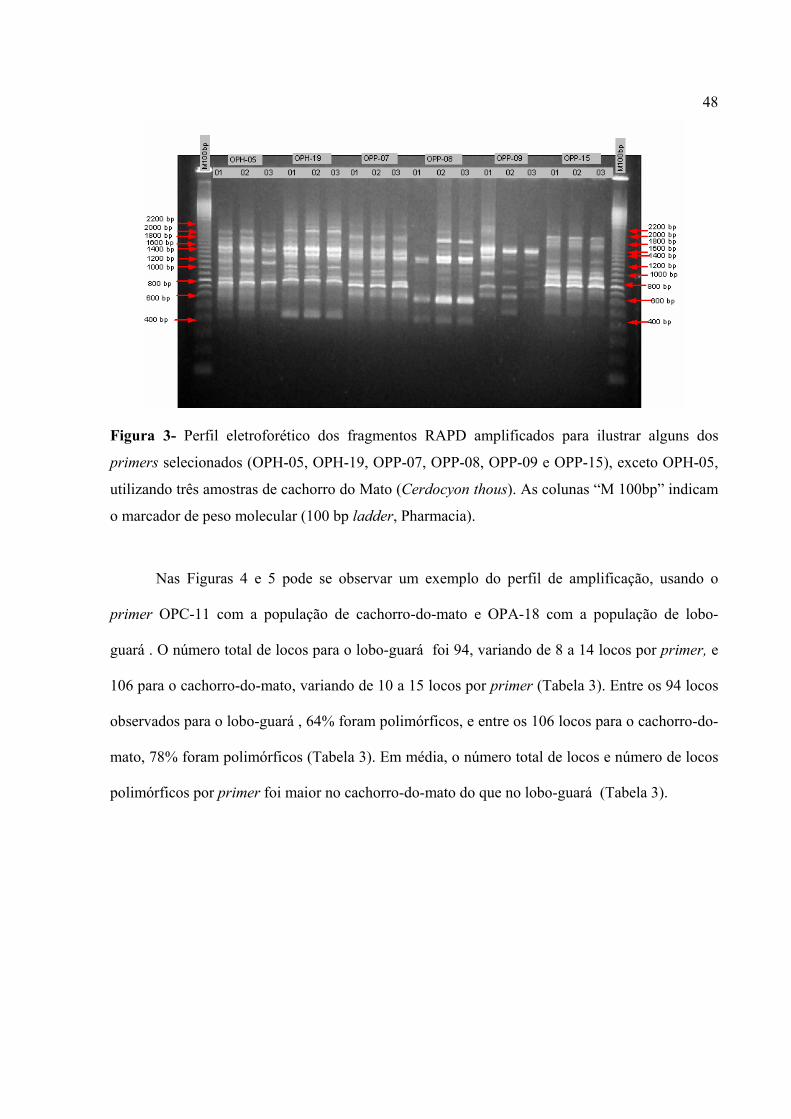
48
Figura 3- Perfil eletroforético dos fragmentos RAPD amplificados para ilustrar alguns dos
primers selecionados (OPH-05, OPH-19, OPP-07, OPP-08, OPP-09 e OPP-15), exceto OPH-05,
utilizando três amostras de cachorro do Mato (Cerdocyon thous). As colunas “M 100bp” indicam
o marcador de peso molecular (100 bp ladder, Pharmacia).
Nas Figuras 4 e 5 pode se observar um exemplo do perfil de amplificação, usando o
primer OPC-11 com a população de cachorro-do-mato e OPA-18 com a população de lobo-
guará . O número total de locos para o lobo-guará foi 94, variando de 8 a 14 locos por primer, e
106 para o cachorro-do-mato, variando de 10 a 15 locos por primer (Tabela 3). Entre os 94 locos
observados para o lobo-guará , 64% foram polimórficos, e entre os 106 locos para o cachorro-do-
mato, 78% foram polimórficos (Tabela 3). Em média, o número total de locos e número de locos
polimórficos por primer foi maior no cachorro-do-mato do que no lobo-guará (Tabela 3).

49
Figura 4- Perfil eletroforético dos fragmentos RAPD amplificados utilizando o primer OPC-11
com alguns indivíduos da população de cachorro-do-mato. As colunas M 100bp, indicam o
marcador de peso molecular (100 bp ladder, Pharmacia).
Figura 5- Perfil eletroforético dos fragmentos RAPD amplificados utilizando o primer OPA-18
com alguns indivíduos da população de lobo-guará . As colunas M 100bp, indicam o marcador de
peso molecular (100 bp ladder, Pharmacia).
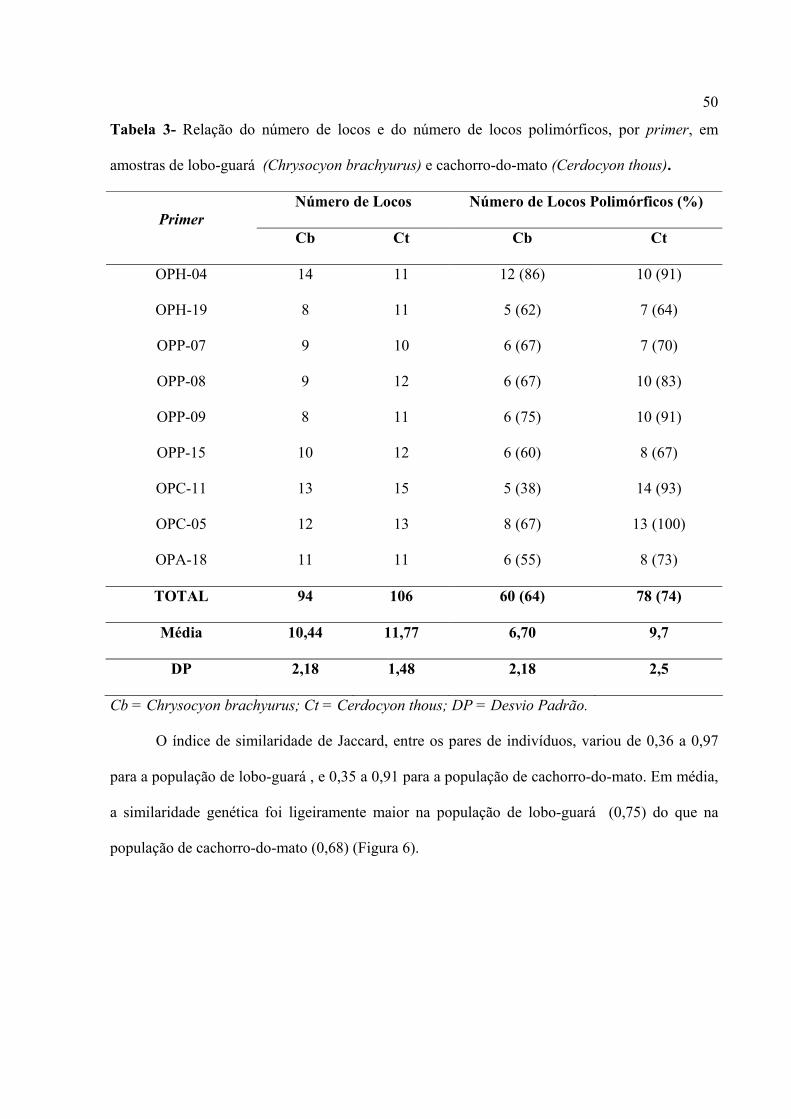
50
Tabela 3- Relação do número de locos e do número de locos polimórficos, por primer, em
amostras de lobo-guará (Chrysocyon brachyurus) e cachorro-do-mato (Cerdocyon thous).
Número de Locos Número de Locos Polimórficos (%) Primer
Cb Ct Cb Ct
OPH-04 14 11 12 (86) 10 (91)
OPH-19 8 11 5 (62) 7 (64)
OPP-07 9 10 6 (67) 7 (70)
OPP-08 9 12 6 (67) 10 (83)
OPP-09 8 11 6 (75) 10 (91)
OPP-15 10 12 6 (60) 8 (67)
OPC-11 13 15 5 (38) 14 (93)
OPC-05 12 13 8 (67) 13 (100)
OPA-18 11 11 6 (55) 8 (73)
TOTAL 94 106 60 (64) 78 (74)
Média 10,44 11,77 6,70 9,7
DP 2,18 1,48 2,18 2,5
Cb = Chrysocyon brachyurus; Ct = Cerdocyon thous; DP = Desvio Padrão.
O índice de similaridade de Jaccard, entre os pares de indivíduos, variou de 0,36 a 0,97
para a população de lobo-guará , e 0,35 a 0,91 para a população de cachorro-do-mato. Em média,
a similaridade genética foi ligeiramente maior na população de lobo-guará (0,75) do que na
população de cachorro-do-mato (0,68) (Figura 6).
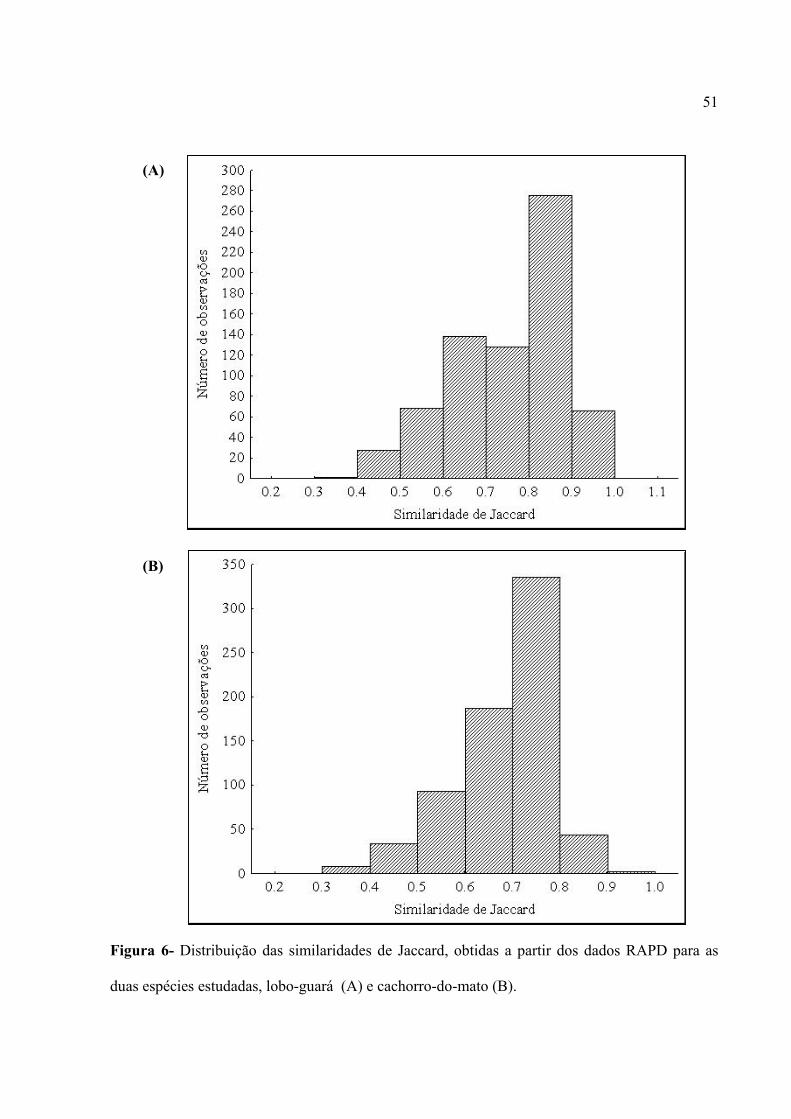
51
Figura 6- Distribuição das similaridades de Jaccard, obtidas a partir dos dados RAPD para as
duas espécies estudadas, lobo-guará (A) e cachorro-do-mato (B).
(A)
(B)

52
Dados STR
Um total de 29 marcadores STR desenvolvidos originalmente para búfalos, bovinos,
cachorro doméstico e gato doméstico (Barker et al., 1997, Cânon et al., 2001, Denise et al., 2004,
Fries et al., 1993, Halverson et al., 1995, Kapes et al., 1997, Menotti-Raymond et al., 1999,
Sunden et al., 1993) foram testados e destes, seis primers (21%), amplificaram nas duas espécies
estudadas (ver Capítulo1).
As freqüências alélicas estimadas nas duas espécies são apresentadas nas Figuras 7 e 8. O
número de alelos encontrados por loco foi semelhante nas duas espécies, com exceção do loco
PEZ-12, para o qual a população de cachorro-do-mato apresentou 12 alelos (Figura 7) enquanto
que, na população de lobo-guará apenas dois alelos foram observados (Figura 8).
A média do número de alelos por loco foi de 7,8 na população de cachorro-do-mato e 4,5
na população de lobo-guará , sendo o máximo de alelos observados igual a 14 (PEZ-13 no
cachorro-do-mato) e o mínimo igual a um (CSSM-038 nas duas populações). Os Locos PEZ-05 e
PEZ-12 representam bem a variabilidade alélica existente entre as duas espécies (Figuras 9 e 10).
Enquanto que o primeiro loco apresentou uma pequena variação (seis alelos no cachorro-do-mato
e cinco alelos no lobo-guará ), no segundo loco, a variação foi maior, tendo sido detectados 12
alelos no cachorro-do-mato e apenas dois alelos na população de lobo-guará .

53
Figura 7- Freqüências alélicas dos seis locos STR, PEZ-05, PEZ-20, PEZ-13 (L-13), PEZ-15 (L-
15), CSSM-038 e PEZ-12, estimadas a partir de 38 indivíduos da população de cachorro-do-mato.
PEZ-05
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
A B C D E F
alelos
freq
. alé
lica
PEZ-20
0
0,050,1
0,15
0,20,25
0,3
0,350,4
0,45
A B C D E F
alelos
freq
. alé
lica
L-13
0
0,05
0,1
0,15
0,2
0,25
0,3
A B C D E F G H I J K L M N
alelos
freq
. alé
lica
L-15
00,050,1
0,150,2
0,250,3
0,350,4
0,45
A B C D E F G
alelos
freq
. alé
lica
CSSM038
0
0,2
0,4
0,6
0,8
1
1,2
A
alelos
freq
. alé
lica
PEZ-12
0
0,05
0,1
0,15
0,2
0,25
0,3
a b c d e f g h i j k l m
alelos
freq
. alé
lica
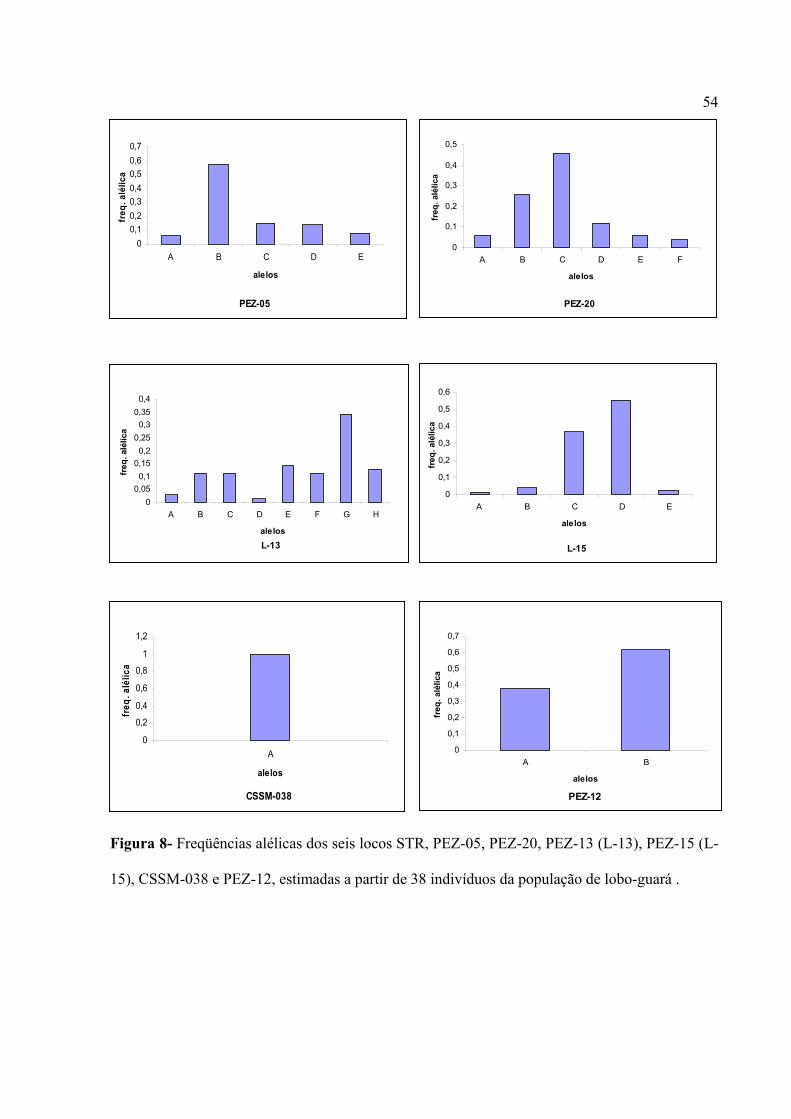
54
Figura 8- Freqüências alélicas dos seis locos STR, PEZ-05, PEZ-20, PEZ-13 (L-13), PEZ-15 (L-
15), CSSM-038 e PEZ-12, estimadas a partir de 38 indivíduos da população de lobo-guará .
PEZ-05
00,10,20,30,40,50,60,7
A B C D E
alelos
freq
. alé
lica
PEZ-20
0
0,1
0,2
0,3
0,4
0,5
A B C D E F
alelos
freq
. alé
lica
L-13
00,050,1
0,150,2
0,250,3
0,350,4
A B C D E F G H
alelos
freq
. alé
lica
L-15
0
0,1
0,2
0,3
0,4
0,5
0,6
A B C D E
alelos
freq
. alé
lica
CSSM-038
0
0,2
0,4
0,6
0,8
1
1,2
A
alelos
freq
. alé
lica
PEZ-12
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
A B
alelos
freq
. alé
lica
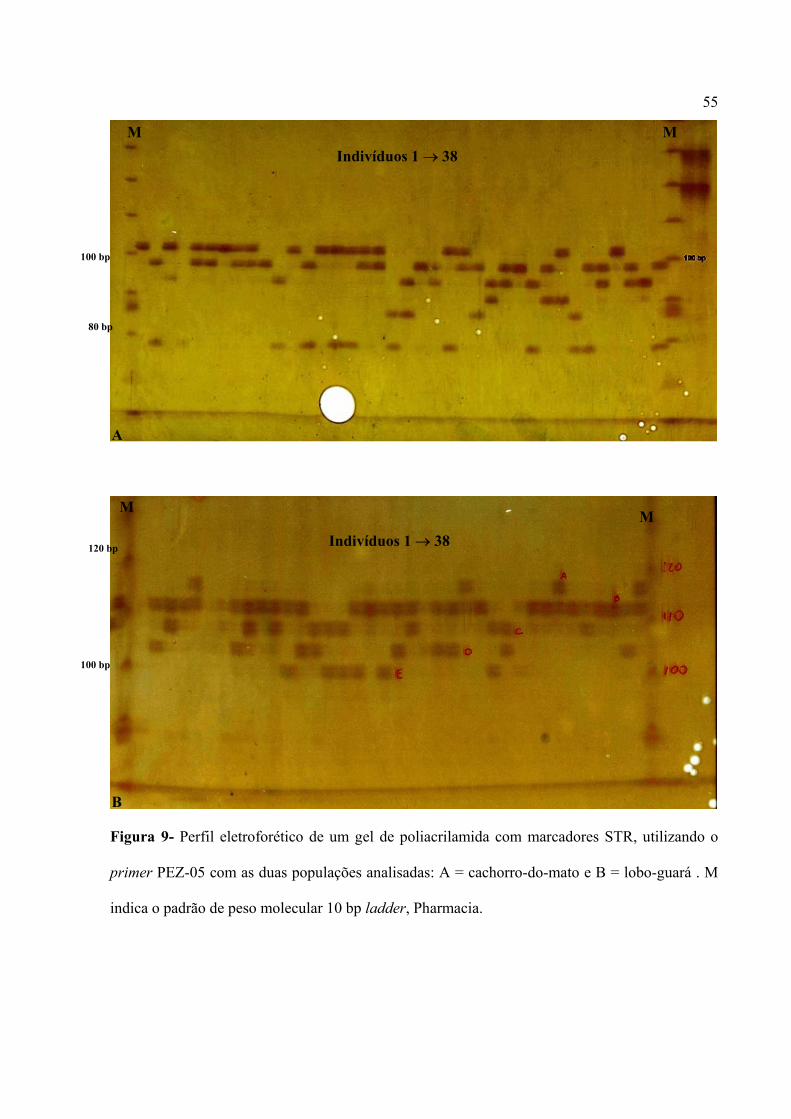
55
Figura 9- Perfil eletroforético de um gel de poliacrilamida com marcadores STR, utilizando o
primer PEZ-05 com as duas populações analisadas: A = cachorro-do-mato e B = lobo-guará . M
indica o padrão de peso molecular 10 bp ladder, Pharmacia.
100 bp
120 bp
M M
M M Indivíduos 1 → 38
Indivíduos 1 → 38
A
B
80 bp
100 bp
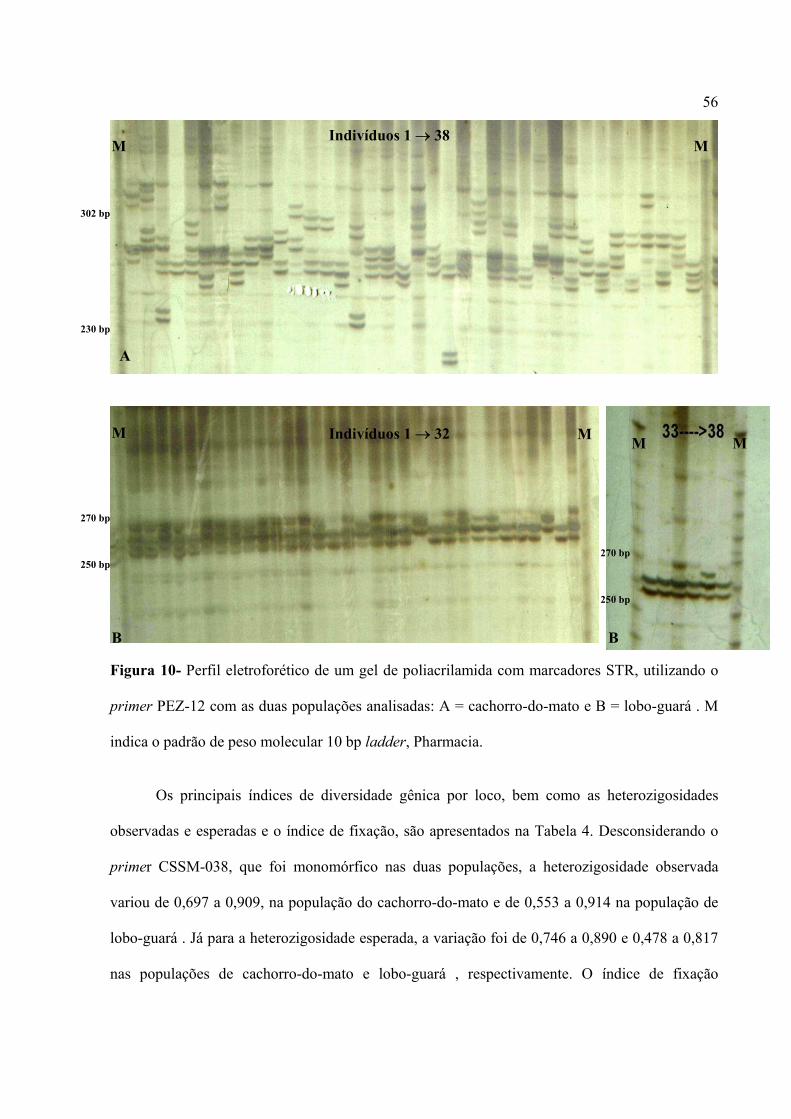
56
Figura 10- Perfil eletroforético de um gel de poliacrilamida com marcadores STR, utilizando o
primer PEZ-12 com as duas populações analisadas: A = cachorro-do-mato e B = lobo-guará . M
indica o padrão de peso molecular 10 bp ladder, Pharmacia.
Os principais índices de diversidade gênica por loco, bem como as heterozigosidades
observadas e esperadas e o índice de fixação, são apresentados na Tabela 4. Desconsiderando o
primer CSSM-038, que foi monomórfico nas duas populações, a heterozigosidade observada
variou de 0,697 a 0,909, na população do cachorro-do-mato e de 0,553 a 0,914 na população de
lobo-guará . Já para a heterozigosidade esperada, a variação foi de 0,746 a 0,890 e 0,478 a 0,817
nas populações de cachorro-do-mato e lobo-guará , respectivamente. O índice de fixação
M M
M M Indivíduos 1 → 38
Indivíduos 1 → 32
A
B
M M
B
302 bp
230 bp
270 bp
250 bp 270 bp
250 bp
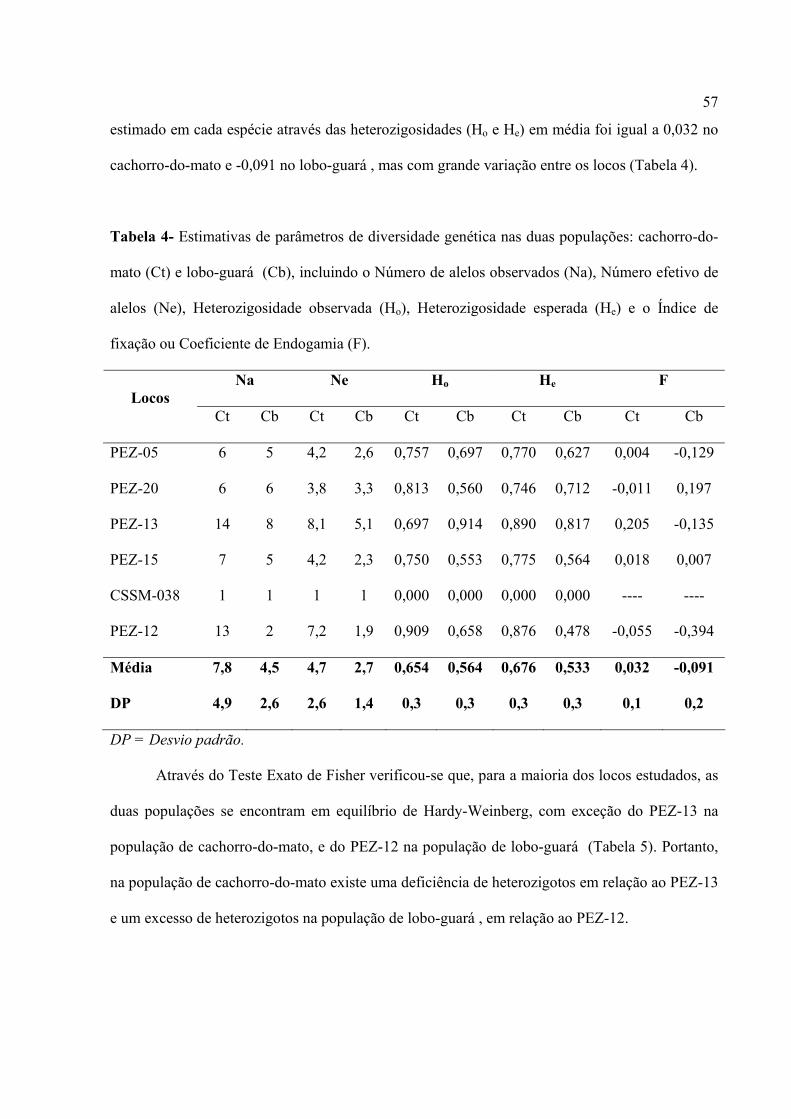
57
estimado em cada espécie através das heterozigosidades (Ho e He) em média foi igual a 0,032 no
cachorro-do-mato e -0,091 no lobo-guará , mas com grande variação entre os locos (Tabela 4).
Tabela 4- Estimativas de parâmetros de diversidade genética nas duas populações: cachorro-do-
mato (Ct) e lobo-guará (Cb), incluindo o Número de alelos observados (Na), Número efetivo de
alelos (Ne), Heterozigosidade observada (Ho), Heterozigosidade esperada (He) e o Índice de
fixação ou Coeficiente de Endogamia (F).
Na Ne Ho He F Locos
Ct Cb Ct Cb Ct Cb Ct Cb Ct Cb
PEZ-05 6 5 4,2 2,6 0,757 0,697 0,770 0,627 0,004 -0,129
PEZ-20 6 6 3,8 3,3 0,813 0,560 0,746 0,712 -0,011 0,197
PEZ-13 14 8 8,1 5,1 0,697 0,914 0,890 0,817 0,205 -0,135
PEZ-15 7 5 4,2 2,3 0,750 0,553 0,775 0,564 0,018 0,007
CSSM-038 1 1 1 1 0,000 0,000 0,000 0,000 ---- ----
PEZ-12 13 2 7,2 1,9 0,909 0,658 0,876 0,478 -0,055 -0,394
Média
DP
7,8
4,9
4,5
2,6
4,7
2,6
2,7
1,4
0,654
0,3
0,564
0,3
0,676
0,3
0,533
0,3
0,032
0,1
-0,091
0,2
DP = Desvio padrão.
Através do Teste Exato de Fisher verificou-se que, para a maioria dos locos estudados, as
duas populações se encontram em equilíbrio de Hardy-Weinberg, com exceção do PEZ-13 na
população de cachorro-do-mato, e do PEZ-12 na população de lobo-guará (Tabela 5). Portanto,
na população de cachorro-do-mato existe uma deficiência de heterozigotos em relação ao PEZ-13
e um excesso de heterozigotos na população de lobo-guará , em relação ao PEZ-12.

58
Tabela 5- Probabilidades do Teste Exato de Fisher para aderência ao equilíbrio de Hardy-
Weinberg.
Locos Cachorro-do-Mato Lobo-Guará
PEZ-05 0,4913 0,4422
PEZ-20 0,5534 0,2679
PEZ-13 0,0000 0,8984
PEZ-15 0,6062 0,9376
CSSM-038 ---- ----
PEZ-12 0,6056 0,0343
Correlação entre as similaridades genéticas obtidas pelo RAPD e STR
O teste de Mantel mostrou que as correlações entre as similaridades genéticas obtidas a
partir dos dados RAPD e dos dados STR foram significativas, porém com valores de r de Pearson
muito baixos, evidenciando que os dois marcadores avaliam as relações existentes entre os
indivíduos de forma diferente (Tabela 6).
Tabela 6- Valores da correlação matricial (r) e o nível de significância (P) obtido a partir do
Teste de Mantel, associando a similaridade genética obtida a partir dos dados RAPD e dados STR
nas duas populações.
População r P
lobo-guará 0,2422 0,0056
cachorro-do-mato 0,2465 0,0126

59
DISCUSSÃO
Variabilidade Genética nas duas espécies estudadas
A interferência do Homem tem levado a um aumento significativo do isolamento de
populações naturais e, conseqüentemente, à perda de diversidade genética (Ceballos e Ehrlich,
2002; Frankhan, 1995). Portanto, a estimativa da diversidade genética em populações naturais,
principalmente naquelas que são consideradas vulneráveis à extinção, como o lobo-guará (IUCN,
2004), é essencial no que diz respeito à adoção de estratégias adequadas de manejo e conservação
de espécies. Vários estudos utilizando marcadores moleculares, como o RAPD e STR, têm
mostrado que estes são ferramentas bastante eficientes para monitorar a variabilidade genética
das populações (Klukowska et al., 2003; Parker et al., 1998; Wandeler et al., 2003).
Neste estudo pode-se observar que a amplitude de variação na similaridade genética
obtida com os dados RAPD foi muito próxima nas duas espécies estudadas, com valores de
similaridade de Jaccard variando de 0,36 a 0,97 no lobo-guará e entre 0,35 a 0,91 no cachorro-
do-mato. Porém, mesmo que a diferença tenha sido pequena, um maior número de indivíduos
com alto índice de similaridade genética (entre 0,8 e 0,9) foi observado na população de lobo-
guará (Figura 6). Este resultado é esperado, uma vez que densidade populacional do lobo-guará
é presumivelmente menor do que a do cachorro-do-mato, o que limita o fluxo gênico entre os
indivíduos, favorecendo a maior semelhança entre os mesmos. Por outro lado, na população de
cachorro-do-mato, que é uma espécie muito comum e abundante em todo o bioma cerrado e no
Parque Nacional das Emas, a proporção de indivíduos com alta similaridade foi menor e foram
observados índices menores do que na população de lobo-guará (Figura 6).
A diversidade genética obtida com os dados de STR, apresentou um nível relativamente
alto de multialelismo nos seis locos analisados, confirmando o elevado conteúdo de informação
genética esperado para este tipo de marcador. Para a população de lobo-guará a média foi igual
4,5 alelos por loco e, se desconsiderarmos o loco CSSM-038, (que foi monomórfico), essa média

60
aumenta para 5,2 alelos por loco. Esse valor foi maior do que o encontrado por De Mattos et al.,
(2004) em um estudo similar com o lobo-guará , onde a média de alelos por loco foi igual a 1,2,
porém utilizando marcadores isoenzimáticos. A média de heterozigosidade observada também foi
maior neste estudo, 56,4% do que a encontrada no estudo de De Mattos et al., (2004) que foi
igual a 5,7% e bem maior do que a encontrada no estudo de Rodrigues (2002), que foi igual a
1,4%, ambos utilizarando marcadores isoenzimáticos. Percebe-se que a escolha do marcador é
bastante crítica para avaliar o nível de variabilidade genética pois, os marcadores isoenzimáticos
evoluem mais lentamente. Portanto, em estudos em nível de população e/ou indivíduos, é
esperada pouca variabilidade e uma saturação de plesimorfias nos dados obtidos com isoenzimas,
ou seja, as semelhanças observadas são devidas a ancestralidade dos alelos e não há eventos
novos o bastante para discriminar os indivíduos estudados (Sole-Cava, 2001).
Quando se compara as estimativas de diversidade genética obtidas neste estudo com
vários estudos com populações de canídeos levantados da literatura, usando diferentes tipos de
marcadores moleculares (Microssatélites, M; Isoenzimas, I e Polimorfismos de Simples
Nucleotídeos, SNP), percebe-se uma grande semelhança entre estes resultados e os obtidos neste
trabalho (Tabela 7). Tanto o número médio de alelos por locos, como as heterozigosidades
esperadas e observadas e o coeficiente de endogamia (F) foram próximos, mesmo levando em
consideração os dados dos marcadores isoenzimáticos que tendem a diminuir a estimativas. O
número médio de alelos por locos obtidos neste estudo foi de 7,8 para o cachorro-do-mato e 4,5
para a população de lobo-guará , valores muito semelhantes aos apresentados na Tabela 7.
Quando considera-se os dados apenas dos marcadores STR, a média de alelos por locos será igual
a 4,3, valor bem próximo ao encontrado no lobo-guará . O mesmo ocorreu para os resultados das
médias das Heterozigosidades esperadas e observadas (He e Ho, respectivamente) e do Índice de
Fixação médio (F). Portanto, existe uma grande aproximação dos resultados obtidos,

61
principalmente para a população de lobo-guará , com os resultados de vários outros estudos com
canídeos na literatura (Tabela 7).
Em canídeos, a diversidade genética parece estar relacionada com o tamanho do corpo das
espécies, ou seja, em espécies menores, (exceto àquelas que estão ameaçadas de extinção), a
diversidade genética tende a ser maior do que em espécies maiores (Wayne, 1996). Esse padrão
pode ser explicado em função da área de ocupação dos indivíduos, onde espécies maiores terão
conseqüentemente uma área de vida mais restrita e portanto, uma menor diversidade genética em
decorrência do menor fluxo gênico. Considerando os resultados apresentados na Tabela 7 e
também os resultados obtidos neste estudo, percebe-se este padrão. Por exemplo, a espécie
Vulpes vulpes na qual em média o tamanho do corpo é de 40 cm, a diversidade genética média
(He = 0,7) foi maior que a observada no Canis lupus (120 cm) que apresentou He = 0,6 e bem
maior que a diversidade genética (He = 0,3) obtida para a espécie Cuon alpinus (90 cm), todos
resultados obtidos com marcadores STR (Sillero-Zubiri et al., 2004) (Tabela 7). Neste trabalho, o
cachorro-do-mato é uma espécie menor (em torno de 66 cm) que o lobo-guará (em torno de
105cm) (Sillero-Zubiri et al., 2004) e apresentou uma maior diversidade genética (He = 0,7).
O valor médio obtido de F pode ser considerado nulo ou muito baixo nas duas espécies
estudadas, indicando que as freqüências alélicas dessas espécies correspondem ao esperado pelo
modelo de equilíbrio de Hardy-Weinberg, neste momento. No entanto, o PEZ-13 e o loco PEZ-12,
no cachorro-do-mato e no lobo-guará , respectivamente, sugerem uma certa endogamia na
primeira e um excesso de heterozigotos na segunda população, significativos no Teste Exato de
Fisher. Como dito anteriormente, o cachorro-do-mato é uma espécie que costuma viver em
bandos, o que provavelmente favorece o cruzamento entre indivíduos aparentados e pode explicar
a endogamia detectada pelo PEZ-13. Já na população de lobo-guará , mesmo sendo uma espécie
vulnerável a extinção, ou seja, com um tamanho populacional mais restrito que a do cachorro-do-
mato, o excesso de heterozigotos detectado pelo primer PEZ-12 pode ser justificados pelo hábito

62
solitário da espécie, ou seja, os encontros ocorrem apenas no período reprodutivo e também
devido às áreas de vida serem grandes, favorecendo encontros aleatórios e não necessariamente
entre indivíduos aparentados geneticamente.
Como pode ser observada na Tabela 7, a maioria dos valores de F das populações de
canídeos são próximos de zero, o que corrobora os resultados obtidos neste estudo. Uma exceção
foi o Canis lupus, cuja população de museu apresentou endogamia elevada (F = 0,183). No
entanto, esse resultado pode ser devido um artefato do próprio processo de amostragem, uma vez
que nas outras populações os valores de F foram próximos de zero (Vilà et al., 2002). E para a
espécie Vulpes vulpes pode se observar um excesso de heterozigotos (F = -0,24) na população
urbana do oeste e endogamia (F = 0,10) na população urbana do leste (Wandeler et al., 2003).
Neste caso, a redução no tamanho das populações devido, por exemplo à fragmentação de
habitats, poderia explicar esses resultados.
Tabela 7- Estimativas de diversidade genética de várias populações de canídeos, usando
diferentes tipos de Marcadores Moleculares, incluindo o número médio de alelos por locos (Na),
Heterozigosidade esperada (He) Heterozigosidade observada (Ho), Índice de Fixação (F), tipo de
marcador e fonte.
População Na He Ho F Marcador Fonte Canis lupus Alaska a Quebec México
4,5 2,7
0,62 0,42
---- ----
---- ----
M Wayne, 1996
Canis lupus Escandinávia Finlândia
---- ----
0,36 0,34
0,35 0,31
0,027 0,088
SNP Seddon et al., 2005
Canis lupus Escandinávia Finlândia e Rússia Amostras de museu
3,8 8,0 5,7
0,55 0,77 0,71
0,59 0,70 0,58
-0,072 0,090 0,183
M Vilà et al., 2002
Canis latrans Alaska
5,9
0,68
----
---- M Wayne, 1996
Canis rufus Cativeiro
5,3
0,55
----
---- M Wayne, 1996
Canis aureus Kenya
4,8
0,52
----
---- M Wayne, 1996
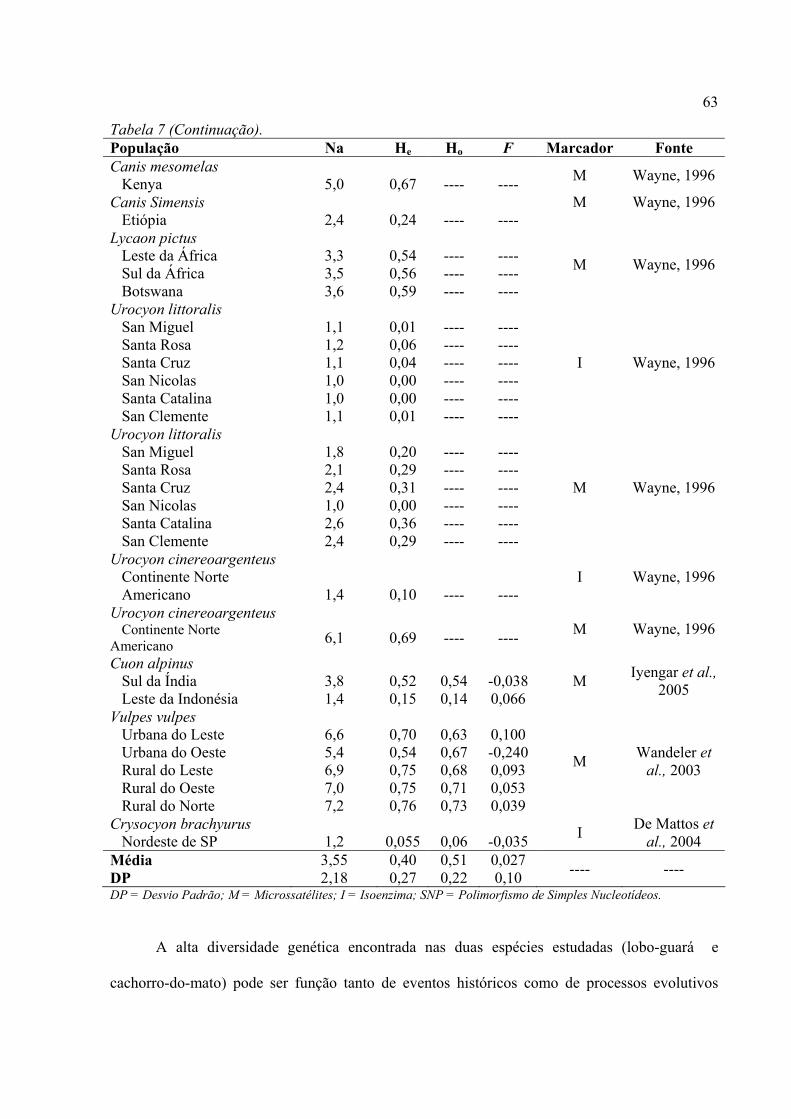
63
Tabela 7 (Continuação). População Na He Ho F Marcador Fonte Canis mesomelas Kenya
5,0
0,67
----
---- M Wayne, 1996
Canis Simensis Etiópia
2,4
0,24
----
----
M Wayne, 1996
Lycaon pictus Leste da África Sul da África Botswana
3,3 3,5 3,6
0,54 0,56 0,59
---- ---- ----
---- ---- ----
M Wayne, 1996
Urocyon littoralis San Miguel Santa Rosa Santa Cruz San Nicolas Santa Catalina San Clemente
1,1 1,2 1,1 1,0 1,0 1,1
0,01 0,06 0,04 0,00 0,00 0,01
---- ---- ---- ---- ---- ----
---- ---- ---- ---- ---- ----
I Wayne, 1996
Urocyon littoralis San Miguel Santa Rosa Santa Cruz San Nicolas Santa Catalina San Clemente
1,8 2,1 2,4 1,0 2,6 2,4
0,20 0,29 0,31 0,00 0,36 0,29
---- ---- ---- ---- ---- ----
---- ---- ---- ---- ---- ----
M Wayne, 1996
Urocyon cinereoargenteus Continente Norte Americano
1,4
0,10
----
---- I Wayne, 1996
Urocyon cinereoargenteus Continente Norte Americano
6,1
0,69
----
---- M Wayne, 1996
Cuon alpinus Sul da Índia Leste da Indonésia
3,8 1,4
0,52 0,15
0,54 0,14
-0,038 0,066
M Iyengar et al., 2005
Vulpes vulpes Urbana do Leste Urbana do Oeste Rural do Leste Rural do Oeste Rural do Norte
6,6 5,4 6,9 7,0 7,2
0,70 0,54 0,75 0,75 0,76
0,63 0,67 0,68 0,71 0,73
0,100 -0,240 0,093 0,053 0,039
M Wandeler et al., 2003
Crysocyon brachyurus Nordeste de SP
1,2
0,055
0,06
-0,035 I De Mattos et
al., 2004 Média DP
3,55 2,18
0,40 0,27
0,51 0,22
0,027 0,10 ---- ----
DP = Desvio Padrão; M = Microssatélites; I = Isoenzima; SNP = Polimorfismo de Simples Nucleotídeos.
A alta diversidade genética encontrada nas duas espécies estudadas (lobo-guará e
cachorro-do-mato) pode ser função tanto de eventos históricos como de processos evolutivos

64
recentes. Como pouco se sabe sobre a história evolutiva das espécies aqui estudadas, as
explicações para os altos níveis de diversidade genética se assentam em inferências. Por exemplo,
uma explicação para a alta diversidade genética tanto no lobo-guará como no cachorro-do-mato
é que ambas são espécies altamente dispersivas, com áreas de vida grandes (ver capítulo 3) e
portanto, com níveis menores de estruturação populacional do que animais sedentários. Uma
outra situação que poderia ser avaliada em estudos futuros nestas duas espécies, principalmente
no lobo-guará , é a possibilidade de que o excesso de heterozigotos seja uma conseqüência do
processo de deriva genética recente, ou seja, em uma população recentemente reduzida, cuja
diversidade genética observada será maior que a esperada em equilíbrio (Luikart e Cornuet,
1998). É importante a identificação de populações que foram submetidas a uma redução drástica
do seu tamanho populacional. Assim sendo, será maior a probabilidade de que os possíveis
efeitos deletérios sejam evitados ou minimizados, através de procedimentos mitigadores de
manejo, como por exemplo a preservação adequada da área de ocupação da espécie, como a
criação de corredores ecológicos favorecendo um maior fluxo gênico entre populações vizinhas
(Luikart et al., 1998).
Outro aspecto importante a ser considerado é que esta alta diversidade genética
encontrada nas duas espécies foi baseada em marcadores considerados neutros (RAPD e STR).
Porém, em termos de variabilidade adaptativa e deletéria, que são difíceis de mensurar, a
variabilidade genética nestas espécies pode ser baixa (Hedrick, 2001). No entanto, seria
interessante em outros estudos quantificar a extensão do efeito evolutivo que se detecta com
marcadores neutros e assim avaliar se é provável que esta alta diversidade tenha conseqüências
biológicas importantes, ou se é devido simplesmente ao alto poder estatístico desses marcadores.
Hedrick (2001), descreve alguns estágios possíveis de associação da significância estatística dos
marcadores moleculares neutros e a importância biológica detectada por estes. Pode acontecer de
não haver diferenças biológicas importantes e não haver significância estatística ou haver uma

65
diferença estatisticamente significante entre os indivíduos, refletindo uma diferença biológica
importante Neste caso, as estatísticas baseadas nestes marcadores moleculares resultam em uma
apropriada avaliação da real situação biológica das espécies (Hedrick, 2001). Porém, os
problemas aparecem quando a significância estatística não reflete a insignificância biológica, ou
vice versa. Por exemplo, pode ocorrer do marcador neutro não detectar significância estatística
entre indivíduos com uma diferença biológica importante, devido à ocorrência de alto fluxo
gênico ou fortes forças seletivas. Portanto, a interpretação dos resultados de marcadores
altamente variáveis, como os estudados neste trabalho (STR’s), deve ser realizada com algumas
precauções, pois as diferenças estatisticamente significantes podem deixar de refletir diferenças
biologicamente importantes (Balloux et al., 2000; Hedrick, 2001; Hedrick, 1999).
Comparação entre os Marcadores RAPD e STR
Existem certas dificuldades em adotar as técnicas moleculares mais eficientes, do ponto
de vista evolutivo, a fim de mensurar a diversidade genética das espécies. Por exemplo, sabe-se
que os marcadores STR e RAPD evoluem rapidamente, portanto são mais úteis para estimativas
de diversidade genética em nível de indivíduos, famílias e populações, enquanto que marcadores
que evoluem mais lentamente, como as Isoenzimas, são melhores utilizados no estudo de
espécies ou táxons supra-espeíficos. No caso dos marcadores RAPD, em termos de custo,
especificidade do primer, quantidade de amostra (DNA) e tempo despendido no procedimento, é
uma técnica mais simples, o que facilita o estudo. No entanto, o caráter dominante do marcador
limita o seu uso na estimativa dos parâmetros genéticos. Há alguns trabalhos que também
discutem a baixa repetência do RAPD, devido a sensibilidade da técnica à fatores como qualidade
e concentração do DNA (Ellsworth et al., 1993).
Os marcadores STR constituem uma poderosa ferramenta para este nível de estudo, ou
seja, a detecção da diversidade genética em nível de populações. No entanto, o alto custo e tempo

66
gastos no desenvolvimento de primers específicos dificultam bastante o uso dos mesmos. Porém,
diversos estudos têm mostrado a possibilidade de amplificação cruzada entre diferentes espécies
e gêneros (Cipriani et al., 1999; Colevati et al., 1999; Roa et al., 2000; Tong et al., 2002;
Williamson et al., 2002; Zucchi et al., 2002, ver capítulo 1). Outro aspecto importante do
marcador STR é a alta taxa de mutação quando comparada com outros marcadores dentro do
mesmo genoma, que são causadas primariamente por alterações no comprimento das unidades de
repetição. Esta alta taxa de mutação constitui, por outro lado, uma limitação do marcador, pois
pode aumentar substancialmente o poder estatístico do marcador e superestimar o nível de
diversidade genética entre os indivíduos de uma população com baixas diferenças biológicas
importantes (Hedrick, 2001). Pode-se, portanto, fazer suposições enganadas de medidas de
conservação das espécies estudadas. Por exemplo, no caso de uma população apresentar alta
diversidade genética (detectada pelos marcadores RAPD ou STR’s) pode-se inferir que esta
população num dado momento não necessita de intervenções para fins de manejo e conservação
adequados. No entanto, pode existir uma alta proporção de alelos deletérios “escondidos” nos
indivíduos heterozigotos ou uma baixa variabilidade adaptativa que não são detectados por estes
marcadores. Portanto, esses resultados podem afetar a tomada de decisões a cerca do manejo e
conservação das espécies.
Neste estudo, foram utilizados os dois marcadores, RAPD e STR, na análise da
variabilidade genética em duas espécies de canídeos do cerrado, para em seguida comparar a
informação gerada por cada um destes marcadores. Estudos que realizaram uma comparação
direta entre dados gerados por RAPD e STR em espécies animais são limitados. Entretanto, pode
ser bastante interessante realizar esta comparação a fim de enriquecer as informações geradas por
cada técnica.
Neste estudo, o teste de Mantel mostrou baixas correlações significativas (em torno de
0,24) entre as similaridades genéticas obtidas com RAPD e STR, o que pode ser parcialmente

67
explicado pela base molecular diferente dos dois marcadores. Alguns eventos genéticos, tais
como recombinação e inserção, não influenciam no polimorfismo gerado pelos marcadores STR
(Mengoni et al., 2000). Ao contrário, a variação observada por meio do RAPD reside em muitos
tipos diferentes de eventos mutacionais, com diferentes implicações evolutivas que ocorrem no
local de anelamento do primer e entre os dois locais adjacentes responsáveis pela amplificação
(Mengoni et al., 2000). Ou seja, os dois marcadores detectaram alta diversidade genética nas duas
espécies estudadas, no entanto de maneiras diferentes, ou seja, baseado em informações genéticas
de regiões diferentes do genoma e que provavelmente terá implicações evolutivas diferentes. No
caso dos marcadores RAPD, as regiões amplificadas são desconhecidas, ou seja, não se sabe se
trata de regiões codificantes ou não do genoma, havendo vários estudos que discutem esta
questão. Mas, de um modo geral esses marcadores são classificados como neutros (Mengoni et
al., 2000). Já para os marcadores STR existem numerosos estudos que demonstram uma
significância funcional dos mesmos, como por exemplo regulação da recombinação e replicação
do DNA, regulação da atividade gênica e do ciclo celular e organização estrutural dos
cromossomos (Gur-Arie et al., 2000; King e Soller, 1999; Li et al., 2002).
Uma outra possível explicação para as diferenças observadas neste estudo está no
procedimento de análise dos dados gerados pelos marcadores STR como caracteres simples, não
permitindo um tratamento de acordo com o modelo mutacional stepwise, usado para estes
marcadores, onde a mutação para um novo alelo de um determinado marcador STR ocorre em
passos, assim alelos com tamanhos mais próximos são considerados mais similares do que alelos
com tamanhos mais distantes (Mengoni et al., 2000).
Vários outros estudos confirmam essa baixa correlação entre os dados gerados por RAPD
e STR (Mengoni et al., 2000; Pejic et al., 1998; Powell et al., 1996; Russell et al., 1997). Porém
alguns estudos mostram que a combinação dos dois marcadores foi eficiente para demonstrar o
relacionamento esperado entre variedades de Alfafa (Medicago sativa) (Mengoni et al., 2000).

68
Portanto, a combinação dessas duas tecnologias pode ser vantajosa para elucidar relacionamentos
genéticos entre populações naturais, principalmente em estudos que consideram escalas menores
de relacionamento entre indivíduos, como este estudo entre variedades de Alfafa.
Estratégias de Conservação e Manejo
A melhor estratégia para proteção em longo prazo da diversidade biológica é a
preservação de populações no ambiente selvagem (preservação in situ) (Primack e Rodrigues,
2001). No entanto, para espécies que exigem grandes áreas de vida, como as estudadas aqui, uma
maneira alternativa para evitar que estas sejam extintas, são programas de criação em cativeiro,
ou ainda o monitoramento intensivo destas espécies em pequenas áreas protegidas conectadas
com corredores de dispersão, que são faixas de habitats que ligam dois ou mais fragmentos
(Rodrigues, 2002). O lobo-guará é uma espécie de difícil manutenção em cativeiro (Primack e
Rodrigues, 2001), de modo que a preservação da área ocupada por esta espécie parece ser a única
alternativa eficiente para sua conservação.
As duas espécies aqui estudadas distribuem-se amplamente pelos cerrados brasileiros
(Sillero-Zubiri et al., 2004). A principal causa do declínio das populações destas espécies,
principalmente do lobo-guará , é a redução drástica do ambiente em que vivem, sendo que na
maioria das vezes suas áreas de distribuição já foi ocupada por empreendimentos agropecuários
(Fonseca et al., 1994). A fragmentação de habitats é hoje uma das maiores ameaças à diversidade
biológica, tanto pela redução dos ambientes naturais como pela divisão dos habitats
remanescentes em fragmentos menores (Meffe e Carroll, 1997; Soulé e Kohm, 1989). No caso do
lobo-guará , além das ameaças inerentes ao pequeno tamanho populacional, devido a fatores
demográficos e genéticos, esta espécie está sujeita a outras ameaças externas, que são importantes
causas de mortalidade e conseqüentemente diminuição do tamanho populacional. Essas ameaças

69
incluem atropelamentos, abates por chacareiros do entorno da área de ocupação da espécie e a
introdução de espécies exóticas, patógenos e parasitas.
Os canídeos estão entre os mamíferos que mais sofrem com atropelamentos (Rodrigues,
2002; Silveira, 1999; Vieira, 1996). Vários estudos identificam os atropelamentos como a
principal causa de mortalidade de Lobos Guarás, e possivelmente do cachorro-do-mato ( Silveira,
1999; Silva et al., 2000; Rocha et al., 2004). A grande mobilidade destas espécies e o
comportamento de comer carcaças de outros animais atropelados, aliado ao excesso de
velocidade dos motoristas, justifica essa alta suscetibilidade. Rodrigues (2002) encontrou em seu
estudo na Estação Ecológica de Águas Emendadas-DF, uma média de 4,5 atropelamentos de
Lobos Guarás por ano, sendo que essa média pode estar subestimada, pois alguns indivíduos
atropelados podem conseguir sair da estrada e mover para dentro da reserva sem serem
detectados. A solução para os problemas de atropelamentos em unidades de conservação envolve
vários segmentos da sociedade, desde o governo à própria população. As principais medidas a
serem adotadas para a redução das mortes por atropelamentos são: redução do limite de
velocidade permitida nos limites com as reservas, instalação de meios de controle desta
velocidade, como radares, limpeza periódica da beira da estrada possibilitando uma melhor
visibilidade dos animais e dos motoristas e campanhas de educação dos motoristas e da
comunidade do entorno, etc (Rodrigues, 2002; Silveira, 1999).
Vários estudos também mostram que os Lobos Guarás são mortos por chacareiros nos
limites das reservas, por predarem animais domésticos, porém parece que esta causa de
mortalidade é menos importante que os atropelamentos (Rodrigues, 2002; Silveira, 1999). No
entanto, é necessário instituir um plano de Educação Ambiental na região a fim de mostrar a
importância destas espécies para os ecossistemas e para o ser humano.
A invasão de animais domésticos, principalmente cachorros, é outra fonte de risco de
mortalidade destas espécies. Segundo Primack e Rodrigues (2001), a introdução de espécies

70
exóticas é uma séria ameaça à vida silvestre e tem levado várias espécies nativas a extinção. No
caso do cachorro doméstico, ele interage com espécies nativas, como o lobo-guará e o cachorro-
do-mato, através da predação, competição por recursos limitados e introdução de doenças
(Primack e Rodrigues, 2001; Rodrigues, 2002). Marinho-Filho et al., (1998), relatou vários casos
de ataques de matilhas a animais silvestres na Estação Ecológica de Águas Emendadas-DF. Em
outros estudos foi observada a competição entre canídeos domésticos e lobos (Canis lupus), que
poderia estar relacionada a causas de extinção destas populações (Boitani e Ciucci, 1995;
Mendelssohn, 1983). Rodrigues (2002), indica que uma possível solução para a minimização dos
problemas causados pela introdução de cachorros domésticos em áreas protegidas seria a
esterilização em massa desses cães domésticos, mas que provavelmente não seria facilmente
aceita pela população humana. Uma outra solução alternativa seria instituir um programa de
vacinação e acompanhamento sanitários dos animais, a fim de minimizar os efeitos da
transmissão de doenças letais que também ocorrem freqüentemente (Primack e Rodrigues, 2001).
Se levado em consideração os parâmetros genéticos estimados no presente trabalho (alta
diversidade genética nas duas espécies estudadas), é interessante considerar a questão da validade
e necessidade de se realizar translocações de animais com o objetivo de elevar a diversidade
genética. Existem estudos (Campbell, 1980) que mostram os efeitos negativos dos translocações,
como por exemplo à impossibilidade de adaptação dos animais ao local que foram introduzidos, a
diluição da freqüência de genes adaptados, introdução de doenças, etc (Hedrick, 2001).
Rodrigues (2002) testou o translocação de três Lobos Guarás na Estação Ecológica de Águas
Emendada-DF e após poucos dias, nenhum dos indivíduos foi registrado na área. Assim sendo, a
translocação de animais em função principalmente de seus pontos negativos parece não ser uma
ferramenta de manejo efetiva. Como os dados obtidos neste trabalho não indicam uma
significativa redução na variabilidade genética e nem endogamia elevada, talvez esta estratégia

71
não seja necessária para estas espécies no PNE, pelo menos no sentido de ampliar a variabilidade
genética existente atualmente.
Contudo, o maior impacto humano nas populações de canídeos silvestres, é a redução e
fragmentação de habitats causados principalmente pelo avanço das fronteiras agrícolas que não
tem demonstrado uma preocupação como o desenvolvimento sustentável da região. São nítidos
os efeitos ecológicos e genéticos da fragmentação de habitats e estes devem ser estudados para
serem incorporados em planejamentos de manejos de espécies ameaçadas (ver capítulo 4). Por
exemplo, a conectividade entre reservas é um fator importante a ser manejada, pois pequenas
populações têm alta probabilidade de extinção. Mas, se este fator for contrabalançado por
recolonização de subpopulações existentes em um sistema de áreas protegidas (ligar o Parque
Nacional das Emas a outras reservas) a metapopulação poderá existir por muito mais tempo
(Meffe e Carroll, 1997). Neste sentido, fica clara a grande necessidade de se desenvolver estudos
mais específicos sobre a ecologia e principalmente sobre o padrão de dispersão destas espécies,
aliado a uma educação ambiental eficiente das populações do entorno do PNE, para que
estratégias de conservação sejam mais bem estabelecidas.

72
AGRADECIMENTOS
Aos colegas do Laboratório de Genética e Melhoramento da Universidade Católica de Goiás –
UCG, Gabriela C. Pádua, Nicole C. L. Dutra, Prof. Dr. Breno de Faria e Vasconcellos, Lorenna L.
de Souza, Luciana M. Bossois e os demais estagiários que nos ajudaram nas análises genéticas. A
todo o pessoal do Parque Nacional das Emas - PNE, principalmente os estagiários que realizaram
os trabalhos de campo. Ao CNPq pelo financiamento de nossas pesquisas dentro do programa
PRONEX para análise de áreas prioritárias para conservação da biodiversidade no Cerrado e em
Goiás. (CNPq/SECTEC-GO, Proc. No. 23234156). JAFD-F recebe uma bolsa de Produtividade
em Pesquisa nível 1B do CNPq.
REFERÊNCIAS BIBLIOGRÁFICAS
Balloux F, Brunner H, et al., (2000) Microsatellites can be misleading: An empirical and
simulation study. Evolution 54: 1414-1422.
Barker JSF, Moore S, Hetzel S, Evan D, Tan SG e Byrne, K (1997) Genetic diversity of Asian
water buffalo (Bubalus bubalis): microsatellite variation and comparison with protein-coding
loci. Animal Genetics 28: 103-115.
Bassan BJ, Anollés GCE e Gresshoff PM (1991) Fast and sensitive silver staining of DNA in
polyacrilamide gels. Analytical Biochemistry 196: 80-83.
Boitani L e Ciucci P (1995) Comparative social ecology of feral dogs and wolves. Ethology, Ecol.
and Evol. 7: 49-72.
Bostein D, White RL, Skolnick MH e Davis RW (1980) Construction of genetic map in man
using restriction fragment length polymorphisms. Am. J. Human Genet. 32: 314–331.

73
Brondani C, Rangel PHN, Borba TCO e Brondani RPV (2003) Transferability of microsatellite
and sequence tagged site markers in Orysa species. Hereditas 138: 187-192.
Campbell S (1980) Is a reintroduction a realistic goal? In: Solé M.E and Wilcox BW (eds)
Conservation Biology: an evolutionary–ecological perspective. Sinauer Association,
Sunderland, 395 pp.
Cañón J, Alexandrino P, Bessa I et al.,(2001) Genetic diversity measures of local European beef
cattle breeds for conservation purposes. Genetics Selection and Evolution 33: 311–332.
Ceballos G e Ehrlich PR (2002) Mammal population losses and the extinction crisis. Science 296:
904-907.
Charlesworth B, Sniegowski P e Stephan W (1994) The evolutionary dynamics of repetitive DNA
in eukaryotes. Nature 371: 215–220
Cipriani G, Lot G, Huang WG et al.,(1999) AC/GT and AG/CT microsatellite repeats in peach
[Prunus persica (L.) Batsch]: isolation, characterization and cross-species amplification in
Prunus . Theoretical Applied Genetics 99: 65-72.
Collevatti RG, Brondani RV e Grattapaglia D (1999) Development and characterization of
microsatellite markers for genetic analysis of a Brazilian endangered tree species of Caryocar
brasiliense. Heredity 83: 748-756.
Crow JF e Kimura MA (1970) An introduction to population genetics theory. London: Harper
Row, 591 pp.
De Mattos PSR, Del Lama MA, Toppa RH e Schwantes AR (2004) Populational Genetic
Structure of free-living maned wolves (Chrysocyon brachyurus) determined by proteic
markers. Braz J Biol 64: 639-644.

74
Denise S, Johnston E, Halverson J, Marshall K, Rosenfeld D, Mckenna S, Sharp T e Edwards J
(2004) Power of exclusion for parentage verification and probability of match for identity in
American kennel club breeds using 17 canine microsatellite markers. Animal Genetics 35:
14–17.
DeSalle R e Amato G (2004) The expansion of conservation genetics. Nature 5: 702-712.
Ellsworth DL, Rittenhouse KR e Honeycutt RL (1993) Artifactual variation in randomly
amplified polymorphic DNA banding patterns. Biotechniques 14: 214-217.
EMBRAPA (2005) Monitoramento por Satélite. Campinas, [S.d.]. Cachorro-do-mato. Disponível
em: <http://www.cnpm.embrapa.br/projetos/fauna/mamifero/cac_mato.html
Farrel LE, Roman J e Sunquist ME (2000) Dietary separation of sympatric carnivores identified
by molecular analysis of scats. Molecular Ecology 9: 1583-1590.
Ferreira ME e Grattapaglia D (1998) Introdução ao Uso de Marcadores Moleculares em Análise
Genética. EMBRAPA - CENARGEN, Brasília, 220pp.
Fonseca GAB, Rylands AB, Costa CRM, Machado RB e Leite YLR (1994) Livro Vermelho Dos
Mamíferos Brasileiros Ameaçados de Extinção. Fundação Biodiversitas, Belo Horizonte.
Francisco LV, Langston AA, Mellersh CS, Neal CL e Ostrander EA (1996) A class of highly
polymorphic tetranucleotide repeats for canine genetic mapping. Mammalian Genome 7:
359–62.
Frankham R (1995) Inbreeding and extinction: a threshold effect. Conservation Biology 9: 792-
799.
Frankham R, Ballou JD e Briscoe DA (2002) Introduction to conservation genetics. Cambridge
University Press. 640pp.

75
Fries R, Eggen A e Womack JE (1993) The bovine genome map. Mammals Genome 4: 405-428.
Gur-Arie R, Cohen CJ, Eitan Y, Shelef L, Hallerman EM e Kashi Y (2000) Simple sequence
repeats in Escherichia coli : abundance, distribution, composition, and polymorphism. Gen.
Res. 10: 62-71.
Halverson J, Dvorak J e Stevenson T (1995) Microsatellite sequences for canine genotyping. US
Patent 05874217.
Hedrick PW (1999) Perspective: Highly variable genetic loci and their interpretation in evolution
and conservation. Evolution 53: 313-318.
Hedrick PW (2001) Conservation genetics: where are we now? Trends in Ecology & Evolution
16: 629-636.
Hillis DM, Mable BK e Moritz C (1996) Molecular systematics. Sinauer Associates, Sunderland,
MA. 655pp.
IBAMA (2003) Lista das Espécies da Fauna Brasileira Ameaçadas de Extinção. Anexo à
Instrução Normativa n° 3, de 27 de maio de 2003, do Ministério do Meio Ambiente.
Disponível em: www.ibama.gov.br.
Isagi Y e Suhandono S (1997) PCR primers amplifying microsatellite loci of Quercus mysinifolia
Blume and their conservation between oak species. Molecular Ecology 6: 897-899.
IUCN (2004) 2004 IUCN Red List of Threatened Species. Disponível em: www.redlist.org.
Iyengar A, Babu VN, Hedges S, Venkataraman AB, Maclean N e Morin PA (2005)
Phylogeography, genetic structure, and diversity in dhole (Cuon alpinus). Molecular Ecology
14: 2281-2297.

76
Kappes SM, Keele JM, Stone RT, Mc Graw RA, Sonstegard TS, Smith TP, Lopes-Corrales NL e
Beattie CW (1997) Second generation linkage map of the bovine genome. Genome Research
7: 235-249.
Karp A, Seberg A e Buiatti M (1996) Molecular techniques in the assessment of botanical
diversity. Annual Botany 78: 143-149.
King DG e Soller M (1999) Variation and fidelity: The evolution of simple sequence repeats as
functional elements in adjustable genes. In: S.P. Wasser, ed., Evolutionary Theory and
Processes: Modern Perspectives. Kluwer Academic Publishers, Dordrecht, The Netherlands,
pp. 65-82.
Klukowska BJ, Strabel, T, Mackowski M e Switonski M (2003) Microsatellite polymorphism and
genetic distances betwen the dog, red fox and artic fox. J. Anim. Breed. Genet. 120: 88-94.
Kosman E e Leonard KJ (2005) Similarity coefficients for molecular markers in studies of genetic
relationships between individuals for haploid, diploid, and polyploid species. Molecular
Ecology 14: 415-424.
Li YC, Korol AB, Fahima T, Beiles A e Nevo E (2002) Microsatellites: genomic distribution,
putative functions and mutational mechanisms: a review. Molecular Ecology 11: 2453-2465.
Luikart G e Cornuet JM (1998) Empirical evaluation of a test for identifying recently
bottlenecked populations from allele frequency data. Conserv. Biol. 12: 228-237.
Luikart G, Allendorf FW, Cornuet JM e Sherwin WB (1998) Distortion of allele frequency
distributions provides a test for recent population bottlenecks. J. Hered. 89: 238-247.
Lynch M e Milligan BG (1994) Analysis of population genetic structure with RAPD markes.
Molecular Ecology 3: 91-99.

77
Manel S, Schwartz M, Luikart G e Taberlet P (2003) Landscape genetics: combining landscape
ecology and population genetics. Trends in Ecology and Evolution 18: 189–197.
Manly BFJ (1991) Randomization and Monte Carlo Methods in Biology. Chapman and Hall,
London.
Marinho-Filho JS, Rodrigues FHG e Guimarães MM (1998) Mamíferos da Estação Ecológica de
Águas Emendadas. 34-63. In: Marinho-Filho, J.S., F.H.G. Rodrigues & M.M. Guimarães
(eds.) Vertebrados da Estação Ecológica de Águas Emendadas. SEMATEC/IEMA, Brasília,
DF.
McDonald DW e Courtenay O (1996) Enduring social relationship in a population of crab-eating
Food habits of two South American canids zorro, Cerdocyon thous, in Amazon Brazil.
Journal of Zoology (London) 239: 329-355.
Meffe GK e Carroll RC (1997) Principles of conservation biology. Sinauer Associates,
Sunderland, Massachusetts, USA.
Mendelssohn H (1983) Conservation of the wolf in Israel. Acta Zoologica Fennica 174: 281-282.
Mengoni A, Gori A e Bazzicalupo M (2000) Use of RAPD and microsatellite (SSR) variation to
assess genetic relationships among populations of tetraploid alfalfa, Medicago sativa. Plant
Breed. 119: 311–317
Menotti-Raymond MV, David A, Lyons LA, Schaffer AA, Tomlin JF, Hutton MK e O' Brien SJ
(1999) A genetic linkage map of microsatellites in the domestic cat (Felis catus). Genomics
57: 9-23.
Miller MP (1997) Tools for population genetics analyses (TFPGA) 1.3: A Windows program for
the analysis of allozyme and molecular population genetic data. Disponível em:
http://herb.bio.nau.edul~miller/tfpga.htm

78
Moran P (2002) Current conservation genetics: building an ecological approach to the synthesis
of molecular and quantitative genetic methods. Northwest Ecology of Freshwater Fish 11:
30–55.
Mullis K e Faloona F (1987) Specific synthesis of DNA in vitro via a polymerase-catalyzed chain
reaction. Methods Enzymol. 155: 335-350.
Neff MW, Broman KW, Mellersh CS, Ray K, Acland GM, Aguirre GD, Ziegle JS, Ostrander EA
e Rine J (1999) A second-generation genetic linkage map of the domestic dog, Canis
familiaris. Genetics 151: 803–820.
Parker PG, Snow AA, Schug MD, Booton GC e Fuerst PA (1998) What molecules can tell us
about populations: choosing and using a molecular marker. Ecology 79: 361–382.
Peakall R, Gilmore S, Keys W, Morgante M e Rafalski A (1998) Cross species amplification of
soybean (Glycine max) simple sequence repeats (SSRs) within the genus and other legume
genera: implications for the transferability of SSRs in plants. Molecular Biology and
Evolution 15: 1275–1287.
Pejic I, Ajmore-Marsan P, Morgante M, Kozumplick V, Castiglioni P, Taramino G e Motto M
(1998) Comparative analysis of genetic similarity among maize inbred lines detected by
RFLPs; RAPDs; SSRs; and AFLPs. Theoretical and Applied Genetics 97: 1248-1255.
Powell W, Morgante M, Andre C, Hanafey M, Vogel J, Tingey P e Rafalsky A (1996) The
comparison of RFLP; RAPD; AFLP and SSR (microsatellite) markers for germplasm
analysis. Molecular Breeding 2: 225-238.
Primack RB e Rodrigues E (2001) Biologia da conservação. E. Rodrigues, Londrina, 328 pp.
Ratnayeke S, Tuskan GA e Pelton MR (2002) Genetic relatedness and female spatial organization
in a solitary carnivore, the raccoon, Procyon lotor. Molecular Ecology 11: 1115-1124.

79
Raymond M e Rousset F (1995) Genepop 3.3, population genetics software for exact tests and
ecomenicism. Journal of Heredity 86: 248–249.
Roa AC, Chavarriaga-Aguirre P, Duque MC et al.,(2000) Cross-species amplification of cassava
(Manhihot esculenta) (Eurphorbiaceae) microsatellites: allelic polymorphism and degree of
realtionship. American Journal of Botany 87: 1647-1655.
Rocha VJ, Reis NR e Sekiama ML (2004) Dieta e dispersão de sementes por Cerdocyon thous
(Linnaeus) (Carnívora, Canidae) em um fragmento florestal no Paraná Brasil. Revista
Brasileira de Zoologia 21: 871-876.
Rodrigues FHG (2002) Biologia e conservação do lobo-guará na Estação Ecológica de Águas
Emendadas, DF. Tese de Doutorado em Ecologia, Universidade Estadual de Campinas,
Campinas.
Rohlf FJ (1991) NTSYS: Numerical taxonomy and multivariate analysis system. Exeter
publishing Co., New York.
Russell RJ, Fuller JD, Macaulay M, Hats BG, Jahoor A, Powell W e Waugh R (1997) Direct
comparison of levels of genetic variation among barley accessions detected by RFLPs,
AFLPs, SSRs and RAPDs. Theor Appl Genet 95: 714-722.
Schonewald-Cox CM, Chambers SM, MacBryde B e Thomas L (1998) Genetics and conservation.
Benjamim/Cummings, Menlo Park, California.
Seddon JM, Parker HG, Ostrander EA e Ellegren H (2005) SNPs in ecological and conservation
studies: a test in the Scandinavian wolf population. Molecular Ecology 14: 503–511.
Sillero-Zubiri C, Hoffmann M e Macdonald DW (eds) (2004) Canids: Foxes, Wolves, Jackals and
Dogs. Status Survey and Conservation Action Plan. IUCN/SSC Canid Specialist Group.
IUCN, Gland, Switzerland and Cambridge, UK.

80
Silva CBX, Nicola PA, Lange RR e Pontes-Filho A (2000) Atropelamentos de Lobos-guarás
(Chrysocyon brachyurus) nas rodovias dos Campos Gerais, Palmeira, Ponta Grossa, Paraná.
8° Reunião Anual da SBPN.
Silveira L (1999) Ecologia e conservação dos mamíferos carnívoros do Parque Nacional das
Emas, Goiás. Dissertação de Mestrado em Ecologia, Universidade Federal de Goiás, Goiânia.
Sneath PHA e Sokal RB (1973) Numerical Taxonomy. The Principles and Practice of Numerical
Classification. San Francisco: W. H. Freeman.
Solé-Cava AM (2001) Biodiversidade molecular e Genética da Conservação. In: Matioli, S.R.
(ed.) Biologia Molecular e Evolução. Holos Editora, Rio de Janeiro, pp. 172-192.
Soulé ME e Kohm KA (1989) Research priorities for conservation biology. Island Press,
Washington.
Soulé ME e Wilcox BA (1980) Conservation biology: An evolutionary-ecological perspective.
Sinauer Associates, Sunderland, Massachusetts.
Sunden SLF, Stone RT, Bishop MD, Kappes SM, Keele JW e Beattie CW (1993) A highly
polymorphic bovine microsatellite locus: BM2113. Animal Genetics 24: 69.
Tautz D (1989) Hypervariability of simple sequences as a general source of polymorphic DNA
markers. Nucleic Acids Res. 17: 6463-6471.
Tong J, Wang Z, Yu X, Wu Q, Chu KH (2002) Cross-species amplification in silver carp and
bighead carp with microsatellite primers of common carp. Molecular Ecology Notes 2: 245-
247.
Vieira EM (1996) Highway mortality of mammals in Central Brazil. Journal of the Brazilian
Association for the Advancement of Science 48: 270-272.

81
Vilà C, Sundquist AK, Flagstad O, Seddon JM, Bjornerfeldt S, Kojola I., Sand, H, Wabakken P e
Ellegren H (2002) Rescue of a severely bottlenecked wolf (Canis lupus) population by a
single inmigrant. Proc. R. Soc. Lond B Biol. Sci. 270: 91–97.
Wandeler P, Funk SM, Largiader CR, Gloor S, Breitenmoser U (2003) The city-fox phenomenon:
genetic consequences of a recent colonization of urban habitat. Molecular Ecology 12: 647-
656.
Wayne RK (1996) Conservation Genetics in the Canidae. In: Avise JC, Hanuick JL (eds)
Conservation genetics: case histories from nature. Chapman & Hall, New York, pp. 75–118.
Welsh J e McClelland M (1990) Fingerprinting genomes using PCR with arbitrary primers.
Nucleic Acids Reaserch 18: 7213-7218.
Williams JGK, Kubelik A, Livak K, Rafalsski J e Tingey SV (1990) DNA polimorphisms
amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Reaserch 18:
6531-6535.
Williamson KS, Cordes JF e May B (2002) Characterization of microsatellite loci in Chinook
salmon (Oncorhynchus tshawytscha) and cross-species amplification in other salmonids.
Molecular Ecology Notes 2: 17-19.
Write G e Powell W (1997) Cross-species amplification of SSR loci in the Meliaceae family.
Molecular Ecology 6: 1195-1197.
Yang G, Saghai Maroof MA, Xu CG, Zhang Q e Biyashev RM (1994) Comparative analysis of
microsatellite DNA polymorphism in landraces and cultivars of rice. Molecular and General
Genetics 245: 187-194

82
Zucchi MI, Brondani RPV, Pinheiro JB, Brondani C e Vencovsky R (2002) Transferability of
microsatellite markers from Eucalyptus spp. to Eugenia dysenterica (Myrtaceae family).
Molecular Ecology Notes 2: 512–513.

83
CAPÍTULO 3

84
Artigo a ser submetido: Genetics and Molecular Biology
Relacionamento Genético e Organização Espacial em uma
População de lobo-guará (Chrysocyon brachyurus, Illiger, 1815) do
Parque Nacional das Emas - Goiás
FLÁVIA MELO RODRIGUES1, 8, JOSÉ ALEXANDRE FELIZOLA DINIZ FILHO2, 3, 5, 8,
MARIANA PIRES DE CAMPOS TELLES4, 5, LUCILEIDE VILELA RESENDE4, NATÁLIA
MUNDIM TÔRRES5, 7, ANAH TEREZA DE ALMEIDA JÁCOMO5, LEANDRO SILVEIRA5,
THANNYA NASCIMENTO SOARES4, CYNTIA K. KASHIVAKURA5, MARIANA M.
FURTADO5, CLAUDIA FERRO5 E SAMUEL ASTETE5 1Departamento de Biologia, Universidade Estadual de Goiás (UEG), Anápolis, GO; 2Departamento de Biologia, Universidade Católica de Goiás (UCG); 3Departamento de Biologia
Geral, ICB, Universidade Federal de Goiás (UFG); 4Laboratório de Genética e Melhoramento,
Departamento de Zootecnia/ITS, Universidade Católica de Goiás (UCG); 5Fundo para a
Conservação da Onça-Pintada (FCOP); 7Programa de Pós-Graduação em Biologia Animal,
Universidade de Brasília (UnB); 8Programa de Pós-Graduação em Ciências Ambientais – UFG.
Resumo
O lobo-guará (Chrysocyon brachyurus) é o maior canídeo sul americano, estando incluído
entre as espécies de mamíferos ameaçadas de extinção desta região, principalmente devido à
transformação da paisagem natural em áreas de sua ocorrência por atividades de desmatamento,
queimadas, agricultura e urbanização. Suas áreas de vida costumam ser grandes e com grandes
variações. O objetivo deste trabalho foi analisar a associação entre a similaridade genética,
mensurada através de marcadores RAPD e STR, com a sobreposição de áreas de vida da
população de lobo-guará do Parque Nacional das Emas, GO (PNE). Entre abril/2002 e
novembro/2003, foram capturados 27 Lobos Guarás, colocados os rádio-colares e coletado
sangue para as análises genéticas. Após as análises genéticas via PCR, os fragmentos
amplificados com os marcadores RAPD foram separados em gel de agarose 1,4% e visualizados
por coloração com brometo de etídio, e aqueles fragmentos obtidos com os marcadores STR
foram separados em gel de poliacrilamida a 4% e visualizado por coloração com nitrato de prata.
Os dados binários foram utilizados para o cálculo do índice de similaridade de Jaccard. Os dados

85
ecológicos foram coletados a partir do monitoramento destes animais por rádiotelemetria no PNE
e as áreas de vida foram estimadas através do método Mínimo Polígono Convexo (MPC),
considerando 80, 90 e 100% dos pontos. Para testar a relação entre a similaridade genética e o
padrão espacial de dispersão dos indivíduos foi utilizado o teste de Mantel, com 5000
permutações. Os nove primers utilizados nas reações RAPD produziram um total de 91 locos,
sendo 42 polimórficos. Um total de 27 locos foi obtido com os marcadores STR e destes 89 %
foram polimórficos. Em média, o tamanho da área de vida dos Lobos Guarás do PNE foi muito
grande, 74,18 ± 44,18 km2 (n = 13) para os machos e 89,70 ± 54,11 km2 (n = 14) para as fêmeas.
A associação entre as similaridades genéticas e espaciais, medidas pelo teste de Mantel, foi
significativa apenas entre os machos (r = 0,247, P = 0,02 para MPC 80% e r = 0,23, P = 0,02 para
MPC 90%), quando considerados os dados gerados pelos marcadores RAPD. Já a associação
entre a similaridade genética, obtida com os marcadores STR, e a similaridade espacial foi
significativa entre os indivíduos de ambos os sexos e entre as fêmeas (r = 0,182, P = 0,03 para
MPC 80%, r = 0,197, P = 0,03 para MPC 90%, r = 0,248, P = 0,002 para MPC 100%
considerando os indivíduos de ambos os sexos e r = 0,236, P = 0,049 para MPC 100%
considerando apenas as fêmeas). Os resultados baseados nos marcadores RAPD, mostraram uma
estruturação espacial da similaridade genética apenas para os machos, concordando com a
hipótese de que a distribuição espacial entre os machos não é aleatória. Isso sugere que este
padrão pode ter sido gerado pela associação da disponibilidade de recursos críticos (i.e. água)
com o nível de parentesco genético. Enquanto, que os resultados obtidos com os marcadores STR
mostraram uma estruturação espacial da variabilidade genética de acordo com o padrão de
filopatria feminina. Como o padrão obtido com os marcadores STR é mais esperado, e possível
que os resultados com os marcadores RAPD sejam artefatos.
Palavaras-Chave: lobo-guará, Área de vida, Rádiotelemetria, Marcadores RAPD, Marcadores
STR.
Enviar correspondência para FM Rodrigues. Universidade Estadual de Goiás, Departamento de
Biologia, Unidade Universitária de Ciências Extas e Tecnológicas, BR 153, Km 3, Campus
Anápolis, CEP: 75110-390, Caixa Postal: 459, Anápolis, GO, Brasil. E-mail: [email protected]

86
Abstract
The Maned wolf (Chrysocyon brachyurus) is the biggest canid of south america included
in the mammal species threatened by extincition list in the region. One of the main reasons is the
transformation of its natural habitat caused by deforestation, fires, agriculture and urbanization.
It’s living areas are usually big and varied. The goal of this paper was to analyse the association
of genetic similarities measured by RAPD and STR markers with the overlap of areas in the life
of the Maned wolves from Ema’s National Park (GO). Between April 2002 and November 2003,
27 individuals were captured, radio-collars were placed on them and their blood was drawn for
genetic analysis. After the analysis via PCR, the amplified fragments with RAPD were separated
in agarosis gel 1.4% and colored with bromite etidium, the fragments obtained with STR were
separated in poliacrilamide gel 4% and colored with silver nitrate. The binary data was used to
find Jaccard’s similarity index. The ecological data was obtained by monitoring these animals
through radiotelemetry and its living areas were estimated by the Minimum Convex Poligon
(MCP) method, considering, 80, 90 and 100% of the points. To test the relation between genatic
similarity and living area deviation standard for the animals, the Mantel test with 5000
permutations was used. The nine primers used in RAPD reactions produced 91 locus, 42 of them
polimorphic. A total of 27 locus was obtained with STR markers and 89% of these were
polimorphic. In average, the size of the area for the Maned wolf in the ENP was of 78,18 +
44,18km2 (n=13) for males and 89,70 + 54,11km2 (n=14) for females. The association between
the genetic and territorial similarities was significant amongst both genders. Between females
( r=0,182, p=0,03 for MPC 80%; r=0,197, p=0,03 for MPC 90%; r=0,248, p=0,002 for MPC
100%). The results using RAPD markers showed a territorial structure based on genetic
similarities only for males, which coincided with the hipothesis that territorial distribution od
males isn´t random. This suggests that this pattern could have been generated by the associaition
of critical resourse avalability (i.e. water) with the level of genetic links. The data from the STR
presented territorial genetic variability according to the females, as the standard obtained by the
STR markers is most expected, it is probable that the results obtained by RAPD are artifacts.
Key words: Maned wolf, living areas, territorial distribution, tadiotelemetry, RAPD markers ,
STR markers.

87
INTRODUÇÃO
O lobo-guará (Chrysocyon brachyurus) é o maior canídeo sul americano, e quando adulto
pode pesar cerca de 25 Kg e medir 90 cm de altura nos ombros e comprimento total entre 145 a
190 cm, sendo a cauda responsável por 45 cm (Mantovani, 2001; Einsenberg e Redford, 1999).
Está incluído entre os mamíferos sul-americanos ameaçados de extinção, principalmente em
função da transformação da paisagem natural, em áreas de sua ocorrência por atividades de
desmatamento, queimadas, agricultura e urbanização (IBAMA, 2003) e também devido à
perseguição dos pecuaristas, principalmente porque acreditam que os Lobos Guarás predam seus
animais domésticos (De Mattos et al., 2004; Fonseca et al., 1994; Silva e Talomi, 2004). A área
de ocorrência original desta espécie compreende a região central do Brasil, o Paraguai, Bolívia,
extremo leste do Peru, nordeste da Argentina e Uruguai, sempre em regiões de vegetação aberta,
como campos e Cerrados. (Dietz, 1985). Está extinto no Uruguai e na Argentina ao sul do Rio da
Prata (Kimberley, 1998). A alimentação consiste principalmente de pequenos animais (roedores,
aves, répteis, etc) e de alguns frutos (fruta do lobo – lobeira e guabiroba) (Motta-Junior, 1996;
Lombardi e Motta-Júnior, 1993; Consorte-McCrea, 1994; Maia e Gouveia, 2002; Mota-Junior,
2002; Jácomo et al., 2004). Esta espécie apresenta funções ecológicas imprescindíveis para a
estabilidade do ecossistema do cerrado, principalmente por ser topo de cadeia alimentar e pela
função como dispersor de sementes, dada a característica onívora de sua dieta (De Mattos, 2003).
O lobo-guará é territorialista, isto é, principalmente na época de reprodução, o casal
demarca uma área geográfica onde não admite a presença de outros indivíduos, e portanto
delimita o seu território. É monógamo facultativo, sendo comum à manutenção do par
reprodutivo, e se apresenta mais ativo ao entardecer e à noite (Dietz, 1984). São animais
solitários e interações entre o casal ocorrem apenas no período reprodutivo que, em cativeiro, se
concentra nos meses de dezembro a junho (Ginsberg e Mcdonald, 1990). Espécies solitárias são
aquelas que têm encontros raros com adultos co-específicos e estão geralmente sozinhos durante

88
períodos de atividade, tal como o forrageamento (Ratnayeke et al., 2002; Waser e Jones, 1983).
No entanto, um estudo realizado com a população de lobo-guará do Parque Nacional das Emas
(PNE) em Goiás, observou alguns casais forrageando juntos, onde sempre era registrado que os
machos defecam sobre as fezes das fêmeas para demarcar território (Silveira, 1999).
A área de vida do lobo-guará costuma ser grande, variando de 21 a 30 km2 na Serra da
Canastra, MG (Dietz, 1984), entre 54 e 115 km2, no município de Águas de Santa Bárbara, SP
(Carvalho e Vasconcellos, 1995) e de 4,67 a 79 km2 no Parque Nacional das Emas, GO (Silveira,
1999). A população do PNE apresenta dois picos de atividade, sendo um entre 8:00 e 10:00h da
manhã e outro entre 20:00 e 22:00h. No entanto, observou-se que o período de atividade da
espécie é mais dependente da umidade relativa do ar e temperatura do que do horário do dia
(Jácomo et al., 2004). Em tempos de clima frio, céu nublado ou após chuva, é possível observar
lobos guará forrageando a qualquer hora do dia (Silveira, 1999).
Geralmente, a área de vida de um animal compreende uma área heterogênea, determinada
por fatores físicos e biológicos, explicando algumas desproporcionalidades no uso de espaço por
algumas espécies (Silva e Talamoni, 2004). A área de uso intensivo tem sido considerada a área
central do home range do animal, que pode estar altamente relacionada com a disponibilidade de
recursos, como o alimento (Silva e Talamoni, 2004). Portanto, são as áreas mais freqüentemente
usadas pelo animal, que são baseadas na média harmônica de uma área de distribuição, também
definidas como centro de atividade (Dixon e Chapman, 1980; Tanaka, 1980). Identificar estas
áreas é de fundamental importância para entendermos a biologia de muitas espécies, incluindo a
interação entre os indivíduos e entre estes com o meio ambiente (Silva e Talamoni, 2004).
Estudos com outros carnívoros solitários, como o guaxinim (Procyon lotor) mostram uma maior
dispersão entre os machos e a filopatria feminina foi uma observação comum. Entretanto,
dispersão entre fêmeas jovens foi também relatado (Cowan, 1973, Ratnayeke, 1997; Ratnayeke et
al., 2002). Para fêmeas de mamíferos que investem muito nos cuidados de seus descendentes,

89
benefícios ecológicos e sociais são fortes fatores a favor da filopatria feminina (Ratnayeke et al.,
2002).
A criação e manutenção de unidades de conservação em áreas de ocorrência do lobo-
guará representam uma estratégia para a conservação da espécie. Embora existam unidades de
conservação implantadas em áreas de cerrado, a área total destas unidades compreende apenas
2% da área total do bioma em questão (Myers, 2000). Ao mesmo tempo, é importante lembrar
que áreas de preservação não oferecem todas as condições necessárias ao estabelecimento de uma
população mínima viável e, portanto não garantem a manutenção da mesma em longo prazo. Um
problema comum é o isolamento das populações em áreas de preservação, que é agravado na
proporção em que estas áreas tornam-se menores e mais distantes entre si, em uma paisagem cujo
uso do solo encontra-se bastante modificado e inadequado para as espécies animais existentes.
Shaffer (1981) definiu que uma população mínima viável para qualquer espécie em um
determinado habitat é a menor população isolada que tenha 99% de chances de continuar
existindo por 1000 anos, a despeito dos efeitos previsíveis de estocasticidadades genética,
ambiental e demográfica, e de catástrofes naturais (Primack e Rodrigues, 2001). No entanto,
pouco se sabe a respeito de informações demográficas e genéticas da maioria das espécies,
dificultando a definição deste número mínimo viável.
Alguns marcadores moleculares, tais como, RAPD (Random Amplified Polymorphic DNA)
e Microssatélites, também conhecidos por marcadores STR (Short Tandem Repeats), têm sido
bastante utilizados para se realizar uma caracterização inicial da variabilidade genética presente
nas populações, a fim de monitorar a variabilidade genética nas espécies, objetivando subsidiar
estratégias de acasalamento e introdução de novos indivíduos na população (Rodrigues et al.,
2004). Assim, a manutenção da diversidade e conseqüentemente o aumento da probabilidade de
viabilidade desta população ao longo do tempo são garantidos, uma vez que pequenas populações
sujeitas à deriva genética são mais suscetíveis a efeitos genéticos deletérios, tais como depressão

90
endogâmica, perda de flexibilidade evolucionária e depressão exogâmica (Ellstrand e Elam, 1993;
Frankham et al., 2003).
O objetivo deste trabalho foi analisar a associação entre a similaridade genética coma a
similaridade espacial (sobreposição de área de vida) e distância espacial (área central) em uma
população de lobo-guará do Parque Nacional das Emas-GO. Deste modo, foram levantadas
informações da espécie no que diz respeito a sua variabilidade genética e estrutura populacional,
fornecendo informações que podem vir a ser relevantes em futuros processos de manejo e
preservação da espécie.
MATERIAL E MÉTODOS
Área de estudo
O estudo foi realizado na maior área de cerrado conservado do sistema de Parques
Nacionais Brasileiros, o Parque Nacional das Emas (PNE). São 132.000 hectares localizados no
sudoeste do Estado de Goiás, próximo ao limite com o Mato Grosso do Sul e Mato Grosso, a
18o19’S e 52o45’E (Figura 1) (Silveira, 1999). Sua altitude varia de 650 a 1000m e possui um
clima seco acentuado, de abril a setembro, com temperaturas podendo atingir a marca de 0o C
onde, é comum a ocorrência de geadas (IBDF/FBCN, 1981). A vegetação do parque corresponde
a aproximadamente 95% de área com predomínio do campo sujo, além de outras vegetações
menos comuns como, mata ciliar, campo úmido, vereda, campo limpo, campo cerrado, cerrado
“Strictu sensu”, cerradão, dentre outras (IBDF/FBCN, 1981).
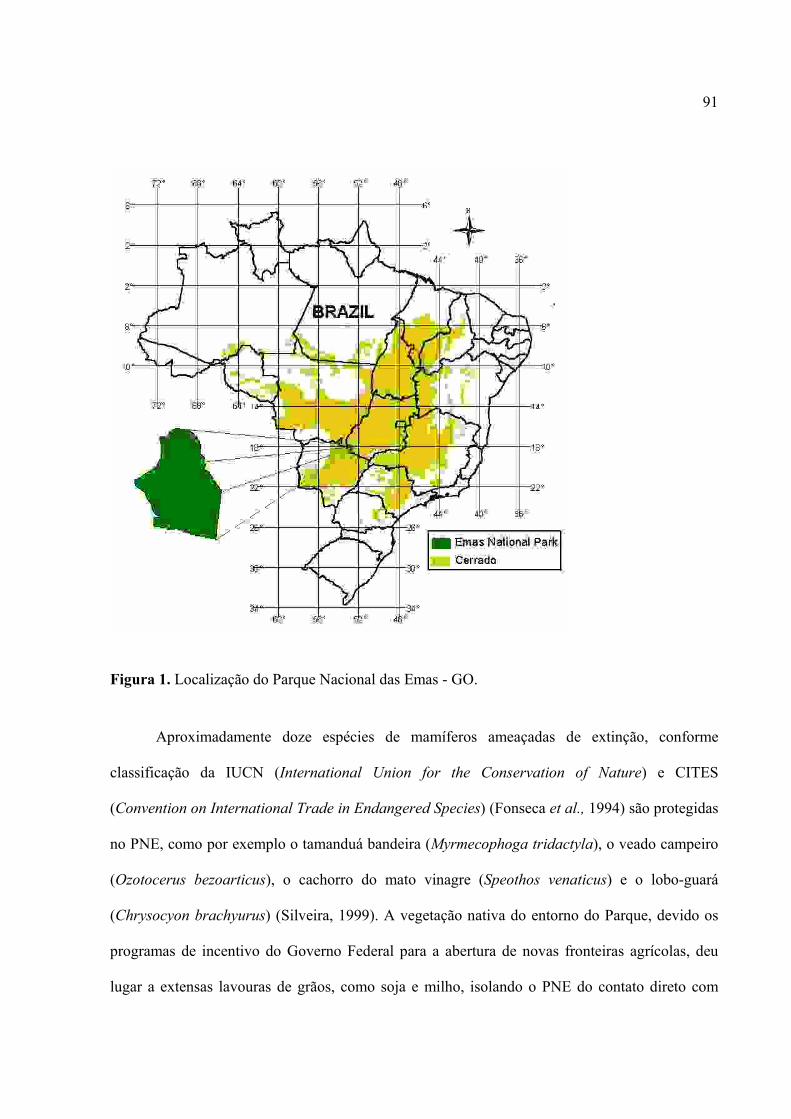
91
Figura 1. Localização do Parque Nacional das Emas - GO.
Aproximadamente doze espécies de mamíferos ameaçadas de extinção, conforme
classificação da IUCN (International Union for the Conservation of Nature) e CITES
(Convention on International Trade in Endangered Species) (Fonseca et al., 1994) são protegidas
no PNE, como por exemplo o tamanduá bandeira (Myrmecophoga tridactyla), o veado campeiro
(Ozotocerus bezoarticus), o cachorro do mato vinagre (Speothos venaticus) e o lobo-guará
(Chrysocyon brachyurus) (Silveira, 1999). A vegetação nativa do entorno do Parque, devido os
programas de incentivo do Governo Federal para a abertura de novas fronteiras agrícolas, deu
lugar a extensas lavouras de grãos, como soja e milho, isolando o PNE do contato direto com

92
outras áreas nativas (Silveira, 1999). Outro problema grave sofrido pelo PNE é o manejo
inadequado de agrotóxicos nas lavouras do seu entorno, nas cabeceiras dos Rios Formoso e
Jacuba, além do atropelamento de animais do Parque na rodovia GO-341, que margeia 25
quilômetros de seu limite oeste (Silveira e Jácomo, 1998).
Dados da Área de Vida
Entre abril de 2002 e novembro de 2003, foram armadas 71 armadilhas em vários pontos
do PNE, sendo 45 do tipo “gaiola” com compartimento para isca viva e 26 do tipo “trampa”. As
armadilhas tipo “gaiola” foram dispostas a cada 1,5 km e iscadas com galinhas ou pombos vivos
e as “trampas” iscadas com um produto comercial feito de extrato de glândulas de cheiro de
carnívoros. Os esforços acumulados foram de 291 armadilhas-noite para as gaiolas e 77
armadilhas-noite para as “trampas”.
Foram capturados 38 Lobos Guarás, dos quais 27 foram considerados nas análises por
apresentarem disponibilidade dos dados de área de vida. A contenção química dos animais foi
feita através da aplicação intramuscular do anestésico Zoletil (250 mg), sendo que a dose
variou de 0,5 a 1,5 ml, de acordo com o peso do animal. Para cada animal capturado foi feita a
biometria, análise clínica, estimativa de idade através de desgaste dentário, coleta de
ectoparasitos, urina, esfregaço vaginal, sangue e colocação de rádio-colares. Foi coletado ao
menos 10 mL de sangue, utilizando tubos vacutainer contendo EDTA, que foram utilizados para
análises de doenças, dosagem hormonal e análise genética. Depois de coletado, o sangue foi
acondicionado em um volume igual de um tampão denominado Easy Blood e armazenado em
geladeira, que auxilia na conservação do material até chegar ao laboratório para a extração do
DNA.
Os rádio-receptores e colares utilizados (Advanced Telemetry Systems, Inc.) nos
indivíduos funcionam através de ondas de rádio VHF (faixa de 150 a 152 MHz), com baterias de

93
duração de 36 a 42 meses. Todos os colares possuem uma freqüência individual e emitem sinais
(pulso), captados pelo rádio-receptor. Os rádios transmissores utilizados são sensíveis às
variações de movimentos dos animais, indicando se o animal está se movimentando (ativo), se
está parado (inativo), ou morto. O monitoramento por rádiotelemetria foi feito de carro, a pé, ou
de avião (Cessna 172), conforme a necessidade e acessibilidade em campo. Foi utilizado o
método da triangulação (White e Garrot, 1990), no qual são tomadas pelo menos duas direções
(azimute) do sinal do transmissor, cada direção tomada de um ponto diferente no terreno, de
modo a formar um triângulo, com o auxílio de uma bússola e GPS (Sistema de Posicionamento
Global) permitindo a identificação da localização. O resultado do estudo de área de vida é
apresentado na forma numérica, geralmente em km2, e na forma geométrica da área sobre um
mapa do local de estudo (Jacob e Rudran, 2003). Para estimar as coordenadas finais da
triangulação foi usado o programa Locate II, versão 2.82 (Nams, 2001).
O cálculo da área de vida dos animais foi feito através do método do Mínimo Polígono
Convexo – MPC (Mohr, 1947), que consiste basicamente na união dos pontos mais extremos da
distribuição de localizações, de forma a fechar o menor polígono possível sem admitir
concavidades (Jacob e Rudran, 2003). Foram considerados 80, 90 e 100% dos pontos, a fim de
estimar o efeito dos outliers, na determinação da área de vida. Foram construídas as matrizes de
sobreposição separadas, de tal modo que uma continha a informação de todos os indivíduos
simultaneamente e outras duas matrizes separadas por sexo. Paras estas análises foi utilizado o
programa Ranges6 versão 1.2 (Kenward et al., 2003).
Dados Genéticos
O DNA dos 27 indivíduos foi extraído a partir das amostras de sangue total, utilizando o
kit de purificação de DNA GFX fornecido pela Amersham Pharmacia Biotech. Após a
quantificação, com o auxílio do marcador de peso molecular Low DNA Mass, e diluição para a

94
concentração ideal de uso, o DNA foi utilizado para a realização de diversas Reações em Cadeia
da Polimerase (PCR), através de reações de amplificação do DNA Polimórfico Amplificado ao
Acaso (RAPD) (Welsh e McClelland 1990; Williams et al. 1990) e reações de amplificação com
os marcadores STR (Tautz, 1989).
As reações de RAPD foram montadas da seguinte forma: 3 µl de DNA (~3 ng/µl); 2,0 µl
de primer (~10 ng/µl); 2,6 µl tampão da enzima (10 X); 0,78 µl de MgCl2 (50 mM); 2,08 µl de
d’NTP (2,5 mM); 0,2 µl da enzima Taq-Polimerase (5 unidades/µl) e completando o volume com
9,34 µl de H2O Milli-Q. Foram utilizadas as seguintes condições nas reações de amplificação:
primeira etapa 2’ a 94°C; segunda etapa 1’30’’ a 92°C, terceira etapa 1’ a 37°C, quarta etapa
2’30’’ a 72°C, quinta etapa 40 ciclos (de 2 a 4) e sexta etapa 5’ a 72°C para a extensão final.
Primeiramente foi realizada uma seleção dos melhores marcadores utilizando o DNA de apenas
três indivíduos para a amplificação com 119 primers (desenvolvidos pela OPERON), seguido
de reações RAPD com todos os 27 indivíduos, usando os marcadores selecionados.
Para as reações com os marcadores STR, o seguinte protocolo foi utilizado: 5 µl de DNA
(~3 ng/µl); 2,2 µl do primers A (forward) (~5 ng/µl); 2,2 µl do primers B (reverse) (~5 ng/µl);
1,3 µl de BSA (10 mg/ml); 1,5 µl tampão da enzima (10 X); 0,5 µl de MgCl2 (50 mM); 1,3 µl de
d’NTP (2,5 mM); 0,2 µl da enzima Taq- Polimerase (5 unidades/µl) e completando o volume
com 0,8 µl de H2O Milli-Q, para um volume final de 15 µl. Para cada reação, realizada no
termociclador MJ Research Inc, utlizamos o seguinte programa: (1º) desnaturação do DNA a
94ºC por 5 minutos e (2º) a 94ºC por 1 minuto; (3º) anelamento dos “primers” a XºC por 1
minuto; (4º) extensão da molécula pela enzima Taq polimerase (fornecida pela Amersham
Pharmacia Biotech) a 72ºC por 1 minuto; (5º) 30 ciclos seguindo do 2º ao 4º passos; (6º) passo
final de extensão de 7 minutos a 72ºC para finalizar os produtos amplificados. Para cada primer
foram testadas temperaturas de anelamento que variaram entre 45ºC e 65ºC, pois este é um dos

95
fatores críticos nas análises com estes marcadores (Hillis et. al, 1996; Ferreira e Grattapaglia,
1998) e após os resultados dos testes foi utilizado a temperatura de anelamento específica de cada
primer (ver capítulo 1).
Os fragmentos de DNA amplificados via marcadores RAPD foram separados através de
eletroforese em gel de agarose 1,5% e em seguida corados com brometo de etídeo. Os fragmentos,
nos géis, foram visualizados através de um transluminador de luz ultravioleta e fotografados com
auxílio do fotodocumentador KODAK EDAS – 120, (Ferreira e Grattapaglia, 1998).
Os produtos de amplificação via marcadores STR foram submetidos à eletroforese em gel
de poliacrilamida a 4% em TBE 1X durante 2 horas, mantendo a corrente elétrica em 90W
constante. Foram aplicados 3µl do produto da PCR, contendo o tampão de carregamento,
previamente desnaturado (94°C por 5 min). O padrão de peso molecular, o marcador Ladder
10bp (Amersham Pharmacia Biotech) foi usado para estimar os tamanhos dos produtos de PCR.
Após a eletroforese, o gel de acrilamida foi corado com nitrato de prata (Bassam et al., 1991).
Após a revelação das bandas e a secagem das placas, estas foram colocadas sobre a luz branca
para a obtenção dos genótipos.
Em seguida foi construída uma matriz de dados binários obtida pela codificação dos géis
em genótipos, para cada conjunto de dados gerados (RAPD e STR). No capítulo 2 é apresentado
um exemplo de como foram transformados os genótipos gerados pelos marcadores STR em
dados binários. Essa matriz binária foi utilizada para estimar a similaridade genética entre os
pares de indivíduos com base no coeficiente de Jaccard (J) (Sneath e Sokal, 1973) que é dado por
PMJ =
onde, M é o número de bandas em comum e P é o número total de bandas, exceto as bandas
ausentes nos dois indivíduos comparadas.

96
Foram obtidas três matrizes de similaridade genética baseadas no índice de Jaccard, para
cada marcador molecular separadamente. A primeira contendo os dados de todos os indivíduos, a
segunda contendo apenas os dados dos machos e a terceira contendo os dados das fêmeas. Essas
matrizes foram construídas utilizando a rotina SIMQUAL (similarity for qualitative data) do
programa NTSYS-PC, versão 1.8 (Rohlf, 1991).
Correlação Genética e Espacial
O teste de Mantel (Manly, 1991) tem sido bastante utilizado para avaliar a correlação
matricial entre duas matrizes simétricas quaisquer E e M comparadas pelo valor de Z, dado por
( )∑ ∑ ⋅=i ijijj
MEZ
onde Eij e Mij são o i-ésimo e j-ésimo elementos das matrizes E e M a serem comparadas. A
significância da relação entre as matrizes pode ser testada recalculando o valor de Z diversas
vezes, permutando aleatoriamente, em cada uma delas, a ordem dos elementos de uma das
matrizes. Deste modo é possível obter uma distribuição de probabilidades à qual o valor de Z
observado pode ser comparado (Manly, 1991). O valor de Z não possui amplitude de variação
definida, mas de fato ele possui uma relação monotônica com o r de Pearson calculado entre as
matrizes. Assim pode-se utilizar o Z de Mantel para testar a significância da correlação matricial,
que varia entre 1,0 e –1,0.
O teste de Mantel requer a construção de duas matrizes de similaridade ou dissimilaridade
simétricas a serem comparadas. Neste estudo, este teste foi utilizado para comparar a similaridade
genética (matriz de Jaccard) com similaridade espacial (matriz de sobreposição), obtidas a partir
dos dados de telemetria para 80, 90 e 100% dos pontos. No entanto, as matrizes de sobreposição
geradas eram assimétricas, portanto estas tiveram que ser previamente transformadas em matrizes

97
simétricas. O modelo é basicamente a estimativa de uma taxa de sobreposição média (Om), que é
dado por
21 OOOm ⋅=
onde O1 é a medida de sobreposição de área do indivíduo A sobre o indivíduo B e O2 a
sobreposição de área do indivíduo B sobre A (Krebs, 1998).
Ainda a partir dos dados de área de vida dos indivíduos, foram construídas também as
matrizes de distâncias espaciais simétricas, usando o índice de distância euclidiana entre os
centros de área, e as mesmas foram usadas para a correlação matricial com as similaridades
genéticas. O cálculo da distância espacial foi realizada no programa NTSYS-PC, versão 1.8, na
rotina SIMINT (Similarity Interval) (Rohlf, 1991).
Foram utilizadas 5000 permutações aleatórias para estabelecer a significância de cada
correlação matricial. Essas análises foram realizadas utilizando a rotina MXCOMP (Matrix
Comparison-Graphics) do mesmo programa, NTSYS-PC, versão 1.8 (Rohlf, 1991).
RESULTADOS
Áreas de Vida
Dos 27 animais analisados, 13 eram machos e 14 eram fêmeas e destes apenas três fêmeas
e um macho eram sub-adultos, sendo os demais adultos. A quantidade total de radiolocalizações
obtidas foi de 2113, com uma média de 78,26 ±94,97 por indivíduo. O número mínimo de
localizações foi 10, para a fêmea no 503 e o máximo foi de 452 localizações para a fêmea no 501.
A área de vida dos indivíduos (machos e fêmeas) calculada com 80% dos pontos variou de 8,8 a
156 km2, para 90% dos pontos, variou de 14,6 a 165,7 km2 e para 100% dos pontos a variação foi
de 23 a 561 km2. As áreas de vida dos machos foram menores que a das fêmeas (Tabelas 1 e 2).
Em média, o tamanho da área de vida dos Lobos Guarás do PNE foi muito grande, 74,18 ± 44,18
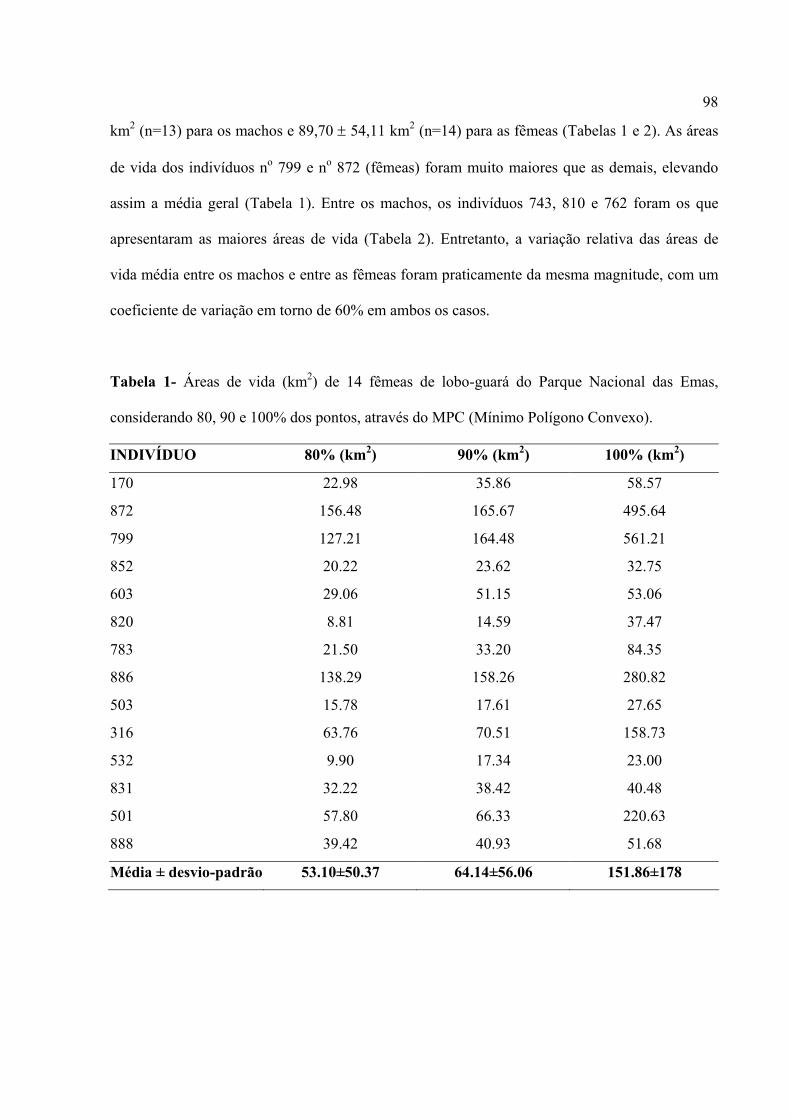
98
km2 (n=13) para os machos e 89,70 ± 54,11 km2 (n=14) para as fêmeas (Tabelas 1 e 2). As áreas
de vida dos indivíduos no 799 e no 872 (fêmeas) foram muito maiores que as demais, elevando
assim a média geral (Tabela 1). Entre os machos, os indivíduos 743, 810 e 762 foram os que
apresentaram as maiores áreas de vida (Tabela 2). Entretanto, a variação relativa das áreas de
vida média entre os machos e entre as fêmeas foram praticamente da mesma magnitude, com um
coeficiente de variação em torno de 60% em ambos os casos.
Tabela 1- Áreas de vida (km2) de 14 fêmeas de lobo-guará do Parque Nacional das Emas,
considerando 80, 90 e 100% dos pontos, através do MPC (Mínimo Polígono Convexo).
INDIVÍDUO 80% (km2) 90% (km2) 100% (km2)
170 22.98 35.86 58.57
872 156.48 165.67 495.64
799 127.21 164.48 561.21
852 20.22 23.62 32.75
603 29.06 51.15 53.06
820 8.81 14.59 37.47
783 21.50 33.20 84.35
886 138.29 158.26 280.82
503 15.78 17.61 27.65
316 63.76 70.51 158.73
532 9.90 17.34 23.00
831 32.22 38.42 40.48
501 57.80 66.33 220.63
888 39.42 40.93 51.68
Média ± desvio-padrão 53.10±50.37 64.14±56.06 151.86±178
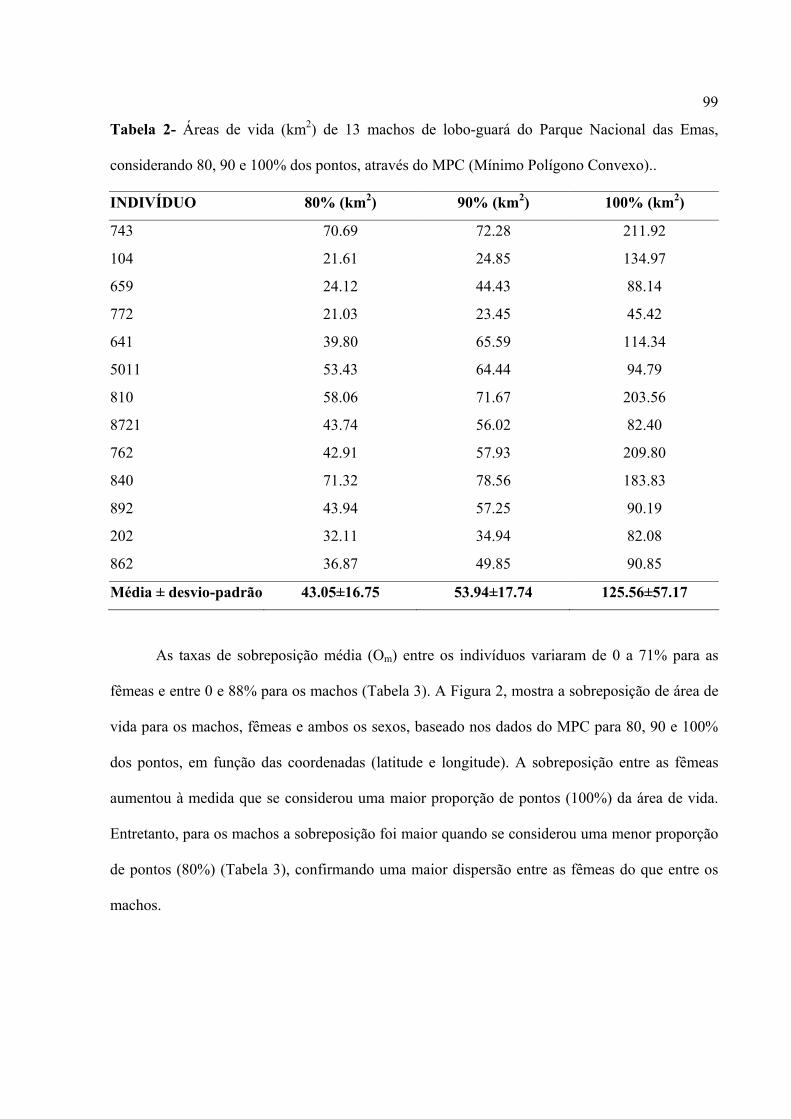
99
Tabela 2- Áreas de vida (km2) de 13 machos de lobo-guará do Parque Nacional das Emas,
considerando 80, 90 e 100% dos pontos, através do MPC (Mínimo Polígono Convexo)..
INDIVÍDUO 80% (km2) 90% (km2) 100% (km2)
743 70.69 72.28 211.92
104 21.61 24.85 134.97
659 24.12 44.43 88.14
772 21.03 23.45 45.42
641 39.80 65.59 114.34
5011 53.43 64.44 94.79
810 58.06 71.67 203.56
8721 43.74 56.02 82.40
762 42.91 57.93 209.80
840 71.32 78.56 183.83
892 43.94 57.25 90.19
202 32.11 34.94 82.08
862 36.87 49.85 90.85
Média ± desvio-padrão 43.05±16.75 53.94±17.74 125.56±57.17
As taxas de sobreposição média (Om) entre os indivíduos variaram de 0 a 71% para as
fêmeas e entre 0 e 88% para os machos (Tabela 3). A Figura 2, mostra a sobreposição de área de
vida para os machos, fêmeas e ambos os sexos, baseado nos dados do MPC para 80, 90 e 100%
dos pontos, em função das coordenadas (latitude e longitude). A sobreposição entre as fêmeas
aumentou à medida que se considerou uma maior proporção de pontos (100%) da área de vida.
Entretanto, para os machos a sobreposição foi maior quando se considerou uma menor proporção
de pontos (80%) (Tabela 3), confirmando uma maior dispersão entre as fêmeas do que entre os
machos.

100
Tabela 3. Taxas de sobreposição média (Om) entre os indivíduos de lobo-guará (Chrysocyon
brachyurus).
No de Pontos (%) Indivíduos Variação (%)
Fêmeas 0 – 61
Machos 0 – 88 80
Machos e Fêmeas 0 – 88
Fêmeas 0 – 68
Machos 0 – 81 90
Machos e Fêmeas 0 – 81
Fêmeas 0 – 71
Machos 0 – 75 100
Machos e Fêmeas 0 – 85

101
8014471
8004324
7994176
7984028
286569 296717 306865 317012
Latitude
Long
itude
a- Sobreposição de machos
MPC 80%
8014847
8004927
7995007
7985087
283712 293632 303552 313472
LatitudeLo
ngitu
de
d- Sobreposição de fêmeas
MPC 80%
8014587
8004418
7994234
7984057
283968 294145 304321 314498
Latitude
Long
itude
g- Sobreposição de machos e fêmeas
MPC 80%
8014471
8004324
7994176
7984028
286199 296347 306495 316642
Latitude
Long
itude
b- Sobreposição de machos
MPC 90%
8014847
8004927
7995007
7985087
281829 291749 301669 311589
Latitude
Long
itude
e- Sobreposição de fêmeas
MPC 90%
8014587
8004410
7994234
7984057
282085 292262 302438 312615
LatitudeLo
ngitu
de
h- Sobreposição de machos e fêmeas
MPC 90%
8025655
8012711
7999768
7986824
285684 298628 311571 324515
Latitude
Long
itude
c- Sobreposição de machos
MPC 100%
8016617
8006254
7995892
7985529
282271 292634 302996 313359
Latitude
Long
itude
f- Sobreposição de fêmeas
MPC 100%
8025655
8012711
7999768
7986824
284852 297796 310739 323683
Latitude
Long
itude
i- Sobreposição de machos e fêmeas
MPC 100%
Figura 2. Áreas de vida entre indivíduos de lobo-guará, considerando machos e fêmeas
separadamente e todos os indivíduos simultaneamente, para 80, 90 e 100% dos pontos do MPC
(Mínimo Polígono Convexo).
102
Variabilidade Genética e Associação com as Áreas de Vida
Dos 119 primers testados nas reações RAPD, nove deles (OPH-04, OPH-19, OPP-07,
OPP-08, OPP-09, OPP-15, OPC-11, OPC-05, OPA-18) mostraram um bom padrão de
amplificação e apresentaram um total de 91 locos, sendo 42 deles polimórficos (46%) (Tabela 4).
O índice de similaridade genética (coeficiente de Jaccard), entre os pares de indivíduos, variou de
0,66 a 0,97 entre as fêmeas, de 0,84 a 0,97 entre os machos e 0,66 a 0,99, quando considerado
ambos os sexos simultâneamente.
Tabela 4. Relação do número de locos e do número de locos polimórficos, por primer, em lobo-
guará (Chrysocyon brachyurus), utilizando marcadores RAPD.
Primer Número de Locos Número de Locos Polimórficos
OPH-04 14 7
OPH-19 8 3
OPP-07 9 4
OPP-08 9 5
OPP-09 7 4
OPP-15 11 4
OPC-11 12 5
OPC-05 11 4
OPA-18 10 4
TOTAL 91 42
Dentre 29 primers de STR testados (ver capítulo 1), desenvolvidos originalmente para
outras espécies, seis (21%) amplificaram com sucesso: PEZ-05, PEZ-20, PEZ-12, LOCO-13,

103
LOCO-15 e CSSM-038 (Denise et al., 2004; Halverson et al., 1995, Barker et al., 1997). Um
total de 27 locos foi obtido e destes 24 (88,88%) foram polimórficos. A similaridade genética
entre os pares de indivíduos variou de 0,09 a 0,80 quando considerado ambos os sexos
simultaneamente.
A associação entre os centros de áreas de vida (distância espacial) e similaridade genética
não foi significativa em nenhuma das condições analisadas (entre machos e fêmeas
separadamente e ambos os sexos simultaneamente), baseado nos dados RAPD (Tabela 5). Já a
associação entre as similaridades genéticas e espaciais foi significativa, apenas entre os machos
quando considerado 80 e 90% dos pontos, com base nos dados RAPD (Tabela 6). Assim,
indivíduos do sexo masculino com áreas mais sobrepostas são mais similares geneticamente.
Com base nos marcadores STR, observou-se uma associação significativa (para 80, 90 e 100%
dos pontos) entre os indivíduos de ambos os sexos simultaneamente e entre as fêmeas (para 100%
dos pontos) (Tabela 6). Portanto, indivíduos (ambos os sexos) mais similares geneticamente
ocupam áreas de vida mais próximas, sendo que existe uma maior tendência de associação entre
as fêmeas, como mostra a Tabela 6.
Tabela 5- Valores da correlação matricial (r ) e o nível de significância (P) obtidos a partir do
Teste de Mantel, comparando a similaridade genética (RAPD) e distância espacial entre os 27
indivíduos da população de lobo-guará (Chrysocyon brachyurus).
Indivíduos R P
Machos e Fêmeas -0,00934 0,41
Machos -0,01514 0,42
Fêmeas 0,05839 0,44
104
Tabela 6- Valores da correlação matricial (r ) e o nível de significância (P) obtidos a partir do
Teste de Mantel, comparando a similaridade genética com a sobreposição espacial (80, 90 e
100% dos pontos) do lobo-guará (Chrysocyon brachyurus), com base nos dados dos marcadores
RAPD e STR.
RAPD STR Indivíduos No de Pontos (%)
r P r P
80 0,015 0,44 0,182 0,003
90 0,013 0,44 0,197 0,003 Machos e Fêmeas
100 0,091 0,20 0,248 0,002
80 0,247 0,02 -0,109 0,199
90 0,230 0,02 -0,104 0,224 Machos
100 0,030 0,44 -0,112 0,267
80 -0,045 0,36 0,172 0,092
90 -0,011 0,45 0,194 0,061 Fêmeas
100 0,160 0,15 0,236 0,049
DISCUSSÃO
Áreas de vida
Os valores de área de vida obtidos para os Lobos Guará no PNE foram maiores do que os
encontrados por Silveira (1999), no mesmo local (PNE) (média de 48,98 ± 31,75 km2, n=5) e do
que os valores obtidos por Rodrigues (2002), na Estação Ecológica de Águas Emendadas - DF
(média 57 km2, n=5) e bem maiores que os dados de área de vida para a população de lobo-guará
do parque Nacional da Serra da Canastra (média 25,21± 4,45 km2, n=3). Os valores encontrados

105
neste estudo se aproximaram mais daqueles encontrados por Carvalho e Vasconcelos (1995), em
fragmentos de cerrado de São Paulo (média 75,03 ± 34,63 km2, n=3).
A variação no tamanho da área de vida de Lobos Guarás pode estar relacionada às
diferenças na disponibilidade alimentar, dentre outros fatores. Uma maior heterogeneidade
espacial e densidade de arbustos e árvores pode refletir uma maior disponibilidade de alimentos.
Como o entorno do Parque Nacional das Emas, área bastante utilizada por esta espécie e muitas
outras que habitam o mesmo, apresenta novas oportunidades de disponibilidade de alimentos,
visto que tratam-se de áreas de lavoura, pastagem e ainda reservas legais, esta podem ser uma
explicação do aumento da área de vida dos indivíduos deste local. Outro fator que poderia
explicar a variação no tamanho da área de vida desta espécie seria a densidade populacional,
onde locais com densidades maiores forçariam uma redução nas áreas individuais. Assim sendo,
grandes reservas são importantes na conservação dos Lobos Guarás, mas estas só serão efetivas
quando há conexão com outras reservas, favorecendo um acréscimo de indivíduos nas populações,
ou disponibilizando recursos alimentares suficientes para a população.
Alguns trabalhos (Rodrigues, 2002) indicam uma alta sobreposição de área de vida entre
indivíduos de diferentes populações de lobo-guará. Os casais de lobo-guará possuem áreas de
vida exclusivas, o que permite a participação do macho no cuidado parental (Dietz, 1984). Os
resultados deste trabalho corroboram parcialmente esta estruturação, mas foi observado uma
maior sobreposição de áreas entre os machos (em média 81%) do que entre as fêmeas (em média
67%). Porém, quando foi estimada a sobreposição considerando os indivíduos de ambos os sexos
simultaneamente, esta foi maior (em média 85%) (Tabela 3). Estas altas taxas de sobreposição
podem justificar a estruturação em pares, que acarreta vantagem ao casal, pois defendem sua área
de outros casais. A troca de parceiros sexuais pode ser facilitada pelas fêmeas por possuírem
áreas de vida maiores, fator que favorece a população por gerar um aumento da variabilidade
genética.

106
Correlações genéticas e espaciais
O grau de polimorfismo genético obtido com os marcadores RAPD (46%) e com os
marcadores STR (89%) foram razoavelmente altos, se comparados com outros estudos usando
marcadores protéicos, como o trabalho de De Mattos et al., (2004), que observou um
polimorfismo médio de 20% para a população de lobo-guará da região nordeste de São Paulo.
Nos trabalhos de Kennedy et al., (1991) e Waine et al., (1991) com lobos cinzentos (Canis lupus),
usando marcadores protéicos, a variabilidade genética mensurada foi de 13,5% e 20%,
respectivamente. Moreira et al., (1998), usando marcadores protéicos, em uma parcela bastante
representativa da população de lobo-guará (Sudeste e Centro-Oeste do Brasil), estimou uma
heterozigosidade média de 1,4 a 8,3%. De Mattos et al., (2004), em seu estudo com 28 Lobos
Guarás do Nordeste de São Paulo, usando também marcadores protéicos, mostrou ausência de
endocruzamento e de estruturação populacional, como conseqüência dos baixos valores de
distância genética. Estes estudos são corroborados pelos resultados deste trabalho no PNE, no que
diz respeito à diversidade genética, que em média foi alta baseando-se no alto polimorfismo
encontrado e na grande variação da similaridade genética observada entre os indivíduos, tanto
para os marcadores STR quanto para os marcadores RAPD. Isto indica que possivelmente uma
medida importante para a conservação do lobo-guará seria a preservação adequada da área que a
espécie ocupa, diminuindo a necessidade de introdução de novos animais no local, visto os
aspectos negativos das translocações e de densidades populacionais muito elevadas. A introdução
de novos animais pode acarretar problemas à população como, por exemplo, a introdução de
doenças infecciosas, diluição da freqüência de genes adaptados, etc (Campbell, 1980; Hedrick,
2001).
Em muitas espécies de mamíferos, a estrutura espacial da variabilidade genética em
pequenas escalas espaciais é determinada pela fidelidade dos indivíduos ao seu local de
nascimento ou grupo social, dependendo de alguns fatores, como sexo e idade (Roff e Fairbairn,

107
2001; Murren et al., 2001; Templeton, 1987; Greenwood, 1980). Em chipanzés (Pan troglotydes),
os grupos familiares tendem a ser patrilineares e as fêmeas têm uma maior dispersão (Wrangham,
1979).
Os resultados obtidos neste estudo, com base nos marcadores RAPD, mostraram uma
estruturação espacial da similaridade genética apenas para os machos, concordando com a
hipótese de que a distribuição espacial entre os machos não é aleatória e está associada com o
grau de parentesco genético. Esta estruturação poderia ser explicada pelo baixo padrão de
dispersão dos machos em relação às fêmeas nesta população, além da distribuição e
disponibilidade de recursos. Portanto, a alta taxa de sobreposição de área de vida entre os machos
aparentados pode estar sendo influenciada pela distribuição de recursos críticos, e não
necessariamente devido o grau de parentesco genético.
Um outro padrão espacial mais comum em mamíferos sociais é a filopatria natal e a
filopatria feminina, que também é um padrão observado em algumas espécies de mamíferos
solitários, como o guaxinim (Procyon lotor) (Ratnayeke et al., 2002). Os jovens freqüentemente
demonstram filopatria natal (Waser e Jones 1983), ou seja, eles permanecem dentro ou próximos
da área natal, dividindo espaço e recursos alimentares com os progenitores, e presumivelmente se
beneficiam desta maior familiaridade, favorecendo o forrageamento, obtenção de melhores
abrigos e maiores chances contra espécies predadoras. Para as fêmeas de mamíferos que investem
muito nos cuidados da prole, os benefícios ecológicos e sociais tendem a favorecer fortemente a
filopatria feminina (Wrangham, 1980). Este padrão espacial tem vantagens e custos. Ao mesmo
tempo em que o grau de parentesco entre as fêmeas aumenta o risco de endogamia,
conseqüentemente da redução da variabilidade genética, a consangüinidade pode favorecer a
seleção de genes que determinam uma boa adaptação a um determinado habitat.
Os resultados obtidos neste estudo com os marcadores STR corroboram a estruturação
espacial da similaridade genética entre os indivíduos de ambos os sexos, com uma maior

108
tendência entre as fêmeas (filopatria feminina), ou seja, as fêmeas mais similares geneticamente
tendem a ocupar a mesma área de vida ou áreas bem próximas. Como esse padrão é mais
esperado do que o observado com os dados RAPD, pode-se levantar a possibilidade de que os
resultados com os marcadores RAPD sejam artefatos. De fato, algumas estimativas de
diversidade genética com marcadores RAPD são questionáveis, devido principalmente a natureza
dominante do marcador.
Alguns estudos mostram que a filopatria não necessariamente é influenciada pelo sexo e
pode ser direcionada pela idade e, neste caso é mais comum entre machos e fêmeas adultos
ocupando a mesma área de vida dos machos (Gompper e Wayne, 1996). No entanto, não está
claro, para mamíferos solitários como o lobo-guará, se indivíduos filopátricos que dividem
espaço têm uma maior tolerância em conseqüência do grau de parentesco genético. Ratnayeke
(1997) observou em outro estudo que a variação no relacionamento genético foi independente da
interação espacial e temporal entre fêmeas de guaxinim. Em algumas espécies, como esquilos
(Van Staaden et al., 1994) a filopatria foi mais fortemente determinada pela sobreposição
espacial do que pela seleção de sociabilidade entre parentes. Neste trabalho, mesmo tendo sido
observada uma associação genética e espacial significativa, entre os indivíduos de ambos os
sexos e entre as fêmeas, com base nos marcadores STR e entre os machos com base nos
marcadores RAPD, os valores de r obtidos foram baixos, em torno de 0,23. Portanto, deve-se
levar em consideração que quando ambos indivíduos são beneficiados com a divisão de espaço,
aumenta-se à chance de tolerância entre eles e, neste caso, a estrutura populacional varia muito
mais em decorrência da demografia local e distribuição de recursos do que devido ao grau de
relacionamento genético entre os indivíduos.

109
AGRADECIMENTOS
Aos colegas do Laboratório de Genética e Melhoramento da Universidade Católica de Goiás -
UCG tais como, Profo. Breno de Faria Vasconcellos, Nicole C. L. Dutra, Gabriela C. Pádua,
Lorenna L. de Souza, Luciana M. Bossois e todos os demais estagiários que nos ajudaram nas
análises genéticas. A todo o pessoal do Parque Nacional das Emas - PNE, principalmente os
estagiários que realizaram os trabalhos de campo. Ao CNPq pelo financiamento de nossas
pesquisas dentro do programa PRONEX para análise de áreas prioritárias para conservação da
biodiversidade no Cerrado e em Goiás. (CNPq/SECTEC-GO, Proc. No. 23234156). JAFD-F
recebe uma bolsa de Produtividade em Pesquisa nível 1B do CNPq.
REFERÊNCIAS BIBLIOGRÁFICAS
Barker JSF, Moore S, Hetzel S, Evan D, Tan SG e Byrne, K (1997) Genetic diversity of Asian
water buffalo (Bubalus bubalis): microsatellite variation and comparison with protein-coding
loci. Animal Genetics 28: 103-115.
Bassan BJ, Anollés GCE e Gresshoff PM (1991) Fast and sensitive silver staining of DNA in
polyacrilamide gels. Analytical Biochemistry 196: 80-83.
Campbell S (1980) Is a reintroduction a realistic goal? In: Soulé M.E and Wilcox BW (eds)
Conservation Biology: an evolutionary–ecological perspective. Sinauer Association,
Sunderland, 395 pp.
Carvalho CT e Vasconcellos LEM (1995) Disease, food and reproduction of the maned wolf
Chrysocyon brachyurus (Illiger) (Carnivora, Canidae) in southeast Brazil. Rev Bras Zoo 12:
627-640.
Consorte-McCrea AG (1994) The maned wolf in captivity. Canid News 2. www.canids.org..

110
Cowan WF (1973) Ecology and life history of the raccoon (Procyon lotor hirsutus, Nelson and
Goldman) in the northern part of its range . PhD Thesis, University of North Dakota, Dakota.
De Mattos PSR, Del Lama M.A, Toppa RH e Schwantes AR (2004) Populational Genetic
Structure of free-living maned wolves (Chrysocyon brachyurus) determined by proteic
markers. Braz J Biol 64: 639-644.
De Mattos PSR. (2003) Epidemiologia e genética populacinal do lobo-guará Chrysocyon
brachyurus (Illiger, 1815) (Carnivora, Canidade) na região nordeste do estado de São Paulo.
Tese de Doutorado em Ecologia e Recursos Naturais, Universidade Federal de São Carlos,
São Paulo.
Denise S, Johnston E, Halverson J, Marshall K, Rosenfeld D, Mckenna S, Sharp T e Edwards J
(2004) Power of exclusion for parentage verification and probability of match for identity in
American kennel club breeds using 17 canine microsatellite markers. Animal Genetics 35:
14–17.
Dietz JM (1984) Ecology and social organization of maned wolf Chrysocyon brachyurus. Tese
Doctor of Philosophy, Michigan State University, Washington.
Dietz JM (1985) Chrysocyon brachyurus. Mammalia Speices 234: 1-4.
Dixon KR e Chapman JA (1980) Harmonic mean measure of animal activity areas. Ecology 61:
1040-1044.
Eisenberg JF e Redford KH (1999) Mammals of the Neotropics - The Central Neotropics:
Ecuador, Peru, Bolivia, Brazil. University of Chicago Press, Chicago, 609pp.
Ellstrand NC e Elam DR (1993) Population genetic consequences os small population size:
Implications for plant conservation. Annual Review of Ecology and Systematics 24: 217-242.

111
Ferreira ME e Grattapaglia D (1998) Introdução ao Uso de Marcadores Moleculares em Análise
Genética. Editora EMBRAPA, Brasília.
Fonseca GAB, Rylands AB, Costa CRM, Machado RB e Leite YLR (1994) Livro Vermelho Dos
Mamíferos Brasileiros Ameaçados de Extinção. Fundação Biodiversitas, Belo Horizonte.
Frankham R, Ballou JD e Briscoe DA (2003) Introduction to Conservation Genetics. Cambridge
University Press, Cambridge.
Ginsberg JR e MacDonald DW (1990) Foxes, wolves, jackals, and dogs: an action plan for the
conservation of canids. Gland: IUCN/SSC Canid Specialist Group & IUCN/SSC Wolf
Specialist Group.
Gompper ME e Wayne RK (1996) Genetic relationships among individuals within carnivore
societies. In: Gittleman JL (ed) Carnivore Behavior Ecology and Evolution. Cornell
University Press, Ithaca, NY, pp 429-452.
Greenwood PJ (1980) Mating systems, philopatry and dispersal in birds and mammals. Animal
Behavior 28: 1140–1162.
Halverson J, Dvorak J e Stevenson T (1995) Microsatellite sequences for canine genotyping. US
Patent 05874217.
Hedrick, PW (2001) Conservation genetics: where are we now? TRENDS in Ecology &
Evolution 16: 629-636.
Hillis DM, Mable BK e Moritz C (1996) Molecular systematics. Sinauer Associates, Sunderland,
MA. 655pp.
IBAMA (2003) Lista das Espécies da Fauna Brasileira Ameaçadas de Extinção. Anexo à
Instrução Normativa n° 3, de 27 de maio de 2003, do Ministério do Meio Ambiente.
Disponível em: www.ibama.gov.br.

112
IBDF/FBCN, (1981) Plano de manejo: Parque Nacional das Emas. Instituto Brasileiro do Meio
Ambiente e Recursos Naturais Renováveis, Brasília.
Illiger K (1815) Uberlick der Saugerthiere nach ihrer Verteilung uber die Welttheile. Akademie
der Wissenshaften Berlin Abhandlungen 1804-1811. pp 39-159.
Jacob AA e Rudran R (2003) Rádiotelemetria em estudos populacionais. In: Cullen Jr L, Rudran
R and Pádua CV (eds) Métodos de Estudos em Biologia da Conservação & Manejo da Vida
Silvestre. Editora da UFPR e Fundação O Boticário de Proteção à Natureza, Curitiba, pp.
285-341.
Jácomo ATA (1999) Nicho alimentar do lobo-guará Chrysocyon brachyurus (Illiger, 1811) no
Parque Nacional das Emas-GO. Dissertação de Mestrado em Ecologia, Universidade Federal
de Goiás, Goiânia.
Jácomo ATA, Silveira L e Diniz-Filho JAF (2004) Niche separation between the maned wolf
(Chrysocyon brachyurus), the crab-eating fox (Dusicyon thous) and the hoary fox (Dusicyon
vetulus) in central Brazil. J. Zool. Lond. 262: 99–106.
Kennedy PK, Kennedy ML, Clarkson PL e Liepins IS (1991) Genetic variability in natural
populations of the gray wolf, Canis lupus. Can. J. Zool 69: 1183-1188.
Kenward RE, South AB e Walls SS (2003) Ranges6 vl.2: For the analysis of tracking and
locartion data. Online Manual. Anatrack Ltd. Walrehan, UK. ISBN 0-9546327-0-2.
Kimberley, AN (1998) Maned wolf (Chrysocyon brachyurus). Disponível em:
http://straylight.tamu.edu/bene/esa/raesg/manedwolf.html.
Krebs JK (1998) Ecological Methodology. 2nd ed. Bemjamin/Cummings imprint. Menlo Park.
Califónia, USA.

113
Lombardi JA e Motta-Jr JC (1993) Seed Dispersal of Solanum lycocarpum St. Hil. (Solanaceae)
by the Maned Wolf, Chrysocyon brachyurus, Illiger (Mammalia, Canidae). Ciência e Cultura
45:126-127.
Maia OB e Gouveia AMG (2002) Birth and mortality of maned wolves Chrysocyon brachyurus
(Illiger, 1815) in captivity. Braz J Biol. 62:25-32.
Manly BFJ (1991) Randomization and Monte Carlo Methods in Biology. Chapman & Hall,
London, 281 pp.
Mantovani JE (2001) Telemetria convencional e via satélite na determinação das áreas de vida de
três espécies de carnívoros da região nordeste do estado de São Paulo. Tese de Doutorado em
Ecologia e Recursos Naturais, Universidade Federal de São Carlos, São Paulo.
Mohr CO (1947) Table of equivalent populations of North American small mammals. American
Midland Naturalist 37:223-249.
Moreira JR, Guimarães NK, Pilla EJS, Contel EPB e Bem ARD (1998) Estudo preliminar da
variabilidade genética do lobo-guará (Chrysocyon brachyurus). Comunicado técnico
EMBRAPA 30:1-10.
Motta-Jr JC (1996) Diet of the maned wolf, Chrysocyon brachyurus, in central Brazil. Journal
Zoology 240: 277-284.
Motta-Jr JC (2002) Fama injusta: novas informações sobre a dieta do lobo-guará podem ajudar a
preservá-lo. Ciência Hoje 31: 71-73.
Murren JC, Julliard R, Schlichting CD e Clobert J (2001) Dispersal, individual phenotype, and
phenotypic plasticity. In: Clobert J, Danchin E, Dhondt, AA and Nichols JD (eds) Dispersal.
Oxford university Press, New York, pp 261-272.

114
Myers N, Mittermeier RA, Mittermeier CG, Fonseca GAB e Kent J (2000) Biodiversity hotspots
for conservation priorities. Nature 403: 853-858.
Nams VO (2001) Locate II, (versão 2.82): A program to triangulate radiotelemetry bearings and
calculate error ellipses. Disponível em www.nsac.ns.ca/envsci/staff/vnams/Locate.htm.
Primack RB e Rodrigues E (2001) Biologia da Conservação. E. Rodrigues, Londrina, 328 pp.
Ratnayeke MS (1997) Female philopatry and spatial organization in the raccoon, Procyon lotor.
PhD Thesis, University of Tennessee, Knoxville.
Ratnayeke S, Tuskan GA e Pelton MR (2002) Genetic relatedness and female spatial organization
in a solitary carnivore, the raccoon, Procyon lotor. Molecular Ecology 11: 1115-1124.
Rodrigues FHG (2002) Biologia e conservação do lobo-guará na Estação Ecológica de Águas
Emendadas, DF. Tese de Doutorado em Ecologia, Universidade Estadual de Campinas,
Campinas.
Rodrigues FM, Pádua GCC, Resende LV, Telles MPC, Jácomo ATA, Silveira L e Bossois LM
(2004) Levantamento do status da variabilidade genética na população de lobo-guará
(Chrysocyon brachyurus - Illiger, 1815) do Parque Nacional das Emas. Resumos do 50o
Congresso Nacional de Genética, Florianópolis, Brazil.
Roff DA e Fairbairn, DJ (2001) The genetic basis of dispersal and migration and its consequences
for the evolution of correlated traits. In: Clobert J, Danchin E, Dhondt, AA and Nichols JD
(eds) Dispersal. Oxford university Press, New York, pp 191-202.
Rohlf FJ (1991) NTSYS: Numerical taxonomy and multivariate analysis system. Exeter
Publishing Co, New York.
Shaffer ML (1981) Minimum population sizes for species conservation. BioScience 31: 131-134.

115
Silva JA e Talamoni AS (2003) Diet adjustments of maned wolves Chrysocyon brachyurus
(Illiger) (Mammalia, Canidade) subjected to supplemental feeding in a private natural reserve,
Southeastern Brazil. Revista Brasileira de Zoologia 20: 339-345.
Silva JA e Talamoni AS (2004) Core area and center of activity of maned wolves Chrysocyon
brachyurus (Illiger) (Mammalia, Canidade), submitted to supplemental feeding. Revista
Brasileira de Zoologia 21: 391-395.
Silveira L (1999) Ecologia e conservação dos mamíferos carnívoros do Parque Nacional das
Emas, Goiás. Dissertação de Mestrado em Ecologia, Universidade Federal de Goiás, Goiânia.
Silveira L e Jácomo ATA (1998) Ecologia e conservação de mamíferos carnívoros do Parque
Nacional das Emas, GO. Relatório Técnico para o IBAMA/DIREC/DEUC.
Sneath PHA e Sokal RR (1973) Numerical taxonomy. W. H. and Freeman, San Francisco.
Tanaka R (1980) Controversial problems in advanced research on estimating population densities
of small rodents. Research Population Ecology Supplement 2: 1-66.
Tautz D (1989) Hypervariability of simple sequences as a general source of polymorphic DNA
markers. Nucleic Acids Res. 17: 6463-6471.
Templeton, AR (1987) Inferences on natural population structure from genetic studies on captive
mammalian population. In: Chepko-Sade DB and Halpin ZT (eds) Mammalian dispersal
patterns: The effects of social structure on population genetics. The University of Chicago
Press, Chicago e London, pp 257-272.
Van Staaden MJ, Chesser RK e Michener GR (1994) Genetic correlation and matrilineal structure
in a population of Spermophilus richardsonii. Journal of Mammalogy 75: 573-582.
Waser PM e Jones WT (1983) Natal philopatry in solitary mammals. Quarterly Review of
Biology 58: 355-390.

116
Wayne RK, Gilbert DA, Lehman N, Hansen K, Eisenhawer A, Girman D, Peterson RO, Mech
LD, Gogan PJP, Seal US e Krumenaker RJ (1991) Conservation genetics of endangered Isle
Royale gray wolf. Conservation Biol 5: 41-51.
Welsh J e McClelland M (1990) Fingerprinting genomes using PCR with arbitrary primers.
Nucleic Acids Reaserch 18: 7213-7218.
Whitte GC e Garrot RA (1990) Analysis of Wildlife Radio-Tracking Data. Academic, San Diego.
Williams JGK, Kubelik Α, Livak K, Rafalsski J e Tingey SV (1990) DNA polimorphisms
amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Reaserch 18:
6531-6535.
Wrangham RW (1979) On the evolution of ape social systems. Social Science Information 18:
334 –368.
Wrangham RW (1980) An ecological model of female-bonded primate groups. Behaviour 75:
262 –300.

117
CAPÍTULO 4

118
Artigo submetido: Genetics and Molecular Biology
Extinção por Depressão Endogâmica sob Estocasticidade Ambiental
em uma População de lobo-guará (Chrysocyon brachyurus) e outra
de cachorro-do-mato (Cerdocyon thous) no Sudoeste do Estado de
Goiás: Um Estudo de Simulação
FLÁVIA MELO RODRIGUES1,2 E JOSÉ ALEXANDRE FELIZOLA DINIZ-FILHO3,4* 1Universidade Estadual de Goiás, Departamento de Biologia; Anápolis-G0; 2Programa de Pós-
Graduação em Ciências Ambientais, UFG; 3Departamento de Biologia Geral, ICB,
Universidade Federal de Goiás (UFG); 4 Departamento de Biologia, MCAS, Universidade
Católica de Goiás (UCG);
Resumo
A perda de variabilidade genética e a depressão endogâmica são os dois principais fatores
genéticos que podem levar uma população à extinção. Populações pequenas têm níveis mais
baixos de diversidade genética do que populações grandes, devido à ocorrência de deriva
genética. A extinção de uma população é uma conseqüência de um feedback positivo entre o
declínio do tamanho populacional e aumento de endogamia, sendo este fenômeno conhecido por
“vortex” de extinção. Flutuações no tamanho populacional devido a estocasticidade ambiental
interagem com a depressão endogâmica e reforçam o vortex de extinção. O Cerrado do sudoeste
goiano, no qual está inserido o Parque Nacional das Emas, está extremamente fragmentado e com
um intenso processo de ocupação humana, principalmente devido à atividade agrícola, que
provoca fortes ameaças à diversidade biológica. O objetivo deste trabalho foi estimar o tempo de
extinção em duas populações de canídeos do Parque Nacional das Emas, o lobo-guará e o
cachorro-do-mato, utilizando um modelo teórico que assume que a combinação dos efeitos da
depressão endogâmica sob estocasticidade ambiental são os possíveis fatores que levam a
extinção dos organismos em populações isoladas. As simulações das dinâmicas populacionais das
duas espécies se basearam em três equações que avaliam a dinâmica da média da freqüência
alélica de genes deletérios (q), o coeficiente de endogamia (F) e o tamanho da população (N). A
heterozigosidade média esperada foi calculada partir dos dados de seis locos STR e usada para
estimar o tamanho efetivo das populações, com base no processo mutacional passo-a-passo

119
(Stepwise). Assim sendo, as simulações para cada espécie foram realizadas usando 12
combinações de três variáveis, i.e., as taxas de redução da capacidade de suporte do ambiente (K),
os coeficientes de endogamia (F) e o desvio-padrão do tamanho da população (sN). Em seguida,
foi realizada uma Análise de Variância Fatorial (ANOVA) para modelar a variação no tempo de
extinção em função dos parâmetros usados e suas interações. A média do tempo de extinção foi
afetado pelos três parâmetros analisados e suas interações, principalmente K e sN. Quando
considerado um pior cenário, ou seja, maior flutuação populacional, alta endogamia e altas taxas
de diminuição da capacidade de suporte, o tempo médio de extinção do lobo-guará foi igual a 363
anos, e para o cachorro-do-mato esse tempo foi igual a 431 anos. Uma análise comparativa,
utilizando uma combinação semelhante de parâmetros para as duas espécies, mostrou que a
diminuição da população de lobo-guará é sempre mais rápida que a da população de cachorro-do-
mato. Este resultado é coerente com expectativas baseadas em parâmetros demográficos e
história de vida diferentes nas duas espécies. Estes resultados mostram que as populações podem
persistir por um tempo razoável mesmo no pior cenário. Porém, deve-se estar atento para estes
resultados, uma vez que este estudo é uma projeção teórica baseada em muitas suposições, e é
bastante liberal principalmente quanto ao tamanho efetivo inicial. Sendo assim, este estudo deve
ser um ponto de partida para futuras avaliações mais precisas da persistência destas populações,
que possam gerar resultados baseados em parâmetros mais realistas da viabilidade destas
populações na região sudoeste do Estado de Goiás.
Palavras-chave: Análise de viabilidade populacional, Extinção, Canídeos, Marcadores STR,
Parque Nacional das Emas.
Enviar correspondência para FM Rodrigues. Universidade Estadual de Goiás, Departamento de
Biologia, Unidade Universitária de Ciências Extas e Tecnológicas, BR 153, Km 3, Campus
Anápolis, CEP: 75110-390, Caixa Postal: 459, Anápolis, GO, Brasil. E-mail: [email protected]

120
Abstract
A typical question addressed by modern programs in conservation biology and the basis
of the so called population viability analysis (PVA) is what is the chance that a population
persists for a given number of years, given certain conditions of habitat and human activities.
Modern PVA analysis must combine factors at genetic, demographic and landscape levels to
create more realistic and precise models predicting population persistence. In this study, we used
Tanaka’s (2000) model of extinction in stochastic environments under inbreeding depression to
mode time to extinction in populations of two canid species living in Emas National Park (ENP)
and surroundings, the maned wolf (Chysocyon brachiurus) and the crab-eating fox (Cerdocyon
thous). Initial estimation of some genetic parameters in the population were based on 6 STR loci,
furnishing estimates of the effective population size, under a stepwise mutation model based on
heterozygosis, and of inbreeding levels of the two species. Our simulations were defined using 12
scenarios, allowing variations in variable rates of habitat loss, magnitude of population
fluctuation and initial inbreeding levels. ANOVA of simulation results showed that times to
extinction were better explained by demographic parameters, and based on large (probably
overestimated) population sizes, the variation in these times for the large-bodied maned wolf
ranged, on average, from 352 to 844, in the worst and best scenario, respectively. For the small-
bodied crab-eating fox, these same estimates are 422 and 974 years. Variations among
simulations are within expectation based on knowledge about species’ life-history, genetics and
demography. Although our simulations show that the populations can persistence through a
reasonable time (i.e., more than 100 years), these results are probably sensitive to unrealistic
assumption on initial population sizes. Despite this, our analyses are a starting point for a more
focused evaluation of persistence in these populations and can drive future research towards
obtained better estimates of parameters that can, in turn, be used to fuel more appropriate and
realist PVA models.
Key words: Population persistence, Population Viability Analysis, Canids, STR markers, Emas
National Park

121
INTRODUÇÃO
A perda de variabilidade genética e a depressão endogâmica são os dois principais fatores
genéticos que podem levar uma população à extinção (Lynch e Lande, 1993; Barret e
Charlesworth, 1991; Soule, 1986). Possivelmente, a contribuição recente mais importante em
relação à genética da conservação seja o reconhecimento da importância da depressão
endogâmica quando se trata de espécies ameaçadas, de modo que a redução da endogamia tem se
tornado uma prioridade em programas de conservação in situ e ex situ (Hedrick, 2001). A
depressão endogâmica resulta no aumento da homozigosidade de mutações deletérias recessivas
em pequenas populações e, portanto, um número menor de descendentes, ou descendentes
estéreis são algumas das características da depressão endogâmica (Charlesworth e Charlesworth,
1987; Ralls et al., 1988). Populações pequenas têm níveis mais baixos de diversidade genética do
que populações grandes. Isto se deve à ocorrência de deriva genética, onde alguns alelos podem
aumentar sua freqüência, outros diminuem e alguns alelos podem ser perdidos completamente
(Frankham et al., 2002). Alguns mecanismos presentes em grandes populações evitam a
endogamia, ou seja, os indivíduos não acasalam com parentes próximos porque eles se dispersam
do seu local de nascimento ou o acasalamento entre parentes é inibido por odores ou outras pistas
sensoriais (Primack e Rodrigues, 2001). Entretanto, em populações com baixas densidades
demográficas, estes mecanismos não conseguem evitar a endogamia. Assim sendo, este fator
(endogamia) pode contribuir para um declínio no tamanho da população e conseqüentemente
favorecer a probabilidade de extinção (Ellstrand e Elam, 1993).
A extinção de uma população é uma conseqüência de um feedback positivo entre o
declínio do tamanho populacional e aumento de endogamia, sendo este fenômeno é conhecido
por “vortex” de extinção (Frankham et al., 2002; Hedrick, 2001). Vortex de extinção é um
sistema de retroalimentação, envolvendo diversos processos genéticos e demográficos. Faz com
que o tamanho populacional, uma vez reduzido abaixo de um dado valor crítico, tenda a se

122
reduzir cada vez mais, levando a população (e, em segundo momento, toda a espécie) à extinção.
Porém, as interações entre fatores genéticos, demográficos e ambientais fazem com que seja
extremamente difícil identificar as causas imediatas para qualquer evento particular de extinção
(Frankham et al., 2002).
Correlações entre aptidão (fitness), heterozigosidade e taxa de endogamia estimadas de
populações naturais fornecem evidências indiretas de depressão endogâmica na natureza (Britten,
1996; Tanaka, 2000a). Não é muito clara a interpretação das estimativas de heterozigosidades
para o processo de extinção. Primeiro, porque existem poucos exemplos de extinção populacional
devido à depressão endogâmica, e segundo porque as investigações das propriedades teóricas do
vortex de extinção por depressão endogâmica tornam difícil avaliar o risco genético de extinção
baseada em dados empíricos (Tanaka, 2000a). Alguns estudos têm considerado o vortex de
extinção usando modelos genéticos e populacionais e sugerem que a rápida extinção devido à
depressão endogâmica é restrita, porém possível com os parâmetros genéticos observados
(Tanaka, 1997, 1998, 2000b).
Se a depressão endogâmica for causada por mutações deletérias recessivas que estão
“escondidas” nos heterozigotos, as grandes populações estão mais sujeitas à depressão
endogâmica do que as pequenas populações. Isso porque, uma grande população que no passado
passou por um efeito “bottleneck” e manteve uma alta heterozigosidade conterá mais mutações
deletérias do que as populações pequenas (Hedrick, 2001). Em pequenas populações, por outro
lado, a maioria das mutações deletérias são rapidamente eliminadas pela seleção, que é mais
eficiente em baixas taxas de heterozigosidades (Tanaka, 1997). Sendo assim, a história
demográfica de uma população pode ter uma importante relação com a extinção, uma vez que a
endogamia pode ser induzida por distúrbios demográficos, genéticos e estocasticidade ambiental
(Tanaka, 1997).

123
As flutuações no tamanho populacional devido a estocasticidade ambiental, tais como
temperatura, disponibilidade de alimento, competição, interagem com a depressão endogâmica e
reforçam o vortex de extinção (Tanaka, 2000b). Segundo Primack e Rodrigues (2001), as maiores
ameaças à diversidade biológica que resultam da atividade humana são: destruição, fragmentação
e degradação do habitat, superexploração das espécies para uso humano, introdução de espécies
exóticas e aumento de ocorrência de doenças. Dentre as principais causas de ameaças de extinção
em mamíferos estão a perda de habitat, responsável por 68%, e a superexploração, que inclui a
caça comercial, esportiva e de subsistência, responsável por 54 % (Reid e Miller, 1989).
O Cerrado da região sudoeste do Estado de Goiás, no qual está inserido o Parque Nacional
das Emas (PNE), está extremamente fragmentado, e com um intenso processo de ocupação
humana. Por exemplo, apenas cerca de 27% do entorno do PNE, em um raio de 30 Km, possui
áreas de remanescentes florestais (Figura 1). Fazem parte desta região extensas lavouras de milho,
soja e pastagem (Pedroso, 2004). Basicamente, o que impulsionou a inserção dos Cerrados nas
áreas produtivas e, conseqüentemente, o desenvolvimento agrícola do Centro-Oeste, foram
alguns programas específicos implementados nessa região ainda nos anos 1950-1960. Dentre os
principais, destacam-se o Programa de Desenvolvimento dos Cerrados (POLOCENTRO) e o
Programa de Cooperação Nipo-Brasileira para Desenvolvimento dos Cerrados (PRODECER)
(Bezerra e Cleps-Júnior, 2004).
O sudoeste goiano teve um desenvolvimento diferenciado das demais regiões do Estado
por ser uma área agropecuária de ocupação bastante antiga, próxima ao Triângulo Mineiro e São
Paulo. Inicialmente, a principal atividade econômica na região era a criação de gado. A partir de
1960, a região passa a vivenciar uma nova fase de desenvolvimento agrícola, incentivada pelo
pacote tecnológico da Revolução Verde e principalmente, pela intervenção do Estado por meio
de políticas desenvolvimentistas (Pedroso, 2004). Com isto ocorre uma desenfreada ocupação
humana no cerrado goiano, em especial na região Sudoeste de Goiás, provocando o

124
desmatamento, afetando dentre outros processos a infiltração da água fluvial e assim o
abastecimento de vários lençóis freáticos, diminui drasticamente. Como conseqüência, vários rios
já secaram nos domínios do cerrado, além de erosões e compactação do solo, devido à pesada
mecanização (Pedroso, 2004; Rodrigues et al., 2001).
Figura 1. Remanescentes florestais no entorno do Parque Nacional das Emas (Adaptado de
Mário Barroso Neto, 2005)
O objetivo deste trabalho foi estimar o tempo de extinção em duas populações de
Canídeos do sudoeste do Estado de Goiás, o lobo-guará (Chrysocyon brachyurus) e o cachorro-
do-mato (Cerdocyon thous), utilizando o modelo de Tanaka (2000a), que considera a depressão
endogâmica e estocasticidade ambiental como possíveis fatores que levam a extinção dos
organismos. O cenário ecológico começou com uma análise genética de populações de cada uma
dessas duas espécies, situadas no Parque Nacional de Emas e usando dados de marcadores

125
moleculares Microssatélites ou STR (Short Tandem Repeats) (Rodrigues et al., 2005; ver capítulo
2). O valor da heterozigosidade esperada (He) foi utilizado para estimar o tamanho efetivo da
população, sob um modelo mutacional stepwise e os marcadores STR foram utilizados para uma
estimativa inicial dos coeficientes de endogamia (F) nas populações. Para a realização das
simulações, foram obtidos outros parâmetros, utilizando abordagens macroecológicas, e os
resultados foram discutidos segundo o conhecimento global da região estudada e a história de
vida das espécies. Nas simulações, assumiu-se que a combinação dos efeitos de isolamento, perda
de hábitat (redução na capacidade de suporte da região) e estocasticidade ambiental podem levar
as populações à extinção.
MATERIAL E MÉTODOS
O modelo genético
As simulações da dinâmica populacional das duas espécies estudadas se basearam em
Tanaka (2000a), que descreve um modelo de extinção por depressão endogâmica sob
estocasticidade ambiental. Assume-se, portanto, que a depressão endogâmica envolve genes
deletérios recessivos, distribuídos entre os n locos autossômicos bi-alélicos, com taxas de
mutação, coeficientes de seleção e dominância semelhantes. O modelo também assumiu que os
efeitos do desequilíbrio de ligação e interações epistáticas são desprezíveis. Os genótipos AA, Aa
e aa, em cada loco, têm um fitness médio igual a 1, 1 e 1 – s, respectivamente, onde s é o
coeficiente de seleção.
O modelo de Tanaka (2000a) se baseia em três equações dinâmicas recorrentes, para a
média da freqüência alélica q de genes deletérios, o coeficiente de endogamia F e o tamanho da
população N. Estas equações são
qt+1 = qt – s[q2 + Ft q (1 – q)] + µ + γ (eq. 1)

126
Ft+1 = ( 1/2Nt + (1 – 1/2Nt)Ft] (1 – 2µ) (1 – sqt) (eq. 2)
Nt+2 = Nt+1 exp[rmax (1 – (Nt+1/ Kt+1) + ε] δ (eq. 3)
Na equação (1), µ é a taxa de mutação e γ é uma variável aleatória com média 0 e desvio
padrão igual a (q (1 - q)/ 2nN)1/2, onde n é o número de locos. A equação (3) mostra que o
tamanho da população N cresce de acordo com um modelo logístico discreto no tempo t, com
capacidade de suporte K e uma flutuação em torno da média de N com o desvio padrão ε. O
parâmetro Maltusiano (taxa de crescimento intrínseca) é dado por rmax, assim sendo um aumento
discreto de N por geração, λ, é determinado por
λ = exp [(rmax)(1 – Nt/Kt)] (eq. 4)
Porém, a população também está sofrendo efeitos de eventos externos, e assim pressupõe-
se que a capacidade de suporte K varia com o tempo e está sendo reduzida com uma taxa h por
unidade de tempo t (em anos, que é a escala da taxa de crescimento λ). Assim, temos que
Kt+1 =Kt - hKt (eq. 5)
O modelo original de Tanaka (2000a) assume uma diminuição exponencial, mas neste
estudo assumiu-se uma diminuição linear e constante através do tempo, gerando uma diminuição
mais rápida em K. Devido os intervalos relativamente pequenos de N (veja abaixo), esta pequena
mudança fará pouca diferença em relação ao modelo original. Além disso, o tamanho da
população é reduzido pelo fator δ, que é uma função das freqüências alélicas q e de como os
genes deletérios afetam a taxa de crescimento da população. Assim, o crescimento da população
está relacionado com a carga genética L e é dado como λδ, onde
δ = (1 – L) = 1 – nsq2 (eq. 6)
Os cálculos descritos acima foram implementados no programa GLOADRUN, escrito em
linguagem Q-BASIC e que será disponibilizado aos interessados mediante solicitação (Apêndice
1).

127
Contexto ecológico e Parametrização do Modelo
O modelo descrito acima foi utilizado para avaliar a dinâmica populacional dentro da
população de lobo-guará e de cachorro-do-mato do Parque Nacional das Emas - GO e regiões do
entorno (Figura 2). A partir das análises de variabilidade genética, utilizando seis locos STR
(Rodrigues et al., 2005; ver capítulo 2) obteve-se a heterozigosidade média esperada para cada
população, iguais a 0,533 e 0,676 para lobo-guará e cachorro-do-mato, respectivamente. Para o
processo mutacional, foi utilizado o modelo Stepwise (SMM – Stepwise Mutation Model, Nei e
Kumar, 2000) a fim de se estimar o tamanho efetivo da população, com base na seguinte equação
He = 1 - [1/(1 + 8Nu)1/2] (eq. 7)
onde He é a heterozigosidade esperada, N é o tamanho efetivo, u é a taxa de mutação por
geração.
O modelo SMM é o mais aceito para marcadores STR e admite que a mutação nos alelos
resultam da perda ou ganho de uma única unidade de repetição, de modo que os alelos poderiam
variar apenas através de estados alélicos anteriormente presentes na população. Assim sendo,
pode-se então aproximar o tamanho efetivo da população de lobo-guará e cachorro-do-mato de
toda região (não necessariamente dentro do PNE) como sendo Ne ≈ 500 e Ne ≈ 1000,
respectivamente. Multiplicando-se estes valores por um fator conservador igual a 2 (Morris &
Doak, 2002), obtem-se uma estimativa inicial razoável do tamanho da população N na região, na
ausência de estimativas empíricas melhores.
O desvio padrão do tamanho da população (sN) foi fixados em 5 ou 10 indivíduos, e assim
este parâmetro pode causar pequenos efeitos em densidades populacionais altas, mas grandes
efeitos em pequenas densidades populacionais, promovendo um vortex de extinção.
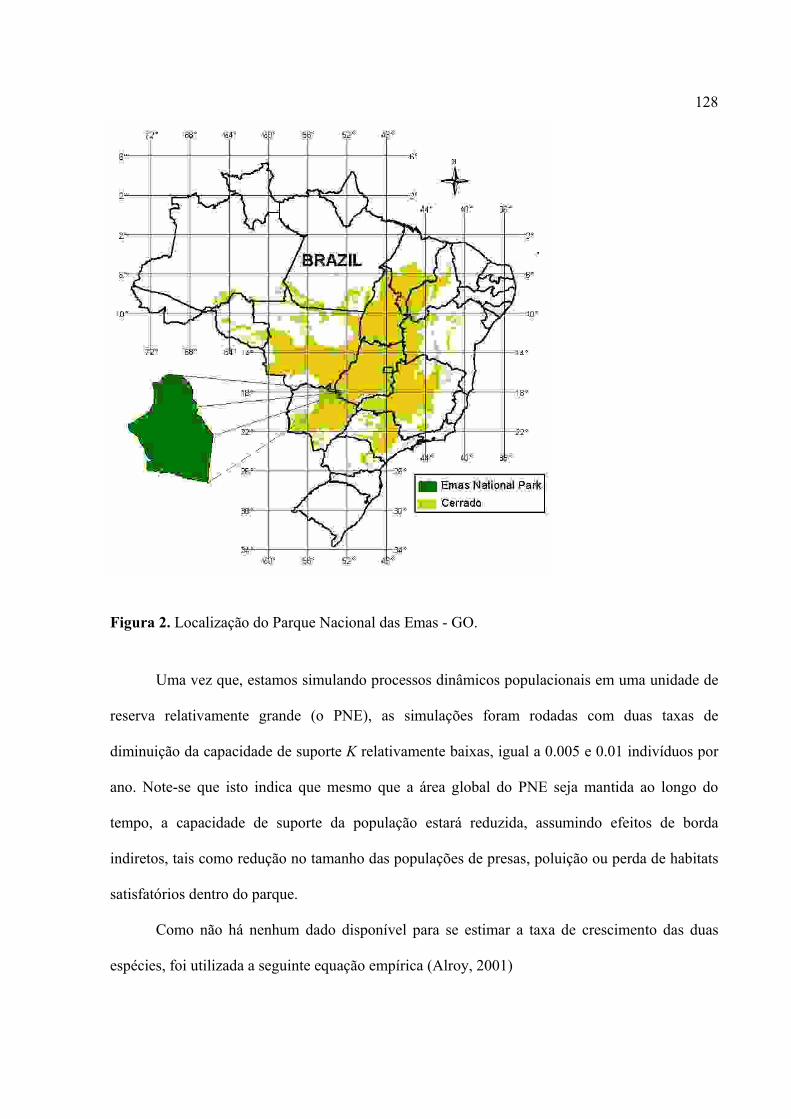
128
Figura 2. Localização do Parque Nacional das Emas - GO.
Uma vez que, estamos simulando processos dinâmicos populacionais em uma unidade de
reserva relativamente grande (o PNE), as simulações foram rodadas com duas taxas de
diminuição da capacidade de suporte K relativamente baixas, igual a 0.005 e 0.01 indivíduos por
ano. Note-se que isto indica que mesmo que a área global do PNE seja mantida ao longo do
tempo, a capacidade de suporte da população estará reduzida, assumindo efeitos de borda
indiretos, tais como redução no tamanho das populações de presas, poluição ou perda de habitats
satisfatórios dentro do parque.
Como não há nenhum dado disponível para se estimar a taxa de crescimento das duas
espécies, foi utilizada a seguinte equação empírica (Alroy, 2001)

129
rmax = exp[ 1.4967 – 0.37 (logM)] (eq. 8)
Onde logM é o logaritmo natural da massa corpórea das espécies, definido como sendo
igual a 24 Kg para o lobo-guará e 6 Kg para o cachorro-do-mato (Einsenberg e Redford, 1999).
Assim, a taxa de crescimento intrínseca máxima para as duas espécies foi igual a 0,10 e 0,17 para
o lobo-guará e cachorro-do-mato, respectivamente.
Devido à grande variação nos coeficientes de endogamia (F) calculados por loco para as
duas espécies (ver capítulo 2) usando os seis locos STR, que variaram de -0,394 a 0,197 (média
igual a -0,091) para o lobo-guará e -0,055 a 0,205 (média igual a 0,032) no cachorro-do-mato,
decidiu-se correr as simulações com três valores iniciais baixos de F, iguais a 0.01, 0.05 e 0.10.
Os outros parâmetros de carga genética foram iguais àqueles utilizados por Tanaka (2000a), de
tal forma que µ = 10-6 , n = 15000 locos e coeficiente de seleção s igual a 1.
Assim sendo, as simulações para cada espécie foram realizadas usando 12 combinações
dos três parâmetros: “baixa” e “alta” taxas de redução da capacidade de suporte K (0.005 e 0.01);
“baixos” “médios” e “altos” coeficientes de endogamia F (0.01, 0.05 e 0.10) e “baixo” e “alto”
desvio padrão do tamanho da população sN (5 e 10 indivíduos) (Tabela 1). Para cada combinação
de parâmetros, foram executadas 500 simulações e o tempo de extinção Te foi obtido.
Para avaliar como a combinação dos parâmetros utilizados afetam Te, uma análise de
variância fatorial (ANOVA) (Sokal & Rohlf, 1985) foi usada para modelar a variação em Te
como uma função dos parâmetros usados e suas interações.

130
Tabela 1- Identificação dos 12 cenários simulados, a partir das combinações dos três parâmetros
utilizados, que incluem variações na capacidade de suporte (K), no desvio padrão no tamanho das
populações (sN ) e no coeficiente de endogamia (F).
Cenários Combinações
1 K = 0,005; sN = 5; F = 0,01
2 K = 0,005; sN = 5; F = 0,05
3 K = 0,005; sN = 5; F = 0,10
4 K = 0,005; sN = 10; F = 0,01
5 K = 0,005; sN = 10; F = 0,05
6 K = 0,005; sN = 10; F = 0,10
7 K = 0,01; sN = 5; F = 0,01
8 K = 0,01; sN = 5; F = 0,05
9 K = 0,01; sN = 5; F = 0,10
10 K = 0,01; sN = 10; F = 0,01
11 K = 0,01; sN = 10; F = 0,05
12 K = 0,01; sN = 10; F = 0,10

131
RESULTADOS
De acordo com a Análise de Variância (ANOVA), a média do tempo de extinção, Te, para
as duas espécies (Tabela 2) foi afetada pelos três parâmetros analisados e suas interações,
principalmente pela taxa da redução na capacidade de suporte e pelo desvio padrão do tamanho
da população.
Tabela 2- Valores de F e nível de significância (P), obtidos a partir da Análise de Variância
(ANOVA), dos três parâmetros analisados: redução na capacidade de suporte (K); desvio padrão
no tamanho das populações (sN) e coeficiente de endogamia (F) e suas interações, em relação ao
tempo de extinção (Te) nas duas espécies estudadas.
F e (P) Fontes de Variação
lobo-guará cachorro-do-mato
K 271470 (0.000) 353855 (0.000)
sN 21113 (0.000) 19756 (0.000)
F 0.680 (0.507) 0.912 (0.402)
K* sN 2241 (0.000) 2128 (0.000)
K*F 0.574 (0.563) 3.239 (0.039)
sN *F 1.586 (0.205) 1.548 (0.213)
K* sN *F 0.299 (0.742) 3.732 (0.024)
Como esperado, os piores cenários (i.e., Te mais curto) ocorreram para ambas as espécies
quando as populações flutuaram mais e apresentaram alta endogamia, vivendo em ambientes com
alta taxa de diminuição na capacidade de suporte (Tabelas 3 e 4).
Para o lobo-guará, os valores de Te variaram de 413 a 291 anos, quando considerado o
pior cenário (cenário 12), e entre 932 a 756 anos, quando considerado o melhor cenário (cenário
1) (Tabela 3). Os principais fatores que afetaram o Te nesta espécie foram a redução na
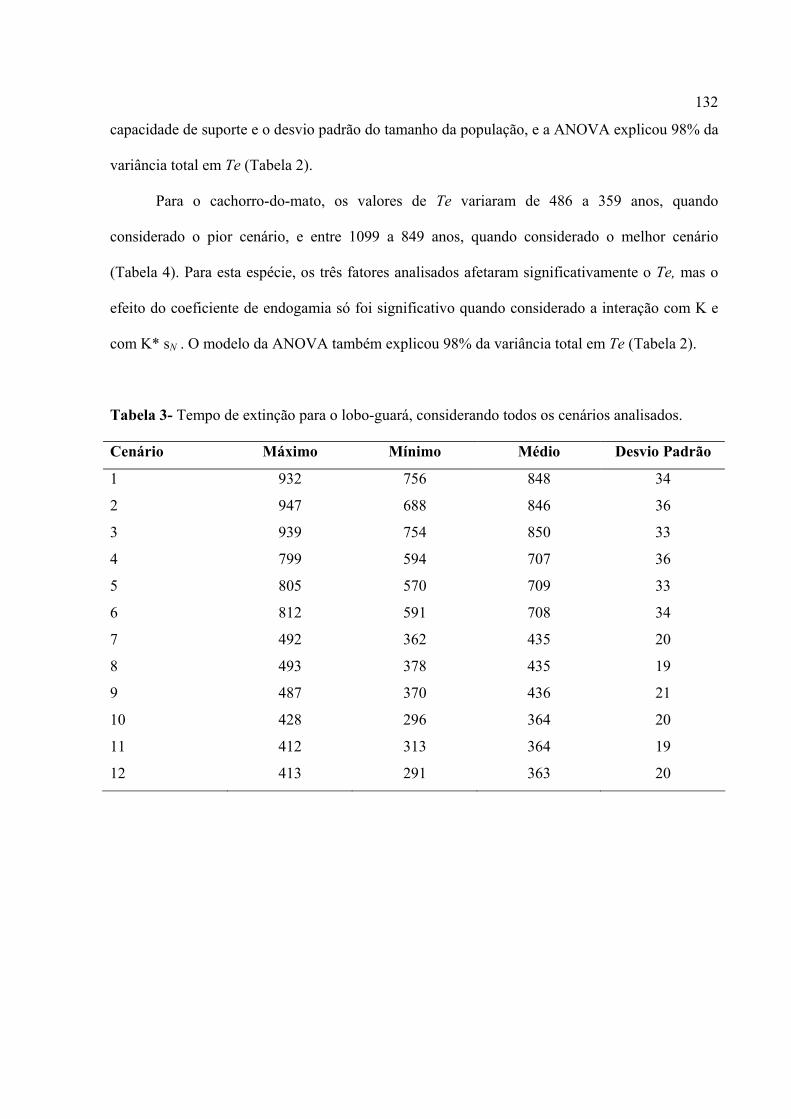
132
capacidade de suporte e o desvio padrão do tamanho da população, e a ANOVA explicou 98% da
variância total em Te (Tabela 2).
Para o cachorro-do-mato, os valores de Te variaram de 486 a 359 anos, quando
considerado o pior cenário, e entre 1099 a 849 anos, quando considerado o melhor cenário
(Tabela 4). Para esta espécie, os três fatores analisados afetaram significativamente o Te, mas o
efeito do coeficiente de endogamia só foi significativo quando considerado a interação com K e
com K* sN . O modelo da ANOVA também explicou 98% da variância total em Te (Tabela 2).
Tabela 3- Tempo de extinção para o lobo-guará, considerando todos os cenários analisados.
Cenário Máximo Mínimo Médio Desvio Padrão
1 932 756 848 34
2 947 688 846 36
3 939 754 850 33
4 799 594 707 36
5 805 570 709 33
6 812 591 708 34
7 492 362 435 20
8 493 378 435 19
9 487 370 436 21
10 428 296 364 20
11 412 313 364 19
12 413 291 363 20
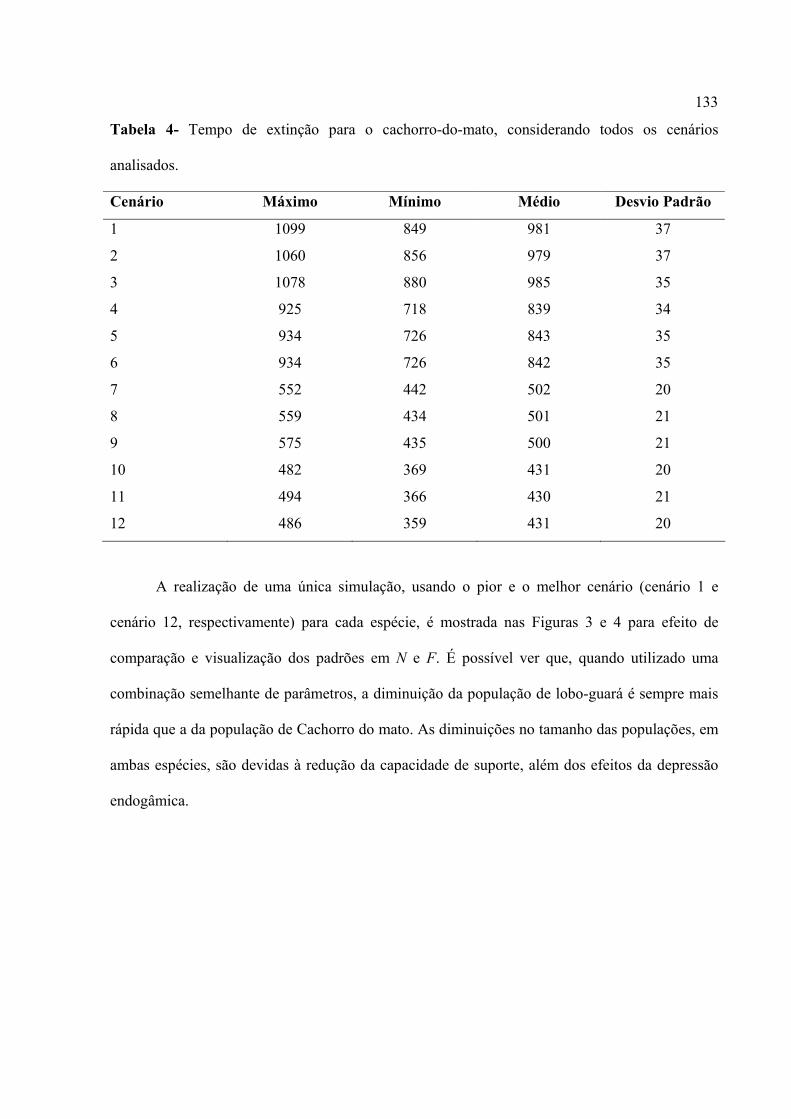
133
Tabela 4- Tempo de extinção para o cachorro-do-mato, considerando todos os cenários
analisados.
Cenário Máximo Mínimo Médio Desvio Padrão
1 1099 849 981 37
2 1060 856 979 37
3 1078 880 985 35
4 925 718 839 34
5 934 726 843 35
6 934 726 842 35
7 552 442 502 20
8 559 434 501 21
9 575 435 500 21
10 482 369 431 20
11 494 366 430 21
12 486 359 431 20
A realização de uma única simulação, usando o pior e o melhor cenário (cenário 1 e
cenário 12, respectivamente) para cada espécie, é mostrada nas Figuras 3 e 4 para efeito de
comparação e visualização dos padrões em N e F. É possível ver que, quando utilizado uma
combinação semelhante de parâmetros, a diminuição da população de lobo-guará é sempre mais
rápida que a da população de Cachorro do mato. As diminuições no tamanho das populações, em
ambas espécies, são devidas à redução da capacidade de suporte, além dos efeitos da depressão
endogâmica.
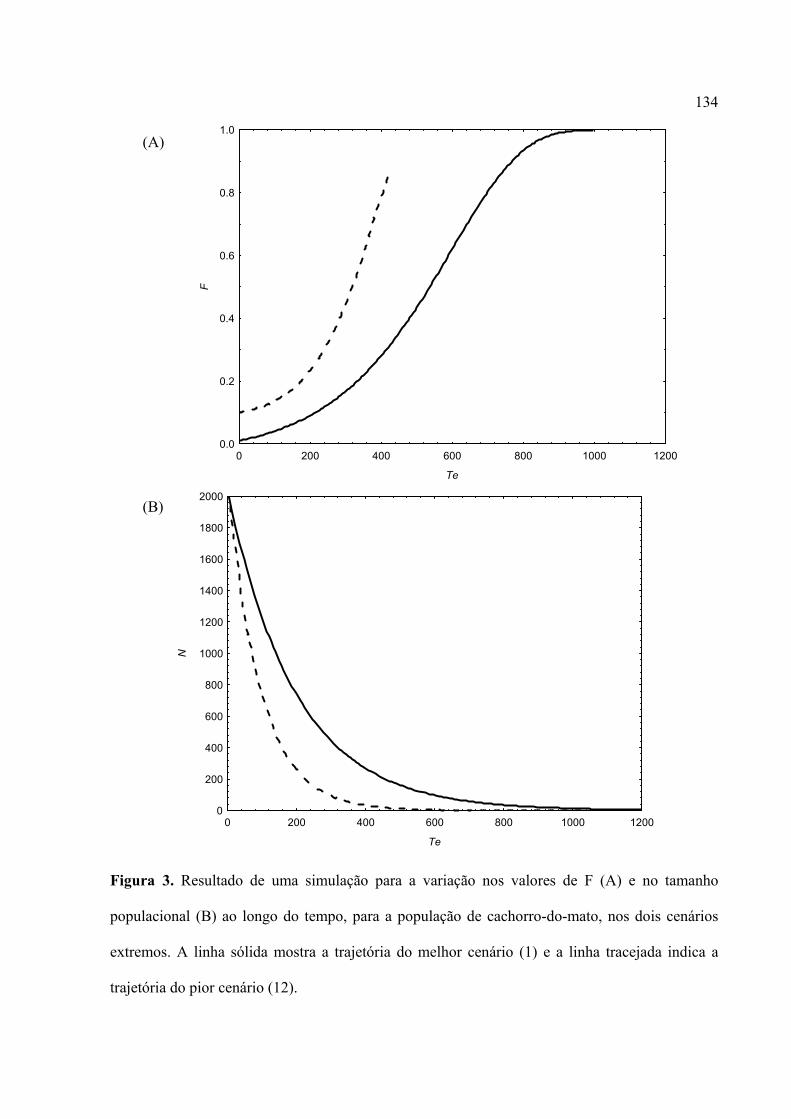
134
0 200 400 600 800 1000 1200
Te
0.0
0.2
0.4
0.6
0.8
1.0
F
0 200 400 600 800 1000 1200
Te
0
200
400
600
800
1000
1200
1400
1600
1800
2000
N
Figura 3. Resultado de uma simulação para a variação nos valores de F (A) e no tamanho
populacional (B) ao longo do tempo, para a população de cachorro-do-mato, nos dois cenários
extremos. A linha sólida mostra a trajetória do melhor cenário (1) e a linha tracejada indica a
trajetória do pior cenário (12).
(A) (A)
(B)
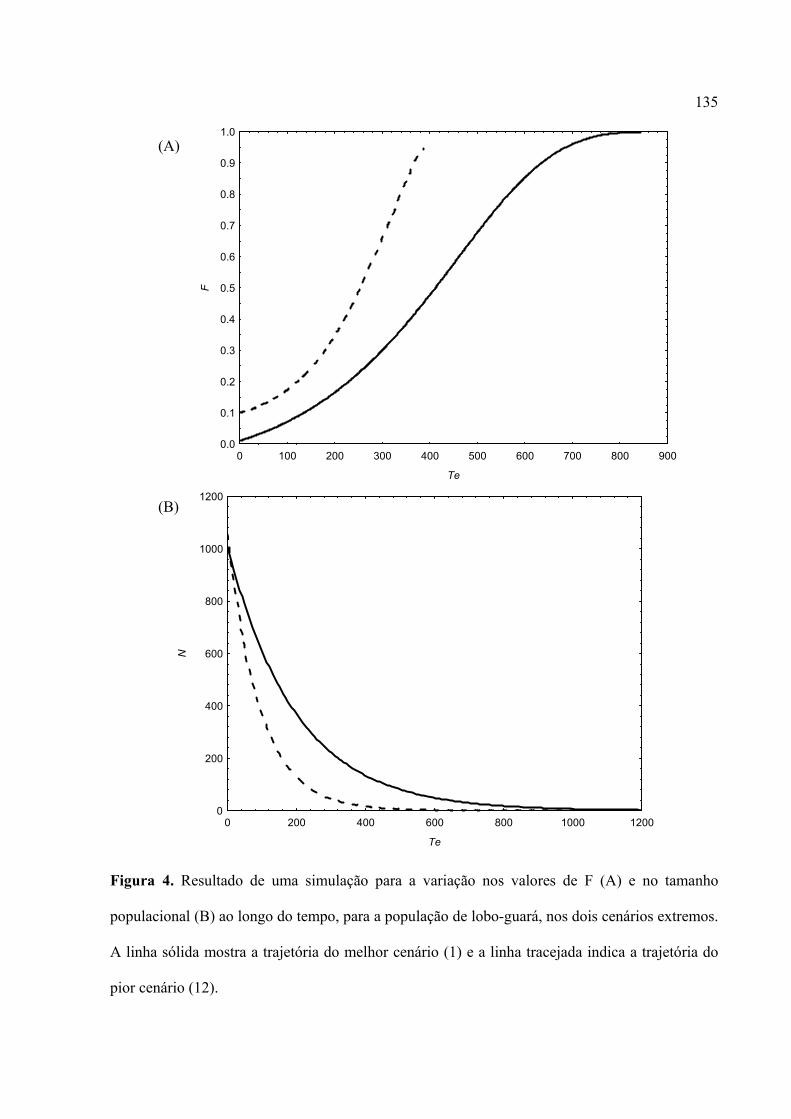
135
0 100 200 300 400 500 600 700 800 900
Te
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
F
0 200 400 600 800 1000 1200
Te
0
200
400
600
800
1000
1200
N
Figura 4. Resultado de uma simulação para a variação nos valores de F (A) e no tamanho
populacional (B) ao longo do tempo, para a população de lobo-guará, nos dois cenários extremos.
A linha sólida mostra a trajetória do melhor cenário (1) e a linha tracejada indica a trajetória do
pior cenário (12).
(A)
(B)

136
DISCUSSÃO
Há um tempo atrás, considerava-se que a contribuição de fatores genéticos para o destino
de espécies em extinção era um fator secundário. Sugeria-se que flutuações demográficas e
estocasticidade ambiental causariam extinção antes que a deterioração genética se tornasse uma
séria ameaça as populações naturais. Porém, as respostas evolutivas de populações frente à
estocasticidade ambiental e demográfica não são independentes da endogamia e da diversidade
genética. A endogamia, em média, reduz as taxas de natalidade e aumentam os índices de
mortalidade e pode influenciar nas relações sexuais entre os indivíduos (Frankham et al., 2002)
Neste estudo, os principais fatores analisados que afetaram o tempo de extinção (Te) nas
duas espécies foram a taxa de redução na capacidade de suporte (K) e o desvio padrão no
tamanho da população (sN). Porém, para a população de cachorro-do-mato, o coeficiente de
endogamia (F) foi um fator importante quando considerado a interação com K e com sN . No
entanto, é importante salientar que a ausência de significância estatística do coeficiente de
endogamia nas analises de extinção para a população de lobo-guará observada neste estudo não
significa que este fator não é importante, e sim que o intervalo de variação utilizado aqui talvez
não tenha sido o bastante para gerar variação no Te final, especialmente considerando uma
variação estocástica ao redor de N. Além disso, alguns estudos com grandes mamíferos mostram
que estas espécies podem não estar sujeitas à depressão endogâmica por conterem baixas taxas de
heterozigosidade, provavelmente devido a um antigo “bottleneck” (Vilà et al., 2002; Merola,
1994; O’Brien et al., 1983; Sage e Wolff, 1986).
Segundo Tanaka (1997), populações muito pequenas provavelmente sofrem grande risco
de extinção devido outros fatores. Alguns estudos mostram que a estocasticidade ambiental e
demográfica colocam em grande risco as populações muito pequenas (Leigh, 1981; Goodman,
1987, Iwasa e Mochizuki, 1988; Lande, 1993). Assim sendo, provavelmente estes efeitos
estocásticos interagem com a depressão endogâmica no estágio final da extinção (Tanaka, 1997).

137
Existem algumas evidências empíricas de que a depressão endogâmica é reforçada em função da
deterioração ambiental, ou seja, o risco de extinção devido à endogamia é mais acelerado em
ambientes deteriorados (habitats fragmentados, devido extensas lavouras, como no PNE) do que
em ambientes ideais (Liberg et al. 2005; Templeton e Read, 1983; Soule e Kohn, 1989; Pray et
al., 1994).
A análise comparativa dos fatores nas duas populações são coerentes com expectativas
baseadas em parâmetros demográficos e história de vida diferentes nas duas espécies (Wayne,
1996). O lobo-guará é uma espécie com o tamanho do corpo maior do que o cachorro-do-mato,
ou seja, ocupam grandes áreas de vida, assim sendo as taxas de crescimento são mais baixas e
estas espécies são mais rapidamente extintas em escalas regionais do que as espécies de pequeno
porte (Henle et al., 2004). Por outro lado, em espécies de porte menor, tais como o cachorro-do-
mato, as taxas de crescimento demográfico são mais elevadas, embora essas sejam usualmente
mais variáveis e estejam mais sujeitas a extinções locais. Entretanto, a extinção total é difícil em
função dos eventos constantes de recolonização (Rocha et al., 2004; Rodrigues et al., 2002;
Beissinger, 2000; Raup, 1991; Silveira e Jácomo, 1998).
Espécies mais generalistas e abundantes, como o cachorro-do-mato, geralmente são
beneficiadas pela fragmentação de habitats, tal como a induzida, por exemplo, pela expansão
agrícola, pois freqüentemente estes animais possuem grande mobilidade, são hábeis para o uso de
diversos habitats e itens alimentares, sendo capazes de explorar também as bordas (Gehring e
Swihart, 2003). Por outro lado, espécies, como o lobo-guará, com alto requerimento individual de
área de vida, baixa abundância, alta flutuação populacional, baixo potencial reprodutivo e certa
especialização de habitat, mostram-se mais sensíveis à fragmentação (Henle et al., 2004).
É claro que as simulações executadas neste estudo só devem ser vistas como avaliações
muito preliminares dos riscos de extinção e viabilidade dessas populações, devido a muitos
fatores. Primeiro, as simulações foram baseadas em algumas suposições que podem ser, em

138
primeira instância, irreais. Por exemplo, não há nenhuma informação detalhada sobre a carga
genética destas populações e como esta pode afetar alguns parâmetros populacionais básicos, tais
como a taxa de mortalidade e fecundidade. Até mesmo uma boa descrição dos parâmetros
populacionais, como taxa de crescimento e flutuação de populacional, não foi realizada. Outra
questão é que foram utilizados diferentes valores de F nas simulações. No entanto, mesmo que
estas estimativas sejam instáveis (mas, normalmente baixas), de um modo geral são obtidos de
fato baixos níveis de endogamia nas duas espécies e em outras espécies de canídeos em
condições semelhantes (De Mattos et al., 2004; Wayne, 1996).
Para este trabalho, nós estimamos o tamanho inicial das populações N com base nas
heterozigosidades esperadas (He), calculadas a partir dos dados de STR. No entanto, a relação
entre o tamanho observado e o tamanho efetivo das espécies é desconhecida. Assumir uma
população de 1000 indivíduos para o lobo-guará e 2000 indivíduos para o cachorro-do-mato em
toda a região (não necessariamente apenas no PNE) não é irrealista, e estas estimativas seriam
muito menores se utilizássemos equações alométricas baseadas na massa corpórea das espécies
(Alroy, 2001, Diniz-Filho, 2004). Mesmo assim, estas estimativas iniciais de 1000 e 2000
indivíduos para as duas espécies podem estar muito superestimadas. Essas estimativas seriam
mais adequadas como um valor inicial da população reprodutiva antes da ocupação humana
acentuada, em equilíbrio genético. Esse valor pode ter sido consideravelmente reduzido nos
últimos 50 anos, especialmente considerando o alto nível de perda de hábitat e ocupação humana
na região sudoeste do Estado de Goiás (apesar da presença do PNE). O modelo utilizado aqui não
assumiu diretamente a fragmentação de hábitat, um componente importante na paisagem da
região, embora este componente possa ser indiretamente capturado pela taxa global de perda de
hábitat (capacidade de suporte).
Uma análise considerando valores mais reais do tamanho destas populações (Leandro
Silveira, comunicação pessoal) no PNE e regiões do entorno foi realizada. Foi estimada uma

139
população com 80 indivíduos para o lobo-guará e 150 indivíduos para o cachorro-do-mato.
Obviamente, o Te foi bem menor neste cenário do que nas análises considerando 1000 e 2000
indivíduos. Para estas estimativas empíricas do tamanho populacional, em média o tempo de
extinção considerando o pior cenário (12) para o lobo-guará foi igual a 116 anos (desvio padrão =
18), com um máximo de 157 anos e mínimo igual a 57 anos, enquanto que para o cachorro-do-
mato o Te médio foi igual a 172 anos (desvio padrão = 19), com um máximo de 231 anos e um
mínimo igual a 106 anos. Quando foi analisado o melhor cenário (1) as estimativas do Te médio
foi igual a 346 anos (desvio padrão = 36), para o lobo-guará e 465 anos (desvio padrão = 35),
para o cachorro-do-mato.
Tanaka (1997), sugere que o tamanho efetivo de uma população pode não reduzir-se
rapidamente, se a heterozigosidade é alta e a taxa intrínseca de crescimento populacional é
suficientemente alta para compensar a endogamia. Uma rápida diminuição no tamanho efetivo da
população pode induzir o vortex de extinção, devido à depressão endogâmica. Vários fatores
podem diminuir o tamanho efetivo de uma população. Alguns estudos mostram que, se a
população está geograficamente subdividida, com restrição na migração entre as subpopulações,
o tamanho efetivo da população pode diminuir rapidamente, sem alterar o tamanho atual da
população (Blanco et al., 2005; Crow e Aoki, 1984).
Assim sendo, os resultados das simulações realizadas aqui mostram que estas populações
podem persistir por um tempo razoável no pior cenário (12), em média 363 anos o lobo-guará e
431 anos o cachorro-do-mato (assumindo 1000 indivíduos de lobo-guará e 2000 indivíduos de
cachorro-do-mato) (Tabelas 3 e 4). Porém, deve-se estar atento para estes resultados, uma vez
que este estudo é uma projeção teórica baseada em muitas suposições, possivelmente irreais dos
processos populacionais que dirigem as populações para a extinção. Sendo assim, este estudo é
um ponto de partida para futuras avaliações mais precisas da persistência destas populações, onde
melhores estimativas dos parâmetros devem ser avaliadas, a fim de se obter resultados mais

140
adequados e modelos mais realistas da viabilidade destas populações na região sudoeste do
Estado de Goiás.

141
AGRADECIMENTOS
Nós agradecemos a Mariana Pires de Campos Telles, Leandro Silveira e Anah T. A. Jácomo
pelas valiosas discussões sobre a genética de populações e conservação de canídeos do Parque
Nacional das Emas - GO. Ao CNPq pelo financiamento de nossas pesquisas dentro do programa
PRONEX para análise de áreas prioritárias para conservação da biodiversidade no Cerrado e em
Goiás. (CNPq/SECTEC-GO, Proc. No. 23234156).
REFERÊNCIAS BIBLIOGRÁFICAS
Alroy J (2001) A multi-species overkill simulation of the end-Pleistocene megafaunal mass
extinction. Science 292: 1893-1896.
Barrett SCH e Charlesworth D (1991) Effects of a change in the level of inbreeding on the genetic
load. Nature 352: 522-524.
Beissinger SR (2000) Ecological mechanisms of extinction. Proceedings of National Academy of
Science USA 97: 11688-11689.
Bezerra LMC e Cleps Júnior J (2004) O desenvolvimento agrícola da região Centro-Oeste e as
transformações no espaço agrário do estado de Goiás. Caminhos da Geografia: 29-49.
Blanco JC, Cortés Y e Virgós E (2005) Wolf response to two kinds of barriers in an agricultural
habitat in Spain. Can. J. Zool. 83: 312–323.
Britten HB (1996) Meta analyses of the association between multi locus heterozygosity and
fitness. Evolution 50: 2158 2164.
Charlesworth D e Charlesworth B (1987) Inbreeding depression and its evolutionary
consequences. Annual Review of Ecology and Systematics 18: 237–268.

142
Crow JF e AOKI K (1984) Group selection for polygenic behavioral trait: estimating the degree
of population subdivision. Proceedings of the National Academy of Sciences of the United
States of America 81:6073-6077.
De Mattos PSR, Del Lama MA, Toppa RH e Schwantes AR (2004) Populational Genetic
Structure of free-living maned wolves (Chrysocyon brachyurus) determined by proteic
markers. Braz J Biol 64: 639-644.
Diniz Filho JAF (2004) Macroecological Analyses Support the Overkill Scenario for the Late
Pleistocene Extinctions. Revista Brasileira de Biologia 64: 407-414.
Eisenberg JF e Redford KH (1999) Mammals of the Neotropics - The Central Neotropics:
Ecuador, Peru, Bolivia, Brazil. University of Chicago Press, Chicago, 609pp.
Ellstrand NC e Elam DR (1993) Population genetic consequences of small population size:
implications for plant conservation. Annual Review of Ecology and Systematics 24: 217-242.
Frankham R, Ballou JD e Briscoe DA (2002) Introduction to conservation genetics. Cambridge
University Press. 640pp.
Gehring TM e Swihart RK (2003) Body size, niche breadth, and ecologically scaled responses to
habitat fragmentation: mammalian predators in an agricultural landscape. Biological
Conservation 109: 283-295.
Goodman D (1987) The demography of chance extinction. In: Soulé ME (ed.) Viable Populations
for Conservation. Cambridge University Press, Cambridge, pp 11-34.
Hedrick PW (2001) Conservation genetics: where are we now? Trends in Ecology & Evolution
16: 629-636.
Henle K, Lindemayer DB, Margules CR, Saunders DA e Wissel C (2004) Species survival in
fragmented landscapes: where are we now? Biodiversity and Conservation 13: 1-8.

143
Iwasa Y e Mochizuki H (1988) Probability of population extinction accompanying a temporary
decrease of population size. Res. Popul. Ecol. 30: 145–164.
Lande R (1993) Risks of population extinction from demographic and environmental stochasticity
and random catastrophes. Am. Nat. 142: 911–927.
Leigh EG (1981) The average lifetime of a population in a varying environment. Journal of
Theoretical Biology 90: 213-239.
Liberg O, Andrén H, Pedersen HC, Sand H, Sejberg D, Wabakken P, Åkesson M e Bensch S
(2005) Severe inbreeding depression in a wild wolf Canis lupus population. Biology Letters 1:
17-20.
Lynch M e Lande R (1993) Evolution and extinction in response to environmental change. In: P
Kareiva, J Kingsolver e R Huey (eds.) Biotic Interactions and Global Change. Sinauer
Assocs., Inc. Sunderland, MA, pp 234-250.
Merola M (1994) A reassessment of homozygosity and the case for inbreeding depression in the
cheetah, Acinonyx jubatus: implications for conservation. Conservation Biology 8: 961–971.
Morris WF e Doak DF (2002) Quantitative Conservation Biology: the Theory and Practice of
Population Viability Analysis. Sinauer Associates, Sunderland, MA, 480 pp.
Nei M e Kumar S (2000) Molecular Evolution and Phylogenetics. Oxford University Press,
Oxford, 333 pp.
O’Brien SJ, Wildt DE, Goldmann D, Merril CR, e Bush M (1983) The cheetah is depauperate in
genetic variation. Science 221: 459–462.
Pedroso ILPB (2004) Meio ambiente, agroindústria e ocupação dos cerrados: o caso do município
do Rio Verde no sudoeste de Goiás. Revista Urutágua (6): 1-10.

144
Pray LA, Schwartz JM, Goodnight GJ e Stevens L (1994) Environmental dependency of
inbreeding depression: implications for conservation biology. Conservation Biology 8: 562–
568.
Primack RB e Rodrigues E (2001) Biologia da conservação. E. Rodrigues, Londrina, 328 pp.
Ralls KJ, Ballou D e Templeton A (1988) Estimates of lethal equivalents and the cost of
inbreeding in mammals. Conserv. Biol. 2: 185-193.
Raup D (1991) Extinction – bad genes or bad luck? W.W. Norton & Company, New York, 210
pp.
Reid WV e Miller KR (1989) Keeping options alive: the scientific basis for conserving
biodiversity. World Resources Institute, Washington, D.C, 185pp.
Rocha VJ, Reis NR e Sekiama ML (2004) Dieta e dispersão de sementes por Cerdocyon thous
(Linnaeus) (Carnívora, Canidae) em um fragmento florestal no Paraná Brasil. Revista
Brasileira de Zoologia, 21 (4), 871-876.
Rodrigues FHG, Silveira L, Jácomo ATA, Bezerra AP, Coelho AMR, Santos DC, Pagnozzi HG e
Hass A (2002) Composição e caracterização da fauna de mamíferos do Parque Nacional das
Emas, Goiás. Revista Brasileira de Zoologia 19: 589-600.
Rodrigues FM, Telles MPC, Resende LV, Soares TN, Diniz-Filho JAF, Jácomo ATA e Silveira L
(2005) Transferibilidade de Marcadores Microssatélite para duas Espécies de Mamíferos
Canídeos do Cerrado Brasileiro. Resumos do 51o Congresso Brasileiro de Genética, Águas de
Lindóia-SP, Brazil.
Rodrigues W, Nogueira J e Imbroisi D (2001) Avaliação Econômica da Agricultura Sustentável:
O Caso dos Cerrados Brasileiros. Cadernos de Ciência & Tecnologia 18: 103-130.

145
Sage RD e Wolff JO (1986) Pleistocene glaciations, fluctuating ranges, and low genetic
variability in a large mammal (Ovis dalli). Evolution 40: 1092–1095.
Silveira L e Jácomo ATA (1998) Ecologia e conservação de mamíferos carnívoros do Parque
Nacional das Emas, GO. Relatório Técnico para o IBAMA/DIREC/DEUC.
Sokal R e Rohlf F (1985) Biometry. W. H. Freeman and Company, USA, 395pp.
Soulé ME (1986) Conservation Biology: The Science of Scarcity and Diversity. Sinauer
Associates, Sunderland, Mass.
Soulé ME e Kohn KA (1989) Research priorities for conservation biology. Washington, DC.
Tanaka Y (1997) Extinction of Populations Due to Inbreeding Depression with Demographic
Disturbances. Researches on Population Ecology 39: 57-66.
Tanaka Y (1998) Theoretical aspects of extinction by inbreeding depression. Researches on
Population Ecology 40: 279–286.
Tanaka Y (2000a) Theoretical Properties of Extinction by Inbreeding Depression Under
Stochastic Environments. In: Ferson S and Burgman M (eds) Quantitative Methods for
Conservation Biology, Springer-Verlag, New York, pp. 274-290.
Tanaka Y (2000b) Extinction of populations by inbreeding depression under stochastic
environments. Population Ecology 42: 55-62
Templeton AR e Read B (1983) The elimination of inbreeding depression in a captive herd of
Speke's gazelle. In: Schonewald-Cox, CM, SM. Chambers, B MacBryde e L Thomas (eds.)
Genetics and Conservation: A Reference for Managing Wild Animal and Plant Populations.
Benjamin/Cummings, Menlo Park, California, pp. 241-261.

146
Vilà C, Sundquist AK, Flagstad O, Seddon JM, Bjornerfeldt S, Kojola I., Sand, H, Wabakken P e
Ellegren H (2002) Rescue of a severely bottlenecked wolf (Canis lupus) population by a
single inmigrant. Proc. R. Soc. Lond B Biol. Sci. 270: 91–97.
Wayne RK (1996) Conservation Genetics in the Canidae. In: Avise JC, Hanuick JL (eds)
Conservation genetics: case histories from nature. New York NY: Chapman & Hall. p 75–118.

147
APÊNDICE 1
CLS PRINT "GLOADRUN - Program for simulating extinction vortex" PRINT "by inbreeding depression (see Tanaka 2000)" PRINT PRINT PRINT "DEMOGRAPHIC PARAMETERS" INPUT "1. Initial population size (K)"; n1 LET k1 = n1 REM INPUT "2. Carrying capacity (K)"; k1 INPUT "2. Rate of decreasing carrying capacity K"; rk INPUT "3. Maximum growth rate (r max)"; rmax INPUT "4. Standard deviation of time variation in population size"; dp2 PRINT PRINT "GENETIC PARAMETERS" REM INPUT "5. Effect of inbreeding on growth rate (INB)"; inb INPUT "5. Initial F value"; F1 REM INPUT "6. Average allele frequency (q)"; q1 REM INPUT "7. Number of loci (nl)"; nl REM INPUT "8. Mutation rate (u)"; mu REM INPUT "9. Selection coeficient (s)"; s LET q1 = .001 LET nl = 15000 LET mu = .000001 LET s = 1 PRINT PRINT "SIMULATION PARAMETERS" INPUT "6. Maximum number of generations (t)"; tm LET sim = 1000 INPUT "7. Number of simulations"; sim INPUT "8. Output file (final step)"; out$ REM INPUT "15. Detailed output file"; outsim$ LET outsim$ = "out" OPEN out$ FOR OUTPUT AS #1 OPEN outsim$ FOR OUTPUT AS #2 PRINT CLS PRINT PRINT "********************************************************" PRINT PRINT "SIMULATING...(1000 times)" PRINT PRINT "********************************************************" PRINT PRINT PRINT PRINT #1, " Simulation time q F N K " PRINT #2, " Simulation time q F N K "

148
APÊNDICE 1 (Continuação) FOR j = 1 TO sim LET K = k1 LET N = n1 LET q = q1 LET F = F1 CONST pi = 3.14159265359# RANDOMIZE (TIMER) FOR i = 1 TO tm IF q > 1 THEN LET q = .999999 md = 0 dp1 = ((q * (1 - q)) / (2 * N * nl)) ^ (1 / 2) u1 = RND u2 = RND x1 = md + SQR(-2 * LOG(u1)) * COS(2 * pi * u2) * dp1 x2 = md + SQR(-2 * LOG(u1)) * COS(2 * pi * u2) * dp2 load = 1 - (q * q * s * nl) LET q = q - (s * ((q * q) + (F * q * (1 - q)))) + mu + x1 IF q < 0 THEN LET q = .00001 LET F = ((1 / (2 * N)) + ((1 - (1 / (2 * N))) * F)) * (1 - (2 * mu)) * (1 - (s * q)) K = (1 - rk) * K REM mort = inb * F IF i >= 2 THEN N = N + (N * (EXP(rmax) * load) * (1 - ((N - 1) / K))) + x2 PRINT #2, USING " ########.### "; j; i; q; F; N; K REM PRINT USING " #######.### "; i; q; F; N; K IF N = 0 THEN GOTO xf: IF N < 0 THEN GOTO xf: NEXT i xf: IF N < 0 THEN LET N = 0 PRINT #1, USING " ########.### "; j; (i - 1); q; F; N; K NEXT j CLS PRINT PRINT "End of simulations" PRINT PRINT "Final simulation, time, q, F, N and K in the file "; out$ PRINT "(Detailed simulation results in the file "; outsim$; ")" CLOSE #1 CLOSE #2 PRINT PRINT PRINT

149
APÊNDICE 1 (Continuação) PRINT INPUT "Do you wish a new simulation? (Y/N)"; g$ IF g$ = "Y" OR g$ = "y" THEN RUN PRINT PRINT "End of data processing..." STOP

150
CONCLUSÕES
- Os marcadores STR (Short Tandem Repeats), são freqüentemente escolhidos para estimar a
variabilidade genética em populações naturais por serem altamente informativos. No entanto, o
uso destes marcadores é limitado, devido, o alto custo e elevado tempo necessário para o
desenvolvimento prévio dos marcadores. Neste estudo, constatou-se um elevado potencial de
transferibilidade de marcadores STR entre canídeos. Do total de 29 locos testados, 21% (6 pares
de primers) foram transferidos com sucesso, sendo que 83% (cinco pares de primers) foram
altamente polimórficos. Estes resultados são importantes uma vez que, permite a caracterização
da variabilidade genética destas populações e conseqüentemente subsidia estratégias de manejo
em populações naturais;
- Utilizando marcadores STR e RAPD (Random Amplified Polymorphic DNA) estimou-se a
diversidade genética das populações de lobo-guará e de cachorro-do-mato do Parque Nacional das
Emas (PNE). Em média, a similaridade genética estabelecida com base no RAPD foi maior entre
os indivíduos da população de lobo-guará do que entre indivíduos da população de cachorro-do-
mato. No entanto, de um modo geral, os resultados do STR não indicaram uma significativa
redução na variabilidade genética e nem endogamia elevada, em nenhuma das duas populações
estudadas, e os parâmetros estão dentro da faixa de variação observada em populações de
canídeos no mundo. Portanto, deve-se pensar em propostas adequadas de conservação e manejo
destas espécies, principalmente do lobo-guará, que é uma espécie ameaçada de extinção, a fim de
aumentar a viabilidade das mesmas em longo prazo. Assim sendo, considerando que as principais
ameaças a estas espécies são a redução drástica do habitat, devido principalmente a ocupação
agropecuária, além dos atropelamentos, abates por chacareiros do entorno do PNE, introdução de

151
espécies exóticas, etc, torna-se necessário uma educação ambiental mais eficiente para que
estratégias de conservação sejam mais bem estabelecidas.
- A associação entre a similaridade genética com a similaridade espacial, baseada nos dados de
área de vida do lobo-guará, permitiu constatar que a estruturação espacial da similaridade
genética foi significativa para os machos, quando considerado os dados obtidos com os
marcadores RAPD. Essa estruturação possivelmente seria explicada pelo baixo padrão de
dispersão entre os machos em relação às fêmeas nesta população. No entanto, os resultados
obtidos com os marcadores STR mostraram uma estruturação espacial da similaridade genética
entre as fêmeas, de acordo com um padrão de filopatria feminina. Uma vez que este padrão é mais
comum nestas espécies, pois para as fêmeas de mamíferos que investem muito no cuidado da
prole os benefícios ecológicos e sociais tendem a favorecer a filopatria feminina, acredita-se que
os resultados obtidos com os marcadores RAPD sejam artefatos. Entretanto, de um modo geral
como os valores das associações foram baixos (correlações matriciais em torno de 0,23), desse
modo acredita-se que quando ambos os indivíduos são beneficiados com a divisão de espaço,
aumenta-se a tolerância entre eles e assim sendo, a estruturação espacial varia muito mais devido
a demografia local e distribuição de recursos do que devido o grau de parentesco genético entre os
indivíduos.
- Quando foi avaliado o tempo de extinção (Te) nestas duas espécies de canídeos do PNE,
assumindo um modelo teórico que combinou os efeitos da depressão endogâmica sob
estocasticidade ambiental, constatou-se que os três parâmetros analisados e suas interações (taxas
de redução da capacidade de suporte, coeficientes de endogamia e flutuação no tamanho da
população), afetaram o Te médio. Uma combinação semelhante de parâmetros para as duas
espécies mostrou que a diminuição da população de lobo-guará é sempre mais rápida que a da

152
população de cachorro-do-mato. Este resultado, é coerente com expectativas baseadas em
parâmetros demográficos e história de vida das duas espécies. Como o lobo-guará ocupa grandes
áreas de vida, possui baixa abundância populacional e certa especialização de habitat em relação
ao cachorro-do-mato, é esperado que esta espécie seja mais sensível a estes parâmetros,
principalmente a fragmentação de habitats. Porém, este estudo é apenas um ponto de partida para
estudos futuros que considerem parâmetros mais realistas da viabilidade destas populações e do
cenário sócio-econômico-ambiental da região sudoeste de Goiás, permitindo assim avaliações
mais precisas da persistência destas populações no PNE e entorno.