estudo da replicaÇÃo do vÍrus dengue-2 sob a …livros01.livrosgratis.com.br/cp071828.pdf ·...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE MEDICINA DE RIBEIRÃO PRETO
PROGRAMA DE PÓS-GRADUAÇÃO EM IMUNOLOGIA BÁSICA E APLICADA
ESTUDO DA REPLICAÇÃO DO VÍRUS DENGUE-2 SOB A
AÇÃO DE DROGAS INIBIDORAS DO METABOLISMO
CELULAR
Kleber Juvenal Silva Farias
Ribeirão Preto
2006

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

KLEBER JUVENAL SILVA FARIAS
ESTUDO DA REPLICAÇÃO DO VÍRUS DENGUE-2 SOB A
AÇÃO DE DROGAS INIBIDORAS DO METABOLISMO
CELULAR
Dissertação de mestrado apresentada à Faculdade de Medicina de Ribeirão Preto da Universidade de São Paulo para obtenção do título de Mestre em Ciências. Área de concentração: Imunologia Básica e Aplicada, opção Bioagentes Patogênicos.
ORIENTADOR: PROF. DR. BENEDITO ANTÔNIO LOPES DA FONSECA
Apoio: Fundação de Amparo à Pesquisa do Estado de São Paulo
-FAPESP-
Ribeirão Preto
2006

FICHA CATALOGRÁFICA
Farias, Kleber Juvenal Silva
Estudo da replicação do vírus dengue-2 sob a ação de drogas inibidoras do metabolismo celular. Ribeirão Preto, 2006.
67 p. : il. ; 30cm
Dissertação de mestrado, apresentada à Faculdade de Medicina de Ribeirão Preto/USP – Programa de pós-graduação em Imunologia Básica e Aplicada – Área de concentração: Bioagentes Patogênicos.
Orientador: Fonseca, Benedito Antônio Lopes.
1. Vírus dengue-2. 2. Cloroquina, Brefeldina A e Citocalasina
B . 3. Quantificação

“Se as coisas são inatingíveis ora...
Não é motivo para não querê-las
Que triste os caminhos
Se não fora a presença
Distante das estrelas”
Mário Quintana.

DEDICATÓRIA
Aos meus pais, Ana Farias e Juvenal Farias (in memorian), que a sua maneira, souberam me
apresentar um mundo de amor, carinho, humildade, amizade, respeito, honestidade, justiça,
sabedoria e razão.
Aos meus irmãos e a minha namorada Paula Renata por estarem sempre ao meu lado.
Amo muito vocês!

AGRADECIMENTOS
Aos meus pais, pela vida, pelo seu amor, pela sua força e por estarem sempre presentes em
minha vida;
Em especial ao Prof. Dr. Benedito Antônio Lopes da Fonseca pela orientação, paciência e
companheirismo.
Ao programa de pós-graduação em Imunologia Básica e Aplicada, na pessoa de seus
Professores pelos ensinamentos e pela convivência;
Aos Professores Luiz Tadeu Morais Figueiredo e Marcos Boulos pelas sugestões e por terem
aceitado participar desta banca;
Ao Nonato, Lurdinha, Sila e Paula, pela amizade, paciência, incentivo e apoio vital nos
momentos finais dessa jornada;
Às secretárias da pós-graduação Ana Cristine, Adriana e Rosângela pela convivência,
amizade, paciência e apoio durante o meu período de mestrado;
Ao Prof. Dr. Victor Hugo Quintana pelos conselhos, amizade, disponibilidade sempre que
precisei, pela orientação no PAE e por sua dedicação em frente aos journals do bioagentes
patogênicos;
Ao Prof. Dr. Francisco Fábio Marques da Silva que muito me ajudou na análise do efeito
citotóxico das drogas;
Aos colegas do laboratório: Patrícia, Mariana, Maira, Camila, Rafael, Emiliana, Luiza,
Teresa, D. Maria, Andréa, Cláudio, Sérgio, Keny e Dani;
Aos colegas da pós-graduação, em especial Janaína, Carlo, Alessandra, Viviane, Juliana,
Veridiana, Marquinhos, Aldo, Lisandra, Nalu, Pink, Patrícia, Deison, Glauciane, Paulinha,
Trança e muitos outros;

Aos colegas do futebol: Benedito, Victor, Cláudio, Carlos, Carlo, Lucas, Pedro, Cacá,
Alexandre, Aldo, Leandro, Pancinha, Round, Gustavo, Felipe, Alexandre, Thiago e muitos
outros pelo companheirismo, apoio e convivência;
Aos técnicos do biotério da Clínica Médica, Roni, Adalberto, Maurício, Áurea, Paulinho,
Pavaneli, Soraia e Sueli pela amizade e auxílio nos experimentos;
A FAPESP pelo indispensável apoio financeiro;
A todos aqueles que direta ou indiretamente colaboraram para que este trabalho fosse
realizado.

SUMÁRIO
RESUMO............................................................................................................................. i
ABSTRACT........................................................................................................................ ii
1. INTRODUÇÃO.............................................................................................................. 02
1.1. Família Flaviviridae..................................................................................................... 02
1.1.1. Vírus dengue............................................................................................................. 02
1.2. Vetor............................................................................................................................. 02
1.3. Biologia do vírus dengue............................................................................................. 03
1.3.1. Aspectos morfológicos.............................................................................................. 03
1.3.2. Estrutura genômica.................................................................................................. 03
1.3.3. Transmissão.............................................................................................................. 04
1.3.4. Ciclo replicativo........................................................................................................ 05
1.4. Manifestações clínicas................................................................................................. 07
1.5. Drogas inibidoras do metabolismo celular................................................................ 09
2. Objetivo........................................................................................................................... 14
2.1. Objetivos específicos.................................................................................................... 14
3. MATERIAIS E MÉTODOS.......................................................................................... 16
3.1. Cultura de células........................................................................................................ 16
3.2. Linhagem viral............................................................................................................. 17
3.3. Estoques virais............................................................................................................. 17
3.3.1. Cultura celular.......................................................................................................... 17
3.3.2. Cérebro de camundongo.......................................................................................... 18
3.4. Extração do RNA viral................................................................................................ 18
3.5. Transcrição Reversa pela Reação em Cadeia da Polimerase (RT-PCR).............. 19

3.6. Titulação do vírus dengue-2....................................................................................... 19
3.6.1. Unidade Formadora de Placa (PFU)...................................................................... 19
3.7. Drogas utilizadas......................................................................................................... 21
3.7.1. Cloroquina................................................................................................................ 21
3.7.2. Brefeldina A.............................................................................................................. 21
3.7.3. Citocalasina B........................................................................................................... 21
3.8. Preparo das drogas...................................................................................................... 21
3.9. Determinação das concentrações de trabalho das drogas para os experimentos.. 22
3.10. Ensaio de acidificação de endossomos (Cloroquina).............................................. 23
3.11. Ensaio de inibição de secreção de proteínas (Brefeldina A).................................. 24
3.12. Ensaio de inibição dos filamentos de actina e microtúbulos (Citocalasina B)..... 25
3.13. Quantificação por PCR em Tempo Real................................................................. 25
3.14. Proteção pós-desafio.................................................................................................. 26
3.15. Análise estatística...................................................................................................... 27
4. RESULTADOS............................................................................................................... 29
4.1. Imunofluorescência indireta....................................................................................... 29
4.2. Titulação dos vírus (PFU)........................................................................................... 29
4.3. Padronização da curva padrão no PCR em tempo real........................................... 30
4.4. Observação de citotoxicidade da cloroquina em células C6/36 e Vero.................. 33
4.5. Efeito de inibidor da acidificação de endossomo (cloroquina) sobre a replicação
do vírus dengue-2................................................................................................................ 33
4.6. Observação de citotoxicidade da Brefeldina A em células Vero............................. 39
4.7. Efeito do bloqueador de secreção de proteínas (brefeldina A) sobre a replicação
do vírus dengue-2................................................................................................................
40
4.8. Observação de citotoxicidade do citocalasina B em células Vero........................... 41

4.9. Efeito do bloqueador de filamentos de actina e microtubulos (citocalasina B)
sobre a replicação do vírus dengue-2................................................................................ 42
4.10. Proteção pós-desafio.................................................................................................. 43
5. DISCUSSÃO................................................................................................................... 49
6. CONCLUSÕES............................................................................................................... 59
7. REFERÊNCIAS BIBLIOGRÁFICAS......................................................................... 60

LISTA DE FIGURAS
Figura 1 – Ciclo replicativo do vírus dengue...................................................................... 7
Figura 2 – Teste de imunofluorescência indireta mostrando presença de vírus DEN-2
em células C6/36 infectadas................................................................................................. 29
Figura 3 – Análise da curva padrão para o sorotipo do vírus dengue-2 através do PCR
em Tempo Real.................................................................................................................... 31
Figura 4 – Perfil de amplificação do vírus dengue-2 por PCR em Tempo Real................. 31
Figura 5 – Análise da curva de dissociação para determinação da especificidade da
reação de amplificação através do PCR em Tempo Real.................................................... 32
Figura 6 – Viabilidade de células Vero e C6/36 submetidas a tratamento com
cloroquina............................................................................................................................. 33
Figura 7 – Ensaio de placa para a titulação do vírus dengue-2........................................... 34
Figura 8 – Ensaio de placa para a titulação do vírus dengue-2 na presença da
cloroquina............................................................................................................................. 35
Figura 9 – Amplificação do fragmento de 419pb da região NS1 do vírus dengue............. 35
Figura 10 – Ação da cloroquina na replicação do vírus dengue-2 em células C6/36......... 36
Figura 11 – Ação da cloroquina sob a replicação do vírus dengue-2 em células Vero...... 37
Figura 12 – Ação da cloroquina adionada em intervalos de 24 horas sob a replicação do
vírus dengue-2 em células Vero........................................................................................... 38
Figura 13 – Ação da cloroquina adicionada em intervalos 12 horas sob a replicação do
vírus dengue-2 em células Vero........................................................................................... 39
Figura 14 – Viabilidade de células Vero submetidas a tratamento com brefeldina A........ 40
Figura 15 – Ação da brefeldina A sob a replicação do vírus dengue-2 em células Vero... 41
Figura 16 – Viabilidade de células Vero submetidas a tratamento com citocalasina B..... 42

Figura 17 – Ação da citocalasina B sob a replicação do vírus dengue-2 em células Vero. 43
Figura 18 – Curvas de sobrevivência dos camundongos infectados via intracerebral com
3X103 PFU/mL do vírus dengue-2 e receberam 30 mg/Kg de cloroquina 2 horas após a
infecção e em intervalos de 24 horas durante 7 dias............................................................ 44
Figura 19 – Curvas de sobrevivência dos camundongos infectados via intracerebral com
3X103 PFU/mL do vírus dengue-2 e receberam 30 mg/Kg de cloroquina 3 dias após a
infecção e em intervalos de 24 horas durante 7 dias............................................................ 45
Figura 20 – Curvas de sobrevivência dos camundongos infectados via intracerebral com
3X103 PFU/mL do vírus dengue-2 e receberam 26,3 mg/Kg de brefeldina A 2 horas
após a infecção..................................................................................................................... 46
Figura 21 – Curvas de sobrevivência dos camundongos infectados via intracerebral com
3X103 PFU/mL do vírus dengue-2 e receberam 1,2mg/Kg de citocalasina B 2 horas após
a infecção............................................................................................................................. 47

i
RESUMO
Os vírus dengue representam o mais importante arbovirus transmissor de doença em humanos, resultando em progressivas manifestações clínicas tais como febre da dengue, febre hemorrágica da dengue e síndrome do choque da dengue. O controle desta infecção é feito apenas pelo controle do vetor, pois não existe até o momento, nenhuma vacina que proteja os indivíduos desta infecção e nenhum antiviral para o controle da replicação viral. Na tentativa de se entender se poderia haver interferência de drogas na replicação do vírus dengue, células C6/36 e Vero foram cultivadas e as monocamadas confluentes foram infectadas com vírus dengue-2, a uma multiplicidade de infecção (MOI) de 0,1. Para confirmação da replicação viral, um ensaio de imunofluorescência indireta foi realizado sobre células infectadas. A análise da inibição da replicação viral pelas drogas que interferem na homeostase celular foi realizada em monocamadas de células Vero e C6/36 tratadas, concomitante e após a infecção viral, com cloroquina (aumenta o pH endossomal), brefeldina A (inibe o fluxo vesicular de proteínas) e citocalasina B (desestabiliza os filamentos de actina). Sobrenadantes de células infectadas foram coletados após períodos de 0, 6, 12, 24, 48, 72, 96, 120, 144 e 168 horas da infecção viral. Foi realizada a extração do RNA total do sobrenadante e a replicação viral foi analisada através da quantificação dos vírus produzidos pela detecção do número de cópias de RNA do vírus dengue-2. Esta quantificação foi realizada através da PCR em tempo real onde a região 3' não-codificadora do genoma dengue-2 foi analisada utilizando primers específicos para o vírus dengue-2 e usando-se o SYBR GREEN® One-Step RT-PCR Master Mix Reagents Kits (Applied Biosystems). Para verificar a ação das drogas in vivo no sentido de investigar o seu possível uso em humanos, grupos de cinco camundongos Swiss de 4 semanas de idade foram desafiados com uma injecção intracerebral de uma cepa virulenta do dengue-2, e tratados ou não 2 horas ou 3 dias após a infecção com as drogas por via intraperitoneal. A cloroquina foi administrada em intervalos de 24 horas durante sete dias, enquanto que a brefeldina A e a citocalasina B foram dose única. Os animais foram observados durante 21 dias. A porcentagem de sobrevivência no grupo de camundongos infectados com vírus e tratados com cloroquina 2 horas após infecção foi melhor do que nos outros grupos (tratadas ou com brefeldina A ou com citocalasina B ou cloroquina 3 dias após infecção) já que houve um desvio da curva de sobrevivência e um deles sobreviveu enquanto nos outros grupos, todos morreram. Todos os animais que não foram tratados morreram por volta do 10° dia. A transcrição reversa por PCR em tempo real mostrou que, comparado as células controle, houve uma diminuição estatisticamente significante na replicação viral em células Vero quando tratadas com cloroquina, brefeldina A e citocalasina B; resultados que foram confirmados por ensaio de placa (PFU). Em células C6/36, a cloroquina induziu um aumento estatisticamente significante na replicação viral a partir de 12 horas quando comparado as células controle, mostrando que este vírus usa uma via diferente de penetração nessas células onde o resultado foi confirmado por ensaio de placa (PFU). A cloroquina mostrou-se eficiente na inibição da replicação do vírus dengue-2 in vivo (camundongos) 2 horas após infecção e in vitro (Vero), enquanto que com a brefeldina A, citocalasina B e cloroquina 3 dias após infecção não tivemos resultados satisfatórios in vivo, porém in vitro essas drogas mostraram-se eficientes. Este trabalho mostra que algumas drogas poderão ter um papel importante no tratamento da dengue em um futuro próximo. Palavras-chave: Dengue-2, Cloroquina, Brefeldina A, Citocalasina B, Quantificação.

ii
ABSTRACT
Dengue viruses represent the most important arboviral disease in humans, resulting in progressively more severe clinical manifestations such as dengue fever, dengue hemorrhagic fever and dengue shock syndrome. The only way to control dengue is vector control since there is no to the moment, no vaccine that protect the individuals of this infection nor an antiviral for the control of the virus replication. In an attempt to understand if drugs would interfere with dengue virus replication, Vero and C6/36 cells were cultivated and confluent monolayers were infected with dengue-2 virus at a multiplicity of infection (MOI) of 0,1. In order to detect viral replication, an indirect immunofluorescence assay was performed on infected cells. To analyse the inhibition of viral replication by drugs that interfere in the homeostase cellular, was performed in monolayers of Vero and C6/36 cells treated, concomitantly and after viral infection, with Chloroquine (increase endosomal pH), brefeldin A (inhibit vesicle flow of proteins) and cytochalasin B (destabilizes actin filamentous). Supernatants of infected cells were collected after defined periods of 0, 6, 12, 24, 48, 72, 96, 120, 144 and 168 hours of viral infection, total RNA was extracted from the supernatant, and viral replication was assessed by Real-time PCR using primers specific to the 3' non-coding region of the dengue-2 genome and SYBR GREEN® One-Step RT-PCR Master Mix Reagents Kits (Applied Biosystems). In order to detect the influence of the drugs in vivo in the sense of investigate his possible use in humans, groups of five 4 week-old Swiss mouse were challenged with an injection intracerebral of a virulent cepa of the dengue-2 and treated either non 2 hours or 3 days after infection with the drugs by intraperitoneal route. The chloroquine was administrated in intervals of 24 hours during 7 days, while the brefeldin A and cytocalasin B were only dose. The animals were observed during 21 days. The percentage of survival in the group of mouse infected with virus and treated with chloroquine 2 hours after infection was better than in the others groups (treated either with brefeldin A or cytocalasin B or chloroquine 3 days after infection) since had a detour of the curve of survival and one of them survived while in the others groups, everybody died. All of the animals that were not treated died by the 10th day. Real-Time PCR results showed that, compared to control cells, there is a statisticatly significant decrease in viral replication in Vero cells when treated with chloroquine, brefeldin A and cytochalasyn B; results that were confirmed by a plaque assay (PFU). In C6/36 cells, chloroquine induced an statisticatly significant increase in viral replication from 12 hours when compared to control cells, showing that this virus uses an unlike road penetration in those cells, results that were also confirmed by the plaque assay (PFU). The chloroquine display an effective inhibition in dengue-2 virus replication in vivo (mouse) 2 hours after infection and in vitro (Vero cells), while brefeldin A, cytocalasin B and chloroquine 3 days after infection did not present a satisfactory result in vivo, but were efficient in vitro. This work shows that some drugs will be able to have an important paper in the handling of the dengue in the near future. Key-Words: Dengue-2, Chloroquine, Brefeldin A, Cytocalasin B, Quantification.

Introdução
1
IINNTTRROODDUUÇÇÃÃOO

Introdução
2
1. INTRODUÇÃO
1.1. Família Flaviviridae
É grande o número de vírus na família Flaviviridae e comum na natureza. Estes são
responsáveis por causar graves doenças em humanos e animais. A família é composta por três
gêneros: Flavivirus, Pestivirus e Hepacivirus. O gênero flavivirus contém mais de 70 vírus
transmitido por artrópodes, entre esses, vírus da Dengue, vírus da Encefalite Japonesa, vírus
da Encefalite transmitido por carrapatos, vírus West Nile e o vírus da Febre Amarela, sendo
considerados patógenos emergentes e reermegentes. Dentro de cada gênero, os vírus podem
ser subdivididos em complexos antigênicos (KUNO et al., 1998; LINDENBACH; RICE,
2001).
1.1.1. Vírus dengue
Os vírus dengue são integrantes da família Flaviviridae e classificados do ponto de
vista epidemiológico como arbovírus, isto é, vírus que é transmitido por artrópodes. Existem 4
sorotipos do vírus do dengue (DEN-1, DEN-2, DEN-3 e DEN-4) e todos podem causar tanto a
forma clássica da doença quanto a dengue hemorrágica. A virulência é diretamente
proporcional à intensidade com que o vírus se multiplica no corpo. As células alvo humanas
para a replicação do vírus dengue são aquelas pertencentes a linhagem monócito-macrófago
(HALSTEAD, 1988).
1.2. Vetor
A dengue é transmitida principalmente pelo Aedes aegypti, vetor também da febre
amarela. É um inseto cosmopolita, encontrado principalmente em locais de grande

Introdução
3
concentração humana. Somente a fêmea transmite a doença, quando pica o homem em busca
de sangue para o desenvolvimento de seus ovos. Ela ataca durante o dia, principalmente ao
amanhecer e no final da tarde. Em média, cada A. aegypti vive em torno de 30 dias e a fêmea
chega a colocar entre 150 e 200 ovos de cada vez. Uma vez infectada com o vírus da dengue,
torna-se vetor permanente da doença e pode também, transmitir a doença para a sua prole
(FONSECA; FIGUEIREDO, 2005).
1.3. Biologia do vírus dengue
1.3.1. Aspectos Morfológicos
Uma partícula viral completa do vírus dengue mede aproximadamente 50nm de
diâmetro e é constituída por um nucleocapsídeo icosaédrico e envelope (KUHN et al., 2002).
O capsídeo viral envolve um core central de RNA formando o nucleocapsídeo. O envelope é
obtido a partir da membrana interna do retículo endoplasmático, quando as partículas virais
brotam do lúmem do retículo endoplasmático para o citoplasma (GUIRAKHOO et al., 1991).
1.3.2. Estrutura Genômica
Os vírus dengue possuem um genoma de RNA de fita simples, polaridade positiva, de
cerca de 11 Kb de comprimento. O RNA genômico consiste de duas regiões não-tradutoras,
uma na porção 5’ e outra na porção 3' , e um fragmento de leitura aberta (RICE et al., 1985).
A ORF codifica uma única poliproteína que é processada por proteases de origem viral e
celular em três proteínas estruturais (capsídeo, premembrana/membrana, e envelope) e sete
não-estruturais (NS1-NS5) na ordem 5' -C-prM-E-NS1-NS2A-NS2B-NS3-NS4A-NS4B-
NS5- 3' (HENCHAL; PUTNAK, 1990; CHAMBERS et al., 1990).

Introdução
4
Dentre as proteínas estruturais, a glicoproteína E do envelope desempenha um papel
dominante na geração de anticorpos neutralizantes e indução da resposta imune (REY, 2003).
A proteína prM é parte dos virions imaturos e sua clivagem proteolítica, por uma protease
celular no complexo de Golgi, remove a porção glicídica e gera virions maduros. Em certos
casos, essa clivagem pode aparentemente ser incompleta, permitindo a proteína prM ser um
alvo adicional para neutralização e para anticorpos protetores (MONATH; BURKE, 2001).
Dentre as sete proteínas não-estruturais, apenas algumas tiveram seus papéis
esclarecidos, ainda que parcialmente. A glicoproteína NS1, com cerca de 430 aminoácidos, é
detectada na superfície celular e secretada para o espaço extracelular, parecendo participar na
fase precoce de replicação viral e estar relacionada à virulência (HENCHAL; PUTNAK,
1990). A proteína NS2 é dividida nas porções NS2A e NS2B, sendo esta última portadora de
atividade proteolítica. As proteínas NS3 e NS5 têm sido associadas com os processos de
replicação e transcrição do RNA viral. Atividades de protease e trifosfatase/helicase foram
atribuídas à proteína NS3 e atividade de RNA polimerase à proteína NS5 (CHAMBERS et
al., 1990; BARTELMA; PADMANABHAN, 2002; ANDREW et al., 2002). A proteína NS4
é clivada nas porções NS4A e NS4B e ambas, juntamente com NS2A e NS2B, associam-se à
membrana da célula infectada durante o processo de maturação viral (CHAMBERS et al.,
1990).
1.3.3. Transmissão
O ciclo de transmissão do vírus dengue começa quando o mosquito fêmea pica uma
pessoa infectada. Dentro do A. aegypti, o vírus multiplica-se no intestino médio do mosquito
e, com o tempo, passa para outros órgãos, chegando finalmente às glândulas salivares
(período de incubação extrínseco, que dura de 7 a 14 dias) e através da picada do mosquito
ocorre a introdução do vírus na pele de outro indivíduo. Assim que penetra na corrente

Introdução
5
sangüínea, o vírus passa a se multiplicar em células mononucleares em órgãos específicos,
como o baço, o fígado e os tecidos linfáticos e também nas células sanguíneas. Esse período é
conhecido como de incubação intrínseco e dura de quatro a sete dias, quando o vírus começa a
circular na corrente sangüínea dos indivíduos e em seguida, ocorrem os primeiros sintomas
(RICHMAN; WHITLEY; HAYDEN, 2002).
1.3.4. Ciclo replicativo
Os vírus dengue entram na célula hospedeira por endocitose mediada por receptor
ainda não esclarecido e replicam-se no citoplasma das células de vertebrados após 12 a 16
horas da infecção (Figura 1). Os receptores de membrana, aos quais se ligam os vírus dengue
na fase inicial da infecção, são desconhecidos, apesar de evidências que o heparan sulfato
seria participante ativo nesta fase da infecção (BARRAGÁN; ANGEL, 2001). Os vírus
dengue entram nas células por passagem direta pela membrana, por pinocitose ou fagocitose
(MONATH; BURKE,2001). Após a penetração no citoplasma ocorre queda do pH intra-
endossomal resultando na fusão do envelope viral com a membrana endossomal. Após a fusão
ter ocorrido, o nucleocapsídeo é liberado para o citoplasma, onde as proteínas do capsídeo e o
RNA se dissociam, iniciando a replicação do RNA genômico e a montagem da partícula viral
(LINDENBACH; RICE, 2003). O RNA viral replica-se em áreas focais perinucleares,
intermediado por um RNA de polaridade negativa que serve como molde na replicação do
RNA do vírus. Este RNA, após uma curta seqüência não codificadora localizada na porção 5’,
possui um longo fragmento de leitura aberta (ORF) que codifica todas as proteínas virais. A
tradução de RNAs ocorre no retículo endoplasmático, próximo à membrana nuclear,
resultando na tradução de uma poliproteína contendo todas as proteínas virais.
Concomitantemente à tradução ou imediatamente após esta, ocorre a clivagem da poliproteína
por uma signal-peptidase e uma serino proteinase contidas na porção N-terminal de NS3, e

Introdução
6
também por proteases celulares. Após o início da tradução e síntese protéica, as proteínas não
estruturais NS3 e NS5 agem como um complexo de replicases e o RNA de polaridade positiva
dos flavivirus é transcrito para um RNA complementar que, por sua vez, origina os RNAs da
progênie viral (CHAMBERS et al., 1990; UCHIL; SATCHIDANANDAM, 2003). Partículas
incompletas são formadas no lúmem do retículo endoplasmático. Essas partículas contém as
glicoproteína E e prM, membrana lipídica e nucleocapsídeo, resultando em partículas virais
imaturas não infecciosas. As partículas virais imaturas são transportadas para o complexo de
Golgi onde ocorre a clivagem da prM pela furina (protease do hospedeiro) removendo a
porção glicídica desta proteína e resultando em partículas virais maturas, que são liberadas da
célula hospedeira por exocitose pelos filamentos de actina (ELSHUBER et al., 2003).
A glicoproteína E do vírus dengue-2 medeia a entrada na célula hospedeira através de
duas funções: ligação ao receptor de membrana plasmática e fusão com membrana
endossomal. Durante a montagem e a formação completa do vírus dengue, a glicoproteína E
se associa com uma segunda proteína (prM), que é o precursor da proteína M de membrana,
em um complexo heterodimérico que protege a proteína E de inativação prematura. A fusão
ocorre entre a membrana viral e uma membrana intracelular seguindo uma mudança
conformacional na glicoproteína E que leva à formação de homotrímeros, sendo esse processo
disparado pela queda do pH endossomal. A formação da partícula viral completa infecciosa
envolve a clivagem da prM e reorientação da glicoproteína E como homodímeros
(HALSTEAD et al., 2005).

Introdução
7
Figura 1 - Ciclo replicativo do vírus dengue - O vírus entra na célula hospedeira por endocitose mediada por receptor; A acidificação da vesícula endossomal dispara uma mudança conformacional da glicoproteína E do vírus, ocorrendo a fusão da membrana viral com a membrana do endossomo e a dissociação do nucleocapsídeo; O genoma é liberado para o citoplasma e o ssRNA de polaridade positiva é traduzido em uma poliproteína; A montagem do vírus ocorre no retículo endoplasmático; A partícula viral imatura é transportada para o complexo de Golgi, onde ocorre a clivagem da proteína prM (RTG - rede trans-Golgi), formando uma partícula matura; Essas partículas são subseqüentemente liberadas por exocitose (Figura adaptada de MUKHOPADHYAY; KUHN; ROSSMANN, 2005).
1.4. Manifestações clínicas
A dengue é hoje a principal doença viral re-emergente no mundo e a mais importante
arbovirose urbana da atualidade e caracteriza-se por ser uma doença febril aguda. É uma
doença endêmica, de caráter urbano, que está presente em praticamente todas as regiões de
clima tropical e subtropical do mundo. Também, as infecções causadas pelos vírus do dengue
assumem grande importância no nosso país (FONSECA; FIGUEIREDO, 2005). Embora se
suspeite que venha causando epidemias há vários séculos, a primeira descrição detalhada da
Infecção viral
Virion maturo
Fusão e dissociação viral
Transporte do vRNA para RER Tradução e processamento da poliproteína
Núcleo
Montagem viral Golgi
RTG
Maturação viral
Replicação do genoma viral

Introdução
8
dengue foi feita por Benjamim Rush, em 1780, durante um surto ocorrido na Philadelphia,
EUA (CAREY, 1971; GUBLER, 1998).
Durante a multiplicação do vírus dengue em células sangüíneas, ocorre intensa resposta
imune com liberação de citocinas e aminas vasoativas que podem agredir as paredes dos
vasos sangüíneos, provocando extravasamento de líquido (plasma) (HALSTEAD, 1988).
Quando isto ocorre muito rapidamente, aliado à diminuição de plaquetas, podem ocorrer
sérios distúrbios no sistema circulatório, hemorragias e queda da pressão arterial (choque) -
este é o quadro da dengue hemorrágico. Clinicamente, as manifestações variam de uma
síndrome viral, inespecífica e benigna, até um quadro grave e fatal, a febre hemorrágica da
dengue e a síndrome do choque da dengue (FHD/SCD) (FONSECA; FIGUEIREDO, 2005)
que são de difícil diagnóstico, sendo confundidas com outras doenças caracterizadas por
distúrbios da permeabilidade capilar (HENCHAL; PUTNAK, 1990).
Durante a segunda guerra mundial, a co-circulação de múltiplos sorotipos dos vírus
dengue, o aumento da atividade epidêmica, aliado ao subseqüente crescimento descontrolado
das cidades, fez emergir a epidemia da febre hemorrágica da dengue (DHF) como o maior
problema de saúde pública em muitos países do sudeste Asiático. O primeiro relato de
epidemia da DHF ocorreu em 1953 em Manila, nas Filipinas, e logo se expandiu para outras
regiões e continentes acarretando graves epidemias, especialmente nas ilhas do Sul do
Pacífico e nas Américas (GUBLER; TRENT, 1994; HALSTEAD, 1970; RIGAU-PÉREZ et
al., 1998; GUBLER, 1998, 2002).
A cada ano é estimado que cerca de 50-100 milhões de casos de dengue ocorram no
mundo todo, incluindo cerca de 500.000 casos de Febre Hemorrágica da Dengue, a mais
grave manifestação da doença (GIBBONS; VAUGHN, 2002). Esta forma da doença tem sido
relatada em cerca de 5% - 10% das infecções secundárias, com uma letalidade de 10%
(RIGAU-PÉREZ et al., 1998).

Introdução
9
Acredita-se que a Febre Hemorrágica da Dengue (FHD) resulte da infecção seqüencial
por diferentes sorotipos, em um processo conhecido como aumento da infecção mediada por
anticorpo (ADE) (TAKEDA; ENNIS, 1990), onde anticorpos adquiridos em infecções prévias
por um determinado sorotipo do vírus do dengue não seriam neutralizantes para o sorotipo
responsável pela infecção atual. Estes anticorpos se ligariam a outros epítopos do sorotipo
responsável pela infecção atual, comuns aos 4 sorotipos virais, mas não o neutralizaria,
formando então, complexos vírus-anticorpos que facilitariam a penetração do vírus
opsonizado em células monocitárias. Células mononucleares expressam na sua superfície
receptores para a porção Fc das imunoglobulinas, região implicada na ligação dos complexos
vírus-anticorpos a estas células, facilitando assim a ligação dos vírus dengue ao seu receptor
bem como a internalização dos complexos vírus-anticorpos durante a reciclagem das
membranas celulares. A facilidade de penetração do vírus nestas células resulta em uma maior
carga viral, maior intensidade da resposta imune a esta infecção com liberação de citocinas e
aminas vasoativas e como conseqüência uma doença mais grave (HALSTEAD, 1988).
Entretanto, há também evidências que outros fatores influem no desenvolvimento desta forma
da doença, sendo um deles, a virulência de cada cepa infectante (HALSTEAD, 1970;
ROTHMAN; ENNIS, 1999). Observou-se que a FHD era mais comum em uma infecção
secundária pelo vírus dengue que em uma infecção primária pelo vírus (THEIN et al., 1997;
HALSTEAD et al., 1970).
1.5. Drogas inibidoras do metabolismo celular
O uso de inibidores metabólicos para estudar aspectos da maturação dos flavivirus não
tem sido investigado extensivamente.

Introdução
10
Alguns vírus entram nas células alvo por endocitose e o baixo pH endossomal, faz
com que ocorra a fusão do envelope viral com a membrana do endossomo, liberando o ácido
nucléico infeccioso para a replicação viral.
Cloroquina, uma 9-aminoquinolina que foi identificada em 1934, é uma base fraca que
aumenta o pH de vesículas ácidas. Extracelularmente a cloroquina está presente
principalmente na forma protonada, devido a sua carga positiva, que é incapaz de atravessar a
membrana plasmática. A porção não-protonada lipofilica da cloroquina entra na célula por
difusão passiva, onde, por sua vez, torna-se protonada em uma maneira inversamente
proporcional ao pH. Assim ela é concentrada dentro de organelas ácidas, tais como,
endossomos, vesículas de Golgi e lisossomos, por um mecanismo que envolve uma bomba de
prótons, que requer energia. A cloroquina exerce efeito antiviral, inibindo passos dependentes
de pH da replicação de diferentes vírus, nos estágios iniciais ou tardios. Para os membros da
família Flaviviridae foi observado a inibição da replicação pela cloroquina in vitro, por afetar
o processamento proteolítico normal da proteína prM para M (RANDOLPH et al., 1990). Há
também descrição da inibição da infectividade do vírus da imunodeficiência humana (TSAI et
al., 1990) e um efetivo inibidor da replicação do coronavirus causador da síndrome
respiratória aguda grave (SARS-CoV) (KEYAERTS et al., 2004).
A brefeldina A foi primeiramente relatada em 1958, como uma droga que exibe uma
extraordinária atividade biológica que inclui efeitos antibióticos, antiviral, antimitótica,
citostático e antitumoral. Tem sido isolada de numerosas espécies de fungos, incluindo
Penicillium decumbens, Penicillium brefeldianum, Penicillium cyaneum e Ascochyta
imperfecta, e embora seja uma molécula pequena possui uma considerável estrutura complexa
(PHILLIPS et al., 1993).
A brefeldina A tem se tornado uma poderosa ferramenta para análise do transporte
vesicular na via secretória em células de mamíferos. Ela inibe a secreção de proteína e induz

Introdução
11
profundas mudanças morfológicas em várias organelas. Em muitos tipos celulares, a droga
leva a uma dissociação do complexo de Golgi e ao transporte retrógrado, dependente de
microtubulos, das proteínas do Golgi de volta ao retículo endoplasmático, com isso
bloqueando a secreção de proteínas. Não somente o complexo de Golgi, mas também outras
organelas nas vias exocitica e endocitica são afetadas pela brefeldina A. Por exemplo, ela
causa a fusão dos compartimentos endossomais iniciais com a rede trans-Golgi (TORII et al.,
1995).
O complexo de Golgi tem um papel crucial no transporte de vesículas secretórias e
proteínas de membrana em células de organismos superiores. Proteínas que se originam no
retículo endoplasmático passam por uma série de modificações no aparelho de Golgi. A
despeito de sua complicada morfologia e função, ele é dinâmico e capaz de rápida dissociação
e reassociação durante mitoses, tratamento com droga, ou em resposta a bloqueios
mutacionais (KASAP et al., 2004).
A secreção de partículas do vírus Kunjin na fase tardia da infecção foi
significantemente afetada pela presença da droga Brefeldina A (BFA) (MACKENZIE;
WESTAWAY, 2001), assim como o vírus minute de camundongo (ROS et al., 2002) e o
vírus West Nile em células de mamíferos (SREENIVASAN et al., 1993).
A citocalasina B tem uma curta história de aplicação na virologia, mas tem sido uma
valiosa ferramenta no estudo sobre a biologia molecular da interação vírus-célula hospedeira.
Tem sido isolada da espécie de fungo Helminthosporium dematoideum, e exerce uma
variedade de alterações nas funções celulares, entre elas a desestabilização do citoesqueleto
(KOCH; KOCH, 1978).
O citoesqueleto é uma estrutura altamente dinâmica que se organiza continuamente
sempre que a célula altera a sua forma, se divide ou responde ao ambiente, participando da
maquinaria necessária para movimentos intracelulares, tais como o transporte de organelas no

Introdução
12
citoplasma e a segregação dos cromossomos na mitose. Há uma grande participação do
citoesqueleto durante o processo de síntese de algumas proteínas celulares, auxiliando no
transporte de vesículas que tem um importante papel na maturação de algumas moléculas
produzidas pela célula. É composto por três diferentes tipos de filamentos protéicos:
filamentos de actina, filamentos intermediários e microtúbulos (ALBERTS et al., 1994).
Os filamentos de actina e os microtubulos estão mais diretamente envolvidos com o
transporte de organelas no ambiente intracelular. O filamento intermediário está mais
relacionado com o processo de sustentação da forma celular (LODISH et al., 2000).
A citocalasina B inibe a replicação viral, como evidenciado em experimentos feitos
com vários vírus. Estudos com o vírus West Nile demonstraram que ocorre uma inibição da
replicação viral em células Vero quando tratadas com a droga (CHU et al., 2003) e com o
vírus Kunjin em células Vero (NG et al., 1983).
Como descrito anteriormente as drogas cloroquina, brefeldina A e citocalasina B
interferem no metabolismo celular e consequentemente na replicação de alguns vírus. Assim,
investigar se essas drogas também têm ação sob o ciclo replicativo do vírus dengue se faz
necessário, pois até o momento não existe um tratamento profilático ou terapêutico para
dengue.

Objetivos
13
OOBBJJEETTIIVVOOSS

Objetivos
14
2. OBJETIVO
Avaliar a eficácia das drogas inibidoras do metabolismo celular que podem interferir
em diferentes estágios do ciclo replicativo do vírus dengue-2, levando em consideração a
produção final de virions pelas células infectadas, tratadas ou não com estas drogas, assim
como a avaliação da taxa de sobrevivência de camundongos infectados e tratados com essas
drogas.
2.1. Objetivos específicos
Avaliar a eficácia da droga cloroquina (aumenta o pH endossomal) sob a replicação do
vírus dengue-2 em células Vero e C6/36.
Avaliar a eficácia da droga citocalasina B (despolimeriza o citoesqueleto) sob a
replicação do vírus dengue-2 em células Vero.
Avaliar a eficácia da droga brefeldina A (inibe o fluxo vesicular de proteínas) sob a
replicação do vírus dengue-2 em células Vero.
Avaliar a porcentagem de sobrevivência após a inoculação das drogas em
camundongos Swiss infectados com vírus dengue-2.

Materiais e Métodos
15
MMAATTEERRIIAAIISS EE MMÉÉTTOODDOOSS

Materiais e Métodos
16
3. MATERIAIS E MÉTODOS
3.1. Cultura de células
Foram utilizadas células C6/36 e Vero. As células C6/36 (linhagem contínua
proveniente do mosquito Aedes albopictus) foram mantidas em frascos de 25cm2 (Corning
Incorporated, Corning, NY, USA) em meio de cultura Leibovitz-15 (L-15) (Invitrogen, New
York, USA) suplementado com 10% Soro Bovino Fetal inativado (SBF), L-glutamina, 1%
antibiótico e 10% triptose fosfato, mantidas em estufa a 28ºC. As células Vero (linhagem
contínua proveniente de rim de macaco verde africano) foram mantidas em frascos de 75cm2
(Corning Incorparated, Corning, NY, USA) em meio Leibovitz-15 (L-15) (Invitrogen, New
York, USA), suplementado como descrito anteriormente, e mantidas em estufa a 37°C, numa
atmosfera úmida contendo 5% de CO2.
O crescimento das células foi acompanhado a cada 24 horas, com o auxilio de
microscópio de fase invertido e o meio de cultura trocado a cada dois dias. O subcultivo foi
realizado ao se observar formação de uma monocamada celular com mais de 80% de
confluência. Para a perpetuação das linhagens celulares, meio dos frascos foi aspirado e a
monocamada celular lavada uma vez com uma solução tampão (PBS). A seguir, as células
foram separadas (Vero) ou não (C6/36) com 2 mL de uma solução de tripsina 0,25% durante
5 minutos. A tripsina foi então inativada com 5mL de meio de cultura (L-15) contendo soro
bovino fetal e as células em suspensão foram subcultivadas em novos frascos, procedimento
esse que deu origem a novas passagens da cultura.

Materiais e Métodos
17
3.2. Linhagem viral
O vírus utilizado neste trabalho foi dengue-2, linhagem New Guinea C. Este vírus foi
fornecido ao nosso laboratório pelo Dr. Robert E. Shope, na universidade de Yale, EUA.
3.3. Estoques virais
3.3.1. Cultura celular
Garrafas de 75cm2 foram semeadas com células C6/36 e crescidas em estufa a 28ºC
em meio L-15 (Invitrogen, New York, USA) suplementado com soro bovino fetal (10%),
triptose fosfato (10%) e antibióticos (penicilina 100 U/ml, estreptomicina 1mg/ml). Após as
monocamadas estarem contínuas, estas foram inoculadas com 25 µl do vírus dengue-2 (New
Guinea C) e mantidas em estufa a 28ºC por 7 dias em meio L-15 (2% SBF) para a replicação
do vírus. Sete dias após a infecção, foi realizado o teste de imunofluorescência indireta para
determinar a presença da infecção viral na cultura. Para tanto, as garrafas de cultura de células
infectadas com o vírus dengue-2 foram agitadas levemente e 15 µl das células do
sobrenadante foram depositadas em uma lâmina própria para a imunofluorescência. Esta foi
seca a 37ºC por aproximadamente 2 horas, em seguida, as células foram fixadas na lâmina
com acetona fria, por 15 minutos, e secas a 37ºC. Logo após, foram adicionados 15 µl de
fluído ascítico de camundongos (MIAF) imunizados contra os vírus do dengue tipo 1, 2, 3 e 4,
e a lâmina foi incubada em câmara úmida a 37ºC por 30 minutos. Em seguida a lâmina foi
lavada em PBS por 10 minutos e seca à temperatura ambiente. Então, foi adicionado 15 µl do
anticorpo secundário (anti-IgG de camundongo ligado à fluoresceína), diluído em PBS-azul
de Evans a 1% e a reação foi novamente incubada em câmara úmida a 37ºC por 30 minutos.
A lâmina foi lavada com PBS por 10 minutos e em seguida com água destilada por 1 minuto,
e após a lavagem, a mesma foi seca em temperatura ambiente. Em seguida, as lâminas foram

Materiais e Métodos
18
analisadas em um microscópio de fluorescência para determinar a presença ou não de
fluorescência. Indicado a positividade da cultura, o sobrenadante foi clarificado, e a ele foi
adicionado 20% de SBF e estocado a -70ºC, para a realização do cálculo do título viral pela
técnica da unidade formadora de placa (PFU).
3.3.2. Cérebro de camundongo
O vírus dengue-2 (New Guinea C) foi amplificado em cérebros de camundongos
recém-nascidos. Cerca de 10µl do estoque de vírus dengue (DEN-2) foi inoculado, pela via
intracerebral, em camundongos Swiss recém nascidos e estes foram observados diariamente.
A partir do quarto dia de infecção, os animais infectados com vírus DEN-2 apresentaram
sinais de paralisia dos membros, principalmente inferiores, e tremores. Os animais foram
sacrificados e tiveram os cérebros removidos, por sucção através de seringa em ambiente
estéril e cada cérebro foi macerado em 900µl de albumina bovina a 7,5%. Em seguida,
centrifugou-se o macerado a 5.000 x g, por 20 minutos a 4ºC e o sobrenadante foi guardado,
em alíquotas, a - 70ºC, para a posterior titulação viral por ensaio de placa (PFU).
3.4. Extração do RNA viral
Para a extração do RNA total do macerado de cérebro de camundongos Swiss recém-
nascidos, infectados com dengue-2, foi utilizado o kit QIAamp® Viral RNA (QIAGEN® ,
USA). Este kit já demonstrou ser eficiente, conforme resultados obtidos em nosso laboratório
(DE PAULA et al., 2001). O kit utiliza uma coluna com sílica gel com alta afinidade para
ácidos nucléicos. Para tanto, 140µl do macerado foram lisados pela solução tampão AVL e,
após lise, a amostra foi aplicada a uma coluna com afinidade para RNA e submetida a uma
centrifugação de 8000 rpm por 1 minuto. Em seguida, a amostra foi lavada duas vezes, uma

Materiais e Métodos
19
com a solução tampão AW1 e outra com a solução tampão AW2. Terminada a lavagem, o
RNA foi eluído da coluna com 60µl da solução tampão AVE e estocado a -70ºC.
3.5. Transcrição Reversa pela Reação em Cadeia da Polimerase (RT-PCR)
Para confirmação da infecção viral do macerado de cérebros de camundongos Swiss
recém-nascidos, foi utilizado o Kit Qiagen® OneStep RT-PCR que é composto de reagentes
que permitem alta eficiência e especificidade da RT-PCR. Essa especificidade é devida à
mistura das enzimas Ominiscript e Sensiscript, desenhadas para a transcrição reversa de
quantidades de RNA acima de 50ng e abaixo de 50ng, respectivamente. Além disso, possui
HotStartTaq DNA Polimerase, que elimina a formação de dímeros de primers. Para tanto, foi
preparada uma mistura contendo 5µl da solução tampão 5X One-Step RT-PCR, 5µl de dNTPs
10mM, 0,5 pmol/µl dos primers AD3 [5' CTGATTTCCAT(A,C,G,T)CC(A,G)TA 3'] e AD4
[5' GA(C,T)ATGGG(A,C,G,T)TA(C,T)TGGATAGA 3'] (HENCHAL et al., 1991), 1µl da
mistura de enzimas e 5µl do RNA, em volume final de 25µl. A reação de amplificação foi
feita a 50ºC por 30’ e a 95ºC por 15’ para a transcrição reversa, seguidos de 40 ciclos a 95ºC
por 1’, 55ºC por 1’, 72ºC por 1’ e um ciclo final de 10’ a 72ºC.
3.6. Titulação do vírus dengue-2
3.6.1. Unidade Formadora de Placa (PFU)
Frascos plásticos de cultivo celular de 75 cm2 (Corning Incorporated, Corning, NY,
USA) contendo monocamada confluente de células Vero (linhagem contínua proveniente de
rim de macaco verde africano) foram incubados com 4mL de tripsina (Invitrogen, New York,
USA) para o deslocamento das células da parede da garrafa que, posteriormente, foram
ressuspensas em 20 mL de meio completo L-15 (Invitrogen, New York, USA) suplementado

Materiais e Métodos
20
com SBF(10%), L- Glutamina 200mM (1%), 1% Antibiótico (penicilina 100 U/mL) e triptose
fosfato (10%). As células foram bem homogeneizadas. Foram semeados 1mL das células
Vero em placas de 24 poços (Corning Incorporated, Corning, NY, USA) em um número
aproximado de 4 x104 células/poço e mantidas em estufa de CO2 (5%) a 37°C por 3 dias, para
formar uma monocamada confluente. As suspensões virais do sobrenadante celular e da
preparação do cérebro dos camundongos foram descongeladas lentamente em gelo e diluições
seriadas decimais foram preparadas até 10-10. O meio de cultura da monocamada celular das
placas de 24 poços foi descartado e adicionou-se 100µl de cada diluição do vírus em cada
poço, em duplicata, e PBS como controle negativo. As placas foram mantidas por 90 minutos
em estufa de CO2 (5%) a 37°C, sendo agitadas de 15 em 15 minutos, para a adsorção viral.
Após a conclusão da adsorção foi retirado o inóculo e adicionou-se 1mL de uma camada de
solução viscosa (meio L-15 2% SBF sem vermelho fenol e carboximetilcelulose a 3% estéril)
(Sigma, Munich, Germany) em cada poço e as placas foram mantidas em estufa de CO2 (5%)
a 37°C por 7 dias. No sétimo dia, a camada viscosa foi descartada, os poços lavados com PBS
e, para visualização das placas virais, foi preparada uma solução de meio L-15 sem vermelho
fenol suplementado a 2% de soro bovino fetal com 2% vermelho neutro (Invitrogen, New
York, USA). Dessa solução, 1mL foi adicionado a cada poço e as placas foram mantidas em
estufa de CO2 (5%) a 37ºC por 5 horas. O cálculo foi realizado baseado na maior diluição dos
títulos virais onde as placas foram visíveis e tendo em conta a diluição e o volume total do
inóculo. Os valores são expressos em unidades formadoras de placa (PFU/mL).

Materiais e Métodos
21
3.7. Drogas utilizadas
3.7.1. Cloroquina – GALENA / SIGMA-ALDRICH
Interfere na bomba de prótons de organelas celulares prevenindo sua acidificação
(SAVARINO et al., 2003).
3.7.2. Brefeldina A – SIGMA-ALDRICH
Bloqueia o transporte de saída de proteínas do retículo endoplasmático para o aparelho de
Golgi (ALBERTS et al., 1994).
3.7.3. Citocalasina B – SIGMA-ALDRICH
Despolimeriza os filamentos de actina (CHU et al., 2003).
3.8. Preparo das drogas
Dentro da capela de fluxo laminar, foi preparada uma solução estoque de cloroquina
em L-15 (2%) na concentração de 97x103 µM (0,5 g em 10 mL), brefeldina A na
concentração de 18x103 µM (0,005g em 1mL de etanol) e citocalasina B na concentração de
2x103 µM (0,001g em 1mL de DMSO). A seguir, essas soluções foram colocadas no agitador
mecânico para a completa diluição da droga no meio. As concentrações utilizadas nos
experimentos foram : (i) cloroquina: 97x102, 97x101, 97, 97x10-1, 97x10-2 µM , em que foram
obtidas por diluições seriadas decimais, a partir da solução de estoque (97x103 µM); (ii)
brefeldina A: 18x102, 18x101, 18, 18x10-1, 18x10-2 µM, em que foram obtidas por diluições
seriadas decimais, a partir da solução de estoque (18x103 µM); (iii) citocalasina B: 2x102,
2x101, 2, 2x10-1, 2x10-2 µM, em que foram obtidas por diluições seriadas decimais, a partir da
solução de estoque (2x103 µM). Somente as diluições a partir de 97, 18, e 20 µM, se

Materiais e Métodos
22
mostraram viáveis para o experimento com cloroquina, brefeldina A e citocalasina B,
respectivamente. Todas as soluções foram preparadas e utilizadas no momento do
experimento.
3.9. Determinação das concentrações de trabalho das drogas para os experimentos
Antes de utilizarmos as drogas foram realizados experimentos preliminares para
determinar níveis citotóxicos destas substâncias em células C6/36 e Vero, e assim determinar
concentrações ideais para os experimentos subseqüentes. Para a realização desta fase do
estudo, células C6/36 e Vero originadas dos frascos de 75cm2 foram plaqueadas em um
número aproximado de 4 x104 células/poço, em placas de cultivo contendo 24 poços, com
uma área aproximada de 2 cm2 cada. Decorrido o período de aproximadamente 72 horas,
quando as culturas celulares já estavam confluentes, as células foram carenciadas de soro,
sendo o meio de cultura substituído para L-15 contendo 2% de SBF com as seguintes drogas
diluídas nas diferentes concentrações descritas no item anterior: (i) cloroquina (inibidor da
acidificação de endossomo); (ii) brefeldina A (bloqueador de secreção de proteínas); (iii)
citocalasina B (despolimerizador do citoesqueleto). Todas as soluções foram preparadas e
utilizadas no momento do experimento. Os tratamentos foram realizados inicialmente
avaliando o efeito de diferentes concentrações das drogas no meio de cultura, ou seja,
realizando uma curva dose-resposta para cada uma das drogas. Após períodos de 1, 6, 12, 24,
48 e 72 horas de contato com as drogas descritas, as células foram removidas por
tripsinização (Vero) e PBS 1X (C6/36), em que a viabilidade celular foi determinada pelo
método de exclusão do azul de Trypan a 0,4% (Invitrogen, New York, USA) e as células
contadas em câmaras de Neubauer, utilizando para isso o microscópio invertido de fase. Este
método consiste no fato de que este corante não sendo lipofílico não consegue entrar
normalmente nas células, o que passa a ocorrer caso haja a ruptura da membrana plasmática,

Materiais e Métodos
23
um típico sinal que antecede a morte celular. Assim, somente as células em morte celular se
apresentam coradas em azul e com limites irregulares. As demais se apresentam
arredondadas, transparentes, com a membrana refringente e com os limites nítidos.
O percentual de viabilidade celular foi obtido pela fórmula a seguir:
Viabilidade celular (%) = N° total de células viáveis x 100 N° total de células
Concluída essa etapa, foram realizadas curvas de porcentagem de viabilidade celular
em função do tempo experimental com as drogas. Os experimentos foram realizados em
duplicata e um número de dois experimentos realizados para cada situação em questão.
3.10. Ensaio de acidificação de endossomos (Cloroquina)
Placas de cultura celular de 24 poços (Corning Incorporated, Corning, New York,
USA) foram semeadas com células C6/36 e Vero, mantidas em estufa de CO2 (5%) a 37ºC
(Vero) e a 28ºC (C6/36). As células foram cultivadas em meio de cultura L-15 (Invitrogen,
New York, USA) modificado, suplementado com soro bovino fetal (10%), triptose fosfato
(10%) e antibióticos (penicilina 100U/mL, estreptomicina 1mg/mL). O monitoramento do
crescimento celular foi feito a cada 24 horas e o meio de cultura trocado a cada dois dias.
Quando as monocamadas apresentavam-se contínuas, estas foram inoculadas com 100µl do
vírus dengue-2 (MOI=0,1) e incubadas novamente a mesma temperatura, durante 1 hora.
Após a adsorção viral, foi realizado a remoção do inóculo e as monocamadas foram lavadas 2
vezes com PBS pH 7,4, e o meio L-15 modificado, suplementado com soro bovino fetal (2%),
triptose fosfato (10%) e antibióticos (1%), juntamente com a droga a uma concentração de
50µg/mL foi adicionado nos seguintes tempos, em diferentes experimentos: (i) juntamente
com o vírus, 1 hora após a adsorção; (ii) juntamente com o vírus, 1 hora após adsorção e a

Materiais e Métodos
24
intervalos de 24-24 horas após adsorção; (iii) juntamente com o vírus, 1 hora após a adsorção
e a intervalos de 12-12 horas após adsorsão. As monocamadas foram mantidas em estufa de
CO2 (5%) a 37ºC (Vero) e 28ºC (C6/36) por 7 dias. Utilizamos três condições: células Vero e
C6/36 infectadas com vírus dengue-2 e não tratadas com a droga; células Vero e C6/36
infectadas com vírus dengue-2 na presença de cloroquina (50µg/mL) e células Vero e C6/36
não infectadas e não tratadas com a droga. Monocamadas infectadas e não infectadas sem a
adição de droga foram usadas como controles. Sobrenadantes foram removidos de duplicatas
de culturas e clarificados por centrifugação em períodos de 0, 6, 12, 24, 48, 72, 96, 120, 144 e
168 horas para a realização do PCR em tempo real e para a determinação do título do vírus
(PFU), em monocamadas de células Vero. O experimento foi repetido três vezes.
3.11. Ensaio de inibição de secreção de proteínas (Brefeldina A)
Monocamadas de células Vero (placas de 24 poços) foram infectadas com vírus
dengue-2. A brefeldina, a uma concentração de 5µg/mL foi adicionada no seguinte tempo:
concomitante e 1 hora após a infecção. As monocamadas foram mantidas em estufa de CO2
(5%) a 37ºC (Vero) por 7 dias. Utilizamos três condições: células Vero infectadas com vírus
dengue-2 e não tratadas com a droga; células Vero infectadas com vírus dengue-2 na presença
de brefeldina A (5µg/mL) e células Vero não infectadas e não tratadas com a droga.
Monocamadas infectadas e não infectadas sem a adição de droga foram usadas como
controles. Os sobrenadantes foram removidos de duplicatas de culturas e clarificados por
centrifugação em intervalos de 0, 6, 12, 24, 48, 72, 96, 120, 144 e 168 horas para a realização
do PCR em tempo real e para a titulação por ensaio de placa (PFU), em monocamadas de
células Vero. O experimento foi repetido três vezes.

Materiais e Métodos
25
3.12. Ensaio de inibição dos filamentos de actina e microtúbulos (Citocalasina B)
Monocamadas de células Vero foram incubadas com citocalasina B (10µg/mL) nos
seguintes tempos: juntamente com o vírus e 1h após a adsorsão. Seguindo os mesmos moldes
dos experimentos do item 3.11.
3.13. Quantificação por PCR em Tempo Real
A curva padrão foi preparada a partir da extração do RNA das cinco diluições
decimais em PBS do estoque do vírus dengue-2 proveniente do macerado de cérebro de
camundongos recém-nascidos com titulação de 3 X 105 PFU/mL. Tanto as amostras de RNA
do padrão quanto do sobrenadante das culturas celulares infectadas com o vírus dengue-2
tratadas ou não com as drogas descritas anteriormente foram analisadas através de reações de
PCR em Tempo Real, utilizando o sistema Sybr Green One-Step RT-PCR Master Mix em um
aparelho ABI5700 (Applied Biosystems, Warrington, Reino Unido). Para as reações, foram
utilizados 12,5µl do reagente Sybr Green Master Mix (Applied Biosystems; que contém o
fluoróforo Sybr Green ; a enzima DNA polimerase AmpliTaq Gold; DNTPs com dUTPs; o
fluoróforo ROX, utilizado como referência passiva para normalização dos níveis de
fluorescência; e as demais soluções tampão, já devidamente otimizadas), 0.5 µl de inibidor de
RNase, 0,13 µl de Multiscribe (50 U/ µl), 0,5 µl dos primers (20nM) DV2U (5’-
AAGGTGAGATGAAGCTGTAGTCTC-3’) e DVL1 (5’-CATTCCATTTTCTGGCGTTCT-
3’) específicos para o vírus dengue-2, que amplifica um fragmento de DNA localizado na
região 3’ não-codificadora (HOUNG et al., 2001), 5,87 µl de água DEPC, e 5 µl de RNA,
totalizando um volume final de 25 µl. As condições de amplificação foram: 48°C por 20 min
e 95°C por 10 min, seguido por 40 ciclos de 95°C por 15 segs e 60°C por 1 min. A curva de
dissociação foi construída com temperatura crescente de 60 a 95ºC, utilizada para a análise da
especificidade de amplificação. O sistema utilizado realiza as reações de amplificação e

Materiais e Métodos
26
detecção, e quantifica as amostras (termociclador ABI5700 associado ao software ABI prism,
Applied Biosystems) através da análise do nível de fluorescência gerado pela incorporação de
nucleases fluorogênicas (Sybr Green) aos produtos de amplificação durante o curso da reação.
Os resultados foram analisados com base no valor de Ct (cicle threshold – ou ciclo limiar),
sendo este o ponto correspondente ao número de ciclos aonde a amplificação das amostras,
atinge um limiar (determinado entre o nível de fluorescência dos controles negativos e a fase
de amplificação exponencial das amostras) que permite a análise quantitativa da expressão do
fator avaliado.
3.14. Proteção pós-desafio
A proteção das drogas inibidoras do metabolismo celular foi avaliada em 9 grupos de
cinco camundongos Swiss fêmeas de 4 semanas. Dessa forma 5 grupos foram infectados via
intracerebral com a inoculação de 100µL de vírus dengue-2 (3X103 PFU/mL) com seringa de
insulina e agulha de 281/2. Após duas horas foram realizadas injeções via intraperitoneal de
cada droga nos respectivos grupos a uma concentração de 30mg/Kg de cloroquina (HONG et
al., 2004), 26,3mg/Kg brefeldina A (PHILLIPS; SUPKO; MALSPEIS, 1993), 1,2mg/Kg de
citocalasina B (AUSTIN; WIND; BROWN, 1982). Sendo que um grupo recebeu cloroquina
somente 3 dias após infecção por via intraperitonial a mesma concentração do grupo anterior
e o outro grupo não foi tratado com nenhuma das drogas (controle positivo). Os grupos
tratados com cloroquina, receberam a droga a intervalos de 24h durante 7 dias. Como controle
negativo um grupo foi inoculado com PBS e os outros 3 grupos foram somente tratados com
cada droga. A análise da sobrevivência foi realizada observando estes animais quanto ao
aparecimento de sinais de doença.

Materiais e Métodos
27
3.15. Análise estatística
A análise estatística foi empregada para avaliar as diferenças na carga viral nos
períodos de coleta entre o experimento (droga) e o controle (sem a droga). Através do
programa de computador GraphPad Prism® (GraphPad Software Inc., EUA) versão 3.0,
foram submetidos ao teste estatístico one-way ANOVA (não-paramétrico), seguido pelo teste
de Bonferroni. Para todas as análises, valores de p< 0,05 foram considerados estatisticamente
significantes.

Resultados
28
RREESSUULLTTAADDOOSS
Resultados
29
4. RESULTADOS
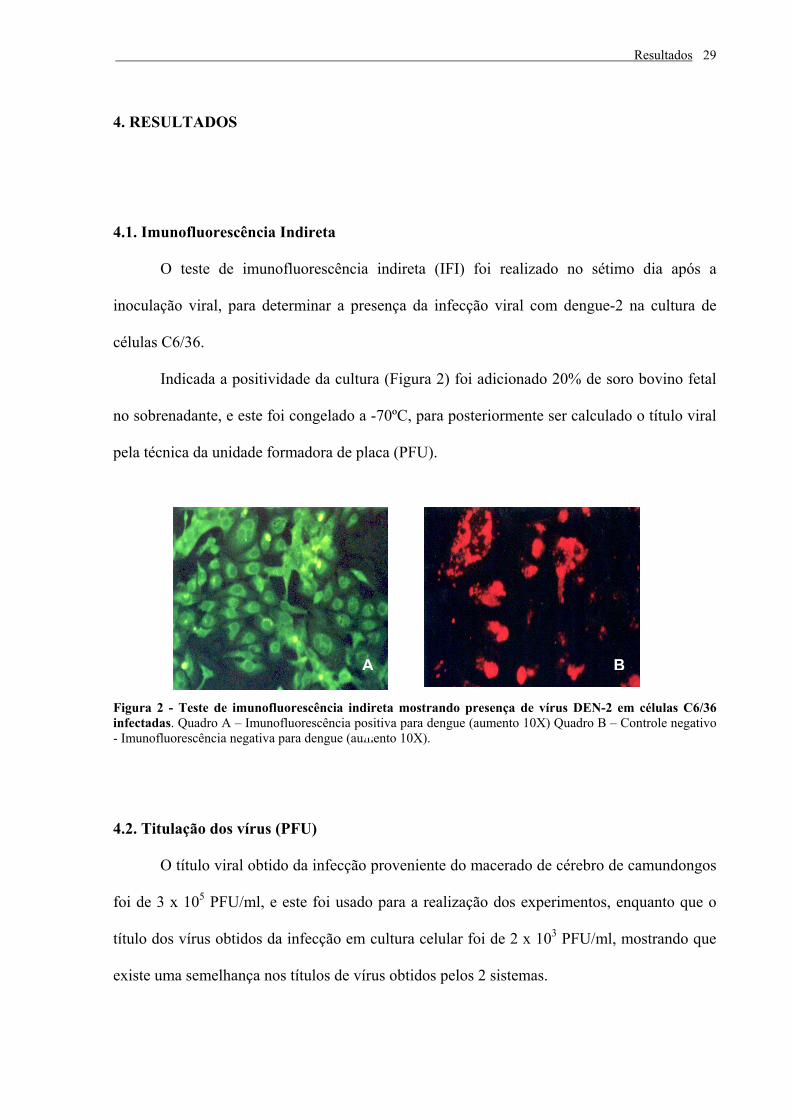
4.1. Imunofluorescência Indireta
O teste de imunofluorescência indireta (IFI) foi realizado no sétimo dia após a
inoculação viral, para determinar a presença da infecção viral com dengue-2 na cultura de
células C6/36.
Indicada a positividade da cultura (Figura 2) foi adicionado 20% de soro bovino fetal
no sobrenadante, e este foi congelado a -70ºC, para posteriormente ser calculado o título viral
pela técnica da unidade formadora de placa (PFU).
Figura 2 - Teste de imunofluorescência indireta mostrando presença de vírus DEN-2 em células C6/36 infectadas. Quadro A – Imunofluorescência positiva para dengue (aumento 10X) Quadro B – Controle negativo - Imunofluorescência negativa para dengue (aumento 10X).
4.2. Titulação dos vírus (PFU)
O título viral obtido da infecção proveniente do macerado de cérebro de camundongos
foi de 3 x 105 PFU/ml, e este foi usado para a realização dos experimentos, enquanto que o
título dos vírus obtidos da infecção em cultura celular foi de 2 x 103 PFU/ml, mostrando que
existe uma semelhança nos títulos de vírus obtidos pelos 2 sistemas.
A
BA

Resultados
30
4.3. Padronização da curva padrão no PCR em tempo real
A partir dos estoques virais de dengue-2 provenientes de cérebros de camundongos
titulados utilizando-se os ensaios de placa (PFU), foram realizadas as cinco diluições seriadas
que tiveram seu RNA extraído para, posteriormente, serem quantificadas por PCR em Tempo
Real utilizando-se o Kit Sybr Green One-Step. A curva padrão no sistema Sybr Green (Figura
3), têm expressados os valores da inclinação, interseção e correlação da reta, no canto inferior
inferior direito, calculados pelo Software GeneAmp 5700 (Applied Biosystems). O
coeficiente de correlação foi excelente (r =0,99), indicando existir uma correlação linear do
ensaio. O limite de detecção mínimo foi de 10 PFU/mL. A análise do perfil da curva de
amplificação é baseado no Ct (ciclo do limiar – início da detecção de fluorescência), em que é
possível visualizar uma relação entre a quantidade de DNA e Ct, pois quanto menor o Ct,
maior é a quantidade de DNA. De acordo com a figura 4, podemos verificar que o início das
amplificações para o vírus dengue-2, teve o Ct de 10. Analisando o perfil da curva de
dissociação, para assegurar que os perfis de amplificação para as diluições dos vírus dengue-2
estavam de fato relacionadas à amplificação do fragmento específico deste vírus, foi possível
validar sua positividade, pois todas as diluições apresentam curvas de dissociação com padrão
específico, como pode ser observada na figura 5.

Resultados
31
Figura 3 – Análise da curva padrão para o sorotipo do vírus dengue-2 através do PCR em Tempo Real. A figura mostra o perfil de amplificação de cada diluição do vírus, aonde podemos verificar que o inicio da amplificação teve um Ct de 10. A curva padrão é expressa com o valor do Ct (Ciclo do limiar) versus log Ct. No lado inferior direito, estão expressos os valores de inclinação da reta, intersecção e correlação.
Figura 4 – Perfil de amplificação do vírus dengue-2 por PCR em Tempo Real. A figura mostra o resultado obtido para a amplificação de cada diluição do vírus (A1-5); exemplificando as fases de amplificação, a determinação de Ct.
A1 A2 A3 A4 A5
Fase de Amplificação
Ct

Resultados
32
Figura 5 – Análise da curva de dissociação para determinação da especificidade da reação de amplificação através do PCR em Tempo Real. (A) demonstra os resultados obtidos pela curva dissociação de cada diluição (A1-5) do vírus dengue-2 a uma mesma concentração de primers (sense e antisense) para a regão 3’ não-codificadora do vírus. A temperatura de “melting” (Tm) de cada diluição do produto específico amplificado do vírus é 81ºC. (B) No controle negativo, a temperatura de “melting” (Tm) do produto inespecífico é de 72,5 ºC.
A1, A2, A3, A4, A5
TM=81ºC Amplificação específica
A
TM=72,5ºC Amplificação inespecífica
B

Resultados
33
4.4. Observação de citotoxicidade da cloroquina em células C6/36 e Vero
Não houve diferença significante entre células C6/36 e Vero com relação à
citotoxicidade. Concentrações iguais ou superiores a 500µg/mL foram acentuadamente
citotóxicas. Concentrações iguais ou inferiores a 50 µg/mL não induziram citotoxicidade
significante (figura 6). Deste modo adotamos a concentração de 50 µg/mL para os
experimentos subseqüentes.
0
10
20
30
40
50
60
70
80
90
100
110
5000 500 50 5 0,5 0,05 0,005
% d
e vi
abili
dade
Células VeroCélulas C6/36
Figura 6 – Viabilidade de células Vero e C6/36 submetidas a tratamento com cloroquina. Concentrações iguais ou inferiores a 50µg/mL não induziram citotoxicidade significante.
4.5. Efeito de inibidor da acidificação de endossomo (cloroquina) sobre a replicação do
vírus dengue-2
A cloroquina não diminuiu a produção do vírus dengue-2 em células C6/36 até o 7°
dia após a infecção quando administrada logo após a adsorção, pois o título viral aumentou de
3 X 105 PFU/mL (sem a droga) para 9 X 107 PFU/mL com a administração da cloroquina
Concentração de cloroquina (µg/mL)
5x103 5x102 5x101 5 5x10-1 5x10-2 5x10-3

Resultados
34
como demonstrado nas figuras 7 e 8. Além disso, no sobrenadante da cultura celular, com e
sem a administração da droga, foi detectada a presença do vírus dengue através da RT-PCR
(Figura 9).
Na tentativa de determinar se a cloroquina agia nas primeiras etapas da inibição da
replicação do vírus dengue-2, monocamadas de células C6/36 foram infectadas com o vírus e
meio de crescimento contendo 50µg/mL de cloroquina foi adicionado. Sobrenadantes foram
coletados a períodos de 6, 12, 24, 48, 72, 96, 120, 144 e 168 horas após a infecção. Esse
resultado (Figura 10) mostrou que a replicação do vírus dengue-2 não foi sensível a droga.
Assim o tratamento com cloroquina resultou em uma redução não significante do número de
cópias de RNA/ml nas primeiras 6 horas e um aumento estatisticamente significante na
replicação viral a partir de 12 até 168 horas (p<0,05), após análise por PCR em tempo real.
Figura 7 – Ensaio de placa para a titulação do vírus dengue-2. O título de 3X 105 PFU/mL foi obtido 7 dias após a infecção, a partir do sobrenadante de cultura de células C6/36 infectadas com vírus dengue-2. O ensaio foi realizado em duplicata e o controle negativo (CN) são células C6/36 não infectadas.
10-4 10-4 10-3 10-3 10-2 10-2
10-6 10-6 10-5 10-5 10-7 10-7
10-10 10-10 10-9 10-9 10-8 10-8
10-1 10-1 CN CN CN CN

Resultados
35
Figura 8 – Ensaio de placa para a titulação do vírus dengue-2 na presença da cloroquina. O título de 9 X 107 PFU/mL foi obtido 7 dias após a infecção, a partir do sobrenadante de cultura de células C6/36 infectadas com vírus dengue-2 e tratadas com cloroquina (50µg/mL) 1 hora após a infecção. O ensaio foi realizado em duplicata e o controle negativo (CN) são células C6/36 não infectadas.
M A B
Figura 9 – Amplificação do fragmento de 419pb da região NS1 do vírus dengue. Eletroforese em gel de agarose 2% corado com brometo de etídio. M (Marcador de peso molecular, 50pb, Invitrogen); A - sobrenadante da cultura de células C6/36 infectadas com vírus DEN-2; B - sobrenadante da cultura de células C6/36 infectadas com vírus dengue-2 na presença de cloroquina (50µg/mL).
← 419 pb 350 pb →
10-4 10-3 10-3 10-2
10-6 10-5 10-5 10-4
10-2 10-1 10-1 CN

Resultados
36
6 12 24 48 72 96 120 144 16810 0
10 1
10 2
10 3
10 4
10 5
sem cloroquina
com cloroquina
Tempo (horas)
Cóp
ias
do R
NA
Vira
l
Figura 10 – Ação da cloroquina na replicação do vírus dengue-2 em células C6/36. Sobrenadante de cultura de células C6/36 infectadas com vírus dengue-2, tratadas ou não com cloroquina, tiveram o RNA total extraído e analisado por PCR em Tempo Real. Os resultados apresentados representam os valores da média do número de cópias de RNA virais de três experimentos (p<0,05).
Em células Vero, quando a cloroquina foi adicionada concomitantemente e 1hora após
infecção, foi realizada a coleta do sobrenadante a 0, 6, 12, 24, 48, 72, 96, 120, 144 e 168 horas
após a infecção para a quantificação do RNA viral por PCR em Tempo Real, levou-se em
consideração que o tempo "zero" é o momento em que a cultura foi incubada a 37ºC, após
adsorção do vírus. Com esta linhagem celular, a droga causou uma redução na replicação viral
nas primeiras horas (6, 12 e 24h), quando comparado com o controle positivo, com p<0,001
(figura 11), mas a partir de então não houve diferença na produção de novos vírus. A análise
por ensaio de placa (PFU), resultou que células Vero infectadas com o vírus e tratadas com a
droga, teve uma carga viral de 10 X menor nas primeiras horas (6, 12 e 24h) quando
comparado com o controle (dados não mostrados).

Resultados
37
0 6 12 24 48 72 96 120 144 16810 0
10 1
10 2
10 3
10 4
sem cloroquina
com cloroquina
Tempo (horas)
Cóp
ias
de R
NA
Vira
l
Figura 11 – Ação da cloroquina sob a replicação do vírus dengue-2 em células Vero. Sobrenadante de cultura de células Vero infectadas com vírus dengue-2, tratadas ou não com cloroquina, tiveram o RNA total extraído e analisados por PCR em Tempo Real. Os resultados apresentados representam os valores da média do número de cópias de RNA virais de três experimentos (p<0,001).
Na tentativa de inibir a replicação viral nas próximas horas, foi adicionada cloroquina
(concomitantemente, 1hora após infecção e em intervalos de 24 horas após infecção),
seguindo os mesmos moldes do experimento anterior. Observou-se uma redução na carga
viral com adição da droga a partir de 6h até 168h quando comparado com o controle positivo
(p<0,05) (figura 12). A análise por ensaio de placa (PFU), resultou que células Vero
infectadas com o vírus e tratadas com a droga (24-24 horas), tiveram uma carga viral 10 a
100X menor quando comparado com o controle (dados não mostrados).

Resultados
38
0 6 12 24 48 72 96 120 144 16810 0
10 1
10 2
10 3
10 4
sem cloroquina
com cloroquina
Tempo (horas)
Cóp
ias
de R
NA
Vira
l
Figura 12 – Ação da cloroquina adionada em intervalos de 24 horas sob a replicação do vírus dengue-2 em células Vero. Sobrenadante de cultura de células Vero infectadas com vírus dengue-2, tratadas (24-24 horas) ou não com cloroquina, tiveram o RNA total extraído e analisados por PCR em Tempo Real. Os resultados apresentados representam os valores da média do número de cópias de RNA virais de três experimentos (p<0,05).
Na tentativa de inibir ainda mais a replicação viral nas próximas horas, foi adicionada
cloroquina, concomitantemente, 1hora após infecção e em intervalos de 12 horas após
infecção (figura 13). Houve uma redução apreciável na carga viral quando comparada com o
uso da cloroquina a intervalos de 24 horas e com o controle positivo, p<0,01. A análise por
ensaio de placa (PFU), resultou que células Vero infectadas com o vírus e tratadas com a
droga (12-12 horas), tiveram uma carga viral 10 a 100 X menor quando comparado com o
controle (dados não mostrados).

Resultados
39
0 6 12 24 48 72 96 120 144 16810 0
10 1
10 2
10 3
10 4
sem cloroquina
com cloroquina
Tempo (horas)
Cóp
ias
de R
NA
Vira
l
Figura 13 – Ação da cloroquina adicionada em intervalos 12 horas sob a replicação do vírus dengue-2 em células Vero. Sobrenadante de cultura de células Vero infectadas com vírus dengue-2, tratadas (12-12 horas) ou não com cloroquina, tiveram o RNA total extraído e analisados por PCR em Tempo Real. Os resultados apresentados representam os valores da média do número de cópias de RNA virais de três experimentos (p<0,01).
4.6. Observação de citotoxicidade da Brefeldina A em células Vero
Concentrações iguais ou inferiores a 5 µg/mL não induziram citotoxicidade
significante para células Vero (figura 14), enquanto que concentrações iguais ou superiores a
50µg/mL foram acentuadamente citotóxicas. Deste modo adotamos a concentração de 5
µg/mL para os experimentos subseqüentes.
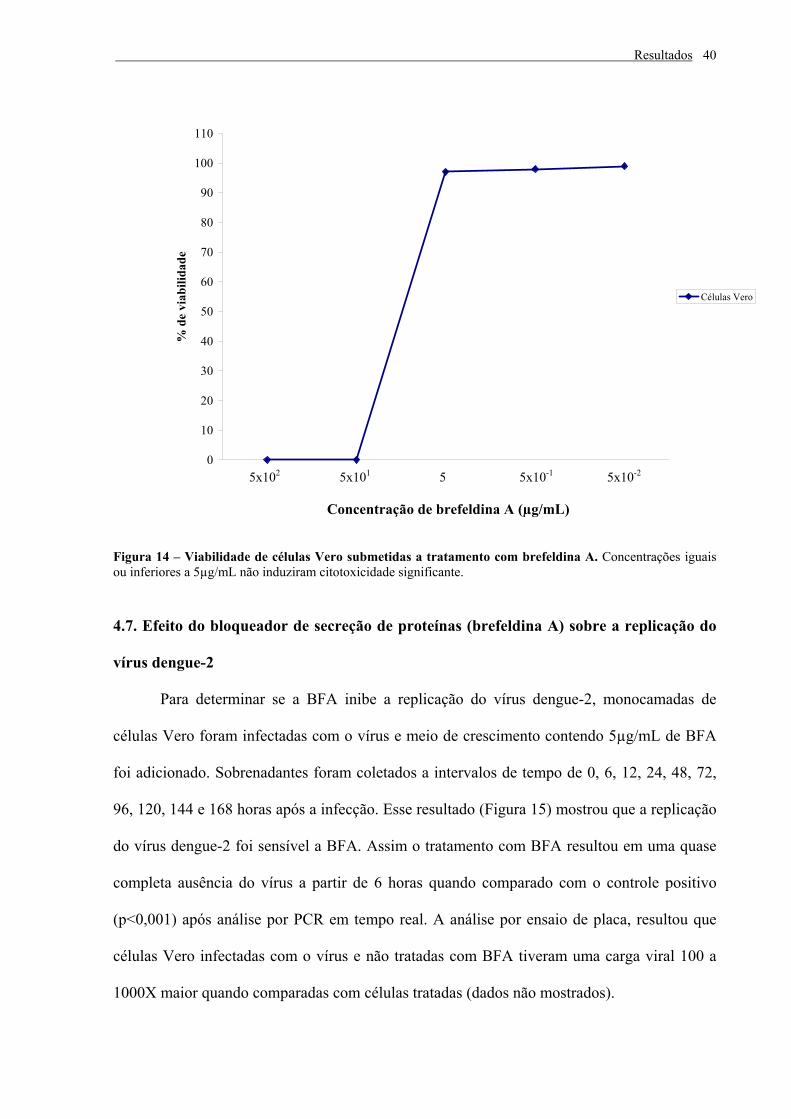
Resultados
40
0
10
20
30
40
50
60
70
80
90
100
110
500 50 5 0,5 0,05
% d
e vi
abili
dade
Células Vero
Figura 14 – Viabilidade de células Vero submetidas a tratamento com brefeldina A. Concentrações iguais ou inferiores a 5µg/mL não induziram citotoxicidade significante.
4.7. Efeito do bloqueador de secreção de proteínas (brefeldina A) sobre a replicação do
vírus dengue-2
Para determinar se a BFA inibe a replicação do vírus dengue-2, monocamadas de
células Vero foram infectadas com o vírus e meio de crescimento contendo 5µg/mL de BFA
foi adicionado. Sobrenadantes foram coletados a intervalos de tempo de 0, 6, 12, 24, 48, 72,
96, 120, 144 e 168 horas após a infecção. Esse resultado (Figura 15) mostrou que a replicação
do vírus dengue-2 foi sensível a BFA. Assim o tratamento com BFA resultou em uma quase
completa ausência do vírus a partir de 6 horas quando comparado com o controle positivo
(p<0,001) após análise por PCR em tempo real. A análise por ensaio de placa, resultou que
células Vero infectadas com o vírus e não tratadas com BFA tiveram uma carga viral 100 a
1000X maior quando comparadas com células tratadas (dados não mostrados).
5x102 5x101 5 5x10-1 5x10-2
Concentração de brefeldina A (µg/mL)
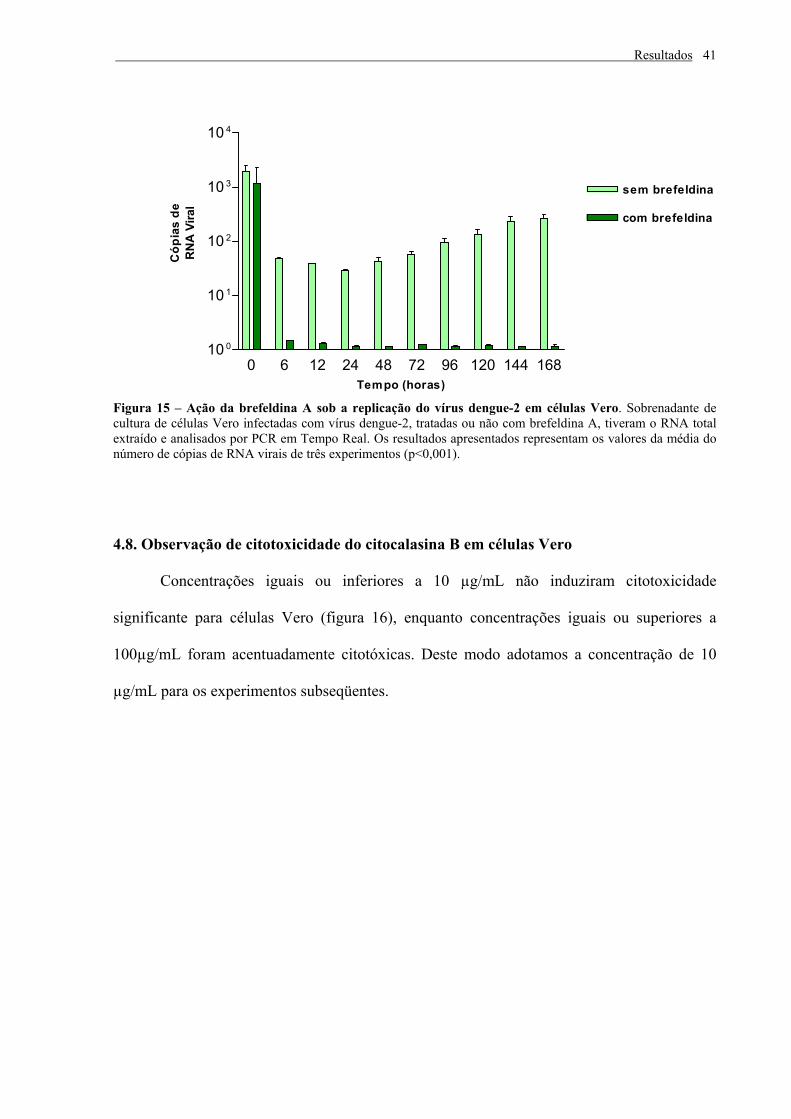
Resultados
41
0 6 12 24 48 72 96 120 144 16810 0
10 1
10 2
10 3
10 4
sem brefeldina
com brefeldina
Tempo (horas)
Cóp
ias
deR
NA
Vira
l
Figura 15 – Ação da brefeldina A sob a replicação do vírus dengue-2 em células Vero. Sobrenadante de cultura de células Vero infectadas com vírus dengue-2, tratadas ou não com brefeldina A, tiveram o RNA total extraído e analisados por PCR em Tempo Real. Os resultados apresentados representam os valores da média do número de cópias de RNA virais de três experimentos (p<0,001).
4.8. Observação de citotoxicidade do citocalasina B em células Vero
Concentrações iguais ou inferiores a 10 µg/mL não induziram citotoxicidade
significante para células Vero (figura 16), enquanto concentrações iguais ou superiores a
100µg/mL foram acentuadamente citotóxicas. Deste modo adotamos a concentração de 10
µg/mL para os experimentos subseqüentes.

Resultados
42
0
10
20
30
40
50
60
70
80
90
100
110
500 50 5 0,5 0,05
% d
e vi
abili
dade
Células Vero
Figura 16 – Viabilidade de células Vero submetidas a tratamento com citocalasina B. Concentrações iguais ou inferiores a 10µg/mL não induziram citotoxicidade significante.
4.9. Efeito do bloqueador de filamentos de actina e microtubulos (citocalasina B) sobre a
replicação do vírus dengue-2.
Para determinar se a citocalasina B inibe a replicação do vírus dengue-2,
monocamadas de células Vero foram infectadas com o vírus e meio de crescimento contendo
10µg/mL da droga foi adicionado. Sobrenadantes foram coletados a intervalos de tempo de 0,
6, 12, 24, 48, 72, 96, 120, 144 e 168 horas após a infecção. Esse resultado (Figura 17)
mostrou que a replicação do vírus dengue-2 foi sensível a citocalasina B. Assim o tratamento
com a droga resultou em uma diminuição da replicação viral a partir de 6 horas que se
manteve durante todo o período observado, quando comparado com o controle positivo
(p<0,05). A análise por ensaio de placa, resultou que células Vero infectadas com o vírus e
103 102 10 10-1 10-2
Concentração de citocalasina B (µg/mL)

Resultados
43
não tratadas com citocalasina B tiveram uma carga viral 10 a 100X maior quando comparada
com células tratadas (dados não mostrados).
0 6 12 24 48 72 96 120 144 16810 0
10 1
10 2
10 3
10 4
sem citocalasina
com citocalasina
Tempo (horas)
Cóp
ias
de R
NA
Vira
l
Figura 17 – Ação da citocalasina B sob a replicação do vírus dengue-2 em células Vero. Sobrenadante de cultura de células Vero infectadas com vírus dengue-2, tratadas ou não com citocalasina B, tiveram o RNA total extraído e analisados por PCR em Tempo Real. Os resultados apresentados representam os valores da média do número de cópias de RNA virais de três experimentos (p<0,05).
4.10. Proteção pós-desafio
Observamos a sobrevivência de todos os camundongos em que foram injetadas apenas
as drogas ou PBS, enquanto que os animais que receberam a inoculação do vírus dengue-2
morreram no 10º dia após o desafio. O grupo que apresentou uma melhor resposta foi o que
recebeu a administração de cloroquina duas horas após a infecção viral e em intervalos de 24
horas durante 7 dias, com uma porcentagem de sobrevivência de 60% 10 dias após a infecção.
Essa porcentagem de sobrevivência se manteve até o 13° dia, caindo para 20% até o 21° dia
(Figura 18). No entanto, os camundongos que receberam cloroquina 3 dias após a infecção
viral e em intervalos de 24 horas durante 7 dias, morreram no 10º dia após o desafio (Figura
19).

Resultados
44
Os grupos de camundongos que receberam as drogas brefeldina A ou citocalasina B
duas horas após a infecção viral tiveram uma porcentagem de sobrevivência de 40% no 10°
dia, no entanto no 11° dia todos os camundongos morreram (Figura 20 e 21).
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Tempo (dias)
% S
obre
vivê
ncia
vírus vírus + cloroquinaPBScloroquina
Figura 18 – Curvas de sobrevivência dos camundongos infectados via intracerebral com 3X103 PFU/mL do vírus dengue-2 e receberam 30 mg/Kg de cloroquina 2 horas após a infecção e em intervalos de 24 horas durante 7 dias. Todos os camundongos do grupo do controle positivo morreram até o 10° dia. Notar que os camundongos que receberam cloroquina + vírus tiveram uma maior sobrevivência comparados àqueles que somente receberam vírus.
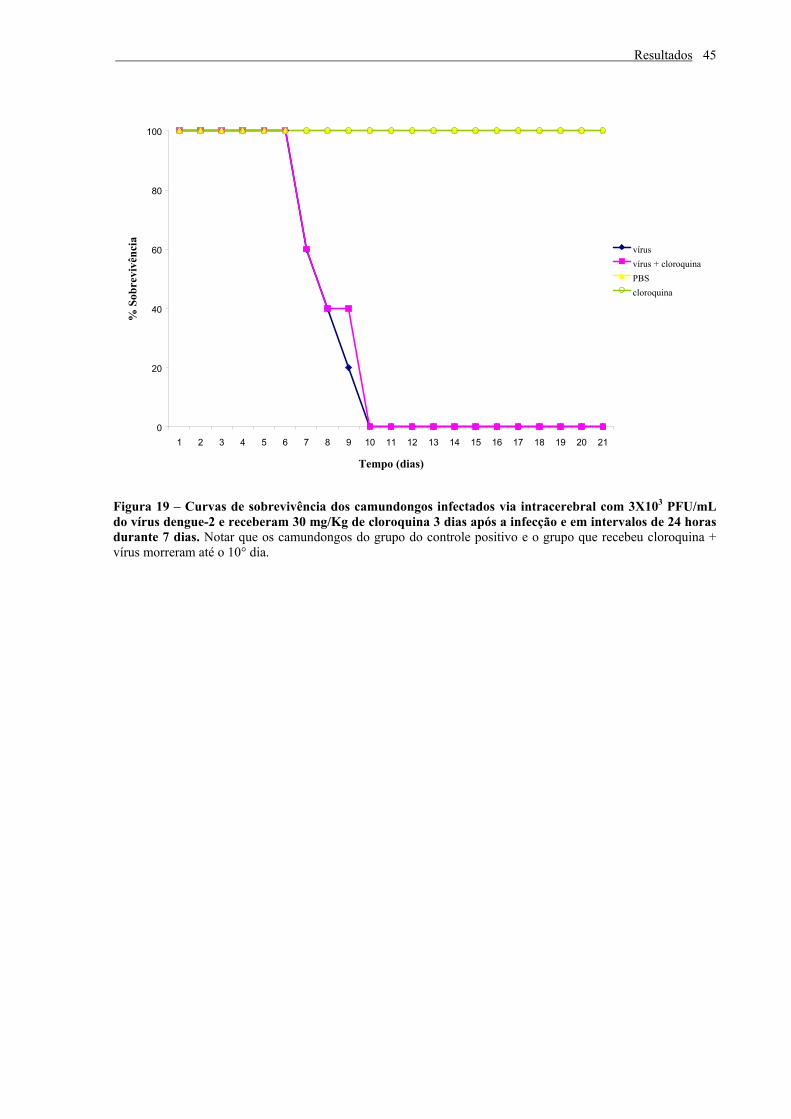
Resultados
45
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Tempo (dias)
% S
obre
vivê
ncia
vírusvírus + cloroquinaPBScloroquina
Figura 19 – Curvas de sobrevivência dos camundongos infectados via intracerebral com 3X103 PFU/mL do vírus dengue-2 e receberam 30 mg/Kg de cloroquina 3 dias após a infecção e em intervalos de 24 horas durante 7 dias. Notar que os camundongos do grupo do controle positivo e o grupo que recebeu cloroquina + vírus morreram até o 10° dia.
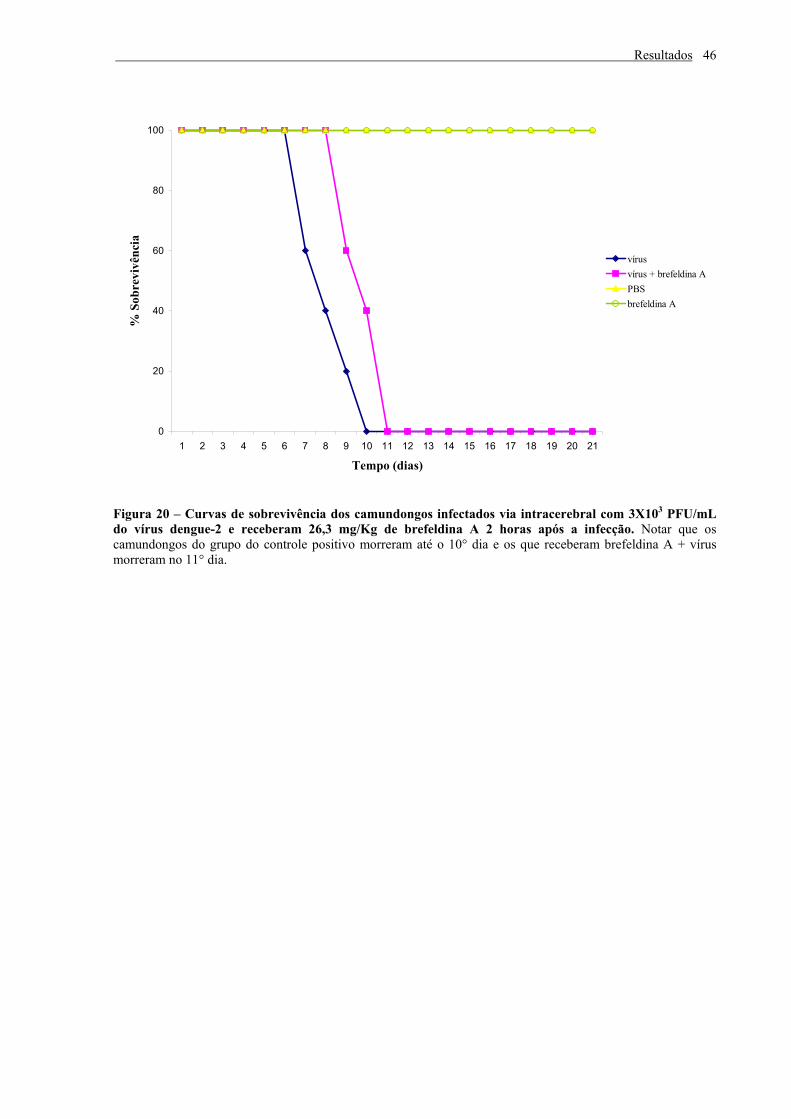
Resultados
46
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Tempo (dias)
% S
obre
vivê
ncia
vírus vírus + brefeldina APBSbrefeldina A
Figura 20 – Curvas de sobrevivência dos camundongos infectados via intracerebral com 3X103 PFU/mL do vírus dengue-2 e receberam 26,3 mg/Kg de brefeldina A 2 horas após a infecção. Notar que os camundongos do grupo do controle positivo morreram até o 10° dia e os que receberam brefeldina A + vírus morreram no 11° dia.

Resultados
47
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Tempo (dias)
% S
obre
vivê
ncia
vírusvírus+ citocalasina BPBScitocalasina B
Figura 21 – Curvas de sobrevivência dos camundongos infectados via intracerebral com 3X103 PFU/mL do vírus dengue-2 e receberam 1,2mg/Kg de citocalasina B 2 horas após a infecção. Notar que os camundongos do grupo do controle positivo morreram até o 10° dia e os que receberam citocalasina B + vírus morreram no 11° dia.

Discussão
48
DDIISSCCUUSSSSÃÃOO

Discussão
49
5. DISCUSSÃO
A dengue é a mais importante arbovirose urbana da atualidade e caracteriza-se por ser
uma doença febril aguda cujos agentes etiológicos, os vírus dengue, são transmitidos ao
homem através de picada da fêmea do mosquito Aedes aegypti. É uma doença endêmica, de
caráter urbano, que está presente em praticamente todas as regiões de clima tropical e
subtropical do mundo. A cada ano é estimado que cerca de 50-100 milhões de casos de
dengue ocorram no mundo, incluindo cerca de 500.000 casos de Febre Hemorrágica da
Dengue, a mais grave manifestação da doença (GIBBONS; VAUGHN, 2002).
Um modo de entrada comum de vírus envelopado na célula hospedeira envolve o
transporte por endocitose seguido pela fusão da membrana viral com a membrana endossomal
(KIELIAN; JUNGERWIRTH, 1990).
O vírus dengue entra na célula hospedeira por endocitose mediada por receptor e
replica-se no citoplasma das células de vertebrados após um período de 12 a 16 horas da
infecção. Os receptores de membrana, aos quais se ligam o vírus dengue na fase inicial da
infecção, são desconhecidos, apesar de haver evidências que o heparan sulfato é participante
ativo nesta fase da infecção (BARRAGÁN; ANGEL, 2001). Após a penetração no citoplasma
ocorre a queda do pH intra-endossomal resultando na fusão do envelope viral com a
membrana endossomal. Após a fusão ter ocorrido, o nucleocapsideo é liberado para o
citoplasma, onde as proteínas do capsídeo e o RNA se dissociam, iniciando a replicação do
genoma RNA e a montagem da partícula viral (LINDENBACH; RICE, 2003).
Apesar de muita pesquisada, ainda não está disponível uma vacina preventiva eficaz,
assim como, drogas antivirais que combatam a infecção pelos vírus dengue. Deste modo a

Discussão
50
investigação de drogas que interferem na homeostase celular e conseqüentemente na
replicação do vírus dengue se faz necessário para o tratamento da dengue.
Drogas que bloqueiam funções celulares específicas e seus efeitos sobre infecções
virais têm sido extensivamente estudadas para explicar diferentes aspectos de infecção dos
vírus.
Até o momento não há estudos publicados sobre a ação de drogas que interferem com
a homeostase celular (Cloroquina, Brefeldina A e Citocalasina B) sob o ciclo replicativo do
vírus dengue. Diante dessa observação, avaliamos a ação dessas três drogas, visando observar
se as mesmas eram capazes de interferir na replicação in vitro e in vivo do vírus dengue-2.
Preliminarmente utilizamos células C6/36 e Vero que são suscetíveis e permissivas à
infecção pelo vírus dengue. Foi necessário investigar a citotoxicidade dessas drogas para as
células Vero, em que diversas diluições de cloroquina, brefeldina A citocalasina B foram
avaliadas, e as concentrações que não induziram citotoxicidade significante foram,
respectivamente, 50µg/mL, 5 µg/mL e 10 µg/mL. Estas concentrações são equivalentes as
utilizadas em experimentos realizados com esta mesma linhagem de células (VINCENTE et
al., 2005; MACKENZIE; WESTAWAY, 2001; CHU et al., 2003). Já a concentração
encontrada no ensaio de citotoxicidade da cloroquina para células C6/36 foi também de 50
µg/mL, essa foi a mesma utilizada por Coombs et al. (1981).
A adsorção do vírus dengue à célula hospedeira ocorre através da interação da
glicoproteína E do envelope do vírus com o receptor da superfície celular, subsequentemente
o vírus penetra na célula por endocitose, e a acidificação da vesícula endossomal faz com que
ocorra a mudança conformacional da glicoproteína E do virion. A mudança conformacional
induz a fusão da membrana viral com a membrana endossomal, o nucleocapsideo é liberado,
ou seja, ocorre o desnudamento, com a posterior liberação do RNA viral para o citoplasma

Discussão
51
(MODIS et al., 2004). Assim a acidificação do endossomo é essencial na replicação do vírus
dengue.
Estudos têm sido realizados para verificar a possível ação da cloroquina, que é uma
base fraca que aumenta o pH de vesículas ácidas, sob a replicação de diversos vírus.
No presente trabalho, observamos que em experimentos realizados com células de
mosquito Aedes albopictus infectadas com vírus dengue-2 e tratadas com cloroquina, ocorreu
a elevação no título final do vírus 7 dias após a infecção. Também se observou o aumento da
carga viral através da RT-PCR realizada a partir do RNA viral extraído da cultura de células
tratadas e não tratadas com cloroquina 7 dias após a infecção, em que a intensidade de banda
de 419pb é mais forte quando foi feito o tratamento com a droga.
Foi observado que a cloroquina agia nas primeiras 6 horas da replicação viral em
células C6/36, onde através da quantificação por PCR em Tempo Real e por ensaio de placa
(PFU), houve um aumento estatisticamente significante na carga viral quando comparada com
o controle após a 2ª hora da infecção.
Em 1981, Coombs, Edwards e Brown. também observaram que a cloroquina não
reduzia a produção viral em experimentos realizados com o vírus Sindbis em células de
mosquito Aedes albopictus.
Hernandez, Luo e Brown (2000) encontraram resultados semelhantes quando usaram a
cloroquina para inibir a replicação do vírus Sindbis. E diante desses achados, duas explicações
eram possíveis: (i) a cloroquina não bloqueia a acidificação dos endossomos das células de
mosquito Aedes albopictus e (ii) a cloroquina bloqueia a acidificação dos endossomos, porém
não a infecção das células de mosquito pelo vírus Sindbis.
Para excluir a explicação que a cloroquina não bloqueava a acidificação dos
endossomos em células de mosquito, foi realizado um teste para verificar se a droga protegia
essas células da intoxicação pela toxina diftérica, a qual requer um pH endossomal menor do

Discussão
52
que 5,5 para entrar no citoplasma da célula. O resultado foi que a droga protegia as células do
mosquito da intoxicação, consequentemente, ela aumentava o pH do endossomo, no entanto
permitia a infecção, assim a exposição ao baixo pH não é essencial para a penetração do vírus
Sindbis à células do mosquito Aedes albopictus (HERNANDEZ; LUO; BROWN, 2000).
Esses dados sugerem que o vírus dengue-2 usado em nossos experimentos obteve um
aumento na replicação em células de mosquito Aedes albopictus, possivelmente por existir
uma outra via de penetração intracitoplasmática que não seja a endossomica, a qual não foi
bloqueada pela cloroquina. Outra possibilidade é que a inibição inicial na replicação viral pela
cloroquina (Figura 10) resultou apenas no acúmulo deste vírus nos compartimentos
endossomais, e que, após a degradação da droga, estes vírus se replicaram em títulos ainda
maiores devido a este acúmulo inicial.
Em experimentos subseqüentes com a cloroquina utilizando células do rim de macacos
verdes africanos (Vero), nós observamos que houve uma diminuição estatisticamente
significante na replicação viral.
A presença da cloroquina provocou um efeito inibitório no ciclo replicativo do vírus
dengue-2 quando comparado com o controle até 24 horas após infecção. Apesar de não haver
a abolição total da replicação viral, houve interferência na liberação de vírus para o meio
extracelular.
Como nós observamos que a droga tinha uma ação limitada, diminuindo a replicação
viral até 24 horas, realizamos experimentos subseqüentes em que a cloroquina foi adicionada
em intervalos de 24 horas na cultura de células Vero infectadas com dengue-2. Observou-se
diminuição no número de cópias do RNA viral no sobrenadante da cultura celular infectada e
tratada com a droga, demonstrando um efeito inibitório da cloroquina no ciclo replicativo do
vírus. Apesar de não haver a abolição total da replicação viral, ocorreu uma redução da carga
viral para o meio extracelular até o 7º dia.

Discussão
53
Com a intenção de diminuir ainda mais a replicação viral, foi adicionada cloroquina
em intervalos de 12 horas. Este foi o experimento em que se conseguiu um melhor resultado
da ação da droga, pois foi observado um efeito inibitório expressivo no ciclo replicativo do
vírus, quando comparado com o controle. Apesar de não haver a abolição total da replicação
viral, houve uma maior interferência na liberação de vírus para o meio extracelular, quando
comparado com o experimento em que se adicionou cloroquina em intervalos de 24 horas.
Nós identificamos neste estudo a cloroquina como um agente antiviral efetivo para o
vírus dengue-2 em condições de cultura celular, como evidenciado pelo seu efeito inibitório
quando a droga foi adicionada após a iniciação e estabelecimento da infecção. O fato que a
cloroquina exerce um efeito antiviral durante condições pós-infecção sugere que ela
provavelmente tem uma ação terapêutica. Quando a cloroquina é adicionada após a infecção,
esse agente pode aumentar rapidamente o pH endossomal e subverter o andamento dos
eventos de fusão entre vírus e endossomo, assim inibindo a infecção. O mecanismo de ação da
cloroquina depende quando ela foi adicionada a célula. Quando adicionada após início da
infecção, essa droga afeta a fusão mediada por endossomo, subseqüentemente a replicação
viral, ou montagem e liberação (VINCENT et al., 2005).
Estudos anteriores com a cloroquina tem demonstrado que ela tem múltiplos efeitos
sobre células de mamíferos além da elevação do pH endossomal, incluindo a prevenção da
glicosilação terminal das imunoglobulinas (THORENS; VASSALLI, 1986). Quando
adicionada em estágios tardios à células infectadas com vírus, a cloroquina inibiu a maturação
do vírus da estomatite vesicular pela inibição da expressão da glicoproteína à superfície
celular (DILLE; JOHNSON, 1982). Essa droga teve ação inibitória até mesmo quando foi
adicionada 3 a 5 horas após a infecção pelo coronavírus (SARS-CV), o agente etiológico da
síndrome respiratória aguda grave (SARS), em células de primata (Vero) (VINCENT et al.,
2005); também inibiu a produção de partículas do vírus da imunodeficiência humana (HIV-1)

Discussão
54
infecciosas pela interferencia com a glicosilação terminal da glicoproteína, em linhagens de
células H9 (TSAI et al., 1990) e linhagens de células monocíticas e linfocíticas (SPERBER et
al., 1993), assim como para o vírus kunjin (flavivirus), em células de primata (Vero)
(MACKENZIE; WESTAWAY, 2001).
Segundo Randolph, Winkler e Stollar (1990) a administração da cloroquina em
momentos tardios da infecção em cultura de células BHK, Vero e KB infectadas com vírus
dengue geravam partículas não infecciosas, pois provavelmente essa droga impedia a
clivagem da proteína prM nos compartimentos vesículares do complexo de Golgi.
Nossos resultados sugerem que a acidificação endossomal deve estar envolvida no
ciclo replicativo do vírus dengue-2 em células de mamíferos, o que poderia ser usado para
considerar a cloroquina como um candidato antiviral para as infecções pelos vírus dengue,
pois ela mostrou-se efetiva na inibição da propagação do vírus dengue-2 em cultura de
células. O fato que a droga teve um efeito antiviral inibitório significante quando as células
susceptíveis (Vero) foram tratadas após a infecção, sugere um possível uso terapêutico, ou
seja, evidencia a possibilidade do uso da cloroquina no tratamento clinico da dengue.
A replicação de alguns vírus ocorre no núcleo ou na região perinuclear do citoplasma,
regiões que se situam, em geral, a alguma distância do local de entrada na célula. Chegar a
esses locais de replicação eficientemente requer transporte organizado. Suporte para a
afirmação de que vírus usam maquinaria de transporte de microtúbulos foi originado a partir
de diversas linhas experimentais, usando drogas despolimerizantes de microtúbulos, que
interferem com a distribuição de proteínas virais no interior da célula infectada (SMITH;
ENQUIST, 2002).
Os vírus podem interagir com o citoesqueleto em múltiplos estágios do seu ciclo
replicativo. Após chegarem aos sítios celulares onde se acumula a progênie viral neoformada,
interações com outros componentes do citoesqueleto podem ocorrer (NEDELLEC et al.,

Discussão
55
1998). Os vírus além de utilizar o citoesqueleto para a sua montagem, podem se utilizar destas
estruturas no momento de sair das células como partículas infecciosas livres (SMITH;
ENQUIST, 2002).
O citoesqueleto da célula hospedeira é frequentemente utilizado pelo vírus em
diferentes estágios do seu ciclo replicativo principalmente, como facilitador do processo
infeccioso. Sabemos que o citoesqueleto é composto por três componentes básicos: filamentos
de actina, microtúbulos e filamentos intermediários, cada um deles com um papel importante
na fisiologia celular (ALBERTS, 1994).
Na tentativa de reduzir a replicação do vírus dengue-2, foi usada a droga citocalasina
B que despolimeriza o citoesqueleto da célula-alvo.
A presença da droga citocalasina B provocou um efeito inibitório no ciclo replicativo
do vírus dengue-2 quando comparado com o controle. Apesar de não haver a abolição total da
replicação viral, houve interferência na liberação de vírus para o meio extracelular.
Experimentos com citocalasina B mostraram que os vírus necessitam da integridade
dos filamentos de actina para a sua replicação, pois a presença da droga provocou um efeito
inibitório no ciclo replicativo do vírus West Nile em células Vero (CHU et al., 2003), assim
como para o vírus Kunjin em células Vero (NG et al., 1983). Somados a estes resultados
obtidos por outros pesquisadores, nossos resultados sugerem que o citoesqueleto de actina
deve estar envolvido no ciclo replicativo do vírus dengue-2, mas o papel destas estruturas na
replicação viral não foi investigado, pois não era o objetivo deste trabalho.
Os vírus de RNA de fita simples e polaridade positiva têm o seu ciclo replicativo
associado à membranas intracelulares. A associação com membranas cria um ambiente que
facilita a replicação viral (BIENZ et al., 1992). Os flavivírus replicam-se na região
perinuclear, em íntima relação com o retículo endoplasmático rugoso.

Discussão
56
Na tentativa de reduzir a replicação do vírus dengue-2, a brefeldina A foi usada como
uma ferramenta para produzir perturbações da rota normal feita pelas glicoproteínas do
retículo endoplasmático para o aparelho de Golgi em infecções de vírus dengue-2 em células
Vero.
A brefeldina A tem se mostrado um potente inibidor do transporte de proteínas do
retículo endoplasmático causando a dissociação do aparelho de Golgi (FUJIWARA et al.,
1988; DOMS; RUSS; YEWDELL, 1989).
A presença da droga brefeldina A provocou um efeito inibitório no ciclo replicativo do
vírus dengue-2 quando comparado com o controle, resultando em uma quase completa
ausência do titulo viral no sobrenadante da cultura de células Vero quando tratadas com a
droga.
Experimentos com brefeldina A mostraram que os vírus necessitam do fluxo vesicular
de proteínas para a sua replicação, pois a presença da droga provocou um efeito inibitório no
ciclo replicativo do vírus Kunjin (MACKENZIE; WESTAWAY, 2001), assim como para o
vírus West Nile em células Vero (SREENIVASAN et al., 1993).
Esses resultados sugerem que o fluxo vesicular de proteínas do retículo
endoplasmático para o complexo de Golgi pode estar envolvido no ciclo replicativo do vírus
dengue-2, mas isto não foi investigado, pois não era o objetivo deste trabalho.
Nos experimentos in vivo, os camundongos tratados com a droga cloroquina 2 horas
após infecção e em intervalos de 24horas durante 7 dias, tiveram a taxa de sobrevivência
considerada satisfatória, pois as manifestações geradas pela encefalite causada pela infecção
viral, com a perda do equilíbrio, tremores, perda de peso, seguida da paralisação das patas
traseiras, somente começaram a ser observadas a partir do 9° dia, enquanto que os animais
infectados não tratados com cloroquina começaram a manifestar estes sinais no 6° dia,
demonstrando que essa droga foi capaz de retardar o início dos sinais de encefalite nos

Discussão
57
camundongos. A porcentagem de sobrevivência observada no 10° dia (mesmo dia em que
todos os camundongos do controle positivo morreram) foi de 60% caindo para 20% no 14°
que se manteve até 21° dia, provavelmente a morte desses camundongos foi devido a eles
ficarem incapacitados de se alimentarem, com a perda do movimento das patas traseiras,
porque notamos que quando foi facilitada a obtenção de água e alimento o camundongo que
sobreviveu apresentou sinais de melhora recuperando o movimento de uma das patas
traseiras. A proteção da droga cloroquina também provavelmente só não foi mais eficiente
devido a administração do vírus pela via intracerebral levando a uma encefalite, pois
comparado a viremia em uma pessoa infectada com vírus dengue, a gravidade da infecção é
bem mais acentuada com o quadro de encefalite, em que está envolvido o sistema nervoso
central. Quando a cloroquina foi administrada 3 dias após infecção viral, não obtivemos um
resultado satisfatório, como foi observado com a administração da droga 2 horas após
infecção, pois todos os camundongos morreram até o 10º dia, provavelmente o intervalo de
tempo prolongado da administração da droga após a infecção viral, fez com que a cloroquina
não tivesse efeito sob a replicação viral.
Os resultados observados para os grupos que receberam a brefeldina A e citocalasina
B não foram satisfatórios, pois as porcentagens de sobrevivência dos camundongos infectados
tratados ou não com as drogas foi similar. Provavelmente a morte dos camundongos pode ter
sido em parte porque os mesmos não receberam reforços dessas drogas.
De acordo com os resultados obtidos, essas drogas poderão ter um papel importante no
tratamento da dengue em um futuro próximo. No entanto, a cloroquina por ser uma droga
relativamente segura, eficaz e de fácil aquisição, usada no tratamento de muitas doenças
incluindo a malária, o seu uso terapêutico mostra-se promissor, pois foi demonstrado que essa
droga possui um efeito antiviral inibitório significante na replicação do vírus dengue.

Conclusões
58
CCOONNCCLLUUSSÕÕEESS

Conclusões
59
6. CONCLUSÕES
1. A acidificação dos endossomos é importante na infecção do vírus dengue em células
Vero, em que houve uma redução significante na replicação desses vírus quando
tratadas com cloroquina.
2. A cloroquina não foi eficaz sob a replicação do vírus dengue-2 em células C6/36.
3. Houve uma quase completa ausência do vírus dengue-2 em cultura de células Vero na
presença da droga brefeldina A.
4. A citocalasina B foi eficaz sob a replicação do vírus dengue-2, em que foi observado
uma redução significante desses vírus em cultura de células Vero.
5. A porcentagem de sobrevivência no grupo de camundongos infectados com vírus
dengue-2 e tratados com cloroquina 2 horas após infecção e em intervalos de 24 horas
durante 7 dias foi satisfatória.
6. Os animais infectados com vírus dengue-2 e tratados com cloroquina 3 dias após a
infecção e em intervalos de 24 horas durante 7 dias não tiveram uma porcentagem de
sobrevivência satisfatória.
7. A porcentagem de sobrevivência no grupo de camundongos infectados com vírus
dengue-2 e tratados com brefeldina A ou citocalasina B não foi satisfatória.

Referências bibliográficas
60
RREEFFEERRÊÊNNCCIIAASS BBIIBBLLIIOOGGRRÁÁFFIICCAASS

Referências Bibliográficas
61
7. REFERÊNCIAS BIBLIOGRÁFICAS
ADELMAN, Z.N.; VARGAS, I.S.; TRAVANTY, E.A.; CARLSON, J.O.; BEATY, B.J.;
BLAIR, C.D.; OLSON, K.E. RNA silencing of Dengue virus type 2 replication in
transformed C6/36 mosquito cells transcribing an inverted-repeat RNA derived from
the virus genome. Journal of Virology, v. 76 (24), p. 12925-12933, 2002.
ALBERTS, B.; BRAY, D.; LEWIS, J.; RAFF, M.; ROBERTS, K.; WATSON, J. D.
Molecular Biology of The Cell. Third edition. Garland Publishing, Inc. New York &
London, Vesicular traffic in the secretory and endocytic pathways, p. 599-609, 1994.
ANDREW, J.B.; MAGNUS, J.; ANNA, V.J.; YIBIN, X.; DAVID, A.J; SUBHASH, G.V.
The Interdomain Region of Dengue NS5 Protein That Binds to the Viral Helicase NS3
Contains Independently Functional Importin β1 and Importin αβ-Recognized Nuclear
Localization Signals. The Journal of Biological Chemistry, v. 277 (39), p. 36399-
36407, 2002.
AUSTIN, W.L.; WIND, M.; BROWN, K.S. Differences in the toxicity and teratogenicity of
cytochalasins D and E in various mouse strains. Teratology, v. 25(1), p. 11-8, 1982.
BARRAGÁN, J.J.M.; ANGEL, R.M. Identification of a putative coreceptor on Vero cells that
participates in dengue 4 virus infection. Journal of Virology, v. 75(17), p. 7818-7827,
2001.
BARTELMA, R.; PADMANABHAN, R. Expression, purification, and characterization of the
RNA 5’- triphosphatase activity of dengue virus type 2 nonstructural protein 3.
Virology, v. 299 (1), p. 122-132, 2002.

Referências Bibliográficas
62
BIENZ, K.; EGGER, D.; PFISTER, T.; TROXLER, M. Structural and functional
characterization of the poliovirus replication complex. Journal Virology, v. 66, p.
2740-2747, 1992.
CAREY, D.E. Chikungunya and dengue: a case of mistaken identy? J. Hist. Med. Allied Sci.,
v. 26(3), p. 243-262, 1971.
CHAMBERS, T.C.; HAHN, C.S.; GALLER, R.; RICE, C.M. Flavivirus genome
organization, expression, and replication. Annu.Rev. Microbiol, v. 44, p. 649-688,
1990.
CHU, J.J.H.; CHOO, B.G.H.; LEE, J.W.M.; NG, M.L. Actin Filaments Participate in West
Nile (Sarafend) Virus Maturation Process. Journal of Medical Virology, v. 71, p. 463-
472, 2003.
COOMBS, K.; MANN, E.; EDWARDS, J.; BROWN, D.T. Effects of chloroquine and
Cytochalasin B on the infection of cells by sindbs virus and vesicular stomatitis virus.
Journal of Virology, v. 37, p. 1060-1065, 1981.
DE PAULA, O.S.; NUNES, C.; MATOS, R.; OLIVEIRA, M.Z.; LIMA, M.D.; FONSECA,
B.A.L. Comparison of techiniques for extracting viral RNA from isolation-negative
serum for dengue diagnosis by the polymerase chain reaction. Journal of Virological
Methods, v. 98, p. 119-125, 2001.
DILLE, B.J.; JOHNSON, T.C. Inhibition of vesicular stomatitis virus glycoprotein expression
by chloroquine. J. Gen. Virol., v. 62, p. 91-103, 1982.
DOMS, R.W.; RUSS, G.; YEWDELL, J.W. Brefeldin A redistributes resident and itinerant
Golgi proteins to the endoplasmic reticulum. J. Cell Biol. v. 109, p. 61-72, 1989.
ELSHUBER, S.; ALLISON, S.L.; HEINZ, F.X.; MANDL, C.W. Cleavage of protein prM is
necessary for infection of BHK-21 cells by tick-borne encephalitis virus. J. Gen.
Virology, v. 84, p. 183-191, 2003.

Referências Bibliográficas
63
FONSECA, B.A.L.; FIGUEIREDO, L.T.M. Dengue. Em “ Tratado de infectologia ” 3ª
edição. Editora Atheneu, Ribeirão Preto. Editor: Roberto Focaccia. p. 343-356, 2005.
FUJIWARA, T.; ODA, K.; YOKOTA, S.; TAKATSUKI, A.; IKEHARA, Y. Brefeldin A
causes disassembly of the Golgi complex and accumulation of secretory proteins in
the endoplasmic reticulum. J. Biol. Chem., v. 263, p. 18545-18552, 1988.
GIBBONS, R.V.; VAUGHN, D.W. Dengue: an escalating problem. Britsh Medical Journal,
v. 324, p. 1563-1566, 2002.
GUBLER, D.J.; TRENT, D.W. Emergence of epidemic dengue/dengue hemorrhagic fever as
a public health problem in the Americas. Infec. Agents Dis., v. 2, p. 283-393, 1994.
GUBLER, D.J. Dengue and Dengue Hemorrhagic Fever. Clinical Microbiology Reviews, v.
3, p. 480-496, 1998.
GUBLER, D.J. The Global Emergence/Ressurgence of Arboviral Diseases As Public Health
Problems. Archives of Medical Research, v. 33, p. 330-342, 2002.
GUIRAKHOO, F.; HEINZ, F.X.; MANDL, C.W.; HOLZMANN, H.; KUNZ, C. Fusion
activity of flaviviruses: comparison of mature and immature (prM-containing) tick-
borne encephalitis virions. J. Gen. Virology, v. 72, p. 1323-1329, 1991.
HALSTEAD, S.B. Observations related to pathogenesis of dengue hemorrhagic fever. VI.
Hypotheses and Discussion. Yale J. Biol. Med., v.42 (5), p. 350-362, 1970.
HALSTEAD, S.B. Pathogenesis of dengue: challenges to molecular biology. Science, v. 239,
p. 476-481, 1988.
HALSTEAD, S.B.; HEINZ, F.X.; BARRETT, A.D.T.; ROEHRIG, J.T. Dengue virus:
molecular basis of cell entry and pathogenesis, 25-27 June 2003, Vienna, Austria.
Vaccine, v. 23, p. 849-856, 2005.
HENCHAL, E.; PUTNAK, J.R. The dengue viruses. Clin. Microbiol. Rev., v. 3, p. 376-396,
1990.

Referências Bibliográficas
64
HENCHAL, E.A.; POLO, S.L.; VORNDAM, V.; YAEMSIRI, C.; INNIS, B.L.; HOKE, C.H.
Sensitivity and specificity of a universal primer set for the rapid diagnosis of dengue
virus infections by polymerase chain reaction and nucleic acid hybridization. Am. J.
Trop. Med. Hyg., v. 45(4), p. 418-428, 1991.
HERNANDEZ, R.H; LUO, T.; BROWN, D.T. Exposure to low pH is not required for
penetration of mosquito cells by Sindbis virus. Journal of Virology, v. 75, p. 2010-
2013, 2000.
HONG, K.; JIANG, Z.; LIANGXI, W.; GUOFU, D; PING, L.; YONGLING, L.;
WENDONG, P.; MINGHAI, W. Chloroquine protects mice from challenge with CpG
ODN and LPS by decreasing proinflammatory cytokine release. International
Immunophamacology, v. 4, p. 223-234, 2004.
HOUNG, H.S.H.; CHEN, R.C.M.; VAUGHN, D.W.; KANESA-THASAN, N. Development
of a fluorogenic RT-PCR system for quantitative identification of dengue virus
serotypes 1-4 using conserved and serotype-specific 3’ noncoding sequences. J. Virol.
Meth., v. 95, p. 19-32, 2001.
KASAP, M.; THOMAS, S.; DANAHER, E.; HOLTON, V.; JIANG, S.; STORRIE, B.
Dynamic nucleation of Golgi apparatus assembly from the endoplasmic reticulum in
interphase HeLa cells. Traffic., v. 5, p. 595-605, 2004.
KEYAERTS, E.; VIJGEN, L.; MAES, P.; NEYTS, J.; RANST, M.V. In vitro inhibition of
severe acute respiratory syndrome coronavirus by chloroquine. Bioch. and Bioph.
Research Communications, v. 323, p. 264-268, 2004.
KIELIAN, M.; JUNGERWIRTH, S. Mechanisms of enveloped virus entry into cells. Mol.
Biol. Med., v. 7, p. 17-31, 1990.
KOCH, G.; KOCH, F. The use of cytochalasins in studies on the molecular biology of virus-
host cell interactions. Front Biol., v. 46, p. 475-498, 1978.

Referências Bibliográficas
65
KUHN, R.J.; ZHANG, W.; ROSSMANN, M.G.; PLETNEV, S.V.; CORVER, J.; LENCHES,
E.; JONES, C.T.; MUKHOPADHYAY, S.; CHIPMAN, P.R.; STRAUSS, E.G.;
BAKER, T.S.; STRAUSS, J.H. Structure of dengue virus: implications for flavivirus
organization, maturation, and fusion. Cell, v. 108, p. 717-725, 2002.
KUNO, G.; CHANG, G.J.; TSUCHIYA, K.R.; KARABATSOS, N.; CROPP, C.B. Phylogeny
of the genus Flavivirus. J. Virology, v. 72, p. 73-83, 1998.
LINDENBACH, B.D.; RICE, C.M. In: Fields Virology (eds Knipe D.M. & Howley P.M.) 4th
edition. Flaviviridae replication, p. 991-1041, 2001. (Lippincott Williams & Wilkins,
Philadelphia, 2001).
LINDENBACH, B.D.; RICE, C.M. Molecular biology of flaviviruses. Adv. Virus Res., v. 59,
p. 23-61, 2003.
LODISH, H.; BERK, A.; ZIPURSKY, S.L.; MATSUDAIRA, P.; BALTIMORE, D.;
DARNELL, J. Cell motility and shape I: microfilaments. Molecular cell biology, 4th
edition. WH Freeman and Company. New York. USA, p. 751-768, 2000.
MACKAY, I.M.; ARDEN, K.E.; NITSCHE, A. Real time PCR in virology. Nucleic Acids
Research, v. 30(6), p. 1292-1305, 2002.
MACKENZIE, J.M.; WESTAWAY, E.G. Assembly and Maturation of the Flavivirus Kunjin
Virus Appear To Occur in the Rough Endoplasmic Reticulum and along the Secretory
Pathway, Respectively. Journal of Virology, v. 75, p. 10787- 10799, 2001.
MODDIS, Y.; OGATA, S.; CLEMENTS, D.; HARRISON, S.C. Structure of the dengue virus
envelope protein after membrane fusion. Nature, v. 427, p. 313-319, 2004.
MONATH, T.P.; BURKE, D.S. Flaviviruses. In: Fields Virology. Philadelphia. Fourth
Edition. Lippincott Willians & Wilkins. Cap. 33, p. 1043-1126, 2001.
MUKHOPADHYAY, S.; KUHN, R.J.; ROSSMANN, M.G. A structural perspective of the
Flavivirus life cycle. Nature, v. 3, p. 13-22, 2005.

Referências Bibliográficas
66
NEDELLEC, P.; VICART, P.; LAURENT-WINTER, C.; MARTINAT, C.; PREVOST,
M.C.; BRAHIC, M. Interaction of Theiler’s virus with intermediate filaments of
infected cells. J. Virol., v. 72, p. 9553-9560, 1998.
NG, M.L.; PEDERSEN, J.S.; TOH, B.H.; WESTAWAY, E.G. Immunofluorescent sites in
Vero cells infected with the flavivirus Kunjin. Arch. Virology, v. 78, p. 177-190,
1983.
NIESTERS, H.G.M. Quantitation of viral load using Real-Time amplification techniques.
Methods, v. 35, p. 419-429, 2001.
PHILLIPS, L.R.; SUPKO, J.G.; MALSPEIS, L. Analysis of brefeldin A in plasma by gas
chromatography with electron capture detection. Analytical Biochemistry, v. 211, p.
16-22, 1993.
RANDOLPH, V.B.; WINKLER, G.; STOLLAR, V. Acidotropic amines inhibit proteolytic
processing of flavivirus prM protein. Virology, v. 174, p. 450-458, 1990.
REY F.A. Dengue virus envelope glycoprotein structure: New insight into its interactions
during viral entry. PNAS, v. 100(12), p. 6899-6901, 2003.
RICE, C.M.; LENCHES, E.M.; EDDY, S.R.; SHIN, S.J.; SHEETS, R.L.; STRAUSS, J.H.
Nucleotide sequence of yellow fever virus: implications for flavivirus gene expression
and evolution. Science, v. 229, p. 726-733, 1985.
RIGAU-PÉREZ, J.G.; CLARK, G.G.; GUBLER, J.D.; REITER, P.; SANDERS, J.E.;
VORNDAM, V. Dengue and dengue haemorrhagic fever. The lancet, v. 353, p. 971-
977, 1998.
RICHMAN, D.D.; WHITLEY, R.J.; HAYDEN, F.G. Flaviviruses. Clinical Virology, 2th
edition. Washington, D.C., p. 1103-1104, 2002.

Referências Bibliográficas
67
ROS, C.; BURCKHARDT, C.J.; KEMPF, C. Cytoplasmic trafficking of minute virus of
mice: Low-pH requeriment, routing to late endosomes, and proteasome interaction.
Journal of Virology, v. 76, p. 12634-12645, 2002.
ROTHMAN, L.A.; ENNIS, A.F. Immunophathogenesis of Dengue Haemorrhagic Fever.
Virology, v. 257, p. 1-6, 1999.
RUIZ, A.C.G. Padronização e utilização da técnica de PCR em tempo real no estudo dos vírus
dengue e da infecção causada pelo dengue-3 em Ribeirão Preto, SP. Tese de
dissertação, 2004.
SAVARINO, A.; BOELAERT, J.R.; CASSONE, A.; MAJORI, G.; CAUDA, R. Effects of
Chloroquine on viral infections: an old drug against today’s diseases? The Lancet, v.
3, p. 722-727, 2003.
SCHMITTGEN, T.D. Real-Time Quantitative PCR. Methods, v. 25, p. 383-385, 2001.
SMITH, G.A.; ENQUIST, L.W. Break ins and break outs: viral interactions with the
cytoskeleton of mammalian cells. Annu Rev. Cell Dev. Biol., v. 18, p. 135-161, 2002.
SPERBER, K.; KALB, T.H.; STECHER, V.J.; BANERJEE, R.; MAYER, L. Inhibition of
human immudeficiency virus type 1 replication by hydroxychloroquine in T cells and
monocytes. AIDS Res. Hum. Retroviruses, v. 9, p. 91-98, 1993.
SREENIVASAN, V.; NG, K.L.; NG, M.L. Brefeldin A affects West Nile virus replication in
vero cells but not C6/36 cells. J. Virol. Methods, v. 45, p. 1-17, 1993.
TAKEDA, A.; ENNIS, F.A. FcR-mediated enhancement of HIV-1 infection by antibody.
AIDS Res. Hum. Retroviruses, v. 6, p. 999-1004, 1990.
TESH, R.B. A method for the isolation and identification of dengue viruses, using mosquito
cell cultures. Am.J.Trop.Med. Hyg., v. 28, p. 1053-1059, 1979.

Referências Bibliográficas
68
THEIN, S.; AUNG, M.M.; SHWE, T.N.; AYE, M.; ZAW, A.; AYE, K.M.; AASKOY, J.
Risk factors in dengue shock syndrome. Am. J. Trop. Med. Hyg., v. 56(5), p. 56-572,
1997.
THORENS, B.; VASSALLI, P. Chloroquine and ammonium chloride prevent terminal
glycosylation of immunoglobulins in plasma cells without affecting secretion. Nature,
v. 321, p. 618-620, 1986.
TORII, S.; BANNO, T.; WATANABE, T.; IKEHARA, Y.; MURAKAMI, K.;
NAKAYAMA, K. Cytotoxicity of brefeldin A correlates with its inhibitory effect on
membrane binding of COP coat proteins. The Journal of Biological Chemistry, v. 270,
p. 11574-11580, 1995.
TSAI, W.P.; NARA, P.L.; KUNG, H.F.; OROSZLAN, S. Inhibition of human
immunodeficiency virus infectivity by chloroquine. AIDS Res. Hum. Retroviruses, v.
6, p. 481-489, 1990.
UCHIL, P.D.; SATCHIDANANDAM, V. Architecture of the Flaviviral Replication
Complex. The Journal of Biological Chemistry, v. 278, p. 24388-24398, 2003.
VINCENT, M.J.; BERGERON, E.; BENJANNET, S.; ERICKSON, B.R.; ROLLIN, P.E.;
KSLAZEK, T.G.; SEIDAH, N.G.; NICHOL, S.T. Chloroquine is a potent inhibitor of
SARS coronavirus infection and spread. Virology journal, v. 2, p. 69, 2005.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo