envolvimento da gsk3 no desenvolvimento embrionÁrio ... · embrionário, ela também se mostrou...
TRANSCRIPT

ENVOLVIMENTO DA GSK3 NO DESENVOLVIMENTO
EMBRIONÁRIO DOCARRAPATO R.(BOOPHILUS) MICROPLUS:
EFEITO DO SILENCIAMENTO POR RNAI EM FÊMEAS
PARCIALMENTE INGURGITADAS E SUA REGULAÇÃO POR CÁLCIO
NAS CÉLULAS EMBRIONÁRIAS BME26
Arianne Fabres de Jesus
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE DARCY
RIBEIRO
CAMPOS DOS GOYTACAZES - RJ
Fevereiro – 2010

II
ENVOLVIMENTO DA GSK3 NO DESENVOLVIMENTO
EMBRIONÁRIO DOCARRAPATO R.(BOOPHILUS) MICROPLUS:
EFEITO DO SILENCIAMENTO POR RNAI EM FÊMEAS
PARCIALMENTE INGURGITADAS E SUA REGULAÇÃO POR CÁLCIO
NAS CÉLULAS EMBRIONÁRIAS BME26
Arianne Fabres de Jesus
“Dissertação apresentada ao Centro de Biociências e
Biotecnologia da Universidade Estadual do Norte
Fluminense Darcy Ribeiro, como parte das
exigências para obtenção do título de Mestre em
Biociências e Biotecnologia”.
Orientador: Prof. Dr. Carlos Logullo
Co-orientadora: Prof. Dr. Anna Okorokova Façanha
Campos dos Goytacazes – RJ
Fevereiro - 2010

III
ENVOLVIMENTO DA GSK3 NO DESENVOLVIMENTO
EMBRIONÁRIO DOCARRAPATO R.(BOOPHILUS) MICROPLUS:
EFEITO DO SILENCIAMENTO POR RNAI EM FÊMEAS
PARCIALMENTE INGURGITADAS E SUA REGULAÇÃO POR CÁLCIO
NAS CÉLULAS EMBRIONÁRIAS BME26
Arianne Fabres de Jesus
Comissão examinadora
_______________________________________ Sirlei Daffre (USP) Membro Externo da Banca
__________________________________________ Profª Antonia Elenir Amâncio Oliveira (LQFPP – CBB – UENF) Membro da Banca
__________________________________________ Prof. Fábio Lopes Olivares (LBCT – CBB – UENF) Membro da Banca
__________________________________________ Prof. Dr. Carlos Logullo (LQFPP - CBB – UENF) Orientador __________________________________________ Prof. Dr. Anna Okorokova (LFBM - CBB – UENF) Co-Orientadora

IV
"Devemos, no entanto, reconhecer,
como me parece, que o homem com
todas as suas nobres qualidades... ainda
sofre em sua prisão corpórea a indelével
marca de sua humilde origem."
Charles Darwin

V
DEDICATÓRIA
Aos meus pais, Neuza e Dulcino, pelo
exemplo, amor e dedicação
incondicional.

VI
Agradecimentos:
Aos meus pais, por sempre estarem do meu lado incondicionalmente;
Ao meu afilhado, João Pedro, que traz alegrias a todo instante;
A toda minha família, pelo apoio;
À amiga Josiana, que sempre fará parte da minha vida;
Aos amigos Érica, Wilson e Rafaela por compartilhar as alegrias e tristezas;
À Paula Pohl por toda a ajuda em meu estagio em Porto Alegre e pela atenção em
São Paulo;
Ao Logullo pela oportunidade;
À galera do laboratório que proporciona um ótimo ambiente de trabalho, com muito
bom humor e amizade (Danielle, Leonardo, Luiz, Renato, Wagner, Barbara, Helga,
Eldo, Evenilton, Jorge, Ojana, Gabriel, Igor, Ronaldinho);
À Adriana, por toda ajuda no lab;
Ao prof. Itabajara por todos os auxílios;

VII
Índice
Resumo ..................................................................................................................... XI
Abstract.................................................................................................................... XII
Introdução.................................................................................................................13
Carrapatos............................................................................................................14
Ciclo de Vida ........................................................................................................16
Embriogênese ......................................................................................................18
Células embrionárias BME26..............................................................................19
Glicogênio Sintase quinase 3(GSK3) ............................................................20
RNAi em carrapatos.............................................................................................25
Objetivos ...................................................................................................................28
Objetivo Geral ......................................................................................................29
Objetivos específicos ..........................................................................................29
Materiais e Métodos .................................................................................................30
Carrapato Rhipicephalus (Boophilus) microplus ....................................................31
Alimentação artificial...........................................................................................31
Síntese do dsRNA................................................................................................32
Injeção do RNAi em partenóginas......................................................................33
Análise da transcrição gênica de GSK3 por qPCR .........................................33
Observação dos embriões por microscopia óptica..........................................34
Manutenção da cultura de células embrionárias ..............................................34
Silenciamento gênico por RNAi da GSK3 em cultura de células ..................35
Dosagem de glicogênio.......................................................................................35
Western blotting...................................................................................................36
Resultados ................................................................................................................37
Silenciamento da GSK3 em fêmeas do carrapato R. microplus ....................38
Silenciamento da GSK3 em células embrionárias BME26 .............................43
Avaliação da regulação da GSK3 por cálcio ...................................................45
Discussão .................................................................................................................50
Conclusões ...............................................................................................................56
Referências ...............................................................................................................58

VIII
Índice de figuras
Fig. 1: O carrapato Rhipicephalus (Boophilus) microplus ........................................16
Fig. 2: O ciclo biológico do Rhipicephalus (Boophilus) microplus .............................19
Fig. 3: Etapas do desenvolvimento do R. microplus .................................................20
Fig. 4: Regulação da GSK3 através da fosforilação de seu resíduo Ser9 ..............23
Fig. 5 - Esquema da GSK3 mostrando os resíduos Ser9 e Tyr216 que podem ser
fosforilados.................................................................................................................24
Fig. 6: Esquema mostrando a provável via de RNAi em carrapatos .........................27
Fig. 7: Alimentação artificial por capilar .....................................................................40
Fig. 8: Ganho de peso das fêmeas depois da alimentação artificial .........................41
Fig. 9: Confirmação do silenciamento da GSK3 em partenóginas ............................42
Fig.10: Efeito do silenciamento gênico por RNAi da
GSK3...........................................................................................................…….....44
Fig. 11: Confirmação do silenciamento da GSK3 na células embrionárias BME26
....................................................................................................................................45
Fig. 12: Avaliação da concentração de glicogênio em células silenciadas ...............46
Fig. 13: Avaliação da fosforilação do resíduo Tyr 216 da GSK3 .................................47
Fig. 14: Avaliação da expressão da GSK3 em células incubadas com CaCl2
....................................................................................................................................48
Fig. 15: Avaliação da expressão da GSK3 em células incubadas com thapsigargina
....................................................................................................................................49
Fig. 16 : Avaliação da transcrição da GSK3b em células incubadas com CaCl2 ou
thapsigargina .............................................................................................................50

IX
Índice de tabelas
Tab. 1: Substratos da GSK3 agrupadas por suas funções celulares ........................22
Tab. 2: Efeito do silenciamento da GSK3 por RNAi na oviposição e eclosão ..........43

X
Abreviaturas
Akt – Proteína quinase B
dsRNA – RNA dupla fita
DAB - Diaminobenzamidina
EDTA- ácido etilenodiaminotetracético
EGTA – ácido etileno glicol tetracético
GS – Glicogênio sintase
GSK3 – Glicogênio sintase quinase 3
PKB/Akt - proteína quinase B
PAGE - gel de poliacrilamida para eletroforese
PI3K - fosfatidil-inositol-3-OH quinase
PMSF - fluoreto de fenil-metil-sulfonila
PVDF - polyvinylidene difluoride
TBS - tampão Tris-HCl 10 mM, NaCl , pH 7,0
TRIS- trishidroximetil aminometano
Wnt – Via de sinalização Wingless

XI
Resumo
O carrapato bovino Rhipicephalus (Boophilus) microplus causa grandes prejuízos a
pecuária de países tropicais e subtropicais de todo o mundo. Neste carrapato, o
ovário da fêmea é o principal sítio de transcrição da enzima Glicogênio sintase
quinase 3 (GSK3). Esta enzima foi originalmente caracterizada como controladora
da síntese de glicogênio em resposta à via de sinalização da insulina, estando
associada a diversos substratos. Esta quinase faz parte de numerosas vias de
sinalização, incluindo metabolismo de glicogênio, transcrição, transporte intracelular
de vesículas, regulação do citoesqueleto, ciclo celular e apoptose. Visando estudar a
função da GSK3 o silenciamento gênico por RNAi em partenóginas (fêmeas
parcialmente alimentadas) foi realizado. O RNAi contra GSK3 reduziu
significativamente a oviposição e eclosão dos ovos dessas fêmeas. Nossos
resultados indicam que a GSK3 é uma proteína essencial envolvida no
desenvolvimento embrionário. Além de estar envolvida no desenvolvimento
embrionário, ela também se mostrou capaz de regular a síntese do glicogênio em
células embrionárias da linhagem BME26, assim como descrito largamente na
literatura para outros organismos. Ainda, fornecemos as evidências que os níveis
celulares de cálcio regulam a atividade da GSK3 já que os níveis de transcrição,
expressão e fosforilação dessa proteína aumentaram em células incubadas com
CaCl2.

XII
Abstract
The cattle ticks, Rhipicephalus (Boophilus) microplus affect cattle production in
tropical and subtropical regions of the world. This tick, the ovary of the female is the
major site of Glicogênio sintase quinase 3 (GSK3) transcription. This This enzyme
was originally characterized as controlling the synthesis of glycogen in response to
the signaling pathway of insulin, standing. associated with various substrates. This
kinase is part of numerous signaling pathways, including glycogen metabolism,
transcription, intracellular transport of vesicles, regulation of the cytoskeleton, cell
cycle and apoptosis. In order to study the function of the GSK3 gene silencing by
RNAi in partenóginas (partially fed females) was performed. RNAi against GSK3
significantly reduced oviposition and hatching of eggs of these females. Our results
indicate that GSK3 is an essential protein involved in embryonic development, it was
also able to regulate the synthesis of glycogen in embryonic stem cell line BME26, as
widely described in the literature for other organisms. More, we provide evidence that
the cellular levels of calcium regulate GSK3 activity because transcription,
expression and phosphorilation levels this protein increased in CaCl2 cells incubated.

13
Introdução

14
Carrapatos
Carrapatos são artrópodes ectoparasitas hematófagos obrigatórios que
infestam um grande número de espécies de vertebrados terrestres tais como
mamíferos, pássaros, répteis e anfíbios. Eles estão distribuídos em três
famílias: Argasídae, Ixodidae e Nuttalliellidae (Keirans 1992; Keirans e
Robbins, 1999). Argasídeos são carrapatos que procuram o hospedeiro para
sugar sangue, abandonando-o em seguida e alimentando-se mais de uma vez.
As fêmeas efetuam várias posturas, alternando ovoposição com alimentação
sangüínea. Cada postura não ultrapassa 150 ovos totalizando um número
realmente pequeno quando comparado aos ixodídeos. Já a família
Nuttalliellidae é representada por apenas uma espécies Nuttalliella namaqua
(Keirans et al., 1976). Pouco se sabe sobre a sua biologia e esta família
aparentemente não tem importância médica ou veterinária. O carrapato
Riphicephalus (Boophilus) microplus pertence à família dos Ixodídeos. Nessa
família a alimentação dura um tempo mais prolongado e é feita uma única vez
em todo o ciclo de vida e a ovoposição resulta num grande número de ovos,
sendo esta a diferença mais marcante entre estas famílias (Flechtmann, 1977).
O carrapato R. microplus (Fig. 1) é originado da Ásia e infesta
preferencialmente bovinos, sendo responsável por grandes perdas econômicas
para a pecuária de regiões tropicais e subtropicais. Sua incidência é maior nos
grandes rebanhos da América, África, Ásia e Austrália, sendo considerado o
carrapato de maior importância econômica nos rebanhos da América do Sul
(Gonzáles, 1995; Nari, 1995). Segundo estimativas realizadas no ano de 2006,

15
os prejuízos econômicos causados por esta espécie no Brasil atingiram a
ordem de dois bilhões de reais (Embrapa, 2005).
Fig. 1: O carrapato Rhipicephalus (Boophilus) microplus parasitando seu hospedeiro, o boi
(Bos taurus).
Na tentativa de minimizar perdas econômicas, têm sido propostas
alternativas que se baseiam na obtenção de antígenos obtidos do próprio
carrapato. Com base neste princípio já foram desenvolvidas duas vacinas já
comercializadas: a TickGARDtm, australiana (Willadsen et al., 1995) e a
GAVACtm, cubana (Rodriguez et al., 1995a, b). Embora estas vacinas
apresentem proteção significativa para os rebanhos de onde elas foram
criadas, essa proteção diminui expressivamente em outras áreas do mundo
dependendo da cepa de carrapato existente (Willadsen et al., 1995; Rodriguez
et al., 1995b). Nosso grupo de trabalho vem realizando testes de vacinação
com outro antígeno, não relacionado à proteína utilizada na GAVAC e
TickGARD. Trata-se de uma proteína de 54 kDa isolada de ovos e
caracterizada como um precursor de proteinase aspártica, denominada BYC
(Pro-catepsina de vitelo de Boophilus) (Logullo et al., 1998, Abreu et al., 2004).
Os testes de vacinação com a BYC demonstraram que ela é capaz de

16
promover uma significativa interferência no processo reprodutivo do carrapato,
com redução no número de fêmeas ingurgitadas, diminiuição da oviposição e
na fertilidade levando a uma proteção de aproximadamente 30% (Vaz Jr. et al.,
1998), comparável as vacinas já comercializadas. Todavia o conhecimento dos
aspectos fisiológicos e bioquímicos que regem a formação dos embriões deste
parasita é fundamental para eleger com acurácia os antígenos a serem
utilizados como vacinas nestes hospedeiros.
Ciclo de Vida
O carrapato R. microplus apresenta duas etapas distintas no seu ciclo de
vida uma fase parasitária durante um período médio de 22 dias sobre um único
hospedeiro e uma fase não parasitária que ocorre no solo e que pode durar de
dois a três meses, dependendo principalmente das condições climáticas
existentes (Fig. 2) (Gonzales et al., 1974).
A fase não parasitária compreende os estágios de fêmea adulta (teleógina),
ovo e larva infestante. A fêmea adulta fecundada, ao desprender-se do bovino
procura um local no solo para efetuar a postura. Em condições adequadas de
temperatura (26-28ºC) e umidade (~80%) a postura se inicia no terceiro dia
após a queda. Após a ovoposição, que é de aproximadamente 3000 ovos, a
fêmea morre. A eclosão dos ovos inicia a partir de 21 dias após o inicio da
postura. As larvas necessitam de um período de maturação médio de uma
semana para estarem aptas a fixarem-se no hospedeiro e continuarem o
desenvolvimento (Gonzales et al., 1974).

17
Na fase parasitária, o carrapato apresenta três variações morfológicas
distintas: larva, ninfa e adulto. A larva alimenta-se e inicia o processo de
desenvolvimento e crescimento tegumentário. Passa por um período de inércia
entre o quarto e quinto dia e atinge a fase de metalarva. Em torno do sexto dia,
adquire uma nova estrutura, com um outro tegumento, mais um par de patas,
ficando agora com 4 pares, e uma fileira de dentição do hipostômio entre outras
alterações: é a fase de ninfa. Esta fase dura em média dois a quatro dias,
sendo que ao continuar seu desenvolvimento, uma nova alteração no
exoesqueleto se processa, havendo um igual período de inatividade,
denominado de metaninfa para que ao final do processo surja o indivíduo
adulto, sexualmente diferenciado. Isto acontece em torno do décimo segundo
dia. A partir dessa fase, inicia-se o processo de maturação dos machos e das
fêmeas, sendo que em torno do décimo sétimo dia os machos já estão aptos à
cópula. As fêmeas, após serem fecundadas, passam de metaninfa para
neógina num período médio de 17 dias. Em seguida, em um período de três
dias, passam a partenógena (parcialmente ingurgitada) e em mais dois dias, a
teleógina (ingurgitamento máximo) (Gonzales et al., 1974). Nota-se um
crescimento mais acentuado do tegumento nessa fase, sendo que nas últimas
horas próximas ao ingurgitamento completo, a alimentação intensifica-se, ao
ponto das fêmeas apresentarem um tamanho cerca de 10 vezes superior ao
dos machos. Aos 22 dias, a maioria das fêmeas cai ao solo. Os machos podem
permanecer no bovino por mais de 38 dias fecundando inúmeras fêmeas
(Gonzales et al., 1974).

18
Fig. 2: O ciclo biológico do Rhipicephalus (Boophilus) microplus. Seu desenvolvimento é
dividido em duas fases: Uma fase de vida livre e outra parasitando seu hospedeiro (Bos
taurus). (Gonzáles et al., 1974)
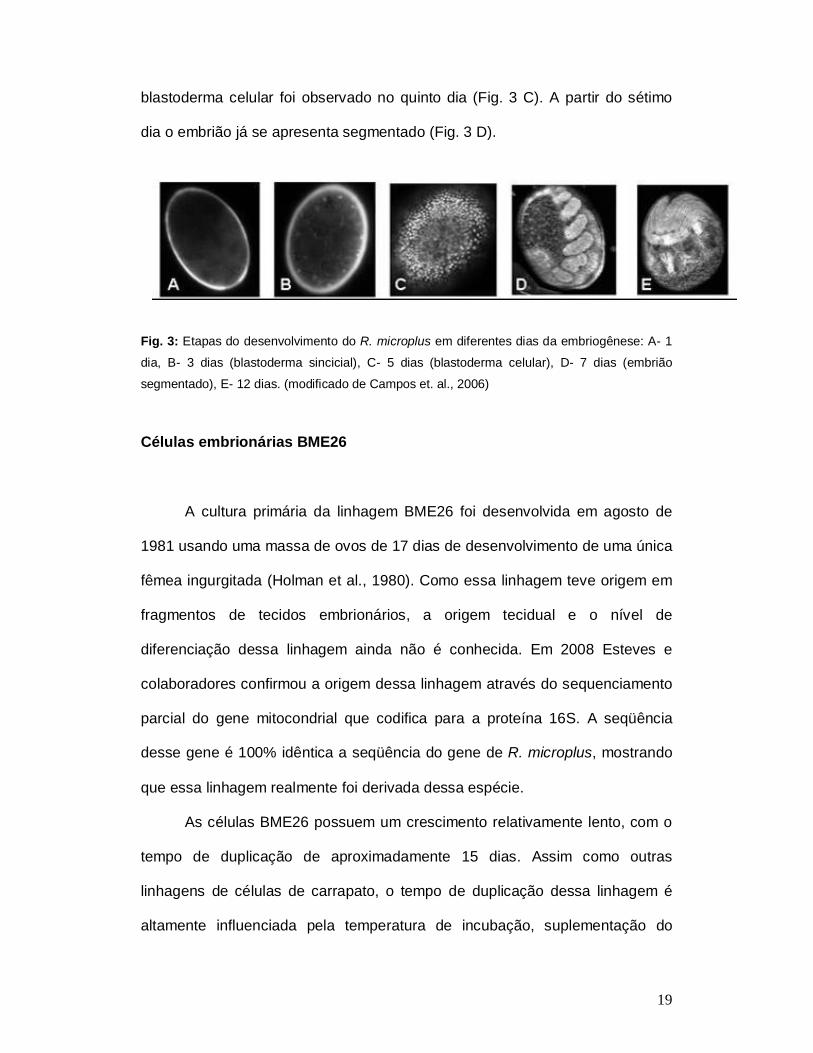
Embriogênese
Campos e colaboradores em 2006 demonstraram algumas etapas da
embriogênese do carrapato R. microplus. O processo ocorre em média de 21
dias, desde a postura dos ovos até a eclosão, em condições controladas de
temperatura e umidade. No desenvolvimento do R. microplus, ocorre a
formação do blastoderma sincicial após a fecundação, na qual muitos núcleos
estão presentes em um citoplasma comum (Fig 3B). A formação do
blastoderma sincicial ocorre no terceiro dia de desenvolvimento. Antes dessa
etapa o ovo, com apenas um dia, apresenta apenas um núcleo (Fig. 3A).
Posteriormente, membranas crescem a partir da superfície envolvendo os
núcleos e formando células, que dão origem ao blastoderma celular. O
19
blastoderma celular foi observado no quinto dia (Fig. 3 C). A partir do sétimo
dia o embrião já se apresenta segmentado (Fig. 3 D).
Fig. 3: Etapas do desenvolvimento do R. microplus em diferentes dias da embriogênese: A- 1
dia, B- 3 dias (blastoderma sincicial), C- 5 dias (blastoderma celular), D- 7 dias (embrião
segmentado), E- 12 dias. (modificado de Campos et. al., 2006)
Células embrionárias BME26
A cultura primária da linhagem BME26 foi desenvolvida em agosto de
1981 usando uma massa de ovos de 17 dias de desenvolvimento de uma única
fêmea ingurgitada (Holman et al., 1980). Como essa linhagem teve origem em
fragmentos de tecidos embrionários, a origem tecidual e o nível de
diferenciação dessa linhagem ainda não é conhecida. Em 2008 Esteves e
colaboradores confirmou a origem dessa linhagem através do sequenciamento
parcial do gene mitocondrial que codifica para a proteína 16S. A seqüência
desse gene é 100% idêntica a seqüência do gene de R. microplus, mostrando
que essa linhagem realmente foi derivada dessa espécie.
As células BME26 possuem um crescimento relativamente lento, com o
tempo de duplicação de aproximadamente 15 dias. Assim como outras
linhagens de células de carrapato, o tempo de duplicação dessa linhagem é
altamente influenciada pela temperatura de incubação, suplementação do

20
meio, histórico de passagens e pode variar de 5 a 15 dias dependendo do meio
utilizado. O tempo de duplicação é altamente contrastante com a rápida
multiplicação celular que ocorre durante a embriogênese (Nuñez et al., 1985).
Essas observações apontam para necessidade de caracterização mais
detalhada de aspectos fisiológicas e nutricionais dessa linhagem.
A linhagem BME26 se adere firmemente ao substrato e é
morfologicamente heterogênea. Culturas iniciais, pouco confluentes, possuem
a aparência de fibroblastos e a medida que aumenta a confluência as células
adotam morfologia arredondada,tornando-se maiores e com uma grande
quantidade de vesículas. O número de vesículas, que contem uma grande
quantidade de lipídeos, varia de célula para célula. Elas também possuem uma
grande quantidade de glicogênio disperso no citoplasma (Esteves et al., 2008).
Glicogênio Sintase quinase 3 (GSK3)
A GSK3 é uma serina/treonina quinase originalmente reconhecida como
um componente da via de sinalização da insulina onde ela é capaz de controlar
a síntese de glicogênio por inibir a glicogênio sintase (Embi et al., 1980). A
enzima está envolvida em diversos processos fundamentais para a célula,
como ciclo celular, transcrição gênica e integridade do citoesqueleto. Isso
ocorre devido ao fato dessa proteína ser capaz de fosforilar enzimas chaves
que coordenam esses processos (Grimes e Jope, 2001). A tabela 1 sumariza
os seus substratos com respectivas funções celulares.

21
Tabela 1: Alguns substratos da GSK3 agrupadas por suas funções celulares (Jope e Johnson, 2004)
Proteínas do metabolismo e sinalização
Proteínas estruturais Fatores de transcrição
AcetilCoA carboxilase Proteína precursora amilóide APC ATP-citrato liase Axina Ciclina D1 Ciclina E elF2B Glicogênio sintase Substrato de receptor de insulina1 Receptor NGF Nucleoporina p62 P21 Presenilina-1 PKA Proteína Fosfatase 1 Piruvato desidrogenase
DF3/MUC1 Dinamina like Kinesina de cadeia leve MAP1B MAP2 NCAM Neurofilamentos tau Telokina
AP-1 b-catenina C/EB CREB GATA4 p53 HIF-1 HSF-1 Mash 1 MITF c-Myb c-Myc NeuroD NFAT NF-kB Notch
A GSK3 se encontra constitutivamente ativa em células não estimuladas
pelas vias de sinalização que ela faz parte sendo inativada por essas vias
quando elas se encontram ativadas (Fig. 4 – Bradley e Woodgett, 2003; Dajani
et al., 2001). Ela é capaz de fosforilar mais de 40 substratos e por causa dessa
ação múltipla deve ser muito bem controlada (Jope e Johnson 2004). Um
desses mecanismos regulatórios consiste na fosforilação do sítio especifico da
GSK3. A atividade da GSK3é aumentada pela fosforilação no resíduo Tyr216,
que pode ser resultado de autofosforilação (Wang et al., 1994) ou da
fosforilação de outras quinases (Lesort et al., 1999; Hartigan e Johnson, 1999).
Inversamente, a fosforilação do resíduo Ser9 resulta na inibição da atividade
dessa quinase.

22
Fig. 4 – Regulação da GSK3 através da fosforilação de seu resíduo Ser9. Em células não estimuladas a GSK3 encontra-se constitutivamente ativa. Tanto substratos pré-fosforilados quanto os não pré-fosforilados podem ser fsoforilados pela GSK3 nessa condição. Quando as quinases que inibem a GSK3 são ativadas pelas vias de sinalização que a que ela participa, a GSK3 é fosforilada por essas quinases em seu resíduo Ser9 sendo inibida. Isso previne a fosforilação de qualquer substrato dessa proteína.
A via fosfatidilinositol 3-quinase (PI3K)/proteína quinase B (AKT) é um
dos sistemas que levam a inativação da GSK3 pelo aumento de sua
fosforilação na Ser9. Nessa via de sinalização a PI3K é ativada por fatores de
crescimento como IGF-1 (insulin-like growth factor-1) que leva a fosforilação de
AKT. Subsequentemente AKT fosforila a Ser9 da GSK3. Adicionalmente a
Ser9 da GSK3 pode ser fosforilada por proteína quinase C (Goode et al.,
1992), proteína quinase A (Fang et al., 2000), p90Rsk e p70 S6 quinase. Assim
múltiplos mecanismos são utilizados para modificar a ativação da GSK3
através da fosforilação. A Fig. 5 mostra o esquema da GSK3 marcando seus
resíduos fosforiláveis (Ser9 e Tyr216).

23
Fig. 5 – Esquema da GSK3 mostrando os resíduos Ser9 e Tyr216 que podem ser fosforilados
(Bradley e Woodgett, 2003)
Além da fosforilação, a atividade da GSK3 pode ser modulada pela sua
distribuição subcelular. Inicialmente a GSK3 foi considerada uma enzima
citoplasmática solúvel (Boyle et al., 1991; Hemming et al., 1981). Contudo, a
GSK3 é capaz de fosforilar proteínas no núcleo. Está capacidade implica na
localização nuclear da GSK3 . De fato, GSK3 foi co-purificada com o
glicogênio de núcleo de fígado de ratos (Caracciolo et al., 1998), também foi
encontrada no núcleo de células NIH-3T3 com o ciclo celular interrompido
(Diehl et al., 1998), em cardiomiócitos estimulados com endotelina-1 (Haq et
al., 2000) e em oocitos de Xenopus laevis submetidas a choque térmico (Xavier
et al., 2000).
Outro modo de regulação dessa proteína é a sua ligação com outras
proteínas, que é tão importante quanto outras modificações que alteram sua
atividade. Esta regulação está bem descrita na via de sinalização Wnt (Fig. 5)
onde a GSK3 se liga a axina fosforilando a -catenina (Dajani et al., 2003). A
ligação específica mostra que existem diferentes domínios de reconhecimento
para cada substrato, mas o conhecimento desses domínios está restrito a via
Wnt (Bax et al., 2001; Fraser et al., 2002). Pouco se sabe sobre estes domínios
de ligação da GSK3com outros substratos
Domínio quinase

24
Existem várias evidências de que o cálcio também regula a ativação da
GSK3. Existem duas hipóteses, uma é o aumento da fosforilação da GSK3 em
seu resíduo Tyr216/279 (Frame e Cohen, 2001; Hartigan e Johnson, 1999;
Bradley e Woodgett, 2003). A outra mostra que a atividade da GSK3 é
aumentada independentemente da fosforilação da Tyr, via aumento da
desfosforilação da Ser9/21 (Song et al., 2002; King et al., 2006).
Estudos com levedura identificaram que Mck1p, proteína kinase da família
GSK3, foi ativada por Ca2+ levando a fosforilação da kinase Hsl1, resultando na
regulação da mitose (Mizunuma et al., 2001). Ainda, Mck1 promove a
fosforilação de RCNs, reguladores negativos de calcineurina (CaN, proteína
fosfatase Ca2+/calmodulina dependente 2B), sendo uma dos componentes
essenciais da via de sinalização calcineurina-dependente (Hilioti et a., 2004)
Apesar das evidências da regulação da GSK3 pelo cálcio, não se sabe
por qual via essa regulação acontece. Song e colaboradores (2002)
demostraram que essa regulação poderia ser via proteína fosfatase 2A que tem
sua atividade aumentada pelo aumento de cálcio que é acompanhado por um
aumento da desfosforilação da GSK3 no resíduo Ser9, o que promove um
aumento de sua atividade. King e colaboradores (2006) demonstraram que o
aumento dessa desfosforilação poderia ser dependente da proteína fosfatase
1A.
Devido ao envolvimento com diversas cascatas de sinalização qualquer
alteração (silenciamento, mutação ou super-expressão) na atividade ou
expressão da GSK3 pode promover mudanças fenotípicas levando a má
formação e/ou letalidade de embriões. A perda da função como conseqüência
da mutação no gene da GSK3 em Drosophila levou a perda das cerdas

25
(Siegfried et al., 1992). Estudos em Xenopus laevis demonstraram que a
expressão de uma versão mutante da GSK3 inativa induziu a duplicação do
eixo embrionário (He et al., 1995). A super-expressão de GSK3 normal ou
mutante em ouriço do mar perturba a formação da padronização do eixo
embrionário (Emily-Fenouil et al., 1998).
RNAi em carrapatos
A técnica de RNA de interferência oferece uma ferramenta alternativa
para a caracterização da função de várias proteínas através do silenciamento
da expressão do mRNA (Aljamali et al., 2002). Nessa técnica as moléculas de
RNA fita dupla são internalizadas pela célula e seu efeito se torna maior nos
próximos estágios do desenvolvimento do carrapato por um mecanismo ainda
não conhecido (Nijhof et al., 2007). Foi demonstrado que em outros organismos
após a internalização os dsRNAs são clivados em fragmentos de 21-25 pb
("small interfering RNA" ou siRNA) por uma enzima chamada Dicer com
atividade de RNAse III. Os siRNAs associam-se no citoplasma ao complexo
multi-protéico induzido de silenciamento por de RNA (RISC) (Hammond et al,
2001) no citoplasma, e o complexo formado e ativado reconhece e cliva RNAm
celulares complementares ao siRNA presente no complexo (Fig. 5). Kurscheid
e colaboradores (2009) forneceram as evidencias da presença da via de RNAi
em R. microplus associada a sequências de ESTs, incluindo Dicer, proteínas
Argonauta, RdRP (RNA polimerase RNA dependente) e proteínas associadas
com o transprote do dsRNA.

26
RISC ativada
RNAm a ser degradado
RISC ativada
RNAm a ser degradado
Fig. 6 – Esquema mostrando a provável via de RNAi em carrapatos. O RNAi sistêmico é capitado pela célula via proteína Sid-1. A dsRNA exógena ou viral entra no citoplasma e é processada em fragmentos de 21 a 23 nucleotídeos de comprimento (siRNA). A proteína chave para essa degradação específica é a proteína Dicer-1. Então o complexo de silenciamento induzido por RNA (RISC) é ativado e degrada o RNAm correspondente à dupla fita (de la Fuent et al., 2007).
A técnica de silenciamento gênico foi utilizada primeiramente em
carrapatos por Aljamali e colaboradores em 2002 e tornou-se rapidamente
aplicada (de la Fuente e Kocan, 2006), sendo útil na caracterização molecular e
funcional de várias proteínas do carrapato (Kocan et al., 2009, de la Fuent et
al., 2006).
Alguns exemplos do uso do RNAi incluem o estudo da proteína ligadora
de histamina (HBP) em fêmeas do carrapato Amblyoma americanum (Aljamali
et al., 2003), o silenciamento de antígenos protetores em I. scapulares (de la
Fuent et al., 2005), o silenciamento das proteínas 4D8 e Rs86, análogas à
Bm86, em R. sanguineus que resultou em diminuição da fixação do carrapato,
ingurgitamento, peso e oviposição (de La Fuent et al., 2006), silenciamento das

27
proteínas v-SNARE e t-SNARE, que mostraram ser importantes no processo
de alimentação do carrapato A. americanum (Karim et al., 2004a; 2004b; 2005).
O silenciamento de uma cistatina salivar de A. americanum promoveu
interferência no processo de alimentação e sobrevida dos carrapatos. Estes
estudos demonstram que a técnica de RNAi pode ser amplamente utilizada no
estudo de função das macromoléculas de carrapatos auxiliando na busca de
alternativas para o controle desse artrópode.

28
Objetivos

29
Objetivo Geral
Avaliar o papel da GSK3 durante a embriogênese do carrapato R.
microplus através da avaliação da oviposição, eclosão e desenvolvimento
embrionário dos ovos de fêmeas silenciadas através de RNAi para a GSK3
Avaliar a interferência do cálcio na transcrição, expressão e fosforilação
do resíduo Tyr216 da GSK3 nas células embrionárias BME26;
Objetivos específicos
Promover o silenciamento da GSK3 em fêmeas parcialmente
ingurgitadas e na linhagem de células embrionárias BME26;
Confirmar o silenciamento da GSK3 nos ovários de fêmeas
parcialmente ingurgitadas e na linhagem de células BME26;
Caracterizar o fenótipo obtido pelo silenciamento através da
avaliação da oviposição, eclosão e desenvolvimento dos ovos das
fêmeas silenciadas;
Analisar a concentração do glicogênio na linhagem de células
BME26 silenciadas para a GSK3;
Avaliar a transcrição, expressão e fosforilação do resíduo Tyr216 da
GSK3nas células embrionárias BME26 incubadas com CaCl2;

30
Materiais e Métodos

31
Carrapato Rhipicephalus (Boophilus) microplus
Os carrapatos da espécie R. microplus foram criados em bovinos na
Faculdade de Veterinária da Universidade Federal do Rio Grande do Sul. Os
carrapatos se desenvolveram em bovinos isolados em estábulos, livres de
qualquer contato com o campo. As larvas foram colocadas no dorso dos
bovinos e após 20 dias as fêmeas no estágio de partenógenas, com
aproximadamente 50 mg, foram coletadas manualmente para os ensaios com
RNAi ou coletadas após a queda (21 dias). Os carrapatos foram mantidos a
28°C e 85% de umidade.
Alimentação artificial
As fêmeas no estágio de partenógenas foram alimentadas com sangue
de um bovino não infestado com carrapato. O sangue foi coletado com sistema
de coleta à vácuo, com o anticoagulante heparina.
As fêmeas foram fixadas em bandejas e separadas em grupos controle,
controle negativo (injetadas com dsRNA de um gene não relacionado) –
galactosidade de E. coli - e injetadas com dsRNA específico para GSK3.
A cada três horas o capilar era trocado e as fêmeas permaneciam
alimentando-se por 28 horas na estufa, a 28ºC, com 80% de umidade relativa.
Os ovos de cada grupo foram coletados e mantidos nas mesmas condições.

32
Síntese do dsRNA
O RNAi foi desenhado para o gene da GSK3 específico para o carrapato
R. microplus. Para a síntese do RNAi, foi selecionada uma região apropriada
do gene. Essa sequência foi submetida a um BLAST no GenBank para
determinar a especificidade do fragmento. O fragmento selecionado contém
798 pb que compreendem os nucleotídeos 697 a 1495 do RNAm da GSK3
(GenBank EF142066).
RNA total foi extraído de ovos de 17º dia de desenvolvimento usando reagente
TRIzol (Invitrogen) como descrito pelo fabricante. O cDNA foi sintetizado a
partir de 2 g de RNA total. A reação foi realizada na presença de RT randon
primer e transcriptase reversa (High-capacity cDNA Reverse Transcripion kit -
Applied Biosystems), de acordo com as instruções do fabricante. Depois da
trascrição reversa, 1 µg do produto foi utilizado para a reação de PCR usando
primers para o gene da GSK3desenhados como descrito acima contendo a
sequência do promotor T7 para a síntese in vitro do dsRNA. Os primers
utilizados foram os seguintes GSK-3T7L: 5’-
TAATACGACTCACTATAGGGTTATGCGACGGCTAGAACACT-3’ and GSK-
3T7R: 5’-TAATACGACTCACTATAGGGGCTCTTGCTCTGTGAAGTTGAA-3’.
Os amplicons gerados foram purificados por gel filtração em coluna spin S-400
(Amersham). A síntese do dsRNA foi realizada utilizando a enzima RNA
polimerase T7 de acordo as especificações do fabricante (T7 RiboMAX™
Express RNAi System - Promega).

33
Injeção do RNAi em partenóginas
Teléoginas foram coletadas logo após a queda e fêmeas no estágio de
partenógenas foram removidas dos bovinos. As fêmeas nos dois estágios
foram usadas para a injeção de aproximadamente 4 g (1,5 l) de dsRNA
específico para GSK3na posição ventral com seringa Hamilton (33 gauge
needle).Carrapatos controle foram injetados com tampão PBS ou com dsRNA
de um gene não relacionado (-galactosidase) de E. coli.
Após a injeção as partenóginas foram alimentadas artificialmente como descrito
acima. Os parâmetros biológicos analisados foram o número de fêmeas que
ingurgitaram e o peso dos carrapatos. Depois do período de oviposição (10
dias) o peso dos ovos postos foi determinado. A eclosão foi verificada 40 dias
após a oviposição.
Análise da transcrição gênica de GSK3 por qPCR
O RNA total foi extraído de 2 ovários de teleóginas 24 e 48 h após a
injeção, 2 ovários de partenóginas 24, 48 e 72 h após a alimentação artificial
ou de células embrionárias BME26 silenciadas para a GSK3 usando reagente
TRIzol (Invitrogen) como descrito pelo fabricante.
O cDNA foi sintetizado a partir de 2 g de RNA total. A reação foi realizada
na presença de RT random primer e transcriptase reversa (High-capacity cDNA
Reverse Transcripion kit - Applied Biosystems), de acordo com as instruções
do fabricante. Depois da reação da transcrição reversa, 200 ng do produto de
cada amostra foi utilizada para a reação de PCR em tempo real usando primers

34
de genes específicos com kit Master SYBR Green (Roche). A amplificação foi
feita usando LightCycler® (Roche). A expressão da proteína ribossomal 40S foi
usada para normalizar e quantificar da expressão relativa de GSK3. Os
primers utilizados foram: 5’-CGAGGTGTACCTGAACCTGGT -3’ (forward) e 5’-
CGATGGCAGATGCCCAGAGAG -3’ (reverse) para GSK-3; 5’-
GGACGACCGATGGCTACCT-3’ (forward) e 5’-
TGAGTTGATTGGCGCACTTCT -3’ (reverse) para mRNA da proteína
ribossomal 40S. A expressão relativa foi expressa em função dos valores
determinados experimentalmente para a eficiência de ambos os pares de
primers, alvo e referência, e de acordo com os valores de Cp, como proposto
por Pfaffl (2001).
Observação dos embriões por microscopia óptica
Amostras de ovos de fêmeas injetadas com ou sem dsRNA foram
coletadas depois de 18 dias do inicio da oviposição, colocadas em lâminas com
óleo de imersão, examinadas na lupa e fotografadas com câmera digital.
Manutenção da cultura de células embrionárias
A cultura de células embrionárias da linhagem BME26 foi mantida a 34º
C em garrafas de cultura de 25 cm2 contendo 5 mL de meio completo (meio L-
15B diluído 3:1 com água estéril e suplementado com 10% de soro fetal bovino,
10% de triptose fosfato, 100 unidades/mL de penicilina e 100 mg/mL de
estreptomicina). As células aderidas foram ressuspendidas em meio completo

35
com seringa de 5 ml e plaqueadas em uma densidade de aproximadamente
5X105 células para os ensaios. A densidade das células foi determinada
através de contagem em câmara de Neubauer e a viabilidade celular foi
avaliada por coloração com azul de trypan (0,4% Sigma)
Silenciamento gênico por RNAi da GSK3 em cultura de células
As células BME26 foram descoladas e plaqueadas em placas de cultura
de células de 24 poços em uma densidade de 5x105 células por poço em 500l
de meio completo. Depois de 24 horas, quando as células se aderem
novamente, o RNA dupla-fita foi adicionado em meio de cultura completo.
Depois de 48 horas o meio foi trocado. Após 72 horas as células foram
utilizadas para extração de RNA para a confirmação do silenciamento ou outros
ensaios. Como controles negativos foram utilizadas células incubadas com
PBS ou RNA dupla-fita contra galactosidase de E. coli .
Dosagem de glicogênio
As células BME26 tratadas com RNA dupla-fita foram lisadas em tampão
Tris-HCL contendo 0,1% Triton X-100, 100 M leupeptina, 1 M pepstatina,
100 M PMSF, 1 mM EDTA, 50 mM fluoreto de sódio, 100 M molibidato de
amônio e 1 mM vanadato de sódio. Depois, 20 µL do sobrenadante foi
incubado com 1 unidade de α-amiloglicosidase (Sigma Chemicals) por 4 h a
40°C. O conteúdo de glicose foi enzimaticamente determinado usando glicose
oxidase (Glucox® Kit enzimatico para dosagem de glicose; Doles, inc.), a partir

36
de incubação por 30 min a 37 °C de acordo com as instruções do fabricante. A
concentração de glicose foi determinada por uma curva padrão e normalizada
pela concentração de proteína de cada amostra.
Western blotting
As células BME26 incubadas na presença de diferentes concentrações
de CaCl2, 1 mM de EGTA ou diferentes concentrações de thapsigargina por 1h
foram lisadas em tampão Tris-HCl 10mM pH 7,4 contendo 0,1% Triton X-100,
100 M leupeptina, 1 M pepstatina, 100 M PMSF, 1 mM EDTA, 50 mM
fluoreto de sódio, 100 M molibidato de amônio e 1 mM vanadato de sódio. A
concentração total de proteína de cada amostra foi determinada pelo méto do
de Bradford (1976). As amostras contendo quantidades equivalentes de
proteína (200 g de proteína) foram aplicadas em SDS-PAGE 10%. As
proteínas foram transferidas para membrana de PVDF por 2 h a 38 mA.
Membranas foram bloqueadas em tampão TBS (Tris–HCl 20 mM pH 7,4, NaCl
150 mM e 0,1% Tween 20 contendo 3% de leite desnatado por 2h a
temperatura ambiente. As membranas foram lavadas e incubadas com
anticorpo policlonal anti-pTyr279/216 GSK3(1:1000 - Sigma) ou anticorpo
monoclonal anti-GSK3(1:1000 - Sigma) em TBS. O anticorpo secundário
utilizado foi o anti IgG de coelho conjugado a peroxidade (1:2000 - Sigma) e a
membrana foi revelada utilizando 0,013% (w/v) de 3,3’-diaminobenzidina
(DAB), 2M de Tris, 0,1M de imidazol e 1% de H2O2 . Imagens foram
digitalmente adquiridas com o scanner e analisadas semi-quantitativamente
usando o programa ImageJ (Sasaki et al., 2006).

37
Resultados

38
Silenciamento da GSK3 em fêmeas do carrapato R. microplus
Primeiramente o silenciamento da expressão da GSK3 por RNAi foi
realizado em fêmeas totalmente alimentadas (teleóginas). A injeção foi
realizada dois dias após a queda da fêmea. Para a confirmação do
silenciamento foi realizada a quantificação da transcrição da GSK3b por PCR
tempo real em ovários das fêmeas retiraods 24 e 48h após a injeção. Não
houve diferença na transcrição da GSK3 entre as fêmeas injetadas com
dsRNA específico para a GSK3 em relação com o dsRNA do controle
negativo (dado não mostrado). Todavia, a expressão gênica de GSK3foi 4
vezes menor após de 48 h de injeção de dsRNA comparando com 24h. A
transcrição dessa proteína já se encontra alta no ovário de teleóginas. Partimos
para o silenciamento da expressão da GSK3 em fêmeas que ainda não
terminaram sua alimentação (partenóginas) e possuem o nível de transcrição
da GSK3 baixo no ovário. As partenóginas foram retiradas do bovino 24h
antes do término da alimentação (em torno de 50 mg) e injetadas com dsRNA
para GSK3, -galactosidase de E. coli ou tampão PBS. Subseqüentemente as
fêmeas foram alimentados artificialmente com capilar por 28 h (Fig. 6).

39
Fig. 7 – Alimentação artificial por capilar. Fêmeas parcialmente ingurgitadas foram
aliementadas com sangue bovino em capilares por 28 h. A – fêmeas antes da alimentação; B –
fêmeas após 3 h de alimentação.
O peso individual das fêmeas de carrapato após a alimentação não foi
significativamente diferente (p> 0,81) entre controle e grupos tratados (média ±
desvio padrão) (grupo controle 70 mg ± 19,74 mg; grupo gene não
relacionado): 75 mg ± 28,05 mg; grupo dsRNA GSK3: 73 mg ± 28,79 mg),
indicando que a presença do dsRNA não interfere o ganho de peso das fêmeas
(Fig. 7).
A B

40
0h 28h0
30
60
90
120 Controle Controle neg.GSK3
Tempo de alimentação artificial
Peso
dos
car
rapa
tos
(mg)
Fig. 8 – Ganho de peso das fêmeas depois da alimentação artificial por capilar. Barra branca:
controle; Barra listrada: Controle negativo; Barra cinza: dsRNA específico para GSK3;
Para a confirmação do silenciamento dois ovários de fêmeas de cada
grupo coletados 24, 48 e 72 h após a alimentação artificial foram utilizados para
quantificação da transcrição da GSK3 por PCR tempo real. Foi observado que
o nível de transcrição da GSK3 nos ovários dos grupos controle aumentou
gradualmente ao longo do tempo analisado. Contudo, em fêmeas injetadas
com dsRNA para a GSK3 os níveis permanecem muito baixos nos ovários,
chegando a 95% de redução depois de 72 h de alimentação artificial quando
comparado ao controle (Fig. 8)
.

41
Fig. 9 – Confirmação do silenciamento da GSK3 em ovários de partenóginas. A transcrição
relativa de GSK3 foi determinada por PCR quantitativo a partir do RNA total extraído de ovários
de fêmeas injetadas com PBS (barra branca), dsRNA de -galactosidase de E. coli (barra
listrada) ou dsRNA de GSK3 de carrapato (barra cinza) coletado 24, 48 ou 72h após o fim da
alimentação artificial.
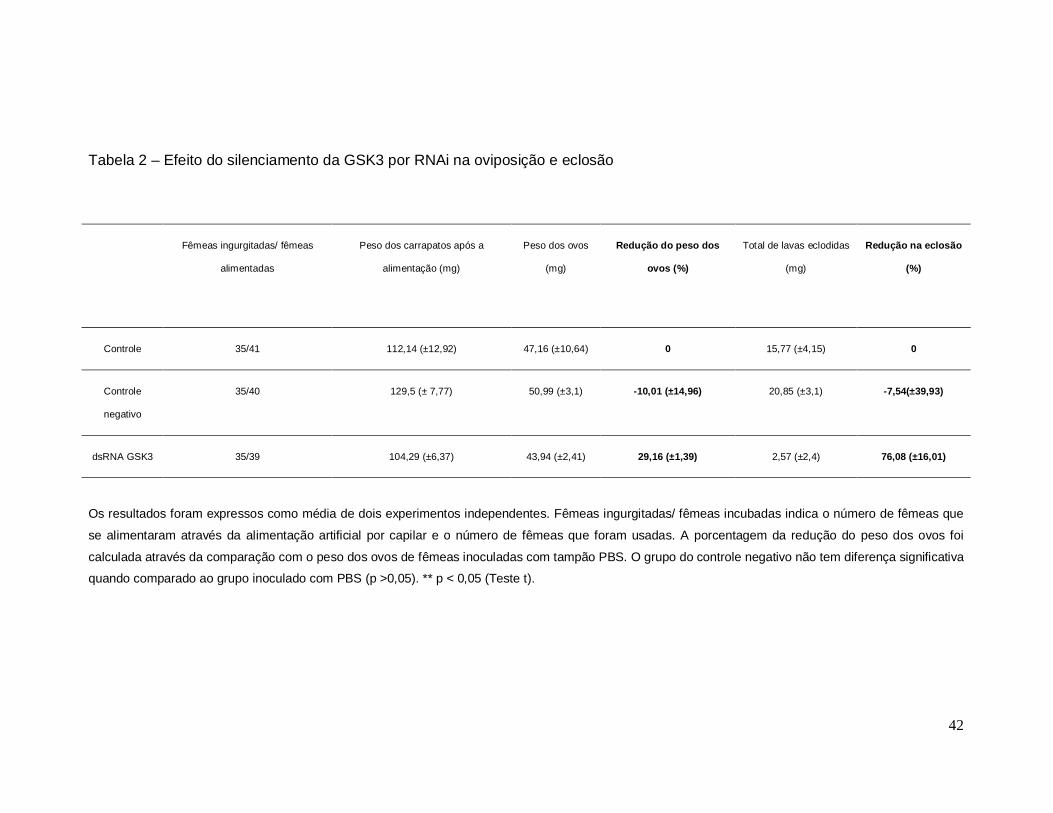
Além disso, as fêmeas tratadas com dsRNA para a GSK3 tiveram uma
redução de 29,1% na oviposição e uma redução na eclosão de seus ovos de
76% quando comparado ao controle negativo (tabela 2).
0.0
0.5
1.0
1.5
2.0
24h 48h 72hTran
scri
ção
Rel
ativ
a (G
SK3/
40S)
42
Tabela 2 – Efeito do silenciamento da GSK3 por RNAi na oviposição e eclosão
Fêmeas ingurgitadas/ fêmeas
alimentadas
Peso dos carrapatos após a
alimentação (mg)
Peso dos ovos
(mg)
Redução do peso dos
ovos (%)
Total de lavas eclodidas
(mg)
Redução na eclosão
(%)
Controle 35/41 112,14 (±12,92) 47,16 (±10,64) 0 15,77 (±4,15) 0
Controle
negativo
35/40 129,5 (± 7,77) 50,99 (±3,1) -10,01 (±14,96) 20,85 (±3,1) -7,54(±39,93)
dsRNA GSK3 35/39 104,29 (±6,37) 43,94 (±2,41) 29,16 (±1,39) 2,57 (±2,4) 76,08 (±16,01)
Os resultados foram expressos como média de dois experimentos independentes. Fêmeas ingurgitadas/ fêmeas incubadas indica o número de fêmeas que
se alimentaram através da alimentação artificial por capilar e o número de fêmeas que foram usadas. A porcentagem da redução do peso dos ovos foi
calculada através da comparação com o peso dos ovos de fêmeas inoculadas com tampão PBS. O grupo do controle negativo não tem diferença significativa
quando comparado ao grupo inoculado com PBS (p >0,05). ** p < 0,05 (Teste t).

43
Os ovos de carrapato de fêmeas injetadas com dsRNA para GSK3
exibiram um fenótipo alterado quando observados em estereomicroscópio e
comparados com os grupos controle (Fig. 9). Enquanto os embriões do grupo
controle foram totalmente visíveis após 18 dias de oviposição, os ovos das
fêmeas injetadas com dsRNA para a GSK3 apresentam um aspecto
indiferenciado.
Fig. 10 – Efeito do silenciamento gênico por RNAi da GSK3 no desenvolvimento embrionário.
(A) Desenvolvimento embrionário de ovos com 18 dias após a oviposição de fêmeas do grupo
controle negativo (B) Desenvolvimento embrionário de ovos inoculadas com dsRNA para
GSK3 com 18 dias de desenvolvimento. Barra de tamanho: 1 mm; barra de tamanho da
amplificação: 0.25 mm; A seta indica o ovo que foi utilizado na ampliação
Silenciamento da GSK3 em células embrionárias BME26
Para minimizar as dificuldades técnicas do silenciamento em
partenóginas partimos para o silenciamento na cultura de células embrionárias
BME26. O silenciamento foi realizado na ausência de agentes usados para
transfecção e a confirmado através de PCR em tempo real após 96 e 120 h de
exposição à dsRNA. A redução na transcrição da GSK3 em células tratadas
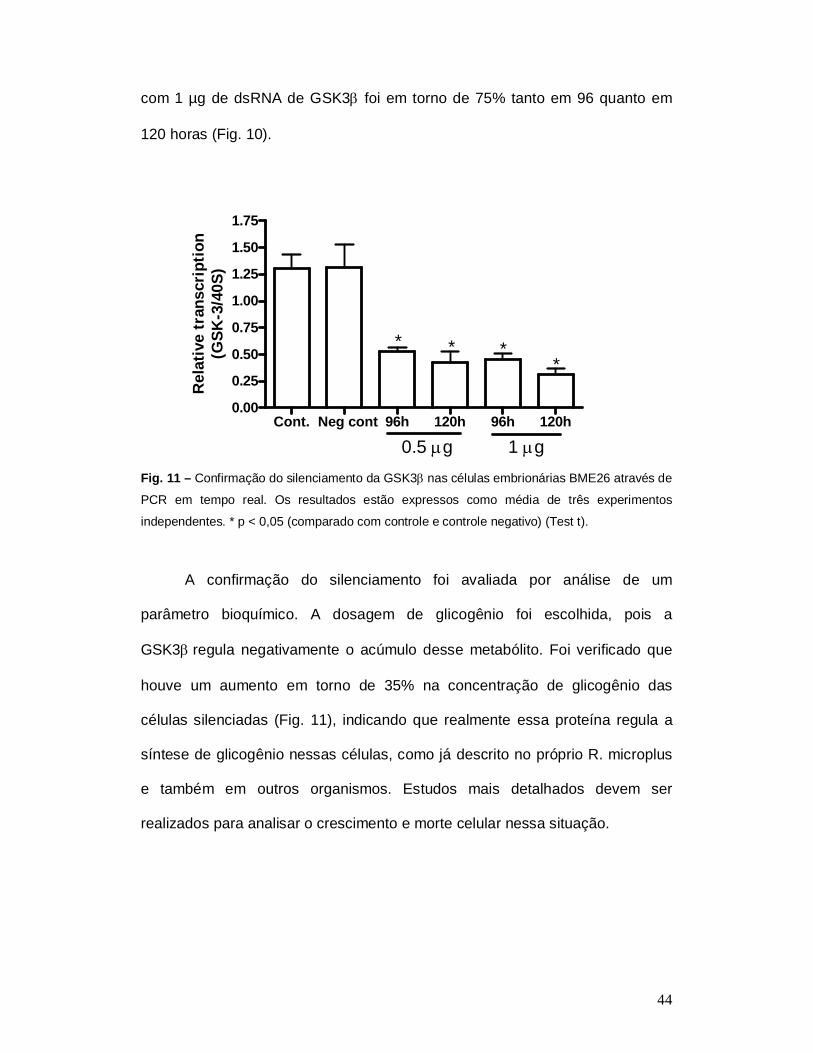
44
Cont. Neg cont 96h 120h 96h 120h0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
0.5 g 1 g
* * **
Rel
ativ
e tr
ansc
ript
ion
(GSK
-3/4
0S)
com 1 µg de dsRNA de GSK3 foi em torno de 75% tanto em 96 quanto em
120 horas (Fig. 10).
Fig. 11 – Confirmação do silenciamento da GSK3 nas células embrionárias BME26 através de
PCR em tempo real. Os resultados estão expressos como média de três experimentos
independentes. * p < 0,05 (comparado com controle e controle negativo) (Test t).
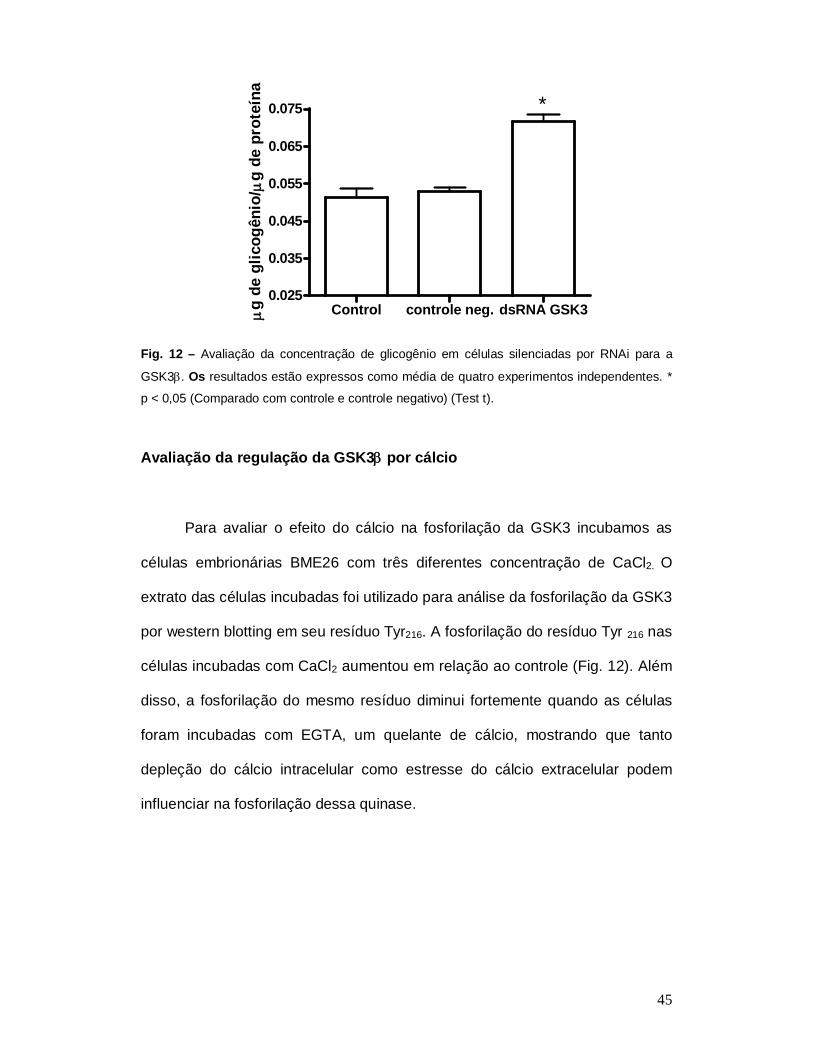
A confirmação do silenciamento foi avaliada por análise de um
parâmetro bioquímico. A dosagem de glicogênio foi escolhida, pois a
GSK3regula negativamente o acúmulo desse metabólito. Foi verificado que
houve um aumento em torno de 35% na concentração de glicogênio das
células silenciadas (Fig. 11), indicando que realmente essa proteína regula a
síntese de glicogênio nessas células, como já descrito no próprio R. microplus
e também em outros organismos. Estudos mais detalhados devem ser
realizados para analisar o crescimento e morte celular nessa situação.
45
Control controle neg. dsRNA GSK30.025
0.035
0.045
0.055
0.065
0.075 *
g
de g
licog
ênio
/ g
de p
rote
ína
Fig. 12 – Avaliação da concentração de glicogênio em células silenciadas por RNAi para a
GSK3. Os resultados estão expressos como média de quatro experimentos independentes. *
p < 0,05 (Comparado com controle e controle negativo) (Test t).
Avaliação da regulação da GSK3 por cálcio
Para avaliar o efeito do cálcio na fosforilação da GSK3 incubamos as
células embrionárias BME26 com três diferentes concentração de CaCl2. O
extrato das células incubadas foi utilizado para análise da fosforilação da GSK3
por western blotting em seu resíduo Tyr216. A fosforilação do resíduo Tyr 216 nas
células incubadas com CaCl2 aumentou em relação ao controle (Fig. 12). Além
disso, a fosforilação do mesmo resíduo diminui fortemente quando as células
foram incubadas com EGTA, um quelante de cálcio, mostrando que tanto
depleção do cálcio intracelular como estresse do cálcio extracelular podem
influenciar na fosforilação dessa quinase.

46
Fig. 13 – Avaliação da fosforilação do resíduo Tyr216 da GSK3através de Western Blotting
utilizando anticorpo policlonal anti-GSK3 (Tyr) em células embrionárias incubadas com
diferentes concentrações de CaCl2 ou 1mM de EGTA.
Além de avaliar a fosforilação da GSK3 também verificamos sua
expressão através de western blotting utilizando anticorpo anti-GSK3. O
extrato de células incubadas com diferentes concentrações de CaCl2 e EGTA
foi utilizado e a expressão da GSK3 aumentou em células incubadas com
CaCl2 (Fig. 13). Não houve diferença entre as células controle e as células
incubadas com 1 mM de EGTA.
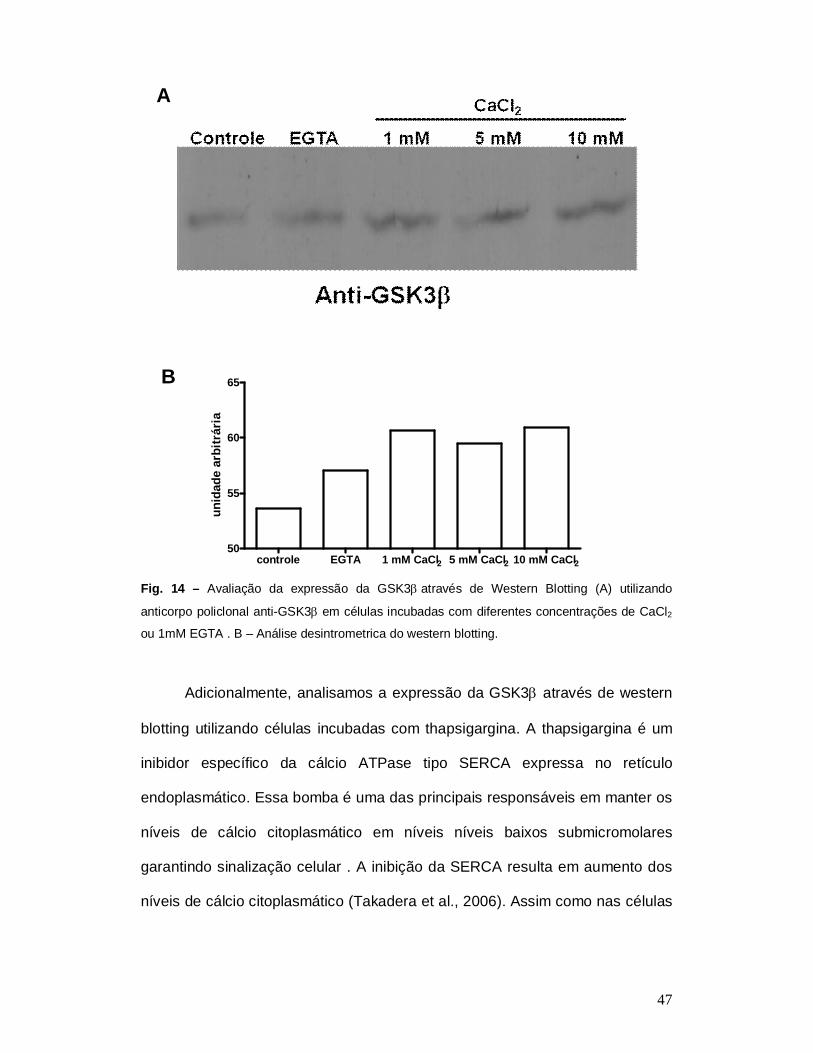
47
controle EGTA 1 mM CaCl2 5 mM CaCl2 10 mM CaCl250
55
60
65
unid
ade
arbi
trár
ia
Fig. 14 – Avaliação da expressão da GSK3através de Western Blotting (A) utilizando
anticorpo policlonal anti-GSK3 em células incubadas com diferentes concentrações de CaCl2
ou 1mM EGTA . B – Análise desintrometrica do western blotting.
Adicionalmente, analisamos a expressão da GSK3 através de western
blotting utilizando células incubadas com thapsigargina. A thapsigargina é um
inibidor específico da cálcio ATPase tipo SERCA expressa no retículo
endoplasmático. Essa bomba é uma das principais responsáveis em manter os
níveis de cálcio citoplasmático em níveis níveis baixos submicromolares
garantindo sinalização celular . A inibição da SERCA resulta em aumento dos
níveis de cálcio citoplasmático (Takadera et al., 2006). Assim como nas células
A
B
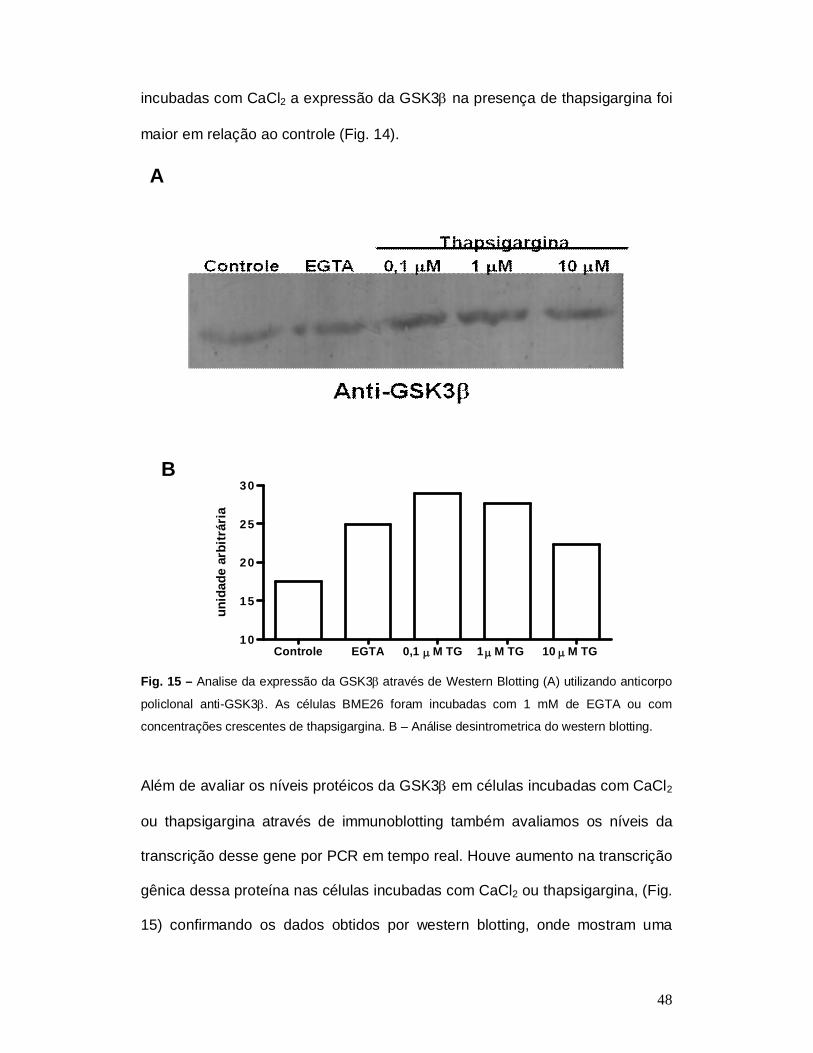
48
Controle EGTA 0,1 M TG 1 M TG 10 M TG10
15
20
25
30
unid
ade
arbi
trár
ia
incubadas com CaCl2 a expressão da GSK3 na presença de thapsigargina foi
maior em relação ao controle (Fig. 14).
Fig. 15 – Analise da expressão da GSK3através de Western Blotting (A) utilizando anticorpo
policlonal anti-GSK3. As células BME26 foram incubadas com 1 mM de EGTA ou com
concentrações crescentes de thapsigargina. B – Análise desintrometrica do western blotting.
Além de avaliar os níveis protéicos da GSK3 em células incubadas com CaCl2
ou thapsigargina através de immunoblotting também avaliamos os níveis da
transcrição desse gene por PCR em tempo real. Houve aumento na transcrição
gênica dessa proteína nas células incubadas com CaCl2 ou thapsigargina, (Fig.
15) confirmando os dados obtidos por western blotting, onde mostram uma
A
B
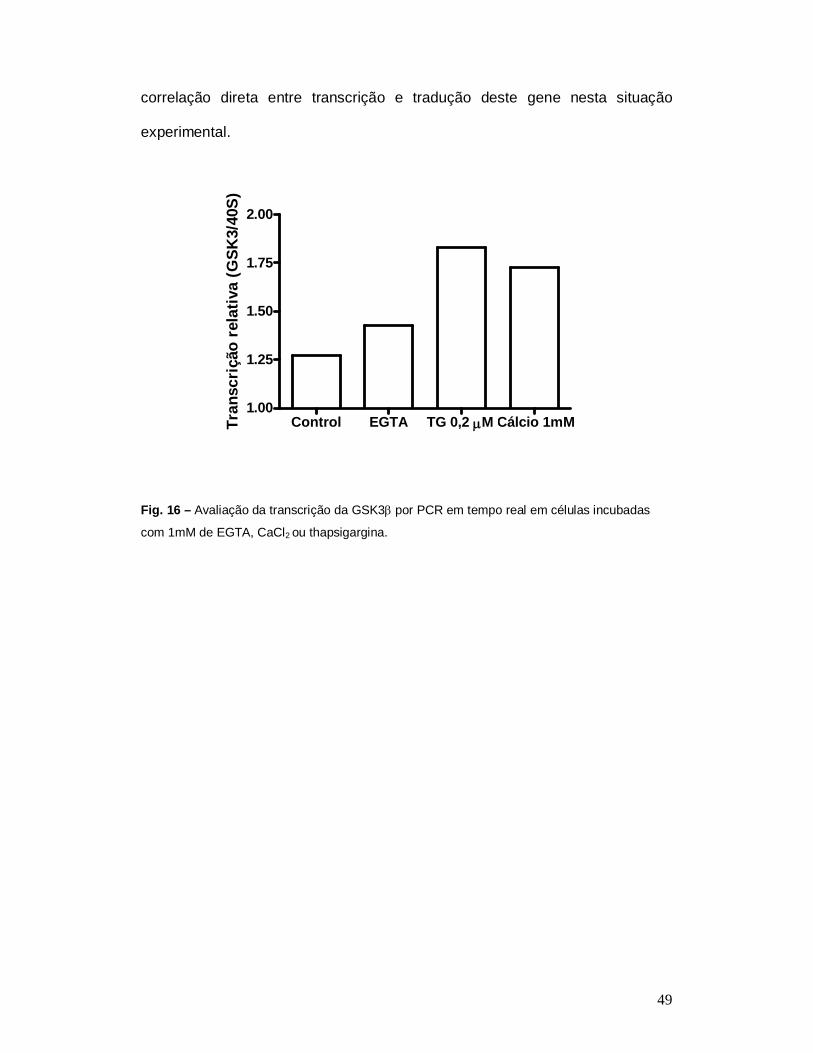
49
Control EGTA TG 0,2 M Cálcio 1mM1.00
1.25
1.50
1.75
2.00Tr
ansc
riçã
o re
lativ
a (G
SK3/
40S)
correlação direta entre transcrição e tradução deste gene nesta situação
experimental.
Fig. 16 – Avaliação da transcrição da GSK3 por PCR em tempo real em células incubadas
com 1mM de EGTA, CaCl2 ou thapsigargina.

50
Discussão

51
Até a presente data, mais de 40 proteínas foram identificados como
substratos de fosforilação medidada pela GSK3. Devido ao seu importante
papel fisiológico, a atividade de GSK3 é alvo de um complexo controle
regulatório (Jope e Johnson, 2004). Existem vários mecanismos que regulam
essa quinase incluindo, fosforilação, formação de complexo protéico,
localização subcelular e proteólise. Ainda há evidências de que sua atividade a
GSK3 pode ser regulada por cálcio (Frame e Cohen, 2001; Hartigan e Johnson,
1999; Bradley e Woodgett, 2003; Song et al., 2002; King et al., 2006). Na
linhagem de células embrionárias BME26 a fosforilação da GSK3 em seu
resíduo Tyr216 foi aumentada quando as células foram incubadas com CaCl2
(Fig. 12). Além disso, a fosforilação neste mesmo resíduo foi bastante
diminuída quando as células foram incubadas com EGTA, um quelante de
cálcio. A mudança na fosforilação dessa quinase pelo CaCl2 e pelo EGTA
sugere que a GSK3 pode ser regulada pelo cálcio. Vale ressaltar que o EGTA
não é um quelante específico para o cálcio, por isso estudos futuros devem ser
realizados, com um quelante específico, para a confirmação desses
dados.Além da fosforilação houve também aumento da expressão e transcrição
da GSK3 quando as células foram incubadas com CaCl2 e thapsigargina (Fig.
13, 14 e 15), um inibidor de cálcio ATPase do retículo endoplasmático que
aumenta os níveis intracelulares de cálcio. Curiosamente o EGTA também foi
capaz de aumentar a fosforilação da GSK3, em níveis menores, provavelmente
por modular os níveis endógenos de cálcio ou outros cátions divalentes.
Já se sabe que a inibição de SERCA pela tapsigargina (Price et al.,
1992) ou depleção de estoque de ER com ionóforos (Drummond et al., 1987)
resultam numa condição fisiológica denominada estresse do retículo

52
endoplasmático. Uma das respostas da célula é ativação de influxo de cálcio
(Putney 1990) e ativação de via de sinalização calcineurina-dependente
(Bonilla et al., 2003) que promovem a sobrevivência celular. A participação da
GSK3 na via calcineurina-dependente foi demonstrada em levedura (Hilioti et
al., 2004). Estudos recentes em animais mostram que durante estresse do
retículo endoplasmático, a GSK3 interage com Hsp105 promovendo a ativação
de caspase-3 (Meares et al., 2008). Desse modo, aumento de níveis de GSK3
e sua fosforilação nas células embrionárias BME26 indica a ocorrência do
estresse do retículo endoplasmático com participação ativa da GSK3. Como
estresse do retículo endoplasmático é envolvido em processo de autofagia e
apoptose, estudos posteriores são necessários para caracterizar esse processo
e sua regulação por cálcio/calcineurina durante embriogênese de carrapato.
Logullo e colaboradores (2009) demostraram que não há, em ovos de R.
microplus uma relação direta entre a transcrição da GSK3b e sua fosforilação.
Quando as células embrionárias BME26 foram incubadas com EGTA foi visto
que existe uma grande diminuição da fosforilação do resíduo Tyr216 da GSK3b
e um pequeno aumento de sua transcrição confirmando os dados mostrados
por Logullo e colaboradores (2009). Ao contrario disso, a transcrição e a
expressão apresentaram uma relação direta, indicando que neste caso o
controle da tradução desta proteína depende da transcrição. As células
incubadas com EGTA, CaCl2 e thapsigargina tiveram um aumento tanto na
transcrição, mostrada por PCR tempo real, quanto na expressão, mostrada por
western blotting.
A GSK3 se encontra constitutivamente ativa em células não estimuladas
e pode ser inibida por uma variedade de estímulos extracelulares como,

53
insulina, fator de crescimento epidermal e proteína Wnt (Grimes e Jope, 2001;
Frame e Cohen, 2001). Estudos em organismos vertebrados e invertebrados
mostram que a GSK3 é uma quinase essencial para a padronização do eixo
embrionário em embriões na via de sinalização Wnt (Ferkey e Kimelman,
2000). Está via determina a polaridade dos segmentos embrionários em
Drosophila e a formação do eixo embrionário em Xenopus. Além disso, essa
via promove o crescimento celular e a diferenciação durante a embriogênese
de mamíferos (Ikeda et al., 1998). Além de seu papel na via Wnt, a GSK3
possui um importante papel no metabolismo do glicogênio (Grimes e Jope,
2001). Na ausência da via da insulina a GSK3 fosforila e inibe a glicogênio
sintase. Quando o nível de glicose sanguínea aumenta o nível de insulina
também aumenta. A Akt é ativada, como resultado da ativação da via da
insulina, e fosforila e inibe a GSK3. Devido a inibição da GSK3 pela ativação da
Akt, a glicogênio sintase pode ser ativada e promover a formação do
glicogênio. A GSK3 é uma quinase altamente conservada presente em
diversas espécies vertebradas e invertebradas (Dornelas et al., 1998). Logullo
e colaboradores (2009) caracterizaram primeiramente a GSK3 em carrapatos.
Este estudo demonstrou que existe somente uma isoforma dessa quinase
presente nos embriões do carrapato R. microplus, a isoforma GSK3. Além
disso, o ovário da fêmea de R. microplus é o principal tecido de transcrição da
GSK3.
O uso do RNAi na caracterização de vias metabólicas em carrapatos foi
primeiramente utilizado por Aljamali e colaboradores em 2002 e rapidamente
se tornou uma poderosa técnica para o estudo de genes (de la Fuente e
Kocan, 2006; Kocan et al. 2009). A redução na oviposição e eclosão de ovos

54
de fêmeas que foram silenciadas (Tabela 2) também foi vista em fêmeas que
tiveram a GSK3 inibida pela injeção de inibidores específicos para essa
quinase (Fabres et al., submetido) sugerindo que a supressão na expressão e
ou função da GSK3 é responsável pelos efeitos biológicos observados nos
ovos de fêmeas tratadas. Estudos anteriores demonstram a resposta ao RNAi
sistêmico em carrapatos não alimentados após a injeção, e o correspondente
gene silenciado ocorre em múltiplos tecidos (de la Fuente et al., 2006a; Nijhof
et al., 2007). Nijhof e colaboradores (2007) e Kocan e colaboradores (2007)
demostraram o efeito do RNAi sistêmico na prole de carrapatos quando a
dsRNA é injetada na hemolinfa de teleóginas causando o silenciamento na
primeira geração de larvas. Adicionalmente, ovos de fêmeas injetadas com
dsRNA para GSK3 apresentam um fenótipo alterado quando comparado ao
controle negativo, demonstrando o efeito do silenciamento da GSK3 durante o
desenvolvimento (Fig. 9). Esses ovos apresentaram um aspecto indiferenciado,
tendo a aparência de ovos de poucos dias de desenvolvimento.
Um problema para a analise morfológica desses embriões é a
impermeabilidade dos ovos de carrapato, que torna inviável o uso de técnicas
de coloração para microscopia. Devido a este problema, ainda não foi possível
avaliar todas as alterações morfológicas causadas pelo silenciamento da
GSK3. Diante desta difilculdade, estamos procurando adaptar um protocolo
de permeabilizacão usado para observar embriões de Tribolium castaneum
para o uso em ovos de carrapato (Wigand et a., 1998). Assim, os embriões
puderam ser observados parcialmente e futuramente acredita-se que será
possível avaliar todas as alterações morfológicas causadas pelo silenciamento
(dados não mostrados). Além disso, por problemas técnicos, ainda não foi

55
possível avaliar por qual via a GSK3 está influenciando o desenvolvimento dos
embriões de carrapato. Por outro lado, preliminarmente, nosso grupo
demonstrou o envolvimento da GSK3 no metabolismo de glicogênio durante a
embriogênese do carrapato R. microplus (Logullo et al. 2009). Uma diminuição
da expressão dessa proteína pode promover uma depleção da energia devido
a diminuição da glicose livre que pode estar sendo mobilizada para a síntese
de glicogênio em excesso, isso pode estar paralizando o desenvolvimento
desses embriões. Além disso, não podemos descartar o envolvimento da GSK3
na via Wnt que possui um grande envolvimento durante a formação
embrionária de organismos vertebrados e invertebrados (Wodarz 1998).
Acredita-se que futuramente será necessário avaliar por qual via esta proteína
esta influenciando o desenvolvimento desses embriões.

56
Conclusões

57
Os resultados desta dissertação permitem concluir que:
A GSK3 tem uma grande importância durante o desenvolvimento
embrionário do carrapato R. microplus, já que o silenciamento
gênico por RNAi dessa proteína causou diminuição na oviposição,
eclosão e desenvolvimento dos ovos de fêmeas injetadas com
dsRNA específico para GSK3;
Foi possível realizar o silenciamento gênico por RNAi nas células
embrionárias BME26 sem o uso de transfecção;
A GSK3 está envolvida no metabolismo de glicogênio das
células embrionárias BME26, como descrito na literatura, já que o
silenciamento dessa proteína alterou os níveis desse metabolito;
O cálcio foi capaz de alterar a fosforilação no resíduo Tyr216,
transcrição e expressão da GSK3 nas células embrionárias
BME26;
Não houve relação entre a fosforilação, a transcrição e expressão
da GSK3 nas células embrionárias incubadas com EGTA,
mostrando que essa proteína é regulada em vários níveis;
Houve relação entre a transcrição e expressão da GSK3 nas
células embrionárias BME26 incubadas com EGTA.

58
Referências
Abreu,L.A., Valle D., Manso P.P., Façanha A.R., Pelajo-Machado M., Masuda
H., Masuda A., Vaz I. Jr., Lenzi H., Oliveira P.L., Logullo C. (2004).
Proteolytic activity of Boophilus microplus Yolk pro-Cathepsin D (BYC) is
coincident with cortical acidification during embryogenesis., Insect
Biochem Mol Biol 34, 443 – 9;
Aljamali, M.N., Sauer, J.R. and Essenberg, R.C. (2002). RNA interference:
applicability in tick research. Experimental and Applied Acarology 28,
89–96;
Aljamali MN, Bior AD, Sauer JR, Essenberg RC. (2003). RNA interference in
ticks: A study using histamine binding protein dsRNA in the female tick
Amblyomma americanum. Insect Mol Biol. 12, 299–305;
Bax B., Carter P.S., Lewis C., Guy A.R., Bridges A., Tanner R., Pettman
G., Mannix C.; Culbert A.A.; Brown M.J.B.; Smith D.G.; Reith A.D.
(2001). The Structure of Phosphorylated GSK-3 Complexed with a
Peptide, FRATtide, that Inhibits -Catenin Phosphorylation. Structure
9,.1143-1152;
Bonilla M, Cunningham KW. Mitogen-activated protein kinase stimulation of
Ca2+ signaling is required for survival of endoplasmic reticulum stress in
yeast. Mol Biol Cell. 2003;14:4296–4305
Boyle WJ, Smeal T, Defize LH, Angel P, Woodgett JR, Karin M, Hunter T.,
(1991). Activation of protein kinase C decreases phosphorylation of c-Jun
at sites that negatively regulate its DNA-binding activity. Cell 64, 573–584;
Bradford, M.M., (1976). A rapid and sensitive method for the quantification of
microgram quantities of protein utilizing the principle of protein-dye binding.
Anal. Biochem. 72, 248–254;
Bradley W. D. and Woodgett (2003). GSK3: tricks of the trade for a multi-
tasking Kinase. Journal of Cell Science 116, 1175 – 1186;
Campos, E., Moraes, J., Façanha, A.R., Moreira, E., Valle, D., Abreu, L.,
Manso, P.P.A., Nascimento, A., Pelajo-Machado, M., Lenzi, H., Masuda,

59
A., Vaz-Jr, I. Logullo, C. (2006), Metabolic activity of Boophilus microplus
during embryonic development. Veterinary Parasitology 138, 349–357;
Caracciolo, M. R., Berlin, W. K., Miller, M. W., Hanover, J.A. (1998) Nuclear
Glycogen Synthase Kinase 3. Biochim. Biophys. Acta 249, 422-427; Cramer, L.G.; Carvalho, L.A.F.; Bridi, A.A.; Amaral, N.K., & Barrick, R.A. (1988).
Efficacy of topically applied ivermectin against Boophilus microplus in
cattle. Vet. Parasitol. 29, 341-349;
Dajani r.,Fraser E., Roe S. M, Young N., Good V., Dale T. C., and Pearl L. H.,
(2001). Crystal Structure of Glycogen Synthase Kinase 3: Structural
Basis for Phosphate-Primed Substrate Specificity and Autoinhibition.
Cell, Vol. 105, 721–732,
Dajani, R., Fraser, E., Roe, S. M., Yeo, M., Good, V. M., Thompson, V., Dale, T.
C., Pearl, L. H. (2003) Structural basis for recruitment of glycogen
synthase kinase 3 beta to the axin-APC scaffold complex. Embo Journal
22, 494-501;
de la Fuente J, Almazan C, Blouin EF, Naranjo V, Kocan KM. (2005). RNA
interference screening n ticks for identification of protective antigens.
Parasitol Res 96, 137-141;
Diehl JA, Cheng M, Roussel MF, Sherr CJ. (1998). Glycogen synthase kinase-
3beta regulates cyclin D1 proteolysis and subcellular localization. Genes
Dev. 15, 3499-511;
de la Fuente J., Almazán C., Blas-Machado U., Naranjo V., Mangold A.J.,
Blouin E.F., Gortazar C. and Kocan K.M. (2006). The tick protective
antigen, 4D8, is a conserved protein involved in modulation of tick blood
digestion and reproduction. Vaccine 24, 4082–4095;
de la Fuente, J. and Kocan, K.M. (2006). Strategies for development of
vaccines for control of ixodid tick species. Parasite Immunology 28, 275–
283;
de la Fuente J., Kocan K. M., Almaza´n C., and Blouin E. F. (2007). RNA
interference for the study and genetic manipulation of ticks. Trends in
Parasitology 23, 427-433;

60
Dornelas, M.C., Lejeune, B., Dron, M. and Kreis, M. (1998). The Arabidopsis
SHAGGY related protein kinase (ASK) gene family: structure,
organization and evolution. Gene 212, 249–257;
Drummond I.A., Lee,A.S., Resendez,E.,Jr and Steinhardt,R.A. (1987) Depletion
of intracellular calcium stores by calcium ionophore A23187 induces the
genes for glucose-regulated proteins in hamster fibroblasts. J. Biol.
Chem., 262, 12801–12805;
Embi, N., Rylatt, D. B. and Cohen, P. (1980). Glycogen synthase kinase-3 from
rabbit skeletal muscle. Separation from cyclic-AMP-dependent protein
kinase and phosphorylase kinase. Eur. J. Biochem. 107, 519-527;
Embrapa (2005). Disponivel em:
http://www.cnpgl.embrapa.br/nova/sala/noticias/jornaldoleite.php?id=163
Emily-Fenouil, F., Ghiglione, C., Lhomond, G., Lepage T. and Gache C. (1998).
GSK-3b/shaggy mediates patterning along the animal- vegetal axis of
the sea urchin embryo. Development 125, 2489-2498;
Esteves E., Lara F. A., Lorenzini D. M., Costa G. H.N., Fukuzawa A. H.,
Pressinotti L. N., Silva J.C., Ferro J. A., Kurtti T. J., Munderloh U. G.,
Daffre S. (2008). Cellular and molecular characterization of an embryonic
cell line (BME26) from the tick Rhipicephalus (Boophilus) microplus.
Insect Biochemistry and Molecular Biology 38, 568–580;
Fabres A., de Andrade C. P., Guizzo M., Sorgine M. H. F., Paiva-Silva G. O.,
Masuda A., da Silva Vaz Jr. I., Logullo C. (2009). Effect of GSK-3 Activity
Enzymatic Inhibition and Gene Silencing by RNAi on Tick Oviposition
and Egg Hatching. Parasitology, submetido;
Fang, X., Yu, S. X., Lu, Y., Bast, R. C., Jr., Woodgett, J. R., and Mills, G. B.
(2000) Proc. Natl. Acad. Sci. U. S. A. 97, 11960–11965;
Ferkey, D.M. and Kimelman, D. (2000). GSK-3: New thoughts on an old
enzyme. Developmental Biology 225, 471–479;
Flechtmann, C. H. W. (1977). Elementos de acarologia. Livraria Nobel, São
Paulo;

61
Frame, S. and Cohen, P (2001). GSK3 takes centre stage more than 20 years
after its discovery. Biochem.J. 359, 1-16;
Fraser E, Young N, Dajani R, Franca-Koh J, Ryves J, Williams RS, Yeo M, Webster MT, Richardson C, Smalley MJ, Pearl LH, Harwood A, Dale TC. (2002) Identification of the Axin and Frat binding region of glycogen synthase kinase-3. J Biol Chem 277, 2176-2185;
Gonzales, J.C., Silva, N.R., Franco, N. (1974), A vida livre do Boophilus
microplus. Arq. Fac. Vet. UFRGS. RG 3, 21-28;
Gonzales, J.C. (1995). O controle do carrapato do boi. 2a ed. Porto Alegre.
Edição do autor;
Goode N, Hughes K, Woodgett JR, Parker PJ. (1992). Differential regulation of
glycogen synthase kinase-3 beta by protein kinase C isotypes. J Biol
Chem 267, 16878–16882;
Grimes, C.A. and Jope, R.S. (2001). The multifaceted roles of glycogen
synthase kinase 3beta in cellular signaling. Progress in Neurobiology 65,
391–426;
Haq, S., Choukroun, G., Kang, Z.B., Ranu, H., Matsui, T., Rosenzweig, A.,
Molkentin, J.D., Alessandrini, A., Woodgett, J., Hajjar, R., Michael, A. and
Force, T. (2000) Glycogen synthase kinase-3beta is a negative regulator
of cardiomyocyte hypertrophy. J. Cell. Biol. 151, 117-130;
Hammond SM, Boettcher S, Caudy AA, Kobayashi R, Hannon GJ. (2001).
Argonaute2, a link between genetic and biochemical analyses of RNAi.
Science.293, 1146-50;
Hartigan J. A. and Johnson G. V. W. (1999). Transient Increases in Intracellular
Calcium Result in Prolonged Site-selective Increases in Tau
Phosphorylation through a Glycogen Synthase Kinase 3-dependent
Pathway. The journal of biological chemistry 274, 21395–21401;
He, X., Saint-Jeannet, J.P., Woodgett, J.R., Varmus, H.E. and Dawid, I.B.
(1995). Glycogen synthase kinase-3 and dorsoventral patterning in
Xenopus embryos. Nature 374, 617-622;
Hemmings BA, Yellowlees D, Kernohan JC, Cohen P. (1981). Purification of
glycogen synthase kinase 3 from rabbit skeletal muscle. Copurification
with the activating factor (FA) of the (Mg-ATP) dependent protein
phosphatase. Eur J Biochem. 119, 443–451;

62
Hilioti Z, Gallagher DA, Low-Nam ST, Ramaswamy P, Gajer P, Kingsbury TJ,
Birchwood CJ, Levchenko A, Cunningham KW. (2004). GSK-3 kinases
enhance calcineurin signaling by phosphorylation of RCNs. Genes Dev.
18, 35-47;
Holman, P.J., Ronald, N.C. (1980). A new tick cell line derived from Boophilus
microplus. Res. Vet. Sci. 29, 383–387;
Hopkins, TJ; Woodley, IR and Blackwell R. (1985). The safety and efficacy of
flumethrin pour-on used to control Boophilus microplus on cattle in
Australia. Vet. Med. Rev. 2, 112-125;
Ikeda, S., Kishida, S., Yamamoto H., Murai, H., Koyama S. and Kikuchi, A.
(1998). Axin, a negative regulator of the Wnt signaling pathway, forms a
complex with GSK-3b and b-catenin and promotes GSK-3b-dependent
phosphorylation of b-catenin. The EMBO Journal 17, 1371–1384;
Jope R. S. and Gail V.W. Johnson (2004). The glamour and gloom of glycogen
synthase kinase-3, Trends in Biochemical Sciences 29, 95 – 102;
Karim S, Miller NJ, Valenzuela J, Sauer JR, Mather TN. (2005). RNAi-mediated
gene silencing to assess the role of synaptobrevin and cystatin in tick
blood feeding. Biochem Biophys Res Commun. 334, 1336-1342;
Karim S, Ramakrishnan VG, Tucker JS, Essenberg RC, Sauer JR. (2004a).
Amblyomma americanum salivary glands: Double-stranded RNA-mediated
gene silencing of synaptobrevin homologue and inhibition of PGE2
stimulated protein secretion. Insect Biochem Mol Biol 34, 407–413;
Karim S, Ramakrishnan VG, Tucker JS, Essenberg RC, Sauer JR. (2004b)
Amblyomma americanum salivary gland homolog of nSec1 is essential for
saliva protein secretion. Biochem Biophys Res Commun 324,1256– 1263;
Keirans, J.E. e Robbins, R.G. (1999). A word checklist of genera, subgenera,
and species o f ticks (Acari: Ixodida) published from 1973-1997. J Vector
Ecol. 24, 115-129;
Keirans, J.E. (1992). Systematics of the Ixodida ( Argasidae, Ixodidae,
Nuttalliellidae): An overviewand some problems. In Tick Vector Biology.
Medical and Veterinary Aspects 1-24;

63
King T. D., Gandy J. C., Bijur G. N. (2006). The protein phosphatase-1/inhibitor-
2 complex differentially regulates GSK3 dephosphorylation and
increases sarcoplasmic/endoplasmic reticulum calcium ATPase 2 levels.
Experimental cell research 312, 3693 – 3700;
Kocan K.M., Manzano-Roman R. and de la Fuente J. (2007). Transovarial
silencing of the subolesin gene in three-host ixodid tick species after
injection of replete females with subolesin dsRNA. Parasitology
Research 100, 1411–1415;
Kocan, K., Zivkovic Z., Blouin E. F., Naranjo V., Almazán C, Mitra R. and de la
Fuente J. (2009). Silencing of genes involved in Anaplasma marginale-
tick interactions affects the pathogen developmental cycle in
Dermacentor variabilis. BMC Developmental Biology 9, 42;
Kurscheid S., Lew-Tabor A. E., Valle M. R., Bruyeres A. G., Doogan V. J.,
Munderloh U. G., Guerrero F. D., Barrero R. A. and Bellgard M. I. (2009).
Evidence of a tick RNAi pathway by comparative genomics and reverse
genetics screen of targets with known loss-of-function phenotypes in
Drosophila. BMC Molecular Biology 26, 1-21;
Lesort M, Jope RS, Johnson GV. (1999). Insulin transiently increases tau
phosphorylation: involvement of glycogen synthase kinase-3beta and
Fyn tyrosine kinase. J Neurochem. 72, 576–584;
Logullo, C., Vaz jr.I.S.,Sorgine, M.H.F.,Paiva-Silva, G.O., Faria, f.s., Zingali, R.,
Lima, ,.F.R., Abreu, l., Oliveira, E.F., Alves, E.W., Masuda, H., Gonzales,
J.C., Masuda,A e Oliveira, P.L. (1998). Isolation of a aspartic proteiase
precursor from the egg of a hard tick, Boophilus microplus. Parasitol.
116, 525-532;
Logullo, C., Witola, W.H., Andrade, C., Abreu, L., Gomes, J., Vaz Jr., I.S.,
Imamura, S., Konnai, S., Ohashi, K. and Onuma, M. (2009). Expression
and activity of glycogen synthase kinase during vitellogenesis and
embryogenesis of Rhipicephalus (Boophilus) microplus. Veterinary
Parasitology 161, 261–269;
McKosker, P.L. (1981). The global importance of Babesiosis. In Babesiosis.
Ristic, M. & J.P. Kreier, Academic Press. New York;

64
Meares GP, Zmijewska AA, Jope RS. 2008. HSP105 interacts with GRP78 and
GSK3 and promotes ER stress-induced caspase-3 activation. Cell
Signal. 20: 347-58
Mizunuma M, Hirata D, Miyaoka R, Miyakawa T., (2001). GSK-3 kinase Mck1
and calcineurin coordinately mediate Hsl1 down-regulation by Ca2+ in
budding yeast. EMBO J. 20, 1074-85;
Nari, A. (1995). Strategies for the control of one-host ticks and relationship with
tick-borne diseases in South America. Vet. Parasitol., 57, 153-165;
Nijhof AM, Taoufik A, de la Fuente J, Kocan KM, de Vries E, Jongejan F.
(2007). Gene silencing of the tick protective antigens, Bm86, Bm91 and
subolesin, in the one-host tick Boophilus microplus by RNA interference.
Int J Parasitol 37, 653-662;
Nolan, J. (1995). Mechanism of resistance to chemical in arthropods parasites
of veterinary importance. Vet. Parasitol. 18, 155-166;
Nuñez, J.L., Muñoz-Cobeñas, M.E., Moltedo, H.L (1985). Boophilus microplus:
The Common Cattle Tick. Springer, Berlin;
Pfaffl MW, (2001). A new mathematical model for relative quantification in real-
time RT-PCR, Nucl. Acids Res.,
Price B.D., Mannheim-Rodman,L.A. and Calderwood,S.K. (1992) Brefeldin A,
thapsigargin and AIF4 stimulate the accumulation of GRP78 mRNA in a
cycloheximide dependent manner, whilst induction by hypoxia is
independent of protein synthesis. J. Cell. Physiol., 152, 545–552;
Putney J.W. (1990) Capacitative calcium entry revisited. Cell Calcium, 11, 611–
624;
Sasaki, Feng Han, Norifumi Shioda, Shigeki Moriguchi, Jiro Kasahara, Koichi
Ishiguro, Kohji Fukunaga (2006). Lithium-induced activation of Akt and
CaM kinase II contributes to its neuroprotective action in a rat microsphere
embolism model. Brain Research
Siegfried, E., Chou, T. and Perrimon, N. (1992). Wingless signaling acts
through zeste-white 3, the Drosophila homolog of glycogen synthase

65
kinase-3, to regulate engrailed and establish cell fate. Cell 71, 1167-
1179;
Solomon, K.R. (1983). Acaricide resistance in ticks. Adv. Vet. Sci. Comp. Med.
27, 341-349;
Song L, De Sarno P, Jope RS (2002). Central role of glycogen synthase kinase-
3beta in endoplasmic reticulum stress-induced caspase-3 activation. J
Biol Chem 277, 44701–44708;
Rodriguez, M.; Penichet, M.L.; Mouris, A.E.; Labarta, V.; Lorenzo Luarces, L;
Rubiera, R.; Cordovés, C.; Sánchez, P.A.; Ramos, E.; Soto, A.; Canales,
M.; Palenzuela, D.; Triguero, A.; Lleonard, R.; Herrera, L. & De la
Fuente, J (1995a). Control of Boophilus microplus population in grazing
cattle vaccinated with a recombinant Bm86 antigen preparation. Vet.
Parasitol. 57, 339-349;
Rodriguez, M; Massard, C.L.; Fonseca, A.H.; Ramos, N.F.; Machado, H.;
Labarta, V. & De la Fuente, J. (1995b). Effect of vaccination with a
recombinant Bm86 antigen preparation on natural infestations of
Boophilus microplus in grazing dairy and beef pure and cross-bred cattle
in Brazil. Vaccine 13, 1804-1808;
Takadera T., Yoshikawa R., Ohyashiki T. (2006). Thapsigargin-induced
apoptosis was prevented by glycogen synthase kinase-3 inhibitors in
PC12 cells. Neuroscience Letters 408 , 124–128;
Vaz Jr., Gonzáles, J. C., Masuda, H., Oliveira, P. L. and Masuda, A. (1998).
Immunization of bovines with an aspartic proteinase precursor isolated
from Boophilus microplus eggs. Vet. Immunol. And Immunopathology
66, 331-341;
Xavier, I. J., Mercier, P. A., McLoughlin, C. M., Ali, A., Woodgett, J. R.,
Ovsenek, N. (2000). Glycogen Synthase Kinase 3beta Negatively
Regulates Both DNA-binding and Transcriptional Activities of Heat Shock
Factor 1. J. Biol. Chem. 275, 29147-29152;
Wang QM, Fiol CJ, DePaoli-Roach AA, Roach PJ. (1994). Glycogen synthase
kinase-3 beta is a dual specificity kinase differentially regulated by
tyrosine and serine/threonine phosphorylation. J Biol Chem. 269, 14566–
14574;

66
Willadsen, P.; Bird, P.; Cobon, G.S. & Hungerford, J. (1995) Commercialization
of a recombinant vaccine against Boophilus microplus. Parasitology, 110,
843-850;
Wigand B., Bucher G. and Klingler M. (1998). A simple whole mount technique
for looking at Tribolium embryos. Electronic reprint from Technical Note
in TIB 38, 281-283;
Wodarz (1998). Mechanisms of Wnt Signaling in Development. Annu. Rev.
Cell Dev. Biol. 14, 59–88.