eliseu marlÔnio pereira de lucena · cv. tommy atkins, da antese até a colheita comercial,...
TRANSCRIPT

ELISEU MARLÔNIO PEREIRA DE LUCENA
DESENVOLVIMENTO E MATURIDADE FISIOLÓGICA DE MANGA ‘TOMMY
ATKINS’ NO VALE DO SÃO FRANCISCO
Tese submetida à Coordenação do Curso de Pós-Graduação em Agronomia, área de Concentração em Fitotecnia, da Universidade Federal do Ceará, como requisito parcial para obtenção do grau de Doutor em Agronomia.
Orientador: Prof. Dr. Joaquim Enéas Filho
FORTALEZA
2006

2
Ficha catalográfica elaborada pela Bibliotecária Ana Cristina A. Ursulino Melo CRB-3/572
L968d Lucena, Eliseu Marlônio Pereira de Desenvolvimento e maturidade fisiológica de manga ‘Tommy
Atkins’ no vale do São Francisco / Eliseu Marlônio Pereira de Lucena. - Fortaleza, 2006. 152 f. il., color. enc.
Orientador: Prof. Dr. Joaquim Enéas Filho Co-Orientador: Dr. Ricardo Elesbão Alves Tese (Doutorado em Agronomia) - Universidade Federal do Ceará, Fortaleza, 2006.
1. Mangifera indica 2. Fisiologia pós-colheita 3. Índice de maturidade 4. Graus-dia I. Enéas Filho, Joaquim (orient.) II. Alves, Ricardo Elesbão (co-orient.) III. Título

3
ELISEU MARLÔNIO PEREIRA DE LUCENA
DESENVOLVIMENTO E MATURIDADE FISIOLÓGICA DE MANGA ‘TOMMY
ATKINS’ NO VALE DO SÃO FRANCISCO
Tese submetida à Coordenação do Curso de Pós-Graduação em Agronomia, área de Concentração em Fitotecnia, da Universidade Federal do Ceará, como requisito parcial para obtenção do grau de Doutor em Agronomia.
Aprovada em 16 de Novembro de 2006.
BANCA EXAMINADORA
__________________________________________________
Prof. Dr. Joaquim Enéas Filho (Orientador)
Universidade Federal do Ceará – UFC
__________________________________________________
Pesq. Dr. Ricardo Elesbão Alves (Co-Orientador)
Embrapa Agroindústria Tropical
__________________________________________________
Pesq. Dr. Joston Simão de Assis
Embrapa Semi-Árido
__________________________________________________
Profª. Dra. Maria Raquel Alcântara de Miranda
Universidade Federal do Ceará – UFC
__________________________________________________
Prof. Dr. Raimundo Wilane de Figueiredo
Universidade Federal do Ceará – UFC

4
A minha mãe,
Antônia (in memoriam),
pelo carinho, compreensão e paciência
nos meus primeiros ensinamentos.

5
AGRADECIMENTOS
À Universidade Federal do Ceará – UFC, em particular, ao Departamento de
Fitotecnia, pela oportunidade concedida para a realização do curso.
À Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico –
FUNCAP, pela concessão da bolsa de estudos durante a realização do curso.
Ao Instituto Centro de Ensino Tecnológico – CENTEC, pela liberação para a
realização do curso.
À Empresa Fruit Fort Agrícola Exportação, por nos ter disponibilizado o seu plantio
de manga para a realização desta pesquisa.
À Embrapa – Semi-Árido, pelo acesso ao laboratório de Fisiologia Pós-Colheita para a
realização das análises preliminares durante a fase de campo e à Embrapa – Agroindústria
Tropical, pela concessão do seu laboratório de Fisiologia e Tecnologia Pós-Colheita para as
análises após a fase de campo.
Ao Professor Joaquim Enéas Filho, pelo apoio, amizade, orientação e confiança
demonstrados.
Ao Pesquisador Joston Simão de Assis, pela eficaz colaboração na condução do
experimento de campo.
Ao Pesquisador Ricardo Elesbão Alves, pela presteza, disponibilidade e apoio durante
as análises laboratoriais deste trabalho.
Ao Pesquisador Lindbergue Araújo Crisóstomo, pela ajuda técnica e gentileza em
ceder o laboratório de Análises de Solos, Água e Plantas, para execução das análises de
nitrogênio.
Ao Professor Raimundo Wilane de Figueiredo pelo apoio e incentivos sempre
presentes durante o curso.
À Professora Maria Raquel Alcântara de Miranda, pela amizade e solidariedade.
Aos Professores do Curso de Pós-Graduação, pela amizade e pelos ensinamentos.
Aos Coordenadores do Curso de Ciências Biológicas da Universidade Estadual do
Ceará – UECE, Professores Germana Costa Paixão e Valberto Barbosa Porto, pela
compreensão e otimização de períodos na distribuição de carga horária.
Ao Diretor Presidente do Instituto Centro de Ensino Tecnológico – CENTEC,
Professor Antônio Amaury Oriá Fernandes, pela atenção, amizade, incentivo e convívio
durante minha permanência no CENTEC Cariri.

6
Ao amigo Diretor Regional do CENTEC Cariri, Cícero de Alencar Leite, pela
colaboração, amizade e excelente convívio durante minha permanência no CENTEC.
Ao Coordenador de Tecnologia de Alimentos do CENTEC Cariri, Antenor Silva
Júnior, pela amizade e apoio incontestável durante a fase inicial desta pesquisa.
Ao Professor do CEFET Petrolina, João Tercio, pela amizade e acolhida no decorrer
do trabalho em campo.
Ao meu ex-aluno e hoje Professor do CENTEC Cariri, Jonas dos Santos Sousa, pela
ajuda na fase preliminar deste experimento.
Ao Gerente de Produção da Fruit Fort e amigo Voltaire Diaz Medina, pela presteza
durante a execução do ensaio em campo.
Ao Sócio Gerente da Projetar Irrigação e companheiro Francisco Fernandes da Costa,
pelo apoio durante a fase preliminar do experimento em campo.
Ao meu colega e hoje pesquisador da Embrapa Agroindústria-Tropical, Carlos Farley
Herbster Moura, pela seriedade e ajuda na realização deste ensaio.
Ao Assistente de Pesquisa do Laboratório de Fisiologia e Pós-Colheita da Embrapa –
Semi-Árido, Joveniano, pela participação nas análises preliminares.
Ao Estagiário da Embrapa – Semi-Árido, Víctor César Macedo da Silva, pela ajuda,
amizade e responsabilidade na condução da pesquisa de campo e análises preliminares.
À Assistente de Pesquisa do Laboratório de Fisiologia e Tecnologia Pós-Colheita da
Embrapa - Agroindústria Tropical, Márcia Régia, pela amizade, atenção e dedicação na
condução dos trabalhos de laboratório.
À Alaís, Edalides, Eliardo, Marcela e Melissa, pela grande ajuda na realização das
análises.
Ao Mestre em Tecnologia de Alimentos, Paolo Germanno, pela amizade e auxilio na
elaboração das planilhas eletrônicas.
Aos companheiros do Laboratório de Fisiologia e Tecnologia Pós-Colheita da
Embrapa - Agroindústria Tropical, pelo maravilhoso convívio durante as análises.
Aos pesquisadores da Embrapa – Agroindústria Tropical, Ebenézer de Oliveira Silva,
Fernando Antônio Souza de Aragão, Heloisa Almeida Cunha Filgueiras e José Luiz Mosca,
pela amizade, apoio e admirável convívio.
Aos funcionários e prestadores de serviço da Embrapa – Agroindústria Tropical, em
especial, D. Maria, pelo entusiasmo e facilidades oferecidas.
Aos funcionários da Fruit Fort: Hélio, Ivanildo, Ivete, Jailson e Wagner, pela atenção e
pela dedicação na condução dos serviços prestados.

7
Aos funcionários do Departamento de Fitotecnia, em especial, Deocleciano Xavier,
secretário do Curso de Pós-Graduação em Agronomia/Fitotecnia da UFC, pela colaboração
durante o curso.
Aos colegas do Curso de Pós-Graduação, pelo incentivo, pela colaboração e pela
solidariedade no decorrer do curso, em especial, Enéas, Fabrício, Gislainy, Orlando e Xavier.
Ao Sansão, pela amizade e eficiente editoração das fotografias.
Aos meus irmãos Macilio e Mardônio, pela incansável, incontestável e sadia
participação nas profícuas discussões técnico-científicas em várias etapas do trabalho.
À Paula, pela atenção, apoio, incentivo e dedicação durante a fase final deste ensaio.
A todos enfim, que de alguma forma contribuíram para a realização desta pesquisa.

8
RESUMO
O presente trabalho objetivou caracterizar as alterações físicas, físico-químicas,
químicas e bioquímicas durante o crescimento dos frutos de mangueira (Mangifera indica),
cv. Tommy Atkins, da antese até a colheita comercial, visando à definição do ponto de
colheita ideal em unidades de calor. Os frutos foram colhidos aos 35, 49, 63, 70, 77, 84, 91,
98, 105 e 112 dias após a antese (DAA), sendo feitas as seguintes determinações: aspectos
morfológicos externos; diâmetros longitudinal, ventral e transversal; produto dos diâmetros;
massas fresca, seca e de água; teor de água; escalas de coloração da casca, de Blush para
coloração da casca e de coloração da polpa; luminosidade, croma e ângulo Hue da polpa;
firmeza; unidades de calor (UC); sólidos solúveis totais (SST); acidez total titulável (ATT);
pH; relação SST/ATT; amido; açúcares solúveis totais, redutores e não redutores; nitrogênio
total, não protéico e protéico; proteínas bruta e verdadeira; vitamina C; clorofila e
carotenóides totais; fenólicos poliméricos, oligoméricos e diméricos; pectinas total, solúvel,
de alta metoxilação e de baixa metoxilação; protopectina; percentagem de solubilização de
pectina; atividade das enzimas pectinametilesterase, poligalacturonase, polifenoloxidase,
peroxidase, amilase total, α- e β-amilases, α- e β-galactosidases extraídas de citosol e de
parede celular; proteínas extraídas de citosol e de parede celular. O trabalho indicou que as
mangas ‘Tommy Atkins’ atingiram a maturidade fisiológica aos 98 DAA, que equivale a
1.685,09 UC. O croma da polpa foi o melhor indicador do estádio de desenvolvimento do
fruto da mangueira cultivada sob irrigação no sub-médio São Francisco, considerando-se o
seu alto coeficiente de determinação, R2=0,9832 (P < 0,01) e seu alto coeficiente de
correlação com pH, açúcares solúveis totais e carotenóides totais, R=0,95; 0,93; e 0,93,
respectivamente (P < 0,01).
Palavras-chave: Mangifera indica, fisiologia pós-colheita, índice de maturidade, graus-dia.

9
ABSTRACT
The objective of this work was to characterize the physical, physicochemical,
chemical and biochemical changes during the development of mango (Mangifera indica), cv.
Tommy Atkins from anthesis to harvest, identifying the optimum harvest maturity stage in
heat units. The fruits were harvested at 35, 49, 63, 70, 77, 84, 91, 98, 105 and 112 days after
the anthesis (DAA), being made the following determinations: fruit external morphology;
longitudinal, ventral e transverse diameters; product of the diameters; fresh and dry mass;
water content and percentage; skin, Blush skin and pulp color scales; pulp luminosity, Hue
angle and chroma; firmness; heat units (HU); total soluble solids (TSS); total titratable acidity
(TTA); pH; TSS/TTA ratio; starch; total, reducing and nonreducing soluble sugars; total,
protein and nonprotein nitrogen; crude and true protein; C vitamin; total chlorophyll and
carotenoids; polymeric, oligomeric and dimeric phenolics; pectin total, soluble and high/low
metoxilation degree; protopectin; solubilization pectin percentage; pectin metyhylesterase,
poligalacturonase, polyphenol oxidase, peroxidase, total amylase, α- and β-amylases, α- and
β-galactosidases enzymes activities; cell wall and cytosol proteins. This work has indicated
that mango ‘Tommy Atkins’ reached the physiological maturity at 98 DAA, that is equivalent
to 1.685,09 HU. The pulp chroma was the best fruit development stage indicator in the
cultivated conditions of this study (São Francisco valley), taking into account the high
coefficient of determination, R2=0,9832 (P < 0,01) and excellent correlation coefficients with
pH, total soluble sugars and total carotenoids, R=0,95; 0,93; e 0,93, respectively (P < 0,01).
Keywords: Mangifera indica, postharvest physiology, maturity indices, degree-days.

10
LISTA DE FIGURAS
Figura Página
1 Temperaturas e umidade relativa do ar na região de Petrolina – PE
durante o ano que antecedeu as colheitas (outubro a dezembro/04) das
mangas ‘Tommy Atkins’............................................................................ 59
2 Radiação solar global e precipitação pluviométrica na região de
Petrolina – PE durante o ano que antecedeu as colheitas (outubro a
dezembro/04) das mangas ‘Tommy Atkins’.............................................. 59
3 Panículas padrões 1 e 2 em mangueira ‘Tommy Atkins’........................... 62
4 Início da diferenciação floral em ramo de mangueira ‘Tommy Atkins’.... 63
5 Formatos do ombro, do nariz e do ápice de mangas ‘Tommy Atkins’ em
desenvolvimento fisiológico e fisiologicamente desenvolvida.................. 64
6 Ilustração dos diâmetros longitudinal, ventral e transversal de mangas
‘Tommy Atkins’......................................................................................... 65
7 Escalas de coloração da casca e da polpa como guia de maturação das
mangas ‘Tommy Atkins’............................................................................ 67
8 Frutos de mangueira ‘Tommy Atkins’ nos estádios chumbinho, bola de
gude, ovo e fruto......................................................................................... 82
9 Diâmetros longitudinal, ventral e transversal de mangas ‘Tommy
Atkins’ durante o desenvolvimento............................................................ 84
10 Produto dos diâmetros de mangas ‘Tommy Atkins’ durante o
desenvolvimento......................................................................................... 84
11 Massas fresca, seca, de água e o teor de água de mangas ‘Tommy
Atkins’ durante o desenvolvimento............................................................ 85
12 Escalas de coloração da casca, de Blush para coloração da casca e de
coloração da polpa de mangas ‘Tommy Atkins’ durante o
desenvolvimento......................................................................................... 87
13 Luminosidade, croma e ângulo Hue da polpa de mangas ‘Tommy
Atkins’ durante o desenvolvimento............................................................ 88
14 Sólidos solúveis totais de mangas ‘Tommy Atkins’ durante o
desenvolvimento......................................................................................... 92

11
15 Acidez total titulável e pH de mangas ‘Tommy Atkins’ durante o
desenvolvimento......................................................................................... 93
16 Relação SST/ATT de mangas ‘Tommy Atkins’ durante o
desenvolvimento......................................................................................... 94
17 Amido, açúcares solúveis totais, redutores e não redutores de mangas
‘Tommy Atkins’ durante o desenvolvimento............................................. 95
18 Nitrogênio total, não protéico e protéico de mangas ‘Tommy Atkins’
durante o desenvolvimento......................................................................... 98
19 Proteína bruta e verdadeira de mangas ‘Tommy Atkins’ durante o
desenvolvimento......................................................................................... 99
20 Vitamina C de mangas ‘Tommy Atkins’ durante o desenvolvimento....... 100
21 Clorofila total da casca de mangas ‘Tommy Atkins’ durante o
desenvolvimento......................................................................................... 101
22 Carotenóides totais de mangas ‘Tommy Atkins’ durante o
desenvolvimento......................................................................................... 102
23 Fenólicos poliméricos, oligoméricos e diméricos de mangas ‘Tommy
Atkins’ durante o desenvolvimento............................................................ 103
24 Pectina total, solúvel e percentagem de solubilização de pectina de
mangas ‘Tommy Atkins’ durante o desenvolvimento............................... 105
25 Pectinas de alta metoxilação, de baixa metoxilação e a protopectina de
mangas ‘Tommy Atkins’ durante o desenvolvimento............................... 106
26 Atividades da pectinametilesterase e poligalacturonase de mangas
‘Tommy Atkins’ durante o desenvolvimento............................................. 107
27 Atividades da polifenoloxidase e peroxidase de mangas ‘Tommy
Atkins’ durante o desenvolvimento............................................................ 109
28 Atividades amilásica total, α-amilásica e β-amilásica de mangas
‘Tommy Atkins’ durante o desenvolvimento............................................. 111
29 Atividades α-galactosidásicas de citosol e de parede celular de mangas
‘Tommy Atkins’ durante o desenvolvimento............................................. 112
30 Atividades β-galactosidásicas de citosol e de parede celular de mangas
‘Tommy Atkins’ durante o desenvolvimento............................................. 113
31 Proteína de citosol e de parede celular de mangas ‘Tommy Atkins’
durante o desenvolvimento......................................................................... 116

12
LISTA DE TABELAS
Tabela Página
1 Análises de fertilidade do solo da rizosfera e de tecido foliar de mangueiras
cv. Tommy Atkins, no inicio do ciclo da cultura em Novembro de 2003....... 60
2 Características das panículas padrões 1 e 2 em mangueira, cv. Tommy
Atkins............................................................................................................... 62
3 Distribuições absoluta e percentual dos frutos por panícula aos 56 DAA em
mangueira, cv. Tommy Atkins......................................................................... 80
4 Valores médios e percentuais da fenologia da floração até o início da
frutificação em mangueira, cv. Tommy Atkins............................................... 81
5 Aspectos morfológicos externos de mangas ‘Tommy Atkins’ durante o
desenvolvimento.............................................................................................. 83
6 Valores médios da firmeza da polpa, em mangas da cv. Tommy Atkins, aos
112 DAA.......................................................................................................... 89
7 Valores de unidades de calor a partir da diferenciação do ramo até a antese
e da antese até o início da frutificação em mangueira, cv. Tommy Atkins..... 90
8 Valores de unidades de calor a partir da antese até o fim da frutificação em
mangueira, cv Tommy Atkins.......................................................................... 91
9 Matriz de correlação entre sólidos solúveis totais e características físicas de
mangas ‘Tommy Atkins’ durante o desenvolvimento..................................... 118
10 Matriz de correlação entre massa seca, colorações da casca e da polpa,
sólidos solúveis totais, acidez, pH e relação SST/ATT de mangas ‘Tommy
Atkins’ durante o desenvolvimento................................................................. 120
11 Matriz de correlação entre croma da polpa, nitrogênio e proteína de mangas
‘Tommy Atkins’ durante o desenvolvimento.................................................. 121
12 Matriz de correlação entre croma da polpa, sólidos solúveis totais,
carboidratos e amilases de mangas ‘Tommy Atkins’ durante o
desenvolvimento.............................................................................................. 122
13 Matriz de correlação entre croma da polpa, vitamina C, clorofila,
carotenóides, fenólicos e oxidases de mangas ‘Tommy Atkins’ durante o
desenvolvimento.............................................................................................. 124

13
14 Matriz de correlação entre croma da polpa, pectinas, pectinases,
galactosidases e proteína de mangas ‘Tommy Atkins’ durante o
desenvolvimento.............................................................................................. 126

14
SUMÁRIO
Página
1 INTRODUÇÃO.................................................................................................. 18
2 REVISÃO DE LITERATURA.......................................................................... 20
2.1 Aspectos gerais da mangueira………………………………………………... 20
2.2 Indução floral………………………………………………………………… 22
2.3 Fenologia da floração à frutificação.................................................................. 24
2.4 Ponto de colheita……………………………………………………………... 27
2.5 Avaliações físicas…………………………………………………………….. 28
2.5.1 Aspectos morfológicos externos…………………………………………… 28
2.5.2 Diâmetros longitudinal, ventral, transversal e produto dos diâmetros........... 29
2.5.3 Massas fresca, seca, de água e o teor de água................................................ 30
2.5.4 Coloração da casca e da polpa........................................................................ 32
2.5.5 Firmeza……………………………………………………………………... 32
2.5.6 Unidades de calor (graus-dia)....................................................................... 33
2.6 Avaliações físico-químicas e químicas............................................................. 36
2.6.1 Sólidos solúveis totais.................................................................................... 36
2.6.2 Acidez total titulável e pH.............................................................................. 36
2.6.3 Relação SST/ATT………………………………………………………….. 37
2.6.4 Carboidratos…………………………………………………………........... 38
2.6.5 Nitrogênio total, não protéico e protéico........................................................ 40
2.6.6 Vitamina C…………………………………………………………………. 42
2.6.7 Clorofila total………………………………………………………………. 43
2.6.8 Carotenóides totais…………………………………………………………. 45
2.6.9 Fenólicos………………………………………………………………….... 47
2.6.10 Pectinas……………………………………………………………………. 49
2.7 Avaliações bioquímicas………………………………………………………. 52
2.7.1 Pectinametilesterase e poligalacturonase…………………………………... 52
2.7.2 Polifenoloxidase e peroxidase……………………………………………… 53
2.7.3 α- e β-Amilases……………………………………………………….......... 55
2.7.4 α- e β-Galactosidases de citosol e de parede celular...................................... 56

15
2.7.5 Proteínas de citosol e de parede celular.......................................................... 56
3 MATERIAL E MÉTODOS............................................................................... 58
3.1 Localização, caracterização edáfica, climática e manejo da área de cultivo..... 58
3.2 Indução floral………………………………………………………………… 61
3.3 Marcação das panículas………………………………………………………. 61
3.4 Fenologia da floração à frutificação.................................................................. 62
3.5 Colheita e manuseio das mangas…………………………………………....... 63
3.6 Avaliações físicas…………………………………………………………...... 64
3.6.1 Aspectos morfológicos externos………………………………………….... 64
3.6.2 Diâmetros longitudinal, ventral, transversal e produto dos diâmetros........... 65
3.6.3 Massas fresca, seca, de água e o teor de água................................................ 65
3.6.4 Escalas de coloração da casca e da polpa....................................................... 66
3.6.5 Coloração da polpa......................................................................................... 67
3.6.6 Firmeza…………………………………………………………………....... 68
3.6.7 Unidades de calor (graus-dia)......................................................................... 68
3.7 Avaliações físico-químicas e químicas............................................................. 69
3.7.1 Sólidos solúveis totais, acidez total titulável, pH e relação SST/ATT........... 69
3.7.2 Determinação dos teores de carboidratos....................................................... 69
3.7.3 Determinação dos teores de nitrogênio total, não protéico e protéico........... 70
3.7.4 Proteína bruta e verdadeira.....................…………………………………… 71
3.7.5 Vitamina C........…………………………………………………………..... 71
3.7.6 Clorofila total……….....…………………………………………………… 72
3.7.7 Carotenóides totais………………………….....…………………………… 72
3.7.8 Fenólicos………….....……………………………………………………... 73
3.7.9 Pectinas…….....…………………………………………………………….. 73
3.8 Avaliações bioquímicas……………...……………………………………….. 74
3.8.1 Atividade da pectinametilesterase……………………………...................... 74
3.8.2 Atividade da poligalacturonase…………...………………………………... 75
3.8.3 Atividade da polifenoloxidase……….……………………………………... 75
3.8.4 Atividade da peroxidase….……………………………………………….... 76
3.8.5 Atividades amilásica total, α- e β-amilásicas................................................ 76
3.8.6 Atividades α- e β-galactosidásicas................................................................ 77
3.8.7 Teor de proteína…………………………………………………………….. 78

16
3.9 Delineamento experimental e análise estatística............................................... 78
4 RESULTADOS E DISCUSSÃO....................................................................... 79
4.1 Indução floral……………………………………………………………….... 79
4.2 Marcação das panículas…………………………………………………...….. 79
4.3 Fenologia da floração à frutificação.................................................................. 80
4.4 Avaliações físicas…...………………………………………………………... 82
4.4.1 Aspectos morfológicos externos………………………………………….... 82
4.4.2 Diâmetros longitudinal, ventral, transversal e produto dos diâmetros........... 83
4.4.3 Massas fresca, seca, de água e o teor de água................................................ 85
4.4.4 Escalas de coloração da casca e da polpa....................................................... 86
4.4.5 Coloração da polpa………...……………………………………………….. 87
4.4.6 Firmeza……………………………………………………………………... 89
4.4.7 Unidades de calor (graus-dia)......................................................................... 90
4.5 Avaliações físico-químicas e químicas............................................................. 91
4.5.1 Sólidos solúveis totais……………………………………………................ 91
4.5.2 Acidez total titulável e pH.............................................................................. 92
4.5.3 Relação SST/ATT………………………………………………………...... 94
4.5.4 Determinação de carboidratos……….........................................…………... 95
4.5.5 Nitrogênio…………………………………………………………………... 97
4.5.6 Proteína bruta e verdadeira............................................................................. 98
4.5.7 Vitamina C........…………………………………………………………..... 99
4.5.8 Clorofila total……………………………………………………………..... 101
4.5.9 Carotenóides totais………………………………………………………..... 102
4.5.10 Fenólicos………………………………………………………………...... 103
4.5.11 Pectinas……………………………………………………………………. 104
4.6 Avaliações bioquímicas……………………………………………………..... 107
4.6.1 Pectinametilesterase e poligalacturonase....................................................... 107
4.6.2 Polifenoloxidase e peroxidase….................................................................... 109
4.6.3 Amilases……………………………………………………………………. 110
4.6.4 Galactosidases…………………………………………………………….... 112
4.6.5 Proteína……………………………………………………………………... 115
4.7 Correlação entre as variáveis estudadas............................................................ 117
4.7.1 Correlação entre sólidos solúveis totais e as variáveis físicas estudadas....... 117

17
4.7.2 Correlação entre as variáveis físicas, físico-químicas e químicas.................. 119
4.7.3 Correlação entre as variáveis físicas, físico-químicas, químicas e
bioquímicas................................................................................................. 121
5 CONCLUSÕES……………………………………………………………….. 128
6 REFERÊNCIAS BIBLIOGRÁFICAS............................................................. 129
APÊNDICE............................................................................................................ 147

18
1 INTRODUÇÃO
O interesse pela fruticultura tem aumentado, nos últimos anos, no Vale do São
Francisco, devido a crescente comercialização exportadora. Entretanto, o desenvolvimento
das técnicas práticas para a determinação do ponto de colheita, não acompanhou o ritmo de
produção.
Quando os frutos são colhidos precocemente, ao serem transportados até o local de
consumo, chegam ainda no estádio verde e muitas vezes murcham e não conseguem atingir a
qualidade fisiológica necessária para o consumo in natura, por outro lado, quando colhidos
tardiamente, sofrem danos mecânicos durante o transporte e chegam ao local de consumo
passados, isto é, já em senescência. Deste modo, ambas as situações resultam em perdas pós-
colheita, as quais refletem em prejuízo para os produtores e exportadores, além de ressaltar
que o erro na determinação do ponto de colheita é o pior cartão de visita deste exportador.
Na determinação do ponto de colheita, pode-se utilizar índices físicos, químicos e
bioquímicos.
Os índices físicos, geralmente são não destrutivos e baseiam-se nas características
físicas do produto ou do ambiente, tais como: morfologia externa, massa, volume, densidade,
tamanho, forma, cor, firmeza, calendário anual e unidades de calor.
Os métodos químicos baseiam-se na composição química do produto, a qual,
dependendo do componente, pode diminuir ou aumentar com o avanço da maturação do fruto.
Já os bioquímicos, fundamentam-se na atividade das enzimas durante a maturação dos
frutos.
Por outro lado, utiliza-se muito da correlação das características físicas, químicas e
bioquímicas, para indicar o número de dias a partir da antese até a colheita, no entanto,
quando o clima é quente, a maturação ocorre mais rapidamente e quando é frio, ocorre mais
lentamente, portanto, quando se determina o ponto de colheita apenas em número de dias, sem
avaliar a temperatura, pode-se estar incorrendo em erro.
Desta forma, sugere-se que a determinação do ponto de colheita seja fixada em dias-
graus centígrados, pois só assim, estará levando-se em conta a variação da temperatura do dia,
pois é determinada somando-se as diferenças entre temperatura média de cada dia e aquela
tida como base, ou mínima em que se registra crescimento da cultura.
Face a essas considerações a presente pesquisa teve como objetivos:

19
i) caracterizar os estádios iniciais de desenvolvimento da manga (Mangifera indica
L.), cv. Tommy Atkins, através das avaliações físicas, físico-químicas, químicas e
bioquímicas de 35 até 112 dias após a antese (DAA);
ii) determinar a maturidade fisiológica da manga cv. Tommy Atkins no vale do São
Francisco, Petrolina, PE;
iii) estimar em unidades de calor (UC) o grau de maturidade da manga ‘Tommy
Atkins’ através da relação com o número de dias após a antese (DAA) e com as
características físicas, físico-químicas, químicas e bioquímicas avaliadas.

20
2 REVISÃO DE LITERATURA
2.1 Aspectos gerais da mangueira
A mangueira (Mangifera indica L.) pertence à classe Dicotiledônea e à família
Anacardiaceae. O gênero Mangifera inclui cerca de 60 espécies, das quais a M. indica é a
mais importante, embora existam outras espécies que produzem frutos comestíveis, como M.
altíssima, M. caesia, M. lagenifera, M. macrocarpa, M. odorata e M. sylvatica. É uma árvore
frondosa, de porte médio a grande, com copa simétrica, de forma arredondada baixa a
piramidal alta, variando de baixa e densa a ereta e aberta, e com folhas sempre verdes. O
sistema radicular é caracterizado por uma raiz pivotante e por raízes de superfície, as quais
apresentam ramificações compostas por raízes finas e fibrosas (SANTOS-SEREJO, 2005).
Mukherjee (1985), seguindo a classificação proposta por Vavilov (1951), para os
centros de origem das plantas cultivadas, relata que a mangueira é originária do segundo
grande centro, o Indiano, e do subcentro Indo-Malaio. Essas regiões distintas deram origem às
duas raças de manga: a indiana, originária do centro Indo-Burma tailandês, que produz frutos
de boa aparência externa, cuja casca é bem colorida, variando de rosa a vermelho intenso,
com sementes monoembriônicas; e a filipina ou indo-chinesa, originária do centro Filipino
Celeste Timor, a qual produz fruto de formato alongado, com casca verde-amarelada quando
maduro, e sementes poliembriônicas.
A manga foi introduzida na América, provavelmente, pelos portugueses no Brasil, no
século XVI. Logo em seguida, foi introduzida no México pelos espanhóis. As primeiras
introduções no Brasil, no entanto, referiam-se às cultivares filipinas, que geralmente
produzem frutos com polpa fibrosa e de baixa qualidade e com semente poliembriônica, com
pequena variação genética. Isso fez com que a cultura da manga ficasse limitada a pequenos
pomares, sem muita expressão, e especificamente para atender ao mercado interno de maneira
bem regionalizada, por quase três séculos (FERREIRA et al., 2002).
Na metade do século XX, no entanto, foram realizadas introduções de cultivares
melhoradas indianas, procedentes da Flórida (USA), portadoras de melhor qualidade com
sementes monoembriônicas, que induzem grande variabilidade quando plantadas de pé
franco. Esse fato modificou sensivelmente a indústria mangícola nacional, dando um novo
alento à cultura, pois essas cultivares americanas, que produzem frutos com pouca fibra, bem

21
coloridos e mais resistentes à antracnose causada pelo fungo Colletotrichum gloeosporioides
são mais comercializáveis, permitindo inicialmente ampliar o excelente mercado interno, e
mais recentemente permitindo conquistar o mercado externo, notadamente dos Estados
Unidos e Japão. A cultivar Haden foi introduzida no Brasil em 1931, mas só a partir da
década de 60 foi plantada comercialmente, e apresenta uma série de limitações,
principalmente com relação à sua suscetibilidade à seca da mangueira e à alternância de
produção. Em 1970, foi introduzida a ‘Tommy Atkins’, junto com muitas outras cultivares
que foram testadas e algumas recomendadas para as condições brasileiras. Com o aumento da
demanda interna e o interesse crescente pelas exportações a partir de 1980, a ‘Tommy Atkins’
se mostrou bastante adequada, principalmente devido a sua maior tolerância à antracnose. A
partir disso, juntamente com a ‘Keitt’ tem sido as cultivares mais plantadas no País (PIZA
JÚNIOR, 1989; DONADIO, 1996).
A cv. Tommy Atkins foi originada nos anos 20 em Fort Lauderdale, Flórida, EUA,
como uma progênie da ‘Haden’, pois devido o alto grau de heterozigose, cada planta, sem a
interferência do homem, é considerada um híbrido F1. Árvore cheia, densa e precoce, com
frutos de tamanho médio a grande, 400 a 700 g, ovalados a oblongos, superfície lisa, com
casca espessa, cor laranja-amarelada coberta com vermelho e púrpura intensa. Polpa amarelo-
escura, firme, suculenta, teor médio de fibra. Semente monoembriônica, pequena, de 6 a 8%
da massa fresca do fruto. O fruto é resistente à antracnose e a danos mecânicos, além de
amadurecer bem se colhido imaturo, propiciando assim, uma maior vida útil pós-colheita.
Apresenta problemas de colapso interno do fruto, alta suscetibilidade ao oídio e malformação
floral. Possui teor de sólidos solúveis totais médio (17ºBrix) e sabor inferior quando
comparada com as cvs. Palmer e Haden. É uma das cultivares de manga mais cultivadas
mundialmente para exportação. Essa cultivar representa 50 e 80% das exportações de manga
no México e no Brasil, respectivamente (CUNHA et al. 1994; DONADIO, 2002; PINTO et
al., 2002b).
De acordo com Almeida et al. (2005), a mangicultura é uma das principais atividades
do agronegócio frutícola do Brasil, apresentando desempenho crescente nos últimos anos,
sendo o nono maior produtor mundial de manga, onde a Índia é o primeiro, e desde 1999, o
segundo maior exportador dessa fruta, perdendo apenas para o México. O Brasil vem
ampliando sua participação nas exportações mundiais e gerando empregos e renda em todo o
território nacional, especialmente no Nordeste, a região que mais exporta manga, destacando-
se os Estados da Bahia e Pernambuco.

22
2.2 Indução floral
De acordo com Albuquerque et al. (2002), o cultivo da mangueira (Mangifera indica,
L) nas condições tropicais semi-áridas permite a produção de frutas durante todo o ano,
inclusive em períodos onde a oferta do produto é escassa, tanto no mercado interno como no
externo, desde que se utilizem técnicas de indução floral.
Os processos naturais de floração em muitas espécies frutíferas lenhosas quase sempre
estão associados à inibição do crescimento vegetativo; essa inibição, em função do tempo e do
estado nutricional da planta, cria condições para floração (BEM-TAL, 1986; FIERRO e
ULLOA, 1991). Sendo assim, conforme Albuquerque et al. (2002), o frio e o estresse hídrico
são condições naturais que induzem a paralisação do crescimento vegetativo da mangueira,
nas condições de clima subtropical e tropical, respectivamente.
Brotos reprodutivos ocorrem geralmente depois de períodos extensos de repouso, nos
trópicos de baixa latitude, ou durante os meses frios de inverno nas regiões tropicais de
latitudes mais altas e nas subtropicais. A inflorescência normal ocorre de janeiro a março no
hemisfério norte e de junho a setembro no hemisfério sul. No entanto, existem variações de
comportamento com relação à floração, dentro da mesma cultivar, dependendo da idade da
planta e do local onde estão plantadas, se nos trópicos secos ou úmidos ou nos subtrópicos
(SINGH, 1960).
Albuquerque et al. (2002) afirmam que técnicas de manejo bem conduzidas podem
melhorar a quantidade e a qualidade dos brotos vegetativos. A poda estimula rapidamente a
brotação em gemas axilares, conseguindo assim multiplicar o número de brotos.
Posteriormente, com técnicas adicionais como manejo nutricional, uso de reguladores e
estresse hídrico, é possível amadurecer os brotos gerados da poda. O nível de estímulo floral
determina a resposta: níveis altos dão origem a panículas normais, níveis intermediários dão
origem a panículas mistas, níveis baixos resultam em crescimento vegetativo.
Schaffer (1994) estudando o efeito do fotoperíodo na cv. Tommy Atkins, constatou
que a mangueira é uma planta neutra.
Segundo Shu e Sheen (1987) estudando o efeito da temperatura no florescimento da
mangueira, observaram que as gemas axilares da cv. Haden, sob temperatura de 19ºC
dia/13ºC noite e 25ºC/19ºC, tiveram 87 e 60% de desenvolvimento floral, respectivamente;
quando a relação temperatura diurna/noturna foi 31ºC/25ºC, foram obtidos apenas ramos
vegetativos das gemas.

23
Na região do sub-médio Vale do São Francisco, a floração natural da mangueira
ocorre durante os meses de maio a agosto, quando o clima está mais seco e as temperaturas
noturnas mais baixas, geralmente menores que 20ºC, e a colheita se completa entre outubro e
janeiro, período que coincide com a safra em outras regiões do País, que é caracterizada por
uma alta oferta do produto no mercado. Porém, a maioria dos produtores procura direcionar
sua produção para janelas de mercado, induzindo a floração, muitas vezes, em condições
adversas (ALBUQUERQUE et al., 2002).
Conforme Chen (1987) durante os períodos de dormência, a disponibilidade de auxina
foliar decresce com a idade do ramo, e os níveis de citocinina se incrementam com o tempo,
sugerindo, portanto, que a auxina age como inibidora e a citocinina como promotora no
processo de quebra da dormência dos ramos.
Albuquerque et al. (2002) relatam que altos níveis de giberelinas inibem a floração e
estimulam o crescimento vegetativo; o declínio dos teores de giberelinas aumentará a
floração, portanto, o principal papel dos reguladores de crescimento vegetal é a supressão da
biossíntese das giberelinas.
Felippe (1979) afirma que entre as várias funções do etileno estão a promoção da
floração em plantas lenhosas e a aceleração de maturação de órgãos das plantas. Neste
sentido, Albuquerque et al. (2002) informam que o uso do etefon (ácido 2-cloroetilfosfônico)
para o amadurecimento de gemas, quando se trabalha em condições adversas, tem sido uma
das principais ferramentas do produtor, embora não tenha bom desempenho quando utilizado
isoladamente, portanto, deve ser combinado com estresse hídrico e/ou paclobutrazol (PBZ).
O impacto primário do estresse hídrico na manga é evitar a emissão de fluxos
vegetativos. A idade acumulada dos brotos é maior em árvores estressadas que em árvores
mantidas sob boas condições de irrigação, as quais podem emitir fluxos vegetativos mais
freqüentemente. Esse retardo no crescimento pode fornecer mais tempo para acumulação do
estímulo floral proposto ou redução no nível do promotor vegetativo; a medida que o ramo
amadurece, diminui o nível de ácido giberélico (GA3) (NUÑEZ-ELISEA e DAVENPORT,
1991).
O PBZ tem sido usado para estimular a floração, promovendo a paralisação do
crescimento vegetativo e reduzindo o alongamento da brotação, sendo a sua ação em função
da inibição da biossíntese das giberelinas, pois bloqueia a oxidação de caureno para ácido
caurenóico. Para a cv. Tommy Atkins recomenda-se aplicar 1 g do princípio ativo do produto
por metro de diâmetro de copa (MANICA, 1996; SILVA, D., 2000; ALBUQUERQUE et al.,
2002).

24
O sulfato de potássio (K2SO4) deve ser utilizado em duas a três aplicações nas
concentrações de 2 a 2,5%. A função do sulfato de potássio está relacionada com o íon
potássio, que vai interferir na relação potássio/nitrogênio (K/N), para impedir que a planta
vegete, além de aumentar o teor de carboidratos (ALBUQUERQUE et al., 2002).
O efeito dos nitratos (KNO3, Ca(NO3)2 e NH4NO3) no processo de indução floral deve
ser interpretado com cautela; os nitratos não induzem a floração, eles estimulam a iniciação
do crescimento. Assim, somente sob condições adequadas da planta e do ambiente à indução
floral, o crescimento será reprodutivo. As dosagens comumente usadas variam de 2 a 4% para
nitrato de potássio (KNO3), de 1,5 a 2% para o nitrato de cálcio (Ca(NO3)2) e até 1% para
nitrato de amônia (NH4NO3) (ALBUQUERQUE et al., 2002; FONSECA e SANTOS-
SEREJO, 2005).
Para Albuquerque et al. (2002), o manejo artificial adequado de floração da mangueira
é delineado de acordo com a época do ano, portanto, utiliza-se dos fatores que mais
influenciam o processo de floração naquele período, tais como: etefon, estresse hídrico,
paclobutrazol, sulfato de potássio, nitrato de potássio e nitrato de cálcio. A indução floral nas
condições climáticas do semi-árido nordestino brasileiro no período de maio a agosto (época
mais fria e de menor precipitação) é mais fácil que no período de outubro a abril (época mais
quente e de maior precipitação).
2.3 Fenologia da floração à frutificação
O florescimento ocorre em ramos com, no mínimo, quatro meses de idade, sob
condições tropicais (SIMÃO, 1971), ou com três meses, sob regime de temperaturas mais
amenas (CUNHA et al., 2002), ou com seis meses, sob condições normais (PINTO et al.,
2002a). É um fenômeno complexo que ocorre durante um período longo que varia de cinco a
seis meses, podendo ter seu início antecipado ou retardado (SIMÃO, 1971), natural ou
artificialmente, em razão das condições climáticas reinantes e da produtividade da safra
anterior ou do uso de determinadas práticas culturais (fitorreguladores de crescimento),
respectivamente (CUNHA et al., 2002). O envolvimento dos fatores climáticos (temperatura,
seca e fotoperíodo) e do etileno no florescimento da mangueira ainda não é plenamente
conhecido (DAVENPORT e NUÑEZ-ELISEA, 1990).

25
De acordo com Pinto et al. (2002a), a mangueira possui inflorescência do tipo
panícula, de forma cônica a piramidal, que se desenvolve de gemas terminais de ramos
maduros, nas quais se encontram flores perfeitas (hermafroditas) e masculinas, portanto, é
polígama. O número de panículas por planta varia de 600 a 6.000, e as flores por panícula
variam de 200 a 4.000 ou, segundo Simão (1971), 400 a 17 mil, as quais são pequenas
(diâmetro de 8 a 11 mm e disco de 2 a 4 mm) e rosadas. Ainda de acordo com esse mesmo
autor, são, em geral, pentâmeras, com androceu composto de quatro a seis estames, dos quais
apenas um é fértil; o ovário é súpero, unilocular e o estigma rudimentar.
A relação sexual em mangueira é a proporção entre flores hermafroditas e
estaminadas, sendo bastante variável dentro de cada panícula da planta e entre cultivares. A
panícula mede de 10 a 60 cm, com o número de flores perfeitas variando de ano para ano e,
dependendo da cultivar e da localização da panícula na planta, pode variar de 2 a 75% (LIMA
FILHO et al., 2002). A cv. Tommy Atkins, em condições de cerrados, chega a alcançar uma
proporção de 52 a 62% de flores masculinas, considerando tanto a posição da flor na raquis
quanto à posição da panícula na planta (PINTO et al., 1987).
A panícula desenvolve-se num período de 35 a 42 dias, e as primeiras flores só abrem
depois de 21 dias de iniciado o desenvolvimento da inflorescência, e a duração do
florescimento varia de 18 a 23 dias, portanto, as flores iniciam a antese antes mesmo que as
panículas atinjam o total comprimento. As flores abrem durante a noite, mas a deiscência só
ocorre no dia seguinte das 12:30 até as 16 horas (SIMÃO, 1971), ou das 6 às 10 horas, como
observado em algumas regiões (CUNHA et al. 2002), pois a deiscência é dependente do
clima. Por outro lado, conforme Pinto et al. (2002a) a maior concentração na abertura das
flores ocorre entre 9 e 11 horas. A receptividade do estigma tem duração de aproximadamente
72 horas após a antese, embora esteja receptivo antes da antese (MUKHERJEE, 1985). O
número de pólen por antera varia de 271 a 648, havendo variação entre cultivares (PINTO et
al. 2002a).
A polinização é feita, geralmente, por insetos, na sua maioria moscas (dípteros), com
pequena participação das abelhas. A baixa população de insetos no período do florescimento e
a presença de outras espécies mais atrativas, entre outros fatores, podem contribuir para
ocorrência de falhas no processo de polinização (FONSECA e SANTOS-SEREJO, 2005).
A inflorescência da mangueira possui centenas a milhares de flores pequenas, porém a
fecundação não é tão eficiente, apenas 3 a 35%, sendo diretamente afetada pelas condições
climáticas, principalmente pela chuva. Assim sendo, a quantidade de frutos que vingam e
amadurecem é pequena em comparação com o número de flores produzidas por árvore. Isso

26
porque o número de estigmas atingidos pelos grãos de pólen não ultrapassa 45% e a
fecundação é ainda menor, provocada pela disposição dos órgãos masculinos e femininos e
pela diferença de altura entre o estilete e o filete, além dos grãos de pólen serem pesados e
agregados, portanto, de difícil soltura, aliados à dicogamia e agentes patogênicos (SIMÃO,
1971; LAROUSSILHE, 1980).
A polinização em mangueira, principalmente naquelas monoembriônicas é
considerada um fator limitante, já que o grande número de flores não corresponde ao muito
pequeno número de frutos vingados. As plantas poliembriônicas produzem embriões
nucelares não sendo, necessariamente, obrigadas a serem polinizadas para ocorrer a
fecundação e vingamento de frutos (PINTO et al., 2002a).
O fenômeno do baixo vingamento de frutos é muito comum em mangueira, uma vez
que, no máximo, 35% do total de flores da mangueira são polinizadas resultando em cerca de
0,01% o número de frutos no stand final (SINGH, 1954). Vários fatores são responsáveis pelo
baixo vingamento de frutos, como, por exemplo, o grande número de flores perfeitas que não
são polinizadas e o alto número de flores masculinas na panícula. Além do pequeno número
de pólen por antera que é um fator genético (cultivar), os fatores nutricional (deficiência de
boro) e ambiental como a temperatura abaixo de 16ºC também afetam a produção e a
viabilidade do pólen, causando um baixo vingamento de frutos (SHARMA e SINGH, 1970).
A abscisão de flores e frutos de mangueira ocorre ao acaso em qualquer posição da
panícula, embora um maior número de frutos se estabeleça ou ocorra o vingamento na porção
terminal da panícula (PINTO et al., 2002a).
Geralmente, apenas uns poucos frutos por panícula (1 a 3) completam seu
desenvolvimento e atingem a maturação, pois muitos sofrem abscisão durante a primeira
semana após a fecundação. Essa é uma característica que pode ser influenciada tanto por
fatores genéticos quanto por fatores climáticos e por manejo inadequado. Baixa temperatura,
chuva, vento forte, plantas mal nutridas e antracnose são alguns dos aspectos que limitam a
produção em várias regiões do mundo (CUNHA et al., 2002).
A frutificação é intensa, entretanto, a mangueira apresenta um elevado índice de queda
de frutos, atingindo cerca de 80%. O fruto é uma drupa com variações na forma, cor e massa,
cuja maturação ocorre entre 3 e 4 meses após a fecundação (LIMA-FILHO et al., 2002).
O fruto da mangueira é uma drupa, com tamanho e massa variando de poucos gramas
a 2 kg, de formas reniforme, ovada, oblonga, arredondada ou cordiforme, e casca com
diferentes variações das cores verde, amarelo e vermelho. Em geral, a cor do fruto está
relacionada à cor da raque. A casca é coriácea e macia e envolve a polpa, de cor amarela

27
(várias tonalidades), mais ou menos fibrosa (de acordo com a cultivar) e de sabor variado. No
interior da polpa encontra-se o caroço ou semente, que é fibroso e apresenta diferentes formas
e tamanhos, de acordo com a cultivar (LAROUSSILHE, 1980).
De acordo com Medina, Voltaire (1996) quando a mangueira é enxertada e conduzida
de acordo com os requisitos técnicos exigidos pela cultura, sua frutificação tem início no
terceiro ano após o plantio, embora a produção econômica só comece a partir do quarto ano,
sendo possível obter rendimentos de 7 a 33 t/ha.
2.4 Ponto de colheita
O grau de maturidade ideal para a colheita, depende do tempo que a manga levará para
ser consumida ou industrializada. Para utilização imediata, colhem-se frutos completamente
maduros e para transporte ou armazenagem por períodos longos, no estádio maturo, a fim de
chegarem ao mercado varejista em bom estado de conservação e maturação (MEDINA,
Valdique, 1996).
De acordo com Alves et al. (2002), os frutos colhidos prematuramente não
amadurecem ou o fazem de forma irregular. Quanto mais imaturo o fruto, maior a
sensibilidade à baixa temperatura, e maior a perda de água por transpiração. As deficiências
no amadurecimento da manga colhida prematuramente manifestam-se, entre outros aspectos,
na cor, na firmeza, no conteúdo de açúcares e na acidez. Por sua vez, uma colheita tardia
reduz a vida útil e torna o fruto mais sensível a danos mecânicos e ao ataque de
microrganismos.
Segundo Cunha et al. (2002), a época de maturação da manga, cujo crescimento
apresenta padrão sigmóide, varia entre as diversas regiões produtoras, e o período de seu
desenvolvimento (da floração à maturidade fisiológica) é, em geral, de 100 a 150 dias. Em
regiões mais quentes, esse período é menor.
Vários parâmetros têm sido sugeridos para determinar a maturidade da manga, com
base no seu aspecto externo, no aspecto físico e na composição química à época da colheita.
Todavia, esses índices variam bastante de cultivar para cultivar, não podendo ser
generalizados para todas as espécies existentes. Quando as mangas ‘Tommy Atkins’ e
‘Haden’ destinam-se a exportação por via marítima, devem se colhidas apenas aquelas que

28
apresentam a espádua elevada na região de inserção do pedúnculo e a espádua dorsal bem
saliente. As mangas que não tiverem essa saliência, em hipótese alguma, devem ser colhidas,
pois não amadurecerão. Para a manga ‘Tommy Atkins’, a colheita tem início após 100 a 105
dias da sua florada, pois após 105 dias inicia-se a mudança na coloração da casca
(BLEINROTH, 1994).
Segundo Srivastava apud Bleinroth (1989), deve-se levar em consideração três fatores
importantes para determinar o ponto de colheita, os quais deverão ser ajustados de acordo
com a cultivar e a região produtora, que são: densidade da fruta entre 1,01 e 1,02 g/cm3;
firmeza entre 17,15 e 19,60 N e sólidos solúveis totais entre 12 e 15ºBrix. Neste mesmo
sentido, Kader (1999) afirma que o teor mínimo de SST necessário para se efetuar a colheita é
de 12 a 14ºBrix.
Para Medlicott e Reynolds (1988) o ponto de colheita da manga deve ser determinado
através da cor da casca e da polpa, associada com outras observações tais como: dias pós-
florada, aspecto das lenticelas, textura, sólidos solúveis totais e acidez.
Conforme Alves et al. (2002), a maturação mínima para colheita da cv. Tommy Atkins
é cor de polpa 1 (creme), cor de casca 2 (verde claro), firmeza de 129,41 N e sólidos solúveis
totais de 7,3ºBrix. Entretanto, o autor adiciona que atualmente recomenda-se que as mangas
que se destinam à Europa e ao Canadá sejam colhidas com cor de polpa correspondente ao
grau entre 2 (até 30% da área amarela e o restante creme) e 3 (amarelo) da escala subjetiva de
coloração da polpa.
2.5 Avaliações físicas
2.5.1 Aspectos morfológicos externos
Conforme Reid (2003) a forma é um índice de maturidade recomendado para:
angulosidade de bananas, faces cheias em mangas, compactação em couves-flor e brócolis.
Este autor também indica a morfologia e a estrutura da superfície como índice de maturidade,
pois pode-se avaliar a formação de cutícula em uvas e tomates, bem como, o brilho de alguns
frutos, o que indicia o desenvolvimento da cera.

29
A forma é estabelecida mediante medições das dimensões ou pelo uso de tabelas com
relação às dimensões, isto é, é avaliada pela relação entre os diâmetros ou por outras
características peculiares da espécie ou cultivar. Como exemplo, tem-se a angulosidade em
banana: de acordo com a cultivar, são colhidas quando apresentam redução na angulosidade,
em geral, quando atinge de 32 a 36 mm de diâmetro; preenchimento das faces em manga,
pêssego e nectarina: são colhidas quando ocorre o preenchimento das faces adjacentes ao
pedúnculo; compactação em brócolis e couve-flor. Para avaliar a morfologia e estrutura
superficial do produto, são observados, visualmente, a formação da cutícula ou o
desenvolvimento de ceras (brilho) na superfície do produto, ou mudanças de estrutura como a
forma reticulada em alguns tipos de melão. Portanto, o conjunto das modificações na
aparência como brilho, cor da casca, formação da cutícula, formação de ceras e
desenvolvimento do ápice, podem, isoladamente ou em conjunto, dar uma idéia do grau de
maturação do produto. Em maçã ocorrem modificações da forma com a evolução do
desenvolvimento e da maturação (CHITARRA e CHITARRA, 2005).
De acordo com Alves et al. (2002), os indicadores físicos como índices de colheita
para manga são baseados, em sua maioria, em características relacionadas à forma e ao
aspecto do fruto, que podem ser percebidas visualmente sem o emprego de métodos
destrutivos, tais como: formato do ombro, textura da casca, brilho da casca, distanciamento
das pontuações brancas e formato do nariz.
Para determinar a maturidade da manga, com base na forma do fruto, observa-se ápice
mais cheio e arredondado, quando o bico começa a aparecer em alguns cultivares. A espádua
(ombro) que na fruta verde está em linha reta com a inserção do pedúnculo, se eleva com a
maturidade (CHITARRA e CHITARRA, 2005).
2.5.2 Diâmetros longitudinal, ventral, transversal e produto dos diâmetros
Conforme Reid (2003), o tamanho é um índice de maturidade recomendado para todas
as frutas e muitas hortaliças. Neste sentido, Kays (1991) afirma que o tamanho pode ser
determinado pela dimensão (comprimento, largura, diâmetro ou circunferência), massa ou
volume.
Por outro lado, Chitarra e Chitarra (2005) explicam que o tamanho é usualmente
limitante como índice de maturidade em frutas, mas é bastante utilizado para hortaliças,

30
especialmente naquelas comercializadas na fase precoce de seu desenvolvimento. As frutas
são, em geral, avaliadas pelo diâmetro.
Miranda (2002), estudando o crescimento dos frutos de sapoti, evidenciou que os
mesmos aumentaram em tamanho durante todo o seu desenvolvimento e que o diâmetro
transversal aumentou linearmente (R² = 0,98), enquanto, o diâmetro longitudinal aumentou
quadraticamente (R² = 0,99). Neste sentido, Leite et al. (2005) pesquisando as características
físicas da manga cv. Tommy Atkins obtiveram os seguintes valores médios, respectivamente,
para diâmetros longitudinal, ventral e transversal: 10,89; 9,55 e 8,67 cm.
Morais (2001) trabalhando com ‘Tommy Atkins’ observou que o volume e as massas
seca e fresca podem ser determinadas a partir do produto dos diâmetros, um método não
destrutivo, baseado em medições fáceis e rápidas. Esta pesquisadora também verificou um
aumento no produto dos diâmetros a partir de 32 dias após floração plena (DAFP), onde foi
constatado 65,38 cm3 até 874,60 cm3 aos 103 DAFP, isto é, com o avanço da maturação.
Resultados semelhantes foram obtidos por Castro Neto e Reinhardt (2003) estudando a cv.
Haden.
2.5.3 Massas fresca, seca, de água e o teor de água
A massa correlaciona-se bem com o tamanho do produto e constitui uma característica
varietal. Ao atingirem o pleno desenvolvimento, as frutas devem apresentar massa variável
dentro dos limites típicos da cultivar, os quais são bastante flexíveis (CHITARRA e
CHITARRA, 2005).
Castro Neto e Reinhardt (2003) pesquisando as relações entre parâmetros de
crescimento do fruto da manga cv. Haden constataram que o acúmulo de massa fresca ocorreu
até 55 dias após a floração, atingindo 436 g. Já Morais (2001) trabalhando com ‘Tommy
Atkins’, verificou acúmulo de massa fresca até 103 DAFP.
Leite et al. (2005) pesquisando as características físicas da manga cv. Tommy Atkins,
confirmaram a ocorrência de uma massa fresca média de 509,19 g ± 23,15, portanto, variou
de 486,04 a 532,34 g. Neste sentido, Morais (2001) constatou uma variação de 36,86 g aos 32
DAFP até 515,58 g aos 103 DAFP, sendo assim, a massa fresca aumenta com o avanço da
maturação.

31
A água contida nos alimentos encontra-se sob as seguintes formas: livre, de estrutura e
de constituição. A água livre é a que não se encontra ligada a nenhuma estrutura molecular
dentro da célula, isto é, encontra-se em estado livre e é relativamente fácil de ser eliminada.
Constitui a fração de água existente nos alimentos. As demais formas de águas existentes nos
alimentos concentrados são denominadas de estrutura (altera a natureza física da substância) e
de constituição (entra na formação da substância), que apesar da importância sob o aspecto
físico-químico não apresentam valores no aspecto prático, pelos baixos teores com que estão
presentes. Na determinação do teor de umidade interessa apenas as formas de água livre e de
estrutura. A determinação da umidade (percentagem de água) pode ser feita por dois
processos: pelo processo indireto e processo direto. No indireto (secagem em estufa), o que se
determina é a matéria seca (massa seca), admitindo-se que a perda de massa corresponda a
massa de água perdida. Na realidade, outras substâncias voláteis além da água são
consideradas, também, como água, ocasionando algum erro, o que vem a ser uma
desvantagem do método. O processo denominado direto (destilação com tolueno), determina-
se a quantidade de água (massa de água) diretamente, embora também esteja sujeito a erros,
uma vez que não é muito fácil distinguir a exata separação da camada de água e do tolueno,
que é também evaporado e condensado, dentro do tubo receptor graduado (ISLABÃO, 1985;
SILVA, 1990).
Conforme Kader (1999), a massa seca é índice objetivo de maturidade estabelecido
para abacate na Califórnia (EUA), devendo variar de 17 a 20,5%, dependendo do cultivar.
Morais (2001) trabalhando com ‘Tommy Atkins’, verificou acúmulo de massa seca até
103 dias após a plena floração (DAPF), variando de 5,33 g (32 DAPF) a 86,65 g (103 DAPF),
desta forma, a massa seca aumenta com o avanço da maturação. Já Castro Neto e Reinhardt
(2003) estudando as relações de crescimento do fruto da manga cv. Haden verificaram que o
padrão de crescimento observado para volume, massa fresca, massa de água e massa seca
foram similares, isto é, padrão sigmoidal, no entanto, o volume, a massa fresca e a massa de
água, atingiram o seu máximo aos 55 dias após a floração (DAF), enquanto a massa seca
atingiu o seu máximo apenas aos 73 DAF, correspondendo, portanto, ao ponto de maturação
fisiológica. Estes autores também evidenciaram que a percentagem de água (teor de água)
manteve-se praticamente constante durante a maturação, no entanto, quando decresce, reduz a
massa fresca.
Leite et al. (2005) avaliando as características físicas da manga cv. Tommy Atkins,
observaram um conteúdo de água (teor de água) médio de 85,24% ± 0,84, portanto, variou de
84,4 a 86,08%.

32
2.5.4 Coloração da casca e da polpa
Embora nem todos os frutos mudem de cor durante o amadurecimento, esta é uma das
características mais associadas ao ponto de colheita e maturidade para consumo (TUCKER,
1993). A época, a velocidade e a intensidade da mudança variam entre espécies e entre
cultivares de uma mesma espécie (KAYS, 1991).
As alterações mais representativas ocorrem em nível de degradação da clorofila.
Apesar do mecanismo exato desta degradação ainda não esteja totalmente entendido, supõe-se
que a molécula de clorofila seja solubilizada das membranas dos tilacóides do cloroplasto
para o estroma, onde é oxidada (TUCKER, 1993).
Simultaneamente à degradação de clorofila, pode haver síntese de outros pigmentos
em alguns frutos (KAYS, 1991; TUCKER, 1993; WILLS et al., 1998). Neste sentido, de
acordo com Medlicott et al. (1992), as mudanças na coloração da manga, se devem à
degradação da clorofila e síntese de carotenóides (carotenos, licopeno e xantofilas) e
flavonóides (antocianinas).
Conforme Saúco (1999), apesar da avaliação da cor da polpa da manga ser um método
destrutivo, é prático e confiável para definição do ponto de colheita, sendo utilizado em
diversos países exportadores.
Rocha et al. (2001), avaliando a cor na cv. Tommy Atkins através de escala subjetiva
indicou que a coloração da casca passou de verde-oliva, no primeiro estádio de maturação,
para vermelha no último estádio, assim como, a coloração da polpa passou de branca para
laranja.
2.5.5 Firmeza
As alterações na firmeza são bastante representativas, podendo afetar a palatabilidade,
a duração do período de armazenamento e a extensão da vida útil do fruto (KAYS, 1991;
YOSHIOKA et al. 1994).
O amolecimento aumenta através de perda de turgescência, degradação do amido ou
degradação das paredes celulares. A perda de turgescência é um processo não-fisiológico
associado á desidratação pós-colheita do fruto, podendo adquirir importância comercial

33
durante o armazenamento. Nos frutos em que o amido constitui uma alta percentagem da
massa fresca, sua degradação resulta em mudanças acentuadas na firmeza. No entanto, em
geral, as alterações na firmeza durante o amadurecimento resultam, predominantemente, da
desestruturação da parede celular (TUCKER, 1993).
Uma vez iniciado o amolecimento, a taxa de mudança na firmeza é função do tipo de
fruto e das condições nas quais é mantido (KAYS, 1991).
É improvável que uma simples enzima seja responsável por mudanças na firmeza. Na
realidade, o processo deve envolver uma interação complexa das atividades das enzimas
amilásicas, pécticas e galactosidases com mudanças físico-químicas na parede. Parece, ainda,
que o amolecimento de diferentes frutos pode ocorrer por diferentes mecanismos (TUCKER,
1993).
Lucena et al. (2000), estudando a caracterização físico-química da manga, cv. Tommy
Atkins, nos estádios 2, 3 e 4 de maturação, constataram que a firmeza média avaliada através
de penetrômetro foi de 95,41, 85,93 e 48,75 N, respectivamente, portanto, é decrescente com
o avanço da maturação.
2.5.6 Unidades de calor (graus-dia)
Os sistemas de unidades de calor (UC) são utilizados para caracterizar o
desenvolvimento da planta, pelo fato da taxa de desenvolvimento variar com a temperatura
(CHITARRA e CHITARRA, 2005).
Segundo Alves et al. (2002), o indicador dias-graus centígrados está relacionado com
o tempo de crescimento, podendo ser útil em regiões onde amplas variações de temperatura
são comuns.
As unidades de calor são utilizadas no estabelecimento do tempo necessário para o
desenvolvimento do produto após a floração, em condições climáticas bem definidas. Para
cada cultura existe um número característico de unidades de calor. Mudanças nas condições
climáticas poderão antecipar (calor) ou retardar (frio) a maturidade. Esse método deve ser
testado durante alguns anos e apresenta o inconveniente de ser necessário o conhecimento da
temperatura mínima requerida, pela cultura, para o seu crescimento (CHITARRA e
CHITARRA, 2005).

34
O conceito de graus-dia ou unidades térmicas pressupõe a existência de uma
temperatura abaixo da qual a planta não se desenvolve ou, se o fizer, o faz a uma taxa muito
reduzida, denominada temperatura base. Esse conceito se baseia no fato de que uma planta
necessita de certa quantidade de energia, representada pela soma de ºC, ou seja, o número de
graus-dia de desenvolvimento (GDD), acima do valor da temperatura-base, para completar
determinada fase fenológica ou, mesmo, seu ciclo total (SILVA et al., 1999).
A predição do estádio de desenvolvimento é, portanto, baseado no acúmulo de graus
(ºC) por unidade de tempo, acima da temperatura base, freqüentemente designada como
“limiar para o crescimento”. A temperatura limite é definida com base no tempo e na
temperatura, desde o plantio até a colheita de uma cultura, desenvolvida em diferentes
localidades ou durante vários anos. Determina-se a temperatura média diária, cuja soma
algébrica é deduzida da temperatura mínima de crescimento. Assim, pode-se predizer a época
provável da maturidade, num determinado ano. À medida que a maturidade se aproxima, pode
ser checada por outros meios (CHITARRA e CHITARRA, 2005).
Os estudos das interações clima-planta foram iniciados, segundo Mota (1986), por
Réaumur, em 1735, sendo este considerado o precursor do sistema de graus-dia (GD) ou
unidades térmicas. Um GD, ou unidade térmica, era definido como a temperatura média do
dia. No método original aplicado por Réaumur, a constante térmica era calculada a partir da
soma das temperaturas médias diárias acima de 0ºC, que podia ser determinada para o ciclo
total ou para cada fase. Esse método foi denominado de método direto, que apresentava o
inconveniente de sofrer variações segundo as localidades consideradas (MOTA, 1986). De
acordo com este autor, para atender ao cálculo de graus-dia para diversas localidades deve-se
usar o método residual, que consiste no somatório das diferenças entre a temperatura média
diária e a temperatura base.
A complexidade fisiológica da planta impede a determinação precisa da temperatura
base. Contudo, é possível encontrar na literatura resultados de pesquisas dando faixas de
valores aproximados das temperaturas ótimas e extremas, nas quais se espera um pleno
desenvolvimento da espécie cultivada. O sucesso do uso de GD para determinar o ponto de
colheita se baseia na informação da temperatura base (SILVA et al., 1999). Chaudhri (1976) e
Silva (1996) sugerem o valor 10ºC como a temperatura base da mangueira.
A exatidão em predizer o estádio de desenvolvimento da planta ou de parte da planta
através de métodos graus-dia, depende da determinação precisa da temperatura base
(HIGLEY et al., 1986). O crescimento e desenvolvimento das plantas tropicais ocorrem mais

35
frequentemente entre 10 e 40ºC. Os relacionamentos de desenvolvimento e temperatura são
usualmente lineares em vez de logarítmicos dentro desses limites (MONTEITH, 1977).
Os sistemas de unidades de calor são utilizados em culturas com uma única colheita
comercial, tais como ervilha e milho. A soma de calor é utilizada na Europa e nos Estados
Unidos, principalmente para produtos cujo teor de amido aumenta consideravelmente com a
maturação. O inconveniente do método é a necessidade do conhecimento da temperatura de
paralisação do crescimento do produto. Em ervilhas, por exemplo, essa temperatura é de
4,4ºC. São necessárias 700 UC para a cultivar industrial e 800 UC para a cultivar de mercado,
para que elas atinjam o ponto ideal de colheita (CHITARRA e CHITARRA, 2005).
Silva et al. (1999) estudando a relação entre coeficientes de cultura e GD da alface,
utilizando 4,4ºC como temperatura base, constataram que a colheita foi realizada com GD
igual a 742ºC e coeficiente de cultura (Kc) médio de 1,2.
Silva (2004) avaliando a fenologia do pequi (Caryocar coriaceum Wittm.) ocorrente
na Chapada do Araripe, Crato-CE, com a temperatura base de 15ºC, afirmou que a somatória
das unidades térmicas no período de janeiro a dezembro de 2002 para a fase vegetativa
revelou a necessidade de um total de 457,8 GD para as áreas de cerradão e cerrado; a floração
um total de 825,0 GD e 829,8 GD para as áreas de cerradão e cerrado, respectivamente,
enquanto que a maturação dos frutos necessitou de 151,6 GD nas quatro áreas do estudo (duas
áreas de cerradão e duas áreas de cerrado).
Conforme Reid (2003), a média de UC durante o desenvolvimento do fruto é um dos
índices de maturidade estabelecidos para pêra, maçã e milho-doce. Neste sentido, Chitarra
(2000) afirma que este método é utilizado para milho, ervilha e tomate, destinados à
industrialização. Já Alves et al. (2002) recomendam este método como indicador físico para
manga.
GD tem sido usado para prever a maturidade do fruto em pessegueiro e nectarina
(MUÑOZ et al., 1986). Entretanto, para plantas tropicais a literatura é escassa; para manga,
por exemplo, 1.000 GD são necessários para se obter fruto de alta qualidade, embora
nenhuma referência seja feita para a temperatura base utilizada nos cálculos
(OPPENHEIMER, 1947 apud SINGN, 1977).
Mosqueda-Vázquez e Ireta-Ojeda (1993) pesquisando GD e temperaturas-base
requeridas para os desenvolvimentos da inflorescência e do fruto da manga ‘Manila’,
verificaram que com a temperatura base de 12ºC são necessários 434,7 GD para elongação da
inflorescência, enquanto com uma temperatura base de 0,33ºC são necessários 2.292,7 GD
para a maturidade do fruto ser atingida.

36
Burondkar et al. (2000) trabalhando com estimativa de UC como índice de maturidade
para diferentes cultivares de manga na região de Konkan de Maharshtra, Índia, observaram
que das três cultivares estudadas com a temperatura base de 17,9ºC, a cv. Alphonso registrou
a menor duração (111 e 93 dias) e unidade de calor (718 e 701 GD) em ambas as localidades
(Vengurla e Deogad), seguido por ‘Kesar’ (118 e 98 dias; 773 e 799 GD) e ‘Ratna’ (127 e 112
dias; 849 e 866 GD) nos respectivos locais.
2.6 Avaliações físico-químicas e químicas
2.6.1 Sólidos solúveis totais
O teor de sólidos solúveis totais (SST), normalmente é feito com objetivo de se ter
uma estimativa da quantidade de açúcares presentes nos frutos, embora, medidos através de
refratômetro, incluem, além dos açúcares, pectinas, sais e ácidos. Os SST aumentam nos
frutos com o amadurecimento, os quais são constituídos principalmente pelos açúcares
solúveis (LIMA, 1997).
A evolução do teor de SST em manga varia de 7,0 a 17,4ºBrix, dependendo da cultivar
e do estádio de maturação do fruto. Algumas cultivares apresentam teores mais baixos como a
Palmer (7,0 a 14,0ºBrix) conforme Melo Neto et al. (1999); a Haden (8,0 a 14,7°Brix) de
acordo com Camillo-López et al. (1995); a Keitt (7,0 a 15,0°Brix) segundo Yamashita (1995)
e a Parvin (7,7 a 15,1°Brix) relatado por Ramos (1994) ou mais elevado como a Tommy
Atkins (8,2 a 17,4°Brix) constatado por Salles e Tavares (1999).
2.6.2 Acidez total titulável e pH
Os dois métodos mais comumente usados para medir a acidez de frutos são a acidez
total titulável (ATT) e o potencial hidrogeniônico (pH), sendo que o primeiro representa todos
os grupamentos ácidos encontrados (ácidos orgânicos livres, na forma de sais e compostos

37
fenólicos), enquanto que o segundo determina a concentração hidrogeniônica da solução
(KRAMER, 1973).
De acordo com Hulme (1974), o conteúdo de ácidos orgânicos diminui com o
amadurecimento na maioria dos frutos tropicais, devido a utilização desses ácidos no ciclo de
Krebs, durante o processo respiratório ou de sua conversão em açúcares (CHITARRA e
CHITARRA, 2005).
Embora algumas cultivares apresentem teor de acidez mais elevado, como por
exemplo, a cv. Alphonso, com aproximadamente 3% de ATT, outras, tais como a cv. Tommy
Atkins apresentam média acidez (0,5 a 1,0%), onde o ácido orgânico predominante é o ácido
cítrico, seguido pelo málico e pelo oxálico (LAKSHMINARAYANA, 1980), por outro lado, a
cv. Parvin tem baixa acidez (0,09 a 0,66%), conforme Ramos (1994).
Quando a maturação é normal e o fruto sadio, o ácido predominante (ácido cítrico)
praticamente desaparece, sendo a acidez remanescente devida aos fenólicos ácidos como o
ácido quínico, ácido clorogênico e ácido neoclorogênico (LIMA, 1997).
Em geral, o sabor ácido está associado, principalmente, ao íon hidrogênio e ao grau de
dissociação. Ácidos fortes (completamente dissociados) apresentam maior acidez do que
soluções de um ácido fraco de normalidade equivalente. Neste sentido, o ácido cítrico tem se
destacado por sua alta acidez relativa (PANGBORN, 1963).
Durante o amadurecimento há diminuição da acidez e conseqüentemente aumento do
pH (MATTOO et al. 1975). A manga é considerada um fruto ácido com a maioria das
cultivares apresentando valores de pH abaixo de 6,0. Dependendo da cultivar e do estádio de
maturação do fruto, algumas cultivares de manga apresentam teores mais baixos como em
Tommy Atkins (3,5 a 3,7) conforme Lucena et al. (2000), médios como em Carabao (3,7 a
4,7) segundo Morga et al. (1979) ou mais elevado como em Bocado (3,2 a 6,0) de acordo com
Castrillo e Bermudez (1992).
2.6.3 Relação SST/ATT
De acordo com Bleinroth (1992), os sólidos solúveis totais (SST) têm tendência de
aumento com o avanço da maturação, enquanto a acidez total titulável (ATT) diminui com o
amadurecimento, sendo assim, a relação SST/ATT é diretamente proporcional aos SST e
inversamente proporcional a ATT.

38
Em muitas frutas, o equivalente entre os ácidos orgânicos e os açúcares é utilizado
como critério de avaliação do “flavor”. Contudo, como são alguns constituintes voláteis, essa
relação é mais indicativa do sabor, porque se utiliza a ATT e não a acidez total (AT), quando
se estabelece essa relação. Além disso, alguns produtos insípidos, contendo ATT e SST muito
baixos, apresentam relação elevada entre esses componentes, o que pode conduzir a
interpretações errôneas da qualidade comestível. Por essa razão, são estabelecidos teores
mínimos de SST e máximos de ATT para alguns frutos, visando à obtenção de um sabor
aceitável. Por exemplo, testes de análise sensorial em laranjas e tangerinas demonstraram que
a relação SST/ATT deve ser de 10:1 e 9:1, respectivamente, desde que o teor de SST seja
igual a 9%. Se, no entanto, o teor de SST for inferior ao mínimo, é necessária uma relação
mais elevada, para obtenção de um sabor aceitável. A relação aumenta com o
amadurecimento devido ao decréscimo na ATT, fato que permite uma relação elevada, em
frutas contendo baixo teor de SST. Do mesmo modo, se o teor de SST for superior a 9%, pode
apresentar uma relação menor que 10:1, ou seja: laranjas com 11% de SST podem apresentar
relação 9:1, adequada ao paladar do consumidor. A relação SST/ATT é mais representativa
que a medição isolada de açúcares ou da acidez, pois essa relação dá uma boa idéia do
equilíbrio entre esses dois componentes (CHITARRA e CHITARRA, 2005).
Para manga, segundo Medlicott e Reynolds (1988), o teor mínimo de SST varia de 7 a
8ºBrix e o teor máximo de ATT varia de 0,65 a 0,70% de ácido cítrico, resultando numa
relação SST/ATT de 10,00 a 12,30.
Lucena et al. (2000) trabalhando com manga cv. Tommy Atkins nos estádios 2, 3 e 4,
obtiveram respectivamente as seguintes relações SST/ATT: 9,20, 16,98 e 20,28.
2.6.4 Carboidratos
A reserva de carbono mais importante nas plantas é o amido (KAYS, 1991; MARTIN
e SMITH, 1995). Ele consiste de diferentes polímeros de glicose arranjados numa estrutura
cristalina tridimensional. Sua biossíntese envolve tanto a produção de glucanas quanto o seu
arranjo no grânulo de amido (MARTIN e SMITH, 1995).
Conforme Dennis e Blakeley (2000) o amido é composto de dois polissacarídeos, a
amilose com longas cadeias não ramificadas de unidades de glicose com ligações α(1→4) e a
amilopectina, a qual é ramificada por ligações α(1→6). Pontos de ramificação na

39
amilopectina são separados por uma média de 20-30 resíduos de glicose. A estrutura da
amilose é uma espiral helicoidal e a da amilopectina é uma rede ramificada, embora ambas
possuam um final redutor e outro não redutor.
De acordo com Evangelista (1999), o amido que se acumula durante o crescimento do
fruto é rapidamente degradado durante o amadurecimento, uma vez que, esta diminuição é
evidente no cloroplasto, onde os grânulos de amido tornam-se menores e praticamente
desaparecem no fruto maduro. Em muitos frutos, a degradação do amido é um evento
característico do amadurecimento (JOHN e DEY, 1986; TUCKER, 1993), que promove a
palatabilidade (KAYS, 1991).
A velocidade e a extensão da degradação do amido durante a maturação variam entre
frutos de diferentes espécies (KAYS, 1991) e pode ocorrer através de duas vias: uma
hidrolítica e outra fosforolítica. A hidrolítica envolve a ação das enzimas α-amilase, β-
amilase, enzima desramificadora e α-glicosidase e a fosforolítica a atividade da enzima
fosforilase do amido (TAIZ e ZEIGER, 2004). A ação cooperativa destas enzimas resulta na
dissolução do amido armazenado, que forma, num primeiro momento, oligossacarídeos e, em
seguida, maltose, glicose ou glicose-1-fosfato (IRVING et al, 1999). Estes produtos podem
ser utilizados na respiração, participando de reações da glicólise, ou na síntese de sacarose
(TUCKER, 1993).
Rocha et al. (2001), trabalhando com a cv. Tommy Atkins, verificou o maior conteúdo
de amido (6,81%), por ocasião da colheita no estádio de maturação 1, polpa branca, e o menor
(3,04%), no estádio 5, polpa laranja. Ainda foi constatado, nesse trabalho, que existe uma
forte correlação entre o índice de degradação de amido, determinado através de teste com
solução de iodo, e a coloração da casca e da polpa, indicando que quanto mais verde a casca e
quanto mais branca a polpa, maior a área do fruto tingida por solução de iodo, pois no estádio
1 de maturação foi observada uma área inferior a 10% sem tingir, enquanto, no estádio 5, essa
área foi superior a 50%. Neste mesmo sentido, Lucena et al. (2000) também trabalhando com
essa cultivar colhida nos estádios 2, 3 e 4 de maturação verificaram conteúdos de 8,50; 7,79 e
7,52% de amido, respectivamente, portanto, decrescente com o avanço da maturação.
O avanço da maturação promove um aumento no conteúdo de açúcares, atribuído
principalmente, à hidrólise de carboidratos de reserva acumulados durante o crescimento do
fruto na planta, resultando na produção de açúcares solúveis totais (AST) (KAYS, 1991;
SIGRIST, 1992; WILLS et al. 1998).

40
Os AST além de terem uma grande participação na composição do flavor, são
utilizados como substratos respiratórios, mas encontram-se nos frutos em quantidades muito
superiores àquelas necessárias à geração de energia (TUCKER, 1993).
Os principais AST presentes nos frutos são a frutose, a glicose e a sacarose. O
dissacarídeo sacarose é o principal açúcar não-redutor enquanto glicose e frutose constituem
os principais açúcares redutores, havendo, na maioria dos frutos, predomínio do primeiro.
Durante o crescimento e a maturação, os teores de açúcares redutores aumentam tanto em
frutos climatéricos quanto nos não-climatéricos (WHITING, 1970).
O conteúdo de açúcares em mangas variam em função da cultivar, condições
nutricionais do solo, condições climáticas, estádio de maturação e temperatura de
armazenamento. Durante o amadurecimento, os teores de glicose e frutose variam, enquanto
que o de sacarose aumenta de três a quatro vezes, sendo, portanto, o principal açúcar
contribuinte para a doçura da manga (LAKSHMINARAYANA, 1980).
A proporção entre os diferentes tipos de açúcares é um importante atributo de
qualidade uma vez que diferem em grau de doçura, sendo assim, a frutose possui grau de
doçura maior que a sacarose e esta por sua vez maior que a glicose (PANGBORN, 1963).
Os AST constituem 91% dos sólidos solúveis totais do mesocarpo de mangas ‘Ngowe’
(BRINSON et al. 1988). Já, Bissoli Júnior (1992) com a cv. Tommy Atkins observou uma
redução gradual nos teores de glicose e frutose e aumento contínuo no de sacarose durante a
maturação e Evangelista (1999) com esta mesma cultivar constatou que os níveis de sacarose
aumentaram de 1,57 para 6,59% durante o período de 35 dias de armazenamento refrigerado.
2.6.5 Nitrogênio total, não protéico e protéico
As proteínas e outros compostos nitrogenados são decompostos na presença do ácido
sulfúrico concentrado à quente produzindo sulfato de amônio. O sulfato de potássio ou de
sódio é adicionado, a fim de aumentar o ponto de ebulição do ácido sulfúrico, apressando a
digestão. Outros compostos como sulfato de cobre e selênio também ajudam a digestão da
matéria orgânica. O sulfato de amônio resultante, na presença da solução concentrada de
hidróxido de sódio, libera NH3 que é recebido na solução de ácido bórico. A amônia, na
solução de ácido bórico, é titulada com ácido sulfúrico ou clorídrico de título conhecido e,

41
assim, determina-se o teor de nitrogênio da amostra. Para o cálculo da proteína bruta, basta
multiplicar o resultado pelo fator 6,25 (SILVA, 1990).
O termo proteína bruta envolve um grande grupo de substâncias com estruturas
semelhantes, porém com funções fisiológicas muito diferentes. Baseado no fato das proteínas
terem porcentagem de nitrogênio quase constante, em torno de 16%, o que se faz é determinar
o nitrogênio e, por meio de um fator de conversão, transformar o resultado em proteína bruta.
No método de Kjeldahl (AOAC, 1995), que é o mais usado, determina-se o nitrogênio contido
na matéria orgânica, incluindo o nitrogênio protéico propriamente dito, ou seja, sob a forma
de proteínas verdadeiras e outros compostos nitrogenados não protéicos, tais como: aminas,
amidas, lecitinas, nitrilas e aminoácidos livres. Isto é, todo o nitrogênio do alimento, com
exceção dos nitratos e nitritos, por isso chama-se proteína bruta (ISLABÃO, 1985; SILVA,
1990).
Quando se quer determinar os nitratos e nitritos, tem-se que reduzi-los com redutores
mais fortes, e depois seguir algum método específico para aqueles compostos (NELSON et
al., 1954; WOOLLEY, et al., 1960).
Quando se quer determinar o nitrogênio contido na amostra, que se apresenta em
forma de proteína verdadeira, isto é, o nitrogênio protéico, faz-se a precipitação da proteína
com reagentes específicos como: ácido wolfrâmico, cloreto de estanho II ou mistura de
CuSO4 a 6% + NaOH 1,25%. A proteína precipitada é então separada por filtração.
Normalmente, faz-se a determinação do nitrogênio não protéico no filtrado, determinando-se,
indiretamente, a proteína verdadeira (RENA e MASCIOTTI, 1976).
Quando se quer estudar a natureza de uma proteína, tem-se que decompô-la por meio
de hidrólise em seus aminoácidos componentes e depois determinar os aminoácidos por
cromatografia (ISLABÃO, 1985).
A maior parte do nitrogênio na planta está em forma de aminoácidos, formando
proteína, a qual é parte do protoplasma. Análises de gramíneas e leguminosas mostram que a
proporção de nitrogênio-amínico varia de 52 a 83% do nitrogênio total, sendo que os valores
maiores foram obtidos em leguminosas. A proporção do nitrogênio total, que está presente na
forma de nitrogênio não protéico, varia de 23 a 30%. Deste nitrogênio não protéico, cerca de
47 a 64% está na forma de peptídeos e aminoácidos livres. Nitrogênio não protéico, bem
como os outros compostos nitrogenados da planta, varia com o estádio de crescimento, nível
de fertilização e cultivar (SILVA, 1990).
As proteínas e aminoácidos livres são constituintes menores de frutos e, como é bem
conhecido, não desempenham nenhum papel na determinação da qualidade comestível.

42
Mudanças nos constituintes nitrogenados indicam, contudo, variações na atividade metabólica
durante diferentes fases do crescimento. Durante a fase climatérica de muitos frutos, existe
uma redução nos aminoácidos livres que geralmente reflete um incremento na síntese
protéica. Durante a senescência, o nível de aminoácidos livres aumenta refletindo uma
degradação de enzimas e redução da atividade metabólica (BOAS, 1999).
Conforme Chitarra e Chitarra (2005) há um número considerável de evidências do
envolvimento da síntese protéica (síntese “de novo” de enzimas) no climatério dos frutos, no
qual ocorre aumento na proporção do nitrogênio protéico em relação ao nitrogênio total.
De acordo com Gortner et al. (1967), avaliando as transformações que ocorrem no
ciclo vital do abacaxi, constataram que o nitrogênio não protéico da casca diminui
continuamente da pré-maturação até a senescência, enquanto o da polpa diminui até a pré-
maturação, mas aumenta da maturação até a senescência.
Lakshminarayana et al. (1970), estudando alguns aspectos do desenvolvimento
fisiológico da manga, constataram uma redução continua nos teores de nitrogênio total da
casca e do fruto completo durante o seu desenvolvimento, no entanto, na polpa, reduziu
inicialmente, depois aumentou e no final reduziu.
2.6.6 Vitamina C
A vitamina C desempenha um papel crucial na nutrição humana na prevenção do
escorbuto. Virtualmente, toda a vitamina C da dieta do homem é obtida a partir dos frutos e
hortaliças. O requerimento diário do homem com relação à vitamina C é de cerca de 50 mg e
muitos frutos contêm esta quantidade em menos que 100 g de tecido (BOAS, 1999).
O teor de vitamina C tende a diminuir com a maturação e com o armazenamento de
muitas hortícolas, devido à atuação da enzima ácido ascórbico oxidase (ascorbinase), ou pela
ação de enzimas oxidantes como a peroxidase. Essa vitamina encontra-se em tecidos vegetais
na forma reduzida como ácido ascórbico (AA), ou na forma oxidada, como ácido
deidroascórbico (DHA), ambos com atividade vitamínica. No entanto, a degradação do DHA
para ácido 2,3-dicetogulônico leva à perda da atividade biológica e esse, através de outras
reações químicas, produz pigmentos escuros que depreciam a aparência do produto. A
vitamina C é um excelente antioxidante e atua nas reações redox como transportador de

43
elétrons para a cadeia respiratória, bem como, regenerando diferentes substratos de sua forma
oxidada para a forma reduzida (CHITARRA e CHITARRA, 2005).
Os frutos constituem a fonte natural mais importante de vitamina C para o ser humano,
e os que se destacam pelo conteúdo dessa vitamina são: caju, mamão, goiaba, frutos cítricos,
morango, manga, caqui, kiwi, maracujá e tomate (AWAD, 1993).
Embora a vitamina C seja considerada como um dos componentes nutricionais mais
importantes nas frutas, o seu teor em geral não ultrapassa o valor de 300 mg/100 g da massa
fresca da polpa. No entanto, alguns produtos são fontes excepcionais como a acerola (1.000 a
1.800 mg/100 g da massa fresca da polpa), a goiaba e o caju (200 a 300 mg/100 g da massa
fresca da polpa). As frutas cítricas, embora não contenham teores elevados (entre 40 e 80
mg/100 g da massa fresca da polpa), são consideradas como boas fontes de vitamina C pelo
alto consumo diário (CHITARRA e CHITARRA, 2005). Neste mesmo sentido, de acordo
com Franco (2003), a manga comum madura possui 43 mg/100 g da massa fresca da polpa,
por outro lado, Wills et al. (1998) afirmam que possui apenas 30 mg/100 g da massa fresca da
polpa.
A vitamina C é acumulada durante o desenvolvimento das frutas na planta. Após a
colheita, o acúmulo é menor ou decresce em alguns produtos como em maçã e manga.
Portanto, quanto mais precoce a colheita, menor o seu teor no produto. Frutas cítricas
imaturas contêm concentração mais elevada de vitamina C que os maduros, mas, a
concentração por fruta tende a aumentar, pelo aumento do volume total de suco e do tamanho
da fruta, com o avanço da maturação na planta (CHITARRA e CHITARRA, 2005).
De acordo com Lakshminarayana et al. (1970) trabalhando com manga, os teores de
vitamina C da polpa e do fruto completo diminuem com o avanço da maturidade e os da casca
aumentam. Porém, conforme Thomas e Oke (1980) independentemente da temperatura de
maturação, na manga madura os teores de vitamina C da casca também diminuem, ainda que
menos acentuadamente do que na polpa, como pôde ser constatado pelas altas porcentagens
de retenção na casca.
2.6.7 Clorofila total
Três tipos principais de pigmentos ocorrem nos produtos vegetais: clorofila,
carotenóides e antocianinas. Portanto, a coloração das frutas e das hortaliças é resultante dos

44
pigmentos clorofila e carotenóides presentes nos cloroplastos e nos cromoplastos, bem como
dos pigmentos fenólicos (antocianinas, flavonóis e proantocianinas) presentes nos vacúolos.
As betalaínas formam um quarto grupo de pigmentos, presentes nos vacúolos e no citosol,
mas são restritas a alguns produtos como, por exemplo, à beterraba. Têm importante papel na
aceitação do produto pelo consumidor (CHITARRA, 2000; CHITARRA e CHITARRA,
2005).
A cor verde dos frutos se deve à presença das clorofilas a e b. A molécula de clorofila
possui duas partes: a primeira é uma estrutura de anel tipo porfirina, contendo Mg2+ no centro,
e a segunda, uma longa cauda de hidrocarbonetos hidrofóbicos denominada fitol. A perda da
cor verde resulta da quebra da estrutura de clorofila, causada principalmente pelas mudanças
de pH, resultantes da presença de ácidos orgânicos provenientes do vacúolo, pela presença de
sistemas oxidantes, pela atividade de clorofilases, que separam o fitol da porfirina na
molécula de clorofila e da ação das enzimas lipoxigenase e peroxidase, as quais parecem estar
indiretamente ligadas ao processo de degradação da clorofila. A ação desses fatores acaba
desorganizando a estrutura interna do cloroplasto (AWAD, 1993; TAIZ e ZEIGER, 2004;
CHITARRA e CHITARRA, 2005).
As clorofilas são encontradas nos cloroplastos, têm coloração que varia de azul-
esverdeado ao verde-amarelado e são os pigmentos predominantes no reino vegetal. São
receptores da luz durante a fotossíntese. As clorofilas a e b são encontradas nas plantas verdes
na proporção aproximada de 3:1 e diferem na substituição do C-3. Durante a maturação dos
frutos, os cromoplastos e suas membranas tilacoidais desintegram-se, ocorrendo uma rápida
degradação da clorofila, com perda da coloração verde dos tecidos, tornando visíveis
pigmentos pré-existentes e/ou síntese de novos pigmentos responsáveis pela coloração
característica de cada espécie, ou de cada cultivar (CHITARRA e CHITARRA, 2005).
A perda da cor verde é utilizada como indicativo ou guia da maturidade. O verde
intenso no fruto jovem muda gradualmente, perdendo a intensidade até tornar-se verde claro.
Em muitas frutas, há perda completa do verde, surgindo então os pigmentos amarelos,
vermelhos ou púrpuros. Embora essas transformações de coloração sejam utilizadas como
guia da maturidade, não são inteiramente confiáveis, porque sofrem a influência de inúmeros
fatores. A exposição à luz solar, por exemplo, pode induzir o desenvolvimento de cor mais
rapidamente em algumas frutas que em outras, na mesma planta, embora ambas possam ter a
mesma época de formação (AWAD, 1993; CHITARRA e CHITARRA, 2005).
Conforme Medlicott et al. (1986) durante a maturação da manga, a cor da casca muda
gradualmente de verde para vermelho-amarelado. Algumas cultivares desenvolvem uma

45
coloração avermelhada brilhante que tem sido atribuída a antocianinas, enquanto outras retêm
uma maior proporção de cor verde, embora estejam completamente maduras. Na cv. Tommy
Atkins observa-se uma rápida destruição de clorofila, sendo a clorofila a preferencialmente
degradada em relação à clorofila b.
De acordo com Mena et al. (1996), ao avaliarem durante 10 dias após a colheita
(DAC) a maturação em frutos de manga cv. Manila armazenadas a temperatura de
comercialização, o teor de clorofila total variou de 4 mg/100 g da massa fresca da casca aos 2
DAC para 1 mg/100 g da massa fresca da casca aos 10 DAC.
2.6.8 Carotenóides totais
Ao mesmo tempo que desaparece a cor verde, podem ser revelados ou sintetizados
pigmentos amarelos, alaranjados e vermelhos, que pertencem ao grupo dos carotenóides. Tais
compostos possuem quarenta átomos de carbono e a presença ou não de átomos de oxigênio
na sua estrutura. Os carotenóides são muito estáveis e permanecem nos tecidos durante a
senescência. Os de cor amarela são bastante comuns e sua presença é um sinal geral por meio
do qual o consumidor julga a maturidade e a qualidade de muitos frutos (AWAD, 1993).
Os pigmentos carotenóides localizam-se nos cromoplastos e também nos cloroplastos
associados com a clorofila. Tem como funções a proteção da clorofila e do aparelho
fotossintético contra a fotodegradação, bem como a absorção de luz em comprimento de onda
diferente do da clorofila, aumentando o potencial energético do sistema. São compostos
terpenóides formados por oito unidades de isopreno divididos em dois subgrupos: os
carotenos e seus derivados oxigenados – as xantofilas. São insolúveis em água e, usualmente,
diferentes compostos encontram-se nos tecidos vegetais em pequenas proporções, resultando
na gama de coloração característica de cada espécie e/ou cultivar. A coloração vermelha pode
ser decorrente de compostos diferentes, como é o caso do pimentão vermelho (que contém
capsantina/capsorubina) e do tomate (licopeno) (CHITARRA e CHITARRA, 2005).
O sistema de duplas ligações conjugadas dos carotenóides serve como cromóforo pela
sua habilidade de absorver luz na região visível e pelo seu poder corante. A cor intensifica-se
com o aumento do número de duplas ligações conjugadas na molécula. Por exemplo, o
licopeno contém 11 duplas ligações conjugadas, já o fitoflueno e o fitoeno, com cinco e três
duplas ligações conjugadas, respectivamente, são incolores. Do mesmo modo, a ciclização

46
nos terminais da molécula também tem relação com a coloração, por colocar as duplas
ligações de dentro dos anéis em plano diferente daquelas da cadeia poliênica. Assim sendo, o
γ-caroteno é laranja-avermelhado, ao passo que o β-caroteno é laranja (CHITARRA e
CHITARRA, 2005). Alguns frutos são também uma boa fonte de β-caroteno (pró-vitamina
A), que será transformado no organismo humano em vitamina A (retinol). Os frutos que se
destacam pelo teor de β-caroteno são: goiaba, manga, caqui, mamão, pêssego, damasco,
melão, maracujá e tomate (AWAD, 1993).
Também, na maturação, a biodegradação dos carotenóides resulta na formação de
alguns compostos voláteis, que podem contribuir para o aroma e o sabor típico de cada
espécie e/ou cultivar (CHITARRA e CHITARRA, 2005).
A diminuição na quantidade de clorofila e o aumento na concentração de carotenóides
podem, em certos casos (tomate), resultar da transformação de cloroplastos em cromoplastos.
Mesmo quando a síntese de clorofila é impedida na ausência da luz, os frutos de tomate
formam cromoplastos e sintetizam carotenóides. Nos frutos cítricos e na banana, a síntese de
carotenóides ocorre durante o desenvolvimento do fruto e bem antes do desaparecimento da
clorofila. Nesse caso, a destruição da clorofila revela a presença dos carotenóides. Os
cromoplastos podem também se desenvolver a partir de amiloplastos. A síntese de
carotenóides pode estar em certos casos (tomate) sob o controle do fitocromo. Nessa hipótese,
a luz vermelha (600 a 700 nm) induz a síntese de licopeno, enquanto a luz vermelha longa
inibe a síntese desse pigmento. Apesar da grande variabilidade de carotenóides nos frutos, os
mais importantes são o β-caroteno e o licopeno (AWAD, 1993).
Em tomates, há uma intensa degradação de clorofila durante o amadurecimento, com
síntese gradual do licopeno. Em banana, a degradação da clorofila é o principal evento, ao
passo que a síntese de outros pigmentos é realizada em níveis relativamente baixos. De modo
oposto, em algumas cultivares de maçã, as modificações na cor da casca são devidas,
preferencialmente, à síntese de carotenos e de antocianinas do que a uma grande degradação
da clorofila (CHITARRA e CHITARRA, 2005).
A oxidação é a principal causa de degradação dos carotenóides e depende da
disponibilidade de oxigênio, luz, calor, metais, enzimas e peróxidos, sendo reduzida pela
presença de antioxidantes como a vitamina C (CHITARRA, 2000).
Segundo Medlicott et al. (1986), o teor de carotenóides em mangas da cv. Tommy
Atkins é superior a 0,5 mg/ 100 g da massa fresca. Neste mesmo sentido, Salunke et al. (1991)
identificaram cerca de dezeseis hidrocarbonetos e oxicarotenóides, sendo que o β-caroteno
predomina no fruto totalmente maduro.

47
De acordo com Salunke e Desai (1984), os teores de pigmentos carotenóides de
mangas maduras variam consideravelmente entre cultivares, sugerindo que as respostas
metabólicas após a colheita podem variar, afetando a longevidade dos frutos das distintas
cultivares, pois constataram 0,9 mg/100 g da massa fresca da polpa para as cvs. Kent e
Peddarasam, 5,7 mg/100 g da massa fresca da polpa para a cv. Pairi, 5,9 mg/100 g da massa
fresca da polpa para a cv. Keitt, 8,3 mg/100 g da massa fresca da polpa para a cv. Alphonso,
9,2 mg/100 g da massa fresca da polpa para a cv. Irwin. Neste mesmo sentido, Medina,
Valdique (1996) afirma que o ponto ótimo de colheita da manga é quando o teor de
carotenóides totais atinge de 3 a 4 mg/100 g da massa fresca da polpa.
2.6.9 Fenólicos
Os compostos fenólicos são encontrados nas plantas em geral e particularmente
notórios nos frutos (VAN BUREN, 1970). Sua distribuição não é uniforme entre as espécies e
nem mesmo entre partes de uma mesma planta (WILLIAMS, 1957). Constituem uma das
principais classes de metabólitos secundários, sendo derivados da via do chiquimato e do
metabolismo dos fenilpropanóides e possuindo funções e estruturas diversas (ROBARDS et
al., 1999).
As principais substâncias classificadas como fenólicos são: os ácidos cinâmicos e seus
derivados, dos quais se destaca o ácido clorogênico; as flavanas; as antocianidinas e
antocianinas; os flavonóis e suas formas glicosídicas; os polifenóis condensados, cujos
precursores possivelmente sejam flavanas; e outros menos comuns, como flavonas,
flavononas e isoflavonas (VAN BUREN, 1970). Deve-se destacar também os taninos,
substâncias que podem formar pontes de hidrogênio entre grupos fenólicos e sítios receptores
de moléculas de colágeno, de maneira a constituir uma estrutura razoavelmente estável
(WHITE, 1957).
A nível subcelular, os fenólicos localizam-se principalmente nos vacúolos com
pequenas quantidades no espaço livre e nenhuma no citoplasma. Em alguns casos, verifica-se
acúmulo de fenólicos do tipo lignina e outras moléculas mais simples, como flavonóides e
ésteres do ácido ferúlico, na parede celular (ROBARDS et al., 1999).
A adstringência dos frutos é, até certo limite, determinada pelos compostos fenólicos
(WILLIAMS, 1957). E resulta da capacidade dos fenólicos de massa molar intermediária

48
(oligoméricos) formar complexos insolúveis com proteínas e mucopolissacarídeos da saliva,
reduzindo sua ação lubrificante (GOLDSTEIN e SWAIN, 1963).
Nos frutos, os compostos fenólicos estão presentes em diferentes graus de
polimerização e podem ser separados em frações, conforme suas solubilidades em solventes
orgânicos puros ou diluídos, portanto, são divididos em três classes com base no número de
anéis aromáticos (fenólicos) presentes. A fração solúvel em metanol absoluto contém
compostos diméricos, são dicíclicos como os flavonóides que possuem dois anéis fenólicos,
de baixa massa molar. A fração solúvel em metanol diluído contém os compostos com mais
de dois anéis fenólicos (oligoméricos), com massa molar intermediária. A fração solúvel em
água, os fenólicos poliméricos, são policíclicos ou polifenóis e contêm flavolanas que estão
firmemente ligadas aos polissacarídeos da parede celular ou a outros polímeros, de massa
molar superior às duas outras frações (GOLDSTEIN e SWAIN, 1963; FILGUEIRAS e
CHITARRA, 1988; LIZADA, 1993).
Comparado a outros tecidos, os níveis de fenólicos nos frutos são relativamente
baixos, mas podem ser significativos na determinação da qualidade (TUCKER, 1993).
Durante a maturação, por exemplo, a adstringência diminui possivelmente por causa da menor
solubilidade das flavanas altamente condensadas, presentes no meio e da sua ligação a outros
componentes celulares (VAN BUREN, 1970). Segundo Aziz e Yusof (1994), o aumento nos
teores de açúcares também contribui para a redução na adstringência.
Em mangas, predominam as formas mais simples (monoméricas), embora outras
formas existam principalmente na fase pré-climatérica (PARK et al., 1980). Os compostos
fenólicos de baixa massa molecular, incluindo os precursores de tanino são aparentemente
muito pequenos para formar ligações cruzadas suficientemente efetivas, sendo, portanto, não
adstringentes.
As mangas quando verdes são adstringentes e algumas cultivares apresentam teores de
fenólicos na faixa de 18,0 a 20,0 mg/100 g da massa fresca da polpa. Estes valores diminuem
com o avanço da maturação atingindo 6,0 a 4,0 mg/100 g da massa fresca da polpa
(GOLDSTEIN e SWAIN, 1963; SELVARAJ e KUMAR, 1989).
Outra importante resposta dos fenólicos na pós-colheita é o escurecimento dos tecidos,
em resposta a injúrias tais como cortes, amassaduras e a desordens fisiológicas. Quando o
escurecimento ocorre, os constituintes fenólicos são oxidados por enzimas específicas,
produzindo quinonas ou produtos semelhantes à quinonas, que polimerizam-se formando
pigmentos marrons (LIMA, 1997).

49
2.6.10 Pectinas
A “American Chemical Society” definiu as substâncias pécticas como sendo
compostas de: protopectina, ácido péctico, ácido pectínico e pectina. Esta classificação é
realizada de acordo com a proporção de grupos carboxílicos das cadeias poligalacturônicas
esterificadas por grupamentos metil-éster, com a presença de cadeias laterais glicosídicas e
com a solubilidade (SAKAI et al., 1993).
A protopectina é a substância péctica que forma a matriz, insolúvel em água e presente
nos tecidos de plantas que, por hidrólise parcial, fornecem pectina; a hidrólise mais completa
produz ácido péctico, ácido galacturônico e álcool metílico. Grande quantidade de
protopectina é encontrada nas frutas imaturas, porém, que já tenham atingido o seu pleno
desenvolvimento, sendo esta, juntamente com os produtos da hidrólise, as substâncias
pécticas das frutas verdes. No amadurecimento, a protopectina, pode ser hidrolisada
enzimaticamente à pectina que, por sua vez, pode ser decomposta formando álcool metílico e
ácido péctico, durante o amadurecimento excessivo ou apodrecimento da fruta. Portanto, a
transformação de protopectina em pectina solúvel é uma das causas do amaciamento das
frutas durante o amadurecimento (CETEC, 1985).
Os ácidos pectínicos e pécticos são constituídos principalmente por unidades de ácido
galacturônico; os primeiros apresentam uma porção considerável dos radicais carboxílicos
esterificados por grupamentos metil, ao passo que as carboxilas dos últimos são
essencialmente livres de metila. Os sais derivados da neutralização desses ácidos por bases
mono ou bivalentes são denominadas pectinato e pectato (SAKAI et al., 1993). Conforme
Raven et al. (2001), sais de cálcio e magnésio do ácido péctico formam a maior parte da
lamela média, uma camada de matéria intercelular que cimenta, unindo as paredes de células
vegetais adjacentes.
Pectinas referem-se aos ácidos pectínicos solúveis em água, com teores variados de
metilação e neutralização, apresentando ligações com cadeias laterais oligo ou
polissacarídicas. São estas as substâncias próprias para a preparação de geléias. Quanto ao
número de grupos metoxila presentes na molécula, as pectinas podem ser classificadas como
de baixo teor de metoxilação, quando for inferior a 7%, ou seja, menos de 50% de
esterificação e como de alto teor de metoxilação, quando superar os 7%, ou mais de 50% de
esterificação. As pectinas de baixa metoxilação não são solúveis em água e não possuem

50
capacidade de formar gel, enquanto as de alta metoxilação são solúveis em água e capazes de
formar gel (MAIA, 1997).
Uma parede celular primária consiste de duas, às vezes, de três camadas
estruturalmente independentes, mas interagindo entre si. A estrutura fundamental de celulose
e de glicanos de ligação cruzada fica embutida em uma segunda camada de polissacarídeos
pécticos da matriz. Uma terceira camada independente consiste de proteínas estruturais, ou de
uma camada de fenilpropanóides. As dicotiledôneas e monocotiledôneas não comelinóides
possuem parede tipo I onde a estrutura xyloglucana-celulose está embutida em uma matriz de
pectina, a qual desempenha muitas funções: determinando a porosidade da parede celular,
fornecendo as cargas das superfícies que modulam o pH da parede e o balanço de íons;
regulando a adesão célula-célula na lamela média; e servindo como moléculas de
reconhecimento que alerta as células da planta para a presença de organismos simbiônticos,
patogênicos e insetos (CARPITA e McCANN, 2000).
As pectinas formam uma fase gel hidratada na qual está implantada a rede celulose-
hemicelulose. Elas atuam como preenchimento hidrofílico, impedindo a agregação e o
colapso da rede de celulose. A parede primária é composta de aproximadamente 35% de
pectinas na base de matéria seca. Como as hemiceluloses, as pectinas incluem vários tipos
diferentes de polissacarídeos, caracteristicamente contendo açúcares ácidos, como ácido
galacturônico, e açúcares neutros, tais com ramnose, galactose e arabinose. As pectinas são os
mais solúveis dos polissacarídeos da parede, podendo ser extraídas com água quente ou com
queladores de cálcio. Na parede, as pectinas podem ter uma estrutura primária simples, tal
como o homogalacturonano e também podem ser grandes e complexas, compostas de tipos
diferentes de polissacarídeos pécticos, assim como os ramnogalacturonanos I e II, que
possuem cadeias laterais de arabinano e galactano. Nos géis pécticos, os grupos carboxila
(COO-) carregados de cadeias de pectinas vizinhas são ligados via Ca²+, que forma um
complexo firme com pectina (TAIZ e ZEIGER, 2004).
De acordo com Carpita e McCann (2000), enzimas específicas da parede celular
podem estar envolvidas com mudanças na rede de pectina, restringindo as suas atividades a
determinadas regiões da parede. Ao limitar a porosidade da parede, as pectinas podem afetar o
crescimento celular, regulando o acesso das enzimas que afrouxam a parede nos substratos
glicanos.
As hemiceluloses e as pectinas podem ser modificadas e quebradas por uma variedade
de enzimas encontradas naturalmente na parede da célula. O processo tem sido estudado

51
detalhadamente em frutos em amadurecimento, cujo amaciamento é considerado resultado da
desagregação da parede (ROSE e BENNETT, 1999).
A maioria dos frutos em que o pericarpo ou endocarpo amolece durante o
amadurecimento desenvolve-se um engrossamento das paredes primárias que são
notadamente enriquecidas com substâncias pécticas, como homogalacturonano e
ramnogalacturonano I. A textura da polpa dos frutos é definida pelo grau de degradação da
parede e perda da adesão celular. Por exemplo, a parede das células do córtex da maçã sofre
poucas mudanças na rigidez e pouca separação celular, enquanto que as paredes celulares das
células do pericarpo do pêssego e tomate amolecem consideravelmente através da expansão
da parede e perda da adesão celular. Em tomate os lóculos que contêm as sementes se
dissolvem completamente em um processo chamado de liquefação. As pectinas
frequentemente constituem mais de 50% da parede celular das frutas (CARPITA e McCANN,
2000).
A determinação da pectina total é a soma das pectinas solúvel e insolúvel em álcool. Já
a determinação da pectina insolúvel em álcool, fração chamada por Esteban et al., (1993)
como sólidos insolúveis em álcool (SIA) deve ser realizada por extração seqüencial
resultando em três classes distintas de substâncias pécticas, sugerida pelo método de
Robertson (1979). O fracionamento envolve a extração por uma série de solventes: água
(ácidos pectínicos ou pectinas de alta metoxilação); solução de oxalato de amônio (ácidos
pécticos ou pectinas de baixa metoxilação) e solução de ácido clorídrico (protopectina ou
pectina ligada covalentemente).
Kapse et al. (1988) verificaram, em seu trabalho, que o desenvolvimento do
amaciamento estava relacionado com a degradação de substâncias pécticas, em mangas das
cvs. Malda e Malgoa, sendo mais acelerado quando os frutos foram armazenados à
temperatura ambiente (25 a 30°C), do que à baixa temperatura (10 ± 1°C), onde a taxa de
conversão foi mais lenta. Lucena et al. (2001), ao estudarem técnicas de conservação pós-
colheita da manga, cv. Tommy Atkins armazenada à temperatura ambiente e refrigerada,
também constataram que os frutos armazenados sob refrigeração tiveram a maturação
retardada.

52
2.7 Avaliações bioquímicas
2.7.1 Pectinametilesterase e poligalacturonase
As enzimas pectinametilesterase (PME) e poligalacturonase (PG), pertencentes ao
grupo das hidrolases, são conhecidas como as enzimas pécticas importantes nos vegetais, pois
podem ocasionar o excessivo amaciamento de frutas e hortaliças. A PME catalisa a remoção
de grupos metoxílicos das moléculas de pectina e de ácido pectínico para formar ácido
péctico. A PG ocasiona a quebra das ligações glicosídicas das substâncias pécticas para
formar finalmente o ácido galacturônico (GAVA, 1984).
Duas poligalacturonases estão geralmente presentes: a endopoligalacturonase que
ataca a molécula de pectina em vários sítios dentro da cadeia e a exopoligalacturonase que
remove seqüencialmente os resíduos de ácido galacturônico a partir da extremidade da
molécula. Das duas, a endopoligalacturonase é muito mais importante com relação às
mudanças físicas na textura, embora, os seus níveis variem de acordo com a espécie e a
cultivar. O processo de solubilização das pectinas contribui para o amaciamento dos tecidos
em decorrência da redução da força de coesão entre as células (KAYS, 1991).
Durante o amaciamento de frutos, estes expressam níveis altos de PME, que hidrolisa
os ésteres metílicos de pectinas. Essa hidrólise torna a pectina mais suscetível à hidrólise
subseqüente por pectinases e enzimas afins, cuja presença, assim como a de enzimas afins na
parede da célula, indica que as paredes são capazes de modificação significativa durante o
desenvolvimento (TAIZ e ZEIGER, 2004).
O processo de amaciamento no tecido parenquimático do tomate está associado com a
perda de metil ésteres do homogalacturonano, em conseqüência da atividade da PME, que
remove os grupos metil éster dos resíduos do α–D-ácido galacturônico das cadeias de
polissacarídeos pécticos. A cadeia de homogalacturonano desesterificado, neste caso, está
susceptível a atividade da PG. A PG I com uma massa molecular de aproximadamente 100
kDa, consiste de PG II de 46 kDa complexada firmemente com uma subunidade β. A
subunidade β é uma proteína rica em aminoácidos aromáticos. Acredita-se que ela atue como
um componente âncora para a subunidade PG II sendo sintetizada precocemente no
desenvolvimento do fruto. A subunidade β pode solubilizar pectinas da parede celular,

53
facilitando progressivamente hidrólises por PG II nas ligações glicosídicas dentro da cadeia
de homogalacturonano não ramificado. Modificação de pectina dentro da parede durante o
amadurecimento é um processo regulado com precisão, o resultado de tal mecanismo é a
modificação do substrato, que restringe o acesso da enzima, ou a presença de enzimas
inibidoras, como na inibição da atividade da PG II por produtos difusíveis da
despolimerização de pectinas (CARPITA e McCANN, 2000).
Em manga, concomitante ao amaciamento, foi observado aumento na atividade das
enzimas pectolíticas, poligalacturonase e pectinametilesterase (AINA e OLADUNJOYE,
1993); decréscimo na atividade da pectinametilesterase e aumento na atividade da
poligalacturonase e celulase (ABU-SARRA e ABU-GOUKH, 1992); e decréscimo nos teores
de açúcares neutros da parede celular (MITCHAM e McDONALD, 1992).
2.7.2 Polifenoloxidase e peroxidase
A polifenoloxidase (PPO) tem duas diferentes atividades catalíticas, ambas
envolvendo o oxigênio. Elas são chamadas de atividades cresolásica e catecolásica. A
cresolase está relacionada com a oxidação de fenóis monoidroxilados, como a tirosina, fenol
ou ortocresol, para formar outro grupo hidroxílico. Os dois elétrons são fornecidos pelo cobre,
sempre associado à enzima. A catecolase envolve a remoção de dois hidrogênios de fenóis
diidroxilados, como o catecol ou diidrofenilalanina, para dar uma ortoquinona
correspondente. As quinonas, por polimerização, produzem melanoidinas. Esta oxidase é
responsável pelo escurecimento enzimático em muitos produtos (GAVA, 1984).
Usualmente, o escurecimento ocorre devido a ferimentos no produto durante as
operações de colheita, armazenamento ou processamento. As quinonas resultantes da
oxidação dos fenólicos apresentam coloração vermelha a marrom-avermelhada; no entanto, a
sua polimerização subseqüente gera macromoléculas mais escuras. Assim, o escurecimento
enzimático dos tecidos vegetais depende não só do tipo de substrato, concentração e
localização, como também do tipo de polímero formado a partir da quinona. Cultivares com
baixa atividade de PPO são desejáveis tanto para o consumo in natura como para o
processamento. No entanto, produtos com elevada atividade dessa enzima podem apresentar
maior resistência ao ataque de patógenos (CHITARRA e CHITARRA, 2005).

54
As PPOs desempenham numerosas funções fisiológicas nas plantas, tais como: na
regulação de potenciais de oxi-redução; em reações do metabolismo intermediário; na
respiração; no funcionamento do fotossistema II da fotossíntese, uma vez que oxidam as
quinonas reduzidas formadas no fotossistema I; na proteção contra patógenos e predadores; na
cicatrização de ferimentos; na formação de pigmentos e na sinalização do amadurecimento
dos frutos (WHITE, 1957; MAYER e HAREL, 1991; SILVA, E., 2000).
Em estudo com sete cultivares de manga (Alphonso, Banganapalli, Dasheri, Fazli,
Langra, Suvarnarekha e Totapuri), em quatro estádios de maturação, foi observado um
aumento da atividade das PPOs desde a maturidade fisiológica até o amadurecimento, seguido
de um declínio da atividade em algumas cultivares (Banganapalli, Dasheri, Fazli e Langra) e
um decréscimo a partir da maturidade fisiológica até alcançar o estádio de maturação
comestível nas demais cultivares estudadas (SELVARAJ e KULMAR, 1989).
Já a peroxidase (POD) decompõe a água oxigenada e um substrato, tal como um fenol
ou ácido ascórbico, produzindo hidróxido e água. Pelo fato de ser facilmente determinada e
por ser uma das enzimas mais resistentes ao calor, a POD é utilizada como indicação da
eficiência do “blanching”, isto é, inativação de enzimas pelo calor, em muitos produtos
(GAVA, 1984).
As isoenzimas de POD atuam sobre diferentes substratos, em reações, tais como:
oxidação de fenólicos e carotenóides, degradação de auxinas, de clorofila e de ácido
ascórbico, bem como na biossíntese da lignina. Portanto, a sua atividade relaciona-se com
modificações nos atributos sensoriais (escurecimento, endurecimento, sabores estranhos) e no
valor nutritivo (perda de atividade vitamínica do ácido ascórbico) dos produtos hortícolas.
Tem função relacionada aos processos de desenvolvimento e de senescência nos tecidos. A
sua atividade aumenta significativamente após a colheita, quando uma gama de compostos
torna-se suscetível à sua ação. O aumento da atividade da POD também está associado com a
biossíntese de compostos das paredes celulares em resposta a danos mecânicos nos tecidos,
resistência a doenças e nos mecanismos de cura ou reparo de ferimento dos tecidos. Elevados
níveis de POD também são associados com a deterioração oxidativa de muitas plantas ou de
frutas que se encontram em estádio avançado de amadurecimento ou senescência
(CHITARRA e CHITARRA, 2005).
Em manga, a atividade da POD aumenta nos primeiros estádios de amadurecimento, e
depois permanece quase constante (ZAUBERMAN et al., 1988; MARIN e CANO, 1992).
Os níveis de fenóis também podem ser muito significativos na determinação da
qualidade dos frutos (TUCKER, 1993). A grande maioria dos fenóis encontrado nos frutos

55
não têm características particulares de sabor quando testados a baixas concentrações na forma
pura. Exceções a esta regra são a acidez associada aos ácidos fenólicos, a adstringência de
flavanas condensadas e o amargor associado a alguns flavonóides de citrus (VAN BUREN,
1970). A perda de adstringência é um dos principais fatores que ocorrem durante o
amadurecimento de muitos frutos e se deve à polimerização de oligômeros adstringentes que
são convertidos em polímeros reativos insolúveis (GOLSTEIN e SWAIN, 1963). Portanto,
neste sentido, a oxidação de fenóis pode resultar da atividade das polifenoloxidases, das
peroxidases ou mesmo sem participação de enzimas (AWAD, 1993).
2.7.3 α- e β-Amilases
Amilases e fosforilases são enzimas que atuam sobre a ligação α(1→4) de polímeros
da glicose como o amido e glicogênio, transformando-os em molécula de menor peso
molecular. Entre as mais importantes temos a α-amilase, a β-amilase e a amido fosforilase. A
α-amilase (enzima dextrinizante) ataca as ligações α(1→4) da amilose ao acaso, produzindo
pequenas dextrinas limite, oligossacarídeos, maltose e glicose. Ao contrário, a β-amilase
(enzima sacarificante) ataca apenas a penúltima ligação, hidrolisando o amido e fornecendo
maltose. A amido fosforilase hidrolisa a ligação terminal α(1→4) para formar glicose-1-
fosfato. Em todos os casos, as enzimas são ativas somente em cadeias glicolíticas de amilose e
amilopectina encontradas no amido. Elas não degradam as ligações α(1→6) encontradas na
amilopectina do amido. Como conseqüência da atividade destas enzimas no amido, por
exemplo, tem-se uma diminuição da viscosidade (por causa do rompimento da cadeia do
polissacarídeo), perda na capacidade do iodo em dar uma coloração azul e no aparecimento de
grupos redutores (GAVA, 1984; AWAD, 1993; TUCKER, 1993).
A α-amilase pode ser responsável por 80% da atividade amilolítica no fruto, portanto,
as atividades β-amilásica e da fosforilase, são bem menores. Na manga ocorre um aumento na
atividade amilásica e na hidrólise do amido durante a maturação, pois este é o principal
carboidrato presente no fruto imaturo (AWAD, 1993).
Tandon e Kalra (1983) trabalhando com mangas ‘Dashehari’, verificaram que a
atividade amilásica aumentou durante o crescimento do fruto, decrescendo quando
amadureceu, constatando assim que provavelmente o decréscimo está associado à redução no
conteúdo de amido que ocorreu nos últimos estádios de desenvolvimento.

56
2.7.4 α- e β-Galactosidases de citosol e de parede celular
Conforme Awad (1993), a β-galactosidase (β-GAL) é uma enzima presente durante a
maturação e o amaciamento de alguns frutos. Ela libera resíduos de galactose da parede
celular da maçã, do tomate, do morango e da pêra. O significado dessa hidrólise no
amaciamento dos frutos não está bem esclarecido. Outra enzima que também libera resíduos
solúveis da parede celular durante a maturação e o amaciamento dos frutos é a α-galactosidase
(α-GAL).
A α- e β-GAL promovem a remoção de galactose na estrutura básica de galactanos e
em cadeias laterais de ramnogalacturonanos I e II, atuando como pectinases, bem como, em
cadeias laterais de xiloglucanos, atuando agora como hemicelulases (TAIZ e ZEIGER, 2004;
CHITARRA e CHITARRA, 2005).
Várias glicosidases também ocorrem nas paredes e algumas aumentam com o
amadurecimento (BRADY, 1987). Ali et al. (1995) registraram as atividades de várias
enzimas deste grupo durante o amadurecimento de manga cv. Harumanis: β-D-glicosidase, α-
L-arabinosidase, α-D-manosidase, α-D-galactosidase e β-D-galactosidase, sendo a última a
predominante. β-galactosidase, α-galactosidase, α-glicosidase e β-glicosidase também têm
alta atividade em laranja ‘Valencia’ (BURNS, 1990). Em melancia, destaca-se a atividade da
α-galactosidase (ITOH et al., 1986). E em maçã, a α-L-arabinofuranosidase pode participar
da perda de resíduos de arabinosil dos poliuronídeos da parede celular (YOSHIOKA et al.,
1995).
2.7.5 Proteínas de citosol e de parede celular
As proteínas estão entre as mais abundantes moléculas orgânicas. Na maioria dos
organismos vivos, as proteínas perfazem 50% ou mais da massa seca. Apenas as plantas, com
o seu alto conteúdo de celulose, têm menos do que 50% de proteína (glicoproteínas e
enzimas). Em sua estrutura, contudo, as proteínas seguem sempre o mesmo esquema: todas
são polímeros de moléculas contendo nitrogênio, conhecidas como aminoácidos, arranjadas
em uma seqüência linear (RAVEN et al., 2001).

57
Embora a constituição estrutural da parede celular seja principalmente composta de
carboidratos, cadeias de proteínas estruturais também são formadas na parede. Existem quatro
grandes classes de proteínas estruturais: glicoproteínas ricas em hidroxiprolina (HRGPs),
proteínas ricas em prolina (PRPs), proteínas ricas em glicina (GRPs) e proteínas
arabinogalactanas (AGPs). As três primeiras são conhecidas pela riqueza em certos
aminoácidos, enquanto a última é denominada de proteoglicanas, pois possui mais de 95% de
carboidratos. A extensina, codificada por uma família multigênica, é uma das mais bem
estudadas HRGPs nas plantas (CARPITA e McCANN, 2000). As glicoproteínas que são
componentes da matriz compreendem cerca de 10% da massa seca de muitas paredes
primárias (RAVEN et al., 2001).
A extensina é uma glicoproteína rica em hidroxiprolina com função de reforço e
proteção, ou seja, proteção do ferimento contra a dessecação, reforço da arquitetura celular e
resistência às doenças. Uma importante característica da extensina é a sua insolubilidade na
parede celular. Na forma monomérica solúvel, pode ser insolubilizada na parede celular por
meio de ligações cruzadas dos monômeros pela ação de peroxidases ligadas às paredes, sendo
um importante componente na formação de barreiras estruturais nos tecidos feridos. Essas
enzimas podem gerar ligações cruzadas nos polímeros das paredes celulares por meio da
formação de ligações bifenil, tais como as do acoplamento de resíduos de tirosina na
extensina (CHITARRA e CHITARRA, 2005).
O teor protéico de frutas, hortaliças (1 a 2%) e legumes (5%) é baixo e, portanto, não é
nutricionalmente significativo, mas tem papel funcional, atuando nos mecanismos
metabólicos como enzimas, compondo a estrutura das paredes celulares ou como parte de
macromoléculas como glicoproteínas, lipoproteínas, com diferentes funções no vegetal.
Entretanto, o consumo intenso de frutos como a banana, abacate e manga, com teores de
proteína entre 1 a 2%, pode contribuir modestamente para a satisfação das necessidades
protéicas do indivíduo (AWAD, 1993; BOAS, 1999; CHITARRA e CHITARRA, 2005).

58
3 MATERIAL E MÉTODOS
Este trabalho foi conduzido na Fazenda Fruit Fort Agrícola Exportação Ltda e no
Laboratório de Fisiologia Pós-Colheita da Embrapa Semi-Árido, situados no município de
Petrolina, PE, durante o período de Fevereiro/2004 a Fevereiro/2005, bem como, nos
Laboratórios de Fisiologia e Tecnologia Pós-Colheita da Embrapa Agroindústria Tropical e de
Fisiologia Vegetal do Departamento de Bioquímica e Biologia Molecular, da Universidade
Federal do Ceará - UFC, em Fortaleza, CE, durante o período de Dezembro de 2004 a Janeiro
de 2006.
3.1 Localização, caracterização edáfica, climática e manejo da área de cultivo
O clima da região é do tipo BSwh’, segundo a classificação de Köeppen, que
corresponde a uma região quente e semi-árida. A área de plantio apresenta as seguintes
características: latitude 9º22’S, longitude 40º33’W, altitude 388 m, temperatura média
26,05ºC, com médias máximas de 32,03ºC e médias mínimas de 20,38ºC, umidade média
mensal é de 66,25%, radiação solar global média de 20,94 MJ/m2/dia, a velocidade média dos
ventos é de 2,05 m/s com direções E e S e precipitação pluvial média anual é de 535,53 mm; o
solo é classificado como latossolo vermelho amarelo-LV (EMBRAPA, 2006).
Nas FIGURAS 1 e 2 encontram-se as características climáticas médias ou totais
referentes ao ano que antecedeu as colheitas, de acordo com os dados fornecidos pelo Centro
de Pesquisa Agropecuária do Tropico Semi-árido – Embrapa. Os dados foram coletados a
partir da Estação Meteorológica Automatizada na Fazenda Fruit Fort Agrícola Exportação
Ltda.

59
,
,
,
, ,,
,,
, ,,
, , ,'
''
' ' ' '' ' ' '
'
' '
&&&
& & &&
&& &
&&
& &
)))
) ) ) )) ) ) )
)) )
nov/0
3jan
/04
mar/04
mai/04
jul/04
set/0
4
nov/0
4
Mês/Ano
15
20
25
30
35
40Te
mpe
ratu
ra (º
C)
35
45
55
65
75
Um
idade relativa (%)
T. média T. máxima T. mínima U. relativa) & ' ,
FIGURA 1 – Temperaturas e umidade relativa do ar na região de Petrolina – PE durante o ano
que antecedeu as colheitas (outubro a dezembro/04) das mangas ‘Tommy Atkins’.
$$$
$
$
$
$ $ $ $ $ $ $ $
*
*
*
* **
** *
*
*
*
*
*
nov/0
3jan
/04
mar/04
mai/04
jul/04
set/0
4
nov/0
4
Mês/Ano
15
20
25
30
Rad
iaçã
o so
lar g
loba
l (M
J/m
²/di a
)
0
100
200
300
400
500
Precipitação (m
m)
Radiação solar global Precipitação* $
FIGURA 2 – Radiação solar global e precipitação pluviométrica na região de Petrolina – PE
durante o ano que antecedeu as colheitas (outubro a dezembro/04) das mangas ‘Tommy Atkins’.

60
No início do ciclo, em Novembro de 2003, foram feitas análises de fertilidade do solo
da rizosfera e foliar das mangueiras. Os resultados destas análises encontram-se listadas na
TABELA 1. De posse dos resultados das análises, foram incorporados na adubação nitrogênio
(nitrato de cálcio-418,3 g/planta e uréia-0,53 g/planta), fósforo (MAP-571,15 g/planta, ácido
fosfórico-0,061 g/planta e nutriox P30-9,47 mL/planta) e potássio (cloreto de potássio-0,61
g/planta, sulfato de potássio-682 g/planta e nitrato de potássio-1.067 g/planta). O magnésio foi
incorporado na forma de sulfato de magnésio (228,03 g/planta). Os micronutrientes (Zn, Fe, B
e Mn) também foram incorporados. Os macro e micronutrientes foram aplicados via
pulverização e fertirrigação em 16 e 6 parcelas respectivamente, distribuídas eqüitativamente
durante todo o ciclo da cultura.
TABELA 1 – Análises de fertilidade do solo da rizosfera e de tecido foliar de mangueiras cv. Tommy Atkins, no início do ciclo da cultura em Novembro de 2003.
Determinações Solo (0-20 cm) Solo (20-40 cm) Folha N - - 12,10 g/kg
M.O. 8,00 g/dm3 2,00 g/dm3 - pH SMP 7,40 7,40 -
pH 6,20 6,20 - P 23,00 mg/dm3 12,00 mg/dm3 1,20 g/kg K 1,30 mmolc/dm3 1,20 mmolc/dm3 7,30 g/kg
Ca2+ 30,00 mmolc/dm3 18,00 mmolc/dm3 20,30 g/kg Mg2+ 4,00 mmolc/dm3 3,00 mmolc/dm3 1,10 g/kg Na 0,20 mmolc/dm3 0,20 mmolc/dm3 185,00 mg/kg
Al3+ < 1,00 mmolc/dm3 < 1,00 mmolc/dm3 280,00 mg/kg H 9,70 mmolc/dm3 9,70 mmolc/dm3 -
H + Al 10,00 mmolc/dm3 10,00 mmolc/dm3 - S.B. 35,30 mmolc/dm3 22,20 mmolc/dm3 -
C.T.C. 45,00 mmolc/dm3 31,90 mmolc/dm3 - V 78,00% 70,00% - S 4,00 mg/dm3 2,00 mg/dm3 1,30 g/kg B 0,33 mg/dm3 0,29 mg/dm3 97,86 mg/kg Cu 10,30 mg/dm3 1,90 mg/dm3 28,00 mg/kg Fe 14,00 mg/dm3 34,00 mg/dm3 175,00 mg/kg Mn 34,50 mg/dm3 16,50 mg/dm3 275,00 mg/kg Zn 145,80 mg/dm3 285,80 mg/dm3 17,00 mg/kg
O sistema de irrigação empregado foi microaspersão, com turno de rega diário,
iniciado após a indução floral com uma lâmina bruta (LB) de 89 litros/planta, sendo ao longo
do ciclo ajustado em função das características climáticas registradas. O controle de plantas
invasoras foi realizado mensalmente, através de capinas com enxada e roço com estrovenga
ou com roçadeira acoplada ao trator. As mangueiras receberam todos os tratos culturais de
acordo com as exigências da cultura.

61
3.2 Indução floral
Utilizou-se para indução floral, mangueiras da cv. Tommy Atkins enxertada sobre a
cv. Espada, com 21 anos de idade, pertencentes ao setor FF1C1 (Lote 642) da Fazenda Fruit
Fort. O lote media 6,09 ha. Era composto de 655 plantas, com altura média de 5,5 m e
estavam dispostas no espaçamento 10 x 10 m.
Para indução floral procedeu-se da seguinte forma:
i) 16/02/04–aplicou-se 34 mL de Cultar (paclobutrazol)/planta, via solo;
ii) 20/04/04–foi feito a 1ª aplicação de sulfato de potássio a 2%, via pulverização;
iii) 30/04/04–foi feito a 2ª aplicação de sulfato de potássio a 2%, via pulverização;
iv) 01/05/04–suspendeu-se a irrigação;
v) 11/05/04–foi feito a 3ª aplicação de sulfato de potássio a 2%, via pulverização;
vi) 20/05/04–aplicou-se 3,74 mL de Ethrel (ethefon)/planta com sulfato de potássio a 2%, via
pulverização;
vii) 31/05/04–aplicou-se 9,1 mL de Ethrel (ethefon)/planta com sulfato de potássio a 2%, via
pulverização;
viii) 02/07/04–aplicou-se nitrato de potássio a 5%, via pulverização;
ix) 09/07/04–aplicou-se nitrato de potássio a 4%, via pulverização;
x) 16/07/04–aplicou-se nitrato de potássio a 3%, via pulverização;
xi) 19/07/04–retomou-se a irrigação com lâmina bruta (LB) = 89 litros/planta; e
xii) 23/07/04–Finalmente aplicou-se nitrato de cálcio a 2,5%, via pulverização.
3.3 Marcação das panículas
Durante a antese, isto é, quando 65% das plantas haviam emitido panículas, foram
selecionadas 211 plantas, onde 797 panículas foram marcadas com fitas de plástico e
monitoradas até 112 dias após a antese (DAA). Em cada planta foi marcada uma panícula por
quadrante quando disponível. As panículas marcadas foram padronizadas em padrão 1 e 2,
conforme as características descritas na TABELA 2 e observadas na FIGURAS 3.

62
TABELA 2 - Características das panículas padrões 1 e 2 em mangueira, cv. Tommy Atkins. Panícula Características Padrão 1 Padrão 2
Comprimento do ápice 10 cm 10 cm Comprimento total 30 cm 30 cm % de Flores visualmente abertas 100% 90% % de Flores visualmente abertas no ápice 100% 0% % de Flores visualmente fecundadas < 5% 0% Formato Pluma Pena
A B
FIGURA 3 – Panículas padrões 1 (A) e 2 (B) em mangueira ‘Tommy Atkins’.
3.4 Fenologia da floração à frutificação
Marcaram-se 10 panículas padrão 1, 10 panículas padrão 2 e 10 ramos com gemas
diferenciando-se para emissão de panícula (FIGURA 4), onde periodicamente, de 3 em 3 dias
foram feitas as seguintes determinações:
• Número de flores em antese nas panículas padrões 1 e 2;
• Número de flores fecundadas nas panículas padrões 1 e 2;

63
• Número de botões nas panículas padrões 1 e 2;
• Número total de flores nas panículas padrões 1 e 2;
• Número de dias para atingir os estádios padrões 1 e 2;
• Número de dias para atingir o estádio chumbinho nas panículas padrões 1 e 2;
• Número de dias para atingir o estádio bola de gude nas panículas padrões 1 e 2;
• Número de dias para atingir o estádio ovo nas panículas padrões 1 e 2; e
• Número de dias para atingir o estádio fruto nas panículas padrões 1 e 2.
FIGURA 4 – Início da diferenciação floral em ramo de mangueira ‘Tommy Atkins’.
3.5 Colheita e manuseio das mangas
Antes da colheita, as mangas foram individualmente numeradas. Colheu-se ao acaso,
na Fazenda Fruit Fort, 24 frutos aos 35, 49, 63, 70, 77, 84, 91, 98, 105 e 112 dias após a
antese (DAA) de Outubro a Dezembro de 2004.
Após a colheita, transportaram-se os frutos para o Laboratório de Fisiologia Pós-
Colheita da Embrapa Semi-Árido, onde foram feitas as avaliações físicas com metade das
mangas colhidas e a outra foi descascada e cortada em cubinhos de 1,5 cm3, sendo uma parte

64
separada para a realização das análises de sólidos solúveis totais, acidez total titulável, pH,
vitamina C e clorofila total e a outra congelada com N2 líquido (-196ºC), acondicionadas em
sacos transparentes de polietileno de baixa densidade (27,0 x 31,0 cm) com fecho hermético,
devidamente identificados e transportadas em caixa de isopor com gelo para os Laboratórios
de Fisiologia e Tecnologia Pós-Colheita da Embrapa Agroindústria Tropical e de Fisiologia
Vegetal do Departamento de Bioquímica e Biologia Molecular, da Universidade Federal do
Ceará - UFC. Então, as amostras foram armazenadas em ultra-freezer a -85ºC até que se
procedesse o restante das análises. Para a realização das análises físico-químicas, químicas e
bioquímicas da polpa os cubinhos de manga foram desintegrados em centrífuga doméstica.
3.6 Avaliações físicas
3.6.1 Aspectos morfológicos externos
Foram determinados o formato do ombro, a textura da casca, o brilho da casca, o
distanciamento das pontuações brancas, o formato do nariz e o formato do ápice. Na FIGURA
5 pode-se observar os tipos de formatos do ombro, do nariz e do ápice da manga durante o
desenvolvimento.
FIGURA 5 – Formatos do ombro, do nariz e do ápice de mangas ‘Tommy Atkins’ em
desenvolvimento fisiológico-A (não deve ser colhida) e fisiologicamente desenvolvida-B (pode ser colhida). Adaptado de Bleinroth, 1994.

65
3.6.2 Diâmetros longitudinal, ventral, transversal e produto dos diâmetros
Os diâmetros foram determinados mediante o uso de um paquímetro digital modelo
SC-6 com escala graduada em milímetros. As leituras, em mm, foram divididas por 10 para
expressar o resultado em centímetros (cm). Veja na FIGURA 6 a ilustração de como foram
feitas estas mensurações.
FIGURA 6 – Ilustração dos diâmetros longitudinal (DL), ventral (DV) e transversal (DT) de
mangas ‘Tommy Atkins’ (adaptado de Campbell, 1992).
O volume dos frutos foi calculado individualmente através do produto entre os
diâmetros longitudinal, ventral e transversal (CASTRO NETO e REINHARDT, 2003). Os
resultados deste método são apresentados como produto dos diâmetros e expressos em
centímetros cúbicos (cm³).
3.6.3 Massas fresca, seca, de água e o teor de água
Para a determinação da massa fresca, os frutos foram pesados individualmente em
balança semi-analítica com precisão de 0,1 g. Todo o processo da colheita do fruto até o final

66
das pesagens foi efetuado em, no máximo, 60 minutos. Este tempo foi rigorosamente mantido
para evitar a perda de água e variação entre as amostragens. Todas as pesagens foram
expressas em gramas (g).
Para a determinação da massa seca, os frutos foram fracionados e colocados para secar
em estufa de circulação forçada de ar a 60 ± 2ºC até a obtenção de massa constante em
balança semi-analítica com precisão de 0,1 g. O tempo de secagem variou de 24 a 72 horas,
de acordo com o tamanho do fruto. Após a secagem, o material foi colocado em dessecador
contendo sílica gel para esfriar de forma a minimizar a sua rehidratação antes da pesagem.
Todas as pesagens foram efetuadas individualmente e expressas em gramas (g).
A massa de água foi determinada por diferença entre a massa fresca (MF) e a massa
seca (MS), sendo, portanto, expresso em gramas (g).
O teor de água foi determinado através da fórmula: Teor de água = massa de água x
MF-1 x 100, sendo, portanto, expresso em percentagem (%).
3.6.4 Escalas de coloração da casca e da polpa
A coloração verde da casca foi obtida através do uso de uma escala subjetiva conforme
GTZ (1992) e Amorim (2002), denominada escala de coloração da casca (ECC): 1-verde
escuro; 2-verde claro; 3-verde-amarelo; 4-amarelo-verde; e 5-amarelo (FIGURA 7). Os
resultados foram expressos em média de notas atribuídas por repetição, através do exame
individual de cada manga por quatro avaliadores treinados.
A coloração vermelha da casca foi obtida através do uso de uma medida subjetiva
onde é avaliado o percentual de cor vermelha na manga (AMORIM, 2002), denominada
escala de Blush para coloração da casca (EBCC). A escala é enumerada de 1 a 5, cada número
correspondendo a uma faixa percentual da cor vermelha: 1-0%; 2->0-25%; 3->25-50%; 4-
>50-75%; e 5->75-100%. Os resultados foram expressos em média de notas atribuídas por
repetição, através do exame individual de cada manga por quatro avaliadores treinados.
A coloração da polpa foi obtida através do uso de uma escala subjetiva conforme GTZ
(1992) e Filgueiras et al. (2000), denominada escala de coloração da polpa (ECP):
1-cor creme: a polpa apresenta a cor creme por completo, podendo variar de creme-
claro a creme-escuro. Não se deve confundir a cor creme com a cor branca;

67
2-mudança da cor creme: há uma mudança em até 30% da área com cor creme para a
cor amarela, partindo do centro do fruto;
3-amarelo: corresponde 30 a 60% da polpa com cor amarela;
4-amarelo-laranja: corresponde a mais de 60% da cor amarela e menos de 30% de cor
laranja; e
5-laranja: corresponde a mais de 90% da cor laranja.
Os resultados foram expressos em média de notas atribuídas por repetição, através do
exame individual de cada manga por quatro avaliadores treinados. A escala de coloração da
polpa para manga pode ser vista na FIGURA 7.
FIGURA 7 – Escalas de coloração da casca e da polpa como guia de maturação das mangas
‘Tommy Atkins’ (adaptado de GTZ, 1992). 3.6.5 Coloração da polpa
Foi realizada com Colorímetro Minolta, modelo CR-300, o qual expressa a cor em três
parâmetros: L, que corresponde a luminosidade (brilho, claridade ou reflectância; 0-
escuro/opaco e 100-branco); C, o croma (saturação ou intensidade da cor; 0-cor impura e 60-

68
cor pura); e H, o ângulo Hue (ângulo da cor; 0º-vermelha; 90º-amarelo; 180º-verde; 270º-azul
e 360º-negro).
3.6.6 Firmeza
Foi determinada em mangas integras, usando-se penetrômetro manual Magness-Taylor
modelo FT 011 com ponta de 8 mm de diâmetro, após a retirada da casca. Foram feitas duas
leituras por manga, em lados opostos da porção equatorial, objetivando efetuar a média. As
leituras, em lbf, foram multiplicadas por 4,4482 para expressar o resultado da força necessária
para romper a resistência da polpa em Newtons (N).
3.6.7 Unidades de calor (graus-dia)
As determinações de unidades de calor (UC) foram feitas de acordo com Chitarra e
Chitarra (2005), onde se calculou a temperatura média diária, cuja soma algébrica foi
deduzida da temperatura mínima de crescimento:
UC = Σ (Tmáx + Tmín/2) – Tbase, onde:
UC = unidades de calor, ºC;
Tmáx = temperatura máxima do ar, ºC;
Tmín = temperatura mínima do ar, ºC; e
Tbase = temperatura base para a planta, ºC.
A temperatura base para a mangueira dada por Chaudhri (1976) e Silva (1996),
representa a temperatura de paralisação do crescimento da planta, neste sentido, estes autores
sugerem o valor de 10ºC.

69
3.7 Avaliações físico-químicas e químicas
3.7.1 Sólidos solúveis totais, acidez total titulável, pH e relação SST/ATT
O teor de sólidos solúveis totais (SST) foi determinado usando-se um refratômetro
digital Atago modelo PR-101, escala de 0 a 45°Brix, com compensação de temperatura
automática (AOAC, 1995).
Determinou-se a acidez total titulável (ATT) por titulometria com solução de NaOH
0,1 N, utilizando titulador automático Mettler DL 12 e expressando os resultados em
percentagem (%) de ácido cítrico na massa fresca (IAL, 1985).
O pH foi medido em potenciômetro digital com eletrodo de membrana de vidro
(AOAC, 1995).
A relação SST/ATT foi obtida através do quociente entre as duas variáveis.
3.7.2 Determinação dos teores de carboidratos
O teor de amido foi determinado conforme metodologia descrita em AOAC (1995),
com algumas adaptações. Amostra de 5 g de polpa foi diluída em 50 mL de água destilada e
centrifugada a 1.000 g, durante 10 min., por três vezes. Ao resíduo, foram adicionados 30 mL
de água destilada mais 5 mL de ácido clorídrico a 37%. Após fervura durante 2 h, sob refluxo,
foi resfriado e neutralizado com solução de carbonato de sódio a 20%. Filtrou-se e o volume
foi completado para 100 mL, com água destilada. A partir do filtrado diluído, determinou-se
os açúcares redutores pelo método do DNS (ácido dinitrosalicílico), conforme Miller (1959),
sendo assim, tomou-se 0,2 mL do filtrado diluído, adicionou-se 1,3 mL de água destilada e 1
mL de ácido dinitrosalicílico (DNS) a 1%, procedendo-se a reação em banho-maria, a 100ºC
por 5 minutos. Após resfriadas em banho de gelo, o volume das amostras foi completado para
10 mL. As leituras foram feitas em espectrofotômetro, a 540 nm. Os resultados obtidos foram
multiplicados pelo fator 0,90 para a obtenção do amido em percentagem (%) da massa fresca.
Os açúcares solúveis totais (AST) foram determinados em amostras de 2 g de polpa,
com extração em 100 mL de álcool etílico a 80% durante 15 minutos e em seguida filtrado.

70
Alíquotas de 10 mL da amostra, diluídas em 100 mL de água destilada, foram usadas nas
determinações, realizadas de acordo com metodologia de Yemn e Willis (1954). Em tubos
contendo alíquota de 0,1 mL do filtrado diluído, adicionou-se 0,9 mL de água destilada e fez-
se reagir com 2 mL do reativo antrona a 0,1%, para depois de agitados, aquecidos em banho-
maria a 100ºC por 8 minutos e resfriados em banho de gelo e a leitura das amostras foi
processada a 620 nm. Os resultados foram expressos em percentagem (%) da massa fresca.
Para a análise dos açúcares redutores (AR), a extração foi feita em água destilada e
determinou-se segundo Miller (1959). A partir de 0,5 g de amostra de polpa diluída para 50
mL e filtrada em papel Wathman qualitativo nº 1, tomou-se 1,5 mL. A este volume,
adicionou-se 1 mL de ácido dinitrosalicílico (DNS) a 1%, procedendo-se a reação em banho-
maria, a 100ºC por 5 minutos. Após resfriadas em banho de gelo, o volume das amostras foi
completado para 10 mL. As leituras foram feitas em espectrofotômetro a 540 nm. Os
resultados foram expressos em percentagem (%) da massa fresca.
Os açúcares não redutores (ANR) foram obtidos por diferença entre os açúcares
solúveis totais (AST) e os redutores (AR), conforme Evangelista (1999), Morais (2001) e
Lima (2002). Os resultados foram expressos em percentagem (%) da massa fresca.
3.7.3 Determinação dos teores de nitrogênio total, não protéico e protéico
O nitrogênio total (NT) foi determinado de acordo com o método de Kjeldahl (AOAC,
1995), com algumas adaptações. Para efetuar a digestão adicionou-se 0,5 g de polpa e 7 mL
da mistura digestora (175 mL de água deionizada + 3,6 g de Na2SeO3.5H2O a 99% + 21,39 g
de Na2SO4 a 99% + 4,0 g de CuSO4.5H2O a 100% + 200 mL de H2SO4 a 98%) em tubos de
digestão, os quais ficaram em bloco digestor a 400ºC até mudarem da cor preta para incolor.
A destilação foi iniciada com a transferência do conteúdo do tubo de digestão para o tubo de
destilação, lavando-o com pequenas porções de água destilada e acrescentando três gotas de
fenolftaleína. Foram colocados 10 mL da solução de ácido bórico a 2% mais indicadores em
Erlenmeyers de 125 mL, sendo em seguida levados para o destilador, onde o tubo de descarga
permaneceu mergulhado na solução. Adicionaram-se 10 a 15 mL de hidróxido de sódio 18 N
ao tubo de destilação e o destilador foi aquecido até quando se destilou 50 mL de amônia.
Finalmente, titulou-se a amônia destilada com H2SO4 0,0525 N até a viragem de azul para
róseo claro, quando se anotou o volume gasto (VA). A prova em branco foi feita sem a adição

71
de polpa, sendo também anotado o volume gasto (VB). Foi feito o cálculo de nitrogênio total
expresso em g/kg da massa fresca da polpa de acordo com a seguinte fórmula:
NT = (VA – VB) x 0,735/Massa da polpa
Para determinação do nitrogênio não protéico (NNP) iniciou-se com a precipitação da
proteína colocando 2 g de polpa e 20 mL de ácido tricloroacético a 15% em um Erlenmeyer
de 125 ml, agitando e deixando em repouso para precipitar por 5 minutos. Após o repouso, foi
filtrado, retirou-se uma alíquota de 5 mL em duplicata, colocando no tubo de digestão e
adicionando 7 mL da mistura digestora. Em seguida, procedeu-se de acordo com o que foi
feito na determinação de nitrogênio total, isto é, digestão, destilação e titulação. Foi feito o
cálculo de nitrogênio não protéico expresso em g/kg da massa fresca da polpa de acordo com
a seguinte fórmula:
NNP = (VA x 4 – VB) x 0,735/Massa da polpa
O nitrogênio protéico (NP) foi obtido pela diferença entre o nitrogênio total e o não
protéico (AOAC, 1995). Os resultados foram expressos em g/kg da massa fresca da polpa.
3.7.4 Proteína bruta e verdadeira
Para o cálculo da proteína bruta (PB), multiplicou-se o nitrogênio total pelo fator 6,25
e para o cálculo da proteína verdadeira (PV), multiplicou-se o nitrogênio protéico pelo fator
6,25 (SILVA, 1990). Os resultados foram expressos em g/kg da massa fresca da polpa.
3.7.5 Vitamina C
A vitamina C foi determinada através da titulometria com solução de DFI (2,6-
diclorofenolindofenol 0,02%) até coloração róseo claro permanente, utilizando 1 g de polpa

72
diluído em 50 mL de ácido oxálico 0,5% de acordo com Strohecker e Henning (1967). Os
resultados foram expressos em mg de ácido ascórbico/100 g da massa fresca da polpa.
3.7.6 Clorofila total
Amostras de epicarpo (espessura de aproximadamente 1 mm) retiradas, ao acaso, das
mangas, foram desintegradas em homogeneizador de tecidos, conforme recomendação de
Bruinsma (1963). Usou-se 1 g do material para 10 mL de uma solução de acetona a 80%. Ao
volume de extrato, após homogeneização, adicionou-se acetona a 80% até completa
descoloração, seguida de filtração. O volume final do extrato foi de 50 mL. A leitura de
absorbância foi efetuada a 652 nm. Os níveis de clorofila total foram expressos em mg/100 g
da massa fresca do epicarpo, segundo a equação adotada por Engel e Poggiani (1991):
Clorofila total = [(A652 x 1000 x V/1000 x W)/34,5] x 100, onde:
A652 = absorbância;
V = volume final do extrato clorofila-acetona em mL; e
W = massa do epicarpo em gramas.
3.7.7 Carotenóides totais
Os carotenóides totais foram determinados pelo método de Higby (1962). Em
Erlenmeyer de 250 mL, foram colocados 10 g de polpa, 30 mL de álcool isopropílico a 99,5%
e 10 mL de hexano a 98,5%, seguido de agitação por 1 minuto. O conteúdo foi transferido
para funil de separação de 125 mL envolvido em alumínio, onde se completou o volume com
água destilada. Deixou-se em repouso por 30 minutos, seguindo-se a lavagem do material.
Repetiu-se esta operação por mais quatro vezes. Filtrou-se o conteúdo com algodão
pulverizado com sulfato de sódio anidro a 99% para um balão volumétrico de 50 mL envolto
com alumínio, onde foram adicionados 5 mL de acetona a 99,5% e completado o volume com

73
hexano a 98,5%. As leituras foram feitas a 450 nm e os resultados expresso em mg/100 g da
massa fresca da polpa, calculados através da fórmula:
Carotenóides totais = (A450 x 100)/(250 x L x W), onde:
A450 = absorbância;
L = largura da cubeta em cm; e
W = quociente entre a massa da amostra original em gramas e o volume final da diluição em
mL.
3.7.8 Fenólicos
A extração dos fenólicos foi realizada de acordo com Swain e Hillis (1959), e o
doseamento segundo metodologia descrita por Reicher et al. (1981). Para a extração de fenóis
diméricos, oligoméricos e poliméricos, pesaram-se 5 g de polpa, utilizando-se
aproximadamente 25 mL do líquido extrator: metanol a 99,5%, metanol a 50% e água
destilada. Para a extração com água destilada, as amostras foram colocadas em banho-maria a
60ºC por 15 minutos. Nos casos de metanol a 99,5% e a 50%, o material foi submetido a
refluxo por 15 minutos. Após a extração, todas as amostras foram agitadas por 15 minutos e,
em seguida, filtradas a vácuo. O filtrado foi evaporado até volume aproximado de 5 mL e
diluído para 50 mL. Alíquotas de 3 mL foram utilizadas para o doseamento, sendo as leituras
realizadas em espectrofotômetro a 720 nm, e os resultados expressos em percentagem (%) da
massa fresca.
3.7.9 Pectinas
As pectinas total e solúvel foram extraídas a partir de 2,5 g da polpa homogeneizada
em etanol 95%, segundo procedimento descrito por McReady e McComb (1952). Após
repouso, a amostra foi lavada por duas vezes com etanol 75%. A extração de pectina solúvel
procedeu-se com filtração e diluição para 50 mL. No caso da pectina total, ajustou-se o pH

74
para 11,5 com solução de NaOH 1 N para posterior repouso por 30 minutos. A seguir, o pH
foi ajustado para 5,0-5,5 com ácido acético glacial para permitir as condições ideais de
hidrólise por meio da pectinase (E.C. 3.2.1.15) de Aspergillus niger, 1,0 U/mg (Merck). As
leituras foram feitas, da mesma forma para as duas variáveis, por colorimetria, a 520 nm,
através da reação de condensação com m-hidroxidifenil, segundo Blumenkrantz e Asboe-
Hansen (1973) e os resultados expressos em mg de pectina por 100 g da massa fresca da
polpa. A percentagem de solubilização de pectina foi obtida através do quociente entre o
conteúdo de pectina solúvel e o da total. Também foram extraídas as frações de pectinas
solúveis em água destilada, oxalato de amônio a 0,75% e ácido clorídrico a 0,05 M, a partir
dos sólidos insolúveis em álcool (SIA), segundo metodologia sugerida por Robertson (1979).
Homogeneizaram-se 2 g da amostra em 30 mL de etanol a 95% e ferveu-se a 85ºC por 15
minutos. Após filtração a vácuo, o resíduo obtido foi lavado com 50 mL de etanol a 95% e 15
mL de acetona a 99,5%, e seco em estufa à temperatura de 40ºC por uma noite. As pectinas
solúveis em água destilada, oxalato de amônio a 0,75% e ácido clorídrico a 0,05 M foram
determinadas pelo método do meta hidroxidifenil (BLUMENKRANTZ e ASBOE-HANSEN,
1973) e os resultados expressos em mg de pectina por 100 g de SIA.
3.8 Avaliações bioquímicas
3.8.1 Atividade da pectinametilesterase
Homogeneizaram-se 10 g da polpa com 20 mL de água destilada a 4ºC. O homogenato
resultante foi centrifugado a 25.000 g, por 10 minutos. O resíduo foi lavado duas vezes com
20 mL de água destilada (4ºC), e em seguida ressuspendido em NaCl 1 M (4ºC) e submetido a
homogeneização por 1 minuto. O pH foi ajustado para 6,0 com NaOH 1 N e o novo
homogenato incubado a 4ºC por 1 hora. Nova centrifugação foi realizada a 25.000 g, por 10
minutos, a 4ºC. O sobrenadante resultante constituiu o extrato enzimático. A este, adicionou-
se pectina cítrica a 1% em NaCl 0,2 N pH 7,0. A taxa de desmetilação do extrato foi medida
por titulação com NaOH 0,01 N, mantendo-se o pH em 7,0 por 10 minutos. Uma unidade de
atividade enzimática (UAE) de pectinametilesterase foi definida como a quantidade de enzima
capaz de catalisar a desmetilação de pectina correspondente ao consumo de 1 nmol de NaOH

75
por 10 minutos. Os resultados foram expressos em UAE por grama da massa fresca por
minuto (JEN e ROBINSON, 1984).
3.8.2 Atividade da poligalacturonase
A extração enzimática para a determinação da atividade poligalacturonásica foi obtida
através da mesma metodologia utilizada para pectinametilesterase. A determinação da
atividade enzimática foi feita conforme Pressey e Avants (1973). O extrato foi incubado com
solução de ácido poligalacturônico 0,25% (lavado com etanol 80% antes do uso) em tampão
acetato de sódio 37,5 mM pH 5,0 a 30ºC por 3 horas. A reação foi interrompida em banho-
maria fervente, e os grupos redutores liberados foram determinados pela técnica do DNS
(ácido dinitrosalicílico), usando glicose anidra como padrão (MILLER, 1959). Uma unidade
de atividade enzimática (UAE) de poligalacturonase foi considerada como sendo a quantidade
de enzima capaz de catalisar a formação de 1 nmol de grupos redutores por minuto. Os
resultados foram expressos em UAE por grama da massa fresca por minuto.
3.8.3 Atividade da polifenoloxidase
O processo de extração da polifenoloxidase foi realizado segundo a técnica proposta
por Wissemann e Lee (1980), com algumas modificações. Homogeneizaram-se 6 g da polpa
em 6 mL de tampão fosfato 0,05 M pH 7,0, contendo 0,1 M de KCl e 1% de
polivinilpirrolidona (PVP). O homogenato foi centrifugado a 11.000 g, por 10 minutos. O
sobrenadante resultante constituiu o extrato enzimático. Até aqui, todo o procedimento foi
realizado a 4ºC. A atividade enzimática foi determinada incubando-se alíquotas de 0,02 a 0,03
do extrato, dependendo do estádio de maturação, e 1,85 mL de tampão fosfato 0,1 M pH 6,0,
contendo 0,1 M de KCl e 0,1 M de catecol, durante 30 minutos, a 30ºC. A reação foi
interrompida pela adição de 0,8 mL de HClO4 2 N. As leituras de absorbância foram
realizadas a 395 nm e considerou-se uma unidade de atividade enzimática (UAE) de
polifenoloxidase como a quantidade de atividade enzimática que produz uma mudança de

76
0,001 unidade de absorbância. Os resultados foram expressos em UAE por grama da massa
fresca por minuto.
3.8.4 Atividade da peroxidase
A extração da peroxidase foi realizada conforme método de Wissemann e Lee (1980)
com as mesmas modificações adotadas para extração da polifenoloxidase. A atividade foi
medida segundo metodologia descrita por Matsuno e Uritani (1972), substituindo-se o
substrato o-fenilenodiamina por guaiacol 1%, adicionado diretamente à solução tampão. As
leituras de absorbância foram realizadas a 470 nm e considerou-se uma unidade de atividade
enzimática (UAE) de peroxidase como a quantidade de atividade enzimática que produz uma
mudança de 0,001 unidade de absorbância. Os resultados foram expressos em UAE por grama
da massa fresca por minuto.
3.8.5 Atividades amilásica total, α- e β-amilásicas
Para atividade amilásica total, a extração e o doseamento seguiram o método descrito
por Bernfeld (1955), modificado por Khader (1992). Extrairam-se 2,5 g da polpa em tampão
fosfato 0,02 M pH 6,9, contendo polivinilpirrolidona (PVP) 2% e 0,5 g de metabissulfito de
sódio a 97%. Após homogeneização, a amostra foi submetida a duas centrifugações a 11.000
g, por 10 minutos. Todo o procedimento de extração foi realizado a 4ºC e o último
sobrenadante obtido constituiu o extrato enzimático. A 0,3 mL do extrato, adicionou-se 0,5
mL de amido solúvel 1% em tampão fosfato 0,02 M pH 6,9, contendo PVP 2% e NaCl 0,007
M, para incubação a 37ºC por 30 minutos. A reação foi interrompida pela adição de 0,5 mL de
NaOH 2 N. A partir daí, procedeu-se às leituras pelo método descrito por Miller (1959),
portanto, determinou-se a atividade desta enzima, pela capacidade de promover a hidrólise do
amido. Os resultados foram expressos em µmol de maltose por grama da massa fresca por
minuto.
A atividade α-amilásica foi feita adicionando-se a 0,3 mL do extrato, 0,5 mL de amido
solúvel 1% em tampão fosfato 0,02 M pH 6,9, contendo polivinilpirrolidona (PVP) 2%, NaCl

77
0,007 M e CaCl2 2,6 mM, para incubação a 70ºC por 30 minutos. A reação foi interrompida
pela adição de 0,5 mL de NaOH 2 N. A etapa de aquecimento a 70ºC na presença de CaCl2
serve para inativar a atividade β-amilásica. A partir daí, procedeu-se às leituras pelo método
descrito por Miller (1959), portanto, determinou-se a atividade desta enzima, pela capacidade
de promover a hidrólise do amido. Os resultados foram expressos em µmol de maltose por
grama da massa fresca por minuto.
A atividade da β-amilase foi obtida por diferença entre amilase total e α-amilase. Os
resultados foram expressos em µmol de maltose por grama da massa fresca por minuto.
3.8.6 Atividades α- e β-galactosidásicas
Para a obtenção das atividades α- e β-galactosidásicas foi realizada a extração
conforme Kitagawa et al. (1995) e a atividade foi determinada segundo Dey e Pridham
(1969). Homogeneizaram-se 10 g da polpa em 5 mL de tampão acetato de sódio 0,1 M pH
5,0, contendo polivinilpirrolidona (PVP) 1%, e centrifugada a 25.000 g, por 15 min, a 4ºC. O
sobrenadante foi utilizado para a determinação da atividade enzimática solúvel no citosol e o
resíduo foi lavado quatro vezes com 5 mL de tampão acetato de sódio 0,02 M pH 5,0,
contendo 2-mercaptoetanol 5 mM, através de centrifugação a 25.000 g, por 5 minutos. Foi,
então, ressuspendido em 10 mL de tampão acetato de sódio 0,02 M pH 5,0, contendo NaCl 3
M, antes de sofrer agitação por 12 horas, mantido a 4ºC. Realizou-se nova centrifugação a
25.000 g, por 20 minutos, a 4ºC. O sobrenadante obtido constituiu o extrato enzimático para
determinação das atividades das enzimas na parede celular. Os extratos foram incubados, por
15 minutos, a 55ºC (para determinação da atividade α-galactosidásica) e a 37ºC (para β-
galactosidásica), em solução dos substratos α- e β-para-nitro-D-galactopiranosídeo 3 mM,
respectivamente, em tampão McIlvaine (citrato a 25 mM + fosfato a 50 mM) pH 5,0 e 4,0,
nesta ordem (McILVAINE, 1921). A reação foi interrompida com carbonato de sódio 0,1 M e
as leituras realizadas em espectrofotômetro, a 400 nm. Considerou-se uma unidade de
atividade enzimática (UAE) de galactosidase como a quantidade de atividade da enzima que
produziu uma mudança de 0,001 unidade de absorbância. Os resultados foram expressos em
UAE por grama da massa fresca por minuto.

78
3.8.7 Teor de proteína
O teor protéico foi determinado segundo o método descrito por Bradford (1976), a
partir dos mesmos extratos de citosol e de parede celular utilizados para determinação das
atividades galactosidásicas, tendo a albumina sérica bovina como padrão. A determinação foi
realizada através da adição de 2,5 mL do reagente Coomassie Brilliant Blue G-250 a 0,05 mL
da amostra e 10 minutos após a adição do reagente realizou-se a leitura em espectrofotômetro,
a 595 nm. Os resultados foram expressos em mg por grama da massa fresca.
3.9 Delineamento experimental e análise estatística
O delineamento experimental utilizado foi o inteiramente casualizado, com 10 ou 8
tratamentos (épocas de colheita), dependendo da análise que foi realizada e 3 repetições,
constituídas por 4 frutos cada. Os dados obtidos em função do tempo de amostragem foram
submetidos à análise de variância observando a significância pelo teste F. Para os casos em
que os tratamentos foram significativos, procedeu-se ajustes através de regressões
polinomiais. O Software ESTAT (Sistema para Análises Estatísticas) foi utilizado nestes
cálculos. Admitiram-se equações polinomiais de até 3º grau, considerando aquelas de grau
superior como desvio de regressão, e coeficientes de determinação maiores que 0,70. Nos
casos em que estes critérios não foram atendidos, optou-se por representar os valores médios
dos tratamentos sem curva de ajuste. Além disso, foram estimados os coeficientes de
correlação linear entre as variáveis estudadas verificando-se a significância pelo teste t,
através do programa estatístico ASSISTAT (Assistência Estatística).

79
4 RESULTADOS E DISCUSSÃO
4.1 Indução floral
As aplicações decrescentes a 5, 4 e 3% de nitrato de potássio (KNO3) no pomar
apresentaram um elevado e uniforme índice de diferenciação floral nos ramos. Este
comportamento corrobora com Albuquerque et al. (2002), quando recomendam de 2 a 4%
KNO3 a fim de antecipar a floração em até 30 dias, além de aumentar significativamente a
frutificação e o rendimento. Mendonça (2000) também conseguiu bons índices de
diferenciação floral apenas com três aplicações de KNO3.
Aos três, dezoito e vinte e seis dias após a aplicação KNO3 a 3% os índices de floração
do pomar foram respectivamente de 20%; 50% e 65%. Esta resposta coincide com a obtida
por Mendonça (2000). O autor concluiu que durante a indução floral de ‘Tommy Atkins’
quando se aplica 30 mL de Paclobutrazol/planta, são necessários apenas 20 dias para atingir
50% de florescimento.
4.2 Marcação das panículas
No pomar foram marcadas 335 panículas no padrão 1 e 462 panículas no padrão 2,
somando um montante de 797 panículas, no entanto, aos 56 DAA observou-se em campo a
existência de apenas 21 frutos no estádio ovo e 340 frutos no estádio fruto, totalizando uma
efetiva produção de 361 frutos, isto é, apenas 35,51% das panículas marcadas tornaram-se
produtivas.
Neste sentido, avaliando o número de frutos por panícula aos 56 DAA, obteve-se a
distribuição absoluta e percentual descrita na TABELA 3. Nota-se que na realidade 64,49%
das panículas marcadas não produziram frutos, enquanto que, 25,85% das panículas marcadas
tiveram apenas um fruto.

80
TABELA 3 - Distribuições absoluta e percentual dos frutos por panícula aos 56 DAA em mangueira, cv. Tommy Atkins.
Número de panículas Distribuição por panícula Absoluto Percentual (%) 0 ovo e 0 fruto 514,00 64,49 1 ovo e 1 fruto 3,00 0,38 1 ovo e 2 frutos 2,00 0,25 2 ovos e 2 frutos 2,00 0,25 1 ovo 12,00 1,50 1 fruto 206,00 25,85 2 frutos 51,00 6,40 3 frutos 7,00 0,88 Total 797,00 100,00
Comportamento semelhante foi relatado por Simão (1971). O autor afirma que a
mangueira fixa no máximo três frutos por panícula. Das panículas formadas apenas 25%
mantêm frutos até a maturação. Dos frutos formados, 60 a 90% caem nos primeiros 30 dias;
94 a 99% aos 60 dias, restando no final apenas de 0,67 a 0,70 dos frutos inicialmente fixados,
isto é, menos de 1% dos frutos atingem o estágio de maturação. Singh (1954) também estudou
este tema e constatou que 35% do total de flores da mangueira são polinizadas resultando em
cerca de 0,01% de frutos no stand final, porém difere daquele relatado por Lima Filho et al.
(2002), quando a queda dos frutos atingiu cerca de 80%. Já Cunha et al. (2002), afirmam que
apenas um a três frutos por panícula completam seu desenvolvimento e atingem a maturação.
4.3 Fenologia da floração à frutificação
Após minuciosa observação das 10 panículas padrão 1, 10 panículas padrão 2 e 10
ramos com gemas se diferenciando para emissão de panícula, constatou-se os valores médios
e percentuais que encontram-se na TABELA 4. Verifica-se uma diferença acentuada entre o
número total de flores dos Padrões 1 e 2, provavelmente esta diferença reflita a ocorrência da
senescência nas flores do Padrão 1, pois constatamos que existe uma diferença de quatro dias
entre os supracitados padrões.

81
TABELA 4 - Valores médios e percentuais da fenologia da floração até o início da frutificação em mangueira, cv. Tommy Atkins.
Panículas Padrão 1 Padrão 2
Determinações
Média (%) Média (%) Nº de flores em antese 3.085,00 57,34 1.082,00 11,74 Nº de flores fecundadas 734,00 13,65 534,00 5,79 Nº de botões 1.561,00 29,01 7.600,00 82,47 Nº total de flores 5.380,00 100,00 9.216,00 100,00 Nº de dias para atingir o estádio padrão 25 DAD1 - 21 DAD - Nº de dias para atingir o estádio chumbinho 2 DAA2 - 5 DAA - Nº de dias para atingir o estádio bola de gude 15 DAA - 19 DAA - Nº de dias para atingir o estádio ovo 33 DAA - 36 DAA - Nº de dias para atingir o estádio fruto 47 DAA - 50 DAA -
¹Dias após a diferenciação. ²Dias após a antese.
Simão (1971) relatou que uma panícula possui de 400 a 17.000 flores, porém difere de
Pinto et al. (2002a), quando constatou apenas de 600 a 6.000 flores. No entanto, o valor
máximo do último autor assemelha-se ao valor mínimo deste estudo, onde se constatou 5.380
flores/panícula (TABELA 4).
Segundo Simão (1971), o período de desenvolvimento da panícula varia de 35 a 42
dias, e as primeiras flores só se abrem a partir de 21 dias do início do desenvolvimento da
inflorescência. Este comportamento assemelha-se com os resultados observados neste
trabalho, pois foram necessários 21 e 25 dias após a diferenciação (DAD) para atingir os
estádios padrão 2 e 1, respectivamente (TABELA 4).
Outro ponto difuso é quanto ao número de dias para atingir a maturação. Lima Filho et
al. (2002) e Medina, Valdique (1996) relatam que são necessários 90 a 120 dias após a
fecundação, no entanto difere daquele constatado por Medlicott e Reynolds (1988) e Bleinroth
(1994), quando registraram de 100 a 105 dias após a florada. Já Cunha et al. (2002) sugere de
100 a 150 dias após a floração. No entanto, a TABELA 4 consta apenas da fenologia da
antese até 47 e 50 DAA, quando os padrões 1 e 2, respectivamente, atingem o estádio de fruto
maturo (FIGURA 8).

82
FIGURA 8 – Frutos de mangueira ‘Tommy Atkins’ nos estádios chumbinho (DLMédio=
0,28cm e DTMédio= 0,29cm), bola de gude (DLMédio= 2,4cm; DV=Médio 2,12cm; e DTMédio= 1,83cm), ovo (DLMédio= 6,35cm; DVMédio= 5,54cm; e DTMédio= 4,48cm) e fruto (DLMédio= 8,06cm; DVMédio= 6,09cm; e DTMédio= 5,22cm).
4.4 Avaliações físicas
As análises de variância (TABELAS 1A a 3A) revelaram efeito significativo do tempo
de colheita para todas as características físicas analisadas.
4.4.1 Aspectos morfológicos externos
Os aspectos morfológicos externos de mangas da cv. Tommy Atkins dos 35 aos 112
DAA encontram-se na TABELA 5. Observa-se que os frutos só apresentaram características
ideais para colheita a partir de 105 DAA. Os resultados aqui encontrados são concordantes
com as recomendações disponíveis na literatura (MEDLICOTT e REYNOLDS, 1988;
ALVES et al., 2002; REID, 2003; CHITARRA e CHITARRA, 2005).

83
TABELA 5 – Aspectos morfológicos externos de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Aspectos morfológicos externos Dias após a antese (DAA) FO1 TC2 BC3 DPB4 FN5 FA6
35 canivete rugosa ausente próximo afilado agudo
49 canivete rugosa ausente próximo afilado agudo
63 canivete rugosa ausente próximo afilado agudo
70 canivete rugosa ausente próximo afilado agudo
77 canivete rugosa ausente próximo afilado agudo
84 canivete rugosa ausente próximo afilado agudo
91 canivete rugosa ausente próximo afilado agudo
98 canivete rugosa ausente próximo afilado agudo
105 cheio lisa presente espaçado achatado obtuso
112 cheio lisa presente espaçado achatado obtuso
¹Formato do ombro. 2Textura da casca. 3Brilho da casca. 4Distanciamento das pontuações brancas. 5Formato do nariz. 6Formato do ápice.
4.4.2 Diâmetros longitudinal, ventral, transversal e produto dos diâmetros
Na FIGURA 9, verifica-se que os diâmetros longitudinal, ventral e transversal
apresentaram comportamento quadrático em resposta à variação de tempo, onde as mangas
aumentaram em tamanho até 70 DAA quando se avalia o diâmetro longitudinal e até 77 DAA
quando se considera os diâmetros ventral e transversal, e após estes períodos praticamente se
mantêm constante. Neste sentido, Leite et al. (2005) pesquisando as características físicas da
manga cv. Tommy Atkins produzida na região do Vale do São Francisco, município de
Petrolina, PE, no período de setembro a outubro de 2004, obtiveram os seguintes valores
médios respectivamente para diâmetros longitudinal, ventral e transversal: 10,89; 9,55 e 8,67
cm. Estes valores são da mesma ordem de grandeza dos obtidos neste trabalho (10,79; 8,51 e
8,05 cm aos 112 DAA). Mendonça (2000) também obteve valores semelhantes para ‘Tommy
Atkins’ cultivada em Jaguaruana-CE para o mesmo período de colheita (11,53; 8,73; e 7,95
cm).

84
,
,
,
,, ,
, , , ,
*
*
*
** *
* * * *
)
)
)
)) )
) ) ) )
35 42 49 56 63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
2
4
6
8
10
12
Diâ
met
ro (c
m)
DL DV DT) * ,
YDL= - 0,5269335 + 0,2690548X - 0,001539007X 2, R2= 0,8342**.YDV= - 2,680375 + 0,2634758X - 0,001474148X 2, R2= 0,9179**.YDT= - 1,818458 + 0,2096275X - 0,001101758X 2, R2= 0,9365**.
FIGURA 9 – Diâmetros longitudinal (DL), ventral (DV) e transversal (DT) de mangas
‘Tommy Atkins’ durante o desenvolvimento.
A FIGURA 10 indica que o produto dos diâmetros apresentou comportamento
quadrático em resposta à variação de tempo. Conforme Morais (2001) e Castro Neto e
Reinhardt (2003), pode-se estimar com precisão as massas fresca e seca das mangas cvs.
Tommy Atkins e Haden, respectivamente, a partir do produto dos diâmetros. Morais (2001)
também constatou que o produto dos diâmetros máximo foi alcançado aos 103 dias após
floração plena (DAFP) com 874,60 cm3, enquanto neste trabalho foi alcançado aos 77 DAA
com 856,84 cm³. Entretanto, ao ser analisada a curva de regressão o ponto máximo do produto
dos diâmetros ocorreu por volta dos 91,17 DAA (812,42 cm³).
)
)
)
)
) )
)) )
)
35 42 49 56 63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
200
400
600
800
1000
Prod
uto
dos d
iâm
etro
s (cm
³)
Y= - 1063,431 + 41,15046X - 0,2256791X 2, R2= 0,8691**.
FIGURA10 – Produto dos diâmetros de mangas ‘Tommy Atkins’ durante o desenvolvimento.

85
4.4.3 Massas fresca, seca, de água e o teor de água
Os efeitos fixos de tratamentos para massa fresca, massa seca e teor de água são
explicados em 90% pela equação de regressão cúbica (R2= 0,9228, R2= 0,9816 e R2= 0,9145
respectivamente), enquanto que para a massa de água, 86% pela equação de regressão
quadrática (FIGURA 11).
&
&
& & &
&
&
&
&
&
,
,
,
,
, , ,, ,
,
**
** * * * * * *)
)
)
)
) ))
) ))
35 42 49 56 63 70 77 84 91 98 105 112Dias após a antese (DAA)
0
100
200
300
400
500
600
Mas
sa (g
)
80
82
84
86
88
90
Teor de água (%)
MF MS MA TA) * , &YMF= 122,1016 - 14,12837X + 0,4161058X2 - 0,002490384X3, R2= 0,9228*.YMS= 113,1149 - 6,707651X + 0,1259424X2 - 0,0006357549X3, R2= 0,9816**.YMA= - 580,5492 + 20,79553X - 0,120173X2, R2= 0,8637**.YTA= 64,57621 + 1,171484X - 0,01735356X2 + 0,00007399974X3, R2= 0,9145**.
FIGURA 11 – Massas fresca (MF), seca (MS), de água (MA) e o teor de água (TA) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Observa-se ainda na Figura 11 que o acúmulo da massa seca ocorreu até 98 dias após a
antese (DAA) coincidindo com a mudança da coloração da casca na escala Blush de 3 para 4
(FIGURA 12). O percentual da cor vermelha evoluiu da faixa de >25-50% para >50-75%. Isto
caracteriza o início da maturação do fruto, quando simultaneamente registrou-se o menor teor
de água. A diferença de acúmulo da massa seca entre 91 e 98 DAA foi de apenas 1,90 g. Este
conjunto de resultados sugere que, para as condições estudadas, a manga cv. Tommy Atkins
atinge a maturação aos 98 DAA.
O período registrado neste trabalho (98 DAA) foi superior aos 75 dias para a cv.
Haden atingir o ponto de colheita (CASTRO NETO e REINHARDT, 2003), na mesma
região, e aos 90 dias relatados para outras cultivares, como Langra, Alphonso, Dashehari e
Krishnabhog, na Índia (SUBRAMANYAN et al., 1975). Estas diferenças podem ser
atribuídas a variações no ciclo de crescimento das plantas e, conseqüentemente, na duração do
período reprodutivo e de crescimento do fruto. No entanto, Morais (2001), estudando o ponto

86
de maturidade para colheita e a vida útil da manga ‘Tommy Atkins’ para o mercado Europeu,
cultivada no Pólo Mossoró-Assú, Rio Grande do Norte, constatou que 96 dias após a floração
plena pode ser considerado como um indicador do ponto de colheita.
A taxa de acúmulo da massa seca no fruto foi maior no período de 49 a 98 DAA.
Conforme Castro Neto e Reinhardt (2003), o acúmulo da massa seca pode ser conseqüência
de maior participação da fotossíntese no período, associada à translocação da matéria seca de
outras partes da planta para o fruto, sendo que a remobilização de fotoassimilados
previamente fotossintetizados é de 40-50% da massa seca do fruto.
Os valores máximos da massa fresca e massa de água do fruto foram, respectivamente,
400,00 g e 335,66 g, aos 84 DAA, havendo em seguida decréscimo, associado à redução no
conteúdo de água do fruto, corroborando assim, com os resultados de Castro Neto e Reinhardt
(2003), que estudando o crescimento de frutos, cv. Haden, constataram que os valores das
massas fresca e de água máximos foram atingidos antes de chegar à maturidade, isto é,
quando a massa seca foi máxima.
4.4.4 Escalas de coloração da casca e da polpa
Embora nem todos os frutos mudem de cor durante o amadurecimento, esta é uma das
características mais associadas ao ponto de colheita e maturidade para consumo (TUCKER,
1993). Esta afirmação é particularmente verdadeira na colheita da manga. As avaliações de
mudança de coloração da casca nas escalas de coloração da casca (ECC), de Blush para
coloração da casca (EBCC) e de coloração da polpa (ECP) mostram claramente esta tendência
na FIGURA 12, quando atingiram a máxima coloração aos 98 DAA.

87
,
,
, , , ,, ,
, ,*
*
* *
* * * *
* *
)
)
) ) ) )) ) ) )
35 42 49 56 63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
1
2
3
4
5
Esca
la d
e c o
lora
ção
(not
as)
ECC EBCC ECP) * ,
YECC= 2,947138 - 0,09818528X + 0,001458854X2 - 0,00000619064X3, R2= 0,9455**.YEBCC= 0,5913978 + 0,03072197X, R2= 0,8602**.YECP= 3,423067 - 0,1195028X + 0,001717478X2 - 0,000006902742X3, R2= 0,9475*.
FIGURA 12 – Escalas de coloração da casca (ECC), de Blush para coloração da casca
(EBCC) e de coloração da polpa (ECP) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Neste trabalho a ECP aumentou de 1,00 aos 35 DAA para 1,83 aos 98 DAA,
mantendo-se estável até os 112 DAA. Por outro lado, Morais (2001) estudando esta mesma
cultivar observou um aumento na ECP de 2,25 aos 82 DAFP para 3,15 aos 103 DAFP. Esta
discrepância talvez seja devido ao fato de que as mangas deste trabalho terem sido marcadas
mais precocemente que as do estudo de Morais (2001), pois floração plena equivale a 80-90%
da emissão de panículas enquanto no presente trabalho foi feito a marcação quando havia
apenas 65% de emissão de panículas.
Alves et al. (2002) recomendam colher a manga com ECC 2,0 e ECP variando de 2,0 a
3,0 e Chitarra e Chitarra (2005) com ECC e ECP 3,0, o que aproxima-se dos valores
detectados no intervalo das colheitas de 98 a 112 DAA para ECC (1,54) e também para ECP
(1,83) neste estudo.
4.4.5 Coloração da polpa
A luminosidade representa o brilho da superfície ou a quantidade de preto. Na manga
‘Tommy Atkins’, houve variação estatisticamente significativa da luminosidade da polpa
durante o período estudado (FIGURA 13), embora o coeficiente de determinação tenha sido

88
inferior a 0,70 (R²= 0,6883). O valor médio obtido foi de 67,26, numa escala em que o
máximo corresponde a 100 e mostra uma tendência de aumento durante o desenvolvimento.
))) )
)) ) )
, ,, , ,
, , ,
*
*
**
**
* *
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
20
40
60
80
100
Lum
inos
idad
e, c
rom
a e
âng u
lo H
ue d
a po
lpa LP CP HP) * ,
YLP= Não ajustado, R2< 0,7000.YCP= 411,0235 - 15,55374X + 0,1981156X2 - 0,000786264X3, R2= 0,9832**.YHP= - 225,0922 + 10,94408X - 0,1219248X2 + 0,0004412443X3, R2= 0,9139**.
FIGURA 13 - Luminosidade (LP), croma (CP) e ângulo Hue da polpa (HP) de mangas
‘Tommy Atkins’ durante o desenvolvimento.
Estes resultados são semelhantes aos de Morais (2001), quando estudando esta mesma
cultivar de 82 a 103 dias após floração plena (DAFP) obteve aumento de luminosidade e um
valor médio de 73,75.
Por outro lado, na FIGURA 13 evidencia-se que as mudanças no croma e no ângulo
Hue asseguram diferenças de cor entre os frutos imaturos e maturos, apoiados por altos
coeficientes de determinação (R²= 0,9832 e R²= 0,9139).
Para o croma, ocorreu um aumento gradual até os 105 DAA (53,56), indicativo de
degradação de clorofila e síntese de carotenóides já que o aumento no croma reflete o
aumento na intensidade da cor amarela. Este resultado difere dos de Morais (2001), que
analisando ‘Tommy Atkins’ detectou o ponto máximo aos 96 DAFP (59,38).
O ângulo Hue, que representa a variação de cor do verde (verde intenso= 180º) para o
amarelo (amarelo intenso= 90º), por sua vez, apresentou resposta menos característica.
Aumentou dos 63 aos 84 DAA (90,96 para 96,83) e depois reduziu até aos 112 DAA (91,03).
Morais (2001) também registrou aumento no valor do croma e redução no ângulo Hue
(passagem do verde para o amarelo) em mangas ‘Tommy Atkins’ colhidas de 82 a 103 DAFP.
Em muitos frutos, a luminosidade é a característica da cor da polpa que varia mais
significativamente durante o amadurecimento. É o que ocorre, por exemplo, nas mangas

89
‘Lippens’ e ‘Smith’, que reduz a luminosidade com o avanço da maturação, conforme
observado por Marin e Cano (1992), embora cultivares como Tommy Atkins desenvolvam
uma cor laranja intensa do mesocarpo, tornando o croma uma variação mais significativa
(MITCHAM e McDONALD, 1992), o que pode ser constatado neste trabalho.
4.4.6 Firmeza
A firmeza da polpa das mangas aos 35, 49, 63, 70, 77, 84, 91, 98 e 105 dias após a
antese (DAA), ultrapassaram a 128,99 N, portanto, não conseguimos determiná-las, uma vez
que, o penetrômetro utilizado nas aferições só registrava até 128,99 N, no entanto, a TABELA
6 apresenta os valores médios da firmeza, em mangas da cv. Tommy Atkins, aos 112 DAA.
Neste sentido, Rocha et al. (2001) também não conseguiram determinar a firmeza da polpa de
mangas ‘Tommy Atkins’ no estádio 1 (1,1 para ECC e 1,3 para ECP), no entanto, obtiveram
no estádio 2 (2 para ECC e 2,1 para ECP; 96,14 N) resultados semelhante aos 112 DAA deste
trabalho (estádio 1,54 para ECC e estádio 1,83 para ECP; 111,57 N). Sendo assim, Morais
(2001) estudando a maturação desta mesma cultivar, aos 82 DAFP também não conseguiu
determinar a firmeza, embora, aos 89 DAFP tenha encontrado um valor médio de 110,78 N, o
que ratifica esta pesquisa. Portanto, a firmeza os 89 DAFP do trabalho de Morais (2001)
equivalem firmeza aos 112 DAA deste trabalho.
TABELA 6 - Valores médios da firmeza da polpa, em mangas da cv. Tommy Atkins, aos 112 DAA.
Textura (N) Repetições 112 DAA 1 100,64 2 112,31 3 121,76
Média 111,57
Medlicott e Reynolds (1988) recomendam colher a manga com a firmeza variando de
107,84 a 127,45 N e Alves et al. (2002) com firmeza de 129,41 N, o que coincide com os
valores observados no intervalo das colheitas de 105 a 112 DAA deste trabalho.

90
4.4.7 Unidades de calor (graus-dia)
A TABELA 7 apresenta os valores de unidades de calor a partir da diferenciação do
ramo até antese e da antese até o início da frutificação em mangueira, cv. Tommy Atkins. O
número de unidades de calor necessário para atingir a panícula padrão 1 (479,82ºC) é maior
que para atingir o padrão 2 (405,02ºC), pois no padrão 1 o percentual de flores visualmente
abertas na panícula é de 100% e o percentual de fecundação visualmente é menos de 5%,
enquanto no padrão 2 o percentual de flores visualmente abertas na panícula é 90% e o
percentual de fecundação visualmente é 0%. Consequentemente, o número de unidades de
calor necessário para atingir os estádios chumbinho, bola de gude, ovo e fruto no padrão 1
será menor que no padrão 2. Mosqueda-Vázquez e Ireta-Ojeda (1993) encontraram resultados
semelhantes para o desenvolvimento da inflorescência de manga ‘Manila’, pois com uma
temperatura base (Tbase) de 12ºC foram necessários 434,7ºC.
TABELA 7 – Valores de unidades de calor a partir da diferenciação do ramo até a antese e da antese até o início da frutificação em mangueira, cv. Tommy Atkins.
Unidades de calor (ºC) Fase fenológica Panícula padrão 1 Panícula padrão 2 Estádio padrão1 479,82 405,02 Estádio chumbinho2 40,29 80,20 Estádio bola de gude2 237,44 304,45 Estádio ovo2 522,12 570,80 Estádio fruto2 751,52 803,81
1A partir da diferenciação. 2A partir da antese.
Os valores de unidades de calor a partir da antese até o fim da frutificação em
mangueira, cv. Tommy Atkins encontram-se na TABELA 8. Observa-se que o número de
unidades de calor variou de 554,61ºC aos 35 DAA a 1.939,70ºC aos 112 DAA. O valor de
unidades de calor calculado para 112 DAA é quase duas vezes o valor relatado por
Oppenheimer, 1947 apud Singn, 1977 (1.000ºC) e mais que duas vezes (866ºC) o valor
relatado por Burondkar et al. (2000) para manga ‘Ratna’ aos 112 DAA com Tbase de 17,9ºC.
Este valor pode ser afetado pela Tbase subestimada. Mais localidades, em épocas diferentes e
durante vários anos deverão ser consideradas em estudos futuros para validar esta nova
metodologia de determinação da maturidade da manga, uma vez que, é necessário a
repetitividade dos resultados para a sua validação.

91
TABELA 8 – Valores de unidades de calor a partir da antese até o fim da frutificação em mangueira, cv. Tommy Atkins. Fase fenológica Unidades de calor (ºC)
35 DAA¹ 554,61 49 DAA 786,27 63 DAA 1.038,89 70 DAA 1.165,97 77 DAA 1.295,96 84 DAA 1.421,97 91 DAA 1.549,60 98 DAA 1.685,09 105 DAA 1.810,88 112 DAA 1.939,70
¹Dias após a antese.
4.5 Avaliações físico-químicas e químicas
A partir das análises de variância, verificou-se efeito significativo do tempo de
colheita sobre as variáveis estudadas, com exceção de pectina solúvel, percentagem de
solubilização de pectina e pectinas de baixa metoxilação (TABELAS 4A a 8A).
4.5.1 Sólidos solúveis totais
O aumento no teor de SST durante o desenvolvimento foi contínuo, partindo-se de
5,6ºBrix aos 35 DAA, para 8,5ºBrix aos 112 DAA, portanto, quase duplicou (FIGURA 14).
Destaca-se que pela curva de regressão o máximo foi atingido aos 106,16 DAA (8,48ºBrix) e
permaneceu constante até os 112 DAA. Este comportamento ratifica alguns estudos
realizados em manga ‘Tommy Atkins’ que têm mostrado valores de SST, respectivamente em
frutos maturos e maduros, de 7,81 e 10,46ºBrix aos 50 e 120 DAF (DUTRA et al., 2005), de
6,41 e 7,72ºBrix aos 82 e 103 DAFP (MORAIS, 2001) e de 7,5 e 10,9ºBrix aos 95 e 125 DAA
(LEDERMAN et al., 1998).

92
)
)
) )
) ) ))
) )
35 42 49 56 63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
1
2
3
4
5
6
7
8
9
10
SST
(ºBrix
)
Y= 9,372455 - 0,2268129X + 0,004038001X 2 - 0,00001864914X 3, R2= 0,9564**.
FIGURA 14 - Sólidos solúveis totais (SST) de mangas ‘Tommy Atkins’ durante o
desenvolvimento.
De acordo com Sigrist (1992), o aumento no teor de SST durante a maturação é
atribuído principalmente à hidrólise dos carboidratos de reserva acumulados durante o
crescimento do fruto na planta. O resultado desta hidrólise é a produção de açúcares solúveis
totais (AST).
Medlicott e Reynolds (1988) recomendam colher a manga com a teor de SST variando
de 7 a 8ºBrix e Alves et al. (2002) com 7,3ºBrix, o que coincide com os valores verificados no
intervalo das colheitas de 70 a 91 DAA (7,36 a 8,03ºBrix) desta pesquisa.
O efeito do tempo de colheita sobre o teor de SST na manga foi altamente
significativo apresentando baixo coeficiente de variação (TABELA 4A). Este comportamento
propicia a adoção do conteúdo de SST como um indicador de maturidade da manga.
4.5.2 Acidez total titulável e pH
Ocorreu diminuição na ATT e aumento no pH com o avanço da maturação (FIGURA
15). As mangas colhidas aos 35 DAA apresentaram menor valor de pH, concordante com a
concentração mais elevada de ATT. Este comportamento decorre do consumo de ácidos
orgânicos no processo respiratório ou de sua conversão em açúcares (HULME, 1974;
CHITARRA e CHITARRA, 2005).

93
*
*
* * ** *
* * *)
)
)
) ) ) ) ) ) )
35 42 49 56 63 70 77 84 91 98 105 112Dias após a antese (DAA)
0
2
4
6
8
10
12
14
16
ATT
(% d
e ác
ido
c ítri
co)
0
1
2
3
4pH
ATT pH) *
YATT= 57,2513 - 1,775669X + 0,01851442X2 - 0,00006386906X3, R2= 0,9804**.YpH= 4,748303 - 0,1004025X + 0,001583358X2 - 0,000006991988X3, R2= 0,9639**.
FIGURA 15 - Acidez total titulável (ATT) e pH de mangas ‘Tommy Atkins’ durante o
desenvolvimento.
A ATT (FIGURA 15) diminuiu durante o desenvolvimento, especificamente de 35 até
112 DAA (14,63 para 0,83% de ácido cítrico). O período em que se verificou maior redução
na concentração de ácidos orgânicos foi de 35 para 63 DAA. Neste período, houve um
decréscimo de 12,96% na ATT. Este resultado assemelha-se com os obtidos por outros
autores em manga: de 1,40 a 0,70% dos 50 aos 120 DAF (DUTRA et al., 2005), de 1,25 a
1,15% dos 82 aos 103 DAFP (MORAIS, 2001) e de 4,5 a 3,0% dos 35 aos 112 DAV
(LAKSHMINARAYANA, 1970).
Comparando-se a ATT aos 112 DAA com aquela observada aos 35 DAA, verificou-se
uma diminuição de aproximadamente dezoito vezes. Comportamento análogo foi obtido por
Rocha (2001), quando trabalhando com manga ‘Tommy Atkins’ registrou uma redução de
cerca de sete vezes.
Medlicott e Reynolds (1988) recomendam colher a manga com a ATT variando de
0,65 a 0,70% de ácido cítrico, o que se aproxima do valor registrado na colheita aos 112 DAA
(0,83%) desta investigação.
Com 35 DAA (FIGURA 15), o pH dos frutos era de 2,84, crescendo a partir daí em
função, principalmente, da redução na ATT. Aos 105 DAA, foi observado o valor máximo do
pH (3,66), decrescendo aos 112 DAA (3,47). Estes resultados são coerentes com os valores
mínimos e máximos de pH disponíveis na literatura para manga ‘Tommy Atkins’: de 3,47 a
4,09 dos 50 aos 120 DAF (DUTRA et al., 2005), de 3,52 a 3,70 dos estádios 2 a 4 (LUCENA
et al., 2000) e de 3,23 a 4,51 dos estádios 1 a 5 (ROCHA, 2001).

94
4.5.3 Relação SST/ATT
Como conseqüência principalmente da redução na ATT, a relação SST/ATT aumenta
ao longo do período avaliado (FIGURA 16). A partir de 35 até 98 DAA (0,38 para 9,14),
verificou-se um incremento acentuado, equivalente a 24 vezes o valor inicial, pois
corresponde ao período de maior redução do ATT, conforme mostra a FIGURA 15. Depois,
até aos 112 DAA (10,13), as mudanças foram pouco representativas uma vez que as duas
variáveis envolvidas na relação tiveram apenas pequenas alterações (FIGURAS 15 e 16). Este
comportamento confirma o estudo de alguns pesquisadores a respeito da marcha da relação
SST/ATT em manga durante o desenvolvimento com os seguintes resultados: de 5,58 a 15,10
dos 50 aos 120 DAF (DUTRA et al., 2005), de 5,13 a 6,74 dos 82 aos 103 DAFP (MORAIS,
2001) e de 9,20 a 20,28 dos estádios 2 a 4 (LUCENA et al., 2000).
)
)
)
)
))
))
))
35 42 49 56 63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
2
4
6
8
10
12
RSA
Y= 2,229463 - 0,2047153X + 0,004912665X 2 - 0,00002190079X3, R2= 0,9805*.
FIGURA 16 - Relação SST/ATT (RSA) de mangas ‘Tommy Atkins’ durante o
desenvolvimento.
A relação SST/ATT é mais representativa que a medição isolada de açúcares ou da
acidez, pois essa relação além de dar uma boa idéia do equilíbrio entre esses dois
componentes, indica o sabor dos frutos (CHITARRA e CHITARRA, 2005).
Para a colheita da manga, Medlicott e Reynolds (1988) recomendam um teor mínimo
de 7 a 8ºBrix e um teor máximo de 0,65 a 0,70% de ATT, visando à obtenção de um sabor
aceitável, resultando numa relação SST/ATT de 10,00 a 12,30, o que coincide com o valor

95
registrado na colheita aos 112 DAA (10,13) nesta pesquisa. Portanto, as mangas oriundas da
colheita aos 112 DAA estavam com um sabor aceitável ao paladar do consumidor.
4.5.4 Determinação de carboidratos
O teor de amido cresceu quadraticamente durante o desenvolvimento, partindo de
2,11% aos 63 DAA para 4,48% aos 112 DAA (FIGURA 17). Resultados equivalentes foram
obtidos por Dutra et al. (2005) com esta mesma cultivar, ao detectarem aumento no teor de
amido a partir de 50 dias após a florada - DAF (2,23%) até 120 DAF (9,77%).
&
&
& & &&
&&
,
,,, , , , ,
*
*
* * **
**
)
)
)) ) )
))
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
1
2
3
4
5
6
7
8
Car
boid
rat o
s (%
da
mas
sa f r
esca
)
AD AST AR ANR) * , &
YAD= 6,716071 - 0,1388549X + 0,001047943X 2, R2= 0,9732*.YAST= 0,3509921 + 0,04772676X, R 2= 0,9640**.YAR= - 5,865496 + 0,2132285X - 0,001169825X 2, R2= 0,8682**.YANR= 3,990437 - 0,1128515X + 0,0008689666X 2, R2= 0,9387*.
FIGURA 17 - Amido (AD), açúcares solúveis totais (AST), redutores (AR) e não redutores
(ANR) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Comportamento também semelhante foi observado por Lakshminarayana et al. (1970),
pois conforme estes autores, durante a permanência da manga na planta, o acúmulo de amido
é a principal atividade nos tecidos da polpa, cujo teor aumenta de 1,5% após o vingamento
para 13% quando atinge o completo desenvolvimento, no entanto, após a colheita esse amido
é totalmente hidrolisado em 8 dias.
Por outro lado, Morais (2001) também estudando a maturação da ‘Tommy Atkins’
constatou um decréscimo do teor de amido de 4,14% aos 82 DAFP para 3,18% aos 103
DAFP. Conforme já comentado anteriormente, possivelmente, a colheita foi realizada em

96
estádio mais avançado do que o usado neste trabalho, o que leva a crer que este é o motivo
porque não se observou a queda do amido, pois a manga aos 112 DAA (4,48%) é equivalente
a manga aos 82 DAFP (4,14%). Portanto, só seria possível constatar esta queda se tivesse sido
feito colheitas após os 112 DAA. Rocha et al. (2001) pesquisando esta mesma cultivar
também verificou um decréscimo no conteúdo de amido do estádio 1 (6,81%) para o estádio 5
(3,04%).
O teor de AST (FIGURA 17), como principal constituinte dos SST, acompanhou o
aumento observado neste durante o desenvolvimento. Os frutos, aos 63 DAA, tinham 3,14%
de AST, que representavam 52,68% do teor de SST naquela data (5,96ºBrix). Com a
maturação, verificou-se ganhos linearmente crescentes nos teores de AST, que, inclusive,
passaram a representar percentuais cada vez maiores dos SST. Aos 112 DAA, os frutos
tinham 5,50% de AST, que equivaliam a 64,70% dos SST (8,50ºBrix). Portanto, durante a
maturação houve um aumento de cerca de 1,75 vezes nos teores de AST.
Resultados semelhantes foram obtidos por Morais (2001) onde, com esta mesma
cultivar, o teor de AST aumentou de 3,07% aos 82 DAFP para 4,72% aos 103 DAFP. Em
estudos realizados com ‘Tommy Atkins’ por Rocha et al. (2001), o teor de AST aumentou de
4,11% no estádio 1 para 13,59% no estádio 5.
O conteúdo de AR (FIGURA 17), principalmente glicose e frutose, aumentou
quadraticamente dos 63 aos 91 DAA (2,77% para 3,84%), depois diminuiu até os 112 DAA
(3,48%). Aos 105 DAA o acúmulo de amido igualou-se à síntese de AR, no entanto, superou
aos 112 DAA, quando a concentração de AR representou cerca de 41% dos SST. Esta
resposta é comparável com a de Dutra et al. (2005), pois com esta mesma cultivar,
constataram aumento no percentual de AR de 50 até 92 DAF (3,05% para 3,11%) e depois
diminuiu até 120 DAF (2,94%).
Resultados coincidentes foram detectados por Rocha (2001), que verificou em manga
‘Tommy Atkins’ a redução no teor de AR do estádio 1 (3,76%) para o estádio 5 (2,75%). Por
outro lado, Morais (2001) observou um aumento no teor de AR a partir dos 82 DAFP (1,57%)
até 103 DAFP (2,34%).
O teor de ANR, onde a sacarose é o principal, aumentou quadraticamente durante o
desenvolvimento da manga (FIGURA 17). Os valores observados deste grupo de açúcares
foram menores que os de AR. Os frutos aos 112 DAA apresentaram valores de ANR iguais a
2,02%, representando cerca de 24% dos SST, que foi aproximadamente seis vezes maior do
que o inicial (0,37%). Esta resposta assemelha-se a de Dutra et al. (2005), pois com esta

97
mesma cultivar, verificaram aumento de cerca de duas vezes no teor de sacarose, principal
ANR, no período de 50 a 120 DAF (3,47% para 5,09%).
Resultados equivalentes foram obtidos por Morais (2001), que registrou em manga
‘Tommy Atkins’ aumento de cerca de duas vezes no conteúdo de ANR, no período de 82 a
103 DAFP (1,51% para 2,38%). Sendo também corroborado por Rocha (2001) que verificou
um aumento no teor de ANR do estádio 1 (0,35%) para o estádio 5 (10,84%), correspondendo
a um incremento de aproximadamente 31 vezes no percentual de ANR.
As proporções observadas entre ANR e AR foram crescentes a partir de 63 até 77
DAA (1:7,69 para 1:11,11) e depois decresceu até os 112 DAA (1:1,72), devido a redução
quadrática dos AR e aumento quadrático dos ANR. Em período equivalente, Dutra et al.
(2005) observaram que a proporção diminuiu de 1:0,87 (50 DAF) para 1:0,57 (120 DAF),
Morais (2001) de 1:1,03 (82 DAFP) para 1:0,98 (103 DAFP) e Rocha (2001) de 1:10,74
(estádio 1) para 1:0,25 (estádio 5), validando a presente pesquisa.
A importância da proporção ANR:AR está diretamente relacionada ao grau de doçura
dos frutos (PANGORB, 1963). Considerando que a frutose possui grau de doçura maior que a
sacarose e esta por sua vez maior que a glicose e sabendo-se que a frutose e a glicose (AR)
estão nas proporções de 2:1 na manga ‘Keitt’ (MEDLICOTT e THOMPSON, 1985), conclui-
se, que os frutos que apresentarem predomínio de AR em relação aos ANR serão mais doces.
Este comportamento foi evidenciado neste estudo (1:1,72 aos 112 DAA).
4.5.5 Nitrogênio
Na FIGURA 18, o conteúdo de nitrogênio total (NT) diminuiu cubicamente dos 63 aos
70 DAA (0,59 para 0,49 g/kg da massa fresca da polpa), depois aumentou até aos 105 DAA
(0,84 g/kg da massa fresca da polpa) e finalmente reduziu até os 112 DAA (0,63 g/kg da
massa fresca da polpa). Resultados equivalentes foram obtidos por Lakshminarayana et al.
(1970), trabalhando com manga, ao detectarem redução no teor de NT do vingamento do fruto
até 42 dias após o vingamento (DAV), depois aumentou até 56 DAV e finalmente reduziu até
112 DAV.

98
,,
,,
,
, , ,
**
* * ** *
*)
)
)
) )
) )
)
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
0,2
0,4
0,6
0,8
1
Nitr
ogên
io (g
/kg
da m
a ssa
fres
c a)
NT NNP NP) * ,
YNT= 12,24107 - 0,4261742X + 0,005047928X 2 - 0,00001933818X 3, R2= 0,7691**.YNNP= Não ajustado, R2< 0,7000.YNP= Não ajustado, R2< 0,7000.
FIGURA 18 - Nitrogênio total (NT), não protéico (NNP) e protéico (NP) de mangas ‘Tommy
Atkins’ durante o desenvolvimento.
Na manga ‘Tommy Atkins’, houve variação estatisticamente significativa do
nitrogênio não protéico (NNP) e do nitrogênio protéico (NP) da polpa durante o período
estudado (FIGURA 18), embora os coeficientes de determinação tenham sido inferiores a
0,70 (R²= 0,6442 e R²= 0,6980). O NNP e o NP mostraram uma tendência de crescimento até
105 DAA, quando suas concentrações representaram 80,24 e 19,76% do NT, respectivamente
e depois decresceram até os 112 DAA, semelhantes ao NT. O valor médio obtido foi de 0,54 e
0,09 g/kg da massa fresca da polpa, respectivamente e o teor de NNP foi superior ao do NP
durante todo o desenvolvimento do fruto, a pesar do incremento no ponto máximo (105 DAA)
em relação ao valor inicial do NP (1,6 vezes) ter sido superior ao do NNP (1,39 vezes).
Estes resultados concordam com a afirmativa de Chitarra e Chitarra (2005) ao
relatarem o papel da síntese do ácido ribonucléico (RNA) e da síntese protéica na indução do
amadurecimento.
4.5.6 Proteína bruta e verdadeira
Na FIGURA 19 o teor de proteína bruta (PB) diminuiu dos 63 aos 70 DAA (3,67 para
3,07 g/kg da massa fresca da polpa), depois aumentou até aos 105 DAA (5,26 g/kg da massa
fresca da polpa) e finalmente reduziu até os 112 DAA (3,96 g/kg da massa fresca da polpa),

99
confirmando o comportamento do NT (FIGURA 18). Este comportamento coincide com
Chitarra e Chitarra (2005), ao afirmarem que as sínteses de ácidos nucléicos e proteína são
mais pronunciadas nas primeiras etapas da maturação.
**
**
*
* **
))
)
))
) )
)
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
1
2
3
4
5
6
Prot
eína
(g/k
g da
mas
sa fr
e sca
)
PB PV) *
YPB= 75,99514 - 2,645288X + 0,03133117X 2 - 0,0001200047X3, R2= 0,7644**.YPV= Não ajustado, R2< 0,7000.
FIGURA 19 - Proteína bruta (PB) e verdadeira (PV) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Para o conteúdo de proteína verdadeira (PV) da polpa houve variação estatisticamente
significativa durante o período estudado (FIGURA 19), embora o coeficiente de determinação
tenha sido inferior a 0,70 (R²= 0,6888). A PV mostrou uma tendência de crescimento até os
105 DAA e depois decresceu até os 112 DAA, semelhante ao NP (FIGURA 18). O valor
médio obtido foi de 0,58 g/kg da massa fresca da polpa e o incremento aos 105 DAA foi de
1,56 vezes em relação ao valor inicial.
4.5.7 Vitamina C
Houve redução no teor de Vitamina C até 98 DAA e a partir de então ficou
praticamente constante (FIGURA 20). As mangas aos 112 DAA apresentaram
aproximadamente metade (12,92 mg/100 g da massa fresca da polpa) do teor inicial aos 35
DAA (21,98 mg/100 g da massa fresca da polpa). Comportamento análogo foi obtido por
Lima (1997), ao detectarem redução no teor de Vitamina C total de 47,9 para 9,3 mg/100 g da
massa fresca da polpa nos frutos normais de ‘Tommy Atkins’.

100
)
)
) )
)) )
)
) )
35 42 49 56 63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
5
10
15
20
25
Vita
min
a C
tot a
l (m
g/1 0
0g d
a m
assa
fre s
ca)
Y= 16,60925 + 0,4786797X - 0,01071689X 2 + 0,00005550309X 3, R2= 0,7265*.
FIGURA 20 - Vitamina C de mangas ‘Tommy Atkins’ durante o desenvolvimento.
A redução na concentração de vitamina C durante o desenvolvimento foi
acompanhada por uma diminuição no conteúdo de açúcar redutor (AR) (FIGURA 17).
Segundo Menezes (1996), o ácido ascórbico é, estruturalmente, um dos componentes
vitamínicos mais simples encontrados em plantas. É uma lactona de açúcar ácido que é
sintetizada em plantas a partir de glicose ou outros carboidratos simples.
O comportamento observado neste experimento para os teores de vitamina C, é
bastante semelhante àquele relatado por Lakshminarayana et al. (1970) em trabalho realizado
com manga, concentrações médias de 175,0 e 87,5 mg/100 g da massa fresca da polpa para os
estádios 35 e 112 DAV, respectivamente. Neste mesmo sentido, de acordo com Franco
(2003), a manga ‘Comum’ madura possui 43 mg/100 g da massa fresca da polpa, por outro
lado, Wills et al. (1998) afirmam que possui 30 mg/100 g da massa fresca da polpa e Salunke
e Desai (1984) em ‘Keitt’ apenas 13,9 mg/100 g da massa fresca da polpa. As diferenças entre
os teores encontrados por estes autores e os deste experimento, podem ser atribuídas à
localização dos plantios, em climas e regiões distintas, diferentes tratos culturais, tipos de
solos, cultivares e uso ou não de irrigação.

101
4.5.8 Clorofila total
Pelos resultados obtidos (FIGURA 21) evidenciou-se um aumento acentuado no
conteúdo de clorofila total da casca de manga dos 35 até os 77 DAA (18,50 para 45,35
mg/100 g da massa fresca da casca), portanto, o valor inicial mais que dobrou e depois
diminuiu até os 112 DAA (30,27 mg/100 g da massa fresca da casca).
)
)
))
)
))
)
)
)
35 42 49 56 63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
10
20
30
40
50
Clo
rofil
a to
tal (
mg/
1 00g
da
mas
sa fr
e sca
)
Y= - 34,04612 + 1,75756X - 0,01056656X 2, R2= 0,7194**.
FIGURA 21 - Clorofila total da casca de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Em trabalho realizado com manga ‘Tommy Atkins’, Medlicott et al. (1986)
observaram uma rápida destruição de clorofila, sendo a clorofila a preferencialmente
degradada em relação à clorofila b. Neste mesmo sentido, Mena et al. (1996) com ‘Manila’
registraram comportamento semelhante ao obtido a partir dos 77 DAA neste experimento,
porém os valores encontrados por aqueles autores não ultrapassaram a 4 mg/100 g da massa
fresca da casca. Isto se deve, provavelmente, devido às mangas estarem em estádio de
maturação mais avançado que o deste experimento.
Tendo em vista que as alterações dessa característica são bem evidentes, constata-se
que a análise de clorofila total é adequada para a avaliação do grau de maturidade da manga.
No período dos 77 aos 112 DAA, enquanto a clorofila diminuiu a escala de coloração da
casca (ECC) aumentou (FIGURAS 12 e 21).

102
4.5.9 Carotenóides totais
O conteúdo de carotenóides totais aumentou quadraticamente com o avanço do
desenvolvimento (FIGURA 22), dos 63 aos 112 DAA (0,26 para 1,14 mg/100 g da massa
fresca da polpa, mais que quadruplicou).
)
)
)
)
))
))
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
0,2
0,4
0,6
0,8
1
1,2
Car
oten
óide
s tot
ais (
mg/
100g
da
mas
sa fr
esca
)
Y= - 3,349373 + 0,07700964X - 0,0003287172X 2, R2= 0,9714*.
FIGURA 22 - Carotenóides totais de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Medlicott et al. (1986), em trabalho realizado com ‘Tommy Atkins’, encontraram um
valor para carotenóides totais superior a 0,5 mg/100 g da massa fresca da polpa. Salunke e
Desai (1984) estudando diversas cultivares de mangas maduras verificaram uma variação no
teor de carotenóides totais de 0,9 a 9,2 mg/100 g da massa fresca da polpa. Mangels et al.
(1993) ressaltam que os níveis desses pigmentos podem ser influenciados pela estação do ano,
localização geográfica, condições de colheita, além de outros fatores.
Neste mesmo sentido, Salunke et al. (1991) identificaram cerca de dezesseis
hidrocarbonetos e oxicarotenóides em manga e constataram que o β-caroteno (pró-vitamina
A), que será transformado no organismo humano em vitamina A (retinol), é predominante no
fruto totalmente maduro.
A distinção marcante de cor constatada pela observação visual, aliada às diferenças
significativas de valores encontrados analiticamente para as medidas de carotenóides totais
durante o desenvolvimento, faz desta variável um parâmetro adequado para a avaliação do
grau de maturidade da manga, concordando com a escala de coloração da polpa (ECP) e

103
croma da polpa (CP), os quais aumentaram com o avanço do desenvolvimento (FIGURAS 12,
13 e 22).
4.5.10 Fenólicos
O conteúdo de fenólicos poliméricos (FIGURA 23) aumentou dos 63 aos 77 DAA
(31,16 para 36,92 mg/100 g da massa fresca da polpa) e depois diminuiu até os 112 DAA
(25,36 mg/100 g da massa fresca da polpa). Resultados equivalentes foram obtidos por
Evangelista et al. (1999), ao detectarem, com o avanço da maturação de ‘Tommy Atkins’,
uma elevação nos teores de compostos fenólicos, seguido de diminuição.
,
,
,
,, ,
,,
*
** * *
* **
)
)
)
)
) )
) )
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
10
20
30
40
50
Fenó
li cos
(mg/
100g
da
mas
sa fr
esca
)
FP FO FD) * ,
YFP= -380,8065 + 14,61817X - 0,1677007X 2 + 0,0006208195X 3, R2= 0,7822**.YFO= Não ajustado, R2< 0,7000.YFD= Não ajustado, R2< 0,7000.
FIGURA 23 - Fenólicos poliméricos (FP), oligoméricos (FO) e diméricos (FD) de mangas
‘Tommy Atkins’ durante o desenvolvimento.
Os teores de fenólicos oligoméricos e diméricos tiveram variação estatisticamente
significativa durante o desenvolvimento da manga (FIGURA 23), embora os coeficientes de
determinação tenham sido inferiores a 0,70 (R²= 0,6263 e R²= 0,6986). A tendência observada
no conteúdo de fenólicos oligoméricos e diméricos durante o desenvolvimento (63 a 112
DAA) foi de redução (34,83 para 28,01 mg/100 g da massa fresca da polpa e 43,14 para 33,58
mg/100 g da massa fresca da polpa, respectivamente). O valor médio obtido foi de 29,19 e
33,91 mg/100 g da massa fresca da polpa, respectivamente.

104
Ao ser considerado apenas os valores iniciais e finais, o teor de fenólicos diméricos foi
superior ao dos oligoméricos e este por sua vez superior ao dos poliméricos. Este
comportamento ratifica o estudo de Park et al. (1980), em trabalho desenvolvido com manga,
segundo qual os fenólicos diméricos predominam sobre os demais.
O somatório dos valores médios dos compostos fenólicos desta pesquisa totaliza em
92,10 mg/100 g da massa fresca da polpa, igualando-se assim ao de Evangelista (1999), o
qual detectou com esta mesma cultivar o valor médio de 93,10 mg/100 g da massa fresca da
polpa no tratamento controle. Por outro lado, os dados obtidos são superiores aos citados por
Lakshminarayana (1980), no qual os compostos fenólicos em mangas maduras variaram de 31
a 75 mg/100 g da massa fresca da polpa e aos de Lima (1997), quando constatou uma
variação de 13,75 a 31,87 mg/100 g da massa fresca em mesocarpo de manga sadia. Esta
diferença pode ser atribuída a diversos fatores que, segundo Lizada (1993) são dependentes da
espécie e/ou cultivar, do estádio fisiológico e da localização.
A perda da adstringência é conseqüência do decréscimo dos compostos fenólicos e do
concomitante acúmulo de açúcares que ocorre durante a maturação (VAN BUREN, 1970;
AZIZ e YUSOF, 1994).
4.5.11 Pectinas
O teor de pectina total (FIGURA 24) diminuiu dos 63 aos 91 DAA (644,62 para
252,90 mg/100 g da massa fresca da polpa) e depois aumentou, muito pouco, até os 112 DAA
(301,96 mg/100 g da massa fresca da polpa). Nota-se que pela curva de regressão o mínimo
do teor de pectina total ocorre aos 97,23 DAA (236,99 mg/100 g da massa fresca da polpa). O
comportamento do teor de pectina total observado em parte coincide com Lima (1997) ao
observar decréscimo (1275 para 375 mg/100 g da massa fresca da polpa) no teor de pectina
total nas mangas ‘Tommy Atkins’ sadias. Por outro lado, Dutra (2005) observou aumento dos
50 aos 85 DAF (1050 para 1150 mg/100 g da massa fresca da polpa), depois redução até os
106 DAF (650 mg/100 g da massa fresca da polpa) e finalmente aumentou até os 120 DAF
(1050 mg/100 g da massa fresca da polpa).
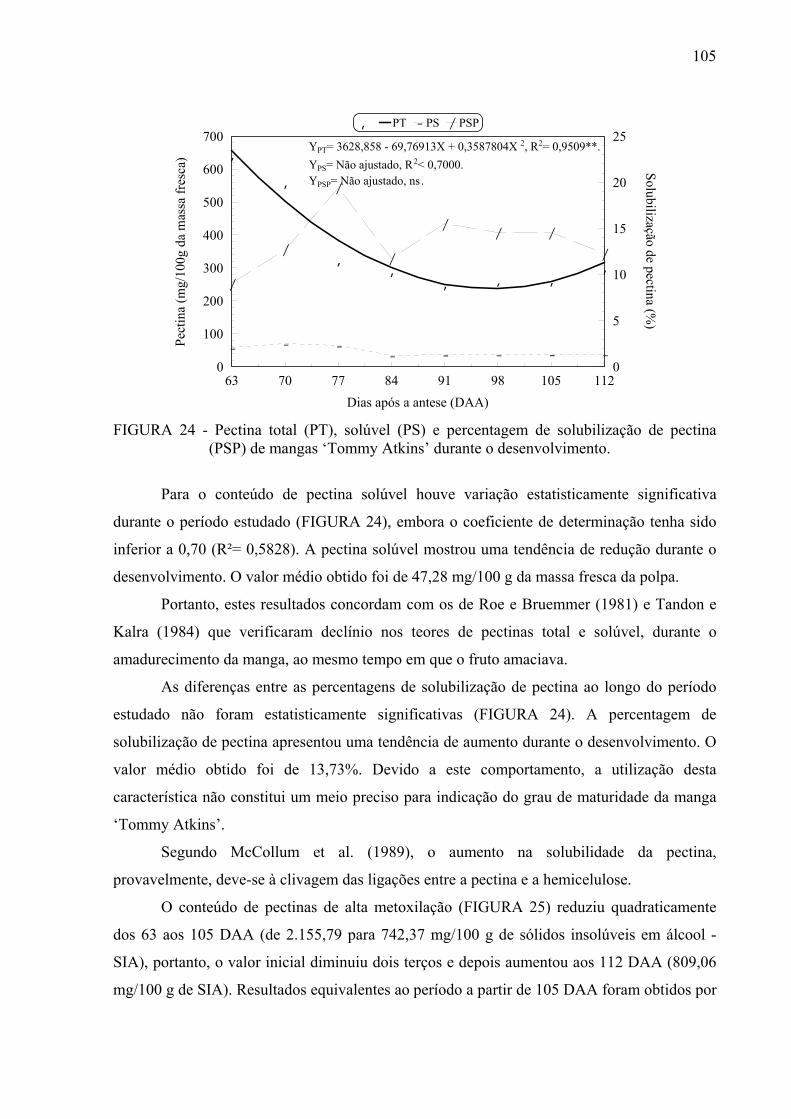
105
,
,,
,
,
,, ,
* ** *
* * * *
)
)
)
))
) ) )
63 70 77 84 91 98 105 112Dias após a antese (DAA)
0
100
200
300
400
500
600
700
Pect
ina
(mg/
100g
da
ma s
sa fr
esc a
)
0
5
10
15
20
25Solubilização de pectina (%
) PT PS PSP) * ,
YPT= 3628,858 - 69,76913X + 0,3587804X 2, R2= 0,9509**.YPS= Não ajustado, R2< 0,7000.YPSP= Não ajustado, ns.
FIGURA 24 - Pectina total (PT), solúvel (PS) e percentagem de solubilização de pectina
(PSP) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Para o conteúdo de pectina solúvel houve variação estatisticamente significativa
durante o período estudado (FIGURA 24), embora o coeficiente de determinação tenha sido
inferior a 0,70 (R²= 0,5828). A pectina solúvel mostrou uma tendência de redução durante o
desenvolvimento. O valor médio obtido foi de 47,28 mg/100 g da massa fresca da polpa.
Portanto, estes resultados concordam com os de Roe e Bruemmer (1981) e Tandon e
Kalra (1984) que verificaram declínio nos teores de pectinas total e solúvel, durante o
amadurecimento da manga, ao mesmo tempo em que o fruto amaciava.
As diferenças entre as percentagens de solubilização de pectina ao longo do período
estudado não foram estatisticamente significativas (FIGURA 24). A percentagem de
solubilização de pectina apresentou uma tendência de aumento durante o desenvolvimento. O
valor médio obtido foi de 13,73%. Devido a este comportamento, a utilização desta
característica não constitui um meio preciso para indicação do grau de maturidade da manga
‘Tommy Atkins’.
Segundo McCollum et al. (1989), o aumento na solubilidade da pectina,
provavelmente, deve-se à clivagem das ligações entre a pectina e a hemicelulose.
O conteúdo de pectinas de alta metoxilação (FIGURA 25) reduziu quadraticamente
dos 63 aos 105 DAA (de 2.155,79 para 742,37 mg/100 g de sólidos insolúveis em álcool -
SIA), portanto, o valor inicial diminuiu dois terços e depois aumentou aos 112 DAA (809,06
mg/100 g de SIA). Resultados equivalentes ao período a partir de 105 DAA foram obtidos por

106
Brinson et al. (1988), ao detectarem aumento no teor de poliuronídeos solúveis em água com
o amadurecimento da manga.
**
**
* * * *
,
,
,, ,
,,
,
)
)
)
)
)
)
)
)
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
500
1000
1500
2000
2500
Pect
inas
(mg/
1 00g
de
S IA
)
PAM PBM PROT) * ,
YPAM= 9356,051 - 160,0225X + 0,7542205X2, R2= 0,7091**.YPBM= Não ajustado, ns.YPROT= - 18486,74 + 700,141X - 8,107922X2 + 0,03000145X3, R2= 0,8973**.
FIGURA 25 - Pectinas de alta metoxilação (PAM), de baixa metoxilação (PBM) e a
protopectina (PROT) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
As diferenças entre os teores de pectinas de baixa metoxilação ao longo do período
estudado não foram estatisticamente significativas (FIGURA 25). As pectinas de baixa
metoxilação apresentaram uma tendência de diminuição durante o desenvolvimento. O valor
médio obtido foi de 1.600,96 mg/100 g de SIA. Este resultado exclui esta característica como
indicador de maturidade em mangas ‘Tommy Atkins’.
O teor de protopectina (FIGURA 25) diminuiu cubicamente dos 63 aos 105 DAA
(1.011,58 para 202,14 mg/100 g de SIA), portanto, o valor inicial reduziu quatro quintos e
depois aumentou até os 112 DAA (445,74 mg/100 g de SIA). Este comportamento coincide
com os resultados de Tandon e Kalra (1984), em manga, ao constatarem que a protopectina
aumenta antes da maturidade fisiológica e decresce, em seguida, com o amadurecimento e o
amaciamento. Esta fração péctica, juntamente com outros materiais cimentantes, é
responsável pela manutenção da firmeza.
O teor das pectinas de baixa metoxilação (1.600,96 mg/100 g de SIA), considerando
apenas os valores médios, foi superior aos de alta metoxilação (1.322,60 mg/100 g de SIA) e
este por sua vez superior ao de protopectina (738,13 mg/100 g de SIA).
Em geral, as alterações nos teores de pectina durante o amadurecimento estão
associadas à degradação enzimática. Porém, a conformação estrutural da molécula, unida,

107
pelo menos parcialmente, por interações não-covalentes, reforça a possibilidade de
degradação não enzimática. Os principais fatores envolvidos neste último tipo são o pH e a
força iônica da parede celular (SEYMOUR e GROSS, 1996).
4.6 Avaliações bioquímicas
Entre as variáveis submetidas à análise de variância representadas nas TABELAS 9A
e 10A, não se verificou efeito significativo do tempo de colheita sobre as atividades da
pectinametilesterase, β-amilásica e proteína de parede celular.
4.6.1 Pectinametilesterase e poligalacturonase
As diferenças entre a atividade da pectinametilesterase (PME) ao longo do período
estudado não foram estatisticamente significativas (FIGURA 26). A atividade da PME
apresentou flutuações ao longo do desenvolvimento, com tendência à estabilidade.
Obviamente este comportamento desclassifica esta característica como indicador do grau de
maturidade.
* **
* * * * *
))
)
)
)
)
)
)
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
50
100
150
200
250
300
Pect
inas
es (U
AE/
g da
mas
s a fr
esca
/ min
)
PME PG) *
YPME= Não ajustado, ns.YPG= Não ajustado, R2< 0,7000.
FIGURA 26 – Atividades da pectinametilesterase (PME) e poligalacturonase (PG) de mangas ‘Tommy Atkins’ durante o desenvolvimento.

108
O valor médio obtido foi de 201,54 UAE/g da massa fresca/min. Este valor médio
assemelha-se com o obtido por Lima (1997) quando encontrou em mesocarpo sadio do fruto
imaturo de ‘Tommy Atkins’ uma atividade da PME de 216,66 UAE/g da massa fresca/min.
Por outro lado, Evangelista (1999) não detectou atividade de PME em nenhum dos
tratamentos nos diferentes estádios de maturação.
O comportamento constatado neste ensaio nem sempre é observado. Segundo Tucker
(1993), a atividade total da PME pode diminuir, permanecer constante ou aumentar durante a
maturação, dependendo do fruto ou do método de extração. Em manga, tem-se obtido
resultados variáveis dependendo da cultivar. Estudos desenvolvidos com as cvs. Keitt (ROE e
BRUEMER, 1981), Dr Knight e Kitchner (ABU-SARRA e ABU-GOUKH, 1992) mostraram
diminuição na atividade da PME durante os primeiros estádios de amadurecimento, com
posterior estabilização. Já nas mangas ‘Carabao’ (LIZADA, 1993) e ‘Abu-Samaka’ (ABU-
SARRA e ABU-GOUKH, 1992), verificou-se aumento inicial e subseqüente decréscimo. Por
outro lado, em algumas cultivares de manga do Paquistão permaneceu constante (ASHRAF et
al., 1981).
Como conseqüência destas variações, a correlação entre o nível de atividade
enzimática da PME e o amaciamento da parede celular ainda não está clara (JOHN e DEY,
1986).
As mudanças na atividade da PME são difíceis de prever devido à presença de
isoformas ou inibidores enzimáticos (TUCKER, 1993). Um destes inibidores, conforme
Bordenave e Goldberg (1993), é o H3O+ que a enzima gera quando desmetila seus substratos.
Segundo Bordenave (1996), devido à alta especificidade das PMEs por metil-ésteres
de galacturonanas, a desesterificação de pectinas que promove nunca é completa. Além disso,
o processo não resulta diretamente no amaciamento, devendo anteceder a efetiva degradação
da pectina pela PG (KAYS, 1991). Assim, Seymour et al. (1987) concluíram que a
solubilização de poliuronídeos em tomates maduros pode ser resultante da atividade conjunta
da PME e da PG.
Para a atividade da poligalacturonase (PG) houve variação estatisticamente
significativa durante o período estudado (FIGURA 26), embora o coeficiente de determinação
tenha sido inferior a 0,70 (R²= 0,6810). A atividade da PG mostrou uma tendência de redução
até os 84 DAA e depois de aumento até os 112 DAA. Este período de redução coincidiu com
a queda brusca no teor de pectina total e com a maior solubilização de pectinas (FIGURA 24),
sugerindo que a maior parte dos substratos da PG foram imediatamente utilizados, quando se
obteve a máxima atividade. Da mesma forma, o período de crescimento coincidiu com a

109
subida no teor de pectina total e com a redução na solubilização de pectinas (FIGURA 24). O
valor médio obtido foi de 31,08 UAE/g da massa fresca/min. Este valor médio é inferior ao
encontrado por Evangelista (1999), mas supera aquele obtido por Lima (1997) que
encontraram em mesocarpo sadio de fruto ‘verde’ de ‘Tommy Atkins’ uma atividade da PG
de 50,0 e de 10,0 UAE/g da massa fresca/min, respectivamente.
Neste sentido, Roe e Bruemmer (1981), Abu-Sarra e Abu-Goukh (1992), Lima (1997)
e Evangelista (1999) registraram aumento na atividade da PG de manga. Abu-Sarra e Abu-
Goukh (1992) observaram, inclusive, uma correlação muito alta (R2>0,96) entre o aumento da
atividade da enzima e a perda de firmeza do fruto.
4.6.2 Polifenoloxidase e peroxidase
Para a atividade de polifenoloxidase (PPO) houve variação estatisticamente
significativa durante o período estudado (FIGURA 27), embora o coeficiente de determinação
tenha sido inferior a 0,70 (R²= 0,6346). A atividade da PPO apresentou flutuações ao longo do
desenvolvimento, com tendência à estabilidade.
) ))
))
)) )
* ** * * * * *
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
100
200
300
400
500
600
Oxi
dase
s (U
AE/
g d a
mas
sa fr
esca
/min
)
PPO POD) *
YPPO= Não ajustado, R2< 0,7000.YPOD= 688,3928 - 21,92311X + 0,232832X 2 - 0,0007982399X3, R2= 0,8151**.
FIGURA 27 – Atividades da polifenoloxidase (PPO) e peroxidase (POD) de mangas ‘Tommy
Atkins’ durante o desenvolvimento.
O valor médio obtido foi de 543,48 UAE/g da massa fresca/min. Este valor médio é
superior aos determinados por Evangelista (1999) e Lima (1997), em mesocarpo sadio de

110
fruto imaturo de ‘Tommy Atkins’ quando encontraram uma atividade da PPO de 31,5 e 15,0
UAE/g da massa fresca/min, respectivamente.
Por outro lado, Selvaraj e Kumar (1989) observaram declínio na atividade desta
enzima durante o amadurecimento dos frutos de varias cultivares indianas, incluindo a
‘Alphonso’, enquanto Lizada (1993) verificou aumento na atividade, durante o
amadurecimento de mangas ‘Malgoa’ e ‘Harumanis’.
Conforme a FIGURA 27, a atividade da peroxidase (POD) diminuiu cubicamente dos
63 aos 84 DAA (32,51 para 13,87 UAE/g da massa fresca/min), o que equivale a uma redução
de quase dois terços do valor inicial e depois aumentou até os 112 DAA (33,58 UAE/g da
massa fresca/min), correspondendo a um incremento de quase três vezes. O período de
diminuição coincidiu com o aumento no conteúdo de fenólicos poliméricos (FIGURA 23),
enquanto o período de aumento coincidiu com a diminuição de fenólicos poliméricos,
sugerindo a associação entre enzima e substrato e, conseqüentemente, à perda de
adstringência aos 112 DAA. Estes resultados são superiores aos de Lima (1997) que em
mesocarpo sadio de ‘Tommy Atkins’ armazenado por 28 dias, verificou uma variação de 6,66
a 10,0 UAE/g da massa fresca/min.
Comparando-se as atividades da PPO e da POD, constata-se que a primeira além dos
níveis mais altos, apresentou variações mais pronunciadas. No entanto, deve-se ressaltar que
mesmo nos casos em que a atividade da POD é alta, sua ação isolada raramente é importante,
pois ela depende da presença da PPO no meio, a fim de gerar o H2O2 necessário à ocorrência
da oxidação (ROBARDS et al., 1999).
4.6.3 Amilases
A atividade amilásica total (FIGURA 28) aumentou linearmente dos 63 aos 112 DAA
(2,53 para 3,00 µmol maltose/g da massa fresca/min). O período de maior atividade coincidiu
com o de maior síntese de amido e de açúcares solúveis totais (FIGURA 17). Este
comportamento em parte coincide com os encontrados por Lima (1997) ao observar
crescimento (0,45 para 1,2 µmol maltose/g da massa fresca/min) na atividade amilásica total
até aos 14 dias nas mangas ‘Tommy Atkins’ sadias armazenadas por 28 dias. Neste sentido,
Awad (1993) relata que em manga ocorre um aumento na atividade amilásica e na hidrólise
do amido durante a maturação, pois este é o principal carboidrato presente no fruto imaturo.

111
, , , ,
* * * *) ) ) )
63 70 77 84
Dias após a
0
0,5
1
1,5
2
2,5
3
3,5
Am
ilase
s (µ
mol
mal
tose
/g d
a m
assa
fres
ca/m
in) AMLT -AML ß-AML) * ,
YAMLT= 1,579683 + 0,01Yα-AML= Não ajustado, Yβ-AML= Não ajustado,
α
FIGURA 28 – Atividades amilásica total (AMLAML) de mangas ‘Tommy Atkin
De acordo com Fuchs et al. (1980), em
atividade amilásica suficiente para iniciar a hid
atividade adicional. Esta atividade adicional p
degradação do amido. Em frutos sobremaduros
substancialmente reduzida.
Neste sentido, a hidrólise do amido, dura
associada com as atividades α- e β-amilásicas e, c
açúcares totais durante o amadurecimento (FUCH
Para a atividade α-amilásica houve varia
período estudado (FIGURA 28), embora o coefic
0,70 (R²= 0,6967). A atividade α-amilásica mos
desenvolvimento. O valor médio obtido foi de
representando cerca de 91% da atividade amil
relatado por Awad (1993), quando afirma que a
atividade amilolítica no fruto.
As diferenças entre a atividade β-amilásic
estatisticamente significativas (FIGURA 28).
tendência de redução durante o desenvolviment
maltose/g da massa fresca/min, equivalendo apr
,,
,
,
**
*
* ))
) )
91 98 105 112
antese (DAA)
323696X , R2= 0,8476**.R2< 0,7000.ns.
T), α-amilásica (α-AML) e β-amilásica (β-s’ durante o desenvolvimento.
manga, imediatamente após a colheita há
rólise do amido e somente depois ocorre
romove um maior incremento na taxa de
, por outro lado, a atividade amilásica é
nte o amadurecimento da manga, tem sido
omo conseqüência, ocorre um aumento dos
S et al., 1980; LIZADA, 1993).
ção estatisticamente significativa durante o
iente de determinação tenha sido inferior a
trou uma tendência de aumento durante o
2,48 µmol maltose/g da massa fresca/min,
ásica total. Este percentual é superior ao
α-amilase pode ser responsável por 80% da
a ao longo do período estudado não foram
A atividade β-amilásica apresentou uma
o. O valor médio obtido foi de 0,25 µmol
oximadamente a 9% da atividade amilásica

112
total. Por outro lado, Sen et al. (1985) observaram que a atividade β-amilásica aumentava com
o desenvolvimento do fruto, declinando com a aproximação da maturidade. Este resultado
exclui esta característica como indicador de maturidade em mangas do tipo ‘Tommy Atkins’.
Lima (1997) estudando as transformações químicas e bioquímicas no mesocarpo com
tecido esponjoso em manga ‘Tommy Atkins’ durante o armazenamento, inferiu que a
atividade amilolítica é uma importante atividade metabólica durante o amadurecimento de
mangas.
4.6.4 Galactosidases
Na FIGURA 29 observa-se que as variações nas atividades α-galactosidásicas,
extraídas do citosol (α-GALc) e da parede celular (α-GALp), ocorreram a partir dos 63 DAA.
A atividade da α-GALc reduziu em cerca de dois terços dos 63 aos 77 DAA (31,50 para 10,85
UAE/g da massa fresca/min), quando alcançou o mínimo. A partir daí, verificou-se um
aumento de aproximadamente cinco vezes até os 112 DAA (57,43 UAE/g da massa
fresca/min). Para a α-GALp, os níveis de atividade foram inferiores e mostraram uma
tendência de crescimento ao longo do desenvolvimento (8,13 para 12,09 UAE/g da massa
fresca/min). O valor médio obtido foi de 11,28 UAE/g da massa fresca/min. Sua importância
para o amaciamento do fruto deve, então, ser secundária.
**
*
* ** *
*
)
)
)
)
)
)
) )
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
10
20
30
40
50
60
-G
AL
(UA
E/g
da m
a ssa
fres
c a/m
in)
-GALc GALp) -*
Yα-GALc= 1162,956 - 39,47594X + 0,441981X 2 - 0,001586173X3, R2= 0,8133*.Yα-GALp= Não ajustado, R2< 0,7000.
α α
α
FIGURA 29 – Atividades α-galactosidásicas de citosol (α-GALc) e de parede celular (α-GALp) de mangas ‘Tommy Atkins’ durante o desenvolvimento.

113
Neste estudo, a atividade da α-GALp representou proporções cada vez menores da
atividade α-galactosidásica total, a partir de 63 DAA, quando representava 20,51% (FIGURA
29). Nas mangas aos 112 DAA, esta percentagem era de apenas 17,39%. Por sua vez,
trabalhos realizados em laranja ‘Valência’ demonstraram que mais de 75% da atividade das
GALs está na fração extraída da parede celular (BURNS, 1990), enquanto, em graviola
‘Crioula’ reduz de 33,94 para 6,92% com o avanço da maturação durante 5 dias após a
colheita (LIMA, 2002).
As atividades das β-GALs (FIGURA 30) durante o desenvolvimento foram diferentes
daquelas observadas para as α-GALs. A atividade da β-GALc reduziu em cerca de metade dos
63 aos 84 DAA (0,22 para 0,12 UAE/g da massa fresca/min), quando alcançou o mínimo. A
partir daí, verificou-se um aumento de aproximadamente dez vezes até os 112 DAA (1,16
UAE/g da massa fresca/min). Este comportamento ratifica o estudo de Evangelista (1999), em
trabalho com esta mesma cultivar, segundo o qual a atividade de β-GALc aumentou à medida
que o fruto tornava-se maduro. Para a β-GALp, os níveis de atividade foram no geral
superiores aos da β-GALc e mostraram uma tendência de crescimento ao longo do
desenvolvimento (0,35 para 0,79 UAE/g da massa fresca/min). O valor médio obtido foi de
0,59 UAE/g da massa fresca/min. Sua importância para o amaciamento do fruto deve, então,
ser primária.
*
*
*
**
**
*
)
)
))
))
)
)
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
ß-G
AL
(UA
E/g
da m
assa
fres
ca/m
in)
ß-GALc ß-GALp) *
Yβ-GALc= - 1,354517 + 0,1062202X - 0,00199009X2 + 0,00001114154X3, R2= 0,9404*.Yβ-GALp= Não ajustado, R2< 0,7000.
FIGURA 30 – Atividades β-galactosidásicas de citosol (β-GALc) e de parede celular (β-
GALp) de mangas ‘Tommy Atkins’ durante o desenvolvimento.

114
A atividade da β-GALc, apesar dos valores mais baixos em relação a β-GALp, durante
quase todo o período de desenvolvimento do fruto, superou a β-GALp aos 112 DAA, quando
apresentou o valor de 1,16 UAE/g da massa fresca/min contra 0,79 UAE/g da massa
fresca/min da β-GALp. Esta resposta sugere que dos 63 aos 105 DAA houve exportação da
enzima para a parede celular, enquanto, aos 112 DAA houve degradação da parede celular.
Podendo ser confirmado pela proporção da atividade da β-GALp em relação à atividade β-
galactosidásica total, que reduziu durante todo o período (61,40 para 50,26%, dos 63 aos 105
DAA), atingindo 40,51% aos 112 DAA. Estes resultados assemelham-se com o de Burns
(1990), que detectou em parede celular de laranja ‘Valência’ mais de 75% da atividade das
GALs.
Levando-se em conta as proporções das atividades α- e β-galactosidásicas totais dos
63 aos 112 DAA, verifica-se que as proporções das GALs citosólicas aumentaram (de 79,49
para 82,61%-α-GALc e de 38,60 para 59,49%-β-GALc) e as proporções das GALs de parede
celular diminuíram (de 20,51 para 17,39%-α-GALp e de 61,40 para 40,51%-β-GALp) com o
avanço da maturação. Este comportamento sugere que nos estádios iniciais de
desenvolvimento dos frutos, as GALs são exportadas do citosol para a parede celular,
enquanto nos estádios finais, após a degradação da parede, as GALs retornam ao citosol.
Portanto, esta atuação é corroborada por Brady (1987) ao afirmar que as GALs são em geral
sintetizadas e exportadas a partir do citoplasma para a parede celular.
As β-GALs citosólicas e de parede celular são mais variáveis que as α-GALs
citosólicas e de parede celular, pois a diferença nas proporções das atividades β- e α-
galactosidásicas totais dos 63 aos 112 DAA, são 20,89 e 3,12%, respectivamente.
Analisando os valores médios deste ensaio, constatou-se que a atividade da α-GALc
(36,70 UAE/g da massa fresca/min) é maior que a da α-GALp (11,28 UAE/g da massa
fresca/min) e esta por sua vez é maior que a da β-GALp (0,59 UAE/g da massa fresca/min), a
qual finalmente é maior que a da β-GALc (0,40 UAE/g da massa fresca/min). Portanto, em
relação a atividade galactosidásica total média as atividades das α-GALc, α-GALp, β-GALp e
β-GALc, representaram: 74,94%, 23,03%, 1,21% e 0,82%, respectivamente. Por outro lado,
Ali (1995) afirma que entre as glicosidases, a β-GAL é a predominante na polpa de mangas
‘Harumanis’.
Mangas maduras podem possuir três isoformas de β-GAL, I, II e III, sendo que a β-
GAL I, aparentemente, é a forma predominante. A perda de galactosil, arabinosil e ramnosil
durante o amadurecimento é atribuída à hidrólise de galactanas e arabinogalactanas pela β-

115
GAL, com atividade de galactanase, pois ela está intimamente relacionada com o decréscimo
na firmeza dos frutos (ALI et al., 1995).
Considerando que a manga sofre amaciamento progressivo, as enzimas que devem
contribuir mais diretamente para o processo são PME, PG (FIGURA 26), α-GAL e β-GAL de
parede celular (FIGURAS 29 e 30). Aos 112 DAA foi caracterizado o início do amaciamento
no fruto (TABELA 6). Sabe-se que a intensidade com que o amaciamento ocorre diminui com
o tempo. Desta forma, espera-se que a atividade das enzimas envolvidas seja reduzida com a
evolução do processo. Em primeiro momento, a PG atuou de modo isolado. Entretanto, a
queda de atividade das GALs de parede celular a partir dos 105 DAA e conseqüente aumento
das GALs citosólicas sugere que as GALs de parede caracterizam melhor esta transformação.
Trabalho realizado por Ketsa et al. (1998) é coerente com esta idéia, pois os autores
encontraram, em manga, uma correlação entre a atividade da β-GAL e a perda de firmeza
bastante superior à da PG.
4.6.5 Proteína
Os teores de proteína mostram uma tendência de decréscimo ao longo do período,
especialmente aquelas quantificadas a partir de extratos do citosol (FIGURA 31). Aos 98
DAA, a queda foi de quase quatro vezes do valor inicial (0,0510 para 0,0150 mg/g da massa
fresca). A partir daí, coincidindo com a fase de maior incremento na atividade da β-GAL
citosólica (quase quatro vezes), o conteúdo de proteína subiu para 0,0598 mg/g da massa
fresca (105 DAA), isto é, um aumento de quatro vezes. Os frutos aos 112 DAA continham
0,0350 mg/g da massa fresca.

116
*
*
*
* *
* * *
)
))
))
)
)
)
63 70 77 84 91 98 105 112
Dias após a antese (DAA)
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
Prot
eína
(mg/
g da
mas
sa fr
e sca
)
Pc Pp) *
YPc= Não ajustado, R2< 0,7000.YPp= Não ajustado, R2< 0,7000.
FIGURA 31 - Proteína de citosol (Pc) e de parede celular (Pp) de mangas ‘Tommy Atkins’
durante o desenvolvimento.
Na parede celular, onde os teores de proteína normalmente são mais baixos, dos 63
aos 112 DAA (0,0294 para 0,0132 mg/g da massa fresca) observou-se uma redução de mais
de duas vezes do valor inicial.
Segundo Kays (1991), durante o amadurecimento, uma proporção considerável das
proteínas é constituída por enzimas, especialmente as hidrolíticas, e estão envolvidas na
degradação de várias moléculas. Além disso, estudos têm verificado que a quantidade de
proteínas extraíveis está relacionada ao estádio de maturação, de forma que nos frutos
maduros os teores são mais baixos (MARIN e CANO, 1992). Naquele estádio, espera-se,
portanto, um incremento na proporção de hidrolases e outras enzimas degradativas em relação
ao total, mesmo com queda característica nos valores absolutos.
Neste sentido, de acordo com Chitarra e Chitarra (2005), há um número considerável
de evidências do envolvimento da síntese protéica (síntese “de novo” de enzimas) no
climatério dos frutos, no qual ocorre aumento na proporção do nitrogênio protéico em relação
ao nitrogênio total. Também existem evidências de um redirecionamento na síntese protéica
durante o amadurecimento. Há aumento no teor das enzimas hidrolíticas, segundo as
mudanças nas suas taxas de síntese e/ou degradação e não como um resultado da ativação de
uma molécula precursora, sintetizada no início do desenvolvimento. Portanto, a síntese dessas
proteínas aumenta de forma acentuada quando a maturação se inicia.

117
4.7 Correlação entre as variáveis estudadas
4.7.1 Correlação entre sólidos solúveis totais e as variáveis físicas estudadas
Os coeficientes de correlação linear determinados com o intuito de medir o grau de
associação entre os sólidos solúveis totais e as variáveis físicas estudadas encontram-se na
TABELA 9. Dentre os coeficientes de correlação linear estimados, para todas as combinações
possíveis, apenas para o teor de água foram observadas correlações negativas e significativas
em relação às demais, com exceção quando correlacionada com o diâmetro longitudinal, que
apesar de continuar negativa foi não significativa. Também foram visualizadas correlações
não significativas da escala de coloração da casca com o diâmetro longitudinal e com o
produto dos diâmetros e da escala de coloração da polpa com o diâmetro longitudinal. As
demais correlações possíveis foram positivas e significativas. Os resultados da correlação
entre o produto dos diâmetros e as massas fresca e seca, apesar de apresentarem menores
valores de coeficientes (R= 0,72 e R= 0,66), assemelham-se àqueles obtidos por Morais
(2001), estudando esta mesma cultivar (R= 0,97 e R= 0,94), e Castro Neto e Reinhardt (2003),
estudando a cv. Haden (R= 0,99 e R= 0,95).
Apesar da massa fresca, massa de água, sólidos solúveis totais, diâmetros transversal e
ventral, escalas de coloração da casca e da polpa terem dado coeficientes de correlação com a
massa seca maiores que o produto dos diâmetros, esse último apresenta a vantagem de ser
uma medida mais confiável (TABELA 9). A massa fresca, massa de água, sólidos solúveis
totais e escalas de coloração da casca são muito sensíveis a variações climáticas (CASTRO
NETO e REINHARDT, 2003) e uma medida de produtos de diâmetros é sempre mais
fidedigna que a avaliação individual de cada diâmetro. Por sua vez, a coloração da polpa é
uma medida subjetiva que depende do observador. Assim, o produto dos diâmetros dos frutos
pode ser usado com segurança para determinar o crescimento da manga cv. Tommy Atkins,
com a vantagem de ser um método não destrutivo, baseado em medições fáceis e rápidas.

118
TABELA 9 - Matriz de correlação entre sólidos solúveis totais e características físicas de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Variáveis DL DV DT PD MF MS MA TA ECC EBCC ECPSólidos solúveis totais (SST) 0,54** 0,66** 0,76** 0,64** 0,71** 0,82** 0,68** -0,79** 0,82** 0,82** 0,79**Diâmetro longitudinal (DL) - 0,95**
0,93** 0,98** 0,69** 0,61** 0,70** -0,30ns 0,24ns 0,45* 0,33ns
Diâmetro ventral (DV) - - 0,97** 0,98** 0,76** 0,70** 0,77** -0,40* 0,38* 0,56** 0,41* Diâmetro transversal (DT) - - - 0,96** 0,80** 0,78** 0,80** -0,53** 0,51** 0,65** 0,52**
Produto dos diâmetros (PD) - - - - 0,72** 0,66** 0,72** -0,40* 0,35ns 0,54** 0,41*Massa fresca (MF) - - - - - 0,94** 0,99** -0,45* 0,50** 0,51** 0,45* Massa seca (MS) - - - - - - 0,91** -0,71** 0,70**
0,62** 0,69**
Massa de água (MA) - - - - - - - -0,39* 0,45* 0,47** 0,39*Teor de água (TA) - - - - - - - - -0,85** -0,71** -0,91**
E. de coloração da casca (ECC) - - - - - - - - - 0,74** 0,88** E. Blush coloração casca (EBCC) - - - - - - - - - - 0,76**
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste t. ns Não-significativo. ECP - Escala de coloração da polpa.

119
Objetivando correlacionar a coloração da polpa feita em colorímetro (LP-CP-HP) com
sólidos solúveis totais e as demais variáveis físicas analisadas na TABELA 9, recalculou-se a
correlação linear com apenas 24 dados/variável, isto é, 8 tratamentos (63, 70, 77, 84, 91, 98,
105 e 112 DAA) e 3 repetições, ao invés de 30 dados/variável, isto é, 10 tratamentos (35, 49,
63, 70, 77, 84, 91, 98, 105 e 112 DAA) e 3 repetições que compõem a TABELA 9. Após esta
análise, constatou-se que a luminosidade e o ângulo Hue da polpa não apresentaram
coeficientes de correlação linear estimados superior a 0,70 quando contrastado com as demais
variáveis analisadas, enquanto o croma da polpa apresentou correlações superiores a 0,70,
positivas e significativas com sólidos solúveis totais (R= 0,83**), escalas de coloração da
casca (R= 0,85**), de Blush para coloração da casca (R= 0,74**) e de coloração da polpa (R=
0,89**) e correlação superior a 0,70, negativa e significativa com teor de água (R= -0,86**).
Portanto, a escala de coloração da polpa possui uma alta correlação com o croma da
polpa. Este resultado é corroborado por Morais (2001) trabalhando com manga ‘Tommy
Atkins’, ao afirmar que os índices obtidos para cor da polpa, através de escala subjetiva são
compatíveis com os obtidos nos parâmetros de avaliação da cor através do colorímetro, apesar
do primeiro ser um método subjetivo, podendo apresentar maior possibilidades de falhas.
Medlicott et al. (1992) também observaram alta correlação entre o método de avaliação da cor
da casca, através de escala subjetiva, e os parâmetros obtidos no colorímetro.
4.7.2 Correlação entre as variáveis físicas, físico-químicas e químicas
Os coeficientes de correlação linear determinados com o intuito de medir o grau de
associação entre massa seca, colorações de casca e da polpa, sólidos solúveis totais, acidez,
pH e relação SST/ATT encontram-se na TABELA 10. Dentre os coeficientes de correlação
linear estimados, para todas as combinações possíveis, apenas para acidez total titulável foram
observadas correlações negativas e significativas. As demais correlações possíveis foram
positivas e significativas.
Objetivando correlacionar a coloração da polpa feita em colorímetro (LP-CP-HP) com
sólidos solúveis totais e as demais variáveis físicas, físico-químicas e químicas analisadas na
TABELA 10, recalculou-se a correlação linear com apenas 24 dados/variável, isto é, 8
tratamentos (63, 70, 77, 84, 91, 98, 105 e 112 DAA) e 3 repetições, ao invés de 30
dados/variável, isto é, 10 tratamentos (35, 49, 63, 70, 77, 84, 91, 98, 105 e 112 DAA) e 3

120
TABELA 10 - Matriz de correlação entre massa seca, colorações da casca e da polpa, sólidos solúveis totais, acidez, pH e relação SST/ATT de
mangas ‘Tommy Atkins’ durante o desenvolvimento. Variáveis ECC EBCC ECP SST ATT pH RSA
Massa seca (MS) 0,70** 0,62** 0,69** 0,82** -0,77** 0,77** 0,80**E. de coloração da casca (ECC) - 0,74** 0,88** 0,82** -0,52** 0,89** 0,82**
E. Blush coloração casca (EBCC) - - 0,76** 0,82** -0,59** 0,81** 0,85** E. de coloração da polpa (ECP) - - - 0,79** -0,49** 0,87** 0,83**
Sólidos solúveis totais (SST) - - - - -0,74** 0,91** 0,93** Acidez total titulável (ATT)
- - - - - -0,67** -0,79**
pH - - - - - - 0,93***, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste t. ns Não-significativo. RSA - Relação SST/ATT.

121
repetições que compõem a TABELA 10. Após esta análise, constatou-se que a luminosidade e
o ângulo Hue da polpa não apresentaram coeficientes de correlação linear estimados superior
a 0,70 quando contrastado com as demais variáveis analisadas, enquanto o croma da polpa
apresentou correlações superiores a 0,70, positivas e significativas com pH (R= 0,95**) e
relação SST/ATT (R= 0,92**) e correlação superior a 0,70, negativa e significativa com
acidez total titulável (R= -0,90**).
Com estes resultados, verifica-se que os índices de maturidade para manga
recomendados por Medlicott e Reynolds (1988) e Alves et al. (2002) possuem altas
correlações entre si. Fica também evidente que o maior coeficiente de correlação linear
estimado observado foi entre o croma da polpa e o pH. Portanto, tendo em vista que o croma
da polpa também apresentou o maior coeficiente de determinação das variáveis estudadas,
este comportamento o credencia como um dos mais precisos e práticos indicadores de
maturidade para manga ‘Tommy Atkins’.
A TABELA 11 apresenta a matriz de correlação entre croma da polpa, nitrogênio e
proteína. Novamente o croma da polpa se destaca por apresentar com o nitrogênio total e a
proteína bruta coeficientes positivos, significativos e iguais a 0,70.
TABELA 11 - Matriz de correlação entre croma da polpa, nitrogênio e proteína de mangas ‘Tommy Atkins’ durante o desenvolvimento. Variáveis NT NNP NP PB PV
Croma da polpa (CP) 0,70** 0,66** 0,46* 0,70** 0,46* Nitrogênio total (NT) - 0,86** 0,76** 0,99** 0,77**
Nitrogênio não protéico (NNP) - - 0,33ns 0,86** 0,33ns
Nitrogênio protéico (NP) - - - 0,77** 0,99** Proteína bruta (PB) - - - - 0,77**
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste t. ns Não-significativo. PV – Proteína verdadeira.
4.7.3 Correlação entre as variáveis físicas, físico-químicas, químicas e bioquímicas
As análises de correlação do amido com croma da polpa, sólidos solúveis totais,
açúcares solúveis totais, açúcares não redutores, amilase total e α-amilase (TABELA 12),
mostraram-se positivas e significativas (P < 0,01), portanto, quanto maior o teor de amido,
maiores serão os valores do croma da polpa, sólidos solúveis totais, açúcares solúveis totais,
açúcares não redutores, amilase total e α-amilase. O maior coeficiente de correlação
encontrado foi entre o croma da polpa e os açúcares solúveis totais, R= 0,93 (P < 0,01). Não

122
TABELA 12 - Matriz de correlação entre croma da polpa, sólidos solúveis totais, carboidratos e amilases de mangas ‘Tommy Atkins’ durante o
desenvolvimento. Variáveis SST AD AST AR ANR AMLT α-AML β-AML
Croma da polpa (CP) 0,83** 0,75** 0,93** 0,29ns 0,86** 0,75** 0,68** -0,02ns
Sólidos solúveis totais (SST) - 0,69** 0,86** 0,56** 0,65** 0,65** 0,60** -0,05ns
Amido (AD) - - 0,80** 0,06ns 0,84**
0,68** 0,62** -0,03ns
Açúcares solúveis totais (AST) - - - 0,41* 0,88** 0,68** 0,65** -0,08ns
Açúcares redutores (AR) - - - - -0,07ns 0,02ns 0,13ns -0,17ns
Açúcares não redutores (ANR) - - - - - 0,73** 0,64** 0,00ns
Amilase total (AMLT) - - - - - - 0,78** 0,16ns
α-amilase (α-AML) - - - - - - - -0,50**, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste t. ns Não-significativo. β-AML - β-amilase.

123
foi encontrado coeficientes de correlação superiores a 0,70 para açúcares redutores e β-
amilase. Dutra et al. (2005) estudando a maturação de ‘Tommy Atkins’ também obtiveram
correlação positiva e significativa entre amido e sólidos solúveis totais, R= 0,82 (P < 0,05),
porém difere daquele relatado por Rocha et al. (2001), quando a coloração da polpa foi a que
apresentou o melhor coeficiente de correlação com o índice de degradação de amido, R= 0,95
(P < 0,01).
De acordo com os coeficientes de correlação do amido com α-amilase e β-amilase e
entre as enzimas α-amilase e β-amilase (TABELA 12), constata-se que estas enzimas não
atuam na degradação do amido simultaneamente, agindo primeiro α-amilase e depois a β-
amilase. Na primeira correlação observa-se que a α-amilase tem uma correlação positiva com
o amido enquanto a β-amilase é negativa. Na segunda correlação verifica-se uma correlação
negativa entre as enzimas, portanto quanto maior a atividade da α-amilase, menor a atividade
da β-amilase. Este comportamento é validado por Hopkins (2000) quando afirma que a α-
amilase é a primeira que “ataca” os grãos de amido intactos, sendo importante nos estágios
iniciais da degradação.
Na TABELA 13 são apresentados os coeficientes de correlação entre croma da polpa,
vitamina C, clorofila, carotenóides, fenólicos e oxidases. Comparando-se os resultados de
croma da polpa com vitamina C total, carotenóides totais, fenólicos poliméricos, oligoméricos
e diméricos, verifica-se que houve altas correlações significativas entre eles. No entanto, a
correlação foi positiva apenas para carotenóides totais, indicando que quanto maior o valor do
croma maior será o conteúdo de carotenóides totais. Por outro lado, as correlações foram
negativas para vitamina C total, fenólicos poliméricos, oligoméricos e diméricos. Isto indica
que as mangas com valores de croma da polpa mais altos possuem em geral menor teor de
vitamina C total e são menos adstringentes. Considerando estas novas correlações, o maior
coeficiente encontrado foi entre o croma da polpa e os carotenóides totais, R= 0,93 (P < 0,01).
Não foi encontrado coeficientes de correlação superiores a 0,70 para vitamina C total,
clorofila total, fenólicos poliméricos e polifenoloxidase.
Ao analisar as correlações da TABELA 13 constatou-se também que entre a PPO e a
POD houve correlação positiva, R= 0,54 (P < 0,01). Este fato concorda com o relato de
Robards et al. (1999) ao afirmarem que a atividade da POD depende da presença da PPO no
meio, a fim de gerar o H2O2 necessário à ocorrência da oxidação.

124
TABELA 13 - Matriz de correlação entre croma da polpa, vitamina C, clorofila, carotenóides, fenólicos e oxidases de mangas ‘Tommy Atkins’
durante o desenvolvimento. Variáveis VCT CLT CAT FP FO FD PPO POD
Croma da polpa (CP) -0,45* -0,13ns 0,93** -0,63** -0,74** -0,77** 0,06ns 0,25ns
Vitamina C total (VCT) - -0,28ns -0,43*
0,24ns 0,26ns 0,23ns 0,30ns 0,32ns
Clorofila total (CLT) - - -0,02ns 0,42* 0,22ns -0,04ns -0,37ns -0,61** Carotenóides totais (CAT) - - - -0,56** -0,69** -0,76** 0,06ns 0,15ns
Fenólicos poliméricos (FP) - - - - 0,55** 0,35ns -0,36ns -0,43* Fenólicos oligoméricos (FO) - - - - - 0,55** -0,03ns -0,19ns
Fenólicos diméricos (FD) - - - - - - 0,01ns -0,06ns
Polifenoloxidase (PPO) - - - - - - - 0,54***, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste t. ns Não-significativo. POD - Peroxidase.

125
Quando se observa o valor do coeficiente de correlação entre croma da polpa,
pectinas, pectinases, galactosidases e proteína (TABELA 14), a associação entre estas
variáveis torna-se mais nítida, evidenciando-se a importância da solubilização da pectina na
diminuição da firmeza de mangas, embora não seja o único fator envolvido. Este
comportamento ratifica o estudo de Malis-Arad et al. (1983), em trabalho desenvolvido com
cultivares mutantes, segundo o qual, já foi constatado que a solubilização de pectinas não é o
único fator que afeta a firmeza, ou que diferencia frutos firmes de macios. Além disso, os
autores citam que a associação entre pectinas e outros polímeros pode afetar sua sensibilidade
à solubilização. Portanto, o estudo da solubilidade de pectinas não deve ser dissociado dos
outros constituintes da parede celular e suas possíveis interações.
Comparando-se os resultados de pectina total com o croma da polpa, pectina solúvel,
pectina de alta metoxilação, protopectina, poligalacturonase, α- e β-galactosidases extraídas
de parede (TABELA 14), verifica-se que houve altas correlações significativas entre eles. No
entanto, a correlação foi positiva para pectina solúvel, pectinas de alta metoxilação,
protopectina e poligalacturonase, indicando que quanto maior o teor da pectina total maior
será o conteúdo de pectina solúvel, pectinas de alta metoxilação, protopectina e
poligalacturonase, portanto, maior será a firmeza do fruto. Por outro lado, as correlações
foram negativas para o croma da polpa, α- e β-galactosidases extraídas de parede. Isto indica
que as mangas com teores de pectina total mais alto possuem em geral menor valor de croma
da polpa e menores atividades de α- e β-galactosidases extraídas de parede, isto é, são mais
firmes. Os maiores coeficientes de correlação encontrados foram entre o croma da polpa e a
protopectina, R= -0,82 (P < 0,01) e entre a pectina total e pectinas de alta metoxilação, R=
0,82 (P < 0,01). Não foram encontrados coeficientes de correlação superiores a 0,70 para
pectina solúvel, percentual de solubilização de pectina, pectinas de baixa metoxilação e
pectinametilesterase.
Associações significativas entre as galactosidases e proteínas só foi detectada entre β-
galactosidase e proteína extraídas de citosol (TABELA 14), R= 0,43 (P < 0,05).
Conforme discutido anteriormente, quando se constatou que no primeiro momento a
PG atua de modo isolado, parece tornar-se mais evidente, ao observar os coeficientes de
correlação negativos entre as atividades de PG com α- e β-GALp e positivo entre as atividades
de PG e a proteína extraída de citosol (TABELA 14). Neste mesmo sentido, detectou-se
também que a PG é a principal responsável pela degradação das pectinas solúvel e de alta
metoxilação, pois a PG possui correlação positiva com estas, enquanto as GALs possuem

126
TABELA 14 - Matriz de correlação entre croma da polpa, pectinas, pectinases, galactosidases e proteína de mangas ‘Tommy Atkins’ durante o
desenvolvimento. Variáveis PT PS PSP PAM PBM PROT PME PG αGALc αGALp βGALc βGALp Pc Pp
Croma da polpa (CP) -0,80** -0,53** 0,19ns -0,71** -0,35ns -0,82** 0,15ns -0,45* 0,72** 0,43* 0,63** 0,64** 0,04ns -0,40ns
Pectina total (PT) - 0,56**
-0,35ns 0,82**
0,38ns 0,46* -0,01ns 0,78** -0,27ns -0,48* -0,32ns -0,63** 0,33ns 0,29ns
Pectina solúvel (PS) - - 0,51* 0,62** 0,41* 0,48* 0,02ns 0,48* -0,38ns -0,30ns -0,33ns -0,43* 0,16ns 0,30ns
% solubilização de pectina (PSP) - - - -0,06ns -0,02ns 0,11ns 0,00ns -0,24ns -0,16ns -0,01ns -0,10ns 0,05ns -0,12ns -0,04ns
Pectinas alta metoxilação (PAM) - - - - 0,46* 0,50* -0,02ns 0,70** -0,32ns -0,62** -0,51* -0,81** 0,10ns 0,17ns
Pectinas baixa metoxilação (PBM)
- - - - - 0,26ns -0,19ns 0,25ns -0,23ns -0,21ns -0,29ns -0,35ns -0,03ns 0,26ns
Protopectina (PROT) - - - - - - -0,16ns 0,08ns -0,71** -0,36ns -0,75** -0,55** -0,41* 0,36ns
Pectinametilesterase (PME) - - - - - - - -0,08ns 0,14ns 0,17ns 0,08ns 0,08ns -0,06ns 0,25ns
Poligalacturonase (PG) - - - - - - - - 0,02ns -0,49* 0,03ns -0,46* 0,50* -0,04ns
α-galactosidase citosol (αGALc) - - - - - - - - - 0,15ns 0,64** 0,29ns 0,32ns -0,37ns
α-galactosidase parede (αGALp) - - - - - - - - - - 0,38ns 0,79** -0,04ns 0,35ns
β-galactosidase citosol (βGALc) - - - - - - - - - - - 0,66** 0,43* -0,26ns
β-galactosidase citosol (βGALp)
- - - - - - - - - - - - 0,03ns 0,05ns
Proteína de citosol (Pc) - - - - - - - - - - - - - -0,06ns
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste t. ns Não-significativo. Pp – Proteína de parede.

127
correlações negativas, portanto, as GALs só atuam na degradação da parede celular após a
ação da PG. Estes resultados coadunam-se com os de Brett e Waldron (1990) ao afirmarem
que o amaciamento dos frutos tem sido associado com o aumento da atividade da PG,
acompanhado de um aumento das pectinas solúveis.
Por outro lado, a PME não apresentou nenhuma correlação significativa com as
pectinas, portanto, isto sugere uma importância secundária no conjunto de processos que
determinam o amadurecimento da manga ‘Tommy Atkins’. Este comportamento é compatível
com o encontrado por Evangelista (1999), quando trabalhando com esta mesma cultivar, não
detectou a presença da PME em nenhum dos tratamentos testados nos diferentes estádios de
maturação, embora as mangas tenham apresentado um amadurecimento normal. Entretanto,
segundo Ahmed e Labavitch (1980), a atividade de PME está presente durante o
desenvolvimento e maturação de frutos e, frequentemente, não mostra correlação com o
processo de amaciamento.

128
5 CONCLUSÕES
Os resultados obtidos permitiram concluir que:
1. Após 26 dias da indução, 65% do pomar estava em floração (antese) e o rendimento
das panículas para a produção efetiva de frutos foi de apenas 35,51% (percentual de
vingamento), das quais 77,03% tiveram apenas 1 fruto;
2. O croma da polpa foi o melhor indicador do estádio de desenvolvimento do fruto da
mangueira ‘Tommy Atkins’ cultivada sob irrigação no sub-médio São Francisco,
considerando-se o seu alto coeficiente de determinação, R2=0,9832 (P < 0,01) e seu alto
coeficiente de correlação com pH, açúcares solúveis totais e carotenóides totais, R=0,95; 0,93;
e 0,93, respectivamente (P < 0,01);
3. Mangas da cv. Tommy Atkins cultivadas no vale do São Francisco, Petrolina, PE,
atingiram a maturidade fisiológica aos 98 dias após a antese (DAA), durante o segundo
semestre de 2004;
4. O calculo acumulado das temperaturas mínimas e máximas e a definição da
temperatura base como sendo 10ºC, indicaram que 1.685,09 unidades de calor (UC)
determinaram a maturidade fisiológica da manga cv. Tommy Atkins, cultivada no vale do São
Francisco, Petrolina, PE.

129
6 REFERÊNCIAS BIBLIOGRÁFICAS
ABU-SARRA, A. F.; ABU-GOUKH, A. A. Changes in pectinesterase, polygalacturonase and cellulase activity during mango fruit ripening. Journal of Horticultural Science, Ashford, v.67, n.4, p. 561-568, July1992. AHMED, A. E.; LABAVITCH, J. M. Cell wall metabolism in ripening fruits: I. Cell wall changes in ripening “Bartlett” pears. Plant Physiology, Baltimore, v. 65, n. 5, p. 1009-1013, May 1980. AINA, J. O.; OLADUNJOYE, O. O. Respiration, pectolytic activity and textural changes in ripening African mango (Irvingia gabonensis) fruits. Journal of the Science of Food and Agriculture, London, v.63, n. 4, p. 451-454, 1993. ALBUQUERQUE, J. A. S. de; MEDINA, V. D.; MOUCO, M. A. do C. Indução floral. In: GENÚ, P. J. de C.; PINTO, A. C. de Q. A cultura da mangueira. 1. ed. Brasília: Embrapa Informação Tecnológica, 2002. cap. 13, p. 259-276. ALI, Z. M.; ARMUGAM, S.; LAZAN, H. β-galactosidase and it’s significance in ripening mango fruit. Phytochemistry, Oxford, v. 38, n. 5, p. 1109-1114, 1995. ALMEIDA, C. O. de; CARDOSO, C. E. L.; SANTANA, M. do A. Comercialização. In: PEREIRA, M. E. C.; FONSECA, N.; SOUZA, F. V. D. (Eds.). Manga: o produtor pergunta, a Embrapa responde. 1. ed. Brasília: Embrapa Informação Tecnológica, 2005. cap. 15, p. 177-184. (Coleção 500 perguntas, 500 respostas). ALVES, R. E. et al. Colheita e pós-colheita. In: GENÚ, P. J. de C.; PINTO, A. C. de Q. A cultura da mangueira. 1. ed. Brasília: Embrapa Informação Tecnológica, 2002. cap. 17, p. 381-405. AMORIM, T. B. F. Colheita e pós-colheita: manejo e conservação da manga. In: SÃO JOSÉ, A. R. (Org.). O agronegócio manga: produção e mercado. Vitória da Conquista: UESB/DFZ, 2002. p. 346-356. 1 CD-ROM. ASHRAF, M. W. et al. Studies on the pectinesterase activity and some chemical constituents of some Pakistani mango varieties during storage ripening. Journal Agricultural Food Chemistry, Columbus, v. 29, n. 3, p. 526-528, May/June 1981.

130
ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS. Official methods of analysis. 16 ed. Arlington: AOAC, 1995. 1141 p. AWAD, M. Fisiologia pós-colheita de frutos. São Paulo: Nobel, 1993. 114 p. AZIZ, P. A.; YUSOF, S. Physico-chemical characteristics of soursop fruit (Anona muricata) during growth and development. ASEAN Food Journal, New York, v. 9, n. 4, p. 147-150, 1994. BEM-TAL, Y. Flowering: its control by vegetative growth inhibition. Acta Horticulturae, Wageninger, n. 179, p. 329-335, 1986. BERNFELD, P. Enzymes of carbohydrate metabolism: amylases, α and β. In: COLWICK, S. P.; KAPLAN, N.O. Methods in enzimology. New York: Academic Press, 1955. v. 1, p. 149-158. BISSOLI JÚNIOR, W. Qualidade de mangas (Mangifera indica L.) cv. Tommy Atkins sob influência da pulverização pré-colheita dos frutos com cálcio e boro. 1999. 86 f. Dissertação (Mestrado em Ciência dos Alimentos)–Escola Superior de Agricultura de Lavras, Lavras, 1992. BLEINROTH, E. W. Manuseio e tratamento de pós-colheita da manga. In: DONADIO, L. C.; FERREIRA, F. R. (Eds.). Anais do II simpósio sobre mangicultura. Jaboticabal: FCAV-FUNEP, 1989. p. 171-184. ______. Determinação do ponto de colheita das frutas. In: BLEINROTH, E. W. (Coord.) et al. Tecnologia de pós-colheita de frutas tropicais. 2. ed. rev. Campinas: ITAL, 1992. cap. 1, p. 1-18. (Manual Técnico, 9). ______. Determinações do ponto de colheita. In: GORGATTI NETTO, A. et al. Manga para exportação: procedimentos de colheita e pós-colheita. Brasília: EMBRAPA-SPI, 1994. cap. 2, p. 11-28. (Série Publicações Técnicas FRUPEX, 4). BLUMENKRANTZ, N.; ASBOE-HANSEN, G. New method for quantitative determination of uronic acids. Analytical Biochemistry, New York, v. 54, p. 484-489, 1973. BOAS, E. V. B. V. Aspectos fisiológicos do desenvolvimento de frutos. Lavras: UFLA/FAEPE, 1999. 75 p.

131
BORDENAVE, M. Analyses of pectin methylesterases. In: LINSKENS, H. F.; JACKSON, J. F. Plant cell wall analysis: modern methods on plant analysis. Berlin: Springer-Verlag, 1996. v. 17, p. 165-180. BORDENAVE, M.; GOLDBERG, R. Purification and characterization of pectin methylesterases from mung bean hypocotyl cell walls. Phytochemestry, Oxford, v. 33, n. 5, p. 999-1003, 1993. BRADFORD, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye-binding. Analytical Biochemistry, New York, v. 72, p. 248-254, 1976. BRADY, J. B. Fruit ripening. Annual Review of Plant Physiology, Palo Alto, v. 38, p. 155-178, 1987. BRETT, C.; WALDRON, K. Physiology and biochemistry of plant cell walls. Londres: Unwin Hyman, 1990. 1993p. BRINSON, K. et al. Post harvest changes in Mangifera indica mesocarp cell walls and cytoplasmic polysaccharides. Phytochemistry, Oxford, v. 27, n. 3, p. 719-723, Feb. 1988. BRUINSMA, J. The quantitative analysis of chlorophylls A and B in plant extracts. Photochemistry and Photobiology, Elmsford, v. 2, p. 241-249, 1963. BURNS, J. α- and β- galactosidase activities in juice vesicles of stored Valencia oranges. Phytochemitry, Oxford, v. 29, n. 8, p. 2425-2429, 1990. BURONDKAR, M. M. et al. Estimation of heat units as maturity indices for different mango varieties in Konkan region of Maharshtra. Acta Horticulturae, Leuven, n. 509, p. 297-299, 2000. CAMILLO-LÓPEZ, A. et al. Ripening and quality of mango fruit as affected by coating with “Semperfresh”. Acta Horticulturae, Leuven, n. 370, p. 203-16, 1995. CAMPBELL, R. J. Mangos: a guide to mangos in Florida. Miami: Fairchild Tropical Garden, 1992. 227 p.

132
CARPITA, N.; McCANN, M. The cell wall. In: BUCHANAN, B. B; GRUISSEM, W.; JONES, R. L. (Eds.). Biochemistry & molecular biology of plants. Rockville: American Society of Plant Physiologists, 2000. ch. 2, p. 52-108. CASTRILLO, M.; BERMUDEZ, A. Post-harvest ripening in wax coated Bocado mango. International Journal of Food Science and Technology, Oxford, v. 27, n. 4, p. 457-463, Aug. 1992. CASTRO NETO, M. T. de; REINHARDT, D. H. Relações entre parâmetros de crescimento do fruto de manga cv. Haden. Revista Brasileira de Fruticultura, Jaboticabal, v. 25, n. 1, p. 35-37, 2003. CETEC – FUNDAÇÃO CENTRO TECNOLÓGICO DE MINAS GERAIS. Manual para fabricação de geléias. Belo Horizonte: CETEC, 1985. 42 p. (Série Publicações Técnicas, 15). CHAUDHRI, S. A. Mango. In: GARDNER, R. S.; CHAUDHRI, S. A. The propagation of tropical fruits trees. England: CAB International, 1976. p. 403-474. CHEN, W. S. Endogenous growth substances in relation to shoot growth and flower bud development of mango. Journal of the American Society Horticulturae of Science, Alexandria, v. 112, n. 2, p. 360-363, 1987. CHITARRA, M. I. F. Tecnologia e qualidade pós-colheita de frutos e hortaliças. Lavras: UFLA/FAEPE, 2000. 68 p. CHITARRA, M. I. F.; CHITARRA, A. B. Pós-colheita de frutas e hortaliças: fisiologia e manuseio. 2. ed. rev. e ampl. Lavras: UFLA, 2005. 785 p. CUNHA, G. A. P. da. et al. A cultura da manga. 1. ed. Brasília: Embrapa-SPI, 1994. 54 p. (Coleção Plantar, 10). CUNHA, G. A. P. da; PINTO, A. C. de Q.; FERREIRA, F. R. Origem, dispersão, taxonomia e botânica. In: GENÚ, P. J. de C.; PINTO, A. C. de Q. A cultura da mangueira. 1. ed. Brasília: Embrapa Informação Tecnológica, 2002. cap. 2, p. 31-36. DAVENPORT, T. L.; NUÑEZ-ELISEA, R. Ethylene and other endogenous factors possibly involved in mango flowering. Acta Horticulturae, Wageninger, v. 2, n. 275, p. 441-447, 1990.

133
DENNIS, D. T.; BLAKELEY, S. D. Carbohydrate metabolism. In: BUCHANAN, B. B; GRUISSEM, W.; JONES, R. L. (Eds.). Biochemistry & molecular biology of plants. Rockville: American Society of Plant Physiologists, 2000. ch. 13, p. 630-675. DEY, P. M.; PRIDHAM, J. B. Purification and properties of α-galactosidases from Vicia faba seeds. Biochemistry Journal, London, v. 113, n. 1, p. 49-55, 1969. DONADIO, L. C. Variedades de mangueira. In: SÃO JOSÉ, A. R. (Coord.) et al. Manga: tecnologia de produção de mercado. Vitória da Conquista: DFZ/UESB, 1996. p. 32-56. ______. Variedades de manga. In: SÃO JOSÉ, A. R. (Org.). O agronegócio manga: produção e mercado. Vitória da Conquista: UESB/DFZ, 2002. p. 119-126. 1 CD-ROM. DUTRA, P. R. S. et al. Indicadores bioquímicos do desenvolvimento de manga Tommy Atkins produzidas no vale do São Francisco. In: SIMPÓSIO BRASILEIRO DE PÓS-COLHEITA DE FRUTOS TROPICAIS, 1., 2005, João Pessoa. Anais... João Pessoa: Embrapa/UFPB/UFS/SBF, 2005. 1 CD-ROM. EMBRAPA. Dados meteorológicos da estação agrometeorológica de Bebedouro. Petrolina: Embrapa Semi-Árido, 2006. Disponível em: <http://www.cpatsa.embrapa.br/serviços/dadosmet/ceb-anual.html>. Acesso em: 22 jun. 2006. ENGEL, V. L.; POGGIANI, F. Estudo da concentração de clorofila nas folhas e seu espectro de absorção de luz em função do sombreamento em mudas de quatro espécies florestais. Revista Brasileira de Fisiologia Vegetal, Brasília, vol. 3, n. 1, p. 39-45, 1991. ESTEBAN, R. M. et al. Pectin changes during the development and ripening of eggplant fruits. Food Chemistry, Oxford, v.46, p. 289-292, 1993. EVANGELISTA, R. M. Qualidade de mangas ‘Tommy Atkins’ armazenadas sob refrigeração e tratadas com cloreto de cálcio pré-colheita. 1992. 129 f. Tese (Doutorado em Ciência dos Alimentos)–Universidade Federal de Lavras, Lavras, 1999. FELIPPE, G. M. Etileno. In: FERRI, M. G. Fisiologia vegetal. 2. ed. São Paulo: EPU, 1979. p. 163-192. FERREIRA, F. R. et al. Germoplasma de manga no Brasil. In: São José, A. R. (Org.). O agronegócio manga: produção e mercado. Vitória da Conquista: UESB/DFZ, 2002. p. 111-118. 1 CD-ROM.

134
FIERRO, C. A; ULLOA, M. A development reference stage for flower induction response to potassium nitrate in mango. Acta Horticulturae, Wageningen, n. 291, p. 71-78, 1991. FILGUEIRAS, H. A. C.; CHITARRA, M. I. F. Influência da embalagem e temperatura de armazenamento sobre os teores de compostos fenólicos em ameixa roxa Delfim Moreira. Pesquisa Agropecuária Brasileira, Brasília, v. 23, n. 1, p. 63-74, jan. 1988. FILGUEIRAS, H. A. C. et al. Colheita e manuseio pós-colheita. In: FILGUEIRAS, H. A. C. (Org.). Manga: pós-colheita. 1. ed. Brasília: Embrapa Comunicação para Transferência de Tecnologia, 2000. cap. 3, p. 22-37. (Série Frutas do Brasil, 2). FONSECA, N.; SANTOS-SEREJO, J. A. dos. Florescimento e frutificação. In: PEREIRA, M. E. C.; FONSECA, N.; SOUZA, F. V. D. (Eds.). Manga: o produtor pergunta, a Embrapa responde. 1. ed. Brasília: Embrapa Informação Tecnológica, 2005. cap. 10, p. 105-122. (Coleção 500 perguntas, 500 respostas). FRANCO, G. Tabela de composição química dos alimentos. 9 ed. São Paulo: Atheneu, 2003. 307 p. FUCHS, Y.; PESIS, E.; ZAUBERMAN, G. Changes in amylase activity, starch and sugars contents in mango fruit pulp. Scientia Horticulturae, Amsterdam, v. 13, n. 2, p. 155-160, 1980. GAVA, A. J. Princípios de tecnologia de alimentos. 1. ed. São Paulo: Nobel, 1984. 284 p. GOLDSTEIN, J. L.; SWAIN, T. Changes in tannins in ripening fruits. Phytochemistry, Oxford, v. 2, n. 4, p. 371-383, 1963.
GORTNER, W. A.; DULL, G. G.; KRAUSS, B. H. Fruit development, maturation, ripening and senescence: a biochemical basis for horticultural terminology. HortScience, Alexandria, v. 2, n. 4, p. 141-144, 1967. GTZ – DEUTSCHE GESELLSCHAFT FÜR TECHNISCHE ZUSAMMENARBEIT. Manual de exportación: frutas tropicales y hortalizas. Eschborn: GTZ, 1992. 34 p.
HIGBY, W. K. A simplified method for determination of some the carotenoid distribution in natural and carotene-fortified orange juice. Journal of Food Science, Chicago, v. 27, p. 42-49, 1962.

135
HIGLEY, L. G.; PEDIGO, L. P.; ASTHIE, K. R. Degday: a program for calculating degree-days, and assumptions behind the degree-day approach. Environmental Entomology, Lanham, v. 15, n. 5, p. 999-1016, 1986. HOPKINS, W. G. Introduction to plant physiology. 2. ed. New York: John Wiley & Sons, Inc., 2000. 512 p. HULME, A. C. The mango. The biochemistry of fruits and their products. 2. ed. London: Academic Press, 1974. v. 2, p. 233-254. INSTITUTO ADOLFO LUTZ. Normas analíticas, métodos químicos e físicos para análise de alimentos. 3. ed. São Paulo: IAL, 1985. v. 1, 533 p. IRVING, D. E.; SHINGLETON, G. J.; HURST, P. L. Starch degradation in buttercup squash (Cucurbita maxima). Journal of the American Society for Horticultural Science, Alexandria, v. 124, n. 6, p. 587-590, 1999. ISLABÃO, N. Manual de cálculo de rações para os animais domésticos. 4. ed. Porto Alegre: SAGRA, 1985. 177 p. ITOH, T.; UDA, Y.; NAKAGAWA, H. Purification and characterization of α-galactosidase from watermelon. Journal of Biochemistry, Tokyo, v. 99, n. 1, p. 243-250, 1986. JEN, J. J.; ROBINSON, M. L. Pectolytic enzymes in sweet bell peppers (Capsicum annuum L.). Journal of Food Science, Chicago, v. 49, n. 4, p. 1085-1087, 1984. JOHN, M. A.; DEY, P. M. Postharvest changes in fruit cell wall. Advances in Food Research, New York, v. 30, p. 139-185, 1986. KADER, A. A. Fruit maturity, ripening, and quality relationships. Acta Horticulturae, Leuven, n. 485, p. 203-208, 1999. KAPSE, B. M.; RANE, D. A.; KHEDKAR, D. M. Correlation between bio-chemical parameters and organoleptic evaluation in mango varieties. Acta Horticulturae, Wageningen, v. 231, n. 2, p. 756-62, 1988. KAYS, S. J. Postharvest physiology of perishable plant products. New York: AVI Book, 1991. 532 p.

136
KETSA, S. et al. Effect of heat treatment on changes in softening, pectic substances and activities of poligalacturonase, pectinesterase and β-galactosidase of ripening mango. Journal of Plant Physiology, Tokyo, v. 153, p. 457-461, 1998. KHADER, S. E. S. A. Effect of gibberellic acid and vapor gard on ripening, amylase and peroxidase activities and quality of mango fruits during storage. Journal of Horticultural Science, Ashford, v. 67, n. 6, p. 855-860, 1992. KITAGAWA, Y; KANAYAMA, Y; YAMAKI, S. Isolation of β-galactosidase fractions from Japanese pear: activity against native cell wall polysaccharides. Physiologia Plantarum, Copenhagen, v. 93, n. 3, p. 545-550, 1995. KRAMER, A. Fruits and Vegetables. In: TWIGG, B. A. Quality control for food industry. Connecticut: AVI Publishing Company, 1973. v. 2, p. 157-227. LAKSHIMINARAYANA, S.; SABRHADRA, N. V.; SUBRAMANYAM, H. Some aspects of developmental physiology of mango fruit. Journal of Horticultural Science, Ashford, v. 45, p. 133-142, 1970. LAKSHIMINARAYANA, S. Mango. In: NAGY, S.; SWAW, P. E. Tropical and subtropical fruits. Westport: AVI Publishing, Inc., 1980. p. 157-184. LAROUSSILHE, F. de. Le manguier. Paris: Maisonneuve et Larose, 1980. 312 p. LEDERMAN, I. E. et al. Determinação do ponto de colheita da manga cv. Tommy Atkins, para a região semi-árida de Pernambuco. Revista Brasileira de Fruticultura, Cruz das Almas, v. 20, n. 2, p. 145-151, 1998. LEITE, J. C. A. et al. Caracterização física da manga variedade Tommy Atkins. In: SIMPÓSIO BRASILEIRO DE PÓS-COLHEITA DE FRUTOS TROPICAIS, 1., 2005, João Pessoa. Anais... João Pessoa: Embrapa/UFPB/UFS/SBF, 2005. 1 CD-ROM. LIMA-FILHO, J. M. P. et al. Ecofisiologia. In: GENÚ, P. J. de C.; PINTO, A. C. de Q. A cultura da mangueira. 1. ed. Brasília: Embrapa Informação Tecnológica, 2002. cap. 3, p.37-50. LIMA, L. C. de O. Tecido esponjoso em manga ‘Tommy Atkins’: transformações químicas e bioquímicas no mesocarpo durante o armazenamento. 1997. 147 f. Tese (Doutorado em Ciência dos Alimentos)–Universidade Federal de Lavras, Lavras, 1997.

137
LIMA, M. A. C. de. Alterações bioquímicas e fisiológicas durante a maturação e o armazenamento de graviola sob refrigeração associada a 1-metilciclopropeno e cera. 2002. 208 f. Tese (Doutorado em Agronomia)-Universidade Federal do Ceará, Fortaleza, 2002. LIZADA, C. Mango. In: SEYMOUR, G. B.; TAYLOR, J. E.; TUCKER, G. A. (ed.). Biochemistry of fruit ripening. London: Chapman & Hall, 1993. ch. 8, p. 255-266. LUCENA, E. M. P. de; SILVA JÚNIOR, A.; CAMPELO, I. K. M. Caracterização físico-química da manga (Mangifera indica L.), variedade Tommy Atkins, em diferentes estádios de maturação. In: CONGRESSO BRASILEIRO DE CIÊNCIA E TECNOLOGIA DE ALIMENTOS, 17., 2000, Fortaleza. Resumos... Fortaleza: Sociedade Brasileira de Ciência e Tecnologia de Alimentos, 2000. v. 2, p. 5.246. LUCENA, E. M. P. de. et al. Técnicas de conservação garantem qualidade e aumento da vida útil da manga Tommy Atkins. Pesquisas FUNCAP: Revista de Ciência e Tecnologia, Fortaleza, ano 3, n. 3, p. 7-8, dez. 2001. MAIA, M. L. L. Geléia e doce em massa. In: TORREZAN, R. (Coord.). Curso de processamento de frutas. Rio de Janeiro: Embrapa-CTAA, 1997. cap. 4, p. 84-99. MALIS-ARAD, S. et al. Pectic substances: changes in soft end firm cultivars and in non-ripening mutants. Journal of Horticultural Science, Ashford, v. 58, n. 1, p. 111-116, Jan. 1983. MANGELS, A. R. et al. Carotenoid content of fruits and vegetables: an evaluation of analytic data. Journal of the American Dietetic Association, Baltimore, v. 93, n. 3, p. 284-296, 1993. MANICA, I. Indução do florescimento em mangueiras. In: SÃO JOSÉ, A. R. (Coord.) et al. Manga: tecnologia de produção de mercado. Vitória da Conquista: DFZ/UESB, 1996. p. 140-144. MARIN, M. A.; CANO, M. P. Patterns of peroxidase in ripening mango (Mangifera indica, L.) fruits. Journal of Food Science, Chicago, v. 57, n. 3, p. 690-692, 1992. MARTIN, C.; SMITH, A. M. Starch biosynthesis. The Plant Cell, Rockville, v. 7, n. 7, p. 971-985, 1995.

138
MATSUNO, H.; URITANI, I. Physiological behavior of peroxidase isozymes in sweet potato root tissue injured by cutting or with black root. Plant and Cell Physiology, Tokyo, v. 13, p. 1091-1101, 1972. MATTOO, A. K. et al. Chemical changes during ripening and senescence. In: PANTASTICO, E. B. Postharvest physiology, handling and utilization of tropical and subtropical fruits and vegetables. Westport: The AVI Publishing, 1975. p. 103-127. MAYER, A. M.; HAREL, E. Phenoloxidases and their significance in fruit and vegetables. In: FOX, P. F. Food Enzimology. London: Elsevier Applied Science, 1991. v. 1, p. 373-398. McCOLLUM, T. G.; HUBER, D. J.; CANTLIFFE, D. J. Modification of polyuronides and hemicelluloses during muskmelon fruit softening. Physiologia Plantarum, Copenhagen, v. 76, n. 3, p. 303-308, 1989. McILVAINE, T. C. A buffer solution for colorimetric comparison. The Journal of Biological Chemistry, Bethesda, v. 49, n. 1, p. 183-186, 1921. McREADY, P. M.; McCOMB, E. A. Extraction and determination of total pectin materials. Analytical Chemistry, Washington, v. 24, n. 12, p. 1586-1588, 1952. MEDINA, Valdique, M. Fisiologia e pós-colheita da manga. In: SÃO JOSÉ, A. R. (Coord.) et al. Manga: tecnologia de produção de mercado. Vitória da Conquista: DFZ/UESB, 1996. p. 202-222. MEDINA, Voltaire, D. Situação da mangicultura no sub-médio São Francisco e perspectivas. In: SÃO JOSÉ, A. R. (Coord.) et al. Manga: tecnologia de produção de mercado. Vitória da Conquista: DFZ/UESB, 1996. p. 285-295. MEDLICOTT, A. P. et al. Measurement of color changes in ripening bananas and mangoes by instrumental, chemical and visual assessments. Tropical Agriculture, Trinidad, v. 69, n. 2, p. 161-166, Apr. 1992. MEDLICOTT, A. P.; REYNOLDS, S. B. Harvest maturity effects on mango fruit ripening. Tropical Agriculture, Trinidad, v. 65, n. 2, p. 153-157, Apr. 1988. MEDLICOTT, A. P.; REYNOLDS, S. B.; THOMPSON, A. K. Effects of temperature on the ripening of mango fruit (Mangifera indica L.) var. Tommy Atkins. Journal of the Science of Food and Agriculture, London, v. 37, n. 5, p. 469-474, May 1986.

139
MEDLICOTT, A. P.; THOMPSON, A. K. Analysis of sugars and organic acids in ripening mango fruits (Mangifera indica var. Keitt) by high performance liquid chromatography. Journal of the Science of Food and Agriculture, London, v. 36, n. 7, p. 561-566, July 1985. MELO NETO, M. L. de. et al. Utilização de embalagens plásticas e refrigeração na conservação da manga (Mangifera indica L.) cv. Palmer. Revista Brasileira de Fruticultura, Jaboticabal, v. 21, n. 2, p. 160-165, ago. 1999. MENA, N. G.; SAUCEDO, V. C.; NIETO, A. D. Evaluacion de tratamientos cuarentenarios en frutos de mango Manila en Mexico. In: SÃO JOSÉ, A. R. (Coord.) et al. Manga: tecnologia de produção de mercado. Vitória da Conquista: DFZ/UESB, 1996. p. 223-240. MENDONÇA, V. Indução floral em mangueira com paclobutrazol, nitrato de potássio e ethephon. 2000. 73 f. Dissertação (Mestrado em Agronomia)–Escola Superior de Agricultura de Mossoró, Mossoró, 2000. MENEZES, J. B. Qualidade pós-colheita de melão tipo galia durante a maturação e o armazenamento. 1996. 157 f. Tese (Doutorado em Ciência dos Alimentos)–Universidade Federal de Lavras, Lavras, 1996. MILLER, G. L. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Analytical Chemistry, Washington, v. 31, p. 426-428, 1959. MIRANDA, M. R. A. de. Alterações fisiológicas e histológicas durante o desenvolvimento, maturação e armazenamento refrigerado do sapoti (Manilkara zapota l. Von Royen). 2002. 135 f. Tese (Doutorado em Agronomia)–Universidade Federal do Ceará, Fortaleza, 2002. MITCHAM, E. J.; McDONALD, R. E. Cell wall modification during ripening of ‘Keitt’ and ‘Tommy Atkins’ mango fruit. Journal of the American Society for Horticultural Science, Alexandria, v. 117, n. 6, p. 919-924, 1992. MONTEITH, J. L. Climate. In: ALVIM, P. de T.; KOZLOWSKI, T. T. (Eds.). Ecophysiology of tropical crops. New York: Academic Press, 1977. ch. 1, p. 1-27. MORAIS, P. L. D. de. Maturidade para colheita e vida útil da manga ‘Tommy Atkins’ para o mercado europeu. 2001. 83 f. Dissertação (Mestrado em Agronomia)–Universidade Federal do Ceará, Fortaleza, 2001.

140
MORGA, N. S. et al. Physicochemical changes in Philippine Carabao mangoes during ripening. Food Chemistry, London, v. 4, n. 2, p. 225-234, Mar. 1979. MOSQUEDA-VÁZQUEZ, R.; IRETA-OJEDA, A. Degree-days and base temperatures required for inflorescence and fruit development in mango ‘Manila’. Acta Horticulturae, Leuven, n. 341, p. 232-237, 1993. MOTA, F. S. Meteorologia agrícola. 7. ed. São Paulo: Nobel, 1986. 376 p. MUKHERJEE, S. K. Systematic and ecogeographic studies of crop genepools: 1. Mangifera. Rome: IBPGR Secretariat, 1985. 86 p. MUÑOZ, C. et al. Determining thermal time and base temperature required for fruit development in low-chilling peaches. HortScience, Alexandria, v. 21, n. 3, p. 520-522, 1986. NELSON, J. L.; KURTZ, L. T.; BRAY, R. H. Rapid determination of nitrates and nitrites. Analytical Chemistry, Washington, v. 26, p. 1081, 1954. NUÑEZ-ELISEA, R.; DAVENPORT, T. L. Florewing of ‘Keitt’ mango in response to deblossoming and gibberellic acid. Proceedings of the Florida State Horticulturae Society, Winter Haven, v. 104, p. 41-43, 1991. PANGBORN, R. M. Relative taste intensities of selected sugars and organic acids. Journal of Food Science, Chicago, v. 28, n. 6, p. 726-733, 1963. PARK, K. Y. et al. Polyphenol oxidase of mango (Mangifera indica L. var. Haden). Journal of Food Science, Chicago, v. 45, n. 6, p. 1619-1621, Dec. 1980. PINTO, A. C. de Q. et al. Melhoramento genético. In: GENÚ, P. J. de C.; PINTO, A. C. de Q. A cultura da mangueira. 1. ed. Brasília: Embrapa Informação Tecnológica, 2002a. cap. 4, p. 51-92. PINTO, A. C. de Q.; COSTA, J. G. da; SANTOS, C. A. F. Principais variedades. In: GENÚ, P. J. de C.; PINTO, A. C. de Q. A cultura da mangueira. 1. ed. Brasília: Embrapa Informação Tecnológica, 2002b. cap. 5, p. 93-116.

141
PINTO, A. C. de Q.; GENÚ, P. J. C.; RAMOS, V. H. V. Avaliação do crescimento e expressão do sexo de cultivares de manga introduzidas na região dos Cerrados. In: CONGRESSO BRASILEIRO DE FRUTICULTURA, 9., 1987, Campinas. Anais... Campinas: Sociedade Brasileira de Fruticultura, 1987. v. 2, p. 567-570. PIZA JÚNIOR, C. de T. A situação da cultura da mangueira em São Paulo. In: DONADIO, L. C.; FERREIRA, F. R. (Eds.). Anais do II simpósio sobre mangicultura. Jaboticabal: FCAV-FUNEP, 1989. p. 31-46. PRESSEY, R.; AVANTS, J. K. Separation and characterization of endopolygalacturonase and exopolygalacturonase from peaches. Plant Physiology, Baltimore, v. 52, n. 3, p. 252-256, 1973. RAMOS, V. H. V. Conservação pós-colheita de manga por meio do tratamento químico, da embalagem plástica e da cera associados à hidrotermia e refrigeração. 1994. 179 f. Tese (Doutorado em Produção Vegetal)-Faculdade de Ciências Agronômicas e Veterinária de Jaboticabal, Jaboticabal, 1994. RAVEN, P. H.; EVERT, R. F.; EICHHORN, S. E. Biologia vegetal. Tradução Jane Elizabeth Kraus et al. 6. ed. Rio de Janeiro: Guanabara Koogan, 2001. 906 p. REICHER, F.; SIERAKOWSKI, M. R.; CORREA, J. B. C. Determinação espectrofotométrica de taninos pelo reativo fosfotúngstico-fosfomolíbdico. Arquivos de Biologia e Tecnologia, Curitiba, v. 24, n. 4, p. 407-411, 1981. REID, M. S. Maturation and maturity indices. In: KADER, A. A. Postharvest technology of horticultural crops. 3. ed. California: University of California, 2003. p. 55-62. (Agriculture and Natural Resources, Publication, 3311). RENA, A. B.; MASCIOTTI, G. Z. Efeito do déficit hídrico sobre o metabolismo de nitrogênio e o crescimento de quatro cultivares de feijão. Revista Ceres, Viçosa, v. 23, n. 128, p. 288-301, 1976. ROBARDS, K. et al. Phenolic compounds and their role in oxidative process in fruits. Food Chemistry, Oxford, v. 66, n. 4, p. 401-436, 1999. ROBERTSON, G. L. The fractional extraction and quantitative determination of pectic substances in grapes and musts. American Journal Enology Viticulture, Davis, v. 30, n. 3, p. 182-186, 1979.

142
ROCHA, R. H. C. et al. Uso do índice de degradação de amido na determinação da maturidade da manga ‘Tommy Atkins’. Revista Brasileira de Fruticultura, Jaboticabal, v. 23, n. 2, p. 302-305, 2001. ROE, B.; BRUEMMER, J. H. Changes in pectic substances and enzymes during ripening and storage of ‘Keitt’ mango. Journal of Food Science, Chicago, v. 46, n. 1, p. 186-189, 1981. ROSE, J. K. C.; BENNETT, A. B. Cooperative disassembly of the cellulose-xyloglucan network of plant cell walls: parallels between cell expansion and fruit ripening. Trends Plant Science, London, v. 4, p. 176-183, 1999. SAKAI, T. et al. Pectin, pectinase and protopectinase: production, properties and applications. Advances in Applied Microbiology, San Diego, v. 39, p. 213-294, 1993. SALLES, J. R. de J.; TAVARES, J. C. Vida útil pós-colheita de manga (Mangifera indica L., cv. Tommy Atkins): influência da temperatura e do estádio de maturação. Revista Brasileira de Fruticultura, Jaboticabal, v. 21, n. 2, p. 171-176, ago. 1999. SALUNKE, D. K.; BOLIN, H. R.; REDDY, N. R. Storage, processing, and nutritional quality of fruits and vegetables. 2. ed. Boston: CRC Press, 1991. v. 1, 323 p. SALUNKE, D. K.; DESAI, B. B. Postharvest biotechnology of fruits. Boca Raton: CRC Press, 1984. v. 1, 168 p. SANTOS-SEREJO, J. A. dos. Classificação e descrição botânica. In: PEREIRA, M. E. C.; FONSECA, N.; SOUZA, F. V. D. (Eds.). Manga: o produtor pergunta, a Embrapa responde. 1. ed. Brasília: Embrapa Informação Tecnológica, 2005. cap. 1, p. 15-17. (Coleção 500 perguntas, 500 respostas). SAÚCO, G. Recolección y postrecolección. In: El cultivo del mango. Madrid: Mundi-Prensa, 1999. p. 267-272. SCHAFFER, B. Mango. In: SCHAFFER, B.; ANDERSEN, P. C. (Eds.). Handbook of environmental physiology of fruit crops: sub-tropical crops. Boca Raton: CRC Press, 1994. SELVARAJ, J. Y.; KUMAR, R. Studies on fruit softening enzymes and polyphenol oxidase activity in ripening mango (Mangifera indica L.) fruit. Journal of Food Science and Technology, Mysore, v. 26, n. 4, p. 218-222, Oct. 1989.

143
SEN, S.; CHATTERJEE, B. K.; ROY, H. B. Amylase activity in ripening mango (Mangifera indica L.) fruit. Indian Biology, Baroda, v. 17, n. 1, p. 25-28, Feb. 1985. SEYMOUR, G. B.; GROSS, K. C. Cell wall disassembly and fruit softening. Postharvest New and Information, London, v. 7, n. 3, p. 45-52, 1996. SEYMOUR, G. B.; LASSLETT, Y.; TUCKER, G. A. Differential effects of pectolytic enzymes on tomato polyuronides in vivo and in vitro. Phytochemistry, Oxford, v. 26, n. 12, p. 3137-3139, 1987. SHARMA, D. K.; SINGH, R. N. Studies on some pollination problems in mango (Mangifera indica L.). Indian Journal Horticultural, New Delhi, v. 27, n. 1, p. 15, 1970. SHU, Z. H.; SHEN, T. F. Floral induction en axillary buds mango (Mangifera indica, L.) as affected by temperature. Scientia Horticulturae, Amsterdam, v. 31, p. 81, 1987. SIGRIST, J. M. M. Respiração. In: BLEINROTH, E. W. (Coord.) et al. Tecnologia de pós-colheita de frutas tropicais. 2. ed. rev. Campinas: ITAL, 1992. cap. 2, p. 19-26. (Manual Técnico, 9). SILVA, D. A. M. da. Mangueira: cultivo sob condição irrigada. 2. ed. Recife: SEBRAE/PE, 2000, 63 p. (Série Agricultura, 9). SILVA, D. J. Análise de alimentos: métodos químicos e biológicos. 2. ed. Viçosa: UFV, 1990, 165 p. SILVA, E. L.; MARTINEZ, L. F.; YITAYEW, M. Relação entre coeficientes de cultura e graus-dia de desenvolvimento da alface. Horticultura Brasileira, Brasília, v. 17, n. 2, p. 134-142, jul. 1999. SILVA, E. M. Mecanismos bioquímicos de fisiopatias importantes de frutas. In: CONGRESSO IBEROAMERICANO DE TECNOLOGIA POSTCOSECHA Y AGROEXPORTACIONES, 2, 2000, Bogotá. Memórias... Bogotá: Universidad Nacional de Colombia, 2000. p. 5-19. SILVA, J. S. O. Produção de manga: manual. Viçosa: CPT, 1996. 34 p. (Série Fruticultura – Manual nº 40).

144
SILVA, M. A. P. da. Morfologia, emergência e ecofisiologia de Caryocar coriaceum Wittm. 2004. 114 f. Tese (Doutorado em Agronomia)-Universidade Federal do Ceará, Fortaleza, 2004. SIMÃO, S. Manual de fruticultura. São Paulo: CERES, 1971. 530 p. SINGH, L. B. The mango: botany, cultivation and utilization. London: Leonard Hill, 1960. ______. Mango. In: ALVIM, P. de T.; KOZLOWSKI, T. T. (Eds.). Ecophysiology of tropical crops. New York: Academic Press, 1977. ch. 18, p. 479-485. SINGH, R. N. Sex ratio and fruit set in mango. Science, Washington, v. 119, p. 389, 1954. STROHECKER, R.; HENNING, H. M. Analisis de vitaminas: métodos comprobados. Madrid: Paz Montalvo, 1967. 428 p. SUBRAMANYAN, H.; SHANTHA, K.; PARPIA, H. A. B. Physiology and biochemistry of mango fruit. Advances in Food Research, San Diego, v. 21, p. 223-305, 1975. SWAIN, T.; HILLIS, E. E. The phenolic constituents of Prunus domestica: II. The analysis of tissues of the ‘victoria’ plum tree. Journal of the Science of Food and Agriculture, London, v. 10, n. 2, p. 135-144, 1959. TAIZ, L.; ZEIGER, E. Fisiologia vegetal. Tradução Eliane Romanato Santarém et al. 3. ed. Porto Alegre: Artmed, 2004. 719 p. TANDON, D. K.; KALRA, S. K. Changes in sugars, starch and amylase activity during development of mango fruit cv. Dashehari. Journal of Horticultural Science, Ashford, v. 58, n. 3, p. 449-453, 1983. ______. Pectin changes during the development of mango fruit cv. Dashehari. Journal of Horticultural Science, Ashford, v. 59, n. 2, p. 283-286, 1984. THOMAS, P.; OKE, M. S. Technical note: vitamin C content and distribution in mangoes during ripening. Journal of Food Science and Technology, Oxford, v. 15, p. 669-672, 1980. TUCKER, G. A. Introduction. In: SEYMOUR, G. B.; TAYLOR, J. E.; TUCKER, G. A. Biochemistry of fruit ripening. London: Chapman & Hall, 1993. p. 1-51.

145
VAN BUREN, J. Fruit phenolics. In: HULME, A. C. (Ed.). The biochemistry of fruits and their products. London: Academic Press, 1970. v. 1, p. 269-304. VAVILOV, N. I. The origin, variation, immunity and breeding of cultivated plants. Translated from Russian by K. Starr Chester. Chronica Botanica, Leiden, v. 13, n. 1, p. 1-366, 1951. WHITE, T. Tannins – their occurrence and significance. Journal of the Science of Food and Agriculture, London, v. 8, n. 7, p. 377-385, 1957. WHITING, G. C. Sugars. In: HULME, A. C. (Ed.). The biochemistry of fruits and their products. London: Academic Press, 1970. v. 1, p. 1-31. WILLIAMS, A. H. The simpler phenolic substances of plants. Journal of the Science of Food and Agriculture, London, v. 8, n. 7, p. 385-389, 1957. WILLS, R. B. H. et al. Postharvest: an introduction to the physiology & handling of fruit, vegetables & ornamentals. 4. ed. Wallingford: New South Wales University Press, 1998. 262 p. WISSEMANN, K. W.; LEE, C. Y. Polyphenoloxidase activity during grape maturation and wine production. American Journal of Enology and Viticulture, Davis, v. 31, n. 3, p. 206-211, 1980. WOOLLEY, J. T.; HICKS, G. P.; HAGEMAN, R. H. Rapid determination of nitrate and nitrite in plant materials. Journal of Agricultural and Food Chemistry, Washington, v. 8, p. 481, 1960. YAMASHITA, F. Armazenagem frigorífica de mangas (Mangifera indica L. cv. Keitt) embaladas sob atmosfera modificada. 1995. 142 f. Tese (Doutorado em Engenharia de Alimentos)-Universidade Estadual de Campinas, Campinas, 1995. YEMN, E. W.; WILLIS, A. J. The estimation of carbohydrate in plant extracts by anthrone. The Biochemical Journal, London, v. 57, p. 508-514, 1954. YOSHIOKA, H.; KASHIMURA, Y.; KANEKO, K. Solubilization and distribution of neutral sugar residues derived from polyuronides during the softening in apple fruit. Journal of Japanese Society of Horticultural Science, Kyoto, v. 63, n. 1, p. 173-182, 1994.

146
______. β-D-galactosidase and α-L-arabinofuranosidase activities during the softening of apples. Journal of Japanese Society of Horticultural Science, Kyoto, v. 63, n. 4, p. 871-878, 1995. ZAUBERMAN, G. et al. Chilling injury, peroxidase, and cellulase activities in the peel of mango fruit at low temperature. HortScience, Alexandria, v. 23, n. 4, p. 732-733, 1988.

147
APÊNDICE

148
Tabela 1A – Resumo das análises de variância dos diâmetros longitudinal (DL), ventral (DV) e transversal (DT), produto dos diâmetros (PD), massas fresca (MF), seca (MS), de água (MA) e o teor de água (TA) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Quadrados médios Causas de variação
Graus de liberdade DL DV DT PD MF MS MA TA
Tempo (9) 6,3518** 6,2472** 5,3781** 179283,9096** 49243,9505** 1498,1663** 34661,1093** 23,3112**1º grau
1 26,7601** 32,4076** 34,6043** 952286,5496** 226775,8645** 9925,6143** 141814,3931** 164,4473** 2º grau 1 20,9284** 19,2016** 10,7257** 450026,3198** 160776,7660** 1914,1430** 127605,3158** 8,5055*
3º grau 1 1,2508ns 0,2341ns 0,0843ns 1519,3753ns 21412,0848* 1395,4231** 11875,1855ns 18,9054** Desvio 6 1,3712ns 0,7303ns 0,4981ns 34953,8236ns 5705,1398ns 41,3860ns 5109,1815ns 2,9904ns
Resíduo 20 0,9786 0,5313 0,2548 27403,2807 4866,1727 144,7487 3411,2841 1,6480CV (%) 9,63 8,98 6,96 25,38 24,02 26,77 23,79 1,50
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste F. ns Não-significativo. Tabela 2A – Resumo das análises de variância das escalas de coloração da casca (ECC), de Blush para coloração da casca (EBCC) e de coloração
da polpa (ECP) de mangas ‘Tommy Atkins’ durante o desenvolvimento. Quadrados médios Causas de
variação Graus de liberdade ECC EBCC ECP
Tempo (9) 0,1887** 2,0000** 0,4222**1º grau
1 1,4032** 15,4839** 3,1162**2º grau 1 0,0703* 0,3871ns 0,3198** 3º grau 1 0,1323** 0,2613ns 0,1645*
Desvio 6 0,0154ns 0,3113ns 0,0333ns
Resíduo 20 0,0125 0,2000 0.0333CV (%) 8,85 14,90 13,35
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste F. ns Não-significativo.

149
Tabela 3A – Resumo das análises de variância de luminosidade (LP), croma (CP) e ângulo Hue da polpa (HP) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Quadrados médios Causas de variação
Graus de liberdade LP CP HP
Tempo (7) 22,4824** 471,2187** 16,6550**1º grau
1 31,6151** 3030,6034** 20,7239**2º grau 1 49,8269** 82,9361** 45,0007**3º grau 1 26,8810* 129,6083** 40,8182**
Desvio 4 12,2635* 13,8458ns 2,5105ns
Resíduo 16
3,2946 8,8996 1,4008CV (%) 2,69 7,84 1,27
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste F. ns Não-significativo. Tabela 4A – Resumo das análises de variância de sólidos solúveis totais (SST), acidez total titulável (ATT), pH, relação SST/ATT (RSA), vitamina
C (VC) e clorofila total (CLT) de mangas ‘Tommy Atkins’ durante o desenvolvimento. Quadrados médios Causas de
variação Graus de liberdade SST ATT pH RSA VC CLT
Tempo (9) 3,8076** 64,1756** 0,2657** 35,8312** 49,6322** 241,5225*1º grau 1 31,5033** 382,4266** 2,1241** 314,4831** 292,3137** 577,2011* 2º grau 1 0,0686ns 169,7758**
0,0117** 0,0398ns 21,5907** 986,5577**3º grau 1 1,2007** 14,0834** 0,1688** 1,6559* 10,6356* 182,2889ns
Desvio 6 0,2493ns 1,8824** 0,0144** 1,0503* 20,3583** 71,2759ns
Resíduo 20 0,1367 0,4309 0,0014 0,3321 2,3127 85,2283CV (%) 5,05 20,14 1,15 9,95 9,48 27,95
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste F. ns Não-significativo.

150
Tabela 5A – Resumo das análises de variância de amido (AD), açúcares solúveis totais (AST), redutores (AR) e não redutores (ANR) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Quadrados médios Causas de variação
Graus de liberdade AD AST AR ANR
Tempo (7) 1,9926** 2,0841** 0,3460** 1,5842**1º grau 1 12,2454** 14,0634** 0,4470* 9,4958** 2º grau 1 1,3289* 0,1095ns 1,6560**
0,9138*3º grau 1 0,1424ns 0,1242 x 10-4ns 0,2913ns 0,2879ns
Desvio 4 0,0578ns 0,1039ns 0,0070ns 0,0980ns
Resíduo 16 0,2231 0,0621 0,0686 0,1215CV (%) 16,51 5,50 7,41 35,11
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste F. ns Não-significativo. Tabela 6A – Resumo das análises de variância de nitrogênio total (NT), não protéico (NNP) e protéico (NP), proteína bruta (PB) e verdadeira (PV)
de mangas ‘Tommy Atkins’ durante o desenvolvimento. Quadrados médios Causas de
variação Graus de liberdade NT NNP NP PB PV
Tempo (7) 0,0357** 0,0135* 0,0086* 1,4067** 0,3326*1º grau 1 0,1128** 0,0607** 0,0087ns 4,4729**
0,3374ns
2º grau 1 0,0010ns 0,0004ns 0,0003ns 0,0350ns 0,0103ns
3º grau 1 0,0784** 0,0100ns 0,0332** 3,0192** 1,2560**Desvio 4 0,0144* 0,0058ns 0,0046ns 0,5800* 0,1811ns
Resíduo 16 0,0043 0,0036 0,0023 0,1727 0,0872CV (%) 10,30 11,08 51,33 10,45 50,85
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste F. ns Não-significativo.

151
Tabela 7A – Resumo das análises de variância de carotenóides totais (CAT), fenólicos poliméricos (FP), oligoméricos (FO) e diméricos (FD) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Quadrados médios Causas de variação
Graus de liberdade CAT FP FO FD
Tempo (7) 0,3639** 59,8327** 58,2995** 174,9624**1º grau 1 2,3438** 219,6586** 255,5895** 595,0196** 2º grau 1 0,1308* 27,1375ns 17,2328ns 260,6030** 3º grau 1 0,0106ns 80,8028**
51,1012ns 58,1674ns
Desvio 4 0,0155ns 22,8075ns 21,0432ns 77,7367* Resíduo 16 0,0159 9,3287 11,6579 20,2477CV (%) 15,99 10,52 11,69 13,26
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste F. ns Não-significativo. Tabela 8A – Resumo das análises de variância de pectina total (PT) e solúvel (PS), percentagem de solubilização de pectina (PSP), pectinas de alta
metoxilação (PAM), de baixa metoxilação (PBM) e a protopectina (PROT) de mangas ‘Tommy Atkins’ durante o desenvolvimento. Quadrados médios Causas de
variação Graus de liberdade PT PS PSP PAM PBM PROT
Tempo (7) 68625,1101** 636,5426ns 28,7021ns 1116272,3714** 94604,2948ns 293709,8901** 1º grau 1 301020,6373** 4852145,8577** 1590706,3628** 2º grau 1 155768,6448** 688365,9603** 65437,1550ns
3º grau 1 1507,2914ns
136297,1220ns 188703,6799**Desvio 4 5519,7994ns 534274,4149**
52780,5082*
Resíduo 16 2529,6703 271,4607 26,0690 59477,8282 84501,1646 17121,6863CV (%) 13,84 34,84 37,16 18,43 18,15 17,72
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste F. ns Não-significativo.

152
Tabela 9A – Resumo das análises de variância das atividades da pectinametilesterase (PME), da poligalacturonase (PG), da polifenoloxidase (PPO), da peroxidase (POD), amilásica total (AMLT), α-amilásica (α-AML) e β-amilásica (β-AML) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Quadrados médios Causas de variação
Graus de liberdade PME PG PPO POD AMLT α-AML β-AML
Tempo (7) 4988,5094ns 163,8706** 2460,6940* 155,6981** 0,1823* 0,2942** 0,0770ns
1º grau 1 197,8011** 519,5786ns 98,2002**
1,0818** 1,4347**2º grau 1 583,3814** 10411,4133** 656,6182** 0,0264ns 0,0765ns 3º grau 1 34,9608ns 534,5928ns 133,5866** 0,1411ns 0,0923ns
Desvio 4 82,7377** 1439,8184ns 50,3704** 0,0067ns 0,1139*Resíduo 16 2274,5710 8,4346 766,9297 8,8731 0,0451 0,0331 0,0323CV (%) 23,66 9,34 5,09 12,42 7,75 7,32 70,29
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste F. ns Não-significativo. Tabela 10A – Resumo das análises de variância das atividades α-galactosidásicas de citosol (α-GALc) e de parede celular (α-GALp), β-
galactosidásicas de citosol (β-GALc) e de parede celular (β-GALp), proteína de citosol (Pc) e de parede celular (Pp) de mangas ‘Tommy Atkins’ durante o desenvolvimento.
Quadrados médios Causas de variação
Graus de liberdade α-GALc α-GALp β-GALc β-GALp Pc Pp
Tempo (7) 793,3285** 29,2356* 0,4999** 0,1640** 0,0008** 0,0002ns
1º grau 1 3195,1607** 60,4033* 2,2081** 0,6916** 0,0004 x 10-4ns 2º grau 1 793,7122** 7,0858ns 1,0569**
0,0034ns 0,0024**3º grau 1 527,4689* 4,4071ns 0,0260* 0,0061ns 0,0005*
Desvio 4 259,2395* 33,1883* 0,0521** 0,1117** 0,0006**Resíduo 16 70,9862 8,9741 0,0046 0,0186 0,0001 0,0001CV (%) 22,95 26,54 16,59 23,01 32,89 53,67
*, ** Significativos, respectivamente, em nível de 5 e de 1% de probabilidade, pelo teste F. ns Não-significativo.