efeitos fisiolÓgicos e metabÓlicos em tomateiro … · companheirismo e imensurável ajuda nesta...
TRANSCRIPT

1
UNIVERSIDADE ESTADUAL DO CENTRO-OESTE, UNICENTRO – PR
EFEITOS FISIOLÓGICOS E METABÓLICOS EM
TOMATEIRO POR ESTROBILURINAS E CARBOXAMIDAS
TESE DE DOUTORADO
JANAINA MAREK
GUARAPUAVA-PR
2018

JANAINA MAREK
EFEITOS FISIOLÓGICOS E METABÓLICOS EM TOMATEIRO POR
ESTROBILURINAS E CARBOXAMIDAS
Tese apresentada à Universidade Estadual do
Centro-Oeste, como parte das exigências do
Programa de Pós-Graduação em Agronomia, área
de concentração em Produção Vegetal, para
obtenção do título de Doutora em Agronomia.
Professora Drª Elizabeth Orika Ono
Orientadora
Professor Dr. João Domingos Rodrigues
Co-Orientador
Professora Drª Cacilda Márcia Duarte Rios Faria
Co-Orientadora
GUARAPUAVA-PR
2018

Catalogação na Publicação
Biblioteca Central da Unicentro, Campus Santa Cruz
Marek, Janaina
M323e Efeitos fisiológicos e metabólicos em tomateiro por estrobilurinas e carboxamidas / Janaina Marek. – – Guarapuava, 2018.
xxii, 149 f. : il. ; 28 cm Tese (doutorado) - Universidade Estadual do Centro-Oeste, Programa de
Pós-Graduação em Agronomia, área de concentração em Produção Vegetal, 2018
Orientadora: Elizabeth Orika Ono Coorientador: João Domingos Rodrigues Coorientadora: Cacilda Márcia Duarte Rios Faria Banca examinadora: Elizabeth Orika Ono, Kátia Regina Freitas Schwan-Estrada, João Domingos Rodrigues, Patricia Carla Giloni de Lima, Cacilda Márcia Duarte Rios Faria
Bibliografia
1. Agronomia. 2. Produção vegetal. 3. Solanum lucopersicum L.. 4.
Alternaria solani. 5. Estresse oxidativo. 6. Proteínas relacionadas à patogênese. 7. Enzimas. I. Título. II. Programa de Pós-Graduação em Agronomia.
CDD 630


“A agricultura é a arte de colher o sol.”
(Provérbio Chinês)

Jamais me envergonharei das minhas raízes...
Sou filha de agricultores humildes,
que com o suor de cada dia e mãos calejadas,
proveram o sustento da família e o estudo de seus filhos.
Pai e Mãe,
Com muito orgulho eu lhes dedico esta tese!

Quero agradecer...
Primeiramente a Deus pelo dom da vida e imensa proteção.
Aos meus pais, José e Juraci, por serem meus exemplos de vida, por todo o amor,
carinho e cumplicidade.
Ao Dione de Azevedo, meu ‘braço direito’, por toda a dedicação, compreensão,
companheirismo e imensurável ajuda nesta caminhada.
A minha irmã Jienefer por toda a ajuda intelectual, física, emocional e espiritual,
minha querida companheira nos momentos de sorrisos e lágrimas.
Ao meu irmão Diogo e minha cunhada Débora, pelo carinho, apoio e pelas palavras
de conforto.
A minha orientadora Drª Elizabeth Orika Ono por todo o ensinamento e exemplo
de profissionalismo, pela paciência e dedicação em me orientar, mesmo com a dificuldade
da distância e, principalmente, pela confiança depositada em mim.
Aos meus co-orientadores Drª Cacilda Márcia Duarte Rios Faria e Dr. João
Domingos Rodrigues pelo apoio incondicional e por toda a atenção e ajuda quando
precisei.
As professoras Drª Kátia Regina Freitas Sckwan-Estrada e Drª Patricia Carla
Giloni de Lima pelas importantes considerações e contribuições para a lapidação deste
trabalho.
A minha querida ‘aprendiz’ Jessica Caroline Miri por ter se dedicado a este trabalho
como se fosse dela, por ter me ajudado do começo ao fim, além da grande amizade em
todas as horas e pelas palavras amigas nos momentos difíceis.
A minha amiga Elaine Pittner, companheira de laboratório, por toda a ajuda e
parceria nas análises enzimáticas e, principalmente, pela amizade construída.

As meninas do “pam”, Regina Lopes, Mariane Dal Comune e Suélen Hellmann,
por toda a ajuda com as análises. A Elisa Adriano e Aline José Maia pela ajuda
com as análises de trocas gasosas.
Aos colegas de laboratório, Laís Gasperotto, Juliane Wolbert, Douglas Bortuli,
Carla Daiane Leite, Leandro Alvarenga Santos, Fábio Telaxca, Letícia Lopes
Dias, Rafael Calixto e Paola Zimmermann por toda a ajuda concedida.
As meninas de Botucatu, Ana Claudia Macedo e Amanda Cristina Esteves Amaro
pelo auxílio com o aporte técnico e científico nas metodologias.
Aos professores Renato e Marcão pela disponibilidade do espaço físico para as
análises de pós-colheita.
A nossa querida secretária Lucilia pela paciência e prontidão em me atender.
Ao pessoal da manutenção pelo empréstimo de ferramentas e aos seguranças pelo
monitoramento prestado em muitas noites e finais de semanas.
A todos os Professores do Programa por auxiliar na construção deste conhecimento.
Aos meus colegas de Pós-Graduação pelas amizades construídas, por estarem comigo
nesse trajeto e por todo o conhecimento que construímos juntos.
Ao Programa de Pós-Graduação em Agronomia e à Universidade Estadual do
Centro-Oeste, Campus Cedeteg pela oportunidade e apoio concedido para a realização
deste trabalho.
A todos os meus familiares e amigos, em especial minha Avó ‘Dona Nena’, Tia
Nega, Andreia, Davina, ‘Dona Zene’, madrinha Ilze, Iara, Ana Paula, Rubia,
Laís, Mayara, Thayna, Jessica (...) obrigada pelas orações, conversas, abraços e
apoio nos momentos de dificuldade.
E aqueles que criticaram, pois só assim pude ver que ainda havia muito para
melhorar.
A todos aqueles que de alguma forma contribuíram para a realização deste trabalho...
Muito obrigada!

SUMÁRIO
ÍNDICE DE FIGURAS ............................................................................................................. i
ÍNDICE DE TABELAS ......................................................................................................... vii
RESUMO .................................................................................................................................. ix
ABSTRACT ............................................................................................................................. xi
1.INTRODUÇÃO ..................................................................................................................... 1
2.OBJETIVOS .......................................................................................................................... 3
2.1Objetivo Geral ....................................................................................................................... 3
2.2Objetivos específicos ............................................................................................................. 3
3. REFERENCIAL TEÓRICO ............................................................................................... 4
3.1 Aspectos gerais do cultivo de tomateiro ............................................................................... 4
3.1.1 Origem ...............................................................................................................................4
3.1.2 Características botânicas e agronômicas ........................................................................... 4
3.1.3 Qualidade pós-colheita de frutos de tomateiro .................................................................. 5
3.2 A importância econômica da cultura do tomateiro ............................................................... 6
3.3 Ocorrência de doenças .......................................................................................................... 7
3.3.1 Pinta preta do tomateiro: aspectos gerais da doença ......................................................... 7
3.3.2 Controle de pinta preta em tomateiro ................................................................................ 9
3.4 Efeitos fisiológicos das estrobilurinas e carboxamidas ...................................................... 11
3.5 Desenvolvimento de plantas ............................................................................................... 12
3.6 Eficiência fotossintética ...................................................................................................... 12
3.6.1 Trocas gasosas ................................................................................................................. 12
3.6.2 Pigmentos fotossintéticos e fluorescência da clorofila a ................................................. 13
3.6.3 Nitrato redutase................................................................................................................ 14
3.7 Estresse oxidativo ............................................................................................................... 15
3.7.1 Enzimas envolvidas na defesa antioxidativa ................................................................... 16
3.7.2 Peroxidação lipídica ........................................................................................................ 18
3.8 Indução de resistência e a defesa vegetal contra patógenos ............................................... 19
3.8.1 Proteínas envolvidas na indução de resistência ............................................................... 23
3.8.2 Metabolismo secundário e defesa vegetal ....................................................................... 24

4. MATERIAL E MÉTODOS ............................................................................................... 27
4.1 Caracterização da área de estudo ........................................................................................ 27
4.2 Material vegetal: semeadura e transplante.......................................................................... 27
4.3 Material vegetal: tratos culturais ........................................................................................ 28
4.4 Delineamento experimental e tratamentos ......................................................................... 29
4.5 Caracterização dos fungicidas ............................................................................................ 30
4.5.1 Boscalida ......................................................................................................................... 30
4.5.2 Boscalida + Cresoxim-metílico ....................................................................................... 30
4.5.3 Piraclostrobina ................................................................................................................. 31
4.5.4 Fluxapiroxade + Piraclostrobina ...................................................................................... 31
4.5.5 Metiram + Piraclostrobina ............................................................................................... 31
4.6 Isolamento e inoculação do patógeno: Alternaria solani ................................................... 32
4.7 Propriedades avaliadas ....................................................................................................... 33
4.7.1 Análise de desenvolvimento ............................................................................................ 33
4.7.2 Trocas gasosas ................................................................................................................. 33
4.7.3 Fluorescência da clorofila a ............................................................................................. 34
4.7.4 Nitrato Redutase .............................................................................................................. 35
4.7.5 Pigmentos fotossintéticos ................................................................................................ 36
4.7.6 Análise de carboidratos ................................................................................................... 37
4.7.7 Avaliação da severidade de Alternaria solani ................................................................. 38
4.7.8 Análises bioquímicas ....................................................................................................... 39
4.7.8.1 Coleta e obtenção dos extratos enzimáticos ................................................................. 39
4.7.8.2 Proteínas totais e atividades enzimáticas ...................................................................... 41
4.7.8.3 Ânion superóxido e peróxido de hidrogênio ................................................................ 43
4.7.8.4 Peroxidação lipídica ..................................................................................................... 45
4.7.8.5 Compostos fenólicos..................................................................................................... 45
4.7.8.6 Deposição de lignina .................................................................................................... 45
4.7.9 Avaliação de produção e pós-colheita de frutos de tomateiro ......................................... 46
4.7.9.1 Produção..... .................................................................................................................. 46
4.7.9.2 Qualidade pós-colheita de frutos .................................................................................. 47
4.7.9.3 Avaliação da perda de massa ........................................................................................ 48
4.8 Análise estatística ............................................................................................................... 49

5. RESULTADOS E DISCUSSÃO ....................................................................................... 50
5.1 Desenvolvimento do tomateiro ........................................................................................... 50
5.2 Eficiência fotossintética ...................................................................................................... 55
5.3 Pigmentos fotossintéticos e atividade da enzima nitrato redutase ...................................... 64
5.4 Síntese e translocação de carboidratos ............................................................................... 69
5.5 Controle de Alternaria solani ............................................................................................. 74
5.6 Estresse oxidativo e enzimas antioxidantes ........................................................................ 78
5.7 Proteínas relacionadas à patogênese e respostas de defesa ................................................ 99
5.7 Produção e Qualidade pós-colheita de frutos de tomateiro .............................................. 115
6. CONSIDERAÇÕES FINAIS ........................................................................................... 125
7. CONCLUSÃO ................................................................................................................... 125
8. Referências Bibliográficas ............................................................................................... 130

i
ÍNDICE DE FIGURAS
Figura 1. Casa de vegetação: vista externa (A) e vista interna (B). Unicentro, Guarapuava/PR.
.................................................................................................................................................. 27
Figura 2. Plantas de tomateiro em casa de vegetação tutoradas verticalmente com fitilho
(Experimento I – 2015) e bambu (Experimento II – 2016). Unicentro, Guarapuava/PR......... 29
Figura 3. Cultura de Alternaria solani antes (A) e após (B) injúria mecânica dos micélios.
Unicentro, Guarapuava/PR. ...................................................................................................... 32
Figura 4. Análise de trocas gasosas em plantas de tomateiro (A e B). Unicentro,
Guarapuava/PR. ........................................................................................................................ 34
Figura 5. Clip para adaptação ao escuro: indicado pela seta (A); Análise de fluorescência da
clorofila a em plantas de tomateiro. Unicentro, Guarapuava/PR. ............................................ 35
Figura 6. Escala diagramática para avaliação de Alternaria solani em tomateiro expressa em
proporção de área foliar lesionada (A). Avaliação da severidade de pinta preta em tomateiro
(B). Unicentro, Guarapuava/PR. .............................................................................................. 38
Figura 7. Ponto de colheita dos frutos de tomateiro: vermelho-claro (A) e pesagem dos frutos
em balança analítica (B). Unicentro, Guarapuava/PR. ............................................................. 46
Figura 8. Colheita de frutos no estádio de maturação verde-salada (A), pesagem dos frutos (B),
frutos no início: 1°dia (C) e final: 21° dia (D) do experimento de avaliação de perda de massa.
Unicentro, Guarapuava/PR. ...................................................................................................... 49
Figura 9. Taxa de assimilação de CO2 (A, μmol m-2 s-1), condutância estomática (gs, mol m-2 s-
1) e concentração interna de CO2 na folha (Ci, μmol mol-1) em plantas de tomateiro híbrido
Conquistador, cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculados com Alternaria solani, aos 34 e 94 dias após o transplantio (DAT),
nos anos de 2015 e 2016. Médias (n = 5) seguidas de letras distintas, na mesma avaliação,
diferem pelo teste Tukey ao nível de 5% de probabilidade (p<0,05). Barras indicam desvio
padrão da média. Unicentro, Guarapuava/PR, 2018. ............................................................... 56
Figura 10. Taxa de transpiração (E, mmol m-2 s-1), eficiência do uso da água (EUA, A/E, μmol
CO2 (mmol H2O)-1) e eficiência de carboxilação (A/Ci) em plantas de tomateiro híbrido

ii
Conquistador, cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculados com Alternaria solani, aos 34 e 94 dias após o transplantio (DAT),
nos anos de 2015 e 2016. Médias (n = 5) seguidas de letras distintas, na mesma avaliação,
diferem pelo teste Tukey ao nível de 5% de probabilidade (p<0,05). Barras indicam desvio
padrão da média. Unicentro, Guarapuava/PR, 2018. ............................................................... 58
Figura 11. Fluorescência variável da clorofila a (Fv), eficiência máxima do PSII (Fv/Fm),
rendimento quântico fotoquímico do PSII, Y(II) e taxa relativa de transferência de elétrons
(ETR) em plantas de tomateiro híbrido Conquistador, cultivadas em casa de vegetação, tratadas
com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, aos 34 e
94 dias após o transplantio (DAT), nos anos de 2015 e 2016. Médias (n = 5) seguidas de letras
distintas, na mesma avaliação, diferem pelo teste Tukey ao nível de 5% de probabilidade
(p<0,05). Barras indicam desvio padrão da média. Unicentro, Guarapuava/PR, 2018. ........... 60
Figura 12. Teor de clorofila a (µg g massa fresca-1), clorofila b (µg g massa fresca-1) e
carotenoides (µg g massa fresca-1) em plantas de tomateiro híbrido Conquistador cultivadas em
casa de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, aos 15, 20, 35, 50, 65, 80, 95 e 110 dias após o transplantio (DAT), nos anos
de 2015 e 2016. Médias (n = 5) seguidas de letras distintas, na mesma avaliação, diferem pelo
teste Tukey ao nível de 5% de probabilidade (p<0,05). Barras indicam desvio padrão da média.
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 65
Figura 13. Atividade da enzima nitrato redutase (NR, μg nitrito min-1 g de massa fresca-1) em
plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com
diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, aos 15, 20, 35,
50, 65, 80, 95 e 110 dias após o transplantio (DAT), nos anos de 2015 e 2016. Médias (n = 5)
seguidas de letras distintas, na mesma avaliação, diferem pelo teste Tukey ao nível de 5% de
probabilidade (p<0,05). Barras indicam desvio padrão da média. Unicentro, Guarapuava/PR,
2018. ......................................................................................................................................... 68
Figura 14. Efeito dos diferentes tratamentos com fungicidas, na área abaixo da curva de
progresso da doença (AACPD), testados no controle de Alternaria solani em plantas de
tomateiro híbrido Conquistador, cultivado em casa de vegetação, nos anos de 2015 e 2016.
Tratamentos: testemunha inoculada (Test+Inoc), testemunha absoluta (Test), boscalida (Bosc),
boscalida mais cresoxim-metílico (Bosc+Cre), piraclostrobina (Pira), fluxapiroxade mais
piraclostrobina (Flux+Pira) e metiram mais piraclostrobina (Me+Pira). Médias (n = 10)
seguidas de letras distintas diferem pelo teste Tukey ao nível de 1% de probabilidade (p<0,05).
Barras indicam e desvio padrão da média. Unicentro, Guarapuava/PR, 2018. ........................ 74
Figura 15. Variação das temperaturas e umidades do ar, máximas e mínimas, no ano de 2015,
avaliadas diariamente durante a condução do experimento, no interior da casa de vegetação

iii
onde foram conduzidas as plantas de tomateiro híbrido Conquistador, tratadas com diferentes
fungicidas de efeitos fisiológicos e inoculados com Alternaria solani. Unicentro,
Guarapuava/PR, 2018. .............................................................................................................. 75
Figura 16. Variação das temperaturas e umidades do ar, máximas e mínimas, no ano de 2016,
avaliadas diariamente durante a condução do experimento, no interior da casa de vegetação
onde foram conduzidas as plantas de tomateiro híbrido Conquistador, tratadas com diferentes
fungicidas de efeitos fisiológicos e inoculados com Alternaria solani. Unicentro,
Guarapuava/PR, 2018. .............................................................................................................. 76
Figura 17. Concentração de ânion superóxido (•O2-, mmol g de massa fresca-1) aos 15, 16, 17,
20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias após
o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 80
Figura 18. Atividade da enzima superóxido dismutase (SOD, U min-1 mg proteína-1) aos 15,
16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias
após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa
de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 83
Figura 19. Concentração de peróxido de hidrogênio (H2O2, µmol g de massa fresca-1) aos 15,
16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias
após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa
de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 86
Figura 20. Atividade da enzima ascorbato peroxidase (APX, U min-1 mg proteína-1) aos 15, 16,
17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias
após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa
de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 88
Figura 21. Atividade da enzima glutationa redutase (GR, U min-1 mg proteína-1) aos 15, 16,
17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias
após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa

iv
de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 89
Figura 22. Atividade da enzima glutationa S-transferase (GST, U min-1 mg proteína-1) aos 15,
16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias
após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa
de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 90
Figura 23. Atividade da enzima glutationa peroxidase (GPX, U min-1 mg proteína-1) aos 15,
16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias
após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa
de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 91
Figura 24. Atividade da enzima catalase (CAT, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29,
30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias após o
transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 94
Figura 25. Peroxidação lipídica (TBARS, µmol g massa fresca-1) aos 15, 16, 17, 20, 29, 30,
31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias após o transplantio
(DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas
com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, nos anos
de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR,
2018. ......................................................................................................................................... 97
Figura 26. Atividade da enzima fenilalanina amônia-liase (PAL, U min-1 mg proteína-1) aos 15,
16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias
após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa
de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. .......................................................................................... 101
Figura 27. Teor de Compostos Fenólicos (mg g massa fresca-1) aos 15, 16, 17, 20, 29, 30, 31,
34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias após o transplantio

v
(DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas
com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, nos anos
de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR,
2018. ....................................................................................................................................... 103
Figura 28. Atividade da enzima peroxidase (POD, U min-1 mg proteína-1) aos 15, 16, 17, 20,
29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias após o
transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. .......................................................................................... 105
Figura 29. Atividade da enzima polifenoloxidase (PPO, U min-1 mg proteína-1) aos 15, 16, 17,
20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias após
o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. .......................................................................................... 106
Figura 30. Deposição de lignina (µg de lignina-1 mg de massa fresca-1) aos 14, 20, 35, 50, 65,
80, 95 e 110 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador
cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e
inoculados com Alternaria solani, nos anos de 2015 e 2016. Médias (n = 10) seguidas de letras
distintas, na mesma avaliação, diferem pelo teste Tukey ao nível de 5% de probabilidade
(p<0,05). Barras indicam desvio padrão da média. Unicentro, Guarapuava/PR, 2018. ......... 108
Figura 31. Atividade da enzima lipoxigenase (LOX, U min-1 mg proteína-1) aos 15, 16, 17, 20,
29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias após o
transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. .......................................................................................... 110
Figura 32. Atividade da enzima quitinase (QUI, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29,
30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias após o
transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. .......................................................................................... 112

vi
Figura 33. Atividade da enzima β-1,3-glucanase (GLU, U min-1 mg proteína-1) aos 15, 16, 17,
20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias após
o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10).
Unicentro, Guarapuava/PR, 2018. .......................................................................................... 113
Figura 34. Atividade das enzimas pectinametilesterase (PME, U min-1 g massa fresca-1) (A.) e
poligalacturonase (PG, U min-1 g massa fresca-1) (B.) em frutos de tomateiro híbrido
Conquistador, cultivadas em casa de vegetação, nos anos de 2015 e 2016. Tratamentos:
testemunha inoculada (Test+Inoc), testemunha absoluta (Test), boscalida (Bosc), boscalida
mais cresoxim-metílico (Bosc+Cre), piraclostrobina (Pira), fluxapiroxade mais piraclostrobina
(Flux+Pira) e metiram mais piraclostrobina (Me+Pira) e inoculadas com Alternaria solani.
Médias (n = 10) seguidas de letras distintas, no mesmo ano, diferem significativamente entre
tratamentos pelo teste Tukey ao nível de 5% de probabilidade (p<0,05). Barras indicam e
desvio padrão da média. Unicentro, Guarapuava/PR, 2018. .................................................. 122
Figura 35. Perda de massa (%) de frutos de tomateiro híbrido Conquistador, cultivadas em casa
de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, durante 21 dias, nos anos de 2015 e 2016. Médias (n = 10) seguidas de letras
diferentes no mesmo dia de avliação diferem significativamente entre si pelo teste Tukey a 5%
de probabilidade (p<0,05). Dados transformados em arco seno de √x(%). Barras indicam desvio
padrão da média. Unicentro, Guarapuava/PR, 2018. ............................................................. 123
Figura 36. Modelo esquemático mostrando os efeitos fisiológicos e metabólicos promovidos
pela aplicação de estrobilurinas: cresoxim-metílico e boscalida e carboxamidas: boscalida e
fluxapiroxade em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação
e inoculadas com Alternaria solani. Fonte: MAREK, 2018. ................................................. 127

vii
ÍNDICE DE TABELAS
Tabela 1. Análise química de solo dos experimentos I e II nos anos de 2015 e 2016,
respectivamente. Unicentro, Guarapuava/PR, 2018. ................................................................ 28
Tabela 2. Altura (cm) aos 14, 21, 28, 35, 42, 49 e 56 dias após o transplantio (DAT) de plantas
de tomateiro híbrido Conquistador, cultivadas em casa de vegetação, tratadas com diferentes
fungicidas de efeitos fisiológicos e inoculadas com Alternaria solani, no ano de 2015.
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 50
Tabela 3. Altura (cm) aos 14, 21, 28, 35, 42, 49 e 56 dias após o transplantio (DAT) de plantas
de tomateiro híbrido Conquistador, cultivadas em casa de vegetação, tratadas com diferentes
fungicidas de efeitos fisiológicos e inoculadas com Alternaria solani, no ano de 2016.
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 51
Tabela 4. Variáveis de desenvolvimento aos 120 dias após o transplantio (DAT) de plantas de
tomateiro híbrido Conquistador, cultivadas em casa de vegetação e tratadas com diferentes
fungicidas de efeitos fisiológicos, inoculados com Alternaria solani, no ano de 2015.
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 53
Tabela 5. Variáveis de desenvolvimento aos 120 dias após o transplantio (DAT) de plantas de
tomateiro híbrido Conquistador, cultivadas em casa de vegetação, tratadas com diferentes
fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, no ano de 2016.
Unicentro, Guarapuava/PR, 2018. ............................................................................................ 54
Tabela 6. Média dos teores de açúcares totais e açúcares redutores (µg g massa fresca-1) em
folha, caule e fruto aos 50 e 120 dias após o transplantio (DAT) de plantas de tomateiro híbrido
Conquistador, cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculadas com Altenaria solani, no ano de 2015. Unicentro, Guarapuava/PR,
2018. ......................................................................................................................................... 70
Tabela 7. Média dos teores de açúcares totais, açúcares redutores (µg g massa fresca-1) em
folha, caule e fruto aos 50 e 120 dias após o transplantio (DAT) de plantas de tomateiro híbrido
Conquistador, cultivadas em casa de vegetação e tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculadas com Alternaria solani, no ano de 2016. Unicentro, Guarapuava/PR,
2018. ......................................................................................................................................... 71
Tabela 8. Valores médios de produção total (PT, kg m-2), número total de frutos (NTF, frutos
m-2), massa fresca de frutos (MFF, g fruto-1) e porcentagem de frutos não comerciais (FNC, %)
de plantas de tomateiro híbrido Conquistador, cultivadas em casa de vegetação, tratadas com

viii
diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, nos anos de
2015 e 2016. Unicentro, Guarapuava/PR, 2018. .................................................................... 116
Tabela 9. Valores médios de firmeza (gf cm-2), pH, teor de sólidos solúveis (SS, °Brix), acidez
titulável (AT, g de ácido cítrico 100 g de polpa-1), relação SS/AT e teor de ácido ascórbico (AA,
mg 100 g de polpa-1) de frutos de tomateiro híbrido Conquistador, cultivados em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, no ano de 2015. Unicentro, Guarapuava/PR, 2018. .................................. 119
Tabela 10. Valores médios de firmeza (gf cm-2), pH, teor de sólidos solúveis (SS, °Brix), acidez
titulável (AT, g de ácido cítrico 100g de polpa-1), relação SS/AT e teor de ácido ascórbico (AA,
mg 100 g de polpa-1) de frutos de tomateiro híbrido Conquistador, cultivados em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani, no ano de 2016. Unicentro, Guarapuava/PR, 2018. .................................. 120

ix
RESUMO
MAREK, Janaina. Efeitos fisiológicos e metabólicos em tomateiro por estrobilurinas e
carboxamidas. Guarapuava: UNICENTRO, 2018. (Tese – Doutorado em Agronomia/Produção
Vegetal).
O objetivo desta pesquisa foi avaliar os efeitos fisiológicos e metabólicos em plantas de
tomateiro híbrido Conquistador, cultivadas em casa de vegetação e inoculadas, com aplicação
de estrobilurinas e carboxamidas. O estudo foi realizado em dois anos (2015 e 2016), nos quais
foram avaliados os efeitos dos diferentes fungicidas no desenvolvimento de plantas de
tomateiro, na eficiência fotossintética, no sistema antioxidativo, no controle da doença e na
ativação de mecanismos de defesa contra o patógeno, bem como na produção e pós-colheita de
frutos. O delineamento experimental utilizado foi blocos ao acaso, com sete tratamentos e cinco
repetições, sendo cada parcela composta por quatro plantas. Os tratamentos utilizados foram:
T1 – testemunha inoculada (água + inóculo); T2 – testemunha absoluta (água); T3 – boscalida
(0,075 g L-1); T4 – boscalida (0,100 g L-1) + cresoxim-metílico (0,050 g L-1); T5 –
piraclostrobina (0,100 g L-1); T6 – fluxapiroxade (0,058 g L-1) + piraclostrobina (0,116 g L-1) e
T7 – metiram (1,100 g L-1) + piraclostrobina (0,100 g L-1). O efeito dos tratamentos no
desenvolvimento foi avaliado pela altura de plantas, área foliar, massa fresca e seca de folhas,
caule e raiz. Para verificar a eficiência fotossintética foram realizadas avaliações de trocas
gasosas, fluorescência da clorofila a, teor de pigmentos, atividade da enzima nitrato redutase e
teor de carboidratos. Para verificar os efeitos dos tratamentos no sistema antioxidativo, no
controle da doença e ativação de mecanismos de defesa ao patógeno, foram avaliadas a área
abaixo da curva de progresso da doença (AACPD), concentração de ânion superóxido e
peróxido de hidrogênio, atividade das enzimas superóxido dismutase, ascorbato peroxidase,
glutationa redutase, glutationa S-transferase, glutationa peroxidase, catalase, fenilalanina
amônia-liase, quitinase, β-1,3-glucanase, peroxidase, polifenoloxidase e lipoxigenase, além do
teor de compostos fenólicos, peroxidação lipídica e deposição de lignina. Para avaliar os efeitos
dos tratamentos na produção e pós-colheita de frutos foram realizadas avaliações de firmeza de
fruto, pH, sólidos solúveis, acidez titulável, relação sólidos solúveis/acidez titulável (SS/AT),
teor de ácido ascórbico, atividade das enzimas pectinametilesterase e poligalacturonase e perda
de massa. Pelos resultados obtidos pode-se concluir que os tratamentos fluxapiroxade mais

x
piraclostrobina, seguido de metiram mais piraclostrobina, se destacaram dos demais
tratamentos, pois apresentaram melhor desenvolvimento das plantas e maior produção de
frutos, em razão da maior eficiência fotossintética, maiores atividades das enzimas de defesa
antioxidativa e ativação de respostas de defesa contra o ataque do patógeno.
Palavras-chave: Solanum lycopersicum L., Alternaria solani, estresse oxidativo, proteínas
relacionadas à patogênese, enzimas.

xi
ABSTRACT
MAREK, Janaina. Physiological and metabolic effects on tomatoes by strobilurins and
carboxamides. Guarapuava: UNICENTRO, 2018. (Thesis - PhD in Agronomy / Plant
Production).
The objective of this research was to evaluate the physiological and metabolic effects of
Conquistador hybrid tomato plants, cultivated in greenhouse and inoculated, with the
application of strobilurins and carboxamides. The study was realized in two years (2015 and
2016), in which the effects of the different fungicides on the development of tomato plants, on
the photosynthetic efficiency, on the antioxidative system, on the control of the disease and on
the activation of defense mechanisms were evaluated the pathogen, as well as in the production
and post-harvest of fruits. The experimental design was a randomized block with seven
treatments and five replicates, each plot being composed of four plants. The treatments used
were: T1 – inoculated control (water + inoculum); T2 – absolute control (water); T3 – boscalide
(0.075 g L-1); T4 – boscalide (0.100 g L-1) + kresoxim-methyl (0.050 g L-1); T5 – pyraclostrobin
(0.100 g L-1); T6 – fluxapiroxade (0.058 g L-1) + pyraclostrobin (0.116 g L-1) and T7 –
methyram (1,100 g L-1) + pyraclostrobin (0.100 g L-1). The effect of treatments on development
was evaluated by plant height, leaf area, fresh and dry mass of leaves, stem and root. To verify
the photosynthetic efficiency, gas exchange, chlorophyll a fluorescence, pigment content,
nitrate reductase enzyme activity and carbohydrate content were evaluated. In order to verify
the effects of treatments in the antioxidative system, in the control of the disease and activation
of defense mechanisms to the pathogen, the area under the disease progress curve (AUDPC),
concentration of superoxide anion and hydrogen peroxide, activity of the enzymes superoxide
dismutase, ascorbate peroxidase, glutathione reductase, glutathione S-transferase, glutathione
peroxidase, catalase, phenylalanine ammonia-lyase, chitinase, β-1,3-glucanase, peroxidase,
polyphenoloxidase and lipoxygenase, in addition to the phenolic compounds content, lipid
peroxidation and deposition of lignin. In order to evaluate the effects of the treatments on fruit
production and post harvesting, fruit firmness, pH, soluble solids, titratable acidity, ratio solid
silts/titratable acidity (SS/AT), ascorbic acid content, pectinametilesterase and
polygalacturonase enzyme activity, and mass loss. Based on the results obtained, it can be
concluded that the treatments fluxapiroxade plus pyraclostrobin, followed by metiram plus

xii
pyraclostrobin stood out from the other treatments, as they presented better plant development
and fruit production, due to the higher photosynthetic efficiency, higher enzyme activities of
antioxidative defense and activation of defense responses against the attack of the pathogen.
Key words: Solanum lycopersicum L., Alternaria solani, oxidative stress, proteins related to
pathogenesis, enzymes.

1
1. INTRODUÇÃO
O tomate está entre as hortaliças de maior relevância econômica e social no mundo e
seu cultivo se destaca como um dos principais geradores de empregos (FONTENELLE et al.,
2011; GRIGOLLI et al., 2011). O tomateiro (Solanum lycopersicum L.) é a segunda solanácea
mais cultivada, com produção estimada de 3,79 milhões de toneladas para a safra 2017 (IBGE,
2017). É cultivado nas mais diversas regiões brasileiras, abrangendo locais com diferentes
características climáticas, além de diversos sistemas de produção. É um dos frutos de hortícolas
mais consumidos no mundo, contribuindo para uma dieta saudável e equilibrada, por serem
ricos em minerais, vitaminas, aminoácidos essenciais, açúcares e fibras dietéticas, além de
conter grandes quantidades de vitaminas B e C, ferro e fósforo (NAIKA et al., 2006).
Em função da crescente expansão do cultivo surgiram vários problemas fitossanitários,
entre eles, muitas doenças fúngicas, incluindo a pinta preta, causada pelo patógeno Alternaria
solani. Esta é uma doença de ocorrência mundial e altamente agressiva, capaz de provocar
danos severos na cultura do tomateiro (DOMINGUES et al., 2009; FONTENELLE et al., 2011;
SONG et al., 2011).
Entre os métodos de controle utilizados a aplicação de fungicidas é a mais frequente
(FREITAS FILHO et al., 2008). Estes produtos químicos protegem a planta contra o ataque do
patógeno (REIS et al., 2007), podendo apresentar ação protetora ou sistêmica. Entre os
fungicidas sistêmicos registrados para o controle da pinta preta em tomateiro, encontram-se as
estrobilurinas e carboxamidas. Entretanto, estas substâncias, além da ação fungitóxica, também
proporcionam efeitos fisiológicos positivos nas plantas.
O efeito fisiológico destas moléculas tem sido estudado em diferentes culturas como
soja, feijão, milho, banana, tomate, pepino, alface, melão e videira (CARRIJO, 2014;
DEMANT; MARINGONI, 2012; LIMA et al., 2009; LIMA et al., 2012; JACOBELIS
JUNIOR, 2015; AMARO et al., 2018; MANSANO, 2014; MACEDO, 2015; MAREK, 2016),
respectivamente. De acordo com os mesmos autores, entre os principais resultados observados,
destacam-se maiores acúmulos de biomassa e produtividade, incremento fotossintético,
aumento da atividade da enzima nitrato redutase, maiores teores de pigmentos fotossintéticos e
redução do estresse oxidativo. Além de melhorar a qualidade pós-colheita de frutos de tomate
e melão (RAMOS et al., 2013; MACEDO et al., 2017).

2
Estes estudos evidenciam os efeitos fisiológicos no crescimento, desenvolvimento e
metabolismo fotossintético da planta, culminando em maior produtividade. No entanto, também
são observados efeitos no sistema antioxidativo da planta e, desta forma, podendo inferir
possíveis ações nas respostas de defesa da planta. Pois, a produção de espécies reativas de
oxigênio (EROs), como o peróxido de hidrogênio (H2O2), tem papel essencial nos mecanismos
de defesa do vegetal, além de atuar como sinalizador no processo de indução de resistência
(LEHMANN et al., 2015).
O fenômeno da indução de resistência em plantas se caracteriza pela ativação do arsenal
de defesa, que inclui barreiras físicas e bioquímicas, que a priori estariam latentes e após
expostas ao agente eliciador ou patógeno são induzidas desencadeando uma sequência de
eventos, ocorrendo maior síntese de compostos de defesa (SONG et al., 2015). Devido a isso,
a planta aumenta sua capacidade de atrasar ou evitar a entrada e, a subsequente, atividade de
um agente agressor em seus tecidos.
Assim, a aplicação de fungicidas formulados a base de estrobilurinas e carboxamidas,
além de controlarem doenças e proporcionarem os efeitos fisiológicos já descritos, também
podem ativar respostas de defesa em plantas de tomateiro. Isto possibilitaria maior controle do
patógeno e de outros possíveis agentes estressores, pois a resistência apresenta sistematicidade,
que conforme Hammerschmidt et al. (2001), a proteção não se limita apenas às partes da planta
que receberam o tratamento, mas se estende para aquelas não tratadas, além de agir de forma
não específica, podendo atuar sobre um ou vários patógenos. Portanto, o uso destes produtos se
torna uma ferramenta importante para o produtor no manejo de controle de pinta preta em
tomateiro, pois auxiliam na melhora do metabolismo das plantas incrementando a
produtividade, além de ativar o arsenal de defesa da planta.

3
2. OBJETIVOS
2.1 Objetivo Geral
Avaliar os efeitos fisiológicos e metabólicos em plantas de tomateiro híbrido
Conquistador, cultivadas em casa de vegetação e inoculadas com Alternaria solani, pela
aplicação de boscalida, boscalida mais cresoxim-metílico, piraclostrobina, fluxapiroxade mais
piraclostrobina e metiram mais piraclostrobina.
2.2 Objetivos específicos
Por meio da aplicação dos diferentes fungicidas já citados, em plantas de tomateiro
inoculadas com A. solani, ao longo do ciclo da cultura objetivou-se:
1 - Avaliar os efeitos no desenvolvimento de plantas de tomateiro, bem como no
acúmulo de biomassa;
2 - Analisar os efeitos na taxa de assimilação de CO2, taxa de transpiração, eficiência
do uso da água, eficiência de carboxilação, fluorescência da clorofila a, teor de pigmentos
fotossintéticos, atividade da enzima nitrato redutase, concentração e translocação de
carboidratos em plantas de tomateiro;
3 - Verificar os efeitos na atividade das enzimas antioxidativas e as respostas na
peroxidação lipídica em plantas de tomateiro;
4 – Avaliar o controle da doença pinta preta em plantas de tomateiro;
5 - Mensurar os efeitos na atividade das proteínas relacionadas à patogênese e a ativação
das respostas de defesa em plantas de tomateiro e
6 - Investigar os efeitos na produção e qualidade pós-colheita de frutos de tomateiro.

4
3. REFERENCIAL TEÓRICO
3.1 Aspectos gerais do cultivo de tomateiro
3.1.1 Origem
O centro de origem do tomateiro é a região andina, desde o Equador, passando pela
Colômbia, Peru, Bolívia, até o norte do Chile. Sua domesticação ocorreu no México, sendo os
espanhóis e portugueses os responsáveis pela difusão do tomate pelo mundo, através de suas
colônias ultramarinas. Chegou ao Brasil no século XIX, por imigrantes europeus que,
difundiram e incrementaram o consumo do tomate após a Primeira Guerra Mundial (NAIKA et
al., 2006; ALVARENGA, 2013).
3.1.2 Características botânicas e agronômicas
O tomateiro, cujo nome científico é Solanum lycopersicon, nomenclatura proposta por
Linnaeus em 1753, pertencente à família Solanaceae, que inclui outras espécies conhecidas,
como a batata, o tabaco, as pimentas e a berinjela (NAIKA et al., 2006). O tomateiro é uma
planta de porte arbustivo cultivado anualmente, podendo apresentar crescimento determinado
ou indeterminado conforme a variedade. Desenvolvem-se bem em diferentes latitudes, tipos de
solo, temperaturas e métodos de cultivos (ALVARENGA, 2013).
Apresenta sistema radicular constituído de raiz principal, raízes secundárias e
adventícias, podendo chegar a profundidades de 1,5 m, porém cerca de 70% das raízes se
encontram a menos de 20 cm da superfície. A parte aérea da planta é constituída por uma haste
principal, as folhas são alternadas, compostas com um grande folíolo terminal e cerca de 6 a 8
folíolos totais (FILGUEIRA, 2005).
A cada três folhas surge uma haste floral. A floração é influenciada por fatores como a
cultivar, temperatura, luminosidade, nutrição mineral e pela relação entre outros órgãos da
planta, assim como o efeito de reguladores vegetais (NAIKA et al., 2006).
O fruto é uma baga carnosa e suculenta, bi, tri ou plurilocular, que irá determinar o
tamanho final do fruto. As sementes são reniformes, pequenas, apresentando pequenos pelos e
de coloração marrom-clara. Um grama contém cerca de 300 sementes, sendo que um fruto pode
ter de 50 a 200 sementes. O desenvolvimento adequado da planta depende de fatores como a
cultivar, luminosidade, temperatura, nutrição, disponibilidade de água e concentração de CO2
(FILGUEIRA, 2005).

5
De acordo com Alvarenga (2013) para o cultivo de tomate encontram-se disponíveis
cultivares ou híbridos com características de hábito de crescimento indeterminado ou
determinado, caracterizados pelos segmentos de mesa e de indústria. As cultivares de hábito de
crescimento indeterminado, se destacam para a produção de frutos de mesa (consumo in natura)
e seu manejo incluem o tutoramento e podas, podendo também ser realizado o raleio de frutos
(NAIKA et al., 2006).
Quanto aos fatores climáticos, o tomateiro apresenta faixas de temperaturas ótimas
conforme o estádio de crescimento e desenvolvimento. Segundo Alvarenga (2013), para a
germinação está entre 18 a 24ºC, para a formação de mudas de 20 a 25ºC, florescimento de 18
a 24ºC, fixação de frutos de 14 a 17ºC durante a noite e de 19 a 24ºC durante o dia, e na fase de
maturação de 20 a 24ºC.
A cultura é muito exigente em água nas fases de desenvolvimento e produção. Quanto
ao fotoperíodo as plantas se desenvolvem bem entre nove e 15 horas de luminosidade diária. A
umidade relativa do ar tem efeitos indiretos, principalmente no que diz respeito à multiplicação
de fungos e bactérias fitopatogênicas (NAIKA et al., 2006).
3.1.3 Qualidade pós-colheita de frutos de tomateiro
A produtividade e a qualidade pós-colheita são reflexos do manejo adotado durante
todo o ciclo da cultura. Frutos com melhor aparência visual influenciam a escolha do
consumidor, quanto ao tamanho, formato e defeitos externos do fruto. De tal forma, que no
supermercado somente os frutos que correspondem às expectativas do consumidor, tais como
cor, brilho e principalmente o tamanho têm especial relevância (HEINE et al., 2015).
A manutenção das características e a qualidade pós-colheita do tomate de mesa, durante
o armazenamento até a comercialização, são fundamentais para preservar a aparência,
principalmente a cor (atributo atrativo) e a qualidade quanto ao teor de açúcares, acidez, pH,
textura, sabor e suculência (FERREIRA et al., 2010).
O tomate caracteriza-se por ser um fruto climatérico. O fruto maduro é o resultado de
inúmeras alterações bioquímicas e fisiológicas que ocorrem nos estádios finais do
desenvolvimento do fruto, resultando em um tomate palatável e atrativo. Entre estas alterações
ocorrem modificações no metabolismo de ácidos orgânicos, açúcares, cor, atividade de enzimas
pectinolíticas e firmeza (TAIZ; ZEIGER, 2013).

6
O fruto do tomateiro é considerado saboroso, quando apresenta a proporção de sólidos
solúveis/acidez titulável (SS/AT) superior a 10 (KADER et al., 1978). Dentre as características
relacionadas à qualidade dos frutos de tomate, os teores de sólidos solúveis, pH e acidez
titulável, são características químicas que conferem o sabor ao fruto e, que também são
perceptíveis pelo consumidor (HEINE et al., 2015).
Em frutos de tomate, os açúcares são os maiores constituintes dos sólidos solúveis e são
medidos em °Brix (medida indireta da quantidade de açúcar no fruto). Desta forma, frutos com
maior conteúdo de °Brix estão relacionados à maior síntese e acúmulo de açúcares durante seu
desenvolvimento, sendo reflexo do manejo adotado na cultura (HEINE et al., 2015). A acidez
em frutos é atribuída à presença dos ácidos orgânicos. Em tomate o ácido orgânico
predominante é o ácido cítrico, correspondendo 90% da acidez titulável (SIMANDLE et al.,
1966).
As modificações na firmeza do fruto são processos desencadeados pela atividade das
enzimas pectinametilesterase (PME) e poligalacturonase (PG), que resultam no amaciamento
do fruto (menor firmeza). A ação da enzima PME é catalisar substâncias pécticas, hidrolizando
grupos metil-éster, produzindo pectinas de menor grau de metilação que servirão de substrato
para a PG. A atuação destas causam mudanças na textura do fruto, pela degradação das pectinas
(SAINZ; VENDRUSCOLO, 2015). No entanto, menor atividade destas enzimas, para frutos de
tomate, permite maior firmeza após o amadurecimento, conferindo ao fruto maior resistência a
impactos físicos sofridos durante o transporte destes até o consumidor final (HEINE et al.,
2015).
3.2 A importância econômica da cultura do tomateiro
O tomateiro é uma das principais hortaliças, sendo a segunda solanácea mais cultivada
no mundo, superada apenas pela produção de batata. Devido a sua flexibilidade na utilização
como alimento, por suas qualidades organolépticas e ao alto teor de vitamina C, tem aceitação
por grande parte dos consumidores (ALVARENGA, 2013).
O aumento do consumo é o principal fator para a expansão da produção de tomate.
Conforme os dados do IBGE (2017), a produção brasileira de tomate possui uma estimativa de
3,79 milhões de toneladas na safra de 2017. A produção está distribuída ao longo de todo país,
sendo a região Sudeste responsável por 45% do total produzido no país, seguida pelas regiões

7
Centro-Oeste, Nordeste e Sul 20,9%, 20,3% e 13,0%, respectivamente, sendo que o Paraná tem
uma participação na produção de 4,5%.
Dentro do agronegócio brasileiro, segundo Rodrigues (2012) o setor da horticultura tem
se destacado como um forte gerador de renda e trabalho para muitas famílias. Estima-se um
total de 700 mil hectares cultivados com geração de cerca de 300 mil empregos em toda a cadeia
produtiva.
Entretanto, a cultura do tomate possui inúmeras doenças fitossanitárias, que podem
acometer a planta ao longo do ciclo, o que a torna uma hortaliça de elevado nível de risco
econômico (ALVARENGA, 2013).
3.3 Ocorrência de doenças
A cultura do tomateiro está sujeita a muitas doenças que causam prejuízos significativos.
Grande parte das doenças é controlada eficientemente a partir de um adequado programa de
manejo, integrando o uso de variedades resistentes e a adoção de medidas de exclusão,
erradicação e proteção da planta contra agentes fitopatogênicos (KUROZAWA; PAVAN,
2005).
Conforme Alvarenga (2013) as doenças que dizimam os tomateiros podem ser causadas
por agentes fúngicos, viróticos, bacterianos, nematoides ou até mesmo doenças fisiológicas.
Entre essas doenças, uma das principais é a pinta preta, causada pelo fungo Alternaria solani,
com ocorrência em todas as regiões onde o tomateiro é cultivado.
3.3.1 Pinta preta do tomateiro: aspectos gerais da doença
O agente causal da pinta preta, Alternaria solani (Ellis; Martin) L.R. Jones; Grout, foi
descrito pela primeira vez em folhas de beterraba em New Jersey em 1882 e a sua
patogenicidade foi estabelecida por volta de 1896. Este agente patológico pertence ao grupo
dos fungos da classe Deuteromycetes, ordem Moniliales e Família Dematiaceae (VALE et al.,
2000).
O fungo se caracteriza por possuir micélio septado e ramificado. Os conidióforos
possuem 12-20 x 120-296 μm, são simples, septados, longos, sub-hialinos a escuros com
conídios terminais. Estes são multicelulares, com septos transversais e longitudinais, clavados
com uma das extremidades pontiagudas, com ou sem apêndice (VALE et al., 2000;
KUROZAWA; PAVAN, 2005).

8
A infecção por A. solani ocorre, principalmente, em condições de temperatura e
umidade elevadas (FILGUEIRA, 2005). As temperaturas mínimas, ótimas e máximas
necessárias para a germinação dos conídios de A. solani são 5, 27 e 32º C, respectivamente
(SALUSTIANO et al., 2006). Desta forma, havendo umidade e calor suficiente, o fungo penetra
diretamente através da cutícula ou via estômatos após a formação de apressório. As lesões
tornam-se visíveis sob condições favoráveis entre 48 a 120 horas após a inoculação. A
ocorrência de epidemias severas desse patógeno está associada a temperaturas diárias de 25 a
32ºC (KUROZAWA; PAVAN, 2005).
O fungo é disseminado facilmente pelo vento e pode sobreviver entre um cultivo e outro,
em restos de culturas infectadas, podendo ainda sobreviver em equipamentos agrícolas, estacas
e caixas usadas para o transporte de frutos. Além do tomateiro, o fungo A. solani infecta outras
solanáceas como batata, berinjela, pimentão e jiló (VALE et al., 2000; KUROZAWA; PAVAN,
2005; ALVARENGA, 2013).
Muitas espécies patogênicas do gênero Alternaria são produtoras de toxinas prolíficas,
que facilitam a sua vida necrotrófica. Estes patógenos necessitam matar as células hospedeiras
antes da colonização e, portanto, são as toxinas secretadas que facilitam a morte da célula
causando danos ao tecido, resultando em necrose da área lesionada (LAWRENCE et al., 2008).
Os fungos fitopatogênicos são capazes de produzir grande número de enzimas
hidrolíticas extracelulares, as quais vêm sendo estudadas em A. solani, sendo que alguns autores
já detectaram atividades aminolíticas, pectinolíiticas e celulolíticas. Resultados obtidos a partir
das atividades aminolítica e pectinolítica de 45 isolados de A. solani, provenientes de diferentes
hospedeiros, revelaram possível envolvimento na patogênese. As diferenças nestas atividades
enzimáticas extracelulares contribuíram para diagnosticar a especificidade de hospedeiro
(MARCHI et al., 2006).
Este perfil enzimático é inter ou intra especificamente variável (MAHMOUD; OMAR,
1994), colaborando para a diferenciação de espécies ou de isolados dentro de uma mesma
espécie. Durante a infecção, segundo Agrios (2004), as enzimas extracelulares parecem
favorecer a penetração, colonização, bem como a obtenção de nutrientes e/ou contribuir nas
reações de defesa da planta.
Devido à forte agressividade com que se desenvolve, a pinta preta provoca perdas diretas
por infectar os frutos e indiretas pela redução do vigor da planta, reduzindo o crescimento

9
devido ao decréscimo da fotossíntese, pelos danos a área fotossinteticamente ativa, além da
ocorrência da desfolha em estágios mais avançados da doença (ALVARENGA, 2013).
A doença apresenta alto potencial destrutivo, incidindo sobre folhas, hastes, pecíolos e
frutos do tomateiro, causando danos em qualquer estádio de desenvolvimento da planta. Toda
parte aérea da planta pode apresentar sintomas da doença, independentemente da idade, mas há
maior tendência de lesões aparecerem, com maior frequência, nas folhas mais velhas
(KUROZAWA; PAVAN, 2005).
Nestas folhas a doença caracteriza-se pela presença de manchas pequenas de cor
marrom-escura, circundadas ou não de um halo amarelado. À medida que a lesão cresce,
formam-se anéis concêntricos, bastante característicos desta doença (FREITAS FILHO et al.,
2008). Quando não é controlada adequadamente há severa destruição da área foliar e, nos
frutos, verifica-se podridão deprimida, circular, próxima ao pedúnculo, coberta por mofo preto
(ALVARENGA, 2013).
Conforme avaliações realizadas por Paula e Oliveira (2003), notou-se que a partir da
quarta semana após o transplante ocorreu a maior incidência de pinta preta, tanto em tomateiros
em sistema rasteiro como no tutorado, acentuando-se gradativamente em algumas cultivares.
Segundo Töfoli e Domingues (2005) o aumento de suscetibilidade à pinta preta está associado
aos tecidos maduros, sendo mais frequente durante a fase de frutificação.
Condições ideais para a ocorrência de epidemias severas também foram observadas por
Salustiano et al. (2006) na região de Viçosa, onde observaram que nas épocas de plantio, em
que predomina temperaturas entre 20 e 30ºC, com presença de molhamento foliar e/ou chuva,
favorece consideravelmente a ocorrência da pinta preta. Entretanto, em épocas de temperaturas
amenas (≤ 20ºC), os mesmos autores relataram que, mesmo na presença de molhamento foliar
e/ou chuvas, a curva de progresso da doença se desenvolveu lentamente, com severidades de
20 a 30% ocorrendo somente ao final do ciclo da cultura, quando a produção já havia atingido
seu máximo.
3.3.2 Controle de pinta preta em tomateiro
Diversos métodos de controle têm sido utilizados no combate das principais doenças
fúngicas do tomateiro. Para o controle de pinta preta são recomendados métodos culturais,
como sementes de boa procedência, rotação de cultura, eliminação de restos culturais,
espaçamento adequado à boa aeração e adubação equilibrada (KUROZAWA; PAVAN, 2005;

10
FREITAS FILHO et al., 2008). Outra opção viável e econômica é a utilização de variedades
resistentes, podendo reduzir a dependência de fungicidas (RAO et al., 2007).
Porém, o controle químico ainda é o método mais utilizado, o qual pode ser feito com
fungicidas protetores e/ou curativos indicados para a cultura (FREITAS FILHO et al., 2008).
De acordo com Reis et al. (2007) fungicidas são produtos químicos aplicados em plantas para
promover a proteção contra a penetração e/ou posterior desenvolvimento dos patógenos,
podendo ser classificados, de maneira geral, como fungicidas de contato ou sistêmico.
Fungicidas sistêmicos, como as estrobilurinas e carboxamidas que são alguns dos
fungicidas registrados para o controle da pinta preta na cultura do tomateiro, são absorvidos
pelas folhas e translocados pelo sistema condutor da planta, proporcionando uma ação protetora
prolongada (REIS et al., 2010). Fungicidas do grupo das estrobilurinas, como a piraclostrobina
e o cresoxim-metílico, atuam na inibição da respiração mitocondrial, que bloqueia a
transferência de elétrons entre o citocromo b e o citocromo c1, interferindo na produção de ATP
e na síntese de energia do patógeno (KANUNGO; JOSHI, 2014; MATOS et al., 2016).
Pertencente ao grupo químico das carboxamidas, o fungicida boscalida vem sendo uma
nova alternativa para o manejo da pinta preta na cultura do tomateiro. Este princípio ativo tem
ação preventiva e curativa, atuando na germinação de esporos, alongamento do tubo
germinativo, crescimento micelial e esporulação de uma ampla variedade de fungos (EFSA,
2015). Sendo classificado como inibidor da respiração na célula fúngica atuando sobre o
complexo II (TÖFOLI, 2004), semelhante ao fluxapiroxade, outra carboxamida. Ambos atuam
na respiração mitocondrial, bloqueando a transferência de elétrons no complexo II, inibindo a
enzima succinato dehidrogenase (SDHI), reduzindo o processo respiratório e bloqueando o
fornecimento de energia das células do fungo (FRAC, 2017).
No entanto, pesquisas têm evidenciado que além da ação fungitóxica destes produtos
formulados a base de estrobilurinas e carboxamidas têm sido relatados efeitos simultâneos na
fisiologia de plantas (DIAZ-ESPEJO et al., 2012; LIMA et al., 2012; DEMANT;
MARINGONI, 2012; RAMOS et al., 2013; COLOMBARI et al., 2015; JADOSKI et al., 2015;
MAREK, 2016; MACEDO et al., 2017; AMARO et al., 2018). A ação extra destes produtos é
importante, pois as práticas agrícolas buscam por manejo que aumentem a eficiência
fotossintética das culturas, e consequentemente, seus rendimentos (BRANDÃO FILHO et al.,
2003).

11
3.4 Efeitos fisiológicos das estrobilurinas e carboxamidas
O uso de fungicidas apenas como agente de controle de fitopatógenos tem mudado.
Segundo Venancio et al. (2003) o lançamento das estrobilurinas mudou o conceito de controle
de doenças, sobretudo pelas vantagens obtidas com a ação destes produtos no metabolismo
vegetal promovendo efeitos fisiológicos positivos nas plantas.
A aplicação de estrobilurinas ativam diferentes rotas metabólicas nos vegetais. Estudos
têm demonstrado que ocorre a ativação da enzima NADH-nitrato redutase, aumento da
assimilação de nitrato e sua posterior incorporação nas moléculas vitais da planta, como a
clorofila. Além disso, ocorre o aumento da eficiência de assimilação de CO2, elevação da taxa
fotossintética e redução da taxa respiratória. Outro efeito promissor é a redução na produção de
etileno, que retarda a senescência das folhas, aumentando o período que a planta permanece
com a fotossíntese ativa, também chamado de ‘efeito verde’ (VENANCIO et al., 2003;
TÖFOLI; DOMINGUES, 2007; FAGAN et al., 2010), repercutindo consequentemente em
aumentos significantes no rendimento das culturas.
A partir dos efeitos fisiológicos observados com o uso das estrobilurinas, estudos com
outras moléculas como boscalida e fluxapiroxade, também têm sido realizados para avaliar seu
potencial no metabolismo e desenvolvimento das plantas. O efeito fisiológico destas moléculas
já foi relatado em diferentes culturas como soja (TSUMANUMA et al., 2010; FAGAN et al.,
2010; SOARES et al., 2011; CARRIJO, 2014), feijão (KOZLOWSKI et al., 2009; DEMANT;
MARINGONI, 2012; JADOSKI et al., 2015), milho (LIMA et al., 2009), banana (LIMA et al.,
2012), tomate (RAMOS et al., 2013; JACOBELIS JUNIOR, 2015), pepino (SIRTOLI et al.,
2011; AMARO, 2011; AMARO et al., 2018), alface (MANSANO, 2014), melão (MACEDO,
2012; 2015; MACEDO et al., 2017), cenoura (COLOMBARI et al., 2015) e videira (MAREK,
2016).
Entre os principais resultados observados se destacam maiores acúmulos de matéria seca
e produtividade (LIMA et al., 2009; KOZLOWSKI et al., 2009; TSUMANUMA et al., 2010;
AMARO, 2011; DEMANT; MARINGONI, 2012; COLOMBARI et al., 2015; JADOSKI et al.,
2015; MAREK, 2016), incremento na taxa fotossintética (FAGAN et al., 2010; CARRIJO,
2014; MANSANO, 2014; AMARO et al., 2018), aumento da atividade da enzima nitrato
redutase e efeito verde (SOARES et al., 2011; MACEDO, 2012; 2015; RAMOS, 2013),
alterações hormonais e atraso na senescência (FAGAN et al., 2010), maior qualidade na pós-
colheita em frutos (RAMOS et al., 2013, MACEDO et al., 2017), formação de óxido nítrico e

12
ativação de respostas de defesa (VENANCIO et al., 2003) e menor estresse oxidativo nas
plantas (AMARO, 2011; MACEDO, 2012; 2015, JACOBELIS JUNIOR, 2015; JADOSKI et
al., 2015; AMARO et al., 2018).
3.5 Desenvolvimento de plantas
As plantas crescem, se desenvolvem e produzem frutos graças ao processo
fotossintético, pelo qual produzem fotoassimilados, a partir do sol, que emite radiação em
diferentes comprimentos de onda (KLUGE et al., 2014). Avaliar o crescimento e
desenvolvimento de plantas é inferir sobre a sua capacidade assimilatória em sintetizar e alocar
os fotoassimilados nos diferentes órgãos. Desta forma, plantas que apresentam maior área
foliar, possuem maior capacidade de aproveitar a energia em prol da fotossíntese, apresentando
melhor desenvolvimento e ganhos de produtividade (REIS et al., 2013).
Unindo maior área fotossintética ativa com alto teor de pigmentos fotossintéticos têm-
se o maior aproveitamento da energia solar para a síntese de compostos orgânicos
(RODRIGUES et al., 2016). Aproximadamente 90% da massa seca de uma planta é formada a
partir dos produtos oriundos da fotossíntese (fotoassimilados), portanto, a planta utiliza uma
parte do que produz para crescer, convertendo em biomassa, e outra parte oxida na respiração
como fonte de energia para seu metabolismo. Sendo assim, a fotossíntese líquida (fotossíntese
bruta menos a respiração) é um indicador da eficiência fotossintética das plantas em assimilar
o CO2 e converter em compostos orgânicos (POPOV et al., 2003).
As plantas realizam fotossíntese durante algumas horas por dia (± 4 horas dia-1) e
respiram durante 24 horas, consumindo considerável parte dos compostos que produziram
durante o período de fotossíntese (informação verbal)1. Desta forma, para se obter maior
produção de frutos em uma planta, é necessário aumentar a fotossíntese líquida através do
aumento da fotossíntese bruta e redução da respiração (KLUGE et al., 2014).
3.6 Eficiência fotossintética
3.6.1 Trocas gasosas
Estudar a fisiologia da planta, no que diz respeito à eficiência fotossintética, visando
ganhos de produtividade tornou-se uma importante estratégia no manejo da cultura. De acordo
1 Rodrigues, J. D. (Professor Titular, Depto. Botânica – IB – UNESP – Botucatu. Comunicação pessoal, 2017.
e-mail: [email protected]

13
com Macedo (2015) as avaliações de trocas gasosas são importantes ferramentas para se avaliar
a capacidade fotossintética das plantas, além da vantagem de ser um método não destrutivo que
permite a análise qualitativa e quantitativa da fotossíntese na planta.
A produção de biomassa depende da eficiência fotossintética dos órgãos fontes, com
forte influência sobre o desenvolvimento do vegetal (FOYER; GALTIER, 1996). A fotossíntese
líquida, de acordo com Popov et al. (2003) indica a eficiência da taxa de assimilação de CO2.
Desta forma, mensurar estes parâmetros fisiológicos em plantas de tomateiro, visando
ganhos de produtividade, é uma ferramenta importante para se avaliar as taxas de assimilação
de CO2, de transpiração, condutância estomática e concentração interna de CO2 nas folhas, além
de determinar as relações entre estas, obtendo a eficiência do uso da água e eficiência de
carboxilação (BRANDÃO FILHO et al., 2003).
3.6.2 Pigmentos fotossintéticos e fluorescência da clorofila a
A eficiência fotossintética de uma planta está fortemente associada à quantidade de
clorofilas presentes no tecido foliar. De acordo com Rodrigues et al. (2016) o aumento nos
teores de clorofilas resulta em maior absorção da luz solar e como resultado, ocorre maior
transporte de elétrons nos fotossistemas durante a fase fotoquímica da fotossíntese. A absorção
da luz ocorre nos fotossistemas I e II (PSI e PSII), sendo estes constituídos de pigmentos
fotossintéticos como as clorofilas a e b que estão presentes nas membranas dos tilacóides
(CASIERRA-POSADA; PEÑA-OLMOS, 2015).
Os pigmentos fotossintéticos são compostos orgânicos que absorvem a energia
proveniente da radiação solar no espectro de luz visível (400 a 700 nm). No entanto, a luz possui
duas importantes características físicas, além de onda eletromagnética ela também transporta
uma quantidade de energia (fóton) (TAIZ; ZEIGER, 2013). A absorção desta energia causa uma
instabilidade na molécula de clorofila, tornando-a excitada. Uma vez na forma instável, a
clorofila precisa retornar ao seu estado basal de energia e para isto precisa dissipar esta energia
na forma de calor e/ou como fluorescência e/ou utilizando nas reações fotoquímicas
(CAMPBELL; FARRELL, 2006).
Pensando em maior eficiência fotossintética na produção vegetal, a via fotoquímica é a
mais importante, pois é nesta que ocorre a fixação do CO2 em uma molécula orgânica pela
enzima Ribulose-1,5-bifosfato carboxilase/oxigenase (Rubisco) que atua no ciclo de Calvin
(TAIZ; ZEIGER, 2013). Para que ocorra a síntese dos compostos orgânicos é necessário

14
primeiramente a produção de ATP e NAPH2, formados durante o transporte de elétrons entre o
PSII e o PSI. Sendo justamente neste processo, que os pigmentos fotossintéticos atuam
eficientemente convertendo a energia solar em energia química nos centros de reação (YEOM
et al., 2014).
No entanto, plantas quando submetidas a condições de estresse abiótico ou biótico,
sofrem alterações no equilíbrio das reações fotossintéticas que acabam causando danos ao
aparato fotossintético. A planta reflete este dano por meio da concentração de pigmentos ou
pela maior emissão de fluorescência da clorofila a (BRESTIC; ZIVCAK, 2013).
A avaliação da fluorescência da clorofila a também é uma técnica não destrutiva que
permite a análise qualitativa e quantitativa da absorção e aproveitamento da energia luminosa
pelo aparato fotossintético, revelando em especial a ‘saúde’ e eficiência dos fotossistemas I e
II (KRAUSE; WEIS, 1991), além de permitir avaliar a taxa de transporte de elétrons (ETR)
entre os centros de reação. Desta forma, evidenciando a qualidade de organização e a eficiência
fotossintética, refletindo no status da planta naquele momento, identificados pelos parâmetros
avaliados (BAKER, 2008; BRESTIC; ZIVCAK, 2013).
3.6.3 Nitrato redutase
Altas atividades da enzima nitrato redutase estão relacionadas a altas produtividades nas
culturas, uma vez que esta é a enzima chave na regulação do metabolismo do nitrogênio
(VIANA; KIEHL, 2010). O nitrogênio participa diretamente do metabolismo da planta, sendo
importante constituinte da molécula de clorofila, ácidos nucleicos, aminoácidos e proteínas
(TAIZ; ZEIGER, 2013).
O nitrogênio, dentre os elementos minerais essenciais para a planta, se destaca como um
dos principais componentes da biomassa. Assim, para suprir suas necessidades, as plantas
absorvem o nitrogênio da solução do solo nas formas de nitrato e amônio, sendo que o nitrato
é a principal forma inorgânica do elemento e sua disponibilidade está ligada diretamente ao pH
do solo e a relação C/N (ERNANI, 2016).
O nitrato (NO3) disponível na solução do solo, após absorvido pelas raízes precisa passar
pelo processo de redução, que ocorre no citoplasma por intermédio da enzima nitrato redutase,
reduzindo nitrato à nitrito (NO2), usando o NADH como doador de elétrons. Na sequência, o
nitrito é transportado até os cloroplastos onde é reduzido à amônia pela enzima nitrito redutase,

15
recebendo elétrons da ferredoxina. Posteriormente a amônia é incorporada em moléculas
orgânicas, entre estas as clorofilas a e b (TAIZ; ZEIGER, 2013).
Entre os efeitos fisiológicos proporcionados pelas estrobilurinas e, recentemente
observados para as carboxamidas, se destaca o aumento da atividade da enzima nitrato redutase.
De acordo com Glaab e Kaiser (1999) estes fungicidas atuam no metabolismo da planta
promovendo a queda nos níveis celulares de ATP, pois eles agem no transporte de elétrons
(como já descrito), desta forma a menor síntese de ATP aumenta a concentração de prótons
(H+) no citosol, este desequilíbrio é a chave propulsora para a ativação da enzima nitrato
redutase. Assim, quanto mais nitrogênio é absorvido e incorporado pela planta, maior é a
produção de compostos nitrogenados como os aminoácidos, que são as bases da biossíntese de
proteínas e, consequentemente, ocorre o aumento na biomassa das plantas.
3.7 Estresse oxidativo
As condições de estresse biótico e abiótico impostas às plantas (alta luminosidade,
extremos de temperatura, seca, salinidade, radiação UV, poluição do ar, herbicidas,
estresses físicos e mecânicos, ou bióticos como o ataque de patógenos) induzem a
superprodução de espécies reativas de oxigênio (EROs ou ROS, do inglês reactive oxygen
species), formadas durante funções metabólicas normais e causam danos oxidativos em
proteínas, lipídios e ácidos nucleicos, caracterizando o estresse oxidativo (SCANDALIOS et
al., 2000; FOYER; NOCTOR, 2003; D´AUTRÉAUX; TOLEDANO, 2007; BARBOSA et al.,
2014).
De acordo com Stangarlin et al. (2011) entre as principais EROs presentes no
metabolismo celular de plantas se destacam o ânion superóxido (•O2-), dotado de baixa
capacidade oxidativa; o peróxido de hidrogênio (H2O2) capaz de romper a membrana do núcleo
e causar danos ao DNA; o radical hidroxila (OH•), com baixa capacidade de difusão, porém alta
reatividade provocando lesões em uma série de moléculas em meio celular; e o oxigênio
singleto (1O2), formado a partir da transferência da energia de ativação para o O2, também
produzindo efeitos deletérios.
As EROs são formadas inevitavelmente nos organismos aeróbicos, em eventos
metabólicos que ocorrem em especial nas mitocôndrias, cloroplastos e peroxissomos. Quando
as plantas estão sob condições de estresse abiótico ou biótico ocorre o aumento na produção
destas espécies (BHATTACHARJEE, 2010; KARUPPANAPANDIAN et al., 2011).

16
A geração das EROs também pode ocorrer a partir de danos no sistema fotossintético
(SOARES; MACHADO, 2007), com a formação de oxigênio singleto (1O2). Quando a energia
armazenada na clorofila não é dissipada para a síntese de carboidratos, é então transferida para
o O2, promovendo o transporte pseudocíclico de elétrons (BHATTACHARJEE, 2010). Isto
ocorre devido à baixa fixação de CO2 e menor poder redutor (NADP+), a ferredoxina acaba
transferindo o elétron para o O2 formando •O2- (AHMAD et al., 2008).
As EROs também atuam na via de sinalização em resposta a situações de estresse nas
plantas, influenciando na expressão de vários genes envolvidos no metabolismo de defesa,
agindo como “moléculas sinalizadoras” ou “mensageiros secundários” (BARBOSA et al.,
2014; RECZEK; CHANDEL, 2015).
3.7.1 Enzimas envolvidas na defesa antioxidativa
De acordo com Moller et al. (2007) para combater os danos oxidativos das EROs, as
plantas dispõem de um sistema de defesa composto por mecanismos antioxidantes
enzimáticos e não enzimáticos, que trabalham em conjunto e em sincronia para
desintoxicar as células.
As defesas não enzimáticas incluem as vitaminas C e E, glutationa, β-caroteno,
compostos fenólicos, tocoferóis e poliaminas, ácido ascórbico, α-tocoferol e os carotenoides. O
sistema defensivo enzimático envolve as enzimas superóxido dismutase (SOD), catalases
(CAT), peroxidases (POD), glutationa peroxidase (GPX), ascorbato peroxidase (APX),
glutationa redutase (GR) e glutationa S-transferase (GST). Todos eles ocorrem em cloroplastos,
mitocôndrias e peroxissomos (SCANDALIOS, 2005; KIM; KWAK, 2010; DINAKAR et al.,
2012; NOCTOR et al., 2015; LEHMANN et al., 2015).
A SOD, uma metalo-enzima, é considerada a primeira linha de defesa contra os danos
causados pelas EROs, pois catalisa a dismutação do radical livre superóxido •O2-, gerando O2 e
H2O2 (GIANNOPOLITIS; RIES, 1977; ALSCHER et al., 2002; MITTLER, 2002;
BHATTACHARJEE, 2010).
A enzima SOD é classificada de acordo com seus cofatores metálicos: cobre e zinco
(Cu/Zn-SOD), manganês (Mn-SOD) e ferro (Fe-SOD) (GILL; TUTEJA, 2010). Geralmente as
plantas contêm uma Mn-SOD localizada na matriz mitocondrial e uma Cu/ZnSOD citosólica,
com Fe-SOD e/ou Cu/Zn-SOD presentes no estroma do cloroplasto. O

17
número de isoenzimas de cada tipo de SOD varia muito de planta para planta, assim
como a quantidade relativa de cada enzima (RESENDE et al., 2003; GILL et al., 2015).
A função principal CAT é prevenir os efeitos potencialmente danosos causados por
mudanças na homeostase de H2O2 (VAN BREUSEGEM et al., 2001; IGAMBERDIEV; LEA,
2002). As CATs são as principais enzimas na eliminação do H2O2 gerado durante a
fotorrespiração e a β-oxidação dos ácidos graxos. Ela converte duas moléculas de H2O2 em
água (H2O) e oxigênio molecular (O2) (DUBEY, 2010).
As plantas possuem várias isoformas de CAT, as quais podem dismutar
diretamente o H2O2 ou oxidar substratos, tais como metanol, etanol, formaldeído e ácido
fórmico (VAN BREUSEGEM et al., 2001; IGAMBERDIEV; LEA, 2002). A atividade da CAT
é efetiva, principalmente em concentrações relativamente altas de H2O2 (mM), por isso são
consideradas indispensáveis para a eliminação das EROs, especialmente em condições de
estresse severo, onde se tem níveis de H2O2 elevados (DUBEY, 2010).
Nas plantas também existem muitas isoformas de PODs, algumas são expressas
enquanto outras são induzidas em resposta ao estresse ambiental. As PODs localizam-se
principalmente na parede celular e no vacúolo, agem utilizando o H2O2 como oxidantes e
compostos de natureza fenólica como doadores de elétrons. São consideradas uma das mais
importantes na eliminação de H2O2 no citosol e nos cloroplastos. Inclusive o H2O2 formado
pela ação da SOD também pode ser eliminado pelas PODs, além da CAT e da APX (LOCATO
et al., 2010).
A enzima APX também é considerada importante na eliminação do H2O2 no citosol e
nos cloroplastos. A degradação do H2O2 ocorre pela utilização do ascorbato como substrato. O
ascorbato é o doador de elétrons específicos para reduzir o H2O2 à água, constituindo parte do
ciclo conhecido como ciclo da ascorbato-glutationa (INZÉ; MONTAGU, 1995; RESENDE et
al., 2003). Esta enzima é importante na defesa de tecidos fotossintéticos contra o estresse
fotoxidativo. A APX também é encontrada no citosol das células não fotossintetizantes atuando
na redução dos níveis de EROs (ASADA, 1999; RESENDE et al., 2003).
Concomitantemente, a enzima GPX catalisa a eliminação de H2O2 e atua na
desintoxicação dos produtos formados na peroxidação lipídica, em decorrência dos danos
oxidativos pela atividade das EROs (DIXON et al., 2010). Nas plantas são encontradas três
isoformas de GPXs, as dependentes de selênio, as hidroperoxidase de fosfolipídios não

18
dependente de selênio (PHGPXs) e glutationa S-transferases com atividade de glutationa
peroxidase (BELA et al., 2015).
Já a GR é uma enzima responsável por catalisar a redução da GSSG (glutationa oxidada)
para GSH (glutationa reduzida). A GR pode ser encontrada tanto em cloroplasto quanto em
mitocôndria, mas sua síntese ocorre no citoplasma da célula de onde é direcionada para as
organelas (HAYES; MCLELLAN, 1999). Esta enzima é encontrada em procariotos e
eucariotos, desde bactérias heterotróficas e fotossintetizantes até plantas e animais superiores
(FOYER et al., 1997).
A GR desempenha papel importante na defesa contra o estresse oxidativo, pois mantém
o equilíbrio entre os níveis de GSSG e GSH na célula. A GSH é uma molécula importante
dentro do sistema celular, participando inclusive do ciclo ascorbato-glutationa (NOCTOR et
al., 2002) ou ainda, da síntese de fitoquelatinas (GRATÃO et al., 2005).
Em parceria com as GRs, as GSTs são enzimas que catalisam a conjugação de GSH à
substratos hidrofóbicos eletrofílicos, como xenobióticos (DIXON et al., 2002). São proteínas
encontradas preferencialmente no citoplasma (MARRS, 1996). Várias classes de GST foram
identificadas em plantas respondendo aos mais diversos tipos de estressores bióticos ou
abióticos (DIXON et al., 2002). De acordo com Ahmad et al. (2010) enquanto a GPX elimina
o H2O2 formando H2O, competindo com a catalase pelo substrato e oxidando GSH, a GST
catalisa a adição de GSH, por isso são diferentes nas suas subunidades e mecanismo catalítico,
no entanto, apresentam funções complementares.
3.7.2 Peroxidação lipídica
As EROs têm funções biológicas definidas quando estão em concentrações fisiológicas
adequadas, atuando como mensageiros e/ou sinalizadores (BARBOSA et al., 2014). Entretanto,
em altas concentrações devem ser evitadas pela planta, considerando que sua reatividade
causará danos oxidativos em proteínas, lipídios e ácidos nucleicos, caracterizando o estresse
oxidativo (SCANDALIOS et al., 2000; FOYER; NOCTOR, 2003; RIBEIRO et al., 2005;
D´AUTRÉAUX; TOLEDANO, 2007; BARBOSA et al., 2014).
Como são moléculas altamente reativas, as EROs irão reagir com os lipídios
formadores de membranas ocorrendo assim, a peroxidação lipídica. A consequência
direta do dano da peroxidação de lipídios sobre as membranas celulares é o
extravasamento do conteúdo celular para o meio, por danificarem irreversivelmente as

19
membranas, além de formar novos radicais lipídicos (DAN HESS, 2000; KRUSE et al.,
2006).
Este efeito deletério das EROs sobre os ácidos graxos poli-insaturados dos
fosfolipídios das membranas celulares, resulta na desintegração destas, permitindo o
livre transporte de substâncias (tanto a entrada quanto a saída) do interior celular. As
EROs ativam as fosfolipases, desintegrando os fosfolipídios, liberando os ácidos
graxos não saturados, resultando em ruptura das membranas celulares (bombas de Na/K e
Ca/Mg), ocasionado danos ao DNA, oxidando lipídios insaturados, formando resíduos
químicos (ex.: malondialdeído) e comprometendo os componentes da matriz extracelular
(RIBEIRO et al., 2005). Além de afetar diretamente diversas moléculas essenciais como
pigmentos fotossintéticos, proteínas e ácidos nucléicos (FOYER et al., 1997), podendo ocorrer
a paralização do metabolismo celular (LIN et al., 2005).
Os lipídios insaturados da mitocôndria, membrana plasmática e de outras organelas são
os principais locais de deterioração oxidativa. Pois, os ácidos graxos insaturados
destas membranas possuem ligações fracas e de fácil rompimento, e tornam-se alvo da presença
excessiva de EROs, ocorrendo à remoção do hidrogênio destes lipídios (MISHRA et al., 2006).
O radical •O2- é um dos principais oxidantes responsáveis pela peroxidação de
lipídios, sua formação causa o aumento na permeabilidade das membranas no ambiente
celular (BREUSEGEM et al., 2001; APEL; HIRT, 2004). O uso de elicitores bióticos
ou abióticos, ou a presença de um patógeno também pode causar estresse oxidativo à
planta, levando a formação de altas concentrações de EROs e a ocorrência de peroxidação de
lipídios (ADAM et al., 1989; KEPPLER; BAKER, 1989).
A peroxidação de lipídios também atua como precursora para a síntese de ácido
jasmônico, por intermédio das enzimas lipoxigenases. O ácido jasmônico ativa a expressão de
vários genes relacionados à defesa durante as reações de hipersensibilidade na planta
(FARMER; RYAN, 1992).
3.8 Indução de resistência e a defesa vegetal contra patógenos
No ambiente de cultivo as plantas estão vulneráveis ao ataque de uma vasta gama de
patógenos. Entretanto, os vegetais dispõem de um sistema de defesa, que inclui barreiras físicas
e bioquímicas pré e pós-formadas, que atuam na proteção contra o ataque de fitopatógenos
(THAKUR; SOHAL, 2013).

20
Quando se refere à indução de resistência, se estuda principalmente as barreiras pós-
formadas, que a priori estariam latentes, e após expostas ao agente eliciador ou patógeno, são
induzidas e desencadeiam uma cascata de eventos que culminam em maior síntese dos
compostos de defesa (SONG et al., 2015).
O fenômeno da indução de resistência em plantas tem sido descrito como resistência
sistêmica adquirida (RSA) e resistência sistêmica induzida (RSI). Conforme Hammerschmidt
et al. (2001) este termo ‘sistêmico’ revela que a proteção não se limita apenas às partes da planta
tratada, mas se estende para as não tratadas e, muitas vezes, até naquelas em desenvolvimento.
Portanto, a resistência do hospedeiro a um patógeno é definida como a capacidade da planta em
atrasar ou evitar a entrada e a subsequente atividade deste em seus tecidos.
A indução de resistência ocorre tanto em nível de plasmalema quanto no interior da
célula da planta hospedeira, podendo ser desencadeada por diferentes agentes elicitores biótico
e abiótico (orgânicos ou sintéticos) (OLIVEIRA et al., 2016). De tal forma, a resistência pode
ocorrer tanto após a infecção (pós-infeccional) ou induzida por um agente ou substância
elicitora (SHAH et al., 2014). No entanto, as substâncias envolvidas nesta defesa serão
sintetizadas somente após a ativação, evitando gasto energético desnecessário (STANGARLIN
et al., 2008).
De acordo com Edreva (2004) quando uma planta é inoculada com um agente
patogênico ("inoculação primária"), e após um intervalo de tempo é submetido a uma
inoculação secundária ("desafio"), observa-se redução no desenvolvimento dos sintomas da
doença. As defesas da planta são pré-condicionadas pela infecção anterior ou tratamento, que
resulta em resistência (ou tolerância) contra o subsequente desafio por um patógeno (RAGAB
et al., 2009).
Como já dito, a indução é a ativação de mecanismos latentes que passam a se expressar
após a exposição da planta ao eliciador. O reconhecimento precoce do patógeno pela planta é
muito importante, uma vez que ela pode mobilizar a tempo as defesas bioquímicas e estruturais
disponíveis para se proteger do patógeno. Quando uma planta é levada ao estado de indução,
seja por eliciadores bióticos ou abióticos, não se trata de um caso de imunidade, pois a doença
pode acontecer, porém em menor intensidade (AGRIOS, 2004).
Durante a interação-planta patógeno ocorre a produção de EROs, que inicia com uma
explosão oxidativa primária, seguida de micro explosões secundárias, culminando no
acionamento dos mecanismos de defesa antioxidante (BHATTACHARJEE, 2010) e atuando

21
na sinalização intracelular de respostas fisiológicas e biológicas (RECZEK; CHANDEL, 2015).
Além disto, o contato entre um eliciador e os tecidos da planta também desencadeia a síntese
destas substâncias, que agem como sinais bioquímicos (BECKERS; SPOEL, 2006).
Entre as EROs, o H2O2 tem papel essencial nos mecanismos de defesa da planta, pois
este agente oxidante tem efeito tóxico agindo diretamente sobre as estruturas do patógeno, além
de se acumular no sítio de infecção restringindo o desenvolvimento da lesão (resposta de
hipersensibilidade – HR). O H2O2 também atua como sinalizador, aumentando a atividade da
enzima benzoico 2-hidrolase, precursora na síntese do ácido salicílico (hormônio crucial na
sinalização da RSA) (LEHMANN et al., 2015).
O H2O2 tem características importantes para o processo de indução de resistência. Esta
molécula apesar de alguns autores não a classificar como um radical livre, por não ter elétrons
desemparelhados na última camada (BARTOSZ, 1997; COTINGUIBA et al., 2015), apresenta
moderada reatividade, meia-vida relativamente longa (1 ms) e tamanho pequeno, características
que permitem ao H2O2 maior facilidade em atravessar membranas celulares e migrar para
diferentes locais, tendo atuação em regiões distantes do local em que foi produzido
(BHATTACHARJEE, 2010; BARBOSA et al., 2014), por isso é tão importante nos processos
de sinalização em condições de estresse.
Na sinalização desencadeada pelo processo de indução de resistência, de acordo com
Shah et al. (2014), podem ser ativadas diferentes rotas metabólicas dependentes do estímulo
inicial. Geralmente ocorre a ativação da rota via ácido salicílico em resposta à infecção por
patógenos biotróficos ou hemibiotróficos, enquanto que para patógenos necrotróficos são
ativadas rotas dependentes de etileno ou ácido jasmônico (THAKUR; SOHAL, 2013). Segundo
Walkvria (1992) a indução de resistência ocorre na planta hospedeira, tendo como efeito
secundário a inibição, bloqueio ou mesmo a morte do patógeno.
Além dos hormônios vegetais, o óxido nítrico (NO) também apresenta importante papel
na difusão da sinalização de respostas de defesa nos vegetais (GAO et al., 2015). Esses sinais
se difundem por toda a planta sistemicamente ou induzindo sua síntese em cadeia. Estudos
demonstram que os diferentes mecanismos de proteção na planta, se iniciam com as HRs, pelo
aumento das EROs e de NO. Os mecanismos regulatórios das EROs funcionam de forma
independente ou em cooperação com o NO, em resposta ao ataque de patógenos (BELLIN et
al., 2013).

22
Quando plantas são previamente expostas a um eliciador, seus tecidos reagem mais
rapidamente e com mais eficiência às tentativas de colonização por um patógeno virulento
(ROMEIRO; GARCIA, 2009). Para ser considerado um eliciador o produto deve conter uma
ou mais moléculas que ativem as respostas de defesa na planta onde foi aplicado (BONALDO
et al., 2005) e seguir os critérios propostos pelos pesquisadores Steiner e Schönbeck (1995).
Estes autores propõem que um agente abiótico ou biótico para ser considerado indutor de
resistência deve obedecer aos seguintes requisitos:
1. Ausência de efeito tóxico sobre o patógeno;
2. Necessidade de intervalo de tempo entre a aplicação do agente indutor e as respostas de
defesa (indução de resistência);
3. Inespecificidade da proteção, ou seja, a indução de resistência deve conferir proteção a uma
ampla gama de patógenos;
4. Ausência de relação entre efeito e doses do indutor.
São exemplos de eliciadores bióticos ou abióticos compostos químicos dos mais
variados grupos, extratos de células ou filtrados de culturas de microrganismos, agentes físicos
ou químicos, moléculas orgânicas, reguladores vegetais e antibióticos (HAMMERSCHMIDT
et al., 2001; OOSTENDORP et al., 2001; AGRIOS, 2004; EDREVA, 2004; FAIZE et al.,
2004; RESENDE et al., 2007; ROMEIRO; GARCIA, 2009).
A utilização de produtos que induzem mecanismos de resistência nas plantas constitui
uma alternativa para o manejo integrado das doenças na cultura do tomateiro. Itako et al. (2012)
avaliaram em casa de vegetação a ação de produtos químicos no controle da mancha bacteriana
(Xanthomonas perforans) do tomateiro e na ativação de proteínas relacionadas à patogênese e,
observaram que os produtos à base de piraclostrobina juntamente com acibenzolar-S-metil
induziram as atividades das enzimas peroxidase, polifenoloxidase e β-1,3-glucanase,
evidenciando que estes produtos podem estar relacionados à indução de resistência a mancha
bacteriana em plantas de tomateiro.
Ao avaliarem a aplicação de ácido abscísico exógeno em plantas de tomateiro, Song et
al. (2011) obtiveram resultados que demonstram a ativação da resistência em plantas de
tomateiro contra A. solani, resultando em redução de áreas de lesão e gravidade da doença.
Ainda, relataram que esse efeito protetor promovido parece estar correlacionado com a
expressão de genes de defesa e com a ativação de enzimas de defesa como a fenilalanina
amônia-liase, polifenoloxidase e peroxidase em plantas de tomateiro.

23
3.8.1 Proteínas envolvidas na indução de resistência
Após o início do processo de indução de resistência ocorre uma cascata de fenômenos
bioquímicos, incluindo as respostas de hipersensibilidade, desencadeadas pelo acúmulo de
H2O2 no tecido vegetal, culminando na ativação da expressão gênica e, subsequente, síntese de
proteínas relacionadas à patogênese (PRPs) (SHAH et al., 2014).
Como já visto, as PRPs são induzidas no hospedeiro em resposta à infecção por um
patógeno ou por estímulos abióticos, e podem estar correlacionadas com a resistência não
específica do hospedeiro ao patógeno (STANGARLIN et al., 2011). Atualmente são conhecidas
17 famílias de PRPs em vegetais com as mais diferenciadas funções, variando de PR-1 até PR-
17 (GORJANOVIĆ, 2009; FILIPENKO et al., 2013). Nos vegetais, segundo Van loon et al.
(2006) as PRPs podem ser encontradas em estômatos, tricomas glandulares, vasos condutores,
epiderme foliar etc., e possuem a capacidade de se acumular em locais distantes do sítio de
infecção (EBRAHIM et al., 2011).
As principais PRPs estudadas no processo de indução de resistência são as peroxidases,
quitinases e β-1,3-glucanases (GAO et al., 2015). As enzimas β-1,3-glucanases, classificadas
como PR-2, são proteínas que hidrolisam ligações glicosídicas do tipo β-1,3 presentes em β-D-
glucanas, liberando glucose como produto principal. Estas enzimas estão presentes em plantas,
algas, fungos, bactérias, cianobactérias, actinomicetos e oomicetos (TAMOI et al., 2007; LIN
et al., 2009).
Em interações planta-patógeno ocorrem a indução da atividade de β-1,3-glucanases
(SELS et al., 2008) as quais degradam as β-glucanas, enzimas multifuncionais que hidrolisam
polissacarídeos como a celulose e outras glucanas, combatendo a invasão do patógeno no tecido
vegetal, uma vez que as β-1,3-1,6- glucanas constituem grande parte da parede celular destes
microrganismos (KUMAR; DEOBAGKAR, 1996; FLEURI; SATO, 2008).
Estas enzimas também depositam β-1,3-glucano após a ocorrência de ferimentos no
tecido vegetal dificultando a penetração do patógeno. Além disto, atuam em processos
fisiológicos como divisão celular, mobilização de moléculas de reserva entre outros
(BUCHNER et al., 2002; EL GHAOUTH et al., 2004).
As quitinases são enzimas amplamente encontradas na natureza, de acordo com a
especificidade do substrato que hidrolisam estão classificadas como PR-8 (atividade sobre a
lisozima) e PR-11 (atividade sobre quitosana) (MARTINS, 2008). As quitinases produzidas por
plantas estão envolvidas na defesa contra microrganismos e contra estresses abióticos. Elas são

24
induzidas durante o processo de infecção por patógenos ou outro tipo de estresse do meio
ambiente (PERRAKIS et al., 1994).
Esta enzima catalisa a hidrólise de ligações glicosídicas da quitina. A quitina é o
segundo biopolímero mais abundante na natureza, perdendo apenas para a celulose, e
encontrado principalmente nas carapaças de crustáceos, cutículas de insetos e parede celular de
fungos (THARANATHAN; KITTER, 2003).
Uma vez induzidas pelo ataque de patógenos as quitinases atuam através de uma série
de eventos degenerativos na célula do patógeno, incluindo a desorganização das células do
fungo fitopatogênico e da sua cutícula. Com isso o fungo sofre desequilíbrio osmótico,
agregação e recolhimento do citoplasma e da membrana plasmática (CASTORIA et al., 2001).
De acordo com Martins (2008) as quitinases e β-1,3-glucanases, por atuarem na hidrólise da
parede celular de fungos, possuem atuação sinérgica potencializando a ação antifúngica.
Além de serem importantes como enzimas antioxidantes, as enzimas peroxidases
(POD), também participam na lignificação, suberização e no metabolismo da parede celular,
por isso são classificadas como PRPs, pertencentes à família PR-9 (VAN LOON; VAN
STREIN, 1999; EBRAHIM et al., 2011).
Estas enzimas catalisam reações envolvendo o H2O2 e compostos fenólicos para reforçar
a parede celular da planta. As macromoléculas polimerizadas pelas PODs são depositadas na
superfície extracelular, fortalecendo a parede celular, restringindo a invasão por patógenos e a
expansão celular, contribuindo para o enrijecimento da estrutura da planta (HIRAGA et al.,
2001).
3.8.2 Metabolismo secundário e defesa vegetal
Os metabólitos secundários são compostos de baixo peso molecular produzidos em
menores concentrações pelas plantas, no entanto, ocorre o aumento da produção destes em
virtude da interação planta-patógeno (THAKUR; SOHAL, 2013). A formação desses
compostos tóxicos é essencial no processo de defesa contra o agente agressor no local de
infecção (SHAH et al., 2014; OLIVEIRA et al., 2016).
A ativação das respostas de defesa ocorre como uma cascata de fenômenos, e
simultaneamente com o aumento da expressão de PRPs, se observa aumento na atividade de
outras enzimas, como a fenilalanina amônia-liase (PAL). O aumento da expressão da enzima
PAL está relacionada a maior síntese e acúmulo de compostos fenólicos, seguido também de

25
maior deposição de lignina no tecido vegetal, aumentando a defesa da planta (OLIVEIRA et
al., 2016).
Entre os produtos oriundos do metabolismo secundário, os compostos fenólicos,
sintetizados no metabolismo dos fenilpropanóides, são importantes substâncias de defesa contra
patógenos (SCHWAN-ESTRADA et al., 2008). O metabolismo dos fenilpropanóides inclui
uma série complexa de caminhos bioquímicos que proporcionam às plantas milhares de
combinações. Com isso, muitos deles são intermediários na síntese de substâncias estruturais
das células, como a lignina. A biossíntese de lignina envolve uma série de enzimas, dentre elas
a enzima PAL (BOATRIGHT et al., 2004).
A PAL está relacionada com a resistência de plantas a patógenos, notadamente, por estar
envolvida no primeiro passo da síntese dos fenilpropanóides, convertendo fenilalanina em
ácido-transcinâmico e síntese de compostos como fitoalexinas, e principalmente, de substâncias
envolvidas na biossíntese de lignina, que confere maior resistência à parede celular das plantas
aos patógenos (NAKAZAWA et al., 2001; ZHAO et al., 2005).
O aumento da atividade da PAL está fortemente relacionado aos diferentes tipos de
estresses, entre eles à invasão por patógenos, no entanto, também pode ser ativada pela presença
de compostos eliciadores. Por ação da maior atividade da PAL, ocorre aumento na síntese dos
compostos fenólicos, que são produzidos rapidamente e se acumulam após a infecção, sendo
tóxicos aos patógenos. Os ácidos clorogênico, caféico e ferúlico são exemplos de alguns desses
compostos (CAMPOS et al., 2004) com forte atuação na defesa contra microrganismos, além
de proteção contra danos por radiação ultravioleta (CAMPOS et al., 2003).
Outras vias do metabolismo secundário também estão envolvidas com a defesa vegetal,
como é o caso da via dos octadecanóides. Danos no tecido foliar desencadeiam vias de
sinalização envolvidas com a ativação das lipases, que liberam ácido linolênico e linoleico da
membrana plasmática. Tais ácidos graxos ativam a cascata de sinalização via ácido jasmônico,
culminado na rota do octadecanóide. Entre as enzimas envolvidas nesta rota, se destacam as
lipoxigenase (LOX) e polifenoloxidase (PPO) (TAIZ; ZEIGER, 2013).
As enzimas LOXs são isoenzimas que estão amplamente distribuídas em plantas
superiores e catalisam a formação de hidroperóxidos a partir dos ácidos linolênico e linoleico
(STENZEL et al., 2003), atuando como enzima chave na biossíntese do ácido jasmônico, sendo
a LOX a primeira enzima desta cascata de sinalização de resposta de defesa vegetal. Os
produtos da via da LOX já foram associados à defesa contra patógenos (RANCÉ et al., 1998;

26
STANGARLIN et al., 2011) e sua atividade é regulada, ente outros fatores, pelo ataque de
patógenos e insetos herbívoros (PORTA et al., 2008).
A PPO é uma enzima encontrada em muitas plantas, bactérias, fungos e algas
(THIPYAPONG et al., 2004). Em plantas superiores se localiza nos tilacóides dos cloroplastos
ou em outros plastídeos. Em sua forma ativa, a PPO catalisa a hidroxilação de monofenóis a o-
dofenóis (cresolase) e a oxidação de o-difenóis a o-diquinonas (catecolase) (SHIN et al., 1997;
BITTNER, 2006).
Pequenas quantidades de PPO podem ser encontradas na parede celular, no entanto, a
maior concentração destas enzimas está localizada nos tilacóides (THIPYAPONG et al., 2004),
são abundantes em tecidos lesionados pela invasão de patógenos ou insetos, sendo importantes
nos mecanismos de defesa (CAMPOS et al., 2003; THIPYAPONG et al., 2004).
A função das PPOs no processo de indução de resistência está relacionada ao aumento
da resistência da parede celular dos tecidos vegetais. Estas enzimas, juntamente com as PODs,
oxidam compostos fenólicos, colaborando com a biossíntese de lignina, aumentando as
barreiras físicas contra o ataque de patógenos (YUAN et al., 2002).
A maior atividade das enzimas PAL, POD e PPO promovem maior síntese e deposição
de lignina nos tecidos vegetais. Em folhas de algodão, Guerra et al. (2013) observaram que altas
concentrações de compostos fenólicos e o aumento da atividade da PPO e POD, aumentaram o
fortalecimento da parede celular, devido a maior deposição de lignina.
O aumento da síntese de compostos fenólicos ocorre por ação da maior atividade da
PAL simultaneamente e, conforme aumenta a lignificação das células, ocorre a redução do
conteúdo de compostos fenólicos nas mesmas, uma vez que estas substâncias são substrato para
a síntese de lignina, por ação da POD e PPO (KUHN; PASCHOLATI, 2010).
A lignina é um composto essencial para as plantas e confere a rigidez das células e
tecidos, sendo fundamental em situações de estresse, conferindo maior resistência, estando
presente na parede celular de folhas, caules e raízes (TAIZ; ZEIGER, 2013). Na indução de
resistência, a maior deposição de lignina constitui importante barreira física e química,
dificultando a penetração pelos patógenos. Por isso, é considerada uma das substâncias mais
importantes presentes na parede celular do hospedeiro (CASTRO et al., 2010).

27
4. MATERIAL E MÉTODOS
4.1 Caracterização da área de estudo
Foram conduzidos dois experimentos: experimento I no ano de 2015 e experimento II
no ano de 2016, em casa de vegetação e laboratório de fitopatologia, pertencentes ao
Departamento de Agronomia, da Universidade Estadual do Centro-Oeste (UNICENTRO),
Campus Cedeteg, no município de Guarapuava/PR, localizada na latitude 25°23’36” S e
longitude 51°27’1’’ W, e 1.025 metros de altitude. O clima da região é classificado como Cfb
subtropical mesotérmico úmido (KÖPPEN, 1948).
Os experimentos foram conduzidos em casa de vegetação (Figura 1), com cobertura de
filme de polietileno, no período de janeiro a maio de 2015 e de 2016 (os dois experimentos
foram conduzidos na mesma época).
Figura 1. Casa de vegetação: vista externa (A) e vista interna (B). Unicentro, Guarapuava/PR.
Fonte: MAREK, 2018.
4.2 Material vegetal: semeadura e transplante
Foi utilizado o híbrido de tomateiro ‘Conquistador’ (Sakata Seeds Sudamérica Ltda.),
tipo salada, de crescimento indeterminado. As mudas foram preparadas em bandejas de
polipropileno com 128 células preenchidas com substrato comercial Carolina Soil® II,
composto de turfa de esfágno, vermiculita expandida, resíduo orgânico agroindustrial classe A,
calcário dolomítico, gesso agrícola e traços de fertilizantes NPK, pH 5,5 ± 0,5, CE 0,7 ± 0,3
mScm-1 e densidade 145 kg m-3, colocando-se uma semente por célula. A semeadura foi
realizada no dia 2 de dezembro de 2015 (experimento I) e de 2016 (experimento II).

28
As mudas foram transplantadas para os vasos de 10 dm3, no dia 6 de janeiro de 2015
(experimento I) e de 2016 (experimento II) e, após, foi realizada a primeira irrigação para
aclimatação das mudas. Os vasos foram preenchidos com solo coletado no campo experimental
na mesma Universidade, classificado como Latossolo Bruno distroférrico típico (EMBRAPA,
2013), para a execução do experimento I (2015) e reutilizado para a execução do experimento
II (2016) e, em ambos, foram realizadas a análise química, conforme Tabela 1.
A distribuição dos vasos foi realizada com espaçamento de 1,0 x 0,5 m, estimando uma
densidade de 20.000 plantas ha-1.
4.3 Material vegetal: tratos culturais
A correção e adubação de base foram realizadas de acordo com a análise química
(Tabela 1) para ambos os experimentos. Ao longo do ciclo, a irrigação foi realizada por
gotejamento. A adubação de cobertura foi realizada conforme a necessidade da cultura, de
acordo com Alvarenga (2013).
Tabela 1. Análise química de solo dos experimentos I e II nos anos de 2015 e 2016,
respectivamente. Unicentro, Guarapuava/PR, 2018.
pH M.O. PMehlich Al H+Al K Ca Mg SB CTC V%
Amostra CaCl2 g dm-3 g dm-3 --------------------------------- mmolc dm-3 -----------------------
2015 5,3 47,1 2,5 0,0 3,6 0,12 2,9 1,5 4,52 8,12 55,7
2016 5,1 46,0 2,3 0,0 3,7 0,15 2,8 1,1 4,05 7,75 52,3
Micronutrientes Enxofre (S) Boro (B) Ferro (Fe) Cobre (Cu) Manganês (Mn) Zinco (Zn)
Amostra -------------------------------------------------------- mg dm-3 ---------------------------------------------------
2015 5,0 0,45 31,3 0,5 28,2 1,9
2016 6,0 0,42 30,2 0,4 26,4 1,4
O controle de plantas daninhas foi efetuado manualmente e o controle de pragas foi
realizado conforme recomendações para a cultura do tomateiro:
• Foi aplicado 15 mL planta-1 via ‘drench’ (esguicho) no transplantio das mudas de 0,6g
L-1 do inseticida formulado com o princípio ativo midacloprido, do grupo químico
neonicotinóide (Evidence®700WG) para o controle de tripes (Thrips palmi), pulgão-verde
(Myzus persicae) e mosca-branca (Bemisia tabaci e B. tabaci raça B);
• Aos 30, 45, 60 e 75 dias após o transplantio foi aplicado via foliar 0,8 mL L-1 do
inseticida formulado com o princípio ativo lufenurom, do grupo químico benzoiluréia (Match

29
EC) para o controle da traça do tomateiro (Tuta absoluta), broca pequena do fruto
(Neoleucinodes elegantalis) e ácaro bronzeado (Aculops lycopersici);
• Aos 25, 35, 45, 55 e 65 dias após o transplantio foi aplicado via foliar 1 mL L-1 do
inseticida formulado com os princípios ativos tiametoxam e lambda-cialotrina, dos grupos
químicos neonicotinóide e piretróide, respectivamente (Engeo Pleno) para o controle de mosca-
branca (Bemisia tabaci).
As plantas foram conduzidas com uma haste ao longo do ciclo, com duas plantas por
vaso e tutoradas na vertical, individualmente, com a utilização de fitilho (experimento I) e de
bambu (experimento 2) (Figura 2). A retirada de brotos foi efetuada quando estes apresentavam
3 a 5 cm de comprimento. As plantas foram conduzidas até o quinto cacho de frutos, quando
foi realizada a poda apical.
Figura 2. Plantas de tomateiro em casa de vegetação tutoradas verticalmente com fitilho (Experimento I – 2015)
e bambu (Experimento II – 2016). Unicentro, Guarapuava/PR.
Fonte: MAREK, 2018.
Durante o período do experimento (janeiro a maio de cada ano), por meio de termo-
higrômetro digital (Incoterm) instalado dentro da casa de vegetação, foram coletados
diariamente dados de temperatura e da umidade relativa do ar (máxima e mínima).
4.4 Delineamento experimental e tratamentos
O delineamento experimental utilizado foi blocos ao acaso, com sete tratamentos e cinco
repetições, sendo cada parcela composta por quatro plantas.
A primeira aplicação dos tratamentos foi realizada 15 dias após o transplantio (DAT) e
as demais em intervalos de quinze dias, totalizando seis aplicações. As aplicações foram

30
realizadas via foliar, na planta toda, com o uso de pulverizador manual de CO2 pressurizado,
com 0,3 kgf/cm2, bico cônico aberto, utilizando-se cortina plástica entre os tratamentos para
evitar deriva.
Os tratamentos utilizados foram:
T1 – testemunha inoculada (água + inóculo);
T2 – testemunha absoluta (água);
T3 – boscalida (0,075 g L-1);
T4 – boscalida (0,100 g L-1) + cresoxim-metílico (0,050 g L-1);
T5 – piraclostrobina (0,100 g L-1);
T6 – fluxapiroxade (0,058 g L-1) + piraclostrobina (0,116 g L-1) e
T7 – metiram (1,100 g L-1) + piraclostrobina (0,100 g L-1).
4.5 Caracterização dos fungicidas
4.5.1 Boscalida
Como fonte de boscalida utilizou-se o fungicida Cantus® contendo 500g kg-1 de
princípio ativo, fabricado pela BASF S.A.
• Nome químico: 2-chloro-N-(4'-chlorobiphenyl-2-yl) nicotinamide;
• Fórmula molecular: C18H12Cl2N2O;
• Classe: Fungicida sistêmico do grupo químico carboxamida (anilida);
• Tipo de formulação: Granulado Dispersível (WG).
4.5.2 Boscalida + Cresoxim-metílico
Como fonte de boscalida + cresoxim-metílico utilizou-se o fungicida Collis® contendo
200g L-1 de princípio ativo boscalida e 100g L-1 de princípio ativo cresoxim-metílico, fabricado
pela BASF S.A.
• Nome químico: 2-chloro-N-(4'-chlorobiphenyl-2-yl) nicotinamide (Boscalida) e methyl(E)-
2-methoxyimino[2-(o-tolyloxymethyl) phenyl]acetate (Cresoxim-metílico);
• Fórmula molecular: C18H12Cl2N2O (Boscalida) e C18H19NO4 (Cresoxim-metílico);
• Classe: Fungicida sistêmico do grupo químico carboxamida (anilida) (Boscalida) e
estrobilurina (Cresoxim-metlico);
• Tipo de formulação: Suspensão Concentrada (SC).

31
4.5.3 Piraclostrobina
Como fonte de piraclostrobina utilizou-se o fungicida Comet® contendo 250g L-1 de
princípio ativo, fabricado pela BASF S.A.
• Nome químico: Methyl N-{2-[1-(4-chlorophenyl) -1H-pyrazol-3-yloxymethyl]phenyl}
(Nmethoxy)carbamate;
• Fórmula molecular: C19H18ClN3O4;
• Classe: Fungicida sistêmico do grupo químico das estrobilurinas;
• Tipo de formulação: Concentrado Emulsionável – EC.
4.5.4 Fluxapiroxade + Piraclostrobina
Como fonte de fluxapiroxade + piraclostrobina utilizou-se o fungicida Orkestra®SC
contendo 167g L-1 do princípio ativo fluxapiroxade e 333g L-1 do princípio ativo
piraclostrobina, fabricado pela BASF S.A.
• Nome químico: 3-(difluoromethyl)-1-methyl-N-(3′,4′,5′-trifluorobiphenyl-2-yl)pyrazole-4-
carboxamide (fluxapiroxade) e MethylN-{2-[1-(4-chlorophenyl)-1H-pyrazol-3-yloxymethyl]phenyl}
(Nmethoxy)carbamate (piraclostrobina);
• Fórmula molecular: C18H12F5N3O (fluxapiroxade) e C19H18ClN3O4 (piraclostrobina);
• Classe: Fungicida de ação protetora e sistêmica, dos grupos químicos estrobilurina
(Piraclostrobina) e carboxamida (Fluxapiroxade);
• Tipo de formulação: Suspensão Concentrada (SC).
4.5.5 Metiram + Piraclostrobina
Como fonte de metiram + piraclostrobina utilizou-se o fungicida Cabrio®Top contendo
550g kg-1 do princípio ativo metiram e 50g kg-1 do princípio ativo piraclostrobina, fabricado
pela BASF S.A.
• Nome químico: Zinc ammoniate ethylenebis(dithiocarbamate)-poly(ethylenethiuram
disulfide) (metiram) e MethylN-{2-[1-(4-chlorophenyl)-1H-pyrazol-3-yloxymethyl]phenyl}
(Nmethoxy) carbamate (piraclostrobina);
• Fórmula molecular: (C16H33N11S16Zn3)x (metiram) e C19H18ClN3O4 (piraclostrobina);
• Classe: Fungicida sistêmico do grupo químico alquilenobis (ditiocarbamato)(Metiram)
e estrobilurina (Piraclostrobina);
• Tipo de formulação: Granulado Dispersível (WG).

32
4.6 Isolamento e inoculação do patógeno: Alternaria solani
O isolado de A. solani inoculado no experimento I (2015) foi obtido a partir de lesões
de folhas de plantas de tomateiro na região de Guarapuava/PR. No experimento II (2016) foi
inoculado o isolado obtido de lesões das plantas testemunhas do experimento I (2015).
Para a manutenção destes isolados, os fungos foram cultivados em meio BDA (batata
ágar dextrose) e incubados em BOD a 25ºC (± 2°C) em fotoperíodo de 12 horas de luz e 12
horas de escuro, por 10 dias. Após, foram guardados discos destas culturas em microtubos
Eppendorf (2 mL) contendo água destilada esterilizada e preservados em BOD, nas mesmas
condições de cultivo.
Para a recuperação destes isolados foram realizados repiques em placas de Petri
contendo meio BDA e incubadas em BOD, nas mesmas condições já citadas, para prover a
suspensão de inóculo inicial para os experimentos. Após o crescimento micelial preencher toda
a placa de Petri (Figura 3A), que ocorreu em média de 8 a 10 dias após a repicagem, foi realizada
a indução da esporulação a partir de injúrias do micélio (estresse físico), utilizando cortes com
bisturi, em toda a extensão da colônia, nos sentidos longitudinal e transversal (Figura 3B).
Realizadas as injúrias, as culturas retornaram às condições de incubação e, após seis
dias, verificou-se a esporulação das culturas. Então foram removidos os esporos com 15 mL de
água destilada esterilizada (com 0,1% de Tween 80), por raspagem com alça de vidro e posterior
filtragem em gaze. A determinação da concentração de esporos na suspensão foi realizada em
câmara de Neubauer ao microscópio de luz, determinando o número de conídios mL-1.
Figura 3. Cultura de Alternaria solani antes (A) e após (B) injúria mecânica dos micélios. Unicentro,
Guarapuava/PR.
Fonte: MAREK, 2018.

33
A concentração da suspensão para a inoculação foi ajustada para 1x104 conídios mL-1.
As plantas, com exceção do tratamento testemunha absoluta (T2) foram inoculadas 24 horas
após a primeira aplicação dos tratamentos (16 DAT). A inoculação foi realizada com
pulverização da suspensão de conídios na face abaxial e adaxial dos folíolos de todas as folhas
da planta, com pulverizador manual spray, até que gotas fossem formadas e começassem a
escorrer e pingar pelas extremidades dos folíolos. Após inoculação das plantas foram
monitoradas a umidade e temperatura da casa de vegetação, para que as condições ideais ao
desenvolvimento do patógeno fossem mantidas.
4.7 Propriedades avaliadas
4.7.1 Análise de desenvolvimento
Foram realizadas avaliações da altura de planta aos 14 (antes da primeira aplicação dos
tratamentos) e aos 21, 28, 35, 42, 49, 56 e 65 DAT (dias após o transplantio). A última avaliação
ocorreu aos 65 DAT, momento no qual foi realizada a poda apical. Para as medições utilizou-
se régua graduada e os resultados foram expressos em cm planta-1.
Aos 120 DAT (término da colheita) foram coletadas cinco plantas de cada tratamento e
divididas em folha, caule e raiz, após foram mensuradas as seguintes características: área foliar,
massa fresca e seca de folhas, caule e raiz.
A área foliar foi mensurada utilizando-se integralizador de área, Area Meter da LiCor®
modelo LI-3100 e seus resultados expressos em centímetros quadrados (cm2). As folhas, caules
e raízes foram secos em estufa de circulação forçada de ar, à temperatura constante de 60°C,
até atingir massa constante (em média 72 horas). Após as amostras foram pesadas em balança
analítica (0,001 g) e os resultados expressos em gramas (g).
4.7.2 Trocas gasosas
As avaliações de trocas gasosas foram realizadas aos 34 e 94 DAT (5º dia após a segunda
e sexta aplicação dos tratamentos, respectivamente), em folhas completamente expandidas,
localizadas no terço médio da planta (Figura 4). Foram realizadas leituras em duplicata, no
período das 9:00 às 11:00 h da manhã.
Estas medidas foram realizadas utilizando-se equipamento com sistema aberto de
fotossíntese com analisador de CO2 e vapor d’água por radiação infravermelha (“Infra Red Gas
Analyser – IRGA”, modelo LI-6400XT, LI-COR) e foram calculadas a partir da diferença entre

34
a concentração de CO2 e o vapor d’água do ar de referência (valor presente na câmara sem a
folha) e da amostra (valor presente na câmara com a folha), obtendo-se as concentrações de
vapor d’água e CO2 que foram liberados (transpiração – vapor d’água) e assimilados
(assimilação de CO2) através dos estômatos foliares.
Figura 4. Análise de trocas gasosas em plantas de tomateiro (A e B). Unicentro, Guarapuava/PR.
Fonte: MAREK, 2018.
As características de trocas gasosas analisadas foram: taxa de assimilação de CO2 (A,
μmol CO2 m-2 s-1), taxa de transpiração (E, mmol vapor d’água m-2 s-1), condutância estomática
(gs, mol m-2 s-1) e concentração interna de CO2 na folha (Ci, μmol CO2 mol-1 ar). Essas
características foram calculadas pelo programa de análise de dados do equipamento medidor de
trocas gasosas, que utiliza a equação geral de trocas gasosas de Von Caemmerer e Farquhar
(1981).
A eficiência do uso da água (EUA, μmol CO2 (mmol H2O)-1) foi determinada através da
relação entre a taxa de assimilação de CO2 e a taxa de transpiração (A/E). A eficiência de
carboxilação da enzima ribulose 1, 5-difosfato carboxilase (Rubisco) foi determinada através
da relação entre taxa de assimilação de CO2 e a concentração interna de CO2 na folha (A/Ci).
4.7.3 Fluorescência da clorofila a
A fluorescência da clorofila a foi avaliada utilizando fluorômetro portátil
(Portable Chlorophyll Fluorometer - PAM-2500). As leituras foram realizadas aos 34 e 94
DAT (5º dia após a segunda e sexta aplicação dos tratamentos, respectivamente), em folhas
completamente expandidas, localizadas no terço médio da planta (Figura 5A). Foram realizadas

35
medições de cinética lenta, com adaptação das folhas ao escuro por 20 minutos (para que todos
os centros de reação estivessem abertos, com capacidade de receber elétrons) (Figura 5B), e as
leituras foram realizadas em duplicata, no período das 9:00 às 11:00 h da manhã,
simultaneamente com as de trocas gasosas.
Figura 5. Clip para adaptação ao escuro: indicado pela seta (A); Análise de fluorescência da clorofila a em plantas
de tomateiro. Unicentro, Guarapuava/PR.
Fonte: MAREK, 2018.
Os parâmetros de fluorescência da clorofila a foram obtidos automaticamente e
armazenados na memória do PAM e, posteriormente, foram descarregados em computador por
meio do programa computacional Wincontrol.
Foram obtidos os parâmetros de fluorescência basal (F0) e fluorescência máxima (Fm),
a partir dos quais foram calculados: a fluorescência variável (Fv) e rendimento quântico máximo
do PSII (Fv/Fm). Também foram obtidas as respostas de rendimento quântico fotoquímico
efetivo do PSII, Y(II) e taxa de transferência relativa de elétrons (ETR).
4.7.4 Nitrato Redutase
Para avaliar a atividade da enzima nitrato redutase (NR, EC 1.6.6.1) foram realizadas
oito coletas de folíolos aos 15, 20, 35, 50, 65, 80, 95 e 110 DAT, que correspondem aos
momentos:
• Tempo 0 hora, antes da aplicação dos tratamentos (15 DAT);
• 5° dia após a primeira aplicação dos tratamentos (20 DAT);
• 5° dia após a segunda aplicação dos tratamentos (35 DAT);
• 5° dia após a terceira aplicação dos tratamentos (50 DAT);

36
• 5° dia após a quarta aplicação dos tratamentos (65 DAT);
• 5° dia após a quinta aplicação dos tratamentos (80 DAT);
• 5° dia após a sexta e última aplicação dos tratamentos (95 DAT);
• 15 dias após a última aplicação dos tratamentos (105 DAT).
Foram coletados folíolos entre as 10 e 11 horas da manhã, período em que as plantas
receberam pelo menos três horas de sol e a enzima nitrato redutase já tinha atingido sua máxima
atividade (HAGEMAN; REED, 1980).
Após coletadas, as folhas foram colocadas em pacotes confeccionado com papel
alumínio e armazenadas em caixa de isopor com gelo (mantendo o material fresco, sem
congelar). Na sequência, o material coletado foi levado para o laboratório e a atividade da
enzima nitrato redutase foi determinada de acordo com metodologia proposta por Streeter e
Bosler (1972), onde foram pesadas 1g da amostra fresca em tubos protegidos da luz (revestidos
de papel alumínio).
Em seguida foram adicionados 8 mL de tampão composto por fosfato (KPi) 0.1M pH
7,5 + KNO3 0,2M, a reação foi conduzida à 37ºC por 1 hora. Depois desse período, foram
adicionados 1 mL de sulfanilamida 1% em HCl 1,5% + 1 mL de α–Naftil 0,2%. Após 5 minutos,
a amostra foi filtrada em algodão e a leitura realizada em espectrofotômetro a 540 nm. As
amostras foram coletadas e analisadas no mesmo dia da coleta. A atividade da enzima nitrato
redutase foi expressa em μg nitrito min-1 g de massa fresca-1.
4.7.5 Pigmentos fotossintéticos
Para a determinação do teor de pigmentos foram realizadas oito coletas de discos de
folhas (2 discos de 200 mm² de diâmetro), realizadasno período entre 9 e 10 horas da manhã,
aos 15, 20, 35, 50, 65, 80, 95 e 110 DAT, que correspondem aos momentos:
• Tempo 0 hora, antes da aplicação dos tratamentos (15 DAT);
• 5° dia após a primeira aplicação dos tratamentos (20 DAT);
• 5° dia após a segunda aplicação dos tratamentos (35 DAT);
• 5° dia após a terceira aplicação dos tratamentos (50 DAT);
• 5° dia após a quarta aplicação dos tratamentos (65 DAT);
• 5° dia após a quinta aplicação dos tratamentos (80 DAT);
• 5° dia após a sexta e última aplicação dos tratamentos (95 DAT);
• 15 dias após a última aplicação dos tratamentos (105 DAT).

37
Imediatamente após a coleta, os discos de folhas foram colocados em pacotes
confeccionado com papel alumínio e, em seguida, congelados em nitrogênio líquido para
paralização imediata de todas as reações. Na sequência, o material coletado foi levado para o
laboratório e a determinação do teor de pigmentos foi realizada pelo método
espectrofotométrico proposto por Lichtenthaler e Wellburn (1983).
Para as análises foram pesadas 0,030 g de material vegetal congelado, após foi triturado
em nitrogênio líquido em almofariz de porcelana. Para a obtenção do extrato, o material vegetal
foi macerado em sala escura, com adição de 8 mL de acetona a 80% e 0,01 g de carbonato de
cálcio (Ca(CO3)2). Esta solução foi centrifugada por 20 minutos a 13500 g.
Para as leituras de absorbância no espectrofotômetro foi utilizado o sobrenadante,
utilizando-se os comprimentos de onda de 470, 646,8 e 663,2nm.
Para os cálculos do teor de pigmentos clorofilianos (chl a, b e total) foram utilizadas as
fórmulas abaixo:
Clorofila a (µg/ml) = 12,21 A663 – 2,81 A646
Clorofila b (µg/ml) = 20,13 A646 – 5,03 A663
Chls a + b = 17,76 A646,6 + 7,34 . A663,6
Para o cálculo do teor de xantofilas e carotenoides (x + c) utilizou-se a fórmula:
Carotenoides (µg/mL) = (1000A470 – 3,27[Chl a] - 104[Chl b])
229
Os teores de pigmentos clorofilianos e carotenoides foram expressos em µg g de massa
fresca-1.
4.7.6 Análise de carboidratos
Para determinar o conteúdo de carboidratos (açúcares redutores e açúcares solúveis)
foram realizadas duas avaliações destrutivas ao longo do desenvolvimento, sendo a primeira
realizada aos 50 DAT e a segunda aos 120 DAT (último dia de colheita). Foram coletadas cinco
plantas por tratamento, as quais foram separadas em folha, caule e frutos. Para cada repetição,
as amostras foram homogeneizadas e alíquotas de 2,0 g foram armazenadas em pacotes
confeccionado com papel alumínio a -20°C até o momento da extração.
O conteúdo de carboidratos foi determinado nos diferentes órgãos vegetais: folhas, caule
e frutos. A extração dos açúcares foi realizada de acordo com metodologia proposta por Garcia
et al. (2006), utilizando 0,1 g de material vegetal fresco e homogeneizado. O extrato obtido foi

38
armazenado, separado em microtubos e armazenado a -20°C até a determinação dos
carboidratos.
Para a quantificação dos açúcares solúveis totais foram utilizadas as metodologias de
Morris (1948) e Yemm e Willis (1954). A leitura foi realizada em espectrofotômetro a
620 nm, utilizando solução de glicose para a confecção da curva padrão. Os açúcares
solúveis totais foram expressos em µg g de massa fresca-1.
A quantificação de açúcares redutores foi realizada, segundo metodologia de Miller
(1959). A leitura foi realizada em espectrofotômetro a 540 nm, utilizando solução de glicose
para a confecção da curva padrão. Os açúcares redutores foram expressos em µg g de massa
fresca-1.
4.7.7 Avaliação da severidade de Alternaria solani
A severidade da doença foi avaliada ao surgimento dos primeiros sintomas, que ocorreu
cinco dias após a inoculação (20 DAT). As avaliações foram realizadas em cinco folíolos
previamente identificados, em 10 plantas por tratamento, por três avaliadores, em intervalos de
7 dias (Figura 6B). Foram realizadas 8 avaliações de severidade no período de 28 de janeiro a
17 de março, em ambos os experimentos. Para a avaliação da severidade foi utilizada a escala
diagramática de Azevedo (1997) (Figura 6A).
Figura 6. Escala diagramática para avaliação de Alternaria solani em tomateiro expressa em proporção de área
foliar lesionada (A). Avaliação da severidade de pinta preta em tomateiro (B). Unicentro, Guarapuava/PR.
Fonte: MAREK, 2018.
Com os dados obtidos das avaliações da severidade da doença foi calculada a área
abaixo da curva de progresso de doença (AACPD) utilizando-se a Eq. 1:

39
𝐴𝐴𝐶𝑃𝐷 = ∑ (𝑌𝑖 + 𝑌𝑖+1
2) (𝑡𝑖+1 − 𝑡𝑖) 𝑒𝑞𝑢𝑎çã𝑜 (1)
Na qual, Y representa a intensidade (severidade) da doença, t o tempo e i o número de
avaliações no tempo (CAMPBELL; MADDEN, 1990).
4.7.8 Análises bioquímicas
4.7.8.1 Coleta e obtenção dos extratos enzimáticos
Foram realizadas 25 coletas de discos de folíolos (200 mm² de diâmetro) para análises
enzimáticas, nos tempos 0, 24, 48 e 120 horas após cada aplicação dos tratamentos e uma coleta
final no 15º dia após a última aplicação, sendo:
• Aos 15 DAT = tempo 0 hora, antes da aplicação dos tratamentos;
• Aos 16 DAT = 24 horas após a primeira aplicação dos tratamentos e antes da
inoculação do patógeno;
• Aos 17 DAT = 48 horas após a primeira aplicação dos tratamentos e 24 horas após a
inoculação do patógeno;
• Aos 20 DAT = 120 horas após a primeira aplicação dos tratamentos e 96 horas após a
inoculação do patógeno;
• Aos 29, 44, 59, 74 e 89 DAT = 0 hora antes da segunda, terceira, quarta, quinta e sexta
aplicação dos tratamentos, respectivamente;
• Aos 30, 45, 60, 75 e 90 DAT = 24 horas após a segunda, terceira, quarta, quinta e sexta
aplicação dos tratamentos, respectivamente;
• Aos 31, 46, 61, 76 e 91 DAT = 48 horas após a segunda, terceira, quarta, quinta e sexta
aplicação dos tratamentos, respectivamente;
• Aos 34, 49, 64, 79 e 94 DAT = 120 horas após a segunda, terceira, quarta, quinta e
sexta aplicação dos tratamentos, respectivamente;
• Aos 105 DAT = 15° dia após a última aplicação dos tratamentos.
Após coletados, os discos de folíolos foram colocados em pacotes confeccionados com
papel alumínio e, em seguida, congeladas em nitrogênio líquido para paralisar todas as reações
imediatamente e armazenado a -20°C para posterior extração enzimática. As coletas foram
realizadas em triplicata, pois foram realizados diferentes tipos de extração.
A primeira extração enzimática foi realizada para determinar o conteúdo proteico e a
atividade das enzimas: superóxido dismutase, ascorbato peroxidase, catalase, peroxidase e

40
polifenoloxidase utilizando 0,3 g de folhas frescas congeladas e trituradas em nitrogênio líquido
com auxílio de almofariz. Em seguida, foi homogeneizada mecanicamente em 4 mL de tampão
fosfato de potássio 50 mM (pH 7,0) contendo 0,1 mM EDTA e 1% (p/p) de PVP (poli-vinil-
pirrolidona), em almofariz. O homogenato foi centrifugado a 15.000 g durante 30 min a 4°C.
Após centrifugação, o sobrenadante obtido foi transferido para microtubos e estes armazenados
a -20°C para posterior determinação (BALBI-PEÑA et al., 2014).
A extração enzimática para a determinação da atividade da enzima glutationa S-
transferase foi realizada segundo Cataneo et al. (2003). Foram macerados 0,3 g de folhas frescas
congeladas e trituradas em nitrogênio líquido com auxílio de almofariz, adicionando-se 10% de
PVPP, tampão de extração gelado na proporção 1:3 (peso/volume), constituído de 50 mM de
Tris-HCl (pH 7,5), 1 mM de EDTA, 1 mM de DTT, 1 mM de GSH, 0,1 mM de Fluoreto de
Fenilmetil-Sulfonil (PMSF) e 10 % (v/v) de glicerol. Os extratos foram centrifugados a 12.000
g por 30 min a 4 ºC, e os sobrenadantes foram armazenados a -20 ºC para posterior utilização
na determinação.
A extração enzimática para a determinação da atividade da enzima glutationa
peroxidase foi realizada pela maceração de 0,5 g de folhas frescas em 2 mL de solução tampão
Tris-HCl (100 mM, pH 7,5) contendo ditiotreitol (1,5 mM), EDTA (1 mM), Triton X-100 (0,1
%), cisteína (5 mM), fluoreto de fenilmetilsulfonilo (PMS, 0,1 mM) e PVPP insolúvel (2%).
Os extratos foram centrifugados a 12.000 g durante 20 min a 4 ºC (ARAVIND; PRASAD,
2005).
Os extratos enzimáticos para determinação da enzima glutationa redutase foram obtidos
por maceração de 0,5 g de folhas frescas em 2 mL de solução tampão Tris-HCl (100 mM, pH
7,5), contendo ditiotreitol (1,5 mM), EDTA (1 mM), Triton X-100 (0,1 %) e PVPP insolúvel
(2%). Os extratos foram centrifugados a 12000 g, durante 20 min a 4 °C (SHANKER et al.,
2004).
Na extração enzimática, para determinar a atividade da enzima fenilalanina amônia-
liase, foram utilizada 0,3 g de folhas frescas em 3,0 mL do tampão TRIS – HCl pH 8,0. O
extrato foi centrifugado a 3200 g, por 10 minutos a 4 ºC, conforme descrito por Umesha (2006).
A extração enzimática para determinar a atividade das enzimas quitinase e β-1,3-
glucanase e o conteúdo total de proteínas para estes ensaios, foi realizada de acordo com
metodologia descrita por Moerschbacher (1988) e citada por Guzzo e Martins (1996). Foram
utilizadas 0,3 g de material vegetal congelado e triturado em nitrogênio líquido com auxílio de

41
almofariz. Em seguida, foram trituradas em almofariz mantido em gelo, contendo 0,3 g de areia
do mar puríssima, 0,1 g de resina Dowex 1-X8 e 0,5 g de PVPP, após foi adicionado 4 mL de
tampão borato de sódio 0,1 M (pH 8,8) contendo EDTA a 1 mM, DTT a 1 mM e ácido ascórbico
a 50 mM, manteve-se em repouso por 5 minutos, centrifugou-se os extratos a 20.000 g, a 4ºC
por 30 minutos. Após centrifugação, o sobrenadante obtido foi transferido para microtubos e
estes armazenados a -20°C para posterior determinação enzimática.
A extração da enzima lipoxigenase foi realizada conforme metodologia proposta por
Axelrod et al. (1981). Foram macerados 0,3 g da amostra em 2 mL de tampão Tris-HCl 50 mM,
CaCl2 20 mM, pH 8,0. Após as amostras forma centrifugadas a 5500 rpm por 10 minutos à 4°C,
e o extrato enzimático obtido foi armazenado a -20°C para posterior determinação.
4.7.8.2 Proteínas totais e atividades enzimáticas
A quantificação do conteúdo total de proteínas nas amostras, em todos os extratos, foi
determinada de acordo com metodologia proposta por Bradford (1976). A concentração de
proteínas foi determinada utilizando-se curva padrão de caseína. A leitura de absorbância foi
realizada em espectrofotômetro a 595 nm, sendo o teor proteico da amostra expresso em mg
proteína g massa fresca-1.
A determinação da atividade da superóxido dismutase (SOD, EC 1.15.1.1) foi
determinada de acordo com metodologia proposta por Giannopolitis e Ries (1997). O ensaio
considerou a capacidade da enzima em inibir a fotorredução do NBT (azul de cloreto de
nitrotetrazólio). A inibição na redução fotoquímica do NBT foi avaliada em espectrofotômetro
a 560 nm. Uma unidade da enzima (U) foi definida como a atividade da SOD necessária para a
iniciação de 50% da fotorredução do NBT. Para o cálculo da atividade específica da enzima
considerou-se a porcentagem de inibição obtida, o volume da amostra e a concentração de
proteína da amostra. A atividade da enzima SOD na amostra foi expressa em U min-1 mg
proteína-1.
A atividade da ascorbato peroxidase (APX, EC 1.11.1.11) foi determinada conforme
metodologia proposta por Nakano e Asada (1981). A atividade enzimática foi calculada
utilizando-se o coeficiente de extinção molar de 2,8 mM cm-1. As leituras foram realizadas em
espectrofotômetro a 290 nm, de 15 em 15 segundos durante 2 minutos. Os valores foram
expressos em U min-1 mg proteína-1.

42
A atividade da glutationa redutase (GR, EC 1.6.4.2) foi determinada de acordo com
metodologia proposta por Shanker et al. (2004). A atividade da GR foi determinada por meio
do aumento da absorvância obtido pela redução de 5,5-ditiobis-2-ácido nitrobenzóico (DTNB)
pela GSH, com a formação de 5-tio-2- ácido nitrobenzóico (TNB). A atividade enzimática foi
determinada pela leitura contínua da absorvância a 412 nm durante 2 min, determinando a
variação na absorção usando o coeficiente de extinção molar de 14,15 mM-1 cm-1. Os valores
da atividade enzimática da GR foram expressos em U min-1 mg proteína-1.
A atividade da glutationa S-transferase (GST, EC 2.5.1.18) foi determinada usando o
método descrito por Wu et al. (1996). A reação foi conduzida a 30ºC por 10 min, com leitura
em espectrofotômetro a 340 nm, determinando a variação na absorção usando o coeficiente de
extinção molar de 9,6 mM cm-1 para GSH-CDNB (GSH = glutationa reduzida e CDNB = 1-
cloro-2,4-dinitrobenzeno). Os valores da atividade enzimática da GST foram expressos em U
min-1 mg proteína-1.
A atividade da glutationa peroxidase (GPX, EC 1.11.1.9) foi determinada conforme
metodologia de Aravind e Prasad (2005). As leituras foram realizadas em espectrofotômetro a
340 nm, durante 2 minutos, a partir da taxa de diminuição da absorbância devido ao consumo
de NADPH. A atividade enzimática foi calculada utilizando-se o coeficiente de extinção molar
de 6,22 mM cm-1. O resultado foi expresso em U min-1 mg proteína-1.
Atividade da catalase (CAT, EC 1.11.1.6) foi determinada de acordo com metodologia
proposta por Peixoto et al. (1999). As leituras de absorbância foram realizadas em
espectrofotômetro a 240 nm aos 0 e 80 segundos, a fim de verificar o decréscimo na
absorbância, monitorado o consumo do H2O2. Para calcular a atividade específica da enzima foi
utilizado o coeficiente de extinção molar do H2O2 (39,4 mmol L-1 cm-1) e a atividade foi
expressa em U min-1 mg proteína-1.
A atividade da peroxidase de pirogalol (POD, EC 1.11.1.7) foi determinada de acordo
com metodologia proposta por Teisseire e Guy (2000). A formação de purpurogalina foi medida
em espectrofotômetro a 430 nm. Para calcular a atividade específica da enzima foi utilizado o
seu coeficiente de extinção molar (2,5 mmol L-1 cm-1) e a atividade foi expressa em U min-1
mg
proteína-1.
A atividade da polifenoloxidase (PPO, EC 1.10.3.1) foi determinada conforme método
descrito por Fujita et al. (1995) e adaptado por Zanatta et al. (2006). O meio de reação foi
incubado por 30 minutos a 30°C em banho-maria, decorrido este período a reação foi paralisada

43
com ácido perclórico (HClO4) 2M. A atividade foi medida em espectrofotômetro a 395 nm,
mensurando a conversão de catecol em quinona. A atividade da PPO foi expressa U min-1 mg
proteína-1.
A atividade da lipoxigenase (LOX, EC 1.13.11.12) sobre o ácido linoléico foi
determinada segundo o método descrito por Axelrod et al. (1981). As leituras foram realizadas
em espectrofotômetro a 234 nm, de 20 em 20 segundos por um período de 60 segundos. A
atividade enzimática foi calculada utilizando-se o coeficiente de extinção molar dos
hidroperóxidos do ácido linoléico a 234 nm (25000 mM cm-1). Os resultados foram expressos
em U min-1 mg proteína-1.
A determinação da atividade da fenilalanina amônia-liase (PAL, EC 4.3.1.5) foi
realizada por quantificação colorimétrica do ácido trans-cinâmico liberado do substrato
fenilalanina, conforme metodologia descrita por Umesha (2006). As leituras foram realizadas
em espectrofotômetro a 290 nm, sendo a atividade determinada em μg de ácido trans-cinâmico
min-1 mg proteína-1, conforme curva padrão de ácido trans-cinâmico. Os resultados foram
expressos em U min-1 mg proteína-1.
A atividade da enzima quitinase (QUI, EC 3.2.1.14) foi determinada segundo Busso e
Mazaro (2015). O meio de reação foi composto por 100 mg de substrato para quitinase, 600 µL
de tampão fosfato de sódio 0,2 M, 300 µL do extrato vegetal, incubado a 50 ºC por 1 hora, após
incubado novamente a 100 ºC por 5 minutos. Após foi centrifugado a 4300 g por 10 minutos, e
o sobrenadante foi retirado para posterior leitura em espectrofotômetro a 595 nm. Os valores
foram expressos em U min-1 mg proteína-1.
A atividade da enzima β-1,3-glucanase (GLU, EC 3.2.1.39) foi determinada segundo
metodologia descrita por Stangarlin et al. (2000), pela quantificação colorimétrica de glicose
liberada de laminarina, por meio do uso da hidrazida do ácido β-hidroxibenzóico (PAHBAH)
a 2,5% conforme metodologia proposta por Lever (1972). As leituras foram realizadas em
espectrofotômetro a 410 nm e a atividade da enzima mensurada de acordo com curva padrão
de glicose. Os valores foram expressos em U min-1 mg proteína-1.
4.7.8.3 Ânion superóxido e peróxido de hidrogênio
Para determinar a concentração do ânion superóxido (•O2-) e de peróxido de hidrogênio
(H2O2), foram realizadas 25 coletas de discos de folíolos (200 mm² de diâmetro), nos tempos

44
0, 24, 48 e 120 horas após cada aplicação dos tratamentos e uma coleta final no 15º dia após a
última aplicação, sendo:
• Aos 15 DAT = tempo 0 hora, antes da aplicação dos tratamentos;
• Aos 16 DAT = 24 horas após a primeira aplicação dos tratamentos e antes da
inoculação do patógeno;
• Aos 17 DAT = 48 horas após a primeira aplicação dos tratamentos e 24 horas após a
inoculação do patógeno;
• Aos 20 DAT = 120 horas após a primeira aplicação dos tratamentos e 96 horas após a
inoculação do patógeno;
• Aos 29, 44, 59, 74 e 89 DAT = 0 hora antes da segunda, terceira, quarta, quinta e sexta
aplicação dos tratamentos, respectivamente;
• Aos 30, 45, 60, 75 e 90 DAT = 24 horas após a segunda, terceira, quarta, quinta e sexta
aplicação dos tratamentos, respectivamente;
• Aos 31, 46, 61, 76 e 91 DAT = 48 horas após a segunda, terceira, quarta, quinta e sexta
aplicação dos tratamentos, respectivamente;
• Aos 34, 49, 64, 79 e 94 DAT = 120 horas após a segunda, terceira, quarta, quinta e
sexta aplicação dos tratamentos, respectivamente;
• Aos 105 DAT = 15° dia após a última aplicação dos tratamentos.
O material vegetal coletado foi colocado em pacotes confeccionados com papel
alumínio e, em seguida, congeladas em nitrogênio líquido, a fim de paralisar todas as reações
imediatamente.
A determinação da concentração de •O2- foi realizada em amostras de 0,1 g de material
vegetal. A produção de •O2 - foi avaliada pela determinação do adenocromo acumulado em
leituras de absorbância a 480 nm (MISRA; FRIDOVICH, 1971). O resultado foi expresso em
mmol g de massa fresca-1.
Para a determinação da concentração de H2O2 foram utilizadas amostras de 0,1 g de
tecido foliar e preparadas conforme metodologia de Kuo e Kao (2003). A concentração de
H2O2, foi estimada com base em curva padrão de H2O2, conforme metodologia de Gay e
Gerbicki (2000). O resultado foi expresso em µmol g de massa fresca-1.

45
4.7.8.4 Peroxidação lipídica
Para a determinação da peroxidação lipídica (TBAR) foram coletados discos de folíolos
(200 mm² de diâmetro) nos mesmos horários descritos para determinação da concentração de
•O2- e H2O2 (subitem 4.6.8.3). A lipoperoxidação foi determinada de acordo com técnica
descrita por Heath e Packer (1968) apud Rama Devi e Prasad (1998). Para a reação, foram
utilizadas 0,3 g de folhas frescas congeladas e trituradas em nitrogênio líquido e
homogeneizadas em 5 mL de solução contendo ácido tiobarbitúrico (TBA) a 0,25% e ácido
tricloroacético (TCA) a 10%.
Após a extração a solução foi incubada em banho-maria a 90°C por 60 minutos.
Decorrido este tempo e após resfriamento, a solução foi centrifugada a 10000 g por 15 minutos,
à temperatura ambiente (25°C). O sobrenadante foi coletado e as leituras foram realizadas em
espectrofotômetro a 560 e 600 nm. Para os cálculos utilizou-se o coeficiente de extinção molar
do malondialdeído (155 mmol L-1 cm-1). Os resultados foram expressos em μmol g de massa
fresca-1.
4.7.8.5 Compostos fenólicos
Da mesma forma, para a determinação do teor de compostos fenólicos foram coletados
discos de folíolos (200 mm² de diâmetro) nos mesmos horários descritos para a determinação
da concentração de •O2- e H2O2 (subitem 4.6.8.3). A reação foi realizada a partir da obtenção
do extrato metanólico de acordo com Moore et al. (2006) e o teor de fenólicos totais foram
determinados como descrito por Yu et al. (2003) e expresso em mg de fenólicos totais em
equivalente de ácido gálico (EAG) por grama de massa fresca das folhas de tomateiro. Os
valores foram expressos em mg g massa fresca-1.
4.7.8.6 Deposição de lignina
Para a determinação da deposição de lignina na parede celular foram coletados folíolos
aos 15, 20, 35, 50, 65, 80, 95 e 110 DAT, que corresponde aos momentos:
• Tempo 0 hora, antes da aplicação dos tratamentos (15 DAT);
• 5° dia após a primeira aplicação dos tratamentos (20 DAT);
• 5° dia após a segunda aplicação dos tratamentos (35 DAT);
• 5° dia após a terceira aplicação dos tratamentos (50 DAT);
• 5° dia após a quarta aplicação dos tratamentos (65 DAT);

46
• 5° dia após a quinta aplicação dos tratamentos (80 DAT);
• 5° dia após a sexta e última aplicação dos tratamentos (95 DAT);
• 15 dias após a última aplicação dos tratamentos (105 DAT).
Após a coleta, os folíolos foram colocados em pacotes confeccionados com papel
alumínio e, em seguida, congeladas em nitrogênio líquido, a fim de paralisar todas as reações
imediatamente. Após a coleta as amostras foram levadas ao laboratório e a deposição de lignina
foi realizada conforme metodologia descrita por Mandal et al. (2011). Foram utilizadas 1,0 g
de folhas frescas congeladas para a reação. As leituras foram realizadas em espectrofotômetro
a 280 nm. Os resultados foram obtidos a partir do aumento de A280nm g de massa fresca-1 de
resíduo seco insolúvel em álcool (RIA) e expressos em µg de lignina-1 mg de massa fresca-1.
4.7.9 Avaliação de produção e pós-colheita de frutos de tomateiro
4.7.9.1 Produção
A colheita dos frutos teve início no dia 27 de março e terminou em 5 de maio de 2015
no experimento I. No experimento II, iniciou em 30 de março e terminou em 7 de maio de 2016.
O período de colheita compreendeu em média 37 dias e foi realizada semanalmente, conforme
os frutos atingiam o estádio de maturação vermelho-claro, conforme descrito por Alvarenga
(2013) (Figura 7A). Todos os frutos foram pesados em balança analítica (Figura 7B), sendo os
resultados expressos em gramas (g), o diâmetro aferido com paquímetro e os valores expressos
em milímetros (mm) (Figura 8B).
Figura 7. Ponto de colheita dos frutos de tomateiro: vermelho-claro (A) e pesagem dos frutos em balança analítica
(B). Unicentro, Guarapuava/PR.
Fonte: MAREK, 2018.

47
Foram avaliadas a produção total e comercial de frutos de tomate (kg m-2), número de
frutos m-2, massa média de frutos (g), diâmetro e comprimento de frutos. Para a avaliação de
produção, os frutos foram selecionados em não comercial e comercial, sendo considerados
frutos com diâmetro menor que 40 mm não comercial e maior que 40 mm e sem defeitos
(sintomas de doenças, pragas, distúrbios fisiológicos e/ou danos físicos) como comercial.
4.7.9.2 Qualidade pós-colheita de frutos
As análises químicas foram realizadas no Laboratório de Fruticultura e Pós-colheita do
Departamento de Agronomia (UNICENTRO) no mesmo dia em que os frutos foram colhidos.
A colheita dos frutos para estas análises foi realizada no dia 9 de abril de 2015 (92 DAT) para
o experimento I e no dia 7 de abril de 2016 (90 DAT) para o experimento II. Os frutos foram
colhidos no estádio de maturação vermelho-claro (ALVARENGA, 2013) (Figura 8A) e retirou-
se o pedúnculo e os cálices para que não houvesse lesões por atrito. No laboratório os frutos
foram lavados em água corrente e secos com papel toalha.
As variáveis analisadas foram: firmeza de polpa, acidez titulável (AT), pH, sólidos
solúveis (SS, oBrix), relação SS/AT, teor de ácido ascórbico, atividade das enzimas
poligalacturonase e pectinametilesterase e perda de massa.
As metodologias são descritas a seguir:
Firmeza de polpa: foi determinada com auxílio de penetrômetro digital (FR-5120
Lutron) empregando-se ponteira de 8 mm. As leituras foram realizadas em dois frutos inteiros
com casca, em dois pontos opostos na região equatorial de cada fruto e os resultados foram
expressos em grama-força (gf cm-2).
Acidez titulável (AT) e pH: foram utilizadas polpa de três tomates, por repetição,
homogeneizadas em triturador doméstico tipo ‘mixer’. Para análise do pH da polpa e da acidez
titulável foram preparadas amostras de 5 g de polpa diluída em 100 mL de água destilada, deste
extrato, uma alíquota de 50 mL foi utilizada para determinação do pH da polpa em peagâmetro
(Marconi, MA 522). Na mesma alíquota foi realizada a titulação com NaOH 0,1 N até pH 8,2
para determinação da acidez titulável. Os dados foram expressos em gramas de ácido cítrico
100 g de polpa-1.
Sólidos solúveis (SS): foi determinado com refratômetro digital automático de
compensação de temperatura (Atago - Pocket refractoneter Pal1). Os resultados foram
expressos em ºBrix.

48
Relação SS/AT: é a relação entre os teores de sólidos solúveis (SS) e a acidez titulável
(AT).
Teor de ácido ascórbico: foi realizado pelo método titulométrico que consiste na redução
de 2,6-diclorofenolindofenol-sódio (DCFI) pelo ácido ascórbico. Foram utilizadas 10 g de
polpa, tituladas em ácido oxálico a 0,5% com DCFI a 0,01 N. O DCFI em meio básico ou neutro
apresenta coloração azul, em meio ácido é rosado e na forma reduzida incolor. O ponto final da
titulação é detectado pela alteração da coloração da solução incolor para rosada, quando a
primeira gota de solução do DCFI é introduzida no sistema e todo ácido ascórbico já foi
consumido (AOAC, 1997). Os dados foram expressos em mg de ácido ascórbico 100 mL de
polpa do fruto-1.
Atividade da enzima poligalacturonase (PG, EC 3.2.1.15): foi determinada de acordo
com metodologia descrita por D’innocenzo e Lajolo (2001) a partir do extrato enzimático
obtido conforme Pressey (1986). Nas condições do ensaio, uma unidade da PG foi considerada
como a quantidade de enzima capaz de catalisar a formação de 1 nmol de grupos redutores min-
1 g de massa fresca-1. Os resultados foram expressos em U g massa fresca-1.
Atividade da enzima pectinametilesterase (PME, EC 3.1.1.11): foi determinada
conforme metodologia descrita por D’innocenzo e Lajolo (2001) a partir do extrato enzimático
obtido conforme Lima (2002). Nas condições do ensaio, uma unidade de PME foi definida
como a quantidade de enzima capaz de catalisar a desmetilação de pectina correspondente ao
consumo de 1 nmol de NaOH min-1 g de massa fresca-1. Os resultados foram expressos em U g
massa fresca-1.
4.7.9.3 Avaliação da perda de massa
Foram selecionados 10 frutos de cada tratamento para a avaliação de perda de massa.
Os frutos foram colhidos no estádio de maturação verde-rosado de acordo com Alvarenga
(2013) (Figura 8A) e mantidos sobre a bancada do laboratório, revestida com papel kraft
(Figuras 8C e 8D). Inicialmente os frutos foram pesados em balança analítica, sendo os
resultados expressos em gramas (g). Para a avaliação da perda de massa ao longo do
experimento, os frutos foram pesados a cada dois dias até o 21º dia.
Neste período, foram monitorados por meio de termo-higrômetro digital (Incoterm), os
dados de temperatura e umidade relativa do ar (máxima e mínima). Para o cálculo da perda de
massa foi utilizado a diferença entre as pesagens de cada intervalo de tempo e os resultados

49
foram expressos em porcentagem (%) (CHITARRA; CHITARRA, 2005), e a partir destes,
foram plotadas curvas de perda de massa ao longo do período avaliado para cada tratamento.
Figura 8. Colheita de frutos no estádio de maturação verde-salada (A), pesagem dos frutos (B), frutos no início:
1°dia (C) e final: 21° dia (D) do experimento de avaliação de perda de massa. Unicentro, Guarapuava/PR.
Fonte: MAREK, 2018.
4.8 Análise estatística
Neste conjunto de dados foram realizados os testes de normalidade por Shapiro-Wilk e
homogeneidade de variância por Cochran C antes de cada teste estatístico paramétrico. A
análise de variância (ANOVA) e as médias também foram compradas pelo teste Tukey, ambos,
ao nível de 1 e 5% de significância.
Os dados de trocas gasosas e fluorescência da clorofila a, não apresentaram normalidade
e foram transformados pelo método Box-Cox. Já os dados de porcentagem de frutos não
comerciais e de perda de massa foram transformados por arco seno de √x (%). Todos os testes
estatísticos foram realizados utilizando o software Ambiente R (R Development Core Team,
2016).
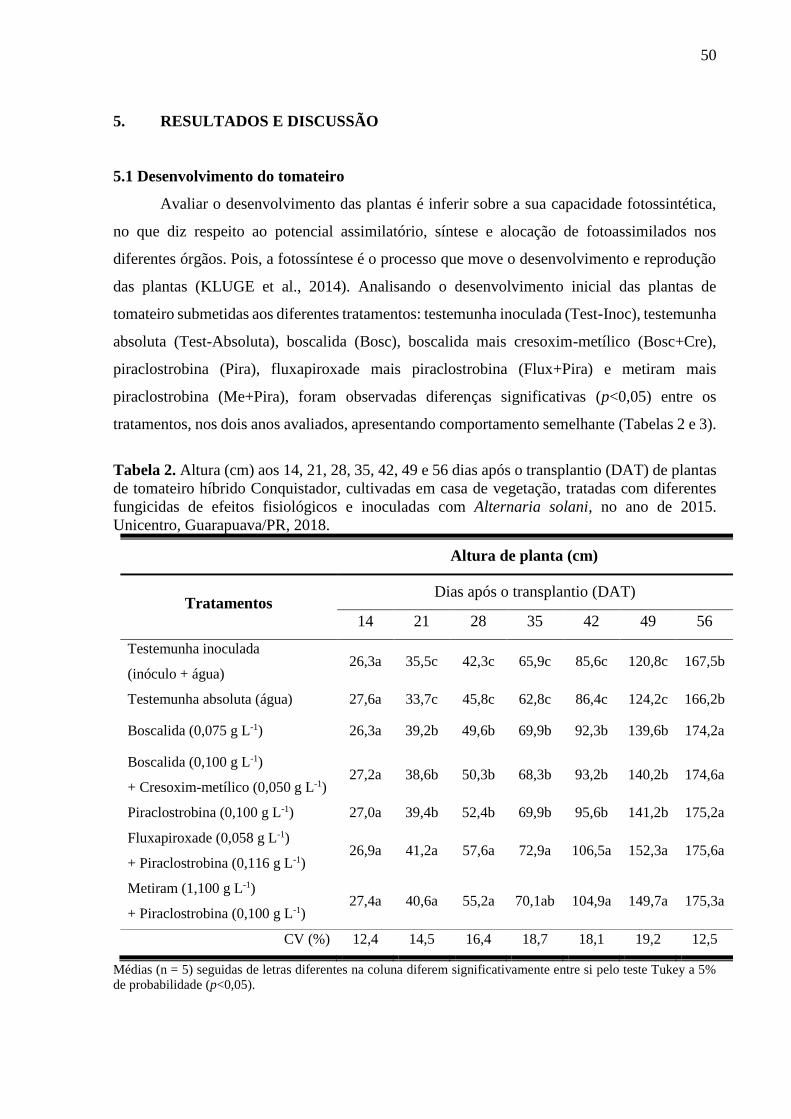
50
5. RESULTADOS E DISCUSSÃO
5.1 Desenvolvimento do tomateiro
Avaliar o desenvolvimento das plantas é inferir sobre a sua capacidade fotossintética,
no que diz respeito ao potencial assimilatório, síntese e alocação de fotoassimilados nos
diferentes órgãos. Pois, a fotossíntese é o processo que move o desenvolvimento e reprodução
das plantas (KLUGE et al., 2014). Analisando o desenvolvimento inicial das plantas de
tomateiro submetidas aos diferentes tratamentos: testemunha inoculada (Test-Inoc), testemunha
absoluta (Test-Absoluta), boscalida (Bosc), boscalida mais cresoxim-metílico (Bosc+Cre),
piraclostrobina (Pira), fluxapiroxade mais piraclostrobina (Flux+Pira) e metiram mais
piraclostrobina (Me+Pira), foram observadas diferenças significativas (p<0,05) entre os
tratamentos, nos dois anos avaliados, apresentando comportamento semelhante (Tabelas 2 e 3).
Tabela 2. Altura (cm) aos 14, 21, 28, 35, 42, 49 e 56 dias após o transplantio (DAT) de plantas
de tomateiro híbrido Conquistador, cultivadas em casa de vegetação, tratadas com diferentes
fungicidas de efeitos fisiológicos e inoculadas com Alternaria solani, no ano de 2015.
Unicentro, Guarapuava/PR, 2018.
Altura de planta (cm)
Tratamentos Dias após o transplantio (DAT)
14 21 28 35 42 49 56
Testemunha inoculada
(inóculo + água) 26,3a 35,5c 42,3c 65,9c 85,6c 120,8c 167,5b
Testemunha absoluta (água) 27,6a 33,7c 45,8c 62,8c 86,4c 124,2c 166,2b
Boscalida (0,075 g L-1) 26,3a 39,2b 49,6b 69,9b 92,3b 139,6b 174,2a
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 27,2a 38,6b 50,3b 68,3b 93,2b 140,2b 174,6a
Piraclostrobina (0,100 g L-1) 27,0a 39,4b 52,4b 69,9b 95,6b 141,2b 175,2a
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 26,9a 41,2a 57,6a 72,9a 106,5a 152,3a 175,6a
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 27,4a 40,6a 55,2a 70,1ab 104,9a 149,7a 175,3a
CV (%) 12,4 14,5 16,4 18,7 18,1 19,2 12,5
Médias (n = 5) seguidas de letras diferentes na coluna diferem significativamente entre si pelo teste Tukey a 5%
de probabilidade (p<0,05).
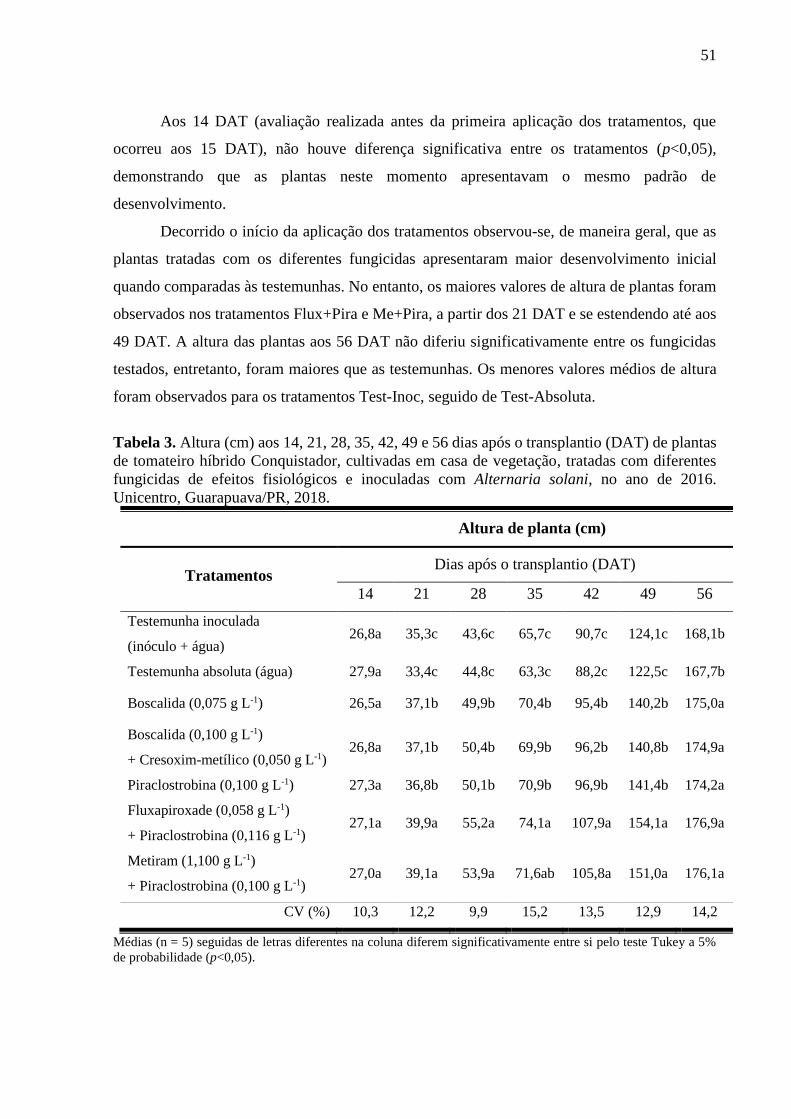
51
Aos 14 DAT (avaliação realizada antes da primeira aplicação dos tratamentos, que
ocorreu aos 15 DAT), não houve diferença significativa entre os tratamentos (p<0,05),
demonstrando que as plantas neste momento apresentavam o mesmo padrão de
desenvolvimento.
Decorrido o início da aplicação dos tratamentos observou-se, de maneira geral, que as
plantas tratadas com os diferentes fungicidas apresentaram maior desenvolvimento inicial
quando comparadas às testemunhas. No entanto, os maiores valores de altura de plantas foram
observados nos tratamentos Flux+Pira e Me+Pira, a partir dos 21 DAT e se estendendo até aos
49 DAT. A altura das plantas aos 56 DAT não diferiu significativamente entre os fungicidas
testados, entretanto, foram maiores que as testemunhas. Os menores valores médios de altura
foram observados para os tratamentos Test-Inoc, seguido de Test-Absoluta.
Tabela 3. Altura (cm) aos 14, 21, 28, 35, 42, 49 e 56 dias após o transplantio (DAT) de plantas
de tomateiro híbrido Conquistador, cultivadas em casa de vegetação, tratadas com diferentes
fungicidas de efeitos fisiológicos e inoculadas com Alternaria solani, no ano de 2016.
Unicentro, Guarapuava/PR, 2018.
Altura de planta (cm)
Tratamentos Dias após o transplantio (DAT)
14 21 28 35 42 49 56
Testemunha inoculada
(inóculo + água) 26,8a 35,3c 43,6c 65,7c 90,7c 124,1c 168,1b
Testemunha absoluta (água) 27,9a 33,4c 44,8c 63,3c 88,2c 122,5c 167,7b
Boscalida (0,075 g L-1) 26,5a 37,1b 49,9b 70,4b 95,4b 140,2b 175,0a
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 26,8a 37,1b 50,4b 69,9b 96,2b 140,8b 174,9a
Piraclostrobina (0,100 g L-1) 27,3a 36,8b 50,1b 70,9b 96,9b 141,4b 174,2a
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 27,1a 39,9a 55,2a 74,1a 107,9a 154,1a 176,9a
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 27,0a 39,1a 53,9a 71,6ab 105,8a 151,0a 176,1a
CV (%) 10,3 12,2 9,9 15,2 13,5 12,9 14,2
Médias (n = 5) seguidas de letras diferentes na coluna diferem significativamente entre si pelo teste Tukey a 5%
de probabilidade (p<0,05).

52
Dentre as características de desenvolvimento mensuradas em plantas de tomateiro,
também foram observadas diferenças significativas (p<0,05) entre os tratamentos para as
variáveis: área foliar (cm2), massas fresca e seca de folhas, caule e raízes (Tabelas 4 e 5).
Respostas semelhantes foram observadas nos dois anos avaliados, nos quais os maiores
valores médios de área foliar foram verificados no tratamento Flux+Pira (10452,1 e 10311,5
cm² de área foliar planta-1, nos anos de 2015 e 2016, respectivamente) seguido do tratamento
Me+Pira (9914,3 e 9889,8 cm² de área foliar planta-1, nos anos de 2015 e 2016,
respectivamente). Tais plantas, por apresentarem maior área foliar, tiveram maior capacidade
de aproveitar a energia em prol da fotossíntese e, assim, apresentaram melhor desenvolvimento
e ganhos de produtividade (REIS et al., 2013).
Ao contrário, os menores valores médios de área foliar foram identificados no
tratamento Test-Inoc (3298,3 e 3098,1 cm² de área foliar planta-1, nos anos de 2015 e 2016,
respectivamente) e Test-absoluta (6247,8 e 6384,2 cm² de área foliar planta-1, nos anos de 2015
e 2016, respectivamente). Como estas avaliações foram realizadas aos 120 DAT (após a última
colheita dos frutos), final do ciclo da cultura, as plantas testemunhas (Test-Inoc) estavam com
maior severidade da doença (A. solani inoculada aos 16 DAT). Devido a sua alta agressividade,
a incidência de A. solani em cultivos de tomateiro, provoca perdas diretas por infectar os frutos
e indiretas pela redução do vigor da planta, reduzindo seu desenvolvimento, devido ao
decréscimo da fotossíntese, pelos danos a área fotossinteticamente ativa, além da ocorrência da
desfolha em estádios mais avançados da doença (TÖFOLI et al., 2014).
Por isso, muitas folhas apresentavam lesões ocasionadas pela colonização do patógeno
e grande parte das folhas do terço inferior destas plantas tinham entrado em senescência e
sofrido abscisão, justificando, parcialmente, a menor área foliar. Parcialmente, porque as
plantas testemunhas (Test-Absoluta) não foram inoculadas no início do experimento e
receberam como tratamento somente a aplicação de água, mesmo assim, apresentaram menor
área foliar (6247,8 e 6384,2 cm² de área foliar planta-1, nos anos de 2015 e 2016,
respectivamente), sendo inferiores aquelas observadas para os tratamentos com os diferentes
fungicidas, diferindo significativamente (p<0,05) destes.
Além do efeito positivo na expansão foliar proporcionada por estas substâncias de
efeitos fisiológicos, também foram observados maiores acúmulos de massa fresca e seca de
folhas, caule e raiz nas plantas tratadas com os diferentes fungicidas. Assim como para área
foliar, os tratamentos Flux+Pira seguido de Me+Pira apresentaram maiores valores médios de

53
Tabela 4. Variáveis de desenvolvimento aos 120 dias após o transplantio (DAT) de plantas de tomateiro híbrido Conquistador, cultivadas
em casa de vegetação e tratadas com diferentes fungicidas de efeitos fisiológicos, inoculados com Alternaria solani, no ano de 2015.
Unicentro, Guarapuava/PR, 2018.
Variáveis de desenvolvimento
Tratamentos Área foliar
(cm²)
Massa fresca
de folhas (g)
Massa seca de
folhas (g)
Massa fresca
de caule (g)
Massa seca de
caule (g)
Massa fresca
de raiz (g)
Massa seca
de raiz (g)
Testemunha inoculada
(inóculo + água) 3298,32e 111,65e 17,26e 418,99c 42,32c 60,18b 9,24b
Testemunha absoluta (água) 6247,84d 211,84d 21,03d 423,56c 44,51c 63,93b 10,41b
Boscalida (0,075 g L-1) 8141,22c 275,37c 26,41c 443,25b 51,47b 81,03a 12,87a
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 8037,21c 270,12c 26,97c 440,24b 48,94b 78,91a 12,04a
Piraclostrobina (0,100 g L-1) 8119,04c 273,98c 26,24c 442,08b 49,12b 79,31a 12,55a
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 10452,11a 330,95a 39,09a 502,34a 56,98a 82,01a 13,02a
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 9914,29b 309,71b 30,42b 495,36a 55,09a 80,96a 12,94a
CV (%) 12,65 13,45 14,72 12,35 10,84 12,72 11,33
Médias (n = 5) seguidas de letras diferentes na coluna diferem significativamente entre si pelo teste Tukey a 5% de probabilidade (p<0,05).
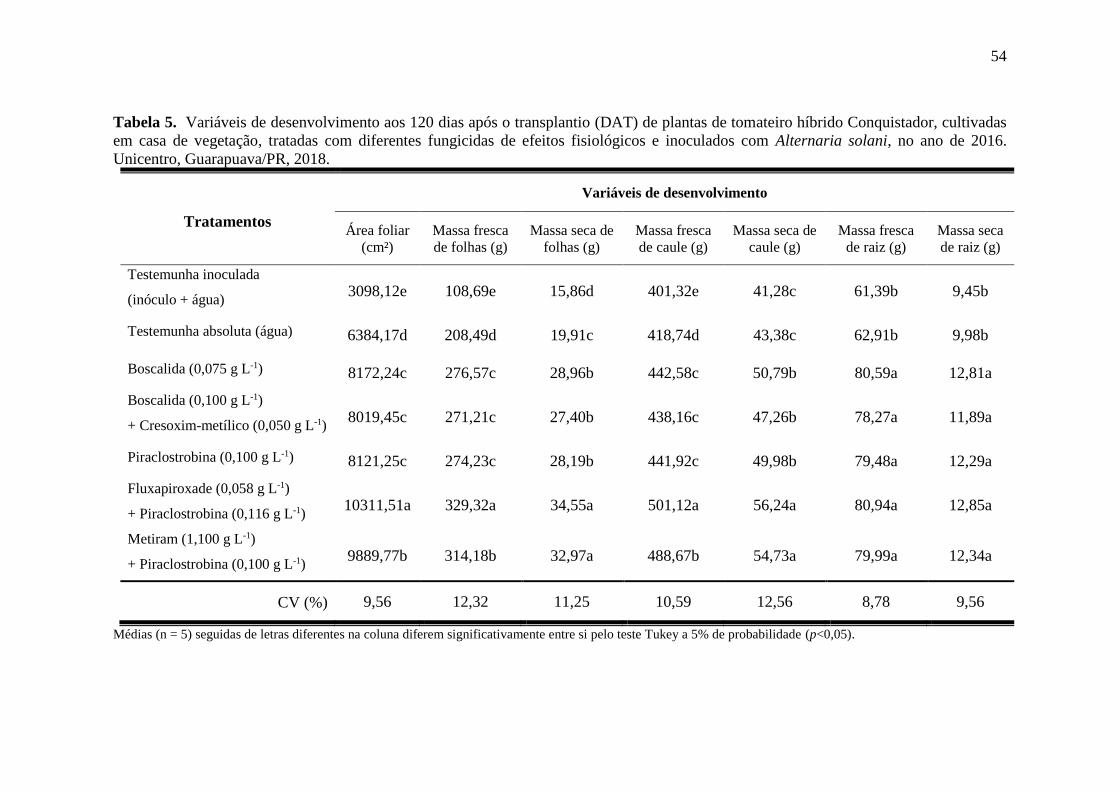
54
Tabela 5. Variáveis de desenvolvimento aos 120 dias após o transplantio (DAT) de plantas de tomateiro híbrido Conquistador, cultivadas
em casa de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, no ano de 2016.
Unicentro, Guarapuava/PR, 2018.
Variáveis de desenvolvimento
Tratamentos Área foliar
(cm²)
Massa fresca
de folhas (g)
Massa seca de
folhas (g)
Massa fresca
de caule (g)
Massa seca de
caule (g)
Massa fresca
de raiz (g)
Massa seca
de raiz (g)
Testemunha inoculada
(inóculo + água) 3098,12e 108,69e 15,86d 401,32e 41,28c 61,39b 9,45b
Testemunha absoluta (água) 6384,17d 208,49d 19,91c 418,74d 43,38c 62,91b 9,98b
Boscalida (0,075 g L-1) 8172,24c 276,57c 28,96b 442,58c 50,79b 80,59a 12,81a
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 8019,45c 271,21c 27,40b 438,16c 47,26b 78,27a 11,89a
Piraclostrobina (0,100 g L-1) 8121,25c 274,23c 28,19b 441,92c 49,98b 79,48a 12,29a
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 10311,51a 329,32a 34,55a 501,12a 56,24a 80,94a 12,85a
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 9889,77b 314,18b 32,97a 488,67b 54,73a 79,99a 12,34a
CV (%) 9,56 12,32 11,25 10,59 12,56 8,78 9,56
Médias (n = 5) seguidas de letras diferentes na coluna diferem significativamente entre si pelo teste Tukey a 5% de probabilidade (p<0,05).

55
massas fresca e seca de folhas e, também, proporcionaram maior acúmulo de massa fresca e
seca de caule, diferindo estatisticamente (p<0,05) dos demais tratamentos. No entanto, não
houve diferença significativa entre os fungicidas para o acúmulo de massa fresca e seca de
raízes, apenas diferindo das testemunhas.
Outros pesquisadores avaliando os efeitos fisiológicos proporcionados por
estrobilurinas e carboxamidas também observaram efeitos positivos no acúmulo de matéria seca
e na produtividade de milho (LIMA et al., 2009), feijão (KOZLOWSKI et al., 2009; DEMANT;
MARINGONI, 2012; JADOSKI et al., 2015), soja (TSUMANUMA et al., 2010), pepino
(AMARO, 2011), cenoura (COLOMBARI et al., 2015) e videira (MAREK, 2016).
Como observado, as plantas tratadas com Flux+Pira, seguidas daquelas que receberam
a aplicação de Me+Pira, se destacaram pelo maior desenvolvimento, apresentando maior área
foliar, configurando desta forma, um maior dossel foliar, consequentemente, tiveram maior
capacidade de absorver a luz incidente e isto influenciou diretamente nas taxas fotossintéticas
e no desenvolvimento das mesmas, como pode ser observado nos resultados obtidos por meio
das análises de trocas gasosas realizadas com o IRGA (“Infra Red Gas Analyser”, Modelo LI-
6400, Li-Cor) (Figuras 9 e 10).
5.2 Eficiência fotossintética
Nas avaliações de trocas gasosas realizadas aos 34 e 94 DAT (Figuras 9 e 10), foram
observadas diferenças significativas (p<0,05) entre os tratamentos, nos dois anos avaliados,
apresentando comportamento semelhante.
Pode ser observado efeito positivo de todos os fungicidas aplicados nos dois momentos
de avaliação (34 e 94 DAT), com aumento da eficiência fotossintética, em relação às
testemunhas. No entanto, plantas tratadas com Flux+Pira obtiveram os maiores valores para
taxa de assimilação de CO2 (A, μmol m-2 s-1) condutância estomática (gs, mol m-2 s-1) e
concentração interna de CO2 na folha (Ci, μmol mol-1) (Figura 9), diferindo estatisticamente
dos demais tratamentos (p<0,05). Em plantas de soja a aplicação de fluxapiroxade (CARRIJO,
2014) e de piraclostrobina (FAGAN et al., 2010) também promoveram aumento da taxa de
assimilação de CO2 e elevação da taxa fotossintética líquida.
A maior eficiência fotossintética observada nas plantas tratadas com Flux+Pira está
relacionada com a maior abertura estomática, através da qual houve maior difusão de CO2 para
a câmara subestomática, que posteriormente, foi utilizado na assimilação líquida de CO2. Uma
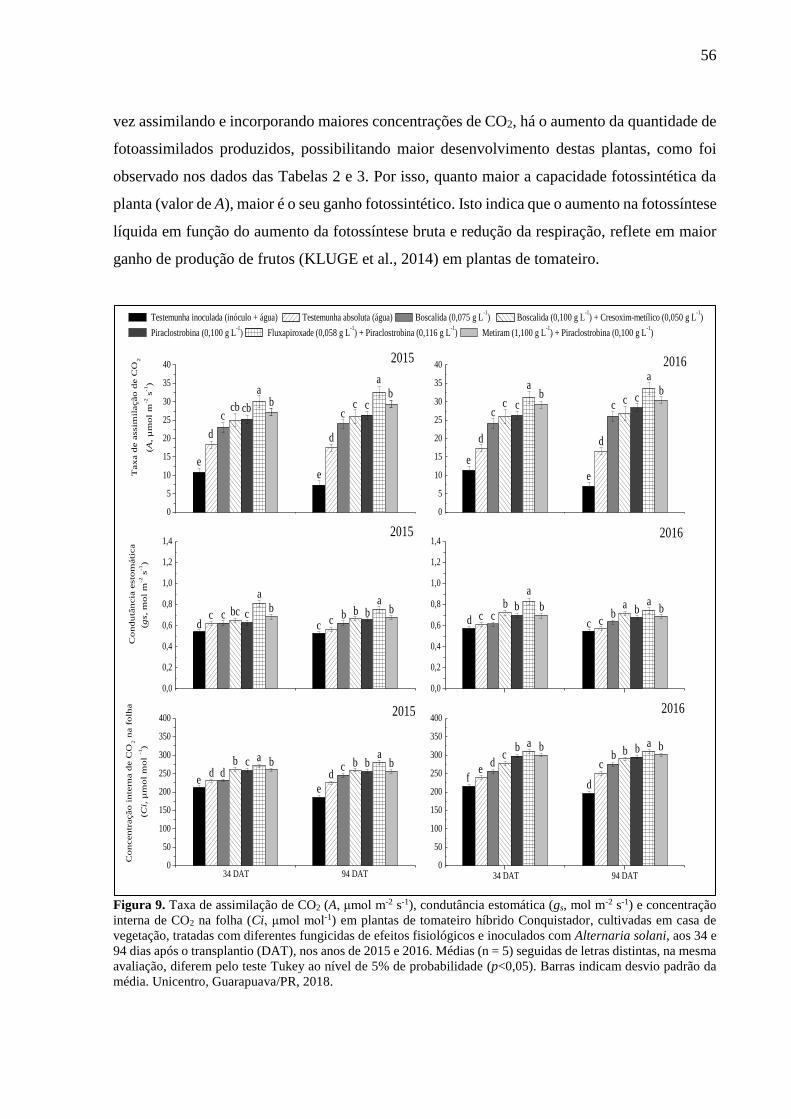
56
vez assimilando e incorporando maiores concentrações de CO2, há o aumento da quantidade de
fotoassimilados produzidos, possibilitando maior desenvolvimento destas plantas, como foi
observado nos dados das Tabelas 2 e 3. Por isso, quanto maior a capacidade fotossintética da
planta (valor de A), maior é o seu ganho fotossintético. Isto indica que o aumento na fotossíntese
líquida em função do aumento da fotossíntese bruta e redução da respiração, reflete em maior
ganho de produção de frutos (KLUGE et al., 2014) em plantas de tomateiro.
Figura 9. Taxa de assimilação de CO2 (A, μmol m-2 s-1), condutância estomática (gs, mol m-2 s-1) e concentração
interna de CO2 na folha (Ci, μmol mol-1) em plantas de tomateiro híbrido Conquistador, cultivadas em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, aos 34 e
94 dias após o transplantio (DAT), nos anos de 2015 e 2016. Médias (n = 5) seguidas de letras distintas, na mesma
avaliação, diferem pelo teste Tukey ao nível de 5% de probabilidade (p<0,05). Barras indicam desvio padrão da
média. Unicentro, Guarapuava/PR, 2018.
0
5
10
15
20
25
30
35
40
a
e
d
cc c
e
d
ccb cb
bb
a
Tax
a d
e a
ssim
ilação
de C
O2
(A
, µ
mo
l m
-2 s
-1)
0
5
10
15
20
25
30
35
40
e
d
c c cb
a
d
e
cc c
ba
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
bbcb
c
ba
dc c bc c
b
Co
nd
utâ
ncia
esto
máti
ca
(g
s,
mo
l m
-2 s
-1)
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1
) Boscalida (0,100 g L-1
) + Cresoxim-metílico (0,050 g L-1
)
Piraclostrobina (0,100 g L-1
) Fluxapiroxade (0,058 g L-1
) + Piraclostrobina (0,116 g L-1
) Metiram (1,100 g L-1
) + Piraclostrobina (0,100 g L-1
)
e
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
d c cb b b
aa
bbcc
a b
34 DAT 94 DAT0
50
100
150
200
250
300
350
400
e
dc b b b
a
ed d
cb ba
2015
2015 2016
2016
2016
Co
ncen
tração
in
tern
a d
e C
O2 n
a f
olh
a
(C
i, µ
mo
l m
ol
-1)
2015
a
34 DAT 94 DAT0
50
100
150
200
250
300
350
400
d
bc
b b ba
fe
dc
bb a

57
As alterações na abertura estomática estão relacionadas ao controle da perda de água
pela planta, que refletem na assimilação de CO2 e manutenção da taxa fotossintética (TAIZ;
ZEIGER, 2013). Desta forma, como observado nos tratamentos Test-Inoc e Test-Absoluta a
redução da abertura estomática restringiu a taxa de fixação de CO2 com consequente redução
de sua concentração nas cavidades subestomáticas e nos espaços intercelulares (concentração
interna de CO2), culminado em menor assimilação de CO2 e menor desenvolvimento destas
plantas (Tabelas 4 e 5). Uma vez que o crescimento e desenvolvimento das plantas dependem
dos processos de divisão e expansão celular, os quais são sensíveis a concentração de CO2
assimilado (TSUTSUMI et al., 2014), o aumento da assimilação de CO2 reflete diretamente na
fotossíntese líquida da planta, potencializando seu crescimento e desenvolvimento e,
consequentemente, o ganho de biomassa (XU et al., 2015).
Ao analisar a taxa de transpiração (E, mmol m-2 s-1) das plantas aos 34 e 94 DAT,
observa-se que não houve diferença estatística entre os fungicidas testados para esta
característica (Figura 10). No entanto, a transpiração das plantas nestes tratamentos é menor
quando comparada às testemunhas, diferindo significativamente destas (p<0,05). Nas plantas,
as variações na transpiração são ocasionadas por alterações na temperatura e no potencial
hídrico foliar, uma vez que os estômatos são sensíveis às mudanças ambientais, por isso quando
há aumento no déficit de pressão de vapor ocorrem limitações no processo fotossintético,
devido ao fechamento estomático e à diminuição de CO2 interno (TAIZ; ZEIGER, 2013).
Avaliando a eficiência do uso da água (EUA, A/E, μmol CO2 mmol H2O-1) e a eficiência
de carboxilação (A/Ci) (Figura 10), novamente o tratamento Flux+Pira se destaca e difere
estatisticamente (p<0,05) dos demais tratamentos, aos 34 e 94 DAT. A alta eficiência de
carboxilação observada neste tratamento reflete o eficiente funcionamento da enzima ribulose
1,5-difosfato carboxilase (Rubisco), enzima chave na fixação do CO2 no processo
fotossintético, em decorrência da maior abertura estomática e dos altos valores de assimilação
de CO2.
As plantas tratadas com Flux+Pira apresentaram maior EUA aos 34 DAT (3,99 e 4,13
μmol CO2 mmol H2O-1 em 2015 e 2016, respectivamente) e aos 94 DAT (4,10 e 4,22 μmol CO2
mmol H2O-1 em 2015 e 2016, respectivamente), devido às altas taxas de assimilação de CO2
observadas. Ao contrário, as plantas Test-Inoc apresentaram as menores EUA, com valores entre
0,70 e 1,14 μmol CO2 mmol H2O-1, nas avaliações realizadas nos dois anos de experimento.

58
Figura 10. Taxa de transpiração (E, mmol m-2 s-1), eficiência do uso da água (EUA, A/E, μmol CO2 (mmol H2O)-
1) e eficiência de carboxilação (A/Ci) em plantas de tomateiro híbrido Conquistador, cultivadas em casa de
vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, aos 34 e
94 dias após o transplantio (DAT), nos anos de 2015 e 2016. Médias (n = 5) seguidas de letras distintas, na mesma
avaliação, diferem pelo teste Tukey ao nível de 5% de probabilidade (p<0,05). Barras indicam desvio padrão da
média. Unicentro, Guarapuava/PR, 2018.
As plantas Test-Absoluta também apresentaram menores valores paras as características
avaliadas de trocas gasosas, quando comparadas aos diferentes fungicidas testados, mas
superiores a Test-Inoc. Mesmo na ausência de doença, as plantas do tratamento Test-Absoluta
apresentaram menor desenvolvimento, em função da menor eficiência fotossintética e,
consequentemente, menor produção de fotoassimilados. Evidenciando assim, o efeito positivo
da aplicação dos diferentes fungicidas no metabolismo fotossintético em plantas de tomateiro.
0
2
4
6
8
10
12
14
16
18
20
cccccb
bbbbb
aa
2015
Tax
a d
e t
ran
sp
iração
(E
, m
mo
l m
-2 s
-1)
2015
0
2
4
6
8
10
12
14
16
18
20
bbbbbb bbbbb
aaa
2016
2016
2016
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
5,0
ef
e
d
cbbc b
dc c
ab
aa
2015
Efi
ciê
ncia
do
uso
da á
gu
a
(A/E
, µ
mo
l C
O2 (
mm
ol
H2O
)-1)
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
5,0
e
d
bcc
b b
e
d
c bc bb
a a
34 DAT 94 DAT0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1
) Boscalida (0,100 g L-1
) + Cresoxim-metílico (0,050 g L-1
)
Piraclostrobina (0,100 g L-1
) Fluxapiroxade (0,058 g L-1
) + Piraclostrobina (0,116 g L-1
) Metiram (1,100 g L-1
) + Piraclostrobina (0,100 g L-1
)
d
c
b b b
e
d
c c cb
ba a
Efi
ciê
ncia
de c
arb
ox
ilação
(A
/Ci)
34 DAT 94 DAT0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
d
c
e
d
b ab c c c bb
a a
ab

59
O incremento na taxa fotossintética também foi relatado após a aplicação de fungicidas
de efeitos fisiológicos nas culturas de soja (FAGAN et al., 2010; CARRIJO, 2014), alface
(MANSANO, 2014), melão (MACEDO, 2012; 2015) e tomate (RAMOS, 2013; JACOBELIS
JUNIOR, 2015).
As plantas sob estresse biótico ou abiótico, apresentam baixa eficiência fotossintética.
Como observado nas análises de desenvolvimento e de trocas gasosas, nas plantas do tratamento
Test-Inoc, o progresso da doença, fator estressante, afetou negativamente o metabolismo
fotossintético. Por terem sido realizadas junto com as análises de trocas gasosas, as análises da
fluorescência da clorofila a, ajudam a entender a proporção da severidade do dano sofrido por
estas plantas sob estresse biótico, neste caso a presença do patógeno A. solani (Figura 11). Pois
os parâmetros obtidos pelas medidas da fluorescência da clorofila a são variáveis importantes
para descrever o estado fisiológico da planta, pois quando há maior dissipação de energia
através do transporte de elétrons, em decorrência de algum fator de estresse, ocorre declínio na
eficiência quântica potencial do fotossistema II (PSII, menor Fv/Fm) e menor taxa de transporte
de elétrons (ETR).
A luz solar ao incidir sobre as plantas é absorvida pelas clorofilas que se tornam
instáveis, isto é, excitadas, pois saem de seu estado basal para um estado de estímulo. No
entanto, para manter a homeostase do metabolismo fotossintético as clorofilas precisam
retornar ao estado basal, mas para isso, é necessário que a energia absorvida seja dissipada
(TAIZ; ZEIGER, 2013). Em plantas há três processos de dissipação da energia luminosa pelas
moléculas de clorofilas: fotoquímico e/ou fluorescência e/ou calor, que são complementares e
ao mesmo tempo competitivos.
As plantas saudáveis e em condições ambientais ótimas, direcionam a maior parte da
energia para a fotossíntese (síntese de moléculas orgânicas). Ao contrário, plantas sob estresse
abiótico ou biótico, apresentam uma maior proporção de energia dissipada de outras formas, ou
seja, a energia se torna não fotoquímica e é perdida (não é convertida em produtos
fotossintéticos). Desta forma, qualquer alteração na taxa fotossintética ou na dissipação de calor
refletirá em maior emissão de fluorescência (CAMPBELL; FARRELL, 2006; TAIZ; ZEIGER,
2013) permitindo assim, identificar a sanidade do aparato fotossintético da planta no momento
da avaliação da fluorescência da clorofila a.
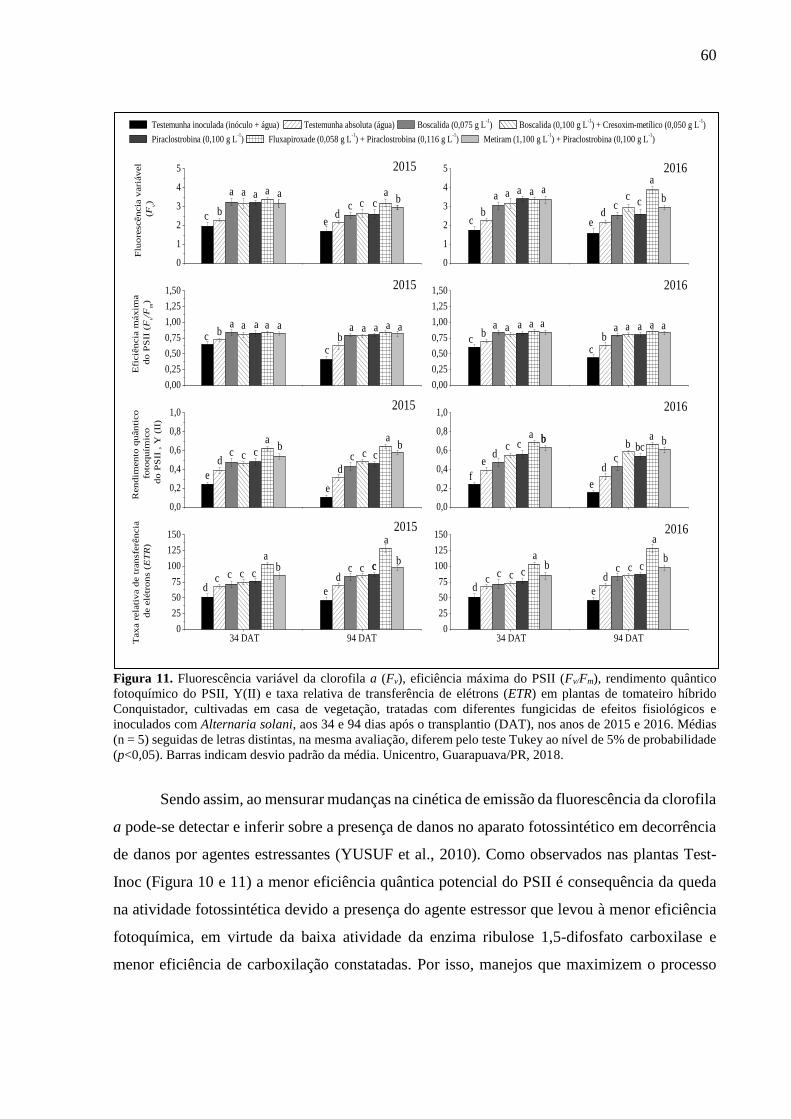
60
Figura 11. Fluorescência variável da clorofila a (Fv), eficiência máxima do PSII (Fv/Fm), rendimento quântico
fotoquímico do PSII, Y(II) e taxa relativa de transferência de elétrons (ETR) em plantas de tomateiro híbrido
Conquistador, cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e
inoculados com Alternaria solani, aos 34 e 94 dias após o transplantio (DAT), nos anos de 2015 e 2016. Médias
(n = 5) seguidas de letras distintas, na mesma avaliação, diferem pelo teste Tukey ao nível de 5% de probabilidade
(p<0,05). Barras indicam desvio padrão da média. Unicentro, Guarapuava/PR, 2018.
Sendo assim, ao mensurar mudanças na cinética de emissão da fluorescência da clorofila
a pode-se detectar e inferir sobre a presença de danos no aparato fotossintético em decorrência
de danos por agentes estressantes (YUSUF et al., 2010). Como observados nas plantas Test-
Inoc (Figura 10 e 11) a menor eficiência quântica potencial do PSII é consequência da queda
na atividade fotossintética devido a presença do agente estressor que levou à menor eficiência
fotoquímica, em virtude da baixa atividade da enzima ribulose 1,5-difosfato carboxilase e
menor eficiência de carboxilação constatadas. Por isso, manejos que maximizem o processo
0
1
2
3
4
5
ed
c c c ba
bc
a a a aa
Flu
ore
scência
vari
ável
(Fv)
0
1
2
3
4
5
ed
cc
c b
a
cb
a a a a a
2016
0,00
0,25
0,50
0,75
1,00
1,25
1,50
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1)
Piraclostrobina (0,100 g L-1) Fluxapiroxade (0,058 g L
-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
cb
a aa aac b
a a aa
Efi
ciê
ncia
máxim
a
do P
SII
(F
v/F
m)
e
0,00
0,25
0,50
0,75
1,00
1,25
1,50
cc
ba a a
ba a aaa aa
2016
0,0
0,2
0,4
0,6
0,8
1,0
e
dc c c
b
e
dc c c
baa
2015
2015
Rendim
ento
quânti
co
foto
quím
ico
do P
SII
, Y
(II
)
0,0
0,2
0,4
0,6
0,8
1,0
b
a
f
e
e
d
dc
c
c bcb bba a
2016
2016
34 DAT 94 DAT0
25
50
75
100
125
150
e
dc
dc c c
c c cbba
a
Taxa r
ela
tiva d
e t
ransfe
rência
de e
létr
ons (
ET
R)
34 DAT 94 DAT0
25
50
75
100
125
150
cd e
dcc cc
c c cbb
a
a
2015
2015

61
fotossintético e dissipem maior quantidade de energia para síntese de carboidratos são
vantajosos.
O rendimento quântico máximo do PSII foi estimado pela razão Fv/Fm e indica a
eficiência da planta na captura da energia de excitação pelos centros de reação abertos do PSII
e sua dissipação no transporte de elétrons do PSII para o fotossistema I (PSI), impulsionando a
síntese de ATP e NADPH2, fontes essenciais de energia para a fixação de CO2. Como pode ser
observado na Figura 11, a aplicação dos diferentes fungicidas de efeitos fisiológicos teve efeito
significativo sobre os parâmetros de fluorescência da clorofila a avaliados aos 34 e 94 DAT. A
fluorescência variável (Fv), eficiência máxima do PSII (Fv/Fm), rendimento quântico
fotoquímico do PSII, Y(II) e a taxa relativa de transferência de elétrons (ETR) apresentaram
comportamento semelhante nos dois anos avaliados (Figura 11).
Estudos realizados com diferentes espécies que apresentam ciclo C3 (como o
tomateiro) demonstram que valores típicos do parâmetro Fv/Fm em folhas saudáveis e, em
condições de iluminação adequadas, variam em torno de 0,83 e valores abaixo deste padrão
indicam que a planta apresenta algum tipo de estresse causando danos ao aparato fotossintético
(BJÖRKMAN; DEMMING, 1987). Neste caso, ocorre a redução na eficiência quântica da
evolução do O2 ou na fixação do CO2, comprometendo o desenvolvimento e produtividade da
planta.
De acordo com os dados apresentados na Figura 11, os valores de Fv/Fm foram
significativamente diferentes (p<0,05) entre os tratamentos, porém não houve diferença
estatística entre as plantas tratadas com os diferentes fungicidas, os quais apresentaram maiores
valores de rendimento quântico potencial do PSII aos 34 e 94 DAT. Os valores variaram entre
0,79 e 0,86, revelando que estas plantas não estavam sob estresse no momento em que as
avaliações foram realizadas. Assim, maiores valores de Fv/Fm indicam maior eficiência de
captura de energia de excitação pelos centros de reação do PSII, reafirmando os resultados
observados nas avaliações de trocas gasosas (Figuras 9 e 10), nas quais a aplicação dos
diferentes fungicidas aumentou, em intensidades diferentes, a eficiência de assimilação e
carboxilação de CO2 pelas plantas.
Ao contrário, o tratamento Test-Inoc, que apresentou menor Fv/Fm teve redução na
eficiência de assimilação e carboxilação de CO2 e, possivelmente, a ocorrência de alterações no
PSII, refletindo em menor capacidade fotossintética e eficiência fotoquímica (CASIERRA-
POSADA et al., 2011). Pois, a presença de condições estressantes leva ao decréscimo na

62
eficiência quântica potencial do PSII, podendo ser detectada pela queda na relação Fv/Fm
(KRAUSE; WEIS, 1991; BAKER; ROSENQVIST, 2004; BAKER, 2008), conforme já
mencionado. Por isso o parâmetro Fv/Fm tem sido amplamente utilizado para identificar e
comparar tecidos saudáveis e danificados por estresses bióticos (ROUSSEAU et al., 2013).
Nas plantas Test-Inoc, foi observado menor Fv/Fm aos 34 e 94 DAT, com queda de
0,64 para 0,43, apresentando declínio acentuado de uma avaliação para outra, indicando
possivelmente a presença de danos no aparato fotossintético. Já no tratamento Test-Absoluta,
os valores de Fv/Fm aos 34 e 94 DAT foram de 0,73 e 0,62, respectivamente.
Uma vez que os valores de Fv/Fm em folhas saudáveis se encontram próximos de 0,83
(BJÖRKMAN; DEMMING, 1987) e como observado, as plantas dos tratamentos Test-Inoc
apresentaram os menores valores deste parâmetro (0,43 e 0,64), pode-se inferir que estas se
encontravam em severas condições de estresse em decorrência do progresso da doença, que
refletiu em perdas da eficiência do PSII. As espécies do gênero Alternaria são produtoras de
toxinas prolíficas, que facilitam a sua vida necrotrófica, provocando a morte da célula, causando
danos ao tecido vegetal (LAWRENCE et al., 2008), resultando em perda de eficiência
fotossintética.
A síntese dos compostos orgânicos é dependente da produção de ATP e NAPH2,
formados durante o transporte de elétrons entre o PSII e o PSI. Assim, a presença do patógeno
A. solani, devido a sua agressividade, afetou a capacidade fotossintética das plantas Test-Inoc
e provocou o desbalanço entre os fotossistemas PSII e PSI, como pode ser observado pela
menor transferência de elétrons (ETR). Em consequência disto, possivelmente, ocorreu o
transporte pseudocíclico de elétrons, também conhecido como Reação de Mehler, onde há
formação de ânion superóxido (•O2-), a partir do oxigênio molecular que recebe o elétron que
seria transferido da ferredoxina para o NADP+ (BHATTACHARJEE, 2010).
Com relação aos parâmetros de fluorescência variável (Fv) e rendimento quântico
efetivo do PSII, Y(II) observou-se diferença significativa entre os tratamentos (Figura 11), com
destaque para o tratamento Flux+Pira que apresentou os maiores valores para Fv aos 94 DAT,
no entanto, sem diferir dos demais fungicidas aos 34 DAT. Tais plantas ao apresentarem
maiores valores de Fv evidenciam maior capacidade de assimilar CO2, uma vez que este
parâmetro é indicativo da capacidade de adaptabilidade do PSII (BAKER; ROSENQVIST,
2004).

63
Plantas que apresentam maiores valores de Fv indicam maior capacidade de
transferência de elétrons entre PSII e PSI para a geração de NADPH2 e ATP, culminando em
maior eficiência fotossintética (BAKER, 2008). Tal fato confirma os resultados verificados para
trocas gasosas, nos quais as plantas submetidas ao tratamento Flux+Pira apresentaram maior
assimilação de CO2, também evidenciado aqui pelos maiores valores de rendimento quântico
efetivo do PSII, sendo aquelas onde se observou os maiores valores de Fv e ETR (Figura 11).
Maiores valores de rendimento quântico efetivo do PSII, Y(II) observados no
tratamento Flux+Pira, revelam maior eficiência fotoquímica do PSII, pois este parâmetro
permite avaliar a capacidade de absorção da luz pelas clorofilas e sua potencial utilização
fotoquimicamente (MAXWELL; JOHNSON, 2000).
Os resultados referentes ao ETR (Figura 11), foram significativamente diferentes entre
os tratamentos (p<0,05), sendo maiores nos tratamentos com aplicação dos diferentes
fungicidas (Flux+Pira > Me+Pira > Pira > Bosc+Cre > Bosc). O transporte de elétrons entre o
PSII e PSI tem como destino final a fixação de CO2, sintetizando carboidratos. Desta forma,
qualquer fator que cause alterações no processo fotossintético refletirá em mudanças na ETR.
De fato, o tratamento Flux+Pira, que apresentou maior rendimento quântico efetivo do PSII
também apresentou maior ETR. Pois, quanto maior o valor de ETR maior é a soma dos destinos
finais dos elétrons no cloroplasto, que serão utilizados principalmente para a fixação de carbono
(MISRA et al., 2011).
Isto comprova a maior eficiência de carboxilação observada no tratamento Flux+Pira,
indicando que estas plantas, tanto aos 34 DAT (que compreendia a fase vegetativa), quanto aos
94 DAT (plena fase reprodutiva), encontravam-se com potencial capacidade fotossintética.
Diante disto, pode-se afirmar que tais plantas produziram maior quantidade de NADPH2 e ATP
(devido ao maior ETR) e utilizaram eficientemente estes produtos no ciclo de Calvin para a
síntese de carboidratos, suprindo as demandas de crescimento, desenvolvimento e produção
que, por consequência, foram maiores neste tratamento.
Ao contrário, as plantas Test-Inoc apresentaram menores valores de Fv, com isto a
eficiência quântica efetiva do PSII da folha diminuiu com o aumento da doença, como já
discutido, em virtude da redução de Fv/Fm em decorrência dos danos causados pelo patógeno
nos tecidos fotossintéticos. Com isso, houve mudanças negativas no processo fotossintético,
prejudicando a síntese e transporte de carboidratos, acarretando em menor utilização de
NADPH2 e ATP e, consequentemente, a eficiência quântica efetiva de PSII diminuiu. Não

64
havendo o consumo de NADPH2 e ATP, há a redução do fluxo de elétrons, como demonstrado
pela baixa ETR nas plantas Test-Inoc, seguidas de Test-Absoluta.
5.3 Pigmentos fotossintéticos e atividade da enzima nitrato redutase
A absorção da luz ocorre nos PSI e PSII, os quais são constituídos de pigmentos
fotossintéticos (clorofila a, clorofila b e carotenoides) (CASIERRA-POSADA; PEÑA-
OLMOS, 2015), por isso a quantidade de clorofilas presentes no tecido foliar reflete
significativamente a eficiência fotossintética de uma planta. Sendo assim o acréscimo nos
teores de clorofilas a e b resulta em maior absorção da luz e consequentemente maior ETR entre
o PSII e PSI (RODRIGUES et al., 2016).
O aumento da eficiência fotossintética, observado nos tratamentos com os diferentes
fungicidas, tem grande relação com o teor de pigmentos fotossintéticos das folhas, analisadas
aos 15, 20, 35, 50, 65, 80, 95 e 110 DAT (Figura 12), que de maneira geral, apresentaram
diferença significativa (p<0,05) entre os tratamentos nos dois anos avaliados.
Na primeira avaliação aos 15 DAT, realizada antes da aplicação dos fungicidas, não se
verificou diferença estatística significativa (p<0,05) entre os tratamentos, no entanto, a partir
dos 20 DAT as plantas tratadas com Flux+Pira apresentaram os maiores teores de clorofila a e
b (µg g massa fresca-1), tendo aumento expressivo com o avanço das avaliações. Este
comportamento está relacionado com o efeito somatório das aplicações, uma vez que as
avalições compreendidas entre 20 e 95 DAT foram realizadas sempre no 5º dia após a aplicação,
portanto, aos 20, 35, 50, 65, 80 e 95 DAT temos: 5º dia após a 1º, 2º, 3º, 4º, 5º e 6º aplicação
dos tratamentos, respectivamente.
Nota-se também que o tratamento Me+Pira com o avançar das avaliações apresentou
efeito semelhante ao Flux+Pira, não diferindo estatisticamente nos teores de clorofila a e b, em
algumas avalições. Estes resultados estão de acordo com os dados observados nas avaliações
de trocas gasosas e fluorescência da clorofila a, onde se constatou maior incremento
fotossintético nas plantas tratadas com Flux+Pira, seguidas de Me+Pira. De acordo com Baker
e Rosenqvist (2004) quanto maior a quantidade de clorofila a, mais elevados estarão os
processos fotoquímicos da fotossíntese, como de fato pode ser observado nos tratamentos
Flux+Pira e Me+Pira, que apresentaram maior concentração de clorofila a a partir dos 20 DAT.
Este start inicial promovido por estes tratamentos refletiu diretamente na fotossíntese líquida e

65
nos demais parâmetros fotossintéticos avaliados, demonstrando ser um dos importantes efeitos
destas moléculas em plantas de tomateiro.
Figura 12. Teor de clorofila a (µg g massa fresca-1), clorofila b (µg g massa fresca-1) e carotenoides (µg g massa
fresca-1) em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes
fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, aos 15, 20, 35, 50, 65, 80, 95 e 110 dias após
o transplantio (DAT), nos anos de 2015 e 2016. Médias (n = 5) seguidas de letras distintas, na mesma avaliação,
diferem pelo teste Tukey ao nível de 5% de probabilidade (p<0,05). Barras indicam desvio padrão da média.
Unicentro, Guarapuava/PR, 2018.
Pelos resultados observados a aplicação destes fungicidas exerce efeitos fisiológicos e
influenciam significativamente a síntese de pigmentos fotossintéticos, em especial de clorofilas
a e b. Incrementos nos teores de clorofilas também foram relatados em meloeiro (MACEDO,
2015) e tomateiro (RAMOS, 2013; JACOBELIS JUNIOR, 2015), após aplicação de fungicidas
a base de estrobilurinas e carboxamidas. Este efeito reflete em maior eficiência fotossintética
300
450
600
750
900
1050
1200
1350
100
150
200
250
300
350
400
450
15 DAT 20 DAT 35 DAT 50 DAT 65 DAT 80 DAT 95 DAT 110 DAT
200
250
300
350
400
450
500
550
15 DAT 20 DAT 35 DAT 50 DAT 65 DAT 80 DAT 95 DAT 110 DAT
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1
) Boscalida (0,100 g L-1
) + Cresoxim-metílico (0,050 g L-1
)
Piraclostrobina (0,100 g L-1
) Fluxapiroxade (0,058 g L-1
) + Piraclostrobina (0,116 g L-1
) Metiram (1,100 g L-1
) + Piraclostrobina (0,100 g L-1
)
ddd
c ccccc
c
bbbbb
bb
aaaaaaa
Clo
rofi
la a
(µ
g g
massa f
resca
-1)
acc
dd
d
dd
ccc
bbbb
b
d
bc
b
aaa
aaaa
a
2016
2016
2016
a
dd
cc
ccccc
c
b b
bc
b
bc
b
bb
aaaaa
a
a
a
2015
Clo
rofi
la b
(µ
g g
massa f
resca
-1)
2015
dddd
ccccc
bbbbbc
dd
c
bb
aaaaa
a
a
ccc
c cc
bb
bb
bb
b
aaa
a
a
aa
a
2015
Caro
ten
oid
es (
µg
g m
assa f
resca
-1)
c
c
cc
c
bb
b
bb
c
b
b
aa
a
a
aa
a
a

66
das plantas, porque a clorofila, em especial a clorofila a, é o elemento essencial no processo
fotossintético e, quanto maior a concentração destes pigmentos, maior é a absorção e transporte
da energia solar pelos fotossistemas, convertendo-a em energia química (YEOM et al., 2014).
Ao contrário, os menores teores de pigmentos fotossintéticos foram observados nos
tratamentos Test-Inoc, seguido de Test-absoluta, em todas as avalições realizadas após os 15
DAT. Ao analisar estes teores de pigmentos e comparar com os parâmetros de emissão de
fluorescência da clorofila a, nota-se que há um avançado grau de estresse nestas plantas
(fotoinibição crônica), pois como já discutido, as plantas Test-Inoc apresentaram níveis mais
reduzidos de Fv/Fm aos 94 DAT, em decorrência do progresso da doença. Sugere-se, portanto,
que a redução dos valores de Fv/Fm pode ter ocorrido devido aos danos causados pelo patógeno
no tecido foliar, pela degradação do conteúdo celular, que acarretou em menores teores de
clorofilas a e b.
Além disto, a presença do patógeno também desencadeou o processo de estresse
oxidativo, todas estas reações em conjunto promoveram, consequentemente, à menor
capacidade de aproveitamento da luz pelas plantas deste tratamento. Pois, quando a energia
armazenada na clorofila não é dissipada para a síntese de carboidratos, é então transferida para
o O2, promovendo o transporte pseudocíclico de elétrons (BHATTACHARJEE, 2010). Isto
ocorre devido à baixa fixação de CO2 e menor poder redutor (NADP+) e, a ferredoxina acaba
transferindo o elétron ao O2 formando •O2- (AHMAD et al., 2008).
As plantas quando submetidas a condições de estresse abiótico ou biótico aumentam a
produção de EROs (BHATTACHARJEE, 2010; KARUPPANAPANDIAN et al., 2011) e,
consequentemente, sofrem alterações no equilíbrio das reações fotoquímicas pelos danos
causados no aparato fotossintético, percebida na cinética de emissão da fluorescência nas
plantas (BAKER; ROSENQVIST, 2004).
Quanto aos teores de carotenoides foram observados maiores valores nas plantas
tratadas com Flux+Pira, seguidas de Me+Pira, diferindo significativamente entre si e dos outros
tratamentos nas avaliações dos 20 aos 110 DAT, nos dois anos avaliados. Ao contrário, os
menores teores de pigmentos foram observados nas plantas Test-Inoc, seguido do tratamento
Test-Absoluta. Supõe-se, portanto, que os menores teores de pigmentos nestes tratamentos,
principalmente nas plantas Test-Inoc, se devam aos danos causados pela presença de A. solani.
Pois, como se trata de um patógeno necrotrófico, a destruição do tecido vegetal reflete em lesões
celulares e causa danos no metabolismo de síntese de pigmentos fotossintéticos. Uma vez que

67
as plantas refletem os danos sofridos no metabolismo fotossintético por meio da concentração
de pigmentos e pela emissão de fluorescência da clorofila a (BRESTIC; ZIVCAK, 2013), como
já observados.
Somado ao efeito fungitóxico, pode-se dizer que os fungicidas de efeitos fisiológicos
como Flux+Pira e Me+Pira também apresentam efeito positivo na proteção do aparato
fotossintético, pois os carotenoides possuem função antioxidativa e protegem a maquinaria
fotoquímica de danos por estresse oxidativo (danos fotoxidativos). Pois, inevitavelmente há
formação de EROs e de seus subprodutos durante o processo da fotossíntese, os quais causam
severos danos oxidativos no aparato fotossintético e diminuem sua eficiência, assim a síntese
de compostos antioxidativos como os carotenoides são importantes fotoprotetores e
antioxidantes (NISAR et al., 2015).
Os teores de pigmentos fotossintéticos nos vegetais são influenciados pela
disponibilidade, absorção e assimilação de nitrogênio em seu metabolismo. O nitrogênio é um
dos principais constituintes da molécula de clorofila (TAIZ; ZEIGER, 2013), por isso a
concentração de clorofilas a e b têm forte relação com a quantidade de nitrogênio que a planta
assimila. A nitrato redutase é a enzima chave na regulação do metabolismo do nitrogênio
(VIANA; KIEHL, 2010) assim, a atividade desta enzima está intrinsicamente ligada ao maior
aporte de pigmentos para o processo fotossintético da planta.
De acordo com a Figura 13 foram avaliados os efeitos fisiológicos na atividade da
enzima nitrato redutase aos 15, 20, 35, 50, 65, 80, 95 e 110 DAT (mesmos dias em que foram
avaliados os teores de pigmentos). De modo geral, plantas tratadas com os diferentes fungicidas
apresentaram maior atividade da enzima nitrato redutase ao longo das avaliações, verificando-
se diferença significativa (p<0,05) entre os tratamentos, em ambos os anos avaliados.
Dentre os tratamentos, as plantas que receberam a aplicação de Flux+Pira e Me+Pira
foram aquelas que apresentaram maior atividade da enzima nitrato redutase, nas coletas
realizadas após os 15 DAT, o que contribuiu para o maior teor de pigmentos fotossintéticos
evidenciados nestes mesmos tratamentos. Justificando a maior eficiência fotossintética e
desenvolvimento destas plantas que, possivelmente, interferiu na produção de frutos.
Aumento da atividade da enzima nitrato redutase e efeito verde também foram relatados
por Soares et al. (2011), Macedo (2015) e Ramos (2013) em plantas de soja, melão e tomate,
respectivamente, tratadas com fungicidas a base de estrobilurinas e carboxamidas.

68
Figura 13. Atividade da enzima nitrato redutase (NR, μg nitrito min-1 g de massa fresca-1) em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação,
tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, aos 15, 20, 35, 50, 65, 80, 95 e 110 dias após o transplantio (DAT), nos
anos de 2015 e 2016. Médias (n = 5) seguidas de letras distintas, na mesma avaliação, diferem pelo teste Tukey ao nível de 5% de probabilidade (p<0,05). Barras indicam
desvio padrão da média. Unicentro, Guarapuava/PR, 2018.
0
1
2
3
4
5
6
7
8
9
10
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
a a aa
b b bb bc
c
c
e
d
dd
cc c
aa
aa
b b bb b
d d
c c
bb b bb b b
aa
a a
a a a a a a
Ati
vid
ade d
a n
itra
to r
eduta
se (
µg n
itri
to m
in-1
g m
ass
a f
resc
a-1
) 2015
ad
c c
b b
aa
15 DAT 20 DAT 35 DAT 50 DAT 65 DAT 80 DAT 95 DAT 110 DAT0
1
2
3
4
5
6
7
8
9
10
e
d
a aa
c bcb
a
a
aa
a
b
c
a a a
bc
aad d d
e
efcc
ddcc
b b b c cc b b
bbcc
d cd cb
aa
a
b
a
aa
2016

69
Pode-se observar na que os maiores picos de atividade da enzima nitrato redutase nos
tratamentos Flux+Pira e Me+Pira (Figura 13) ocorreram nas avaliações realizadas aos 50, 65 e
80 DAT, período no qual as plantas de tomateiro se encontravam em transição entre a fase
vegetativa e início da fase reprodutiva, momento de maior necessidade de nitrogênio para suprir
a demanda de fotoassimilados para o desenvolvimento dos frutos. Aos 95 e 110 DAT, fase em
que foi observado maior crescimento e maturação de frutos, as plantas tratadas com Flux+Pira
novamente apresentaram maior atividade da enzima nitrato redutase, seguidas do tratamento
Me+Pira, evidenciando assim os efeitos positivos destas substâncias na fisiologia de plantas de
tomateiro.
A aplicação de fungicidas que promovem efeitos fisiológicos, como as estrobilurinas e
mais recentemente as carboxamidas, aumentam a biomassa e a produção, mesmo em plantas
que não são afetadas por fungos, por isso são práticas agrícolas interessantes no cenário
agrícola. Tais fungicidas, em especial a piraclostrobina, aumentam a assimilação de nitrogênio
por elevarem a atividade de NADH-nitrato, por isso a aplicação de piraclostrobina pode ser
considerada uma alternativa importante para o manejo da adubação nitrogenada (KANUNGO;
JOSHI, 2014).
Plantas que assimilam maior quantidade de nitrogênio também sintetizam maiores
concentrações de compostos nitrogenados essenciais para o seu crescimento (VIANA; KIEHL,
2010). De fato, as plantas Test-Inoc e Test-Absoluta apresentaram a menor atividade da enzima
nitrato redutase, a qual culminou em menor teor de pigmentos, baixa atividade fotossintética,
acarretando menor altura de planta e menor acúmulo de massa fresca e seca de plantas, como
observado nas análises de desenvolvimento.
5.4 Síntese e translocação de carboidratos
A maior obra da fotossíntese é permitir o crescimento e desenvolvimento do vegetal, a
partir de um complexo processo de reações de oxi-redução, fixando o CO2 em compostos
orgânicos. Uma vez que a produção de carboidratos é o produto final da fotossíntese, relações
entre órgãos fonte-dreno também são influenciados pela eficiência fotossintética. Desta forma,
foi avaliada a translocação de fotoassimilados nas plantas de tomateiro aos 50 e 120 DAT
(Tabelas 6 e 7).
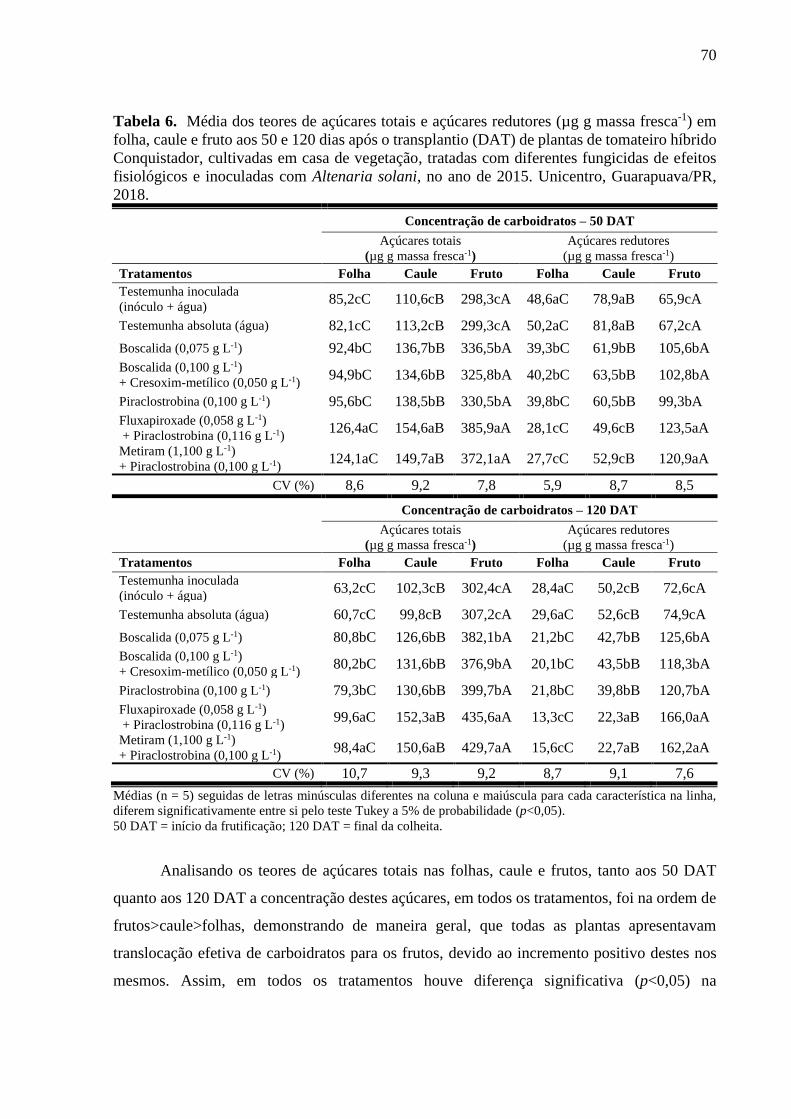
70
Tabela 6. Média dos teores de açúcares totais e açúcares redutores (µg g massa fresca-1) em
folha, caule e fruto aos 50 e 120 dias após o transplantio (DAT) de plantas de tomateiro híbrido
Conquistador, cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculadas com Altenaria solani, no ano de 2015. Unicentro, Guarapuava/PR,
2018.
Concentração de carboidratos – 50 DAT
Açúcares totais
(µg g massa fresca-1)
Açúcares redutores
(µg g massa fresca-1)
Tratamentos Folha Caule Fruto Folha Caule Fruto
Testemunha inoculada
(inóculo + água) 85,2cC 110,6cB 298,3cA 48,6aC 78,9aB 65,9cA
Testemunha absoluta (água) 82,1cC 113,2cB 299,3cA 50,2aC 81,8aB 67,2cA
Boscalida (0,075 g L-1) 92,4bC 136,7bB 336,5bA 39,3bC 61,9bB 105,6bA
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 94,9bC 134,6bB 325,8bA 40,2bC 63,5bB 102,8bA
Piraclostrobina (0,100 g L-1) 95,6bC 138,5bB 330,5bA 39,8bC 60,5bB 99,3bA
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 126,4aC 154,6aB 385,9aA 28,1cC 49,6cB 123,5aA
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 124,1aC 149,7aB 372,1aA 27,7cC 52,9cB 120,9aA
CV (%) 8,6 9,2 7,8 5,9 8,7 8,5
Concentração de carboidratos – 120 DAT
Açúcares totais
(µg g massa fresca-1)
Açúcares redutores
(µg g massa fresca-1)
Tratamentos Folha Caule Fruto Folha Caule Fruto
Testemunha inoculada
(inóculo + água) 63,2cC 102,3cB 302,4cA 28,4aC 50,2cB 72,6cA
Testemunha absoluta (água) 60,7cC 99,8cB 307,2cA 29,6aC 52,6cB 74,9cA
Boscalida (0,075 g L-1) 80,8bC 126,6bB 382,1bA 21,2bC 42,7bB 125,6bA
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 80,2bC 131,6bB 376,9bA 20,1bC 43,5bB 118,3bA
Piraclostrobina (0,100 g L-1) 79,3bC 130,6bB 399,7bA 21,8bC 39,8bB 120,7bA
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 99,6aC 152,3aB 435,6aA 13,3cC 22,3aB 166,0aA
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 98,4aC 150,6aB 429,7aA 15,6cC 22,7aB 162,2aA
CV (%) 10,7 9,3 9,2 8,7 9,1 7,6
Médias (n = 5) seguidas de letras minúsculas diferentes na coluna e maiúscula para cada característica na linha,
diferem significativamente entre si pelo teste Tukey a 5% de probabilidade (p<0,05).
50 DAT = início da frutificação; 120 DAT = final da colheita.
Analisando os teores de açúcares totais nas folhas, caule e frutos, tanto aos 50 DAT
quanto aos 120 DAT a concentração destes açúcares, em todos os tratamentos, foi na ordem de
frutos>caule>folhas, demonstrando de maneira geral, que todas as plantas apresentavam
translocação efetiva de carboidratos para os frutos, devido ao incremento positivo destes nos
mesmos. Assim, em todos os tratamentos houve diferença significativa (p<0,05) na

71
concentração de açúcares totais entre os diferentes órgãos avaliados, sendo o mesmo observado
para os teores de açúcares redutores.
Tabela 7. Média dos teores de açúcares totais, açúcares redutores (µg g massa fresca-1) em
folha, caule e fruto aos 50 e 120 dias após o transplantio (DAT) de plantas de tomateiro híbrido
Conquistador, cultivadas em casa de vegetação e tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculadas com Alternaria solani, no ano de 2016. Unicentro, Guarapuava/PR,
2018.
Concentração de carboidratos – 50 DAT
Açúcares totais
(µg g massa fresca-1)
Açúcares redutores
(µg g massa fresca-1)
Tratamentos Folha Caule Fruto Folha Caule Fruto
Testemunha inoculada
(inóculo + água) 89,5cC 121,9cB 299,9cA 45,5aC 84,6aB 60,3cA
Testemunha absoluta (água) 86,3cC 124,2cB 303,1cA 48,2aC 85,7aB 62,4cA
Boscalida (0,075 g L-1) 96,2bC 143,1bB 342,5bA 37,3bC 66,2bB 95,3bA
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 98,8bC 147,8bB 338,3bA 36,4bC 65,6bB 96,8bA
Piraclostrobina (0,100 g L-1) 99,1bC 146,4bB 337,4bA 37,9bC 68,2bB 93,4bA
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 129,1aC 160,2aB 388,9aA 25,5cC 55,6cB 121,3aA
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 126,9aC 159,9aB 389,0aA 24,9cC 54,8cB 118,9aA
CV (%) 9,6 8,7 7,2 6,4 7,4 8,4
Concentração de carboidratos – 120 DAT
Açúcares totais
(µg g massa fresca-1)
Açúcares redutores
(µg g massa fresca-1)
Tratamentos Folha Caule Fruto Folha Caule Fruto
Testemunha inoculada
(inóculo + água) 68,0cC 99,3cB 322,9cA 31,9aC 56,2cB 78,1cA
Testemunha absoluta (água) 66,8cC 103,6cB 318,9cA 32,6aC 55,0cB 79,4cA
Boscalida (0,075 g L-1) 80,2bC 126,6bB 384,1bA 25,1bC 40,7bB 123,7bA
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 78,4bC 123,2bB 385,9bA 23,9bC 44,6bB 126,9bA
Piraclostrobina (0,100 g L-1) 79,9bC 124,0bB 387,2bA 26,5bC 41,2bB 125,7bA
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 89,7aC 139,6aB 447,2aA 14,4cC 27,4aB 158,1aA
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 91,9aC 141,3aB 446,1aA 11,9cC 28,6aB 162,0aA
CV (%) 9,4 8,7 10,4 9,7 8,4 6,8
Médias (n = 5) seguidas de letras minúsculas diferentes na coluna e maiúscula para cada característica na linha,
diferem significativamente entre si pelo teste Tukey a 5% de probabilidade (p<0,05).
50 DAT = início da frutificação; 120 DAT = final da colheita.
No entanto, os tratamentos Flux+Pira e Me+Pira foram aqueles que apresentaram as
maiores concentrações destes carboidratos nas folhas, caules e frutos, nas duas avaliações

72
realizadas (50 e 120 DAT), diferindo significativamente (p<0,05) dos demais tratamentos, com
comportamento semelhante nos dois anos avaliados.
Isto indica que estas substâncias promoveram maior síntese e acúmulo de carboidratos
em plantas de tomateiro, estando de acordo com os dados obtidos para as análises de eficiência
fotossintética. Verificando os teores destes carboidratos nos tratamentos Bosc, Bosc+Cre e Pira
aos 50 e 120 DAT observou-se que não houve diferença estatística entre eles para os teores de
açúcares totais e redutores nos órgãos avaliados. Porém, estes três tratamentos diferem
significativamente (p<0,05) dos demais, apresentando valores médios destas características
superiores aos das testemunhas (Test-Inoc e Test-Absoluta) e inferiores aos observados para
Flux+Pira e Me+Pira.
Nos dois anos avaliados (Tabelas 6 e 7), tanto aos 50 DAT quanto aos 120 DAT, as
menores concentrações de açúcares totais e redutores foram observados nas plantas Test-Inoc
e Test-Absoluta, evidenciando que os diferentes fungicidas avaliados influenciaram na
concentração e partição de carboidratos em plantas de tomateiro.
Avaliando a aplicação de piraclostrobina e boscalida, isolados e combinados com
outros produtos químicos em plantas de tomateiro, Ramos et al. (2015) também observaram
que em todos os tratamentos as concentrações de açúcares totais e redutores foram superiores
à testemunha, além de verificarem que nos frutos foi estatisticamente superior às folhas e caule,
evidenciando que estes tratamentos são efetivos na maior translocação destes açúcares, por
promoverem maior desenvolvimento do tecido, aumentando a capacidade de dreno do órgão
em crescimento, neste caso o fruto.
Quanto à concentração de açúcares redutores os menores valores foram encontrados nas
folhas das plantas tratadas com Flux+Pira e Me+Pira, diferindo significativamente dos demais
tratamentos. Isto condiz com o observado nos dados obtidos para as trocas gasosas,
apresentando feedback positivo, uma vez que estas plantas ao possuírem menor concentração
destes açúcares na fonte, não apresentaram limitações no processo fotossintético (devido ao
acúmulo de fotoassimilados) e com isso, as taxas de assimilação de CO2 se mantiveram altas.
Os açúcares são os produtos primários da fotossíntese, por isso quando estão acima da
concentração normal nas folhas (fonte) estimulam rotas de sinalização que bloqueiam a
expressão de genes associados à fotossíntese, caracterizando um feedback negativo do produto
final e os processos fotossintéticos descontinuam até que esta fase se reverta (QUIRINO et al.,
2000; TAIZ; ZEIGER, 2013).

73
Por serem menos reativos, os carboidratos translocados a longas distâncias no floema
são os açúcares não redutores (DINANT; LEMOINE, 2010), assim, a sacarose é o açúcar
predominantemente translocado via floema, mas outros açúcares móveis também estão
presentes como a rafinose e estaquiose, além dos açúcares-álcoois, como o sorbitol e manitol
(VAN BEL; HESS, 2008).
Altas taxas fotossintéticas promovem abundante síntese de açúcares que reflete em
maior crescimento e armazenamento de carboidratos nos drenos (TAIZ; ZEIGER, 2013). O
principal dreno em plantas de tomateiro a partir da floração são os frutos, como observado os
tratamentos Flux+Pira e Me+Pira apresentaram as maiores concentrações de açúcares totais e
redutores aos 120 DAT nos frutos, diferindo significativamente (p<0,05) dos demais
tratamentos.
A maior síntese e acúmulo destes carboidratos nos frutos pode ser explicada pela maior
eficiência fotossintética já relatada, entretanto, também se observou eficiente translocação
destes açúcares no sentido fonte-dreno. Estudos têm demonstrado que as estrobilurinas, como
a piraclostrobina alteram o balanço hormonal da planta, promovendo a síntese de auxinas e
ácido abscísico (ABA) e diminuindo a produção de etileno (KANUNGO; JOSHI, 2014). As
citocininas, por promoverem maior crescimento do dreno e expansão foliar, aumentam a taxa
fotossintética e a capacidade do dreno no tecido. Já o etileno inibe o crescimento dos drenos e
diminui a atividade das invertases (ALBACETE et al., 2013). Diante disto, é possível inferir
que tenha ocorrido um melhor balanço hormonal influenciando a regulação das relações fonte-
dreno com a aplicação destas substâncias, promovendo maior crescimento do tecido dreno e
aumentando a translocação de fotoassimilados para estes órgãos.
Aos 120 DAT os tratamentos Flux+Pira e Me+Pira apresentaram as maiores
concentrações de açúcares totais em todos os órgãos avaliados, evidenciando eficiente acúmulo
de reservas no decorrer do tempo. Isto condiz com o observado nas análises de desenvolvimento
para estas mesmas plantas, uma vez que 90% da massa seca da planta é formada por
fotoassimilados que foram convertidos em biomassa ao longo do desenvolviemnto da planta
(POPOV et al., 2003). Portanto, de acordo com os efeitos observados, os tratamentos Flux+Pira
e Me+Pira promoveram alterações nas relações entre fonte-dreno e, consequentemente, na
produtividade das plantas de tomateiro.
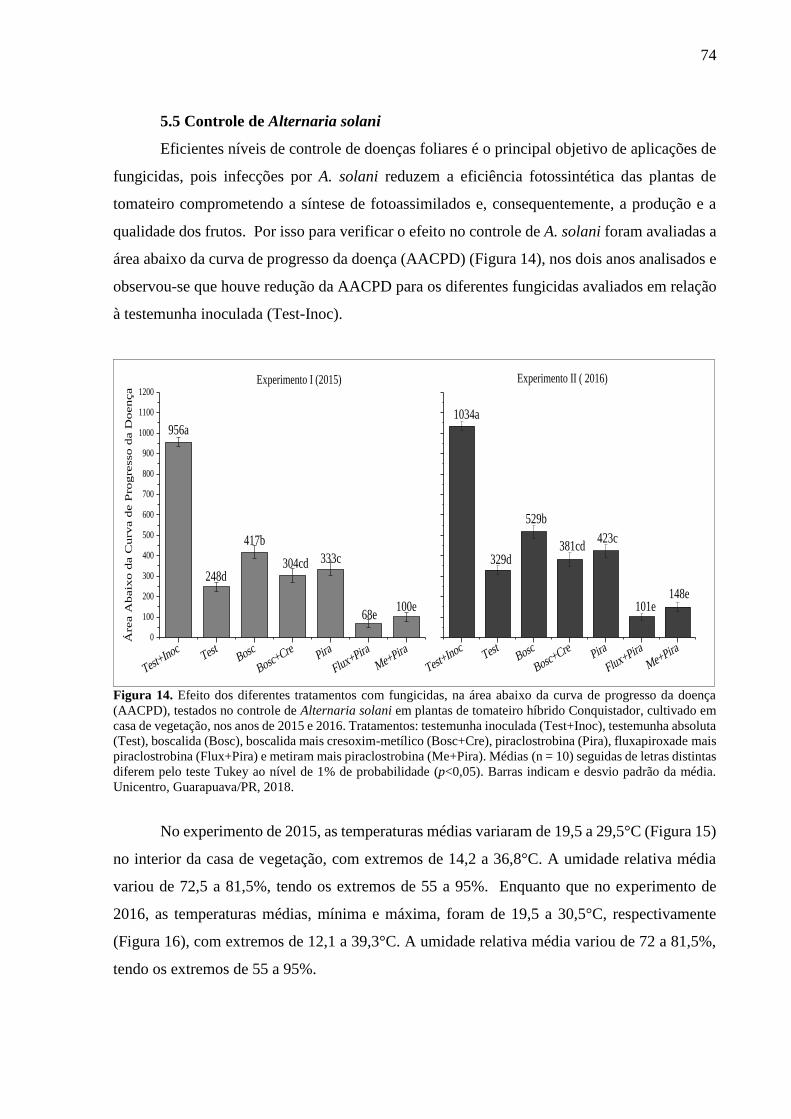
74
5.5 Controle de Alternaria solani
Eficientes níveis de controle de doenças foliares é o principal objetivo de aplicações de
fungicidas, pois infecções por A. solani reduzem a eficiência fotossintética das plantas de
tomateiro comprometendo a síntese de fotoassimilados e, consequentemente, a produção e a
qualidade dos frutos. Por isso para verificar o efeito no controle de A. solani foram avaliadas a
área abaixo da curva de progresso da doença (AACPD) (Figura 14), nos dois anos analisados e
observou-se que houve redução da AACPD para os diferentes fungicidas avaliados em relação
à testemunha inoculada (Test-Inoc).
Test+Inoc
Test Bosc
Bosc+Cre Pira
Flux+Pira
Me+Pira0
100
200
300
400
500
600
700
800
900
1000
1100
1200
Experimento II ( 2016)
100e68e
333c304cd
417b
248d
Áre
a A
baix
o d
a C
urv
a d
e P
rog
resso
da D
oen
ça
956a
Experimento I (2015)
Test+Inoc
Test Bosc
Bosc+Cre Pira
Flux+Pira
Me+Pira
329d381cd
423c
529b
101e148e
1034a
Figura 14. Efeito dos diferentes tratamentos com fungicidas, na área abaixo da curva de progresso da doença
(AACPD), testados no controle de Alternaria solani em plantas de tomateiro híbrido Conquistador, cultivado em
casa de vegetação, nos anos de 2015 e 2016. Tratamentos: testemunha inoculada (Test+Inoc), testemunha absoluta
(Test), boscalida (Bosc), boscalida mais cresoxim-metílico (Bosc+Cre), piraclostrobina (Pira), fluxapiroxade mais
piraclostrobina (Flux+Pira) e metiram mais piraclostrobina (Me+Pira). Médias (n = 10) seguidas de letras distintas
diferem pelo teste Tukey ao nível de 1% de probabilidade (p<0,05). Barras indicam e desvio padrão da média.
Unicentro, Guarapuava/PR, 2018.
No experimento de 2015, as temperaturas médias variaram de 19,5 a 29,5°C (Figura 15)
no interior da casa de vegetação, com extremos de 14,2 a 36,8°C. A umidade relativa média
variou de 72,5 a 81,5%, tendo os extremos de 55 a 95%. Enquanto que no experimento de
2016, as temperaturas médias, mínima e máxima, foram de 19,5 a 30,5°C, respectivamente
(Figura 16), com extremos de 12,1 a 39,3°C. A umidade relativa média variou de 72 a 81,5%,
tendo os extremos de 55 a 95%.
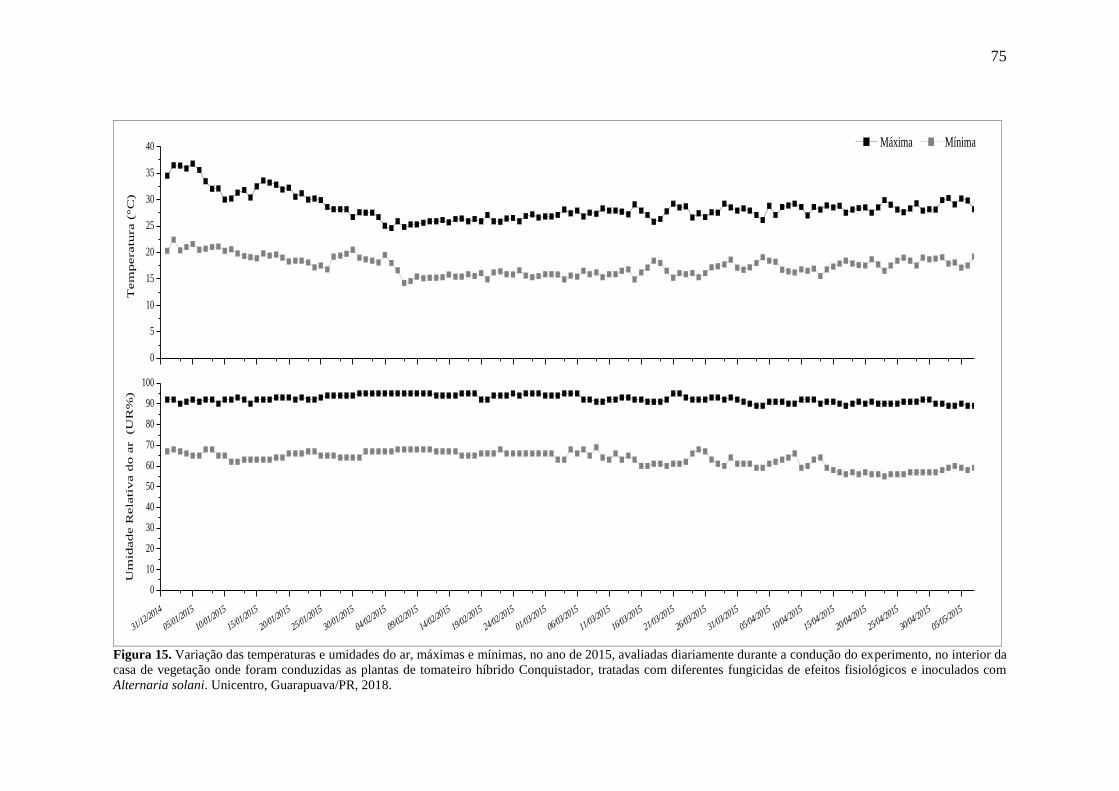
75
0
5
10
15
20
25
30
35
40
31/12/2014
05/01/2015
10/01/2015
15/01/2015
20/01/2015
25/01/2015
30/01/2015
04/02/2015
09/02/2015
14/02/2015
19/02/2015
24/02/2015
01/03/2015
06/03/2015
11/03/2015
16/03/2015
21/03/2015
26/03/2015
31/03/2015
05/04/2015
10/04/2015
15/04/2015
20/04/2015
25/04/2015
30/04/2015
05/05/2015
0
10
20
30
40
50
60
70
80
90
100
Tem
peratu
ra (
°C
)
Máxima Mínima
Um
idad
e R
ela
tiv
a d
o a
r
(U
R%
)
Figura 15. Variação das temperaturas e umidades do ar, máximas e mínimas, no ano de 2015, avaliadas diariamente durante a condução do experimento, no interior da
casa de vegetação onde foram conduzidas as plantas de tomateiro híbrido Conquistador, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani. Unicentro, Guarapuava/PR, 2018.
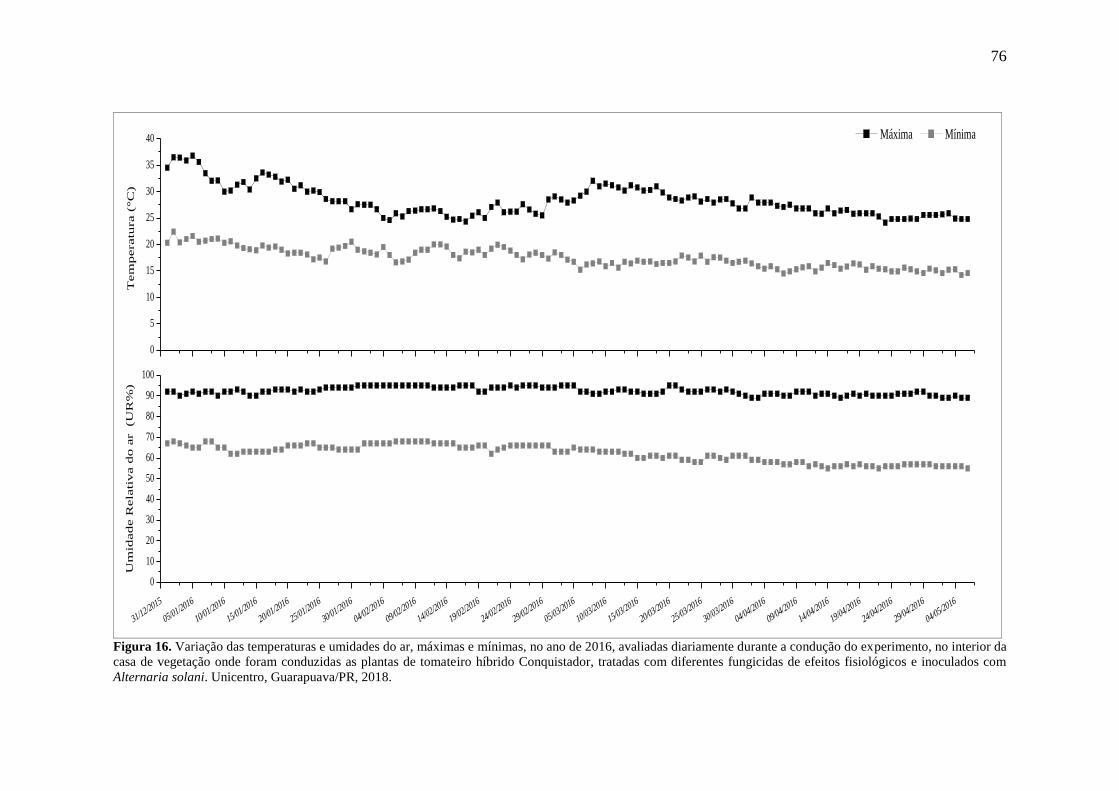
76
0
5
10
15
20
25
30
35
40
31/12/2015
05/01/2016
10/01/2016
15/01/2016
20/01/2016
25/01/2016
30/01/2016
04/02/2016
09/02/2016
14/02/2016
19/02/2016
24/02/2016
29/02/2016
05/03/2016
10/03/2016
15/03/2016
20/03/2016
25/03/2016
30/03/2016
04/04/2016
09/04/2016
14/04/2016
19/04/2016
24/04/2016
29/04/2016
04/05/2016
0
10
20
30
40
50
60
70
80
90
100
Tem
peratu
ra (
°C
) Máxima Mínima
Um
idad
e R
ela
tiv
a d
o a
r
(U
R%
)
Figura 16. Variação das temperaturas e umidades do ar, máximas e mínimas, no ano de 2016, avaliadas diariamente durante a condução do experimento, no interior da
casa de vegetação onde foram conduzidas as plantas de tomateiro híbrido Conquistador, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com
Alternaria solani. Unicentro, Guarapuava/PR, 2018.

77
O aumento da severidade da pinta preta é proporcional ao aumento da umidade relativa
e de temperaturas na faixa de 25 a 32ºC, onde ocorre intensa redução da área foliar, queda do
vigor das plantas, danos em caules e depreciação de frutos (TÖFOLI; DOMINGUES, 2005).
De fato, a ausência de controle e as condições ambientais ideais (Figuras 15 e 16) para o
desenvolvimento da doença acarretou em maior intensidade desta na testemunha inoculada
(Test-Inoc), verificado pelas maiores AACPDs, nos dois anos avaliados.
Devido às condições ideais observadas para o desenvolvimento da doença, altas
temperaturas e umidade relativa do ar, somadas a presença de inóculo (plantas doentes: Test-
Inoc) as plantas Test-Absoluta, que não foram inoculadas no início do experimento, também
manifestaram certo percentual de severidade de doença nas últimas avaliações, por isso
apresentaram índices de AACPD de 248 e 329 nos anos de 2015 e 2016, respectivamente.
As lesões de A. solani são desenvolvidas a partir de penetrações que ocorrem após a
formação do apressório e, no caso deste patógeno, esta estrutura é essencial para o processo de
infecção. Por isso, quando se tem menor número de apressórios formados se observa menor
frequência de lesões no hospedeiro (BALBI-PEÑA et al., 2014). Pode-se observar que os
fungicidas Flux+Pira, Me+Pira, Bosc, Pira e Bosc+Cre reduziram os valores em 92,88, 89,54,
56,38, 65,16 e 68,20% para AACPD no ano de 2015, e em 90,23, 85,68, 48,84, 59,09 e 63,15%
no ano de 2016, respectivamente, em relação à testemunha inoculada.
As menores AACPDs nas plantas tratadas com os fungicidas avaliados em contraste
com a testemunha Test-Inoc indicam que estas moléculas apresentam eficiente controle durante
a germinação dos conídios de A. solani devido ao seu efeito sobre a cadeia de transporte de
elétrons na mitocôndria, afetando a respiração do fungo. As estrobilurinas atuam no complexo
III da respiração mitocondrial, sendo que os princípios ativos piraclostrobina (Pira) e cresoxim-
metílico (Cre) bloqueiam a transferência de elétrons entre o citocromo b e o citocromo c1,
interferindo no transporte de elétrons e, consequentemente, na formação de ATP para as células
do fungo (KANUNGO; JOSHI, 2014; MATOS et al., 2016).
Já as carboxamidas, fluxapiroxade (Flux) e boscalida (Bosc), atuam na respiração
mitocondrial bloqueando a transferência de elétrons no complexo II, inibindo a enzima
succinato dehidrogenase (SDHI), reduzindo o processo respiratório e bloqueando o
fornecimento de energia das células do fungo (FRAC, 2017). São fungicidas de ação preventiva
e curativa, atuando na germinação de esporos, alongamento do tubo germinativo, crescimento
micelial e esporulação de uma ampla variedade de fungos (EFSA, 2015).

78
No entanto, os tratamentos Flux+Pira e Me+Pira apresentaram as menores AACPDs,
diferindo significativamente (p<0,01) dos demais tratamentos, com resultados semelhantes nos
dois anos avaliados. Os fungicidas Bosc, Bosc+Cre e Pira também foram eficientes em reduzir
a severidade da doença, quando comparados à testemunha Test-Inoc (Figura 14). Assim, o uso
destas estrobilurinas e carboxamidas são eficientes tanto para o controle de A. solani, além de
promoverem maior rendimento fotossintético de plantas de tomateiro. Honorato Júnior et al.
(2015) também verificaram eficiente controle de Hemileia vastatrix em Coffea arabica e
melhor desempenho fotossintético das plantas após a aplicação foliar de piraclostrobina,
comprovando a ação ‘extra’ destes produtos.
5.6 Estresse oxidativo e enzimas antioxidantes
Por consequência da presença da doença nas plantas, decorrentes da inoculação
realizada aos 16 DAT, houve exposição destas a um fator estressante (estresse biótico). Como
observados nos dados de eficiência fotossintética (trocas gasosas e fluorescência da clorofila
a), síntese e translocação de carboidratos, desenvolvimento e acúmulo de biomassa nas plantas
de tomateiro, nota-se que a presença deste fator estressante também teve influência no
metabolismo fotossintético, no desenvolvimento e, possivelmente, na produtividade destas
plantas.
O estresse dispara várias respostas bioquímicas no metabolismo da planta, entre elas a
biossíntese de componentes oxidativos e, consequentemente, a atividade de enzimas
antioxidativas, configurando uma cascata de eventos. Em plantas sob condições de crescimento
normais a produção de EROs é baixa. No entanto, quando submetidas à algum tipo de estresse
biótico ou abiótico, a primeira resposta observada na planta é o aumento drástico da produção
de espécies reativas de oxigênio (EROs ou ROS, do inglês reactive oxygen species) que causam
danos oxidativos em proteínas, lipídios e ácidos nucleicos, caracterizando o estresse oxidativo
(BARBOSA et al., 2014).
Quando se trata de estresse por agente fitopatogênico, uma das primeiras respostas
celulares após o reconhecimento planta-patógeno é a explosão oxidativa, caracterizada por uma
superprodução de ânion superóxido (•O2-) e do produto de sua dismutação, o peróxido de
hidrogênio (H2O2), principalmente no apoplasto, mas também podendo ser produzidos em
outros compartimentos celulares, como nas mitocôndrias e cloroplastos (SHARMA et al.,
2012). No entanto, Resende et al. (2003) relatam que primeiro ocorre uma pequena e rápida

79
explosão oxidativa, e após ocorre uma segunda explosão, mais forte e prolongada que
caracteriza o processo de resistência, culminando em respostas de defesa como a HR.
O ânion •O2- (radical moderadamente reativo e instável) é gerado por meio de diferentes
vias metabólicas na planta a partir da redução do O2 por um único elétron (RIBEIRO et al.,
2005). Por ser a primeira espécie reativa formada, o •O2- dá origem às outras espécies reativas.
O superóxido •O2-, por intermédio da enzima SOD é dismutado a H2O2 (BHATTACHARJEE,
2010). Uma vez protonado, o •O2- dá origem ao radical peroxila (HO2
•), altamente reativo,
porém presente em pequenas concentrações em pH fisiológico (BARBOSA et al., 2014).
Desta maneira, também foi avaliada a influência dos tratamentos no sistema
antioxidativo enzimático ao longo do ciclo de crescimento e desenvolvimento das plantas de
tomateiro. Nas avaliações da concentração do ânion •O2- em folhas de tomateiro (Figura 17),
aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e
105 DAT, observou-se comportamento similar nos dois anos avaliados. Pode-se notar que
houve aumento da produção desta espécie reativa de oxigênio aos 17 DAT (coleta realizada 24
horas após a inoculação do patógeno) nas plantas Test-Inoc. Isto ocorreu porque durante o
contato inicial entre o patógeno e a célula vegetal, dá-se início a uma cascata de eventos, tendo
como uma das primeiras respostas a explosão oxidativa formando as EROs, e como já relatado,
o •O2- é a primeira molécula reativa a ser formada.
As EROs, iniciando com o •O2-, são formadas pelo vazamento inevitável de elétrons que
acabam sendo ‘aceitos’ pelo O2. Tais elétrons são oriundos dos processos de transporte de
elétrons nos cloroplastos, mitocôndrias e membranas plasmáticas, ou até mesmo, como um
subproduto de várias vias metabólicas localizadas em diferentes compartimentos celulares. Por
isso, as EROs são produzidas em diferentes locais na célula, tais como: cloroplastos,
mitocôndrias, plasmalema, peroxissomas, apoplasto, retículo endoplasmático e paredes
celulares (SHARMA et al., 2012). A transferência de elétrons na mitocôndria ou no cloroplasto
são importantes vias de formação de •O2-, pois ativam as NADPH-oxidases/sintases ligadas à
membrana, além de peroxidases da parede celular e lipoxigenases (RESENDE et al., 2003;
RIBEIRO et al., 2005).
Estudos têm demonstrados que as EROs também agem como “moléculas sinalizadoras”
ou “mensageiros secundários”, atuando na via de sinalização em resposta a situações de
estresses nas plantas, influenciando na expressão de vários genes envolvidos no metabolismo
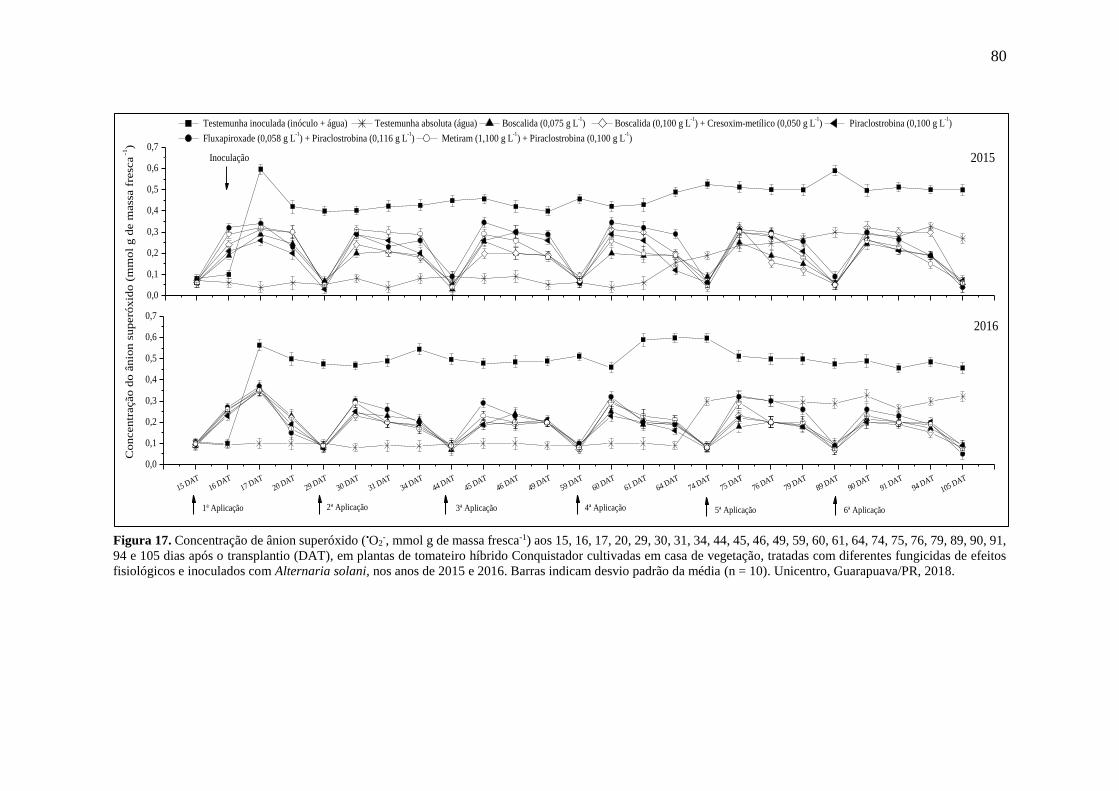
80
Figura 17. Concentração de ânion superóxido (•O2
-, mmol g de massa fresca-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91,
94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
15 DAT16 DAT
17 DAT20 DAT
29 DAT30 DAT
31 DAT34 DAT
44 DAT45 DAT
46 DAT49 DAT
59 DAT60 DAT
61 DAT64 DAT
74 DAT75 DAT
76 DAT79 DAT
89 DAT90 DAT
91 DAT94 DAT
105 DAT
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
2016
Co
ncen
tração
do
ân
ion
su
peró
xid
o (
mm
ol
g d
e m
ass
a f
resc
a-1
)
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

81
de defesa da planta (BARBOSA et al., 2014; RECZEK; CHANDEL, 2015). Por isso, as EROs
também são produzidas nos estádios posteriores da patogênese, como de fato ocorreu.
Pode-se observar que a concentração de •O2- permaneceu alta ao longo de todo o
crescimento e desenvolvimento das plantas Test-Inoc, em função do progresso da doença.
Porém, nas plantas Test-Absoluta o aumento da concentração de •O2- só ocorreu aos 64 e 74
DAT, nos anos de 2015 e 2016, respectivamente (Figura 17). Como a concentração de •O2-
manteve-se baixa, quando comparada aos demais tratamentos até estas datas, pode-se aferir que
o aumento significativo da produção de •O2- aos 64 e 74 DAT, ocorreu em função do início do
desenvolvimento da doença nestas plantas, que se confirma pela presença dos primeiros
sintomas observados nesta fase do experimento.
Apesar de todas as plantas, exceto as Test-Aboluta, terem sido inoculadas aos 16 DAT,
ou seja, 24 horas após a aplicação dos fungicidas, o comportamento observado nas plantas
tratadas com os diferentes fungicidas foi totalmente diferente do que se observou nas plantas
Test-Inoc, porém apresentaram comportamento similar entre eles.
Nota-se que a concentração de •O2- nas plantas tratadas com Flux+Pira, Me+Pira, Bosc,
Bosc+Cre e Pira (Figura 17) apresentou picos de produção sempre 24 horas após cada
aplicação, permanecendo alta, porém com pequenas oscilações pelas próximas 48 e 120 horas,
voltando a uma concentração baixa após 15 dias, que compreende o tempo de ‘zero hora’ antes
da próxima aplicação. Como já discutido, tanto as estrobilurinas, quanto as carboxamidas,
atuam na respiração mitocondrial do fungo. As estrobilurinas se ligam ao centro da oxidação
ubi-hidroquinona do complexo bc 1 mitocondrial (complexo III), já as carboxamidas inibem a
enzima succinato dehidrogenase (SDHI) no complexo II da respiração mitocondrial (FRAC,
2017). O efeito destas moléculas, portanto, é no transporte de elétrons que reduz a síntese de
NADH e ATP, bloqueando a produção de energia para a sobrevivência do fungo (KANUNGO;
JOSHI, 2014).
Isto indica que, possivelmente, a geração de •O2- nestes tratamentos, tenha relação com
o modo de ação destas moléculas, uma vez que estes fungicidas testados (estrobilurinas e
carboxamidas), atuam na cadeia de transporte de elétrons do patógeno irreversivelmente, mas
reversivelmente na planta. De fato, Diaz-Espejo et al. (2012) sugerem que há uma possível
evidência de que a inibição do transporte de elétrons, que somente ocorreria a nível
mitocondrial, também possa ocorrer em cloroplastos, após a aplicação de estrobilurinas em
plantas. Desta forma, a geração de •O2- possivelmente esteja ocorrendo durante a fase deste

82
‘bloqueio transitório’ no transporte de elétrons nos cloroplastos e mitocôndrias, pela ação dos
fungicidas, gerando elétrons ‘perdidos’ que acabam sendo capturados pelo O2 formando o •O2.
As EROs, dependendo da sua concentração nas plantas, podem gerar efeitos benéficos
ou deletérios. Em baixas ou moderadas concentrações, as EROs atuam como mensageiros
secundários em cascatas de sinalização intracelular que medeiam várias respostas em plantas.
No entanto, em altas concentrações elas causam danos em diversas biomoléculas (SHARMA
et al., 2012). Para evitar os danos deletérios das EROs as plantas possuem um sistema de defesa
composto por mecanismos antioxidantes enzimáticos e não enzimáticos, que trabalham em
conjunto e em sincronia para desintoxicar as células (MOLLER et al., 2007) em nível de
cloroplastos, mitocôndrias e peroxissomos (SCANDALIOS, 2005; KIM; KWAK, 2010;
DINAKAR et al., 2012; NOCTOR et al., 2015; LEHMANN et al., 2015).
Entre o arsenal de defesa enzimático, a enzima superóxido dismutase (SOD), uma
metalo-enzima, é a primeira enzima na linha de defesa contra os danos causados pelos ânions
•O2- (BHATTACHARJEE, 2010). Ao analisar a atividade da enzima SOD em folhas de
tomateiro (Figura 18), aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75,
76, 79, 89, 90, 91, 94 e 105 DAT, observa-se que houve um padrão de resposta similar nos dois
anos avaliados. Os resultados revelam que a atividade da enzima SOD encontrava-se alta nas
plantas tratadas com os diferentes fungicidas de efeitos fisiológicos, nas avalições que
compreendiam 24, 48 e 120 horas após cada aplicação, tendo forte relação com o observado
nas respostas obtidas para a concentração de •O2- (Figuras 17). De maneira geral, todos os
tratamentos com fungicidas de efeitos fisiológicos apresentaram alta atividade da SOD.
No entanto, as maiores atividades da enzima SOD foram observadas nos tratamentos
Flux+Pira, seguido de Me+Pira, demonstrando que houve maior defesa antioxidativa nestas
plantas, a fim de desintoxicar as células pela eliminação do •O2-, minimizando os danos ao
metabolismo. Isto fica evidente quando se observa os dados de eficiência fotossintética nestes
tratamentos, os quais apresentaram maior eficiência de carboxilação e rendimento quântico
efetivo do PSII, além de maior teor de pigmentos fotossintéticos.
Pois a SOD, por ser a primeira enzima na linha de defesa contra as EROs a nível celular,
atua dismutando o •O2- em H2O2 (BHATTACHARJEE, 2010), apresentando uma conversão
altamente eficiente numa velocidade de 1010 vezes maior que na reação espontânea (RESENDE
et al., 2003). Estudos demonstram que o •O2- também pode atuar na redução de metais como o
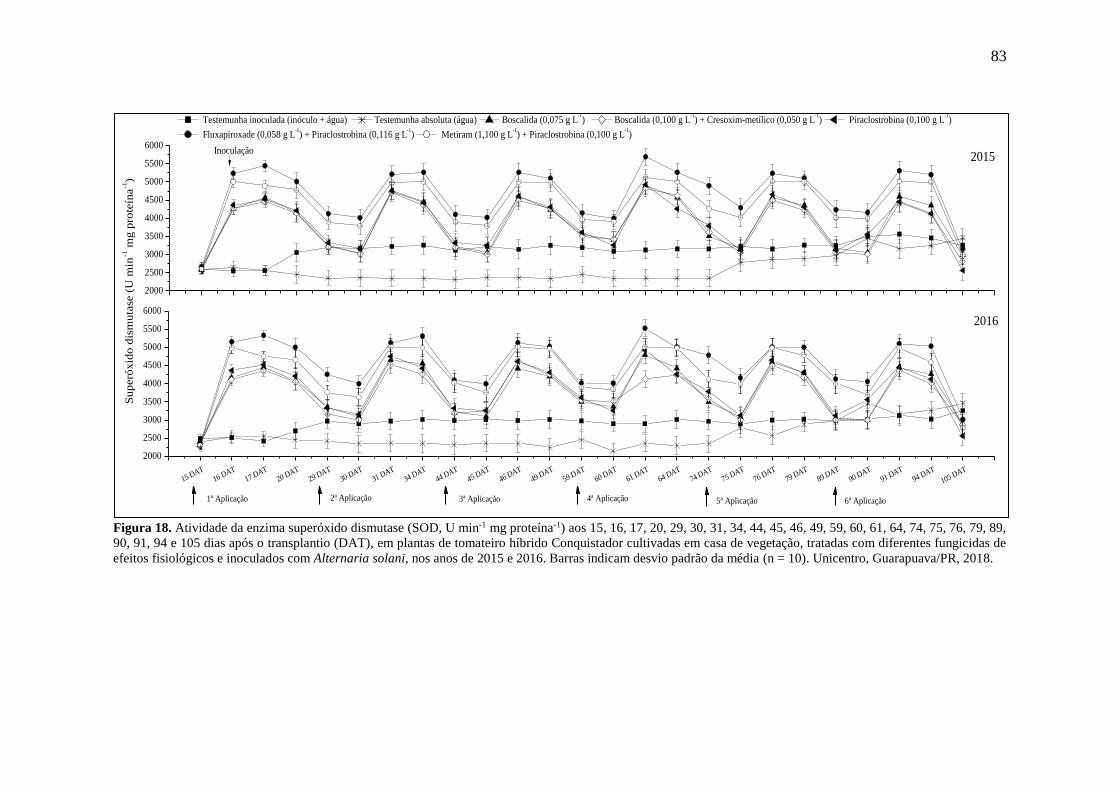
83
Figura 18. Atividade da enzima superóxido dismutase (SOD, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89,
90, 91, 94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de
efeitos fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
2000
2500
3000
3500
4000
4500
5000
5500
6000
15 DAT16 DAT
17 DAT20 DAT
29 DAT30 DAT
31 DAT34 DAT
44 DAT45 DAT
46 DAT49 DAT
59 DAT60 DAT
61 DAT64 DAT
74 DAT75 DAT
76 DAT79 DAT
89 DAT90 DAT
91 DAT94 DAT
105 DAT
2000
2500
3000
3500
4000
4500
5000
5500
6000
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Su
peró
xid
o d
ism
uta
se (
U m
in-1
mg
pro
teín
a-1
)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

84
Fe3+, nas reações de Haber-Weiss ou Fenton, sendo a principal via de geração do radical
hidroxila (OH•) (GILL; TUTEJA, 2010), o radical mais reativo da família das EROs
(KARUPPANAPANDIAN et al., 2011).
Assim, a enzima SOD por estar presente na maioria dos compartimentos subcelulares
que geram •O2- (SHARMA et al., 2012) e, devido a sua velocidade de ação em dismutar o •O2
-,
acabam indiretamente contribuindo também para a redução de OH• gerado a partir de reações
com o ânion •O2- (DINAKAR et al., 2012).
Nas plantas Test-Inoc houve aumento da atividade da enzima SOD após os 20 DAT,
enquanto que nas plantas Test-Absoluta ocorreu apenas aos 74 DAT, tendo relação com o
observado para a concentração de •O2-. Porém, nas duas testemunhas a atividade da SOD foi
inferior aos tratamentos com fungicidas indicando menor desintoxicação oxidativa nestas
plantas.
A menor atividade da enzima SOD nestas plantas tem relação com a alta agressividade
do patógeno, visto que o processo de infecção de A. solani é muito rápido, pois estudos
demonstram que entre 6 a 12 horas após a inoculação, cerca de 98,9% e 97,1% dos conídios
haviam germinado em folhas de plantas suscetível e resistente de tomateiro, respectivamente,
iniciando o processo de infecção sobre o hospedeiro (ARAÚJO; MATSUOKA, 2004). De fato,
o maior acúmulo de •O2- foi detectado logo após a inoculação (aos 17 DAT, que compreende
24 horas após inoculação) nas plantas Test-Inoc (Figura 17) em resposta ao estresse decorrente.
No entanto, estudos realizados por Balbi-Peña et al. (2014) verificaram que o aumento
de EROs e as respostas de hipersensibilidade (HR) não contribuem para a resistência de plantas
de tomateiro contra A. solani. Os autores descrevem que esta ineficiência das respostas de HR
está relacionado ao fato de A. solani ser um patógeno necrotrófico e, por isso, na interação A.
solani-tomateiro tais respostas de defesa não se fazem eficientes para conter as lesões, pelo
contrário, provavelmente facilitam a colonização dos tecidos do hospedeiro, uma vez que a
morte da célula hospedeira não limita o crescimento do patógeno. Portanto, a alta agressividade
do patógeno somada a baixa eficiência das EROs em combater o seu progresso, resultou em
maior percentual de tecido foliar danificado, isto interferiu na eficiência do arsenal de defesa.
Por isso se observou menor atividade da SOD e, consequentemente, das outras enzimas
antioxidantes, o que comprometeu sanidade dos fotossistemas e, consequentemente, a
eficiência fotossintética destas plantas.

85
Como visto, a maior concentração de H2O2 celular surge da dismutação do •O2-
catalisada pela SOD, onde dois radicais •O2- formam um H2O2 (BHATTACHARJEE, 2010).
Apesar de ser ausente de carga, o H2O2, é um oxidante relativamente estável e de pequeno
tamanho, o que facilita a passagem através da bicamada lipídica da membrana celular e sua
migração em compartimentos diferentes e distante do local de formação, difundindo os danos,
mas também atuando como importante mensageiro secundário em condições de estresse
(RESENDE et al., 2003; RIBEIRO et al., 2005; BARBOSA et al., 2014). Desta forma,
subsequentemente, as vias geradoras de •O2- formam o H2O2, no entanto, é em condições de
estresse severo que se nota as maiores gerações de H2O2 (SHARMA et al., 2012).
Ao analisarmos as concentrações de H2O2 em folhas de tomateiro (Figura 19) aos 15,
16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105
DAT, observou-se que houve novamente um padrão de resposta similar nos dois anos avaliados,
complementando o que foi observado para concentração de •O2- e na atividade da enzima SOD.
Nos tratamentos em que a atividade da SOD estava alta, se observa maiores concentrações de
H2O2, ou seja, maiores quantidades de ânions superóxidos foram dismutados à peróxido de
hidrogênio, desintoxicando o metabolismo celular.
As menores concentrações de H2O2 foram observadas nas plantas Test-Absoluta, no
entanto, estas também apresentaram baixas concentrações de •O2- e da atividade da SOD. Já as
plantas Test-Inoc apresentaram baixas concentrações de H2O2, quando comparadas às plantas
tratadas com os diferentes fungicidas. Entretanto, como já discutido, foram observadas para
estas mesmas plantas altas concentrações de •O2- e baixa atividade da SOD ao longo do seu
ciclo, fato que acarretou danos oxidativos no metabolismo celular, indicado pela baixa
eficiência fotossintética observada neste tratamento.
Na célula vegetal a regulação dos níveis intracelulares de H2O2 também se faz
necessário. Pois o peróxido de hidrogênio também é prejudicial à célula, por ter elevada ação
deletéria, pois é capaz de inativar enzimas, além de participar nas reações de Haber-Weiss ou
Fenton, gerando OH•, o oxidante mais reativo da família das EROs (KARUPPANAPANDIAN
et al., 2011). Sendo assim, o H2O2, quando em concentrações deletérias, também precisa ser
eliminado do metabolismo celular. Por isso, as enzimas antioxidantes trabalham em conjunto,
buscando diminuir a concentração destas EROs originadas por condições de estresses sofridos
pelo vegetal ao longo do ciclo de desenvolvimento.
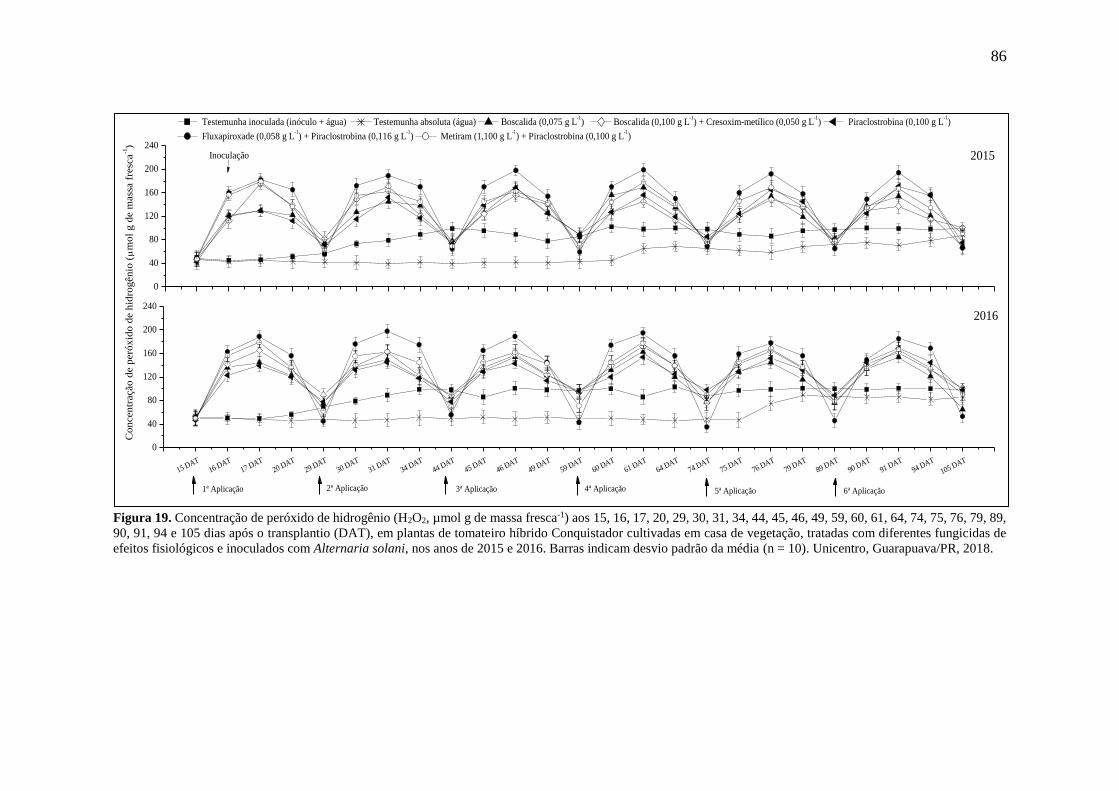
86
Figura 19. Concentração de peróxido de hidrogênio (H2O2, µmol g de massa fresca-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89,
90, 91, 94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de
efeitos fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0
40
80
120
160
200
240
15 DAT16 DAT
17 DAT20 DAT
29 DAT30 DAT
31 DAT34 DAT
44 DAT45 DAT
46 DAT49 DAT
59 DAT60 DAT
61 DAT64 DAT
74 DAT75 DAT
76 DAT79 DAT
89 DAT90 DAT
91 DAT94 DAT
105 DAT
0
40
80
120
160
200
240
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Concentr
ação d
e p
eró
xid
o d
e h
idro
gênio
(
mol
g d
e m
ass
a f
resc
a-1
)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

87
A regulação dos níveis de H2O2 ocorre por meio de uma grande variedade de enzimas,
tais como ascorbato peroxidase (APX), catalase (CAT), peroxidase (POX), glutationa S-
transferase (GST), glutationa redutase (GR) e a glutationa peroxidase (GPX) (SCANDALIOS,
2005; KIM; KWAK, 2010; DINAKAR et al., 2012; NOCTOR et al., 2015; LEHMANN et al.,
2015), que estão comumente envolvidas na defesa vegetal contra o estresse oxidativo.
Estas enzimas são proteínas que catalisam diferentes reações, controlando os níveis de
EROs, atuando em diferentes organelas para manter a homeostase celular (BARBOSA et al.,
2014). Diante disto, a atividade da enzima APX foi avaliada em folhas de tomateiro (Figura
20), aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91,
94 e 105 DAT, submetidas aos diferentes tratamentos, nos dois experimentos avaliados,
apresentando efeitos similares. Para os diferentes fungicidas testados observou-se maior
atividade da enzima APX em relação às testemunhas, apresentando picos de atividade sempre
às 48 horas após cada aplicação (Figura 20).
A enzima APX é responsável por degradar o H2O2 utilizando o ascorbato como
substrato, tendo alta afinidade com o H2O2, permitindo eliminá-lo em pequenas concentrações,
pois esta enzima apresenta constante de Michaelis-Menten (Km) na ordem µM para seu
substrato (SHARMA et al., 2012). Sua atuação principal é na remoção de H2O2, nos
cloroplastos (RESENDE et al., 2003), defendendo os tecidos fotossintéticos contra o estresse
fotoxidativo. De fato, a alta atividade das APXs colaborou para a manutenção da integridade
do aparato fotossintético, resultando em maior eficiência fotossintética, conforme foi observado
nos tratamentos com os diferentes fungicidas (Figuras 9, 10 e 11).
As menores atividades da APX foram observadas nas plantas Test-Inoc e Test-Absoluta
com pequenas oscilações ao longo do ciclo da cultura. Ao contrário do que foi observado para
as plantas tratadas com fungicidas de efeitos fisiológicos, as testemunhas por apresentarem
baixa atividade da APX, ficaram mais vulneráveis aos danos oxidativos causados pela presença
de H2O2.
Juntamente com a APX, as enzimas GR, GPX e GST compõem o arsenal de defesa ao
estresse oxidativo, atuando na eliminação do H2O2. Semelhante ao observado para as enzimas
SOD e APX, houve diferença na atividade das enzimas GR (Figura 21), GPX (Figura 22) e
GST (Figura 23) em folhas de tomateiro aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59,
60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 DAT, submetidas aos diferentes tratamentos, nos
dois experimentos avaliados, apresentando efeitos semelhantes.

88
Figura 20. Atividade da enzima ascorbato peroxidase (APX, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89,
90, 91, 94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de
efeitos fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0
10
20
30
40
50
15 DAT16 DAT
17 DAT20 DAT
29 DAT30 DAT
31 DAT34 DAT
44 DAT45 DAT
46 DAT49 DAT
59 DAT60 DAT
61 DAT64 DAT
74 DAT75 DAT
76 DAT79 DAT
89 DAT90 DAT
91 DAT94 DAT
105 DAT
0
10
20
30
40
50
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Asc
orb
ato
pero
xid
ase
(U
min
-1 m
g p
rote
ína
-1)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

89
Figura 21. Atividade da enzima glutationa redutase (GR, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90,
91, 94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de
efeitos fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
15 DAT16 DAT
17 DAT19 DAT
29 DAT30 DAT
31 DAT34 DAT
44 DAT45 DAT
46 DAT49 DAT
59 DAT60 DAT
61 DAT64 DAT
74 DAT75 DAT
76 DAT79 DAT
89 DAT90 DAT
91 DAT94 DAT
105 DAT
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Glu
tati
ona r
eduta
se (
U m
in-1
mg p
rote
ína
-1)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

90
Figura 22. Atividade da enzima glutationa S-transferase (GST, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89,
90, 91, 94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de
efeitos fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
15 DAT16 DAT
17 DAT20 DAT
29 DAT30 DAT
31 DAT34 DAT
44 DAT45 DAT
46 DAT49 DAT
59 DAT60 DAT
61 DAT64 DAT
74 DAT75 DAT
76 DAT79 DAT
89 DAT90 DAT
91 DAT94 DAT
105 DAT
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Glu
tath
ion
e-S
-tra
nsf
era
se (
U m
in-1
mg
pro
teín
a-1
)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

91
Figura 23. Atividade da enzima glutationa peroxidase (GPX, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89,
90, 91, 94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de
efeitos fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0
30
60
90
120
150
15 DAT
16 DAT
17 DAT
20 DAT
29 DAT
30 DAT
31 DAT
34 DAT
44 DAT
45 DAT
46 DAT
49 DAT
59 DAT
60 DAT
61 DAT
64 DAT
74 DAT
75 DAT
76 DAT
79 DAT
89 DAT
90 DAT
91 DAT
94 DAT
105 DAT
0
30
60
90
120
150
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Glu
tati
on
a p
ero
xid
ase
(U
min
-1 m
g p
rote
ína
-1)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

92
Nota-se que as maiores atividades da enzima GR (Figura 21) foram proporcionadas pela
aplicação de Flux+Pira, seguido de Me+Pira. Os outros fungicidas promoveram aumento na
atividade da enzima, porém em menor intensidade. De maneira geral, é possível observar um
pico de atividade da enzima GR 24 horas após cada aplicação dos fungicidas, com acentuado
declínio após 48 e 120 horas.
A enzima GR também atua na defesa contra o estresse oxidativo, pois mantém o
equilíbrio entre os níveis de GSSG (glutationa oxidada) e GSH (glutationa reduzida) na célula,
em nível de cloroplasto, mitocôndrias, citosol e peroxissomos, no entanto, cerca de 80% da
atividade da GR ocorre em tecidos fotossintéticos (SHARMA et al., 2012). Por isso, juntamente
com a APX, a GR protege o aparato fotossintético dos efeitos nocivos das EROs, confirmando
mais uma vez o efeito positivo destes fungicidas na sanidade do sistema fotossintético.
Como visto, o mecanismo de ação da glutationa ocorre por meio da redução do H2O2 e
de hidropeptídeos orgânicos e envolve simultaneamente a oxidação da GSH para GSSG, por
intermédio da GR, que oxida o NADPH e regenera a GPX. Já a GST, por sua vez, auxilia na
detoxificação do organismo, catalisando a conjugação da GSH a compostos endógenos ou
exógenos como xenobióticos (COGO et al., 2009). De fato, a atividade da enzima GST (Figura
22), apresentou comportamento similar ao da GR em todos os tratamentos, observando-se
maiores atividades para as plantas tratadas com Flux+Pira, seguidas de Me+Pira. No entanto,
as plantas tratadas com Bosc, Bosc+Cre e Pira quando comparadas as testemunhas, também
apresentaram maiores atividades da enzima GST, porém em menor intensidade do que o
observado para aquelas tratadas com Flux+Pira e Me+Pira. O oposto foi observado nas
testemunhas, Test-Absoluta e Test-Inoc, as quais apresentaram as menores atividades das
enzimas GR e GST, elucidando novamente a baixa ativação do sistema de defesa antioxidativo
nestas plantas.
Para a atividade da enzima GPX (Figura 23) constatou-se maior atividade nos
tratamentos Flux+Pira, seguidos de Me+Pira, Bosc, Bosc+Cre e Pira, com aumento expressivo
nas avaliações às 24 horas e mantendo-se alta às 48 e 120 horas após cada aplicação. De maneira
geral, estes tratamentos também foram eficientes em ativar a atividade da enzima GPX
auxiliando na eliminação de H2O2 do metabolismo celular. Como já mencionado, as enzimas
GPXs também atuam na desintoxicação dos produtos formados na peroxidação lipídica, em
decorrência dos danos oxidativos pela atividade das EROs (DIXON et al., 2010). A atividade
da enzima GPX é muito eficiente, pois assim como relatado para a APX, possui KM na ordem

93
de 1µM para o H2O2 (RIBEIRO et al., 2005) e, por isso, quando atuam em conjunto, agem
eficientemente em níveis baixíssimos de H2O2, realizando o ajuste ‘fino’ na remoração destas
EROs do sistema celular.
Além das enzimas já descritas, a enzima catalase (CAT) também compõe o arsenal de
defesa contra o estresse oxidativo, cooperando na eliminação das EROs dos tecidos vegetais, a
partir da conversão de duas moléculas de H2O2 em H2O e O2 (DUBEY, 2010; SHARMA et al.,
2012). A atividade da enzima CAT, avaliada aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49,
59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 DAT em folhas de tomateiro (Figura 24), foi
maior nos tratamentos Flux+Pira e Me+Pira, apresentando pico de atividade às 24 horas,
seguido de declínio da atividade ás 48 e 120 horas, após cada aplicação.
As plantas tratadas com os demais fungicidas apresentaram baixa atividade da enzima
CAT, porém nota-se pequenos picos de atividade 24 horas após cada aplicação, assim como
observado para os tratamentos Flux+Pira e Me+Pira. Diferentemente, o tratamento Test-Inoc
apresentou pico de atividade da enzima CAT somente aos 20 DAT, período que corresponde
às 96 horas após a inoculação do patógeno, depois ocorreu queda na atividade permanecendo
baixa nas demais avaliações. Nas plantas Test-Absoluta observou-se baixa atividade em todas
as avaliações realizadas.
A enzima CAT também previne os efeitos potencialmente danosos causados por
mudanças na homeostase de H2O2, atuando em conjunto com as enzimas do ciclo ascorbato-
glutationa (APX, GR, GST e GPX), apesar de suas características serem diferentes podem
funcionar efetivamente em paralelo e são importantes na eliminação do H2O2 nas diferentes
organelas celulares. Pois a CAT não necessita de agente redutor, sendo energeticamente
eficiente para remoção do H2O2 (SHARMA et al., 2012) e, principalmente, é efetiva em
concentrações relativamente altas de H2O2 (mM), sendo consideradas indispensáveis para a
desintoxicação de EROs, especialmente, em condições de estresse severo, quando os níveis de
H2O2 estão elevados (DUBEY, 2010).
O aumento da atividade das enzimas SOD, APX, GR, GST, GPX e CAT após a
aplicação dos diferentes fungicidas evidenciou que estas substâncias promovem a ativação do
sistema de defesa antioxidativo, preparando a planta para resistir a um possível agente
estressante. No entanto, nota-se que o efeito não perdura por muitos dias, em alguns casos,
como no observado para a enzima APX, o pico de atividade ocorre 48 horas após cada
aplicação, depois a atividade diminui, enquanto que para a enzima GST o efeito é mais
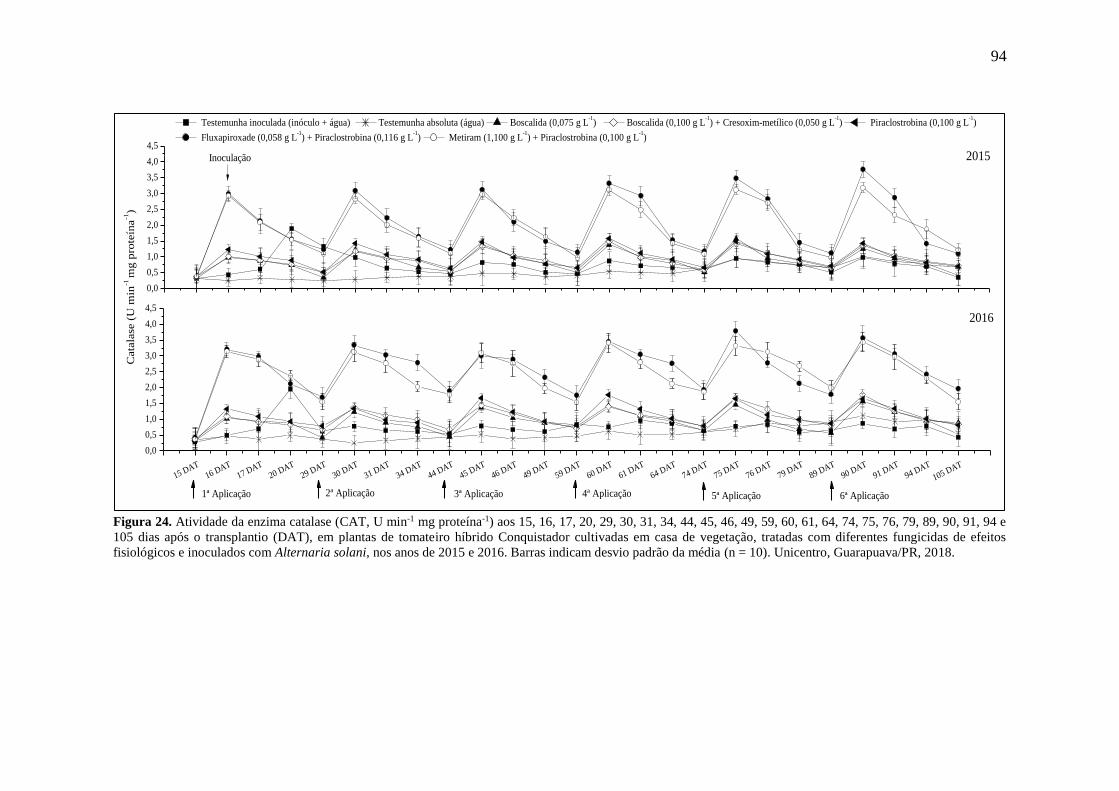
94
Figura 24. Atividade da enzima catalase (CAT, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e
105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
15 DAT
16 DAT
17 DAT
20 DAT
29 DAT
30 DAT
31 DAT
34 DAT
44 DAT
45 DAT
46 DAT
49 DAT
59 DAT
60 DAT
61 DAT
64 DAT
74 DAT
75 DAT
76 DAT
79 DAT
89 DAT
90 DAT
91 DAT
94 DAT
105 DAT
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Cata
lase
(U
min
-1 m
g p
rote
ína
-1)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

95
prolongado, apresentando maior atividade da enzima até o quinto dia após a aplicação (120
horas). A diferença percebida na expressão das enzimas antioxidativas ocorre porque elas atuam
em momentos e locais diferentes para conter o acúmulo de EROs e, evitando assim, os danos
oxidativos e distúrbios metabólicos, interrompendo e reparando os danos desta cascata de
oxidação (SHARMA et al., 2012), que consequentemente levam à morte celular.
Outro fator interessante observado, é que o efeito da aplicação dos diferentes fungicidas
se repete a cada aplicação, isto comprova o potencial destas substâncias como promotoras de
defesa ao estresse oxidativo em plantas de tomateiro. No entanto, verificou-se que os fungicidas
que afetaram em maior intensidade a atividade das enzimas SOD, APX, GR, GST, GPX e CAT
em plantas de tomateiro foram Flux+Pira e Me+Pira. O aumento da atividade destas enzimas
foi importante para manter a integridade celular, garantindo sua homeostase, pois como
relatado, elas atuam em organelas diferentes, assim potencializando a defesa antioxidativa da
célula. A ação das enzimas antioxidantes mantém a concentração de EROs no organismo dentro
dos limites fisiológicos (RIBEIRO et al., 2005) e de acordo com Sharma et al. (2012) a
expressão simultânea de várias enzimas antioxidantes tem demonstrado ser mais eficiente do
que ações isoladas em plantas.
Todos esses resultados estão de acordo com os dados observados nos parâmetros
fotossintéticos, uma vez que as plantas tratadas com estes fungicidas apresentaram as maiores
eficiências fotossintéticas, indicando que essas plantas estavam mais saudáveis que as
testemunhas. Alterações positivas nos mecanismos antioxidativos proporcionado pela aplicação
de fungicidas de efeitos fisiológicos também já foram relatados em plantas de pepineiro
(AMARO, 2011), meloeiro (MACEDO, 2012; 2015), tomateiro (JACOBELIS JUNIOR, 2015),
feijoeiro (JADOSKI et al., 2015), cafeeiro (HONORATO JÚNIOR et al., 2015), pepineiro
japonês (AMARO et al., 2018).
Contrariamente, as plantas Test-Inoc, as quais apresentaram elevado grau de estresse
oxidativo observado pela alta concentração de EROs (•O2- e H2O2) e baixa atividade das
enzimas antioxidativas avaliadas, acabaram sofrendo com a menor supressão dos níveis tóxicos
de EROs na célula, que acarretou danos irreversíveis. Uma vez que as EROs em altas
concentrações são extremamente prejudiciais para o metabolismo celular, e quando o nível de
EROs excede os mecanismos de defesa, tem-se o fenômeno celular caracterizado como estado
de ‘estresse oxidativo’ (SHARMA et al., 2012), causando consequentemente a peroxidação de
lipídios. Tais danos afetam negativamente a eficiência quântica potencial do PSII, detectada

96
pela menor relação Fv/Fm, conforme já mencionado. Concomitantemente, à queda da eficiência
fotoquímica, ocorreu redução no desenvolvimento destas plantas, bem como o menor acúmulo
de carboidratos e biomassa.
Nota-se, portanto, que a capacidade fotossintética das plantas, nos diferentes
tratamentos diminuiu em proporção a severidade da doença e a intensidade das respostas de
defesa ao estresse oxidativo, decorrentes da interação planta-patógeno. Além dos danos
ocasionados pelo processo de infecção, penetração e colonização pelo patógeno no tecido foliar
do hospedeiro (síntese de enzimas hidrolíticas extracelulares), somam-se a eles, os danos
ocasionados pela alta concentração de EROs, em níveis tóxicos à célula vegetal, causando a
degradação do tecido vegetal devido a peroxidação lipídica.
O processo de peroxidação lipídica, determinada a partir do acúmulo de substâncias
reativas ao ácido tiobarbitúrico (TBARS) é um dos parâmetros utilizados para avaliar a
intensidade do estresse oxidativo em plantas. Pois, durante o processo de lipoperoxidação, a
reação desencadeada pelas EROs ocasiona a degradação de ácidos graxos poli-insaturados das
membranas celulares, como os ácidos linolênico e linolênico, que têm como subprodutos
principais os íons peróxido e o malondialdeíodo (MDA), os quais são mensurados por meio da
reação com os ácidos tiobarbitúrico e tricloroacético.
Diante disto, a peroxidação lipídica avaliada aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45,
46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 DAT em folhas de tomateiro (Figura
25), demonstrou que os danos oxidativos às membranas celulares foram superiores no
tratamento Test-Inoc, apresentando aumento da intensidade com o decorrer do tempo, seguido
do tratamento Test-Absoluta, quando comparados aos demais tratamentos. As plantas Test-
Absoluta também apresentaram significativa peroxidação lipídica, porém em menor
intensidade que as plantas Test-Inoc.
A alta lipoperoxidação observada nas plantas testemunhas foi reflexo do nível de
estresse elevado nestas plantas, em decorrência do progresso da doença e, concomitantemente,
pela alta produção de EROs que não foram eliminadas devido a ineficiência do sistema
antioxidativo, ou seja, pela baixa atividade das enzimas SOD, APX, GR, GST, GPX e CAT.
Pois, para conter os efeitos deletérios da alta reatividade das EROs as plantas dependem de um
sistema antioxidativo eficaz, quando isto não ocorre, as EROs em altas concentrações
fisiológicas reagem com os lipídios das membranas, ocorrendo assim, a peroxidação lipídica.
A consequência direta do dano da peroxidação de lipídios sobre as membranas celulares é o

97
Figura 25. Peroxidação lipídica (TBARS, µmol g massa fresca-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105
dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos
e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0
30
60
90
120
150
180
210
15 DAT16 DAT
17 DAT20 DAT
29 DAT30 DAT
31 DAT34 DAT
44 DAT45 DAT
46 DAT49 DAT
59 DAT60 DAT
61 DAT64 DAT
74 DAT75 DAT
76 DAT79 DAT
89 DAT90 DAT
91 DAT94 DAT
105 DAT
0
30
60
90
120
150
180
210
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Pero
xid
ação L
ipíd
ica (
TB
AR
S,
mol
g m
ass
a f
resc
a-1
)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

98
extravasamento do conteúdo celular para o meio, por danificarem irreversivelmente as
membranas, além de formar novos radicais lipídicos e intensificando os danos (DAN HESS,
2000; KRUSE et al., 2006).
Além das membranas, outras moléculas essenciais como pigmentos fotossintéticos,
proteínas e ácidos nucléicos também são alvos das EROs (FOYER et al., 1997; SHARMA et
al., 2012), por isso quando em altas concentrações as EROs podem desencadear a paralização
do metabolismo celular (LIN et al., 2005). Logo, estes efeitos do estresse oxidativo interferiram
fortemente no processo fotossintético, causando danos aos pigmentos e aos fotossistemas, bem
como na expressão das enzimas envolvidas no processo de fixação de CO2, como pode ser
observado nos tratamentos Test-Inoc e Test-Absoluta, que apresentaram menores teores de
clorofilas a e b, menor assimilação de CO2, eficiência de carboxilação e rendimento quântico
do PSII (Figuras 9, 10, 11 e 12) e com isto o menor desenvolvimento das plantas.
Ao contrário, a alta atividade das enzimas antioxidativas resultou em menores níveis de
peroxidação lipídica nas plantas tratadas com Flux+Pira, Me+Pira, Bosc, Bosc+Cre e Pira, ao
longo do ciclo da cultura e possibilitaram maior sanidade do aparato fotossintético. Somado a
este efeito, o que possivelmente também pode ter contribuído para a menor peroxidação
lipídica, é a maior concentração de carotenoides observada nestes tratamentos (Figura 12), pois
estes pigmentos são importantes na eliminação do •O2- , além de possuírem a capacidade de
suprimir, prevenir e minimizar o estado tripleto da clorofila devido ao excesso de energia de
excitação, assim prevenindo a formação de •O2- e protegendo o aparato fotossintético
(SHARMA et al., 2012).
A eficiente eliminação das EROs, principalmente do •O2- por intermédio da SOD, é
muito importante para a homeostase celular, porque o radical •O2- é um dos principais oxidantes
responsáveis pela peroxidação de lipídios e sua formação causa o aumento na permeabilidade
das membranas no ambiente celular (BREUSEGEM et al., 2001; APEL; HIRT, 2004). Além
disso, a SOD também contribui indiretamente na redução de OH• (DINAKAR et al., 2012), que
segundo Sharma et al. (2012), uma única molécula de OH• pode resultar na peroxidação de
diversos ácidos graxos poli-insaturados, pois as reações envolvidas neste processo são cíclicas.
De modo geral, pode-se dizer que a aplicação destes fungicidas proporcionou adequado
controle do estresse oxidativo, que refletiu em menores danos celulares e, consequentemente,
em menor perda de área fotossinteticamente ativa, que contribuiu para o maior desenvolvimento
e acúmulo de matéria seca e, possivelmente maior produção de frutos.

99
5.7 Proteínas relacionadas à patogênese e respostas de defesa
De acordo com os resultados já apresentados, a aplicação de diferentes fungicidas
também interferiu na concentração das EROs: •O2- e H2O2 (Figuras 17 e 19), por isso tais
resultados inferem a possível atuação destas moléculas como sinalizadoras de respostas de
defesa em plantas de tomateiro.
Os altos níveis de EROs, como descrito anteriormente, causam sérios danos celulares
por aumentarem a lipoperoxidação. No entanto, as EROs desempenham papéis diferentes nas
plantas, dependendo do equilíbrio entre sua produção e eliminação nos tecidos: em baixas
concentrações atuam como sinalizadores bioquímicos na cascata de transdução de sinais para a
ativação de genes de defesa e de proteção celular, enquanto que em altas concentrações causam
exacerbados danos aos componentes celulares (SHARMA et al., 2012). Assim, as EROs têm
funções biológicas definidas quando estão em concentrações fisiológicas adequadas
(BARBOSA et al., 2014).
Desta forma, uma das funções de grande importância destas espécies reativas de
oxigênio na interação planta-patógeno, é atuarem como mediadores de sinalização intracelular
em respostas fisiológicas e biológicas, nos processos de defesa da planta, caracterizada como
indução de resistência. Os radicais livres são moléculas altamente reativas, devido à
instabilidade da molécula (possuem elétrons desemparelhados), no entanto, o H2O2, mesmo não
sendo classificado como um radical livre, por não ter elétrons desemparelhados na última
camada (BARTOSZ, 1997; COTINGUIBA et al., 2015) é a principal EROs envolvida na
sinalização de respostas de defesa ao ataque por patógenos.
O H2O2 aumenta a atividade da enzima benzóico 2-hidrolase, precursora na síntese do
ácido salicílico (hormônio vegetal crucial na sinalização da resistência sistêmica adquirida –
RSA). Além disso, o H2O2, no apoplasto da célula hospedeira, tem papel essencial nos
mecanismos de defesa da planta por apresentar efeito tóxico ao patógeno, agindo diretamente
sobre as estruturas do fungo e se acumulando no sítio de infecção, desta forma, restringindo o
desenvolvimento da lesão pelas reações de respostas de hipersensibilidade – HR (RESENDE
et al., 2003; LEHMANN et al., 2015).
Esta sequência de eventos culmina no aumento da expressão gênica de proteínas de
defesa, conhecidas como proteínas relacionadas à patogênese (PRPs) tais como a peroxidase
(POD), quitinase e β-1,3-glucanase. Somada a estas respostas também ocorre ativação da
expressão de genes que codificam enzimas da via dos fenilpropanóides, como a enzima

100
fenilalanina amônia-liase (PAL) e da via dos octadecanóides, como as enzimas
polifenoloxidase (PPO) e lipoxigenase (LOX). A sinalização desencadeada pelo processo de
indução de resistência, de acordo com Shah et al. (2014), pode ativar diferentes rotas
metabólicas. Na maioria das interações entre plantas e patógenos biotróficos ou
hemibiotróficos ocorre a ativação da rota via ácido salicílico em resposta à infecção, enquanto
que para patógenos necrotróficos são ativadas rotas dependentes de etileno ou ácido jasmônico
(THAKUR; SOHAL, 2013).
No entanto, a própria peroxidação de lipídios, por intermédio das enzimas LOXs pode
ser precursora da síntese de ácido jasmônico, e ativar a expressão de vários genes relacionados
à defesa durante as reações de HR na planta (DEUNER et al., 2015). Segundo Pinto et al.
(2011), a HR resulta na indução da morte celular programada (MCP) no sítio da infecção e por
consequência, na ativação de diferentes sinais, os quais resultam na expressão de uma variedade
de genes de defesa, no aparecimento de lesões necróticas no local da infecção, impedindo o
desenvolvimento da infecção pelo patógeno.
Esta cascata de fenômenos bioquímicos, normalmente observada após a indução,
resultará em aumento da defesa do vegetal ao ataque do patógeno, reforçando também as
barreiras físicas, por aumentar a atividade de outras enzimas, como a PAL. O aumento da
atividade da PAL está fortemente relacionado com a maior síntese e acúmulo de compostos
fenólicos, seguido também de maior deposição de lignina no tecido vegetal, aumentando a
defesa da planta (OLIVEIRA et al., 2016).
A PAL é considerada a enzima chave no metabolismo secundário das plantas e atua
sobre um substrato específico, a fenilalanina. Portanto, a atividade da enzima PAL avaliada aos
15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105
DAT em folhas de tomateiro (Figura 26) demonstrou que houve diferença na atividade desta
enzima entre os tratamentos, ao longo das avaliações. As maiores atividades da enzima PAL
foram observadas para as plantas tratadas com os diferentes fungicidas às 24 e 48 horas após
cada aplicação. No entanto, os tratamentos Flux+Pira, seguido de Me+Pira foram aqueles que
apresentaram os maiores picos de atividade desta enzima.
Para as plantas Test-Inoc nota-se que ocorreu maior pico de atividade da PAL aos 20
DAT, ou seja, 96 horas após a inoculação do patógeno, mantendo-se alta aos 29 DAT e após,
apresentou pequena redução da atividade, com pequenas oscilações no decorrer das demais
avaliações, sem apresentar queda significativa. Este comportamento observado demonstra que

101
Figura 26. Atividade da enzima fenilalanina amônia-liase (PAL, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79,
89, 90, 91, 94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas
de efeitos fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
15 DAT
16 DAT
17 DAT
20 DAT
29 DAT
30 DAT
31 DAT
34 DAT
44 DAT
45 DAT
46 DAT
49 DAT
59 DAT
60 DAT
61 DAT
64 DAT
74 DAT
75 DAT
76 DAT
79 DAT
89 DAT
90 DAT
91 DAT
94 DAT
105 DAT
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Fen
ilala
nin
a a
mô
nia
lia
se (
U m
in-1
mg
pro
teín
a-1
)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

102
o aumento da atividade da PAL ocorreu em resposta ao estresse ocasionado pela infecção do
patógeno.
Nas plantas tratadas com os fungicidas ocorreu aumento da atividade da PAL 24 e 48
horas após cada aplicação, e após este pico, houve redução da expressão aos níveis normais
(próximo do observado aos 15 DAT, momento antes da aplicação dos tratamentos). Já na Test-
Absoluta a atividade da enzima PAL permaneceu baixa até os 64 e 74 DAT, nos anos de 2015
e 2017, respectivamente, onde ocorreu pico da atividade da enzima PAL, apresentando queda
gradativa no decorrer das demais avaliações.
Verificou-se, portanto, que os tratamentos com fungicidas de efeitos fisiológicos
influenciaram na atividade da enzima PAL. Esta é a enzima chave na via metabólica da
fenilalanina, atuando no metabolismo dos fenilpropanóides e apresenta relação direta com a
síntese de compostos fenilpropanóides, como compostos fenólicos e lignina, importantes
compostos de defesa contra patógenos (SCHWAN-ESTRADA et al., 2008), pois a presença de
compostos fenólicos inibem metabólitos do patógeno, dificultando seu desenvolvimento
(STANGARLIN et al., 2011) e a maior deposição de lignina dificulta e restringe a penetração
do patógeno nos tecidos do hospedeiro (STANGARLIN et al., 2011).
A formação desses compostos tóxicos é essencial no processo de defesa contra o agente
agressor no local de infecção (SHAH et al., 2014; OLIVEIRA et al., 2016). Para constatar o
efeito do aumento da atividade da enzima PAL na síntese de compostos fenólicos foram
avaliados os teores destes aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74,
75, 76, 79, 89, 90, 91, 94 e 105 DAT em folhas de tomateiro (Figura 27), demonstrando que
houve diferença na concentração destas substâncias, entre os tratamentos, ao longo das
avaliações.
Observou-se maior concentração de compostos fenólicos 24 horas após cada aplicação
dos tratamentos Flux+Pira, Me+Pira, Bosc+Cre, Bosc e Pira (picos de teores), tendo declínio
acentuado nas avaliações às 48 e 120 horas após cada aplicação. No entanto, o tratamento
Flux+Pira apresentou os maiores picos de concentração tendo relação com as maiores
atividades da enzima PAL, observadas neste mesmo tratamento. Neste caso, pode-se dizer que
as maiores concentrações de compostos fenólicos foram influenciadas pela atividade da enzima
PAL, pois os picos são observados nos mesmos horários para estas plantas.
Comportamento semelhante ocorreu para as plantas Test-Inoc que apresentaram
maiores concentrações dos compostos fenólicos após os 20 DAT, mesmo momento em que
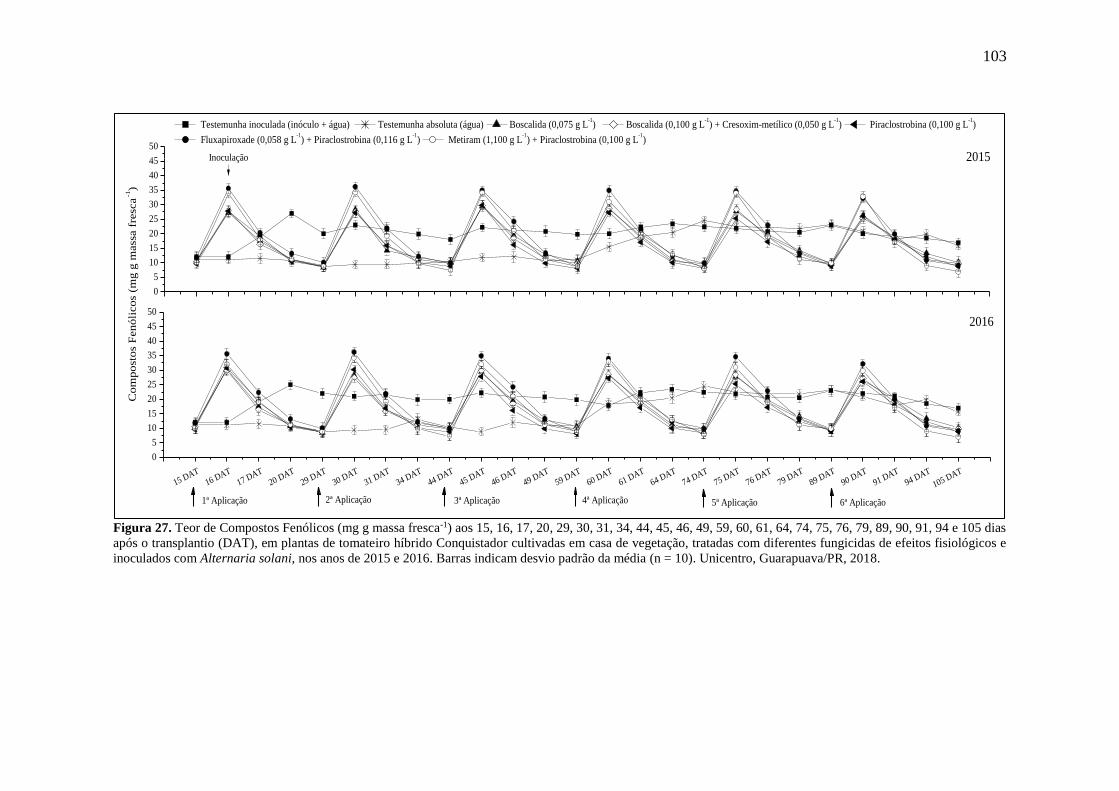
103
Figura 27. Teor de Compostos Fenólicos (mg g massa fresca-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 dias
após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e
inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0
5
10
15
20
25
30
35
40
45
50
15 DAT
16 DAT
17 DAT
20 DAT
29 DAT
30 DAT
31 DAT
34 DAT
44 DAT
45 DAT
46 DAT
49 DAT
59 DAT
60 DAT
61 DAT
64 DAT
74 DAT
75 DAT
76 DAT
79 DAT
89 DAT
90 DAT
91 DAT
94 DAT
105 DAT
0
5
10
15
20
25
30
35
40
45
50
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Co
mp
ost
os
Fen
óli
co
s (m
g g
mass
a f
resc
a-1
)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

104
houve maior atividade da enzima PAL. Isto indica forte relação entre a atividade da PAL e a
síntese de compostos fenólicos observados em plantas de tomateiro, nas condições destes
experimentos.
Os compostos fenólicos são metabólitos secundários das plantas e, geralmente, estão
envolvidos na defesa do vegetal contra organismos patogênicos (substâncias antinutritivas)
(CAMPOS et al., 2004) e na proteção aos raios ultravioletas (CAMPOS et al., 2003). Os
compostos fenólicos são importantes antioxidantes por possuírem propriedade redutora, devido
sua estrutura química, que permite o sequestro ou neutralização dos radicais livres, formando
intermediários instáveis (SILVA et al., 2010). Desta forma, altos níveis de compostos fenólicos
no metabolismo celular contribuem para a eliminação das EROs, além de protegerem o sistema
de defesa antioxidante.
Portanto, pode-se inferir que os tratamentos com os diferentes fungicidas promoveram
maior defesa antioxidante em plantas de tomateiro, constatada pelo maior teor de compostos
fenólicos nos tratamentos Flux+Pira, Me+Pira, Bosc+Cre, Bosc e Pira, nos quais se observou a
menor peroxidação lipídica. Além destas funções, os compostos fenólicos também são
substratos para a síntese de lignina pela ação de PODs e PPOs. Estas enzimas oxidam
compostos fenólicos, colaborando com a biossíntese de lignina, aumentando as barreiras físicas
contra o ataque de patógenos (YUAN et al., 2002).
De fato, ao analisarmos a atividade das enzimas POD e PPO aos 15, 16, 17, 20, 29, 30,
31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 DAT em folhas de
tomateiro (Figura 28 e 29, respectivamente) pode-se observar a sequência dos picos de atividade
nos mesmos horários em que houve maior concentração de compostos fenólicos (24 horas após
cada aplicação dos fungicidas). Portanto, tanto o teor de compostos fenólicos, quanto a
atividade das enzimas POD e PPO foram afetados pela aplicação de Flux+Pira, Me+Pira,
Bosc+Cre, Bosc e Pira em plantas de tomateiro ao longo do ciclo da cultura.
No entanto, nas plantas Test-Inoc notou-se que ocorreu aumento na atividade das
enzimas POD e PPO aos 20 DAT, que corresponde às 96 horas após a inoculação do patógeno.
Observou-se que a atividade destas enzimas se manteve constante durante as demais avaliações.
Nas plantas Test-Absoluta pode-se verificar que a atividade das enzimas POD e PPO manteve-
se baixa até os 64 DAT, onde ocorreu pequeno aumento da atividade da POD e aos 74 DAT
para a enzima PPO. Comportamento semelhante foi observado para a atividade da PAL e dos
teores de compostos fenólicos nestes mesmos tratamentos.

105
Figura 28. Atividade da enzima peroxidase (POD, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94
e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0
50
100
150
200
250
300
350
400
15 DAT
16 DAT
17 DAT
20 DAT
29 DAT
30 DAT
31 DAT
34 DAT
44 DAT
45 DAT
46 DAT
49 DAT
59 DAT
60 DAT
61 DAT
64 DAT
74 DAT
75 DAT
76 DAT
79 DAT
89 DAT
90 DAT
91 DAT
94 DAT
105 DAT
0
50
100
150
200
250
300
350
400
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Pero
xid
ase
(U
min
-1 m
g d
e p
rote
ína
-1)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

106
Figura 29. Atividade da enzima polifenoloxidase (PPO, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90,
91, 94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0,0
0,2
0,4
0,6
0,8
1,0
1,2
15 DAT16 DAT
17 DAT20 DAT
29 DAT30 DAT
31 DAT34 DAT
44 DAT45 DAT
46 DAT49 DAT
59 DAT60 DAT
61 DAT64 DAT
74 DAT75 DAT
76 DAT79 DAT
89 DAT90 DAT
91 DAT94 DAT
105 DAT
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Poli
fen
olo
xid
ase
(U
min
-1 m
g p
rote
ína
-1)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

107
Isto indica que a atividade da POD foi influenciada pela presença do patógeno, pois a
POD é uma das enzimas correlacionadas às respostas de resistência em vegetais nas interações
planta-patógeno, por isso, classificada como PRP. As enzimas PODs estão localizadas
principalmente na parede celular e no vacúolo (LOCATO et al., 2010) e atuam em reações de
polimerização, participando da lignificação, suberização e metabolismo da parede celular, a
partir da oxidação de compostos fenólicos, além de estarem relacionadas ao aumento de H2O2
(ação antimicrobiana).
Portanto, uma das funções e, neste caso a mais importante, é sua atuação nas respostas
de defesa da planta ao ataque de patógenos, por participarem de diversos processos fisiológicos
que reforçam a estrutura da parede celular restringindo a entrada do patógeno (BALBI-PEÑA
et al., 2014). As PODs também são importantes enzimas antioxidantes por catalisarem reações
envolvendo o H2O2 (EBRAHIM et al., 2011) e o aumento de sua expressão pode ser indicativo
bioquímico de estresse resultante de fatores bióticos e abióticos (KIM; KWAN, 2010).
Assim como as PODs, as PPOs também estão relacionadas com respostas de defesa.
Estas enzimas aumentam sua atividade após a ocorrência de danos na célula (CAMPOS et al.,
2003; THIPYAPONG et al., 2004) por estarem presentes nos tilacóides, por isso sua ação contra
fitopatógenos ocorre, principalmente após a ruptura da célula pelo processo de penetração do
patógeno. Quando a enzima é liberada destas organelas, simultaneamente inicia o processo de
oxidação de compostos fenólicos que foram simultaneamente liberados do vacúolo, produzindo
quinonas que são substâncias antimicrobianas (THIPYAPONG et al., 2004).
Além disto, as PPOs também atuam no processo de lignificação, juntamente com a POD
e a PAL, aumentando as barreiras de defesa à invasão pelo patógeno. A lignificação proporciona
maior resistência às paredes celulares, exigindo do patógeno, no momento da penetração no
tecido hospedeiro, maior atividade e gasto metabólico para degradar a lignina e avançar com a
colonização (STANGARLIN et al., 2011).
Conforme os resultados observados, a expressão das enzimas POD e PPO está
fortemente correlacionada com a ocorrência de infecção por patógenos e, também, pela
presença de um agente elicitor, neste caso os fungicidas de efeitos fisiológicos. Como a PPO é
uma enzima que se encontra latente e torna-se ativa quando liberada da membrana dos tilacóides
(devido ao rompimento destas) pode-se relacionar os picos de atividade da PPO com os picos
de produção das EROs, •O2- e H2O2 (Figuras 17 e 19), observados após cada aplicação dos
diferentes fungicidas. De fato, uma vez que aumentava a concentração de EROs na célula
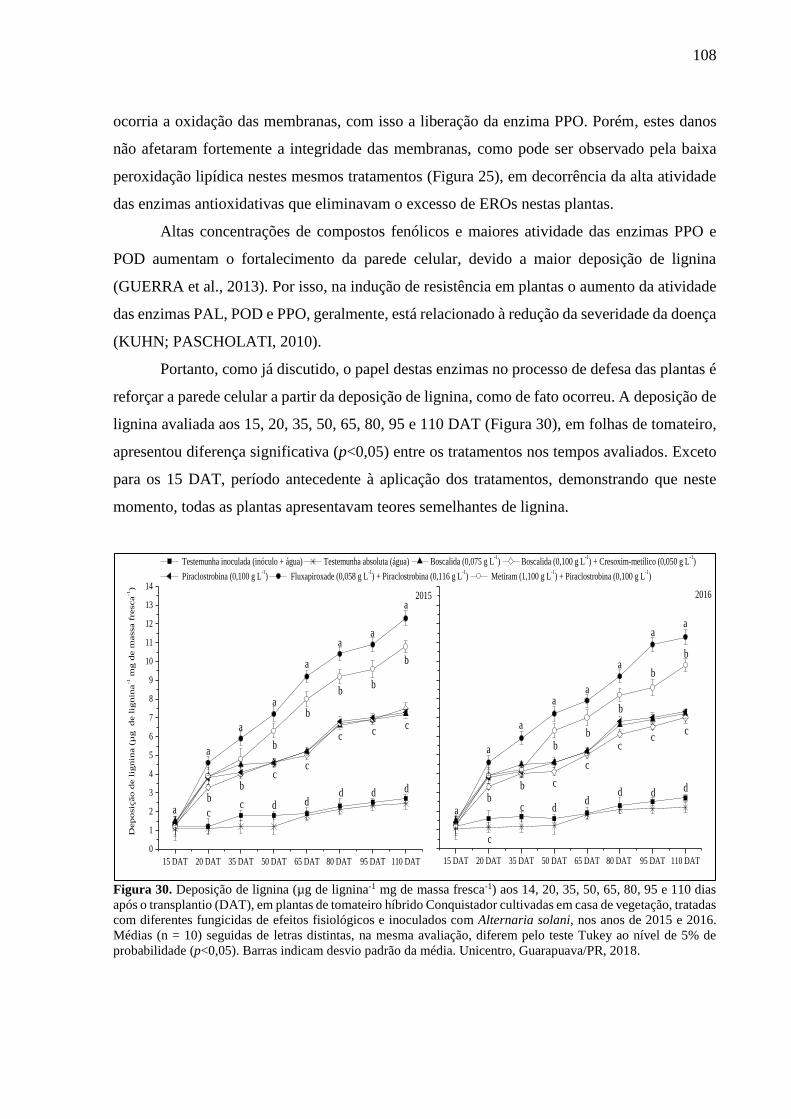
108
ocorria a oxidação das membranas, com isso a liberação da enzima PPO. Porém, estes danos
não afetaram fortemente a integridade das membranas, como pode ser observado pela baixa
peroxidação lipídica nestes mesmos tratamentos (Figura 25), em decorrência da alta atividade
das enzimas antioxidativas que eliminavam o excesso de EROs nestas plantas.
Altas concentrações de compostos fenólicos e maiores atividade das enzimas PPO e
POD aumentam o fortalecimento da parede celular, devido a maior deposição de lignina
(GUERRA et al., 2013). Por isso, na indução de resistência em plantas o aumento da atividade
das enzimas PAL, POD e PPO, geralmente, está relacionado à redução da severidade da doença
(KUHN; PASCHOLATI, 2010).
Portanto, como já discutido, o papel destas enzimas no processo de defesa das plantas é
reforçar a parede celular a partir da deposição de lignina, como de fato ocorreu. A deposição de
lignina avaliada aos 15, 20, 35, 50, 65, 80, 95 e 110 DAT (Figura 30), em folhas de tomateiro,
apresentou diferença significativa (p<0,05) entre os tratamentos nos tempos avaliados. Exceto
para os 15 DAT, período antecedente à aplicação dos tratamentos, demonstrando que neste
momento, todas as plantas apresentavam teores semelhantes de lignina.
Figura 30. Deposição de lignina (µg de lignina-1 mg de massa fresca-1) aos 14, 20, 35, 50, 65, 80, 95 e 110 dias
após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas
com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016.
Médias (n = 10) seguidas de letras distintas, na mesma avaliação, diferem pelo teste Tukey ao nível de 5% de
probabilidade (p<0,05). Barras indicam desvio padrão da média. Unicentro, Guarapuava/PR, 2018.
15 DAT 20 DAT 35 DAT 50 DAT 65 DAT 80 DAT 95 DAT 110 DAT
0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15 DAT 20 DAT 35 DAT 50 DAT 65 DAT 80 DAT 95 DAT 110 DAT
a
ddddd
ccc
c
cc
c
bb
b
b
bb
b
a
aa
a
a
a
a
Deposiç
ão d
e l
ignin
a (
µg de l
ignin
a-1
mg d
e m
assa f
resca
-1)
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1
) Boscalida (0,100 g L-1) + Cresoxim-metílico (0,050 g L
-1)
Piraclostrobina (0,100 g L-1) Fluxapiroxade (0,058 g L
-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
dddd
d
cc
c
c
c
c
c
b
b
b
bb
bb
a
a
aa
a
a
a
20162015
a

109
As plantas tratadas com os diferentes fungicidas, de maneira geral, apresentaram maior
síntese e deposição de lignina em folhas de tomateiro ao longo do ciclo da cultura, em
comparação com as testemunhas Test-Inoc e Test-Absoluta. No entanto, os maiores teores de
lignina foram observados para o tratamento Flux+Pira, seguido de Me+Pira.
A lignina é um polímero que, juntamente com a celulose e outros polissacarídeos que
constituem a parede celular das plantas, funciona como uma barreira física à penetração do
patógeno e por isso, é considerada uma das substâncias mais importantes presentes na parede
celular do hospedeiro (CASTRO et al., 2010). Assim, plantas que apresentam maiores teores
de lignina são mais resistentes ao ataque por micro-organismo. Isto indica, portanto, que a
aplicação de fungicidas de efeitos fisiológicos como estrobilurinas e carboxamidas, também
atuam como eliciadores e induzem respostas de defesa da planta como observado nos resultados
até aqui demonstrados.
Além das enzimas PAL, POD e PPO podem-se adicionar à lista das enzimas
sinalizadoras de respostas de indução de resistência, a enzima LOX e as PRPs, quitinase e β-
1,3-glucanase. A enzima LOX, assim como a PPO, faz parte das enzimas da via sinalizadora
dos octadecanóides na qual ocorre a biossíntese de ácido jasmônico, importante hormônio
vegetal relacionado ao estresse vegetal que ativa várias respostas de defesa. As LOXs estão
entre as enzimas chave nesse metabolismo. De acordo com Pinto et al. (2011) diversas vias de
sinalização estão interligadas e interagem com a formação de ácido jasmônico.
Cerca de 90% da membrana plasmática é composta de ácidos graxos insaturados,
principalmente, ácido linolênico e ácido linoleico, os quais são precursores para a atividade da
LOX (TAIZ; ZEIGER, 2013). Desta forma, sua atividade está associada com o metabolismo
dos lipídios, principalmente, no que diz respeito à lipoperoxidação. Observou-se que a atividade
da enzima LOX avaliada aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74,
75, 76, 79, 89, 90, 91, 94 e 105 DAT em folhas de tomateiro (Figura 31) apresentou diferença
entre os tratamentos no decorrer do tempo, com expressão semelhante nos dois anos avaliados.
Para a aplicação dos diferentes fungicidas pode-se observar que houve comportamento
semelhante ao observado nas demais enzimas avaliadas, com picos de atividades após cada
aplicação. No entanto, é possível verificar que os picos de atividade vão reduzindo no decorrer
das aplicações. Ao contrário, as plantas Test-Inoc apresentaram aumento da atividade da LOX
aos 16 DAT, compreendendo 24 horas após a inoculação do patógeno, permanecendo nas
demais avaliações.
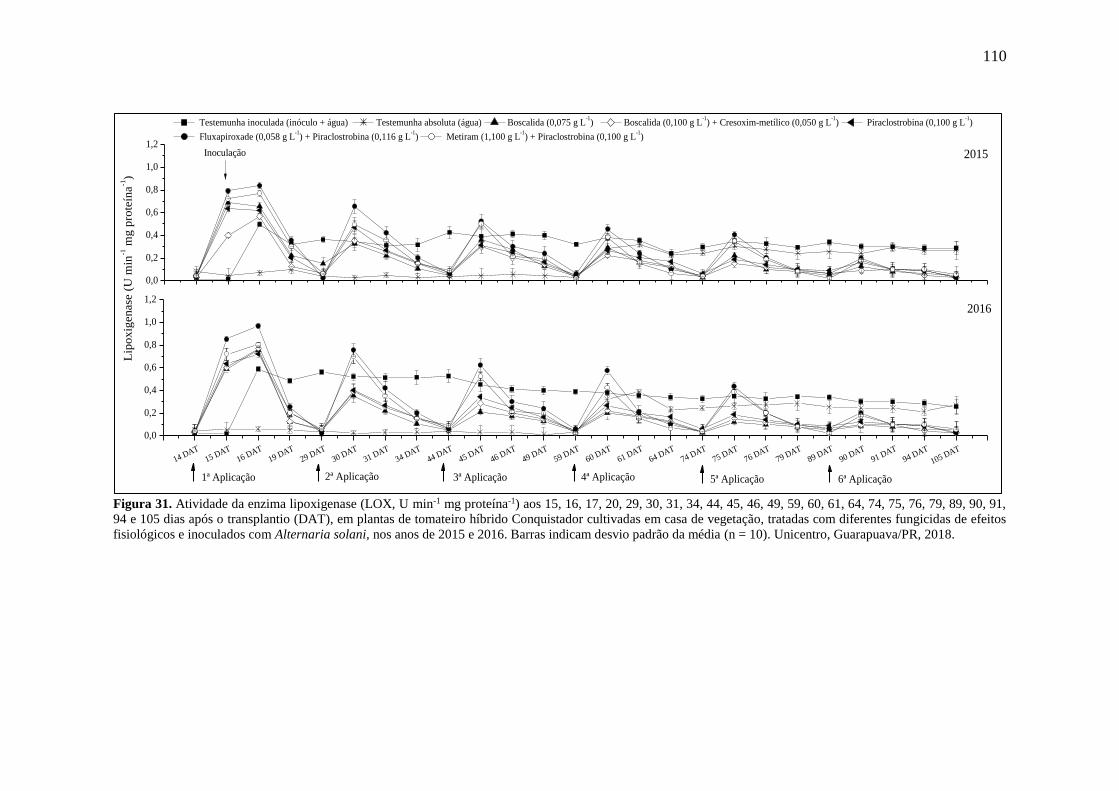
110
Figura 31. Atividade da enzima lipoxigenase (LOX, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91,
94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0,0
0,2
0,4
0,6
0,8
1,0
1,2
14 DAT15 DAT
16 DAT19 DAT
29 DAT30 DAT
31 DAT34 DAT
44 DAT45 DAT
46 DAT49 DAT
59 DAT60 DAT
61 DAT64 DAT
74 DAT75 DAT
76 DAT79 DAT
89 DAT90 DAT
91 DAT94 DAT
105 DAT
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Lip
oxig
enase
(U
min
-1 m
g p
rote
ína
-1)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

111
Possivelmente, o padrão de resposta observado tem relação com os níveis de
peroxidação lipídica, uma vez que, essas enzimas catalisam os ácidos graxos poli-insaturados
para hidroxiperóxidos, originando moléculas reativas (como H2O2), resultando na síntese de
ácido jasmônico e, por conseguinte, na sinalização de respostas de defesa contra patógenos,
como relatado por Stangarlin et al. (2011). De fato, nas plantas tratadas com Flux+Pira,
Me+Pira, Bosc+Cre, Bosc e Pira houve menor peroxidação lipídica ao longo das avaliações,
devido à ativação do aparato de defesa antioxidante. Como a via das LOXs dá origem a radicais
reativos, a presença simultânea das enzimas antioxidativas, auxiliou na proteção da célula
contra possíveis danos causados pelos radicais formados.
A ativação da expressão das enzimas PPO e LOX em resposta à aplicação dos diferentes
fungicidas demonstrou que ocorreu a ativação da rota dos octadecanóides, gerando moléculas
sinalizadoras e ativadoras de genes envolvidos com a defesa vegetal. Portanto, além da ativação
da rota metabólica dos fenilpropanóides e via do ácido salicílico, a aplicação destas moléculas
também induziu a ativação da via do ácido jasmônico, evidenciando o potencial destes
fungicidas como indutores de resistência em plantas de tomateiro.
O efeito aqui observado é muito importante pois, como já discutido, na interação A.
solani-tomateiro, por se tratar de patógeno necrotrófico, a morte celular não dificulta e, sim,
facilita a colonização do tecido pelo fungo, por isso a resistência gene-a-gene via sinalização
dependente de ácido salicílico não se faz tão eficiente. Portanto, nesse tipo de interação, as
respostas de defesa dependem da via de sinalização por intermédio do ácido jasmônico e/ou
etileno (GLAZEBROOK, 2005).
Como já visto, as PRPs são induzidas no hospedeiro em resposta à infecção por um
patógeno ou por estímulos abióticos (STANGARLIN et al., 2011). Entre as principais PRPs
estudadas no processo de indução de resistência estão as PODs (já discutidas), as quitinases e
β-1,3-glucanases (GAO et al., 2015). As enzimas quitinases e β-1,3-glucanases, por atuarem na
hidrólise da parede celular de fungos, possuem atuação sinérgica, potencializando a ação
antifúngica (MARTINS, 2008), por isso, o aumento na expressão destas enzimas são
importantes na defesa contra micro-organismos.
Quanto à atividade das enzimas quitinase e β-1,3-glucanase avaliadas aos 15, 16, 17,
20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e 105 DAT em
folhas de tomateiro (Figura 32 e 33, respectivamente) pode-se observar que os tratamentos
também interferiram na expressão destas enzimas. De maneira geral, é possível observar picos
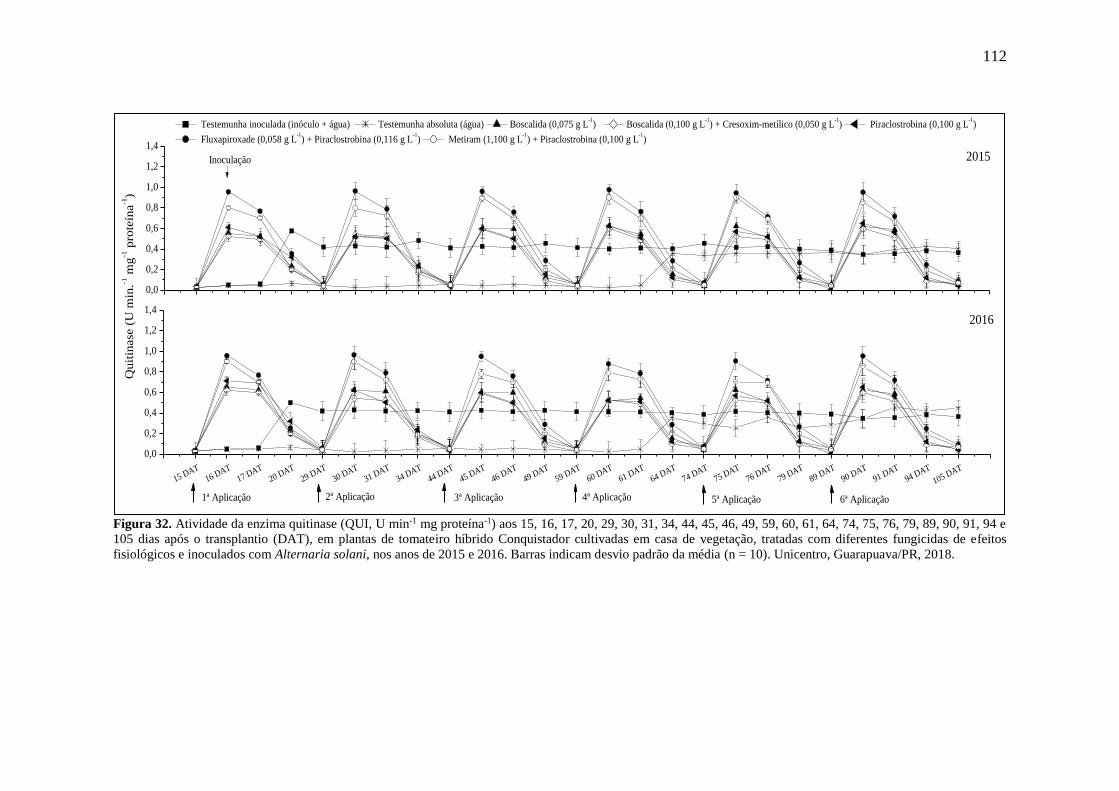
112
Figura 32. Atividade da enzima quitinase (QUI, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90, 91, 94 e
105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
15 DAT
16 DAT
17 DAT
20 DAT
29 DAT
30 DAT
31 DAT
34 DAT
44 DAT
45 DAT
46 DAT
49 DAT
59 DAT
60 DAT
61 DAT
64 DAT
74 DAT
75 DAT
76 DAT
79 DAT
89 DAT
90 DAT
91 DAT
94 DAT
105 DAT
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
Quit
inase
(U
min
.-1 m
g-1
pro
teín
a-1
)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

113
Figura 33. Atividade da enzima β-1,3-glucanase (GLU, U min-1 mg proteína-1) aos 15, 16, 17, 20, 29, 30, 31, 34, 44, 45, 46, 49, 59, 60, 61, 64, 74, 75, 76, 79, 89, 90,
91, 94 e 105 dias após o transplantio (DAT), em plantas de tomateiro híbrido Conquistador cultivadas em casa de vegetação, tratadas com diferentes fungicidas de efeitos
fisiológicos e inoculados com Alternaria solani, nos anos de 2015 e 2016. Barras indicam desvio padrão da média (n = 10). Unicentro, Guarapuava/PR, 2018.
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
15 DAT16 DAT
17 DAT20 DAT
29 DAT30 DAT
31 DAT34 DAT
44 DAT45 DAT
46 DAT49 DAT
59 DAT60 DAT
61 DAT64 DAT
74 DAT75 DAT
76 DAT79 DAT
89 DAT90 DAT
91 DAT94 DAT
105 DAT
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1) Piraclostrobina (0,100 g L
-1)
Fluxapiroxade (0,058 g L-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
1,3
-Glu
can
ase
(U
min
-1 m
g p
rote
ína
-1)
2016
Inoculação
6ª Aplicação4ª Aplicação 5ª Aplicação3ª Aplicação2ª Aplicação1ª Aplicação
2015

114
de atividade 24 horas após cada aplicação dos fungicidas, ocorrendo declínio após 48 e 120
horas. No entanto, as maiores atividades destas enzimas foram proporcionadas pela aplicação
de Flux+Pira, seguido de Me+Pira. Os outros fungicidas testados promoveram aumento na
atividade da enzima, porém em menor intensidade.
Para as plantas Test-Inoc nota-se que ocorreu aumento da atividade, tanto para quitinase
quanto para β-1,3-glucanase, aos 20 DAT (96 horas após a inoculação do patógeno) e manteve-
se durante as demais avaliações. Nas plantas Test-Absoluta pode-se verificar que a atividade
destas enzimas se manteve baixa até os 64 DAT, após houve um pequeno aumento.
Diferentes pesquisas relatam que a alta atividade de quitinase e de β-1,3-glucanase estão
relacionadas com o aumento da resistência das plantas aos patógenos. Pois a enzima quitinase,
hidrolisa a quitina da parede celular de fungos (THARANATHAN; KITTER, 2003) e, assim,
prejudica o desenvolvimento do patógeno. Já a enzima β-1,3-glucanase hidroliza polímeros de
β-1,3glucanas (LIN et al., 2009), que são componentes abundantes na parede celular de muitos
patógenos, juntamente com a quitina. Desta forma, no processo de indução de resistência, as
enzimas quitinase e β-1,3-glucanase, agem de forma conjunta (MARTINS, 2008).
Na interação planta-patógeno, a atuação conjunta de quitinase e de β-1,3-glucanase,
segundo Mauch e Staehelin (1989), inicia com a síntese de pequena quantidade de
β-1,3-glucanase, que atua em nível de lamela média e, conforme ocorre o crescimento do fungo,
as hifas vão sendo degradadas por ação desta enzima. Então, os subprodutos desta hidrólise
atuam como sinalizadores da invasão e induzem a síntese em maior concentração de quitinases
e β-1,3-glucanases que são armazenadas nos vacúolos. À medida que o patógeno consegue
romper as primeiras barreiras de defesa e adentra à célula, o conteúdo dos vacúolos é liberado
e as enzimas agem sobre as estruturas do patógeno, contendo a invasão.
Deste modo, as plantas tratadas com Flux+Pira, Me+Pira, Bosc+Cre, Bosc e Pira ao
promoverem o aumento nos níveis de quitinase e de β-1,3-glucanase comprovam sua eficiência
na ativação de respostas de defesa, pois desencadearam a síntese e acúmulo destas PRPs,
aumentando a capacidade de defesa aos patógenos em plantas de tomateiro. Na planta a ação
destas PRPs contra os micro-organismos patogênicos pode ser tanto direto como indireto. Pode
atuar, por exemplo, de forma direta inibindo a germinação de esporos ou o crescimento do
patógeno, por apresentarem atividade antimicrobiana. No entanto, a ação indireta é a mais
importante pela ação inibitória na atividade hidrolítica sobre a parede celular do fungo
(STANGARLIN et al., 2011).

115
De modo geral, pode-se dizer que a aplicação destes fungicidas proporcionou a ativação
de respostas de defesa em plantas de tomateiro, pois aumentou a expressão de importantes
enzimas de diferentes vias bioquímicas como PAL, POD, PPO, LOX, quitinase e
β-1,3-glucanase, além de maior acúmulo de compostos fenólicos e deposição de lignina. Este
efeito das estrobilurnas e, mais recentemente das carboxamidas, ainda é pouco conhecido.
Alguns trabalhos têm verificado que a aplicação de estrobilurinas interferem positivamente na
defesa de plantas contra agentes patogênicos virais e bacterianos em feijoeiro
(UDAYASHANKAR et al., 2012) e tomateiro (SKANDALIS et al., 2016). Estes estudos
revelam que a aplicação de piraclostrobina aumenta a expressão de genes envolvidos na
iniciação de respostas ao estresse biótico.
Portanto, com base nos resultados apresentados, todas as enzimas antioxidativas, assim
como as relacionadas com a indução de resistência, expressaram maiores atividades, após cada
aplicação dos diferentes fungicidas testados. Isto indica que, mesmo atuando em organelas
distintas e pertencendo a famílias diferentes, todas as enzimas apresentam em comum o fato de
estarem relacionadas à defesa da planta em condições de estresse biótico.
5.7 Produção e Qualidade pós-colheita de frutos de tomateiro
Em todos os parâmetros avaliados neste trabalho até o momento constatou-se efeitos
positivos em decorrência da aplicação dos diferentes fungicidas. No entanto, como o objetivo
final do cultivo de tomateiro é a produção de frutos com qualidade, observou-se que o efeito
dos benefícios fisiológicos e metabólicos também foi positivo na produção e qualidade pós-
colheita de frutos de tomateiro.
No experimento conduzido em 2015, a colheita dos frutos iniciou-se no dia 27 de março
de 2015 (79 DAT) e estendeu-se até o dia 5 de maio de 2015 (118 DAT). Enquanto que no
experimento de 2016, a colheita dos frutos iniciou-se no dia 30 de março de 2016 (82 DAT) e
estendeu-se até dia 7 de maio de 2016 (120 DAT).
De acordo com a Tabela 8 pode-se observar que houve diferença significativa (p<0,05)
entre os tratamentos nas características de produção e produtividade de frutos. Os resultados
encontrados foram similares para os dois experimentos (anos 2015 e 2016). De maneira geral,
pode-se observar que a aplicação de fungicidas influenciou positivamente na produção total de
frutos (kg m-2), com incremento de 104,2, 98,5, 63,4, 64,7 e 56,2% em relação à Test-Inoc, para
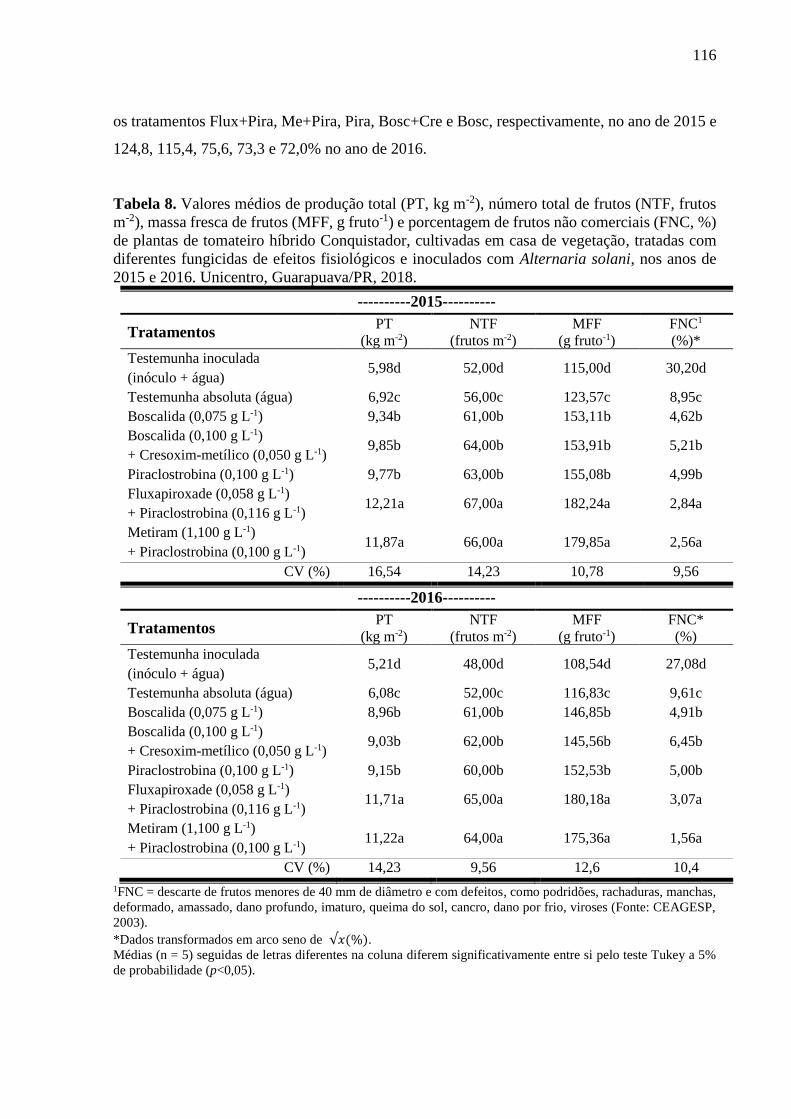
116
os tratamentos Flux+Pira, Me+Pira, Pira, Bosc+Cre e Bosc, respectivamente, no ano de 2015 e
124,8, 115,4, 75,6, 73,3 e 72,0% no ano de 2016.
Tabela 8. Valores médios de produção total (PT, kg m-2), número total de frutos (NTF, frutos
m-2), massa fresca de frutos (MFF, g fruto-1) e porcentagem de frutos não comerciais (FNC, %)
de plantas de tomateiro híbrido Conquistador, cultivadas em casa de vegetação, tratadas com
diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, nos anos de
2015 e 2016. Unicentro, Guarapuava/PR, 2018.
----------2015----------
Tratamentos PT
(kg m-2)
NTF
(frutos m-2)
MFF
(g fruto-1)
FNC1
(%)*
Testemunha inoculada
(inóculo + água) 5,98d 52,00d 115,00d 30,20d
Testemunha absoluta (água) 6,92c 56,00c 123,57c 8,95c
Boscalida (0,075 g L-1) 9,34b 61,00b 153,11b 4,62b
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 9,85b 64,00b 153,91b 5,21b
Piraclostrobina (0,100 g L-1) 9,77b 63,00b 155,08b 4,99b
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 12,21a 67,00a 182,24a 2,84a
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 11,87a 66,00a 179,85a 2,56a
CV (%) 16,54 14,23 10,78 9,56
----------2016----------
Tratamentos PT
(kg m-2)
NTF
(frutos m-2)
MFF
(g fruto-1)
FNC*
(%)
Testemunha inoculada
(inóculo + água) 5,21d 48,00d 108,54d 27,08d
Testemunha absoluta (água) 6,08c 52,00c 116,83c 9,61c
Boscalida (0,075 g L-1) 8,96b 61,00b 146,85b 4,91b
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 9,03b 62,00b 145,56b 6,45b
Piraclostrobina (0,100 g L-1) 9,15b 60,00b 152,53b 5,00b
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 11,71a 65,00a 180,18a 3,07a
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 11,22a 64,00a 175,36a 1,56a
CV (%) 14,23 9,56 12,6 10,4
1FNC = descarte de frutos menores de 40 mm de diâmetro e com defeitos, como podridões, rachaduras, manchas,
deformado, amassado, dano profundo, imaturo, queima do sol, cancro, dano por frio, viroses (Fonte: CEAGESP,
2003).
*Dados transformados em arco seno de √𝑥(%).
Médias (n = 5) seguidas de letras diferentes na coluna diferem significativamente entre si pelo teste Tukey a 5%
de probabilidade (p<0,05).

117
Também se observou incremento de 76,4, 71,5, 41,2, 42,3 e 35,0% em relação à Test-
Absoluta para os tratamentos Flux+Pira, Me+Pira, Pira, Bosc+Cre e Bosc, respectivamente, no
ano de 2015 e de 92,6, 84,5, 50,5, 48,5 e 47,4% no ano de 2016. Os maiores incrementos de
produção observados no ano de 2016, em comparação com 2015, devem-se ao fato das
testemunhas terem apresentado menor produção no ano de 2016, em decorrência da maior
severidade da doença como comprovado pelas maiores AACPDs (Figura 14).
O tratamento que apresentou maior produção de frutos foi o Flux+Pira (12,2 e 11,7 kg
m-2 em 2015 e 2016, respectivamente) seguido do tratamento Me+Pira (11,9 e 11,2 7 kg m-2 em
2015 e 2016, respectivamente), não diferindo significativamente entre si. Estes mesmos
tratamentos também apresentaram o maior número de frutos totais m-2, com maior massa fresca
de frutos (g fruto-1) e, consequentemente, menor porcentagem de frutos não comerciais.
Estes resultados evidenciam que a aplicação destas estrobilurinas e carboxamidas
proporcionaram incrementos de produtividade na cultura do tomateiro e são consistentes com
Giuliani et al. (2010) e Cantore et al. (2016), que relataram que a aplicação de azoxistrobina e
piraclostrobina, respectivamente, em plantas de tomateiro promoveram maior rendimento de
frutos comercializáveis. Pesquisas anteriores também demonstraram incrementos de
produtividade por estes fungicidas em outras culturas como videira (DIAZ-ESPEJO et al.,
2012), soja (JOSHI et al., 2014), feijão (JADOSKI et al., 2015), milho (SHETLEY et al., 2015),
cenoura (COLOMBARI et al., 2015) e pepino japonês (AMARO et al., 2018).
A maior produção de frutos e, concomitantemente, maior número de frutos comerciais,
nos tratamentos Flux+Pira e Me+Pira está associada aos efeitos colaterais sobre a fisiologia da
planta adicionados a melhor eficácia de controle de doenças (DIAZ-ESPEJO et al., 2012). De
fato, a maior eficiência fotossintética foi observada para estes tratamentos, evidenciada pelas
altas taxas de assimilação de CO2 e eficiência de carboxilação, demonstradas pelo maior
rendimento quântico do PSII, aliados a maior atividade da enzima nitrato redutase que
influenciou no teor de pigmentos fotossintéticos e na assimilação e disponibilidade de
nitrogênio durante o ciclo da cultura.
Somados a estes efeitos também se observou maior atividade das enzimas antioxidantes
que minimizaram os danos oxidativos, além do aumento nas respostas de defesa da planta.
Estudos demonstram que a síntese de compostos de defesa no processo de indução de
resistência, como proteínas/enzimas específicas utilizam como substratos, por exemplo,
eritrose-4-fosfato do ciclo das pentoses ou fosfoenolpiruvato da glicólise na rota dos

118
fenilpropanoides. Assim, acabam competindo pela energia necessária ao metabolismo primário,
podendo comprometer o crescimento, desenvolvimento e produção do vegetal (MAZARO et
al., 2009). Isto indica que a ativação de respostas de defesa observadas em plantas de tomateiro,
gerou baixo custo energético, ou seja, o efeito positivo no aumento da síntese de fotoassimilados
compensou a perda metabólica gasta na ativação das diferentes rotas de defesa.
Analisando os parâmetros de qualidade pós-colheita de frutos tratados com diferentes
fungicidas pode-se observar nas Tabelas 9 e 10 que houve diferença significativa (p<0,05) entre
os tratamentos para a as características de firmeza de polpa, teor de sólidos solúveis, relação
SS/AT e teor de ácido ascórbico, nos dois anos avaliados.
Os valores de pH dos frutos de tomate variaram entre 4,38 a 4,44 e entre 4,38 e 4,45 em
2015 e 2016, respectivamente, não sendo influenciados significativamente pelos tratamentos
aplicados nas plantas durante os estádios vegetativo e reprodutivo (Tabelas 9 e 10). Esta
característica também não foi influenciada pela aplicação de estrobilurinas e carboxamidas em
tomateiro (RAMOS et al., 2013) e meloeiro (MACEDO et al., 2017). Comportamento
semelhante foi observado nos valores de acidez titulável, onde os valores variaram entre 0,10 a
0,13 e 0,10 a 0,14 em 2015 e 2016, respectivamente, e não houve diferença significativa entre
os tratamentos.
O teor de sólidos solúveis, determinado em °Brix, apresentou diferença significativa em
função dos tratamentos. Os maiores valores foram observados nos tratamentos Flux+Pira (3,78
e 3,82 em 2015 e 2016, respectivamente) e Me+Pira (3,76 e 3,79 em 2015 e 2016,
respectivamente), diferindo dos demais tratamentos. Em frutos de tomateiro, os açúcares são os
maiores constituintes sólidos, assim, frutos que apresentam maior °Brix refletem maior síntese
e acúmulo de açúcares durante seu desenvolvimento (HEINE et al., 2015), aumentando
conforme a maturação se aproxima (CHITARRA; CHITARRA, 2005).
De fato, as maiores concentrações de açúcares totais e redutores aos 120 DAT foram
observados nos frutos de plantas tratadas com Flux+Pira e Me+Pira (Tabelas 6 e 7),
evidenciando que estes tratamentos aplicados durante o desenvolvimento da planta promovem
maior qualidade dos frutos, influenciando principalmente no sabor. A aplicação de
estrobilurinas e carboxamidas também influenciaram positivamente as características físico-
químicas dos frutos de tomateiro (RAMOS et al., 2013) e meloeiro (MACEDO et al., 2017).

119
Tabela 9. Valores médios de firmeza (gf cm-2), pH, teor de sólidos solúveis (SS, °Brix), acidez titulável (AT, g de ácido cítrico 100 g de
polpa-1), relação SS/AT e teor de ácido ascórbico (AA, mg 100 g de polpa-1) de frutos de tomateiro híbrido Conquistador, cultivados em casa
de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, no ano de 2015. Unicentro,
Guarapuava/PR, 2018.
Análises Pós-Colheita - 2015
Tratamentos Firmeza
(gf cm-2) pH
SS
(°Brix)
AT
(g ác. cítrico) SS/AT
AA
(mg 100 g-1)
Testemunha inoculada
(inóculo + água) 91,12c 4,42a 3,16c 0,12a 26,33c 43,25c
Testemunha absoluta (água) 94,85c 4,44a 3,34c 0,12a 27,83c 45,75c
Boscalida (0,075 g L-1) 99,47b 4,39a 3,46b 0,13a 26,62c 54,50b
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 98,12b 4,42a 3,58b 0,11a 32,55b 53,25b
Piraclostrobina (0,100 g L-1) 101,26b 4,38a 3,53b 0,12a 29,42b 54,75b
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 132,59a 4,41a 3,78a 0,10a 37,80a 59,25a
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 129,84a 4,38a 3,76a 0,12a 31,33b 57,25a
CV (%) 10,25 9,52 6,89 5,89 9,47 8,95
Médias (n = 10) seguidas de letras diferentes na coluna diferem significativamente entre si pelo teste Tukey a 5% de probabilidade (p<0,05).
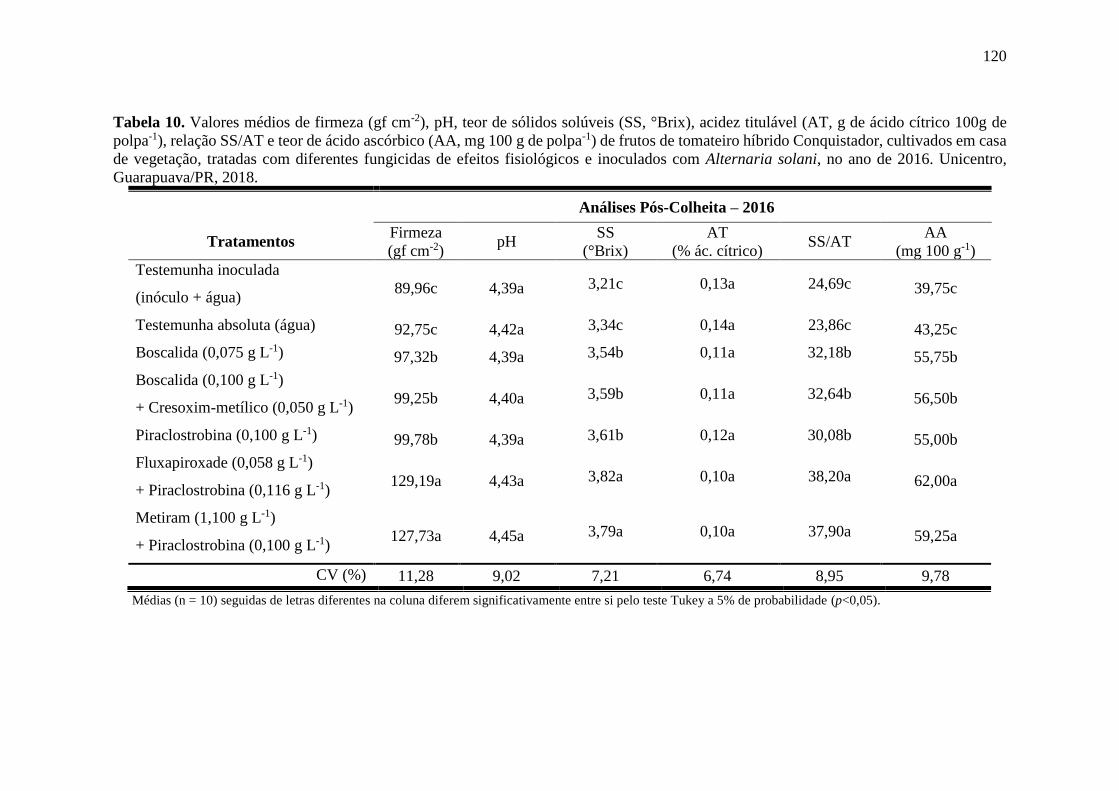
120
Tabela 10. Valores médios de firmeza (gf cm-2), pH, teor de sólidos solúveis (SS, °Brix), acidez titulável (AT, g de ácido cítrico 100g de
polpa-1), relação SS/AT e teor de ácido ascórbico (AA, mg 100 g de polpa-1) de frutos de tomateiro híbrido Conquistador, cultivados em casa
de vegetação, tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, no ano de 2016. Unicentro,
Guarapuava/PR, 2018.
Análises Pós-Colheita – 2016
Tratamentos Firmeza
(gf cm-2) pH
SS
(°Brix)
AT
(% ác. cítrico) SS/AT
AA
(mg 100 g-1)
Testemunha inoculada
(inóculo + água) 89,96c 4,39a 3,21c 0,13a 24,69c 39,75c
Testemunha absoluta (água) 92,75c 4,42a 3,34c 0,14a 23,86c 43,25c
Boscalida (0,075 g L-1) 97,32b 4,39a 3,54b 0,11a 32,18b 55,75b
Boscalida (0,100 g L-1)
+ Cresoxim-metílico (0,050 g L-1) 99,25b 4,40a 3,59b 0,11a 32,64b 56,50b
Piraclostrobina (0,100 g L-1) 99,78b 4,39a 3,61b 0,12a 30,08b 55,00b
Fluxapiroxade (0,058 g L-1)
+ Piraclostrobina (0,116 g L-1) 129,19a 4,43a 3,82a 0,10a 38,20a 62,00a
Metiram (1,100 g L-1)
+ Piraclostrobina (0,100 g L-1) 127,73a 4,45a 3,79a 0,10a 37,90a 59,25a
CV (%) 11,28 9,02 7,21 6,74 8,95 9,78
Médias (n = 10) seguidas de letras diferentes na coluna diferem significativamente entre si pelo teste Tukey a 5% de probabilidade (p<0,05).

121
As características sensoriais como doçura, acidez, aroma, textura, etc., determinam o
sabor de frutos maduros (ANTUNES et al., 2014) e podem ser avaliadas pela relação SS/AT,
analisando o equilíbrio entre o conteúdo de açúcares e ácidos no fruto (CHITARRA;
CHITARRA, 2005). Pode-se observar que o tratamento que apresentou os maiores valores para
a relação SS/AT foi Flux+Pira (37,8 e 38,8 em 2015 e 2016, respectivamente), seguido de
Me+Pira (31,3 e 37,8 em 2015 e 2016, respectivamente). No entanto, não houve diferença
estatística entre eles no ano de 2016, diferindo dos demais tratamentos.
O tomate é considerado saboroso, quando apresenta a proporção SS/AT superior a 10
(KADER et al., 1978). De maneira geral, todos os frutos apresentaram excelente qualidade,
estando adequados para o consumo in natura. No entanto, quanto maior o valor da relação
SS/AT mais suave é o sabor do fruto, devido a excelente combinação entre açúcar e ácido
(SHIRAHIGE et al., 2010).
Na análise do teor de ácido ascórbico os tratamentos Flux+Pira e Me+Pira apresentaram
os melhores resultados. A aplicação de Flux+Pira em plantas de tomateiro ao longo do ciclo
proporcionou incremento de 36,9 e 55,9% em relação à Test-Inoc (em 2015 e 2016,
respectivamente) e 29,5 e 43,4% (em 2015 e 2016, respectivamente), enquanto que para
Me+Pira foi de 32,4 e 49,1% (em 2015 e 2016, respectivamente) e 25,1 e 36,9% (em 2015 e
2016, respectivamente) de ácido ascórbico nos frutos.
O ácido ascórbico, também conhecido como vitamina C, é um composto antioxidante
essencial para a saúde, por estar envolvido nas respostas fisiológicas ao estresse (ORNELAS-
PAZ et al., 2013) e, por isso, frutos com altos teores de vitamina C são mais atrativos aos olhos
dos consumidores (SHIRAHIGE et al., 2010). Além de ser essencial, a preservação das
características físico-químicas e a qualidade pós-colheita dos frutos de tomate de mesa
(FERREIRA et al., 2010) conferem melhor sabor ao fruto, que são perceptíveis pelo
consumidor e definem sua escolha no momento da aquisição (HEINE et al., 2015).
Quanto à firmeza dos frutos (gf cm-2), os tratamentos Flux+Pira e Me+Pira também
apresentaram valores superiores de firmeza, isto indica que estes tratamentos proporcionaram
o desenvolvimento de frutos mais firmes e com maior conservação pós-colheita. Tais resultados
têm correlação com as respostas observadas para as atividades das enzimas pectinametilesterase
(PME) e poligalacturonase (PG) e com a perda de massa (Figuras 34 e 35) nestes mesmos
tratamentos, pois o amaciamento dos frutos decorre de processos desencadeados pela atividade
das enzimas PME e PG (SAINZ; VENDRUSCOLO, 2015). Tratamentos que interferem na
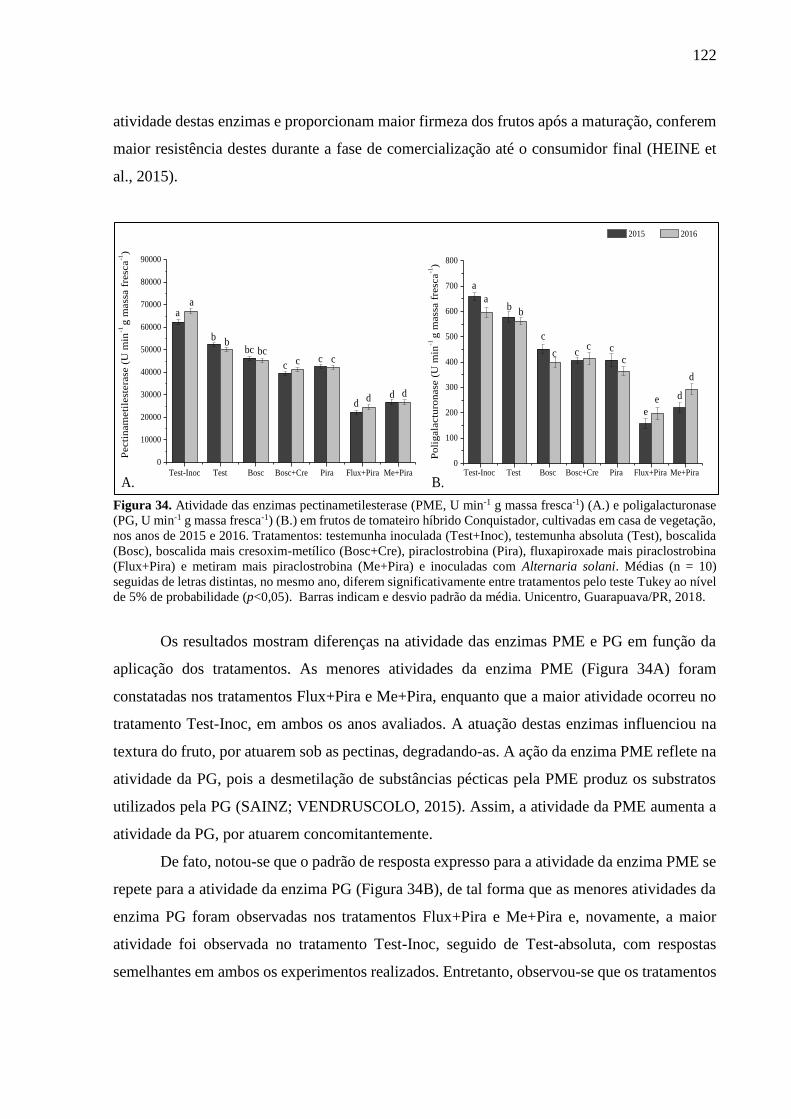
122
atividade destas enzimas e proporcionam maior firmeza dos frutos após a maturação, conferem
maior resistência destes durante a fase de comercialização até o consumidor final (HEINE et
al., 2015).
Figura 34. Atividade das enzimas pectinametilesterase (PME, U min-1 g massa fresca-1) (A.) e poligalacturonase
(PG, U min-1 g massa fresca-1) (B.) em frutos de tomateiro híbrido Conquistador, cultivadas em casa de vegetação,
nos anos de 2015 e 2016. Tratamentos: testemunha inoculada (Test+Inoc), testemunha absoluta (Test), boscalida
(Bosc), boscalida mais cresoxim-metílico (Bosc+Cre), piraclostrobina (Pira), fluxapiroxade mais piraclostrobina
(Flux+Pira) e metiram mais piraclostrobina (Me+Pira) e inoculadas com Alternaria solani. Médias (n = 10)
seguidas de letras distintas, no mesmo ano, diferem significativamente entre tratamentos pelo teste Tukey ao nível
de 5% de probabilidade (p<0,05). Barras indicam e desvio padrão da média. Unicentro, Guarapuava/PR, 2018.
Os resultados mostram diferenças na atividade das enzimas PME e PG em função da
aplicação dos tratamentos. As menores atividades da enzima PME (Figura 34A) foram
constatadas nos tratamentos Flux+Pira e Me+Pira, enquanto que a maior atividade ocorreu no
tratamento Test-Inoc, em ambos os anos avaliados. A atuação destas enzimas influenciou na
textura do fruto, por atuarem sob as pectinas, degradando-as. A ação da enzima PME reflete na
atividade da PG, pois a desmetilação de substâncias pécticas pela PME produz os substratos
utilizados pela PG (SAINZ; VENDRUSCOLO, 2015). Assim, a atividade da PME aumenta a
atividade da PG, por atuarem concomitantemente.
De fato, notou-se que o padrão de resposta expresso para a atividade da enzima PME se
repete para a atividade da enzima PG (Figura 34B), de tal forma que as menores atividades da
enzima PG foram observadas nos tratamentos Flux+Pira e Me+Pira e, novamente, a maior
atividade foi observada no tratamento Test-Inoc, seguido de Test-absoluta, com respostas
semelhantes em ambos os experimentos realizados. Entretanto, observou-se que os tratamentos
Test-Inoc Test Bosc Bosc+Cre Pira Flux+Pira Me+Pira0
100
200
300
400
500
600
700
800
e
de
d
ccc
cc
c
bb
a
a
B.
Poli
gala
ctu
ronase
(U
min
-1 g
mass
a f
resc
a-1
)
2015 2016
A.Test-Inoc Test Bosc Bosc+Cre Pira Flux+Pira Me+Pira
0
10000
20000
30000
40000
50000
60000
70000
80000
90000
ddd
d
cc
bccc
bcb
b
a
Pecti
nam
eti
lest
era
se (
U m
in-1
g m
ass
a f
resc
a-1
)
a
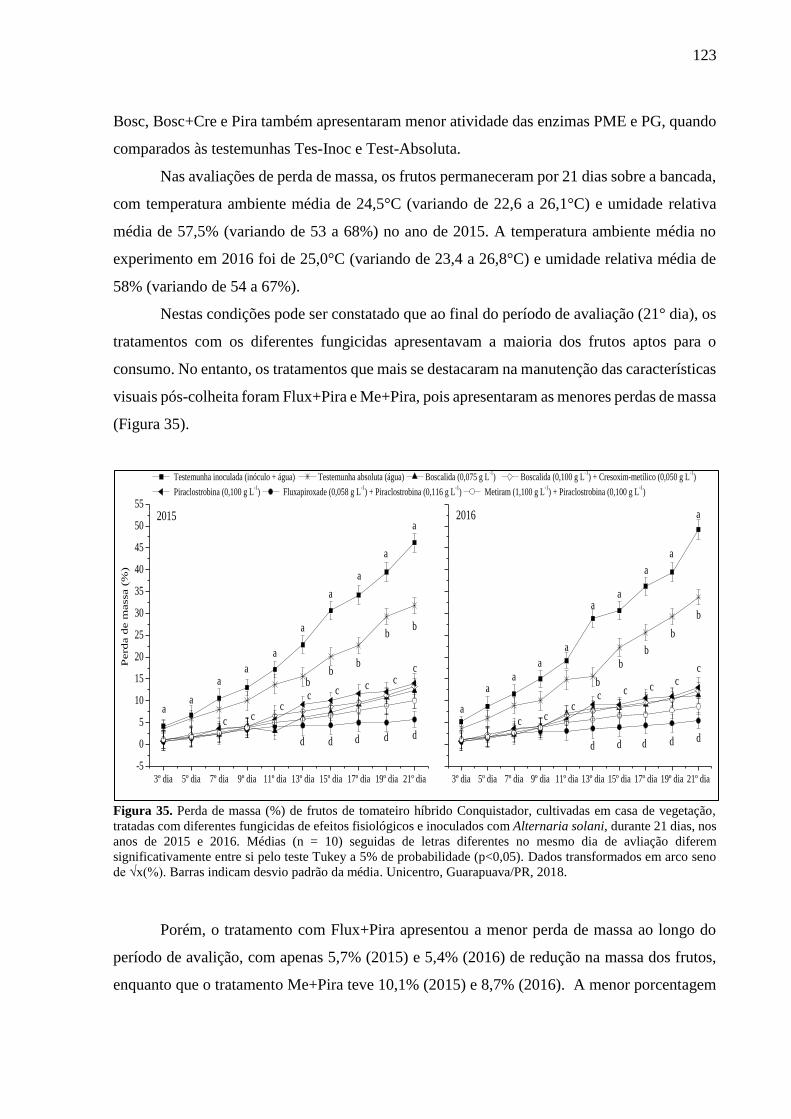
123
Bosc, Bosc+Cre e Pira também apresentaram menor atividade das enzimas PME e PG, quando
comparados às testemunhas Tes-Inoc e Test-Absoluta.
Nas avaliações de perda de massa, os frutos permaneceram por 21 dias sobre a bancada,
com temperatura ambiente média de 24,5°C (variando de 22,6 a 26,1°C) e umidade relativa
média de 57,5% (variando de 53 a 68%) no ano de 2015. A temperatura ambiente média no
experimento em 2016 foi de 25,0°C (variando de 23,4 a 26,8°C) e umidade relativa média de
58% (variando de 54 a 67%).
Nestas condições pode ser constatado que ao final do período de avaliação (21° dia), os
tratamentos com os diferentes fungicidas apresentavam a maioria dos frutos aptos para o
consumo. No entanto, os tratamentos que mais se destacaram na manutenção das características
visuais pós-colheita foram Flux+Pira e Me+Pira, pois apresentaram as menores perdas de massa
(Figura 35).
Figura 35. Perda de massa (%) de frutos de tomateiro híbrido Conquistador, cultivadas em casa de vegetação,
tratadas com diferentes fungicidas de efeitos fisiológicos e inoculados com Alternaria solani, durante 21 dias, nos
anos de 2015 e 2016. Médias (n = 10) seguidas de letras diferentes no mesmo dia de avliação diferem
significativamente entre si pelo teste Tukey a 5% de probabilidade (p<0,05). Dados transformados em arco seno
de √x(%). Barras indicam desvio padrão da média. Unicentro, Guarapuava/PR, 2018.
Porém, o tratamento com Flux+Pira apresentou a menor perda de massa ao longo do
período de avalição, com apenas 5,7% (2015) e 5,4% (2016) de redução na massa dos frutos,
enquanto que o tratamento Me+Pira teve 10,1% (2015) e 8,7% (2016). A menor porcentagem
3º dia 5º dia 7º dia 9º dia 11º dia 13º dia 15º dia 17º dia 19º dia 21º dia
-5
0
5
10
15
20
25
30
35
40
45
50
55
3º dia 5º dia 7º dia 9º dia 11º dia 13º dia 15º dia 17º dia 19º dia 21º dia
ddddd
cc
cccc
cc
bb
b
bb
a
a
a
a
a
a
aa
a
2015
Perd
a d
e m
assa (
%)
a
Testemunha inoculada (inóculo + água) Testemunha absoluta (água) Boscalida (0,075 g L-1) Boscalida (0,100 g L
-1) + Cresoxim-metílico (0,050 g L
-1)
Piraclostrobina (0,100 g L-1) Fluxapiroxade (0,058 g L
-1) + Piraclostrobina (0,116 g L
-1) Metiram (1,100 g L
-1) + Piraclostrobina (0,100 g L
-1)
2016
ddddd
cc
cccc
cc
b
bb
b
b
a
a
a
aa
a
a
aa
a

124
de perda de massa ao longo do período de avaliação está relacionada à firmeza dos frutos, que
de fato foram maiores para estes dois tratamentos. A perda de massa pode ser atribuída,
principalmente, à perda de umidade e de material de reserva pela transpiração e respiração,
respectivamente, sendo um dos principais fatores limitantes da vida útil pós-colheita de frutos.
O maior período de conservação pós-colheita do fruto, permite ter uma perspectiva de quanto
tempo ele resiste ao transporte e/ou comercialização sem que perca suas características e
qualidades externas e internas dos frutos.
Observou-se que as testemunhas, Tes-Inoc e Test-Absoluta, apresentaram as maiores
perdas de massa, quando comparados aos tratamentos Flux+Pira, Me+Pira, Bosc, Bosc+Cre e
Pira. Melhor qualidade pós-colheita de frutos de tomateiro também foi observada por Ramos et
al. (2013), relatando que o tratamento com a mistura de piraclostrobina e boscalida
proporcionaram frutos mais firmes com uma melhor conservação pós-colheita, uma vez que
frutos mais firmes poderão ser comercializados por períodos mais longos.
Portanto, indicando que, de maneira geral, estes fungicidas também apresentam efeito
positivo na qualidade pós-colheita de frutos de tomateiro, desacelerando o processo de
degradação do fruto, proporcionando maior vida de prateleira e conservação da qualidade no
armazenamento. Em última análise, pode-se inferir que a produtividade e a qualidade pós-
colheita de frutos de tomateiro são reflexos do manejo adotado durante o ciclo da cultura e que
a aplicação dos fungicidas testados promoveram efeitos positivos em todas as características
avaliadas.

125
6. CONSIDERAÇÕES FINAIS
Considerando os resultados obtidos nas condições destes experimentos foi possível
verificar por meio dos efeitos fisiológicos proporcionados pelos diferentes fungicidas, que em
todas as características avaliadas as plantas testemunhas, Test-Inoc e Test-Absoluta, foram
aquelas que apresentaram os menores resultados, nos dois anos avaliados, comprovando o efeito
fisiológico dos fungicidas nas plantas de tomateiro híbrido Conquistador cultivadas em casa de
vegetação.
É importante destacar que as temperaturas médias (mínima e máxima) observadas
durante os dois anos de condução do experimento encontravam-se entre 19,5 a 30,5°C e,
sabendo-se que para plantas C3 a temperatura ótima para todas as reações bioquímicas da
fotossíntese é de aproximadamente 25°C, nota-se que as temperaturas foram ótimas para os
processos fotossintéticos durante todo o ciclo de desenvolvimento das plantas de tomateiro.
A partir dos dados observados nas avaliações de trocas gasosas e fluorescência da
clorofila a comprovou-se que plantas de tomateiro sob estresse biótico (Test-Inoc) em
decorrência da presença da doença pinta preta sofreram danos no aparato fotossintético. Este
fato foi verificado pela baixa eficiência fotossintética, que refletiu diretamente no
desenvolvimento e produtividade destas plantas.
Constatou-se também que, os diferentes fungicidas testados, além de serem eficientes
no controle de pinta preta em tomateiro foram capazes de promover a ativação de mecanismos
de defesa, induzindo respostas bioquímicas, observadas pelo aumento da atividade das enzimas
antioxidantes e das enzimas relacionadas à patogênese com baixo custo metabólico. Pois, nestes
mesmos tratamentos observou-se maior acúmulo de biomassa e produção de frutos, quando
comparados às testemunhas. Isto indica que há um importante mecanismo de resistência sendo
induzido por estas moléculas, que até o momento era pouco conhecido.
Para a atividade das enzimas antioxidativas e PRPs, avaliadas ao longo do ciclo da
cultura, notou-se que o efeito se repetiu, apresentando picos de atividades entre 24 e 48 horas
após cada aplicação dos tratamentos, comprovando o potencial destas substâncias como
promotoras de respostas de defesa ao estresse biótico em plantas de tomateiro. Este efeito
protetor ao estresse oxidativo somado a maior eficiência fotossintética resultou nos maiores
índices produtivos e qualidade de frutos observados nas plantas tratadas com estas
estrobilurinas e carboxamidas, pois incrementar a eficiência fotossintética e controlar o estresse
são fatores cruciais para aumentar a produtividade da cultura.

126
Os maiores valores da atividade da enzima nitrato redutase observados nas plantas
tratadas com os diferentes fungicidas, refletiram em maiores teores de pigmentos
fotossintéticos, em especial das clorofilas a e b, pois o teor de clorofila tem estreita relação com
a quantidade de nitrogênio absorvido e assimilado pela planta. Também, o aumento na síntese
de pigmentos incrementou a eficiência fotossintética nestes tratamentos, demonstrando que há
relação entre o aumento da atividade da enzima nitrato redutase e a concentração de clorofilas
interferindo diretamente na fotossíntese líquida e nos demais parâmetros fotossintéticos
avaliados em plantas tratadas com estas estrobilurinas e carboxamidas.
Outra característica importante é que a enzima nitrato redutase pode produzir óxido
nítrico a partir da redução de um elétron do nitrito, utilizando NADPH como doador de elétrons.
Por sua vez, o óxido nítrico, além de outras funções, apresenta importante papel na difusão da
sinalização de respostas de defesa nos vegetais. Assim, há a hipótese de que a maior atividade
da enzima nitrato redutase também esteja relacionada, pela maior biossíntese de óxido nítrico,
com a cascata de eventos, observadas nas respostas de defesas em plantas tratadas com estas
estrobilurinas e carboxamidas. Portanto, diante desta inferência novos estudos com estas
moléculas no patossistema tomateiro-A. solani são necessários para elucidar esta hipótese.
Durante a condução dos experimentos foram realizadas aplicações quinzenais,
totalizando 6 aplicações de fungicidas, que permitiram excelente controle da doença. No
entanto, em condições de campo, onde os fatores climáticos não podem ser controlados, são
necessárias aplicações semanais ou em menor intervalo, dependendo da severidade da doença
e das condições climáticas. Por isso, é comum encontrar sistemas de cultivo que realizam mais
de 12 aplicações de fungicidas durante o ciclo da cultura. Sabendo dos efeitos positivos que os
fungicidas aqui testados proporcionam na fisiologia e no metabolismo das plantas, recomenda-
se que estas moléculas sejam utilizadas em um manejo racional (rotação de modos de ação)
para o controle de A. solani em tomateiro.
Em última análise, tem-se como desafio compreender como todos os efeitos fisiológicos
e metabólicos promovidos pelas estrobilurinas e carboxamidas avaliadas se interligam. A
Figura 36 ilustra, de modo simplificado, a interação entre todos os efeitos fisiológicos e
metabólicos observados em plantas de tomateiro híbrido Conquistador, cultivadas em casa de
vegetação e inoculadas com A. solani, pela aplicação de boscalida, boscalida mais cresoxim-
metílico, piraclostrobina, fluxapiroxade mais piraclostrobina e metiram mais piraclostrobina.

> Firmeza dos frutosAtividade enzimas PME e PG
Perda de massa
Qualidade pós colheita
Sólidos solúveisRelação SS/ATÁcido ascórbico
Melhora o sabor
N frutos m²Massa fresca frutos g% frutos comerciais
> Produtividade
o
> E
ficiê
nci
a foto
ssin
tétic
a
{Taxa de assimilação de COCondutância estomática[ ] interna de COTaxa transpiraçãoEficiência do uso da águaEficiência de carboxilaçãoEficiência máxima de PS IIRendimento quântico fotoquímico do PS IITaxa relativa de transporte de elétrons
[ ] de clorofilas a e b
Crescimento vegetalÁrea foliarBiomassa de folha, caule e raízes[ ] Açúcares totais e açúcaresredutores nas folhas, caule e frutos
> D
ese
nvo
lvim
ento
{2
2
Inibe o transporte de e temporariamente-
[ERO ] = O e H O 2S 2 2-.
Danos oxidativos Defesa antioxidativa
Peroxidação lipídica O 2
2 2
-.
H O
[ ]
[ ]
SODAPXGRGSTGPXCAT
{ {Peroxidaçãolipídica
Tóxico ao patógeno
Respostas dehipersensibilidade
Assimilaçãode nitrogênio
NO NO- -23
Respostasde defesa
Ativação de defesas
NO
?
?
Ativa enzima NR
2 2H O
Via fenilpropanóides Via Octadecanóides
Biossíntese de AS Biossíntese de AJ
PAL
Comp. fenóicos
POD
LOX
PPO
{+
{ Quitinase
B-1, 3-Glucanase
Biossíntesede lignina {?
Eficiente controlede Alternaria solani
Alternaria solani
> PRPs
{
FungicidasEstrobilurinas
e Carboxamidas
BOSBOS + CREPIRAFLUX + PIRAME + PIRA{
{
nível de ATP celular
[H ] no citosol+
Inibe o transporte de e -*
e/ou
{ {
{*
Box 1
Box 2
Box 4
Box 3
Box 5
Box 6
Box 7
Box 8
Box 9
Box 10
Legenda
*Mensageiro celular - metabolismo secundário
Proteção do aparatofotossintético
EROs
MA
RE
K, 2018

128
Figura 36. Modelo esquemático mostrando os efeitos fisiológicos e metabólicos promovidos pela aplicação de
estrobilurinas: cresoxim-metílico e boscalida e carboxamidas: boscalida e fluxapiroxade em plantas de tomateiro
híbrido Conquistador cultivadas em casa de vegetação e inoculadas com Alternaria solani. Fonte: MAREK, 2018.
A eficiência de controle de pinta preta (A. solani) em plantas de tomateiro submetidas à
aplicação de estrobilurinas e carboxamidas está representado no BOX 1: a ação transitória das
moléculas no metabolismo celular vegetal – inibição temporária do transporte de elétrons e
ativação da atividade da enzima nitrato redutase (NR) e o possível envolvimento de moléculas
sinalizadoras como o óxido nítrico (NO). Com a ativação da enzima NR há o aumento da
assimilação de nitrogênio (BOX 2), que culmina na maior síntese de pigmentos fotossintéticos
e, consequentemente, aumenta a eficiência fotossintética da planta (BOX 3), refletindo em
maior desenvolvimento (BOX 4). Em função da ação dos fungicidas no metabolismo celular
(BOX 5) há a superprodução de espécies reativas de oxigênio (EROs). As EROs quando estão
em altas concentrações podem causar danos celulares, no entanto, quando o arsenal de defesa
enzimático antioxidativo é ativado (BOX 7), ocorre menores danos em função da menor
peroxidação lipídica (BOX 6) e, consequentemente, maior proteção do aparato fotossintético.
Entretanto, a superprodução de EROs (BOX 5) também desencadeia respostas de
defesa, além de serem tóxicas ao patógeno (BOX 8), por isso, em concentrações fisiológicas
adequadas, o H2O2 também atua como mensageiro secundário, ativando importantes vias de
defesa vegetal no metabolismo secundário (BOX 9) e a possível biossíntese de hormônios
vegetais como ácido salicílico e ácido jasmônico (importantes sinalizadores de respostas de
defesa). Unindo todos os efeitos fisiológicos e metabólicos a resposta final é a maior
produtividade e qualidade pós-colheita (BOX 10).

129
7. CONCLUSÃO
Com base nos resultados obtidos e nas condições destes experimentos pode-se concluir
que as aplicações de boscalida, boscalida mais cresoxim-metílico, piraclostrobina,
fluxapiroxade mais piraclostrobina e metiram mais piraclostrobina apresentaram eficiente
controle da doença, promoveram efeitos fisiológicos positivos e ativaram os mecanismos de
defesa em plantas de tomateiro.
Os fungicidas fluxapiroxade mais piraclostrobina, seguido de metiram mais
piraclostrobina, foram os tratamentos que proporcionaram os melhores controles de A. solani e
as melhores respostas fisiológicas e metabólicas em plantas de tomateiro híbrido Conquistador,
cultivado em casa de vegetação.

130
8. Referências Bibliográficas
ADAM, A.; FARKAS, T.; SOMLYAI, G.; HEVESI, M.; KIRALY, Z. Consequence of •O2-
generation during a bacterially induced hypersensitive reaction in tobacco: deterioration of
membrane lipids. Physiological and Molecular Plant Pathology, v. 34, n. 1, p. 13-26, 1989.
AGRIOS, G. N. Plant pathology. 5. ed. San Diego, Califórnia (EUA): Elsevier Academi Press,
2004. 922 p.
AHMAD, P. SARWAT, M. SHARMA, S. Reactive oxygen species, antioxidants and signaling
in plants. Journal of Plant Biology, v.51, n.3, p.167-173, 2008.
AHMAD, P.; JALEEL, C. A.; SALEM, M. A.; NABI, G.; SHARMA, S. Roles of enzymatic
and nonenzymatic antioxidants in plants during abiotic stress. Critical reviews in
biotechnology, v. 30, n. 3, p. 161-175, 2010.
ALBACETE, A. A.; MARTÍNEZ-ANDÚJAR, C.; PÉREZ-ALFOCEA, F. Hormonal and
metabolic regulation of source-sink relations under salinity and drought: From plant survival to
crop yield stability. Biotechnology Advances, v. 32, n. 1, p. 12-30, 2014.
ALSCHER, R. G.; ERTURK, N.; HEATH L, S. Role of superoxide dismutases (SODs) in
controlling oxidative stress in plants. Journal of Experimental Botany, v. 53, n. 372, p. 1331–
1341, 2002.
ALVARENGA, M. A. R. Tomate: produção em campo, casa de vegetação e
hidroponia. 2ed., Lavras: UFLA, 2013. 455p.
AMARO, A. C. E. Efeitos fisiológicos de fungicidas no desenvolvimento de plantas de pepino
japonês enxertadas e não enxertadas, cultivadas em ambiente protegido. 2011. 86f. Dissertação
(Mestrado em Agronomia) – Programa de Pós-Graduação em Agronomia, área de concentração
em Horticultura, Faculdade de Ciências Agronômicas, Universidade Estadual Paulista “Júlio
de Mesquita Filho”, Botucatu.
AMARO, A. C. E.; RAMOS, A. R. P.; MACEDO, A. C.; ONO, E. O.; RODRIGUES, J.
D. Effects of the fungicides azoxystrobin, pyraclostrobin and boscalid on the physiology of
Japanese cucumber. Scientia Horticulturae, v. 228, n. 26, p. 66-75, 2018.
ANTUNES, M. C.; CUQUEL, F. L.; ZAWADNEAK, M. A. C.; MOGOR, A. F.; RESENDE,
J. T. V. Postharvest quality of strawberry produced during two consecutive seasons.
Horticultura Brasileira, v. 32, n. 2, p. 168-173, 2014.
AOAC - ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTRY. Official methods of
analysis of AOAC international. 16. ed. USA, Association of Analytical Communities, 1997.
APEL, K.; HIRT, H. Reactive oxygen species: Metabolism, oxidative stress, and signal
transduction. Annual Review of Plant Biology, v. 55, p. 373-399, 2004.
ARAÚJO, J. C. A.; MATSUOKA, K. Histopatologia da interação Alternaria solani e
tomateiros resistente e suscetível. Fitopatologia Brasileira v. 29, n. 3, p. 268-275. 2004.

131
ARAVIND, P.; PRASAD, M. Modulation of cadmium-induced oxidative stress in
Ceratophyllum demersum by zinc involves ascorbato-glutathione cycle and glutathione
metabolism. Plant Physiology and Biochemistry, v. 43, n. 2, p. 107-116, 2005.
ASADA, K. The water-water cycle in chloroplasts: scavenging of active oxygens and
dissipation of excess photons. Annual Review of Plant Biolog, v.50, n. 1, p. 601-639, 1999.
AXELROD, B.; CHEESBROUGH, T. M.; LAAKSO, S. [53] Lipoxygenase from soybeans:
EC 1.13. 11.12 Linoleate: oxygen oxidoreductase. Methods in enzymology, New York. v.71,
p.441-451, 1981.
AZEVEDO, L.A.S.de. Manual de quantificação de doenças de Plantas. São Paulo: Luiz
Azevedo. 1997, p. 51-102.
BAKER, N.R. Chlorophyll fluorescence as a probe of photosynthesis In Vivo. Annual Review
Plant Biology, v. 59, p. 89-113, 2008.
BAKER, N.R.; ROSENQVIST, E. Applications of chlorophyll fluorescence can improve crop
production strategies: an examination of future possibilities. Journal of Experimental Botany,
v. 55, n. 403, p. 1607-1621, 2004.
BALBI-PEÑA, M. I.; SCHWAN-ESTRADA, K. R. F.; STANGARLIN, J. R. Oxidative burst
and the activity of defense-related enzymes in compatible and incompatible tomato-Alternaria
solani interactions. Semina: Ciências Agrárias, v. 35, n. 5, p. 2399-2414, 2014.
BARBOSA, M. R.; SILVA, M. M. A.; WILLADINO, L.; ULISSES, C.; CAMARA,
T. R. Geração e desintoxicação enzimática de espécies reativas de oxigênio em plantas.
Ciência Rural, v. 44, n. 3, p. 453-460, 2014.
BARTOSZ, G. Oxidative stress in plants. Acta Physiologiae Plantarum, v. 19, n. 1, p. 47-64,
1997.
BECKERS, G. J. M.; SPOEL, S. H. Fine-tuning plant defence signalling: salicylate versus
jasmonate. Plant Biology, v. 8, n. 1, p. 1-10, 2006.
BELA, K.; HORVÁTH, E.; GALLÉ, Á.; SZABADOS, L.; TARI, I.; CSISZÁR, J. Plant
glutathione peroxidases: emerging role of the antioxidant enzymes in plant development and
stress responses. Journal of plant physiology, v. 176, p. 192-201, 2015.
BELLIN, D.; ASAI, S.; DELLEDONNE, M.; YOSHIOKA, H. Nitric oxide as a mediator for
defense responses. Molecular plant-microbe interactions, v. 26, n. 3, p. 271-277, 2013.
BHATTACHARJEE, S. Sites of generation and physicochemical basis of formation of reactive
oxygen species in plant cell. In: GUPTA, S. D. (Ed.) Reactive oxygen species and
antioxidants in higher plants. Enfield: Science Publishers, p.178-203. 2010.
BITTNER, S. When quinones meet amino acids: chemical, physical and biological
consequences. Amino acids, v. 30, n. 3, p. 205-224, 2006.

132
BJÖRKMAN, O.; DEMMING, B. Photon yield of O2 evolution and chlorophyll fluorescence
characteristics at 77k among vasculas plants of diverse origins. Planta, v. 170, n. 4, p. 489-504,
1987.
BOATRIGHT, J.; NEGRE, F.; CHEN, X.; KISH, C. M.; WOOD, B.; PEEL, G.; DUDAREVA,
N. Understanding in vivo benzenoid metabolism in petunia petal tissue. Plant Physiology, v.
135, n. 4, p. 1993-2011, 2004.
BONALDO, S. M.; PASCHOLATI, S. F. ROMEIRO, R. Indução de resistência: noções
básicas e perspectivas. In: CAVALCANTI, L. S.; DI PIERO, R. M.; CIA, P.; PASCHOLATI,
S. F.; RESENDE, M. L.V.; ROMEIRO, R. S. Indução de resistência em plantas a patógenos
e insetos. Piracicaba: FEALQ, p. 11-28. 2005.
BRADFORD, M. M. A rapid and sensitive method for the quantitation of microgram quantities
of protein utilizing the principle of protein-dye binding. Analytical biochemistry, v. 72, n. 1-
2, p. 248-254, 1976.
BRANDÃO FILHO, J. U. T.; GOTO, R.; GUIMARÃES, V. F.; HABERMANN, G.;
RODRIGUES, J. D.; CALLEGARI, O. Influência da enxertia nas trocas gasosas de dois
híbridos de berinjela cultivados em ambiente protegido. Horticultura Brasileira, p. 474-477,
2003.
BRESTIC, M.; ZIRVCAK, M. PSII fluorescence techniques for measurement of drought and
high temperature stress signal in crop plants: protocols and applications. In: ROUT, G. R.; DAS,
A. B. Molecular Stress Physiology of Plants. Bhubaneswar, India: Springer Dordrecht, p 87-
133, 2013.
BREUSEGEM, F. V.; VRANOVA, E.; DAT, J. F.; INZE, D. The role of active oxygen species
in plant signal transduction. Plant Science, v. 161, n. 3, p. 405-414, 2001.
BUCHNER, P.; ROCHAT, C.; WUILLEME, S.; BOUTIN, J. P. Characterization of a tissue-
specific and developmentally regulated β-1, 3-glucanase gene in pea (Pisum sativum). Plant
molecular biology, v. 49, n. 2, p. 171-186, 2002.
BUSSO, C.; MAZARO, S. M. Utilizaton of coloidal chitin stained with remazol brilliant blue
RSA a sensitive, rapid and economical alternative to detect chitinase in plants. In: 23rd
Congress of the International Union for Biochemistry and Molecular Biology (IUBMB),
Abstracts Book, v. 1. p. 239, 2015.
CAMPBELL, C. L.; MADDEN, L. V. Introduction to plant disease epidemiology. John
Wiley & Sons. 1990. 560p.
CAMPBELL, M. K.; FARRELL, S. O. Bioquímica. 5. ed. São Paulo: Thomson, p.845, 2006.
CAMPOS, A. D.; BRANCÃO, N.; FERREIRA, A. G.; SILVEIRA, E. P. D.; HAMPE, M. M.
V.; OSORIO, V. A.; ANTUNES, I. F.; AUGUSTIN, E. Atividade de peroxidase e
polifenoloxidase na resistência do feijão à antracnose. Pesquisa Agropecuária Brasileira, v.
39, n. 7, p. 637-643, 2004.

133
CAMPOS, A. D.; FERREIRA, A. G.; HAMPE, M. M. V.; ANTUNES, I. F.; RANCAO, N.;
SILVEIRA, E. P.; SILVA, J. B.; OSORIO, V. A. Induction of chalcone synthase and
phenylalanine ammonia-lyase by salicylic acid and Colletotrichum lindemuthianum in common
bean. Brazilian Journal of Plant Physiology, v. 15, n. 3, p. 129-134, 2003.
CANTORE, V.; LECHKAR, O.; KARABULUT, E.; SELLAMI, M. H.; ALBRIZIO, R.;
BOARI, F.; STELLACCI, A. M.; TODOROVIC, M. Combined effect of deficit irrigation and
strobilurin application on yield, fruit quality and water use efficiency of “cherry” tomato
(Solanum lycopersicum L.). Agricultural Water Management, v. 167, n. 31, p. 53-61, 2016.
CARRIJO, D. R. Efeitos fisiológicos provocados pelo fungicida Fluxapiroxade, isolado e em
mistura com a Piraclostrobina, na cultura de soja. 2014. Dissertação (Mestrado em Agronomia)
– Programa de Pós-Graduação em Agronomia, área de concentração em Fitotecnia, Faculdade
de Ciências Agronômicas, Universidade Estadual Paulista “Júlio de Mesquita Filho”, Botucatu.
CASIERRA-POSADA, F.; PEÑA-OLMOS, J. E. Modificaciones fotomorfogénicas inducidas
por la calidad de la luz en plantas cultivadas. Revista de la Academia Colombiana de Ciencias
Exactas, Físicas y Naturales, v. 39, p. 84-92, 2015.
CASIERRA-POSADA, F.; PEÑA-OLMOS, J.E.; ULRICHS, C. Growth and photochemical
efficiency of Photosystem II in strawberry plants (Fragaria sp.) affected by the light quality:
agronomic implications Revista Actualidad & Divulgación Científica, v. 14, n. 2, p. 43-53,
2011.
CASTORIA, R.; CURTIS, F.; LIMA, G.; CAPUTO, L.; PACIFICO, S.; CICCO, V.
Aureobasidium pullulans (LS-30) an antagonist of postharvest pathogens of fruits: study on its
modes of action. Postharvest Biology and Technology, v. 22, n. 1, p. 7-17, 2001.
CASTRO, N. R.; COÊLHO, R. S. B.; LARANJEIRA, D.; PIMENTEL, R. M. D. M.;
FERREIRA, C. P. Murcha de fusário em helicônia: fontes de resistência, método alternativo de
detecção e defesa estrutural. Summa Phytopathologica, v. 36, n. 1, p. 30-34, 2010.
CATANEO, A. C.; DESTRO, G. F. G.; FERREIRA, L. C.; CHAMMA, K. L.; SOUSA, D. C.
F. Glutathione S-transferase activity on the degradation of the herbicide glyphosate in maize
(Zea mays) plants. Planta Daninha, v. 21, p 307-312. 2003.
CEAGESP – COMPANHIA DE ENTREPOSTOS E ARMAZÉNS GERAIS DE SÃO PAULO.
Centro de Qualidade em Horticultura. Programa Brasileiro para Modernização da
Horticultura: Normas de classificação de tomates. São Paulo, 2003. 3 p. (CQH Doc. 26).
CHITARRA, M. I.; CHITARRA, A. B. Pós-colheita de frutas e hortaliças: fisiologia e
manuseio. 2.ed. Lavras, UFLA, 2005. 783p.
COGO, A. J. D.; SIQUEIRA, A. F.; RAMOS, A. C.; CRUZ, Z. M.; SILVA, A. G. Utilização
de enzimas do estresse oxidativo como biomarcadoras de impactos ambientais, Natureza on
line, v. 7, n. 1, p. 37-42, 2009.

134
COLOMBARI, L. F.; BALDINI, L. F. G.; BALDINI, V.; CARDOSO, A. I. I.; GOTO, R. Efeito
fisiológico de fungicidas sistémicos em parâmetros agronómicos da cenoura. Revista de
Ciências Agrárias, v. 38, n. 3, p. 366-371, 2015.
COTINGUIBA, G. G.; SILVA, J. R. D. N.; AZEVEDO, R. R. D. S.; ROCHA, T. J. M.,
SANTOS, A. F. D. Método de avaliação da defesa antioxidante: uma revisão de
literatura. Journal of Health Sciences, v. 15, n. 3, p. 231-237, 2015.
D´AUTRÉAUX, B.; TOLEDANO, M. B. ROS as signalling molecules: mechanisms that
generate specificity in ROS homeostasis. Nature reviews Molecular cell biology, v. 8, n. 10,
p. 813, 2007.
DAN HESS, F. Light-dependent herbicides: an overview. Weed Science, v. 48, n. 2, p.
160-170, 2000.
DEMANT, L. A. R.; MARINGONI, A. C. Controle da mancha angular do feijoeiro com uso
de fungicidas e seu efeito na produção das plantas. Idesia (Arica), v. 30, n. 2, p. 93-100, 2012.
DEUNER, C.; BORGES, C. T.; ALMEIDA, A. S.; MENEGHELLO, G. E.; TUNES, L.
V. Ácido jasmónico como promotor de resistência em plantas. Revista de Ciências Agrárias,
v. 38, n. 3, p. 275-281, 2015.
DIAZ-ESPEJO, A.; CUEVAS, M. V.; RIBAS-CARBO, M.; FLEXAS, J., MARTORELL, S.;
FERNÁNDEZ, J. E. The effect of strobilurins on leaf gas exchange, water use efficiency and
ABA content in grapevine under field conditions. Journal of plant physiology, v. 169, n. 4, p.
379-386, 2012.
DINAKAR, C.; DJILIANOV, D.; BARTELS, D. Photosynthesis in desiccation tolerant plants:
energy metabolism and antioxidative stress defense. Plant Science, v. 182, p. 29-41, 2012.
DINANT, S.; LEMOINE, R. The phloem pathway: New issues and old debates. Comptes
Rendus Biologies, v. 333, n. 4, p. 307-319, 2010.
D'INNOCENZO, M.; LAJOLO, F. Effect of gamma irradiation on softening changes and
enzyme activities during ripening of papaya fruit. Journal of Food Biochemistry, v. 25, n. 5,
p. 425-438, 2001.
DIXON, D. P.; LAPTHORN, A.; EDWARDS, R. Plant glutathione transferases. Genome
biology, v. 3, n. 3, p. 3004.1-3004.10, 2002.
DIXON, D. P.; SKIPSEY, M.; EDWARDS, R. Roles for glutathione transferases in plant
secondary metabolism. Phytochemistry, v. 71, n. 4, p. 338-350, 2010.
DOMINGUES, R. J.; SOUZA, J. D. F. D.; TÖFOLI, J. G.; MATHEUS, D. R. Ação “in vitro”
de extratos vegetais sobre Colletotrichum acutatum, Alternaria solani e Sclerotium rolfsii.
Arquivos do Instituto Biológico, v. 76, n. 4, p. 643-649, 2009.
DUBEY, R. S. Metal toxicity, oxidative stress and antioxidative defense system in
plants. In: GUPTA, S. D. (Ed.) Reactive oxygen species and antioxidants in higher plants.
Enfield: Science Publishers, p.178-203. 2010.

135
EBRAHIM, S.; USHA, K.; SINGH, B. Pathogenesis related (PR) proteins in plant defense
mechanism. In: MÉNDEZ-VILAS, A. (Ed.) Science Against Microbial Pathogens:
Communicating Current Research and Technological Advances. Formatec, p. 1043–1054,
2011.
EDREVA, A. A novel strategy for plant protection: Induced resistance. Journal of Cell and
Molecular Biology, v. 3, n. 2, p. 61-69, 2004.
EFSA: European Food Safety Authority. Reasoned opinion on the modification of the existing
maximum residue levels for propamocarb in onions, garlic, shallots and leeks. EFSA Journal,
v. 13, n. 4, 2015.
EL GHAOUTH, A.; WILSON, C. L.; CALLAHAN, A. M. A novel strategy for plant
protection: Induced resistance. Journal of Cell and Molecular Biology, v. 3, n. 2, p. 61-69,
2004.
EMBRAPA - Empresa Brasileira de Pesquisa Agropecuária. Sistema brasileiro de
classificação de solos. 3ed. Rio de Janeiro: Embrapa Solos, 2013. 353 p.
ERNANI, P. R. Química do solo e disponibilidade de nutrientes. 2 ed. Lages: O autor, 2016.
256p.
FAGAN, E. B.; DOURADO NETO, D.; VIVIAN, R.; FRANCO, R. B.; YEDA, M. P.;
MASSIGNAM, L. F.; MARTINS, K. V. Efeito da aplicação de piraclostrobina na taxa
fotossintética, respiração, atividade da enzima nitrato redutase e produtividade de grãos de
soja. Bragantia, v. 69, n. 4, 2010.
FAIZE, M.; FAIZE, L.; KOIKE, N.; ISHIZAKA, M.; ISHII, H. Acibenzolar-S-methyl-induced
resistance to Japanese pear scab is associated with potentiation of multiple defense
responses. Phytopathology, v. 94, n. 6, p. 604-612, 2004.
FARMER, E. E.; RYAN, C. A. Octadecanoid precursors of jasmonic acid activate the synthesis
of wound-inducible proteinase inhibitors. The Plant Cell Online, v. 4, n. 2, p. 129-134, 1992.
FERREIRA, S. M. R.; QUADROS, D. A. D.; KARKLE, E. N. L.; LIMA, J. J. D.; TULLIO, L.
T.; FREITAS, R. J. S. D. Qualidade pós-colheita do tomate de mesa convencional e
orgânico. Ciência e Tecnologia de Alimentos, v. 30, n. 4, p. 858-869, 2010.
FILGUEIRA, F. A. R. Novo manual de olericultura: agrotecnologia moderna na produção
e comercialização de hortaliças. 2. ed. UFV: Viçosa - Minas Gerais, 2005. 412 p.
FILIPENKO, E. A.; KOCHETOV, A. V.; KANAYAMA, Y.; MALINOVSKY, V. I.;
SHUMNY, V. K. PR-proteins with ribonuclease activity and plant resistance against
pathogenic fungi. Russian Journal of Genetics: Applied Research, v. 3, n. 6, p. 474-480,
2013.
FLEURI, L. F.; SATO, H. H. β-1, 3 Glucanases e quitinases: aplicação na lise de leveduras e
inibição de fungos. Ciência e Agrotecnologia, v. 32, p. 1224- 1231, 2008.

136
FONTENELLE, A. D. B.; GUZZO, S. D.; LUCON, C. M. M.; HARAKAVA, R. Growth
promotion and induction of resistance in tomato plant against Xanthomonas euvesicatoria and
Alternaria solani by Trichoderma spp. Crop Protection, v. 30, n. 11, p. 1492-1500, 2011.
FOYER, C. H.; GALTIER, N. Source-sink interaction and communication in leaves. In:
ZAMSKI, E.; SCHAFFER, A. A. Photoassimilate distribution in plants and crops: Sourcesink
relationships. New York: Marcel Dekker, INC., 1996. p.331-340.
FOYER, C. H.; LOPEZ‐DELGADO, H.; DAT, J. F.; SCOTT, I. M. Hydrogen peroxide‐and
glutathione‐associated mechanisms of acclimatory stress tolerance and signalling. Physiologia
Plantarum, v. 100, n. 2, p. 241-254, 1997.
FOYER, C. H.; NOCTOR, G. Redox sensing and signalling associated with reactive oxygen in
chloroplasts, peroxisomes and mitochondria. Physiologia plantarum, v. 119, n. 3, p. 355-364,
2003.
FRAC Code List ©*2017: Fungicides sorted by mode of action (including FRAC Code
numbering), p. 1-12, 2017.
FREITAS FILHO, A. M. D. ; SANTOS, S. S.; BRANDÃO FILHO, J. U. T.; SANTOS, H. S.;
VIDA, J. B.; HORA, R. C. D. Controle da pinta preta (Alternaria solani) por meio de fungicidas
na cultura do tomate. Horticultura Brasileira, v. 26, n. 2, p. 2922-2926, 2008.
FUJITA, S.; SAARI, N. B.; MAEGAWA, M.; TETSUKA, T.; HAYASHI, N.; TONO, T.
Purification and properties of polyphenol oxidase from cabbage (Brassica oleracea
L.). Journal of Agricultural and Food Chemistry, v. 43, n. 5, p. 1138-1142, 1995.
GAO, Q. M.; ZHU, S.; KACHROO, P.; KACHROO, A. Signal regulators of systemic acquired
resistance. Frontiers in plant science, v. 6, n. 228, p. 1-12, 2015.
GARCIA, I. S.; SOUZA, A.; BARBEDO, C. J.; DIETRICH, S. M. C.; FIGUEIREDO-
RIBEIRO, R. C. L. Changes in soluble carbohydrates during storage of Caesalpinia echinata
LAM. (Brazilwood) seeds, an endangered leguminous tree from the Brazilian Atlantic
Forest. Brazilian Journal of Biology, v. 66, n. 2B, p. 739-745, 2006.
GAY, C.; GEBICKI, J. M. A critical evaluation of the effect of sorbitol on the ferric–xylenol
orange hydroperoxide assay. Analytical biochemistry, v. 284, n. 2, p. 217-220, 2000.
GIANNOPOLITIS, C. N.; REIS, S. K. Superoxide dismutases: I. Occurrence in higher
plants. Plant physiology, v. 59, n. 2, p. 309-314, 1977.
GILL, S. S.; ANJUM, N. A.; GILL, R.; YADAV, S.; HASANUZZAMAN, M.; FUJITA, M.;
TUTEJA, N. Superoxide dismutase-mentor of abiotic stress tolerance in crop
plants. Environmental Science and Pollution Research, v. 22, n. 14, p. 10375-10394, 2015.
GILL, S. S.; TUTEJA, N. Reactive oxygen species and antioxidant machinery in abiotic stress
tolerance in crop plants. Plant physiology and biochemistry, v. 48, n. 12, p. 909-930, 2010.

137
GIULIANI, M. M.; NARDELLA, E.; GATTA, G.; DE CARO, A.; QUITADAMO, M.
Processing tomato cultivated under water deficit conditions: the effect of azoxystrobin. Acta
Horticulturae: III International Symposium on Tomato Diseases, n. 914, p. 287-294, 2010.
GLAAB, J.; KAISER, W. M. Increased nitrate reductase activity in leaf tissue after application
of the fungicide Kresoxim-methyl. Planta, v. 207, n. 3, p. 442-448, 1999.
GLAZEBROOK, J. Contrasting mechanisms of defense against biotrophic and necrotrophic
pathogens. Annual Review of Phytopathology, v. 43, p. 205-227. 2005.
GORJANOVIĆ, S. A review: biological and technological functions of barley seed
Pathogenesis‐related proteins (PRs). Journal of the Institute of Brewing, v. 115, n. 4, p. 334-
360, 2009.
GRATÃO, P. L.; POLLE, A.; LEA, P. J.; AZEVEDO, R. A. Making the life of heavy metal-
stressed plants a little easier. Functional Plant Biology, v. 32, n. 6, p. 481-494, 2005.
GRIGOLLI, J. F. J.; KUBOTA, M. M.; ALVES, D. P.; RODRIGUES, G. B.; CARDOSO, C.
R.; SILVA, D. J. H. D.; MIZUBUTI, E. S. G. Characterization of tomato accessions for
resistance to early blight. Crop Breeding and Applied Biotechnology, v. 11, n. 2, p. 174-180,
2011.
GUERRA, A. M. N. M.; RODRIGUES, F. Á.; BERGER, P. G.; BARROS, A. F.;
RODRIGUES, Y. C.; LIMA, T. C. Resistência do algodoeiro à ferrugem tropical
potencializada pelo silício. Bragantia, v. 72, n. 3, p. 279-291, 2013.
GUZZO, S. D.; MARTINS, E. M. F. Local and systemic induction of β‐1, 3‐glucanase and
chitinase in coffee leaves protected against Hemileia vastatrix by Bacillus
thuringiensis. Journal of phytopathology, v. 144, n. 9‐10, p. 449-454, 1996.
HAGEMAN, R. H.; REED, A. J. [24] Nitrate reductase from higher plants. Methods in
enzymology, v. 69, p. 270-280, 1980.
HAMMERSCHMIDT, R.; MÉTRAUX, J. P.; VAN LOON, L. C. Inducing resistance: a
summary of papers presented at the First International Symposium on Induced Resistance to
Plant Diseases, Corfu, May 2000. European Journal of Plant Pathology, v. 107, n. 1, p. 1-6,
2001.
HAYES, J. D.; MCLELLAN, L. I. Glutathione and glutathione-dependent enzymes represent
a co-ordinately regulated defence against oxidative stress. Free radical research, v. 31, n. 4,
p. 273-300, 1999.
HEINE, A. J. M.; MORAES, M. O. B.; PORTO, J. S.; SOUZA, J. R.; REBOUÇAS, T. N. H.;
SANTOS, B. S. R. Número de haste e espaçamento na produção e qualidade do
tomate. Scientia Plena, v. 11, n. 9, p. 1-7, 2015.
HIRAGA, S.; SASAKI, K.; ITO, H.; OHASHI, Y.; MATSUI, H. A large family of class III
plant peroxidases. Plant and Cell Physiology, v. 42, n. 5, p. 462-468, 2001.

138
HONORATO JÚNIOR, J.; ZAMBOLIM, L.; AUCIQUE-PÉREZ, C. E.; RESENDE, R. S.;
RODRIGUES, F. A. Photosynthetic and antioxidative alterations in coffee leaves caused by
epoxiconazole and pyraclostrobin sprays and Hemileia vastatrix infection. Pesticide
biochemistry and physiology, v. 123, p. 31-39, 2015.
HULTIN, H. O.; SUN, B.; BULGER, J. Pectin methyl esterases of the banana: purification
and properties. Journal of Food Science, v. 31, n. 3, p. 320-327, 1966.
IBGE, Instituto Brasileiro de Geografia e Estatística. LSPA - Levantamento sistemático da
produção agrícola: pesquisa mensal de previsão e acompanhamento das safras agrícolas
no ano civil. AGROPECUÁRIA, D. D. P. C. D. Rio de Janeiro, p.25-130, 2017.
IGAMBERDIEV, A. U.; LEA, P. J. The role of peroxisomes in the integration of metabolism
and evolutionary diversity of photosynthetic organisms. Phytochemistry, v. 60, n. 7, p. 651-
674, 2002.
INZÉ, D.; VAN MONTAGU, M. Oxidative stress in plants. Current Opinion in
Biotechnology, v. 6, n. 2, p. 153-158, 1995.
ITAKO, A.T.; TOLENTINO JÚNIOR, J. B.; SILVA JÚNIOR, T. A. F. D. S.; SOMAN, J. M.;
MARINGONI, A. C. Efeito de produtos químicos sobre a mancha bacteriana (Xanthomonas
perforans) e na ativação de proteínas relacionadas à patogênese em tomateiro. Idesia (Arica),
v. 30, n. 2, p. 85-92, 2012.
JACOBELIS JUNIOR, W. Ação de fungicidas no desenvolvimento de plantas de tomate, em
condições de ambiente protegido. 2015. Dissertação (Mestrado em Agronomia) – Programa de
Pós-Graduação em Agronomia, área de concentração em Horticultura, Faculdade de Ciências
Agronômicas, Universidade Estadual Paulista “Júlio de Mesquita Filho”, Botucatu.
JADOSKI, C. J.; RODRIGUES, J. D.; SORATTO, R. P.; DOS SANTOS, C. M.; RIBEIRO,
E. Ação fisiológica da piraclostrobina na assimilação de CO2 e enzimas antioxidantes em
plantas de feijão condicionado em diferentes tensões de água no solo. Irriga, v. 20, n. 2, p. 319-
333, 2015.
JOSHI, J.; SHARMA, S.; GURUPRASAD, K. N. Foliar application of pyraclostrobin fungicide
enhances the growth, rhizobial-nodule formation and nitrogenase activity in soybean (var. JS-
335). Pesticide biochemistry and physiology, v. 114, p. 61-66, 2014.
KADER, A. A.; MORRIS, L. L.; STEVENS, M. A.; HOLTON, M. A. Composition and flavour
quality of fresh market tomatoes as influenced by some postharvest handling
procedures. Journal of American Horticultural Science, v. 103, n. 1, p. 6-13.1978, 1978.
KANUNGO, M.; JOSHI, J. Impact of Pyraclostrobin (F-500) on crop plants. Plant Science
Today, v. 1, n. 3, p. 174-178, 2014.
KARUPPANAPANDIAN, T.; MOON, J. C.; KIM, C.; MANOHARAN, K.; KIM, W.
Reactive oxygen species in plants: their generation, signal transduction, and scavenging
mechanisms. Australian Journal of Crop Science, v. 5, n. 6, p. 709-725, 2011.

139
KEPPLER, L. D.; BAKER, C. J. O2- initiated lipid peroxidation in a bacteria-induced
hypersensitive reaction in tobacco cell suspensions. Phytopathology, v. 79, n. 5, p. 555-562,
1989.
KIM, Y. H.; KWAK, S. S. The role of antioxidant enzymes during leaf development. In:
GUPTA, S. D. (Ed.) Reactive oxygen species and antioxidants in higher plants. Enfield:
Science Publishers, p.178-203. 2010.
KLUGE, R. A.; TEZOTTO-ULIANA, J. V.; SILVA, P. P. Aspectos fisiológicos e ambientais
da fotossíntese. Revista Virtual de Química, v. 7, n. 1, p. 56-73, 2014.
KÖPPEN, W. Climatologia: com um estudio de los climas de La tierra. México: Fondo de
Cultura Econômica, 1948. 478p.
KOZLOWSKI, L. A.; SIMÕES, D. F. M.; DE SOUZA, C. D.; TRENTO, M. Efeito fisiológico
de estrobilurina F 500® no crescimento e rendimento do feijoeiro. Revista Acadêmica:
Ciência Animal, v. 7, n. 1, p. 41-54, 2009.
KRAUSE, G.; WEIS, E. Chlorophyll fluorescence and photosynthesis: The basics. Annual
Review Plant Physiology and Plant Molecular Biology, v. 42, n. 1, p. 313-349, 1991.
KRUSE, N. D.; VIDAL, R. A.; TREZZI, M. M.; SIQUEIRA, I. R. Estresse oxidativo em
girassol (Helianthus annuus) indica sinergismo para a mistura dos herbicidas metribuzin e
clomazone. Planta daninha, v. 24, n. 2, p. 379-390, 2006.
KUHN, O. J.; PASCHOLATI, S. F. Custo adaptativo da indução de resistência em feijoeiro
mediada pela rizobactéria Bacillus cereus ou acibenzolar-S-metil: atividade de enzimas, síntese
de fenóis e lignina e biomassa. Summa Phytopathologica, v. 36, n. 2, p. 107-114, 2010.
KUMAR, N. N.; DEOBAGKAR, D. N. Multifactional glucanases. Biotechnology advances,
v. 14, n. 1, p. 1-15, 1996.
KUO, M. C.; KAO, C. H. Aluminum effects on lipid peroxidation and antioxidative enzyme
activities in rice leaves. Biologia plantarum, v. 46, n. 1, p. 149-152, 2003.
KUROZAWA, C.; PAVAN, M. A. Doenças do tomateiro. In: KIMATI, H.; AMORIM, L.,
REZENDE, J. A. M.; BERGAMIN FILHO, A.; CAMARGO, L. E. A. (Eds.). Manual de
fitopatologia: doenças de plantas cultivadas. 4 ed. São Paulo: Agronômica Ceres, v. 2, 2005,
607-626 p.
LAWRENCE, C. B.; MITCHELL, T. K.; CRAVEN, K. D.; CHO, Y. R.; CRAMER, R. A.;
KIM, K. H. At death's door: Alternaria pathogenicity mechanisms. The Plant Pathology
Journal, v. 24, n. 2, p. 101-111, 2008.
LEHMANN, S.; SERRANO, M.; L’HARIDON, F.; TJAMOS, S. E.; METRAUX, J. P.
Reactive oxygen species and plant resistance to fungal pathogens. Phytochemistry, v. 112, p.
54-62, 2015.
LEVER, M. A new reaction for colorimetric determination of carbohydrates. Analytical
biochemistry, v. 47, n. 1, p. 273-279, 1972.

140
LICHTENTHALER, H. K.; WELLBURN, A. R. Determinations of total carotenoids and
chlorophylls a and b of leaf extracts in different solvents. Biochemical Society Transactions,
v. 11, p. 591-592, 1983.
LIMA, A. D. S.; REFFATTI, M. T. N.; JUNCOS, M. C.; BURBULHAN, T.; MARTIKOSKI,
L. Efeito fisiológico de fungicida pyraclostrobin e tratamento de sementes na cultura do milho.
Pesquisa Aplicada & Agrotecnologia, v. 2, n. 3, p. 113-120, 2009.
LIMA, H.C. Modificações de carboidratos estruturais e de enzimas pécticas em jabuticaba.
61p.Tese (doutorado) - Universidade Federal de Lavras. Lavras, 2002.
LIMA, J. D.; MORAES, W. D. S.; SILVA, S. H. M. G. D. Respostas fisiológicas em mudas de
banananeira tratadas com estrobilurinas. Semina: Ciências Agrárias, v. 33, n. 1, p. 77-86,
2012.
LIN, C. W; CHANG, H. B; HUANG, H. J. Zinc induces mitogen-activated protein kinase
activation mediated by reactive oxygen species in rice roots. Plant Physiology and
Biochemistry, v.43, n. 10, p. 963–968, 2005.
LIN, L.; MENG, X.; LIU, P.; HONG, Y.; WU, G.; HUANG, X.; LI, C.; DONG, J.; XIAO, L.;
LIU, Z. Improved catalytic efficiency of Endo-β-1, 4-glucanase from Bacillus subtilis BME-15
by directed evolution. Applied microbiology and biotechnology, v. 82, n. 4, p. 671-679, 2009.
LOCATO, V.; PINTO, M. C.; PARADISO, A.; GARA, L. Reactive oxygen
species and ascorbate glutathione interplay in signaling and stress responses. In: GUPTA, S. D.
(Ed.) Reactive oxygen species and antioxidants in higher plants. Enfield: Science
Publishers, p.178-203. 2010.
MACEDO, A. C. Efeitos fisiológicos de fungicidas no desenvolvimento de plantas de melão
rendilhado, cultivadas em ambiente protegido. 2012. 66f. Dissertação (Mestrado em
Agronomia) – Programa de Pós-Graduação em Agronomia, área de concentração em
Horticultura, Faculdade de Ciências Agronômicas, Universidade Estadual Paulista “Júlio de
Mesquita Filho”, Botucatu.
MACEDO, A. C. Efeitos fisiológicos de fungicidas no desenvolvimento de plantas de melão
rendilhado, cultivadas em ambiente protegido. 2015. 88f. Tese (Doutorado em Agronomia) –
Programa de Pós-Graduação em Agronomia, área de concentração em Horticultura, Faculdade
de Ciências Agronômicas, Universidade Estadual Paulista “Júlio de Mesquita Filho”, Botucatu.
MACEDO, A. C.; AMARO, A. C. E.; RAMOS, A. R. P.; ONO, E. O.; RODRIGUES, J. D.
Strobilurin and boscalid in the quality of net melon fruits. Semina: Ciências Agrárias, v. 38,
n. 2, p. 543-550, 2017.
MAHMOUD, A. L.; OMAR, S. A. Enzymatic activity and mycotoxin-producing potential of
fungi isolated from rotted lemons. Cryptogamie Mycologie, v. 15, n. 2, p. 117-124, 1994.
MANDAL, S.; DAS, R. K.; MISHRA, S. Differential occurrence of oxidative burst and
antioxidative mechanism in compatible and incompatible interactions of Solanum lycopersicum

141
and Ralstonia solanacearum. Plant Physiology and Biochemistry, v. 49, n. 2, p. 117-123,
2011.
MANSANO, A. R. Fungicidas e reguladores vegetais nas características fisiológicas e
produtivas de alface ‘vera’. 2014. 48f. Dissertação (Mestrado em Agronomia) – Programa de
Pós-Graduação em Agronomia, área de concentração em Horticultura, Faculdade de Ciências
Agronômicas, Universidade Estadual Paulista “Júlio de Mesquita Filho”, Botucatu.
MARCHI, C. E.; BORGES, M. D. F.; MIZUBUTI, E. S. G. Atividades amilolítica e
pectinolítica de Alternaria solani e a relação com a agressividade em tomateiro. Summa
Phytopathologica, v. 32, n. 4, p. 345-352, 2006.
MAREK, J. Fungicidas de efeitos fisiológicos no controle do míldio e no desenvolvimento de
mudas de videira. 2016. 57f. Dissertação (Mestrado em Agronomia) – Programa de Pós-
Graduação em Agronomia, área de concentração em Produção Vegetal, Universidade Estadual
do Centro-Oeste, Guarapuava.
MARRS, K. A. The functions and regulation of glutathione S-transferases in plants. Annual
review of plant biology, v. 47, n. 1, p. 127-158, 1996.
MARTINS, E. M. F. Proteínas relacionadas à patogênese. In: PASCHOLATI. S. F.; LEITE.
B.; STANGARLIN, J. R.; CIA, P. Interação Planta-Patógeno: Fisiologia, Bioquímica e
Biologia Molecular. Piracicaba: FEALQ. 2008. 627 p.
MATOS, G. A.; SOUSA, F. A.; PAULO JÚNIOR, J.; LIMA, L. M. Avaliação da mistura de
fungicidas no controle de doenças do cafeeiro. Revista Getec, v. 5, n. 9, p. 90-103, 2016.
MAUCH, F.; STAEHELIN, L. A. Functional implications of the subcellular localization of
ethylene-induced chitinase and [beta]-1, 3-glucanase in bean leaves. The Plant Cell, v. 1, n. 4,
p. 447-457, 1989.
MAXWELL, K.; JOHNSON, G. N. Chlorophyll fluorescence – a pratical guide. Journal of
Experimental Botany, v. 51, n. 345, p. 659-668, 2000.
MAZARO, S. M.; WAGNER JÚNIOR, A.; SANTOS, I. D.; CITADIN, I.; POSSENTI, J. C.;
GOUVÊA, A. D. Controle do tombamento de plântulas de beterraba e tomate pelo tratamento
de sementes com quitosana. Pesquisa Agropecuária Brasileira, v. 44, n. 11, p. 1424-1430,
2009.
MILLER, G. L. Use of dinitrosalicylic acid reagent for determination of reducing sugar.
Analytical Chemistry, v. 31, n. 3, p. 426-428, 1959.
MISHRA, S.; SRIVASTAVA, S.; TRIPATHI, R. D.; GOVINDARAJAN, R.;
KURIAKOSE, S. V.; PRASAD, M. N. V. Phytochelatin synthesis and resopsne of
antioxidants during cadmium stress in Bacopa monnieri L. Plant Physiology and
Biochemistry, v. 44, n. 1, p. 25-37, 2006.

142
MISRA, A.N.; LATOWSKI, D.; STRZALKA, K. Violaxanthin de-epoxidation in aging
cabbage (Brassica oleracea L.) leaves play as a sensor for photosynthetic excitation pressure.
Journal of Life Sciences, v.5, p. 182-191, 2011.
MISRA, H. P.; FRIDOVICH, I. The generation of superoxide radical during the autoxidation
of hemoglobin. Journal of Biological Chemistry, v. 247, n. 21, p. 6960-6962, 1972.
MITTLER, R. Oxidative stress, antioxidants and stress tolerance. Trends in Plant in
Science, v. 7, n. 9, p. 405-410, 2002.
MOLLER, I. M.; JENSEN, P. E.; HANSSON, A. Oxidative modifications to cellular
components in plants. Annual Review of Plant Biology, v 58, p. 459-481, 2007.
MOORE, J.; LIU, J. G.; ZHOU, K.; YU, L. Effects of genotype and environment on the
antioxidant properties of hard winter wheat bran. Journal of Agricultural and Food
Chemistry, v. 54, n. 15, p. 5313-5322, 2006.
MORRIS, D. L. Quantitative determination of carbohydrates with dreywood's anthrone
reagent. Science, v. 107, p. 254-255, 1948.
NAGALAKSHMI, N.; PRASAD, M. N. V. Responses of glutathione cycle enzymes and
glutathione metabolism to copper stress in Scenedesmus bijugatus. Plant Science, v. 160, n. 2,
p. 291-299, 2001.
NAIKA, S.; JEUDE, J. V. L. D.; GOFFAU, M. D.; HILMI, M.; DAM, B. V. A cultura do
tomate: produção, processamento e comercialização. Wageningen, Países Baixos: Digigrafi,
2006. 104 p.
NAKANO, Y.; ASADA, K. Hydrogen peroxide is scavenged by ascobate specific peroxidase
in spinach cloroplasts. Plant and Cell Physiology, v. 22, p. 867-880, 1981.
NAKAZAWA, A.; NOZUE, M.; YASUDA, H. Expression pattern and gene structure of
phenylalanine ammonia-lyase in Phabitis nil. Journal of Plant Reseasch, v. 114, n. 2, p. 323-
328, 2001.
NISAR, N.; LI, L.; LU, S.; KHIN, N. C.; POGSON, B. J. Carotenoid metabolism in plants.
Molecular plant, v. 8, n. 1, p. 68-82, 2015.
NOCTOR, G.; GOMEZ, L.; VANACKER, H.; FOYER, C. H. Interactions between
biosynthesis, compartmentation and transport in the control of gluthatione homeostasis and
signalling. Journal of Experimental Botany , v. 53, n. 372, p. 1283-1304, 2002.
NOCTOR, G.; LELARGE-TROUVERIE, C.; MHAMDI, A.; The metabolomics of oxidative
stress. Phytochemistry, v. 112, p. 33-53, 2015.
OLIVEIRA, M. D. M.; VARANDA, C. M. R.; FÉLIX, M. R. F. Induced resistance during the
interaction pathogen x plant and the use of resistance inducers. Phytochemistry letters, v. 15,
p. 152-158, 2016.
OOSTENDORP, M.; DIETRICH, B.; STAUB, T. Induced disease resistance in plants by
chemicals. European Journal of Plant Pathology, v. 107, n. 1, p. 19-28, 2001.

143
ORNELAS-PAZ, J. J.; YAHIA, E. M.; RAMIREZ-BUSTAMANTE, N.; PEREZ-
MARTINEZ, J. D.; ESCALANTE-MINAKATA, M. P.; IBARRA-JUNQUERA, V.;
ACOSTA-MUNIZ, C.; GUERRERO-PRIETO, V.; OCHOA-REYES, E. Physical attributes
and chemical composition of organic strawberry fruit (Fragaria x ananassa Duch, cv. Albion)
at six stages of ripening. Food Chemistry, v. 138, n. 1, p. 372–381, 2013.
PAULA, R. S. D.; OLIVEIRA, W. F. D. Resistência de tomateiro (Lycopersicon esculentum)
ao patógeno Alternaria solani. Pesquisa Agropecuária Tropical, v. 33, n. 2, p. 89-95, 2003.
PEIXOTO, P. H. P.; CAMBRAIA, J.; SANT'ANNA, R.; MOSQUIM, P. R.; MOREIRA, M.
A. Aluminum effects on lipid peroxidation and on the activities of enzymes of oxidative
metabolism in sorghum. Revista Brasileira de Fisiologia Vegetal, v. 11, n. 3, p. 137-143,
1999.
PERRAKIS, A.; TEWS, I.; DAUTER, Z.; OPPENHEIM, A. B.; CHET, I.; WILSON, K. S.;
VORGIAS, C. E. Crystal structure of a bacterial chitinase at 2.3 A resolution. Structure, v. 2,
n. 12, p. 1169-1180, 1994.
PINTO, M. D. S. T.; RIBEIRO, J.M.; DE OLIVEIRA, E. A. G. O estudo de genes e proteínas
de defesa em plantas. Revista Brasileira de Biociências, v. 9, n. 2, p. 241-248, 2011.
POPOV, E. G.; TALANOV, A. V.; KURETS, V. K.; DROZDOV, S. N. Effect of temperature
on diurnal changes in CO2 Exchange in intact cucumber plants. Russian Journal of Plant
Physiology, v. 50, n. 2, p. 178-182, 2003.
PORTA, H.; FIGUEROA-BALDERAS, R. E.; ROCHA-SOSA, M. Wounding and pathogen
infection induce a chloroplast-targeted lipoxygenase in the common bean (Phaseolus vulgaris
L.). Planta, v. 227, n. 2, p. 363-373, 2008.
PRESSEY, R. Extraction and assay of tomato polygalactorunases. Horticultural Science, v.
21. n.3. p. 490-492. 1986.
PRESSEY, R.; AVANTS, J.K. Solubilization of cell walls by tomato polygalacturonase: effects
of pectinesterases. Journal of Food Biochemistry, v. 1, n. 6, p. 57-74, 1982.
QUIRINO, B. F.; NOH, Y. S.; HIMELBLAU, E.; AMASINO, R. M. Molecular aspects of leaf
senescence. Trends in Plant Science, v. 5, n. 7, p. 278-282, 2000.
R Development Core Team (2016). Softwaer R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0,
URL <http://www.R-project.org/.>
RAGAB, M. M. M.; SABER, M. M.; EL-MORSY, S. A.; ABDEL-AZIZ, A. R. M. Induction
of systemic resistance against root rot of basil using some chemical inducers. Egyptian Journal
of Phytopathology, v. 37, n. 1, p. 59-70, 2009.
RAMA DEVI, S.; PRASAD, M. N. V. Copper toxicity in Ceratophyllum demersum L.
(Coontail), a free floating macrophyte: Response of antioxidant enzymes and antioxidants.
Plant Science, v. 138, n. 2, p. 157-165, 1998.

144
RAMOS, A. C. Produtos de efeitos fisiológicos no desenvolvimento de plantas de tomate
‘Giuliana’, na produção e pós-colheita de frutos. 2013. 143f. Tese (Doutorado em Agronomia)
– Programa de Pós-Graduação em Agronomia, área de concentração em Horticultura,
Faculdade de Ciências Agronômicas, Universidade Estadual Paulista “Júlio de Mesquita
Filho”, Botucatu.
RAMOS, A. R. P.; AMARO, A. C. E.; MACEDO, A. C.; SUGAWARA, G. S. D. A.;
EVANGELISTA, R. M.; RODRIGUES, J. D.; ONO, E. O. Qualidade de frutos de tomate
‘giuliana’ tratados com produtos de efeitos fisiológicos. Semina: Ciências Agrárias, v. 34, n.
6, p. 35-43, 2013.
RAMOS, R. P.; AMARO, A. C. E.; MACEDO, A. C., SOUZA, E. R.; RODRIGUES, J. D.;
ONO, E. O. Acúmulo de carboidratos no desenvolvimento de tomateiro tratado com produtos
químicos. Semina: Ciências Agrárias, v. 36, n. 2, p. 705-718, 2015.
RANCÉ, I.; FOURNIER, J.; ESQUERRÉ-TUGAYÉ, M. The incompatible interaction between
Phytophthora parasitica var. nicotianae race and tobacco is suppressed in transgenic plants
expressing antisense lipoxygenase sequences. Proceedings of the National Academy of
Sciences, v. 95, n. 11, p. 6554-6559, 1998.
RAO, E. S.; MUNSHI, A. D.; SINHA, P.; RAJKUMAR. Genetics of rate limiting disease
reaction to Alternaria solani in tomato. Euphytica, v. 159, n. 1-2, p. 123-134, 2007.
RECZEK, C. R.; CHANDEL, N. S. ROS-dependent signal transduction. Current opinion in
cell biology, v. 33, p. 8-13, 2015.
REIS, E. M.; REIS, A. C.; CARMONA, M. A. Manual de fungicidas: guia para o controle
químico de doenças de plantas. 6. ed., Passo Fundo: Ed. Universidade de Passo Fundo, 2010.
226p.
REIS, E. M.; REIS, A. C.; FORCELINI, C. A. Manual de fungicidas: guia para o controle
químico de doenças de plantas. 5. ed., Passo Fundo: Ed. Universidade de Passo Fundo, 2007.
153p.
REIS, L. S.; AZEVEDO, C. A. V.; ALBUQUERQUE, A. W.; JOSUÉ JUNIOR, F. S. Índice
de área foliar e produtividade do tomate sob condições de ambiente protegido. Revista
Brasileira de Engenharia Agrícola e Ambiental, v. 17, n.4, p. 386-391, 2013.
RESENDE, M. L. V.; BARRETI, P. B.; MEDEIROS, F. C. L.; SILVA, D.; PEREIRA, R. B.;
LINS, S. R. O.; PEREIRA, L. M.; CAMPOS, M. A. Percepção e transdução de sinais para a
ativação de respostas de defesa em plantas contra patógenos. Revisão Anual de Patologia de
Plantas, Passo Fundo, v. 15, p. 173-242, 2007.
RESENDE, M. L.V.; SALGADO, S. M. L.; CHAVES, Z. M. Espécies ativas de oxigênio na
resposta de defesa de plantas a patógenos. Fitopatologia Brasileira, v. 28, n. 2, p. 123-130,
2003.

145
RIBEIRO, S. M. R.; QUEIROZ, J. H.; PELUZIO, M. D. C. G.; COSTA, N. M. B.; MATTA,
S. L. P.; QUEIROZ, M. E. L. R. A formação e os efeitos das espécies reativas de oxigênio
no meio biológico. Bioscience journal, v. 21, n. 3, p. 133-149, 2005.
RODRIGUES, C. R.; RODRIGUES, T. M.; LUZ, J. M. Q.; SOUSA, V. B. F.; SOUSA, J. B.;
NUNES, A. C. P.; TRINDADE, P. R. Clorofila a e b de tomateiro tratado com silicato de
potássio e fungicida. Global Science and Technology, v. 9, n. 2, p. 54-64, 2016.
RODRIGUES, L. E. ABCSEM - Associação Brasileira do Comércio de Sementes e Mudas.
Tomaticultura: valioso segmento do agronegócio nacional, 2012. Disponível em: <
http://www.abcsem.com.br/noticia.php?cod=2420 >. Acesso em: 26/09/2017.
ROMEIRO, R. D. S.; GARCIA, F. A. O. Indução de resistência em plantas a patógenos por
eliciadores de natureza bacteriana. In: BETTIOL, W. e MORANDI, M. A. B. (Eds.).
Biocontrole de Doenças de Plantas: uso e perspectivas. Jaguariúna, SP: Embrapa Meio
Ambiente, 2009. 85-99 p.
ROUSSEAU, C.; BELIN, E.; BOVE, E.; ROUSSEAU, D.; FABRE, F.; BERRUYER, R.;
GUILLAUMÉS, J.; MANCEAU, C.; JACQUES, M.A.; BOUREAU, T. High throughput
quantitative phenotyping of plant resistance using chlorophyll fluorescence image analysis.
Plant Methods, v. 9, n. 17, p.1-13, 2013.
SAINZ, R. L.; VENDRUSCOLO, J. L. S. Propriedades da poligalacturonase e
pectinametilesterase em pêssegos [Prunus persica (L.) Batsch] de cultivares
brasileiras. Revista Brasileira de Tecnologia Agroindustrial, v. 9, n. 1, p. 1724-1743, 2015.
SALUSTIANO, M. E.; VALE, F. X. R. D.; ZAMBOLIM, L.; FONTES, P. C. R. O manejo da
pinta-preta do tomateiro em épocas de temperaturas baixas. Summa Phytopathologica, v. 32,
n. 4, p. 353-359, 2006.
SCANDALIOS J. G.; ACEVEDO A.; RUZSA S. Catalase gene expression in response
to chronic high temperature stress in maize. Plant Science, v. 156, n. 1, p. 103-110,
2000.
SCANDALIOS, J. G. Oxidative stress: molecular perception and transduction of signals
triggering antioxidant gene defenses. Brazilian Journal of Medical and Biological
Research, v. 38, n. 7, p. 995-1014, 2005.
SCHWAN-ESTRADA, K. R. F.; STANGARLIN, J. R.; PASCHOLATI, S. F. Mecanismos
bioquímicos e defesa vegetal. In: PASCHOLATI. S. F.; LEITE. B.; STANGARLIN, J. R.; CIA,
P. Interação Planta-Patógeno: Fisiologia, Bioquímica e Biologia Molecular. Piracicaba:
FEALQ, Cap. 6, 2008, 227-248 p.
SELS, J.; MATHYS, J.; CONINCK, B. M. A.; CAMMUE, B. P. A.; DE BOLLE, M. F.
C. Plant pathogenesis-related (PR) proteins: a focus on PR peptides. Plant Physiology and
Biochemistry, v. 46, n. 11, p. 941-950, 2008.

146
SHAH, J.; CHATURVEDI, R.; CHOWDHURY, Z.; VENABLES, B.; PETROS, R.A.
Signaling by small metabolites in systemic acquired resistance. The Plant Journal, v. 79, n. 4,
p. 645-658, 2014.
SHANKER, A. K.; DJANAGUIRAMAN, M.; SUDHAGAR, R.; CHANDRASHEKAR, C. N.;
PATHMANABHAN, G. Differential antioxidative response of ascorbate glutathione pathway
enzymes and metabolites to chromium speciation stress in green gram (Vigna radiata (L.)
R.Wilczek. cv CO 4) roots. Plant Science, v. 166, n. 4, p. 1035-1043, 2004.
SHARMA, P.; JHA, A. B.; DUBEY, R. S.; PESSARAKLI, M. Reactive oxygen species,
oxidative damage, and antioxidative defense mechanism in plants under stressful
conditions. Journal of botany, v. 2012, p. 1-26, 2012.
SHETLEY, J.; NELSON, K. A.; STEVENS, W. G.; DUNN, D.; BURDICK, B.;
MOTAVALLI, P. P.; ENGLISH, J. T.; DUDENHOEFFER, C. J. Corn yield response to
pyraclostrobin with foliar fertilizers. Journal of Agricultural Science, v. 7, n. 7, p. 18-34,
2015.
SHIN, R.; FRODERMAN, T.; FLURKEY, W. H. Isolation and characterization of a mung bean
leaf polyfhenol oxidase. Phytochemistry, v. 45, n. 1, p. 15-21, 1997.
SHIRAHIGE, F. H.; MELO A. M. T.; PURQUERIO L. F. V.; CARVALHO C. R. L.; MELO
P. C. T. Produtividade e qualidade de tomates Santa Cruz e Italiano em função do raleio de
frutos. Horticultura Brasileira, v. 28, n. 3, p. 292-298, 2010.
SILVA, M. L. C.; COSTA, R. S., SANTANA, A. D. S.; KOBLITZ, M. G. B. Compostos
fenólicos, carotenóides e atividade antioxidante em produtos vegetais. Semina: Ciências
Agrárias, v. 31, n. 3, p. 275-281, 2010.
SIMANDLE, P. A.; BROGDON, J. L.; SWEENEY, J. P.; MOBLEY, E. O.; DAVIS, D. W.
Quality of six tomato varieties as affected by some compositional factors. Proceedings of the
American for Horticultural Science, v. 89, p. 532-538, 1966.
SIRTOLI, L. F.; RODRIGUES, J. D.; GOTO, R. Efeito fisiológico do fungicida boscalida na
atividade da nitrato redutase e nas características fitotécnias de pepineiro japonês enxertado e
não enxertado. Scientia Agraria Paranaensis, v. 10, n. 3, p. 58-69, 2011.
SKANDALIS, N.; DIMOPOULOU, A.; BERI, D.; TZIMA, A.; MALANDRAKI, I.
THEOLOGIDIS, I. Effect of pyraclostrobin application on viral and bacterial diseases of
tomato. Plant Disease, v. 100, n. 7, p. 1321-1330, 2016.
SOARES, A. M. S.; MACHADO, O. L. T. Defesa de plantas: sinalização química e
espécies reativas de oxigênio. Revista Trópica – Ciências Agrárias e Biológicas, v.1,
n. 1, p. 9-19, 2007.
SOARES, L. H.; FAGAN, E. B.; CASAROLI, D.; ANDRADE, D. M. D.; SOARES, A. L.;
MARTINS, K. V.; ROCHA, F. J. D. Aplicação de diferentes estrobilurinas na cultura da soja.
Revista da FZVA, v. 18, n. 1, p. 78-97, 2011.

147
SONG, W.; MA, X.; TAN, H.; ZHOU, J. Abscisic acid enhances resistance to Alternaria solani
in tomato seedlings. Plant Physiology and Biochemistry, v. 49, n. 7, p. 693-700, 2011.
SONG, Y.; CHEN, D.; LE, K.; SUN, Z.; ZENG, R. Enhanced tomato disease resistance primed
by arbuscular mycorrhizal fungus. Frontiers in plant science, v. 6, n. 786, p. 1-13, 2015.
STANGARLIN, J. R.; KUHN, O. J.; TOLEDO, M. V.; PORTZ, R. L.; SCHWAN-ESTRADA,
K. R. F.; PASCHOLATI, S. F. A defesa vegetal contra fitopatógenos. Scientia Agraria
Paranaenis, v. 10, n. 1, p. 18-46, 2011.
STANGARLIN, J. R.; LEITE, B. Alterações fisiológicas na suscetibilidade. In:
PASCHOLATI. S. F.; LEITE. B.; STANGARLIN, J.R.; CIA, P. Interação Planta-Patógeno:
Fisiologia, Bioquímica e Biologia Molecular. Piracicaba: FEALQ, 2008, 627 p.
STANGARLIN, J. R.; PASCHOLATI, S. F; LABATE, C. A. Efeito de Phaeoisariopsis griseola
na atividade de ribulose-1,5-bifosfato carboxilase-oxigenase (rubisco), clorofilase, β-1,3-
glucanase e quitinase em cultivares de Phaseolus vulgaris. Fitopatologia Brasileira, v. 24, p.
59-66, 2000.
STEINER, U.; SCHÖNBECK, F. induced disease resistance in monocots. In:
HAMERSCHIMIDT, R.; KUC, J. (Ed.). Induced resistance to disease in plants:
developments in plant pathology. Dordrecht: Kluwer Academic Publishers, 1995, p. 86-110.
STENZEL. I.; HAUSE, B.; MAUCHER, H.; PITZSCHKE, A.; MIERSCH, O.; ZIEGLER, J.
Allene oxide cyclase dependence of the wound response and vascular bundle‐specific
generation of jasmonates in tomato amplification in wound signalling. The Plant Journal, v.
33, n. 3, p. 577-589, 2003.
STREETER, J. G.; BOSLER, M. E. Comparison of in vitro and in vivo assays for nitrate
reductase in soybean leaves. Plant Physiology, v. 49, n. 3, p. 448-450, 1972.
TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 4. ed. Porto Alegre: Artmed, 2013. 820 p.
TAMOI, M.; KUROTAKI, H.; FUKAMIZO, T. β-1,4-glucanase-like protein from the
cyanobacterium Synechocystis PCC6803 is a β-1,3-1,4-glucanase and functions in salt stress
tolerance. Biochemical Journal, London, v. 405, n. 1, p. 139-146, 2007.
TEISSEIRE, H.; GUY, V. Copper-induced changes in antioxidant enzymes activities in fronds
of duckweed (Lemna minor). Plant science, v. 153, n. 1, p. 65-72, 2000.
THAKUR, M.; SOHAL, B. S. Role of elicitors in inducing resistance in plants against pathogen
infection: a review. ISRN biochemistry, v. 2013, p. 1-10, 2013.
THARANATHAN, N. R.; KITTER, F. S. Chitin: the undisputed biomolecule of great potential.
Critical Reviews in Food Science and Nutrition, v. 43, n. 1, p. 61-87, 2003.
THIPYAPONG, P.; HUNT, M. D.; STEFFENS, J. C. Antisense downregulation of polyphenol
oxidase results in enhanced disease susceptibility. Planta, v. 220, n. 1, p. 105-117, 2004.

148
TÖFOLI, J. G. Pinta preta - uma ameaça constante aos cultivos da batata e do tomate.
2004. Disponível em: < http://www.biologico.sp.gov.br/artigos_ok.php?id_artigo=13 >.
Acesso em: 03/10/2017.
TÖFOLI, J. G.; DOMINGUES, R. J. Controle da pinta preta do tomateiro com o uso de
acibenzolar-s-metil isolado, em mistura com fungicidas e em programas de aplicação.
Arquivos do Instituto Biológico, v. 72, n. 4, p. 481-487, 2005.
TÖFOLI, J. G.; DOMINGUES, R. J. Severa pinta preta. 2007. Disponível em: <
http://www.biologico.sp.gov.br/artigos_ok.php?id_artigo=59 >. Acesso em: 03/10/2017.
TÖFOLI, J. G.; DOMINGUES, R. J.; FERRARI, J. T. Requeima e mancha de alternaria nas
culturas da batata e tomate: Divulgação técnica. Biológico, São Paulo, v. 76, n. 1, p. 41-50,
2014.
TSUMANUMA, G. M.; CARVALHO, S. J. P. D.; FANCELLI, A. L.; BERNARDES, M. S.;
RODRIGUES, M. A. T.; BEGLIOMINI, E. Crescimento de dois cultivares de soja submetidos
a aplicações de herbicidas e fungicidas. Revista Ceres, v. 57, n. 6, p. 742-750, 2010.
TSUTSUMI, K.; KONNO, M.; MIYAZAWA, S.I.; MIYAO, M. Sites of action of elevated
CO2 on leaf development in rice: Discrimination between the effects of elevated CO2 and
nitrogen deficiency. Plant and Cell Physiology, v. 55, n. 2, p. 258-268, 2014.
UDAYASHANKAR, A. C.; NAYAKA, C. S.; ARCHANA, B.; NAYAK, U.; NIRANJANA,
S. R.; PRAKASH, H. S. Strobilurins seed treatment enhances resistance of common bean
against bean common mosaic virus. Journal of Phytopathology, v. 160, n. 11-12, p. 710-716,
2012.
UMESHA, S. Phenylalanine ammonia lyase activity in tomato seedlings and its relationship to
bacterial canker disease resistance. Phytoparasitica, v. 34, n. 1, p. 68-71, 2006.
VALE, F. X. R. D.; ZAMBOLIM, L.; PAUL, P. A.; COSTA, H. Doenças causadas por fungos
de tomate. In: ZAMBOLIM, L.;VALE, F. X. R. D., et al. Controle de doenças de plantas -
hortaliças. Viçosa: Minas Gerais, v.2, 2000, 699-755 p.
VAN BEL, A. J. E.; HESS, P. H. Hexoses as phloem transport sugars: the end of a dogma?
Journal of Experimental Botany, v. 59, n. 2, p. 261-272, 2008.
VAN BREUSEGEM, F.; VRANOVA, E.; DAT, J. F.; INZE, D. The role of active
oxygen species in plant signal transduction. Plant Science, v. 161, n. 3, p. 405-414,
2001.
VAN LOON, L. C.; REP, M.; PIETERSE, C. M. J. Significance of inducible defense-related
proteins in infected plants. Annual Review of Phytopathology, v. 44, p. 135-162, 2006.
VAN LOON, L. C.; VAN STREIN, E. A. The families of pathogenesis-related proteins, their
activities, and comparative analysis of PR-1 type proteins. Physiological and molecular plant
pathology, v. 55, n. 2, p. 85-97, 1999.

149
VENANCIO, W. S.; RODRIGUES, M. A. T.; BEGLIOMINI, E.; SOUZA, N. L. D.
Physiological effects of strobilurin fungicides on plants. Publicatio UEPG: Ciências Exatas
e da Terra, Agrárias e Engenharias, v. 9, n. 3, p. 59-68, 2003.
VIANA, E. M.; KIEHL, J. C. Doses de nitrogênio e potássio no crescimento do
trigo. Bragantia, v. 69, n. 4, p. 975-982, 2010.
VON CAEMMERER, S.; FARQUHAR, G. D. Some relationships between the biochemistry
of photosynthesis and the gas exchange of leaves. Planta, v. 153, n. 4, p. 376-387, 1981.
WALKVRIA, B. C. M. Controle alternativo de fitopatógenos. Pesquisa Agropecuária
Brasileira, v. 27, n. 13, p. 175-190, 1992.
WU, J.; OMOKAWA, H.; HATZIOS, K. K. Glutathione S-transferase activity in unsafened
and fenclorim-safened rice (Oryza sativa). Pesticide Biochemistry and Physiology, v. 54, n.
3, p. 220-229, 1996.
XU, Z.; JIANG, Y.; ZHOU, G. Response and adaptation of photosynthesis, respiration, and
antioxidant systems to elevated CO2 with environmental stress in plants. Frontiers in Plant
Science, v. 6, n. 70, p. 1-17, 2015.
YEMM, E. W.; WILLIS, A. J. The estimation of carbohydrates in plant extracts by anthrone.
Biochem Journal, v.57, n.3, p.508-514, 1954.
YEOM, M.; KIM, H.; LIM, J.; SHIN, A. Y.; HONG, S.; KIM, J.; NAM, H. G. How do
phytochromes transmit the light quality information to the circadian clock in Arabidopsis.
Molecular Plant, v. 7, n. 11, p. 1701-1704, 2014.
YU, L.; PERRET, J.; HARRIS, M.; WILSON, J.; HALEY, S. Antioxidant properties of bran
extracts from “Akron” wheat grown at different locations. Journal of Agricultural and Food
Chemistry, v. 51, n. 6, p. 1566-1570, 2003.
YUAN, Y. J.; LI, C.; HU, Z. D.; WU, J. C. A double oxidative burst for taxol production in
suspension cultures of Taxus chinensis var. mairei induced by oligosaccharide from Fusarium
oxysprum. Enzyme and Microbial Technology, v. 30, n. 6, p. 774-778, 2002.
YUSUF, M. A.; KUMAR, D.; RAJWANSHI, R.; STRASSER, R.J.; TSIMILLIMICHAEL,
M.; GOVINDJEE; SARIN, N. B. Overexpression of y-totopherol methyl transferase gene in
transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a
fluorescence measurements. Biochimica et Biophysica Acta (BBA)-Bioenergetics, v. 1797,
n. 8, p. 1428-1438, 2010.
ZANATTA, C. L.; ZOTARELLI, M. F.; CLEMENTE, E. Peroxidase (POD) e polifenoloxidase
(PPO) em polpa de goiaba (Psidium guajava R.). Ciência e Tecnologia dos Alimentos, v. 26,
p. 705-708, 2006.
ZHAO, H.; WANG, B. C.; ZHAO, H. C.; WANG, J. B. Stress stimulus induced resistance to
Cladosporium cucumerinum in cucumber seeding. Colloids and surfaces B: Biointerfaces, v.
44, n. 1, p. 36-40, 2005.