efeitos da farinha da casca de maracujÁ passiflora …siaibib01.univali.br/pdf/aline de faveri...
TRANSCRIPT

ALINE DE FAVERI
EFEITOS DA FARINHA DA CASCA DE MARACUJÁ
(Passiflora edulis Var. flavicarpa) SOBRE A SÍNDROME
METABÓLICA INDUZIDA POR DIETA DE
CAFETERIA EM CAMUNDONGOS
Itajaí (SC)
2019


UNIVERSIDADE DO VALE DO ITAJAÍ
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS
FARMACÊUTICAS
ÁREA DE CONCENTRAÇÃO EM PRODUTOS NATURAIS E
SUBSTÂNCIAS SINTÉTICAS BIOATIVAS
ALINE DE FAVERI
EFEITOS DA FARINHA DA CASCA DE MARACUJÁ
(Passiflora edulis Var. flavicarpa) SOBRE A SÍNDROME
METABÓLICA INDUZIDA POR DIETA DE
CAFETERIA EM CAMUNDONGOS
Dissertação submetida à Universidade do
Vale do Itajaí como parte dos requisitos para a obtenção do grau de Mestre em Ciências Farmacêuticas.
Orientador: Prof. Dr. José Roberto Santin
Itajaí (SC)
Março de 2019




“Dedico este trabalho a minha família, em especial aos meus pais, irmã e
noivo, e a todos que contribuíram para
minha formação acadêmica”.


AGRADECIMENTOS
Agradeço aos meus pais Anísio De Faveri e Rosangela Farias De
Faveri, pela dedicação à minha formação pessoal e profissional, amor e
incentivo;
A minha irmã e aluna do Curso de Biomedicina da UNIVALI, Renata
De Faveri pelo companheirismo e dedicação durante a execução deste
projeto;
Ao meu noivo Rodrigo Escobar, pelo apoio e compreensão nos
momentos de ausência;
Ao meu orientador Prof. Dr. José Roberto Santin, pelos ensinamentos,
paciência e imensurável contribuição à minha formação acadêmica;
A Profª. Drª Nara Lins Meira Quintão pela disponibilidade e
colaboração na execução deste projeto;
A Profª. Drª. Ana Mara de Oliveira Silva e suas alunas, por me receber
na Universidade Federal de Sergipe e proporcionar inestimável contribuição
na realização dos ensaios antioxidantes;
A amiga e nutricionista Marina Jagielski Goss, pelo incentivo à
ingressar no programa de mestrado;
A todos colegas do programa de pós-graduação, em especial as amigas
que levarei do mestrado para a vida, Milena Fronza Broering, Izabel
Bousfield e Roberta Nunes;
Aos membros da banca, que gentilmente aceitaram o convite para
examinarem esta dissertação de mestrado;
A todos os professores e colaboradores do Curso de Pós-Graduação
em Ciências Farmacêuticas da UNIVALI que de alguma forma contribuíram
para a realização deste trabalho;
Ao apoio da Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior – Brasil (CAPES, cod. 001).
A todos, meus sinceros agradecimentos!


“Suba o primeiro degrau com fé. Não
é necessário que você veja toda a
escada. Apenas dê o primeiro passo.”
Martin Luther King


EFEITOS DA FARINHA DA CASCA DE MARACUJÁ
(Passiflora edulis Var. flavicarpa) SOBRE A SÍNDROME
METABÓLICA INDUZIDA POR DIETA DE
CAFETERIA EM CAMUNDONGOS
Aline De Faveri
Março/2019
Orientador: José Roberto Santin, Doutor.
Área de Concentração: Produtos naturais e substâncias sintéticas bioativas
Número de Páginas: 112.
Os hábitos alimentares vem mudando continuamente, o qual é um dos fatores mais fortes que explica a obesidade mundial. O aumento na ingestão de produtos processados e ultraprocessados altamente calóricos e com baixo valor nutricional, tem sido relacionado à diversas desordens metabólicas, como a resistência à insulina, obesidade central, alterações no metabolismo lipídico e glicídico e indução de estresse
oxidativo, responsável pelas disfunções características da síndrome metabólica. Em modelos animais, a dieta de cafeteria visa mimetizar estas alterações metabólicas, sendo muito utilizada para estudos de obesidade e síndrome metabólica. Diante da escassez de publicações acerca dos benefícios da farinha da casca do fruto de Passiflora edulis (FCM) e das modificações no padrão alimentar, o presente projeto teve como objetivo avaliar os efeitos da suplementação com farinha da casca de maracujá na ração de camundongos submetidos a dieta de cafeteria, bem como avaliar suas implicações no processo metabólico. Os ensaios FRAP, atividade sequestradora
do NO2 e sistema β Caroteno/Ácido Linoleico foram empregados para determinação da atividade antioxidante da FCM. Camundongos C57BL/6 machos foram divididos em diferentes grupos: 1) controle (ração padrão AIN-93M); 2) Dieta de cafeteria (Dieta hipercalórica) e 3) Dieta de cafeteria acrescida de Farinha de casca de maracujá (FCM) (Dieta hipercalórica + FCM 15%). Os animais receberam os diferentes tratamentos durante 16 semanas. Durante este período, foram avaliados o peso dos animais, consumo de ração e de água. Na última semana foi avaliada a resistência à insulina e tolerância oral a glicose. Ao final do experimento, os animais foram
anestesiados, o sangue foi coletado para avaliação de parâmetros bioquímicos (glicose, insulina, AST, ALT, creatinina, colesterol total e triglicerídeos). O tecido hepático foi coletado para determinação das enzimas antioxidantes e quantificação de lipídios. Os tecidos hepático, pancreático e renal foram submetidos a avaliação histológica. Os dados obtidos demonstraram que a dieta de cafeteria foi capaz de induzir importantes alterações metabólicas como: aumento significativo no índice de adiposidade, glicemia de jejum, colesterol e triglicerídeos séricos e hepáticos, TNF e IL-6, e infiltração de lipídeos no fígado, resistência a insulina e intolerância oral a glicose, hiperplasia das Ilhotas de Langherhans bem como atrofia dos glomérulos
renais. A suplementação com FCM foi capaz de atenuar os efeitos da dieta de

cafeteria, melhorando todos os parâmetros avaliados, seja pela ação de fibras ou devido a presença de compostos bioativos.
Palavras chave: Passiflora edulis. Farinha da casca do maracujá. Dieta de cafeteria.
Síndrome metabólica. Resistência a insulina.

PASSION FRUIT PEEL FLOUR (Passiflora edulis Var.
Flavicarpa) EFFECTS IN METABOLIC SYNDROME
INDUCED BY CAFETERIA DIETIN MICE
Aline De Faveri
March/2019
Professor: José Roberto Santin, Doutor.
Concentration area: Natural products and synthetic bioactive
substances
Number of pages: 112.
Changes in the eating habits are one of the strongest factors that explains obesity worldwide. The higher consumption of processed and ultraprocessed foods, high calorie and with low nutritional value has been related to several metabolic disorders, such as insulin resistance, central obesity, changes in lipid and glucose metabolism and oxidative stress, which is responsible for the characteristic dysfunctions of the metabolic syndrome. In animal models, cafeteria diet aims to mimic these metabolic alterations, being widely used on studies of obesity and metabolic syndrome.
Considering the few publications about the benefits of Passiflora edulis peel flour (PEPF), the aim of this study was to evaluate the effects of the supplementation with PEPF in mice on a cafeteria diet. To determinate the antioxidant activity of PEPF, FRAP assay, NO2 sequestering activity and β Carotene/Linoleic Acid system were used. C57Bl/6 black male mice were divided into different groups: 1) control (fed on a standard AIN-93M diet); 2) Cafeteria diet (Hypercaloric diet) and 3) Cafeteria diet plus Passionfruit Peel Flour (PFPF) (Hypercaloric diet associated to 15% of PEPF). Mice were treated for 16 weeks. During the period, the weight of the animals, food
and water intake were mesuared. In the last week, insulin resistance and glucose tolerance tests were applied. In the end of the experimentation, animals were anesthetized, blood was collected to evaluate biochemical parameters (glucose, insulin, AST, ALT, creatinine, total cholesterol and triglycerides), as well as pancreas, liver and adipose tissues. Liver tissue was also used to determinate the antioxidant enzymes and lipid quantification. Data demonstrated that supplementation with PEPF was able to attenuate metabolic changes induced by cafeteria diet such as: a significant increase in the rate of adiposity, fasting glucose, serum and hepatic concentration of cholesterol, triglycerides, TNF and IL-6, lipid infiltration in the liver, insulin
resistance and oral glucose intolerance, hyperplasia of the Langerhans Islets as well as renal glomerulus atrophy. In conclusion, cafeteria diet induces important metabolic disorders in mice, that were attenuated with PEPF supplementation. This effect could be by the actionpresence of fibers or due to the presence of bioactive compounds presents in the PFPF.
Key-words: Passiflora edulis. Passion fruit peel flour. Cafeteria diet.
Metabolic Syndrome. Insulin resistance.


LISTA DE FIGURAS
Figura 1 Principais alterações metabólicas induzidas pela dieta de
cafeteria...........................................................................
42
Figura 2 Plantação de
Maracujá..........................................................................
44
Figura 3 Partes do
maracujá..........................................................................
45
Figura 4 Estrutura química do ζ-caroteno (a), β-caroteno (b), pró-
licopeno (c), β-criptoxantina (d), violaxantina (e)
presentes no maracujá amarelo.......................................
46
Figura 5 Subdivisão dos grupos experimentais de acordo com o
tratamento dietético e cronograma de realização do
experimento.....................................................................
57
Figura 6 Atividade antioxidante do extrato da farinha da casca do
maracujá..........................................................................
66
Figura 7 Percentual de ganho de peso e área sobre a curva dos
diferentes grupos após 16 semanas de
experimentação................................................................
67
Figura 8 Consumo alimentar em gramas e calorias, ganho de peso
por consumo calórico e ingestão hídrica dos diferentes
grupos experimentais.......................................................
68
Figura 9 Perfil glicêmico dos diferentes grupos
experimentais..................................................................
70
Figura 10 Histologia, pesos e área das ilhotas de Langerhans do
pâncreas...........................................................................
72
Figura 11 Colesterol total e triglicerídeos
séricos..............................................................................
73
Figura 12 Parâmetros de tecido adiposo dos diferentes grupos
experimentais..................................................................
75
Figura 13 Análise histológica, peso e concentração de colesterol e
triglicerídeos no fígado....................................................
78
Figura 14 Concentração de IL-6 e TNF no tecido
hepático...........................................................................
79
Figura 15 Parâmetros antioxidantes no fígado dos diferentes
grupos experimentais após o período de intervenção......
80

Figura 16 Perfil renal dos diferentes grupos
experimentais..................................................................
81
Figura 17 Resumo gráfico de resultados da pesquisa....................... 94

LISTA DE TABELAS
Tabela 1 Critérios para diagnóstico de síndrome
metabólica.........................................................................
33
Tabela 2 Teor de nutrientes em cascas in natura de maracujá
amarelo (P. edulis var. flavicarpa) (Gondim et al.,
2005).................................................................................
49
Tabela 3 Teor de cianogênicos na casca do maracujá (in natura)
(Nascimento et al.,2013)...................................................
52
Tabela 4 Informação nutricional da FCM (Passiflora edulis Var.
flavicapa) empregada no presente estudo. Valores
referente a 1 porção do produto (50g = 7 colheres de
sopa)..................................................................................
53
Tabela 5 Concentração das enzimas hepáticas aspartato
aminotransferase (AST) e alanina aminotransferase
(ALT) no soro dos animais...............................................
76


LISTA DE ABREVIATURAS
ADA - American Dietetic Association
AGEs - Produtos finais de glicação avançada
AGL - Ácidos Graxos Livres
AIN-93M - American Institute of Nutrition (Instituto Americano de Nutrição)
ALT - Alanina Aminotransferase
ANVISA – Agência Nacional de Vigilância Sanitária
AST - Aspartato Aminotransferase
C – Grupo dieta controle
CAF - Grupo Dieta de Cafeteria
CAT – Catalase
CCD - Cromatografia em camada delgada
CEUA - Comitê de Ética no Uso de Animais
CGPCC - Coeficiente de Ganho de Peso por Consumo Calórico
CLAE- Cromatografia Líquida de Alta Eficiência
CONCEA - Conselho Nacional de Controle de Experimentação Animal
DCV - Doenças Cardiovasculares
DHGNA – Doença hepática gordurosa não alcoólica
DM – Diabetes Mellitus
DRIs - Dietary Reference Intakes (Ingestão dietética de referência)
EMBRAPA – Empresa Brasileira de Pesquisa Agropecuária
ERO - Espécies Reativas de Oxigênio
FAO - Food and Agriculture Organization of the United Nations
(Organização das Nações Unidas para Alimentação e Agricultura)
FCM – Farinha da Casca do Maracujá
FRAP - Ferric Reducing Antioxidant Power (Poder antioxidante de redução
do ferro)
GLUT – Transportador de glicose 4
GPx - Glutationa Peroxidase
GSH - Glutationa Reduzida
GSSG - Glutationa Oxidada
HAS - Hipertensão Arterial Sistêmica
HDL - High Density Lipoprotein-cholesterol
HOMA - Homeostatic model assessment (Modelo de Avaliação da
Homeostase)

IBGE - Instituto Brasileiro de Geografia e Estatística
IDF - International Diabetes Federation (Federação Internacional de
Diabetes)
IG – Índice Glicêmico
IL-1β – Interleucina 1β
IL-6 - Interleucina 6
IRS1 – Insulin Receptor Substrate 1 – Substrato Receptor de Insulina 1
LDL-c - Low Density Lipoprotein-cholesterol (Lipoproteína-Colesterol de
baixa densidade)
NCEP-ATPIII - Third Report of the National Cholesterol Education
Program Expert Panel on Detection, Evaluation, and Treatment of High
Blood Cholesterol in Adults (Terceiro Relatório do Painel de Especialistas do
Programa Nacional de Educação sobre Colesterol na Detecção, Avaliação e
Tratamento de Hipercolesterolemia em Adultos)
NF-kB - Fator Nuclear kB
OMS – Organização Mundial de Saúde
PA – Pressão Arterial
PCR – Proteína C Reativa
RI - Resistência à Insulina
SM – Síndrome metabólica
SOD - Superóxido Dismutase
TG - Triglicerídeos
TLR2 – Toll Like Receptor (Receptor tipo “Toll”)
TNF – Fator de Necrose Tumoral
VIGITEL - Pesquisa de Vigilância de Fatores de Risco e Proteção para
Doenças Crônicas por Inquérito Telefônico
VLDL-c - Very Low Density Lipoprotein-cholesterol (Lipoproteína-
Colesterol de muito baixa densidade)

SUMÁRIO
1 INTRODUÇÃO................................................................................ 255
2 OBJETIVOS .................................................................................... 299
2.1 Objetivo Geral ..............................................................................299
2.2 Objetivos Específicos ...................................................................299
3 REVISÃO BIBLIOGRÁFICA........................................................... 31
3.1 Transição nutricional e suas repercussões ....................................... 31
3.2 Síndrome Metabólica ..................................................................... 32
3.2.1 Dieta de Cafeteria ................................................................... 38
3.3 Maracujá amarelo (Passiflora edulis Var. flavicarpa) ..................... 43
3.3.1 Farinha da Casca do Maracujá amarelo (Passiflora edulis Var.
flavicarpa) ....................................................................................... 47
4 MATERIAL E MÉTODOS ............................................................... 53
4.1 Farinha da casca de maracujá (FCM).............................................. 53
4.1.1 Obtenção do produto ............................................................... 53
4.2 Ensaios in vitro .............................................................................. 51
4.2.1 Obtenção do extrato de FCM ................................................... 53
4.2.1.1 Avaliação do potencial antioxidante da FCM in vitro ............ 54
4.2.1.2 Método FRAP ....................................................................... 54
4.2.1.3 Sistema de cooxidação do β-caroteno/ácido linoléico ............ 54
4.2.1.4 Capacidade antioxidante por eliminação do Óxido Nítrico (NO)
........................................................................................................ 55
4.3 Ensaios in vivo .............................................................................566
4.3.1 Animais ..................................................................................566
4.3.2 Tratamento e parâmetros murinométricos ...............................566
4.3.3 Elaboração das dietas experimentais ......................................577
4.3.4 Consumo energético ...............................................................588
4.3.5 Eficiência Alimentar ...............................................................588
4.3.6 Curva de tolerância oral a glicose e teste de resistência à insulina
..........................................................................................................599
4.3.7 Eutanásia e coleta do material biológico ................................599
4.3.8 Determinação do peso relativo dos órgãos e tecidos ................ 60
4.3.9 Parâmetros bioquímicos no soro .............................................. 60
4.3.10 Avaliação histológica ............................................................ 60
4.3.11 Determinação de lipídeos no fígado ....................................... 61

4.3.12 Marcadores de estresse oxidativo ........................................... 61
4.3.12.1 Preparo dos homogeneizados no tecido hepático ................. 61
4.3.12.2 Atividade antioxidante total do tecido pelo método FRAP .... 62
4.3.12.3 Quantificação de glutationa reduzida (GSH) ........................ 62
4.3.13 Determinação da atividade das enzimas antioxidantes ............ 62
4.3.13.1 Superóxido Dismutase (SOD) .............................................. 62
4.3.13.2 Catalase (CAT) ................................................................... 63
4.3.13.3 Glutationa Peroxidase (GPx) ............................................... 63
4.3.14 Quantificação de proteínas no tecido hepático ........................ 64
4.3.15 Determinação das citocinas inflamatórias no fígado ............... 64
4.4 Análise estatística ........................................................................... 64
5 RESULTADOS .................................................................................. 65
5.1 Avaliação da atividade antioxidante in vitro do extrato hidroalcoólico
obtido a partir da FCM ......................................................................... 65
5.2 Ensaios in vivo ............................................................................... 66
6 DISCUSSÃO ...................................................................................... 83
7 CONCLUSÕES.................................................................................. 95
REFERÊNCIAS ................................................................................... 97

25
1 INTRODUÇÃO
Nas últimas décadas, o Brasil, assim como diversos países em
desenvolvimento, passou por mudanças numa acelerada transição
demográfica, epidemiológica e nutricional. As altas taxas de mortalidade
infantil e desnutrição deram lugar a um crescente número de distúrbios
metabólicos e doenças crônicas não transmissíveis, como diabetes,
hipertensão arterial, hipercolesterolemia, doença hepática gordurosa não
alcóolica, obesidade e, consequentemente, síndrome metabólica (SM), tendo
sua origem na maioria das vezes, na alimentação inadequada (BATISTA
FILHO; BATISTA, 2010; MAYER JR et al., 2018).
As mudanças nas práticas alimentares contemporâneas, principalmente
o aumento do consumo de gorduras, açúcares, alimentos açucarados e ultra
processados, e redução da ingestão de carboidratos complexos e de fibras,
presentes principalmente em alimentos como frutas, verduras e legumes,
contribuem para o acometimento por estas doenças. Este novo padrão
alimentar tornou-se objeto de preocupação das ciências da saúde, uma vez
que, estudos epidemiológicos passaram a demonstrar correlações entre a dieta
e doenças crônicas (COUTINHO; GENTIL; TORAL, 2008; PROENÇA,
2010; GIL-CARDOSO et al., 2017, VIGITEL, 2018).
Estudos com modelos animais de obesidade são relatados, porém,
muitos não reproduzem a ingestão de alimentos regularmente consumidos
por humanos. Com o objetivo de compreender o perfil nutricional atual,
mimetizar seus efeitos a longo prazo, e proporcionar o entendimento da
fisiologia do organismo, têm sido empregadas as chamadas dietas de cafeteria
(FAGUNDES; TAHA, 2004; DAMATTA, 2010; GIL-CARDOSO et al.,
2017).
A dieta de cafeteria consiste em oferecer um aporte calórico elevado,
por meio do aumento dos carboidratos e/ou lipídeos presentes na
alimentação, assemelhando-se, portanto, aos hábitos alimentares modernos.
Por esta analogia, este tipo de dieta vem sendo utilizada em muitas pesquisas
relacionadas à obesidade e distúrbios metabólicos (NASCIMENTO et al.,
2008; GIL-CARDOSO et al., 2017). Alimentos como chocolate, amendoim,
biscoito, bacon, leite condensado, bolo, refrigerante e entre outros, podem
compor essa dieta (LOUZADA et al., 2013).
Autores apontam que esta dieta, administrada em ratos, promove um
padrão alimentar hiperfágico e hipercalórico, com rápido ganho de peso, e

26
aumento da massa de tecido adiposo, intolerância à glicose, resistência à
insulina e hiperinsulinemia (ESTADELLA et al., 2004; SAMPEY et al.,
2011; GOULARTE; FERREIRA; SANVITTO, 2012; BALLESTRERI;
MARCON; TAVARES1, 2015), podendo induzir o desenvolvimento de
distúrbios metabólicos, não apenas por intermédio de desequilíbrios entre a
distribuição dos macro nutrientes, como também, ao tipo de carboidratos e
gorduras consumidas, e não ao valor energético atribuído a estes nutrientes
(PAWLAK; KUSHNER; LUDWIG, 2004).
Em contrapartida estudos epidemiológicos demonstram que, dietas ricas
em frutas e vegetais parecem ter efeito protetor, podendo reduzir a incidência
de síndrome metabólica, sendo estes efeitos atribuídos sobretudo pela
presença de fibras nestes alimentos. Como um nutriente essencial, as fibras
têm apresentado um papel crucial na manutenção da saúde, com ações de
prevenção e proteção para várias doenças humanas, sendo complementares
no tratamento de diversas doenças relacionadas a alimentação como a
diabetes mellitus (DM), dislipidemias e obesidade (MATTOS; MARTINS,
2000; BERNAUD; RODRIGUES, 2013; SLAVIN, 2013; ZHANG et al.,
2016).
Neste contexto, visando a prevenção e tratamento destas desordens
metabólicas, a utilização de compostos bioativos derivados de produtos
naturais estão ganhando cada vez mais destaque, uma vez que podem
proporcionar a melhora da saúde, além de apresentarem baixa toxicidade e
poucos efeitos adversos. O uso do gênero Passiflora, e suas aplicações na
farmacologia, já é bem descrito na literatura em estudos clínicos e
experimentais, sendo atribuídos efeitos sedativos, antiespasmódico e
ansiolítico às folhas, e atividade antioxidante à polpa (DHAWAN;
DHAWAN; SHARMA, 2004) além da grande quantidade de fibras
alimentares, principalmente as solúveis como a pectina, presentes na casca
do maracujá (YAPO; KOFFI, 2006; GOSS et al., 2018).
Tendo em vista a influência do comportamento alimentar sobre o
acometimento da população por doenças crônicas e distúrbios metabólicos,
e, sendo muitos mecanismos fisiopatológicos interligados entre si, se faz
necessário estudos que abranjam os aspectos relacionados ao surgimento
desses quadros clínicos. É possível que o sucesso desse modelo se dê devido
ao fato de se assemelhar a forma como a obesidade é adquirida pela
população humana, podendo assim reproduzir suas causas e consequências.

27
Assim, o objetivo do presente projeto foi avaliar os efeitos metabólicos
da suplementação com farinha da casca de maracujá sobre a síndrome
metabólica induzida por dieta de cafeteria em camundongos.

28

29
2 OBJETIVOS
2.1 Objetivo Geral
Avaliar os efeitos da farinha da casca de maracujá (Passiflora edulis
Var. Flavicarpa) sobre a síndrome metabólica induzida por dieta de cafeteria
em camundongos.
2.2 Objetivos Específicos
a) Avaliar o potencial antioxidante da farinha de casca de P. edulis Var.
flavicarpa;
b) Avaliar o efeito da incorporação de farinha de casca de P. edulis
Var. flavicarpa na síndrome metabólica induzida por dieta de
cafeteria sobre:
Parâmetros de consumo alimentar, energético e hídrico dos animais;
Parâmetros murinométricos;
Tolerância oral a glicose e resistência à insulina;
Marcadores bioquímicos: perfil lipídico, glicemia, insulina, função
renal, enzimas hepáticas;
Capacidade antioxidante (enzimas, FRAP) no tecido hepático dos
diferentes grupos experimentais;
Concentração das citocinas inflamatórias no tecido hepático (TNF e
IL- 6);
Morfologia dos diferentes tecidos (fígado, pâncreas, rins e tecido
adiposo).

30

31
3 REVISÃO BIBLIOGRÁFICA
3.1 Transição nutricional e suas repercussões
A obesidade é definida pela Organização Mundial da Saúde (OMS)
pelo índice de massa corporal acima de 30 kg/m2, podendo ser desencadeada
por diversos fatores. A elevação do índice de massa corporal é o maior fator
de risco para complicações que incluem o diabetes, doenças cardiovasculares,
doenças do trato gastrointestinal e câncer (WHO, 2015; BORTOLIN et al.,
2018).
Em 2014, segundo dados da OMS, mais de 1,9 bilhão da população
mundial de adultos, apresentavam excesso de peso. Dentre estes indivíduos,
mais de 600 milhões estavam obesos. Estes dados sugerem que, naquele ano,
aproximadamente 13% da população mundial adulta estava obesa e 39%
estava com sobrepeso, evidenciando que a prevalência mundial de obesidade
duplicou entre os anos de 1980 e 2014 (WHO, 2015).
A prevalência de obesidade tem aumentado em taxas alarmantes. Dados
preocupantes vem de estudos bem recentes que mostram que mais de 90%
dos adultos dos EUA, Nova Zelândia, Grécia e Islândia são “overfat”, um
novo termo que se refere a um acúmulo excessivo de gordura corporal que se
torna suficiente para prejudicar a saúde (MAFFETONE; RIVERA-
DOMINGUEZ; LAURSEN, 2017). Essa taxa extremamente alta é explicada
pelo fato de pessoas com peso normal, mas que tenham uma proporção de
gordura suficiente para prejudicar a saúde entram na conta, mostrando que o
problema de excesso de gordura é bem maior do que se imagina.
No Brasil, a prevalência de sobrepeso e obesidade é elevada. Segundo
levantamento da Pesquisa de Vigilância de Fatores de Risco e Proteção para
Doenças Crônicas por Inquérito Telefônico - Vigitel (2018), realizada em
todas as capitais brasileiras entre fevereiro e dezembro de 2016, uma em cada
cinco pessoas no País está acima do peso. A prevalência de obesidade passou
de 11,8%, em 2006, para 18,9%, em 2016, e de excesso de peso também
cresceu, passou de 42,6% para 53,8% em 10 anos.
Ainda segundo a pesquisa, dados demonstram, que este crescimento
pode ter colaborado para o aumento da prevalência de doenças crônicas,
como diabetes e hipertensão. O diagnóstico médico de diabetes passou de
5,5% em 2006, para 8,9% em 2016. Já o de hipertensão, no mesmo período,

32
saiu de 22,5% para 25,7%. E em ambos os casos, o diagnóstico é mais
prevalente em mulheres (VIGITEL, 2018).
As causas da obesidade na população são múltiplas e complexas. O
sedentarismo e a mudança nos hábitos alimentares têm sido apontados como
os dois principais fatores por trás desta epidemia global (DAVIS;
PLAISANCE; ALLISON, 2018). Estudos epidemiológicos apontam que o
hábito alimentar dos indivíduos vem sofrendo importantes mudanças, o que
indica ser um dos fatores mais relevantes que explicam a obesidade mundial.
Uma destas modificações, é o tipo de macronutriente ingerido. Consome-se
cada vez mais produtos processados contendo açúcar refinado, xarope de
milho, óleos refinados, sal (sódio). Outra mudança, é o consumo de alimentos
ultra processados, o qual vem aumentando progressivamente, como por
exemplo, alimentos pré-cozidos, biscoitos recheados, salgadinhos, todos com
alta densidade calórica e baixo valor nutricional (BORTOLIN et al., 2018).
Concomitante a isso, observa-se a redução da ingestão de frutas, verduras e
legumes, consequentemente de fibras, vitaminas e minerais, além de
compostos bioativos e substâncias antioxidantes. De um modo geral, as
mudanças nos hábitos alimentares são cada vez mais em direção ao aumento
no consumo calórico, que, a longo prazo, pode estar relacionado com a
epidemia global de obesidade.
Porém, este acúmulo excessivo de gordura corporal traz prejuízos ao
organismo, podendo desencadear “co-morbidades”, como hipertensão,
dislipidemia, diabetes, doença hepática gordurosa, doenças cardiovasculares
e a resistência à insulina (RI), todas condições metabólicas que compreendem
a síndrome metabólica, sendo a última, a que parece ser comum e
correlacionar-se com todas outras.
3.2 Síndrome Metabólica
O conceito da síndrome metabólica existe há pelo menos 90 anos. Foi
descrita pela primeira vez na década de 1920 por Kylin, um médico sueco,
conceituando-a como uma condição metabólica resultante do agrupamento
de fatores de risco para doenças cardiovasculares (DCV), como a hipertensão
arterial sistêmica (HAS) e hiperglicemia. Em 1947, Vague correlacionou-a
também com a obesidade andróide, caracterizada pela adiposidade corporal
central, fenômeno comumente associado a anormalidades metabólicas

33
associadas à DM tipo 2 (KYLIN, 1923; CAMERON; SHAW; ZIMMET,
2004; ECKEL; GRUNDY; ZIMMET, 2005).
Em 1988, Reaven descreveu a síndrome metabólica como uma série de
anormalidades metabólicas e hemodinâmicas, comumente presentes no
indivíduo obeso (MOTTILLO, 2010). Mais tarde, ao final da década de 1990,
a Organização Mundial da Saúde estabeleceu o termo unificado de síndrome
metabólica, para o conjunto de fatores de risco para DCV, tais como a
obesidade, hiperglicemia, HAS e dislipidemia, visando caracterizar uma
possível predisposição para complicações do aparelho circulatório, como
insuficiência cardíaca, doença arterial coronária, infarto agudo do miocárdio
e acidente vascular encefálico. Em 2001, o “Third Report of the National
Cholesterol Education Program Expert Panel on Detection, Evaluation, and
Treatment of High Blood Cholesterol in Adults” (NCEP-ATPIII) definiu
critérios clínicos e laboratoriais para o diagnóstico de síndrome metabólica.
Posteriormente, a “International Diabetes Federation” redefiniu alguns
critérios, enfatizando a importância da obesidade abdominal central, e
considerando valores distintos de acordo com o grupo racial do indivíduo
(NAKAZONE et al., 2007).
Para fins de diagnóstico, a I Diretriz Brasileira no Diagnóstico e
Tratamento da Síndrome Metabólica adotou os critérios do NCEP-ATPIII
(do inglês, National Cholesterol Education Program - Adult Treatment Panel
III) como referência para a população brasileira devido sua simplicidade,
definindo o diagnóstico da síndrome metabólica como a presença de três ou
mais critérios dentre os 5 estabelecidos (SOCIEDADE BRASILEIRA DE
CARDIOLOGIA, 2005) (Tabela 1).
Tabela 1. Critérios para diagnóstico de síndrome metabólica.
Componentes Valores de referência
Circunferência da cintura Homens > 102 cm
Mulheres > 88 cm
Pressão arterial PAD ≥ 130 e/ou PAS ≥85 mmHg
Glicemia de jejum ≥ 100 mg/dL
Triglicerídeos ≥ 150 mg/dL
HDL-colesterol Homens < 40 mg/dL
Mulheres < 50mg/dL
Fonte: National Cholesterol Education Program (2002). Legenda: NCEP-ATPIII- National Cholesterol Education Program -Adult Treatment Panel III. PAD: Pressão

34
Arterial Diastólica. PAS: Pressão Arterial Sistólica. HDL-colesterol: Lipoproteína de alta densidade (do inglês, High density lipoprotein).
No entanto, considerando as evidências, e relacionando a conexão entre
obesidade central e risco de DCV e síndrome metabólica, a International
Diabetes Federation (IDF) publicou em 2005 um novo critério, propondo a
redução nos valores de referência de circunferência abdominal para ≥94 cm
e ≥80cm para homens e para mulheres, respectivamente (ALBERTI, 2006),
uma vez que os estudos demonstram que o aumento do risco, está diretamente
e progressivamente associado ao aumento da circunferência da cintura
(SABOYA et al., 2016).
Além destas disfunções metabólicas, a literatura discute a relação da
resistência à insulina na fisiopatologia da síndrome metabólica, uma vez que
até 70% dos indivíduos acometidos apresentam RI, sugerindo assim, uma
menor sensibilidade à ação deste hormônio (FORD, 2005; MAYER JR,
2018). Atualmente, vem sendo descrita como fator de ligação entre a
adiposidade central, intolerância à glicose, HAS, dislipidemia, distúrbios da
coagulação, hiperuricemia e microalbuminúria, comorbidades integrantes da
síndrome metabólica, desempenhando um papel importante na sua
patogênese (RIBEIRO FILHO, 2006; SABOYA et al., 2016, RANASINGHE
et al., 2017).
A RI é uma condição complexa, definida como um estado de menor
resposta metabólica a níveis circulantes de insulina em tecido periféricos
(músculo, fígado e tecido adiposo), sistema nervoso central e células beta-
pancreáticas. Também é definida por uma menor capacidade de supressão da
gliconeogênese hepática (CZECH, 2017), tendo relação direta com a
adiposidade e com a alta prevalência de obesidade central (LOPES, 2005). O
fígado e tecido adiposo branco, também se tornam menos sensíveis às suas
ações metabólicas, promovendo alteração nas concentrações séricas de
glicose, podendo evoluir para DM tipo 2 (CHAWLA; NGUYEN; GOH,
2011).
A insulina é um hormônio anabólico, tendo sua sinalização iniciada
com sua ligação a um receptor específico da membrana, uma proteína com
atividade quinase intrínseca, denominada receptor de insulina. A ativação do
receptor de insulina resulta na fosforilação do substrato receptor de insulina
(IRS) 1 e 2 em tirosina, criando sítios de ligação para uma outra proteína
citosólica, denominada fosfatidilinositol 3-quinase (PI3k), que quando

35
ativada aumenta a fosforilação em serina da proteína quinase B (Akt) e isso
permite o transporte da glicose no músculo e no tecido adiposo, através da
translocação do transportador de glicose (GLUT4) para a membrana,
permitindo a captação da glicose (VÁZQUEZ-JIMÉNEZ et al., 2017).
Na obesidade ocorrem alterações em diversos pontos da via de
transdução do sinal de insulina. Ocorre redução na concentração e atividade
quinase do receptor de insulina, na concentração e fosforilação dos IRS 1 e
2, na atividade PI3k, na translocação dos GLUT´s e na atividade de enzimas
intramusculares, consequentemente isso atenua a captação da glicose nos
tecidos insulino-dependentes (FOSTER; KLIP, 2000).
A existência de uma relação entre a adiposidade e a resistência à
insulina já está bem esclarecida, contudo ainda não se pode afirmar com
exatidão se a resistência à insulina é um fator promotor ou uma consequência
do ganho de tecido adiposo (LOPES, 2005). O entendimento da estrutura e
funcionalidade do tecido adiposo vem sendo aprimorado ao longo dos anos.
Atualmente, além de um depósito de lipídeos, este tecido já é reconhecido
como um órgão, com diversas funcionalidades autócrinas e parácrinas,
implicações no sistema endócrino, e em processos metabólicos,
principalmente em processos inflamatórios (TRAYHURN; WOOD, 2004;
GOLLISCH et al., 2009).
Outros fatores também devem ser considerados na patogênese da
síndrome metabólica, como a ativação crônica do sistema imunológico,
distúrbios do eixo hipotálamo-hipófise-adrenal, alteração da ação dos
hormônios glicocorticoides, estresse crônico e fatores genéticos que possam
estar envolvidos. E ainda, a contribuição das citocinas, hormônios, e outras
moléculas sintetizadas e secretadas pelos adipócitos, são relacionadas a
síndrome metabólica (FORD, 2005; PICON et al., 2006).
Com o acúmulo de gordura corporal, os adipócitos secretam maiores
concentrações de Fator de Necrose Tumoral (TNF) e Interleucina 6 (IL-6),
que são antagonistas à ação da insulina. O TNF favorece a fosforilação em
serina do substrato do receptor da insulina-1 (IRS1). O IRS1 fosforilado em
serina não é capaz de ativar fosfatidilinositol 3 quinase (PI3-q) o que resulta
em transporte diminuído de glicose, o que tem sido associado ao aumento da
resistência à insulina (CAMPOS et al., 2006; FURUKAWA et al., 2017).
O NF-kB também é um fator que possui relação com o surgimento e
manutenção da inflamação na síndrome metabólica, já que, quando ativado

36
no adipócito, leva a produção de citocinas pró-inflamatórias, e como
consequência, irá contribuir para a resistência à insulina (LIMA et al., 2012).
Em condições normais, a insulina participa da regulação do
metabolismo lipídico por meio de várias ações, todavia, em indivíduos com
síndrome metabólica essa ação não é observada, favorecendo as
dislipidemias. A dislipidemia presente na síndrome metabólica é chamada de
aterogênica e consiste em três irregularidades lipídicas: hipertrigliceridemia,
HDL-c diminuído, do inglês High Density Lipoprotein-cholesterol, e valores
do LDL-c, do inglês Low Density Lipoprotein-cholesterol, entre a
normalidade e leve a moderada elevação (GUIMARÃES, 2006).
Esse fenômeno está relacionado a resistência à insulina, na qual, devido
ao menor metabolismo das VLDL-c (Very Low Density Lipoprotein-
cholesterol), decorrente da hiperinsulinemia, a concentração plasmática de
triglicerídeos (TG) se eleva, ao mesmo tempo em que a de HDL-c sofre
decréscimo, promovendo o aumento do nível de VLDL-c, redução dos níveis
e do tamanho da HDL-c e aumento de partículas de LDL-c, as quais são ricas
em apolipoproteína B (GUIMARÃES, 2006; OLIVEIRA et al., 2004).
De forma simplificada, a hipertrigliceridemia observada no indivíduo
com síndrome metabólica, pode ser atribuída ao aumento da síntese de
apolipoproteína C-III (APO c-III), que interfere na ação da lipoproteína
lipase, envolvida no processo de hidrólise dos TG da partícula de VLDL-c.
Na cascata do metabolismo endógeno das lipoproteínas, a ação das lipases,
também sobre as VLDL-c, cria remanescentes mais aterogênicos. A alta
concentração de APO c-III interfere na captação de remanescentes de VLDL-
c pelos receptores de LDL-c nas células hepáticas, gerando aumento de TG
na circulação. Ademais, a APO c-III liga-se ao receptor do tipo Toll (TLR2),
um mediador pró-inflamatório que intensifica o processo aterosclerótico
(GRUNDY et al., 2004).
Corroborando, autores demonstraram que os ácidos graxos saturados de
cadeia longa, são os lipídeos mais nocivos no que tange acúmulo de massa
adiposa. Nesse estudo, os pesquisadores verificaram que essas moléculas
ligam-se aos receptores do tipo Toll (TLR2 e TLR4) das micróglias, células
protetoras do hipotálamo, estimulando a produção de citocinas pró-
inflamatórias (TNF, IL-1β e IL-6) e, consequentemente, a destruição dos
neurônios responsáveis pelo controle do apetite e da termogênese
(MILANSKI et al., 2009), demonstrando a importante participação do
sistema imune e hormonal na gênese da síndrome metabólica.

37
Outros eventos também podem ser observados no quadro de síndrome
metabólica, tais como aumento da aderência de células mononucleares,
diminuição da vasodilatação endotélio-dependente, secreção aumentada de
citocinas pró-inflamatórias como proteína C reativa (PCR), IL-6 e IL-1,
elevadas concentrações séricas de ácido úrico e microalbuminúria
(MOZAFFARIAN et al., 2004; GOMES et al., 2010; FURUKAWA et al.,
2017).
Não incluso nos critérios de diagnóstico, mas característica nesta
síndrome é a doença hepática gordurosa não alcoólica (DHGNA), a qual é
considerada a manifestação hepática da síndrome (LONARDO, 1999;
MUSSO et al., 2008; YKI-JÄRVINEN, 2014). A DHGNA corresponde ao
acúmulo anormal de lipídeos nos hepatócitos, e engloba um espectro de
doenças hepáticas crônicas na ausência de uso excessivo de álcool. Estima-
se que a DHGNA afete cerca de 1 bilhão de indivíduos em todo o mundo
(LOOMBA; SANYAL, 2013).
A resistência à insulina também está diretamente correlacionada a
DHGNA (MAVROGIANNAKI; MIGDALIS, 2013). Conforme
mencionado, o efeito antilipolítico da insulina está diminuído, favorecendo a
síntese hepática de triglicerídeos. Além disso, quando há dificuldade na
sinalização celular de insulina, ocorre aumento na liberação de ácidos graxos
livres do tecido adiposo, os quais podem se depositar no tecido adiposo
subcutâneo sob a forma de gordura ectópica ou em órgãos como o coração e
o fígado (GAGGINI et al., 2013).
Sabe-se que, em resposta à resistência tecidual provocada pela gordura
excessiva, a secreção de insulina é aumentada, culminando em
hiperinsulinemia. Estudos sugerem que a hiperinsulinemia, poderia elevar a
pressão arterial, por meio da ativação do sistema nervoso simpático, do
comprometimento da vasodilatação periférica, da maior resposta a
angiotensina e do aumento da reabsorção renal de sódio e água, com
consequente aumento de volume sanguíneo (BARROSO; ABREU;
FRANCISCHETTI, 2002). A HAS está relacionada com alterações
funcionais e/ou estruturais dos órgãos-alvo (rins, coração, encéfalo e vasos
sanguíneos) e, portanto, a um maior risco de acidentes cardiovasculares
(SOCIEDADE BRASILEIRA DE CARDIOLOGIA, 2010). A associação da
síndrome metabólica com a DCV atua aumentando a mortalidade geral em
aproximadamente 1,5 vezes e a mortalidade cardiovascular em cerca de 2
vezes (LAKKA et al., 2002; MOTTILLO, 2010).

38
A prevalência da síndrome metabólica é alarmante, e está aumentando
à nível mundial, tanto em países desenvolvidos como em desenvolvimento
(ALBERTI et al., 2009). Estudos apontam que nas últimas décadas, a
incidência aumentou progressivamente, estimando uma prevalência de até
23,7%, de acordo com os critérios ATP III, em um estudo realizado nos EUA
com uma amostra de 8.814 adultos (FORD; GILES; DIETZ, 2002; SABOYA
et al., 2016).
Dados epidemiológicos demonstram que de 20 à 25% da população
mundial adulta apresenta síndrome metabólica (KASSI et al., 2011) sendo
um indicador de risco de vida a longo prazo (MAMIKUTTY et al., 2014).
Este rápido aumento tem sido acompanhado pela crescente epidemia de DM,
HAS, DCV e obesidade (RANASINGHE et al., 2017), a qual vem crescendo
em proporções epidêmicas em todo o mundo, aumentando a predisposição
para todas as causas específicas ou não de morbidade e mortalidade (WHO,
2011).
Apesar dos avanços na compreensão e tratamento, a síndrome
metabólica é um importante problema de saúde pública (SABOYA et al.,
2016). Predisposição genética, idade, estilo de vida e perfil socioeconômico
podem estar envolvidos no desenvolvimento desta condição metabólica
(CORNIER et al., 2008; RANASINGHE et al., 2017), todavia, observa-se
que um dos principais fatores que contribuem para o seu desenvolvimento,
são dietas ricas em carboidratos e gorduras (MAMIKUTTY et al., 2014). O
aumento do consumo de gorduras, principalmente saturadas, açúcar, sódio,
alimentos refinados e redução da ingestão diária de carboidratos complexos
e de fibras, vem sendo o novo padrão dietético da população, sendo este, não
saudável, e um importante gatilho desencadeador dos quadros de obesidade
e síndrome metabólica (CARVALHO; ALFENAS, 2008).
3.2.1 Dieta de Cafeteria
Alimentos ultraprocessados, são componentes cada vez mais
frequentes na alimentação diária da população. Por serem industrializados,
apresentam uma alta densidade energética, em função da presença de altos
teores de açúcares, sódio, gorduras saturadas e trans, além de um teor de
fibras diminuído, tendo impacto negativo na qualidade da alimentação. Esse
perfil alimentar tem um grande impacto à saúde, podendo ser responsável por

39
comprometer o balanço energético do organismo, favorecendo o acúmulo de
gordura, contribuindo para a instalação e perpetuação de quadros de DCV,
DM, HAS, obesidade e câncer (LOUZADA et al., 2015).
Assemelhando-se aos hábitos alimentares da sociedade ocidental,
estudos com animais têm sido realizados, utilizando as chamadas dietas de
cafeteria, que consistem em oferecer um aporte calórico elevado, por meio
do aumento dos carboidratos e/ou lipídeos presentes na alimentação. Este
desequilíbrio dos macro nutrientes é induzido através da inclusão de
alimentos comuns à população humana na dieta dos animais, entretanto, não
há um protocolo que padronize este tipo de dieta, e assim os autores podem
optar pelos alimentos hipercalóricos que desejarem utilizar em sua pesquisa.
Alimentos como chocolate, amendoim, bacon, leite condensado, bolo, patê,
refrigerante e outros podem compor essa dieta (LOUZADA et al., 2013).
A composição da dieta de cafeteria empregada no estudo, será uma
dieta hipercalórica (normoproteica e hiperlipídica) proposta por Estadella et
al. (2004), a qual consiste na associação da dieta AIN-93 M com alimentos
hipercalóricos, ricos em carboidratos e lipídeos, sendo: amendoim, chocolate
ao leite e biscoito de maisena. Estes autores têm seu modelo reproduzido em
diversos estudos, com bons resultados na indução da síndrome metabólica em
modelos animais (EGUCHI et al., 2008; OLIVEIRA et al., 2011;
BALLESTRERI; MARCON, TAVARES, 2015).
Sampey e colaboradores (2011) demonstraram que o modelo de indução
da síndrome metabólica por meio de dietas de cafeteria, parece ser o melhor
modelo, por sua robustez, refletindo a fisiopatologia da doença de forma mais
próxima a condição a qual é adquirida pela população humana, tornando esse
modelo viável, podendo contribuir nos progressos de compreensão da
etiologia, patogenia e intervenções terapêuticas da síndrome metabólica.
Outros autores também reforçam que a utilização deste tipo dieta em animais
mostra-se eficiente para o estudo da fisiopatologia das complicações
associadas à obesidade, como é o caso das doenças cardiovasculares e, mais
especificamente, da função endotelial, visto que é o modelo mais próximo da
gênese da obesidade em humanos (ROSINI; SILVA; MORAES, 2012).
Autores demonstram que essa dieta, induz o desenvolvimento de
distúrbios metabólicos envolvidos na gênese da síndrome metabólica, como
alteração do metabolismo da glicose, hiperglicemia, adiposidade, DHGNA,
alterações do perfil lipídico e resistência à insulina (EGUCHI et al., 2008;
CASTELL-AUVI et al., 2012; ANDRADE et al., 2013; BALLESTRERI;

40
MARCON; TAVARES1, 2015) não só por intermédio de desequilíbrios entre
a distribuição dos macro nutrientes, como também, devido a qualidade do
carboidrato consumido, não apenas pelo seu valor energético a ele atribuído.
Ressalva-se o papel do índice glicêmico (IG), ou seja, a capacidade que o
carboidrato contido em um alimento tem em aumentar a glicemia, o que
parece exercer maior influência sobre o surgimento de doenças metabólicas
como a resistência à insulina, tanto em modelos experimentais como em
humanos (PAWLAK; KUSHNER; LUDWIG, 2004; SAMPEY et al., 2011).
A resistência à insulina pode ser desencadeada por consumo crônico de
dietas ricas em gorduras. Estas dietas elevam os ácidos graxos livres (AGL)
circulantes fornecidos pelo tecido adiposo na lipólise. No meio intracelular,
os AGL exercem ação inibitória na secreção de insulina pelas células b-
pancreáticas e efeitos sistêmicos sobre a sensibilidade à insulina na utilização
periférica da glicose, referidos como lipotoxicidade (GAUR et al., 2014).
Esta condição está associada a produção de espécies reativas de oxigênio
(ERO), a diminuição do sistema de defesa antioxidante endógeno e aumento
do estresse oxidativo, que resulta em hiperinsulinemia compensatória,
hiperglicemia e armazenamento de lipídeos no tecido adiposo (STYSKAL et
al., 2012).
A produção de radicais livres consiste em um processo contínuo e
fisiológico, associado a importantes funções metabólicas como a
transferência de elétrons em células aeróbicas, especialmente pela cadeia
transportadora de elétrons mitocondrial (LAMBETH; EISH, 2014). Porém, a
produção contínua de radicais livres precisa ser balanceada por mecanismos
de defesa antioxidante. Portanto, o estresse oxidativo resulta em um
desequilíbrio entre a formação de ERO e a competência antioxidante
endógena, como forma de combater o excedente de radicais livres. Os
radicais livres em excesso podem provocar oxidação de biomoléculas com
consequente perda de suas funções biológicas e/ou desequilíbrio
homeostático, o que causa um dano oxidativo potencial contra células e
tecidos (SABITHA et al., 2014). As enzimas antioxidantes endógenas,
associadas aos antioxidantes provenientes da dieta são capazes de minimizar
essa desordem metabólica e, consequentemente, o desenvolvimento de dano
celular (MIGLIO et al., 2014).
O acúmulo de lípideos tóxicos em tecidos não adiposos é associado a
resistência à insulina. Ao mesmo tempo, o aumento da demanda de insulina
promove uma compensação das células beta, que envolve aumento da função

41
do pâncreas e/ou aumento da massa celular. Assim, a resistência à insulina
pode levar a DM tipo 2, concomitante com a condução da falha das células
beta. Avaliando a função pancreática após 17 semanas de tratamento com
dieta de cafeteria, estudos demonstraram um estado pré-diabético like em
animais, com acúmulo ectópico de lipídeos no pâncreas, conteúdo de insulina
e expressão gênica elevadas, condição de hiperinsulinemia, e ainda, sinais
iniciais de apoptose no pâncres (CASTELL-AUVI et al., 2012).
Outro achado é a correlação dos níveis aumentados de triglicerídeos por
este tipo de dieta, com a redução da função endotelial. Rosini, Silva e Moraes
(2012) apresentam em sua revisão que, em modelo de obesidade, os estudos
mostram que o consumo de dieta enriquecida de gorduras e sacarose por três
dias e 12 semanas provoca diminuição tempo-dependente da resposta a
agentes vasodilatadores com ação mediada pelo endotélio (carbacol) ou ação
direta sobre o músculo liso vascular (nitroprussiato de sódio) em artéria
mesentérica de ratos Wistar, mostrando que reduções na função endotelial
podem estar mais relacionadas com os níveis aumentados de triglicerídeos.
Todos estes achados comprovam os efeitos sistêmicos deletérios das
dietas de cafeteria, reforçando a importância de estudar mecanismos
envolvidos na síndrome metabólica. A figura 1 ilustra as principais
alterações metabólicas induzidas pela dieta de cafeteria.

42
Figura 1. Principais alterações metabólicas induzidas pela dieta de cafeteria.
Fonte: o autor.
As frutas e os vegetais, fazem parte do grupo de alimentos mais
importantes no controle do peso e prevenção de doenças. O consumo
adequado deste grupo de alimentos, tem sido associado com risco diminuído
para o acometimento por doenças crônicas, incluindo a síndrome metabólica.
Porém, o padrão dietético atual da população, é caracterizado pelo consumo
insuficiente de frutas, verduras e legumes. A OMS estima que
aproximadamente 2,7 milhões de mortes por ano em todo mundo podem ser
atribuídas a esse consumo inadequado, estando entre os dez principais fatores
de risco para a carga total global de doença em todo o mundo. A Organização
das Nações Unidas sobre Alimentos e Agricultura (Food and Agriculture
Organization of the United Nations - FAO) e a OMS recomendam um
consumo mínimo de frutas, verduras e legumes de 400 g/dia ou o equivalente
a cinco porções desses alimentos (WHO, 2011), ressaltando a importância da
inclusão deste grupo de alimentos a alimentação diária, bem como de
produtos à base destes alimentos.

43
3.3 Maracujá amarelo (P. edulis Var. flavicarpa)
Maracujá é uma denominação geral dada ao fruto e à planta de várias
espécies do gênero Passiflora. O nome maracujá é de origem tupi-guarani e
significa ”alimento que se toma de sorvo” ou “alimento em forma de cuia”,
uma referência a sua forma após cortado transversalmente. As primeiras
referências às plantas do gênero Passiflora foram feitas no século 16,
incluindo sua citação na obra Tratado descritivo do Brasil, de 1587, na qual
o português Gabriel Soares de Sousa fez referência ao maracujá, como uma
planta exótica com múltiplas potencialidades alimentares, ornamentais e
medicinais (EMBRAPA, 2016).
Apesar de pouco usual no Brasil, outra designação é "fruto-da-paixão"
que tem origem na correlação da morfologia da sua flor chamada "flor-da-
paixão", fazendo alusão aos símbolos da Paixão de Cristo. Em 1610, o
historiador italiano Giacomo Bosio associou as estruturas das flores, da
gavinha e das folhas do maracujazeiro com vários elementos da paixão
(crucificação) de Jesus Cristo, o que foi publicado na obra La trionfante e
gloriosa croce, ou a triunfante e gloriosa cruz. Entre tais associações, os três
pistilos representam a Santíssima Trindade (Pai, Filho e Espírito Santo), os
três estiletes representam os cravos usados na crucificação, as cinco anteras
representam as cinco chagas de Cristo; os filamentos da corona representam
a coroa de espinhos; as gavinhas os chicotes, e as folhas, as lanças dos
soldados que açoitaram Jesus Cristo. Essa associação da Paixão de Cristo às
estruturas das flores do maracujá deu origem ao nome do gênero Passiflora,
vindo do latim passio (Paixão) e floris (flor) de forma que o maracujá também
é conhecido como Flor da Paixão. Em inglês o fruto é conhecido como
“passion fruit”, ou fruto da paixão (EMBRAPA, 2016).
Segundo a mesma publicação da Empresa Brasileira de Pesquisa
Agropecuária (2016), as espécies de maracujazeiro são plantas trepadeiras
herbáceas ou lenhosas, podendo atingir de 5 m a 10 m de comprimento,
conforme demonstra a figura 2. Dependendo da espécie, as folhas, flores e
frutos apresentam vários formatos, cores e tamanhos. Estima-se que o gênero
Passiflora é composto por mais de 500 espécies diferentes, das quais mais de
150 são nativas do Brasil, considerado um dos maiores centros de
diversidade. A cada ano, novas espécies de maracujá têm sido identificadas e
descritas no País (EMBRAPA, 2016). As espécies desse gênero são bem
distribuídas em clima temperado, quente e regiões tropicais. O Instituto

44
Brasileiro de Geografia e Estatística (IBGE) (2012) apresenta dados que
demonstram que a produção anual de maracujá equivale a 923 mil toneladas,
com destaque para as regiões produtoras nordeste e sudeste, sendo o Brasil o
maior produtor mundial de maracujá, e também o maior consumidor,
chegando a atingir aproximadamente 1 milhão de toneladas por ano, sendo
uma das principais frutas do consumo rotineiro da população brasileira.
Figura 2. Plantação de Maracujá.
Fonte: Banco de Imagens da Embrapa, 2015 (https://www.embrapa.br/busca-de-
imagens/-/midia/2572001/plantacao-de-maracuja).
O maracujá do tipo Passiflora edulis flavicarpa, conhecido como
maracujá azedo ou amarelo, é formado de pedúnculo, epicarpo (a casca
amarela mais exterior), mesocarpo (a parte branca da casca também
denominada albedo), endocarpo ou polpa (que contém o arilo carnoso) e as
sementes (NASCIMENTO et al., 2013) representado na figura 3. É o mais
produzido e comercializado representando 95% dos pomares, valorizado
quanto a produção devido ao seu melhor rendimento. Seu cultivo está
basicamente voltado para a indústria de sucos e polpas (ZERAIK et al., 2010;
NASCIMENTO et al., 2013).

45
Figura 3. Partes do maracujá.
Fonte: Adaptada de Banco de Imagens da Embrapa, 2016
(https://www.embrapa.br/busca-de-imagens/-/midia/2823002/maracuja-brs-sol-do-
cerrado-brs-sc1)
As propriedades medicinais e funcionais do fruto maracujá são
conhecidas mundialmente. É uma planta bastante estudada, tendo seus efeitos
elucidados na literatura (EMBRAPA, 2016). A cor amarela intensa do suco
de maracujá deve-se aos pigmentos carotenóides. Mercadante, Britton e
Rodriguez-Amaya (1998) identificaram por meio de técnicas
espectroscópicas (EM, UV-Vis, RMN 1H e 13C) e cromatográficas (CCD e
CLAE-cromatografia líquida de alta eficiência) treze carotenoides do
maracujá amarelo: ζ-caroteno (principal carotenoide), fitoeno, fitoflueno,
neurosporeno, β-caroteno, licopeno, pró-licopeno, monoepóxi-β-caroteno, β-

46
criptoxantina, β-citraurina, anteraxantina, violaxantina e neoxantina. O
principal papel dos carotenoides na dieta humana é de serem precursores de
vitamina A. Poucos carotenoides possuem esta atividade vitamínica, que é
atribuída à estrutura retinóide (com anel β-ionona). O β-caroteno é o que
possui maior atividade como provitamina A, podendo atuar como
antioxidantes na prevenção do câncer, catarata, arteriosclerose e processos de
envelhecimento em geral (ZERAIK et al., 2010).
Figura 4. Estrutura química do ζ-caroteno (a), β-caroteno (b), pró-licopeno
(c), β-criptoxantina (d), violaxantina (e) presentes no maracujá amarelo.
Fonte: Wondracek e colaboradores, 2012.
Já os flavonoides encontrados nos frutos das espécies de Passiflora,
são principalmente do tipo C-glicosídeos. Os flavonoides apresentam vários
efeitos biológicos e farmacológicos, incluindo atividade antibacteriana,
antiviral, anti-inflamatória, antialérgica e vasodilatadora. Além disso, estas
substâncias inibem a peroxidação lipídica e reduzem o risco de doenças

47
cardiovasculares, efeitos estes relacionados à sua atividade antioxidante,
caracterizada pela capacidade de sequestrar radicais livres em organismos
vivos (ZERAIK et al., 2010).
São atribuídos efeitos sedativos, antiespasmódico e ansiolítico às folhas
do maracujá, confirmados em experimentos em animais (DHAWAN;
DHAWAN; SHARMA, 2004). Já a polpa apresenta em sua composição
elevado teor de carotenoides (15,36 a 27,14 mg/g), compostos como a
provitamina A, ácido ascórbico, assim como flavonoides e alcaloides,
responsáveis por uma importante atividade antioxidante (SILVA;
MERCADANTE, 2002; DHAWAN; DHAWAN; SHARMA, 2004;
CAZARIN et al., 2014). E as sementes são ricas em ácidos poliinsaturados
como os ω-3 e ω-6 (ZERAIK et al., 2010).
Na casca do fruto encontram-se cianidina-3-O-glicosídeo,
quercetina-3-O-glicosídeo e ácido edúlico, além de pectina, a qual
corresponde a 19,1% de sua composição (YAPO; KOFFI, 2006). Apesar de
apresentar ações farmacológicas e de interesse nutricional, ainda são poucos
os estudos existentes com enfoque sobre os constituintes químicos das
diversas partes dos frutos do maracujá. Destaca-se que, a casca e semente
do maracujá, subprodutos da indústria de alimentos, sendo que mais de 75%
deste resíduo poderia ser transformado em ingredientes com propriedades
bioativas para promoção de saúde (MARTINEZ et al., 2012; CAZARIN et
al., 2014).
3.3.1 Farinha da Casca do Maracujá amarelo (P. edulis Var. flavicarpa)
Cerca de 52% da composição mássica do fruto maracujá é constituída
pela casca (CAZARIN et. al., 2014). Nas indústrias alimentícias, menos de
30% do peso do maracujá é aproveitado, sendo descartados cerca de 60 à 70%
o que constitui a maior parte do peso do fruto. O destino dos resíduos vegetais
representa um crescente problema, devido ao aumento da produção a cada
ano (MELETTI, 2011) tornando-se tema de pesquisa a busca do uso desses
resíduos no desenvolvimento de produtos de maior valor agregado como, por
exemplo, farinhas com alto teor de fibras (NASCIMENTO et al., 2013).
As fibras alimentares são definidas como a porção de hidrato de carbono
comestível das plantas, resistentes à digestão enzimática, são constituídas por
fibras solúveis (pectinas) e insolúveis (lignina). Como um nutriente essencial,

48
as fibras têm apresentado um papel crucial na manutenção da saúde, com
ações de prevenção e proteção para várias doenças humanas (MATTOS;
MARTINS, 2000; BERNAUD; RODRIGUES, 2013; SLAVIN, 2013;
ZHANG et al., 2016).
Segundo recomendações da OMS, o consumo de fibras acima de 25
gramas ao dia está associado à prevenção de doenças crônicas. Já a American
Dietetic Association (ADA) indica um consumo diário de 14 g de fibras por
1000 kcal, ou 25 g para mulheres e 38 g para homens adultos. As Dietary
Reference Intakes (DRIs) recomendam um consumo médio de 30 g/dia de
fibra total para adultos sadios (INSTITUTE OF MEDICINE, FOOD AND
NUTRITION BOARD, 2002).
O consumo humano das fibras dietéticas promove diversos efeitos
fisiológicos benéficos, dentre os quais pode-se citar o retardo no
esvaziamento gástrico e, por conseguinte, reduzir a velocidade de absorção
dos carboidratos e liberação dos nutrientes para a circulação, reduzindo o pico
hiperglicêmico pós-prandial imediato (SANTOS, 2013), auxiliando na
prevenção e tratamento do DM e síndrome metabólica.
Outro efeito da ingestão de fibras é o aumento do volume e da
viscosidade fecal, estimulando os movimentos peristálticos do intestino, e,
consequentemente, melhorando o trânsito intestinal, o que diminui o tempo
de contato de substâncias potencialmente carcinogênicas com a mucosa
(CAZARIN et al., 2014).
A metabolização ou fermentação das fibras pelo microbioma no
intestino grosso geram como produtos finais os ácidos graxos de cadeia curta,
que promovem diversos efeitos benéficos ao indivíduo. O efeito das fibras
nas doenças metabólicas e inflamatórias, parecem ter associação com os
ácidos graxos de cadeia curta, particularmente o propionatato e butirato.
Estudos sugerem que os ácidos graxos de cadeia curta reduzem a inflamação
de baixo grau observada na obesidade (GALISTEO et al., 2008; TOPPING,
2001).
A casca do maracujá é rica em fibras, sendo composta pelo flavedo, que
corresponde à camada externa de coloração verde a amarela, rica em fibras
insolúveis e o albedo, que corresponde à camada interna branca, que é rica
em fibra solúvel, em especial a pectina, com pequenas quantidades de
mucilagens (JANEBRO et al., 2008).
Ademais, a casca do maracujá é também rica em niacina (vitamina B3),
ferro, cálcio e fósforo (Tabela 2) (GONDIM et al., 2005).
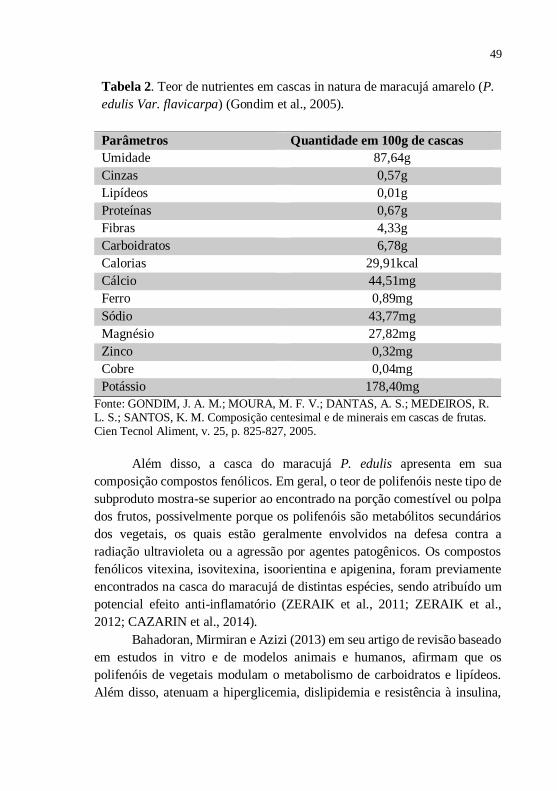
49
Tabela 2. Teor de nutrientes em cascas in natura de maracujá amarelo (P.
edulis Var. flavicarpa) (Gondim et al., 2005).
Parâmetros Quantidade em 100g de cascas
Umidade 87,64g
Cinzas 0,57g
Lipídeos 0,01g
Proteínas 0,67g
Fibras 4,33g
Carboidratos 6,78g
Calorias 29,91kcal
Cálcio 44,51mg
Ferro 0,89mg
Sódio 43,77mg
Magnésio 27,82mg
Zinco 0,32mg
Cobre 0,04mg
Potássio 178,40mg
Fonte: GONDIM, J. A. M.; MOURA, M. F. V.; DANTAS, A. S.; MEDEIROS, R. L. S.; SANTOS, K. M. Composição centesimal e de minerais em cascas de frutas. Cien Tecnol Aliment, v. 25, p. 825-827, 2005.
Além disso, a casca do maracujá P. edulis apresenta em sua
composição compostos fenólicos. Em geral, o teor de polifenóis neste tipo de
subproduto mostra-se superior ao encontrado na porção comestível ou polpa
dos frutos, possivelmente porque os polifenóis são metabólitos secundários
dos vegetais, os quais estão geralmente envolvidos na defesa contra a
radiação ultravioleta ou a agressão por agentes patogênicos. Os compostos
fenólicos vitexina, isovitexina, isoorientina e apigenina, foram previamente
encontrados na casca do maracujá de distintas espécies, sendo atribuído um
potencial efeito anti-inflamatório (ZERAIK et al., 2011; ZERAIK et al.,
2012; CAZARIN et al., 2014).
Bahadoran, Mirmiran e Azizi (2013) em seu artigo de revisão baseado
em estudos in vitro e de modelos animais e humanos, afirmam que os
polifenóis de vegetais modulam o metabolismo de carboidratos e lipídeos.
Além disso, atenuam a hiperglicemia, dislipidemia e resistência à insulina,

50
melhoram o metabolismo do tecido adiposo, aliviam vias de sinalização
sensíveis ao estresse oxidativo e processo inflamatórios.
As fibras alimentares presentes na casca do maracujá (P. edulis Var.
flavicapa), representam 19,1% de sua composição, formam géis em meio
aquoso cuja consistência viscosa altera o tempo de esvaziamento gástrico,
aumentam a saciedade e retardam o tempo de absorção de carboidratos
simples. Este gel é capaz de formar complexos com os sais biliares
aumentando a excreção de colesterol, podendo a farinha ser usada no
tratamento dietoterápico de diversas doenças (YAPO; KOFFI, 2006; GOSS
et al., 2018).
Para analisar o efeito da farinha na redução do colesterol, pesquisadores
avaliaram o efeito da suplementação de 30g da farinha da casca de maracujá
por 60 dias por 19 mulheres hipercolesterolêmicas. Após o período de
intervenção, foi observada uma redução estatisticamente significante nos
níveis colesterol total e colesterol LDL, porém não houve alteração dos
valores de colesterol HDL (RAMOS et al., 2007).
Janebro et al. (2008) realizaram um estudo para avaliar o efeito do
consumo regular de farinha da casca de maracujá amarelo sobre a glicemia
em jejum de indivíduos saudáveis e com diabetes. Após 60 dias de
suplementação da dieta desses indivíduos com 30g da farinha da casca de
maracujá, obtida do albedo e do flavedo (casca), a qual correspondia a 17,4g
de fibras totais, sendo 6,3 g de fibras solúveis e 11,1g de fibra insolúvel, para
ser ingerida ao longo do dia juntamente com os alimentos, demonstrou uma
diminuição significativa dos valores de glicemia do grupo teste em relação
ao grupo controle, após a suplementação.
Corroborando, Queiroz e colaboradores (2012) também avaliaram o
efeito da suplementação de 30g de farinha da casca do maracujá em 43
indivíduos diabéticos por 60 dias, e evidenciou-se que o consumo desta
farinha pode ajudar no tratamento coadjuvante do controle glicêmico, com
diminuição significativa na glicemia de jejum, hemoglobina glicada, índice
HOMA IR e aumento no índice HOMA.
Embora os estudos apresentem resultados promissores, os
pesquisadores referem que, em decorrência ao forte sabor residual e
distúrbios gastrointestinais provocados pelo uso da farinha, alguns
voluntários tendem a interromper sua participação nas pesquisas (RAMOS et
al., 2007; COQUEIRO; PEREIRA; GALANTE, 2016).

51
O aproveitamento de resíduos vegetais como fonte de fibras alimentares
por meio de sua incorporação como ingrediente em produtos alimentícios tem
grande potencial de viabilidade do ponto de vista nutricional, econômico e
também ecológico. Porém, o produto final deve proporcionar segurança
toxicológica para o consumidor. Apesar de produtos à base de plantas sejam
geralmente considerados seguros, sua toxicidade não deve ser descartada
(NASCIMENTO et al., 2013). A casca do maracujá possui compostos
considerados altamente tóxicos, que são os glicosídeos cianogênicos,
presentes em quantidades significativas em todas as partes da P. edulis, como
no albedo e polpa do fruto (MEDEIROS et al., 2009; MELETTI, 2011).
O ácido cianídrico é um líquido volátil e incolor, considerado uma das
substâncias mais tóxicas existentes. Normalmente está associado a um
aldeído ou uma cetona e a um ou mais açucares, formando compostos
denominados glicosídeos cianogênicos. Embora seja considerado um sistema
próprio de defesa das plantas, quando ocorre rompimento do tecido vegetal,
há um desdobramento de reações enzimáticas entre os glicosídeos até o ácido
cianídrico. Na corrente sanguínea, este ácido libera íon cianeto, o qual se liga
ao citocromo mitocondrial da hemoglobina. Esse processo é altamente tóxico
e acarreta em asfixia celular que, dependendo do teor de cianeto no sangue,
pode levar à morte (NASCIMENTO et al., 2013).
Medeiros e colaboradores (2009) avaliaram a segurança do uso da
farinha do maracujá-amarelo (P. edulis var. flavicarpa) como complemento
alimentar, através de ensaios toxicológicos clínicos de fase 1 em 36
voluntários saudáveis. O protocolo consistiu na administração de 10g da
farinha três vezes ao dia, totalizando 30g ao dia, durante 8 semanas. Os
resultados da pesquisa não evidenciaram sinais de toxicidade nos diversos
órgãos e sistemas avaliados, assim, os autores concluíram que a
administração, na dose preconizada, mostrou-se segura, sem apresentar
alterações que pudessem comprometer seu uso como alimento com
propriedade de saúde. Entretanto, não é possível afirmar a ausência de efeitos
tóxicos com a administração crônica, uma vez que o estudo apresentou
duração de somente oito semanas (COQUEIRO; PEREIRA; GALANTE,
2016).
Alguns estudos quantificaram o teor de cianogênicos na casca do
maracujá, conforme demonstra a tabela 3.
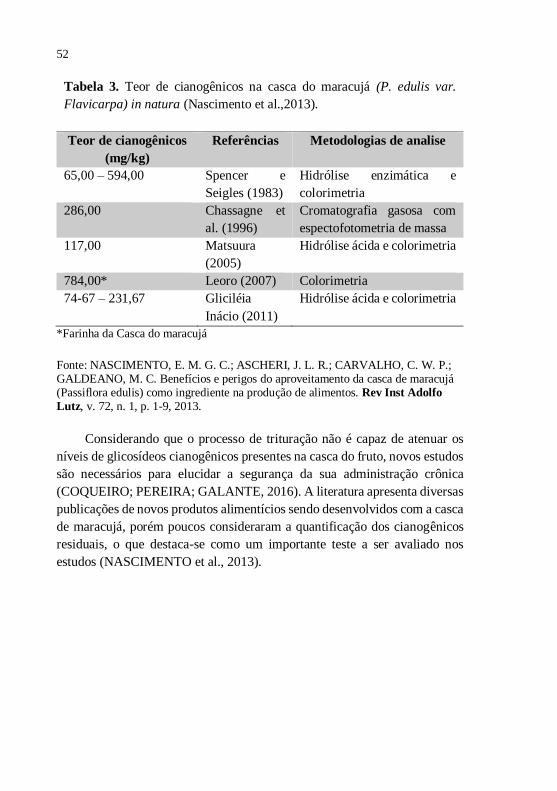
52
Tabela 3. Teor de cianogênicos na casca do maracujá (P. edulis var.
Flavicarpa) in natura (Nascimento et al.,2013).
Teor de cianogênicos
(mg/kg)
Referências Metodologias de analise
65,00 – 594,00 Spencer e
Seigles (1983)
Hidrólise enzimática e
colorimetria
286,00 Chassagne et
al. (1996)
Cromatografia gasosa com
espectofotometria de massa
117,00 Matsuura
(2005)
Hidrólise ácida e colorimetria
784,00* Leoro (2007) Colorimetria
74-67 – 231,67 Gliciléia
Inácio (2011)
Hidrólise ácida e colorimetria
*Farinha da Casca do maracujá
Fonte: NASCIMENTO, E. M. G. C.; ASCHERI, J. L. R.; CARVALHO, C. W. P.; GALDEANO, M. C. Benefícios e perigos do aproveitamento da casca de maracujá (Passiflora edulis) como ingrediente na produção de alimentos. Rev Inst Adolfo
Lutz, v. 72, n. 1, p. 1-9, 2013.
Considerando que o processo de trituração não é capaz de atenuar os
níveis de glicosídeos cianogênicos presentes na casca do fruto, novos estudos
são necessários para elucidar a segurança da sua administração crônica
(COQUEIRO; PEREIRA; GALANTE, 2016). A literatura apresenta diversas
publicações de novos produtos alimentícios sendo desenvolvidos com a casca
de maracujá, porém poucos consideraram a quantificação dos cianogênicos
residuais, o que destaca-se como um importante teste a ser avaliado nos
estudos (NASCIMENTO et al., 2013).

53
4 MATERIAL E MÉTODOS
4.1 Farinha da casca de maracujá (FCM)
4.1.1 Obtenção do produto
A FCM (P. edulis Var. flavicapa) industrializada e embalada, foi
adquirida no comércio local da cidade de Balneário Camboriú – Santa
Catarina no mês de abril de 2017, e armazenada sob refrigeração à 4 ºC. Os
dados nutricionais encontram-se na tabela 4.
Tabela 4. Informação nutricional da FCM (P. edulis Var. flavicapa)
empregada no presente estudo. Valores referente a 1 porção do produto (50g
= 7 colheres de sopa).
Quantidade por porção %VD*
Valor energético 67kcal = 281kJ 3%
Carboidratos 13g 4%
Proteínas 3g 4%
Gorduras totais 0g 0%
Gorduras saturadas 0g 0%
Gorduras trans 0g **
Fibra alimentar 29g 117%
Sódio 41mg 2%
*% Valores Diários de referência com base em uma dieta de 2.000kcal ou
8.400kJ.
** Valor Diário não estabelecido.
Fonte: o autor.
4.2 Ensaios in vitro
4.2.1 Obtenção do extrato de FCM
O material vegetal foi submetido a extração em solvente
hidroalcoólico. Este material foi pesado e depositado em recipiente de vidro
com adição de solução hidroalcoólica a 70% na proporção de 1:3 do pó, como
líquido extrator. A mistura resultante permaneceu em maceração estática por

54
7 dias, passando pelo processo de filtração para obtenção do extrato bruto. O
extrato bruto foi concentrado em evaporador rotativo sob pressão reduzida,
com temperatura entre 55 ºC e 60 ºC, para eliminação total do solvente
(CECHINEL-FILHO; YUNES, 1998).
4.2.1.1 Avaliação do potencial antioxidante da FCM in vitro
Para a determinação da atividade antioxidante da FCM, foram
realizados os métodos FRAP, sistema de cooxidação β-caroteno/ácido
linoleico e atividade antioxidante por eliminação do Óxido Nítrico a partir do
extrato obtido.
4.2.1.2 Método FRAP
A determinação do poder antioxidante de redução do ferro (FRAP),
do inglês (ferric reducing antioxidant power), a qual consiste da capacidade
dos fenóis em reduzir o Fe3+ em Fe2+, foi primeiramente descrita por Benzie
e Strain (1996) e modificada por Pulido e colaboradores (2000). Em triplicata,
9 μL do extrato bruto da FCM ou solução Trolox, foram adicionados a 27 μL
de água destilada e 270 μL de solução FRAP em uma placa de 96 poços. A
solução FRAP foi preparada no momento do seu uso, a qual consiste em 10
mL de Tampão Acetato pH 3.6 0,3 mM, 1,0 mL de TPTZ 10mM em HCl 40
mM e 1,0 mL de Cloreto Férrico 20 mM. As amostras foram incubadas à
37°C, por 30 minutos e a leitura realizada a 595 nm, utilizando
espectrofotômetro de microplacas Sinergy (Biotek). O sulfato ferroso
(FeSO4) foi utilizado como padrão, e utilizado para fins de cálculo do poder
de redução do extrato. Os resultados foram expressos em µM de equivalentes
de sulfato ferroso produzido.
4.2.1.3 Sistema de cooxidação do β-caroteno/ácido linoléico
A capacidade antioxidante do extrato foi avaliada em sistema modelo
de cooxidação de substratos: β-caroteno/ácido linoleico por radicais de
peróxido. O método foi originalmente descrito por Marco (1968), e
posteriormente modificado por Miller (1971). Em triplicata, 10 μL do extrato
bruto de FCM, água ou solução Trolox (padrão) foram adicionados em uma
placa de 96 poços. Paralelamente, foram dissolvidos 2,0 mg de β-caroteno

55
em 1,0 mL de clorofórmio. Para o preparo da solução reativa, foram
homogeneizados em um Erlenmeyer 150 µL desta mistura resultante, 50 µL
de ácido linoléico, 200 µL de Tween 80 e 500 µL de clorofórmio. Logo após,
a mistura foi submetida à completa evaporação do clorofórmio sob
nitrogênio. Adicionou-se a esta mistura isenta de clorofórmio, cerca de 25
mL de água destilada previamente saturada com oxigênio e agitou-se
vigorosamente durante 30 min. Imediatamente, foram transferidos 240 µL da
solução reativa para a placa, para mensuração da absorbância do tempo zero
em 470 nm, utilizando espectrofotômetro de microplacas Sinergy (Biotek). O
sistema de emulsão foi incubado a 50 ºC por 2 horas para acelerar as reações
de oxidação e iniciar o descoramento do β-caroteno, sendo a leitura da
absorbância final realizada após 120 min. Todas as determinações foram
acompanhadas de um controle sem antioxidante. Os resultados foram
expressos em percentual de proteção da oxidação. O decaimento da densidade
ótica do controle (Absinicial – Absfinal) foi considerado como 100% de oxidação
(MILLER, 1971; MOREIRA; MANCINI-FILHO, 2004). A partir desta
relação, o decréscimo na leitura da absorbância das amostras foi
correlacionado com o controle, estabelecendo desta forma, o percentual de
proteção da oxidação das amostras pela equação:
% de proteção da oxidação = 100 – [(Abs amostra x 100)/ Abs controle]
4.2.1.4 Capacidade antioxidante por eliminação do Óxido Nítrico (NO)
Seguindo o método de avaliação descrito por Griess (1879),
modificado por Basu e Hazra (2006), este consiste em determinar a ação
sequestradora do íon NO2-, por meio dos nitritos que são quantificados
espectrofotometricamente pela reação com o Reagente de Griess
(sulfanilamida à 2% + cloridrato de N-(naftil)etilenodiamina, à 0,2%). Em
uma microplaca de 96 poços, o meio reacional consistiu em 50 μL do extrato
de FCM ou solução Trolox, adicionados de 50 μL de nitroprussiato de sódio
20 mM em tampão fosfato (pH 7,4) e incubados a 37°C por 1 hora.
Posteriormente, 100 μL do reagente de Griess foi acrescido e a absorbância
mensurada em espectrofotômetro a 540 nm. Uma curva padrão de nitrito de
sódio foi construída, e os resultados foram expressos como μmol de nitrito
produzido/L.

56
4.3 Ensaios in vivo
4.3.1 Animais
A amostra foi composta de 30 camundongos da espécie Black,
linhagem C57BL/6, machos, adultos jovens com idade entre 2 a 3 meses, com
peso médio de 25 a 35 gramas, provenientes do Biotério da Universidade do
Vale do Itajaí. Os animais foram mantidos em condições normais de biotério,
com ciclo de luz, temperatura e umidade controlados, bem como acesso a
alimento e água ad libitum. Todos os procedimentos foram realizados de
acordo com os princípios éticos de experimentação animal recomendados
pelo Conselho Nacional de Controle de Experimentação Animal (CONCEA),
e de acordo com a Lei nº 11.794 de 2008 da Constituição Federal. No que
tange a questões éticas, o projeto foi submetido para apreciação do Comitê
de Ética no Uso de Animais (CEUA) da Universidade do Vale do Itajaí, e
aprovado sob o parecer nº016/17 (Anexo I).
4.3.2 Tratamento e parâmetros murinométricos
Para o ensaio biológico, com duração de 16 semanas (112 dias), os
animais foram distribuídos em blocos ao acaso, em três grupos de 10 animais,
divididos de acordo com a dieta e tratamento ofertado: grupo dieta controle
(C); grupo dieta de cafeteria (CAF); grupo dieta de cafeteria acrescida de 15%
de FCM. Durante o período, foi realizado o controle semanal do consumo
alimentar, hídrico e peso corporal dos animais (Figura 4).

57
Figura 5. Subdivisão dos grupos experimentais de acordo com o tratamento
dietético e cronograma de realização do experimento.
Fonte: o autor.
4.3.3 Elaboração das dietas experimentais
A dieta adotada na pesquisa para o grupo controle (C), seguiu as
recomendações do Instituto Americano de Nutrição para uma dieta de
manutenção de ratos adultos (AIN-93M) (REEVES; NIELSEN; FAHEY,
1993). Para elaboração da dieta de cafeteria, ofertada para os grupos tratados
(CAF e FCM) foram associados à dieta AIN-93 alimentos hipercalóricos,
ricos em carboidratos e lipídeos, sendo: amendoim moído, chocolate em pó e
biscoito de maisena triturado, nas proporções de 3:2:2:1, previamente
padronizada por Estadella et al. (2004) e analisada por Oliveira et al. (2011).
Destaca-se que, ao grupo tratado, foram acrescidos 15% da FCM à dieta de
cafeteria.
No preparo das dietas experimentais, os ingredientes foram pesados
em balança (Gehaka®) com capacidade de 4 kg e sensibilidade de 0,001g.
Após a pesagem dos ingredientes de cada dieta, os mesmos foram

58
homogeneizados, e a mistura umedecida com água destilada para modelagem
manual. As diferentes dietas experimentais foram mantidas em estufa com
circulação de ar a 50 ºC por 48 horas para secagem, e permaneceram
armazenadas em recipientes plásticos, herméticos, sob refrigeração (4 ºC)
durante todo o período experimental.
4.3.4 Consumo energético
Com base na informação nutricional fornecida pelos fabricantes dos
produtos (amendoim, chocolate, biscoito maizena e FCM), foram calculados
o valor energético por grama de dieta, e ao final do experimento foi calculada
a média de calorias consumidas por animal. Adotou-se os valores de 4
(quatro) calorias por grama de carboidratos e proteínas e 9 (nove) calorias por
grama de lipídeos.
4.3.5 Eficiência Alimentar
A eficiência alimentar foi avaliada segundo Nery et al. (2011). A
capacidade do animal em converter energia alimentar consumida em peso
corporal, se dá através do Coeficiente de Ganho de Peso por Consumo
Calórico (CGPCC). Durante o experimento os animais foram pesados e
mensurada a ingesta calórica para cada grupo. O CGPCC é a relação entre o
ganho de peso por quantidade de calorias do alimento ingerido.
𝐶𝐺𝑃𝐶𝐶 = (𝑃𝐹 − 𝑃𝐼)
𝐾𝑐𝑎𝑙
PF: peso final em gramas do animal durante o período de acompanhamento
PI: peso corporal do animal no início do experimento em gramas
Kcal: valor calórico da dieta ingerida.

59
4.3.6 Curva de tolerância oral a glicose e teste de resistência à insulina
Após o período de intervenção, os animais foram submetidos aos testes
para determinação da tolerância oral a glicose e resistência à insulina,
segundo as recomendações de Wang e Liao (2012). Os animais foram
submetidos a um período de jejum de seis horas para ambos experimentos,
entre as 07:00 e 13:00 horas, uma vez que murinos são ativos no ciclo escuro,
sendo o horário escolhido mais fisiológico quando comparado comparados a
humanos.
Para realização da curva de tolerância oral a glicose, após o período de
jejum, foi mensurada a glicemia capilar basal (tempo 0) através de uma
pequena incisura na cauda para coleta do sangue, sendo imediatamente após
administrado glicose por via oral (2 g/kg), e, mensurada a glicemia nos
tempos 30, 60, 90 e 120 minutos. Os níveis de glicose foram mensurados com
glicosímetro (ACCU-CHEK Active) em cada tempo, sendo os resultados
apresentados em forma de curva.
Para o teste de resistência à insulina em outro momento, o mesmo
protocolo de jejum foi adotado, e em seguida, foi mensurada a glicemia
capilar basal (tempo 0), e administrou-se por via intraperitoneal insulina
humana na concentração de 2 U/kg. Posteriormente, a glicemia capilar foi
mensurada nos tempos 30, 60, 90 e 120 minutos, sendo os resultados
apresentados em forma de curva.
4.3.7 Eutanásia e coleta do material biológico
Ao final das 16 semanas, os animais receberam anestésicos via
intraperitoneal para completa sedação antes do procedimento de eutanásia,
sendo administrados uma associação de 80 mg/kg de quetamina e 8 mg/kg de
xilazina. Foi realizada a coleta de sangue circulante através da veia braquial,
e posteriormente, foi realizado o deslocamento cervical. Para retirada dos
órgãos, a cavidade abdominal foi aberta, seguida da dissecação do tecido
adiposo das regiões mesentérica, retroperitoneal, e epididimal. A soma do
peso das três gorduras forneceu o total de gordura visceral. Fígado, pâncreas
e rins também foram coletados para análise morfológica e realização de
ensaios in vitro.

60
4.3.8 Determinação do peso relativo dos órgãos e tecidos
Os órgãos e tecidos foram imediatamente pesados após removidos, em
balança (Gehaka®) com precisão de 0,005 gramas. Os valores foram
expressos em percentual obtidos através do seguinte cálculo:
𝑷𝒆𝒔𝒐 𝒓𝒆𝒍𝒂𝒕𝒊𝒗𝒐 (%) = 𝑃𝑒𝑠𝑜 𝑑𝑜 𝑡𝑒𝑐𝑖𝑑𝑜 𝑜𝑢 ó𝑟𝑔ã𝑜 𝑋 100
𝑃𝑒𝑠𝑜 𝑓𝑖𝑛𝑎𝑙 𝑑𝑜 𝑎𝑛𝑖𝑚𝑎𝑙
4.3.9 Parâmetros bioquímicos no soro
O sangue coletado dos animais submetidos as diferentes intervenções,
foi utilizado para determinação da glicose, perfil lipídico: colesterol total,
HDL colesterol, LDL colesterol, triglicerídeos; creatinina; enzimas
hepáticas: fosfatase alcalina, aspartato aminotransferase (AST), alanina
aminotransferase (ALT), os quais foram avaliados por meio de kits
enzimáticos específicos. A insulina foi dosada por meio de
eletroquimioluminescência.
4.3.10 Avaliação histológica
Amostras biológicas de fígado, tecido adiposo, rins e pâncreas dos
animais, foram fixados em solução de formol, onde ficaram imersos por 24
horas, e após este período, imergidos em uma solução de álcool a 70%.
Posteriormente, os órgãos foram emblocados em parafina e cortados (7 μm
de espessura) em micrótomo de maneira semi-seriada. As lâminas obtidas
segundo esse processo foram submetidas à coloração por hematoxilina-
eosina para análise morfológica. Foram tomadas microfotografias dos cortes histológicos utilizando
microscópio óptico Olympus CBA®. Para análise do pâncreas foram
capturadas 15 imagens para cada grupo experimental utilizando-se objetiva
de 10x. Para análise dos rins, foram capturadas 20 imagens para cada grupo
experimental, utilizando-se uma objetiva de 10x. As análises das imagens
foram determinadas com auxílio do software Image J® que quantificou as
áreas das respectivas unidades funcionais de cada tecido (RASBAND, 2014).
Os resultados foram expressos em micrometros/campo. Para o tecido
adiposo, foram capturadas 10 imagens de cada grupo experimental

61
utilizando-se a objetiva de 10x, e as imagens analisadas com auxílio do
software Adiposoft®.
4.3.11 Determinação de lipídeos no fígado
Foi utilizado o método de extração proposto por FOLCH et al. (1957)
para extração da fração lipídica no fígado dos animais. Neste método, 100
mg de fígado foram triturados e homogeinizados em disruptor de tecidos
durante 3 minutos com 1900 μL de solução clorofórmio-metanol 2:1. Depois
deste procedimento, o material triturado foi adicionado de 400 μL de metanol.
Este material foi então centrifugado a 3.000 rpm, durante 10 minutos. O
sobrenadante foi transferido para outro tubo e adicionado de 800 μL de
clorofórmio e 640 μL de solução NaCl a 0,73%. Após este procedimento,
houve nova centrifugação a 3.000 rpm durante 10 minutos. Desta vez,
descartou-se o sobrenadante e o precipitado foi lavado por três vezes com a
solução de Folch (3% de clorofórmio, 48% de metanol, 47% de água, 2% de
NaCl 0,2%). Os extratos lipídicos foram secos em estufa a 37 ºC. O extrato
lipídico seco foi ressuspenso com 1 mL de isopropanol e procedeu-se a
determinação de colesterol total e triglicerídeos utilizando kit enzimático.
4.3.12 Marcadores de estresse oxidativo
Para averiguação do estresse oxidativo no tecido hepático, foram
empregadas metodologias: método de FRAP e quantificação da glutationa
reduzida (GSH), ambas descritas a seguir.
4.3.12.1 Preparo dos homogeneizados no tecido hepático
Fragmentos do fígado foram homogeneizados com tampão fosfato de
potássio 0,1 M pH 7,0, igual a três vezes o valor absoluto da massa da amostra
em homogeneizador rotativo de tecidos. O homogeneizado foi centrifugado
a 13.000 rotações por minuto à temperatura de 4 ºC por 30 minutos. Em
seguida, o sobrenadante foi coletado, obtendo-se a fração citosólica para a
determinação da atividade antioxidante do tecido: FRAP e GSH; além da
determinação das enzimas antioxidantes: SOD, CAT e GPx.

62
4.3.12.2 Atividade antioxidante total do tecido pelo método FRAP
Seguiu-se a metodologia descrita no sub item 4.2.1.2, com algumas
modificações. A reação se deu com 9 µL de cada homogeneizado de fígado,
27 µL de água destilada e 270 µL do reagente FRAP. Os resultados foram
expressos em µmol de Fe2SO4/mg de proteína.
4.3.12.3 Quantificação de glutationa reduzida (GSH)
O ensaio foi realizado de acordo com o protocolo descrito por Tietze
(1969). Os homogeinatos de fígado foram ressuspensos numa solução de
EDTA em TCA 5%. Posteriormente, foram centrifugados a 17000g, a 2 ºC
por 15 min, e coletou-se o sobrenadante. Em microplacas, foram adicionados
270 µL de tampão fosfato pH 8,0 e 20 µL do sobrenadante contendo GSH.
Em seguida, adicionou-se 10 µL de 5’5 ditiobis-22 ácido nitrobenzoico
10mM (DTNB), depois de 15 minutos a absorbância foi lida em
espectofotômetro a 412 nm. A concentração de GSH foi quantificada
utilizando curva padrão de GSH sintética e os resultados, expressos em µmol
de GSH/mg de tecido.
4.3.13 Determinação da atividade das enzimas antioxidantes
As atividades das enzimas antioxidantes foram determinadas
conforme as metodologias descritas nas próximas subseções.
4.3.13.1 Superóxido Dismutase (SOD)
A atividade da SOD citoplasmática foi avaliada de acordo com a
metodologia de McCord e Fridovich (1969), que verifica a produção dos
ânios peróxidos produzidos pela xantina oxidase em presença de xantina. O
volume da xantina oxidase utilizado na reação foi determinado em um
branco, na ausência de SOD, obtendo-se uma variação na absorbância, a 550
nm, entre 0,0250 e 0,0300/min. O meio de reação foi composto por citocromo
c 100 μM diluído em H2O mQ, xantina 500 μM, EDTA 1 mM e KCN 200
μM em tampão fosfato de potássio 0,05 M pH 7,8. Em duplicata, 250 μL do
meio de reação foi pipetado em uma placa de 96 poços, sendo adicionados
xantina oxidase (volume encontrado no branco) e 15 μL da fração citosólica

63
do tecido. Os resultados foram expressos em U/mg de proteína. Entendeu-se
por uma unidade (U) a atividade da enzima que que promoveu 50% de
inibição da reação da xantina a 25 ºC em pH 7,8.
4.3.13.2 Catalase (CAT)
A CAT propicia a oxidação do peróxido de hidrogênio a H2Oe O2. A
metodologia empregada foi descrita por Beutler (1975), a qual quantifica a
velocidade de decomposição do peróxido de hidrogênio pela enzima,
mediante o decréscimo da densidade óptica a 230 nm à 37 ºC. O meio de
reação foi composto de H2O2 10mM (10 μL de peridrol 30% em 10mL de
H2O mQ) e tampão Tris HCl 1M EDTA 5 mM pH8,0. Para a reação, foi
utilizada a diluição de 10 μL do homogeneizado adicionados a 2990 μL do
meio. Em duplicata, 250 μL do meio de reação foi pipetado em uma placa de
96 poços, sendo adicionado 10 μL das amostras (frações citosólicas), e
incubadas a 37 ºC para realizada a leitura da absorbância a cada minuto
durante 6 minutos. Os resultados foram expressos em U/mg de proteína.
Uma unidade (U) da catalase correspondeu a atividade da enzima que
realizou a hidrólise de 1 μmol de H2O2 por minuto a 37 ºC em pH 8,0.
4.3.13.3 Glutationa Peroxidase (GPx)
A atividade da GPx na fração citosólica foi determinada pela
metodologia padronizada por Sies (1979). Este método fundamenta-se na
aferição do decaimento da densidade óptica, a 340 nm, promovido pela
oxidação do NADPH a 30 ºC durante a redução da glutationa oxidada
(GSSG) catalisada pela glutationa redutase. O meio de reação continha
glutationa 1 mM, GR 0,1 U/mL, NADPH 20 mM, EDTA 5 mM pH 7,0 e
tampão fosfato de potássio 0,1 M pH 7,0. Em duplicata, 250 μL deste meio
de reação foi pipetado em uma placa de 96 poços, sendo adicionado 10 μL
das amostras (frações citosólicas) e 5 μL de t-butil 0,5 mM em H2O mQ, e
incubadas a 30 ºC para a leitura da absorbância a cada minuto durante 6
minutos. Os resultados foram expressos em U/mg de proteína.
Uma unidade (U) da enzima foi definida como atividade da enzima
que oxidou 1 μmol de NADPH por minuto a 30 ºC em pH 7,0.

64
4.3.14 Quantificação de proteínas no tecido hepático
A determinação do conteúdo de proteínas presente no tecido hepático
foi realizada segundo o método de Bradford (1985). Uma curva padrão de
proteína com solução padrão de albumina foi adotada.
4.3.15 Determinação das citocinas inflamatórias no fígado
Fragmentos do fígado foram homogeneizados com uma solução
tampão PBS para ELISA pH 7,2, igual a cinco vezes o valor absoluto da
massa da amostra em homogeneizador rotativo de tecidos. O homogeneizado
foi centrifugado a 6.900 rotações por minuto à temperatura de 4 ºC por 10
minutos. Em seguida, o sobrenadante foi coletado, para a determinação da
concentração das citocinas inflamatórias: TNF e IL-6. Ambas foram
determinadas pelo método de ELISA de acordo com as instruções do
fabricante (P&D System).
4.4 Análise estatística
Os resultados foram expressos como média ± erro padrão da média
(E.P.M.) e analisados estatisticamente pela análise de variância com
comparações múltiplas (ANOVA) e, quando necessário, utilizado como pós
teste o teste de Tukey ou teste t. Os dados foram analisados no programa
GraphPad Prism 5, admitindo como significativo p < 0,05.

65
5 RESULTADOS
5.1 Avaliação da atividade antioxidante in vitro do extrato hidroalcoólico
obtido a partir da FCM
Os dados apresentados na figura 6 demonstram os resultados in vitro
do extrato de FCM evidenciando seu potencial na redução do ferro por meio
do método FRAP nas maiores concentrações de extrato (300 e 1000 μg/mL).
O extrato hidroalcoólico de FCM promoveu proteção do β-caroteno/ácido
linoleico na concentração de 1000 μg/mL e também apresentou capacidade
de sequestro do NO nas concentrações de 30 à 1000 μg/mL.
Juntos, estes dados demonstram que o extrato de FCM apresenta
atividade antioxidante, a qual pode auxiliar na modulação dos radicais livres
e estresse oxidativo, ambos envolvidos em doenças metabólicas.

66
Figura 6. Atividade antioxidante do extrato da farinha da casca do maracujá.
A) Resultados do ensaio FRAP do extrato da FCM; B) Capacidade antioxidante do extrato por sequestro do NO2; C) Resultados do ensaio sistema β Caroteno/Ácido Linoleico. Valores expressam a média ± e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de Tukey. * p <0,05 vs. grupo sistema
empregado.
5.2 Ensaios in vivo
Os experimentos realizados in vivo evidenciaram que a dieta de
cafeteria administrada pelo período de 16 semanas promoveu importantes
alterações no metabolismo lipídico e glicídico, mimetizando um perfil
semelhante a síndrome metabólica.
O ganho de peso foi acompanhado semanalmente. Os dados obtidos
estão apresentados na figura 7. Animais do grupo CAF não apresentaram
diferenças no ganho de peso em relação à dieta controle ao final das 16
semanas de experimento. No entanto, o grupo que consumiu a dieta de
cafeteria acrescida de FCM apresentou ganho de peso inferior ao grupo CAF,
conforme pode ser visualizado na figura 7A e 7B.
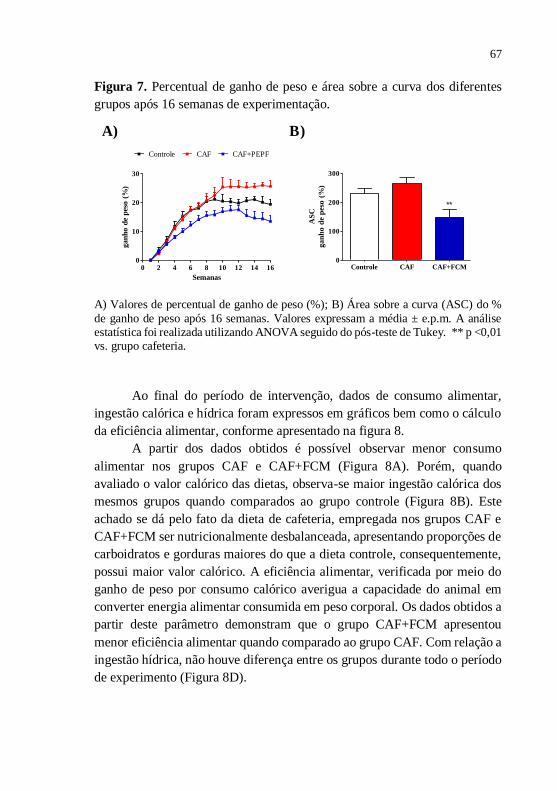
67
Figura 7. Percentual de ganho de peso e área sobre a curva dos diferentes
grupos após 16 semanas de experimentação.
0 2 4 6 8 10 12 14 160
10
20
30
Controle CAF CAF+PEPF
Semanas
ga
nh
o d
e p
eso
(%
)
Controle CAF CAF+FCM0
100
200
300
**
AS
C
ga
nh
o d
e p
eso
(%
)
A) B)
A) Valores de percentual de ganho de peso (%); B) Área sobre a curva (ASC) do %
de ganho de peso após 16 semanas. Valores expressam a média ± e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de Tukey. ** p <0,01 vs. grupo cafeteria.
Ao final do período de intervenção, dados de consumo alimentar,
ingestão calórica e hídrica foram expressos em gráficos bem como o cálculo
da eficiência alimentar, conforme apresentado na figura 8.
A partir dos dados obtidos é possível observar menor consumo
alimentar nos grupos CAF e CAF+FCM (Figura 8A). Porém, quando
avaliado o valor calórico das dietas, observa-se maior ingestão calórica dos
mesmos grupos quando comparados ao grupo controle (Figura 8B). Este
achado se dá pelo fato da dieta de cafeteria, empregada nos grupos CAF e
CAF+FCM ser nutricionalmente desbalanceada, apresentando proporções de
carboidratos e gorduras maiores do que a dieta controle, consequentemente,
possui maior valor calórico. A eficiência alimentar, verificada por meio do
ganho de peso por consumo calórico averigua a capacidade do animal em
converter energia alimentar consumida em peso corporal. Os dados obtidos a
partir deste parâmetro demonstram que o grupo CAF+FCM apresentou
menor eficiência alimentar quando comparado ao grupo CAF. Com relação a
ingestão hídrica, não houve diferença entre os grupos durante todo o período
de experimento (Figura 8D).
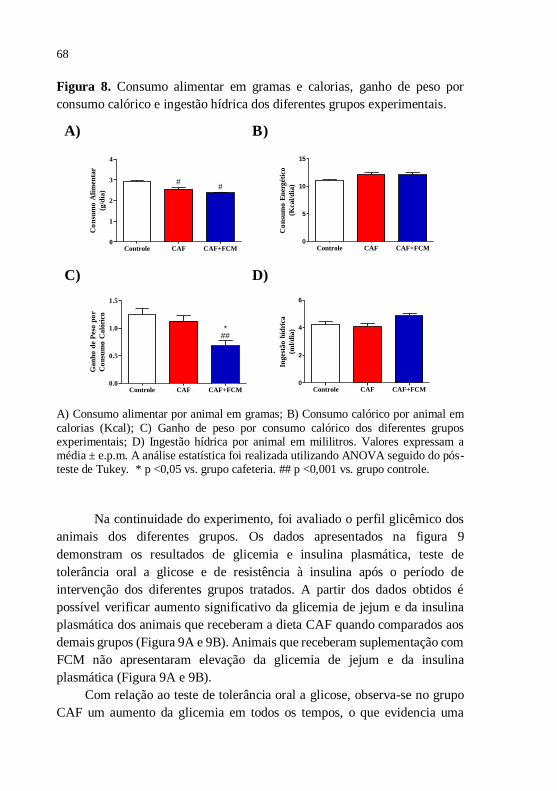
68
Figura 8. Consumo alimentar em gramas e calorias, ganho de peso por
consumo calórico e ingestão hídrica dos diferentes grupos experimentais.
Controle CAF CAF+FCM0
1
2
3
4
##
Co
nsu
mo
Ali
men
tar
(g/d
ia)
Controle CAF CAF+FCM0
5
10
15
Co
nsu
mo
En
ergét
ico
(Kca
l/d
ia)
Controle CAF CAF+FCM0.0
0.5
1.0
1.5
*##
Gan
ho
de
Pes
o p
or
Co
nsu
mo
Caló
rico
Controle CAF CAF+FCM0
2
4
6
Inges
tão
híd
rica
(ml/
dia
)
A) B)
C) D)
A) Consumo alimentar por animal em gramas; B) Consumo calórico por animal em
calorias (Kcal); C) Ganho de peso por consumo calórico dos diferentes grupos experimentais; D) Ingestão hídrica por animal em mililitros. Valores expressam a média ± e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de Tukey. * p <0,05 vs. grupo cafeteria. ## p <0,001 vs. grupo controle.
Na continuidade do experimento, foi avaliado o perfil glicêmico dos
animais dos diferentes grupos. Os dados apresentados na figura 9
demonstram os resultados de glicemia e insulina plasmática, teste de
tolerância oral a glicose e de resistência à insulina após o período de
intervenção dos diferentes grupos tratados. A partir dos dados obtidos é
possível verificar aumento significativo da glicemia de jejum e da insulina
plasmática dos animais que receberam a dieta CAF quando comparados aos
demais grupos (Figura 9A e 9B). Animais que receberam suplementação com
FCM não apresentaram elevação da glicemia de jejum e da insulina
plasmática (Figura 9A e 9B).
Com relação ao teste de tolerância oral a glicose, observa-se no grupo
CAF um aumento da glicemia em todos os tempos, o que evidencia uma

69
tendência a intolerância à glicose neste grupo quando comparado ao grupo
controle (Figura 9C e 9D). Animais provenientes do grupo CAF+FCM
apresentaram resultados de concentração de glicemia inferiores em todos os
tempos avaliados (Figura 9C), apresentando redução global no valor de
glicemia (Figura 9D). Estes dados corroboram a afirmativa de que a dieta de
cafeteria gera alterações no metabolismo glicídico, alterando o
comportamento da glicemia após a administração de uma dose de 2 mg/kg de
peso de glicose por via oral.
Após a observação do perfil glicêmico, foi realizado ensaio de
resistência à insulina, com intuito de observar o perfil insulinêmico dos
animais. O ensaio de resistência à insulina, demonstra o comportamento do
hormônio após a administração intraperitoneal. Os dados obtidos
demonstram que o grupo CAF apresenta maiores concentrações de glicose
em todos os tempos avaliados, quando comparados ao grupo controle
conforme pode ser visualizado na figura 9E e F. Estes dados demonstram que
neste modelo, o grupo CAF desenvolveu resistência a insulina, enquanto o
grupo tratado com FCM e o grupo controle apresentaram comportamento
glicêmico esperado em resposta à administração de insulina exógena (Figura
9E e 9F).
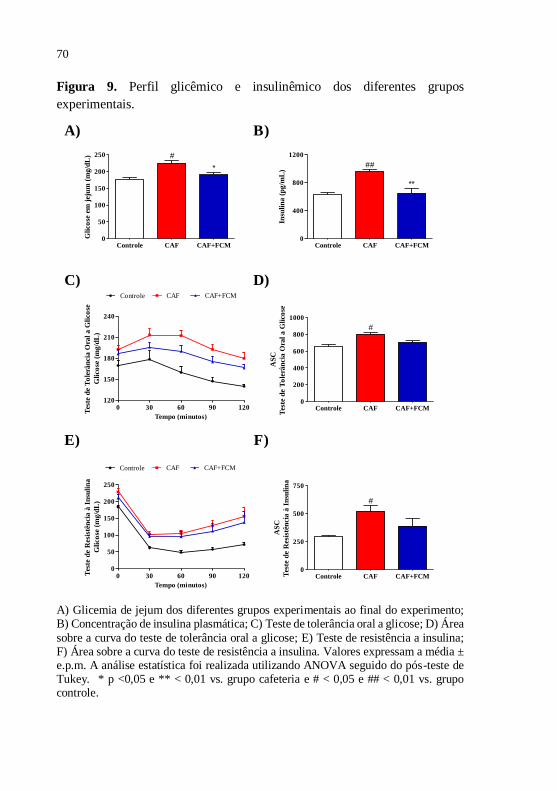
70
Figura 9. Perfil glicêmico e insulinêmico dos diferentes grupos
experimentais.
Controle CAF CAF+FCM0
50
100
150
200
250 #
*
Gli
cose
em
jej
um
(m
g/d
L)
Controle CAF CAF+FCM0
400
800
1200
##
**
Insu
lin
a (
pg
/mL
)
0 30 60 90 120120
150
180
210
240
Controle CAF CAF+FCM
Tempo (minutos)
Tes
te d
e T
ole
rân
cia
Ora
l a
Gli
cose
Gli
cose
(m
g/d
L)
Controle CAF CAF+FCM0
200
400
600
800
1000
#
AS
C
Tes
te d
e T
ole
rân
cia
Ora
l a
Gli
cose
0 30 60 90 1200
50
100
150
200
250
Controle CAF CAF+FCM
Tempo (minutos)
Tes
te d
e R
esis
tên
cia
à I
nsu
lin
a
Gli
cose
(m
g/d
L)
Controle CAF CAF+FCM0
250
500
750
#
AS
C
Tes
te d
e R
esis
tên
cia
à I
nsu
lin
a
A) B)
C) D)
E) F)
A) Glicemia de jejum dos diferentes grupos experimentais ao final do experimento; B) Concentração de insulina plasmática; C) Teste de tolerância oral a glicose; D) Área
sobre a curva do teste de tolerância oral a glicose; E) Teste de resistência a insulina; F) Área sobre a curva do teste de resistência a insulina. Valores expressam a média ± e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de Tukey. * p <0,05 e ** < 0,01 vs. grupo cafeteria e # < 0,05 e ## < 0,01 vs. grupo controle.

71
Adicionalmente, foram realizadas análises histológicas no tecido
pancreático, com o intuito de observar a morfologia do tecido, em especial
das ilhotas de Langerhans, que são as unidades funcionais deste órgão,
responsáveis pela produção de insulina. Os dados obtidos na avaliação
histológica (Figura 10A) demonstram um aumento no diâmetro das ilhotas
de Langerhans do grupo CAF, quando comparados aos grupos controle e
CAF+FCM. Esta hiperplasia foi confirmada pelo do cálculo de área das
ilhotas de Langerhans no pâncreas, onde foi possível observar aumento das
ilhotas de animais do grupo CAF quando comparado a animais do grupo
controle (Figura 10B). Estas alterações não foram evidenciadas no grupo
CAF+FCM, como apresentado na figura 10A e B. Com relação ao peso
absoluto e relativo, não foram observadas diferenças significativas (Figura 10
C e D).
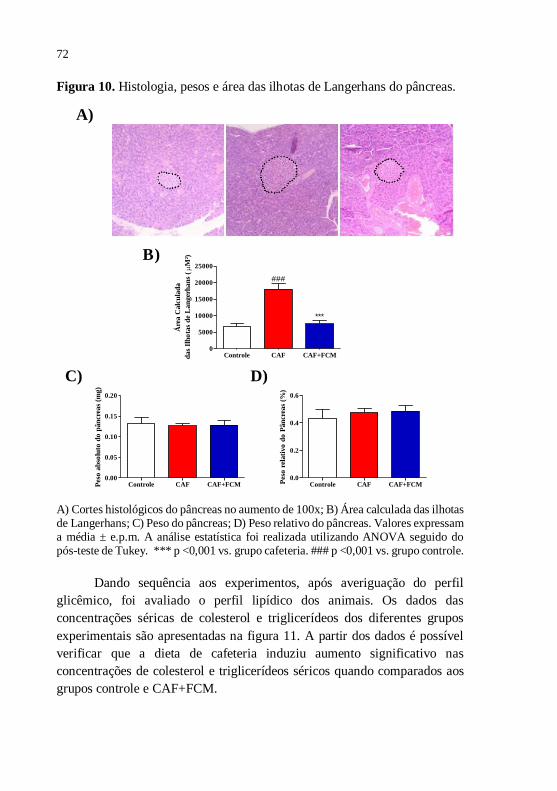
72
Figura 10. Histologia, pesos e área das ilhotas de Langerhans do pâncreas.
Controle CAF CAF+FCM0
5000
10000
15000
20000
25000
###
***
Áre
a C
alc
ula
da
das
Ilh
ota
s d
e L
an
ger
han
s (
M²)
Controle CAF CAF+FCM0.00
0.05
0.10
0.15
0.20
Pes
o a
bso
luto
do
pân
crea
s (m
g)
Controle CAF CAF+FCM0.0
0.2
0.4
0.6
Pes
o r
elati
vo
do
Pân
crea
s (%
)
A)
B)
C) D)
A) Cortes histológicos do pâncreas no aumento de 100x; B) Área calculada das ilhotas de Langerhans; C) Peso do pâncreas; D) Peso relativo do pâncreas. Valores expressam a média ± e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de Tukey. *** p <0,001 vs. grupo cafeteria. ### p <0,001 vs. grupo controle.
Dando sequência aos experimentos, após averiguação do perfil
glicêmico, foi avaliado o perfil lipídico dos animais. Os dados das
concentrações séricas de colesterol e triglicerídeos dos diferentes grupos
experimentais são apresentadas na figura 11. A partir dos dados é possível
verificar que a dieta de cafeteria induziu aumento significativo nas
concentrações de colesterol e triglicerídeos séricos quando comparados aos
grupos controle e CAF+FCM.
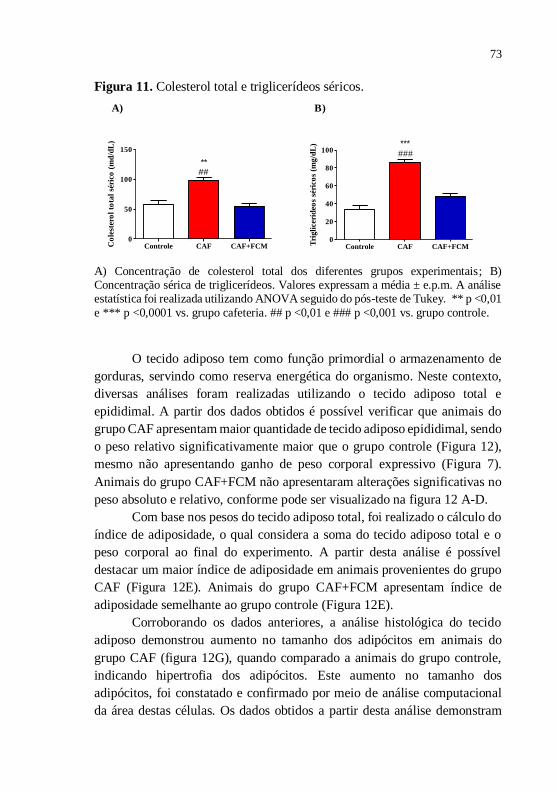
73
Figura 11. Colesterol total e triglicerídeos séricos.
Controle CAF CAF+FCM0
50
100
150
**##
Co
lest
ero
l to
tal
séri
co (
md
/dL
)
Controle CAF CAF+FCM0
20
40
60
80
100***
###
Tri
gli
cerí
deo
s sé
rico
s (m
g/d
L)
A) B)
A) Concentração de colesterol total dos diferentes grupos experimentais; B) Concentração sérica de triglicerídeos. Valores expressam a média ± e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de Tukey. ** p <0,01
e *** p <0,0001 vs. grupo cafeteria. ## p <0,01 e ### p <0,001 vs. grupo controle.
O tecido adiposo tem como função primordial o armazenamento de
gorduras, servindo como reserva energética do organismo. Neste contexto,
diversas análises foram realizadas utilizando o tecido adiposo total e
epididimal. A partir dos dados obtidos é possível verificar que animais do
grupo CAF apresentam maior quantidade de tecido adiposo epididimal, sendo
o peso relativo significativamente maior que o grupo controle (Figura 12),
mesmo não apresentando ganho de peso corporal expressivo (Figura 7).
Animais do grupo CAF+FCM não apresentaram alterações significativas no
peso absoluto e relativo, conforme pode ser visualizado na figura 12 A-D.
Com base nos pesos do tecido adiposo total, foi realizado o cálculo do
índice de adiposidade, o qual considera a soma do tecido adiposo total e o
peso corporal ao final do experimento. A partir desta análise é possível
destacar um maior índice de adiposidade em animais provenientes do grupo
CAF (Figura 12E). Animais do grupo CAF+FCM apresentam índice de
adiposidade semelhante ao grupo controle (Figura 12E).
Corroborando os dados anteriores, a análise histológica do tecido
adiposo demonstrou aumento no tamanho dos adipócitos em animais do
grupo CAF (figura 12G), quando comparado a animais do grupo controle,
indicando hipertrofia dos adipócitos. Este aumento no tamanho dos
adipócitos, foi constatado e confirmado por meio de análise computacional
da área destas células. Os dados obtidos a partir desta análise demonstram

74
que animais do grupo CAF apresentam aumento da área de adipócitos,
quando comparado a animais controle (Figura 12F). Não foram observadas
alterações significativas no tamanho de adipócitos de animais do grupo FCM
(Figura 12F e G).

75
Figura 12. Parâmetros de tecido adiposo dos diferentes grupos
experimentais.
Controle CAF CAF+FCM0.00
0.05
0.10
0.15
0.20
Pes
o a
bso
luto
do
teci
do
ad
ipo
so a
bd
om
inal
(mg)
Controle CAF CAF+FCM0.0
0.2
0.4
0.6
0.8
Pes
o r
elati
vo
do
teci
do
ad
ipo
so a
bd
om
inal
(%)
Controle CAF CAF+FCM0.0
0.2
0.4
0.6
0.8
Pes
o a
bso
luto
do
teci
do
ad
ipo
so e
pid
idim
al
(mg)
Controle CAF CAF+FCM0.0
0.5
1.0
1.5
2.0
2.5
Pes
o r
elati
vo
do
teci
do
ad
ipo
so e
pid
idim
al
(%)
Controle CAF CAF+FCM0
1
2
3
4
*
#
Índ
ice
de
Ad
ipo
sid
ad
e (%
)
Controle CAF CAF+FCM0
5000
10000
15000
20000
#
*
Áre
a C
alc
ula
da
de
Ad
ipóci
tos
(M
²)
Control CAF CAF+PEPF
A) B)
C) D)
E) F)
G)

76
A) Peso absoluto do tecido adiposo abdominal; B) Peso relativo do tecido adiposo abdominal; C) Peso absoluto do tecido adiposo epididimal; D) Peso relativo do tecido adiposo epididimal; E) Índice de adiposidade calculado; F) Cortes histológicos do
tecido adiposo no aumento de 100x; G) Área calculada dos adipócitos. Valores expressam a média ± e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de Tukey. * p <0,05 vs. grupo cafeteria. # p <0,05 vs. grupo controle.
A dieta de cafeteria ocasionou aumento da concentração de colesterol
e triglicerídeos séricos, como exposto anteriormente (figura 11). Esta
alteração no metabolismo de lipídeos, pode induzir disfunções no fígado,
relacionadas ao aumento de enzimas hepáticas, como aspartato
aminotransferase (AST) e alanina aminotransferase (ALT). Neste contexto,
análises para avaliação do perfil hepático foram realizadas.
Dados obtidos no presente estudo, demonstram que a dieta de cafeteria
nas condições estudadas não ocasiona alterações significativas na
determinação de AST e ALT. Animais que receberam suplementação com
FCM, apresentaram redução significativa de AST e ALT (Tabela 5).
Tabela 5. Concentração das enzimas hepáticas aspartato aminotransferase
(AST) e alanina aminotransferase (ALT) no soro dos animais.
Grupos AST ALT
Controle 63,28±1,19U/L 22,4±0,67U/L
CAF 72,87±4,06U/L 21,33±1,05U/L
CAF+FCM 52,2±0,96U/L* 18,2±0,73U/L*
Valores expressam a média ± e.p.m. A análise estatística foi realizada utilizando
ANOVA seguido do pós-teste de Tukey. * p <0,05 vs. grupo cafeteria.
Apesar do padrão normal das enzimas hepáticas (Tabela 4), os dados
obtidos na análise histológica do tecido hepático demonstraram que a dieta
de cafeteria provoca importantes alterações no tecido hepático, apresentando
áreas de esteatose hepática multifocal, relacionada com aumento no infiltrado
de lipídeos (Figura 13A) e, sem evidências da mesma anormalidade no grupo
FCM. Com relação aos dados de peso absoluto e relativo do fígado, não
houver diferenças significativas entre os grupos (Figura 13B e C).

77
A maior deposição de lipídeos no tecido hepático foi determinada, e
os resultados obtidos demonstraram aumento de colesterol e triglicerídeos no
tecido hepático no grupo CAF quando comparado ao grupo controle,
corroborando assim com os dados histológicos (13A). O grupo tratado com
FCM não apresentou aumento na infiltração de triglicerídeos quando
comparado ao grupo controle, conforme demonstrado na figura 13.
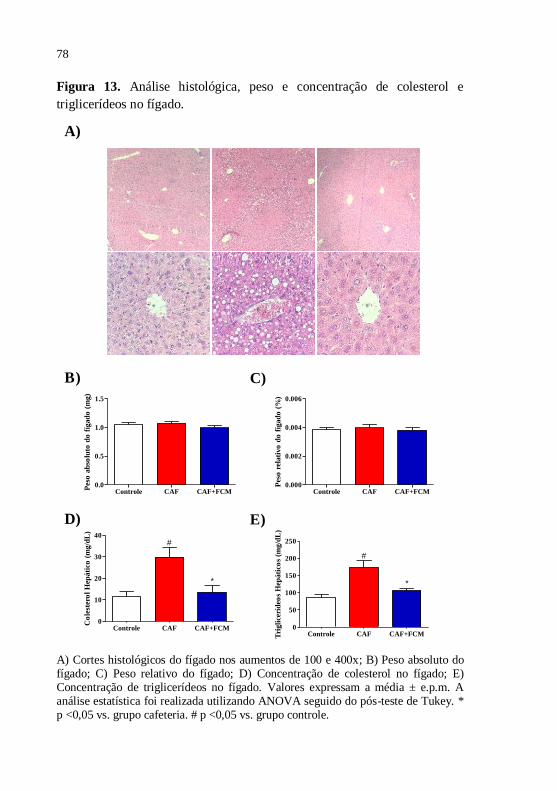
78
Figura 13. Análise histológica, peso e concentração de colesterol e
triglicerídeos no fígado.
Controle CAF CAF+FCM0.0
0.5
1.0
1.5
Pes
o a
bso
luto
do
fíg
ad
o (
mg)
Controle CAF CAF+FCM0.000
0.002
0.004
0.006
Pes
o r
elati
vo
do
fíg
ad
o (
%)
Controle CAF CAF+FCM0
10
20
30
40#
*
Co
lest
ero
l H
epáti
co (
mg/d
L)
Controle CAF CAF+FCM0
50
100
150
200
250
#
*
Tri
gli
cerí
deo
s H
epáti
cos
(mg/d
L)
A)
B) C)
D) E)
A) Cortes histológicos do fígado nos aumentos de 100 e 400x; B) Peso absoluto do fígado; C) Peso relativo do fígado; D) Concentração de colesterol no fígado; E) Concentração de triglicerídeos no fígado. Valores expressam a média ± e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de Tukey. * p <0,05 vs. grupo cafeteria. # p <0,05 vs. grupo controle.

79
As alterações na morfologia do tecido hepático, associado a um
aumento na infiltração de lipídeos, pode ocasionar no tecido hepático a
indução de um processo inflamatório. Neste contexto, foram avaliadas as
concentrações de IL-6 e TNF, no tecido hepático. Os dados obtidos nesta
avaliação demonstraram que a dieta de cafeteria após 16 semanas promoveu
aumento nos níveis de IL-6 no tecido hepático (Figura 14). Animais do grupo
CAF+FCM apresentaram alterações significativas nos níveis de IL-6 (Figura
14).
Figura 14. Concentração de IL-6 e TNF no tecido hepático.
Controle CAF CAF+FCM0
5000
10000
15000
20000
25000
TF
N (
pg/m
g d
e t
eci
do
)
Controle CAF CAF+FCM0
200
400
600
800
*
#
IL-6
(p
g/m
g d
e t
eci
do
)
A) B)
a) Concentração de interleucina 6 no homogeinato de fígado dos diferentes grupos experimentais; b) Concentração de fator de necrose tumoral no homogeinato de fígado. Valores expressam a média ± e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de Tukey. * p <0,05 vs. grupo cafeteria. # p <0,01 vs. grupo controle.
Além disso, foi realizada avaliação de FRAP, GSH, SOD, CAT e GPx
no fígado. Os dados obtidos demonstram que a dieta de cafeteria não foi
capaz de modificar significativamente nenhum dos parâmetros avaliados
(Figura 15A-E).

80
Figura 15. Parâmetros antioxidantes no fígado dos diferentes grupos
experimentais após o período de intervenção.
Controle CAF CAF+FCM0
5000
10000
15000
FR
AP
do
Fíg
ad
o
(m
M/m
g d
e p
rote
ína)
Controle CAF CAF+FCM0
1000
2000
3000
4000
GS
H
(mM
/mg d
e p
rote
ína)
Controle CAF CAF+FCM0
500
1000
1500
SO
D
(U/m
g d
e p
rote
ína)
Controle CAF CAF+FCM0
500
1000
1500
2000
CA
T
(U/m
g d
e p
rote
ína)
Controle CAF CAF+FCM0
2
4
6
GP
x
(U/m
g d
e p
rote
ína)
A) B)
C) D)
E)
A) FRAP do fígado; B) GSH; C) SOD; D) CAT; E) GPx. Valores expressam a média
± e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de
Tukey.
Adicionalmente também foram realizadas avaliação da função renal
dos animais submetidos ao experimento. Os rins são órgãos filtradores do
sistema urinário, tendo como unidades funcionais os glomérulos renais. A
figura 16 apresenta as lâminas histológicas do tecido renal, onde foram
observadas alterações morfológicas deste órgão. É possível perceber discreta
atrofia dos glomérulos renais (Figura 16A), constatada e confirmada por meio
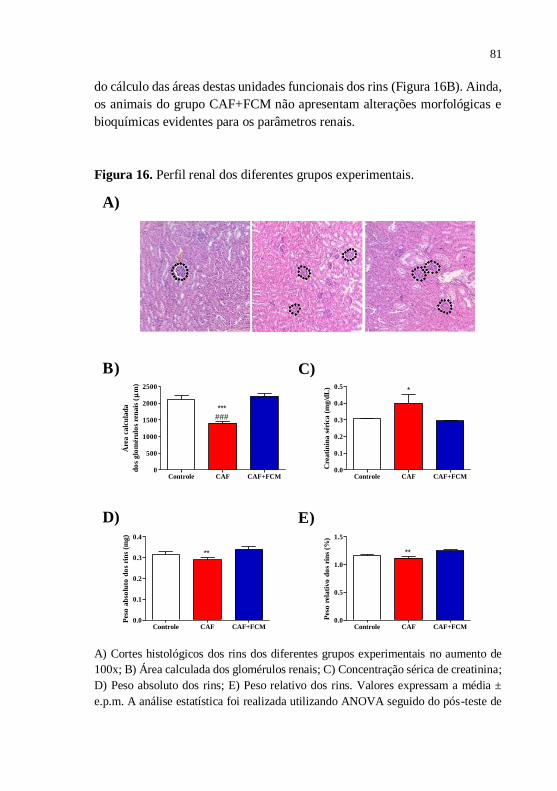
81
do cálculo das áreas destas unidades funcionais dos rins (Figura 16B). Ainda,
os animais do grupo CAF+FCM não apresentam alterações morfológicas e
bioquímicas evidentes para os parâmetros renais.
Figura 16. Perfil renal dos diferentes grupos experimentais.
Controle CAF CAF+FCM0
500
1000
1500
2000
2500
***###
Áre
a c
alc
ula
da
do
s glo
méru
los
ren
ais
(
m)
Controle CAF CAF+FCM0.0
0.1
0.2
0.3
0.4
0.5 *
Cre
ati
nin
a s
éri
ca (
mg/d
L)
Controle CAF CAF+FCM0.0
0.1
0.2
0.3
0.4
**
Peso
ab
solu
to d
os
rin
s (m
g)
Controle CAF CAF+FCM0.0
0.5
1.0
1.5
**
Peso
rela
tivo
do
s ri
ns
(%)
A)
B) C)
D) E)
A) Cortes histológicos dos rins dos diferentes grupos experimentais no aumento de
100x; B) Área calculada dos glomérulos renais; C) Concentração sérica de creatinina;
D) Peso absoluto dos rins; E) Peso relativo dos rins. Valores expressam a média ±
e.p.m. A análise estatística foi realizada utilizando ANOVA seguido do pós-teste de

82
Tukey. * p <0,05, ** p <0,01 e *** p <0,001 vs. grupo cafeteria. ### p <0,001 vs.
grupo controle.

83
6 DISCUSSÃO
O maracujá está entre os frutos mais produzidos e consumidos no
Brasil, e seu principal resíduo, a sua casca, além da grande quantidade de
fibras, é rica em compostos fenólicos que apresentam propriedades
funcionais, como antioxidante, podendo desta forma desempenhar papel
importante na saúde (GOSS et al., 2018). Destaca-se que em geral, o teor de
polifenóis mostra-se superior na casca dos frutos do que na sua porção
comestível ou polpa (TEHRANIFAR et al., 2011), salientado a possibilidade
de utilização da FCM como um alimento funcional para o tratamento de
doenças metabólicas.
No estudo de Goss et al. (2018), foram identificados os compostos
fenólicos como compostos majoritários deste material vegetal, sendo assim,
foi elaborado um extrato hidroalcoólico (70%) para avaliação in vitro dos
efeitos antioxidantes. Neste contexto, segundo Silva (2012), as metodologias
para determinação da capacidade antioxidante são numerosas e podem estar
sujeitas a interferências, além de se basearem em fundamentos diversos.
Assim, a atividade antioxidante de extratos e frações pode ser determinada
por diversos métodos em diferentes sistemas: lipídico e aquoso. No entanto,
as metodologias mais utilizadas estão normalmente relacionadas com os
mecanismos antioxidantes que os compostos podem apresentar. Dessa forma,
preconizou-se a utilização de três técnicas, já que nenhum ensaio utilizado
isoladamente para determinar a capacidade antioxidante irá refletir
exatamente a “capacidade antioxidante total” de uma amostra. A capacidade antioxidante do extrato hidroalcoólico de FCM pela
redução do ferro férrico à ferroso foi avaliado por meio do método FRAP,
sendo este efeito dose dependente. O ensaio FRAP (ferric reducing
antioxidant power) (BENZIE; STRAIN, 1996) é baseado na capacidade dos
fenóis em reduzir o Fe3+ em Fe2+. Quando isto ocorre, na presença de 2,4,6-
tripiridil-s-triazina (TPTZ), a redução é acompanhada pela formação de um
complexo corado com Fe2+. Apesar de ser um método relativamente simples,
é muito utilizado, podendo ser aplicado não somente para estudos da
atividade antioxidante em extratos de alimentos, mas também, para o estudo
da eficiência antioxidante de substâncias puras, órgãos e tecidos (PULIDO;
BRAVO; SAURA-CALIXTO, 2000).
Outra metodologia empregada na determinação do potencial
antioxidante do extrato da FCM foi a determinação da ação sequestradora do

84
íon NO2, baseada na reação com o reagente de Griess, na qual o nitrito reage
com a sulfanilamida em meio ácido (RAMOS; CAVALHEIRO;
CAVALHEIRO, 2006). Neste ensaio, o extrato hidroalcoólico de FCM
apresentou atividade sequestradora do íon NO2 nas concentrações de 30 à
1000 μg/mL.
Outro método distinto, o sistema β Caroteno/Ácido Linoleico tem
como função avaliar a capacidade de um antioxidante inibir a peroxidação do
ácido linoleico a radicais livres, os quais são responsáveis pela descoloração
do β-caroteno. O β-caroteno é extremamente sensível à oxidação mediada por
radicais livres, podendo os antioxidantes doar átomos de hidrogênio para
eliminar os radicais e impedir a descoloração de carotenoides (SILVA, 2012).
Utilizando este modelo, foi possível verificar que o extrato de FCM apresenta
capacidade de inibir a peroxidação do ácido linoleico somente na
concentração de 1000 μg/mL.
Juntos, os resultados demonstram que a FCM apresenta atividade
antioxidante, a qual pode auxiliar na modulação dos radicais livres e estresse
oxidativo. Ainda, os dados obtidos corroboram com dados recentes
publicados por Goss et al. (2018), que demonstraram o efeito antioxidante do
extrato hidroalcoólico de FCM utilizando o método DPPH. No mesmo
trabalho, os autores reportam a presença de flavonoides e compostos
fenólicos, sendo identificados o ácido cafeico e a isoorientina no extrato
hidroalcoólico de FCM. Além destes, outros compostos fenólicos como a
vitexina, isovitexina, isoorientina e apigenina também já foram reportados
em extratos obtidos a partir da casca do maracujá. A estes compostos, têm
sido atribuídos efeitos anti-inflamatório (ZERAIK et al., 2011; ZERAIK et
al., 2012) anti-hipertensivo (LEWIS et al., 2013) e antioxidante (CAZARIN
et al., 2014).
A presença de flavonoides e compostos fenólicos identificados na
farinha da casca do maracujá, pode ser a razão pela qual lhe é conferida
atividade antioxidante. As substâncias antioxidantes possuem papel
importante na saúde, visto que há correlação positiva entre a prevenção de
doenças crônicas e o consumo de vegetais e frutas, as quais naturalmente
contém substâncias antioxidantes (CAZARIN et. al., 2014).
Por outro lado, o hábito alimentar dos seres humanos vem mudando
continuamente, o qual é um dos fatores mais fortes que explica a epidemia
mundial de obesidade. Atualmente, se consome cada vez menos alimentos
frescos e in natura como frutas e vegetais, em contrapartida, vem aumentando

85
o consumo de alimentos processados e ultraprocessados, como biscoitos,
salgadinhos, bolos, alimentos pré-cozidos, etc., os quais são altamente
calóricos, com baixo valor nutricional, ricos em açúcar refinado, xarope de
milho e óleos refinados (BARRETT et al., 2016). De um modo geral, as
mudanças nos hábitos alimentares são cada vez mais direcionadas ao
aumento no consumo de calorias.
Nos estudos experimentais em animais, vários modelos de dietas
indutoras de obesidade e síndrome metabólica tem sido utilizadas para
estudar a etiologia destas doenças, com o objetivo de obter melhor
compreensão dos mecanismos moleculares e possíveis tratamentos. Destaca-
se que as dietas de cafeteria são modelos muito relevantes de indução de
síndrome metabólica e obesidade, por abrangerem alimentos palatáveis e de
alta densidade energética, uma vez que as metodologias costumam empregar
alimentos consumidos pela sociedade ocidental, diretamente relacionados à
obesidade em humanos (LOUZADA et al., 2013; NEIA et al., 2018).
A dieta de cafeteria, empregada neste estudo, é caracterizada pela alta
densidade calórico energética, com elevadas quantidades de carboidratos e
gorduras, sendo responsáveis por aumentar a massa de tecido adiposo e
induzir a obesidade. Além disso, a dieta de cafeteria é acompanhada de
importantes alterações funcionais e metabólicas que vão muito além da
deposição de gordura corporal (OLIVEIRA, 2015). A alta ingestão de
gorduras e açúcares, promovida por meio desta dieta, são oxidados,
produzindo substâncias tóxicas, tais como produtos finais de glicação
avançada (AGEs), gerando desequilíbrio do sistema redox, associado à
inflamação de baixo grau e ao estresse oxidativo, bem como disfunções
metabólicas e danos inflamatórios em diversos órgãos (IANTORNO et al.,
2014; NAVARRO et al., 2016).
No presente estudo, conforme esperado, a dieta de cafeteria foi capaz
de induzir desordens no metabolismo glicídico e lipídico, nos tecidos
adiposo, hepático, pancreático e renal. Os resultados obtidos são similares a
condição de síndrome metabólica, segundo parâmetros preconizados pelo
NCEP ATP III, demonstrando a eficácia do modelo testado na indução da
síndrome metabólica em animais. Outros estudos que empregaram a mesma
metodologia de dieta de cafeteria padronizada também obtiveram resultados
semelhantes (ESTADELLA et al., 2004; SAMPEY et al., 2011; HIGA et al.,
2014; BALLESTRERI et al., 2015; SOARES et al., 2017).

86
Além das alterações “esperadas” causadas pela dieta de cafeteria na
indução da síndrome metabólica e obesidade, como: aumento da adiposidade
visceral, hiperinsulinemia, intolerância à glicose, dislipidemia com níveis
séricos de triglicerídeos elevados e redução do HDL-colesterol (GOMEZ-
SMITH, 2016), doença hepática gordurosa não alcoólica (PARAFATI et al.,
2015), entre outras alterações, estudos recentes em animais, que também
empregaram esta metodologia, demonstram outros prejuízos causados pela
exposição a dietas de cafeteria, destacando-se: elevados níveis plasmáticos
de produtos tóxicos como os AGEs (NAVARRO et al., 2016); o aumento da
resposta metabólica ao estresse oxidativo e danos oxidativos no DNA, no
plasma e principais tecidos envolvidos na defesa antioxidante (MELLOUK
et al., 2016); danos no coração (fibrose perivascular coronária e esteatose),
nos rins (inflamação intersticial crónica e esclerose glomerular) e no fígado
(fígado, fibrose portal, apoptose e esteatose) (ZEENI et al., 2015); alteração
na expressão dos genes envolvidos na permeabilidade intestinal, estresse
oxidativo e inflamação intestinal (GIL-CARDOSO et al., 2017); o aumento
da ansiedade, prejuízos na aprendizagem espacial, memória e na neurogênese
(FERREIRA, et. al., 2018); alterações neurobiológicas, incluindo alterações
neuroplásticas e funcionais, ativação da microglia e neuroinflamação
(GUTIÉRREZ-MARTOS et al., 2018); alterações morfológicas e
moleculares associadas à hiperplasia uterina associada a carcinogênese
endometrial (GASTIAZORO et al., 2018); entre outros.
Todavia, as dietas de cafeteria podem apresentar desvantagens.
Autores criticam a reprodutibilidade deste modelo, pelo fato de não ser um
protocolo padronizado. De maneira geral, cada pesquisador faz sua própria
composição de alimentos industrializados, de acordo com a realidade de sua
região, como por exemplo: salgadinhos tipo chips, biscoitos recheados, bolos,
salame, bacon, batata frita, queijo, chocolates, refrigerantes a base de cola,
etc. Outra desvantagem, são as deficiências nutricionais que podem ser
causadas, de nutrientes como proteínas, vitaminas e minerais, uma vez que
alimentos industrializados são pobres nestes nutrientes. Estas deficiências
nutricionais podem interferir positiva ou negativamente com os resultados
das pesquisas. E ainda, por apresentar vários aditivos alimentares que não
estariam presentes nas rações padrão de manutenção dos animais, sendo:
corantes, flavorizantes, conservantes, emulsificantes, que podem gerar outras
alterações no organismo, dificultando a interpretação dos resultados
(BORTOLIN et al., 2017). Desta forma, ressalta-se a importância do cuidado

87
na escolha da metodologia empregada neste tipo de modelo de indução de
síndrome metabólica, bem como a seleção dos estudos que seguiram
metodologias semelhantes a adotada, para fins de comparação dos resultados
obtidos.
Um resultado importante, foi com relação ao ganho de peso. No
presente estudo, animais que consumiram a dieta CAF, não ganharam peso
em relação a dieta controle, porém, o grupo que recebeu FCM apresentou
menor ganho de peso quando comparado aos demais grupos. Assim como os
resultados obtidos neste experimento, outros autores também demonstraram
a ineficiência da dieta de cafeteria em induzir ganho de peso expressivo
(MUNTZEL et al., 2012; HIGA et al., 2014; MARQUES et al., 2015;
BORTOLIN et al. 2017).
Apesar de alguns estudos apresentarem resultados significativos na
avaliação deste parâmetro (ESTADELLA et al., 2004; SAMPEY et al., 2011;
BALLESTRERI et al., 2015; DALBY et al., 2017; SOARES et al., 2017)
destaca-se o viés da não padronização das metodologias de dieta de cafeteria,
o que pode gerar resultados de ganho de peso diferentes conforme a escolha
dos alimentos (BORTOLIN et al., 2017).
Segundos os mesmos autores, essa diferença também pode se dar
devido a escolha da dieta do grupo controle. Existem duas dietas comumente
utilizadas como controle, uma destas é uma ração padrão feita a base de grãos
(produtos não refinados) (conhecido em inglês como Standard Chow,
Unpurified Diet ou Grain-based Diet). A outra, é uma ração com baixo
conteúdo de gordura feita com produtos refinados, geralmente contendo a
mesma base nutricional da dieta obesogênica utilizada (conhecida em inglês
como refined low-fat diet – rLFD). As rações purificadas são feitas com
ingredientes refinados tais como caseína, proteína isolada de soja, amido de
milho, sacarose, celulose e óleos refinados, os quais contém um nutriente
principal e pouquíssimo de outros nutrientes e não-nutrientes. Diferente de
rações padrão a base de grãos, as quais são compostas, geralmente, por três
grãos integrais: milho, aveia e trigo, podendo incluir ainda farelo de alfafa,
ração para peixe, levedura de cerveja, conservantes, e um adicional de
vitaminas e minerais que, na maioria dos casos, não leva em consideração o
que já está presente nos grãos integrais acrescentados (BORTOLIN et al.,
2017; DALBY et al., 2017; PELLIZZON; RICCI, 2018).
Dalby e colaboradores (2017) observaram que animais que consomem
uma rLFD (empregada neste trabalho) ganham bem mais peso do que os

88
animais que consomem uma ração padrão baseada em grãos. Reforça-se a
grande diferença entre estes tipos de dietas, e por tal diferença na composição,
apesar de comum, torna-se impossível autores compararem e tirar conclusões
sobre as diferenças fenotípicas conduzidas por dietas completamente
diferentes (BORTOLIN et al., 2017).
Com relação ao consumo alimentar dos grupos experimentais, foi
observada diferença no consumo de ração dos grupos CAF e FCM quando
comparados o grupo controle, entretanto, não houve diferença no valor
calórico consumido ao dia entre os grupos. Quanto a ingestão hídrica, animais
que receberam a dieta CAF ingeriram menor quantidade de água quando
comparado ao grupo FCM.
Apesar do parâmetro ganho de peso do grupo CAF não ter sido
expressivo neste estudo, quando avaliada a eficiência alimentar através do
ganho de peso por consumo calórico, cálculo proposto por Nery et al. (2011)
para relacionar o ganho de peso com o valor calórico ingerido, observou-se
que o grupo cafeteria apresentou maior ganho de peso quando comparado ao
grupo FCM. Sabe-se que as fibras solúveis como a pectina, presentes na
FCM, apresentam alta capacidade em reter água e diminuir a absorção de
macronutrientes, principalmente ácidos graxos no intestino (CATALANI et
al., 2003), mecanismo que pode explicar o menor ganho de peso do grupo
FCM.
Ademais, a dieta de cafeteria ocasionou desordens metabólicas
importantes. Quanto aos parâmetros bioquímicos, foi observado no grupo
CAF intolerância oral a glicose, resistência a ação da insulina, maiores
concentrações de glicose em jejum, colesterol e triglicerídeos séricos.
Todavia, o grupo que recebeu a suplementação com a FCM, não apresentou
estas alterações metabólicas, atenuando os efeitos da exposição a dieta de
cafeteria, o que demonstra um efeito protetor a estes animais no modelo de
dieta de cafeteria.
Outros autores, que também estudaram o modelo de dieta de cafeteria
por período de tempo semelhante, obtiveram resultados parecidos. Sampey et
al. (2011) observaram que animais que receberam dieta de cafeteria por 15
semanas, apresentaram hiperglicemia, hiperinsulinemia, intolerância a
glicose, e severas alterações nos tecidos hepático e adiposo. Ballestreri et al.
(2015) constataram após 120 dias de intervenção com dieta de cafeteria,
aumento significativo nos níveis de glicemia, e triglicerídeos. Castell-Auvı et
al., (2012) observaram que a administração de dieta de cafeteria por 17

89
semanas mimetizou um estado pré-diabético, com aumento da glicemia e
secreção de insulina pelo pâncreas. Contudo, destaca-se que estudos com
período mais curtos de intervenção apresentam resultados idênticos. Dalby et
al. (2017) após 8 semanas de dieta de cafeteria constataram intolerância oral
a glicose, e Soares et al. (2017), após o mesmo período de 8 semanas
observaram concentrações aumentadas de glicose, colesterol e triglicerídeos.
Corroborando aos parâmetros bioquímicos, foram evidenciadas
alterações na morfologia das ilhotas de Langerhans do pâncreas dos animais
do grupo CAF, evidenciando hiperplasia destas células. Esta alteração no
tecido pancreático foi observada por Gao, Ma e Lio (2015) após
administração de dieta hiperlipídica por 12 semanas. Os autores sugerem que
a resistência à insulina e hiperglicemia também apresentada no estudo em
questão, desencadeou a aumento do volume celular das ilhotas pancreáticas,
mimetizando estágios iniciais da patogênese do DM tipo 2.
Estudos com a FCM demonstraram sua ação hipoglicemiante, tanto
em modelos animais (LIMA et al., 2012) quanto em pacientes com
diagnóstico de diabetes ou hiperglicemia (JANEBRO et al., 2008).
Outrossim, a FCM apresenta efeitos no metabolismo lipídico. Estudo de
pouco mais de 8 semanas com ratos diabéticos, verificou o efeito da
suplementação de 15 e 30% de FCM sob os parâmetros metabólicos,
demostrando redução nos níveis séricos de triglicerídeos em ambos os grupos
suplementados com a farinha (CORRÊA et al., 2014).
Os efeitos da FCM também são evidenciados em ensaios com
humanos, na melhora do metabolismo lipídico e glicídico, melhorando a
resistência a insulina de pacientes diabéticos tipo 2 (QUEIROZ et. al., 2012),
na redução do colesterol total e LDL em pacientes hipercolesterolêmicas
(RAMOS et al. 2007) e também na redução dos triglicerídeos e aumento do
colesterol HDL em pacientes portadores de diabetes tipo 2 (JANEBRO et al.,
2008).
Estes resultados podem estar relacionados ao efeito das fibras
presentes nesse alimento, principalmente a pectina. A hidratação da fibra
ocorre pela adsorção de água à sua superfície ou pela incorporação ao
interstício macromolecular. Na mucosa intestinal, há formação de uma
camada viscosa (gel) que altera a difusão e absorção de nutrientes. Em função
dessa maior viscosidade do conteúdo entérico, efeitos críticos regulam a
resposta metabólica à carga de nutrientes, como por exemplo, o decréscimo
na absorção de carboidratos pelo organismo, mecanismo que pode explicar

90
sua ação hipoglicemiante (MEDEIROS et al., 2009). Além disso, esse gel é
capaz de formar complexos com sais biliares, aumentando a excreção de
colesterol, podendo ser útil na prevenção e tratamento de doenças
metabólicas como: dislipidemias, DM tipo 2 e obesidade (SALLES;
BROCH; CARDOSO, 2004).
O aumento excessivo do tecido adiposo (hipertrofia e hiperplasia) é a
principal característica da obesidade. No presente estudo, observou-se
aumento da massa de tecido adiposo abdominal e epididimal dos animais do
grupo CAF, bem como no índice de adiposidade deste grupo, o que não foi
evidenciado nos animais que receberam a dieta acrescida de FCM. Nos
achados histológicos, foi evidenciada hipertrofia dos adipócitos no grupo
CAF. Animais tratados com a FCM apresentaram resultados semelhantes aos
animais do grupo controle.
No que tange ao aumento da gordura corporal, Sampey et al. (2011)
compararam o efeito da dieta de cafeteria e uma dieta rica em gordura na
indução da síndrome metabólica em ratos. Os autores demonstraram que a
dieta de cafeteria induziu um aumento de gordura epididimal. E ainda, estudo
conduzido por Navarro et al. (2016) em sua investigação sobre os efeitos da
dieta de cafeteria, observaram que animais submetidos a esta dieta
apresentaram índice de adiposidade significativamente maior em comparação
com animais do grupo controle, após a 6ª semana de experimento.
As anormalidades no tecido adiposo parecem estar diretamente
relacionadas a condição de resistência a insulina. O tecido adiposo já foi
considerado um tecido inerte servindo principalmente como estocagem de
energia. Porém, atualmente sabe-se que ele é metabolicamente ativo, grande
fonte geradora de citocinas pró e anti-inflamatórias, conhecidas como
adipocinas, tais como TNF, IL-6, leptina, resistina e adiponectina. Sabe-se
que na obesidade há um desequilíbrio entre estas adipocinas, levando ao
favorecimento de um estado pró-inflamatório. Por esse motivo, a obesidade
está associada a um quadro pró-inflamatório crônico de baixo grau, o qual
pode levar a uma série de disfunções metabólicas (LEE et al., 2013; GAO;
MA; LIU, 2015; GONZÁLEZ-MUNIESA et al., 2017).
Adipócitos hipertrofiados são menos responsivos à insulina, secretam
mais leptina, citocinas pró-inflamatórias e menos adiponectina, reproduzindo
um eterno estado inflamatório crônico, com repercussões no centro de
saciedade cerebral, aumentando o apetite e sustentando a resistência a

91
insulina e síndrome metabólica (GUSTAFSON et al, 2015; PAN; MYERS,
2018).
Este quadro pró-inflamatório crônico pode levar a um desequilíbrio
redox conhecido como estresse oxidativo, contribuindo para as disfunções
metabólicas associadas à obesidade. Adicionalmente, o estresse oxidativo
pode levar a danos celulares ou teciduais e disparar uma resposta
inflamatória, configurando um ciclo vicioso (BONDIA-PONS et al., 2012).
A inflamação crônica e o estresse oxidativo são duas importantes condições
para o desenvolvimento de síndrome metabólica, bem como de muitas
doenças degenerativas, como por exemplo DM tipo 2, doenças
cardiovasculares e hepatopatias, as quais tem seu aparecimento favorecido
pela obesidade (GONZÁLEZ-MUNIESA et al., 2017; BORTOLIN et al.,
2018).
Alterações no metabolismo dos ácidos graxos pode originar acúmulo
inapropriado de lipídeos no músculo e no fígado, prejudicando,
consequentemente suas funções (DEFRONZO, 2010). Neste modelo de
indução de síndrome metabólica através da dieta de cafeteria, o fígado parece
ser o órgão mais prejudicado pela dieta CAF. Nos achados histológicos pôde-
se verificar áreas de esteatose hepática multifocal, compatível com DHGNA,
acúmulo de colesterol e triglicerídeos no fígado, indicando que a
administração de dieta de cafeteria favorece o desenvolvimento de
anormalidades no tecido intra-hepático, com aumento de IL-6 e tendência de
elevação do TNF, mesmo sem alterações nas transaminases hepáticas,
conforme constatado neste estudo. Esse acúmulo de lipídeos hepáticos está
diretamente relacionado ao desenvolvimento de esteatose hepática e, quando
associado à FCM parece não causar tal dano.
A DHGNA é reconhecida como a manifestação hepática da síndrome
metabólica, acredita-se que assim como no tecido adiposo a DHGNA
promove uma resposta inflamatória do tipo Th1, onde as células de Kupffer
podem ser polarizadas para um fenótipo pró-inflamatório, o que contribui
com a resistência a insulina. As células de Kupffer são os macrófagos
residentes no tecido hepático, e a principal fonte de citocinas inflamatórias
no fígado. Diversos autores consolidam a teoria de que a ativação destas
células é necessária na indução da resistência a insulina hepática (KREMER
et al., 2006; LANTHIER et al., 2010; SAMPEY et al., 2011). Esta ativação
diminui a concentração ou fosforilação dos substratos de receptor de insulina,
IRS 1 e 2 e atividade da PI3K através da fosforilação em serina, translocando

92
menos o GLUT-4 e perpetuando a resistência insulínica (LANTHIER et al.,
2010).
Os mesmos autores demonstram que em resposta a alimentação com
alto teor de gordura, a sobrecarga lipídica hepática, os metabólitos lipídicos
e os mediadores pró-inflamatórios alteram a sensibilidade à insulina hepática,
consolidando que as alterações metabólicas e inflamatórias ocorrem
primeiramente no fígado, e posteriormente nos tecidos periféricos, sendo a
resistência a insulina hepática crucial para o desenvolvimento da DHGNA.
Segundo Lazo et al. (2013), o diagnóstico de DHGNA pode ser feito
mesmo com valores normais de transaminases, utilizando a presença de
infiltrado lipídico no fígado. Os autores referem que a mensuração das
enzimas pode potencialmente subestimar muitos casos da doença, já que a
definição para a presença de esteatose hepática é baseada em exames de
imagem, como a ultrassonografia (análise do quadrante hepático avaliado) ou
por meio de biópsia do tecido hepático. Todavia, estudos mostram correlação
positiva entre os níveis aumentados de insulina e ALT com risco de
desenvolvimento de esteatose hepática não alcoólica.
No entanto, até o presente momento, a associação dos níveis séricos
de ALT e AST com a incidência de síndrome metabólica não foi confirmada
em estudos transversais (ELIZONDO-MONTEMAYOR et al., 2014). Um
estudo de coorte demonstra associação entre os níveis de ALT e a incidência
tanto de síndrome metabólica e DM num período de vinte anos de
acompanhamento. Porém os autores advertem que os níveis de ALT dentro
dos parâmetros normais também podem estar associados com resultados
adversos à saúde, já que podem apresentar alterações histológicas de
DHGNA (GOESSLING et al., 2008). Autores reforçam que associação entre
os valores de ALT e a síndrome metabólica e DM são consistentes, com o
fato de que ALT é uma enzima altamente específica do fígado, já que se
encontra limitada a um componente citosólico de hepatócitos (KEW, 2000).
Por outro lado, AST é menos discriminativa ao tecido hepático, uma vez que
se encontra aumentada também por dano no coração, músculo esquelético,
rins, cérebro, pâncreas e eritrócitos (GOESSLING et al., 2008).
Além do mais, a administração de dieta de cafeteria induziu aumento
nos níveis hepáticos de IL-6, reforçando a evidência de dano hepático. A IL-
6 é uma citocina pró-inflamatória que é elevada em pacientes com DHGNA.
Neste contexto, os níveis de IL-6 indicam claramente a intensidade da lesão
do hepatócito (THIESEN et al., 2018). Juntos, o aumento nas concentrações

93
de TNF e IL-6 reforçam o grau de dano hepático provocado pela dieta CAF,
e atenuado pela dieta acrescida de FCM.
A DHGNA é a causa mais comum de enfermidade crônica do fígado
em todo o mundo (VERNON; BARANOVA; YOUNOSSI, 2011). É definida
pelo acúmulo de gordura (>5%) nas células hepáticas, sem ingestão excessiva
de álcool ou originário de outras causas de doença hepática como fatores
autoimunes, hepatotoxicidade por uso de drogas ou hepatites virais. O
espectro histológico da DHGNA alinha-se a partir de uma simples esteatose
à esteatohepatite não alcoólica, fibrose hepática, cirrose (CHALASANI et al.,
2012) e ainda está associada ao desenvolvimento de carcinoma hepatocelular.
Essa doença parece afetar aproximadamente 30% da população de países
ocidentais, especialmente em pacientes com síndrome metabólica, obesidade
e diabetes tipo 2. A DHGNA é considerada um fator de risco independente
para o desenvolvimento de doença cardiovascular e as evidências suportam
sua relação também como causa do desenvolvimento de doença renal crônica
(LOOMBA; SANYAL, 2013).
No presente estudo, foram evidenciadas alterações no tecido e
marcadores renais. Animais do grupo CAF apresentaram discreta atrofia dos
glomérulos renais, menor área destas células e peso do órgão, igualmente,
maiores concentrações séricas de creatinina. Em contrapartida, o grupo FCM
não demonstrou as mesmas alterações, apresentando resultados semelhantes
ao grupo controle.
Segundo Navarro e colaboradores (2016), os mecanismos pelos quais
a obesidade predispõe as pessoas a danos renais são desconhecidos,
entretanto, sabe-se que esta condição provoca aumento da inflamação e lesão
no tecido renal, principalmente causadas pelo estresse oxidativo. Os mesmos
autores não observaram alterações estruturais nos rins após a administração
de dieta CAF por 6 semanas, porém, observaram aumento de citocinas
inflamatórias nos rins e níveis plasmáticos de AGEs elevados.
Os resultados apresentados indicam que o uso da FCM traz efeitos
positivos em animais com síndrome metabólica induzida por dieta de
cafeteria, podendo ser utilizado como estratégia no tratamento da obesidade
e síndrome metabólica.
Medeiros et al. (2009) com o objetivo de avaliar, não somente o efeito
terapêutico, mas também o potencial tóxico da FCM, submeteram indivíduos
saudáveis a 10 g do produto três vezes ao dia, totalizando 30g durante oito
semanas. O tratamento promoveu redução significativa na glicemia e peso
94
corporal (2% de redução) dos indivíduos, e constatou que o seu uso foi bem
tolerado pelos voluntários, não sendo relatadas reações adversas ou sinais de
toxicidade que pudessem comprometer sua utilização como alimento com
propriedade de saúde, nesta dosagem e período experimental.
Embora os resultados com humanos sejam interessantes no tratamento
de doenças metabólicas como o diabetes e hipercolesterolemia, reforça-se a
necessidade de estudos mais aprofundados que visem avaliar seu efeito
toxicológico, bem como o teor de substâncias designadas glicosídeos
cianogênicos, que são tóxicos e apresentam-se em quantidades significativas
em todas as partes da Passiflora edulis (COQUEIRO; PEREIRA;
GALANTE, 2016).
Figura 17. Resumo gráfico de resultados da pesquisa.
Fonte: o autor.

95
7 CONCLUSÕES
Os achados do presente trabalho demonstram que a dieta de cafeteria
é um modelo robusto para estudar os efeitos da Síndrome Metabólica,
gerando importantes alterações após o período de 16 semanas, tais alterações
que tem seu início a partir da resistência à insulina e um processo de
deposição de gorduras, tendo o fígado e pâncreas como principais órgãos
comprometidos.
Animais tratados com FCM apresentaram diminuição nos parâmetros
que estabelecem e caracterizam a Síndrome Metabólica, principalmente o
quadro de resistência à insulina, tolerância a glicose, deposição de gordura
corporal e acúmulo de lipídeos no fígado, bem como parâmetros
pancreáticos e renais. Desta forma, a FCM pode ser estabelecida como uma
fonte importante de fibras e metabólitos secundários que ajudam a evitar os
danos ocasionados pelo consumo da dieta de cafeteria. Tendo em vista a
necessidade de encontrar novas alternativas capazes de melhorar a saúde e
qualidade de vida da população, além de considerar que a FCM se constitui
de um resíduo agroindustrial, apresentando baixo custo e pouca
industrialização, destaca-se seu potencial uso como coadjuvante nas doenças
metabólicas. Novos estudos acerca dos benefícios desse produto necessitam
ser realizados a fim de definir dose ideal por dia, assim como, outras formas
de consumo.

96

97
REFERÊNCIAS ALBERTI, K. G. M. M.; ZIMMET, P.; SHAW, J.. Metabolic syndrome-a new
world-wide definition. A Consensus Statement from the International Diabetes Federation. Diabetic Medicine, v. 23, n. 5, p.469-480, 2006. ALBERTI, K. G.; ECKEL, R. H.; GRUNDY, S. M.; ZIMMET, P. Z.; CLEEMAN, J. I.; DONATO, K. A.; FRUCHART, J. C.; JAMES, W. P.; LORIA, C. M.; SMITH JR, S. C. Harmonizing the metabolic syndrome: a joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World
Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation, v. 120, n. 16, p. 1640–1645, 2009. ANDRADE, G. F.; ALMEIDA, C. G.; ESPESCHIT, A. C. R.; DANTAS, M. I. S.; BENJAMIN, L. A.; RIBEIRO, S. M. R.; MARTINO, H. S. D. The addition of whole soy flour to cafeteria diet reduces metabolic risk markers in wistar rats. Lipids in Health and Disease, v. 12, n. 145, 2013.
BAHADORAN, Z.; MIRMIRAN, P.; AZIZI, F. Dietary polyphenols as potential nutraceuticals in management of diabetes: a review. Journal of diabetes &
metabolic disorders, v. 12, n. 1, p. 43, 2013.
BALLESTRERI, E.; MARCON, I. F.; TAVARES, R. G. Comparação de modelos de indução da síndrome metabólica: dieta com excesso de frutose e dieta hiperlipidêmica. Revista Brasileira de Obesidade, Nutrição e Emagrecimento, v. 9, n. 51, p. 96-104, 2015.
BARRETT, P.; MERCER, J. G.; MORGAN, P. J. Preclinical models for obesity research. Disease Models and Mechanisms, v. 9, p.1245–55, 2016. BARROSO, S. R.; ABREU, V. G.; FRANCISCHETTI, E. A. A participação do tecido adiposo visceral na gênese da hipertensão e doença cardiovascular aterogênica: um conceito emergente. Arquivos Brasileiros de Cardiologia, v. 78, n. 6, p. 618-630, 2002.
BASU, S.; HAZRA, B. Evaluation of nitric oxide scavenging activity, in vitro and ex vivo, of selected medicinal plants traditionally used in inflammatory diseases. Phytotherapy research, v. 20, n. 10, p. 896-900, 2006. BATISTA FILHO, M.; BATISTA, L. V. Transição alimentar/nutricional ou mutação antropológica? Ciencia e Cultura, v. 62, n. 4, São Paulo, 2010.
BENZIE, I. F. F.; STRAIN, J. J. The ferric reducing ability of plasma (frap) as a measure of “antioxidant power”: the frap assay. Analytical Biochemistry, v. 239, p. 70-76, 1996.

98
BERNAUD, F. S. R.; RODRIGUES, T. C. Fibra alimentar: ingestão adequada e efeitos sobre a saúde do metabolismo. Arquivos brasileiros de endocrinologia &
metabologia, v. 57, n. 6, p. 397-405, 2013. BEUTLER, E. Red cell metabolism: a manual of biochemical methods. 2.ed. New York, London: Grune & Stratton, p. 89-90, 1975. BORTOLIN, R. C.; VARGAS, A. R ; GASPAROTTO, J.; CHAVES, P. R.; SCHNORRL, C. E.; MARTINELLO, K. B.; SILVEIRA, A. K.; RABELO, T. K.; GELAIN, D. P.; MOREIRA, J. C. F. A new animal diet based on human Western
diet is a robust diet-induced obesity model: comparison to high-fat and cafeteria diets in term of metabolic and gut microbiota disruption. International Journal of
Obesity, v. 42, n. 3, p. 525, 2018. BRADFORD, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, v. 72, p. 677-85, 1985. BRASIL. Ministério da Saúde. ANVISA. Agência Nacional de Vigilância Sanitária .
Resolução RDC n° 259 de 20 de setembro de 2002. Regulamento técnico para
rotulagem de alimentos embalados. Disponível em: < http://elegis. anvisa. gov. br/leisref/public/showAct. php?id=1261 &word= > Acesso em: 28 Julho de 2018. CAMERON, A. J.; SHAW, J. E.; ZIMMET, P. Z. The metabolic syndrome: prevalence in worldwide populations. Endocrinology and Metabolism Clinics
North America, v. 33, p. 351–75, 2004.
CAMPOS, K. E.; SINZATO, Y. K.; DAMASCENO, D. C.; RUDGE, M. V. C. Obesidade e Resistencia à insulina. FEMINA, v. 34, n. 9, 2006. CARVALHO, G. Q.; ALFENAS, R. C. G. Índice glicêmico: uma abordagem crítica acerca de sua utilização na prevenção e no tratamento de fatores de risco cardiovasculares. Revista de Nutrição, v. 21, n. 5, p. 577-587, 2008. CARVALHO, Melicia Cintia GALDEANO. Benefícios e perigos do aproveitamento
da casca de maracujá (Passiflora edulis) como ingrediente na produção de alimentos. Rev Inst Adolfo Lutz, v. 72, n. 1, p. 1-9, 2013. CATALANI, L. A.; KANG, E. M. S.; DIAS, M. C. G.; MACULEVICIUS, J. Fibras alimentares. Rev Bras Nutr Clin, v. 18, n. 4, p. 178-82, 2003.
CASTELL-AUVI, A.; CEDO, L.; PALLARES, V.; BLAY, M.; ARDEVOL, A.; PINENT, M. The effects of a cafeteria diet on insulin production and clearance in rats. British Journal of Nutrition, v. 108, n. 7, p. 1155-1162, 2012.

99
CAZARIN, C. B. B.; SILVA, J. K. S.; COLOMEU, T. C.; ZOLLNER, R. L.; MARÓSTICA JUNIOR, M. R. Capacidade antioxidante e composição química da casca de maracujá (Passiflora edulis). Ciência Rural, Santa Maria, v. 44, n. 9, p.
1699-1704, set, 2014. CECHINEL-FILHO, V.; YUNES, R. A. Estratégias para a obtenção de compostos farmacologicamente ativos a partir de plantas medicinais. Conceitos sobre modificação estrutural para otimização da atividade. Química nova, v. 21, n. 1, p. 99-105, 1998. CHALASANI, N.; YOUNOSSI, Z.; LAVINE, J.E.; DIEHL, A.M.; BRUNT, E.M.;
CUSI, K.; CHARLTON, M.; SANYAL, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology, v. 55, p. 2005–2023, 2012. CHAWLA, A.; NGUYEN, K. D.; GOH, Y. P. S. Macrophage mediated inflammation in metabolic disease. Nat. Rev. Immunol., v. 11, n. 11, p. 738–749, 2011.
COQUEIRO, A. Y.; PEREIRA, J. R. R.; GALANTE, F. Farinha da casca do fruto de Passiflora edulis f. flavicarpa Deg (maracujá-amarelo): do potencial terapêutico aos efeitos adversos. Rev. Bras. Pl. Med., Campinas, v. 18, n. 2, p. 563-569, 2016. CORNIER, M. A.; DABELEA, D.; HERNANDEZ, T. L.; LINDSTROM, R. C.; STEIG, A. J.; STOB, N. R.; VAN PELT, R.. E.; WANG, H.; ECKEL, R. H. The metabolic syndrome. Endocr Rev, v. 29, n. 7, p. 777-822, 2008.
CORRÊA, E. M.; MEDINA, L.; BARROS-MONTEIRO, J.; VALLE, N. O.; SALES, R.; MAGALÃES, A.; SOUZA, F. C. A.; CARVALHO, T. B.; LEMOS, J. R.; LIRA, E. F.; LIMA, E. S.; GALENO, D. M. L.; MORALES, L.; ORTIZ, C.; CARVALHO, R. P. The intake of fiber mesocarp passionfruit (Passiflora edulis) lower of triglyceride and cholesterol decreasing principally insulin and leptin. J
Aging Res Clin Pract., v. 3, n.1, p. 31-35, 2014. COUTINHO, J. G.; GENTIL, P. C.; TORAL, N. A desnutrição e obesidade no
Brasil: o enfrentamento com base na agenda única da nutrição. Cad. Saúde Pública, v. 24, n. 2, p. 332-340, 2008. CZECH, M. P. Insulin action and resistance in obesity and type 2 diabetes. Nature
medicine, v. 23, n. 7, p. 804, 2017.
DALBY, M. J.; ROSS, A. W.; WALKER, A. W.; MORGAN, P. J. Dietary
Uncoupling of Gut Microbiota and Energy Harvesting from Obesity and Glucose Tolerance in Mice. Cell Rep, v. 21, p. 1521–33, 2017.

100
DAMATTA, R. A. Modelos animais na pesquisa biomédica. Scientia Medica, v. 20, n. 3, p. 210-211, 2010.
DAVIS, R. A. H.; PLAISANCE, E. P.; ALLISON, D. B. Complementary Hypotheses on Contributors to the Obesity Epidemic. Obesity (Silver Spring), v. 26, n. 1, p. 17–21, 2018. DEFRONZO, R. A. Insulin resistance, lipotoxicity, type 2 diabetes and atherosclerosis: the missing links. The Claude Bernard Lecture 2009. Diabetologia, v. 53, n. 7, p. 1270-1287, 2010.
DHAWAN, K.; DHAWAN, S.; SHARMA, A. Passiflora: a rewiew update. J.
Ethnopharmacol., v. 94, p. 1-23, 2004. ECKEL, R. H.; GRUNDY, S. M.; ZIMMET, P. Z. The metabolic syndrome Lancet, v. 365, p. 1415–28, 2005. EGUCHI, R.; CHEIK, N. C.; OYAMA, L. M.; NASCIMENTO, C. M. O. MELLO,
M. T.; TUFIK, S.; DAMASO, A. Efeitos do Exercício Crônico Sobre a Concentração Circulante da Leptina e Grelina em Ratos com Obesidade Induzida por Dieta. Rev Bras Med Esporte, v. 14, n. 3, 2008. ELIZONDO-MONTEMAYOR, L.; UGALDE-CASAS, P. A.; LAM-FRANCO, L., et al. Association of ALT and the metabolic syndrome among Mexican children. Obes Res Clin Pract, v. 8, p. 79-87, 2014.
EMBRAPA. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Maracujá: o produtor pergunta, a Embrapa responde - Fábio Gelape Faleiro, Nilton Tadeu Vilela Junqueira, editores técnicos. Brasília, DF: Embrapa, 2016. ESTADELLA, D.; OYAMA, L. M.; BUENO, A. A.; HABITANTE, C. A.; SOUZA, G. I.; RIBEIRO, E. B.; MOTOYAMA, C. S. M.; NASCIMENTO, C. M. O. Effect of palatable hyperlipidic diet on lipid metabolism of sedentary and exercised rats. Nutrition, v. 20, p. 218-24, 2004.
FAGUNDES, D. J.; TAHA, M. O. Modelo animal de doença: critérios de escolha e espécies de animais de uso corrente. Acta Cirúrgica Brasileira, v. 19, n. 1, p. 59-65, 2004. FERREIRA, A.; CASTRO, J. P.; ANDRADE, J. P.; DULCE MADEIRA, M.; CARDOSO, A. Cafeteria-diet effects on cognitive functions, anxiety, fear and neurogenesis in the juvenile rat. Neurobiol Learn Mem., v. 31, p. 197-207, 2018.
FOLCH, J.; LEES, M.; SLOANE STANLEY, G. H. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem., v. 226, n. 1, p. 497–509, 1957.

101
FORD, E. S. Risks for all-causemortality, cardiovascular disease,and diabetes associated with the metabolic syndrome: a summary of the evidence. Diabetes
Care, v. 28, n. 7, p. 1769–1778, 2005.
FORD, E. S.; GILES, W. H.; DIETZ, W. H. Prevalence of the metabolic syndrome among US adults: findings from the third National Health and Nutrition Examination Survey. Jama, v. 287, n. 3, p. 356-359, 2002. FOSTER, L. J; KLIP, A. Mechanism and regulation of GLUT-4 vesicle fusion in muscle and fat cells. Am J Physiol Cell Physiol, v. 279, n. 4, p. 877-90, 2000.
FURUKAWA, S.; FUJITA, T.; SHIMABUKURO, M.; IWAKI, M.; YAMADA, Y.; NAKAJIMA, Y.; NAKAYAMA, O.; MAKISHIMA, M.; MATSUDA, M.; SHIMOMURA1, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J Clin Invest., v. 114, n. 12, p. 1752-1761, 2017. GAGGINI, M.; MORELLI, M.; BUZZIGOLI, E.; DEFRONZO, R. A.; BUGIANESI, E.; GASTALDELLI, A. Non-alcoholic fatty liver disease (NAFLD) and its connection with insulin resistance, dyslipidemia, atherosclerosis and coronary heart disease. Nutrients, v. 5, n. 5, p. 1544–1560, 2013.
GALISTEO, M.; DUARTE, J.; ZARZUELO, A. Effects of dietary fibers on disturbances clustered in the metabolic syndrome. J. Nutr. Biochem., v. 19, p. 71–84, 2008. GAO, M.; MA, Y.; LIU, D. High-Fat Diet-Induced Adiposity, Adipose Inflammation, Hepatic Steatosis and Hyperinsulinemia in Outbred CD-1 Mice. Plos
one, v. 13, 2015.
GASTIAZORO, M. P.; GUERRERO-SCHIMPF, M.; DURANDO, M.; LAZZARINO, G. P.; ANDREOLI, M. F.; ZIERAU, O.; LUQUE, E. H.; RAMOS, J. G.; VARAYOUD, J. Induction of uterine hyperplasia after cafeteria diet exposure. Molecular and Cellular Endocrinology, v. 18, 2018. GAUR, A.; PAL, G. K.; ANANTHANARAYANAN, P. H.; PAL, P. Role of ventromedial hypothalamus in high fat diet induced obesity in male rats:association
with lipid profile, thyroid profile and insulin resistance. Anna neurosci, v. 21, n. 3, p. 104-7, 2014. GIL-CARDOSO, K.; GINÉS, I.; PINENT, M.; ARDÉVOL, A.; TERRA, X.; BLAY, M. A cafeteria diet triggers intestinal inflammation and oxidative stress in obese rats. British Journal of Nutrition, v. 117, p. 218–229, 2017. GOESSLING, W. M. J.; WASAN, R. S.; D'AGOSTINO, R. B., et al.
Aminotransferase levels and 20-year risk of metabolic syndrome, diabetes, and cardiovascular disease. Gastroenterology, v. 135, p. 1935-44, 2008.

102
GOLLISCH, K. S. C.; BRANDAUER, J.; JESSEN, N.; TOYODA, T.; NEYER, A.; HIRSHMAN, M. F.; GOODYEAR, L. J. Effects of exercise training on subcutâneos and visceral adipose tissue in normal and high-fat diet-fed rats. Am. J. Physiol.
Endocrinol. Metab, 2009. GOMES, F.; TELO, D. F.; SOUZA, H. P.; NICOLAU, J. C.; HALPERN, A.; SERRANO JR, C. V. Obesidade e doença arterial coronariana: papel da inflamação vascular. Arquivos Brasileiros de Cardiologia, v. 94, n. 2, p. 273-279, 2010. GOMEZ-SMITH, M.; KARTHIKEYAN, S.; JEFFERS, M. S.; JANIK, R.; THOMASON, L. A.; STEFANOVIC, B.; CORBETT, D. A physiological
characterization of the Cafeteria diet model of metabolic syndrome in the rat. Physiology & Behavior, v. 167, n. 1, p. 382-91, 2016. GONDIM, J. A. M.; MOURA, M. F. V.; DANTAS, A. S.; MEDEIROS, R. L. S.; SANTOS, K. M. Composição centesimal e de minerais em cascas de frutas, Cien
Tecnol Aliment, v. 25, p. 825-827, 2005. GONZÁLEZ-MUNIESA, P.; MÁRTINEZ-GONZÁLEZ, M. A.; HU, F. B.; DESPRÉS, J. P.; MATSUZAWA, Y.; LOOS, R. J. F.; MORENO, L. A.; BRAY, G.
A.; MARTINEZ, J. A. Obesity. Nat. Rev. Dis. Prim, v. 3, 2017. GOSS, M. J.; NUNES, M. L. O.; MACHADO, I. D.; MERLIN, L.; MACEDO, N. B.; SILVA, A. M. O.; BRESOLIN, T. M. B.; SANTIN, J. R. Peel flour of Passiflora edulis Var. Flavicarpa supplementation prevents the insulin resistance and hepatic steatosis induced by low-fructose-diet in young rats. Biomedicine &
Pharmacotherapy, v. 102, p. 848–54, 2018.
GOULARTE, J. F.; FERREIRA, M. B. C.; SANVITTO, G. Effects of food patern change and physical exercise on cafeteria diet-induced obesity in female rats. British Journal of Nutrition, p. 1-8, 2012. GRIESS, P. Bemerkungen zu der abhandlung der H.H. Weselsky und Benedikt .Ueber einige azoverbindungen. Chem Ber, v. 12 p. 426-428, 1879. GRUNDY, S. M.; BREWER, H. B. Jr; CLEEMAN,J. I.; SMITH, S. C. Jr;
LENFANT, C.; AMERICAN HEART ASSOCIATION; NACIONAL HEART, LUNG, AND BLOOD INSTITUTE. Definition of metabolic syndrome report of the National Heart, Lung, and Blood Institute/American Heart Association Conference on scientific issues related to definition. Circulation, v. 109, n. 3, p. 433-438, 2004. GUIMARÃES, I. C. B.; GUIMARÃES, A. C. Síndrome Metabólica na infância e adolescência. Um fator maior de risco cardiovascular. Revista Baiana de Saúde
Pública, v. 30, n. 2, p. 349, 2006.

103
GUSTAFSON, B.; HEDJAZIFAR, S.; GOGG, S.; HAMMARSTEDT, A.; SMITH, U. Insulin resistance and impaired adipogenesis. Trends in Endocrinology &
Metabolism, v. 26, n. 4, p. 193-200, 2015
GUTIÉRREZ‐MARTOS, M.; GIRARD, B.; MENDONÇA‐NETTO, S.; PERROY, J.; VALJENT, E.; MALDONADO, R.; MARTIN, M. Cafeteria diet induces neuroplastic modifications in the nucleus accumbens mediated by microglia activation. Addiction Biology, v. 23, n. 2, p. 735-49, 2018. HIGA, T. S.; SPINOLA, A. V.; FONSECA-ALANIZ, M. H.; EVANGELISTA, F. S. Comparison between cafeteria and high-fat diets in the induction of metabolic
dysfunction in mice. Int J Physiol Pathophysiol Pharmacol, v. 6, n. 1, p. 47–54, 2014. IANTORNO, M.; CAMPIA, U.; DI DANIELE, N.; NISTICO, S.; FORLEO, G. B.; CARDILLO, C.; TESAURO, M. Obesity, inflammation and endothelial dysfunction. J Biol Regul Homeost Agents, v. 28, p. 169-76, 2014. IBGE. INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Anuário
Estatístico do Brasil. Brasil. Rio de Janeiro, p. 458, 2012.
IBGE. INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Pesquisa
Nacional de Saúde, 2013. Brasil. Rio de Janeiro; 2013. INSTITUTE OF MEDICINE, FOOD AND NUTRITION BOARD. Dietary
Reference Intakes: Energy,Carbohydrates, Fiber, Fat, Fatty Acids, Cholesterol, Protein and Amino Acids. Washington: National Academies Press, 2002.
JANEBRO, D. I.; QUEIROZ, M. S. R.; RAMOS, A. T.; SABAA-SRUR, A. U. O. CUNHA, M. A. L. Efeito da farinha da casca do maracujá amarelo (Passiflora edulis flavicarpa) nos níveis glicêmicos e lipídicos de pacientes diabéticos tipo 2. Rev. Bras. Farmacogn., v. 2, p. 724-732, 2008. KASSI, E.; PERVANIDOU, P.; KALTSAS, G.; CHROUSOS, G. Metabolic syndrome: definitions and controversies. BMC Med, v. 9, n. 48, 2011.
KEW, M. C. Serum aminotransferase concentration as evidence of hepatocellular damage. Lancet, v. 355, p. 591–2, 2000. KREMER, M.; HINES, I. N.; MILTON, R. J.; WHEELER, M. D. Favored T helper 1 response in a mouse model of hepatosteatosis is associated with enhanced T cellmediated hepatitis. Hepatology, v. 44, p. 216-27, 2006. KYLIN, E. S. Hypertonie-Hyperglykamie-Hyperurikamie syndrome. Zentralblatt
fur innere Medizin, v.44, 1923.

104
LAKKA, H. M.; LAAKSONEN, D. E.; LAKKA, T. A.; NISKANEN, L. K.; KUMPUSALO, E.; TUOMILEHTO, J.; SALONEN, J. T. The metabolic syndrome and total and cardiovascular disease mortality in middle-aged men. The Journal of
American Medical Association, v. 288, n. 21, p. 2709- 2716, 2002. LAMBETH, J. D.; NEISH, A. S. Nox enzymes and new thinking on reactive oxygen: a double-edged sword revisited. Annu Rev Pathol, v. 9, p. 119-45, 2014. LANTHIER, N.; MOLENDI-COSTE, O.; HORSMANS, Y.; VAN ROOIJEN, N.; CANI, P. D.; LECLERCQ, I. A. Kupffer cell activation is a causal factor for hepatic insulin resistance. Am J Physiol Gastrointest Liver Physiol, v. 298, p.107–16,
2010. LAZO, M.; HERNAEZ, R.; EBERHARDT, M. S.; BONEKAMP, S.; KAMEL, I.; GUALLAR, E.; KOTEISH, A.; BRANCATI, F. L.; CLARK, J. M. Prevalence of nonalcoholic fatty liver disease in the United States: the Third National Health and Nutrition Examination Survey, 1988-1994. Am J Epidemiol., v. 178, n. 1, p. 38-45, 2013 LEE, E. J.; PARK, J. H. Receptor for advanced glycation endproducts (RAGE), its
ligands, and soluble RAGE: Potential biomarkers for diagnosis and therapeutic targets for human renal diseases. Genomics Inform, v. 11, n. 4, p. 224–9, 2013. LIMA, E. S.; SCHWERTZ, M. C.; SOBREIRA, C. R. C.; BORRAS, M. R. L. Efeito hipoglicemiante da farinha do fruto de maracujá-do-mato (Passiflora nítida Kunth) em ratos normais e diabéticos. Rev. Bras. Pl. Med, v.14, n.2, p.383-388, 2012.
LIU, F. F.; ANG, C. Y. W.; SPRINGER, D. Optimization of extraction conditions for active components in Hypericum perforatum using surface methodology. Journal of Agriculture and Food Chemistry, v. 48, p. 3.364-3.371, 2000. LONARDO, A. Fatty liver and nonalcoholic steatohepatitis. Where do we stand and where are we going? Digestive Diseases, v. 17, n. 2, p. 80-89, 1999. LOOMBA, R.; SANYAL, A.J. The global NAFLD epidemic. Nat. Rev.
Gastroenterol. Hepatol., v. 10, 686–690, 2013. LOPES, H. F. Hipertensão, obesidade, resistência à insulina e síndrome metabólica. Rev. Bras. Hipertens, v. 12, p. 154-158, 2005. LOUZADA, M. L. C.; MARTINS, A. P. B.; CANELLA, D. S.; BARALDI, L. G.; LEVY, R. B.; CLARO, R. M.; MOUBARAC, J.; CANNON, G.; MONTEIRO, C. A. Alimentos ultraprocessados e perfil nutricional da dieta no Brasil. Rev. Saúde
Pública, 2015.

105
LOUZADA, S. M.; GUILHERME, C.; MACHADO, F. S.; ARAÚJO, A. S.; SANVITTO, G. L.; FUNCHAL, C. Efeito da obesidade induzida pela dieta da cafeteria sobre os parâmetros oxidativos no ovário de ratas. Revista da Sociedade
Brasileira de Ciência em Animais de Laboratório, v. 2, n. 2, p. 92-102, 2013. MAFFETONE, P. B.; RIVERA-DOMINGUEZ, I.; LAURSEN, P. B.; Overfat adults and children in developed countries: The public health importance of identifying excess body fat. Front. Public Heal, v. 5, p. 1–11, 2017. MAMIKUTTY, N.; THENT, Z. C.; SAPRI, S. R.; SAHRUDDIN, N. N.; MOHD YUSOF, M. R.; HAJI SUHAMI, F. The establishment of metabolic syndrome
model by induction of fructose drinking water in male Wistar rats. Biomed. Res.
Int., 2014. MARCO, G.J. A rapid method for evaluation of antioxidants. J. Am. Oil Society, v. 45, p. 594-598, 1968 MARQUES, A. C. R.; GABBIATTI, G. C.; GRAVENA, Â. A. F.; AMARAL, V. Influência das dietas hipercalóricas sobre os parâmetros de obesidade, dislipidemia e hiperglicemia em ratos. Revista Saúde e Pesquisa, v. 8, n. 1, p. 55-62, 2015.
MARTINEZ, R.; TORRES, P.; MENESES, M. A.; FIGUEROA, J. G.; PEREZ-ALVAREZ, J. A.; VIUDA-MARTOS, M. Chemical, technological and in vitro antioxidant properties of mango, guava, pineapple and passion fruit dietary fibre concentrate. Food Chemistry, v.135, n.3, p.1520-1526, 2012. MATSUURA, F. C. A. U. Estudo do albedo de maracujá e de seu aproveitamento em barra de cereais. 2005. 157 f. Tese (Doutorado em Tecnologia de Alimentos) -
Universidade Estadual de Campinas, Campinas, 2005. MATTOS, L. L.; MARTINS, I. S. Consumo de fibras alimentares em população adulta. Rev Saúde Pública, v. 34, n. 1, p. 50-55, 2000. MAVROGIANNAKI, A. N.; MIGDALIS, I. N. Nonalcoholic fatty liver disease, diabetes mellitus and cardiovascular disease: newer data. Int J Endocrinol, 2013.
MAYER JR, O.; BRUTHANS, J.; SEIDLEROVÁ, J.; KARNOSOVÁ, P.; VANĚK, J.; HRONOVÁ,M.; GELŽINSKÝ, J.; CVÍČELA, M.; WOHLFAHRT, P.; CÍFKOVÁ, R.; FILIPOVSKÝ, J. Prospective study of metabolic syndrome as a mortality marker in chronic coronary heart disease patients. European Journal of
Internal Medicine, v. 47, p. 55-61, 2018. MC CORD, J. M.; FRIDOVICH, I. Superoxide dismutase, an enzyme function for erythrrhrocuprein (hemocuprein). Journal of Biological Chemistry, v. 244, p.
6049-55, 1969.

106
MEDEIROS, J. S.; DINIZ, M. F. F. M.; SRUR, A. U. O. S.; PESSOA, M. B.; CARDOSO, M. A. A.; CARVALHO, D. F. Ensaios toxicológicos clínicos da casca do maracujá-amarelo (Passiflora edulis, f.flavicarpa), como alimento com
propriedade de saúde. Brazilian Journal of Pharmacognosy. v.19, n.2, p. 394-399, 2009. MELETTI, L. M. M. Avanços na cultura do maracujá no Brasil. Rev Bras Frutic, p. 83-91, 2011. MELLOUK, Z.; AGUSTINA, M.; RAMIREZ, M.; PENA, K.; ARIVALO, J. The therapeutic effects of dietary krill oil (Euphausia superba) supplementation on
oxidative stress and DNA damages markers in cafeteria diet-overfed rats. Annales
de Cardiologie et D'angeiologie, v. 65, v. 3, p. 223-8, 2016. MERCADANTE, A. Z.; BRITTON, G.; RODRIGUEZ-AMAYA, D. B. Carotenoids from Yellow Passion Fruit (Passiflora e dulis). Journal of Agricultural and Food
Chemistry, v. 46, n. 10, p. 4102-4106, 1998. MIGLIO, C.; PELUSO, I.; RAGUZZINI, A.; VILLANO, D. V.; CESQUI, E.; CATASTA, G.; TOTI, E.; SERAFINI, M. Fruit juice drinks prevent endogenous
antioxidant response to high-fat meal ingestion. British Journal of Nutrition, v. 111, n. 2, p. 294–300, 2014. MILANSKI, M.; DEGASPERI, G.; COOPE, A.; MORARI, J.; DENIS, R.; CINTRA, D. E.; TSUKUMO, D. M. L.; ANHE, G.; AMARAL, M. E.; TAKAHASHI, H. K.; CURI, R.; OLIVEIRA, H. C.; CARVALHEIRA, J. B. C.; BORDIN, S.; SAAD, M. J.; VELLOSO, L. A. Saturated fatty acids produce an inflammatory response predominantly through the activation of TLR4 signaling in
hypothalamus: implications for the pathogenesis of obesity. J Neurosci, v. 29, n. 2, p. 359-70, 2009. MILLER, H. E. A simplified method for the evaluation of antioxidants. Journal of
the American Oil Chemists' Society, v. 48, n. 2, p. 91-91, 1971. MOREIRA, A. V. B.; MANCINI FILHO, J. Influência dos compostos fenólicos de especiarias sobre a lipoperoxidação e o perfil lipídico de tecidos de ratos. Revista de
Nutrição da PUCCAMP, v. 17, n. 4, p. 411-24, 2004. MOTTILLO, S. FILION, K. B.; GENEST, J.; JOSEPH, L.; PILOTE, L.; POIRIER,P.; RINFRET,S.; SCHIFFRIN, E. L.; EISENBERG, M. J. The metabolic syndrome and cardiovascular risk: a systematic review and meta-analysis. Journal
of the American College of Cardiology, v. 56, n. 14, p. 1113- 1132, 2010. MOZAFFARIAN, D.; PISCHON, T.; HANKINSON, S. E.; RIFAI, N.;
JOSHIPURA, K.; WILLET, W. C.; RIMM, E. B. Dietary intake of trans fatty acids and systemic inflammation in women. The American Journal of Clinical
Nutrition, v. 79, n. 4, p. 606-612, 2004.

107
MUNTZEL, M. S.; AL-NAIMI, O. A.; BARCLAY, A.; AJASIN, D. Cafeteria Diet Increases Fat Mass and Chronically Elevates Lumbar Sympathetic Nerve Activity in
Rats. Hypertension, p. 1498–1502, 2012. MUSSO, G.; GAMBINO; R., BO, S.; UBERTI, B.; BIROLI, G.; PAGANO, G.; CASSADER, M. Should nonalcoholic fatty liver disease be included in the definition of metabolic syndrome? A cross-sectional comparison with Adult Treatment Panel III criteria in nonobese nondiabetic subjects. Diabetes care, v. 31, n. 3, p. 562-568, 2008.
NAKAZONE, M. A.; PINHEIRO, A.; BRAILE, M. C. V. B.; PINHEL, M. A. S.; SOUSA, G. F.; PINHEIRO JÚNIOR, S.; BRANDÃO, A. C. TOLEDO, J. C. Y.; BRAILE, D. M.; SOUZA, D. R. S. Prevalência de síndrome metabólica em indivíduos brasileiros pelos critérios de NCEP-ATPIII e IDF. Rev Assoc Med Bras, v. 53, n. 5, p. 407-13, 2007. NASCIMENTO, A. F.; SUGIZAKI, M. M.; LEOPOLDO, A. S.; LIMA-LEOPOLDO, A. P.; LUVIZOTTO, R. A. M.; NOGUEIRA, C. R.; CICOGA, A. C. A hypercaloric pellet-diet cycle induces obesity and co-morbidities in Wistar
rats. Arquivos Brasileiros de Endocrinologia & Metabologia, v. 52, n. 6, p. 968-974, 2008. NASCIMENTO, E. M. G. C.; ASCHERI, J. L. R.; CARVALHO, C. W. P.; GALDEANO, M. C. Benefícios e perigos do aproveitamento da casca de maracujá (Passiflora edulis) como ingrediente na produção de alimentos. Rev Inst Adolfo
Lutz, v. 72, n. 1, p. 1-9, 2013.
NAVARRO, M. E. L.; SANTOS, K. C.; NASCIMENTO, A. F.; FRANCISQUETI, F. V.; MINATEL, I. O.; PIERINE, D. T.; LUVIZOTTO, R. A. M.; FERREIRA, A. L.; CAMPOS, D. H. S.; CORRÊA, C. R. Inflamação renal, alterações metabólicas e oxidativas após 6 semanas de dieta de cafeteria em ratos. Brazilian Journal of
Nephrology, p. 9-14, 2016. NÉIA, V. B. M. J. C.; AMBROSIO-ALBUQUERQUE, E. P.; FIGUEIREDO, I. L.; BOEING, J. S.; SILVA, T. C.; LEWANDOWSKI, V.; RIBEIRO, R. P.;
VISENTAINERE, J. E. L.; VISENTAINER, J. V. Impact of cafeteria diet on the composition of fatty acids in zebrafish (Danio rerio) fillets. Journal of the
Brazilian Chemical Society, v. 29, n. 6, p. 1183-1188, 2018. NERY, C. D. S.; PINHEIRO, I. L.; MUNIZ, G. D. S.; VASCONCELOS, D. A. A. D.; FRANÇA, S. P. D.; NASCIMENTO, E. D. Medidas murinométricas e eficiência alimentar em ratos provenientes de ninhadas reduzidas na lactação e submetidos ou não ao exercício de natação. Revista Brasileira de Medicina do Esporte, v. 17, n.
1, p. 49-55, 2011.

108
OLIVEIRA, C. L.; MELLO, M. T.; CINTRA, I. P.; FISBERG, M. Obesidade e síndrome metabólica na infância e na adolescência. Revista de nutrição, v. 17, n. 2, p. 237-245, 2004.
OLIVEIRA, T. W. S.; LEANDRO, C. G.; DEIRO, T. C. B. J.; PEREZ, G. S.; SILVA, D. F.; DRUZIAN, J. I.; COUTO, R. D.; BARRETO-MEDEIROS, J. M. A perinatal palatable high-fat diet increases food intake and promotes hypercholesterolemia in adult rats. Lipids. v. 46, 1071-4, 2011. OLIVEIRA, V.; MARINHO, R.; VITORINO, D.; SANTOS, G. A.; MORAES, J. C.; DRAGANO, N.; SARTORI-CINTRA, A.; PEREIRA, L.; CATHARINO, R. R.;
SILVA, A. S.; ROPELLE, E. R.; PAULI, J. R.; SOUZA, C. T.; VELLOSO, L. A.; CINTRA, D. E. Diets containing α-linolenic (ω3) or oleic (ω9) fatty acids rescues obese mice from insulin resistance. Endocrinology, v. 156, p. 4033–46, 2015. PAN, W. W.; MYERS, J. R. M. G. Leptin and the maintenance of elevated body weight. Nature Reviews Neuroscience, v. 19, n. 2, p. 95, 2018 PARAFATI, M.; LASCALA, A.; MORITTU, V. M.; TRIMBOLI, F.; RIZZUTO, A.; BRUNELLI, E.; COSCARELLI, F.; COSTA, N.; BRITTI, D.; EHRLICH, J.;
ISIDORO, C.; MOLLACE, V.; JANDA, E. Bergamot polyphenol fraction prevents nonalcoholic fatty liver disease via stimulation of lipophagy in cafeteria diet-induced rat modelo of metabolic syndrome. The Journal of Nutritional Biochemistry, v. 26, n. 9, p. 938-48, 2015. PAWLAK, D. B.; KUSHNER, J. A.; LUDWIG, D. S. Effects of dietary glycaemic index on adiposity, glucose homoeostasis, and plasma lipids in animals. The
Lancet, v. 364, n. 9436, p. 778-785, 2004.
PELLIZZON, M. A.; RICCI, M. R. The common use of improper control diets in diet-induced metabolic disease research confounds data interpretation: The fiber factor. Nutrition & metabolism, v. 15, n. 1, p. 3, 2018. PICON, P. X.; ZANATTA, C. M.; GERCHMAN, F.; ZELMANOVITZ, T.; GROSS, J. L.; CANANI, L. H. Analysis of the criteria used for the definition of metabolic syndrome in patients with type 2 diabetes mellitus. Arquivo Brasileiro
de Endocrinologia e Metabologia, São Paulo, v. 50, n. 2, p. 264-270, 2006. PROENÇA, R. P. C. Alimentação e globalização: algumas reflexões. Ciencia e
Cultura, v. 62, n. 4, p. 43-47, 2010. PULIDO, R.; BRAVO, L.; SAURA-CALIXTO, F. Antioxidant activity of dietary as determined by a modified ferric reducing/ antioxidant power assay. Journal
Agriculture and Food Chemistry, v. 48, p. 3396-3402, 2000.
QUEIROZ, M. S. R.; JANEBRO, D. I.; CUNHA, M. A. L.; MEDEIROS, J. S.; SABAA-SRUR, A. U.; DINIZ, M. F. F. M.; SANTOS, S. C. Effect of the yellow

109
passion fruit peel flour (Passiflora edulis f. flavicarpa deg.) in insulin sensitivity in type 2 diabetes mellitus patients. Nutrition Journal, v. 11, n. 89, 2012.
RAMOS, A. T.; CUNHA, M. A. L.; SABAA-SRUR, A. U. O.; PIRES, V. C. F.; CARDOSO, M. A. A.; DINIZ, M. F. M.; MEDEIROS, C. C. M. Use of Passiflora edulis f. flavicarpa on cholesterol reduction. Revista Brasileira de Farmacognosia, v. 17, n.4, p. 592-597, 2007. RAMOS, L. A.; CAVALHEIRO, C. C. S.; CAVALHEIRO, E. T. G.; Determinação de nitrito em águas utilizando extrato de flores. Química Nova, v. 29, n. 5, 2006.
RANASINGHE, P.; MATHANGASINGHE, Y.; JAYAWARDENA, R.; HILLS, A. P.; MISRA, A. Prevalence and trends of metabolic syndrome among adults in the asia-pacific region: a systematic review. BMC Public Health, v. 17, n. 101, p. 1-9, 2017. RASBAND, W. S. National Institutes If Health, Bethesda, Maryland, USA. Disponível em: <https://imagej.nih.gov/ij/>, Acesso em: 10 jun. 2018. REEVES, P. G.; NIELSEN, F. H.; FAHEY, G. C. AIN-93 purified diets for
laboratory rodents: final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. Journal of Nutrition,
v. 123, p. 1939 – 1951, 1993. RIBEIRO FILHO, F. F.; MARIOSA, L. S.; FERREIRA, S. R. G.; ZANELLA, M. T. Gordura visceral e síndrome metabólica: mais que uma simples associação. Arquivos Brasileiros de Endocrinologia e Metabologia, v. 50, n. 2, p. 230-238, 2006.
ROSINI, T. C.; SILVA, A. S. R, MORAES, C. Diet-induced obesity: rodent model for the study of obesity-related disorders. Revista da Associação Médica
Brasileira (English Edition), v. 58, n. 3, p. 383-387, 2012. SABITHA, K.; VENUGOPAL, B.; RAFI, M. D.; RAMANA, K. V. Role os Antioxidant Enzymes in Glucose and Lipid Metabolism in Association with Obesity and Type 2 Diabetes. American Journal of Medical Sciences and Medicine, v. 2,
n. 1, p. 21-24, 2014. SABOYA, P. P.; BODANESE, L. C.; ZIMMERMANN, P. R.; GUSTAVO, A.S.; ASSUMPÇÃO, C. M.; LONDERO, F. Metabolic syndrome and quality of life: a systematic review. Revista latino-americana de enfermagem, v. 24, 2016. SALLES, G. F.; BLOCH, K. V.; CARDOSO, C. R. L. Mortality and predictors of mortality in a cohort of Brazilian type 2 diabetic patients. Diabetes Care, v. 27, p.
1299–1305 2004

110
SAMPEY, B. P.; VANHOOSE, A. M.; WINFIELD, H. M.; FREEMERMAN, A. J.; MUEHLBAUER, M. J.; FUEGER, P. T.; NEWGARD, C. B.; MAKOWSKI, L. Cafeteria diet is a robust model of human metabolic syndrome with liver and
adipose inflammation: comparison to higt-fat diet. Obesity, v. 19, n. 6, p. 1109-17, 2011. SANTOS, R. D.; GAGLIARDI, A. C. M.; XAVIER, H. T.; MAGNONI, C. D.; CASSANI, R.; LOTTENBERG, A. M. P.; CASELLA FILHO, A.; ARAUJO, D. B.; CESENA, F. Y.; ALVES, R. J.; FENELON, G.; NISHIOKA,S. A. D.; FALUDI, A. A.; GELONEZE, B.; SCHERR, C.; KOVACS, C.; TOMAZZELA, C.; CARLA, C.; BARRERA-ARELLANO, D.; CINTRA, D.; QUINTÃO, E.; NAKANDAKARE, E.
R.; FONSECA, F. A. H.; PIMENTEL, I.; SANTOS, J. E.; BERTOLAMI, M. C.; ROGERO, M.; IZAR, M. C. O.; NAKASATO, M.; DAMASCENO, N. R. T.; MARANHÃO, R.; CASSANI, R. S. L.; PERIM, R.; RAMOS, S. I Diretriz sobre o consumo de Gorduras e Saúde Cardiovascular. Arquivos Brasileiros de
Cardiologia, v. 100, n. 1, p. 1-40, 2013. SIES, H.; KOCH, O. R.; MARTINO, E.; BOVERIS, A. Increased biliary glutathione disulfide release in chronically ethanol treated rats. FEBS Letters, v. 103, p. 287-90, 1979.
SILVA, A. M. O. Efeito dos compostos fenólicos do alecrim (Rosmarinus
officinalis L.) na inflamação aguda e sobre os marcadores de estresse oxidativo em ratos diabéticos. São Paulo, 2012. Tese de Doutorado – Faculdade de Ciências Farmacêuticas – Universidade de São Paulo. SILVA, A. M. O. Efeito dos compostos fenólicos presentes no alecrim
(Rosmarinus officinalis L.) sobre as enzimas antioxidantes e os parâmetros
bioquímicos de ratos diabéticos induzidos por estreptozotocina. São Paulo, 2008. Dissertação de Mestrado – Faculdade de Ciências Farmacêuticas – Universidade de São Paulo. SILVA, S. R. D.; MERCADANTE, A. Z. Composição de carotenóides de maracujá-amarelo (Passiflora edulis fl avicarpa) in natura. Food Science and Technology, v.22, p.254-258, 2002.
SLAVIN, J. Fiber and Prebiotics: Mechanisms and Health Benefits. Nutrients, p. 1417-1435, 2013. SOARES, M. B.; RAMALHO, J. B.; IZAGUIRRY, A. P.; PAVIN, N. F.; SPIAZZI, C. C.; SCHIMIDT, H. L.; MELLO-CARPES, P. B.; SANTOS, F. W. Comparative effect of Camellia sinensis teas on object recognition test deficit and metabolic changes induced by cafeteria diet. Nutrition Neuroscience, p. 1–10, 2017.
SOCIEDADE BRASILEIRA DE CARDIOLOGIA. I Diretriz brasileira de diagnóstico e tratamento da síndrome metabólica. Arquivos Brasileiros de
Cardiologia, v. 84, n. 1, p. 3- 28, 2005.

111
SOCIEDADE BRASILEIRA DE CARDIOLOGIA. VI Diretrizes Brasileiras de Hipertensão. Arquivos Brasileiros de Cardiologia, v. 95, n.1, 2010.
STYSKAL, J.; REMMEN, H. V.; RICHARDSON, A.; SALMON, A. B. Oxidative stress and diabetes: What can we learn about insulin resistance from antioxidant mutant mouse models? Free Radical Biology & Medicine, v. 52, p. 46–58, 2012. TEHRANIFAR, A.; SELAHVARZI, Y.; KHARRAZI, M.; BAKHSHB, V. J. High potential of agro-industrial by-products of pomegranate (Punica granatum L.) as the powerful antifungal and antioxidant substances. Industrial Crops and Products,
v.34, n.3, p.1523-1527, 2011. THALLAS‐BONKE, V.; COUGHLAN, M. T.; TAN, A. L.; HARCOURT, B. E.; MORGAN, P. E.; DAVIES, M. J.; BACH, L. A.; COOPER, M. E.; FORBES, J. M. Targeting the AGE-RAGE axis improves renal function in the context of a healthy diet low in advanced glycation end-product content. Nephrology, v. 18, p. 47-56, 2013. THIESEN, L. C.; NUNES, M. L. O.; MEYRE-SILVA, C.; PASTOR, V. D.;
ANDRADE, S. F.; COUTO, A. G.; SILVA, L. M.; BRESOLIN, T. M. B.; SANTIN, J. R. The hydroethanolic Litchi chinensis leaf extract alleviate hepatic injury induced by carbon tetrachloride (CCl4) through inhibition of hepatic inflammation. Biomedicine & Pharmacotherapy, v. 107, p. 929–936, 2018. TIETZE, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: applications to mammalian blood and other tissues. Analythical Biochemistry, v. 27, n. 3, p. 502-22, 1969.
TOPPING, D. L.; CLIFTON, P. M. Short-chain fatty acids and human colonic function: roles of resistant starch and nonstarch polysaccharides. Physiol. Rev., v. 81, p. 1031–1064. 2001. TRAYHURN, P. WOOD, I. S. Adipokines: inflammation and pleiotropic role of white adipose tissue. British Journal of Nutrition. 2004
VERNON G.; BARANOVA A.; YOUNOSSI Z. M. Systematic review: The epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in adults. Alimentary Pharmacology and Therapeutics, v. 34, p. 274–285, 2011. VIGITEL. Vigitel Brazil 2017: surveillance of risk and protective factors for chronic diseases by telephone survey: estimates of Frequency and sociodemographic distribution of risk and protective factors for chronic diseases in the capitals of the
26 Brazilian states and the Federal District in 2017. 1 ed. Brasilia, 2018.

112
WANG, C.; LIAO, J. K. A mouse model of diet-induced obesity and insulin resistance. Methods Mol Biol, v. 821, p. 421-33, 2012.
WHO, World Health Organization. Health Topics. Obesity: World Health Organization, disponível em < http://www.who.int/topics/obesity/en/> 2015. WHO, World Health Organization. Obesity and overweight, disponível em <http://www.who.int/mediacentre/factsheets/fs311/en/index.html> 2011.
WONDRACEK, D. C.; VIEIRA, R. F.; SILVA, D. B.; AGOSTINI-COSTA, T. S.; SANO, S. M.; FALEIRO, F. G. Influência da saponificação na determinação de
carotenoides em maracujás do cerrado. Química Nova, v. 35, n. 1, 2012.
YAPO, B. M.; KOFFI, K. L.; Yellow passion fruit rinds a potencial source of low-methoxyl pectin. Journal of Agricultural and Food Chemistry, v. 54, p. 2738-2744, 2006. YKI-JÄRVINEN, H. Non-alcoholic fatty liver disease as a cause and a consequence of metabolic syndrome. The lancet Diabetes & endocrinology, v. 2, n. 11, p. 901-910, 2014.
ZEENI, N.; DAGHER-HAMALIAN, C.; DIMASSI, H.; FAOUR, W. H. Cafeteria diet-fed mice is a pertinente modelo of obesity-induced organ damage: a potential role of inflammation. Inflammation Research, v. 64, n. 7, p. 501-12, 2015. ZERAIK, M. L.; PEREIRA, C. A. M.; ZUIN, V. G.; YARIWAKE, J. H. Passion
fruit: a functional food?. Revista Brasileira De Farmacognosia-Brazilian Journal
of Pharmacognosy, v. 20, n. 3, p. 459-471, 2010. ZERAIK, M. L.; SERTEYN, D.; DEBY-DUPONT, G.; WAUTERS, J. N.; TITS, M.; YARIWAKE, J. H.; ANGENOT, L.; FRANCK, T. Evaluation of the antioxidant activity of passion fruit (Passifl ora edulis and Passifl ora alata) extracts on stimulated neutrophils and myeloperoxidase activity assays. Food Chemistry, v.128, n.2, p.259-265, 2011.
ZERAIK, M. L.; YARIWAKE, J. H.; WAUTERS, J. N.; TITS, M. ANGENOT, L. Analysis of passion fruit rinds (Passifl ora edulis): isoorientin quantification by HPTLC and evaluation of antioxidant (radical scavenging) capacity. Química Nova, v.35, p.541-545, 2012. ZHANG, Z.; SHI, L.; PANG, W.; LIU, W.; LI, J.; WANG, H.; SHI, G. Dietary Fiber Intake Regulates Intestinal Microflora and Inhibits Ovalbumin-Induced Allergic Airway Inflammation in a Mouse Model. Plos One, v. 11, n. 2, p. 1-16,
2016.