ecologia e comportamento do mico-leÃo da-cara · universidade estadual de santa cruz - uesc ......
TRANSCRIPT

i
UNIVERSIDADE ESTADUAL DE SANTA CRUZ - UESC
Programa de Pós-Graduação em Zoologia
CARLOS EDUARDO GUIDORIZZI
ECOLOGIA E COMPORTAMENTO DO MICO-LEÃO-DA-CARA-
DOURADA, LEONTOPITHECUS CHRYSOMELAS (KUHL, 1820)
(PRIMATES, CALLITRICHIDAE), EM UM FRAGMENTO DE
FLORESTA SEMIDECIDUAL EM ITORORÓ, BAHIA, BRASIL
ILHÉUS – BA 2008

ii
UNIVERSIDADE ESTADUAL DE SANTA CRUZ - UESC
Programa de Pós-Graduação em Zoologia
CARLOS EDUARDO GUIDORIZZI
ECOLOGIA E COMPORTAMENTO DO MICO-LEÃO-DA-CARA-
DOURADA, LEONTOPITHECUS CHRYSOMELAS (KUHL, 1820)
(PRIMATES, CALLITRICHIDAE), EM UM FRAGMENTO DE
FLORESTA SEMIDECIDUAL EM ITORORÓ, BAHIA, BRASIL
Dissertação apresentada para a obtenção do título de Mestre em Zoologia, à Universidade Estadual de Santa Cruz. Área de Concentração: Zoologia Aplicada
Orientador: Drª. Maria Cecília Kierulff
Co-Orientador: Prof. Martín R. Alvarez
ILHÉUS – BA 2008

iii
UNIVERSIDADE ESTADUAL DE SANTA CRUZ - UESC
Programa de Pós-Graduação em Zoologia
CARLOS EDUARDO GUIDORIZZI
ECOLOGIA E COMPORTAMENTO DO MICO-LEÃO-DA-CARA-DOURADA,
LEONTOPITHECUS CHRYSOMELAS (KUHL, 1820) (PRIMATES, CALLITRICHIDAE), EM UM
FRAGMENTO DE FLORESTA SEMIDECIDUAL EM ITORORÓ, BAHIA, BRASIL
Ilhéus – Bahia, 25/03/2008
________________________________________ Maria Cecília Martins Kierulff – Drª.
Fundação Parque Zoológico de São Paulo – FPZSP (Orientadora)
________________________________________ Martín Roberto Del Valle Alvarez – Dr.
UESC/DCB (Co-Orientador)
________________________________________ Yvonnick Le Pendu – Dr.
UESC/DCB
________________________________________ Paula Procópio de Oliveira – Drª.
Instituto Pri-Matas

iv
DEDICO ESTE TRABALHO
AOS MEUS PAIS, PLÍNIO E VERA,
POR TODO APOIO E INCENTIVO
EM TODOS ESSES ANOS
LONGE DE CASA

v
“MAS O MATO MUDAVA BRUTO, NO ESCONSO, MAIS MATO
SE FECHANDO. TINHAM MEXIDO NO GALHO, MAS NÃO ERA
OUTRO SERELEPE, NÃO.
SUSTO QUE UNS ESTAVAM CONVERSANDO, COCHICHO,
DEPRESSA, FERVIDO, DAVAM BICOTAS. (...) ESCORREGOU
PELAS FOLHAGENS, DE SONSAGATO, QUERENDO MAIS
ESCONDIDO. DESORDEM DE AMEAÇA, QUE DISSE-DISSE,
ERA LÁ EM CIMA: (...) MUITOS OLHOS ESTALAVAM, NO
MIOLOSO”
JOÃO GUIMARÃES ROSA

vi
Agradecimentos
À Cecília, pela orientação e incentivo durante todo o trabalho e por compartilhar todo seu
conhecimento sobre micos-leões, contribuindo, em muito, para minha formação.
Ao Martin, pela co-orientação, pela ajuda, sempre presente quando necessária, e pelas
proveitosas discussões.
À Dra. Kristel De Vleeschouwer e à Dra. Dehorah Faria, pelos comentários para melhora
do trabalho no exame de qualificação. À Dra. Paula Procópio-de-Oliveira e ao Dr.
Yvonick Le Pendu, por aceitarem participar da banca de defesa.
À minha família, Plínio, Vera, Lu e Má, por todo o apoio, mesma à distância. Em especial
à minha mãe, pela revisão ortográfica do texto.
Ao Instituto de Estudos Socioambientais do Sul da Bahia – IESB, pela administração do
projeto, e a todos que fazem parte da Instituição.
Em especial, agradeço ao Gabriel Rodrigues dos Santos, pelo incentivo desde que cheguei
em Ilhéus e por acreditar em meu trabalho.
Ao Sr. “Julho”, meu assistente de campo, pela grande ajuda, imprescindível em muitos
momentos do trabalho. E por estar sempre bem humorado e disposto a fazer o que fosse
necessário. E à D. Maria, pela companhia e pela comida.
Ao Buguinha, o Marisburgo, da fazenda Sta. Rita, vizinha à Barro Branco, pelo apoio
logístico, pela ajuda sempre que necessária (quando o carro quebrava!) e pela companhia.
Enfim, por ter feito meus dias de campo mais fáceis.
Ao proprietário e ao gerente da Fazenda Barro Branco, Sr. Antônio Fernandes e Jurandir,
por permitirem a realização do trabalho e pelo apoio logístico. E também por manterem a
floresta em pé.

vii
Aos funcionários da fazenda Barro Branco, principalmente ao Demar e ao Márcio, pela
ajuda quando necessária, pela companhia e por compartilhar os conhecimentos sobre a
região.
Ao pessoal da fazenda Cabana da Ponte, principalmente ao Mário Elmo, por ceder os
dados pluviométricos durante o período de estudo.
A todos que participaram, em algum momento, das capturas dos micos: Lílian, a
veterinária responsável pelo projeto, Leo, Kristel, Adriana, Vinícius e Neca,
À Michaelle Pessoa, pelo treinamento e ajuda imprescindível na parte de fenologia.
Ao Zé Lima e ao Mardel M. Lopes, pela ajuda na coleta e identificação do material
botânico, e à Thalita Fontoura, pela identificação das bromélias.
A todos que leram e contribuíram com comentários valiosos em versões não finalizadas
da dissertação: Nayara Cardoso, Fábio Falcão, Gustavo Canale e Leonardo Oliveira.
A todos os amigos do mestrado e de Ilhéus, em especial a Camila, Cassiano, Fábio,
Gustavo, Leo, e Priscila, pela companhia, amizade e conversas sobre conservação e outras
coisas da vida.
À Nayara, pelo apoio nos momentos difíceis, estímulo, idéias compartilhadas e pela
amizade e carinho.
Este trabalho não seria possível, desta forma, sem a participação dessas pessoas.
Em tempo: à Ana Carla e à Carol, pela ajuda na formatação.
Por fim, agradeço ao IBAMA, pela licença de pesquisa , e aos financiadores do projeto: a
FAPESB, que outorgou minha bolsa de Mestrado, o Rufford Small Grants, o Primate
Action Fund, o Lion Tamarin Fund e o Zoológico da Antuérpia, Bélgica.

viii
SUMÁRIO ÍNDICE DE FIGURAS ..............................................................................................X ÍNDICE DE TABELAS ........................................................................................... XI RESUMO.................................................................................................................XII ABSTRACT........................................................................................................... XIII CAPÍTULO I - INTRODUÇÃO GERAL ..................................................................1
1. INTRODUÇÃO GERAL .....................................................................................................................2 1.1 O gênero Leontopithecus Lesson, 1840.........................................................................................2 1.2 As Florestas Semideciduais do Sul da Bahia ................................................................................8 1.3 Diferenças ambientais: ecologia e comportamento ......................................................................9
2. OBJETIVOS ......................................................................................................................................11 2.1 Geral ...........................................................................................................................................11 2.2 Específicos...................................................................................................................................11
3. MÉTODOS ........................................................................................................................................12 3.1 Área de Estudo ............................................................................................................................12
3.1.1 Informações climáticas ........................................................................................................................15 3.2 Grupos monitorados....................................................................................................................16
CAPÍTULO II - PADRÃO DE ATIVIDADES E DIETA DO MICO-LEÃO-DA-CARA-DOURADA (LEONTOPITHECUS CHRYSOMELAS) EM UM FRAGMENTO DE FLORESTA SEMIDECIDUAL...............................................18
1. INTRODUÇÃO .................................................................................................................................19 2. MÉTODOS ........................................................................................................................................21
2.1 Área de Estudo e Grupos.............................................................................................................21 2.2 Disponibilidade de frutos ............................................................................................................21 2.3 Composição e variação sazonal da dieta ....................................................................................23 2.4 Padrão de Atividades ..................................................................................................................24
3. RESULTADOS..................................................................................................................................26 3.1 Grupos monitorados....................................................................................................................26 3.2 Disponibilidade de frutos e hábitats............................................................................................27 3.3 Composição e variação sazonal da dieta. ...................................................................................28 3.4 Padrão de atividade ....................................................................................................................34
4. DISCUSSÃO......................................................................................................................................36 5. CONCLUSÕES..................................................................................................................................44
CAPÍTULO III - ÁREA DE VIDA E USO DO ESPAÇO DO MICO-LEÃO-DA-CARA-DOURADA (LEONTOPITHECUS CHRYSOMELAS) EM UM FRAGMENTO DE FLORESTA SEMIDECIDUAL...............................................46
1. INTRODUÇÃO .................................................................................................................................47 2. MÉTODOS ........................................................................................................................................48
2.1 Área de Estudo e Grupos.............................................................................................................48 2.2 Período de Atividade ...................................................................................................................48 2.3 Área de Vida, Uso do Espaço e Deslocamento Diário................................................................48 2.4 Densidade....................................................................................................................................50
3. RESULTADOS..................................................................................................................................51 3.1 Grupos monitorados....................................................................................................................51 3.2 Período de atividade ...................................................................................................................51 3.3 Área de Vida e Uso do Espaço ....................................................................................................51
3.3.1 Uso da borda e tipos de mata ...............................................................................................................53 3.3.2 Deslocamento diário e locais de dormida ............................................................................................57 3.3.3 Fruteiras visitadas ................................................................................................................................59
3.4 Densidade....................................................................................................................................63 4. DISCUSSÃO......................................................................................................................................65 5. CONCLUSÕES..................................................................................................................................72

ix
CAPÍTULO IV - A CONSERVAÇÃO DO MICO-LEÃO-DA-CARA-DOURADA (LEONTOPITHECUS CHRYSOMELAS) NAS FLORESTAS SEMIDECIDUAIS DO SUL DA BAHIA .................................................................................................73
1. SITUAÇÃO ATUAL DE L. CHRYSOMELAS NAS FLORESTAS SEMIDECIDUAIS.............................................74 2. PRIORIDADES DE PESQUISA E AÇÕES PARA A CONSERVAÇÃO DO MLCD EM FRAGMENTOS SEMIDECIDUAIS.......................................................................................................................................82 3. CONCLUSÕES ......................................................................................................................................86
REFERÊNCIAS........................................................................................................87 ANEXO I: ESPÉCIES MONITORADAS NAS TRILHAS DE FENOLOGIA E NÚMERO DE INDIVÍDUOS DE CADA ESPÉCIE. ..............................................96 ANEXO II: PADRÃO DE TIVIDADES DO GRUPO GRA EM CADA MÊS DE OBSERVAÇÃO E NO TOTAL................................................................................98

x
ÍNDICE DE FIGURAS FIGURA 1. DISTRIBUIÇÃO HISTÓRICA E ATUAL DAS QUATRO ESPÉCIES DE MICOS-LEÕES. (MODIFICADO DE
KLEIMAN E RYLANDS, 2002).........................................................................................................3 FIGURA 2. O MICO-LEÃO-DA-CARA-DOURADA, LEONTOPITHECUS CHRYSOMELAS, COM RADIO COLAR. ..........6 FIGURA 3. DISTRIBUIÇÃO DE LEONTOPITHECUS CHRYSOMELAS, DESTACANDO A RESERVA BIOLÓGICA DE
UNA. (MODIFICADO DE RYLANDS ET AL., 2002)...............................................................................6 FIGURA 4. TIPOS DE VEGETAÇÃO NO SUL E EXTREMO SUL DO ESTADO DA BAHIA, E LOCALIZAÇÃO DA
FAZENDA BARRO BRANCO, ONDE FOI REALIZADO ESTE ESTUDO, E DA RESERVA BIOLÓGICA DE UNA. BASE DE DADOS RETIRADA DE ARQUIVO DIGITAL (PRADO ET AL., 2003). ..........................................7
FIGURA 5. ACIMA: IMAGEM DE SATÉLITE (CBERS, 2007) DA ÁREA DE ESTUDO, MOSTRANDO O CENTRO URBANO DO MUNICÍPIO DE ITORORÓ E FRAGMENTOS FLORESTAIS DA REGIÃO. ABAIXO, MAPA DO FRAGMENTO ESTUDADO (FAZENDA BARRO BRANCO, MUNICÍPIO DE ITORORÓ, BAHIA)....................14
FIGURA 6. ACIMA: PLUVIOSIDADE DURANTE O PERÍODO DE ESTUDO. DADOS COLETADOS NA FAZENDA CABANA DA PONTE. ABAIXO: PLUVIOSIDADE, TEMPERATURA MÁXIMA E MÍNIMA MÉDIAS NO PERÍODO DE 1974 A 1978. DADOS OBTIDOS NA ESTAÇÃO METEOROLÓGICA DA CEPLAC LOCALIZADA, NA ÉPOCA, NA FAZENDA CABANA DA PONTE...............................................................15
FIGURA 7. A FLORESTA DA FAZENDA BARRO BRANCO NA ESTAÇÃO CHUVOSA (FEVEREIRO/2007) E NO FINAL DA ESTAÇÃO SECA (SETEMBRO/2007). .....................................................................................16
FIGURA 8: LOCALIZAÇÃO DOS TRANSECTOS UTILIZADOS PARA OBSERVAÇÕES FENOLÓGICAS. ...................22 FIGURA 9. ESQUEMA MOSTRANDO A METODOLOGIA DOS PONTOS QUADRANTES PARA MARCAÇÃO DAS
ÁRVORES MONITORADAS PARA FENOLOGIA. EM CADA PONTO, ERAM MARCADAS AS QUATRO ÁRVORES MAIS PRÓXIMAS AO PONTO CENTRAL, UMA EM CADA QUADRANTE. FORAM MARCADAS ÁRVORES COM DAP ≥ 7,5 CM.............................................................................................................22
FIGURA 10: COBERTURA FOLIAR DAS ÁRVORES MONITORADAS NO FRAGMENTO DA FAZENDA BARRO BRANCO DURANTE O PERÍODO DE OBSERVAÇÕES FENOLÓGICAS........................................................27
FIGURA 11: PLUVIOSIDADE E QUANTIDADE DE FRUTOS VERDES, MADUROS E DE FLORES E BOTÕES DAS ÁRVORES MONITORADAS DURANTE O PERÍODO DE OBSERVAÇÕES FENOLÓGICAS. .............................28
FIGURA 12. PRODUÇÃO DE FRUTOS DE TRÊS ESPÉCIES ARBÓREAS MONITORADAS NA FENOLOGIA E CUJOS FRUTOS FORAM CONSUMIDOS PELOS MICOS. ......................................................................................34
FIGURA 13. PADRÃO DE ATIVIDADES DO GRUPO GRA NAS ESTAÇÕES SECA, CHUVOSA E NO PERÍODO TOTAL. AS ATIVIDADES FORRAGEAR POR ANIMAL E COMER ANIMAL FORAM AGRUPADAS.............................34
FIGURA 14. PADRÃO DE ATIVIDADES DO GRUPO PRINCIPAL GRA EM CADA MÊS DE ESTUDO NO FRAGMENTO DA FAZENDA BARRO BRANCO. ..........................................................................................................35
FIGURA 15. CICLO DIÁRIO DE ATIVIDADES DO GRUPO PRINCIPAL GRA, MOSTRANDO A PROPORÇÃO DE CADA COMPORTAMENTO EM CADA HORA DO DIA. DADOS REFERENTES A TODO O PERÍODO DE ESTUDO. .....36
FIGURA 16. ÁREA DE VIDA DOS GRUPOS ESTUDOS, CALCULADO PELO MÉTODO DO MÍNIMO POLÍGONO CONVEXO (MPC), ADAPTADO PARA ABRANGER SOMENTE ÁREA DE FLORESTA. A ÁREA DE VIDA DO GRUPO GRA FOI DE 71 HA E A DO GRUPO MAN, 68 HA......................................................................52
FIGURA 17. ÁREA DE VIDA DO GRUPO GRA, CALCULADA PELO MÉTODO DO KERNEL-FIXO. AS ÁREAS MAIS ESCURAS REPRESENTAM AS REGIÕES COM MAIOR PROBABILIDADE DE ENCONTRAR O GRUPO. A ÁREA DE VIDA, DEFINIDA COMO O CONTORNO DE 95%, FOI DE 58 HA. .........................................................53
FIGURA 18. REGISTROS DAS LOCALIZAÇÕES DO GRUPO E MAPA MOSTRANDO OS CONTORNOS DAS BORDAS DE 10, 50 E 100 METROS.....................................................................................................................54
FIGURA 19. REGISTROS DAS LOCALIZAÇÕES DO GRUPO E MAPA MOSTRANDO OS TIPOS FLORESTAIS: FLORESTA SECA E RIACHOS/BREJOS....................................................................................................55
FIGURA 20. PRODUTIVIDADE DE FRUTOS MADUROS, FRUTOS VERDES E FLORES NOS AMBIENTES DE INTERIOR DA FLORESTA, RIACHOS E NOS CEM PRIMEIROS METROS DE BORDA. ...................................56
FIGURA 21. PRODUTIVIDADE DE FRUTOS MADUROS E VERDES NOS AMBIENTES DE INTERIOR DA FLORESTA, RIACHOS E NOS CEM PRIMEIROS METROS DE BORDA, EM CADA MÊS DE OBSERVAÇÃO........................57
FIGURA 22. DESLOCAMENTO DIÁRIO MÉDIO DO GRUPO GRA E PLUVIOSIDADE EM CADA MÊS DE OBSERVAÇÃO. ....................................................................................................................................57
FIGURA 23. FREQÜÊNCIA DE UTILIZAÇÃO DA ÁREA E LOCALIZAÇÕES DOS OCOS UTILIZADOS COMO LOCAIS DE DORMIDA. OS SÍMBOLOS MAIORES REPRESENTAM OS OCOS MAIS UTILIZADOS. .............................59
FIGURA 24. FREQÜÊNCIA DE UTILIZAÇÃO DA ÁREA E LOCALIZAÇÕES DAS FRUTEIRAS UTILIZADAS PELO GRUPO GRA.......................................................................................................................................60
FIGURA 25. FREQÜÊNCIA DE UTILIZAÇÃO DA ÁREA E LOCALIZAÇÕES DAS BROMÉLIAS UTILIZADAS PELO GRUPO GRA.......................................................................................................................................61

xi
FIGURA 26. ÁREA DE VIDA DO GRUPO GRA, CALCULADA PELO MÉTODO DO KERNEL-FIXO, ENTRE OS MESES DE AGOSTO/2006 E SETEMBRO/2007. OS PONTOS REPRESENTAM AS FRUTEIRAS UTILIZADAS EM CADA MÊS. ...................................................................................................................................................62
FIGURA 27. MPC DOS GRUPOS MONITORADOS E LOCALIZAÇÕES DAS ÁREAS DE SOBREPOSIÇÃO COM OUTROS GRUPOS, A, B E C, ONDE HAVIA ENCONTROS INTERGRUPAIS. ...............................................64
FIGURA 28. MICO-LEÃO-DA-CARA-DOURADA OBSERVADO NO FRAGMENTO DA FAZENDA BARRO BRANCO, COM PADRÃO DE COLORAÇÃO DIFERENTE DO DA ESPÉCIE (INDICADA PELA ESPÉCIE). PROVAVELMENTE UMA FÊMEA...........................................................................................................79
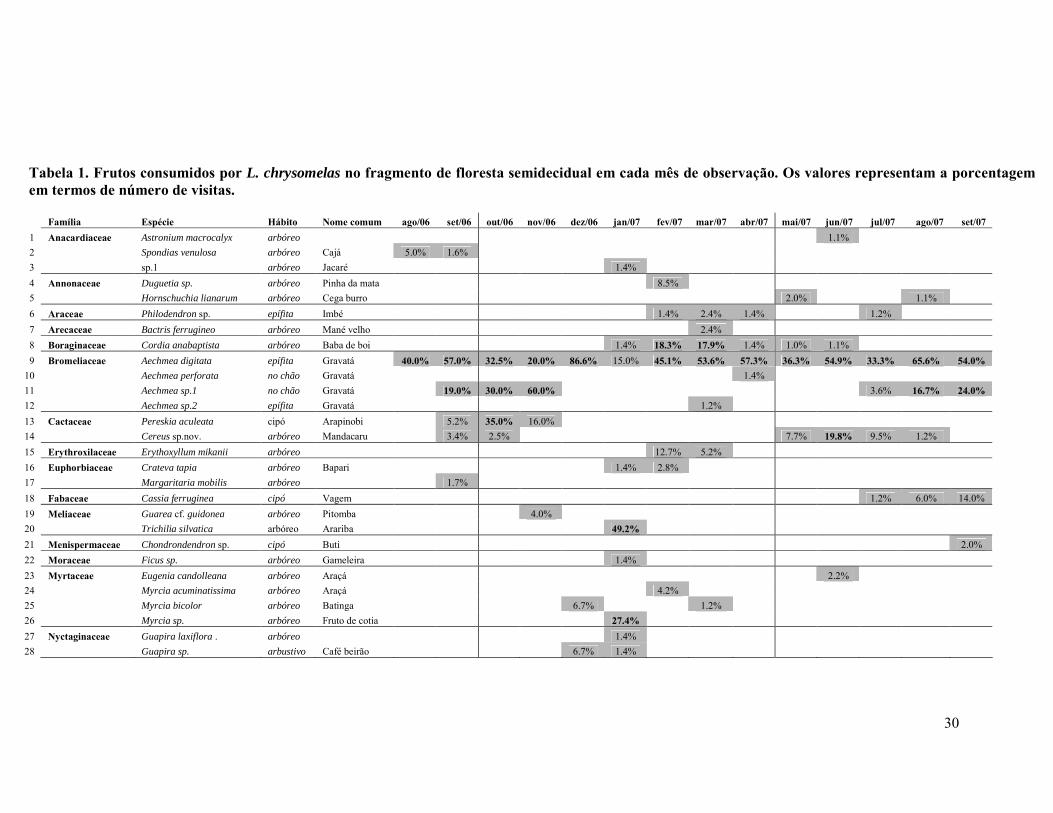
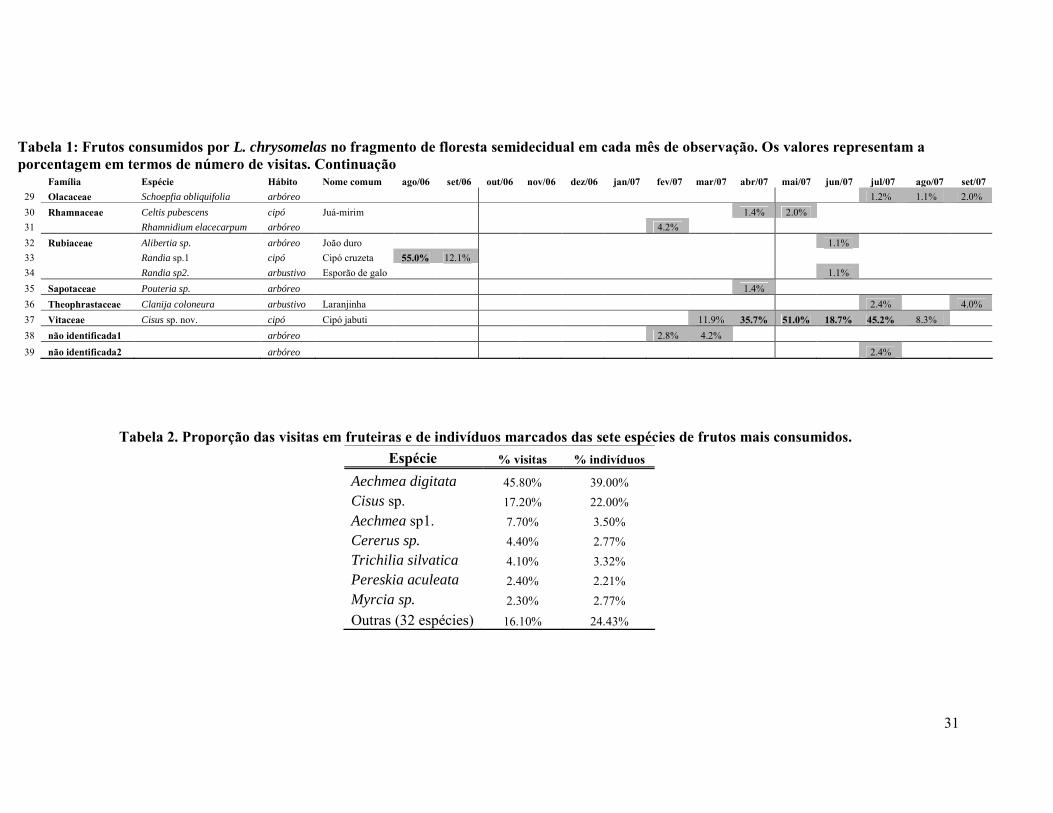
ÍNDICE DE TABELAS TABELA 1. FRUTOS CONSUMIDOS POR L. CHRYSOMELAS NO FRAGMENTO DE FLORESTA SEMIDECIDUAL EM CADA MÊS DE OBSERVAÇÃO. OS VALORES REPRESENTAM A A PORCENTAGEM EM TERMOS DE NÚMERO DE VISITAS. ..............................................................30
TABELA 2. PROPORÇÃO DAS VISITAS EM FRUTEIRAS E DE INDIVÍDUOS MARCADOS DAS SETE ESPÉCIES DE FRUTOS MAIS CONSUMIDOS. ..........................................................................31
TABELA 3. NÚMERO DE ESPÉCIES CONSUMIDAS, NÚMERO DE FRUTEIRAS VISITADAS, TEMPO TOTAL EM FRUTEIRAS, NÚMERO DE VISITAS POR DIA (MÉDIA) E O TEMPO EM FRUTEIRAS POR DIA (MÉDIA). DADOS DO GRUPO GRA, EM CADA MÊS DE OBSERVAÇÃO32
TABELA 4. COMPARAÇÃO DO PADRÃO DE ATIVIDADES EM TRÊS ESTUDOS COM L. CHRYSOMELAS.......................................................................................................................................41
TABELA 5. PERÍODO DE ATIVIDADE DO GRUPO GRA DURANTE O PERÍODO DE ESTUDO, DE AGOSTO/2006 A SETEMBRO/2007..................................................................................................51
TABELA 6. COMPARAÇÃO DA ALTURA E DIÂMETRO NA ALTURA DO PEITO (DAP) DAS ÁRVORES MONITORADAS PARA FENOLOGIA NOS AMBIENTES DE FLORESTA SECA E RIACHO/BREJO........................................................................................................................................56
TABELA 7. NÚMERO DE INDIVÍDUOS OBSERVADOS NOS GRUPOS MONITORADOS (MAN E GRA) E EM OUTROS TRÊS GRUPOS AVISTADOS FREQUENTEMENTE NO FRAGMENTO. .....64

xii
Resumo
Conhecer a ecologia e o comportamento de espécies ameaçadas é essencial para
nortear ações visando a sua conservação. O mico-leão-da-cara-dourada (Leontopithecus
chrysomelas) é um calitriquídeo endêmico da Mata Atlântica do Sul da Bahia e
ameaçado de extinção. A espécie ocupa dois tipos de hábitats: floresta ombrófila e
semidecidual. Todas as informações sobre a ecologia e comportamento provêm da
floresta ombrófila, na Reserva Biológica de Una. Dadas as diferenças florísticas, de
estrutura de vegetação, de sazonalidade climática, além das diferentes pressões
antrópicas, é esperado que haja diferenças ecológicas e comportamentais entre as
populações. Esse trabalho teve como objetivo obter as primeiras informações sobre
dieta, padrão de atividades e uso do espaço de L. chrysomelas em floresta semidecidual.
Dois grupos foram monitorados, entre janeiro de 2006 e setembro de 2007, em um
fragmento de 450 ha da Fazenda Barro Branco, Itororó, Bahia. Foram realizadas 169
horas de observação de um grupo e 683 do outro. A dieta foi semelhante à de outras
populações de Leontopithecus, em termos de itens consumidos, sendo composta por
frutos (geralmente maduros), insetos, pequenos vertebrados (anfíbios e lagartos) e
exsudatos. Foram consumidos frutos de 39 espécies vegetais, pertencentes a 21 famílias,
número bem menor do que o observado para a população da Rebio-Una. As famílias
Bromeliaceae e Myrtaceae foram as mais consumidas, em número de espécies. Frutos
da bromélia Aechmea digitata foram os mais consumidos e bromélias foram os locais
mais utilizados para forrageio por animais, representando um recurso essencial para a
espécie no fragmento semidecidual. Frutos de cipós foram muito consumidos, o que
pode representar uma adaptação às condições do fragmento florestal. O consumo de
exsudatos só foi observado na estação seca, quando também houve aumento do tempo
dedicado à procura/consumo de animais e aumento do consumo de frutos de bromélias.
Os grupos utilizaram 70 ha de floresta cada. O tamanho do território foi limitado pelo
formato irregular do fragmento. O grupo utilizou a periferia mais frequentemente do
que o centro do território, porém, evitou os 50 primeiros metros da borda (contato da
floresta com pasto). O uso do espaço e o deslocamento diário estiveram relacionados às
localizações dos locais de dormida (todos ocos de árvores), das fruteiras utilizadas e da
presença de grupos vizinhos (defesa de território). Os resultados evidenciaram a grande
flexibilidade ecológica e comportamental da espécie, o que não garante, no entanto, sua
sobrevivência a longo prazo em florestas semideciduais. Esforços devem ser feitos para
aumentar a conectividade entre os fragmentos e o fluxo gênico da população.

xiii
Abstract
Knowledge on a given species ecology and behavior is an essential step for its
management and conservation. The golden-headed lion tamarin (Leontopithecus
chrysomelas) is an endemic and threatened Callithrichidae from Atlantic Forest of
Southern Bahia, inhabiting two forest types: wet and semideciduous forest. Only
populations living in wet forest have been monitored (at Una Biological Reserve).
Given the differences between these two habitats (floristic, climatic and anthropogenic
pressures), it is expected differences in the ecology and behavior of populations
inhabiting these forest types. The objective of this study was to understand the diet,
activity patterns and use of space of L. chrysomelas in a semideciduous forest. Two
groups were monitored, between Jan/2006 and Sep/2007, in a 450 ha forest fragment at
Fazenda Barro Branco, Itororó, Bahia. One group had 169 hrs, and the other 683 hrs of
observation. Items of their diet were similar to the described for others Leontopithecus
populations (fruits, insects, small vertebrates, such as lizards and frogs, and exudates).
Fruits of 39 plant species (in 21 families) were consumed, lesser than was reported in
Rebio-Una. Bromeliaceae and Myrtaceae were the families with more species
consumed. Fruits of Aechmea digitata (Bromeliaceae) were the most consumed during
the study, and bromeliads were also the main foraging site for animal prey, being an
essential resource for L. chrysomelas in semideciduous forest fragment. Fruits from
lianas were also used often, which may be an adaptation to the local conditions of the
fragment. Exudates were only eaten during the dry season, when the time dedicated to
search/eating of animals increased. During this season, the proportion of bromeliads
fruits in the diet also increased. Both groups had a home range of 70 ha. The size of the
territory may have been limited by the irregular shape of the fragment. The periphery of
the territory was used more often then the center, but the group avoided the 50 meters
from the edge (contact of the forest with the pasture area). The use of space and dairy
movements were associated with the locations of sleeping trees (tree holes), the fruit
trees and the presence of neighboring groups. Results showed the great ecological and
behavioral flexibility of the species, which, however, does not guarantee the survival of
the species in semideciduous forest. Efforts should be made towards increasing
fragments connections and gene flow for L. chrysomelas population.

1
CAPÍTULO I - Introdução Geral

Capítulo 1
2
1. INTRODUÇÃO GERAL
Esta dissertação está dividida em quatro capítulos: (1) uma introdução geral,
aqui apresentada, com descrição da área de estudo; (2) um capítulo sobre dieta e padrão
de atividade; (3) um capítulo sobre uso do espaço; e (4) um capítulo final com
considerações a respeito da conservação do mico-leão-da-cara-dourada em florestas
semideciduais. Metodologias específicas são apresentadas e cada capítulo e todas as
referências estão agrupadas no final.
1.1 O gênero Leontopithecus Lesson, 1840
A família Callitrichidae inclui os menores primatas do Novo Mundo, com massa
corporal variando de 100g a 700g (FLEAGLE, 1999) sendo, também, a mais diversa em
termos de espécies e subespécies, com 60 taxa reconhecidos (RYLANDS et al., 2000).
Seis gêneros estão incluídos na família: Cebuella, Callithrix, Mico, Saguinus,
Leontopithecus e Callimico. Os três primeiros possuem adaptações para utilização de
exsudatos vegetais: os dentes incisivos são de tamanho semelhante aos caninos e
voltados para frente, e são usados para abrir buracos em troncos (STEVENSON E
RYLANDS, 1988). Saguinus, Leontopithecus e Callimico não possuem tal
especialização e os caninos são bem maiores do que os incisivos (KLEIMAN et al.,
1988; FLEAGLE, 1999).
Os micos-leões, gênero Leontopithecus (Lesson, 1840), são os maiores
Callitrichidae, com os adultos podendo atingir mais de 650g. Quatro espécies de
distribuição alopátrica são reconhecidas, todas endêmicas da Mata Atlântica (Fig. 1):
mico-leão-da-cara-dourada, L. chrysomelas (Kuhl, 1820), que ocorre no sul da Bahia e
extremo norte de Minas Gerais; mico-leão-dourado, L. rosalia (Linnaeus, 1766),
encontrado na região costeira do Rio de Janeiro; mico-leão-preto, L. chrysopygus
(Mikan, 1823), que ocorre no oeste de São Paulo; e mico-leão-da-cara-preta, L. caissara
Lorini e Persson, 1990, com distribuição no litoral norte do Paraná e litoral sul de São
Paulo (RYLANDS et al., 2002). Todas as quatro espécies de Leontopithecus estão
incluídas em listas brasileiras e internacionais de espécies ameaçadas (MMA, 2005;
IUCN, 2007), sendo a destruição da Mata Atlântica a principal ameaça (RYLANDS et
al., 2002).

Capítulo 1
3
Figura 1. Distribuição histórica e atual das quatro espécies de micos-leões.
(Modificado de KLEIMAN E RYLANDS, 2002).
Os micos-leões possuem um sistema de reprodução cooperativo, no qual todos
os indivíduos do grupo ajudam no cuidado dos filhotes, seja carregando ou provendo
alimentos (DIETZ E BAKER, 1993; BAKER et al., 2002) e, normalmente, há apenas
um casal reprodutor (BAKER et al., 2002).
A ecologia comportamental das diferentes espécies e populações do gênero foi
revisada e comparada por Kierulff et al. (2002). A dieta dos micos-leões é composta
principalmente por frutos e insetos, consumindo também flores, néctar, exsudatos,
fungos e vertebrados de pequeno porte, como rãs, lagartos e filhotes de aves
(KLEIMAN et al., 1988; KIERULFF et al., 2002). Possuem os dedos alongados, o que
facilita o forrageio por presas animais em locais específicos como bromélias, buracos
em troncos e outras epífitas (COIMBRA-FILHO E MITTERMEIER, 1973; KLEIMAN

Capítulo 1
4
et al., 1988; RYLANDS, 1996). Os grupos utilizam ocos de árvore como locais de
dormida (COIMBRA-FILHO, 1978) e usam principalmente os estratos intermediários
da floresta, subindo ocasionalmente à copa para se alimentarem de frutos (COIMBRA-
FILHO E MITTERMEIER, 1973). A área de vida é relativamente grande, variando de
cerca de 40 ha, em algumas populações de L. rosalia, a mais de 300 ha, em L. caissara
(DIETZ et al., 1997; KIERULFF et al., 2002).
Apesar de serem espécies essencialmente de florestas primárias (RYLANDS,
1996), devido às necessidades de ocos para dormida e de bromélias e epífitas para
forrageio – características mais comuns em florestas não alteradas – os micos-leões
utilizam áreas secundárias com diferentes graus de alteração (KIERULFF et al., 2002;
RABOY et al., 2004). Além disso, as quatro espécies de micos-leões ocupam hábitats
bem distintos no que se refere à composição florística, sazonalidade e disponibilidade
de alimentos, evidenciando a grande flexibilidade ecológica e comportamental do
gênero (KIERULFF et al., 2002).
1.1.1 O Mico-Leão-da-Cara-Dourada, Leontopithecus chrysomelas (Kuhl, 1820)
O mico-leão-da-cara-dourada (MLCD) (Fig. 2) é classificado como “Em Perigo”
na Lista Brasileira de Espécies Ameaçadas e na lista vermelha da IUCN (MMA, 2005;
IUCN, 2007), e é endêmico da Mata Atlântica do sul da Bahia e extremo norte de Minas
Gerais (RYLANDS et al., 2002). Essa região destaca-se por apresentar elevadas taxas
de biodiversidade e endemismo, tanto de plantas como de animais (FONSECA et al.,
2002). Os primatas representam um bom exemplo dessa diversidade: a região é
considerada um centro de endemismo para esta ordem (KINZEY, 1982) e é no sul da
Bahia que encontramos, originalmente, a taxocenose de primatas mais diversificada em
toda a Mata Atlântica, com representantes dos seis gêneros que ocorrem no Bioma
(RYLANDS et al., 1996) – Callithrix, Leontopithecus, Callicebus, Cebus, Alouatta e
Brachyteles.
Como na maior parte do bioma Mata Atlântica, grandes blocos de floresta não
existem mais na região, e os remanescentes encontram-se isolados ou conectados
parcialmente por sistemas agroflorestais, como as cabrucas – sistema no qual o cacau
(Theobroma cacao) é plantado à sombra de árvores da copa da floresta (ALGER E
CALDAS, 1996).

Capítulo 1
5
A espécie ocorre na área limitada ao sul pelo Rio Jequitinhonha e ao norte pelo
Rio de Contas (Fig. 3); a oeste, a distribuição de L. chrysomelas é limitada por mudança
vegetacional, de floresta semidecidual para “mata de cipó”, formação florestal de
transição entre a Mata Atlântica e a Caatinga, associada a um aumento de altitude em
decorrência do Planalto de Vitória da Conquista (PINTO E RYLANDS, 1997;
RYLANDS et al., 2002). O extremo oeste da distribuição localiza-se a cerca de 150 km
da costa (RYLANDS et al., 2002). A área de distribuição total da espécie é de cerca de
19.000 Km2 e a população selvagem é estimada entre 6.000 e 15.000 indivíduos
(PINTO E RYLANDS, 1997).
Duas formações florestais são encontradas na área de distribuição do MLCD
(PINTO 1994; RYLANDS et al., 2002): Floresta Higrófila, ou ombrófila, sem estação
seca definida (GOUVEA et al., 1976), ocorrendo em uma faixa paralela à linha da costa
de aproximadamente 70 km de largura (VINHA et al., 1976); e Floresta Mesófila, ou
semidecidual (GOUVEA et al., 1976), na região mais oeste da distribuição (Fig. 4).
Além das diferenças na vegetação, essas regiões também apresentam realidades sócio-
econômicas bastante distintas. No sul da Bahia, as florestas semideciduais tiveram
desmatamento anterior ao das florestas ombrófilas e com maior intensidade
(MENDONÇA et al., 1994). Na região de floresta ombrófila, há predomínio de
plantações de cacau, na forma de cabrucas, o que permite certa conectividade entre os
fragmentos. É nessa região que podem ser encontrados os maiores fragmentos habitados
pelo MLCD (PINTO E RYLANDS, 1997). Já na região de floresta semidecidual, as
condições edafo-climáticas são propícias à pecuária, a atividade predominante (VINHA
et al., 1976), e os fragmentos florestais remanescentes são menores e encontram-se
bastante isolados por extensas áreas de pastagens.
Apesar de a floresta semidecidual ocupar mais da metade da área de distribuição
do MLCD, a maior parte das populações da espécie habita regiões de floresta ombrófila,
e as populações remanescentes em florestas semideciduais são encontradas em
numerosos fragmentos pequenos e isolados (PINTO, 1994; PINTO E RYLANDS,
1997).

Capítulo 1
6
Figura 2. O mico-leão-da-cara-dourada, Leontopithecus chrysomelas, com radio
colar.
Figura 3. Distribuição de Leontopithecus chrysomelas, destacando a Reserva
Biológica de Una. (Modificado de RYLANDS et al., 2002).

Capítulo 1
7
Figura 4. Tipos de vegetação no sul e extremo sul do Estado da Bahia, e localização
da Fazenda Barro Branco, onde foi realizado este estudo, e da Reserva Biológica
de Una. Base de dados retirada de arquivo digital (PRADO et al., 2003).

Capítulo 1
8
1.2 As Florestas Semideciduais do Sul da Bahia
A região oeste da distribuição de L. chrysomelas está inserida na região de
“Floresta Latifoliada Subcaducifólia Pluvial”, ou Mata Mesófila ou semidecidual, de
acordo com Gouvêa et al. (1976), e encontra-se bastante fragmentada. Ameaças
causadas pela ação humana, como corte seletivo, queimadas e entrada de gado nos
fragmentos, além dos efeitos negativos causados pelo efeito de borda, ainda ameaçam
os fragmentos florestais remanescentes (MARIANO-NETO, 2004).
Inicia-se a cerca de 50 km da costa marinha e ocupa uma faixa de
aproximadamente 50 km de largura, no sentido norte-sul, e sua composição florística
varia de acordo com a altitude, solo e drenagens (GOUVÊA et al., 1976). Os poucos
estudos florísticos ou ecológicos no sul da Bahia foram realizados em florestas
ombrófilas (MARIANO-NETO, 2004; AMORIM et al., 2005), e pouca informação
existe para as florestas semideciduais. Um levantamento florístico de algumas florestas
semideciduais da região, que incluíram os Municípios de Itororó, Caatiba, Ibicuí e Nova
Canaã, reportou uma estrutura de vegetação variável e com árvores altas, porém, de
diâmetros menores do que as encontradas na região de floresta ombrófila (VINHA et
al., 1976). No estrato arbóreo, foram encontradas espécies tanto de áreas mais úmidas
como de áreas mais secas, destacando-se as famílias Leguminosae, Moraceae,
Myrtaceae e Sapotaceae (VINHA et al., 1976). O substrato arbustivo era denso, com
grande quantidade de cipós, e rico em Rutáceas, Bromeliáceas e Ciperáceas (GOUVÊA
et al., 1976; VINHA et al., 1976). Este estudo, porém, teve foco em espécies com
potencial de exploração.
Mais recentemente, Amorim et al. (2005) fizeram um levantamento de plantas
vasculares na Serra do Teimoso, Município de Jussari, região de transição entre floresta
ombrófila e semidecidual, e que é habitada por micos-leões (OLIVER E SANTOS,
1991). De modo geral, os autores encontraram similaridade florística com as florestas
ombrófilas do sul da Bahia, porém muitos gêneros de plantas encontrados estão
presentes apenas em regiões mais secas, não ocorrendo nas florestas ombrófilas. As
famílias mais comuns foram Fabaceae (Leguminosae), Myrtaceae e Rubiaceae;
Bromeliaceae foi bastante comum no sub-bosque. As bordas das florestas e clareiras, no
entanto, apresentaram maior diferença em relação a esses mesmos ambientes de
florestas ombrófilas (AMORIM et al., 2005).

Capítulo 1
9
Mariano-Neto (2004) estudou a estrutura de duas florestas semideciduais nos
Municípios de Itambé e Itororó. Apesar desses locais não serem habitados por micos-
leões, pelo menos atualmente, suas estruturas e composições podem ser semelhantes a
outras florestas semideciduais localizadas mais a leste, onde ocorre L. chrysomelas. Os
resultados revelaram alta diversidade e riqueza nas florestas semideciduais, porém,
inferiores às observadas nas florestas ombrófilas do sul da Bahia. A grande diversidade
encontrada pode ser explicada pela presença de espécies tanto de áreas mais secas como
de florestas mais úmidas (MARIANO-NETO, 2004). A quantidade de bromélias e
lianas foi bastante alta, o que pode estar relacionado às perturbações antrópicas, como o
efeito de borda e o corte seletivo. Essas perturbações alteram condições
microclimáticas, como intensidade de luz, o que pode ser favorável ao crescimento de
bromélias (MARIANO-NETO, 2004).
1.3 Diferenças ambientais: ecologia e comportamento
Diferenças ecológicas e comportamentais em diferentes populações de uma
mesma espécie de primata já foram relatadas e são comumente atribuídas às diferenças
nos hábitats dessas populações (KIERULFF et al., 2002). A disponibilidade de
alimentos, estrutura da vegetação, composição florística do local e pressão de
predadores são características do ambiente que podem afetar os aspectos ecológicos e
comportamentais de uma espécie, como o tamanho e uso da área de vida, padrão de
atividades e dieta (OATES, 1987).
A destruição de hábitat e a fragmentação são uma das principais causas da perda
de biodiversidade (LAURENCE E COCHRANE, 2001). Além da diminuição da
variabilidade genética, decorrente do isolamento de pequenas populações (RALLS et
al., 1986), a fragmentação potencializa efeitos negativos como a caça, incidência de
doenças, ação de predadores e eventos estocásticos (LAURENCE E COCHRANE,
2001). A fragmentação florestal também pode afetar a qualidade do hábitat,
modificando a composição florística, densidade e distribuição de espécies vegetais
(MENON E POIRIER, 1996), o que pode ter efeito na ecologia e comportamento de
primatas (BICCA-MARQUES, 2005).
A fragmentação florestal pode afetar os primatas de diversas formas, tanto em
aspectos demográficos, como tamanho de grupos e densidade de indivíduos, como em

Capítulo 1
10
aspectos comportamentais, como a estratégia alimentar, além de afetar o fluxo gênico da
população (MARSH, 2003).
Todas as informações sobre a ecologia e o comportamento do MLCD existentes
provêm de trabalhos realizados na região de floresta ombrófila: um estudo de três meses
realizado na Estação Experimental Lemos Maia, da CEPLAC (RYLANDS, 1982;
RYLANDS, 1989) e um estudo de longa duração realizado na Reserva Biológica de
Una (DIETZ et al., 1994; DIETZ et al., 1996; RABOY, 2002; RABOY e DIETZ, 2004;
RABOY et al., 2004; HANKERSON et al., 2006). Nesses locais, não ocorre
sazonalidade pluviométrica, que é de cerca de 2000 mm anuais (RYLANDS, 1989;
RABOY, 2002). Outros estudos de monitoramento a longo prazo estão em andamento
na Rebio-Una - um em uma área com grande proporção de floresta primária
(coordenado por Dr. James Dietz e Dra. Becky Raboy) e outro em área da Reserva mais
degradada (coordenado pela Dra. Kristel De Vleeschouwer).
Alterações ecológicas e comportamentais, causadas por diferenças nos hábitats,
já foram relatadas para as diferentes espécies de Leontopithecus, e mesmo para
diferentes populações de uma mesma espécie do gênero (KIERULFF et al., 2002).
Essas alterações permitem a sobrevivência de micos-leões em florestas de diferentes
estruturas, idades e composições florísticas, características que afetam a quantidade e
qualidade de recursos disponíveis (KIERULFF et al., 2002).
Nada se sabe sobre a ecologia e o comportamento de L. chrysomelas no
ambiente de floresta semidecidual, e mesmo informações demográficas são bastante
escassas (ver PINTO, 1994). Não existem informações sobre a genética dessas
populações, ou mesmo o quanto são distintas geneticamente da população da Rebio-Una
(HOLST et al., 2006), única área de proteção integral em que a espécie pode ser
encontrada (RYLANDS et al., 2002). As populações habitando florestas semideciduais
são pequenas, embora ainda sejam numerosas, podendo servir como reservatório de
diversidade genética da espécie, e não existe nenhuma população de L. chrysomelas
protegida na região da floresta semidecidual.
Diferentemente de outros calitriquídeos, como Callithrix, Mico e Saguinus, que
são gêneros fortemente associados às florestas secundárias e bordas de mata, o gênero
Leontopithecus presumivelmente adaptou-se a utilização de florestas maduras,
especializando-se em forrageio por presas animais em locais específicos e na utilização
de ocos como locais de dormida (RYLANDS, 1996). Atualmente, poucas regiões da
Mata Atlântica ainda retêm características originais e os micos-leões habitam florestas

Capítulo 1
11
extremamente alteradas pela ação humana, com condições ecológicas muito diferentes
das que a espécie evoluiu. Essa é a situação encontrada nas florestas semideciduais
habitadas por L. chrysomelas no sul da Bahia, onde os fragmentos florestais são
menores do que na região de floresta ombrófila, com grande efeito de borda e tendo
sofrido intenso corte seletivo.
Dadas as diferenças florísticas, estruturais e sazonais, além das diferentes
pressões antrópicas, entre as florestas semidecidual e ombrófila, é esperado que haja
diferenças ecológicas e comportamentais entre as populações de L. chrysomelas
habitando esses ambientes. O conhecimento de tais diferenças permite avaliar quais
condições ambientais afetam a espécie (ZHOU et al., 2007), além de oferecer
informação essencial para o manejo e a formulação de estratégias visando a conservação
da espécie (PRIMACK E RODRIGUES, 2001; VALLADARES-PADUA et al., 2002).
Além disso, entender como uma espécie sobrevive em florestas degradadas nos permite
compreender a flexibilidade ecológica e comportamental da espécie (RYLANDS,
1996), avaliando as condições mínimas que permitem sua sobrevivência e permitindo a
elaboração de ações para conservação ou mesmo enriquecimento dessas florestas
(WONG E SICOTTE, 2007).
2. OBJETIVOS
2.1 Geral
Estudar a ecologia e o comportamento do mico-leão-da-cara-dourada,
Leontopithecus chrysomelas, em um fragmento de floresta semidecidual.
2.2 Específicos
1) Descrever o padrão de atividades do grupo em uma área de floresta semidecidual
(Cap. II);
2) Listar as espécies de plantas utilizadas na alimentação (Cap. II);
3) Avaliar a disponibilidade temporal e espacial de frutos na floresta (Caps. II e III);
4) Estimar a área de vida e avaliar o padrão de utilização da área por um grupo (Cap.
III);
5) Avaliar como a disponibilidade de frutos nas estações seca e chuvosa ao longo do
ano afeta os parâmetros ecológico-comportamentais estudados (Cap. II e III);

Capítulo 1
12
6) Comparar os resultados com as informações sobre a ecologia da espécie existentes
para a Reserva de Una (Floresta Ombrófila) e com dados de outras espécies de
Leontopithecus, para entender como diferenças nos hábitats afetam o comportamento e
a ecologia da espécie (Cap. II e III);
7) Discorrer sobre a conservação do mico-leão-da-cara-dourada na região de floresta
semidecidual (Cap. IV).
3. MÉTODOS
3.1 Área de Estudo
A pesquisa foi realizada em um fragmento de floresta semidecidual na Fazenda
Barro Branco (15° 08’ 25” S, 39° 57’ 21” W), localizada a aproximadamente 15 km da
zona urbana do Município de Itororó, Bahia, e distante cerca de 100 km, em linha reta,
da Reserva Biológica de Una (Fig. 4).
O fragmento é cercado por áreas de pastagens e possui aproximadamente 450
ha, com formato muito irregular, aumentando bastante a proporção de bordas, além de
ser atravessado por uma estrada de terra (Fig. 5). Encontra-se isolado e com seu formato
atual há pelo menos 50 anos, segundo informações dos proprietários e moradores
antigos da região, e é composto por um mosaico de floresta secundária em diferentes
estágios de regeneração. A floresta sofreu intenso corte seletivo, que ainda acontece,
porém em escala reduzida. A exploração da floresta ocorreu, principalmente, para a
retirada de madeira para ser usada na própria fazenda, na construção de cercas e currais,
e como lenha. Alguns riachos temporários, que ficam cheios apenas na época de mais
chuva, formando brejos, cortam a mata.
A região na qual está localizado o fragmento foi apontada por Pinto (1994)
como uma localidade de interesse para a conservação de L. chrysomelas. O autor
destacou esta e outras áreas pela quantidade e qualidade da floresta, interesse
conservacionista dos proprietários ou pela infra-estrutura e logística para a realização de
pesquisas. Considerando os fragmentos vizinhos, a região da Fazenda Barro Branco
apresenta uma das maiores áreas de floresta da região.
O sub-bosque é bastante denso e com muitos cipós, e as árvores emergentes
atingem até 30 metros de altura. Bromélias são comuns, tanto epífitas como no chão
(principalmente as espécies do gênero Aechmea).

Capítulo 1
13
Além de L. chrysomelas, outras três espécies de primatas ocorrem na área:
Callithrix kuhlii, Callicebus melanochir e Cebus xanthosternos. Moradores antigos
indicaram a presença de Alouatta guariba no passado. Possíveis predadores de L.
chrysomelas que podem ser encontrados no fragmento são felinos (Puma concolor e
Leopardus sp.), irara (Eira barbara) e possivelmente aves de rapina. Embora inserido
em uma área particular, a caça não é comum no fragmento, não existindo, portanto,
pressão de caça significativa.
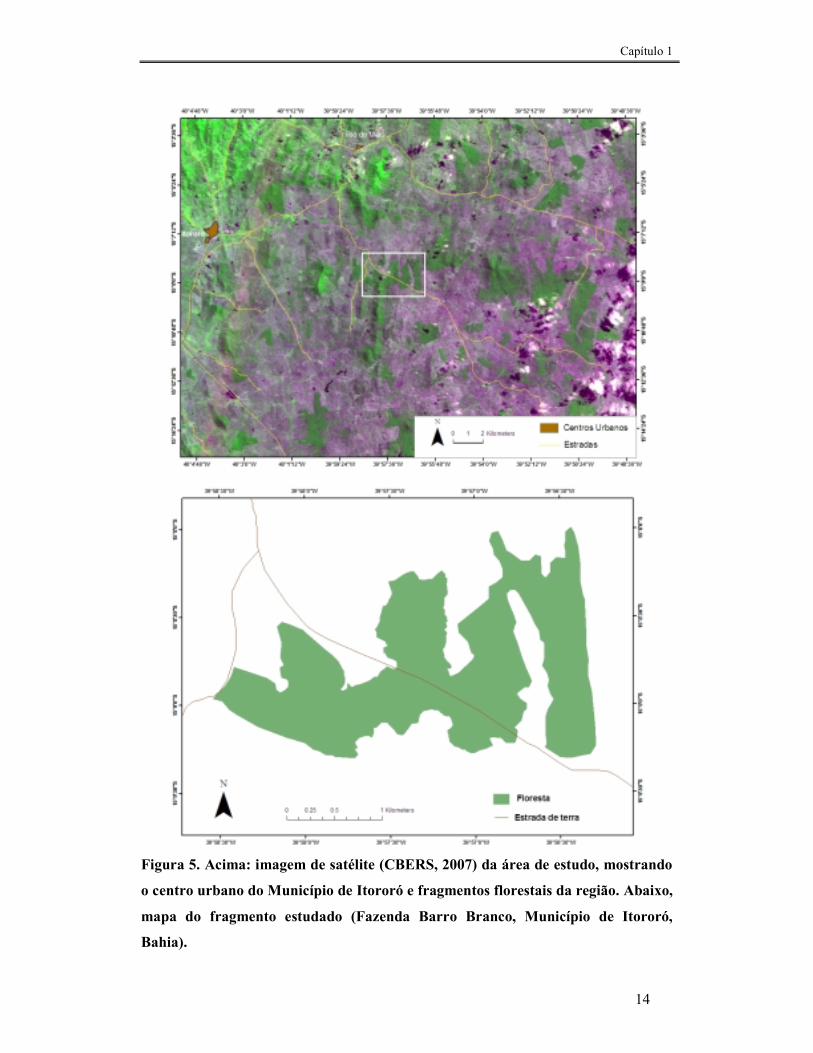
Capítulo 1
14
Figura 5. Acima: imagem de satélite (CBERS, 2007) da área de estudo, mostrando
o centro urbano do Município de Itororó e fragmentos florestais da região. Abaixo,
mapa do fragmento estudado (Fazenda Barro Branco, Município de Itororó,
Bahia).
Capítulo 1
15
3.1.1 Informações climáticas
A região de floresta semidecidual na Bahia ocorre em áreas com precipitação
por volta de 1000 mm anuais e com uma estação seca bem definida com duração de 3 a
4 meses, de junho a setembro, quando algumas árvores perdem as folhas (GOUVÊA et
al., 1976).
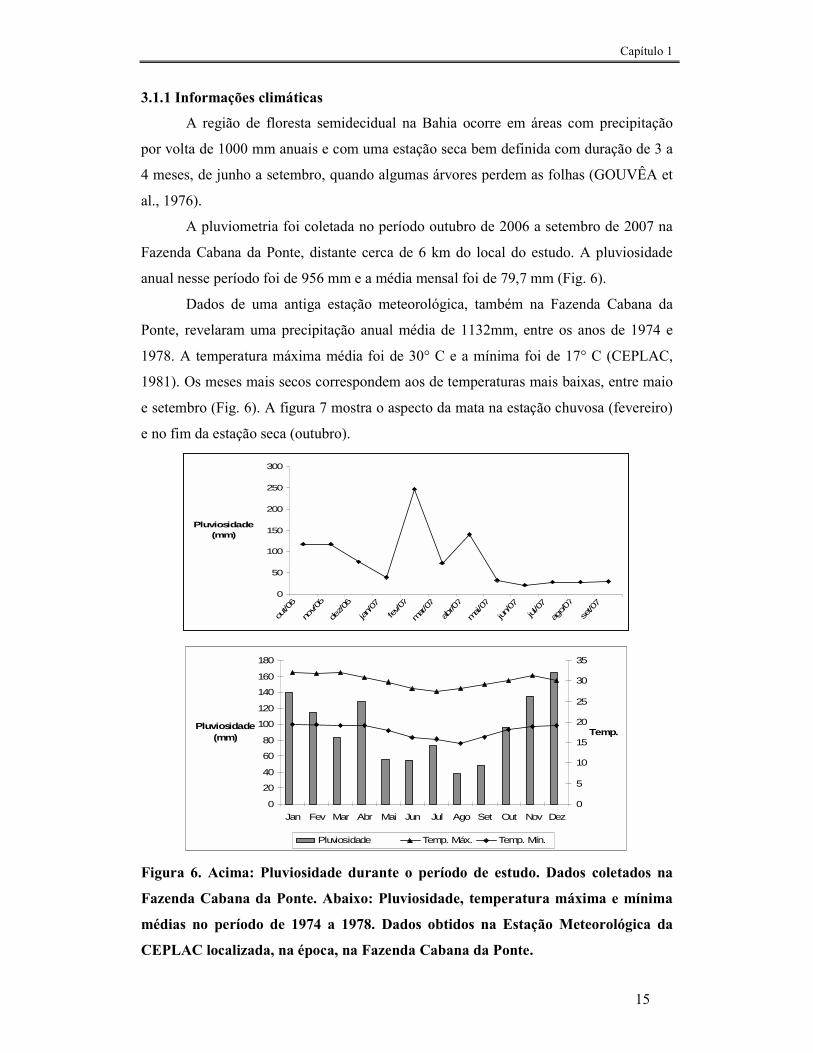
A pluviometria foi coletada no período outubro de 2006 a setembro de 2007 na
Fazenda Cabana da Ponte, distante cerca de 6 km do local do estudo. A pluviosidade
anual nesse período foi de 956 mm e a média mensal foi de 79,7 mm (Fig. 6).
Dados de uma antiga estação meteorológica, também na Fazenda Cabana da
Ponte, revelaram uma precipitação anual média de 1132mm, entre os anos de 1974 e
1978. A temperatura máxima média foi de 30° C e a mínima foi de 17° C (CEPLAC,
1981). Os meses mais secos correspondem aos de temperaturas mais baixas, entre maio
e setembro (Fig. 6). A figura 7 mostra o aspecto da mata na estação chuvosa (fevereiro)
e no fim da estação seca (outubro).
0
50
100
150
200
250
300
out/0
6
nov/0
6
dez/0
6
jan/07
fev/07
mar/07
abr/0
7
mai/07
jun/07
jul/07
ago/0
7
set/0
7
Pluviosidade (mm)
0
20
40
60
80
100
120
140
160
180
Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
Pluviosidade (mm)
0
5
10
15
20
25
30
35
Temp.
Pluviosidade Temp. Máx. Temp. Mín.
Figura 6. Acima: Pluviosidade durante o período de estudo. Dados coletados na
Fazenda Cabana da Ponte. Abaixo: Pluviosidade, temperatura máxima e mínima
médias no período de 1974 a 1978. Dados obtidos na Estação Meteorológica da
CEPLAC localizada, na época, na Fazenda Cabana da Ponte.

Capítulo 1
16
Figura 7. A floresta da Fazenda Barro Branco na estação chuvosa (fevereiro/2007)
e no final da estação seca (setembro/2007).
3.2 Grupos monitorados
A pesquisa foi realizada no período de janeiro/2006 e agosto/2007. Dois grupos
foram monitorados: MAN e GRA. Foi considerado um dia completo de observação
quando esta ocorreu desde o momento em que os animais deixaram o local de dormida
até o momento em que eles chegaram ao local de dormida (“oco a oco”).
Inicialmente, o grupo MAN foi monitorado com observação direta, sem a
utilização de rádios-transmissores. Posteriormente, foi realizada a captura dos dois
grupos para colocação de rádios-transmissores (Holohil Systems®). O monitoramento
foi feito com receptor e antena da marca Telonics®. A técnica de captura foi a mesma
utilizada em outros estudos com L. chrysomelas e L. rosalia (KIERULFF, 2000;
RABOY E DIETZ, 2004). Os grupos foram cevados com bananas e capturados em
armadilhas tipo Tomahawk. Durante a captura e manipulação dos animais, eram
registrados o sexo, o peso e medidas biométricas, além de uma rápida análise do estado
de saúde dos animais. Os animais foram marcados com tinta preta nyanzol nas partes
douradas para reconhecimento individual. Todo o procedimento foi realizado de acordo
com as normas para captura e manipulação de animais e tinha autorização do IBAMA.

Capítulo 1
17
Antes de o monitoramento ser iniciado, os grupos passaram por um processo de
habituação ao observador, que durou cerca de um mês para cada grupo. O processo de
habituação foi necessário para permitir uma maior aproximação e um melhor contato
visual com os grupos.

18
CAPÍTULO II - Padrão de atividades e dieta do
mico-leão-da-cara-dourada (Leontopithecus
chrysomelas) em um fragmento de floresta
semidecidual

Capítulo 2
19
1. INTRODUÇÃO
O padrão de atividades representa o modo como os indivíduos distribuem as
diferentes atividades ao logo do tempo (DEFLER, 1995). Diversas variáveis afetam o
padrão de atividade de primatas, tanto ambientais, como temperatura e disponibilidade
espacial e temporal de recursos alimentares, como variáveis intrínsecas da espécie ou
dos grupos, como a composição da dieta, o tamanho corporal, o tamanho e a
composição dos grupos, a época reprodutiva e a densidade populacional (OATES, 1987;
STRIER, 1987; DORAN, 1997; HANYA, 2004; GURSKY, 2000; LI E ROGERS,
2004; VASEY, 2005; ZHOU et al., 2007).
O padrão de atividades está relacionado às restrições energéticas e metabólicas
(OATES, 1987) e para consumir a quantidade de energia necessária, os primatas
ajustam suas atividades de acordo com o ambiente (ZHOU et al., 2007). Assim, o
padrão de atividades de uma espécie representa um balanço entre as diferentes
atividades e pode variar ao longo do ano ou da vida do indivíduo, refletindo o
investimento em energia e tempo necessários para sua sobrevivência e reprodução
(DEFLER, 1995). Por exemplo, grupos com mais filhotes podem apresentar maior taxa
de comportamentos sociais, como catação e brincadeira (LI E ROGERS, 2004), e a
presença de fêmeas grávidas ou em lactação, com filhotes que exigem cuidados, pode
aumentar o requerimento energético do grupo, aumentando o tempo dedicado à
alimentação (VASEY, 2005).
A procura por alimentos é parte essencial da vida de qualquer primata, afetando
todas as suas outras atividades (OATES, 1987) Os primatas respondem de diversas
maneiras às variações temporais na oferta de alimentos, e variações sazonais no padrão
de atividades de um mesmo grupo ou população já foram amplamente relatadas. De
modo geral, espécies com dieta predominantemente folívora (por exemplo, as do gênero
Alouatta) tendem a diminuir a atividade e repousar mais nas épocas de escassez de
alimentos, enquanto que primatas frugívoros e insetívoros tendem a locomover-se mais
e a aumentar o tempo de forrageio por esses itens durante a escassez. (OATES, 1987).
Gursky (2000), estudando Tarsius spectrum, um prosímio insetívoro, observou que
quando a abundância de alimentos diminuía, o tempo dedicado à sua procura
aumentava. Doran (1997) observou que chimpanzés gastaram mais tempo procurando e
alimentando-se de frutos durante a época em que estes eram mais escassos. Isso ocorre
porque na época de menor disponibilidade de frutos, as fontes de alimentos encontram-

Capítulo 2
20
se mais espalhadas e em menor quantidade e/ou qualidade, como também observou
Terborgh (1983) ao estudar Cebus apella. Essa relação, porém, pode ser mais complexa.
Muitos primatas frugívoros também podem apresentar diminuição de atividade durante
períodos de escassez de alimentos, uma resposta apresentada como estratégia para
diminuir o gasto energético (VASEY, 2005).
Variações sazonais na dieta de diversos primatas também já foram amplamente
relatadas. Por exemplo, Lophocebus albigenia, espécie onívora, alimentou-se de mais
frutos durante a época em que estes eram mais abundantes e aumentou a proporção de
flores, sementes e folhas na época de escassez de frutos (POULSEN et al., 2001).
Callithrix geoffroyi, um calitriquídeo da Mata Atlântica, também aumentou a proporção
de frutos durante a época de chuvas, quando havia mais frutos disponíveis. Estas
variações estão associadas à disponibilidade sazonal de alimentos no ambiente e às
variações das necessidades energéticas e metabólicas dos indivíduos.
As quatro espécies do gênero Leontopithecus (Callitrichidae) são frugívoro-
insetívoras, sendo frutos os principais componentes da dieta, seguidos de insetos
(RYLANDS, 1989; DIETZ et al., 1997; PASSOS, 1999, KIERULFF et al., 2002,
RABOY E DIETZ, 2004). Há preferência por frutos maduros, na maioria pequenos,
macios e doces; frutos maiores são consumidos apenas ocasionalmente (DIETZ et al.,
1997; PASSOS, 1999; LAPENTA et al., 2003; RABOY E DIETZ, 2004) e frutos de
árvores são mais consumidos do que os de cipós (KIERULFF et al., 2002). Presas
animais representam o segundo item na dieta dos micos-leões, que forrageiam por
presas ativamente, manipulando substratos (RYLANDS, 1996; KIERULFF et al.,
2002). Insetos da Ordem Orthoptera são os animais mais consumidos, seguidos de
larvas de Coleoptera e Lepidoptera (DIETZ et al., 1997; KIERULFF et al., 2002).
Flores, néctar, fungos, exsudatos e vertebrados de pequeno porte também são
consumidos, em diferentes proporções, dependendo da população estudada.
O comportamento alimentar e o padrão de atividades de L. chrysomelas foram
mais detalhadamente estudados por Raboy e Dietz (2004) em área de floresta ombrófila
bem preservada da Reserva Biológica de Una, com mais de 7.000 ha de floresta, e por
Rylands (1989), também em floresta ombrófila da região de Una. Este último estudo foi
realizado em um fragmento florestal bem menor, com cerca de 400 ha. Entretanto, até o
momento, nada se conhece sobre a dieta e o padrão de atividades de populações
habitando florestas semideciduais.

Capítulo 2
21
Variações na dieta e no padrão de atividades, decorrentes de diferenças
ambientais, também já foram relatadas para as diferentes espécies de Leontopithecus e
mesmo para diferentes populações de uma mesma espécie (KIERULFF et al., 2002).
Assim, é esperado que populações de L. chrysomelas em floresta semidecidual e a
população de floresta ombrófila da Rebio-Una apresentem diferenças de dieta e padrão
de atividades. Além disso, comparar o padrão de atividades em diferentes condições
ecológicas permite avaliar as influências ambientais e sazonais no comportamento das
espécies (ZHOU et al., 2007). O entendimento dessas variações é essencial para o
manejo e a formulação de estratégias visando a conservação das espécies (PRIMACK E
RODRIGUES, 2001; VALLADARES-PADUA et al., 2002).
Este capítulo tem como objetivo descrever o padrão de atividades de um grupo
de L. chrysomelas habitando um fragmento de floresta semidecidual, apresentar a
composição da dieta do grupo e avaliar como o padrão de atividades e o consumo dos
diferentes itens alimentares variam ao longo do ano, de acordo com a disponibilidade de
frutos na floresta. Os resultados foram comparados com as informações disponíveis para
a Rebio-Una e para as outras espécies de Leontopithecus.
2. MÉTODOS
2.1 Área de Estudo e Grupos
O trabalho foi realizado em um fragmento de floresta semidecidual da fazenda
Barro Branco, Município de Itororó, Bahia. Uma descrição detalhada da área de estudo
e informações sobre os grupos estudados (MAN e GRA), encontram-se no Capítulo I.
2.2 Disponibilidade de frutos
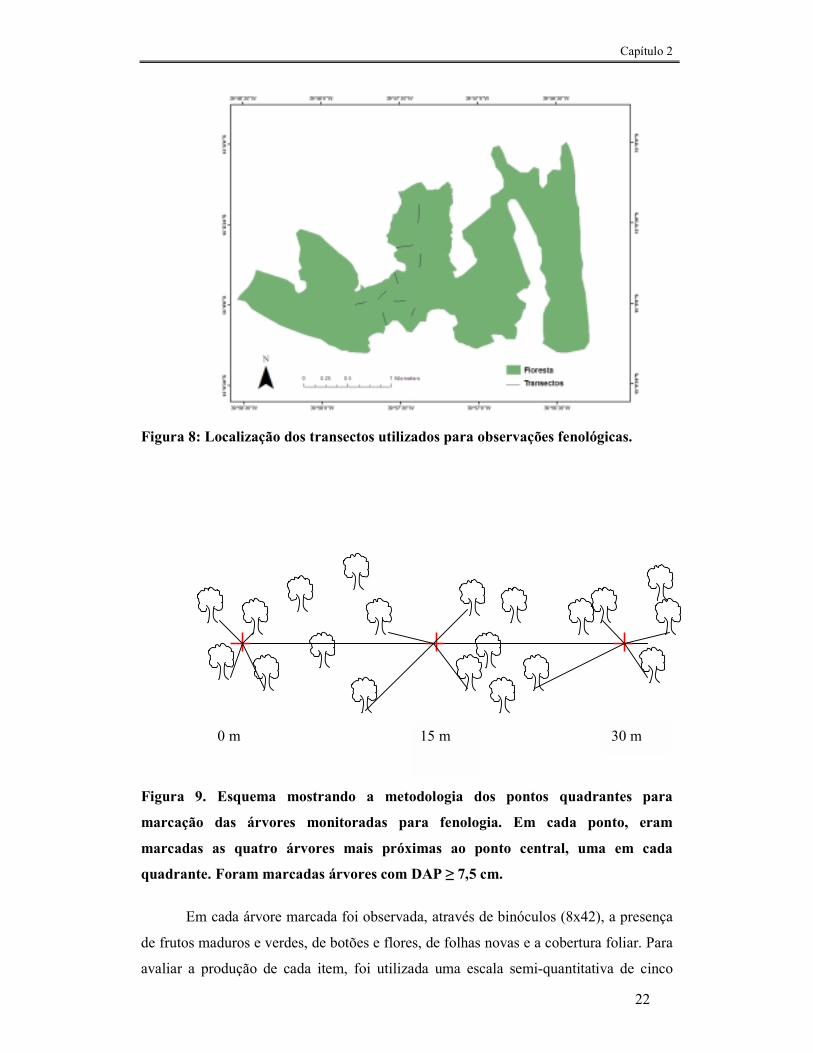
Foi realizada uma avaliação da disponibilidade de frutos de espécies arbóreas na
mata através do método do ponto quadrante (COTTAN E CURTIS, 1956). Transectos
com tamanho entre 150 a 210 metros foram abertos (Fig. 8). A cada 15 metros foram
marcadas as quatro árvores mais próximas do ponto central, uma em cada quadrante
(Fig. 9). Foram consideradas apenas as árvores com DAP (diâmetro na altura do peito)
≥ 7,5cm. As observações foram mensais, realizadas durante um período de 12 meses
consecutivos (outubro/2006 – setembro/2007).
Capítulo 2
22
Figura 8: Localização dos transectos utilizados para observações fenológicas.
Figura 9. Esquema mostrando a metodologia dos pontos quadrantes para
marcação das árvores monitoradas para fenologia. Em cada ponto, eram
marcadas as quatro árvores mais próximas ao ponto central, uma em cada
quadrante. Foram marcadas árvores com DAP ≥ 7,5 cm.
Em cada árvore marcada foi observada, através de binóculos (8x42), a presença
de frutos maduros e verdes, de botões e flores, de folhas novas e a cobertura foliar. Para
avaliar a produção de cada item, foi utilizada uma escala semi-quantitativa de cinco
0 m 15 m 30 m

Capítulo 2
23
categorias (de 0 a 4), conforme Fournier (1974), onde: zero representa ausência; 1
indica presença com quantidade variando de 1 a 25%; 2, presença com quantidade de 26
a 50%; 3, presença com quantidade de 51 a 75%; e 4, presença com quantidade de 76 a
100%. O índice calculado representa a proporção do item observado em relação ao total,
sendo calculado pela seguinte fórmula: I = Σ Fournier x 100/ 4 x N; onde, Σ Fournier é a
somatória dos valores de cada item e N é o número de indivíduos observados.
Dois ambientes foram analisados:
(1) Margem de Riacho/Brejo. No caso dos riachos, que só ficam cheios na
época de chuvas, os transectos acompanharam o seu leito, de fundo arenoso. Os brejos
eram regiões que ficavam alagadas durante a época de chuvas, porém sem água
corrente. Os pontos dos transectos foram marcados no centro do riacho, que pode atingir
até cerca de três metros de largura e, em alguns locais, até um metro de profundidade, e
no meio dos brejos;
(2) Floresta seca. Foram abertos três transectos na borda mata, que se iniciavam
no contato da mata com o pasto e seguiam no sentido borda para o interior da floresta.
Outros três transectos foram abertos no interior da floresta. Esses transectos estavam
distantes pelo menos 100 metros da borda e dos transectos dos riachos. Essa disposição
dos transectos foi escolhida para se ter uma representação de diferentes ambientes da
floresta
Os ambientes também foram comparados em termos de altura das árvores e
diâmetro na altura do peito (DAP), através do teste T de Student. No total foram
monitoradas 440 árvores, sendo 148 em riachos e 292 em floresta seca. Todas as árvores
monitoradas foram identificadas taxonomicamente por especialistas e o material foi
depositado no Herbário da Universidade Estadual de Santa Cruz - UESC.
2.3 Composição e variação sazonal da dieta
Todos os itens alimentares consumidos pelos grupos foram anotados. Os animais
consumidos foram identificados até o nível de Ordem. As espécies vegetais utilizadas
na alimentação tiveram ramos (com ou sem flores) coletados e foram identificadas até o
nível de espécie, sempre que possível. O material foi identificado taxonomicamente por
especialistas e depositado no Herbário da UESC.
Todas as visitas em fruteiras do grupo GRA foram anotadas. Uma visita em
fruteira ocorria quando pelo menos um indivíduo do grupo consumia frutos por pelo
menos um minuto. Nestas ocasiões, a fruteira utilizada foi marcada e numerada. Foi

Capítulo 2
24
anotado o tempo em que o grupo ficou se alimentando, marcado a partir da entrada do
primeiro indivíduo na fruteira até a saída do último, e o número de micos que se
alimentou de seus frutos.
No caso de alimentação de frutos de bromélias, o registro desse tempo nem
sempre era possível, já que a visibilidade do grupo era muitas vezes prejudicada quando
eles entravam dentro da bromélia. Em muitos casos o fruto da bromélia não estava
visível, e só era possível saber que um mico alimentou-se de frutos quando estes caíam
no chão, já comidos. Quando isso acontecia, era anotada a hora em que o primeiro
indivíduo entrou na bromélia e a hora em que o último saiu, o que não necessariamente
representava o tempo em que o grupo se alimentou daquela bromélia. Além disso,
diversas atividades, além de comer frutos, são realizadas dentro da bromélia,
principalmente forrageio por insetos, o que torna difícil o registro do tempo exato que o
animal se dedicou a cada atividade, quando estava dentro da bromélia.
As espécies de frutos consumidas em cada mês foram listadas e localizadas em
um mapa. Para verificar se houve diferença entre as estações seca e chuvosa, foram
comparadas as seguintes medidas, através do teste T de Student: média de fruteiras
visitadas por dia e tempo médio de permanência em fruteiras. Essas análises foram
feitas para todas as fruteiras, somente para bromélias e somente para fruteiras diferentes
de bromélias. No caso de bromélias, pelo motivo apresentado no parágrafo anterior, não
foi considerado o tempo de permanência, apenas o número de bromélias visitadas para
alimentação de frutos.
2.4 Padrão de Atividades
Para permitir uma comparação mais acurada com os estudos realizados na
Reserva Biológica de Una, a metodologia usada para a coleta dos dados
comportamentais foi a mesma descrita por Raboy e Dietz (2004). Os registros dos
comportamentos de cada indivíduo do grupo foram coletados pelo método de varredura
instantânea, ou scan sampling (ALTMANN, 1974), com duração de cinco minutos, a
cada 20 minutos. Assim, foram coletados três registros de todo o grupo a cada hora.
Para o cálculo do padrão de atividades, foram utilizados apenas os dias
completos de observação. Apenas os dados referentes ao grupo GRA foram analisados,
já que o grupo MAN teve poucos dias completos de observação.
Os comportamentos dos indivíduos foram classificados nas seguintes categorias,
adaptado do etograma descrito por Raboy e Dietz (2004):

Capítulo 2
25
1. Locomovendo - indivíduo se deslocando de um ponto a outro;
2. Parado/Repouso – indivíduo não realiza nenhuma atividade ou deitado em
substrato, dormindo ou não;
3. Forrageando ou comendo partes de plantas – indivíduo procurando,
manipulando ou ingerindo alimento vegetal;
4. Forrageando ou comendo presa animal – indivíduo comendo e procurando
ativamente por presa animal, de forma manipulativa ou visual;
5. Atividade Social – indivíduo em atividade social, como catação, brincadeira,
vocalização e marcação com glândula;
6. Outras – quaisquer atividades que não se enquadram nas categorias acima –
bebendo água, esfregando corpo em seiva.
Forragear e comer partes vegetais foram agrupados devido à dificuldade de
separar esses comportamentos durante a observação em campo. Como os eventos de
comer inseto ou outros animais são relativamente rápidos e nem sempre visíveis, o
método de varredura utilizado pode não ter sido eficaz para registrar este
comportamento. Assim, a categoria “comendo presas” foi tratada juntamente com
“forrageando por presas” na análise do tempo dedicado à alimentação.
Os itens ingeridos durante a alimentação (fruto, flor, néctar ou exsudato) e o
micro-hábitat de forrageio (bromélias, folhas secas de bromélias, outras epífitas, folhas
de palmeira, buraco em troncos, galhos ou outros) foram anotados. O monitoramento
dos indivíduos que nasceram durante a pesquisa começou a ser anotado apenas quando
estes estavam independentes, isto, é, quando deixavam de ser carregados pelos adultos,
o que ocorre com cerca de três meses de idade (BAKER et al., 2002).
Para o padrão de atividades, foi calculada a proporção de cada atividade em
relação ao número total de registros, tanto para o período total do estudo como para
cada mês separadamente. O ciclo diário das atividades foi realizado do mesmo modo,
levando-se em conta cada hora do dia.
Comer exsudatos foi agrupado na categoria “outros comportamentos” nas
análises estatísticas, devido ao baixo número de registros.
Para verificar se houve diferenças na proporção dos comportamentos entre as
estações, os dados foram agrupados em estação seca e chuvosa e foi realizado o teste do
Qui-quadrado. A porcentagem dos registros de cada classe comportamental foi

Capítulo 2
26
comparada entre as estações pelo teste de Mann-Whitney para verificar quais
comportamentos apresentaram variação sazonal. Foi realizado o teste de correlação de
Spearman entre a proporção de cada comportamento e a (1) pluviosidade; e (2) índice
de Fournier para frutos maduros, frutos verdes e flores.
Todos os testes estatísticos foram feitos com o programa SPSS 10.1 e todos
foram bilaterais, considerando o nível de significância < 0,05 (ZAR, 1996).
3. RESULTADOS
3.1 Grupos monitorados
O grupo MAN foi monitorado por oito meses, entre janeiro/2006 e agosto/2006.
O grupo teve 169,5 horas de observação, porém foram apenas cinco dias completos, que
representaram 56,5 horas. A composição desse grupo variou de dois a seis indivíduos –
chegando a ter dois machos adultos, uma fêmea adulta, um adulto de sexo não
identificado e dois filhotes. Estes nasceram em fevereiro/2006, porém, desapareceram
duas semanas depois. Dois indivíduos adultos desapareceram em fevereiro e
junho/2006, restando apenas dois machos adultos, que continuaram juntos e ocupando a
mesma área que o grupo utilizava anteriormente. Em março de 2007 um indivíduo
juntou-se aos dois, provavelmente uma fêmea, embora falte confirmação. Em junho de
2007, o macho que carregava o rádio transmissor foi encontrado morto, com sinais de
predação - não foi possível, porém, identificar o predador.
O grupo GRA foi monitorado por 14 meses consecutivos, entre agosto/2006 e
setembro/2007, com observações de dois a seis dias por mês e teve 682,79 horas de
observação, com 64 dias completos, que corresponderam a 654,29 horas. Ao todo,
foram realizados 1.959 scans e 12.310 registros individuais, dos quais 9.833 (79,9%)
foram válidos, isto é, a atividade do indivíduo foi observada. Os restantes representaram
registros “não visíveis”.
A composição desse grupo variou de quatro a nove indivíduos – chegando a ser
composta por dois machos adultos, duas fêmeas adultas, dois machos jovens, uma
fêmea jovem e dois filhotes. Quatro filhotes, dois gêmeos de cada fêmea, nasceram em
janeiro de 2006 e mais quatro, novamente dois gêmeos de cada fêmea, em
setembro/outubro de 2006. Houve nascimento de mais um par de gêmeos em fevereiro
de 2007. Dos 10 nascimentos, sete filhotes desapareceram antes de completarem sete
meses de idade; desses, cinco desapareceram antes dos três meses.

Capítulo 2
27
3.2 Disponibilidade de frutos e hábitats
O acompanhamento fenológico teve início no começo da estação chuvosa de
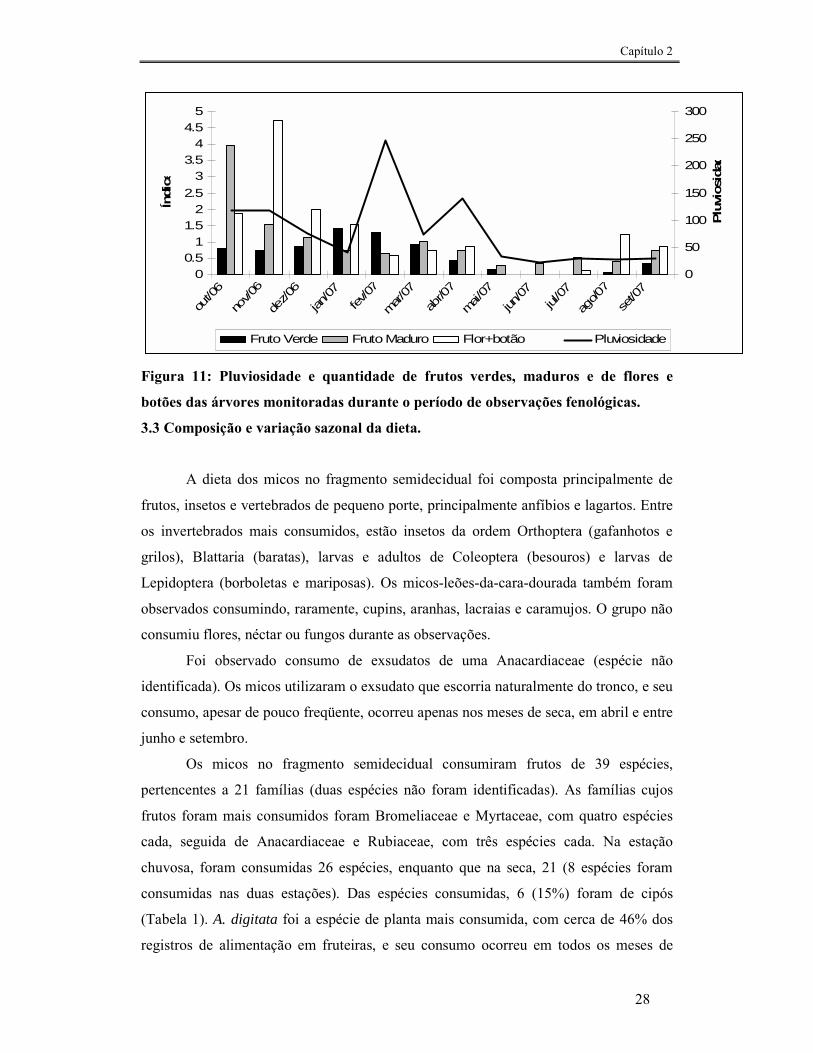
2006, no mês de outubro. As observações mostraram queda acentuada da cobertura
foliar nos meses de agosto, setembro e outubro (Fig. 10). A quantidade de frutos na
mata variou bastante ao longo do ano, com um pico de frutos no mês de outubro. Os
meses que apresentaram menor quantidade de frutos, tanto maduros como verdes, foram
os mais secos e mais frios (maio – agosto). Houve um pico de floração no mês de
novembro, e nos meses seguintes a quantidade de flores e botões diminuiu, até estarem
totalmente ausentes nos meses de junho e julho. (Fig. 11).
Nas 440 árvores monitoradas, foram identificadas 28 famílias botânicas (ver
lista de espécies no Anexo I). A família mais comum foi a das leguminosas (Fabaceae),
com 46% das árvores monitoradas, seguida de Euphorbiaceae (11%), Anacardiaceae
(8,5%) e Meliaceae e Myrtaceae (6,7%). As espécies mais comuns foram Piptadenia sp.
(Fabaceae), Gonirrachis marginata (Fabaceae) e uma Euphorbiaceae não identificada,
que, juntas, representaram 39% de todas as árvores identificadas (23 indivíduos, do
total, não foram identificados).
Cobertura Foliar
40.0
50.0
60.0
70.0
80.0
90.0
100.0
out/0
6no
v/06
dez/0
6jan
/07fev
/07
mar/07
abr/0
7
mai/07
jun/07
jul/07
ago/0
7se
t/07
Índice
Figura 10: Cobertura foliar das árvores monitoradas no fragmento da Fazenda
Barro Branco durante o período de observações fenológicas.
Capítulo 2
28
00.5
11.5
22.5
33.5
44.5
5
out/0
6
nov/0
6
dez/0
6jan
/07fev
/07
mar/07
abr/0
7
mai/07
jun/07
jul/07
ago/0
7
set/0
7
Índi
ce
0
50
100
150
200
250
300
Plu
vios
idad
Fruto Verde Fruto Maduro Flor+botão Pluviosidade
Figura 11: Pluviosidade e quantidade de frutos verdes, maduros e de flores e
botões das árvores monitoradas durante o período de observações fenológicas.
3.3 Composição e variação sazonal da dieta.
A dieta dos micos no fragmento semidecidual foi composta principalmente de
frutos, insetos e vertebrados de pequeno porte, principalmente anfíbios e lagartos. Entre
os invertebrados mais consumidos, estão insetos da ordem Orthoptera (gafanhotos e
grilos), Blattaria (baratas), larvas e adultos de Coleoptera (besouros) e larvas de
Lepidoptera (borboletas e mariposas). Os micos-leões-da-cara-dourada também foram
observados consumindo, raramente, cupins, aranhas, lacraias e caramujos. O grupo não
consumiu flores, néctar ou fungos durante as observações.
Foi observado consumo de exsudatos de uma Anacardiaceae (espécie não
identificada). Os micos utilizaram o exsudato que escorria naturalmente do tronco, e seu
consumo, apesar de pouco freqüente, ocorreu apenas nos meses de seca, em abril e entre
junho e setembro.
Os micos no fragmento semidecidual consumiram frutos de 39 espécies,
pertencentes a 21 famílias (duas espécies não foram identificadas). As famílias cujos
frutos foram mais consumidos foram Bromeliaceae e Myrtaceae, com quatro espécies
cada, seguida de Anacardiaceae e Rubiaceae, com três espécies cada. Na estação
chuvosa, foram consumidas 26 espécies, enquanto que na seca, 21 (8 espécies foram
consumidas nas duas estações). Das espécies consumidas, 6 (15%) foram de cipós
(Tabela 1). A. digitata foi a espécie de planta mais consumida, com cerca de 46% dos
registros de alimentação em fruteiras, e seu consumo ocorreu em todos os meses de

Capítulo 2
29
observação O cipó Cisus sp. foi a segunda espécie mais consumida, com 17,2% das
visitas (Tabela 2).
Foram registradas 881 visitas do grupo em fruteiras (considerando apenas os
dias completos de observação). Bromélias estão entre os frutos mais consumidos, com
475 (54%) das visitas (Tabela 3).
30
Tabela 1. Frutos consumidos por L. chrysomelas no fragmento de floresta semidecidual em cada mês de observação. Os valores representam a porcentagem em termos de número de visitas.
Família Espécie Hábito Nome comum ago/06 set/06 out/06 nov/06 dez/06 jan/07 fev/07 mar/07 abr/07 mai/07 jun/07 jul/07 ago/07 set/07 1 Anacardiaceae Astronium macrocalyx arbóreo 1.1% 2 Spondias venulosa arbóreo Cajá 5.0% 1.6% 3 sp.1 arbóreo Jacaré 1.4% 4 Annonaceae Duguetia sp. arbóreo Pinha da mata 8.5% 5 Hornschuchia lianarum arbóreo Cega burro 2.0% 1.1% 6 Araceae Philodendron sp. epífita Imbé 1.4% 2.4% 1.4% 1.2% 7 Arecaceae Bactris ferrugineo arbóreo Mané velho 2.4% 8 Boraginaceae Cordia anabaptista arbóreo Baba de boi 1.4% 18.3% 17.9% 1.4% 1.0% 1.1% 9 Bromeliaceae Aechmea digitata epífita Gravatá 40.0% 57.0% 32.5% 20.0% 86.6% 15.0% 45.1% 53.6% 57.3% 36.3% 54.9% 33.3% 65.6% 54.0%
10 Aechmea perforata no chão Gravatá 1.4% 11 Aechmea sp.1 no chão Gravatá 19.0% 30.0% 60.0% 3.6% 16.7% 24.0% 12 Aechmea sp.2 epífita Gravatá 1.2% 13 Cactaceae Pereskia aculeata cipó Arapinobi 5.2% 35.0% 16.0% 14 Cereus sp.nov. arbóreo Mandacaru 3.4% 2.5% 7.7% 19.8% 9.5% 1.2% 15 Erythroxilaceae Erythoxyllum mikanii arbóreo 12.7% 5.2% 16 Euphorbiaceae Crateva tapia arbóreo Bapari 1.4% 2.8% 17 Margaritaria mobilis arbóreo 1.7% 18 Fabaceae Cassia ferruginea cipó Vagem 1.2% 6.0% 14.0% 19 Meliaceae Guarea cf. guidonea arbóreo Pitomba 4.0% 20 Trichilia silvatica arbóreo Arariba 49.2% 21 Menispermaceae Chondrondendron sp. cipó Buti 2.0% 22 Moraceae Ficus sp. arbóreo Gameleira 1.4% 23 Myrtaceae Eugenia candolleana arbóreo Araçá 2.2% 24 Myrcia acuminatissima arbóreo Araçá 4.2% 25 Myrcia bicolor arbóreo Batinga 6.7% 1.2% 26 Myrcia sp. arbóreo Fruto de cotia 27.4% 27 Nyctaginaceae Guapira laxiflora . arbóreo 1.4% 28 Guapira sp. arbustivo Café beirão 6.7% 1.4%
31
Tabela 1: Frutos consumidos por L. chrysomelas no fragmento de floresta semidecidual em cada mês de observação. Os valores representam a porcentagem em termos de número de visitas. Continuação
Família Espécie Hábito Nome comum ago/06 set/06 out/06 nov/06 dez/06 jan/07 fev/07 mar/07 abr/07 mai/07 jun/07 jul/07 ago/07 set/07 29 Olacaceae Schoepfia obliquifolia arbóreo 1.2% 1.1% 2.0% 30 Rhamnaceae Celtis pubescens cipó Juá-mirim 1.4% 2.0% 31 Rhamnidium elacecarpum arbóreo 4.2% 32 Rubiaceae Alibertia sp. arbóreo João duro 1.1% 33 Randia sp.1 cipó Cipó cruzeta 55.0% 12.1% 34 Randia sp2. arbustivo Esporão de galo 1.1% 35 Sapotaceae Pouteria sp. arbóreo 1.4% 36 Theophrastaceae Clanija coloneura arbustivo Laranjinha 2.4% 4.0% 37 Vitaceae Cisus sp. nov. cipó Cipó jabuti 11.9% 35.7% 51.0% 18.7% 45.2% 8.3% 38 não identificada1 arbóreo 2.8% 4.2%
39 não identificada2 arbóreo 2.4%
Tabela 2. Proporção das visitas em fruteiras e de indivíduos marcados das sete espécies de frutos mais consumidos.
Espécie % visitas % indivíduos
Aechmea digitata 45.80% 39.00% Cisus sp. 17.20% 22.00% Aechmea sp1. 7.70% 3.50% Cererus sp. 4.40% 2.77% Trichilia silvatica 4.10% 3.32% Pereskia aculeata 2.40% 2.21% Myrcia sp. 2.30% 2.77% Outras (32 espécies) 16.10% 24.43%
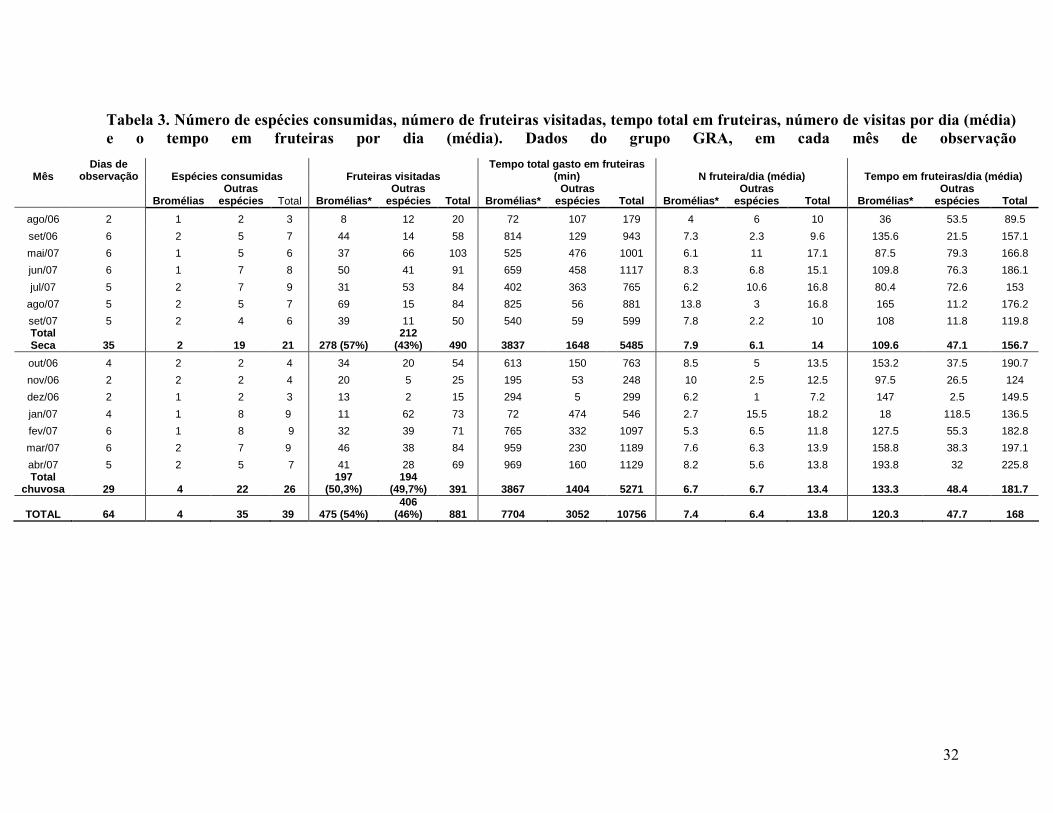
32
Tabela 3. Número de espécies consumidas, número de fruteiras visitadas, tempo total em fruteiras, número de visitas por dia (média) e o tempo em fruteiras por dia (média). Dados do grupo GRA, em cada mês de observação
Mês Dias de
observação Espécies consumidas Fruteiras visitadas Tempo total gasto em fruteiras
(min) N fruteira/dia (média) Tempo em fruteiras/dia (média)
Bromélias Outras
espécies Total Bromélias* Outras
espécies Total Bromélias* Outras
espécies Total Bromélias* Outras
espécies Total Bromélias* Outras
espécies Total ago/06 2 1 2 3 8 12 20 72 107 179 4 6 10 36 53.5 89.5 set/06 6 2 5 7 44 14 58 814 129 943 7.3 2.3 9.6 135.6 21.5 157.1 mai/07 6 1 5 6 37 66 103 525 476 1001 6.1 11 17.1 87.5 79.3 166.8 jun/07 6 1 7 8 50 41 91 659 458 1117 8.3 6.8 15.1 109.8 76.3 186.1 jul/07 5 2 7 9 31 53 84 402 363 765 6.2 10.6 16.8 80.4 72.6 153
ago/07 5 2 5 7 69 15 84 825 56 881 13.8 3 16.8 165 11.2 176.2 set/07 5 2 4 6 39 11 50 540 59 599 7.8 2.2 10 108 11.8 119.8 Total Seca 35 2 19 21 278 (57%)
212 (43%) 490 3837 1648 5485 7.9 6.1 14 109.6 47.1 156.7
out/06 4 2 2 4 34 20 54 613 150 763 8.5 5 13.5 153.2 37.5 190.7 nov/06 2 2 2 4 20 5 25 195 53 248 10 2.5 12.5 97.5 26.5 124 dez/06 2 1 2 3 13 2 15 294 5 299 6.2 1 7.2 147 2.5 149.5 jan/07 4 1 8 9 11 62 73 72 474 546 2.7 15.5 18.2 18 118.5 136.5 fev/07 6 1 8 9 32 39 71 765 332 1097 5.3 6.5 11.8 127.5 55.3 182.8 mar/07 6 2 7 9 46 38 84 959 230 1189 7.6 6.3 13.9 158.8 38.3 197.1 abr/07 5 2 5 7 41 28 69 969 160 1129 8.2 5.6 13.8 193.8 32 225.8 Total
chuvosa 29 4 22 26 197
(50,3%) 194
(49,7%) 391 3867 1404 5271 6.7 6.7 13.4 133.3 48.4 181.7
TOTAL 64 4 35 39 475 (54%) 406
(46%) 881 7704 3052 10756 7.4 6.4 13.8 120.3 47.7 168

Capítulo 2
33
Na estação seca, a proporção de bromélias na dieta, em termos de visitas, foi
significativamente maior (X2 = 4,042; gl = 1; p = 0,044).
Apesar de a proporção de bromélias, em relação a todas as fruteiras visitadas, ter
sido superior na estação seca, o número médio de bromélias visitadas para alimentação
de frutos por dia não diferiu entre as estações seca e chuvosa (t = 1,208; gl = 62; p =
0,232). O tempo que os micos passaram em bromélias também não diferiu
significativamente entre as estações (t = -1,344; gl = 62; p = 0,184). Vale ressaltar,
porém, que, devido à impossibilidade de observação do grupo quando estavam dentro
das bromélias, esse valor pode estar subestimado, e não representa o tempo real de
alimentação de frutos, nem o tempo total que o grupo passou dentro de bromélias, já
que o tempo não era registrado quando não havia alimentação de frutos de bromélias.
Foi registrado o tempo em bromélias apenas nas ocasiões em que foi possível observar
consumo de frutos.
Também não houve diferença significativa entre as duas estações em relação ao
número médio de visitas diárias em outras fruteiras (não bromélias) (t = -0,542; gl = 62;
p = 0,590), assim como o tempo total gasto nessas fruteiras (t = -0,135; gl = 62; p =
0,893).
Das espécies arbóreas cujos frutos foram consumidos, nove foram amostradas na
fenologia. Dessas, apenas três (Trichilia silvatica, Cordia anabaptista e Spondias
velunosa) apresentaram produção de frutos, verdes ou maduros, ao longo do
monitoramento. T. silvatica teve frutificação rápida, com um pico em janeiro. C.
anabaptista teve pico em fevereiro, porém continuou apresentando frutos, em menor
quantidade, até o mês de junho. S. velunosa apresentou padrão diferente. Seus
amadureceram de forma lenta, a partir de dezembro, atingindo um pico no mês de abril
(Fig. 12).
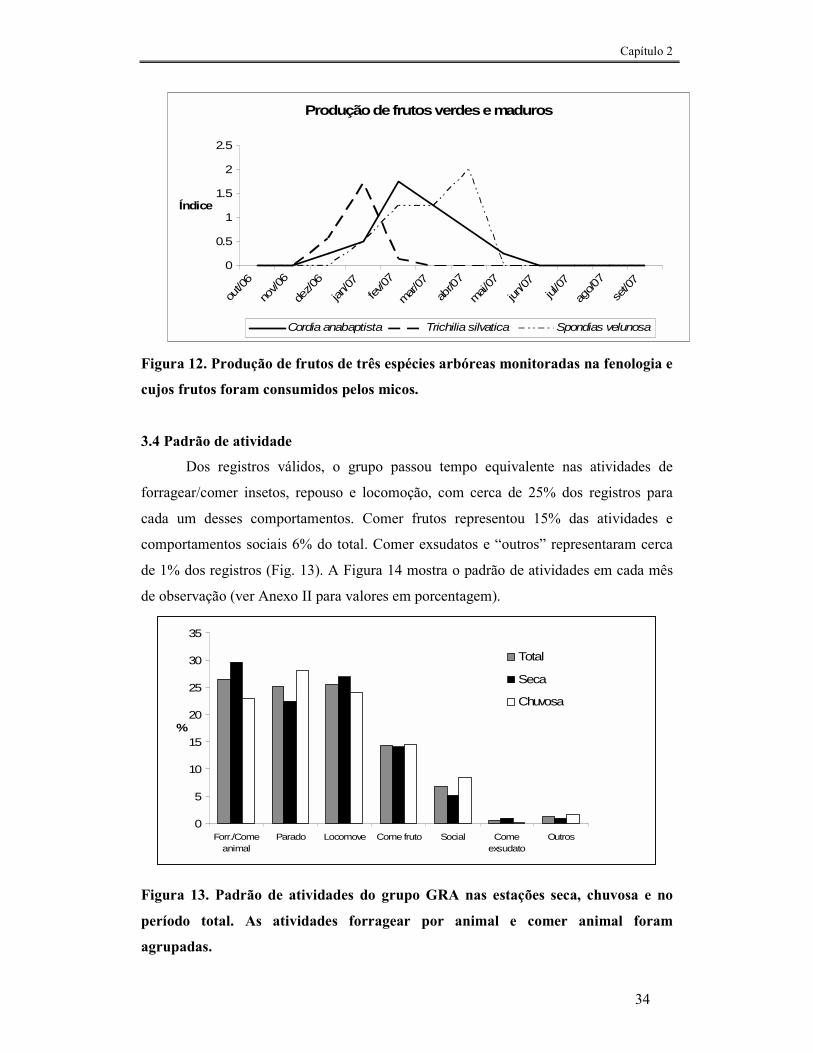
Capítulo 2
34
Produção de frutos verdes e maduros
0
0.5
1
1.5
2
2.5
out/0
6no
v/06
dez/0
6jan
/07fev
/07
mar/07
abr/0
7
mai/07
jun/07
jul/07
ago/0
7se
t/07
Índice
Cordia anabaptista Trichilia silvatica Spondias velunosa
Figura 12. Produção de frutos de três espécies arbóreas monitoradas na fenologia e
cujos frutos foram consumidos pelos micos.
3.4 Padrão de atividade
Dos registros válidos, o grupo passou tempo equivalente nas atividades de
forragear/comer insetos, repouso e locomoção, com cerca de 25% dos registros para
cada um desses comportamentos. Comer frutos representou 15% das atividades e
comportamentos sociais 6% do total. Comer exsudatos e “outros” representaram cerca
de 1% dos registros (Fig. 13). A Figura 14 mostra o padrão de atividades em cada mês
de observação (ver Anexo II para valores em porcentagem).
0
5
10
15
20
25
30
35
Forr./Comeanimal
Parado Locomove Come fruto Social Comeexsudato
Outros
%
Total
Seca
Chuvosa
Figura 13. Padrão de atividades do grupo GRA nas estações seca, chuvosa e no
período total. As atividades forragear por animal e comer animal foram
agrupadas.

Capítulo 2
35
0%10%20%30%40%50%60%70%80%90%
100%
ago/
06
set/0
6
out/0
6
nov/
06
dez/
06
jan/
07
fev/
07
mar
/07
abr/0
7
mai
/07
jun/
07
jul/0
7
ago/
07
set/0
7
ParadoSocialLocomoveCome exsudatoCome frutoForr/Come animalOutros
Figura 14. Padrão de atividades do grupo principal GRA em cada mês de estudo
no fragmento da Fazenda Barro Branco.
Considerando apenas os registros de alimentação, o grupo dedicou 64% do
tempo de alimentação para forrageio/consumo de presas, 35% para consumo de frutos e
apenas 1% para consumos de exsudatos.
A maior parte do forrageio por animais foi manipulativo, com 90.2% dos
registros, enquanto que o forrageio visual ocupou apenas 9.8% dos registros. Em
relação aos locais preferidos para forrageio, bromélias ou folhas secas de bromélias
foram os mais utilizados (86%), enquanto que troncos (incluindo casca e buracos em
troncos) foram utilizados em 10% dos registros. Outros locais, como folhas no chão,
folhas de palmeiras e outras epífitas, corresponderam a apenas 4% dos registros.
Houve diferenças entre as proporções dos registros de cada atividade nas
estações seca e chuvosa (X2 = 121,5; gl = 5; p < 0,001). Os comportamentos de
comer/forragear insetos, parado e social apresentaram diferenças significativas entre as
estações seca e chuvosa (U = 254,0; Z = -3,42; p = 0,001; U = 237,5; Z = -3,64; p <
0,001 e U = 301,0; Z = -2,78; p = 0,005, respectivamente).
Apenas o comportamento “parado” apresentou correlação significativa com a
pluviosidade (r = 0,657; p = 0,011; N = 14). Houve correlação significativa entre a
abundância de frutos maduros e o tempo que o grupo passou locomovendo-se (r = -
0,628; p = 0,029; N = 12) e em atividade social (r = 0,688; p = 0,013; N = 12).
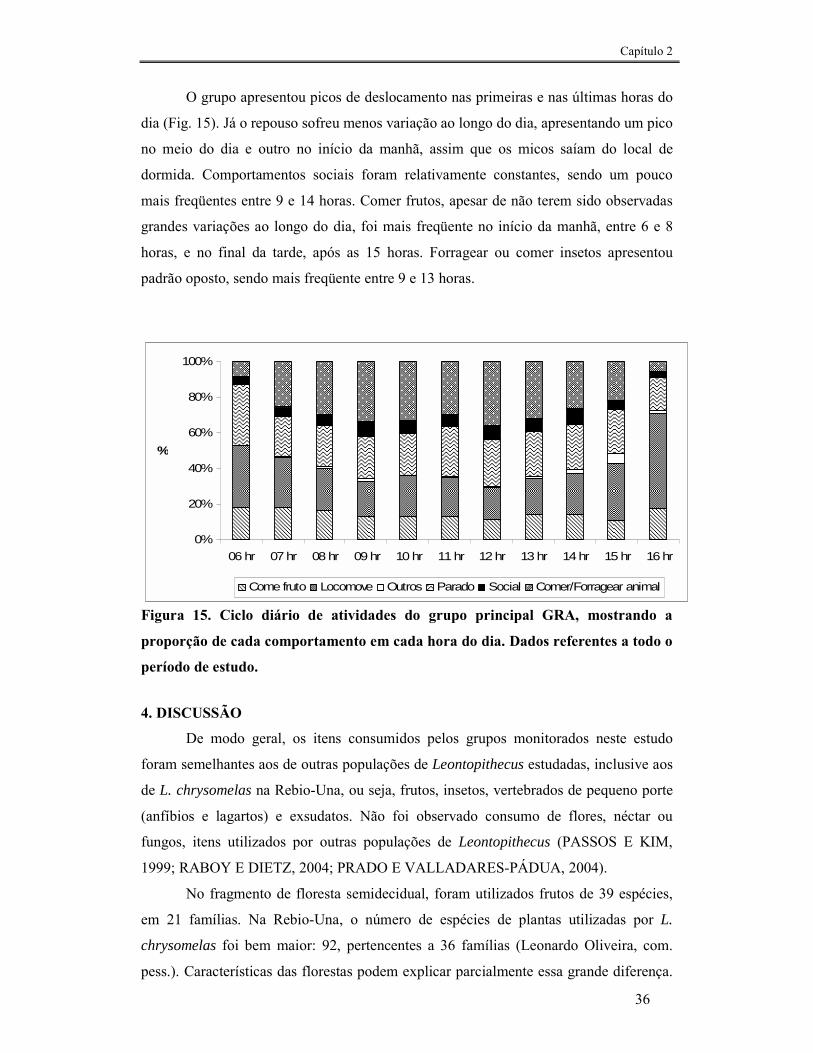
Capítulo 2
36
O grupo apresentou picos de deslocamento nas primeiras e nas últimas horas do
dia (Fig. 15). Já o repouso sofreu menos variação ao longo do dia, apresentando um pico
no meio do dia e outro no início da manhã, assim que os micos saíam do local de
dormida. Comportamentos sociais foram relativamente constantes, sendo um pouco
mais freqüentes entre 9 e 14 horas. Comer frutos, apesar de não terem sido observadas
grandes variações ao longo do dia, foi mais freqüente no início da manhã, entre 6 e 8
horas, e no final da tarde, após as 15 horas. Forragear ou comer insetos apresentou
padrão oposto, sendo mais freqüente entre 9 e 13 horas.
0%
20%
40%
60%
80%
100%
06 hr 07 hr 08 hr 09 hr 10 hr 11 hr 12 hr 13 hr 14 hr 15 hr 16 hr
%
Come fruto Locomove Outros Parado Social Comer/Forragear animal
Figura 15. Ciclo diário de atividades do grupo principal GRA, mostrando a
proporção de cada comportamento em cada hora do dia. Dados referentes a todo o
período de estudo.
4. DISCUSSÃO
De modo geral, os itens consumidos pelos grupos monitorados neste estudo
foram semelhantes aos de outras populações de Leontopithecus estudadas, inclusive aos
de L. chrysomelas na Rebio-Una, ou seja, frutos, insetos, vertebrados de pequeno porte
(anfíbios e lagartos) e exsudatos. Não foi observado consumo de flores, néctar ou
fungos, itens utilizados por outras populações de Leontopithecus (PASSOS E KIM,
1999; RABOY E DIETZ, 2004; PRADO E VALLADARES-PÁDUA, 2004).
No fragmento de floresta semidecidual, foram utilizados frutos de 39 espécies,
em 21 famílias. Na Rebio-Una, o número de espécies de plantas utilizadas por L.
chrysomelas foi bem maior: 92, pertencentes a 36 famílias (Leonardo Oliveira, com.
pess.). Características das florestas podem explicar parcialmente essa grande diferença.

Capítulo 2
37
As florestas ombrófilas do sul da Bahia apresentam elevados índices de diversidade de
plantas (FONSECA et al., 2003), maiores do que os da floresta semidecidual (VINHA
et al., 1976; GOUVEA et al.,1976; MARIANO-NETO, 2004), o que pode refletir em
maior número de espécies utilizadas por micos na Rebio-Una. Na Rebio-Una também
existe maior diversidade de hábitats, como florestas maduras, florestas secundárias,
brejos e cabrucas, todos utilizados, em maior ou menor grau, pelos micos (RABOY et
al., 2004). Já o fragmento de floresta semidecidual estudado é mais homogêneo. Assim,
a diversidade de hábitats e de espécies na Rebio-Una pode aumentar a gama de frutos
consumidos.
A fragmentação da floresta semidecidual também pode afetar negativamente a
diversidade de plantas e, como conseqüência, diminuir as espécies que podem ser
utilizadas pelos micos-leões na alimentação. A fragmentação pode levar a extinções
locais e diminuir a diversidade de plantas (TABARELLI et al., 2004). O efeito negativo
da fragmentação no número de plantas utilizadas já foi demonstrado em primatas.
Bicca-Marques (2003) observou correlação positiva entre o tamanho do fragmento e o
número de espécies de plantas utilizadas pelo gênero Alouatta, que é essencialmente
folívoro.
Por fim, o tempo dos estudos e o número de grupos monitorados podem explicar
a diferença observada no número de espécies de plantas consumidas. Raboy et al.
(2004), analisando dados de várias populações de Leontopithecus, observou correlação
positiva entre a duração do estudo e o número de espécies consumidas. Os dados sobre a
dieta de L. chrysomelas na Rebio-Una foram baseados na observação de três grupos
que, somados, tiveram mais de 2.200 horas de observação em um período de mais de
dois anos (RABOY E DIETZ, 2004), enquanto que o presente trabalho teve dois grupos
monitorados em cerca de 850 horas.
Myrtaceae e Bromeliaceae foram as famílias de plantas com maior número de
espécies consumidas no fragmento de floresta semidecidual. Myrtaceae também é a
família mais consumida por L. chrysopygus (VALLADARES-PADUA, 1993) e L.
caissara (PRADO E VALLADARES-PADUA, 2004). Na Rebio-Una, Myrtaceae e
Sapotaceae foram as famílias mais utilizadas (RABOY et al., 2004). Melastomataceae
foi a mais utilizada por L. chrysomelas em um fragmento próximo à Rebio-Una
(RYLANDS, 1989). Essa família é mais comum em áreas degradas e também foi
bastante consumida por micos na Rebio-Una que habitavam florestas secundárias
(RABOY E DIETZ, 2004). No fragmento semidecidual, apesar da floresta ser

Capítulo 2
38
degradada devido ao corte seletivo e ao efeito borda, a família Melastomataceae não é
comum, e não foi encontrado nenhum indivíduo arbóreo dessa família nas árvores
monitoradas nos transectos ou mesmo fora destes.
Apenas duas espécies (Myrcia acuminatissima e Schoepfia obliquifolia)
consumidas no fragmento semidecidual também estão presentes na dieta dos micos-
leões-da-cara-dourada na Rebio-Una. Outros seis gêneros apresentaram espécies em
comum (Aechmea, Duguetia, Eugenia, Guapira, Philodendron e Pouteria). Essa
diferença mostra grande flexibilidade ecológica da espécie, e parece ser reflexo das
diferenças florísticas existentes entre as florestas ombrófila e semidecidual. Amorim et
al. (2005), estudando um fragmento de transição entre floresta ombrófila e floresta
semidecidual no sul da Bahia, encontrou considerável diferença na composição
florísticas em comparação com a floresta ombrófila de Una. É de se esperar que na
Fazenda Barro Branco, localizada mais a oeste do que a região de transição, a diferença
em relação à floresta de Una seja ainda maior.
Apesar de não terem sido incluídas no acompanhamento fenológico, os frutos de
A. digitata estavam disponíveis o ano todo, e os micos alimentaram-se de seus frutos em
todos os meses durante a pesquisa.. Mais da metade das visitas em fruteiras foi em
bromélias (cerca de 54%). Em apenas um dos meses de observação (janeiro/2007),
frutos de bromélias não estavam entre os mais consumidos.
A. digitata foi o fruto mais consumido. Aparentemente, a maturação desses
frutos se dá de forma lenta, já que, em diversas ocasiões, uma mesma infrutescência foi
visitada diversas vezes ao longo dos meses. Em muitas ocasiões, os grupos estavam nas
bromélias, porém não era possível observar a atividade, nem se a bromélia estava com
frutos maduros. Em outras ocasiões, foram observadas sementes de bromélias nas fezes,
sem que anteriormente tenha ocorrido registro de alimentação de frutos de bromélia.
Assim, a proporção observada de visitas em bromélias em relação ao total de visitas em
fruteiras foi subestimada, e a relevância de frutos de bromélias na alimentação dos
micos no fragmento semidecidual foi ainda maior do que a observada, principalmente
durante a estação seca.
Desse modo, bromélias foram recursos essenciais para os micos no fragmento de
floresta semidecidual estudado, servido não apenas como local preferido de forrageio
por presas animais, mas também como fonte de frutos, principalmente durante a estação
seca. A disponibilidade de um recurso alimentar essencial pode ajudar na sobrevivência
de primatas em fragmentos florestais (MARSH, 2003). Esse parece ser o caso das

Capítulo 2
39
bromélias para os micos nos fragmentos semideciduais, apesar de não ser possível
avaliar quanto os micos dependem das bromélias para sua sobrevivência.
Embora nem todos os trabalhos com Leontopithecus indiquem o número de
cipós utilizados na alimentação, a proporção de frutos de cipós consumidos por micos-
leões-da-cara-dourada no fragmento de floresta semidecidual parece ser maior do que
na Rebio-Una. De todas as espécies consumidas de frutos, cerca de 15% foram de cipós.
Em agosto de 2006 um cipó (Randia sp1.) foi a espécie mais consumida e em outubro
do mesmo ano o cipó Pereskia aculeata foi a mais consumida. Cisus sp., outra espécie
de cipó, foi a segunda espécie mais utilizada no total e esteve entre as mais consumidas
entre abril e julho de 2007 (Tabela 1). O fragmento possui grande quantidade de cipós
(observação pessoal), o que também foi observado por Mariano-Neto (2004) em outros
fragmentos de florestas semideciduais no sul da Bahia. A abundância de cipós é
característica de florestas alteradas e com forte influência do efeito borda (LAURANCE
et al., 1997; MARIANO-NETO, 2004). O elevado consumo de frutos de cipós pode
representar uma adaptação dos micos-leões-da-cara-dourada às condições locais, isto é,
à grande quantidade de lianas decorrente da degradação do fragmento, evidenciando a
grande flexibilidade ecológica da espécie.
Trichilia silvatica apresentou frutificação apenas em janeiro. Naquele mês, essa
espécie foi a mais consumida, com cerca de 50% das visitas em fruteiras. Cordia
anabaptista apresentou pico de frutificação em fevereiro, e continuou apresentando
frutos, em menor quantidade, até junho. Fevereiro e março foram os meses em que essa
espécie foi mais consumida, com cerca de 18% das visitas em fruteiras, mas seus frutos
continuaram sendo consumidos até junho. Nesses meses em que essas espécies foram
mais consumidas, houve também uma diminuição proporcional no consumo de
bromélias, que presumivelmente estavam disponíveis o ano todo, mostrando uma
preferência dos micos por Trichilia silvatica e Cordia anabaptista.
Já o cajá (Spondias velunosa) apresentou um padrão diferente de frutificação e
utilização pelos micos. Essa espécie apresentou frutificação lenta ao longo dos meses,
entre fevereiro e maio. Os micos consumiram cajá em agosto e setembro de 2006,
quando o monitoramento fenológico ainda não havia se iniciado, porém não
consumiram no período em que foi registrada maior frutificação nas trilhas de
fenologia. Durante esse período, houve muito consumo de Cordia anabaptista e Cisus
sp., além de bromélias. Assim, houve uma preferência por esses frutos, em detrimento
do cajá. Análises nutricionais poderiam explicar essas preferências, ou mesmo as

Capítulo 2
40
propriedades organolépticas dos frutos. Micos-leões preferem frutos pequenos, macios e
doces (DIETZ et al., 1997; LAPENTA et al., 2003), e os cajás no fragmento
semidecidual tinham pouca polpa e eram bastante azedos (observação pessoal), além de
serem relativamente grandes.
Exsudatos e néctar representam a segunda parte de plantas mais consumidas na
alimentação de outras populações de Leontopithecus (KIERULFF et al., 2002) e todas
as espécies do gênero as consomem, principalmente como recursos alternativos durante
a época de menor disponibilidade de frutos (RYLANDS, 1989; DIETZ et al., 1997;
PASSOS E KIM, 1999; RABOYE DIETZ, 2004). Uma diferença em relação aos grupos
da Rebio-Una e de outras populações de Leontopithecus foi que os micos no fragmento
semidecidual não se alimentaram de flores, néctar ou fungos. Flores foram recursos
alimentares bastante utilizados por grupos de L. chrysomelas na Rebio-Una (RABOY E
DIETZ, 2004) e na Estação Experimental Lemos Maia, chegando a 20% dos registros
de alimentação de plantas (RYLANDS, 1989). Néctar de cinco espécies (Symphonia
globulifera e quatro espécies de Manilkara) e flores de duas (Hortia arborea e uma não
identificada) foram consumidos por L. chrysomelas na Rebio-Una (RABOY E DIETZ,
2004). No entanto, a espécie mais utilizada para consumo de néctar, S. globulifera,
assim como as demais, não foram observadas no fragmento semidecidual.
Na Rebio-Una, L. chrysomelas consome exsudatos de Parkia pendula (juerana)
a partir de buracos abertos por Callithrix kuhlii (RYLANDS, 1989; RABOY E DIETZ,
2004). No fragmento de floresta semidecidual não existe P. pendula, porém os micos
alimentam-se de exsudatos que escorrem naturalmente do tronco de uma
Anacardiaceae. No fragmento também ocorre C. kuhlii e eles roem tronco para
alimentação de exsudatos (observação pessoal), porém os micos-leões não foram
observados utilizando os buracos abertos por C. kuhlii.
De modo geral, os locais usados para forrageio por presas animais no fragmento
semidecidual foram os mesmos de outras populações de Leontopithecus. No entanto, há
diferenças quanto às proporções dos locais utilizados, o que parece refletir a
disponibilidade desses micro-hábitat nos ambientes (KIERULFF et al., 2002). L. rosalia
utilizou mais freqüentemente bromélias ou fendas e cascas de tronco, dependendo da
população estudada (KIERULFF, 2000; KIERULFF et al., 2002), enquanto que L.
chrysopygus, que ocupa um ambiente onde não há bromélias, utilizou mais cavidades de
tronco e folhas de palmeiras (KEUROGHLIAN E PASSOS, 2001; PASSOS, 1999),

Capítulo 2
41
mesmos micro-hábitats usados por L. caissara, que também utilizou bromélias
(PRADO E VALLADARES-PADUA, 2004).
Assim como nos estudos de Rylands (1989) e Raboy e Dietz (2004), bromélias
também foram os locais preferidos de forrageio de L. chrysomelas por presas animais no
fragmento semidecidual, confirmando a relevância deste recurso para diferentes
populações da espécie. Para o grupo estudado no presente trabalho, a utilização de
bromélias ou folhas secas de bromélias como locais de forrageio foi ainda maior: 86%
dos registros de forrageio, contra cerca de 50% no estudo de Rylands (1989) e, em
média, 76,5% no estudo de Raboy e Dietz (2004).
Comparando os resultados do padrão de atividades com as informações
existentes para Rebio-Una, o grupo no fragmento semidecidual passou mais tempo
forrageando por animal e menos tempo locomovendo-se (Tabela 4). No grupo estudado
por Rylands (1989) o padrão observado foi bastante diferente, com 43% dos registros
locomovendo, 20% parado, 24% comendo planta e apenas 13% forrageando por
animais. Este estudo, porém, teve apenas três meses de duração, o que limita a
comparação. Raboy e Dietz (2004) observaram cerca de 20% dos registros de forrageio
por animais (Tabela 4). O fato de o presente estudo ter mostrado um tempo maior de
forrageio pode indicar que a quantidade de frutos na floresta semidecidual é menor do
que na floresta ombrófila. A hipótese de que as necessidades nutricionais das duas
populações são diferentes também não pode ser descartada, já que estão sujeitas à
diferentes condições ambientais.
Tabela 4. Comparação do padrão de atividades em três estudos com L. chrysomelas. Este estudo Raboy (2002) Rylands (1989)
Locomovendo 26% 33% 43%
Parado/Social 32% 33% 20%
Comendo/Forrageando planta 15% 12% 24%
Comendo/Forrageando animal 26% 20% 13%
Outros 1% 2% 0%
Número de grupos estudados 1 3 1
Duração do estudo (meses) 14 30 3
Foi observada maior taxa de alimentação de frutos nas primeiras e nas últimas
horas do dia. Esse padrão está associado à necessidade de alimento rico em energia e de

Capítulo 2
42
fácil digestão antes e imediatamente após um período prolongado de jejum durante a
noite (OATES, 1987). Esse padrão é semelhante ao encontrado em outras espécies de
primatas onívoros e outros calitriquídeos, inclusive outras populações de Leontopithecus
(RYLANDS, 1982; TERBORGH, 1983; FERRARI, 1988; RABOY E DIETZ, 2004).
Repouso e comportamentos sociais apresentaram curvas semelhantes, com a
diferença que repouso apresentou um pico na primeira hora do dia. Essa semelhança é
esperada, já que essas atividades estão normalmente associadas (FERRARI, 1988). O
padrão observado para o comportamento de locomoção, com picos no início e final do
dia, também corresponde ao descrito para outros calitriquídeos (FERRRARI, 1988;
RABOY E DIETZ, 2004). Raboy e Dietz (2004) sugerem que esse padrão pode estar
relacionado à localização e ao número limitado de locais de dormida. Assim, os micos
precisariam de um grande deslocamento pela manhã para chegarem à fonte de alimento
e um grande deslocamento no fim do dia, para chegarem ao local de dormida.
Os resultados de fenologia mostraram que houve grande variação na
disponibilidade de frutos das árvores ao longo do ano, com um pico de frutificação e
floração (meses de outubro e novembro) e uma época em que a oferta de frutos nas
árvores era muito baixa (a estação seca, de maio a agosto). No entanto, o tempo que os
indivíduos passaram alimentando-se de frutos não variou ao longo dos meses e não
apresentou correlação significativa com a quantidade de frutos das árvores monitoradas
pela fenologia, nem com a pluviosidade. Poucas espécies que foram amostradas na
fenologia, porém, foram utilizadas pelos micos para alimentação e grande parte dos
frutos consumidos foi de bromélias e cipós, que não tiveram acompanhamento
fenológico. Além disso, muitas árvores cujos frutos foram utilizados não entraram na
amostragem fenológica. Portanto, não é possível dizer, a partir dos resultados da
fenologia, que durante o ano, houve queda na disponibilidade dos frutos utilizados pelos
micos.
Por outro lado, o consumo de exsudatos só foi observado nos meses entre abril e
setembro, quando a disponibilidade de frutos de árvores na mata estava baixa. Em
outras populações estudadas, os micos complementam a dieta com néctar, fungos ou
exsudatos em época de escassez de alimento (KIERULFF et al., 2002). A proporção de
exsudatos em relação à dieta total (cerca de 1%) foi bastante baixa e semelhante à
observada por Raboy e Dietz (2004) na Rebio-Una (0,75%) e por Dietz et al. (1997)
para L. rosalia (1,5%). No entanto, foi muito inferior à observada em outras espécies do
gênero, principalmente em relação à L. chrysopygus na Estação Ecológica Caetetus,

Capítulo 2
43
cuja dieta na época seca foi constituída por até 55% de exsudatos (PASSOS, 1999).
Características do ambiente podem explicar essa grande diferença. L. chrysopygus
habita floresta semidecidual, com alguns meses de seca, período em que há forte
diminuição da disponibilidade de frutos, além de não haver bromélias no local
(KIERULFF et al., 2002). Já L. chrysomelas na Rebio-Una habita floresta ombrófila
densa, com chuvas mais regulares no período de um ano, indicando que provavelmente
não há diminuição acentuada da disponibilidade de frutos ou que os frutos têm melhor
qualidade nutricional do que nos outros lugares (RABOY E DIETZ, 2004).
Assim, é possível que os micos tenham utilizado exsudatos como fonte
complementar de alimentos durante a estação seca devido a uma queda na
disponibilidade de frutos, ainda que essa queda tenha sido pouco pronunciada. A
presença de frutos de bromélias durante o ano todo pode ter contribuído para que essa
queda não tenha sido significativa.
Durante a época seca, foi consumido um menor número de espécies e houve
aumento proporcional no consumo de frutos de bromélias, o que pode ser conseqüência
da queda na disponibilidade de outros frutos, diferentes de bromélias, durante a época
seca.
Houve um aumento do tempo dedicado ao forrageio por presas animais na
estação seca, o que também pode estar relacionado à diminuição na disponibilidade de
frutos. Kierulff (2000) também observou aumento no tempo de forrageio por animais
durante a estação seca em L. rosalia na Reserva Biológica União, e micos-leões-pretos,
que habitam florestas semideciduais no interior de São Paulo, também aumentam o
tempo de forrageio por presas durante a estação seca, quando há queda na
disponibilidade de frutos (KEUROGHLIAN E PASSOS, 2001).
O tempo que o grupo passou parado e em atividade social foi maior na estação
chuvosa. Assim, houve uma transferência do tempo dedicado ao repouso e atividade
social na estação chuvosa para a procura/consumo de animais na estação seca.
Procópio-de-Oliveira (2002) também observou maior taxa de descanso durante a
estação chuvosa para L. rosalia, relacionando esse aumento às altas temperaturas e
umidade relativa durante a época chuvosa, o que desencoraja atividades com alto custo
energético. O aumento de repouso e atividades sociais observado no fragmento
semidecidual também pode estar relacionado à época de nascimentos, que ocorreram no
final da estação seca ou meio da chuvosa. Assim, durante estação chuvosa, havia no

Capítulo 2
44
grupo filhotes e jovens. Grupos com mais filhotes normalmente apresentam maiores
taxas de comportamentos sociais (LI E ROGERS, 2004), principalmente brincadeiras.
O fato de o tempo de alimentação de frutos não ter sofrido variação entre as
estações, somado ao aumento do tempo dedicado ao forrageio na estação seca, pode
indicar (1) que a qualidade dos alimentos durante a estação seca sofreu uma queda,
obrigando os micos a passarem mais tempo na procura de insetos ou (2) que a
disponibilidade de insetos diminuiu na estação seca e, portanto, os micos tiveram que
passar mais tempo em forrageio para capturar uma mesma quantidade de presas. Isto é,
diminuíram a taxa de captura de presas animais. Essas duas possibilidades não são
mutuamente exclusivas. Durante a estação seca, foi utilizado um número bem menor de
espécies de frutos para a alimentação, o que pode refletir negativamente na qualidade
nutricional da dieta. Além disso, estudos já mostraram que a abundância de insetos é
maior em épocas de chuva (FERRARI, 1988). Apenas estudos sobre a qualidade
nutricional dos frutos e o monitoramento da disponibilidade de insetos ao longo do ano,
avaliando a taxa de captura de presas animais, podem responder claramente esta
questão.
5. CONCLUSÕES
• Os itens da dieta do mico-leão-da-cara-dourada em fragmento de floresta
semidecidual foram semelhantes ao observado em outras populações do gênero,
inclusive à da população de L. chrysomelas da Rebio-Una. Foram consumidos
frutos (na maioria maduros), insetos (principalmente gafanhotos), vertebrados de
pequeno porte (rãs e lagartos) e exsudatos.
• Frutos de cipós foram mais utilizados do que na Rebio-Una, o que pode
representar uma adaptação às condições locais.
• O número de espécies utilizadas na alimentação foi bem menor do que na Rebio-
Una, o que pode ser explicado tanto pelas características das florestas – menos
diversificada na semidecidual – como pelo tamanho reduzido do fragmento e
pelo menor tempo de duração do estudo.
• Bromeliaceae e Myrtaceae foram as famílias mais importantes em termos de
número de espécies. Em relação ao número de indivíduos visitados para
consumo de frutos, Bromeliaceae foi a mais utilizada.

Capítulo 2
45
• Bromélias foram recursos muito utilizados, tanto como fonte de frutos como
local de forrageio por presas animais, confirmando sua relevância para
diferentes populações de L. chrysomelas.
• Houve queda acentuada na disponibilidade de frutos na floresta durante a
estação seca. Essa queda, porém, não refletiu na taxa de consumo de frutos pelos
micos, já que bromélias estavam presentes o ano todo e houve aumento do
consumo de seus frutos durante a estação seca.
• O consumo de exsudatos, no entanto, apesar de pouco freqüente, ocorreu
somente nos meses de seca, o que pode indicar uma queda, ainda que pouco
acentuada, na disponibilidade de frutos durante esta estação.

46
CAPÍTULO III - Área de vida e uso do espaço do
mico-leão-da-cara-dourada (Leontopithecus
chrysomelas) em um fragmento de floresta
semidecidual

Capítulo 3
47
1. INTRODUÇÃO
A área de vida dos primatas, de forma simplificada, representa o espaço onde os
indivíduos do grupo exercem suas atividades ao longo do tempo (OATES, 1987) e seu
tamanho está relacionado tanto a características do ambiente, como a fatores sociais. A
localização e o padrão de distribuição temporal e espacial dos recursos utilizados, que
podem ser alimentos, água, locais de descanso ou de proteção contra predadores afetam
o tamanho da área de vida (HAMILTON, 1982; TERBORGH, 1983; BARTON et al.,
1992; COWLISHAW, 1997). O tamanho e composição dos grupos também influenciam
o tamanho da área de vida, assim como a densidade de grupos (BICCA-MARQUES,
2003). De modo geral, uma densidade populacional maior pode levar a uma diminuição
da área de vida dos grupos (KIERULFF et al., 2002). A densidade, por sua vez, está
relacionada à qualidade do hábitat (KIERULFF et al., 2002). Assim, o uso do espaço
por primatas está fortemente relacionado à abundância de recursos, principalmente
alimentares, características do grupo como tamanho e composição, e densidade e
interações entre grupos vizinhos (CROCKETT E EISENBERG, 1987).
O mico-leão-da-cara-dourada, Leontopithecus chrysomelas, é um calitriquídeo
endêmico da Mata Atlântica do sul da Bahia e ameaçado de extinção, sendo classificado
na categoria “Em Perigo” (MMA, 1995, IUCN, 2007). A espécie é frugívora-insetívora
e os grupos defendem o território de grupos vizinhos nos chamados “encontros inter-
grupais” (RABOY E DIETZ, 2004). Esses encontros ocorrem normalmente na borda
dos territórios e envolve vocalizações e, em certas ocasiões, perseguições e agressões.
Todas as informações existentes sobre área de vida, uso de hábitat e densidade
da espécie provêm de estudos de monitoramento realizados na região de floresta
ombrófila. Rylands (1989) monitorou um grupo por três meses em fragmento no
entorno da Reserva Biológica de Una e Raboy e Dietz (2004) e Raboy et al. (2004)
realizaram o estudo mais detalhado com a espécie, monitorando grupos em floresta
madura na Rebio-Una. Já informações ecológicas e populacionais sobre a espécie em
florestas semideciduais são bastante escassas. O único estudo realizado foi de Pinto
(1994), que realizou rápido levantamento populacional com uso de play-back em alguns
fragmentos de floresta semidecidual.
Enquanto que alguns calitriquídeos são especialistas em utilizar florestas
secundárias e áreas nas bordas das matas (Callithrix, Mico e Saguinus), principalmente
como locais de procura por presas animais, Leontopithecus adaptou-se a utilização de

Capítulo 3
48
florestas maduras, especializando-se em forrageio por presas animais em locais
específicos e na utilização de ocos como locais de dormida - recursos mais comuns em
florestas maduras (RYLANDS, 1996). Ainda assim, utilizam bordas e florestas
secundárias em diferentes graus de degradação (KIERULFF et al., 2002).
Estudar a espécie em um ambiente fragmentado e degradado nos permite
entender como a espécie de adapta a essas condições, e revela sua flexibilidade
ecológica e comportamental, além de orientar ações específicas para a conservação da
espécie em florestas semideciduais. Este trabalho tem como objetivo apresentar as
primeiras informações sobre área de vida, uso do espaço e aspectos demográficos de
grupos de L. chrysomelas habitando um fragmento de floresta semidecidual,
comparando os resultados com informações existentes para a espécie na Rebio-Una.
Serão avaliados se a localização de recursos (fruteiras e locais de dormida) e outras
variáveis (tipo de hábitat e distância da borda do território) afetam o padrão de
movimentação de um grupo.
2. MÉTODOS
2.1 Área de Estudo e Grupos
O trabalho foi realizado em um fragmento de floresta semidecidual de 450 ha na
fazenda Barro Branco, Itororó, Bahia. Dois grupos foram monitorados (MAN e GRA).
A descrição mais detalhada do local de estudo e mais informações sobre os grupos
encontram-se no Capítulo I.
2.2 Período de Atividade
O período de atividade foi calculado apenas para o grupo GRA, nos dias de
observação completa. O período de atividade foi o tempo da saída do primeiro mico do
local de dormida pela manhã até a entrada do último mico no local de dormida no
período da tarde.
2.3 Área de Vida, Uso do Espaço e Deslocamento Diário
A posição do grupo era registrada com GPS (E-trex, Garmin) em intervalos
regulares de 20 minutos. A área de vida foi calculada usando-se dois métodos: mínimo
polígono convexo (MPC), no qual os pontos mais externos são ligados formando o
polígono (JACOB E RUDRAN, 2004); e Kernel-fixo, que descreve a probabilidade do

Capítulo 3
49
grupo ser encontrado em determinada área (WORTON, 1989). Para o cálculo do MPC
foram utilizados todos os pontos dos dois grupos. O Kernel-fixo foi calculado apenas
para o grupo GRA, e foram utilizados apenas os dias completos de observação e
registros a cada duas horas, para diminuir a dependência entre registros consecutivos.
Para a análise do Kernel, o valor do fator h, ou parâmetro de “suavização”
(bandwidth ou smoothing parameter) tem grande influência no resultado (KERNOHAN
et al., 2001), pois irá determinar o grau de detalhamento da estimativa de densidade
(JACOB E RUDRAN, 2004). O fator h pode ser visto como o raio de percepção da
espécie (POWEL, 2003) e quanto maior seu valor, mais suaves são os contornos
gerados e menor é o detalhamento (WORTON, 1989). O cálculo do fator h foi realizado
através do processo LSCV (Least Squares Cross Validation). Essa é a forma sugerida
por diversos autores como a mais adequada para o cálculo do fator h, já que diminui a
subjetividade na escolha do valor e faz o cálculo a partir dos próprios dados
(KERNOHAN et al., 2001; JACOB E RUDRAN, 2004). O método foi o mesmo
utilizado por Raboy e Dietz. (2004) para calcular a área de vida dos grupos de L.
chrysomelas na Rebio-Una. Foram realizados os contornos de 25%, 50%, 75% e 95%,
que representam a probabilidade do grupo ser encontrado nessas áreas. A área de vida
foi definida como o contorno de 95%, conforme Raboy et al. (2004).
Para o grupo GRA, foi avaliado o uso das bordas do fragmento, considerando
bordas com 100, 50 e 10 metros de largura. Foi realizado o teste do Qui-quadrado para
avaliar se o número observado de registros em cada largura de borda e no interior do
fragmento correspondeu à quantidade de registros esperados de acordo com a área
disponível, isto é, se o uso da área era aleatório.
A cada localização do grupo, era registrado também o tipo de mata: (1) riachos
ou brejos (que ficam alagados na época de chuvas); e (2) floresta seca. Os dois
ambientes foram comparados em temos de disponibilidade de frutos, altura das árvores
e diâmetro das árvores na altura do peito (DAP). Para avaliar a disponibilidade de
frutos, foi realizada fenologia do estrato arbóreo a partir do método dos quadrantes,
monitorando árvores de DAP ≥ 7,5 cm (ver capítulo II para mais detalhes sobre a
metodologia). A partir dos pontos registrados e do conhecimento da área, foi
confeccionado um mapa com a disponibilidade desses ambientes na área de vida do
grupo GRA. Do mesmo modo realizado com as bordas, foi avaliado se o uso desses
ambientes correspondeu à quantidade disponível desses ambientes.

Capítulo 3
50
Todas as visitas em fruteiras do grupo GRA foram anotadas. Uma visita era
registrada quando pelo menos um indivíduo do grupo consumia fruto por pelo menos
um minuto. Nestas ocasiões, a fruteira utilizada foi marcada, numerada e teve a posição
registrada com GPS. A localização de todas as fruteiras foi sobreposta em mapa
produzido pelo método do Kernel.
O deslocamento diário foi calculado apenas para o grupo GRA, já que o grupo
MAN teve poucos dias completos de observação, e foram utilizados apenas os pontos
obtidos em dias completos de observação. A distância diária percorrida foi calculada a
partir da soma das distâncias entre os pontos consecutivos registrados a cada 20
minutos. As posições de todos os locais de dormida foram registradas com GPS e as
localizações sobrepostas em mapa de área de uso produzido pelo método do Kernel.
Os cálculos da área de vida e deslocamento diário foram realizados utilizando-se
a extensão “Home Range Tools” do software ArcGis 9.2
Para verificar se houve diferença na média de deslocamento diário entre as
estações seca e chuvosa foi realizado o teste T de Student. Para verificar se o
deslocamento diário estava relacionado com período de atividade, foi realizado o teste
de Correlação de Spearman. O mesmo teste foi realizado entre o deslocamento diário e
as distâncias entre o local de dormida e a primeira e última fruteira visitada pelos micos,
com o objetivo de se verificar a influência dessas fruteiras na escolha do local de
dormida.
Para verificar se a presença de grupos vizinhos e a defesa do território afetam o
uso do espaço e o deslocamento diário, foram comparadas as médias de deslocamento
diário entre dias em que ocorreram encontros intergrupais nas bordas das áreas de vida e
dias em que não ocorreram, através do teste não paramétrico de Mann-Whitney.
Foi realizado o teste de Correlação de Spearman entre o deslocamento diário e o
número de fruteiras visitadas por dia. Os testes estatísticos foram feitos com o programa
SPSS 10.1 e todos foram bilaterais, considerando o nível de significância < 0,05 (ZAR,
1996).
2.4 Densidade
O cálculo da densidade e do número de micos no fragmento foi realizado a
partir da densidade conhecida na área ocupada pelos dois grupos monitorados,
extrapolando o resultado para todo o fragmento . Também são apresentados dados sobe
o tamanho de outros grupos presentes no fragmento e que eram frequentemente

Capítulo 3
51
avistados, cujas áreas eram vizinhas aos grupos monitorados. Sempre que eram
avistados, era anotado o número de indivíduos no grupo e se havia ou não filhotes.
3. RESULTADOS
3.1 Grupos monitorados
O tamanho do grupo MAN variou de dois a seis indivíduos durante o período de
estudo, e o grupo GRA, de quatro a nove. Informações mais detalhadas sobre a
composição dos grupos e tempo de observação foram apresentados no Capítulo II.
3.2 Período de atividade
O período médio de atividade do grupo GRA foi de 9,89 (± 1,5) horas (N = 64
dias). O tempo mínimo de atividade foi de 8,53 horas, no mês de junho, e o máximo foi
de 12,01 horas, em janeiro (Tabela 5). Houve diferença significativa entre os períodos
de atividade das estações, sendo maior durante a estação chuvosa, que corresponde aos
meses de verão (T = -7,874; df = 62; p < 0,001).
Tabela 5. Período de atividade do grupo GRA durante o período de estudo, de agosto/2006 a setembro/2007.
Horas/Tempo ATIVIDADE Estação seca Estação chuvosa
Início da atividade (mín. e máx.) 5:24 (Set) – 6:51 (Ago) 5:01 (Nov) - 6:36 (Abr)
Retirada para o local de dormida (mín. e máx.) 14:45 (Jun) - 16:44 (Set) 15:55 (Abr)– 17:38 (Jan)
Tempo Máximo de Atividade (horas) 11,52 (Set) 12,01 (Jan)
Tempo Mínimo de Atividade (horas) 8,53 (Jun/2007) 9,32 (Abr)
Média do Tempo de Atividade (horas) 9,5 10,9
3.3 Área de Vida e Uso do Espaço
O grupo GRA utilizou uma área de vida de 81 ha, calculada pelo método do
Mínimo Polígono Convexo. Devido ao formato irregular do fragmento, o polígono
formado abrange áreas de pastagem, que obviamente não são usadas pelos micos.
Desconsiderando essa área de pasto, a área de mata utilizada pelo grupo foi de 71 ha. A
área de vida do grupo MAN apresentou um MPC de 71 ha e 68 ha considerando apenas
a área de floresta (Fig. 16).
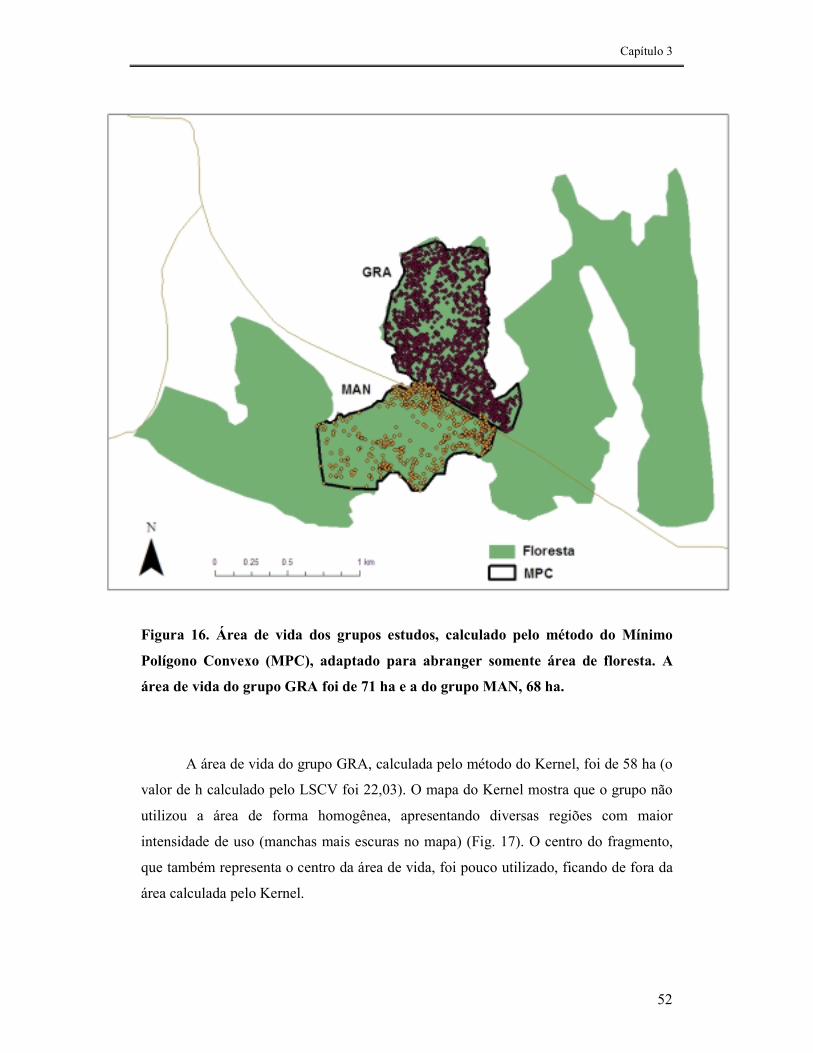
Capítulo 3
52
Figura 16. Área de vida dos grupos estudos, calculado pelo método do Mínimo
Polígono Convexo (MPC), adaptado para abranger somente área de floresta. A
área de vida do grupo GRA foi de 71 ha e a do grupo MAN, 68 ha.
A área de vida do grupo GRA, calculada pelo método do Kernel, foi de 58 ha (o
valor de h calculado pelo LSCV foi 22,03). O mapa do Kernel mostra que o grupo não
utilizou a área de forma homogênea, apresentando diversas regiões com maior
intensidade de uso (manchas mais escuras no mapa) (Fig. 17). O centro do fragmento,
que também representa o centro da área de vida, foi pouco utilizado, ficando de fora da
área calculada pelo Kernel.
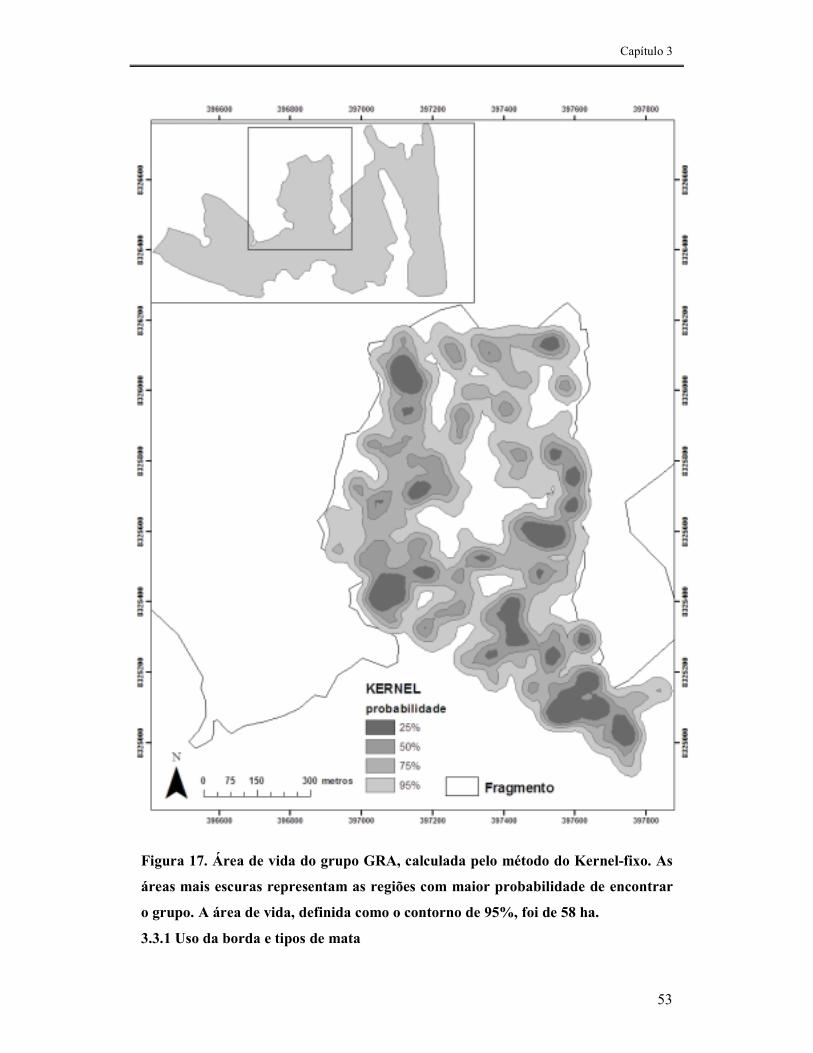
Capítulo 3
53
Figura 17. Área de vida do grupo GRA, calculada pelo método do Kernel-fixo. As
áreas mais escuras representam as regiões com maior probabilidade de encontrar
o grupo. A área de vida, definida como o contorno de 95%, foi de 58 ha.
3.3.1 Uso da borda e tipos de mata
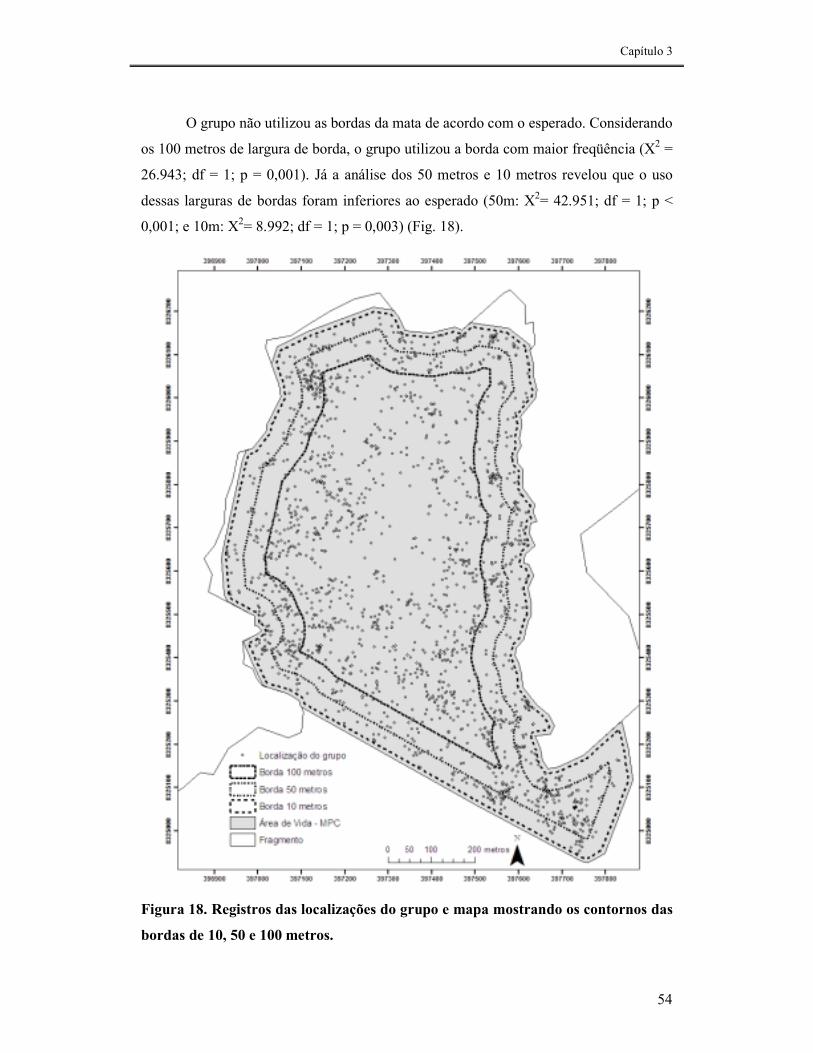
Capítulo 3
54
O grupo não utilizou as bordas da mata de acordo com o esperado. Considerando
os 100 metros de largura de borda, o grupo utilizou a borda com maior freqüência (X2 =
26.943; df = 1; p = 0,001). Já a análise dos 50 metros e 10 metros revelou que o uso
dessas larguras de bordas foram inferiores ao esperado (50m: X2= 42.951; df = 1; p <
0,001; e 10m: X2= 8.992; df = 1; p = 0,003) (Fig. 18).
Figura 18. Registros das localizações do grupo e mapa mostrando os contornos das
bordas de 10, 50 e 100 metros.

Capítulo 3
55
Em relação aos diferentes tipos de matas - floresta seca e brejo - , o uso também
não foi de acordo com o disponível. Na área de vida do grupo GRA, 20% eram cobertos
por brejos, e o uso deste ambiente foi menor do que o esperado (X2 = 55,70; df = 1; p <
0,001) (Fig. 19).
Figura 19. Registros das localizações do grupo e mapa mostrando os tipos
florestais: floresta seca e riachos/brejos.

Capítulo 3
56
A caracterização desses dois ambientes não revelou diferenças em relação à
altura das árvores (t = -0.79; df = 438; p = 0.94). Já o DAP apresentou diferença
significativa (t = -3.02; df = 438; p = 0.003), sendo maior nos riachos (Tabela 6). A
produção de frutos foi maior no ambiente riacho/brejo e menor nos cem metros de borda
(Figs. 20 e 21).
Tabela 6. Comparação da altura e diâmetro na altura do peito (DAP) das árvores monitoradas para fenologia nos ambientes de floresta seca e riacho/brejo.
Altura (m) DAP (cm) média desvio média desvio
Floresta seca (N = 292) 15,57 7,09 16,6 8,4
Riacho/brejo (N = 148) 15,62 7,16 19,6 12,0
0
0.2
0.4
0.6
0.8
1
1.2
Fruto maduro Fruto verde Flores
ÍndiceInteriorRiachoBorda (100m)
Figura 20. Produtividade de frutos maduros, frutos verdes e flores nos ambientes
de interior da floresta, riachos e nos cem primeiros metros de borda.
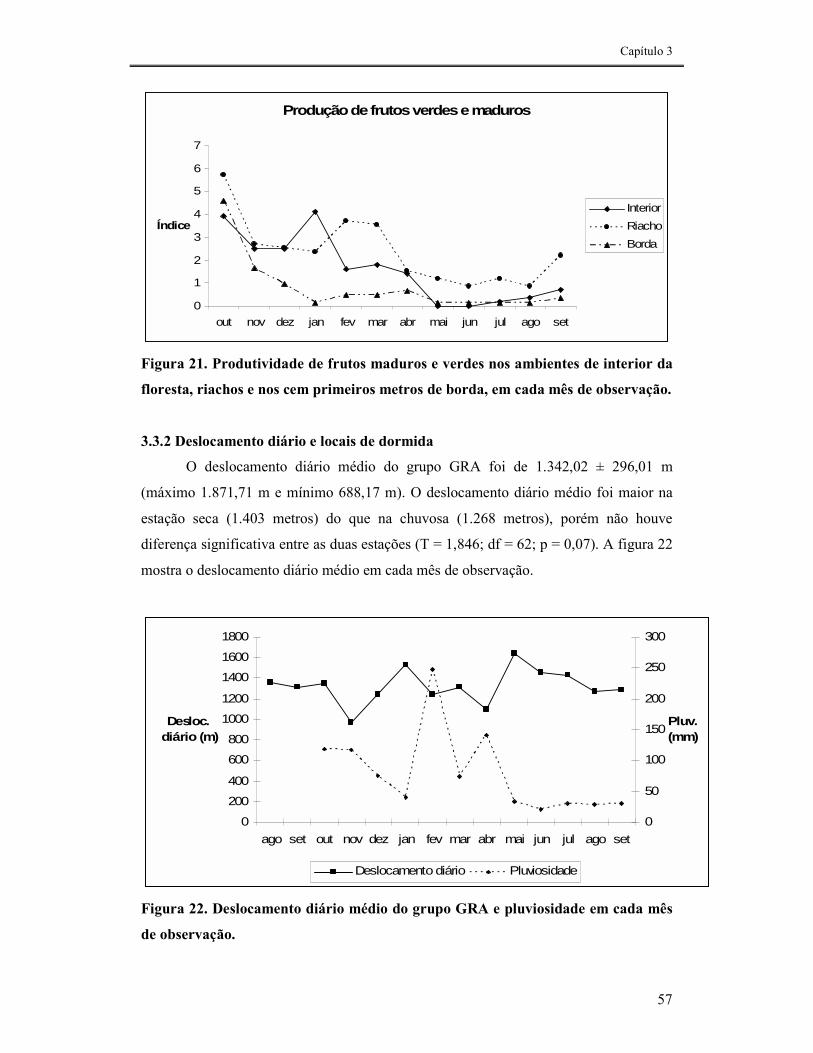
Capítulo 3
57
Produção de frutos verdes e maduros
0
1
2
3
4
5
6
7
out nov dez jan fev mar abr mai jun jul ago set
ÍndiceInteriorRiachoBorda
Figura 21. Produtividade de frutos maduros e verdes nos ambientes de interior da
floresta, riachos e nos cem primeiros metros de borda, em cada mês de observação.
3.3.2 Deslocamento diário e locais de dormida
O deslocamento diário médio do grupo GRA foi de 1.342,02 ± 296,01 m
(máximo 1.871,71 m e mínimo 688,17 m). O deslocamento diário médio foi maior na
estação seca (1.403 metros) do que na chuvosa (1.268 metros), porém não houve
diferença significativa entre as duas estações (T = 1,846; df = 62; p = 0,07). A figura 22
mostra o deslocamento diário médio em cada mês de observação.
0
200
400
600
800
1000
1200
1400
1600
1800
ago set out nov dez jan fev mar abr mai jun jul ago set
Desloc. diário (m)
0
50
100
150
200
250
300
Pluv. (mm)
Deslocamento diário Pluviosidade
Figura 22. Deslocamento diário médio do grupo GRA e pluviosidade em cada mês
de observação.

Capítulo 3
58
O deslocamento diário não apresentou correlação significativa com o período de
atividade (r = - 0,021; p = 0,872; N = 64), nem com a distância da primeira e última
fruteira visitada do dia em relação ao local de dormida (r = 0,219; p = 0,082; N = 64; r =
0,211; p = 0,097; N = 64). Comparando os dias em que houve encontro (N = 16) com os
dias em que não houve (N = 48), a diferença do deslocamento diário foi significativa,
sendo maior nos dias de encontro (U = 186.000; p = 0,02). O deslocamento também
apresentou correlação significativa com o número de fruteiras visitadas por dia (r =
0,355; p = 0,004; N = 64).
Durante o período de estudo, o grupo principal GRA utilizou 35 árvores como
local de dormida (N = 96 dias de observação). Todos foram ocos naturais de árvores e
alguns foram mais utilizados do que outros (Fig. 23). Três ocos foram utilizados em seis
ocasiões e outros três em cinco ocasiões; apenas dois ocos foram utilizados quatro vezes
e oito, três vezes. A maioria dos ocos (12) foi usada duas vezes e sete ocos foram
usados apenas uma vez. Os três ocos mais utilizados foram responsáveis por 18,7% das
vezes em que foi observado o local de dormida.
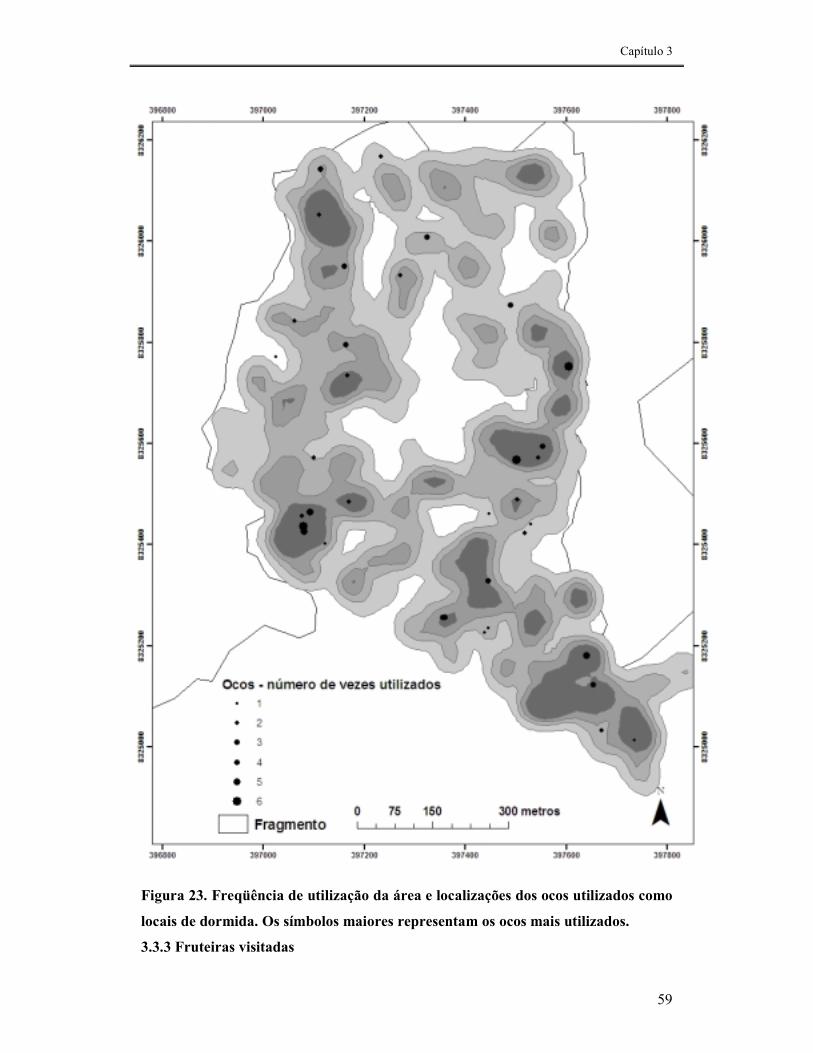
Capítulo 3
59
Figura 23. Freqüência de utilização da área e localizações dos ocos utilizados como
locais de dormida. Os símbolos maiores representam os ocos mais utilizados.
3.3.3 Fruteiras visitadas
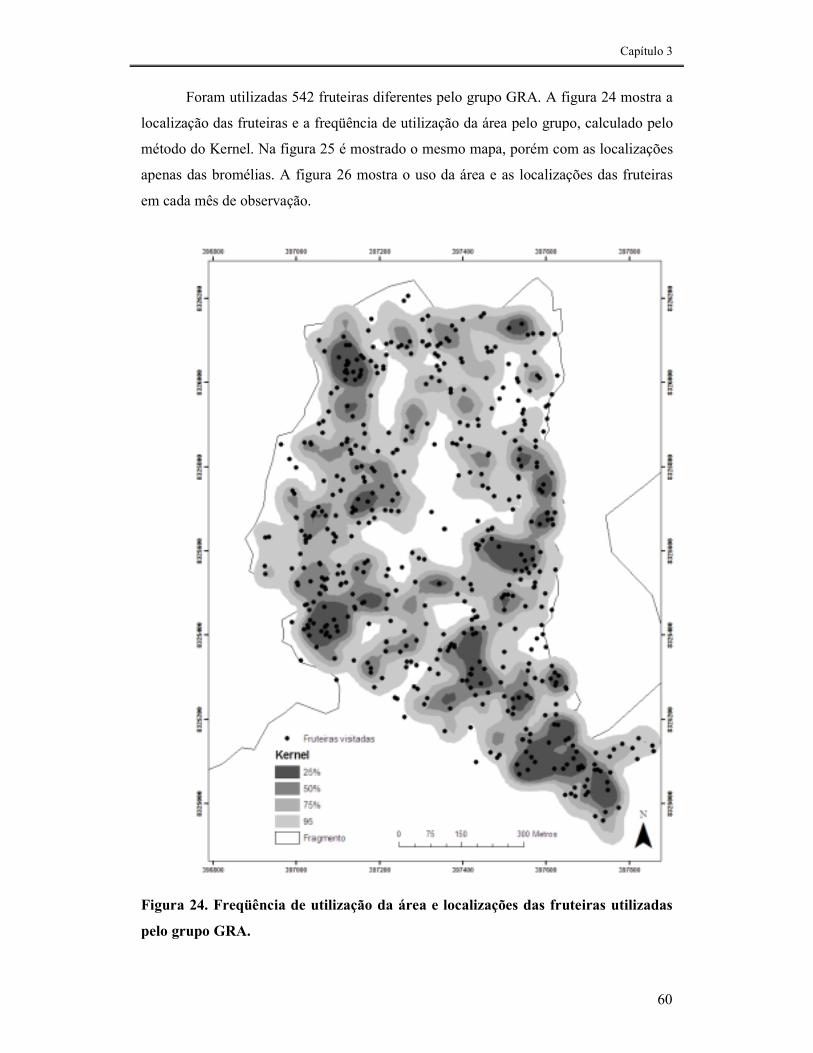
Capítulo 3
60
Foram utilizadas 542 fruteiras diferentes pelo grupo GRA. A figura 24 mostra a
localização das fruteiras e a freqüência de utilização da área pelo grupo, calculado pelo
método do Kernel. Na figura 25 é mostrado o mesmo mapa, porém com as localizações
apenas das bromélias. A figura 26 mostra o uso da área e as localizações das fruteiras
em cada mês de observação.
Figura 24. Freqüência de utilização da área e localizações das fruteiras utilizadas
pelo grupo GRA.
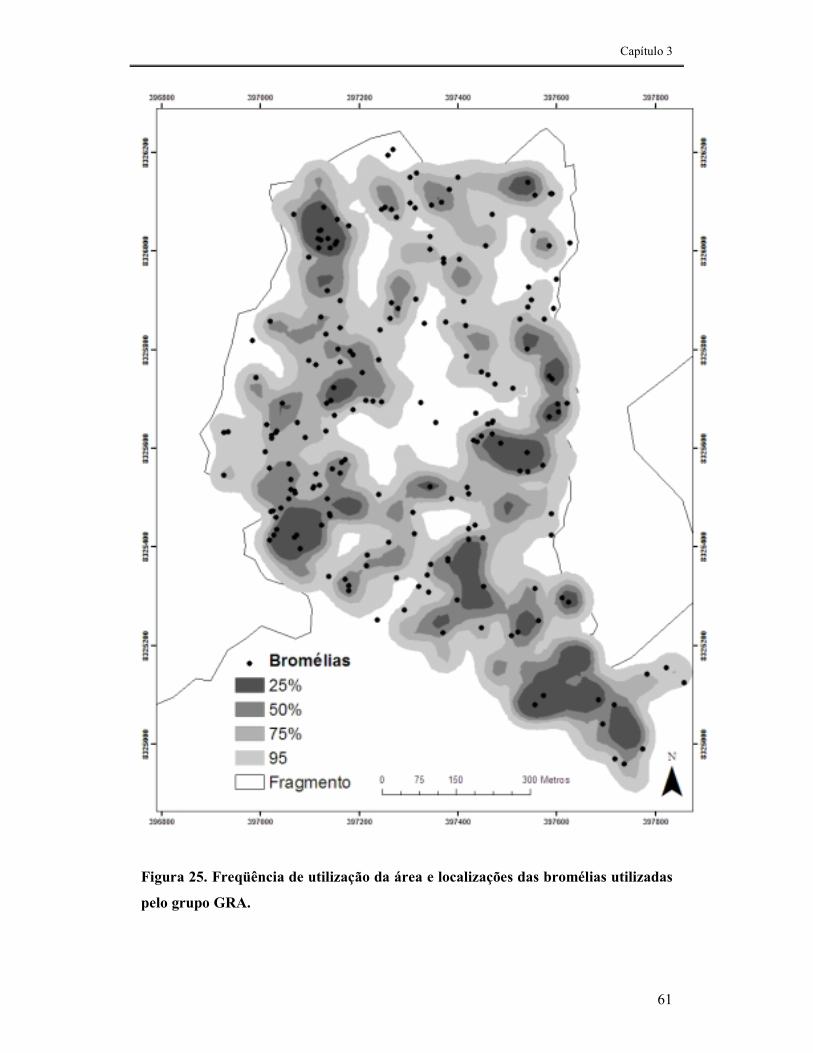
Capítulo 3
61
Figura 25. Freqüência de utilização da área e localizações das bromélias utilizadas
pelo grupo GRA.

Capítulo 3
62
Figura 26. Área de vida do grupo GRA, calculada pelo método do Kernel-fixo,
entre os meses de agosto/2006 e setembro/2007. Os pontos representam as fruteiras
utilizadas em cada mês.
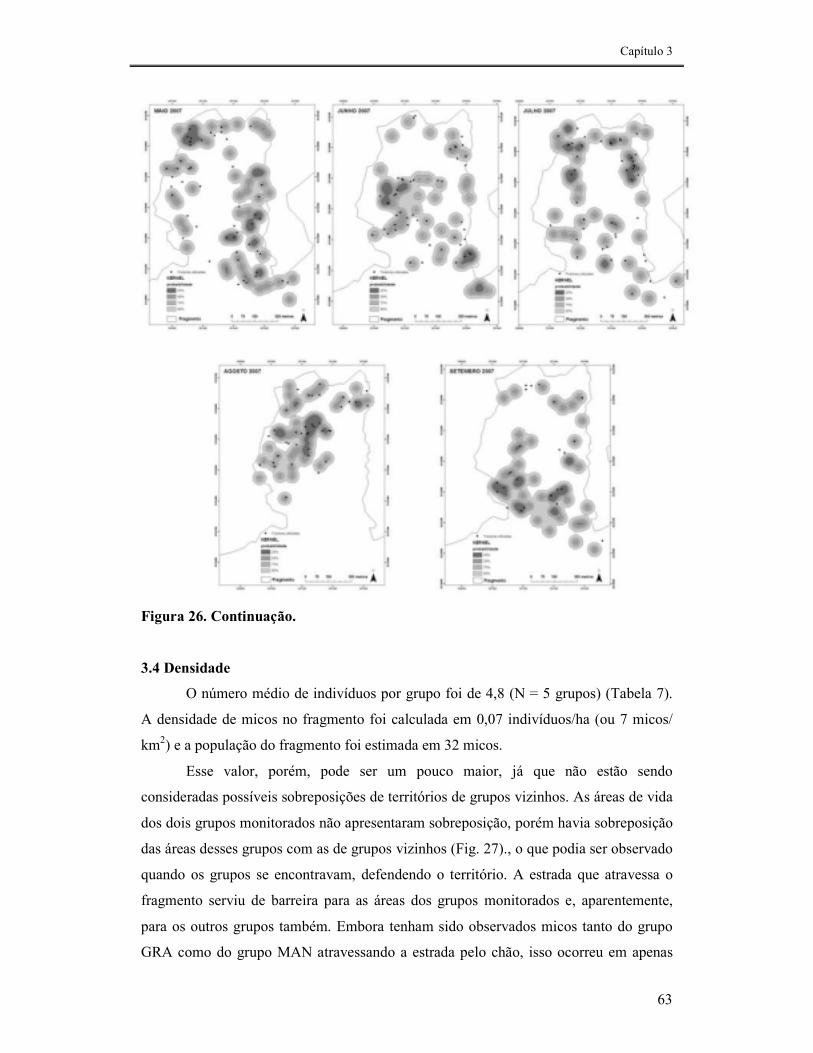
Capítulo 3
63
Figura 26. Continuação.
3.4 Densidade
O número médio de indivíduos por grupo foi de 4,8 (N = 5 grupos) (Tabela 7).
A densidade de micos no fragmento foi calculada em 0,07 indivíduos/ha (ou 7 micos/
km2) e a população do fragmento foi estimada em 32 micos.
Esse valor, porém, pode ser um pouco maior, já que não estão sendo
consideradas possíveis sobreposições de territórios de grupos vizinhos. As áreas de vida
dos dois grupos monitorados não apresentaram sobreposição, porém havia sobreposição
das áreas desses grupos com as de grupos vizinhos (Fig. 27)., o que podia ser observado
quando os grupos se encontravam, defendendo o território. A estrada que atravessa o
fragmento serviu de barreira para as áreas dos grupos monitorados e, aparentemente,
para os outros grupos também. Embora tenham sido observados micos tanto do grupo
GRA como do grupo MAN atravessando a estrada pelo chão, isso ocorreu em apenas
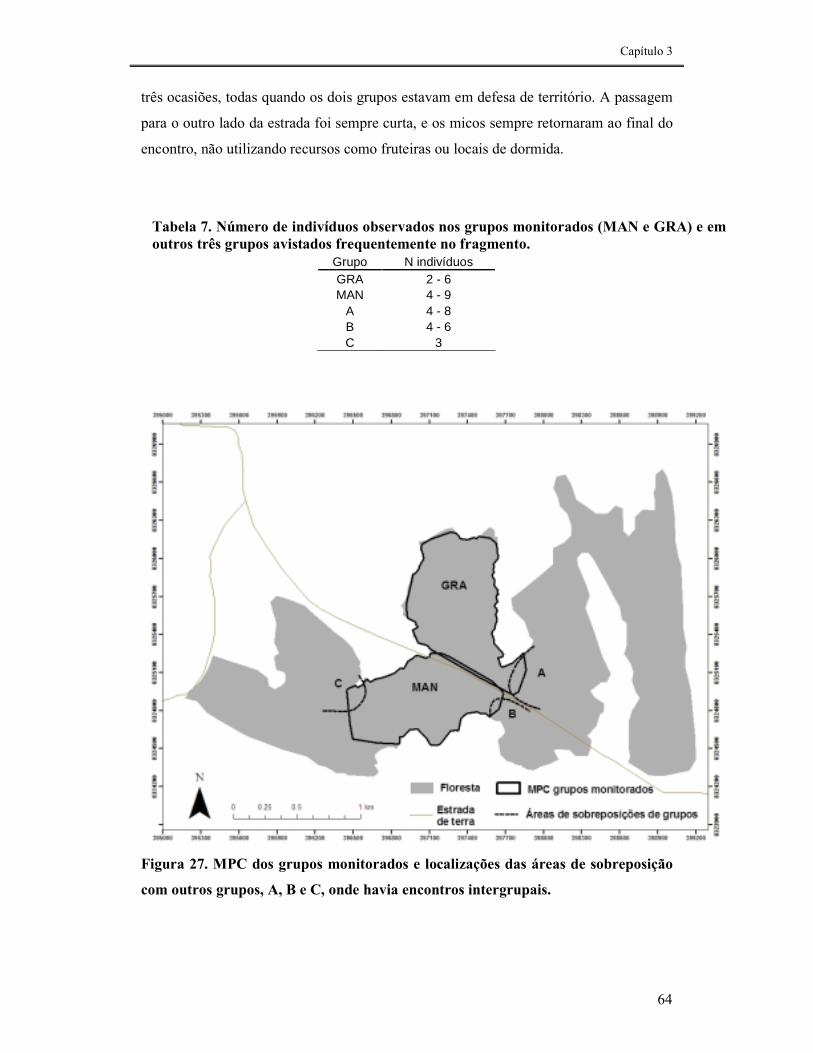
Capítulo 3
64
três ocasiões, todas quando os dois grupos estavam em defesa de território. A passagem
para o outro lado da estrada foi sempre curta, e os micos sempre retornaram ao final do
encontro, não utilizando recursos como fruteiras ou locais de dormida.
Tabela 7. Número de indivíduos observados nos grupos monitorados (MAN e GRA) e em outros três grupos avistados frequentemente no fragmento.
Grupo N indivíduos GRA 2 - 6 MAN 4 - 9
A 4 - 8 B 4 - 6 C 3
Figura 27. MPC dos grupos monitorados e localizações das áreas de sobreposição
com outros grupos, A, B e C, onde havia encontros intergrupais.

Capítulo 3
65
4. DISCUSSÃO
O período de atividade observado foi similar ao já descrito para outras espécies
do gênero, entre 8 e 12 horas diárias, durante a fase diurna (KIERULFF, 2000;
PROCÓPIO-DE-OLIVEIRA, 2002, LAPENTA, 2006). O período foi mais longo na
estação de chuva, que representa os meses de verão, quando o fotoperíodo e a
temperatura são maiores. Esse padrão já foi observado para outras populações de
Leontopithecus (PASSOS, 1997; PROCÓPIO-DE-OLIVEIRA, 2002; LAPENTA,
2006).
Todos os locais de dormida utilizados pelos grupos estudados foram ocos de
árvores, assim como em outras populações de Leontopithecus. Outros estudos já
mostraram que locais alternativos, como, bromélias, palmeiras e emaranhados de
vegetação, também podem ser usados (RYLANDS, 1989; RABOY, 2002; PROCÓPIO-
DE-OLIVEIRA, 2002; KIERULFF et al., 2002). Assim, o uso preferencial de ocos
como locais de dormida parece ser uma característica de todas as populações de L.
chrysomelas.
As espécies do gênero Leontopithecus possuem área de vida relativamente
grande (DIETZ et al., 1997), que podem passar de 300 ha. Junto com Saguinus,
possuem as maiores áreas em relação à massa de um grupo dentre todos os primatas do
Novo Mundo (DIETZ et al., 1997). O grande tamanho da área pode estar relacionado
aos hábitos alimentares da espécie, que se alimenta de frutos e insetos, recursos que
estão distribuídos esparsamente no ambiente.
O tamanho e a composição dos grupos são fatores que podem afetar o tamanho
da área de vida (OATES, 1987). As áreas de vida de ambos os grupo no fragmento
semidecidual, calculada pelo MPC, foram similares, desconsiderando-se a área de pasto.
Nesse caso, os tamanhos e composições dos grupos, que eram diferentes, não afetaram o
tamanho da área de vida.
A área de vida do grupo GRA, calculada pelo MPC e considerando apenas as
áreas de floresta, foi de71 ha, enquanto que o cálculo pelo Kernel (95%) resultou em 58
ha. Essa diferença era esperada, já que, normalmente, o método do MPC resulta em
áreas maiores do que o método do Kernel (PIMLEY et al., 2005). O MPC pode incluir
áreas que nunca foram usadas pelo grupo. No caso do grupo GRA, a diferença deveu-se
principalmente à exclusão, pelo método do Kernel, do centro do fragmento, região que
foi muito pouco utilizada pelo grupo. Essa região central também não é utilizada por
outros grupos do fragmento, já que o grupo GRA ocupa todo seu entorno. Assim, o

Capítulo 3
66
MPC resulta em um valor mais próximo da real quantidade de floresta disponível ao
grupo, e é mais eficiente para o cálculo de densidade populacional (PROCÓPIO-DE-
OLIVEIRA, 2002). O método do MPC, porém, não mostra uma representação da
intensidade de uso dentro da área de vida, característica que é levada em conta no
método do Kernel (WORTON, 1989).
Em primatas, além do tamanho e composição dos grupos, características da
floresta, como idade, estrutura da vegetação, disponibilidade e distribuição de recursos,
também afetam o tamanho da área de vida e a densidade de indivíduos (KIERULFF et
al., 2002). Diferenças entre tamanhos de áreas de vida nas diferentes populações de
Leontopithecus parecem relacionadas às diferenças de hábitats e à quantidade de
recursos disponíveis (KIERULFF et al., 2002). Quanto menor a qualidade do hábitat,
maiores são as áreas de vida. Por exemplo, L. chrysopygus possui área de vida de até
mais de 200 ha (VALLADARES-PADUA, 1993; KEUROGHLIAN E PASSOS, 2001),
relativamente grande quando comparada às áreas de L. chrysomelas e L. rosalia. L.
chrysopygus ocupa florestas semideciduais no interior de São Paulo, e seu grande
tamanho da área de vida está relacionado à forte sazonalidade climática da região, à
baixa densidade de bromélias e ao uso de uma variedade de micro-hábitats para
forrageio por animais, distribuídos esparsamente no ambiente (VALLADARES-
PADUA, 1993; KEUROGHLIAN E PASSOS, 2001).
Para L. chrysomelas, a área de vida no fragmento semidecidual foi bastante
inferior ao observado por Raboy e Dietz (2004) na floresta ombrófila da Rebio-Una, que
teve média de 123 ha, calculada pelo método do Kernel. O tamanho e composição do
grupo GRA foram semelhantes ao observado na Rebio-Una - média de oito indivíduos
(RABOY, 2002). Apesar de não poder ser descartada a hipótese de que o tamanho
menor da área de vida encontrado no fragmento semidecidual seja devido a uma melhor
qualidade do hábitat (por exemplo, maior disponibilidade de frutos ou maior quantidade
de bromélias), os resultados podem ser explicados, também, pela própria conformação
do fragmento.
Já foi observado para outras espécies que o tamanho da área de vida está
relacionado ao tamanho do fragmento florestal. Wong e Sicotte (2007), estudando
Colobus vellerosus em Gana, observaram que as áreas de vida de grupos habitando
fragmentos tendiam a ser menores do que a de grupos em floresta contínua. Em uma
análise de diversas espécies de Alouatta, Bicca-Marques (2003) encontrou correlação
positiva entre o tamanho da área de vida e o tamanho do fragmento.

Capítulo 3
67
Rylands (1989) estudando um grupo de L. chrysomelas na Estação Ecológica
Lemos Maia, próxima à Rebio-Una, encontrou área de vida de cerca de 40 ha. Esse
local é bastante alterado, com predominância de vegetação secundária e representa um
fragmento de floresta isolado de blocos maiores.
Assim, o tamanho relativamente pequeno da área de vida do grupo no fragmento
de floresta semidecidual, em relação às dos grupos da Rebio-Una, pode ser
conseqüência do tamanho e da conformação do fragmento florestal.
No fragmento onde foi realizado o estudo, o seu formato bastante irregular
limitou o tamanho da área que pode ser utilizada pelo grupo. No Caso do grupo GRA,
as fronteiras norte, leste e oeste da área de vida fazem contato com pastos e, portanto, a
área do grupo não poderia crescer nestes sentidos. Na fronteira sul da área, há uma
estrada de terra, criando um vão de cerca de 10 metros e não permitindo que as copas
das árvores dos lados opostos da estrada se toquem. A estrada serviu como barreira para
a área de vida dos grupos estudados, apesar dos micos poderem, potencialmente,
atravessá-la pelo chão. Os grupos, no entanto, não utilizavam frequentemente a área do
lado oposto da estrada, e só atravessaram em ocasiões de encontro. A área de vida do
grupo GRA ainda era sobreposta à área de outro grupo, na parte sudeste de seu
território. No lado oposto da estrada ainda havia dois grupos. Se fosse um bloco de
floresta maior e contínuo, a área utilizada poderia ser maior. Portanto, por se tratar de
um fragmento de formato irregular, atravessado por uma estrada e com outros grupos
presentes, o grupo permaneceu confinado em uma parte da mata de cerca de 70 ha, sem
possibilidade de expansão do território.
É difícil afirmar se o grupo utilizaria uma área maior se estivesse em uma
floresta contínua, ou se sobreviveria se estivesse confinado em uma parte ainda menor
de floresta. Os resultados só permitem dizer que o grupo, que chegou a ter nove micos,
estava sobrevivendo em uma área com cerca de 70 ha de floresta e, portanto, podem
sobreviver em fragmentos deste tamanho.
Vale ressaltar que o tamanho da área de vida e a densidade de indivíduos são
dinâmicos, podendo sofrer variações ao longo do tempo. Por exemplo, a população
estudada por Raboy e Dietz (2004), na região de Maruim da Rebio-Una, apresentou área
de vida média de 123 ha. Em 2005, essa mesma população apresentava média de 58 ha,
menor ainda do que o observado no fragmento semidecidual (HOLST, 2006). A
densidade de micos nesse local praticamente dobrou em quase dez anos, de 0,053
micos/ha para 0,10 micos/ha (HOLST, 2006). Variação na densidade de indivíduos

Capítulo 3
68
também foi observada para L. rosalia na Reserva Biológica Poço das Antas, onde houve
queda pronunciada da densidade devido a um aumento de predação de micos
(FRANKLIN et al., 2007).
A densidade de micos calculada para o fragmento estudado, 0,07 micos/ha, foi
pouco menor do que o valor mais atual constatado para a Rebio-Una, que é de 0,10
indivíduos/ha para a região de Maruim, onde a floresta está melhor preservada, e 0,11
indivíduos/ha na região de Piedade, onde a floresta é mais degradada e há maior
proporção de florestas secundárias (HOLST, 2006). Esse último cálculo, assim como no
presente estudo, não leva em consideração possíveis sobreposições de territórios e,
portanto, esses valores podem ser ainda pouco maiores. A menor densidade pode ser
parcialmente explicada pelo tamanho médio dos grupos, que foi menor no fragmento
semidecidual (4,8 indivíduos). Na Rebio-Una, Dietz et al. (1996) encontrou média de
cinco indivíduos por grupo, sem contar os filhotes, e os grupos estudados por Raboy
(2002) tinham, em média, oito indivíduos. Comparando com outras espécies de
Leontopithecus, a densidade no fragmento semidecidual é semelhante às de L. rosalia
(0,035 a 0,12 micos/ha) e maiores do que as de L. chrysopygus (0,01 a 0,037 micos/ha)
e L. caissara (0,015 micos/ha) (KIERULFF et al., 2002).
Considerando o padrão de utilização da área de vida do grupo GRA em todo o
período de estudo, o mapa produzido aplicando o método do Kernel-fixo revelou que o
centro da área de uso foi menos utilizado do que as bordas e que houve várias regiões
dentro da área de vida que foram mais intensamente utilizadas. O padrão de utilização
da área de vida de L. chrysomelas na Rebio-Una foi diferente do encontrado no
fragmento semidecidual. Raboy e Dietz (2004) observaram que os grupos de micos-
leões utilizaram as bordas do território menos frequentemente do que o interior, além de
terem apresentado apenas uma região de uso mais intenso (área núcleo).
Micos-leões são territorialistas e defendem o território ativamente de grupos
vizinhos (PERES, 1989; KIERULFF et al., 2002). Já foi observado um uso maior das
bordas do território em L. rosalia na Reserva Biológica Poço das Antas (PERES, 1989).
A movimentação dos grupos esteve fortemente associada ao patrulhamento das bordas,
defesa do território e à competição exploratória. Raboy e Dietz (2004) sugerem que a
defesa de território é mais freqüente em locais onde há alta densidade de grupos e áreas
de vida menores. Diferentemente do observado em Poço das Antas, no Rio de Janeiro,
esses autores encontraram que, para L. chrysomelas na Rebio-Una, a procura por
alimentos e o número limitado de locais de dormida são fatores mais importantes do que

Capítulo 3
69
a defesa de recursos para determinar o padrão de movimentação dos grupos. Assim, o
tamanho relativamente grande das áreas de vida e a baixa densidade de grupos nesse
local diminuem os comportamentos de defesa de território (RABOY E DIETZ, 2004) e,
conseqüentemente, o uso da periferia do território também diminui. O mesmo foi
observado para L. rosalia na Reserva Biológica União (KIERULFF et al., 2002). Nesse
local, a densidade de micos é baixa, e os grupos passaram a maior parte do tempo no
centro do território e apresentaram pouco comportamento de defesa de território.
No fragmento de floresta semidecidual, o grupo GRA mantinha contato com
outros três grupos apenas nas partes sul/sudeste do território. Todo o restante do
território era limitado por áreas de pasto, onde obviamente não há grupos para serem
repelidos. Portanto, o padrão observado - maior uso das bordas do que do interior – não
pode ser totalmente explicado pela movimentação para defesa de território e competição
com grupos vizinhos, como foi descrito para L. rosalia em Poço das Antas (PERES,
1989).
O uso preferencial de certas áreas pode estar relacionado, também, a variações
na qualidade do hábitat e na distribuição de recursos, como alimento e locais de
dormida. No mapa produzido pelo método do Kernel, a maioria das manchas mais
escuras (áreas mais utilizadas) são também áreas de concentração de locais de dormida
e, em alguns casos, de fruteiras visitadas. Todas as árvores que foram mais utilizadas
como locais de dormida (5 ou 6 vezes) estão localizadas em regiões que foram mais
frequentemente usadas. A região mais utilizada na parte noroeste da área de vida
apresenta grande concentração de fruteiras. A maior mancha escura do mapa, localizada
na parte sudeste da área de vida, além de ter uma concentração de fruteiras e de locais
de dormida, também era local de encontro com pelo menos outros dois grupos.
Procópio-de-Oliveira (2002) observou que a utilização da área por L. rosalia estava
relacionada à presença de recursos alimentares e locais de forrageio. Resultados
semelhantes foram observados para L. chrysopygus (PASSOS, 1997; VALLADARES-
PADUA, 1993).
Os mapas de utilização da área de vida ao longo do período de monitoramento
revelaram que o uso da área em cada mês não foi concentrado, mas espalhado por todo
o território. Se em alguns meses o mapa revelou um padrão que sugere uma
concentração do uso em determinada parte, isso pode ter ocorrido devido ao baixo
número de observações nesses meses, como ocorreu em novembro e dezembro de 2006.
Ainda assim, o uso do espaço esteve associado à localização das fruteiras utilizadas. De

Capítulo 3
70
modo geral, as fruteiras estão dispersas em toda a área de vida do grupo, inclusive as
bromélias cujos frutos foram utilizados (Figs. 24 e 25). A distribuição dispersa e não
concentrada das fruteiras, principalmente das bromélias, que foram utilizadas o ano
todo, pode explicar o uso não concentrado da área de vida pelos micos ao longo dos
meses de observação.
Assim, os resultados sugerem que o padrão de utilização da área de vida dos
micos-leões-da-cara-dourada no fragmento de floresta semidecidual está relacionado
tanto à localização de grupos vizinhos, como à localização de recursos alimentares e de
locais de dormida, conforme observado em outras espécies de Leontopithecus.
Raboy (2002), estudando três grupos de L. chrysomelas na Rebio-Una,
encontrou uma média de deslocamento diário de 1753 metros, maior, portanto, do que a
observada no fragmento de floresta semidecidual, que foi de 1342 metros. O
deslocamento diário também foi maior para L. rosalia na Reserva Biológica União,
variando de 1502 a 2134 metros (KIERULFF, 2000; PROCOPIO DE OLIVEIRA,
2002; LAPENTA, 2006) Tendo em vista que a área de vida do grupo no fragmento
semidecidual é menor, era esperado um deslocamento diário também menor (OATES,
1987, KIERULFF et al., 2002). Procópio-de-Oliveira (2002), estudando L. rosalia
observou que o deslocamento diário de L. rosalia foi positivamente relacionado com a
área utilizada mensalmente. Lapenta (2006), no entanto, observou resultado contrário
em um grupo da mesma população estudada por Procópio-de-Oliveira (2002). A autora
explica o resultado pela utilização constante de um mesmo oco de dormida. Assim, o
grupo percorria grandes distâncias para chegar a esse oco, mesmo tendo área de vida
relativamente pequena.
O deslocamento diário não teve correlação com o período de atividade e,
portanto, com o comprimento do dia. Resultado semelhante foi encontrado por Lapenta
(2006) para L. rosalia. Outros fatores, no entanto, como aspectos demográficos e a
localização das fontes de alimento, afetam o deslocamento diário.
No presente estudo, o deslocamento foi maior em dias em que houve encontro
intergrupal. Nestas ocasiões, o grupo GRA deslocava-se do local em que estava até o a
borda do território, principalmente na parte sudeste, onde havia o contato com os grupos
vizinhos. Muitas vezes, o grupo atravessava todo seu território, sem parar para se
alimentar ou forragear por presas animais. Esses dias foram os de maiores
deslocamentos do grupo.

Capítulo 3
71
Também foi encontrada correlação positiva entre o deslocamento diário e o
número de fruteiras visitadas. Procópio-de-Oliveira (2002) observou resultado
semelhante: o deslocamento diário de L. rosalia foi positivamente relacionado com o
número de espécies utilizadas. Esses resultados sugerem que a localização das fruteiras
influencia fortemente o deslocamento diário e o padrão de utilização do espaço pelos
grupos.
Apesar de o grupo ter utilizado a periferia do território mais do que seu interior,
as análises de uso da borda mostraram que o grupo utilizou com freqüência maior do
que a esperada a faixa entre 100 e 50 metros da borda, porém, evitou a parte mais
externa do território. No caso do grupo GRA, a borda do território também representa a
borda do fragmento. Assim, os micos evitaram a região mais próxima às áreas de pasto.
Micos-leões utilizam florestas secundárias com diferentes graus de alteração
(KIERULFF, et al., 2002; RABOY et al., 2004) e a pesquisa realizada mostrou que os
micos-leões podem sobreviver em um fragmento bastante alterado. Ainda assim, eles
evitaram a região mais próxima da borda, que é mais degradada devido ao efeito de
borda, bastante evidente no fragmento estudado (observação pessoal).
A análise de disponibilidade de frutos mostrou que as árvores dos primeiros 100
metros da borda apresentaram menor produtividade do que os ambientes de interior da
floresta ou riacho. Os primeiros 50 metros da borda podem ter sido menos utilizados
pelos micos simplesmente por apresentarem menor quantidade de frutos. No entanto,
não foi avaliada a disponibilidade de frutos de cipós e de bromélias na borda da floresta.
O efeito borda também produz alterações que também podem afetar a utilização
das bordas pelos micos. Nessa região, os micos podem estar mais expostos à predação,
principalmente por aves de rapina. Assim, é possível que os micos tenham evitado essa
região como forma de diminuir o risco de predação. O efeito borda também altera de
maneira acentuada as condições microclimáticas, por exemplo aumentando a
temperatura e diminuindo a umidade. Normalmente, esses efeitos são mais evidentes
nos primeiros 60 metros de borda (KAPOS et al., 1997; TURTON E FREIBURGER,
1997). Os micos podem ter utilizado menos os primeiros metros da borda para evitar as
condições microclimáticas mais adversas dessa região. Todos esses fatores podem ter
contribuído em conjunto para a menor utilização das bordas pelos micos.
A constatação de que os micos evitaram os primeiros 50 metros de borda indica
que esse hábitat – a borda – não é o mais adequado para a espécie. Esse fator deve ser
levado em consideração em análises populacionais da espécie, principalmente na região

Capítulo 3
72
de floresta semidecidual, que é extremamente fragmentada e, portanto, a proporção de
ambientes de borda é elevada. Normalmente, as análises consideram os fragmentos de
forma homogênea, o que, no caso dos micos em florestas semideciduais, pode
superestimar a densidade.
O grupo também utilizou com menor freqüência do que o esperado as áreas de
riachos/brejos, apesar de essas áreas terem apresentado maior produtividade de frutos de
árvores. A presença de outros recursos, como bromélias e cipós, recursos muito
utilizados pelos micos no fragmento (Capítulo II), também podem influenciar o padrão
de utilização da área pelos micos. Apenas estudos mais detalhados, que avaliem a
disponibilidade de frutos tanto de árvores como de cipós, a quantidade de bromélias e as
condições microclimáticas nos diferentes ambientes e em diferentes distâncias da borda,
poderiam elucidar melhor o padrão de utilização da área pelos micos.
5. CONCLUSÕES
• Grupos de L. chrysomelas podem viver em uma área bastante alterada de cerca
de 70 ha de floresta semidecidual, porém o formato irregular do fragmento
limitou o tamanho da área de vida;
• Todos os locais de dormida foram ocos de árvores, o que parece ser
característica de todas as populações de L. chrysomelas;
• O padrão de utilização da área de vida pelo grupo não apresentou uma região
mais utilizada (área núcleo), mas várias regiões apresentaram maior freqüência
de utilização, que se concentraram na periferia do território;
• O grupo evitou os 50 primeiros metros da borda, o que pode estar relacionado à
menor produtividade de frutos na borda, proteção contra predadores ou proteção
contra condições microclimáticas mais adversas;
• O deslocamento diário foi menor do que o observado na Rebio-Una e foi afetado
pela localização das fruteiras utilizadas e pela presença de grupos vizinhos e
defesa do território;
• O padrão de utilização da área de vida foi fortemente influenciado pela
localização dos locais de dormida, das fontes de frutos, e pela presença de
grupos vizinhos e defesa do território.

73
CAPÍTULO IV - A conservação do mico-leão-da-
cara-dourada (Leontopithecus chrysomelas) nas
florestas semideciduais do sul da Bahia

Capítulo 4
74
1. Situação atual de L. chrysomelas nas florestas semideciduais
O mico-leão-da-cara-dourada, L. chrysomelas, é classificado como “Em perigo”,
de acordo com a Lista Nacional das Espécies da Fauna Brasileira Ameaçadas de
Extinção e a lista da IUCN (MMA, 2005; IUCN, 2007). A maior população de L.
chrysomelas está localizada na Reserva Biológica de Una. Na última Análise de
Viabilidade Populacional e de Hábitat (PHVA) para a espécie, realizada em 2005, foi
identificado que a população da Rebio-Una é viável a longo prazo e capaz de manter a
diversidade genética em níveis considerados elevados, isto é, maior de 98% em 100
anos (HOLST et al., 2006). Esta, porém, é a única população da espécie protegida em
Unidade de Conservação Integral.
Na porção leste da distribuição da espécie, onde está localizada a Rebio-Una, há
predomínio de plantações de cacau na forma de cabrucas, onde o cacau é plantado à
sombra de árvores nativas. Essa prática ajudou a preservar boa parte da biodiversidade
da fauna das florestas ombrófilas do sul da Bahia (FARIA et al., 2006; FARIA E
BAUMGARTEN, 2007). Apesar de não sabermos se grupos de micos-leões podem
viver exclusivamente em cabrucas, suas áreas de vida podem incluí-las. Desse modo,
cabrucas também podem servir de corredores entre fragmentos florestais para grupos de
micos-leões (RABOY et al., 2004).
Na parte oeste da distribuição do mico-leão-da-cara-dourada, por outro lado, a
situação é bem diferente. A atividade econômica predominante é a pecuária extensiva,
que foi responsável pelo desmatamento mais intenso e mais antigo do que o ocorrido
nas florestas ombrófilas (MENDONÇA et al., 1994). Embora mais da metade da área de
distribuição da espécie esteja inserida na região de floresta semidecidual, a maior parte
das populações encontra-se nas florestas ombrófilas (PINTO E RYLANDS, 1997). As
populações habitando florestas semideciduais encontram-se em fragmentos pequenos e
isolados por extensas áreas de pastagens, colocando-as em situação crítica de
conservação.
Não existe Unidade de Conservação, seja de uso direto ou indireto, em toda a
região de floresta semidecidual e, portanto, nenhuma população de L. chrysomelas
habitando esse tipo florestal encontra-se protegida. Apenas a RPPN Serra do Teimoso
possui população de MLCD, porém, situa-se em uma região de transição entre floresta
ombrófila e semidecidual (AMORIM et al., 2005).

Capítulo 4
75
O estudo da ecologia e do comportamento de L. chrysomelas em um fragmento
de floresta semidecidual com forte pressão antrópica (ambiente no qual nada se sabia
sobre a espécie), evidenciou a grande flexibilidade ecológica/comportamental e
adaptabilidade da espécie, tal como descrito para L. chrysopygus, que habita florestas
com diferentes características no interior de São Paulo (VALLADARES-PADUA et al.,
2002).
A dieta do mico-leão-da-cara-dourada no fragmento semidecidual foi bastante
distinta do descrito para a Rebio-Una em termos de espécies consumidas, apesar dos
itens serem basicamente os mesmos: frutos (na maioria maduros), insetos, anfíbios,
lagartos e exsudatos. Apenas duas espécies foram consumidas por micos tanto na
Rebio-Una e no fragmento semidecidual, reflexo da diferença florística das duas áreas.
Frutos de cipós foram bastante consumidos, o que pode representar uma adaptação às
condições ecológicas locais (um fragmento alterado e com forte efeito borda), propícias
ao crescimento de cipós (Capítulo II).
O tempo dedicado à procura e consumo de animais foi maior no fragmento
semidecidual, e o tamanho da área de vida e a distância diária percorrida também
apresentaram grande variação nos dois locais, sendo menores no fragmento
semidecidual. As semelhanças foram a utilização de ocos como locais de dormida e o
freqüente uso de bromélias, tanto para a alimentação de frutos como para procura por
presas animais, o que sugere que essas são características comuns a diversas populações
de L. chrysomelas (Capítulos II e III).
Em relação às bromélias, a freqüência de sua utilização foi maior no fragmento
semidecidual do que na Rebio-Una e áreas adjacentes, tanto para consumo de seus
frutos como para procura por presas animais, indicando que esse recurso pode ser mais
essencial para as populações da parte oeste da distribuição do que para a população da
Rebio-Una. Quatro espécies foram consumidas, sendo que A. digitata foi utilizada em
todos os meses. Durante a estação seca, os frutos de bromélias foram ainda mais
consumidos e os micos alimentaram-se de poucas espécies de outros frutos (Capítulo
II).
Embora seja difícil afirmar até que ponto a sobrevivência dos micos nos
fragmentos semideciduais depende das bromélias, sem dúvida elas representaram um
recurso essencial para eles. A disponibilidade de um recurso alimentar essencial pode
ajudar na sobrevivência de primatas em fragmentos florestais (MARSH, 2003), o que
parece ser o caso das bromélias para os micos na região de floresta semidecidual. A

Capítulo 4
76
presença de bromélias deve ser uma variável de peso a ser considerada para a avaliação
da qualidade de hábitat para a espécie.
Os grupos no fragmento semidecidual utilizaram uma área de 70 ha, calculada
pelo método do Mínimo Polígono Convexo (MPC). O Kernel resultou em uma área
menor (58 ha), por excluir o interior do território, que foi pouco utilizado pelo grupo.
Essa região, no entanto, não está disponível para outros grupos. Assim, o MPC dá uma
representação melhor da área de floresta que pode ser utilizada pelo grupo, e deve ser
usado em cálculos de avaliação populacional (Capítulo III).
O formato irregular do fragmento limitou o tamanho da área que poderia ser
utilizada pelo grupo, já que o território do grupo estava quase todo cercado por áreas de
pastagens. Na parte sul do território, uma estrada de terra serviu como barreira do
território (apesar de os micos poderem, potencialmente, atravessá-la pelo chão). É difícil
afirmar se o grupo utilizaria uma área maior se estivesse em uma floresta contínua, ou
se sobreviveria se estivesse confinado em uma parte ainda menor de floresta. Os
resultados só permitem dizer que o grupo, que chegou a ter nove micos, estava
sobrevivendo em uma área com cerca de 70 ha de floresta (Capítulo III).
Diferenças ecológicas e comportamentais já foram relatadas para diferentes
espécies do gênero Leontopithecus e mesmo para diferentes populações de uma mesma
espécie (KIERULFF et al., 2002). Essas diferenças estão relacionadas a diferenças no
hábitat, como idade da floresta, estrutura da vegetação, composição florística e
sazonalidade climática (KIERULFF et al., 2002). Portanto, as diferenças ecológicas e
comportamentais em L. chrysomelas observadas entre a populações da Rebio-Una e do
fragmento semidecidual, provavelmente são decorrentes das diferenças ambientais –
diferenças em composição florística, estrutura, além das pressões antrópicas (ver
Capítulo I). Efeitos oriundos de diferenças genéticas, no entanto, não podem ser
descartados, apesar de ainda não ser possível determinar o quanto as diferenças
genéticas entre as populações são responsáveis pelas diferenças comportamentais
observadas.
Essa grande flexibilidade ecológica e comportamental pode aumentar as chances
de sobrevivência da espécie em diferentes hábitats, inclusive ambientes fortemente
alterados pela ação humana. Essa flexibilidade, no entanto, não garante sua
sobrevivência a longo prazo nos fragmentos de florestas semideciduais.
Diversos fragmentos de floresta semidecidual ainda possuem populações de L.
chrysomelas, porém os fragmentos são pequenos e encontram-se isolados por uma

Capítulo 4
77
matriz de pastagens. A fragmentação florestal pode afetar os primatas de diversas
formas, tanto em aspectos demográficos, como tamanho de grupos e densidade de
indivíduos, como em aspectos comportamentais, como estratégia alimentar (MARSH,
2003). Além disso, o isolamento dos fragmentos pode afetar o fluxo gênico da
população.
Sem dúvida, a principal ameaça à espécie na região de florestas semideciduais é
o tamanho reduzido e o isolamento das populações restritas aos fragmentos. Esse
isolamento pode afetar a viabilidade das populações a longo prazo. Populações
relativamente pequenas e isoladas são mais susceptíveis à extinção devido a fatores
genéticos e demográficos, que podem atuar em conjunto. As ameaças vêm
principalmente da perda da variabilidade genética e da depressão endogâmica (RALLS
et al., 1986; GONÇALVES et al., 2003). Além disso, a fragmentação também
potencializa outros efeitos negativos, como a caça, incidência de doenças, ação de
predadores e eventos estocásticos (LAURENCE E COCHRANE, 2001).
Pode-se argumentar que as populações de L. chrysomelas habitando florestas
semideciduais são pequenas e, por isso, não viáveis a longo prazo. No entanto, essas
populações podem ser reservatórios para a diversidade genética da espécie como um
todo. Não existem informações sobre a genética das populações habitando florestas
semideciduais e não se sabe o quanto essas populações são distintas geneticamente da
população da Rebio-Una (HOLST et al., 2006). Também não sabemos até que ponto os
efeitos negativos decorrentes da depressão endogâmica estão afetando essas populações.
Obter tais informações é essencial para avaliar o valor das populações das florestas
semideciduais na manutenção da diversidade genética da espécie e na orientação de
possíveis ações de manejo. Mesmo pequenas, essas populações podem representar um
reservatório genético que viabilize a conservação da espécie.
Os efeitos da fragmentação na genética de primatas é assunto que deve ser mais
explorado. Gonçalves et al. (2003) não observou efeito da fragmentação na diversidade
genética do calitriquídeo Mico argentatus, na Amazônia, já que mesmo populações
habitando fragmentos pequenos apresentaram alta diversidade genética. O isolamento
dessas populações, no entanto, pode ser recente, e, portanto, os efeitos negativos
decorrentes do isolamento podem ainda não ser visíveis.
Para o mico-leão-dourado, L. rosalia, no entanto, já foi demonstrada perda de
diversidade genética em populações pequenas (GRATIVOL et al., 2001). Essa perda foi
maior nas menores populações. Dietz et al. (2000), trabalhando com a mesma espécie,

Capítulo 4
78
encontrou evidências de depressão endogâmica na Reserva Biológica Poço das Antas,
no Rio de Janeiro.. Nesse estudo, os autores observaram que a mortalidade de infantes
era mais alta para os indivíduos cujos pais já tinham algum grau de parentesco. Assim, é
bastante provável que a população do fragmento estudado, estimada em cerca de 30
indivíduos, apresente altas taxas de endogamia, perda de diversidade genética e
depressão endogâmica.
Durante o período do presente estudo, foi constatada alta mortalidade de micos-
leões no fragmento semidecidual. Houve nascimentos de 12 filhotes (seis pares de
gêmeos) nos dois grupos monitorados. Desses, sete (60%) desapareceram antes de
completarem três meses de vida (tempo aproximado do desmame) e outros dois, entre
três e sete meses. Esses valores são bem maiores do que o observado na Rebio-Una,
onde apenas 10% desapareceram antes dos três meses e 23% antes de completarem um
ano (RABOY et al., 2001). Mesmo sendo uma amostra pequena (um ano e meio de
observação de dois grupos), esse fato gera preocupação a respeito da conservação dos
micos-leões nesses fragmentos. Apesar de não ser possível indicar a causa das mortes
desses infantes, é possível que tenham sido decorrentes da endogamia e da depressão
endogâmica. Outras explicações, no entanto, como alta taxa de predação e deficiência
nutricional, não podem ser descartadas.
Durante o monitoramento, foi observada a entrada de um indivíduo,
provavelmente uma fêmea, em um dos grupos. Esse indivíduo apresentava coloração
diferente do padrão de L. chrysomelas. A face, os antebraços e parte dorsal da cauda
eram dourados, como os demais, porém o resto do corpo não era preto, mas levemente
dourado (Fig. 28). Padrão muito semelhante já foi descrito para L. chrysomelas em um
indivíduo em cativeiro, e está associado à deficiência nutricional, principalmente de
proteínas (COIMBRA-FILHO E ROCHA, 1978). Esse indivíduo cativo recuperou a
coloração padrão após alimentação adequada. Embora tenha sido apenas um caso, no
qual nem se tinha monitoramento antes da sua entrada no grupo, a hipótese de que esse
indivíduo apresentava deficiência nutricional não pode ser descartada. Ele permaneceu
com o grupo até o fim de seu monitoramento, três meses depois, quando o macho com
rádio-colar foi encontrado morto, e o restante do grupo não foi mais observado.

Capítulo 4
79
Figura 28. Mico-leão-da-cara-dourada observado no fragmento da Fazenda Barro
Branco, com padrão de coloração diferente do da espécie (indicada pela seta).
Provavelmente uma fêmea.
Vale ressaltar que essa população não se encontra totalmente isolada, já que a
espécie habita outros fragmentos vizinhos, a menos de um quilômetro de distância (ver
Capítulo I para descrição e imagem da região). Já foram observados micos-leões-
dourados atravessando áreas de pasto de mais de um quilômetro (OLIVEIRA et al.,
2004). Assim, é possível que exista dispersão entre esses fragmentos, embora durante o
período de estudo não tenha sido observada. Essa mesma situação, de diversos
fragmentos com possível migração de micos entre eles, é encontrada em outras regiões
de floresta semidecidual.
Essa disposição de fragmentos, alguns isolados parcialmente por pastos sujos e a
maioria habitada por micos-leões, com provável dispersão entre eles, sugere uma
estrutura de metapopulação - um conjunto de sub-populações com certa migração entre
elas, podendo haver extinções locais e novas recolonizações (HANSKI, 1998).
Estratégias de manejo que envolvam a estrutura da metapopulação podem ser usadas
para aumentar as chances de sobrevivência de uma espécie (HANSKI, 1998), e
conhecer a estrutura da metapopulação permite que os esforços de manejo e
conservação sejam maximizados (RODRIGUES-VARGAS et al., 2003).
Também é de interesse a conservação das adaptações comportamentais e
ecológicas do mico-leão-da-cara-dourada. A diversidade ecológica da espécie pode ter
papel essencial na sua sobrevivência a longo prazo, e o conhecimento das adaptações às
condições ecológicas específicas, como em fragmentos degradados, pode ser útil na
orientação de ações de manejo.

Capítulo 4
80
Além disso, primatas influem na ecologia das florestas tropicais e também são
responsáveis por grande parte da biomassa de vertebrados nessas florestas (CHAPMAN
E ONDERDONK, 1998). Desse modo, é essencial compreender o papel ecológico de
primatas em fragmentos florestais e avaliar como eles podem auxiliar na regeneração de
florestas (MARSH, 2003). Outras espécies do gênero Leontopithecus são dispersoras de
sementes (L. chrysopygus – PASSOS, 1997b; L. rosalia – LAPENTA, 2006). A maioria
dos frutos consumidos pelos micos no fragmento semidecidual tinham as sementes
engolidas, e estas passavam aparentemente intactas pelo trato digestivo (observação
pessoal). Assim, é possível que L. chrysomelas nos fragmentos semideciduais também
tenham papel na dispersão e regeneração das florestas.
Os fragmentos de floresta semidecidual no sul da Bahia, apesar de não serem
muito grandes, são numerosos, e sem dúvida possuem uma relevância local para a
conservação não só de L. chrysomelas, mas de outras espécies também. A presença de
primatas nesses fragmentos é um indicativo de sua viabilidade como reservatórios de
fauna. Por exemplo, no local onde foi realizado o presente estudo, além de L.
chrysomelas, também estavam presentes Cebus xanthosternos e Callicebus melanochir,
espécies consideradas criticamente ameaçada e vulnerável, respectivamente (IUCN,
2007). Em outros fragmentos da região ainda é possível encontrar Alouatta guariba
(Leonardo G. Neves, com. pess.), espécie também considerada criticamente ameaçada
(IUCN, 2007).
Se estudos vegetacionais na região de floresta semidecidual são raros
(AMORIM et al., 2005; MARIANO-NETO, 2004), estudos referentes à fauna são ainda
menos comuns. Silveira (2003) realizou um rápido levantamento de aves em alguns
fragmentos de floresta semideciduais em Itambé, município localizado cerca de 50 km a
oeste de Itororó. Nesse estudo, o autor encontrou algumas espécies ameaçadas de
extinção como Amazona rodocorytha, Aratinga auricapilla e Myrmeciza ruficauda.
Apesar dos fragmentos de Itambé não possuírem L. chrysomelas, a presença dessas
espécies é indício de que os fragmentos mais a leste, ocupados por micos-leões, também
podem abrigar essas ou outras espécies ameaçadas de aves, dada a proximidade e
semelhança entre as florestas. Por exemplo, foi observado no fragmento em Itororó,
onde foi realizado o presente trabalho, Rhopornis ardesiaca, espécie ameaçada que na
Bahia é conhecida apenas na região de Boa Nova (SILVEIRA, 2003). Levantamentos
de diversos grupos de fauna são necessários para um melhor conhecimento das florestas
semideciduais e para subsidiar ações de conservação na região.

Capítulo 4
81
Além das ameaças decorrentes do isolamento das populações relativamente
pequenas, os próprios fragmentos ainda sofrem ameaça. A fragmentação florestal
aumenta a proporção de borda da floresta. O próprio tamanho dos fragmentos, na
imensa maioria menor do que 1000 ha (embora uma análise minuciosa não tenha sido
feita), e os seus formatos bastante irregulares, contribuem para o aumento do efeito
borda.
O efeito borda é extremamente variável em diferentes florestas (LAURANCE et
al., 1997), porém algumas tendências parecem existir, tanto físicas como bióticas. As
mudanças climáticas associadas ao efeito borda geralmente atingem cerca de 60 metros
da borda (KAPOS et al., 1997; TURTON E FREIBURGER, 1997), e há mudança na
composição das comunidades e abundâncias de diversas espécies animais e vegetais
(TERBORGH et al., 1997; LAURANCE et al., 1997; DIDHAM, 1997).
Embora não tão comum como antigamente, o corte seletivo ainda existe na
região, principalmente para sustentar as próprias atividades da fazenda. A madeira
cortada é utilizada em construção de cercas e currais e como combustível, na forma de
lenha.
A maioria dos fragmentos não é cercada, o que permite a entrada do gado no
interior da floresta. Isso ocorre mais frequentemente na época da seca, quando o capim
está mais seco e disponível em menor quantidade nas áreas de pasto. O gado alimenta-
se de brotos no interior da floresta, além de pisotear e matar plântulas, o que desacelera
ou até mesmo inibe a regeneração da floresta.
Outra prática da região que coloca em risco os fragmentos é a queimada,
realizada principalmente para limpeza de pastos sujos, mas que pode, eventualmente,
entrar na floresta. Esse risco obviamente é bem maior durante a estação seca.
Tanto o efeito de borda como a entrada de gado nos fragmentos e as queimadas,
provocam fortes alterações na estrutura dos fragmentos e dificultam a regeneração
natural da floresta. Um dos efeitos é o aumento da quantidade de lianas, o que foi
observado por Mariano-Neto (2004) em alguns fragmentos de florestas semideciduais
da região. O aumento de cipós prejudica o estrato arbóreo, aumentando a mortalidade e
o sombreamento, diminuindo a massa foliar das árvores e afetando o crescimento e a
produção de frutos e sementes das árvores. (LAURANCE et al., 2007). Os micos
utilizaram frutos de muitos cipós em sua dieta (Capítulo II), mais do que o observado na
Rebio-Una (RABOY E DIETZ, 2004), o que pode ser simplesmente uma adaptação às
condições locais, isto é, à grande quantidade de lianas decorrente da perturbação do

Capítulo 4
82
fragmento. Mais uma vez observa-se a flexibilidade da espécie, porém não é possível
dizer se essas alterações terão conseqüências para a sobrevivência dos grupos no
fragmento a longo prazo.
Vale ressaltar, ainda, que a caça e captura de animais vivos ainda pode
representar uma ameaça. Embora exista caça na região (observação pessoal), a captura
de micos-leões é eventual, já que não é um animal visado. As caças mais visadas são
tatus, veados e caititus. Primatas são caçados de forma oportunística, e normalmente
para captura de filhotes para serem criados como animais de estimação. Foram
observados na região de Itororó Cebus xanthosternos e Callithrix kuhlii mantidos em
cativeiro. Apesar de não ter sido observado nenhum mico-leão, eles sem dúvida estão
sujeitos à mesma ameaça.
A densidade de indivíduos encontrada no fragmento semidecidual foi menor do
que a reportada para a Rebio-Una, em parte devido ao tamanho médio dos grupos serem
um pouco menor (Capítulo III). Isso trás maior preocupação em relação à conservação
da espécie na região de floresta semidecidual. Esse resultado, no entanto, deve ser visto
com cautela. Primeiro, não foram consideradas possíveis sobreposições de territórios de
grupos vizinhos, o que pode aumentar a densidade de indivíduos no fragmento.
Segundo, o fragmento é bastante recortado, e os resultados podem ser verdadeiros
apenas para o esse fragmento. Ainda assim, o valor encontrado para área de vida dos
grupos (70 ha) pode ser usado em análises de densidade populacional para a espécie na
região , já que a situação da maioria dos fragmentos semideciduais na área de
distribuição do mico-leão-da-cara-dourada é parecida com a da fazenda Barro Branco,
isto é, bastante irregulares, com grande proporção de bordas (observação pessoal).
Fragmentos menores, no entanto podem conter grupos, e não devem ser, em princípio,
descartados de análises.
2. Prioridades de pesquisa e ações para a conservação do MLCD em fragmentos
semideciduais
A abordagem em nível metapopulacional tem sido uma ferramenta útil no
delineamento de estratégias visando à conservação de diversas espécies de primatas que
habitam ambientes fragmentados, inclusive outras espécies de Leontopithecus (L.
chrysopygus -VALLADARES-PADUA et al., 2002; Saimiri oerstedii - RODRIGUEZ-
VARGAS, 2003). No caso de L. chrysopygus, a população cativa tem sido considerada
como parte da metapopulação e ações de manejo, como translocação e reintrodução,

Capítulo 4
83
foram usadas para aumentar as chances de sobrevivência da espécie a longo prazo
(VALLADARES-PADUA et al., 2002). O manejo metapopulacional pode aumentar o
fluxo gênico entre as populações de micos-leões-da-cara-dourada habitando fragmentos
de florestas semideciduais, elevando, assim, as chances de sobrevivência dessas
populações.
Apesar de ações de manejo como translocações – retirada de animais de um
local e soltura em outro local em que a espécie ocorre ou já ocorreu - não terem sido
apontadas como prioritárias para a conservação da espécie na última Análise de
Viabilidade Populacional e de Hábitat (PHVA) realizado (HOLST et al., 2006), a
sobrevivência de muitas populações em fragmentos de floresta semidecidual talvez
dependa de tal técnica. Muitos grupos de micos-leões estão confinados em fragmentos
muito pequenos e totalmente isolados, e apenas a retirada desses grupos garantiriam sua
sobrevivência. Translocações aumentam o fluxo gênico entre as populações e a técnica
já foi usada, com sucesso, em ações de manejo para L. rosalia e L. chrysopygus
(KIERULFF, 2000; VALLADARES-PADUA et al., 2002).
Para tais ações, no entanto, é necessário um melhor conhecimento da estrutura
da metapopulação dos micos-leões-da-cara-dourada nos fragmentos semideciduais. Um
melhor detalhamento sobre a presença ou ausência da espécie em fragmentos de
diferentes tamanhos e características, além de avaliações de hábitats adequados para a
espécie, são essenciais para a identificação de grupos a serem translocados e possíveis
locais para a soltura. Esforços no sentido de identificar populações em fragmentos de
diferentes tamanhos estão em andamento (Becky Raboy, com. pess.).
Pesquisas futuras na região também deveriam priorizar o conhecimento da
diversidade genética das populações habitando diferentes fragmentos de florestas
semideciduais, e entre essas populações e a da Rebio-Una. Se as populações da parte
oeste realmente apresentarem diversidade genética substancialmente diferente da
população da Rebio-Una, esta unidade de conservação estaria protegendo apenas parte
da diversidade genética total da espécie (HOLST et al., 2006).
Estudos sobre tamanho, composição de grupos e densidade em outros
fragmentos também são necessários para uma melhor perspectiva sobre demografia do
mico-leão-da-cara-dourada em fragmentos de floresta semideciduais.
Um problema que certamente surgiria no caso de ações como translocação, é em
relação ao local para possíveis solturas, já que fragmentos grandes o suficientes para a
sobreviv6encia de grupos e que já não contenham a espécie talvez não existam mais na

Capítulo 4
84
região. Ainda assim, com um conhecimento demográfico e genético dessas populações,
seria possível aumentar o fluxo gênico através de tais técnicas.
Alguns aspectos da biologia e história natural da espécie em florestas
semideciduais ainda precisam ser mais bem compreendidos, de modo a orientar ações
de manejo e aprimorar os modelos de viabilidade populacionais. Por exemplo,
informações sobre aspectos reprodutivos, mortalidade e migração entre grupos e entre
fragmentos ajudam a refinar esses modelos. Existem alguns dados para a região de
floresta ombrófila (HOLST et al., 2006), no entanto, dadas as diferenças ambientais,
esses aspectos podem ser bem diferentes nos fragmentos semideciduais (Capítulo I). O
presente estudo obteve algumas informações, porém, estudos em outros fragmentos da
região e estudos de longa duração podem prover respostas mais precisas.
Estudos mais detalhados sobre uso do hábitat também são necessários. Por
exemplo, o uso das bordas pode ser assunto mais explorado. As bordas podem ser
bastante utilizadas por muitas espécies de primatas (MARSH, 2003), inclusive alguns
calitriquídeos, como Callithrix e Saguinus (RYLANDS, 1996), porém não parece ser
um hábitat muito utilizado por Leontopithecus. Se atualmente vemos espécies de micos-
leões usando hábitats degradados e florestas em regeneração (KIERULFF et al., 2002;
RABOY E DIETZ, 2004), é devido à condição atual da Mata Atlântica, praticamente
sem florestas maduras na área de distribuição das quatro espécies (RYLANDS, 1996).
Os resultados do presente trabalho confirmam que a borda parece não ser um
hábitat adequado para a espécie. Apesar dos micos utilizarem os 100 primeiros metros
da borda com certa freqüência, foi observado que a borda mais próxima do contato com
a pastagem (os 50 primeiros metros) foi evitada (Capítulo III). Não se sabe até que
ponto grupos de micos poderiam sobreviver em florestas ainda mais degradadas do que
o local onde foi realizado este estudo, ou locais com efeito de borda ainda mais
pronunciado. Esse fato é mais um ponto de preocupação para a conservação da espécie
na região de floresta semidecidual, dados os tamanhos e condições dos fragmentos
nessa região, já que nem toda a floresta pode representar um hábitat ideal. Nas análises
de disponibilidade de hábitat e viabilidade populacional, normalmente não há
diferenciação entre o interior da floresta e as bordas. Sem dúvida, estudos adicionais
que verifiquem como os micos-leões utilizam diferentes tipos de borda e quais variáveis
afetam a utilização das bordas, são necessários para uma melhor compreensão sobre
quais tipos de hábitat são adequados para a espécie. Essa informação pode orientar e
refinar análises de viabilidade populacional.

Capítulo 4
85
Um estudo mais minucioso da relação dos micos-leões com as bromeliáceas
pode revelar o quanto os micos dependem dessa família de plantas para sua
sobrevivência, e se bromélias realmente representam um recurso essencial. Esse
conhecimento também permitiria avaliar qual hábitat é mais adequado para a espécie na
região de floresta semidecidual.
Esforços também deveriam ser feitos no sentido de aumentar a conectividade
entre os fragmentos ou mesmo a qualidade do hábitat. Além de aumentar a quantidade
de hábitat, elevaria também o fluxo gênico entre diferentes grupos. Espécies vegetais
utilizadas pelos micos (Capítulo II) poderiam ser usadas na formação de corredores
entre fragmentos, aumentando as chances de esses corredores serem utilizados pela
espécie.
Outra preocupação é a manutenção dos fragmentos florestais. Como mencionado
anteriormente, o corte seletivo, o fogo e a entrada do gado nas florestas são as principais
ameaças e não existe unidade de conservação na região de floresta semidecidual no sul
da Bahia. A maioria dos fragmentos da região está em apenas uma ou poucas
propriedades
Apesar de a população de micos-leões-da-cara-dourada na Rebio-Una ser viável
a longo prazo, ela é a única população da espécie em área de proteção integral. A
proteção de outras populações daria maior segurança para a conservação da espécie no
futuro, e esforços para a criação de reservas na região de floresta semidecidual, onde
não há unidades de conservação, são necessários. Para isso, inventários de diversos
grupos de fauna e flora são essências. Conforme dito anteriormente, outras espécies de
primatas e aves ameaçados de extinção podem ser encontradas nos fragmentos. O
incentivo à criação de RPPN’s pode ser uma alternativa à falta de unidades de
conservação no local.
Por fim, programas de educação/informação ambiental e envolvimento de
moradores e fazendeiros locais, são essenciais para a conservação dos fragmentos e de
sua fauna. Durante a estada na região, foi detectada uma grande falta de conhecimento a
respeito da fauna, inclusive do mico-leão, por parte dos moradores. Os donos das
fazendas ou gerentes são os tomadores de decisão a respeito do que é feito com o
fragmento e, portanto, deveriam ser o público alvo. Informações a respeito de práticas
de manejo menos nocivas aos fragmentos e sobre a presença de espécies ameaçadas,
além de informações sobre criação de reservas particulares deveriam ser disseminadas
através desses programas de educação/informação ambiental.

Capítulo 4
86
3. Conclusões
• A grande flexibilidade ecológico-comportamental de L. chrysomelas não garante
a sobrevivência da espécie a longo prazo nos fragmentos semideciduais;
• A principal ameaça é o isolamento de pequenas populações. A alta taxa de
mortalidade observada pode ser conseqüência desse isolamento;
• O manejo metapopulacional na região de floresta semidecidual pode aumentar as
chances de sobrevivência da espécie, porém estudos sobre demografia e análise
de hábitats, além de estudos genéticos, são necessários para tal manejo.
• Pesquisas futuras na região podem priorizar aspectos como dispersão entre
fragmentos, a relação dos micos-leões com as bromélias e o uso de diferentes
hábitats, como as bordas da floresta.
• Aumentar a conectividade entre os fragmentos de florestas semideciduais e
garantir a integridade dos fragmentos existentes são ações essenciais para a
conservação de L. chrysomelas na região.

Referências
87
REFERÊNCIAS ALGER, K.; CALDAS, M. Cacau na Bahia: decadência e ameaça à Mata Atlântica. Ciência
Hoje. Vol. 20, 1996, p. 28-35. ALTMANN, J. Observational study of behavior: sampling methods. Behavior. Vol. 49,
1974, pgs. 227-267. AMORIM, A.M; FIASCHI, P.; JARDIM, J.G.; THOMAS W.W.; CLIFTON, B.C.; DE
CARVALHO, A.M.V. The vascular plants of a forest fragment in southern Bahia, Brazil. Sida. Vol 21 (3). 2005. pgs. 1726-1752.
BAKER, A.J.; BALES, K.; DIETZ, J.M. Mating system and group dynamics in lion tamarins.
In: Lion Tamarins Biology and Conservation D.G. Kleiman and A.B. Rylands (eds.). 2002, pgs. 1188-212.
BARTON, R.A.; Whiten, A. ;Strum, S.C.; Byrne, R.W.; Simpson, A.J. Habitat use and
resource availability in baboons. Animal Behaviour. Vol 43 (5). 1992. pgs. 831-844. BICCA-MARQUES, J.C. How do howler monkeys cope with habitat fragmentation?
Primates in fragments: ecology and conservation. Marsh, L. (ed.). New York: Kluwer Academic/Plenum 2003. pgs. 455-475.
CEPLAC. Centro de Pesquisas do cacao. Resumos dos dados climatológicos. Posto: Fazenda Cabana da Ponte. Município: Itororó-BA. Período: 1973/1978. 1981
CHAPMAN, C.A.; ONDERDONK, D.A. Forests without primates: Primate/plant
codependency. American Journal of Primatology. Vol 45 (1). 1998. pgs. 127-141. COIMBRA-FILHO, A.F. Natural Shelter of Leontopithecus rosalia and Some Ecological
Implications. In Biology and Conservation of the Callitrichdae. D.G. Kleiman (ed). Smithsonian Institution. 1978, p. 79-89.
COIMBRA-FILHO, A.F.; METTERMEIER, R.A. Distribution and ecology of the genus
Leontopithecus in Brasil. Primates. Vol. 14 (1), 1973, pgs. 47-66. COIMBRA-FILHO, A.F.; ROCHA, N.C. A loss of pigment in Leontopithecus rosalia
chrysomelas (Kuhl, 1820), its treatment and reversal (Callitrichidae, Primates). Revista Brasileira de Biologia. Vol. 38, 1978, Pgs. 165-170.
COTTAN, G. e CURTIS J.T. The use of distance measures in phytsociological samplimg. Ecology. Vol. 37 (3), 1965, p. 451-460.
COWLISHAW, G. Trade-offs between foraging and predation risk determine habitat use in a
desert baboon population. Animal Behaviour. Vol 53 (4). 1997. pgs. 667-686 CROCKETT, C.M.; EISENBERG, J.F. Howlers: Variations in group size and demography.
In: Primates Societies. B.B. Smuts D.L. Cheney R.M. Seyfarth R.W. Wrangham T.T. Struhsaker, (Ed). University of Chicago Press., Chicago. 1987, Pgs.54-68.

Referências
88
DEFLER, T.R. The time budget of a group of wild woolly monkeys (Lagothrix lagotricha).
International Journal of Primatology. Vol. 16, 1995, pgs. 107-120. DIDHAM, R.K. The influence of edge effects and forest frsgmentation on leaf litter
invertebrates in Central Amazonia. In: Tropical forest remnants. Ecology, management and conservation of fragmented communities. Laurance, W.F. and Bierregaard, R.O.Jr. (editors). The University of Chicago Press. 1997. pgs. 55-70
DIETZ, J.M..; BAKER, A.J. Polygyny and female reproductive success in golden lion
tamarins, Leontopithecus rosalia. Animal Behaviour. Vol. 46 (6), 1993, pgs. 1067-1078.
DIETZ, J.M.; SOUZA, S.N.F.; SILVA, J.R.O. Population structure and territory size in
golden-headed lion tamarins, Leontopithecus chrysomelas. Neotropical Primates. Vol. 2 (suppl), 1994, pgs. 21-23.
DIETZ, J.M.; SOUSA, S.N.; BILLERBECK, R. Populations dynamics of golden-headed lion
tamarins Leontopithecus chrysomelas in Una Reserve, Brazil. Dodo (Journal of Wildlife Preservation Trusts). Vol 32. 1996. pgs. 115-122
DIETZ, J.M.; PERES, C.; PINDER, L. Foraging ecology and use of space in wild golden lion
tamarins (Leontopithecus rosalia). American Journal of Primatology. Vol. 41 (4), 1997, pgs. 289-305.
DIETZ, J.M.; BAKER, A.J.; BALLOU, J.D. Demographic evidence of inbreeding depression
in golden lion tamarin. In: Genetics, demography and population viability. Young, A.G. and Clarke, G.M. (editors).Cambridge University Press. 2000. pgs. 203-211.
DORAN, D. Influence of seasonality on activity patterns, feeding behavior, ranging, and
grouping patterns in Tai chimpanzees. International Journal of Primatology. Vol. 18 (2), 1997, pgs. 183-206.
FARIA D.; LAPS R. R.; BAUMGARTEN, J.; CETRA, M. Bat and bird assemblages from
forests and shade cacao plantations in two contrasting landscapes in the Atlantic Forest of southern Bahia, Brazil. Biodiversity and Conservation, n. 15, 2006, p. 587–612.
FARIA, D.; BAUMGARTEN, J. Shade cacao plantations (Theobroma cacao) and bat conservation in southern Bahia, Brazil. Biodiversity and Conservation. vol.16, 2007, p.291-312.
FERRARI, S.F. The behaviour and ecology of the buffy-headed marmoset, Callithrix flaviceps (O. Thomas, 1903). Tese (Doutorado). University College of London. 1988.
FLEAGLE, L.G. Primate Adaptation and Evolution, 2ª ed. San Diego Academic Press. 596
pg. 1999.
FONSECA, G.A.B., et al. Abordagens Ecológicas e Instrumentos Econômicos para o Estabelecimento do Corredor do Descobrimento: uma Estratégia para Reverter a Fragmentação Florestal da mata atlântica no Sul da Bahia. Instituto de Estudos

Referências
89
Sócio Ambientais do Sul da Bahia (IESB), Consevation International do Brasil, PROBIO/MMA, Universidade Federal de Minas Gerais, Núcleo de Estudos e Pesquisas Ambientais da UNICAMP, Departamento de Desenvolvimento Florestal do Estado da Bahia, Brasília. Relatório técnico não publicado. 2002.
FOURNIER, L.A. Um método cuantitativo para la medición de características fenológicas em
árboles. Turrialba. Vol. 24, 1974, pgs. 422-423. FRANKLIN, S.P.; HANKERSON, S.J.; BAKER, A.J.; DIETZ, J.M. Golden lion tamarin
sleeping-site use and pre-retirement behavior during intense predation. American Journal of Primatology. Vol. 69. 2007. Pgs. 325-335.
GALETTI, M.; PIZO, M.A.; MORELLATO, P.C. Fenologia, frugivoria e dispersão de
sementes. In: Métodos de estudos em Biologia da Conservação e manejo da vida silvestre. Cullen Jr., L., Rudran, R., Valladares-Pádua, C. (Ed). Editora da universidade Federal do Paraná, Curitiba. 2004, Pgs. 395-422
GONÇALVES, E.C.; FERRARI, S.F.; SILVA, A.; COUTINHO, P.E.G.; MENEZES, E.V.;
SCHNEIDER, M.P.C. Effects of habitat fragmentation on the genetic variability of silvery marmosets, Mico argentatus. In: Primates in fragments: ecology and conservation. Marsh, L. (ed.). New York: Kluwer Academic/Plenum 2003. pgs. 17-28.
GOUVÊA, J.B.S., MATTOS, L.A. a HORI, M.. Fitogeografia In: Diagnóstico Socioeconômico da Região Cacaueira, vol 7: Recursos Florestais. Comissão Executiva do Plano da Lavoura Cacaueira (CEPLAC) e Instituto Interamericano de Ciências Agrícolas – OEA. Ilhéus, Bahia, Brasil. 1976.
GRATIVOL, A.D.; BALLOU, J.D.; FLEISCHER, R.C. Microsatellite variation within and
among recently fragmented populations of the golden lion tamarin (Leontopithecus rosalia). Conservation genetics. Vol. 2. 2001. pgs. 1-9.
GURSKY, S.L. Effect of seasonality on the behavior of an insectivorous primate, Tarsius
spectrum. International Journal of Primatology. Vol. 20 (3), 2000, pgs. 477-495. HAMILTON, W.J. Baboon sleeping site preferences and relationships to primate grouping
patterns. American Journal of Primatology. Vol. 3 (1-4). 1982. pgs. 41-53. HANKERSON, S.J; DIETZ, J.; RABOY, B.E. Associations between golden-headed lion
tamarins and the bird community in the Atlantic Forest of southern Bahia. International Journal of Primatology. Vol. 27 (2), 2006, pgs. 487-495.
HANSKI, I. Metapopulation dynamics. Nature. Vol. 396, 1998. pgs. 41-49. HANYA, G. Seasonal variation in the activity budget of Japanese macaques in the coniferous
forest of Yakushima: effects of food and temperature. American Journal of Primatology. Vol. 63, 2004, pgs. 165-177.
HOLST, B.; MEDICE, O.J.; MARINHO-FILHO, O.J.; KLEIMAN, D.; LEUS, K.,; PISSINATTI, A., VIVEKANDA, G..; BALLOU, J.D.; TRAYLOR-HOLZER, K.; RABOY, B.; VLEESCHOUWER, K.; MONTENEGRO, M.M. (eds.). Lion Tamarin

Referências
90
population and habitat viability assessment workshop 2005, final report. IUCN/SSC Conservation Breeding Specialist Group, Apple Valley, MN, USA. 2006.
IUCN. 2007 IUCN Red List of Threatened Species. Disponível em
http://www.iucnredlist.org Acessado em 10 set. 2007. JACOB, A.A.; RUDRAN, R. Radiotelemetria em estudos populacionais. In: Métodos de
estudos em Biologia da Conservação e manejo da vida silvestre. Cullen Jr., L., Rudran, R., Valladares-Pádua, C. (Ed). Editora da universidade Federal do Paraná, Curitiba. 2004, Pgs. 285-341.
KAPOS, V.; WANDELLI, E.; CAMARGO, J.L.; GANADE, G. Edge-related changes in
environment and plant responses due to forest fragmentation in Central Amazônia. In: Tropical forest remnants. Ecology, management and conservation of fragmented communities. Laurance, W.F. and Bierregaard, R.O.Jr. (editors). The University of Chicago Press. 1997. pgs. 33-44.
KERNOHAN, B.J; GITZEN, R.A.; MILLSPAUGH, J. Analysis of animal space use and
movements. In: Radio tracking and animal populations. Millspaugh, J. and Marzluff, J.M. Academic press, San Diego. 2001. pgs. 125-166.
KEUROGHLIAN, A.; PASSOS, F.C. Prey foraging behaviour, seasonality and time-budgets in black lion tamarins (Leontopithecus chrysopygus) (Mikan 1823) (Mammalia, Callitrichidae). Brazilian Journal of Biology. Vol. 61 (3), 2001, pgs. 455-459.
KIERULFF, M.C.M.. Ecology and Behaviour of translocated groups of golden lion
tamarins (Leontopithecus rosalia). 2000. Tese (Doutorado), University of Cambridge, UK.
KIERULFF, M.C.M; RABOY, B.E.; PROCÓPIO-DE-OLIVEIRA, P.; MILLER, K.;
PASSOS, F.; PRADO, F. Behavioral Ecology of Lion Tamarins. In: Lion Tamarins Biology and Conservation D.G. Kleiman and A.B. Rylands (eds.). 2002, pgs. 157-187.
KINZEY, W.G. Distribution of primates and forest refuges. In: Biological Diversification in
the Tropics. G. T. Prance, Editors. New York: Columbia University Press. 1982, Pgs. 455-482.
KLEIMAN D.G., HOAGE R.J., GREEN K.M. The lion tamarins, genus Leontopithecus. In: Ecology and behavior of neotropical primates. Mittermeier A.B. Rylands A.F. Coimbra-Filho G.A.B. da Fonseca, Editors. Washington, DC: World Wildlife Fund. 1988, Pgs. 299-347.
LAPENTA, M.N. Frugivoria, Dispersão Primária e Secundária de Sementes Consumidas por
Micos-Leões-Dourados (Leontopithecus rosalia) na Reserva Biológica União, RJ. Tese (doutorado). Universidade de São Paulo, São Paulo. 2006.
LAPENTA, M.J.; PROCÓPIO-DE-OLIVEIRA, P.; KIERULFF, M.C.M., MOTTA-JUNIOR,
J.C. Fruit explotation by golden-headed lion tamarins (Leontopithecus rosalia) in the União Biológica Reserve, Rio das Ostras, RJ-Brasil. Mammalia. Vol. 57 (1), 2003, 41-45.

Referências
91
LAURENCE, W.F et al. Tropical forest fragmentation: synthesis of a diverse and dynamic
discipline. In Tropical forest remnants. Ecology, management and conservation of fragmented communities. Laurance, W.F. and Bierregaard, R.O.Jr. (editors). The University of Chicago Press. 1997.pgs. 502-504.
LAURENCE, W.F.; COCHRANE, M.A. Synergystic effects in fragmented landscapes. Conservation Biology. Vol. 15, 2001, pgs. 1488-1489.
LI, Z.; ROGERS, E. Habitat quality and activity budgets of white-headed langurs in Fusui, China. International Journal of Primatology. Vol. 25 (1), 2004, pgs. 41-54.
MARIANO-NETO, E. As florestas do Planalto da Conquista e regiões adjacentes. Relatório técnico não publicado. 2004.
MARSH., L. The nature of fragmentation. In: Primates in fragments: ecology and conservation. Marsh, L. (ed.). New York: Kluwer Academic/Plenum 2003. pgs. 1-10.
MENDONÇA, J.R.; DE CARVALHO, A.M.; MATTOS SILVA L.A.; THOMAS, W.W. 45 Anos de Desmatamento no Sul da Bahia, Remanescentes da Mata Atlântica -1945, 1960, 1974, 1990. Projeto Mata Atlântica Nordeste, CEPEC, Ilhéus, Bahia, Brasil. 1994.
MENON, S.; POIRIER, F.E. Lion-tailed macaques (Macaca silenus) in a disturbed forest fragment: activity patterns and time budget. International Journal of Primatology. Vol. 17, 1996, pgs. 969-985.
MMA – MINISTÉRIO DO MEIO AMBIENTE. Lista Nacional das Espécies da Fauna Brasileira Ameaçadas de Extinção. Disponível em www.mma.gov.br/port/sbf/fauna/index, acesso em 14 set. 2007.
OATES, J.F. Food distribution and primate behaviour. In: Primates Societies. B.B. Smuts D.L. Cheney R.M. Seyfarth R.W. Wrangham T.T. Struhsaker, (Ed). University of Chicago Press., Chicago. 1987, Pgs. 197-209
OLIVEIRA, L.C.; FERNANDEZ, F.A.S.; SCHITTINI, G.M; PASSAMANI, M. Uso de
fragmentos pequenos de Mata Atlântica pelo mico-leão-dourado, Leontopithecus rosalia. In: A primatologia no Brasil Volume 8. Mendes, S.L e Chiarello, A.D. (editors). Vitória, ES. 2004.
OLIVER, W.L.R.; SANTOS I.B. Threatened endemic mammals of the Atlantic Forest region
of south-east Brazil. Wildlife Preservation Trust, Special Scientific report n°. 4. 1991.
PASSOS, F.C. Padrão de atividade, dieta e uso do espaço em um grupo de mico-leão preto (Leontopithecus chrysopygus) na Estação Ecológica dos Caetetus, SP. Tese (doutorado). Universidade Federal de São Carlos. 1997.
PASSOS, F.C. Seed dispersal by black lion tamarin, Leontopithecus chrysopygus (Primates, Callitrichidae), in southeastern Brazil. Mammalia. Vol. 61 (1), 1997b. Pgs. 109-111.

Referências
92
PASSOS, F.C. Dieta de um Grupo de Mico-Leão-Preto, Leontopithecus chrysopygus
(Mammalia, Callitrichidae), na Estação Ecolégica dos Caetetus, São Paulo. Revista Brasileira de Zoologia. Vol. 16, 1999, pgs. 269-278.
PASSOS,F.C E KIM, A.C. Néctar feeding on Mabbea fistulifera Mart. (Euphorbiaceae) by
black lio tamarins, Leontopithecus chrysopygus Mikan, 1983 (Callitrichidae, during the dry season of southeastern Brazil. Mammalia. Vol. 63 (4), 1999, pgs 519-521.
PIMLEY, E.R.; BEARDER, S.K.; DIXSON, A.F. Home range analysis of Perodicticus potto
edwardsi and Sciurocheirus cameronensis. International Journal of Primatology. Vol. 26 (1), 2005, pgs. 191-206.
PERES, C.A. Costs and benefits of territorial defense in wild golden lion tamarins,
Leontopithecus rosalia. Behavior Ecology and Sociobiology. Vol 25. 1989. pgs 227-223.
PINTO, L.P. de S.. Distribuição Geográfica, População e Estado de Conservação do Mico-Leão-da-Cara-Dourada, Leontopithecus chrysomelas (Callitrichidae, Primates). Dissertação de mestrado, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais. 1994.
PINTO, L.P. de S. and RYLANDS, A.B.. Geographic distribution of the golden-headed lion
tamarim, Leontopithecus chrysomelas: Implications for its management and conservation. Folia Primatologica. Vol. 68, 1997, pgs. 161-180.
POULSEN, J.R.; CLARK, C.J.; SMITH, T.B. Seasonal variation in the feeding ecology of the
grey-cheeked mangabey (Lophocebus albigena) in Cameroon. American Journal of Primatology. Vol. 54 (2), 2001, pgs. 91-105.
POWEL, R.A. Animal home ranges and territories and home ranges estimator. In: Research
techniques in animal ecology: controversies and consequences. Bioitani, L., Fuller, T.K. (Orgs.). New York, Columbia University Press. 2003. Pgs 65-110.
PRADO, P.I. et al. (Orgs.). Corredor de Biodiversidade da Mata Atlântica do Sul da
Bahia. Publicação em CD room. Ilhéus. IESB/CI/CABS/UFMG/UNICAMP. 2003. PRADO, F.; VALLADRES-PÁDUA, C. Ecologia alimentar de um grupo de mico-leão-da-
cara-preta, Leontopithecus caissara (Primates: Callithrichidae), no Parque Nacional de superagui, guaraqueçaba – PR, Brasil. In: A Primatologiua no Brasil, Vol. 8. Mendes, S.L. e Chiarrello, A.G. (Org.). Vitória, 2004. Pgs. 145-154.
PRIMACK, R.B.; RODRIGUES, E. Biologia da Conservação. Londrina. 2005. 325p. PROCÓPIO-DE-OLIVEIRA, P. Ecologia alimentar, dieta e área de uso d micos-leões
dourados (Leontopithecus chrysomelas) translocados e a sua relação com a distribuição espacial e temporal de recursos alimentares na Reserva Biológica União, RJ. Tese (Doutorado). Universidade Federal de Minas Gerais, Belo Horizonte. 2002.

Referências
93
RABOY, B.E.; BACH, A.; DIETZ, J.M. Birth seasonality and infant survival in GHLT. Tamarin Tales. Vol. 5. 2001. Pgs.3-4.
RABOY, B.E.. The Ecology and Behavior of Wild Golden-Headed-Lion-Tamarins
(Leontopithecus crysomelas). Tese (doutorado), University o Maryland. 2002, 160p. RABOY, B.E.; DIETZ, J.M.. Diet, foraging, and use of space in wild golden-headed lion
tamarins American Journal of Primatology. Vol. 63(1), 2004, Pgs. 1-15. RABOY, B.E., CHRISTMAN, M.C., DIETZ, J.M. The use of degraded and shade cocoa
forests by endangered golden-headed lion tamarins Leontopithecus chrysomelas Oryx. Vol. 38(1), 2004, Pgs. 75-83.
RALLS, K.; HARVEY, P.H.; LYLES, A.M. Inbreeding in natural populations of birds and
mammals. In: Conservation Biology: the science of scarcity and diversity. Soulé, M.E. (Ed.). Sinauer, Sunderland, 1986, pgs. 35-56.
RODRIGUES-VARGAS, A.R. Analysis of the hypothetical population structure of the squirrel monkey (Saimiri oerstedii) in Panama. In: In: Primates in fragments: ecology and conservation. Marsh, L. (ed.). New York: Kluwer Academic/Plenum 2003. pgs. 53-62.
RYLANS, A.B. The Behaviour and Ecology of three Species of Marmosets and tamarins (Callitrichidae, Primates) in Brazil. Tese de Doutorado, University of Cambridge, UK. 1982.
RYLANDS, A.B. Sympatric brazilian callitrhchids: the black tufted-ear marmoset, Callithrix
kuhli, and the golden-headed lion tamarin, Leontopithecus chrysomelas. Journal of Human evolution. Vol 18, 1989, 679-695.
RYLANS, A.B. Habitat and the evolution of social and reproductive behavior in Callitrichidae. American Journal of Primatology. Vol. 38(1)., 1996, Pgs: 5-18.
RYLANDS, A.B.; DA FONSECA, G.A.B.; LEITE, Y.L.R.; MITTERMEIER, R.A. Primates
of the Atlantic forest: Origin, distribution, endemism, and communities. In: Adaptative radiation of the neotropical primates. Norconk, M.A., Editors. New York: Plenum Press. 1996. Pgs: 21-51.
RYLANDS, A.B, SCHNEIDER, H., LANGGUTH, A., MITTERMEIER, R.A, GROVES,
C.P., RODRIGUES-LUNA, E. An assessment of the diversity os New World Primates. Neotropical Primates. Vol. 8 (2), 2000, Pgs. 61-93.
RYLANDS, A.B., KIERULFF, M.C.M. e PINTO, L.P.de S. Distribution and status of lion
tamarins. In D.G. Kleiman and A.B. Rylands (eds.), Lion Tamarins Biology and Conservation, 2002, Pgs. 42-70.
SILVEIRA, L.F. Espécies de aves ameaçadas e quase ameaçadas de extinção registradas
nos fragmentos entre Boa Nova e Itambé, Bahia. Relatório técnico não publicado. 2003.

Referências
94
STEVENSON, M.F. AND RYLANDS, A.B. The marmosets, genus Callithrix. In: Ecology and behavior of neotropical primates. Mittermeier A.B. Rylands A.F. Coimbra-Filho G.A.B. da Fonseca, Editors. Washington, DC: World Wildlife Fund. 1988 Pgs. 131-222.
STRIER, K.B. Activity budgets of woolly spider monkeys, or muriquis (Brachyteles
arachnoides). American Journal of Primatology. Vol 1, 1987, pgs.385-395. TABARELLI, M.; SILVA, J.M.C.; GASCON, C. Forest fragmentation and the
impovershment of neotropical forests. Biodiversity and Conservation. Vol. 13, 2004, pgs. 1419-1425,
TERBORGH, J. Five New World Monkeys: a study in comparative ecology. Princeton
University Press, Princeton, New Jersey. 1983, 260 pgs. TERBORGH, J.; LOPES, L.; TELLO, J.; YU, D.; BRUNI, A.R. Transitory states in relaxing
ecosystems of land bridge islands. In Tropical forest remnants. Ecology, management and conservation of fragmented communities. Laurance, W.F. and Bierregaard, R.O.Jr. (editors). The University of Chicago Press. 1997.pgs. 256-274.
TURTON, S.M.; FREIBURGER, H.J. Edge and aspects effects on the microclimate of a
small tropical forest remnant on the Atherton Tableland, Northeastern Australia. In Tropical forest remnants. Ecology, management and conservation of fragmented communities. Laurance, W.F. and Bierregaard, R.O.Jr. (editors). The University of Chicago Press. 1997.pgs. 45-54.
VALLADARES-PÁDUA, C. The ecology, behaviour and conservation of the black lion
tamarin (Leontopithecus chrysopygus, Mikan, 1823). Tese (doutorado). University of Florida, Gainesville, Florida. 1993.
VALLADARES-PÁDUA, C.; BALLOU, J.D.; MARTINS, C.S.; CULLEN, L.JR.
Metapopulation management for the conservation of black lion tamarins. In D.G. Kleiman and A.B. Rylands (eds.), Lion Tamarins Biology and Conservation, 2002, Pgs. 301-314.
VASEY, N. Activity budgets and activity rhythms in red ruffed lemurs (Varecia rubra) on the
Masaola península, Madagascar: seasonality and reproductive energetics. American Journal of Primatology. Vol. 63, 2005, pgs. 23-44.
VINHA, S.G. da, RAMOS, T.J.S. e HORI, M. Inventário Florestal. In: Diagnóstico Socioeconômico da Região Cacaueira, vol 7: Recursos Florestais. Comissão Executiva do Plano da Lavoura Cacaueira e Instituto Interamericano de Ciências Agrícolas – OEA. Ilhéus, Bahia, Brasil. 1976.
WONG, N.P.; SICOTTE, P. Activity budgets and ranging patterns ofColobus vellerosus in forest fragments in Central Ghana. Folia Primatologica. Vol. 78, 2007, pgs. 245-254.
WORTON, B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology. Vol. 70, 1989, pgs. 164-168.

Referências
95
ZAR, J. Biostatiscal Analisys. USA: Prentice Hall, 1966.
ZHOU, Q.H.; WEI, F.W.; HUANG, C.M.; LI, M.; REN, B.P.; LUO, B. Seasonal variation in the activity patterns and time budgets of Trachypithecus francoisi in the Nonggang Nature Reserve, China. International Journal of Primatology. Vol. 28 (3), 2007, 657-671.

96
ANEXO I: Espécies monitoradas nas trilhas de fenologia e número de indivíduos de cada espécie.
FAMÍLIA (APG) FAMÍLIA (Cronquist) Espécie* N
ANACARDIACEAE ANACARDIACEAE Loxopterygium sp. 5 Myracrodruon macrocalyx Engl. 27 Spondias venulosa Mart. ex Engl. 1 sp.1 4 BIGNONIACEAE BIGNONIACEAE Handroanthus chrysotrichus (Mart. ex DC.) Mattos 2 Tabebuia elliptica (DC.) Sandwith 4 Tabebuia roseo-alba (Ridl.) Sandwith 1 BORAGINACEAE BORAGINACEAE Cordia anabaptista Cham. 4 CAPPARACEAE CAPPARACEAE Capparis flexuosa L. S. L. 4 Capparis nectares Vell. 5 CELASTRACEAE CELASTRACEAE Maytenus sp. 6 CLUSIACEAE CLUSIACEAE Garcinia gardneriana (Planch. & Triana) Zappi 1 COMBRETACEAE COMBRETACEAE Terminalia sp. 9 ERYTHROXILACEAE ERYTHROXILACEAE Erythoxyllum mikanii Peyri: Vel aff. 1 EUPHORBIACEAE EUPHORBIACEAE cf. Croton 1 Sebastiania brasiliensis 1 sp. 1 2 sp. 2 1 sp. 3 39 FABACEAE CAESALPINIACEAE Caesalpinia ferrea Mart. 2 Dialium sp. 2 Goniorrachis marginata Taub. 54 Macrolobium sp. 1 Melanoxylon brauna Schott 1 Peltogyne pauciflora Benth. 11 FABACEAE FABACEAE Andira sp. 1 Centrolobium microchaete 2 Dalbergia sp. 6 Lonchocarpus cultratus (Vell.) M. c. Lima 5 Machaerium acutifolium 4 Platmyscium sp. 1 Zollernia latifolia Benth. 2 FABACEAE MIMOSACEAE Acacia sp. 2 cf. Piptadenia sp. 71 Inga sp. 1 3 Inga sp. 2 1 Piptadenia cf. moniliformis Benth. 26 Stryphnodendron sp. 1 LECYTHIDACEAE LECYTHIDACEAE Cariniana sp. 4 MALVACEAE BOMBACACEAE Ceiba ventricosa 10 Chorisia insignis 1 MALVACEAE STERCULIACEAE Guazuma ulmiflora Lam. 2 MELIACEAE MELIACEAE Guarea cf. guidonea (L.) Sleumer. 3 Guarea macrophylla Vell. 10 sp. 1 2 Trichilia casaretti C.DC. 1 Trichilia cf. hirta 3 Trichilia silvatica C. DC. 7

97
ANEXO I: Continuação MORACEAE MORACEAE Ficus bahiensis 1 MYRTACEAE MYRTACEAE Campomanesia sp. 1 Eugenia candolleana Dc. 10 Eugenia platyphylla O. Berg. 2 Myrcia acuminatissima O. Berg. 5 Myrcia falax (Rich.) DC. 1 sp. 1 1 sp. 2 1 sp. 3 3 NYCTAGINACEAE NYCTAGINACEAE Guapira venosa (Choisy) Lundll. 6 OLACACEAE OLACACEAE Heisteria sp. 2 PHYLLANTACEAE EUPHORBIACEAE Margaritaria nobilis L.f. 1 POLYGONACEAE POLYGONACEAE Coccoloba sp. 2 PUTRANJIVACEAE EUPHORBIACEAE Drypetes sp. 3 RUBIACEAE RUBIACEAE Alseis floribunda Schott. 6 Coutaria hexandra 2 Tocoyena sp. 1 Galipia rimiplicilelis 1 Zanthoxylum nemorale Mart. 1 SALICACEAE FLACOURTIACEAE Banara kuhlmanii 4 Casearia melliodora Eichler 2 SAPINDACEAE SAPINDACEAE Cupania cff. racemosa 1 SAPOTACEAE SAPOTACEAE Micropholis guianensisis 3
* 23 indivíduos não foram identificado
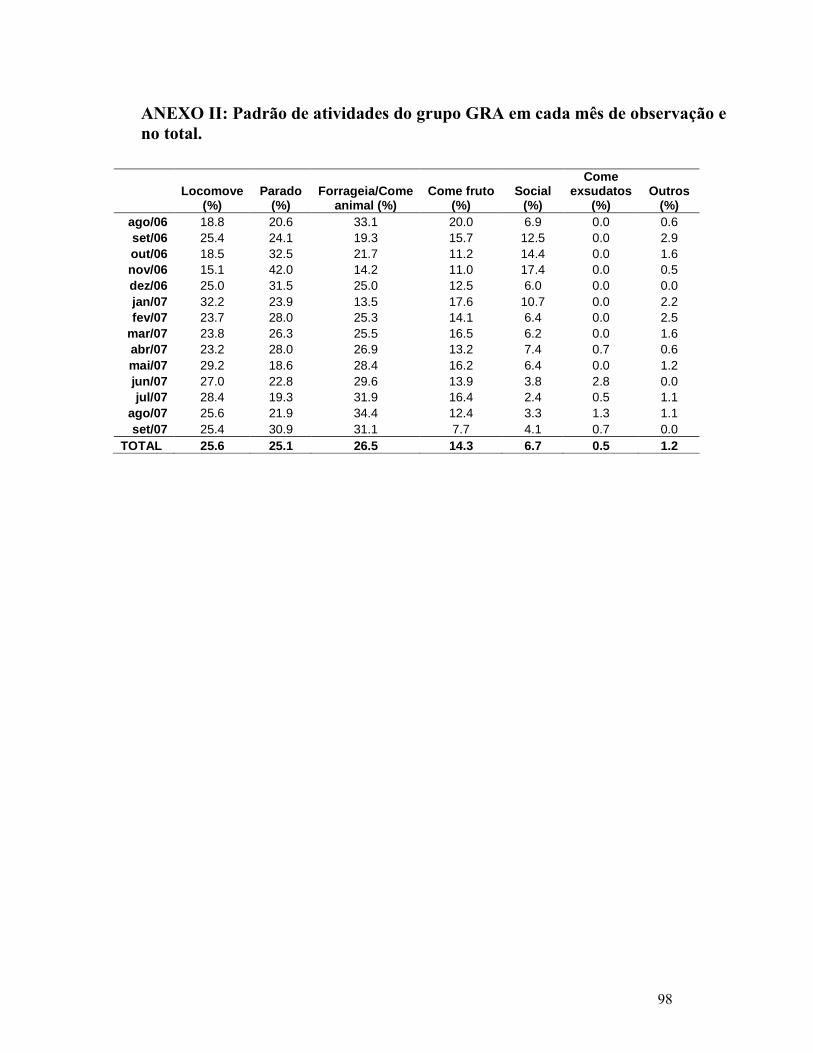
98
ANEXO II: Padrão de atividades do grupo GRA em cada mês de observação e no total.
Locomove
(%) Parado
(%) Forrageia/Come
animal (%) Come fruto
(%) Social
(%)
Come exsudatos
(%) Outros
(%) ago/06 18.8 20.6 33.1 20.0 6.9 0.0 0.6 set/06 25.4 24.1 19.3 15.7 12.5 0.0 2.9 out/06 18.5 32.5 21.7 11.2 14.4 0.0 1.6 nov/06 15.1 42.0 14.2 11.0 17.4 0.0 0.5 dez/06 25.0 31.5 25.0 12.5 6.0 0.0 0.0 jan/07 32.2 23.9 13.5 17.6 10.7 0.0 2.2 fev/07 23.7 28.0 25.3 14.1 6.4 0.0 2.5
mar/07 23.8 26.3 25.5 16.5 6.2 0.0 1.6 abr/07 23.2 28.0 26.9 13.2 7.4 0.7 0.6 mai/07 29.2 18.6 28.4 16.2 6.4 0.0 1.2 jun/07 27.0 22.8 29.6 13.9 3.8 2.8 0.0 jul/07 28.4 19.3 31.9 16.4 2.4 0.5 1.1
ago/07 25.6 21.9 34.4 12.4 3.3 1.3 1.1 set/07 25.4 30.9 31.1 7.7 4.1 0.7 0.0
TOTAL 25.6 25.1 26.5 14.3 6.7 0.5 1.2