associaÇÃo de polimorfismos genÉticos com a...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DE CAMPINAS
FACULDADE DE CIÊNCIAS MÉDICAS
EMILIA ANGELA SIPPERT
ASSOCIAÇÃO DE POLIMORFISMOS GENÉTICOS COM A ALOIMUNIZAÇÃO ERITROCITÁRIA EM PACIENTES PORTADORES DE ANEMIA FALCIFORME
CAMPINAS
2016

EMILIA ANGELA SIPPERT
ASSOCIAÇÃO DE POLIMORFISMOS GENÉTICOS COM A ALOIMUNIZAÇÃO ERITROCITÁRIA EM PACIENTES PORTADORES DE ANEMIA FALCIFORME
Tese apresentada à Faculdade de Ciências Médicas da Universidade Estadual de Campinas como parte dos requisitos exigidos para a obtenção do título de Doutora em Ciências na área de concentração Clínica Médica.
ORIENTADORA: LILIAN MARIA DE CASTILHO ESTE EXEMPLAR CORRESPONDE À VERSÃO FINAL DA TESE DEFENDIDA PELA ALUNA EMILIA ANGELA SIPPERT, ORIENTADA PELA PROFa. DRa. LILIAN MARIA DE CASTILHO.
CAMPINAS
2016


BANCA EXAMINADORA DA DEFESA DE DOUTORADO
EMILIA ANGELA SIPPERT
ORIENTADORA: LILIAN MARIA DE CASTILHO
MEMBROS:
1. PROF(A). DR(A). LILIAN MARIA DE CASTILHO
2. PROF(A). DR(A). ANA MARIA SELL
3. PROF(A). DR(A). CARINE PRISCO ARNONI
4. PROF(A). DR(A). NICOLA AMANDA CONRAN ZORZETTO
5. PROF. DR. FERNANDO FERREIRA COSTA
Programa de Pós-Graduação em Clínica Médica da Faculdade de Ciências Médicas
da Universidade Estadual de Campinas
A ata de defesa com as respectivas assinaturas dos membros da banca examinadora
encontra-se no processo de vida acadêmica do aluno.
Data: 21/01/2016

DEDICATÓRIA
A minha mãe por todo amor, compreensão e
incentivo, pelos esforços em todas as etapas da
minha vida! Por acreditar e confiar em mim!

AGRADECIMENTOS
Agradeço a Deus, pelas infinitas bênçãos concedidas e pela força nos
momentos difíceis.
Agradeço à minha orientadora Professora Doutora Lilian Castilho por
acreditar em minha capacidade, por me incentivar e me dar força em todos os
momentos! Obrigada pela contribuição com o meu crescimento científico, acadêmico
e pessoal! Agradeço a sua dedicação como orientadora, a maneira compreensiva e
amiga com a qual sempre me atendeu e orientou. O seu direcionamento foi
essencial para a realização deste trabalho. Agradeço por todas as oportunidades de
crescimento que me proporcionou.
À FAPESP (Fundação de amparo a pesquisa do Estado de São Paulo)
pela concessão da bolsa de doutorado Processo nº 2013/01426-1 (EAS) e pelo
auxílio financeiro para o desenvolvimento deste projeto Processos nº 2012/04651-3
(LMC) e nº 2014/00984-3 (FFC).
Ao INCTs (Instituto de Ciência e Tecnologia do Sangue) pelo apoio
financeiro e ao Hemocentro da Unicamp, pela estrutura adequada para a realização
do trabalho.
Aos membros titulares da banca: Dra. Ana Maria Sell, Dra. Carine Arnoni,
Dra. Nicola Amanda Conran Zorzetto, Dr. Fernando Costa e aos membros suplentes:
Dra. Flavia Roche Moreira Latini, Dr. José Mauro Kutner e Dra. Andrana Karla
Calgarotto pelo tempo dedicado, para a leitura e discussão deste trabalho.
À minha mãe pelo seu apoio e palavras de coragem nos momentos que
mais precisei, por confiar em mim e por estar sempre presente. Obrigada Mãe! TE
AMO!
Ao meu grande amor Henrique, que sempre me apoiou e compreendeu
minha ausência! Obrigada pelo carinho que sempre teve comigo e pela sua
paciência.

Às minhas irmãs Luciane e Fabiane que muito me incentivaram durante o
desenvolvimento deste trabalho, obrigada pelas boas energias transmitidas e pelas
orações!
À Sueli, Mario, Sâmia e Ary que acompanharam todos os meus passos
durante esses anos, obrigada pelo incentivo, força, carinho e atenção!
À minha amiga Josielle que sempre esteve do meu lado me apoiando e
me dando coragem para enfrentar todos os desafios que apareceram durante o
doutorado.
Aos amigos da Pós-graduação e do laboratório: Ane, Beatriz, Carol,
Mayra e Dr. Vagner pela convivência harmoniosa, pelo aprendizado e incentivo.
À Marcela pela ajuda na seleção dos pacientes e nas análises
laboratoriais. Muito Obrigada!
Aos amigos Andrana, Matheus, Irene e Ana Leda que me ajudaram a
entender o mundo da citometria. Muito obrigada pela atenção e ajuda!
À Dra. Simone Gilli e ao Dr. Marcelo Addas que ajudaram no
delineamento do projeto e sempre contribuíram com valiosas sugestões para a
melhoria do mesmo.
Ao Dr. Jordão pela amizade, atenção, incentivo e contribuições científicas
prestadas durante o desenvolvimento deste trabalho.
À Ucha e à Dulcineia que compartilharam gentilmente seus ensinamentos
e sempre estiveram prontas em ajudar, sempre me apoiando e incentivando durante
o trabalho.
Aos pacientes que com muita paciência e boa vontade colaboraram com o
desenvolvimento deste trabalho.
À todos do Laboratório de Imunogenética da Universidade Estadual de
Maringá por todo apoio que me foram prestados desde o mestrado e, especialmente
à Professora Doutora Jeane L. Visentainer, à Camila Rodrigues e ao Hugo Vicentin,
pela colaboração para realização desse trabalho.

Agradeço a Professora Doutora Ana Maria Sell que me introduziu nos
estudos de Imunohematologia e Imunogenética, muito obrigada por compartilhar seu
conhecimento comigo desde o Mestrado e pelas valiosas discussões que
engrandeceram este trabalho.
À todos os funcionários do Hemocentro e a todos aqueles que
indiretamente contribuíram para a realização deste trabalho. Muito obrigada!

Trabalho realizado no Laboratório de Biologia Molecular de Grupos
Sanguíneos Eritrocitários do Hemocentro da UNICAMP, com auxílio financeiro da
FAPESP Processos nº 2012/04651-3 (LMC), nº 2013/01426-1 (EAS) e nº
2014/00984-3 (FFC), e do Instituto Nacional de Ciências e Tecnologia do Sangue
(INCTs).

RESUMO
A aloimunização eritrocitária é uma das sérias complicações da terapia
transfusional em pacientes com Anemia Falciforme (AF). A presença de
aloanticorpos além de dificultar a obtenção de sangue compatível para esses
pacientes pode resultar a reações transfusionais hemolíticas tardias (RTHTs) e na
formação de autoanticorpos. A maioria dos anticorpos desenvolvidos por estes
pacientes são direcionados aos antígenos do sistema Rh. No entanto, diferenças na
resposta imunológica de pacientes transfundidos que desenvolvem aloanticorpos
(respondedores) e dos que não desenvolvem anticorpos (não respondedores) não
são totalmente conhecidas. A hipótese é de que a susceptibilidade a aloimunização
seja governada por vários fatores inclusive pela diversidade genética entre os
indivíduos. O objetivo geral deste estudo foi identificar o perfil genético de grupos
sanguíneos de 161 pacientes com doença falciforme em regime de transfusão
crônica (67 aloimunizados e 94 não aloimunizados) e as possíveis associações de
polimorfismos presentes em genes imunologicamente relevantes (genes HLA e de
citocinas) com a aloimunização eritrocitária. A genotipagem eritrocitária foi realizada
pela técnica de microarray com os Kits HEA BeadChipTM, RHD BeadChipTM e RHCE
BeadChipTM (BioArray Solutions). Os polimorfismos em genes de citocinas foram
analisados pelas técnicas de PCR, PCR-RFLP e pelo ensaio de genotipagem
TaqMan (Applied Biosystems), enquanto a genotipagem HLA de classe II foi
realizada por PCR-SSO (One Lambda). Para a determinação do equilíbrio de Hardy-
Weinberg e das frequências alélicas e genotípicas foi utilizado o software Arlequin e
a comparação das frequências dos polimorfismos foi analisada pelo teste exato de
Fisher. A determinação do perfil eritrocitário dos pacientes revelou que o fenótipo C–
E–, K–, Fy(a–), Jk(b–), S– é prevalente. Também constatou-se que 65% dos
pacientes com anticorpos anti-Rh possuíam antígenos Rh variantes que levaram ao
desenvolvimento de anticorpos clinicamente significantes. Além disso, a análise dos
polimorfismos em genes de citocinas mostrou que o alelo A e o genótipo GA do
polimorfismo TNFA–308G/A foram mais frequentemente encontrados em pacientes
aloimunizados do que em não aloimunizados (alelo A: 16,4% vs 6,8%; P=0,004;
genótipo GA: 32,8% vs 11,7%; P=0,0021). As frequências do alelo IL1B–511T e
dos genótipos IL1B–511TT e CT foram significativamente maiores em aloimunizados
quando comparado com os pacientes não aloimunizados (alelo T: 53,0% vs 37,5%;

P= 0,0085; genótipos CT + TT: 81,82% vs 60,87%; P=0,0071). Com relação aos
alelos HLA classe II, encontrou-se uma alta frequência do alelo HLA-DRB1*15 em
pacientes aloimunizados aos antígenos Rh comparados aos não aloimunizados
(15,63% vs 6,98%; P=0,044). Os resultados demonstraram que polimorfismos nos
genes TNFA, IL1B e HLA-DRB1 estão associados com um maior risco de
desenvolvimento de anticorpos anti-eritrocitários em pacientes com doença
falciforme. Os achados deste trabalho podem contribuir futuramente para melhorar
as estratégias transfusionais e terapêuticas para os pacientes com doença
falciforme.
Palavras-chave: Anemia Falciforme, antígenos HLA, citocinas, antígenos de grupos
sanguíneos, anticorpos.

ABSTRACT
Red blood cell (RBC) alloimmunization is one of the serious complications
associated with transfusion therapy in patients with sickle cell disease (SCD). The
presence of the alloantibodies also hinders the provision of compatible blood for
these patients and can lead to delayed hemolytic transfusion reactions (DHTR) and
autoantibody formation. Most developed antibodies are directed to the Rh antigens.
However, differences in the immune response of transfused patients who develop
alloantibodies (responders) and who do not develop antibodies (non-responders) are
not completely known. The hypothesis is that susceptibility to RBC alloimmunization
is governed by many factors including genetic diversity among individuals. The aim of
this study was to identify blood group alleles of 161 patients with SCD in chronic
transfusion therapy (67 alloimmunized and 94 non-alloimmunized) and associations
of polymorphisms on immunologically relevant genes (HLA genes and cytokines) with
RBC alloimmunization. RBC genotyping was performed by microarray analysis with
HEA BeadChipTM, RHD BeadChipTM and RHCE BeadChipTM (Immucor). Cytokine
gene polymorphisms were analysed by PCR, PCR-RFLP and TaqMan genotyping
assay (Applied Biosystems) while, HLA class II typing was performed using PCR-
SSO (One Lambda). The Hardy-Weinberg equilibrium and the allelic and genotypic
frequencies were obtained by Arlequin software. Allele and genotype frequencies
were compared using the Fisher’s exact test. The RBC antigen profile determination
revealed that the C–E–, K–, Fy(a–), Jk(b–), S– phenotype is prevalent. Also it was
also found that 65% of patients with alloantibodies anti-Rh had Rh antigens variants
which led to the development of clinically significant antibodies. Furthermore,
analysis of cytokine gene polymorphisms revealed increased percentage of A allele
and GA genotype of the TNFA–308G/A in alloimmunized compared to non-
alloimmunized patients (A allele: 16.4% vs 6.8%; P=0.004; GA genotype: 32.8% vs
11.7%; P=0.0021). Moreover, IL1B–511T allele and IL1B–511TT and CT genotype
frequencies were significantly higher in alloimmunized than in non-alloimmunized
patients (T allele: 53.0% vs 37.5%; P=0.0085; CT + TT genotypes: 81.82% vs
60.87%; P=0.0071). In relation to HLA class II, we found a higher frequency of HLA-
DRB1*15 in alloimmunized patients to Rh antigens compared to non-alloimmunized
(15.63% vs 6.98%; P=0.044). Results showed that TNFA, IL1B and HLA-DRB1 gene
polymorphisms are associated with an increased risk of developing RBC antibodies

in Brazilian patients with SCD. These findings can contribute, in the future, to improve
transfusion and therapeutic strategies for patients with SCD.
Key words: Sickle Cell Disease, HLA antigens, cytokines, blood group antigens,
antibodies.

LISTA DE FIGURAS
Figura 1 - Fisiopatologia da Doença Falciforme.. ..................................................... 22
Figura 2 - Mecanismo de aloimunização eritrocitária.. ............................................. 25
Figura 3 - Complexo gênico HLA presente no cromossomo 6.. ............................... 30
Figura 4 - Estrutura das moléculas HLA classe I e II.. .............................................. 31
Figura 5 - Etapas do protocolo de genotipagem de alelos de grupos sanguíneos em larga escala pela plataforma HEA BeadChipTM.. ....................................................... 49
Figura 6 - Etapas da tipificação HLA pela técnica de PCR-SSO.. ............................ 52
Figura 7 - Géis de eletroforese evidenciando os padrões polimórficos dos genes estudados.. ................................................................................................................ 55
Figura 8 - Discriminação alélica do SNP IL10–819T/C obtido por PCR em Tempo real.. .......................................................................................................................... 57
Figura 9 - Frequências e especificidades dos aloanticorpos desenvolvidos pelos pacientes com Anemia Falciforme politranfundidos.. ................................................ 61

LISTA DE TABELAS
Tabela 1 - Sequências das sondas utilizadas para determinação da zigozidade RHD. .................................................................................................................................. 42
Tabela 2 - Alelos e SNPs presentes no HEA BeadChipTM ........................................ 48
Tabela 3 - Variantes RHD analisadas pelo RHD BeadChipTM .................................. 50
Tabela 4 - Sequência de primers para detecção dos SNPs IL17F 7488A/G, IL17A –197G/A, IL1B–511C/T, IL10(–1082A/G, –592 A/C), TNFA–308G/A, condições de ciclagem da PCR, produto da PCR, enzima de restrição utilizada e tamanho dos fragmentos obtidos. ................................................................................................... 54
Tabela 5 - Características da população estudada ................................................... 60
Tabela 6 - Variantes RH encontradas em 13 pacientes com Anemia Falciforme associados com anticorpos clinicamente significativos envolvidos em RTHT ou diminuição da sobrevivência de hemácias transfundidas. ......................................... 65
Tabela 7 - Frequências alélicas e genotípicas dos SNPs TNFA–308G/A (rs1800629) em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles. .......................................................................................................... 68
Tabela 8 - Frequências alélicas e genotípicas do SNP IL1B –511C/T (rs16944) em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles. ............................................................................................................. 69
Tabela 9 - Frequências alélicas e genotípicas do SNP IL10 –1082G/A (rs1800896) em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles. .......................................................................................................... 70
Tabela 10 - Frequências dos diplótipos formados pelos polimorfismos IL10 (–1082G/A, –819T/C, –592C/A) em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles. ................................................ 71
Tabela 11 - Frequências dos diplótipos formados pelos polimorfismos IL4-590C/T e IL4 íntron 3 VNTR em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles. ........................................................................... 72
Tabela 12 - Distribuição das frequências dos grupos alélicos HLA-DRB1 entre pacientes portadores de Anemia Falciforme aloimunizados a antígenos eritrocitários e pacientes não aloimunizados. ................................................................................ 74
Tabela 13 - Frequências dos grupos alélicos DRB1 em pacientes com aloanticorpos contra antígenos Rh (D, C, c, E e/ou e), em pacientes com aloanticorpos não Rh e pacientes não aloimunizados. ................................................................................... 75

LISTA DE ABREVIATURAS E SIGLAS
AF Anemia Falciforme
RTHTs Reações transfusionais hemolíticas tardias
HbS Hemoglobina S
Hb Hemoglobina A
TRALI Lesão pulmonar aguda relacionada à transfusão
APCs Células apresentadoras de antígenos
TCR Receptor de célula T
Treg Células T reguladoras
IFN-γ Interferon gama
IL-10 Interleucina-10
HLA Antígenos Leucocitários Humanos
TNF-α Fator de necrose tumoral-α
MHC Complexo Principal de Histocompatibilidade
IL-2 Interleucina-2
IL-4 Interleucina-4
IL-5 Interleucina-5
IL-6 Interleucina-6
IL-9 Interleucina-9
IL-13 Interleucina-13
IL-17A Interleucina-17A
IL-17F Interleucina-17F
IL-21 Interleucina-21
IL-22 Interleucina-22
IL-8 Interleucina-8
IL-1β Interleucina-1β

TGF-β Fator de transformação do crescimento beta
NFAT Fator nuclear de células T ativadas
Células NK Células natural killer
IL-3 Interleucina-3
GM-CSF Fator estimulador de colônias de macrófagos e granulócitos
FcεRI Receptor de IgE de alta afinidade
IL-15 Interleucina-15
VNTR Número variável de repetições in tanden, do inglês: variable number of tanden repet
DNA Ácido desoxirribonucléico
DO Densidade óptica
eMAP Elongation-mediated multiplex analysis of polymorphisms
FY Sistema de grupo sanguíneo Duffy
HEA BeadChipTM
Plataforma de microarray
PCR Reação em cadeia da polimerase
PCR-AS Reação em cadeia da polimerase alelo específica
PCR-RFLP Análise dos fragmentos da digestão enzimática dos produtos da reação em cadeia da polimerase
PCR-SSO Reação em cadeia da polimerase sequência específica de oligonucleotídeos, do inglês: Sequence-Specific Oligonucleotide
EDTA Ácido etilenodiamino tetra-acético
RHD*Ѱ RHD*pseudogene
dNTP Desoxinucleotídeos
μL Microlitro
mL Mililitro
CTLA-4 Cytotoxic T-Lymphocyte Antigen 4
CCR7 CC-chemokine receptor 7
ng Nanograma

μg Micrograma
pb Pares de bases

SUMÁRIO
1 INTRODUÇÃO ....................................................................................................... 22
1.1 ANEMIA FALCIFORME E ALOIMUNIZAÇÃO ERITROCITÁRIA ........................ 22
1.2 FATORES DE RISCO ASSOCIADOS À ALOIMUNIZAÇÃO ERITROCITÁRIA .. 26
1.2.1 Gênero ...................................................................................................... 26
1.2.2 Imunogenicidade do antígeno ............................................................... 26
1.2.3 Inflamação ............................................................................................... 27
1.2.4 Desregulação Imune ............................................................................... 28
1.2.5 Genes de resposta imune ....................................................................... 28
1.2.5.1 Sistema HLA ....................................................................................... 29
1.2.5.2 Citocinas ............................................................................................. 31
2 OBJETIVOS ........................................................................................................... 38
2.1 OBJETIVO GERAL ............................................................................................. 38
2.2 OBJETIVOS ESPECÍFICOS ............................................................................... 38
3 MATERIAIS E MÉTODOS ..................................................................................... 39
3.1 CASUÍSTICA ....................................................................................................... 39
3.1.1 Grupo de pacientes com Anemia Falciforme ....................................... 39
3.1.2 Grupo controle ........................................................................................ 39
3.1.3 Aspectos éticos da pesquisa ................................................................. 40
3.2 MATERIAIS ......................................................................................................... 40
3.2.1 Kit de extração de DNA ........................................................................... 40
3.2.2 Água livre de nuclease ........................................................................... 40
3.2.3 Etanol absoluto (CH3CH2OH) .................................................................. 40
3.2.4 Taq DNA Polimerase ............................................................................... 40
3.2.5 dNTP 10Mm .............................................................................................. 41
3.2.6 Primers ..................................................................................................... 41
3.2.7 Sondas TaqMan para determinação da zigozidade RHD ..................... 41
3.2.8 Marcador de peso molecular .................................................................. 42
3.2.9 Tampão Tris-EDTA-Borato (TEB) 10X ................................................... 42
3.2.10 Enzimas de restrição ............................................................................ 42
3.2.11 Gel de agarose ...................................................................................... 42
3.2.12 Gel de poliacrilamida 12% .................................................................... 42

3.2.13 Plataformas HEA BeadChipTM, RHCE BeadChipTM e RHD BeadChipTM
........................................................................................................................... 43
3.2.14 Kit de extração Qiaex II ......................................................................... 43
3.2.15 Trizol ....................................................................................................... 43
3.2.16 Superscript II Kit.................................................................................... 43
3.2.17 Big Dye ................................................................................................... 44
3.2.18 Marcador de peso Molecular Low Mass .............................................. 44
3.2.19 Kit LABType® SSO................................................................................ 44
3.2.20 Ensaios TaqMan® para genotipagem de SNPs em genes de
citocinas............................................................................................................ 44
3.2.21 Mastermix de genotipagem Taqman® ................................................. 44
3.3 MÉTODOS .......................................................................................................... 45
3.3.1 Dados transfusionais .............................................................................. 45
3.3.2 Avaliação sorológica .............................................................................. 45
3.3.3 Coleta da amostra e Extração de DNA .................................................. 45
3.3.4 Análise molecular de antígenos eritrocitários ...................................... 46
3.3.4.1 Genotipagem de grupos sanguíneos em larga escala ........................ 46
3.3.4.2 Genotipagem RHD e RHD*Ѱ ............................................................. 49
3.3.4.3 Determinação da zigozidade do gene RHD ........................................ 49
3.3.4.4 Identificação de variantes RHD e RHCE ............................................ 49
3.3.4.5 Sequenciamento do DNA e Clonagem ............................................... 50
3.3.5 Avaliação do significado clínico dos anticorpos anti-Rh em pacientes
portadores de variantes Rh ............................................................................. 51
3.3.6 Genotipagem HLA de classe II (-DRB1, -DQA1, -DQB1) ...................... 51
3.3.7 Genotipagem de polimorfismos em genes de citocinas ..................... 52
3.3.8 Análise estatística ................................................................................... 57
4 RESULTADOS ....................................................................................................... 59
4.1 CARACTERÍSTICAS DA POPULAÇÃO ESTUDADA ......................................... 59
4.2 PERFIL DE ALOIMUNIZAÇÃO ERITROCITÁRIA DOS PACIENTES COM
ANEMIA FALCIFORME ...................................................................................... 60
4.3 PERFIL GENÉTICO DOS GRUPOS SANGUÍNEOS EM PACIENTES COM
ANEMIA FALCIFORME ...................................................................................... 61

4.4 VARIANTES RH E IMUNIZAÇÃO RH EM PACIENTES COM ANEMIA
FALCIFORME ..................................................................................................... 62
4.4.1 Análise sorológica dos antígenos e anticorpos Rh ............................. 62
4.4.2 Análise molecular de variantes RHD e RHCE ....................................... 62
4.4.3 Imunização Rh e significado clínico dos anticorpos Rh ..................... 63
4.5 FREQUÊNCIA DOS POLIMORFISMOS EM GENES DE CITOCINAS E A
ASSOCIAÇÃO COM A ALOIMUNIZAÇÃO ERITROCITÁRIA EM PACIENTES
COM ANEMIA FALCIFORME ............................................................................. 66
4.6 FREQUÊNCIAS DOS GRUPOS ALÉLICOS HLA DE CLASSE II E A
ASSOCIAÇÃO COM A ALOIMUNIZAÇÃO ERITROCITÁRIA EM PACIENTES
COM ANEMIA FALCIFORME ............................................................................. 73
5 DISCUSSÃO .......................................................................................................... 76
5.1 CARACTERÍSTICAS DA POPULAÇÃO ESTUDADA ......................................... 76
5.2 PERFIL DE ALOIMUNIZAÇÃO ERITROCITÁRIA DOS PACIENTES COM
ANEMIA FALCIFORME ...................................................................................... 76
5.3 PERFIL GENÉTICO DOS GRUPOS SANGUÍNEOS EM PACIENTES COM
ANEMIA FALCIFORME ...................................................................................... 77
5.4 VARIANTES RH E IMUNIZAÇÃO RH EM PACIENTES COM ANEMIA
FALCIFORME ..................................................................................................... 78
5.5 INFLUÊNCIA DOS POLIMORFISMOS EM GENES DE CITOCINAS NA
ALOIMUNIZAÇÃO ERITROCITÁRIA EM PACIENTES PORTADORES DE
ANEMIA FALCIFORME ...................................................................................... 79
5.6 INFLUÊNCIA DOS GRUPOS ALÉLICOS HLA DE CLASSE II NA
ALOIMUNIZAÇÃO ERITROCITÁRIA EM PACIENTES COM ANEMIA
FALCIFORME ..................................................................................................... 82
5.7 APLICAÇÃO CLÍNICA DOS RESULTADOS OBTIDOS NA ANÁLISE DOS
POLIMORFISMOS EM GENES DE CITOCINAS E DOS ALELOS HLA............. 84
6 CONCLUSÃO ........................................................................................................ 86
7 REFERÊNCIAS ...................................................................................................... 88
8 APÊNDICE ........................................................................................................... 103
9 ANEXOS .............................................................................................................. 114

22
___________________________________________________________________
1 INTRODUÇÃO 1.1 ANEMIA FALCIFORME E ALOIMUNIZAÇÃO ERITROCITÁRIA
A Anemia Falciforme é um distúrbio genético das hemoglobinas que
ocorre devido a uma mutação de ponto envolvendo a troca do nucleotídeo adenina
pela timina (GAG>GTG), no gene da β-globina, presente no cromossomo 11.
Consequentemente a esta troca ocorre a substituição de um ácido glutâmico por
uma valina na sexta posição da cadeia polipeptídica (glu6val; rs334) e a
subsequente formação de uma hemoglobina anômala – a hemoglobina S (HbS -
α2β2S) (1).
Como resultado desta mutação pontual, a HbS, em seu estado
desoxigenado (desoxi-HbS) e em concentração elevada, é capaz de estabelecer
interações intermoleculares hidrofóbicas que promovem a sua polimerização no
interior da célula, causando a deformação, enrijecimento, diminuição da flexibilidade
(hemácias falcizadas) e diminuição da sobrevida das hemácias levando à hemólise
intravascular crônica e à oclusão da microvasculatura (2,3), como demonstrado na
Figura 1.
Figura 1 - Fisiopatologia da Doença Falciforme. A síntese de HbS é resultado de uma mutação de ponto; quando está desoxigenada, a HbS, forma polímeros no interior da hemácia, deformando-a e tornando-a rígida (formato de foice). O processo de vaso-oclusão, em que participam hemácias, neutrófilos e reticulócitos, e a hemólise intravascular de hemácias são eventos que caracterizam a Anemia Falciforme. Fonte: Adaptado de Steinberg (3).

23
Quando os níveis de oxigênio são restabelecidos ocorre o retorno da
hemácia a sua forma bicôncava original, porém, eventos sucessivos de falcização
podem alterar a estrutura da membrana eritrocitária, tornando as hemácias
irreversivelmente falcizadas (4,5). Estas células apresentam uma alta concentração
de HbS, alto conteúdo de cálcio e pouco potássio, não retornando à forma original
mesmo quando há um aumento da tensão de oxigênio (6).
O quadro clínico dos indivíduos com AF é heterogêneo, com gravidade
elevada, podendo apresentar retardo no crescimento e desenvolvimento, bem como
alterações em vários órgãos, sempre em decorrência da hemólise contínua e de
fenômenos vaso-oclusivos (4). Outras complicações associadas à doença incluem: o
acidente vascular cerebral, a retinopatia, a síndrome torácica aguda, úlceras de
perna, sequestro esplênico, alterações pulmonares, priapismo, infecções, entre
outras (5). Infecções e crises vaso-oclusivas estão entre as principais causas de
hospitalização entre esses indivíduos (4,5).
A transfusão de concentrado de hemácias é uma das terapias utilizada
para prevenir e tratar as complicações agudas e crônicas associadas à doença
falciforme (7) e tem por objetivo aumentar o hematócrito e desta forma aumentar a
oxigenação sanguínea, e assim diminuir a hipóxia tecidual (7).
Entretanto, apesar das transfusões de concentrados de hemácias
diminuírem significativamente a morbidade e mortalidade nesta população de
pacientes (8,9), sua utilização é complicada devido à alta incidência de
aloimunização, uma vez que 18% a 52% dos pacientes tornam-se imunizados (10–
12). A aloimunização eritrocitária pode causar diminuição da sobrevida das
hemácias transfundidas, reação transfusional hemolítica (aguda ou tardia) e
consequentemente um aumento dos riscos inerentes à transfusão como a lesão
pulmonar aguda relacionada à transfusão (TRALI), doenças infecciosas e a
síndrome de hiperhemólise que pode por a vida do paciente em risco (13,14). Além
disso, a presença de aloanticorpos, que muitas vezes está associada com a
presença concomitante de autoanticorpos, pode levar a atrasos na liberação da
transfusão devido à complexidade dos testes para identificação dos anticorpos
formados e a dificuldade de encontrar unidades compatíveis (15).
Adicionalmente, pacientes que se tornam aloimunizados estão 20 vezes
mais propensos a formar anticorpos adicionais após um ou mais eventos de

24
transfusões (16). Reações transfusionais hemolíticas são mais comuns em pacientes
falciformes do que na população geral, mas são difíceis de serem diagnosticadas,
pois os sintomas da doença e os dados laboratoriais que geralmente encontram-se
alterados nos pacientes com hemólise crônica podem mascarar este diagnóstico
(17–19).
Com o objetivo de prevenir a formação de aloanticorpos contra antígenos
de grupos sanguíneos e as consequências negativas subsequentes às RTHTs,
muitos serviços tem adotado o protocolo transfusional de compatibilidade fenotípica
para os antígenos ABO, Rh (D, C, c, E, e) e K (20,21), enquanto outros tem
implementado um protocolo de compatibilidade mais estendida envolvendo os
antígenos Fya/Fyb, Jka/Jkb e Ss (22). A transfusão fenótipo compatível com base no
perfil genotípico dos pacientes falciformes demonstra ser ainda mais eficiente na
prevenção da aloimunização e de RTHTs em pacientes em regime de transfusão
crônica (23–25). No entanto, embora estes programas tenham contribuído
significativamente para a redução da aloimunização, o custo-benefício ainda é
controverso porque nem todos os pacientes desenvolvem anticorpos, e outros
continuam ainda a desenvolver anticorpos contra antígenos de outros sistemas e
principalmente contra variantes do sistema Rh (26,27). Alelos RH alterados que
codificam antígenos D, C e e variantes em grupos Afrodescendentes são
responsáveis pela produção de autoanticorpos e aloanticorpos complexos muitas
vezes dirigidos a antígenos de alta frequência populacional.
A alta frequência de alelos RH alterados devido a grande diversidade
genética do locus RH em Afrodescendentes e as limitações dos métodos sorológicos
para distinguir os antígenos variantes contribuem para a alta taxa de aloimunização
Rh em pacientes com AF (28–30). A genotipagem RH revelou que muitos pacientes
com AF apresentam alelos que codificam antígenos Rh parciais, mas poucas
evidências clínicas ou biológicas relacionadas com aloimunização e reações
transfusionais hemolíticas foram relatadas para todos os alelos RH variantes.
A aloimunização eritrocitária envolve vários passos, tais como: o
reconhecimento do antígeno alogênico pelas células apresentadoras de antígeno
(APCs); o processamento e a apresentação de antígenos por moléculas HLA de
classe II para o receptor de célula T CD4+ (TCR); a ativação de células T CD4+; a
interação de células T e B e a liberação de citocinas que ativam as células B a se

25
diferenciarem em células produtoras de anticorpos (plasmócitos) (13), que secretam
anticorpos específicos para os determinantes antigênicos (Figura 2).
Figura 2 - Mecanismo de aloimunização eritrocitária. I: Reconhecimento do antígeno pelas células dendríticas, II: apresentação do peptídeo pelas moléculas HLA de classe II ao receptor de linfócitos T CD4
+ (TCR), III: ativação de linfócitos T CD4
+, IV: interação de células T e B e
liberação de citocinas, V: Citocinas ativam os linfócitos B a diferenciarem-se em plasmócitos que secretam anticorpos específicos aos determinantes antigênicos. Fonte: Adaptado de Yazdanbakhsh et al. (13).
Este processo de aloimunização eritrocitária é desencadeado pela
transfusão de concentrados de hemácias contendo antígenos de grupos sanguíneos
que estão ausentes no receptor (31). No entanto, tem se verificado que alguns
pacientes, mesmo recebendo transfusões antígeno incompatível, não produzem
aloanticorpos eritrocitários, e são chamados de não respondedores, enquanto outros
apesar de receberem transfusões fenótipo compatível para os principais antígenos
clinicamente significantes desenvolvem anticorpos adicionais contra antígenos de
outros sistemas inclusive contra variantes dos sistema Rh e são considerados como
indivíduos respondedores (22,27,31,32).
Devido a estas diferenças na resposta imunológica de pacientes
transfundidos que desenvolvem aloanticorpos (respondedores) e dos que não
desenvolvem anticorpos (não respondedores), diversos estudos têm sido realizados
na tentativa de esclarecer quais são os fatores de risco associados à aloimunização
eritrocitária. Entre os principais fatores estudados estão: o gênero (33–36), a

26
imunogenicidade do antígeno (37), a inflamação (38–41), fatores imunes (42–45) e
fatores genéticos (46,47).
1.2 FATORES DE RISCO ASSOCIADOS À ALOIMUNIZAÇÃO ERITROCITÁRIA 1.2.1 Gênero
As investigações com relação à influência do gênero feminino e masculino
na aloimunização eritrocitária têm mostrado resultados conflitantes. Alguns estudos
têm relatado um risco elevado para as mulheres (32–34), enquanto outros não
mostraram diferença de risco entre homens e mulheres (35). Em uma revisão
sistemática realizada por Verduin et al. (36) em que foram analisados 30 estudos
com relação à influência do gênero na aloimunização eritrocitária observou-se que
as mulheres com doença falciforme apresentaram um aumento do risco relativo para
a produção de aloanticorpos em comparação com os homens. De acordo com os
autores, isso pode ser explicado pela maior exposição das mulheres com doença
falciforme a eventos que podem levar à imunização, como por exemplo, a gravidez
e/ou transfusões sanguíneas (36).
1.2.2 Imunogenicidade do antígeno
Os antígenos eritrocitários variam na sua capacidade de induzir uma
resposta humoral. A imunogenicidade de um antígeno é, em parte, determinada pelo
processamento dos antígenos, o número de peptídeos gerados e a medida em que
estes peptídeos eficientemente estimulam as células T alorreativas (37).
O antígeno D é significativamente mais imunogênico do que os outros
antígenos de grupos sanguíneos e também o mais imunogênico dentro do sistema
Rh. Em parte isso ocorre devido às diferenças de aminoácidos entre a proteína D e a
proteína RhCE. A proteína RhD difere em cerca de 35 aminoácidos da proteína
RhCE e, portanto, a possibilidade de geração de epitopos imunogênicos é muito
maior do que para os alelos caracterizados por uma única diferença de aminoácido
como por exemplo os antígenos Fya e Fyb (37).
Os antígenos K, E, c, Jka e Fya que consistem de apenas um polimorfismo
de nucleotídeo único podem não ser tão imunogênicos quanto o antígeno D, mas
possuem capacidade de imunizar um número significativo de receptores (48). Os

27
outros antígenos menos imunogênicos possuem uma capacidade de imunização
menor que 1% (48,49).
1.2.3 Inflamação
A hipótese de que o aumento da inflamação sistêmica predisponha a
aloimunização eritrocitária foi estudada em modelos animais. Hendrickson et al. (38)
verificaram que o estado inflamatório desencadeia uma melhor apresentação
antigênica por células dendríticas, levando consequentemente a uma maior resposta
proliferativa dos linfócitos T CD4+. Adicionalmente, Smith et al. (41) demonstraram
que a transfusão de concentrados de hemácias, na ausência de inflamação, pode
levar a uma tolerância antigênica. Contudo, em um estudo com animais homozigotos
para o gene da globina βs, não se verificou a associação de inflamação e
aloimunização eritrocitária (40).
Adicionalmente aos modelos em animais, vários estudos em humanos
têm demonstrado que a condição inflamatória encontrada em algumas doenças
predispõe o indivíduo ao desenvolvimento de aloanticorpos. Por exemplo, Ramsey et
al. (50) verificaram uma alta taxa de formação de anticorpos contra antígenos de
baixa incidência e baixa imunogenicidade entre pacientes com doenças autoimunes.
Outro estudo desenvolvido por Papay et al. (51) encontrou que pacientes com
doença inflamatória intestinal apresentaram 2-3 vezes mais chance de desenvolver
aloanticorpos quando comparado com um grupo controle saudável. Um terceiro
grupo (52) encontrou que a taxa de aloimunização em receptores que apresentaram
reação transfusional febril à transfusão de plaquetas era maior do que no grupo
controle que também havia recebido transfusões. Recentemente, um estudo que
avaliou a relação entre a presença de manifestações clínicas do paciente com AF no
momento da transfusão de concentrados de hemácias e a aloimunização eritrocitária
verificou que pacientes com síndrome torácica aguda e crises vaso oclusivas
apresentavam maior risco de aloimunização demonstrando, portanto, uma relação
positiva entre a inflamação do receptor e a aloimunização subsequente a transfusão
de hemácias (53).

28
1.2.4 Desregulação Imune
Além do estado inflamatório do receptor, outros fatores imunes podem
afetar a aloimunização. As células T reguladoras (Tregs) são conhecidas por
suprimir a ativação e as funções efetoras de vários tipos de células e em diferentes
situações (54). Estudos mostraram uma menor atividade de células Tregs, altos
níveis de interferon gama (IFN-γ) e baixos níveis de Interleucina-10 (IL-10) em
indivíduos respondedores quando comparados com indivíduos não respondedores,
especulando-se que a IL-10 pode ter um papel na desativação da apresentação de
antígenos/ativação das células T nestes pacientes (42,43). Além disso, verificou-se
que as células B reguladoras de pacientes com AF aloimunizados e não
aloimunizados diferem na sua capacidade de produzir IL-10 e atenuar a ativação de
monócitos (44).
Outro grupo de pesquisa, no entanto, não encontrou diferenças funcionais
das células Tregs de pacientes aloimunizados e não aloimunizados com AF (45),
mas verificou que pacientes com AF exibem um fenótipo especial de Treg, ou seja,
têm uma proporção elevada de Treg naive (nTreg) expressando CTLA-4 e CD39
(marcadores relacionados com a atenuação da ativação de células T), e uma alta
proporção de Treg ativada (aTreg) com baixos níveis de expressão HLA-DR (menor
atividade supressora). Os autores também sugerem que tanto as células Treg naives
quanto as Tregs ativadas podem apresentar defeitos de migração em pacientes com
AF devido aos seus baixos níveis de expressão CCR7 (45).
1.2.5 Genes de resposta imune
Nos últimos anos, diversos estudos demostraram que fatores genéticos
também podem estar envolvidos na susceptibilidade à aloimunização eritrocitária. Na
AF, foi demonstrado que o polimorfismo rs660 no gene TRIM21 (Ro52) pode ser um
marcador da eficiência da indução de tolerância na primeira infância e
desenvolvimento de competência imunitária a antígenos eritrocitários em pacientes
com AF pré-adolescentes / adolescentes. Neste estudo, foi verificado que a maioria
dos pacientes com genótipo rs660CT desenvolveram anticorpos nos primeiros anos
de vida, enquanto pacientes com genótipo rs660TT o fizeram predominantemente
após os cinco anos de idade (46). Posteriormente, o mesmo grupo de pesquisa

29
verificou que dois SNPs presentes no gene que codifica a molécula envolvida no
sinal de modulação de linfócitos B (gene CD81), podem conferir um maior risco à
aloimunização eritrocitária (47).
Estudos mais recentes envolvendo a pesquisa de um amplo número de
polimorfismos no genoma humano (GWAS - do inglês genome wide association
studies) têm demonstrado diferentes resultados, por exemplo, Milton et al. (55)
encontraram que vários genes de resposta imune podem estar relacionados com a
susceptibilidade à aloimunização eritrocitária, seja a nível antigênico específico ou a
nível global, enquanto Hanchard et al. (56) não encontraram associações
significantes.
Outros genes importantes no contexto da aloimunização eritrocitária
incluem os genes de citocinas e genes do sistema HLA que ainda não foram
completamente estudados em pacientes com AF politransfundidos e que, portanto,
tornaram-se objetos deste estudo.
1.2.5.1 Sistema HLA
O sistema HLA (antígenos leucocitários humanos, do inglês: Human
Leukocyte Antigen) representa um conjunto de genes altamente polimórficos que
desempenham um papel de grande importância no sistema imunológico. As
proteínas codificadas por estes genes são responsáveis pela apresentação de
antígenos aos receptores de linfócitos T e B e estão situadas na membrana celular
de quase todas as células do sistema imune (57).
Os genes responsáveis pela codificação dos antígenos HLA estão
localizados no complexo HLA, que corresponde a um pequeno segmento
cromossômico, no braço curto do cromossomo 6, na região p21.3. O complexo
contém mais de 220 genes que codificam três classes de moléculas, denominadas
classe I, II e III (57).
A região classe I (mais telomérica) contém genes que codificam as
moléculas clássicas de histocompatibilidade HLA-A, -B e -C. A região de classe II
(mais centromérica) contém os genes que codificam as moléculas clássicas HLA
-DR, -DQ e -DP e a região de Classe III que é telomérica à região classe II não
contém genes que codificam moléculas de histocompatibilidade, e sim outras

30
moléculas, tais como os componentes do complemento (C2, Fator B e C4), 21-
hidroxilase, linfotoxina e Fator de necrose tumoral-α (TNF-α) (57–59). A localização
do complexo gênico HLA está representada na Figura 3.
Figura 3 - Complexo gênico HLA presente no cromossomo 6. Esquema da localização dos genes HLA no cromossomo 6, as três regiões e a disposição dos genes de classe I, II e III. Fonte: Adaptado de Westover et al. (60).
As moléculas HLA de classe I são expressadas constitutivamente em
todas as células nucleadas e plaquetas, e são responsáveis pela apresentação de
peptídeos endógenos aos linfócitos T CD8+, enquanto que as moléculas de classe II
têm uma distribuição mais restrita, sendo encontradas na superfície das APCs
(células dendríticas, linfócitos B e macrófagos) com a função de apresentação de
peptídeos exógenos aos linfócitos T CD4+ (57,58).
As moléculas de classe I e II são estruturalmente semelhantes. Os
resíduos de aminoácidos polimórficos que caracterizam as diversas moléculas estão
localizados nas fendas de ligação com o peptídeo, como pode ser visualizado na
Figura 4. Na molécula de classe I, o sulco de ligação ao peptídeo é formado pela
interação dos segmentos α1 e α2 e na molécula de classe II, pela interação dos
segmentos α1 e β1 (57,58).

31
Figura 4 - Estrutura das moléculas HLA classe I e II. O diagrama da esquerda ilustra as diversas regiões da molécula de classe I. Os domínios α1, α2 e α3 da cadeia α. Cadeia polipeptídica β2-microglobulina não codificada pelo complexo principal de histocompatibilidade (MHC), ligada não-covalentemente à cadeia α. A estrutura da porção extracelular de uma molécula HLA classe l ligada a um peptídeo, é exibida pela cristalografia de raios X. O diagrama da direita mostra a estrutura das moléculas HLA classe II que consistem de duas cadeias peptídicas que dobram-se em dois domínios: α1 e α2 ou β1 e β2. Ao seu lado direito é exibido pela cristalografia de raios X a estrutura da porção extracelular da molécula ligada a um peptídeo. Fonte: Abbas et al. (57).
O grande polimorfismo apresentado pelos genes HLA tem sido utilizado
para obtenção de histocompatibilidade entre doadores e receptores de órgãos e
tecidos, em estudos que tentam verificar a existência de associações de genes HLA
com doenças e em estudos de associação de moléculas HLA com a aloimunização
eritrocitária e plaquetária em populações transfundidas.
Devido à importância das moléculas HLA no mecanismo de
aloimunização eritrocitária, estudos em populações transfundidas têm documentado
uma possível relação entre alelos HLA e a resposta imune a uma variedade de
antígenos eritrocitários, incluindo os antígenos D (61), Jka (62,63), K (64,65), Fya
(65,66), Mia (67) e Dia (68). No entanto, até o momento não foram demonstradas
associações consistentes entre moléculas HLA e o risco de aloimunização em
pacientes com AF (69,70).
1.2.5.2 Citocinas
As citocinas são glicoproteínas de baixo peso molecular que são
secretadas pelas células da imunidade natural e imunidade adquirida em resposta a

32
um estímulo que modula o comportamento da célula alvo. A atuação dessas
proteínas pode ser parácrina, autócrina ou endócrina, alcançando células alvo
distantes via corrente sanguínea (58). Essas moléculas agem mediante ligação a
receptores específicos presentes na superfície de células alvo e participam tanto dos
processos inflamatórios quanto na resposta imune (71).
Na fase de ativação das respostas imunes adquiridas, as citocinas
estimulam o crescimento e a diferenciação de linfócitos, e nas fases efetoras da
imunidade natural e imunidade adquirida, elas ativam diferentes células efetoras
para eliminar microrganismos e outros antígenos (57).
As citocinas Th1 típicas incluem IFN-γ, TNF-α e IL-2 e direcionam a
imunidade celular na defesa, principalmente contra vírus e células cancerosas,
enquanto que as citocinas produzidas por células Th2 tais como: IL-4, IL-5, IL-6, IL-
9, IL-10 e IL-13 favorecem ativação de células B, a produção de imunoglobulinas e
inibição da resposta Th1. Citocinas Th1 inibem citocinas Th2 e vice-versa (57). Tal
como as células Th1 e Th2, as células Th17 produzem um grupo de citocinas
distintas: a interleucina-17 (também chamado de interleucina-17A), IL-17F, IL-22 e
IL-21, todas participam na produção de um tipo específico de resposta inflamatória
(72).
As citocinas podem ser também divididas em dois grupos de acordo com
a função: citocinas anti-inflamatórias (IL-4 e IL-10) e pró-inflamatórias (TNF-α, IL-8,
IL-1, IL-6, IFN-γ e IL-17). A falta de equilíbrio entre citocinas pró e anti-inflamatórias
desativa a função adequada do sistema imunológico (73).
Indivíduos portadores de AF, até mesmo no estado estável, têm como
principal característica uma produção exacerbada de citocinas inflamatórias (IL-
1,TNF-α, IL-6) (74–76). No entanto, o padrão de citocinas anti-inflamatórias como IL-
4 (77,78) e IL-10 (74,79) são divergentes entre os estudos.
Como a produção de citocinas é controlada geneticamente, variações
devido a polimorfismos na região promotora ou outras regiões reguladoras dos
genes de citocinas, podem afetar a produção dessas moléculas e, o equilíbrio da
resposta imune (80). Os polimorfismos podem ser de 3 formas principais:
polimorfismo de único nucleotídeo (do Inglês: single nucleotide polymorphisms - sigla
SNPs), o de número variável de repetições in tanden (do inglês variable number of

33
tanden repet – sigla VNTR) ou microssatélites. Alguns polimorfismos levam à
substituição de um aminoácido na cadeia proteica, porém a maioria, encontram-se
na região promotora do gene, na região intrônica ou na região 3’. Assim, um
indivíduo pode ter diferentes fenótipos de produção de citocinas dependendo do seu
genótipo ou haplótipo para cada citocina (58,80).
Os avanços nas áreas de genética e da biologia molecular permitiram a
identificação dos genes que codificam a produção de diferentes citocinas, bem
como, a identificação dos polimorfismos relacionados a estes genes. Estas
descobertas trouxeram novas perspectivas, permitindo avaliação da distribuição, o
perfil genético de produção e a associação destes polimorfismos com diversas
condições clínicas em que as citocinas tenham um papel importante.
Algumas citocinas e genes de citocinas que parecem possuir um papel
importante na aloimunização eritrocitária incluem o fator de necrose tumoral-α (TNF-
α) e o gene TNFA; Interleucina-6 (IL-6) e o gene IL6; Interleucinas17A e 17F(IL-17A,
IL-17F) e os genes IL17A e IL17F; Interleucina-1β (IL-1β) e gene IL1B; Interleucina-
10 (IL-10) e o gene IL10; Interleucina-4 (IL-4) e o gene IL4.
O TNF-α é uma citocina produzida principalmente por macrófagos, células
natural killer (células NK), linfócitos T e B (57,58). Esta citocina contribui para o
processo inflamatório por meio da ativação dos leucócitos, do estímulo ao aumento
da expressão de moléculas de adesão nas células endoteliais contribuindo para a
adesão leucocitária. Além disso, o TNF-α induz a degranulação de neutrófilos e o
aumento da permeabilidade capilar, inibe a proliferação celular e induz a célula à
morte (57,81). O TNF-α contribui ainda para o estado vascular inflamatório presente
em várias doenças, incluindo a AF (74,82).
O gene que codifica a citocina TNF-α (gene TNFA) está situado no
cromossomo 6, na região p21.3. O SNP mais estudado neste gene é o –308G/A
(rs1800629) que está presente no sítio de ligação para o fator de transcrição AP1 na
região promotora do gene (83) sendo o alelo TNFA-308A associado à alta produção
desta citocina (83). Em estudos com doenças, foi encontrado associações entre o
SNP TNFA–308G/A e o prognóstico por infecção pelo vírus da Hepatite B (84), a
proteção ao acidente vascular cerebral em pacientes com AF (85,86), ao lúpus
eritematoso sistêmico, a diabetes e suas complicações (87–91).

34
A Interleucina-6 é uma citocina pró-inflamatória, produzida por uma
variedade de células, tais como: linfócitos B e T, fibroblastos, monócitos,
queratinócitos, células endoteliais, entre outras células (92). IL-6 não é expressa
constitutivamente, mas pode ser expressa em resposta a estímulos inflamatórios
diversos tais como IL-1 e TNF-α (93). É uma citocina que atua em vários processos
fisiológicos incluindo: hematopoese, crescimento celular, indução da produção de
proteínas de fase aguda, manutenção do processo inflamatório, indução a
diferenciação de células T auxiliares em células Th2 e diferenciação de células B em
plasmócitos, estimulando a secreção de imunoglobulinas e controle do equilíbrio
entre as células T regs e células Th17 (94–97).
O gene IL6 está localizado no cromossomo 7, na região p21, possui cinco
éxons e quatro íntrons distribuídos em uma região de aproximadamente 5kb. Na
posição –174 do gene IL6 foi descrito o polimorfismo IL6–174G/C rs1800795 que
resulta em três possíveis genótipos: G/G, G/C e C/C. Estas variações podem
influenciar a ligação de fatores de transcrição influenciando a taxa de transcrição e
de expressão da citocina. Estudos anteriores encontraram o alelo C associado a
menor expressão de IL-6 (93,98). Este polimorfismo tem sido estudado em muitas
doenças onde o componente inflamatório é importante (99–102)
As citocinas IL-17A e IL-17F são secretadas por células Th17, que são
geradas a partir da diferenciação de células CD4+ efetoras. A diferenciação destas
células é induzida por TGF-β + IL-6 e também na presença de IL-1 e IL-21. Além
disso, IL-17 atua como uma citocina pró-inflamatória que induz o recrutamento de
neutrófilos durante a resposta inflamatória e a secreção de citocinas, entre as quais
IL-1, IL-6 e TNF-α (72,103,104).
IL-17A e IL-17F compartilham uma forte homologia na sequência, e os
genes que as codificam estão localizados próximos, no cromossomo 6, na região
p12 (105). O polimorfismo no gene IL17A−197G/A (rs2275913) está localizado
dentro do motivo de ligação para o fator nuclear de células T do gene IL17, sendo o
alelo A, relacionado com a alta produção de IL-17 (106,107) e também com a
produção mais eficiente, isto porque possui alta afinidade pelo regulador essencial
do promotor de IL17 (NFAT). Este polimorfismo tem sido associado com um maior
risco ao desenvolvimento e severidade de doenças inflamatórias como doença do

35
enxerto contra o hospedeiro (106), colite ulcerativa (107,108) e câncer gástrico
(109).
O gene IL17F contém 2 éxons, o segundo éxon compartilha uma
homologia significante com o éxon 3 do gene IL17. O polimorfismo IL17F (7488T/C)
que leva a substituição do aminoácido histidina por arginina na posição 161 da
proteína (His161Arg, rs763780) (110) tem sido associado com a proteção a doenças
autoimunes (107,110,111), risco de câncer gástrico (112), colite ulcerativa (107,108)
e susceptibilidade a leucemia mielóide aguda (113).
IL-1β é uma citocina pro-inflamatória, fortemente expressa por monócitos,
macrófagos teciduais, células dendríticas, linfócitos B, células NK e células epiteliais
(114). IL-1β afeta quase todos os tipos de células, é importante na imunidade inata e
adaptativa, na patogênese de doenças infecciosas, autoimunes e doenças auto
inflamatórias, bem como na mediação de respostas inflamatórias sistêmicas
compreendendo febre, proteínas hepáticas de fase aguda, e ativação de leucócitos
(114–116). IL-1β também possui um papel adjuvante na expansão e diferenciação
de células T CD4+ e produção de anticorpos (117).
O gene IL1B está localizado no cromossomo 2, na região q14. O
polimorfismo IL1B–511C/T rs16944 localiza-se na região promotora do gene. O alelo
menos comum (IL1B–511T) foi associado com o fenótipo de alta capacidade
produtora e tem sido associado com doenças que tem a inflamação como
componente na sua fisiopatologia (118–120).
IL-10, membro da família de citocinas Th2, é produzida por quase todos
os tipos de células do sistema imune inato (macrófagos, monócitos, células
dendríticas, mastócitos, neutrófilos, eosinófilos e células natural Killer) e adaptativo
(incluindo células T CD4+, T CD8+ e células B). Ao contrário das células da
imunidade inata, que podem produzir IL-10 imediatamente após um estímulo
ambiental, as células T CD4+ naives necessitam reconhecer o peptídeo apresentado
por APCs, após a interação de moléculas HLA, peptídeo e TCR, para posteriormente
se diferenciarem em Th1, Th2 e Th17 e ocorrer a produção de IL-10 (121,122).
IL-10 está envolvida no controle das reações da imunidade natural e da
imunidade mediada por células (57,122,123), mas possui também atividade anti-
inflamatória, como inibidor da síntese de IFN-γ, IL-2, IL-3, TNF-α e GM-CSF

36
produzidas por macrófagos ativados e células T helper. Ela regula negativamente a
expressão de citocinas Th1, a expressão de antígenos HLA de classe II e moléculas
CD54 (ICAM-1), CD80 (B7), e CD86 (B7.2) em monócitos (124). A expressão
reduzida destas moléculas pode afetar significativamente a capacidade de ativação
de células T por monócitos (APCs)(124–126).
O gene IL10, localizado no cromossomo 1q31-32, contém cinco éxons e
ocupa aproximadamente 5,1 kb. A produção celular de IL-10 parece ser influenciada
por três SNPs localizados na região promotora do gene IL10: –1082G/A
(rs1800896), –819T/C (rs1800871) e –592C/A (rs1800872) por resultarem em
alteração de sítios específicos de reconhecimento de fatores transcricionais
(127,128). Os estudos mostram que os SNPs –1082G/A, –819C/T e –592C/A estão
em desequilíbrio de ligação e podem formar 8 possíveis haplótipos (GCC, GCA,
GTC, GTA, ACC, ACA, ATC e ATA), sendo apenas 3 encontrados em populações
caucasóides: GCC, ACC e ATA os quais podem determinar 3 fenótipos possíveis:
alto produtor (GCC/GCC), médio produtor (GCC/ACC e GCC/ATA) e baixo produtor
(ACC/ACC, ACC/ATA e ATA/ATA) (127–129).
A citocina IL-4 é membro da família de citocinas Th2, sendo produzida
principalmente por linfócitos T CD4+, mas também por mastócitos e basófilos (57).
Ela estimula o desenvolvimento de células Th2 a partir de células Th naives,
aumenta a expressão de HLA-II, possibilitando maior ativação de Th2, aumenta
ainda a expressão de receptores de alta afinidade para IgE (FcεRI) em mastócitos e
basófilos e de baixa afinidade para IgE (FcεRII) em células B não-ativadas. Nas
células B ativadas a IL-4 estimula a síntese principalmente de IgE e de IgG1, sendo
seu efeito antagonizado por IFN-γ (57).
O gene IL4 está localizado no cromossomo 5q31-q33 em um cluster de
genes de citocinas (IL-3, IL-5, IL-9, IL-13, IL-15, GM-CSF e fator regulatório de IFN).
O polimorfismo –590C/T (rs2243250), descrito na região promotora do gene IL4,
está localizado próximo a região de controle do gene e pode afetar a atividade
transcricional da proteína (130). Outro polimorfismo localizado no terceiro íntron é o
VNTR (número variável de repetições in tanden, do inglês variable number of tanden
repet). Tanto o alelo T na posição –590 quanto o alelo RP1 do polimorfismo íntron 3
VNTR foram associados com maior produção de IL-4 ou atividade de células T (130–
132). Estes polimorfismos presentes no gene IL4, foram associados a determinadas

37
condições autoimunes (133,134) e inflamatórias (135–137).
Considerando que as citocinas tem um importante papel na inflamação e
na regulação da resposta imune, que os níveis de produção de citocinas frente a um
estímulo podem variar entre os indivíduos e ainda considerando que moléculas HLA
possuem diferentes capacidades para o encaixe de peptídeos e sua consequente
apresentação ao sistema imune, acreditamos que os polimorfismos em genes de
citocinas, acima descritos e alelos HLA possam estar relacionados com a
susceptibilidade a aloimunização eritrocitária em pacientes falciformes.
Ao identificar marcadores genéticos de aloimunização eritrocitária, como
os polimorfismos em genes de citocinas e alelos HLA, pode ser possível prever com
antecedência respondedores e não-respondedores. Este conhecimento pode auxiliar
futuramente no direcionamento de unidades fenotipadas ou genotipadas, inclusive
em relação aos haplótipos RH, apenas para os pacientes respondedores, evitando-
se assim o alto custo da utilização de unidades fenótipo compatível para não-
respondedores.

38
___________________________________________________________________
2 OBJETIVOS 2.1 OBJETIVO GERAL
Este trabalho teve como objetivo geral identificar o perfil genético de
grupos sanguíneos, os haplótipos RH em pacientes que desenvolveram anticorpos
contra antígenos Rh e as possíveis associações de polimorfismos presentes em
genes imunologicamente relevantes (genes HLA e genes de citocinas) com a
aloimunização eritrocitária em pacientes portadores de AF politransfundidos.
2.2 OBJETIVOS ESPECÍFICOS
– Identificar a taxa de aloimunização eritrocitária e a frequência dos
aloanticorpos e autoanticorpos encontrados na população de pacientes falciformes
politransfundidos.
– Identificar a frequência dos principais antígenos de grupos sanguíneos
entre os pacientes falciformes através da análise molecular.
– Realizar a genotipagem de alelos RHD e RHCE em pacientes com AF
que desenvolveram anticorpos contra antígenos Rh e avaliar a significância clínica
dos anticorpos produzidos.
– Determinar as frequências de alelos, genótipos e haplótipos dos
polimorfismos IL17F (7488A/G), IL17A–197G/A, IL4 (–590C/T e íntron 3 VNTR), IL10
(–1082A/G, –819T/C, –592A/C), TNFA–308G/A, IL1B–511C/T e IL6–174C/G e as
possíveis associações com a aloimunização eritrocitária em pacientes portadores de
AF com múltiplas transfusões.
– Determinar a frequência dos alelos HLA classe II (HLA-DRB1, -DQA1, -
DQB1) e as possíveis associações com a aloimunização eritrocitária em pacientes
portadores de AF com múltiplas transfusões.
– Determinar as frequências dos haplótipos formados por HLA-DRB1 e o
polimorfismo TNFA–308G/A e as possíveis associações com a aloimunização
eritrocitária em pacientes portadores de AF com múltiplas transfusões.

39
__________________________________________________________________
3 MATERIAIS E MÉTODOS 3.1 CASUÍSTICA 3.1.1 Grupo de pacientes com Anemia Falciforme
Participaram deste estudo 161 pacientes portadores de AF
politransfundidos que foram atendidos no Hemocentro da UNICAMP, Centro Infantil
Boldrini e Hemonúcleo de Taubaté durante o período de setembro/2012 a
maio/2015. Foram incluídos pacientes homozigotos para HbS, de ambos os sexos
independentemente da idade. Cada voluntário concordou em participar deste estudo
mediante entrevista e esclarecimentos verbais, sendo submetidos à coleta
sanguínea, segundo proposto no projeto aprovado pelo Comitê de Ética em
Pesquisa Institucional. A coleta das amostras foi realizada no dia da consulta de
rotina dos pacientes que apresentavam condição clínica estável (steady-state). Os
dados demográficos de cada paciente foram obtidos por meio de entrevista com os
pacientes e pais.
Foram incluídos pacientes que apresentavam histórico de transfusão
sanguínea nos últimos 6 meses antes da data de inclusão do paciente. Todos os
pacientes transfundidos receberam unidades de sangue HbS-negativas,
leucoreduzidas e não irradiadas.
Foram excluídos da casuística pacientes aparentados, pacientes que
haviam recebido menos de 5 transfusões sanguíneas ou que somente haviam
recebido transfusões compatíveis para os principais antígenos clinicamente
significantes.
3.1.2 Grupo controle
O grupo controle foi constituído de 288 doadores de sangue e medula
óssea autodeclarados como Afro-Brasileiros, que foram atendidos no Hemocentro da
Universidade Estadual de Campinas (UNICAMP). Esses indivíduos possuíam nível
normal de Hb, ausência de anemia ou HbS, ausência de condições inflamatórias e
doenças hematológicas. Este grupo foi utilizado para determinação das frequências

40
dos polimorfismos em genes de citocinas e alelos HLA em uma população saudável
de etnia similar aos pacientes.
3.1.3 Aspectos éticos da pesquisa
Este estudo foi aprovado em 10/08/2012 pelo Comitê de Ética em
Pesquisa (CEP) da Faculdade de Ciências Médicas da UNICAMP, Parecer no
68683/2012 (Anexo I), conforme previsto na resolução no 196/96 do Conselho
Nacional de Saúde (CNS). Todos os participantes assinaram o termo de
consentimento livre e esclarecido, previamente aprovado pelo CEP.
3.2 MATERIAIS
3.2.1 Kit de extração de DNA
Para a extração do DNA do sangue periférico foi utilizado o kit comercial
QIAamp DNA Blood Mini Kit da empresa Qiagen® (Qiagen, Valencia, CA, USA).
Estão inclusos neste kit os reagentes: Buffer AL, Buffer AW1, Buffer AW2, proteinase
K e os materiais: colunas de sílica e tubos de lise.
3.2.2 Água livre de nuclease
A água livre de nuclease (Applied Biosystems®, Foster City, CA, EUA) foi
utilizada na etapa final da extração de DNA, na reconstituição dos primers e nas
reações de PCR.
3.2.3 Etanol absoluto (CH3CH2OH)
O etanol absoluto (P.A) (Sigma-Aldrich, St. Louis, MO, EUA) foi utilizado
na preparação dos reagentes de extração de DNA e no protocolo de
sequenciamento de DNA.
3.2.4 Taq DNA Polimerase
A enzima Taq DNA polimerase (Invitrogen®, Carisbad, CA, EUA) foi
utilizada nas Reações em Cadeia da Polimerase Alelo Específica (PCR-AS), PCR-
multiplex, Análise dos Fragmentos da Digestão Enzimática dos Produtos da Reação

41
em Cadeia da Polimerase (do inglês - restriction fragment length polymorphism –
sigla: PCR-RFPL), e Reação em Cadeia da Polimerase Sequência Específica de
Oligonucleotídeos (do inglês - sequence specific oligonucleotide – sigla: PCR-SSO).
O kit incluía 5 unidades/μL da enzima, 1mL de tampão 10X (200mM Tris-HCL
(pH8.4) e 500mM de KCl e 1mL de MgCl2 50mM.
Para a técnica de microarray HEA BeadChipTM (Human Erythrocyte
Antigen, BioArray Solutions, Immucor, Warren, NJ, EUA) foi utilizada a enzima Taq
polimerase Platinum (Invitrogen®). O kit incluía 5 unidades/μL da enzima, 1mL de
tampão 10X (200mM Tris-HCL (pH8.4) e 500mM de KCl e 1mL de MgCl2 50mM.
3.2.5 dNTP 10Mm
As dNTPs (dATP, dTTP, dCTP, dGTP) adquiridas da empresa
Invitrogen®, foram utilizadas nas técnicas de PCR convencional na concentração de
10mM.
3.2.6 Primers
Os primers utilizados na análise molecular dos grupos sanguíneos estão
descritos nas referências (30,138–141). Os primers utilizados na genotipagem dos
polimorfismos IL17F (7488A/G), IL17A–197G/A, IL4–590C/T, IL10 (–1082A/G, –
819T/C, –592A/C), TNFA–308G/A, IL1B–511C/T e IL6–174C/G estão descritos na
Tabela 4 e os primers utilizados para determinação do polimorfismo IL4 íntron 3
VNTR está descrito na referência (142). Todos os primers foram adquiridos da
empresa Invitrogen®.
3.2.7 Sondas TaqMan para determinação da zigozidade RHD
Sondas fluorescentes duplamente marcadas foram sintetizadas pela
Applied Biosystems® para determinação da zigozidade RHD por PCR em tempo real
pela metodologia de genotipagem TaqMan®. Os fluorocromos utilizados foram: FAM
(6-carboxyfluorescein), TAMRA (6-carboxytetramethylrhodamine) e JOE (2,7-
dimethoxy-4,5-dichloro-6-carboxyfluorescein). As sequências das sondas estão
descritas na Tabela 1.

42
Tabela 1 - Sequências das sondas utilizadas para determinação da zigozidade RHD.
Técnica Sequência das sondas
PCR em tempo real
(genotipagem
TaqMan®)
RD-T 5'(FAM)TACGTGAGAAACGCTCATGACAGCAAAGTCT(TAMRA)3'
Alb-probe
5’(JOE)-CCTGTCATGCCCACACAAATCTCTCC-(TAMRA)3’
3.2.8 Marcador de peso molecular
Marcador de peso molecular de 100pb (Invitrogen®) foi utilizado nas
corridas de eletroforese para análise do tamanho dos fragmentos de DNA.
3.2.9 Tampão Tris-EDTA-Borato (TEB) 10X
Tampão utilizado no preparo do gel de agarose e na corrida de
eletroforese. Foi preparado dissolvendo-se 108g de Tris (Sigma-Aldrich®), 55g de
ácido bórico (Sigma-Aldrich®) e 40mL de EDTA 0,5M pH 8.0 (Sigma-Aldrich®) em
quantidade de água destilada (dH2O) suficiente para 1000mL.
3.2.10 Enzimas de restrição
As enzimas de restrição utilizadas neste trabalho foram adquiridas das
empresas New England Biolab® (Beverly, MA, EUA) e MBI Fermentas® (Amherst,
NY, EUA) e foram utilizadas para digestão dos produtos de PCR de acordo com o
polimorfismo estudado, como pode ser visualizado na Tabela 4.
3.2.11 Gel de agarose
A solução de agarose foi preparada em TEB 1X de acordo com a
concentração específica de cada protocolo (1%, 2% ou 3%). Esta solução era
aquecida em forno micro-ondas, durante 1 minuto e, após resfriamento, adicionou-se
50µL de brometo de etidio (Invitrogen®).
3.2.12 Gel de poliacrilamida 12%
A solução de poliacrilamida foi preparada misturando 23,3mL de
acrilamida 40% (Gibco BRL®, Gaithersburg, MD, EUA); 8,8mL de TEB 10X, 55mL
de água destilada, 363μL de persulfato de amônio 10% (Gibco BRL®) e 16μL de

43
TEMED (Gibco BRL®). Esta solução foi dispensada em uma placa de vidro,
previamente preparada, para que ocorresse a sua polimerização.
3.2.13 Plataformas HEA BeadChipTM, RHCE BeadChipTM e RHD BeadChipTM
As plataformas de microarray HEA BeadChipTM, RHCE BeadChipTM e
RHD BeadChipTM foram adquiridas da empresa BioArray Solutions (Immucor. NJ,
USA) e foram utilizadas para a genotipagem eritrocitária estendida dos grupos
sanguíneos e para a pesquisa de variantes nos genes RHCE e RHD,
respectivamente.
Estas plataformas utilizam kits que contém todos os reagentes
necessários para a execução da técnica, incluindo: lâminas de vidro com chips de
DNA (sondas de oligonucleotídeos específicas ao polimorfismo estudado) com
capacidade para 8 ou 96 amostras, PCR-Mix (contendo primers, dNTPs e tampão) e
as enzima ExoSAPIT, Lambda Exonuclease e Thermo Sequenase, provenientes da
Empresa Amersham Pharmacia Biotech (Piscataway, NJ, EUA).
Nos Kits de RHCE e RHD também estão inclusos a enzima Hot Start Taq
DNA polimerase (BioArray Solutions).
3.2.14 Kit de extração Qiaex II
Este Kit foi adquirido da Empresa Qiagen® e foi utilizado na extração e
purificação do DNA contido em gel de agarose para subsequente reação de
sequenciamento de DNA.
3.2.15 Trizol
Este regente foi utilizado para extração de RNA segundo as instruções do
fabricante (Invitrogen®).
3.2.16 Superscript II Kit
Este Kit foi utilizado na transcrição do mRNA e foi adquirido da Empresa
Invitrogen®.

44
3.2.17 Big Dye
O reagente Big Dye (BD Half-term, GenPark, Perkin Elmer Biosystems,
Foster City, CA, EUA) foi utilizado nas reações de sequenciamento de DNA.
3.2.18 Marcador de peso Molecular Low Mass
O marcador de peso molecular Low Mass (Invitrogen®) foi utilizado na
análise do peso molecular dos fragmentos designados para sequenciamento.
3.2.19 Kit LABType® SSO
O kit LABType® SSO (One Lambda Inc., Canoga Park, CA, EUA),
tecnologia Luminex xMap (Luminex Corporation, Austin, EUA) foi utilizado para
genotipagem de grupos alelicos HLA de classe II. Foram utilizado os kits LABType®
SSO HLA DRB1 e SSO HLA DQA1/DQB1 (One Lambda Inc.) para genotipagem dos
alelos HLA dos loci HLA-DRB1 e -DQA1/-DQB1, respectivamente.
Os kits eram compostos por reagentes de pré-PCR: solução de primers
(específico para cada loci HLA) e solução D-Mix para primers; e reagentes de pós-
PCR (hibridização): tampão de hibridização, tampão de lavagem, solução de
desnaturação, tampão de neutralização, tampão de coloração SAPE e mistura de
pérolas (microesferas ligadas a sondas de oligonucleotídeos específicos para cada
alelo).
3.2.20 Ensaios TaqMan® para genotipagem de SNPs em genes de citocinas
Os ensaios de genotipagem TaqMan® foram confeccionados e
sintetizados pela Applied Biosystems®. Os ensaios utilizados foram: rs1800871
(C_1747362_10), rs2243250 (C_16176216_10) e rs1800795 (C_1839697_20).
Cada ensaio possuía um par de iniciadores que flanqueiam o SNP e um
par de sondas: uma homóloga do tipo selvagem, marcadas com VIC®, e outra
marcada com 6-carboxifluoresceína (FAM®), homóloga ao tipo mutante.
3.2.21 Mastermix de genotipagem Taqman®
O mastermix de genotipagem TaqMan® (Applied Biosystems®) foi
utilizado para a genotipagem de polimorfismos por PCR em tempo real. O mix incluía

45
a enzima DNA polimerase AmpliTaq Gold® UP (Ultra Pura), dNTPs e tampão de
forma otimizada.
3.3 MÉTODOS 3.3.1 Dados transfusionais
Dados de fenotipagem eritrocitária das bolsas de sangue recebidas,
fenotipagem dos pacientes e anticorpos identificados foram obtidos por meio do
banco de dados do serviço de transfusão.
O número de transfusões sanguíneas recebidas por paciente foi
identificado pela análise do prontuário, banco de dados do serviço da transfusão e
relato do paciente, visto que algumas vezes os mesmos se submetiam a transfusão
sanguínea em outra instituição e este dado poderia não ter sido registrado pelo
clínico no prontuário.
3.3.2 Avaliação sorológica
Nas amostras de hemácias dos pacientes foram realizadas fenotipagens
para os antígenos D, C, c, E, e, utilizando-se dois antissoros comerciais diferentes
(Immucor, Norcross, GA, EUA; Bio-Rad, Lagoa Santa, Brasil). Nas amostras de
soros foram realizadas a pesquisa e identificação de anticorpos por hemaglutinação
em gel (Bio-Rad, Cressier sur Morat, Suíça). O Teste de antiglobulina direta e
controle autólogo foram realizados em gel para todas as amostras antígeno positivas
para a especificidade correspondente do anticorpo Rh. Estudos de eluato foram
realizados em todas as amostras com um teste direto de antiglobulina positivo
utilizando um método de eluição ácida (DiaCidel, Bio-Rad, Brasil). Adsorções com
hemácias autólogas também foram efetuadas para auxiliar a diferenciação de auto-
e aloanticorpos.
Os anticorpos foram classificados como auto- ou aloanticorpos com base
nos resultados dos testes sorológicos, fenotipagem e genotipagem eritrocitária.
3.3.3 Coleta da amostra e Extração de DNA
Amostras de sangue de todos os pacientes e controles foram coletadas
em tubos contendo anticoagulante (EDTA) e centrifugadas a 210g durante 15

46
minutos, posteriormente os DNAs foram extraídos a partir dos leucócitos do sangue
periférico utilizando-se o Kit comercial QIAamp DNA Blood Mini Kit (Qiagen®). Após
a extração, a qualidade e a quantidade do DNA foi determinada pela leitura óptica da
absorbância em um equipamento de espectrofotometria (NanoDrop Lite, Thermo
Scientific, Wilmington, USA). Os DNAs foram armazenados em freezer -20C para
posterior realização dos testes moleculares.
3.3.4 Análise molecular de antígenos eritrocitários 3.3.4.1 Genotipagem de grupos sanguíneos em larga escala
A genotipagem para os principais alelos de grupos sanguíneos foi
realizada pela técnica de microarray utilizando-se a plataforma HEA BeadChipTM
(BioArray Solutions, Immucor) de acordo com o protocolo do fabricante. Os
polimorfismos avaliados estão descritos na Tabela 2.
A amplificação das sequências alvo foi realizada por PCR-multiplex, com
a utilização do PCR-Mix presente no Kit e a amostra de DNA do paciente. Em todas
as reações foram incluídos uma amostra controle positivo (amostra com genótipo e
fenótipo conhecido) e um controle negativo (água ao invés de DNA). As
amplificações foram feitas no termociclador Veriti (Applied Biosystems®).
Após a PCR-multiplex realizou-se a remoção de todo o excesso de
primers e dNTPs através da adição da enzima ExoSAPIT (Amersham Pharmacia
Biotech) ao produto de PCR amplificado. A obtenção de fita simples de DNA foi
realizada posteriormente através da digestão enzimática com a enzima Lambda
Exonuclease (Amersham Pharmacia Biotech).
Em seguida, adicionou-se ao produto da PCR a solução composta pela
enzima Thermo Sequenase, tampão (1X) e 1mmol de cada dNTP (dCTP marcada
com fluoróforos). Esta mistura foi dispensada em cada BeadChipTM. As lâminas
contendo os microchips foram então incubadas para que houvesse ou não a
hibridização do DNA alvo amplificado às sondas pré-existentes no BeadChipTM.
Completado o período de incubação, todos os BeadChipsTM foram
lavados 3 vezes com água destilada, a fim de remover todo e qualquer excesso de
dNTP, para minimizar a emissão indesejada de sinal de fluorescência.

47
Ao término da hibridização, as lâminas contendo os BeadChipsTM foram
levadas ao sistema de captura de imagem, o AIS Data Acquisition BasisTM (BioArray
Solutions Information System), responsável pela captura e decodificação da
fluorescência. Este sistema é composto por um leitor óptico de código de barras
(lâminas) e um microscópio de fluorescência acoplado a uma câmera fotográfica
interligada a um software, o wHEATM (Web-Based Human Erythrocyte antigen and
Hemoglobinopathy analysis – BioArray Solutions), que possibilitou a identificação e
interpretação do alelo hibridizado através da comparação dos valores de
fluorescência obtidos com os valores de fluorescência dos polimorfismos
depositados no Banco de Dados BasisTM (BioArray Solutions). A emissão dos
resultados foi feita na forma de tabelas e gráficos. Um resumo de todos os
procedimentos que compõe a técnica de genotipagem em larga escala da
plataforma de microarray HEA BeadChipTM estão descritos na Figura 5.

48
Tabela 2 - Alelos e SNPs presentes no HEA BeadChipTM
Sistema de grupo sanguíneo Alelos SNPs
Rh
C/c 307 C>T
109 Ins
E/e 676 G>C
VS 733 C>G
V 1006 G>T
Kell
K/k 698 T>C
Jsa/Jsb 1910 C>T
Kpa/Kpb 961 C>T
Duffy
Fya/Fyb 125 G>A
GATA (Silencing FY) -33 T>C
Fyx[Fy(b+w)] 265 C>T
Kidd Jka/Jkb 838 G>A
MNS
M/N 59 C>T
S/s 143 T>C
Silencing S Ex5 230 C>T
Silencing S Int5 g>t
Diego Dib/Dia 2561 C>T
Lutheran Lua/Lub 230 A>G
Dombrock
Doa/Dob 793 A>G
Hy+/Hy- 323 G>T
Jo(a+)/Jo(a-) 350 C>T
Landsteiner-Wiener LWa/LWb 308 A>G
Colton Coa/Cob 134 C>T
Scianna Sc1/Sc2 169 G>A
Hemoglobin S HgbS 173 A>T

49
Figura 5 - Etapas do protocolo de genotipagem de alelos de grupos sanguíneos em larga escala pela plataforma HEA BeadChip
TM. Fonte: Adaptado de Da Costa (143).
3.3.4.2 Genotipagem RHD e RHD*Ѱ
A determinação da presença do gene RHD (íntron 4 e éxon 7) e a
detecção da inserção de 37pb no gene RHD, conhecida como pseudogene RHD ou
RHD*Ѱ, foi realizada conforme protocolo descrito por Singleton et al. (138).
3.3.4.3 Determinação da zigozidade do gene RHD
A determinação da zigozidade do gene RHD foi realizada pela técnica de
PCR-RFLP, de acordo com o protocolo descrito por Wagner et al. (144) e por PCR
quantitativo em tempo real como previamente descrito por Chiu et al. (139).
3.3.4.4 Identificação de variantes RHD e RHCE
A identificação de alelos RHD variantes foi realizada utilizando-se a
plataforma RHD BeadChipTM (BioArray Solutions) que contém 34 marcadores

50
associados com expressão alterada do gene RHD. Os alelos e SNPs presentes na
plataforma RHD BeadChipTM estão descritos na Tabela 3.
Tabela 3 - Variantes RHD analisadas pelo RHD BeadChipTM
D fraco tipo: 1, 1.1, 2, 3, 4.0, 4.1, 4.2/DAR, 4.3, 5, 11, 14, 15, 17, 25, 29, 34, 40, 47, 51
D negativo: RHDΨ; W16X; D-CE(3-7)-D; D-CE(4-7)-D; (C)dceS; DCE(3-9)-D; CE(1-3)-D(4-
10); rG; RHD(Y269X)
Del: 1227 G>A; IVS3+1G>A; M295I
D Parcial: DAR/ D fraco 4.2; DIIIa (DIII tipo 5), DIII tipo 4,6; DIIIc; DIVa; DIVa -2; DIV tipo 3,
4, 5; DIVb; DV tipo 1,2,3 (DBS-0),4,5(DHK),6,7,8,9; DVI tipo 1,2,3,4; DNB; DHMi; DUC-2;
DAU 1,2,3,4,5; DBT 1,2; DCS 1, 2; DOL; DOL-3; DFR; DFR -2; DTO; DBS-0,1,2.
A identificação dos alelos RHCE variantes foi realizada com a utilização da
plataforma RHCE BeadChipTM (BioArray Solutions, Immucor) que permite a
detecção de 25 polimorfismos associados com alterações dos antígenos C (RH2), c
(RH4), E (RH3), e (RH5), CW (RH8), CX (RH9), V (RH10), VS (RH20), e Crawford
(RH43). Com este método podemos identificar os alelos RHCE variantes: ceRT,
ceAR, ceMO, ceRA, ceVG, ceEK, ceBI, CeMA, ceSL, CeVA, ceTI, ceFV, DHAR, E
tipo I/II/III/IV, EKH, ceS, (C)ceS, ceS(340,378) e 48C.
Para a determinação das variantes RHD e RHCE pelas plataformas de
microarray, RHD e RHCE BeadChip, realizou-se o mesmo procedimento técnico da
plataforma HEA BeadChipTM, descrito anteriormente. As plataformas diferem apenas
no programa de ciclagem da reação de PCR e nos tempo e temperatura de
incubação das reações de purificação, desnaturação e hibridização.
3.3.4.5 Sequenciamento do DNA e Clonagem
A análise da sequência de DNA foi realizada em produtos de PCR
amplificados a partir de DNA genômico em todas as amostras que não foram
identificadas pelas plataformas BeadChip utilizando-se pares de iniciadores RHD e
RHCE específicos descritos previamente (30,140).

51
Os produtos de PCR foram purificados por eluição a partir de gel agarose
1%, utilizando um kit de extração de gel Qiaex II (Qiagen) e sequenciados
diretamente, sem subclonagem, em um sequenciador ABI 373XL Perkin Elmer
Biosystems, com o reagente Big Dye BD Half-term (GenPak, Perkin Elmer
Biosystems, Foster City, CA, EUA).
A fim de determinar as combinações alélicas RH das amostras
identificadas com variantes RHCE, foi realizada a clonagem e sequenciamento de
Rh-cDNA. O RNA foi isolado dos reticulócitos com Trizol (Invitrogen). A transcrição
reversa foi realizada com Superscript First Strand Synthesis (Invitrogen), utilizando
iniciadores específicos para o gene (141). Os produtos de PCR amplificado a partir
do cDNA foram purificados com ExoSAP-IT (USB, Cleveland, OH, EUA), clonado em
um vetor TA (Invitrogen) e sequenciados utilizando primers previamente descritos
(141).
3.3.5 Avaliação do significado clínico dos anticorpos anti-Rh em pacientes portadores de variantes Rh
O significado clínico dos anticorpos Rh em pacientes portadores de
variantes Rh foi avaliado por análise retrospectiva dos níveis de HbS e Hb antes e
após as transfusões de concentrados de hemácias antígeno positivo. O
aproveitamento transfusional dos pacientes foi determinado por alterações dos
valores basais de Hb e porcentagem (%) de HbS de cada paciente.
3.3.6 Genotipagem HLA de classe II (-DRB1, -DQA1, -DQB1)
A tipagem HLA classe II das amostras de DNA de pacientes portadores
de AF foi realizada pela técnica de PCR-SSO (One Lambda Inc.) com tecnologia
Luminex. A reação de amplificação do DNA foi feita com oligonucleotídeos
biotinilados (primers) específicos para o éxon 2 (tipificações de HLA-DRB1, -DQA1 e
-DQB1). Após amplificação, os fragmentos de DNA obtidos foram separados por
eletroforese, em cuba micro SSP gel System (One Lambda Inc.), em gel de agarose
a 2,0%, a 150 volts por 10 minutos. A visualização das bandas foi realizada com
Syber Green (Invitrogen) em luz azul, e a interpretação dos resultados foi baseada
na presença ou ausência do fragmento específico de DNA amplificado.

52
Em seguida, realizou-se a desnaturação da dupla fita de DNA do produto
já amplificado, a neutralização química da fita aberta e posteriormente a hibridização
com sondas oligoespecíficas. Tais sondas já estão pré-conjugadas em microesferas
(“pérolas”), de forma que cada microesfera possui um tipo de sonda específico para
cada alelo de HLA. O produto hibridizado foi marcado com um produto fluorescente,
o SAPE (Estreptavidina conjugada com R-Ficoeritrina, do inglês: R-Phycoerythrin-
conjugated Strepavidin). A leitura das microesferas marcadas foi realizada em um
citômetro de fluxo (LABScanTM
), neste equipamento os lasers com diferentes
comprimentos de onda reconhecem as sondas marcadas ou não com o SAPE.
A leitura no citômetro de fluxo baseia-se na percentagem de valores
atribuídos para a sonda, que são superiores ou inferiores ao valor de cut-off (corte)
pré-definido para a sonda. Após leitura no citômetro de fluxo, a interpretação dos
resultados foi feita pelo software HLA FusionTM Research da One Lambda versão
3.0. O software analisa as combinações de sondas nas microesferas detectadas
pelo aparelho, consulta um banco de dados interno e define quais são os genes ou
alelos presentes. As etapas de tipificação HLA estão resumidas na Figura 6.
Figura 6 - Etapas da tipificação HLA pela técnica de PCR-SSO. Fonte: Adaptado de www.onelambda.com acesso em agosto, 2015 (145).
3.3.7 Genotipagem de polimorfismos em genes de citocinas
As reações de PCR para a determinação dos polimorfismos IL17F
7488A/G, IL17A–197G/A, IL10 (–1082A/G, –592A/C), TNFA–308G/A, IL1B–511C/T
foram compostas de 100ng de DNA, 0,1μM de cada primer (10uM) (Invitrogen),

53
0,3mM de cada dNTP (10 mM) (Invitrogen), 1,0U de Taq DNA polimerase Platinum
(5,0 U), 2,5mM de MgCl2 (50mM) e tampão (1x) (Invitrogen) em um volume final de
30L. Os ciclos de amplificação foram realizados no termociclador Veriti (Applied
Biosystems®). Após a amplificação, cada produto de PCR foi digerido com a
respectiva enzima de restrição para obtenção do genótipo do polimorfismo. Os
produtos da digestão enzimática do polimorfismo TNFA–308G/A foram separados
por eletroforese em gel de acrilamida 12%. Os produtos da digestão enzimática dos
polimorfismos IL17F 7488A/G, IL17A–197G/A, IL10 (–1082A/G, –592A/C) e IL1B
–511C/T foram separados por eletroforese em gel de agarose 3% (Figura 7). A
sequência dos primers utilizados para detecção dos SNPs, as condições de
ciclagem da PCR, produto da PCR, enzima de restrição utilizada, e tamanho dos
fragmentos obtidos estão descritos na Tabela 4.
O polimorfismo IL4 íntron 3 VNTR foi determinado por um ensaio de PCR
com primers previamente descritos por Cantagrel et al. (142). A reação de
amplificação foi realizada em um volume final de 20 𝜇L contendo 100ng de DNA
genômico 0,1𝜇M de cada primer (10𝜇M), 0,3mM de dNTP (10mM) (Invitrogen),
2,0mM de MgCl2 (50mM), 3𝜇L 10x PCR buffer, e 1,5U Taq DNA polymerase
(Invitrogen). As condições de ciclagem da PCR foram: desnaturação inicial 96°C por
5 min, 35 ciclos de 96°C-1 min, 60°C-1 min and 72°C-1min e incubação final a 72°C
por 7 min. Os produtos de PCR foram separados por eletroforese em gel de agarose
2,5%. Os fragmentos de DNA obtidos de 254bp (três repetições de 70-pb), 183pb
(duas repetições de 70-pb) e 114bp (ausência de repetição) foram identificados
como Alelo 2(RP2), alelo 1(RP1) e alelo 3(RP3), respectivamente (Figura 7).

54
Tabela 4 - Sequência de primers para detecção dos SNPs IL17F 7488A/G, IL17A –197G/A, IL1B–511C/T, IL10(–1082A/G, –592 A/C), TNFA–308G/A, condições de ciclagem da PCR, produto da PCR, enzima de restrição utilizada e tamanho dos
fragmentos obtidos.
SNP Sequência de primer,
condições de ciclagem da PCR
Produto da PCR
Enzima de restrição,
fragmentos obtidos (pb)
Metódo de referência
IL17F
7488A/G
(rs763780)
5-ACCAAGGCTGCTCTGTTTCT-3
5-GGTAAGGAGTGGCATTTCTA-3
96°C-5 min, 35 ciclos (95°C-40s, 58°C-1 min e
72°C-1 min) 72°C-7min.
143pb
NlaIII
(A) 80+63
(G) 143
(112)
IL17A
–197G/A
(rs2275913)
5-AACAAGTAAGAATGAAAAGAGGACATGGT-3
5-CCCCCAATGAGGTCATAGAAGAATC-3
96°C-5 min, 35 ciclos (95°C-40s, 58°C-1 min e
72°C-1 min) 72°C-7min.
102pb
EcoNI
(G) 68+34
(A) 102
(112)
IL1B
–511C/T
(rs16944)
5´- TGGCATTGATCTGGTTCATC – 3´
5´- GTTTAGGAATCTTCCCACTT – 3´
95°C-5 min, 35 ciclos (95°C-1min, 55°C-1 min e
72°C-1 min) 72°C-7min
305pb
AvaI
(C) 190 + 115
(T) 305
(146)
IL10
–1082 A/G
(rs1800896)
5’-CCAAGACAACACTACTAAGGCTCCTTT-3’ 5’-
GCTTCTTATATGCTAGTCAGGTA-3’
95°C-5 min, 35 ciclos (95°C-1min, 60°C-1 min e
72°C-1 min) 72°C-7min.
377pb
EcoNI
(A) 280 + 97
(G) 252 + 97+ 28
(147)
IL10
–592 A/C
(rs1800872)
5’-GGTGAGCACTACCTGACTAGC-3´
5’-CCTAGGTCACAGTGACGTGG-3’
95°C-5 min, 35 ciclos (95°C-1min, 61°C-1 min e
72°C-1 min) 72°C-7min.
412pb
RsaI
(A)236+176
(C) 412
(148,149)
TNFA
–308G/A
(rs1800629)
5’-AGGCAATAGGTTTTGAGGGCCAT-3’
5’-TCCTCCCTGCTCCGATTCCG-3’
95°C-5 min, 35 ciclos (95°C-1min, 61°C-1 min e
72°C-1 min) 72°C-7min.
107pb
NcoI
(G) 87 + 20
(A) 107
(150)

55
Figura 7 - Géis de eletroforese evidenciando os padrões polimórficos dos genes estudados. A) IL4 íntron 3 VNTR, B) TNFA –308G/A, C) IL1B–511C/T, D) IL10–592A/C, E) IL10–1082G/A, F) IL17F (7488A/G), G) IL17A–197G/A. Fonte: autora.

56
Os SNPs IL10–819T/C(rs1800871), IL4–590C/T(rs2243250) e IL6–
174C/G (rs1800795) foram investigados por PCR em Tempo Real com utilização do
sistema TaqMan® genotyping Assay (Applied Biosystems®).
Os ensaios de genotipagem utilizados foram: rs1800871
(C_1747362_10), rs2243250 (C_16176216_10) e rs1800795 (C_1839697_20).
A reação de amplificação foi realizada em um volume final de 5 μL
contendo 2,5μL de TaqMan Universal PCR Master Mix, 0,125μL do mix de primers e
sondas (20✕) e 1μL de DNA (20ng/ μL). As condições de ciclagem foram: 95°C por
10 minutos seguidos de 50 ciclos de 92°C por 15 segundos e 60°C durante 1 minuto.
Os ensaios foram realizados utilizando o sistema de PCR em tempo real
StepOnePlus™ (Applied Biosystems®).
Após a ciclagem os dados de fluorescência alélica específica de cada
amostra foram analisados utilizando o software SDS v1.3.1 (Applied Biosystems®,
2005) para determinar o genótipo de cada amostra. A Figura 8, mostra um gráfico de
discriminação alélica para o polimorfismo IL10–819T/C. A determinação dos alelos
dos SNPs IL4–590C/T e IL6–174C/G foi realizada utilizando-se a mesma estratégia
descrita na Figura 8.
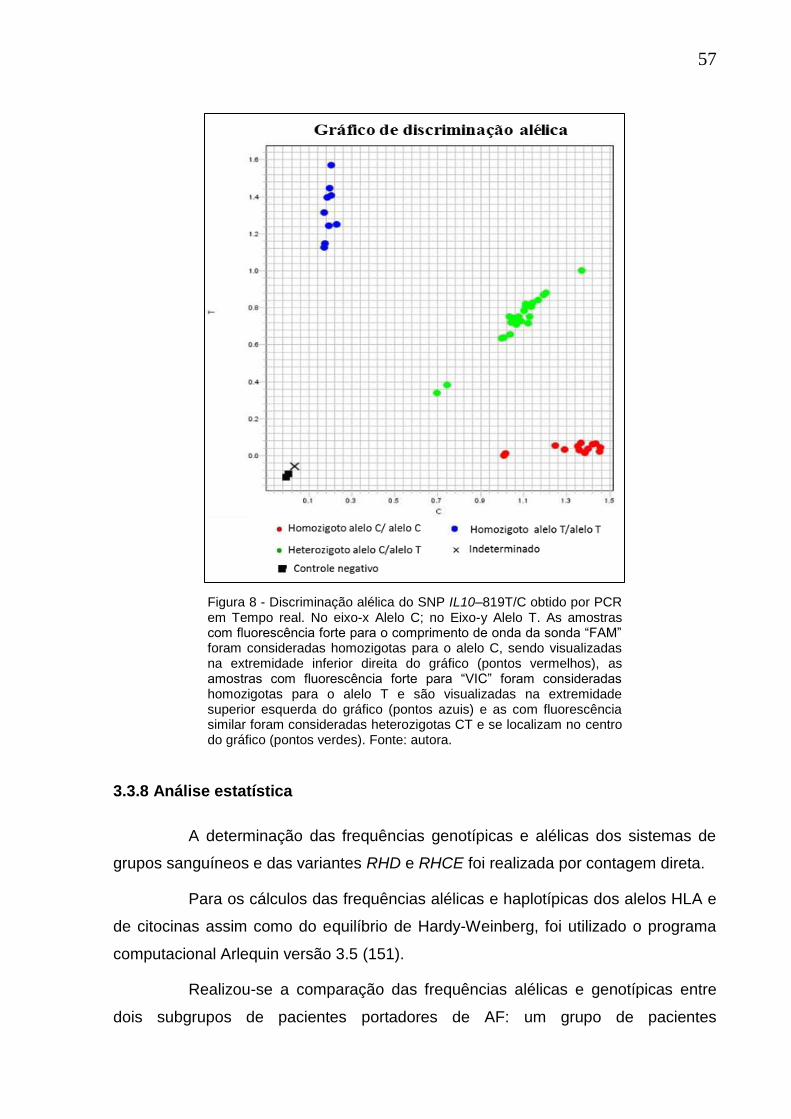
57
Figura 8 - Discriminação alélica do SNP IL10–819T/C obtido por PCR em Tempo real. No eixo-x Alelo C; no Eixo-y Alelo T. As amostras com fluorescência forte para o comprimento de onda da sonda “FAM” foram consideradas homozigotas para o alelo C, sendo visualizadas na extremidade inferior direita do gráfico (pontos vermelhos), as amostras com fluorescência forte para “VIC” foram consideradas homozigotas para o alelo T e são visualizadas na extremidade superior esquerda do gráfico (pontos azuis) e as com fluorescência similar foram consideradas heterozigotas CT e se localizam no centro do gráfico (pontos verdes). Fonte: autora.
3.3.8 Análise estatística
A determinação das frequências genotípicas e alélicas dos sistemas de
grupos sanguíneos e das variantes RHD e RHCE foi realizada por contagem direta.
Para os cálculos das frequências alélicas e haplotípicas dos alelos HLA e
de citocinas assim como do equilíbrio de Hardy-Weinberg, foi utilizado o programa
computacional Arlequin versão 3.5 (151).
Realizou-se a comparação das frequências alélicas e genotípicas entre
dois subgrupos de pacientes portadores de AF: um grupo de pacientes

58
aloimunizados a antígenos eritrocitários e o outro grupo de pacientes não-
aloimunizados e também a comparação do grupo de pacientes com o grupo
controle. A comparação das frequências foi realizada pelo Teste Exato de Fisher e a
correção de Bonferroni foi aplicada quando adequada. A comparação das variáveis
numéricas entre os grupos foi realizada pelo Teste de Mann Whitney. O nível de
significância adotado para os testes estatísticos foi de 5%, ou seja, p<0,05.
O cálculo de desequilíbrio de ligação entre os genes HLA e o polimorfismo
TNFA–308G/A foi realizado segundo Imanish et al. (152).

59
___________________________________________________________________
4 RESULTADOS 4.1 CARACTERÍSTICAS DA POPULAÇÃO ESTUDADA
O presente estudo envolveu 161 pacientes com AF, autodeclarados como
brasileiros com descendência Africana, que receberam múltiplas transfusões de
concentrados de hemácias. Sessenta e sete pacientes (41,6%) estavam
aloimunizados a antígenos eritrocitários e 94 (58,4%) não estavam aloimunizados.
Entre os 67 pacientes aloimunizados 60% eram do sexo feminino e a
mediana de idade entre eles foi de 36 anos (idade mínima: 4 anos, idade máxima:
68 anos). A média de unidades de concentrados de hemácias recebidos por estes
pacientes foi de 83 unidades/paciente (mínimo de 5 unidades e máximo de 449
unidades).
No grupo de pacientes não aloimunizados o sexo feminino também foi
prevalente entre os pacientes (62%). A mediana de idade foi de 34 anos (idade
mínima: 2 anos, idade máxima: 70 anos) e a média de concentrados de hemácias
recebidos por paciente foi de 46 unidades (mínimo de 5 unidades e máximo de 289
unidades).
Foram estudados também 288 doadores de sangue e de medula óssea,
auto identificados como brasileiros com descendência Africana. A maioria destes
doadores eram do sexo feminino (57%) e o grupo apresentou uma mediana de idade
de 33 anos (idade mínima: 18 anos e a idade máxima: 63 anos). Estes doadores não
receberam transfusões sanguíneas.
Na Tabela 5 encontra-se detalhadamente a distribuição das variáveis:
idade, gênero e número de transfusões nos grupos estudados.

60
Tabela 5 - Características da população estudada
Categoria Pacientes com AF
aloimunizados n=67
Pacientes com AF não aloimunizados
n=94
Controles n=288
Gênero
Feminino 40(60%) 58(62%) 165(57%)
Masculino 27(40%) 36(38%) 123(43%)
Idade (em anos)
Mediana (mínimo-máximo) 36(4-68) 34(2-70) 33(18-63)
Transfusões
Média (mínimo-máximo) 83(5-449) 46(5-289) 0(0%)
4.2 PERFIL DE ALOIMUNIZAÇÃO ERITROCITÁRIA DOS PACIENTES COM
ANEMIA FALCIFORME
Entre os 67 pacientes aloimunizados contra antígenos eritrocitários 35
(52,2%) desenvolveram um único aloanticorpo e 32 (47,8%) desenvolveram dois ou
mais aloanticorpos.
Dezesseis especificidades de aloanticorpos foram identificadas e as mais
frequentes foram contra os antígenos RhE (38,8%); RhC (32,8%); RhD (31,34%) e K
(20,9%) (Figura 9). Verificou-se também que quarenta e nove pacientes (73%)
apresentaram pelo menos um anticorpo contra antígenos do sistema Rh (D, C, c, E,
e, CW) e que 37 (55,22%) pacientes aloimunizados apresentavam autoanticorpos.
Entre os pacientes não aloimunizados apenas 11 (11,70%) apresentaram
autoanticorpos (P<0,0001; OR=7,57; IC95%=3,21-19,24).
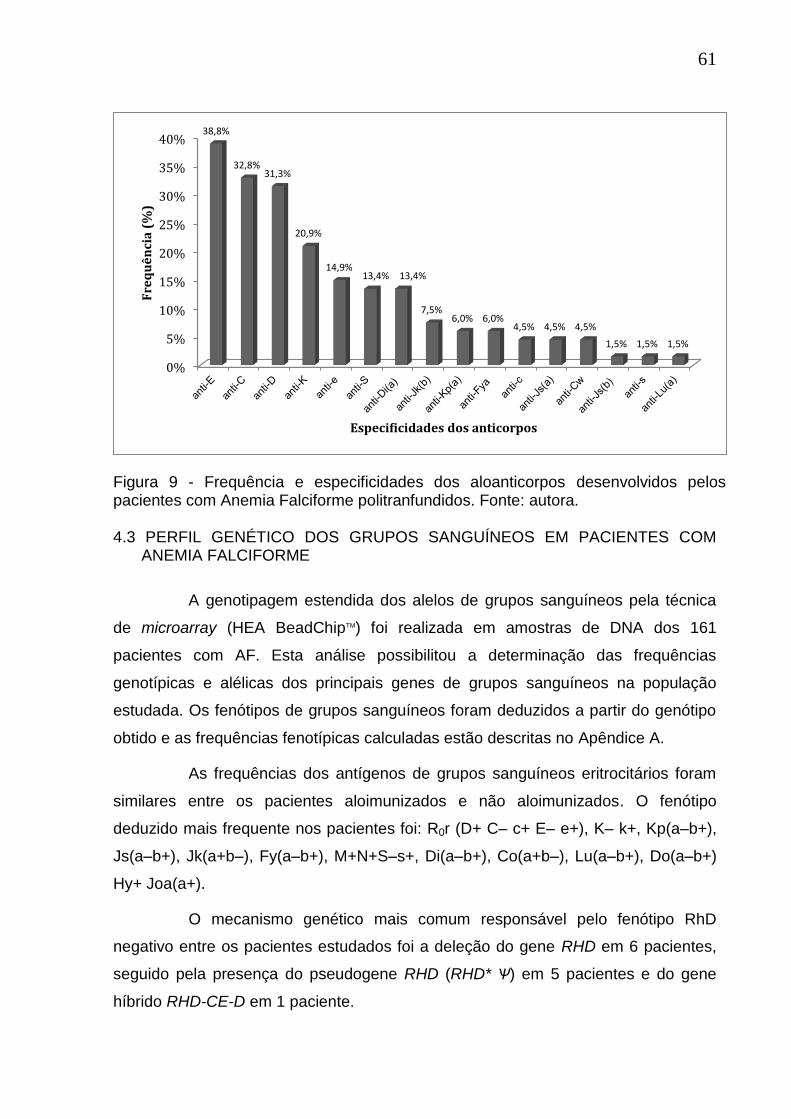
61
Figura 9 - Frequência e especificidades dos aloanticorpos desenvolvidos pelos pacientes com Anemia Falciforme politranfundidos. Fonte: autora. 4.3 PERFIL GENÉTICO DOS GRUPOS SANGUÍNEOS EM PACIENTES COM
ANEMIA FALCIFORME
A genotipagem estendida dos alelos de grupos sanguíneos pela técnica
de microarray (HEA BeadChipTM) foi realizada em amostras de DNA dos 161
pacientes com AF. Esta análise possibilitou a determinação das frequências
genotípicas e alélicas dos principais genes de grupos sanguíneos na população
estudada. Os fenótipos de grupos sanguíneos foram deduzidos a partir do genótipo
obtido e as frequências fenotípicas calculadas estão descritas no Apêndice A.
As frequências dos antígenos de grupos sanguíneos eritrocitários foram
similares entre os pacientes aloimunizados e não aloimunizados. O fenótipo
deduzido mais frequente nos pacientes foi: R0r (D+ C– c+ E– e+), K– k+, Kp(a–b+),
Js(a–b+), Jk(a+b–), Fy(a–b+), M+N+S–s+, Di(a–b+), Co(a+b–), Lu(a–b+), Do(a–b+)
Hy+ Joa(a+).
O mecanismo genético mais comum responsável pelo fenótipo RhD
negativo entre os pacientes estudados foi a deleção do gene RHD em 6 pacientes,
seguido pela presença do pseudogene RHD (RHD* Ѱ) em 5 pacientes e do gene
híbrido RHD-CE-D em 1 paciente.
0%
5%
10%
15%
20%
25%
30%
35%
40%38,8%
32,8% 31,3%
20,9%
14,9% 13,4% 13,4%
7,5% 6,0% 6,0%
4,5% 4,5% 4,5%
1,5% 1,5% 1,5%
Fre
qu
ên
cia
(%
)
Especificidades dos anticorpos

62
Na análise do sistema Duffy, observamos a presença do fenótipo
Fy(a–b–) em 21,12% dos pacientes, cujo mecanismo molecular responsável pela
supressão da expressão do antígeno Fyb foi a presença da mutação –67T/C no
promotor eritroide GATA1 do gene FY em homozigose (FY*02N01/FY*02N01). O
fenótipo Fy(a+b–) decorrente desta mutação em heterozigose foi encontrado em
21,74% dos pacientes, os quais possuíam o genótipo FY*01/FY*02N01.
Identificamos também um paciente com o fenótipo Fy(a+bx) devido a presença da
mutação 265C/T em heterozigose (genótipo FY*01/FY02W01) e um paciente Fy(a–
bx) que possuía tanto a mutação –67T/C quanto a mutação 265C/T (genótipo
FY*02N.01/ FY*02W.01) em heterozigoze.
4.4 VARIANTES RH E IMUNIZAÇÃO RH EM PACIENTES COM ANEMIA
FALCIFORME
Com a finalidade de caracterizarmos os anticorpos Rh desenvolvidos por
pacientes falciformes realizamos um estudo sorológico, molecular e clínico em 48
pacientes aloimunizados contra os antígenos Rh. Os resultados estão descritos a
seguir.
4.4.1 Análise sorológica dos antígenos e anticorpos Rh
Entre os 48 pacientes com AF que desenvolveram apenas anticorpos anti-
Rh 9 não apresentavam o antígeno para a especificidade do anticorpo
correspondente (3 anti-D em pacientes D– e 6 anti-C em pacientes C–) e 39
apresentavam o antígeno Rh correspondente (Quadro 1). Controles autólogos, teste
direto da antiglobulina (TDA) e eluato foram positivos em 8 dos 39 pacientes com
anticorpos Rh e antígenos correspondentes. Adsorções em hemácias autólogas
demonstraram que estes 8 pacientes tinham auto-anticorpos.
4.4.2 Análise molecular de variantes RHD e RHCE
A análise molecular revelou alelos RHD e RHCE associados com
variantes dos antígenos RhD, RhC e Rhe em 31 dos 48 pacientes com anticorpos
anti-Rh. Variantes RHD foram encontradas em 23/31 pacientes, variantes RHCE em
28/31 e combinações alélicas RHD e RHCE em 20/31 pacientes. Os alelos RHD

63
mais frequentemente encontrados em nosso grupo de pacientes foram RHD*DAR,
RHD*fraco parcial 4.0, RHD*DIIIa e RHD*DAU0. Com relação aos alelos RHCE, os
mais frequentes foram RHCE*ceS, RHCE*ce48C, RHCE*ce733G, RHCE*ceAR e
RHCE*ceTI. Alelos RHD variantes estavam presentes em homozigose e em
heterozigose em 44% dos casos e no estado hemizigoto em 12% dos pacientes.
Alelos variantes RHCE em homozigose foram encontrados em 25% dos casos,
heterozigoto composto em 43% e em heterozigose com um alelo convencional em
29% dos pacientes. Associações alélicas mostraram que 5 pacientes eram
heterozigotos compostos para variantes RHD e RHCE (Quadro 1).
4.4.3 Imunização Rh e significado clínico dos anticorpos Rh
A análise retrospectiva mostrou que 13 pacientes com variantes Rh que
desenvolveram aloanticorpos (4 anti-D, 4 anti-C e 5 anti-e) apresentaram RTHT ou
diminuição da sobrevivência das hemácias transfundidas, verificado pela mudança
dos valores basais de Hb pré-transfusional e porcentagem (%) de HbS no momento
da detecção dos anticorpos. Esses anticorpos foram associados com piora da
anemia e/ou aumento de HbS (Tabela 6).

64
Quadro 1 - Alelos RH em 39 pacientes portadores de Anemia Falciforme que desenvolveram anticorpos anti-Rh e que apresentavam tipagem eritrocitária positiva
para o antígeno correspondente.
n Genótipo RH e Fenótipo deduzido
RHD RHCE
Pacientes D+ com anti-D considerados aloanticorpos
Genótipo Fenótipo deduzido
Genótipo Fenótipo deduzido
3 DIIIa/DIIIa D parcial ceS/ce
S c+, e parcial
3 DAR/DAR D parcial ce/ceAR c+e+
1 DAR/D deletado D parcial ce/ceEK c+e+
2 DAU0/D deletado D parcial ce/ce c+e+
1 DAU3/DAU5 D parcial cE/ce c+E+e+
1 DAU4/DAU4 D parcial ce48C/ce48C c+e+w
3 Fraco parcial 4.0/Fraco parcial 4.0 D parcial ce733G/ce733G c+, e parcial
1 Fraco parcial 4.0/DIIIa-CE(4-7)-D D parcial ce/ceS c+e+
Pacientes e+ com anti-e considerados autoanticorpos
5 RHD/RHD D+ ce/ce c+e+
1 RHD/D deletado D+ Ce/ce C+c+e+
2 RHD/D deletado D+ ce/cE c+E+e+
Pacientes e+ com anti-e considerados aloanticorpos
2 RHD/RHD D+ Ce/ce48C,733G e parcial
3 D Deletado D– ceS/ce733G e parcial
2 RHD/RHD D+ ce48C/ceS e parcial
1 D Deletado D– ce48C,733G/ce733G e parcial
Pacientes com D+ e e+ com anti-D e anti-e considerados aloanticorpos
1 DAR/DAR D parcial ceS/ceAR e parcial
1 DAU5/DAU0 D parcial ce48C/ceMO e parcial
Pacientes com C+ com anti-C considerados aloanticorpos
1 DIIIa-CE(4-7)-D/DAU0 D parcial (C)ces/ce C parcial
3 DIIIa-CE(4-7)-D/ DIVa-2 D parcial (C)ces/ceTI C parcial, e parcial
Pacientes com D+ e C+ com anti-D e anti-C considerados aloanticorpos
2 DIIIa-CE(4-7)-D/ DIVa-2 D parcial (C)ces/ceTI C parcial, e parcial

65
Tabela 6 - Variantes RH encontradas em 13 pacientes com Anemia Falciforme associados com anticorpos clinicamente significativos envolvidos em RTHT ou
diminuição da sobrevivência de hemácias transfundidas. ID
Especificidade do anticorpo
variante RH Transfusões de hemácias
Hemoglobina S (%) na detecção do
anticorpo
Hemoglobina (g/dL) na detecção do
anticorpo
n Pré-transfusão
Pós-transfusão
Pré-transfusão
Pós-transfusão
375 Anti-e ceS/ce733G 64 23,8 45,8 7,8 6,9
509 Anti-e ceS/ce733G 38 24,3 58,1 8,5 6,4
510 Anti-e ce48C/ceS 142 29,6 42,1 9,8 8,1
516 Anti-e ce733G/ce733G 94 25,3 40,8 8,9 7,5
517 Anti-e ce733G/ce733G 8 28,9 46,1 9,5 8,2
534 Anti-D DIIIa/DIIIa 103 19,6 44,6 8,4 7,2
536 Anti-D DIIIa/DIIIa 32 29,3 47,2 9,0 7,8
538 Anti-D DAR/DAR 4 26,1 34,6 7,9 6,8
562 Anti-D DAU4/DAU4 6 23,1 43,6 8,6 8,0
711 Anti-C DIIIa-CE(4-7)-D/DIVa-2
84 24,1 45,3 7,9 6,8
722 Anti-C DIIIa-CE(4-7)-D/DIVa-2
114 44,3 54,8 6,9 5,9
738 Anti-C DIIIa-CE(4-7)-D/DIVa-2
62 27,6 58,1 8,6 7,6
752 Anti-C DIIIa-CE(4-7)-D/DAU0
321 26,4 50,7 7,3 6,5
Anticorpos Rh detectados nestes pacientes foram considerados aloanticorpos. RTHT: Reação Transfusional Hemolítica Tardia.

66
4.5 FREQUÊNCIA DOS POLIMORFISMOS EM GENES DE CITOCINAS E A ASSOCIAÇÃO COM A ALOIMUNIZAÇÃO ERITROCITÁRIA EM PACIENTES COM ANEMIA FALCIFORME
Em todos os grupos estudados, as distribuições genotípicas dos
polimorfismos IL10–1082G/C, IL10–819T/C, IL10–592C/A, IL17A–197A/G, IL17F
7488A/G, IL6–174C/T, TNFA–308G/A, IL1B–511C/T, IL4-590C/T e IL4 íntron 3
VNTR encontraram-se em equilíbrio de Hardy-Weinberg (P≥0,05 nos pacientes e
nos controles). As associações significativas encontradas na análise dos
polimorfismos nos genes das citocinas analisadas com a aloimunização eritrocitária
estão resumidas nas Tabela 7 e 8.
A análise dos polimorfismos em genes de citocinas revelou que o alelo
TNFA–308A foi prevalente nos pacientes aloimunizados quando comparado com os
não aloimunizados (16,42% vs 5,85% P=0,004; OR=3,14; IC=1,4-7,49; Tabela 7).
Uma diferença significativa também foi encontrada na distribuição dos genótipos
deste polimorfismo, sendo o genótipo TNFA–308GA encontrado em maior
frequência em pacientes aloimunizados do que em pacientes não-aloimunizados
(32,84% vs 11,70%; P=0,0021; OR=3,65; IC=1,53-9,16; Tabela 7). Estes resultados
sugerem uma forte associação do alelo TNFA-308A e do genótipo TNFA–308GA
com a susceptibilidade à aloimunização eritrocitária.
Diferenças significativas foram encontradas na distribuição dos alelos e
genótipos do SNP IL1B–511C/T em pacientes aloimunizados e não aloimunizados.
A frequência do alelo T foi significativamente maior em aloimunizados do que em
não aloimunizados (aloimunizados: 53,03% vs não aloimunizados: 37,50%;
P=0.0085; OR=1.87; IC=1.16-3.04; Tabela 8). Além disso, os modelos estatísticos
dominate e codominante demonstraram que ambos os genótipos IL1B–511TT e
IL1B–511TC foram significativamente mais frequentes em pacientes aloimunizados
do que em não aloimunizados indicando uma associação positiva destes genótipos
com a aloimunização eritrocitária em pacientes com AF (Tabela 8).
A análise dos polimorfismos –1082G/A, –819T/C e –592C/A no gene IL10
não mostrou associações significantes com a aloimunização eritrocitária (Tabela 9
Apêndices B e C). No entanto, as comparações das frequências alélicas e
genotípicas do SNP –1082G/A entre pacientes e controles demonstraram que o
alelo G e o genótipo GG foram significativamente menos frequente em pacientes

67
com AF do que em controles saudáveis (alelo G: 27,64% vs 34,2%, P= 0,05;
Genótipo GG: 6,21% vs 13%, P=0,02; Tabela 9). Além disso, menor frequência do
genótipo –1082GG também foi encontrado em pacientes aloimunizados quando
comparado com controles saudáveis (genótipo GG: 4,48% vs 13%; P= 0,05).
Vários possíveis diplótipos foram encontrados na análise das
combinações entre os alelos dos polimorfismos IL10 (–1082G/A, –819T/C e
–592C/A) (Tabela 10). Nos pacientes com AF os mais comumente encontrados
foram: ATA/ACC (25,79%), ATA/GCC (20,75%) e ACC/GCC (20,13%). O diplótipo
GCC/GCC foi encontrado em menor frequência nos pacientes falciformes do que
nos controles (6,29% vs 13,2%; P=0,031).
Dois polimorfismos foram estudados no gene IL4, a inserção de
fragmentos de 70pb no íntron 3 e o SNP –590C/T que se localiza na região
promotora do gene. Embora as frequências alélicas e genotípicas de ambos os
polimorfismos não diferiram estatisticamente entre os grupos (Apêndices D e E), a
análise haplotípica destes polimorfismos mostrou que, entre os pacientes, os
diplótipos mais frequentes foram: C,RP2/T,RP1 (37,58%), C,RP2/C,RP2 (24,84%),
T,RP1/T,RP1 (15,29%) e T,RP2/T,RP1 (8,28%). Diferença estatística significativa foi
encontrada nas frequências dos diplótipos T,RP2/T,RP1 e T,RP2/ T,RP2 entre
pacientes com AF e controles, como pode ser verificado na Tabela 11. O diplótipo
T,RP1/T,RP1 foi significativamente mais frequente em pacientes aloimunizados
quando comparados aos controles (22,22% vs 10,8%; P=0,031), mas não alcançou
significância estatística quando comparado aos não-aloimunizados (15,29% vs 10,64
%, P>0,05).
As frequências alélicas e genotípicas dos SNPs IL17A–197A/G, IL17F
7488A/G, IL6–174C/T e haplótipos de IL17, não diferiram estatisticamente entre
pacientes aloimunizados e não aloimunizados e entre pacientes e controles
(Apêndices F - I).

68
Tabela 7 - Frequências alélicas e genotípicas dos SNPs TNFA–308G/A (rs1800629) em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
SNP Modelo
Estatístico Genótipos /
alelos
PA PNA Pacientes
totais Controles Análise Estatística
n=67(%) n=94(%) n=161(%) n=288(%) PA vs PNA PA vs
Controles
Pacientes Totais vs Controles
TNFA –308G/A (rs1800629)
Genótipos
Codominante
GG 45(67,16) 83(88,30) 128(79,50) 222(77,10) ref ref ref
GA 22(32,84) 11(11,70) 33(20,50) 62(21,50) P=0,0021; OR=3,65; IC=1,53-9,16 ns ns
AA 0(0,00) 0(0,00) 0(0,00) 4(1,40) ns ns ns
Dominante
GG 45(67,16) 83(88,30) 128(79,50) 222(77,10) ref ref ref
GA+AA 22(32,84) 11(11,70) 33(20,50) 66(22,90) P=0,0021; OR=3,65; IC=1,53-9,16 ns ns
Recessivo
GG+GA 67(100,00) 94(100,00) 161(100,00) 284(98,60) ref ref ref
AA 0(0,00) 0(0,00) 0(0,00) 4(1,40) ns ns ns
Over dominante
GG+AA 45(67,16) 83(88,30) 128(79,5) 226(78,50) ref ref ref
GA 22(32,84) 11(11,70) 33(20,5) 62(21,50) ns ns ns
Alelos
G 112(83,58) 177(94,15) 289(89,75) 506(87,80) ref ref ref
A 22(16,42) 11(5,85) 33(10,25) 70(12,20) P=0,004; OR=3,14; IC=1,4-7,49 ns ns
PA: pacientes aloimunizados portadores de Anemia Falciforme; PNA: pacientes não aloimunizados portadores de Anemia Falciforme; Pacientes Totais: pacientes aloimunizados + não aloimunizados; OR: odds ratio; IC: intervalo de 95% de confiança; ref: grupo de referência; ns: não significativo.

69
Tabela 8 - Frequências alélicas e genotípicas do SNP IL1B –511C/T (rs16944) em pacientes portadores de Anemia Falciforme
aloimunizados e não aloimunizados e em controles.
SNP Modelo Estatístico Genótipos / alelos
PA PNA Pacientes
totais Controles Análise Estatística
n=66(%) n=92(%) n=158(%) n=288(%) PA vs PNA PA vs
Controles
Pacientes Totais vs Controles
IL1B–511C/T (rs16944)
Genótipos
Codominante
CC 12(18,18) 36(39,13) 48(30,38) 81(28,12) ref ref ref
CT 38(57,58) 43(46,74) 81(51,27) 151(52,44) P=0,02; OR=2,63; IC=1,14-6,39 ns ns
TT 16(24,24) 13(14,13) 29(18,35) 56(19,44) P=0,015; OR=3,6; IC=1,24-11,05 ns ns
Dominante
CC 12(18,18) 36(39,13) 48(30,38) 81(28,10) ref ref ref
CT+TT 54(81,82) 56(60,87) 110(69,62) 207(71,90) P=0,0071; OR=2,87; IC=1,29-6,73 ns ns
Recessivo
CC+CT 50(75,80) 79(85,90) 129(81,70) 232(80,60) ref ref ref
TT 16(24,20) 13(14,10) 29(18,40) 56(19,40) ns ns ns
Over dominante
CC+TT 28(42,4) 49(53,3) 77(48,7) 137(47,60) ref ns ns
CT 38(57,6) 43(46,7) 81(51,3) 151(52,40) ns ns ns
Alelos
C 62(46,97) 115(62,50) 177(56,01) 313(54,34) ref ref ref
T 70(53,03) 69(37,50) 139(43,99) 263(45,65) P=0,0085; OR=1,87; IC=1,16-3,04 ns ns
PA: pacientes aloimunizados portadores de Anemia Falciforme; PNA: pacientes não aloimunizados portadores de Anemia Falciforme; Pacientes Totais: pacientes aloimunizados + não aloimunizados; OR: odds ratio; IC: intervalo de 95% de confiança; ref: grupo de referência; ns: não significativo.

70
Tabela 9 - Frequências alélicas e genotípicas do SNP IL10 –1082G/A (rs1800896) em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
SNP Modelo
Estatístico Genótipos /
alelos
PA PNA Pacientes
totais Controles Análise Estatística
n=67(%) n=94(%) n=161(%) n=288(%) PA vs PNA PA vs Controles Pacientes Totais vs Controles
IL10 –1082G/A
(rs1800896)
Genótipo
Codominante
AA 35(52,24) 47(50,00) 82(50,93) 130(45,10) ref ref ref
AG 29(43,28) 40(42,55) 69(42,86) 119(41,30) ns ns ns
GG 3(4,48) 7(7,45) 10(6,21) 39(13,60) ns P=0,05; OR=0,29; IC=0,053-0,99 P=0,02; OR=0,41; IC=0,17-0,89
Dominante AA 35(52,24) 47(50) 82(50,93) 130(45,10) ref ref ref
AG+GG 32(47,76) 47(50) 79(49,07) 158(54,90) ns ns ns
Recessivo AG+AA 64(95,52) 87(92,55) 151(93,79) 249(86,50) ref ref ref
GG 3(4,48) 7(7,45) 10(6,21) 39(13,50) ns ns P=0,021; OR=0,42; IC=0,183-0,895
Over dominante
AA+GG 38(56,72) 54(57,45) 92(57,10) 169(58,70) ref ref ref
AG 29(43,28) 40(42,55) 69(42,90) 119(41,30) ns ns ns
Alelo
A 99(73,88) 134(71,28) 233(72,36) 379(65,80) ref ref ref
G 35(26,12) 54(28,72) 89(27,64) 197(34,20) ns ns P=0,050; OR=0,74; IC=0,54-1,00
PA: pacientes aloimunizados portadores de Anemia Falciforme; PNA: pacientes não aloimunizados portadores de Anemia Falciforme; Pacientes Totais: pacientes aloimunizados + não aloimunizados; OR: odds ratio; IC: intervalo de 95% de confiança; ref: grupo de referência; ns: não significativo.

71
Tabela 10 - Frequências dos diplótipos formados pelos polimorfismos IL10 (–1082G/A,–819T/C, –592C/A) em pacientes portadores de Anemia Falciforme
aloimunizados e não aloimunizados e em controles.
Diplótipos PA
n=66(%) PNA
n=93(%) Pacientes Totais
n=159(%) Controles n=288(%)
ATA/ACC 20(30,30) 21(22,58) 41(25,79) 69(23,90)
ATA/GCC 15(22,73) 18(19,35) 33(20,75) 64(22,20)
ACC/GCC 12(18,18) 20(21,51) 32(20,13) 52(18,00)
ATA/ATA 6(9,09) 12(12,90) 18(11,32) 37(12,80)
ACC/ACC 6(9,09) 12(12,90) 18(11,32) 18(6,60)
GCC/GCC 3(4,55) 7(7,53) 10(6,29)a 38(13,20)
a
ATA/ATC 2(3,03) 0(0,00) 2(1,26) 1(0,35)
ATC/ATC 1(1,52) 0(0,00) 1(0,63) 3(1,00)
ACA/GCA 1(1,52) 0(0,00) 1(0,63) 1(0,35)
ACA/ACC 0(0,00) 2(2,15) 2(1,26) 1(0,35)
GCC/ATC 0(0,00) 1(1,08) 1(0,63) 1(0,35)
GTA/ATA 0(0,00) 0(0,00) 0(0,00) 1(0,35)
ACA/ACA 0(0,00) 0(0,00) 0(0,00) 1(0,35)
GCA/GCC 0(0,00) 0(0,00) 0(0,00) 1(0,35)
PA: pacientes aloimunizados portadores de Anemia Falciforme; PNA: pacientes não aloimunizados portadores de Anemia Falciforme; Pacientes Totais: pacientes aloimunizados + não aloimunizados; OR: odds ratio; IC: intervalo de 95% de confiança;
aP=0,031; OR=0,44; IC=0,19-0,93.

72
Tabela 11 - Frequências dos diplótipos formados pelos polimorfismos IL4-590C/T e IL4 íntron 3 VNTR em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
Diplótipos PA
n=63(%) PNA
n=94(%)
Pacientes Totais
n=157(%)
Controles n=288(%)
Análise Estatística
PA vs PNA PA vs controles Pacientes vs controles
C, RP2/C, RP2 13(20,63) 26(27,66) 39(24,84) 93(32,29) ns ns ns
C, RP2/T, RP2 6(9,52) 3(3,20) 9(5,73) 32(11,11) ns ns ns
C, RP2/C, RP1 1(1,59) 0(0,00) 1(0,64) 2(0,70) ns ns ns
C, RP2/T, RP1 22(34,92) 37(39,36) 59(37,58) 96(33,33) ns ns ns
T, RP2/T, RP2 1(1,59) 3(3,19) 4(2,55) 24(8,33) ns ns P=0,021; OR=0,28; IC=0,071-0,86
T, RP2/T, RP1 4(6,35) 9(9,57) 13(8,28) 7(2,43) ns ns P=0,010; OR=3,61; IC=1,30-10,94
T, RP1/T, RP1 14(22,22) 10(10,64) 24(15,29) 31(10,76) ns P=0,031; OR=2,36; IC=1,07-4,97 ns
T, RP1/T, RP3 1(1,59) 2(2,13) 3(1,91) 1(0,35) ns ns ns
C, RP2/T, RP3 0(0,00) 3(3,19) 3(1,91) 0(0,00) ns ns ns
C, RP1/C, RP1 1(1,59) 0(0,00) 1(0,64) 2(0,70) ns ns ns
C, RP1/T, RP1 0(0,00) 1(1,06) 1(0,64) 0(0,00) ns ns ns
PA: pacientes aloimunizados portadores de Anemia Falciforme; PNA: pacientes não aloimunizados portadores de Anemia Falciforme; Pacientes Totais: pacientes aloimunizados + não aloimunizados; OR: odds ratio; IC: intervalo de 95% de confiança; ns: não significativo.

73
4.6 FREQUÊNCIAS DOS GRUPOS ALÉLICOS HLA DE CLASSE II E A ASSOCIAÇÃO COM A ALOIMUNIZAÇÃO ERITROCITÁRIA EM PACIENTES COM ANEMIA FALCIFORME
Em todos os grupos analisados a distribuição das frequências genotípicas
dos grupos alélicos HLA mostrou-se em equilíbrio de Hardy-Weinberg (p≥0,05).
Com relação ao locus HLA-DRB1, a análise das frequências dos grupos
alélicos entre os pacientes portadores de AF e o grupo de indivíduos controles
saudáveis não mostrou diferença estatística significativa (Tabela 12). Quando
analisamos pacientes aloimunizados e não aloimunizados observamos uma maior
frequência do HLA-DRB1*15 no grupo de pacientes com aloanticorpos do que no
grupo de não aloimunizados, no entanto, apenas uma tendência à significância
estatística foi encontrada (13,85% vs 6,98%; P=0,068; OR=2,17; CI=0,94-5,16).
Além disso, a frequência do HLA-DRB1*04 em pacientes aloimunizados foi menor do
que em não aloimunizados, mas novamente apenas uma tendência a significância
estatística foi verificada (7,69% vs 15,79%; P=0,057; OR=0,456; CI=0,18-1,02).
Devido à alta porcentagem de pacientes aloimunizados que
desenvolveram um ou múltiplos aloanticorpos dirigidos à antígenos do sistema Rh
(D, E, C, c, e), analisamos comparativamente as frequências dos grupos alélicos
HLA-DRB1 entre o grupo de pacientes com aloanticorpos anti-Rh versus o grupo de
pacientes não aloimunizados. Além disso, comparamos o grupo de pacientes com
aloanticorpos anti-Rh versus o grupo de pacientes aloimunizados por outros
antígenos (anticorpos não Rh). Observamos que o HLA-DRB1*15 foi mais frequente
em pacientes com anticorpos Rh do que em pacientes não aloimunizados, não
diferindo do grupo de pacientes com outros aloanticorpos, indicando que este grupo
alélico pode ser um fator de suscetibilidade à aloimunização Rh (15,63% vs 6,98%,
P= 0,044, OR= 2,46; IC95%: 1,021-6,05; Tabela 13).
Na análise dos possíveis haplótipos formados pelos grupos alélicos HLA-
DRB1 e o polimorfismo TNF–308G/A encontramos uma maior frequência do
haplótipo DRB1*15/TNF-308G em pacientes aloimunizados contra antígenos Rh do
que em pacientes não aloimunizados (14,6% vs 5,81%; P=0,031; OR=2,75; IC=1,08-
7,25). O cálculo do desequilíbrio de ligação do haplótipo DRB1*15/TNF-308G em
pacientes aloimunizados e não aloimunizados não mostrou diferença estatística
significante (P>0.05).

74
As frequências dos grupos alélicos -DQA1 e -DQB1 foram semelhantes
entre os grupos de pacientes com AF aloimunizados e o grupo de pacientes não
aloimunizados e também não foi encontrada associação significante destes locus
com a aloimunização eritrocitária (Apêndices J, K e L).
Tabela 12 - Distribuição das frequências dos grupos alélicos HLA-DRB1 entre pacientes portadores de Anemia Falciforme aloimunizados a antígenos eritrocitários e
pacientes não aloimunizados.
HLA-DRB1 PA
n=130(%) PNA
n=172(%) Pacientes totais
n=302(%) Controles n=576(%)
DRB1*01 8(6,15) 16(9,30) 24(7,95) 54(9,38)
DRB1*10 3(2,31) 3(1,74) 6(1,99) 8(1,39)
DRB1*11 23(17,69) 24(13,95) 47(15,56) 73(12,67)
DRB1*12 5(3,85) 5(2,91) 10(3,31) 19(3,30)
DRB1*13 15(11,54) 19(11,05) 34(11,26) 90(15,63)
DRB1*14 3(2,31) 8(4,65) 11(3,64) 17(2,95)
DRB1*15 18(13,85)a 12(6,98)
a 30(9,93) 69(11,98)
DRB1*16 3(2,31) 10(5,81) 13(4,30) 20(3,47)
DRB1*03 13(10,00) 17(9,88) 30(9,93) 76(13,19)
DRB1*04 10(7,69)b 27(15,70)
b 37(12,25) 43(7,47)
DRB1*07 18(13,85) 17(9,88) 35(11,59) 61(10,59)
DRB1*08 8(6,15) 11(6,40) 19(6,29) 36(6,25)
DRB1*09 3(2,31) 3(1,74) 6(1,99) 10(1,74)
PA: pacientes aloimunizados portadores de Anemia Falciforme; PNA: pacientes não aloimunizados portadores de Anemia Falciforme; Pacientes Totais: pacientes aloimunizados+ não aloimunizados; OR: odds ratio; IC: intervalo de 95% de confiança;
aPA versus PNA, P=0,068 (OR=2,17; CI=0,94-
5,16); bPA versus PNA, P=0,057 (OR=0,456; CI=0,18-1,02).

75
Tabela 13 - Frequências dos grupos alélicos DRB1 em pacientes com aloanticorpos contra antígenos Rh (D, C, c, E e/ou e), em pacientes com aloanticorpos não Rh e
pacientes não aloimunizados.
HLA-DRB1
Pacientes com aloanticorpos anti- Rh
(A) n=96(%)
Pacientes com aloanticorpos não Rh
(B) n=34(%)
Pacientes não aloimunizados (C)
n=172(%)
DRB1*01 7(7,29) 1(2,94) 16(9,30)
DRB1*10 3(3,13) 0(0,00) 3(1,74)
DRB1*11 16(16,67) 7(20,59) 24(13,95)
DRB1*12 3(3,13) 2(5,88) 5(2,91)
DRB1*13 10(10,42) 4(14,71) 19(11,05)
DRB1*14 2(2,08) 1(2,94) 8(4,65)
DRB1*15 15(15,63)a
2(8,82) 12(6,98)a
DRB1*16 3(3,13) 0(0,00) 10(5,81)
DRB1*03 7(7,29) 6(17,65) 17(9,88)
DRB1*04 8(8,33) 2(5,88) 27(15,70)
DRB1*07 12(12,50) 6(17,65) 17(9,88)
DRB1*08 7(7,29) 1(2,94) 11(6,40)
DRB1*09 3(3,13) 0(0,00) 3(1,74) a A versus C, P=0,044 (OR=2,46;CI=1,021-6,05).

76
___________________________________________________________________
5 DISCUSSÃO 5.1 CARACTERÍSTICAS DA POPULAÇÃO ESTUDADA
As variáveis idade, sexo e etnia não diferiram estatisticamente entre os
pacientes aloimunizados, pacientes não aloimunizados e indivíduos controles,
demonstrando que houve homogeneidade entre os grupos em relação a estas
variáveis.
Com respeito ao número de transfusões observou-se que a média de
unidades de concentrado de hemácias transfundidas foi maior em aloimunizados,
porém não diferiu estatisticamente do número de bolsas transfundidas aos pacientes
não aloimunizados, o que demonstra que o número de transfusões não foi um fator
que interferiu em nossa análise.
5.2 PERFIL DE ALOIMUNIZAÇÃO ERITROCITÁRIA DOS PACIENTES COM
ANEMIA FALCIFORME
A prevenção da aloimunização continua a ser um grande desafio para
todos os serviços que tratam pacientes com AF. Apesar das inúmeras estratégias
transfusionais propostas ao longo dos anos (20,27) as taxas de aloimunização neste
grupo de pacientes têm se mantido entre 18% (10) e 47%(11). Em nosso estudo, a
taxa de aloimunização eritrocitária encontrada foi de 41,6%; enquanto que, um
estudo realizado no estado da Bahia encontrou uma taxa de aloimunização ainda
maior (52%), com alta prevalência de anticorpos anti-E, anti-K e anti-C (39,3%,
21,4% e 16,1%), respectivamente (12).
Entre os pacientes aloimunizados da nossa casuística, 73% possuíam
anticorpos dirigidos aos antígenos do sistema Rh, sendo anti-E (38,8%) e anti-C
(32,8%) os aloanticorpos mais frequentemente encontrados, ocorrendo sozinhos ou
em combinação com outros aloanticorpos. Outros estudos também mostraram que a
maioria dos anticorpos encontrados em pacientes com AF são dirigidos a esses
antígenos (10,12,32) e isto pode ser parcialmente explicado pelo fato de muitos
pacientes receberem transfusões Rh incompatíveis.

77
Nossos resultados mostraram ainda uma alta frequência de
autoanticorpos entre os pacientes aloimunizados quando comparados com os não
aloimunizados, demonstrando que pacientes aloimunizados com AF possuem 7,5
vezes mais chances de apresentar autoanticorpos do que os pacientes não
aloimunizados, o que está de acordo com estudos anteriores (11,21,22).
5.3 PERFIL GENÉTICO DOS GRUPOS SANGUÍNEOS EM PACIENTES COM
ANEMIA FALCIFORME
Em nossa casuística de pacientes com AF, o fenótipo eritrocitário mais
frequente foi C–, E–, K–, Fy(a–), Jk(b–), S–, Di(a–). De acordo com a literatura este
fenótipo é característico de populações Africanas e reflete, portanto a população
estudada (13). A alta frequência deste fenótipo eritrocitário nestes pacientes pode
explicar em parte a alta taxa de aloimunização encontrada em nosso estudo para os
antígenos E (38,8%), C(32,8%), K (20,9%), S e Dia (13,4%).
De acordo com estudos prévios, o fenótipo eritrocitário C–, E–, S–, K–,
Fy(a–), Jk(b–) é encontrado em 1 a cada 6 doadores de sangue Afro-americanos,
mas em apenas 1 a cada 131 doadores Caucasianos (153). Assim, pacientes
falciformes transfundidos com sangue de doadores Caucasianos possuem grande
probabilidade de receber hemácias positivas para esses antígenos e se aloimunizar.
Por isso, com o objetivo de aumentar o número de doadores Afrodescendentes
muitos serviços americanos têm incentivado o recrutamento de doadores com
descendência Africana para suprir a necessidade transfusional de pacientes com AF
e reduzir a taxa de aloimunização (154).
Em nossa casuística, identificamos 69 pacientes com o fenótipo Fy(b–)
que poderiam receber sangue Fy(b+), pois apresentam a mutação –67T>C no
promotor eritroide GATA1. Essa mutação frequente em descendentes de Africanos
leva a não expressão do antígeno Fyb nas hemácias, no entanto, o gene FY*B é
expresso em outros tecidos, como por exemplo, nas células do endotélio dos vasos
sanguíneos, não havendo risco de desenvolvimento de anticorpos anti-Fyb (155–
157). Isto significa que a disponibilidade de sangue compatível para estes pacientes
pode aumentar consideravelmente se este polimorfismo for analisado (158).
Ainda com relação ao Sistema Duffy, verificamos que 2 pacientes foram
genotipados como: FY*01/FY*02W.01 e FY*02N.01/ FY*02W.01, ou seja, portadores

78
do alelo FY*B–265T, que leva a fraca expressão do antígeno Fyb nas hemácias de
Caucasianos. É interessante notar que o segundo paciente possui tanto a mutação
GATA (–67T/C) quanto a mutação 265C/T, caracterizando a miscigenação da
população brasileira (158).
O fenótipo RhD negativo, está associado a vários mecanismos genéticos,
dentre os quais, a presença do gene hibrido RHD-CE-D e a presença de uma
inserção de 37pb que resulta em um stop códon prematuro e caracteriza o
pseudogene RHD (RHD*Ѱ), frequentemente encontrados em Africanos RhD
negativos, e a deleção do gene RHD que é mais comum em Caucasianos mas
também ocorre em Afrodescendentes (138,144). Em nossos 12 pacientes tipados
como RhD-negativo o mecanismo molecular mais frequente foi a deleção do gene
RHD (50%) seguido pela presença do pseudogene RHD (41,7%) e pela formação de
gene hibrido RHD-CE-D (8,3%). Quatro destes pacientes estavam aloimunizados ao
antígeno D, demonstrando assim a importância da análise destes três mecanismos
genéticos quando se realiza a genotipagem RHD em pacientes portadores de AF.
5.4 VARIANTES RH E IMUNIZAÇÃO RH EM PACIENTES COM ANEMIA
FALCIFORME
A presença de alelos RH variantes nos 48 pacientes que desenvolveram
anticorpos Rh foi verificada, assim como o significado clínico dos anticorpos
produzidos.
As variantes RH caracterizadas neste trabalho foram associadas
anteriormente com aloimunização Rh em pacientes com AF (27,29,30,159) e foi
demonstrado que anticorpos anti-Rh produzidos por pacientes com as variantes
encontradas entre nossos pacientes ou anticorpos contra antígenos de alta
prevalência podem estar envolvidos na ocorrência de RTH tardias ou agudas
(27,30). Em nosso estudo, 13 dos anticorpos anti-Rh desenvolvidos contra antígenos
D, C e e alterados demonstraram ser clinicamente significantes, o que foi
evidenciado por uma piora na anemia e/ou um aumento da HbS quando os
pacientes foram transfundidos com os antígenos correspondentes.
Além disso, em pacientes com AF, existe uma forte associação entre o
desenvolvimento de autoanticorpos e aloimunização eritrocitária (27,29,159). Em
nosso trabalho, a análise do gene RHCE em conjunto com a análise sorológica

79
auxiliou a identificar autoanticorpos contra o antígeno Rhe em 8 dos 48 pacientes
falciformes (17%). O conhecimento da prevalência de variantes RH e estudos como
este que demonstram o significado clínico dos anticorpos produzidos em pacientes
portadores destas variantes fornecem suporte ao desenvolvimento de estratégias
para compatibilidade RH com objetivo de evitar a aloimunização Rh e o risco de
reação transfusional hemolítica.
A tipagem molecular dos antígenos Rh tem sido utilizada para identificar
alelos RH alterados, para auxiliar a diferenciar autoanticorpos de aloanticorpos e
para realizar transfusões RH haplótipo compatível em pacientes com AF (27,29,160).
A possibilidade de realizar genotipagem em larga escala em doadores de sangue
pelas plataformas de microarray pode melhorar a terapia transfusional destes
pacientes possibilitando a compatibilidade mais exata (161). No entanto, o custo
desta estratégia tem dificultado sua implementação para todos os pacientes
falciformes com fenótipos eritrocitários mais raros e os portadores de variantes RH.
Ao identificar diferenças na resposta imunológica de pacientes falciformes
transfundidos que desenvolvem aloanticorpos (respondedores) e dos que não
desenvolvem anticorpos (não respondedores), pode ser possível prever com
antecedência respondedores e não respondedores, evitando assim o alto custo da
utilização de unidades fenótipo compatível para não-respondedores e, selecionando
unidades fenotipadas ou genotipadas inclusive em relação aos haplótipos RH
apenas para os respondedores com a finalidade de reduzir a aloimunização
associada a morbidade e mortalidade.
5.5 INFLUÊNCIA DOS POLIMORFISMOS EM GENES DE CITOCINAS NA
ALOIMUNIZAÇÃO ERITROCITÁRIA EM PACIENTES PORTADORES DE ANEMIA FALCIFORME
Em nossa análise dos polimorfismos em genes de citocinas encontramos
associações importantes entre polimorfismos em genes de citocinas inflamatórias
IL1B e TNFA e um maior risco a aloimunização eritrocitária em pacientes
politransfundidos com AF.
TNF-α, é uma citocina pró-inflamatória potente que tem sido implicada em
vários processos patológicos. Altos níveis de TNF-α podem alterar a resposta imune
e aumentar o estado inflamatório do indivíduo (84–87,89,91). Um número grande de

80
polimorfismos foram descritos no gene que codifica esta citocina, mas o
polimorfismo com potencial relevância funcional está na região promotora na posição
–308 do gene TNFA, onde ocorre a transição de uma guanina (G) por uma adenina
(A) (83). A presença do alelo A pode afetar a ligação de fatores de transcrição e
consequentemente pode aumentar a transcrição do gene de seis a sete vezes,
aumentando significativamente o nível de TNF-α produzido (83,150,162–164).
Interessantemente, em nosso estudo observou-se uma associação positiva do alelo
que é alto produtor de TNF-α (TNFA–308A) e do genótipo heterozigoto TNFA–
308GA com a aloimunização eritrocitária. De acordo com estes resultados pacientes
falciformes que recebem transfusões de concentrados de hemácias e apresentam o
genótipo TNFA-308GA possuem 3 vezes mais risco de se aloimunizar contra os
antígenos eritrocitários.
Existem evidências de que um estado inflamatório no receptor favorece a
ocorrência de aloimunização em animais (38,39,41) e em humanos (51–53,165), e
que alguns receptores respondem facilmente a uma estimulação imune enquanto
outros não respondem. Assim, nossa hipótese é que o genótipo TNFA–308GA
contribui para o aumento do estado inflamatório do paciente com AF por meio de
uma maior produção de TNF-α do que o genótipo TNFA–308GG (mais frequente em
não aloimunizados) e, consequentemente este estado inflamatório elevado
predispõe a um maior risco de aloimunização. Portanto, a análise deste SNP TNFA
–308G/A pode ajudar a diferenciar pacientes respondedores e não respondedores.
A IL-1β é uma citocina pró-inflamatória que tem um papel adjuvante na
produção de anticorpos(166). Nakae et al. (166) demonstraram em um estudo in vivo
que não houve produção de anticorpos contra antígenos de eritrócitos de carneiro
(uma resposta T dependente) em camundongos deficientes em IL-1β. Nos
camundongos do tipo selvagem, a IL-1β aumentou a expansão de células T CD4+
naives e de memória em resposta a um estímulo antigênico.
Vários polimorfismos genéticos foram identificados no gene que codifica a
expressão de IL-1, incluindo o polimorfismo de único nucleotídeo rs16944 que ocorre
devido a uma troca de base C/T na posição –511 da região promotora do gene IL1B
(167). Este polimorfismo IL1B–511C/T tem sido associado com alterações na
transcrição gênica (168) e na produção da citocina IL-1β (168–170). No presente
estudo, houve prevalência do alelo IL1B–511T e dos genótipos IL1B–511TT e CT no

81
grupo de pacientes aloimunizados quando comparado com o grupo de não
aloimunizados mostrando uma possível correlação deste SNP com a
susceptibilidade a aloimunização eritrocitária em pacientes com AF. Este achado
demonstra uma correlação entre a suscetibilidade genética e condição inflamatória
com a aloimunização eritrocitária.
A presença de polimorfismos no gene IL1B pode ter efeitos combinados
na expressão de IL-1β, uma vez que este SNP pode estar em desequilíbrio de
ligação com outros SNPs na região promotora do gene (170). Portanto, novos
estudos incluindo outros SNPs funcionais no promotor do gene IL1B devem ser
realizados para aumentar nossa compreensão de como os haplótipos de IL1B
podem estar relacionados com o fenótipo respondedor.
A interleucina-10 (IL-10) é uma importante citocina imunoreguladora
(124). A IL-10 exerce função anti-inflamatória pela limitação de secreção de citocinas
pró-inflamatórias, tais como IL-1, IFN-γ e TNF-α (125,171) e pela regulação negativa
da função de APCs, regulando assim o desenvolvimento de células Th1 (172).
Em estudo anterior realizado por Bao et al. (43) pacientes não
aloimunizados apresentaram baixa atividade de células Treg, altos níveis de IL-10 e
menor produção de IFN-γ em comparação com pacientes aloimunizados. Em nosso
estudo, não encontramos associações significativas dos haplótipos associados com
baixa/alta produção de IL-10 com a suscetibilidade à aloimunização. No entanto, o
genótipo IL10–1082GG e o diplótipo GCC/GCC, que é formado pelos alelos dos
polimorfismos de IL10 (–1082G,–819C,–592C) em homozigose, que são
relacionados com alta produção de IL-10, foram encontrados em menor frequência
em pacientes com AF (aloimunizados e não aloimunizados) do que em controles.
Este resultado demostra que em nossa coorte de pacientes com AF transfundidos a
maioria dos pacientes codifica níveis baixos de IL-10.
A interleucina-4 (IL-4), produzida por células T CD4+ ativadas tipo Th2, é
uma citocina chave na diferenciação que induz o desenvolvimento do subconjunto
de linfócitos Th2 naives (57). Tanto o alelo T na posição –590 quanto o alelo RP1 do
polimorfismo íntron 3 VNTR foram associados com maior produção de IL-4 ou
atividade de células T (130–132). Estes polimorfismos, foram associados
anteriormente a determinadas condições autoimunes (133,134) e inflamatórias (135–
137).

82
Estudo prévio conduzido por Bao et al. (43) mostrou que metade dos
pacientes com AF aloimunizados tinham maior expressão de IL-4 em comparação
com os não-respondedores, indicando um desvio para a resposta imune humoral
(Th2) neste subgrupo de respondedores. Em nosso estudo, o diplótipo que
possivelmente codifica uma maior produção de IL-4 (T, RP1/T, RP1) foi mais
frequente em aloimunizados do que em não aloimunizados e controles, mas
diferença estatística significativa só foi observada em comparação com o grupo
controle. Assim, não podemos confirmar a associação entre o diplótipo de maior
produção de IL-4 (T, RP1/T, RP1) e a aloimunização eritrocitária.
Alta frequência do diplótipo T, RP2/T, RP1 (possível alto produtor de IL-4)
e menor frequência do diplótipo T, RP2/T RP2 (possível intermediário ou baixo
produtor de IL-4) foram encontrados em nossos pacientes politransfundidos com AF,
independentemente do status de aloimunização quando comparados com indivíduos
controles. Assim, podemos concluir que nossos pacientes falciformes podem
produzir níveis mais elevados de IL-4 do que os indivíduos controles, o que já foi
previamente demonstrado em estudos anteriores (78,173) o que consequentemente
pode resultar em um maior risco de morbidade para estes pacientes (173).
5.6 INFLUÊNCIA DOS GRUPOS ALÉLICOS HLA DE CLASSE II NA
ALOIMUNIZAÇÃO ERITROCITÁRIA EM PACIENTES COM ANEMIA FALCIFORME
Antígenos leucocitários humanos (HLA) modulam a resposta imune,
porque são responsáveis por apresentar o peptídeo às células T CD4+. A indução da
resposta imune depende de uma apresentação adequada por moléculas HLA de
classe II e das sequências de aminoácidos lineares curtas derivadas de
aloantígenos contendo o epitopo alvo, para ser reconhecido por células T CD4+. O
loci HLA classe II mais envolvido na resposta a alopeptídeos é o HLA-DR. (57).
Estudos prévios sugerem que alelos do locus HLA-DRB1 podem
influenciar o aloimunização eritrocitária, devido à ligação seletiva de peptídeos por
sua molécula (63–65,67,68,174–176); HLA-DRB1*01 e DRB1*10 foram associados
com a imunização pelo antígeno Jka (62,63), DRB1*04 e DRB1*15 com
aloimunização por Fya (65), HLA-DRB1*07:01 foi associado com um maior risco de
aloimunização pelo antígeno Dia (68) e HLA-DRB1*09:01 pelo antígeno Mia (67).

83
Chiaroni et al. (2006) encontraram os grupos alélicos HLA-DRB1*11 e DRB1*13
associados com imunização ao antígeno K (64), enquanto que Noizat-Pirenne et al.
(2006) mostraram que várias moléculas DRB1* poderiam apresentar o antígeno K
(65). Em nosso estudo a análise específica de fatores de risco para os antígenos
Jka, Fya, Dia e K não foi realizada devido ao pequeno número de pacientes com
estes anticorpos. A análise das frequências dos alelos HLA-DRB1 em pacientes
aloimunizados em geral versus pacientes não aloimunizados não mostrou
associação significativa entre os alelos HLA e o risco de aloimunização global.
Resultados similares foram encontrados por Hanchard et al. (56); Nickel et al. (177)
e Tatari-Calderone et al. (178).
Considerando a alta frequência de anticorpos contra antígenos do sistema
RH encontrada em nossos pacientes, realizamos a comparação entre as frequências
dos alelos HLA presentes nos pacientes aloimunizados contra antígenos Rh com a
frequência dos alelos HLA identificados no grupo de pacientes não aloimunizados.
Nesta análise, verificamos que o alelo HLA-DRB1*15 foi significativamente mais
frequente nos pacientes aloimunizados conferindo portanto um maior risco de
aloimunização contra os antígenos Rh.
Devido ao forte desequilíbrio de ligação entre alelos presentes em todo o
Complexo Principal de Histocompatibilidade (MHC) (83), é difícil determinar se um
determinado polimorfismo pode ser responsável por um fenótipo associado com a
aloimunização eritrocitária quando o mesmo encontra-se dentro da região de MHC.
Como o polimorfismo –308G/A no gene TNFA e o grupo alélico HLA-DRB1*15
mostraram-se associados com a aloimunização eritrocitária e estes genes estão
situados na região do MHC, realizamos análise comparativa das frequências dos
haplótipos HLA-DRB1/TNFA–308G/A em pacientes aloimunizados e não-
aloimunizados. Interessantemente, o haplótipo DRB1*15/TNF–308G mostrou-se
significativamente mais frequente nos pacientes aloimunizados para antígenos Rh
quando comparados com pacientes não aloimunizados. Mesmo com a força de
oposição do TNFA–308G, que foi associado à proteção para aloimunização, o grupo
alélico DRB1*15 manteve a associação de susceptibilidade, o que reforça a
associação entre DRB1*15 e a aloimunização Rh. Uma vez que nenhum
desequilíbrio de ligação significativo foi encontrado para este haplótipo nos grupos
analisados, os nossos resultados indicam que a ação de DRB1*15 e TNFA–308G é

84
independente.
Estudos anteriores já haviam demonstrado que o HLA-DRB1*15 está
envolvido com a aloimunização eritrocitária. Hall et al. (179) mostraram que uma
resposta imune ao antígeno D, pelo HLA DRB1*15, pôde ser induzida em
camundongos transgênicos e Urbaniak (180) verificaram que o alelo HLA-
DRB1*15:01 foi altamente frequente em doadores D– que produziram anticorpos
anti-D em resposta à exposição ao antígeno D por transfusão sanguínea . Além
disso, estudos com pacientes portadores de AF desenvolvidos por Hoppe et al. (70)
e Castilho et al. (181) mostraram DRB1*15 associado com a suscetibilidade a
aloimunização eritrocitária independente da especificidade do anticorpo. No entanto,
observamos que em ambos os estudos, apesar da alta frequência de anticorpos
contra antígenos do sistema RH, uma análise específica para estes antígenos não
foi realizada.
Por outro lado, os nossos resultados não são concordantes com os
achados de Schonewille et al. (182) que encontrou DRB1*01 associado com anti-D e
HLA-DRB1*09 associada com risco de aloimunização por anti-E. No entanto, este
resultado foi obtido em uma população de pacientes sem especificação de doença e
em controles normais predominantemente Caucasianos, e, não foi realizada
comparação entre pacientes aloimunizados e não aloimunizados. A existência de
discrepâncias observadas entre os estudos envolvendo o polimorfismo HLA e o risco
de aloimunização pode ocorrer (64,65,86,179,182) principalmente porque foi
demonstrado que um determinado alelo HLA pode apresentar comportamento
diferente, isto é, conferir resistência ou susceptibilidade, de acordo com o ambiente
e/ou grupo étnico da população.
5.7 APLICAÇÃO CLÍNICA DOS RESULTADOS OBTIDOS NA ANÁLISE DOS
POLIMORFISMOS EM GENES DE CITOCINAS E DOS ALELOS HLA.
Após identificação que o alelo DRB1*15 e que os polimorfismos TNFA–
308G/A e IL1B–511C/T podem conferir maior suscetibilidade ao desenvolvimento
de anticorpos contra antígenos eritrocitários, verificamos em nossa casuística o
número de pacientes que eram portadores de um ou mais destes fatores (alelos
TNFA–308A, IL1B–511T e/ou DRB1*15). Esta análise mostrou que 121 dos 161
pacientes com AF (75%) analisados apresentavam pelo menos um dos fatores de

85
risco à aloimunização eritrocitária. Assim, com base em nossos resultados 75% dos
pacientes com AF deveriam receber unidades de sangue compatíveis para um
maior número de antígenos eritrocitários, incluindo variantes RH como uma forma
de profilaxia da aloimunização eritrocitária.
A análise destes polimorfismos como fatores de risco a aloimunização
eritrocitária em pacientes portadores de AF pode contribuir futuramente na seleção
dos pacientes respondedores que devem receber concentrado de hemácias com
compatibilidade genética estendida, com o objetivo de redução da aloimunização
eritrocitária e dos efeitos adversos às transfusões.

86
___________________________________________________________
6 CONCLUSÃO
Com base nos resultados obtidos podemos concluir que:
- A frequência de aloanticorpos eritrocitários em pacientes portadores de
AF foi 41,6%.
- A aloimunização eritrocitária não foi associada com o número de
transfusões, sexo ou idade.
- A frequência de autoanticorpos em pacientes aloimunizados foi de
55,22%.
- Não foi observado diferença significativa entre os fenótipos eritrocitários
dos pacientes aloimunizados e não aloimunizados.
-O fenótipo eritrocitário mais frequente entre os pacientes foi C– E–, K–,
Fy(a–), Jk(b-), S–, Di(a-).
- A caracterização molecular de variantes RHD e RHCE contribuiu para a
diferenciação de auto e aloanticorpos Rh.
- Os alelos RHD variantes mais frequentemente encontrados entre os
pacientes com anticorpos Rh foram RHD*DAR, RHD*fraco parcial 4.0, RHD*DIIIa e
RHD*DAU0. Os alelos RHCE variantes mais frequentes foram RHCE*ceS,
RHCE*ce48C, RHCE*ce733G, RHCE*ceAR e RHCE*ceTI.
- Aloimunização Rh associada com a presença de alelos RH variantes é
alta e os anticorpos podem ser clinicamente significantes.
- O alelo A do SNP TNFA–308G/A, o alelo T do SNP IL1B–511T/C e os
genótipos TNFA–308GA e IL1B–511TT e CT podem conferir susceptibilidade a
aloimunização eritrocitária em pacientes portadores de AF.
- Os polimorfismos IL10–1082G/C, IL10–819T/C, IL10–592C/A, IL17A–
197A/G, IL17F 7488A/G, IL6–174C/T, IL4-590C/T e IL4 íntron 3 VNTR não
mostraram associação com a aloimunização eritrocitária em pacientes com AF de
nossa casuística.

87
- A molécula HLA-DRB1*15 e o haplótipo HLA-DRB1*15/TNFA–308G
podem conferir suscetibilidade a aloimunização Rh em pacientes com AF
politransfundidos.
- A detecção de polimorfismos em genes de citocinas pró-inflamatórias e
alelos HLA podem ajudar a prever pacientes respondedores e não-respondedores.
- Em nosso trabalho, 75% dos pacientes com AF deveriam receber
unidades de sangue compatíveis para um maior número de antígenos eritrocitários,
incluindo variantes RH como uma forma de profilaxia da aloimunização eritrocitária.
- Os resultados obtidos podem contribuir futuramente na seleção de
unidades de sangue compatíveis para pacientes com AF em risco de aloimunização.

88
___________________________________________________________________
7 REFERÊNCIAS
1. Ingram VM. Gene mutations in human haemoglobin: the chemical difference between normal and sickle cell haemoglobin. Nature. 1957;180(4581):326–8.
2. Madigan C, Malik P. Pathophysiology and therapy for haemoglobinopathies. Part I: sickle cell disease. Expert Rev Mol Med. 2006;8(9):1–23.
3. Steinberg MH. Management of sickle cell disease. N Engl J Med. 1999;340(13):1021–30.
4. Steinberg MH. Sickle cell anemia, the first molecular disease: overview of molecular etiology, pathophysiology, and therapeutic approaches. Sci World J. 2008;8:1295–324.
5. Stuart MJ, Nagel RL. Sickle-cell disease. Lancet. 2004;364(9442):1343–60.
6. Naoum PC. Interferentes eritrocitários e ambientais na anemia falciforme. Rev Bras Hematol Hemoter. 2000;22(1):5–22.
7. Josephson CD, Su LL, Hillyer KL, Hillyer CD. Transfusion in the patient with sickle cell disease: a critical review of the literature and transfusion guidelines. Transfus Med Rev. 2007;21(2):118–33.
8. Adams RJ, Brambilla D. Discontinuing prophylactic transfusions used to prevent stroke in sickle cell disease. N Engl J Med. 2005;353(26):2769–78.
9. Wang WC, Dwan K. Blood transfusion for preventing primary and secondary stroke in people with sickle cell disease. Cochrane database Syst Rev. 2013;11:CD003146.
10. Rosse WF, Gallagher D, Kinney TR, Castro O, Dosik H, Moohr J, et al. Transfusion and alloimmunization in sickle cell disease. The Cooperative Study of Sickle Cell Disease. Blood. 1990;76(7):1431–7.
11. Aygun B, Padmanabhan S, Paley C, Chandrasekaran V. Clinical significance of RBC alloantibodies and autoantibodies in sickle cell patients who received transfusions. Transfusion. 2002;42(1):37–43.
12. Dias Zanette AM, de Souza Gonçalves M, Vilasboas Schettini L, Magalhães Aguiar L, Santos Bahia RC, Vasconcelos Nogueira LA, et al. Alloimmunization and clinical profile of sickle cell disease patients from Salvador-Brazil. Ethn Dis. 2010;20(2):136–41.
13. Yazdanbakhsh K, Ware RE, Noizat-Pirenne F. Red blood cell alloimmunization in sickle cell disease: pathophysiology, risk factors, and transfusion management. Blood. 2012;120(3):528–37.

89
14. Silliman CC, Ambruso DR, Boshkov LK. Transfusion-related acute lung injury. Blood. 2005;105(6):2266–73.
15. Chou ST. Transfusion therapy for sickle cell disease: a balancing act. Hematology Am Soc Hematol Educ Program. 2013;2013:439–46.
16. Schonewille H, van de Watering LMG, Brand A. Additional red blood cell alloantibodies after blood transfusions in a nonhematologic alloimmunized patient cohort: is it time to take precautionary measures? Transfusion. 2006;46(4):630–5.
17. Cox J V, Steane E, Cunningham G, Frenkel EP. Risk of alloimmunization and delayed hemolytic transfusion reactions in patients with sickle cell disease. Arch Intern Med. 1988;148(11):2485–9.
18. Petz LD, Calhoun L, Shulman IA, Johnson C, Herron RM. The sickle cell hemolytic transfusion reaction syndrome. Transfusion. 1997;37(4):382–92.
19. Vidler JB, Gardner K, Amenyah K, Mijovic A, Thein SL. Delayed haemolytic transfusion reaction in adults with sickle cell disease: a 5-year experience. Br J Haematol. 2015;169(5):746–53.
20. Vichinsky EP, Luban NL, Wright E, Olivieri N, Driscoll C, Pegelow CH, et al. Prospective RBC phenotype matching in a stroke-prevention trial in sickle cell anemia: a multicenter transfusion trial. Transfusion. 2001;41(9):1086–92.
21. Sakhalkar VS, Roberts K, Hawthorne LM, McCaskill DM, Veillon DM, Caldito GC, et al. Allosensitization in patients receiving multiple blood transfusions. Ann N Y Acad Sci. 2005;1054:495–9.
22. Lasalle-Williams M, Nuss R, Le T, Cole L, Hassell K, Murphy JR, et al. Extended red blood cell antigen matching for transfusions in sickle cell disease: a review of a 14-year experience from a single center (CME). Transfusion. 2011;51(8):1732–9.
23. Castilho L, Rios M, Bianco C, Pellegrino J, Alberto FL, Saad STO, et al. DNA-based typing of blood groups for the management of multiply-transfused sickle cell disease patients. Transfusion. 2002;42(2):232–8.
24. Ribeiro KR, Guarnieri MH, da Costa DC, Costa FF, Pellegrino J, Castilho L. DNA array analysis for red blood cell antigens facilitates the transfusion support with antigen-matched blood in patients with sickle cell disease. Vox Sang. 2009;97(2):147–52.
25. Da Costa DC, Pellegrino J, Guelsin GAS, Ribeiro KAR, Gilli SCO, Castilho L. Molecular matching of red blood cells is superior to serological matching in sickle cell disease patients. Rev Bras Hematol Hemoter. 2013;35(1):35–8.

90
26. Chou ST, Westhoff CM. Molecular biology of the Rh system: clinical considerations for transfusion in sickle cell disease. Hematology Am Soc Hematol Educ Program. 2009;178–84.
27. Chou ST, Jackson T, Vege S, Smith-Whitley K, Friedman DF, Westhoff CM. High prevalence of red blood cell alloimmunization in sickle cell disease despite transfusion from Rh-matched minority donors. Blood. 2013;122(6):1062–71.
28. Chou ST, Liem RI, Thompson AA. Challenges of alloimmunization in patients with haemoglobinopathies. Br J Haematol. 2012;159(4):394–404.
29. Noizat-Pirenne F, Tournamille C. Relevance of RH variants in transfusion of sickle cell patients. Transfus Clin Biol. 2011;18(5-6):527–35.
30. Noizat-Pirenne F, Lee K, Pennec P-Y Le, Simon P, Kazup P, Bachir D, et al. Rare RHCE phenotypes in black individuals of Afro-Caribbean origin: identification and transfusion safety. Blood. 2002;100(12):4223–31.
31. Vichinsky EP, Earles A, Johnson RA, Hoag MS, Williams A, Lubin B. Alloimmunization in sickle cell anemia and transfusion of racially unmatched blood. N Engl J Med. 1990;322(23):1617–21.
32. Ameen R, Al Shemmari S, Al-Bashir A. Red blood cell alloimmunization among sickle cell Kuwaiti Arab patients who received red blood cell transfusion. Transfusion. 2009;49(8):1649–54.
33. Spielmann W, Seidl S. Prevalence of irregular red cell antibodies and their significance in blood transfusion and antenatal care. Vox Sang. 1974;26(6):551–9.
34. Hoeltge GA, Domen RE, Rybicki LA, Schaffer PA. Multiple red cell transfusions and alloimmunization. Experience with 6996 antibodies detected in a total of 159,262 patients from 1985 to 1993. Arch Pathol Lab Med. 1995;119(1):42–5.
35. Saverimuttu J, Greenfield T, Rotenko I, Crozier J, Jalaludin B, Harvey M. Implications for urgent transfusion of uncrossmatched blood in the emergency department: The prevalence of clinically significant red cell antibodies within different patient groups. Emerg Med (Fremantle). 2003;15(3):239–43.
36. Verduin EP, Brand A, Schonewille H. Is female sex a risk factor for red blood cell alloimmunization after transfusion? A systematic review. Transfus Med Rev. 2012;26(4):342–53.e5.
37. Urbaniak SJ. Alloimmunity to RhD in humans. Transfus Clin Biol. 2006;13(1-2):19–22.
38. Hendrickson JE, Chadwick TE, Roback JD, Hillyer CD, Zimring JC. Inflammation enhances consumption and presentation of transfused RBC antigens by dendritic cells. Blood. 2007;110(7):2736–43.

91
39. Zimring JC, Hendrickson JE. The role of inflammation in alloimmunization to antigens on transfused red blood cells. Curr Opin Hematol. 2008;15(6):631–5.
40. Hendrickson JE, Hod EA, Perry JR, Ghosh S, Chappa P, Adisa O, et al. Alloimmunization to transfused HOD red blood cells is not increased in mice with sickle cell disease. Transfusion. 2012;52(2):231–40.
41. Smith NH, Hod EA, Spitalnik SL, Zimring JC, Hendrickson JE. Transfusion in the absence of inflammation induces antigen-specific tolerance to murine RBCs. Blood. 2012;119(6):1566–9.
42. Bao W, Yu J, Heck S, Yazdanbakhsh K. Regulatory T-cell status in red cell alloimmunized responder and nonresponder mice. Blood. 2009;113(22):5624–7.
43. Bao W, Zhong H, Li X, Lee MT, Schwartz J, Sheth S, et al. Immune regulation in chronically transfused allo-antibody responder and nonresponder patients with sickle cell disease and β-thalassemia major. Am J Hematol. 2011;86(12):1001–6.
44. Bao W, Zhong H, Manwani D, Vasovic L, Uehlinger J, Lee MT, et al. Regulatory B-cell compartment in transfused alloimmunized and non-alloimmunized patients with sickle cell disease. Am J Hematol. 2013;88(9):736–40.
45. Vingert B, Tamagne M, Desmarets M, Pakdaman S, Elayeb R, Habibi A, et al. Partial dysfunction of Treg activation in sickle cell disease. Am J Hematol. 2014;89(3):261–6.
46. Tatari-Calderone Z, Minniti CP, Kratovil T, Stojakovic M, Vollmer A, Barjaktarevic I, et al. rs660 polymorphism in Ro52 (SSA1; TRIM21) is a marker for age-dependent tolerance induction and efficiency of alloimmunization in sickle cell disease. Mol Immunol. 2009;47(1):64–70.
47. Tatari-Calderone Z, Tamouza R, Le Bouder GP, Dewan R, Luban NLC, Lasserre J, et al. The association of CD81 polymorphisms with alloimmunization in sickle cell disease. Clin Dev Immunol. 2013;2013:937846.
48. Prigent A, Maillard N, Absi L, Aloui C, Cognasse F, Laradi S, et al. From Donor to Recipient: Current Questions Relating to Humoral Alloimmunization. Antibodies. 2014;3(1):130–52.
49. Schonewille H, Brand A. Alloimmunization to red blood cell antigens after universal leucodepletion. A regional multicentre retrospective study. Br J Haematol. 2005;129(1):151–6.
50. Ramsey G, Smietana SJ. Multiple or uncommon red cell alloantibodies in women: association with autoimmune disease. Transfusion. 1995;35(7):582–6.

92
51. Papay P, Hackner K, Vogelsang H, Novacek G, Primas C, Reinisch W, et al. High risk of transfusion-induced alloimmunization of patients with inflammatory bowel disease. Am J Med. 2012;125(7):717.e1–8.
52. Yazer MH, Triulzi DJ, Shaz B, Kraus T, Zimring JC. Does a febrile reaction to platelets predispose recipients to red blood cell alloimmunization? Transfusion. 2009;49(6):1070–5.
53. Fasano RM, Booth GS, Miles M, Du L, Koyama T, Meier ER, et al. Red blood cell alloimmunization is influenced by recipient inflammatory state at time of transfusion in patients with sickle cell disease. Br J Haematol. 2015;168(2):291–300.
54. Vignali DAA, Collison LW, Workman CJ. How regulatory T cells work. Nat Rev Immunol. 2008;8(7):523–32.
55. Milton JN, Ashley-Koch AE, Garrett ME, Soldano KL, Orringer EP, Sebastiani P, et al. Genes Associated with Alloimmunization to Blood Group Antigens in Sickle Cell Disease. Blood. 2014;124(21):762.
56. Hanchard NA, Moulds JM, Belmont JW, Chen A. A Genome-Wide Screen for Large-Effect Alloimmunization Susceptibility Loci among Red Blood Cell Transfusion Recipients with Sickle Cell Disease. Transfus Med hemotherapy. 2014;41(6):453–61.
57. Abbas AK, Lichtman AHH, Pillai S. Imunologia Celular e Molecular. 7th ed. Rio de Janeiro: Elsevier; 2012. 560 p.
58. Murphy K. Imunobiologia de Janeway. 8th ed. Artmed Editora; 2014. 888 p.
59. Klein J, Sato A. The HLA system. First of two parts. N Engl J Med. 2000;343(10):702–9.
60. Jonna B. Westover, Thayne L. Sweeten, Michael Benson PB-W and ART. Immune Dysfunction in Autism Spectrum Disorder. Autism - A Neurodevelopmental Journey from Genes to Behaviour [Internet]. In Tech; 2011. p. 498. Available from: http://www.intechopen.com/books/autism-a-neurodevelopmental-journey-from-genes-to-behaviour/immune-dysfunction-in-autism-spectrum-disorder
61. Brain P, Hammond MG. Association between histocompatibility type and the ability to make anti-Rh antibodies. Eur J Immunol. 1974;4(3):223–5.
62. Ansart-Pirenne H, Zeliszewski D, Lee K, Martin-Blanc S, Rouger P, Noizat-Pirenne F. Identification of immunodominant alloreactive T-cell epitopes on the Jka red blood cell protein inducing either Th1 or Th2 cytokine expression. Blood. 2004;104(10):3409–10.
63. Reviron D, Dettori I, Ferrera V, Legrand D, Touinssi M, Mercier P, et al. HLA-DRB1 alleles and Jk(a) immunization. Transfusion. 2005 Jun;45(6):956–9.

93
64. Chiaroni J, Dettori I, Ferrera V, Legrand D, Touinssi M, Mercier P, et al. HLA-DRB1 polymorphism is associated with Kell immunisation. Br J Haematol. 2006;132(3):374–8.
65. Noizat-Pirenne F, Tournamille C, Bierling P, Roudot-Thoraval F, Le Pennec P-Y, Rouger P, et al. Relative immunogenicity of Fya and K antigens in a Caucasian population, based on HLA class II restriction analysis. Transfusion. 2006;46(8):1328–33.
66. Picard C, Frassati C, Basire A, Buhler S, Galicher V, Ferrera V, et al. Positive association of DRB1 04 and DRB1 15 alleles with Fya immunization in a Southern European population. Transfusion. 2009;49(11):2412–7.
67. Chu C-C, Ho H-T, Lee H-L, Chan Y-S, Chang F-J, Wang C-L, et al. Anti-“Mi(a)” immunization is associated with HLA-DRB1*0901. Transfusion. 2009;49(3):472–8.
68. Baleotti W, Ruiz MO, Fabron A, Castilho L, Giuliatti S, Donadi EA. HLA-DRB1*07:01 allele is primarily associated with the Diego a alloimmunization in a Brazilian population. Transfusion. 2014;54(10):2468–76.
69. Alarif L, Castro O, Ofosu M, Dunston G, Scott RB. HLA-B35 is associated with red cell alloimmunization in sickle cell disease. Clin Immunol Immunopathol. 1986;38(2):178–83.
70. Hoppe C, Klitz W, Vichinsky E, Styles L. HLA type and risk of alloimmunization in sickle cell disease. Am J Hematol. 2009;84(7):462–4.
71. Miyajima A, Kitamura T, Harada N, Yokota T, Arai K. Cytokine receptors and signal transduction. Annu Rev Immunol. 1992;10:295–331.
72. Miossec P, Korn T, Kuchroo VK. Interleukin-17 and type 17 helper T cells. N Engl J Med. 2009;361(9):888–98.
73. Hauser SL. Tumor necrosis factor: immunogenetics and disease. Ann Neurol. 1995;38(5):702–4.
74. Lanaro C, Franco-Penteado CF, Albuqueque DM, Saad STO, Conran N, Costa FF. Altered levels of cytokines and inflammatory mediators in plasma and leukocytes of sickle cell anemia patients and effects of hydroxyurea therapy. J Leukoc Biol. 2009;85(2):235–42.
75. Taylor SC, Shacks SJ, Qu Z. In vivo production of type 1 cytokines in healthy sickle cell disease patients. J Natl Med Assoc. 1999;91(11):619–24.
76. Jison ML, Munson PJ, Barb JJ, Suffredini AF, Talwar S, Logun C, et al. Blood mononuclear cell gene expression profiles characterize the oxidant, hemolytic, and inflammatory stress of sickle cell disease. Blood. 2004;104(1):270–80.

94
77. Musa BOP, Onyemelukwe GC, Hambolu JO, Mamman AI, Isa AH. Pattern of serum cytokine expression and T-cell subsets in sickle cell disease patients in vaso-occlusive crisis. Clin Vaccine Immunol. 2010 Apr;17(4):602–8.
78. Raghupathy R, Haider MZ, Azizieh F, Abdelsalam R, D’Souza TM, Adekile AD. Th1 and Th2 cytokine profiles in sickle cell disease. Acta Haematol. 2000;103(4):197–202.
79. Hassan DA, Marques C, Santos-Gomes GM, do Rosario VE, Mohamed HS, Elhussein AM, et al. Differential expression of cytokine genes among sickle-cell-trait (HbAS) and normal (HbAA) children infected with Plasmodium falciparum. Ann Trop Med Parasitol. 2009;103(4):283–95.
80. Jin P, Wang E. Polymorphism in clinical immunology - From HLA typing to immunogenetic profiling. J Transl Med. 2003;1(1):8.
81. Pathare A, Al Kindi S, Alnaqdy AA, Daar S, Knox-Macaulay H, Dennison D. Cytokine profile of sickle cell disease in Oman. Am J Hematol. 2004;77(4):323–8.
82. Keikhaei B, Mohseni AR, Norouzirad R, Alinejadi M, Ghanbari S, Shiravi F, et al. Altered levels of pro-inflammatory cytokines in sickle cell disease patients during vaso-occlusive crises and the steady state condition. Eur Cytokine Netw. 2013;24(1):45–52.
83. Wilson AG, Symons JA, McDowell TL, McDevitt HO, Duff GW. Effects of a polymorphism in the human tumor necrosis factor alpha promoter on transcriptional activation. Proc Natl Acad Sci U S A. 1997;94(7):3195–9.
84. Du T, Guo X-H, Zhu X-L, Li J-H, Lu L-P, Gao J-R, et al. Association of TNF-alpha promoter polymorphisms with the outcomes of hepatitis B virus infection in Chinese Han population. J Viral Hepat. 2006;13(9):618–24.
85. Hoppe C, Klitz W, Cheng S, Apple R, Steiner L, Robles L, et al. Gene interactions and stroke risk in children with sickle cell anemia. Blood. 2004;103(6):2391–6.
86. Hoppe C, Klitz W, D’Harlingue K, Cheng S, Grow M, Steiner L, et al. Confirmation of an association between the TNF(-308) promoter polymorphism and stroke risk in children with sickle cell anemia. Stroke. 2007;38(8):2241–6.
87. Wang Y, Ng MCY, So W-Y, Ma R, Ko GTC, Tong PCY, et al. Association between tumour necrosis factor-alpha G-308A polymorphism and risk of nephropathy in obese Chinese type 2 diabetic patients. Nephrol Dial Transplant. 2005;20(12):2733–8.
88. Liu Z, Ding Y, Xiu L, Pan H, Liang Y, Zhong S, et al. A meta-analysis of the association between TNF-A -308G>A polymorphism and type 2 diabetes mellitus in Han Chinese population. PLoS One. 2013;8(3):e59421.

95
89. Meng N, Zhang Y, Li H, Ma J, Qu Y. Association of tumor necrosis factor alpha promoter polymorphism (TNF-α 238 G/A and TNF-α 308 G/A) with diabetic mellitus, diabetic retinopathy and diabetic nephropathy: a meta-analysis. Curr Eye Res. 2014;39(2):194–203.
90. Sesti LFC, Crispim D, Canani LH, Polina ER, Rheinheimer J, Carvalho PS, et al. The -308G>A polymorphism of the TNF gene is associated with proliferative diabetic retinopathy in Caucasian Brazilians with type 2 diabetes. Invest Ophthalmol Vis Sci. 2015;56(2):1184–90.
91. Dhamodharan U, Viswanathan V, Krishnamoorthy E, Rajaram R, Aravindhan V. Genetic association of IL-6, TNF-α and SDF-1 polymorphisms with serum cytokine levels in diabetic foot ulcer. Gene. 2015;565(1):62–7.
92. Ataie-Kachoie P, Pourgholami MH, Morris DL. Inhibition of the IL-6 signaling pathway: a strategy to combat chronic inflammatory diseases and cancer. Cytokine Growth Factor Rev. 2013;24(2):163–73.
93. Terry CF, Loukaci V, Green FR. Cooperative influence of genetic polymorphisms on interleukin 6 transcriptional regulation. J Biol Chem. 2000;275(24):18138–44.
94. Rincón M, Anguita J, Nakamura T, Fikrig E, Flavell RA. Interleukin (IL)-6 directs the differentiation of IL-4-producing CD4+ T cells. J Exp Med. 1997;185(3):461–9.
95. Kishimoto T. Interleukin-6: discovery of a pleiotropic cytokine. Arthritis Res Ther. 2006;8 Suppl 2:S2.
96. Neurath MF, Finotto S. IL-6 signaling in autoimmunity, chronic inflammation and inflammation-associated cancer. Cytokine Growth Factor Rev. 2011;22(2):83–9.
97. Ataie-Kachoie P, Pourgholami MH, Richardson DR, Morris DL. Gene of the month: Interleukin 6 (IL-6). J Clin Pathol. 2014;67(11):932–7.
98. Fishman D, Faulds G, Jeffery R, Mohamed-Ali V, Yudkin JS, Humphries S, et al. The effect of novel polymorphisms in the interleukin-6 (IL-6) gene on IL-6 transcription and plasma IL-6 levels, and an association with systemic-onset juvenile chronic arthritis. J Clin Invest. 1998;102(7):1369–76.
99. Vozarova B, Fernández-Real J-M, Knowler WC, Gallart L, Hanson RL, Gruber JD, et al. The interleukin-6 (-174) G/C promoter polymorphism is associated with type-2 diabetes mellitus in Native Americans and Caucasians. Hum Genet. 2003;112(4):409–13.
100. Chumaeva N, Hintsanen M, Pulkki-Råback L, Jokela M, Juonala M, Lehtimäki T, et al. Interleukin-6 gene polymorphism, chronic stress and atherosclerosis: interleukin-6-174G>C polymorphism, chronic stress and risk of early

96
atherosclerosis in the Cardiovascular Risk in Young Finns Study. J Psychosom Res. 2014;76(4):333–8.
101. Biswas S, Ghoshal PK, Mandal N. Synergistic effect of anti and pro-inflammatory cytokine genes and their promoter polymorphism with ST-elevation of myocardial infarction. Gene. 2014;544(2):145–51.
102. Humphries SE, Luong LA, Ogg MS, Hawe E, Miller GJ. The interleukin-6 -174 G/C promoter polymorphism is associated with risk of coronary heart disease and systolic blood pressure in healthy men. Eur Heart J. 2001;22(24):2243–52.
103. Kawaguchi M, Adachi M, Oda N, Kokubu F, Huang S-K. IL-17 cytokine family. J Allergy Clin Immunol. 2004;114(6):1265–73; quiz 1274.
104. Isailovic N, Daigo K, Mantovani A, Selmi C. Interleukin-17 and innate immunity in infections and chronic inflammation. J Autoimmun. 2015;60:1–11.
105. Wang X, Zhang Y, Yang XO, Nurieva RI, Chang SH, Ojeda SS, et al. Transcription of IL17 and IL17F is controlled by conserved noncoding sequence 2. Immunity. 2012;36(1):23–31.
106. Espinoza JL, Takami A, Nakata K, Onizuka M, Kawase T, Akiyama H, et al. A genetic variant in the IL-17 promoter is functionally associated with acute graft-versus-host disease after unrelated bone marrow transplantation. PLoS One. 2011;6(10):e26229.
107. Arisawa T, Tahara T, Shibata T, Nagasaka M, Nakamura M, Kamiya Y, et al. The influence of polymorphisms of interleukin-17A and interleukin-17F genes on the susceptibility to ulcerative colitis. J Clin Immunol. 2008;28(1):44–9.
108. Hayashi R, Tahara T, Shiroeda H, Saito T, Nakamura M, Tsutsumi M, et al. Influence of IL17A polymorphisms (rs2275913 and rs3748067) on the susceptibility to ulcerative colitis. Clin Exp Med. 2013;13(4):239–44.
109. Shibata T, Tahara T, Hirata I, Arisawa T. Genetic polymorphism of interleukin-17A and -17F genes in gastric carcinogenesis. Hum Immunol. 2009;70(7):547–51.
110. Kawaguchi M, Takahashi D, Hizawa N, Suzuki S, Matsukura S, Kokubu F, et al. IL-17F sequence variant (His161Arg) is associated with protection against asthma and antagonizes wild-type IL-17F activity. J Allergy Clin Immunol. 2006;117(4):795–801.
111. Chen B, Zeng Z, Hou J, Chen M, Gao X, Hu P. Association of interleukin-17F 7488 single nucleotide polymorphism and inflammatory bowel disease in the Chinese population. Scand J Gastroenterol. 2009;44(6):720–6.
112. Wu X, Zeng Z, Chen B, Yu J, Xue L, Hao Y, et al. Association between polymorphisms in interleukin-17A and interleukin-17F genes and risks of gastric cancer. Int J Cancer. 2010;127(1):86–92.

97
113. Wróbel T, Gębura K, Wysoczańska B, Jaźwiec B, Dobrzyńska O, Mazur G, et al. IL-17F gene polymorphism is associated with susceptibility to acute myeloid leukemia. J Cancer Res Clin Oncol. 2014;140(9):1551–5.
114. Dinarello CA. Interleukin-1 in the pathogenesis and treatment of inflammatory diseases. Blood. 2011;117(14):3720–32.
115. Dinarello CA. Immunological and inflammatory functions of the interleukin-1 family. Annu Rev Immunol. 2009;27:519–50.
116. Contassot E, Beer H-D, French LE. Interleukin-1, inflammasomes, autoinflammation and the skin. Swiss Med Wkly. 2012;142:w13590.
117. Ben-Sasson SZ, Hu-Li J, Quiel J, Cauchetaux S, Ratner M, Shapira I, et al. IL-1 acts directly on CD4 T cells to enhance their antigen-driven expansion and differentiation. Proc Natl Acad Sci U S A. 2009;106(17):7119–24.
118. El-Omar EM, Carrington M, Chow WH, McColl KE, Bream JH, Young HA, et al. Interleukin-1 polymorphisms associated with increased risk of gastric cancer. Nature. 2000;404(6776):398–402.
119. Trevilatto PC, de Souza Pardo AP, Scarel-Caminaga RM, de Brito RB, Alvim-Pereira F, Alvim-Pereira CC, et al. Association of IL1 gene polymorphisms with chronic periodontitis in Brazilians. Arch Oral Biol. 2011;56(1):54–62.
120. Stefanidis I, Kreuer K, Dardiotis E, Arampatzis S, Eleftheriadis T, Hadjigeorgiou GM, et al. Association Between the Interleukin-1β Gene ( IL1B ) C−511T Polymorphism and the Risk of Diabetic Nephropathy in Type 2 Diabetes: A Candidate–Gene Association Study. DNA Cell Biol. 2014;33(7):463–8.
121. Zhu J, Yamane H, Paul WE. Differentiation of effector CD4 T cell populations (*). Annu Rev Immunol. 2010;28:445–89.
122. Saraiva M, O’Garra A. The regulation of IL-10 production by immune cells. Nat Rev Immunol. 2010;10(3):170–81.
123. Gabryšová L, Howes A, Saraiva M, O’Garra A. The regulation of IL-10 expression. Curr Top Microbiol Immunol. 2014;380:157–90.
124. Moore KW, de Waal Malefyt R, Coffman RL, O’Garra A. Interleukin-10 and the interleukin-10 receptor. Annu Rev Immunol. 2001;19:683–765.
125. Fiorentino DF, Zlotnik A, Vieira P, Mosmann TR, Howard M, Moore KW, et al. IL-10 acts on the antigen-presenting cell to inhibit cytokine production by Th1 cells. J Immunol. 1991;146(10):3444–51.
126. Ding L, Shevach EM. IL-10 inhibits mitogen-induced T cell proliferation by selectively inhibiting macrophage costimulatory function. J Immunol. 1992;148(10):3133–9.

98
127. Turner DM, Williams DM, Sankaran D, Lazarus M, Sinnott PJ, Hutchinson I V. An investigation of polymorphism in the interleukin-10 gene promoter. Eur J Immunogenet. 1997;24(1):1–8.
128. Suárez A, Castro P, Alonso R, Mozo L, Gutiérrez C. Interindividual variations in constitutive interleukin-10 messenger RNA and protein levels and their association with genetic polymorphisms. Transplantation. 2003;75(5):711–7.
129. Oleksyk TK, Shrestha S, Truelove AL, Goedert JJ, Donfield SM, Phair J, et al. Extended IL10 haplotypes and their association with HIV progression to AIDS. Genes Immun. 2009;10(4):309–22.
130. Wu MC, Huang CM, Tsai JJP, Chen HY, Tsai FJ. Polymorphisms of the interleukin-4 gene in chinese patients with systemic lupus erythematosus in Taiwan. Lupus. 2003;12(1):21–5.
131. Vandenbroeck K, Martino G, Marrosu M, Consiglio A, Zaffaroni M, Vaccargiu S, et al. Occurrence and clinical relevance of an interleukin-4 gene polymorphism in patients with multiple sclerosis. J Neuroimmunol. 1997;76(1-2):189–92.
132. Nakashima H, Miyake K, Inoue Y, Shimizu S, Akahoshi M, Tanaka Y, et al. Association between IL-4 genotype and IL-4 production in the Japanese population. Genes Immun. 2002;3(2):107–9.
133. Maksymowych WP, Suarez-Almazor ME, Buenviaje H, Cooper B-L, Degeus C, Thompson M, et al. HLA and cytokine gene polymorphisms in relation to occurrence of palindromic rheumatism and its progression to rheumatoid arthritis. J Rheumatol. 2002;29(11):2319–26.
134. Mahmoudi M, Tahghighi F, Ziaee V, Harsini S, Rezaei A, Soltani S, et al. Interleukin-4 single nucleotide polymorphisms in juvenile systemic lupus erythematosus. Int J Immunogenet. 2014;41(6):512–7.
135. Mishra A, Jha AN, van Tong H, Singh VK, Gomes CEM, Singh L, et al. Analysis of genetic variants in the IL4 promoter and VNTR loci in Indian patients with Visceral Leishmaniasis. Hum Immunol. 2014;75(12):1177–81.
136. Birbian N, Singh J, Jindal SK, Sobti RC. High risk association of IL-4 VNTR polymorphism with asthma in a North Indian population. Cytokine. 2014;66(1):87–94.
137. Cabantous S, Ranque S, Poudiougou B, Traore A, Berbache S, Vitte J, et al. Genotype combinations of two IL4 polymorphisms influencing IL-4 plasma levels are associated with different risks of severe malaria in the Malian population. Immunogenetics. 2015;67(5-6):283–8.
138. Singleton BK, Green CA, Avent ND, Martin PG, Smart E, Daka A, et al. The presence of an RHD pseudogene containing a 37 base pair duplication and a nonsense mutation in africans with the Rh D-negative blood group phenotype. Blood. 2000;95(1):12–8.

99
139. Chiu RW, Murphy MF, Fidler C, Zee BC, Wainscoat JS, Lo YM. Determination of RhD zygosity: comparison of a double amplification refractory mutation system approach and a multiplex real-time quantitative PCR approach. Clin Chem. 2001;47(4):667–72.
140. Legler TJ, Eber SW, Lakomek M, Lynen R, Maas JH, Pekrun A, et al. Application of RHD and RHCE genotyping for correct blood group determination in chronically transfused patients. Transfusion. 1999;39(8):852–5.
141. Hipsky CH, Lomas-Francis C, Fuchisawa A, Reid ME, Moulds M, Christensen J, et al. RHCE*ceCF encodes partial c and partial e but not CELO, an antigen antithetical to Crawford. Transfusion. 2011;51(1):25–31.
142. Cantagrel A, Navaux F, Loubet-Lescoulié P, Nourhashemi F, Enault G, Abbal M, et al. Interleukin-1beta, interleukin-1 receptor antagonist, interleukin-4, and interleukin-10 gene polymorphisms: relationship to occurrence and severity of rheumatoid arthritis. Arthritis Rheum. 1999 Jun;42(6):1093–100.
143. Da Costa DC. Genotipagem de grupos sanguíneos no suporte transfusional para pacientes com anemia falciforme [dissertação]. Campinas (SP): Universidade Estadual de Campinas, Faculdade de Ciências Médicas, Departamento de Clínica Médica; 2011.
144. Wagner FF, Flegel WA. RHD gene deletion occurred in the Rhesus box. Blood. 2000;95(12):3662–8.
145. One Lambda. 2016. Acesso em agosto 2015. Disponível em: www.onelambda.com
146. López NJ, Jara L, Valenzuela CY. Association of interleukin-1 polymorphisms with periodontal disease. J Periodontol. 2005 Feb;76(2):234–43.
147. Salhi A, Rodrigues V, Santoro F, Dessein H, Romano A, Castellano LR, et al. Immunological and genetic evidence for a crucial role of IL-10 in cutaneous lesions in humans infected with Leishmania braziliensis. J Immunol. 2008;180(9):6139–48.
148. Yao C-J, Du W, Chen H-B, Xiao S, Wang C-H, Fan Z-L. Associations of IL-10 gene polymorphisms with acute myeloid leukemia in Hunan, China. Asian Pac J Cancer Prev. 2013;14(4):2439–42.
149. Scarel-Caminaga RM, Trevilatto PC, Souza AP, Brito RB, Camargo LEA, Line SRP. Interleukin 10 gene promoter polymorphisms are associated with chronic periodontitis. J Clin Periodontol. 2004 Jun;31(6):443–8.
150. Lin H-J, Tsai F-J, Chen W-C, Shi Y-R, Hsu Y, Tsai S-W. Association of tumour necrosis factor alpha -308 gene polymorphism with primary open-angle glaucoma in Chinese. Eye (Lond). 2003;17(1):31–4.

100
151. Excoffier L, Lischer HEL. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour. 2010;10(3):564–7.
152. Imanishi T, Akaza T, Kimura A TK& GT. Allele and haplotype frequencies for HLA and complement loci in various ethnic groups. In: Tsuji K, Aizawa M, Sasazuki T, eds. HLA-1991. Proceedings of the Eleventh International Histocompatibility Workshop and Conference. Oxford: Oxford University Press; 1992.
153. Sosler SD, Jilly BJ, Saporito C, Koshy M. A simple, practical model for reducing alloimmunization in patients with sickle cell disease. Am J Hematol. 1993;43(2):103–6.
154. Frye V, Caltabiano M, Kessler DA, Schaffler H, Reboza M, Hillyer CD, et al. Evaluating a program to increase blood donation among racial and ethnic minority communities in New York City. Transfusion. 2014;54(12):3061–7.
155. Tournamille C, Colin Y, Cartron JP, Le Van Kim C. Disruption of a GATA motif in the Duffy gene promoter abolishes erythroid gene expression in Duffy-negative individuals. Nat Genet. 1995;10(2):224–8.
156. Parasol N, Reid M, Rios M, Castilho L, Harari I, Kosower NS. A novel mutation in the coding sequence of the FY*B allele of the Duffy chemokine receptor gene is associated with an altered erythrocyte phenotype. Blood. 1998;92(7):2237–43.
157. Tournamille C, Le Van Kim C, Gane P, Le Pennec PY, Roubinet F, Babinet J, et al. Arg89Cys substitution results in very low membrane expression of the Duffy antigen/receptor for chemokines in Fy(x) individuals. Blood. 1998;92(6):2147–56.
158. Yazdanbakhsh K, Rios M, Storry JR, Kosower N, Parasol N, Chaudhuri A, et al. Molecular mechanisms that lead to reduced expression of duffy antigens. Transfusion. 2000;40(3):310–20.
159. Pham B-N, Peyrard T, Juszczak G, Beolet M, Deram G, Martin-Blanc S, et al. Analysis of RhCE variants among 806 individuals in France: considerations for transfusion safety, with emphasis on patients with sickle cell disease. Transfusion. 2011;51(6):1249–60.
160. Flegel WA. Molecular genetics and clinical applications for RH. Transfus Apher Sci. 2011;44(1):81–91.
161. Keller MA, Horn T, Crowley J et al. Abstracts of the AABB Annual Meeting and CTTXPO. Denver, Colorado, USA. October 12-15, 2013. Transfusion. 2013;53 Suppl 2:174A (SP304).
162. Kroeger KM, Carville KS, Abraham LJ. The -308 tumor necrosis factor-alpha promoter polymorphism effects transcription. Mol Immunol. 1997;34(5):391–9.

101
163. Wu WS, McClain KL. DNA polymorphisms and mutations of the tumor necrosis factor-alpha (TNF-alpha) promoter in Langerhans cell histiocytosis (LCH). J Interferon Cytokine Res. 1997;17(10):631–5.
164. Kroeger KM, Steer JH, Joyce DA, Abraham LJ. Effects of stimulus and cell type on the expression of the -308 tumour necrosis factor promoter polymorphism. Cytokine. 2000;12(2):110–9.
165. Tyler LN, Harville TO, Blackall DP. Multiple alloantibodies after transfusion in an infant treated with infliximab. N Engl J Med. 2007;357(20):2092–3; discussion 2093.
166. Nakae S, Asano M, Horai R, Iwakura Y. Interleukin-1 beta, but not interleukin-1 alpha, is required for T-cell-dependent antibody production. Immunology. 2001;104(4):402–9.
167. Di Giovine FS, Takhsh E, Blakemore AI, Duff GW. Single base polymorphism at -511 in the human interleukin-1 beta gene (IL1 beta). Hum Mol Genet. 1992;1(6):450.
168. Rogus J, Beck JD, Offenbacher S, Huttner K, Iacoviello L, Latella MC, et al. IL1B gene promoter haplotype pairs predict clinical levels of interleukin-1beta and C-reactive protein. Hum Genet. 2008;123(4):387–98.
169. Iacoviello L, Di Castelnuovo A, Gattone M, Pezzini A, Assanelli D, Lorenzet R, et al. Polymorphisms of the interleukin-1beta gene affect the risk of myocardial infarction and ischemic stroke at young age and the response of mononuclear cells to stimulation in vitro. Arterioscler Thromb Vasc Biol. 2005;25(1):222–7.
170. Hall SK, Perregaux DG, Gabel CA, Woodworth T, Durham LK, Huizinga TWF, et al. Correlation of polymorphic variation in the promoter region of the interleukin-1 beta gene with secretion of interleukin-1 beta protein. Arthritis Rheum. 2004;50(6):1976–83.
171. Fiorentino DF, Zlotnik A, Mosmann TR, Howard M, O’Garra A. IL-10 inhibits cytokine production by activated macrophages. J Immunol. 1991;147(11):3815–22.
172. O’Garra A, Murphy KM. From IL-10 to IL-12: how pathogens and their products stimulate APCs to induce T(H)1 development. Nat Immunol. 2009;10(9):929–32.
173. Taylor SC, Shacks SJ, Qu Z, Wiley P. Type 2 cytokine serum levels in healthy sickle cell disease patients. J Natl Med Assoc. 1997 Nov;89(11):753–7.
174. Wojtulewicz-Kurkus J, Zupańska B, Podobińska I, Brojer E. Preliminary evaluation of DR antigens in responders and nonresponders to antigen D from the Rh blood group system. Arch Immunol Ther Exp (Warsz). 1981;29(4):447–50.

102
175. Kumar US, Ghosh K, Gupte SS, Gupte SC, Mohanty D. Role of HLA antigens in Rh (D) alloimmunized pregnant women from Mumbai, Maharashtra, India. J Biosci. 2002;27(2):135–41.
176. Verhagen OJHM, Valle L Della, Dohmen SE, Swelsen W, Vidarsson G, Haas M De, et al. HLA-DRB1null15 is not strongly linked to rhd immunization risk, but associated with high anti-D titers after hyperimmunization. Vox Sang. 2013;105:235–6.
177. Nickel RS, Horan JT, Fasano RM, Meyer E, Josephson CD, Winkler AM, et al. Immunophenotypic Parameters and RBC Alloimmunization in Children with Sickle Cell Disease on Chronic Transfusion. Am J Hematol. 2015;
178. Tatari-Calderone Z, Gordish-Dressman H, Fasano R, Riggs M, Fortier C, Campbell AD, et al. Protective effect of HLA-DQB1 alleles against alloimmunization in patients with sickle cell disease. Hum Immunol. 2015;
179. Hall AM, Cairns LS, Altmann DM, Barker RN, Urbaniak SJ. Immune responses and tolerance to the RhD blood group protein in HLA-transgenic mice. Blood. 2005;105(5):2175–9.
180. J.Urbaniak S. Alloimmunity to human red blood cell antigens. Vox Sang. 2002;83 S1:293–7.
181. Castilho SL, Vianna TVC, Frauches TF CF. Abstracts of the 25th Regional Congress of the International Society of Blood Transfusion in conjunction with the 33rd Annual Conference of the British Blood Transfusion Society, London, UK, June 27-July 1, 2015. Vox Sang. 2015;109 Suppl:1–379.
182. Schonewille H, Doxiadis IIN, Levering WHBM, Roelen DL, Claas FHJ, Brand A. HLA-DRB1 associations in individuals with single and multiple clinically relevant red blood cell antibodies. Transfusion. 2014;54(8):1971–80.

103
___________________________________________________________________
8 APÊNDICE
Apêndice A - Frequências fenotípicas dos antígenos de grupos sanguíneos deduzidas a partir dos genótipos em pacientes com Anemia Falciforme
Sistema Fenótipo deduzido do
genótipo
Pacientes Aloimunizados com
AF
Pacientes Não aloimunizados com
AF
Pacientes totais
RH RhD – 4(5,97) 8(8,51) 12(7,45)
RhD+ 63(94,03) 86(91,49) 149(92,55)
n(%) 67(100,00) 94(100,00) 161(100,00)
C– E– c+ e+ 28(41,79) 31(32,98) 59(36,65)
C+ E– c+ e+ 22(32,84) 32(34,04) 54(33,54)
C– E+ c+ e+ 10(14,93) 15(15,96) 25(15,53)
C+ E– c- e+ 2(2,99) 6(6,38) 8(4,97)
C- E+ c+ e– 1(1,49) 3(3,19) 4(2,48)
C+ E+ c+ e+ 3(4,48) 5(5,32) 8(4,97)
C+ E+ c+ e– 1(1,49) 2(2,13) 3(1,86)
n(%) 67(100,00) 94(100,00) 161(100,00)
Kell K–k+ 63(94,03) 89(94,68) 152(94,41)
K+k+ 4(5,97) 5(5,32) 9(5,59)
K+ k– 0(0,00) 0(0,00) 0(0,00)
n(%) 67(100,00) 94(100,00) 161(100,00)
Kp(a+b–) 0(0,00) 0(0,00) 0(0,00)
Kp(a+b+) 0(0,00) 2(2,15) 2(1,26)
Kp(a–b+) 66(100,00) 91(97,85) 157(98,74)
n(%) 66(100,00) 93(100,00) 159(100,00)
Js(a+b–) 1(1,49) 0(0,00) 1(0,63)
Js(a+b+) 5(7,46) 12(12,90) 17(10,62)
Js(a–b+) 61(91,04) 81(87,10) 142(88,75)
n(%) 67(100,00) 93(100,00) 160(100,00)
Kidd Jk(a+b–) 30(45,45) 39(41,94) 69(43,40)
Jk(a+b+) 31(46,97) 45(48,39) 76(47,80)
Jk(a–b+) 5(7,58) 9(9,68) 14(8,80)
n(%) 66(100,00) 93(100,00) 159(100,00)
Duffy Fy(a+b–) 2(2,99) 9(9,57) 11(6,83)
Fy(a+b–)a 15(22,39) 20(21,28) 35(21,74)
Fy(a+b+) 10(14,93) 13(13,83) 23(14,29)
Fy(a+bx)c 1(1,49) 0(0,00) 1(0,62)
Fy(a-b+) 6(8,96) 11(11,70) 17(10,56)
Fy(a–b+)a 16(23,88) 23(24,47) 39(24,22)
Fy(a–bx)c 0(0,00) 1(1,06) 1(0,62)
Fy(a–b–)b 17(25,37) 17(18,09) 34(21,12)
n(%) 67(100,00) 94(100,00) 161(100,00)
.

104
Apêndice A continuação - Frequências fenotípicas dos antígenos de grupos sanguíneos deduzidas a partir dos genótipos em pacientes com Anemia Falciforme
Sistema Fenótipo deduzido do
genótipo
Pacientes Aloimunizados com
AF
Pacientes Não aloimunizados com
AF Pacientes totais
MNSs M+N– 16(24,24) 38(41,76) 54(34,39)
M+N+ 35(53,03) 42(46,15) 77(49,04)
M–N+ 15(22,73) 11(12,09) 26(16,56)
n(%) 66(100,00) 91(100,00) 157(100,00)
S+s– 4(6,06) 10(10,64) 14(8,75)
S+s+ 26(39,39) 37(39,36) 63(39,38)
S–s+ 36(54,55) 47(50,00) 83(51,88)
n(%) 66(100,00) 94(100,00) 160(100,00)
Lutheran Lu(a+b–) 0(0,00) 0(0,00) 0(0,00)
Lu(a+b+) 5(7,58) 8(8,79) 13(8,28)
Lu(a–b+) 61(92,42) 83(91,21) 144(91,72)
n(%) 66(100,00) 91(100,00) 157(100,00)
Diego Di(a+b–) 0(0,00) 0(0,00) 0(0,00)
Di(a+b+) 2(3,03) 6(6,52) 8(5,06)
Di(a–b+) 64(96,97) 86(93,48) 150(94,94)
n(%) 66(100,00) 92(100,00) 158(100,00)
Colton Co(a+b–) 65(98,48) 88(95,65) 153(96,84)
Co(a+b+) 1(1,52) 4(4,35) 5(3,16)
Co(a–b+) 0(0,00) 0(0,00) 0(0,00)
n(%) 66(100,00) 92(100,00) 158(100,00)
Dombrock Do(a+b–) Hy+ Joa(a+) 4(6,35) 9(9,68) 13(8,33)
Do(a+b+) Hy+ Joa(a+) 28(44,44) 37(39,78) 65(41,67)
Do(a–b+) Hy+ Joa(a+) 31(49,21) 47(50,54) 78(50,00)
n(%) 63(100) 93(100) 156(100)
a Presença da mutação gata (–67T/C) em heterozigose;
b Presença da mutação gata (–67T/C) em homozigose;
c Presença da
mutação 265C/T e –67T/C.

105
Apêndice B - Frequências alélicas e genotípicas do SNP IL10–819T/C em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
SNP Modelo
Estatístico Genótipo / alelo
PA PNA Pacientes Totais Controles Teste Estatístico
n=67(%) n=94(%) n=161(%) n=288(%) PA vs PNA PA vs controle Pacientes totais
vs controles
IL10-819T/C (rs1800871)
Genótipos
Codominante
CC 22(33,33) 41(44,09) 63(39,62) 112(38,90) ref ref ref
TC 35(53,03) 40(43,01) 75(47,17) 134(46,50) ns ns ns
TT 9(13,64) 12(12,90) 21(13,21) 42(14,60) ns ns ns
Dominante
CC 22(33,33) 41(44,09) 63(39,62) 112(38,90) ref ref ref
TT + TC 44(66,67) 52(55,91) 96(60,38) 176(61,10) ns ns ns
Recessivo
CC + TC 57(86,36) 81(87,10) 138(86,79) 246(85,40) ref ref ref
TT 9(13,64) 12(12,90) 21(13,21) 42(14,60) ns ns ns
Over dominante
CC + TT 31(47,00) 53(57,00) 84(52,80) 154(53,50) ref ref ref
TC 35(53,00) 40(43,00) 75(47,20) 134(46,50) ns ns ns
Alelos
C 79(59,85) 124(65,96) 203(63,44) 358(62,20) ref ref ref
T 53(40,15) 64(34,04) 117(36,56) 218(37,80) ns ns ns
PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF, Pacientes totais: Pacientes aloimunizados + não aloimunizados, Controle: doadores de sangue e medula óssea, ref: grupo de referência, ns: não significante.

106
Apêndice C - Frequências alélicas e genotípicas do SNP IL10–592C/A em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
SNP
Modelo Estatístico
Genótipo / alelo
PA PNA Pacientes Totais Controles Teste Estatístico
n=67(%) n=94(%) n=161(%) n=288(%) PA vs PNA
PA vs controle
Pacientes totais vs controles
IL10–592C/A
(rs1800872)
Genótipos
Codominante
CC 22(32,84) 41(43,62) 63(39,13) 115(39,90) ref ref ref
CA 38(56,72) 41(43,62) 79(49,07) 133(46,20) ns ns ns
AA 7(10,45) 12(12,77) 19(11,80) 40(13,90) ns ns ns
Dominante
CC 22(32,84) 41(43,62) 63(39,13) 115(39,90) ref ref ref
AA + CA 45(67,16) 53(56,38) 98(60,87) 173(60,10) ns ns ns
Recessivo
CC + CA 60(89,55) 82(87,23) 142(88,20) 248(86,10) ref ref ref
AA 7(10,45) 12(12,77) 19(11,80) 40(13,90) ns ns ns
Over dominante
CC + AA 29(43,30) 53(56,40) 82(50,90) 155(53,80) ref ref ref
CA 38(56,70) 41(43,60) 79(49,10) 133(46,20) ns ns ns
Alelos
C 82(61,19) 122(64,89) 204(63,35) 363(63,00) ref ref ref
A 52(38,81) 65(34,57) 117(36,34) 213(37,00) ns ns ns
PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF, Pacientes totais: Pacientes aloimunizados + não aloimunizados, Controle: doadores de sangue e medula óssea, ref: grupo de referência, ns: não significante.

107
Apêndice D - Frequências alélicas e genotípicas do polimorfismo IL4 íntron 3 VNTR em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
Polimorfismo Genótipo / alelo
PA PNA Pacientes Totais Controles Teste Estatístico
n=67(%) n=94(%) n=161(%) n=288(%) PA vs PNA PA vs controle Pacientes totais
vs controles
IL4 intron 3 VNTR
Genótipos
RP2/RP2 22(32,84) 32(34,04) 54(33,54) 131(45,50) ref ref ref
RP1/RP1 15(22,39) 12(12,77) 27(16,77) 34(11,80) ns ns ns
RP2/RP1 29(43,28) 46(48,94) 75(46,58) 122(42,40) ns ns ns
RP1/RP3 1(1,49) 2(2,13) 3(1,86) 1(0,30) ns ns ns
RP2/RP3 0(0,00) 2(2,13) 2(1,24) 0(0,00) ns ns ns
Sem RP1 22(32,84) 34(36,17) 56(34,78) 131(45,50) ref ref ref
Com RP1 45(67,16) 60(63,83) 105(65,22) 157(54,50) ns ns ns
outros 52(77,61) 82(87,23) 134(83,23) 254(88,20) ref ref ref
somente RP1 15(22,39) 12(12,77) 27(16,77) 34(11,80) ns ns ns
Alelos
RP2 73(54,48) 112(59,57) 185(57,45) 384(66,70) ref ref ref
RP1 60(44,78) 72(38,30) 132(41,00) 191(33,20) ns ns ns
RP3 1(0,75) 4(2,13) 5(1,55) 1(0,10) ns ns ns
Sem RP1: RP2/RP2 + RP2/RP3; Com RP1: RP2/RP1+ RP1/RP1+ RP1/RP3; somente RP1: RP1/RP1. PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF, Pacientes totais: Pacientes aloimunizados + não aloimunizados, Controle: doadores de sangue e medula óssea, ref: grupo de referência, ns: não significante.

108
Apêndice E - Frequências alélicas e genotípicas do SNP IL4–590C/T (rs2243250) em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
SNP Modelo
Estatístico Genótipo / alelo
PA PNA Pacientes Totais Controles Teste Estatístico
n=64(%) n=93(%) n=157(%) n=288(%) PA vs PNA
PA vs controle
Pacientes totais vs controles
IL4–590C/T (rs2243250)
Genótipos
Codominante
CC 15(23,4) 26(27,96) 41(26,28) 94(32,60) ref ref ref
CT 28(43,80) 43(46,24) 71(45,51) 131(45,50) ns ns ns
TT 21(32,80) 24(25,81) 45(28,21) 63(21,90) ns ns ns
Dominante
CC 15(23,40) 26(27,96) 41(26,10) 94(32,60) ref ref ref
CT+TT 48(76,60) 67(72,04) 116(73,90) 194(67,40) ns ns ns
Recessivo
CC + CT 43(67,20) 69(74,19) 112(71,30) 225(78,10) ref ref ref
TT 20(32,80) 24(25,81) 45(28,70) 63(21,90) ns ns ns
Over dominante
CC + TT 36 (56,20) 50 (53,80) 86 (54,80) 157(54,50) ref ref ref
CT 28(43,80) 43 (46,20) 71 (45,20) 131(45,50) ns ns ns
Alelos
C 58(44,40) 95(51,08) 153(48,40) 319(55,40) ref ref ref
T 70(55,60) 91(48,92) 161(51,60) 257(44,60) ns ns ns
PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF, Pacientes totais: Pacientes aloimunizados + não aloimunizados, Controle: doadores de sangue e medula óssea, ref: grupo de referências, ns: não significante.

109
Apêndice F - Frequências alélicas e genotípicas do SNP IL17A–197G/A (rs2275913) em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
SNP Modelo
Estatístico Genótipo / alelo
PA PNA Pacientes Totais Controles Teste Estatístico
n=65(%) n=91(%) n=156(%) n=288(%) PA vs PNA PA vs controle Pacientes totais vs controles
IL17A –197G/A (rs2275913)
Genótipos
Codominante
GG 42(64,62) 62(68,13) 104(66,67) 195(67,70) ref ref ref
AA 2(3,08) 3(3,30) 5(3,21) 15(5,20) ns ns ns
GA 21(32,31) 26(28,57) 47(30,13) 78(27,00) ns ns ns
Dominante
GG 42(64,62) 62(68,13) 104(66,67) 195(67,70) ref ref ref
AA+GA 23(35,38) 29(31,87) 52(33,33) 93(32,30) ns ns ns
Recessivo
GG + GA 63(96,92) 88(96,70) 151(96,79) 273(94,80) ref ref ref
AA 2(3,08) 3(3,30) 5(3,21) 15(5,20) ns ns ns
Over dominante
GG + AA 44 (67,70) 65 (71,40) 109(69,90) 210(72,90) ref ref ref
GA 21 (32,30) 26 (28,60) 47(30,10) 78(27,10) ns ns ns
Alelos
G 105(80,77) 150(82,42) 255(81,73) 469(81,10) ref ref ref
A 25(19,23) 32(17,58) 57(18,27) 109(18,90) ns ns ns
PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF, Pacientes totais: Pacientes aloimunizados + não aloimunizados, Controle: doadores de sangue e medula óssea, ref: grupo de referência, ns: não significante.

110
Apêndice G - Frequências alélicas e genotípicas do SNP IL17F 7488A/G (rs763780) em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
SNP Modelo
Estatístico Genótipo / alelo
PA PNA Pacientes Totais Controles Teste Estatístico
n=65(%) n=91(%) n=156(%) n=288(%) PA vs PNA PA vs controle Pacientes totais
vs controles
IL17F 7488A/G (rs763780)
Genótipos
Codominante
AA 56(86,15) 78(85,71) 134(85,90) 254(88,20) ref ref ref
GG 1(1,54) 0(0,00) 1(0,64) 0(0,00) ns ns ns
AG 8(12,31) 13(14,29) 21(13,46) 34(11,80) ns ns ns
Dominante
AA 56(86,15) 78(85,71) 134(85,90) 254(88,20) ref ref ref
AG+GG 9(13,85) 13(14,29) 22(14,10) 34(11,80) ns ns ns
Recessivo
AG+AA 64(98,46) 91(100,00) 155(99,36) 288(100,00) ref ref ref
GG 1(1,54) 0(0,00) 1(0,64) 0(0,00) ns ns ns
Over dominante
AA + GG 57 (87,70) 78 (85,70) 135(86,50) 254(88,20) ref ref ref
GA 8 (12,30) 13 (14,30) 21(13,50) 34(11,80) ns ns ns
Alelos
A 120(92,31) 169(92,86) 289(92,63) 542(94,10) ref ref ref
G 10(7,69) 13(7,14) 23(7,37) 34(5,90) ns ns ns
PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF, Pacientes totais: Pacientes aloimunizados + não aloimunizados, Controle: doadores de sangue e medula óssea, ref: grupo de referência, ns: não significante.

111
Apêndice H - Frequências alélicas e genotípicas do SNP IL6–174C/G (rs1800795) em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
SNP Modelo
Estatístico Genótipo /alelo
PA PNA Pacientes Totais Controles Teste Estatístico
n=64(%) n=93(%) n=157(%) n=288(%) PA vs PNA PA vs
controle Pacientes totais vs
controles
IL6–174C/G (rs1800795)
Genótipos
Codominante
GG 40(62,50) 67(72,04) 107(68,15) 207(71,90) ref ref ref
CG 20(31,25) 25(26,88) 45(28,66) 75(26,00) ns ns ns
CC 4(6,25) 1(1,08) 5(3,18) 6(2,10) ns ns ns
Dominante
GG 40(62,50) 67(72,04) 107(68,15) 207(71,90) ref ref ref
GC + CC 24(37,50) 26(27,96) 50(31,85) 81(28,10) ns ns ns
Recessivo
GC + GG 60(93,75) 92(98,92) 152(96,82) 282(97,90) ref ref ref
CC 4(6,25) 1(1,08) 5(3,18) 6(2,10) ns ns ns
Over dominante
GG + CC 44 (68,75) 68(73,12) 112(371,30) 213(74,00) ref ref ref
GC 20(31,25) 25(26,88) 45(28,7) 75(26,00) ns ns ns
Alelos
G 100(79,37) 159(85,48) 259(82,48) 489(84,90) ref ref ref
C 28(22,22) 27(14,52) 55(17,52) 87(15,10) ns ns ns
PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF, Pacientes totais: Pacientes aloimunizados + não aloimunizados, Controle: doadores de sangue e medula óssea, ref: grupo de referência, ns: não significante.

112
Apêndice I - Frequências dos diplótipos formados pelos SNPs IL17A–197G/A e IL17F 7488A/G em pacientes portadores de Anemia Falciforme aloimunizados e não aloimunizados e em controles.
SNPs diplótipos
PA PNA Pacientes Totais Controles Teste Estatístico
n=65(%) n=91(%) n=156(%) n=288(%) PA vs PNA PA vs controle Pacientes totais
vs controles
IL17A–197G/A/ IL17F 7488A/G
AA/AA 1(1,54) 3(3,30) 4(2,55) 13(4,50) ns ns ns
AG/AA 1(1,54) 0(0,00) 1(0,64) 2(0,70) ns ns ns
GA/AA 17(26,15) 22(24,18) 39(24,84) 71(24,70) ns ns ns
GA/GA 38(58,46) 53(58,24) 91(57,96) 170(59,00) ns ns ns
GG/GA 3(4,62) 9(9,89) 12(7,64) 24(8,30) ns ns ns
AA/GG 4(6,15) 4(4,40) 0(0,00) 8(2,80) ns ns ns
GG/GG 1(1,54) 0(0,00) 1(0,64) 0(0,00) ns ns ns
PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF, Pacientes totais: Pacientes aloimunizados + não aloimunizados, Controle: doadores de sangue e medula óssea, ns: não significante.

113
Apêndice J - Distribuição das frequências dos grupos alélicos HLA-DQA1 e -DQB1
em pacientes portadores de Anemia Falciforme aloimunizados a antígenos eritrocitários e pacientes não aloimunizados.
HLA-DQA1 PA PNA
HLA-DQB1 PA PNA
n=132(%) n=174(%) n=132(%) n=174(%)
DQA1*01 56(41,79) 68(39,08) DQB1*01 0(0,00) 1(0,57)
DQA1*02 18(13,43) 16(9,20) DQB1*02 30(22,73) 28(16,09)
DQA1*03 16(11,94) 34(19,54) DQB1*03 44(33,33) 58(33,33)
DQA1*04 7(5,22) 17(9,77) DQB1*04 4(3,03) 15(8,62)
DQA1*05 36(26,87) 37(21,26) DQB1*05 25(18,94) 41(23,56)
DQA1*06 1(0,75) 2(1,15) DQB1*06 29(21,97) 31(17,82)
PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF.
Apêndice K- Distribuição das frequências dos grupos alélicos HLA-DQA1 em pacientes portadores de Anemia Falciforme com anticorpos anti-Rh, pacientes com
outros anticorpos não Rh e pacientes não aloimunizados.
HLA-DQA1 PA com anti-Rh PA com anticorpos não Rh PNA
n=96(%) n=32(%) n=174(%)
DQA1*01 41(42,71) 12(37,5) 68(39,08)
DQA1*02 12(12,5) 6(18,75) 16(9,20)
DQA1*03 13(13,54) 2(6,25) 34(19,54)
DQA1*04 7(7,29) 0(0) 17(9,77)
DQA1*05 22(22,92) 12(37,5) 37(21,26)
DQA1*06 1(1,04) 0(0) 2(1,15)
PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF.
Apêndice L - Distribuição das frequências dos grupos alélicos HLA-DQB1 em pacientes portadores de Anemia Falciforme com anticorpos anti-Rh, pacientes com
anticorpos não Rh e pacientes não aloimunizados.
HLA-DQB1 PA com anti-Rh
Pacientes aloimunizados por outros anticorpos
PNA
n=96(%) n=32(%) n=174(%)
DQB1*01 0(0) 0(0) 1(0,57)
DQB1*02 19(19,79) 11(34,38) 28(16,09)
DQB1*03 33(34,38) 9(28,13) 58(33,33)
DQB1*04 4(4,17) 0(0) 15(8,62)
DQB1*05 19(19,79) 5(15,63) 41(23,56)
DQB1*06 21(21,88) 7(21,86) 31(17,82)
PA: Pacientes aloimunizados com AF, PNA: Pacientes não aloimunizados com AF.

114
__________________________________________________________________
9 ANEXOS
Anexo I. Parecer do Comitê de Ética em Pesquisa

115

116
Anexo II. Dados parciais publicados

117

118

119

120

121

122
Anexo III. Permissão da Editora para utilização do artigo publicado na Tese