anÁlise de biomarcadores bioquÍmicos no … · resumo os estuários são ambientes de grande...
TRANSCRIPT

ANÁLISE DE BIOMARCADORES BIOQUÍMICOS NO CARANGUEJO UCA
MORDAX EXPOSTO A DIFERENTES CONDIÇÕES DE SALINIDADE E
FONTES DE CONTAMINAÇÃO NA BACIA DO MARAJÓi
Carla Carolina Miranda dos Santosii
Lílian Lund Amadoiii
Universidade Federal do Pará
Resumo
Os estuários são ambientes de grande relevância ecológica e biológica importantes para
o desenvolvimento humano através do crescimento populacional e de atividades
econômicas em seu entorno. Dentre os fatores abióticos deste ambiente destaca-se a
salinidade em virtude de suas constantes variações. O estuário da Bacia de Marajó, PA
recebe a descarga de uma grande variedade de compostos tóxicos oriundos dos rios que
compõem sua bacia de drenagem, além dos contaminantes liberados durante operações
portuárias.Os biomarcadores são variações induzidas por xenobióticos em moléculas,
componentes celulares, estruturas ou funções que configuram-se ferramentas de
diagnóstico precoce da saúde ambiental. O objetivo deste trabalho é avaliar a qualidade
ambiental do ponto de vista biológico, utilizando biomarcadores bioquímicos em
caranguejos Uca mordax oriundos de diferentes pontos da bacia de Marajó levando em
conta o gradiente de salinidade e a influência antrópica. Neste contexto foram
selecionados quatro pontos: Abaetetuba, PA (referência dulciaquícola), Barcarena, PA
(área portuária dulciaquícola), Vigia, PA (referência estuarina) e Baía do Guajará-Belém,
PA (área urbana estuarina). Foram analisados três biomarcadores nas brânquias e
músculo: capacidade antioxidante contra radicais peroxil (ACAP- biomarcador de
exposição), atividade da glutationa-s-transferase (GST- biomarcador de exposição) e a
lipoperoxidação (LPO- biomarcador de efeito). No período de transição a capacidade
antioxidante em brânquias apresentou maiores valores de indução diferenciando-se
(p<0,05) do período seco e chuvoso. No músculo os animais coletados no período
chuvoso apresentaramvalores superiores (p<0,05) em relação aos coletados no período
seco. Percebe-se o mesmo padrão de atividade da GST para ambos os órgãos com maior
atividade no período chuvoso e menor no período de transição. Nas brânquias
oscaranguejos coletados no período de transição diferenciaram-se (p<0,05) doscoletados
no período chuvoso apresentando os menores valores de lipoperoxidação enquanto no
músculo no período chuvoso não houve diferenças entre os caranguejos dos distintos
pontos (p>0,05). Conclui-se que os animais de todos os pontos amostrados demonstraram
que suas defesas antioxidantes variam com sazonalidade e que o período de transição
parece configurar um maior desafio metabólico para os animais induzindo suas defesas
antioxidantes.Não houve diferenças expressivas entre os ambientes com e sem histórico
de poluição.
Palavras-chave: Poluição aquática, Salinidade, Uca mordax.

Abstract
Estuaries are environments of great ecological and biological relevance and important for
human development through the population growth and economic activities in its
surroundings. Among the abiotic factors of this environment, salinity stands out because
of its constant variations. The estuary of Marajó Bay, PA receives the discharge of a
variety of toxic compounds from the rivers that compose its drainage basin, beside the
contaminants released during port operations. Biomarkers are changes induced by
xenobiotics in molecules, cellular components, structures or features that works as tools
for early diagnosis of environmental health. The objective is to evaluate the
environmental quality from a biological perspective, using biochemical biomarkers in
crabs Uca mordax from different point of Marajó Bay considering the salinity gradient
and the anthropogenic influence. In this context, four sampling sites were
selected:Abaetetuba, PA (freshwater reference); Barcarena, PA (freshwater portuary
area); Vigia, PA (estuarine reference) and the Guajará Bay-Belém, PA (estuarine urban
area). Three biomarkers were analyzed in gills and muscle: antioxidant capacity against
peroxyl radicals (ACAP- biomarker of exposure), glutathione-s-transferase activity
(GST- biomarker of exposure) and lipid peroxidation (LPO – effect biomarker). During
the transition period the antioxidant capacity in gills showed higher induction values
differing (p < 0.05) of dry and rainy season while muscle showed higher values on rainy
season (p < 0.05) when compared to dry season. There is the same pattern of GST activity
for both tissues with greater activity in the rainy season and shorter during the transition
season. Gills in the transition season differed (p< 0.05) of the rainy season presenting the
lowest lipid peroxidation values while in the muscle during the rainy season there were
no differences between the sampling sites (p> 0.05). We conclude that animals of all
sampling sites showed that their antioxidant defenses vary with seasonality and the
transition period seems to be an important metabolic challenge for animals inducing its
antioxidant defenses. There were no significant differences between environments with
and without history of pollution.
Keywords: Aquatic Pollution, Salinity, Uca mordax.

Os estuários são áreas de grande
importância ecológica, econômica e
social, por serem ambientes de produção,
criação e reprodução de várias espécies
biológicas incluindo espécies de
importância comercial (ASMUS,1996)
dando suporte ao estabelecimento de
cadeias alimentares complexas
(BRAGA, 2000).
O estuário da Bacia de Marajó recebe a
descarga de uma grande variedade de
compostos tóxicos oriundos dos rios que
compõem sua bacia de drenagem, além
dos contaminantes liberados durante
operações portuárias (PEREIRA FILHO
et al., 2001).
A poluição de ambientes estuarinos é
considerada como um problema
ambiental crítico devido à alta variação
dos fatores abióticos, que não só impõem
severas restrições à vida dos organismos
nessas áreas como também alteram
constantemente a biodisponibilidade de
contaminantes (AMADO et al.,2006).
Dentre os fatores abióticos, destaca-se a
salinidade como parâmetro ambiental
importante em virtude de suas altas e
constantes variações, as quais afetam a
fisiologia e a ecologia dos organismos
(ANGER, 2001).Como uma das
adaptações fisiológicas fundamentais de
organismos estuarinos podemos destacar
a osmorregulação que é a habilidade de
manter ativamente a concentração
osmótica dos fluidos extracelulares,
apesar das variações na osmolaridade
do ambiente (MONSERRAT et al.,
2007).
O efeito combinado das atividades
antrópicas com as flutuações marcantes
de salinidade podem gerar condições de
estresse ambiental para os organismos
uma vez que contribuem
significativamente com a
biodisponibilidade de poluentes e
promove a realocação da energia que
seria utilizada para detoxificação para a
iono e osmorregulação, ou vice-versa
(MONSERRAT et al., 2007).
Neste contexto, variações induzidas por
xenobióticos em moléculas,
componentes celulares, estruturas ou
funções são utilizadas como
biomarcadores (DEPLEDGE et al.,
1995). Biomarcadores são capazes de
informar se um organismo foi exposto à
um xenobiótico (biomarcador de
exposição),as consequências danosas
dessa exposição no organismo
(biomarcadores de efeito) ou a
susceptibilidade do organismo ao
xenobiótico (SOGORB et al, 2014).
Essas alterações em respostas biológicas
quando um determinado organismo está
fora de suas condições fisiológicas
normais, seja devido à presença de
poluentes ou devido a modificações
físico-químicas de seu habitat
(AMORIM, 2003) configuram-se como
excelentes ferramentas de diagnóstico
precoce na saúde ambiental por
representarem respostas em baixo nível
de organização biológica
(CARAJAVILLE et al., 2000),
permitindo a identificação precoce de
mudanças antes que alcancem níveis
mais complexos de organização
(MONSERRAT et al., 2003a) e de
maior relevância ecológica.
A análise de biomarcadores é feita em
órgãos de organismos residentes nos
ambientes em que se pretende avaliar sua
qualidade do ponto de vista biológico,
tais organismos são chamados de
biomonitores.
Estudos têm demonstrado que as
populações de caranguejos chama-marés
podem chegar à extinção local devido à
degradação crescente dos hábitats
intertidais por poluição, dragagem e
aterro o que demonstra a sensibilidade
destes organismos a estresses ambientais
(BARNWELL, 1986)

O presente estudo tem como objetivo
fazer uma abordagem do ambiente
estuarino utilizando biomarcadores
bioquímicos em caranguejos daespécie
Uca mordax sujeitos à diferentes
condições de salinidade e de influência
antrópica. Estes resultados serão
comparados em tempo e espaço com
fatores ambientais como salinidade, pH e
oxigênio dissolvido além de identificar
possíveis alterações devido ao impacto
antropogênico.
Materiais e Métodos
1.1 Área de estudo
A área escolhida para o estudo é a Bacia
do Marajó formada pela junção dos rios
Pará/Tocantins e, posteriormente
somada as águas da Baía de Guajará, que
circunda a cidade de Belém, e todos os
demais afluentes que irão formar a Baía
de Marajó antes de atingir o oceano.
Assim, os pontos de coleta são (Figura
1):
A) Referência dulciaquícola: localizado
no município de Abaetetuba, em uma
ilha às margens do rio Campumpema,
consiste de um local sem histórico de
contaminação e de água doce ao longo de
todo o ano.
B) Área portuária dulciaquícola:
localizado no município de Barcarena,
consiste em uma área portuária com
ambientes semelhantes ao ponto “a” nas
suas mediações, com a diferença de ter
um grande histórico de impacto
antropogênico.
C) Área urbana estuarina: nas margens
da Baía de Guajará, próximo ao canal do
Tucunduba na cidade de Belém, área
com leve influência de águas estuarinas
e com alto grau de impacto urbano.
D) Referência estuarina: localizado no
município de Vigia, próximo ao mercado
de pescados do município, área sob forte
influência estuarina e baixo histórico de
poluição,
1.2 Atividades de campo
Para coleta dos caranguejos (apenas
machos) foi utilizada a catação manual,
logo em seguida a maré de baixa-mar.
Foram realizadas 3 coletas em cada
ambiente: uma no período de transição
(Julho/2014), uma no período menos
chuvoso(Outubro/2014) e outra no
período chuvoso (Fevereiro/2015). No
momento da coleta foram registrados os
dados ambientais de cada ponto, junto
com uma descrição do ecossistema. Os
dados abióticos mensurados foram:
salinidade, pH, oxigênio dissolvido,
temperatura da superfície da água,
condutividade.
Os organismos coletados, foram
devidamente etiquetados em relação ao
local e dados ambientais obtidos e
imediatamente crioanestesiados, mortos
e armazenados em gelo (0°C) até a
dissecação, o que ocorreu no mesmo dia
da captura.
1.3 Atividades Laboratoriais
-Biometria e dissecação dos caranguejos
Em laboratório, no mesmo dia da coleta,
os animais foram biometrados com o
auxílio de um paquímetro (0,01 mm)
sendo medidas a maior largura da
carapaça em vista dorsal, alturae
comprimento. Os organismos também
foram pesados em balança semianalítica
(0,01g) para que em seguida as brânquias
e porções do músculo fossem retirados e
acondicionados em tubos tipo eppendorf
para armazenamento em ultra-freezer (-
80ºC) até as análises bioquímicas.
-Preparo dos homogeneizados para
dosagens bioquímicas
As amostras de músculo e brânquias são
homogeneizadas (1:4, p/v) em tampão

contendo 20 mM Tris–Base, 1 mM
EDTA, 1 mM ditiotreitol (Sigma, St.
Louis, MO), 500 mM sacarose, 150 mM
KCl, e 0,1 mM fenilmetilsufonil fluoride
(PMSF; Sigma), com pH ajustado para
7,6 (BAINY et al., 1996). A seguir os
homogeneizados foram centrifugados a
20.000 x g por 20 min a 4ºC. O
sobrenadante foi retirado, aliquotado e
acondicionado a -80ºC até o momento
das dosagens.
-Determinação de Proteínas Totais
A análise de proteínas totais foi realizada
com kit comercial (Doles Ltda, Brazil)
baseado no teste Biureto (citrato
trissódico 0,114M, carbonato de sódio
0,21M e sulfato de cobre 0,01M) para
proteínas, as leituras foram realizadas em
leitor de microplacasmultimodal (Victor
X3, Perkin Elmer) a 550 nm. Os
resultados foram expressos em
miligramas de proteínas/mL.
-Determinação da atividade da
glutationa-S-transferase (GST)
A dosagem de GST foi baseada nos
trabalhos de Habig et al. (1974) e Habig
& Jakoby (1981). A metodologia
consiste em avaliar a conjugação de
1mM de GSH (Sigma) com 1mM do
reagente 1-cloro-2,4 dinitrobenzeno
(CDNB,Sigma),processo catalisado pela
GST. O complexo conjugado formado
possui absorbância máxima em 340 nm
(ε = 9,6 mM -1.cm-1). Como meio de
reação é utilizado tampão fosfato 0,1 M,
pH 7,00. As leituras foram realizadas em
espectrofluorímetro (Victor 2, Perkin
Elmer) com leitora de microplacas. Os
resultados são expressos em UGST/ mg
de proteína que representa a quantidade
Figura 1: Figura1: Pontos de coleta (círculos escuros) em cada município para a Bacia de Marajó, Pará: A) referência dulciaquícola, Abaetetuba; B) área portuária dulciaquícola, Barcarena; C) área urbana estuarina, Belém e D) referência estuarina,Vigia.

necessária da enzima para conjugar 1
µMol de CDNB/ min/mg de proteína, a
25ºC e pH 7,0.
-Determinação da capacidade
antioxidante total contra radicais peroxil
A capacidade antioxidante total contra
radicais peroxil foi analisada através da
determinação da concentração de
espécies ativas de oxigênio (EAO) nas
amostras com e sem exposição a um
gerador de radicais peroxil. O princípio
deste método é que amostras com maior
capacidade antioxidante terão menor
formação de radicais.
Os radicais peroxil são produzidos por
decomposição termal (35°) de 2’2’-
azobis-2-metilpropiamidina
dihidrocloreto (ABAP, 4mM; Aldrich)
(Winston et al., 1998). As leituras
foram realizadas em leitora de
microplaca de fluorescência (485 e 530
para excitação e emissão
respectivamente) durante uma hora
(Victor 2, Perkin Elmer), em um meio
contendo 30 mM de HEPES (pH 7,2),
200 mM KCl, 1 mM MgCl2, 40 µM
H2DCF-DA com e sem a adição de
ABAP. O incremento de fluorescência
de cada amostra ao longo de uma hora de
leitura foi plotado e área sob a reta
formada foi calculada. A diferença
relativa entre as áreas de uma mesma
amostra exposta e não exposta ao ABAP
foram consideradas como medida da
capacidade antioxidante total. Os
resultados são expressos como inverso
da área relativa.
-Lipoperoxidação (LPO) - Ensaio FOX
modificado
A lipoperoxidação foi determinada de
acordo com Hermes-Lima et al. (1995)
adaptada para microplacas segundo
Monserrat et al. (2003). Esse método se
baseia na oxidação do Fe2+ a Fe 3+ por
hidroperóxidos lipídicos presentes na
amostra. Sob condições de pH ácido o
Fe3+ formado se complexa ao corante
laranja de xilenol. As amostras foram
homogeneizadas (1:6 m/v) em metanol
100% à frio (4ºC). Os homogeneizados
foram centrifugados a 1000x g, durante
10 min à 4°C. A presença de
hidroperóxidos lipídicos nas amostras é
avaliada espectrofotometricamente em
550 nm. O hidroperóxido de cumeno
(CHP) foi utilizado como padrão. O
conteúdo de peróxidos lipídicos foi
expresso como equivalentes de 1nM
CHP/g de tecido úmido.
-Integração dos dados e análise
estatística
Foi montada uma base de dados, com os
resultados das variáveis nas áreas
selecionadas para estudo. Após
avaliação da distribuição normal e da
homocedasticidade destas variáveis
foram empregados testes estatísticos
paramétricos para comparação das
médias (ANOVA). Havendo diferença
significativa estas são determinadas pelo
teste post-hoc de Tukey. Para os dados
de biometria foi realizado o teste não
paramétrico de Kruskal-Wallis (ZAR,
1984). O nível de significância aceito é
de 5%.
1.4 Resultados
- Dados de precipitação
Observa-se na Figura 2 o padrão de
variação de precipitação para a região de
Belém ao longo de 2 anos. Com base
nesses padrões que foram selecionados
os meses de coleta. Dentre os meses em
que as coletas ocorreram o maior valor
de precipitação foi no mês de
Fevereiro/15 com 397mm, enquanto
Outubro/14 apresentou o menor valor 78
mm. Julho/14 apresentou valor
intermediário de 188 mm.
-Parâmetros Físico-Químicos

Os parâmetros físico-químicos foram
medidos na maré baixa e maré alta
respectivamente como mostra a Tabela
1.
Não houve diferenças expressivas
quanto a temperatura entre os pontos e
nem entre as estações. O pH manteve-se
na faixa de 5-7 em todos os pontos para
todos os períodos.
Percebe-se grande diferença na faixa de
oxigênio dissolvido entre a maré-baixa
com predominância de baixos valores e
maré-alta com predominância de valores
maiores para todos os pontos ao longo
dos três períodos (com exceção de
Abaetetuba e Vigia no período chuvoso).
Em ambos os períodos de estiagem e
transição percebe-se que os ambientes
dulciaquícolas apresentam valores de
OD superiores aos ambientes estuarinos.
Nos períodos de estiagem e chuvoso
observa-se que os ambientes estuarinos
Belém e Vigia possuem condutividades
maiores em relação aos ambientes
dulciaquícolas. No período de transição
Vigia, Belém e Abaetetuba apresentaram
altos valores de condutividade.
Abaetetuba, Barcarena e Belém não
apresentaram variação na salinidade para
nenhum dos períodos amostrados. Vigia
apresentou alta variação de salinidade
entre maré alta e baixa no entanto, sem
diferenças marcantes entre os períodos
amostrados.
-Biometria
Como mostra a Tabela 2 Os animais de
Abaetetuba não diferenciaram-se
(p>0,05) entre os períodos amostrados
para peso, comprimento total e largura.
Para o peso os organismos de Vigia e
Belém tiveram diferenças (p<0,05) entre
os três períodos amostrados com maior
valor no período de estiagem (3,54±0,07
cm e 3,37±0,07 cm respectivamente) e
menor no período de transição
(1,36±0,01 cm e 1,35±0,002 cm).
No período de estiagem e de transição os
animais de Abaetetuba e Barcarena não
diferenciaram-se (p>0,05) para o peso e
largura. Organismos de Barcarena e
Belém apresentaram diferenças (p>0,05)
para o comprimento total entre os
períodos de estiagem (1,42±0,008 cm e
1,41±0,009 cm) e transição, onde neste
último apresentaram menores valores
(1,35±0,002 cm e 1,35±0,002 cm) não
diferenciando-se (Tabela 2).
Caranguejos de Vigia, Belém e
Barcarena apresentaram diferenças
(p<0,05) para a largura entre os períodos
de estiagem (1,99±0,01 cm, 1,97±0,01
cm e 1,94±0,01 cm) e transição onde
apresentaram valores menores
(1,87±0,06 cm, 1,83±0,003 cm e
1,80±0,004 cm) (Tabela 2).
No período de estiagem os caranguejos
de Abaetetuba diferenciaram-se dos
animais dos demais pontos para largura
apresentando o menor valor (1,84±0,01
cm). No período de transição os animais
dos ambientes estuarinos diferenciaram-
se (p<0,05) dos animais dos ambientes
dulciaquícolas. (Tabela 2)
Não houve diferença entre os
organismos de Belém e Vigia em
nenhum dos períodos amostrados e nem
entre os períodos para altura (p>0,05).
Os animais de Barcarena diferenciaram-
se dos animais dos demais pontos
amostrados para altura nos períodos de
transição e chuvoso apresentando os
maiores valores (1,23±0,003 cm e
1,25±0,11 cm) (Tabela 2).

Figura 2: Precipitação registrada para Belém do período de Janeiro/13 à Junho/2015 (dados obtidos do site
do INMET)
Tabela 1: Parâmetros físico-químicos dos ambientes amostrados ao longo de três períodos sazonais. Os
dados são apresentados como os valores registrados na maré baixa –maré alta.
Tabela 2: Biometria de Uca mordax para todos os pontos amostrados ao longo de três períodos sazonais.
Os dados estão expressos em mediana ± quartil. As letras maiúsculas diferentes representam diferenças
entre períodos para um mesmo local e as letras minúsculas diferentes representam diferenças entre locais
em um mesmo período.
PERÍODO LOCAL TEMPERATURA DA ÁGUA(ºC) pH OXIGÊNIO DISSOLVIDO(mg/L) CONDUTIVIDADE (µS/cm) SALINIDADE
ABAETETUBA 25,1 - 30,7 7,2 - 6,99 4,08 - 7,15 378,4 - 34,1 0 - 0
VIGIA 27,4 - 28,3 7,1 - 7,56 2,4 - 5,54 136,6 - 90,2 0 - 12
BARCARENA 28,6 - 29,8 6,95 - 7,33 5,2 - 9,0 44,1 - 60,9 0 - 0
BELÉM 26,6 - 28,5 7,17 - 6,7 2,71 - 6,8 219,9 - 25,2 0 - 0
ABAETETUBA 28,9 - 30,4 7,92 - 6,87 4,72 - 7,18 40,9- 34,1 0 - 0
VIGIA 27,6 - 30,8 6,9 - 7,28 2,58 – 6,93 58,6 – 4,4 0 - 13
BARCARENA 28,9 - 29,4 6,81 - 7,28 4,99 - 8,7 46,7 – 59,6 0 - 0
BELÉM 26,3 - 29,5 7,12 – 6,99 3,17 - 6,47 296,6 – 37,1 0 - 0
ABAETETUBA 28,1 - 38,5 5,49 - 6,5 5,8 - 1,38 143,8 - 201 0 - 0
VIGIA 24,9 - 26,8 7,38 - 7,5 6,2 - 5 992 - 2520 4--13
BARCARENA 27,6 - 30 7,2 - 6,84 5,65 - 6,71 66,5 - 75,8 0 - 0
BELÉM 27,4 - 28,5 7,42 - 7,22 1,3 - 6,32 303 - 64,4 0 - 0
TRANSIÇÃO
ESTIAGEM
CHUVOSO
PERÍODO LOCAL PESO(cm) COMPRIMENTO TOTAL(cm) LARGURA(cm) ALTURA(cm)
ABAETETUBA 2,33±1,12 Aab 1,33±0,02 Aa 1,82±0,03 Aa 1,01±0,01 Aa
VIGIA 2,63±0,07Ba 1,36±0,01 Ba 1,87±0,06 Ba 1,00±0,008 Aa
BELÉM 2,44±0,01 Bab 1,35±0,002 Ba 1,83±0,003 Ba 1,01±0,01 Aa
BARCARENA 2,31±0,01 Bb 1,35±0,002 Ba 1,80±0,004 Ba 1,23±0,003 Bb
ABAETETUBA 2,49±0,04 Aa 1,36±0,01 Ab 1,84±0,01 Aa 1,13±0,09 Bb
VIGIA 3,54±0,07 Ab 1,44±0,007 Aa 1,99±0,01 Ab 1,00±0,008 Aa
BELÉM 3,37±0,07 Ab 1,41±0,009 Aab 1,97±0,01 Ab 1,00±0,008 Aa
BARCARENA 3,11±0,06 Aa 1,42±0,008 Aa 1,94±0,01 Ab 1,00±0,007 Aa
ABAETETUBA 2,48±0,03 Ab 1,34±0,08 Aa 1,84±0,007 Aa 1,05±0,04 Aa
VIGIA 3,09±0,07 Ca 1,40±0,009 Ab 1,94±0,01 Ab 1,04±0,007 Aa
BELÉM 3,04±0,06 Ca 1,39±0,009 Bb 1,92±0,01 Ab 1,01±0,008 Aa
BARCARENA 2,78±0,04 Aba 1,38±0,007 ABb 1,88±0,008 Ba 1,25±0,11 Bb
TRANSIÇÃO
ESTIAGEM
CHUVOSO

-Biomarcadores
Brânquias
Capacidade antioxidante total-
Biomarcador de exposição.
Os animais de Barcarena diferenciaram-
se (p<0,05) dos animais dos demais
pontos no período de estiagem
apresentando a menor capacidade
antioxidante (0,24 ± 0,05), enquanto no
período chuvoso não houve diferença
entre os organismos dos pontos
amostrados (p>0,05). No período de
transição os organismos de Belém
(3,16±0,28) diferenciaram-se dos de
Abaetetuba (0,74±0,13) apresentando
maior indução das defesas antioxidantes
(Figura 3).
Organismos de Barcarena, Belém e
Vigia apresentaram no período de
transição maior capacidade antioxidante
(2,53±0,39; 3,16±0,28 e 2,16 ±0,06
respectivamente) quando comparados
aos períodos chuvoso (0,38±0,09; 0,68 ±
0,06 e 0,41±0,03) e de estiagem
(0,24±0,05; 1,03±0,26 e 0,65±0,16) que
apresentam-se iguais com capacidades
antioxidantes menores (Figura 3).
Atividade da glutationa-s-transferase-
Biomarcador de exposição.
A atividade da GST nas brânquias de
caranguejos de Abaetetuba (140,25±8,32
UGST/ mg proteína) diferenciou-se das
de organismos de Belém (102,01±7,05
UGST/ mg proteína) no período de
estiagem apresentando maior atividade
desta enzima, não houve diferenças
(p>0,05) entre os pontos amostrados
para o período de transição. No período
chuvoso os animais de Belém
(147,08±9,66 UGST/ mg proteína)
diferenciaram-se dos de Vigia
(108,94±6,32 UGST/ mg proteína) e
Barcarena (102,57±5,80 UGST/ mg
proteína) com maiores níveis de
atividade (Figura 4).
As brânquias de caranguejo de
Abaetetuba, Barcarena e Vigia
apresentaram no período de transição
menores atividades da GST (40,50 ±2,05
UGST/ mg de proteína; 51,73 ± 1,85
UGST/ mg de proteína e 49,93±2,51
UGST/ mg de proteína respectivamente)
diferenciando-se de ambos os períodos
de estiagem (140,25±8,32 UGST/ mg de
proteína; 121,12 ± 7,04 UGST/ mg de
proteína e 128,24±8,55 UGST/ mg de
proteína) e chuvoso (122,41±3,93
UGST/ mg de proteína; 102,57±5,80
UGST/ mg de proteína e 108,94±6,32
UGST/ mg de proteína) que por sua vez
apresentaram-se iguais e com níveis
maiores de atividade.(Figura 4)
Houve diferenças (p<0,05) entre todos
os períodos climáticos para caranguejos
de Belém, com maior atividade da
enzima no período chuvoso
(147,08±9,66 UGST/ mg de proteína) e
menor no período de transição
(39,01±2,64 UGST/ mg de proteína
(Figura 4).
Figura 3: Capacidade Antioxidante Total em brânquias de Uca mordax
(n=5-10).Valores expressos em média±erro padrão. As letras maiúsculas
diferentes representam diferenças (p<0,05) entre períodos para um
mesmo local e letras minúsculas diferentes representam diferenças (p<
0,05) entre os locais no mesmo período.

Lipoperoxidação- Biomarcador de
efeito.
A lipoperoxidação nas brânquias de
caranguejos de Vigia (25,01±5,10 nM
CHP/ g de tecido úmido) diferenciou-se
das de animais de Belém (16,03 ± 2,73
nM CHP/ g de tecido úmido) e
Abaetetuba (19,56 ± 1,67 nM CHP/ g de
tecido úmido) no período de estiagem
apresentando maiores quantidades de
lipídeos peroxidados. Não houve
diferença (p>0,05) entre os pontos
amostrados nos períodos de transição e
chuvoso (Figura 5).
Os animais de Abaetetuba, Barcarena e
Belém apresentaram diferenças entre o
período chuvoso (27,65±0,94nM CHP/ g
de tecido úmido; 29,39±1,05 nM CHP/ g
de tecido úmido e 26,82±1,44 nM CHP/
g de tecido úmido respectivamente) e o
período de transição (15,30±1,50 nM
CHP/ g de tecido úmido; 19,5±1,78 nM
CHP/ g de tecido úmido e 14,56±0,96
nM CHP/ g de tecido úmido) onde o
primeiro apresentou maiores
quantidades de lipoperoxidação. O
período de estiagem não diferenciou-se
dos demais períodos para os organismos
de todos os pontos amostrados (p>0,05)
mas percebe-se quantidades de lipídeos
peroxidados superiores ao período de
transição (Figura 5).
O período de estiagem para os animais
de Vigia (25,01±5,10nM CHP/ g de
tecido úmido) diferenciou-se do período
de transição (18,88±1,31 nM CHP/ g de
tecido úmido) com maiores níveis de
LPO, enquanto o período chuvoso não
diferenciou-se (p>0,05) dos demais. No
entanto, suas quantidades de lipídeos
peroxidados mostram-se maiores que a
do período de transição (Figura 5).
Músculo
Capacidade antioxidante total-
Biomarcador de exposição.
Não houve diferenças (p>0,05) entre os
organismos de todos os pontos
amostrados no período de estiagem
enquanto no período chuvoso
organismos dos pontos estuarinos Belém
(0,30±0,04) e Vigia (0,30±0,05)
apresentaram maior indução das defesas
antioxidantes diferenciaram-se dos
organismos dos pontos dulciaquícolas
Abaetetuba (0,11±0,01) e Barcarena
(0,14±0,01) (Figura 6)
A capacidade antioxidante total em
músculo de caranguejos de
Barcarena,Vigia e Belém apresentaram
diferenças entre os períodos chuvoso
(0,14±0,01; 0,30±0,05 e 0,30±0,04
respectivamente) e de estiagem
(0,04±0,01; 0,04±0,02 e 0,03±0,01) onde
o primeiro demonstra maiores níveis de
Figura 4: Atividade da Glutationa-s-transferase em brânquias de Uca
mordax (n=5-10). Valores expressos em média ± erro padrão. As
letras maiúsculas diferentes representam diferenças (p<0,05) entre
períodos para um mesmo local e letras minúsculas diferentes
representam diferenças (p< 0,05) entre os locais no mesmo período.
Figura 5: Lipoperoxidação em brânquias de Uca mordax (n=5-10) Valores
expressos em média±erro padrão. As letras maiúsculas diferentes
representam diferenças (p<0,05) entre períodos para um mesmo local e
letras minúsculas diferentes representam diferenças (p< 0,05) entre os
locais no mesmo período.
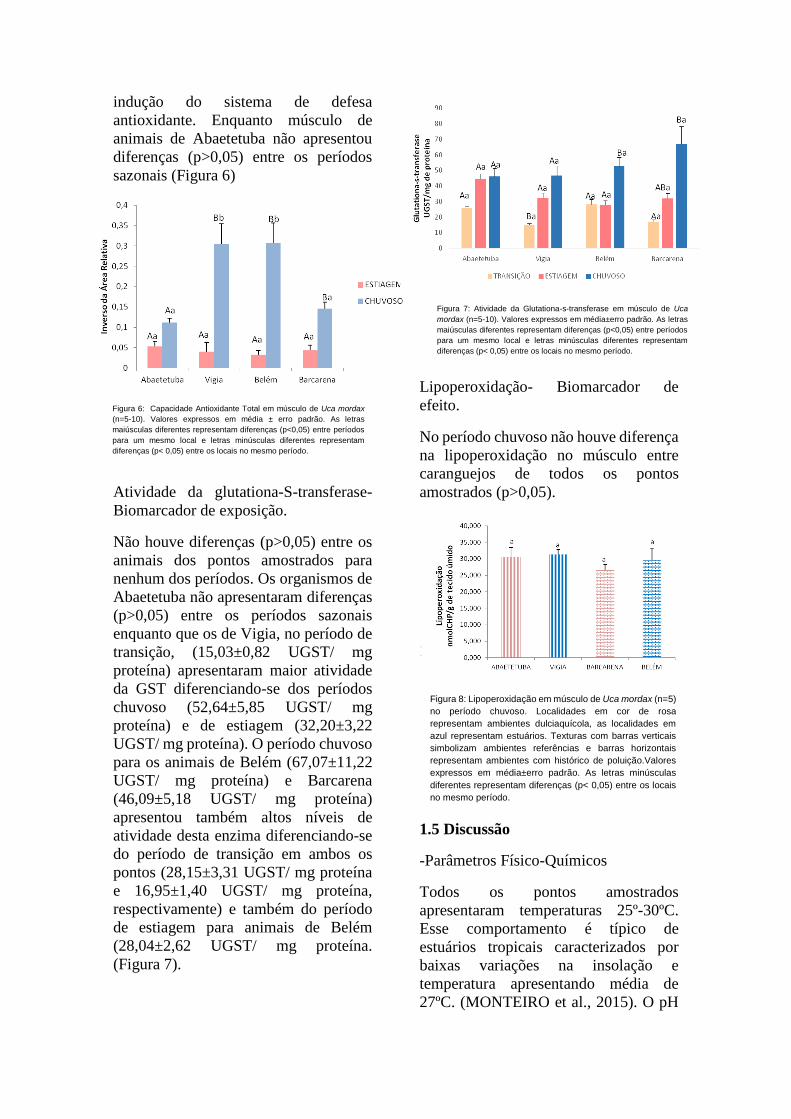
indução do sistema de defesa
antioxidante. Enquanto músculo de
animais de Abaetetuba não apresentou
diferenças (p>0,05) entre os períodos
sazonais (Figura 6)
Atividade da glutationa-S-transferase-
Biomarcador de exposição.
Não houve diferenças (p>0,05) entre os
animais dos pontos amostrados para
nenhum dos períodos. Os organismos de
Abaetetuba não apresentaram diferenças
(p>0,05) entre os períodos sazonais
enquanto que os de Vigia, no período de
transição, (15,03±0,82 UGST/ mg
proteína) apresentaram maior atividade
da GST diferenciando-se dos períodos
chuvoso (52,64±5,85 UGST/ mg
proteína) e de estiagem (32,20±3,22
UGST/ mg proteína). O período chuvoso
para os animais de Belém (67,07±11,22
UGST/ mg proteína) e Barcarena
(46,09±5,18 UGST/ mg proteína)
apresentou também altos níveis de
atividade desta enzima diferenciando-se
do período de transição em ambos os
pontos (28,15±3,31 UGST/ mg proteína
e 16,95±1,40 UGST/ mg proteína,
respectivamente) e também do período
de estiagem para animais de Belém
(28,04±2,62 UGST/ mg proteína.
(Figura 7).
Lipoperoxidação- Biomarcador de
efeito.
No período chuvoso não houve diferença
na lipoperoxidação no músculo entre
caranguejos de todos os pontos
amostrados (p>0,05).
1.5 Discussão
1.5 Discussão
-Parâmetros Físico-Químicos
Todos os pontos amostrados
apresentaram temperaturas 25º-30ºC.
Esse comportamento é típico de
estuários tropicais caracterizados por
baixas variações na insolação e
temperatura apresentando média de
27ºC. (MONTEIRO et al., 2015). O pH
Figura 6: Capacidade Antioxidante Total em músculo de Uca mordax
(n=5-10). Valores expressos em média ± erro padrão. As letras
maiúsculas diferentes representam diferenças (p<0,05) entre períodos
para um mesmo local e letras minúsculas diferentes representam
diferenças (p< 0,05) entre os locais no mesmo período.
Figura 7: Atividade da Glutationa-s-transferase em músculo de Uca
mordax (n=5-10). Valores expressos em média±erro padrão. As letras
maiúsculas diferentes representam diferenças (p<0,05) entre períodos
para um mesmo local e letras minúsculas diferentes representam
diferenças (p< 0,05) entre os locais no mesmo período.
Figura 8: Lipoperoxidação em músculo de Uca mordax (n=5)
no período chuvoso. Localidades em cor de rosa
representam ambientes dulciaquícola, as localidades em
azul representam estuários. Texturas com barras verticais
simbolizam ambientes referências e barras horizontais
representam ambientes com histórico de poluição.Valores
expressos em média±erro padrão. As letras minúsculas
diferentes representam diferenças (p< 0,05) entre os locais
no mesmo período.

de levemente ácido ao neutro é típico das é típico de águas brancas da Amazônia
(GAILLADERT et al., 1997).
Abaetetuba e Barcarena apresentaram
pH em torno de 6,0, sendo este valor
esperado em ambientes com alta taxa de
matéria orgânica (CONAMA, 2005) Os
valores mais elevados de oxigênio
dissolvido durante a maré alta se dá pelo
maior influxo que promove maior
agitamento e aeração das águas.
Os ambientes estuarinos naturalmente
apresentam altas quantidade de oxigênio
dissolvido devido à grande
produtividade primária destes ambientes
costeiros. Assim, os valores encontrados
no presente estudo estão na faixa normal,
superiores a 5mg/L (CONAMA, 2005) e
podem auxiliar na elucidação de
mudanças da qualidade do estuário
(CHAPMAN, 1992). Os ambientes
dulciaquícolas apresentam faixa normal
para os valores de OD não inferiores a
6mg/L (CONAMA,2005).
A condutividade maior nos ambientes
estuarinos pode ser atribuída às
variações mais expressivas de salinidade
(principalmente em Vigia) que leva à
variação da composição iônica
(MILLERO, 1984).
A variação dos parâmetros abióticos
parecem ter dinâmicas mais relacionadas
à natureza dos ambientes sem apresentar
variações expressivas entre as estações,
uma vez que, na Amazônia as estações
não são bem delimitadas e apresenta
pluviosidade regular durante o ano todo
(SOUZA et al, 2009).
-Biometria
No período chuvoso os animais de todos
os pontos amostrados mostram maiores
valores de peso em relação aos animais
coletados nos outros períodos, isso pode
ocorrer devido maiores demandas
alimentares que são intensificadas em
principalmente em ambientes costeiros
com o aumento da pluviosidade
(MARTINS, 2006).
Os animais de Vigia diferenciaram-se de
Abaetetuba apresentando maiores
valores para peso, comprimento e largura
em todos os períodos amostrados
podendo demonstrar que animais de
ambientes estuarinos apresentam
melhores condições sugerindo assim
menor impacto antrópico e/ou um
ambiente mais favorável ao
desenvolvimento desta espécie. Segundo
AMARAL & JABLONSKI (2005) em
estudo com caranguejos Ucides
chordatus alterações no tamanho das
populações e dos espécimes podem ser
atribuídas a destruição das áreas de
manguezal.
As diferenças nas dimensões corporais
de uma mesma espécie estão
relacionadas a variados fatores, dentre os
quais pode-se citar a quantidade de
matéria orgânica presente no substrato
(MASUNARI & DISSENHA, 2005)
maior em ambientes de manguezais,
assim, animais estuarinos apresentam
maior porte físico devido à alta
produtividade de seu habitat.
Os animais de Barcarena, Belém e Vigia
mostraram aumento dos valores de quase
todas as medidas (exceto altura para
Belém e Vigia) do período de transição
para o período de estiagem. Uma vez que
o período de transição pode apresentar
oscilações diárias de salinidade menos
regulares em relação aos outros períodos
os animais podem estar redirecionando
seu investimento energético em proteção
do organismo e não mais em alimentação
e crescimento como evidenciados nos
períodos chuvoso e seco que
apresentaram maiores valores para todas
as medidas.
Não percebe-se influência direta dos
históricos de contaminação no
desenvolvimento dos animais, isso pode
ser explicado pela dinâmica das águas

amazônicas que de um modo geral são o
ricas em nutrientes e íons, o que se sabe,
interfere na biodisponibilidade para
muitos compostos tóxicos
(MONSERRAT et al., 2007). Todos os
animais amostrados em cada um dos
pontos já estavam em maturação sexual
uma vez que todos apresentaram valores
superiores a 1,19 cm de largura da
carapaça (FRANSOZO et al., 2009).
-Biomarcadores
Brânquias:
A capacidade antioxidante total
apresentou-se maior para o período de
transição em relação às estações de
estiagem e chuvosa. No período de
transição, que equivale a diminuição
gradual das chuvas e início de estiagem,
os organismos enfrentam uma menor
constância nas condições do meio.
Assim, necessitam constantemente
adaptarem-se á essas variações, o que
demanda grande gasto energético. Sabe-
se que o aumento do gasto energético
está diretamente relacionado com o
aumento da geração de espécies reativas
de oxigênio (SILVEIRA, 2008) o que
induz um aumento das defesas
antioxidantes (HOLMGREN, 1985).
Concomitantemente, nota-se menor
valores de peso, comprimento e largura
nesta estação sugerindo novamente
alocação energética também para a
defesa do organismo através da indução
das defesas antioxidantes. Segundo
CHEN et al. (1997) os crustáceos
decápodos apresentam desajuste no
balanço osmótico quando submetidos à
variações na salinidade do meio uma vez
que o reajuste da concentração osmótica
requer muita energia. Durante os
períodos chuvoso e de estiagem onde
existe maior constância nos valores de
salinidade e demais características
abióticas, nos ambientes estuarinos
(Belém e Vigia) observou-se maior
indução da GST, uma enzima de
detoxificação. Isso pode estar associado
ao aumento da biodisponibilidade de
compostos xenobióticos na água que
mudam sua dinâmica de complexação
conforme a variação na composição
iônica e de matéria orgânica no meio, o
que pode levar à indução desta enzima.
Este fato é corroborado por uma maior
atividade da GST encontrada em Belém
(ambiente estuarino com histórico de
contaminação) quando comparada a
Vigia (ambiente estuarino referência).
Além disso, a realocação de energia para
crescimento (elucidada por maiores
valores de peso, comprimento, largura e
altura para todos os ambientes no
período de estiagem e chuvoso) podem
fragilizar os sistema de defesa
antioxidante potencializando a geração
de espécies reativas de oxigênio.
MYUNG et al(2010) em experimento
com o bivalve Scapharca broughtonii
submetidos à estresse térmico e osmótico
observou que ambos estresse hipo e
hiperosmótico aumentam a expressão e
atividades de enzimas antioxidantes
como catalase e superóxido dismutase
bem como o tempo de exposição
prolongado às salinidades experimentais
(25, 35 e 45) levava também ao aumento
da atividade destas enzimas. Assim, no
período de transição os altos níveis de
defesa antioxidante parecem ter caráter
compensatório não levando à indução da
GST, isso se expressa nos baixos valores
de lipoperoxidação encontrados para
esta estação.
Os altos valores de lipoperoxidação para
o período chuvoso podem estar
associados ao processo de ressuspensão
de poluentes com a maior hidrodinâmica
somados à investimento energético em
alimentação e reprodução. Segundo
DIAS-FILHO (2006) sobretudo no
período chuvoso há aumento da
comunidade planctônica, principalmente
de algas Clorofíceas, que levam a
maiores níveis de produtividade neste
período aumentando assim a demanda

alimentar para os consumidores da teia
alimentar.
Músculo:
No músculo visualiza-se aumento da
capacidade antioxidante no período
chuvoso e o mesmo padrão na atividade
para GST, sendo o menor valor
registrado para o período de transição.
Observa-se que os níveis de atuação da
enzima foram muito menores no
músculo quando comparados ás
brânquias isso se dá principalmente pelas
brânquias serem o principal órgão de
contato com o meio aquático estando
exposta às suas variações abióticas
inerentes (PAITAL et al., 2010).
A lipoperoxidação não mostrou
diferenças entre os pontos no período
chuvoso. Assim, o aumento da atividade
da GST no período chuvoso para todos
os ambientes pode ser um mecanismo
inerente dos animais que se preparam
para maiores demandas metabólicas e
consequentemente maiores danos
oxidativos.
Assim, tanto os biomarcadores
analisados quanto os valores biométricos
apontam que o período de transição
impõe condições mais severas aos
animais do que os períodos de estiagem
e chuvoso havendo investimento em
defesa do organismo. Os períodos
chuvoso e estiagem parecem não
oferecer adversidades metabólicas e os
animais parecem encontrar condições de
alimentação e desenvolvimento
adequado. Não se observa relação direta
entre a natureza dos ambientes
(estuarinos ou dulciaquícolas) e nem a
presença de xenobióticos no
comportamento metabólico dos animais.
A sazonalidade se configura como fator
determinante no direcionamento do
metabolismo dos animais.
1.6 Referências
AMADO, L.L., DA ROSA, C.E.,
MEIRELLES LEITE, A., MORAES, L.,
VAZ PIRES, W., PINHO, G.L.L.,
MARTINS, C.M.G., ROBALDO, R.B.,
NERY, L.E.M., MONSERRAT,
J.M.,BIANCHINI, A., MARTÍNEZ,
P.E., GERACITANO, L.A. Biomarkers
in croakers Micropogonias furnieri
(Teleostei, Scianidae) from polluted
and non-Polluted areas from the Patos
Lagoon estuary (Southern Brazil):
evidences of genotoxic and
immunological effects. Marine
Pollution Bulletin. 52, 199–206, 2006.
AMARAL, A.C.Z. & JABLONSKI, S.
Conservação da biodiversidade
marinha e costeira no Brasil.
Megadiversidade, Brasília, DF, Brasil,
1(1):43-51, 2005..
AMORIM LC. Os biomarcadores e
sua aplicação na avaliação da
exposição aos agentes químicos
ambientais. Revista Brasileira de
Epidemiologia 6: 158-170. 2003.
ANGER, K.The Biology of Decapod
Crustacean Larvae. Crustacean
Issues, 14. AA Balkema, pp. 420,2001.
ASMUS, M. L. Análise e Usos do
Sistema Estuário da Lagoa dos
Patos. In: Reunião especial da
SBPC: Ecossistemas costeiros, do
conhecimento à gestão, 3, 1996,
Florianópolis. Anais. Florianópolis,
1996, p.105-108.
BAINY,A.C.D.;SAITO,E.;CARVALH
O,P. S. M. & JUNQUEIRA, V. B. C.
Oxidative stress in gill, erythrocytes,
liver and kidney of Nile tilapia
(Oreochromis niloticus) from a

polluted site. Aquatic Toxicology,
34:151-162, 1996.
BARNWELL, F.H; THURMAN, C.L.
Taxonomy and Biogeography of the
fiddler crabs (Ocypodidae, Genus
Uca) of the Atlantic and Gulf Coasts of
Eastern North-America. Zoological
Journal of the Linnean 81: 23-87. 1984.
BRAGA, R. A. P. Caracterização
das Zonas Estuarinas de
Pernambuco. In: Seminário
Internacional: Perspectivas e
implicações da carcinicultura estuarina
de estado de pernambuco, 1, 2000,
Recife. Anais. Editora Bagaço, Recife,
2000, p.13-20.
CAJARAVILLE, M.P., BEBIANNO,
M.J., BLASCO, J., PORTE, C.,
SARASQUETE, C.,VIARENGO, A.
The use of biomarkers to assess the
impact of pollution in costal
environments of the Iberian
Peninsula: a practical approach.
Science of Total Environment 247, 295–
311, 2000.
CHAPMAN, D. Water Quality
Assessments. Chapman and Hall, New
York, NY, 1992.
CHEN, J-C, CHIA P-G. Osmotic and
ionic concentrations of Scylla
serrata(Forska °1) subjected to
different salinity levels. Comparative
Biochemistry and Physiology part A
227:239-244, 1997.
CONSELHO NACIONAL DO MEIO
AMBIENTE. Resolução n°357, de 17
de março de 2005. DOU nº 053, p. 58-
63, 2005.
DEPLEDGE, M. H.; AAGAARD, A.
& GYÖRKÖS, R. Assessment of
trace metal toxicity using molecular,
physiological and behavioural
biomarkers. Marine Pollution Bulletin,
31:19-27, 1995.
DIAS-FILHO, M.B. A fotossíntese e o
aquecimento global. Belém:
EmbrapaAmazônia Oriental, 2006b.
24p. Embrapa Amazônia Oriental.
Documentos, 234.
FRANSOZO, V.C.; MORTARI, R. C.;
BENETTI, A. S. Population biology of
Uca mordax (Smith, 1870)
(Crustacea,Decapoda, Ocypodidae)
from the southeastern coast of
Brazil. Estudos de Biologia(UCP.
Impresso), v. 31, p. 23-31. 2009.
GAILLARDET, J.; DUPRÉ, B.;
ALLÈGRE, C.; NÉGREL, P. Chemical
and Physical Denudation in the
Amazon River Basin. Chemical
Geology, 142: 141-173, 1997.
HABIG, W. H. & JAKOBY, W. B. 1981.
Assay for differentiation of
glutathione-S-transferases. Methods in
Enzymology, 77:398-405.
HABIG, W. H.; PABST, M. J. &
JAKOBY, W. B. 1974. Glutathione-
S-trasferases: The first enzymatic
step in mercapturic acid formation.
Biological Chemistry, 249:7130-7139.
HERMES-LIMA, M.; WILLMORE, W.
G. & STOREY, K.B. Quantification of
the lipid peroxidation in tissue
extracts based on Fe(III) xylenol
orange complex formation. Free
Radical Biology & Medicine, 19:271-
280, 1995.

HOLMGREN, A. Thioredoxin.Annual
Review of Biochemistry.v.54, p.237-271,
1985.
MARTINS, A. A.V; COSTA, R.A.M;
PEREIRA, L. C.C. Space-time
distribution of zooplankton
community in an Amazonian artificial
coastal lagoon in Bragança, Pará,
Brazil. Boletim Museu Parense Emílio
Goeldi, Ciências Naturais, Belém, v. 1,
n. 3, p. 103-111, 2006.
MASUNARI, S.; DISSENHA, N. &
FALCÃO, R. C. Crescimento relativo e
destreza dos quelípodos de Uca
maracoani (Latreille) (Crustacea,
Decapoda, Ocypodidae) no Baixio
Mirim, Baía de Guaratuba, Paraná,
Brasil. Revista Brasileira de Zoologia
22(4):974-983, 2005.
MONSERRAT, J.M., GERACITANO,
L.A., BIANCHINI, A. Current and
future perspectives using biomarkers
to assess pollution in aquatic
ecosystems.Comments on Toxicology 9,
255–269, 2003a.
MONSERRAT, J.M.; MARTÍNEZ, P.E;
GERACITANO, L.A.; AMADO, L.L.;
MARTINS, C.M.; PINHO, GT.L.;
CHAVES, I.S,; FERREIRA-CRAVO,
M.; VENTURA-LIMA, J. &
BIANCHINI, A.. Pollution biomarkers
in estuarine animals: critical review
and new perspectives. Comparative
Biochemistry and Physiology part
C:Toxicology and Pharmacology,
146(1-2): 221-34, 2007.
MILLERO, F.J. The conductivity-
density-salinity-chlorinity
relationship for estuarine waters.
Limnology and Oceangraphy, 29(6),
p.1317-1321, 1984.
MONTEIRO, S.M. EL-ROBRINI, M.;
ALVES, I.C.C. Seasonal dynamics of
nutrients in an Amazon estuary.
Mercator, Fortaleza, v. 14, n. 1, p.151-
162, 2015.
MYUNG, I.A; CHEOL, Y.C. Activity of
antioxidant enzymes and physiological
responses in ark shell,Scapharca
broughtonii, exposed to thermal and
osmotic stress: Effects on hemolymph
and biochemical parameters.
Comparative Biochemistry and
Physiology part B, 155, 34-42, 2010.
PAITAL, B., CHAINY, G.B.N., 2010.
Antioxidant defenses and oxidative
stress parameters in tissues of mud
crab (Scylla serrata) with reference to
changing salinity. Comparative
Biochemistry and Physiology part
C:Toxicology and Pharmacology,151,
142–151.
PEREIRA FILHO, J.; SCHETTINI, C.
A. F.; RÖRIG, L. & SIEGLE, E.
Intratidal variation and net transport
of dissolved inorganic nutrients, POC
and chlorophyll a in the Camboriú
river estuary, Brazil. Estuarine,
Coastal and Shelf Science, 53:249-257,
2001.
SILVEIRA, R.L; HIRABARA, S.N;
LAMBERTUCCI, R.H; LEANDRO,
C.V; FIAMONCINI, J; PINHEIRO,
C.H.J; D’ANGELO., A.C.A; BASSIT,
R.A; PITHON-CURI, T.C; CURI, R.
Metabolic regulation and production
of oxygen reactive species during
muscule contraction: effect of
glycogen on intracellular redox state
Revista Brasileira de Medicina e
Esporte vol.14 no.1 Niterói Jan./Feb,
2008.

SOGORB, M.A; ESTÉVEZ, J.;
VILANOVA, E. Biomarkers in
Toxicology.Academic Press, p.965–973,
2014.
SOUZA, B.E; LOPES M.N.G; ROCHA,
E.J.P; SOUZA, J.R.S; CUNHA, A.C;
SILVA, R.R; FERREIRA, D.B.S;
SANTOS, D.M; DO CARMO, A.M.C;
SOUZA, J.R.A; GUIMARÃES, P.L; DA
MOTA, M.A.S; MAKINO, M; SENNA,
R.C; SOUSA, A.M.L; MOTA, G.V;
KUHN, P.A.F; SOUZA, P.F.S;
VITORINO,M.I. Precipitação sazonal
sobre a amazônia oriental no período
chuvoso:observações e simulações
regionais com o REGCM3. Revista
Brasileira de Meteorologia, v.24, n.2,
111-124, 2009.
WITTERS, H.E. Chemical speciation
dynamics and toxicity assessment in
aquatic ecosystems. Ecotoxicoly and
Environmental Safety. 41, 90–95.1998.
ZAR, J. H. Biostatistical analysis. 2º ed.
Prentice Hall: New Jersey, 1984.
____________________
ii Graduando do curso de Licenciatura
em Biologia da Universidade Federal do
Pará. Bolsista PRODOUTOR. E-mail:
iiiDocente do Instituto de Ciências
Biológicas, Universidade Federal do
Pará. E-mail: [email protected].
Parecer do orientador
A aluna apresenta perfil de pesquisadora, apresentando rendimento extremamente satisfatório ao longo do desenvolvimento de suas atividades relacionadas ao plano de trabalho. Neste período atuou conjuntamente em outros projetos de pesquisa em desenvolvimento no grupo BIOPAQ sem prejuízo algum para com seu plano PIBIC.
Belém, 09 de agosto de 2015
Profa. Dra. Lílian Lund Amado
iTrabalho desenvolvido com o apoio do
Programa PRODOUTOR.