alguns fungos presentes em plantas nativas e … · 2017. 11. 22. · romero, guilherme lapidus,...
TRANSCRIPT

I
ALGUNS FUNGOS PRESENTES EM PLANTAS NATIVAS E
EXÓTICAS DO CERRADO
Erica Santos do Carmo de Souza
Brasília-DF 2011
Universidade de Brasília Instituto de Ciências Biológicas Departamento de Fitopatologia Programa de Pós-Graduação em Fitopatologia

II
ALGUNS FUNGOS PRESENTES EM PLANTAS NATIVAS E
EXÓTICAS DO CERRADO
Erica Santos do Carmo de Souza
Brasília-DF 2011
Universidade de Brasília Instituto de Ciências Biológicas Departamento de Fitopatologia Programa de Pós-Graduação em Fitopatologia

III
ALGUNS FUNGOS PRESENTES EM PLANTAS NATIVAS E
EXÓTICAS DO CERRADO
Erica Santos do Carmo de Souza
Brasília-DF 2011
Universidade de Brasília Instituto de Ciências Biológicas Departamento de Fitopatologia Programa de Pós-Graduação em Fitopatologia
Dissertação apresentada ao Departamento de Fitopatologia, Instituto de Ciências Biológicas da Universidade de Brasília, como requisito parcial para a obtenção de grau de Mestre em Fitopatologia.

IV
Trabalho realizado junto ao Departamento de Fitopatologia, Instituto de
Ciências Biológicas da Universidade de Brasíla, sob a orientação do
Professor José Carmine Dianese, e com o apoio financeiro concedido pela
Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Dissertação aprovada em:_____,_____,________ por:
____________________________________________
Prof. José Carmine Dianese (Orientador)
Universidade de Brasília
____________________________________________
Dra. Arailde Fontes Urben (Examinadora)
Embrapa Cenargen
____________________________________________
Prof. Carlos Antonio Inácio (Examinador)
Universidade de Brasília

V
Dedico este trabalho à minha mãe América
pelo seu amor incondicional,
cuidado e confiança.

VI
AGRADECIMENTOS
Agradeço primeiramente a Deus pela minha vida, por ter me proporcionado
integridade física e mental, por Ele abrir as portas para as minhas conquistas e por
ser o condutor do meu caminho me dando uma vivência abençoada.
Ao meu orientador, Professor José Carmine Dianese, pela confiança em meu
trabalho, pelos seus valiosos ensinamentos e por ser espelho de motivação e
determinação pela carreira científica.
À professora Rita de Cássia (mais precisamente à amiga Rita), por ser tão
maravilhosa comigo, compartilhando seus conhecimentos e me dando suporte
sempre, tanto na vida profissional como também na pessoal.
Ao Professor Helson Mário por todo o seu apoio e consideração e pela
parceria na execução das medidas e documentações fotográficas em vários
trabalhos.
À Professora Mariza Sanchez, curadora da Coleção Micológica de Referência
Herbário UB, pessoa que me acolheu com cuidados de mãe, a qual tenho muito
carinho. Agradeço pela ajuda constante nas atividades rotineiras do laboratório e
principalmente na identificação dos espécimes botânicos.
Ao Professor Carlos Antonio Inácio, pela prontidão instantânea em cooperar
nos estudos micológicos.
Ao nosso grupo de estudos em Micologia, Professores: Dianese, Rita, Helson,
Carlos Inácio e Mariza Sanchez. Aos colegas Leila, Eugênio, William, Marcelo, Maria
do Desterro e Eliane, pela compartilhação mútua de conhecimentos.
Ao Professor José Elias de Paula do Departamento de Botânica, pela ajuda
na identificação das plantas hospedeiras.
À Professora Sônia Nair Baó, responsável pelo laboratório de Microscopia
Eletrônica de Varredura. Agradeço ao Felipe Guimarães que, com toda atenção,
ajudou-me prontamente na execução das fotos.
À professora Rosana Tindon, pela simpatia e disponibilidade no uso de
equipamentos em seu laboratório.
Aos Professores do Programa de Pós-graduação em Fitopatologia, pela arte
de ensinar e transferir experiências construtoras do caráter profissional. Adalberto
Corrêa Café Filho, Alice Kazuko Inoue Nagata, Carlos Antonio Inácio, Carlos Hidemi
Uesugi, Cláudio Lúcio Costa, Cleber Furlanetto, Denise Vilela de Resende Santiago,

VII
Helson Mário Martins do Vale, José Carmine Dianese, Juvenil Enrique Cares, Luiz
Eduardo Bassay Blum, Marisa Álvares Ferreira, Renato Resende de Oliveira, Rita de
Cássia Pereira Carvalho e Robert Neil Gerard Miller. A vocês, a minha admiração
pela competência acadêmica.
À Universidade de Brasília, especialmente ao Departamento de Fitopatologia,
pela oportunidade da execução deste trabalho.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
pelo financiamento da bolsa de estudos.
Aos membros da banca examinadora: Prof. José Carmine Dianese, Dra.
Arailde Fontes Urben, Prof. Carlos Antonio Inácio e Prof. Adalberto Corrêa Café
Filho, por terem aceitado o convite com grande satisfação e contribuir para a
melhora deste trabalho.
Aos colegas de curso: Ana Cristina, Bruno Barros, Cecília Rodrigues, Daniel
Lage, Eder Marques, Eliane Armando, Esdras Henrique, Eugenio Sperandio, Flávio
Romero, Guilherme Lapidus, Jessica da Mata, Kamila Araújo, Leonardo Braúna,
Liamar dos Anjos, Maria do Desterro, Mariana Severo, Maíra Teixeira, Marcelo
Tavares, Mikhail Leastro, Mônica Alves, Pablo Melo, William Soares, em especial a
Andressa Quintana, Ana Paula Pagani, Claudênia Ferreira e Justino Dias. A todos o
meu muito obrigada por termos compartilhado momentos difíceis e muitas vezes de
sufoco, e também de muita descontração, mas sempre de mãos unidas em prol das
vitórias em comum.
Aos funcionários do Departamento de Fitopatologia: Arenildo Soares, Arlindo
Epaminondas, Kamila Araújo, Leila Santos, Maria Dias, Marivaldo Farias, Ribamar
Frazão e em especial, José Cézar de Castro, por proporcionarem condições
favoráveis ao desenvolvimento deste trabalho.
Ao Milton Luiz da Paz Lima, professor e amigo, por ser um dos maiores
incentivadores e por me despertar para o mundo de uma pós-graduação. Obrigada
por acreditar em mim e por me fazer conhecer a “Corrente do Bem”.
Aos meus amigos do Laboratório de Cogumelos da Embrapa Cenargen,
Vanessa Alves, Thaís Torquato, Eduardo Lima e Márcia Motta, por toda amizade
verdadeira, bom humor e companheirismo.
À minha mãe América e aos meus irmãos, Eduardo, Samuel, Simeão e
Sizaltino, pelo apoio e confiança.

VIII
A toda a minha família pela admiração e incentivo constante.
À minha prima Edvânia por sempre me dar força e me apoiar, sobretudo nos
momentos mais difíceis.
À minha amiga Raquel Alves, por compartilhar desafios, incertezas, e mais
que isto, pelo companheirismo nas alegrias e tristezas, cumplicidade e crédito.
As amigas Eliane Lagasse e Daiane Aparecida, as quais tenho muita
satisfação em compartilhar uma amizade intensa concretizada na confiança. Muito
obrigada por torcerem e fazerem parte da minha história.
Aos meus padrinhos Izabel Lagasse e Edson Ferreira e a toda a nossa
família, por todo cuidado, afeto e consideração.
À Waléria e Alfredo pela valiosa ajuda nos momentos de sufoco.
A todos que de alguma maneira contribuíram para o êxito deste trabalho.

IX
“As pessoas que vencem neste mundo, são as que procuram as
circunstâncias de que precisam e, quando não as encontram, as criam.”
(Bernard Shaw)

X
ÍNDICE GERAL
AGRADECIMENTOS ................................................................................................ VI
ÍNDICE DE TABELAS .............................................................................................. XI
ÍNDICE DE FIGURAS ............................................................................................... XI
RESUMO................................................................................................................. XIII
ABSTRACT ............................................................................................................. XIV
INTRODUÇÃO ............................................................................................................ 1
MATERIAIS E MÉTODOS .......................................................................................... 6
RESULTADOS E DISCUSSÃO .................................................................................. 8
CAPÍTULO I ................................................................................................................ 8
DESCRIÇÃO E TAXONOMIA DE FUNGOS DO CERRADO ..................................... 8
1 HIFOMICETO ............................................................................................................ 8 1.1 Janetia sp.(provável espécie nova nº 1) ....................................................... 8
2 CELOMICETOS ....................................................................................................... 14 2.1 Acrostroma sp. (provável espécie nova nº 2) ............................................. 14 2.2 Pestalotiopsis sp. (provável espécie nova nº 3) ......................................... 17 2.3 Stigmopeltis sp. (provável espécie nova nº 4) ........................................... 22 2.4 Celomiceto (provável gênero novo nº 1) .................................................... .26
3 ASCOMICETOS ...................................................................................................... 29 3.1 Staibia connari Bat. & Peres ...................................................................... 29 3.2 Phyllachora davillae F. Stevens ................................................................. 32 3.3 Asterinella sp. (provável espécie nova nº 5)............................................... 35 3.4 Coccodiella sp. (provável espécie nova nº 6) ............................................. 39 3.5 Johansonia sp. (provável espécie nova nº 7) ............................................. 43 3.6 Limacinia sp. (provável espécie nova nº 8) ................................................ 47 3.7 Phillipsiella sp. (provável espécie nova nº 9) ............................................. 51 3.8 Phyllachora sp. (provável espécie nova nº 10)........................................... 55 3.9 Stomatogene sp. (provável espécie nova nº 11) ........................................ 58 3.10 Stomiopeltis sp. (provável espécie nova nº 12) ....................................... 62 3.11 Vonarxella sp. (provável espécie nova nº 13) ........................................... 66 3.12 Ascomiceto (provável gênero novo nº 2) .................................................. 70 3.13 Ascomiceto (provável gênero novo nº 3) .................................................. 70
4 BASIDIOMICETO ..................................................................................................... 75 4.1 Puccinia malvacearum Bertero ex Montagne ............................................ 75
CAPÍTULO II ............................................................................................................. 78
FUNGOS EM PLANTAS EXÓTICAS PRESENTES NO CERRADO ....................... 78
1 HIFOMICETOS ........................................................................................................ 78 1.1 Cercospora sp. (provável espécie nova nº 14) ........................................... 78 1.2 Cercospora sp. (provável espécie nova nº 15) ........................................... 78 1.3 Pseudocercospora sp. (provável espécie nova nº 16) ............................... 85
CONCLUSÃO ........................................................................................................... 88
REFERÊNCIAS BIBLIOGRÁFICAS ......................................................................... 89

XI
ÍNDICE DE TABELAS
TABELA 1. Comparação morfométrica entre as espécies de Janetia sp. e o
espécime encontrado em Ocotea sp ..................................................................... ...12
TABELA 2. Comparação entre as espécies de Pestalotiopsis encontradas em
Arecaceae no Brasil. ............................................................................................. ...19
TABELA 3. Comparação entre P. tecomicola e o espécime encontrado sobre
Mauritia sp.. ........................................................................................................... ...20
TABELA 4. Comparação entre as espécies de Stigmopeltis com o espécime
encontrado sobre Micropholis venulosa ................................................................ ...24
TABELA 5. Comparação entre as espécies de Asterinella sp. encontradas em
Myrtaceae.............................................................................................................. ...37
TABELA 6. Comparação entre as espécies de Coccodiella sp. encontradas em
Arecaceae. ............................................................................................................ ...41
TABELA 7. Comparação entre as espécies de Johansonia sp.com a espécie
encontrada em Lauraceae. ................................................................................... ...45
TABELA 8. Comparação entre as espécies de Limacinia da América do Sul com a
encontrada em Lantana camara. ........................................................................... ...49
TABELA 9. Comparacão entre as espécies de Phillipsiella e a espécie encontrada
em Ocotea paniculata. .......................................................................................... ...53
TABELA 10. Comparação morfométrica entre o espécime em estudo e as espécies
conhecidas de Stomatogene sp. ........................................................................... ...60
TABELA 11. Comparação das espécies de Stomiopeltis encontradas no Brasil.. ...64
TABELA 12. Comparação entre a espécie monotípica de Vonarxella e o espécime
encontrado em Ocotea sp. .................................................................................... ...68
TABELA 13. Comparação entre as espécies de Cercospora encontradas em Piper
sp. e o espécime em estudo. ................................................................................. ...81
TABELA 14. Comparação entre as espécies de Cercospora encontradas em
Jatropha sp. e o espécime em estudo. .................................................................. ...82
TABELA 15. Comparação entre as espécies de Pseudocercospora encontradas em
Arecaceae com o espécime sobre Licuala jeanencyi. ............................................ ...86

XII
ÍNDICE DE FIGURAS
FIGURA 1. Distribuição geográgica do Cerrado em comparação com outros Biomas
Brasileiros.....................................................................................................................1
FIGURA 2. Professor Ezechias Paulo Heringer...........................................................3
FIGURA 3. A-D. Janetia sp. em tricomas de Ocotea sp............................................13
FIGURA 4. A-E. Acrostroma sp. em Roessleria brasiliana........................................16
FIGURA 5. A-H. Pestalotiopsis sp. em Mauritia sp...................................................21
FIGURA 6. A-G. Stigmopeltis em Micropholis venulosa............................................25
FIGURA 7. A-E. Provável gênero novo nº 1 parasitando uma espécie de
Asteraceae.................................................................................................................28
FIGURA 7. A-F. Staibia connari em folhas de Ocotea sp..........................................31
FIGURA 8. A-G. Phyllachora davillae em Davilla rugosa..........................................34
FIGURA 9. A-H. Asterinella sp. em folhas de Eugenia sp.........................................38
FIGURA 10. A-H. Coccodiella sp. sobre Butia sp......................................................42
FIGURA 11. A-G. Johansonia sp. em Ocotea paniculata..........................................46
FIGURA 12. A-H. Limacinia sp. em Lantana camara................................................50
FIGURA 13. A-H. Phillipsiella sp. em Ocotea sp.......................................................54
FIGURA 14. A-G. Phyllachora sp. em Ilex affinis......................................................57
FIGURA 15. A-I. Stomatogene em folhas de Rapanea guianensis...........................61
FIGURA 16. A-F. Stomiopeltis sp. em folhas de Ilex affinis......................................65
FIGURA 17. A-G. Vonarxella em folhas de Ocotea sp..............................................69
FIGURA 18. A-G. Novo gênero (nº 2) de ascomiceto setoso em tricomas foliares de
membro de uma Lauraceae.......................................................................................73
FIGURA 19. A-J. Provável gênero novo (nº 3) de ascomiceto sobre tricomas de
membro de uma Lauraceae.......................................................................................74
FIGURA 20. A-I. Puccinia malvacearum em Triumffeta rhomboidae........................77
FIGURA 21. A-D. Cercospora sp. em Piper nigrum..................................................83
FIGURA 22. A-D. Cercospora sp. em Jatropha curcas.............................................84
FIGURA 23. A-F. Pseudocercospora sp. em folhas de Licuala jeanencyi.................87

XIII
RESUMO
De um total de 22 fungos estudados, 19 foram encontrados em plantas
nativas do Cerrado e três em plantas exóticas. Espécies possivelmente novas de
fungos foram estudadas em folhas de Ocotea sp. (Janetia relatada pela primeira vez
em Lauraceae e espécies de Vonarxella, Johansonia, Phillipsiella e Staibia connari);
em folhas de uma Asteraceae não identificada (um novo celomiceto); em Roessleria
brasiliana (Acrostroma sp.); e em Mauritia sp. (Pestalotiopsis sp.) e Micropholis
venulosa (Stigmopeltis sp.), além disso, em Lauraceae não identificada foram
estudados dois gêneros novos de ascomicetos ambos associados a tricomas
foliares. Outras prováveis espécies novas foram descritas em Ilex affinis
(Phyllachora sp. e Stomiopeltis sp.); em Eugenia sp. (Asterinella sp.); em Davilla
rugosa (Phyllachora davilae); em Butia sp. (Coccodiella sp.); em Lantana camara
(Limacinia sp.), em Triunfetta rhomboidea (Puccinia malvacearum). Em plantas
exóticas no cerrado resgistraram-se duas novas espécies de Cercospora em
pimenta-do-reino (Piper nigrum) e pinhão-manso (Jatropha curcas) e uma
Pseudocercospora em Licuala jeanencyi (Arecaceae).

XIV
ABSTRACT
A total of 22 fungal species were studied; nineteen of them on native Cerrado
plants and three on exotic species grown in the Cerrado. Fungal species considered
as probably new taxa were studied on leaves of Ocotea sp. (Janetia sp., a first
record on Lauraceae, and Vonarxella sp., Johansonia sp., Phillipsiella sp., and
Staibia connari); on leaves of a non-identified Asteraceae (a new coelomycete); on
Roessleria brasiliana (Acrostroma sp.); on Mauritia sp. (Pestalotiopsis sp.); and on
Micropholis venulosa (Stigmopeltis sp.). On a non-identified species belonging in the
Lauraceae, two new trichome-associated ascomycete genera were studied. Other
probably new species were described and illustrated on Ilex affinis (Phyllachora sp.
and Stomiopeltis sp.); on Eugenia sp. (Asterinella sp); on Davilla rugosa (Phyllachora
davilae); on Butia sp. (Coccodiella sp.); on Lantana camara (Limacinia sp.), on
Triunfetta rhomboidea (Puccinia malvacearum).Two new Cercospora species were
detected on exotic host species found in Brasilia (Piper nigrum and Jatropha
curcas), and a Pseudocercospora species on Licuala jeanencyi (Arecaceae)
collected in Recife, Pernambuco.

1
INTRODUÇÃO
O Cerrado constitui o segundo maior bioma do Brasil e da América do Sul,
superado apenas pela Floresta Amazônica, ocupando originalmente mais de dois
milhões de km2 (23 % de toda a área do país) e abrangendo uma área contínua do
Distrito Federal e dos estados de Minas Gerais, Goiás, São Paulo, Mato Grosso do
Sul, Mato Grosso e Tocantins; uma parte dos estados de Rondônia, Bahia, Ceará,
sul do Maranhão e do Piauí, áreas isoladas ao norte do Amapá, Amazonas, Pará e
Roraima, apresentando ainda pontos isolados no Paraná (Ribeiro & Walter, 1998) [Fig.1].
Este importante bioma abriga um rico patrimônio de recursos naturais
adaptados às condições climáticas e edáficas que determinam a sua existência. O
Cerrado é uma savana tropical na qual uma vegetação rasteira, formada principalmente por
gramíneas, coexiste com árvores e arbustos esparsos (Klink et al., 1995).
Os solos do Cerrado são antigos, profundos e bem drenados,
nutricionalmente são ácidos e de baixa fertilidade, com altos níveis de ferro e
alumínio.
Mata Atlântica
Figura 01. Distribuição geográfica do Cerrado em comparação com outros Biomas Brasileiros.

2
O clima mostra duas estações bem definidas, uma seca e uma chuvosa. A
precipitação média anual é de 1500 mm, com grandes variações intra-regionais
(Klink et al., 1995).
A região do Cerrado exibe uma enorme heterogeneidade espacial,
estendendo-se por mais de 20 graus de latitude, com altitudes variando de quase 0
a 1800 metros. A área do Cerrado ocupa diferentes bacias hidrográficas (Amazonas,
Tocantins, Paraná, Paraguai, São Francisco e Parnaíba) e exibe uma grande
diversidade de solos e climas que se refletem numa vasta biota (Cole, 1986; Ribeiro
& Walter, 2008).
Esse bioma constitui a maior savana tropical do mundo em termos de
diversidade apresentando 44% de suas espécies de plantas lenhosas endêmicas
(Klink & Machado, 2005).
A vegetação do Cerrado ocupa cerca de 12% de toda a flora brasileira
(Ribeiro & Walter, 1998) e possui sua diversidade estimada em cerca de 10.000
espécies de angiospermas, 1.000 pterófitas, 2.000 briófitas e 2.000 algas,
totalizando em torno de 15.000 espécies de plantas sendo 4.400 endêmicas
(IBAMA, 2011). O Distrito Federal com extensão territorial de 54 Km2 apresenta
cerca de 2.000 espécies de plantas conhecidas (Filgueiras & Pereira, 1993).
Considerando a grande biodiversidade existente nos trópicos, de acordo com
Hawksworth (1991, 2001) para cada espécie de planta vascular, estão associados
de seis a oito fungos. Dianese (2000) estimou a existência de 70 a 100 mil espécies
de fungos (incluindo os liquenizados) no Cerrado.
Estudos recentes na Universidade de Brasília (UnB) mostram dados que
extrapolam a estimativa de Hawksworth (1991, 2001), constando a associação de
mais de 25 fungos por espécie (Dianese et al., 1997; Chaves,1998; Dornelo-Silva,
1999; Castro, 2009; Santos, 2011) confirmando a estimativa feita por Dianese (2000).
No geral, a micobiota do Cerrado é pouco estudada. A diversidade
microbiana, incluindo os microfungos, não tem recebido a devida atenção por parte
dos países neotropicais apesar das nações desenvolvidas alertarem para o potencial
biotecnológico destes microorganismos (Dianese, 2000).
Dentre as mais de 10.000 espécies de plantas do Cerrado, um grande
número delas se destaca pelo grande potencial econômico, dados as possibilidades
reais de virem a ser domesticadas e cultivadas para produção de alimentos

3
(principalmente frutas como: pequi, maracujás diversos, cagaita, pêra-do-cerrado,
gabiroba, araçá e jatobá) e também o possível uso como plantas medicinais, há
muito tempo conhecido por leigos e cientistas (Guarim-Neto & Morais, 2003). Em
contrapartida plantas exóticas de interesse agronômico e ornamental, são
comumente introduzidas no Cerrado tornando-se hospedeiras alternativas de
vários fungos.
Os estudos da micodiversidade do Cerrado inciaram-se na UnB em meados
de 1993, com apoio da Fundação Banco do Brasil e CNPq, resultando no que é hoje
a Coleção Micológica de Referência Herbário UB (CMR-UB) com aproximadamente
22.000 exsicatas. A grande maioria do acervo da CMR-UB está ainda em fase inicial
de estudo, com a maioria do material classificado apenas em termos de grandes
grupos (basidiomicetos, ascomicetos e seus anamorfos). Nos últimos 12 anos, foram
descritas e publicadas 104 novas espécies e cerca de 20 novos gêneros de fungos
associados a plantas do Cerrado (Dianese et al., 1997; Dianese, 2000; Dornelo-Silva
& Dianese, 2004; Dornelo-Silva et al., 2007; Gutiérrez & Dianese, 2008, 2009;
Dianese et al., 2008; Souza et al., 2008; Pereira-Carvalho et al., 2009a,b).
Um dos mais importantes coletores de fungos do Cerrado foi seguramente o
Professor Ezechias Paulo Heringer (Fig.2).
Figura 02. Professor Ezechias Paulo Heringer.

4
O mineiro Heringer veio para Brasília no ano de 1960, a convite do Presidente
Juscelino Kubitschek e foi pioneiro no estudo do Cerrado e suas orquídeas.
Em 1962, trabalhou na implantação do Parque Zoobotânico de Brasília. De
1963 a 1977, atuou como professor da UnB e responsável pela Fazenda Água
Limpa (FAL) tornando-se, após sua aposentadoria, Professor Emérito da UnB. Em
1964 doou seu herbário particular à UnB.
Quando Heringer chegou a Brasília ainda havia muito Cerrado intocado. Seu
trabalho abriu caminho para pesquisas no Cerrado e serviu como ponto de partida
para muitos outros estudos. Além do Parque Nacional de Brasília, criou a Reserva
Biológica de Águas Emendadas, a Estação Experimental de Agricultura Cabeça de
Veado e o Parque Municipal do Gama.
O professor Heringer foi responsável pelo fornecimento de muitas amostras
de fungos do Cerrado aos micólogos Amhés Pinto Viégas e Augusto Chaves Batista,
do Instituto Agronômico de Campinas e da Universidade do Recife, hoje
Universidade Federal de Pernambuco, respectivamente (Bezerra, 2005; Dianese,
2005).
Outra grande contribuição na identificação dos fungos das plantas do Cerrado
coletadas pelo Professor Heringer na década de 70 foi a colaboração da micologista
Dra. Arailde Fontes Urben, pesquisadora da Embrapa Recursos Genéticos e
Biotecnologia.
Uma parte das amostras coletadas por ele constitui hoje o que aqui se designou
como Coleção Micológica E. P. Heringer (CMEPH), a qual está incorporada à Coleção
Micológica do Herbário UB (CMHUB), pertencente à Universidade de Brasília.
A Coleção contém cerca de 1.600 espécimes de plantas herborizadas
contendo fungos coletados principalmente no Cerrado abrangendo o Distrito Federal
e os estados de Minas Gerais (principalmente em Paraopeba), Goiás, Mato Grosso,
Pernambuco, Alagoas, Pará, Amapá, Rio de Janeiro, Bahia e São Paulo.
Diante da considerável quantidade e qualidade do material micológico
coletado pelo Professor Heringer e tendo em vista que os fungos compõem hoje
uma coleção, com identificações apenas parciais (a maioria em níveis de grandes
grupos como ascomicetos, basidiomicetos e seus anamorfos), torna-se necessária a
revisão taxonômica desse material. Assim, apenas alguns de seus espécimes foram
incluídos nesse trabalho, como um passo inicial de uma futura revisão da CMEPH.

5
Objetivo Geral
O trabalho visa descrever e identificar parte da micobiota do Cerrado contida
na CMHUB, incluindo-se também alguns materiais da CMEPH.
Objetivos específicos
Ampliar o conhecimento da biodiversidade fúngica associada a diferentes
famílias botânicas representadas no Cerrado.
Revisar taxonomicamente e ilustrar alguns fungos da CMEPH, incluindo
material previamente identificado.
Descrever e publicar possíveis novos táxons ou ocorrências da micobiota
presente na CMR-UB e CMEPH, com isso ampliando o conhecimento sobre os
fungos oriundos do Cerrado.

6
MATERIAIS E MÉTODOS
Um total de 300 exsicatas foi analisado. Deste, aproximadamente 90 %
pertencente à Coleção Micológica Ezechias Paulo Heringer e 10 % à Coleção
Micológica de Referência Herbário UB da Universidade de Brasília.
Algumas hospedeiras foram identificadas pelo próprio Prof. Heringer e
confirmadas pelo Prof. José Elias de Paula, do Departamento de Botânica e pela
curadora da CMHUB Profa. Mariza Sanchez, do Departamento de Fitopatologia da
Universidade de Brasília. Algumas hospedeiras não foram identificadas em nível de
gênero ou mesmo de espécie devido às condições inadequadas de armazenamento
ou quantidade insuficiente de material botânico.
Alguns fungos aqui estudados foram previamente identificados por Amhés
Pinto Viégas e Augusto Chaves Batista e foram agora submetidos à atualização
taxonômica/nomenclatural.
A identificação preliminar dos fungos foi feita com a utilização de microscópio
estereoscópico e microscópio de luz, observando-se sintomas como manchas e
áreas escurecidas e, nelas, sinais do fungo como micélio superficial e frutificações
diversas, bem como evidência de colonização do tecido por fungos e a presença de
fungos hiperparasitas.
Para o manuseio do material e a retirada de amostras dos fungos, foram
utilizados estiletes metálicos de vários calibres. Os cortes histológicos foram
realizados em crio–micrótomo (MICRON, HM 505 E, Micron Laborgerate, Walldorf,
Alemanha) com espessura variando de 10 a 30 µm, a fim de observar as estruturas
imersas nas plantas hospedeiras e a morfologia da relação dessas com os fungos.
Amostras fúngicas removidas por estiletes ou agulhas, como também os
cortes, foram montadas em lâminas semi–permanentes utilizando corante lacto–
glicerol com e sem azul de algodão para se observar as estruturas do fungo em
geral. Utilizou-se o corante glicerol-KOH/floxina básica para se estudar a
conidiogênese em hifomicetos e celomicetos. Também foi utilizado somente água
para se verificar a coloração das estruturas do fungo.
As lâminas foram examinadas em microscópio de luz Leica DM 2500 e as
análises morfométricas foram feitas através do programa Leica QWin–Plus.

7
A documentação microfotográfica foi realizada com câmara fotográfica (Leica
DFC 490) acoplada a computador. Foram realizadas documentações fotográficas
com o Microscópio digital Dino-Lite Pro e em Microscopia de varredura (JEOL JSM-
700 1 F).
As medidas referentes a cada estrutura fúngica foram realizadas com 20 a 50
repetições e foram consideradas as dimensões extremas (ordem crescente) e as de
maior frequência (entre colchetes) mostradas nas medições. As medidas dispostas
entre parêntesis referem-se aos valores ocasionalmente encontrados.
As identificações dos fungos foram feitas ou confirmadas com base em
chaves contidas em literatura de cada grupo fúngico tais como: Ellis (1971 e 1976) e
Carmichael et al. (1980) para hifomicetos; Sutton (1980) e Nag–Raj (1993) para
celomicetos; Kiffer & Morelet (2000) para celomicetos e hifomicetos; Müller & Von
Arx (1962, 1973), Arx & Müller (1954,1975), Batista & Ciferri (1963a, 1963b), Batista
& Ciferri (1962) e Rossman (1987) para ascomicetos e Cummins & Haratsuka
(1983, 2003) para ferrugens (basidiomicetos).
A verificação de nomes científicos, nomes de autores, validade e sinonímias
foram feitas em portais on line: o Index Fungorum (www.indexfungorum.org) do CABI
Bioscience Database, elaborado por Paul Kirk e colaboradores, o Cyberliber
(http://www.cybertruffle.org.uk/cyberliber/) mantido por David W. Minter, o portal do
United State Department of Agriculture sob a responsabilidade de Farr & Rossman
(http://nt.ars–grin.gov/fungaldatabases/fungushost/fungushost.cfm), e o banco de
dados da coleção micológica da Embrapa Recursos Genéticos e Biotecnologia
(http://pragawall.cenargen.embrapa.br/aiqweb/michtml/fgbanco01.asp).
Alguns novos táxons determinados e ocorrências inéditas referentes a tipo de
hospedeira e local já foram publicados em forma de resumo e apresentação oral em
Congressos Nacionais e Internacionais (Dianese et al., 2010; Dianese et al.; Vale et
al., 2010a, b; Vale et al., 2011a, b, c; Souza et al., 2011a, b, c; Sperandio et al.,
2011).

8
RESULTADOS E DISCUSSÃO
CAPÍTULO I
DESCRIÇÃO E TAXONOMIA DE FUNGOS DO CERRADO
1 Hifomiceto
1.1 Janetia sp. (provável espécie nova nº 1)
Colônias efusas, hipófilas, marrons a marrom-escuras, (Fig.3. A). Micélio
marrom-claro, superficial sobre tricomas. Hifas 2-4 [2,5] µm, septadas, ramificadas,
lisas, marrons claras (Fig.3. B). Conidióforos micronemáticos, mononemáticos.
Células conidiogênicas 3-7 [4,5] × 4-7 [5,5] µm, integradas, intercalares,
holoblásticas, denticuladas e marrons (Fig.3. B e C). Conídios 12-21 [16,5] x 3-6 [4]
µm, solitários, secos, obclavados, marrons, lisos e de base truncada, 3-5 [4] septos
constritos (Fig.3. B, C e D).
Espécime examinado: Brasil. Distrito Federal, Brasília, Jardim Botânico de Brasília
em folhas de Ocotea sp. (Lauraceae), 20 Ago 1994, leg. J.C. Dianese, holótipo (UB
Micol. Coll. 6902).
O gênero Janetia M.B Ellis (1976) foi caracterizado inicialmente por possuir
micélio superficial com hifas sem apressórios (ou hifopódios), conidióforos
micronemáticos e mononemáticos, células conodiogênicas poliblásticas, integradas,
intercalares, denticuladas com paredes finas e coloração escura, conídios solitários,
secos, obclavados, multisseptados, marrons e lisos (Ellis, 1976).
O conceito do gênero foi ampliado para incluir também espécies com conídios
distosseptados, obovóides, elipsóides e conídios com ápice curvado, uma espécie
com conidióforo sinemático e até micélio imerso, além de outras duas espécies
apresentando associação com outros fungos, sendo que uma destas possui ainda
micélio interno (Hughes, 1983; Silvanesan & Hsieh, 1990; Goh & Hyde, 1996).

9
Assim, foram incluídas quatro espécies com conídios distosseptados: Janetia
bacilliformes Gamundí, Aramb. & Giaiotti, Janetia refugia e Janetia canescens,
descritas por B. Sutton & Pascoe, além de Janetia synnematosa Sivan. & W.H.
Hsieh. Além disso, em J. canescens, os conídios e hifas vegetativas são verrugosos,
à semelhança de outras duas espécies que também apresentam verrugosidade nos
conídios, J. refugia e Janetia faureae (Piroz.) M.B. Ellis. Entretanto, J. canescens,
apresenta conídios cilíndricos, enquanto as outras duas espécies possuem conídios
obclavados. Ainda, com relação ao formato dos conídios, a maioria das espécies
apresenta conídios obclavados ou cilíndricos, ou ainda cilíndrico-obclavados. Porém,
Janetia leprosa (Piroz.) S. Hughes, Janetia obovata M. Calduch, Gené, Abdullah &
Guarro e Janetia curviapicis Goh & K.D. Hyde representam exceções, uma vez que
na primeira espécie os conídios são elipsóides, na segunda, estes são obovóides e
na última espécie os conídios se apresentam curvados no ápice.
Outra característica importante incluída entre as espécies do gênero, além
das acima mencionadas (conídios com pseudosseptos, formato e verrugosidade),
refere-se à presença de conidióforos sinemáticos apenas em J. synnematosa. Ellis
(1976) ao estabelecer o gênero, recombinou Sporidesmium faureae, espécie
proposta por Pirozynski, para J. faureae, considerando nesta espécie a presença de
conídios finamente verrugosos (Goh & Hyde, 1996; Calduch et al., 2002).
Características de conídios com pseudosseptos e outros formatos, além de
obclavados não foram considerados por Ellis (1976).
Este conceito ampliado de Janetia foi formalizado por Goh & Hyde (1996) os
quais, por outro lado, consideraram J. tetracentri Y.L. Guo como sinônimo de J.
faureae. Logo, Janetia cubensis Matsush com células conidiogênicas monoblásticas
originando conídios cilíndrico-clavados e apresentando secessão rexolítica, foi
excluída do gênero.
As espécies acrescidas recentemente ao gênero foram, J. obovata e Janetia
salvertia Dornelo-Silva & Dianese. Janetia obovata foi descrita na Espanha e o
epíteto específico refere-se ao formato dos conídios, que neste aspecto, difere
radicalmente de todas as demais espécies do gênero por apresentar conídios com
três a cinco septos (predominando conídios com quatro septos), caracteristicamente
obovóides escuros na porção apical e progressivamente mais claros na base, ao
contrário das outras espécies do gênero que apresentam uma coloração mais

10
escura nas células basais, e mais clara no ápice. Janetia salvertia encontrada em
Salvertia convallariaeodora A. St-Hil, apresenta células conidiogênicas poliblásticas
denticuladas com 1-4 dentículos diferenciando-se das demais espécies,
principalmente pelo formato e dimensões dos conídios (Calduch et al., 2002;
Dornelo-Silva & Dianese, 2003).
A maioria das espécies do gênero apresenta células conidiogênicas
poliblásticas, porém J. capnophila S. Hughes, J. matsushimae Subram., J. refugia e
J. obovata apresentam células conidiogênicas monoblásticas. De acordo com
Hughes (1983) J. capnophila e J. bacilliformis podem ser poliblásticas, mas isto não
é comum para estas duas espécies.
As duas espécies (J. euphorbiae e J. faureae) propostas por Ellis (1976) ao
estabelecer o gênero Janetia foram caracterizadas pela presença de micélio
superficial, conforme mencionado anteriormente. No entanto, Kirk (1985) descreveu
J. longispora P.M. Kirk com micélio parcialmente superficial e parcialmente imerso,
neste aspecto idêntico a J. curviapicis e J. obovata (Goh & Hyde, 1996; Calduch et
al., 2002).
Entre as espécies adicionadas ao gênero, três apresentam associação com
outros fungos. Colônias de J. capnophila foram encontradas em associação com
espécies de Euantennaria Speg. e Acrogenotheca elegans (Fraser) Cif. & Bat. e
Janetia interna H.J. Swart (1985) foi descrita como possível micoparasita sobre
acérvulos e ascomas de Discostromopsis stoneae H.J. Swart. Janetia mangiferae foi
encontrada em associação com uma espécie de Stomiopeltis Theiss., entretanto a
relação de micoparasitismo não foi determinada, assim como ocorre em J.
curviapicis que apresenta micélio superficial ou imerso no substrato e cresce em
associação ou junto com outros hifomicetos (Hughes & Cavalcanti, 1983; Swart,
1985).
A permanência das espécies J. obovata e J. synnematosa no gênero é aqui
questionada, uma vez que estas duas espécies apresentam características muito
distintas quando comparadas às demais. A segregação de espécies no gênero é
baseada principalmente na morfometria dos conídios. Este argumento foi usado para
acomodar J. obovata dentro de Janetia. Entretanto, a forma divergente do conídio de
J. obovata (três a cinco septos e versicolor) forçaria sua inclusão em outro gênero,

11
bem como a presença de conidióforos formando sinêmio poderia também, excluir J.
synnematosa de Janetia.
O fato de o gênero abrigar espécies com diferentes hábitos (parasíticas ou
saprofíticas de plantas ou fungos) poderia ser questionado em uma revisão futura do
táxon. Entretanto, optou-se por aceitar critérios já usados para outros gêneros como
Cercospora Fresen. e Mycovellosiella Rangel que, além das espécies
fitopatogênicas, incluem também micoparasíticas como, Cercospora uromycestri
Pollack, hiperparasita de écios de Uromyces cestri Mont. (Pollack, 1987) e
Mycovellosiella raveneliae M.D. Mehrotra & R.K. Verma (hiperparasita de
urediniósporos e teliósporos de Ravenelia clemensiae Syd.) (Mehrotra & Verma,
1993). A presença de conídios pseudosseptados, pode também ser objeto de
discussão. Dentre os hifomicetos graminícolas, anteriormente englobados pelo
gênero Helmintosporium Link, Curvularia Boedjin, é separada de Bipolaris
Shoemaker, por possuir conídios eusseptados (Silvanesan, 1987). Por analogia a
este grupo fúngico, questiona-se a inclusão, dentro de Janetia, de espécies com
conídios pseudosseptados. Para resolver estes questionamentos seria necessário
um estudo mais minucioso do gênero.
O gênero Janetia tem 20 espécies (Tabela 1) encontradas em várias famílias
botânicas (Kirk, 2011). No Brasil são publicadas apenas Janetia salvertiae
(Vochysiaceae) e J. mangiferae (Anacardiaceae), porém há relatos de ocorrência no
Brasil de J. euphorbiae, encontrada em Gomidesia sp. (Myrtaceae) e em Caryocar
brasiliense (Caryocaraceae), e de J. mangiferae encontrada também em folhas de
Caryocar brasiliense (Pereira-Carvalho, 2004; Armando, 2004).
Dentre as espécies conhecidas para o gênero, o espécime em estudo possui
suas dimensões conidiais vagamente próximas das espécies J. leprosa e J.
mangiferae. A septação dos conídios é correspondente a da espécie J. obovata. O
espécime tem suas células conidiogênicas e micélio com medições muito próximas
de J. euphorbiae UB 506 (Tabela 1).
Apesar de algumas medidas aproximarem de algumas espécies, estas não
são suficientes para acomodar o espécime em estudo em alguma das espécies já
conhecidas.
Trata-se do primeiro relato de Janetia sp. sobre Ocotea sp., e também para
membro da família Lauraceae, sendo uma provável nova espécie.
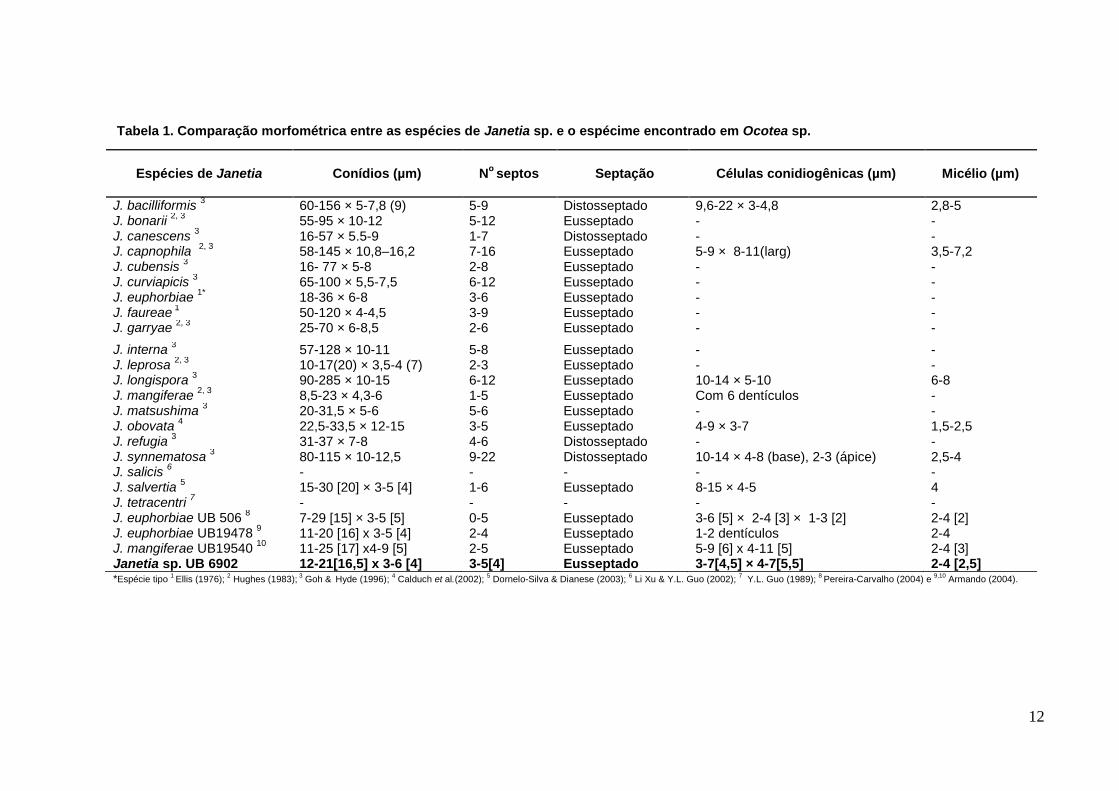
12
Tabela 1. Comparação morfométrica entre as espécies de Janetia sp. e o espécime encontrado em Ocotea sp.
*Espécie tipo 1 Ellis (1976);
2 Hughes (1983);
3 Goh & Hyde (1996);
4 Calduch et al.(2002);
5 Dornelo-Silva & Dianese (2003);
6 Li Xu & Y.L. Guo (2002);
7 Y.L. Guo (1989);
8 Pereira-Carvalho (2004) e
9,10 Armando (2004).
Espécies de Janetia Conídios (µm) No
septos
Septação
Células conidiogênicas (µm)
Micélio (µm)
J. bacilliformis 3 60-156 × 5-7,8 (9) 5-9 Distosseptado 9,6-22 × 3-4,8 2,8-5
J. bonarii 2, 3
55-95 × 10-12 5-12 Eusseptado - - J. canescens
3 16-57 × 5.5-9 1-7 Distosseptado - -
J. capnophila 2, 3
58-145 × 10,8–16,2 7-16 Eusseptado 5-9 × 8-11(larg) 3,5-7,2 J. cubensis
3 16- 77 × 5-8 2-8 Eusseptado - -
J. curviapicis 3 65-100 × 5,5-7,5 6-12 Eusseptado - -
J. euphorbiae 1*
18-36 × 6-8 3-6 Eusseptado - - J. faureae
1 50-120 × 4-4,5 3-9 Eusseptado - -
J. garryae 2, 3
25-70 × 6-8,5 2-6 Eusseptado - -
J. interna 3 57-128 × 10-11 5-8 Eusseptado - -
J. leprosa 2, 3
10-17(20) × 3,5-4 (7) 2-3 Eusseptado - - J. longispora
3 90-285 × 10-15 6-12 Eusseptado 10-14 × 5-10 6-8
J. mangiferae 2, 3
8,5-23 × 4,3-6 1-5 Eusseptado Com 6 dentículos - J. matsushima
3 20-31,5 × 5-6 5-6 Eusseptado - -
J. obovata 4 22,5-33,5 × 12-15 3-5 Eusseptado 4-9 × 3-7 1,5-2,5
J. refugia 3 31-37 × 7-8 4-6 Distosseptado - -
J. synnematosa 3
J. salicis 6
80-115 × 10-12,5 -
9-22 -
Distosseptado -
10-14 × 4-8 (base), 2-3 (ápice) -
2,5-4 -
J. salvertia 5
J. tetracentri 7
15-30 [20] × 3-5 [4] -
1-6 -
Eusseptado -
8-15 × 4-5 -
4 -
J. euphorbiae UB 506 8
J. euphorbiae UB19478 9
J. mangiferae UB19540 10
Janetia sp. UB 6902
7-29 [15] × 3-5 [5] 11-20 [16] x 3-5 [4] 11-25 [17] x4-9 [5] 12-21[16,5] x 3-6 [4]
0-5 2-4 2-5 3-5[4]
Eusseptado Eusseptado Eusseptado Eusseptado
3-6 [5] × 2-4 [3] × 1-3 [2] 1-2 dentículos 5-9 [6] x 4-11 [5] 3-7[4,5] × 4-7[5,5]
2-4 [2] 2-4 2-4 [3] 2-4 [2,5]

13
10 µm
F
50 µm 20 µm
A B C
D
10 µm
10 µm
Figura 03. A-D. Janetia sp. em tricomas de Ocotea sp. A, B e C. Crescimento do micélio superficial esporulado ao longo de tricomas. D. Conídio obclavado, marrom e com constrição nos septos.

14
2 Celomicetos
2.1 Acrostroma sp. (provável espécie nova nº 2)
Conidióforos em sinêmio 341-1588 [1255] x 68-412 [175] µm, escuros e
rígidos, ramificados ou não com ápice mostrando massa esférica de conídios 74-295
[187] x 95-342 [215] µm (Fig.4. A e B) Células conidiogênicas enteroblásticas
fialídicas, marrons. Conídios 5-9 [7] x 2,5-6 [3] µm, elipsóides, amerosseptados,
hialinos e gutulados quando imaturos e marrons claros quando atingem a
maturidade (Fig.4. C, D e E).
Espécime examinado: Brasil. Minas Gerais, em caule de Roessleria brasiliana
(Amaranthaceae), leg. E. P. Heringer, (CMEPH M29 408).
A Família Batistiaceae Samuels & K. F. Rodrigues é composta por membros
com ascoma não ostiolado, negro, parede espessa e cephalotecóide. Tecido
interascal ausente. Asco diminuto deliquescente. Ascósporos translúcidos,marrons,
simples, sem poro germinativo. Os amamorfos são hifomicetos sinemáticos
pertencentes ao gênero Acrostroma.
Esta Família foi estabelecida com base em um único gênero monotípico
(espécie-tipo: Batistia annulipes) e originalmente colocada na ordem Sordariales
com base na conidiogênese fialídica do anamorfo (Samuels & Rodrigues, 1989).
Anteriormente o gênero foi indevidamente colocado em Xylariaceae (Ciferri, 1958) e
a família Cephalothecaceae com base no seu perídio cephalotecóide (Samuels &
Rodrigues, 1989). Kirk et al. (2001) removeu Batistiaceae da ordem Sordariales e a
colocou como família incertae sedis em Sordariomycetidae.
Para Huhndorf et al. (2004), baseados em dados moleculares, família
Batistiaceae não pertence à ordem Sordariales, nem tampouco pode ser alocada
como família incertae sedis em Sordariomycetidae. Finalmente, na falta de dados
confiáveis, decidiram tratar Batistiaceae como família incertae sedis pertencente ao
filo Ascomycota.
Atualmente de acordo com Lumbsch & Huhndorf (2010) a família
Batistiaceae permanece conforme estabelecido anteriormente por Kirk et al. (2001).

15
O gênero Acrostroma possui três espécies registradas: Acrostroma
annellosynnema Seifert, Acrostroma madhucae R.K. Verma, N. Sharma & Soni e
Acrostroma sterculiae R.K. Verma, N. Sharma & Soni (Kirk, 2011).
O espécime se difere das espécies já descritas e, além disso, este é o
primeiro relato deste fungo na famíla Amaranthaceae sendo, portanto uma provável
espécie nova.
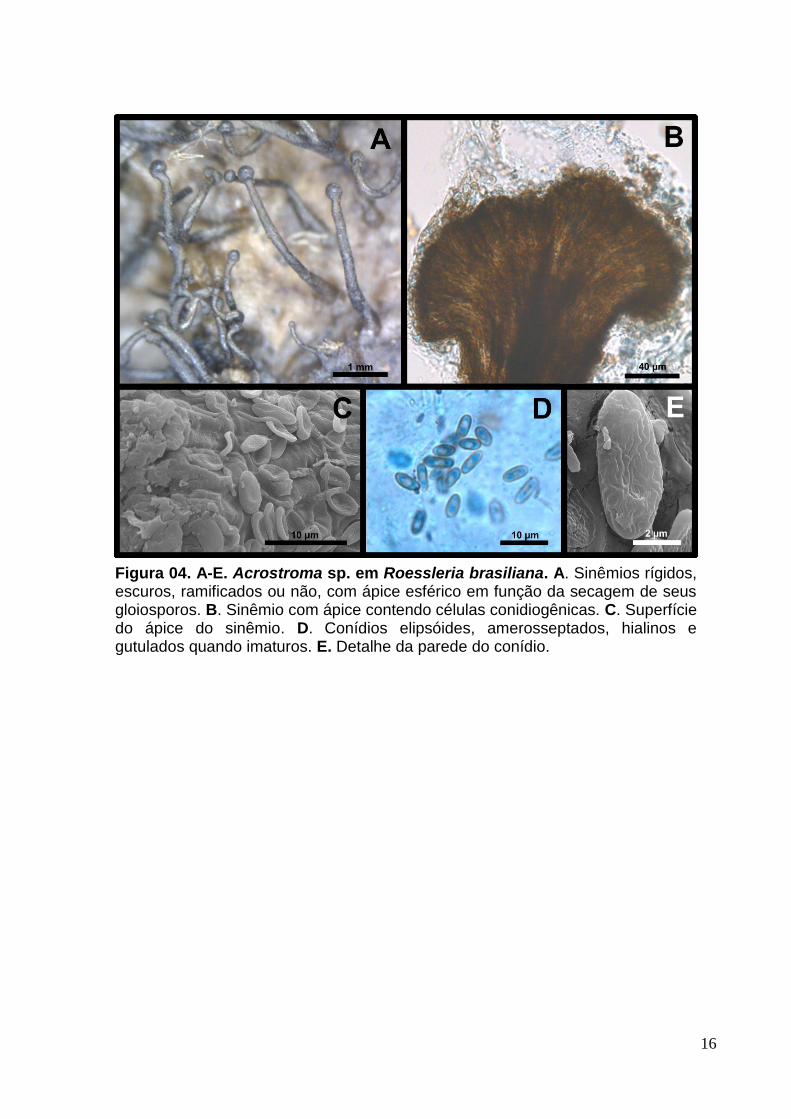
16
Figura 04. A-E. Acrostroma sp. em Roessleria brasiliana. A. Sinêmios rígidos, escuros, ramificados ou não, com ápice esférico em função da secagem de seus gloiosporos. B. Sinêmio com ápice contendo células conidiogênicas. C. Superfície do ápice do sinêmio. D. Conídios elipsóides, amerosseptados, hialinos e gutulados quando imaturos. E. Detalhe da parede do conídio.

17
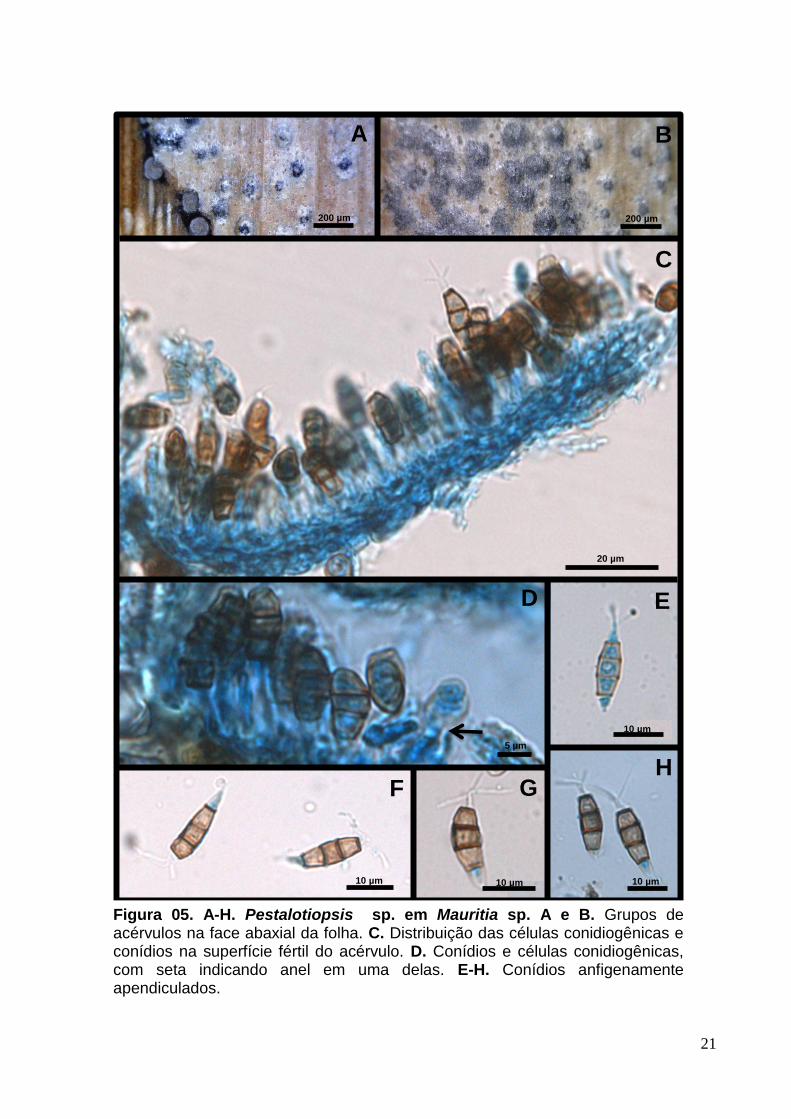
2.2 Pestalotiopsis sp. (provável espécie nova nº 3)
Conidioma 84-175 x 120-186 µm, acervular, subepidérmico, solitário e
formado de parede escura com textura angularis (Fig.5. A e B). Deiscência irregular.
Conidióforos micronemáticos, hialinos e cilíndricos (Fig.5. C e D). Células
conidiogênicas 4-6 [6] x 3-5 [3] µm holoblásticas, anelídicas, indeterminadas,
cilíndricas e hialinas (Fig.5. D). Conídios 9-18,5 [14] x 5-6,5 [5] µm, fusiformes, retos
ou levemente curvados, quatro septos, célula basal hialina, simples, célula apical
cônica, hialina, com três apêndices, simples, filiformes, células medianas
escurecidas, com paredes mais espessas (Fig.5. E-H).
Espécime examinado: Brasil. Distrito Federal, Gama, em folhas de Mauritia sp.
(Arecaceae), 02 Jun 1962, leg. E. P. Heringer (CMEPH M39 806).
Segundo Sutton (1980) o gênero Pestalotiopsis é caracterizado por possuir
micélio imerso, ramificado, septado, hialino a marrom-claro. Conidioma acervular,
epidérmico a subepidérmico, separado ou confluente formado de parede escura com
textura angularis. Deiscência irregular. Conidióforos hialinos, ramificados e septados
na base e na porção apical, cilíndricos ou lageniformes. Células conidiogênicas
holoblásticas, anelídicas, indeterminadas, integradas, cilíndricas, hialinas, suaves,
com proliferações percorrentes. Conídios fusiformes retos ou levemente curvados,
quatro septos, célula basal hialina, truncada, simples; célula apical cônica, hialina,
com dois ou mais apêndices, simples ou ramificados, filiformes, células medianas
escurecidas, com paredes mais espessas, suaves ou verruculosas.
De acordo com Farr & Rossman (2011) há cinco registros do gênero
Pestalotiopsis na Família Arecaceae no Brasil. São elas: P. acrocomiarum em
Acrocomia intumescens, P. leucothoes em Bactris sp., Pestalotiopsis sp. em Elaeis
guineensis e P. versicolor em Cocos sp. O espécime em estudo apresenta
dimensões muito menores do que em todas as espécies encontradas em
arecáceaes relatadas no Brasil (Tabela 2). Apenas as dimensões referentes à
largura dos conídios [5-(5)-6,5 µm] são próximas as da espécie P. leucothoes (4,9 -
6,9 µm).
De acordo com a chave para o gênero em Nag Raj (1993) o espécime se
enquadra na espécie P. tecomicola, encontrada em Tecoma radicans (Bignoniaceae),

18
por apresentar conídios com quatro septos, célula apical com apêndices típicos e
uniformidade na coloração das células. Apesar de o espécime possuir essas
características morfológicas semelhantes, a espécie P. tecomicola apresenta
dimensões superiores com relação a acérvulo, suas células conidiogênicas são mais
cumpridas, porém mais estreitas e conídios maiores e mais largos (Tabela 3).
Esta é a primeira vez que Pestalotiopsis sp. é encontrado em Mauritia sp. no
Brasil.
O espécime é uma provável espécie nova a ser posteriormente publicada de
acordo com o Código Internacional de Nomenclatura para algas, fungos e plantas (ICN).
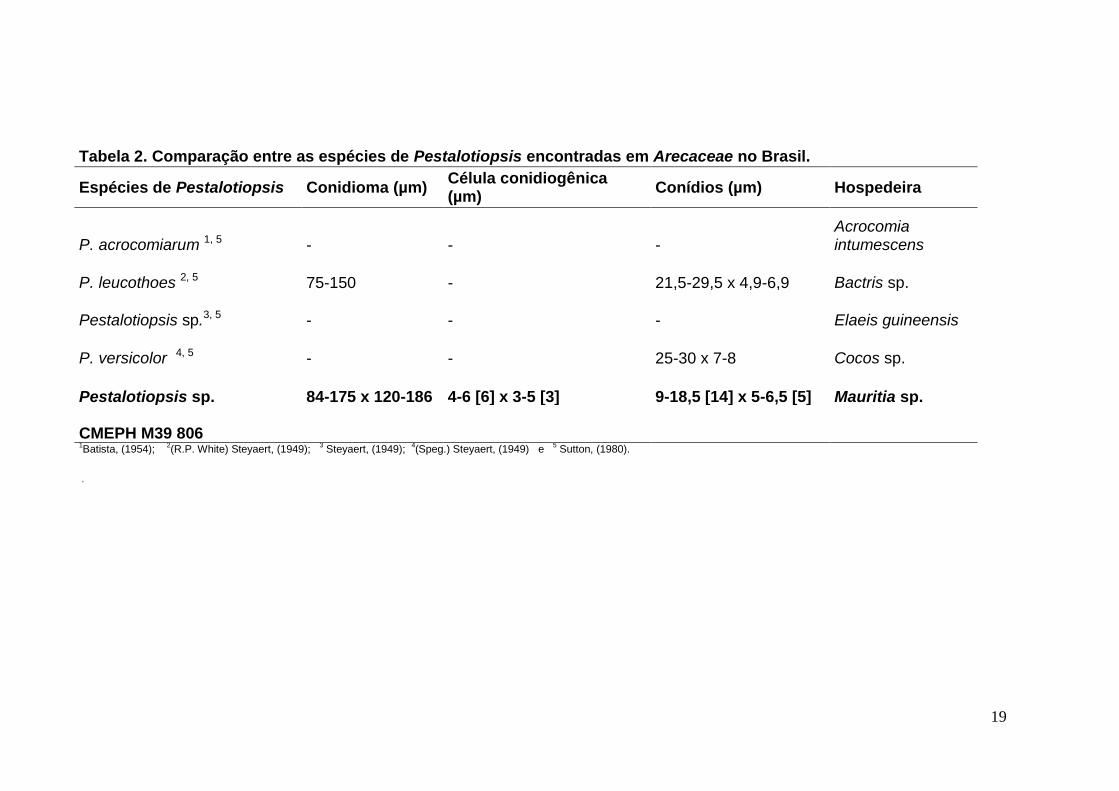
19
Tabela 2. Comparação entre as espécies de Pestalotiopsis encontradas em Arecaceae no Brasil.
Espécies de Pestalotiopsis Conidioma (µm) Célula conidiogênica (µm)
Conídios (µm) Hospedeira
P. acrocomiarum 1, 5 - - - Acrocomia intumescens
P. leucothoes 2, 5 75-150 - 21,5-29,5 x 4,9-6,9 Bactris sp.
Pestalotiopsis sp.3, 5 - - - Elaeis guineensis
P. versicolor 4, 5 - - 25-30 x 7-8 Cocos sp.
Pestalotiopsis sp. 84-175 x 120-186 4-6 [6] x 3-5 [3] 9-18,5 [14] x 5-6,5 [5] Mauritia sp.
CMEPH M39 806
1Batista, (1954);
2(R.P. White) Steyaert, (1949);
3 Steyaert, (1949);
4(Speg.) Steyaert, (1949) e
5 Sutton, (1980).
.
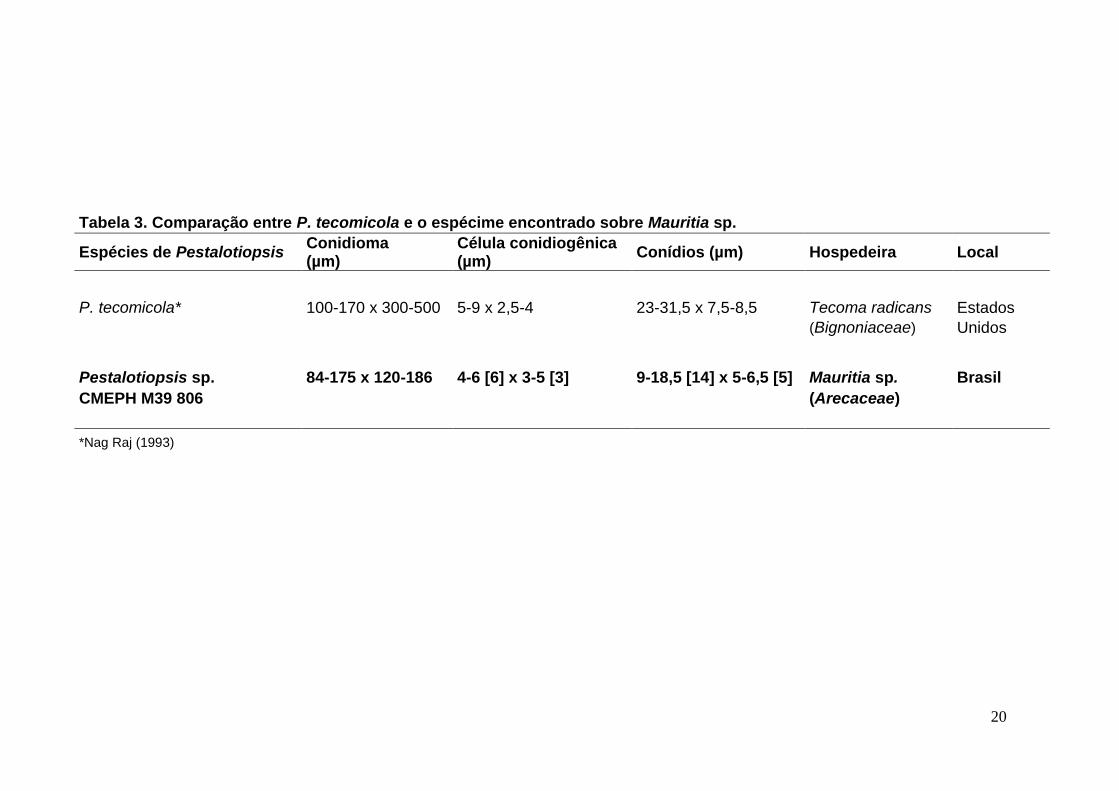
20
Tabela 3. Comparação entre P. tecomicola e o espécime encontrado sobre Mauritia sp.
Espécies de Pestalotiopsis Conidioma (µm)
Célula conidiogênica (µm)
Conídios (µm) Hospedeira Local
P. tecomicola* 100-170 x 300-500 5-9 x 2,5-4 23-31,5 x 7,5-8,5 Tecoma radicans Estados
(Bignoniaceae) Unidos
Pestalotiopsis sp. 84-175 x 120-186 4-6 [6] x 3-5 [3] 9-18,5 [14] x 5-6,5 [5] Mauritia sp. Brasil
CMEPH M39 806
(Arecaceae)
*Nag Raj (1993)
21
1.1. Stigmopeltis sp. (provável espécie nova nº 4)
H
E
F G
C
BA
10 µm
50 µm
0,5 µm
C
D
200 µm 200 µm
20 µm
10 µm
10 µm 10 µm
5 µm
D
Figura 05. A-H. Pestalotiopsis sp. em Mauritia sp. A e B. Grupos de acérvulos na face abaxial da folha. C. Distribuição das células conidiogênicas e conídios na superfície fértil do acérvulo. D. Conídios e células conidiogênicas, com seta indicando anel em uma delas. E-H. Conídios anfigenamente apendiculados.

22
2.3 Stigmopeltis sp. (provável espécie nova nº 4)
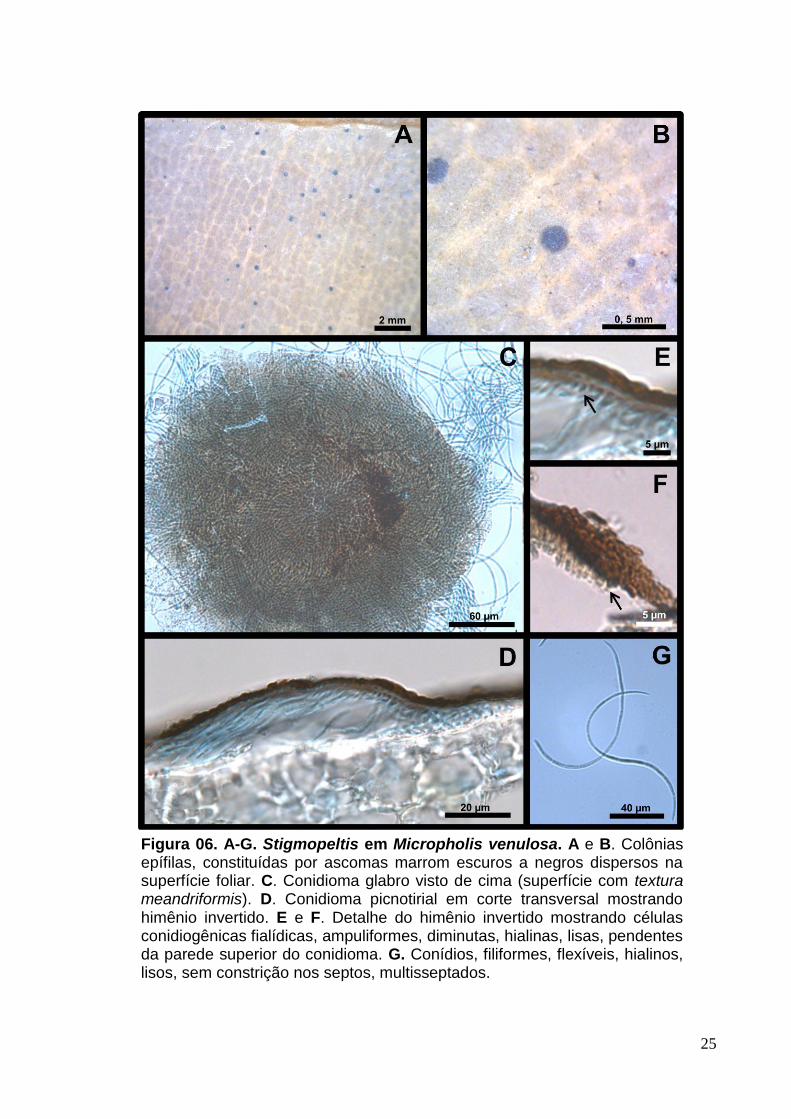
Colônias epífilas, marrom-escuras a negras (Fig.6. A e B). Micélio imerso,
limitado a hifas finas sob os conidiomas. Conidiomas 91-250 [205] x 13-42 [21] µm,
superficiais, glabros, isolados ou agregados, escutelares, dimidiados, marrons, não
ostiolados, deiscência irregular, superfície mostrando textura meandriformis, sem
parede basal, fixando-se no espécime vegetal através de hifas finas e hialinas (Fig.6.
C e D). Células conidiogênicas 1-3,5 [1,5] x 0,5-2 [1,0] µm, fialídicas, ampuliformes,
diminutas, hialinas, lisas, pendentes da parede de um conidioma de himênio
invertido (Fig.6. E e F).Conídios 43-182 [135] x 1,5-3 [2] µm, filiformes, aciculares,
hialinos, lisos, multisseptados (7-19 [16] septos) sem constrição nos septos (Fig.6. G).
Espécime examinado: Brasil. Em folhas de Micropholis venulosa (Sapotaceae). 03
Jul 1962; leg. E.P Heringer, holótipo (CMEPH M39 825).
Batista & Peres (1963), decreveram o gênero monotípico Scoliotidium e este
apresenta semelhanças morfológicas com a espécie tipo do gênero Stigmopeltis (S.
roupalae Syd.), mas diferem em relação à posição do himênio, pois apenas o gênero
Stigmopeltis possui himênio invertido, ou seja, suas células conidiogênicas são
presas à parede superior do conidioma ao contrário do gênero Scoliotidium.
O gênero Stigmopeltis é caracterizado por apresentar conidiomas superficiais,
escudados, parede com textura meandriforme, himênio invertido, deiscência
irregular e conídios longos, multisseptados e filiformes (Sydow, 1927). O espécime
em estudo é um membro desse gênero.
Atualmente existem quatro espécies de Stigmopeltis publicadas (Kirk, 2011)
sendo elas: S. roupalae Syd. (1927), S. graminicola Wehm. (1952), S.illicis Bat. &
Peres (1960) e S. phoebes Syd. (1927). Segundo Pereira-Carvalho (2004) e Santos
(2011) duas prováveis espécies novas foram encontradas em folhas de Elymus
glaucus (Myrtaceae) e em folhas de Salacia crassifolia (Celastraceae) [Tabela 4].
O espécime em estudo apresenta dimensões do conidioma próximas de S.
graminicola e ambos são epífilos, entretanto seus conídios são bem maiores do que
S. graminicola.

23
Com relação às medidas dos conídios o espécime se aproxima relativamente
da espécie S. roupaleae, porém seus conidiomas são distintos sendo S. roupaleae
hipófilo e com dimensões maiores.
Portanto, trata-se provavelmente de uma nova espécie, relatada pela primeira
vez em plantas da família Sapotaceae.
24
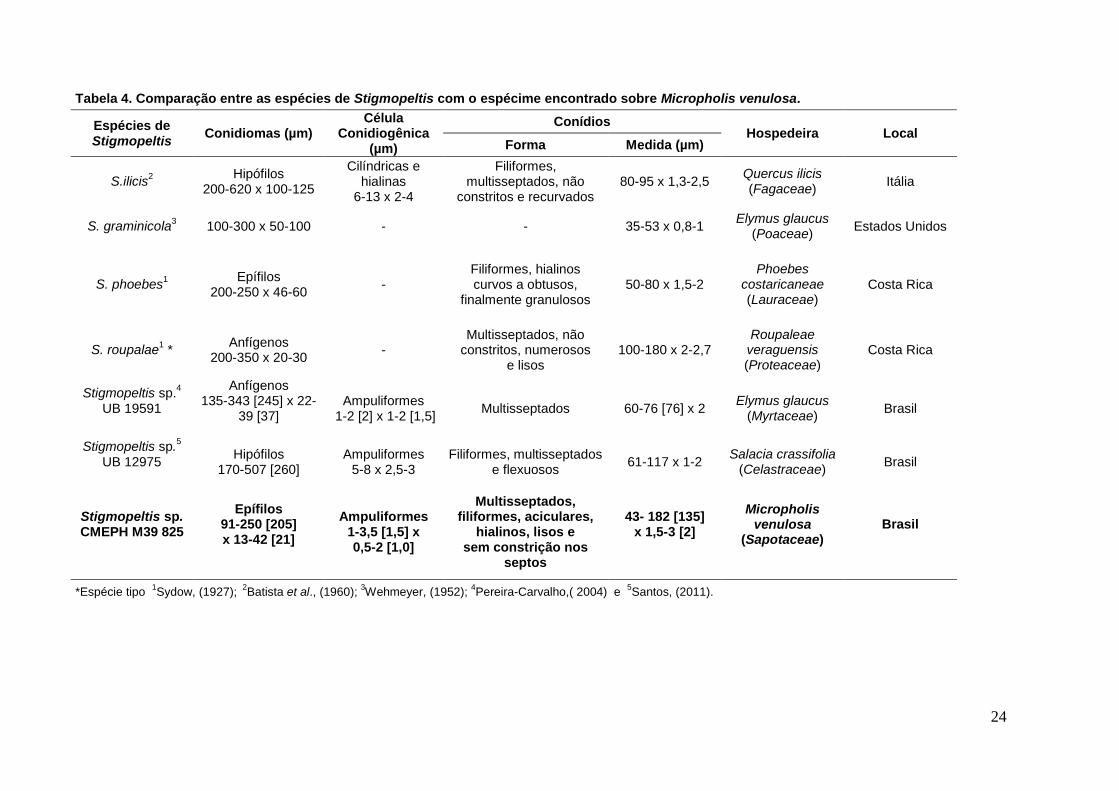
Tabela 4. Comparação entre as espécies de Stigmopeltis com o espécime encontrado sobre Micropholis venulosa.
Espécies de Stigmopeltis
Conidiomas (µm) Célula
Conidiogênica (µm)
Conídios Hospedeira Local
Forma Medida (µm)
S.ilicis2
Hipófilos 200-620 x 100-125
Cilíndricas e hialinas
6-13 x 2-4
Filiformes, multisseptados, não
constritos e recurvados 80-95 x 1,3-2,5
Quercus ilicis (Fagaceae)
Itália
S. graminicola3 100-300 x 50-100 - - 35-53 x 0,8-1
Elymus glaucus (Poaceae)
Estados Unidos
S. phoebes1
Epífilos 200-250 x 46-60
- Filiformes, hialinos curvos a obtusos,
finalmente granulosos 50-80 x 1,5-2
Phoebes costaricaneae (Lauraceae)
Costa Rica
S. roupalae1 *
Anfígenos 200-350 x 20-30
- Multisseptados, não
constritos, numerosos e lisos
100-180 x 2-2,7 Roupaleae
veraguensis (Proteaceae)
Costa Rica
Stigmopeltis sp.4
UB 19591
Anfígenos 135-343 [245] x 22-
39 [37]
Ampuliformes 1-2 [2] x 1-2 [1,5]
Multisseptados 60-76 [76] x 2 Elymus glaucus
(Myrtaceae) Brasil
Stigmopeltis sp.5
UB 12975
Hipófilos 170-507 [260]
Ampuliformes 5-8 x 2,5-3
Filiformes, multisseptados e flexuosos
61-117 x 1-2 Salacia crassifolia
(Celastraceae) Brasil
Stigmopeltis sp. CMEPH M39 825
Epífilos 91-250 [205] x 13-42 [21]
Ampuliformes 1-3,5 [1,5] x 0,5-2 [1,0]
Multisseptados, filiformes, aciculares,
hialinos, lisos e sem constrição nos
septos
43- 182 [135] x 1,5-3 [2]
Micropholis venulosa
(Sapotaceae)
Brasil
*Espécie tipo 1
Sydow, (1927); 2
Batista et al., (1960); 3Wehmeyer, (1952);
4Pereira-Carvalho,( 2004) e
5Santos, (2011).
25
Figura 06. A-G. Stigmopeltis em Micropholis venulosa. A e B. Colônias epífilas, constituídas por ascomas marrom escuros a negros dispersos na superfície foliar. C. Conidioma glabro visto de cima (superfície com textura meandriformis). D. Conidioma picnotirial em corte transversal mostrando himênio invertido. E e F. Detalhe do himênio invertido mostrando células conidiogênicas fialídicas, ampuliformes, diminutas, hialinas, lisas, pendentes da parede superior do conidioma. G. Conídios, filiformes, flexíveis, hialinos, lisos, sem constrição nos septos, multisseptados.

26
2.4 Celomiceto (provável gênero novo nº 1)
Conidiomas picnidiais 141-358 [260] x 87-304 [206] µm, epífilos, superficiais,
estromáticos, agregados, globosos, parede com textura globulosa, escuros e
uniloculares (Fig.7. A e B). Conidióforos ausentes. Células conidiogênicas 6,5-13
[10] x 4-6 [5], enteroblásticas fialídicas, discretas e marrom-claras (Fig.7. C).
Conídios 23-35 [28] x 7-10,5 [9] µm cilíndricos a clavados, marrom-claros e
bicelulares, ocasionalmente com uma leve constrição no septo (Fig.7. D e E).
Espécime examinado: Brasil. Em membro de Asteraceae, leg. E.P Heringer,
holótipo (CMEPH M16 446-50).
De acordo com Sutton (1980) e Kiffer & Morelet (2000) o espécime em estudo
se aproxima do gênero Didymochaeta por este possuir conidiomas picnidiais,
desenvolvendo-se superficialmente, solitários ou agregados, globosos, marrom-
escuros, uniloculares, de paredes espessas, escuras, textura globulosa inicialmente,
tornando-se hialinas com textura angularis; ostíolo central, circular com papila pouco
desenvolvida; conidióforos ausentes; células conidiogênicas enteroblásticas,
fialídicas, discretas, determinadas, hialinas, com canal proeminente e parede
periclinal espessa, formado a partir de células internas da parede picnidial; porém
seus conídios são cilíndricos, de 1 a 2 septos, hialinos e obtusos.
Ainda utilizando a chave de Sutton (1980) o celomiceto aqui estudado
também foi comparado com outros gêneros picnidiais e de conídios hialinos. Os
gêneros comparativos foram: Chaetodiplodia sp., Ascochytulina sp., Pseudodiplodia
sp., Stenocarpella sp. e Tunicago sp.
Os gêneros em comparação se diferem do celomiceto em questão por
apresentar as seguintes características.
Em Chaetodiplodia sp. os picnídios são setosos, em Ascochytulina sp., os
picnídios são imersos e clipeados e apresentam conídios verruculosos, em
Pseudodiplodia sp., os picnídios são imersos e de parede marrom-clara,
Stenocarpella sp., possui picnídios imersos e células conidiogênicas variando de
ampuliformes a lageniformes, e em Tunicago sp. os picnídios são imersos, possuem
conidióforos irregulares, ramificados e septados, além de seus conídios possuírem
uma bainha rígida e granular.

27
Diante tais informações, o celomicelo aqui estudado é um provável gênero
novo a ser publicado de acordo com as normas do Código Internacional de
Nomenclatura para algas, fungos e plantas (ICN).

28
1 mm
10 µm
20 µm20 µm100 µm
A
B C
D E
Figura 07. A-E. Provável gênero novo nº 1 parasitando uma espécie de Asteraceae. A. Aspecto do agregado estromático negro na superfície adaxial da folha, contendo conidiomas cujos ostíolos podem ser vistos (seta). B. Corte transversal de dois conidiomas superficiais, estromáticos, agregados, globosos, escuros e uniloculares. C. Células conidiogênicas ampuliformes a lageniformes, fialídicas, discretas, marrom-claras a marrons. D e E. Conídios, oblongos a clavados, marrom-claros, bicelulares.

29
3 Ascomicetos
3.1 Staibia connari Bat. & Peres
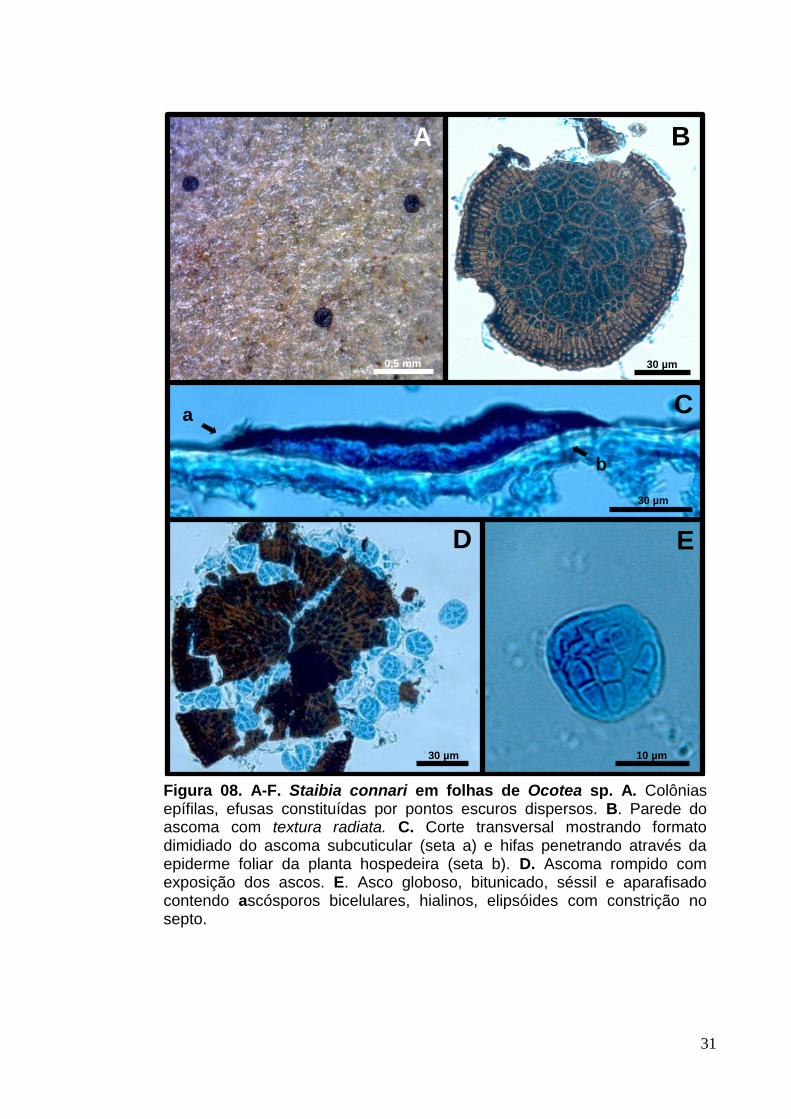
Colônias epífilas, efusas e escuras (Fig.8. A). Micélio superficial inconspícuo,
marrom-claro. Ascomas 78-155 [153] x 13,5-17 µm, subcuticulares, dispersos,
dimidiados, escuros, glabros não ostiolados, textura radiata (Fig.8. B e C). Ascos
11,7-19,5 [16,5] x 11-20 [14,5] µm, numerosos, globosos a subglobosos, bitunicados
contendo oito esporos, sésseis e aparafisados (Fig.8. D e E). Ascósporos 6,5-10,5
[8,5] x 2,5-4 [3,5] µm, hialinos, elipsóides com um septo mediano sem constrição no
septo (Fig.8. F).
Espécime examinado: Brasil. Em folhas de Ocotea paniculata (Lauraceae), 24 Jun
1962, leg. E.P. Heringer, holótipo (CMEPH M39 823 B).
De acordo com Arx & Müller (1975) a família Leptopeltidaceae é composta por
fungos saprofíticos, com micélio conspícuo, imerso no hospedeiro, subcuticular ou
superficial, com a parede do ascoma podendo ser de textura radiata ou
epidermoidea, sem ostíolo e abrindo-se por deiscência irregular na maturidade.
Ascos paralelos, clavados, bitunicados, parafisados, contendo oito esporos podendo
ser hialinos ou marrom-claros, variando de unicelular a multicelular.
O gênero Staibia, monotípico (espécie-tipo: S. connari Bat. & Peres) possui
registro apenas no Cerrado Brasileiro. A espécie tipo foi encontrada sobre Connarus
suberosus Planch. (Connaraceae) coletada no estado de Minas Gerais, descrito e
alocado por Batista & Peres em Leptopeltidaceae Höhn. (Batista et. al., 1966).
Staibia connari já foi relatada em várias plantas do Cerrado como: Andira sp.
e Cassia sp. (Fabaceae), Anemopaegma sp. (Bignoniaceae), Bombax sp.
(Bombacaceae), Diospyros sp. (Ebenaceae), Kielmeyera sp. (Clusiaceae), Plumeria
sp. (Apocynaceae), Salacia sp. (Celastraceae), Salvertia convallariaeodora
(Vochysiaceae) e Connarus suberosus (Connaraceae), enquanto Staibia sp. já foi
relatada em Anacardium sp. (Anacardiaceae), e em Salacia crassifolia (Dornelo-
Silva, 1999; Farr & Rossman, 2011; Santos, 2011). Uma provável espécie nova

30
pertencente ao gênero foi encontrada pela primeira vez em folhas de Eugenia
dysenterica (Myrtaceae) [Pereira-Carvalho, 2004].
A espécie tipo (S. connari) apresentou ascomas 130-141 x 11-16 µm; ascos
13-31 x 15-20 µm e ascósporos 9-12 x 3,5-6 µm. Apesar do fungo estudado, encontrado pela primeira vez em folhas de uma
espécie da família Lauraceae, apresentar medidas relativamente menores do que as
da espécie-tipo, essa diferença não é suficiente para propor uma nova espécie.
31
A
0,5 mm
E
B
30 µm
D
30 µm
C
30 µm
10 µm
a
b
Figura 08. A-F. Staibia connari em folhas de Ocotea sp. A. Colônias epífilas, efusas constituídas por pontos escuros dispersos. B. Parede do ascoma com textura radiata. C. Corte transversal mostrando formato dimidiado do ascoma subcuticular (seta a) e hifas penetrando através da epiderme foliar da planta hospedeira (seta b). D. Ascoma rompido com exposição dos ascos. E. Asco globoso, bitunicado, séssil e aparafisado contendo ascósporos bicelulares, hialinos, elipsóides com constrição no septo.

32
3.2 Phyllachora davillae F. Stevens
Colônias hipófilas e dispersas, parede negra e brilhante (Fig.9. A e B).
Ascomas 116-270 [226] x 121-411 [234] µm, multiloculares (3-5 por estroma),
imersos, subepidérmicos, globosos a periformes (Fig.9. C). Clípeos presentes e
bem desenvolvidos (Fig.9. C). Ascos 70-132 [129] x 10-17 [15] µm, numerosos,
cilíndricos, unitunicados contendo oito esporos (Fig.9. D, E e F). Paráfises 1,5-4
[2,5] µm, hialinas e não ramificadas (Fig.9. D, E e F). Ascósporos 10-17,5 [13] x 4-
10 [6] µm, elípticos, amerosseptados, gutulados e hialinos (Fig.9. G).
Espécime examinado: Brasil. Em folhas de Davilla rugosa (Dilleniaceae), leg. E.P.
Heringer, holótipo (CMEPH M16 456-45).
Segundo Farr & Rossman (2011), o gênero Phyllachora possui três registros de
ocorrência em Davilla rugosa, sendo um no Brasil (Mendes et al., 1998) e dois no
Panamá (Dennis, 1970; Piepenbring, 2006) .
A espécie Phyllachora davillae descrita em literatura por Stevens (1930)
apresentou lesões consideravelmente pequenas, 1-3 mm, circulares, anfígenas.
Estroma central, solitário, diminuto, 0,5 - 1 mm, preto fosco e ligeiramente levantado.
Contém lóculos no mesófilo de 1-3 em cada estroma, alongados verticalmente com
170 µm de profundidade, 110 µm de largura ou subgloboso, 170 µm ou oval 380 µm
de largura e 230 µm de profundidade. Clípeos superiores bem desenvolvidos,
negros e espessos, 60 µm, muitas vezes estendendo muito, 200 µm, lateralmente a
partir dos lóculos, paredes laterais e basais 30 µm considerado espesso. Estroma
lateral bem desenvolvido e de células paralelas longas. Ascos clavados, 8 esporos,
oblíquos, unitunicados 126 x 14 µm. Paráfises filiformes e gelatinosas. Ascósporos
ovais, obtusos de 15 x 7-8 µm. As estruturas fúngicas foram encontradas associadas
às folhas de Davilla rugosa.
O espécime ora reencontrado em D. rugosa apresenta características
dimensionais similares da espécie já publicada, entretanto diferenças morfológicas
foram encontradas. O espécime em estudo apresentou entre 3-5 lóculos por
estroma, ascos cilíndricos e ascósporos elípticos. Já na espécie publicada foi
observado um número entre 1-3 de lóculos por estroma, ascos clavados e
ascósporos variando de ovais a obtusos.

33
Apesar dessas diferenças encontradas, estas não são suficientes para
acomodar o espécime em estudo em outra espécie, até porque as espécies dentro
do gênero Phyllachora são determinadas de acordo com o hospedeiro. Sendo
assim, o filacoráceo encontrado aqui em D. rugosa corresponde a mesma espécie
estabelecida por Stevens (1930).

34
Figura 09. A-G. Phylachora davillae em Davilla rugosa. A e B. Ascomas hipófilos e dispersos, parede negra e brilhante. C. Ascomas em estromas multiloculares, globosos a ovóides, imersos, subepidérmicos. D, E e F. Ascos numerosos, parafisados, cilíndricos, unitunicados contendo oito esporos. G. Ascósporo, hialino, elíptico, amerosseptado e gutulado.

35
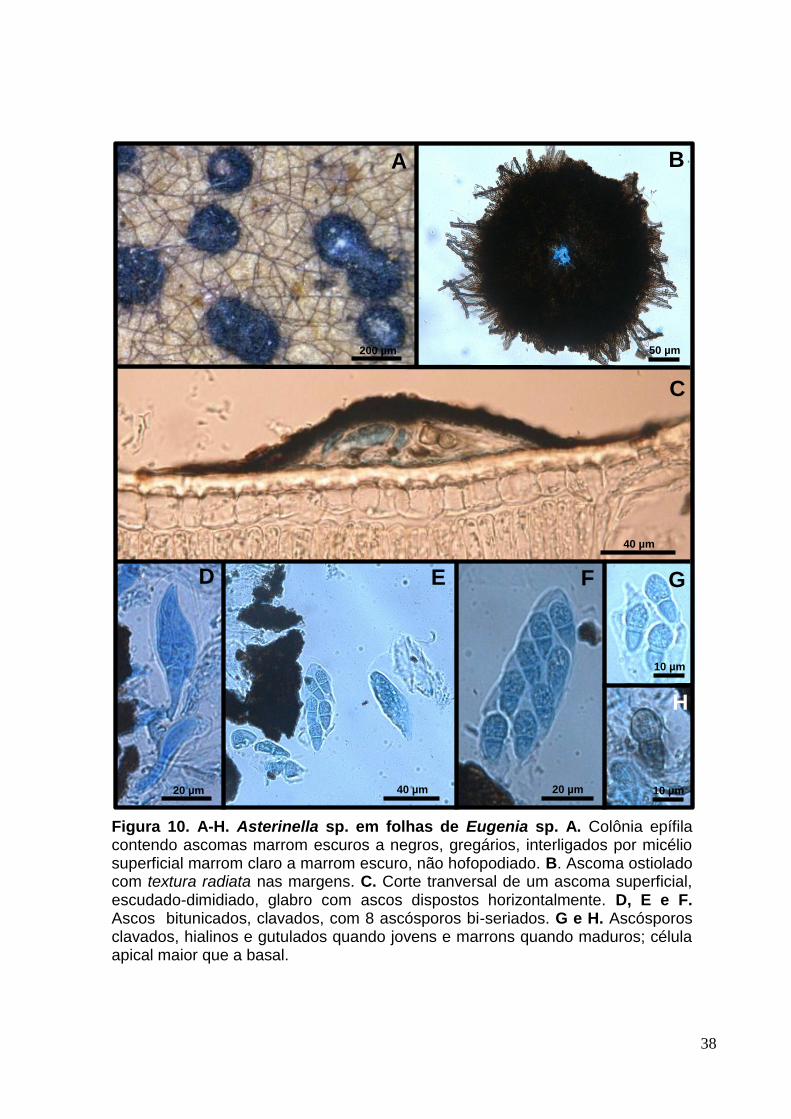
3.3 Asterinella sp. (provável espécie nova nº 5)
Colônias epífilas, gregárias, circulares e escuras (Fig.10. A). Lesões epífilas,
esbranquiçadas a marrons, sob os ascomas (Fig.10. A). Micélio superficial sub-
hialino a marrom-claro com hifas não hifopodiadas (Fig.10. A). Células miceliais 2-5
[3,5] µm de diâmetro. Ascomas 153-361 [250] x 21-69 [50] µm, superficiais,
escudado-dimidiados, glabros, ostiolados, textura meandriforme e radiata (Fig.10. B
e C). Ascos 47-87 [67] x 14-27 [20] µm, bitunicados, clavados, com 8 ascósporos,
parafisados (Fig.10. D, E e F). Paráfises 8-12 µm, hialinas, ramificadas, filiformes
septadas. Ascósporos 17-25 [21] × 6-10 [8] µm clavados, hialinos e gutulados
quando jovens, tornando-se marrons com a maturidade, constritos no septo (Fig.10.
G e H).
Espécime examinado: Brasil. Distrito Federal, Brasília, em folhas de Eugenia sp.
(Myrtaceae), 10 Jun 1962, leg. E. P. Heringer, (CMEPH M39 809).
De acordo com Arx & Müller (1975) o espécime em estudo pertence à família
Microtyriaceae (por apresentar características que o alocam nesta família e também
por ser parasítico) e ao gênero Asterinella por conter ascomas escudados,
ostiolados, micélio superficial com hifas não hifopodiadas, hipostroma ausente,
ascos bitunicados e ascósporos bicelulares, hialinos quando imaturos e marrons na
maturidade.
Comparações com a espécie tipo, Asterinella puiggarii (Speg.) Theiss,
confirmam a identificação com o espécime em questão, uma vez que ambos
possuem em princípio a formação da parede do ascoma com textura meandriforme
e com o desenvolvimento assumem uma textura radiata propriamente dita.
Membros da família Asterinaceae, por exemplo, Asterina sp., podem ser
confundidos com Asterinella sp., graças a semelhanças na textura da parede do
ascoma (textura radiata), entretanto em Asterinella as hifas não são hifopodiadas e a
deiscência do ascoma ocorre através de ostíolo diferente da deiscência estelar do
ascoma em espécies de Asterina.
Existem aproximadamente 101 registros de ocorrência de espécies de
Asterinella encontradas em diversas hospedeiras (Kirk, 2011). Além disso, Farr &
Rossman (2011) relatam que apenas duas espécies (A. cylindrotheca e A. puiggarii)

36
ocorrem em Myrtaceae, porém Batista & Peres (1961a), mencionaram cinco
espécies, incluindo também A. caaguazensis, a A. puiggarii var. minor e a A.
sublibera. A espécie A. sublibera também foi encontrada sobre folhas de Eugenia
bimarginata (Pereira-Carvalho, 2004).
O espécime em estudo pode ser separado das demais espécies registradas
em Myrtaceae (Tabela 5), por possuir ascomas de dimensões próximas às de A.
caaguazensis; seus ascos apresentam medidas parecidas com as de A.
cylindrotheca, porém, as mensurações dos ascóporos diferem claramente das de
todas as espécies sobre Myrtaceae. Os demais dados obtidos permitem considerar
o espécime aqui estudado como pertencente a uma provável espécie nova, a ser
posteriormente publicada conforme as normas vigentes no Código Internacional de
Nomenclatura para algas, fungos e plantas (ICN).
37
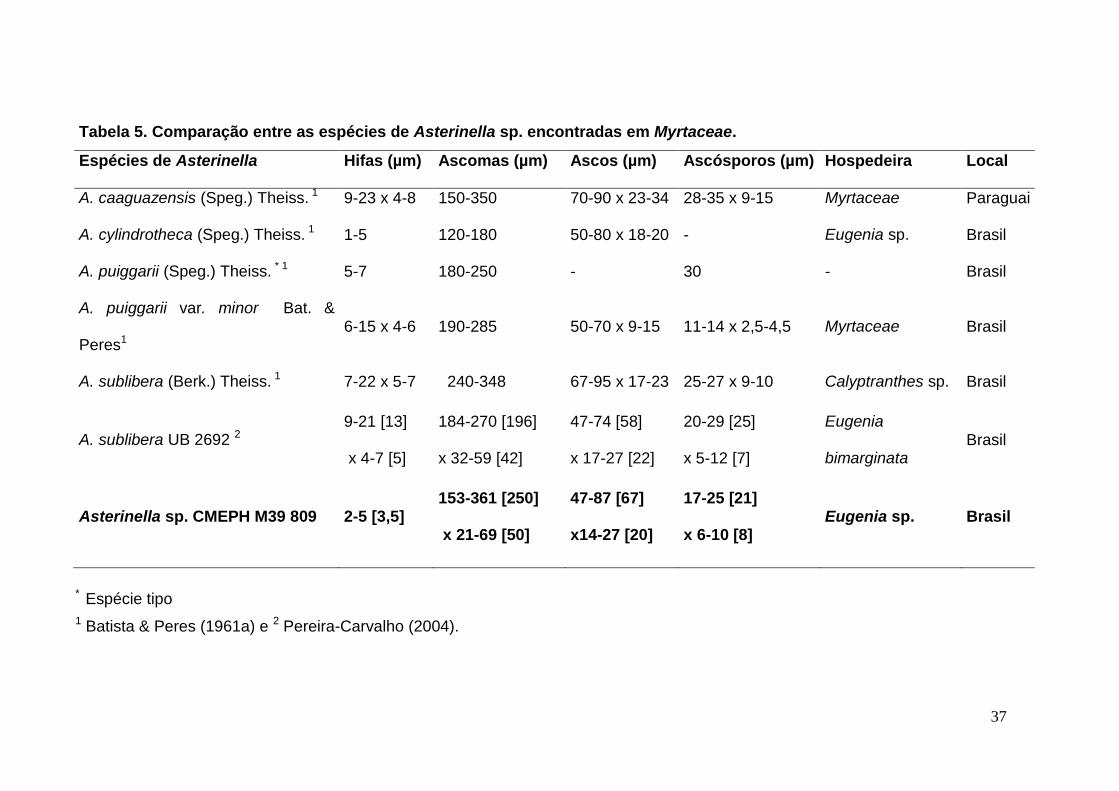
Tabela 5. Comparação entre as espécies de Asterinella sp. encontradas em Myrtaceae.
Espécies de Asterinella Hifas (µm) Ascomas (µm) Ascos (µm) Ascósporos (µm) Hospedeira Local
A. caaguazensis (Speg.) Theiss. 1 9-23 x 4-8 150-350 70-90 x 23-34 28-35 x 9-15 Myrtaceae Paraguai
A. cylindrotheca (Speg.) Theiss. 1 1-5 120-180 50-80 x 18-20 - Eugenia sp. Brasil
A. puiggarii (Speg.) Theiss. * 1 5-7 180-250 - 30 - Brasil
A. puiggarii var. minor Bat. &
Peres1 6-15 x 4-6 190-285 50-70 x 9-15 11-14 x 2,5-4,5 Myrtaceae Brasil
A. sublibera (Berk.) Theiss. 1 7-22 x 5-7 240-348 67-95 x 17-23 25-27 x 9-10 Calyptranthes sp. Brasil
A. sublibera UB 2692 2 9-21 [13]
x 4-7 [5]
184-270 [196]
x 32-59 [42]
47-74 [58]
x 17-27 [22]
20-29 [25]
x 5-12 [7]
Eugenia
bimarginata Brasil
Asterinella sp. CMEPH M39 809 2-5 [3,5] 153-361 [250]
x 21-69 [50]
47-87 [67]
x14-27 [20]
17-25 [21]
x 6-10 [8] Eugenia sp. Brasil
* Espécie tipo
1 Batista & Peres (1961a) e 2 Pereira-Carvalho (2004).
38
A B
200 µm
40 µm
40 µm 20 µm20 µm 10 µm
10 µm
C
E
H
FD G
50 µm
Figura 10. A-H. Asterinella sp. em folhas de Eugenia sp. A. Colônia epífila contendo ascomas marrom escuros a negros, gregários, interligados por micélio superficial marrom claro a marrom escuro, não hofopodiado. B. Ascoma ostiolado com textura radiata nas margens. C. Corte tranversal de um ascoma superficial, escudado-dimidiado, glabro com ascos dispostos horizontalmente. D, E e F. Ascos bitunicados, clavados, com 8 ascósporos bi-seriados. G e H. Ascósporos clavados, hialinos e gutulados quando jovens e marrons quando maduros; célula apical maior que a basal.

39
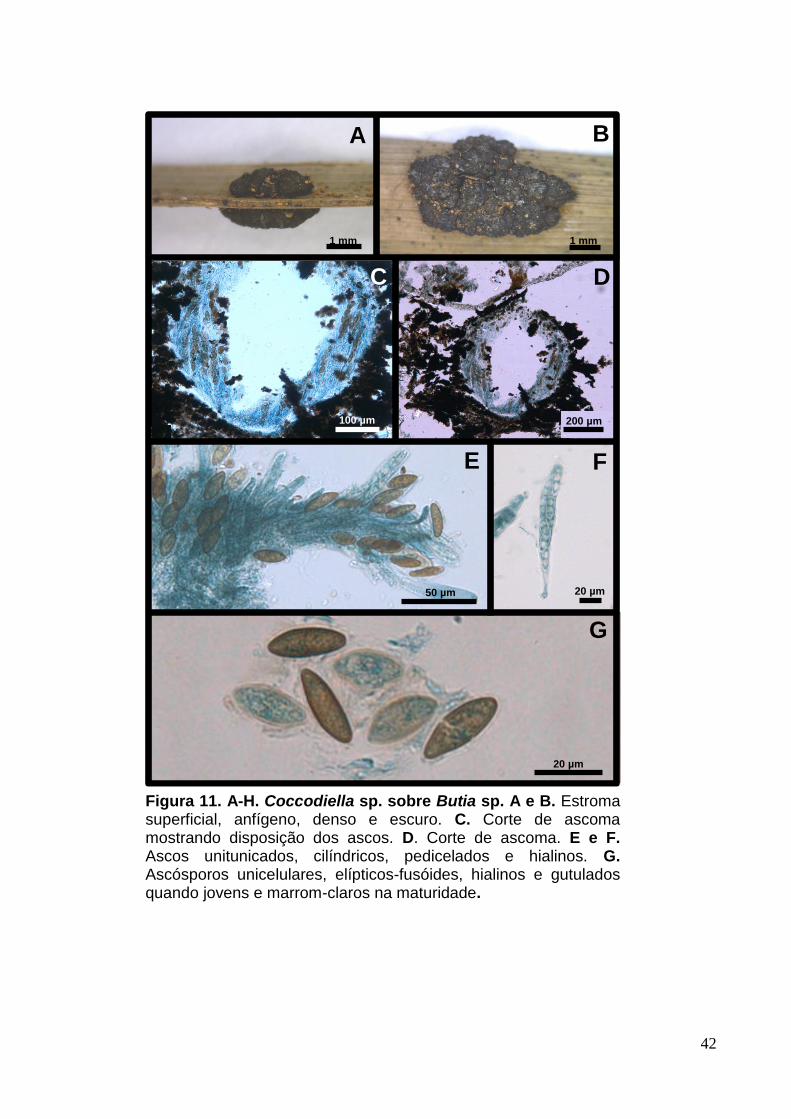
3.4 Coccodiella sp. (provável espécie nova nº 6)
Estromas 2-6 µm, superficiais, multiloculares, anfígenos com crescimento
através do limbo foliar e projetando-se para fora da folha, multiloculares, densos,
escuros (Fig.11. A e B). Peritécios 205-687 [361] x 276-555 [509] µm, superficiais,
globosos (Fig.11. C e D). Ascos 57-123 [82] x 12-21 [12] µm, numerosos,
unitunicados, cilíndricos, pedicelados, hialinos e parafisados (Fig.11. E e F).
Paráfises septadas, ramificadas e hialinas. Ascósporos 20-28 [25] x 6-11 [9] µm,
unicelulares, elíptico-fusóides, hialinos e gutulados quando jovens e marrom-claros
na maturidade (Fig.11. G).
Espécime examinado: Brasil. Em folhas de Butia sp.(Arecaceae), leg. E. P.
Heringer, (CMEPH M39 813).
O Gênero Coccodiella pertence à família Phyllachoraceae que é representada
por membros que possuem estroma bem desenvolvido, imerso no tecido do
hospedeiro, geralmente com a presença de clípeo, negros e brilhantes ou raramente
coloridos, contendo de um a vários ascomas. Ascoma peritecial, parede fina, ostíolo
composto por perífises. Perídio composto de tecido escuro ou sem cor. Hamatécio
constituído por paráfises, algumas vezes deliqüescentes. Ascos cilíndricos,
persistentes. Ascósporos frequentemente hialinos podendo ser marrons,
amerosseptados, raramente ornamentados, muitas vezes com uma bainha
mucilaginosa. Possuem sua fase anamórfica em celomicetos. Conidioma
usualmente formados em lóculos dentro do estroma. Conidioma varia bastante
quanto a forma por ser frequentemente bastante achatado ou comprimido por
ascomas adjacentes, ocasionalmente são picnidiais, as paredes do conidioma são
frequentemente indiferenciadas. Células conidiogênicas são cilíndricas,
frequentemente aglomeradas em pequenos conidióforos com proliferação
percorrente. Conídios pequenos, asseptados, hialinos, filiformes (geralmente
curvados) ou bacilar em geral com função espermacial (Cannon & Kirk, 2007).
Coccodiella Hara [sin. Coccostroma sp.Theiss. & Syd.] é um gênero que inclui
espécies com estroma pulvinado, irrompente, tornando-se superficial, com centro
monolocular com ascos em fascículos, parcialmente imersos em associação com um
hipostroma. Ascos com oito esporos unicelulares e marrons.

40
De acordo com Farr & Rossman (2011), quatro espécies são relatadas na
família Arecaceae (Tabela 6).
O fungo em estudo apresenta dimensões dos ascos mais largos do que os de
C. petrakii (Cif.) Arx & E. Müll. e ascósporos mais largos do que os de C. bactridis
(Rehm) Arx & E. Müll.
Os ascomas e ascósporos do espécime em estudo superam (vagamente) em
tamanho os das demais espécies do gênero que ocorrem sobre espécies da família
Arecaceae (Tabela 6). Essas características sugerem que o fungo estudado
pertence provavelmente a uma nova espécie.

41
Tabela 63. Comparação entre as espécies de Coccodiella sp. encontradas em Arecaceae.
Espécies de Estroma (mm)
Peritécios (µm) Ascos Ascósporos (µm)
Hospedeira Local Coccodiella sp.
C. bactridis (Rehm) Arx & E. Müll 1-1,5 - 70-75 x 12-15 µm 20-25 x 6-7 Bactris sp. Brasil
Clavados
C. nervisequens (Petr. & Cif.) Arx E. Müll
0,5-3 180-300 x 350 75-95 x 14-18 µm 14-20 x 5-7 Euterpes globosa
América Central
cilíndrico-clavados
C. palmicola (Speg.) Arx & E. Müll 1-4 150-250 x 300-400 75-100 x 20-25 µm 19-26 x 7-10 Cocos yatai Brasil
C. petrakii (Cif.) Arx & E. Müll 1-3 150-400 x 200-300 60-100 x 8,5-13 µm 11-18 x 8-11 Oreodoxa regia
América Central
cilíndricos
Coccodiella sp. CMEPH M39 813
2-6 205-687 [361] x 276-555 [509]
57-123 [82] x 12-21 [12] µm cilíndricos
20-28 [25] x 6-11 [9]
Butia sp. Brasil
Arx & Müller, (1954).
42
100 µm
20 µm
D
E F
1 mm1 mm
A B
C
50 µm 20 µm
200 µm
G
Figura 11. A-H. Coccodiella sp. sobre Butia sp. A e B. Estroma superficial, anfígeno, denso e escuro. C. Corte de ascoma mostrando disposição dos ascos. D. Corte de ascoma. E e F. Ascos unitunicados, cilíndricos, pedicelados e hialinos. G. Ascósporos unicelulares, elípticos-fusóides, hialinos e gutulados quando jovens e marrom-claros na maturidade.

43
3.5 Johansonia sp. (provável espécie nova nº 7)
Colônias hipófilas, dispersas, setosas, marrom-escuras (Fig.12. A e B).
Micélio superficial, setoso. Setas miceliais 77,5-157,5 [143,5] x 3-6 [6] µm, eretas,
septadas, marrom-escuras, filiformes (Fig.12. C, D e E). Hifas ramificadas, septadas,
hialinas a marrom-claras, setosas. Ascomas 156-360 [218,5] µm, superficiais,
discóides, marrom-escuros a negros (Fig.12. D e E). Ascos 27-38 [30] x 7-11,5 [9]
µm, numerosos, cilíndricos a elipsóides, bitunicados, contendo oito esporos (Fig.12.
F). Ascósporos 5,5-11 [7] x 1-3,5 [3] µm, hialinos, bicelulares, clavados, e sem
constrição nos septos (Fig.12. G).
Espécime examinado: Brasil. Em folhas de Ocotea paniculata (Lauraceae), 24 Jun
1962, leg. E. P. Heringer, (CMEPH M39 823 B).
A família Saccardiaceae abriga os ascomicetos saprofíticos (ou parasíticos)
com desenvolvimento superficial e micélio presente, frequentemente inconspícuo,
ocasionalmente setoso, composto por hifas hialinas ou escuras, septadas. Estroma
ausente ou pequeno na região basal dos ascomas. Ascomas superficiais, discóides,
de paredes finas com a parede superior gelatinosa, composto por células
isodiamétricas ou irregulares, não ostiolados e ascos paralelos em camada única,
bitunicados, clavados ou obovóides com hamatécio interascal filamentoso,
constituído por paráfises apicalmente enegrecidas, ramificadas ou não, formando
epitécio. Ascósporos bicelulares ou multicelulares, hialinos ou escuros com bainha
ausente (Arx & Müller, 1975; Kirk et al, 2001; Cannon & Kirk, 2007).
O gênero Johansonia é caracterizado por formar os ascomas sobre um
pequeno subículo. Ascos numerosos, bitunicados e parafisados. As paráfises são
inter-ascais, hialinas, septadas, ramificam e tornam-se escurecidas formando um
epitécio. Ascósporos são bicelulares e hialinos. Ascomas são cercados por micélio
setoso que se espalha pela superfície foliar. Setas eretas marrons e septadas.
Johansonia se difere de Phillipsiella pela presença de seta e por apresentar subículo
mais desenvolvido (Müller & von Arx,1962; Arx & Müller, 1975).
A descrição do gênero Johansonia é baseada na espécie tipo J. setosa Sacc. (1889), uma espécie encontrada em folhas de membro da família Sapindaceae
coletado na América do Sul (Müller & von Arx, 1962).

44
Devido o gênero possuir ascomas superficiais e discóides, ascos bitunicados
e ascósporos hialinos, Müller & von Arx (1962) alocaram o gênero na família
Schizothyriaceae. Em um estudo posterior, no entanto, von Arx & Müller (1975) o
transferiram para Saccardiaceae em Dothideales, pelo fato de formar epitécio
agregando paráfises ramificadas. Barr (1993) aceitou Johansonia em
Phillipsiellaceae, enquanto Lumbsch e Huhndorf (2007) concluíram que era um
membro da família Saccardiaceae, uma família que consideravam como incertae
sedis em Ascomycota. Segundo Lumbsch e Huhndorf (2010) Johansonia sp. ainda
permanece na família Saccardiaceae. Como atualmente não existem dados de
sequencia de DNA representada para qualquer espécie de Johansonia no GenBank,
a sua posição taxonômica permanece obscura (Crous et al., 2010).
De acordo com Kirk (2011) existem 13 espécies publicadas no gênero
Johansonia e, além disso, Dornelo-Silva (1999) e Pereira-Carvalho (2004) relataram
prováveis espécies novas de Johansonia encontradas em Qualea grandiflora e em
Eugenia dysenterica (Tabela 7).
O espécime em questão possui suas dimensões próximas de J. pandani
quanto ao tamanho das setas e comprimento dos ascos, entretanto seu ascoma é
consideravelmente maior aproximando-se de J. brasiliensis. Os ascósporos são
notavelmente menores entre todas as espécies do gênero relacionadas.
Portanto pode-se inferir que o espécime é uma provável espécie nova. Este é
o primeiro relato de Johansonia em membro da família Lauraceae.

45
Tabela 7. Comparação entre as espécies de Johansonia sp. com a espécie encontrada em Lauraceae.
Espécies de Johansonia Seta (µm) Ascoma (µm) Ascos (µm) Ascósporos (µm) Hospedeira Local
J. amadelfa (Syd.) Arx1 170 x 3-5 250-400 x 25- 25-33 x 11-16 9-12 x 3-4 Roupalae veraguensis (Proteaceae) Costa Rica
J. arxii Bose & Müll. 2 - - - - Quercus dilatata (Fagaceae) Índia
J. brasiliensis Arx1 200-300 x 7-9 151-280 x 65-80 40-54 x 18-23 19-24 x 6-7,5 Inga sp. (Leguminosae) Brasill
J. chapadiensis Crous et al. 3 130-260 x 4-5 300 x 200 32-45 x 11-19
(13-)15-19(-44) x (5-)6-7
Dimorphandra mollis (Leguminosae) Brasil
J. consociata (Syd.) Arx1 - 180-220 - 10-13 x 3,5-4,5 Roupalae veraguensis (Proteaceae) Costa Rica
J. cupulata (Ellis Mart.) Barr.1 210 x 6 300 48 x 12 12-13 x 4-4,5 Quercus laurifoliae (Fagaceae) Estados Unidos
J. formosa (Ellis Mart.) Barr.1 160-300
180-280 x 80-100
33-60 x 14,5-28 21-25,5 x 7,5-9 Olea americana (Oleaceae) Estados Unidos
J. guazumae P. Henn1 - 250 40-50 x 5-6 6-8 x3-3,5 Guazuma rosea (Sterculiaceae) Brasil
J. micheliae Bose & Müll. 2 - - - - Michelia kisopa (Magnoliaceae) Índia
J. microglossae (Hansf.) Arx1 140 120-180 x 60-80 - 17-20 x 6-8 Microglossa volubilis (Asteraceae) África
J. nigro-capitata (Wint.) Sacc1 - 83-128 25-27 x 12,5-16 11-13 x 4 Solanaceae Brasil
J. pandani E.Müll.4 100 150-200 x 50-60 27-36 x 11-13 11-13 x 3,5-5 Pandani sp. (Pandanaceae) Indonésia
J. setosa (Wint.) Sacc. *1 Maior que 500 190-28- x 52-74 40-52 x 12-14 11-14 x 3-4 Sapindaceae América do Sul
Johansonia sp. 5
UB 10041 135-620 x 4-6 [5] 230-450 x 45-60 30-57 [45] x 10-18 [15] 10-22 [17] x 4-6 [5] Qualea grandiflora (Vochysiaceae) Brasil
Johansonia sp.6
UB 7744 77-130 x 1-3 [1] 60-139 [80] x 50-57 35-45 [39] x 8-13 [10] 8-14 [11] x 2-4 [3] Eugenia dysenterica (Myrtaceae) Brasil
Johansonia sp. CMEPH M39 823B
77,5-157,5 [143,5] x 3-6 [6]
156-360 [218,5] 27-38[30] x 7-11,5 [9]
5,5-11 [7] x 1-3,5 [3]
Ocotea paniculata (Lauraceae) Brasil
* Espécie tipo
1 Müller & Arx (1962);
2 Bose & Müll (1964)
3 Crous et al.( 2010);
4 E. Müll. (1954);
5 Dornelo-Silva (1999) e
6 Pereira-Carvalho (2004)
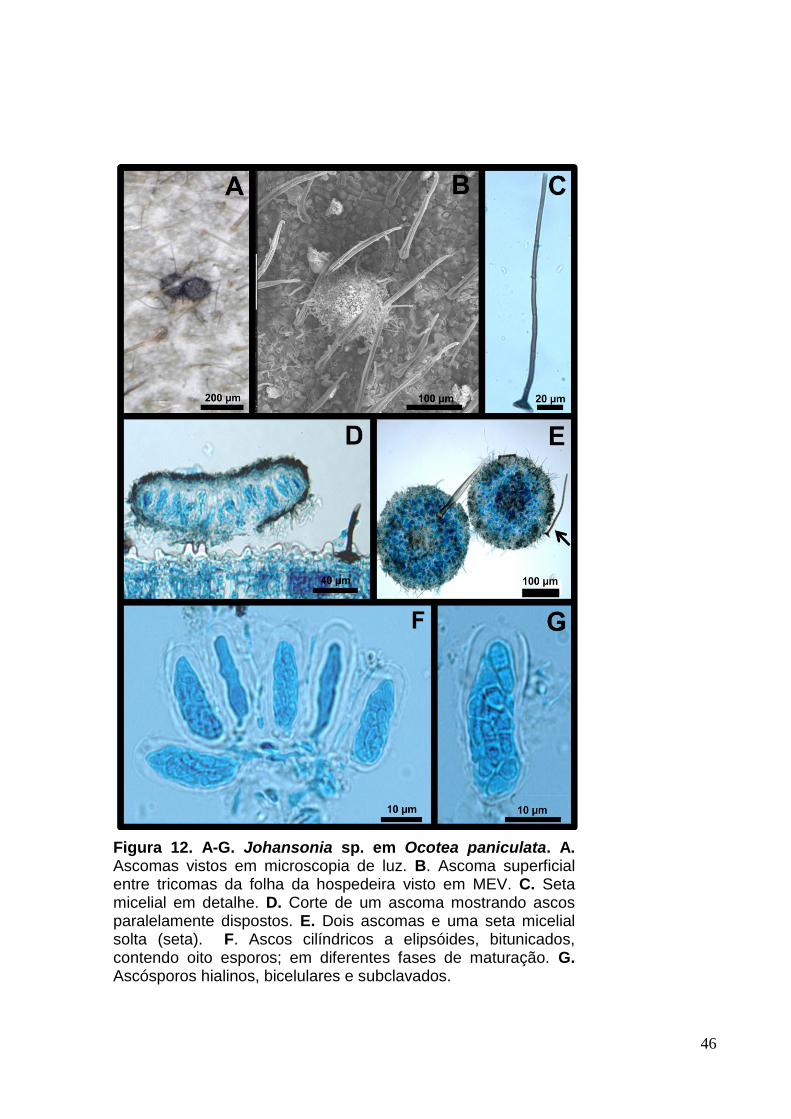
46
Figura 12. A-G. Johansonia sp. em Ocotea paniculata. A. Ascomas vistos em microscopia de luz. B. Ascoma superficial entre tricomas da folha da hospedeira visto em MEV. C. Seta micelial em detalhe. D. Corte de um ascoma mostrando ascos paralelamente dispostos. E. Dois ascomas e uma seta micelial solta (seta). F. Ascos cilíndricos a elipsóides, bitunicados, contendo oito esporos; em diferentes fases de maturação. G. Ascósporos hialinos, bicelulares e subclavados.

47
3.6 Limacinia sp. (provável espécie nova nº 8)
Colônias hipófilas e efusas (Fig.13. A). Ascomas 97,5-209 [130] x 88-188
[132] µm, superficiais, periteciais, sub-globosos, solitários, glabros, escuros, textura
angularis (Fig.13. B e C). Ascos 38-65 [60] x 9-25 [19] µm, 8 esporos, bitunicados,
hialinos, clavados e pedicelados (Fig.13. D e E). Ascósporos 33-59 [48] x 2-4,5 [3]
µm, filiformes a fusiformes, hialinos, multisseptados (6-9 septos), não constritos,
levemente curvados, célula apical com proeminência papilada (Fig.13. F).
Espécime examinado: Brasil. Em folhas de Lantana camara (Verbenaceae), leg. E.
P. Heringer, (CMEPH M16 438-54).
O gênero Limacinia Neger (Capnodiaceae), apresenta micélio superficial,
hialino ascomas globosos, glabros, ostiolados. Ascos bitunicados, fissitunicados,
ovóides a clavados. Ascósporos alongados e fusiformes, hialinos ou pigmentados,
multisseptados, geralmente com septos transversais (Arx & Müller, 1975). O gênero
Limacinia possui como tipo a espécie L. fenandeziana Neger encontrada sobre
Myrceugenia fernandeziana (Myrtaceae) no Chile.
Em Limacinia sp. são incluídos alguns ascomicetos causadores de fumagina
com esporos hialinos e pigmentados e com ascomas glabros ou setosos.
Saccardo & Sydow (1899) incluíram no gênero Limacinia 14 recombinações
em adição à espécie-tipo de Neger.
Lindau (1897) incluiu Limacinia em Perisporiales como tendo ascósporos
marrons e mais de duas células. Ele caracterizou o gênero Limacinia por ascomas
glabros diferindo-se do gênero Morfea de ascomas setosos.
Arnaud (1910) aceitou o trabalho feito por Saccardo como um ponto em seu
novo conceito de Limacinia, incluiu espécies com ascósporos hialinos a escuros e
ascomas glabros ou setosos. Ele alocou quatro espécies de regiões temperadas,
cinco tropicais e uma espécie questionável da Austrália dentro de Limacinia.
Theissen & Sydow (1917) concordaram com uma maior amplitude para o
gênero Limacinia estabelecida por Saccardo. Eles consideraram para Limacinia as
espécies de capnodiáceos com ascósporos hialinos com septos transversais. Este

48
conceito foi mantido por vários autores (Clements & Shear, 1931; Fraser, 1935;
Batista & Ciferri,1963; Arx & Muller, 1975).
Atualmente constam aproximadamente 81 espécies no gênero, dessas
apenas seis (Tabela 8) são encontradas na América do Sul (Kirk, 2011; Farr &
Rossmam, 2011).
O espécime aqui estudado, quando comparado com as demais espécies
encontradas na América do Sul, possui medidas de ascomas e ascos próximas das
da espécie Limacinia quitensis, porém seus ascósporos são muito maiores que os
de todas as outras espécies analisadas do continente exceto L. melioloides. Além
disso, em L. penziguii o formato dos ascos é obovado e os conídios são muriformes
e apresentam três septos com constrição, ao contrário do espécime estudado que
possui ascos clavados e conídio com mais de três septos (apenas transversais) e
não constritos.
Este é o primeiro relato de Limacinia em planta da família Verbenaceae.
Assim, o fungo ora analisado deve tratar-se de uma espécie nova para o
gênero a ser publicada de acordo com as normas vigentes no Código Internacional
de Nomenclatura para algas, fungos e plantas (ICN).
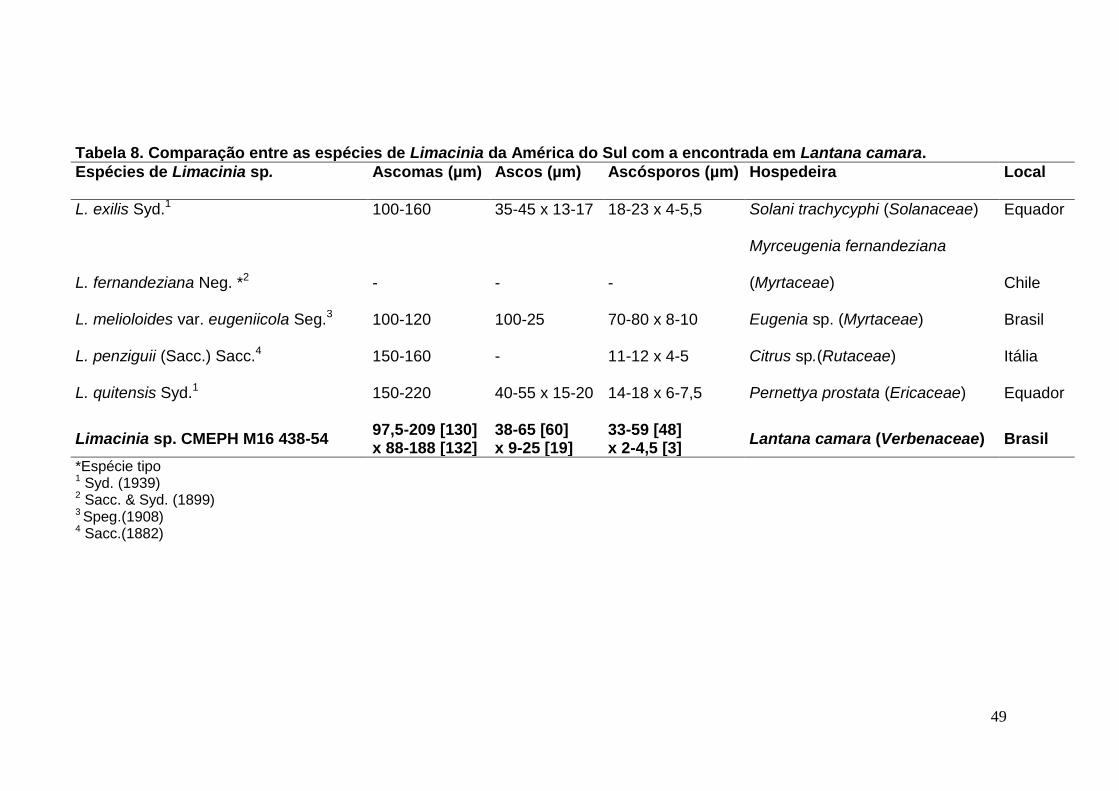
49
Tabela 8. Comparação entre as espécies de Limacinia da América do Sul com a encontrada em Lantana camara.
Espécies de Limacinia sp. Ascomas (µm) Ascos (µm) Ascósporos (µm) Hospedeira Local
L. exilis Syd.1 100-160 35-45 x 13-17 18-23 x 4-5,5 Solani trachycyphi (Solanaceae) Equador
L. fernandeziana Neg. *2 - - -
Myrceugenia fernandeziana
(Myrtaceae) Chile
L. melioloides var. eugeniicola Seg.3 100-120 100-25 70-80 x 8-10 Eugenia sp. (Myrtaceae) Brasil
L. penziguii (Sacc.) Sacc.4 150-160 - 11-12 x 4-5 Citrus sp.(Rutaceae) Itália
L. quitensis Syd.1 150-220 40-55 x 15-20 14-18 x 6-7,5 Pernettya prostata (Ericaceae) Equador
Limacinia sp. CMEPH M16 438-54 97,5-209 [130] x 88-188 [132]
38-65 [60] x 9-25 [19]
33-59 [48] x 2-4,5 [3]
Lantana camara (Verbenaceae) Brasil
*Espécie tipo 1 Syd. (1939) 2 Sacc. & Syd. (1899) 3 Speg.(1908) 4 Sacc.(1882)
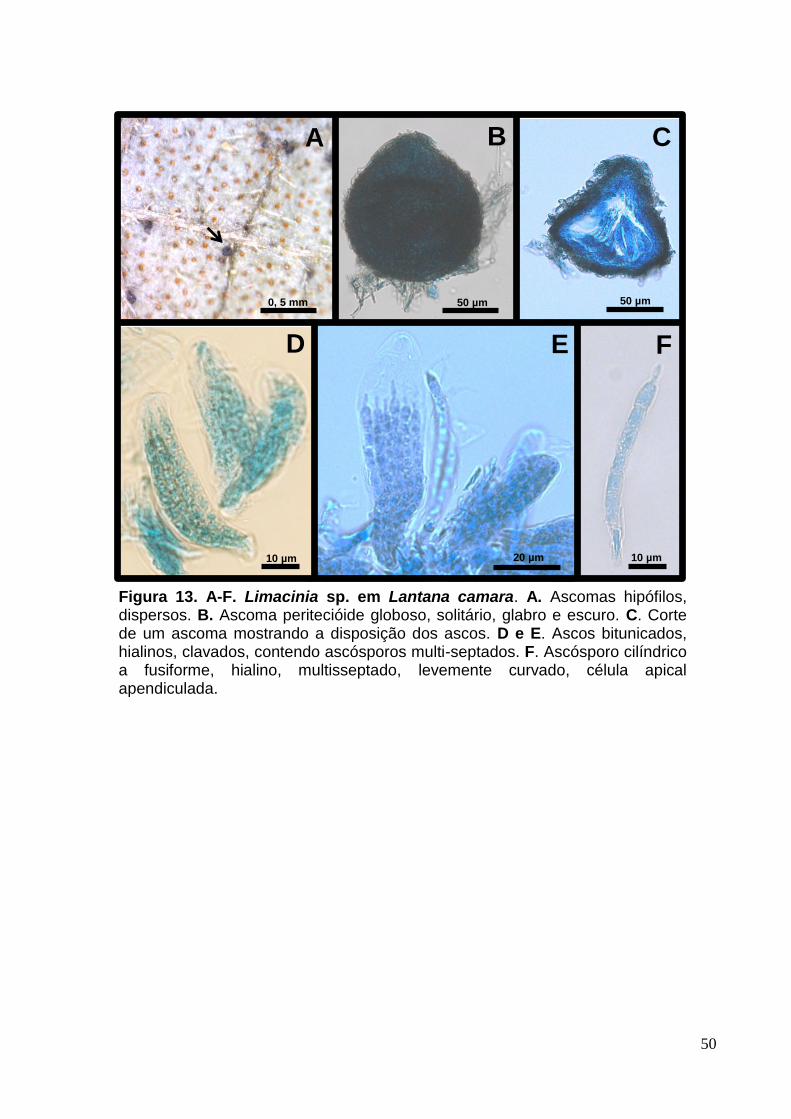
50
50 µm
A
FD E
B
0, 5 mm 50 µm
20 µm
C
50 µm
10 µm 10 µm
Figura 13. A-F. Limacinia sp. em Lantana camara. A. Ascomas hipófilos, dispersos. B. Ascoma peritecióide globoso, solitário, glabro e escuro. C. Corte de um ascoma mostrando a disposição dos ascos. D e E. Ascos bitunicados, hialinos, clavados, contendo ascósporos multi-septados. F. Ascósporo cilíndrico a fusiforme, hialino, multisseptado, levemente curvado, célula apical apendiculada.

51
3.7 Phillipsiella sp. (provável espécie nova nº 9)
Colônias hipófilas, dispersas, marrom-escuras. Micélio superficial. Hifas
ramificadas, septadas, hialinas a marrom-claras. Setas ausentes. Ascomas 212-427
[237] x 214-408 [356,5] µm, superficiais, discóides, marrom-escuros a negros
(Fig.14. A-D). Ascos 23,5-43 [32,5] x 7-12 [9,5] µm, numerosos, cilíndricos a
elipsóides, bitunicados, contendo oito esporos (Fig.14. E, F e G). Ascósporos 6,5-9,5
[7] x 2-4 [2,5] µm, hialinos, bicelulares, clavados e sem constrição no septo (Fig.14.
H).
Espécime examinado: Brasil. Em folhas de Ocotea paniculata (Lauraceae), 24 Jun
1962, leg. E.P. Heringer, holótipo (CMEPH M39 823 B).
A família Phillipsiellaceae compreende os fungos saprofíticos e epifíticos
muitas vezes associados a folhas pilosas. Estroma ausente. Ascomas pequenos,
escudados para pulvinados possivelmente apoteciais, escuros, às vezes micélio
pouco desenvolvido. Possuem perídio pouco desenvolvido com células
pseudoparenquimatosas. Tecido inter-ascal composto por pseudoparáfises
alargadas no ápice para formar uma camada de epitécio. Ascos sacados, sésseis
com paredes grossas pelo menos no ápice, não fissitunicados. Ascósporos de
hialinos a escuros, simples, septos transversais ou muriformes. (Cannon & Kirk,
2007). De acordo com Lumbsch e Huhndorf (2010) o gênero Phillipsiella permanece
em Saccardiaceace.
O gênero é caracterizado por ascoma superficial, séssil, discóide situado em
um emaranhado de hifas que formam excípulo. Ascos numerosos, bitunicados e
parafisados. As paráfises são inter-ascais, hialinas, septadas, ramificam e tornam-se
escurecidas na região do epitécio. Ascósporos são bicelulares e hialinos. Ascomas
são cercados por micélio superficial.
Existem 11 espécies de Phillipsiella (Kirk, 2011) sendo P. atra Cooke a
espécie tipo. Uma provável espécie nova de Phillipsiella foi encontrada em folhas de
Qualea sp. (Dornelo-Silva, 1999). Essas espécies foram comparadas com o
espécime estudado (Tabela 9).

52
Os valores das medidas dos ascomas do espécime em estudo se
aproximaram da espécie P. ellisii, entretanto possui ascomas mais largos. As
espécies: P. atra, P. nigella e P. puiggani apresentaram ascos com dimensões iguais
e estas são relativamente semelhantes ao fungo ora estudado. Quanto aos
ascósporos as medidas se aproximam mais de P. ellisii. Em termos métricos gerais
nota-se que o espécime em estudo está mais próximo da espécie P. illisii, porém
apresenta ascos maiores.
Este é o primeiro relato de Phillipsiella em membro da família Lauraceae. Sendo
assim, infere-se que o espécime em questão é provavelmente uma espécie nova.
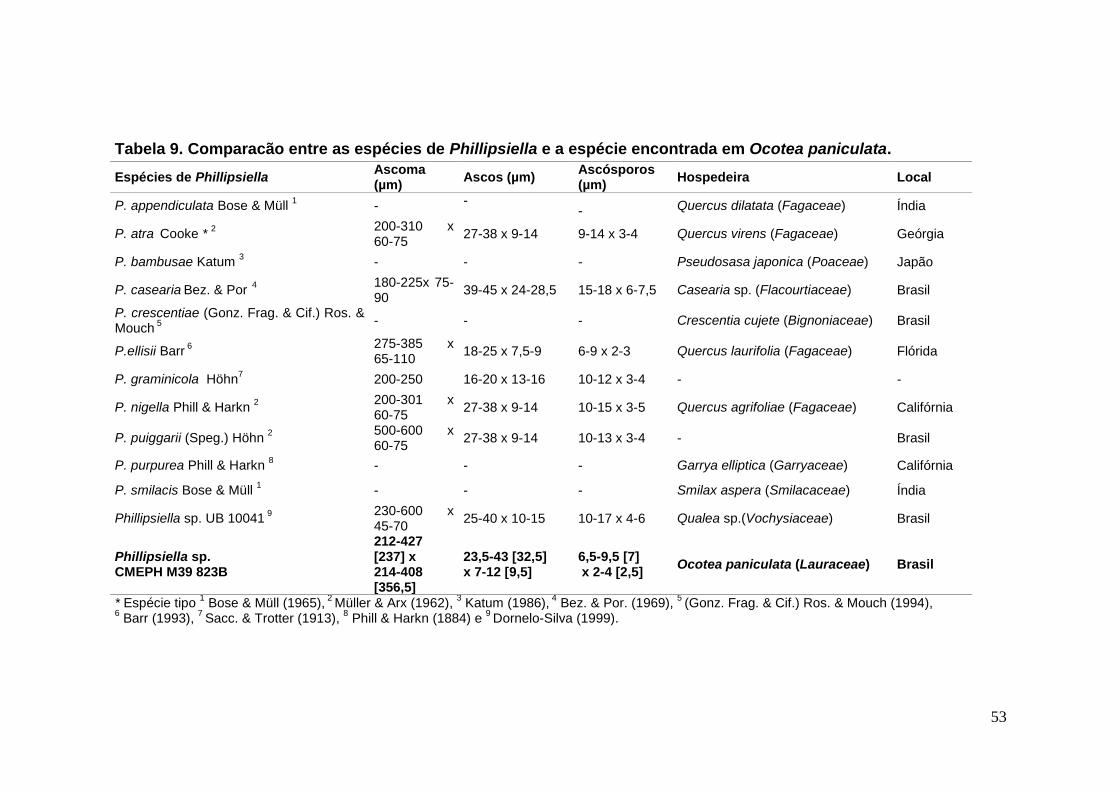
53
Tabela 9. Comparacão entre as espécies de Phillipsiella e a espécie encontrada em Ocotea paniculata.
Espécies de Phillipsiella Ascoma (µm)
Ascos (µm) Ascósporos (µm)
Hospedeira Local
P. appendiculata Bose & Müll 1 - -
- Quercus dilatata (Fagaceae) Índia
P. atra Cooke *
2
200-310 x 60-75
27-38 x 9-14 9-14 x 3-4 Quercus virens (Fagaceae) Geórgia
P. bambusae Katum 3 - - - Pseudosasa japonica (Poaceae) Japão
P. casearia Bez. & Por
4
180-225x 75-90
39-45 x 24-28,5 15-18 x 6-7,5 Casearia sp. (Flacourtiaceae) Brasil
P. crescentiae (Gonz. Frag. & Cif.) Ros. & Mouch
5
- - - Crescentia cujete (Bignoniaceae) Brasil
P.ellisii Barr 6
275-385 x 65-110
18-25 x 7,5-9 6-9 x 2-3 Quercus laurifolia (Fagaceae) Flórida
P. graminicola Höhn7 200-250 16-20 x 13-16 10-12 x 3-4 - -
P. nigella Phill & Harkn 2
200-301 x 60-75
27-38 x 9-14 10-15 x 3-5 Quercus agrifoliae (Fagaceae) Califórnia
P. puiggarii (Speg.) Höhn 2
500-600 x 60-75
27-38 x 9-14 10-13 x 3-4 - Brasil
P. purpurea Phill & Harkn 8 - - - Garrya elliptica (Garryaceae) Califórnia
P. smilacis Bose & Müll 1 - - - Smilax aspera (Smilacaceae) Índia
Phillipsiella sp. UB 10041 9
230-600 x 45-70
25-40 x 10-15 10-17 x 4-6 Qualea sp.(Vochysiaceae) Brasil
Phillipsiella sp. CMEPH M39 823B
212-427 [237] x 214-408 [356,5]
23,5-43 [32,5] x 7-12 [9,5]
6,5-9,5 [7] x 2-4 [2,5]
Ocotea paniculata (Lauraceae) Brasil
* Espécie tipo 1 Bose & Müll (1965),
2 Müller & Arx (1962),
3 Katum (1986),
4 Bez. & Por. (1969),
5 (Gonz. Frag. & Cif.) Ros. & Mouch (1994),
6 Barr (1993),
7 Sacc. & Trotter (1913),
8 Phill & Harkn (1884) e
9 Dornelo-Silva (1999).
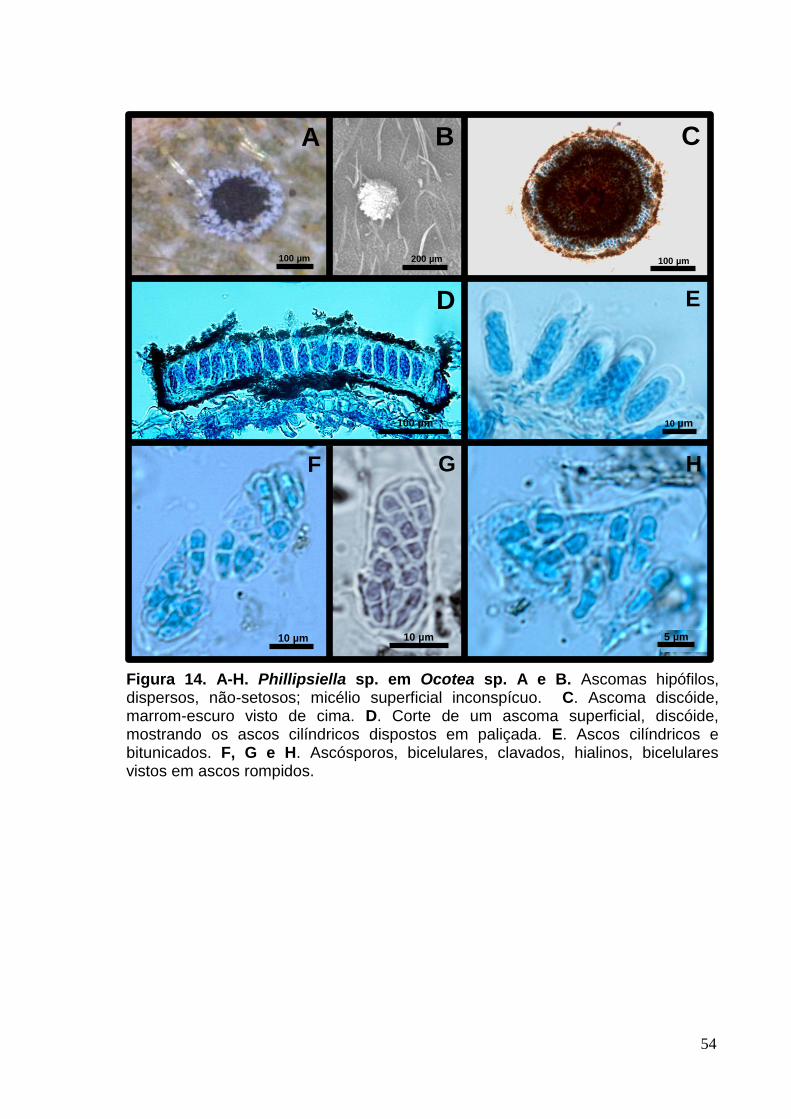
54
100 µm 10 µm
A
E
F G
200 µm100 µm 100 µm
10 µm10 µm 5 µm
B C
D
H
Figura 14. A-H. Phillipsiella sp. em Ocotea sp. A e B. Ascomas hipófilos, dispersos, não-setosos; micélio superficial inconspícuo. C. Ascoma discóide, marrom-escuro visto de cima. D. Corte de um ascoma superficial, discóide, mostrando os ascos cilíndricos dispostos em paliçada. E. Ascos cilíndricos e bitunicados. F, G e H. Ascósporos, bicelulares, clavados, hialinos, bicelulares vistos em ascos rompidos.

55
3.8 Phyllachora sp. (provável espécie nova nº 10)
Teleomorfo - Colônias hipófilas e dispersas, parede negra e brilhante. (Fig.15.
A e B). Ascomas 101-420 [250] µm, multiloculares, imersos, subepidérmicos,
periformes (Fig.15. C, D e E). Ostíolo conspícuo (Fig.15. D e E). Clípeo pouco
desenvolvido (Fig.15. C, D e E). Perífises 1-2,5 [1,5] µm, hialinas e não ramificadas
(Fig.15. F e G). Ascos 35-70 [56] x 7-11 [11] µm, numerosos, cilíndricos,
unitunicados contendo oito esporos (Fig.15. F). Paráfises 1,5-3 [2] µm, hialinas e não
ramificadas (Fig.15. F). Ascósporos 12-25 [22] x 2-5,5 [4] µm, hialinos, fusóides,
amerosseptados e gutulados (Fig.15. F e G). Anamorfo - Colônias hipófilas e
dispersas, parede negra e brilhante (Fig.15. A e B). Conidioma picnidial 174-232
[185] x 107-163 [156,5] µm, imersos, subepidérmicos. Ostíolo conspícuo (Fig.15. C).
Células conidiogênicas, fialídicas e hialinas. Conídios 4-7 [6] x 1-2 [2] µm, gutiformes
a fusóides, hialinos, amerosseptados e gutulados.
Espécime examinado: Brasil. Em folhas de Ilex affinis (Verbenaceae), 06 Jun.
1962, leg. E.P. Heringer, holótipo (CMEPH M39 808).
A Família Phyllachoraceae é caracterizada por possuir estroma bem
desenvolvido, imerso no tecido do hospedeiro, geralmente com a presença de
clípeo, negros e brilhantes ou raramente coloridos, contendo de um a vários
ascomas. Ascoma peritecial, parede fina, ostíolo composto por perífises. Perídio
composto de tecido escuro ou sem cor. Hamatécio constituído por paráfises,
algumas vezes deliqüescentes. Ascos cilíndricos, persistentes. Ascósporos
frequentemente hialinos podendo ser marrons, amerosseptados, raramente
ornamentados, muitas vezes com uma bainha mucilaginosa. Possuem sua fase
anamórfica em celomicetos. Conidioma usualmente formados em lóculos dentro do
estroma. Conidioma varia bastante quanto à forma por ser frequentemente bastante
achatado ou comprimido por ascomas adjacentes, ocasionalmente são picnidiais, as
paredes do conidioma são frequentemente indiferenciadas. Células conidiogênicas
são cilíndricas, frequentemente aglomeradas em pequenos conidióforos com
proliferação percorrente. Conídios pequenos, asseptados, hialinos, filiformes (geralmente
curvados) ou bacilar em geral com função espermacial (Cannon & Kirk, 2007).

56
O gênero Phyllachora tem como característica principal a presença de clípeo
e sintoma semelhante à mancha de “pixe”, brilhante e negra. Possui
aproximadamente 1504 registros entre espécies, sub-espécies e variedades,
ocorrentes em hospedeiras diversas (Kirk, 2011)
As espécies do gênero são determinadas de acordo com o hospedeiro devido
a sua especificidade. Sendo assim, por se tratar do primeiro relato no gênero Ilex, o
espécime em questão trata-se de uma provável nova espécie.
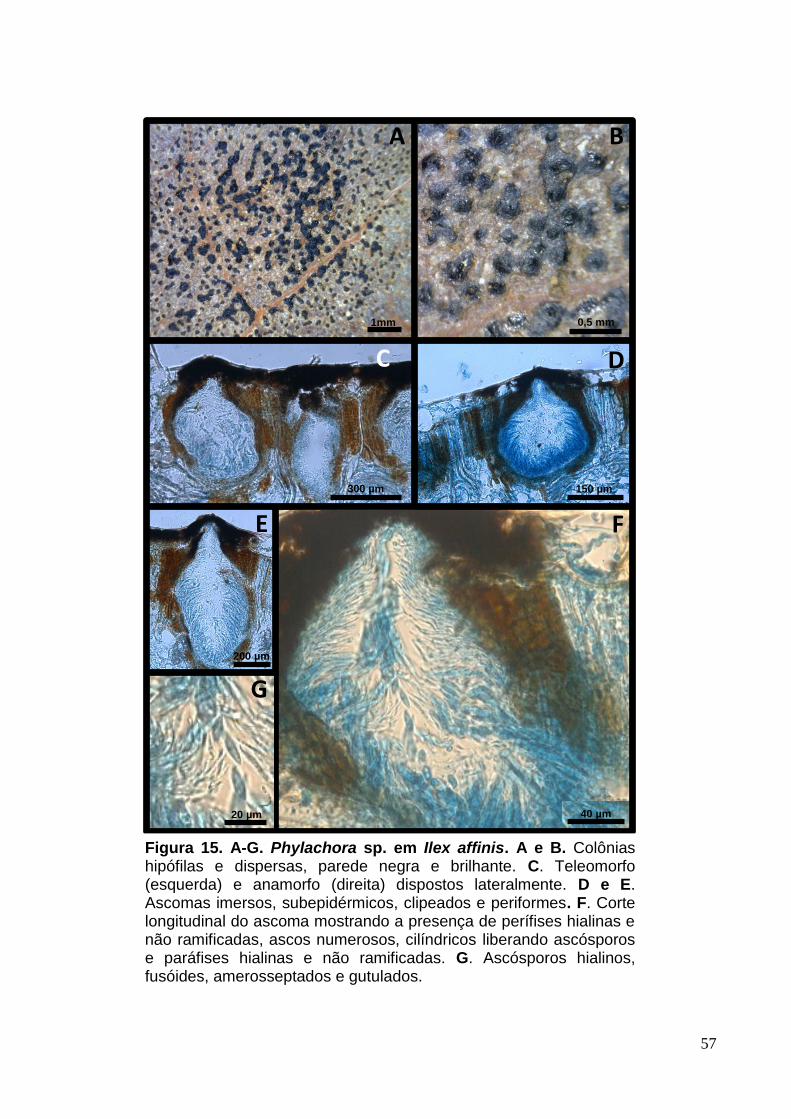
57
C
A B
FE
D
1mm 0,5 mm
300 µm 150 µm
G
40 µm20 µm
200 µm
Figura 15. A-G. Phylachora sp. em Ilex affinis. A e B. Colônias hipófilas e dispersas, parede negra e brilhante. C. Teleomorfo (esquerda) e anamorfo (direita) dispostos lateralmente. D e E. Ascomas imersos, subepidérmicos, clipeados e periformes. F. Corte longitudinal do ascoma mostrando a presença de perífises hialinas e não ramificadas, ascos numerosos, cilíndricos liberando ascósporos e paráfises hialinas e não ramificadas. G. Ascósporos hialinos, fusóides, amerosseptados e gutulados.

58
3.9 Stomatogene sp. (provável espécie nova nº 11)
Colônias hipófilas esparsas, marrons escuras (Fig.16. A). Ascomas 67-101
[90] µm de diâmetro, multiloculares, estromáticos (5-6 lóculos por ascoma)
desenvolvido sobre estomatopódio, marrons escuros, parede superior com textura
angularis (Fig.16. B, D e E). Estomatopódio escuro composto por células
pseudoparênquimatosas (Fig.16. E). Ascos 6-11 [10] x 30-40 [34] µm bitunicados,
inicialmente clavados tornando-se alargados na porção mediana, pedicelados, 8
esporos (Fig.16. F e G). Ascósporos 9,5-14 [10] x 3-4 [3] µm, clavados, hialinos,
bicelulares, célula apical 4,5-6,5 [5] x 2,5-4,5 [3] μm e célula basal 4,5-8,5 [6,5] x 1,5-
3 [2] μm, septo no terço superior, envolvidos por bainha mucilaginosa (Fig.16. H e I).
Espécime examinado: Brasil. Distrito Federal, Brasília, Embrapa Recursos
Genéticos e Biotecnologia em folhas de Myrsine guianensis (=Rapanea guianensis)
(Myrsinaceae), 20 Ago 1994, leg. T.S.L. Barros e J.C. Dianese, holótipo (UB Micol.
Coll. 06915).
O gênero Stomatogene foi estabelecido por Theissen baseado em Asterina
agaves Ellis & Everth (Ellis & Everth, 1990) que mostrou micélio hifopodiado, no
entanto, a característica principal do gênero foram os ascomas estromáticos que
apresentavam desenvolvimento sobre estomatopódios (Theissen, 1918).
Segundo Barr (1987), o gênero Stomatogene Theiss. pertence à família
Pseudosphaericeae que agrupa fungos com ascomas imersos, irrompentes ou
superficiais, minúsculos a pequenos, globosos ou esféricos, com hifas penetrando a
cavidade estomatal e o tecido foliar. Ascos em forma de saco, ovóides,
relativamente poucos, formados na camada basal do ascoma. Resíduos de tecido
intersticial frequentemente permanecem entre e sobre os ascos. Ascósporos
hialinos, amarelados ou marrom-claros, raramente marrom-escuros, obovóides ou
fusóides alongados, um ou vários septos ou ainda muriformes, envolvidos por uma
capa gelatinosa, com o tempo a parede torna-se verrugosa.
Outros autores alocaram o gênero em famílias diferentes. Segundo Müller &
Arx (1962) o gênero Stomatogene Theiss. faz parte da família Perisporiopsidaceae
(Dothiorales); Lutrell (1973) considera o gênero como pertencente à família
Parodiopsidaceae (Dothideales); Arx & Müller (1975) colocou o gênero na família

59
Parodiellinaceae. Barr (1987) manteve o gênero em Pseudosphaeriaceae pela
similaridade do perídio e do tecido intersticial com os outros membros da família.
Lumbsch & Huhndorf (2010) acomodaram o gênero Stomatogene na família
Parodiopsidaceae, concordando com Luttrell (1973).
Atualmente são aceitas três espécies de Stomatogene: S. yuccae Hansford
(espécie tipo) encontrado em Yucca sp. (Agavaceae), S. agaves (Ellis & Everth.)
Theiss, em Agave sp. (Agavaceae), e S. lycopodii M.L. Farr em Lycopodium
cernuum pertencente à família Lycopodiaceae (Kirk, 2011).
Nota-se que duas espécies neste gênero foram descritas em Agavaceae, a
hospedeira do espécime em estudo é uma espécie endêmica do Cerrado, membro
da família Myrsinaceae, uma família de dicotiledônea filogeneticamente distante da
família Agavaceae, e mais ainda da outra hospedeira, Lycopodium sp. (família
Lycopodiaceae), um licopódio, planta vascular primitiva que se reproduz por meio de
esporos.
O espécime em estudo, embora mostrando ascoma com diâmetro semelhante
aos de S. agaves, ainda difere de ambas as espécies conhecidas em Agavaceae por
ter ascos (30-40 [34] x 6-11 [10] µm), e ascósporos (9,5-14 [10] x 3-4 [3] µm)
significativamente menores, quando comparados com S. agaves (ascos: 50-60 x 20
µm; ascósporos: 15-27 x 6-8 µm), e S. yuccae (ascos: 30-60 µm; ascósporos 30-35
x 8-10 µm) [Tabela 10].
Finalmente, o espécime é facilmente separado de S. lycopodii, pois este
possui ascósporos marrons. Além disso, o espécime foi encontrado sobre uma
Myrsinaceae. Mediante as evidências discutidas, o espécime em estudo será
preparado para publicação como uma espécie nova conforme as normas do Código
Internacional de Nomenclatura para algas, fungos e plantas (ICN).
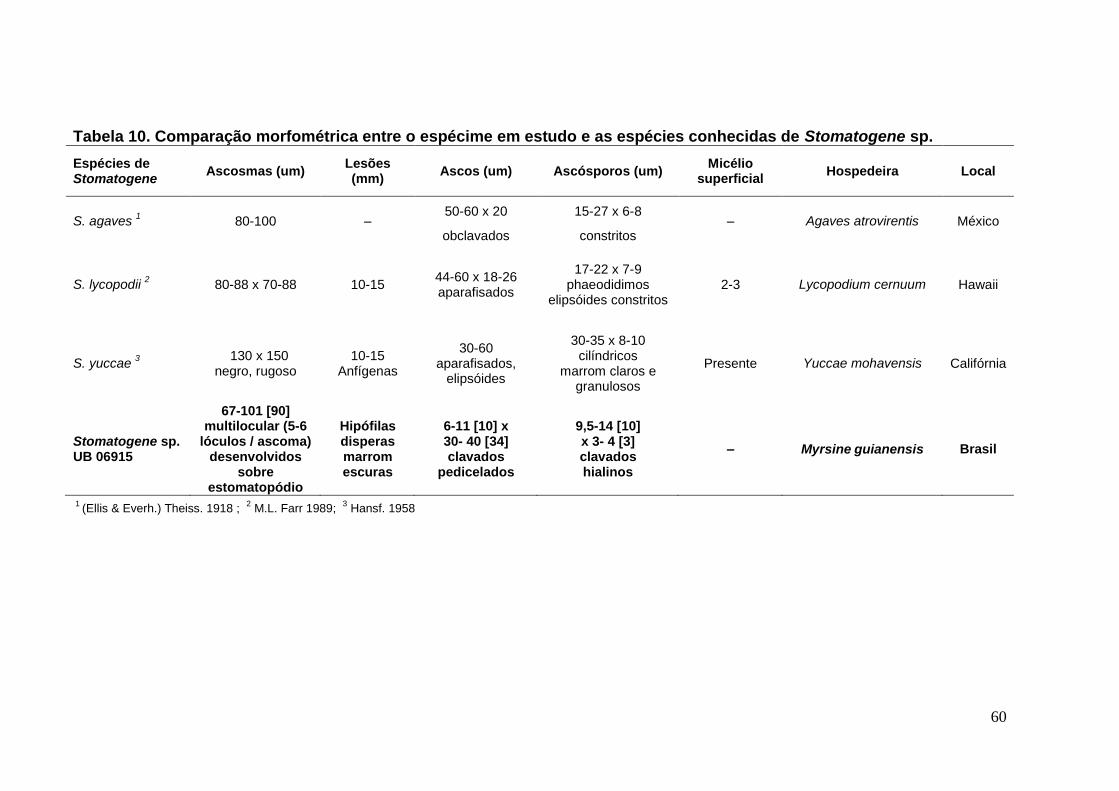
60
Tabela 10. Comparação morfométrica entre o espécime em estudo e as espécies conhecidas de Stomatogene sp.
Espécies de Stomatogene
Ascosmas (um) Lesões (mm)
Ascos (um) Ascósporos (um) Micélio
superficial Hospedeira Local
S. agaves 1 80-100 –
50-60 x 20 15-27 x 6-8 – Agaves atrovirentis México
obclavados constritos
S. lycopodii 2 80-88 x 70-88 10-15
44-60 x 18-26 aparafisados
17-22 x 7-9 phaeodidimos
elipsóides constritos 2-3 Lycopodium cernuum Hawaii
S. yuccae 3
130 x 150 negro, rugoso
10-15 Anfígenas
30-60 aparafisados,
elipsóides
30-35 x 8-10 cilíndricos
marrom claros e granulosos
Presente Yuccae mohavensis Califórnia
Stomatogene sp. UB 06915
67-101 [90] multilocular (5-6
lóculos / ascoma) desenvolvidos
sobre estomatopódio
Hipófilas disperas marrom escuras
6-11 [10] x 30- 40 [34] clavados
pedicelados
9,5-14 [10] x 3- 4 [3] clavados hialinos
– Myrsine guianensis Brasil
1 (Ellis & Everh.) Theiss. 1918 ;
2 M.L. Farr 1989;
3 Hansf. 1958
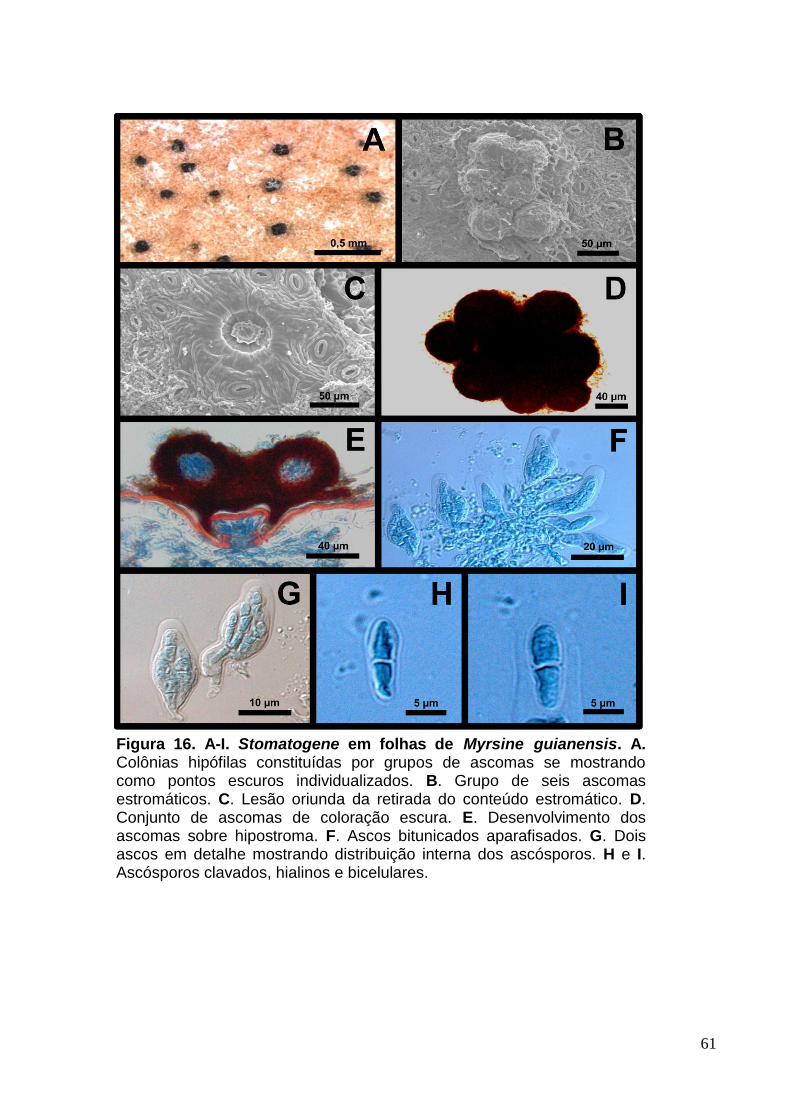
61
Figura 16. A-I. Stomatogene em folhas de Myrsine guianensis. A. Colônias hipófilas constituídas por grupos de ascomas se mostrando como pontos escuros individualizados. B. Grupo de seis ascomas estromáticos. C. Lesão oriunda da retirada do conteúdo estromático. D. Conjunto de ascomas de coloração escura. E. Desenvolvimento dos ascomas sobre hipostroma. F. Ascos bitunicados aparafisados. G. Dois ascos em detalhe mostrando distribuição interna dos ascósporos. H e I. Ascósporos clavados, hialinos e bicelulares.

62
3.10 Stomiopeltis sp. (provável espécie nova nº 12)
Colônias hipófilas, gregárias, marrom-escuras a pretas (Fig.17. A). Micélio
superficial, hipófilo, glabro, constituído de hifas marrons, não hifopodiadas,
flexuosas, ramificadas, septadas, apresentando uma leve constrição. Ascomas 70-
129 [106] × 15-27 [15] µm, superficiais, glabros, dispersos, plano-orbiculares, parede
de textura epidermoidea, parede basal inconspícua (Fig.17. B e C). Ascos 23,5-33
[33] × 6,5-9 [8,5] µm, cilíndrico-clavados, pedicelados, bitunicados, contendo oito
ascósporos (Fig.17. D e E). Paráfises 0,5-1,5 [1] µm de diâmetro, filiformes, hialinas,
septadas e ramificadas. Ascósporos 5,5-10,5 [7] × 2-3 [2] µm, clavados, hialinos,
bicelulares, sem constrição nos septos (Fig.17. D e E).
Espécime examinado: Brasil. Distrito Federal, Brasília, em folhas de Ilex affinis
(Aquifoliaceae), 06 Jun 1962, leg. E. P. Heringer (CMEPH M39 808).
O gênero Stomiopeltis foi estabelecido por Theissen em 1914, baseado em
uma espécie encontrada em uma laurácea. Posteriormente o gênero foi revisado por
Luttrell em 1946, que acrescentou uma espécie com peritécios multiloculares (S.
polyloculata), ampliando-se assim o conceito do gênero.
De acordo com Batista (1959) o gênero é alocado em Micropeltidaceae. A
família Micropeltidaceae é composta por fungos que apresentam micélio
parcialmente superficial. Ascomas escutelares, ostiolados; perídio composto de uma
ou várias camadas de células pseudoparenquimáticas, algumas vezes
epidermóideas. Tecido interascal composto de pseudoparáfises. Ascos clavados,
fissitunicados. Ascósporos hialinos ou marron-claros, com septos transversais a
muriformes, algumas vezes atenuados na base (Kirk et al., 2001).
Batista considerou quatro subfamílias para a Família Micropeltidaceae:
Dictyopeltoideae, a Haplopeltoideae, a Gymnopeltoideae e a Stomiopeltoideae (com
Stomiopeltis como gênero tipo). Stomiopeltis é representado por membros que
apresentam micélio superficial, reticulado, marrom, ascomas superficiais, dimidiados,
escudados, uni ou multiloculares, textura meandriformis, ascos bitunicados e
ascósporos bicelulares e hialinos.
Arx & Müller (1975), Barr (1987) e Ericksson (1981), também alocaram
Stomiopeltis em Micropeltidadeae, Dothideomycetidae (incert sedis). Esta

63
classificação foi aceita por Kirk et al. (2001) e atualmente por Lumbsch &
Huhndorf (2010).
O gênero Stomiopeltis possiu 47 espécies descritas (Kirk, 2011). Do total de
espécies descritas 12 delas ocorrem no Brasil. Ainda no país há o relato de
Stomiopeltis em Qualea sp. e em Salacia sp. (Dornelo-Silva, 1999; Santos, 2011).
Essas espécies incluem: Stomiopeltis allophyli, S. aspersa (espécie tipo), S. batistae,
S. cassiae, S. citri, S. japônica, S. minor, S. myrciae, S. rubi, S. suttoniae, S.
tephrosiae e S. tetrasperma (Farr & Rossman, 2011; Kirk, 2011).
Comparando-se o espécime estudado com as espécies descritas, esse
mostra seus ascomas semelhantes à espécie S.cassiae, apresenta dimensões
relativamente parecidas quanto aos ascos de S.citri e principalmente quanto aos
ascósporos, se aproxima da espécie S. minor (Tabela 11).
Quanto aos ascomas de S. minor (50-80 µm), esses são muito menores
quando comparados com o fungo em estudo.
Mediante tais diferenças infere-se que o espécime em estudo é uma provável
espécie nova. Além disso, este é o primeiro relato do gênero Stomiopeltis na família
Aquifoliaceae.
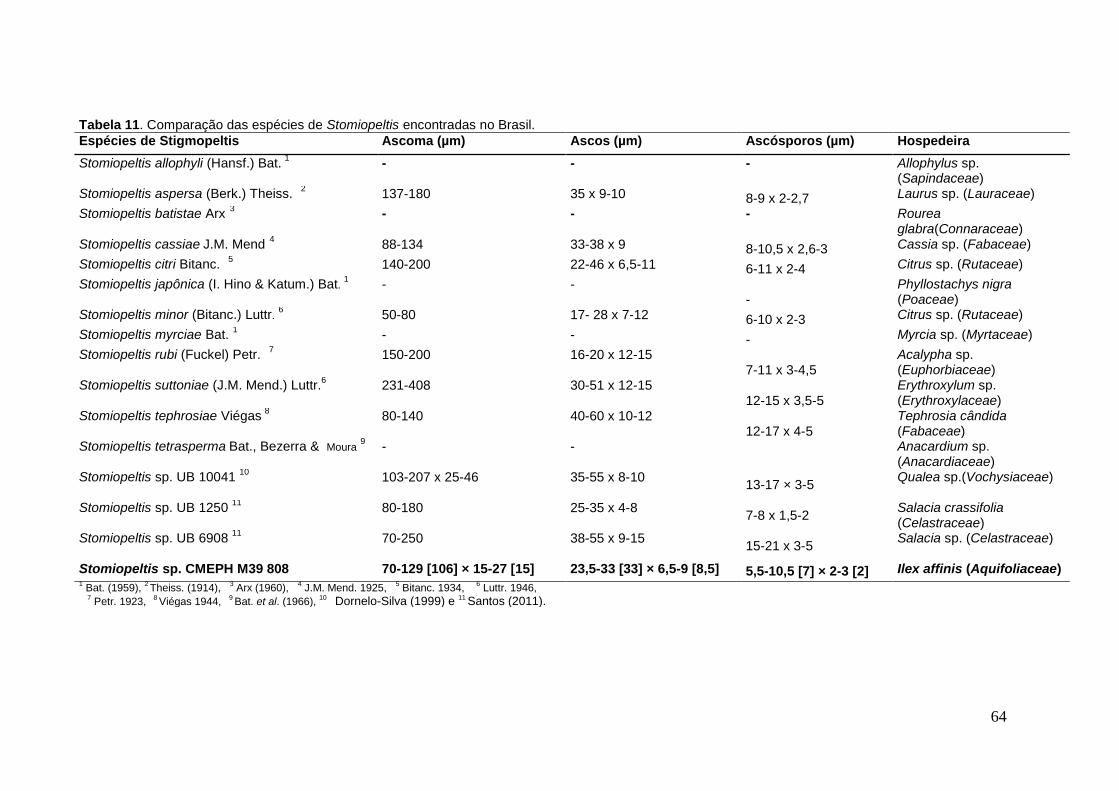
64
Tabela 11. Comparação das espécies de Stomiopeltis encontradas no Brasil. Espécies de Stigmopeltis Ascoma (µm) Ascos (µm) Ascósporos (µm) Hospedeira
Stomiopeltis allophyli (Hansf.) Bat. 1
- - - Allophylus sp. (Sapindaceae)
Stomiopeltis aspersa (Berk.) Theiss. 2 137-180 35 x 9-10 8-9 x 2-2,7 Laurus sp. (Lauraceae)
Stomiopeltis batistae Arx 3 - - - Rourea
glabra(Connaraceae) Stomiopeltis cassiae J.M. Mend 4
88-134 33-38 x 9 8-10,5 x 2,6-3 Cassia sp. (Fabaceae)
Stomiopeltis citri Bitanc. 5 140-200 22-46 x 6,5-11 6-11 x 2-4 Citrus sp. (Rutaceae)
Stomiopeltis japônica (I. Hino & Katum.) Bat. 1 - -
- Phyllostachys nigra (Poaceae)
Stomiopeltis minor (Bitanc.) Luttr. 6 50-80 17- 28 x 7-12 6-10 x 2-3 Citrus sp. (Rutaceae)
Stomiopeltis myrciae Bat. 1 - - - Myrcia sp. (Myrtaceae)
Stomiopeltis rubi (Fuckel) Petr. 7 150-200 16-20 x 12-15
7-11 x 3-4,5 Acalypha sp. (Euphorbiaceae)
Stomiopeltis suttoniae (J.M. Mend.) Luttr.6 231-408 30-51 x 12-15
12-15 x 3,5-5 Erythroxylum sp. (Erythroxylaceae)
Stomiopeltis tephrosiae Viégas 8 80-140 40-60 x 10-12
12-17 x 4-5 Tephrosia cândida (Fabaceae)
Stomiopeltis tetrasperma Bat., Bezerra & Moura 9 - -
Anacardium sp. (Anacardiaceae)
Stomiopeltis sp. UB 10041 10
103-207 x 25-46 35-55 x 8-10
13-17 × 3-5 Qualea sp.(Vochysiaceae)
Stomiopeltis sp. UB 1250 11
80-180 25-35 x 4-8
7-8 x 1,5-2 Salacia crassifolia (Celastraceae)
Stomiopeltis sp. UB 6908 11
70-250 38-55 x 9-15
15-21 x 3-5 Salacia sp. (Celastraceae)
Stomiopeltis sp. CMEPH M39 808 70-129 [106] × 15-27 [15] 23,5-33 [33] × 6,5-9 [8,5] 5,5-10,5 [7] × 2-3 [2] Ilex affinis (Aquifoliaceae) 1 Bat. (1959),
2 Theiss. (1914),
3 Arx (1960),
4 J.M. Mend. 1925,
5 Bitanc. 1934,
6 Luttr. 1946,
7 Petr. 1923,
8 Viégas 1944,
9 Bat. et al. (1966),
10 Dornelo-Silva (1999) e 11 Santos (2011).

65
A B
ED
1 mm 20 µm
10 µm 10 µm
40 µm
C
Figura 17. A-F. Stomiopeltis sp. em folhas de Ilex affinis. A. Colônia hipófila, composta por ascomas dispersos. B. Parede do ascoma com textura epidermoidea. C. Corte de ascoma superficial, com parede basal inconspícua. D e E. Ascos cilíndrico-clavados, pedicelados, bitunicados, contendo ascósporos clavados, hialinos, bicelulares, sem constrição nos septos.

66
3.11 Vonarxella sp. (provável espécie nova nº 13)
Colônias hipófilas, esparsas, cobrindo grande parte do limbo foliar (Fig. 18. A).
Micélio superficial marrom-claro que ocorre principalmente em torno e sob o ascoma
(Fig. 18. B e C). Ascomas 183-248 [215] x 160-290 [186] μm de diâmetro,
superficiais, marrom-escuros, discóides orbiculares com a superfície irregular,
apotecióides, levemente convexos, sobre subículo com hifas hialinas (Fig. 18. B e
C). Ascos 28-53 [50] x 11-25 [11,5] μm, obovóides a clavados, bitunicados,
pedicelados, com 8 esporos (Fig. 18. D e E). Ascósporos 12-21 [16,5] x 4-6,5 [5] μm,
clavados, septados, bicelulares (célula apical 7-9 [7,5] x 4-7 [6] μm e célula basal 7-
10,5 [7,5] x 3,5-6 [5,5] μm), hialinos e constritos no septo (Fig. 18. F e G).
Espécime examinado: Brasil. Distrito Federal, Brasília, Jardim Botânico de Brasília
em folhas de Ocotea sp. (Myrsinaceae), 20 Ago 1994; leg. M.L. Nishijima e J.C.
Dianese, holótipo (UB Micol. Coll. 06902).
Segundo Batista et al., (1965) o gênero Vonarxella é caracterizado por
apresentar subículo peliculoso, marrom, com hifas pouco ramificadas, septadas.
Apotécios elevados sobre subículo, superficiais, plano-orbiculares, glabros, marrom-
escuros, membranosos, discóides, planos, não pruinosos, excípula prosenquimática,
epitécio marrom, hipotécio marrom e himênio hialino. Ascos bitunicados,
inoperculados, com 8 esporos, subsésseis, parafisados. Ascósporos oblongos,
bicelulares e hialinos.
O espécime estudado pertence ao gênero Vonarxella Bat. por apresentar
características morfológicas próximas, porém pouco definidas com relação a forma
do excípulo, do epitécio e do hipotécio, como na descrição da espécie tipo.
Trata-se de um gênero monotípico tendo como espécie tipo V. dipterygis Bat.
Peres & Bez. encontrada sobre folhas de Dipteryx alata Vog. (Fabaceae).
A espécie tipo foi determinada como V. dipterygis e possui subículo miceliano,
delgado, glabro, peliculoso, formado de hifas flexuosas, marrom septadas, pouco
constritas, reticuladas, 2-3-5 μm de diâmetro. Apotécios superficiais, elevados sobre
o subículo, glabros, membranosos, plano orbiculares, levemente constritos, na base,
185-420 μm de diâmetro e 63-77 μm de altura, sobre hifas suportes bem
características, de ápice dilatado, marrom–escuros, 4-5 μm de diâmetro, bordos

67
abruptos, discóides, planos, não pruinosos, excípulo prosenquimático, 7-17 μm de
espessura; epitécio marrom, 5-10,5 μm de espessura, hipotécio marrom,
prosenquimático, 9-13 μm de espessura. Himênio basal, plano, hialino, 245-385 μm
de diâmetro e 47,55 μm de altura. Ascos oblongos – clavados, bitunicados,
inoperculados, 8 esporos, subsésseis, paralelos, 28-43 x 10-13 μm, paráfises
hialinas, filiformes, septadas e ramificadas, 1-1,5 μm de diâmetro. Ascósporos
oblongos, hialinos, bicelulares, pouco constritos, polísticos de pólos rotundos, 10-13
x 3.5-4 μm.
O espécime diferencia- se de V. dipterygis por apresentar subículo com hifas
hialinas; ascomas orbiculares, levemente convexos e de menor diâmetro; ascos
mais largos; ascósporos maiores e mais largos (Tabela 12).
O espécime-tipo de V. dipterygis, depositado no Instituto de Micologia de
Recife em Pernambuco, foi examinado e nele não existe mais vestígio do fungo.
Diante disso será possível ampliar o conceito do gênero incluindo a característica
hialina das hifas e neotipificar o gênero usando como espécime-tipo o material ora
estudado.
Mediante as evidências discutidas e por ser o primeiro relato do gênero na
família Lauraceae, propõe-se a criação de um novo táxon específico para acomodar
o espécime, que será devidamente documentado e submetido à publicação.

68
1 Bat., J.L. Bezerra & Peres 1965
Tabela 12. Comparação entre a espécie monotípica de Vonarxella e o espécime encontrado em Ocotea sp.
Espécies de Vonarxella
Ascomas (µm) Colônias Ascos (µm) Ascósporos
(µm) Hospedeira Local
V. dipterygis 1 185-420 x 63-77 –
28-43 x 10-13 oblongos clavados
bitunicados
10-13 x 3,5-4 bicelulares oblongos
pouco constritos hialinos
Dipteryx alata Brasil
Vonarxella sp. UB 06902
183-248 [215] x 160-290 [186]
Hipófilas dispersas
28-53 [50] x 11-25 [11,5] obovóides a
clavados pedicelados
12-21 [16,5] x 4-6,5 [5]
bicelulares clavados
constritos hialinos
Ocotea sp. Brasil
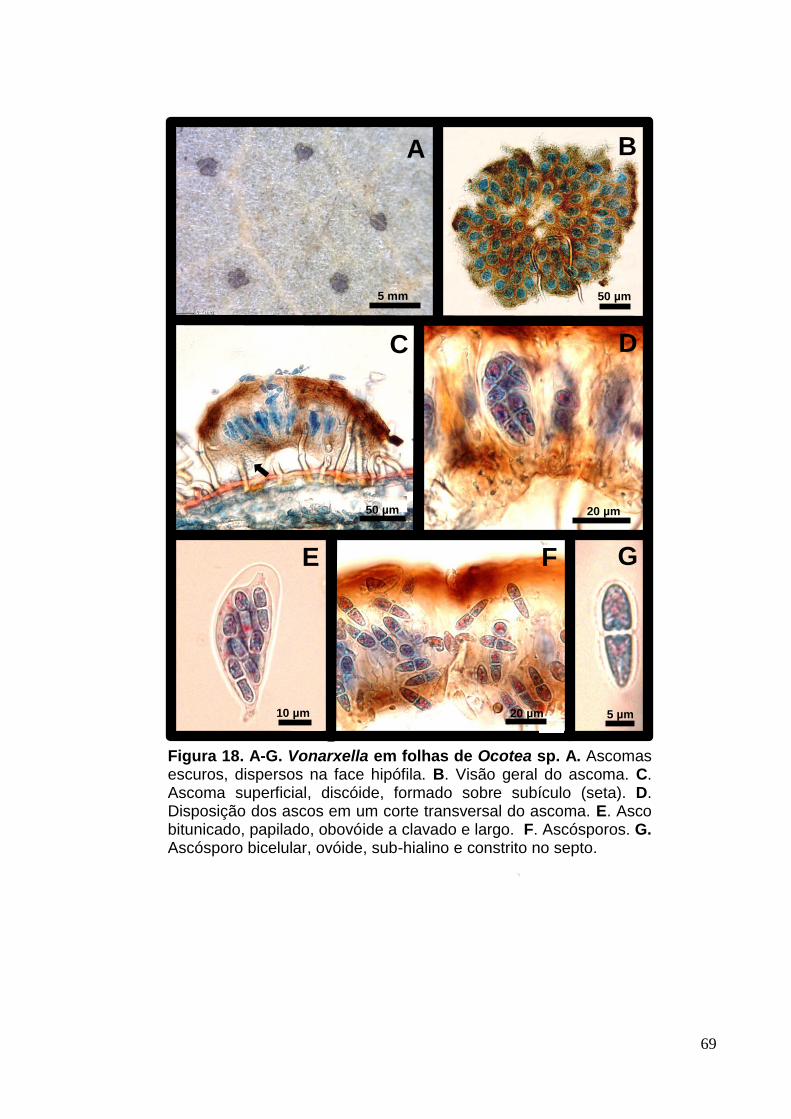
69
BA
DC
GFE
5 mm
5 µm
50 µm 20 µm
50 µm
10 µm 20 µm
Figura 18. A-G. Vonarxella em folhas de Ocotea sp. A. Ascomas escuros, dispersos na face hipófila. B. Visão geral do ascoma. C. Ascoma superficial, discóide, formado sobre subículo (seta). D. Disposição dos ascos em um corte transversal do ascoma. E. Asco bitunicado, papilado, obovóide a clavado e largo. F. Ascósporos. G. Ascósporo bicelular, ovóide, sub-hialino e constrito no septo.

70
3.12 Ascomiceto (provável gênero novo nº 2)
Colônias efusas desenvolvendo-se superficialmente na face abaxial da folha e
envolvendo o tricoma em toda a sua extensão, a partir de micélio disperso na
superfície do limbo foliar (Fig.19. A). Micélio superficial, semi-conspícuo, hialino a
marrom-claro, crescendo nos tricomas a partir do micélio superficial (Fig.19. C).
Hifas septadas, hialinas a marrom-claras e se aglomeram no ápice, ocasionalmente,
na região mediana ou basal do tricoma para formação dos ascomas (Fig.19. B).
Ascomas 71-95 [88,5] × 59-88 [80] µm, distribuídos ao longo dos tricomas mas
predominantemente nas pontas, globosos, ostiolados, parede de textura
epidermoidea, setosos (Fig.19. B e D). Setas septadas, eretas, rígidas, afiladas a
pontiagudas (Fig.19. B e E). Ascos 43,5-93 [60,5] x 9,5-20 [15] µm, bitunicados,
clavados a cilíndricos, aparafisados, 8 esporos (Fig.19. F). Ascósporos 17-26 [22,5]
x 3-6 [4,5] µm bicelulares, hialinos, clavados (Fig.19. G).
3.13 Ascomiceto (provável gênero novo nº 3)
Colônias efusas desenvolvendo-se superficialmente na face abaxial da folha e
envolvendo o tricoma em toda a sua extensão (Fig. 20. A e B), a partir de micélio
disperso na superfície do limbo foliar (Fig. 20. C). Hifas septadas, hialinas a marrom-
claras e se aglomeram no ápice, ocasionalmente, na região mediana ou basal do
tricoma para formação dos ascomas (Fig. 20. D e E). Ascomas 125-179 [146] x 121-
157 [149] µm, ovóides, ostiolados, paredes rígidas, localizados em sua maioria nas
extremidades dos tricomas, setosos (Fig. 20. D e E). Setas 18-69 [58,5] x 2-5 [3,5]
µm, fortemente recurvadas, uncinadas, rígidas, septadas, afinando em direção ao
ápice, cobrindo completamente a superfície do ascoma (Fig. 20. D e F). Ascos 58,5-
82 [71,5] x 10,5-21 [15,5] µm, bitunicados, abundantes, cilíndrico-elipsóides,
aparafisados, 8 esporos (Fig.20. G e H). Ascósporos 18-29 [22,5] x 4-6 [5] µm,
bicelulares, hialinos, cilíndricos a clavados (Fig.20. I e J).
Espécimes examinados: Brasil. Distrito Federal, Brasília. Em folhas de membro da
família Lauraceae, leg. E.P. Heringer, holótipo (CMEPH M24 1756).

71
Estudos preliminares sugerem que os espécimes (mencionados acima como
prováveis gêneros novos números 2 e 3) pertencem a gêneros novos pertencentes à
Classe Dothideomycetes.
A Classe Dothideomycetes acomoda gêneros com ascos bitunicados, com
ascomas de formato variável, podendo ser apoteciais, periteciais e cleistoteciais,
formado como lóculos dentro de um tecido estromático, himenio às vezes gelatinoso.
Tecido interascal geralmente presente, usualmente composto por parafisóides ou
pseudoparáfises ramificadas ou anastomosadas, pelo menos inicialmente,
conectadas em ambas as partes basal e apical. Ascos geralmente clavados ou
cilíndricos, de parede espessa, fissitunicados, raramente com estruturas apicais.
Ascósporos quase sempre septados, constritos ou não no primeiro septo, às vezes
muriformes, hialinos ou escuros, muitas vezes não ornamentados (Kirk et al., 2008)
Segundo Kirk et al. (2008) dentro desta Classe estão duas Subclasses:
Dothideomycetidae e Pleosporomycetidae.
Dothideomycetidae compõe três Ordens: Capnodiales, Myriangiales e
Dothideales (Kirk et al., 2008).
A Ordem Capnodiales abriga membros com micélio superficial bem
desenvolvido e escuro. Ascomas globosos ou elongados, setosos, ou com
apêndices. Apêndices ausentes ou inconspícuos (Kirk et al., 2008).
Membros que apresentam, em termos gerais, estroma crustoso ou pulvinado
de coloração hialina ou escura e parede espessa, imersos, pertencem a Ordem
Myriangiales (Kirk et al., 2008).
Já a Ordem Dothideales é representada por ascomicetos que possuem
ascomas periteciais, lóculos estromáticos ou não, escuros e com paráfises ausentes
(Arx & Müller, 1975; Kirk et al., 2008).
A Subclasse Pleosporomycetidae é composta por apenas uma Ordem a
Pleosporales. Esta ordem é definida por membros que raramente são contituídos por
estromas ou elementos filamentosos. Ascoma peritecial ou raramente cleistotecial,
às vezes clipeados, às vezes globosos, parede espessa, imersos ou erumpentes,
escuros, ostiolados, às vezes setosos ou com apêndices. Perídio usualmente
espesso composto por células pseudoparemquimatosas. Tecido interascal composto
por pseudoparáfises (Kirk et al. 2008).

72
Diante de tais definições, é coerente alocar os prováveis gêneros novos
propostos na Subclasse Dothideomycetidae, Ordem Dothideales, Família
Pseudoperisporiaceae.
Em Pseudoperisporiaceae incluem membros sapróbios ou biotróficos no
tecido da planta ou de outros fungos, possuem micélio superficial, ascomas
superficiais, periteciais, pequenos mais ou menos globosos, com ostíolo definido, às
vezes setosos, perídio de paredes finas compostas por células
pseudoparenquimatosas. Tecido interascal composto de pseudoparáfises
deliqüescentes na maturidade. Ascos sacados e fissitunicados. Ascósporos hialinos
a escuros de város formatos com septos transversais (Cannon & Kirk, 2007).
De acordo com Arx & Müller (1975), o gênero Wentiomyces, um membro
típico desta família, apresenta ascomas com dimensões menores do que 200 µm,
setosos, aparafisados, ostiolados e apendiculados. Características estas
semelhantes aos espécimes aqui estudados. A diferença encontrada em
Wentiomyces é que este possui, além de setas rígidas, apêndices bifurcados
dispostos no ascoma, possui hábito parasídico, contrapondo com os fungos em
questão em que um apresenta ascomas somente com setas rígidas e outro com
ascomas contendo apêndices sinuosos sem bifurcação, mas ambos com hábitos de
crescimento sobre tricomas. Para se estabelecer uma taxonomia correta e precisa
faz-se nessário recorrer a análises moleculares e filogenéticas que indique grau de
similaridade entre os organismos propostos.
Os dois espécimes aqui estudados são facilmente separados com base em
diferenças no formato e quantidade das setas bem como em diferenças
dimensionais, sendo cada um deles prováveis gêneros novos.
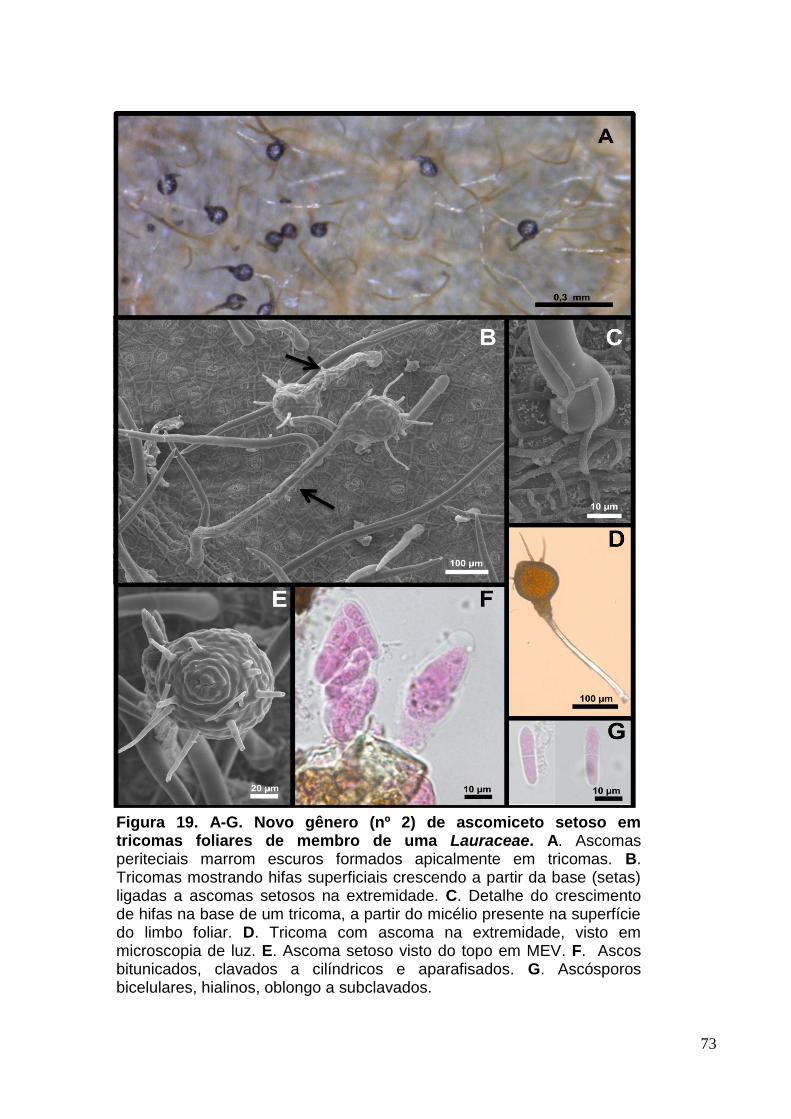
73
Figura 19. A-G. Novo gênero (nº 2) de ascomiceto setoso em tricomas foliares de membro de uma Lauraceae. A. Ascomas periteciais marrom escuros formados apicalmente em tricomas. B. Tricomas mostrando hifas superficiais crescendo a partir da base (setas) ligadas a ascomas setosos na extremidade. C. Detalhe do crescimento de hifas na base de um tricoma, a partir do micélio presente na superfície do limbo foliar. D. Tricoma com ascoma na extremidade, visto em microscopia de luz. E. Ascoma setoso visto do topo em MEV. F. Ascos bitunicados, clavados a cilíndricos e aparafisados. G. Ascósporos bicelulares, hialinos, oblongo a subclavados.
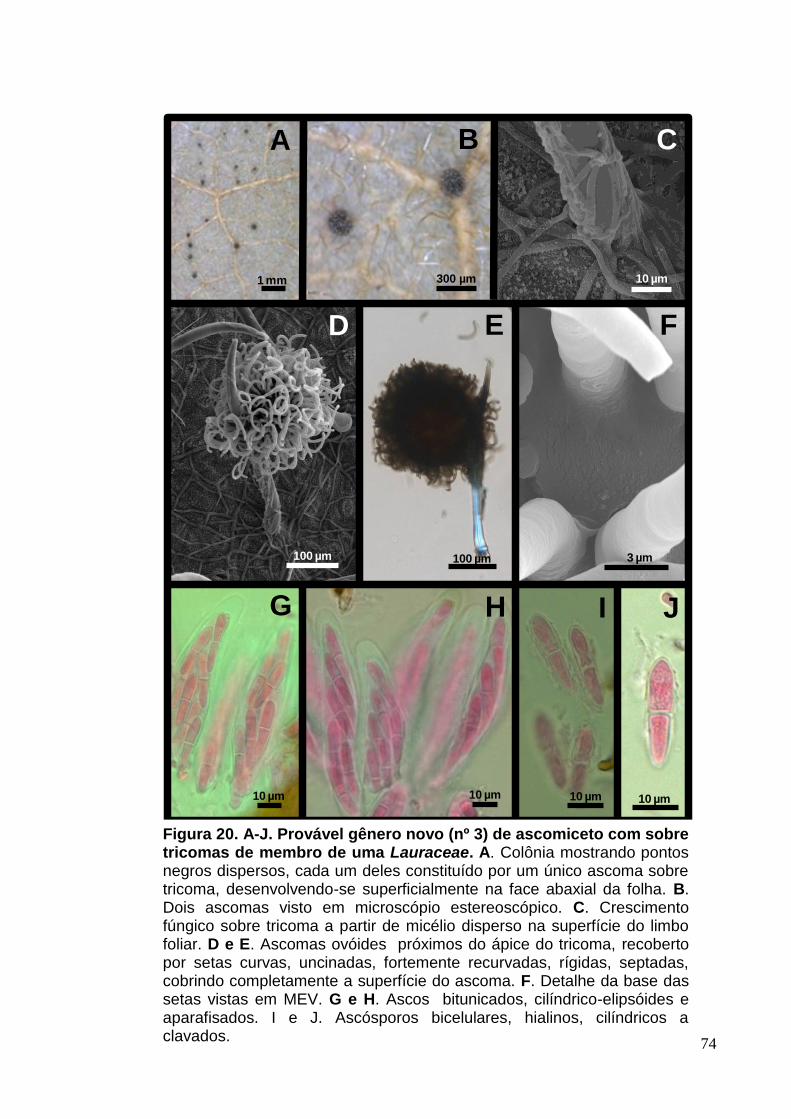
74
A
G H
F
10 µm10 µm10 µm 10 µm
100 µm 100 µm 3 µm
10 µm1 mm
ED
I J
B
300 µm
C
Figura 20. A-J. Provável gênero novo (nº 3) de ascomiceto com sobre tricomas de membro de uma Lauraceae. A. Colônia mostrando pontos negros dispersos, cada um deles constituído por um único ascoma sobre tricoma, desenvolvendo-se superficialmente na face abaxial da folha. B. Dois ascomas visto em microscópio estereoscópico. C. Crescimento fúngico sobre tricoma a partir de micélio disperso na superfície do limbo foliar. D e E. Ascomas ovóides próximos do ápice do tricoma, recoberto por setas curvas, uncinadas, fortemente recurvadas, rígidas, septadas, cobrindo completamente a superfície do ascoma. F. Detalhe da base das setas vistas em MEV. G e H. Ascos bitunicados, cilíndrico-elipsóides e aparafisados. I e J. Ascósporos bicelulares, hialinos, cilíndricos a clavados.

75
4 Basidiomiceto
4.1 Puccinia malvacearum Bertero ex Montagne
Espermogônias, picnios, écios e urédias ausentes. Télios 449-1640 [1300] x
296-441[408] µm, adaxiais, irrompentes, subepidérmicos, aparafisados, marrom-
claros a alaranjados, circulares e dispersas (Fig.21. A e B). Teliósporos 40-79 [54] x
15-23 [18] µm, bicelulares (ocasionalmente com mais de um septo), ligeira
constrição nos septos, elipsóides a oblongo-fusóides, parede lisa, marrom-claros,
célula apical geralmente papiladas, pedicelados (Fig.21. C-I). Pedicelos 69-157 [118]
µm hialinos, lisos e persistentes (Fig.21.D-I).
Espécime examinado: Brasil. Em folhas de Triumfetta romboidae (Malvaceae), leg.
E. P. Heringer, (CMEPH M16 463-39).
Vinte e seis espécies de Puccinia foram reportadas em membros da família
Malvaceae (Farr & Rossman, 2011). Apenas Puccinia heterospora ocorre em
Triumfetta sp. (Hennen et al., 2005). Nesta hospedeira também são registradas a
ocorrência de: Didymopsora triumfettae e Pucciniosira pallidula. O espécime
encontrado não pertence aos gêneros mencionados acima, pois em Didymopsora
sp. os teliósporos são bicelulares, oblongos, hialinos, e vagamente verrugosos e em
Pucciniosira sp. são restritamente unicelulares, ovóides a oblongos e ligeiramente
verrugosos.
De acordo com Hennen et al. (2005), a espécie Puccinia heterospora é
semelhante a várias espécies microcíclicas do gênero Puccinia em diferentes
hospedeiros da família Malvaceae.
Em 1982, Lindquist modificou uma chave para melhor alocar e identificar as
espécies encontradas em membros de Malvaceae. O espécime aqui descrito deve
ser alocado em P. malvacearum por possuir teliósporos 41-70 x 14-25 μm
bicelulares (ocasionalmente com dois septos) e com formato elipsóide a oblongo-
fusiformes (Lidquist, 1982). Puccinia heterospora se difere de P. malvacearum por
conter teliósporos unicelulares de formato estritamente elipsóide além de serem um
pouco mais escuros.

76
De acordo com Hennen et al. (2005), esta espécie já foi reportada em outras
hospedeiras da família Malvaceae, tais como: Alcea rosea, Anoda sp., Hibiscus
esculentos, Malva parviflora, Malva rotundifolia, Malva sylvestris, Malva sp.,
Malvastrum coromandelianum e Sida sp., todas no Brasil.
Este é o primeiro relato de P. malvacearum incidente em Triumfetta sp.
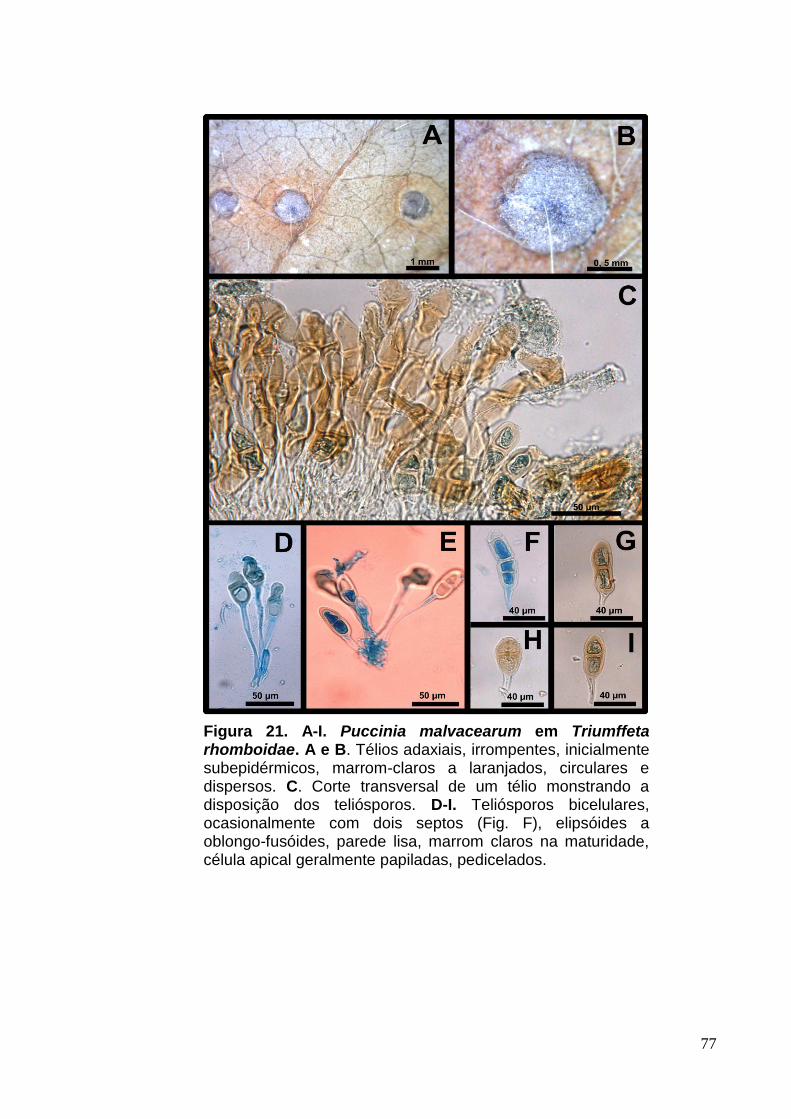
77
Figura 21. A-I. Puccinia malvacearum em Triumffeta rhomboidae. A e B. Télios adaxiais, irrompentes, inicialmente subepidérmicos, marrom-claros a laranjados, circulares e dispersos. C. Corte transversal de um télio monstrando a disposição dos teliósporos. D-I. Teliósporos bicelulares, ocasionalmente com dois septos (Fig. F), elipsóides a oblongo-fusóides, parede lisa, marrom claros na maturidade, célula apical geralmente papiladas, pedicelados.

78
CAPÍTULO II
FUNGOS EM PLANTAS EXÓTICAS PRESENTES NO CERRADO
1 Hifomicetos
1.1 Cercospora sp. (provável espécie nova nº 14)
Manchas foliares hipófilas, marrons com bordos escurecidos (Fig.22. A).
Conidióforos 91-154 [118] x 2,5-6,5 [4] fasciculados, simples, macronemáticos,
geniculados, simpodiais, lisos e marrons (Fig.22. B). Células conidiogênicas 9-56
[34] x 2,5-6 [4] poliblásticas, conspícuas, com cicatrizes planas (Fig.22. C).
Conídios 25-350 [122] x 2-4 solitários, aciculares, flexíveis, filiformes, curvados,
hialinos, lisos, septados (até 25 septos) e com base truncada (Fig.22. D).
Espécime examinado: Brasil. Distrito Federal, Brasília, Estação Biológica da
Universidade de Brasília em folhas de Piper nigrum (Piperaceae), 01 Nov 2009, leg.
R.C Pereira-Carvalho, holótipo (UB Micol. Coll. 21134).
1.2 Cercospora sp. (provável espécie nova nº 15)
Manchas foliares hipófilas, circulares ou irregulares, marrons com bordos
escurecidos arroxeados e centro claro de coloração acinzentada (Fig.23. A).
Conidióforos 79-223 x 3-4 fasciculados, simples, macronemáticos, geniculados,
simpodiais, lisos e marrons (Fig.23. B). Células conidiogênicas 1,5-3 poliblásticas,
com cicatrizes conspícuas e planas (Fig.23. C). Conídios 49-272 x 2-4 solitários,
aciculares, flexíveis, filiformes, curvados, hialinos, lisos, septados (até 25 septos) e
com base truncada (Fig.23. D).
Espécime examinado: Brasil. Distrito Federal, Planaltina, Embrapa Cerrados em
folhas de Jatropha curcas (Euphorbiaceae), 18 Set. 2009, leg. A.C Dianese, holótipo
(UB Micol. Coll. 21035).

79
Ambas as espécies estudadas, em pimenta do reino (Piper nigrum) e pinhão
manso (Jatropha curcas) são espécies recentemente detectadas no Cerrado, em
Brasília que, em campo produziram nas folhas das hospedeiras, manchas foliares
típicas de cercosporóides.
Os hifomicetos cercosporóides são anamorfos de espécies de ascomicetos da
família Mycosphaerellaceae. Esses fungos são causadores de manchas foliares ou
lesões necróticas, delimitadas por bordas geralmente marrons e centros
acinzentados, variando em tamanho e forma. Esses organismos também podem ser
encontrados em pecíolos, frutos e sementes (Chupp, 1954; Crous & Braun, 2003).
Dentre os cercosporóides, o gênero Passalora foi o primeiro a ser descrito,
entretanto o gênero Cercospora é o mais conhecido e com maior número de
espécies.
A classificação taxonômica dos fungos cercosporóides sofreu mudanças
recentes com o advento das técnicas moleculares. Assim, Crous & Braun (2003),
baseados em sequencias das regiões ITS1 e ITS2 do rDNA, designaram como
fungos cercosporóides verdadeiros apenas quatro gêneros: Pseudocercospora
Speg., Stenella Syd., Cercospora Fresen. e Passalora Fr. emend. Crous & Braun.
Em alguns casos, os gêneros tiveram seus conceitos ampliados passando a
incluir espécies até então pertencentes a outros gêneros, como no caso de
Passalora sp. que no novo conceito englobou os gêneros Mycovellosiella Rangel,
Phaeoramularia Munt.-Cvetk e parte do gênero Phaeoisariopsis Ferraris (Crous &
Braun, 2003).
Os espécimes ora estudados pertencem tipicamente ao gênero Cercospora
(Crous & Braun 2003).
A espécie encontrada em P. nigrum é claramente separada das outras
espécies de Cercospora anteriormente detectadas em hospedeiras do gênero Piper,
a saber: C. piperis-betle Sawada & Katsuki e C. piperata Asthana & Mahmud (Hsieh
& Goh, 1990; Chupp, 1954). A espécie em P. nigrum apresenta conidióforos e
conídios muito maiores quando comparados aos das outras duas espécies de
Cercospora presentes em Piper spp. (Tabela 13).
Já no caso de J. curcas, duas espécies de Cercospora são conhecidas,
Cercospora jatrophicola (Speg.) Chupp, e C. jatrophigena Braun (Chupp, 1954;
Braun, 2001).

80
A espécie C. jatrophicola apresenta conidióforos e conídios significativamente
menores quando comparados com o espécime estudado; ao contrário, a espécie C.
jatrophigena possui conidióforos e conídios maiores (Tabela 14).
Embora diferentes um do outro, como mostrado nas figuras 22 e 23, tanto o
espécime encontrado sobre P. nigrum quanto aquele em J. curcas, com base nos
dados morfológicos obtidos, foram alocados em C. apii Fresen. com base na
amplitude morfológica estabelecida por Crous & Braun (2003) para a espécie. Para
eles as características morfométricas de C. apii mostraram uma amplitude
extraordinária, principalmente no que diz respeito às dimensões dos conídios. Com
isso, limitou-se e muito a possibilidade de segregação de espécies obviamente
distintas em termos morfológicos. Sendo assim, muitos espécimes morfologicamente
diferentes são englobados em C. apii, podendo ser separadas somente através de
análises moleculares (Hsieh & Goh, 1990).
Não há registro de C. apii em ambas as espécies de plantas aqui estudadas.
Apesar das diferenças morfológicas evidentes entre elas em termos de condióforos
e forma dos locos conidiogênicos, apenas a análise molecular irá permitir uma
efetiva segregação de espécies, bem como a retirada de ambas de dentro do
complexo C. apii estabelecido por Crous & Braun (2003).
Apesar de a identificação estar incompleta esses registros são importantes
porque os fungos são patogênicos e inéditos em ambos os hospedeiros. As
informações obtidas e analisadas até o presente momento indicam que ambos os
espécimes são prováveis espécies novas do gênero Cercospora.

81
Tabela 13. Comparação entre as espécies de Cercospora encontradas em Piper sp. e o espécime em estudo.
Espécies de Cercospora Conidióforo (µm) Célula Conidiogênica (µm) Conídio (µm) Hospedeira Local
C. pireris-betle Sawada & Katsuki 1 30-80 x 4-5 – 30-90 x 2-3 Piper betle Taiwan
C. piperata Asthana & Mahmud 2 10-50 x 2-4 – 15-75 x 2-3,5 Piper longum Índia
Cercospora sp. CMHUB 21134 91-154 [118] x 2,5-6,5 [4] 9-56 [34] x 2,5-6 [4]
25-350 [122] x 2-4 Piper nigrum Brasil
1 Hsieh & Goh, 1990 2 Chupp, 1954
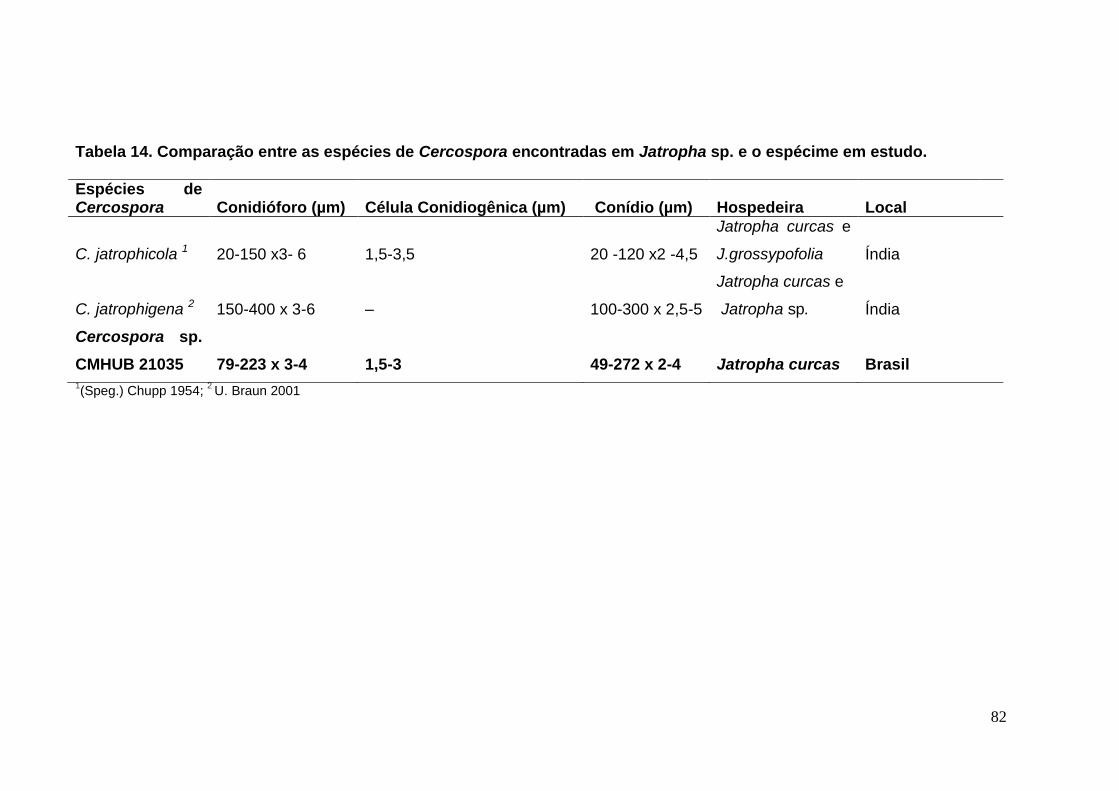
82
1(Speg.) Chupp 1954;
2 U. Braun 2001
Tabela 14. Comparação entre as espécies de Cercospora encontradas em Jatropha sp. e o espécime em estudo.
Espécies de Cercospora Conidióforo (µm) Célula Conidiogênica (µm) Conídio (µm) Hospedeira Local
C. jatrophicola 1 20-150 x3- 6 1,5-3,5 20 -120 x2 -4,5
Jatropha curcas e
J.grossypofolia Índia
C. jatrophigena 2 150-400 x 3-6 – 100-300 x 2,5-5
Jatropha curcas e
Jatropha sp. Índia
Cercospora sp.
CMHUB 21035 79-223 x 3-4 1,5-3 49-272 x 2-4 Jatropha curcas Brasil
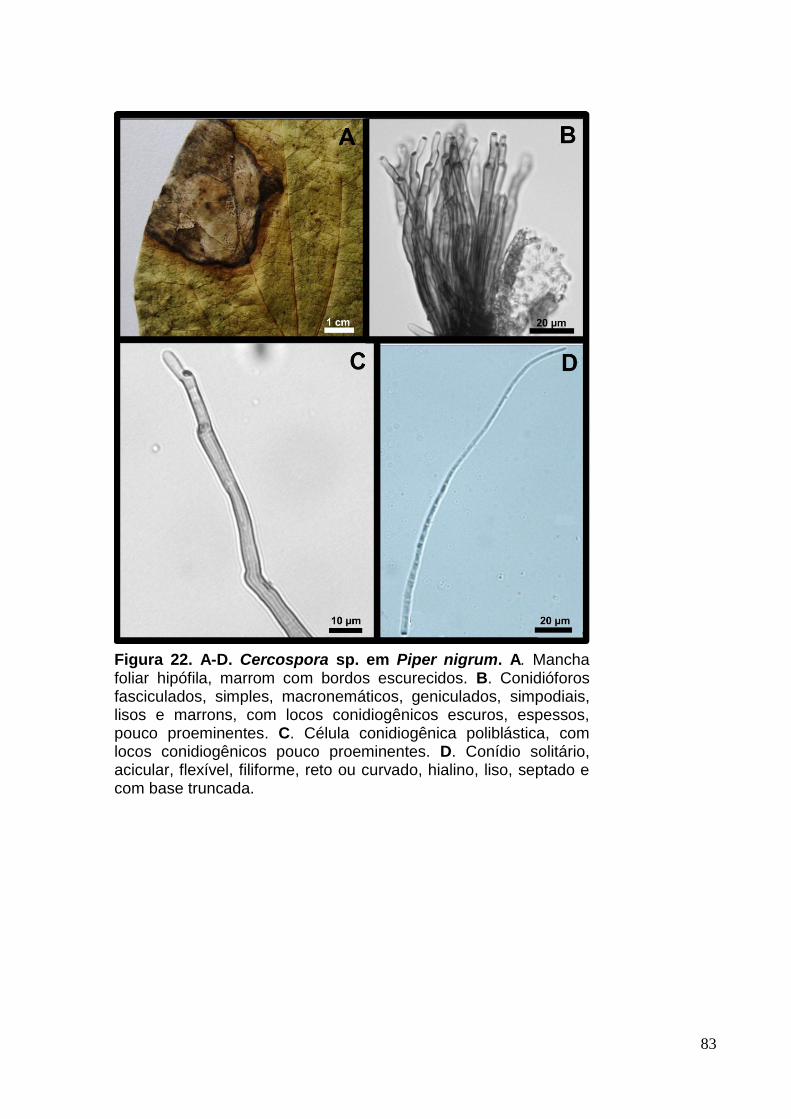
83
Figura 22. A-D. Cercospora sp. em Piper nigrum. A. Mancha foliar hipófila, marrom com bordos escurecidos. B. Conidióforos fasciculados, simples, macronemáticos, geniculados, simpodiais, lisos e marrons, com locos conidiogênicos escuros, espessos, pouco proeminentes. C. Célula conidiogênica poliblástica, com locos conidiogênicos pouco proeminentes. D. Conídio solitário, acicular, flexível, filiforme, reto ou curvado, hialino, liso, septado e com base truncada.
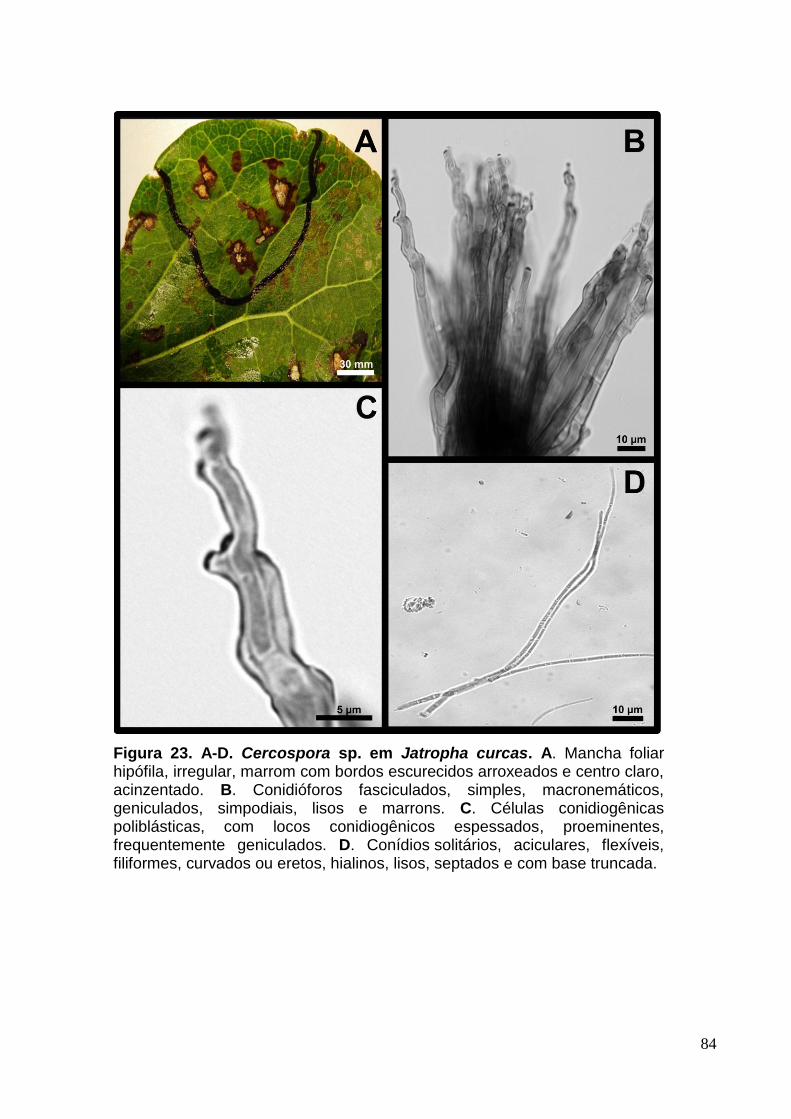
84
Figura 23. A-D. Cercospora sp. em Jatropha curcas. A. Mancha foliar hipófila, irregular, marrom com bordos escurecidos arroxeados e centro claro, acinzentado. B. Conidióforos fasciculados, simples, macronemáticos, geniculados, simpodiais, lisos e marrons. C. Células conidiogênicas poliblásticas, com locos conidiogênicos espessados, proeminentes, frequentemente geniculados. D. Conídios solitários, aciculares, flexíveis, filiformes, curvados ou eretos, hialinos, lisos, septados e com base truncada.

85
1.3. Pseudocercospora sp. (provável espécie nova nº 16)
Manchas necróticas irregulares, epífilas e marrons (Fig. 24. A). Estromas 11-
45,5 [20,5] x 10,5-33 [23,5] µm, marrons e irrompentes (Fig. 24. C). Conidióforos
21,5-45 [31,5] x 2-4 [2,5] µm, com 1 a 2 septos, fasciculados, simpodiais não
ramificados e marrons (Fig. 24. D). Células conidiogênicas 3,5-20,5 [8,5] x 1,5-3,5
[2,5] µm, com 2-6 [3], geniculadas, poliblásticas, com cicatrizes planas, inconspícuas
(Fig. 24. F). Conídios 18,5-93,5 [76,5] x 1-3 [2,5] µm, 3-6 [4] septos, hialinos, cilíndricos a
aciculares, curvados, flexíveis, base truncada, cicatriz plana (Fig. 24. E e F).
Espécime examinado: Brasil. Pernambuco, Recife, Cerâmica Francisco Brennand
em folhas de Licuala jeanencyi (Arecaceae), 07 Mar 2011, leg. J.C. Dianese,
holótipo (UB Micol. Coll. 21489).
De acordo com Farr & Rossman (2011) existem quatro registros de espécies
de Pseudocercospora sp. na família Arecaceae: P. arecacearum U. Braun & C.F. Hill
(2006), P. coperniciae U. Braun & F.O. Freire (2003), P. rhapisicola (Tominaga) Goh
& W.H. Hsieh (1989) e P. roystoneae U. Braun & Crous (2003).
O espécime em estudo difere dos membros das espécies conhecidas por ser
o único que apresenta lesões epífilas. Além disso, seus conidióforos apresentam
dimensões distintas das demais espécies, suas células conidiogênicas são próximas
das de P. roystoneae; seus conídios são menores e mais estreitos que os conídios
de P. arecacearum e a septação dos conídios é semelhante à de P. rhapisicola
(Tabela 15).
Este é o primeiro registro de ocorrência do gênero Pseudocercospora em
Licuala sp.
Assim, os dados mostrados indicam tratar-se de uma nova espécie de
Pseudocercospora, sobre um membro da família Arecaceae.
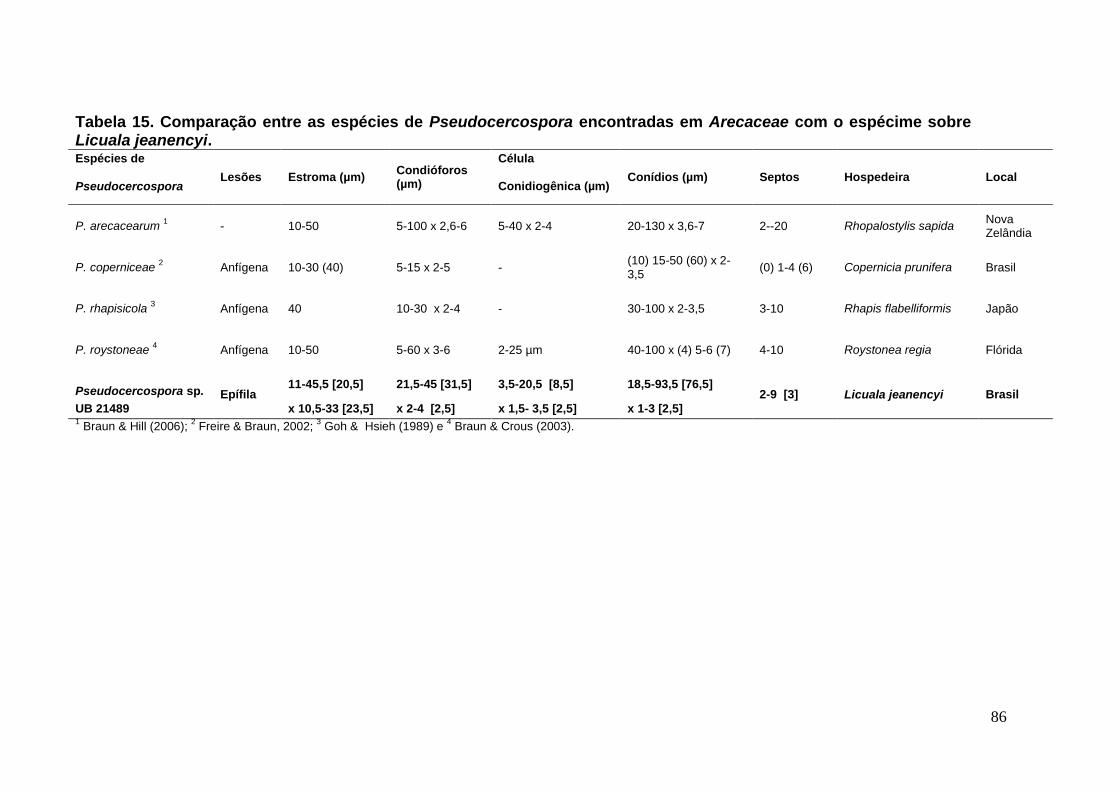
86
Tabela 15. Comparação entre as espécies de Pseudocercospora encontradas em Arecaceae com o espécime sobre Licuala jeanencyi. Espécies de
Lesões Estroma (µm) Condióforos (µm)
Célula
Conídios (µm) Septos Hospedeira Local Pseudocercospora Conidiogênica (µm)
P. arecacearum
1
- 10-50 5-100 x 2,6-6 5-40 x 2-4 20-130 x 3,6-7 2--20 Rhopalostylis sapida
Nova Zelândia
P. coperniceae
2
Anfígena
10-30 (40) 5-15 x 2-5 - (10) 15-50 (60) x 2-3,5
(0) 1-4 (6) Copernicia prunifera Brasil
P. rhapisicola
3
Anfígena
40 10-30 x 2-4 - 30-100 x 2-3,5 3-10 Rhapis flabelliformis Japão
P. roystoneae
4
Anfígena 10-50 5-60 x 3-6 2-25 µm 40-100 x (4) 5-6 (7) 4-10 Roystonea regia Flórida
Pseudocercospora sp. Epífila
11-45,5 [20,5] 21,5-45 [31,5] 3,5-20,5 [8,5] 18,5-93,5 [76,5] 2-9 [3] Licuala jeanencyi Brasil
UB 21489 x 10,5-33 [23,5] x 2-4 [2,5] x 1,5- 3,5 [2,5] x 1-3 [2,5] 1 Braun & Hill (2006);
2 Freire & Braun, 2002;
3 Goh & Hsieh (1989) e
4 Braun & Crous (2003).

87
H
A C
E
I
2,5 cm
20 µm
10 µm
D
B
E
100 µm
10 µm
F
10 µm
Figura 24. A-F. Pseudocercospora sp. em folhas de Licuala jeanencyi. A. Dois folíolos mostrando manchas marrom-escuras a negras, em maior quantidade na parte basal. B. Sinais do fungo representados pelos conidióforos sustentando massa de conídios. C. Fascículo de conidióforos sobre estroma intradérmico de textura prismático-angularis (seta). D. Detalhe dos conidióforos
fasciculados, simpodiais não ramificados e marrons. E. Conídio solitário
hialino, septado, curvo e flexível. F. Células conidiogênicas poliblásticas (seta) e conídios aciculares.

88
CONCLUSÃO
Foram selecionados e estudados sistematicamente 22 fungos associados à
famílias botânicas nativas e exóticas do Cerrado, a saber: Aquifoliaceae, Arecaceae,
Asteraceae, Dilleniaceae, Euphorbiaceae, Sapotaceae, Lauraceae, Malvaceae,
Myrsinaceae, Myrtaceae, Piperaceae e Verbenaceae.
Dentre os fungos analisados, três correspondem a espécies já conhecidas:
Staibia connari, um ascomiceto inédito em Ocotea paniculata, Phyllachora davilae,
um ascomiceto relatado pela primeira vez para a espécie hospedeira, Davilla rugosa
e por último a espécie Puccinia malvacearum, uma ferrugem já reportada para o
gênero da hospedeira (Triumfetta rhomboidea).
Dezesseis prováveis espécies novas foram encontradas, dentre elas, quatro
hifomicetos (Janetia, pela primeira vez em Ocotea sp. duas em Cercospora sp.
encontradas em Piper nigrum e em Jatropha curcas, ambas inéditas para as
hospedeiras e Pseudocercospora, pela primeira vez em folhas de Licuala jeanencyi),
três celomicetos (Acrostroma, Pestalotiopsis e Stigmopeltis em Roessleria brasiliana,
Mauritia sp. e Micropholis venulosa, respectivamente), e nove ascomicetos
(Asterinella em Eugenia, Coccodiella em Butia sp., Johansonia e Phillipsiella em
Ocotea paniculata, Limacinia em Lantana camara, Phyllachora e Stomiopeltis em
Ilex affinis, Stomatogene em Myrsine guianensis e Vonarxella em Ocotea sp.; com
excessão de Asterinella, todos possuem os primeiros registros de ocorrência em
seus hospedeiros correspondentes).
Também foram encontrados três prováveis gêneros novos: um celomiceto
sobre uma asterácea, e dois ascomicetos associados a tricomas foliares de membro
da família Lauraceae.

89
REFERÊNCIAS BIBLIOGRÁFICAS
ARMANDO, E.A.S. Micobiota associada à família Caryocaraceae. Dissertação de
Mestrado. Universidade de Brasília. Brasília. 2004.
ARNAUD, G. Contribution a l’étude des fumagines. 1 Partie (Limacinia, Seuratia,
Pleosphaeria, etc.). Annales de l’École National d’Agriculture de Montpellier. Série 2.
9: 239–277. 1910.
ARX, J.A. VON & MÜLLER, E. A Re-evaluation of the bitunicate ascomycetes with
keys to families and genera. Centraalbureau voor Schimmelcultures, Baarn. Studies
in Mycologia. 1-159. 1975.
ARX, J.A. VON & MÜLLER, E. Die Gattungen der amerosporen Pyrenomyceten.
Beiträge zur Kryptogamenflora der Schweiz. 11 (1): 434. 1954.
BARR, M.E. Prodomus to the Loculoascomycetes. Printed USA. Amherst,
Massachusetts. 1987.
BARR, M.E. Redisposition of some taxa described by JB Ellis. Mycotaxon. 46: 45–
76. 1993.
BATISTA, A.C. & CIFERRI, R. Capnodiales. Saccardoa. 2: 296. 1963a.
BATISTA, A.C. & CIFERRI, R. The Chaetothyriales. Sydowia, Beihefte. 3: 129.
1962.
BATISTA, A.C. & CIFERRI, R. The sooty-molds of the family Asbolisiaceae.
Laboratorio Crittogamico, Istituto Botanico della Università di Pavia. 31:229. 23
plates. 1963b.
BATISTA, A.C. & PERES. G.E.P. Asterinella Theiss. Revisão de algumas espécies.
Atas do Instituto de Micologia da Universidade do Recife. 305:9-22.1961a.

90
BATISTA, A.C. & PERES, G.E.P. Alguns novos micropeltis da Amazônia. UFPe.
Instituto de Micologia (Universidade do Recife. Instituto de Micologia. Publicação
386. 115. Recife-PE. 1963.
BATISTA, A.C. BEZERRA, J.L. & PERES, G.E.P. Vonarxella e outros novos
ascomicetos. Rivista di Patologia Vegetale, Pavia. Série 4.1:61. 1965.
BATISTA, A.C. Monografia dos fungos Micropeltaceae. Publicações do Instituto de
Micologia da Universidade do Recife. 56:519. 1959.
BATISTA, A.C. PERES, G.E.P. & TORRES, S.V. Staibia - Um novo gênero de
Leptopeltaceae. Atas do Instituto de Micologia da Universidade do Recife 478:140-
144. 1966a.
BEZERRA, J.L. A História de Augusto Chaves Batista. Fitopatologia Brasileira.
(Suplemento). 30: S4-S6. 2005.
BEZERRA, J.L.; POROCA, D.J.M. Alguns Deuteromycotina do Brasil. Publicações.
Instituto de Micologia da Universidade Federal de Pernambuco 639: 1-10. 1969.
BOSE, S.K.; MÜLLER, E. Central Himalayan fungi. I. Indian Phytopathology 17 (1):
3-22. 1964.
BRAUN, U., CROUS P. W. & KAMAL: New species of Pseudocercospora,
Pseudocercosporella, Ramularia and Stenella (cercosporoid hyphomycetes). Mycol.
Progress 2(3): 197-208. 2003.
FREIRE, F.C.O.; BRAUN, U. Some cercosporoid hyphomycetes fron Brazil-II.
Cryptogamie Mycologie 23: 295-328. 2002.
BRAUN, U., HILL, C.F. & SCHUBERT, K. New species and new records of biotrophic
micromycetes from Australia. Fungal Diversity 22: 13-35. 2006.

91
BRAUN, U. Revision of Cercospora species described by K.B. Boedijn. Nova
Hedwigia. 73: 419-436. 2001.
CALDUCH, M. GENE, J. ABDULLAH, S.K & GUARRO, J. Janetia obovata and
Stachybotryna excentrica, two new hyphomycetes from submerged plant material in
Spain. Mycologia 94:355-361. 2002.
CANNON, P.F. & P.F. KIRK. Fungal Families of the World. CAB International.
Wallingford. 2007.
CARMICHAEL, J.W. KENDRICK, W.B. CONNERS, I.L. & SIGLER, L. Genera of
Hyphomycetes. University of Alberta Press. Edmonton, Canadá. 1980.
CASTRO, M.T. Fungos associados à Copaifera. Monografia. Universidade de
Brasília. Brasília. 2009.
CHAVES, Z.M. Fungos associados à Mauritia flexuosa (Buriti) e Mauritiella armata
(Buritirana). Tese de Mestrado. Universidade de Brasília. Brasília. 1998.
CHUPP, C.A. Monograph of the Fungus Genus Cercospora. Ithaca. New York.
1954.
CIFERRI, R. Batistia, a new genus of the Cephalothecaceae family. Atti Ist Bot Univ
Pavia. Serie 5. 15:160–170. 1958.
CLEMENTS, F.E. & SHEAR, C.L. The genera of fungi. H.W. Wilson Company. New
York. 1931.
COLE, M.M. The Savannas - Biogeography and Geobotany, Academic Press,
London. 1986.
CROUS, P.W. & BRAUN, U. Mycosphaerella and its anamorphs: 1. Names published
in Cercospora and Passalora. CBS Biodiversity. Series 1. Ultrecht. 2003.

92
CROUS, P.W. BARRETO, R.W. ALFENAS, A.C. ALFENAS, R.F. & GROENEWALD,
J.Z. What is Johansonia? IMA Fungus 1: 117–122. 2010.
CUMMINS, G.E. & HIRATSUKA, Y. llustrated Genera Rust Fungi. Revision Edition.
Minnesota: Ed. The Am. Phytopath. Soc. St. Paul. 152. 1983.
CUMMINS, G.E. & HIRATSUKA, Y. llustrated Genera Rust Fungi. Third Edition.
Minnesota: Ed. The Am. Phytopath. Soc. St. Paul. 225. 2003.
CYBERLIBER, an Electronic Library for Mycology. Disponível em:
<http://www.cybertruffle.org.uk/cyberliber/>, Acesso em: Agosto de 2011.
DENNIS, R.W.G. Kew Bulletin Additional Series III. Fungus Flora of Venezuela and
Adjacent Countries. Verlag von J. Cramer. 531 pages. 1970.
DIANESE, A.C. COSTA, A.M. DIANESE, J.C. A new Pseudocercospora species on
Passiflora setacea. Mycotaxon. 105:1-5. 2008.
DIANESE, J.C. MEDEIROS, R.B. & SANTOS, L.T. Biodiversity of Microfungi Found
on Native Plants of the Brazilian Cerrado. In: Hyde, K. D. Biodiversity of Tropical
Microfungi. Hong Kong University Press. 367-417. 1997.
DIANESE, J.C. Micodiversidade associada a plantas do Cerrado. In: Tópicos atuais
em Botânica. Embrapa. 109-115. Brasília. 2000.
DIANESE, J.C. Trinta Anos de Fitopatologia na Universidade de Brasília.
Fitopatologia Brasileira. (Suplemento). 30: S18-S20. 2005.
DIANESE. J.C. DIANESE A.C. SOUZA, E.S.C. & PEREIRA-CARVALHO R.C. New
record of Cercospora apii s. lato on Piper nigrum and Jatropha curcas in the Cerrado.
In: Joint Meeting of the Mycological Society of America and The International

93
Symposium on Fungal Endophytes of Grasses. Kentucky. MSA & ISFEG. Lexington.
25. 2010.
DORNELO-SILVA, D. & DIANESE, J.C. Hyphomycetes on the Vochysiaceae from
the Brazilian Cerrado. Mycologia. 95:1239-1251. 2003.
DORNELO-SILVA, D. & DIANESE, J.C. New hyphomycete genera on Qualea
species from the Brazilian Cerrado. Mycologia. 96: 879-884. 2004.
DORNELO-SILVA, D. Fungos associados a plantas da família Vochysiaceae
presentes no Cerrado. Dissertação de Mestrado. Universidade de Brasília. Brasília.
1999.
DORNELO-SILVA, D. PEREIRA-CARVALHO, R.C. & DIANESE, J.C. New Stenella
and Parastenella species from the Brazilian Cerrado. Mycologia. 99: 753-764. 2007.
ELLIS, J.B. & EVERHART, B.M. New species of fungi from various localities. Bulletin
of the Torrey Botanical Club. 27: 571. 1900.
ELLIS, M.B. Dematiaceous Hyphomycetes. Commonwealth Mycological
Institute,Kew, Surrey. 608. England. 1971.
ELLIS, M.B. More Dematiaceous Hyphomycetes. Commonwealth Mycological
Institute, Kew, Surrey. 507. England. 1976.
ERIKSSON, O. The familes of bitunicate ascomycetes. In: editor (Ed) Ópera
Botânica. 60:1-220. 1981.
FARR, D.F. & ROSSMAN, A.Y. Fungal Databases, Systematic Mycology and
Microbiology Laboratory, ARS, USDA. Disponível em: <http://nt.ars-
grin.gov/fungaldatabases/fungushost/fungushost.cfm> Acesso em: Janeiro-Agosto
de 2011.

94
FARR, M.L. Two new species of tropical fungi. Mem. N. Y. bot. Gdn. New York
Botanical Garden. 49:70-73. 1989.
FILGUEIRAS, T.S. & PEREIRA, B.S. Flora do Distrito Federal. In Cerrado:
caracterização, ocupação e perspectivas. (M. Novaes Pinto, coord.). Editora da
Universidade de Brasília. 345-405. Brasília 1993.
FRASER, L.R. An investigation of the sooty moulds of New South Wales.
Proceedings of the Linnean Society of New South Wales. 60(3-4): 171. 1935.
GOH, T.K & HSIEH, W.H. Studies on Cercospora and allied genera of Taiwan.
Transactions of the Mycological Society of the Republic of China. 4(2-3): 46. 1989.
GOH, T.K & HYDE, K.D. Janetia with Key. Mycologia 88:1014-1021.1996.
GUARIM-NETO, G. & MORAIS, R.G. Recursos medicinais de espécies do Cerrado
de Mato Grosso: um estudo bibliográfico. Acta Botanica Brasileira 17(4): 561-584.
2003.
GUTIÉRREZ, A.H. & DIANESE, J.C. New cercosporoid fungi from the Brazilian
Cerrado 1. Species on hosts of the families Anacardiaceae, Araliaceae,
Bombacaceae, Burseraceae and Celastraceae. Mycotaxon 106: 41-63. 2008.
GUTIÉRREZ, A.H. & DIANESE, J.C. New cercosporoid fungi from the Brazilian
Cerrado 2. Species on hosts of the subfamilies Caesalpinioideae, Faboideae and
Mimosoideae (Leguminosae s. lat.). Mycotaxon.107: 1-24. 2009.
HANSFORD, C.G. Tropical fungi. VIII. Sydowia 11(1-6): 68. 1958.
HAWKSWORTH, D.L. The fungal dimension of biodiversity: magnitude, significance,
and conservation. Mycological Research 95: 641–655. 199. 1991.

95
HAWKSWORTH, D.L. The magnitude of fungal diversity: the 1.5 million species
estimate revisited. Mycological Research 105: 1422–1432. 2001.
HENNEN, J.F. FIGUEIREDO, M.B. CARVALHO J.R, & HENNEN, P.G. Catalogue of
the species of plant rust fungi (Uredinales) of Brazil. Rio de Janeiro: Instituto de
pesquisas. Jardim Botânico do Rio de Janeiro, 2005. Disponível em:
<http://www.jbrj.gov.br/publica/uredinales/index.htm>, Acesso em: Maio de 2011.
HSIEH, W. H. & GOH, T. K. Cercospora and similar fungi from Taiwan. Maw Chang
Book. Co., Taipei. 1990.
HUGHES, S.J & CAVALCANTI, W.A. Janetia mangiferae n. sp. and the
Johncourchian. gen. anamorph of Septobasidium pilosum from Brazil. Canadian
Journal of Botanic. 61: 2224-2230. 1983.
HUGHES, S.J. New Zealand Fungi 32. Janetia capnophila sp. nov. and some allies.
New Zealand Journal of Botany. 21:177-182. 1983.
HUHNDORF, S.M. MILLER, A.N. & FERNÁNDEZ F.A. Molecular systematics of the
Sordariales: the order and the family Lasiosphaeriaceae redefined. Mycologia. 96
(2):368–387. 2004.
IBAMA. Ecossistemas Brasileiros. Disponível em: <http://www.ibama.gov.br/
ecossistemas/cerrado.htm> Acesso em Julho de 2011.
KIFFER, W. & MORELET, M. The Deuteromycetes Mitosporic Fungi Classification
and Generic Keyz. Science Publishers Inc. 1a ed. France. 2000.
KIRK, P. Index Fungorum. CABI Bioscience, CBS and Landcare Research, available
online, ed. 2011. Disponível em: < www.indexfungorum.org>, Acesso em: Janeiro –
Setembro de 2011.

96
KIRK, P.M. CANNON, P.F. & DAVID, J.C. Ainsworth and Bisby’s Dictionary of the
Fungi. 9 ed. CABI. UK. 2001.
KIRK, P.M. CANNON, P.F. MINTER, D.W. & STALPERS, J.A. Dictionary of the
fungi. 10 ed. CAB International. Wallingford. 2008.
KIRK, P.M. New or interesting microfungi XIV. Dematiaceous hyphomycetes from Mt.
Kenya. Mycotaxon. 23:305-352.1985.
KLINK, C.A. MACEDO, R.F. & MUELLER, C.C. De Grão em Grão, o Cerrado Perde
Espaço (Cerrado - Impactos do Processo de Ocupação). WWF- Fundo Mundial para
a Natureza. Brasília. 1995.
KLINK, C.A. MACHADO, R.B. A conservação do Cerrado Brasileiro.
Megadiversidade 1: 147-155. Belo Horizonte. 2005.
LINDAU, G. Pyrenomycetinae. In: Engler, A. & Prantl, K. Die natürlichen
Pflanzenfamilien. 1: 1: 178-278, 321-505. 1897.
LINDQUIST, J.C. Royas de la republica Argentina y zonas limitrofes. Instituto
Nacional de Tecnología Agropecuária. Buenos Aires. 1982.
LUMBSCH, H.T. & HUHNDORF, S.M. Myconet.14. Part One. Outline of Ascomycota
– 2009. Fieldiana, Life and Earth Sciences. 1: 1–42. 2010.
LUMBSCH, H.T. & HUHNDORF, S.M. Outline of Ascomycota. Myconet. 13: 1- 58.
2007.
LUTTRELL, E.S. The genus Stomiopeltis. Mycologia. 38: 565-585.1946.
LUTTRELL, E.S. The Loculoascomycetes. In: Ainsworth, C.G., Sparroe, F.K. &
Sussman, A.S. The Fungi. (Ed.) Academic Press, New York, London. Chapter 7. 4 A:
135-219. 1973.

97
MEHROTRA, M.D. & VERMA, R.K. Mycovellosiella raveneliae, a hyperparasite of
Ravenelia clemensiae from India. Mycological Research. 97: 1020. 1993.
MENDES, M.A.S. SILVA, V.L. DIANESE, J.C. FERREIRA, M.A.S.V. SANTOS,
C.E.N. NETO, E.G. URBEN, A.F. & CASTRO, C. Fungos em plantas no Brasil.
Brasília-DF. Embrapa. 555. 1998. Disponível em:
http://pragawall.cenargen.embrapa.br/aiqweb/michtml/fgbanco01.asp>, Acesso em:
Agosto de 2011.
MÜLLER, E. & ARX, J.A. VON. Pyrenomycetes: Meliolales, Coronophorales,
Sphaeriales. In: Ainsworth, G. C. & al. The Fungi IV A: 87-132. 1973.
MÜLLER, E. Ein neuer Discomycet aus Java: Johansonia pandani. Sydowia 8 (1-6):
54-56. 1954.
MÜLLER, E. & ARX, J.A. von. Die Gattungen der didymosporen Pyrenomyceten.
Beitrage zur Kryptogamenflora der Schweiz 11: 1-922. 1962.
NAG RAJ, T.R. Coelomycetous anamorphs with appendage-bearing conidia.
University of Waterloo. Printed Edwards Brothers, Michigan USA. Canada. 1993.
NEGER, F.W. Enfermedades de los cereales, nuevamente introducidas en
Chile. Bol. Soc. Nac. Agric. Chile 27: 398. 1896.
PEREIRA-CARVALHO, R.C. DORNELO-SILVA, D. INACIO, C.A., & DIANESE, J.C.
Chaetothyriomyces: a new genus in family Chaetothyriaceae. Mycotaxon 107: 483-
488. 2009a.
PEREIRA-CARVALHO, R.C. SEPÚLVEDA-CHAVERA, G. ARMANDO, E.A.
DIANESE, J.C. An overlooked source of fungal diversity: novel hyphomycete genera
on trichomes of Cerrado plants. Mycological Research 113: 261-274. 2009b.

98
PIEPENBRING, M. Checklist of fungi in Panama. Preliminary version. Puente Biol. 1:
1-190. 2006.
POLLACK, F.G. An annotated compilation of Cercospora names. Mycologia Memoirs
12:1-212. 1987.
RIBEIRO, J. F. & WALTER, B. M. T. Fisionomias do bioma Cerrado. In: Cerrado:
ambiente e flora. Sano, S.M. & Almeida, S.P. (Ed.), Planaltina-DF. 1998.
RIBEIRO, J.F. & WALTER, B.M.T. As principais fitofisionomias do bioma Cerrado. In:
SANO, S.M. ALMEIDA, S.P. & RIBEIRO, J.F. (orgs.). Cerrado: ecologia e flora. 151-
212. EMBRAPA. Brasília. 2008.
ROSSMAN, A.Y. The Tubeufiaceae and similar Loculoascomycetes. Mycological
Papers 157: 1–71. 1987.
SACCARDO, P. A. & SYDOW, P. Hymenomycetae. Sylloge Fungorum 14: 474.
Pavia. 1899.
SACCARDO, P.A. Sylloge Fungorum omnium hucusque cognitorum. Pavia 1. 1882.
SAMUELS, G.J. & RODRIGUES, K.F. Batistia annulipes and its anamorph,
Acrostroma annellosynnema. Mycologia 81: 52-56. 1989.
SANTOS, L.T.P. Micobiota foliícola de Salacia crassifolia. Dissertação de Mestrado.
Universidade de Brasília. Brasília. 2011.
SILVANESAN, A. & HSIEH, W.H. A synnematous species of Janetia from Taiwan.
Mycological Research 94: 566-569. 1990.
SILVANESAN, A. Graminicolous species of Bipolaris, Curvularia, Drechslera,
Exserohilum and their teleomorphs. Mycological Papers 158: 1-261. 1987.

99
SPEGAZZINI, C.L. Fungi aliquot Paulistani. Revista del Museo de La Plata 15: 7-48.
1908. Disponível em: <http://www.cybertruffle.org.uk/spegazzini/eng/005039a_.htm>,
Acesso em: Abril de 2011.
SOUZA, C.A.P. VITÓRIA, N. S. BEZERRA, J.L. LUZ, E.D.M.N. INÁCIO, C.A. &
DIANESE, J.C. Camarotella brasiliensis sp. nov. (Phyllachoraceae) on Syagrus
schizophylla (Arecaceae) from Brazil. Mycotaxon 103: 313-317. 2008.
SOUZA, E.S.C. PEREIRA-CARVALHO, R.C. SOARES, W.R.O. & DIANESE J.C.
Staibia conari em Ocotea sp. (Lauraceae). In: VII Congresso Latino Americano de
Micologia. Resumenes y Programa del VII Congresso Latino Americano de
Micologia. San José. Costa Rica. 2011b.
SOUZA, E.S.C. SPERANDIO, E.M. VALE, H.M.M. SANCHEZ, M. & DIANESE, J.C.
Pseudocercospora sp. em folhas de Licuala jeanencyi. In: VII Congresso Latino
Americano de Micologia. Resumenes y Programa del VII Congresso Latino
Americano de Micologia. San José. Costa Rica. 2011c.
SOUZA, E.S.C. VALE, H.M.M. SOARES, W.R.O. PEREIRA-CARVALHO, R.C. &
DIANESE, J.C. Revisitando a Coleção Micológica de Ezechias Paulo Heringer:
dados iniciais. In: 26 Congresso Brasileiro de Microbiologia. Microbiologia in foco.
São Paulo: Vox Editora Ltda. 4: 245-245. Foz do Iguaçu- PR. 2011a.
SPERANDIO, E. M. SOUZA, E.S.C. VALE, H.M.M. PEREIRA-CARVALHO, R.C.
SOARES, W.R.O. & DIANESE, J.C. Primeiro relato de Johansonia sp. e Phillipsiella
sp. em Ocotea sp. In: 44 Congresso Brasileiro de Fitopatologia. Tropical Plant
Pathology - Fitopatologia Brasileira. 36: 667-667. Bento Gonçalves-RS. 2011.
STEVENS, F.L. Parasitic fungi of British Guiana, Trinidad and Costa Rica. Annales
Mycologici. 28 (3/4): 282. 1930.
SUTTON, B.C. The Coelomycetes. Fungi Imperfecti with Pycnidia Acervuli and
Stromata. Kew. Commonwealth Mycological Insitute. UK. 1980.

100
SWART, H.J. Australian leaf-inhabiting fungi. XVII. Janetia interna sp. nov.
Transactions of the British Mycological Society. 84:174-175.1985.
SYDOW, H. Fungi aequatorienses (series prima). Annales Mycologici 37: 275-438.
1939.
SYDOW, H. Fungi in itinere costaricensi collecti. Annales Mycologici 25 (1/2): 125-
160. 1927.
THEISSEN, F. & SYDOW, H. "Synoptische Tafeln" in Annales Mycologici. Annals of
Mycology 15:6. 389 – 491. 1917.
THEISSEN, F. Annotaçoes à mycoflora Brasileira. Brotéria, Série Botânica 12: 85.
1914.
THEISSEN, F. Mycologische Mitteilungen. Annales Mycologici 14(6): 406. 1918.
VALE, H.M.M. SOUZA, E.S.C. ARMANDO, E.A.S. SALES, H.B. PEREIRA-
CARVALHO, R.C. & DIANESE, J.C. Provável espécie nova de Vonarxella em folhas
de Ocotea sp. In: VI Congresso Brasileiro de Micologia. Anais do VI Congresso
Brasileiro de Micologia. Brasília-DF. 2010b.
VALE, H.M.M. SOUZA, E.S.C. ARMANDO, E.A.S. SALES, H.B. PEREIRA-
CARVALHO, R.C. & DIANESE, J.C. Provável espécie nova de Stomatogene em
folhas de Rapanea guianensis. In: VI Congressso Brasileiro de Micologia. Anais do
VI Congresso Brasileiro de Micologia. Brasília-DF. 2010a.
VALE, H.M.M. SOUZA, E.S.C. PEREIRA-CARVALHO, R.C. & DIANESE, J.C.
Janetia sp. em tricomas de Ocotea sp. no Brasil. In: 44 Congresso Brasileiro de
Fitopatologia. Tropical Plant Pathology - Fitopatologia Brasileira. 36: 1002-1002.
Bento Gonçalves- RS. 2011a.

101
VALE, H.M.M. SOUZA, E.S.C. PEREIRA-CARVALHO, R.C. & DIANESE, J.C.
Provável espécie nova de Stigmopeltis sp. em folhas de Micropholis venulosa
(Sapotaceae). In: 26 Congresso Brasileiro de Microbiologia, Microbiologia in foco -
Programa e Resumos. São Paulo: Vox Editora Ltda. 4: 243-243. Foz do Iguaçu- PR.
2011b.
VALE, H.M.M. SOUZA, E.S.C. PEREIRA-CARVALHO, R.C. & DIANESE, J.C. Dois
novos ascomicetos tricomáticos em planta da família Lauraceae. In: 26 Congresso
Brasileiro de Microbiologia. Microbiologia in foco - Programa e Resumos. São Paulo:
Vox Editora Ltda. 4: 244-244. Foz do Iguaçu- PR. 2011c.
WEHMEYER, L.E. Some fungi imperfecti from Mt. Rainer National Park. Sydowia
6:964: 433-442. 1952.
Y.L. GUO. Fungi and Lichens of Shennongjia. Mycological and Lichenological
Expedition to Shennongjia (Beijing). 356. 1989.