343o graciela s. centenaro.doc) - ppgalimentos.furg.br do grau de hidrlise... · capita/ano), mais...
TRANSCRIPT

1 INTRODUÇÃO
As proteínas são macromoléculas complexas, compostas de aminoácidos, e
necessárias para os processos químicos que ocorrem nos organismos vivos. Estas
moléculas especiais realizam as mais variadas funções no nosso organismo, desde o
transporte de nutrientes e metabólitos à catálise de reações biológicas. São os
constituintes básicos da vida: tanto que seu nome deriva da palavra grega "proteios", que
significa “em primeiro lugar”.
As proteínas têm muitas funções importantes no organismo humano. São os
principais componentes dos tecidos estruturais como a pele e o colágeno, encontrados
em tecidos conjuntivos como tendões e ligamentos. O sangue necessita das proteínas
para os glóbulos vermelhos, glóbulos brancos e numerosos compostos do plasma. A
imunidade do seu corpo também depende das proteínas, que são necessárias para a
formação dos anticorpos e dos glóbulos brancos que combatem as doenças. As enzimas
e os hormônios (por exemplo, a insulina) também são proteínas (ISTOÉ, 2002).
A deficiência de proteínas na dieta já foi um sério problema no passado,
principalmente nos países em desenvolvimento, onde havia grande escassez de
alimentos protéicos. Uma provável solução que tem diminuído esse fato é a utilização de
fontes alternativas de proteínas e o desenvolvimento de tecnologias que permitem o
melhor aproveitamento dos produtos protéicos ou das proteínas tradicionais.
A proteína de origem animal, pela sua qualidade nutricional, pode ser o foco
para o desenvolvimento de tecnologias que permitam seu melhor aproveitamento. Nesse
sentido, os concentrados de proteínas, a silagem enzimática e os isolados e hidrolisados
de proteínas de pescado parecem ser promissores (KRISTINSSON e RASCO, 2000a). A
produção mundial de pescado é de aproximadamente 130 milhões de toneladas por ano,
sendo que 970 mil toneladas são capturadas no Brasil, das quais mais de 50% são
considerados de baixo valor comercial ou subprodutos. Na região de Rio Grande, no sul
do Brasil, cerca de 45 a 60 mil toneladas de pescado são capturadas por ano,
correspondendo a 4,5 - 6% do total nacional. (FAO, 2005).
A aplicação de tecnologia enzimática para a transformação da carne de
pescado em uma nova fonte protéica é um método alternativo para o melhor
aproveitamento de espécies sem valor comercial, a fim de recuperar e modificar as
proteínas para a produção de diversos ingredientes alimentícios e produtos de uso
industrial. Para que esses produtos possam ser aplicados com êxito, é necessário
conhecer as propriedades funcionais, que estão diretamente relacionadas com as
características sensoriais e de textura dos alimentos adicionados de hidrolisados. O
conhecimento das propriedades funcionais específicas dos hidrolisados tende a facilitar o

2
direcionamento da sua aplicação, contribuindo para um melhor aproveitamento,
resultando em produtos de maior qualidade tecnológica e nutricional (KRISTINSSON e
RASCO, 2000a).
A hidrólise de proteínas pode ser realizada com enzimas, ácidos ou álcali, mas a
hidrólise enzimática é mais indicada que métodos químicos para a produção de
hidrolisados com aplicações nutricionais. A modificação enzimática das proteínas, usando
preparados de enzimas proteolíticas selecionadas para clivar ligações peptídicas
específicas, é amplamente utilizada na indústria de alimentos (LAHL e BRAUN, 1994).
As propriedades funcionais de maior interesse para a indústria de alimentos são
a solubilidade, a capacidade de retenção de água, a capacidade de emulsificação e de
formação de espuma e ainda as propriedades sensoriais. Pela modificação das
condições de reação enzimática das proteínas, é possível produzir hidrolisados com
diferentes características funcionais (KRISTINSSON e RASCO, 1998).
Através da hidrólise da proteína de pescado, empregando proporções
enzima/substrato e tempos de reação adequados, é possível produzir hidrolisados com
estruturas moleculares e propriedades funcionais diferentes que podem encontrar
aplicações em vários produtos alimentícios. (KRISTINSSON e RASCO, 2000a),
Com o crescimento da população mundial e a pesca perto de exceder os limites
sustentáveis de mais de 100 milhões de toneladas a cada ano, é extremamente
importante que a utilização dos recursos marinhos se dê com mais precaução e
inteligência, considerando que a demanda de produtos alimentícios será cada vez maior,
principalmente por aqueles com proteína de alto valor nutricional e valor tecnológico
agregado (KRISTINSSON e RASCO, 2000a).
O município de Rio Grande é considerado uma área tradicional de captura de
pescado e abriga diferentes indústrias pesqueiras que são responsáveis por parte do
descarte de subprodutos da pesca (BRASIL, 2005).
Alguns dos subprodutos do processamento são espécies de baixo valor
comercial, destinadas, na sua maioria, para fabricação de farinha de pescado juntamente
com os demais resíduos da indústria, ou então são descartados no meio ambiente.
Muitas indústrias processadoras de pescado investem parte do que arrecadam tratando
seus subprodutos antes de descartá-los no oceano, por exemplo (KRISTINSSON e
RASCO, 2000a). Porém, para que a indústria desenvolva processos para recuperar e
utilizar subprodutos, é necessário que os mesmos sejam viáveis e melhores que o seu
descarte (KRISTINSSON e RASCO, 2000b).
A possibilidade de recuperação e modificação das proteínas do músculo de
pescado mediante hidrólise enzimática justifica o estudo das características físico-
químicas que permitem o uso destes materiais como ingredientes funcionais. A utilização

3
de enzimas para hidrolisar proteínas alimentares é um processo empregado para
aperfeiçoar ou modificar as propriedades funcionais, físico-químicas e sensoriais das
proteínas nativas, sem comprometer seu valor nutritivo, melhorando sua absorção.
O conhecimento da relação do grau de hidrólise com alguma característica
funcional específica do hidrolisado, permite elaborar produtos com propriedades
funcionais previamente definidas. (SHAHIDI e KAMIL, 2001).
Diante disso, este estudo buscou oferecer uma alternativa à indústria em relação
ao aproveitamento de biomassa de pescado, agregando valor à mesma através de
processos economicamente acessíveis, visando sua aplicação em produtos amplamente
consumidos, através do melhoramento de suas propriedades funcionais.
A partir destas considerações este trabalho teve como objetivo geral estudar a
influência do grau de hidrólise enzimática sobre as propriedades funcionais de
hidrolisados protéicos obtidos a partir de corvina (Micropogonias furnieri) com a utilização
da enzima proteolítica Alcalase. Para isso, os objetivos específicos foram:
- Definir as condições da reação enzimática, (pH, tempo e temperatura),
utilizando corvina como substrato e alcalase como enzima;
- Realizar reações enzimáticas com variação de fatores de maior influência para
obtenção de hidrolisados com diferentes graus de hidrólise;
- Obter hidrolisados de corvina com diferentes graus de hidrólise para a
avaliação das propriedades funcionais;
- Avaliar as propriedades funcionais de interesse tecnológico: solubilidade,
capacidade de formação de espuma, capacidade emulsificante e capacidade de retenção
de água e de óleo dos diferentes hidrolisados obtidos;
- Verificar como o grau de hidrólise influi sobre as propriedades funcionais
medidas.

4
2 REVISÃO BIBLIOGRÁFICA
2.1 Pescado
Entende-se como pescado fresco, o produto obtido de espécies saudáveis e de
qualidade adequada ao consumo humano, convenientemente lavado e que seja
conservado somente pelo resfriamento a uma temperatura próxima a do ponto de fusão
do gelo. O pescado é um importante elemento da dieta humana, pois é uma fonte de
diversos componentes com elevado valor nutricional, como as proteínas, e os ácidos
graxos da série ômega-3, que trazem inúmeros benefícios à saúde (BRASIL, 1997).
2.1.1 Consumo e produção de pescado
Recentemente estudos científicos veiculados em vários periódicos internacionais
apontam para um colapso na pesca de recursos marinhos costeiros, em nível
internacional. Na costa do Brasil, a tendência a diminuição da disponibilidade de pescado
acompanha a restante do planeta. O Brasil apresenta um dos mais baixos índices de
consumo de pescado. Este índice, dentre outros fatores, provalvemente deve-se à falta
de conhecimento da importância do pescado na alimentação. As perspectivas da
demanda mundial de pescado para consumo humano direto são determinadas pelo
crescimento da população, mudanças de ingresso per capita e ritmo da urbanização
(NEIVA, 2005).
Segundo a FAO (2005), a proteína de origem animal mais consumida
mundialmente é o pescado, no entanto o Brasil apresenta um baixo consumo (6,7kg/per
capita/ano), sendo que no Rio Grande do Sul este índice é ainda menor (2kg/per
capita/ano), mais especificamente no sul do Rio Grande do Sul, o consumo é de 0,5
kg/per capita/ano, tendo-se o tradicionalismo e o nível cultural da população da região
como os principais motivos do baixo consumo. Este índice torna-se um pouco mais
elevado nas capitais (em torno de 15kg/per capita/ano), mas ainda baixo se comparado
com outros países, onde o consumo de pescado pode chegar a mais de 40kg/per
capita/ano.
Espécies de pescado de baixo valor comercial vêm sendo amplamente
estudadas com a finalidade de transformar essa matéria-prima em produtos aceitáveis
pela população, conduzindo a um maior consumo.

5
2.1.2 Corvina (Micropogonias furnieri)
A corvina (Micropogonias furnieri), é considerada um dos mais importantes
recursos costeiros da plataforma sul do Brasil, podendo atingir 70 cm de comprimento.
Apresenta ampla distribuição geográfica ocorrendo no Atlântico desde o México até a
Argentina (ELSDON e GILLANDERS, 2002). O tamanho mínimo para captura da corvina
é de 25cm medido da extremidade da cabeça até a nadadeira caudal (BRASIL, 2003).
Possui grande tolerância às variações de salinidade, o que facilita a alimentação e
melhores condições para proteger-se de predadores (CASTELLO, 1986). É um peixe
onívoro e prefere uma dieta baseada em pequenos crustáceos como caranguejos e
camarões (COSTA e ARAÚJO, 2003).
No ciclo de vida desta espécie os indivíduos jovens migram para as áreas
estuarinas e os adultos alcançam o litoral para reproduzir. Dessa forma, o tamanho da
população de corvina varia de um ano para outro em conseqüência da migração, a qual
também ocorre devido a disponibilidade de alimento (COSTA e ARAUJO, 2003)
Segundo Badolato et al. (1994), a composição proximal da corvina varia em
função das estações do ano, apresentando uma oscilação de 77,2 a 83,8% para o teor de
umidade, 14,5 a 20,7% para proteína, 0,8 a 1% de gordura e 1 a 1,2% de cinzas. Estas
variações também podem ocorrer devido a fatores como sexo, tamanho, ciclo reprodutor
e alimentação. Yeannes e Almandos (2003), avaliando a composição química de
diferentes espécies de peixes nacionais encontraram para a corvina valores médios de
79,1% para umidade, 19,2% de proteína, 1% de cinza e 0,8% de lipídios.
Esta espécie apresenta baixo rendimento no que se refere ao filé. Gonçalves e
Passos (2003), utilizando a corvina (Micropogonias furnieri) na elaboração de produtos
reestruturados encontraram rendimento de 32% para o filé, valor próximo ao encontrado
por Stori (2000), que foi de 34,8%. Enquanto que Bruschi (2001), estudando o
rendimento, composição química e perfil de ácidos graxos de pescados e seus resíduos
obteve rendimento de 36% para o filé da corvina, valor este baixo considerando outras
espécies como a pescada, sardinha e atum que apresentaram rendimentos 44%, 51% e
52% respectivamente. O rendimento do filé de corvina é baixo restando o resíduo que
contém cerca de 10% de carne.
A utilização da corvina como matéria-prima se deve pela mesma ser considerada
de baixo valor comercial. A espécie apresenta duas fases no seu desenvolvimento. Na
primeira fase, quando jovem, possui uma carne branca e macia, mas na segunda fase,
quando adulta, possui carne amarelada e mais rígida, e contaminada por cestóides que
migram do intestino do pescado para o músculo, causando uma proteólise indesejada no

6
mesmo, o que leva a corvina a ser pouco aproveitada (GONÇALVES e PASSOS, 2003).
O aspecto da corvina pode ser observado na Figura 1.
Figura 1: Corvina (Micropogonias furnieri)
2.2 Proteínas
Todas as proteínas são compostas de aproximadamente 20 aminoácidos, sendo
que a proporção dos mesmos varia conforme a característica da fonte de proteína. A
qualidade nutricional de qualquer proteína está ligada à sua composição de aminoácidos,
digestibilidade e capacidade de fornecer aminoácidos essenciais nas quantidades
requeridas pela espécie que consome a proteína. As proteínas são nutrientes essenciais
aos organismos animal e humano, devendo estar presentes na alimentação em
quantidades adequadas. Além do aspecto quantitativo, deve-se levar em conta o aspecto
qualitativo das proteínas, ou seja, seu valor nutritivo (SGARBIERI, 1996).
Sabe-se que as proteínas fornecem aminoácidos, que são os elementos que as
compõem. São usados para a formação de músculos e outros componentes do corpo
que contêm proteínas, incluindo imunoglobulinas, albuminas, enzimas e hormônios. O
organismo sintetiza aminoácidos não essenciais, enquanto outros que são essenciais
precisam ser fornecidos por fontes alimentares. As proteínas e outros compostos
contendo nitrogênio são constantemente degradados e reconstruídos. Todas essas
perdas devem ser repostas por um suprimento contínuo de aminoácidos fornecidos pela
dieta.
Cada animal, incluindo o homem, deve ter uma fonte adequada de proteínas
para crescer e se manter. Como as proteínas são os constituintes principais de tecidos
ativos do organismo, e este por sua vez depende das proteínas alimentares, a qualidade
e quantidade destas substâncias indispensáveis são de grande importância para a dieta
diária (MITCHELL et al., 1978). Em muitas partes do mundo, sobretudo nos países em
desenvolvimento, as fontes alimentares de proteína especialmente proteínas de boa
qualidade, são muito escassas. As proteínas há muito tempo foram reconhecidas como
elementos estruturais indispensáveis de cada célula do organismo. No seu papel como
enzimas, as proteínas controlam a degradação dos alimentos, para fornecerem energia e
para síntese de novos compostos a fim de manter e reparar tecidos do corpo

7
(ROBINSON, 1991). Cada tipo de proteína possui uma estrutura molecular única que
determina suas propriedades funcionais, em conjunto com as condições ambientais nas
quais se apresentam tais propriedades (DEMETRIADES et al., 1997).
2.2.1 Proteínas do Pescado
O músculo de pescado, assim como o de outras espécies, é composto de
proteínas de elevado valor nutritivo por conter alta proporção de aminoácidos essenciais,
principalmente aqueles que são limitantes em proteínas de origem vegetal, como por
exemplo, a lisina, um aminoácido limitante em cereais e farinha de trigo. (OGAWA e
MAIA, 1999; SGARBIERI, 1996). O músculo do pescado contém dois grupos principais
de proteínas: as proteínas solúveis do sarcoplasma e as proteínas estruturais das
miofibrilas. Os principais componentes das proteínas estruturais são: actomiosina,
tropomiosina, miosina e actina. As proteínas miofibrilares representam de 66 a 77 % das
proteínas totais do músculo do pescado e apresentam alta funcionalidade quando
comparadas com as proteínas sarcoplasmáticas (SIMÕES et al.,1998; CONTRERAS-
GUZMÁN, 1994). As proteínas presentes em vísceras e cartilagens apresentam em geral,
balanço de aminoácidos inferior ao das proteínas miofibrilares. No entanto, em produtos
de pescado, a qualidade da proteína irá variar dependendo, da proporção de proteínas
musculares neles contida (OGAWA e MAIA, 1999; SGARBIERI, 1996).
Segundo Ogawa e Maia (1999), a composição protéica da carne de pescado
pode variar em função da espécie, tamanho, sexo, época do ano, entre outros. Porém,
geralmente o músculo contém cerca de 20% de proteína. Ressalte-se que este
percentual é um pouco menor na carne escura do que na carne branca, verificando-se o
contrário em relação aos lipídeos. Bioquimicamente sabe-se que o músculo escuro tem
maior proporção de proteína sarcoplasmática e do estroma que o músculo branco e que
o conteúdo de glicogênio é mais alto em carnes escuras, e por isso, no rigor mortis, o pH
atinge valores menores, na faixa de 5,6 a 6,0, comparado com o músculo branco com
valores de 6,0 a 6,4.
Também o músculo escuro é mais rico em lipídeos, em taurina e em Fe,
contendo mais nitrogênio extrativo, sobretudo histidina (OGAWA e MAIA, 1999). As
proteínas sarcoplasmáticas representam aproximadamente 20-25 % da proteína total do
músculo, são solúveis em água e como principais características estão a sua capacidade
de adesão às proteínas miofibrilares, impedindo a formação de gel de alta elasticidade,
baixa viscosidade, baixa capacidade de retenção de água e baixa capacidade de
absorção de sabores e corantes (SIMÕES et al.,1998). Segundo Sgarbieri (1996),

8
particular atenção deve ser dada a preservação de aminoácidos sulfurados, uma vez que
a concentração destes aminoácidos não é muito alta nas proteínas do pescado.
As proteínas miofibrilares são as proteínas contidas nas células musculares,
formadoras dos tecidos esqueléticos e em grande parte, responsáveis pelo fenômeno de
contração muscular. Do ponto de vista alimentar, as proteínas miofibrilares são as
principais proteínas do músculo do peixe, e representam 10% do peso do músculo
estriado e 60 a 75% do conteúdo de proteína total do músculo. A miosina e actina, que
contribuem com cerca de 75% do total das proteínas miofibrilares, são responsáveis pela
contração e relaxamento muscular (OGAWA e MAIA, 1999).
Segundo este mesmo autor, particular atenção deve ser dada na preservação de
aminoácidos sulfurados, uma vez que a concentração destes aminoácidos não é muito
alta nas proteínas do pescado. Conforme Shahidi e Kamil (2001), as proteínas de
pescado podem ser utilizadas para diversos fins:
- fortificar o valor nutritivo de alimentos à base de cereais;
- preparação de pós e hidrolisados como agentes dietéticos e hipoalergênicos
e como ração animal;
- fornecer de maneira adequada a utilização de proteínas não-lácteas em
substitutos do leite para indivíduos jovens;
- contribuir para as propriedades atrativas de ração animal.
2.3 Enzimas
Enzimas são catalisadores bioquímicos essenciais para sobrevivência, pois
aceleram reações químicas entre constituintes orgânicos das células. Existe uma grande
variedade de enzimas disponíveis comercialmente que diferem em relação à fonte
biológica, atividade, pureza, forma física e características tais como pH e temperatura
ótima, além de seu preço. As enzimas utilizadas pela indústria de alimentos e em
pesquisas são predominantemente as hidrolases, a maioria carboidrases, seguido das
proteases e lípases. (WHITAKER, 1994).
Na ciência e tecnologia de alimentos, as enzimas são exploradas para realizar
determinadas funções em processos e análises facilitando a conversão de matérias-
primas em alimentos de alta qualidade. As enzimas tornam isso possível porque seu sítio
ativo é altamente específico para certos substratos. As enzimas agem de maneira
específica, formando um complexo com o substrato catalisando sua transformação
(KRISTINSSON e RASCO, 2000b).
A hidrólise de proteínas alimentares se dá por várias razões, incluindo
características de melhoramento nutricional, retardamento da deterioração, aumento ou

9
diminuição da solubilidade, aumento de propriedades espumantes ou coagulantes,
aumento da capacidade emulsificante, prevenção de interações indesejáveis, remoção de
sabores ou odores e ingredientes tóxicos ou inibidores (LAHL e BRAUN, 1994).
2.3.1 Enzimas proteolíticas
As enzimas proteolíticas, proteases ou proteinases pertencem ao grupo das
hidrolases as quais têm em comum o envolvimento da água na formação do produto. As
proteinases catalisam a reação de hidrólise das ligações peptídicas das proteínas,
ocorrendo a transferência de componentes do substrato para a água. Estas enzimas
podem ter origem animal, vegetal ou microbiana (WHITAKER, 1994).
As proteases estão presentes entre as enzimas mais estudadas. Preparados de
enzimas proteolíticas são economicamente, o mais importante grupo de enzimas e seu
uso está bem estabelecido na indústria de alimentos. As proteases são caracterizadas de
acordo com a especificidade de ligações peptídicas atacadas e o seu mecanismo de
ação (KRISTINSSON e RASCO, 2000a).
As enzimas proteolíticas derivadas de plantas e microrganismos são as mais
apropriadas para o desenvolvimento de hidrolisados protéicos de pescado. As enzimas
utilizadas na hidrólise da proteína de pescado possuem pelo menos uma característica
em comum que é fazerem parte da categoria dos alimentos, e, se forem de origem
microbiana, o organismo produtor tem que ser não-patogênico. A escolha da enzima é
normalmente determinada pela combinação entre eficácia e economia (KRISTINSSON e
RASCO, 2000a). A hidrólise de proteínas de pescado com enzimas proteolíticas
selecionadas, possibilita o controle do grau de hidrólise da proteína do substrato. Usando
proporções enzima-substrato e tempos de reação adequados é possível produzir
hidrolisados com diferentes estruturas moleculares e diferentes propriedades funcionais
que podem encontrar aplicações em vários produtos alimentícios (ONODENALORE e
SHAHIDI, 1996).
Quatro principais classes de proteases são conhecidas, sendo denominadas
pelo grupo funcional principal no seu sitio ativo: serina, tiol, carboxil e metalo. As
proteases são caracterizadas em endoproteinases e exopeptidases. As endoproteinases
hidrolisam as ligações peptídicas nas moléculas de proteínas, geralmente em resíduos
específicos a fim de produzir peptídeos relativamente grandes. As exopeptidases
removem sistematicamente aminoácidos do nitrogênio terminal chamadas
aminopeptidases ou do carbono terminal chamadas carboxipeptidases. Para hidrolisar
proteínas alimentares, geralmente são usadas endoproteinases, ocasionalmente
combinadas com exopeptidases para realizar uma degradação completa (KRISTINSSON
10
e RASCO, 2000a). No processo de hidrólise, as enzimas proteolíticas são utilizadas para
solubilizar a proteína muscular do pescado, resultando em duas fases distintas, a solúvel
e a insolúvel. A fração insolúvel é utilizada como ração animal, enquanto que a solúvel
pode ser convertida em ingredientes e incorporada em alimentos. O hidrolisado solúvel
pode ser submetido à desidratação, resultando numa forma mais estável, em pó, com
alta concentração protéica. Tal produto é conhecido como hidrolisado protéico de
pescado (DINIZ e MARTIN, 1996).
A Alcalase, uma enzima alcalina produzida por fermentação submersa de uma
espécie selecionada de Bacillus licheniformis, desenvolvida pela Novo Nordisk
(Bagsvaerd, Dinamarca) para indústria de detergentes, tem sido estudada repetidamente
por vários pesquisadores por ser uma das melhores enzimas usadas para preparar
hidrolisados protéicos de pescado funcionais e outros hidrolisados protéicos (BENJAKUL
e MORRISSEY, 1997; DINIZ e MARTIN, 1996; SHAHIDI et al., 1995). Apresenta
atividade ótima à temperatura entre 50ºC e 70ºC, a valores de pH entre 6,0 e 10,0 (DINIZ
e MARTIN, 1996).
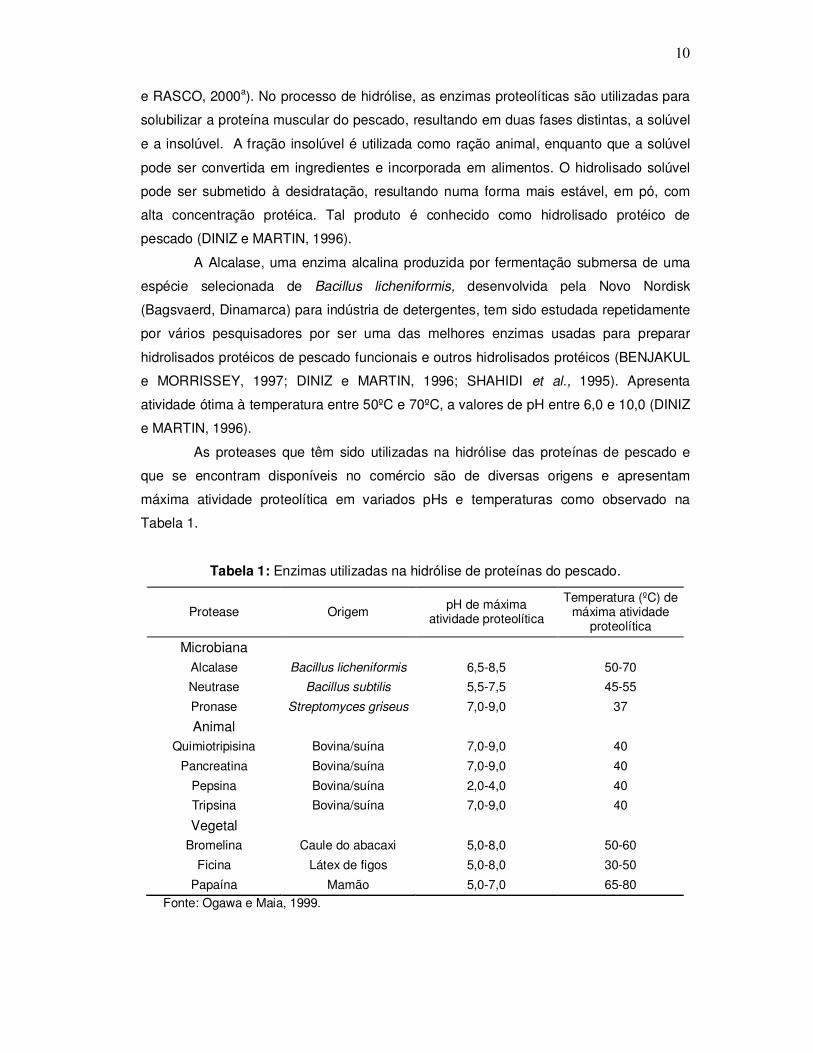
As proteases que têm sido utilizadas na hidrólise das proteínas de pescado e
que se encontram disponíveis no comércio são de diversas origens e apresentam
máxima atividade proteolítica em variados pHs e temperaturas como observado na
Tabela 1.
Tabela 1: Enzimas utilizadas na hidrólise de proteínas do pescado.
Protease Origem pH de máxima
atividade proteolítica
Temperatura (ºC) de máxima atividade
proteolítica
Microbiana
Alcalase Bacillus licheniformis 6,5-8,5 50-70
Neutrase Bacillus subtilis 5,5-7,5 45-55
Pronase Streptomyces griseus 7,0-9,0 37
Animal
Quimiotripisina Bovina/suína 7,0-9,0 40
Pancreatina Bovina/suína 7,0-9,0 40
Pepsina Bovina/suína 2,0-4,0 40
Tripsina Bovina/suína 7,0-9,0 40
Vegetal
Bromelina Caule do abacaxi 5,0-8,0 50-60
Ficina Látex de figos 5,0-8,0 30-50
Papaína Mamão 5,0-7,0 65-80 Fonte: Ogawa e Maia, 1999.

11
2.3.2 Fatores relacionados com a hidrólise enzimática
O processo hidrolítico e as condições de reação diferem entre diferentes
enzimas e substratos utilizados e também depende das propriedades desejadas para o
hidrolisado. A temperatura e o pH do processo são normalmente ajustados para otimizar
a cinética da enzima ou mistura de enzimas selecionadas. Uma protease comercial é
adicionada em variadas concentrações dependendo da taxa de hidrólise necessária.
Dada uma determinada enzima e um substrato em particular, qualquer processo de
hidrólise envolve ao menos 5 variáveis independentes. Estas são, pH, temperatura,
proporção enzima-substrato, concentração do substrato protéico em % ou unidades de
atividade por Kg, e tempo (KRISTINSSON e RASCO, 2000a).
2.3.2.1 pH
O pH do meio da reação é um fator importante na hidrólise enzimática. Este fator
está em função da natureza dos grupos funcionais da enzima, principalmente os grupos
dos sítios ativos da enzima, entretanto sofre influência da temperatura, pois influi no grau
de associação/dissociação dos grupos funcionais. A força iônica do meio é outro fator
que pode influir no pH em função da atividade iônica desse meio (WHITAKER, 1994).
A inativação química pode se dar tanto pela diminuição ou aumento do pH do
meio, até um ponto onde as enzimas forem inativadas. Algumas enzimas são mais
sensíveis para mudanças de pH do que para mudanças na temperatura. A alcalase é
uma enzima relativamente termoestável, mas é muito sensível em pH ácido. A completa
inativação da alcalase, portanto, é obtida pela redução do pH para o valor de 4 (SHAHIDI
et al., 1995). Extremos de pH podem também, assim como temperaturas elevadas, ter
efeitos negativos nas proteínas e peptídeos. Muitas proteínas se desdobram a valores de
pH menores que 5 e maiores que 10 (KRISTINSSON e RASCO, 2000a).
2.3.2.2 Temperatura
A temperatura é um dos fatores mais estudados na reação enzimática, e verifica-
se que a temperatura da reação de hidrólise determina a atividade enzimática, portanto,
recomenda-se utilizar temperaturas para máxima atividade das enzimas, ou utilizar um
intervalo de temperatura em que a estabilidade das enzimas e dos substratos não seja
comprometida (WHITAKER, 1994).

12
A temperatura na qual várias proteínas desnaturam e desdobram-se varia
enormemente. Como as proteínas do músculo de pescado se adaptam a baixas
temperaturas, elas não são estáveis ao calor. A desnaturação é normalmente indesejada
porque resulta na alteração das propriedades físico-químicas, particularmente na perda
de solubilidade e funcionalidade da proteína (KRISTINSSON e RASCO, 2000a).
Os mesmos autores relatam que o término da reação enzimática pode ser um
grande obstáculo na preparação de um hidrolisado protéico de pescado. A escolha do
método de inativação deve ser cuidadosamente baseado na sensibilidade da enzima que
está sendo estudada, em relação ao pH ou temperatura. Em alguns casos são utilizados
uma combinação de temperatura elevada e pH baixo.
A inativação das enzimas é alcançada por meios químicos ou térmicos.
Normalmente a pasta fluida do hidrolisado e as enzimas são transferidas para um banho
quente, onde as enzimas são inativadas através da sua exposição a temperaturas entre
75 e 100°C por 5 à 30 minutos, dependendo do tipo de enzima (HOYLE e MERRITT,
1994).
2.3.2.3 Concentração enzima versus concentração de substrato
A relação enzima/substrato (E/S) é um fator importante na cinética da reação e
influi também na recuperação do produto. A especificidade da enzima determina
principalmente a natureza do produto, podendo resultar frações de diferentes pesos
moleculares e conseqüentemente diferentes características funcionais. A natureza e o
estado de desnaturação das proteínas do substrato são fatores determinantes na taxa de
conversão da enzima influenciando na natureza do produto. Assim, podem resultar
peptídeos de diferentes tamanhos ou, até mesmo, aminoácidos livres em função da
natureza e do estado do substrato (WHITAKER, 1994).
Pelo aumento da concentração de proteases, até certo ponto e, desta forma,
aumento do grau de hidrólise, a recuperação de nitrogênio solúvel aumenta. A
concentração de substrato também possui efeitos negativos na recuperação das
proteínas. Linder et al. (1995), citados por Kristinsson e Rasco (2000a), encontraram que
mais de 8% de concentração protéica no sistema, sem considerar a enzima, pareceu ter
um efeito inibidor na recuperação das proteínas.
Espécies de pescado magras, ou materiais derivados destas, são o substrato
adequado para hidrólises enzimáticas, pois o problema de oxidação dos lipídios pode ser
reduzido. O preparo de hidrolisado protéico de pescado com espécies gordas como
arenque, sardinha e anchova, requererá tratamentos adicionais como centrifugação para
remover o excesso de gordura (KRISTINSSON e RASCO, 2000a).

13
2.3.3 Grau de Hidrólise da reação enzimática
Segundo Panyam e Kilara (1996), a hidrólise enzimática da proteína resulta na
diminuição do peso molecular, aumento do número de grupos ionizáveis e exposição de
grupos hidrofóbicos que estavam protegidos na estrutura original da proteína.
Para acompanhar a cinética de reação e encontrar a medida da hidrólise, aplica-
se o parâmetro chamado Grau de Hidrólise (GH). O critério quantitativo da reação de
proteólise é o grau de hidrólise (NIELSEN e OLSEN, 2002), definido como a porcentagem
de ligações peptídicas clivadas em relação ao total de ligações peptídicas (PILOSOF,
2000).
A vantagem de se usar o GH como parâmetro do processo é que o mesmo
parece determinar, sem ambigüidade, as propriedades de um hidrolisado protéico para
um dado sistema proteína-enzima. O GH demonstra tanto teórica como empiricamente
que 4 variáveis do processo, substrato (S), proporção enzima/substrato (E/S),
temperatura (T) e tempo (t), não precisam ser controladas, contanto que %GH seja
controlado. (ADLER-NISSEN, 1984, citado por KRISTINSSON e RASCO, 2000a). Assim,
fica claro que o GH é um método simples e rápido para medir a extensão da quebra das
proteínas (KRISTINSSON e RASCO, 2000a).
Quando o grau de hidrólise é atingido, é necessário terminar a reação
enzimática. Isto é muito importante, caso contrário as enzimas continuam ativas no
substrato hidrolisando as proteínas e es seguida os peptídeos (HOYLE e MERRITT,
1994).
Existem vários métodos de controle, mas sob condições industriais práticas
existem poucos. O grau de hidrólise é mais comumente empregado para descrever a
hidrólise de proteínas alimentícias.
2.4 Hidrolisados enzimáticos
Modificações químicas e enzimáticas têm sido usadas para melhorar as
propriedades funcionais das proteínas (POMERANZ, 1991), no entanto, a hidrólise
enzimática apresenta maiores benefícios e vantagens sobre a modificação química
porque inclui a especificidade da enzima em relação ao substrato, havendo pouca
probabilidade de ocorrer reações indesejáveis, que resultem na formação de produtos
tóxicos, além de se processar sob condições mais brandas (PATEL et al., 1988).
Os hidrolisados protéicos são obtidos por ação enzimática ou por processo
químico (ácido ou alcalino), obtendo-se uma mistura de peptídeos e aminoácidos. A
hidrólise com enzimas é mais indicada para a produção de hidrolisados com aplicações

14
nutricionais, que métodos químicos. A hidrólise enzimática pode produzir hidrolisados
com perfil de peptídeos bem definido, enquanto que as hidrólises ácida e alcalina podem
destruir L-aminoácidos, produzindo D-aminoácidos e ainda formar substâncias tóxicas
como lisino-alanina. Isso é especialmente crítico em hidrolisados protéicos, onde as
proporções de aminoácidos, dipeptídeos e tripeptídeos são importantes para a absorção
pelo organismo (LAHL e BRAUN, 1994). A hidrólise enzimática das proteínas musculares
de pescado é caracterizada por uma rápida fase inicial, durante a qual, várias ligações
peptídicas são hidrolisadas. Após, esta taxa de hidrólise enzimática decresce e atinge
uma fase estacionária, na qual nenhuma hidrólise aparente ocorre (KRISTINSSON e
RASCO, 2000a).
A hidrólise enzimática de proteínas de pescado é importante para o preparo de
produtos com propriedades funcionais únicas e características benéficas relacionadas à
saúde (SHAHIDI e KAMIL, 2001). Ela tem sido empregada para converter biomassa de
pescado subutilizada, a qual é geralmente destinada à produção de ração animal ou
fertilizantes, em produtos protéicos. Logo, a aplicação de tecnologia enzimática é um
método alternativo para um melhor aproveitamento de espécies sem valor comercial, o
que torna o custo baixo e eleva o valor protéico dos produtos disponíveis, sendo mais
aceitáveis para o consumo humano, na forma de uma nova fonte alimentar (DINIZ e
MARTIN, 1996).
Sampedro et al. (1986) consideraram que no processo de hidrólise enzimática se
obtém duas frações, uma solúvel e outra insolúvel, sendo que a fração insolúvel pode ser
usada como ingrediente para ração animal, enquanto o hidrolisado solúvel pode ser
submetido à desodorização, concentração e desidratação, resultando em um pó mais
estável, com alta concentração de proteína, normalmente denominado de hidrolisado
protéico de pescado que pode ser utilizado como ingrediente para produtos alimentícios
de consumo humano. Shahidi et al. (1995), descobriram que uma alta concentração de
peptídeos solúveis de pescado na mistura reacional, liberados durante a fase inicial de
hidrólise, reduz tanto a taxa de hidrólise quanto à recuperação das proteínas solúveis.
Assim, a remoção de hidrolisado da mistura reacional deveria acentuar a taxa de hidrólise
e a recuperação protéica.
Kristinsson e Rasco (2000a), informaram que a hidrólise protéica de pescado,
com enzimas proteolíticas selecionadas, fornece a possibilidade de controlar o grau de
quebra da proteína no substrato, indicando como vantagens que a utilização de
proporções enzima/substrato e tempos de reação adequados, permite a produção de
hidrolisados com estruturas moleculares e propriedades funcionais diferentes,
encontrando aplicações em várias formulações de alimentos.

15
A Figura 2 representa um processo típico de obtenção de hidrolisado protéico de
pescado sugerido por Kristinsson e Rasco (2000a).
Figura 2: Fluxograma do processo de obtenção de hidrolisado protéico de pescado.
2.5 Hidrolisados enzimáticos de pescado
A hidrólise de proteínas pode ser produzida com dois objetivos, alteração de
propriedades funcionais e produção de pequenos peptídeos e aminoácidos para serem
utilizados em alimentos dietéticos ou como agente flavorizante (KILARA, 1985). Shahidi
et al. (1995), usaram Alcalase com sucesso para otimizar as condições de processo para
produzir um hidrolisado protéico de capelin (Mallotus villosus). O hidrolisado produzido
com alcalase exibiu uma recuperação superior da proteína comparada com as enzimas
neutrase e papaína e também apresentou o menor conteúdo lipídico e propriedades
funcionais excelentes.
Quaglia e Orban (1990), citados por Kristinsson e Rasco (2000b), estudaram
essas três enzimas sob condições ótimas de solubilização enzimática de proteína de
sardinha. Os hidrolisados produzidos usando alcalase e papaína foram quase idênticos
na recuperação de nitrogênio, que aumentou proporcionalmente com a concentração da
Matéria-prima
Homogeneização (ajuste pH e Temp.)
Fração Solúvel
Inativação (química/termicamente)
Desidratação
Hidrolisado Protéico de Pescado
Centrifugação Proteína não hidrolisada e
outros materiais insolúveis
Enzimas
Reação Enzimática (tempos diferentes)

16
enzima. Os hidrolisados de alcalase e papaína apresentaram propriedades funcionais
melhores e com maior valor nutricional do que com a neutrase.
As enzimas alcalase e neutrase foram estudadas por Benjakul e Morrissey
(1997), em resíduo sólido branco de merluza em condições de pH 9,5 e 60°C e de pH
7,0, e 55°C. A alcalase teve uma atividade consideravelmente maior que a neutrase e
produziu uma hidrólise mais eficiente. Condições ótimas para alcalase foram 20 unidades
Anson (AU)/Kg, 1h de tempo de reação à 60°C e pH 9,5. O hidrolisado resultante teve um
alto conteúdo protéico com excelente recuperação de nitrogênio superior a 70%, e uma
composição de aminoácidos comparável ao músculo de pescado.
Novas enzimas têm mostrado um excelente potencial para hidrólise de proteínas
de pescado para produção de hidrolisados protéicos de alta funcionalidade, incluindo a
flavourzyme 1000L, fabricada pela Novo Nordisk, Bagsvaerd, Denmark e a corolase
7089, da indústria Rohm Enzymes, Somerset, NJ. (KRISTINSSON e RASCO, 2000b).
Estudo de Hale (1969) reporta os efeitos de várias condições de processamento
com enzimas proteolíticas comerciais avaliando o rendimento e a composição de
hidrolisados protéicos de pescado solúveis em água. Ele concluiu que a hidrólise de
abrótea crua com alcalase em pH 8,5 ou superior gerou o melhor balanço de
aminoácidos essenciais e uma porcentagem maior de produto solúvel, seguido pelo
hidrolisado obtido com pancreatina.
2.6 Efeito do grau de hidrólise nas propriedades funcionais de hidrolisados de
pescado.
Uma das principais vantagens e objetivos da hidrólise enzimática das proteínas
de pescado é modificar e melhorar suas propriedades funcionais. Essas propriedades
dos hidrolisados protéicos de pescado são importantes especialmente se eles vão ser
usados como ingredientes de produtos alimentícios (GILDGERG, 1993). Existem
dois métodos para produzir hidrolisados a partir de proteínas de pescado, o processo
autolítico e o processo de hidrólise acelerada. Em ambos os métodos as enzimas
proteolíticas catalisam a ruptura hidrolítica das ligações peptídicas das proteínas e dos
polipeptídios, produzindo peptídios de baixo peso molecular e/ou aminoácidos (MOHR,
1980; SIMPSON et al., 1991).
Estes dois métodos consistem em misturar o pescado moído com enzimas
endógenas ou com proteases microbianas ou de vegetais incubadas em reatores sob
condições ótimas de atividade enzimática. A escolha da enzima e das condições de
processamento determinam as propriedades do produto, sendo que condições brandas
como, períodos curtos de hidrólise e temperaturas moderadas são importantes para obter

17
propriedades emulsificantes e geleificantes dos produtos (MOHR, 1980; OWENS e
MENDOZA, 1985).
A hidrólise prolongada e não controlada pode produzir peptídeos de cadeia curta
com perda das propriedades funcionais das proteínas nativas, podendo contribuir desta
forma com surgimento de um gosto amargo nos produtos obtidos. Por isso, o grau de
hidrólise se torna importante na otimização dos parâmetros de processo no
desenvolvimento do produto (VENUGOPAL e SHAHIDI, 1995). Em geral, as
endopeptidases menos específicas das bactérias e dos vegetais, são mais eficientes na
solubilização do músculo de pescado que enzimas mais específicas como pepsina e
tripsina (MACKIE, 1982). Lahl e Braum (1994), verificaram que papaína, ficina e
bromelina não foram seletivas quando comparadas com enzimas microbianas, que
tendem a serem mais seletivas. Tanto a tripsina como a quimiotripsina apresentaram
atividade de endo e exopeptidases com especificidade para clivar a ligação arginina-
lisina.
Os mesmos autores citam como desvantagens que a hidrólise de proteína de
lote comercial pode apresentar um alto custo pela utilização de grandes quantidades de
enzimas, dificuldade no controle da extensão da reação (que pode resultar em produtos
não-homogêneos, consistindo de frações de peso molecular variável), baixo rendimento,
a necessidade de inativar enzimas pelo pH ou tratamento térmico no final das reações, (o
que eleva os custos do processo) e ainda que as enzimas empregadas no processo não
podem ser reutilizadas. Finalmente, eles consideraram que as operações do processo
enzimático podem ser bem dimensionadas, desde que a especificidade da enzima, as
condições da reação enzimática (temperatura, pH, meio reacional, tempo) e as condições
da inativação da enzima sejam conhecidas.
2.7 Propriedades funcionais das proteínas
A funcionalidade das proteínas dos alimentos é definida como “aquelas
propriedades funcionais e químicas que afetam o comportamento das proteínas nos
alimentos durante o processamento, armazenamento, preparo e consumo” (KINSELLA,
1976). As propriedades físicas e químicas que governam a funcionalidade das proteínas
incluem tamanho, forma, composição e seqüência de aminoácidos, carga líquida,
distribuição das cargas, relação hidrofobicidade/hidrofilicidade a e capacidade de reagir
com outros componentes. Tem se verificado, por exemplo, que a hidrólise de proteínas
de pescado com proteases, resulta em produtos com melhores propriedades de
solubilidade e melhores propriedades funcionais que dependem da solubilidade
(DAMODARAM, 1996).

18
Sgarbieri (1998), explicou que o termo “funcionalidade” tem sido aplicado para se
referir às propriedades não-nutritivas que conferem aos alimentos maior conveniência de
manuseio, melhor aparência na apresentação e melhor aceitação pelo consumidor e que
a maioria das propriedades funcionais influenciam o caráter sensorial de um alimento,
em especial a textura. De acordo com este autor, a manifestação de funcionalidade, por
certos componentes dos alimentos, irá depender de propriedades físicas e químicas
desses componentes que são muito importantes para o preparo de determinados tipos de
alimentos e sua aceitação pelo consumidor, como:
- Propriedades de hidratação, dependentes das interações com água como
solubilidade, dispersibilidade, suculência, adsorção e retenção de água.
- Propriedades dependentes da interação entre macromoléculas e a água, como
geleificação, textura, formação de estruturas viscoelásticas.
- Propriedades que dependem do tamanho e da forma das moléculas ou de
partículas em suspensão como viscosidade, sensação tátil, modificada em função da
transformação de material sólido em micropartículas.
- Propriedades que resultam da interação de um grande número de constituintes
dos alimentos, tais como, a água, as proteínas, polissacarídeos como amido e
componentes solúveis e insolúveis da fibra alimentar.
Mullally et al. (1995), informaram que a hidrólise enzimática da proteína de
pescado gera uma mistura de aminoácidos livres, di-, tri-, e oligopeptídios, aumenta o
número de grupos polares e a solubilidade do hidrolisado e, portanto, modifica as
características funcionais das proteínas, melhorando suas qualidades funcionais e
biodisponibilidade. A escolha do substrato e das proteases empregadas e o grau de
hidrólise da proteína afetam as propriedades físico-químicas do hidrolisado resultante. A
hidrólise de produtos com enzimas é um meio atraente de oferecer melhores
propriedades funcionais e nutricionais às proteínas de alimentos. A Tabela 2 mostra
algumas das mais importantes propriedades das proteínas e suas aplicações nos
alimentos.
As propriedades funcionais das proteínas também são influenciadas por diversos
parâmetros intrínsecos, além de fatores relacionados a metodologias utilizadas no seu
estudo. Os dados disponíveis são bastante variáveis e muitas vezes conflitantes, devido
a fatores interferentes como pH, força iônica, concentração de proteína e temperatura do
meio (DUARTE et al,. 1998).

19
Tabela 2: Propriedades funcionais de proteínas nos alimentos e suas aplicações
Propriedade Aplicação
Emulsificação Carnes, molhos de salada
Hidratação Massas, carnes
Viscosidade Bebidas, massas
Geleificação Salsichas, sobremesas de geléia, queijos
Espumosidade Coberturas, merengues, pães-de-ló
Propriedades texturais Alimentos texturizados
Solubilidade Bebidas
Fonte: Sgarbieri,1998.
2.7.1 Solubilidade
A solubilidade é uma propriedade física e também funcional muito importante,
além do que influi em outras propriedades funcionais, em graus diferentes (SGARBIERI,
1998). A solubilidade é provavelmente a propriedade funcional mais importante das
proteínas e dos hidrolisados protéicos. Muitas das outras propriedades funcionais, como
emulsificação e a formação de espuma, são afetadas pela solubilidade, sendo, portanto
um excelente indicador da funcionalidade do hidrolisado protéico, do seu potencial e da
limitação de sua aplicação (KRISTINSSON e RASCO, 2000a).
Segundo Sgarbieri (1998), alguns dos fatores que afetam a solubilidade das
proteínas são: pH, distribuição de cargas, força iônica ou concentração de sais,
temperatura e natureza do solvente. O pH afeta a densidade e a distribuição das cargas
na molécula da proteína de tal forma que no pH do ponto isoelétrico (pI), a proteína
apresentará um mínimo de solubilidade, devido ao número de cargas positivas e
negativas ser igual, portanto, elas se neutralizam intramolecularmente. Nessas
condições, as moléculas de proteína apresentam menor afinidade pela água, atraindo-se
mutuamente e formando uma massa insolúvel que precipita.
De acordo com o autor, à medida que o pH torna-se mais ácido ou mais alcalino,
há predominância, respectivamente, de cargas positivas ou negativas, havendo interação
mais forte das moléculas de proteína com a água e também uma maior repulsão entre
moléculas de proteína, aumentando significativamente a solubilidade. A diferença de
comportamento na solubilidade das proteínas se deve as propriedades físicas, químicas
e estruturais dessas proteínas. As proteínas miofibrilares de pescado possuem pI entre
pH 5 e 6 e são praticamente insolúveis em pHs ácidos neutros e ligeiramente alcalinos.

20
Ele relata ainda, que a solubilidade aumenta abruptamente a partir de pH 11,
sugerindo que essas proteínas poderão estar formando complexos insolúveis e que
somente a predominância de cargas negativas em pHs fortemente alcalinos geram forças
de repulsão eletrostática capazes de promover a dissociação dos complexos e
solubilização.
A temperatura aumenta a solubilidade das proteínas até o ponto em que começa
a ocorrer desnaturação térmica. A desnaturação da molécula da proteína coloca grupos
hidrofóbicos em contato com o solvente, ocasionando diminuição de solubilidade,
podendo haver coagulação e precipitação. Temperaturas acima de 50ºC podem provocar
desnaturação e diminuição de solubilidade nas proteínas de acordo com o autor.
2.7.2 Capacidade de formação de espuma
As espumas alimentares consistem em gotas de ar dispersas e envolvidas em
líquido que contém um surfactante que diminui a tensão interfacial e de superfície do
líquido (KINSELLA, 1976). As propriedades de formação de espuma se relacionam com
as propriedades de superfície que as proteínas apresentam (KRISTINSSON e RASCO,
2000a). A capacidade de formar espuma estável em presença de ar é uma propriedade
funcional importante das proteínas em muitos produtos alimentícios do tipo bolos de anjo,
suspiros, suflês, vários tipos de coberturas de bolos e sobremesas, além de bebidas
como cervejas. Várias substâncias tensoativas como detergentes e saponinas são
formadoras de espumas, mas em alimentos, as proteínas são as mais importantes
(SGARBIERI, 1998).
A qualidade espumante de uma proteína deve ser estudada quanto a
estabilidade e densidade de sua espuma e quanto ao volume formado da mesma sob
certas condições. A capacidade de formação de espuma é medida pelo aumento do
volume de uma dispersão de proteína como resultado da incorporação de ar por
batimento, agitação ou aeração. Proteínas ideais para formação de espuma têm a
propriedade de formar membranas extensíveis em torno das bolhas de ar passando por
um certo grau de desnaturação que parece ser crítico para a estabilização das espumas
e de sua estrutura tridimensional (SGARBIERI, 1998).
Um bom critério para a avaliação da estabilidade da espuma é a medida do
tempo em que ocorre algum evento, como o tempo decorrido desde a formação até a
ruptura total das bolhas. Obtém-se, neste caso, o tempo de vida da espuma, determinado
por dois processos que são o afinamento e o colapso dos filmes. Soluções diversas
podem formar espumas cujo tempo de vida pode ser de apenas alguns segundos ou de
até centenas de meses. A evolução dos processos vai depender da natureza química do

21
agente espumante, considerando-se que seu papel consiste basicamente em estabilizar
os filmes de espuma, e do controle de efeitos externos (FIGUEREDO et al., 1999).
As propriedades da espuma de hidrolisados protéicos de pescado tem sido
pouco estudadas. Shahidi et al. (1995), e Onodenalore e Shahidi (1996), citados por
Kristinsson e Rasco (2000a) estudaram a formação de espuma e a estabilidade da
espuma de hidrolisados protéicos de tubarão e capelin. Em ambos processos, a Alcalase
foi a protease utilizada para hidrólise. O hidrolisado protéico de capelin obteve uma boa
formação de espuma, de 90%, com 12% de grau de hidrólise. O hidrolisado protéico de
carne de tubarão apresentou uma maior porcentagem de formação de espuma, 106%, e
o hidrolisado de polpa lavada de tubarão mostrou uma capacidade de formação de
espuma de 130%. O hidrolisado protéico de capelin teve uma estabilidade de espuma
maior após 30 segundos quando comparada com a proteína hidrolisada de tubarão.
2.7.3 Capacidade emulsificante
Emulsões são definidas como misturas de pelo menos dois líquidos imiscíveis,
estando um deles disperso no outro em forma de minúsculas gotículas. Os materiais
imiscíveis são usualmente a água e um óleo ou gordura líquida. Como a água e o óleo
são imiscíveis, há necessidade de agitação mecânica para que se forme uma dispersão
homogênea dos dois líquidos. Contudo, essa dispersão não constituirá uma emulsão em
virtude da tendência natural de separação dos dois líquidos. Uma emulsão alimentícia
deverá ser estável por, pelo menos, alguns meses. O leite é um exemplo de emulsão
alimentícia estável, em que as partículas de gordura estão dispersas em solução aquosa.
O líquido que aparece disperso em forma de gotícula constitui a fase dispersa,
descontínua ou interna (SGARBIERI, 1998).
Para este autor, a capacidade de emulsificação é normalmente definida como o
volume de óleo (mL) que pode ser emulsionado pelo hidrolisado protéico (g), antes que
ocorra a inversão da fase ou colapso da emulsão. Como os hidrolisados protéicos de
pescado produzem emulsões de baixa viscosidade, a maneira mais apropriada de medir
esta propriedade é através do método de titulação, medindo a resistência elétrica durante
a titulação do óleo. O repentino aumento da resistência é observado quando o máximo da
capacidade de emulsificação é alcançado.
A estabilidade da emulsão refere-se à habilidade de uma emulsão de resistir a
mudanças em suas propriedades ao longo do tempo e depende das características físico-
químicas das moléculas das proteínas e das propriedades hidrodinâmicas e
viscoelásticas do filme de proteína formada (SGARBIERI, 1998). A medida envolve uma
mistura de hidrolisado com o óleo e água, centrifugação, e medida total do volume da

22
emulsão, onde a estabilidade é expressa como a diferença entre o volume total da
emulsão e o volume de água com relação ao volume total (KRISTINSSON e RASCO,
2000a).
As propriedades de emulsificação estão diretamente ligadas às propriedades de
superfície, o que demonstra o quão eficiente são os hidrolisados na sua ação de diminuir
a tensão interfacial entre componentes hidrofóbicos e hidrofílicos. As proteínas adsorvem
na sua superfície as gotas de óleo formadas durante a homogeneização transformando-
se em uma membrana protetora que previne a coalescência das gotas (DEMETRIADES
et al., 1997). Os hidrolisados apresentam esta “atividade superficial” promovendo a
formação de emulsões óleo em água, pois eles possuem grupos funcionais hidrofílicos e
hidrofóbicos sendo também solúveis em água (WILDING et al., 1984).
As propriedades de emulsificação das proteínas hidrolisadas são melhoradas
quando a extensão da hidrólise é cuidadosamente controlada, já que uma hidrólise
extensiva resulta numa drástica perda das propriedades emulsificantes (MAHMOUD,
1994). A distribuição dos pesos moleculares dos hidrolisados mostra que altos conteúdos
de frações protéicas de elevado peso molecular desempenham um papel importante na
estabilidade das emulsões. Os hidrolisados com grau de hidrólise baixo possuem
hidrofobicidade superficial maior, como foi demonstrado experimentalmente com
hidrolisados de sardinha, obtidos por ação da alcalase, de baixo grau de hidrólise,
apresentando melhor capacidade emulsificante que caseinato de sódio comercial
(QUAGLIA e ORBAN, 1990). Estes autores relatam que há uma relação entre o grau de
hidrólise e as propriedades de emulsificação dos hidrolisados protéicos de pescado. A
hidrólise enzimática possui uma influência negativa na capacidade de formar uma
emulsão estável à medida que o grau de hidrólise aumenta. Hidrolisados com diferentes
distribuições de pesos moleculares demonstraram que um conteúdo maior de frações
protéicas de alto peso molecular desempenha um papel importante no que diz respeito à
estabilização da emulsão.
A hidrólise parcial das proteínas geralmente aumenta o número de grupos
polares e hidrofílicos, diminui o peso molecular, altera a estrutura globular das proteínas
podendo expor maior número de grupos hidrofóbicos. Estas alterações podem afetar as
propriedades emulsificantes (NIELSEN, 2002).
2.7.4 Capacidade de retenção de água
A capacidade de retenção de água é um fenômeno importante na tecnologia de
alimentos, pois a água absorvida em pequenas quantidades não atua como solvente,
mas contribui para dar corpo e aumentar a viscosidade (Cândido et al., 1998), ou seja,

23
refere-se à habilidade das proteínas de embeber água e retê-la dentro da matriz protéica
como acontece com os géis protéicos de músculo bovino e de pescado, sendo possível
correlacioná-la com a capacidade de ligar a água (DAMODARAN, 1996). A capacidade
de retenção de água das proteínas é de grande importância na indústria de alimentos
devido a que a retenção de água em um alimento normalmente melhora sua textura.
Deve-se observar que o estado de desnaturação das proteínas também influencia na
capacidade de retenção de água.
Para Kristinsson e Rasco (2000a), as propriedades funcionais das proteínas em
um sistema alimentício dependem, em parte, da interação proteína-água dependendo o
resultado final de como e quanto às proteínas ligam e mantém água no sistema. Os
hidrolisados protéicos de pescado possuem excelente capacidade de retenção de água,
sendo adequados para certas formulações de alimentos.
Em um estudo de Kristinsson e Rasco (1998), hidrolisados protéicos de salmão
com diferentes graus de hidrólise (5, 10 e 15%) foram adicionados a pastas de salmão
moído, sendo que esses produtos apresentaram perdas de água de 1% após o
congelamento e descongelamento, armazenados durante 48 dias, quando comparados
com o controle que apresentou perdas de 3%.
2.7.5 Capacidade de retenção de óleo
A absorção de gordura é uma propriedade funcional muito considerada. No caso
dos hidrolisados, estes são misturados com uma quantidade específica de excesso de
gordura por um tempo determinado, e em seguida centrifugados com força centrífuga
baixa, sendo a quantidade de gordura adsorvida, expressa em mililitros de gordura ligada
por grama de proteína do hidrolisado (SHAHIDI et al.,1995; KRISTINSSON e RASCO,
1998).
De acordo com Donadel e Prudencio-Ferreira (1999), a absorção de gordura
varia em função do número de grupos hidrofóbicos expostos da proteína e,
provavelmente, as cadeias laterais não polares das proteínas tem afinidade com as
cadeias hidrofóbicas da molécula de óleo, contribuindo para a absorção.
O mecanismo de absorção de gordura comprovada por este método, é atribuído
principalmente à captura física do óleo sendo maior a gordura absorvida para maior
densidade protéica (KINSELLA, 1976). A capacidade de ligação de gordura das proteínas
se correlaciona com a hidrofobicidade superficial. A absorção de gordura de hidrolisado
protéico de salmão, obtidos com diferentes graus de hidrólise foi estudada por Kristinsson
e Rasco no ano de 1998, verificando que os hidrolisados protéicos de pescado em todos

24
os casos apresentaram uma excelente absorção de gordura, maior que os concentrados
protéicos de albumina de ovo e de proteína de soja.
Em estudo de CHO et al. (2004), foram avaliadas as propriedades funcionais de
gelatina de tubarão (Isurus oxyrinchus). As propriedades analisadas foram CRA e CRO,
onde foram comparados três tipos de gelatina, sendo elas, gelatina da cartilagem,
gelatina de grau analítico e gelatina obtida em aditivos alimentares. Os autores
verificaram que a CRO da gelatina obtida da cartilagem de tubarão foi maior do que as
outras duas gelatinas. Para eles, a CRA e a CRO são propriedades funcionais que estão
relacionadas com textura pela interação entre componentes como água, óleo e outros
compostos.
2.8 Propriedades Nutricionais
Os métodos de preparação dos hidrolisados protéicos podem ter um profundo
efeito nas suas propriedades funcionais, assim como nas suas propriedades nutricionais.
Eles podem aumentar o potencial dos pescados de baixo custo comercial agregando
valor ao produto, apresentando bom perfil de aminoácidos essenciais e boas
propriedades funcionais (KRISTINSSON e RASCO, 2000a). As proteínas de pescado
podem ser usadas para fortificar o valor nutricional dos alimentos a base de cereais, para
preparar produtos em pó e hidrolisados com finalidades dietéticas e hipoalergênicas,
assim como para a alimentação animal (SHAHIDI e VENUGOPAL, 1994), e também para
proporcionar uma maneira promissora de usar proteínas não lácteas como substitutos
lácteos para animais jovens (REBECA et al., 1991), e por contribuir nas propriedades
sensoriais atrativas para ração animal. Onodenalore e Shahidi (1996), informaram que a
hidrólise enzimática de proteínas de pescado é importante para o preparo de produtos
com propriedades funcionais únicas e com características benéficas a saúde.
A hidrólise enzimática controlada de proteínas tem atingido considerável
importância na ciência dos alimentos e nutrição, muitas vezes melhorando as
propriedades funcionais e nutricionais dos alimentos. Ela também melhora a
digestibilidade sem afetar o valor nutritivo das proteínas (KRISTINSSON e RASCO,
2000b). Cândido e Sgarbieri (2003), elaboraram um concentrado protéico de carne
mecanicamente separada de pescado eviscerado e o compararam com um hidrolisado
protéico de pescado obtido com a enzima flavourzyme, o qual foi preparado com um grau
de hidrólise abrangendo de 3,5 a 4,5%. A parte insolúvel dos hidrolisados apresentou
melhor perfil de aminoácidos que a fração solúvel. O valor nutricional determinou que a
fração insolúvel não diferiu do hidrolisado total, e foi consideravelmente maior que o da
fração solúvel.

25
A qualidade nutricional de hidrolisado protéico em pó de tilápia foi analisada por
Abdul-Hamid et al. (2002), utilizando duas temperaturas diferentes de secagem, 150°C e
180°C. A temperatura mais alta utilizada resultou num decréscimo no conteúdo de todos
os aminoácidos analisados para o hidrolisado. Todavia, a qualidade protéica dos dois
hidrolisados foi alta, com valores de digestibilidade in vitro de 88,4 e 92%
respectivamente. Dentre os métodos de avaliação da qualidade nutricional das proteínas
tem-se o poder de proteólise ou digestibilidade da proteína in vitro. Segundo Sgarbieri
(1996), a digestibilidade de uma proteína deve ser entendida como sendo a parte ou
porção da proteína que pode ser hidrolisada pelas enzimas digestivas até aminoácidos e
que portanto, estaria disponível biologicamente, desde que não houvesse nenhuma
interferência na absorção dos aminoácidos pelo organismo humano.

26
3 MATERIAL E MÉTODOS
3.1 Material
3.1.1 Matéria-prima
A matéria-prima utilizada para a obtenção dos hidrolisados foi a corvina
(Micropogonias furnieri), espécie de baixo valor comercial, doada pela indústria PESCAL
S.A, localizada na cidade de Rio Grande, RS.
3.1.2 Reagentes
Os reagentes químicos utilizados durante o processamento assim como nas
análises químicas formam todos de qualidade analítica (P.A).
3.1.3 Enzima
A enzima proteolítica utilizada foi a Alcalase 2.4L, fornecida pela Novozymes
Latin América Ltda.
3.1.4 Infra-estrutura
O trabalho experimental foi desenvolvido nos laboratórios de Tecnologia de
Alimentos e de Bioquímica Tecnológica, localizados no Campus Cidade da Fundação
Universidade Federal do Rio Grande (FURG), RS.
3.2 Métodos
3.2.1 Obtenção dos filés e da polpa de corvina
O pescado foi transportado em recipiente com gelo até o Laboratório de
Tecnologia de Alimentos da FURG. Foi realizada uma pré-lavagem com o intuito de
retirar as sujidades superficiais antes de se iniciar a etapa de descabeçamento e
evisceração, feita de modo que não houvesse contaminação do pescado pelas vísceras,
seguida da filetagem. Os filés limpos foram acondicionados em embalagens de polietileno
e armazenados sob congelamento em freezer horizontal em temperatura de -18 ºC.

27
3.2.2 Determinação da atividade específica da enzima
A atividade específica da enzima foi determinada conforme Gildberg et al.
(2002), Rebeca et al., (1991), e Gildberg e Raa (1983), usando caseína como substrato.
Realizou-se uma hidrólise com caseína (2%) e outra com o filé de pescado como
substrato (2% proteína) a uma temperatura de 40ºC, pH 7,0 (tampão fosfato 0,2M, vide
Anexo E) e 1% de enzima (p/p), durante um intervalo de duas horas. A atividade
enzimática foi expressa como unidade de atividade (U) que corresponde à quantidade em
gramas de enzima capaz de liberar ou produzir 1µmol de tirosina por minuto, nas
condições do experimento.
3.2.3 Caracterização fisico-química
3.2.3.1 Da matéria-prima
Os filés foram descongelados em refrigerador por 14 horas, homogeneizados em
triturador de facas duplas (Arno, modelo PL, “pic-liq”) e caracterizados fisicamente
mediante a determinação de pH e rendimento. O pH foi medido através da utilização de
potenciômetro de bancada e o rendimento foi calculado a partir do peso dos filés obtidos
em relação ao peso inicial do pescado. A composição proximal do filé de corvina foi
determinada de acordo com a metodologia oficial da AOAC (1995), onde proteínas foram
determinadas pelo método de Kjeldahl (N x 6,25), lipídios pelo método de Soxhlet, cinzas
por método gravimétrico em mufla 550-600ºC e umidade por método gravimétrico em
estufa 105ºC. Todas as determinações foram realizadas em triplicata.
3.2.3.2 Dos hidrolisados
Os hidrolisados foram caracterizados fisicamente mediante a determinação do
rendimento. Quanto à caracterização química determinaram-se os maiores componentes,
de acordo com a metodologia oficial da AOAC (1995), onde proteína foi determinada pelo
método de Kjeldahl (N x 6,25), cinzas por método gravimétrico em mufla 550-600ºC e
umidade por método gravimétrico em estufa 105ºC. Todas as determinações foram
realizadas em triplicata.

28
3.2.4 Obtenção do hidrolisado protéico de pescado
Para a determinação das faixas de trabalho da reação enzimática, para atingir
graus de hidrólise adequados sem prejudicar as características funcionais e nutricionais
dos hidrolisados consultou-se a literatura científica (GBOGOURI et al., 2004, DINIZ E
MARTIN, 1996). Foram realizados inicialmente, testes para obtenção de hidrolisados com
a protease alcalase, nas faixas ótimas de pH e temperatura da enzima.
3.2.4.1 Testes preliminares e definição das condições de hidrólise enzimática
Para determinar o tempo de hidrólise, foram realizados testes preliminares.
Inicialmente foram realizados dois experimentos para obter hidrolisados em diferentes
graus de hidrólise, que se denominou teste preliminar 1 (T1). As condições de reação
empregadas foram: pH 8,0, concentração de enzima 2,5% (p/p), concentração de
substrato 1,5% (p/v), e tempo de 60 minutos em temperaturas de 50 e 55ºC. Foram
tomadas amostras de hidrolisados nos tempos de 15, 30, 45 e 60 minutos, determinando-
se o grau de hidrólise em cada tempo. Testes posteriores foram realizados com o objetivo
de obter valores maiores de grau de hidrólise, que se denominou teste preliminar 2 (T2)
onde foram pesquisadas novas condições de reações, alterando a concentração de
enzima [E] e a concentração de proteínas do substrato [S], com base na literatura
(GBOGOURI et al., 2004). As condições da reação foram: pH 8,0, temperatura 50ºC, [E]:
4%, 6% e 8% (p/p) e [S]: 3%, 4% e 5% (p/v).
3.2.4.2 Fluxograma geral dos hidrolisados protéicos
O fluxograma geral utilizado no processo de obtenção dos hidrolisados protéicos
de pescado pode ser visualizado na Figura 3. As reações enzimáticas foram conduzidas
com a enzima em estudo, em valores ótimos de pH e temperatura estabelecidos com
base na literatura e através dos testes preliminares, variando-se as concentrações de
substrato e de enzima.

29
Figura 3: Fluxograma da produção de hidrolisado enzimático de pescado.
Na Figura 4, observa-se o sistema utilizado para as reações de hidrólise,
composto por um reator de vidro encamisado (A), aberto com capacidade de 1000mL,
conectado a um banho ultratermostático (Quimis, modelo 214. D2) (B) para controle de
temperatura e um agitador eixo-élice (Quimis, modelo Q-215 D2K) (C).
Figura 4: Sistema utilizado para as reações de hidrólise enzimática.
Vísceras, pele, espinhos, escamas
Pescado inteiro
Pré-Lavagem
Descabeçamento e Evisceração
Lavagem
Água
Polpa
Filetagem
Resíduo
Resíduo
Adição de solvente /Ajuste do pH
Homogeneização
Reação Enzimática
Inativação
Desidratação
Adição da enzima
Térmica TCA
Hidrolisado Protéico de Pescado
Controle de pH e T
A
B C
(A) reator de vidro encamisado (B) banho ultratermostático (C) agitador eixo-élice

30
Foram realizados ensaios com um volume total de 800mL, em reator
encamisado mediante agitação de 600rpm (GBOGOURI et al., 2004), com agitador eixo-
élice a uma temperatura de 50ºC, por 60 minutos. O pH foi ajustado utilizando tampão
fosfato pH 8,0 (MORITA, 1972). Tomaram-se amostras no final da reação para a medida
do grau de hidrólise de cada ensaio. Após o término da reação, a enzima foi inativada
termicamente a 85ºC por um tempo de 15 minutos.
Os hidrolisados inativados foram desidratados em estufa com circulação forçada
de ar (modelo Q314M, Quimis) em temperatura de 60°C por um tempo de 10 horas. Os
hidrolisados secos foram moídos em triturador de facas duplas (Arno, modelo PL “pic-
liq”), peneirados e acondicionados em recipientes herméticos em temperatura de
refrigeração.
3.2.4.3 Medida do grau de hidrólise
Previamente foram determinados intervalos de tempos onde amostras de
hidrolisado seriam retiradas. A reação foi interrompida com adição de solução de ácido
tricloroacético (TCA 6,25%) conforme Pezoa e Salas-Mellado (1979). Tomaram-se 6mL
de amostra e adicionaram-se 4mL de TCA (6,25%). Após o repouso de 15 minutos a
amostra foi filtrada em papel filtro (Whatman, nº1) e o teor de proteínas solúveis foi
determinado no filtrado pelo método de Lowry et al. (1951), expressando a concentração
de proteínas como mg de albumina, através de leitura em espectrofotômetro visível
(Biospectro, modelo SP-22) à absorbância de 750 ηm. Para tal, foi necessária a
construção de uma curva padrão de albumina. O grau de hidrólise foi expresso como a
relação entre as proteínas solubilizadas e as proteínas totais presentes no substrato
inicial, em % de acordo com a equação 1.
% GH = mg proteína hidrolisada (solubilizadas em TCA 2,5%) x 100 (1)
mg proteína total
3.2.4.4 Determinação da funcionalidade dos hidrolisados enzimáticos de corvina
Os hidrolisados enzimáticos secos em pó foram analisados pelas propriedades
funcionais de solubilidade, capacidade de retenção de água, capacidade de retenção de
óleo, capacidade emulsificante e capacidade de formação de espuma.

31
3.2.4.4.1 Solubilidade
A solubilidade (%S) dos hidrolisados protéicos de corvina foi determinada de
acordo com o método descrito por Morr et al. (1985), com variação de pH na faixa de 3 a
11. Pesou-se 500 mg de proteína em um béquer de 50 mL, adicionou-se 2 mL de NaCl
0,1M obtendo-se uma pasta homogênea. A seguir foi adicionada solução tampão
(MORITA, 1972), no pH correspondente até o volume de 40 mL.
A dispersão protéica foi mantida sob agitação por 45 minutos em agitador
magnético (QUIMIS, modelo 261-2) e após transferida para um balão volumétrico de 50
mL completando o volume com solução tampão correspondente. Em seguida,
centrifugou-se a dispersão por 30 minutos a 9000xg em centrífuga de tubos (Biosystems,
tipo MPW-350/350R) e tomando-se alíquotas para determinação do conteúdo de proteína
solúvel pelo método descrito por Lowry et al. (1951). Os teores de proteína total e solúvel
no sobrenadante foram determinados pelo método de Kjeldahl, (NT x 6,25) e pelo método
de Folin-Ciocalteau de acordo com Lowry et al. (1951), respectivamente.
A solubilidade da proteína foi calculada como a quantidade de proteína no
sobrenadante dividida pela quantidade de proteína total x 100. Determinou-se a %
solubilidade conforme a equação 2.
%S = A x 50 x 100 (2)
W x P
100
Onde: A = Concentração de proteína no sobrenadante (mg/mL);
W = Peso da amostra (mg);
P = Quantidade de proteína na amostra (determinada pelo método de Kjeldhal);
3.2.4.4.2 Capacidade de retenção de água (CRA)
A capacidade de retenção de água (CRA) dos hidrolisados foi medida segundo o
método de Regenstein et al. (1979), adaptado às condições do laboratório. Foram
preparadas dispersões protéicas a 1%, com variação de pH na faixa de 3 a 11.
Adicionou-se à dispersão 2 mL de solução de NaCl 0,1 M até a obtenção de uma
pasta homogênea. Em seguida adicionou-se solução tampão de acordo com o pH
correspondente até o volume de 40 mL. A dispersão foi mantida sob agitação por 15
minutos e centrifugado a 3000xg por 25 minutos em centrífuga de tubos (Biosystems, tipo
MPW-350/350R). As proteínas solúveis no sobrenadante foram quantificadas pelo
método de Lowry et al. (1951), e descontadas do total de proteínas da amostra original. A

32
capacidade de retenção de água foi determinada como a quantidade de água retida pela
proteína contida na amostra, expressa em g de água retida por grama de proteína, de
acordo com a equação 3.
CRA = quantidade de água retida (g) x 100 (3)
massa proteína original (g)
3.2.4.4.3 Capacidade de retenção de óleo (CRO)
A capacidade de retenção de óleo (CRO) dos hidrolisados obtidos foi
determinada segundo metodologia descrita por Fonkwe e Singh (1996), adaptado as
condições do laboratório, onde foi pesado 1 grama de amostra protéica em tubos e
adicionados 20 mL de óleo vegetal. A amostra foi agitada em agitador de tubos
(Certomet, MU) para que ocorresse a homogeneização da mistura durante 10 minutos.
Em seguida centrifugaram-se as amostras a 9000xg em centrífuga de tubos (Biosystems,
tipo MPW-350/350R) por 15 minutos. A diferença entre o óleo adicionado e o óleo não
retido foi considerada como a quantidade de óleo retido pelos hidrolisados. A capacidade
de retenção de óleo foi expressa como a quantidade de óleo retido por grama de proteína
de acordo com a equação 4.
CRO = óleo retido (mL) (4)
massa proteína (g)
3.2.4.4.4 Capacidade Emulsificante (CE)
A capacidade emulsificante (CE) dos hidrolisados foi determinada pelo método
descrito por Okezie e Bello (1988), que consistiu em preparar uma suspensão contendo
1g de proteína com 34 mL de solução de NaCl 3%, em liquidificador doméstico (Arno,
modelo LIR-P) por 30 segundos em velocidade média. Em seguida foram adicionados,
com o auxílio de uma bureta, 30 mL de óleo vegetal a uma vazão de 10 mL/min sob
agitação.
A emulsão formada foi transferida para um tubo graduado de centrífuga e levado
a um banho de água (modelo Q-215-2, Quimis) a 85ºC por 15 minutos. Após as
amostras foram centrifugadas a 3000xg por 40 minutos em centrifuga de tubos
(Biosystems, tipo MPW-350/350R). O volume de óleo separado em cada amostra após a
centrifugação foi medido diretamente no tubo. A diferença entre a camada de óleo
remanescente e a quantidade de óleo adicionado foi expressa como a quantidade de óleo

33
emulsificado e retido por grama de proteína contida na amostra, de acordo com a
equação 5.
CE = quantidade de óleo emulsificado (mL) (5)
massa proteína (g)
3.2.4.4.5 Propriedades espumantes
A capacidade de formação de espuma (CFE) foi medida de acordo com um
método combinado, apresentado por Phillips et al. (1987), e Dipak e Kumar (1986),
descrito por Glória e Reginato-d’Arce (2000), adaptado as condições do laboratório, onde
se preparou uma suspensão de 5g de amostra protéica com 100mL de água. A
suspensão foi agitada em liquidificador doméstico (Arno, modelo LIR-P), por 5 minutos
com agitação média, transferindo-se a dispersão para uma proveta graduada de 500mL.
A capacidade de formação de espuma foi calculada como a % de aumento de
volume baseando-se no volume inicial e após a formação de espuma, conforme a
equação 6.
CFE = (B-A) x 100 (6)
A
Onde: A = volume antes da agitação (mL);
B = volume após batimento (mL);
A determinação da estabilidade de espuma (EEs) dos hidrolisados foi medida
através do repouso da amostra à temperatura ambiente (20-25ºC) com leitura do volume
após intervalos 1, 5, 10, 30 e 60 minutos, sendo a estabilidade calculada pela equação 7.
EEs = V x 100 (7)
Vo
Sendo V o volume final de espuma após cada intervalo de tempo e Vo,
corresponde ao volume inicial da espuma formada (mL). Todas as medidas dos valores
das propriedades funcionais foram realizadas em triplicata.
3.2.4.5 Determinação da digestibilidade dos hidrolisados
A avaliação nutricional dos hidrolisados de corvina foi realizada mediante a
determinação da digestibilidade in vitro através de hidrólise enzimática de acordo com o
método de Sgarbieri (1996). Inicialmente pesou-se uma quantidade de amostra que

34
correspondesse ao equivalente a 0,5 g de proteína, preparou-se uma solução de 1,5mg
de pepsina por mL de HCl 0,10N e adicionou-se 15 mL dessa solução à amostra, após
adicionou-se 0,5 mL de solução de merthiolate incolor, homogemeizou-se bem e levou-se
ao banho-maria a 37º C por 3 horas, com agitação periódica. Decorrido este tempo os
tubos contendo as amostras foram resfriados e a solução foi neutralizada com NaOH
0,2N em pHmetro, e em seguida ajustou-se a pH 8,0 com tampão fosfato.
Foi preparada uma solução de 0,5 mg de pancreatina por mL de tampão fosfato
pH 8,0. Adicionou-se 10 mL da solução de pancreatina ao produto hidrolisado pela
pepsina e levou-se novamente a banho-maria por 37º C, porém por 24 horas com
agitação periódica. Após o tempo de hidrólise foi adicionado 5 mL de TCA 5% ao
hidrolisado, centrifugou-se por 15 minutos a 5000 xg para separar o material insolúvel. O
hidrolisado foi então filtrado e o precipitado redissolvido pela adição de 5 mL de NaOH
0,2 N. O filtrado foi recolhido para que fosse determinado o nitrogênio digerido, pelo
método de Kjeldhal. Seguiu-se o mesmo procedimento para a solução branco.
3.2.5 Planejamento experimental e tratamento estatístico
Realizou-se um planejamento experimental fatorial completo 22 com três
repetições no ponto central, duas variáveis de entrada estudadas em dois níves,
totalizando onze ensaios (NETO et al., 1996). A Tabela 3 mostra os níveis para cada
variável independente estudada e a Tabela 4 apresenta a matriz do planejamento.
Como resposta, avaliou-se o grau de hidrólise (GH), o rendimento em proteína
bem como as propriedades funcionais, (Solubilidade, CRA, CRO, CE, CFE). As variáveis
independentes foram: concentração de substrato, sendo considerada a quantidade de
proteína em relação ao volume de tampão (p/v) expresso como porcentagem e a
concentração de enzima, considerando a massa de enzima em relação à massa em
proteínas presente no substrato, expresso em porcentagem (p/p).
Tabela 3: Valores das variáveis do planejamento experimental e seus
respectivos níveis codificados e reais.
níveis
Variáveis - α -1 0 +1 + α
[S] (p/v) 2,18 3 5 7 7,82
[E] (p/p) 0,0318 0,04 0,06 0,08 0,0882
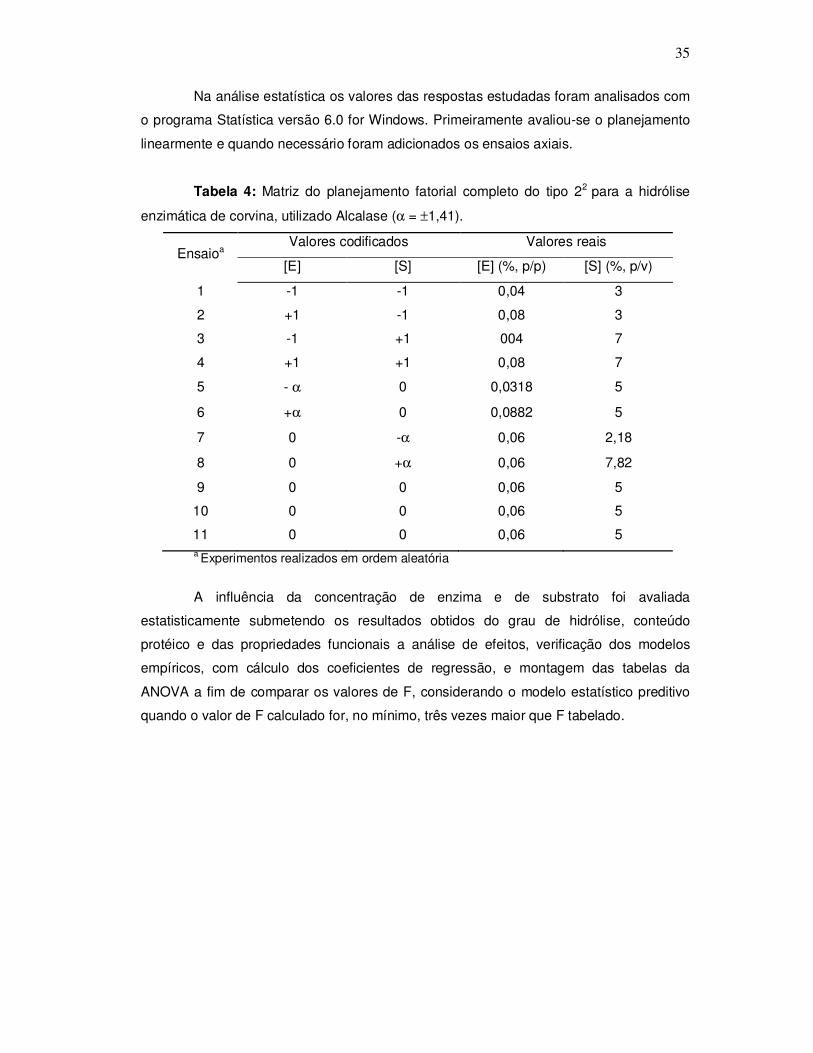
35
Na análise estatística os valores das respostas estudadas foram analisados com
o programa Statística versão 6.0 for Windows. Primeiramente avaliou-se o planejamento
linearmente e quando necessário foram adicionados os ensaios axiais.
Tabela 4: Matriz do planejamento fatorial completo do tipo 22 para a hidrólise
enzimática de corvina, utilizado Alcalase (α = ±1,41).
Ensaioa Valores codificados Valores reais
[E] [S] [E] (%, p/p) [S] (%, p/v)
1 -1 -1 0,04 3
2 +1 -1 0,08 3
3 -1 +1 004 7
4 +1 +1 0,08 7
5 - α 0 0,0318 5
6 +α 0 0,0882 5
7 0 -α 0,06 2,18
8 0 +α 0,06 7,82
9 0 0 0,06 5
10 0 0 0,06 5
11 0 0 0,06 5
a Experimentos realizados em ordem aleatória
A influência da concentração de enzima e de substrato foi avaliada
estatisticamente submetendo os resultados obtidos do grau de hidrólise, conteúdo
protéico e das propriedades funcionais a análise de efeitos, verificação dos modelos
empíricos, com cálculo dos coeficientes de regressão, e montagem das tabelas da
ANOVA a fim de comparar os valores de F, considerando o modelo estatístico preditivo
quando o valor de F calculado for, no mínimo, três vezes maior que F tabelado.

36
4 RESULTADOS E DISCUSSÃO
4.1 Atividade específica da enzima
A Figura 5 representa a atividade dos preparados enzimáticos com caseína
(padrão) e filé de corvina como substrato. A caseína é o substrato protéico mais
freqüentemente utilizado para a determinação da atividade específica sob condições
neutras e alcalinas. Assim, de acordo com Gildberg et al. (2002), como o filé de corvina é
o substrato protéico utilizado no experimento, torna-se relevante a comparação entre
ambos os substratos.
0
0,5
1
1,5
2
2,5
3
3,5
4
Ati
vid
ade
esp
ecíf
ica
(u
mo
l T
iro
sin
a/m
in/g
)
Corvina Caseína
Substratro
Figura 5: Atividade específica da Alcalase determinada com padrão caseína e
músculo de corvina como substrato
A Alcalase apresentou valor de atividade de 1,68µmol de tirosina por minuto por
grama utilizando corvina como substrato. Comparando com os resultados de Gildberg et
al. (2002), que trabalharam com proteína de bacalhau e Santos (2006), com proteína de
cabrinha, os mesmos encontraram valores de atividade enzimática em torno de 1µmol e
1,56µmol de tirosina por minuto por grama, respectivamente, utilizando esta mesma
enzima. Assim, observou-se que o substrato no qual a enzima atua é de fundamental
importância para a determinação do grau de hidrólise final em relação às demais
variáveis do processo de hidrólise como afirmado por Kristinsson e Rasco (2000a,b).

37
4.2 Caracterização fisico-química da matéria-prima
4.2.1 pH
Os filés de corvina, utilizados como matéria-prima, apresentaram valor de pH de
6,7 (±0,02) indicando bom estado de conservação. Neste sentido, os resultados
mostraram que o produto se encontrava dentro de uma faixa normal de qualidade para
este parâmetro, considerando que os valores de pH para o pescado “in natura”, não
ultrapassaram o indicado pela legislação brasileira, que propõe um máximo de 6,8
(BRASIL, 1997). De acordo com Martin (1982), o pH é um parâmetro que indica a
qualidade do alimento, pois é afetado por reações que ocorrem após a morte do animal e
indica a presença de microrganismos que através de seu metabolismo causam o
acúmulo de material metabólico alcalino, que eleva o valor do pH, diminuindo a qualidade
do produto.
4.2.2 Rendimento
Na Tabela 5 estão apresentados os valores do rendimento de três exemplares
de corvina, coletados para essa determinação.
Tabela 5: Rendimento do processo de obtenção de filés de corvina
(Micropogonias furnieri)
Porção Exemplar 1 (g) Exemplar 2 (g) Exemplar 3 (g) Média (g)
pescado inteiro 1236,6 1573,1 1296,4 1322
s/ cabeça 841,2 1027,7 888,4 919,1
s/ vísceras 766,3 950,7 811,3 842,77
Filé 363,3 542,7 400,8 435,6
Rendimento 29,4 34,5 30,9 31,6
A corvina apresentou um rendimento médio de filés de 31,6% ±2,62. Observou-
se que esta espécie apresentou menor rendimento quando comparado com outras como
o atum, que de acordo com o trabalho desenvolvido por Bruschi (2001), apresentou um
rendimento em filés de 51,1%. Durante o processamento no laboratório constatou-se
dificuldade para a filetagem de corvina, devido aos ossos, espinhos e ao tamanho do
pescado. Em função disso, parte do filé ficou aderida à pele diminuindo o rendimento final
do processo. Dessa forma, ficou claro que o rendimento de filé depende, além da
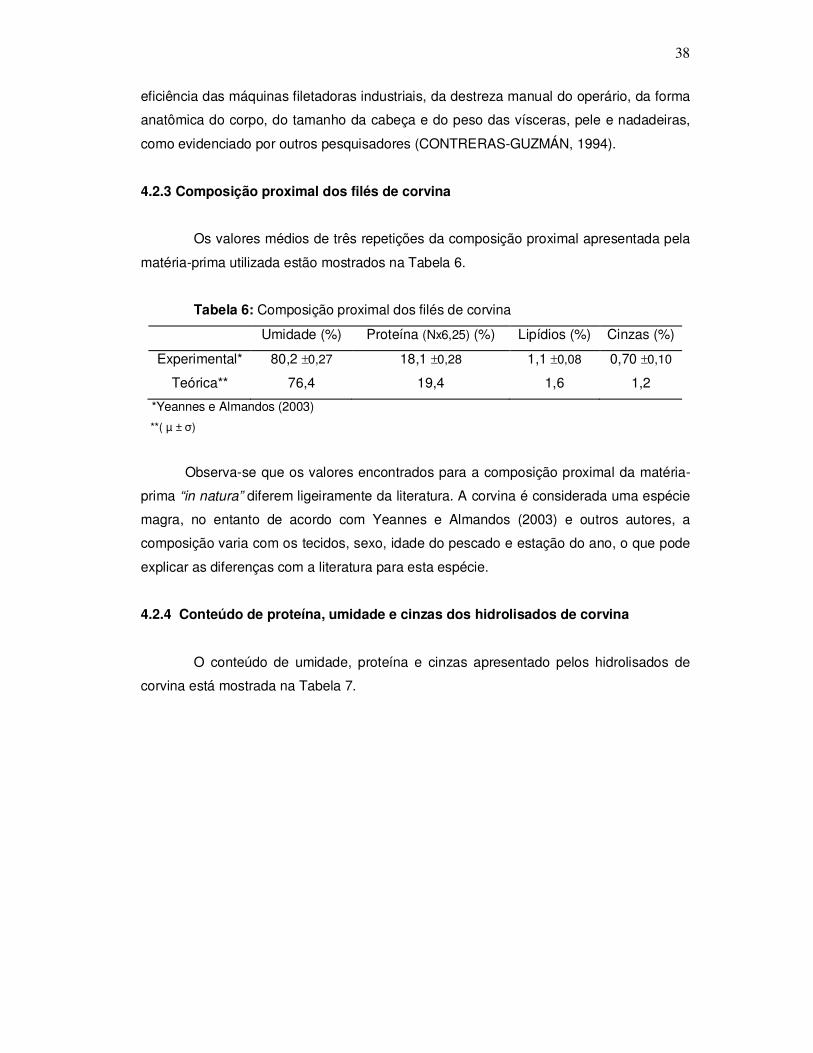
38
eficiência das máquinas filetadoras industriais, da destreza manual do operário, da forma
anatômica do corpo, do tamanho da cabeça e do peso das vísceras, pele e nadadeiras,
como evidenciado por outros pesquisadores (CONTRERAS-GUZMÁN, 1994).
4.2.3 Composição proximal dos filés de corvina
Os valores médios de três repetições da composição proximal apresentada pela
matéria-prima utilizada estão mostrados na Tabela 6.
Tabela 6: Composição proximal dos filés de corvina
Umidade (%) Proteína (Nx6,25) (%) Lipídios (%) Cinzas (%)
Experimental* 80,2 ±0,27 18,1 ±0,28 1,1 ±0,08 0,70 ±0,10
Teórica** 76,4 19,4 1,6 1,2
*Yeannes e Almandos (2003)
**( µ ± σ)
Observa-se que os valores encontrados para a composição proximal da matéria-
prima “in natura” diferem ligeiramente da literatura. A corvina é considerada uma espécie
magra, no entanto de acordo com Yeannes e Almandos (2003) e outros autores, a
composição varia com os tecidos, sexo, idade do pescado e estação do ano, o que pode
explicar as diferenças com a literatura para esta espécie.
4.2.4 Conteúdo de proteína, umidade e cinzas dos hidrolisados de corvina
O conteúdo de umidade, proteína e cinzas apresentado pelos hidrolisados de
corvina está mostrada na Tabela 7.
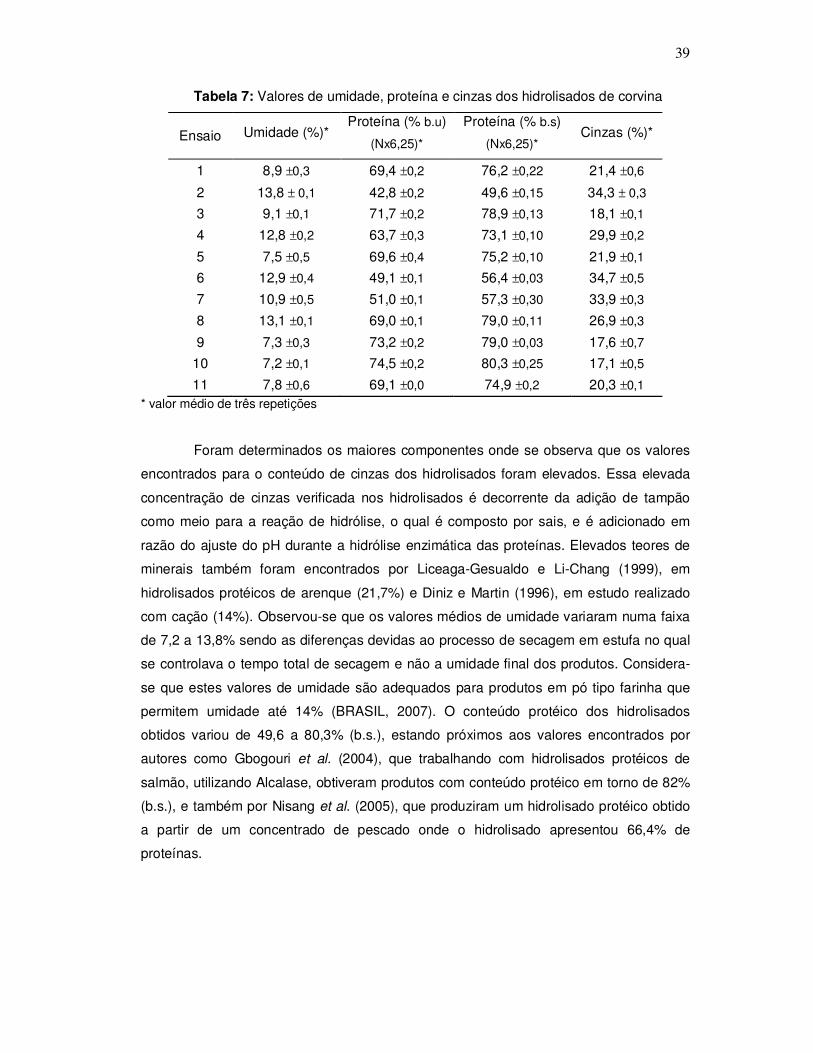
39
Tabela 7: Valores de umidade, proteína e cinzas dos hidrolisados de corvina
Ensaio Umidade (%)* Proteína (% b.u)
(Nx6,25)*
Proteína (% b.s)
(Nx6,25)* Cinzas (%)*
1 8,9 ±0,3 69,4 ±0,2 76,2 ±0,22 21,4 ±0,6
2 13,8 ± 0,1 42,8 ±0,2 49,6 ±0,15 34,3 ± 0,3
3 9,1 ±0,1 71,7 ±0,2 78,9 ±0,13 18,1 ±0,1
4 12,8 ±0,2 63,7 ±0,3 73,1 ±0,10 29,9 ±0,2
5 7,5 ±0,5 69,6 ±0,4 75,2 ±0,10 21,9 ±0,1
6 12,9 ±0,4 49,1 ±0,1 56,4 ±0,03 34,7 ±0,5
7 10,9 ±0,5 51,0 ±0,1 57,3 ±0,30 33,9 ±0,3
8 13,1 ±0,1 69,0 ±0,1 79,0 ±0,11 26,9 ±0,3
9 7,3 ±0,3 73,2 ±0,2 79,0 ±0,03 17,6 ±0,7
10 7,2 ±0,1 74,5 ±0,2 80,3 ±0,25 17,1 ±0,5
11 7,8 ±0,6 69,1 ±0,0 74,9 ±0,2 20,3 ±0,1 * valor médio de três repetições
Foram determinados os maiores componentes onde se observa que os valores
encontrados para o conteúdo de cinzas dos hidrolisados foram elevados. Essa elevada
concentração de cinzas verificada nos hidrolisados é decorrente da adição de tampão
como meio para a reação de hidrólise, o qual é composto por sais, e é adicionado em
razão do ajuste do pH durante a hidrólise enzimática das proteínas. Elevados teores de
minerais também foram encontrados por Liceaga-Gesualdo e Li-Chang (1999), em
hidrolisados protéicos de arenque (21,7%) e Diniz e Martin (1996), em estudo realizado
com cação (14%). Observou-se que os valores médios de umidade variaram numa faixa
de 7,2 a 13,8% sendo as diferenças devidas ao processo de secagem em estufa no qual
se controlava o tempo total de secagem e não a umidade final dos produtos. Considera-
se que estes valores de umidade são adequados para produtos em pó tipo farinha que
permitem umidade até 14% (BRASIL, 2007). O conteúdo protéico dos hidrolisados
obtidos variou de 49,6 a 80,3% (b.s.), estando próximos aos valores encontrados por
autores como Gbogouri et al. (2004), que trabalhando com hidrolisados protéicos de
salmão, utilizando Alcalase, obtiveram produtos com conteúdo protéico em torno de 82%
(b.s.), e também por Nisang et al. (2005), que produziram um hidrolisado protéico obtido
a partir de um concentrado de pescado onde o hidrolisado apresentou 66,4% de
proteínas.

40
4.3 Testes preliminares
4.3.1 Testes preliminares T1
Nos testes preliminares foram determinados valores de grau de hidrólise,
medidos em diferentes tempos, com a finalidade de se obter graus de hidrólise variados,
em um tempo total de 60 minutos. Foram obtidos valores de GH na faixa de 7,12 a 19,9%
(vide Anexo A, Tabela A.1). Os valores de GH nos tempos de 15, 30, 45 e 60 minutos
foram analisados estatisticamente a um nível de confiança de 95%. A análise de
variância (Anexo A, Tabela A.2), mostrou que não houve diferença significativa entre os
valores de GH (Figura 6), para um tempo de reação de 60 minutos nas temperaturas
estudadas.
55ºC 50ºC
0 15 30 45 60
Tempo (min)
0
10
20
30
% G
H
Figura 6: Grau de hidrólise versus tempo em dois valores de temperatura
Condições de hidrólise: [E] 2,5% (g enzima/g proteína); [S] 1,5% (g proteína/ vol. tampão); pH 8,0.
Os valores de GH, tanto para a temperatura de 50 como para a temperatura de
55ºC aumentaram progressivamente até 30 minutos de reação, apresentando um novo
aumento em 60 minutos, onde os valores de GH para estas duas temperatura
mostraram-se semelhantes. Então, decidiu-se trabalhar com temperatura de 50ºC, afim
de não propiciar a desnaturação térmica, visto que se pretendia obter hidrolisados em
faixas distintas de GH para verificar a influência do mesmo nas propriedades funcionais
dos produtos obtidos a partir da corvina.
4.3.2 Testes preliminares T2
Através da análise de variância dos valores obtidos no teste preliminar T2,
observou-se que houve diferença significativa (α=5%), em relação ao GH, nos tempos de

41
reação de 30 e 60 minutos para as diferentes condições de concentração de substrato e
enzima estudadas (vide Anexo B, Tabela B.1).
Analisando as condições de reação para o tempo de 30 minutos observou-se
que somente as condições de [S] 3% e [E] 6% e [S] 4% e [E] 6% não apresentaram
diferença significativa (Anexo B, Tabela B.2), no entanto todas as outras condições
estudadas no tempo de 30 minutos diferiram significativamente.
A análise de variância para o tempo de 60 minutos mostrou que somente as
condições das combinações [S] 5% e [E] 6% e [S] 4% e [E] 8% não apresentaram
diferença significativa entre si (Anexo B, Tabela B.3). Todas as condições estudadas em
um tempo de 60 minutos diferiram significativamente entre si, exceto as citadas
anteriormente. Verificou-se que, com um maior tempo de reação foi possível obter uma
ampla faixa de valores GH, juntamente com concentrações de enzima e de substrato
maiores.
Os resultados mostraram que houve um aumento considerável no grau de
hidrólise em relação aos testes anteriores, com valores na faixa de 5,8 a 38,2%, (Figura
7) quando foram aumentadas as concentrações de enzima e substrato. A partir destes
resultados realizou-se o planejamento experimental definitivo, descrito no item 3.2.5
S 3% E 4% S 4% E 4% S 5% E 4% S 3% E 6% S 4% E 6% S 5% E 6% S 3% E 8% S 4% E 8% S 5% E 8%0 10 20 30 40 50 60 70
Tempo (min)
0
5
10
15
20
25
30
35
40
% G
H
Figura 7: Grau de hidrólise da reação enzimática em função do tempo em
diferentes condições de [E] e [S]
Condições de hidrólise: pH=8,0; T=50ºC
O tempo estabelecido para as reações enzimáticas do planejamento
experimental foi de 60 minutos, visto que seria possível obter uma faixa ampla de grau de
hidrólise neste intervalo, considerando que um período de tempo longo poderia prejudicar
algumas propriedades funcionais relacionadas com a hidrofobicidade de superfície das
proteínas (KRISTINSSON E RASCO, 2000a).
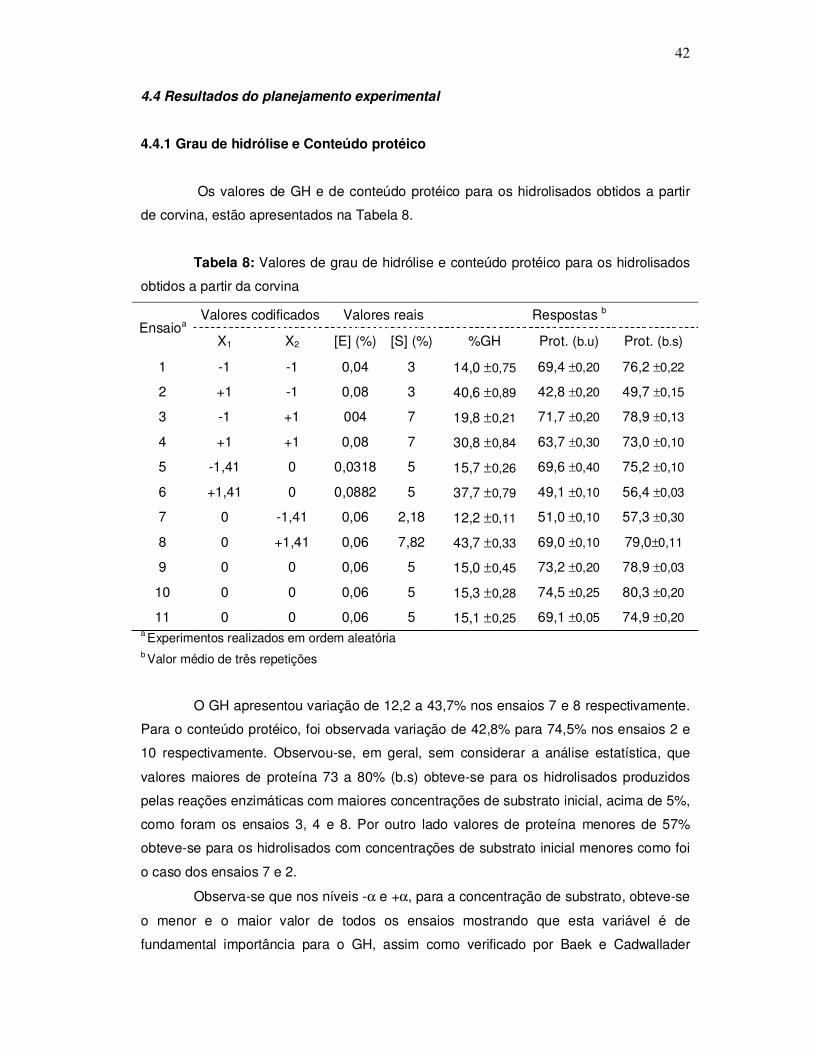
42
4.4 Resultados do planejamento experimental
4.4.1 Grau de hidrólise e Conteúdo protéico
Os valores de GH e de conteúdo protéico para os hidrolisados obtidos a partir
de corvina, estão apresentados na Tabela 8.
Tabela 8: Valores de grau de hidrólise e conteúdo protéico para os hidrolisados
obtidos a partir da corvina
Ensaioa Valores codificados Valores reais Respostas b
X1 X2 [E] (%) [S] (%) %GH Prot. (b.u) Prot. (b.s)
1 -1 -1 0,04 3 14,0 ±0,75 69,4 ±0,20 76,2 ±0,22
2 +1 -1 0,08 3 40,6 ±0,89 42,8 ±0,20 49,7 ±0,15
3 -1 +1 004 7 19,8 ±0,21 71,7 ±0,20 78,9 ±0,13
4 +1 +1 0,08 7 30,8 ±0,84 63,7 ±0,30 73,0 ±0,10
5 -1,41 0 0,0318 5 15,7 ±0,26 69,6 ±0,40 75,2 ±0,10
6 +1,41 0 0,0882 5 37,7 ±0,79 49,1 ±0,10 56,4 ±0,03
7 0 -1,41 0,06 2,18 12,2 ±0,11 51,0 ±0,10 57,3 ±0,30
8 0 +1,41 0,06 7,82 43,7 ±0,33 69,0 ±0,10 79,0±0,11
9 0 0 0,06 5 15,0 ±0,45 73,2 ±0,20 78,9 ±0,03
10 0 0 0,06 5 15,3 ±0,28 74,5 ±0,25 80,3 ±0,20
11 0 0 0,06 5 15,1 ±0,25 69,1 ±0,05 74,9 ±0,20 a Experimentos realizados em ordem aleatória b Valor médio de três repetições
O GH apresentou variação de 12,2 a 43,7% nos ensaios 7 e 8 respectivamente.
Para o conteúdo protéico, foi observada variação de 42,8% para 74,5% nos ensaios 2 e
10 respectivamente. Observou-se, em geral, sem considerar a análise estatística, que
valores maiores de proteína 73 a 80% (b.s) obteve-se para os hidrolisados produzidos
pelas reações enzimáticas com maiores concentrações de substrato inicial, acima de 5%,
como foram os ensaios 3, 4 e 8. Por outro lado valores de proteína menores de 57%
obteve-se para os hidrolisados com concentrações de substrato inicial menores como foi
o caso dos ensaios 7 e 2.
Observa-se que nos níveis -α e +α, para a concentração de substrato, obteve-se
o menor e o maior valor de todos os ensaios mostrando que esta variável é de
fundamental importância para o GH, assim como verificado por Baek e Cadwallader
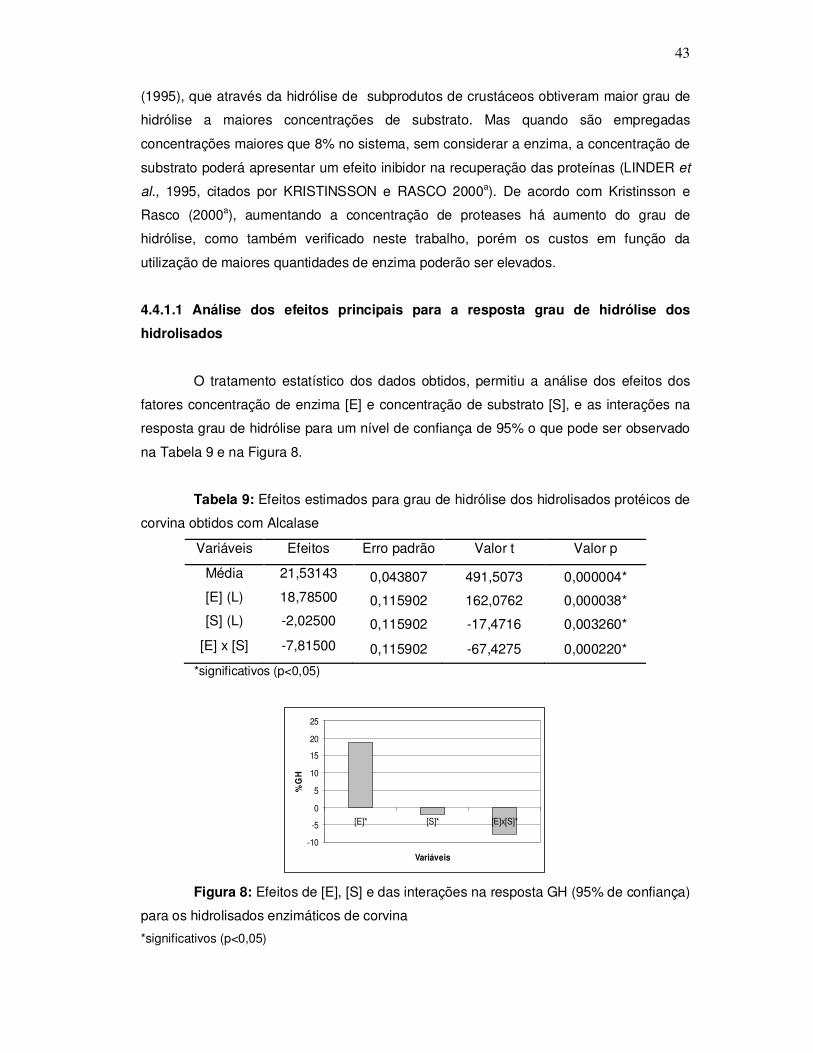
43
(1995), que através da hidrólise de subprodutos de crustáceos obtiveram maior grau de
hidrólise a maiores concentrações de substrato. Mas quando são empregadas
concentrações maiores que 8% no sistema, sem considerar a enzima, a concentração de
substrato poderá apresentar um efeito inibidor na recuperação das proteínas (LINDER et
al., 1995, citados por KRISTINSSON e RASCO 2000a). De acordo com Kristinsson e
Rasco (2000a), aumentando a concentração de proteases há aumento do grau de
hidrólise, como também verificado neste trabalho, porém os custos em função da
utilização de maiores quantidades de enzima poderão ser elevados.
4.4.1.1 Análise dos efeitos principais para a resposta grau de hidrólise dos
hidrolisados
O tratamento estatístico dos dados obtidos, permitiu a análise dos efeitos dos
fatores concentração de enzima [E] e concentração de substrato [S], e as interações na
resposta grau de hidrólise para um nível de confiança de 95% o que pode ser observado
na Tabela 9 e na Figura 8.
Tabela 9: Efeitos estimados para grau de hidrólise dos hidrolisados protéicos de
corvina obtidos com Alcalase
Variáveis Efeitos Erro padrão Valor t Valor p
Média 21,53143 0,043807 491,5073 0,000004*
[E] (L) 18,78500 0,115902 162,0762 0,000038*
[S] (L) -2,02500 0,115902 -17,4716 0,003260*
[E] x [S] -7,81500 0,115902 -67,4275 0,000220*
*significativos (p<0,05)
-10
-5
0
5
10
15
20
25
[E]* [S]* [E]x[S]*
Variáveis
%G
H
Figura 8: Efeitos de [E], [S] e das interações na resposta GH (95% de confiança)
para os hidrolisados enzimáticos de corvina
*significativos (p<0,05)

44
Tanto a [E] como a [S] foram significativas no processo bem como a interação,
sendo que a variável com maior efeito foi concentração de enzima, onde se pode
observar um efeito positivo, quando se aumenta a concentração de Alcalase no meio de
0,04 para 0,08%, significando um aumento de 18,78% na variável resposta. Esta relação
direta a positiva de [E] com o GH era esperado, já que o aumento da enzima permitirá
uma maior quebra (cisão) das cadeias protéicas, aumentando a quantidade de proteínas
de menor tamanho molecular e o grau de hidrólise do produto obtido.
Para a concentração de substrato quando a concentração no meio passou de 3%
para 7%, ocorreu efeito negativo no GH significando uma diminuição de 2,02% no valor
da resposta. A interação das variáveis de entrada, [E] e [S], influenciou negativamente o
GH causando uma diminuição de 7,8%. Levando em conta o discutido anteriormente, que
a [E] teve um efeito positivo no GH e a [S] um efeito negativo, pode-se dizer que a [S]
nestas condições de estudo, teve um efeito maior quando se considerou o efeito das
duas variáveis em conjunto sobre o GH.
4.4.1.2 Verificação do modelo matemático preditivo
A análise dos dados permitiu concluir que o planejamento experimental
realizado para obtenção dos hidrolisados protéicos de corvina, com a enzima alcalase,
não apresentou um modelo preditivo para o GH, visto que o valor de F calculado não foi
suficientemente maior que o tabelado, como mostram as Tabelas C.1 e C.2 (Anexo C).
Isto não torna os resultados inválidos, pois se constatou que aumentando a concentração
de enzima é possível elevar os valores de GH quando isto for desejado, do contrário,
diminuindo-se a concentração dessa variável, podem-se obter menores GH. Como se
pretendia obter faixas distintas de GH, para verificar a influência do mesmo nas
propriedades funcionais dos produtos obtidos a partir da corvina, os resultados
encontrados foram satisfatórios para este tipo de estudo visto que não se pretendia uma
maximização na resposta dessa variável.
4.4.1.3 Análise dos efeitos principais para a resposta Conteúdo Protéico dos
hidrolisados
O tratamento estatístico dos dados obtidos, permitiu a análise dos efeitos
principais da concentração de enzima e concentração de substrato, na variável conteúdo
protéico para um nível de confiança de 90% o que pode ser observado na Tabela 10 e na
Figura 9.

45
Tabela 10: Efeitos estimados para o conteúdo protéico dos hidrolisados
protéicos de corvina obtidos com alcalase
Variáveis Efeitos Erro padrão Valor t Valor p
Média 66,3429 1,065252 62,27902 0,000258*
[E] -17,3000 2,818392 -6,13825 0,025529*
[S] 11,6000 2,818392 4,11582 0,054271*
Interação 9,3000 2,818392 3,29975 0,080858*
* significativos (p<0,10)
Verificou-se que quando a concentração de enzima aumentou de 0,04 para
0,08%, houve um efeito negativo no rendimento do conteúdo protéico dos hidrolisados,
significando uma diminuição de 17,3%, no entanto quando a concentração de substrato
do meio passou de 3% para 7% observou-se um efeito positivo, visto que houve um
aumento de 11,6% no conteúdo protéico dos hidrolisados. A interação entre as
variáveis [E] e [S], influenciou positivamente, causando um aumento de 9,3% no
conteúdo protéico.
-20
-15
-10
-5
0
5
10
15
[E]* [S]* [E]x[S]*
Variáveis
Co
nte
úd
o P
roté
ico
(%
)
Figura 9: Efeitos da [E] e da [S] no conteúdo protéico (90% de confiança) para
os hidrolisados enzimáticos de corvina
*significativos (p<0,10)
4.4.1.4 Verificação do modelo matemático preditivo
Considerando-se somente os efeitos significativos prosseguiu-se a análise
estatística (Tabela 11), obtendo-se o coeficiente de determinação do modelo matemático
(R2 = 0,9744) e a respectiva equação desenvolvida para predizer o conteúdo protéico dos
hidrolisados obtidos.

46
Tabela 11: Análise de variância dos valores da resposta conteúdo protéico dos
hidrolisados a um nível p de confiança de 90%.
Fonte de variação SQ GL MQ F
Regressão 1175,291 5 235,05 30,09
Resíduo 30,885 5 6,177
Falta de Ajuste 14,998 34
Erro Puro 15,887 2
Total 1206,176 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tabelado = 3,45
Coeficiente de determinação R2 = 0,9744
A análise de variância (teste F) mostrou que o modelo quadrático se ajustou bem
aos dados experimentais quanto ao conteúdo protéico, o que pode ser observado na
Tabela 11. Obteve-se um modelo preditivo, visto que F calculado foi suficientemente
superior que o F tabelado. Os coeficientes de regressão utilizados na construção do
modelo podem ser visualizados na Tabela 12.
Tabela 12: Coeficientes de regressão do conteúdo protéico dos hidrolisados de
corvina obtidos com alcalase
Variáveis Coef.de
Regressão
Erro
padrão t(2) p
Lim. Conf.
- 90%
Lim. Conf.
+ 90%
Média 72,25754 1,627180 44,40661 0,000507* 65,2564 79,25873 [E] (L) -7,96181 0,997938 -7,97827 0,015349* -12,2556 -3,66803 [E] (Q) -5,91389 1,190799 -4,96632 0,038234* -11,0375 -0,79029
[S] (L) 6,09062 0,997938 6,10321 0,025811* 1,7968 10,38440
[S] (Q) -5,58695 1,190799 -4,69176 0,042550* -10,7105 -0,46335
[E] x [S] 4,65000 1,409196 3,29975 0,080858* -1,4133 10,71328
O modelo com as variáveis codificadas que representa o conteúdo protéico em
%, em função da concentração de enzima e de substrato, está representado pela
equação 8.
Conteúdo Protéico = 72,25 – 7,96[E] – 5,91[E]2 + 6,09[S] (8)
– 5,58[S] 2 + 4,56[E].[S]
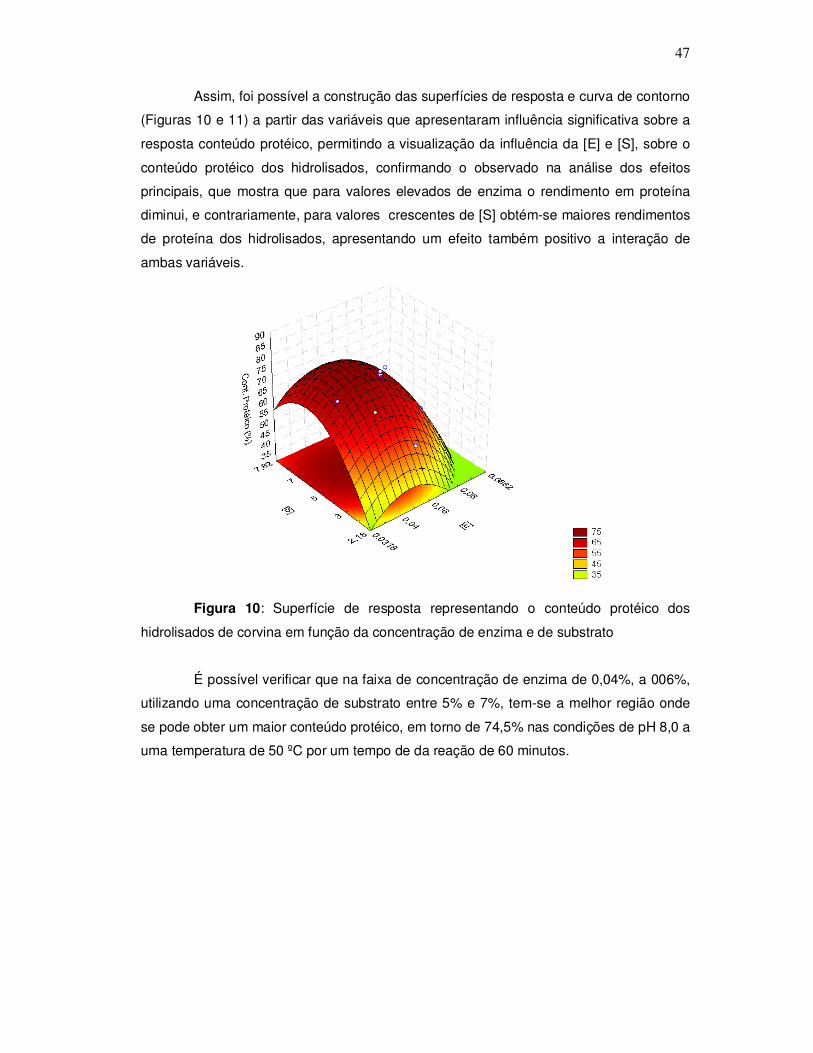
47
Assim, foi possível a construção das superfícies de resposta e curva de contorno
(Figuras 10 e 11) a partir das variáveis que apresentaram influência significativa sobre a
resposta conteúdo protéico, permitindo a visualização da influência da [E] e [S], sobre o
conteúdo protéico dos hidrolisados, confirmando o observado na análise dos efeitos
principais, que mostra que para valores elevados de enzima o rendimento em proteína
diminui, e contrariamente, para valores crescentes de [S] obtém-se maiores rendimentos
de proteína dos hidrolisados, apresentando um efeito também positivo a interação de
ambas variáveis.
Figura 10: Superfície de resposta representando o conteúdo protéico dos
hidrolisados de corvina em função da concentração de enzima e de substrato
É possível verificar que na faixa de concentração de enzima de 0,04%, a 006%,
utilizando uma concentração de substrato entre 5% e 7%, tem-se a melhor região onde
se pode obter um maior conteúdo protéico, em torno de 74,5% nas condições de pH 8,0 a
uma temperatura de 50 ºC por um tempo de da reação de 60 minutos.
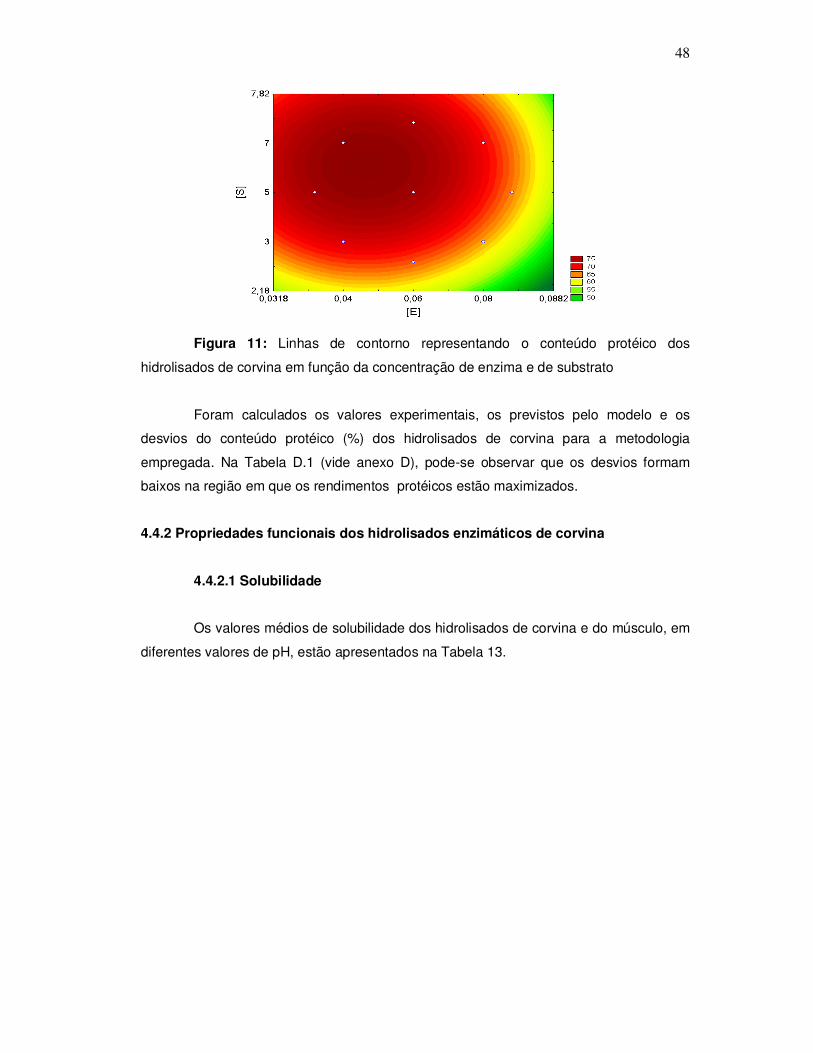
48
Figura 11: Linhas de contorno representando o conteúdo protéico dos
hidrolisados de corvina em função da concentração de enzima e de substrato
Foram calculados os valores experimentais, os previstos pelo modelo e os
desvios do conteúdo protéico (%) dos hidrolisados de corvina para a metodologia
empregada. Na Tabela D.1 (vide anexo D), pode-se observar que os desvios formam
baixos na região em que os rendimentos protéicos estão maximizados.
4.4.2 Propriedades funcionais dos hidrolisados enzimáticos de corvina
4.4.2.1 Solubilidade
Os valores médios de solubilidade dos hidrolisados de corvina e do músculo, em
diferentes valores de pH, estão apresentados na Tabela 13.

49
Tabela 13: Valores médios de solubilidade (%) dos hidrolisados protéicos e do
músculo de corvina em diferentes valores de pH.
Ensaio [E] (%) [S] (%) Solubilidade (%) *
pH 3 pH 5 pH 7 pH 9 pH 11
1 0,04 3 32,87 30,97 36,11 38,18 38,21
2 0,08 3 47,46 41,42 47,56 50,61 51,90
3 004 7 33,02 29,81 30,75 31,17 35,85
4 0,08 7 40,90 38,81 39,33 39,59 41,54
5 0,0318 5 31,82 28,75 31,22 31,77 33,05
6 0,0882 5 41,20 37,52 43,62 44,42 44,80
7 0,06 2,18 25,69 20,26 21,30 23,18 26,82
8 0,06 7,82 38,80 37,10 38,91 39,69 46,79
9 0,06 5 31,40 29,82 32,58 35,79 36,62
10 0,06 5 32,10 30,05 33,24 35,80 35,96
11 0,06 5 31,08 29,97 33,13 33,24 34,90
músculo - - 12,47 8,97 10,19 12,92 15,36 * valor médio de três repetições
Na Figura 12 pode-se verificar que a hidrólise enzimática contribuiu para
aumentar a solubilidade dos hidrolisados protéicos comparados com o músculo in natura
como pode ser observado através das curvas de solubilidade obtidas em diferentes
valores de pH.
0
5
10
15
20
25
30
35
40
45
50
55
0 1 2 3 4 5 6 7 8 9 10 11 12
pH
So
lub
ilid
ade
(%)
Ensaio1
Ensaio 2
Ensaio 3
Ensaio 4
Ensaio 5
Ensaio 6
Ensaio 7
Ensaio 8
Ensaio 9
Ensaio 10
Ensaio 11
in natura
Figura 12: Solubilidade dos hidrolisados de corvina e do músculo in natura em
diferentes valores de pH

50
Para Das e Kinsella (1990), citados por Duarte et al. (1998), a hidrólise
enzimática leva a uma maior exposição de grupos carregados, diminui o tamanho
molecular e aumenta a hidrofilicidade das proteínas. Os resultados mostraram que as
curvas de solubilidade para todos os hidrolisados, apresentaram o menor valor no pH 5,0,
pois este pH encontra-se próximo ao ponto isoelétrico das proteínas, onde o número de
cargas negativas e positivas tende a se neutralizar intramolecularmente, apresentando
uma menor afinidade pela água.
Verificou-se que houve um pequeno aumento dessa propriedade no valor de pH
ácido, e um aumento progressivo do pH neutro para os pHs alcalinos, sendo que os
maiores valores foram obtidos no pH 11,0. Segundo Cheftel et al. (1993), esse aumento
ocorre devido ao número de resíduos de cargas negativas em pH acima do ponto
isoelétrico, ser maior do que os resíduos de cargas negativas abaixo do ponto isoelétrico.
Para Sgarbieri (1998), as proteínas miofibrilares podem estar formando complexos
insolúveis o que somente a predominância de cargas negativas em pHs fortemente
alcalinos geram forças de repulsão eletrostática capazes de promover a dissociação dos
complexos e solubilização. Comparado o músculo de corvina com os hidrolisados, o
músculo obteve valores de solubilidade inferiores. Estes resultados estão em
concordância com Rebeca et al. (1991), pois segundo estes autores as proteínas
miofibrilares do pescado são muito insolúveis devido ao maior tamanho das moléculas,
visto que os hidrolisados apresentam diminuição no tamanho dos peptídeos comparado
com a proteína intacta.
Também foi estudada a relação entre solubilidade e GH. Os hidrolisados com
maior GH mostraram maior solubilidade que os hidrolisados com menores valores de GH,
concordando com os resultados obtidos por Gbogouri et al. (2004), que estudaram a
influência do GH nas propriedades funcionais de hidrolisados de cabeça de salmão. Eles
encontraram que para o menor GH encontrado (11,5%), os valores de solubilidade
variaram de 75 a 85% aproximadamente em diferentes valores de pH. Para o maior valor
de GH obtido pelos autores (17,3%), os valores de solubilidade foram maiores variando
de 85 a 95% aproximadamente, para diferentes pHs. Segundo Panyam e Kilara (1996),
uma das principais conseqüências da hidrolise enzimática é o aumento da solubilidade e,
normalmente este aumento está associado ao aumento do grau de hidrólise. O aumento
da solubilidade dos hidrolisados é provavelmente devido à diminuição do tamanho das
moléculas e correspondente aumento da exposição de grupos hidrofílicos amino e
carboxil ionizáveis. O hidrolisado obtido a partir das condições do ensaio 2 foi o que
apresentou os maiores valores de solubilidade em todos os pHs, e também um dos
maiores valores de GH (40,64%) obtidos deste estudo, sendo o mesmo observado para
os ensaios 4, 6 e 8 que apresentaram respectivamente valores de GH de 30,8, 37,73 e

51
43,68%. O hidrolisado do ensaio 7 apresentou a menor solubilidade em todos os valores
de pH sendo este fato justificado pelo baixo GH apresentado por esse produto (12,8%).
Os hidrolisados obtidos através dos ensaios dos pontos centrais 9, 10 e 11 apresentaram
perfis de solubilidade semelhantes dos hidrolisados 1, 3 e 5 bem como seus valores de
GH. Nas Figuras 13 e 14 pode ser observada a relação entre o GH e a solubilidade dos
hidrolisados de corvina, medida nos pHs 5,0 e 11,0 respectivamente.
S pH 5,0 GH (%)
1 2 3 4 5 6 7 8 9 10 11
Ensaios
0
5
10
15
20
25
30
35
40
45
50S
olub
ilida
de (
%)
GH
(%
)
Figura 13: Solubilidade e GH dos hidrolisados de corvina e do músculo in natura
em pH 5,0
Observa-se que a solubilidade em pH 5,0 de todos os hidrolisados é menor que
a solubilidade em pH 11,0 e que os valores de GH apresentam uma relação direta com a
solubilidade.
S pH 11,0 GH (%)
1 2 3 4 5 6 7 8 9 10 11
Ensaios
0
10
20
30
40
50
60
Sol
ubili
dade
(%
)
GH
(%
)
Figura 14: Solubilidade e GH dos hidrolisados de corvina e do músculo in natura
em pH 11,0

52
4.4.2.1.1 Análise dos efeitos principais para a resposta solubilidade dos
hidrolisados
Na Tabela 14 encontram-se os efeitos da solubilidade dos hidrolisados de
corvina em diferentes valores de pH.
Tabela 14: Efeitos estimados dos valores da solubilidade dos hidrolisados de
corvina em diferentes valores de pH
Propriedade
Funcional Variáveis Efeitos Erro padrão Valor t(2) Valor p
Média 35,54714 0,197170 180,2864 0,000031*
Solubilidade [E] (L) 11,23500 0,521664 21,5369 0,002149*
pH 3,0 [S] (L) -3,20500 0,521664 -6,1438 0,025484* (p<0,05)* Interação -3,35500 0,521664 -6,4313 0,023334*
Solubilidade Média 32,97857 0,044132 747,2739 0,000002*
pH 5,0 [E] (L) 9,72500 0,116762 83,2892 0,000144* (p<0,05)* [S] (L) -1,88500 0,116762 -16,1440 0,003815*
Interação -0,72500 0,116762 -6,2092 0,024970*
Solubilidade Média 36,10000 0,133648 270,1117 0,000014*
pH 7,0 [E] (L) 10,01500 0,353601 28,3229 0,001244* (p<0,05)* [S] (L) -6,79500 0,353601 -19,2166 0,002697*
Interação -1,43500 0,353601 -4,0583 0,055695
Solubilidade Média 37,76857 0,557550 67,74025 0,000218*
pH 9,0 [E] (L) 10,42500 1,475138 7,06713 0,019440* (p<0,05)* [S] (L) -9,01500 1,475138 -6,11129 0,025746*
Interação -2,00500 1,475138 -1,35919 0,307057
Solubilidade Média 39,28286 0,327966 119,7771 0,000070*
pH 11,0 [E] (L) 9,69000 0,867717 11,1672 0,007924* (p<0,05)* [S] (L) -6,36000 0,867717 -7,3296 0,018110*
Interação -4,00000 0,867717 -4,6098 0,043977*
* significativas
As representações gráficas da análise de efeitos da solubilidade em diferentes
valores de pH, dos hidrolisados protéicos de corvina obtidos a partir da alcalase, estão
apresentados na Figura 15.
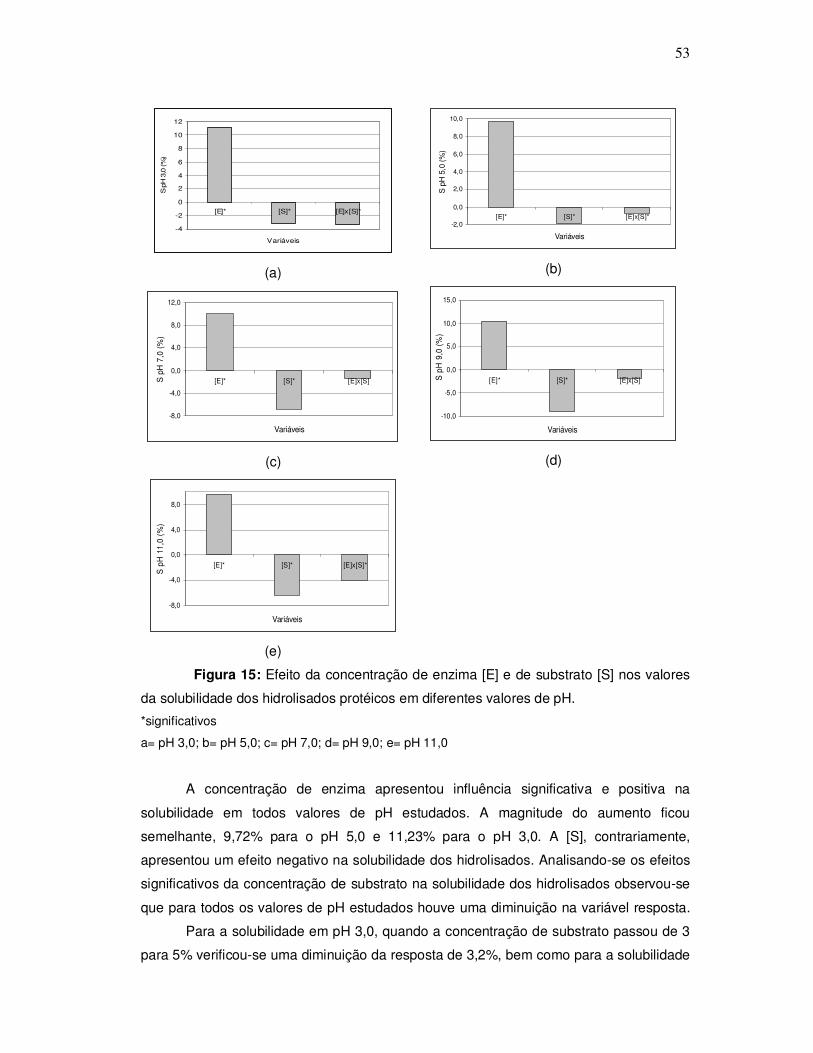
53
-4
-2
0
2
4
6
8
10
12
[E]* [S]* [E]x[S]*
Variáveis
S p
H 3
,0 (%
)
(a)
-8,0
-4,0
0,0
4,0
8,0
12,0
[E]* [S]* [E]x[S]
Variáveis
S p
H 7
,0 (
%)
(c)
-8,0
-4,0
0,0
4,0
8,0
[E]* [S]* [E]x[S]*
Variáveis
S p
H 1
1,0
(%)
(e)
-2,0
0,0
2,0
4,0
6,0
8,0
10,0
[E]* [S]* [E]x[S]*
Variáveis
S p
H 5
,0 (
%)
(b)
-10,0
-5,0
0,0
5,0
10,0
15,0
[E]* [S]* [E]x[S]
VariáveisS
pH
9,0
(%)
(d)
Figura 15: Efeito da concentração de enzima [E] e de substrato [S] nos valores
da solubilidade dos hidrolisados protéicos em diferentes valores de pH.
*significativos
a= pH 3,0; b= pH 5,0; c= pH 7,0; d= pH 9,0; e= pH 11,0
A concentração de enzima apresentou influência significativa e positiva na
solubilidade em todos valores de pH estudados. A magnitude do aumento ficou
semelhante, 9,72% para o pH 5,0 e 11,23% para o pH 3,0. A [S], contrariamente,
apresentou um efeito negativo na solubilidade dos hidrolisados. Analisando-se os efeitos
significativos da concentração de substrato na solubilidade dos hidrolisados observou-se
que para todos os valores de pH estudados houve uma diminuição na variável resposta.
Para a solubilidade em pH 3,0, quando a concentração de substrato passou de 3
para 5% verificou-se uma diminuição da resposta de 3,2%, bem como para a solubilidade

102
54
em pHs 5,0, 7,0, 9,0 e 11,0 que apresentou um decréscimo de 1,88, 6,79, 9,01 e 6,36%
respectivamente. A interação entre as variáveis [E] e [S] não apresentou efeito
significativo na solubilidade dos hidrolisados nos pHs 7,0 e 9,0. Para a solubilidade nos
valores de pH 3,0, 5,0 e 11,0 a interação entre as variáveis mostrou efeito significativo,
porém negativo, de 3,35, 0,72 e 4,0 % respectivamente na solubilidade.
4.4.2.1.2 Verificação do modelo matemático preditivo
Elaboraram-se as tabelas da análise de variância (ANOVA) onde foi verificado se
os modelos eram preditivos ou não. Observou-se que para a solubilidade nenhum modelo
foi preditivo. As tabelas das análises dos modelos não preditivos para a resposta
solubilidade encontram-se no Anexo C.
4.4.2.2 Capacidade de retenção de água (CRA)
Os valores médios de três repetições da capacidade de retenção de água (CRA)
dos hidrolisados de corvina, e do músculo in natura, em diferentes valores de pH, estão
apresentados na Tabela 15.
Tabela 15: Valores médios de CRA (g água/g proteína) dos hidrolisados
protéicos em diferentes valores de pH
Ensaio [E] (%) [S] (%) CRA (g água/ g proteína) *
pH 3 pH 5 pH 7 pH 9 pH 11
1 0,04 3 6,5 4,5 5,9 6 6,2
2 0,08 3 4,6 3,6 3,9 4 4,2
3 004 7 5,9 3,9 4 4,2 5,1
4 0,08 7 5 4 4,3 4,6 4,8
5 0,0318 5 6,6 4,9 5,5 6,1 6,3
6 0,0882 5 4,7 3,8 4,1 4,2 4,6
7 0,06 2,18 7 5,2 5,8 6,2 6,5
8 0,06 7,82 4,2 3,1 3,3 3,7 3,9
9 0,06 5 6,3 4,8 5,6 6,2 6,3
10 0,06 5 6 5 5,5 6,1 6,3
11 0,06 5 6,4 4,9 5,6 6,2 6,3
músculo - - 3,5 2,9 3,1 3,2 3,2
*valor médio de três repetições
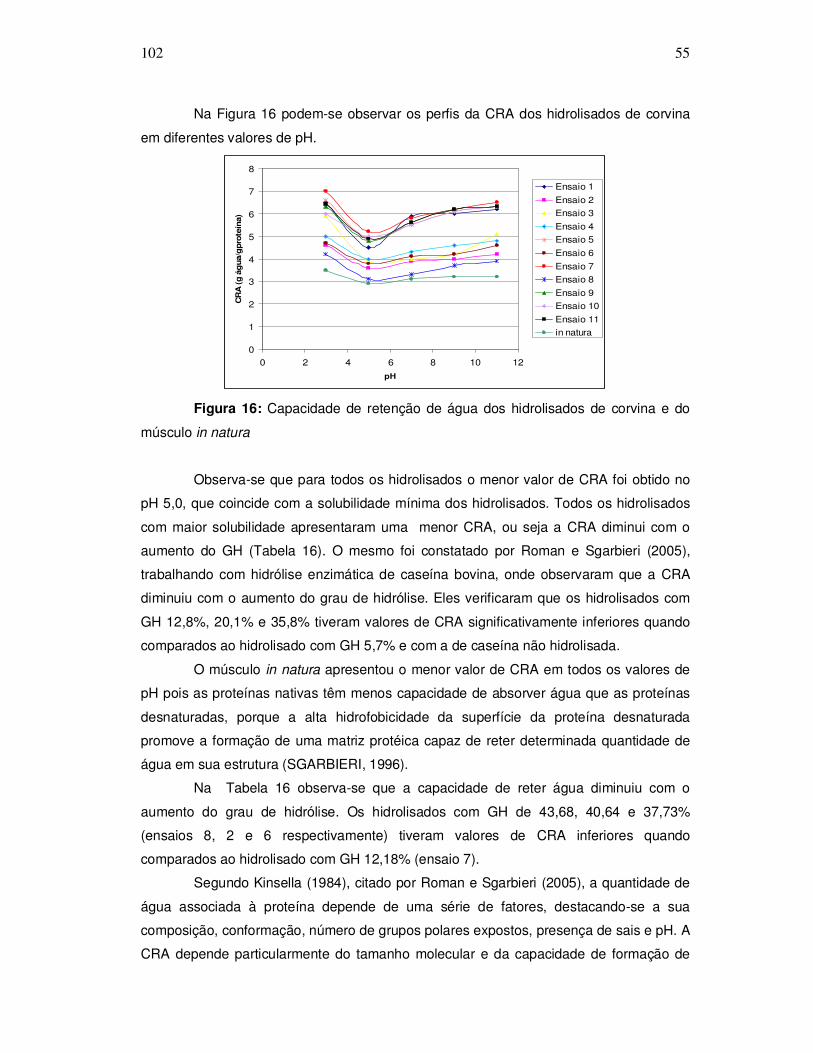
102
55
Na Figura 16 podem-se observar os perfis da CRA dos hidrolisados de corvina
em diferentes valores de pH.
0
1
2
3
4
5
6
7
8
0 2 4 6 8 10 12
pH
CR
A (
g á
gua/
gpro
teín
a)
Ensaio 1
Ensaio 2
Ensaio 3
Ensaio 4
Ensaio 5
Ensaio 6
Ensaio 7
Ensaio 8
Ensaio 9
Ensaio 10
Ensaio 11
in natura
Figura 16: Capacidade de retenção de água dos hidrolisados de corvina e do
músculo in natura
Observa-se que para todos os hidrolisados o menor valor de CRA foi obtido no
pH 5,0, que coincide com a solubilidade mínima dos hidrolisados. Todos os hidrolisados
com maior solubilidade apresentaram uma menor CRA, ou seja a CRA diminui com o
aumento do GH (Tabela 16). O mesmo foi constatado por Roman e Sgarbieri (2005),
trabalhando com hidrólise enzimática de caseína bovina, onde observaram que a CRA
diminuiu com o aumento do grau de hidrólise. Eles verificaram que os hidrolisados com
GH 12,8%, 20,1% e 35,8% tiveram valores de CRA significativamente inferiores quando
comparados ao hidrolisado com GH 5,7% e com a de caseína não hidrolisada.
O músculo in natura apresentou o menor valor de CRA em todos os valores de
pH pois as proteínas nativas têm menos capacidade de absorver água que as proteínas
desnaturadas, porque a alta hidrofobicidade da superfície da proteína desnaturada
promove a formação de uma matriz protéica capaz de reter determinada quantidade de
água em sua estrutura (SGARBIERI, 1996).
Na Tabela 16 observa-se que a capacidade de reter água diminuiu com o
aumento do grau de hidrólise. Os hidrolisados com GH de 43,68, 40,64 e 37,73%
(ensaios 8, 2 e 6 respectivamente) tiveram valores de CRA inferiores quando
comparados ao hidrolisado com GH 12,18% (ensaio 7).
Segundo Kinsella (1984), citado por Roman e Sgarbieri (2005), a quantidade de
água associada à proteína depende de uma série de fatores, destacando-se a sua
composição, conformação, número de grupos polares expostos, presença de sais e pH. A
CRA depende particularmente do tamanho molecular e da capacidade de formação de
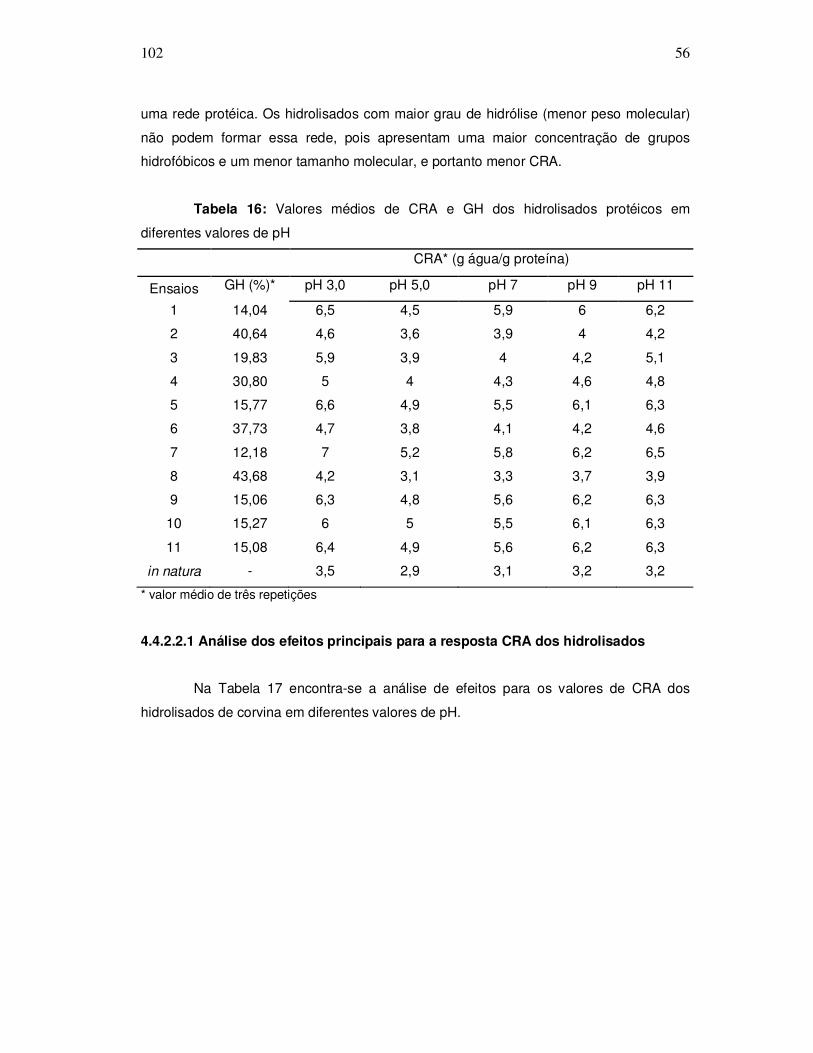
102
56
uma rede protéica. Os hidrolisados com maior grau de hidrólise (menor peso molecular)
não podem formar essa rede, pois apresentam uma maior concentração de grupos
hidrofóbicos e um menor tamanho molecular, e portanto menor CRA.
Tabela 16: Valores médios de CRA e GH dos hidrolisados protéicos em
diferentes valores de pH
CRA* (g água/g proteína)
Ensaios GH (%)* pH 3,0 pH 5,0 pH 7 pH 9 pH 11
1
2
3
4
5
6
7
8
9
10
11
in natura
14,04
40,64
19,83
30,80
15,77
37,73
12,18
43,68
15,06
15,27
15,08
-
6,5
4,6
5,9
5
6,6
4,7
7
4,2
6,3
6
6,4
3,5
4,5
3,6
3,9
4
4,9
3,8
5,2
3,1
4,8
5
4,9
2,9
5,9
3,9
4
4,3
5,5
4,1
5,8
3,3
5,6
5,5
5,6
3,1
6
4
4,2
4,6
6,1
4,2
6,2
3,7
6,2
6,1
6,2
3,2
6,2
4,2
5,1
4,8
6,3
4,6
6,5
3,9
6,3
6,3
6,3
3,2
* valor médio de três repetições
4.4.2.2.1 Análise dos efeitos principais para a resposta CRA dos hidrolisados
Na Tabela 17 encontra-se a análise de efeitos para os valores de CRA dos
hidrolisados de corvina em diferentes valores de pH.
102
57
Tabela 17: Efeitos estimados para os valores de CRA em pH 3,0 a 11,0 dos
hidrolisados protéicos de corvina obtidos com alcalase
Propriedade
Funcional Variáveis Efeitos Erro padrão Valor t Valor p
Média 5,81429 0,078680 73,89828 0,000183*
CRA [E] -1,40000 0,208167 -6,72538 0,021402*
pH 3,0 [S] -0,10000 0,208167 -0,48038 0,678366 p(<0,05)* Interação 0,50000 0,208167 2,40192 0,138273
Média 4,385714 0,037796 116,0351 0,000074*
CRA [E] -0,400000 0,100000 -4,0000 0,057191*
pH 5,0 [S] -0,100000 0,100000 -1,0000 0,422650 p(<0,10)* Interação 0,500000 0,100000 5,0000 0,037750*
Média 4,971429 0,021822 227,8195 0,000019*
CRA [E] -0,850000 0,057735 -14,7224 0,004582*
pH 7,0 [S] -0,750000 0,057735 -12,9904 0,005874*
(p<0,10) preditivo* Interação 1,150000 0,057735 19,9186 0,002511*
Média 5,328571 0,021822 244,1858 0,000017*
CRA [E] -0,800000 0,057735 -13,8564 0,005168*
pH 9,0 [S] -0,600000 0,057735 -10,3923 0,009133* (p<0,20) preditivo* Interação 1,200000 0,057735 20,7846 0,002307*
Média 5,60000 0,350000 16,00000 0,000531*
CRA [E] -1,15000 0,926013 -1,24188 0,302521
pH 11,0 [S] -0,25000 0,926013 -0,26997 0,804686 p(<0,10)* Interação 0,85000 0,926013 0,91791 0,426356
* significativas
Na Figura 17 é possível visualizar as representações gráficas dos efeitos em
relação a concentração de enzima e de substrato, e suas interações na capacidade de
retenção de água dos hidrolisados, nos diferentes valores de pH estudados.
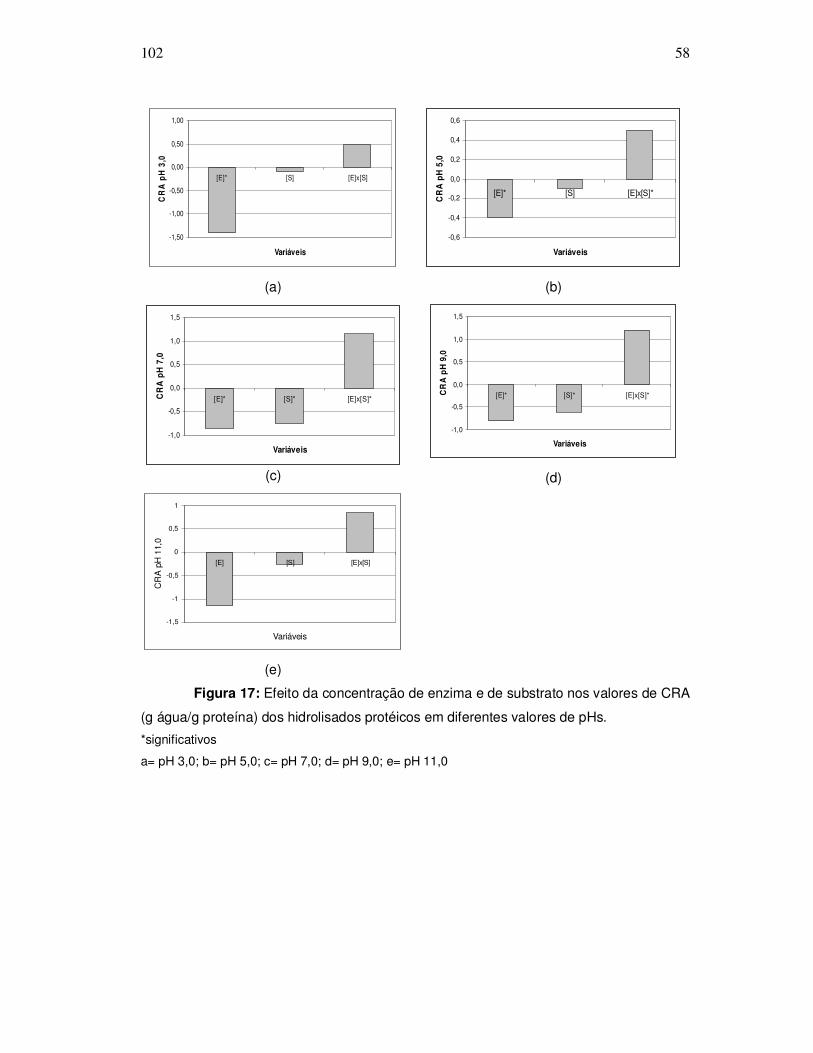
102
58
-1,50
-1,00
-0,50
0,00
0,50
1,00
[E]* [S] [E]x[S]
Variáveis
CR
A p
H 3
,0
(a)
-1,0
-0,5
0,0
0,5
1,0
1,5
[E]* [S]* [E]x[S]*
Variáveis
CR
A p
H 7
,0
(c)
-1,5
-1
-0,5
0
0,5
1
[E] [S] [E]x[S]
Variáveis
CR
A p
H 1
1,0
(e)
-0,6
-0,4
-0,2
0,0
0,2
0,4
0,6
[E]* [S] [E]x[S]*
Variáveis
CR
A p
H 5
,0
(b)
-1,0
-0,5
0,0
0,5
1,0
1,5
[E]* [S]* [E]x[S]*
VariáveisC
RA
pH
9,0
(d)
Figura 17: Efeito da concentração de enzima e de substrato nos valores de CRA
(g água/g proteína) dos hidrolisados protéicos em diferentes valores de pHs.
*significativos
a= pH 3,0; b= pH 5,0; c= pH 7,0; d= pH 9,0; e= pH 11,0

59
Analisando a Figura 17 observa-se que [E] foi significativa e negativa em todos
os pHs exceto para o pH 11,0. A [S] foi significativa e negativa somente nos valores de
pH 7,0 e 9,0, mas a [E] foi mais influente que a [S]. No entanto a interação de ambas
variáveis foi significativa nos pHs 5,0, 7,0 e 9,0 e positiva nos 3 casos. Observa-se que a
interação entre as variáveis (Figura 17), concentração de enzima e concentração de
substrato, exerceu um efeito positivo na capacidade de retenção de água dos
hidrolisados de corvina em todos os valores de pH.
4.4.2.2.2 Verificação do modelo matemático preditivo
Foram elaboradas as tabelas da ANOVA para verificar se os modelos
quadráticos eram preditivos e verificou-se que somente a CRA nos valores de pH 7,0 e
9,0 apresentaram modelos preditivos. Nos valores de pH 3,0, 5,0, e 11,0 os modelos não
foram preditivos e as tabelas da ANOVA encontram-se no Anexo C (Tabelas C.17 a
C.21). Na Tabela 18 pode ser observada a ANOVA da CRA medida em pH 7,0.
Tabela 18: Análise de variância dos valores da CRA dos hidrolisados protéicos
de corvina em pH 7,0 a um nível p de confiança de 90%
Fonte de variação SQ GL MQ F
Regressão 8,2852 5 1,657 14,28
Resíduo 0,580206 5 0,11604
Falta de Ajuste 0,5735439 3
Erro Puro 0,00667 2
Total 8,86545 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 3,45
Coeficiente de determinação R2 =0,93455
Os coeficientes de regressão utilizados na construção do modelo podem ser
visualizados na Tabela 19.

60
Tabela 19: Coeficientes de regressão da CRA em pH 7,0 dos hidrolisados de
corvina obtidos com alcalase.
Variáveis Coef.de
Regressão
Erro
padrão t(2) p
Lim. Conf.
- 90%
Lim. Conf.
+ 90%
Média 5,567240 0,033333 167,0192 0,000036* 5,469909 5,664572
[E] (L) -0,460620 0,020443 -22,5321 0,001964* -0,520313 -0,400928
[E] (Q) -0,422297 0,024394 -17,3118 0,003320* -0,493526 -0,351068
[S] (L) -0,629999 0,020443 -30,8176 0,001051* -0,689692 -0,570306
[S] (Q) -0,548045 0,024394 -22,4667 0,001975* -0,619274 -0,476816
[E] x [S] 0,575000 0,028868 19,9186 0,002511* 0,490707 0,659293 *significativas (p<0,10)
O modelo com as variáveis codificadas que representa o valor da CRA em pH
7,0 em função da concentração de enzima e concentração de substrato, na faixa
estudada, está representado pela equação 9.
CRA (pH 7,0) = 5,56 -0,46[E] - 0,42[E]2 -0,629[S] - 0,548[S]2 - 0,575[E].[S] (9)
As superfícies de resposta geradas pelo modelo podem ser observadas nas
Figuras 18 e 19.
Figura 18: Superfície de resposta representando a CRA em pH 7,0 dos
hidrolisados de corvina em função da concentração de enzima e de substrato
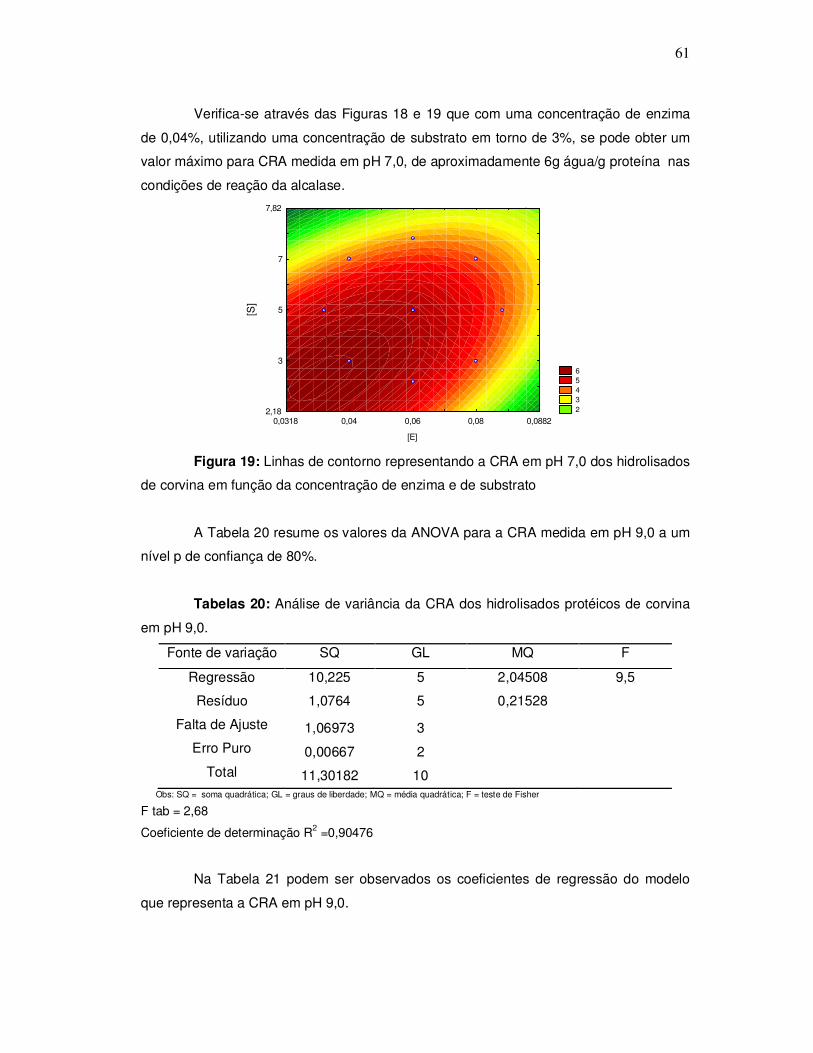
61
Verifica-se através das Figuras 18 e 19 que com uma concentração de enzima
de 0,04%, utilizando uma concentração de substrato em torno de 3%, se pode obter um
valor máximo para CRA medida em pH 7,0, de aproximadamente 6g água/g proteína nas
condições de reação da alcalase.
6 5 4 3 2
0,0318 0,04 0,06 0,08 0,0882
[E]
2,18
3
5
7
7,82
[S]
Figura 19: Linhas de contorno representando a CRA em pH 7,0 dos hidrolisados
de corvina em função da concentração de enzima e de substrato
A Tabela 20 resume os valores da ANOVA para a CRA medida em pH 9,0 a um
nível p de confiança de 80%.
Tabelas 20: Análise de variância da CRA dos hidrolisados protéicos de corvina
em pH 9,0.
Fonte de variação SQ GL MQ F
Regressão 10,225 5 2,04508 9,5
Resíduo 1,0764 5 0,21528
Falta de Ajuste 1,06973 3
Erro Puro 0,00667 2
Total 11,30182 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 2,68
Coeficiente de determinação R2 =0,90476
Na Tabela 21 podem ser observados os coeficientes de regressão do modelo
que representa a CRA em pH 9,0.
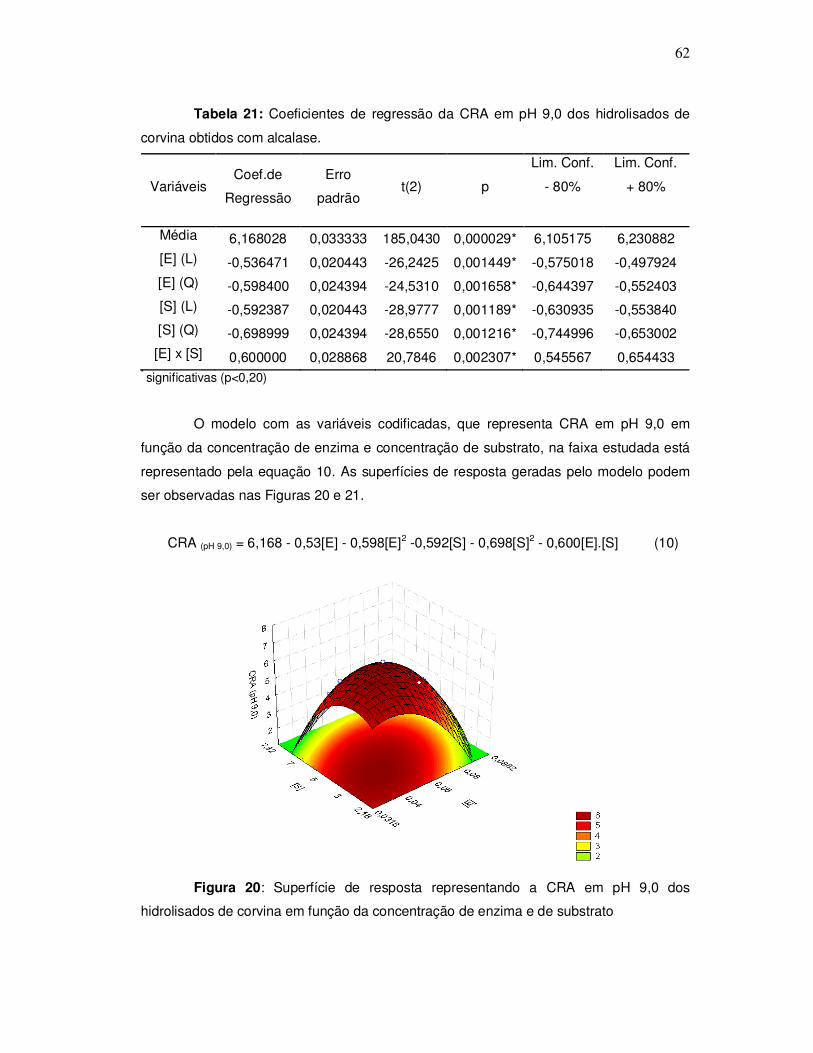
62
Tabela 21: Coeficientes de regressão da CRA em pH 9,0 dos hidrolisados de
corvina obtidos com alcalase.
Variáveis Coef.de
Regressão
Erro
padrão t(2) p
Lim. Conf.
- 80%
Lim. Conf.
+ 80%
Média 6,168028 0,033333 185,0430 0,000029* 6,105175 6,230882
[E] (L) -0,536471 0,020443 -26,2425 0,001449* -0,575018 -0,497924
[E] (Q) -0,598400 0,024394 -24,5310 0,001658* -0,644397 -0,552403
[S] (L) -0,592387 0,020443 -28,9777 0,001189* -0,630935 -0,553840
[S] (Q) -0,698999 0,024394 -28,6550 0,001216* -0,744996 -0,653002
[E] x [S] 0,600000 0,028868 20,7846 0,002307* 0,545567 0,654433 * significativas (p<0,20)
O modelo com as variáveis codificadas, que representa CRA em pH 9,0 em
função da concentração de enzima e concentração de substrato, na faixa estudada está
representado pela equação 10. As superfícies de resposta geradas pelo modelo podem
ser observadas nas Figuras 20 e 21.
CRA (pH 9,0) = 6,168 - 0,53[E] - 0,598[E]2 -0,592[S] - 0,698[S]2 - 0,600[E].[S] (10)
Figura 20: Superfície de resposta representando a CRA em pH 9,0 dos
hidrolisados de corvina em função da concentração de enzima e de substrato
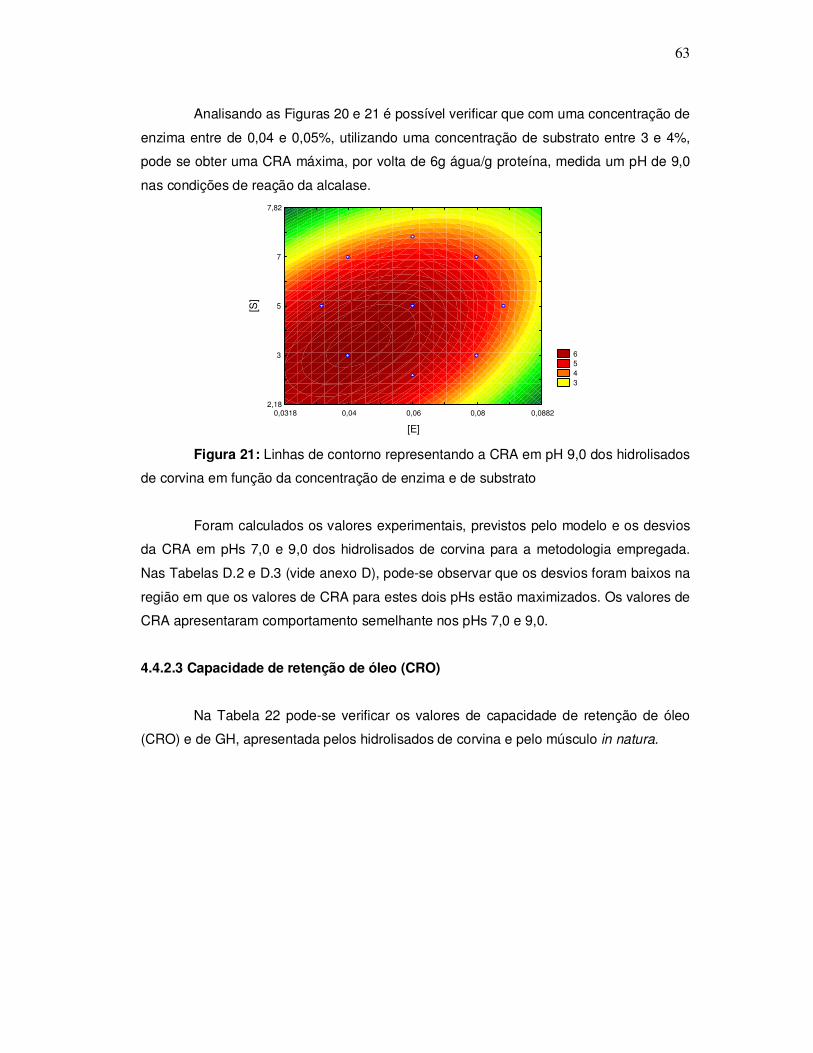
63
Analisando as Figuras 20 e 21 é possível verificar que com uma concentração de
enzima entre de 0,04 e 0,05%, utilizando uma concentração de substrato entre 3 e 4%,
pode se obter uma CRA máxima, por volta de 6g água/g proteína, medida um pH de 9,0
nas condições de reação da alcalase.
6 5 4 3
0,0318 0,04 0,06 0,08 0,0882
[E]
2,18
3
5
7
7,82
[S]
Figura 21: Linhas de contorno representando a CRA em pH 9,0 dos hidrolisados
de corvina em função da concentração de enzima e de substrato
Foram calculados os valores experimentais, previstos pelo modelo e os desvios
da CRA em pHs 7,0 e 9,0 dos hidrolisados de corvina para a metodologia empregada.
Nas Tabelas D.2 e D.3 (vide anexo D), pode-se observar que os desvios foram baixos na
região em que os valores de CRA para estes dois pHs estão maximizados. Os valores de
CRA apresentaram comportamento semelhante nos pHs 7,0 e 9,0.
4.4.2.3 Capacidade de retenção de óleo (CRO)
Na Tabela 22 pode-se verificar os valores de capacidade de retenção de óleo
(CRO) e de GH, apresentada pelos hidrolisados de corvina e pelo músculo in natura.
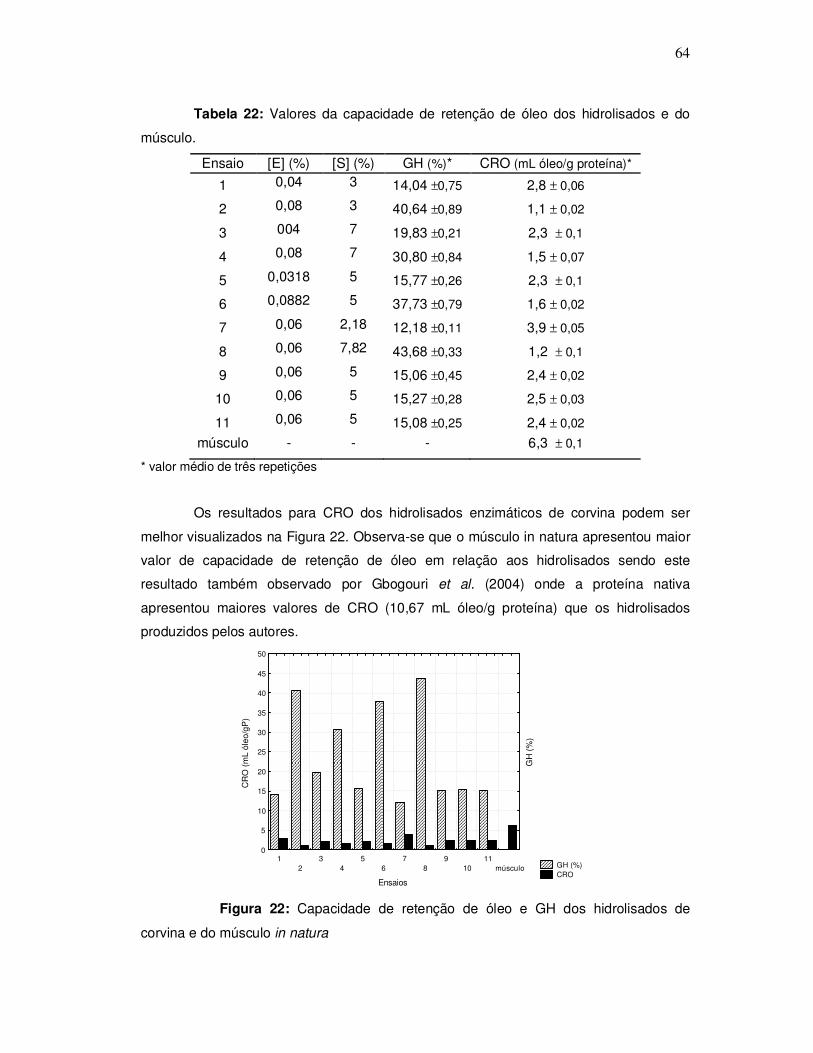
64
Tabela 22: Valores da capacidade de retenção de óleo dos hidrolisados e do
músculo.
Ensaio [E] (%) [S] (%) GH (%)* CRO (mL óleo/g proteína)*
1 0,04 3 14,04 ±0,75 2,8 ± 0,06
2 0,08 3 40,64 ±0,89 1,1 ± 0,02
3 004 7 19,83 ±0,21 2,3 ± 0,1
4 0,08 7 30,80 ±0,84 1,5 ± 0,07
5 0,0318 5 15,77 ±0,26 2,3 ± 0,1
6 0,0882 5 37,73 ±0,79 1,6 ± 0,02
7 0,06 2,18 12,18 ±0,11 3,9 ± 0,05
8 0,06 7,82 43,68 ±0,33 1,2 ± 0,1
9 0,06 5 15,06 ±0,45 2,4 ± 0,02
10 0,06 5 15,27 ±0,28 2,5 ± 0,03
11 0,06 5 15,08 ±0,25 2,4 ± 0,02
músculo - - - 6,3 ± 0,1
* valor médio de três repetições
Os resultados para CRO dos hidrolisados enzimáticos de corvina podem ser
melhor visualizados na Figura 22. Observa-se que o músculo in natura apresentou maior
valor de capacidade de retenção de óleo em relação aos hidrolisados sendo este
resultado também observado por Gbogouri et al. (2004) onde a proteína nativa
apresentou maiores valores de CRO (10,67 mL óleo/g proteína) que os hidrolisados
produzidos pelos autores.
GH (%) CRO
12
34
56
78
910
11músculo
Ensaios
0
5
10
15
20
25
30
35
40
45
50
CR
O (
mL
óleo
/gP
)
GH
(%
)
Figura 22: Capacidade de retenção de óleo e GH dos hidrolisados de
corvina e do músculo in natura

65
Estes valores indicam uma baixa hidrofobicidade dos hidrolisados protéicos, pois
quando há grande quantidade de regiões hidrofóbicas nas proteínas, há favorecimento de
interação destas com o óleo (CHEFTEL et. al., 1993). Isso explica porque a CRO do
músculo foi maior que a dos hidrolisados. Nota-se que os hidrolisados com valores
menores de GH, apresentaram valores de CRO maiores. Os hidrolisados obtidos nas
condições dos ensaios 7 e 8, que tiveram o menor e o maior valor de GH (12,18 e
43,68%), apresentaram o maior e o menor valor de CRO (3,9 e 1,2 mL óleo/ g proteína)
respectivamente.
Estes resultados concordam com estudos de Gbogouri et al. (2004), que
trabalhando com hidrolisados protéicos de subprodutos de salmão, verificaram que a
capacidade de retenção de óleo era menor nos hidrolisados com GH maior. Eles
concluíram que para um GH de 11,5% a CRO apresentou um valor de 3,55 mL óleo/ g
proteína, já em hidrolisados com valores de GH de 17, 8% a CRO diminuiu para 2,8 (mL
óleo/g proteína). Kristinsson (1998), citado por Kristinsson e Rasco (2000ª), estudou a
absorção de óleo em hidrolisados de proteína de salmão e verificou que o hidrolisado
com GH de 5% apresentou uma absorção de óleo significativamente maior que os
hidrolisados com valores de 10 e 15% de GH. Hidrolisados de salmão vermelho
produzidos por Sathivel et al. (2005), em um tempo de 25 minutos tiveram uma maior
habilidade para reter óleo de soja que hidrolisados produzidos em um tempo de 75
minutos. De acordo com Kinsella (1976), altos valores de CRO são desejáveis em
produtos como extensores de carne para melhorar a sua sensação na boca, assim como
em produtos viscosos como sopas, queijos processados e massas.
4.4.2.3.1 Análise dos efeitos principais para a resposta CRO dos hidrolisados
Na Tabela 23 encontram-se os efeitos da CRO dos hidrolisados de corvina.
Tabela 23: Efeitos estimados para a CRO dos hidrolisados protéicos de corvina
obtidos com alcalase.
Propriedade Funcional
Variáveis Efeitos Erro padrão
Valor t(2) Valor p
Média 2,14286 0,021822 98,1981 0,000104*
CRO [E] (L) -1,25000 0,057735 -21,6506 0,002127*
[S] (L) -0,05000 0,057735 -0,8660 0,477767
Interação 0,45000 0,057735 7,7942 0,016065* * significativas (p<0,05)

66
Na Figura 23 observa-se a representações gráfica dos efeitos estimados em
relação à concentração de enzima [E] e de substrato [S], e suas interações na
capacidade de retenção de óleo dos hidrolisados protéicos.
-1,50
-1,20
-0,90
-0,60
-0,30
0,00
0,30
0,60
[E]* [S] [E]x[S]*
Variáveis
CR
O (
mL
óleo
/g P
)
Figura 23: Efeito da concentração de enzima e de substrato na CRO dos
hidrolisados protéicos
* significativas (p<0,05)
A concentração de enzima [E] apresentou um efeito significativo na capacidade
de retenção de óleo dos hidrolisados, porém negativo, onde se observou que quando a
concentração de enzima do meio aumentou de 0,04 para 0,08%, houve uma diminuição
de 1,25% nessa propriedade. A concentração de substrato [S] não foi significativa para a
capacidade de retenção de óleo neste estudo. A interação entre as variáveis [E] e [S] foi
significativa para o processo e influenciou positivamente a CRO dos hidrolisados.
4.4.2.3.2 Verificação do modelo matemático preditivo
Elaboraram-se as tabelas da ANOVA onde foi verificado se os modelos se
apresentavam preditivos ou não. Observou-se que para a CRO os modelos não foram
preditivos. As tabelas das análises dos modelos linear e quadrático não preditivos
encontram-se no Anexo C (Tabelas C.3 e C.4).
4.4.2.4 Capacidade Emulsificante (CE)
Na Tabela 24 encontram-se os valores da CE dos hidrolisados de corvina e do
músculo in natura e na Figura 24, estão apresentados os perfis da capacidade
emulsificante juntamente com os valores de GH dos hidrolisados de corvina e do músculo
in natura.
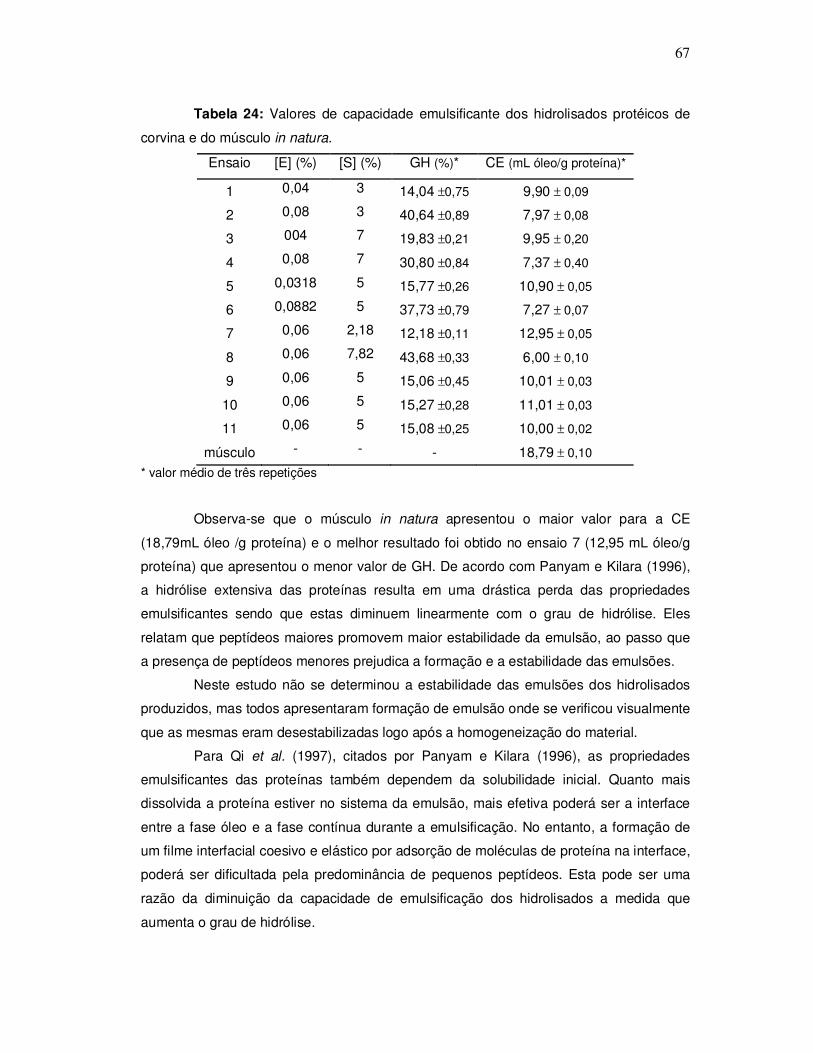
67
Tabela 24: Valores de capacidade emulsificante dos hidrolisados protéicos de
corvina e do músculo in natura.
Ensaio [E] (%) [S] (%) GH (%)* CE (mL óleo/g proteína)*
1 0,04 3 14,04 ±0,75 9,90 ± 0,09
2 0,08 3 40,64 ±0,89 7,97 ± 0,08
3 004 7 19,83 ±0,21 9,95 ± 0,20
4 0,08 7 30,80 ±0,84 7,37 ± 0,40
5 0,0318 5 15,77 ±0,26 10,90 ± 0,05
6 0,0882 5 37,73 ±0,79 7,27 ± 0,07
7 0,06 2,18 12,18 ±0,11 12,95 ± 0,05
8 0,06 7,82 43,68 ±0,33 6,00 ± 0,10
9 0,06 5 15,06 ±0,45 10,01 ± 0,03
10 0,06 5 15,27 ±0,28 11,01 ± 0,03
11 0,06 5 15,08 ±0,25 10,00 ± 0,02
músculo - - - 18,79 ± 0,10 * valor médio de três repetições
Observa-se que o músculo in natura apresentou o maior valor para a CE
(18,79mL óleo /g proteína) e o melhor resultado foi obtido no ensaio 7 (12,95 mL óleo/g
proteína) que apresentou o menor valor de GH. De acordo com Panyam e Kilara (1996),
a hidrólise extensiva das proteínas resulta em uma drástica perda das propriedades
emulsificantes sendo que estas diminuem linearmente com o grau de hidrólise. Eles
relatam que peptídeos maiores promovem maior estabilidade da emulsão, ao passo que
a presença de peptídeos menores prejudica a formação e a estabilidade das emulsões.
Neste estudo não se determinou a estabilidade das emulsões dos hidrolisados
produzidos, mas todos apresentaram formação de emulsão onde se verificou visualmente
que as mesmas eram desestabilizadas logo após a homogeneização do material.
Para Qi et al. (1997), citados por Panyam e Kilara (1996), as propriedades
emulsificantes das proteínas também dependem da solubilidade inicial. Quanto mais
dissolvida a proteína estiver no sistema da emulsão, mais efetiva poderá ser a interface
entre a fase óleo e a fase contínua durante a emulsificação. No entanto, a formação de
um filme interfacial coesivo e elástico por adsorção de moléculas de proteína na interface,
poderá ser dificultada pela predominância de pequenos peptídeos. Esta pode ser uma
razão da diminuição da capacidade de emulsificação dos hidrolisados a medida que
aumenta o grau de hidrólise.
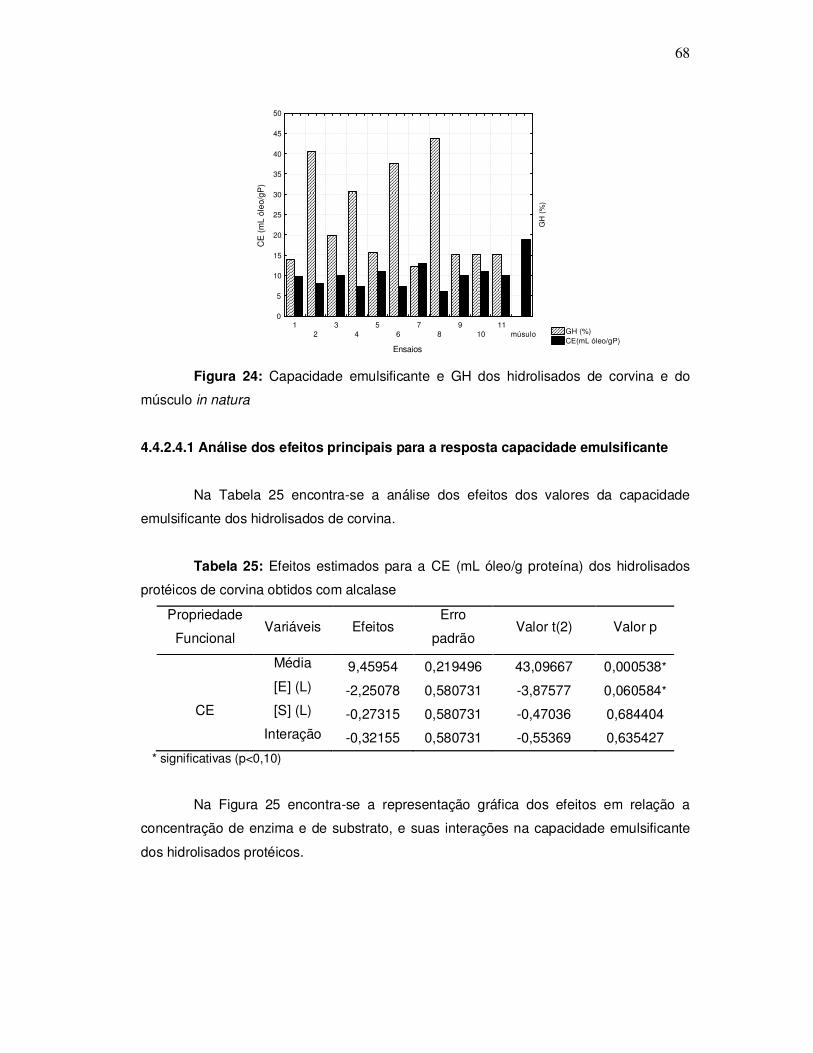
68
GH (%)CE(mL óleo/gP)
12
34
56
78
910
11músulo
Ensaios
0
5
10
15
20
25
30
35
40
45
50
CE
(m
L ól
eo/g
P)
GH
(%
)
Figura 24: Capacidade emulsificante e GH dos hidrolisados de corvina e do
músculo in natura
4.4.2.4.1 Análise dos efeitos principais para a resposta capacidade emulsificante
Na Tabela 25 encontra-se a análise dos efeitos dos valores da capacidade
emulsificante dos hidrolisados de corvina.
Tabela 25: Efeitos estimados para a CE (mL óleo/g proteína) dos hidrolisados
protéicos de corvina obtidos com alcalase
Propriedade
Funcional Variáveis Efeitos
Erro
padrão Valor t(2) Valor p
Média 9,45954 0,219496 43,09667 0,000538*
[E] (L) -2,25078 0,580731 -3,87577 0,060584* CE [S] (L) -0,27315 0,580731 -0,47036 0,684404
Interação -0,32155 0,580731 -0,55369 0,635427 * significativas (p<0,10)
Na Figura 25 encontra-se a representação gráfica dos efeitos em relação a
concentração de enzima e de substrato, e suas interações na capacidade emulsificante
dos hidrolisados protéicos.

69
-3,0
-2,0
-1,0
0,0
1,0
2,0
[E]* [S] [E]x[S]
VariáveisC
E (
mL
óleo
/gP
)
Figura 25: Efeito da concentração de enzima e de substrato na CE dos
hidrolisados protéicos
* significativas (p<0,10)
A análise dos efeitos permitiu concluir que somente a [E] foi significativa no
processo, no entanto apresentou um efeito negativo, onde se observa através da Tabela
25 que quando o conteúdo de enzima no meio aumenta, há uma diminuição na
capacidade emulsificante em aproximadamente 2,25%, o que comprova o discutido
anteriormente, que com o aumento do GH, há uma diminuição desta propriedade, então
se a concentração de enzima aumenta, também aumentará o GH e o efeito sobre a
capacidade emulsificante será o mesmo.
4.4.2.4.2 Verificação do modelo matemático preditivo
Elaboraram-se as tabelas da ANOVA onde foi verificado se os modelo se
apresentavam preditivos ou não. Observou-se que para a capacidade emulsificante os
modelos não foram preditivos. As tabelas das análises dos modelos não preditivos
encontram-se no Anexo C (Tabelas C.5 e C.6).
4.4.2.5 Propriedades espumantes
4.4.2.5.1 Capacidade de formação de espuma (CFE)
Na Tabela 26 estão reunidos os valores de CFE apresentados pelos hidrolisados
de corvina, e pelo músculo in natura.
70
Tabela 26: Valores de capacidade de formação de espuma dos hidrolisados
Ensaio GH (%)* CFE (%)*
1 14,04 35 2 40,64 5 3 19,83 21 4 30,8 9 5 15,77 32 6 37,73 4 7 12,18 38 8 43,68 10 9 15,06 30 10 15,27 30 11 15,08 28
músculo in natura - 39 * valor médio de três repetições
Os resultados de CFE dos hidrolisados e do músculo apresentaram diferença
entre si, onde se pode observar que o músculo de corvina in natura apresentou o maior
valor para esta propriedade. Os maiores valores entre os hidrolisados foram obtidos pelos
ensaios 7, 1, 5, 9, 10 e 11.
Os perfis de capacidade de formação de espuma e o GH (%) dos hidrolisados
protéicos de corvina estão apresentados na Figura 26.
GH (%) CFE (%)
12
34
56
78
910
11músculo
Ensaios
0
5
10
15
20
25
30
35
40
45
50
CF
E
GH
(%
)
Figura 26: Capacidade de formação de espuma versus GH dos hidrolisados de
corvina.
Observa-se que a maior expansão de espuma entre os hidrolisados foi
conseguida com o ensaio 7 que tem um valor baixo de GH. O ensaio 6 resultou no
hidrolisados com a menor CFE, seguido dos ensaios 2, 4 e 8, sendo que os mesmos
apresentam os maiores valores de GH, concordando com Turgeon et al. (1992), citados
por Roman e Sgarbieri (2005), onde relataram que, apesar de pequenos peptídeos se

71
difundirem rapidamente na interface e serem adsorvidos, eles serão menos eficientes na
redução da tensão superficial, por dificuldade de se reorientarem na interface água-ar e
formarem membrana contínua viscoelástica, dificultando a formação de espuma.
A formação e a estabilidade de espumas em sistemas protéicos é dependente
de uma série de fatores que, no presente trabalho, não foram avaliados. Segundo
Kinsella (1984), citado por Roman e Sgarbieri (2005), o ideal para que uma proteína
apresente o melhor desempenho na formação e estabilidade de espuma, é que possua
um peso molecular maior que 20Kda, um mínimo de carga liquida, presença de sítios de
ligações hidrofóbicas, além de uma conformação flexível. Para o autor, não somente
fatores relacionados ao grau de hidrólise podem influir no desempenho das propriedades
de espuma, mas também o pH e outros aspectos relacionados à composição dos
hidrolisados. Neste estudo, a capacidade de formação de espuma aumentou com a
diminuição do GH.
4.4.2.5.2 Estabilidade de espuma (EEs)
Na Tabela 27 estão reunidos os valores de estabilidade de espuma, em
intervalos de tempo por um período de 60 minutos, apresentados pelos hidrolisados de
corvina, e pelo músculo in natura. Os valores de volume de espuma utilizados para o
cálculo dos valores de EEs encontram-se no Anexo C (Tabela C. 22)
Tabela 27: Valores de estabilidade de espuma dos hidrolisados de corvina em
diferentes tempos.
Estabilidade de Espuma (%)*
Ensaio 5 minutos 15 minutos 30 minutos 60 minutos
1 98,53 97,79 97,79 97,79 2 97,27 97,27 96,36 96,36 3 99,18 98,36 98,36 98,36 4 99,06 99,06 98,11 97,17 5 100,00 97,74 97,74 97,74 6 99,05 99,05 97,14 96,19 7 98,56 98,56 98,56 98,56 8 99,10 96,40 96,40 95,50 9 99,24 98,47 98,47 98,47 10 99,24 97,71 97,71 97,71 11 100,00 98,45 98,45 98,45
in natura 97,14 97,14 97,14 97,14 * valor médio de três repetições
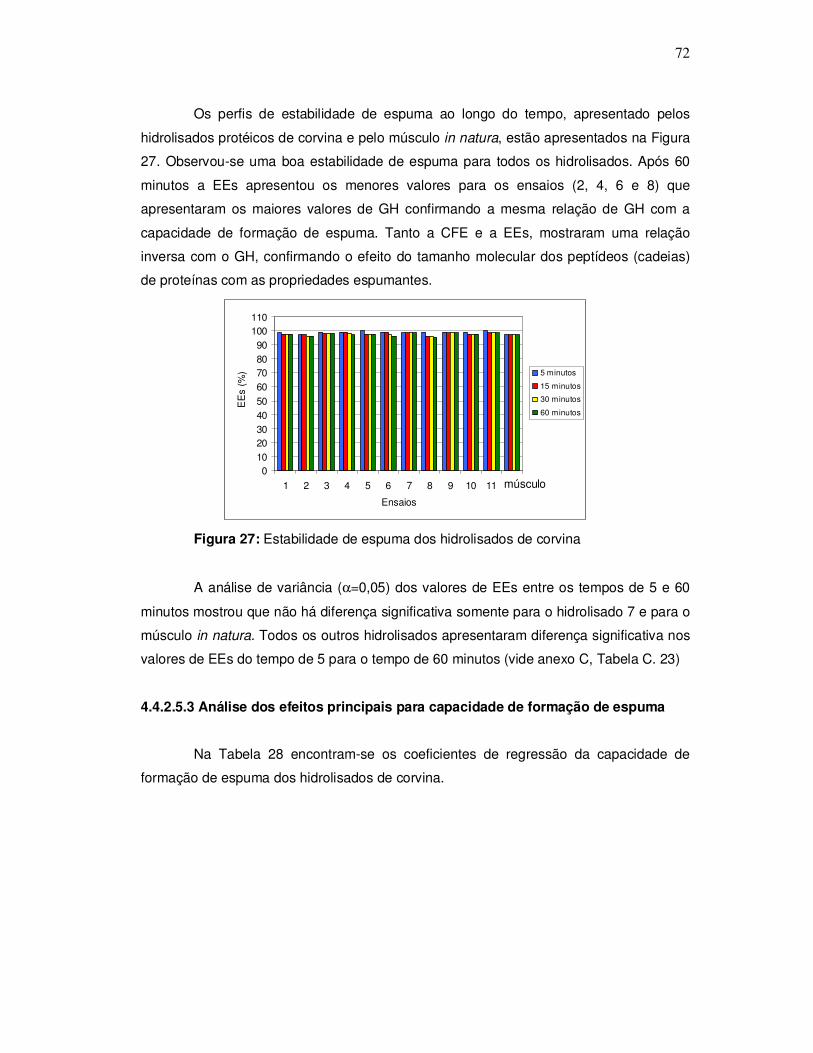
72
Os perfis de estabilidade de espuma ao longo do tempo, apresentado pelos
hidrolisados protéicos de corvina e pelo músculo in natura, estão apresentados na Figura
27. Observou-se uma boa estabilidade de espuma para todos os hidrolisados. Após 60
minutos a EEs apresentou os menores valores para os ensaios (2, 4, 6 e 8) que
apresentaram os maiores valores de GH confirmando a mesma relação de GH com a
capacidade de formação de espuma. Tanto a CFE e a EEs, mostraram uma relação
inversa com o GH, confirmando o efeito do tamanho molecular dos peptídeos (cadeias)
de proteínas com as propriedades espumantes.
0102030405060708090
100110
1 2 3 4 5 6 7 8 9 10 11 12
Ensaios
EE
s (%
) 5 minutos
15 minutos
30 minutos
60 minutos
Figura 27: Estabilidade de espuma dos hidrolisados de corvina
A análise de variância (α=0,05) dos valores de EEs entre os tempos de 5 e 60
minutos mostrou que não há diferença significativa somente para o hidrolisado 7 e para o
músculo in natura. Todos os outros hidrolisados apresentaram diferença significativa nos
valores de EEs do tempo de 5 para o tempo de 60 minutos (vide anexo C, Tabela C. 23)
4.4.2.5.3 Análise dos efeitos principais para capacidade de formação de espuma
Na Tabela 28 encontram-se os coeficientes de regressão da capacidade de
formação de espuma dos hidrolisados de corvina.
músculo

73
Tabela 28: Efeitos estimados para a CFE (%) dos hidrolisados de corvina.
Propriedade
Funcional Variáveis Efeitos
Erro
padrão Valor t(2) Valor p
Média 22,5714 0,436436 51,7176 0,000374 *
[E] (L) -21,0000 1,154701 -18,1865 0,003010*
CFE [S] (L) -9,0000 1,154701 -7,7942 0,016065 *
Interação 5,0000 1,154701 4,3301 0,049414 * * significativas (p<0,05)
Na Figura 28 encontra-se a representação gráfica dos efeitos que foram
significativos em relação a concentração de enzima e de substrato, e suas interações na
capacidade de formação de espuma dos hidrolisados protéicos. Observa-se que tanto a
[E] como a [S] foram significativas na CFE, porém esses efeitos foram negativos visto que
quando a [E] do meio aumentou, a capacidade de formação de espuma apresentou uma
diminuição de 21%. O mesmo ocorreu para a [S], pois quando a mesma aumentou houve
uma diminuição na CFE de 9%. Levando em conta a magnitude do efeito, a [E] mostrou-
se uma variável com maior influência do que a [S]. Observa-se que apesar das duas
variáveis terem apresentado um efeito negativo sobre a CFE, a interação das variáveis
apresentou um efeito positivo.
-25
-20
-15
-10
-5
0
5
10
[E]* [S]* [E]x[S]*
Variáveis
CFE
(%
)
Figura 28: Efeito da concentração de enzima e de substrato na CFE dos
hidrolisados protéicos
*significativas (p<0,05)
4.4.2.5.4 Verificação do modelo matemático preditivo
A análise de variância (teste F) mostrou que o modelo quadrático se ajustou bem
aos dados experimentais quanto à capacidade de formação de espuma, o que pode ser
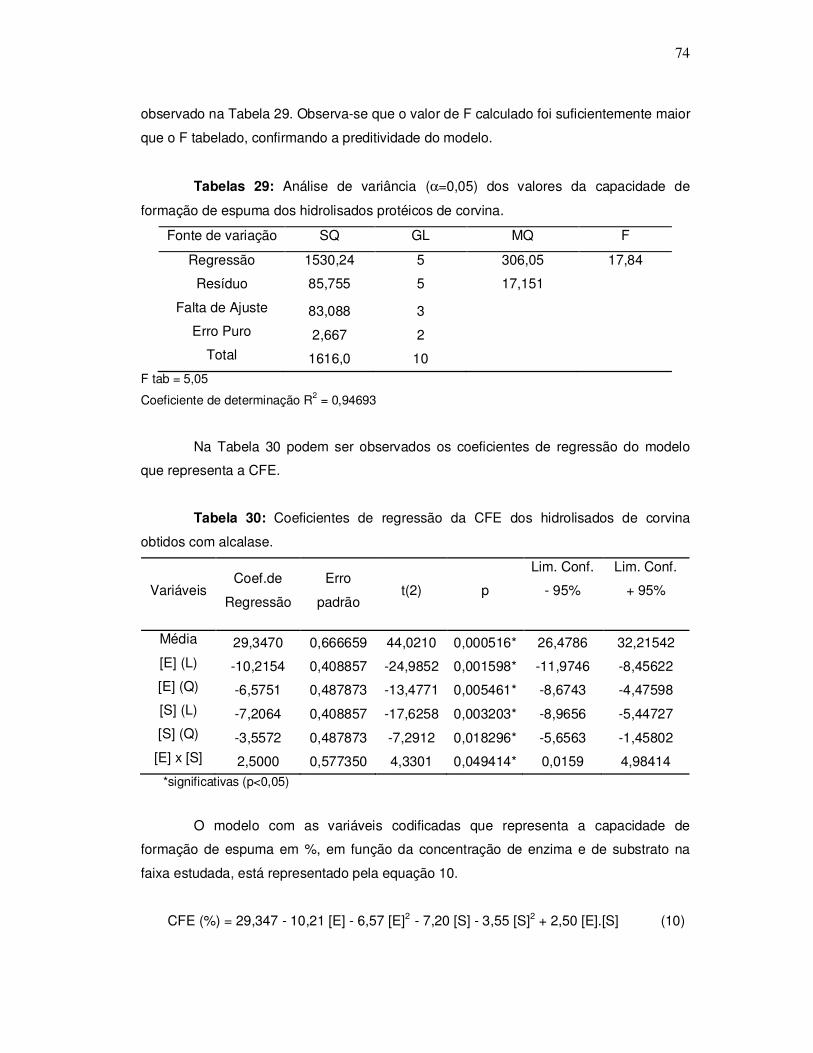
74
observado na Tabela 29. Observa-se que o valor de F calculado foi suficientemente maior
que o F tabelado, confirmando a preditividade do modelo.
Tabelas 29: Análise de variância (α=0,05) dos valores da capacidade de
formação de espuma dos hidrolisados protéicos de corvina.
Fonte de variação SQ GL MQ F
Regressão 1530,24 5 306,05 17,84
Resíduo 85,755 5 17,151
Falta de Ajuste 83,088 3
Erro Puro 2,667 2
Total 1616,0 10
F tab = 5,05
Coeficiente de determinação R2 = 0,94693
Na Tabela 30 podem ser observados os coeficientes de regressão do modelo
que representa a CFE.
Tabela 30: Coeficientes de regressão da CFE dos hidrolisados de corvina
obtidos com alcalase.
Variáveis Coef.de
Regressão
Erro
padrão t(2) p
Lim. Conf.
- 95%
Lim. Conf.
+ 95%
Média 29,3470 0,666659 44,0210 0,000516* 26,4786 32,21542
[E] (L) -10,2154 0,408857 -24,9852 0,001598* -11,9746 -8,45622
[E] (Q) -6,5751 0,487873 -13,4771 0,005461* -8,6743 -4,47598
[S] (L) -7,2064 0,408857 -17,6258 0,003203* -8,9656 -5,44727
[S] (Q) -3,5572 0,487873 -7,2912 0,018296* -5,6563 -1,45802
[E] x [S] 2,5000 0,577350 4,3301 0,049414* 0,0159 4,98414
*significativas (p<0,05)
O modelo com as variáveis codificadas que representa a capacidade de
formação de espuma em %, em função da concentração de enzima e de substrato na
faixa estudada, está representado pela equação 10.
CFE (%) = 29,347 - 10,21 [E] - 6,57 [E]2 - 7,20 [S] - 3,55 [S]2 + 2,50 [E].[S] (10)

75
Assim, o modelo foi utilizado na construção da superfície de resposta e curvas
de contorno, permitindo a visualização do comportamento dos hidrolisados enzimáticos
de corvina, obtidos com a enzima alcalase em função das variáveis que apresentaram
influência significativa sobre a variável resposta capacidade de formação de espuma,
demonstrado nas Figuras 29 e 30.
Figura 29: Superfície de resposta representando a capacidade de formação de
espuma dos hidrolisados de corvina em função da concentração de enzima e de
substrato.
Analisando as Figuras 29 e 30 é possível verificar que com uma concentração de
enzima entre de 0,04 %, utilizando uma concentração de substrato até 3%, pode se obter
uma CFE máxima, nas condições de pH 8,0 a uma temperatura de 50 ºC por um tempo
de reação de 60 minutos.

76
Figura 30: Linhas de contorno representando a capacidade de formação de
espuma dos hidrolisados de corvina em função da concentração de enzima e de
substrato.
Calcularam-se os valores experimentais, os previstos pelo modelo e os desvios
da CFE dos hidrolisados de corvina para a metodologia empregada. Na Tabela D.4 (vide
anexo D), pode-se observar que os desvios formam baixos na região em que os valores
de CFE apresentam-se mais elevados.
4.4.3 Digestibilidade in vitro da proteína dos hidrolisados de corvina
Na Tabela 31, estão apresentados os valores de digestibilidade in vitro dos
hidrolisados enzimáticos de corvina. Observa-se que todos os hidrolisados apresentaram
valores de digestibilidade acima de 90% o que demonstra a alta qualidade protéica dos
produtos obtidos. Estes dados foram obtidos por uma simulação da ação das enzimas no
organismo animal necessitando de complementação com um teste in vivo.
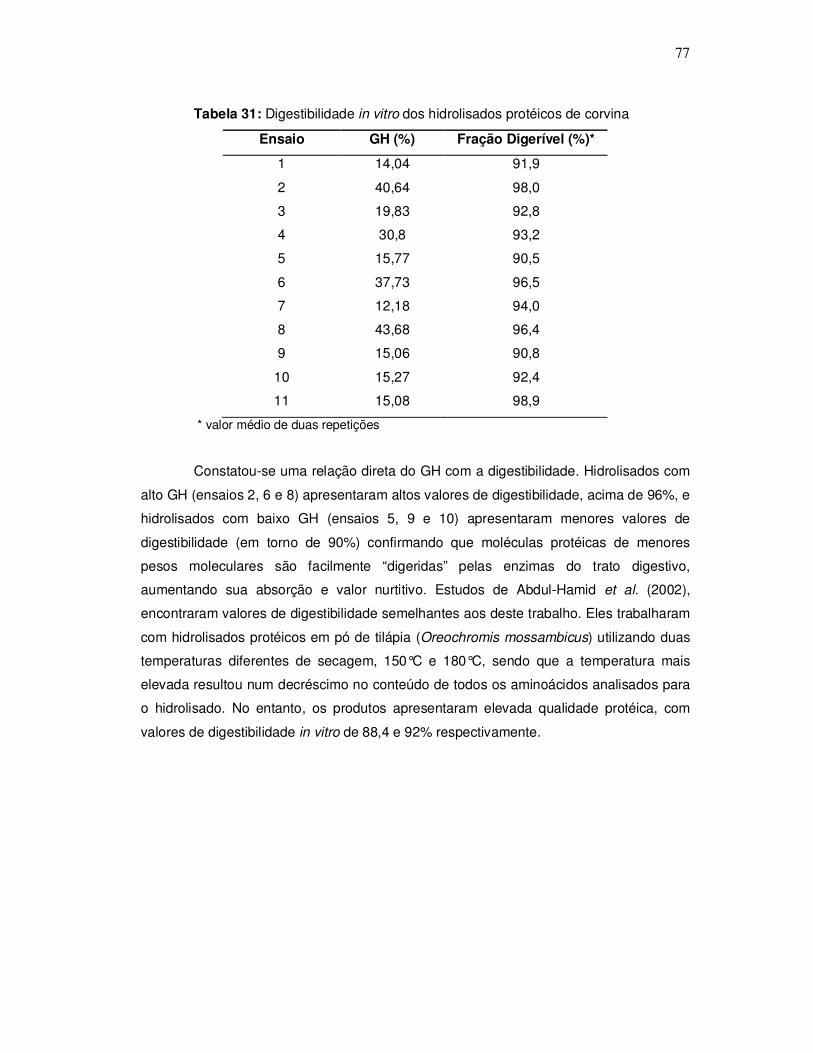
77
Tabela 31: Digestibilidade in vitro dos hidrolisados protéicos de corvina
Ensaio GH (%) Fração Digerível (%)*
1 14,04 91,9
2 40,64 98,0
3 19,83 92,8
4 30,8 93,2
5 15,77 90,5
6 37,73 96,5
7 12,18 94,0
8 43,68 96,4
9 15,06 90,8
10 15,27 92,4
11 15,08 98,9
* valor médio de duas repetições
Constatou-se uma relação direta do GH com a digestibilidade. Hidrolisados com
alto GH (ensaios 2, 6 e 8) apresentaram altos valores de digestibilidade, acima de 96%, e
hidrolisados com baixo GH (ensaios 5, 9 e 10) apresentaram menores valores de
digestibilidade (em torno de 90%) confirmando que moléculas protéicas de menores
pesos moleculares são facilmente “digeridas” pelas enzimas do trato digestivo,
aumentando sua absorção e valor nurtitivo. Estudos de Abdul-Hamid et al. (2002),
encontraram valores de digestibilidade semelhantes aos deste trabalho. Eles trabalharam
com hidrolisados protéicos em pó de tilápia (Oreochromis mossambicus) utilizando duas
temperaturas diferentes de secagem, 150°C e 180°C, sendo que a temperatura mais
elevada resultou num decréscimo no conteúdo de todos os aminoácidos analisados para
o hidrolisado. No entanto, os produtos apresentaram elevada qualidade protéica, com
valores de digestibilidade in vitro de 88,4 e 92% respectivamente.

78
5 CONCLUSÃO
Foi possível obter hidrolisados enzimáticos de corvina com diferentes valores de
grau de hidrólise.
O grau de hidrólise mostrou uma relação direta com a solubilidade e indireta com
a capacidade de retenção de água, capacidade de retenção de óleo, capacidade
emulsificante e capacidade de formação de espuma.
As concentrações de enzima e de substrato influenciaram diferentemente as
variáveis resposta estudadas.
A análise estatística dos valores experimentais permitiu obter modelos
matemáticos que relacionaram a concentração de enzima e a concentração de substrato
com o conteúdo protéico, com a capacidade de formação de espuma, e com a
capacidade de retenção de água em pH 7,0 e 9,0.
A obtenção de hidrolisados com diferentes graus de hidrólise e diferentes
propriedades funcionais e de digestibilidade, permitiu cumprir os objetivos deste estudo,
para disponibilizar hidrolisados que possam ser aplicados com diferentes finalidades
tecnológicas.

79
6 REFERÊNCIAS BIBLIOGRÁFICAS
AOAC, OFFICIAL METHODS OF ANALYSIS Association of Official Analytical
Chemists., 16ª ed, v. 1-2, USA, 1995.
ABDUL-HAMID, A.; BAKAR, J.; BEE, G. H. Nutritional quality of spray dried protein
hydrolysate from Black Tilapia (Oreochromis mossambicus). Food Chemistry. v. 78,
p.69, 2002.
BADOLATO, E. S. G.; CARVALHO, J. B.; AMARAL MELLO, M. R. P.; TAVARES, M.;
CAMPOS, N.C.; AUED-PIMENTEL,S.; MORAIS,C. Composição centesimal, de ácidos
graxos e valor calórico de cinco espécies de peixes marinhos nas diferentes estações
do ano. Revista do Instituto Adolfo Lutz, v. 54, n. 1, p. 27-35, 1994.
BAEK, H. H.; CADWALLADER, K.R. Enzimatic hydrolysis of crayfish processing by-
products. Journal of Food Science. v. 60, n. 5, p.929-935, 1995.
BENJAKUL, B.; MORRISSEY, M. T. Protein hydrolysates from Pacific whiting solid waste.
Journal of Agricultural and Food Chemistry. v. 45, p. 3424, 1997.
BRASIL. Métodos Analíticos Oficiais para Controle de Produtos de Origem Animal e
seus Ingredientes: Métodos Físico-químicos. Brasília 1981.
BRASIL, Ministério da Agricultura. Secretária Municipal do Instituto Brasileiro do Meio
Ambiente e dos Recursos Naturais Renováveis. Tamanhos mínimos para captura
de peixes marinhos. IBAMA, Portaria nº 73 de 24 de novembro de 2003.
BRASIL. Ministério da Agricultura. Secretária Municipal do Instituto Brasileiro do Meio
Ambiente e dos Recursos Naturais Renováveis. Desembarque de Pescado no Rio
Grande do Sul em 2004. Rio Grande, IBAMA, 2005.
BRASIL. Ministério da Agricultura e do Abastecimento. Regulamento Técnico de
Identidade e Qualidade de Peixe Fresco. Portaria nº 185 de 13 de maio de 1997.
Disponível em: <http://www.engetecno.com.br>. Acesso em 08 de março de 2006.
BRUSCHI, F. L. F. Rendimento, composição química e perfil de ácidos graxos de
pescados e seus resíduos: uma comparação. Trabalho de Conclusão apresentado
ao Curso de Oceanografia. Universidade do Vale do Itajaí, Centro de Ciências
Tecnológicas, da Terra e do Mar, Curso De Oceanografia, 2001.
CASTELLO, J. P. Distribucion, crecimiento y maduracion sexual de la corvina juvenil
(Micropogonias furnieri) en el estuario de la “Lagoa dos Patos”, Brasil. PHYSIS v. 44,
p. 21-26, 1986.
CÂNDIDO, L. M. B.; NOGUEIRA, A. K.; SGARBIERI, V. C. Propriedades funcionais de
concentrados protéicos de pescado preparados por vários métodos. Brazilian
Journal of Food Technology. 1(1,2): 77- 89, 1998.

80
CÂNDIDO, L. M. B.; SGARBIERI, V. C. Enzymatic hydrolysis of Nile tilapia (Oreochromus
niloticus) myofibrillar proteins: effects on nutritional and hydrophilic properties. Journal
of the Science of Food and Agriculture. v. 83, p. 937, 2003.
CHEFTEL, J. C.; CUQ, J. L.; LORIENT, D. Aminoácidos, peptidos y proteínas. In:
FENNEMA, O. Química de los Alimentos. Zaragoza: Ed. Acribia, p. 275, 1993.
CHO, S. M.; KWAK, K. S.; PARK, D. C. GU, Y. S.; JI, C. I.; JANG, D. H.; LEE, Y. B.; KIM,
S. B. Processing optimization an functional properties of gelatin from shark (Isurus
oxyrinchus) cartilage. Food Hydrocolloids, v. 18, p. 573-579, 2004.
CONTRERAS-GUZMÁN, E. S. Bioquímica de pescados e derivados. Jaboticabal:
FUNEP, 1994.
COSTA, M. R., ARAÚJO, F. Use of a tropical bay in southeastern Brazil by juvenile and
subadult Micropogonias furnieri (Perciformes, Sciaenidae) Journal of Marine
Science V. 60, p. 268-277, 2003.
DAMODARAN, S. Amino acids, peptides and proteins. Food Chemistry. Ed. Marcel
Deker Inc. New York, 1996.
DEMETRIADES, K.; COUPLAND, J. N.; McCLEMENTS, D. J. Physical properties of whey
protein stabilized emulsions as related to pH and NaCl. Journal of Food Science. v.
62, p.342, 1997.
DINIZ, F. M.; MARTIN, A. M. Use of response surface methodology to describe the
combined effects of pH, temperature and E/S ratio on the hydrolysis of dogfish
(Squalus acanthias) muscle. International Journal of Food Science and
Technology. v. 31, p.419, 1996.
DONADEL, M. E.; PRUDENCIO-FERREIRA, S. H. Propriedades funcionais de
concentrado protéico de feijão envelhecido. Ciência e Tecnologia de Alimentos. 19
(3): Campinas, set/dez, 1999.
DUARTE, A. J.; CARREIRA, R. L.; JUNQUEIRA, R. G. Propriedades emulsificantes e
solubilidade da caseína bovina e de seus hidrolisados trípticos: 1. Efeito do pH e do
tempo de hidrólise. Ciência e Tecnologia de Alimentos. 18 (3): 295-302, 1998.
ELSDON, T.S. GILLANDERS B.M. Interactive effects of temperature and salinity on
otolith chemistry: challenges for determining environmental histories of fish. Fish
Aquatic Science. v. 59, p. 1796-1808, 2002.
FAO, Food and Agriculture Organization of the United Nations, Fishery Statistics 2003.
Disponível em: <http://www.fao.org/>. Acesso em: 24 de outubro de 2005.
FIGUEREDO, R. C. R.; SABADINI, E.; RIBEIRO, F. A. L. Ciência de Espumas- Aplicação
na Extinção de Incêndios. Química Nova. v.22, n.1, p.126-130, 1999.

81
FONKWE, L. G.; SINGH, R. K. Protein Recovery from Mechanically Deboned Turkey
Residue by Enzymic Hydrolysis. Process Biochemistry. v. 31, n. 6, p. 605-616,
1996.
GBOGOURI, G.A.; LINDER, M.; FANNI, J.; PARMENTIER, M. Influence of Hydrolysis
Degree on the Functional Properties of Salmon Byproducts Hydrolysates. Journal of
Food Science: Food Chemistry and Toxicology. v. 69, n 8, 2004.
GILDBERG, A.; RAA, J. Purification and characterization of pepdins from the artic fish
capelin (Mallotus villosus). Comp. Biochem. Physiol. v. 75 (3), p.337, 1983.
GILDBERG, A. Enzymatic Processing of Marine Raw Materials. Process Biochemistry.
28, p. 1-15, 1993.
GILDBERG, A.; ARNESEN, J. A.; CARLEOG, M. Utilization of cod backbone by
biochemical fractionation. Process Biochemistry. v. 38, p. 475, 2002.
GLÓRIA, M. M.; REGINATO-D’ARCE, M. A. B. Concentrado e Isolado Protéico de Torta
de Castanha do Pará: Obtenção e Caracterização Química e Funcional. Ciência e
Tecnologia de Alimentos. 20(2): 240-245, 2000.
GONÇALVES A. A.; PASSOS, M. G. Uso da enzima transglutaminase na elaboração
de um produto reestruturado à base de peixe. Revista Nacional da Carne. v. 317,
2003.
HALE, M. B. Relative activities of commercial-available enzymes in the hydrolysis of fish
proteins. Food Technology. v. 23, p.107, 1969.
HOYLE, N.; MERRITT, J. H. Quality of fish protein hydrolysates from herring (Clupea
harengus). Journal of Food Science. v. 59, p.76, 1994.
GUIA DA SAÚDE FAMILIAR, REVISTA ISTOÉ, volume 16, março de 2002.
KILARA, A. Enzyme-modified protein foods ingredients. Process Biochemistry.
Wattford, v.20, n.5, p.149-157, 1985.
KINSELLA, J.E. Functional properties of proteins in foods: a survey. Critical Reviews in
Food Science and Nutrition. 8(4): 219, 1976.
KRISTINSSON, H.G. Reaction kinetics biochemical and functional properties of salmon
muscle proteins hydrolysed by different alkaline proteases. Master’s Thesis.
University of Washington, Seattle W.A., 1998.
KRISTINSSON, H.G.; RASCO, B.A. Fish Protein Hydrolysates: Production, Biochemical
and Functional Properties. Critical Reviews in Food Science and Nutrition.
40(1):43-81, Washington, 2000a.
KRISTINSSON, H. G.; RASCO, B. A. Kinetics of the hydrolysis of Atlantic salmon
(Salmon solar) muscle proteins by alkaline proteases and a visceral serine protease
mixture. Process Biochemistry. v. 36, p. 131-139, 2000b.

82
LAHL, W.J.; BRAUN, S.D.; Enzymatic Production of Protein Hydrolysates for Food
Use. Food Technology, 48 (10): 68, 1994.
LICEAGA-GESUALDO, A. M.; LI-CHAN, E. C. Y.; Functional properties of fish protein
hydrolysate from herring (clupea harengus). Journal of Food Science, v. 64, n.6, p.
1000-1004, 1999.
LOWRY, O. H.; ROSEBROUGH, N. J.; FARR, A. L.; RANDALL, R. J. Protein
measurement whit the Folin phenol reagent. Journal Biol. Chem. 193, p 265-275,
1951.
MACKIE, L.M. Fish protein hydrolysates. Process Biochemistry. 1-2, 26, 1982.
MAHMOUD, M. L. Physicochemical and functional properties of protein hydrolysates in
nutritional products. Food Technology. 58(10): 89, 1994.
MARTIN, R. E. Chemistry and Biochemistry of Marine Food Products. AVI Publishing
Company, Westport: 1982.
MITCHELL, H. S.; RYNBERGEN, H. J.; ANDERSON, L.; DIBBLE, M. V. Nutrição.
Interamericana, Rio de Janeiro, RJ, ed. 16, 1978.
MOHR, V. Enzymes technology in the meat and fish industries. Process Biochemistry.
8-9, 18, 1980.
MORR, V.; GERMAN, B.; KINSELA, J. E.; REGENSTEIN, J. M.; VAN BUREN, J. P.;
KILARA, A.; LEWIS, B. A.; MANGINA, M. E. A collaborative study to develop a
standarized food protein solubility procedure. Journal of Food Science, v. 50, p.
1715, 1985.
MORITA, T; ASSUMPÇÃO, R.M.V.; Manual de soluções, reagentes e solventes. Ed.
Edgard Blücher Ltda, p. 272, 1972.
MULLALLY, M. M.; O’CALLAGHAN, D. M.; FITZGERALD, R. J.; DONNELLY, W. J.;
DALTON, J. P. Zymogen activation in pancreatic endoproteolytic preparations and
influence on some whey protein characteristics. Journal of Food Science, v. 60 (2),
p. 227, 1995.
NEIVA, G. S. Sumário sobre a pesca mundial. Disponível em: < http://www.pescabrasil.
com.br/comercial/artigo2.asp > Acesso em: 03 de novembro de 2005.
NETO, B. B.; SCARMINIO, I. S.; BRUNS, R. E. Planejamento e Otimização de
Experimentos. 2ª edição. São Paulo: Editora da Unicamp, 1996.
NIELSEN, P. M.; OLSEN, H. S. Enzymic modification of food protein. "Enzymes in food
technology". Sheffield Academic Press, Cap. 6, p. 109-143, 2002.
OGAWA, M; MAIA, E. L. Manual de pesca – Ciência e Tecnologia do Pescado, v 1.
Livraria Varela, São Paulo, SP, 1999.

83
OKEZIE, B. O.; BELLO, A. B. Physicochemical and Functional Properties of Winged bean
Flour and Isolate compared with Soy Isolate. Journal of Food Science, v. 53 (2), p.
540-454, 1988.
OWENS, J. D., MENDOZA, L. S. Enzymically hydrolysed and bacterially fermented
fishery products. Journal of Food Technology, v. 20, 273-293, 1985.
ONODENALORE, A.C.; SHAHIDI, F. Protein dispersions and hydrolysates from shark
(Isurus oxyrinchus), Journal of Aquatic Food Production Technology, v. 5, p. 43,
1996.
PANYAM, D., KILARA, A. Enchanting the functionality of food proteins by enzymatic
modification. Trends in Food Science & Technology. Cambridge, v.7, n.4, p.120-
125, 1996.
PATEL, P. D.; STRIPP, A. M.; FRY, J. C. Whipping test for the determination of foaming
capacity of protein: a collaborative study. International Journal of Food Science and
Technology, v.23, n.1, p. 57-63, 1988.
PEZOA, V. G.; MELLADO, M. S. Estudo das condições ótimas de pH e temperatura para
atividade enzimática de estratos dos componentes do aparelho digestivo da corvina
(Micropogon furnieri) e da castanha (Umbrina canosai). Obtenção de um
concentrado de proteínas de pescado para alimentos pelo método enzimático,
utilizando as próprias enzimas do pescado, p. 5-38, FURG, Rio Grande, 1979.
PILOSOF, A. M. R. Propiedades de Hidratación. In: "Caracterización funcional y
estructural de proteínas". Ed. Pilosof, A.M.R & Bartholomai G. B. Buenos Aires
Eudeba, v.1, p.17-28, 2000.
POMERANZ, Y. Functional properties of food components. 2º ed. San Diego:
Academic Press, p. 569, 1991.
QUAGLIA, G. B.; ORBAN, E. Influence of enzymatic hydrolysis on structure and
emulsifying properties of sardine (Sardina pilchardus) protein hydrolysates. Journal of
Food Science. v. 55, p.1571, 1990.
REBECA, B. D.; PEÑA-VERA, M. T.; DÍAZ-CASTAÑEDA, M. Production of fish protein
hydrolysates with bacterial proteases; Yield and nutritional value. Journal of Food
Science. v. 56, p. 309, 1991.
REGENSTEIN, J. M.; GORIMAR, T. S.; SHERBON, J. M. Measuring the water capacity of
natural actomyosin from chicken breast muscle in the presence of pyrophosphate and
divalent cation. Journal of Food Biochemistry. v. 3, p. 205-211, 1979.
ROBINSON, D.S. Bioquímica y valor nutritivo de los alimentos. Ed. Acribia, S.A.
Zaragoza, 1991.

84
ROMAN, J. A.; SGARBIERI, V. C. Efeito da Hidrólise Enzimática sobre Propriedades
Funcionais de Caseína Bovina coagulada pela ação da Quimiosina. Ciência e
Tecnologia de Alimentos. 25(3): 468-474, 2005.
SAMPEDRO, G., LÓPEZ-BENITO, M., PASTORIZA, L. Cabezas y vísceras de túnidos:
Su aprovechamiento para alimentación animal. Informe técnico nº 137 del Instituto
de Investigaciones Pesqueras. Barcelona, 1986.
SANTOS, S. D. Obtenção e avaliação de hidrolisado enzimático obtido a partir de
pescado de baixo valor comercial. Dissertação de mestrado. Fundação
Universidade Federal do Rio Grande - FURG, Rio Grande, RS, 2006.
SATHIVEL, S.; SMILEY, S.; PRINYAWIWATKUL, W.; BECHTEL, P J. Functional and
Nutritional Properties of Red Salmon (Oncorhynchus nerka) Enzymatic Hydrolysates.
Food Chemistry and Toxicology. v. 70, n. 6, 2005.
SGARBIERI, V.C. Proteínas em alimentos protéicos. Propriedades – Degradações –
Modificações. Livraria Varela, São Paulo, SP, 1996.
SGARBIERI, V.C. Propriedades funcionais de proteínas em alimentos. Boletim da
Sociedade Brasileira de Ciência e Tecnologia de Alimentos. 32(1):105-126, 1998.
SHAHIDI, F.; VENUGOPAL, V. Solubilization and thermostability of water dispersions of
muscle structural proteins of Atlantic Herring (Clupea harengus). Journal of
Agricultural and Food Chemistry. v. 42, p. 1440, 1994.
SHAHIDI, F.; HAN, X-Q.; SYNOWIECKI, J. Production and characteristics of protein
hydrolysates from capelin (Mallotus villosus). Food Chemistry. v. 53, p.285, 1995.
SHAHIDI, F.; KAMIL, Y.V.A.J. Enzymes from fish aquatic invertebrates and their
application in the food industry. Food Science & Technology. V.12, p. 435-464,
2001.
SIMÕES, D.R.S.; AUGUSTO-RUIZ, W.; PEDROSO, M. A. ALMEIDA, T.L. Hambúrgueres
formulados com base protéica de pescado. Ciência e Tecnologia de Alimentos.
18(4):4-14, 1998.
SIMPSON, B. K.; SMITH, J. P.; HAARD, N.F. Marine enzymes. In. Y. H. Hui (Ed.)
Encyclopedia of Food Science and Technology. p. 1645-1653, New York N. Y.
Jonh Wiley and Sons, 1991.
STORI, F.T. Avaliação dos resíduos da industrialização do pescado em Itajaí e
Navegantes (SC), como subsídio à implementação de um sistema gerencial de bolsa
de resíduos. Trabalho de Conclusão de Curso, Universidade do Vale do Itajaí -
UNIVALI, SC, p. 145, 2000.
VENUGOPAL, V.; SHAHIDI, F. Value-added products from underutilized fish species.
Critical Reviews in Food Science and Nutrition. v. 35, 431-453, 1995.

85
YEANNES, M. I.; ALMANDOS, M.E. Estimation of Fish Proximate Composition Starting
From Water Contend Journal of Food Composition and Analysis v. 16, p. 81-92,
2003.
WHITAKER, R. Principles of Enzymology for Food Science. Marcel Decker, N.Y.,
1994.
WILDING, P.; LILLIFORD, P.J.; RUGENSTEIN, J.M. Functional properties of proteins in
foods. Journal of Chemistry, Technology and Biotechnology. 34B, p.182, 1984.
86
ANEXO A
Tabela A.1: Valores médios de GH em diferentes temperaturas do teste preliminar 1 (T1)
Tempo (min) GH (T = 50ºC)* Tempo (min) GH (T = 55ºC)*
0 8,75 0 7,12
15 10,10 15 14,95
30 15,31 30 17,91
45 15,43 45 17,97
60 19,87 60 19,90
* valores médios de três repetições
Tabela A.2: Análise de Variância do teste preliminar 1 (T1)
Tabela A.3: Teste de Tukey para o teste preliminar 1 (T1)
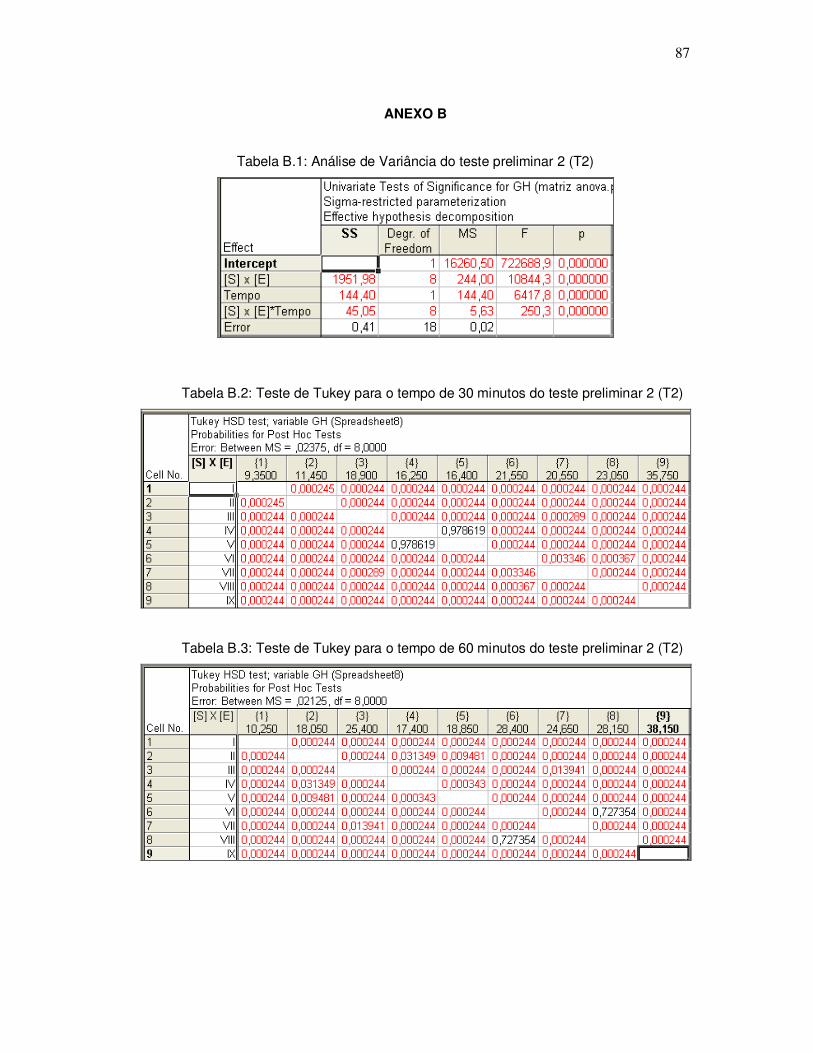
87
ANEXO B
Tabela B.1: Análise de Variância do teste preliminar 2 (T2)
Tabela B.2: Teste de Tukey para o tempo de 30 minutos do teste preliminar 2 (T2)
Tabela B.3: Teste de Tukey para o tempo de 60 minutos do teste preliminar 2 (T2)
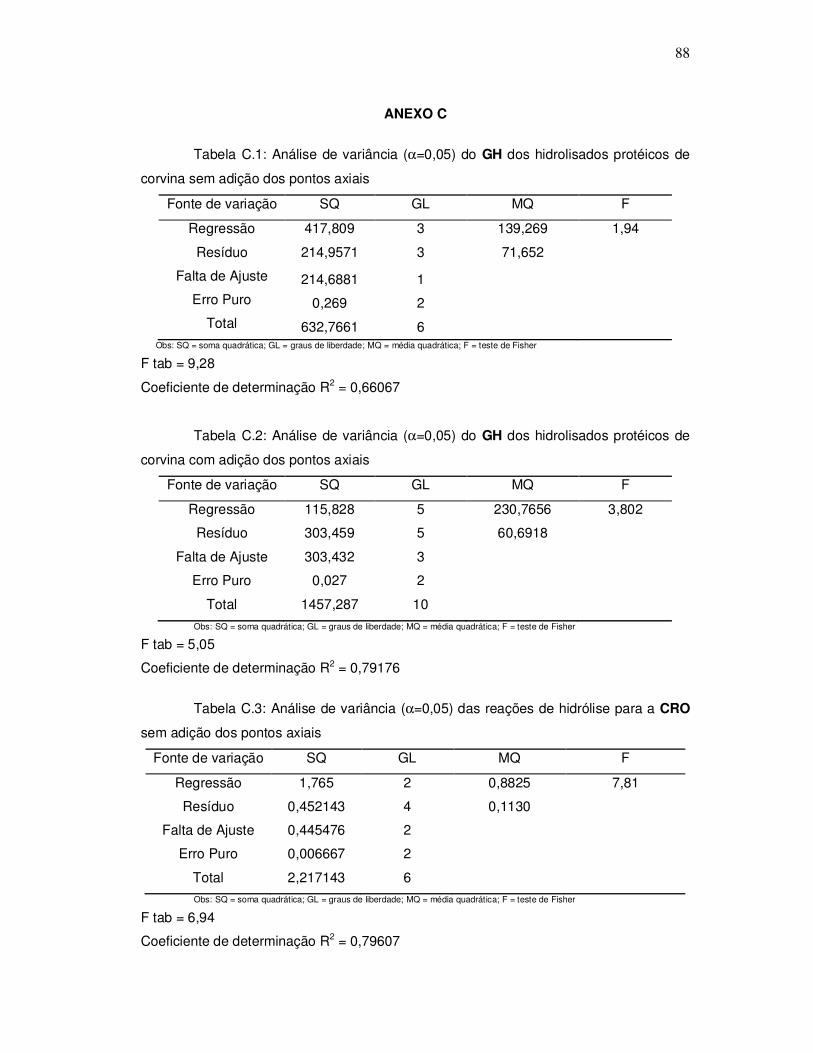
88
ANEXO C
Tabela C.1: Análise de variância (α=0,05) do GH dos hidrolisados protéicos de
corvina sem adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 417,809 3 139,269 1,94
Resíduo 214,9571 3 71,652
Falta de Ajuste 214,6881 1
Erro Puro 0,269 2
Total 632,7661 6
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 9,28
Coeficiente de determinação R2 = 0,66067
Tabela C.2: Análise de variância (α=0,05) do GH dos hidrolisados protéicos de
corvina com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 115,828 5 230,7656 3,802
Resíduo 303,459 5 60,6918
Falta de Ajuste 303,432 3
Erro Puro 0,027 2
Total 1457,287 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 5,05
Coeficiente de determinação R2 = 0,79176
Tabela C.3: Análise de variância (α=0,05) das reações de hidrólise para a CRO
sem adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 1,765 2 0,8825 7,81
Resíduo 0,452143 4 0,1130
Falta de Ajuste 0,445476 2
Erro Puro 0,006667 2
Total 2,217143 6
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 6,94
Coeficiente de determinação R2 = 0,79607
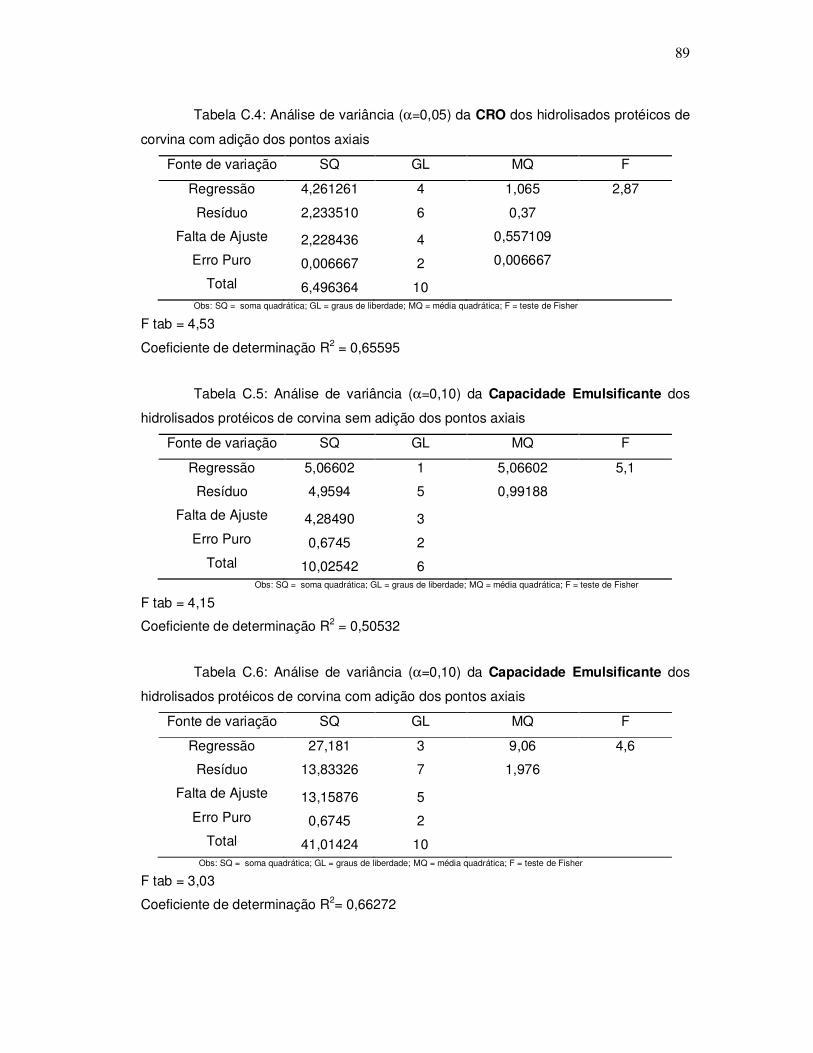
89
Tabela C.4: Análise de variância (α=0,05) da CRO dos hidrolisados protéicos de
corvina com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 4,261261 4 1,065 2,87
Resíduo 2,233510 6 0,37
Falta de Ajuste 2,228436 4 0,557109
Erro Puro 0,006667 2 0,006667
Total 6,496364 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 4,53
Coeficiente de determinação R2 = 0,65595
Tabela C.5: Análise de variância (α=0,10) da Capacidade Emulsificante dos
hidrolisados protéicos de corvina sem adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 5,06602 1 5,06602 5,1
Resíduo 4,9594 5 0,99188
Falta de Ajuste 4,28490 3
Erro Puro 0,6745 2
Total 10,02542 6
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 4,15
Coeficiente de determinação R2 = 0,50532 Tabela C.6: Análise de variância (α=0,10) da Capacidade Emulsificante dos
hidrolisados protéicos de corvina com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 27,181 3 9,06 4,6
Resíduo 13,83326 7 1,976
Falta de Ajuste 13,15876 5
Erro Puro 0,6745 2
Total 41,01424 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 3,03
Coeficiente de determinação R2= 0,66272
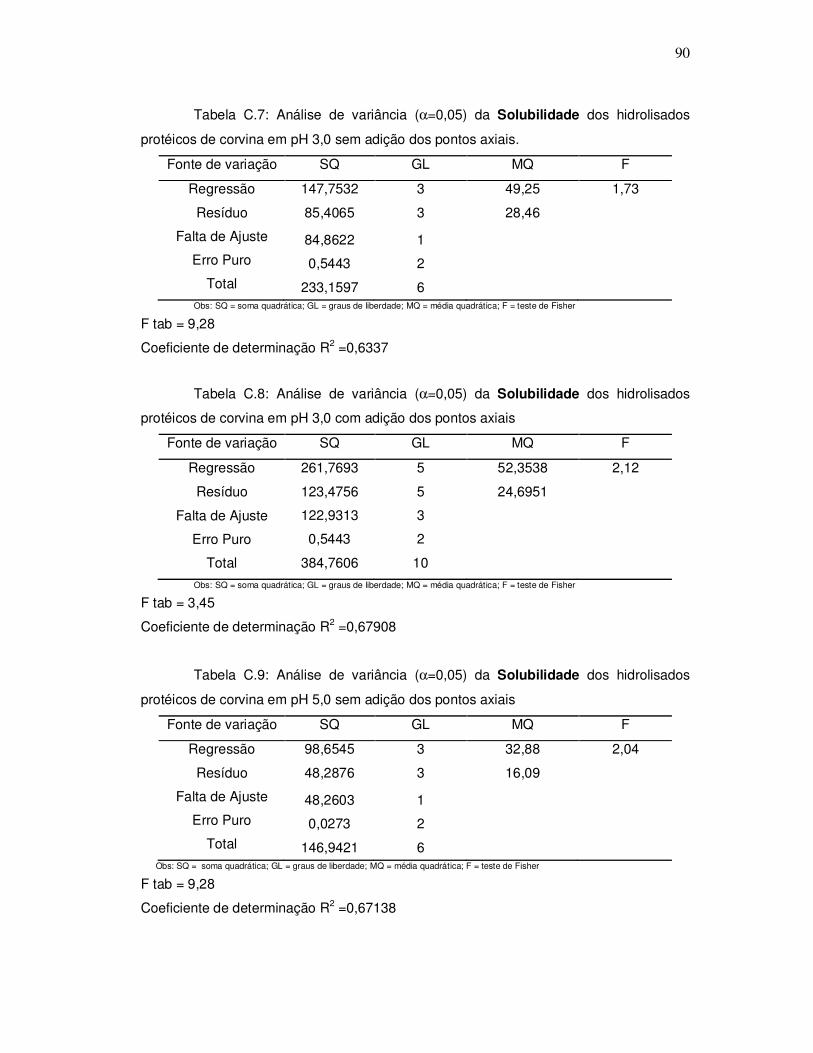
90
Tabela C.7: Análise de variância (α=0,05) da Solubilidade dos hidrolisados
protéicos de corvina em pH 3,0 sem adição dos pontos axiais.
Fonte de variação SQ GL MQ F
Regressão 147,7532 3 49,25 1,73
Resíduo 85,4065 3 28,46
Falta de Ajuste 84,8622 1
Erro Puro 0,5443 2
Total 233,1597 6
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 9,28
Coeficiente de determinação R2 =0,6337
Tabela C.8: Análise de variância (α=0,05) da Solubilidade dos hidrolisados
protéicos de corvina em pH 3,0 com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 261,7693 5 52,3538 2,12
Resíduo 123,4756 5 24,6951
Falta de Ajuste 122,9313 3
Erro Puro 0,5443 2
Total 384,7606 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 3,45
Coeficiente de determinação R2 =0,67908
Tabela C.9: Análise de variância (α=0,05) da Solubilidade dos hidrolisados
protéicos de corvina em pH 5,0 sem adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 98,6545 3 32,88 2,04
Resíduo 48,2876 3 16,09
Falta de Ajuste 48,2603 1
Erro Puro 0,0273 2
Total 146,9421 6
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 9,28
Coeficiente de determinação R2 =0,67138

91
Tabela C.10: Análise de variância (α=0,05) da Solubilidade dos hidrolisados
protéicos de corvina em pH 5,0 com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 219,04 5 43,81 1,57
Resíduo 138,901 5 27,78
Falta de Ajuste 138,8823 3
Erro Puro 0,02763 2
Total 357,9567 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab=3,45
Coeficiente de determinação R2= 0,61164
Tabela C.11: Análise de variância (α=0,05) da Solubilidade dos hidrolisados
protéicos de corvina em pH 7,0 sem adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 146,4722 2 73,23 5,49
Resíduo 53,3058 4 13,32
Falta de Ajuste 53,0557 2
Erro Puro 0,2501 2
Total 199,7780 6
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
Ftab = 6,94
Coeficiente de determinação R2= 0,73318
Tabela C.12: Análise de variância (α=0,05) da Solubilidade dos hidrolisados
protéicos de corvina em pH 7,0 com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 266,9378 3 88,97 2,68
Resíduo 232,1936 7 33,17
Falta de Ajuste 231,9435 5
Erro Puro 0,2501 2
Total 499,13 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab= 4,35
Coeficiente de determinação = R2= 0,5348

92
Tabela C.13: Análise de variância (α=0,05) da Solubilidade dos hidrolisados
protéicos de corvina em pH 9,0 sem adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 183,9508 2 94,97 7,55
Resíduo 50,2775 4 12,57
Falta de Ajuste 45,9254 2
Erro Puro 4,3521 2
Total 240,2283 6
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab =6,94
Coeficiente de determinação R2 =0,79071
Tabela C.14: Análise de variância (α=0,05) da Solubilidade dos hidrolisados
protéicos de corvina em pH 9,0 com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 243,46 2 121,73 3,47
Resíduo 280,389 8 35,048
Falta de Ajuste 276,0362 6
Erro Puro 4,3521 2
Total 523,849 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 4,46
Coeficiente de determinação R2 =0,46475
Tabela C.15: Análise de variância (α=0,05) da Solubilidade dos hidrolisados
protéicos de corvina em pH 11,0 sem adição dos pontos axiais.
Fonte de variação SQ GL MQ F
Regressão 150,3456 3 50,11 2,34
Resíduo 64,2185 3 21,40
Falta de Ajuste 62,7126 1
Erro Puro 1,5059 2
Total 214,5641 6
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 9,28
Coeficiente de determinação R2 =0,7007
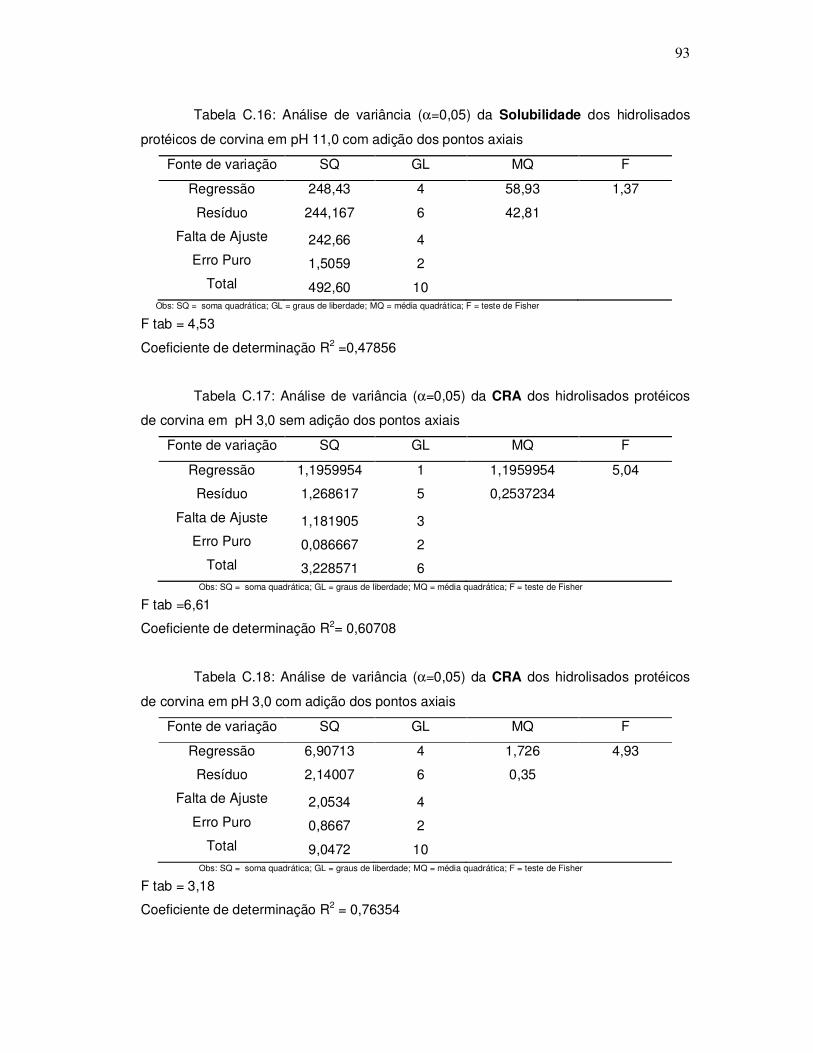
93
Tabela C.16: Análise de variância (α=0,05) da Solubilidade dos hidrolisados
protéicos de corvina em pH 11,0 com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 248,43 4 58,93 1,37
Resíduo 244,167 6 42,81
Falta de Ajuste 242,66 4
Erro Puro 1,5059 2
Total 492,60 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 4,53
Coeficiente de determinação R2 =0,47856
Tabela C.17: Análise de variância (α=0,05) da CRA dos hidrolisados protéicos
de corvina em pH 3,0 sem adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 1,1959954 1 1,1959954 5,04
Resíduo 1,268617 5 0,2537234
Falta de Ajuste 1,181905 3
Erro Puro 0,086667 2
Total 3,228571 6
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab =6,61
Coeficiente de determinação R2= 0,60708
Tabela C.18: Análise de variância (α=0,05) da CRA dos hidrolisados protéicos
de corvina em pH 3,0 com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 6,90713 4 1,726 4,93
Resíduo 2,14007 6 0,35
Falta de Ajuste 2,0534 4
Erro Puro 0,8667 2
Total 9,0472 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 3,18
Coeficiente de determinação R2 = 0,76354
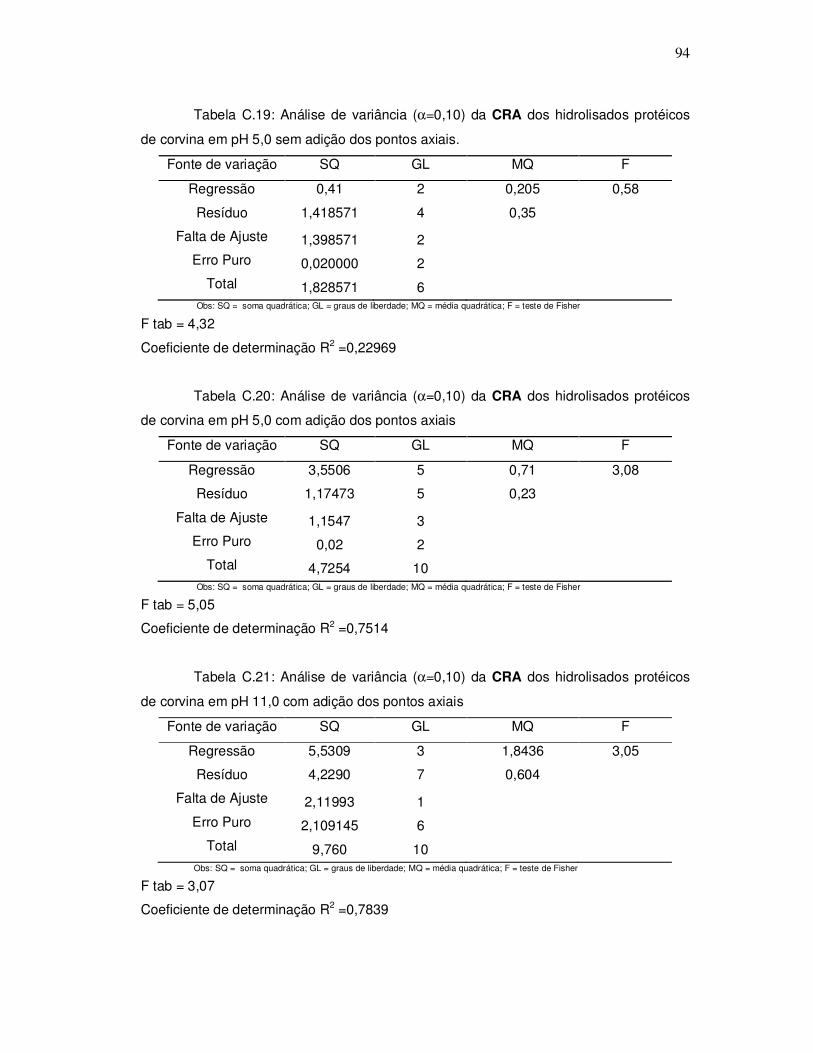
94
Tabela C.19: Análise de variância (α=0,10) da CRA dos hidrolisados protéicos
de corvina em pH 5,0 sem adição dos pontos axiais.
Fonte de variação SQ GL MQ F
Regressão 0,41 2 0,205 0,58
Resíduo 1,418571 4 0,35
Falta de Ajuste 1,398571 2
Erro Puro 0,020000 2
Total 1,828571 6
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 4,32
Coeficiente de determinação R2 =0,22969
Tabela C.20: Análise de variância (α=0,10) da CRA dos hidrolisados protéicos
de corvina em pH 5,0 com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 3,5506 5 0,71 3,08
Resíduo 1,17473 5 0,23
Falta de Ajuste 1,1547 3
Erro Puro 0,02 2
Total 4,7254 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 5,05
Coeficiente de determinação R2 =0,7514
Tabela C.21: Análise de variância (α=0,10) da CRA dos hidrolisados protéicos
de corvina em pH 11,0 com adição dos pontos axiais
Fonte de variação SQ GL MQ F
Regressão 5,5309 3 1,8436 3,05
Resíduo 4,2290 7 0,604
Falta de Ajuste 2,11993 1
Erro Puro 2,109145 6
Total 9,760 10
Obs: SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática; F = teste de Fisher
F tab = 3,07
Coeficiente de determinação R2 =0,7839
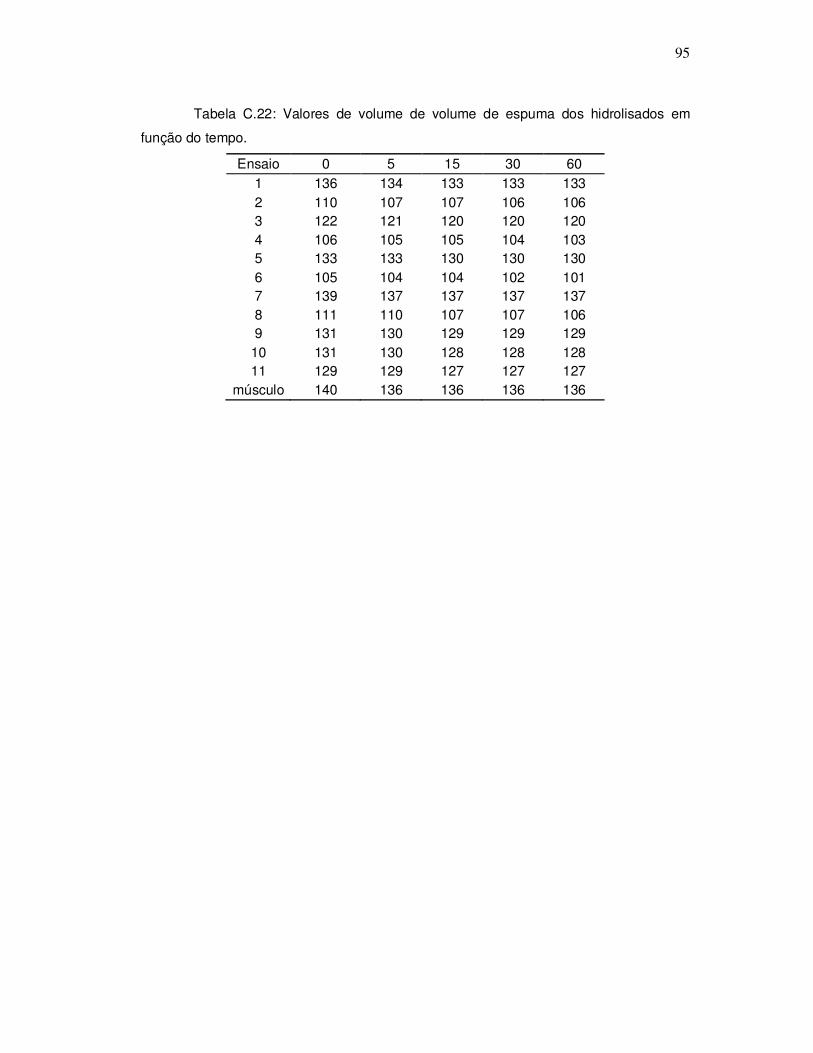
95
Tabela C.22: Valores de volume de volume de espuma dos hidrolisados em
função do tempo.
Ensaio 0 5 15 30 60 1 136 134 133 133 133 2 110 107 107 106 106 3 122 121 120 120 120 4 106 105 105 104 103 5 133 133 130 130 130 6 105 104 104 102 101 7 139 137 137 137 137 8 111 110 107 107 106 9 131 130 129 129 129 10 131 130 128 128 128 11 129 129 127 127 127
músculo 140 136 136 136 136
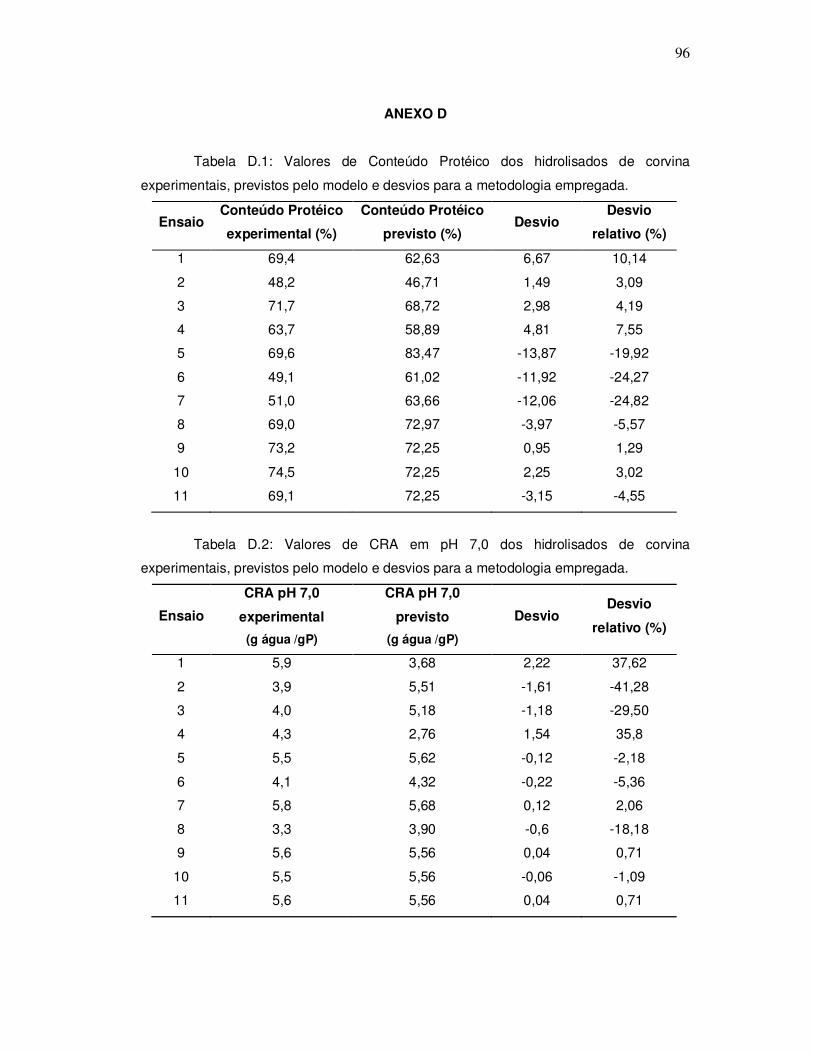
96
ANEXO D
Tabela D.1: Valores de Conteúdo Protéico dos hidrolisados de corvina
experimentais, previstos pelo modelo e desvios para a metodologia empregada.
Ensaio Conteúdo Protéico
experimental (%)
Conteúdo Protéico
previsto (%) Desvio
Desvio
relativo (%)
1 69,4 62,63 6,67 10,14
2 48,2 46,71 1,49 3,09
3 71,7 68,72 2,98 4,19
4 63,7 58,89 4,81 7,55
5 69,6 83,47 -13,87 -19,92
6 49,1 61,02 -11,92 -24,27
7 51,0 63,66 -12,06 -24,82
8 69,0 72,97 -3,97 -5,57
9 73,2 72,25 0,95 1,29
10 74,5 72,25 2,25 3,02
11 69,1 72,25 -3,15 -4,55
Tabela D.2: Valores de CRA em pH 7,0 dos hidrolisados de corvina
experimentais, previstos pelo modelo e desvios para a metodologia empregada.
Ensaio
CRA pH 7,0
experimental
(g água /gP)
CRA pH 7,0
previsto
(g água /gP)
Desvio Desvio
relativo (%)
1 5,9 3,68 2,22 37,62
2 3,9 5,51 -1,61 -41,28
3 4,0 5,18 -1,18 -29,50
4 4,3 2,76 1,54 35,8
5 5,5 5,62 -0,12 -2,18
6 4,1 4,32 -0,22 -5,36
7 5,8 5,68 0,12 2,06
8 3,3 3,90 -0,6 -18,18
9 5,6 5,56 0,04 0,71
10 5,5 5,56 -0,06 -1,09
11 5,6 5,56 0,04 0,71
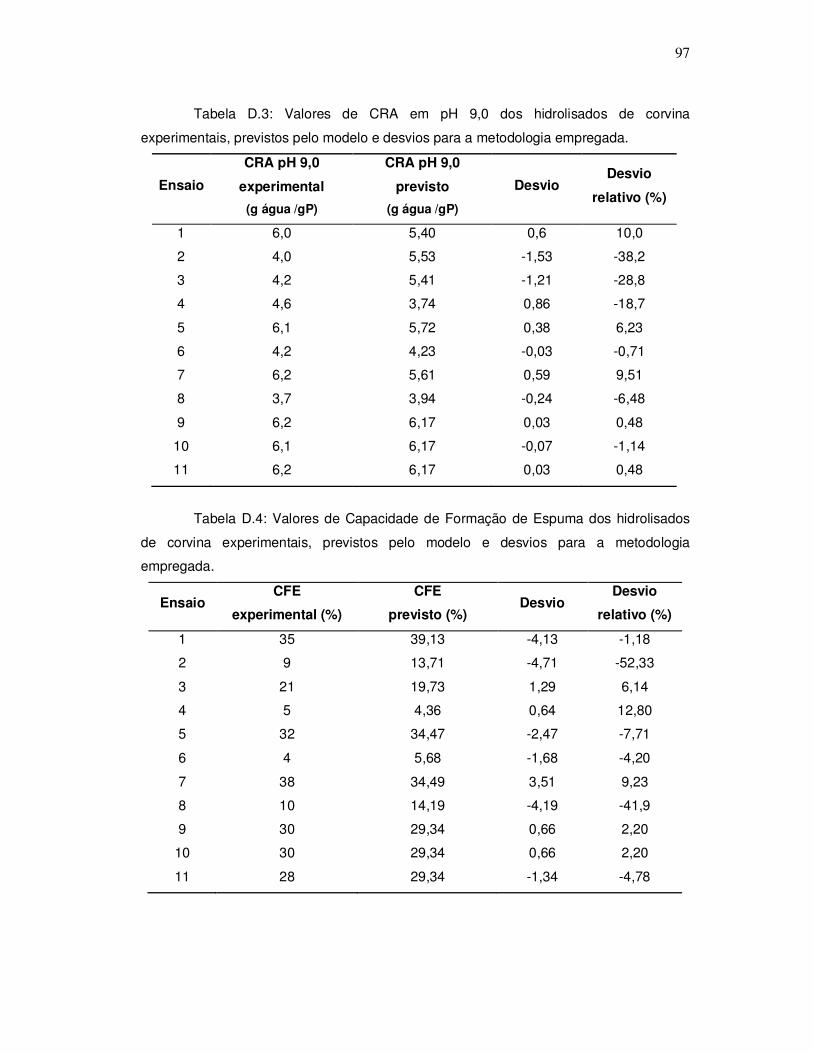
97
Tabela D.3: Valores de CRA em pH 9,0 dos hidrolisados de corvina
experimentais, previstos pelo modelo e desvios para a metodologia empregada.
Ensaio
CRA pH 9,0
experimental
(g água /gP)
CRA pH 9,0
previsto
(g água /gP)
Desvio Desvio
relativo (%)
1 6,0 5,40 0,6 10,0
2 4,0 5,53 -1,53 -38,2
3 4,2 5,41 -1,21 -28,8
4 4,6 3,74 0,86 -18,7
5 6,1 5,72 0,38 6,23
6 4,2 4,23 -0,03 -0,71
7 6,2 5,61 0,59 9,51
8 3,7 3,94 -0,24 -6,48
9 6,2 6,17 0,03 0,48
10 6,1 6,17 -0,07 -1,14
11 6,2 6,17 0,03 0,48
Tabela D.4: Valores de Capacidade de Formação de Espuma dos hidrolisados
de corvina experimentais, previstos pelo modelo e desvios para a metodologia
empregada.
Ensaio CFE
experimental (%)
CFE
previsto (%) Desvio
Desvio
relativo (%)
1 35 39,13 -4,13 -1,18
2 9 13,71 -4,71 -52,33
3 21 19,73 1,29 6,14
4 5 4,36 0,64 12,80
5 32 34,47 -2,47 -7,71
6 4 5,68 -1,68 -4,20
7 38 34,49 3,51 9,23
8 10 14,19 -4,19 -41,9
9 30 29,34 0,66 2,20
10 30 29,34 0,66 2,20
11 28 29,34 -1,34 -4,78

98
ANEXO E
Procedimento de preparo da Solução Tampão Fosfato pH 8,0 utilizada nas
reações de hidrólise de acordo com Morita (1972).
Os cálculos para o preparo de um volume de 2000mL estão descritos abaixo:
- preparo da solução de ácido cítrico::
M = m 0,1M = m
Mol.v 210,14 x 2(litros)
m = 42,028g de ácido cítrico (pesar e elevar a 2000 mL)
1375mL ----------500mL
x------------------2000mL
x = 55mL (ácido cítrico 0,1M) (volume necessário para preparar 2 litros de tampão pH 8,0)
- preparo da solução de fosfato de sódio (Na2HPO4):
M = m 0,2M = m
Mol.v 141,965 x 2(litros)
m = 56,786g de fosfato de sódio (pesar e elevar a 2000 mL)
486,25mL--------500mL
y---------------2000mL
y = 1945mL (fosfato de sódio 0,2M) (volume necessário para preparar 2 litros de tampão pH 8,0)
Tomar 55mL de ácido cítrico mais 1945mL de fosfato de sódio e misturar,
formando 2 litros de tampão. Ajustar o pH adicionando ácido cítrico ou fosfato de sódio
até ph 8,0.

99
RESUMO
O objetivo deste trabalho foi estudar a influência do grau de hidrólise nas propriedades funcionais de hidrolisados protéicos de corvina (Micropogonias furnieri). Os hidrolisados enzimáticos foram obtidos utilizando músculo de corvina como substrato, a enzima foi Alcalase 2.4L, o pH 8,0 e a temperatura da reação foi 50ºC. Após 60 minutos de reação, a enzima foi inativada por aquecimento do hidrolisado a 85ºC por 15 minutos. Os hidrolisados inativados foram desidratados em temperatura de 60ºC por 10 horas, moídos, peneirados e acondicionados em recipientes herméticos. Após a realização de testes preliminares, foram definidas as concentrações de enzima [E] a de substrato [S] que foram utilizadas na elaboração de um planejamento fatorial completo 22, com três repetições nos pontos centrais, tendo como variáveis respostas o grau de hidrólise (%GH), o conteúdo protéico (%), e as propriedades funcionais, solubilidade, capacidade de retenção de água (CRA), capacidade de retenção de óleo (CRO), capacidade emulsificante (CE) e capacidade de formação de espuma (CFE). Obteve-se hidrolisados com diferentes valores de GH e conteúdo protéico variando de 12,2 a 43,7% e 51,0 a 74,5%, respectivamente. A análise estatística dos resultados permitiu avaliar o efeito dos variáveis [E] e [S] sobre o GH e conteúdo protéico e as propriedades funcionais dos hidrolisados enzimáticos de corvina. Os resultados mostraram que a concentração de enzima influenciou significativamente o GH e o conteúdo protéico dos hidrolisados e a concentração de substrato apresentou influencia significativa somente para os valores do conteúdo protéico dos mesmos. Os perfis apresentados pelas propriedades funcionais de solubilidade e CRA foram semelhantes em todos os casos, com valores mínimos no pH do ponto isoelétrico, com leve aumento para a faixa de pH ácido e alcalino. A [E] mostrou influência significativa no aumento dos valores de solubilidade para todos os pHs analisados, já a [S] não apresentou efeito benéfico na resposta solubilidade em todos os pHs estudados. O efeito da [S] não foi significativo para as respostas CRO e CE e a [E] influenciou negativamente essas respostas. O grau de hidrólise mostrou uma relação direta com a solubilidade e indireta com a capacidade de retenção de água, capacidade de retenção de óleo, capacidade emulsificante e capacidade de formação de espuma. Constatou-se que o conteúdo protéico, CFE e CRA nos pHs 7,0 e 9,0, permitiram obter modelos preditivos. Os hidrolisados mostraram digestibilidade in vitro variando de 90,8 a 98,8% de fração digerível. Estes resultados indicam a potencialidade dos hidrolisados enzimáticos de corvina para a utilização em uma ampla variedade de formulações alimentícias para consumo humano. Palavras-chave: corvina; enzima; hidrolisados enzimáticos; grau de hidrólise; propriedades funcionais

100
ABSTRACT
The objective of this work was to study the influence of the hydrolysis degree in the functional properties of hydrolysates protein croaker (Micropogonias furnieri). The enzymatic hydrolysates were obtained using croaker muscle as substrate, the enzyme was Alcalase 2.4L, the pH 8.0 and the reaction temperature was 50ºC. After 60 minutes of reaction the enzyme was inactivated by heating of the hydrolysates at 85ºC for 15 minutes. After the accomplishment of preliminary tests were defined the enzyme [E] and substrate [S] concentration that used in the elaboration of complete factorial design 22, with triplicate of the central points. The variables answers were the hydrolysis degree (%GH), protein content (%), and the functional properties: solubility, capacity of retention of water (CRA), capacity of oil retention (CRO), capacity emulsifying (CE) and capacity foaming (CFE). Were obtained hydrolysates with different values of GH and protein content varying from 12.18 to 43.68% and 51.0 to 74.5%, respectively. The statistical analysis of the results allowed to evaluate the effect of the variables [E] and [S] on GH and protein content and the functional properties of the enzymatic hydrolysates of croaker. The results showed that the enzyme concentration influenced the GH and protein content significantly and the substrate concentration presented influence significant only for the values of the protein content of the same ones. The profiles presented by the functional properties of solubility and CRA they were similar in all the cases, with minimum values in the pH of the isoeletric point, with increase light for the range of acid and alkaline pH. The [E] showed significant influences in the increase of the solubility values for all the analyzed pHs, already the [S] it didn't present beneficial effect in the answer solubility in all the studied pHs. The effect of the [S] was not significant for the answers CRO and CE and the [E] influenced those answers negatively. The hydrolysis degree showed a direct relationship with the solubility and indirect with the capacity of retention of water, capacity of oil retention, capacity emulsifying and capacity foaming. Was verified that the protein content, CFE and CRA in the pHs 7.0 and 9.0, allowed to obtain models preditivos. The hydrolysates showed a quality protein good with values of digestibility in vitro from 90.8 to 98.8%. These results indicate the potentiality of the enzymatic hydrolysates of croaker for the use in a wide variety of nutritious formulations for human consumption.
Keywords: croaker; enzyme; enzymatic hydrolysates; hydrolysis degree; functional properties

101
SUMÁRIO LISTA DE TABELAS............................................................................................................i
LISTA DE FIGURAS...........................................................................................................iii
RESUMO.............................................................................................................................v
ABSTRACT........................................................................................................................vi
1 INTRODUÇÃO ............................................................................................................... 1
2 REVISÃO BIBLIOGRÁFICA........................................................................................... 4
2.1 Pescado ....................................................................................................................... 4
2.1.1 Consumo e produção de pescado ............................................................................................... 4
2.1.2 Corvina (Micropogonias furnieri) .............................................................................................. 5
2.2 Proteínas ..................................................................................................................... 6
2.2.1 Proteínas do Pescado................................................................................................................. 7
2.3 Enzimas ....................................................................................................................... 8
2.3.1 Enzimas proteolíticas ................................................................................................................. 9
2.3.2 Fatores relacionados com a hidrólise enzimática ..................................................................... 11
2.3.2.1 pH .................................................................................................................................................. 11 2.3.2.2 Temperatura ................................................................................................................................. 11 2.3.2.3 Concentração enzima versus concentração de substrato ......................................................... 12
2.3.3 Grau de Hidrólise da reação enzimática .................................................................................. 13
2.4 Hidrolisados enzimáticos ........................................................................................... 13
2.5 Hidrolisados enzimáticos de pescado ........................................................................ 15
2.6 Efeito do grau de hidrólise nas propriedades funcionais de hidrolisados de pescado. 16
2.7 Propriedades funcionais das proteínas ...................................................................... 17
2.7.1 Solubilidade ............................................................................................................................. 19
2.7.2 Capacidade de formação de espuma ........................................................................................ 20
2.7.3 Capacidade emulsificante ........................................................................................................ 21
2.7.4 Capacidade de retenção de água .............................................................................................. 22
2.7.5 Capacidade de retenção de óleo ............................................................................................... 23
2.8 Propriedades Nutricionais .......................................................................................... 24
3 MATERIAL E MÉTODOS ............................................................................................. 26
3.1 Material ...................................................................................................................... 26
3.1.1 Matéria-prima ......................................................................................................................... 26
3.1.2 Reagentes ................................................................................................................................ 26
3.1.3 Enzima .................................................................................................................................... 26
3.1.4 Infra-estrutura ......................................................................................................................... 26
3.2 Métodos ..................................................................................................................... 26
3.2.1 Obtenção dos filés e da polpa de corvina.................................................................................. 26
3.2.2 Determinação da atividade específica da enzima ..................................................................... 27
3.2.3 Caracterização fisico-química.................................................................................................. 27 3.2.3.1 Da matéria-prima .......................................................................................................................... 27
3.2.3.2 Dos hidrolisados ........................................................................................................................... 27
3.2.4 Obtenção do hidrolisado protéico de pescado .......................................................................... 28 3.2.4.1 Testes preliminares e definição das condições de hidrólise enzimática................................... 28 3.2.4.2 Fluxograma geral dos hidrolisados protéicos ............................................................................. 28 3.2.4.3 Medida do grau de hidrólise ....................................................................................................... 30
3.2.4.4 Determinação da funcionalidade dos hidrolisados enzimáticos de corvina.............................. 30 3.2.4.4.1 Solubilidade ........................................................................................................................... 31 3.2.4.4.2 Capacidade de retenção de água (CRA) ............................................................................. 31
3.2.4.4.3 Capacidade de retenção de óleo (CRO) .............................................................................. 32

102
3.2.4.4.4 Capacidade Emulsificante (CE) ............................................................................................ 32 3.2.4.4.5 Propriedades espumantes .................................................................................................... 33
3.2.4.5 Determinação da digestibilidade dos hidrolisados .................................................................... 33
3.2.5 Planejamento experimental e tratamento estatístico ................................................................. 34
4 RESULTADOS E DISCUSSÃO ................................................................................. 36
4.1 Atividade específica da enzima .................................................................................. 36
4.2 Caracterização fisico-química da matéria-prima ......................................................... 37
4.2.1 pH ...................................................................................................................................... 37
4.2.2 Rendimento .............................................................................................................................. 37
4.2.3 Composição proximal dos filés de corvina ................................................................................ 38
4.2.4 Conteúdo de proteína, umidade e cinzas dos hidrolisados de corvina ....................................... 38
4.3 Testes preliminares .................................................................................................... 40
4.3.1 Testes preliminares T1 ............................................................................................................. 40
4.3.2 Testes preliminares T2 ............................................................................................................. 40
4.4 Resultados do planejamento experimental ................................................................. 42
4.4.1 Grau de hidrólise e Conteúdo protéico ..................................................................................... 42 4.4.1.1 Análise dos efeitos principais para a resposta grau de hidrólise dos hidrolisados .................. 43 4.4.1.2 Verificação do modelo matemático preditivo .............................................................................. 44 4.4.1.3 Análise dos efeitos principais para a resposta Conteúdo Protéico dos hidrolisados ............... 44 4.4.1.4 Verificação do modelo matemático preditivo .............................................................................. 45
4.4.2 Propriedades funcionais dos hidrolisados enzimáticos de corvina ............................................ 48 4.4.2.1 Solubilidade .................................................................................................................................. 48
4.4.2.1.1 Análise dos efeitos principais para a resposta solubilidade dos hidrolisados ................... 52 4.4.2.1.2 Verificação do modelo matemático preditivo ....................................................................... 54
4.4.2.2 Capacidade de retenção de água (CRA) .................................................................................... 54 4.4.2.2.1 Análise dos efeitos principais para a resposta CRA dos hidrolisados ............................... 56 4.4.2.2.2 Verificação do modelo matemático preditivo ....................................................................... 59
4.4.2.3 Capacidade de retenção de óleo (CRO)..................................................................................... 63 4.4.2.3.1 Análise dos efeitos principais para a resposta CRO dos hidrolisados ............................... 65 4.4.2.3.2 Verificação do modelo matemático preditivo ....................................................................... 66
4.4.2.4 Capacidade Emulsificante (CE)................................................................................................... 66 4.4.2.4.1 Análise dos efeitos principais para a resposta capacidade emulsificante ......................... 68 4.4.2.4.2 Verificação do modelo matemático preditivo ....................................................................... 69
4.4.2.5 Propriedades espumantes ........................................................................................................... 69 4.4.2.5.1 Capacidade de formação de espuma (CFE) ....................................................................... 69 4.4.2.5.2 Estabilidade de espuma (EEs) ............................................................................................. 71 4.4.2.5.3 Análise dos efeitos principais para capacidade de formação de espuma ......................... 72
4.4.2.5.4 Verificação do modelo matemático preditivo ........................................................................... 73
4.4.3 Digestibilidade in vitro da proteína dos hidrolisados de corvina ............................................... 76
5 CONCLUSÃO ............................................................................................................... 78
6 REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................. 79
ANEXO A ........................................................................................................................ 86
ANEXO B ........................................................................................................................ 87
ANEXO C ........................................................................................................................ 88
ANEXO D ........................................................................................................................ 96
ANEXO E ........................................................................................................................ 98