universidade federal do rio grande do norte centro … · 2019-01-29 · felicidade, o da amizade...
TRANSCRIPT

AADDRRIIAANNEE TTRRIINNDDAADDEE MMEEDDEEIIRROOSS LLIIMMAA
AAVVAALLIIAAÇÇÃÃOO DDOO PPOOTTEENNCCIIAALL AANNTTIIOOXXIIDDAANNTTEE EE
AANNTTIIIINNFFLLAAMMAATTÓÓRRIIOO DDEE GGAALLAACCTTOOMMAANNAANNAA DDOO FFUUNNGGOO
TTyyllooppiilluuss bbaalllloouuiiii
NATAL
2009
UUNNIIVVEERRSSIIDDAADDEE FFEEDDEERRAALL DDOO RRIIOO GGRRAANNDDEE DDOO NNOORRTTEE
CCEENNTTRROO DDEE BBIIOOCCIIÊÊNNCCIIAASS
DDEEPPAARRTTAAMMEENNTTOO DDEE BBIIOOQQUUÍÍMMIICCAA

AADDRRIIAANNEE TTRRIINNDDAADDEE MMEEDDEEIIRROOSS LLIIMMAA
AAVVAALLIIAAÇÇÃÃOO DDOO PPOOTTEENNCCIIAALL AANNTTIIOOXXIIDDAANNTTEE EE
AANNTTIIIINNFFLLAAMMAATTÓÓRRIIOO DDEE GGAALLAACCTTOOMMAANNAANNAA DDOO FFUUNNGGOO
TTyyllooppiilluuss bbaalllloouuiiii
NATAL
Dissertação apresentada ao Departamento de Bioquímica da Universidade Federal do Rio Grande do Norte como requisito parcial para obtenção do título de Mestre em Bioquímica.
Orientadora: Profa. Dra. Edda Lisboa Leite



2009
Agradeço à excelente mestre, mãe, amiga, e principalmente a Mulher Edda Lisboa Leite, minha orientadora, por ter me aceitado e me incentivado sempre nesta caminhada, pela
simplicidade, generosidade e ética com qual transmite todos os seus conhecimentos e por expressá-los com tamanha humildade.
Aprendi com a primavera a deixar-me cortar e voltar sempre inteira”. Cecília Meireles

À minha família. Em especial a minha mãe, pelo seu incentivo para que eu voltasse
aos estudos, pela educação transmitida com carinho e amor, pelos valores ensinados durante todos esses anos e pela sabedoria da luz divina que lhes foi dada para orientar e encaminhar seus filhos para a luz da sabedoria humana. A minha existência não teria sentido algum sem a tua presença constante na minha vida.
A minha vozinha Expedita pelas suas constantes orações em minha intenção e ao meu avô Beto por suas palavras carinhosas sempre que ligo para saber como eles estão.
A minha alma sempre fica alimentada quando falo com vocês. Ao meu filho Anthony motivo pelo qual busco sempre o melhor para poder oferecê-
lo de maneira incondicional. A minha mãe de Natal (RN). Dona Fátima que cuida de mim desde que comecei a
minha graduação por ter dedicado seu valioso tempo na minha recuperação. Amo muito você. E claro a Denise por aceitar sem ciúmes (raridade) tamanha dedicação de sua mãe para comigo.
A Alfredo Luis “In memória” pelo grande homem, médico, amigo e mestre brilhante que me inspirou na escolha da minha profissão. Mestre querido tua grandeza e elevação espiritual foram percebidas e sentidas por todos aqueles que tiveram a oportunidade de estarem ao seu lado.
“A Biologia é pura arte, nós somos seus admiradores e DEUS o maior dos artistas”. Alfredo Luis- -Fred -Dr. Formiga.

A Marília por todos os momentos bons e difíceis que passamos nestes dois anos de
mestrado e principalmente pela imensa força que me deu há um ano, momento que mais precisei e até pensei em desistir. Obrigada nêga, tu és uma pessoa de grande personalidade e que estimo muito.
A Monique, companheira de experimentos. Tenho muito orgulho de ter conhecido uma pessoa tão especial como você. Torço pelas suas vitorias que serão muitas.
A Almino pela tranqüilidade e sensatez que sempre fez questão de nós passar. E a sua valiosa ajuda com seus conhecimentos histológicos. Obrigada, boa sorte no seu projeto e vê se não erra meu nome nos agradecimentos.
A Alisson, pequeno grande homem. Obrigada pela paciência nestes últimos momentos da realização deste trabalho.
A Thuane pela sua amizade e excelentes ajudas na realização dos experimentos in
vivo. A Luiza (lulu), Karhen e sua mãe Dona Raimunda pelas conversas prazerosas e
grandes conselhos passados nas várias caronas que me deram no trajeto de volta para casa. Amei conhecé-las!
A Leonardo, o cara. Obrigada por toda ajuda com as formatações, onde sou lenta demais, e em especial por ter me dado a oportunidade de conhecê-lo melhor fazendo com que eu aprendesse que neste mundo não devemos nunca pré-julgar os outros pelas opiniões de outrem. Sua autenticidade é maravilhosa.
A Thiago, Kahena, Lissandra, Maria Emília e Celina que apesar de não termos compartilhado muito tempo juntos no dia-dia do laboratório, não deixaram de ter sua importância nesta jornada.
A todos do departamento que contribuíram em alguma parte neste trabalho, em especial a Cleisyvan, Norberto, Mariana, Nedinaldo e Ana Katarina.
A Virginia e a Sheila que com palavras de incentivo para minha amiga Carol, também me ajudaram a continuar lutando pelos meus objetivos. Muitas vezes quando agimos com pureza no coração, mesmo que indiretamente podemos acrescentar muito aos nossos semelhantes.
A Carol Perez amiga que quero levar para sempre em meu coração, pois pessoas como você neste mundo estão em extinção. Valeu Carol tudo que passamos foi para o nosso engrandecimento tenha certeza, pois nada acontece por acaso. Adoloo ocê!
A turma da limpeza Jonas, Ângela e seu Raimundo pela ajuda no biotério e no laboratório tornando o ambiente mais agradável de trabalhar.
A turma da secretaria, Ricardo, Rogério e Olívia, por sempre nos atenderem de maneira educada e prestativa.
A todos os meus amigos fora da universidade pela compreensão das ausências do nosso convívio. É bom saber que fazemos falta! Márcia, Marina, Camila, Vanda, Jaíra, Eruska, obrigada pelo carinho.
Ao casal mais lindo e especial do Jiqui, Tereza e Junior e sua irmã Mariney e netinho Gabriel pelos momentos de descontração nas caminhadas pelos quarteirões com nossos amigos LayKa, fofinha,Duque e Princesinha.
Ao meu eterno e mais fiel amigo Lion pelo amor sem cobranças. Os cincos anos que passei com a sua existência foram muitos felizes, principalmente quando chegava à casa arrasada e o primeiro que ia me receber era você sempre com muita felicidade.

A Roberta Godone por me apresentar a umas das pessoas mais ilustres do Departamento da Bioquímica, a professora Dilma Lima, exemplo de determinação, competência e sabedoria. Professora Dilma, minha eterna gratidão por todo apoio que me deste.
A Thiago Dk pelo auxilio técnico no meu computador que insistia em quebrar nos momentos em que eu mais precisava.
A minha cunhada Ellen Fabiana por me proporcionar momentos maravilhosos na leitura de livros, fontes não científica, os quais foram também muito importantes para escrita deste projeto.
A professora Luciana Da Matta por ter me despertado o interesse pela Bioquímica nas suas excelentes aulas da graduação.
A professora Selma Jerônimo pela compreensão, oportunidade que me foi dada e principalmente pela ética com a qual conduz os seus atos.
Aos vários amigos do Departamento de Botânica, Ecologia e Zoologia, em especial a Bianca e ao professor Iure que me auxiliaram na coleta e na pesquisa dos fungos.
A Creuza, uma página seria pouco para agradecer-te. És uma pessoal de beleza única, com infinitas qualidades, obrigada por estar sempre ao meu lado, me incentivando, se preocupado, me aconselhando, sentirei saudades da nossa convivência e dos nossos banquetes.
A Capes pela concessão de bolsa em parte do meu trabalho e a UFRN pela disponibilidade do espaço físico.
Pelo simples bom dia, pelas conversas informais ou de ajuda no trabalho de rotina ou nas fases preliminares do projeto ou ajudas em formas de “dicas” para os experimentos. Enfim, por tudo e por não ter estado sozinho, venho agradecer a todos que contribuíram direta ou indiretamente para este trabalho.
E em especial ao meu GRANDE amigo JESUS CRISTO por proporcionar sempre na minha vida a oportunidade de conviver com pessoas iluminadas. Por estar sempre ao meu lado nos bons e maus momentos e sempre usar as pessoas certas para representá-lo em corpo. A ti Senhor todo Honra e toda Glória.
“De todos os bens que a sabedoria nos faculta como meio de obter a nossa felicidade, o da amizade é de longe o maior”.
Epicuro, 341-270 a.C
"É melhor tentar, ao invés de sentar-se e nada fazer; É melhor falhar, mas não deixar
a vida passar; Eu prefiro na chuva caminhar, do que em dias tristes em casa me esconder; Prefiro ser feliz, embora louco, do que viver infeliz em são conformismo".
Martin Luther King

RESUMO
Polímeros da parede celular de fungos são conhecidos por possuírem muitas atividades biológicas como suas ações antiinflamatórias, antioxidante e antitumoral. Polissacarídeos de diferentes pesos moleculares obtidos de cogumelos podem ativar os leucócitos, estimular a atividade fagocítica, citotóxica e antimicrobiana, incluindo a produção de espécies reativas de oxigênio. No presente estudo, foi investigada a característica química dos polissacarídeos extraídos de Tylopilus ballouii sua atividade antioxidante in vitro e a sua atividade antiinflamatória no modelo de inflamação aguda. Os resultados mostraram que foram extraídos predominantemente polissacarídeos e esses consistiram primariamente de manose e galactose e possui quantidades variáveis de xilose e fucose. As análises de infravermelho mostraram a possível interação entre estes polissacarídeos e proteínas. Além disso, seu peso molecular é de cerca de 140 kDa. A atividade antioxidante foi testada com relação ao “seqüestro” sobre os radicais superóxido e hidroxila, atividade antioxidante total e peroxidação lipídica. Com relação à inibição da formação dos radicais superóxido e hidroxila, os polissacarídeos atingiram um IC50 de 1,64 e 1,25 mg/ml, respectivamente. Os resultados do ensaio de peroxidação lipídica mostraram que os polissacarídeos de Tylopilus ballouii apresentam um IC50 de 1,65 mg/ml. Além disso, a atividade antiinflamatória mostrou que eles agem reduzindo o edema em 32,8, 42 e 56% nas doses de 30, 50 e 70 mg/kg, respectivamente. Assim, estes resultados podem indicar o possível uso dos polissacarídeos de Tylopilus ballouii como antiinflamatório e antioxidante. Palavras-chave: Polissacarídeos; Cogumelo; radicais livres; inflamação.

ABSTRACT
Polymers of mushroom cellular wall are recognized for presenting a lot of biological activities such as anti-inflammatory, antioxidant and anti-tumoral action. Polysaccharides from mushrooms of different molecular mass obtained mushrooms can activate leucocytes, stimulate fagocitic, citotoxic and antimicrobial activity including oxygen reactive species production. In this study were investigated chemical characteristics, in vitro antioxidant activity and anti-inflammatory action in an acute inflammation model of the polysaccharides extracted from Tylopilus ballouii. Results showed that were mainly extracted polysaccharides and that it primarily consisted of mannose and galactose with variable amounts of xylose and fucose. Infrared analysis showed a possible interation between this polysaccharides and proteins. In addition, molecular mass was about 140KDa. Antioxidant activity was tested by superoxide and hydroxyl radical scavenging assay, total antioxidant activity and lipid peroxidation assay. For superoxide and hydroxyl radical generation inhibition, polysaccharides have an IC50 of 2.36 and 0.36 mg/mL, respectively. Lipid peroxidation assay results showed that polysaccharides from Tylopilus ballouii present an IC50 of 3.42 mg/mL. Futhermore, anti-inflammatory assay showed that polysaccharides cause an paw edema decreasing in 32.8, 42 and 56% in 30, 50 and 70 mg/Kg dose, respectively. Thus, these results can indicate a possible use for these polysaccharides from Tylopilus ballouii as an anti-inflammatory and antioxidant.
Keywords: Polysaccharides; Mushroom; free radicals, inflammation.

LISTA DE FIGURAS
FIGURA 1 Mecanismo de interação entre as células sanguíneas a partir de
um estímulo tecidual (injúria vascular).
19
FIGURA 2 Processo de formação de espécie reativa de oxigênio (EROS) 20
FIGURA 3 Representação esquemática da conformação induzida por
temperatura da solução de lentinana a 137 °C
29
FIGURA 4 Estrutura parcial da galactomana e suas respectivas unidades
monossacarídicas.
30
FIGURA 5 Fungo Tylopilus ballouii 34
FIGURA 6 Organograma da metodologia 37
FIGURA 7 Esquema de obtenção do composto do fungo T. ballouii 42
FIGURA 8 Percentual dos constituintes açúcares totais e proteínas após
extração de T. balloui
52
FIGURA 9 Cromatografia descendente em papel do hidrolisado dos
polissacarídeos de T. ballouii utilizando como padrão
monossacarídeos. A: Sistema solvente Acetato de etila: Piridina:
água. B: Sistema solvente Butanol: Ácido acético: água
53
FIGURA 10 Cromatografia em HPLC da fração polissacarídica do fungo T. ballouii e eluição em gradiente de água e acetonitrila.
54
FIGURA 11 Perfil dos polissacarídeos do fungo T. ballouii em cromatografia
gel filtração.
55
FIGURA 12 Espectro de Infravermelho dos polissacarídeos do fungo T.
ballouii.
56
FIGURA 13 Espectros de RMN de 1H obtidos das frações de
polissacarídeos de T. balloui.
58
FIGURA 14 Efeito antiinflamatório dos polissacarídeos de T. ballouii no
modelo de edema plantar induzido por carragenana.
60
FIGURA 15 Efeito antiinflamatório dos polissacarídeos de T. ballouii
(70 mg/kg) e antiinflamatórios como diclofenaco de sódio e
61

indometacina no modelo de edema plantar induzido por
carragenana.
FIGURA 16 Análises histológicas (HEX200) da pata de animais com edema
plantar induzido por carragenana. A- Controle positivo; B –
Tylopilus SP (30 mg/kg); C – Tylopilus SP (50 mg/kg); D –
Tylopilus SP (70 mg/kg); E- Diclofenaco de sódio; F-
Indometacina; G- Controle negativo
62
FIGURA 17 Efeito dos polissacarídeos de Tylopilus ballouii sobre a
viabilidade celular no lavado peritoneal de camundongos Swiss
63
FIGURA 18 Efeito inibitório dos radicais superóxido pelos polissacarídeos do
fungo T. balloui
64
FIGURA 19 Efeito inibitório dos radicais hidroxilas pelos polissacarídeos do
fungo T. ballouii
65
FIGURA 20 Capacidade antioxidante total dos polissacarídeos do fungo T.
ballouii
66
FIGURA 21 Inibição da Peroxidação Lipídica da membrana da gema de
ovos pelos polissacarídeos do fungo T. balloui
67

LISTA DE TABELAS
TABELA 1 Fontes usuais de alguns polissacarídeos 25
TABELA 2 Homo- e heteropolissacarídeos produzidos por fungos
filamentosos
27
TABELA 3 Análise da composição centesimal do fungo T .balloui 51
TABELA 4 Composição monossacarídica dos polissacarídeos do fungo T.
ballouii
54
TABELA 5 Sinais de hidrogênio (H) nos espectros de RMN de 1H
encontrados na literatura e na fração de polissacarídeos de T.
ballouii.
57
TABELA 6 Valores de IC50 de polissacarídeos de T. balloui 68

LISTA DE ABREVIATURAS
µg Micrograma
OH • Radical hidroxila
ARA Arabinose
CDP Cromatrografia descendente em papel
COX Ciclooxigenase
COX- 1 Ciclooxigenase 1
COX-2 Ciclooxigenase 2
EDTA Ácido etilenodiamino tetra-acético
EPS Exopolissacarídeos
EROS Espécies Reativas do Oxigênio
FUC Fucose
g Grama
GAL Galactose
GL Glicose
h Hora
HPLC Cromatografia líquida de alta eficiência
i.p Via intraperitoneal
IFN-α Interferon α
IFN-γ Interferon β
IL-1 Interleucina 1
IL-1ß Interleucina 1ß
IL-6 Interleucina 6
IL-8 Interleucina 8
IV infravermelho
MAN Manose
MDA Malonildialdeíldo
ml Mililitro
mM Milimolar
N Normal
NADH Nicotinamina adenina dinucleotídeo

NBT Nitroblue tetrazolium
O2•− Radical ânion superóxido
ºC Graus centígrados
PBS Tampão salina fosfato
PG Prostaglandina
RMN Ressonância magnética nuclear
TNFα Fator de Necrose Tumoral α
TR Tempo de retenção
Vf Volume final
Vi. Volume inicial
Vp Volume da pata
XIL Xilose
α Alfa
β Beta
δ Deslocamento químico em PPM

ÍNDICE
Página
RESUMO
ABSTRACT
LISTA DE FIGURAS
LISTA DE TABELAS
LISTA DE ABREVIATURAS...
1- INTRODUÇÃO 19
1.1 INFLAMAÇÃO. 19
1.2 RADICAIS LIVRES. 20
1.3 ASPECTOS GERAIS DOS FUNGOS 22
1.4 ATIVIDADES BIOLÓGICAS DOS POLISSACARÍDEOS DOS FUNGOS 23
1.5 GALACTOMANANAS 29
1.6..GÊNERO TYLOPILUS 31
2 - OBJETIVOS. 33
2.1 – Objetivo Geral. 33
2.2 – Objetivo Específicos. 33
3 – MATERIAIS E MÉTODOS 34
3.1 - Materiais 34
3.1.1 - Fungo. 34
3.1.2 – Animais 35
3.1.3 - Outros materiais 35
3.1.4 - Aparelhos 36
3.2 - Metodologia 37
3.2.1 – Composição centesimal 38
3.2.1.1 – Determinação da fração cinza. 38
3.2.1.2 – Determinação de lipídeos 38
3.2.1.3 – Determinação de proteínas 39
3.2.1.4 – Determinação de fibra 40
3.2.1.5. – Determinação de Umidade 40
3.2.1.6 – Determinação de Carboidratos. 41

3.2.2 – Obtenção dos polissacarídeos do fungo T.balloui 41
3.2.3 – Caracterização química 43
3.2.3.1- Análises Colorimétricas. 43
3.2.3.2 – Cromatografia Descendente em Papel
3.2.3.3 - Cromatografia Líquida de Alta Eficiência
43
43
3.2.3.4 – Determinação da massa molecular 44
3.2.3.5 – Análise de Infravermelho 44
3.2.3.6 – Ressonância Magnética Nuclear (RMN)
3.2.4 – Atividade Antiinflamatória
44
3.2.4.1 – Avaliação do efeito do composto de T.ballouii em um modelo de
edema induzido por carragenana
44
3.2.4.2 – Análises Histológicas 46
3.2.5 – Atividade Citotóxica 46
3.2.5.1 – Determinação da viabilidade celular ex vivo
3.2.6 – Atividade antioxidante
46
47
3.2.6.1 – Radicais superóxidos. 47
3.2.6.2 – Radicais hidroxilas 47
3.2.6.3 – Atividade antioxidante total 47
3.2.6.4 – Peroxidação lipídica
3.2.7. Cálculo do IC50
48
48
3.2.8 - Análise Estatística
49
4 - RESULTADOS 50
4.1 – Extração 50
4.2 – composição monossacarídica
4.3 - Análises Químicas.
51
52
4.4 - Composição monossacarídica do precipitado 2 52
4.5– Determinação da massa molecular. 55
4.6- Espectroscopia de Infravermelho. 55
4.10 - Determinação da viabilidade celular ex vivo
4.11 - Atividade Antioxidante
4.12 - Seqüestro de Radicais Superóxidos
63
63
63
4.13 - Radicais hidroxilas 64

4.14 - Atividade antioxidante total 66
4.15 - Peroxidação lipídica
5- DISCUSSÃO
6-CONCLUSÕES
REFERÊNCIAS
67
69
74

17
1. INTRODUÇÃO
1.1. INFLAMAÇÃO
A inflamação é uma resposta à infecção, antígenos ou estímulo lesivo e tem
por função destruir os agentes irritantes e potencializar o reparo tecidual (NATHAN,
2002; SHERWOOD; TOLIVER, 2004). Diferentes tipos de respostas podem ser
mediados por este processo inflamatório, entretanto os quatros sinais clínicos da
inflamação aguda são sempre observados como: dor, rubor, calor e tumor (BRUNE;
HINZ, 2004).
Apesar de ser um mecanismo de defesa, a inflamação pode ser prejudicial
por reações de hipersensibilidades fatais ou de lesionar os órgãos de maneira
progressiva e permanente (MITCHELL et al., 2006).
No processo inflamatório, duas fases podem ser caracterizadas, são elas:
- A fase aguda que é de curta duração sendo caracterizada por formação do
edema e respostas vasculares e celulares. As respostas vasculares ocorrem com a
inflamação após a lesão. A vasoconstricção é seguida imediatamente pela
vasodilatação das arteríolas e vênulas que suprem à área. Como resultado, a área
torna-se congesta causando o rubor (eritema) e calor associado com a inflamação
aguda. Estas respostas são acompanhadas do aumento da permeabilidade capilar
e extravasamento dos fluidos para dentro dos tecidos (edema). A resposta celular da
inflamação aguda é marcada pelo movimento dos leucócitos para a área da lesão ou
trauma (MITCHELL et al., 2006).
- A fase crônica de longa duração sendo caracterizada por infiltrado de células
mononucleares, incluindo macrófagos, linfócitos e plasmócitos, por destruição
tecidual induzida pela persistência do agente nocivo ou pelas células inflamatórias, e
pelas tentativas de cicatrização com a substituição do tecido lesado por tecido
conjuntivo, efetuado através da proliferação de pequenos vasos (ALLER et al., 2006;
KUMAR et al., 2005).
Dos mediadores envolvidos no processo inflamatório podem ser citadas a
ação do sistema complemento, sistema de coagulação, sistema cinina, hormônios,
com efeito, na angiogênese, processos de reparo e resposta imunológica humoral e
celular (HAVSTEEN, 2002). O recrutamento de células durante a inflamação
depende da liberação de mediadores locais, a histamina e a serotonina que estão

18
entre os primeiros mediadores liberados durante o processo inflamatório (LUSTER et
al., 2005).
Posteriormente a esta resposta fisiológica ocorre à formação das
prostaglandinas, leucotrienos dentre outros. As prostaglandinas e os leucotrienos
são metabólitos do ácido araquidônico produzidos pela ciclooxigenase (COX) 1 e
ciclooxigenase (COX) 2 bem como pela lipooxigenase. A enzima COX-1 é expressa
de forma constitutiva em células e tecidos de mamíferos enquanto a COX-2
geralmente está presente em tecidos de mamíferos em baixos níveis, mas é
altamente induzível e têm seus níveis aumentados perante um ou mais estímulos,
tais como citocinas e fatores de crescimento. As prostaglandinas são importantes
mediadores inflamatórios com diversas propriedades vasoativas, de contração de
músculo liso e quimiotáxicas (CHANDRASEKHARAN; SIMMONS, 2004).
A quimioatração de leucócitos no sítio lesado, por exemplo, é uma das
principais características do processo inflamatório (FIGURA 1). A migração
adequada e em tempo hábil de leucócitos da microcirculação para o foco
inflamatório é uma das etapas fundamentais deste processo.
Nesta etapa, os leucócitos sanguíneos são ativados, se aderem à superfície
das células endoteliais através de receptores de superfície como as selectinas e os
carboidratos, produzindo uma grande quantidade de espécies reativas de oxigênio
(EROS) no tecido inflamatório.
Os macrófagos e neutrófilos produzem EROS como primeira linha de defesa
contra bactérias, vírus e outras partículas estranhas ao organismo. Essa produção
também acelera a formação de eicosanóides e citocinas, que podem estender a
resposta inflamatória (MITCHELL et al., 2006).
Esta reação pode ser importante como mecanismo de defesa, entretanto, um
excesso na formação de EROS pode levar à oxidação e perda da função de
moléculas essenciais como, ácidos nucléicos, lipídeos de membrana, grupos
sulfidrílicos de proteínas e enzimas (HADDAD, 2002).

19
FIGURA 1. Mecanismo de interação entre as células sanguíneas a partir de um estímulo tecidual (injúria vascular). Por diapedese, os leucócitos migram para os sítios da inflamação e são ativados aderindo às células endoteliais que são induzidas imediatamente a liberarem os conteúdos dos seus grânulos estocados (corpos de Weibel-Palace) compostos de P-selectinas e Fatores de Von Willebrand (FVW) os quais desempenham um papel importante na adesão plaquetária. O acúmulo de fibrina é determinado por este processo inflamatório formando um envoltório ao redor do local, evitando a progressão da infecção. Fonte: http://www.medstudents.com.br/original/revisao/selectin/selectin1.jpg
1.2. RADICAIS LIVRES
Os radicais livres podem ser definidos como qualquer espécie química capaz
de existência independente, que possui um ou mais elétrons desemparelhados em
seu orbital, sendo altamente reativos e capazes de atacar biomoléculas. (MARRONI,
MARRONI, 2002). A formação de EROS é uma conseqüência natural do
metabolismo aeróbico e é parte integrante para a manutenção da homeostase do
oxigênio nos tecidos (CASTRO; FREEMAN, 2001).
Cerca de 1-3% do oxigênio consumido pelos organismos aeróbicos são
convertidos em EROS (SOHAL; WEINDRUCH, 1996). Desta forma, a maioria dos
radicais livres é derivado do metabolismo do oxigênio (BANERJEE; DASGUPTA,
2005).
20
Os EROS mais comumente formado são o radical ânion superóxido (O2•−) e o
radical hidroxila (OH•) (WILSON et al., 2001). O O2•− é formado quando um elétron
é adicionado a uma molécula de oxigênio, e é considerado o tipo menos reativo de
EROS. Depois do O2•− ser produzido, ele dispara uma cascata de eventos rápidos
que criam outros radicais livres, finalmente encerra na formação de água (KOHEN;
NYSKA, 2002).
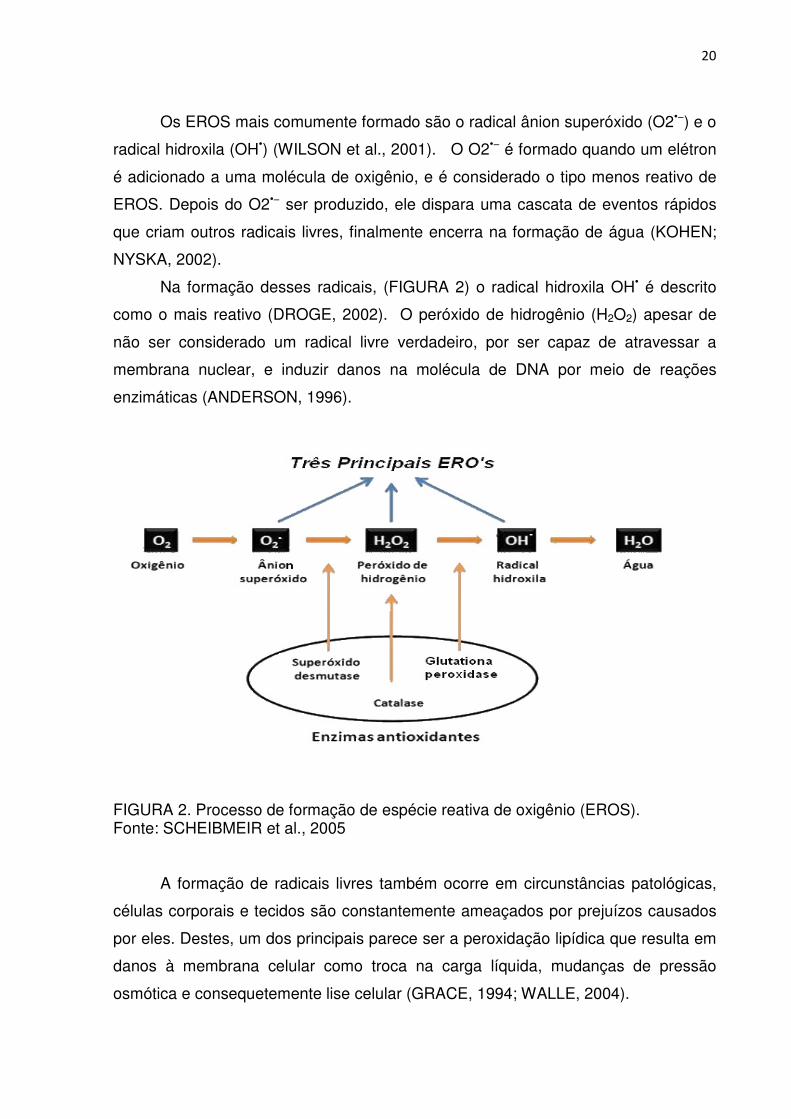
Na formação desses radicais, (FIGURA 2) o radical hidroxila OH• é descrito
como o mais reativo (DROGE, 2002). O peróxido de hidrogênio (H2O2) apesar de
não ser considerado um radical livre verdadeiro, por ser capaz de atravessar a
membrana nuclear, e induzir danos na molécula de DNA por meio de reações
enzimáticas (ANDERSON, 1996).
FIGURA 2. Processo de formação de espécie reativa de oxigênio (EROS). Fonte: SCHEIBMEIR et al., 2005
A formação de radicais livres também ocorre em circunstâncias patológicas,
células corporais e tecidos são constantemente ameaçados por prejuízos causados
por eles. Destes, um dos principais parece ser a peroxidação lipídica que resulta em
danos à membrana celular como troca na carga líquida, mudanças de pressão
osmótica e consequetemente lise celular (GRACE, 1994; WALLE, 2004).

21
Radicais livres podem agir sobre vários mediadores inflamatórios
contribuindo para a inflamação comprometendo a função celular (NIJVELDT et al.,
2001), além de estarem ligados com processos de envelhecimento corporal
(DEGÁSPARI; WASZCZYNSKYJ, 2004).
O excesso de radicais é potencialmente prejudicial para a manutenção
celular, quando a lesão é pronunciada ou persistente, a resposta compensatória
torna-se insuficiente para corrigir o desequilíbrio do estado de oxi-redução, dando
origem ao estresse oxidativo. Mecanismos efetivos foram desenvolvidos pelos
organismos vivos para evitar este dano, como os antioxidantes enzimáticos e não
enzimáticos (MANSON, 2003). O mecanismo enzimático inclui as enzimas
superóxido desmutase que elimina o radical superóxido, bem como catalase e
glutationa peroxidase para a remoção de H2O2. Enzimas antioxidantes
desempenham um papel fundamental no equilíbrio do corpo e são essenciais na
manutenção das funções fisiológicas.
A maioria dessas enzimas antioxidantes é induzida sob a inflamação, trauma
ou outras condições de estresse, e essa indução representam o mecanismo-chave
para o corpo em resposta a uma variedade de estressores (CASTRO; FREEMAN,
2001; MCCORD, 2000). Os antioxidantes não enzimáticos incluem algumas
vitaminas como ácido ascórbico, vitamina E, carotenóides, compostos fenólicos e
polifenólicos e outros produtos naturais e sintéticos. Antioxidantes são compostos
que podem retardar ou inibir a oxidação de lipídios ou outras moléculas, evitando o
início ou propagação das reações de oxidação em cadeia (DEGÁSPARI;
WASZCZYNSKYJ, 2004).
A busca por compostos bioativos para desenvolver novos medicamentos vem
atraindo nos últimos anos, a atenção da comunidade científica. O emprego de
antioxidantes sintéticos na indústria de alimentos tem sido alvo de questionamentos
quanto à sua inocuidade e, devido a isso, pesquisas estão voltadas para a
descoberta de compostos naturais que exibam esta propriedade funcional, atuando
sozinhos ou sinergisticamente com outros aditivos (YANISHLIEVA et al., 2006).
Dentre as fontes de antioxidantes naturais podem ser citados os cereais, os
cogumelos, as ervas e as sementes de frutas cítricas (YANISHLIEVA et al., 2006;
ELMASTAS et al., 2007).

22
1.3. ASPECTOS GERAIS DOS FUNGOS
Os fungos são organismos eucariontes heterotróficos que obtêm sua energia
pela ruptura de moléculas orgânicas, podendo ocupar diversos nichos ecológicos,
atuando tanto como parasitas, sapróbios ou então estabelecendo relações
simbióticas, por exemplo, com algas, formando os líquens (TORTORA et al., 2000).
Os fungos constituem um reino diferenciado das plantas e animais por várias
características, dentre as quais, a mais marcante é o modo de nutrição absortiva.
Entre os demais caracteres está incluída a formação de estruturas vegetativas
filamentosas, as hifas, que constituem o micélio e estruturas especializadas para
reprodução sexuada e assexuada próprias para cada grupo taxonômico. Embora a
maioria dos fungos forme micélios bem desenvolvidos, alguns como as leveduras
são unicelulares e por isso não formam micélios (ALEXOPOULOS et al., 1996).
Com relação ao tamanho, grande parte dos fungos é microscópica e por
habitarem os mais diversos substratos, eles são na sua maioria desconhecidos
(HYDE, 1997). Essa falta de conhecimento se acentua mais nas regiões tropicais.
Rossman (1997) considera que os microfungos tropicais representam um
universo praticamente inexplorado da biodiversidade. Dos macrofungos conhecidos
grande parte está incluída no filo Basidiomycota. Entre as 30.000 espécies desse
filo estão presentes várias espécies de cogumelos comestíveis e os venenosos, os
gasteromicetos, as orelhas - de - pau, bem como dois grupos fitopatogênicos
importantes, as ferrugens e os carvões. Os membros desse filo têm papel
preponderante na decomposição de substratos vegetais. (RAVEN et al., 2001).
Os corpos de frutificação são denominados cogumelos e são conhecidos
desde a antiguidade, quando o homem já os utilizava com finalidades terapêuticas,
alimentar e religiosa (MELITO, 1999). Vários relatos na literatura têm comprovado
que o valor nutritivo dos cogumelos é considerado de qualidade para uma dieta
balanceada (MANZI et al., 1999).
O crescente interesse no estudo dos fungos está relacionado à essencial
importância destes nas diversas cadeias tróficas e na possibilidade de aplicação
comercial em distintas áreas como na nutrição, na saúde humana e animal, no
tratamento de águas residuais, dentre outras (KIRK et al. 2001). Substâncias como

23
antibióticos, antioxidantes, antiinflamatórios, esteróides, ácidos orgânicos produzidos
por algumas espécies de fungos têm um importante destaque na indústria
farmacêutica. É o caso dos fungos do gênero Penicillium, que são produtores de
antibióticos; da cravagem do centeio, Claviceps purpurea, que contém vários
alcalóides, entre eles ergonovina e ergotamina, que atuam como vasoconstritores e
dos produtores de β-D-glucanas, Botryosphaeria rhodina, Sclerotium glucanicum,
entre vários outros (ROSADO et al., 2003; WISBECK; FURLAN; NINOW, 2003;
MAZIERO; CAVAZZONI; BONONI, 1999).
A utilização desses organismos pode ser através do seu material celular
(biomassa) ou então de macromoléculas isoladas. Há uma forte tendência em se
explorar comercialmente a biomassa fúngica para isolamento de seus componentes
celulares e consequentemente de seus principais constituintes, tais como enzimas,
nucleotídeos, proteínas, lipídeos e principalmente polissacarídeos (BELEM; LEE,
1998; BEROVIČ et al., 2003; CAMERON et al., 1988; KOLLÁR et al., 1992;
PAVLOVA et al., 2005).
A biomassa excedente da produção industrial de antibióticos por Penicillium
chrysogenum, além de ser aproveitada para ração e ou no preparo de fertilizantes
(MUZARELI et al., 2000), também vem se mostrando uma fonte viável de
polissacarídeos do tipo glucanas já que estas apresentam atividades antitumorais e
antiinflamatórias (WANG et al., 2007).
Trabalhos recentes têm dado grande ênfase a compostos bioativos de
fungos, além de seus polissacarídeos, compostos fenólicos e proteínas com ação
sobre o sistema imune e como potente antioxidante (SANTOS et al., 2004; CUI et
al., 2005).
1.4. ATIVIDADE BIOLOGICA DOS POLISSACARÍDEOS DE FUNGOS
Os polissacarídeos são os biopolímeros mais amplamente distribuídos na
natureza (ZAIKOV, 2005). São constituídos por resíduos de monossacarídeos
unidos uns aos outros por ligações glucosídicas e em comparação com outros
biopolímeros, como proteínas e ácidos nucléicos, oferecem uma maior capacidade
para transportar informação biológica, devido a sua variabilidade estrutural

24
(SHARON; LIS, 1993). Polissacarídeos bioativos isolados de fontes naturais estão
demonstrados na TABELA 1.
Os polissacarídeos bioativos de fungos podem ser estruturais, excretados ao
meio como exopolissacarídeos (EPS) ou localizarem-se no citoplasma, exercendo a
função de reserva energética (KRCMAR et al., 1999; NELSON; COX, 2002). Os
polissacarídeos estruturais isolados da parede celular fúngica têm sido
caracterizados como homopolímeros, heteropolímeros ou, ainda, encontram-se
ligados aos resíduos de proteínas formando complexos polissacarídeo-proteína (KIM
et al., 2003; ZHANG et al., 2002).
A parede celular é uma camada relativamente rígida que fornece suporte
osmótico e determina o formato da célula, além de estar relacionada a eventos de
sinalização celular, adesão e digestão extracelular (MAGNELLI et al., 2005).
Estruturalmente, a parede celular dos fungos é muito semelhante à parede
celular de vegetais, entretanto, sua composição química é distinta. Os
polissacarídeos correspondem a cerca de 80% a 90% da composição química da
parede celular dos fungos, sendo os outros componentes: proteínas, lipídios,
polifosfatos e íons inorgânicos que constituem a matriz de sustentação (GORIN;
BARRETO-BERGTER, 1983; MADIGAN et al., 1997; LIPKE; OVALLE, 1998; CABIB
et al., 2001).
Quitina, glucanas, galactomananas e proteínas são compostos mais
freqüentes, embora sua quantidade varie entre as diferentes espécies de fungos
(ADAMS, 2004). Baseados nas interações entre as diversas moléculas constituintes
da parede celular dos fungos, especialmente a composição dos polissacarídeos,
diferentes associações foram relatadas, como a de celulose-glicogênio, celulose-
glucana, celulose-quitina, quitosana-quitina, quitina-glucana, manana-glucana (ou
polissacarídeos contendo manose-glucana), manana-quitina e poligalactosamina-
galactana (GORIN; BARRETO-BERGTER, 1983).
Diante dos resultados provenientes da caracterização química dessas
macromoléculas, alguns autores propõem a sua utilização como marcadores para a
classificação taxonômica de fungos filamentosos, leveduras e liquens
(CARBONERO et al., 2003,2005; PESSONI et al. 2005; TEIXEIRA et al., 1995).

25
TABELA 1. Fontes usuais de alguns polissacarídeos. Fonte: DA CUNHA et al., 2009.
Os EPS são definidos como polissacarídeos extracelulares, produzidos por
alguns fungos e bactérias, os quais são encontrados ligados à superfície das células
ou são excretados para o meio extracelular, na forma de limo (SUTHERLAND,
1998). Nos fungos, os EPS constituem uma importante percentagem da biomassa,
participando com mais de 75% dos polissacarídeos constituintes da parede da hifa
(BARBOSA et al., 2004).
Fungos produtores de EPS desempenham papel importante no processo de
degradação de xenobióticos, uma vez que imobilizam as enzimas extracelulares. O
gel formado por estes biopolímeros impede a desidratação da hifa e permite adesão
entre as células ou a adesão destas às superfícies, além de selecionar moléculas do
meio (MAZIERO et al., 1999).

26
Os EPS produzidos por fungos também podem ser heteropolissacarídeos ou
homopolissacarídeos. Em relação aos heteropolissacarídeos de EPS, foi descrita
uma galactomanana com cadeia principal encontrada em Collybia maculata (LIM et
al., 2005).
Considerando-se o grande número de fungos encontrados na natureza e a
possibilidade de terem em sua composição endopolissacaríedos e
exopolissacarídeos com diferentes aplicações medicinais, estudos estão sendo
realizados para verificar o potencial dessas macromoléculas como antioxidantes,
antimutagênicos, anticoagulantes, antitrombóticos e imunomoduladores (FREIMUND
et al., 2003; CORRADI DA SILVA et al., 2006; ZHANG et al., 2005).
Alguns fungos produtores de exopolissacarídeos, sua composição
monomérica e o tipo de ligação glucosídica estão sumarizados na (TABELA 2), onde
pode se observar que as principais estruturas descritas são de glucanas e mananas.

27
TABELA 2. Homo- e heteropolissacarídeos produzidos por fungos filamentosos. Fonte: BARBOSA et al., 2004.

28
Dos polissacarídeos biologicamente ativos encontrados nos fungos as
glucanas são os homopolissacarídeos mais comuns. Glucanas são polímeros de
glucose amplamente distribuídos na natureza e classificadas conforme o tipo de
ligação glucosídica [α, β] da cadeia principal. As β-D-glucanas dos fungos são
polissacarídeos com função estrutural na parede celular do micélio, das frutificações,
ou também podem ser exopolissacarídeos. (BROWN; GORDON, 2001; PELOSI et
al., 2003).
A característica estrutural é considerada um fator fundamental para a
atividade das β-glucanas, principalmente no sistema imunológico (BROWN;
GORDON, 2001). Esta, por sua vez, pode atuar por meio do aumento da atividade
de células T e células T helper, indução da atividade citotóxica de macrófagos
peritoneais, aumento das proteínas de fase aguda e da sensibilidade à histamina,
estimulação da atividade da interleucina IL-1 em monócitos humanos, dentre outras
ações (OKA et al., 1996; POUTSIAKA et al., 2003; FORTES; NOVAES, 2006;
CHEN; SEVIOUR, 2007; MORADALI et al, 2007).
As β-glucanas são reconhecidas pelo sistema imune inato de vertebrados
através de receptores de superfície celular, designados primariamente para o
controle de patógenos fúngicos (ADEREM; ULEVITCH, 2000). Três receptores
celulares de β-glucanas (CR3, dectin-1 e lactosilceramida) foram identificados e
caracterizados quanto a sua função (ZIMMERMAN et al., 1998; ROSS et al., 1999;
BROWN ; GORDON, 2001, 2003).
Os CR3 são receptores celulares responsáveis por diversas atividades in
vitro e in vivo, estimulando a secreção de citocinas (TNF-α, IFN-α, IFN-γ e IL-6) em
células natural killer, principalmente, na presença de patógenos (ROSS et al., 1999).
Os receptores do tipo dectin-1, de monócitos e macrófagos, possuem um
ligante para polissacarídeos exógenos e outro ligante co-estimulatório para células
T. Alguns estudos demonstraram que esses receptores celulares e também o
lactosilceramida de linfócitos reconhecem β-glucanas, principalmente com estrutura
do tipo β-(1→3) e β-(1→6) (ZIMMERMAN et al., 1998; BROWN; GORDON, 2003).
Muitos medicamentos utilizados em neoplasias apresentam como efeito
colateral uma ação imunossupressora, que pode culminar em infecção e septicemia.
Devido à capacidade das β-D-glucanas em estimular o sistema imunológico,
elas podem ser utilizadas como adjuvantes desses quimioterápicos no tratamento de

29
pacientes imunocomprometidos (BOHN; BEMILLER, 1995; LEE et al., 2001; LEUNG
et al., 2006; SUGAWARA et al., 2004; ZHANG et al., 2005).
Além das glucanas outros polissacarídeos bioativos, como os
heteropolissacarídeos também estão sendo investigados com o intuito de determinar
suas atividades biológicas. De acordo com Zhang et al. (2007), manoglucanas,
galactomananas e glucomananas, obtidas da biomassa de basidiomicetos, também
podem apresentar atividades importantes para a indústria farmacêutica e alimentícia.
As atividades desses polissacarídeos fungicos parecem estar relacionadas à
estrutura química da cadeia principal da glucana na forma de hélice tríplice (FIGURA
3), à freqüência e complexidade das cadeias laterais e à sua massa molecular
(SCHMID et al., 2007).
FIGURA 3. Representação esquemática da conformação induzida por temperatura da solução de lentinana a 137 °C. Fonte: WANG et al., 2008
1.5. GALACTOMANANAS
Galactomananas são polissacarídeos de reserva de parede celular
encontradas em maior quantidade na natureza. Além disso, é um dos sistemas
sobre o qual se tem maior conhecimento acerca da biossíntese (BUCKERIDGE;
DIETRICH, 1996; LIMA, 2000). As galactomananas são polissacarídeos neutros,
comumente encontrados no endosperma das sementes de algumas leguminosas,
porém também podem ser obtidas a partir de outras fontes como leveduras e fungos
(ANDRADE et al., 1999).

30
As galactomananas são ramificadas, pouco densas e solúveis em solução
aquosa, são compostas de uma cadeia principal formada por unidades
monossacarídicas de β-D-manopiranose unidas por ligações glicosídicas (1→4),
substituídas em O-6, por unidades monossacarídicas de α-D-galactopiranose
(FIGURA 4), em graus variados, dependendo da espécie (DAAS et al., 2000;
SIERAKOWSKI et al., 2000; CUNHA et al., 2009).
Na natureza, o grau de ramificação das galactomananas varia de polímeros
em que quase todas as unidades de manose apresentam ramificação com galactose
a polímeros em que nenhuma ramificação é detectada. O nível e a distribuição de
galactose sobre a cadeia principal ocupam um papel importante nas propriedades
físico-químicas do polímero (BUCKERIDGE; DIETRICH, 1990; BUCKERIDGE;
DIETRICH, 1996; LIMA, 2000).
A solubilidade em água tende a aumentar com o aumento de galactose, mas
devido às grandes interações internas (pontes de hidrogênio), isso normalmente não
é verificado, sendo observado um aumento da viscosidade da solução contendo a
galactomanana que é considerado um bom espessante (SIERAKOWSKI et al.
2000).
FIGURA 4. Estrutura parcial da galactomana e suas respectivas unidades monossacarídicas. Legenda: Man: manose, Gal: galactose. Fonte: SIERAKOWAKI et al., 2000.
As galactomananas são muito empregadas pelas indústrias farmacêutica,
cosmética e alimentícia. Estudos relatam que galactomananas podem interferir na
inflamação (CRAY et al., 2009), na geração de radicais livres (NAKAMURA et al.,

31
1998) e na coagulação (PIRES et al., 2001). Também são usados no tratamento de
diabetes mellitus reduzindo o pico de concentração sanguínea de glicose que ocorre
após a ingestão de alimentos (REYNOLDS, 1989; WANDE et al., 1994;
BRUNETON, 1991).
1.6. GÊNERO TYLOPILUS
Os representantes do gênero Tylopilus pertence à ordem Boletales, a qual é
bem conhecida na Europa, América do Norte e várias regiões tropicais, mas apenas
nas últimas décadas têm registros de estudos na América do Sul. Esses registros
são concentrados com base em documentação de fungos em florestas de altitude
(HALLING; MUELLER, 1999; HALLING; MUELLER, 2002). A diversidade de
Boletales no Brasil é muito pouco conhecida, em comparação com outros países,
onde este grupo tem sido mais intensamente estudado (HALLING, 1989; HALLING;
FRANCO-MOLANO, 1996; HALLING; MUELLER, 2002; WATLING, 2001).
Alguns registros foram realizados para esse gênero durante a primeira
metade do século XX (RICK, 1907, 1960), e outros foram feitos na região amazônica
na década de 1980 (SINGER et al., 1983). Publicações sobre Boletales estão sendo
adicionadas ao conhecimento desse grupo no Brasil (SINGER; ARAUJO, 1986;
PUTZKE; PEREIRA, 1994; WATLING; MEIJER, 1997; GIACHINI et al., 2000;
BASEIA; ADAUTO, 2002).
Tylopilus é um gênero de fungos micorrízicos e seu membro mais conhecido
são os boletos Bitter (Tylopilus felleus), única espécie encontrada na Europa e
Tylopilus chromapes, espécie comestível encontrada na América do Norte (SINGER;
ARAUJO, 1979).
Este grupo de fungos foi descritos pela primeira vez por Singer e Araujo na
América Central e ainda são pouco conhecido nas florestas tropicais da América do
Sul (SINGER, 1984). Tylopilus ballouii caracteriza-se por apresentar a superfície do
píleo alaranjada quando fresca e os esporos elipsóides. No entanto, a coloração se
destaca apenas nos espécimes jovens e desaparece com a idade e se torna um
marrom pálido. O seu registro de ocorrência é relatado na América do Norte,
América Central (Costa Rica) e do Sul (Guiana) (HENKEL, 1999, 2001). Como a

32
maioria dos boletos é simbiontes ectomicorrízicos obrigatórios, a sua distribuição
biogeográfica depende da distribuição de suas plantas hospedeiras. A invasão de
novas áreas geográficas de uma planta hospedeira e seus fungos simbiontes deve
ocorrer simultaneamente (RICHARDSON; RUNDE, 1998).
Considerando a carência de levantamento sobre a diversidade dos fungos no
Brasil, especialmente para o Nordeste e a importância destes como fonte de
moléculas bioativas, além da rápida degradação das formações vegetais nativas que
abrigam ampla variedade destes organismos, estudos intensivos são necessários
para o conhecimento dessa biodiversidade com possíveis potenciais
biotecnológicos. Tylopilus ballouii é um fungo nativo da América do Norte e não têm
registro em regiões tropicais (HEINEMANN, 1954), e está ausente da lista de
espécies de gênero Tylopilus na Amazônia (SINGER; IVORY 1983).

33
2. OBJETIVOS
2.1. OBJETIVO GERAL
Este trabalho tem como objetivo geral extrair, caracterizar e avaliar o
efeito dos polissacarídeos presentes no corpo de frutificação do fungo Tylopilus
ballouii na inflamação e no sequestro de radicais livres.
2.2. OBJETIVOS ESPECÍFICOS
Para alcançar o objetivo geral, as seguintes estratégias foram propostas:
• Extrair os polissacarídeos presentes no corpo de frutificação do fungo
Tylopilus ballouii;
• Determinar a composição centesimal e caracterizar a estrutura dos
polissacarídeos por métodos químicos e espectroscópicos;
• Realizar bioensaio utilizando os polissacarídeos extraídos de Tylopilus
ballouii em ratos Wistar para avaliar sua ação na inflamação, bem como,
analisar histologicamente o efeito deste composto neste modelo;
• Avaliar o potencial citotóxico dos polissacarídeos do fungo Tylopilus
ballouii;
• Analisar o efeito in vitro dos polissacarídeos sobre a geração de radicais
livres.

34
3. MATERIAS E MÉTODOS
3.1. MATERIAIS
3.1.1. Fungo
O composto polissacarídico utilizado nesse estudo foi obtido dos corpos de
frutificação do fungo Tylopilus ballouii (Figura 5). A coleta do material foi realizada
durante o período chuvoso de janeiro/ 2008 a junho/ 2008, no Parque Estadual das
Dunas, Reserva da Biosfera da Mata Atlântica Brasileira (05º46'S - 35º12'O),
localizado no Estado do Rio Grande do Norte, no município de Natal. A análise e
coleta do material foram realizadas pelo Prof. Dr. Iuri Goulart Baseia do
Departamento de Botânica, Ecologia e Zoologia da UFRN.
Classificação taxonômica:
Reino: Fungi
Filo: Basidiomycota
Classe: Agaricomycetes
Ordem: Boletales
Família: Boletaceae
Gênero: Tylopilus.
Espécie: Tylopilus ballouii
FIGURA 5. Fungo Tylopilus ballouii

35
3.1.2. Animais
A atividade antiinflamatória dos polissacarídeos do fungo T. ballouii foi
avaliada no modelo de edema plantar utilizando-se ratos da linhagem Wistar com
aproximadamente dois meses de idade, pesando entre 150 – 200 g, obtidos do
biotério do Departamento de Bioquímica - UFRN. Para a atividade citotóxica através
do ensaio de viabilidade celular ex vivo, foram utilizados camundongos da linhagem
Swiss, do biotério do Departamento de Bioquímica - UFRN. Todos os animais
utilizados na fase experimental foram mantidos em gaiolas individuais, submetidos à
água e dieta ad libitum em condições controladas de iluminação (ciclo 12h
claro/escuro) e temperatura constante a 25º C. Os animais foram acondicionados no
laboratório, por no mínimo, 2 h antes dos testes e utilizados uma única vez para os
experimentos. Os ensaios foram desenvolvidos de acordo com as normas vigentes.
3.1.3. Outros Materiais
• Ácido acético, acetona, metanol, etanol, n-butanol, cloreto de sódio, tiossulfato de
sódio da Reagen Quimibrás Indústrias Químicas S.A. (Rio de Janeiro, RJ, Brasil).
• Hidroximetilamino metano, carragenana, coomasie brilliant blue R 250 (Sigma
Chemical Company, St. Louis, MO, EUA).
• NADH, azul de tripan, nitroazul de tetrazolio, fenazim metossulfato, molibdato de
amônio, ácido tricloroacético, ácido 2-tiobarbitúrico 98%, ácido ascórbico 99%,
1,2-diaminoetano, ácido isobutírico, piridina, metil e tolueno da Aldrich Chemical
Co. Inc. (Millwaukee, WI, EUA).
• Papéis Whatman n.º 1 e 3 MM foram obtidos da W & R Balston Ltd. (Maidstone,
Inglaterra).
• Cloridrato de xilazina a 2% e cloridrato de ketamina 5% (Köning do Brasil, Ltda)
• Ácido sulfúrico, ácido clorídrico, ácido fosfórico da Merck (Darmstadt, Alemanha).

36
3.1.4. Aparelhos
Além dos aparelhos usuais de laboratório podemos destacar:
• Agitador orbital mod. 255-B da FANEM Ltda. (São Paulo, SP, Brasil).
• Banhos e estufas de temperatura constante da FANEM Ltda. (São Paulo, SP,
Brasil).
• Centrífuga refrigerada RC 2-B da Ivan Sorvall Inc. (Norwalk, CO, EUA).
• Espectrofotômetros Varian - Series 634 da Varian Techtron PPTY Ltd.
(Springvale, Vico, Austrália) e Hitachi U-2000 (Tóquio, Japão).
• Evaporador rotatório Evapo-Mix da Buchler Instruments (Fort Lee, NJ, EUA).
• Espectrômetro de infravermelho modelo FT1 6PC da Perkin Elmer (EUA).

37
3.2. MÉTODOS
Organograma geral
O estudo detalhado da estrutura e atividade de biomoléculas requer muitas
vezes o seu isolamento. A partir da obtenção dos corpos de frutificação do fungo T.
ballouii foi realizada a primeira etapa desse processo (extração). O polissacarídeo
resultante foi usado para estudo das atividades biológicas e caracterizações
químicas. No esquema abaixo temos representado uma visão geral dos
experimentos realizados neste trabalho.
Corpos de frutificação de Tylopilus ballouii
PolissacarídeosPolissacarídeos
Caracterização químicaCaracterização química
Atividade antiinflamatória, antioxidante e citotoxicidade
celular
Atividade antiinflamatória, antioxidante e citotoxicidade
celular
Dosagens químicasAnálise de infravermelho
Ressonância Magnética nuclear do H1
Composição monossacarídicaMassa molecular
Dosagens químicasAnálise de infravermelho
Ressonância Magnética nuclear do H1
Composição monossacarídicaMassa molecular
Composição centesimalComposição centesimal
Atividades biológicasAtividades biológicas
FIGURA 6. Organograma da metodologia

38
3.2.1. Composição Centesimal
Para a determinação dos teores de cinza, proteínas, fibras e umidades foram
utilizados os métodos descritos pela AOAC (1984). Para a determinação de lipídeos
utilizou-se o método descrito por BLIGH e DYER (1959). O conteúdo de carboidratos
do fungo foi calculado por diferença. Todas as determinações foram realizadas em
triplicata.
3.2.1.1. Determinação da fração cinza
A fração cinza foi determinada de acordo com o método gravimétrico através
da incineração da matéria orgânica total. Amostras de 2 g do fungo pulverizado
foram pesadas e colocadas em cadinhos de porcelana previamente tarados e o
material foi incinerado em bico de Busen até ponto de carvão; a seguir os cadinhos
foram colocados em mufla a 550ºC até adquirir cor uniforme (esbranquiçada) que vai
do cinza ao branco, sem pontos de carvão. Os cadinhos contendo a amostra foram
retirados da mufla quando a temperatura atinge 50ºC e colocados em dessecador
para esfriar e, posteriormente, foram pesados. A diferença de peso encontrada nas
amostras utilizadas após incineração fornece o teor de cinzas.
3.2.1.2. Determinação de Lipídeos
Para quantificação dos lipídeos foram pesadas 3,0 g do fungo pulverizado e
colocadas em erlenmeyer com rolha, sendo adicionados 10 mL de clorofórmio, 20
mL de metanol e 8 mL de água destilada. Em seguida, os erlenmeyers foram
arrolhados hermeticamente e colocados em agitador rotativo (rumbeira) durante 30
minutos. Após esse período foram adicionados 10 mL de clorofórmio e 10 mL de
sulfato de sódio a 1,5%, os tubos foram arrolhados novamente e agitados durante 2
minutos e deixados em repouso para separar as camadas. Foram retirados 13 mL
da camada inferior que continha clorofórmio e lipídeos para um tubo cônico de 30
mL com rolha esmerilhada e adicionou-se 1 g de sulfato de sódio anidro. O tubo foi
arrolhado e agitado para remover os traços de água, o material foi filtrado e 5 mL
transferidos para um becker de 50 mL previamente pesado e levado à estufa a 105º

39
C até completa evaporação do solvente, depois foi resfriado em dessecador. Os
beckers foram pesados e o percentual de lipídeos foi calculado a partir do teor
encontrado nos 5 mL, relacionando com o peso da amostra.
3.2.1.3. Determinação de Proteínas
Os teores de proteínas das amostras foram determinados pelo método
Semimicro Kjeldahl, através da dosagem do nitrogênio total. Amostras de 200 mg do
fungo previamente delipidado com auxílio de solventes orgânicos foram dessecadas
e colocadas em balão de Kjeldahl de 50 mL onde foram adicionados 0,5 g de sulfato
de cobre, 1,0 g de sulfato de sódio e 5 mL de ácido sulfúrico concentrado. Os balões
foram levados ao digestor de Kjeldahl até completar mineralização durante
aproximadamente 4 horas, fato este caracterizado pela formação de um líquido
translúcido de cor verde-azulada. Após digestão e resfriamento foram adicionados
10 mL de água destilada ao balão de Kjeldahl, o qual foi acoplado ao aparelho de
destilação. Na mangueira do destilador foi conectado um erlenmeyer contendo 10
mL de solução de ácido bórico saturado com três gotas do indicador misto. Através
do funil do destilador, foram adicionados ao balão de Kjeldahl, 25 mL de hidróxido de
sódio a 40%. Foi destilado aproximadamente um volume de 50 mL, o qual foi titulado
com ácido sulfúrico 0,02 N, previamente padronizado. O teor de proteína foi
calculado de acordo com a seguinte fórmula:
N = V x Normalidade x Fator do ácido x 14,01
g da amostra
P = 6,25 x N

40
3.2.1.4. Determinação de Fibras
O método utilizado foi o da digestão ácida e alcalina. Amostras de 2 g do
fungo, previamente delipidada, foram pesadas e colocadas em béquer de 600 mL,
contendo 200 mL de H2SO4 a 1,25% fervente e mantido em ebulição por 30 minutos;
após esse período o material ainda quente foi filtrado a vácuo sobre papel de filtro
comum; o béquer foi lavado com água destilada e a seguir, o material foi recolocado
no mesmo béquer sendo adicionados 200 mL de hidróxido de sódio a 1,25% e
mantido em ebulição por 30 minutos. Decorrido esse tempo, o material ainda quente
foi filtrado a vácuo em cadinhos de Gooch com lã de vidro, previamente pesados, e
o material lavado sucessivamente com água destilada para remover o excesso de
base existente no filtrado, sendo este procedimento monitorado com um papel
indicador, até completa neutralização. Em seguida, o resíduo foi lavado com álcool e
éter etílico por duas vezes. Após a secagem à temperatura ambiente, o cadinho foi
colocado em estufa a 105° C até peso constante. A variação do peso encontrado
fornece a quantidade de fibras da amostra.
3.2.1.5. Determinação da umidade
A umidade foi determinada pelo método gravimétrico, o qual quantifica o teor
de água da amostra através da diferença de peso da amostra hidratada e seca. Para
este procedimento foram pesadas amostras de 10 g em cápsulas de porcelana
previamente taradas e levadas à estufa a 105º C para dessecar até peso constante.
O teor de umidade foi determinado através da diferença do peso inicial e final.

41
3.2.1.6. Determinação de Carboidratos
O teor de carboidratos presente no fungo T. ballouii foi obtido por estimativa,
através da diferença dos componentes já determinados para um total de 100%,
conforme mostra a fórmula abaixo:
% Lipídeos + % Proteínas + % Fibras + % Cinzas = X%
100% - X% = % de Carboidratos
3.2.2. Obtenção dos polissacarídeos do fungo T. ballouii
Antes do processo de extração para obtenção dos polissacarídeos, os corpos
de frutificação do fungo foram lavados, dessecados a 40º C e posteriormente
pulverizados. O método de extração dos polissacarídeos foi realizado a partir de
modificações no método de Gonzaga et al., (2005). De um total de 70 g do fungo
seco, 20 g foi destinado para análise da composição centesimal e 50 g à extração de
polissacarídeos. O fungo foi suspenso em 5% de água destilada (w/w)
permanecendo a mistura em banho-maria a 100º C por 4 h. O filtrado foi
neutralizado para pH 7.0 por adição de solução de hidróxido de sódio e adicionado
etanol/solução do extrato, 5:1, para obter as precipitações, as quais foram isoladas
por centrifugação, lavadas com acetona e dessecadas. Peróxido de hidrogênio
(30%) 1:1 etanol/peróxido foi adicionado por aproximadamente 15 h em temperatura
ambiente para a clarificação do material extraído. Devido ao seu material
parcialmente solubilizado foram submetidos à segunda extração usando
etanol/volume de extrato (4:1). As precipitações foram isoladas por centrifugação,
lavadas com etanol, depois com acetona e liofilizadas. Para purificar as amostras de
possíveis substancias insolúveis, os materiais isolados foram suspensos em água
destilada com agitação em 40°C por 12 h. Depois desse procedimento as
suspensões foram centrifugadas e a fase solúvel foi removida, submetida à
liofilização e acondicionados de forma adequada para manter suas características
físico-químicas (FIGURA 7).

42
Corpos de frutificação de Tylopilus ballouii
70 g
Precipitado 1Precipitado 1Sobrenadante
5% em água destilada, 100°C/ 4hPrecipitação com etanol (5:1 v/v)Centrifugação (8000g, 20min, 4°C)
Precipitado 2Precipitado 2Sobrenadante
H2O2 a 30% (1:1 v/v)Precipitação com etanol (4:1 v/v)Centrifugação (8000g, 20min, 4°C)
Sobrenadante Precipitado
Ressuspenso em água/12h a 40°CCentrifugação (8000g, 20min, 4°C
Liofilização
FIGURA 7. Esquema de obtenção dos polissacarídeos do fungo T. ballouii.
FFrraaççããoo ssoollúúvveell

43
3.2.3. Caracterização química
3.2.3.1. Análises Colorimétricas
3.2.3.1.1. Açúcares totais
Açúcares totais foram determinados pelo método do fenol/ácido sulfúrico de
acordo com Dubois e colaboradores (1956), empregando-se como padrão L-
galactose, sendo as leituras realizadas a 490 nm.
3.2.3.1.2. Proteínas
O conteúdo de proteínas foi determinado segundo o método de Bradford
(1976) utilizando o corante Coomassie Brilliant Blue G-250 empregando-se como
padrão albumina sérica bovina.
3.2.3.2. Cromatografia Descendente em Papel
O composto polissacarídico foi hidrolisado (HCl 2N, 2 h, 100º C), neutralizado,
seco, ressuspenso em água destilada e aplicado em papel Whatman N°1. O qual foi
submetido a dois sistemas de solventes: butanol: ácido acético: Água (v/v/v, 2/ 3/
1,5) e acetato de etila: piridina: água (v/v/v, 8/ 2/ 1). Os monossacarídeos foram
revelados após redução do nitrato de prata (TREVELYAN, 1950).
3.2.3.3. Cromatografia Líquida de Alta Eficiência
Os polímeros foram hidrolisados (HCl 2N, 2 h, 100° C) e a composição
monossacarídica foi determinada utilizando-se um sistema de cromatografia liquida
de alta eficiência acoplado a um detector de índice de refração, usou-se uma coluna
LiChroCART ® 250-4. Como padrão para a análise foram utilizados os seguintes
açúcares: glicose, galactose, arabinose, fucose, manose, ramnose e xilose.

44
3.2.3.4. Determinação da massa molecular
As frações de polissacarídeos foram submetidas à cromatografia de gel
filtração em Sephadex G-100 (140 × 1 cm) com 0,2 M de ácido acético e 0,15 M de
cloreto de sódio como eluente. A eluição foi monitorada pela análise de açúcares
totais (DUBOIS et al., 1956). Para estimar a massa molecular dos polissacarídeos,
foram utilizadas dextranas de diferentes tamanhos como padrões (Pharmacia).
3.2.3.5. Análise de Infravermelho
A espectroscopia de infravermelho foi realizada em espectômetro FT-IR ABB
Bomen modelo MB 104, de 4000 a 400 cm-1. O polissacarídeo foi analisado após
secagem sob a forma de pastilha de KBr.
3.2.3.6. Ressonância Magnética Nuclear (RMN)
Os espectros de RMN foram obtidos a 60ºC utilizando amostras dissolvidas
em D2O. As análises dos espectros de H1 foram realizadas em espectrômetro Varian
Mercury 200 Magneto Oxford.
3.2.4. Atividade Antiinflamatória
3.2.4.1. Avaliação do efeito do composto de Tylopilus ballouii em um
modelo de edema induzido por carragenana.
Neste experimento foram utilizados ratos da linhagem Wistar para verificação
do efeito do polissacarídeo de T. ballouii estimulado por um agente pró-inflamatório
denominado carragenana, na face plantar da pata direita. Os animais foram
previamente anestesiados utilizando-se cloridrato de xilazina e cloridrato de
ketamina por via intraperitoneal (i.p), antes da aplicação do agente inflamatório no
intuito de aliviar sua sensação dolorosa. Todos os animais foram pesados e um
cronômetro digital foi utilizado para verificação do tempo decorrido desde a
administração.

45
Sete grupos foram formados com n = 5
• Grupo 1: Controle negativo: solução salina
• Grupo 2: Controle positivo: 1% de carragenana (sem tratamento)
• Grupo 3: Animais tratados com polissacarídeos de T.ballouii a 30 mg/kg i.p e
1% de carragenana r.p.
• Grupo 4: Animais tratados com polissacarídeos de T. ballouii a 50mg/kg i.p e 1%
de carragenana r.p.
• Grupo 5: Animais tratados com polissacarídeos de T. ballouii a 70mg/kg i.p e 1%
de carragenana r.p
• Grupo 6: Animais tratados com Indometacina a 50 mg/kg i.p e 1% de
carragenana r.p
• Grupo 7: Animais tratados com Diclofenaco de Sódio a 50mg/kg i.p e 1%
carragenana r.p
Nos grupos 3, 4 e 5 foram administrados o composto polissacarídico de T.
ballouii nas concentrações de 30, 50 e 70 mg/kg de animal por i.p 30 minutos antes
da injeção da carragenana. Nos grupos 1 e 2 foram administrados i.p solução
fisiológica. Nos grupos 6 e 7 aplicou-se indometacina e diclofenaco de sódio i.p,
respectivamente. Após o intervalo de 30 minutos, realizou-se a medida da espessura
da pata direita com o uso de um paquímetro, em seguida a inflamação foi induzida
através da aplicação da injeção plantar de carragenana 1% (0,1 mL) na pata direita
de todos os ratos. Os grupos controles foram submetidos ao mesmo procedimento
sendo que os animais receberam solução salina estéri i.p. Este procedimento foi
repetido após o intervalo de 1ª, 2ª, 3ª e 4ª h. Os resultados da medição foram
expressos como a diferença entre o volume final da pata a cada tempo (Vf) e o
volume inicial (Vi), representados pela fórmula: Vp = Vf-Vi. Para a realização das
análises histológicas, após 4ª h das injeções i.p, r.p e aferição da espessura da pata
por paquímetro, os animais foram eutanisiados e amostras da pele e tecido celular
subcutâneo da região plantar foram colhidas para análise histológica.

46
3.2.4.2. Análises histológicas
Para os exames histológicos, foram retiradas amostras da pele com tecido
celular subcutâneo da pata dos animais. O tecido foi fixado em formaldeído 10% por
48hs, embebido em parafina, cortados em micrótomo rotativo na espessura de 5
mm. Seqüencialmente, os cortes foram submetidos à técnica de coloração pela
hematoxilina-eosina (H.E.) e analisados em microscópio de luz e fotografados em
fotomicroscópio (JUNQUEIRA; CARNEIRO, 1995).
3.2.5. Atividade Citotóxica
3.2.5.1. Determinação da Viabilidade Celular ex vivo
A viabilidade celular foi determinada utilizando-se o método de exclusão por
azul de tripan descrito por Phillips (1973). Doze camundongos Swiss foram reunidos
em 4 grupos, anestesiados e o polissacarídeo foi administrado i.p em diferentes
doses (50 e 100 mg/kg). Os animais foram eutanasiados após 4 h e outros após 24
h. O grupo controle recebeu apenas PBS (pH 7,4) no lugar da amostra. O lavado
peritoneal foi coletado e uma alíquota de 20 µL da suspensão de células foi retirada
e diluída em 20 µL de azul de tripan. A porcentagem de células viáveis foi
determinada após contagem diferencial em Câmara de Neubauer. A taxa de
viabilidade foi calculada conforme a equação: Células Viáveis (%) = 100 x células
não coradas/ células totais (não coradas+ coradas). A viabilidade dos controles foi
normatizada para 100%.

47
3.2.6. Atividade Antioxidante
3.2.6.1. Radicais Superóxidos
Os radicais superóxidos foram gerados em 3 mL de tris-HCl (16mM, pH 8,0),
que continham 78 µM de NADH (forma reduzida), 50 µM de nitroazul de tetrazolio,
10 µM de fenazina metossulfato e concentrações variadas de polissacarídeos. A
reação foi detectada por monitoramento da absorbância a 560 nm. Foram realizados
controles para cada amostra. O branco não continha NADH, sendo este substituído
pelo tris-HCl (ZHANG et al., 2003).
3.2.6.2. Radicais Hidroxilas
Os radicais hidroxilas foram gerados por um método modificado de Smirnoff e
Cumners (1989) em 3 mL de fosfato de sódio (150 mM, pH 7,4), que continha 10
mM de sulfato ferroso heptahidratado (FeSO4.7H2O), 10 mM de
etilenodiaminotetraacético (EDTA), 2m M de salicilato de sódio, 200 µL de peróxido
de hidrogênio a 30 % e concentrações variadas de polissacarídeos. A solução foi
incubada a 37°C por 1 h, e a atividade do radical hidroxila sobre o salicilato foi
detectada a absorbância de 510 nm. No branco o peróxido foi substituído pelo
tampão fosfato (ZHANG et al., 2003).
3.2.6.3. Capacidade Antioxidante Total
A capacidade antioxidante total foi determinada pelo método do
fosfomolibdênio, que se baseia na determinação espectrofotométrica da redução do
Mo+5 a Mo+4 com formação subseqüente de fosfato de Mo+5, que apresenta
absorção máxima a 695 nm. Alíquotas de 0,1 mL da solução da amostra
(concentração variada de polissacarídeos) foi misturada em um tubo Eppendorf com
1 mL da solução reagente (0,6 M de ácido sulfúrico, 28 mM fosfato de sódio e
molibdato de amônio 4 mM).Os tubos foram fechados e incubados a 95 °C por 90

48
min. Após resfriamento, à temperatura ambiente, foi determinada a absorbância a
695 nm. Foi utilizado ácido ascórbico como padrão e a capacidade antioxidante total
foram expressos em equivalentes de ácido ascórbico (PRIETO et al. 1999).
3.2.6.4. Peroxidação Lipídica
A inibição da peroxidação lipídica foi determinada pela quantificação do
produto de decomposição de peróxido lipídico MDA baseada na reação ao ácido
tiobarbitúrico com gema de ovo como substrato oxidável. Resumidamente, 0,5 mL
de homogeneizado de ovo (10% em 0,2 MPBS (pH 7,4), v / v) e 0,5 mL de amostra
em diferentes concentrações foram misturadas, em seguida, 0,05 mL de sulfato de
ferro (0,07 M) foi adicionado ao para iniciar a peroxidação lipídica. Após a
incubação a 37 ° por 30 min, 1,5 mL de tricloroacético ácido (20%) (w / v) e 1,5 mL
de ácido tiobarbitúrico 0,8% (w / v) foram adicionados para saciar a reação, a
mistura resultante foi agitada e aquecida a 100 ° C por 15 min, e em seguida,
centrifugado a 3000 rpm por 10 min. A absorbância da camada superior foi medido
em 532 nm (ZHANG ;YU, 1997). A inibição da peroxidação lipídica foi calculada da
seguinte forma: taxa de inibição (%) = [1 - (A Amostra - Branco Amostra) / AControle] x 100,
O α-tocoferol foi utilizado com padrão.
3.2.7. Cálculo do IC50
O IC50, ou seja, a quantidade em mg/mL da amostra necessária para inibir em
50% os radicais antioxidantes foi determinado para seqüestro dos radicais
superóxido e hidroxila, bem como para inibição da peroxidação lipídica. O cálculo do
IC50 de cada atividade foi realizado por regressões não lineares.

49
3.2.8. Análises Estatísticas
Para análises estatísticas dos modelos experimentais de inflamação e
citotoxicidade foi utilizado o teste de análise de variância ANOVA com nível de
significância de p<0,001 e p>0,5 para determinar que grupos diferem entre os
valores obtido para os grupos controle e experimental.

50
4. RESULTADOS
4.1. Extração
A extração foi desenvolvida conforme descrito anteriormente em materiais e
métodos, apresentando um rendimento de 0, 997 g de polissacarídeos o qual foi
caracterizado quimicamente e analisadas suas propriedades farmacológicas.
4.2. Composição centesimal
O conhecimento da composição centesimal é realizado através da análise
química determinando-se o teor de umidade, cinzas, lipídeos, proteínas, fibras e
carboidratos (MORETTO et al., 2002). Pelo fato desta pesquisa estar direcionada
no estudo das propriedades antioxidantes e antiinflamatória do fungo T. ballouii,
outros parâmetros nutricionais como minerais e vitaminas não foram quantificados
individualmente neste trabalho. Os resultados obtidos na análise da composição
centesimal do fungo T. ballouii podem ser observados na TABELA 3. Estas análises
demonstraram que os carboidratos (32,5%) representam o principal constituinte do
corpo de frutificação do fungo T. ballouii. Outros constituintes, proteínas (27,0%) e
fibras (18,2%) também se destacaram com altos percentuais na composição desse
fungo.

51
TABELA 3: Análise da composição centesimal do fungo T. ballouii.
Componentes
Teor (%)
Umidade
Proteína
Lipídeos
Fibras
Cinzas
Carboidrato
11,3
27,0
5,2
18,2
5,8
32,5
4.3. Análises químicas
Após extração e fracionamento dos componentes dos corpos de frutificação
do fungo T. ballouii por procedimentos já referidos em materiais e métodos as
analises químicas foram realizadas no sentido de verificar os constituintes
resultantes do processo de fracionamento. Neste trabalho, realizou-se a extração
com água quente seguida de posteriores precipitações com etanol (precipitado 1) e
(precipitado 2). Essa metodologia é largamente utilizada por ser uma forma eficiente
de precipitação de polissacarídeos e por isto, bastante relatada pela literatura
(PEREYRA et al., 2003; ALQUINI, et al., 2004). Os resultados das análises do
precipitado 2 demonstraram conter altos teores de carboidratos (80,5%) e razoável
teor de proteínas (19.5 %) (FIGURA 8).

52
FIGURA 8. Percentual dos constituintes açúcares totais e proteínas após extração de T. ballouii.
4.4. Composição monossacarídica do precipitado 2
Para analise estrutural de um polissacarídeo faz-se necessário primeiro,
determinar os tipos de monossacarídeos que o constituem (NELSON et al., 2000).
Para isso foram realizados dois métodos cromatográficos, cromatografia
descendente em papel (CDP) e cromatografia líquida de alta eficiência (HPLC).
4.4.1. Cromatografia descendente em papel
A CDP realizada no sistema de solvente acetato de etila: piridina: água (v/v/v,
8: 2: 1) demonstra, de forma qualitativa, a presença dos monossacarídeos: xilose e/
ou fucose, manose, e galactose (FIGURA 9A). A CDP realizada no sistema de
solvente butanol: ácido acético: água (v/v/v, 2: 3: 1,5) apresentou como esperado,
diferenças na migração entre os monossacarídeos xilose e fucose caracterizando a
presença destes dois açúcares (FIGURA 9B).

53
Acetato de etila Butanol
FIGURA 9. Cromatografia descendente em papel do hidrolisado dos polissacarídeos de T. ballouii, utilizando como padrão monossacarídeos. A: Sistema solvente Acetato de etila: Piridina: água. B: Sistema solvente Butanol: Ácido acético: água. Legenda das siglas na figura: FUC: fucose; XIL: xilose; GAL: galactose; GLI: glicose; MAN: manose; ARA: arabinose; P: polissacarídeo de T. ballouii.
4.4.2. Análise da fração de polissacarídeos por cromatografia líquida de
alta eficiência (HPLC)
Os dados obtidos por HPLC mostraram de forma quantitativa a presença de
xilose, fucose, arabinose (traços) manose e galactose, demonstrando predominância
dos dois últimos, galactose (26,88%) e manose (54,20%) comprovando a presença
de galactomananas neste composto (TABELA 4). Os perfis cromatográficos foram
inicialmente obtidos com gradiente exploratório de água e acetonitrila (5 a 90% do
modificador orgânico em 60 min), em coluna de LiChroCART 250-4 usando-se como
referencial diversos padrões monossacarídicos. Os perfis cromatográficos obtidos
por HPLC resultaram na presença de quatro unidades monossacarídicas (FIGURA
10).

54
TABELA 4. Composição monossacarídica dos polissacarídeos do fungo T. ballouii.
Galactose Manose
Fucose Arabinose Xilose
Relação molar
0,50 1 0,13 0,04 0,30
Composição (%)
26,88 54,20 6,57 1,91 12,40
.
FIGURA 10. Cromatografia em HPLC da fração polissacarídica do fungo T. ballouii e eluição em gradiente de água e acetonitrila.

55
4.5. Determinação da massa molecular
A determinação da massa molecular do composto polissacarídico foi
analisada por cromatografia em gel filtração em Sephadex G-100. Esta, mostrou
uma simetria no pico de eluição indicando homogeneidade no polímero e massa
molecular de aproximadamente 147 kDa quando comparado com os padrões da
curva de calibração da coluna (FIGURA 11).
FIGURA 11. Perfil dos polissacarídeos do fungo T. ballouii em cromatografia gel filtração. 4.6. Espectroscopia de Infravermelho
O espectro de infravermelho dos polissacarídeos do fungo T. ballouii
apresentou similaridade para os padrões de carboidratos e proteínas. As atribuições
das principais absorções são características das estruturas glucosídicas e são
relacionadas com o estiramento de OH (3000- 3500cm-1), e carbono anomérico
(1070 cm-1).
O sinal existente na faixa entre as bandas (1630 e 1400 cm-1) estão
relacionados com as vibrações de grupo amida o que confirma a presença de

56
proteínas nesse composto, sugerindo uma associação entre carboidrato e proteínas
(FIGURA 12).
Figura 12. Espectro de Infravermelho dos polissacarídeos do fungo T. ballouii.
4.7. Ressonância magnética nuclear do hidrogênio
A ressonância magnética nuclear (RMN) é uma das mais poderosas técnicas
espectroscópicas no estudo de aspectos estruturais e dinâmicos de moléculas
orgânicas em solução (KAISER, 2000). Para a química de carboidratos se tornou
uma ferramenta largamente utilizada, portanto torna-se imprescindível sua utilização,
principalmente na elucidação estrutural de polissacarídeos com estruturas
complexas. Pelas análises de RMN várias informações podem ser obtidas a respeito
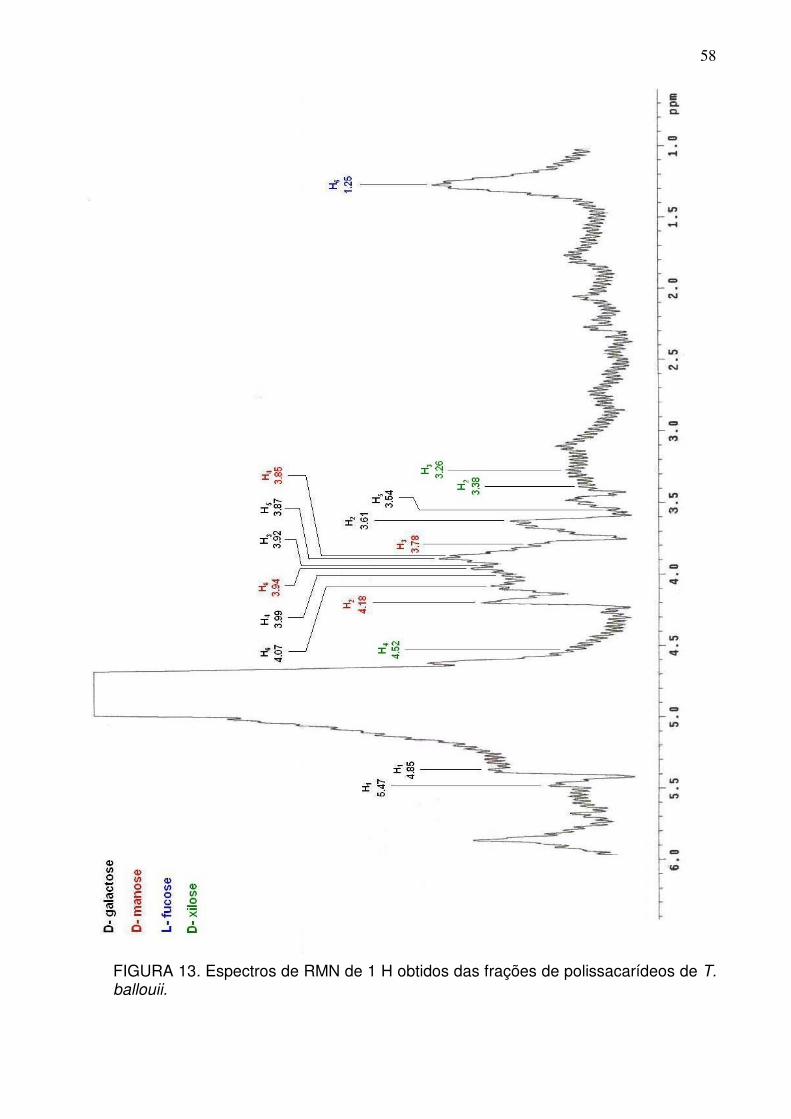
da estrutura do polissacarídeo. O espectro de RMN do H1 do composto
polissacarídico contém sinais característicos de uma galactomanano. Apresenta
sinais nos prótons anoméricos (δ 5.4 e 5.1) para D-galactose e D-manose,
57
respectivamente. Também, foi possível evidenciar a presença de outros
monossacarídeos como fucose e xilose conforme demontrado na FIGURA 13. Na
fucose observou-se forte ressonância do H6 (δ 1.25) e sinais referentes à presença
de xilose (TABELA 5).
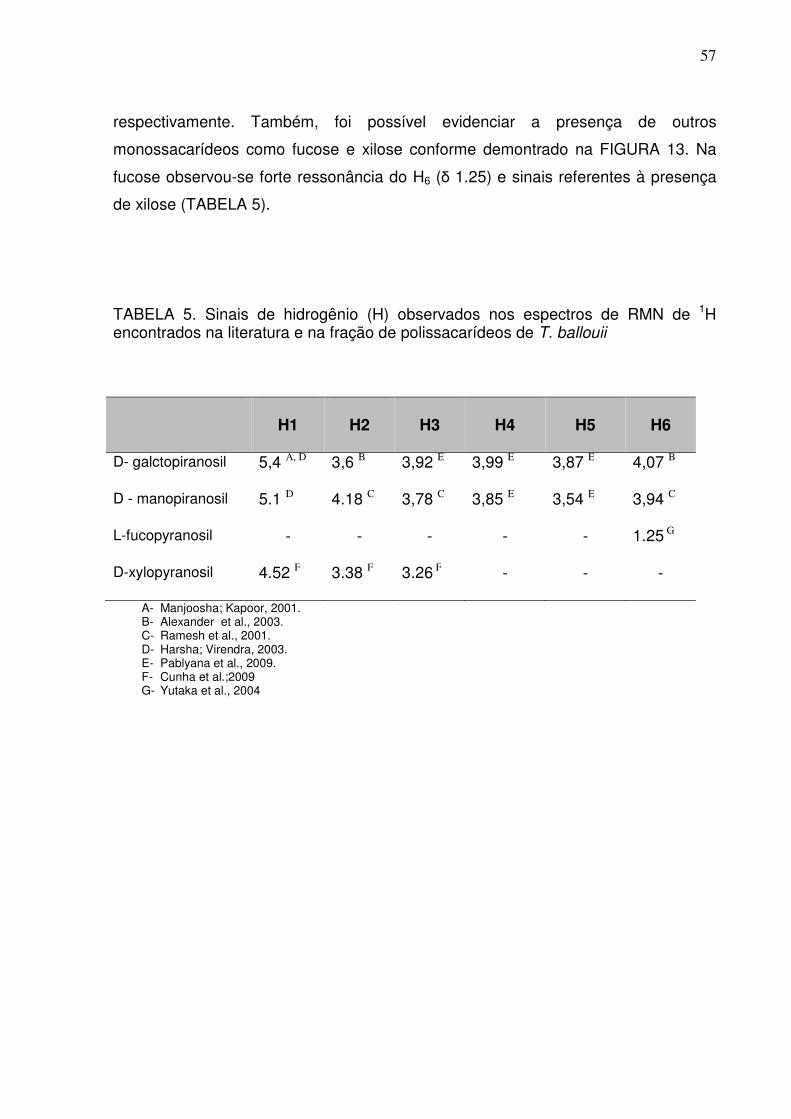
TABELA 5. Sinais de hidrogênio (H) observados nos espectros de RMN de 1H encontrados na literatura e na fração de polissacarídeos de T. ballouii
H1
H2
H3
H4
H5
H6
D- galctopiranosil 5,4 A, D 3,6 B 3,92 E 3,99 E 3,87 E 4,07 B
D - manopiranosil 5.1 D 4.18 C 3,78 C 3,85 E 3,54 E 3,94 C
L-fucopyranosil - - - - - 1.25 G
D-xylopyranosil 4.52 F 3.38 F 3.26 F - - -
A- Manjoosha; Kapoor, 2001. B- Alexander et al., 2003. C- Ramesh et al., 2001. D- Harsha; Virendra, 2003. E- Pablyana et al., 2009. F- Cunha et al.;2009 G- Yutaka et al., 2004
58
FIGURA 13. Espectros de RMN de 1 H obtidos das frações de polissacarídeos de T. ballouii.

59
4.8. Avaliação do efeito do polissacarídeo de Tylopilus ballouii em um modelo de
edema induzido por carragenana
Para se conhecer melhor o processo inflamatório, alguns modelos foram
desenvolvidos na tentativa de reproduzi-lo experimentalmente. O presente trabalho
utilizou como modelo de inflamação o edema de pata induzido por carragenana. A
ação do polissacarídeo rico em galactomanana foi testada nas doses de 30, 50 e 70
mg/kg de peso corporal em diferentes grupos de animais, conforme já descrito. A
FIGURA 14 representa os resultados obtidos deste pré-tratamento em relação aos
controles positivos (carragenana 1%) e controle negativo (solução salina). Esta
demonstra uma discreta redução do edema na 1ª hora para as doses de 30 e 50
mg/kg. Após a 2ª, 3ª e 4ª horas observa-se uma redução de 33% e 55% do volume
da pata, respectivamente. O polissacarídeo rico em galactomanana reduziu 56% do
edema quando aplicado na concentração de 70 mg/kg do peso do animal, logo após
há 1ª hora e na 3ª e 4ª hora, pico da caragenana, houve uma redução de 83% do
edema. Com relação à dose de 70 mg/kg, pode-se inferir que apresentou melhor
efeito antiinflamatório quando comparadas com as demais concentrações mesmo
nas primeiras horas e quando comparada ao grupo controle observa-se uma
redução significativa (p >0,05) na evolução do edema.

60
FIGURA 14. Efeito antiinflamatório do polissacarídeo de T. ballouii no modelo de edema plantar induzido por carragenana. Os dados foram expressos em média ± DP onde *** corresponde a p< 0.001.
Para fins comparativos do efeito antiinflamatório do polissacarídeo em estudo,
o mecanismo de ação deste biopolímero foi analisado com substâncias padrões cujo
mecanismo de ação fosse conhecido e amplamente comprovado. Para tal, foi
administrada a concentração que apresentou melhor efeito inibitório (70 mg/kg) e os
antiinflamatórios não esteroidas (AINES) diclofenaco de sódio e indometacina. Os
resultados observados na FIGURA 15 demonstraram que na 1ª h após aplicação do
polissacarídeo rico em galactomanana já foi possível verificar a redução do edema
de pata. O composto a partir da 2ª h apresentou uma ação similar à indometacina e
o diclofenaco. Na 4ª h a redução do volume do edema foi significante para as
substâncias analisadas.
Os resultados obtidos da avaliação antiinflamatória indicaram que o
polissacarídeo rico em galactomanana apresentou redução no edema induzido por
carragenana. Segundo os testes ANOVA todas as doses testadas com os
polissacarídeos e os grupos tratados com os fármacos indometacina e diclofenaco
de sódio, quando comparados ao controle positivo apresentaram resultados
significativos estatisticamente, principalmente a partir da segunda hora (p< 0.001).

61
FIGURA 15. Efeito antiinflamatório dos polissacarídeos de T. ballouii (70 mg/kg) e antiinflamatórios como diclofenaco de sódio e indometacina no modelo de edema plantar induzido por carragenana. Os dados foram expressos em média ± DP onde * corresponde a p< 0.05; ** p<0.01; ***p<0.001.
4.9. Análises Histológica do edema de pata em ratos induzidos por carragenana
Através de observações microscópicas e da utilização de técnicas de biologia
molecular é possível caracterizar melhor a participação das células sanguíneas e
substâncias químicas envolvidas no processo inflamatório (SOUZA et al., 2003). Nas
análises histológicas do edema de pata em ratos induzidos por carragenana usou-se
controles positivo e negativo (salina) assim como diferentes concentrações do
composto polissacarídico de T. ballouii. No grupo destes animais observou-se um
intenso infiltrado de células polimorfonucleares (Figura 16 A). Por outro lado, foi
observado que as análises das patas dos animais tratados com galactomanas do
citado fungo apresentaram uma diminuição do infiltrado (Figura 16 C, D e E).
*** ***
**
*** *
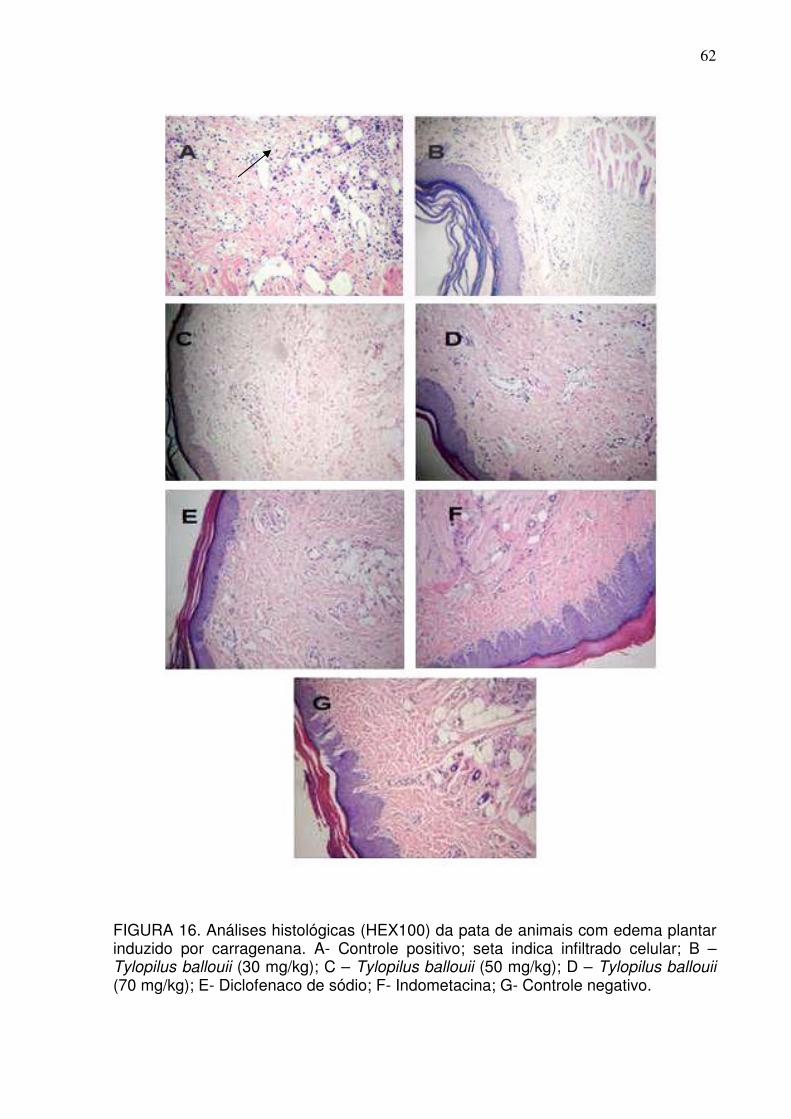
62
FIGURA 16. Análises histológicas (HEX100) da pata de animais com edema plantar induzido por carragenana. A- Controle positivo; seta indica infiltrado celular; B – Tylopilus ballouii (30 mg/kg); C – Tylopilus ballouii (50 mg/kg); D – Tylopilus ballouii (70 mg/kg); E- Diclofenaco de sódio; F- Indometacina; G- Controle negativo.

63
4.10. Determinação da Viabilidade Celular ex vivo
Uma das dificuldades de se estabelecer a ação imunomoduladora de um
fungo é a presença de citotoxicidade em algumas espécies de fungos. Observamos
que os polissacarídeos desse fungo têm ação antiinflamatória e que não
apresentam, neste experimento, citotoxicidade frente a macrófagos peritoneais no
modelo de Viabilidade celular ex vivo (FIGURA 17).
FIGURA 17. Efeito dos polissacarídeos de T. ballouii sobre a viabilidade celular no lavado peritoneal de camundongos Swiss.
4.11. Atividade Antioxidante
4.12. Seqüestro de Radicais Superóxidos
Foi testada a atividade seqüestrante de radicais superóxido do composto
polissacarídico de T. ballouii utilizando o modelo de sistema in vitro, onde o radical
superóxido é gerado - fenazina metassulfato / NADH - com redução do NBT. A
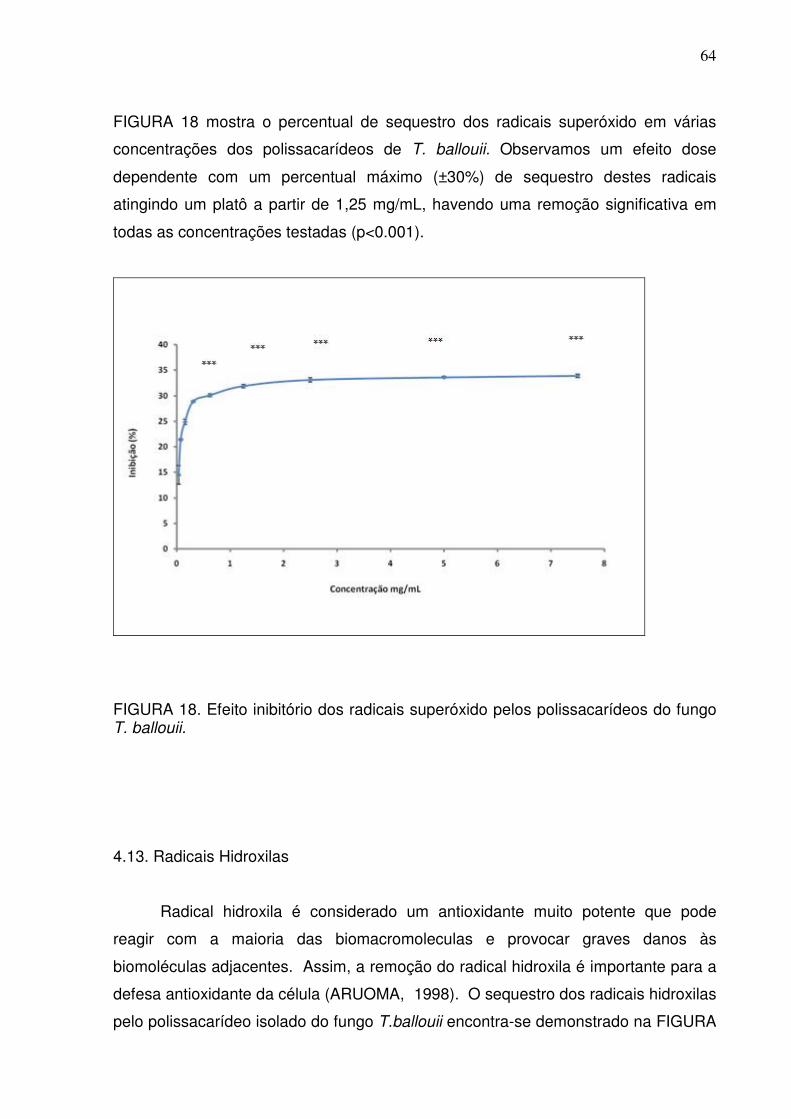
64
FIGURA 18 mostra o percentual de sequestro dos radicais superóxido em várias
concentrações dos polissacarídeos de T. ballouii. Observamos um efeito dose
dependente com um percentual máximo (±30%) de sequestro destes radicais
atingindo um platô a partir de 1,25 mg/mL, havendo uma remoção significativa em
todas as concentrações testadas (p<0.001).
FIGURA 18. Efeito inibitório dos radicais superóxido pelos polissacarídeos do fungo T. ballouii.
4.13. Radicais Hidroxilas
Radical hidroxila é considerado um antioxidante muito potente que pode
reagir com a maioria das biomacromoleculas e provocar graves danos às
biomoléculas adjacentes. Assim, a remoção do radical hidroxila é importante para a
defesa antioxidante da célula (ARUOMA, 1998). O sequestro dos radicais hidroxilas
pelo polissacarídeo isolado do fungo T.ballouii encontra-se demonstrado na FIGURA

65
19. Observamos que este polissacarídeo apresentou no inicio uma atividade mais
discreta e depois mais potencializada na concentração 0, 625 mg/mL, atingindo
inibição máxima em torno de 60% (p<0.001). A capacidade dos polissacarídeos no
seqüestro de radicais hidroxilas foi em concentração dose-dependente.
FIGURA 19. Efeito inibitório dos radicais hidroxilas pelos polissacarídeos do fungo T. ballouii.

66
4.14. Atividade Antioxidante Total
Os polissacarídeos do fungo T. ballouii apresentaram uma atividade
antioxidante total frente à molibidato de amônia com aumento rápido da inibição na
concentração de 0, 039 mg/mL (3,8%) para 0, 078 mg/mL (19,3%). O máximo de
atividade atingida pelos polissacarídeos deste fungo pode ser comparado ao
sequestro realizado por 1,22 mg de ácido ascórbico. Observou-se que em
concentrações a partir de 0, 250 mg/mL de polissacarídeos ocorria uma saturação
do seqüestro dos radicais livres (FIGURA 20).
FIGURA 20. Capacidade antioxidante total dos polissacarídeos do fungo T. ballouii.
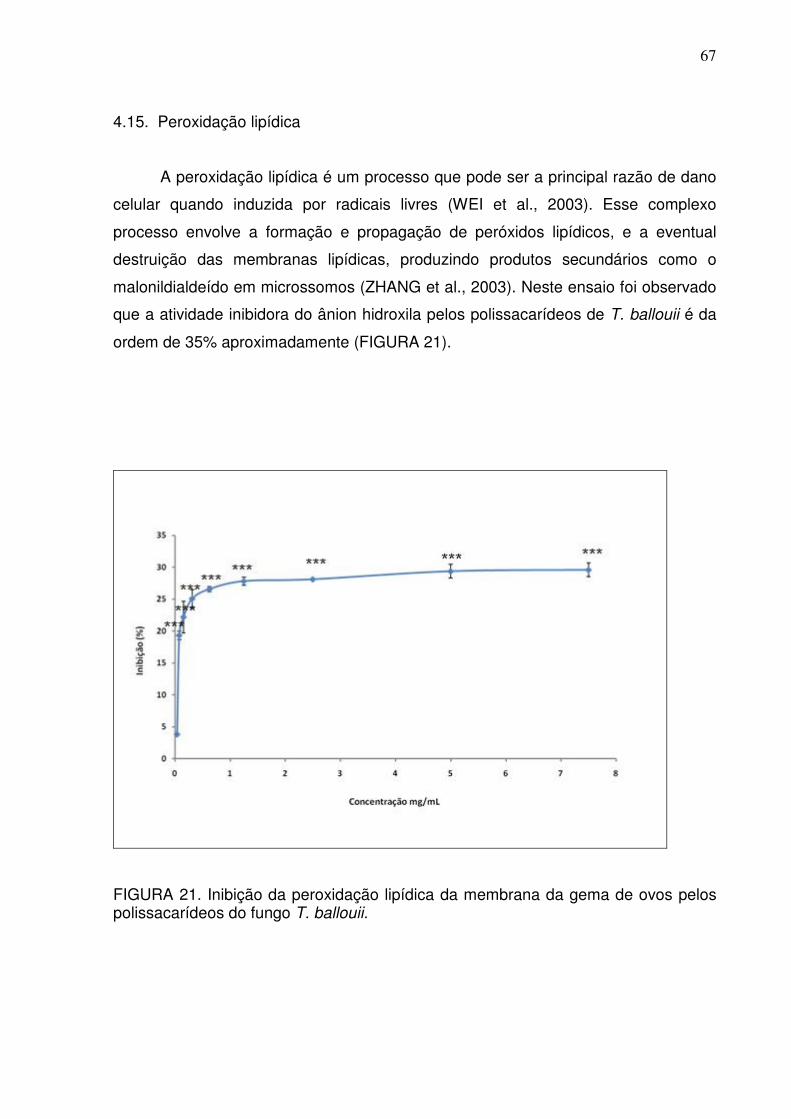
67
4.15. Peroxidação lipídica
A peroxidação lipídica é um processo que pode ser a principal razão de dano
celular quando induzida por radicais livres (WEI et al., 2003). Esse complexo
processo envolve a formação e propagação de peróxidos lipídicos, e a eventual
destruição das membranas lipídicas, produzindo produtos secundários como o
malonildialdeído em microssomos (ZHANG et al., 2003). Neste ensaio foi observado
que a atividade inibidora do ânion hidroxila pelos polissacarídeos de T. ballouii é da
ordem de 35% aproximadamente (FIGURA 21).
FIGURA 21. Inibição da peroxidação lipídica da membrana da gema de ovos pelos polissacarídeos do fungo T. ballouii.

68
4.16. IC50
O IC50 representa a concentração da amostra necessária para proporcionar
50% da atividade inibitória (seqüestro) de radicais livres. Este parâmetro foi
calculado a partir das concentrações dos compostos analisados e da taxa de
inibição da cada amostra. A formação de radicais livres nestes sistemas foi inibida
pelos polissacarídeos do fungo T. ballouii o qual mostrou valores de IC50 de 1,25
mg/ml, 1,64 e 1,70 mg/mL, para os testes de radicais hidroxila, superóxido e
peroxidação lipídica, respectivamente (TABELA 6).
TABELA 6. Valores de IC50 de polissacarídeos de T. ballouii
Tylopilus ballouii
IC50 (mg /ml)
Radicais hidroxilas
1,25
Radicais Superoxido 1,64
Peroxidação Lipídica 1,65

69
5. DISCUSSÃO
Os fungos têm sido utilizados durante milênios devido às suas propriedades
medicinais e nutricionais (FORTES; NOVAES, 2006; TAVEIRA et al, 2007,
FIRENZUOLI et al., 2008). Trabalhos científicos apontando essas propriedades têm
levado a um maior interesse no consumo destes no Brasil (COLAUTO et al., 1998). O
corpo de frutificação do fungo T. ballouii foi utilizado para obtenção dos polissacarídeos
e análise de composição centesimal.
A avaliação da composição centesimal do fungo T. ballouii foi realizada, tendo
em vista que o cogumelo em estudo pertence ao gênero onde são encontradas
algumas espécies comestíveis. Esta avaliação demonstrou que esse fungo é composto
principalmente por carboidratos (32,5%) e proteínas (27,0%). Teores semelhantes
foram publicados por Chang e Milles (1989) para os fungos Agaricus bisporus
(champignon de Paris), Lentinus edodis (shiitake) e Pleurotus ostreatus (shimeji), os
quais contêm como principais constituintes: carboidratos com teor médio para as
espécies analisadas da ordem de 63,17% e proteínas 28%, 19% e 22%,
respectivamente.
A extração deste fungo resultou em um composto cujo teor de carboidratos foi de
80,5%, valores semelhantes foram encontrados por Dore et al (2007) que citou valores
da ordem de 84% para o fungo Geastrum saccatum. O composto extraído do fungo T.
ballouii apresentou também razoável teor de proteínas (19,5%) em sua composição.
Este resultado difere de outros analisados por Queiroz et al (2009) para o fungo Caripia
montagnei que apresentou baixos teores de proteinas (1,3%) e Dore et al (2007) para
os fungos G. saccatum e Polyporos dermoporus que apresentaram teores de proteínas
de 7,7% e 1%, respectivamente. Relatos da literatura descrevem vários polissacarídeos
e complexo polissacarídeo-proteína isolados de fungos sendo usados como fonte de
agentes terapêuticos (CHAKRABORTY et al, 2004).
É de fundamental importância nos estudos de polissacarídeos bioativos a
determinação de sua composição monossacarídica para uma interpretação bem
sucedida das suas atividades biológicas. Dois métodos cromatográficos, portanto, foram
utilizados neste estudo. O primeiro, mais tradicional que avalia de forma qualitativa a

70
cromatografia descendente em papel (CDP), e um segundo, um método de
quantificação, sendo este mais sensível e rápido (HPLC). Os dois métodos mostraram
que os polissacarídeos do fungo estudado são formados principalmente de manose e
galactose, bem como xilose e fucose, com vestígios de arabinose. No entanto, as
análises de HPLC demonstram teores maiores de galactose (26,88%) e manose
(54,2%), sugerindo se tratar de um composto rico em galactomanana.
Polissacarídeos contendo galactose, manose e fucose também foram
caracterizados para Ganoderma applanatum (USUI et al., 1981), Fomitella fraxinea
(CHO et al., 1998), Laetiporus sulphureus (ALQUINI et al., 2004) e Flammulina velutipes
(SMIDERLE et al., 2008).
Polissacarídeos são polímeros de média a alta massa molecular, constituídos
por monossacarídeos conectados por ligações glucosídicas. Esses polímeros
diferenciam-se, principalmente, pelos monômeros constituintes, tipo de ligação,
comprimento da cadeia e número de ramificações (FREIMUND et al., 2003). Análises
feitas por cromatografia em gel filtração utilizando coluna Sephadex G-100 mostraram
que o polímero em estudo apresentou um alto peso molecular (147 KDa).
A espectroscopia de infravermelho (IV) tem sido largamente usada na
caracterização de diversos compostos. Torna-se de fundamental importância não só na
caracterização de alguns grupos funcionais, como nas modificações químicas induzidas
em alguns polímeros. As atribuições das principais absorções do espectro do IV (4000–
400 cm-1) são característicos de estruturas glucosídicas e estão relacionados ao
estiramento OH (3000-3500 cm-1); vibração do grupo anomérico C1H (1070 cm-1)
(GONZAGA et al., 2005), sendo esta mesma banda relacionada a polissacarídeos com
ligações α-(1�6) e α-(1�4), (SHINGEL, 2002). Da mesma forma os padrões de
proteínas também foram evidenciados com absorções característica 1630 e 1400 cm-1
(CAREY, 1992). As bandas relacionadas a proteínas confirmam mais uma vez a sua
presença, o que e possível uma associação entre carboidrato e proteínas.
Outro método espectroscópico que pode ser utilizado na caracterização química
de moléculas é a ressonância magnética nuclear. Para a química de carboidratos se
tornou uma ferramenta largamente utilizada, portanto torna-se imprescindível sua
utilização, principalmente na elucidação estrutural de polissacarídeos com estruturas

71
complexas. Diversos trabalhos na literatura relatam aspectos de galactomananas de
diferentes fontes em relação aos resultados obtidos por RMN, indicando similaridade
nos deslocamentos químicos desses polissacarídeos. Dessa forma, as ressonâncias do
hidrogênio (1H) e do carbono (13C) podem ser atribuídas baseadas em dados
reportados na literatura (JOSHI e KAPOOR, 2003; VIEIRA et al., 2007; CUNHA et al.,
2009). O espectro do hidrogênio para as amostras dissolvidas em água deuterada
confirmou mais uma vez a presença dos monossacarídeos galactose, manose, xilose e
fucose. Em relação aos heteropolissacarídeos foi descrita uma galactomanana
encontrada em Collybia maculata (LIM et al., 2005) e Lasallia pustulata (PEREYRA et
al., 2003).
Polissacarídeos fúngicos vêm sendo muito estudados e relatados na literatura
por possuírem diferentes aplicações medicinais. Dentre esses diversos estudos muitos
estão sendo realizados para verificar o potencial dessas macromoléculas como
antioxidantes, antimutagênicos, anticoagulantes, antiinflamatórios, antitrombóticos e
imunomoduladores (FREIMUND et al., 2003; CORRADI et al., 2006; ZHANG et al.,
2005). Dessa forma, o alvo deste estudo foi caracterizar os polissacarídeos obtidos do
corpo de frutificação do fungo T. ballouii e verificar seu potencial antioxidante e
antiinflamatório. Para avaliar a possível atividade antiinflamatória do composto
polissacarídico do fungo T. ballouii foi utilizado o modelo de edema plantar induzido por
carragenana em ratos Wistar. Segundo Lapa et al (2003) este modelo induz aumento
agudo e progressivo do volume da pata injetada que é proporcional à intensidade da
resposta inflamatória, constituindo um parâmetro útil na avaliação da atividade
antiinflamatória de novos compostos. Este modelo de inflamação envolve a liberação
de vários mediadores que induzem a reação inflamatória em duas fases distintas
(SULEYMAN; BUYUKOKUROGLU, 2001). A fase inicial (0-1h) onde ocorre
principalmente à liberação de histaminas e a fase posterior (1-4h) onde há uma elevada
e destacada produção de prostaglandinas, produtos da cicloxigenase
(PEPIANAYAGAM et al., 2006; SILVA et al., 2005). Na segunda fase a ação das
prostaglandinas é caracterizada principalmente por migração de leucócitos e edema
pronuciado, com pico entre a terceira e quarta hora (RANGEL, 2005).

72
Polissacarídeos obtidos a partir do fungo T. ballouii reduziram o edema
provocado pela injeção s.p de carragenana de forma dose dependente e com perfil de
redução semelhante ao diclofenaco de sódio e indometacina, fármacos comerciais que
atuam sobre a COX (cicloooxigenase). Polissacarídeos obtidos de Ganoderma tsugae,
Ganoderma lucidum e Grifola frondosa apresentaram efeito antiinflamatório
semelhantes reduzindo a migração de leucócitos e a liberação de mediadores pró-
inflamatórios (LIN et al., 2006 HONG et al., 2004; ZHANG et al, 2002, 2003). Para
confirmar os resultados obtidos no modelo de edema plantar induzido por carragenana
bem como verificar o efeito dos polissacarídeos do fungo T. ballouii sobre o infiltrado
celular foram realizadas análises histológicas das patas de ratos que foram submetidos
à indução por carragenana.
As análises histológicas também exibiram um perfil de dose-dependência na
ação dos polissacarídeos em questão sobre o recrutamento de leucócitos. A melhor
concentração utilizada para inibição da inflamação foi a de 70 mg/kg por peso corporal.
Polissacarídeos de T. felleus testados pelo modelo de edema de pata induzido por
carragenana apresentam uma inibição semelhante da inflamação em doses superiores
a 50 mg / kg (KOHLMUNZER et al., 1977). É possível que a ação antiinflamatória desse
polímero seja atribuída à ação de sequestro sobre os radicais livres e sobre a COX
(ciclooxigenase) ou à inibição de fatores como citocinas que pode mediar à lesão
tecidual relacionada à resposta inflamatória (GORDON, 2001; HERMANN et al., 2001).
Técnicas de coloração são usualmente utilizadas para avaliar a citotoxicidade de
substâncias. Neste trabalho foi utilizado o teste de exclusão com o azul de tripan, que é
um corante que só penetra nas células mortas, cujas membranas não podem mais
excluí-lo. Este fenômeno de permeabilidade da membrana celular permite estimar
indiretamente o grau de integridade da mesma (WALUM et al., 1990). Através desta
técnica foi observado que o composto polissacarídico do fungo T. ballouii não
apresentou ação citotóxica no período de 4 h e 24 h. Os efeitos adversos dos fungos
com propriedades medicinais não são descritos detalhadamente na literatura, mas
estudos in vivo comprovam inexistência de toxicidade quando utilizados em diferentes
concentrações (KUROIWA et al, 2005; NOVAES et al, 2005).

73
Vários compostos fúngicos já foram descritos na literatura por possuírem
atividade antioxidante (BARROS et al., 2008; JAYAKUMAR et al., 2006; CHEUNG;
CHEUNG, 2005). Entretanto, a maioria dos antioxidantes utilizados ainda são os
sintéticos, os quais têm sido suspeitos de ser responsável por danos no fígado e
carcinogênese (GRICE, 1988; QI et al., 2005). Assim, pesquisas direcionadas para a
utilização de eficazes antioxidantes naturais são essenciais para que possam proteger
o corpo humano dos radicais livres e retardar o progresso de muitas doenças crônicas
(LUO, 2008; NANDITA; RAJINI, 2004).
O composto polissacarídico extraído de T. ballouii demonstrou capacidade de
sequestrar radicais livres nas metodologias testadas, principalmente no seqüestro do
radical hidroxila onde atingiu o IC50 com 1,25 mg/ mL. Compostos polissacarídicos
derivados de diferentes espécies de fungos já foram descritas na literatura por
apresentarem potencial atividade “sequestradora” de radicais superóxido e hidroxila
(LIU; OOI; CHANG, 1997). A metodologia realizada para peroxidação lipídica envolveu
a observação de produtos secundários a partir da gema do ovo no qual a reação com o
tiobarbitúrico produz um composto de coloração rosa, que tem forte absorção a 532 nm
(WEI et al., 2003).
Comparado com outros sistemas geradores de radicais realizados neste
trabalho, o seqüestro de peróxidos lipídico pelo composto polissacarídico do fungo T.
ballouii foi o que apresentou menor efeito antioxidante. Polissacarídeos derivados de
cogumelos podem ser efetivos na inibição de radicais superóxidos e hidroxilas,
entretanto, não exibir efeito no processo de peroxidação lipídica (LIU et al., 1997). Por
outro lado, Sun et al (2009) conseguiram inibir em cerca de 64% a peroxidação lipídica
em gema de ovo.

74
6. CONCLUSÕES
� A composição centesimal do tecido do fungo Tylopilus ballouii mostrou um teor
de 32,5% de carboidratos, seguido de 27,0% de proteínas;
� As análises químicas, a espectrofotometria de infravermelho e a composição
monossacarídica demonstraram que os polissacarídeos obtidos do fungo são
compostos, principalmente, por galactomananas associado a porções protéicas,
comuns em fungos;
� No modelo de inflamação testado, o edema plantar induzido por carragenana, os
polissacarídeos de Tylopilus ballouii mostrou um efeito antiinflamatório significativo na
concentração de 70 mg/Kg;
� O perfil de redução do edema dos polissacarídeos de Tylopilus ballouii é
semelhante à indometacina e ao diclofenaco sódico, dois fármacos antiinflamatórios,
indicando que esses polissacarídeos em estudo podem possuir atividade
antiinflamatória por inibir a COX;
� As combinações das análises realizadas sugerem que este fungo apresenta um
potencial efeito antiinflamatório e antioxidante podendo assim contribuir para a
prevenção ou redução do desenvolvimento de patologias;
� Os ensaios de viabilidade celular mostraram que os polissacarídeos do fungo
Tylopilus ballouii não promovem citotoxicidade significativa no período de 4 e 24h
após administração. Estas características são bastante relevantes, portanto mais
estudos precisam ser realizados para que este fungo possa ser validado
cientificamente, podendo assim apresentar uma maior utilidade pela população tanto
na área de nutricêuticos com farmacológica.

ADAMS, D. Fungal cell wall chitinases and glucanases.Microbiology, Reading, 150(7): 2029-2035, 2004. ADEREM, A.; ULEVITCH, R. J. Toll-like receptors in the induction of the innate immune response. Nature, v. 406, p. 782-787, 2000. ALEXANDER, S.; SHASHKOV, A., GALINA, M.; STRESHINSKAYA, B.; LARISA, N.; KOSMACHEVSKAYA, B.; SOF’YA, N.; SENCHENKOVA, A.; LYUDMILA, I.; EVTUSHENKO, C.; IRINA, B.; NAUMOVA, B. NMR-based identification of cell wall galactomannan of Streptomyces sp. VKM Ac-2125. Carbohydrate Research, 338 2021-/2024 ALEXOPOULOS, C.J.; MIMS, C. W.; BLACKWELL, M. Introductory mycology, 4ed. John Wiley & Sons, New York, 1996. ALLER, M. A.; ARIAS, J.L; ARIAS, J.I.; SÁNCHEZ-PATÁN, F.; ARIAS, J. The inflammatory response: An efficient way of life. Medical Science Monitor, v. 12 (10), p. 225 – 234, 2006. ALQUINI, G.; CARBONERO, E. R.; ROSADO, F. R.; COSENTINO, C.; IACOMINI, M. Polysaccharides from the fruit bodies of the basidiomycete Laetiporus sulphureus (Bull.: Fr.) Murr. FEMS Microbiology Letters, 230, 47-52, 2004. ANDERSON, D. Antioxidant defences against reactive oxygen species causing genetic and other damage. Mutation Research, Amsterdam, v.350, n.1, p.103-108, 1996. ANDRADE, C.T.; AZERO, E. G.; LUCIANO, L.; GONÇALVES, M. P. Solution properties of the galactomannans Extracted from the seeds of Caesalpinia pulcherrima and Cassia javanica: comparison with locust bean gum. International Journal of Biological Macromolecules, 26: 181-185, 1999. A.O.A.C. ASSOCIATION OF OFFICIAL ANALITICAL CHEMISTS. Official methods of analysis, 14ed. Washington, p988-1141, 1984. BANERJEE, A.; DASGUPTA, N.; DE, B. In vitro study of antioxidant activity os Syzygium cumini fruit. Food Chemistry, vol. 20, p. 727-733, 2005. BARBOSA, A. M; CUNHA, P. D. T.; PIGATTO, M. M.; CORRADI da SILVA, M. L. Produção e Aplicações de Exopolissacarídeos Fúngicos. Semina: Ciências Exatas e Tecnológicas, Londrina, 25(1): 29-42, 2004. BASEIA, I. G.; ADAUTO, I. M.. Geastrum setiferum (Gasteromycetes): a new species with a setose endoperidium. Mycotaxon 84:135-139, 2002.

BELEM, M. A. F.; LEE, B. H. Production of bioingredients from Kluyveromyces marxianus grown on whey: an alternative. Critical Reviews in Food Science and Nutrition, 38(7): 565-598, 1998. BEROVI Č, M.; HABIJANI Č, J.; ZORE , I.; WRABER ,B.; HOD ŽAR , D.; BOH , B.; POHLEVEN , F. Submerged cultivation of Ganoderma lucidum biomass andimmunostimulatory effects of fungal polysaccharides. Journal of Biotechnology, 103(1): 77-86, 2003. BLIGH, E.G.; DYER, W.J.A. A rapid method of total lipid extraction and purification. Can J. Biochem. Physiol., 37: 911-917, 1959. BOHN, J. A.; BE MILLER, J. N. (1→3)-β-D-Glucans as biological response modifers: a review of structure-functional activity relationships. Carbohydrate Polymers, 28(1): 3-14, 1995. BRADFORD, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem., 7(72):248-54, 1976. BROWN, G. D.; GORDON, S. A. New receptor for β-glucans. Nature, 413: 36–37,2001 BROW, G.D.; TAYLOR, P.R.; REID, D. M.; WILLMENT, J.A.; WILLIAMS, D.L.; MARTINEZ-POMARES, L.; WONG, S.Y.C.; GORDON, S. Dectin-1 is a major β-glucan receptor on macrophages. J. Exp. Med, 2002. BROWN, G.D.; GORDON, S. Fungal β-glucans and mammalian imunity. Immunity, 19: 311- 315, 2003. BRUNE, K., HINZ, B. Selective cyclooxygenase-2 inhibitors: similarities and differences, Scand Journal of Rheumatology, 33, p. 1-6, 2004. BRUNETON, J. Elementos de fitoquímica e farmacognosia. Espanha: Acribia, 1991. BUCKERIDGE, M.S.; DIETRICH, S.M.C. Mobilisation of the raffinose family oligosaccharides and galactomannan in germinating seeds of Sesbania marginata Benth. (Leguminosae-Faboideae). Plant Science, 117(1): 33-43, 1996. BUCKERIDGE, M.S.; DIETRICH, S.M.C. Galactomannan from Brazilian native legume seeds. Revista Brasileira de Botânica,13(2):109-112, 1990.

CABIB, E.; ROH, D. H.; SCHMIDT, M.; CROTTI, L. B.; VARMA, A. The Yeast Cell Wall and Septum as Paradigms of Cell Growth and Morphogenesis. Journal of Biological Chemistry, 276(23): 19679-19682, 2001. CAMERON, D. R.; COOPER, D. G.; NEUFELD, R. J. The mannoprotein of Saccharomyces cerevisiae is effective bioemulsifier. Applied and Environmental Microbiology, 54(6): 1420-1425, 1988 CARBONERO , A. R.; TISCHER , C.A.; COSENTINO ,C.; GORIN , P. A. J.; IACOMINI , M. Structural characterization of a galactomannan from the cyanolichen Leptogium azureum. Carbohydrate Polymers, 53(4): 469–473, 2003. CARBONERO , A. R.; MONTAI , A. V.; MELINGER ,C. G.; ELIASARO , S.; SASAKI , G. L.; GORIN , P. A. J.; IACOMINI , M. Glucans of lichenized fungi:significance for taxonomy of the genera Parmotrema and Rimelia. Phytochemistry, 66(8): 929- 934, 2005. CASTRO, L.; FREEMAN, B. A. Reactive oxygen species in human health and disease. Nutrition, 17:161,163–1, 165, 2001. Carey, P. R.. (1992). Biochemcal application of Roman and resonance Raman spectroscopies. New York: Academic Press. CAREY, P. R... Biochemcal application of Roman and resonance Raman spectroscopies. Academic Press, New York, 1992 CHAKRABORTY, I.; MONDAL, S.; PRAMANIK, M.; ISLAM, D. R. Structural investigação of a water soluble glucan from an a edible mushroom, Astraeus hygrometricus. Carboydrate Research. 339 2249-2254, 2004 CHANDRASEKHARAN, N.V.; SIMMONS, D. L. The cyclooxygenases. Genome Biology, London, v.5, n.241, 2004. CHANG, S. T.; MILLES, P. G. Edible mushrooms and their cultivation. boca raton: crc press, p. 345, 1989. CHEN, J.; SERVIOUR, R. Medicinal importance of fungal beta (1-->3), (1-->6)-glucans . Mycol Res. v.111, n.6, p.635-652, 2007.
CHO, S. M.; KOSHINO, H.; YU, S. H.; YOO, I. D. A mannofucogalactan, fomitellan A,with mitogenic effect from fruit bodies of Fomitella fraxinea (Imaz.). Carbohydrate Polymers, 37, 13-18, 1998 CHO, J.H.; CHO, S.D.; HU, H.; KIM, S.H.; LEE, S.K.; LEE, Y.S.; KANG, K.S. The roles of ERK 1/2 and p38 MAP kinases in the prevention mechanism of mushroom Phellinus linteus against the inhibiton of gap junctional intercellular commuinication by hydrogen peroxide. Carcinogenesis, 23: 1164- 1169; 2002.

COLAUTO, N. B.; EIRA, A. F.; MINHONI, M. T. A. Fatores físicos que afetam a produtividade do cogumelo comestível Pleurotus sajor-caju (Fr.) Singer. Científica, São Paulo, v. 26, p. 25-43, 1998. CORRADI DA SILVA, M. L.; MARTINEZ, P. F.; IZELI, N. L.; SILVA, I. R.; VASCONCELOS, A. F. D.; CARDOSO, M. S. Caracterização química de glucanas fúngicas e suas aplicações biotecnológicas. Química Nova, 29(1): 85-92, 2006. CRAY, C.; WATSON, T.; RODRIGUEZ, M.; ARHEART, K. L. Application of Galactomannan Analysis and Protein Electrophoresis in the Diagnosis of Aspergillosis in Avian Species. Journal of Zoo and Wildlife Medicine, 40(1): 64-70, 2009. CUNHA, P.L.R.; VIEIRA, Í.G.P.; ARRIAGA, Â.M.C.; DE PAULA, R.C.M.; FEITOSA, J.P.A. Isolation and characterization of galactomannan from Dimorphandra gardneriana Tul. seeds as a potential guar gum substitute, Food Hydrocolloids, 23: 880-885, 2009. CUI, Y.; KIM, D.S.; PARK, K.C. Antioxidant effect of Inonotus obliquus. Journal of Ethno Pharmacology,p. 96: 79-85, 2005. DAAS, P.J. H.; SCHOLS, H.A.; DE JONGH, H. H. On the galactosyl distribution of commercial galactomannans. Carbohydrate Research, 329: 609–619, 2000. DANTAS,J. C. M.; SILVA, F. R. F.; ROCHA, H. A. O.; BASEIA, I. G.; LEITE, E. L. Antiinflammatory, antioxidant and cytotoxic actions of b-glucan-rich extract from Geastrum saccatum mushroom. International Immunopharmacology, 7, 1160- 1169, 2007. DORE, C. M. P. G.; AZEVEDO, T. C. G.; SOUZA, M. C. R.; REGO, L. A.; DEGÁSPARI, C.H., WASZCZYNSKYJ, N. Propriedades antioxidantes de compostos fenólicos. Visão Acadêmica, Curitiba, v. 5, n. 1, p. 33-40, 2004. DROGE W. Free radicals in the physiological control of cell function Physiol Rev, 82:47—95, 2002. DUBOIS, M.; GILLES, K. A.; HAMILTON, J. K.; REBERS, P. A. & SIMTH, F. Colorimetric method for determination of sugars, and related substances. Anal. Chem., 28: 350-356, 1956 ELMASTAS, M.; ISILDAK, O.; TURKEKUL, I.; TEMUR, N.; J. Food Compos. Anal, p. 20- 345, 2007. FIRENZUOLI, F.; GORI, L.; LOMBARDO, G. The medicinal mushroom Agaricus blazei Murrill: review of literature and pharmaco-toxicological problems. Evid. Based. Complement. Alternat. Med. v.5, n.1, p.3-15, 2008.

FORTES, R.C.; NOVAES, M.R.C.G. Efeitos da suplementação dietética com cogumelos Agaricales e outros fungos medicinais na terapia contra o câncer. Rev. Bras. Cancerol. v.52, n.4, p.363-371, 2006. FREIMUND, S.; SAUTER, M.; KÄPPELI, O.; DUTLER H. A new non-degrading isolation process for 1,3--D-glucan of high purity from baker´s yeast Saccharomyces cerevisiae. Carbohydrate Polymers, 54, 159-171, 2003. GIACHINI A. J.; OLIVEIRA, V. L.; CASTELLANO, M. A.; RAPPE, J. M .T. Ectomycorrhizal fungi in Eucalyptus and Pinus plantations in Southern Brazil. Mycologia 92(6): 1166-1177, 2000. GORIN, P. A. J., BARRETO-BERGTER, E. M. The chemistry of polysaccharides of fungi and lichens. In G. O. Aspinall (Ed.), The polysaccharides, 2: 365–409, 1983. GONZAGA, M. L. C.; RICARDO, N. M. P. S.; HEATLEY, F. SOARES, S. A. Isolation and characterization of polysaccharides from Agaricus blazei Murill. Carbohydrate Polymers. v. 60, p.43-49, 2005. GRACE, P.A. Ischaemia-reperfusion injury. Br. J. Surg, v. 81, p. 637–647, 1994. GUTTERIDGE JMC, MITCHELL J. Redox imbalance in the critically ill. Br Med Bull, 55(1):49—75, 1999. HADDAD, J.J. Pharmaco-redox regulation of cytokine-related pathways: from receptor signaling to pharmacogenomics. Free Rad. Biol. Med, p. 33: 907-926, 2002. HALLIWELL, B.; GUTTERIDGE, J. C. Free Radicals in Biology and Medicine. 3.ed. Oxford, New York, 2000. HALLING , R.E .; MUELLER, G. M. New boletes from Costa Rica.Mycologia 91: 893-899, 1999. HALLING, R.E.; MUELLER, G. M. Agarics and boletes of Neotropical oakwoods, p. 1-10. In: R WATLING, J. C.; FRANKLAND, A.M.; AINSWOR T.H. S.; ISAAC; ROBINSON, C.H.(eds.). Tropical mycology. United Kingdom: CABI Publishing, 2002. HALLING, R. A Synopsis of Colombian Boletes. Mycotaxon 34:93 – 113, 1989. HALLING ,R.E.; FRANCO-MOLANO, E. Agaricales from Costa Rica:New taxa with ornamented spores. Mycologia 88:666-670, 1996.

HARSHA, J.; VIRENDRA, P. K. Cassia grandis Linn. f. seed galactomannan: structural and crystallographical studies. Carbohydrate Research, 338 19071912, 2003. HAVSTEEN, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol Ther, p. 96: 67-202, 2002. HEINEMANN P. Flore iconographique des champignons du Congo. In: Fascicle 3. Boletineae. Brussels: Le jardin Botanique de 1'Etat. p 51-78, 1954. HENKEL, T. W. New taxa and distribution records for Tylopilus from Dicymbe forest of Guyana. Mycologia 91: 655-665, 1999. HENKEL, T. W. Tylopilus pakaraimensis, a new species of Tylopilus section Potamogetones from Guyana. Mycotaxon 78: 105-1 14, 2001. HYDE, K.D. Biodiversity of tropical microfungi. Hong Kong, University Press, Hong Kong, 1997. JOSHI, H.; KAPOOR, V.P. Cassia grandis Linn. f. seed galactomannan: structural and crystallographical studies. Carbohydrate Research, v.338, p.1907–1912, 2003. JUNQUEIRA, L.C ; CARNEIRO, José. Histologia Básica. 8ªed. Rio de Janeiro: Guanabara Koogan S.A.,1995 KAISER, C. R. RMN 2D: Detecção inversa e gradientes de campo na composição estrutura de compostos orgânicos. Química Nova, v.23, p.231-236, 2000. KIM, S. W.; XU, C. P.; HWANG, H. J.; CHOI, J. W.; KIM, C. W.; YUN, J.W. Production and characterization of exopolyasaccharides from an entomopathogenic fungus Cordyceps militaris Ng3. Biotechnology Progress, 19( 2): 428-435, 2003. KIRK, P.M.; CANNON, P.F.; DAVID, J.C.; STALPERS, J.A. Ainsworth & Bisby´s Dictionary of the Fungi. 9 th ed.CABI Bioscience, Egham, 2001. KOHEN R, NYSKA A. Oxidation of biological systems: oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicol Pathol, 30(6):620—50, 2002. KOHLMUNZER S, QUINN ML, BENOIT PS, FARNSWORTH N.R. Antiinflammatory activity of Tylopilus felleus (Bull. ex Fr.) P. Karst. Pol J Pharmacol Pharm. 1977 29(5):539-41. KOLLÁR, R.; STURDIK, E.; SAJBIDOR, J. Complete fractionation of Saccharomyces cerevisiae biomass. Food Biothecnology, 6(3): 225-237, 1992.

KRCMAR, P.; NOVOTNY, C.; MARAIAS, M. F.; JOSELEAU, J. P. Structure of extracellular polysaccharide produced by lignin-degrading fungus Phlebia radiate in liquid culture. International Journal of Biologica Macromolecules, 24(1): 61-64, 1999. KUMAR, V.; ABBAS, A. K.; FAUSTO, N. ROBBINS , C. Bases patológicas das doenças. 7ed. Rio de Janeiro: Elsevier, p. 49 – 79, 2005. LEE, J. N.; LEE, D. Y.; JI, I. H.; KIM, G. E.; KIM, H. N.; SOHN, J.; KIM, S.; KIM, C. W. Purifcation of soluble β-glucan with immune-enhancing activity from the cell wall of yeast. Bioscience Biotechnology and Biochemistry, 65(4): 837-841, 2001. LEUNG, M. Y. K.; LIUB, C.; KOON, J. C. M.; FUNG, K. P. Polysaccharide biological response modifers. Immunology Letters, 105(2): 101–114, 2006. LIM, M. J.; JOO, J. H.; KIM, H. O.; KIM, H. M.; KIM, S. W.; HWANG, H. .J.; YUN, J. W. Structural analysis and molecular characterization of exopolysaccharides 76 produced by submerged mycelial culture of Collybia maculata TG-1. Carbohydr. Polym, 61: 296-303, 2005. LIMA, D.U. Galactomannanans as the reserve carbohydrate in legume seeds. In: GUPTA, A.K.; KAUR, N. Carbohydrate reserves in plants: synthesis and regulation. Amsterdam: p.283-316, Elsevier, 2000. LIPKE, P. N.; OVALLE, R. Cell wall architecture in yeast: new structure and new chalenges. Journal of Bacteriology, 180(15): 3735-3740, 1998. LIU, F., OOI, V.C.E., CHANG, S.T. Free radical scavenging activities of mushroom polysaccharide extracts. Life Sciences, 60(10): 763-771, 1997. LIU, W.K.; NG, T.B.; SZE, S.F.; & TSUI, K.W. Activation of peritoneal macrophage by polysaccharides from the mushroom Coriolus versicolor. Immunopharmacology, 26: 139-146, 1993. LUSTER, A.D., ALON, R., VON ANDRIAN, U.H. Immune cell migration in inflammation: present and future therapeutic targets. Nature Immunology, p.6, 1182–1190, 2005. MADIGAN, M. T.; MARTINKO, J. M.; PARKER, J. Brock Biology of Microorganisms. New Jersey: Prentice Hall International, Inc. p. 774, 1997. MAGNELLI, P.; CIPOLLO, E.J.; ROBBINS, F. P. W. Glucanase-driven fractioation allows redefnition of Shizoccharomyces pombe cell wall compositon and structure:assigment of diglucan. Analytical Biochemistry, 336(2): 202-212, 2005

MANJOOSHA, C, VIRENDRA, P. K. Structure of a galactomannan from the seeds of Cassia angustifolia Vahl. Carbohydrate Research, 332 439–444, 2001.
MANSON, M. Cancer prevention The potencial for diet to modulate molecular signaling. Trends in Molecular Medicine, v.9, p. 11-18, 2003. MANZI, P.; GAMBELLI, L.; MARCONI, S.; VIVANTI, V.; PIZZOFERRATO, L. Nutrients in edible mushrooms: an inter-species comparative study. Food Chemistry, 65: 477-482, 1999. . MARRONI, N. P., MARRONI, C. A. Estresse Oxidativo e Antioxidante. Porto Alegre: Editora Ulbra, p.33-48, 2002. MAZIERO, R.; CAVAZZONI, V.; BONONI, V. L. R. Screening of Basidiomycetes for the production of exopolysaccharide and biomass in submerged culture. Revista de Microbiologia, 30: 77-84, 1999. MCCORD JM. The evolution of free radicals and oxidative stress. Am J Med, p. 9-108-652, 2000 . MELITO, F.C. COGUMELO DO SOL. In: http://educar.sc.usp.br/licenciatura/1999/cogumelo.html; 1999. MITCHELL, R. N.; KUMAR, U.; ABBAS, A. K; FAUSTO, N. Fundamentos de patologia da doenças. 7 ed. Rio de janeiro: Elsevier, p.800, 2006. MORADALI, M.F.; MOSTAFAVI, H.; GHODS, S.; HEDJAROUDE, G. Immunomodulating and anticancer agents in the realm of macromycetes fungi. Int Immunopharmacol. v.7, n.6, p.701-724, 2007.
MORETTO, E. FETT R.; GONZAGA, L.V.; KUSKOSKI, E.M. Introdução à ciência de alimentos. Editora da UFSC, 255p., 2002. MUZARELI , R. A. A.; MILIANI , M.; CARTOLARI, M.; TARSI , R.; TOSI , G.; MUZARELI, C. Polyuronans obtained by regiospecific oxidation of polysaccharides from Aspergillus niger, Trichoderma reesei and Saprolegnia sp. Carbohydrate Polymers, 43(1): 55-61, 2000. NAKAMURA, S., OGAWA, M.; NAKAI, S.; KATO, A.; KITTS, D. D. Antioxidant activity of a Maillard-type phosvitin-galactomannan conjugate with emulsifying properties and heat stability. Journal of agricultural and food chemistry, 46(10): 3958-3963, 1998. NATHAN, C. Points of control in inflammation. Nature insight review articles, v. 420 (19/26), p. 846 – 852, 2002.

NELSON, D. L.; COX, M. M. Lehninger, princípios de bioquímica. 3. ed. São Paulo: Sarvier, 2002. NIJVELDT, J.R. Flavonoids: a review of probable mechanisms of action and potential applications. Am J Clin Nutr, v. 74, p. 418–425, 2001. OKA, M.; HAZAMA,S.; SUZUKI, M.; WANG, F.; WADAMORI, K.; IIZUKA, N.; TAKEDA, S.; AKITOMI, Y.; OHBA, Y.; KAJIWARA, K.; SUGA, T.; SUZUKI, T. In vitro and in vivo analyses of human leucocyte binding the antitumor polysaccharide lentinan. Int. J. Immunopharmacology. v.18, n.3, p.211-216, 1996.
PABLYANA, L. R.; CUNHA, A.; I´CARO, G. P.; VIEIRA, B.; AˆNGELA, M. C.; ARRIAGA, A. R. C. M..; JUDITH, P. A.; FEITOSA, P. A. Isolation and characterization of galactomannan from Dimorphandra gardneriana Tul. seeds as a potential guar gum substitute. Food Hydrocolloids, 23 880–885, 2009 PAVLOVA, K.; PANCHEV, I.; HRISTOZOVA , T. S. Physico-chemical characterization of exomannan from Rhodotorula acheniorum MC. World Journal of Microbiology and Biotechnology, 21(3): 279-283, 2005. PELOSI, L.; IMAI, T., CHANZY, H.; HEUX, L.; BUHLER, E.; BULONE, V. Structural and morphological diversity of (1→3)-β-D-glucans synthesized in vitro by enzymes from Saprolegnia monoïca: comparison with a corresponding in vitro product from blackberry (Rubus fruticosus). Biochemistry, 42: 6274-6274, 2003. PEREYRA, M. T.; PRIETO, A.; BERNABÉ, M.; LEAL, J. A. Studies of new polysaccharide from Lasallia pustulata (L.) Hoffm. The Lichenologist, v. 35, p. 177-185, 2003. PERIANAYAGAM, J. B.; SHARMA, S. K.; PILLAI, K.K. Anti-inflammatory activity of trichodesma indicum root extract in experimental animals. J. of Ethnopharmacology. v.104, p.410-414, 2006. PESSONI, R. A. B.; FRESHOUR, G.; FIGUEIREDO, R. L.; HAHN, M. G.; BRAGA, M. R. Cellwall structure and composition of Penicillium janczewskii as affected inulin. Mycologia, 97(2): 304-311, 2005. PHILLIPS, H.J. Dye exclusion test for viability. In: KRUSE Jr, P.F.; PATTERSON, Jr.M.H. Tissue Culture: methods and application. New York, Academic Press, p. 406-408, 1973. PIRES, L.; GORIN, P. A. J.; REICHER, F.; SIERAKOWSKI, M. R. An active heparinoid obtained by sulphation of a galactomannan extracted from the endosperm of Senna macranthera seeds. Carbohydrate Polymers, 46(2):165-169, 2001.

POUTSIAKA D. D. ; MENGOZZI M. ; VANNIER E. ; BHANU SINHA ; DINARELLO C. A. ;. Cross-Linking of the β-glucan receptor on human monocytes results in interleukin-1 receptor antagonist but not interleukin-1 production. Blood. v.82, n.12, p.3695-3700, 2003.
PRIETO, P.; PINEDA, M.; AGUILAR, M. Spectrophotometric Quantitation of Antioxidant Capacity through the Formation of a Phosphomolybdenum Complex: Specific Application to the Determination of Vitamin E1. Analytical Biochemistry, 269: 337–341, 1999. PUTZKE, J. ; PEREIRA, A. B. Os fungos da família Boletaceae conhecidos do Rio Grande do Sul (Fungi, Basidiomycota).Caderno de Pesquisa, Série Botânica 6 (1):75-100, 1994. RAMESH, H. P.; YAMAKI, K.; ONO, H.; TSUSHIDA, T. Two-dimensional NMR spectroscopic studies of fenugreek (Trigonella foenum-graecum L.) galactomannan without chemical fragmentation. Carbohydrate Polymers 45 69-77, 2001. RANGEL, E. T. Triagem antinflamatória dos extratos brutos de Echinodorus macrophyllus Michell (chapéu-de-couro) e estudo farmacológico antinflamatório pré-clinico do extrato hidroalcoolico. 2005. 130f. Dissertação (mestrado em ciências da saúde) – Universidade Federal de Mato Grosso, Cuiabá, Mato grosso. RAVEN, P. H.; EVERT, R. F.; EICHHORN, S. E. Biologia Vegetal, 6.ed. Rio de Janeiro. Guanabara Koogan, 2001. REYNOLDS, J.E.F. Guar gun. In: REYNOLDS, J.F.E. Martindale : the Extra pharmacopeia. 29. Ed., London: The Pharmaceutical Press.p.391, 1989. RICK, J. Contributio ad monographiam Agaricearum Brasiliensium I. Broteria 6: 65-92, 1907. RICK, J. Basidiomycetes eubasidii in Rio Grande do Sul - Brasilia.4. Meruliaceae, Polyporaceae, Boletaceae. Iheringia série Botânica 7: 193-295, 1960. RICHARDSON, D .M.; RUNDE, P. W. Ecology and biogeography of Pinus. In: Ecology and Biogeography of Pinus (ed. D.M. Richardson). Cambridge University Press, 1998. ROSADO, F. R. et al. Biomass and exopolysaccharide production in submerged cultures Pleurotus ostreatoroseus sing and Pleurotus ostreatus florida (Jack: Fr) kummer. Journal Basic Microbiol, 43(3): 230-237, 2003.

ROSS, G.D.; VETVICKA, V.; YAN, J.; XIA, Y.; VETVICKOVA, J. Therapeutic intervention with complement and beta-glucan in cancer. Immunopharmacology, 42: 61-74, 1999. ROSSMAN, A.Y. Biodiversity of tropical microfungi: an overview. Pp-1-10. In: K.D. Hyde (Ed.). Biodiversity of Tropical Microfungi. University Press, Hong Kong, 1997. SANTOS, L.C.; HONDA, N.K.; CARLOS, I.Z.; VILEGAS, W. Intermediate reactive oxygen and nitrogen from macrophages induced by Brazilian lichens. Fitoterapia, 75: 473-479, 2004. SCHMID, F.; SEPAROVIC, F.; RESCK, I. S.; MCDOUGALL, B. M.; STONE, B. A.; BROWNLEE, R. T. C. Characterisation of the extracellular polysaccharides produced by isolates of the fungus Acremonium. Carbohydrate Research, 342, 2481–2483, 2007. SHARON, N.; LIS, H. Carbohydrates in cell recognition. Sci Am Jan:74–81,1993. SHERWOOD, E. R.; TOLIVER-KINSKY, T. Mechanisms of the inflammatory response. Best Practice & Research Clinical Anaesthesiology, v.18, p. 385–405, 2004. SHINGUEL, K. I. Determination of structural peculiarities of dexran, pullulan and γ-irradiated pullulan by Fourier-transform IR spectroscopy. Carbohydrate Research. 337: 1445–1451, 2002. SIERAKOWSKI, M. R.; MILAS, M.; DESBRIE`RES, J.; RINAUDO, M. Specific modifications of galactomannans. Carbohydrate Polymers, 42: 51-57, 2000. SINGER, R.; ARAUJO, I.; IVORY, M. H. The ectotrophically mycorrhizal fungi of the neotropical lowlands, especiallycentral Amazonia. Beih. Nova Hedwigia 77: 9352, 1983. SINGER, R.; ARAUJO, I. Litter decomposing and ectomycorrhizal Basidiomycetes in an Igapó forest. Pl. Syst. Evol. 153:107-117, 1986. SINGER, R.; ARAUJO, I. Litter decomposition and ectomycorrhizae in Amazonian forests 1.Composition of litter decomposing and ectomycorrhizal basidiomycetes in latosol-terra firme rainforest and in white podzol campinarana. Acta Amazonica 9:23-41, 1979. SINGER R. The role of fungi in Amazonian forests and in reforestation. In: Sioli H, ed. The Amazon. Limnology and landscape ecology of a mighty tropical river and its basin. Boston, Massachusetts: W. Junk Publishers. p .603-614, 1984.

SINGER, R.; IVORY M. H. The ectotrophically mycorrhizal fungi of the neotropical lowlands, especially Central Amazonia. Beih Nova Hedwigia 77: l-352, 1983. SMIDERLE, F. R.; CARBONERO, E. R.; SASSAKI, G. L.; GORIN, P. A. J.; IACOMINI, M. Characterization of a heterogalactan: Some nutritional values of the edible mushroom Flammulina velutipes. Food Chemistry, 108, 1, 329-333, 2008. SOHAL RS, WEINDRUCH R. Oxidative stress, caloric restriction, and aging. Science, 273:59–63, 1996. SOUZA, L. A. R. et al. Caracterização química e atividades farmacológicas de polissacarídeos sulfatados extraidos da alga marinha Amansia multifida. Infarma. Pharmácia Brasileira, v.19, p. 1-6, 2007. SOUZA, M. M.; BELLA CRUZ, A.; SCHUMACHER, M.B. ; KRUGER, M. R. O. ; FREITAS, R.A. ; BELLA CRUZ, R. C. ; Métodos de avaliação biológica de produtos naturais e sintéticos. In: BRESOLIN, T. M. B.; CHERCHINEL FILHO, V. Ciências químico-farmacêuticas: Contribuição ao desenvolvimento de novos fármacos e medicamentos. Itajaí: UNIVALI, 2003. SUGAWARA, T.; TAKAHASHI, S.; OSUMI, M.; OHNO, N. Refnement of the structures of cell-wall glucans of Schizosaccharomyces pombe by chemical modifcation and NMR spectroscopy. Carbohydrate Research, 339(13): 2255–2265, 2004. SULEYMAN, H, Buyukokuroglu ME. The effects of newly synthesized pyrazole derivatives on formaldehyde-, carrageenan-, and dextran-induced acute paw edema in rats. Biol Pharm Bull 2001;24: 1133–6. SUTHERLAND, J. W. Novel and established applications of microbial polysaccharides. Trends in Biotechnology, 16:41-46, 1998. TAVEIRA, V.C.; NOVAES, M.R.C.G. Consumo de cogumelos na nutrição humana: uma revisão da literatura. Com. Ciências Saúde. v.18, n.4, p.315-322, 2007. TEIXEIRA, A. Z..; IACOMINI , A. M.; GORIN , P. A. J. Chemotypes of mannose-containing polysaccharides of lichen mycobionts: a possible aid in classification and identification. Carbohydrate Research, 266(2): 309-314, 1995. TORTORA, G. J.; FUNKE, B. R.; CASE, C. L. Microbiologia. 6. ed. Porto Alegre Artmed, 2000. TREVELYAN, W.E.; PROCTER, D.P.; HARRISON, J.S. Detection of sugars on paper chromatograms. Nature, 166: 444 - 445, 1950. URBEN, A.F.; SIQUEIRA, P. Cogumelos e suas Dehcias. Brasilia, D.F., EMBRAPA Informação Tecnológica ,2003.

USUI, T.; IWASAKI, Y.; MIZUNO, T. Isolation and characterization of two kinds of heterogalactan from the fruit bodies of Ganoderma applanatum by employing a column of concanavalin A-sepharose 4B. Carbohydrate Research, 92, 103-114, 1981. VIEIRA, I. G. P.; MENDES, F.N.P.; GALLÃO, M. I.; BRITO, E. S. NMR study of galactomannans from the seeds of mesquite tree (Prosopis juliflora (Sw) DC). Food Chemistry, v. 101, p. 70-73, 2007. YANISHLIEVA, N. V.; MARINOVA, E.; POKORNÝ, J. Eur. J. Lipid. Sci. Tech, p. 108- 793, 2006. YEUNG, J.H.K.; CHIU, L.C.M. & OOI, V.E.C. Effect of polysaccharide peptide on glutathione and protection against paracertanol induced hepatotoxicity in the rat. Methods and Findings in Experimental and Clinical Pharmacology, 16: 723-729, 1994. YUTAKA, K.; BARBARA. M.; KYOKO, I.; AKIHIRO, T.; TAKUJI, K.; AKIRA, A.; IYOSHI, S.; HIROYUKI, M.; MAMORU, K.; TADASHI, I. Isolation and partial characterization of fucan sulfates from the body wall of sea cucumber Stichopus japonicus and their ability to inhibit osteoclastogenesis. Carbohydrate Research, 339 1339–1346, 2004. WALLE, T. Flavonoids and isoflavones (phytoestrogens): absorption, metabolism and bioactivity. Free radical biology & medicine, v.36, n.7, p.829-837, 2004. WANDE, A. Handbook of Pharmaceutical Excipients. 2.ed., Washington: American Phamaceutical Association, 1994. WANG, T.; DENG, L.; LI, S.; TAN, T. Structural characterization of a water-insoluble (1→3)-α-D-glucanisolated from the Penicillium chrysogenum. Carbohydrate Polymers, 67(1): 133–137, 2007. WANG, X.; XU, X.; ZHANG, L. Thermally induced conformation transition of triple-helical Lentinan in NaCl aqueous solution. The Journal of Physical Chemistry B, 112, 10343–10351, 2008. WILSON JN, PIERCE JD, CLANCY RL. Reactive oxygen species in acute respiratory distress syndrome. Heart Lung, 30:370—5, 2001. WATLING, R. Australian boletes: their diversity and possible origins. Austr. Syst. Bot. 14:407-416, 2001. WATLING, R.; MEIJER, A. Macromycetes from the state of Paraná, Brazil. Edinburgh J. Bot. 54(2): 243, 1997

WISBECK, E; FURLAN, S. A; NINOW, J. Influencia do pH e do Kla na produção de biomassa de Pleurotus ostreatus DSM 1833 em cultivo submerso. In: XIV Simpósio Nacional de Fermentação- SINAFERME. Florianópolis SC, 5 a 8/08/2003. ZAIKOV, G. E. Chemistry of Polyssacharides. Leiden: Ridderprint BV, Ridderkerk. p. 15-18, 2005 ZHANG, J.; TANG, O.; ZIMMERMAN-KORDMANN, M.; REUTTER, W.; FAN, H. Activation of B lymphocytes by GLIS, a bioactive proteoglycan from Ganoderma lucidum. Life Sciences, 71(6): 623–638, 2002. ZHANG, L.; LI, X.; XU, X.; ZENG, F. Correlation between antitumor activity, molecular weight, and confromation of lentin. Carbohydrate Research, 340(8): 1515-1521, 2005. ZHANG, Q.; YU, P.; LI, Z.; ZHANG, H.; XU, Z.; LI, P. Antioxidant activities of sulfated polysaccharide fractions from Porphyra haitanesis. J. Applied Phycology, 15: 305-310, 2003. ZHANG, E. X.; YU, L. J. Studies on polysaccharide from Sargassum thunberg for its ability to scavenge active oxygen species. Chinese Journal of Marine Drugs, 3: 1–4, 1997 ZHANG, M.; CUI, S. W.; CHEUNG, P. C. K.; WANG, Q. Antitumor polysaccharide from mushrooms: a review on their isolation process, structural characteristics and antitumor activity. Trends in Food Science & Technology, 18: 4-19, 2007. ZHOU, Y-C e ZHENG, R-L. Phenolic compounds and analog as superoxide anion scavengers and antioxidants. Biochemical Pharmacology, 42(6): 1177-1179, 1991. ZIMMERMAN, J. W.; LINDERMUTH, J.; FISH, P. A.; PALACE, G. P.; STEVENSON, T. T.; DEMONG D. E. A novel carbohydrate-glycosphingolipid interaction between a beta-(1-3)-glucan immunomodulator, PGG-glucan, and lactosylceramide of human leukocytes, J Biol Chem, 273(34):22014-20, 1998.