universidade federal do piauí - leg.ufpi.br§ão... · avaliou a diversidade genética de ovinos...
TRANSCRIPT

Universidade Federal do Piauí
Caracterização da Diversidade Genética de Ovinos Santa Inês no Estado do
Piauí
Iolly Tábata Oliveira Marques
Teresina
2014
Dissertação apresentada à Universidade Federal
do Piauí como parte das exigências do Programa
de Pós-graduação em Genética e Melhoramento,
área de concentração em Genética e
Melhoramento, para obtenção do título de
“Mestre”.

Iolly Tábata Oliveira Marques
Bióloga
Caracterização da Diversidade Genética de Ovinos Santa Inês no Estado do
Piauí
Teresina
2014
Dissertação apresentada à Universidade
Federal do Piauí como parte das exigências do
Programa de Pós-graduação em Genética e
Melhoramento, área de concentração em
Genética e Melhoramento, para obtenção do
título de “Mestre”.
Orientador:
Prof. Dr. Fábio Barros Britto

Caracterização da Diversidade Genética de Ovinos Santa Inês no Estado do
Piauí
Iolly Tábata Oliveira Marques
Aprovada em 30 / 05 / 2014
Comissão julgadora:
__________________________________________________________________
Profa. Dra. Cintia de Souza Clementino – UESPI
__________________________________________________________________
Prof. Dr. Marcos Jacob de Oliveira Almeida – EMBRAPA
__________________________________________________________________
Prof. Dr. Fábio Barros Britto - UFPI
(Orientador)

Dedico ao meu marido, Rodrigo Alves de Souza Galvão,
por ser meu principal incentivador para seguir a carreira acadêmica.
Aos meus pais, Eudes Gomes Marques e
Mª do Perpétuo Socorro de Oliveira Marques
pelo apoio incondicional.

AGRADECIMENTOS
À Deus, a Ele todas as honras e glórias, por ser minha fortaleza, meu refúgio,
minha redenção, minha verdade e minha vida.
À Universidade Federal do Piauí e ao Programa de Pós Graduação em
Genética e Melhoramento, por nos proporcionar um ensino de qualidade com
professores de alto gabarito.
À CAPES, pela bolsa concedida durante parte da realização do curso.
À Coordenadora Profa. Dra. Ângela Celis, por ser uma mãe para todos nós,
auxiliando, aconselhando e consolando nos momentos difíceis, e sempre nos
estimulando para alcançarmos nossos objetivos.
Ao meu orientador Dr. Fábio Barros Britto (o melhor de todos!) pela paciência
infinita, calma, atenção e confiança dedicadas a mim. Obrigada por aguentar minhas
paranoias, reclamações e prantos de maneira tranquila e sempre encontrar a melhor
saída para tudo.
Ao professor do Campus Cinobelina Elvas – Bom Jesus, José Lindenberg
Rocha Sarmento e ao aluno de pós graduação em Ciência Animal, Daniel Biagiotti
por fornecer o material biológico e parte dos reagentes fundamentais para a
realização desse trabalho.
Aos professores Dr. Fábio Diniz e Dra. Regina Lúcia Ferreira Gomes e ao
Márcio Costa, por fornecer reagentes necessários e em especial ao professor Dr.
Evando Aguiar, que além de material, disponibilizou tempo e atenção para
esclarecer as mais diversas dúvidas.
Às amigas de mestrado, Kátia Carvalho e Kaline Gonzalez, por me ensinarem
as técnicas que desenvolvi nesse trabalho e por me ajudarem na adaptação em
Bom Jesus.

Aos companheiros de Bom Jesus: Rafael Lino e Renan pela ajuda no
laboratório e Sarah Albuquerque, por me abrigar em sua casa, fazendo que com a
temporada que passei na cidade fosse mais fácil.
Aos colegas de turma, pela ajuda mútua nas dificuldades das disciplinas, em
especial aos amigos: Rafael Almeida, Rosimeire Xavier, Joseane Inácio, Polyanna
Bacelar e Larrone Silva.
Ao amigo que esse curso me deu, Sérgio Ewerton Menezes, pela amizade
sincera, pela ajuda abnegada, por todos os momentos em que dividimos as muitas
dificuldades e alegrias, dentro e fora do laboratório. Somos irmãos em Cristo e em
Maria, e nossa amizade durará para sempre.
Aos meus amigos do Encontro de Jovens com Cristo (EJC), pela força e
incentivo, que mesmo com a minha ausência no grupo, nunca deixaram de torcer e
rezar por mim.
Aos meus amigos e família pelo apoio.

“Eu vou e quem me impedirá?
Se ao meu lado está o autor da minha fé;
Eu vou! A força ele me dá, coragem pra enfrentar o que vier;
Eu vou e crendo levarei a salvação aos meus, à minha família;
E já não importa o grau a que eu cheguei;
Eu vou seguir decididamente o amado Rei.”
Decididamente - Walmir Alencar

SUMÁRIO
RESUMO..................................................................................................................VIII
ABSTRACT.................................................................................................................IX
LISTA DE FIGURAS....................................................................................................X
LISTA DE TABELAS...................................................................................................XI
1. INTRODUÇÃO.......................................................................................................12
2. REVISÃO DE LITERATURA..................................................................................14
2.1. Ovinocultura no Brasil.........................................................................................14
2.2. Ovinos deslanados e a raça Santa Inês..............................................................16
2.3. A raça Santa Inês como um recurso genético.....................................................19
2.4. A biologia molecular como ferramenta na conservação de recursos
genéticos....................................................................................................................20
2.5. Caracterização molecular de populações...........................................................24
3. MATERIAL E MÉTODO.........................................................................................26
3.1. Amostragem........................................................................................................26
3.2. Extração de DNA................................................................................................27
3.3. Seleção e amplificação de Marcadores RAPD....................................................28
3.4. Genotipagem.......................................................................................................30
3.5. Análise dos dados...............................................................................................30
4. RESULTADOS.....................................................................................................33
4.1. Marcadores RAPD selecionados.........................................................................33
4.2. Diversidade genética...........................................................................................34
4.3. Análise de variância molecular (AMOVA) e Estrutura populacional....................35
5. DISCUSSÃO..........................................................................................................39
6. CONCLUSÕES E RECOMENDAÇÕES.............................................................43
7. REFERÊNCIAS BIBLIOGRÁFICAS.......................................................................44

VIII
RESUMO
MARQUES. I.T.O. Caracterização da Diversidade Genética de Ovinos Santa Inês no
Estado do Piauí. 2014. 53p. Dissertação para obtenção do título de Mestre na área
de concentração de Genética e Melhoramento - Universidade Federal do Piauí,
Teresina, Piauí, 2014.
A raça Santa Inês está entre os ovinos deslanados de maior destaque existentes no
Brasil. Esta raça apresenta vantagens em qualidade da carne, carcaça de
desenvolvimento considerável e o sistema de criação relativamente simples. Devido
a estas razões, uma crescente comercialização da raça foi observada entre os
produtores. No entanto, o manejo reprodutivo sem controle com cruzamentos
intercorrentes e poucos reprodutores pode levar à perda da diversidade genética.
Assim, são necessários estudos genéticos de populações, considerando à
conservação de germoplasma. Este estudo realizado com marcadores RAPD
avaliou a diversidade genética de ovinos Santa Inês em rebanhos do Piauí, Brasil.
Seis populações localizadas nos municípios de Cristino Castro, Bom Jesus,
Redenção, São Raimundo Nonato, Teresina e Campo Maior foram analisadas com
cinco primers. Trinta e seis locos polimórficos foram encontrados e um índice de
heterozigosidade (Hs) de 0,289. O coeficiente de endogamia (f) das populações foi
de 0,172, mostrando um nível médio de diferenciação entre as populações. Os
valores de Hs observados foram, respectivamente, 0,132 e 0,199, que revelam a
diferenciação genética significativa entre os locais pesquisados. A Análise de
Variância Moleucular mostrou que 80% da variabilidade é explicada pelo
componente "dentro das populações", e 8% pelas diferenças "entre os grupos", no
entanto a análise de cluster e teste de Mantel não mostrou relação entre as
diferenças genéticas e localização geográfica das amostras. Análise no software
Structure estabeleceu que as amostras poderiam ser organizadas em três grupos,
com certo fluxo gênico entre as localidades. A origem dos animais utilizados neste
estudo e a troca de reprodutores entre os locais podem ser responsáveis pelos
resultados encontrados neste trabalho.
Palavras-chave: Caracterização molecular, marcadores moleculares, ovinos
deslanados, estrutura populacional, RAPD.

IX
ABSTRACT
MARQUES. I.T.O. Characterization of Genetic Diversity of Santa Ines Sheep in the
state of Piaui. 2014. 53p. Dissertação para obtenção do título de Mestre na área de
concentração de Genética e Melhoramento - Universidade Federal do Piauí,
Teresina, Piauí, 2014.
Santa Inês is one of the most prominent hair sheep found in Brazil. This breed
present good quality on meat, considerable carcass development and a relative
simple rearing system. Due to these reasons, an increasing on the marketing
considering this breeding was observed among producers. However, the
uncontrolled reproductive management, with intercurrent crossing using few sires,
may lead to the loss of genetic diversity. Thus, studies on population genetics
considering the germoplasm conservation are necessary. This study was carried out
with RAPD makers and evaluated the genetic diversity of Santa Inês sheep in herds
located in Piaui state, Brazil. Six populations located in the municipalities of Cristino
Castro, Bom Jesus, Redenção, São Raimundo Nonato, Teresina and Campo Maior
were analyzed with five markers. Thirty-six polymorphic loci were found and showed
heterozygosity index (Hs) of 0.289. The mean inbreeding coefficient (f) of the
populations was 0.172, showing an intermediate level of inbreeding; θII and ΦST index
show the level of differentiation among populations. The values observed were
respectively 0.132 and 0.199, which reveal significant genetic differentiation among
locations surveyed. The Analysis of Molecular Variance showed that 80 % of the
variability is explained by the “within populations” component, and 8% by the
differences “among groups”, however cluster analysis and Mantel test showed no
relationship between genetic differences and geographical location of samples.
Analysis on the software Structure established that samples could be organized in
three groups, with certain gene flow among locations. The origin of the animals used
in this study and the exchange of reproducers among locations may be responsible
for the results found in this work.
Keywords: Molecular characterization, molecular markers, hair sheep, population
structure, RAPD.

X
LISTA DE FIGURAS
Figura 1 - Fenótipo de representante da raça Santa Inês........................................18
Figura 2 - Variação de cor de pelagem entre animais da raça Santa Inês................18
Figura 3 - Mapa do Estado do Piauí destacando municípios onde foram realizadas
as coletas de pelos em ovinos Santa Inês.................................................................26
Figura 4 - Gel de agarose 0,8% mostrado resultados de extração de DNA de pelos
de ovinos Santa Inês com amostras numeradas do município de Teresina – PI......28
Figura 5 - Gel de Agarose a 1,5% mostrando padrão de bandas obtido pelo primer
RAPD UBC 180, com amostras numerdas de DNA de ovinos Santa Inês
provenientes de São Raimundo Nonato, PI. As setas indicam locos com presença de
polimorfismos.............................................................................................................34
Figura 6 - Dendrograma gerado pelos de valores de ΦST par a par entre as
populações, através do método UPGMA. SRN: São Raimundo Nonato, CM: Campo
Maior, RE: Redenção do Gurguéia, CC: Cristino Castro, BJ: Bom Jesus, TE:
Teresina......................................................................................................................36
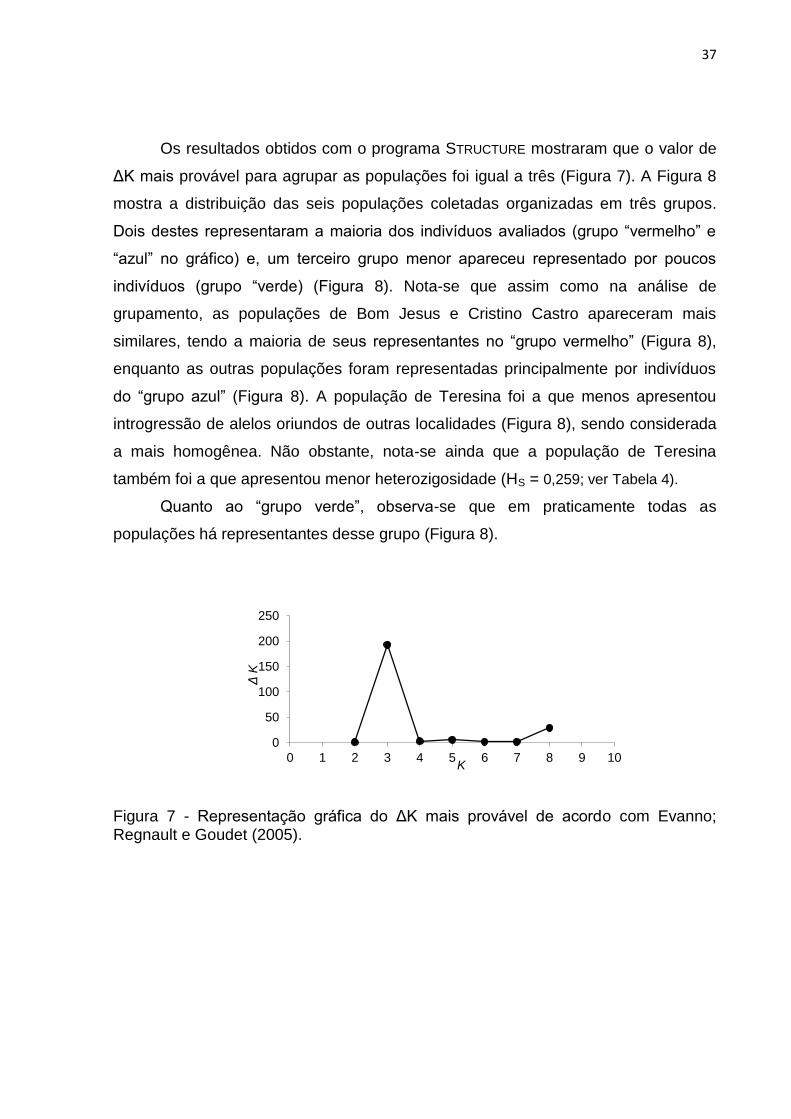
Figura 7 - Representação gráfica do ΔK mais provável de acordo com Evanno;
Regnault e Goudet (2005).........................................................................................37
Figura 8 - Designação das populações de Ovinos Santa Inês usando a estrutura
baseada na alocação das amostras para K=3. Em A: Cristino Castro, B: Bom Jesus,
C: Redenção do Gurguéia, D: São Raimundo Nonato, E: Teresina e F: Campo
Maior......................................................................................................................... 38

XI
LISTA DE TABELAS
Tabela 1 - Relação de municípios e quantidade de animais amostrados..................27
Tabela 2 - Lista de marcadores RAPD testados no experimento e suas
sequências................................................................................................................30
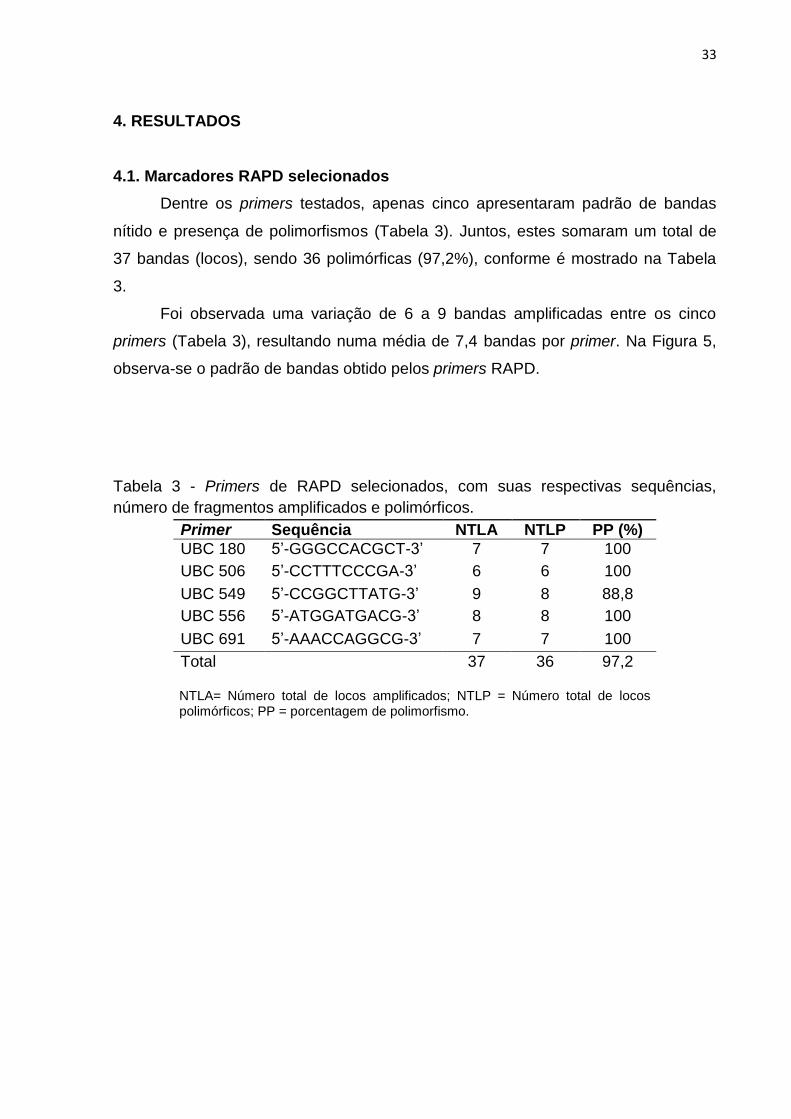
Tabela 3 - Primers de RAPD selecionados, com suas respectivas sequências,
número de fragmentos amplificados e polimórficos...................................................33
Tabela 4 - Estimativa da diversidade genética para cada uma das seis populações
de ovinos Santa Inês no estado do Piauí analisadas no presente estudo.................33
Tabela 5 - Análise de Variância Molecular (AMOVA) para as populações de ovinos
Santa Inês no estado do Piauí...................................................................................35
Tabela 6 - Distâncias genéticas par-a-par entre cada localidade estudada (ΦST; diagonal
superior) e distâncias geográficas em quilômetros (diagonal inferior). CC: Cristino castro, BJ:
Bom Jesus, RE: Redenção do Gurguéia, SRN: São Raimundo Nonato, TE: Teresina, CM:
Campo Maior..........................................................................................................................36

12
1. INTRODUÇÃO
As raças de ovinos trazidas para América do Sul tiveram procedência tanto
europeia quanto africana e, com o passar dos anos, estas foram se adaptando às
condições locais, adquirindo características próprias. Esse fato levou ao surgimento
de raças nativas que apresentam grande rusticidade, tendo como principal aptidão a
produção de carne, leite e peles para subsistência (EGITO et al., 2002; PAIVA et al.,
2005a).
No Brasil, a ovinocultura teve destaque inicialmente na produção de lã na
região Sul do país, onde durante décadas acumulou um contingente de maior
significado nacional. Já na região Nordeste encontrava-se um segundo agrupamento
de ovinos, característico da pecuária de corte de subsistência, formada por raças
nativas e mestiças (MORAIS, 2000). Com a crise do mercado da lã na década de
90, a ovinocultura de corte brasileira ganhou destaque (VIANA, 2008).
Os rebanhos do Nordeste têm como finalidade a produção de carne, com raças
deslanadas (IBGE, 2011), que representam uma importante fonte de proteína para
as populações locais, principalmente nos locais que enfrentam longos períodos de
estiagem. Tais raças deslanadas são adaptadas à região Nordeste, porém
apresentam baixos índices de produtividade (SOUSA e LEITE, 2000). Este fato está
diretamente relacionado à falta de um sistema de produção adequado, de modo que
se utilizem raças apropriadas para cada objetivo de produção (CARNEIRO et al.,
2007). Segundo Vieira et al. (2000) as raças que mais se destacam no Nordeste são
as naturalizadas Somalis Brasileira, Bergamácia Brasileira; e as nativas Morada
Nova Rabo Largo, Cariri e Santa Inês.
No estado do Piauí existe um efetivo de aproximadamente 1,4 milhões de
ovinos, formados tanto por raças nativas como raças exóticos, onde a raça Santa
Inês é a que mais se destaca, devido ao potencial genético, porte e adaptabilidade
(BIAGIOTTI et al., 2012). Os animais são rústicos, precoces e se ajustam a qualquer
sistema de criação e pastagem, em diversas regiões do país. Atualmente, encontra-
se em fase de expansão por ser o destaque entre as raças de ovinos (ABSI, 2012;
ARCO, 2012).
Em busca do aumento da produtividade, muitos produtores lançam mão de
cruzamentos de suas raças nativas com raças comerciais (ARANDAS et al., 2012;
GUIMARÃES et al., 2012), o que pode ocasionar à perda de variabilidade de

13
germoplasma nativo. Diante deste fato, a caracterização das populações da raça
Santa Inês torna-se importante para evitar a erosão genética e direcionar estratégias
de preservação (FARID et al., 2000; MENEZES et al., 2006). Com o aumento da
taxa de perda de raças e a limitação nos recursos disponíveis, o abandono dos
rebanhos nordestinos tem se tornado mais significativo, levando, em último caso, ao
risco de extinção (RODRIGUES, 2009).
A utilização de marcadores moleculares como técnica para caracterização
genética de raças tem ganhado espaço, servindo como uma ferramenta útil para a
identificação dos animais e a estimativa da distância genética (TENEVA, 2009;
SILVA et al., 2011).
Objetivou-se avaliar a diversidade genética de ovinos da raça Santa Inês em
rebanhos do Piauí, utilizando marcadores RAPD. Devido à velocidade e baixo custo
de obtenção de dados com marcadores RAPD, o presente trabalho apresenta os
resultados de uma análise preliminar da diversidade genética e da estrutura de
populações de ovinos da raça Santa Inês. Neste Estado, os dados técnicos são
escassos e os resultados gerados nortearão futuros estudos de conservação da
raça, visando o melhoramento da mesma sem perder o foco na diversidade genética
presente na mesma.

14
2. REVISÃO DE LITERATURA
2.1. Ovinocultura no Brasil
Evidências mostram que a domesticação dos animais consistiu num processo
lento e complexo, no qual envolveu não só a modificação do comportamento das
espécies, mas também sua morfologia e fisiologia. As ovelhas constam no primeiro
grupo de animais domesticados para fins de abate, datando por volta de 7000 anos
a.C. Naquele período, os animais não representavam somente alimento (carne e
leite), mas também fonte de matéria prima para agasalho, força de tração e
companhia (MARIANTE e CAVALCANTE, 2006).
Ovinos estão presente na América do Sul desde a época da colonização,
quando espanhóis e portugueses os trouxeram em seus navios, juntamente com
bovinos, caprinos, aves, cavalos e suínos; fazendo deste evento o ponto inicial de
disseminação de animais domesticados no continente (PRIMO, 2004; McMANUS et
al., 2010). Há indícios que as raças ovinas chegadas ao Brasil tenham origem tanto
europeia quanto africana, e que ao longo dos anos esses animais passaram por
processos de adaptação e seleção natural resultando em várias raças localmente
adaptadas em toda área territorial do país (PAIVA et al., 2005a). Tais raças ficaram
conhecidas como “locais”, “crioulas” ou “nativas” por terem agregado valores como a
rusticidade, o que promoveu a criação de ovinos para fins de subsistência, na
produção de carne, leite e peles (EGITO et al., 2002).
No Brasil, a ovinocultura teve destaque inicialmente na região Sul do país,
mais precisamente no estado de Rio Grande do Sul, onde a demanda de ovinos
melhorados advindos da Austrália, Europa e Estados Unidos foi bem significativa,
levando a essa região posição de destaque na exploração comercial (ARAÚJO,
2000). Apesar de algumas raças serem de corte, o forte da ovinocultura sulista foi o
comércio de lã, tanto que em 1942 foi criada a primeira associação do ramo, a
Associação Riograndense de Criadores de Ovinos (ARCO), que após alguns anos
veio a ser Associação Brasileira de Criadores de Ovinos (sem mudar a sigla), a qual
iniciou as primeiras avaliações objetivas para seleção de ovinos, no final da década
de 80, visando melhorar a produtividade e a qualidade da lã (OJEDA, 1999).
O declínio do promissor mercado de lã nacional se deu no início dos anos
1990, quando o mercado foi atingindo por uma profunda crise internacional (devido
ao início da comercialização de tecidos sintéticos no mercado) a qual atingiu

15
fortemente os produtores gaúchos, que ainda tentaram continuar com a manutenção
da raça Corriedale como uma forma de voltar para a produção de carne. Porém,
depois de uma ligeira recuperação, a crise voltou a um profundo agravamento,
concomitante ao aumento de áreas cultivadas com grãos, cultura que se mostrou
competitiva e lucrativa. A soma desses fatores fez com que muitos produtores
desistissem da ovinocultura (MORAIS, 2000; VIANA, 2008).
Foi nesse cenário que a ovinocultura de corte da região Nordeste, que até
então se tratava somente de subsistência, ganhou destaque no mercado brasileiro.
O número de rebanhos efetivos tem aumentado como um todo a cada ano no país,
sendo que dados da Produção da Pecuária Municipal de 2011 estimaram mais de 17
milhões de cabeças, apontando um aumento de 1,6% em relação aos dados
apresentados em 2010, porém a seca prolongada no Nordeste no ano de 2012
acarretou uma redução de 5% no efetivo de cabeças em relação ao ano anterior,
somando 16,789 milhões de cabeças (IBGE 2012). Desse total, mais de 9 milhões
estão concentrados no Nordeste brasileiro, representando 55,5% do total nacional.
Destacam-se os estados da Bahia, Ceará, Pernambuco e Piauí, os quais estão
respectivamente em 2º, 3º, 4º e 5º lugar na listagem nacional (IBGE, 2012). Esse
efetivo nordestino é composto de raças deslanadas, especializadas para corte e com
grande capacidade adaptativa às condições peculiares da região, além de
representar uma importante fonte de proteína para a população (SILVA, 2002).
Com o objetivo de incrementar a ovinocultura de corte, alguns produtores
importaram raças exóticas, que foram usadas em cruzamentos para produção de
animais “meio-sangue” (MORAIS, 2000). Porém, como essas raças foram
selecionadas em climas temperados, não obtiveram o mesmo sucesso adaptativo
das raças locais (EGITO, 2002; CARNEIRO, 2010) fazendo com que todas as
atenções se voltassem às raças deslanadas nativas.
Sinais de que a ovinocultura de corte tem ganhado espaço ao longo dos anos
no país são notados através das exigências dos consumidores por carcaças de
melhor qualidade, da regulamentação do Governo Federal na comercialização de
carnes e na constante ampliação de pesquisas no ramo; criando uma situação
favorável para expansão do comércio, o que consequentemente desperta o
interesse de produtores visando o aumento da qualidade dos animais (ROSANOVA,
2005; SIMPLÍCIO E SIMPLÍCIO, 2006; FARIA, 2008).

16
Diante disso, Faria e Silva (2006) relataram a crescente demanda por carne
de ovinos em cortes padronizados ou produtos processados, de maneira mais
industrializada (embalados e comercializados de forma resfriada ou congelada)
principalmente em áreas habitadas pelo segmento populacional detentor de maior
poder aquisitivo. Além disso, Carvalho e Souza (2008) destacam a modificação do
mercado consumidor nos centros urbanos do Nordeste, onde se observou um
aumento da procura por carne ovina e caprina, em consequência das campanhas e
propagandas relacionadas ao consumo de alimentos mais saudáveis, com baixos
teores de gordura, além do resgate da valorização dos hábitos alimentares
regionais, estimulada pelo turismo da região.
2.2. Ovinos deslanados e a raça Santa Inês
Com a crescente demanda de carne ovina, as raças deslanadas mostram-se
claramente como a melhor opção, tanto pelo tamanho da carcaça e qualidade da
carne (ZAPATA et al., 2001), quanto pelas características atribuídas aos animais
que são adaptados a altas temperaturas, resistentes a parasitas, além da rusticidade
e prolificidade (EGITO et al., 2002). Porém, por outro lado, estas raças apresentam
baixos índices de produtividade (SOUSA e LEITE, 2000). Este fato pode estar
diretamente relacionado à falta um sistema de produção adequado, de modo que se
utilizem raças apropriadas para cada objetivo de produção (CARNEIRO et al., 2007).
Pinheiro Jr. et al. (2010) destacam as exigências de uma exploração racional de
ovinos de corte, tais como: planejamento, infra-estrutura, mão-de-obra qualificada e
conhecimento de mercado. Já Simplício et al. (2007) falam da importância que a
nutrição, a saúde e o ambiente exercem sobre os animais e como isso reflete no
desempenho produtivo. Dessa maneira, uma relação custo/benefício rentável é
obtida através de animais geneticamente superiores, os quais podem ser
trabalhados em sistemas modernos e eficazes de produção, agregados aos manejos
corretos de nutrição, reprodução e sanitário, culminando no melhoramento
zootécnico (ROSANOVA et al., 2005).
Segundo Vieira et al. (2000) as raças mais presentes no Nordeste são:
Somalis Brasileira, Bergamácia Brasileira, Morada Nova,
Rabo Largo, Cariri e Santa Inês. Os representantes da raça Santa Inês
ganharam destaque em detrimento aos animais de outras raças, por apresentar alto
valor adaptativo e reprodutivo, potencial genético, resistência a parasitas

17
gastrointestinais, excelente qualidade de pele, além de um bom desenvolvimento
ponderal (SOUSA et al., 2003). A raça teve provável origem no estado da Bahia
(PAIVA et al., 2005b), sendo desenvolvida a partir de cruzamentos intercorrentes
das raças Bergamácia, Morada Nova, Somalis e outros ovinos sem raça definida.
Algumas características morfológicas corroboram com esta teoria, tais como: tipo de
orelhas, formato da cabeça e os vestígios de lã que correspondem à Bergamácia, a
condição deslanada e a pelagem se dá pela participação da raça Morada Nova. Já a
participação da Somalis é caracterizada pela presença de alguma gordura em torno
da implantação da cauda, quando o animal está muito gordo (ABSI, 2012; ARCO,
2012; VERÍSSIMO et al., 2009). Em contrapartida, Paiva et al. (2003) concluíram em
seus trabalhos de caracterização genética de Santa Inês que a raça é mais próxima
da raça Rabo Largo ao invés da raça Morada Nova, o que aumenta a dúvida em
relação à sua origem.
O fenótipo apresentado pelos animais pertencentes à raça é bem
determinado (Figura 1), podendo-se citar a presença de pelos curtos e sedosos, com
pelagens que podem variar entre castanha, vermelha, preta, malhada de preto e
branco ou malhada de vermelho e branco (Figura 2).
Os machos são de porte médio a grande, com peso variando de 80 a 120 Kg
e fêmeas pesando de 60 a 90 Kg. A cabeça tem tamanho médio, focinho alongado,
as orelhas são carnudas, em forma de lança; a fronte apresenta-se curta, larga e
reta. O pescoço é alongado nas fêmeas e curto e forte nos machos, o peito é largo,
arredondado e um pouco proeminente. O tronco dos animais é forte, com os quartos
dianteiro e traseiro grandes, com a garupa levemente inclinada; a cauda tem base
larga, estreitando ao longo do comprimento.
A raça também apresenta pele de altíssima qualidade com grande
elasticidade e excelente flexibilidade, maciez, resistência e textura fina, podendo
alcançar até 30% do valor total do animal, utilizada na indústria de vestiário e
calçados finos. Os animais são rústicos, precoces e se adaptam a qualquer sistema
de criação e pastagem nativa, em diversas regiões do país e estiveram em fase de
expansão por serem o destaque entre as raças de ovinos, sendo encontrados em
grade escala em fazendas do Centro-oeste, Sudeste e Sul (ABSI, 2012; ARCO,
2012; SANTOS, 2003; SOUSA et al., 2003).

18
Figura 1 - Fenótipo de representante da raça Santa Inês. Fonte: http://ovinossantainesedorper.blogspot.com.br/
Figura 2 - Variação de cor de pelagem entre animais da raça Santa Inês. Fonte: http://www.revistaberro.com.br/?materias/ler,1100

19
2.3. A raça Santa Inês como um recurso genético
Vários autores relatam que as principais raças nativas nordestinas estão
sofrendo cada vez mais a ação de cruzamentos indiscriminados com raças exóticas
(ARANDAS et al., 2012; GUIMARÃES et al., 2012; PAIVA et al., 2005c), o que
ocasionam perda de variabilidade genética destes animais localmente adaptados.
Sendo assim, torna-se essencial sua conservação, promovendo a manutenção das
variabilidades inter-racial, a qual evita a extinção das raças, e intra-racial, que evita a
erosão genética (MENEZES et al., 2006).
Recentes dados de polimorfismo genômico apontam que a população de
ovinos ao redor do mundo manteve um efetivo populacional grande mesmo após
gerações de domesticação e/ou seleção, conservando altos níveis de diversidade
genética entre raças (KIJAS et al., 2012). Mesmo nessas condições, raças ovinas do
Nordeste brasileiro sofrem com o abandono da variedade local em favor de raças
exóticas e seus cruzamentos. Este evento promove o aumento da taxa de perda de
raças e a limitação nos recursos disponíveis, fornecendo evidências de que podem
estar sob risco de erosão da diversidade genética e, em último caso, de extinção
(RODRIGUES, 2009).
Paiva e Pimentel (2008) afirmaram que a raça Santa Inês é uma forte
candidata a perder grande parte de suas características de rusticidade e adaptação
em detrimento de ganhos de heterose obtidos por cruzamentos com várias raças
comerciais, o que pode, ao longo dos anos, levar à formação de ecótipos. McManus
et al. (2010) reforçaram essa tese, quando diz que a "nova Inês Santa" pode perder
características importantes herdadas ao longo dos anos, como: qualidade da pele,
resistência a parasitas gastro-intestinais; assim como adquirir características não-
desejáveis como a sensibilidade à scrapie (IANELLA et al., 2009). A raça Suffolk,
suspeita de ser utilizada para a criação deste novo animal, tem um histórico de
susceptibilidade a esta doença (HEGGEBØ et al., 2002).
Como uma maneira de conservação de material genético ameaçado de
extinção, ovinos nativos foram incluídos no Programa de Conservação de Recursos
Genéticos, da Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA).
Rebanhos de conservação in vivo são encontrados em Centros de Pesquisa da
EMBRAPA (em deles localizado em Campo Maior – PI), Universidades, Empresas

20
Estaduais de Pesquisa, assim como por criadores particulares, sendo esta rede
coordenada pelo Centro Nacional de Pesquisa de Recursos Genéticos e
Biotecnologia (Cenargen, Brasília - DF). O centro de conservação de germoplasma
in situ da raça Santa Inês fica localizado no estado de Sergipe (EGITO et al., 2002),
sendo proposto por Sousa et al. (2003) o “Programa de melhoramento genético para
ovinos deslanados e caprinos de corte no Brasil”. Este, por sua vez, deu origem ao
Programa de Melhoramento Genético de Ovinos da Raça Santa Inês (PROMOSI)
gerido pela Empresa Estadual de Pesquisa Agropecuária da Paraíba (EMEPA)
(LOBO e LOBO, 2007; SOUSA et al., 2008).
Vários estudos envolvendo a raça Santa Inês foram realizados nos últimos
anos, como uma forma de aumentar o conhecimento acerca da raça, o que justifica
a variedade dos temas abordados, tais como: reprodução (SILVA e ARAÚJO, 2000;
QUESADA et al., 2002; NETO, 2008), alimentação (SILVA, 2002), resistência à
parasitas (AMARANTE et al., 2004), tolerância à temperatura (CEZAR et al., 2003;
SANTOS et al., 2006; VERÍSSIMO et al., 2009), desenvolvimento ponderal
(OLIVEIRA, et al., 2010; SILVA et al., 2012), morfometria (COSTA JÚNIOR et al.,
2006; BIAGIOTTI et al., 2013; TORRES, et al., 2013), manejo (MORAIS e
MADALENA, 2006) e principalmente estudos genéticos (PAIVA et al., 2003; PAIVA,
et al., 2005d; ROCHA et al., 2009; PETROLI et al., 2009; OLIVEIRA et al., 2010;
SANTOS et al., 2012; SOUZA et al., 2012).
Essas informações vêm agregar conhecimento e reforçar a importância de
conservação da raça, além de sugerir uma forma mais eficiente de realizar a
manutenção da mesma, contribuindo para melhores estratégias futuras de
melhoramento e seleção. Com isso, pode-se aumentar a produtividade da raça,
despertando o interesse por parte dos produtores com a percepção destes de um
melhor retorno financeiro.
2.4. A biologia molecular como ferramenta na conservação de recursos
genéticos
Até os anos 60, estudos de genética e melhoramento eram realizados apenas
por meio de marcadores morfológicos, baseados em caracteres fenotípicos. Com o
avanço da tecnologia uma nova ferramenta passou a ser utilizada, fazendo com que
as informações dos marcadores morfológicos passassem a ser complementadas por
marcadores isoenzimáticos. Porém para o uso de uma investigação mais detalhada

21
do genoma, as isoenzimas apresentaram algumas limitações como, por exemplo, o
baixo número de locos que podem ser detectados no genoma e o baixo número de
alelos por loco (FERREIRA e GRATTAPAGLIA, 1998; CASTIGLIONI e BICUDO,
2003).
A descoberta da Reação em Cadeia da Polimerase (PCR), desenvolvida por
Kary Mullis em 1983 (MULLIS et al., 1986), inaugurou uma nova era em estudos
genéticos. A PCR é uma técnica bioquímica e molecular para amplificar
exponencialmente um fragmento de DNA, através de replicação enzimática, sem o
uso de um organismo vivo (PAVLOV et al., 2004); tornou-se uma ferramenta
essencial na biologia molecular e desempenha um papel fundamental em quase
todas as técnicas que estão atualmente aplicáveis à análise e caracterização de
genomas (TENEVA, 2009).
Após o advendo da PCR, surgiram diversos métodos de detecção de
polimorfismos genéticos diretamente ao nível de DNA, que permitem a obtenção de
uma gama de marcadores, cobrindo todo o genoma do organismo em questão. Tais
marcadores, denominados “marcadores moleculares” têm sua funcionalidade
baseada no dogma central da Biologia Molecular e na pressuposição do que as
diferenças genéticas significam (CASTIGLIONI e BICUDO, 2003).
Com o uso de marcadores moleculares pode-se obter um número ilimitado de
polimorfismos genéticos, como também a identificação direta de genótipos sem
influência do ambiente; e como são capazes de detectar a variação genética ao nível
da sequência de DNA, foram removidas as limitações dos marcadores morfológicos,
cromossômicos e proteínas. Podem ser utilizados em estudos ecológicos,
filogenéticos e taxonômicos, como também em estudos evolutivos, de diversidade
genética inter e intra-específica, de identidade, origem genética e identificação de
novas variantes; gerando informações importantes para subsidiar diferentes ações
de pesquisa desde a coleta até o uso dos recursos genéticos em programas de
melhoramento (CASTIGLIONI e BICUDO, 2003; FALEIRO, 2007).
Na genética animal, os marcadores moleculares são peças-chave, pois atuam
revelando o polimorfismo repetitivo em nível de DNA. Servem como uma ferramenta
útil para identificar animais, estimar a distância genética, assim como avaliar a
variabilidade genética inter e intrapopulacional, tornando a seleção de acordo com o
genótipo uma ferramenta importante em animais silvestres, domésticos e na criação
de animais de fazenda (ROSA e PAIVA, 2009; TENEVA, 2009). Tais polimorfismos

22
podem ser utilizados para construir mapas genéticos e avaliar as diferenças entre os
marcadores na expressão de características particulares (VIGNAL et al., 2002).
Foram construídos mapas genéticos extensos em uma variedade de espécies
animais, tais como: bovinos, ovinos e suínos, usados para o marcador de seleção
assistida, análise segregação e para a detecção de genes principais (KINGHORN et
al., 1993. ; ROHER et al., 1994; KINGHORN, 1997; VIGNAL et al., 2002).
No caso de animais de rebanho intensamente selecionados (rebanhos
comerciais), esse conhecimento molecular pode auxiliar na otimização de
acasalamentos visando um maior controle sobre os níveis de endogamia, já que
esse controle é crucial para planejamento de programas de conservação de
espécies ou raças adaptadas ameaçadas de extinção, além de nortear programas
de melhoramento animal que visem ganho genético a logo prazo (ROSA e PAIVA,
2009).
Entre os marcadores disponíveis, os RADP (Polimorfismo de DNA
amplificado ao acaso) mostraram resultados importantes em diversas pesquisas com
mamíferos (TELLES, et al., 2001; SPRITZE et al., 2003; ALBUQUERQUE et al.,
2006), inclusive de ovinos (PAIVA et al., 2003), para caracterização e estimativa de
variabilidade, com o objetivo de investigação da diversidade genética e similaridade
entre e dentro de raças. Esses dados gerados são necessários para fornecer
informação genética útil essencial para o desenvolvimento de planos de gestão
eficazes para a conservação e melhoria dos recursos genéticos (KHALDI et al.,
2010).
Os RADP são fragmentos de DNA amplificados através da técnica de PCR
utilizando primers curtos e aleatórios (geralmente 8-10 nucleotídeos), funcionando
de tal maneira que não há necessidade do conhecimento prévio da sequência de
DNA a ser amplificada. Esses primers quando submetidos a temperaturas ideais se
hibridizam com sequências genômicas complementares (WILLIAMS et al., 1990). A
técnica pode ser utilizada para produzir os perfis de DNA relativamente detalhados e
complexos para a detecção de polimorfismos do fragmento amplificado entre os
organismos (TENEVA, 2009). A amplificação de um fragmento RAPD no genoma em
questão é atrelada à existência de duas sequências de DNA complementares ao
primer aleatório, que devem estar posicionadas em orientação oposta e adjacentes o
suficiente para permitir a amplificação exponencial de um segmento de DNA pela
enzima DNA polimerase. Cada primer utilizado, em diferentes amplificações, dirige a

23
síntese de vários segmentos de DNA simultaneamente em diversos pontos do
genoma, resultando em várias bandas (FERREIRA e GRATTAPAGLIA, 1998;
CASTIGLIONI e BICUDO, 2003).
A visualização das bandas é realizada através de eletroforese horizontal em
gel de agarose, seguida de coloração (geralmente realizada com brometo de etídio)
e fotodocumentação sob luz ultravioleta. Os polimorfismos são detectados como a
presença ou ausência de bandas de um tamanho específico. Em teoria, o número de
bandas visualizadas no gel depende exclusivamente do comprimento do primer
utlizado e do tamanho e complexidade do genoma analisado. Porém, dados
experimentais têm mostrado que o número de bandas obtido é independente da
complexidade do genoma (CASTIGLIONI e BICUDO, 2003; TENEVA, 2009).
RAPD caracterizam-se como marcadores dominantes, isto é, não é possível
diferenciar homozigotos dominantes de heterozigotos. Como consequência, ao se
observar uma banda no gel, não é possível informar se aquele segmento originou-se
a partir de uma ou duas cópias da sequência amplificada (considerando um
organismo diploide). Assim, o genótipo recessivo é identificado pela ausência de
banda no gel e os genótipos dominantes e heterozigotos são identificados de igual
forma, com a presença de banda (CASTIGLIONI e BICUDO, 2003; FERREIRA e
GRATTAPAGLIA, 1998).
As vantagens que se podem destacar do uso do marcador em questão, de
acordo com Vignal et al. (2002) e Mburu e Hanotte (2005), são: baixo custo,
simplicidade e rapidez da técnica, produção de grande número de bandas (cada
uma correspondendo a um loco do genoma), sem necessidade de conhecimento
prévio da sequência, podendo ser utilizada em qualquer organismo e pequena
quantidade de amostras de DNA requerida, já que será amplificado pela técnica de
PCR. Existem também algumas desvantagens a respeito da técnica, principalmente
a sensibilidade às condições de PCR, como a temperatura de anelamento dos
primers, a quantidade de íons de magnésio, a concentração e qualidade do DNA e a
presença de possíveis contaminantes na mesma (CASTIGLIONI e BICUDO, 2003;
TENEVA, 2009).
A utilização de marcadores RAPD pode ser observada em recentes estudos
de várias espécies, como por exemplo: diversidade em plantas (VALLE et al., 2013;
SANTOS et al., 2013), caracterização molecular de populações e diversidade
genética em peixes (USMAN et al., 2013; MASSAGO et al., 2009), diversidade

24
genética e estudos populacionais em insetos (RAMPELOTTI et al., 2008; SUZUKI et
al., 2006; HIRAGI et al., 2009). Em ovinos, um dos pioneiros em estudos
moleculares de raças brasileiras foi Paiva et al. (2005b), que utilizou marcadores
RAPD para estimar a variabilidade genética de cinco raças brasileiras deslanadas
em quatro estados mais o Distrito Federal e identificar a origem racial de ovinos
Santa Inês.
2.5. Caracterização molecular de populações
A variabilidade genética se mostra útil na investigação das relações genéticas
entre subpopulações de uma espécie onde os alelos são compartilhados entre
subpopulações por ocasião da migração, como também são compartilhados dentro
das populações em virtude da ancestralidade em comum (HARTL e CLARK, 2010).
A variabilidade genética de um indivíduo ou de uma população é medida
através da heterozigosidade (Hs). O índice Hs representa o nível de heterozigose
que pode ser encontrado em uma população se a mesma estiver sendo submetida a
acasalamentos ao acaso. Ou seja, o Hs estima heterozigosidade média esperada
dentro de subpopulações que tem o sistema de acasalamento ao acaso como modo
reprodutivo (McMANUS et al., 2011).
Em 1951 Wright introduziu o coeficiente de endogamia como um parâmetro
que afeta diretamente a diversidade genética de uma população, sendo que o dividiu
em três índices: FIS, FST, FIT.
O efeito da endogamia em população subdividida é dado através do índice de
fixação (f), o qual mede a falta de heterozigotos dentro de uma população. Como é
sabido, a subdivisão ou fragmentação de uma população causa carência de
heterozigotos, portanto, o aumento da endogamia ao longo das gerações. O índice
de fixação é análogo ao FIS de Wright (endogamia do indivíduo em relação à
subpopulação) e se mostra como uma ferramenta útil para indicar a diferenciação
genética, pois permite uma comparação objetiva do efeito geral da estrutura
populacional entre diferentes organismos (HEDRICK, 2009).
O índice FST de Wright mede a carência de heterozigotos entre
subpopulações e é a medida mais inclusiva da subdivisão populacional, pois mostra
a proporção da diversidade total (HEDRICK, 2009). Essa redução de
heterozigosidade pode ser usada para quantificar o nível de diferenciação genética

25
entre as subpopulações. Análogos modernos de FST incluem GST (NEI, 1973; 1987),
θ (COCKERHAM 1969; 1973; WEIR e COCKERHAM 1984), e ФST (EXCOFFIER et
al. 1992). Os parâmetros para interpretação do FST variam de 0 a 1, e segue a
seguinte escala: de 0 a 0,05 considera-se baixo, de 0,05 a 0,15 considera-se
moderado, de 0,15 a 0,25 considera-se alto e acima de 0,25 considera-se valores
muito altos de diferenciação genética (HARTL e CLARK, 2010).
A redução da heterozigosidade em relação à população total é dada pelo
coeficiente FIT de Wright, o qual leva em consideração os efeitos do acasalamento
não aleatório e os efeitos da subdivisão da população. Weir e Cockerham (1984)
definiram F como índice similar ao FIT de Wright. Os três coeficientes de endogamia
se relacionam da seguinte maneira:

26
3. MATERIAL E MÉTODOS
3.1. Amostragem
Foram coletadas amostras de pelos de 164 ovinos da raça Santa Inês
oriundos de seis fazendas no estado do Piauí, localizadas em seis municípios:
Cristino Castro, Bom Jesus, Redenção do Gurguéia, São Raimundo Nonato,
Teresina e Campo Maior (Figura 3, Tabela 1). Os pelos foram coletados próximo à
região caudal do animal, colocados em envelopes identificados individualmente por
fazenda/localidade, transportados em caixa térmica até o Laboratório de Genética e
Conservação de Germoplasma da Universidade Federal do Piauí, Campus Profª.
Cinobelina Elvas. O material coletado foi acondicionado a aproximadamente 4°C até
o momento da extração de DNA. As coletas foram realizadas entre os anos de 2010
e 2013, por membros do Programa de Pós-Graduação em Ciência Animal da
Universidade Federal do Piauí, Campus Profª. Cinobelina Elvas, os quais
gentilmente cederam parte de suas amostras para a realização deste trabalho.
Figura 3 – Mapa do Estado do Piauí destacando municípios onde foram realizadas
as coletas de pelos em ovinos Santa Inês.

27
Tabela 1 – Relação de municípios e quantidade de animais amostrados.
Município N° de Indivíduos
Cristino Castro 33
Bom Jesus 36
Redenção do Gurguéia 13
São Raimundo Nonato 22
Teresina 30
Campo Maior 30
Total 164
3.2. Extração de DNA
O DNA dos bulbos pilosos foi extraído no Laboratório de Genética e
Conservação de Germoplasma da Universidade Federal do Piauí, Campus Profª.
Cinobelina Elvas, por meio do kit de extração Qiagen QIAamp® DNA Tissue Kit com
as modificações descritas abaixo.
Foram utilizados aproximadamente 40 pelos de cada indivíduo, dos quais
foram individualmente retirados os bulbos pilosos com auxílio de bisturi, para logo
em seguida serem armazenados em microtubos de 1,5 mL devidamente
identificados. Em cada tubo foi adicionado 45 µL de tampão ATL1 e, em seguida, o
material foi rapidamente centrifugado para concentrar o conteúdo no fundo do tubo.
Dando prosseguimento, 10 µL de Proteinase K foram adicionados a cada amostra,
sendo estas agitadas em vórtex e incubadas a 56°C por quatro horas. Durante a
incubação o material foi homogeneizado a cada 30 minutos. Após a incubação
adicionou-se 50 µL de tampão AL1 sendo o material agitado em vórtex e incubado a
70°C por 10 minutos. Após essa incubação, foram adicionados 50 µL de álcool
absoluto, levando os microtubos ao vórtex e, em seguida, a uma rápida
centrifugação. O sobrenadante foi transferido para um novo tubo contendo um filtro
de membrana de sílica, o qual foi centrifugado por 1 minuto a 8.000 rpm. O filtrado
foi descartado e 250 µL de tampão AW11 foi adicionado à cada membrana, sendo as
1 Nome do Tampão fornecido pelo fabricante. Não há descrição da sigla no manual do mesmo.

28
mesmas centrifugadas por 1 minuto a 8.000 rpm. O filtrado foi descartado e
adicionou-se 500 µL de tampão AW21. Novamente o material foi centrifugado por 3
minutos a 14.000 rpm. O tampão foi despejado e os tubos novamente centrifugados
por 1 minuto a 14.000 rpm. Os filtros contendo as membranas foram colocados em
outros microtubos de 1,5mL, nos quais foram adicionados 50 µL de tampão AE2 (10
mM Tris·Cl; 0.5 mM EDTA; pH 9.0) para conservação do DNA. Nesta fase as
amostras ficaram 5 minutos em temperatura ambiente e em seguida foram
centrífugadas por 1 minuto a 8.000 rpm para a recuperação do DNA. Todos os
tampões e a Proteinase K foram fornecidos no Kit.
Os resultados das extrações foram verificados através de gel de agarose a
0,8% e a concentração das amostras foi ajustada para ~10ng/µL utilizando marcador
DNA nas concentrações de 10 e 25 ng/µL (Figura 4). Em seguida as mesmas
foram estocadas em freezer a -20°C para procedimentos posteriores.
Figura 4 – Gel de agarose 0,8% mostrado resultados de extração de DNA de
pelos de ovinos Santa Inês com amostras numeradas do município de Teresina – PI.
3.3. Seleção e Amplificação de Marcadores RAPD
Foram avaliados 34 primers RAPD, desenhados pela University of Britsh
Columbia (Tabela 2) os quais foram testados em amostras de DNA escolhidas
aleatoriamente. Para cada um dos primers foi realizada Reação em Cadeia da
Polimerase (PCR) em volume final de 10 µL contendo os seguintes componentes:
2 Nome do Tampão fornecido pelo fabricante. Não há descrição da sigla no manual do mesmo.

29
tampão de PCR a 1; MgCl2 a 3,4 mM; dNTP a 0,8 mM; primer a 0,8 µM, 1 U de Taq
DNA polimerase; 0,5 µL de DNA genômico (~5 ng) e água ultrapura q.s.p. 10 µL.
A amplificação do DNA foi realizada em Termociclador Maxygene Gradiente
(Axygen) e Termocilador Veriti 96 Well Thermal Cycler 9902 (Applied Biosystems)3
com desnaturação inicial de 5 minutos a 94ºC, seguida de 36 ciclos com
desnaturação de 40 segundos a 94ºC, 1 minuto a 36ºC para anelamento e 2 minutos
a 72ºC para extensão. Foi realizada uma etapa de extensão final de 10 minutos a
72ºC. Ao final da reação, os produtos das amplificações foram congelados até o
momento da genotipagem.
3Cada primer foi processado em apenas um termociclador.

30
Tabela 2 – Lista de marcadores RAPD testados no experimento e suas sequências.
Marcador Sequência
UBC-001 5’-CCTGGGCTTC-3’ UBC-013 5’-CCTGGGTGGA-3’ UBC-040 5’-TTACCTGGGC-3’ UBC-088 5’-CGGGGGATGG-3’ UBC-097 5’-ATCTGCGAGC-3’ UBC-125 5’-GCGGTTGAGG-3’ UBC-158 5’-TAGCCGTGGC-3’ UBC-180 5’-GGGCCACGCT-3’ UBC-197 5’-TCCCCGTTCC-3’ UBC-243 5’-GGGTGAACCG-3’ UBC-247 5’-TACCGACGGA-3’ UBC-248 5’-GAGTAAGCGG-3’ UBC-270 5’-TGCGCGCGGG-3’ UBC-272 5’-AGCGGGCCAA-3’ UBC-321 5’-ATCTAGGGAC-3’ UBC-358 5’-GGTCAGGCCC-3’ UBC-378 5’-GACAACAGGA-3’ UBC-386 5’-TGTAAGCTCG-3’ UBC-409 5’-TAGGCGGCGG-3’ UBC-436 5’-GAGGGGGCCA-3’ UBC-386 5’-TGTAAGCTCG-3’ UBC-474 5’-AGGCGGGAAC-3’ UBC-494 5’-TGATGCTGTC-3’ UBC-506 5’-CCTTTCCCGA-3’ UBC-529 5’-CACTCCTACA-3’ UBC-549 5’-CCGGCTTATG-3’ UBC-556 5’-ATGGATGACG-3’ UBC-564 5’-CGGCGTTACG-3’ UBC-573 5’-CCCTAATCAG-3’ UBC-626 5’-CCAAGCCCGG-3’ UBC-643 5’-ATAAGCGGTG-3” UBC-663 5’-CGTATAGCCG-3’ UBC-686 5’-CGTGACAGGA-3’ UBC-691 5’-AAACCAGGCG-3’
3.4. Genotipagem
A genotipagem foi realizada através de eletroforese em gel de agarose a
1,5%, corados com SYBR SAFE ou banho em brometo de etídio e visualização em
Transiluminador L.PIX (Loccus Biotecnologia).
As informações moleculares obtidas nos géis foram convertidas em dados
binários, onde a presença ou ausência de bandas foram codificadas,
respectivamente, com os valores 1 e 0.

31
3.5. Análise dos dados
A partir dos dados gerados em análise direta no gel, foi possível verificar a
taxa de polimorfismo após a contagem de locos polimórficos entre o número total de
locos amplificados. Além disso, a taxa de polimorfismo foi avaliada para cada
população isoladamente.
O programa HICKORY 1.1, que implementa o método Bayesiano descrito por
Holsinger; Lewis e Dey, (2002), foi usado para estimar as heterozigosidades (Hs).
Também foram estimados os índices θII (análogo do FST de Weir e Cockerham) e f
(análogo do FIS). Foram utilizadas 25.000 simulações de Cadeias de Markov Monte
Carlo com burn in de 5.000. A confiabilidade dos dados foi estimada a partir do
intervalo de confiança entre 2,5% a 97,5% (IC=95%).
A análise de variância molecular (AMOVA) para as seis populações foi
realizado para determinar ΦST (outro análogo FST) utilizando ARLEQUIN 3.0
(EXCOFFIER; LAVAL; SCHNEIDER, 2005). A AMOVA foi configurada agrupando-se
as populações em duas regiões, de acordo com sua proximidade geográfica, como
segue:
Grupo 1 (Região Sul): Cristino Castro, Bom Jesus, Redenção do
Gurguéia e São Raimundo Nonato;
Grupo 2 (Região Centro-Norte): Teresina e Campo Maior.
Também foram calculados os valores de ΦST par-a-par entre cada localidade
para a realização do teste de Mantel, que mostra a proporção da variação genética
explicada pela proporção da variação geográfica. O nível de significância do teste foi
calculado após 1.000 permutações dos dados. A distância geográfica entre cada
localidade foi calculada com o auxílio do Google Maps
(https://www.google.com/maps/). Estes dados também foram utilizados para a
construção de um dendrograma utilizando o método Ligação Média entre Grupos
(UPGMA) realizado no programa MEGA 5.02 (KUMAR et al., 2008).
O programa STRUCTURE versão 2.2 (PRITCHARD; STEPHENS; DONNELLY,
2000) foi utilizado para definir o número de grupos (K) mais provável nas amostras
coletadas, por meio de métodos Bayesianos sem informações a “priori” sobre a
origem das amostras. Foram utilizadas 100.000 simulações de Cadeias de Markov
Monte Carlo com burn in de 50.000 e modelo de ancestralidade admixture. Foram

32
testados valores de K variando de 1 a 8. A determinação do K mais provável em
relação aos propostos foi realizada utilizando valores de ΔK (EVANNO; REGNAULT;
GOUDET, 2005), como segue:
| |
[ ]
onde, m = média, s = desvio padrão e K = número de grupos proposto. E L’ sendo:
[ ]
33
4. RESULTADOS
4.1. Marcadores RAPD selecionados
Dentre os primers testados, apenas cinco apresentaram padrão de bandas
nítido e presença de polimorfismos (Tabela 3). Juntos, estes somaram um total de
37 bandas (locos), sendo 36 polimórficas (97,2%), conforme é mostrado na Tabela
3.
Foi observada uma variação de 6 a 9 bandas amplificadas entre os cinco
primers (Tabela 3), resultando numa média de 7,4 bandas por primer. Na Figura 5,
observa-se o padrão de bandas obtido pelos primers RAPD.
Tabela 3 - Primers de RAPD selecionados, com suas respectivas sequências,
número de fragmentos amplificados e polimórficos.
NTLA= Número total de locos amplificados; NTLP = Número total de locos polimórficos; PP = porcentagem de polimorfismo.
Primer Sequência NTLA NTLP PP (%)
UBC 180 5’-GGGCCACGCT-3’ 7 7 100
UBC 506 5’-CCTTTCCCGA-3’ 6 6 100
UBC 549 5’-CCGGCTTATG-3’ 9 8 88,8
UBC 556 5’-ATGGATGACG-3’ 8 8 100
UBC 691 5’-AAACCAGGCG-3’ 7 7 100
Total 37 36 97,2
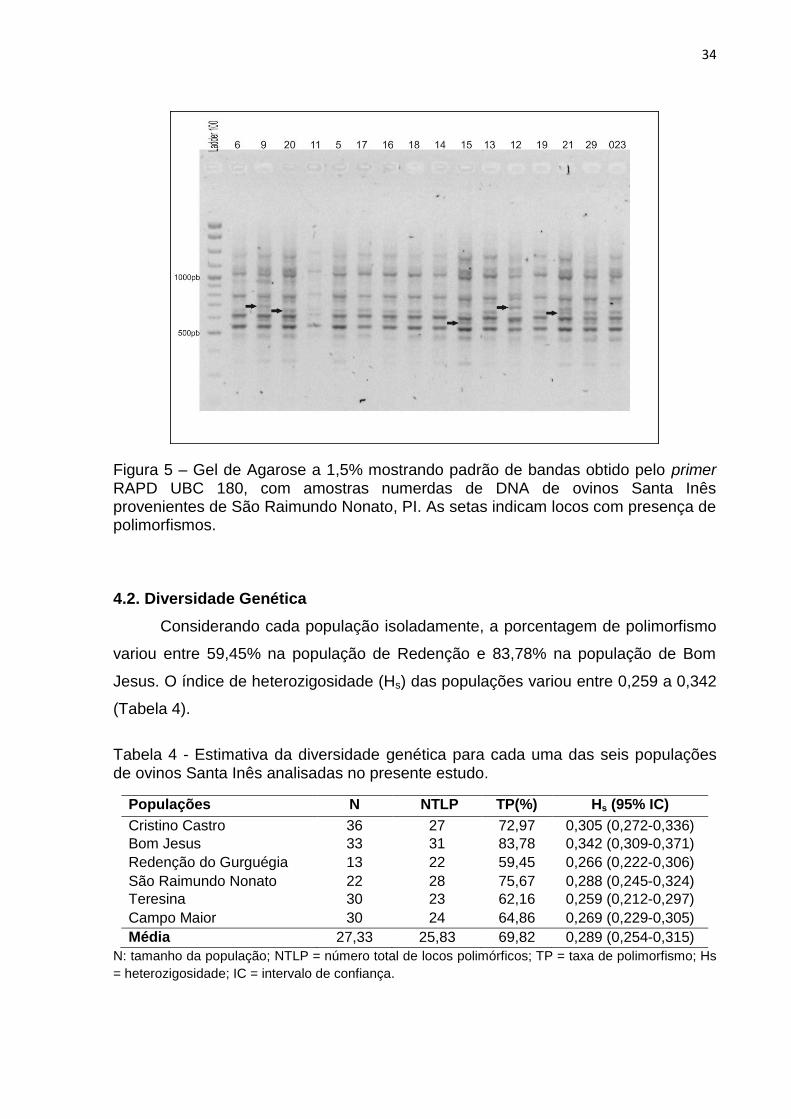
34
Figura 5 – Gel de Agarose a 1,5% mostrando padrão de bandas obtido pelo primer RAPD UBC 180, com amostras numerdas de DNA de ovinos Santa Inês provenientes de São Raimundo Nonato, PI. As setas indicam locos com presença de polimorfismos.
4.2. Diversidade Genética
Considerando cada população isoladamente, a porcentagem de polimorfismo
variou entre 59,45% na população de Redenção e 83,78% na população de Bom
Jesus. O índice de heterozigosidade (Hs) das populações variou entre 0,259 a 0,342
(Tabela 4).
Tabela 4 - Estimativa da diversidade genética para cada uma das seis populações de ovinos Santa Inês analisadas no presente estudo.
Populações N NTLP TP(%) Hs (95% IC)
Cristino Castro 36 27 72,97 0,305 (0,272-0,336)
Bom Jesus 33 31 83,78 0,342 (0,309-0,371)
Redenção do Gurguégia 13 22 59,45 0,266 (0,222-0,306)
São Raimundo Nonato 22 28 75,67 0,288 (0,245-0,324)
Teresina 30 23 62,16 0,259 (0,212-0,297)
Campo Maior 30 24 64,86 0,269 (0,229-0,305)
Média 27,33 25,83 69,82 0,289 (0,254-0,315)
N: tamanho da população; NTLP = número total de locos polimórficos; TP = taxa de polimorfismo; Hs
= heterozigosidade; IC = intervalo de confiança.

35
Foi obtido o coeficiente de endogamia f que mostrou média de 0,172 (IC =
0,148-0,493). O coeficiente diferenciação entre populações θII apresentou valor de
0,132 (IC = 0,098-0,175).
4.3. Análise de Variância Molecular (AMOVA) e Estrutura Populacional
A análise revelou que a maior parte da variância foi encontrada dentro das
populações de forma geral (80,03%); alta variabilidade essa pode ser atribuída às
diferenças genéticas entre as fêmeas do rebanho. Porém as diferenças entre
populações pode ser considerada significativa, de acordo com os valores de ΦST
(0,199; P < 0,001). Considerando a organização das amostras entre região Centro-
Norte e Sul do Piauí, a variação encontrada nos dados mostra que 8,42% desta
pode ser explicada pela variação entre regiões (ΦCT = 0,084; P < 0,001). Por fim,
observou-se que 11,55% da variação é explicada pelas diferenças encontradas
entre populações dentro das regiões (ΦSC = 0,126; P < 0,001).
Tabela 5 - Análise de Variância Molecular (AMOVA) para as populações de ovinos Santa Inês no estado do Piauí.
Fonte de Variação GL SQ Componente de Variância
% Total Φ P*
Entre Regiões 1 47,201 0,374 8,42% 0,084a <0,001
Entre populações
dentro das regiões
4 67,815 0,514 11,55% 0,126b <0,001
Dentro das
populações
157 559,437 3,563 80.03% 0,199c <0,001
Total 162 674,454 4,45247 100%
GL: grau de liberdade; SQ: soma dos quadrados; Φ: estatísticas F para marcadores
dominantes; P*: significância a 0,001 calculado após 1000 permutações; a = ΦCT; b =
ΦSC, c = ΦST
Apesar dos valores obtidos na AMOVA, não foi observada relação entre
diferenciação genética e distância geográfica. O dendrograma obtido através das
análises de agrupamento com valores de ΦST par-a-par mostra este fato (Figura 5),
bem como os resultados do teste de Mantel, que não mostrou-se significativo (r2 =
0,30; P = 0,106). A única correlação positiva foi observada entre as populações de
Teresina e Cristino Castro, com a maior distância genética (ΦST =0,314), as quais
encontram-se a 561 km de distância (Tabela 6). Porém, altos valores de distância
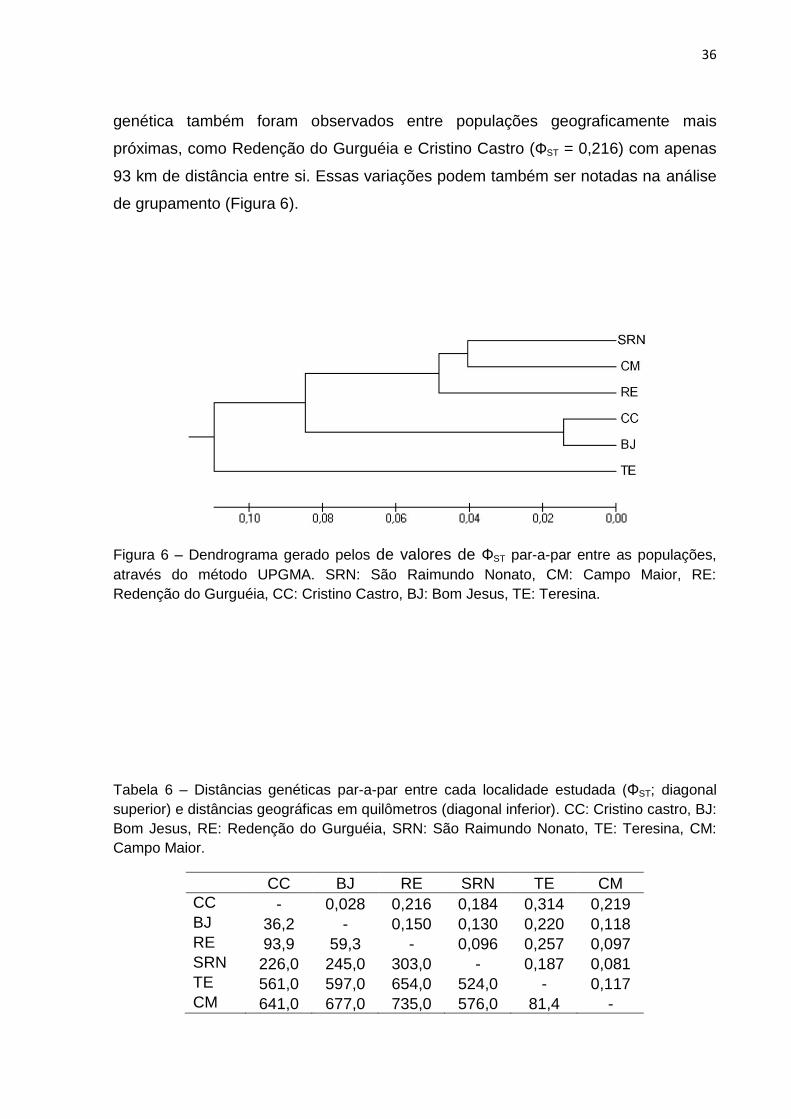
36
genética também foram observados entre populações geograficamente mais
próximas, como Redenção do Gurguéia e Cristino Castro (ΦST = 0,216) com apenas
93 km de distância entre si. Essas variações podem também ser notadas na análise
de grupamento (Figura 6).
Figura 6 – Dendrograma gerado pelos de valores de ΦST par-a-par entre as populações,
através do método UPGMA. SRN: São Raimundo Nonato, CM: Campo Maior, RE:
Redenção do Gurguéia, CC: Cristino Castro, BJ: Bom Jesus, TE: Teresina.
Tabela 6 – Distâncias genéticas par-a-par entre cada localidade estudada (ΦST; diagonal
superior) e distâncias geográficas em quilômetros (diagonal inferior). CC: Cristino castro, BJ:
Bom Jesus, RE: Redenção do Gurguéia, SRN: São Raimundo Nonato, TE: Teresina, CM:
Campo Maior.
CC BJ RE SRN TE CM
CC - 0,028 0,216 0,184 0,314 0,219 BJ 36,2 - 0,150 0,130 0,220 0,118 RE 93,9 59,3 - 0,096 0,257 0,097 SRN 226,0 245,0 303,0 - 0,187 0,081 TE 561,0 597,0 654,0 524,0 - 0,117 CM 641,0 677,0 735,0 576,0 81,4 -
37
Os resultados obtidos com o programa STRUCTURE mostraram que o valor de
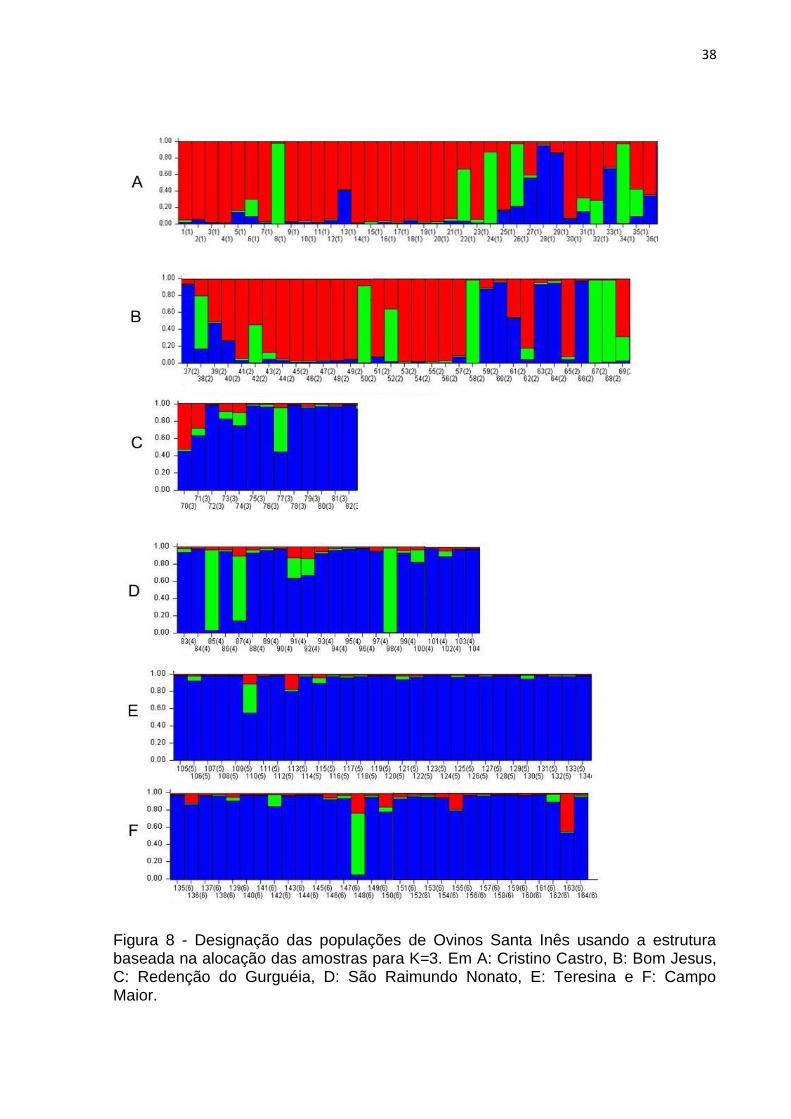
ΔK mais provável para agrupar as populações foi igual a três (Figura 7). A Figura 8
mostra a distribuição das seis populações coletadas organizadas em três grupos.
Dois destes representaram a maioria dos indivíduos avaliados (grupo “vermelho” e
“azul” no gráfico) e, um terceiro grupo menor apareceu representado por poucos
indivíduos (grupo “verde) (Figura 8). Nota-se que assim como na análise de
grupamento, as populações de Bom Jesus e Cristino Castro apareceram mais
similares, tendo a maioria de seus representantes no “grupo vermelho” (Figura 8),
enquanto as outras populações foram representadas principalmente por indivíduos
do “grupo azul” (Figura 8). A população de Teresina foi a que menos apresentou
introgressão de alelos oriundos de outras localidades (Figura 8), sendo considerada
a mais homogênea. Não obstante, nota-se ainda que a população de Teresina
também foi a que apresentou menor heterozigosidade (HS = 0,259; ver Tabela 4).
Quanto ao “grupo verde”, observa-se que em praticamente todas as
populações há representantes desse grupo (Figura 8).
Figura 7 - Representação gráfica do ΔK mais provável de acordo com Evanno; Regnault e Goudet (2005).
0
50
100
150
200
250
0 1 2 3 4 5 6 7 8 9 10
Δ K
K
38
Figura 8 - Designação das populações de Ovinos Santa Inês usando a estrutura baseada na alocação das amostras para K=3. Em A: Cristino Castro, B: Bom Jesus, C: Redenção do Gurguéia, D: São Raimundo Nonato, E: Teresina e F: Campo Maior.

39
5. DISCUSSÃO
O alto polimorfismo encontrado (97,2%) nas amostras de DNA de ovinos
Santa Inês através do uso de marcadores RAPD pode representar uma variabilidade
genética alta das populações, porém é preciso levar em consideração que se obteve
somente de 6 a 8 bandas polimórficas por primer. Outros estudos sobre
caracterização genética de ovinos com RAPD (KHALDI et al., 2010) e outras
espécies de ruminantes (ALBUQUERQUE et al., 2006) mostraram resultados
semelhantes, evidenciando que, mesmo com um pequeno número de marcadores é
possível mensurar a variabilidade genética devido ao alto grau de polimorfismo
destes marcadores. Considerando a análise para cada população, isoladamente, a
taxa de locos polimórficos variou de 59,45% a 83,78%, sendo a menor taxa
encontrada na população de Redenção do Gurguéia. No entanto, este resultado
pode refletir o baixo número de amostras analisadas neste município.
O valor encontrado de diversidade genética (Hs), medido pelo índice de
heterozigosidade, revela variação entre populações. A estimativa apresentou menor
valor para Teresina (0,289) maior para Bom Jesus (0,342). Esta característica
também ficou representada pela análise realizada no programa Structure, onde a
população de Teresina não mostrou grande variação de cor (“grupo azul”). No
entanto, mesmo com esta constatação, esses valores podem ser considerados
baixos, revelando a predominância de indivíduos homozigotos. Este fato pode estar
atrelado ao uso de poucos reprodutores e/ou cruzamentos entre meio-irmãos nas
localidades, os quais levam ao aumento da endogamia. Esta prática pode levar a um
processo conhecido como “Depressão Endogâmica” que pode reduzir sensivelmente
a produtividade de um rebanho (BREDA et al.,2004; CARNEIRO et al., 2007).
Os valores de Hs encontrados neste estudo (média de 0,289) foram um pouco
abaixo dos valores encontrados por Paiva et al. (2003), que obtiveram uma média de
Hs = 0,367 também estudando a raça Santa Inês (populações de Sergipe, Goiás e
Distrito Federal), e de Hs = 0,322 para a raça ovina Somalis usando marcadores
RAPD. Esses resultados podem ser um indicativo da ocorrência de deriva genética
e/ou endogamia. Segundo Eding (2001), a perda contínua de diversidade genética
de uma raça reduz a possibilidade de melhoramento genético no futuro.
Corroborando com estes resultados, o índice de endogamia estimado (f =
0,172) indicou presença de endogamia dentro das populações, porém, a medida

40
deve ser interpretada com cautela já que o intervalo de confiança obtido foi
considerado alto (0,010 a 0,492; a 95%). Entretanto, mesmo nessas condições, as
populações usadas no presente estudo eram formadas na sua maioria por fêmeas,
indicando o pequeno número de machos reprodutores dentro dos rebanhos
estudados. Como consequência direta, pode-se esperar o aumento da endogamia
observada pela presença constate de meios-irmãos dentro da população. Rego Neto
(2013) em seu trabalho de estrutura genética de populações de ovinos Santa Inês
no Estado do Piauí através de dados de pedigree observou que, na população
estudada, o nível de endogamia aumentou entre os anos de 2010 e 2012, por
consequência da diminuição do tamanho efetivo das populações. Diante disso a
raça encontra-se em estado de monitoramento no Estado, pois populações com
tamanho efetivo reduzido têm maior probabilidade de perda da diversidade genética
e consequentemente, aumento no coeficiente de endogamia.
Teixeira Neto et al. (2013) obtiveram um valor de Fis = -0,005 através de
dados de pedigree em rebanhos Santa Inês dos estados da Bahia, Ceará, Paraíba,
Pernambuco, Piauí, Rio Grande do Norte, Sergipe e São Paulo. O valor negativo foi
justificado pelos autores como um provável resultado do consórcio de reprodutores,
prática utilizada por produtores dos rebanhos avaliados, para redução de custo na
aquisição de animais melhoradores. Dados moleculares, como os do presente
estudo, são mais precisos na estimativa de índice de endogamia, pois dados de
pedigree podem conter erros ou estar incompletos. Entretanto a prática de consórcio
de reprodutores, se bem conduzida, pode ser estratégia para diminuir os índices de
endogamia acima do limite em rebanhos e favorecer o melhoramento genético da
raça.
Outro fator resultante da endogamia dentro dos rebanhos é a justamente o
acúmulo de diferenças interpopulacionais. Essas diferenças foram avaliadas pelos
índices θII e ΦST, que apresentaram valores considerados de moderados a altos. Hartl
e Clark (2010) afirmam que quando há subdivisão populacional, quase não se pode
evitar a ocorrência de alguma diferenciação genética entre as populações, visto que
as frequências alélicas entre as subpopulações se tornam diferentes ao longo do
tempo. As diferenças encontradas foram avaliadas com a AMOVA que revelou que
cerca de 8% da variação pode ser explicada pela divisão entre as regiões Centro-
Norte e Sul do Piauí. Já as diferenças entre populações explicam cerca de 20% da
variação. Porém, as análises realizadas no programa STRUCTURE mostraram que,

41
mesmo com a subdivisão populacional sendo significativa e com a baixa
heterozigosidade encontrada, há compartilhamento de alelos entre as populações. A
única localidade que mostrou baixa introgressão de alelos foi Teresina, onde
também foi observada a menor heterozigosidade, fato que pode ser justificado pelo
uso dos animais, na sua maioria, para fins de exposição; o que é uma prática
comum entre os produtores do município.
De acordo com o Φst par a par de cada localidade, as populações de Bom
Jesus e Cristino Castro são mais assemelhadas, podendo esse ser um reflexo da
proximidade entre os municípios (36,2 Km), e consequentemente o uso do(s)
mesmo(s) reprodutor(es). Os demais grupamentos formados não seguem nenhum
padrão específico de estruturação que esteja relacionado com a posição geográfica
das amostras, sendo este fato confirmado pelo teste de Mantel. Como não se sabe
da ancestralidade dos animais em questão, pode-se supor que o comércio de ovinos
existente entre os municípios colabora com esses resultados. Outro ponto a ser
destacado é entre os municípios de Campo Maior e Teresina, que além da
proximidade geográfica, compartilham alguns produtores e reprodutores. A falta de
relação positiva encontrada no presente trabalho pode ser devido a fazendas sem
relacionamento ou até mesmo aquisição de reprodutores da “velha Santa Inês” (ver
próximo parágrafo). Assim, para ovinos onde há manipulação antrópica, padrões de
isolamento genético por distância não são tão óbvios quanto em populações naturais
(SUN et al., 2007).
A estrutura das amostras nos três grupos indicados pelas análises do
STRUCTURE também pode estar relacionada com a presença de animais
pertencentes ao “Novo Santa Inês”, citado por Carneiro et al. (2010) e Paiva et al.
(2005b). Estes autores concluíram que raças de ovinos nativos no Brasil eram
intimamente relacionadas por ter ocorrido cruzamentos aleatórios no passado, além
da ação da deriva genética e mesmo com esse fato, as raças nativas apresentam
maior diversidade genética do que raças comerciais. Em alguns casos, as distâncias
entre as populações de Santa Inês eram maiores que a distância entre as raças, o
que reforça a hipótese da existência de uma “velha raça Santa Inês” versus a “nova
raça Santa Inês”. Criadores e técnicos classificam a “velha Santa Inês” como
animais mais rústicos e menores, que eram predominantes nas décadas de 1980 e
1990. A “nova Santa Inês” é descrita com o fenótipo que temos atualmente, como
um animal de grande porte e bem desenvolvido, que tem justificativa mais provável

42
os cruzamentos com a raça Suffolk do que a seleção dentro da própria raça
(McMANUS et al., 2010; CARNEIRO et al., 2010). Entretanto, informações
fenotípicas são necessárias para afirmar a presença desses duas linhagens entre os
animais analisados no presente estudo.
Assim, este trabalho traz resultados preliminares sobre a estruturação e a
diversidade genética de importantes rebanhos da raça Santa Inês localizados no
Estado do Piauí. Estes dados serão importantes para nortear futuras pesquisas onde
serão considerados marcadores com maior poder de resolução e populações mais
representativas.

43
6. CONCLUSÕES E RECOMENDAÇÕES
Constatou-se a prática de endocruzamentos nos rebanhos estudados através
dos altos índices de f, θII e Φst, porém a variabilidade ainda encontrada dentro
das populações revela que há diversidade genética entre os indivíduos dentro
das populações.
As seis populações estudadas formaram apenas três grupos genéticos, sendo
que a população de Teresina se mostrou a mais endogâmica.
Recomenda-se aos produtores a aquisição de mais animais reprodutores ou
de origens diferentes e/ou o rodízio destes entre as fazendas, para que assim
os efeitos da endogamia sejam diminuídos e a diversidade genética possa
aumentar nos rebanhos de Santa Inês no Estado do Piauí.

44
7. REFERÊNCIAS BIBLIOGRÁFICAS
ABSI. Associação Brasileira de Santa Inês: A Raça. Disponível em: www.absantaines.com.br. Acesso em 29.08.2012.
ALBUQUERQUE, M. S. M.; EGITO, A. A.; MARQUES, J. R. F.; CIAMPI, A. Y.; MARIANTE, A. S.; CASTRO, S.T. R.; COSTA,M. R.; PAIVA, S. R.; SILVA, A. M.; CONTEL, E. P. B. Variabilidade genética em búfalos estimada por marcadores RAPD. Pesquisa Agropecuária Brasileira, Brasília, v.41, n.4, p.623-628, 2006.
AMARANTE, A. F. T; BRICARELLO, P. A.; ROCHA, R. A.; GENNARI, S. M. Resistance of Santa Inês, Suffolk and Lle de France sheep to naturally acquired gastrointestinal nematode infections. Veterinary Parasitology, ed. 120, p. 91-106, 2004.
ARANDAS, J. K. G.; RIBEIRO, M. N.; PIMENTA FILHO, E. C.; DA SILVA, R. C. B.;
FACÓ, O.; ESTEVES, S. N. Estrutura Populacional de Ovinos da Raça Morada
Nova. In: IX SIMPÓSIO BRASILEIRO DE MELHORAMENTO ANIMAL, Anais... João
Pessoa. 2012.
ARAÚJO, A. M.; SIMPLÍCIO, A. A. Melhoramento Genético em Caprinos e Ovinos no
Brasil: Importância do padrão racial. In: SIMPÓSIO NACIONAL DE
MELHORAMENTO ANIMAL, Anais... Belo Horizonte, 2000.
ARCO. Associação Brasileira de Criadores de Ovinos: Padrões raciais. Disponível em: www.arcoovinos.com.br. Acesso em 19.07.2012. BIAGIOTTI, D.; NERI, V. S.; DO Ó, A.O.; TORRES, T.S.; GUIMARÃES, F. F.; PIRES, L.C.; SARMENTO, J.L.R. Diversidade biométrica entre ovinos criados no Estado do Piauí utilizando analise multivariada. In: IX SIMPÓSIO BRASILEIRO DE MELHORAMENTO ANIMAL, Anais... João Pessoa, 2012. BIAGIOTTI, D.; SARMENTO, J.L.R; Ó, A.O. do; RÊGO NETO, A. de A.; SANTOS; G. V. dos; SANTOS, N. P. da S.; TORRES, T.S.; NERI, V. S. Caracterização fenotípica de ovinos da raça Santa Inês no Estado do Piauí. Revista Brasileira de Saúde e Produção Animal, v.14, n.1, p.29-42, 2013. BREDA F.C., EUCLYDES R.F., PEREIRA C.S., TORRES R. de A., CARNEIRO
P.L.S., SARMENTO R.L., FILHO R. de A. T., MOITA A.K.F. Endogamia e limite de
seleção em populações selecionadas obtidas por simulação. Revista Brasileira de
Zootecnia, v.33, p. 2017-2025, 2004.
CARNEIRO, H.; LOUVANDINI, H.; PAIVA, S.R.; MACEDO, F.; MERNIES, B.;
McMANUS, C. Morphological characterization of sheep breeds in Brazil, Uruguay
and Colombia. Small Ruminant Research, v.94, p.58-65, 2010.
CARNEIRO, P.L.S.; MALHADO, C.H.M.; SOUZA JÚNIOR, A. A. O.; SILVA, A. G. S.;
SANTOS, P.F. et al. Desenvolvimento ponderal e diversidade fenotípica entre

45
cruzamentos de ovinos Dorper com raças locais. Pesquisa Agropecuária
Brasileira, v. 42, p. 991-998, 2007.
CARNEIRO, T.X.; GONÇALVES, E.C.; SCHNEIDER, M.P.C.; SILVA, A. Diversidade
genética e eficiência de DNA microssatélites para o controle genealógico da raça
Nelore. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.59, n.5, p.1257-
1262, 2007.
CARVALHO, D. M; SOUZA, J.P. Análise da cadeia produtiva de caprino-ovinocultura em Garanhuns. In: XLVI Congresso da Sociedade Brasileira de Economia, Administração e Sociologia Rural. Anais... Acre, 2008.
CASTIGLIONI, L.; BICUDO, H. E. M. C. A Técnica de RAPD (Random Amplified
Polymorphic DNA) e suas aplicações para estudos em genética molecular. Revista
UNORP, v. 3, n.2, p. 63-77, 2003.
CEZAR, M. F.; SOUZA, B. B. de; SOUZA, W. H. de; PIMENTA FILHO, E. C.; TAVARES, G.de P.; MEDEIROS, G. X. Avaliação de parâmetros fisiológicos de ovinos Dorper, Santa Inês e seus mestiços perante condições climáticas do trópico semi-árido nordestino. Ciência e Agrotecnologia, v.28, n.3, p. 614 – 620, 2003. COCKERHAM, C.C. Variance of gene frequencies. Evolution, v. 23, p.72–84, 1969. COCKERHAM, C.C. Analyses of gene frequencies. Genetics, v. 74, p.679–700, 1973.
COSTA JÚNIOR, G. da S.; CAMPELO, J. E. G.; AZEVÊDO, D. M. M. R.; MARTINS FILHO, R.; CAVALCANTE, R. R.; LOPES, J. B.; OLIVEIRA, M. E. Caracterização morfométrica de ovinos da raça Santa Inês criados nas microrregiões de Teresina e Campo Maior, Piauí. Revista Brasileira de Zootecnia, v. 35, n.6, p. 2260-2267, 2006. EDING, H. Conservation of genetic diversity: assessing genetic variation using
marker estimated kinships. Thesis (Ph.D. in Animal Science). Wageningen,
Wageningen University, 79p, 2001.
EGITO, A. A.; MARIANTE, A. S.; ALBUQUERQUE, M. S. M. Programa brasileiro de conservação de recursos genéticos animais. Archivos de Zootecnia, v. 51, p. 39-52, 2002.
EVANNO, G.; REGNAULT, S.; GOUDET, J. Detecting the number of clusters of
individuals using the software STRUCTURE: a simulation study. Molecular Ecology.
v. 14, p. 2611-2620, 2005.
EXCOFFIER, L.; LAVAL, G.; SCHNEIDER, S. Arlequin (version 3.0): An integrated
software package for population genetics data analysis. Evolutionary
Bioinformatics Online, v. 1, p. 47-50, 2005.

46
EXCOFFIER, L.; SMOUSE, P. E.; QUATTRO, J. M. Analysis of molecular variance
inferred from metric distances among DNA haplotypes: application to human
mitochondrial DNA restriction data. Genetics, v.131, p. 479–491, 1992.
FALEIRO, F.G. Marcadores genético moleculares aplicados a programas de
conservação e uso de recursos genéticos. Embrapa Cerrados, Planaltina, DF,
107p., 2007.
FARIA, L. A. Estratégias de coordenação da cadeia da carne ovina e a qualidade de carne, 2008. Disponível em: www.farmpoint.com.br. Acesso em 09.02.14.
FARIA, L.A.; SILVA, D.J. Segmentos de distribuição e consumo na cadeia da ovinocultura brasileira. Instituto Didatus de Ensino e Qualificação. 2006. Disponível em: http://www.didatus.com.br/noticias/noticia.asp?cod=42. Acesso em 09.02.14.
FARID, A.; O’REILLY, E.; DOLLARD, C.; KELSEY JR, C.R. Genetic analysis of ten sheep breeds using microsatellite markers. Canadian Journal of Animal Science, v. 80, p. 9–17, 2000.
FERREIRA, M. E.; GRATTAPAGLIA, D. Introdução ao uso de marcadores
moleculares em análise genética. 3 ed., Embrapa CENARGEN, Brasília, DF. 1998
GUIMARÃES, F. F.; REGO NETO, A. A.; ROCHA, L. M. C.; DO Ó, A.O.; BIAGIOTTI, D.; SARMENTO, J.L.R. Parâmetros populacionais de ovinos Santa Inês na região sul do Piauí. In: IX SIMPÓSIO BRASILEIRO DE MELHORAMENTO ANIMAL, Anais... João Pessoa. 2012.
HARTL, D. L. e A. G., CLARK. Princípios de genética de populações. Editora Artmed, 4ª edição, 2010.
HEDRICK, PHILIP. Genetics of Populations. Jones e Bartlett Learning, 4th Edition,
2009.
HEGGEBØ, R.; PRESS, C.M.; GUNNES, G.; GONZÁLEZ, L.; JEFFREY, M. Distribution and accumulation of PrP in gut-associated and peripheral lymphoid tissue of scrapie-affected Suffolk sheep. Journal of General Virology, v.83, n.2, p.479-489, 2002.
HIRAGI, C.; SIMÕES, K.; MARTINS, E.; QUEIROZ, P.; LIMA, LUZIA; MONNERAT, R. Variabilidade Genética em Populações de Aedes aegypti (L.) (Diptera:Culicidae) Utilizando Marcadores de RAPD. Neotropical Entomology, v.38, n.4, p.542-547, 2009.
HOLSINGER, K.E.; LEWIS, P.O.; DEY, D.K. A Bayesian approach to inferring
population structure from dominant markers. Molecular Ecology, v.11, p.1157-
1164, 2002.

47
IANELLA, P.; McMANUS, C.; CAETANO, A.R.; MARTINS, C. F.; SOUZA, C. J.H. de; FACÓ, O.; AZEVEDO, H.C.; CARNEIRO, P.S.; PAIVA, S.R. Avaliação dos polimorfismos do gene PRNP ligados a scrapie clássica em núcleos de conservação de ovinos do Brasil. In: SIMPOSIO DE RECURSOS GENETICOS DA AMERICA LATINA Y EL CARIBE, 7., 2009, Pucon. Proceedings... Pucon: v.2, p.265-266. 2009.
IBGE. Produção da Pecuária Municipal, volume 39, 2011.
IBGE. Produção da Pecuária Municipal, volume 40, 2012.
KHALDI, Z.; REKIK, B.; HADDAD, B.; ZOURGUI, L.; SOUID, S. Genetic
characterization of the three ovine breeds in Tunisia using randomly amplified
polymorphic DNA markers. Livestock Research for Rural Development, v. 22, n.
3, 2010.
KIJAS, J.W.; LENSTRA, J. A.; HAYES, B.; BOITARD, S.; NETO, L. R. P.;
CRISTOBAL, M. S.; SERVIN, B.; McCULLOCH, R.; WHAN, V.; GIETZEN, K.;
PAIVA, S.; BARENDSE, W.; CIANI, E.; RAADSMA, H.; McEWAN, J.; DALRYMPLE,
B. Genome-wide analysis of the world´s sheep breeds reveals high levels of historic
mixture and strong recent selection. PLoS Biology, v.10 n.2, 2012.
KINGHORN, B.P. An index of information content for genotype probabilities derived from segregation analysis. Genetics, v. 145, p. 479-483, 1997.
KINGHORN, B.P.; KENNEDY, B.W.; SMITH, C. A method for screening for genes of major effect. Genetics, v.134, p. 351-360, 1993. KUMAR, S., NEI, M., DUDLEY, J., TAMURA, K. MEGA: a biologist-centric
softwarefor evolutionary analysis of DNA and protein sequences. Briefings in
Bioinformatics, v. 9, p. 299-306. 2008.
LÔBO, R. N. B.; LÔBO, A. M. B. O. Melhoramento genético como ferramenta para o
crescimento e o desenvolvimento da ovinocultura de corte. Revista Brasileira de
Reprodução Animal, v.31, n.2, p.247-253, 2007.
MARIANTE, A. S.; CAVALCANTE, N. Animais do Descobrimento: Raças
domésticas da história do Brasil. Brasília, 2ª ed. Embrapa. 2006.
MASSAGO, H.; RIBEIRO, R. P.; BARRERO, N. M. L.; POVH, J. A.; CASTAGNOLLI,
N.; GOMES, P.C. Diversidade genética de quatro linhagens de Oreochromis niloticus
utilizando o marcador RAPD. Bioscience Journal, v. 25, n.4, p.150-159, 2009.
MBURU, D.; HANOTTE, O. A practical approach to microsatellite genotyping with
special reference to livestock population genetics. ILRI Biodiversity project, Nairobi,
Kenya, 2005.

48
McMANUS, C.; PAIVA, S. R.; ARAÚJO, R.O. Genetics and breeding of sheep in
Brazil. Revista Brasileira de Zootecnia, v. 39, p. 236-246, 2010.
MCMANUS, C.; PAIVA, S. Estatísticas para descrever genética de populações. Informação Genético-Sanitária da Pecuária Brasileira. Publicado on-line em www.animal.unb.br em 07.01.2011.
MENEZES, M. P. C.; MARTINEZ, A. M.; RIBEIRO, M. N.; PIMENTA FILHO, E. C.;
BERMEJO, J. V. D. Caracterização genética de raças caprinas nativas brasileiras
utilizando-se 27 marcadores microssatélites. Revista Brasileira de Zootecnia, v.35,
n.4, p.1336-1341, 2006.
MORAIS, O.R. O melhoramento genético dos ovinos no Brasil: situação atual e
perspectivas para o futuro. In: SIMPÓSIO NACIONAL DE MELHORAMENTO
ANIMAL, 2000, Belo Horizonte. Anais... Belo Horizonte: FEPMVZ, p.266-272, 2000.
MORAIS, O. R.; MADALENA, F. E. Economic values for production traits in Santa
Inês sheep. In: 8th World Congress on Genetics Applied to Livestock Production.
Anais… Belo Horizonte, 2006.
MULLIS, K.; FACOMA, F.; SCHARF, S.; SNIKL, R.; HORN, G.; ERLISH, H. Specific amplification of DNA in vitro: the polymerase chain reaction. Cold Sping Harbor Symposium Quantitative Biology, v. 51, p.260, 1986.
NEI, M. Analysis of gene diversity in subdivided populations. Proceedings of the
National Academy of Sciences, v. 70, p. 3321–3323, 1973.
NEI, M. Molecular Evolutionary Genetics. New York, NY: Columbia University
Press, 1987.
NETO, A. C. B. Avaliação de cruzamentos de ovinos das raças Dorper, Poll
Dorset, Santa Inês, e Somalis Brasileira. 45 f. Dissertação de Mestrado em
Zootecnia - CCA, Universidade Federal do Ceará, Fortaleza, 2008.
OJEDA, D.B. Participação do melhoramento genético na produção ovina. Revista
Brasileira de Reprodução Animal, v.23, n.2, p.146-149, 1999.
OLIVEIRA, K. A. P. de; LÔBO, R. N. B.; FACÓ, O. Genetic evaluation of partial
growth trajectory of Santa Inês breed using random regression models. Revista
Brasileira de Zootecnia, v. 39, n.5, p. 1029 – 1036, 2010.
PAIVA, S. R.; PIMENTEL, C. McM. Uso de marcadores moleculares como
ferramenta adicional para conservação e melhoramento da raça Santa Inês. In: V
Congresso Nordestino de produção animal, 2008, Aracajú. Anais... Aracajú:
Sociedade Nordestina de Produção Animal, 2008.
PAIVA, S. R.; FARIA, D. A.; SILVÉRIO, V. C.; McMANUS, C.; EGITO, A. A.;
DERGAM, J. A.; GUIMARÃES, S. E. F.; CASTRO, S. T. R.; ALBUQUERQUE, M. S.

49
M.; MARIANTE, A. S. Genetic variability among Brazilian sheep using microsatellites.
The role of biotechnology for the characterization and conservation of crop,
forestry, animal and fishery genetic resources. Turim, Itália: Rome, Itália : FAO
Working Group on Biotechnology, p.195-196, 2005c.
PAIVA, S. R.; FARIA, D. A.; SILVÉRIO, V. C.; McMANUS, C.; EGITO, A. A.;
DERGAM, J. A.; GUIMARÃES, S. E. F.; CASTRO, S. T. R.; ALBUQUERQUE, M. S.
M.; MARIANTE, A. S. Phylogenetic relationships among brazilian sheep breeds. The
role of biotechnology, 2005d.
PAIVA, S. R.; SILVÉRIO, V. C.; EGITO, A. A.; MCMANUS, C.; FARIA, D. A.;
MARIANTE, A. S.; CASTRO, S.R.; ALBUQUERQUE, M.S.M.; DERGAM, J. Genetic
variability of the brazilian hair sheep breeds. Pesquisa Agropecuária Brasileira,
v.40, n.9, p.887-893, 2005b.
PAIVA, S. R.; SILVÉRIO, V. C.; EGITO, A. A.; MCMANUS, C.; FARIA, D. A.;
MARIANTE, A. S.; CASTRO, S.R.; ALBUQUERQUE, M.S.M.; DERGAM, J.
Caracterização Genética da raça Santa Inês. In: 2ND International Symposium on
Sheep and Goat Production, 2003, João Pessoa. Anais… João Pessoa: Empresa
Estadual de Pesquisa Agropecuária da Paraíba, p. 487- 499, 2003.
PAIVA, S. R.; SILVÉRIO, V. C.; PAIVA, D. A. de F.; McMANUS, C.; EGITO, A. A.;
MARIANTE, A. da S.; CASTRO, S. R.; ALBUQUERQUE, M.S.M.; DERGRAM, J. A.
Origin of the main locally adapted sheep breeds of Brazil: a RFLP-PCR molecular
analysis. Archivos de Zootecnia, v. 54, p. 395-399, 2005a.
PAVLOV, A.R.; PAVLOVA, N.V.; KOZYAKIN, S.A.; SLESAREV, A.I. Recent development in the optimization of termostable SNA polymerases for efficient applications. Trends Biotechnology, v. 22, p. 253-260, 2004.
PETROLI, C. D. ; PAIVA, S. R. ; CORREA, M. P. C. ; McMANUS, C. Genetic
monitoring of a Santa Ines herd using microsatellite markers near or linked to the
sheep MHC. Revista Brasileira de Zootecnia, v.38, n.4, p. 670-675, 2009.
PINHEIRO JÚNIOR, J. W.; OLIVEIRA, A. A. Da F.; ANDERLINI, G. A.; ABREU, S. R. De O.; VALENÇA, R. M. B.; MOTA, R. A. Aspectos sociais, higiênico-sanitários e reprodutivos da ovinocultura de corte do Estado de Alagoas, Brasil. Revista Brasileira de Ciências Agrárias, v. 5, n. 4, p. 600-605, 2010.
PRITCHARD, J. K.; STEPHENS, M.; DONNELLY, P. Inference of population
structure using multilocos genotype data. Genetics, v. 115, p. 945-959, 2000.
PRIMO, AT. América: conquista e colonização. Porto Alegre: Movimento, 2004.
QUESADA, M.; McMANUS, C.; COUTO, F. A. D’A. Efeitos genéticos e fenotípicos
sobre características de produção e reprodução de ovinos deslanados no Distrito
Federal. Revista Brasileira de Zootecnia, v.31, n.1, p. 342-349, 2002 (suplemento).

50
RAMPELOTTI, F. T.; FERREIRA, A.; TCACENCO, F. A.; MARTINS, J. F. DA S.; GRÜTZMACHER, A. D. E PRANDO, H. F. Diversidade genética de Tibraca limbativentris Stål (Hemiptera:Pentatomidae) de Santa Catarina e do Rio Grande do Sul, usando marcadores RAPD. Neotropical Entomology, v.37, n.1, 2008. REGO NETO, A. A. Estrutura genética da população de ovinos Santa Inês no
Estado do Piauí. Dissertação de Mestrado em Ciência Animal - CCA, Universidade
Federal do Piauí, Teresina, 2013.
ROCHA, S. S. N. da; SILVA, D. M. F. da; LEAL, C. A. de S.; ROCHA NETO, M. N.
da; OLIVEIRA, A. E. T.; ADRIÃO, M.; WISCHRAL, A. Caracterização gênica de
ovinos nativos, da raça Santa Inês, para o gene da BMP15. Anais... IX Jepex,
Recife, 2009.
RODRIGUES, D. S. Estrutura Populacional de um Rebanho Morada Nova
Variedade Branca no Estado do Ceará. 2009. 45 f. Dissertação de Mestrado em
Zootecnia - CCA, Universidade Federal do Ceará, Fortaleza, 2009.
ROHRER, G.A.; ALEXANDER, L.J.; KEELE, J.W.; SMITH, T.P.; BEATTIE, C.W. A microsatellite linkage map of the porcine genome. Genetics, n.136, v.1, p. 231-245, 1994.
ROSA, A. J. M., PAIVA, S. R. Marcadores moleculares e suas aplicações em estudos populacionais de espécies de interesse zootécnico. Planaltina, DF: Embrapa Cerrados, 35p, 2009.
ROSANOVA, C.; SOBRINHO, A. G. Da S.; NETO, S. G. A raça Dorper e sua
caracterização produtiva e reprodutiva. Veterinária Notícias, Uberlândia, v. 11, n. 1,
p. 127-135, 2005.
SANTOS, C. R.; MORI, E.; LEÃO, D. A.; MAIORKA, P. C. Genotipagem de
polimorfismos no gene prnp em ovinos da raça Santa Inês no Estado de São Paulo.
Pesquisa Veterinária Brasileira, v. 32, n.3, p. 221-226, 2012.
SANTOS, J. R.S. dos; SOUZA, B. B. de; SOUZA, W. H. de; CEZAR, M. FONTES;
TAVARES, G. de P. Respostas fisiológicas e gradientes térmicos de ovinos das
raças Santa Inês, Morada Nova e de seus cruzamentos com a raça Dorper às
condições do semi-árido nordestino. Ciência e Agrotecnologia, v.30, n.5, p.995-
1001, 2006.
SANTOS, P. S.; BLANK, M. F. A.; BLANK, A. F.; SILVA, S. A.; GOIS, I. G.; MANN,
R. S. Caracterização molecular de germoplasma de patchouli (Pogostemon sp.) por
marcadores RAPD. Scientia plena, v.9, n.5, 2013.
SANTOS, R. A cabra e a ovelha no Brasil. Uberaba: Agropecuária Tropical, 2003.

51
SILVA, F. L. R. da. Desempenho de ovinhos deslanados e mestiços criados em
pastagem nativa, na região semi-árida do Nordeste do Brasil. Revista Científica de
Produção Animal, v.4, n. 12, p. 71-76, 2002.
SILVA, F. L. R. da; ARAÚJO, A. M. Características de reprodução e de crescimento
de ovinos mestiços Santa Inês, no Ceará. Revista Brasileira de Zootecnia, v.29,
n.6, p.1712-1720, 2000.
SILVA, L. S. A. da; FRAGA, A. B., SILVA, F. de L. da; BEELEN, P. M. G.; SILVA, R.
M. de O. Growth curve in Santa Inês sheep. Small Ruminant Research, v. 105, p.
182-185, 2012.
SILVA, R. C. B. DA; PIMENTA FILHO, E. C.; RIBEIRO, M. N.; SILVA, E. C. DA;
FACÓ, O.; PAIVA, S. R. Diversidade genética de ovinos Morada Nova no estado do
Ceará, Brasil. In: 48ª Reunião Anual da Sociedade Brasileira de Zootecnia, Belém,
2011, Anais... Reunião Anual da Sociedade Brasileira de Zootecnia, 2011.
SIMPLÍCIO, A.A.; FREITAS, V.J.F.; FONSECA, J.F. Biotécnicas da reprodução
como técnicas de manejo reprodutivo em ovinos. Revista Brasileira de
Reprodução Animal, v.31, n.2, p.234-246, 2007.
SIMPLÍCIO, A.A.; SIMPLÍCIO, K.M.M.G. Caprinocultura e ovinocultura de corte: desafios e oportunidades. Revista CFMV. Brasília, DF, p 7-18, 2006.
SOUSA, W.H.; LEITE, P.R.M. Ovinos de corte: a raça Dorper. João Pessoa:
Emepa-PB, 76p, 2000.
SOUSA, W.H.de; LÔBO, R.N.B.; MORAIS, O. R. Ovinos Santa Inês: estado da arte
e perspectivas. In: Simpósio Internacional sobre caprinos e ovinos de corte, 2003.
João Pessoa. Anais... João Pessoa: Empresa Estadual de Pesquisa Agropecuária
da Paraíba, 2003.
SOUSA ,W.H. de; OJEDA, M. D.B.; MORAIS, O. R.; ROCHA, J. L. Programa de melhoramento genético para ovinos deslanados e caprinos de corte do Brasil: uma proposta. Tecnologia e Ciência Agropecuária, João Pessoa, v.2, n.3, p.41-46, 2008. SOUZA, C. A.; PAIVA, S. R.; McMANUS, C.M.; AZEVEDO, H. C.; MARIANTE, A.S.;
GRATTAPAGLIA, D. Genetic diversity and assessment of 23 microsatellite markers
for parentage testing of Santa Inês hair sheep in Brazil. Genetic and Molecular
Research, v. 11, n.2, p.1217-1229, 2012.
SPRITZE, A.; EGITO, A. A. DE; MARIANTE, A. DA S.; MCMANUS, C.
Caracterização genética da raça bovina Crioulo Lageano por marcadores
moleculares RAPD. Pesquisa Agropecuária Brasileira, v. 38, n. 10, p. 1157-1164,
2003.

52
SUN, W.; CHANG, H.; JI, D.; LIAO, X.; DU, L.; LU, S.; KENJI, T. Distance of different
ecological type sheep breeds in Mongolia sheep group. Journal of Genetics and
Genomics, v.34, n. 11, p. 1001-1009, 2007.
SUZUKI, K. M.; ALMEIDA, F.S.; SODRÉ, L. M. K.; PASCUAL, A. N.T.; SOFIA, S. H.
Genetic similarity among male bees of Euglossa truncate Rebêlo e Moure
(Hymenoptera: Apidae) revealed by RADP markers. Neotropical Entomology, v.
35, n.4, p. 477-482, 2006.
TEIXEIRA NETO, M. R.; DA CRUZ,J.F.; CARNEIRO, P. L. S.; MALHADO, C. H. M.;
FARIA, H. H. N. Parâmetros populacionais da raça ovina Santa Inês no Brasil.
Pesquisa Agropecuária Brasileira, v.48, n.12, p.1589-1595, 2013.
TELLES, M. P. DE C.; MONTEIRO, M. S. R.; RODRIGUES, F. M.; SOARES, T. N.;
RESENDE, L. V.; AMARAL, A. DAS G.; MARRA, P. R. Marcadores RAPD na análise
da divergência genética entre raças de bovinos e número de locos necessários para
a estabilidade da divergência estimada. Ciência Animal Brasileira, v. 2, n. 2, p. 87-
95, 2001.
TENEVA, A. Molecular markers in animal genome analysis. Biotechnology in Animal Husbandry, v. 25 (5-6), p.1267-1284, 2009.
TORRES, T. S.; NERI, V. dos S.; BIAGIOTTI, D.; FIGUEIREDO FILHO, L. A. S.;
SENA, L. S.; SARMENTO, J. L. R. Uso do método de agrupamento da ligação média
(UPGMA) associado à caracterização fenotípica de ovinos Santa Inês. In: X
Simpósio Brasileiro de Melhoramento Animal. Anais... Uberaba, 2013.
USMAN, B. A.; AGBEBI, O.T.; BANKOLE, M. O.; OGUNTADE, O .R.; POPOOLA, M.
O. Molecular characterication of two cichlids populations (Tilapia guineenses and
Sarotherodon melanotheron) from diferente water bodies in Lagos State, Nigeria.
International Journal for Biotechnology and Molecular Biology Research, v. 4,
n.5, p. 71-77, 2013.
VALLE, J. S.; FONSECA, B. K. D.; NAKAMURA, S.S.; LINDE, G.A.; MATTANA,
R.S.; MING, L.C.; COLAUTO, N.B. Diversidade genética de populações naturais de
pariparoba [Pothomorphe umbellata (L.) Miq.] por RAPD. Revista Brasileira de
Plantas Medicinais, v. 15, n.1. p.47-53, 2013.
VERÍSSIMO, C. J.; TITTO, C. G.; KATIKI, L. M.; BUENO, M. S.; CUNHA, E. A. da;
MOURÃO, G. B.; OTSUK, I. P.; PEREIRA, A. M. F.; NOGUEIRA FILHO, J. C. M.;
TITTO, E. A. L. Tolerância ao calor em ovelhas Santa Inês de pelagem clara e
escura. Revista Brasileira de Saúde e Produção Animal, v. 10, n.1, p. 159-167,
2009.
VIANA, J G. A. Panorama geral da ovinocultura no mundo e no Brasil. Revista
Ovinos, ano 4, n.12, p. 44 - 47, 2008.

53
VIEIRA, O. R.; SIMPLICIO, A. A.; LEITE, E. R.; CIRIACO, A. L. T. Padrão Racial no
melhoramento genético de caprinos e ovinos no Brasil. In: SIMPÓSIO NACIONAL
DE MELHORAMENTO ANIMAL, Anais... Belo Horizonte, 2000.
VIGNAL, A.; MILAN, D.; SANCRISTOBAL, M.; EGGEN, A. A review on SNP and other types of molecular markers and their use in animal genetics. Genetics Selection Evolution, v. 34, p. 275-305., 2002. WEIR, B.S. e COCKERHAM, C.C. Estimating F-estatistics for the analysis of population structure. Evolution, v. 38, n.6, p. 1358-1370, 1984.
WILLIAMS, J. G. K.; KUBELIK, A. R.; LIVAK, K. J.; RAFASLKI, J. A. e TINGEY, S. V.
DNA polymorphisms amplified by arbitrary primers are useful as genetic markers.
Nucleic Acids Research, v. 18, n. 22, p. 6531-5, 1990.
WISCHRAL, A.; SILVA, D. M. F.; LEAL, C. A. S.; ROCHA S. S. N.; ADRIÃO, M.
Caracterização genotípica de ovinos das raças Santa Inês e Morada Nova para o
marcador microssatélite SSR-GHR. Anais... IX Jepex, Recife, 2009.
ZAPATA, J. F. F.; SEABRA, L. M. A . J.; NOGUEIRA, C. M.; BEZERRA, L. C.;
BESERRA, F. J. Características de carcaça de pequenos ruminantes do nordeste do
Brasil. Ciência Animal, v. 11, p. 79-86, 2001.