universidade estadual do sudoeste da bahia uesb … · 11 citogeneticamente, das quais 7 apresentam...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DO SUDOESTE DA BAHIA – UESB
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, BIODIVERSIDADE E
CONSERVAÇÃO
ESTUDOS CITOGENÉTICOS DOS GÊNEROS Phyllodytes E Hypsiboas (AMPHIBIA,
ANURA)
MARCELLE AMORIM CARVALHO
Jequié-BA
2012
PPGGBC

MARCELLE AMORIM CARVALHO
ESTUDOS CITOGENÉTICOS DOS GÊNEROS Phyllodytes E Hypsiboas (AMPHIBIA,
ANURA)
Dissertação de mestrado apresentada ao
programa de Pós-Graduação em Genética,
Biodiversidade e Conservação da Universidade
Estadual do Sudoeste da Bahia, como requisito
para obtenção do título de Mestre em Genética,
Biodiversidade e Conservação.
Orientador: Prof. Dr. Sérgio Siqueira Júnior.
Co-orientadora: Profa. Dra. Caroline Garcia
Jequié-BA
2012
PPGGBC

Jequié, 23 de Março de 2012
BANCA EXAMINADORA
Prof. Dr. Sérgio Siqueira Júnior (Orientador) – UESB / Jequié
________________________________
Prof. Dr. Marco Antônio Costa – UESC / Ilhéus
________________________________
Prof. Dr. Paulo Roberto Antunes de Mello Affonso – UESB / Jequié
________________________________
SUPLENTE
Profa. Dra. Débora Diniz – UESB / Jequié
________________________________
PPGGBC

Dedico meu trabalho aos professores Dra.
Caroline Garcia e Dr. Sérgio Siqueira Jr.; e
a minha família amada (Weliton, Jane,
Denise, Jamille, Vinícius, Pedrinho...)
PPGGBC

AGRADECIMENTOS
Agradeço,
À UESB e ao Programa de Pós Graduação em Genética, Biodiversidade e Conservação,
pela oportunidade;
À Fapesb pela concessão da bolsa, muito importante nessa caminhada;
Ao prof. Dr. Sérgio Siqueira Jr. pelo convite à orientação, pelas coletas, por me ensinar
a buscar, pela confiança, compreensão, paciência, conselhos, incentivo, e por tudo que pôde
me ensinar.
A profa. Dra. Caroline Garcia, pela incessante ajuda, mesmo antes de se tornar minha
co-orientadora; pelo exemplo de professora/orientadora; por ter persistido junto comigo nas
técnicas da cito, por se mostrar sempre acessível e feliz por ajudar, por ter tornado possível
muito desse trabalho, por ter sido um anjo no meu caminho... faltam palavras pra dizer o
quanto foi importante nessa caminhada. Por sua leveza, amizade... por vibrar comigo a cada
conquista! Nunca vou conseguir te agradecer o suficiente!
Aos meus pais, que sempre me incentivaram, fizeram e fazem tudo o que podem para
me proporcionar a melhor educação/formação possível. Por me acudir sempre que precisei,
pelas palavras e gestos de força e carinho, por serem responsáveis pela melhor parte do que eu
sou; pelo amor incondicional, que me da segurança pra tanto nessa vida;
Às minhas irmãs, companheiras, cada uma a sua maneira; sempre torcendo por mim;
Ao meu namorido, ou noivorido por me compreender, me fortalecer, incentivar... por
sempre ter uma palavra otimista pra me oferecer, por me cuidar nos momentos mais difíceis e
me reconhecer como batalhadora no caminho que escolhi. Pelo carinho e amor todos os dias...
À Doce, minha princesa linda, por ser tão companheira, e me fazer rir sempre ao chegar
em casa, mesmo depois dos dias mais cansativos;
Aos meus colegas de laboratório, em especial: Lívia, Isabel, Roque, Oziel, Aline,
Jamille, Naiana, Milena por tornarem essa jornada mais alegre, mais leve. E pela ajuda, de
uma forma ou outra no decorrer do trabalho; ao colega Paulo Borges, pela ajuda com coletas e
fotos;
Aos outros professores do programa, em especial aos da Genética, por tudo que me
ensinaram, pelos bons exemplos;
À Deus, acima de tudo, pois Ele, mais que qualquer pessoa, me permitiu, me fortaleceu,
me ajudou a vencer os obstáculos;
Enfim, a todos que participaram direta ou indiretamente desse trabalho, muito obrigada!
PPGGBC

BIOGRAFIA
Marcelle Amorim Carvalho, filha de Weliton Santana Carvalho e Janísia Antunes
Amorim Carvalho, nasceu em Itapetinga-Bahia aos 12 dias do mês de Janeiro de 1985.
Estudou nas escolas O sossego da Mamãe e na Cooperativa Educacional de Itapetinga, onde
concluiu o ensino médio, no ano de 2002, aos 17 anos. Desde criança dizia querer ser
cientista, sonho que começou a se realizar aos 20, quando foi aprovada na Faculdade de
Ciências Biológicas na Universidade Estadual do Sudoeste da Bahia. Desde então, passou a
morar no município de Jequié-Bahia. Já no ensino superior, participou do Grupo Ecofau, no
laboratório de Invertebrados/Ecologia da UESB, onde obteve auxílio para a construção dos
primeiros alicerces científicos, e para a realização dos trabalhos e escritos da monografia
intitulada: Inventário da Espongiofauna da Baía de Camamu e seu entorno-BA, orientada pelo
Prof. Dr. Ulisses Pinheiro. Em 2009 formou-se Bacharel Ciências Biológicas com Ênfase em
Ecologia de Águas continentais. No ano seguinte, foi aprovada no Programa de Pós
Graduação em Genética, Biodiversidade e Conservação, onde, em conjunto com o Prof. Dr.
Sérgio Siqueira e a Profa. Dra. Caroline Garcia, desenvolveu projeto de pesquisa relacionada
à citogenética de anuros. Está concluindo, em 2012, o mestrado, aos 27 anos de idade.
PPGGBC

"Bom mesmo é ir a luta com determinação,
abraçar a vida e viver com paixão, perder
com classe e vencer com ousadia, porque o
mundo pertence a quem se atreve" (Charles
Chaplin)
PPGGBC

RESUMO 1
A família Hylidae é constituída por aproximadamente 900 espécies agrupadas em três 2
subfamílias: Phyllomedusinae, Pelodryadinae e Hylinae, sendo a última a maior, 3
compreendendo 37 gêneros, incluindo Hypsiboas e Phyllodytes. Apresentam 2n=18 a 52 4
cromossomos, sendo que, em sua maioria, apresentam número cromossômico 2n=24. O 5
gênero Hypsiboas, possui 84 espécies, distribuídas principalmente em áreas de Mata 6
Atlântica, Cerrado adjacente à Caatinga e formações de Cerrado, ocorrendo também na 7
Floresta Amazônica. Os indivíduos deste gênero variam em tamanho e coloração e são 8
organizados em grupos, tais como H. benitezi, H. punctatus, H. similineatus, H. pellucens, H. 9
albopunctatus, H. faber e H. pulchellus. Destas espécies, apenas 21 foram analisadas 10
citogeneticamente, das quais 7 apresentam apenas dados de número diplóide. O gênero 11
Phyllodytes, composto por animais de pequeno porte e endêmicos da Mata Atlântica, é 12
também dividido em grupos (P. auratus, P. luteolos e P. tuberculosus). Das 11 espécies 13
descritas para o gênero, apenas três foram submetidas a estudos moleculares e nenhuma 14
apresenta dados cromossômicos. De acordo estudos de dados moleculares, morfológicos e 15
número diplóide, as espécies de ambos os gêneros apresentam incertezas em suas relações 16
filogenéticas e encontram-se ameaçadas principalmente pela perda de habitat, pelas mudanças 17
ambientais globais e doenças infecciosas emergentes, estando inseridos em biomas 18
considerados hotspots. Desta forma torna-se importante a sua caracterização genética, tanto 19
para sanar as dúvidas em relação a sua filogenia, quanto para, em conjunto com estudos 20
ecológicos, ser utilizado em programas de manejo e conservação. No presente estudo foram 21
analisados citogeneticamente três espécies do gênero Phyllodytes: P. luteolos, P. 22
melanomystax e P. tuberculosus, e quatro espécies do gênero Hypsiboas: H. atlanticus, H. 23
crepitans, H. semilineatus e H. pombali; todas provenientes de fragmentos de Mata Atlântica, 24
utilizando coloração convencional, bandamento C e impregnação por nitrato de prata. Todas 25
as espécies em questão apresentaram o mesmo número diplóide e número fundamental 26
(2n=24 e NF=48), com pequenas diferenças no que diz respeito à morfologia cromossômica e 27
a quantidade e localização das regiões de heterocromatina constitutiva e das RONs, 28
possibilitando inferências sobre os possíveis rearranjos cromossômicos envolvidos na 29
evolução cariotípica desses grupos. 30
Palavras-chave: Ag-RONs, bandamento C, cromossomos, evolução cariotípica, inversões 31
pericêntricas. 32
33
PPGGBC

ABSTRACT 1
The Hylidae family comprises nearly 900 species grouped into three subfamilies: 2
Phyllomedusinae, Pelodryadinae and Hylinae, the last being the largest, comprising 37 3
genera, including Hypsiboas and Phyllodytes. They present 2n=18 to 52 chromosomes, and, 4
mostly, have a chromosome number 2n=24. The genus Hypsiboas has 84 species, distributed 5
mainly in the Atlantic Forest areas, Cerrado adjacent to the Caatinga, and Cerrado domain, 6
also occurring in the Amazon Rainforest. The individuals of this genus vary in size and color 7
and are organized into groups, such as: H. benitezi, H. punctatus, H. similineatus, H. 8
pellucens, H. albopunctatus, H. faber and H. pulchellus. Among these species, only 21 were 9
analyzed cytogenetically, of which 7 present only diploid number data. The genus 10
Phyllodytes, composed of small animals and endemic of the Atlantic Forest, is also divided 11
into the groups P. auratus, P. luteolos and P. tuberculosus. Among the 11 species described 12
for the genus, only 3 were subjected to molecular studies and none presents chromosomal 13
studies. According to molecular data, morphological, and diploid number studies, the species 14
of both genera present uncertainties in their phylogenetic relationships and also are threatened 15
primarily by habitat loss, the global environmental changes, and emerging infectious diseases, 16
being inserted in biomes considered hotspots. Thus, it becomes important to their genetic 17
characterization, both to address the questions regarding their phylogeny, and for, in 18
conjunction with ecological studies to be used in management and conservation programs. In 19
the present study were analyzed cytogenetically 3 species of the genus Phyllodytes: P. 20
luteolos, P. melanomystax and P. tuberculosus, and four species of the genus Hypsiboas: H. 21
atlanticus, H. crepitans, H. semilineatus, and H. pombali, all from Tropical Forest fragments, 22
using conventional stain, C-banding, and silver nitrate impregnation. All the mentioned 23
species have presented the same diploid number and fundamental number (2n=24 and 24
NF=48), with minor differences regarding chromosome morphology and the amount and 25
location of constitutive heterochromatic regions and the RONs, allowing inferences about 26
possible chromosomal rearrangements involved in the karyotype evolution of these groups. 27
Keywords: Ag-NORs, C-banding, chromosomes, karyotype evolution, pericentric inversions. 28
29
PPGGBC

LISTA DE FIGURAS 1
Figura 1. Fotos dos indivíduos de Phyllodytes analisados no presente estudo: a) Phyllodytes 2
luteolos, b) Phyllodytes melanomystax e c) Phyllodytes tuberculosus. 3
Figura 2. Cariótipos submetidos a coloração convencional (1) e bandamento C (2) de: a) 4
Phyllodytes luteolos, b) Phyllodytes melanomystax e c) Phyllodytes tuberculosus. Em 5
destaque os cromossomos nucleolares submetidos à impregnação por nitrato de prata e 6
bandamento C. 7
Figura 3. Fotos dos indivíduos de Hypsiboas analisados no presente estudo: a) Hypsiboas 8
atlanticus, b) Hypsiboas crepitans, c) Hypsiboas pombali, d) Hypsiboas semilineatus. (Fonte: 9
a) Sérgio Siqueira Jr.; b, c, d) Mauro Teixeira Jr.) 10
Figura 4. Cariótipos submetidos a coloração convencional (1) e bandamento C (2) de: a) H. 11
atlanticus; b) H. crepitans; c) H. pombali e d) H. semilineatus. Em destaque os cromossomos 12
nucleolares submetidos à impregnação por nitrato de prata e bandamento C. 13
Figura 5. Idiogramas das espécies de Hypsiboas analisadas citogeneticamente. 14
LISTA DE TABELAS 15
Tabela 1 – Localização dos blocos de heterocromatina e das RONs nas espécies de Phyllodytes 16
analisadas no presente trabalho 17
Tabela 2 – Fórmulas cariotípicas das espécies de Phyllodytes analisadas no presente trabalho 18
Tabela 3 – Fórmulas cariotípicas descritas para espécies de Hypsiboas baseadas em trabalhos 19
previamente publicados. 20
Tabela 4 - Fórmulas cariotípicas descritas para espécies de Hypsiboas baseadas em trabalhos 21
previamente publicados após padronização da medidas cromossômicas segundo Levan (1964). 22
Tabela 5 – Resumo dos dados citogenéticos disponíveis para espécies de Hypsiboas. 23
24
PPGGBC

LISTA DE ABREVIATURAS, SIGLAS, SÍMBOLOS 1
2n – Número diplóide 2
Ag-RON – Impregnação por nitrato de prata em região organizadora de nucléolo 3
Ba(OH)2 – Hidróxido de bário 4
BI- Banda intersticial 5
BP – Banda pericentromérica 6
BT – Banda terminal 7
DNA – Ácido desoxirribonucléico 8
DNAr – Ácido desoxirribonucléico ribossomal 9
EEWG – Estação Ecológica Wenceslau Guimarães 10
GC – Guanina-Citosina 11
HCl – Ácido clorídrico 12
M - Molaridade 13
m - metacêntrico 14
Mp – Megapixels 15
N – Normalidade 16
NF – Número fundamental 17
p - braço cromossômico curto 18
q - braço cromossômico longo 19
RPMI – Meio de Cultura 20
RNA – Ácido ribonucléico 21
RNAr - Ácido ribonucléico ribossomal 22
RON – Região organizadora de nucléolo 23
sm - submetacêntrico 24
SSC – Solução citrato-salina 25
st - subtelocêntrico 26
- ou * - dado ausente 27
PPGGBC

SUMÁRIO
RESUMO ................................................................................................................................................ 1
ABSTRACT ............................................................................................................................................ 2
LISTA DE FIGURAS ............................................................................................................................. 3
LISTA DE TABELAS ............................................................................................................................ 3
LISTA DE ABREVIATURAS, SIGLAS, SÍMBOLOS ......................................................................... 4
1. INTRODUÇÃO ................................................................................................................................ 14
1.1 A Mata Atlântica e os Anfíbios ................................................................................................... 14
1.2 História Natural dos Anfíbios ..................................................................................................... 15
1.3 Citogenética de Anfíbios ............................................................................................................. 15
1.4 A Família Hylidae e os Gêneros Hypsiboas e Phyllodytes ......................................................... 16
2 REVISÃO DE LITERATURA .......................................................................................................... 18
2.1 A Classe Amphibia e a Ordem Anura - Panorama Geral ............................................................ 18
2.1.2 História Natural .................................................................................................................... 18
2.1.3 Sistemática e Taxonomia...................................................................................................... 19
2.2 Citogenética em Anuros .............................................................................................................. 20
2.2.1 Região Organizadora do Nucléolo (RON) ........................................................................... 22
2.2.2 Heterocromatina e Bandamento C ....................................................................................... 24
2.3 A Família Hylidae ....................................................................................................................... 25
2.4 Os Gêneros Phyllodytes e Hypsiboas (Hylidae, Hylinae) ........................................................... 26
3. OBJETIVOS ..................................................................................................................................... 29
3.1 Objetivos Específicos .................................................................................................................. 29
4. MATERIAIS E MÉTODOS ............................................................................................................. 30
4.1 Coletas ......................................................................................................................................... 30
4.2 Procedimentos Metodológicos .................................................................................................... 30
4.2.1 Estudos Citogenéticos .......................................................................................................... 30
4.2.2 Preparação de Cromossomos Mitóticos - Intestino (King & Rofe, 1976)............................ 30
4.2.3 Detecção das Regiões Organizadoras de Nucléolo (Ag-RONs) (Howell e Black, 1980) .... 31
4.2.4 Detecção da Distribuição da Heterocromatina Constitutiva (Sumner, 1972) ...................... 31
4.3 Captura de Imagens ..................................................................................................................... 32
4.4 Montagem de Cariótipos e Idiogramas ....................................................................................... 32
5. REFERÊNCIAS ................................................................................................................................ 33
CAPÍTULO I - Estudos citogenéticos no gênero Phyllodytes (Anura, Hylidae) .................................. 40
CAPÍTULO II - Dinâmica da Evolução Cromossômica no Gênero Hypsiboas (Anura: Hylidae) ....... 51
6 CONCLUSÕES GERAIS .................................................................................................................. 71
PPGGBC

14
1. INTRODUÇÃO 1
1.1 A Mata Atlântica e os Anfíbios 2
Classificada como um conjunto de fisionomias e formações florestais, a Mata Atlântica 3
se distribui na faixa litorânea, florestas de baixada, matas interioranas e campos de altitude do 4
Rio Grande do Sul até o Amapá. 5
É considerada um dos principais biomas no que se refere a biodiversidade, pois sua 6
heterogeneidade de hábitats e micro-hábitats, favorece espécies especialistas em determinado 7
tipo de ambiente e, conseqüentemente, o alto número de endemismos. 8
Atualmente, possui menos de 7% de seu tamanho original, e, apresenta-se sob a forma 9
de fragmentos descontínuos, como resultado de processos de uso e ocupação do solo. 10
Levando em conta, portanto, estes dados sobre biodiversidade, endemismo e status de 11
ameaçado, este bioma é considerado um hotspot prioritário para a conservação. 12
Das aproximadamente 400 espécies de anfíbios descritas na Mata Atlântica, cerca de 90 13
são endêmicas e, poucos vertebrados dependem tanto das condições ambientais quanto eles. 14
Sua distribuição, ecologia e comportamento são influenciados especialmente pela vegetação, 15
umidade, espessura de folhiço, variedade de microambientes e disponibilidade de água. 16
Assim, os efeitos do processo de fragmentação têm trazido prejuízos para as espécies de 17
anfíbios, acarretando no empobrecimento dessas comunidades e em riscos de extinção 18
associados às mudanças climáticas, perda de hábitat e introdução patógenos. 19
Na última revisão da lista da fauna brasileira ameaçada de extinção realizada em 2002 20
(Ibama, 2003), constam 15 espécies de anfíbios ameaçados e uma espécie considerada extinta, 21
todas da Mata Atlântica. Recentes estudos indicam que estes números estão subestimados. O 22
rápido declínio das espécies de anfíbios tem mobilizado a comunidade científica em relação à 23
importância da expansão dos programas de pesquisa de genética e biodiversidade e da prática 24
de estratégias de conservação especialmente em regiões onde existem poucos dados sobre 25
diversidade, abundância e distribuição das espécies, como no Brasil. 26
27
PPGGBC

15
1.2 História Natural dos Anfíbios 1
Atualmente na classe Amphibia existem 6771 espécies descritas classificadas em três 2
ordens: Caudata, que reúne as salamandras com aproximadamente 619 espécies, 3
Gymnophiona que representa as cobras-cegas, com 186 espécies descritas, e a ordem Anura, 4
representada pelas rãs, sapos e pererecas, constituída atualmente por 5966 espécies. 5
Os anuros são conhecidos popularmente por serem animais que têm uma parte do seu 6
ciclo de vida na água, sob forma de larvas, e outra parte terrestre, quando transformam-se em 7
adultos. Entretanto, algumas espécies não apresentam a fase larval aquática, possuindo 8
desenvolvimento direto, enquanto outras, mesmo em estágio adulto, vivem em ambientes 9
aquáticos. São vertebrados com pele úmida, permeável e rica em glândulas; apresentam 10
respiração pulmonar, cutânea ou branquial e a temperatura do corpo desses animais varia 11
conforme a temperatura do ambiente (heterotérmicos). Por estes motivos a grande maioria das 12
espécies possui hábitos noturnos e/ou crepusculares, sendo que algumas podem ficar em 13
atividade durante todo o dia se estiverem no interior de matas fechadas, com pouca 14
luminosidade. Este grupo apresenta como importante característica a vocalização, que é 15
distinta para cada espécie e emitida apenas pelo macho para aproximar fêmeas para o 16
acasalamento, e para marcação de território. 17
Os Anura, pertencente à classe dos anfíbios, distribui-se por quase todos os continentes, 18
exceto a Antártica e algumas ilhas oceânicas, estando 77% de suas famílias em regiões 19
tropicais. 20
Os indivíduos dessa ordem são elementos importantes nas cadeias e teias ecológicas, 21
agindo como controladores de insetos e outros vertebrados. São também considerados 22
indicadores biológicos e ambientais, pois necessitam de um ecossistema equilibrado e 23
dependem de microclima para a sua sobrevivência. 24
1.3 Citogenética de Anfíbios 25
Os Anura são relativamente conservados em sua morfologia, o que dificulta resolver 26
questões relativas à sua taxonomia e sistemática através destes caracteres. Dessa forma, 27
principalmente a partir do ano 2000 estudos baseados em análise de quantidades significativas 28
de caracteres, como dados moleculares, passaram a contribuir para o entendimento das 29
relações filogenéticas desse grupo. Da mesma forma dados citogenéticos têm sido utilizados 30
para estudos citotaxonômicos e de evolução cromossômica. 31
PPGGBC

16
Até a década de 1980, a citogenética em Anura baseava-se apenas em análises de 1
cariótipos corados convencionalmente com Giemsa, permitindo apenas a caracterização 2
numérica e morfológica dos cromossomos, sendo, ainda assim, possível detectar certa 3
variabilidade no grupo, assim como a possível existência de rearranjos. 4
As primeiras descrições cariotípicas sugeriram que um maior número cromossômico era 5
característico de anuros morfologicamente mais primitivos, e que o número reduzido de 6
cromossomos das famílias mais derivadas teria surgido a partir da redução de um cariótipo de 7
26 cromossomos. 8
A partir do desenvolvimento de técnicas como bandamento C, Ag-RON, colorações 9
com fluorocromos e hibridação in situ, houve uma facilitação do reconhecimento de 10
homologias dos cromossomos, permitido uma melhor compreensão das relações entre gêneros 11
e espécies, através da indicação dos possíveis rearranjos cromossômicos envolvidos na 12
evolução cariotípica e especiação dos Anura. 13
1.4 A Família Hylidae e os Gêneros Hypsiboas e Phyllodytes 14
A família Hylidae, constituída por aproximadamente 901 espécies, é agrupada em três 15
subfamílias: Phyllomedusinae, Pelodryadinae e Hylinae, e em geral, são arborícolas, possuem 16
cabeça e olhos grandes, cintura delgada, mãos e pés longos, discos digitais aumentados e 17
tamanho variável entre 17-140 mm. 18
O cariótipo com 2n=24 cromossomos é o mais comumente encontrado na família 19
Hylidae, com ocorrência de números menores ou maiores, como 2n=18, 20 e 22, e 2n=26, 30, 20
34 e indivíduos triplóides 3n=48 e tetraplóides 4n=52. Para o gênero Hypsiboas foram 21
caracterizadas citogeneticamente 21 das 84 espécies do gênero, das quais 7 tem publicação 22
apenas do número diplóide. Até o momento, não existem dados citogenéticos para 23
Phyllodytes. 24
Até a década de 90, a família Hylidae possuía o maior número de espécies até então 25
cariotipadas (129), sendo que o maior número de espécies analisadas pertence a subfamília 26
Hylinae. Contudo, a maioria das espécies dessa família ainda não possui cariótipo conhecido, 27
e a maior parte das informações citogenéticas existentes baseia-se somente em estudos feitos 28
com coloração convencional. 29
Recentemente, o refinamento das técnicas de marcações cromossômicas permitiu a 30
realização de trabalhos que revelaram a ocorrência de alterações cariotípicas em muitas 31
espécies de Hylidae. Entre estes, a presença de cromossomos sexuais morfologicamente 32
PPGGBC

17
diferenciados, cromossomos B, variação na quantidade e distribuição de heterocromatina e 1
diferenças em relação ao número e localização da Ag-RON. 2
Ressalta-se também a importância dos trabalhos que utilizam técnicas mais avançadas, 3
como a coloração com os fluorocromos base-específicos, a hibridação in situ fluorescente em 4
análises cromossômicas de representantes dessa família. Tais estudos vêm fornecendo dados 5
importantes para a compreensão da evolução cariotípica nos Hylidae. 6
O gênero Hypsiboas é organizado em grupos, tais como H. benitezi, H. punctatus, H. 7
similineatus, H. pellucens, H. albopunctatus, H. faber e H. pulchellus; relevantes para a 8
definição de unidades monofiléticas em estudos moleculares. No entanto, algumas espécies 9
têm caracteres que tornam difícil alocá-las em qualquer um dos grupos atualmente 10
reconhecidos. 11
Os Phyllodytes, por sua vez, com base em dados morfológicos possuem três grupos (P. 12
auratus; P.luteolos e P.tuberculosus), e apresentam 11 espécies, sendo que algumas destas 13
não possuem nenhum grupamento. Este é um grupo pouco estudado tanto taxonomicamente 14
quanto a partir de dados moleculares. Além disso, trabalhos recentes divergem quanto a sua 15
filogenia, utilizando dados moleculares de apenas 3 espécies. 16
O presente trabalho, portanto, considerando o escasso conhecimento acerca das espécies 17
de Phyllodytes e Hypsiboas analisadas, e o fato de estarem inseridos em hotspots, tem como 18
objetivo realizar estudos utilizando dados citogenéticos, produzindo uma importante 19
ferramenta para estudos citotaxonômicos e de evolução cromossômica, além de tentar 20
esclarecer as relações filogenéticas destes gêneros. 21
22
PPGGBC

18
2 REVISÃO DE LITERATURA 1
2.1 A Classe Amphibia e a Ordem Anura - Panorama Geral 2
2.1.2 História Natural 3
A classe Amphibia constitui o grupo basal dos vertebrados terrestres, e sua 4
nomenclatura indica que a maioria das espécies que compõe esta classe vive parcialmente na 5
água e parcialmente na terra (Storer et al., 1989; Romer & Parsons, 1985). 6
Para atender as necessidades tanto terrestres quanto aquáticas, apresentam e importantes 7
estruturas adaptativas, como pele úmida e glandular, extremidades para nadar e andar (exceto 8
para Ordem Gymnophiona), respiração pulmonar, cutânea, bucal e branquial, e membros 9
adaptados para o salto em alguns representantes (Romer & Parson, 1985; Pough, 2002). 10
De acordo com Pough (2002) e Frost (2012), os anuros ocorrem em todos os 11
continentes, exceto na Antártida e algumas ilhas oceânicas, apresentando 77% de suas 12
famílias com distribuição tropical e 14% em áreas subtropicais do hemisfério Norte. 13
Aproximadamente 1600 das espécies dessa ordem vivem na América do Sul e, 14
principalmente, na Amazônia e nas florestas tropicais úmidas, não havendo em nenhuma 15
outra parte do mundo tamanha diversidade (Zug, 1993). No Brasil, o número de espécies de 16
anuros é bastante elevado, tendo sido encontradas 20 das 48 famílias existentes (Frost, 2012). 17
São organismos que possuem pele úmida, permeável e com presença de glândulas; 18
apresentam respiração pulmonar, cutânea ou branquial e são heterotérmicos. O que explica os 19
hábitos noturnos e/ou crepusculares da maioria das espécies, sendo que algumas podem ficar 20
em atividade durante todo o dia em locais úmidos e de pouca luminosidade (Pough, 2002). 21
Diferentemente de muitos animais terrestres, os anuros utilizam o som tanto para marcar 22
território, quanto para identificar possíveis companheiros. De maneira geral, o macho vocaliza 23
com o intuito de atrair a fêmea e, ao mesmo tempo, afastar outros machos da mesma espécie. 24
O canto é bastante diversificado entre as espécies, sendo essa uma das características que 25
auxiliam na identificação da espécie e determinação do sexo do indivíduo (Lutz, 1973). O 26
canto específico das espécies é considerado um caráter evolutivo conservado e, táxons 27
aparentados podem apresentar padrões audioespectográficos similares (Pough, 2002). 28
Os anfíbios anuros apresentam também variados modos de reprodução, como ovos e 29
larvas aquáticas, deposição de ovos fora d’água, ovoviviparidade e viviparidade, sendo a 30
fertilização externa o mais freqüente (Duellman & Rodrigues, 1994). 31
PPGGBC

19
Muitas dessas espécies possuem ainda, cuidado parental, desempenhando funções como 1
vigiar os ninhos, ou transportar ovos e girinos para a água. Este papel pode ser maternal, 2
paternal ou biparental (Beck, 1998). A maioria dos representantes dessa ordem não possui 3
órgão copulador introdutório. As fêmeas depositam ovos em local apropriado, ou os armazena 4
em parte do corpo. Até 10.000 ovos podem ser postos em uma única desova (Pough, et al., 5
2002). 6
2.1.3 Sistemática e Taxonomia 7
Atualmente na classe Amphibia existem três ordens e 6771 espécies descritas: ordem 8
Caudata, que reúne as salamandras com 619 espécies, Gymnophiona que representa as 9
cobras-cegas, com 186 espécies, e a ordem Anura, representada pelas rãs, sapos e pererecas, 10
constituída atualmente por 5966 espécies descritas (Frost, 2012). 11
Os Anura são ainda subdivididos em três subordens classificadas com base em 12
características como o número de vértebras, estrutura do cinturão peitoral, e morfologia dos 13
girinos (Frost et al., 2006). Subordem Archaeobatrachia, apresentando quatro famílias de rãs 14
consideradas mais primitivas; Mesobatrachia, que inclui seis famílias de rãs em um nível 15
evolutivo intermediário, e subordem Neobatrachia, o maior grupo, englobando cerca de 95% 16
de todas as espécies de Anura (Ford & Cannatella, 1993). 17
Em um grupo tão diversificado e de morfologia conservada como os Anura, são comuns 18
questões relativas à taxonomia e sistemática (Hillis & Dixon, 1991), o que tem motivado a 19
realização de extensas revisões, utilizando, além dos critérios tradicionais, àqueles obtidos 20
através da análise do seqüenciamento de DNA, reunidos ou não a dados citogenéticos 21
(Faivovich, 2002; Faivovich et al., 2004; Faivovich et al., 2005; Frost et al., 2006, Grant et 22
al., 2006). 23
Os principais avanços na taxonomia de anuros nos anos 80 e 90 foram baseados na 24
classificação fenotípica. Entretanto, até o fim da década de 90, apenas as relações entre as 25
famílias basais estavam bem estabelecidas (Frost et al., 2006). A partir do ano 2000, estudos 26
baseados em análises de grande quantidade de caracteres, como dados moleculares, tiveram 27
contribuição significativa no entendimento das relações filogenéticas entre os neobatráquios 28
(exemplo, Faivovich, 2002; Faivovich et al., 2004). Uma das mais importantes contribuições 29
foi realizada por Frost et al. (2006) e Grant et al. (2006) que fizeram extensos estudos 30
englobando todas ordens de anfíbios e praticamente todas as famílias de anuros, fornecendo 31
um mapeamento geral dos relacionamentos filogenéticos e embasamentos para estudos 32
PPGGBC

20
subsequentes de grupos de anfíbios não amostrados anteriormente por dados moleculares, 1
como é o caso dos trabalhos de Heinicke et al. (2007) e Hedges et al. (2008). 2
2.2 Citogenética em Anuros 3
Os anuros apresentam-se como um grupo de grande interesse para pesquisas sobre 4
mecanismos de evolução cromossômica, por sua abundância, extensa distribuição e 5
diversidade cariotípica. Evidencia-se, assim, a importância da citogenética como ferramenta 6
auxiliar na taxonomia e no esclarecimento das relações filogenéticas entre as espécies de 7
anuros. Esse papel torna-se mais relevante, a medida em que as técnicas empregadas nas 8
análises dos cromossomos se aperfeiçoam ao longo do tempo (Catroli, 2008). 9
A citogenética vem contribuindo enormemente para as investigações em sistemática de 10
vários grupos de organismos. Analisando as variações cromossômicas é possível formular 11
hipóteses sobre a filogenia de diferentes grupos taxonômicos e sobre rearranjos 12
cromossômicos ocorridos durante a evolução desses grupos, possibilitando, inclusive, 13
inferências sobre eventos populacionais (Lourenço, 1996). 14
A filogenia sugerida pelos dados de citogenética muitas vezes coincide com as 15
provenientes de análises morfo-anatômicas, moleculares e bioquímicas, contudo, existem 16
casos de controvérsia entre as relações filogenéticas entre esses tipos de análise. Tal resultado 17
pode decorrer da superficialidade de alguma das análises feitas, levando a interpretações 18
incorretas da ocorrência de convergência durante a evolução de algum desses grupos de 19
caracteres, ou de diferenças de taxas evolutivas nos diferentes conjuntos de caracteres 20
(Futuyma, 1992). 21
A análise conjunta de dados de diferentes naturezas é utilizada para maximizar o 22
entendimento correto da filogenia do grupo de interesse. Assim, estudos citogenéticos 23
fornecem dados que devem ser analisados em conjunto com dados os anatômicos, 24
moleculares e bioquímicos para a escolha da árvore filogenética mais provável (ex. Frost et 25
al., 2006). 26
Até a década de 80 muitos estudos citogenéticos em Anura foram desenvolvidos com 27
base na análise de cariótipos corados convencionalmente com Giemsa onde a maior 28
preocupação dessas investigações era a caracterização numérica e morfológica dos 29
cromossomos. Este dados permitiram detectar a ocorrência de variabilidade cariotípica e 30
possível existência de rearranjos cromossômicos (Bogart, 1973; 1991 Beçak et al., 1970; 31
Morescalchi, 1973; Miura 1995). 32
PPGGBC

21
As primeiras descrições cariotípicas sugeriram que anuros morfologicamente mais 1
primitivos possuem cariótipos com maior número cromossômico e que, os cariótipos com 2
poucos cromossomos, das espécies mais derivadas, sejam resultantes de um longo processo 3
evolutivo (Bogart, 1973). Ainda com base em dados citogenéticos, Bogart (1973) inferiu que 4
cariótipos dos indivíduos de famílias mais derivadas (como Dendrobatidae, Hylidae, 5
Leptodactilydae e Ranidae) possam ter surgido a partir de um cariótipo com 26 cromossomos 6
que provavelmente sofreu redução, originando cariótipos com 24 e 22 cromossomos. 7
As reduções do número de cromossomos ocorridas ao longo do processo evolutivo em 8
Anura parecem ser resultantes de fusões cêntricas, ou de fusão “in tandem”, sendo que 9
existem casos em que os dados são insuficientes para apontar a polaridade da transformação 10
cariotípica em análise, sendo impossível determinar se houve redução ou aumento do número 11
cromossômico (Bogart, 1973, 1991; Beçak et al., 1970; e Morescalchi, 1973; Miura et al., 12
1995). 13
Bogart (1973, 1991) e Miura (1995) consideram ainda que translocações, inversões, 14
adições, deleções e amplificações, mesmo não estando envolvidas em mudanças de números 15
cromossômicos, são também prováveis rearranjos ocorridos na evolução cariotípica da 16
Ordem. É possível inferir também que os estudos citogenéticos em Anura auxiliem no 17
esclarecimento dos processos de especiação, fornecendo exemplos em que os rearranjos 18
cromossômicos tenham sido fundamentais para a especiação (White, 1978) ou em que essas 19
alterações tenham ocorrido de forma independente ou subsequente à especiação (como 20
sugerido por Imai, 1983 e Coyne, 1984). 21
Uma melhor compreensão das relações cariotípicas passou a ocorrer após o uso de 22
técnicas citogenéticas mais avançadas, como o bandamento-C, o método de Ag-RON, e as 23
colorações com fluorocromos e hibridação in situ, que facilitaram o reconhecimento de 24
homologias dos cromossomos e possibilitaram uma melhor compreensão das relações entre 25
gêneros e espécies, através da indicação dos rearranjos cromossômicos que podem estar 26
envolvidos na evolução cariotípica dos anuros (exemplos, Bogart 1981; King, 1990; Siqueira 27
et al., 2004). 28
Apesar dessas técnicas evidenciarem vários tipos de rearranjos cromossômicos, estes 29
foram descritos para poucas espécies até agora e evidenciaram rearranjos envolvendo 30
principalmente as RONs (exemplos, Lourenço et al., 1998, Siqueira et al., 2004). Em células 31
meióticas foram observados anéis multivalentes possivelmente resultantes da presença de 32
translocações cromossômicas múltiplas e inversões pericêntricas e paracêntricas (exemplos, 33
Lourenço et al., 2000, Siqueira et al., 2004). 34
PPGGBC

22
Com o uso de tais técnicas, foi possível diferenciar espécies morfologicamente 1
semelhantes e que apresentaram mesmo número e morfologia cromossômica, além de mostrar 2
variações intra-específicas de localização e número de RONs e de bandas heterocromáticas 3
(ver em Quinderé, 2007). 4
Cromossomos sexuais heteromórficos são raros em anuros e foram observados em 5
pouco mais de 20 espécies já analisadas citogeneticamente, e apresentaram os sistemas 6
XY/XX, ZZ/ZW, XAAY/XXAA, 00/W0 (ver, entre outros, Schmid et al., 1991, 1992, 1993; 7
Miura, 1994; Lourenço et al., 1998; Ananias et al., 2007; Busin et al., 2007). 8
Apesar de se mostrar como uma ferramenta valiosa no estudo dos anuros, até o 9
momento, aproximadamente 1000 espécies foram estudadas citogeneticamente sendo que, 10
menos de um terço destas foram analisadas com técnicas diferenciais como, bandamento-C, 11
Ag-RON, coloração com fluorocromos base-espécificos e hibridação in situ fluorescente (ver 12
referências, King 1990; Schmid et al., 2003; Siqueira et al., 2004). 13
2.2.1 Região Organizadora do Nucléolo (RON) 14
As RONs são sítios cromossômicos ricos em seqüências GC, formados por numerosas 15
cópias de genes ribossomais que codificam os RNAs ribossomais (RNAr) 18S, 5,8S e 28S, 16
arranjadas in tandem e separadas por seqüências espaçadoras. Esses RNAr tornam-se parte 17
integrante do ribossomo (Galls & Pardue, 1969; Sumner, 1990). Também compõem as RONs, 18
um grupo peculiar das proteínas ácidas altamente argirofílicas, permitindo assim que elas 19
sejam muito clara e rapidamente visualizadas por procedimentos de impregnação por nitrato 20
de prata (Goodpasture & Bloom, 1975). Geralmente são regiões de constrições secundárias, 21
inicialmente observadas por Henderson et al. (1972). No entanto, nem todas as RONs 22
aparecem como constrições secundárias e em alguns casos contêm heterocromatina (Schmid, 23
1982). Em alguns casos ainda, a RON localiza-se adjacente a regiões de heterocromatina 24
(Schmid, 1982). 25
A impregnação pelo íon prata é o método mais usado para a identificação de RONs, 26
embora detecte apenas as RONs que estiveram ativas na última intérfase. Essa técnica foi 27
proposta inicialmente por Howell et al. (1975) Variações da técnica de impregnação pela 28
prata foram oferecidas por outros pesquisadores, como por exemplo, Orlet (1979), que 29
misturou uma solução de prata a 50% com ácido fórmico diluído e incubou a lâmina nesta 30
solução. Este procedimento foi modificado por Howell & Black (1980), que obteve uma 31
solução coloidal misturando ácido fórmico à solução de gelatina diluída e posteriormente este 32
PPGGBC

23
revelador coloidal foi misturado com uma solução de nitrato de prata imediatamente antes do 1
uso, sendo então as lâminas incubadas em estufa. 2
A impregnação por prata só é possível de ser observada em RONs que estão ou 3
estiveram associadas à síntese de RNA (Miller et al., 1976a, b), sendo que a especificidade 4
pela prata se dá em razão da reação desta com as proteínas acídicas, associadas ao RNAr 5
recém-transcrito (Schwarzacher & Wachtler, 1993). Portanto, apenas as RONs ativas na 6
intérfase anterior serão evidenciadas pelo método em cromossomos metafásicos. 7
A análise RON e suas características, como número, tamanho e posição no genoma, tem 8
se mostrado importante para o entendimento da evolução cromossômica do grupo em estudo, 9
podendo ser um caráter informativo para a distinção de espécies e populações. 10
Schmid (1982) analisou cromossomos de 260 espécimes de anuros entre 23 diferentes 11
gêneros, e pôde confirmar um alto grau de fixação de heteromorfismo de tamanho de RON 12
entre os espécimes, e também que a maioria dos anuros, tanto de famílias tidas como basais 13
como derivadas, apresenta um par de RONs por genoma diplóide (Schmid, 1982; King et al., 14
1990; Schmid et al., 1990), sugerindo que possivelmente esta seja uma condição 15
plesiomórfica em Anura. De acordo com King et al. (1990), as RONs em Anura tendem a 16
coincidir com bandas C positivas indicando a presença de heterocromatina nesses sítios. 17
Alguns mecanismos de dispersão de RONs, possivelmente envolvidos no surgimento de 18
RONs múltiplas, têm sido considerados por diversos autores (Wiley et al., 1989; King et al., 19
1990; Lourenço et al., 1998; Silva et al., 1999). Estes autores sugerem que essas variações 20
possam ser um resultado de diferenças estruturais das RONs, causadas por rearranjos como 21
transposição e amplificação de genes do DNAr, e erros de reinserção durante a amplificação 22
extra-cromossômica de genes ribossomais na oogênese. 23
A análise das regiões organizadoras de nucléolos (RONs) representa uma importante 24
ferramenta para o estudo de cariótipos em anfíbios (exemplos, Wiley et al., 1989, Lourenço et 25
al., 1998, Silva et al., 1999, Siqueira et al., 2004). Segundo Schmid (1982), o número e a 26
localização das RONs costumam ser características de espécies e populações, que em geral 27
apresentam apenas um par cromossômico portador dessas regiões. A origem da variação intra-28
específica no número e na localização das RONs, tem sido explicada pela ocorrência de 29
translocações e inversões, transposição de segmentos de DNA por meio de elementos 30
genéticos móveis, amplificações de genes de DNAr fita simples formando RONs 31
identificáveis e funcionais, além de erros de reinserções durante a amplificação do DNAr no 32
curso da oogênese (Schmid et al.,2003). 33
PPGGBC

24
2.2.2 Heterocromatina e Bandamento C 1
A heterocromatina foi distinguida da eucromatina pela primeira vez em 1928, por Heitz 2
com base na compactação diferencial na intérfase. A primeira é altamente condensada e 3
possui arranjos nucleossômicos altamente compactados juntamente com a cromatina, 4
enquanto a última apresenta-se menos condensada, mais acessível e geralmente transcreve 5
mais facilmente (Huisinga et al., 2006). As regiões cromossômicas de alta densidade e de 6
elementos repetitivos do DNA tais como seqüências satélites e elementos transponíveis, que 7
são encontrados nos centrômeros e telômeros são os alvos principais da formação de 8
heterocromatina (ver revisão de Grewal & Jia 2007). 9
A heterocromatina pode ser classificada em heterocromatina facultativa e 10
heterocromatina constitutiva; a depender do seu tipo de empacotamento ou condensação. As 11
regiões de heterocromatina constitutiva permanecem condensadas durante todo o ciclo celular 12
em locais específicos do genoma, enquanto a heterocromatina facultativa é encontrada 13
também nos loci de regulação, onde o estado da cromatina pode mudar em resposta aos sinais 14
celulares e às atividades genéticas (ver revisão de Grewal & Jia 2007). 15
Hoje já se conhecem diversas funções adicionais para a heterocromatina. Segundo a 16
recente revisão realizada por Grewal & Jia (2007), a heterocromatina, dentre outras funções 17
que exerce, se porta como uma “plataforma dinâmica” no recrutamento e extensão de 18
proteínas regulatórias ao logo dos seus domínios, facilitando diversos processos, tais como, 19
transcrição, segregação cromossomal e interações cromatínicas. 20
Normalmente, a maior parte do DNA que é compactado em heterocromatina não 21
contém genes, porém os genes que se tornam compactados em heterocromatina são, em geral, 22
resistentes à expressão ou silenciados. Isso não significa que a heterocromatina tenha que ser 23
desprezada, muito pelo contrário, pois ela é responsável, dentre outros fatores, pelo 24
funcionamento apropriado dos telômeros e centrômeros, auxiliando na proteção do genoma 25
(Alberts et al., 2002). 26
A heterocromatina constitutiva é geralmente detectada citogeneticamente através da 27
técnica de bandamento C (Sumner, 1990), apresentando três etapas básicas: tratamento com 28
ácido com HCl seguido de um tratamento alcalino com solução aquosa de hidróxido de bário 29
[Ba2 (OH)] saturado, e depois com solução salina de cloreto de sódio e citrato de sódio (2x 30
SSC). 31
PPGGBC

25
O tratamento com hidróxido de bário resulta na extração de até 50% do DNA dos 1
cromossomos. Esse DNA é mais facilmente extraído das regiões banda C negativas, 2
resultando em uma fraca coloração quando comparada com regiões de heterocromatina 3
constitutiva (banda C positivas) (Schmid, 1990). 4
O bandamento C vem sendo empregado no estudo de um grande número de espécies de 5
anuros, com diversas finalidades, dentre as quais: identificação de espécies parentais com a 6
identificação de padrões em determinadas espécies ou populações, procura de possíveis 7
polimorfismos e homologias, além da caracterização de gêneros e famílias e identificação de 8
pares heteromórficos entre machos e fêmeas (Schmid 1980; Sumner 1990; Siqueira et al., 9
2008). 10
Freqüentemente, o padrão de banda C varia entre as diferentes espécies, podendo ser um 11
importante caráter sistemático, que permite a sugestão de rearranjos genéticos ocorridos 12
durante a evolução do grupo em estudo (Sumner, 1990). Além de variações interespecíficas, 13
variações intra-específicas no padrão de banda C, que embora pouco freqüentes, já foram 14
reportadas em anuros, mostrando principalmente polimorfismo de tamanho dessas regiões. 15
Diferenças nos padrões heterocromáticos entre espécies são encontradas tanto nos anuros 16
tidos como basais quanto nos mais derivados (King, 1991). 17
O padrão de distribuição e quantidade de heterocromatina, detectado pela técnica de 18
bandamento C, assim como a caracterização da composição das bandas heterocromáticas, 19
obtidas a partir do uso de fluorocromos, possibilita também a observação das transformações 20
que ocorreram em grupos de espécies próximas que apresentam cariótipos muito semelhantes 21
(Schmid, 1980; Guerra, 1988). Essas características da heterocromatina banda-C positiva 22
mencionadas acima evidenciam que o seu estudo tem sido um importante caráter citogenético 23
a ser analisado na busca de uma melhor compreensão das questões sistemáticas, bem como da 24
evolução cariotípica nesta Ordem (Schmid, 1980; Odierna et al., 2001; Kasahara et al., 2003). 25
2.3 A Família Hylidae 26
A família Hylidae é constituída por aproximadamente 900 espécies, agrupadas em três 27
subfamílias: Phyllomedusinae, Pelodryadinae e Hylinae, sendo a última a maior, 28
compreendendo 37 gêneros e 646 espécies (Faivovich et al., 2005; Frost, 2012). No Brasil, os 29
representantes dessa família correspondem a cerca de 40% do total de espécies de anuros aqui 30
descritas (SBH, 2007). 31
PPGGBC

26
Essa grande diversidade de Hylinae, associada a sua ampla distribuição reflete na 1
existência de conflitos taxonômicos e sistemáticos, o que vem despertando o interesse dos 2
pesquisadores em estudos filogenéticos que buscam testar o monofiletismo e a validade do 3
seu status taxonômico (Faivovich et al., 2005; Salducci et al., 2005; Wiens et al., 2010). 4
Os hilídeos, em geral, são arborícolas, possuem cabeça e olhos grandes, cintura delgada, 5
patas longas, discos digitais aumentados (Pough, et al., 2002) e tamanho variável entre 17-6
140mm (Duellman & Trueb, 1986). O cariótipo com 2n=24 cromossomos é o mais comum 7
em Hylidae, ocorrendo números menores 2n=18, 20 e 22, ou maiores, 2n=26, 30, 34, 3n=48 8
(triplóide) e 4n=52 (tetraplóide) (Catroli, 2008). É provável que o cariótipo da todas ou quase 9
todas as espécies com número diplóide menor ou igual a que 24 seja derivado de um ancestral 10
com 2n=26 (Bogart, 1973), o que é corroborado nos resultados dos estudos filogenéticos de 11
Faivovich et al., (2005). 12
De acordo com os dados das revisões de King (1990) e Kuramoto (1990), os hilídeos se 13
apresentavam como a família com o maior número de espécies até então cariotipadas, sendo a 14
maior representatividade em espécies analisadas pertencentes a subfamília Hylinae, e ao 15
gênero Hyla. Ainda assim, a maioria das espécies dessa família não tem cariótipo conhecido, 16
restando muitas lacunas quanto à caracterização cromossômica de seus representantes. A 17
maior parte das informações citogenéticas nos hilídeos era baseada somente em estudos feitos 18
com coloração convencional. 19
Com o refinamento das técnicas de bandamento cromossômico puderam ser realizados 20
trabalhos capazes de revelar a ocorrência de alterações cariotípicas em muitas espécies de 21
Hylidae, como a presença de cromossomos sexuais morfologicamente diferenciados, 22
cromossomos B, variação na quantidade e distribuição de heterocromatina, e diferenças em 23
relação ao número e localização da Ag-RON. É importante ressaltar os trabalhos que utilizam 24
técnicas mais avançadas, como a coloração com os fluorocromos base-específicos, a 25
Hibridação in situ fluorescente em análises cromossômicas de representantes da família 26
Hylidae. Esses estudos, têm fornecido dados importantes para compreender da evolução 27
cariotípica nos Hylidae (Kasahara et al., 1998; Kasahara et al., 2003; Schmid e Steinlein, 28
2003). 29
2.4 Os Gêneros Phyllodytes e Hypsiboas (Hylidae, Hylinae) 30
O gênero Phyllodytes pertence a família Hylidae, subfamília Hylinae e tribo 31
Lophiohylini (Frost, 2012). Esses animais são popularmente conhecidos como pererecas das 32
PPGGBC

27
bromélias (Musso & Lima, 2002), apresentando pequeno porte, onde o adulto pode variar 1
entre 1,7 a 3,5 cm de comprimento, e ventosas nas pontas dos dedos, essenciais para aderência 2
na vegetação (Duellman & Trueb, 1994). 3
Possuem hábito bromelícola e são endêmicos de regiões de Mata Atlântica, um dos 4
biomas mais ameaçados, que vem sofrendo drástica redução devido ação antrópica, restando 5
apenas 7% de sua cobertura original. (Fundação SOS Mata Atlântica, 2012). 6
Apresenta 11 espécies (Frost, 2012), das quais 9 (exceto P. gynaethes e P. maculosus) 7
são pertencentes aos grupos: grupo P. auratus (P. autratus e P. wuchereri), grupo P. luteolos 8
(P. acuminatus, P. brevirostris, P. edelmoi, P. kautskyi, P. luteolos e P. melanomistax) e 9
grupo P. tuberculosus (P. punctatus e P. tuberculosus). 10
Análise de dados moleculares de Faivovich et al. (2005) reconheceram três grupos de 11
espécies e uma espécie não atribuída ao grupo demonstrando o polifiletismo de Phyllodytes. 12
Entretanto a última revisão do gênero, feita por Wiens et al. (2010) indica que Phyllodytes é 13
monofilético e o aloca próximo de grupos como, Trachycephalus, Osteocephalus, 14
Aparasphenodon e Itapotihyla, que morfologicamente são muito distintos deste gênero 15
gênero. 16
Faivovich et al. (2005) baseando-se em análises moleculares de 45 espécies 17
demonstraram o monofiletismo do grupo de Hylas da América do Sul, nesse sentido 18
sinonimizou 15 gênero e alocou as espécies brasileiras no gênero (Hypsiboas). 19
Este gênero constitui-se de 84 espécies distribuídas principalmente em áreas de Mata 20
Atlântica, Cerrado adjacente à Caatinga e formações de Cerrado, ocorrendo também na 21
Floresta Amazônica. O gênero Hypsiboas é organizados em grupos tais como, H. benitezi, H. 22
punctatus, H. similineatus, H. pellucens, H. albopunctatus, H. faber e H. pulchellus; de 23
acordo com suas semelhanças morfológicas. Estes grupos são relevantes para a definição de 24
unidades monofiléticas, no entanto, algumas espécies têm caracteres que tornam difícil alocá-25
las em qualquer um dos grupos atualmente reconhecidos (Faivovich et al., 2005). 26
Até então, foram caracterizadas citogeneticamente 21 espécies do gênero, das quais 7 27
apresentam somente dados do número cromosssômico (Carvalho et al., 2009; Nunes & 28
Fagundes, 2008; Gruber et al., 2006; Raber et al., 2004; Ananias et al., 2004), Essas 29
caracterizações mostram números diplóides que variam entre 2n=22 a 24 cromossomos 30
(Gruber et al., 2006), sendo 2n=24 o número cromossômico mais comum (Faivovich, 2005). 31
Estudos citogenéticos em anuros, ao contrário do observado em nível morfológico, têm 32
mostrado grande variabilidade em Anura e contribuído bastante em investigações de ordem 33
sistemática e relações filogenéticas de vários grupos (Toledo et al., 2007). Até o momento 34
PPGGBC

28
cerca de 1000 espécies de Anura foram estudadas citogeneticamente dentre as 5966 espécies 1
existentes nessa ordem. Apesar da importância dada a estudos citogenéticos ainda não há 2
registro de tais dados para o gênero Phyllodytes, nem para algumas espécies do gênero 3
Hypsiboas, como H. atlanticus e H. pombali. 4
5
PPGGBC

29
3. OBJETIVOS 1
Considerando escasso conhecimento das espécies de Phyllodytes e de algumas espécies 2
de Hypsiboas (Anura, Hylidae), e o fato de esses gêneros estarem inseridos em hotspots, o 3
presente estudo tem como objetivo realizar estudos taxônomicos destes gêneros, utilizando 4
dados citogenéticos, produzindo uma importante ferramenta citotaxonômica que, em conjunto 5
com dados ecológicos, podem auxiliar em programas de manejo e conservação. 6
3.1 Objetivos Específicos 7
A. Coleta e identificação de espécies do gênero Phyllodytes e Hypsiboas, preferencialmente 8
de suas localidades tipo; 9
B. Definição do número diplóide e fórmula cariotípica das populações amostradas a partir de 10
técnicas de coloração convencional; 11
C. Estabelecimento de padrões de distribuição da heterocromatina constitutiva e localização 12
de sítios ativos de NORs, eficientes marcadores cromossômicos, a partir, respectivamente, de 13
técnicas de bandamento-C e impregnação por nitrato de prata nas espécies analisadas; 14
D. Analisar diversidade inter e intraespecífica através de estudos comparativos entre os 15
espécimes; 16
E. Analisar os resultados obtidos comparando com os encontrados na literatura para outros 17
anuros, visando estudos de evolução cromossômica no grupo; 18
19 PPGGBC

30
4. MATERIAIS E MÉTODOS 1
4.1 Coletas 2
Os espécimes do gênero Phyllodytes utilizados provêm de três localidades do estado da 3
Bahia/ Brasil: o Parque Estadual da Serra do Conduru, localizado entre os municípios de 4
Ilhéus, Uruçuca e Itacaré, reserva da Natura, localizada no município de Serra Grande e da 5
cidade de Ilhéus. Os indivíduos do gênero Hypsiboas foram coletados de um fragmento de 6
Mata Atlântica do município de Itacaré-BA e da Estação Ecológica Wenceslau Guimarães, no 7
município que da nome à reserva-BA. As coletas foram realizadas através de busca ativa e 8
vocalização. 9
Foram analisados 13 indivíduos do gênero Phyllodytes, identificados como P. 10
melanomistacs, P. luteolos e P. tuberculosus; e 8 indivíduos do gênero Hypsiboas, 11
identificados como H. atlanticus, H. crepitans, H. pombali e H. semilineatus. 12
Os espécimes vivos coletados foram encaminhados ao Laboratório de Citogenética da 13
Universidade Estadual do Sudoeste da Bahia – Campus de Jequié onde foram processados, 14
recebendo numeração correspondente ao registro de tombo. Os exemplares foram fixados em 15
formol a 4% e posteriormente mantidos em álcool 70%, identificados e depositados também 16
no Laboratório de Citogenética da Universidade Estadual do Sudoeste da Bahia – Campus de 17
Jequié. 18
4.2 Procedimentos Metodológicos 19
4.2.1 Estudos Citogenéticos 20
4.2.2 Preparação de Cromossomos Mitóticos - Intestino (King & Rofe, 1976) 21
A. Foi injetado intraperitonealmente uma solução aquosa de colchicina 0,5%, na proporção de 22
0,02 ml por grama de peso do animal; 23
B. Após quatro a seis horas, o animal previamente anestesiado, foi sacrificado sendo 24
removidos coração, fígado, intestino e testículos; 25
C. O coração e os testículos foram acodicionados em eppendorf contendo álcool 100% para 26
posterior análise molecular; 27
PPGGBC

31
D. O intestino foi deixado imerso em água destilada por 20 minutos; sendo posteriormente 1
acondicionado em tubo falcon contendo fixador Carnoy (3:1) por 24 horas em geladeira. Após 2
esse período procedeu-se a raspagem da parte interna para obtenção das células a serem 3
analisadas; 4
E. O material obtido do intestino, foi homogeneizado com auxílio de pipetas pasteur e, em 5
seguida, centrifugdo por duas vezes a 950 rpm por dez minutos cada vez; 6
F. Seguiu-se então o armazenamento do material em eppendorfs contendo fixador Carnoy 7
(3:1), em congelador; 8
G. As suspensões obtidas foram pingadas em lâminas, e coradas com uma solução de Giemsa 9
5% em tampão fosfato 0,1M a pH 6,8 por aproximadamente 10 minutos. Após coloração, 10
foram lavadas em água corrente e secas ao ar. 11
4.2.3 Detecção das Regiões Organizadoras de Nucléolo (Ag-RONs) (Howell e Black, 12
1980) 13
A. Foram pingadas sobre uma lamínula, 2 gotas de solução aquosa de gelatina a 2%, acrescida 14
de ácido fórmico na proporção de 2ml/100 ml de solução, e 4 gotas de solução aquosa de 15
nitrato de prata a 50%; 16
B. Cobriu-se lamínula com a lâmina previamente preparada; 17
C. O conjunto lâmina-lamínula foi incubado em estufa ou banho-maria a 60º C até que a 18
lâmina adquirisse coloração de caramelo a marro claro; 19
D. Lavou-se a lâmina em água destilada, com o auxilio de uma piceta, para que a lamínula 20
escorresse naturalmente com a corrente d’água; 21
E. Posterior à retirada da lamínula, a lâmina foi lavada em água corrente (torneira) e seca ao 22
ar; 23
F. Quando se fez necessário, a lâmina foi corada com Giemsa diluído em tampão fosfato pH 24
6,8 por cerca de 20 segundos. 25
4.2.4 Detecção da Distribuição da Heterocromatina Constitutiva (Sumner, 1972) 26
A. Inicialmente a lâmina foi tratada com HCl 0,2N por 13 minutos em temperatura ambiente; 27
B. Lavou-se a lâmina em água corrente e, depois, seca ao ar; 28
C. Depois de seca, a lâmina foi tratada em solução aquosa e Ba(OH)2 5% a 60ºC durante 60 29
segundos; 30
PPGGBC

32
D. Imediatamente após retirada da solução de bário, imergiu-se a lâmina rapidamente em HCl 1
0,2N; 2
E. A lâmina foi então deixada em solução salina 2XSSC a 60ºC por 30 minutos; 3
F. Lavou-se a lâmina em água corrente, deixando secar ao ar. 4
G. Após todo esse procedimento, corou-se a lâmina Giemsa a 5% diluído em tampão fosfato 5
pH 6,8 durante 10 minutos. 6
4.3 Captura de Imagens 7
As preparações foram analisadas em microscópio óptico ou microscópio de epifluorescência e 8
capturadas com 5 Mp de definição através do sistema de análise de imagens CoolSnap Pro, 9
utilizando-se para tal, o programa Image Pro Plus 6.0 (Media Cybernetics). 10
4.4 Montagem de Cariótipos e Idiogramas 11
Os cariótipos foram montados utilizando Adobe Photoshop 7.0.1, e a morfologia 12
cromossômica foi determinada segundo a razão de braços proposta por Levan, 1964. 13
14
PPGGBC

33
5. REFERÊNCIAS 1
ALBERTS, B.; JOHSON, A.; LEWIS, J.; RAFF; ROBERTS, K. & WALTER, P. 2003. 2
Molecular Biology of the Cell, 4º ed. Garland Science, New York, NY, 352-4, 378-80. 3
ANANIAS, F.; GARCIA, P. C. A. & RECCO-PIMENTEL, S.M. 2004. Conserved 4
karyotypes in the Hyla pulchella species group (Anura, Hylidae). Hereditas 140: 42-48. 5
ANANIAS, F.; MODESTO, A. D. S.; MENDES, S. C. & NAPOLI, M. F. (2007) Unusual 6
primitive heteromorphic ZZ/ZW sex chromosomes in Proceratophrys boiei (Anura 7
Cycloramphidae Alsodinae) with description of C-Band interpopulational 8
polymorphism. Hereditas 144:206-212 9
BEÇAK, M. L.; DENARO, L. & BEÇAK, W. 1970. Polyploid and mechanisms of karyotypic 10
diversification in Amphibia. Cytogenetics 9:225-238. 11
BECK, C. W. 1998. Mode of fertilization and parental care in anurans. Animal Behavior 55: 12
439-449. 13
BOGART, J. P. Evolution of anuram karyotypes. 1973. In Vial, J. L. (ed.). Evolutionary 14
Biology of the Anurans: contemporary research on major problems. University of 15
Missuru Press, Columbia, pp. 337-349. 16
BOGART, J.P. 1981. Chromosomes studies in Sminthillus from Cuba and Eleutherodactylus 17
from Cuba and Puerto Rico (Amphibia: Leptodactylidae). Life Sci. Cont. R. Ont. Mus. 18
129:1-22. 19
BOGART, J.P. 1991. The influence of life history on karyotypic evolution in frogs. In: 20
Green, D. M. and Sessions SK (eds), Amphibian Cytogenetics and Evolution, Acad 21
Press, New York, pp 233-258. 22
BUSIN, C. S.; ANDRADE, G. V.; BERTOLDO, J.; DEL GRANDE, M. L.; 23
UETANABARO, M. & RECCO-PIMENTEL, S. M. (2007) Cytogenetic analysis of 24
four species of Pseudis (Anura Hylidae) with the description of ZZ/ZW sex 25
chromosomes in P Tocantins. Genetica 133:119-127. 26
CATROLI, G. F. 2008. Cariótipo se seis espécies de Bokermannohyla dos grupos de B. 27
circumdata e B. pseudopseudis (Anura, Hylidae). Rio Claro: Universidade Estadual 28
Paulista. Dissertação de Mestrado em Ciências Biológicas. 29
CARVALHO, K. A.; GARCIA, P. C. A. & RECCO-PIMENTEL, S. M. 2009. Cytogenetic 30
comparison of tree frogs of the genus Aplastodiscus and the Hypsiboas faber group 31
(Anura, Hylidae). Genetics and molecular Research 8 (4): 1498-1508. 32
PPGGBC

34
COYNE, J. A. 1984. Correlation between heterozygosity and rate of chromosome evolution 1
in animals. Arm. At. 123:725-729. 2
DUELLMAN, W. E. & RODRIGUES, L. O. 1994. Guide to the frogs of Iquitos region, 3
Amazonian Peru. University of Kansas Printing Service, Lawrence, Kansas. 4
DUELLMAN, W. &.; TRUEB, L. 1994. In: Biology of Amphibians. The Johnns Hopkins 5
University Press, Baltimore and Lodon. 6
DUELLMAN, W. E. & TRUEB, L. 1986. In Biology of Amphibians. Mc Graw Hill. New 7
York, Caps 5 and 6, pp. 109-112. E pp. 407-408. 8
FAIVOVICH, J. 2002. A cladistic analysis of Scinax Anura: (Hylidae). Cladistics 18: 367–9
393. 10
FAIVOVICH, J., GARCIA, P. C. A., ANANIAS, F., LANARI, L., BASSO, N. G. & 11
WHEELER, W. C. 2004. A molecular perspective on the phylogeny of the Hyla 12
pulchella species group (Anura, Hylidae). Mol. Phylogenetics Evol. 32: 938-950. 13
FAIVOVICH, J.; HADDAD, C. F. B.; GARCIA, P. C. A.; FROST, D. R.; CAMPBELL, J.A. 14
& WHEELER, W. C. 2005. Systematic review of the frog family Hylidae, with special 15
reference to Hylinae: phylogenetic analysis and taxonomic revision. Bull. Am. Museum 16
Nat. Hist. 294:1-240 17
FORD, L. & CANNATELLA, D. C. 1993. The major clades of frogs. Hepertol. Monigr. 18
7:94-117. 19
FROST, D. R., GRANT, T., FAIVOVICH, J., BAIN, R. H., HAAS, A., HADDAD, C. F. B., 20
SÁ, R. O., CHANNING, A., WILKINSON, M., DONNELLAN, S. C., RAXWORTHY, 21
C. J., CAMPBELL, J. A., BLOTTO, B. L., MOLER, P., DREWES, R. C., 22
NUSSBAUM, R. A., LYNCH, J. D., GREEN, D. M. & WHEELER, W. C. 2006. The 23
amphibian tree of life. Bull. Am. Museum Nat. Hist. 297: 1-370. 24
FROST, D.R. Amphibian Species of theWorld: An Online Reference, version 5.3. Electronic 25
database accessible at http://research.amnh.org/herpetology/amphibia American 26
Museum of Natural History, New York, USA., 2012. Acesso em: 20/02/2012. 27
FUNDAÇÃO SOS MATA ATLÂNTICA. 2012. Mata Atlântica. Portal SOS Mata Atlântica, 28
disponível em: 29
http://www.sosmatatlantica.org.br/index.php?section=info&action=mata. Acesso em: 30
14/11/11. 31
FUTUYMA, D. J. 1992. Biologia Evolutiva. 2 º edição, Ribeirão Preto. Sociedade Brasileira 32
de Genética/CNPq, 631pp. 33
PPGGBC

35
GALLS, J.G. & PARDUE, M.L. 1969. Molecular hybridization of radioactive DNA to the 1
DNA of cytological preparation Proc Natl Acad Sci USA 64 pp 600-604. 2
GOODPASTURE, C. & BLOOM S.E. 1975. Visualization of nucleolar organizer regions in 3
mammalian chromosomes using silver staining. Chromosoma, 53: 37-50. 4
GRANT, T., FROST, D. R., CALDWELL, J. P., GAGLIARDO, R., HADDAD, C. F. B., 5
KOK, P. J. R., MEANS, D. B., NOONAN, B. P., SCHARGEL, W. E. & WHEELER, 6
W. C. 2006. Phylogenetic systematics of dart-poison frogs and their relatives 7
(Amphibia: Athesphatanura: Dendrobatidae). Bull. Am. Museum Nat. Hist. 269: 1-262. 8
GREWAL, S.I.S. & JIA, S.T. 2007. Heterochromatin revisited. Nature Rev Gen 8:35-46. 9
GRUBER, S. L.; HADDAD, C. F. B. & KASAHARA, S. 2006. Chromosome banding in 10
three species of Hypsiboas (Hylidae, Hylinae), with special reference to a new case of 11
B-chromosome in anuran frogs and to the reduction of the diploid number of 2n=24 to 12
2n=22 in the genus. Genetica 130:281-291. 13
GUERRA, M.S. 1988. Introdução à Citogenética Geral. (ed) Guanabara Rio de Janeiro RJ pp 14
97-98. 15
HEDGES, S.B.; DUELLMAN, W.E. & HEINICKE, M.P. 2008. New World direct-16
developing frogs (Anura: Terrarana): Molecular phylogeny classification biogeography 17
and conservation Zootaxa 3-182. 18
HEINICKE, M.P.; DUELLMAN, W.E. & HEDGES, S.B. 2007. Major Caribbean and Central 19
American frog faunas originated by ancient oceanic dispersal. Proc Natl Acad Sci 20
104:10092-10097. 21
HEITZ, E. D. 1928. As heterochromatin der moose. L. Jabet. Wiss. Bot. 69:762-818. 22
HENDERSON, A. S.; WARBURTON, D. & ATWOOD, K.C.; 1972. Location of ribosomal 23
DNA in the human chromosome complement. Proc. Natl. Acad. Sci, USA. 69:3394-24
3398. 25
HILLIS, D. M. & DIXON, M. T. 1991.Ribosomal DNA: molecular evolution and 26
phylogenetic inference. Q Rev Biol, 66, 411 -53. 27
HOWELL, W. M. & BLACK, D. A. 1980. Controlled silver staining of nucleolar organizer 28
regions with a protective colloidal developer; a 1-step method. Experientia 36:1014-29
1015. 30
HUISINGA, K.L.; BROWER-TOLAND, B. & ELGIN, S.C. 2007. The contradictory 31
definitions of heterochromatin:transcription and silencing. Chromosoma 115 110-122. 32
PPGGBC

36
IBAMA. Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis. Lista 1
de Reservas Biológicas. Disponível em: 2
http://www.ibama.gov.br/siucweb/mostraUc.php?seqUc=811. Acesso 05/01/2012. 3
IMAI, H. T. 1983. Quantitative analyses of karyotype alteration an differentiation in 4
mammals. Evolution 37:1154-1161. 5
KASAHARA, S., SILVA, A. P. Z. & GRUBER, S. L. 1998. Use of lymphocyte cultures for 6
BrdU replication banding patterns in anuran species (Amphibia). Genet. Mol. Biol. 21: 7
471-467. 8
KASAHARA, S.; ZAMPIERI, A. P. S.; GRUBER S.L. & HADDAD, C.F.B. 2003. 9
Comparative cytogenetic four tree frog species (Anura, Hylidae, Hylinae) from Brazil. 10
Cytogenet. Gen. res. 103:155-167. 11
KING, M. & ROFE, R. 1976. Karyotype variation in the Australian gekko Phyllodactylus 12
marmoratus (Gray) (Gekkonidae: Reptilia). – Chromosoma 54: 75-87. 13
KING, M. 1990. Amphibia. In John B, Gwent C (Eds) Animal Cytogenetics. Amphibia 4 14
Chordata 2. Gebruder Borntraeger, Berlin, pp. 1-241. 15
KING, M.; COTRERAS, N. & HONEUCUTT, R. L. 1990.Variation within and between 16
nucleolar regions in Australian hylids frogs (Anura) shown by 18S ad 28S I situ 17
hybridization. Genetica, 80:17-29. 18
KING, M. 1991. The evolution of the heterochromatin in the amphibian genome. In Green, 19
M. G. & Sessions S. K. (eds.). Amphibians Cytogenetic and Evolution. Academic 20
Prees, San Diego, pp.359-391. 21
KURAMOTO, M. 1990. A list of chromosome number of anuran amphibians. Bull. Fukuoka 22
Univ. Educ., 39:83-127. 23
LEVAN, A.; FREDGA, K. & SANDBERG, A. A. 1964. Nomenclature for centromeric 24
position on chromosomes. Hereditas 52:201-220. 25
LOURENÇO, L. B.; RECCO-PIMENTEL, S. M. & CARDOSO, A. J. 1998. Polymorphism 26
of the nucleolus organizer regions (NORs) Physalaemus petersi (Amphibia, Anura, 27
leptodactylidae) detected by silver staining and fluorescent in situ hybridization. Chrom. 28
Res., 6:621-328. 29
LOURENÇO, L. B. 1996. Estudo citogenético comparativo de Edalorhina perezi e 30
Physalaemus petersi (Amphibia, Anura, Leptodactylidae). Campinas: Universidade 31
Estadual de Campinas. Dissertação de Mestrado em Ciências Biológicas. 32
PPGGBC

37
LOURENÇO, L.B., GARCIA, P.C.A. & RECCO-PIMENTEL, S. M. 2000. Cytogenetics of 1
two species of Paratelmatobius (Anura, Leptodactylidae), with phylogenetic comment. 2
Hereditas, 133: 201-210. 3
LUTZ, B. 1973. Brazilian species of Hyla. University of Texas press. Austin & London. 4
MILLER, D.A.; DEV, V.G.; TANTRAVAHI, R. & MILLER, O.J. 1976 a. Suppression of 5
human nucleolus organizer activity in mouse-human somatic hybrid cells. Exp Cell 6
Res 101:235-243. 7
MILLER, O.J.; MILLER, D.A.; DEV, V.G.; TANTRAVAHI, R & CROCE, C.M. 1976 b. 8
Expression of human and suppression of mouse nucleolus organizer activity in mouse-9
human somatic cell hybrids. Proc Natl Acad Sci 73:4531-4535. 10
MIURA, I. 1994. Sex chromosome differentiation in the Japanese brown frog Rana japonica. 11
Sex-related heteromorphism of the distribution pattern of constitutive heterochromatin 12
in chromosome nº 4 of the Nakuya population. Zool. Sci. 11:797-806. 13
MIURA, I. 1995. The late replication banding patterns of chromosomes are higly conserved 14
in the genera Rana, Hyla and Bufo (Amphibia, Anura). Chromosoma, 103:567-574. 15
MORESCALCHI, A. 1972. Amphibia. In Chiareli, A.B. & Capana, E.. (eds.). Cytotaxonomy 16
and Vertebrate Evolution. Academic Press, New York, pp. 233-348. 17
MORESCALCHI, A. 1973. Amphibia. In Chiareli, A. B. & Capana, E. (eds.). Cytotaxonomy 18
and Vertebrate Evolution. Academic Press, New York, pp. 233-348. 19
MUSSO, C. M. & LIMA, R. N. 2002. Diagnóstico Ambiental – Fauna. Zoneamento 20
Ambiental Reserva Ecológica de Jacarenema, Vila Velha – ES. Disponível em: 21
http://www.avidepa.org.br 22
NUNES, R. D. R. A. & FAGUNDES, V. 2008. Patterns of ribosomal DNA distribution in 23
hylid frogs from the Hypsiboas faber an H. semilineatus species groups. Genetics and 24
molecular biology, 31, 4, 982-987. 25
ODIERNA, G.; VENCES, M. & APREA, G. 2001. Chromosome data for Malagasy poison 26
frogs (Amphibia, Ranidae, Mantella) and their bearing o taxonomy and phylogeny, 27
Zool. Sci., 18:505-514. 28
OLERT, J. 1979. Interphase studies with simplified methods of silver staining of nucleoli. 29
Experientia, 35: 283-385. 30
POUGH, F. H.; HEISER, J. B. & MCFARLAND, W. N. 2002. Salamandras, Anuros e 31
Cecílias. In A vida dos vetebrados. Atheneu, São Paulo. 32
PPGGBC

38
QUINDERÉ, Y. R. S. D. 2007. Citogenética de populações e espécies de Physalaemus do 1
grupo "cuvieri" (Anura, Leiuperidae).” Campinas: Universidade Estadual de Campinas. 2
Dissertação de Mestrado em Biologia Celular e Estrutural. 3
RABER, S.C., CARVALHO, K.A., GARCIA, P.C., VINCIPROVA, G. & RECCO-4
PIMENTEL, S.M. 2004. Chromosomal characterization of H. bischoffi bischoffi and 5
Hyla guentheri (Anura, Hylidae). Phyllomedusa, 3: 43-49. 6
ROMER, A.S. & PARSON, T.S. 1985. Quem é quem entre os vertebrados. In: Anatomia 7
Comparada dos Vertebrados, Atheneu Ed., São Paulo, pp. 29-80. 8
SALDUCCI, M. D.; MARTY, C.; FOUQUET, A. & GILLES, A. 2005. Phylogenetic 9
relationships and biodiversity in Hylids (Anura: Hylidae) from French Guiana. C R 10
Biol. 328 (10-11): 1009-1024. 11
SBH. 2007. Lista de espécies de anfíbios do Brasil. Sociedade Brasileira de Herpetologia 12
(SBH). Disponível em: <http://sbherpetologia.or.br/checklist/anfibios.htm>. Acesso em: 13
09/12/10. 14
SCHMID, M. 1980. Chromosome banding in Amphibia V. Highly differentiated ZW/ZZ sex 15
chromosomes and exceptional genome size in Pyxicephalus adspersus (Anura, 16
Ranidae). Chromosoma 80:69-96. 17
SCHMID, M. 1982. Chromosome banding in Amphibia. VII. Analysis of structure and 18
variability of NORs in Anura. Chromosoma, 87:327-344. 19
SCHMID, M.; STEILEIN, C.; NANDA, I. & EPPLEN, J. T. 1990. In: E. Olmo ed. 20
Chromosome banding in Amphibia. Cytogenetics of Amphibians and Reptiles, 21
Birkhauser Verlag, Basel, p. 21-45. 22
SCHMID, M., NANDA, I., STEINLEIN, C., KAUSCH, K., EPPLEN, J. T. & HAAF, T. 23
1991. Sex-determining mechanisms and sex chromosomes in Amphibia. In: D. M. 24
Green e S. K. Sessions eds. Amphibian Cytogenetics and Evolution. Academic Press, 25
San Diego, p. 393-430. 26
SCHMID, M.; STEILEIN, C. & FEICHTINGER, W. 1992. Chromosome banding in 27
Amphibia. XVII. First demonstration of multiple sex chromosomes in amphibians: 28
Eleutherodactylus maussi (Anura, Leptodactylidae). Cytogenet. Cell. Genet, 62: 284-29
292. 30
SCHMID, M.; OHTA, S.; STEILEIN, C. & GUTTENBACH, M. 1993. Chromosome banding 31
in Amphibia. XIX. Primitive ZW/ZZ Sex chromosome I Burgeria buergeri (Anura, 32
Rhacophoridae). Cytogenet. Cell Genet., 62: 238-246. 33
PPGGBC

39
SCHMID, M. & STEINLEIN, C. 2003. Chromosome banding in Amphibia. XXIX. The 1
primitive XY/XX sex chromosome of Hyla femoralis (Anura, Hylidae). Cytogenet. 2
Genome Res 101:74-79. 3
SCHWARZACHER, H. G. & WACHTLER, F. 1993. The nucleolus. Anat. Embriolo., 4
188:515-536. 5
SILVA, A. P. Z., HADDAD, C. F. B. & KASAHARA, S. 1999. Nucleolus organizer region 6
in Physalaemus cuvieri (Anura, Leptodactylidae), with evidence of a unique case of Ag-7
NOR variability. Hereditas 131: 135-141. 8
SIQUEIRA, S.; ANANIAS, F. & RECCO-PIMENTEL, S.M. 2004. Cytogenetic analysis of 9
three species of Eleutherodactylus (Anura, Leptodactylidae) from southeastern Brazil. 10
Genet Mol Biol 27:363-372. 11
SIQUEIRA, S.; AGUIAR, O. JR.; TRÜSSMANN,C.; DEL-GRANDE, M. L. & RECCO-12
PIMENTEL, S. M. 2008. Chromosomal analysis of three Brazilian “eleutherodactyline” 13
frogs (Anura: Terrarana), with suggestion of a new species. Zootaxa 1860: 51–59. 14
STORER, T. I.; USINGER, R. L.; STEBBINS, R. C. & NYBAKKEN, J. W. 1989. Classe 15
Amphibia – Anfíbios. In Zoologia Geral. Companhia Ed. Nacional, pp. 618- 41. 16
SUMNER, A.T. 1972. A simple technique for demonstrating centromeric heterocromatio. 17
Exp Cell 75:304-306. 18
SUMNER, A. T. 1990. C-banding and related methods. In: Chromosome Banding. Unwin 19
Hyman. Ed. London. 39-90. 20
TOLEDO, L. F., BRITO, F. B., ARAÚJO, O. G. S., GIASSON, L. O. M. & HADDAD, C. F. 21
B. 2007. The occurrence of Batrachochytrium dendrobatidis in Brazil and the inclusion 22
of 17 new cases of infection. Sou. Am J Herpet 1:185-191. 23
WIENS, J. J.; KUCZYNSKI, C. A.; HUA, X. & MOEN, D. S. 2010. An expanded phylogeny 24
of treefrogs (Hylidae) based on nuclear and mitochondrial sequence data. Molecular 25
Phylogenetics and Evolution 55 871–882. 26
WHITE, M. J. D. 1978. Modes of speciation. WH freeman, São Francisco. 27
WILEY, J.E.; LITTLE, M.L.; ROMANO, M.A.; BLOUNT, D.A. & CLINE, G.R. 1989. 28
Polymorphism in the location of the 18S and 28S rDNA genes on the chromosomes of 29
the diploid-tetraploid treefrogs Hyla chrysoscelis and H. versicolor. Chromosoma 97: 30
481–487. 31
ZUG, G. R. 1993. Amphibians. In: Herpetology – Na Introductory Biology of Amphibians 32
and Reptiles. Academic Press, Inc. Washington, DC. 33
34
PPGGBC

40
CAPÍTULO I - Estudos citogenéticos no gênero Phyllodytes (Anura, Hylidae) 1
Marcelle Amorim Carvalho1, Caroline Garcia
1 e Sérgio Siqueira
1 2
1 Departamento de Ciências Biológicas, Laboratório de Citogenética, Universidade Estadual do Sudoeste da 3
Bahia. Jequié, BA, Brasil. 4
RESUMO 5
Estudos de natureza sistemática e filogenética dentro da ordem Anura foram, em sua 6
maioria, baseados em dados morfológicos, os quais, em muitos grupos, apresentam-se 7
altamente conservados, constituindo caracteres diagnósticos fracos. A partir de 2002, estudos 8
genéticos mostraram-se como ferramentas promissoras na resolução de conflitos taxonômicos 9
e na elucidação de relações evolutivas, sendo uma delas a Citogenética. Dentre as 5966 10
espécies de anfíbios anuros descritas, em torno de 1000 espécies já apresentam algum tipo de 11
informação cariotípica. Estes dados tem se mostrado extremamente úteis em inferências sobre 12
os mecanismos de evolução cromossômica e estudos de taxonomia, e quando associados a 13
dados morfológicos, ecológicos e moleculares permitem um melhor entendimento de suas 14
relações filogenéticas. O gênero Phyllodytes é composto por animais de pequeno porte e 15
endêmicos da Mata Atlântica, e das 11 espécies descritas para o gênero, apenas três destas 16
foram estudadas molecularmente e nenhuma apresenta dados cromossômicos. No presente 17
estudo foram analisados citogeneticamente três espécies deste gênero - Phyllodytes luteolos, 18
P. melanomystax e P. tuberculosus, utilizando coloração convencional, bandamento C e 19
impregnação por nitrato de prata. As três espécies apresentaram o mesmo número diplóide e 20
número fundamental (2n=24 e NF=48), com pequenas diferenças no que diz respeito à 21
morfologia cromossômica e a quantidade e localização das regiões de heterocromatina 22
constitutiva e das RONs. 23
Palavras-chave: Ag-RONs, bandamento C, cromossomos, evolução cariotípica, inversões 24
pericêntricas 25
26
PPGGBC

41
ABSTRACT 1
Cytogenetic studies of genus Phyllodytes (Anura, Hylidae) 2
Phylogenetic and systematics studies within the Anura order were mostly based on 3
morphological data, which in many groups, present themselves as highly conserved, 4
constituting poor diagnostic characters. However, since 2002, genetic studies showed to be 5
promising tools for resolving conflicts regarding taxonomy and in elucidating evolutionary 6
relationships, one of which the Cytogenetics. Among the approximately 5966 species of 7
amphibians described, about 1000 species present some karyotype information. This 8
information has been extremely useful in inferences about the mechanisms of chromosomal 9
evolution and taxonomy studies, and when associated with morphological, ecological and 10
molecular techniques allow a better understanding of their phylogenetic relationships. The 11
genus Phyllodytes consists of small and endemic animals from the Atlantic Forest, and from 12
the 11 species described of the genus, only 3 of these were molecularly studied and they had 13
none chromosomal data presented. In the present study 3 species of the genus Phyllodytes 14
were analyzed cytogenetically; P. luteolos, P. melanomystax and P. tuberculosus, using 15
conventional stain, C-banding, and silver nitrate impregnation. These species have the same 16
diploid and fundamental number (2n=24 and NF=48), with minor differences regarding 17
chromosome morphology, amount and location of constitutive heterochromatic regions and 18
the RONs. 19
Keywods: Ag-NORs, C-banding, chromosomes, karyotype evolution, pericentric inversions 20
21 PPGGBC

42
INTRODUÇÃO 1
O gênero Phyllodytes, composto por 11 espécies, pertence a família Hylidae, subfamília 2
Hylinae e tribo Lophiohylini (Frost, 2012). Esses animais, popularmente conhecidos como 3
pererecas das bromélias (Musso & Lima, 2002), apresentam pequeno porte, onde o indivíduo 4
adulto pode variar entre 1,7 a 3,5 cm de comprimento, apresentando ventosas nas pontas dos 5
dedos, essenciais para aderência na vegetação (Duellman & Trueb, 1994). 6
Esses animais apresentam hábito bromelícola e são exclusivos de ambientes de Mata 7
Atlântica, um dos biomas mais ameaçados e que vem sofrendo drástica redução devido ação 8
antrópica, contando atualmente, com apenas 7% de sua cobertura original. (Fundação SOS 9
Mata Atlântica, 2012). 10
O fato deste bioma marcado por alto grau de endemismo de sua fauna encontrar-se sob 11
severa ameaça, atinge, inevitavelmente, a riqueza de espécies encontradas nessas áreas. 12
Atualmente, reconhece-se a existência de cerca de 340 espécies de anfíbios vivendo em 13
remanescentes de Mata Atlântica, sendo que 90 dessas são consideradas endêmicas (Fundação 14
SOS Mata Atlântica, 2012; Lagos & Muller, 2007). Esses fatos vêm despertando o interesse 15
de pesquisadores em melhor caracterizar os anfíbios desse bioma visando estudos de 16
biodiversidade e projetos de manejo e conservação. Nesse sentido, marcadores 17
cromossômicos e moleculares apresentam grande aplicabilidade em sua caracterização 18
genética, permitindo a identificação de variações intra e interespecífica e o monitoramento da 19
variação genética das populações. 20
O gênero Phyllodytes é um dos menos estudados dentro de Hylinae, tanto do ponto de 21
vista morfológico quanto genético. Os poucos estudos disponíveis sobre as relações 22
filogenéticas dentro da família Hylidae utilizam dados morfológicos e alguns poucos dados 23
moleculares (Wiens, et al., 2010; Faivovich et al., 2005). 24
Faivovich et al. (2005) utilizando dados morfológicos e seqüenciamento de genes 25
mitocondriais (12S, 16S e citocromo b), propõem o monofiletismo do gênero, recuperando-o 26
como grupo mais basal para a tribo Lophiohylini. Wiens et al. (2010) em estudos mais 27
recentes, baseados apenas em dados moleculares (12S e 16S) não confirmou o monofiletismo 28
de Phyllodytes, alocando P. auratus mais próximo filogeneticamente de Itapotihyla do que 29
das demais espécies do gênero. Levando-se em consideração o pequeno número de espécies 30
analisadas no estudo de Wiens (2010) ou Faivovich (2005) (P. auratus, P, luteolos e 31
Phyllodytes sp.) as relações evolutivas dentro do gênero permanecem pouco esclarecidas. 32
PPGGBC

43
Em grupos com espécies que apresentam ampla distribuição, ou para aquelas que 1
possuem diversidade críptica, os caracteres morfológicos nem sempre são suficientemente 2
fortes para permitir a correta identificação de espécies. Nesses casos, onde a morfologia 3
mostra-se conservada ou limitada outras ferramentas podem ser utilizadas. Uma dessas 4
ferramentas é a citogenética (Hillis & Dixon, 1991; Toledo et al., 2010). 5
O presente estudo teve por objetivo analisar citogeneticamente três espécies de 6
Phyllodytes gerando informações que possam ser úteis para estudos de evolução 7
cromossômica e citotaxonomia que poderão ser utilizadas também para promover o sucesso 8
de programas de conservação. 9
MATERIAIS E MÉTODOS 10
Foram analisados 11 indivíduos provenientes de três localidades do estado da Bahia/ 11
Brasil: o Parque Estadual da Serra do Conduru, localizado entre os municípios de Ilhéus, 12
Uruçuca e Itacaré; da reserva da Natura, localizada no município de Serra Grande e da cidade 13
de Ilhéus. Os espécimes foram identificados como Phyllodytes luetolos, P. melanomistax e P. 14
tuberculosus e tombados no LAGOA (Laboratório de Genética de Organismos Aquáticos), 15
Universidade Estadual do Sudoeste da Bahia, Jequié, sob os números 330, 331, 332, 333, 334, 16
431, 432, 434, 435, 444 e 445. (Fig. 1 e Tabela 1). 17
Para a obtenção dos cromossomos, os animais foram tratados com injeção 18
intraperitoneal com uma solução aquosa de colchicina 0,5%, na proporção de 0,02 ml por 19
grama de peso do animal por um período de 4 a 6 horas antes de serem anestesiados e 20
processados. Os cromossomos mitóticos foram obtidos através da raspagem do epitélio 21
intestinal, conforme protocolo descrito por King & Rofe, (1976) com modificações. 22
Para a determinação do número e morfologia dos cromossomos foi utilizada a coloração 23
convencional por Giemsa e a classificação cromossômica seguiu a proposta de Levan et al. 24
(1964). 25
Para o cálculo do número de braços cromossômicos (NF), assumiu-se os cromossomos 26
metacêntricos, submetacêntricos e subtelocêntricos como portadores de dois braços 27
cromossômicos, e cromossomos acrocêntricos como portadores de apenas um braço. As 28
medições cromossômicas foram realizadas com o auxílio de um programa computacional que 29
simula um paquímetro (Bressane e Bressane, em preparação). 30
PPGGBC

44
O bandamento C, seguiu o protocolo proposto por Sumner (1972) com modificações; e 1
para a detecção das regiões organizadoras de nucléolo utilizou-se a impregnação por nitrato 2
de prata, com pequenas alterações segundo o Howell & Black (1980). 3
RESULTADOS 4
As três espécies de Phyllodytes apresentaram número diplóide 2n=24 cromossomos e 5
NF=48. Os cariótipos obtidos mostraram-se bastante semelhantes, diferindo quanto a 6
morfologia de alguns pares cromossômicos. Para todas as espécies os pares 1, 9, 10, 11 e 12 7
mostraram-se metacêntricos, e o par 4 subtelocêntrico. Por outro lado, os pares 2, 3, 6 e 8 8
variaram entre metacêntricos e submetacêntricos e os pares 5 e 7 variaram entre 9
submetacêntricos e subtelocêntricos. Pode-se ainda observar a ocorrência de uma constrição 10
secundária em um dos cromossomos do par 7 de Phyllodytes melanomystax, responsável por 11
um heteromorfismo de tamanho entre os homólogos (Fig. 2 A, B, C e Tabela 2). 12
O bandamento C revelou a presença de blocos de heterocromatina constitutiva, 13
localizados pericentromericamente em todos os cromossomos de P. tuberculosus e na maioria 14
dos cromossomos de P. melanomystax e P. luteolos. Blocos banda C positivos também 15
puderam ser observados nas regiões terminais de alguns cromossomos nas três espécies e 16
intersticialmente apenas em P. melanomystax e P. luteolos (Figura 3 D, E, F e Tabela 1). 17
Com relação ao numero e localização das RONs, Phyllodytes melanomystax apresentou 18
uma marcação de Ag-RONs na posição terminal dos braços longos do par 7, coincidente com 19
um bloco de heterocromatina e com a constrição secundária em um dos homólogos do par. 20
Phyllodytes luteolos apresentou RONs terminais localizadas nos braços longos do par 8. Não 21
foi possível obter dados sobre o número e localização das RONS para P. tuberculosus (Fig. 2 22
– boxes). 23
DISCUSSÃO 24
Os cariótipos dos espécimes analisados deste estudo apresentam número diplóide 25
equivalente à 2n=24 cromossomos. Esse número diplóide é uma das características 26
citogenéticas mais marcantes da família Hylidae, apesar de já haverem sido descritas 27
variações neste que vão de 2n=18 cromossomos (ex.: Aplastodiscus leucopygius) até 2n=52 28
cromossomos (ex.: Phyllomedusa tretraploidea) (Catroli, 2008). 29
PPGGBC

45
De acordo Bogart (1973) os cariótipos de todas ou quase todas as espécies com 1
número diplóide diferente do número diplóide modal para a família são, provavelmente, 2
derivados de um ancestral comum que apresentava 2n=26 cromossomos. Essa proposta foi 3
corroborada pelos resultados dos estudos de Faivovich et al. (2005), que propõem filogenia 4
para os hilídeos utilizando dados moleculares, porém não deixando de levar em consideração 5
o número cromossômico das espécies. 6
A macroestrutura cromossômica de Phyllodytes apresenta seis pares cromossômicos 7
(1, 4, 9, 10, 11 e 12) com morfologia conservada e outros seis (2, 3, 5, 6, 7 e 8) de morfologia 8
variada. Analisando essas fórmulas cariotípicas, pode-se inferir que as semelhanças entre 9
essas três espécies é resultado de sua ancestralidade comum, já as diferenças na 10
macroestrutura resultantes do posicionamento do centrômero devem-se, provavelmente à 11
inversões pericêntricas como sugerido para espécies de Dendropsohus (Medeiros, 2005), 12
Hypsiboas (Carvalho et al., 2008) e Phyllomedusa (Barth, 2008), as quais possivelmente 13
constituam o principal mecanismo de evolução cariotípica para o grupo. 14
O bandamento C permitiu revelar uma maior diferenciação entre as espécies 15
analisadas, identificando três padrões de distribuição e quantidade dos blocos de 16
heterocromatina. A diminuição do número e do tamanho de blocos heterocromáticos em 17
regiões pericentroméricas, em muitos casos está associada a presença de blocos banda C 18
positiva em regiões intesticiais e/ou terminais, como pode ser observado para P. 19
melanomystax e P. luteolos. Esse fato parece indicar que a ocorrência de heterocromatina 20
intersticial e/ou terminal possivelmente resulta de eventos de inversão pericêntrica 21
envolvendo os blocos proximais à região centromérica, resultando no novo posicionamento 22
(Medeiros, 2005; Carvalho et al., 2008; Barth, 2008). 23
Considerando as RONs, estas são simples e localizadas terminalmente nos braços 24
longos dos pares 7 e 8, em P. melanomystax e P. luteolos, respectivamente. Esses pares 25
cromossômicos apresentam a mesma morfologia nas duas espécies indicando que eles possam 26
ter uma ancestralidade comum e sua localização esteja dependente de rearranjos que tenham 27
ocorrido em outros pares adjacentes durante a evolução cromossômica do grupo. A existência 28
de homeologias entre pares de diferentes espécies pode ser confirmado em estudos dos 29
gêneros Physalaemus (Quinderé, 2007) e Phyllomedusa (Bruschi, 2010). 30
Ainda não foi possível obter resultados de Ag-RON para P. tuberculosus, mas, devido 31
a sua natureza heterocromática e das outras duas espécies estudadas, pode-se supor que, a 32
impregnação pelo nitrato de prata revele a RON coincidente com os blocos heterocromáticos 33
PPGGBC

46
existentes na região terminal dos cromossomos do par 8, o qual também mostra-se semelhante 1
em morfologia ao par nucleolar das demais espécies em estudo. 2
Embora translocações/transposições e inversões pericêntricas sejam os eventos mais 3
prováveis de terem ocorrido no grupo em questão, acarretando nas diferenças entre as 4
espécies, não podemos afirmar que estes foram os únicos eventos envolvidos na evolução 5
cromossômica do grupo, pois sabe-se que esses podem ser mais complexos, como observado 6
por Ventura (2009), através de pintura cromossômica, em roedores do gênero Akodon. 7
O presente estudo mostrou que existem importantes diferenças citogenéticas entre as espécies 8
estudadas, o que contribui com os estudos de evolução cariótipica e citotaxonomia, além de 9
poderem ser utilizadas em programas de conservação, visto que, ações bem sucedidas de 10
reintrodução e/ou a permanência das espécies, dependem de uma compatibilidade genética 11
entre elas. Contudo, para auxiliar o entendimento das relações filogenéticas, se faz necessário 12
a coleta e análise de um maior número de espécies utilizando uma maior variedade de técnicas 13
genéticas. 14
AGRADECIMENTOS: 15
Os autores agradecem ao Prof. Dr. Mirco Solé pela identificação dos exemplares 16
Phyllodytes e auxílio nas coletas. Este trabalho contou com o apoio financeiro da Fundação de 17
Amparo à Pesquisa do Estado da Bahia (FAPESB – processo BOL 1399/2010). 18
REFERÊNCIAS 19
BARTH, A. 2008. Estudos citogenéticos nas espécies do gênero Phyllomedusa (Anura, 20
Hylidae) encontradas na região sul do estado da Bahia. Resumo de Dissertação de 21
Mestrado. 2p. 22
BRESSANE & BRESSANE em preparação. 23
BRUSCHI, D. P. 2010. Relações inter e intraespecíficas no grupo de Phyllomedusa 24
hypochondrialis (Anura, Hylidae): Estudos citogenéticos e de DNA mitocondrial. 25
Campinas: Universidade Estadual de Campinas. Dissertação de Mestrado em Biologia 26
Celular e Estrutural. 27
BOGART, J. P. 1973. Evolution of anuram karyotypes. In Vial, J. L. (ed.). Evolutionary 28
Biology of the Anurans: contemporary research on major problems. University of 29
Missuru Press, Columbia, pp. 337-349. 30
PPGGBC

47
CARVALHO, F. K, DE; FERNANDES, A.; BARTH, A. & CUSTÓDIO, R. J. 2008. Tempo 1
ou antropismo como fatores causadores de alterações cromossômicas para uma 2
população de Hypsiboas raniceps (Anura: Hylidae) (Cope, 1862). 3
CATROLI, G. F. 2008. Cariótipo se seis espécies de Bokermannohyla dos grupos de B. 4
circumdata e B. pseudopseudis (Anura, Hylidae). Dissertação apresentada ao Instituto 5
de Biociências da Universidade Estadual Paulista, UNESP, Campus de Rio Claro, para 6
a obtenção do título de Mestre em Ciências Biológicas (Biologia Celular e Molecular) 7
Rio Claro – SP. 8
DUELLMAN, W. E. & TRUEB, L. 1994. Biology of Amphibians, The John Hopkins Univ 9
Press. Baltimore, 670 pp. 10
FAIVOVICH, J.; HADDAD, C. F. B.; GARCIA, P. C. A.; FROST, D. R.; CAMPBELL, J.A. 11
& WHEELER, W. C. 2005. Systematic review of the frog family Hylidae, with special 12
reference to Hylinae: phylogenetic analysis and taxonomic revision. Bull. Am. Museum 13
Nat. Hist. 294:1-240. 14
FROST, D.R. 2012 Amphibian Species of the World: An Online Reference, version 5.3. 15
Electronic database accessible at http://research.amnh.org/herpetology/amphibia 16
American Museum of Natural History, New York, USA. 17
FUNDAÇÃO SOS MATA ATLÂNTICA. 2012. Mata Atlântica. Portal SOS Mata Atlântica, 18
disponível em: http://www.sosmatatlantica.org.br/index.php?section=info&action=mata 19
. Acessado em 07/01/2012. 20
HILLIS, D. M. & DIXON, M. T. 1991.Ribosomal DNA: molecular evolution and 21
phylogenetic inference. Q Rev Biol, 66, 411 -53. 22
HOWELL, W. M. & BLACK, D. A. 1980.Controlled silver staining of nucleolar organizer 23
regions with a protective colloidal developer; a 1-step method. Experientia 36:1014-24
1015. 25
KING, M. & ROFE, R. 1976. Karyotype variation in the Australian gekko Phyllodactylus 26
marmoratus (Gray) (Gekkonidae: Reptilia). – Chromosoma 54: 75-87. 27
LAGOS, A. R. & MULLER, B. L. A. 2007. Hotspot brasileiro – Mata Atlântica. Saúde e 28
Ambiente em Revista. Universidade do Grande Rio – UNIGRANRIO 29
LEVAN, A.; FREDGA, K. & SANDBERG, A. A. 1964. Nomenclature for centromeric 30
position on chromosomes. Hereditas 52:201-220. 31
MEDEIROS, L. R. 2005. Citogenética de Dendropsophus (Anura, Hylidae): Caracterizações 32
e comparações cromossômicas. Tese de Doutorado em Biologia Celular e Estrutural 33
110p. 34
PPGGBC

48
MUSSO, C. M. & LIMA, R. N. 2002. Diagnóstico Ambiental – Fauna. Zoneamento 1
Ambiental Reserva Ecológica de Jacarenema, Vila Velha – ES. Disponível em: 2
http://www.avidepa.org.br 3
QUINDERÉ, Y. R. S. D. 2007. Citogenética de populações e espécies de Physalaemus do 4
grupo "cuvieri" (Anura, Leiuperidae).” Campinas: Universidade Estadual de Campinas. 5
Dissertação de Mestrado em Biologia Celular e Estrutural. 6
SUMNER, A.T. 1972. A simple technique for demonstrating centromeric heterocromatio. 7
Exp Cell 75:304-306. 8
TOLEDO, L. F., SIQUEIRA, S., DUARTE, T. C., VEIGA-MENONCELLO, A. P. C. & 9
RECCO-PIMETEL, S. M. 2010. Description of a new species of Pseudopaludicola 10
Miranda-Ribeiro, 1926 from the state of São Paulo, Southeastern Brazil (Anura, 11
Leiuperidae). Sou. Am J Herpet 1:185-191. 12
VENTURA, K. 2009. Estudos citogenéticos e filogenia molecular em roedores da tribo 13
Akodontini. Tese apresentada ao Departamento de Genética e Biologia Evolutiva do 14
Instituto de Biociências da Universidade de São Paulo para a obtenção do título de 15
Doutor em Ciências na área de Biologia/Genética. São Paulo.WIENS, J. J.; 16
KUCZYNSKI, C. A.; HUA, X.; MOEN, D. S .2010. An expanded phylogeny of 17
treefrogs (Hylidae) based on nuclear and mitochondrial sequence data. Molecular 18
Phylogenetics and Evolution 55 871–882. 19
PPGGBC

49
Tabela 1 – Localização dos blocos de heterocromatina e das RONs nas espécies de
Phyllodytes analisadas no presente trabalho
Espécie Nº de tombo Localidade Sexo Bandamento C RONs
P. luteolos 444 e 4445 Ilhéus/BA 2 Machos BP= maioria
BI=1q, 1p, 5p,
7q, 10p
BT= 1q, 2p, 3q,
4q
B
T=8q
P. melanomystax 331, 431, 432,
434 e 435
PESC/BA e
Reserva da
Natura/BA
4 Machos e
1 Fêmea
BP= 1, 4, 6, 7, 8,
9, 10, 11 e 12
BI=1q
BT=1p, 2q, 7q
B
T =7q
P. tuberculosus 330, 332, 333 e
334
PESC/BA 4 Machos BP=todos os
cromossomos
BT= 8q
_
Tabela 2 – Morfologia cromossômica das espécies de Phyllodytes analisadas no presente trabalho
Espécie 1 2 3 4 5 6 7 8 9 10 11 12
P. luteolos m m m st st sm st sm* m m m m
P. melanomistacs m m sm st sm m sm* m m m m m
P. tuberculosus m sm sm st st sm st sm m m m m
* par onde houve marcação das RONs
PPGGBC

50
Fig. 1 - Fotos dos indivíduos de Phyllodytes analisados no presente estudo:
a) Phyllodytes luteolos, b) Phyllodytes melanomystax e c) Phyllodytes
tuberculosus.
Fig. 2 - Cariótipos submetidos a coloração convencional (1) e bandamento C (2) de: a) Phyllodytes
luteolos, b) Phyllodytes melanomystax e c) Phyllodytes tuberculosus. Em destaque os cromossomos
nucleolares submetidos à impregnação por nitrato de prata e bandamento C.
1
a) b) c)
PPGGBC

51
CAPÍTULO II - Dinâmica da Evolução Cromossômica no Gênero Hypsiboas (Anura: 1
Hylidae) 2
Marcelle Amorim Carvalho1, Miguel Trefault Rodrigues
2, Caroline Garcia
1 e Sérgio Siqueira
1 3
1 Departamento de Ciências Biológicas, Laboratório de Citogenética, Universidade Estadual do Sudoeste da 4
Bahia. Jequié, BA, Brasil. 5
2 Departamento de Zoologia, Instituto de Biociências, Universidade de São Paulo. São Paulo, SP, Brasil. 6
RESUMO 7
Hylidae é uma das famílias mais numerosas dos anuros, representando no Brasil 40% da 8
diversidade dessa Ordem. Essa grande diversidade, somada a ampla distribuição desses 9
animais se reflete em conflitos taxonômicos e sistemáticos. Os hilídeos apresentam número 10
diplóide 2n=24 cromossomos, embora já tenham sido relatadas variações. Entre os Hypsiboas, 11
apenas 16% de suas espécies possuem dados citogenéticos, sendo a maioria resultado apenas 12
de coloração convencional. O presente trabalho analisa dados citogenéticos de diferentes 13
espécies e populações de Hypsiboas e discute a dinâmica evolutiva da macro e microestrura 14
cromossômica desses anfíbios. O número diplóide 2n=24 cromossomos é o mesmo nas 15
espécies analisadas, no entanto o grupo apresentou grande variabilidade na morfologia e 16
marcações cromossômicas. A existência de possíveis marcadores cromossômicos com 17
potencial citotaxônomico e a existência de uma possível relação filogeografica dos padrões 18
cromossômicos são discutidos. 19
Palavras-chave: Ag-RONs, bandamento C, cromossomos, evolução cariotípica, inversões 20
pericêntricas 21
ABSTRACT 22
Chromosomal Evolution Dynamics in the genus Hypsiboas (Anura: Hylidae) 23
Hylidae is one of the larger families of the anurans, representing in Brazil 40% of the 24
diversity of this order. This large diversity, added to the wide distribution of these animals is 25
reflected in taxonomic and systematic conflicts. The hylids present the diploid number of 26
PPGGBC

52
2n=24 chromosomes, although variations have already been reported. Among the Hypsiboas, 1
only 16% of its species have cytogenetic data, and most only results from conventional 2
staining. This paper analyzes cytogenetic data of different species and Hypsiboas populations 3
and discusses the evolutionary dynamics of both, macro and micro chromosomal structure of 4
these amphibians. The diploid number 2n=24 chromosomes is the same in the analysed 5
species, however the group showed great variability in morphology and chromosomal 6
markers. The presence of possible chromosomal markers with cytotaxonomic potential and 7
the existence of a possible phylogeographic relationship of the chromosomal patterns are 8
discussed. 9
INTRODUÇÃO 10
Hylidae é reconhecida como uma das famílias mais numerosas da ordem Anura. 11
Representantes desse grupo de anfíbios, caracterizados principalmente pela presença de discos 12
adesivos nas bases dos dedos e hábitos arbóreos, podem ser encontrados por todos os 13
continentes do globo, com exceção do continente Antártico, sendo sua maior diversidade 14
relatada para a região Neotropical (Frost, 2012). 15
Os hilídeos totalizam 907 espécies e 47 gêneros (Amphibiaweb, 2012), comumente 16
subdivididos em três subfamílias (Faivovich et al., 2005, Wiens et al., 2010), dentre essas 17
destaca-se a subfamília Hylinae por ser a mais diversa, abrigando cerca de 650 espécies 18
(Frost, 2012). 19
Toda a diversidade de espécies de Hylinae, associada à sua ampla distribuição 20
geográfica refletem na existência de inúmeros conflitos taxonômicos e sistemáticos. Na 21
última década essa subfamília vem sendo foco de estudos filogenéticos que buscaram testar 22
seu monofiletismo e a validade do status taxonômico de seus representantes (Faivovich 23
et al., 2005; Salducci et al., 2005; Wiens et al., 2010). Um dos estudos pioneiros nessa 24
abordagem e que trouxe grandes modificações à taxonomia vigente para Hylinae foi realizado 25
por Faivovich et al. (2005), utilizando o sequenciamento de genes mitocondriais e nucleares 26
associado a dados morfológicos para 226 espécies de hilídeos. Com base neste trabalho 2/3 27
das espécies atribuídas ao gênero Hyla foram alocadas em outros 15 gêneros, sendo um desses 28
o gênero Hypsiboas. 29
Os representantes do gênero Hypsiboas são popularmente conhecidos como pererecas, 30
sapos martelo, sapos ferreiro, entre outros, e são bastante freqüentes nas regiões úmidas da 31
mata atlântica, floresta amazônica, caatinga e formações de cerrado, totalizando 86 espécies 32
PPGGBC

53
(Amphibiaweb, 2012). Com base em dados genéticos, morfológicos, alguns dados 1
comportamentais e ecológicos os Hypsiboas estão alocados em sete grupos de espécies: H. 2
albopunctatus, H. benitizi, H. faber, H. pellucens, H. puchellus, H. punctatus e H. 3
semilineatus. 4
Estes grupos não embasados em sinapomorfias morfológicas são relevantes para a 5
definição de unidades monofiléticas, no entanto, algumas espécies têm caracteres que tornam 6
difícil alocá-las em qualquer um dos grupos atualmente reconhecidos (Faivovich et al., 2005). 7
Wiens et al. (2010), através de seqüenciamento gênico também reconheceu a existência 8
destes sete grupos monofiléticos entretanto, infere relações evolutivas entre os grupos, 9
ligeiramente diferentes das propostas por Faivovich et al. (2005). 10
Embora muitos esforços venham sendo empregados no entendimento das relações 11
sistemáticas do grupo, Hylidae permanece pouco estudada do ponto de vista citogenético. 12
Menos de 17% das espécies de Anura apresentam algum dado citogenético disponível, sendo 13
a maioria destes estudos restrita a caracterização da macroestrutura cariotípica utilizando 14
coloração convencional (King, 1990). 15
Durante muito tempo assumiu-se que os anfíbios constituíam um dos poucos grupos de 16
vertebrados que apresentavam cariótipos extremamente conservados. Entretanto, com o 17
emprego de técnicas de coloração diferencial e de citogenética molecular, descobriu-se uma 18
grande variabilidade na microestrutura cromossômica que permite uma melhor identificação 19
de espécies e suas variantes geográficas, constituindo uma importante ferramenta auxiliar em 20
estudos taxonômicos (Kasahara et al., 2003; Siqueira et al., 2004). 21
Os Hylidae, em sua maioria, apresentam número diplóide 2n=24 cromossomos, com 22
cariótipo constituído predominantemente por cromossomos de dois braços, enquanto 23
variações no número cromossômico têm sido consideradas apomorfias para o grupo (King, 24
1990; Catroli, 2008). 25
Com relação ao gênero Hypsiboas, apenas 16% apresentam algum dado citogenético, 26
sendo a maioria dos trabalhos focada apenas na descrição do cariótipo de diferentes espécies e 27
sem importantes inferências sobre evolução cromossômica do grupo com base em marcadores 28
heterocromáticos ou RONs (Raber et al., 2004; Ananias et al., 2004; Gruber et al., 2006; 29
Nunes & Fagundes, 2008 a,b; Carvalho et al., 2009). 30
O presente trabalho teve como objetivo realizar uma análise citogenética de diferentes 31
espécies e/ou populações de Hypsiboas visando identificar possíveis marcadores 32
cromossômicos que auxiliem no melhor entendimento da dinâmica evolutiva da macro e 33
microestrura cromossômica desses anfíbios. 34
PPGGBC

54
MATERIAIS E MÉTODOS 1
Foram analisados espécimes de H. crepitans, H. pombali e H. semilineatus provenientes 2
da Estação Ecológica Wenceslau Guimarães - EEWG (Wenceslau Guimarães/BA) e H. 3
atlanticus, proveniente de um fragmento de Mata Atlântica do município de Itacaré-BA (Fig. 4
3). Estes encontram-se depositados no Laboratório de Citogenética da UESB com devidos 5
números de tombo: 438, 439, L115, 22260, 22265, 22266, 22087 e 22088. 6
Para a preparação da suspensão de cromossomos mitóticos, utilizaram-se a raspagem do 7
epitélio intestinal, conforme protocolo descrito por King e Rofe, (1976) com modificações, ou 8
a dissociação do tecido hepático segundo protocolo de Garcia, ainda não publicado. 9
Para a determinação do número e morfologia dos cromossomos foi utilizada a coloração 10
convencional por Giemsa e o número de braços cromossômicos (NF) foi calculado segundo 11
Levan (1964). 12
Na elaboração dos idiogramas foram utilizados tanto os resultados obtidos nesse 13
trabalho, como os disponíveis em artigos da área. As figuras dos cariótipos foram 14
digitalizadas e submetidas à medição cromossômica no programa “Paquimetro” (Bressane e 15
Bressane, em preparação) tendo como base a proposta de Levan et al. (1964). As medidas 16
cromossômicas obtidas foram utilizadas para a construção de idiogramas através do programa 17
para cosntrução de idiogramas “Idiograma” (Bressane e Bressane, em preparação). A 18
padronização da classificação cromossômica se fez necessária para permitir uma melhor 19
comparação das fórmulas cariotípicas e da proporção de tamanho entre os cromossomos, uma 20
vez que muitos autores seguiram padrões de medidas cromossômicas diferenciados. 21
O bandamento C, deu-se segundo protocolo proposto por Sumner (1972) com 22
modificações; e para a detecção das regiões organizadoras de nucléolo utilizou-se a 23
impregnação por nitrato de prata, com pequenas alterações, segundo o Howell & Black 24
(1980). 25
RESULTADOS 26
Todas as espécies de Hypsiboas analisadas neste estudo apresentaram 2n=24 27
cromossomos e NF=48 com diferentes fórmulas cariotípicas (Fig. 4 - Tabela 4). 28
A impregnação por nitrato de prata permitiu evidenciar a ocorrência de RONs simples 29
localizadas intersticialmente nos braços longos do par cromossômico 7 para H. crepitans, H. 30
pombali e H. semilineatus, sendo que para H. pombali e H.semilineatus em todas as metáfases 31
PPGGBC

55
analisadas apenas um dos homólogos evidenciou marcações Ag-RONs. Hypsiboas atlanticus, 1
apresentou RONs múltiplas localizadas terminalmente nos braços longos de dois pares de 2
cromossomos (10 e 12) (Fig 5 - boxes). 3
O bandamento C revelou a existência de blocos de heterocromatina constitutiva 4
adjacente aos centrômeros de todos os cromossomos de H. crepitans, H. semilineatus e H. 5
pombali e nos pares 1, 2, 3, 4 e 5 de H. atlanticus. Puderam ainda ser evidenciadas regiões 6
Banda C+ localizadas intersticialmente nos braços longos de seis pares de cromossomos de H. 7
atlanticus (1, 2, 3, 6, 7 e 8) e nos braços curtos do par 3 de H. crepitans. Blocos 8
heterocromáticos terminais foram observados apenas para H. atlanticus e mostraram-se 9
coincidentes com as RONs. Para as demais espécies analisadas não havia heterocromatina 10
associada à RON (Fig 5). 11
A associação dos dados obtidos nesse trabalho com os dados citogenéticos disponíveis 12
na literatura para espécies de Hypsiboas permitiu uma melhor comparação das fórmulas 13
cartiotípicas já evidenciadas para estes animais (Tabela 3). Após a padronização das medidas 14
cromossômicas (Tabela 4) foram identificados oito padrões de composição cromossômica, 15
muitos desses compartilhados por diferentes espécies (Tabela 4 e Figura 6). 16
Com relação à macroestrutura cariotípica, pode-se observar que os pares 17
cromossômicos 1 e 5, são os únicos que mantém morfologia constante (metacêntrico e 18
submetacêntrico, respectivamente) para todas as espécies estudadas. Dentre os demais pares 19
cromossômicos, os pares 2, 10, 11 e 12 mostram-se mais conservados, enquanto os demais 20
pares apresentaram uma maior variação morfológica devido ao posicionamento diferencial da 21
constrição primária (Tabela 4). 22
A comparação entre os padrões de distribuição de heterocromatina constitutiva e das 23
Ag-RONs permitiu evidenciar alguns marcadores cromossômico para H. albomarginatus 24
(RONs interstiticiais nos braços curtos do par 2), H. crepitans (bloco heterocromático 25
interstitical nos braços curtos do par 3) e espécies do grupo H. pulchellus (bloco 26
heterocromático terminal nos braços longos do par 1) (Fig. 5). 27
DISCUSSÃO 28
Dentre as 86 espécies reconhecidas para o gênero Hypsiboas, apenas 13 já foram 29
analisadas até o momento, incluindo os dados do presente trabalho, este número aumenta para 30
15. Todas as espécies apresentaram 2n=24 cromossomos e NF=48, com exceção de H. 31
albopunctatus com 2n=22 cromossomos (Tabela 3). A manutenção do número diplóide em 24 32
PPGGBC

56
cromossomos é uma das principais características citogenéticas da família Hylidae, apesar de 1
números diplóides maiores ou menores já terem sido observados (2n=18 em Aplastodiscus 2
leucopygius até 2n=52 em Phyllomedusa tretraploidea) (Catroli, 2008). Acredita-se que o 3
cariótipo de todas, ou quase todas as espécies, com número diplóide menor ou igual a 24 4
cromossomos e pertencentes às famílias de anuros consideradas mais derivadas, como 5
Dendrobatidae, Hylidae, Leptodactilydae e Ranidae, tenham surgido por um evento de 6
redução a partir de um ancestral comum com 2n=26 cromossomos (Bogart, 1973), o que é 7
corroborado pela filogenia proposta por Faivovich et al. (2005), 8
Segundo Gruber et al. (2006) dois possíveis mecanismos poderiam estar envolvidos na 9
redução de número diplóide de H. albopunctatus, fusões em tandem ou translocações 10
envolvendo os pequenos cromossomos do cariótipo, o que poderia ter resultado também na 11
mudança na posição das RONs. Os autores também consideram a possibilidade de que a 12
ocorrência de um cromossomo supranumerário, observada apenas nessa espécie, represente 13
um vestígio de que, no passado, H. albopunctatus também tenha apresentado 2n=24 14
cromossomos. 15
Com relação à morfologia dos cromossomos, observa-se que há predomínio de 16
cromossomos de dois braços no cariótipo e que alguns pares cromossômicos apresentam uma 17
evolução mais dinâmica do que outros. Os pares 1 e 5 mantém morfologia e tamanho 18
conservados para todas as espécies/populações já estudadas, enquanto que os pares 3, 4, 6 e 7 19
variam mais sua morfologia com relação ao posicionamento do centrômero, podendo ser 20
metacêntricos, submetacêntricos ou subtelocêntricos (Tabela 4). Esse fato, associado à 21
manutenção do tamanho dos pares cromossômicos entre os cariótipos são um forte indício de 22
que as inversões pericêntricas seriam o principal mecanismo envolvido na diferenciação da 23
macroesturura cariotípica do grupo. 24
Ainda com relação macroestrurura cromossômica foi possível distribuir as espécies e/ou 25
populações já analisadas em oito grupos de acordo com sua fórmula cariotípica (o que 26
chamaremos de grupos cariotípicos) (Fig. 5 e Tabela 4). Pode-se ainda observar que estes 27
grupos cariotípicos não são compostos necessariamente por populações de uma mesma 28
espécie ou por espécies que pertençam aos mesmos grupos de espécie propostos por 29
Faivovitch et al. (2005) e Wiens et al. (2010). Para o grupo H. faber foram encontradas cinco 30
fórmulas cariotípicas (grupos cariotípicos 1, 2, 3, 5 e 6). O grupo H. semilineatus apresentou 31
dois cariótipos distintos (grupos cariotípicos 4 e 5), assim como o grupo H. pulchellus (grupos 32
cariotípicos 3 e 7). Os grupos H. albopunctatus e H. punctatus apresentaram apenas um 33
padrão cromossômico (grupos cariotípicos 2 e 8, respectivamente). Apesar do 34
PPGGBC

57
compartilhamento de um mesmo padrão cariotípico entre as espécies de Hypsiboas, observa-1
se que a microestrutura cromossômica é muito mais variável permitindo a diferenciação de 2
espécies e suas variantes geográficas. 3
De modo geral, as RONs são tidas como um importante marcador citogenético para o 4
estudo de anfíbios, isso porque, para maioria dos anuros, o número e a localização desses 5
sítios mostram-se conservados dentro das espécies estudadas (Lourenço et al., 1998). 6
Para as espécies e populações de Hypsiboas analisadas, o estudo das RONs demonstrou 7
uma grande variabilidade, onde a maioria das espécies apresentou RONs simples localizadas 8
intersticialmente nos braços longos de cromossomos médio/pequenos (pares 7, 10 e 11) 9
(presente estudo, Ananias et al., 2004; Gruber et al., 2006; Nunes & Fagundes, 2008a). Essa 10
variação não está restrita apenas à espécies diferentes, o que pôde ser observado nas duas 11
populações de H. crepitans (presente estudo; Gruber et al., 2006) e H. faber (Nunes & 12
Fagundes, 2008a; Carvalho et al., 2009) estudadas que diferiram com relação ao par 13
cromossômico nucleolar. Esses dados indicam que as RONs são marcadores cromossômicos 14
fracos para identificar as possíveis espécies do gênero e que outros marcadores devem ser 15
aplicados conjuntamente para que os dados apresentem potencial citotaxonômico. Entretanto, 16
em alguns casos, as RONs terminais e/ou RONs localizadas nos maiores cromossomos do 17
complemento (pares 1 e 2) mostraram-se conservadas e restritas a grupos específicos, 18
podendo ser consideradas, até o momento, marcadores cromossômicos espécie-específico 19
para H. albomarginatus e H. semiguttatus (Ananias et al., 2004; Nunes & Fagundes, 2008a). 20
O primeiro relato de RONs múltiplas para o gênero é descrito no presente estudo para 21
H. atlanticus, que apresentou marcações terminais discretas nos braços longos dos pares 10 e 22
12. De modo geral os anfíbios anuros apresentam RONs simples terminais (King et al., 1990), 23
entretanto para Hypsiboas, quando leva-se em conta as filogenias moleculares disponíveis 24
para o grupo, RONs simples intersticiais (Nunes & Fagundes, 2008a), podem ser 25
consideradas como uma característica plesiomórfica, e condições variantes como apomorfias. 26
A variação de localização das RONs poderia ser mais facilmente explicada por 27
possíveis eventos de translocação ou transposição cromossômica, uma vez que há manutenção 28
do número e do tamanho cromossômico entre os diferentes cariótipos analisados. Esses 29
mesmos mecanismos também poderiam explicar o surgimento das RONs múltiplas em H. 30
altlanticus. Possivelmente a espécie ancestral possuía RONs terminais localizadas no par 31
cromossômico 10 e por translocação ou transposição, parte dos sítios de rDNA foram 32
transferidos para a região terminal do par 12, resultando no pequeno tamanho das marcações 33
evidenciadas pelo nitrato de prata nessa espécie. Eventos de transposição e/ou translocação 34
PPGGBC

58
também são considerados como os possíveis mecanismos de dispersão de sítios ribossomais 1
45S em muitas espécies de anuros que apresentam RONs múltiplas (Schmid et al., 1995). 2
A aplicação da FISH com sondas de rDNA 45S foi realizada apenas para quatro 3
espécies de Hypsiboas (H. albomarginatus, H. faber, H. pardalis e H. semilineatus) 4
confirmando a existência de RONs simples para estas (Nunes & Fagundes, 2008a). 5
Ressaltamos aqui a importância da realização da técnica de FISH com genes ribossomais para 6
confirmar o numero e a localização dos sítios de rDNA 45S nas espécies estudadas nesse 7
trabalho, uma vez que para H. pombalis e H. semilineatus a prata só evidenciou um dos 8
homólogos do par nucleolar, o que poderia indicar uma ativação preferencial dos sítios 9
ribossomais durante a interfase precedente. Além disso, essa técnica também seria útil para 10
verificar a real existência de RONs múltiplas para H. atlanticus, pois existem relatos para 11
outros grupos animais de marcações inespecíficas obtidas pela impregnação por nitrato de 12
prata e que se mostraram coincidentes com blocos heterocromáticos de natureza ácida 13
(Sumner, 1990; Sánchez et al., 1995). 14
O padrão de distribuição de heterocromatina é uma das características mais utilizadas 15
para a diferenciação de espécies/populações dentre os anuros, sendo um dos marcadores 16
cromossômicos mais empregados em estudos citotaxonômicos, como observado por 17
Anderson (1991) em espécies de Hyla, em espécies de Telmatobufo (Formas et al., 2000) e 18
em Colostethus (Veiga-Menoncello et al., 2003). 19
Os poucos dados disponíveis para o gênero Hypsiboas mostram que a maioria das 20
espécies apresenta grande quantidade de heterocromatina pericentromérica distribuída por 21
todo o complemento cromossômico. Blocos BC+ interstiticiais são observados em apenas 22
alguns cromossomos de poucas espécies e blocos terminais são encontrados com menos 23
freqüência (Tabela 3 e Figura 5). Uma melhor comparação do padrão de distribuição da 24
heterocromatina entre populações fica dificultada, pois a maioria dos trabalhos disponível não 25
aplicou esta metodologia. 26
Tendo como base as relações evolutivas entre as espécies estudadas na filogenia 27
molecular de Faivovich et al. (2005) associadas aos dados reunidos na figura 2 e dados 28
disponíveis para espécies próximas, observa-se que cariótipos contendo apenas 29
heterocromatina constitutiva pericentromérica constituem uma característica plesiomórfica 30
para o grupo. Blocos heterocromáticos intersticiais ou terminais consistiriam em variações do 31
padrão ancestral. 32
Pode-se observar que as espécies de Hypsiboas que possuem heterocromatina em 33
posição intersticial/terminal apresentam uma diminuição na quantidade de heterocromatina 34
PPGGBC

59
pericentromérica ou no tamanho desses blocos, como pode ser visto para H.albomarginatus, 1
H. albopunctatus, H. crepitans, H. raniceps e H atlanticus (Gruber et al., 2006, Carvalho et 2
al., 2009, presente trabalho). Esse fato poderia ser facilmente explicado como resultado de 3
eventos de inversões cromossômicas que resultariam na transferência dessas regiões ao longo 4
dos braços cromossômicos, reforçando que as inversões seriam o principal mecanismo de 5
evolução cariotípica em Hypsiboas. Essas inversões envolvendo blocos heterocromáticos 6
também poderiam explicar o fato de algumas espécies apresentarem RONs heterocromáticas, 7
enquanto a maioria apresenta RONs eucromáticas, possivelmente estas regiões tornaram-se 8
intercaladas ou associadas à heterocromatina após esses eventos de reorganização estrutural 9
dos cromossomos. 10
Com base nos padrões observados, nota-se que o bloco heterocromático intersticial 11
localizado nos braços curtos do par cromossômico 3 de H. crepitans apresenta um grande 12
potencial para ser considerado um marcador espécie específico, pois está presente apenas nas 13
populações dessa espécie, mesmo que essas apresentem diferenças que as caracterizem como 14
variantes geográficas. O mesmo ocorre para o bloco terminal localizado nos braços longos do 15
par cromossômico 1 das espécies pertencentes ao grupo H. pulchellus e o bloco terminal nos 16
braços curtos do par 6 de H. bischoffi (Fig. 4). 17
Seria de se esperar que populações de uma espécie ou espécies pertencentes a um 18
mesmo grupo taxonômico compartilhassem mais semelhanças com relação ao padrão de 19
distribuição de heterocromatina e/ou número e localização das RONs do que com outras 20
espécies, entretanto, com base nos dados reunidos na figura 2 e no local de origem das 21
populações analisadas pode-se inferir que a evolução da microestrutura cariotípica apresenta-22
se fortemente associada com a distribuição geográfica desses animais. Por exemplo, as 23
espécies do presente estudo provenientes da Estação Ecológica de Wenceslau Guimarães, 24
apesar de diferirem com relação à fórmula cariotípica, apresentam o mesmo padrão de 25
Bandamento C e RONs, assim como ocorre para H. marginatus, H. semigutattus e H. aff. 26
semigutattus (Ananias et al., 2004) e H. bischoffi e H. guentheri (Ruber et al., 2004). Essa 27
possível relação entre a dinâmica evolutiva da microestrutura cromossômica e a localização 28
geográfica dessas espécies poderia ser resultado de características ambientais únicas que 29
favorecem uma organização cariotípica ótima, situação semelhante é proposta para variantes 30
geográficas de lagartos do gênero Tropidurus (Pellegrino et al., 1996) e para grupos de 31
peixes, como o gênero Leporinos (Galetti et al., 1981, 1984), entretanto, esta hipótese só 32
poderá ser confirmada com o aumento dos estudos citogenéticos no gênero. 33
PPGGBC

60
Com base nos dados reunidos, o gênero Hypsiboas mostra-se um interessante material 1
para estudos de evolução cromossômica em anuros, uma vez que, mesmo com os poucos 2
trabalhos já desenvolvidos, nota-se uma acentuada variabilidade cariotípica para o grupo. A 3
presença de marcadores citotaxonômicos e possível relação filogeográfica da microestrutura 4
cariotípica só poderão ser confirmados com o aumento dos estudos de natureza cromossômica 5
para o gênero. Devido a complexidade taxonômica do grupo, estudos moleculares e 6
citogenéticos populacionais também mostram-se necessários para confirmar se há 7
diversidade críptica em algumas espécies, como, por exemplo, H. crepitans e H. faber, como 8
sugerido pelos dados apresentados. 9
AGRADECIMENTOS: 10
Os autores agradecem à equipe do Prof. Dr. Miguel T. Rodrigues pela coleta e 11
identificação dos animais. À Mauro Teixeira Júnior, pelas fotos dos exemplares analisados. 12
Ao Prof. Dr. Mirco Solé pela identificação dos exemplares de H. atlanticus. Aos 13
coordenadores da Estação Ecológica de Wenceslau Guimarães pelo auxílio durante as 14
expedições de campo. Este trabalho contou com o apoio financeiro da Fundação de Amparo à 15
Pesquisa do Estado da Bahia (FAPESB – processo BOL 1399/2010). 16
REFERÊNCIAS 17
AMPHIBIAWEB. 2012. Disponível em <amphibiaweb.org>. Acessado em 02/03/2010. 18
ANANIAS, F.; GARCIA, P. C. A. & RECCO-PIMENTEL, S.M. 2004. Conserved 19
karyotypes in the Hyla pulchella species group (Anura, Hylidae). Hereditas 140: 42-48. 20
ANDERSON, K., 1991.Chromosome evolution in Holoartic treefrogs. In: Green, D.M. & 21
Sessions, S.K. Amphibians cytogenetics and evolution. Academic Press pp. 299-331. 22
BOGART, J. P. 1973. Evolution of anuram karyotypes. In Vial, J. L. (ed.). Evolutionary 23
Biology of the Anurans: contemporary research on major problems. University of 24
Missuru Press, Columbia, pp.. 337-349. 25
BRESSANE & BRESSANE. Em preparação. 26
CARVALHO, K. A., GARCIA, P. C. A. & RECCO-PIMENTEL, S. M. 2009. Cytogenetic 27
comparison of tree frogs of the genus Aplastodiscus and the Hypsiboas faber group 28
(Anura, Hylidae). Genetic and Molecular Research 8 (4): 1498-1508. 29
PPGGBC

61
CATROLI, G. F. 2008. Cariótipo se seis espécies de Bokermannohyla dos grupos de B. 1
circumdata e B. pseudopseudis (Anura, Hylidae). Dissertação apresentada ao Instituto 2
de Biociências da Universidade Estadual Paulista, UNESP, Campus de Rio Claro, para 3
a obtenção do título de Mestre em Ciências Biológicas (Biologia Celular e Molecular) 4
Rio Claro – SP. 5
FAIVOVICH, J.; HADDAD, C. F. B.; GARCIA, P. C. A.; FROST, D. R.; CAMPBELL, J.A. 6
& WHEELER, W. C. 2005. Systematic review of the frog family Hylidae, with special 7
reference to Hylinae: phylogenetic analysis and taxonomic revision. Bull. Am. Museum 8
Nat. Hist. 294:1-240. 9
FORMAS, J.R. & CUEVAS, C.C. 2000. Comparative cytogenetic analysis of the Chilean 10
leptodactylid frog genus Telmatobufo, with description of T. venustosus. P. Biol. Soc. 11
Wash. 113:890-899. 12
FROST, D.R. 2012 Amphibian Species of theWorld: An Online Reference, version 5.3. 13
Electronic database accessible at http://research.amnh.org/herpetology/amphibia 14
American Museum of Natural History, New York, USA. 15
GALETTI JUNIOR, P. M.; FORESTI, F.; BERTOLLO, L. A. C. & MOREIRA-FILHO, O. 16
1984. Characterization of eight species of Anostomidae fish on the basis of the 17
Nucleolar Organizing Regions. Caryologia (Firenze), v. 37, n. 4, p. 401-406. 18
GALETTI JUNIOR, P. M.; FORESTI, F.; BERTOLLO, L. A. C. & MOREIRA-FILHO, O. 19
1981. Karyotipic similarity in three genera (Leporinus, Leporellus and Schizodon) of the 20
family Anostomidae (Teleostei). Brazilian Journal of Genetics, v. IV, n. 1, p. 11-15. 21
GARCIA, C. Melhoria na metodologia da obtenção de cromossomos mitóticos em 22
representantes da herpetofauna. Em preparação. 23
GRUBER, S. L.; HADDAD, C. F. B, & KASAHARA, S. 2006. Chromosome banding in 24
three species of Hypsiboas (Hylidae,Hylinae), with special reference to a new case of B-25
chromosome in anuran frogs and to the reduction of the diploid number of 2n = 24 to 26
2n=22 in the genus. Springer Science+Business Media B.V. 27
HOWELL, W. M. & BLACK, D. A. 1980.Controlled silver staining of nucleolar organizer 28
regions with a protective colloidal developer; a 1-step method. Experientia 36:1014-29
1015. 30
KASAHARA, S.; ZAMPIERI, A. P. S.; GRUBER S.L. & HADDAD, C.F.B. 2003. 31
Comparative cytogenetic four tree frog species (Anura, Hylidae, Hylinae) from Brazil. 32
Cytogenet. Gen. res. 103:155-167. 33
PPGGBC

62
KING, M. 1990. Amphibia. In John B, Gwent C (Eds) Animal Cytogenetics. Amphibia 4 1
Chordata 2. Gebruder Borntraeger, Berlin, pp. 1-241. 2
KING, M. & ROFE, R. 1976. Karyotype variation in the Australian gekko Phyllodactylus 3
marmoratus (Gray) (Gekkonidae: Reptilia). – Chromosoma 54: 75-87. 4
LEVAN, A.; FREDGA, K. & SANDBERG, A. A. 1964. Nomenclature for centromeric 5
position on chromosomes. Hereditas 52:201-220. 6
LOURENÇO, L. B.; RECCO-PIMENTEL, S. M. & CARDOSO, A. J. 1998. Polymorphism 7
of the nucleolus organizer regions (NORs) Physalaemus petersi (Amphibia, Anura, 8
leptodactylidae) detected by silver staining and fluorescent in situ hybridization. Chrom. 9
Res., 6:621-328. 10
NUNES, R. D. R. A. & FAGUNDES, V. 2008. Patterns of ribosomal DNA distribution in 11
hylid frogs from the Hypsiboas faber an H. semilineatus species groups. Genetics and 12
molecular biology, 31, 4, 982-987. 13
PELLEGRINO, K. C. M.; RODRIGUES, M. T. & YASSUDA, Y. Y. 1996. Comparative 14
cytogenetic studies of eleven species of the Tropidurus torquatus group (Sauria, 15
Tropiduridae) with banding patterns. Hereditas (Lund), Suécia, v. 125, p. 37-46. 16
RABER, S.C., CARVALHO, K.A., GARCIA, P.C., VINCIPROVA, G. & RECCO-17
PIMENTEL, S.M. 2004. Chromosomal characterization of H. bischoffi and Hyla 18
guentheri (Anura, Hylidae). Phyllomedusa, 3: 43-49. 19
SALDUCCI, M. D.; MARTY, C.; FOUQUET, A. & GILLES, A. 2005. Phylogenetic 20
relationships and biodiversity in Hylids (Anura: Hylidae) from French Guiana. C R 21
Biol. 328 (10-11): 1009-1024. 22
SÁNCHEZ, A.; JIMENEZ, R.; BURGOS, M.; STITOU, S.; ZURITA, F. & DIAZ DE LA 23
GUARDIA, R. 1995. Cytogenetic peculiarities in the Algerian hedgehog: silver stains 24
not only NORs but also heterochromatic blocks. Heredity, vol. 75, p. 10-6. 25
SCHMID, M., FEICHTINGER, W., WEIMER, R., MAIS, C., BOLAÑOS, F. & LEÓN, P. 26
Chromosome banding in Amphibia XXI. Inversion polymorphism and multiple 27
nucleolus organizer regions in Agalychnis callidryas (Anura, Hylidae). Cytogenet. Cell 28
Genet. 69: 18-26, 1995. 29
SIQUEIRA, S.; ANANIAS, F. & RECCO-PIMENTEL, S.M. 2004. Cytogenetic analysis of 30
three species of Eleutherodactylus (Anura, Leptodactylidae) from southeastern Brazil. 31
Genet Mol Biol 27:363-372. 32
SUMNER, A.T. 1972. A simple technique for demonstrating centromeric heterocromatin. 33
Exp Cell 75:304-306. 34
PPGGBC

63
SUMNER, A. T. 1990. Chromosome banding. Uniwin Hyman, Londres, UK. P.434. 1
WIENS, J. J.; KUCZYNSKI, C. A.; HUA, X. & MOEN, D. S. 2010. An expanded phylogeny 2
of treefrogs (Hylidae) based on nuclear and mitochondrial sequence data. Molecular 3
Phylogenetics and Evolution 55 871–882. 4
PPGGBC
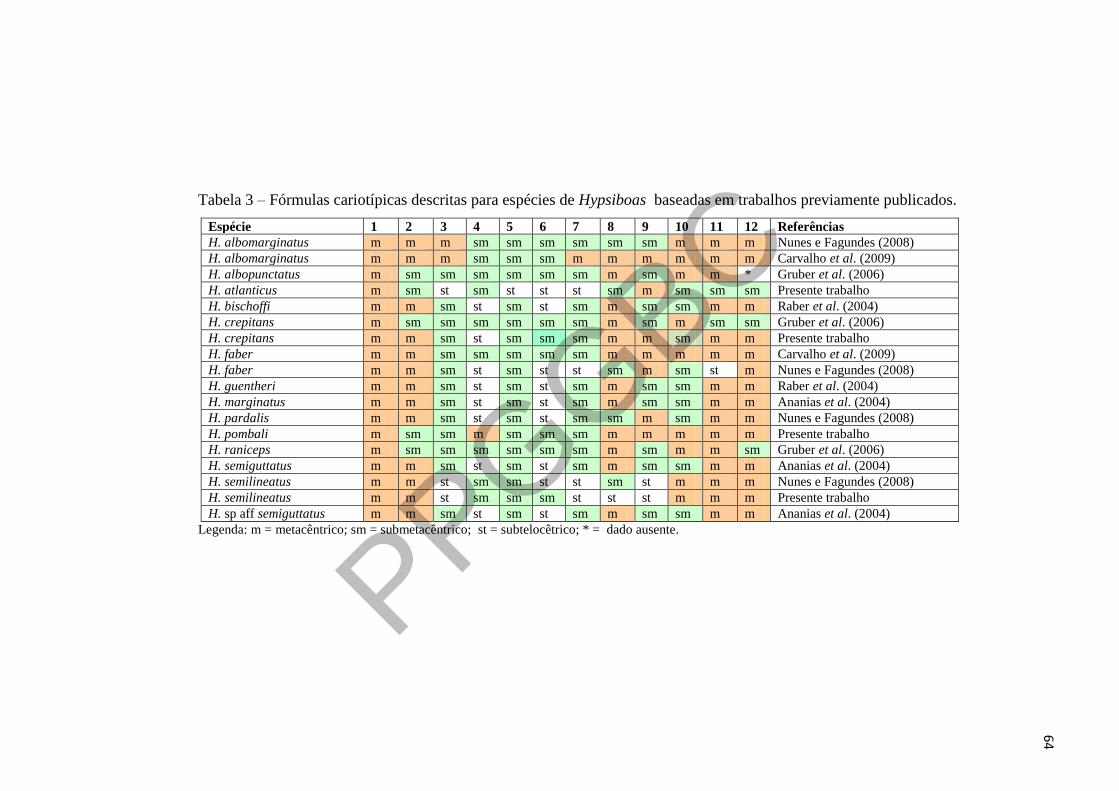
6
4
Tabela 3 – Fórmulas cariotípicas descritas para espécies de Hypsiboas baseadas em trabalhos previamente publicados.
Espécie 1 2 3 4 5 6 7 8 9 10 11 12 Referências
H. albomarginatus m m m sm sm sm sm sm sm m m m Nunes e Fagundes (2008)
H. albomarginatus m m m sm sm sm m m m m m m Carvalho et al. (2009)
H. albopunctatus m sm sm sm sm sm sm m sm m m * Gruber et al. (2006)
H. atlanticus m sm st sm st st st sm m sm sm sm Presente trabalho
H. bischoffi m m sm st sm st sm m sm sm m m Raber et al. (2004)
H. crepitans m sm sm sm sm sm sm m sm m sm sm Gruber et al. (2006)
H. crepitans m m sm st sm sm sm m m sm m m Presente trabalho
H. faber m m sm sm sm sm sm m m m m m Carvalho et al. (2009)
H. faber m m sm st sm st st sm m sm st m Nunes e Fagundes (2008)
H. guentheri m m sm st sm st sm m sm sm m m Raber et al. (2004)
H. marginatus m m sm st sm st sm m sm sm m m Ananias et al. (2004)
H. pardalis m m sm st sm st sm sm m sm m m Nunes e Fagundes (2008)
H. pombali m sm sm m sm sm sm m m m m m Presente trabalho
H. raniceps m sm sm sm sm sm sm m sm m m sm Gruber et al. (2006)
H. semiguttatus m m sm st sm st sm m sm sm m m Ananias et al. (2004)
H. semilineatus m m st sm sm st st sm st m m m Nunes e Fagundes (2008)
H. semilineatus m m st sm sm sm st st st m m m Presente trabalho
H. sp aff semiguttatus m m sm st sm st sm m sm sm m m Ananias et al. (2004)
Legenda: m = metacêntrico; sm = submetacêntrico; st = subtelocêtrico; * = dado ausente.
PPGGBC

6
5
Tabela 4 – Fórmulas cariotípicas descritas para espécies de Hypsiboas baseadas em trabalhos previamente publicados
após padronização da medidas cromossômicas segundo Levan (1964).
Espécie 1 2 3 4 5 6 7 8 9 10 11 12 Referências
Grupo cariotípico 1
H. albomarginatus m m m sm sm sm m m m m m m Carvalho et al. (2009)
H. albomarginatus m m m sm sm st m m m m m m Nunes e Fagundes (2008)
Grupo cariotípico 2
H. albopunctatus m m sm st sm sm sm m sm m m - Gruber et al. (2006)
H. crepitans m m sm st sm sm sm m m sm m m Gruber et al. (2006)
H. crepitans m m sm st sm sm sm m sm m m m Presente trabalho
H. raniceps m m sm st sm sm sm m sm m m m Gruber et al. (2006)
Grupo cariotípico 3
H. bischoffi m m sm st sm st sm m m m m m Raber et al. (2004)
H. guentheri m m sm st sm st sm m m m m m Raber et al. (2004)
H. marginatus m m sm st sm st sm m m m m m Ananias et al. (2004)
H. pardalis m m sm st sm st sm m m m m m Nunes e Fagundes (2008a)
H. semiguttatus m m sm st sm st sm m m m m m Ananias et al. (2004)
H. sp aff semiguttatus m m sm st sm st sm m m m m m Ananias et al. (2004)
Grupo cariotípico 4
H. semilineatus m m st sm sm st sm sm sm m m m Nunes e Fagundes (2008a)
H. semilineatus m m st sm sm st sm sm sm m m m Presente trabalho
Grupo cariotípico 5
H. pombali m m sm m sm sm sm m m m m m Presente trabalho
H. faber m m sm sm sm sm sm m m m m m Carvalho et al. (2009)
Grupo cariotípico 6
H. faber m m sm st sm st st sm m m sm m Nunes e Fagundes (2008a)
Grupo cariotípico 7
H. polytaenius m sm sm sm sm st sm m m m m m Nunes e Fagundes (2008b)
Grupo cariotípico 8
H. atlanticus m m st st sm st m sm m sm sm sm Presente trabalho
Legenda: m = metacêntrico; sm = submetacêntrico; st = subtelocêtrico. PPGGBC
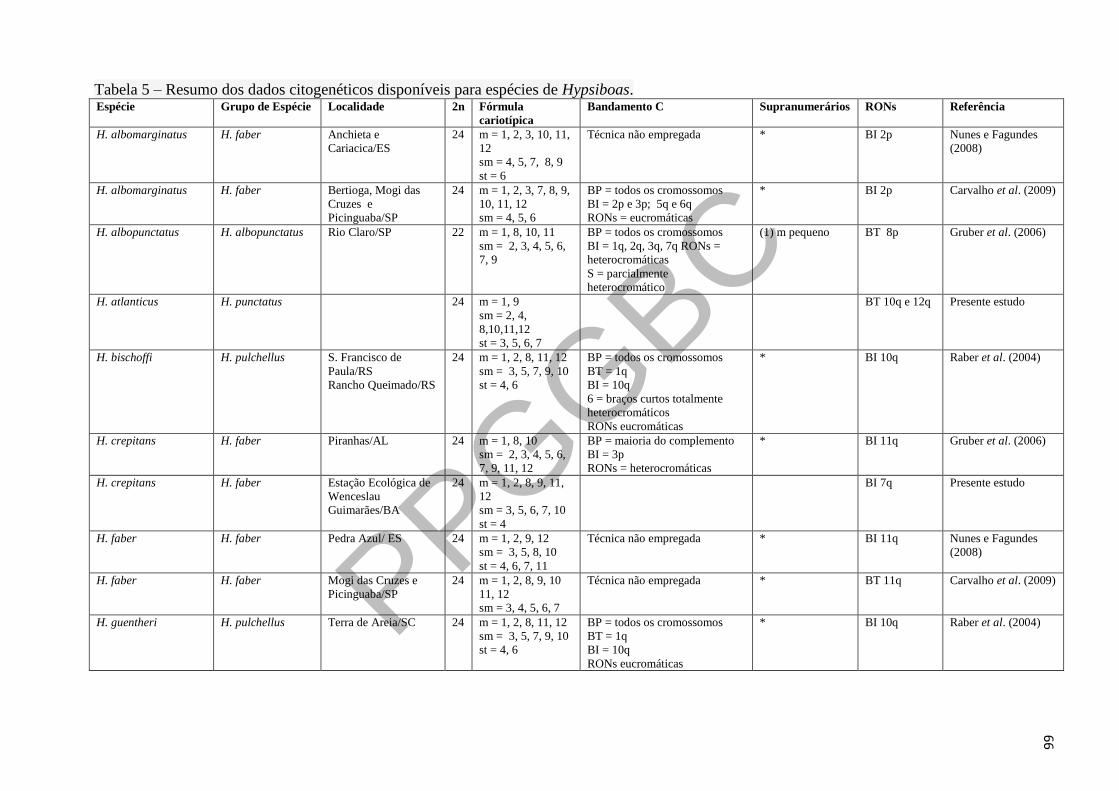
6
6
Tabela 5 – Resumo dos dados citogenéticos disponíveis para espécies de Hypsiboas. Espécie Grupo de Espécie Localidade 2n Fórmula
cariotípica
Bandamento C Supranumerários RONs Referência
H. albomarginatus H. faber Anchieta e
Cariacica/ES
24 m = 1, 2, 3, 10, 11,
12
sm = 4, 5, 7, 8, 9
st = 6
Técnica não empregada * BI 2p Nunes e Fagundes
(2008)
H. albomarginatus H. faber Bertioga, Mogi das
Cruzes e
Picinguaba/SP
24 m = 1, 2, 3, 7, 8, 9,
10, 11, 12
sm = 4, 5, 6
BP = todos os cromossomos
BI = 2p e 3p; 5q e 6q
RONs = eucromáticas
* BI 2p Carvalho et al. (2009)
H. albopunctatus H. albopunctatus Rio Claro/SP 22 m = 1, 8, 10, 11
sm = 2, 3, 4, 5, 6,
7, 9
BP = todos os cromossomos
BI = 1q, 2q, 3q, 7q RONs =
heterocromáticas
S = parcialmente
heterocromático
(1) m pequeno BT 8p Gruber et al. (2006)
H. atlanticus H. punctatus 24 m = 1, 9
sm = 2, 4,
8,10,11,12
st = 3, 5, 6, 7
BT 10q e 12q Presente estudo
H. bischoffi H. pulchellus S. Francisco de
Paula/RS
Rancho Queimado/RS
24 m = 1, 2, 8, 11, 12
sm = 3, 5, 7, 9, 10
st = 4, 6
BP = todos os cromossomos
BT = 1q
BI = 10q
6 = braços curtos totalmente
heterocromáticos
RONs eucromáticas
* BI 10q Raber et al. (2004)
H. crepitans H. faber Piranhas/AL 24 m = 1, 8, 10
sm = 2, 3, 4, 5, 6,
7, 9, 11, 12
BP = maioria do complemento
BI = 3p
RONs = heterocromáticas
* BI 11q Gruber et al. (2006)
H. crepitans H. faber Estação Ecológica de
Wenceslau
Guimarães/BA
24 m = 1, 2, 8, 9, 11,
12
sm = 3, 5, 6, 7, 10
st = 4
BI 7q Presente estudo
H. faber H. faber Pedra Azul/ ES 24 m = 1, 2, 9, 12
sm = 3, 5, 8, 10
st = 4, 6, 7, 11
Técnica não empregada * BI 11q Nunes e Fagundes
(2008)
H. faber H. faber Mogi das Cruzes e
Picinguaba/SP
24 m = 1, 2, 8, 9, 10
11, 12
sm = 3, 4, 5, 6, 7
Técnica não empregada * BT 11q Carvalho et al. (2009)
H. guentheri H. pulchellus Terra de Areia/SC 24 m = 1, 2, 8, 11, 12
sm = 3, 5, 7, 9, 10
st = 4, 6
BP = todos os cromossomos
BT = 1q
BI = 10q
RONs eucromáticas
* BI 10q Raber et al. (2004) PPGGBC

6
7
Espécie Grupo de Espécie Localidade 2n Fórmula
cariotípica
Bandamento C Supranumerários RONs Referência
H. marginatus H. pulchellus S. Francisco de
Paula/RS
24 m = 1, 2, 8, 11, 12
sm = 3, 5, 7, 9, 10
st = 4, 6
BP = todos os cromossomos
10q = totalmente
heterocromáticos
BT = 1q
RONs = heterocromáticas
* BI 10q Ananias et al. (2004)
H. p. joaquini H. pulchellus - 24 m = 1, 2, 8, 11, 12
sm = 3, 5, 7, 9, 10
st = 4, 6
BP = todos os cromossomos
10q = totalmente
heterocromáticos
BT = 1q
RONs = eucromáticas
* BT 1p Ananias et al. (2004)
H. pardalis H. faber Cariacica/ES 24 m = 1, 2, 9, 11, 12
sm = 3, 5, 7, 8, 10
st = 4, 6
Técnica não empregada * BT 11p Nunes e Fagundes
(2008)
H. pombalis H. semilineatus Estação Ecológica de
Wenceslau
Guimarães/BA
24
m = 1, 4, 8, 9, 10,
11, 12
sm = 2, 3, 5, 6, 7
BI 7q Presente estudo
H. raniceps H. albopunctatus Brasilândia/MS 24 m = 1, 8, 10, 11
sm = 2, 3, 4, 5, 6,
7, 9, 12
BP = 11q
BI = 6,p e 7p
RONs = heterocromáticas
* BT 11q Gruber et al. (2006)
H. semiguttatus H. pulchellus Carambá do Sul/SC
S. Francisco de
Paula/RS
Piraquara/PR
24 m = 1, 2, 8, 11, 12
sm = 3, 5, 7, 9, 10
st = 4, 6
BP = todos os cromossomos
10q = totalmente
heterocromáticos
BT = 1q
RONs = eucromáticas
* BT 1 p Ananias et al. (2004)
H. semilineatus H. semilineatus Sta Teresa/ES 24 m = 1, 2, 10, 11,
12
sm = 4, 5, 8
st = 3, 6, 7, 9
Técnica não empregada * BI 7q Nunes e Fagundes
(2008)
H. semilineatus H. semilineatus Estação Ecológica de
Wenceslau
Guimarães/BA
24 m = BI 7q Presente estudo
H. sp aff semiguttatus H. pulchellus Misiones/Argentina 24 m = 1, 2, 8, 11, 12
sm = 3, 5, 7, 9, 10
st = 4, 6
BP = todos os cromossomos
10q = totalmente
heterocromáticos
BT = 1q
RONs = eucromáticas
* BT 1p Ananias et al. (2004)
Legenda: m = metacêntrico; sm = submetacêntrico; st = subtelocêtrico; BP = banda pericentromérica; BT = banda terminal; BI = banda intersticial; q = braço longo; p = braço
curto; * = dado ausente. PPGGBC

68
Fig.3 Fotos dos indivíduos de Hypsiboas analisados no presente estudo: a)
Hypsiboas atlanticus, b) Hypsiboas crepitans, c) Hypsiboas pombali, d)
Hypsiboas semilineatus. (Fonte: a) Sérgio Siqueira Jr.; b, c d) Mauro Teixeira Jr.)
a) b)
c) d)
PPGGBC

69
Fig. 4 Cariótipos submetidos a coloração convencional (1) e bandamento C (2) de: a) H. atlanticus; b) H.
crepitans; c) H. pombali e d) H. semilineatus. Em destaque os cromossomos nucleolares submetidos à
impregnação por nitrato de prata e bandamento C.
1
PPGGBC
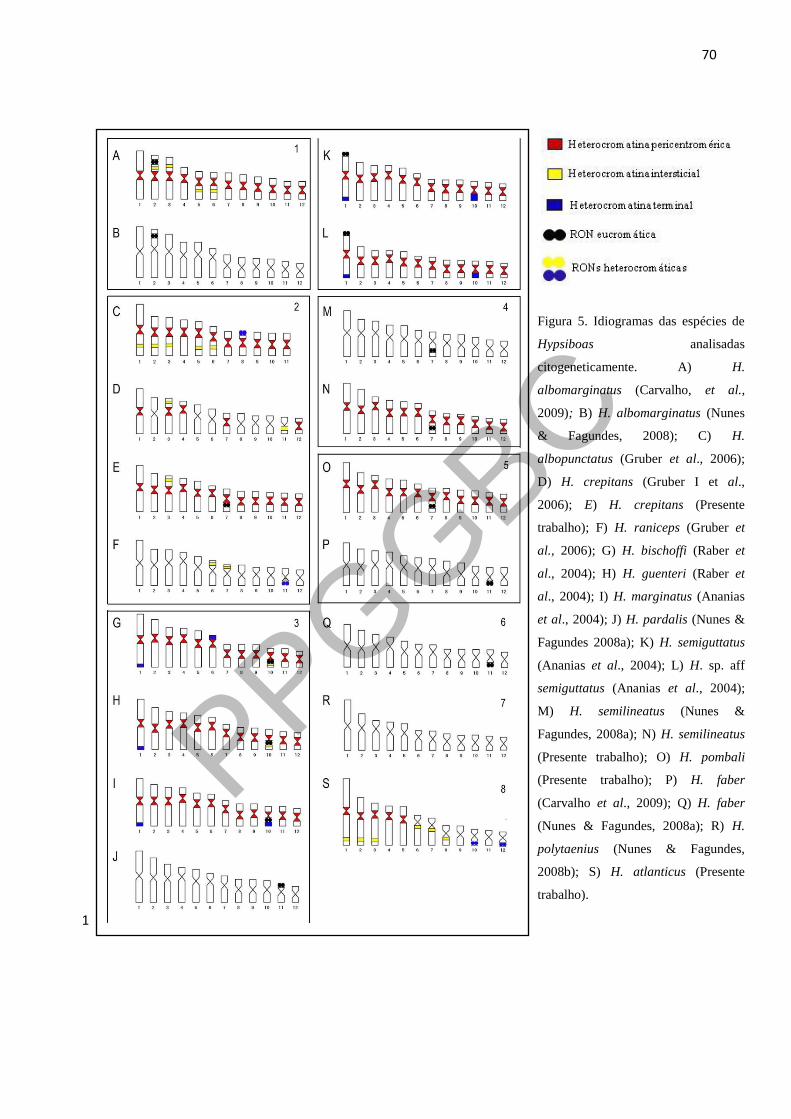
70
Figura 5. Idiogramas das espécies de
Hypsiboas analisadas
citogeneticamente. A) H.
albomarginatus (Carvalho, et al.,
2009); B) H. albomarginatus (Nunes
& Fagundes, 2008); C) H.
albopunctatus (Gruber et al., 2006);
D) H. crepitans (Gruber I et al.,
2006); E) H. crepitans (Presente
trabalho); F) H. raniceps (Gruber et
al., 2006); G) H. bischoffi (Raber et
al., 2004); H) H. guenteri (Raber et
al., 2004); I) H. marginatus (Ananias
et al., 2004); J) H. pardalis (Nunes &
Fagundes 2008a); K) H. semiguttatus
(Ananias et al., 2004); L) H. sp. aff
semiguttatus (Ananias et al., 2004);
M) H. semilineatus (Nunes &
Fagundes, 2008a); N) H. semilineatus
(Presente trabalho); O) H. pombali
(Presente trabalho); P) H. faber
(Carvalho et al., 2009); Q) H. faber
(Nunes & Fagundes, 2008a); R) H.
polytaenius (Nunes & Fagundes,
2008b); S) H. atlanticus (Presente
trabalho).
1
PPGGBC

71
6 CONCLUSÕES GERAIS 1
A partir das análises citogenéticas em Phyllodytes, pode-se inferir que: 2
As semelhanças cariotípicas entre as três espécies são provavelmente resultado de uma 3
ancestralidade comum, e que as diferenças são resultantes do posicionamento do 4
centrômero, provavelmente devido à inversões pericêntricas; 5
A diminuição do número e do tamanho de blocos heterocromáticos em regiões 6
pericentroméricas, está possivelmente associada à presença de blocos banda C positiva 7
em regiões intersticiais e/ou terminais, sendo essas bandas resultantes de eventos de 8
inversão pericêntrica envolvendo os blocos proximais à região centromérica; 9
As RONs em P. melanomystax e P. luteolos apareceram no pares 7 e 8 respectivamente, 10
indicando uma provável homeologia desses pares, que apresentam a mesma morfologia 11
nessas duas espécies; 12
Mesmo ainda não tendo obtido resultados de Ag-RONs para P. tuberculosus, pode-se 13
supor que, devido a sua natureza heterocromática e das outras duas espécies estudadas, a 14
impregnação pelo nitrato de prata revele a RON coincidente com os blocos 15
heterocromáticos existentes na região terminal dos cromossomos do par 8, o qual também 16
mostra-se semelhante em morfologia ao par nucleolar das demais espécies em estudo. 17
18
Em se tratando do gênero Hypsiboas, observou-se que: 19
A manutenção do tamanho dos pares cromossômicos entre os cariótipos são um forte 20
indício de que as inversões pericêntricas seriam o principal mecanismo envolvido na 21
diferenciação da macroestrutura cariotípica do grupo; 22
Para espécies estudadas é possível distinguir oito grupos cariotípicos que não são 23
compostos necessariamente por populações de uma mesma espécie ou por espécies que 24
pertençam aos mesmos grupos taxonômicos de espécie; 25
Há a presença de marcadores citotaxonômicos e uma possível relação filogeográfica da 26
microestrutura dos cromossomos – contudo, ainda não se sabe se essa relação é uma 27
constante no gênero. Para confirmar tal fato faz-se necessário o estudo de outras espécies 28
e populações, além do emprego de técnicas moleculares e citogenéticas mais apuradas, 29
como hibridação in situ. 30
PPGGBC

72
O estudo mostra indícios de que as RONs são marcadores cromossômicos fracos para 1
identificar as espécies do gênero neste estudo, já que as duas populações de H. crepitans 2
e H. faber estudadas diferiram com relação ao par cromossômico nucleolar. 3
Levando-se em conta as filogenias moleculares disponíveis para o grupo, e que, de modo 4
geral os anfíbios anuros apresentam RONs simples terminais, as RONs simples 5
intersticiais em Hypsiboas podem ser consideradas como uma característica 6
plesiomórfica, e condições variantes como apomorfias. 7
A variação de localização das RONs poderia ser explicada por possíveis eventos de 8
translocação ou transposição cromossômica, uma vez que há manutenção do número e do 9
tamanho cromossômico entre os diferentes cariótipos analisados. Esses mesmos 10
mecanismos também poderiam explicar o surgimento das RONs múltiplas em H. 11
atlanticus. 12
Pode-se observar que as espécies de Hypsiboas que possuem heterocromatina em posição 13
intersticial/terminal apresentam uma diminuição na quantidade de heterocromatina 14
pericentromérica ou no tamanho desses blocos, o que poderia ser resultado de eventos de 15
inversões cromossômicas que resultariam na transferência dessas regiões ao longo dos 16
braços cromossômicos. 17
As inversões envolvendo blocos heterocromáticos também poderiam explicar o fato de 18
algumas espécies apresentarem RONs heterocromáticas, enquanto a maioria apresenta 19
RONs eucromáticas, possivelmente estas regiões tornaram-se intercaladas ou associadas 20
à heterocromatina após esses eventos de reorganização estrutural dos cromossomos. 21
Esses dados mostram-se capazes de contribuir para os estudos de evolução cromossômica 22
e citotaxonomia, podendo, inclusive, ser uma ferramenta útil em programas de 23
conservação. 24
PPGGBC