universidade estadual do cearÁ faculdade de …§ão.pdfuniversidade estadual do cearÁ faculdade...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DO CEARÁ
FACULDADE DE VETERINÁRIA
PROGRAMA DE PÓS GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
SUZAN VITÓRIA GIRÃO LIMA
PESQUISA MICROBIOLÓGICA EM CALOPSITAS (Nymphicus hollandicus)
ORIUNDAS DE PET SHOPS E RESIDÊNCIAS DE FORTALEZA, CEARÁ
FORTALEZA - CEARÁ
2016

SUZAN VITÓRIA GIRÃO LIMA
PESQUISA MICROBIOLÓGICA EM CALOPSITAS (NYMPHICUS HOLLANDICUS)
ORIUNDAS DE PET SHOPS E RESIDÊNCIAS DE FORTALEZA, CEARÁ
Dissertação apresentada ao Programa de Pós-
Graduação em Ciências Veterinárias da
Faculdade de Veterinária da Universidade
Estadual do Ceará, como requisito parcial para
obtenção do grau de Mestre em Ciências
Veterinárias.
Orientador: Prof. Dr. William Cardoso Maciel
Co-orientador: Dr. Régis Siqueira de Castro
Teixeira
FORTALEZA - CEARÁ
2016



Ao meu pai, pelo cuidado especial nesses anos
de mestrado e de vida
À minha mãe, pela esperança que sempre
ilumina meus dias e horas
Aos meus melhores amigos, Débora Nishi e
Anderson Vieira, por me lembrarem que ainda
existem pessoas sãs em meio à loucura
Aos animais, pela jornada que enfrentamos
juntos

AGRADECIMENTOS
A Deus, por tudo e cada parte.
À Universidade Estadual do Ceará e ao Programa de Pós-Graduação em Ciências Veterinárias
(PPGCV), pela oportunidade de cumprir essa jornada.
Ao Laboratório de Estudos Ornitológicos - LABEO, pela oportunidade de conhecer o mundo
das aves e da pesquisa científica.
Ao meu orientador Prof. Dr. William Cardoso Maciel, pelo incentivo ao trabalho científico no
início da minha vida acadêmica, e contínuo apoio durante minha carreira científica.
Ao Dr. Régis Siqueira de Castro Teixeira, Super Régis, meu co-orientador, pelo apoio
científico, pelas conversas filosóficas, pelos conselhos espirituais, e tantas outras pequenas
coisas que fizeram grande diferença ao longo de toda essa jornada.
À doutoranda Elisângela de Sousa Lopes, que encerra comigo essa caminhada árdua, por todo
o companheirismo e ajuda em todo o meu experimento.
À mestranda Fernanda Gaio, uma surpresa bem-vinda nesses últimos momentos tensos que a
vida traz, pela amizade que eu tanto aprendi a apreciar.
À IC Ana Paula, pelo apoio inestimável ao meu trabalho, e essa alma genuína que possui.
À toda equipe do LABEO: Jackson Beleza, Ruben Horn, Anne Bezerra, Felipe Rebouças,
Bruno Pessoa, Neilton Monteiro, Cecília Casimiro e Marcel Lucena, por todo e cada
momento vivido em equipe, marcados por alegrias e atribulações.
À mestranda Mariana Duarte, do Laboratório de Toxinologia Molecular da UFC, por todo o
cuidado durante a análise de PCR, e paciência com as nossas várias mensagens e e-mails.
E, finalmente, agradeço a todos aqueles que me ajudaram de alguma forma durante esses
anos, quer tenham sido inspiração, ou apenas exemplos a não ser seguidos.

RESUMO
As calopsitas (Nymphicus hollandicus) pertencem à ordem Psittaciformes, sendo uma das
aves de companhia mais populares no Brasil e em todo o mundo. Sendo nativas da Austrália,
essas aves são encontradas no Brasil obtidas em lojas pet shop ou feiras livres. O contato
direto entre psitaciformes e seres humanos é bastante abrangente e a análise da flora
bacteriana dessas aves é pouco conhecida, e infecções bacterianas representam uma das
principais causas de doenças entéricas em aves. A presença de bactérias Gram-negativas tais
como Escherichia coli e Pseudomonas sp. em psitacídeos já foi relatada na literatura, embora
a realidade dos plantéis de criadores de calopsitas seja desconhecida. Assim sendo, o objetivo
desse estudo foi identificar e avaliar os perfis de resistência de cepas de bactérias Gram-
negativas isoladas de calopsitas oriundas de residências e pet shops em Fortaleza – CE, a
analisar a presença de cepas de E. coli diarreiogênicas (DEC) nesses animais. Para a
realização desse estudo foram coletadas 182 amostras de swabs de arrasto e suabes cloacais de
calopsitas provenientes de 8 domicílios, 7 Pet shops e 3 aviários. Este estudo foi aprovado
pelo Comitê de Ética para o Uso de Animais da Universidade Estadual do Ceará (Protocolo
647396-2015). O procedimento microbiológico para isolamento e identificação das cepas foi
composto pelas seguintes etapas: pré-enriquecimento com Água Peptonada, seguido pelo
enriquecimento seletivo em caldos Rappaport-Vassialidis, Seletino-Cistina e Brain Heart
Infusion (BHI), plaqueamento em Ágar Verde Brilhante, MacConkey e Salmonella-Shigella,
e identificação bioquímica presuntiva. Adicionalmente, os perfis de resistência antimicrobiana
das cepas foram determinados pelo método de discodifusão, e a detecção de oito genes de
virulência nas cepas de E. coli isoladas foi realizada por PCR. Foi possível o isolamento de
bactérias pertencentes aos gêneros Pantoea, Citrobacter, Serratia, Escherichia coli,
Providencia spp., Enterobacter sp, Shigella, Proteus e Pseudomonas, sendo Pantoea
agglomerans a mais prevalente. O teste sensibilidade evidenciou maior resistência à ácido
nalidíxico, azitromicina, sulfonamida e cotrimoxazol. Apenas uma cepa de E. coli foi positiva
para os genes eaeA e bfpA, sendo classificada como EPEC típica. Os resultados indicam que
calopsitas podem albergar bactérias Gram-negativas, e que as cepas apresentam alto índice de
resistência a antimicrobianos.
Palavras-chave: Enterobactérias. Antibiograma. EPEC. Pseudomonas sp. Calopsitas.

ABSTRACT
Cockatiels (Nymphicus hollandicus) belong to the order Psittaciformes, being one of the most
popular company birds in Brazil and around the world. Being native to Australia, these birds
are found in Brazil obtained at a pet shop or free trade shows. Direct contact between
psittaciformes and humans is quite comprehensive and the analysis of bacterial flora of these
birds is little known, although bacterial infections represent a major cause of enteric diseases
in birds. The presence of Gram negative bacteria such as Escherichia coli and Pseudomonas
sp. in psittacines has been reported in the literature, but the reality of the breeding grounds of
calopsitas breeders is unknown. Thus, the objective of this study was to identify and evaluate
the resistance profiles of strains of Gram-negative bacteria isolated from calopsitas from
residences and pet shops in Fortaleza, CE, to analyze the presence of diarrheogenic E. coli
strains Animals. For this study, 182 samples were collected from drag swabs and cloacal
swabs of cockatiels from 8 households, 7 pet shops, and 3 aviaries. This work was approved
by the Ethics Committee for the use of animals of the State University of Ceará (UECE):
647396-2015. The microbiological procedure for the isolation and identification of
Salmonella was composed of the following steps: pre-enrichment with Peptone Water,
followed by selective enrichment in Rappaport-Vassiliadis, Selenito-Cystine and Brain Heart
Infusion (BHI), plating in Bright Green, MacConkey and Salmonella-Shigella Agar, and
presumptive biochemical identification. Additionally, the antimicrobial resistance of the
strains was determined by the disc-diffusion method, and the detection of eight virulence
genes in strains of E. coli isolated was performed by PCR. It was possible to isolate bacteria
belonging to the genus Pantoea, Citrobacter, Serratia, Escherichia coli, Providencia spp.,
Enterobacter sp, Shigella, Proteus, and Pseudomonas, being Pantoea agglomerans the most
prevalent. The sensitivity test showed greater resistance to nalidixic acid, azithromycin,
sulfonamide and cotrimoxazole. Only one strain of E. coli was positive for the eaeA and bfpA
genes and was classified as typical EPEC. The results indicate that cockatiels can harbor
Gram-negative bacteria and that the strains present a high rate of antimicrobial resistance.
Key words: Enterobacteria. Antibiogram. EPEC. Pseudomonas sp. Cockatiels.

LISTA DE FIGURAS
Figura 1 - Lutino Macho .............................................................................................. 19
Figura 2 - Canela Macho e Fêmea .............................................................................. 20
Figura 3 - Pérola Macho e Fêmea ............................................................................... 20
Figura 4 - Cara amarela Macho .................................................................................. 21
Figura 5 - Cara branca Arlequim cinza reverso ........................................................ 22
Figura 6 - Cara branca Macho e Fêmea ..................................................................... 22
Figura 7 - Macho fulvo arlequim (esq) e Fêmea fulvo pérola (dir) .......................... 23
Figura 8 - Pastel Macho e Fêmea ................................................................................ 23
Figura 9 - Oliva Macho e Fêmea ................................................................................. 23
Figura 10 - Macho Cara Branca pérola prata recessivo ........................................... 24
Figura 11 - Silvestre Macho e Fêmea .......................................................................... 24
Figura 12 - Macho cara branca arlequim prata dominante ..................................... 25
Figura 13 - Platinum Macho e Fêmea ......................................................................... 26
Figura 14. Albino Macho ............................................................................................ 26
CAPÍTULO 1
Figura 1 - Eletroforese em gel de agarose de produtos de PCR .............................. 52

LISTA DE TABELAS
Tabela 1 Nomenclatura oficial segundo a Ordem Brasileira de Juízes de Ornitologia:
CALOPSITA LINHA CARA AMARELA (FUNDO AMARELO) ........... 19
Tabela 2 Nomenclatura oficial segundo a Ordem Brasileira de Juízes de Ornitologia:
CALOPSITA LINHA CARA-BRANCA (FUNDO BRANCO) .................. 20
Tabela 3 Nomenclatura oficial segundo a Ordem Brasileira de Juízes de Ornitologia:
CALOPSITA LINHA PÉROLA (FUNDO AMARELO E BRANCO) ..... 25
CAPÍTULO 1
Tabela 1 Frequências absolutas e relativas de enterobactérias isoladas de suabes
cloacais e de arrasto de calopsitas (Nymphicus hollandicus) de Fortaleza,
Brasil ................................................................................................................ 50
Tabela 2 Porcentagem de cepas de Enterobacteriaceae isoladas de calopsitas
(Nymphicus hollandicus) resistentes a antimicrobianos .............................. 51
Tabela 3 Enterobactérias Multirresistentes (MDR) isoladas de calopsitas
(Nymphicus hollandicus) ................................................................................. 52
CAPÍTULO 2
Table 1 Percentage of Pseudomonas sp. strains isolated from cockatiels (Nymphicus
hollandicus) resistant to antimicrobials …………………………………… 62
Table 2 Multidrug-resistant (MDR) Pseudomonas sp. strains isolated from
cockatiels (Nymphicus hollandicus) ……………………………………… 63

LISTA DE ABREVIATURAS E SIGLAS
APEC: Escherichia coli patogênica para aves
DAEC: Escherichia coli difusamente aderente
DEC: Escherichia coli diarreiogênica
EAEC: Escherichia coli enteroagregativa
EHEC: Escherichia coli enterohemorrágica
EIEC: Escherichia coli enteroinvasiva
EPEC: Escherichia coli enteropatogênica
ETEC: Escherichia coli enterotoxigênica
ExPEC: Escherichia coli extraintestinal
NMEC: Escherichia coli causadora de meningite neonatal
PCR: Polimerase chain reaction
STEC: Escherichia coli produtora de toxina de shiga
UPEC: Escherichia coli uropatogênica

SUMÁRIO
1 INTRODUÇÃO ....................................................................................................... 13
2 REVISÃO DE LITERATURA ............................................................................... 14
2.1 ORDEM PSITTACIFORMES .................................................................................. 14
2.2 CALOPSITAS (Nymphicus hollandicus) .................................................................. 14
2.2.2 Tipos de mutações .................................................................................................... 18
2.2.3 Características gerais .............................................................................................. 26
2.3 FAMÍLIA ENTEROBACTERIACEA ...................................................................... 28
2.3.1 Escherichia coli ........................................................................................................ 28
2.3.2 Salmonella spp. ........................................................................................................ 34
2.3.3 Proteus spp. .............................................................................................................. 34
2.3.4 Citrobacter spp. ........................................................................................................ 35
2.3.5 Pantoea sp. ................................................................................................................ 35
2.3.6 Providencia spp. ....................................................................................................... 36
2.4 Pseudomonas aeruginosa .......................................................................................... 37
2.5 RESISTÊNCIA ANTIMICROBIANA ..................................................................... 38
3 JUSTIFICATIVA .................................................................................................... 40
4 HIPÓTESE CIENTÍFICA ...................................................................................... 41
5 OBJETIVOS ............................................................................................................ 42
5.1 OBJETIVO GERAL .................................................................................................. 42
5.2 OBJETIVOS ESPECÍFICOS .................................................................................... 42
6 CAPÍTULO 1 ........................................................................................................... 43
7 CAPÍTULO 2 ........................................................................................................... 58
8 CONCLUSÃO .......................................................................................................... 70
REFERÊNCIAS BIBLIOGRÁFICAS .................................................................. 71

13
1 INTRODUÇÃO
A calopsita (Nymphicus hollandicus) pertence à ordem Psittaciformes, sendo
considerada a menor cacatua existente. Essas aves se destacam no comércio pet em
decorrência do baixo custo de aquisição e manutenção, bem como por sua beleza, canto e
docilidade (CÔRREA, 2012). A criação dessas aves não exige nenhuma autorização especial,
visto que a legislação vigente as considera como animais domésticos, possibilitando sua
criação em cativeiro (IBAMA, 1998). Por conta disso, o número de criadores de calopsitas no
Brasil vem se intensificando, sendo seus exemplares encontrados, em sua grande maioria, em
criadouros comerciais e em residências.
As aves da ordem Psittaciformes são passíveis de albergarem inúmeros agentes
etiológicos, e várias doenças são transmitidas de aves em cativeiro para os seres humanos
através do contato direto ou indireto com aves doentes ou portadoras assintomáticas. Dentre
estas doenças, destacam-se as de origem bacteriana, considerados os agentes zoonóticos mais
comuns (AKHTER et al., 2010). Inúmeros são os micro-organismos que afetam as aves
ornamentais, sendo as bactérias Gram-negativas consideradas as mais patogênicas nesta
espécie, merecendo destaque Escherichia coli, Salmonella sp, Klebsiella sp, Pseudomonas sp,
Proteus sp, Bordetella sp, Citrobacter freundii, e Yersinia sp.
Desse modo, além da possibilidade de carrearem agentes de potencial
patogenicidade, aptos a serem transmitidos para outros animais, inclusive ao homem, os
psitacídeos de vida livre ou de cativeiro podem albergar bactérias com inúmeros mecanismos
de resistência a antimicrobianos (HIDASI et al., 2013). Com o crescente interesse pela
comercialização dessas aves, torna-se de fundamental importância um estudo da microbiota
desses animais com o intuito de auxiliar no diagnóstico das possíveis doenças que acometem
as calopsitas, possibilitando a instituição de uma terapêutica adequada para o tratamento dos
animais debilitados, assim como uma maior compreensão da colonização da microbiota
entérica dessas aves, fornecendo dados relevantes para os ajustes necessários de manejo e de
acondicionamento dessas aves nos criadouros (CÔRREA, 2012).
Dessa forma, o objetivo do presente trabalho foi identificar e avaliar os perfis de
resistência de cepas de enterobactérias isoladas de calopsitas de residências e pet shops em
Fortaleza – CE, e analisar a presença de cepas de E. coli diarreiogênicas (DEC) nesses
animais.

14
2 REVISÃO DE LITERATURA
2.1 ORDEM PSITTACIFORMES
A ordem Psittaciformes é bastante diversificada, constituída por cerca de 78
gêneros e 332 espécies, sendo classificada atualmente em três famílias: Psittacidae,
representada pelas araras (Ara spp., Anodorhynchus spp.), papagaios (Amazona spp.),
maracanãs (Propyrrhura spp) e periquitos (Brotogeris spp).; Loridae, formada pelos Lóris
(Trichoglossus spp., Eos spp.); e Cacatuidae, composta pelas cacatuas (Cacatua spp) e
calopsitas (Nymphicus spp.) (COLLAR, 1997; CHRISTIDIS & BOLES, 2008; GODOY &
CUBAS, 2009; HARCOURT-BROWN, 2010).
Os Psittaciformes caracterizam-se pelo bico forte, grosso e recurvo, língua
carnuda e grossa, e pela mandíbula superior recurvada sobre a inferior, adaptada à
alimentação a base de sementes e frutos. As patas são zigodáctilas, sendo o primeiro e o
quarto dedos em posição posterior enquanto o segundo e o terceiro dedos se dispõem na
dianteira, facilitando em uma maior agilidade para captura de alimentos e de posicionamento
(SICK, 1997; GODOY, 2007). Modificações antrópicas de habitat, a caça furtiva e o
comércio ilegal são ameaças significativas: 85 espécies estão listadas como crítico, em perigo
ou vulnerável e 19 espécies como extinto pela União Internacional para a Conservação da
Natureza (IUCN, 2015).
Quanto ao dimorfismo sexual, verifica-se que a maioria das espécies são
monomórficas, necessitando de testes moleculares ou endoscópicos para a determinação do
sexo das mesmas (GODOY, 2007). Mediante algumas espécies serem capazes de
mimetizarem a voz humana, como devido à variedade de coloração e exuberância em suas
plumagens, os psitacídeos são amplamente visados para atuar como animais de companhia ou
de exibição (SIGRIST, 2014).
2.2 CALOPSITAS (Nymphicus hollandicus)
A calopsita pertence à ordem Psittaciformes, família Psittacidae, subfamília
Cacatuinae (BIRDLIFE INTERNATIONAL, 2012). Inicialmente descrita pelo escritor e
naturalista escocês Robert Kerr em 1793, como Hollandicus psittacuse, foi transferida para
seu próprio gênero, Nymphicus, por Wagler em 1832 (SANTOS, 2013). Em 1864, se tornou

15
bastante conhecida dos ingleses e, por volta de 1884, já se encontrava bem estabelecida nos
aviários europeus. Entretanto, a disseminação maciça dessa ave somente ocorreu a partir do
surgimento da primeira mutação de cor, o arlequim, pouco antes de 1949, no estado da
Califórnia (Estados Unidos). A partir daí outros padrões de cores foram sendo fixados,
ganhando então grande popularidade, igualando-se, praticamente, àquela do periquito
australiano, e a introdução destas aves no Brasil se deu a partir da década de 1970 (BENEZ,
2001; AUSTRALIAN MUSEUM, 2006).
Segundo pesquisa realizada pelo painel online CONECTAí em todo o país, em
setembro e outubro de 2013 mostra que 80% dos internautas brasileiros possuem animal de
estimação, dos quais as aves ocupam o terceiro lugar (12%). Entre as aves, a mais comum é o
periquito, citado por 28%, seguido do canário (26%), calopsita (21%) e papagaio (14%). O
gasto médio mensal com animais é de R$ 99,25. 32% dos respondentes gastam entre R$ 30 e
75 com os animais e 30% gastam entre R$ 75 e R$ 150. Nos extremos, 15% gastam menos do
que R$ 30 por mês e 15% gastam mais do que R$ 300 mensais. A maioria compra os
produtos para os animais em petshops (61%), 42% compram em supermercados e 20% em
mercadinhos. Na hora da compra, 76% levam em consideração a qualidade do produto, 45%
consideram o preço e 26% a indicação do veterinário. Cerca de três em cada 10 internautas
(34%) levam seu animal ao veterinário somente quando ele apresenta algum problema de
saúde. 15% levam seus animais ao veterinário pelo menos uma vez por mês, 17% entre dois a
três meses e 18% levam uma vez entre três e seis meses. Um em cada 10 (9%) leva o animal
ao veterinário uma vez por ano (IBOPE, 2013).
2.2.1 Cores e Mutações
Os diversos padrões de cores existentes, originadas de padrões silvestres e
mutações, podem se mesclar originando uma grande gama de cores, tornando cada calopsita
única.
De forma geral, as aves possuem duas cores básicas:
1) Melânica (melanina): pigmentos melânicos e podem variar do negro ao cinza
azulado, do marrom (castanho) escuro ao bege claro ou mesmo estar ausente, se distribuem
principalmente no eixo das penas e na periferia;

16
2) Lipocrômica (lipocromo): pode se apresentar em várias gradações de
vermelho, amarelo ou simplesmente branca.
A cor que vemos nas aves é resultante da combinação dos efeitos dessas duas
cores básicas. São transmitidas em genes diferentes de forma independente. As diversas
possíveis mutações podem atuar tanto nas melaninas como nos lipocromos, reduzindo sua
expressão em graus variáveis, inibindo seu aparecimento ou aumentando seu efeito.
Mutação é toda alteração espontânea dos caracteres hereditários (genes)
transmissíveis à descendência. Chamamos de pássaros mutantes aqueles que diferem dos seus
ancestrais.
As mutações não afetam a posição do gene no cromossoma, mas o gene mutante
produz um efeito diferente do gene original. Muitas mutações passam despercebidas pois seus
efeitos são de pouca expressão, há outras onde o efeito é percebido sensivelmente. Toda
mutação é progressiva e só se manifesta completamente com o passar do tempo. Quando
ocorre uma mutação o gene passa a existir de duas formas, a original que produz um efeito
conhecido, e a mutante que produz um novo efeito. As formas diferentes de um gene são
denominadas de alelos. Existem genes que apresentam mais de uma mutação, neste caso,
teremos uma série alomórfica ou alelos múltiplos. Embora possam haver três ou mais versões
de cada gene, cada indivíduo possui apenas dois alelos.
As mutações podem se manifestar de diferentes maneiras e recebem as seguintes
denominações:
1) Dominante: Para que se manifeste, basta que o gene esteja presente uma vez
no par de cromossomos correspondentes (heterozigótico) e pode ser herdado do pai ou da
mãe;
2) Recessiva autossômica: Só se manifesta em quem herda um gene recessivo de
cada progenitor (homozigoto), ou seja, sempre se manifesta aos pares. Caso esteja presente
em um único cromossomo, dizemos que o pássaro é portador;
3) Ligada ao sexo: Transmitida por genes contidos no cromossomo que transmite
as características do sexo. Como todas as mutações sexo-ligadas são recessivas em relação ao
gene original, nos machos o efeito delas só se fará sentir no fenótipo quando em homozigose.

17
Assim somente os machos poderão portar genes recessivos sexo-ligados. As fêmeas jamais
serão portadoras, serão puras ou normais.
São infinitas as possibilidades de tonalidades de cores nos pássaros. Quando
combinadas com fatores mutantes a variedade cresce mais ainda. Estas variações podem tanto
atuar nos lipocromos como nas melaninas.
Variedades das cores lipocrômicas
As cores lipocrômicas podem se apresentar em várias tonalidades de amarelo,
passando pelo laranja, podendo chegar ao vermelho vivo, ou mesmo ser inibida por alguma
mutação, chegando ao branco total ou parcial. O pássaro que tem a melanina inibida é
chamado de lipocrômico puro. São as seguintes possibilidades de cores lipocrômicas:
1 - Branco Recessivo - A inibição do lipocromo é total, o pássaro apresenta uma
brancura imaculada e a cor da pele fica com tom violeta. A transmissão hereditária é recessiva
não ligada ao cromossomo do sexo (autossômica). Só se exterioriza no fenótipo em estado de
homozigoto.
2 - Branco dominante - O lipocromo se apresenta de maneira latente e sua inibição
é parcial, vestígios de lipocromos serão perceptíveis, principalmente nos encontros. Se
manifesta estando apenas em um dos cromossomos do par hereditário (heterozigótico),
quando em estado de homozigoto pode ser letal.
3 – Amarelo - Nessas cores o lipocromo pode se apresentar de maneira intensa ou
com nevoas brancas espalhadas uniformemente pela plumagem na forma de escamas. O fator
intenso é dominante em relação ao nevado, ou seja, os intensos podem portar nevado, sendo
heterozigoto para o esse fator, ou ainda serem homozigotos com duplo fator intenso. Os
nevados só se exteriorizam quando os pássaros são homozigotos para o nevadismo. Tanto o
lipocromo intenso, quando o nevado, podem ser inibidos pelo branco recessivo ou dominante.
4 – Inos - Albinos (brancos com olhos vermelhos) e Lutinos (amarelos com olhos
vermelhos). São os chamados Inos lipocrômicos (sem melanina). Na verdade, existem duas
diferentes mutações que causam o mesmo efeito no fenótipo. Uma de transmissão recessiva
autossômica e outra sexo-ligada, podem atuar em todas as cores lipocrômicas.

18
Variedades das cores melânicas
As melaninas se apresentam em várias tonalidades de cor, partindo do negro
imaculado podendo chegar ao bege claro, em algumas vezes é quase imperceptível. Pode até
em algumas circunstâncias ser totalmente inibidas. Quando o pássaro tem os lipocromos
inibidos e apresentam alguma melanina, são chamados de melânicos puros.
1 – Negro - O negro é a oxidação máxima das possíveis melaninas e é dominante
em relação a todas outras manifestações melânicas.
2 – Canela - A melanina canela tem a mesma característica da negra oxidada, a
única diferença é a tonalidade da cor que passa a ser marrom escuro oxidado. Sua transmissão
é de natureza sexo-ligada, é recessiva em relação a melanina negra e dominante a todos
fatores de diluição melânica.
3 – Pastel - A mutação pastel causa mudança na tonalidade das melaninas negras
que passam a se apresentar com uma coloração cinza escuro (quase negra). Já nas melaninas
marrons o efeito é de diluição e dispersão. Sua transmissão é sexo-ligada recessiva e pode
atuar em todas as cores melânicas clássicas.
2.2.2 Tipos de mutações
As mutações básicas ou primárias são Lutino, Arlequim, Canela, Pérola, Cara
Branca, e Prata. As mutações combinadas ou cruzadas são Pérola-Arlequim, Canela-
Arlequim, Canela-Pérola-Arlequim, Lutino-Canela, Lutino-Pérola, Lutino-Pérola Arlequim,
Cara Branca-Lutino (albino), Cara Branca-Arlequim, Cara Branca-Pérola, Cara Branca-
Pérola-Arlequim, Cara Branca-Canela, Cara Branca-Canela-Arlequim, e Cara Branca-Canela-
Pérola-Arlequim.
Diversas das mutações existentes são raras, das quais muitas ainda não são
encontradas no Brasil. Essas mutações são classificadas da seguinte forma:

19
1) Sexo-ligado: Lutino, Canela, Pérola, Cara Amarela
Lutino (Lutino): Sem dúvida alguma, é o padrão mais popular e apreciado, tendo surgido
nos EUA em 1958. Essa mutação causa perda do
pigmento melanina (que confere o tom cinza à ave).
Desse modo, são aves de cor dominante branca, com
olhos vermelhos, pés rosados, crista amarela, bico
marfim, cabeça amarelada com bochechas vermelhas.
Nas asas e cauda, também está presente o amarelo. Na
realidade, os lutinos não podem ser considerados como
brancos ou albinos, pois não são inteiramente brancos,
em razão da presença das cores amarela e vermelha
(dadas pelo pigmento psitacina). Os indivíduos podem
apresentar desde um amarelo forte até um branco quase
total no corpo. Neste padrão ocorre um defeito de
origem genética, caracterizado pela existência de uma
área sem penas localizada atrás da cabeça. As fêmeas têm estrias amarelas na face inferior
da cauda e pontos amarelos embaixo da asa (RUPLEY, 1999).
Tabela 1. Nomenclatura oficial segundo a Ordem Brasileira de Juízes de Ornitologia
CALOPSITA LINHA CARA AMARELA (FUNDO AMARELO)
CÓD. COR
PS070101 Calopsita cinza fêmea
PS070102 Calopsita cinza macho
PS070103 Calopsita canela fêmea
PS070104 Calopsita canela macho
PS070105 Calopsita arlequim cinza (padrão 50% e reversos)
PS070106 Calopsita arlequim canela (padrão 50% e reversos)
PS070107 Calopsita lutino
PS070108 Calopsita fulvo
PS070120 Calopsita outras mutações fundo amarelo
PS070150 Calopsita combinações fundo amarelo
Figura 1. Lutino Macho
Fonte: calopsitasmansas.net

20
Tabela 2. Nomenclatura oficial segundo a Ordem Brasileira de Juízes de Ornitologia
CALOPSITA LINHA CARA-BRANCA (FUNDO BRANCO)
CÓD. COR
PS070201 Calopsita cara-branca cinza fêmea
PS070202 Calopsita cara-branca cinza macho
PS070203 Calopsita cara-branca canela fêmea
PS070204 Calopsita cara-branca canela macho
PS070205 Calopsita cara-branca arlequim cinza (padrão 50% e reversos)
PS070206 Calopsita cara-branca arlequim canela (padrão 50% e reversos)
PS070207 Calopsita cara-branca ino (albino)
PS070208 Calopsita cara-branca fulvo
PS070209 Calopsita cara-branca outras mutações fundo branco
PS070250 Calopsita cara-branca combinações fundo branco
Canela (Cinnamon): Também conhecida como Isabelino, surgiu pela primeira vez na
Bélgica, em 1960. As aves são semelhantes ao padrão normal, com exceção da alteração
na coloração da melanina, produzindo uma coloração marrom-claro (ou canela). Também
as patas e os olhos são de coloração mais clara. Os machos adultos são um pouco mais
escuros que as fêmeas (em razão da maior presença de melanina). Algumas fêmeas podem
ter mais amarelo na face do que os machos, além de apresentarem o barramento típico sob
as penas da cauda (RUPLEY, 1999).
Pérola (Pearl): Surgiu pela primeira vez na Alemanha
Ocidental em 1967. É uma
mutação que afeta as penas
individualmente (há uma
falta de melanina no centro
de cada pena,
individualmente), fazendo
com que haja uma falta de
coloração uniforme,
resultando em penas com
Figura 2 Canela Macho e Fêmea Fonte: calopsitasmansas.net
Figura 3. Pérola Macho e Fêmea
Fonte: calopsitasmansas.net

21
coloração em forma de "concha". São aves extremamente vistosas, sendo que o padrão
básico pode variar bastante. De modo geral, mostram as duas manchas laterais à cabeça,
as faces são amarelas salpicadas de cinza, e a crista amarela riscada de cinza. As penas das
costas exibem um padrão “escamado”, resultante da ausência de melanina no seu centro,
podendo a cor desta parte das penas variar do branco ao amarelo. As penas das asas são
cinza, com faixas amarelas. A cauda é amarela, e o peito e a barriga, listrados de amarelo
e cinza. As fêmeas carregam o perolado nas costas, asas, nuca e cabeça, com uma
concentração maior nas costas. Os machos adultos podem perder totalmente o perolado,
principalmente na cabeça e na nuca (RUPLEY, 1999).
Cara amarela (Yellow Face ou Yellow Cheek): É uma das mais recentes mutações,
surgida na Europa na década de 80, e ainda não há
notícia de sua existência no Brasil. Os primeiros
exemplares deste padrão chegaram aos EUA por
volta de 1992. São em tudo, semelhantes aos demais
padrões, diferindo apenas na cor das bochechas, que,
ao invés de serem vermelhas, mostram-se amarelas.
A principal diferença entre os sexos é o amarelo da
bochecha, que é mais forte no macho. Há três
formas dessa mutação (como ocorre com o padrão
prata): a dominante simples-fator, a dominante
duplo-fator e a recessiva (RUPLEY, 1999).
2) Recessivo: Arlequim, Cara Branca, Fulvo, Pastel, Esmeralda, Prata Recessivo
Arlequim (Pied): É a mutação mais antiga, surgida nos EUA em 1949; causa alteração da
coloração normal em extremidades. Esse padrão é extremamente variável, podendo
apresentar aves bastante semelhantes ao normal até aquelas com poucas áreas de cor
cinza, predominando o amarelo claro e apenas algumas penas de coloração cinza. A
Figura 4. Cara amarela Macho
Fonte: calopsitasmansas.net

22
cabeça exibe um amarelo forte, faces vermelhas e crista amarela. Idealmente, um arlequim
deve mostrar 75% de penas com ausência de melanina e 25% com presença. Um arlequim
puro possui, idealmente, uma máscara “limpa”, livre de manchas cinzas, uma cauda limpa
e asas de voos com um balanço igual de marcas, com simetria perfeita. Nesse padrão, é
virtualmente impossível a distinção de sexo (uma vez que a marcação arlequim obscurece
as diferenças de plumagem), só sendo possível no caso em que a fêmea apresente barras
na parte inferior do rabo. São reconhecidas quatro classificações de arlequim: escuro (ou
heavy, com 75% ou mais de melanina), claro (ou light, com apenas 25% de melanina),
reverso (ou reverse, com marcações apenas nas asas de voos, tendo o restante do corpo
sem melanina) e limpo (ou clear, um pássaro totalmente amarelo ou branco; é também
chamado de lutino com olhos pretos) (RUPLEY, 1999).
Cara branca (White Face): O padrão Cara
Branca surgiu na
Holanda por volta de
1969. No final da
década de 1970,
passou a ser produzido
na Alemanha e
Inglaterra. Essa mutação
causa perda do pigmento psitacina (que confere tons
amarelo e laranja), causando a falta da pigmentação laranja
e amarela nas bochechas e no corpo. A fêmea tem o corpo
cinza, bordas das asas brancas e face inferior da cauda com
estrias pretas e brancas, tornando a face inteiramente cinza. O macho segue um padrão
parecido com o normal, porém com a face totalmente branca e as cores cinzas com um
tom mais escuro, crista cinza e bordas das asas brancas (RUPLEY, 1999).
Figura 5. Cara branca Arlequim cinza reverso Fonte: Arquivo pessoal
Figura 6. Cara branca Macho e Fêmea
Fonte: calopsitasmansas.net

23
Figura 7. Macho fulvo arlequim (esq) e Fêmea fulvo pérola (dir)
Fonte: calopsitasmansas.net
Fulvo (Fallow): Surgidos em 1971, nos EUA. Semelhante ao Canela (também há
mudança da coloração da melanina de preto para marrom), mas aqui também ocorre uma
diminuição da densidade da melanina, fazendo com que pareçam um Canela pálido. O
amarelo é mais pronunciado (principalmente embaixo do corpo e crista), olhos são
vermelhos e peito é de coloração mostarda ou creme. As fêmeas costumam ser mais
bonitas que os machos, por apresentarem cores mais brilhantes. Os sexos são praticamente
iguais, tornando-se mais difícil a identificação (RUPLEY, 1999).
Pastel (Pastelface): A ave
possui a coloração
das bochechas no tom
pastel. Apesar de
conferir a mesma
coloração, o padrão
Pastel não deve ser
confundido com o cara amarela. Essa é uma mutação
sutil, que promove um tom mais brando de todas as
cores. Externamente é em tudo semelhante ao cara
amarela, mas tem herança genética autossômica
recessiva, o que facilita e acelera as combinações entre os padrões, principalmente com
aqueles de herança ligada ao sexo. É dominante apenas para o padrão cara branca.
Também aqui ocorre duas formas: fator-simples e fator-duplo.
Oliva ou Esmeralda (Olive ou Spangle ou Esmerald
Green): Mutação extremamente nova, surgida nos EUA,
de padrão ainda não muito definido. Caracteriza-se,
basicamente, por uma coloração canela-esverdeada,
podendo variar de claro a escuro, e um padrão de
marcação das penas muito característico (que as pessoas
denominam padrão de "lantejoulas", ou spangled no
inglês) (RUPLEY, 1999).
Figura 8. Pastel Macho e Fêmea Fonte: calopsitasmansas.net
Figura 9. Oliva Macho e Fêmea
Fonte: calopsitasmansas.net

24
Prata Recessivo:
Mutação estabilizada na década de 60, na Europa (apesar
de ter surgido na Nova Zelândia, em 1950). Difere do
padrão normal pelo fato de os olhos serem de cor
vermelha e o cinza global do corpo ter passado à cor
prateada, ocorrendo uma grande flutuação de tonalidades
entre os indivíduos. As demais características de cor são as
mesmas do padrão normal, inclusive quanto à
identificação do macho e da fêmea (RUPLEY, 1999).
3) Dominante: Cinza Silvestre, Prata Dominante, Bochecha Amarela Dominante
Cinza ou Normal (Normal Grey): A variedade original, encontrada na natureza, tem o
corpo cinza com as bordas das asas brancas. A crista do macho é amarela sobre uma
cabeça amarela e, na fêmea, cinza-amarelada com a
cabeça cinza. Ambos têm as faces formadas por uma
mancha vermelha, circular, em cada lateral da face, de
tom mais suave na fêmea. A cauda é totalmente negra no
macho e na fêmea intercalada de negro com amarelo na
parte de baixo. No macho adulto, a cabeça é amarela,
com duas manchas circulares laterais (bochechas) de cor
vermelha, crista amarela, corpo revestido com penas de
cor cinza, com o dorso mais escuro, bordas das asas
brancas e cauda negra. A fêmea adulta apresenta a
mesma coloração dos filhotes. O corpo é de cor cinza,
cabeça também cinza com as bochechas de cor vermelha mais suave, crista cinza-
amarelada, bordas das asas brancas e face interior da cauda estriada de amarelo e preto,
Figura 10. Macho Cara Branca pérola prata recessivo
Fonte: calopsitasmansas.net
Figura 11. Silvestre Macho e Fêmea
Fonte: calopsitasmansas.net

25
com penas laterais externas amarelas. Em ambos os sexos, os olhos são marrons, o bico
cinza escuro e as patas, cinza escurecido (RUPLEY, 1999).
Tabela 3. Nomenclatura oficial segundo a Ordem Brasileira de Juízes de Ornitologia
CALOPSITA LINHA PÉROLA (FUNDO AMARELO E BRANCO)
CÓD. COR
PS070301 Calopsita pérola cinza
PS070302 Calopsita pérola canela
PS070303 Calopsita pérola arlequim cinza (padrão 50% e reversos)
PS070304 Calopsita pérola arlequim canela (padrão 50% e reversos)
PS070305 Calopsita pérola cara-branca cinza
PS070306 Calopsita pérola cara-branca canela
PS070307 Calopsita pérola cara-branca arlequim cinza (padrão 50% e reversos)
PS070308 Calopsita pérola cara-branca arlequim canela (padrão 50% e reversos)
PS070309 Calopsita pérola lutina
PS070310 Calopsita pérola fulvo (fundo amarelo e branco)
PS070311 Calopsita pérola outras mutações
Prata Dominante: É um padrão de cor obtido
recentemente, tendo sido fixado por volta de 1979.
São aves que apresentam a cor cinza do padrão
normal diluída, mostrando um tom pastel prateado. Os
olhos e patas são pretos, as penas cinzas, mantendo o
amarelo forte das faces e da crista e o vermelho das
bochechas, com um prateado mais escuro na região do
pescoço. A graduação do prateado varia de ave para
ave, sendo a cor dos machos mais brilhante e intensa.
A diferenciação entre os sexos pode ser feita do
mesmo modo que o padrão normal. Nesta mutação,
os genes produzem dois efeitos visuais diferentes,
caso ocorram como fator simples ou duplo. Aves fator-duplo são mais claras que as aves
fator-simples, parecendo lutinos, mas com um tom acinzentado; eles retêm a marcação
mais escura na cabeça, olhos e pés escuros (RUPLEY, 1999).
Bochecha Amarela Dominante: É muito similar ao Bochecha Amarela Sexo Ligado, com
diferenças sutis quanto à intensidade na coloração da cor amarela e laranja.
Figura 12. Macho cara branca arlequim prata dominante
Fonte: calopsitasmansas.net

26
Outras mutações existentes são:
Albino (White Face Lutino): Ave inteiramente branca,
com os olhos vermelhos e pés rosados, com ausência
total de qualquer pigmentação (na realidade, resultam
da combinação de duas mutações: lutino e cara branca).
As fêmeas são mais fáceis de serem encontradas, por
ser um padrão com herança ligada ao sexo (RUPLEY,
1999).
Platinum: Mutação
só existente na Austrália.
Há uma confusão com relação a esse nome, uma vez que na
América do Norte chamam de Platinum aves prata
dominante. Essa mutação se caracteriza por uma coloração
"cinza-fumaça" clara (smokey-grey), com asas e cauda cinza
mais escuro. Bico, pés e patas são bege claro. Os olhos são
vermelhos ao nascer, mas escurecem logo em seguida
(RUPLEY, 1999).
2.2.3 Características gerais
Consideradas a menor espécie de cacatua existente, apresentam, em média, 30 a
33 cm de comprimento, 45 cm de envergadura e o peso varia de 80 a 150 gramas. São aves
dóceis, que podem ser utilizadas como animal de estimação. Resistentes quando adultas, com
uma alimentação balanceada e cuidado adequado podem viver até 25 anos (BENEZ, 2001;
FORSHAW, 2010; GORMAN, 2010).
Figura 13. Albino Macho
Fonte: calopsitasmansas.net
Figura 14. Platinum Macho e Fêmea Fonte: calopsitasmansas.net

27
Na natureza, essas aves habitam quase todas as partes do continente australiano,
vivendo tanto em áreas úmidas quanto em regiões secas. São comumente encontrados em
áreas agrícolas, savanas, jardins, área de vegetação arbustiva, e até em parque de cidades,
embora dificilmente sejam vistos nas zonas costeiras. Em áreas secas onde água torna-se um
fator limitante, calopsitas tendem a disputar por comida, o que gera a necessidade de
migração, quando essas aves viajam em grupos se movendo para áreas com fontes de água e
alimentos (TORLONI, 1991).
Em cativeiro, vivem com uma dieta básica de ração e mistura de sementes
apropriadas, com um complemento de milho, pequenas quantidades de frutas e legumes
(VERHOEF-VERHALLEN, 2004). O tamanho e comportamento dos grupos formados em
cativeiro variam de acordo com o habitat e sazonalidade. Animais jovens e adultos de ambos
os sexos podem formar grandes grupos ou ainda subgrupos tanto em vida livre quanto em
cativeiro (RUPLEY, 1999).
Existem diferentes e variadas colorações de plumagem devido a mutações, sendo
a plumagem de adulto desenvolvida entre três e seis meses de idade. Segundo alguns autores,
a calopsita atinge a maturidade sexual em torno dos seis meses de idade (HARPER & LOWE,
1998; WATSON & HURLEY, 2003; FORSHAW, 2010), enquanto que para HICKMAN
(2001) a maturidade ocorre a partir de um ano de idade. Os indivíduos adultos, durante o
período reprodutivo, são divididos em pares, em gaiolas destinadas à reprodução. O macho
adulto tem ainda um comportamento de defesa de seu território, enquanto a fêmea se limita
exclusivamente à postura e sobrevivência da ninhada (BENEZ, 2001). O período médio de
incubação é de 18 dias, onde os pais se revezam chocando os ovos de uma ninhada,
normalmente, formada por seis ovos (WATSON & HURLEY, 2003; VERHOEF-
VERHALLEN, 2004).
A criação dessas aves não exige nenhuma autorização especial, visto que a
portaria Nº 93, de 07 de junho de 1998 do Instituto Brasileiro do Meio Ambiente e dos
Recursos Naturais Renováveis - IBAMA (1998) as considera como animais domésticos,
possibilitando sua criação em cativeiro. Recentemente, a criação de calopsitas no Brasil vem
se intensificando, sendo seus exemplares encontrados, em sua grande maioria, em criadouros
comerciais particulares e em estabelecimentos comerciais (BENEZ, 2001).

28
2.3 FAMÍLIA ENTEROBACTERIACEA
Entre as bactérias apontadas como importantes patógenos aviários relacionados às
falhas de manejo em aves de cativeiro, destacam-se as pertencentes à família
Enterobacteriaceae, que apresenta cerca de 42 gêneros e mais de 142 espécies bacterianas
(JANDA e ABBOT, 2008). Os membros da família Enterobacteriaceae são bacilos Gram
negativos de até 3 µm de comprimento, classificados em aproximadamente 28 gêneros e 80
espécies, que se distinguem bioquimicamente (QUINN et al., 2005); são capazes de fazerem a
redução de nitrato a nitrito e a não formação de esporos, apresentando um ótimo crescimento
a uma temperatura de 37 °C (BERGEY & HOLT, 1994; QUINN et al., 2005), e de fermentar
a glicose e uma ampla variedade de carboidratos, sendo oxidase-negativos e catalase-
positivos, com exceção da Shigella dysenteriae.
Por afetarem o trato intestinal de seus hospedeiros, sejam como habitantes
naturais ou parasitários, as enterobactérias de um modo geral são responsáveis por quadros de
diarreia, sonolência e perda de apetite (SEGABINAZI, 2004). As enterobactérias mais
problemáticas aos psitaciformes são: Escherichia coli, Yersinia sp, e Salmonella sp, advindo
daí a importância dos psitacídeos em relação à saúde pública por conta de seu potencial de
albergar patógenos zoonóticos (SIMPSON, 1996; IKUNO et al, 2008). O grau de infecção e
de invasão bacteriana varia quanto ao tipo de enterobactéria, porém, de um modo geral, a sua
proliferação e disseminação no organismo do homem ou do animal que atinge, depende da
idade do hospedeiro, das espécies e de suas condições fisiológicas.
2.3.1 Escherichia coli
Escherichia coli é um bacilo Gram negativo, fermentativo, geralmente móvel,
com flagelos peritríquios e fímbrias (QUINN et al., 2005). Apresentam hastes restas, cujo
comprimento se estendem de 1,1 – 1,5 μm a 2,0-6,0 μm. Suas cepas podem ser circundadas
por cápsulas ou microcápsulas, sendo anaeróbias facultativas (BERGEY & HOLT, 1994). São
capazes de fermentarem, bioquimicamente, os açúcares glicose, ramnose, maltose, manose,
manitol, xilose, sorbitol e arabinose. Em contrapartida, a fermentação de adonitol, sacarose,
salicina, rafinose, ornitina, dulcitol e arginina é variável (KONEMAN et al., 2010). Cresce
rapidamente em meios bacterianos como ágar MacConkey, onde forma colônias lactose
positiva grandes. E. coli é a principal bactéria anaeróbica facultativa, presente no trato
intestinal da maioria das espécies animais e é geralmente recuperada da cultura de fezes, mas

29
em aves de estimação saudáveis é recuperada em apenas uma pequena porcentagem de
indivíduos (GYLES; FAIRBROTHER, 2010).
Logo após o nascimento, sem necessariamente causar infecção, a Escherichia coli
coloniza o intestino dos indivíduos, porém o papel dela como membro da microbiota de aves
não é completamente elucidado, podendo atuar como fonte de vitaminas e ocupando os sítios
na mucosa intestinal, impedindo assim a colonização do epitélio por microrganismos
patogênicos (FERREIRA & KNÖBL, 2000). Diversos estudos já realizados em aves
demonstraram que E. coli é um dos patógenos mais prevalentes entre as outras enterobactérias
isoladas (AKHTER et al., 2010; HIDASI et al., 2013; HORN et al., 2015; LOPES et al.,
2015). GORDON & COWLING (2003), argumentaram que a associação com os humanos, as
alterações de dieta e a massa corporal são fatores predisponentes significativos na prevalência
dessa bactéria nos animais, ao realizarem um estudo de isolamento de E. coli em mamíferos e
aves silvestres na Austrália.
A patogenia das infecções por E. coli em aves ainda é pouco conhecida. As cepas
de E. coli patogênicas para aves são menos toxigênicas do que as cepas que infectam
mamíferos (BARNES; VAILLANCOURT; GROSS, 2003). As E. coli de aves podem
produzir enterotoxinas que induzem a hipersecreção de fluidos no lume intestinal causando
diarreia (GERLACH, 1994).
Nos psitacídeos, tanto em infecções primárias ou secundárias de colibacilose, os
sinais clínicos geralmente se apresentam de forma inespecífica, sendo as aves passíveis de
desenvolverem reações de letargia, incoordenação motora, penas arrepiadas, rinite, dispneia,
presença de sons estertores, anorexia, diarreia, políúria, dor abdominal, inchaço articular,
edema subcutâneo, conjuntivite, perda de peso, vômito e morte súbita. Foi observada também
a presença de sinais como morte embrionária, onfalite, coligranuloma, sinovite, doença
respiratória crônica complicada, colisepticemia, salpingite, síndrome da cabeça inchada e
celulite aviária (FERREIRA, KNÖBL, 2000; GODOY, 2007; MARIETTO-GONÇALVES et
al, 2010).
Quanto ao aspecto necroscópico são inclusos atrofia da musculatura peitoral,
penas descoloridas, hepatomegalia com a disposição de pontos amarelados ao longo do órgão,
pneumonia, aerossaculite, opacificação dos sacos aéreos, pericardite, congestão hepática e
renal, hiperemia da mucosa intestinal, peritonite, esplenomegalia e polisserosite fibrinosa. Nas

30
septicemias bacterianas nas aves são descritas lesões esplênicas caracterizadas por presença
de exsudato fibrinoso ao redor dos capilares embainhados, depleção linfoide, hiperemia e
infiltrado de heterofilos e trombócitos (RIDELL, 1987).
A existência de cepas patogênicas de E. coli causadoras de colibacilose está
relacionada aos fatores de virulência que podem ser avaliados para diferenciar amostras
patogênicas de não patogênicas (JOHNSON, 1991; SAIDENBERG et al., 2012). Mediante a
esses diversos fatores de virulência, associados com uma série de mecanismos de
patogenicidade, essa espécie bacteriana foi classificada em patotipos, divididos em duas
categorias: os patotipos associados a infecções intestinais, denominados de E. coli
diarreagênica (DEC), e aqueles integrantes causadores de infecções extraintestinais,
pertencente a Escherichia coli patogênica extraintestinal (ExPEC) (MARTINEZ e
TRABULSI, 2008).
As DEC são geralmente agrupadas em pelo menos seis categorias: E. coli
enterotoxigênica (ETEC), E. coli enteropatogênica (EPEC), E. coli enteroinvasiva (EIEC), E.
coli entero-hemorrágica (EHEC), E. coli enteroagregativa (EAEC) e E. coli difusamente
aderente (DAEC) (KAPER; NATARO; MOBLEY, 2004; KONEMAN et al., 2010). Já as
ExPEC, comumente, são divididas em E. coli uropatogênica (UPEC), E. coli associada à
meningite neonatal (MNEC) e E. coli patogênica aviária (APEC) (SANTOS et al., 2009).
Em humanos e animais, cepas de EPEC causam diarreia aquosa contendo muco
acompanhada de vômitos e febre, sendo uma das principais causas de diarreia infantil em
países em desenvolvimento (LEVINE, 1987; DONNENBERG, KAPER, 1992). Produzem
uma lesão característica de ligação ou desaparecimento nas bordas da microvilosidade,
causando diarreia crônica, que leva a sequelas como má absorção, má nutrição, perda de peso
e retardo no crescimento (MINAGAWA, 2007). O gene eaeA, localizado no loci de
desaparecimento dos enterócitos (LEE) em uma ilha de patogenicidade (PAI), tem sido
utilizado para identificação de EPEC. A presença desse gene determina um padrão de
aderência de bactérias ao epitélio intestinal em enterócitos com lesão em forma de pedestal
conhecida por attaching and effacing (A/E), que é o mecanismo principal da patogênese da
EPEC (A/E) (VALLANCE, FINLAY, 2000; SYDOW, 2005). O contato com as células
epiteliais resulta na secreção de diversas proteínas disparando resposta na célula hospedeira,
incluindo a ativação de um sinal de rotas de transdução, a despolarização das células e a

31
ligação da proteína intimina da membrana externa. A intimina é codificada pelo gene eaeA.
Não produzem nenhuma enterotoxina ou citotoxina (CROXEN & FINLAY, 2010). Tal como
acontece com outros patotipos de E. coli, a transmissão de EPEC é através da via fecal-oral, e
diversos estudos mostraram que este patotipo causa grande parte das infecções de origem
alimentar (DILIELO, 1982; PRASAD, 1998; DE BUYSER et al., 2001; EVANCHO et al.,
2001; SOOMRO et al., 2002).
E. coli entero-hemorrágica (EHEC) possui a capacidade de destruir células
epiteliais e produzir uma citotoxina potente, a toxina Shiga, que provoca diarreia com ou sem
a presença de sangue, síndrome urêmico-hemolítica, e é fatal para crianças. Existem dois
grupos de toxina Shiga, denominados Stx1 e Stx2. Pela presença do gene eaeA ligam-se
fortemente às células dos mamíferos e produzem o mesmo fenômeno que as linhagens de
EPEC. (ROCHA, 2008; CROXEN & FINLAY, 2010; SILVA et al., 2011).
Em humanos e animais, as E. coli enterotoxigênicas (ETEC) colonizam as
proximidades do intestino delgado causando diarreia aquosa e hipotermia. Este patotipo se
parece com o Vibrio cholerae no fato de aderirem-se à mucosa do intestino delgado e
causarem diarreia sem invadir a mucosa, porém produzindo toxinas que agem nas células da
mucosa. Possui fatores de colonização específicos (CFA/ I a IV) (ROCHA, 2008; CROXEN
& FINLAY, 2010). Produzem dois tipos de enterotoxinas, uma semelhante à toxina da cólera,
denominada toxina termolábil (LT), e outra do tipo diarreica, chamada de toxina termoestável
(ST). Existem dois tipos de toxinas LT: LT-I e LT-II. A ST é uma família de pequenas
toxinas, às quais podem ser divididas em dois grupos: as solúveis (STa) e as insolúveis em
metanol (STb), ambas são codificadas por plasmídeos (CAMPOS & TRABULSI, 2002;
BERCHIERI JUNIOR et al., 2009)
Em humanos e animais cepas de E. coli enteroinvasiva (EIEC) causam distúrbios
no intestino grosso, provocam febre e diarreias profusas contendo muco e sangue. O
microrganismo coloniza o cólon e contém plasmídeos de 120 a 140 MDa necessário para a
invasibilidade, o qual carrega todos os genes necessários para a virulência (BERCHIERI
JUNIOR et al., 2009). Causam um distúrbio que é indistinguível dos sintomas da disenteria
causada pelas espécies de Shigella. Invadem e proliferam em células epiteliais in vivo e in
vitro (CAMPOS & TRABULSI, 2002; SYDOW, 2005). As linhagens EIEC invadem
ativamente as células do cólon e propagam-se lateralmente para as células adjacentes,

32
virtualmente idênticas às espécies de Shigella. No entanto, as EIEC não produzem
shigatoxinas. Quando a infecção é severa, pode levar a uma forte reação inflamatória com
grande ulceração.
Cepas de E. coli enteroagregativa (EAEC) podem colonizar e causar doença no
intestino de humanos e animais, causando diarreia aquosa persistente por mais de 14 dias. O
padrão de aderência é em forma de agregados de cultura em célula de cólon humano. Esta
bactéria estimula a secreção mucoide e se liga a ela, formando um biofilme, causando assim
uma colonização persistente e diarreia (MINAGAWA, 2007). Produzem uma toxina
termolábil relacionada antigenicamente à hemolisina, mas que não é hemolítica, e uma toxina
(citotoxina) termoestável enteroagregativa codificada por um plasmídeo (EAST1). Produzem
uma toxina do tipo ST e uma do tipo hemolisina. Algumas cepas são conhecidas por
produzirem uma toxina do tipo shigatoxina (verotoxina) (CAMPOS & TRABULSI, 2002;
SYDOW, 2005).
E. coli uropatogênica (UPEC) é responsável por infecções urinárias em humanos e
animais (cistite e pielonefrite). A bactéria penetra o trato urinário e invade o epitélio da
bexiga, causando cistite, e quando não é tratada corretamente, pode ascender aos rins causar
pielonefrite, lesão renal irreversível, insuficiência renal e septicemia. Acredita-se que o
reservatório para esta linhagem de E. coli seja o próprio trato gastrointestinal (SIDOW, 2005;
BERCHIERI JUNIOR, et al., 2009).
E. coli causadora de meningite neonatal (NMEC) afeta crianças recém-nascidas.
Este tipo se caracteriza por atravessar a barreira hematoencefálica e colonizar o sistema
nervoso central causando meningite. A proteína IbeA presente na NMEC possui a habilidade
de invadir células endoteliais da microvasculatura cerebral, causando meningite neonatal em
humanos (SAVIOLLI, 2010).
Escherichia coli que adere difusamente (DAEC) está associado em alguns estudos
com diarreia, embora sua patogenia não seja definida com consistência. O termo DAEC foi
inicialmente utilizado para se referir a qualquer cepa que se adere às células HEp-2 e He-La
que não forme microcolônias típicas de EPEC. Com a descoberta da EAEC, alguns autores
reconhecem a DAEC como uma categoria independente, potencialmente causadora de
diarreia. Como se trata de uma categoria ainda não muito estudada, pouco se sabe sobre sua
patogênese (MINAGAWA, 2007).

33
E. coli patogênica aviária (APEC) apresentam fatores de virulência como a
expressão de adesinas, a produção de sideróforos e a capacidade de resistir aos efeitos
microbicidas do soro como agentes fundamentais da manutenção e sobrevivência do
microrganismo no hospedeiro, os quais são responsáveis pelo desenvolvimento da
colibacilose aviária, termo que se refere a qualquer infecção, localizada ou sistêmica, causada
por APEC (DELICATO et al., 2003; PRIOSTE et al., 2013). A maior parte das APEC
isoladas de aves de produção é patogênica apenas para aves e apresentam um baixo risco de
doença para humanos ou outros animais (BARNES et al., 2003; BARCELOS, 2005), embora
alguns trabalhos frequentemente comparem cepas APEC a cepas isoladas de casos humanos
de infecções extra intestinais (RODRIGUEZ-SIEK et al., 2005; EWERS et al., 2007; MORA
et al., 2012; MORA et al., 2013).
O diagnóstico da colibacilose é feito pela associação do histórico, dos sinais
clínicos, duração da doença, juntamente com práticas laboratoriais baseadas no isolamento e
identificação do agente microbiano. O exame citológico por coloração de Gram, quando
associado ao processamento microbiológico padrão, tem sido evidenciado como uma boa
técnica para detecção bacteriana (MACHADO, 2000; QUINN et al., 2005; CATÃO-DIAS &
CARVALHO, 2007; EVANS et al., 2014). Os clínicos de aves, geralmente, não têm acesso a
testes de sorotipagem, ensaio de toxicidade, sondas de DNA e técnica de PCR, e sem estes
exames complementares não é possível obter informações sobre a patogenicidade da cepa
isolada, dificultando o estabelecimento de protocolo terapêutico e prognóstico da ave avaliada
(FUDGE, 2001).
O tratamento é feito com antibióticos de amplo espectro de ação, suplementos
vitamínicos, dieta balanceada e melhoras nas condições sanitárias e de manejo, sendo
importante ressaltar o emprego prévio de testes de sensibilidade dos isolados para a promoção
terapêutica adequada, sendo necessário acrescentar suplementos vitamínicos, dieta balanceada
e um manejo adequado ao tratamento terapêutico (QUINN et al., 2005; GODOY, 2007;
HIRSH, 2009).
Práticas de desinfecção das habitações das aves, o manejo alimentar adequado aos
animais, o controle de roedores e insetos, assim como a realização rotineira do monitoramento
sanitário e microbiológico do ambiente e dos animais é recomendada por MARIETTO-
GONÇALVES et al. (2010) para garantir a sanidade animal.

34
2.3.2 Salmonella spp.
As salmonelas são bactérias da família Enterobactereaceae que incluem mais de
2500 sorotipos (CAMPOS, 2002). O gênero Salmonella é dividido em duas espécies S.
enterica, no qual são incluídas seis subespécies (enterica, salamae, arizonae, diarizonae,
houtenae e indica) e S. bongori (SÁNCHEZ-VARGAS et al.,2011). São bastonetes curtos,
Gram negativos, aeróbios e anaeróbios facultativos de fácil crescimento em meios comuns
(TORTORA, 2012).
Salmonella spp tem como habitat primário o trato intestinal do homem e de
diversos animais, incluindo insetos, aves, répteis e mamíferos, de forma que sua excreção
resulta na contaminação da água, alimentos e o próprio ambiente (BAÚ et al., 2001). A
ocorrência de Salmonella tem sido muito comum em aves (SILVA & DUARTE, 2002).
Em passeriformes e psitacídeos, a salmonelose é uma causa frequente de doença e
mortalidade causada por uma grande variedade de sorotipos, sendo S. Typhimurium o sorotipo
mais isolado destas aves (PASMANS et al, 2013). Outras espécies isoladas incluem, S.
arizonae, S. houtenae, S. Enteritidis, S. Rissen e S. Pullorum (MARIETTO-GONÇALVES et
al, 2010). O resultado da infecção pode variar desde o desenvolvimento de portadores
assintomáticos a surtos de doenças com mortalidade elevada. Nos casos subclínicos, as aves
podem tornar-se portadores persistentes ou temporários, eliminando contínua ou
intermitentemente o agente através das fezes (FLAMER, 1999; PASMANS et al., 2013).
2.3.3 Proteus spp.
As bactérias do gênero Proteus sp. são bacilos Gram negativos anaeróbicos
facultativos, móveis, catalase positivos e oxidase negativos. Hidrolisam ureia, produzem H2S,
e são lisina descarboxilase e lactose negativos. Para a diferenciação entre as espécies (Proteus
vulgaris e Proteus mirabilis) devem ser analisadas as variações que ocorrem nos testes de
indol, Voges-Proskauer e citrato de Simmons (HOLT et al., 1994; OLIVEIRA, 1995).
Habitualmente encontradas no solo, na água e em materiais contaminados com
fezes, as bactérias desse gênero são constituídas por cinco espécies: P. vulgaris, P. mirabilis,
P. myxofaciens, P. penneri e P. hauseri (KONEMAN et al., 2010). P. mirabilis e P. vulgaris
correspondem às duas espécies mais estudadas desse gênero, nos quais ambas já foram
isoladas do trato intestinal de mamíferos, aves e répteis (MANOS & BELAS, 2006).

35
Proteus spp. fazem parte da microbiota do intestino grosso. Potencialmente
patogênicos para aves, apontados como causa de lesões nos pés e doenças respiratórias como
aerosaculite e pneumonia caseosa, quando o animal tem imunodepressão, esta bactéria pode
infectar os ovos através da penetração do microrganismo pela casca, que é facilitada pela
contaminação fecal. Apesar de Proteus spp. estar frequentemente relacionado a infecções no
trato urinário, estas bactérias podem ser isoladas de muitas partes infectadas do corpo
(GODOY; CUBAS, 2009; BROOKS et al., 2014), e quadros de septicemia foram observados
em codornas e faisões infectados com vírus apatogênico da influenza aviária. Em aves
aquáticas é capaz de causar salpingite, artrite, aerossaculite e septicemia (BARNES, 2003).
2.3.4 Citrobacter spp.
As espécies do gênero Citrobacter apresentam potencial zoonótico, atuando como
agente de infecções secundárias ao atingir as aves (GODOY, 2007; RITCHIE et al., 1994).
Segundo GERLACH (1994) as três espécies mais comuns do gênero Citrobacter são C.
freundii, C. amalonaticus e C. diversus, que são menos frequentemente isoladas de aves do
que outros membros da família Enterobacteriaceae. Destas, a espécie C. freundii parece ser a
mais patogênica das três, proprocionando o desenvolvimento de sinais como depressão,
diarreia e óbito (GODOY, 2007), enquanto que C. diversus é raramente isolada de aves.
Adicionalmente, é de conhecimento científico que as aves que sobrevivem a uma infecção por
esse patógeno podem se tornar carreadoras. Este gênero causa infecções secundárias graves,
ocorrendo rápida bacteremia seguida por morte aguda quando o microrganismo penetra a
mucosa intestinal. Contudo, não há casos de infecções humanas derivadas de exposição a aves
infectadas (GERLACH, 1994).
2.3.5 Pantoea sp.
Pertencente à familia Enterobacteriaceae, Pantoea agglomerans (previamente
classificada como Bacillus agglomerans e Enterobacter agglomerans) é um bacilo Gram
negativo, comumente encontrado em nichos ecológicos como água, solo, esgoto, sementes,
vegetais, material feculento e alimentos, bem como relatado como patógenos oportunistas de
animais e humanos (SHARMA et al.2012). KIRZINGER et al. (2015) relata que várias cepas
de Pantoea, incluindo uma de ambiente clínico, realizam intercâmbio genético de
mecanismos metabólicos importantes na colonização de hospedeiros vegetais e animais com
outros membros da família Enterobacteriaceae, incluindo espécies patógenas para humanos.

36
GIBBS et al. (2007) relata que a bactéria P. agglomerans foi a mais isolada de
amostras de fezes de Graúnas de Cabeça Amarela (Xanthocephalus xanthocephalus). Estes
autores afirmaram que esta espécie pode ser estudada como possível organismo de exclusão
competitiva para a microbiota de aves de produção, devido à habilidade que este
microrganismo possui em inibir a colonização por patógenos em plantas e a aparente
adaptabilidade ao sistema gastrointestinal destas aves.
2.3.6 Providencia spp.
O gênero Providencia pertence à família Enterobacteriaceae e apresenta
semelhanças fenotípicas com os gêneros Proteus e Morganella, também incluídos nessa
família. A taxonomia do gênero Providencia tem sido caracterizada por uma grande
instabilidade. Atualmente, este gênero consiste de cinco espécies: Providencia alcalifaciens,
Providencia stuartii, Providencia rettgeri, Providencia rustigianii e Providencia heimbachae
(PIGNATO et al, 1999).
São bastonetes Gram-negativos e anaeróbios facultativos, com motilidade devido
à presença de flagelo peritríqueo, apresentando as seguintes características bioquímicas:
citrato de Simmons positivo, não produz H2S, negativo no teste de lisina e ornitina
descarboxilase, fermenta manose e a hidrólise da ureia é variável (O'HARA et al., 2000).
Apesar de considerada uma bactéria da flora fisiológica de algumas espécies de
aves, Providencia spp. já foi associada a nefrite devido a ascensão da bactéria a partir da
cloaca (LUMEIJ, 1994).
2.3.7 Outras enterobactérias
Klebsiella possui células capsuladas e alta capacidade de fermentação de diversos
carboidratos, sendo um dos poucos membros da família Enterobacteriaceae que é imóvel. A
principal espécie do gênero é K. pneumoniae que, assim como K. oxytoca e algumas outras
espécies do gênero menos frequentes, atuam como patógenos oportunistas em humanos,
sendo encontrados no ambiente e muitas vezes causando infecções nosocomiais (BERGEY et
al., 1994). As infecções por Klebsiella spp. em psitacídeos, mais especificamente por K.
pneumoniae e K. oxytoca, envolvem o trato respiratório e intestinal, acarretando em
problemáticas como sinusite, aerossaculite, pneumonia caseosa e, eventualmente, hepatite,
nefrite, afecções do sistema nervoso e desenvolvimento de uma resposta septicêmica

37
(GODOY, 2007). Não são conhecidas informações especificas sobre as vias de transmissão,
período de incubação e patogenia de Klebsiella spp nas aves (BROWN, 2000), mas de acordo
com GERLACH (1994), as bactérias desse gênero podem desencadear um quadro de
bacteremia, sendo passíveis de afetarem os rins, promovendo problemáticas relacionadas a
falhas renais, bem como nas infecções crônicas, os pulmões podem ser apresentar como
órgãos alvo. Desse modo, embora, a infecção sistêmica possa ser mais comum que a local, em
psitaciformes afecções no pele, cavidade oral e papo podem ser desenvolvidas.
As bactérias do gênero Enterobacter fazem parte da microbiota entérica comensal,
sendo associadas a uma variedade de infecções oportunistas que afetam as vias urinarias e o
trato respiratório (KONEMAN et al., 2010). O gênero possui 16 espécies, todavia as espécies
E. aerogenes e E. cloacae são as mais comumente isoladas de amostras biológicas.
Encontram-se distribuídas amplamente na água, esgoto, solo e vegetais. O grau de infecção e
de invasão bacteriana varia quanto ao tipo de enterobactéria, porém, de um modo geral, a sua
proliferação e disseminação no organismo do homem ou do animal que atinge depende da
idade do hospedeiro, das espécies e de suas condições fisiológicas, estando associadas uma
variedade de infecções oportunistas humanas que afetam as vias urinárias, o trato respiratório,
as feridas cutâneas e, em ocasiões, causam septicemia e meningite (KONEMAN et al, 2010).
As outras enterobactérias, tais como as espécies dos gêneros Edwarsiella,
Morganella, e Serratia atuam como patógenos oportunistas para os animais, suscitando
infecções em diferentes regiões anatômicas, além de depender de circunstâncias como a
imunidade do hospedeiro, a presença de infecções primárias, tecidos danificados e a própria
virulência bacteriana para o desenvolvimento de uma atividade patogênica (QUINN et al.,
2005; TIZARD, 2009). Essas enterobactérias já foram previamente isoladas em estudos
realizados com psitacídeos de cativeiro e canários (HIDASI et al., 2013; HORN et al., 2015;
LOPES et al., 2015).
2.4 Pseudomonas aeruginosa
Este microrganismo da família Pseudomonadaceae (PALLERONI et al., 1973),
caracteriza-se como bastonete Gram-negativo reto ou ligeiramente curvo, aeróbio estrito,
podendo ser observado como células isoladas, aos pares, ou em cadeias curtas, revelando
mobilidade através de flagelo polar monotríquio (POLLACK, 2000). P. aeruginosa é não
fermentadora de carboidratos, produtora de citocromo-oxidase, utiliza o nitrato em

38
substituição ao oxigênio como aceptor final de elétrons, produzindo também Arginina
dehidrolase e Ornitina-descarboxilase. A pioverdina e a piocianina são pigmentos
fluorescentes difusíveis no meio de cultura produzidos por este microrganismo. Algumas
cepas produzem um pigmento avermelhado (piorrubina) ou preto (piomelanina) (POLLACK,
1983).
É um patógeno oportunista que normalmente habita o solo e superfícies em
ambientes aquosos. Sua adaptabilidade e alta resistência intrínseca aos antibióticos permitem
sobreviver em uma ampla gama de outros ambientes naturais e artificiais, incluindo
superfícies em instalações médicas. As infecções graves de P. aeruginosa são frequentemente
nosocomiais e quase todas estão associadas a defesas do hospedeiro comprometidas, como em
neutropenia, queimaduras graves ou fibrose cística (LYCZAK et al., 2000). O espectro de
doenças causadas por este agente compreende desde infecções superficiais da pele a sepse
fulminante (MURRAY, 1995). P. aeruginosa pode causar infecção aguda pela produção de
toxinas e infecção crônica pela ação da camada espessa que consiste no seu biofilme, e ainda,
pode resultar no somatório dos tipos de infecção pela ação concomitante desses componentes
(PALLERONI, 1998).
Devido ao surgimento e disseminação contínua de cepas resistentes aos
antimicrobianos, as opções terapêuticas são cada vez mais limitadas; como resultado, as
infecções por P. aeruginosa demonstram alta morbidade e mortalidade. Nos Estados Unidos,
está entre os patógenos hospitalares mais comuns, e é o segundo patógeno mais isolado de
pacientes com pneumonia associada ao ventilador (HIDRON et al., 2008).
2.5 RESISTÊNCIA ANTIMICROBIANA
A resistência bacteriana pode ser classificada em dois tipos: resistência adquirida,
na qual microrganismos que são originalmente susceptíveis aos antimicrobianos se tornam
resistentes a eles pela presença de mutação cromossômica ou pela aquisição de plasmídeos ou
transposons que carreiam marcadores de resistência após prolongada exposição; e a
resistência natural, onde os microrganismos apresentam resistência intrínseca a uma variedade
de agentes antimicrobianos (NIKAIDO, 1994).
Segundo Gutiérrez et al. (1990) a resistência bacteriana a antibióticos é um sério
problema do ponto de vista clínico e de saúde pública. Há evidências que o tratamento

39
indiscriminado de animais com antibióticos torne seus produtos e derivados fonte para
resistência aos antibióticos em patógenos da espécie humana. Portanto, é de extrema
importância o isolamento e identificação desses agentes em laboratório, como prova definitiva
no diagnóstico das enfermidades a análise in vitro da sensibilidade antimicrobiana se faz
necessário nas amostras isoladas. Desta forma, contribui-se para um melhor controle, com a
utilização de terapêutica adequada, além de promover um decréscimo na resistência aos
antibióticos.
Caracterizado como uma problemática de alarde mundial, tem se evidenciado
cada vez mais o surgimento de cepas resistentes a antimicrobianos ou resistentes a múltiplas
drogas (MDR) carreados por aves (SHOBRAK e ABO-AMER, 2014). Essa resistência
adquirida é uma das principais causas de insucesso da terapia antimicrobiana, uma vez que
prejudica os tratamentos ao limitar as opções viáveis de antimicrobianos (SPINOSA, 2006).
A transferência de genes por membros da família Enterobacteriaceae já foi
demonstrada in vitro por Okamoto et al. (2011), os quais demonstraram que E. coli pode
transferir genes plasmidiais de resistência de forma horizontal para Salmonella Enteritidis,
todavia os mesmos afirmam que in vivo outras condições podem ser desfavoráveis para que
este fenômeno ocorra.
Resistência a múltiplos antimicrobianos em cepas de enterobactérias já foram
evidenciados em diversos estudos com aves de vida livre (SANTOS et al., 2013; SMITH et
al., 2014; STEDT et al., 2014), como também em aves silvestres de cativeiro (HIDASI et al.,
2013; LOPES et al., 2015).
Existem dois métodos de avaliação da sensibilidade bacteriana: o método
quantitativo e o qualitativo. O método quantitativo determina a concentração inibitória
mínima (CIM), definida como a menor concentração do antibiótico capaz de inibir o
crescimento bacteriano, enquanto o método qualitativo mais usual consiste no método de
discodifusão, no qual emprega discos de papel de filtro impregnados com concentrações fixas
de antibióticos (FREITAS et al., 2013).
Os testes in vitro não são capazes de mimetizar a concentração de antimicrobianos
e a sua real biodisponibilidade a nível intestinal, tendo a sua confiabilidade reduzida, o que
representa desvantagens no emprego dessas técnicas de avaliação do perfil de sensibilidade.

40
Além disso, a ausência das condições naturais no método in vivo tais como a relação
hospedeiro e droga empregada, assim como a própria reação bacteriana ante o ambiente
apresentada pelo hospedeiro (PRESCOTT, 2009) são outras desvantagens. Apesar disso, os
métodos in vitro ainda consistem no melhor exame para orientação ante uma terapia anti-
infecciosa (FREITAS et al., 2013).
3 JUSTIFICATIVA
Com a popularização da criação de calopsitas como animais de companhia tem se
verificado seu crescente contato com o homem, gerando o risco da transmissão de micro-
organismos potencialmente patogênicos para outros animais, incluindo o homem. Nesse
contexto, essas aves são passíveis de albergarem diversas bactérias, dentre elas destacando-se as
pertencentes ao gênero Escherichia coli, agente de inúmeras patologias.
Desse modo, o conhecimento da presença, comportamento patogênico e
resistência antimicrobiana desses patógenos são de grande importância para o controle de
possíveis problemas sanitários relacionados a esses micro-organismos, permitindo o
fornecimento de dados essenciais para o estabelecimento de diagnóstico de enfermidades, como
também de medidas preventivas de controle a serem adotadas pelos criadores.

41
4 HIPÓTESE CIENTÍFICA
É possível isolar cepas de diversas bactérias Gram-negativas em Psittaciformes, portanto
estas podem ser disseminadas ao meio ambiente pelas calopsitas. Os microrganismos presentes
nas calopsitas podem apresentar resistência a antibióticos e assim promover um risco de
transmissão de resistência a patógenos humanos.

42
5 OBJETIVOS
5.1 OBJETIVO GERAL
Investigar a presença de bactérias Gram-negativas em calopsitas (Nymphicus
hollandicus) obtidas de pet shops e residências e avaliar seus perfis de resistência a
antimicrobianos.
5.2 OBJETIVOS ESPECÍFICOS
5.2.1 Isolar e tipificar as bactérias Gram-negativas de amostras de suabes cloacais
e fezes de calopsitas presentes em cativeiro domiciliar e Pet shops, assim como de aves
clinicamente doentes e destinadas à avaliação clínica;
5.2.2 Avaliar a resistência a antimicrobianos das cepas de bactérias Gram-
negativas isoladas de calopsitas;

43
5.2.3 Identificar genes diarreiogênicos presentes nas cepas de Escherichia coli
isoladas das aves;
5.2.5 Verificar os riscos de calopsitas serem albergadoras de microrganismos de
interesse na saúde humana.
6 CAPÍTULO 1
Detection of enterobacteria and antimicrobial sensibility in cockatiels (Nymphicus
hollandicus) from house-captive and pet shops
Detecção de enterobactérias e sensibilidade antimicrobiana em calopsitas (Nymphicus
hollandicus) de cativeiro domiciliar e Pet shops

44
Periódico submetido: Arquivo Brasileiro de Medicina Veterinária e Zootecnia
(Submetido em Novembro – QUALIS A2)
DETECÇÃO DE ENTEROBACTÉRIAS, SENSIBILIDADE ANTIMICROBIANA E
ANÁLISE DE PCR DE Escherichia coli DIARREIOGÊNICAS EM CALOPSITAS
(Nymphicus Hollandicus) DE PET SHOPS E RESIDÊNCIAS DE FORTALEZA - CE
[Enterobacterial Detection, Antimicrobial Sensitivity and PCR Analysis of Escherichia coli
Diarreiogenic in Pet shops and Household Cockatiels]
S.V.G. Lima*1, A.P.A da Silva
1, E.S. Lopes
1, M.D. Bona
2, R.V. Horn
1, P.H.Q.S. de
Medeiros2, R.S.C. Teixeira
1, A.H. Bindá
2, W.M. Cardoso
1
1Laboratório de Estudos Ornitológicos (LABEO). Programa de Pós-Graduação em Ciências
Veterinárias (PPGCV), Faculdade de Veterinária, Universidade Estadual do Ceará (UECE),
1700, Campus do Itaperi, Fortaleza, CE, Brasil.
2Laboratório de Toxinologia Molecular. Escola de Medicina da Universidade Federal do
Ceará (UFC), Campus do Porangabussu, Fortaleza, CE, Brazil
*Autor para correspondência: email: [email protected]

45
RESUMO
O objetivo do presente trabalho foi identificar e avaliar os perfis de resistência de cepas de
enterobactérias isoladas de calopsitas e analisar a presença de cepas de E. coli diarreiogênicas
(DEC) nesses animais. 182 amostras foram coletadas e submetidas a pré-enriquecimento
utilizando água peptonada; enriquecimento seletivo com os meios Rappaport-Vassiliadis,
Selenito-Cistina e Brain Heart Infusion (BHI); e triagem em placas contendo os ágares Verde
Brilhante, MacConkey e Salmonella-Shigella. As colônias foram selecionadas e submetidas à
identificação bioquímica e susceptibilidade antimicrobiana. Foram avaliadas 21 cepas de E.
coli quanto à presença de oito genes de virulência de DEC por reação em cadeia da
polimerase (PCR). As enterobactérias encontrada com maior frequência foi Pantoea
agglomerans (28%), Citrobacter freundii (11%) e Serratia liquefaciens (10%). Dois
antibióticos apresentaram baixa eficácia contra as bactérias testadas: Sulfonamida (22%) e
Cotrimoxazol (sulfametoxazol + trimetoprim) (21%). O total de bactérias multirresistentes
(MDR) foi de 158 (50%). Apenas uma cepa de E. coli foi positiva para os genes eaeA e bfpA,
que foi classificada como EPEC típica. Calopsitas apresentam enterobactérias
multirresistentes e cepas de E. coli contendo genes de virulência relacionados a EPEC, que são
potencialmente patogênicas para animais e humanos.
Palavras-chave: Enterobactérias, antibiograma, Escherichia coli diarreiogênicas, EPEC,
calopsitas
SUMMARY
The objective of the present work was to identify and evaluate the resistance profiles of
strains of enterobacteria isolated from calopsites and to analyze the presence of strains of
diarrheogenic E. coli (DEC) in these animals. 182 samples were collected and submitted to
pre-enrichment using peptone water; selective enrichment with Rappaport-Vassiliadis,
Selenito-Cystine and Brain Heart Infusion (BHI) media; and screening on plaques containing
the Green Bright, MacConkey and Salmonella-Shigella agar. The colonies were selected and
submitted to biochemical identification and antimicrobial susceptibility. Twenty-one E. coli
strains were evaluated for the presence of eight DEC virulence genes by polymerase chain
reaction (PCR). The enterobacteria found most frequently were Pantoea agglomerans (28%),

46
Citrobacter freundii (11%) and Serratia liquefaciens (10%). Two antibiotics showed low
efficacy against the tested bacteria: Sulfonamide (22%) and Cotrimoxazole (sulfamethoxazole
+ trimethoprim) (21%). Total multiresistant bacteria (MDR) was 158 (50%). Only one E. coli
strain was positive for the eaeA and bfpA genes, which was classified as typical EPEC.
Cockatiels present multiresistant enterobacteria and E. coli strains containing EPEC-related
virulence genes, that are potentially pathogenic to animals and humans
Key words: Enterobacteria, antibiogram, diarrheogenic Escherichia coli, EPEC, cockatiels
INTRODUÇÃO
Calopsitas (Nymphicus hollandicus) são atualmente um dos psitacídeos mais
populares mantidos como animais de companhia, devido à sua inteligência e habilidades, e à
adaptabilidade da espécie a pequenos ambientes. Contudo, erros de manejo associados ao
stress do confinamento predispõem esses animais à diversas doenças, e devido à proximidade
dessas aves com seus proprietários, o risco da ocorrência de doenças relacionadas a patógenos
capazes de infectar diferentes hospedeiros se torna relevante (DLUGOSZ et al, 2015.)
Entre as bactérias apontadas como importantes patógenos aviários, os principais
agentes envolvidos nas doenças gastrintestinais são as enterobactérias, particularmente
Salmonella sp., Escherichia coli e Enterobacter sp., que podem causar tanto infecções
intestinais como extraintestinais, afetando também outras espécies de aves e mamíferos
(MEYER JÚNIOR, et.al, 2006; QUEIROZ, 2008).
E. coli é uma das principais causas de mortalidade em aves e é responsável por
perdas econômicas significativas em todo o mundo (SCHOULER et al., 2012). Além disso, E.
coli diarreiogênicas (DEC) estão relacionadas com doenças intestinais/diarreicas, infecções do
trato urinário, sepse e meningite em humanos. Em relação as E. coli diarreiogênicas, existem

47
seis tipos bem definidos: EPEC – E. coli enteropatogênica, EHEC - E. coli enterohemorrágica,
EAEC – E. coli enteroagregativa, EIEC - E. coli enteroinvasiva, ETEC - E. coli
enterotoxigênica e DAEC - E. coli difusamente aderente (KAPER; NATARO; MOBLEY,
2004). Dessa forma, o objetivo do presente trabalho foi identificar e avaliar os perfis de
resistência de cepas de enterobactérias isoladas de calopsitas provenientes de residências e pet
shops em Fortaleza – CE, e analisar a presença de cepas de E. coli diarreiogênicas nesses
animais.
MATERIAL E MÉTODOS
Este estudo foi aprovado pelo Comitê de Ética para o Uso de Animais da
Universidade Estadual do Ceará (Protocolo 647396-2015). De setembro de 2015 a maio de
2016, foram coletadas 182 amostras, das quais 110 foram suabes cloacais individuais e 72
suabes de arrasto, de 18 criatórios. As aves analisadas foram obtidas de 8 domicílios, 7 pet
shops e 3 aviários localizados na cidade de Fortaleza, no Ceará, Brasil. As amostras foram
coletadas em tubos contendo água peptonada tamponada (Himedia, Mumbai, India) e
transportadas para o Laboratório de Estudos Ornitológicos (LABEO) da Universidade
Estadual do Ceará (UECE) para processamento microbiológico. Posteriormente, alíquotas das
amostras foram transferidas para tubos contendo Brain Heart Infusion (Himedia, Mumbai,
India) e tubos com caldo Selenito Cistina (Himedia, Mumbai, India) e Rappaport Vassiliadis
(Merck). Sequencialmente, as amostras foram estriadas em placas com ágar MacConkey
(Himedia, Mumbai, India) e em ágar Hektoen (Himedia, Mumbai, India). Colônias com
características morfológicas de enterobactérias foram selecionadas e identificadas por meio de
bateria de provas bioquímicas. A temperatura e o período de incubação para cada etapa foram
padronizados respectivamente em 37° C e 24 horas.
O teste de sensibilidade antimicrobiana foi realizado utilizando o método de
difusão em disco em ágar Mueller-Hinton (Himedia®) seguindo os padrões fornecidos pelo
Clinical and Laboratory Standards Institute (CLSI, 2013). Foram utilizados os seguintes
discos nas respectivas concentrações: Azitromicina (15 μg), ácido nalidíxico (30 μg),
Ciprofloxacina (5 μg), Enrofloxacina (5 μg), Cloranfenicol (30 μg), Estreptomicina (10 μg),
Gentamicina (10 μg), Tetraciclina (30 μg), Ceftiofur (30 μg), Polimixina B (300 μg),
Cotrimoxazol (sulfametoxazol + trimetoprim) (25 μg) e Sulfonamida (300 μg) (Laborclin,
Pinhais, PR, Brasil). Os resultados foram interpretados medindo as zonas de inibição ao redor

48
dos discos (CLSI, 2015), e as zonas intermediárias foram interpretadas como resistência ao
respectivo antibacteriano; as cepas que mostraram resistência fenotípica a três ou mais
antibióticos foram consideradas multirresistentes. A amostra de controle utilizada para este
teste foi a estirpe de E. coli ATCC 25922.
Adicionalmente, realizou-se pesquisa de genes de virulência nas cepas de E. coli
isoladas. As cepas mantidas em ágar nutriente foram reativadas em BHI e plaqueadas em
MacConkey e incubadas durante 24h a 37°C. A partir de cada placa, foram recolhidas duas a
três colônias e colocadas em tubos contendo 1 mL de Triton X-100 a 0,5%, que foram
agitados em vórtice durante 15s e fervidos durante 20min a 94ºC. Os tubos foram então
centrifugados a 10.000 rpm durante 10 min. O sobrenadante contendo DNA foi quantificado e
qualificado por espectrofotometria usando NanoDrop Spectrophotometer 2000 (Thermo
Scientific, Wilmington, EUA). Para diagnóstico molecular de DEC as amostras de DNA
foram submetidas a reações em cadeia da polimerase (PCR). A presença de oito genes de
genes de virulência de cinco patotipos foi avaliada como se segue: genes stx1 (348pb) e stx2
(584pb) para a identificação de E. coli produtoras de Shiga-Toxina (STEC); eltB (508 pb) e
estA (147 pb) para E. coli enterotoxigênica (ETEC); eaeA (881 pb) e bfpA (300 pb) para E.
coli enteropatogênica (EPEC); ipaH (483 pb) para E. coli enteroinvasiva (EIEC); aatA (630
pb) e aaiC (215 pb) para a E. coli enteroagregativa (EAEC) (TANIUCHI ET AL., 2012). As
cepas positivas para os genes eaeA e bfpA foram consideradas EPEC típicas, enquanto que as
positivas exclusivamente para o gene eaeA foram classificadas como EPEC atípicas.
(DONNENBERG & FINLAY, 2013). As estirpes EAEC 042, EHEC O157: H7, EIEC O124,
EPEC 2348/69 e ETEC H10407 foram utilizadas como controles positivos para as reações. A
PCR foi realizada utilizando GoTaqGreen kit (Promega) e iniciadores a 0,4 uM em MyCycler
Thermal Cycler (Biorad, CA, EUA), com o seguinte protocolo: 95°C durante 15 min; 40
ciclos de 95°C durante 30 s, 57°C durante 30 s e 72°C durante 110 min; seguido de 72°C
durante 10 min. Os produtos amplificados foram submetidos a eletroforese em gel de agarose
corado com brometo de etídio a 2% e fotografados pelo sistema transiluminador ChemicDoc
XRS 112 (Biorad, CA, EUA).
RESULTADOS E DISCUSSÃO
O total de enterobactérias encontrada em suabes cloacais das calopsitas analisadas
foi de 67,9% e de 32,3% em suabes de arrasto (Tab. 1). Dos 315 isolados obtidos, verificou-se
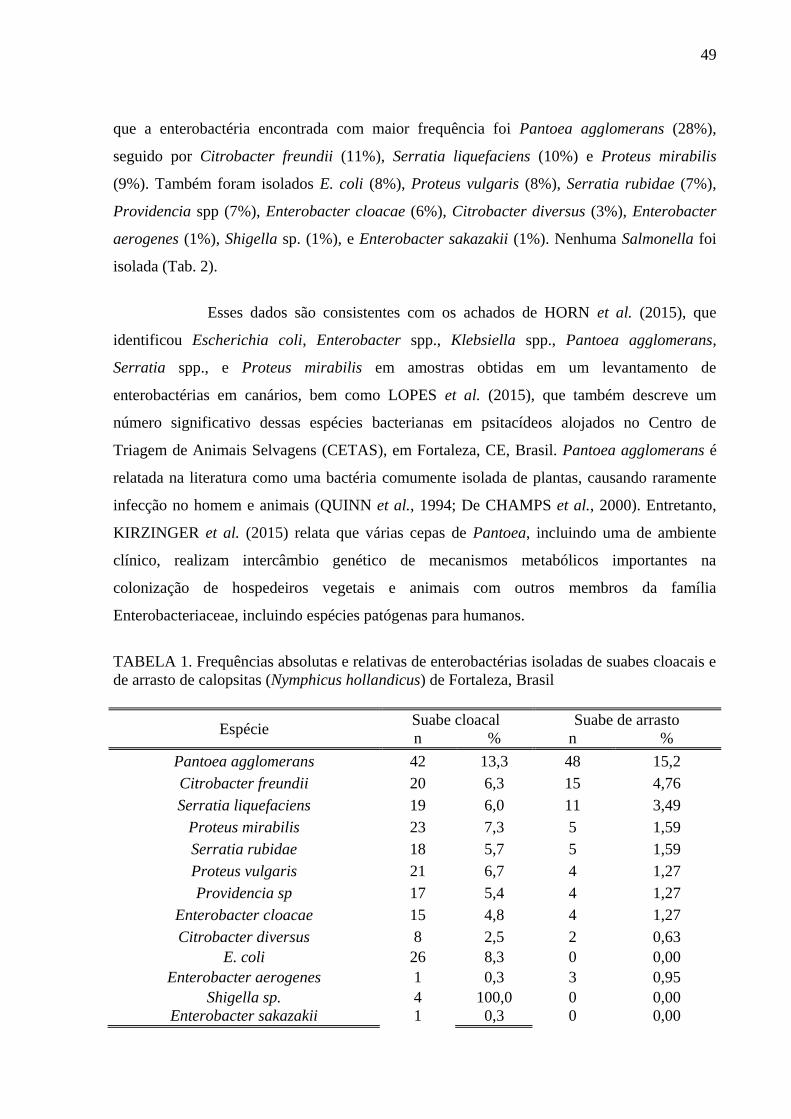
49
que a enterobactéria encontrada com maior frequência foi Pantoea agglomerans (28%),
seguido por Citrobacter freundii (11%), Serratia liquefaciens (10%) e Proteus mirabilis
(9%). Também foram isolados E. coli (8%), Proteus vulgaris (8%), Serratia rubidae (7%),
Providencia spp (7%), Enterobacter cloacae (6%), Citrobacter diversus (3%), Enterobacter
aerogenes (1%), Shigella sp. (1%), e Enterobacter sakazakii (1%). Nenhuma Salmonella foi
isolada (Tab. 2).
Esses dados são consistentes com os achados de HORN et al. (2015), que
identificou Escherichia coli, Enterobacter spp., Klebsiella spp., Pantoea agglomerans,
Serratia spp., e Proteus mirabilis em amostras obtidas em um levantamento de
enterobactérias em canários, bem como LOPES et al. (2015), que também descreve um
número significativo dessas espécies bacterianas em psitacídeos alojados no Centro de
Triagem de Animais Selvagens (CETAS), em Fortaleza, CE, Brasil. Pantoea agglomerans é
relatada na literatura como uma bactéria comumente isolada de plantas, causando raramente
infecção no homem e animais (QUINN et al., 1994; De CHAMPS et al., 2000). Entretanto,
KIRZINGER et al. (2015) relata que várias cepas de Pantoea, incluindo uma de ambiente
clínico, realizam intercâmbio genético de mecanismos metabólicos importantes na
colonização de hospedeiros vegetais e animais com outros membros da família
Enterobacteriaceae, incluindo espécies patógenas para humanos.
TABELA 1. Frequências absolutas e relativas de enterobactérias isoladas de suabes cloacais e
de arrasto de calopsitas (Nymphicus hollandicus) de Fortaleza, Brasil
Espécie Suabe cloacal Suabe de arrasto
n % n %
Pantoea agglomerans 42 13,3 48 15,2
Citrobacter freundii 20 6,3 15 4,76
Serratia liquefaciens 19 6,0 11 3,49
Proteus mirabilis 23 7,3 5 1,59
Serratia rubidae 18 5,7 5 1,59
Proteus vulgaris 21 6,7 4 1,27
Providencia sp 17 5,4 4 1,27
Enterobacter cloacae 15 4,8 4 1,27
Citrobacter diversus 8 2,5 2 0,63
E. coli 26 8,3 0 0,00
Enterobacter aerogenes 1 0,3 3 0,95
Shigella sp. 4 100,0 0 0,00
Enterobacter sakazakii 1 0,3 0 0,00

50
Total
214,0 67,9 101 32,38
Espécies dos gêneros Proteus, Hafnia e Serratia podem atuar como patógenos
oportunistas em certas circunstâncias e resultar em um perigo para a saúde aviária. Na maioria
dos estudos, no entanto, os gêneros Enterobacter spp, Proteus spp., Providencia spp. e
Citrobacter são suspeitas de causarem doenças em homens, e muitas vezes estão associadas a
infecções nosocomiais (STEELE et al., 2005). Citrobacter freundii é considerada uma
bactéria oportunista e ubíqua (FERNANDEZ et al., 2011), foi detectada como agente primário
em casos de morte súbita em papagaios cativos (CHURRIA et al., 2014). Proteus já foi
relatado em aves jovens, sendo identificado como uma população microbiana que tende a
diminuir à medida que as aves envelhecem (NALDO et al., 1988). É considerado um
microrganismo oportunista e têm sido associados a distúrbios do sistema respiratório superior
e pododermatite em aves selvagens (OLINDA et al., 2012). Enterobacter sakazakii tem sido
correlacionada com a ocorrência de surtos de alta mortalidade em rebanhos de canários, e
estes microorganismos têm sido considerados tão importantes como Salmonella e Escherichia
coli (CATTAROSSI et al. 2013).
A ausência de Salmonella tem sido relatada em vários estudos brasileiros recentes,
com baixa ou nenhuma detecção desta bactéria em aves aparentemente saudáveis (BEZERRA
et al. 2013; HORN et al, 2015; LOPES et al, 2015). DLUGOSZ et al. (2105) relatou que não
houve isolamento desse patógeno em um estudo em que se avaliou a presença de Salmonella
sp. em amostras fecais de 86 calopsitas mantidas em cativeiro na região metropolitana de
Curiti
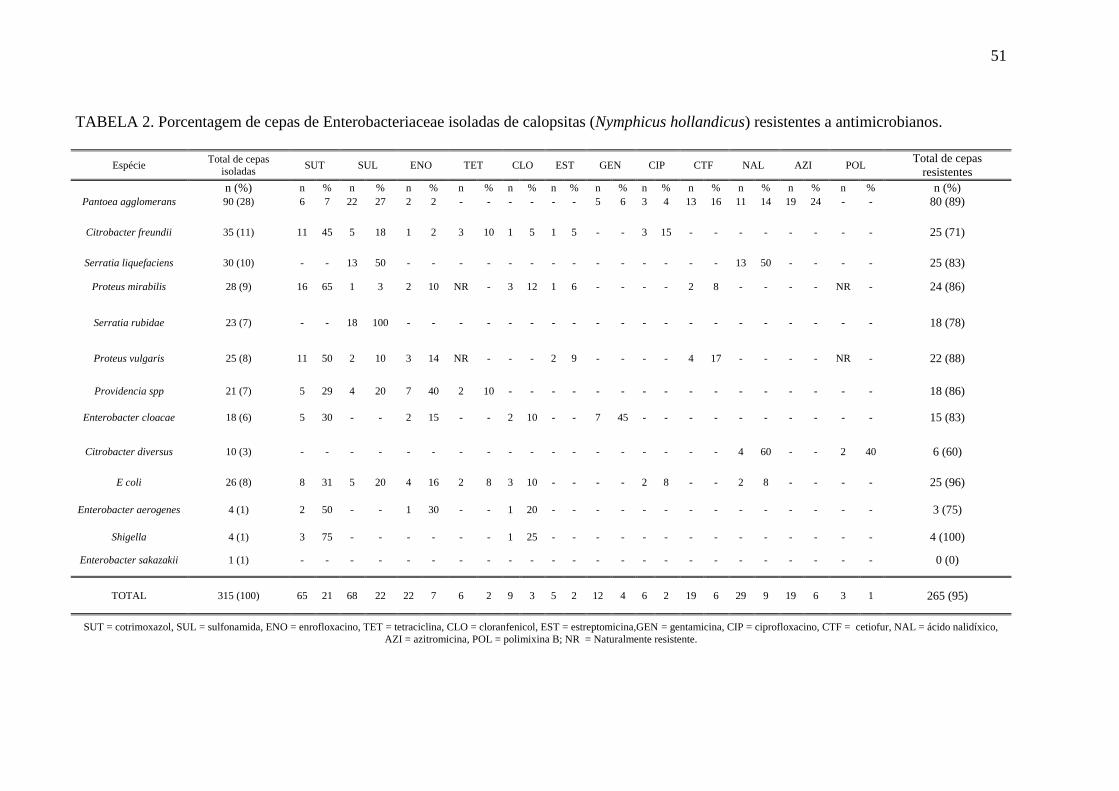
51
TABELA 2. Porcentagem de cepas de Enterobacteriaceae isoladas de calopsitas (Nymphicus hollandicus) resistentes a antimicrobianos.
Espécie Total de cepas
isoladas SUT SUL ENO TET CLO EST GEN CIP CTF NAL AZI POL
Total de cepas
resistentes
n (%) n % n % n % n % n % n % n % n % n % n % n % n % n (%)
Pantoea agglomerans 90 (28) 6 7 22 27 2 2 - - - - - - 5 6 3 4 13 16 11 14 19 24 - - 80 (89)
Citrobacter freundii 35 (11) 11 45 5 18 1 2 3 10 1 5 1 5 - - 3 15 - - - - - - - - 25 (71)
Serratia liquefaciens 30 (10) - - 13 50 - - - - - - - - - - - - - - 13 50 - - - - 25 (83)
Proteus mirabilis 28 (9) 16 65 1 3 2 10 NR - 3 12 1 6 - - - - 2 8 - - - - NR - 24 (86)
Serratia rubidae 23 (7) - - 18 100 - - - - - - - - - - - - - - - - - - - - 18 (78)
Proteus vulgaris 25 (8) 11 50 2 10 3 14 NR - - - 2 9 - - - - 4 17 - - - - NR - 22 (88)
Providencia spp 21 (7) 5 29 4 20 7 40 2 10 - - - - - - - - - - - - - - - - 18 (86)
Enterobacter cloacae 18 (6) 5 30 - - 2 15 - - 2 10 - - 7 45 - - - - - - - - - - 15 (83)
Citrobacter diversus 10 (3) - - - - - - - - - - - - - - - - - - 4 60 - - 2 40 6 (60)
E coli 26 (8) 8 31 5 20 4 16 2 8 3 10 - - - - 2 8 - - 2 8 - - - - 25 (96)
Enterobacter aerogenes 4 (1) 2 50 - - 1 30 - - 1 20 - - - - - - - - - - - - - - 3 (75)
Shigella 4 (1) 3 75 - - - - - - 1 25 - - - - - - - - - - - - - - 4 (100)
Enterobacter sakazakii 1 (1) - - - - - - - - - - - - - - - - - - - - - - - - 0 (0)
TOTAL 315 (100) 65 21 68 22 22 7 6 2 9 3 5 2 12 4 6 2 19 6 29 9 19 6 3 1 265 (95)
SUT = cotrimoxazol, SUL = sulfonamida, ENO = enrofloxacino, TET = tetraciclina, CLO = cloranfenicol, EST = estreptomicina,GEN = gentamicina, CIP = ciprofloxacino, CTF = cetiofur, NAL = ácido nalidíxico,
AZI = azitromicina, POL = polimixina B; NR = Naturalmente resistente.

52
Curitiba-PR. GOPEE et al. (2000) citam que aves de vida livre são apontadas como potenciais
carreadoras de patógenos para o interior de plantéis de animais, mas que a frequência de
Salmonella sp. em aves de cativeiro é relativamente baixa, quando comparada a mamíferos e
répteis.
Os resultados dos testes de sensibilidade aos antimicrobianos estão demonstrados
na tab. 2. Dois antibióticos apresentaram baixa eficácia contra as bactérias testadas:
Sulfonamida (22%) e Cotrimoxazol (sulfametoxazol + trimetoprim) (21%). Todas as
diferentes espécies de enterobactérias isoladas de calopsitas apresentaram taxas de resistência
variada para os antibióticos testados. Apenas a única Enterobacter sakazakii isolada foi
sensível a todos.
As cepas de Serratia liquefaciens apresentaram resistência para Sulfonamida
(50%) e Ácido Nalidíxico (50%). Citrobacter diversus apresentou alta resistência para Ácido
Nalidíxico (60%) e Polimixina B (40%). As cepas de Shigella (75%), Proteus mirabilis
(65%), Proteus vulgaris (50%), Enterobacter aerogenes (50%), e E. coli (30%) apresentaram
as maiores taxas de resistência à Cotrimoxazol (sulfametoxazol + trimetoprim). O gênero
Providencia apresentou resistência de 40% à Enrofloxacina, enquanto 45% das cepas de
Enterobacter cloacae foram resistentes à Gentamicina. Todas as cepas de Serratia rubidae
apresentaram alta resistência apenas a Sulfonamida (100%).
Entre as 315 enterobactérias isoladas neste estudo, 265 (95,1%) apresentaram
resistência a pelo menos um dos antibióticos testados, e o total de bactérias multirresistentes
(MDR) foi de 158 (50%). O máximo de antimicrobianos para o qual 17 isolados foram
resistentes foi de sete, seguidos por 50 isolados resistentes a seis antimicrobianos (Tab. 3).
TABELA 3. Enterobactérias Multirresistentes (MDR) isoladas de calopsitas (Nymphicus
hollandicus)
Nº de antibióticos Nº de cepas resistentes %
Pelo menos 1
265
84,2
>1
207
65,8
>2
158
50
>3
158
50
>4
149
47,4
>5
149
47,4
>6
50
15,8
>7
17
5,3

53
>8
0
0
Resultados semelhantes já foram obtidos anteriormente por HORN et al. (2015),
que relatou resistência de 55,7% para sulfonamidas. Várias das espécies estudadas por esses
autores também apresentaram resistência para sulfazotrim (sulfametoxazol + trimetoprim) e
ácido nalidíxico. Além disso, 55,7% das bactérias apresentaram multirresistência, e o máximo
de antimicrobianos para o qual um único isolado foi resistente foi de oito, seguidos por três
isolados resistentes a sete antimicrobianos, o que corrobora com os achados deste estudo.
LOPES et al. (2015) relata que 95,1% das cepas analisadas por esses autores mostraram
resistência a pelo menos um dos antibióticos testados, e apenas uma única cepa de Proteus
mirabilis (0,6%) apresentou resistência a 11 dos 12 antibióticos utilizados.
21 cepas de E. coli foram submetidas a PCR apenas 1 (4,7%) foi positiva para os
genes eaeA e bfpA (fig. 1). Considerando a presença desses genes, essa cepa foi classificada
como EPEC típica (tEPEC). KNÖBL e MENÃO (2010) investigou a presença de EPEC típica
e atípica em psitacídeos em um estudo com 80 suabes de fezes frescas coletadas de
psitacídeos no Estado de São Paulo. Vinte e quatro cepas (30%) foram isoladas, mas apenas
duas foram positivas para o gene eae.
Fig. 1 Eletroforese em gel de agarose de produtos de PCR. Coluna 1: Marcador Molecular. Coluna 2: Controle positivo para
o gene eaeA. Coluna 3: amostra negativa. Coluna 4: amostra isolada de calopsita positiva para o gene eaeA. Colunas 5, 6 e
7: amostras negativas. Coluna 8: Controle negativo. Coluna 9: Controle positivo para o gene bfpA. Coluna 10: amostra
negativa. Coluna 11: amostra isolada de calopsita positiva para o gene bfpA. Colunas 12, 13 e 14: amostras negativas.
Coluna 15: Controle negativo.
EPEC foi o primeiro patotipo a ser descrito e continua sendo um importante
agente etiológico de diarreia infantil em todo mundo, com elevada prevalência em ambientes
hospitalares e comunidades carentes (WHO, 2002), bem como também é relatado ser uma das
principais causas de diarreia persistente (ABBA et al., 2009).

54
As cepas de EPEC são classificadas em típicas quando abrigam o plasmídeo para
o fator de aderência de EPEC (EAF), que codifica bfp (bundle forming pili), enquanto as
cepas atípicas são negativas para o gene bfp (TRABULSI et al, 2002). Em aEPEC, a presença
de certos marcadores genômicos adicionais está fortemente associada com diarreia crônica
(AFSET, J.E. et al.,2006).
REPLE et al. (2015) investigou a presença de grupos filogenéticos de EPEC e
STEC em cepas isoladas de fezes de 448 psitacídeos, obtendo elevada frequência de
ocorrência de E. coli positiva para os genes eae e bfp em calopsitas (39,3%). Destas, 14,58%
foram classifcadas como EPEC típicas e como EPEC atípicas (5/48). TRABULSI et al.
(2002) relata que EPEC típicas são raramente isoladas de animais, sendo este patotipo
encontrado majoritariamente em humanos. A presença de EPEC típicas nesses psitacídeos
sugere a transmissão de caráter antropozoonótico como a mais provável.
CONSIDERAÇÕES FINAIS
Verificou-se a presença de enterobactérias nas calopsitas estudadas, além da
detecção de uma amostra de E. coli positiva para genes de virulência relacionados a EPEC.
Além disso, os resultados mostraram que essas bactérias possuíam elevado percentual de
resistência aos antimicrobianos estudados. Estes resultados indicam a necessidade de medidas
preventivas que visem melhorar as condições sanitárias, como por exemplo, reduzir a
contaminação ambiental e dos alimentos ofertados aos animais.
REFERÊNCIAS
AFSET, J. E.; BRUANT, G.; BROUSSEAU, R. et al. Identification of virulence genes linked
with diarrhea due to atypical enteropathogenic Escherichia coli by DNA microarray analysis
and PCR. Journal of Clinical Microbiology, v. 44, n. 10, p. 3703-3711, 2006.
BEZERRA, W. G. A.; CARDOSO, W. M.; DE CASTRO TEIXEIRA; R. S., et al. Survey of
Salmonella sp. in budgerigars (Melopsittacus undulatus) in Fortaleza, Brazil. Acta Scientiae
Veterinariae, v. 41, p. 01-07, 2013.
CATTAROSSI, D; AZZARA, E; CATANIA, S. Clinical and laboratory practice for canaries
and true finches. Veterinary Clinics of North America: Exotic Animal Practice, v. 16, n. 1,
p. 31-46, 2013.

55
CHURRIA, C.D.G.; ARIAS, N.; ORIGLIA, J. et al. Citrobacter freundii infection in two
captive Australian king parrots (Alisterus scapularis). Journal of zoo and Aquarium
Research, v.2, n. 2, p. 52-53, 2014
DE CHAMPS, C., LE SEAUX, S., DUBOST, J. J. et al. Isolation of Pantoea agglomerans in
two cases of septic monoarthritis after plant thorn and wood sliver injuries. Journal of
Clinical Microbiology, v. 38, n. 1, p. 460-461, 2000.
DE SOUZA LOPES, E.; MACIEL, W. C.; DE ALBUQUERQUE, Á. H et al. Prevalence and
Antimicrobial Resistance Profile of Enterobacteria Isolated from Psittaciformes of Illegal
Wildlife Trade. Acta Scientiae Veterinariae, v. 43, p. 1313, 2015.
DLUGOSZ, A. P. "Prevalência de Salmonella Sp. em calopsitas (Nymphicus Hollandicus)
mantidas em cativeiro comercial." Archives of Veterinary Science 20.2. 2015.
DONNENBERG, M. S.; FINLAY, B. B. Combating enteropathogenic Escherichia coli
(EPEC) infections: the way forward. Trends in Microbiology, v. 21, n. 7, p. 317-319, 2013.
FERNÁNDEZ, A.; VELA, A.; ANDRADA, M. et al. Citrobacter freundii septicemia in a
stranded newborn Cuvier's beaked whale (Ziphius cavirostris). Journal of Wildlife Diseases,
v. 47, n. 4, p. 1043-1046, 2011.
GOPEE, N.V.; ADESIYUN, A.A.; CAESAR. K. Retrospective and longitudinal study of
salmonellosis in captive wildlife in Trinidad. Journal of Zoo and Wildlife Medicine, v.36,
p.284-293, 2000
HORN, R. V.; CARDOSO, W. M.; LOPES, E. S. et al. Identification and antimicrobial
resistance of members from the Enterobacteriaceae family isolated from canaries (Serinus
canaria). Pesquisa Veterinária Brasileira, v. 35, n. 6, p. 552-556, 2015.
KAPER, J. B.; NATARO, J. P.; MOBLEY, H. L. T. Pathogenic Escherichia coli. Nature
Reviews; v. 2, p. 123-140, 2004.
KIRZINGER, M. WB; BUTZ, C. J.; STAVRINIDES, J. Inheritance of Pantoea type III
secretion systems through both vertical and horizontal transfer. Molecular Genetics and
Genomics, v. 290, n. 6, p. 2075-2088, 2015.

56
KNÖBL, T; MENÃO, M. C. Escherichia coli Enteropatogênica (EPEC) Isoladas De
Psitacídeos. FIEP BULLETIN - volume 80 - Special Edition - ARTICLE I –2010.
Disponível em:
<http://www.fiepbulletin.net/index.php/fiepbulletin/article/viewFile/1112/2155> Acessado
em: 29 outubro de 2016.
MEYER JÚNIOR, J.C.; DIAS, H.L., ARAÚJO; J.B. da C. Determinação qualitativa das
enterobactérias presentes no trato digestivo de muçuãs (Kinosternon scorpioides) mantidas
em cativeiro. In: Anais do VII Congresso Internacional sobre Manejo de Fauna Silvestre
na Amazônia e America Latina. Ilhéus. p.15. 2006.
NALDO, J. L.; SILVANOSE, C. D.; SAMOUR, J. H.; & BAILEY, T. A. et al.
Developmental intestinal aerobic microflora in the kori bustard (Ardeotis kori). Avian
Pathology, v. 27, n. 4, p. 359-365, 1998.
OLINDA, R. G.; SOUZA, M. C. A.; FIGUEIREDO, J. N.; SILVA, J. M. C. et al. Diagnosis
of Proteus spp. in wild birds raised under captivity in Rio Grande do Norte, Brazil. Arquivos
do Instituto Biológico, v. 79, n. 2, p. 301-303, 2012.
QUEIROZ, B. D. Principais doenças respiratórias que acometem Psitacídeos e Passeriformes
criados como animais de estimação. Qualittas Documentos. Rio de Janeiro: UCB. 98 p.
2008.
QUINN, P. J.; CARTER, M. E.; MARKEY, B. et al. Enterobacteriaceae. Clinical
Veterinary Microbiology. Wolfe Publishing, London, p. 209-236, 1994.
REPLE, J. N.; OLIVEIRA, M. C. V.; OLIVEIRA, M. G. X. et al. Análise filogenética de
Escherichia coli Enteropatogenica (EPEC) e Produtoras de Toxina Shiga (STEC) isoladas de
fezes de psitacídeos: avaliação do potencial zoonótico. Revista de Educação Continuada em
Medicina Veterinária e Zootecnia, v. 13, n. 1, p. 81-83. 2015.
RITCHIE, B.W.; HARRISON, G.J.; HARRISON, L.R. 1994. Avian Medicine: principles
and application. Lake Worth: Wingers Publishing, 1384p.

57
SCHOULER, C; SCHAEFFER; B; BRÉE, A. et al. Diagnostic strategy for identifying avian
pathogenic Escherichia coli based on four patterns of virulence genes. J. Clin. Microbiol.
50(5): 1673-1678. 2012.
STEELE, C. M.; BROWN, R. N.; BOTZLER, R. G. Prevalences of zoonotic bacteria among
seabirds in rehabilitation centers along the Pacific Coast of California and Washington,
USA. Journal of Wildlife Diseases, v. 41, n. 4, p. 735-744, 2005.
TANIUCHI, M.; WALTERS, C. C.; GRATZ, J., M. et al. Development of a multiplex
polymerase chain reaction assay for diarrheagenic Escherichia coli and Shigella spp. and its
evaluation on colonies, culture broths, and stool. Diagnostic microbiology and infectious
disease, v. 73, n. 2, p. 121-128, 2012.
TRABULSI, L. R.; KELLER, R.; GOMES, T. A. T. Typical and atypical Enteropathogenic
Escherichia coli. (Synopsis). Emerging infectious diseases, v. 8, n. 5, p. 508-514, 2002.
WORLD HEALTH ORGANIZATION. Improving diarrhoea estimates. Geneva, 2002.
Disponível em: <http:www.who.int/child_adolescent_health/documents/pdfs/improving_
diorrhoea_estimates.pdf> Acessado em: 11 de novembro de 2016.

58
7 CAPÍTULO 2
Prevalence and antibiogram analysis of Pseudomonas spp. in cockatiels (Nymphicus
hollandicus) from Fortaleza, CE
Prevalência e análise de antibiograma de Pseudomonas spp. em calopsitas (Nymphicus
hollandicus) de Fortaleza, CE

59
Periódico submetido: Arquivo Brasileiro de Medicina Veterinária e Zootecnia
(Submetido em Novembro – QUALIS A2)
PREVALENCE AND ANTIBIOGRAM ANALYSIS OF Pseudomonas spp. IN
COCKATIELS (Nymphicus hollandicus) FROM FORTALEZA, CE
[Pseudomonas spp. in cockatiels from Fortaleza]
Suzan Vitória Girão Lima1,2
, William Cardoso Maciel1, Windleyanne Gonçalves Amorim
Bezerra1, Jackson Forte Beleza
1, Fernanda Gaio Conceição
1, Régis Siqueira de Castro
Teixeira1
1Laboratory of Ornithological Studies (LABEO). Graduate Program in Veterinary Science
(PPGCV), Faculty of Veterinary Medicine, State University of Ceará (UECE), Itaperi
Campus, Fortaleza, CE, Brazil. Fax +55 (85) 3101-9860]. Faculty of Veterinary Medicine,
PPGCV, UECE. Av. Dr. Silas Munguba, n. 1700, Itaperi Neighborhood. CEP 60.714.903
Fortaleza-CE. Brazil.
2Correspondence: S.V.G. Lima [[email protected]]
Abstract
Cockatiels (Nymphicus hollandicus) are among the most common pet bird, but little is known
about the zoonotic risks of these animals. Thus, the objective of the present work was to
identify and evaluate the resistance profiles of Pseudomonas aeruginosa strains isolated from
cockatiels (Nymphicus hollandicus) from residences and pet shops in Fortaleza - CE. A total
of 182 samples (110 individual cloacal suabes and 72 drag suabes) were collected from
eighteen domiciliary breeding location. Twenty-nine (Pseudomonas aeruginosa) strains were
identified, and the antimicrobial resistance profiles of the strains were determined using the
disk diffusion method. All the 29 strains tested presented resistance to at least one antibiotic.
Furthermore, 21 isolates showed a multidrug resistance profile, corresponding to 72.4 % of
the total, and the maximum number of antibiotics to which a strain presented resistance was 7.
The results indicate that common pet birds such as cockatiels can harbor strains of
Pseudomonas sp., a pathogen harmful to both animal and human health. In addition, a high
rate of antimicrobial resistance was verified.

60
Keywords: Antimicrobial resistance, Pseudomonas aeruginosa, Cockatiel
INTRODUCTION
Pseudomonas aeruginosa is a ubiquitous and metabolically versatile Gram-negative
bacterium notable for its ability to thrive in highly diverse ecological niches, and due to
numerous virulence factors that contribute to its pathogenesis, can cause significant morbidity
and mortality among compromised humans1,2
. Moreover, P. aeruginosa possesses an intrinsic
resistance to many antimicrobials because of the bacterium’s outer membrane barrier, the
presence of multidrug efflux transporters, and endogenous antimicrobial inactivation3.
Due to the frequency with which it is involved in infections in man, it represents a serious
problem, particularly in hospitalized patients, since most infections with Pseudomonas
species occur in compromised hosts4,5,6
. The pathogenicity of these organisms is based on its
ability to produce a variety of toxins and proteases and on its ability to resist phagocytosis4.
The clinical picture is extremely severe, especially in patients with immune system
deficiencies or chronic infections and in patients with acquired immunodeficiency syndrome,
with severe burns, cancer, and mortality rates may reach 50%5,6
.
Pseudomonas aeruginosa causes diseases in plants and animals, being a common avian
pathogen7,8,9
, principally affecting the upper respiratory tract, causing respiratory tract
infections8,10,11
, being also frequently isolated from hemorrhagic enteritis, corneal ulcers, and
septicemia in birds12
.
P. aeruginosa resistance to different antimicrobials may be intrinsic due to the low
permeability of its membrane and the ability to form a biofilm, or readily acquired by the
association with microorganisms commonly resistant to antibiotics via chromosomal
mutations and lateral gene transfer, becoming a dangerous and dreaded pathogen3,13,14
. The
only antibiotic agents to which strains are regularly sensitive are cephalosporins,
5carbenicillin, colistin, gentamicin, polymyxin, quinolones, and streptomycin; however,
degrees of cross-resistance between these agents have been reported13
.

61
Therefore, the aim of this study was to identify and evaluate the resistance profiles of
Pseudomonas aeruginosa strains isolated from cockatiels (Nymphicus hollandicus) from
residences and pet shops in Fortaleza - CE.
MATERIAL AND METHODS
Sampling
This study was approved by the Local Ethics Committee for the Use of Animals from the
State University of Ceará (Protocol 647396-2015). From September 2015 to May 2016 a total
of 182 samples were collected, from which 110 were individual cloacal suabes and 72 were
drag suabes samples from eighteen domiciliary breeding location. The birds analyzed were
from households, pet shops and aviaries located in the city of Fortaleza in Ceará State, Brazil.
The cloacal suabe samples were placed in tubes containing buffered peptone water broth
(Himedia, Mumbai, India), which were maintained in a box containing ice in order to preserve
their intrinsic properties. The collected material was transported to the Laboratory of
Ornithological Studies (LABEO), located at the State University of Ceará, for the
microbiological procedure. When present, clinical signs from each sampled bird were taken
note.
Laboratory method
The samples stored in tubes containing buffered peptone water were removed from the ice
box and incubated in a bacteriological incubator for 24 h at 37ºC, which was the
standard duration and the temperature used in all procedures performed in this study. In the
following step, aliquots of 1 mL of the initial samples were transferred to tubes containing
Brain Heart Infusion broth (Himedia, Mumbai, India), and then placed again in the incubator.
In the third step, the colonies were removed aseptically with a nickel-chrome handle and
plated on MacConkey agar (Himedia, Mumbai, India), and posteriorly incubated.
For each sample, one to three bacterial colonies were selected based on morphology and
pigmentation of the colonies and submitted to the following biochemical tests for bacterial
identification: investigation of classic fermentation characteristics in TSI agar (Triple– Sugar-
Iron), decarboxylation reactions in the LIA (Lysine Iron agar), production of sulfide, indole
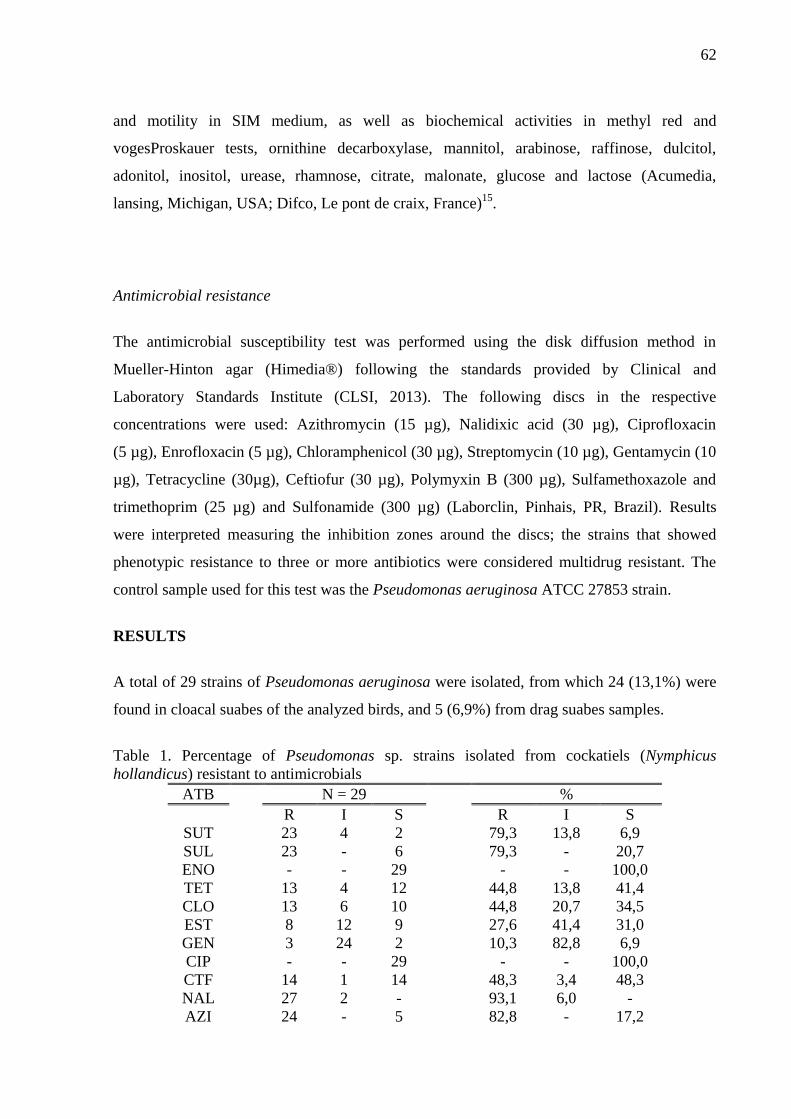
62
and motility in SIM medium, as well as biochemical activities in methyl red and
vogesProskauer tests, ornithine decarboxylase, mannitol, arabinose, raffinose, dulcitol,
adonitol, inositol, urease, rhamnose, citrate, malonate, glucose and lactose (Acumedia,
lansing, Michigan, USA; Difco, Le pont de craix, France)15
.
Antimicrobial resistance
The antimicrobial susceptibility test was performed using the disk diffusion method in
Mueller-Hinton agar (Himedia®) following the standards provided by Clinical and
Laboratory Standards Institute (CLSI, 2013). The following discs in the respective
concentrations were used: Azithromycin (15 µg), Nalidixic acid (30 µg), Ciprofloxacin
(5 µg), Enrofloxacin (5 µg), Chloramphenicol (30 µg), Streptomycin (10 µg), Gentamycin (10
µg), Tetracycline (30µg), Ceftiofur (30 µg), Polymyxin B (300 µg), Sulfamethoxazole and
trimethoprim (25 µg) and Sulfonamide (300 µg) (Laborclin, Pinhais, PR, Brazil). Results
were interpreted measuring the inhibition zones around the discs; the strains that showed
phenotypic resistance to three or more antibiotics were considered multidrug resistant. The
control sample used for this test was the Pseudomonas aeruginosa ATCC 27853 strain.
RESULTS
A total of 29 strains of Pseudomonas aeruginosa were isolated, from which 24 (13,1%) were
found in cloacal suabes of the analyzed birds, and 5 (6,9%) from drag suabes samples.
Table 1. Percentage of Pseudomonas sp. strains isolated from cockatiels (Nymphicus
hollandicus) resistant to antimicrobials
ATB N = 29 %
R I S R I S
SUT 23 4 2 79,3 13,8 6,9
SUL 23 - 6 79,3 - 20,7
ENO - - 29 - - 100,0
TET 13 4 12 44,8 13,8 41,4
CLO 13 6 10 44,8 20,7 34,5
EST 8 12 9 27,6 41,4 31,0
GEN 3 24 2 10,3 82,8 6,9
CIP - - 29 - - 100,0
CTF 14 1 14 48,3 3,4 48,3
NAL 27 2 - 93,1 6,0 -
AZI 24 - 5 82,8 - 17,2

63
POL 2 - 27 6,9 - 93,1 ATB = antimicrobial; SUT = Sulfazotrim (Sulfamethoxazole+trimethoprim; SUL = sulfonamides;
ENO = enrofloxacin; TET = tetracycline; CLO = chloramphenicol; EST = streptomycin; GEN =
gentamycin; CIP = ciprofloxacin; CTF = cetiofur; NAL = nalidixic acid; AZI = azithromycin; POL
= polimyxin B.
Table 1 shows the variable resistance levels exhibited by the strains to 10 of the 12
tested antibiotics, where only enrofloxacin and ciprofloxacin were fully effective against all of
the tested strains. The antibiotic to which strains presented the most frequent resistance was
Nalidixic acid (93,1%), followed by azithromycin (82,8%). All the 29 (100,0%) strains tested
presented resistance to at least one antibiotic. Furthermore, 21 isolates showed a multidrug
resistance profile, corresponding 72.4 % of the total. The maximum number of antibiotics to
which a strain presented resistance was 7 (7 strains), followed by 6 (8 strains) (Table 2).
Table 2. Multidrug-resistant (MDR) Pseudomonas sp. strains isolated from cockatiels
(Nymphicus hollandicus)
Nº of antimicrobials Nº of resistant strains (%)
At least 1 29 (100)
>1 29 (100)
>2 28 (96,6)
>3 21 (72,4)
>4 13 (44,8)
>5 8 (27,6)
>6 7 (24,1)
>7 0 (0)
DISCUSSION
The analysis of the samples allowed the identification of 29 strains of P. aeruginosa.
24 (21,8%) were found in cloacal suabes of the analyzed birds. A study from 2015 in nestling
red-tailed amazon indicated a prevalence of Pseudomonas spp. in 31.57% of the samples16
.
The authors regarded the finding as common since these bacteria are widely distributed in the
world and are frequently found in cultures with samples of coana and cloaca17
. However,
several studies in broiler chicks and egg layers demonstrated the presence of systemic
symptoms such as congested liver, heart, lungs and kidneys, enlargement of the yolk sac,
embryonic mortality in hatcheries, septicemic infections in newly hatched chickens.
Moreover, the pathogenicity of P. aeruginosa was associated with localized and systemic
hemorragyc lesions both in young and growing birds18,19
.

64
5 (6,9%) strains were obtained from drag swabs samples. this P. aeruginosa causes
diseases in plants and animals. Thus, P. aeruginosa is known to be highly adaptable and to
have diverse phenotypic characteristics, which enable it to subsist on a variety of different
carbon sources for energy, is able to utilize nitrogen as a terminal electron acceptor to respire
under anaerobic conditions, has minimal nutrient requirements, and grows at temperatures of
up to 42°C20,21
. Also, this pathogen naturally forms polysaccharide-encased, surface attached
communities known as biofilms, and resists protozoan predation by directly injecting
cytotoxic effector proteins into the cytosol of eukaryotic cells through a type III secretion
system20,21,22
. These mechanisms act on opportunistic infections in humans as well. Due to its
metabolic diversity, P. aeruginosa can even multiply in certain disinfectants and metabolize
many antibiotics19,24
. While antimicrobials were originally developed and used to kill bacteria,
recent work reveals that the biological functions of antibiotics are not limited to bactericidal
(killing) or bacteriostatic (growth inhibition) effects. Pseudomonas aeruginosa possesses
multifactorial mechanisms of responses and resistance to antimicrobials, such as altered target
sites, bacterial efflux pumps, enzyme production or inhibition and loss of membrane
protein24,25,26.
The higher antibiotic resistance was to Nalidixic acid (93,1%). This result is
corroborated by a study performed in different species of psittacines in 2015, where the
authors observed that the Pseudomonas strains analyzed were generally resistant to nalidixic
acid with a 94,12% prevalence. All 29 strains were susceptible to enrofloxacyn, although the
literature relates that enrofloxacyn resistance is most frequently reported for Pseudomonas
aeruginosa27.
Similarly, all strains were susceptible to Ciprofloxacin. A prevalence study
done in clinical specimens collected from different hospitals of Karachi, Pakistan28
, revealed
75% sensitivity against Ciprofloxacin and Nalidixic acid. Other studies performed in human
samples, however, demonstrated a varied percentage of resistance to Ciprofloxacin, and
Nalidixic acid ranging from 87.8% to 100%29,30,31.
Azithromycin had a resistance prevalence of 82,8%. This might be explained by the
P. aeruginosa cells being intrinsically resistant to macrolides when tested in standard broth
culture32
. However, macrolides such as azithromycin are still widely used antibiotics, since it
causes a reduced P. aeruginosa virulence, delayed bactericidal effects, reduced bacterial
adherence to the respiratory epithelium, decreased bacterial motility, and impaired biofilm
production32,33,34
.

65
21 isolates showed a multidrug resistance profile, corresponding to 72.4 % of the
total. The maximum number of antibiotics to which a strain presented resistance was 7 (7
strains), followed by 6 (8 strains). Multidrug-resistant (MDR) P. aeruginosa phenotype is
defined as resistant to one anti-microbial agent in three or more anti-pseudomonal anti-
microbial classes (carbapenems, fluoroquinolones, penicillins /cephalosporins and
aminoglycosides)35.
A report describing the clinical and laboratory findings of kori bustards
(Ardeotis kori) infected with Pseudomonas aeruginosa showed an antibacterial resistance
pattern to 6 antibiotics. Similarly, in a study performed in different species of psittacines in
2015, the authors observed that the Pseudomonas strains analyzed had an MDR pattern to 5
antibiotics24
.
In the light of the considerable increase in the prevalence of multidrug resistance in
P. aeruginosa, which complicates decisions on treatment with antibiotics and its effects on
high morbidity and mortality in humans36
, some authors state that the volume of antibiotics
used and further released directly or through the microbiota into municipal effluents and
sewage creates a selective pressure that results, for the environment, in a "reservoir Natural
"strains of multidrug-resistant strains37
. There has also been observed that resistance of
Pseudomonas aeruginosa species towards the antibiotic action has a genetic correspondent,
although there is no full correspondence between the “in vitro” sensitivity of Pseudomonas
aeruginosa and the one “in vivo”25.
CONCLUSION
The results indicate that common pet birds such as cockatiels can harbor strains of
Pseudomonas sp., and a high resistance rate to antimicrobials was detected. Due to the health
importance of these bacteria, the emergence of multidrug resistant strains is concerning, and
greater prevention and control efforts must be imposed to avoid the presence of this pathogen
in the environment where these birds are held.
CONFLICT OF INTEREST
The authors declare no conflicts of interest in this study.
REFERENCES

66
1. Schurek, K. N., Breidenstein, E. B. M., and Hancock, R. E. W. “Pseudomonas aeruginosa:
a persistent pathogen in cystic fibrosis and hospital-associated infections,” in Antibiotic
Discovery and Development, Vol. 1, eds T. J. Dougherty and M. J. Pucci (New York:
Springer), 2012;679–715.
2. Kung, Vanderlene L., Egon A. Ozer, and Alan R. Hauser. "The accessory genome of
Pseudomonas aeruginosa." Microbiology and Molecular Biology Reviews 74.4 2010: 621-
641.
3. Poole, K. Pseudomonas aeruginosa: resistance to the max. Front. Microbiol. 2011;2:65.
doi:10.3389/fmicb.2011.00065
4 R. S. Baltimore, “Pseudomonas,” in Nelson Textbook of Pediatrics, pp. 862–864, 2000.
5. Molina-cabrillana, J. et al. E. Incidence of nosocomial infections at a neonatal intensive
care unit: a six-year surveillance study. Enfermedades Infecciosas y Microbiología Clínica,
v. 24, n. 5, p. 307-312, 2006.
6. Peterson, D. L. The epidemiological profile of infections with multidrug-resistant
Pseudomonas aeruginosa and Acinetobacter species-Review. Clinical Infectious Diseases,
suppl. 2, p. S43-48, 2006.
7. Brittingham, M. C., Temple, S. A. & Duncan, R. M. A survey of the prevalence of selected
bacteria in wild birds. Journal of Wildlife Diseases 24, 299–307, 2008.
8. Gerlach, H. Bacteria. In B.W. Ritchie, G.J. Harrison & L.R.Harrison (Eds.) Avian
medicine: principles and application. Lake Worth, FL: Wingers Publishing Inc. 1994; pp.
950–983.
9. Walker, S. E., Sander, J. E., Cline, J. L. & Helton, J. S. Characterization of Pseudomonas
aeruginosa isolates associated with mortality in broiler chicks. Avian Diseases 46, 1045–
1050, 2002.
10. BAILEY, T. A., SILVANOSE, C. D., NALDO, J. N. & HOWLETT, J. H. Pseudomonas
aeruginosa infections in kori bustards (Ardeotis kori). Avian Pathology 29, 41–44, 2000.

67
11. Momotani, E., Kiryu, M., Ohshiro, M., Murakami, Y., Ashida, Y., Watanabe, S. &
Matsubara, Y. Granulomatous lesions caused by Pseudomonas aeruginosa in the Ostrich
(Struthio camelus). Journal of Comparative Pathology 112, 273–28; 1995.
12. Benskin, C. M. H.; Wilson, K.; Jones, K.; Hartley, I. R. Bacterial pathogens in wild birds:
a review of the frequency and effects of infection. Biological Reviews, v.89, p. 349-373,
2009. doi: http://dx.doi.org/10.1111/j.1469-185X.2008.00076.x.
13. M. Toda, S. Okubo, R. Hiyoshi, and T. Shimamura, “The bactericidal activity of tea and
coffee,” Letters in Applied Microbiology, vol. 8, no. 4, pp. 123–125, 1989.
14. Crespo, M. P. La lectura interpretativa del antibiograma: Uma herramienta para predecir
la resistencia bacteriana en el laboratorio de microbiología de rutina. Colombia Médica, v.
33, n. 4, p. 179-193, 2002.
14. Koneman E, Winn WJR, Allen S, Janda W, Procop G, Schreckenberger P, Woods, G.
Diagnóstico microbiológico: texto e atlas colorido, 6 ed, Ed. Guanabara Koogan, Rio de
Janeiro. 2010.
15. Serafini, P. P.; Meurer, R.; Biesdorf, S. M.; Sipinski, E. A. B. O uso da microbiologia
como ferramenta para a conservação de aves ameaçadas: dados preliminares para o papagaio-
de-cara-roxa, Amazona brasiliensis (Aves: Psittacidae) no Paraná. Arq. Ciênc. Vet. Zool.
UNIPAR, Umuarama, v. 18, n. 1, p. 65-69, jan./mar. 2015.
15. Pier, G. B., and R. Ramphal. Pseudomonas aeruginosa, p. 2587–2615. In G. Mandell, J.
Bennett, and R. Dolin (ed.), Principles and practice of infectious diseases. Elsevier
Churchill Livingstone, Philadelphia, PA. 2005.
16. Qeuiroz, B. D. Principais doenças respiratórias que acometem psitacídeos e passeriformes
criados como animais de estimação. Rio de Janeiro, 2008.78 f. Monografa (Trabalho de
Conclusão de Curso) - Universidade Castelo Branco, São Paulo, 2008.
17. Dinev I, S Denev and G Beev. Clinical and morphological studies on spontaneous cases of
Pseudomonas aeruginosa infections in birds. Pak Vet J, 33(3): 398-400. 2013.
18. Satish and Priti, 2015. International J Ext Res. 6:37-39. 2015.

68
19. Abd, H., B. Wretlind, A. Saeed, E. Idsund, K. Hultenby, and G. Sandstrom. Pseudomonas
aeruginosa utilises its type III secretion system to kill the free-living amoeba Acanthamoeba
castellanii. J. Eukaryot. Microbiol. 55:235–243, 2008.
20. Tashiro, Yosuke, et al. "Interspecies interaction between Pseudomonas aeruginosa and
other microorganisms." Microbes and Environments 28.1 2013; 13-24.
21. Matz, C., A. M. Moreno, M. Alhede, M. Manefield, A. R. Hauser, M. Givskov, and S.
Kjelleberg. 2008. Pseudomonas aeruginosa uses type III secretion system to kill biofilm-
associated amoebae. ISME J. 2:843–852, 2008.
22. Bodey, G. P., R. Bolivar, V. Fainstein, and L. Jadeja. Infections caused by Pseudomonas
aeruginosa. Rev. Infect. Dis. 5:279–313, 1983.
23. Dantas, G., M. O. Sommer, R. D. Oluwasegun, and G. M. Church. Bacteria subsisting on
antibiotics. Science 320:100–103, 2008.
24. Linares, J. F., Gustafsson, I., Baquero, F., and Martinez, J. L. Antibiotics as intermicrobial
signaling agents instead of weapons. Proc. Natl. Acad. Sci. U.S.A. 2006;103, 19484–19489.
doi: 10.1073/pnas.0608949103
24. Oprea, Anca, and Maria Crivineanu. "The Sensitivity of Psittacine Isolated Pseudomonas
Strains towards Antibiotics." Bulletin of University of Agricultural Sciences and Veterinary
Medicine Cluj-Napoca. Veterinary Medicine 66.1 2009.
25. Aminov, R. I. Biotic acts of antibiotics. Front. Microbiol. 2013;4:241.
doi:10.3389/fmicb.2013.00241
26. Elizabeth BH, Vincent HT. Impact of multi-drug resistant Pseudomonas aeruginosa
infection on patient outcomes. Expert Rev Pharmacoeconomics Outcomes Res.
2010;10(4):441–451.
27 Flammer, Keven, and E. U. A. Carolina do Norte. "USO DA ENROFLOXACINA
(BAYTRIL®) EM PSITACÍDEOS." 1999.

69
28 Khan F, Khan A, Kazmi SU. Prevalence and susceptibility pattern of multi drug resistant
clinical isolates of Pseudomonas aeruginosa in Karachi. Pak J Med Sci 2014;30(5):951-954.
doi: http://dx.doi.org/10.12669/pjms.305.5400
29. Gill MM, Usman J, Kaleem F, Hassan A, Khalid A, Anjum R, Fahim Q. Frequency and
Antibiogram of Multi-drug Resistant Pseudomonas aeruginosa. J Coll Physicians Surg Pak.
2011;21(9):531-534. doi: 09.2011/JCPSP.531534
30. Tam VH, Chang KT, Abdelraouf K, Brioso CG, Ameka M, McCaskey LA, et al.
Prevalence, mechanism and susceptibility of multidrug resistant bloodstream isolates of
Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2010;54:1160-1164
31. Abdallah IA, Abdulla ME. Antibiotic resistance in Pseudomonas aeruginosa isolated
from various clinical specimens in Ibn e Sina Hospital-Sirte-Libya. Bull Alex Fac Med.
2009;45(3):771-775
32. Morita, Yuji, Junko Tomida, and Yoshiaki Kawamura. "Responses of Pseudomonas
aeruginosa to antimicrobials." Low-dose antibiotics: current status and outlook for the future.
2014;30.
33. Shinkai M, Rubin BK. Macrolides and airway inflammation in children. Paediatr Respir
Rev. 2005;6(3):227-35. http://dx.doi.org/10.1016/j.prrv.2005.06.005 PMid:16153572
34. McArdle JR, Talwalkar JS. Macrolides in cystic fibrosis. Clin Chest Med.
2007;28(2):347-60. http://dx.doi.org/10.1016/j.ccm.2007.02.005 PMid:17467553.
35 Bailey, T. A., et al. "Pseudomonas aeruginosa infections in kori bustards (Ardeotis kori)."
Avian Pathology 29.1 2000; 41-44.
36. Ansari, Ayesha, S. M. Salman, and Shadma Yaqoob. Antibiotic Resistance Pattern in
Pseudomonas aeruginosa Strains Isolated at Era’s Lucknow Medical College and Hospital,
Lucknow, India. 2015.
37 Mahmoud, Ahmed Bakr, et al. "Prevalence of multidrug-resistant Pseudomonas
aeruginosa in patients with nosocomial infections at a University Hospital in Egypt, with
special reference to typing methods." J Virol Microbiol 13, 2013.

70
8 CONCLUSÃO
Os resultados indicam que as calopsitas criadas em ambiente doméstico e comercial
podem abrigar cepas de bactérias Gram-negativas prejudiciais para a saúde animal e humana e
animais, tais como Pseudomonas sp. e Escherichia coli.
Adicionalmente, foi possível evidenciar que as cepas isoladas apresentam alto índice
de resistência, apresentando cepas resistentes a múltiplas drogas, o que denota um possível
risco
para a saúde de proprietários e demais pessoas envolvidas na criação destas aves.
Além disso, o estudo dos fatores de virulência de E. coli revelou a presença de cepas
patogênicas de cepas contendo genes relacionados a EPEC. A investigação destas questões é
importante para avaliar a patogenicidade, o potencial zoonótico e os riscos envolvidos tanto
para a saúde animal quanto para a saúde pública.

71
REFERÊNCIAS BIBLIOGRÁFICAS
AKHTER, J.M.; HOSSAIN.; ISLAM, M.T. et al. Isolation and identification of microflora
from apparently healthy caged parrots of Dhaka Zoo of Bangladesh. Bangl. J. Vet. Med., v.8,
p.5-10, 2010.
AUSTRALIAN MUSEUM. In BIRDS IN BACKYARDS. Cockatiel (Nymphicus
hollandicus). 2006. Disponível em: <http://www.birdsinbackyards.net/bird/49>. Acesso em:
27 julho de 2015.
AWADALLAH, M.A.; MERWAD, A.M.; MOHAMED, R.E. Prevalence of zoonotic
Escherichia coli and Salmonella in wild birds and humans in Egypt with emphasis on
RAPDPCR fingerprinting of E. coli. Global Veterinaria, v. 11, n. 6, p. 781-788, 2013.
BARCELOS, A. S. Avaliação macroscópica, histopatológica e bacteriológica
de fígados de frangos (Gallus gallus) condenados no abate pela inspeção
sanitária. 2005. 83 f. Dissertação de Mestrado - Universidade Federal de Santa
Maria, Santa Maria.
BARNES, H. J. Other bacterial diseases. In: SAIF, Y. M.; BARNES, H. J.; GLISSON, J. R.;
FADLY, A. M.; McDOUGALD, L. R.; SWAYNE, D. E. (Org.) Diseases of Poultry. Ames:
Iowa State Press, 2003. p. 797-862.
BARNES, H. J.; VAILLANCOURT, J. P.; GROSS, W. B. Colibacillosis In:
SAIF W. M. Diseases of poultry. (11ª ed.). Iowa, p. 138-144, 2003
BAÚ, A. C.; CARVALHAL, J. B.; ALEIXO, J. A. G. Prevalência de Salmonella em produtos
de frango e ovos de galinha comercializados em Pelotas, RS, Brasil. Ciência Rural, Santa
Catarina, RS, v. 31, n.2, p.303-307, 2001.
BENEZ, S.M. Aves: Criação, Clínica, Teoria, Prática. São Paulo – SP: Robe Editorial, 522 p,
2001.
BERCHIERI JUNIOR, A.; MACARI, M. Doenças das aves. Campinas:
FACTA, p.455-469. 2009

72
BIRDLIFE INTERNATIONAL. 2012. Nymphicus hollandicus. In: IUCN Red List of
Threatened Species. Versão 2013.1. Disponível em:<http://www.iucnredlist.org.>. Acesso
em 30 de julho de 2015.
BROOKS, G.F.; BUTEL, J.S.; ORNSTON, L.N. et al. Microbiologia Médica de Jawetz,
Melnick & Adelberg-26. AMGH Editora, 2014.
CAMPOS, L.C.; TRABULSI, L.R. Escherichia. In.: TRABULSI, L.R. et al.
Microbiologia. 3 ed. São Paulo: Atheneu, 2002, p.215-228.
CATÃO-DIAS, J. L.; CARVALHO, V. M. Tuberculose. In: CUBAS, Z. S.; SILVA, J. C. R.;
CATÃO-DIAS, J. L. Tratado de animais selvagens. Medicina Veterinária. 1. ed. São Paulo:
Roca, 2007. cap. 43, p. 726-735
CHRISTIDIS, L., BOLES, W.E. Systematics and Taxonomy of Australian Birds.
CSIRO Publishing, Australia, 2008.
COLLAR, N.J. Family Psittacidae (Parrots). In: DEL HOYO, J.; ELLIOTT, A.; SARGATAL,
J. Handbook of the Birds of the World, v.4, Barcelona: Lynx Edicions, 1997, p. 280- 477.
CÔRREA, F.A.F. Características dos patótipos de E. coli e implicações de
E. coli patogênica para aves em achados de abatedouros frigoríficos.
2012. 37 f. Seminário (Mestrado em Ciência Animal). Escola de Veterinária e
Zootecnia. Universidade Federal de Goiás, Goiânia.
CROXEN, M. A. & FINLAY, B. B. Molecular mechanisms of Escherichia coli
pathogenicity. Nature, 8: 20-38, 2010.
DE BUYSER M.L.; DUFOUR, B.; MAIRE M et al. Implication of milk and milk products in
food-borne diseases in France and in different industrialised countries. International Journal
of Food Microbiology, v. 67, n. 1, p. 1-17, 2001.
DELICATO, E.R.; BRITO, B.G.; GAZIRI, L.C.J.; VIDOTTO, M.C. Virulence-associated
genes in Escherichia coli isolates from poultry with colibacillosis. Veterinary Microbiology,
v.94; p.97-103, 2003.
DILIELLO, Leo R. et al. Methods in food and dairy microbiology. AVI Publishing Co. Inc.,
1982.
DONNENBERG, M. S.; KAPER, J. B. Enteropathogenic Escherichia coli. Infection and
immunity, v. 60, n. 10, p. 3953, 1992.
EVANCHO, G. M.; SVEUM, W. H.; MOBERG, L. J et al. Compendium methods for the
microbiological examination of foods. Compendium of methods for the microbiological
examination of foods, 2001.
EVANS, E.E.; MITCHEKK, M.A.; WHITTINGTON, J.K.; TULLY JR.; T.N. Measuring the
level of agreement between clocal Gram’s stains and bacterial cultures in Hispaniolan
Amazon Parrots (Amazona ventralis). Journal of Avian Medicine and Surgery, v. 28, p.
290-296, 2014.

73
EWERS, C.; LIA, G.; WILKING, H. et al. Avian pathogenic, uropathogenic, and newborn
meningitis-causing Escherichia coli: How closely related are they? International Journal of
Medical Microbiology, v.297, p. 163-176, 2007.
FERREIRA, A.J.P.; KNÖBL. Colibacilose aviária. In: BERCHIERI, A.J.; MACARI, M.
Doença das aves. Campinas: FACTA, 2000, p. 197-2005.
FORSHAW, Joseph M. Parrots of the World. Princeton University Press, 2010.
FREITAS, A.N.; LUTZ, L.; MACHADO, A.; ZAVASZKI, A.P.; BARTH, A.L. Antibiograma:
metodologia e interpretação clínica. In: BARROS, E.; MACHADO, A.; SPRINZ, E.
Antimicrobianos. Porto Alegre: Artmed, 2013, p. 93- 101.
FUDGE, A. M. Diagnosis and treatment of avian bacterial diseases. Seminars in Avian and
Exotic Pet Medicine, v.10, ed.1, p.3-11, 2001.
GERLACH H. Bacteria. In: RITCHIE, B.W.; HARRINSON, G.J.; HARRINSON, L.R. Avian
Medicine: Principles and application. Florida: Wingers Publishing, 1994, p. 949-983.
GIBBS, P. S.; KASA, R.; NEWBREY, J. L. et al. Identification, Antimicrobial Resistance
Profiles, and Virulence of Members from the Family Enterobacteriaceae from the Feces of
Yellow-Headed Blackbirds (Xanthocephalus xanthocephalus) in North Dakota. Avian
Diseases, v.51, n.3, p.649-655, 2007.
GYLES, C. L.; FAIRBROTHER, J. M. Escherichia coli. In: GYLES, C. L.; PRESCOTT, J. F.;
SONGER, J. G.; THOEN, C. O. (Org.) Pathogenesis of bacterial infections in animals.
Ames: Blackwell Publishing, 2010. p. 267-308.
GODOY, S. N.; CUBAS, Z. S. Principais doenças bacterianas e fúngicas em Psittaciformes–
revisão. Clínica Veterinária, v. 14, n. 81, p. 88-98, 2009.
GODOY, S.N. Psittaciformes (arara, papagaio, periquito). In: CUBAS, Z.S.; SILVA, J.C.R.;
CATÃO-DIAS, J.L. Tratado de animais selvagens. São Paulo: ROCCA Ltda, 2007, p. 222-
251.
GORDON, D.M.; COWLING, A. The distribution and genetic structure of Escherichia coli in
Australian vertebrates: host and geographic effects. Microbiology, v. 149, p. 3575-3586,
2003.
GORMAN, M. The cockatiel handbook. Barron's Educational Series, 2010.
GUTIÉRREZ, L.M. et al. Incidence of Staphylococci in ovine mastitic milk and antibiotic
susceptibility of the strains. Milchwissenschaft, v.45, p.778-781, 1990.
HARCOURT-BROWN, N.H. Aves Psitaciformes. In: TULLY T.; DORRESTEIN, G. M.;
JONES A. K. Clínica de aves. 323p. Rio de Janeiro, Elsevier, 2010. p. 122 – 149.
HARPER, E.J.; LOWE, B. Hematology values in a colony of budgerigars (Melopsittacus
undulatus) and changes associated with aging. The Journal of Nutrition, v.128, n.12,
p.2639-2640, 1998.

74
HIDASI, H.W.; NETO, J.H.; MORAES, D.M.C.; LINHARES, G.F.C.; JAYME, V.S.
ANDRADE, M.A. Enterobacterial detection and Escherichia coli antimicrobial resistant in
parrots seized from the illegal wildlife trade. Journal of Zoo and Wildlife Medicine, v.44, p.
1-7, 2013.
HIDRON, A. I.; EDWARDS, J. R.; PATEL, J. et al. Antimicrobial-resistant pathogens
associated with healthcare-associated infections: annual summary of data reported to the
National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2006–
2007. Infection Control & Hospital Epidemiology, v. 29, n. 11, p. 996-1011, 2008.
HIRSH, D.C. Família Enterobacteriaceae. In: HIRSH, D.W.; ZEE, Y.C. Microbiologia
Veterinária. Rio de Janeiro: Guanabara Koogan, 2009, p. 59-76.
BERGEY, D.H.; HOLT, J. Bergey’s Manual of Determinative Bacteriology. 9 ed. Ed.:
Baltimore: William & Wilkis, 1994, 787p.
FLAMMER, K.; E. U. A. CAROLINA DO NORTE. "USO DA ENROFLOXACINA
(BAYTRIL®) EM PSITACÍDEOS." 1999.
IBAMA. Manual do Sistema - Serviços On-Line – IBAMA Portaria IBAMA Nº 93, de 7 de
julho de 1998. Disponível em <https://servicos.ibama.gov.br/ctf/manual/html/0 42200.htm>.
Acesso em 14 de maio de 2015.
IBOPE. Oito em cada dez internautas possuem animal de estimação no Brasil. Disponível
em <http://www.ibope.com.br/pt-br/noticias/Paginas/Oito-em-cada-dez-internautas-possuem-
animal-de-estimacao-no-Brasil.aspx>. Acesso em 26 de novembro de 2016.
IKUNO A.A., GAMA N.M.S.Q, GUASTALLI E.A.L., GUIMARÃES M.B. e FERREIRA
V.C.A. Características de isolados de Escherichia coli provenientes de aves silvestres quanto
a genes de virulência e resistência a antibióticos. Anais 35º Combravet, Gramado, RS.
(Resumo). 2008.
IUCN. Red List of Threatened Species. Version 2015-3. Disponível em
<www.iucnredlist.org>. Acesso em 30 de julho de 2015.
JANDA, J.M. e ABBOT, S.L. The Family Enterobacteriaceae. In: GOLDMAN, E.; GREEN,
L.M. Pratical Handbook of Microbiology. 2ed. Ed.: ACRC title, 2008, p. 217-219.
JOHNSON, J.R. Virulence factors in Escherichia coli urinary tract infection. Clin Microbiol
Rev, v.4, n.1, p.80-128, 1991.
KONEMAN, E.; WINN, W.JR.; ALLEN, S.; et al. As Enterobacteriaceae. In: ______.
Diagnóstico microbiológico: texto e atlas colorido. 6 ed. Rio de Janeiro.: Guanabara
Koogan, 2010, p.208-299.
LEVINE, M. M. Escherichia coli that cause diarrhea: enterotoxigenic, enteropathogenic,
enteroinvasive, enterohemorrhagic, and enteroadherent. Journal of Infectious Diseases, v.
155, n. 3, p. 377-389, 1987.

75
LOPES, E.S.; MACIEL, W.C.; ALBUQUERQUE, A.H.; MACHADO, D.N.; BEZERRA,
W.G.A.; VASCONCELOS, R.H.; LIMA, B.P.; MARIETTO-GONÇALVES, G.A.;
TEIXEIRA, R.S.C. Prevalence and Antimicrobial resistance profile of Enterobacteria isolated
from psittaciformes of illegal wildlife trade. Acta Scientiae Veterinariae, v. 43, n. 1313, p. 1-
9, 2015.
LUMEIJ, J. T. Nephrology. In: RITCHIE, B. W.; HARRISON, G. J.; HARRISON, L. R. (Org.)
Avian medicine: principles and application. Lake Worth: Wingers Publishing, 1994. p.
538-555.
LYCZAK, J. B.; CANNON, C. L.; PIER, G. B. Establishment of Pseudomonas aeruginosa
infection: lessons from a versatile opportunist. Microbes and Infection, v. 2, n. 9, p. 1051-
1060, 2000.
MACHADO, A. A. C. Prospecção sorológica de Salmonella Pullorum em pombos (Columbia
livia) capturados em granjas avícolas de Fortaleza. 2000. 45f. Dissertação (Mestrado) -
Programa de Pós-graduação em Ciências Veterinária, Universidade Estadual do Ceará,
Fortaleza, 2000.
MANOS, J.; BELAS, R. The genera Proteus, Providencia and Morganella. Prokaryotes, v.6,
p. 245-269, 2006.
MARIETTO-GONÇALVES, ALMEIDA, S.M.; LIMA, E.T.; ANDREATTI-FILHO, R.L.
Detecção de Escherichia coli e Salmonella spp. em microbiota intestinal de Psittaciformes em
fase de reabilitação para soltura. Brazilian Journal Research of Animal Science, v.47, n.3,
p.185-189, 2010.
MARTINEZ, M.B.; TRABULSI, L.R. Enterobacteriaceae. In: TRABULSI, L.R.;
ALTHERTHUM, F. Microbiologia. 5ed. São Paulo: Atheneu, 2008, p.267-279.
MINAGAWA, C. W. Estudo microbiológico fecal de linhagens de camundongos, de estirpe de
E. coli e do meio ambiente em biotérios. 2007. 108 f. Dissertação de Mestrado.
Universidade de São Paulo, São Paulo.
MORA, A.; LÓPEZ, C.; HERRERA, A.; VISO, S.; MAMANI, R.; DHABI, G.; ALONSO, M.
P.; BLANCO, M.; BLANCO, J. E.; BLANCO J. Emerging avian pathogenic Escherichia coli
strains belonging to clonal groups O111:H4-D-ST2085 and O111:H4-D-ST117 with high
virulence-gene content and zoonotic potential. Veterinary Microbiology, v.156, p.347-352,
2012.
MORA, A.; VISO, S.; LÓPEZ, C.; ALONSO, M. P.; GARCÍA-GARROTE, F.; DABHI, G.;
MAMANI, R.; HERRERA, A.; MARZOA, J.; BLANCO, M.; BLANCO, J. E.;
MOULINSCHOULEUR, M.; SCHOULEUR, C.; BLANCO, J. Poultry as reservoir for
extraintestinal pathogenic Escherichia coli O45:K1:H7-B2-ST95 in humans. Veterinary
Microbiolology, 2013. Disponível em <http://dx.doi.org/10.1016/j.vetmic.201 3.08.007>
MURRAY, P. R. Laboratory Procedures for Epidemiologic Analysis. Manual of Clinical
Microbiology, v. 6, 1995.

76
NIKAIDO, H. Prevention of drug access to bacterial targets: permeability barriers and active
efflux. Science-AAAS-Weekly Paper Edition-including Guide to Scientific Information, v.
264, n. 5157, p. 382-387, 1994.
O’HARA, C. M.; BRENNER, F. W.; MILLER, J. M. Classification, identification, and clinical
significance of Proteus, Providencia, and Morganella. Clinical Microbiology Reviews, v.13,
p. 534–546, 2000.
OKAMOTO, A. S.; ANDREATTI FILHO, R. L.; ROCHA, MILBRADT, E. L. Transference in
vitro of the resistance to the antimicrobials between Escherichia coli, Lactobacillus spp. and
Salmonella Enteritidis isolated from chickens. Arquivo Brasileiro de Medicina Veterinária
e Zootecnia, v.63, n.5, p.1149-1153, 2011.
OLIVEIRA, S. J. Guia bacteriológico prático: Microbiologia Veterinária. Canoas: ULBRA,
1995. 142p.
PALLERONI, N. J., KUNISAWA, R., CONTOPOULOU, R., & DOUDOROFF, M. Nucleic
acid homologies in the genus Pseudomonas. International Journal of Systematic and
Evolutionary Microbiology, v. 23, n. 4, p. 333-339, 1973.
PASMANS, F; BOYEN, F; HAESEBROUCK, F. Salmonella Infections in Exotic Pets. In:
BARROW, P. A.; Methner, U., Ed(s). Salmonella in Domestic Animals. CABI, 2ª Edição. p.
337 – 350. 2013.
PIGNATO, S.; GIAMMANCO, G. M.; GRIMONT, F. et al. Molecular Characterization of the
Genera Proteus, Morganella, and Providencia by Ribotyping. Journal of Clinical
Microbiology, v. 37, n. 9, p. 2840-2847, 1999.
POLLACK, Matthew. Pseudomonas aeruginosa. Principles and practice of infectious
diseases, v. 5, p. 1980-2003, 2000.
POLLACK, Matthew. The role of exotoxin A in Pseudomonas disease and immunity. Review
of Infectious Diseases, v. 5, n. Supplement 5, p. S979-S984, 1983.
PRASAD, D. N. Hygienic and safety aspects of dairy products. Indian Dairyman, v. 50, p.
112-118, 1998.
PRESCOTT, J.F. Quimioterapia Antimicrobiana. In: HIRSH, D.W.; ZEE, Y.C.
Microbiologia Veterinária. Rio de Janeiro: Guanabara Koogan, 2009, p. 27-46.
PRIOSTE, F.E.S.; CUNHA, M.P.V.; TEIXEIRA, R.H.F.; ZWARGG, T.; DI-CHIACCHIO, G.;
MELVILLE, P.A.; BENITES, N.R.; SINHORINI, J.; MATUSHIMA, E.R.; KNÖBL, T.
Genetic similarity between APEC and Escherichia coli strains isolated from Guaruba
guarouba in a survey on healthy captive psittacine birds. Brazilian Journal of Veterinary
and Research Animal Science, v.50, n.2, p.145-151, 2013.
QUINN, P.J; MARKEU, B.K.; CARTER, N.E.; DONNELY, W.J.; LEONARD, E.C.
Microbiologia veterinária e doenças infecciosas. Editora Artmed, Porto Alegre. 512p. 2005.

77
RIDELL, C. Lymphoid system. In: RIDELL, C. Avian histopathology. 1. ed.
Pennsylvania: American Association of Avian Pathologists, 1987. cap. 2, p. 7-17.
ROCHA, S.L.S. Detecção de fatores de virulência de amostras de
Escherichia coli isoladas de granjas avicolas do RS através do Multiplex PCR. Dissertação
de Mestrado. 2008. 68 f. Universidade do Rio Grande do Sul.
RODRIGUEZ-SIEK, K. E.; GIDDINGS, C. W.; DOETKOTT, C.; JOHNSON, T. J.; FAKHR,
M. K.; NOLAN, L. K. Comparison of Escherichia coli isolates implicated in human urinary
tract infection and avian colibacillosis. Microbiology, v.151, p.2097-2110, 2005.
RUPLEY, A. E. Manual de Clínica Aviária. São Paulo: Roca, 1999. p.301.
SAIDENBERG, A.B.; TEIXEIRA, R.H.F.; GUEDES, N.M.R.; ALLGAYER, M.C.;
MELVILLE, P.A.; BENITES, N.R. Molecular detection of enteropathogenic Escherichia coli
in asymptomatic captive psittacines. Pesquisa Veterinária Brasileira, v. 32, p. 922-926,
2012.
SÁNCHEZ-VARGAS, F.M.; ABU-EL-HAIJA, M.A.; GÓMEZ-DUARTE, O.G. Salmonella
infections: An update on epidemiology, management, and prevention. Travel Medicine and
Infectious Disease, v. 9, p. 263-277, 2011
SANTOS, A.C.M.; ZIDKO, A.C.M.; PIGNATARI, A.C.C.; GALES, A.C.; SILVA, R.M. A
virulência de Escherichia coli patogênica extra-intestinal (ExPEC) em relação à idade e ao
sexo do hospedeiro. O Mundo da Saúde, v.33, p.392-400, 2009.
SANTOS, T.; SILVA, N.; IGREJAS, G.; RODRIGUES, P.; MICAEL, J.; RODRIGUES, T.;
RESENDES, R.; GONÇALVES, A.; MARINHO, C.; GONÇALVES, D.; CUNHA, R.;
POETA, P. Dissemination of antibiotic resistant Enterococcus spp. and Escherichia coli from
wild birds of Azores Archipelago. Anaerobe, v. 24, p. 25-31, 2013.
SAVIOLLI, J.Y. Pesquisa e caracterização de Escherichia coli
patogênica (E. coli produtora de toxina Shiga – STEC; E. coli aviária
patogênica - APEC) de fragatas (Fregata magnificens) da Costa do Estado
de São Paulo. 2010. 84 f. Dissertação de Mestrado. Universidade de São Paulo.
SEGABINAZI, S.D. Presença de bactérias de família Enterobacteriaceae nas superfícies
externa e interna de Alphitobius diapenerius (panzer) oriundos de granjas avícolas dos
Estados do rio Grande do Sul e Santa Catarina. Santa Maria, 2004. Dissertação de Mestrado.
Universidade Federal de Santa Maria. 105 p.
SHARMA, M.; DOGRA, B.B.; MISRA, R. et al. Multidrug resistant Pantoea agglomerans in a
patient with septic arthritis-a rare report from India. Int J Microbiol Res 2012; 4: 263-265.
SHOBRAK, M.Y.; ABO-AMER, A.E. Role of wild birds as carriers of multi-drug resistant
Escherichia coli and Escherichia vulneris. Brazilian Journal of Microbiology, v.45, n.4, p.
1199-1209, 2014.
SICK, H. Ordem Psittaciformes. In: ______. Ornitologia brasileira. Rio de Janeiro: Nova
Frontera, 1997, p. 351-382.

78
SIGRIST, T. Guia de Campo Avis Brasilis – Avifauna Brasileira. São Paulo: Avis Brasilis,
2014, 608 p.
SILVA, R. M. Caracterização fenotípica e genotípica de Escherichia coli
proveniente de lesões de celulite de frangos de corte. 2011. 63 f. Dissertação
de Mestrado. Universidade Federal da Bahia.
SIMPSON, V. R. Post-mortem examination. In: Manual of psittacine birds. Cheltenham:
BSAVA, p. 69-86, 1996.
SMITH, S.; WANG, J.; FANNING, S.; MCMAHON, B.J. Antimicrobial resistant bacteria in
wild mammals and birds: a coincidence or cause for concern? Irish Veterinary Journal,
v.67, n.8, p. 1-3, 2014.
SOOMRO, A. H. et al. Isolation of Escherichia coli from raw milk and milk products in
relation to public health sold under market conditions at Tandojam. Pakistan Journal of
Nutrition, v. 1, n. 3, p. 151-152, 2002.
SPINOSA, H. S. Considerações Gerais sobre os Antimicrobianos. In: SPINOSA, H. S.;
GÓRNIAK, S. L.; BERNARDI, M. M. Farmacologia Aplicada à Medicina Veterinária,
ed. 4, 2006.
STEDT, J.; BONNEDAHL, J.; HERNANDEZ, J.; MCMAHON, B.J.; HASAN, B.; OLSEN,
B.; DROBNI, M.; WALDESNTRO, J. Antibiotic resistance patterns in Escherichia coli from
gulls in nine European countries. Infection Ecology and Epidemiology, v. 4, n. 215, p.1-10,
2014.
SYDOW, A. C. M. D.G. V. Avaliação da ocorrência de fatores de
virulência em estirpes de Escherichia coli em fezes de cães errantes. 2005. 89 f. Dissertação de Mestrado. Universidade de São Paulo.
TIZARD, I.A. Imunidade adquirida a bactérias e fungos. In: TIZARD, I.R. Imunologia
Veterinária: Uma Introdução. 8ed. São Paulo: ROCCA, 2009, p. 295-306
TORLONI, CEC. Criação de calopsitas. Guarulhos: LIS Gráfica e Editora LTDA, 1991.
TORTORA, G.J.; FUNKE, B.R.; CASE, C.L. Microbiologia. 10 ed. Porto Alegre: Artmed,
2012, 934p.
VALLANCE, B. A.; FINLAY, B. B. Exploitation of host cells by enteropathogenic
Escherichia coli. Proceedings of the National Academy of Sciences, v. 97, n. 16, p. 8799-
8806, 2000.
VERHOEF-VERHALLEN, E.J.J. The Complete Encyclopedia of cage e aviary birds. Rebo
Publishers, 2004, 312p.
WATSON, B.; HURLEY, M. Gran guía ilustrada de los loros. Tikal ediciones, 2003, 288p.