universidade de são paulo escola superior de agricultura ... · vitor antonio corrêa pavinato...
TRANSCRIPT

Universidade de São Paulo Escola Superior de Agricultura “Luiz de Queiroz”
Variabilidade e estrutura genética de populações de Alabama argillacea (Hüeb.) (Lepidoptera: Noctuidae) no Brasil: subsídios para o
manejo da resistência à toxina Cry1Ac em algodão geneticamente modificado
Vitor Antonio Corrêa Pavinato
Dissertação apresentada para a obtenção do título de Mestre em Ciências. Área de concentração: Entomologia
Piracicaba 2010

Vitor Antonio Corrêa Pavinato
Bacharel e Licenciado em Ciências Biológicas
Variabilidade e estrutura genética de populações de Alabama argillacea (Hüeb.) (Lepidoptera: Noctuidae) no Brasil: subsídios para o manejo da resistência à
toxina Cry1Ac em algodão geneticamente modificado
Orientador: Prof. Dr. CELSO OMOTO
Dissertação apresentada para a obtenção do título de Mestre em Ciências. Área de concentração: Entomologia
Piracicaba 2010

Dados Internacionais de Catalogação na Publicação
DIVISÃO DE BIBLIOTECA E DOCUMENTAÇÃO - ESALQ/USP
Pavinato, Vitor Antonio Corrêa Variabilidade e estrutura genética de populações de Alabama argillacea (Hüeb.) (Lepidoptera:
Noctuidae) no Brasil: subsídios para o manejo da resistência à toxina Cry1Ac em algodão geneticamente modificado / Vitor Antonio Corrêa Pavinato. - - Piracicaba, 2010.
121 p. : il.
Dissertação (Mestrado) - - Escola Superior de Agricultura “Luiz de Queiroz”, 2010. Bibliografia.
1. Algodão 2. Curuquerê - Genética - Resistência 3. Genética ecológica 4. Marcador molecular 5Organismos geneticamente modificados 6. Variação genética I. Título
CDD 633.51 P338v
“Permitida a cópia total ou parcial deste documento, desde que citada a fonte – O autor”

3
Aos meus pais José Alberto Pavinato
e Eliete Fátima Corrêa Pavinato
DEDICO
À minha avó Zoraide Sinicato Corrêa
OFEREÇO

4

5
AGRADECIMENTOS
Ao Prof. Dr. Celso Omoto pela orientação, pelos importantes ensinamentos, pela
motivação e pela oportunidade concedida.
À Dra. Maria Imaculada Zucchi pelo importante ajuda e orientação e
ensinamentos na área de genética de populações.
Ao Prof. Dr. José Baldin Pinheiro pela oportunidade, apoio e incentivo para a
realização de uma etapa desse trabalho no Laboratório de Diversidade Genética e
Melhoramento da ESALQ/USP.
Ao Dr. Samuel Martinelli pela construção da biblioteca enriquecida em SSR
Ao Biólogo e colega Miklos Maximiliano Bajay pela importante ajuda no desenho
dos primers e durante meu aprendizado das técnicas moleculares utilizadas no trabalho.
Ao Engenheiro Agrônomo Marcelo Mattos Cavallari e à Bióloga Aluana Abreu
pela ajuda com análise dos dados de genética de populações.
Ao Biólogo Felipe Antonio Domingues pela ajuda com o trabalho de linha – básica
de suscetibilidade, durante seu estágio no Lab. de Resistência de Artrópodes a
Pesticidas.
Aos professores do Programa de Pós-Graduação em Entomologia pelos
conhecimentos transmitidos.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pelo
financiamento do projeto de pesquisa.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela
concessão de bolsa de estudos.
À Monsanto do Brasil pelo suporte técnico na coleta de populações
Aos amigos Nádia F. Bertan Casarin, Eloisa Salmeron, Felipe Antonio
Domingues, Gislaine O. Campos, Danielle Thomazoni, Edgar Francisco Gaona Mena,
Oderlei Bernardi, Karina Cordeiro Albernaz, Oscar A. Batista Neto e Silva, Miklos
Maximiliano Bajay, Thiago Luiz da Mata, Carlos Eduardo de Araujo Batista, Giuliana
Etore do Valle, Fatima Bosetti, Glauber Campacci Pavan, Milene Möller pelo agradável
convívio durante os últimos anos.
Às bibliotecárias Eliana M. Garcia e Silvia M. Zinsly da ESALQ/USP pelo auxílio
na formatação das referências bibliográficas.

6

7
“Há um tempo em que é preciso abandonar as roupas usadas, que já tem a forma do
nosso corpo, e esquecer os nossos caminhos, que nos levam sempre aos mesmos
lugares. É tempo da travessia: e, se não ousarmos fazê-la, teremos ficado, para sempre,
à margem de nós mesmos.”
Fernando Pessoa

8

9
SUMÁRIO
RESUMO ....................................................................................................................... 11
ABSTRACT ................................................................................................................... 13
LISTA DE FIGURAS...................................................................................................... 15
LISTA DE TABELAS ..................................................................................................... 17
1 INTRODUÇÃO ........................................................................................................ 19
2 DESENVOLVIMENTO ............................................................................................ 23
2.1 Revisão bibliográfica ............................................................................................. 23
2.1.1 Características bioecológicas de Alabama argillacea (Hüebner) ........................ 23
2.1.2 Resistência de insetos a plantas geneticamente modificadas ............................ 25
2.1.3 Marcadores moleculares ..................................................................................... 29
2.1.4 Marcadores moleculares microssatélites e genética de populações .................. 32
2.2 Material e métodos ............................................................................................... 37
2.2.1 Coleta das populações ....................................................................................... 37
2.2.2 Criação em laboratório........................................................................................ 39
2.2.3 Caracterização da suscetibilidade de Alabama argillacea à toxina Cry1Ac. ....... 40
2.2.3.1 Linha-básica de suscetibilidade ...................................................................... 40
2.2.3.2 Determinação e validação da concentração diagnóstica para o monitoramento
da suscetibilidade .......................................................................................................... 43
2.2.4 Desenvolvimento e caracterização dos microssatélites ...................................... 43
2.2.4.1 Extração e quantificação de DNA ................................................................... 43
2.2.4.2 Construção de biblioteca genômica enriquecidas em microssatélites ............ 45
2.2.4.3 Seleção, seqüenciamento dos clones positivos e desenho de primers .......... 48
2.2.4.4 Amplificações iniciais e otimização dos primers ............................................. 49
2.2.4.5 Caracterização dos locos microssatélites ....................................................... 50
2.2.5 Estudo da variabilidade genética e estrutura populacional ............................. 50
2.2.5.1 Genotipagem dos locos microssatélites ......................................................... 50
2.2.5.2 Variabilidade genética intrapopulacional ........................................................ 51
2.2.5.3 Variabilidade interpopulacional e estrutura genética ...................................... 51
2.3 Resultados e discussão ........................................................................................ 53
2.3.1 Caracterização da suscetibilidade de Alabama argillacea à toxina Cry1Ac. ....... 53

10
2.3.1.1 Linha-básica de suscetibilidade ...................................................................... 53
2.3.1.2 Caracterização da curva de concentração–resposta de inibição de
crescimento... ................................................................................................................ 59
2.3.1.3 Determinação e validação da concentração diagnóstica para o monitoramento
da suscetibilidade .......................................................................................................... 65
2.3.2 Desenvolvimento e caracterização dos microssatélites ...................................... 70
2.3.2.1 Construção de biblioteca genômica enriquecidas em Microssatélites ............ 70
2.3.2.2 Seleção, seqüenciamento dos clones positivos e desenho de primers .......... 70
2.3.2.3 Amplificações iniciais e otimização dos primers ............................................. 76
2.3.2.4 Caracterização dos locos microssatélites ....................................................... 80
2.3.3 Estudo da variabilidade genética e estrutura populacional... ............................... 84
2.3.3.1 Variabilidade genética intrapopulacional ......................................................... 84
2.3.3.2 Variabilidade interpopulacional e estrutura genética populacional .................. 89
3 CONCLUSÕES ..................................................................................................... 109
REFERÊNCIAS.... ....................................................................................................... 111

11
RESUMO
Variabilidade e estrutura genética de populações de Alabama argillacea (Hüeb.) (Lepidoptera: Noctuidae) no Brasil: Subsídios para o manejo da resistência à
toxina Cry1Ac em algodão geneticamente modificado
Algodão geneticamente modificado que expressa a toxina Cry1Ac de Bacillus thuringiensis Berliner tem sido plantado no Brasil desde 2006. Entre as pragas-alvo da tecnologia, Alabama argillacea (Hüeb.) é uma espécie monófoga e apresenta alto potencial de risco de evolução da resistência. Para a implantação de um programa de manejo da resistência de A. argillacea à toxina Cry1Ac no Brasil, os principais objetivos do trabalho foram: a) estabelecer a linhas-básicas de suscetibilidade à toxina Cry1Ac em populações de A. argillacea e definir concentrações diagnósticas para o monitoramento da resistência e b) isolar e caracterizar locos microssatélites para avaliar a variabilidade e estruturação genética de populações de A. argillacea no Brasil. As linhas-básicas de suscetibilidade foram estimadas por meio de bioensaio de imersão de discos de folhas em soluções contendo a toxina Cry1Ac para populações de A. argillacea coletadas nos estados da Bahia, Goiás, Mato Grosso e Mato Grosso do Sul, durante as safras agrícolas de 2008 e 2009. Foram isolados e caracterizados dez locos microssatélites. Para avaliar a variabilidade genética foram estimadas as heterozigosidades observadas e esperadas. Para o estudo da estruturação genética foram estimadas as estatísticas F e feita a análise de agrupamento (distância de Nei) e análise Bayesiana. Baseado na estimativa da CL50 foram encontradas variações naturais de até seis vezes na suscetibilidade à toxina Cry1Ac entre as populações testadas. A partir da análise conjunta dos dados de concentração-mortalidade das populações testadas, foram definidas as concentrações diagnósticas de 10 e 32 µg de Cry1Ac/ml de água para futuros programas de monitoramento da resistência. O número médio de alelos por loco foi de 7,1 (variando de dois a 23 alelos). As heterozigosidades observada e esperada médias foram de 0,532 e 0,329. O índice de fixação intrapopulacional médio (f≈ FIS) foi de 0,268, com variação entre os locos de -0,008 a 0,736. O índice de fixação da espécie (FIS) estimado através da análise de variância foi de 0,244 (IC 95% de 0,093 a 0,418). O valor de FST estimado foi de 0,036 (IC 95% de 0,007 a 0,080). Esse valor de FST não diferiu significativamente de zero, indicando a ausência de estruturação genética. Contudo foi detectado certo grau de endogamia intrapopulacional. A estruturação espacial da variabilidade genética não foi detectada, pois as populações avaliadas apresentaram uma coesão que é mantida pela alta taxa de migração (6,7 migrantes por geração). Entretanto, foi identificada indícios de estruturação genética determinada pelo tempo, uma vez que tanto o agrupamento baseado em distâncias genéticas quanto à análise Bayesiana identificaram grupos que são formados por populações coletadas em safras agrícolas diferentes. As causas ligadas a essa mudança na variabilidade genética não puderam ser identificadas, entretanto pode se inferir que possivelmente causas naturais ou práticas de manejo estejam determinando eventos de gargalo genético. Devido ao intenso fluxo gênico entre populações de A. argillacea no Brasil, estratégias de manejo da resistência devem ser implantadas no âmbito nacional. Palavras-chave: Alabama argillacea; Algodão geneticamente modificado; Marcadores
microssatélites; Variabilidade genética; Estrutura genética

12

13
ABSTRACT
Variability and genetic structure of Alabama argillacea (Hüeb.) (Lepidoptera: Noctuidae) populations in Brazil: Basis for managing resistance to Cry1Ac toxin in
genetically modified cotton
Genetically modified cotton expressing Cry1Ac toxin of Bacillus thuringiensis Berliner has been planted in Brazil since 2006. Among target pests of this technology, Alabama argillacea (Hüeb.) is a monophagous species and offers a high potential risk of resistance evolution. In order to implement a resistance management program of A. argillacea to Cry1Ac toxin in Brazil, the objectives of this research were: a) to establish baseline susceptibility to Cry1Ac toxin in A. argillacea populations and define diagnostic concentrations for resistance monitoring and b) to isolate and characterize microsatellite loci to evaluate the variability and genetic structure of A. argillacea populations in Brazil. The baseline susceptibility data were estimated with leaf-disc bioassays by dipping into different concentration of Cry1Ac solution. Populations of A. argillacea were collected in Bahia, Goiás, Mato Grosso and Mato Grosso do Sul States, during 2008 and 2009 cotton-growing seasons. Ten microsatellite loci were isolated and characterized. The genetic variability was evaluated estimating observed and expected heterozygosities. For the studied of genetic structure, the F statistics was estimated, and Cluster analysis (Nei´s distance) and Bayesian analysis were performed. Based on estimation of LC50, natural variation up to 6-fold was detected in the susceptibility to Cry1Ac among tested populations. Based on analysis of concentration-mortality data by combining all populations, diagnostic concentrations of 10 and 32 µg of Cry1Ac/ml of water were defined for monitoring resistance. The mean number of alleles per loci was 7.1 (varying from 2 to 23 alleles). The observed and expected heterozigosities was 0,523 e 0, 395. The mean intrapopulation fixation index (f ≈ FIS) 0,268, varying from -0.008 to 0.736 between loci. The species fixation index (FIS) estimated by analysis of variance was 0.244(95% CI of 0.093 to 0.418). The estimated value of FST was 0.036 (95% CI of 0.007 to 0.080). The FST value was not significantly different from zero, indicating absence of genetic structure However, some degree of intrapopulational inbreeding was detected. Spatial structure of genetic variability was not detected because tested populations showed cohesion kept by high migration rate (6.7 migrants per generation). However, evidence of genetic structure across time was detected by Cluster analysis of genetic distance as well as by Bayesian analysis with group formation by population collection seasons. Factors affecting changes in genetic variability were not identified; however, natural factors or management practices may be determining some genetic bottleneck events. Due to intense gene flow among A. argillacea populations in Brazil, resistance management strategies must be implemented in a national basis. Keywords: Alabama argillacea; Genetically modified cotton; Microsatellite markers;
Genetic variability; Genetic structure

14

15
LISTA DE FIGURAS
Figura 1 – A) Preparo das placas para bioensaio de imersão de discos de folhas: células contendo 500 µL de solução de ágar a 2,5%; camada de papel filtro sobre a camada de ágar; discos de folhas que foram imersos em soluções de Cry1Ac (µg de I.A. / ml de água); B) transferência de lagartas de A. argillacea para cada célula após a secagem da solução contendo toxina Cry1Ac; e C) material utilizado para a realização do bioensaio de imersão de discos de folhas: a) pincel para a transferência das lagartas; b) filme plástico para selagem das placas ................................................................................... 42
Figura 2 – Caracterização da linha-básica de suscetibilidade das populações de Alabama argillacea coletadas nos estados da Bahia (BA-1, BA-3 e BA-4), Mato Grosso (MT-1), Mato Grosso do Sul (MS-1) e a suscetível de referência (LAB) à toxina Cry1Ac ........................................................................................................................................ 57
Figura 3 – Caracterização da resposta de inibição de crescimento para a população de A. argillacea do estado de Bahia (BA-3) à toxina Cry1Ac ............................................... 63
Figura 4 – Caracterização da resposta de inibição de crescimento para a população de A. argillacea do estado de Mato Grosso (MT-1) à toxina Cry1Ac .................................... 64
Figura 5 – Caracterização da linha-básica de suscetibilidade à toxina Cry1Ac mediante agrupamento de dados obtidos com as caracterizações de curvas de concentração-resposta de quatro populações de A. argillacea coletadas nas safras 2007/2008. ......... 68
Figura 6 – Freqüências de motivos microssatélites perfeitos, interrompidos, compostos e compostos interrompidos isolados em A. argillacea. ....................................................... 72
Figura 7 – Porcentagem de motivos microssatélites encontrados nos 117 insertos de A. argillacea seqüenciados que apresentaram pelo menos uma região microssatélite. ...... 72
Figura 8 – Porcentagem de motivos microssatélites encontrados nos primers obtidos para A. argillacea. ........................................................................................................... 75
Figura 9 – Freqüência dos motivos dinucleotídeos que são acessados pelos primers obtidos para A. argillacea ................................................................................................ 75
Figura 10 – Teste inicial utilizando os primers 13, 14, 15, 16, 17, 18, 19, 20, 21, 22 e 23; e três indivíduos de A. argillacea escolhidos ao acaso nas populações diferentes à temperatura de 60°C ....................................................................................................... 79
Figura 11 – Gel de acrilamida 7% com os testes dos primers isolados na primeira triagem. Primers que acessam os locos Aar14, Aar15, Aar16, Aar19, Aar21 e Aar23 em A. argillacea. .................................................................................................................... 79
Figura 12 – Histograma das freqüências alélicas dos dez locos microssatélites, estimados para ≈20 indivíduos de A. argillacea. O eixo Y indica a freqüência alélica e o eixo X indica o tamanho do alelo, em pares de base ...................................................... 83

16
Figura 13 – Padrão de divergência genética entre as populações de A. argillacea, definido pelo agrupamento UPGMA, a partir das distâncias genéticas de Nei (1978). Correlação cofenética igual a 0,99%. Para testar a consistência dos nós foram realizadas 1000 reamostragens por bootstrap. ............................................................... 99
Figura 14 – Resultado da análise obtida para a determinação do valor de agrupamento que melhor representa a variabilidade genética das populações de A. argillacea estudadas: K variando de 1 a 15, com cinco repetições cada K, burn in de 50.000 permutações e seguidas por 500.000 permutações MCMC ......................................... 100
Figura 15 – Resultado obtido para K=2, através do programa Structure 2.2. O numero de K que melhor representa o conjunto de dados determinou a formação de dois grupos, que se distinguem apenas pelo ano em que as populações desses grupos foram coletadas; a) as populações foram ordenadas segundo suas proximidades geográficas, e b) foram ordenadas segundo ano de coleta. Essas informações não foram determinadas a priori para a análise bayesiana ........................................................... 101

17
LISTA DE TABELAS
Tabela 1 – Caracterização da linha-básica de suscetibilidade à toxina Cry1Ac em populações de Alabama argillacea .................................................................................. 56
Tabela 2 – Modelo de regressão não-linear para estimativa dos parâmetros EC50 e EC99
para a população de A. argillacea da Bahia (BA-3) ......................................................... 61
Tabela 3 – Modelo de regressão não – linear para estimativa dos parâmetros EC50 e EC99 para a população de A. argillacea Mato Grosso (MT-1) .......................................... 62
Tabela 4 – Caracterização da linha-básica de suscetibilidade à toxina Cry1Ac mediante agrupamento de dados obtidos com as caracterizações de curvas de concentração-resposta de quatro populações de A. argillacea coletadas nas safras 2007/2008 .......... 69
Tabela 5 – Primers desenhados para A. argillacea a partir das regiões microssatélites . 73
Tabela 6 – Característica dos dez locos microssatélites obtidos para Alabama argillacea: Sequência do primer forward (F) e reverse (R) ............................................................... 82
Tabela 7 – Estimativas de variabilidade genética para os locos microssatélites utilizados em A. argillacea ............................................................................................................... 85
Tabela 8 – Estimativa de parâmetros descritivos de variabilidade genética nas onze populações de A. argillacea ............................................................................................ 86
Tabela 9 – Probabilidade do Teste Exato de Fisher para aderência ao equilíbrio de Hardy-Weinberg .............................................................................................................. 87
Tabela 10 – Freqüência alélica encontrada em cada loco microssatélite encontrada em cada população de A. argillacea ..................................................................................... 90
Tabela 11 – Alelos privados obtidos em 70 alelos de marcadores microssatélites. Loco, alelo, freqüência que ocorrem na população que são encontrados e população de origem ............................................................................................................................. 92
Tabela 12 – Estimativa das estatísticas F de Wright, através da análise de variância das freqüências alélicas e do numero de migrantes por geração (Nem) em onze populações naturais de A. argillacea. Intervalo de confiança (IC) de 95% de probabilidade .............. 94
Tabela 13 – Estimativa do parâmetro θ par-a-par entre as populações .......................... 95
Tabela 14 – Distâncias genéticas de NEI (1978), calculadas entre as populações ........ 98

18

19
1 INTRODUÇÃO
As plantas geneticamente modificadas (GM) resistentes a insetos são uma das
novas tecnologias para controle de pragas. Desde meados dos anos 90 tem sido
crescente a utilização de plantas GM contendo genes da bactéria entomopatogênica
Bacillus thuringiensis Berliner que codificam proteínas com ação inseticida, na
agricultura mundial. O Brasil ocupa a terceira posição no quadro mundial em relação à
área plantada com cultivares transgênicos, ficando atrás dos EUA e da Argentina
(INTERNATIONAL SERVICE FOR THE ACQUISITION OF AGRI-BIOTECH
APPLICATIONS - ISAAA, 2008). Das variedades transgênicas para controle de insetos
plantadas no Brasil, destacam-se as de algodão e de milho. O algodão Bollgard®
(Monsanto) foi aprovado pela Comissão Técnica Nacional de Biossegurança (CTNBio)
no Brasil em 2005. Essa variedade possui o gene proveniente de Bacillus thuringiensis
(Bt) que codifica a proteína inseticida Cry1Ac. Essa proteína tem como pragas-alvos, os
seguintes insetos da ordem Lepidoptera na cultura do algodão no Brasil: Alabama
argillacea (Hüeb.), Heliothis virescens (Fabr.) e Pectinophora gossypiella (Saund.).
Recentemente, em 2009, a CTNBio aprovou a liberação de mais duas variedades de
algodão GM resistentes a insetos, o algodão Bollgard® II (Monsanto) que expressa as
toxinas Cry1Ac e Cry2Ab2 e o algodão Widestrike® (Dow AgroSciences Industrial Ltda)
que expressa as toxinas Cry1Ac e Cry1F de B. thuringiensis. Além de controlarem as
pragas alvos do algodão Bollgard®, outras espécies são controladas por essas novas
variedades, tais como: Helicoperva zea (Hüeb.), Spodoptera exigua (Hüeb.), Spodoptera
eridania (Cramer), Pseudoplusia includens (Walker), Trichoplusia ni (Hüeb.)
(Widestrike®) e Spodoptera frugiperda (J.E. Smith) (Bollgard® II e Widestrike®).
Um dos problemas da utilização indiscriminada de uma tecnologia para o manejo
de pragas é a evolução da resistência. Com isso, esforços devem ser tomados para
frear ou mitigar a evolução da resistência, a fim de manter o uso sustentável de uma
tecnologia e permitir que não ocorra o seu desuso. Sendo assim, as primeiras ações
que devem ser tomadas, dentro de programas de manejo da resistência é a
caracterização de linhas–básicas de suscetibilidade, a definição de concentrações
diagnósticas e a implementação de programas de monitoramento da evolução da
resistência (ROUSH; MILLER, 1986; MARÇON et al,. 2000; WU et al., 2002; ALI et al.,

20
2006). O monitoramento é eficiente de acordo com a habilidade de detectar alelos raros
que conferem resistência quando estes estão em baixa freqüência na população, pois
assim é possível direcionar as estratégias de manejo antes da resistência se tornar um
problema (ROUSH; MILLER, 1986; ANDOW; HUTCHINSON,1998)
A principal estratégia de manejo da resistência a proteínas inseticidas expressas
por plantas GM tem sido baseada na manutenção de uma área de refúgio, onde são
plantadas variedades convencionais não transgênicas da mesma cultura próximas à
área de plantio da variedade GM que expressa uma ou mais toxinas em alta dose
(GOULD, 1998). Contudo, para o sucesso dessa estratégia é necessário que ocorra às
seguintes condições: a) gene da resistência seja de caráter recessivo, b) a concentração
da toxina expressa seja alta o suficiente para matar os indivíduos heterozigotos da
população e c) que o cruzamento entre indivíduos que carregam o gene da resistência e
indivíduos suscetíveis ocorra ao acaso (ALSTAD; ANDOW, 1995).
Estudos de variabilidade genética, estrutura populacional e fluxo gênico são de
grande importância dentro do contexto de manejo pró-ativo da resistência de insetos a
plantas GM (CAPRIO; TABASHNIK, 1992; CAPRIO et al., 2000). Como o fluxo gênico é
uma força coesiva que mantém a uniformidade da variabilidade genética entre as
populações de uma espécie e é responsável em trazer variação nas populações, é por
meio desse fluxo que os alelos que conferem resistência são espalhados para outras
populações. Além disso, como a estrutura populacional e variabilidade genética variam
entre populações, a freqüência dos alelos que conferem resistência pode estar também
distribuída desigualmente. Portanto, entendimento da estrutura genética e do fluxo
gênico intra-específico entre as populações de uma espécie-praga permite o
delineamento de práticas de manejo mais eficientes com o objetivo de se retardar a
evolução da resistência (CAPRIO; TABASHNIK, 1992) e identificar unidades evolutivas
distintas que são áreas focos para o manejo da resistência.
Conhecimentos de processos ecológicos que influenciam a variabilidade genética
entre populações são de grande relevância para refinar as estratégias de manejo da
resistência (GOULD, 1998). Esses estudos permitem entender a estruturação
hierárquica da diversidade genética, padrões e processos que determinam a
diferenciação genética e o fluxo gênico entre populações de insetos. Marcadores

21
moleculares têm sido utilizados para os estudos reportados para os principais alvos de
controle da primeira geração de plantas GM, e envolvem, por exemplo, insetos-pragas
como Ostrinia nubilalis (Hüeb.) (BOURGUET et al., 2000; COATES et al., 2004; 2005),
Helicoverpa armigera (Hüeb.) (SCOTT et al., 2006), e Spodoptera frugiperda (J.E.
Smith) (MARTINELLI et al., 2006; 2007).
Dentre os insetos-alvo do controle das plantas de algodão Bt, A. argillacea é uma
das espécies que apresenta o mais elevado risco para evolução da resistência (FITT et
al., 2006). A. argillacea é uma espécie monófaga que se alimenta exclusivamente de
algodão (Gossypium spp.). Características como a sua alimentação apenas em plantas
de algodão e a alta mobilidade das lagartas e dos adultos desta praga elevam a
probabilidade da exposição continuada deste inseto ao longo do tempo às toxinas de Bt
e deste modo aumentam o risco de evolução da resistência (FITT et al., 2006).
Portanto, o objetivo do trabalho foi de realizar um estudo genético ecológico em
populações de A. argillacea no Brasil para obter subsídios para o manejo pró-ativo da
resistência à toxina Cry1Ac expressa em algodão GM. Para tanto, foram conduzidos (a)
estudos para caracterização das linhas-básicas de suscetibilidade à proteína Cry1Ac de
Bt para verificar se existe variabilidade natural na resposta à toxina dependente da
localidade geográfica e (b) estudos da variabilidade e estrutura genética de populações
de A. argillacea mediante uso de marcadores microssatélites para avaliar se a
variabilidade genética está estruturada dentro ou entre populações no Brasil e testar se
ocorre fluxo gênico entre as populações de A. argillacea.

22

23
2 DESENVOLVIMENTO
2.1 Revisão bibliográfica
2.1.1 Características bioecológicas de Alabama argillacea (Hüebner)
Alabama argillacea (Hüebner) é uma espécie nativa das Américas do Sul e
Central, e é encontrada em todos os países do continente. A distribuição geográfica
dessa espécie abrange o extremo meridional da Argentina e toda região algodoeira dos
Estados Unidos (CALGAGNOLO, 1965). No Brasil, A. argillacea é o principal inseto-
praga da cultura do algodão, podendo reduzir em até 67% a produtividade da cultura
(RAMALHO, 1994).
As lagartas de A. argillacea, também conhecida como curuqurê-do-algodoeiro,
atacam o limbo das folhas, devorando-as quase totalmente, e quando o ataque se dá
por ocasião abertura das maçãs, ocorre à maturação forçada das mesmas, diminuindo a
resistência das fibras. Na América Tropical, o seu ataque pode ocorrer em qualquer
época do ano ou durante todo o ciclo da planta dependendo das condições climáticas de
cada região (SILVA et al., 1980). As lagartas alimentam-se das folhas e quando em altas
densidades populacionais, podem desfolhar completamente o algodoeiro, reduzindo
assim consideravelmente a produtividade da planta (GRAVENA; CUNHA 1991;
JÁCOME et al., 2001; QUIRINO; SOARES 2001). A produção do algodão, peso do
capulho e das sementes, e a porcentagem de uniformidade das fibras foram
influenciadas pelos níveis de população das pragas até os 135 dias, não proporcionando
mais danos econômicos após este período (MARCHINI et al., 1976). Em algodão mocó
(Gossipium hirsutum var. Maria Galante), o ataque de A. argillacea é responsável pelo
retardo do ciclo floral da planta em aproximadamente um mês (SEARA, 1970). A.
argillacea é uma espécie monófoga durante o estágio larval, sendo encontrada apenas
se alimentando de folhas de algodão, por essa razão, esta espécie-praga tem grande
importância para essa cultura (AZEVEDO et al., 2002).
A. argillacea é uma espécie que possui hábito migratório. Essa praga surge, na
cultura do algodão, em épocas distintas de ocorrência entre estados e países, devido ao

24
fato dessa espécie realizar migrações cíclicas (SILVEIRA NETO, 1972; HENDRICKS,
1975). Mediante estudos de flutuação populacional, foi observado que A. argillacea
ocorre na cultura do algodão no Brasil, nos meses de janeiro a julho. Essa espécie
ocorre a partir de dezembro, na região Centro-Sul do país (LADEIRA, 1957). No estado
de São Paulo, A. argillacea começa a surgir também no mês de dezembro e permanece
até julho, apresentando pico populacional no mês de abril (SILVEIRA NETO, 1972;
BOTELHO, 1976). Nos estado de Minas Gerais e Goiás infestações dessa praga podem
ocorrer a partir de novembro (GALLO et al., 2002). Na região Nordeste, no município de
Canindé, Ceará, A. argillacea surge na cultura do algodão Mocó nos primeiros dias do
mês de janeiro, permanecendo na região até o início do mês de maio (SANTOS, 1980).
As infestações no estado de São Paulo iniciam-se com adultos provenientes da região
Centro-Norte que em seguida se dirigem para a região Centro-Sul do país e norte da
Argentina, que partem por fim para a região Norte do Brasil. Ocorrem de três a cinco
gerações de A. argillacea em todo o ciclo do algodoeiro, sendo que cada geração
ocorrem em um período médio de 24 dias. Com isso as primeiras gerações que ocorrem
na cultura em determinado ano são de adultos migrantes provenientes de outra região
produtora adjacente, as gerações intermediárias são de indivíduos que estabelecem a
população na área, e as ultimas gerações são de indivíduos migrantes responsáveis
pela colonização de outras regiões (CALCAGNOLO, 1965). Com a expansão da cultura
do algodão para a região dos Cerrados nos últimos anos, a ocorrência dessa praga está
em sincronia com as épocas de cultivo nas diferentes regiões produtoras de algodão no
Brasil.
O controle populacional de A. argillacea tem sido feito principalmente mediante
a utilização de inseticidas sintéticos. Contudo, métodos alternativos de controle têm sido
sugeridos. A utilização de agentes microbianos, como Beauveria bassiana (Bals.) Vuill.,
Metharizium anisopliae (Metsch.) Sorok. e Bacillus thuringiensis, mostraram ser
eficientes para o controle de A. argillacea (MARTINS et al., 2007). Além disso, a
utilização de variedades de algodão que apresentam algum tipo de resistência ao
ataque de A. argillacea também tem se mostrado eficientes no controle da praga
(SANTOS; BOIÇA JUNIOR, 2001). Dentro desse contexto, a utilização de plantas
geneticamente modificadas que expressam toxinas Cry, provenientes de B.

25
thuringiensis, são as mais novas alternativas para o controle desse inseto na cultura do
algodão no Brasil. Entretanto, as características bioecológicas de A. argillacea como
hábito alimentar monófago e grande capacidade de dispersão, aumentam a exposição
do inseto à proteína inseticida de B. thuringiensis, aumentando assim os riscos de
evolução de resistência a toxinas expressas por plantas GM (FITT et al., 2006).
2.1.2 Resistência de insetos a plantas geneticamente modificadas
Plantas geneticamente modificadas que expressam toxinas provenientes de
Bacillus thuringiensis são as mais novas tecnologias para o controle de pragas
agrícolas. Nos EUA são plantadas desde 1996, com a liberação comercial do algodão Bt
Bollgard® (Monsanto) que expressa à toxina Cry1Ac. No Brasil a liberação comercial da
primeira variedade ocorreu em 2005, com a aprovação do algodão Bollgard®. Mais
recentemente, em 2009, houve a liberação comercial de mais duas variedades, o
algodão Bollgard® II (Monsanto) que expressa às toxinas Cry1Ac e Cry2Ab2 e o algodão
Widestrike® (Dow AgroSciences) que expressa às toxinas Cry1Ac e Cry1F. Apesar de
essas novas tecnologias trazerem uma maior variedade de toxinas para o sistema de
produção do algodão, todas expressam a toxina Cry1Ac. Portanto essa toxina ainda irá
impor uma pressão de seleção para as pragas alvos de seu controle (ALI et al., 2006).
As plantas GM provaram ter sucesso em termos de controle de pragas,
permitindo a redução do uso de inseticidas químicos (BETZ et al., 2000; FERRÉ; VAN
RIE, 2002). Os cristais protéicos (também chamados de δ-endotoxinas) de Bt são
extremamente tóxicos para determinadas pragas, contudo são pouco ou não-tóxicos
para a maioria dos insetos benéficos e outros organismos não-alvos (CROFT, 1990).
As toxinas de Bt são protoxinas que necessitam ser ativadas pelas proteases do
sistema digestivo de insetos para se tornarem tóxicas. As toxinas ativadas se ligam a
receptores específicos nas microvilosidades das membranas celulares das células
epiteliais do mesêntero (HOFFMANN et al., 1988; VAN RIE et al., 1989). Ao serem
ativados, esses receptores específicos desencadeiam a formação de poros no epitélio
do mesêntero, e essa formação modifica o equilíbrio osmótico que leva a lise celular e

26
ruptura do sistema digestivo, causando a morte do inseto (KNOWLES; ELLAR, 1987;
GILL et al.,1992; SCHNEPF et al.,1998). A ligação da toxina nos receptores é específica
por isso sua atividade depende da presença de receptores específicos no mesêntero
dos insetos (VAN RIE et al., 1989).
Devido à expressão contínua das toxinas inseticidas, as plantas GM que contém
os genes de Bt exercem uma elevada pressão de seleção, na direção da sobrevivência
dos genótipos resistentes nas populações de insetos-pragas alvos do controle. O
potencial de evolução de resistência de populações de insetos às toxinas de Bt é uma
das principais ameaças e limitações ao emprego dessa tecnologia no manejo de pragas
agrícolas (GOULD, 1998; BATES et al., 2005; FITT et al., 2006).
A resistência envolve a sobrevivência e reprodução diferencial de organismos
expostos à pressão de seleção que resultaria na mortalidade da maioria dos indivíduos
de uma população. A resistência a toxinas (inseticidas naturais e sintéticos) provêm do
estresse imposto por essas substâncias que agem na variabilidade genética existente
de populações naturais. Segundo Dobzhansky (1951), a resistência é uma característica
“pré-adaptativa”, genética e hereditária. Na evolução da resistência ocorre uma
substituição de alelos: o alelo para a suscetibilidade é substituído, ao longo das
gerações, pelo alelo que confere resistência. Os alelos que conferem resistência surgem
ao acaso por mutação e ocorrem em baixas freqüências. Com a pressão de seleção,
esses alelos passam a ocorrer com maior freqüência, alterando assim a proporção entre
os alelos existentes na população. Esta substituição pode ocorrer em um ou em vários
locos e é favorecida pela forte seleção que o pesticida promove na variação genética
inicial (ROUSH; McKENZIE, 1986).
Em princípio, o mecanismo de resistência ao Bt poderia ser localizado em cada
um das diferentes etapas do modo de ação das proteínas Cry, tais como: solubilização,
transformação proteolítica, passagem através da membrana peritrófica, sítio de ação na
membrana plasmática das células do mesêntero, formação dos poros, e lise osmótica
das células (FERRÉ; VAN RIE, 2002). Considerando que diversos mecanismos têm sido
observados em linhagens resistentes selecionadas em laboratório, apenas dois
mecanismos de resistência têm sido relatados. O mecanismo de resistência mais
importante tem sido aquele ligado à diminuição da sensibilidade dos sítios de ação da

27
toxina. Em segundo lugar está o mecanismo relacionado à pro-ativação das δ-
endotoxinas pelas proteases do sistema digestivo (TABASHNIK, 1994).
Pelo fato de Bt ter sido utilizado comercialmente por mais de três décadas sem
relatos de desenvolvimento de resistência nas populações de campo, alguns autores
presumiram que a evolução da resistência a essas toxinas seria improvável
(TABASHNIK, 1994). Entretanto, foram observados casos de resistência a biopesticidas
formulados à base de Bt em populações de campo da traça-das-crucíferas, Plutella
xylostella L. (TABASHNIK 1991, 1994). Além disso, foram detectadas mudança e
diminuição da suscetibilidade de populações naturais de Heliothis virescens e
Helicoverpa zea às proteínas purificadas Cry1Ac e Cry1Ab (STONE; SIMS, 1993; ALI et
al., 2006). Já em relação a plantas GM, já foram reportados a capacidade de
Helicoverpa armigera (Hüeb.) em desenvolver resistência, quando submetida à pressão
de seleção com algodão Bt, que expressa à toxina Cry1Ac, em condições de laboratório
na China (FENGXIA et al., 2003; LU et al., 2004). Contudo o único caso de resistência a
Cry1Ac, presente em planta GM foi de H. zea, nos estados de Arizona e Mississipi,
EUA, onde a área de refúgio plantada é menor que 20% (TABASHNIK et al., 2008)
Nos países em que se tem utilizado com sucesso a tecnologia das plantas GM
resistentes a insetos, os programas de manejo da resistência tem se baseado na
expressão das toxinas inseticidas em altas doses associadas à manutenção de áreas
refúgio e monitoramento da suscetibilidade a estas proteínas nas populações de insetos.
A área de refugio consiste no plantio de variedade convencionas que não expressam
a(s) toxina(s), juntamente com as plantas GM (GOULD, 1998; BATES et al., 2005;
TABASHNIK, 2008).O uso de concentrações diagnósticas em um programa de
monitoramento da resistência, permite que um número maior de insetos seja testado
apenas nas concentrações com maior capacidade de informar sobre a suscetibilidade a
um determinado inseticida e/ou biopesticida na população da praga, e
conseqüentemente, os resultados podem ser obtidos de uma forma mais rápida, precisa
e prática (ROUSH; MILLER, 1986) Contudo, para um programa monitoramento da
resistência ser eficiente, esse necessita de métodos de bioensaios sensíveis, capazes
de identificar a presença de indivíduos resistentes quando esses estão em baixa
freqüência na população. Sims et al. (1996) compararam a sensibilidade relativa de dois

28
métodos de bioensaio de ingestão com a avaliação da mortalidade e da inibição de
crescimento como critérios de resposta e concluíram que os ensaios onde foi avaliada a
inibição de crescimento larval utilizando concentrações subletais da proteina Cry1Ac
foram mais sensíveis que os ensaios de concentração-resposta.
Muitas vezes os programas de monitoramento da resistência são negligenciados,
pois a ao invés de ocorrer a sua implementação concomitante com o inicio do uso de
uma determinada tecnologia para o controle de pragas, essa ocorre quando a
resistência de insetos já é um problema no campo (SIMS et al., 1996). Sobre estratégias
de manejo e programas de monitoramento da resistência, existe uma discussão a
respeito de quando se deve introduzir uma variedade GM e desenvolver áreas de
refúgio: a primeira abordagem diz respeito a uma prática de manejo pró-ativa da
resistência, no qual a uma interpretação conservativa dos dados disponíveis é utilizada
para o desenvolvimento de estratégias de manejo da resistência. Essa abordagem
empregaria uma área de refúgio relativamente grande associada ao plantio inicial de
plantas transgênicas, além de envolver uma série de estudos envolvendo a planta
transgênica, ecologia dos insetos e outros aspectos antes de haver a liberação da planta
para o plantio comercial; a segunda abordagem seria a introdução rápida das plantas
transgênicas combinada com práticas de manejo adaptativa da resistência. Essa prática
adotaria uma interpretação menos conservativa dos dados disponíveis, mas utilizaria
uma abordagem contínua de monitoramento e avaliação dos dados para estar sempre
adotando práticas de manejo condizentes com a situação. Essa prática de manejo
depende da nossa habilidade de identificar alelos raros que conferem resistência
quando esses ainda estão em baixa freqüência nas populações (ANDOW;
HUTCHINSON,1998).
Sendo assim, percebe-se que tanto as práticas de utilização conservativa quanto
as táticas de manejo adaptativo depende de geração de conhecimento sobre o
complexo de pragas que são alvos de controle de determinada planta GM e informações
sobre os aspectos agroecológicos do ambiente. Portanto, conhecer a diversidade
genética dos insetos pragas, e como essa está estruturada em unidades populacionais
menores dentro do complexo ecossistema permite entender os processos que
determinam essa distribuição espacial além de permitir isolar locais e populações com

29
maior potencial para a evolução da resistência, uma vez que a estrutura populacional e
a variabilidade genética divergirem entre populações, da mesma forma os alelos que
conferem resistência também podem estar distribuídos desigualmente. Dessa forma
percebe-se a importância de se entender os padrões de distribuição da variabilidade
genética e fluxo gênico para se determinar áreas foco para o monitoramento e
desenvolver estratégias para mitigar a evolução da resistência nesses locais. (CAPRIO
et al., 2000).
2.1.3 Marcadores moleculares
A genética de populações sofreu uma revolução nas últimas décadas, após o
advento dos marcadores moleculares. Essa revolução se deve ao fato de hoje, ser
possível estudar genética de qualquer organismo existente, sem a necessidade de se
ter um organismo mutante que manifesta diferenças visíveis, como ervilhas lisas e
rugosas, ou Drosophila spp. com olhos vermelhos ou brancos.
E essa revolução veio com a abundância de dados experimentais gerados pelo
uso de métodos moleculares nos estudos dos polimorfismos genéticos. Presente em
todos os organismos, os polimorfismos moleculares permitem que as populações sejam
estudadas independentemente da espécie ou do hábitat, e sem a necessidade de
cruzamentos controlados, genes mutantes ou de quaisquer estudos genéticos anteriores
(HARTL, 2008)
Sendo assim, marcador molecular é todo e qualquer fenótipo molecular
proveniente de um gene expresso, como no caso de isoenzimas, ou de um segmento
específico de DNA (correspondendo a regiões expressas ou não do genoma). A
revolução neste plano se iniciou com o descobrimento e utilização de marcadores
isoenzimáticos a partir do ano de 1959, ampliando vastamente o número de marcadores
genéticos e possibilitando a aplicação da técnica praticamente todas as espécies de
plantas e animais (FERREIRA; GRATTAPAGLIA, 1998).
Os marcadores isoenzimáticos são formas distintas de uma mesma enzima, que
não precisam estar num mesmo indivíduo, com função catalítica idêntica ou similar que
diferem entre si pela a seqüência de aminoácidos. O princípio básico da técnica consiste

30
na utilização de eletroforese em gel e na visualização da atividade enzimática por meio
de métodos histoquímicos. Trata-se de uma técnica de fácil manipulação, ainda muito
utilizada, custo relativamente baixo, e que permite analisar vários sistemas
isoenzimáticos e grande número de indivíduos. Os marcadores isoenzimáticos são do
tipo co-dominantes. Apresentam como propriedade mais expressiva, a base genética
simples envolvida ligada a essas enzimas (SOLTIS; SOLTIS, 1989), entretanto com
limitações na cobertura reduzida dos genomas investigados, devido ao pequeno número
de locos que podem ser detectados, e o baixo nível de polimorfismo identificado por loco
(ALFENAS, 1998; BORÉM; CAIXETA, 2006).
Outro marcador molecular desenvolvido foi o RFLP – “Restriction Fragment
Lenght Polymorphism” (polimorfismo no tamanho de fragmentos de restrição). O
desenvolvimento dessa técnica só foi possível a partir de descobertas das enzimas de
restrição na década de 60. Esse marcador possibilitou detectar diferenças entre
indivíduos por meio de cortes no DNA estudado. Com esta técnica, o DNA genômico de
um indivíduo pode ser isolado e digerido com enzimas de restrição, os fragmentos
obtidos são separados em um gel num processo denominado eletroforese, gerando
fragmentos de tamanho e número variáveis que evidenciam o polimorfismo genético. Os
fragmentos são transferidos para uma membrana de náilon ou nitrocelulose – técnica
denominada “Southern blot”. A próxima etapa consiste na hibridização do DNA das
amostras já imobilizadas em membranas, com uma sonda radioativa de DNA
complementar ao fragmento de interesse. A última etapa é a auto-radiografia, ou seja,
exposição da membrana hibridizada com a sonda radioativa a um filme de raios-X,
revelando a presença de bandas, que são os marcadores RFLP (HELENTJARIS et al.,
1986). Uma das principais vantagens desse marcador é a sua expressão co-dominante,
que permite distinguir indivíduos homozigóticos de indivíduos heterozigóticos, outra
vantagem que merece destaque é a possibilidade de utilização de sondas heterólogas,
permitindo o mapeamento comparativo entre as espécies. As principais desvantagens
são o trabalho que envolve a execução das várias etapas, o alto custo e o uso de
radioatividade (FERREIRA; GRATTAPAGLIA, 1998).
Com o desenvolvimento de uma tecnologia simples e eficiente que automatizou
a reação em cadeia da polimerase – PCR “Polymerase Chain Reaction” pelo

31
pesquisador Kary Mullis permitiu uma revolução na biologia molecular e
conseqüentemente no desenvolvimento e aplicação de marcadores moleculares, tais
como RAPD, STS, SCARS, AFLP, SNPs e microssatélites. Através deste processo é
possível multiplicar “in vitro” e em escala exponencial cópias de um segmento específico
de DNA, na presença da enzima DNA polimerase (MULLIS; FALOONA, 1987).
O RAPD – “Random Amplified Polymorphic” DNA (DNA polimórfico amplificado
arbitrariamente) – é basicamente uma variação do protocolo de PCR, difere em duas
características, utiliza um primer único e menor ao invés de um par de primers e este
tem seqüência alvo desconhecida. Esta técnica permite a detecção de fragmentos
aleatórios de DNA. Em relação aos RFLPs, os RAPDs são mais baratos, requerem
pouco tempo e não necessitam de radioisótopos, entretanto são marcadores
dominantes, ou seja, não é possível distinguir os genótipos dos heterozigotos, e
conseqüentemente não é possível se estimar a freqüências alélicas e genotípicas de um
determinado loco, a não ser que se assuma equilíbrio de Hardy-Weinberg para aquele
loco e população. Outra desvantagem deste marcador é que se trata de uma técnica
muito sensível e que, portanto, não possui reprodutibilidade entre laboratórios
(WILLIAMS, 1990).
AFLP – “Amplified Fragment Length Polymorphisms” (polimorfismos de
comprimento de fragmentos amplificados) são marcadores moleculares desenvolvidos
por Zabeu (1993); cuja técnica combina a digestão de fragmentos de DNA com enzimas
de restrição de corte raro e enzimas de corte freqüente e a amplificação desses
fragmentos por PCR (VANTOAL et al., 1996). Assim sendo, o DNA de um indivíduo é
clivada com uma combinação de duas enzimas, uma de corte raro, outra de corte
freqüente e esses fragmentos após serem clivados são ligados às extremidades dos
adaptadores específicos de cada enzima de restrição, que servem de sítios de ligação
dos primers para posterior reação de PCR; os produtos dessa amplificação são
separados em gel de poliacrilamida desnaturante e o polimorfismo é identificado pela
presença ou ausência de bandas (VOS et al., 1995). Os marcadores AFLP também são
dominantes. Contudo, apresenta como vantagens a visualização em um único gel de um
grande número de fragmentos gerados, proporcionando uma amostragem ampla e
simultânea de um genoma, grande poder de detecção de variabilidade genética, e

32
apresentam maior robustez e reprodutibilidade quando comparado com marcador
RAPD.
Os marcadores moleculares os SNPs – “Single Nucleotide Polymorphisms”
(polimorfismo de um único nucleotídeo) são mais recentes. Os SNPs são alterações de
uma única base no genoma, ocorrendo a cada 600 pares de bases aproximadamente,
quando se compara segmentos correspondentes do genoma entre indivíduos da mesma
espécie. Esses polimorfismos ocorrem quando há substituições de um único nucleotídeo
A, C, G, T em uma determinada posição do genoma com freqüência alélica mínima de
1% em uma população (BROOKES, 1999). Os SNPs são, portanto, marcadores
genéticos baseados na detecção de polimorfismos resultantes de alterações de
nucleotídeos, que devido à freqüência em que ocorrem são considerados importantes
ferramentas para obtenção de mapas genéticos de alta resolução. A via direta para
identificação de SNP é o seqüenciamento de fragmentos de DNA, amplificados por
PCR, com primers desenhados para este fim. A identificação também pode ser feita por
análise eletrônica de variação pontual (KWOK, 2001).
2.1.4 Marcadores moleculares microssatélites e genética de populações
Microssatélites, também conhecidos como sequências de repetição simples –
“Simple Sequence Repeats” (SSR), ou repetições curtas em “tandem” – “Short Tandem
Repeats” (STR), são regiões de DNA repetitivo não codificantes compostos por
pequenos motivos de um a seis nucleotídeos repetidos em “tandem”, que são
encontrados em quase todos os genomas de procariotos e eucariotos (FERREIRA;
GRATTAPAGLIA, 1998; TOTH; GASPARI; JURKA, 2000; ZANE; BARGRLLONI;
PATARNELLO, 2002). Estão presentes em regiões codificantes e não-codificantes e
geralmente são caracterizados por um alto grau de polimorfismo. São muito utilizados
como marcadores moleculares por possuírem um atributo particular que é ter uma alta
taxa de mutação quando comparados com o restante do genoma (JARNE; LAGODA,
1996).
A forma como se originam e a dinâmica de mutação dos microssatélites no
genoma dos organismos ainda é incerta (SCHLOTTERER, 2000), entretanto sabe se

33
que a taxa de mutação dos microssatélites é muito alta quando comparada com outras
partes do genoma, variando de 10-6 a 10-2 nucleotídeos por loco por geração (SIA et. al.,
2000). Muitos mecanismos já foram sugeridos para explicar as altas taxas de mutação
dos microssatélites, incluindo erros durante a recombinação, “crossing-over” desigual e
“slippage” da DNA polimerase durante a replicação ou reparo do DNA (STRAND et al.,
1993). Sobre as teorias da dinâmica de mutação dos microssatélites duas são mais
aceitas e dizem que essa pode estar associada ao sistema de reparo pela DNA
polimerase ou ainda, ser conseqüência do processo de recombinação (SCHLOTTERER;
TAUTZ, 1992; FIELD; WILLS, 1996). Durante o processo de recombinação o “crossing-
over” desigual pode ser responsável pela alta taxa de polimorfismo destes marcadores,
que por problemas no pareamento dessas seqüências durante o quiasma, aumenta a
taxa de mutação das regiões microssatélites e são estas mutações que tornam estes
marcadores tão informativos (STRAND et al., 1993; SCHLÖTTERER et al., 1998). Já
sobre mutações ligadas a eventos de “slippage”, ou deslizamento da DNA polimerase
durante a replicação, acredita-se que na replicação de uma região repetitiva, as fitas de
DNA separam-se e une-se novamente de forma incorreta, o que geraria cópias de
trechos de DNA (alelos) com diferentes tamanhos ou números de repetições de um
determinado motivo no próximo ciclo de replicação, por meio da inserção ou deleção de
uma unidade de repetição. Esse tipo de evento possivelmente gera alta taxa de mutação
dos microssatélites, numa freqüência de 10-4 e 10-3 por loco e por gameta em cada
geração (SCHLOTTERER; TAUTZ 1992; SCHLOTTERER, 2004).
O polimorfismo de um loco microssatélite é resultado do número de repetições do
motivo que o caracteriza. Portanto, os diferentes alelos que um loco microssatélite pode
apresentar distinguem-se uns dos outros pelo número de repetições dos motivos
microssatélites. Um loco microssatélite homozigoto tem o mesmo número de repetições
em ambos os cromossomos homólogos, entretanto um loco heterozigoto tem um
número de repetições do motivo para cada alelo de cada cromossomo homólogo. Por
exemplo, um alelo pode ter nove repetições e outro dez (OLIVEIRA et al., 2006). O
número de repetições é um fator crucial para determinar a dinâmica evolutiva do DNA
microssatélite, e por isso é importante conhecer quais parâmetros influenciam o
comprimento das repetições.

34
Os microssatélites são classificados conforme a composição das seqüências
repetidas: (a) repetições perfeitas, quando não apresentam nenhuma interrupção, por
exemplo: GTGTGTGTGTGTGT; (b) repetições imperfeitas, quando são interrompidas
por bases que não correspondem ao motivo, por exemplo: GTGTGTGTAGTGTGTT; (c)
repetições compostas, quando duas ou mais repetições (classes) de microssatélites
estão dispostas adjacentes, por exemplo: GTGTGTGTGTGTCACACACACACA
(BORÉM; CAIXETA, 2006).
Apesar das seqüências microssatélites variarem de um indivíduo para outro, as
seqüências de DNA que as flanqueiam são bastante conservadas entre indivíduos da
mesma espécie. Conhecendo essas regiões é possível desenhar primers (iniciadores)
capazes de amplificar, através de PCR, as regiões adjacentes e as que contêm os
microssatélites. Os produtos de amplificação podem ser visualizados em gel de
poliacrilamida desnaturante ou não desnaturante, em gel de agarose de alta resolução
ou por meio de primers fluorescentes em seqüenciador automático. O polimorfismo
entre bandas é decorrente dos tamanhos diferentes entre as seqüências repetitivas
(SOUZA, 2001; BORÉM; CAIXETA, 2006). Esses marcadores são, portanto, facilmente
detectados através de PCR, produzindo resultados altamente reprodutíveis se
comparado a outros tipos de marcadores moleculares (TAUTZ; TRICK; DOVER, 1986;
WRIGHT; BENTZEN, 1994; O’CONNELL; WRIGHT, 1997).
Os microssatélites são marcadores moleculares úteis em um grande número de
análises. Sua aplicação vai desde o mapeamento gênico a estudos forenses,
abrangendo áreas de estudo como a da genética de populações e conservação e
manejo de recursos naturais (JARNE; LAGODA, 1996). São marcadores ideais para
estudos de genética e evolução de populações devido ao alto poder discriminatório, à
robustez, confiabilidade, praticidade operacional e por serem mais informativos
geneticamente (SLATKIN, 1995).
Além disso, apresentam as seguintes características, que fazem com que seja
preferencialmente escolhido como marcador molecular: são marcadores co-dominantes;
são amplamente distribuídos no genoma dos eucariotos; são multialélicos; são
facilmente amplificados via PCR; e os primers uma vez desenvolvidos, podem ser
facilmente compartilhados entre laboratório (SCHLÖTTERER, 2004).

35
Contudo o emprego da tecnologia envolve algumas limitações, como o grande
trabalho para o desenvolvimento dos primers específicos para os locos microssatélites
de cada espécie. Entretanto, pelo fato das sequências que flanqueiam os microssatélites
serem altamente conservadas, os mesmos primers obtidos para uma espécie podem ser
utilizados para outras espécies relacionadas (FERREIRA; GRATTAPAGLIA, 1998).
O desenvolvimento de marcadores microssatélites para espécies do grupo dos
lepidópteros é extremamente difícil por razões não compreendidas. Entretanto novas
descobertas demonstraram que muitas seqüências de microssatélites de lepidópteros
existem em múltiplas cópias no genoma, e possuem regiões flanqueadoras iguais ou
quase semelhantes. Essas descobertas forneceram uma convincente explicação para a
baixa eficiência de isolamento de microssatélites em lepidópteros, e também deram luz
para o entendimento da dinâmica evolutiva das sequências microssatélites nesse grupo
de insetos (ZHANG, 2004). A abundância de microssatélites encontrada em genomas
de lepidópteros é considerada baixa, quando comparada com outros grupos
taxonômicos. Dentre 20 trabalhos reportados em literatura que foram feitos o isolamento
de locos microssatélites, em 80% dos casos não mais do que cinco locos polimórficos
foram encontrados (ZHANG, 2004; JI; ZHANG, 2004). Além disso, é importante o fato
de que a eficiência de clonagem também é baixa. Enquanto que a eficiência foi de 14%
para a clonagem de sequências contendo microssatélies em Locusta migratória (L.),
para H. armigera essa eficiência foi em torno de 2,5% (JI, 2003; ZHANG, 2003). Esses
resultados mostram a baixa freqüência de microssatélites nos genomas de lepidópteros.
O isolamento de locos microssatélites em lepidópteros também é complicado por causa
da ocorrência de famílias de DNA microssatélites. Essas famílias possuem as regiões
flanqueadoras semelhantes (NEVE; MENGLÉZ, 2000), e na maioria dos casos essas
regiões possuem DNA repetitivo. A associação de microssatélites com regiões
flanqueadoras que possuem DNA repetitivo parece ser característica dos microssatélites
de lepidópteros (JI; ZHANG, 2004). Contudo, em estudo de genética de populações,
apenas a ocorrência de microssatélites cópia-única é usada para acessar o polimorfismo
de populações naturais. Isso determina conseqüentemente, que as regiões
flanqueadoras dos microssatélites isolados devem ser únicas no genoma do organismo
estudado. Sendo assim, os microssatélites de lepidópteros são considerados

36
redundantes, pois as várias regiões flanqueadoras semelhantes podem ser amplificadas
utilizando um mesmo conjunto de primers desenvolvidos para acessar apenas um loco
microssatélite. As conclusões a respeito das características dos locos microssatélites de
lepidópteros (baixa freqüência e alta redundância) são as de que os microssatélites
estão em estágios iniciais de evolução, e que muitas dessas seqüências
experimentaram eventos recentes de propagação e multiplicação. Portanto, as regiões
flanqueadoras dos microssatélites de lepidópteros ainda não acumularam mutações
suficientes para promover a diferenciação dessas regiões, resultando baixa proporção
de microssatélites únicos (ZHANG, 2004).
Em estudos de genéticas de populações, a escolha do marcador determina o
grau de resolução das suas análises de variabilidade genética e estrutura populacional.
Dentre as possibilidades de marcadores moleculares, hoje os microssatélites são
amplamente utilizados por permitir esse grau de resolução, e tem sido amplamente
utilizados em estudos de populações de pragas-alvo do controle de plantas transgênicas
e no contexto do manejo integrado de pragas (COATES; HELLMICH; LEWIS, 2005; JI,
2003; SCOTT et al., 2006).
Uma população, sob o ponto de vista genético, é definida como um conjunto de
indivíduos de uma mesma espécie, que compartilham o mesmo pool gênico, isto é,
indivíduos que compartilham seus genes através de reprodução (FALCONER;
MACKAY, 1989). Em genética de populações os parâmetros utilizados para descrever
uma população são: as freqüências alélicas e genotípicas, que se referem,
respectivamente, à quantidade observada de um determinado alelo frente ao total de
alelos da população e a quantidade observada de indivíduos com um determinado
genótipo frente ao total de indivíduos da população, as heterozigosidades esperada (HE)
e observada (HO) e a diversidade genética, que é medida da variabilidade genética da
população, que permitem identificar os alelos e identificar o número de indivíduos
heterozigotos, bem como a quantidade de indivíduos homozigotos diferentes (WEIR,
1996). Esses parâmetros além de descrever populações individuais são necessários
para se estudar a variabilidade genética entre populações e como essa variabilidade
está estruturada no espaço. A alteração genética de uma população pode ser avaliada
pela mudança nas suas freqüências gênicas, o que torna a estimativa da freqüência de

37
heterozigotos fundamental nos estudos evolutivos (NEI, 1978). Além disso, as
mudanças e distribuição das freqüências alélicas entre as populações de uma espécie
determinam o grau de estruturação da variabilidade genética existente. Sendo assim os
fatores que determinam a estruturação são: deriva genética, migração, mutação,
seleção, sistema reprodutivo e número de gerações (HARTL, 2008).
O entendimento da estrutura genética e do fluxo gênico intra-específico entre
populações e subpopulações de uma determinada espécie de inseto-praga é essencial
para se delinear melhores de práticas de manejo com o objetivo de retardar a evolução
da resistência a qualquer tática de controle de insetos, uma vez que é por meio dos
eventos de migração que os alelos que conferem resistência são rapidamente
distribuídos entre as populações de um inseto-praga (CAPRIO; TABASHNIK, 1992;
TABASHNIK, 1991). Portanto a escolha do marcador molecular microssatélite foi feita
baseada nas suas características únicas e no seu grau de resolução que melhor irá
responder os questionamentos que irão auxiliar nos estudos sobre evolução da
resistência.
2.2 Material e métodos
2.2.1 Coleta das populações
As populações de A. argillacea foram coletadas nas regiões produtoras de
algodão no Brasil, em quatro estados sobre a variedade comercial cultivada (Gossipium
hirsutum L.), e também foram coletados indivíduos em G. hirsutum L. var. Maria
Galante, que é uma variedade selvagem de algodão, comumente conhecido como
algodão arbóreo ou mocó. Nas coletas, preferencialmente foram amostrados formas
larvais ou então pupas do inseto. As coletas foram realizadas buscando a máxima
variabilidade possível. Os pontos de amostragem da praga foram distribuídos dentro de
cada região de coleta. Esses indivíduos foram armazenados adequadamente,
juntamente com o substrato para alimentação para lagartas, e foram trazidos para o
Laboratório de Resistência de Artrópodes a Pesticidas, Departamento de Entomologia e
Acarologia, ESALQ-USP. Os insetos provenientes de cada localidade foram criados em

38
laboratório de acordo com a metodologia descrita a seguir (item 2.2.2) e foram utilizados
tanto para a caracterização da linha-básica de suscetibilidade à toxina Cry1Ac quanto
para o estudo de variabilidade, estrutura genética e fluxo gênico da espécie no Brasil. As
regiões de coleta, a identificação de cada população, o local (município) de origem e a
data de coleta estão expressos no Quadro 1.
Região População Município Coordenadas Geográficas
Data de Coleta
Bahia BA-1 Barreiras 44°59’32,9”O 12°08’53,4”S
13/02/2008
BA-2 Luis Eduardo Magalhães 45°47’53,7”O 12°05’57,9”S
24/02/2008
BA-3 Roda Velha 45°49’23,2”O 12°41’36,9”S
17/03/2008
Goiás GO-1 Montevidiu 51°08'27,9"S 17°29'11,9”O
17/03/2008
Mato Grosso MT-1 Campo Verde 55°09’58,6”S 15°32’44,0”O
26/06/2008
Mato Grosso do Sul MS-1 Chapadão do Sul 52°36’59,4”S 18°46’44,0”O
17/03/2008
Bahia BA-4 São Desidério 44°59'03,2”S 12°21'07,6”O
05/03/2009
Goiás GO-2 Cristalina 47°36'45,4"S 16°17'00,2"O
16/03/2009
Mato Grosso MT-2 Rondonópolis 54°40’09,3”S 16°42’07,5”O
12/03/2009
MT-3 Primavera do Leste 54°17’58,5”S 15°33’34,9”O
18/03/2009
Mato Grosso do Sul MS-2 Chapadão do Sul 52°38’34,7”S 18°46’24,0”O
25/03/2009
Paraíba PB-1 Campina Grande 35°52’56,3”S 07°13’51,6”O
13/09/2009
Quadro 1 – Identificação das populações de Alabama argillacea amostradas nas diferentes regiões, municípios, coordenadas geográficas e datas de coleta

39
2.2.2 Criação em laboratório
Os adultos de A. argillacea foram criados em laboratório, em gaiolas de PVC
revestidas com papel sulfite, que foi utilizado com substrato para postura, e alimentados
com solução de mel 30%. As gaiolas permaneceram em sala climatizada regulada a 25
± 1ºC, umidade relativa (UR) acima de 60% e fotofase de 14h. A cada dois dias foi feita
a troca das gaiolas, da solução de mel, e a separação das posturas. Essas posturas
foram colocadas em recipientes plásticos de 10 ml com tampa juntamente com papel
filtro umedecidos e acondicionados em câmara climatizada regulada a 25 ± 1ºC,
umidade relativa (UR) 60% e fotoperíodo de 14:10 (claro:escuro) até a eclosão das
lagartas. As lagartas neonatas foram transferidas para placas de Petri de vidro,
juntamente folhas novas de algodão (cultivar Delta Opa)l e permaneceram nesses
recipientes até a fase de pupa. Para cada população foram separadas em torno de dez
placas de Petri com aproximadamente 10 lagartas por placa, sendo que nos estádios
larvais mais próximos do estágio de pupa, foram deixadas em torno de cinco lagartas
por placa. As folhas de algodão foram trocadas e as placas limpas a cada dois dias nos
primeiros instares, e todos os dias após o terceiro instar até a fase de pupa. Essas
placas foram acondicionadas em sala climatizada regulada a 25 ± 1ºC, umidade relativa
(UR) 30% e fotofase de 14h até as lagartas se tornarem pupas. As pupas foram
separadas do restante das lagartas e colocadas sobre vermiculita, em placas de Petri, e
foram acondicionadas em sala climatizada, regulada para umidade relativa (UR) para
60%, sendo que a vermiculite foi umedecida a cada dois dias. Uma parte das lagartas
(10%) foi utilizada para a manutenção das populações, e o restante e foi utilizada para a
realização dos bioensaios de caracterização da linha-básica de suscetibilidade e
validação da concentração diagnóstica.

40
2.2.3 Caracterização da suscetibilidade de Alabama argillacea à toxina Cry1Ac.
2.2.3.1 Linha-básica de suscetibilidade
As caracterizações da linha-básica de suscetibilidade das populações de A.
argillacea (Quadro 1) foram realizadas por meio de bioensaios de imersão de discos de
folhas de algodão em soluções contendo toxina Cry1Ac, obtida a partir do biopesticida
MVP II (Dow Agrosciences, San Diego, EUA), e 0,1% do agente surfactante Triton X-
100 . Os discos, após a imersão nas diferentes concentrações, foram colocados em
placas de acrílico de 12 células de 2,0 cm de diâmetro (Costar, Cambridge,
Massachusetts, EUA) contendo no fundo 500 µL de solução de ágar 2,5% e sobre essa
camada, disco de papel de filtro. Após a secagem da toxina foi realizada a transferência
de uma lagarta neonata (24 h após eclosão) de A. argillacea em cada célula. As lagartas
do tratamento testemunha foram dispostas sob o mesmo substrato sem a presença da
toxina. Para cada concentração foram utilizadas aproximadamente 96 lagartas
neonatas, divididas em oito repetições. Essas placas foram fechadas com filme plástico
e esse foi perfurado com alfinete entomológico para haver trocas gasosas entre as
células e o ambiente da câmara climatizada, a fim de evitar o acúmulo de água dentro
de cada célula (Figura 1). Essas placas foram mantidas em câmara climatizada regulada
a 27 ± 1ºC e fotofase de 14 h. Os bioensaios foram avaliados cinco dias após a
transferência das lagartas. Os critérios de resposta avaliados foram a mortalidade e a
inibição de crescimento.
A mortalidade foi observada pela constatação visual das condições diagnóstico-
toxicológicas características da toxina, que são: não desenvolvimento das lagartas, ou
seja, não ultrapassam o primeiro instar, as lagartas apresentam sinais de dessecação e
as folhas contendo a toxina não apresentam sinais de consumo. As concentrações
utilizadas foram definidas a fim de compreender as porcentagens de mortalidade entre o
intervalo de 5 a 95%. Os dados de mortalidade obtidos com os bioensaios foram
submetidos à análise de regressão utilizando o modelo de Probit, por meio do programa
PoloPlus (LeOra Software, 2002) e as estimativas dos seguintes parâmetros foram
obtidas: CL50 e CL95 (Concentração Letal capaz de causar mortalidade em 50% e 95%

41
da população, respectivamente); intervalo de confiança a 95% (IC 95%) das
concentrações letais; o coeficiente angular (± erro padrão) da regressão de Probit. Para
a comparação das linhas–básicas obtidas foram comparados os limites dos intervalos
de confiança a 95% (IC 95%) das CL50s e CL95s estimadas para cada população. As
estimativas foram consideradas significativamente diferentes (p < 0,05) quando os IC
95% não se sobrepuseram. O teste de igualdade e paralelismo de retas, que compara
os valores de coeficiente angular (parâmetro β) e os valores de intercepto das retas no
eixo Y (parâmetro α) entre as retas obtidas com a regressão de Probit, foi utilizado para
confirmar as diferenças estatisticamente com um nível de significância. As retas foram
consideradas não paralelas se os valores de β apresentaram diferenças significativas (p
< 0,05). Já as retas foram consideradas diferentes se os valores de α e de β foram
significativamente diferentes (p < 0,05). Contudo as retas foram consideradas
significativamente diferentes se o teste de hipótese H0 tanto para o paralelismo, quanto
para o de similaridade foram rejeitados.
Para a avaliação da inibição de crescimento, foram obtidos os valores de peso
agrupando as lagartas sobreviventes em cada tratamento, de cada repetição. Para os
dados de peso médio foi realizado análise de regressão não-linear utilizando o modelo
de regressão logístico não-linear seguinte: peso=W0/ [(1+(concentração/EC50)B], onde
W0 é o peso esperado da testemunha, concentração é a quantidade de toxina Cry1Ac
por ml de água, EC50 é a concentração efetiva capaz de reduzir 50% do peso da lagarta
e B parâmetro de angulação da função logística (SIMS; BERBERICH, 1996). Para o
calculo da EC99 e seu respectivo intervalo de confiança (IC 95%) a seguinte equação
modificada foi utilizada: peso=W0 / [(1+(100-1)(concentração / EC99)B]. Através desse
modelo foi possível obter as estimativas dos parâmetros EC50 e EC99 e seus respectivos
intervalos de confiança a 95% de probabilidade (IC95).

42
Figura 1 – A) Preparo das placas para bioensaio de imersão de discos de folhas: células contendo 500 µL de solução de ágar a 2,5%; camada de papel filtro sobre a camada de ágar; discos de folhas que foram imersos em soluções de Cry1Ac (µg de I.A. / ml de água); B) transferência de lagartas de A. argillacea para cada célula após a secagem da solução contendo toxina Cry1Ac; e C) material utilizado para a realização do bioensaio de imersão de discos de folhas: a) pincel para a transferência das lagartas; b) filme plástico para selagem das placas
A
B
C

43
2.2.3.2 Determinação e validação da concentração diagnóstica para o
monitoramento da suscetibilidade
As duas abordagens para a caracterização da suscetibilidade permitiram
determinar as concentrações diagnósticas que foram utilizadas no monitoramento da
suscetibilidade de A. argillacea à toxina Cry1Ac. Foi feita uma regressão de Probit
agrupando todos os dados obtidos com as caracterizações das linhas-básicas de
suscetibilidade das populações da safra agrícola de 2007/2008. As estimativas dos
parâmetros: CL50, CL95 e CL99 (concentração letal de 50, 95 e 99%, respectivamente),
intervalo de confiança a 95% (IC 95%) para cada concentração letal e o coeficiente
angular (± erro padrão) da regressão de Probit também foram obtidas (item 2.2.3.1). Foi
feita a comparação dos resultados obtidos com a caracterização da linha–básica de
suscetibilidade com os dados obtidos com a regressão para o modelo de resposta de
inibição de crescimento.
Os procedimentos de bioensaio foram os mesmos utilizado para a
caracterização da linha-básica de suscetibilidade, Foram testadas aproximadamente
500 lagartas neonatas, divididas em repetições de 12 lagartas. Foram estimados a
porcentagem de mortalidade em cada repetição e a média dessa estimativa e seu
respectivo erro padrão em cada população estudada.
2.2.4 Desenvolvimento e caracterização dos microssatélites
2.2.4.1 Extração e quantificação de DNA
A extração de DNA foi realizada com adultos provenientes de lagartas coletadas
no campo. Esses adultos foram armazenados a -20ºC até o momento da extração de
DNA. Para a extração do DNA foi utilizado o protocolo CTAB descrito por Doyle e Doyle
(1990) com modificações. O DNA foi extraído a partir do tórax de indivíduos adultos de
A. argillacea macerados da seguinte maneira: (1) Em tubos plásticos de tipo

44
“Eppendorff’s” de 1,5 mL identificados foi adicionado um tórax, devidamente separado
do restante do corpo; (2) foram adicionados 500 µL de tampão de extração CTAB pré-
aquecido, os tubos foram fechados e agitados para ressuspender o tecido no tampão;
(3) foram levados ao banho-maria (65ºC) por 60 minutos, agitando-os manualmente a
cada 10 minutos; (5) após serem retirados do banho-maria e com a mistura em
temperatura ambiente foram adicionados 700 µL de clorofórmio-álcool isoamílico (24:1)
e os tubos foram agitados por 10 minutos invertendo-os no mínimo 20 vezes até fazer
uma emulsão homogênea; (6) os tubos foram centrifugados por 5 minutos a 12000 rpm;
(7) o sobrenadante foi transferido para novos tubos também marcados; (8) foram
adicionados 2,5x do volume do volume da solução (≈ 1500 µl) de isopropanol gelado,
que foi misturado suavemente ao sobrenadante que estava no tubo e esse foi deixado
em repouso, por 60 minutos, a -20°C, para formar um precipitado; (9) os tubos foram
centrifugados durante 10 minutos a 12000 rpm; (10) suavemente foi descartado o
máximo de sobrenadante invertendo os tubos sem perder o pélete; e após a secagem
do precipitado à temperatura ambiente; (11) foram adicionados 300 µL de etanol 70%
(v/v) para lavar o precipitado, deixando-o imerso por 5 a 10 minutos, invertendo-o
suavemente; (12) os tubos foram centrifugados durante 5 minutos a 12000 rpm; (13) foi
retirado o máximo do etanol sem perder o pélete; (14) foi adicionado novamente 300 µL
de etanol 70% (v/v) para lavar o precipitado, invertendo-o suavemente; (15) os tubos
foram centrifugados durante 5 minutos a 12000 rpm; (16) foi retirado o máximo do
etanol e o pellet secou em temperatura ambiente; (17) cada precipitado foi dissolvido
em 50 µL de tampão TE (10 mM Tris-HCl, 1 mM de EDTA, pH 8,0) acrescido de 1%
RNAse (10mg/mL) e deixando-se sobre a bancada no decorrer da noite. As amostras
foram então armazenadas a –20ºC.
A quantidade de DNA presente em cada amostra foi estimada pela intensidade
de fluorescência emitida pelo corante Sybr Safe® (Invitrogen) sob UV em géis de
agarose a 0,8% (p/v). Essa intensidade foi comparada a de padrões com pesos
moleculares e concentrações específicas e conhecidas de DNA (DNA do fago λ).

45
2.2.4.2 Construção de biblioteca genômica enriquecidas em microssatélites
Para a construção da biblioteca genômica enriquecida em microssatélites foram
utilizados DNA do tórax de dois indivíduos adultos de Alabama argillacea que foi
extraído utilizando o protocolo descrito anteriormente. O protocolo utilizado para a
construção da biblioteca foi o descrito por Billotte et al. (1999), com modificações.
Inicialmente foi feita a digestão do DNA genômico utilizando enzima de restrição. A
digestão de aproximadamente 30,0 μg de DNA (1000 ng/μL) foi feita utilizando 5,0 μL
enzima RsaI (10 U/ μL). Foram adicionados na reação 10,0 μL de tampão de reação (50
mM Tris-HCL, p H7,8; 10 mM MgCl2; 10 mM DTT; 25 μg/ml BSA), 10,0 μL de
espermidina (40mM) e água milliQ autoclavada para completar 100 μL. O material foi
incubado a 37º C por 6 horas.
Em seguida foi realizada a ligação dos adaptadores. Os adaptadores Rsa21 (5’
– CTC TTG CTT ACG CGT GGA CTA – 3’) e Rsa25 (5’ – TAG TCC ACG CGT AAG
CAA GAG CAC A – 3’) foram ligados aos fragmentos digeridos de extremidade abrupta
com a enzima T4 DNA ligase. Para isso foram utilizados 1,5 μL do adaptador Rsa21 (10
μM) e 1,5 μL do adaptador Rsa 25 (10 μM), 5,0 μg do DNA genômico digerido, 5,0 μL do
tampão 5X (50 mM Tris-HCL, pH7,8; 10 mM MgCl2; 10 mM DTT; 25 μg/ml BSA), 2,0 μL
de T4 DNA ligase (1U / μL) e água milliQ autoclavada para completar 25,0 μL. Esta
reação foi incubada a 20°C por 2 horas.
Após a ligação dos adaptadores foi realizada uma reação de amplificação para
gerar uma maior quantidade de DNA para a seleção. Os fragmentos foram amplificados
utilizando 5,0 μL do produto da ligação; 2,0 μL do primer Rsa21 (10 μM), 5 μL de
tampão 10 X (50 mM KCl; 10 mM de Tris-HCl pH 8,9); 1,5 μL de MgCl2 (50, mM); 4 μL
(2,5 mM) de dNTP) 5,0 μL (3U) de Taq polimerase e água milliQ autoclavada para
completar 50,0 μL. Esta reação foi submetida a uma etapa inicial de desnaturação a
95°C por 4 minutos, seguida de 20 ciclos de 94°C por 30 segundos, 60°C por 1 minuto e
72°C por 1 minuto, e uma etapa de extensão de 72°C por 8 minutos foi adicionado após
o último ciclo. Os fragmentos amplificados foram purificados utilizando o Kit “Qiaquick
PCR purification kit” (Qiagem).

46
A seleção dos fragmentos contendo seqüências ricas em GA/CT, AT/TA e
CA/GT foi feita por meio de hibridização com sondas biotiniladas, biotina - III(CT)8 e
biotina - III(GT)8 e recuperados com estreptavidina ligadas a “beads” magnéticas
utilizando o kit Streptavidine-Magnesphere da Promega (Promega, USA). Para
realização das lavagens, as esferas magnetizadas foram atraídas por um imã
posicionado lateralmente ao tubo em um suporte. A fração enriquecida contendo os
fragmentos de DNA previamente ligados aos adaptadores foi incubada a 95°C por 15
minutos para a desnaturação da dupla fita. Ao DNA desnaturado foram adicionados 3 µL
de cada oligonucleotídeo marcado com as biotinas. Esta solução de hibridização foi
incubada a temperatura ambiente sob constante agitação e após 20 minutos foi
adicionada às esferas magnetizadas previamente lavadas seguindo as recomendações
do fabricante. A suspensão contendo as esferas magnetizadas e o complexo DNA-
sonda foi incubada por 10 minutos à temperatura ambiente sob suave agitação. Após a
incubação, foram realizadas lavagens conforme descrito por Billotte et al. (1999) e os
fragmentos selecionados foram recuperados através de lavagem com 250 µL de água
milliQ autoclavada.
Após a seleção dos fragmentos enriquecidos, estes foram submetidos a uma
PCR utilizando como primer o adaptador Rsa21. A amplificação foi realizada utilizando
20 µL do produto da ligação; 4,0 µL do primer Rsa21 (20 µM); 10 µL de tampão 10 X (50
mM KCl; 10 mM de Tris-HCl pH 8,9); 4,0 µL de MgCl2 (2,0 mM); 8 µL de dNTP (2,5
mM), 5U de Taq polimerase (Invitrogen) e água milliQ autoclavada para completar 100,0
µL. Esta reação foi submetida a uma etapa inicial de desnaturação a 95°C por 1 minuto,
seguida de 25 ciclos de 94°C por 40 segundos, 60°C por 1 minuto e 72°C por 2 minutos.
Um passo prolongado de extensão de 72°C por 5 minutos foi adicionado após o último
ciclo.
Em seguida, 3 µL dos produtos da PCR foram ligados a 1 µL do vetor pGEM-T
(kit Promega), segundo o protocolo fornecido com o vetor plasmidial, e foram
transformados em XL1-Blue SuperCompetent. As células transformadas foram
plaqueadas em meio LB sólido contendo ampicilina (100 µg/ml), 60 µL IPTG (24 mg/ml)
e 60 µL X-Gal (20 mg/ml). As placas foram incubadas invertidas a 37 °C por 18 horas
em estufa para o crescimento de colônias.

47
Os clones positivos (colônias brancas) foram selecionados com a ajuda de
palitos estéreis. Cada colônia selecionada foi colocada em dos poços de uma placa de
ELISA com fundo em U com 80 µL de meio LB contendo ampicilina (100µg/ml). As
placas foram incubadas a 37 °C overnight em estufa para o crescimento de colônias.
Após esse período as placas foram armazenadas em freezer -20°C por 30 minutos e
então foram armazenadas em refrigerador -80°C com glicerol 50% (p/v).
Após o crescimento das colônias foi realizada uma reação de amplificação dos
insertos diretamente das colônias obtidas na etapa anterior, utilizando o primer Rsa21,
com o objetivo e verificar a eficiência do procedimento de enriquecimento e clonagem.
Colônias individuais foram transferidas com um palito estéril para o tubo de PCR
contendo 30,5 µL de água milliQ autoclavada. Esta suspensão de células foi utilizada
em uma reação com volume final de 45,0 µL contendo as seguintes reagentes: 5,0 µL
de tampão 10X (50 mM KCl; 10mM de Tris-HCl, pH 8,9), 4,0 µL de MgCl2 (2,5 mM), 4,0
µL de dNTP (2,5 mM), 2,5 µL de Rsa21 (10 mM), 1,0 µL de Taq DNA polimerase (5
U/µL). Esta reação foi submetida a uma etapa inicial de desnaturação a 95ºC por 4
minutos, seguido de 30 ciclos de 94ºC por 30segundos, 52ºC por 45segundos, 72ºC por
1 minuto e 30 segundos e uma etapa final de extensão a 72ºC por 8 minutos. Para
controle, 10 µL do volume da reação foram utilizados na eletroforese em gel de agarose
1,5% (p/v).
Com o objetivo de isolar o DNA plasmidial das colônias recombinantes para
posterior sequenciamento foi colocado 1 ml de meio Circle Grow contendo 100 µg/ml de
ampicilina em cada pocinho da microplaca; foram inoculadas colônias individuais com o
auxílio de pipeta multicanal; a placa foi selada com adesivo e furada com o auxilio de
uma agulha para aeração durante o crescimento; esta placa foi incubada a 37ºC em
agitador a 300 rpm durante 22h; o adesivo foi trocado e a placa foi centrifugada por 6
minutos a 3000 rpm, para sedimentar as células; o sobrenadante foi descartado e a
placa foi mantida invertida sobre papel absorvente por 1 minuto; foi adicionado a cada
pocinho 240 µL de solução GTE [Glicose 20%, Tris 1 M (pH 7,4), EDTA 0,5 M (pH 8,0)],
a placa foi selada com adesivo e feita a ressuspensão das células agitando-as no vortex
por 2 minutos; a placa foi centrifugada por 6 minutos a 4000 rpm e o sobrenadante foi
descartado; foi adicionado a cada pocinho 80 µL de GTE, a placa foi selada com

48
adesivo para etanol e agitada no vortex por 5 minutos; foi transferido 60 µL de cada
suspensão de células para placa com fundo em U contendo 5 µL de RNAse (10 mg/mL);
foram adicionados a cada pocinho 60 µL de NaOH 0,2M – SDS 1% (p/v), a placa foi
selada com adesivo e a solução foi misturada 10 vezes por inversão e incubada 10
minutos à temperatura ambiente; após esse período a placa foi centrifugada
brevemente; foram adicionados a cada pocinho 60 µL de KOAc 3 M, a placa foi selada
com adesivo e a solução foi misturada 10 vezes por inversão, centrifugada brevemente;
o adesivo foi removido e a placa incubada em estufa a 90ºC por exatos 30 minutos; a
placa foi esfriada em gelo por 10 minutos e centrifugada por 4 minutos a 4000 rpm; foi
fixada com fita adesiva uma placa filtro no topo de uma placa de fundo em U, foi
transferido todo o volume para a placa filtro, e centrifugada por 4 minutos a 4000 rpm, a
20ºC; foi removida a placa filtro e adicionado ao filtrado 110 µL de isopropanol; a placa
foi selada com adesivo e a solução foi misturada 20 vezes por inversão e centrifugada
por 45 minutos a 4000 rpm a 20ºC; o sobrenadante foi cuidadosamente descartado e
foram adicionados 200 µL de etanol 70% (v/v) gelado; a placa foi centrifugada por 5
minutos a 4000 rpm a 20ºC, e o sobrenadante descartado; a placa foi invertida sobre
papel absorvente, centrifugada invertida por 600 rpm. A placa secou por 60 minutos à
temperatura ambiente e o pellet foi ressuspendido em 60 µL de água milliQ (overnight).
2.2.4.3 Seleção, seqüenciamento dos clones positivos e desenho de primers
Os fragmentos contendo clones positivos para SSR foram extraídos das
colônias que apresentaram o incerto (colônias brancas) e os plasmídeos foram
seqüenciados em seqüenciador automático. Foi realizada uma reação com 1μg do
plasmídeo, 8 μL do Big-Dye v.3.1 (Applied Biosystem) e água milliQ para completar
20μL. Os cromatogramas, produzidos pelo sequenciador, foram analisados, a fim de
checar a qualidade do seqüenciamento, utilizando o programa CHROMAS 2.2.1
(http://www.technelysium.com.au/chromas.html).
Os pares de primers que flaqueam as regiões de microssatélites foram
desenhados com o auxilio dos seguintes programas: WEBTROLL
(http://www.bioinformatica.ucb.br/troll.html), utilizado para identificar as regiões de
sequências repetitivas, ou seja, regiões que contem os motivos microssatélites repetidos

49
em tandem nos fragmentos seqüenciados; PRIMER3 (http://fokker.wi.mit.edu/cgi-
bin/primer3/primer3_www_slow.cgi), que foi utilizado para localizar as regiões que
flanqueiam as sequências microssatélites e posteriormente a obtenção de sugestões de
primers que mais se adéquam aos critérios como: temperatura de anelamento dos
oligonucleotídeos, variando entre 55 e 70 ºC, a diferença de temperatura de anelamento
entre os pares de primers de no máximo de 3 ºC e conteúdo GC no mínimo de 50% e
máximo de 60%; GENE RUNNER (http://www.generunner.net/), utilizado para testar a
qualidade dos primers pré-selecionados; e novamente o programa CHROMAS 2.2.1 que
também foi utilizado para testar a qualidade dos primers, contudo este teste é realizado
na própria sequência genômica seqüenciada, a fim de observar se os primers que foram
desenhados acessam uma região que foi bem seqüenciada, permitindo assim ter uma
maior certeza sobre a qualidade e precisão desses primers obtidos.
2.2.4.4 Amplificações iniciais e otimização dos primers
Primeiramente foram feitos testes para se determinar as melhores condições de
amplificação dos primers desenhados. Esses testes iniciais foram necessários para se
determinar a reação e o ciclo de amplificação que puderam ser utilizadas nas
otimizações dos primers. As amplificações iniciais foram feitas com cinco indivíduos
provenientes das populações naturais e 3 primers escolhidos ao acaso Em uma reação
de amplificação utilizando um dos primers selecionados, para um volume final de 20µL,
foram utilizados cerca de 10 ηg do DNA genômico; 0,4 µL do primer foward (10 µM) e
0,4 µL do primer reverse (10 µM); 2,5 µM de cada desoxirribonucleosídeo trifosfato
(dNTP); 2,0 µL solução tampão Buffer 10 X (50 mM de KCl; 10 mM de Tris-HCl, pH 8,9);
2,0 µL de MgCl2 (2,5 mM); 5,0 µL de BSA (2,5µg / ml) e 1 U de Taq DNA polimerase.
As reações de amplificação foram realizadas em um termociclador MyCyclerTM (BIO-
RAD, Inc.) programado para uma desnaturação inicial de 2 minutos a 95ºC, seguido de
45 ciclos com 1 minuto de desnaturação a 95ºC, 1 minuto de anelamento a 60ºC e 1
minuto de extensão a 72ºC, finalizando com uma etapa de extensão final de 5 minutos a
72ºC.
Para a determinação das condições ideais de amplificação de cada primer
sintetizado, as reações foram submetidas a diferentes temperaturas de anelamento. Foi

50
realizado um teste inicial utilizando quatro temperaturas (48, 52, 56 e 60°C), o mesmo
ciclo, para os 23 pares de primers, em três indivíduos das populações naturais
escolhidos ao acaso. Outros testes também foram realizados para otimização dos
primers que não apresentaram amplificação no teste com as quatro temperaturas. Esses
testes foram realizados submetendo as reações de PCRs com variação de temperatura
em gradiente, variação nas concentrações de MgCl2, DNA e BSA. O objetivo dessas
otimizações foi de obter um mínimo de 10 primers que acessam locos microssatélies
polimórficos. Os produtos da PCR foram observados em géis de agarose a 1,5%. A
partir desses resultados foi possível determinar a temperatura de anelamento dos
primers utilizados para a genotipagem dos locos microssatélites.
2.2.4.5 Caracterização dos locos microssatélites
A caracterização dos locos obtidos nas otimizações foi feita em uma população
de campo de A. argillacea com aproximadamente 20 indivíduos, escolhida ao acaso,
buscando locos altamente polimórficos. Essa caracterização foi feita em géis de
poliacrilamida 7% revelados com nitrato de prata, segundo procedimento descrito por
Creste e colaboradores (2001). Os dados obtidos após leitura dos géis foram
submetidos à análise utilizando o programa MStools, que é um suplemento
desenvolvido para Microsoft Excel®, e foi utilizado para estimar as estatísticas
descritivas de cada loco como: HO, HE, número de alelos por loco, amplitude alélica e o
PIC (Polymorphism Information Content).
2.2.5 Estudo da variabilidade genética e estrutura populacional
2.2.5.1 Genotipagem dos locos microssatélites
Os locos obtidos por meio da otimização e caracterização foram utilizados para
a genotipagem dos indivíduos das populações de A. argillacea coletadas, para o estudo
de genética de populações, em géis de poliacrilamida 7% desnaturante, corado com
nitrato de prata (CRESTE; TULMANN NETO; FIGUEIRA, 2001). O tamanho da amostra
(N) utilizado foi de aproximadamente 20 adultos de cada população natural coletada. Os

51
dados de freqüência alélica e genotípica de cada loco por população, obtida através das
leituras dos géis, foram analisados utilizando os programas computacionais
desenvolvido para estudo de genética de populações.
Após a leitura dos géis foi feito um teste utilizando o programa MICRO-CHEKER
(VAN OOSTERHOUT et al., 2004) a fim de observar erros de genotipagem e a presença
de alelos nulos nos locos utilizados. A constatação de presença de alelos nulos também
foi feita através do teste para Equilíbrio de Hardy-Weinberg, onde foi utilizado como
critério para definir que um loco possui alelos nulos, quando esse não aderiu às
proporções de Equilíbrio de Hardy-Weinberg em todas as populações.
2.2.5.2 Variabilidade genética intrapopulacional
Para as estimativas dos parâmetros descritivos populacionais como: de número
médio de alelos por população (nA), número médio de alelos por loco (A),
heterozigosidade observada (HO) e esperada (HE) por loco e as heterozigosidades
observadas (HO) e esperadas (HE) médias por população, além das estimativas do
índice de fixação intrapopulacional médio por loco e por população, foi utilizado o
programa GDA (LEWIS; ZAYKIN, 2006). Foi feito o teste para equilíbrio Hardy-Weinberg
utilizando o teste de aderência (teste exato de Fisher), conforme definido por Weir
(1996) utilizando o programa TFPGA (MILLER, 1997). O valor de significância foi
ajustado realizando a correção de Bonferroni, e para isso foi dividido o valor de alpha a
95% de probabilidade (α = 0,05) pelo número de locos (10 locos). Foi realizado o teste
de desequilíbrio de ligação entre os locos, ajustando o valor de significância (p-value)
para o nível nominal 5%, através do programa FSTAT (GOUDET, 2009).
2.2.5.3 Variabilidade interpopulacional e estrutura genética populacional
Para o estudo da variabilidade interpopulacional e estruturação genética de A.
argillacea no Brasil, inicialmente foram obtidos as estimativas de diversidade e
freqüência alélica por loco e por população, utilizando o programa FSTAT (GOUDET,

52
2001). Além disso, foram obtidas as estimativas de alelos exclusivos (ou privados) de
cada loco em cada população, utilizado o programa GDA, uma vez que a distribuição
desses alelos entre as populações pode indicar a similaridade e determinar o grau de
isolamento entre as populações.
As estimativas das estatísticas F de Wright (FIS, FST e FIT) foram obtidas através
do método proposto por Weir e Cockerham (1984) (FIS ≈ f; e FST ≈ θ), utilizando o
programa GDA que assume o modelo aleatório de acordo com Weir (1996), em que as
populações amostradas são consideradas como representativas da espécie e com uma
história evolutiva comum, e que assume também o modelo de alelos infinitos. Além
disso, foram obtidas as estimativas de RST, que é uma estatística análoga ao FST de
Wright, desenvolvida por Slatkin (1995), mas que assume o modelo de mutação
stepwise para os locos microssatélites. Também foram obtidas as estimativas de FST
par-a-par, entre as 11 populações (onde FST ≈ θ), utilizando também o programa FSTAT,
com o objetivo de observar quais são as contribuições de cada pareamento de
populações para a estimativa de FST total.
Para o cálculo das distâncias genéticas de Nei (1978) e da consistência dos nós
dos agrupamentos empregando o procedimento de reamostragem por 10000 bootstrap
foi utilizado o programa TFPGA. Para a construção dos dendrogramas, a partir das
distâncias de Nei (1978) e dos valores de bootstrap, e utilizando o critério de
agrupamento UPGMA foi empregado o programa NTSYSpc 2.1.
Para aumentar a robustez das análises, também foi utilizada a estatística
bayesiana para verificar a estruturação da variabilidade genética e o fluxo gênico entre
as populações. Para isso foi utilizado o programa STRUCTURE 2.2 (PRITCHARD et al.,
2002). Para a estimativa do número de populações (K) foi utilizado o modelo de
ancestralidade “admixture”, com alelos correlacionados (“correleted”) e K variando de
um a 16, foi definido a variação de K para cinco agrupamentos a mais do número
original de populações. Para cada valor de K foram realizadas cinco interações
independentes com 50.000 permutações burn in e 500.000 permutações que utilizam o
algoritmo Monte Carlo cadeia de Markov (MCMC). Para estimar qual valore de K que se
ajusta aos dados, foi utilizado o procedimento descrito por Evanno e colaboradores

53
(2005). Os gráficos dos resultados do STRUCTURE 2.2, foram editados utilizando o
programa DISTRUCT 1.1 (ROSENBERG, 2004).
2.3 Resultados e discussão
2.3.1 Caracterização da suscetibilidade de Alabama argillacea à toxina Cry1Ac.
2.3.1.1 Linha-básica de suscetibilidade
A CL50 estimada para a toxina Cry1Ac para a população de laboratório (LAB) foi
de 0,16 µg de Cry1Ac / mL de água, com intervalo de confiança de 95% (IC 95%) de
0,09 – 0,28 e coeficiente angular (± erro padrão) de 1,49 (± 0,19). A CL50 estimada para
essa população se deve ao aumento na proporção de indivíduos resistentes, uma vez
que essa população não foi exposta à toxina. Isso se deve ao fato de que a população
LAB apresenta baixa variabilidade genética e tenha sofrido eventos de depressão
endogâmica durante o tempo que está sendo criada em laboratório. Dessa forma, a
amplitude de resposta à toxina dessa população foi baixa. Portanto, a caracterização da
resposta dessa população ao aumento da toxina serviu de controle permitindo realizar a
comparação das respostas das outras populações em relação a uma população de
laboratório que não tinha sido exposta à toxina Cry1Ac.
A caracterização da suscetibilidade através das curvas de concentração-
resposta permitiu observar a existência de variabilidade natural de até seis vezes na
resposta à toxina nas populações coletadas em diferentes locais e estados produtores
de algodão (Tabela 1). A CL50 estimada para a toxina Cry1Ac na população coletada na
Bahia, município de Barreiras (BA-1), foi de 0,96 µg de Cry1Ac / mL de água, IC 95% de
0,58 – 1,63 e coeficiente angular (± erro padrão) de 1,62 (± 0,21). Já a CL50 estimada na
população coletada na Bahia, município de Roda Velha (BA-3), foi de 0,45 µg de Cry1Ac
/ mL de água, com IC 95% de 0,37 - 0,56 e coeficiente angular (± erro padrão) de 1,40
(± 0,10). Para a população coletada em Mato Grosso, município de Campo Verde (MT-
1), a CL50 estimada para a toxina Cry1Ac foi de 0,50 µg de Cry1Ac / mL de água, com IC
95% de 0,38 - 0,64 e coeficiente angular (± erro padrão) de 1,38 (± 0,10). Para a

54
população coletada em Mato Grosso do Sul, município de Chapadão do Sul (MS-1), a
CL50 estimada para a toxina Cry1Ac foi de 0,53 µg de Cry1Ac / mL de água, com IC 95%
de 0,25 – 0,92 e coeficiente angular (± erro padrão) de 1,16 (± 0,17). E por fim para a
população coletada na Bahia, município de São Desidério (BA-4), a CL50 estimada para
a toxina Cry1Ac foi de 0,22 µg de Cry1Ac / mL de água, com IC 95% de 0,08 – 0,39 e
coeficiente angular (± erro padrão) de 1,72 (± 0,23).
Comparando os valores de CL50 percebe-se que não houve diferenças
significativas na resposta ao aumento da concentração da toxina Cry1Ac entre as
populações BA-3, MT-1 e MS-1, pois os valores de CL50 são semelhantes e os
intervalos de confiança estimados para esse parâmetro se sobrepuseram. Além disso,
quando comparado a razão relativa de suscetibilidade entre essas populações, foi
possível observar que as respostas dessas populações foram semelhantes. A
população BA-1 se assemelhou as populações MT-1 e MS-1 pela sobreposição dos
intervalos de confiança, mas foi a que apresentou os maiores valores das concentrações
letais de 50 e 95%, e a que apresentou a maior razão relativa de suscetibilidade.
Percebe-se nitidamente que essa população respondeu significativamente a toxina.
Comparando-se os intervalos de confiança das CL50s e CL95s estimadas, a resposta da
população BA-4 se comportou igualmente a resposta das populações BA-3, MT-1 e MS-
1, e diferiu da população BA-1. Interessante notar que a resposta dessa população foi à
única mais próxima da resposta da população de laboratório (Tabela 1).
Já através do teste de paralelismo de retas foi possível confirmar a hipótese que
as retas são paralelas (p >0,05). Entretanto as retas não são semelhantes (p < 0,05).
Foram testadas então as retas de cada população par-a-par, a fim de se determinar
quais delas são semelhantes e quais divergem significativamente pela comparação dos
valores de intercepto (parâmetro β) e do coeficiente angular (parâmetro α). Foram
consideradas conclusivamente semelhantes aquelas que diferiram significativamente no
teste de similaridade de retas. As populações BA-3 MT-1 e MS-1 foram
significativamente semelhantes entre si (p > 0,05). A população BA-1, apesar da análise
dos IC 95% CL50s e CL95s estimadas ter apresentado semelhança a população MS-1,
através desse teste pode se confirmar que são significativamente semelhantes(p >
0,05). Contudo BA-1 diferiu significativamente de BA-3 e MT-1 (p < 0,05). Todas as

55
populações apresentaram respostas distintas em relação à população de laboratório,
exceto a população BA-4 (p > 0,05).
As linhas-básicas de suscetibilidade das populações BA-3, MT-1 e MS-1 foram
bastante próximas. A resposta da população BA-4 foi mais próxima da população LAB
(Tabela 1, Figura 2).

Tabela 1 – Caracterização da linha-básica de suscetibilidade à toxina Cry1Ac em populações de Alabama argillacea
População N1
Coeficiente
angular
(± erro padrão)
2 g.l.2
CL50
(µg de I.A.3/ml de
água)
(95% I.C.)
CL95
(µg de I.A.c/ml de água)
(95% I.C.)
RR4
LABc 171 1,49 ± 0,19 4,31 3 0,16 (0,09 – 0,28) 1,98 (0,68 – 33,32) ---
BA-1b 176 1,62 ± 0,21 2,62 3 0,96 (0,58 – 1,63) 19,38 (7,97 – 105,21) 6,00
BA-3ª 631 1,40 ± 0,10 2,78 5 0,45 (0,37 – 0,56) 6,66 (4,75 – 10,15) 2,81
BA-4c 399 1,72 ± 0,23 6,23 5 0,22 (0,08 – 0,39) 1,28 (1,15 – 5,30) 1,37
MT-1a 588 1,38 ± 0,10 1,00 5 0,50 (0,38 – 0,64) 7,70 (5,31 – 12,47) 3,12
MS-1ab 261 1,16 ± 0,17 3,85 3 0,53 (0,25 – 0,92) 13,60 (4,04 – 466,55) 3,31 1 Número de lagartas testadas 2 graus de liberdade
3 µg de Cry1Ac / mL de água
4 Razão relativa de suscetibilidade: CL50 da população de campo / CL50 da população de laboratório
* significância ao teste de paralelismo de retas: letras iguais não apresentaram diferenças significativas à α = 0,05.

57
Figura 2 – Caracterização da linha-básica de suscetibilidade das populações de Alabama argillacea coletadas nos estados da Bahia (BA-1, BA-3 e BA-4), Mato Grosso (MT-1), Mato Grosso do Sul (MS-1) e a suscetível de referência (LAB) à toxina Cry1Ac
O conhecimento da suscetibilidade de uma espécie de inseto-praga a
inseticidas entre regiões geográficas é importante para se avaliar o risco potencial de
evolução de resistência, entender se a variação na suscetibilidade entre diferentes
populações está associada com o histórico de uso de um inseticida e para mensurar o
quanto programas de manejo da resistência estão sendo eficientes (US EPA, 2001).
Variações significativas na suscetibilidade a um inseticida entre regiões geográficas de
uma espécie geralmente indica alto riso para evolução de resistência (KINSINGER;
McGAUGHEY, 1979).
Diferenças geográficas na suscetibilidade à toxina Bt em várias espécies –
alvos do controle de plantas GM já foram reportadas, incluindo, Ostrinia nubilalis nos

58 Estados Unidos (MARÇON et al., 1999; REED; HALLIDAY, 2001) na Espanha
(GONZALEZ-NUÑEZ et al., 2000), e Alemanha (SAEGLITZ et al., 2006); Helicoverpa
zea nos Estados Unidos (SIEGFRIED et al. ,2000); e Diabrotica virgifera virgifera
LeConte no Centro-Oeste dos EUA no cinturão do milho (SIEGFRIED et al., 2005).
Contudo a variação na suscetibilidade entre populações de regiões geográficas dessas
espécies estudadas, em geral, foram relativamente baixas, com diferenças entre as
CL50s menores que sete vezes. Contudo, a variação da suscetibilidade de populações
de Diatraea saccharalis a toxina Cry1Ab presente em milho transgênico foi ligeiramente
maior (≤ 10 vezes a diferença entre CL50s) que a variação na suscetibilidade reportada
nos estudos anteriores (HUANG et al., 2008).
Já para a principal praga-alvo de controle do algodão transgênico nos Estados
Unidos, os resultados para as caracterizações das linhas-básicas de populações de
diferentes regiões geográficas se assemelharam aos resultados obtidos com as pragas
de milho. Antes da introdução do algodão Bt, uma notável variação, acima de oito
vezes, foi encontrada na suscetibilidade de Heliothis virescens de regiões geográficas
diferentes nos Estados Unidos (STONE; SIMS, 1993; LUTTRELL et al., 1999), contudo
essas populações de campo apresentaram suscetibilidade semelhante as populações
de laboratório, que nunca tiveram contato com a toxina Bt (LUTTRELL et al., 1999).
Após vários anos de uso comercial do algodão Bt, as populações de campo de Heliothis
virescens apresentaram a mesma suscetibilidade que as populações de laboratório
(HARDEE et al., 2001; ALI; LUTRELL; YOUNG, 2006).
Comparando os dados obtidos para A. argillacea no Brasil, as populações de
campo não apresentaram diferenças na suscetibilidade maiores que seis vezes. Dentre
as populações estudadas, apenas as resposta das populações BA-1 e BA-4 se
diferenciaram das respostas das outras populações. A população BA-1 foi a que
apresentou a maior diferença em relação à população suscetível de laboratório, sendo
essa diferença maior que seis vezes. Contudo, comparando essa população com as
outras, a diferença na suscetibilidade não ultrapassa cinco vezes. Já a população BA-4,
que foi coletada no estado da Bahia, em uma região próxima a da população BA-1, foi a
que apresentou a suscetibilidade mais próxima da população de laboratório. Curioso,
que essa população diferiu das outras em relação à safra de coleta. Enquanto que as

59
outras foram coletadas na safra agrícola de 2007/2008, essa ultima foi coletada na safra
agrícola de 2008/2009, mais precisamente no inicio do ano de 2009.
As diferenças na suscetibilidade a toxina Bt entre populações de A. argillacea
observadas aqui estão mais ligadas a variações naturais entre as populações do que
causada por pressões seletivas causada pela exposição à toxina Bt. Como reportado
para outras espécies de insetos-pragas, essas diferenças podem estar relacionadas a
diferenças no valor adaptativo (crescimento e desenvolvimento) de cada população
(ROSSITER; YENDOL; DUBOIS, 1990; MARÇON et al., 1999), variações incontroláveis
nas condições dos bioensaios, ou outros fatores não genéticos (SIMS et al., 1996).
Portanto, os resultados obtidos com as caracterizações das linhas-básicas das
populações de campo de A. argillacea sugerem que estão ainda permanecem
suscetíveis à toxina Cry1Ac, mesmo após quatro anos do início da utilização da
tecnologia do algodão GM. Os dados obtidos com as caracterizações das
suscetibilidades foram utilizados para se estabelecer as concentrações diagnósticas
que poderão ser utilizadas em programas de monitoramento da evolução da resistência
de A. argillacea a toxina Bt. Além disso, as linhas-básicas obtidas também poderão ser
utilizadas como referencia para se compara as alterações na suscetibilidade de outras
populações da espécie, coletadas futuramente, como um indicativo de potencial de
evolução de resistência da espécie a toxina Cry1Ac.
2.3.1.2 Caracterização da curva de concentração–resposta de inibição de
crescimento
Para as populações BA-3 e MT-1 os valores de peso médios obtidos se
ajustaram ao modelo de regressão logístico não–linear e assim permitiu estimar as
EC50 e EC99 (concentração capaz de inibir o crescimento em 50% e 99%,
respectivamente) para cada população (Tabelas 2 e 3).
Para a população coletada na Bahia, município de Roda Velha (BA-3), a EC50
estimada foi de 0.093, IC 95% de 0,061 – 0,125. Já a EC99 estimada foi de 29,65 com
IC 95% de -4,82 – 64,12 (tabela 2). Já para a população proveniente do município de
Campo Verde, Mato Grosso (MT-1), a EC50 estimada foi de 0,067 com IC 95% de 0,048

60 – 0,086. A EC99 estimada para essa população foi de 17,27 com IC 95% de -0,15 –
34,70 (tabela 3). É possível observar que existe variabilidade na resposta de inibição de
crescimento entre as duas populações. Mesmo havendo sobreposição dos intervalos de
confiança (IC 95%) da estimativa da EC50 entre as populações, os valores de cada
concentração foram significativamente diferentes e, além disso, o IC 95% da EC50 da
população MT-1 não abrange o valor de EC50 estimado para a população BA-3. Já em
relação a EC99, apesar de o IC 95% da estimativa de EC99 da população BA-3 abranger
todo o IC 95% dessa concentração letal da população MT-1, esse intervalo é muito
extenso, não permitindo afirmar com precisão a semelhança dessa estimativa entre as
populações.
Apesar de haver variabilidade na resposta entre as duas populações avaliadas, o
comportamento das curvas obtidas pela regressão linear não-logística foi semelhante.
Isso pode ser visto na extremidade final das curvas de ambas as populações, onde a
concentração de 10 µg de Cry1Ac / mL foi capaz de reduzir o desenvolvimento em
99%. (Figuras 3 e 4)..

61
Tabela 2 – Modelo de regressão não-linear para estimativa dos parâmetros EC50 e EC99 para a população de A. argillacea da Bahia (BA-3)
Fonte g.l.a Soma de
Quadrados Quadrado
Médio p-valueb
Modelo 3 34159,9 11386,6 < 0,0001
Erro 5 90,5659 18,1132
Total não corrigido
8 34250,4
Parâmetro Estimativa Erro padrão Intervalo de Confiança 95%
(IC 95%) W0c 136,9 4,2381 126,0 – 147,8
Bd 0,7972 0,0673 0,624 – 0,970
EC50e 0,0931 0,0125 0,061 – 0,125
EC99f 29,649 13,409 -4,818 – 64,116
a graus de liberdade
b valor de significância ao ajuste ao modelo de regressão logístico não - linear
c peso esperado da testemunha d parâmetro de angulação da função logística e concentração efetiva capaz de reduzir 50% do peso da lagarta f concentração efetiva capaz de reduzir 99% do peso da lagarta

62
Tabela 3 – Modelo de regressão não – linear para estimativa dos parâmetros EC50 e EC99 para a população de A. argillacea Mato Grosso (MT-1)
Fonte g.l. Soma de
Quadrados Quadrado
Médio p-value
Modelo 3 18165,9 6055,3 < 0,0001
Erro 5 37,0692 7,4138
Total não
corrigido 8 18202,9
Parâmetro Estimativa Erro padrão Intervalo de Confiança 95%
(IC 95%) W0 104,8 2,7225 97,782 – 111,8
b 0,8285 0,0634 0,665 – 0,991
EC50 0,0674 0,00748 0,048 – 0,086
EC99 17,2770 6,7799 -0,151 – 34,705 a graus de liberdade b valor de significância ao ajuste ao modelo de regressão logístico não - linear c peso esperado da testemunha d parâmetro de angulação da função logística e concentração efetiva capaz de reduzir 50% do peso da lagarta
f concentração efetiva capaz de reduzir 99% do peso da lagarta

63
Figura 3 – Caracterização da resposta de inibição de crescimento para a população de A. argillacea do estado de Bahia (BA-3) à toxina Cry1Ac

64
Figura 4 – Caracterização da resposta de inibição de crescimento para a população de A. argillacea do estado de Mato Grosso (MT-1) à toxina Cry1Ac
As caracterizações das respostas de inibição de crescimento para as
populações BA-3 e MT-1 permitiram estimar com maior precisão as concentrações
diagnósticas para o monitoramento da evolução da resistência de A. argillacea a
proteína inseticida Cry1Ac. Apesar de haver variabilidade na respostas de inibição de
crescimento das duas populações, as estimativas da EC99 juntamente com os
respectivos IC 95% para cada população serviram de confirmação para as
concentrações letais determinadas como concentrações diagnósticas. A confiabilidade
desse método de bioensaio em identificar genótipos resistentes em populações
naturais quando esses se encontram em baixa freqüência foi levada em consideração
na utilização das estimativas dos parâmetros EC50 e EC99 para auxiliar a escolhas das
concentrações diagnósticas. Além disso, esse método de bioensaio permite uma
adequada discriminação entre lagartas suscetíveis de resistentes utilizando uma
concentração muito menor da toxina (SIMS et al., 1996)

65
2.3.1.3 Determinação e validação da concentração diagnóstica para o
monitoramento da suscetibilidade
A caracterização da linha-básica de suscetibilidade e da resposta de inibição de
crescimento permitiu se determinar com maior precisão as concentrações diagnósticas
com potencial para serem utilizadas no monitoramento da alteração da suscetibilidade
de A. argillacea ao uso de plantas GM que expressão a toxina Cry1Ac. Entretanto,
percebe-se que existe uma grande amplitude de variação da resposta à toxina entre as
populações. Por isso, foi então feita caracterização da linha-básica agrupando em um
mesmo conjunto de dados todos os valores de mortalidade obtidos com as
caracterizações das linhas-básicas das quatro populações de campo da safra 2007-
2008 (BA-1, BA-3, MS-1, MT-1) (Figura 5). Isso foi feito objetivando aumentar a
sensibilidade da análise de regressão e da estimativa da dos parâmetros: CL95 e CL99;
coeficiente angular e IC 95% das concentrações letais (Tabela 4).
Com as caracterizações das linhas-básicas das populações coletadas na safra
agrícola de 2007/2008 pode se estimar a CL95 que poderia ser escolhida como
concentração diagnóstica. Observando os valores obtidos, essa concentração está
próximo a de 10 µg de Cry1Ac / mL de água. Os limites superiores dos IC 95% de das
CL95 e CL99 foram escolhidos como ponto de partida para determinação das
concentrações diagnósticas. Para aumentar a probabilidade de escolha de
concentrações diagnósticas ideais, valores totais de EC99 (concentração que
efetivamente causa redução em 99% no desenvolvimento larval) para a toxina Cry1Ac
também foram estimados. Os valores estimados para a EC99 das populações BA-3 e
MT-1 e a análise das curvas obtidas com o modelo de regressão logístico não-linear,
permitiram definir as concentrações diagnósticas com mais acuidade, uma vez que os
bioensaios de resposta de inibição de crescimento são mais sensíveis em se detectar
genótipos resistentes (SIMS et al., 1996). As concentrações diagnósticas definidas
foram as que determinam os menores valores de peso médio das lagartas e estão
próximo aos valores de EC99 determinados.
Sendo assim, com base na análise de Probit dos dados agrupados foi possível
determinar as concentrações de 10 e de 32 µg de Cry1Ac / mL de água como

66 concentrações diagnósticas, que correspondem respectivamente as CL95 e CL99 da
caracterização da linha-básica dos dados agrupados.
Um dos componentes mais importantes da estratégia de manejo da resistência é
a habilidade de monitorar efetivamente a evolução da resistência em populações da
praga alvo de controle (DENNEHEY, 1987). A habilidade em se detectar a resistência é
necessária para (1) se determinar quando as falhas no controle são determinadas pela
presença de indivíduos resistentes na população, (2) avaliar a extensão e a distribuição
das populações resistentes, e (3) testar a efetividade dos programas de manejo em
reduzir a freqüência dos indivíduos resistentes (HALLIDAY; BURNHAM, 1990).
A resistência a inseticidas químicos e microbianos tem sido tradicionalmente
monitorada e comparada utilizando caracterizações de linhas-básicas de populações de
campo obtidas através da regressão de Probit. Essa abordagem permite calcular a
razão de resistência (CL50 da testada dividida pela CL50 da população suscetível de
referência) e a comparação estatística das CL50s e do coeficiente angular obtido com a
regressão de Probit (ROBERTSON; PREISLER, 1992). Similarmente, estudos para se
caracterizar linha- básica de insetos alvos de controle de plantas GM gerou estimativas
de CL50s e coeficiente angulares de diferentes populações expostas à toxina Bt.
Contudo as estimativas de CL50s e de coeficiente angulares permitem distinguir
fenótipos resistentes em populações naturais quando estes estão já em alta freqüência,
e essa abordagem não é suficientemente sensível para detectar a presença de
indivíduos resistentes quanto são raro nas populações (freqüência entre 10-4 e 10-3)
(ROUSH; MILLER, 1986; HALLIDAY; BURNHAM, 1990). Uma alternativa a
caracterização e comparação de linhas-básicas de suscetibilidade envolvem a
determinação e o uso de concentrações diagnósticas ou discriminatórias em programas
de monitoramento (FFRENCH-CONSTANT; ROUSH, 1990). Com o uso de
concentrações diagnósticas, um número maior de insetos é testado apenas nas
concentrações mais informativas sobre as mudanças nas freqüências dos fenótipos
resistentes em populações naturais, conseqüentemente os resultados podem ser
obtidos de uma forma rápida de precisa (ROUSH; MILLER, 1986)
Após a determinação das concentrações diagnósticas deveriam ter sido feitas a
validação das mesmas, realizando bioensaios de imersão de discos de folhas contendo

67
toxina Cry1Ac apenas nas concentrações determinadas. Contudo, por problemas
ocorridos durante a criação em laboratório das populações de A. argillacea obtidas de
campo, na safra agrícola de 2008/2009, foi possível apenas validar a concentração de
10 µg de Cry1Ac / mL de água.
Essa validação foi feita apenas em uma população, utilizando em torno de 500
indivíduos da população da Bahia, coletada no município de São Desidério. A
porcentagem de mortalidade 97,08% (± 1,00) na concentração diagnóstica. Contudo,
diferenças nos valores de mortalidade em populações naturais, utilizando essa
metodologia são esperadas (MARÇON et al., 2000). Entretanto, em programas de
manejo da resistência onde os objetivos são identificar os genótipos resistentes quando
esses são menos raros na população, a determinação das concentrações diagnósticas
para o monitoramento das mudanças na suscetibilidade deve ser criteriosa e mais
precisa possível. Pois, mesmo pequenos erros na estimativa de CL99 e EC99 podem
causar erros consideráveis na estimativa da freqüência de indivíduos resistentes
quando essas concentrações são utilizadas como concentrações diagnósticas (ROUSH;
MILLER, 1986). Portanto, a concentração diagnóstica final pode apenas ser designada
após a validação empírica feita testando em várias populações da praga (MARÇON,
2000). Sendo assim, para uma melhor determinação dessas concentrações há a
necessidade de validar essas em outras populações naturais de A. argillacea.

68
Figura 5 – Caracterização da linha-básica de suscetibilidade à toxina Cry1Ac mediante agrupamento de dados obtidos com as caracterizações de curvas de concentração-resposta de quatro populações de A. argillacea coletadas nas safras 2007/2008.

Tabela 4 – Caracterização da linha-básica de suscetibilidade à toxina Cry1Ac mediante agrupamento de dados obtidos com as caracterizações de curvas de concentração-resposta de quatro populações de A. argillacea coletadas nas safras 2007/2008
População Na
Coeficiente
angular
(± erro padrão)
2 g.l.b
CL50
(µg de I.A.c/ml de água)
(95% I.C.)
CL95
(µg de I.A.c/ml de
água)
(95% I.C.)
CL99
(µg de I.A.c/ml de água)
(95% I.C.)
LAB 171 1,49 ± 0,19 4,31 3 0,16 (0,09 – 0,28) 1,98 (0,68 – 33,32) 5,66 ( 1,43 – 253,42)
Conjunto 1668 1,34 ± 0,63 2,29 5 0,52 (0,45 – 0,60) 8,77 (6,88 – 11,64) 28,22 (20,31 – 41,65) a Número de lagartas testada
b graus de liberdade c µg de Cry1Ac / mL de água

70 2.3.2 Desenvolvimento e caracterização dos microssatélites
2.3.2.1 Construção de biblioteca genômica enriquecidas em Microssatélites
A extração do DNA genômico a partir do tórax de indivíduos adultos de A.
argillacea forneceu uma concentração de aproximadamente 250 ηg/µL de DNA e foram
utilizados aproximadamente 5 μg na reação. Na digestão com a enzima Rsal o DNA
genômico foi digerido não foi gerado nenhum tamanho preferencial de fragmento, assim
foi produzido o perfil adequado para a construção da biblioteca. A ligação dos
adaptadores foi importante para garantir que cada fragmento tivesse uma terminação
comum e conhecida. A pré-amplificação foi realizada com sucesso, confirmando a
ligação dos adaptadores aos fragmentos e gerando uma maior quantidade de DNA para
a seleção. Não houve a amplificação aparente de fragmentos preferenciais, o que
poderia ser prejudicial nas etapas posteriores da construção da biblioteca, pois levaria a
clonagem e seqüenciamento de fragmentos preferenciais, obtenção de microssatélites
redundantes e aumentando o tempo e os custos para a identificação de microssatélites.
A eficácia do enriquecimento foi verificada através dos produtos de PCR obtidos
diretamente de colônias individuais e foi confirmada com a técnica de seqüenciamento.
Cerca de 90% dos clones analisados apresentaram presença de motivos de sequências
repetitivas, sendo que muitas das seqüências apresentavam mais de uma região
microssatélite.
2.3.2.2 Seleção, seqüenciamento dos clones positivos e desenho de primers
Dos 117 insertos que foram seqüenciados, 109 apresentaram regiões
microssatélites, sendo que muitos apresentaram mais de uma região. Foram
encontrados 236 motivos microssatélites no total e os motivos encontrados
apresentaram as seguintes freqüências: 89% perfeitos, 6% interrompidos, 2%
compostos perfeitos e 3% compostos interrompidos (Figura 6). Dentre os motivos
encontrados nas 109 sequências, os pentanucleotídeos foram os mais abundantes,
sendo 48%, seguidos pelos motivos dinucleotídeos e mononucleotídeos com 35% e

71
13% respectivamente (Figura 7). Vinte e três dessas sequências puderam ser utilizadas
para o desenho dos primers das regiões flanqueadoras dos motivos microssatélites
(Tabela 5). As seqüências restantes não foram utilizadas, pois a região flanqueadora
era também um microssatélite ou estava muito próximo de uma das extremidades da
seqüência impossibilitando o desenho dos primers. Dentre as 23 primers obtidos, 87 %
apresentaram motivos dinucleotídeos e 13 % apresentaram motivos trinucleotídeos
(Figura 8). Dentre os motivos dinucleotídeos os mais abundantes, nos microssatélites
isolados, foram às repetições AC/CA, seguida das repetições TG/GT, AG/GA TC/CT
(Figura 9).
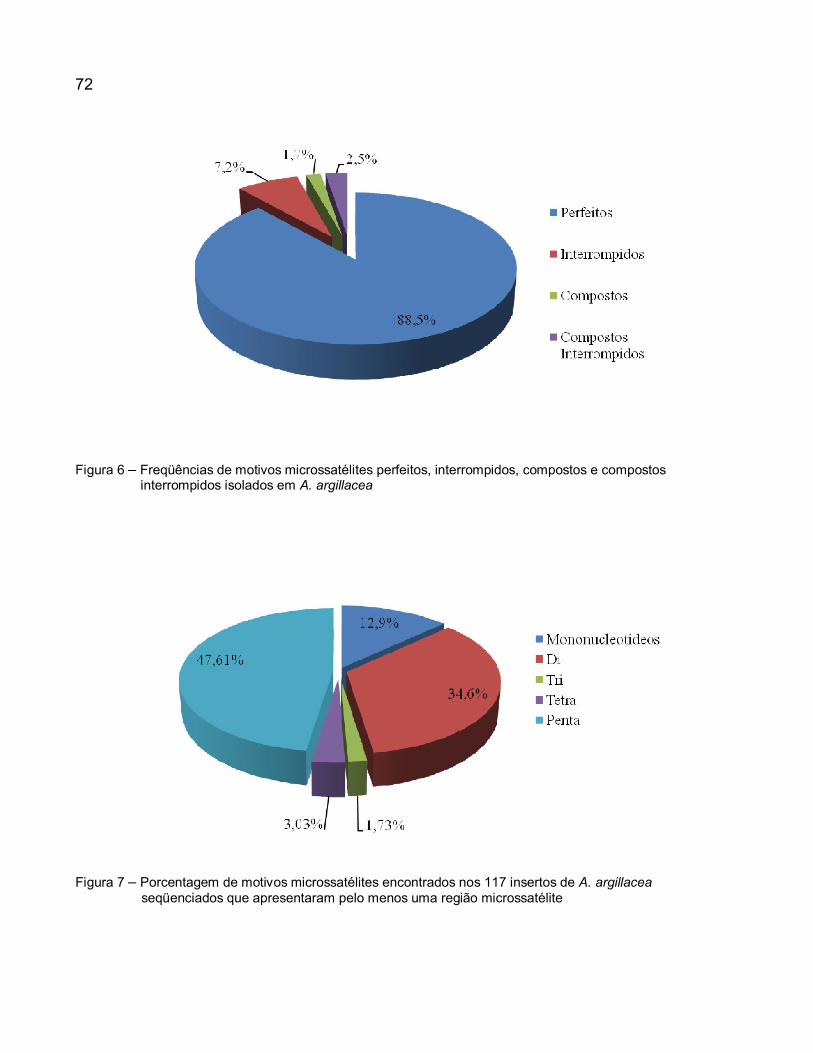
72
Figura 6 – Freqüências de motivos microssatélites perfeitos, interrompidos, compostos e compostos interrompidos isolados em A. argillacea
Figura 7 – Porcentagem de motivos microssatélites encontrados nos 117 insertos de A. argillacea seqüenciados que apresentaram pelo menos uma região microssatélite
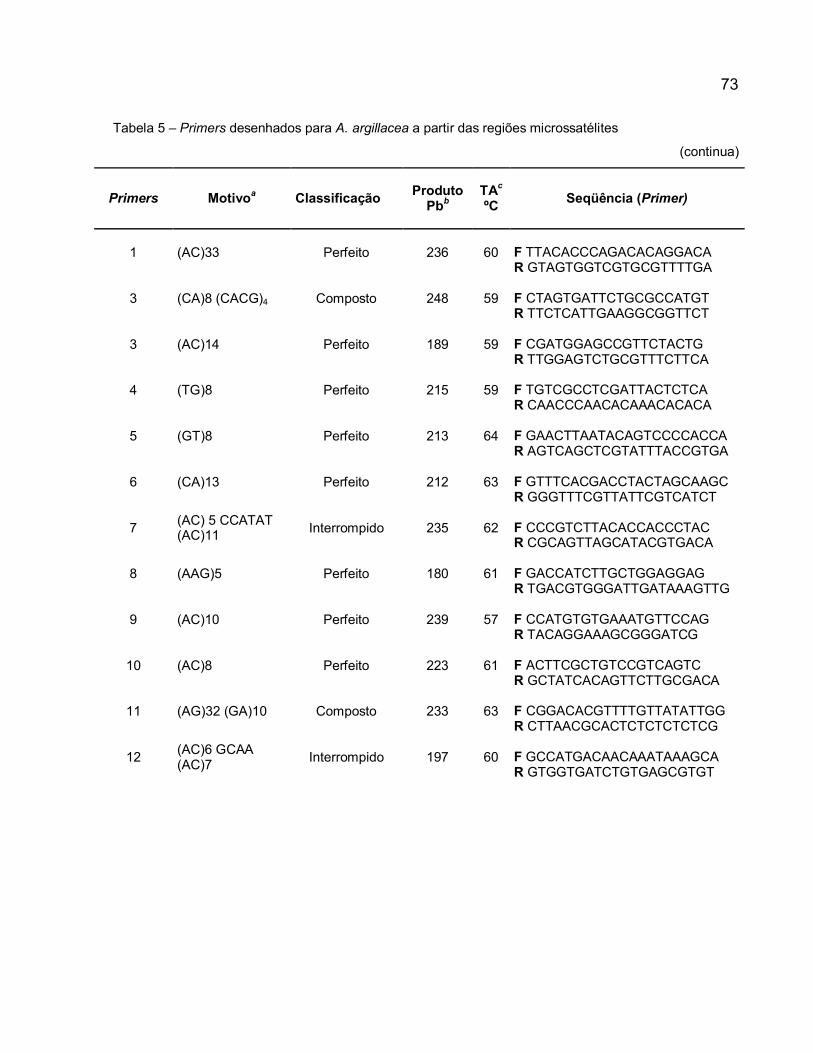
73
Tabela 5 – Primers desenhados para A. argillacea a partir das regiões microssatélites
(continua)
Primers Motivoa Classificação Produto
Pbb
TAc ºC
Seqüência (Primer)
1 (AC)33 Perfeito 236 60 F TTACACCCAGACACAGGACA R GTAGTGGTCGTGCGTTTTGA
3 (CA)8 (CACG)4 Composto 248 59 F CTAGTGATTCTGCGCCATGT R TTCTCATTGAAGGCGGTTCT
3 (AC)14 Perfeito 189 59 F CGATGGAGCCGTTCTACTG R TTGGAGTCTGCGTTTCTTCA
4 (TG)8 Perfeito 215 59 F TGTCGCCTCGATTACTCTCA R CAACCCAACACAAACACACA
5 (GT)8 Perfeito 213 64 F GAACTTAATACAGTCCCCACCA R AGTCAGCTCGTATTTACCGTGA
6 (CA)13 Perfeito 212 63 F GTTTCACGACCTACTAGCAAGC R GGGTTTCGTTATTCGTCATCT
7 (AC) 5 CCATAT (AC)11
Interrompido 235 62 F CCCGTCTTACACCACCCTAC R CGCAGTTAGCATACGTGACA
8 (AAG)5 Perfeito 180 61 F GACCATCTTGCTGGAGGAG R TGACGTGGGATTGATAAAGTTG
9 (AC)10 Perfeito 239 57 F CCATGTGTGAAATGTTCCAG R TACAGGAAAGCGGGATCG
10 (AC)8 Perfeito 223 61 F ACTTCGCTGTCCGTCAGTC R GCTATCACAGTTCTTGCGACA
11 (AG)32 (GA)10 Composto 233 63 F CGGACACGTTTTGTTATATTGG R CTTAACGCACTCTCTCTCTCG
12 (AC)6 GCAA (AC)7
Interrompido 197 60 F GCCATGACAACAAATAAAGCA R GTGGTGATCTGTGAGCGTGT

74 Tabela 5 – Primers desenhados para A. argillacea a partir das regiões microssatélites
(conclusão)
Primers Motivoa Classificação Produto
Pbb
TAc ºC
Seqüência (Primer)
13 (ATA)4 Perfeito 250 62 F TTTGTTCCATCTAAGGCTACG R GCGGACTATGAGAGTGAAGGA
14 (GT)9 Perfeito 185 58 F TCACACTGATGCGTGATTGA R TCGATTGAAAACCTCCTCCT
15 (GT)6 Perfeito 245 62 F TCCATAGAAGACAGCGATTTTG R CCCACAAACCACACTAAAGGA
16 (CT)10 CACT (ACTC)3
Interrompido 194 62 F GTCCCTATGCGTGGACAGTA R TTCTGGCTAGTTTTGTTCCCTA
17 (CA)9 A (AC)5
Imperfeito 170 56 F CTTGCGGTTTGAGGCTCT R TCCATTAAAGCGATCAATGC
18 (AC)10 Perfeito 242 56 F AGCCATGTGTGAAATGTTCC R AAGAAAAGCGGGGATCGT
19 (TG)7 (TC)5 (TG)5
Composto 166 62 F GCCAGGTTTCTCTCTCTCGAT R TGACGCACACACACAAAGAG
20 (AC)22 Perfeito 193 62 F AGTCCAAATCCAACCACGAG R GAGCACTTACTGTGTGCCAGA
21 (GAA)6 Perfeito 177 61 F CACGAAAATGCTTACAACACG R CAAAGGGTCAAAAGTCAAAACC
22 (TG)8 Perfeito 167 59 F GCACACACCGGGTCTTTAT R CTCCAAAGTCACATCGTCCA
23 (CA)7 Perfeito 239 62 F TTGACTATGGGTGGAGTGAGG R TGGTCAGACTTCCTGCACAT
a Repetição de nucleotídeos em tandem que caracterizam uma região microssatélite.
b Pares de bases.
c Temperatura de Anelamento do primer.

75
Figura 8 – Porcentagem de motivos microssatélites encontrados nos primers obtidos para A. argillacea
Figura 9 – Freqüência dos motivos dinucleotídeos que são acessados pelos primers obtidos para A. argillacea

76
O grau de eficiência, da metodologia utilizada para isolar locos microssatélites
presentes no genoma de A. argillacea pôde ser avaliada pela razão entre eficiência de
clonagem vezes a razão número de sequências com microssatélites utilizada dividido
pelo número total de seqüências clonadas com microssatélites. O resultado dessa
estimativa é de uma eficiência de 19,53%.
Já a eficiência de isolamento de locos de microssatélites através dos primers
desenhados, pode ser medida através da razão entre número total de microssatélites
em condições de serem utilizados, sobre o número total de microssatélites isolados. A
razão de eficiência foi de 9,74%.
2.3.2.3 Amplificações iniciais e otimização dos primers
Dos 23 primers sintetizados, apenas dez foram utilizados no estudo de genética
de populações. Através da primeira triagem, utilizando quatro temperaturas de
anelamento (48, 52, 56 e 60ºC), com o objetivo de abranger um espectro de condições
maior, foi possível isolar inicialmente seis pares de primers (Figura 10). As condições de
PCR utilizadas para esta séria de triagens foram às mesmas determinadas nos testes
iniciais para os primers sintetizados para A. argillacea. Contudo, desses seis, um não
apresentou polimorfismo (primer 15) em gel de poliacrilamida 7%, sendo assim, esse foi
descartados. Apenas os primers 14, 16, 19, 21 e 23 foram isolados. Pode – se ver que
esses primers foram os únicos que acessaram locos polimórficos nesse teste (Figura
11).
Na tentativa de se isolar dez pares de primers dos 23 sintetizados, uma série de
testes foi realizada. Foram então testados seis pares de primers, utilizando a mesma
reação, mas submetendo as reações em gradiente de temperatura. Três foram
posteriormente descartados, pois não apresentaram polimorfismo e as amplificações
geraram produtos de amplificação inespecíficos (05 e 20), ou não apresentaram produto
de amplificação (18) Apenas os primers 13 e 17 apresentaram polimorfismo e produtos
de amplificação satisfatório.
Após esse teste, uma série de outros foram realizados alterando as condições da
reação de PCR, como: alteração na concentração de primers, MgCl2, dNTP’s, de DNA,

77
temperatura de anelamento e número de ciclos. Desses testes foram isolados três
pares de primers: 06, 10 e 11. As condições de amplificação utilizadas para o par de
primers 06 e 11foram: para um volume final de 20µL, foram utilizados cerca de 6 ηg do
DNA genômico; 0,4 µL do primer foward (10 µM) e 0,4 µL do primer reverse (10 µM);
2,5 µM de cada desoxirribonucleosídeo trifosfato (dNTP); 2,0 µL solução tampão Buffer
10 X (50 mM de KCl; 10 mM de Tris-HCl, pH 8,9); 0,5 µL de MgCl2 (0,625 mM); 5,0 µL
de BSA (2,5µg / ml) e 3 U de Taq DNA polimerase. As reações de amplificação foram
realizadas em um termociclador MyCyclerTM (BIO-RAD, Inc.) programado para uma
desnaturação inicial de 2 minutos a 95ºC, seguido de 35 ciclos com 1 minuto de
desnaturação a 95ºC, 1 minuto de anelamento a 50ºC, para o primer 06; 1 minuto de
anelamento a 56ºC para o primers 11, e 1 minuto de extensão a 72ºC, finalizando com
uma etapa de extensão final de 5 minutos a 72ºC.
Já para o primers 10, as condições utilizadas foram: para um volume final de
20µL, foram utilizados cerca de 6 ηg do DNA genômico; 0,4 µL do primer foward (10
µM) e 0,4 µL do primer reverse (10 µM); 2,5 µM de cada desoxirribonucleosídeo
trifosfato (dNTP); 2,0 µL solução tampão Buffer 10 X (50 mM de KCl; 10 mM de Tris-
HCl, pH 8,9); 0,5 µL de MgCl2 (2,5 mM); 3,0 µL de BSA (2,5µg / ml) e 3 U de Taq DNA
polimerase. As reações de amplificação foram realizados em um termociclador
MyCyclerTM (BIO-RAD, Inc.) programado para uma desnaturação inicial de 2 minutos a
95ºC, seguido de 35 ciclos com 1 minuto de desnaturação a 95ºC, 1 minuto de
anelamento a 56ºC e 1 minuto de extensão a 72ºC, finalizando com uma etapa de
extensão final de 5 minutos a 72ºC.
Essa etapa do trabalho foi a mais demorada e a que foi mais trabalhosa. Muitos
dos testes realizados não serviram para separar pares de primers, outros apenas
serviram para se chegar próximo de uma condição ideal para a amplificação. Outros
testes determinaram as condições idéias, mas quando foi feita a caracterização dos
primers otimizados, esses não apresentaram polimorfismo ou apresentaram produtos
de amplificação inespecíficos.
Dos 23 pares de primers, dez foram apresentaram condições satisfatórias para
amplificação, quatro não apresentaram polimorfismo, e nove não apresentaram
produtos de amplificação satisfatórios e foram descartados. Portanto os primers

78 escolhidos para a genotipagem e avaliação da variabilidade e estrutura genética foram
aqueles que apresentaram maior especificidade, melhores amplificações e
polimorfísmo.

79
Figura 10 – Teste inicial utilizando os primers 13, 14, 15, 16, 17, 18, 19, 20, 21, 22 e 23; e três indivíduos de A. argillacea escolhidos ao acaso nas populações diferentes à temperatura de 60°C
Figura 11 – Gel de acrilamida 7% com os testes dos primers isolados na primeira triagem. Primers que acessam os locos Aar14, Aar15, Aar16, Aar19, Aar21 e Aar23 em A. argillacea
13 14 15 16 17 18 19 20 21 22 23
14
15
16
19 r21
23

80 2.3.2.4 Caracterização dos locos microssatélites
Os locos Aar06, Aar10, Aar11, Aar13, Aar14, Aar16, Aar17, Aar19, Aar21 e
Aar23, que são acessados pelos primers: 06, 10, 11, 13, 14, 16, 17, 19, 21 e 23,
respectivamente, foram caracterizados quanto à amplitude de variação alélica, número
de alelos, HO, HE, conteúdo de informação polimórfica (PIC) e se o loco se ajusta as
proporções de equilíbrio de Hardy – Weinberg. Para isso, foram feitas as genotipagens
de uma população (≈ 20 indivíduos) escolhida ao acaso, em gel de acrilamida 7% e
posterior leitura desses géis.
Dos dez locos caracterizados, três desses apresentaram alto grau de
polimorfismo, possuindo cada um deles de cinco a 11 alelos (Aar16, Aar19 e Aar11),
sendo o loco Aar11, que apresentou o maior valor de PIC. Cinco desses locos
apresentaram grau de polimorfismo intermediário com número de alelos variando de
três a quatro. E por fim dois locos apresentaram baixo grau de polimorfismo (locos
Aar13 e Aar23). A heterozigosidade observada (HO) variou de 0,000 a 0,630, sendo que
os menores valores pertencem aos locos Aar06 e Aar13 e o maior valor pertence ao
loco Aar19. Já a heterozigosidade esperada (HE) variou de 0,114 a 0,920, com o menor
valor atribuído ao loco Aar13, e o maior valor ao loco Aar11. Os valores de PIC
variaram de 0,104 a 0,870, sendo o menor determinado pelo loco Aar13 e o maior pelo
loco Aar11. Apenas o loco Aar06 não se ajustou as proporções de equilíbrio de Hardy-
Weinberg (p < 0,005) (Tabela 6). Os baixos valores de HO dos locos Aar06 e Aar13
foram determinados pela baixa proporção de heterozigotos nesses locos na população
estudada. Já os valores altos de HO foram determinados pela grande porcentagem de
indivíduos heterozigotos. Os valores de HE foram determinados pelo número de alelos e
pela distribuição da freqüência dos alelos em cada loco. Pode-se notar que os locos
que apresentaram os maiores valores de HE são aqueles que possuem um maior
número de alelos e uma distribuição homogênea das freqüências desses alelos na
população. Já os que possuem os menores valores de HE são os que apresentaram
menor número de alelos.
No total, foram encontrados 42 alelos distribuídos entre os dez locos
caracterizados. Os alelos encontrados em cada loco e as freqüências relativas desses

81
alelos estimados para ≈20 indivíduos de A. argillacea estão representadas graficamente
na Figura 12.

Tabela 6 – Característica dos dez locos microssatélites obtidos para Alabama argillacea: Sequência do primer forward (F) e reverse (R)
Loco Seqüências dos pares
de primers (5´- 3´) Motivo
T (°C)
Amplitude alélica (bp)
Na HOb HE
c PICd P value EHWe
Aar06 F: GTTTCACGACCTACTAGCAAGC R: GGGTTTCGTTATTCGTCATCT
(CA)13 60 196 – 200 3 0,000 0,420 0,363 0,0016*
Aar10 F: ACTTCGCTGTCCGTCAGTC R: GCTATCACAGTTCTTGCGACA
(AC)8 56 269 – 273 3 0,300 0,347 0,311 1,0000
Aar11 F: CGGACACGTTTTGTTATATTGG R: CTTAACGCACTCTCTCTCTCG
(AG)32(GA)10 56 196 – 270 11 0,333 0,920 0,870 0,0062
Aar13 F: TTTGTTCCATCTAAGGCTACG R: GCGGACTATGAGAGTGAAGGA
(ATA)4 60 250 – 259 2 0,000 0,114 0,104 0,0303
Aar14 F: TCACACTGATGCGTGATTGA R: TCGATTGAAAACCTCCTCCT
(GT)9 60 206 – 212 4 0,412 0,586 0,484 0,3280
Aar16 F:GTCCCTATGCGTGGACAGTA R:TTCTGGCTAGTTTTGTTCCCTA
(CT)10CACT(ACTC)3 62 230 – 240 5 0,579 0,815 0,760 0,6478
Aar17 F: CTTGCGGTTTGAGGCTCT R: TCCATTAAAGCGATCAATGC
(CA)9A(AC)5 60 178 – 182 3 0,350 0,304 0,265 1,0000
Aar19 F: GCCAGGTTTCTCTCTCTCGAT R: TGACGCACACACACAAAGAG
(TG)7(TC)5(TG)5 60 194 – 242 6 0,632 0,735 0,668 0,1682
Aar21 F: CACGAAAATGCTTACAACACG R: CAAAGGGTCAAAAGTCAAAACC
(GAA)6 60 180 – 186 3 0,500 0,422 0,345 1,0000
Aar23 F: TTGACTATGGGTGGAGTGAGG R: TGGTCAGACTTCCTGCACAT
(CA)7 60 310 – 312 2 0,400 0,467 0,351 0,6331 a Número de alelos encontrados no loco.
b Heterozigosidade observada.
c Heterozigosidade esperada. d Conteúdo de informação polimórfica. e Grau de significância ao teste exato de Fisher para Equilíbrio de Hardy-Weinberg – Correção de Bonferroni α=0,005. *
Não se ajusta as proporções do Equilíbrio de Hardy – Weinberg.

83
Figura 12 – Histograma das freqüências alélicas dos dez locos microssatélites, estimados para ≈20 indivíduos de A. argillacea. O eixo Y indica a freqüência alélica e o eixo X indica o tamanho do alelo, em pares de base

84 2.3.3 Estudo da variabilidade genética e estrutura populacional
2.3.3.1 Variabilidade genética intrapopulacional
Através do uso de marcadores moleculares microssatélites foi possível observar
variabilidade genética nas populações de A. argillacea estudadas. O número de alelos
encontrados nos dez locos microssatélites isolados variou de dois a 23. No total foram
identificados 71 alelos. A heterozigosidade observada média entre os locos foi de 0,541,
variando de 0,205 a 0,938. Já a heterozigosidade esperada média entre os locos foi de
0,396, com variação de 0,173 a 0,717. O índice de fixação intrapopulacional, também
chamado de coeficiente de endogamia foi de 0,268, com variação de -0,039 a 0,736.
(Tabela 7)
Já em relação a estimativas de variabilidade genética obtidas para cada
população, o número médio de alelos encontrados, foi de 4,281, variando de 3,70 alelos
ao máximo de cinco alelos. A heterozigosidade observada e esperada média foram
0,523 e 0,395, respectivamente. A maior heterozigosidade observada foi a da
população MT-3 e a menor foi a da população MS-1. Já em relação à heterozigosidade
espera, o maior valor foi o da população GO-1 e o menor valor foi da população GO-2.
Os índices de fixação intrapopulacional (f) estimados para cada população através das
heterozigosidades (HO e HE), estão apresentados na tabela 8. O valor médio foi de
0,017, com variação de 0,098 a 0,405 (Tabela 8).
Através do teste exato de Fisher foi possível verificar quais populações não se
ajustaram as proporções de equilíbrio de Hardy-Weinberg para cada loco estudado
Podemos ver que todas as populações estão em EHW para a maioria dos locos.
Contudo podemos notar que o teste Exato de Fischer deu significativo (p < 0,005) para
a maioria das populações no loco Aar06, na população MT-1 para o loco Aar11, nas
populações MS-2 e MT-3 para o loco Aar14 e apenas na população MT-3 para o loco
Aar16 (Tabela 9). O teste para verificar se existe desequilíbrio de ligação entre os locos,
nas populações estudadas, foi feito com o valor de significância (p-value) para o nível
nominal de 5% de 0,0001. Nenhum desequilíbrio foi detectado entre todos os locos (p >
0,0001).

85
Tabela 7 – Estimativas de variabilidade genética para os locos microssatélites utilizados em A. argillacea
Loco Aa HOb HE
c fd
Aar06 7,0 0,657 0,173 0,736
Aar10 7,0 0,429 0,369 0,140
Aar11 23,0 0,938 0,451 0,519
Aar13 3,0 0,205 0,178 0,130
Aar14 5,0 0,590 0,369 0,376
Aar16 6,0 0,735 0,575 0,218
Aar17 5,0 0,255 0,265 -0,039
Aar19 9,0 0,740 0,717 0,031
Aar21 4,0 0,370 0,364 0,016
Aar23 2,0 0,491 0,495 -0,008
Média 7,1 0,541 0,396 0,268 a Número de alelos total por loco polimórfico b Heterozigosidade observada por loco
c Heterozigosidade esperada por loco
d Ìndice de fixação intrapopulacional estimado para cada loco.

86
Tabela 8 – Estimativa de parâmetros descritivos de variabilidade genética nas onze populações de A. argillacea
População Na nAb HO
c HEd fe
BA-2 11 4,300 0,436 0,548 0,211
BA-3 20 4,200 0,350 0,513 0,321
BA-4 22 4,100 0,441 0,489 0,098
GO-1 20 4,500 0,434 0,559 0,225
GO-2 24 4,200 0,355 0,474 0,255
MS-1 20 4,500 0,326 0,547 0,405
MS-2 24 4,300 0,389 0,513 0,245
MT-1 20 3,700 0,376 0,523 0,281
MT-2 24 4,400 0,367 0,531 0,314
MT-3 24 5,000 0,471 0,540 0,131
PB-1 24 3,900 0,395 0,511 0,230
Média 4,281 0,523 0,395 0,248 a Número de indivíduos amostrados. bNúmero médio de alelos por população. c Heterozigosidade observada média.
d Heterozigosidade esperada média.
e Índice de fixação intrapopulacional.

87 Tabela 9 – Probabilidade do Teste Exato de Fisher para aderência ao equilíbrio de Hardy-Weinberg
População Aar06 Aar10 Aar11 Aar13 Aar14 Aar16 Aar17 Aar19 Aar21 Aar23
BA-2 0,015 0,537 0,157 1,000 0,520 0,537 1,000 0,554 1,000 0,554
BA-3 0,001 1,000 0,006 0,030 0,328 0,648 1,000 0,168 1,000 0,633
BA-4 0,130 0,353 0,076 0,025 0,659 0,034 1,000 0,213 1,000 1,000
GO-1 0,003 0,149 0,058 1,000 0,281 0,644 0,538 0,149 0,276 1,000
GO-2 0,000 0,365 0,019 1,000 0,695 1,000 1,000 1,000 0,164 0,391
MS-1 0,002 0,149 0,007 0,151 0,021 0,645 1,000 0,597 1,000 0,543
MS-2 0,000 1,000 0,012 1,000 0,005* 0,328 0,272 1,000 0,235 0,038
MT-1 0,004 0,619 0,002 1,000 0,656 0,603 1,000 0,647 1,000 0,367
MT-2 0,040 0,113 0,438 1,000 0,048 0,199 0,525 0,684 0,151 0,150
MT-3 0,089 0,538 1,000 0,056 0,002 0,002 0,000 0,387 0,538 1,000
PB-1 0,000 0,501 0,075 0,501 0,015 0,684 0,398 1,000 1,000 1,000
Em negrito: não se ajusta as proporções do Equilíbrio de Hardy – Weinberg com grau de significância α = 0,005.
Esses valores representam as estimativas médias desses parâmetros nas
populações e servem para mostrar a variabilidade genética encontrada nos locos
microssatélites isolados. As estimativas de índice de fixação intrapopulacional obtidas
para cada loco representam as diferenças entre as freqüências de heterozigotos reais
encontradas nas populações, em relação à freqüência de heterozigotos esperadas,
baseadas no número de alelos e nas freqüências desses, por loco e por população.
Diferenças significativas entre HO e HE são influenciadas por eventos que alteram a
freqüência dos genótipos homozigotos em relação à freqüência dos genótipos
heterozigotos, como a endogamia e a seleção natural (HARTL; CLARK, 2007). Como
não podemos ter certeza sobre a ligação desses locos microssatélites com alelos de
genes que possam estar aumentando o valor adaptativo de determinados indivíduos na
população, pela forma que esses microssatélites foram isolados, apenas podemos
então inferir que os valores de índice de fixação intrapopulacional podem ter sido
influenciados por eventos de endogamia. Entretanto, pelos dados apresentados na
tabela 7, podemos descartar essa possibilidade, pois existe uma grande variação
nesses valores. Podemos então aceitar que o número de indivíduos utilizados não foi

88 suficiente para amostrar toda variabilidade genética dos locos Aar06, Aar11, Aar14 e
Aar16.
O índice de fixação intrapopulacional é determinado pela variação das
estimativas dos parâmetros HO e HE. Quanto maior as diferenças entre essas
estimativas na população, maior será o valor de f estimado, uma vez que as estimativas
de HE são obtidas através das freqüências alélicas e informa sobre a quantidade de
heterozigotos esperados na população em relação à freqüência encontrada dos alelos,
e o parâmetro HO informa sobre o número total de heterozigotos contados na
população. Havendo então uma grande diferença entre HE em relação à HO, essa deve
estar relacionada à endogamia, pois essa aumenta a freqüência de genótipos
homozigotos em detrimento aos heterozigotos na população. Nesse caso, como os
valores de índice de fixação intrapopulacional são obtidos através desses valores
estimados para cada loco e calculado então o valor médio para cada população,
podemos considerar então que valores altos indicam então haver certo grau de
endogamia na população. Entretanto, não podemos afirmar com certeza, pois não
podemos descartar a possibilidade de erro amostral.
Para os locos Aar14 e Aar16 os desvios no equilíbrio de Hardy-Weinberg
podem estar relacionados a problemas de amostragem, não estando relacionados com
desvios causados pela endogamia, uma vez que para os outros locos essas populações
não apresentaram desvios significativos. Já em relação ao loco Aar06 em que a maioria
das populações apresentou desvios no EHW, esse fato deve estar relacionado com a
presença de alelos nulos, que podem ter determinado um excesso de genótipos
homozigotos levando assim os desvios no EHW observados. Alelos nulos ocorrem
quando a região que flanqueia os microssatélites sofre mutação na porção de
anelamento do primer, não permitindo assim que esse se ligue corretamente (DAKIN;
AVISE, 2004). Com isso não ocorre à amplificação do segmento que contem o
microssatélite. A conseqüência disso pode ser vista em uma amostra de uma
população, onde uma parte dos indivíduos tem seus alelos amplificados e outra parte
não. Outra possível causa da ocorrência de alelos nulos seria a amplificação diferencial
dos alelos de tamanhos diferentes (WATTIER et al., 1998), e a erros de genotipagem,
que por motivos relacionados às condições da eletroforese, os dois alelos por

89
possuírem tamanhos poucos discrepantes acabam sendo lidos como sendo apenas um
só. Em ambos os casos ocorrem erros de genotipagem que levam a um excesso de
homozigotos na amostra, determinando os desvios no EHW.
Para testar a presença de alelos nulos nos locos e nas populações foi então
utilizado o programa Micro-Checker. Esse programa testa a presença de alelos nulos
comparando as freqüências observadas dos alelos, com a esperada sobre equilíbrio de
Hardy-Weinberg. Foram então detectados a presença de alelos nulos, em todas as
populações, para os locos Aar06 e Aar11. Contudo, esses não apresentaram desvios
no EHW em todas as populações e, portanto, foram mantidos nas análises, por serem
locos informativos.
2.3.3.2 Variabilidade interpopulacional e estrutura genética populacional
Através das estimativas das freqüências alélicas foi possível observar a
diversidade existente em cada loco estudado, nas onze populações de A. argillacea.
Podemos afirmar que A. argillacea apresenta grande variabilidade genética, que pode
ser confirmada pela diversidade alélica encontrada. Foram identificados 71 alelos totais
existentes nas populações estudadas sendo que 90,15% são compartilhados entre
todas as populações (Tabela 10). Através estimativa de alelos exclusivos, pode se
verificar que apenas 9,85%, ou seja, apenas sete alelos são restritos a algumas
populações (Tabela 11). Das 11 populações, seis possuem alelos exclusivos. Contudo,
dentre essas populações, o número de alelos exclusivos é baixo, variando de um ao
máximo de dois (GO-2). Portanto a maioria dos alelos é compartilhada entre todas as
populações estudadas indicando assim, de forma indireta, que existe fluxo gênico entre
essas populações. As estimativas de alelos exclusivos indicam que apenas o loco
Aar06 possui 14,3% de seus alelos restritos a população MS-1. O loco Aar11 possui
8,7% de seus alelos restritos a duas populações, GO-1 e GO-2. Os locos Aar14 e Aar16
possuem cada um 20% de alelos de seus exclusivos nas populações MT-1 e MS-2,
respectivamente. Já os locos Aar19 e Aar21, possuem 11,1% e 25% de alelos
exclusivos, respectivamente.

90
Tabela 10 – Freqüência alélica encontrada em cada loco microssatélite encontrada em cada população de A. argillacea
(continua)
Loco BA-2 BA-3 BA-4 GO-1 GO-2 MS-1 MS-2 MT-1 MT-2 MT-3 PB-1
Aar06
N 8 12 22 10 23 9 13 9 18 13 24
196 - 0,083 0,045 - 0,022 - 0,077 - 0,139 0,231 - 0,125 0,167 - 0,100 - - - 0,111 0,167 0,115 -
200 0,750 0,750 0,182 0,700 0,152 0,444 0,423 0,333 0,389 0,115 0,292
202 - - 0,682 0,100 0,804 0,444 0,462 0,278 0,306 0,500 0,604
208 0,125 - 0,045 - 0 - - - - 0,038 0,104
210 - - 0,045 0,100 0,022 - 0,038 0,278 - - -
212 - - - - - 0,111 - - - - -
Aar10
N 11 20 20 20 23 20 18 20 24 22 24
213 0,091 - - 0,025 - 0,025 - - 0,042 - -
217 - - - 0,050 - 0,025 - - 0,021 - -
265 - - 0,050 0,150 0,065 0,050 0,028 0,025 0,083 0,045 0,063
269 0,182 0,125 0,050 0,100 0,152 0,100 0,111 0,100 0,042 0,091 0,042
271 0,636 0,800 0,850 0,650 0,696 0,650 0,750 0,700 0,750 0,795 0,833
273 0,091 0,075 0,050 0,025 0,087 0,150 0,083 0,175 0,063 0,045 0,063
275 - - - - - - 0,028 - - 0,023 -
Aar11
N 10 12 8 9 15 11 9 11 11 15 13
192 - - - - - 0,091 0,111 - 0,091 - -
196 - 0,083 - - - - 0,167 - 0,091 - -
218 - - - 0,056 - 0,091 0,056 - - 0,033 -
220 0,100 0,125 0,063 0,167 - 0,091 0,056 0,182 0,136 0,067 0,192
224 0,100 0,125 0,125 - 0,067 - - - - 0,033 -
228 0,100 0,167 0,313 - - 0,182 - - 0,045 0,100 0,231
230 - - - 0,056 0,067 - 0,167 0,182 - 0,033 0,154
236 0,100 0,042 - - 0,100 - 0,111 0,091 0,227 0,133 -
238 0,100 - - - - - - - - 0,033 -
240 - 0,042 - 0,278 - 0,182 - - - - -
242 0,05 0,042 - 0,056 - - - - - - -
244 0,15 0,042 0,063 - - - - - 0,091 - -
253 - - - 0,056 - - - - - - -
254 - 0,167 - 0,111 0,033 0,091 - - - 0,033 0,038
258 0,100 - 0,063 - 0,067 - 0,111 0,091 0,045 - 0,077
260 0,100 0,042 0,125 0,056 0,167 0,091 0,222 0,273 0,045 0,167 -
270 - 0,125 - - 0,067 0,091 - - 0,091 0,033 -
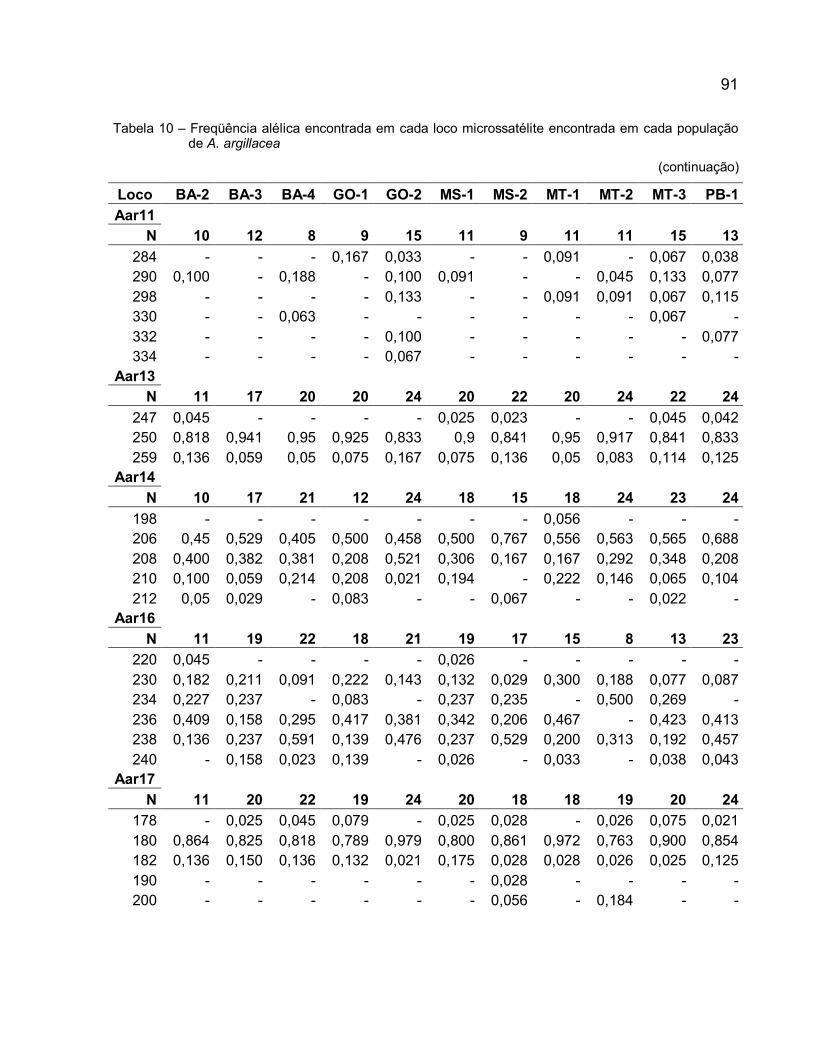
91
Tabela 10 – Freqüência alélica encontrada em cada loco microssatélite encontrada em cada população de A. argillacea
(continuação)
Loco BA-2 BA-3 BA-4 GO-1 GO-2 MS-1 MS-2 MT-1 MT-2 MT-3 PB-1
Aar11
N 10 12 8 9 15 11 9 11 11 15 13
284 - - - 0,167 0,033 - - 0,091 - 0,067 0,038
290 0,100 - 0,188 - 0,100 0,091 - - 0,045 0,133 0,077
298 - - - - 0,133 - - 0,091 0,091 0,067 0,115
330 - - 0,063 - - - - - - 0,067 -
332 - - - - 0,100 - - - - - 0,077
334 - - - - 0,067 - - - - - -
Aar13
N 11 17 20 20 24 20 22 20 24 22 24
247 0,045 - - - - 0,025 0,023 - - 0,045 0,042
250 0,818 0,941 0,95 0,925 0,833 0,9 0,841 0,95 0,917 0,841 0,833
259 0,136 0,059 0,05 0,075 0,167 0,075 0,136 0,05 0,083 0,114 0,125
Aar14
N 10 17 21 12 24 18 15 18 24 23 24
198 - - - - - - - 0,056 - - -
206 0,45 0,529 0,405 0,500 0,458 0,500 0,767 0,556 0,563 0,565 0,688
208 0,400 0,382 0,381 0,208 0,521 0,306 0,167 0,167 0,292 0,348 0,208
210 0,100 0,059 0,214 0,208 0,021 0,194 - 0,222 0,146 0,065 0,104
212 0,05 0,029 - 0,083 - - 0,067 - - 0,022 -
Aar16
N 11 19 22 18 21 19 17 15 8 13 23
220 0,045 - - - - 0,026 - - - - -
230 0,182 0,211 0,091 0,222 0,143 0,132 0,029 0,300 0,188 0,077 0,087
234 0,227 0,237 - 0,083 - 0,237 0,235 - 0,500 0,269 -
236 0,409 0,158 0,295 0,417 0,381 0,342 0,206 0,467 - 0,423 0,413
238 0,136 0,237 0,591 0,139 0,476 0,237 0,529 0,200 0,313 0,192 0,457
240 - 0,158 0,023 0,139 - 0,026 - 0,033 - 0,038 0,043
Aar17
N 11 20 22 19 24 20 18 18 19 20 24
178 - 0,025 0,045 0,079 - 0,025 0,028 - 0,026 0,075 0,021
180 0,864 0,825 0,818 0,789 0,979 0,800 0,861 0,972 0,763 0,900 0,854
182 0,136 0,150 0,136 0,132 0,021 0,175 0,028 0,028 0,026 0,025 0,125
190 - - - - - - 0,028 - - - -
200 - - - - - - 0,056 - 0,184 - -
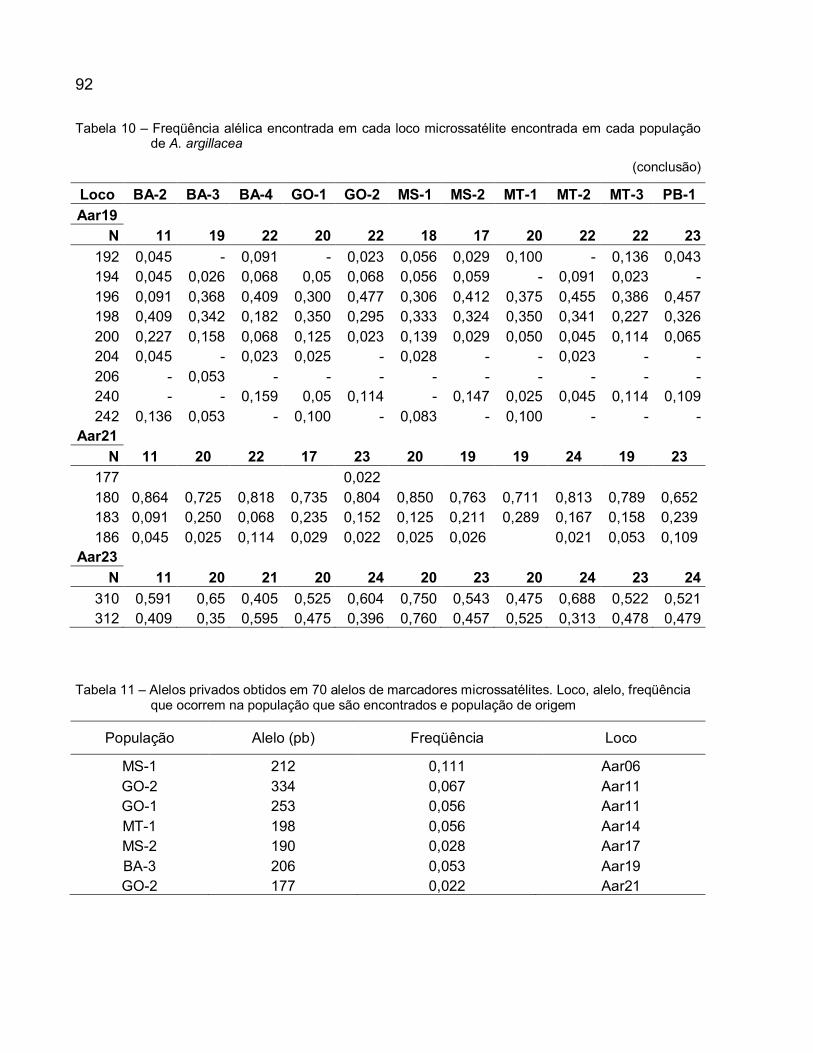
92
Tabela 10 – Freqüência alélica encontrada em cada loco microssatélite encontrada em cada população de A. argillacea
(conclusão)
Loco BA-2 BA-3 BA-4 GO-1 GO-2 MS-1 MS-2 MT-1 MT-2 MT-3 PB-1
Aar19
N 11 19 22 20 22 18 17 20 22 22 23
192 0,045 - 0,091 - 0,023 0,056 0,029 0,100 - 0,136 0,043
194 0,045 0,026 0,068 0,05 0,068 0,056 0,059 - 0,091 0,023 -
196 0,091 0,368 0,409 0,300 0,477 0,306 0,412 0,375 0,455 0,386 0,457
198 0,409 0,342 0,182 0,350 0,295 0,333 0,324 0,350 0,341 0,227 0,326
200 0,227 0,158 0,068 0,125 0,023 0,139 0,029 0,050 0,045 0,114 0,065
204 0,045 - 0,023 0,025 - 0,028 - - 0,023 - -
206 - 0,053 - - - - - - - - -
240 - - 0,159 0,05 0,114 - 0,147 0,025 0,045 0,114 0,109
242 0,136 0,053 - 0,100 - 0,083 - 0,100 - - -
Aar21
N 11 20 22 17 23 20 19 19 24 19 23
177 0,022
180 0,864 0,725 0,818 0,735 0,804 0,850 0,763 0,711 0,813 0,789 0,652
183 0,091 0,250 0,068 0,235 0,152 0,125 0,211 0,289 0,167 0,158 0,239
186 0,045 0,025 0,114 0,029 0,022 0,025 0,026 0,021 0,053 0,109
Aar23
N 11 20 21 20 24 20 23 20 24 23 24
310 0,591 0,65 0,405 0,525 0,604 0,750 0,543 0,475 0,688 0,522 0,521
312 0,409 0,35 0,595 0,475 0,396 0,760 0,457 0,525 0,313 0,478 0,479
Tabela 11 – Alelos privados obtidos em 70 alelos de marcadores microssatélites. Loco, alelo, freqüência que ocorrem na população que são encontrados e população de origem
População Alelo (pb) Freqüência Loco
MS-1 212 0,111 Aar06
GO-2 334 0,067 Aar11
GO-1 253 0,056 Aar11
MT-1 198 0,056 Aar14
MS-2 190 0,028 Aar17
BA-3 206 0,053 Aar19
GO-2 177 0,022 Aar21

93
O índice de fixação intrapopulacional da espécie, estimado pelo processo da
análise de variância, com base nos dez locos microssatélites, 71 alelos e nas onze
populações apresentou valor de 0,244 (IC 95% de 0,093 a 0,418). A estimativa de f não
foi significativamente igual a zero, mostrando haver certo grau de endogamia
intrapopulacional. Esse valor de endogamia intrapopulacional médio indica haver certo
grau de cruzamento entre parentes, e também é o componente que mais influência a
endogamia total entre as populações estudadas. Já o FST estimado a partir do
parâmetro theta (θ) (WEIR; COCKERHAM, 1984) foi de 0,036, com (IC 95% de 0.007 a
0,080). A estimativa de RST foi de 0,040. A estimativa de θ total foi significativamente
igual a zero, e indicam ausência de estruturação genética. O parâmetro Nem foi
estimado com base na estimativa de θ, e resultou no valor de 6,694 indivíduos adultos
migrantes em condições de se acasalar por geração (Tabela 12).
Para refinar as estimativas de θ com o objetivo de observar o grau de
diferenciação entre pares de populações foi então estimado o θ par-a-par. Podemos
observar que os valores dessa estimativa variaram de -0,0019 a 0,1082. As diferenças
calculadas entre pares de populações estimativas não foram significativamente
diferentes de zero para a maioria das interações, indicando não haver diferenciação
genética entre as populações, mesmo entre as que estão mais distantes
geograficamente. Entretanto apenas dois valores θ par-a-par foram superiores a 0,1 e
ambos foram obtidos entre a população GO-2 e as duas populações da Bahia, BA-2 e
BA-3, e todas as estimativas com a população GO-1 não foram possíveis calcular o
valor de significância, apesar das não haver diferenciação significativas, que podem ser
observadas pelos baixos valores de θ estimados. (Tabela 13).

94
Tabela 12 – Estimativa das estatísticas F de Wright, através da análise de variância das freqüências alélicas e do numero de migrantes por geração (Nem) em onze populações naturais de A. argillacea. Intervalo de confiança (IC) de 95% de probabilidade
f F θ RST *Nem
Estimativa 0,244 0,271 0,036 0,040 6,694
Limite Superior (IC 95%) 0,400 0,435 0,074 -
Limite Inferior (IC 95%) 0,073 0,090 0,008 -
* Número médio de migrantes por geração calculada através da estimativa de FST.

Tabela 13 – Estimativa do parâmetro θ par-a-par entre as populações
BA-2 BA-3 BA-4 GO-1 GO-2 MS-1 MS-2 MT-1 MT-2 MT-3 PB-1
BA-2 BA-3 -0,0095
BA-4 0,0978 0,0971 GO-1 -0,0068 -0,0019 0,0878
GO-2 0,1045 0,1082 0,0182 0,0974 MS-1 0,0029 0,0105 0,0380 0,0022 0,0362
MS-2 0,0526 0,0421 0,0394 0,0422 0,0308 0,0142 MT-1 0,0320 0,0440 0,0571 0,0036 0,0522 0,0196 0,0201
MT-2 0,0347 0,0140 0,0737 0,0443 0,0680 0,0059 0,0086 0,0484 MT-3 0,0500 0,0570 0,0231 0,0495 0,0171 0,0108 0,0136 0,0193 0,0226
PB-1 0,0752 0,0669 0,0104 0,0501 0,0202 0,0211 0,0065 0,0231 0,0550 0,0147 NS: Não significativo (P < 0,05). ** não foi possível calcular.

96
Os valores de FST obtidos através do estimador θ (WEIR; COCKERHAM, 1984)
tanto o total, quanto os par-a-par, ficaram entre o intervalo de 0 e 0,10, com
predominância de valores abaixo de 0,5, indicando que a estruturação genética entre as
populações de A. argillacea estudadas ficou entre baixa e moderada, sendo a baixa
estruturação predominante. Segundo Hartl e Clark (2007) valores de FST entre 0 e 0,05
indicam baixa estruturação genética; valores entre 0,05 e 0,15 indicam estruturação
genética moderada, valores entre 0,15 e 0,25 estão relacionados à estruturação alta e
valores acima de 0,25 indicam uma forte estruturação genética.
A estimativa de RST, que assume outro modelo mutacional para estimar o grau
de diferenciação entre as populações corroborou com o resultado estimado para FST. As
estimativas de FST geralmente diferem das estimativas de RST, uma vez que cada uma
assume um modelo diferente de mutação dos alelos. O parâmetro FST assume o
modelo de Alelos Infinitos (IAM), enquanto que o parâmetro RST baseia – se no modelo
de mutação stepwise (SMM), que também é o modelo mais indicado para estimar
diferenciação genética entre populações, uma vez que é o modelo que mais próximo
chega da explicação da forma como os microssatélites evoluem. Sendo assim, espera –
se que a estimativa de RST seja maior que a do FST (SLATIKIN, 1995). Nos resultados
obtidos, pode – se notar que a estimativa de RST foi maior que a do FST, entretanto não
podemos afirmar que foram diferentes significativamente. Isso se deve ao fato de que, o
modelo de SMM ser capaz de explicar a evolução de todos os locos microssatélites,
uma vez que outras formas de evolução dos mesmos podem ocorrem (BALLOUX;
LUGON-MOULIN, 2002). Sendo assim, para o calculo de fluxo gênico médio entre as
populações, foi utilizado o a estimativa do parâmetro FST.
Já a estimativa de fluxo gênico, indicou haver uma taxa alta de migração total
entre as populações. Govindajaru (1989) distinguiu três níveis de fluxo gênico: alto Nem
> 1, Intermediário (0,25 < Nem < 0,99) e baixo Nem < 0,25. Em relação à ocorrência de
deriva genética, essa causa diferenciação populacional se Nem <1, mas não resultará
se Nem > 1 (SLATKIN; BARTON, 1989). A estimativa obtida nesse estudo foi de
(6,694), indicando haver alto fluxo gênico entre as populações sendo capaz, portanto,
de impedir que haja perdas significativas na variabilidade genética determinada por
deriva entre as populações.

97
Os baixos valores das estatísticas F de Wright, da estimativa de e RST, das
estimativas de diferenciação entre cada par de populações e o alto valor de Nem podem
ser indicativos de que a espécie no Brasil possui estrutura de metapopulação com certo
alto grau de panmixia (acasalamentos ao acaso). Isso sem se deve ao fato de fluxo
gênico alta ser capaz de manter a coesão entre todas as populações, evitando assim as
suas diferenciações. Para testar se existe algum grau de estruturação genética
relacionado com a distância geográfica, foram então utilizadas duas abordagens para
análise de dados. Primeiramente foram estimadas as distâncias genéticas segundo
modelo de Nei (NEI, 1978) e construído o dendrograma utilizando o critério de
agrupamento UPGMA, e assim inferir sobre possível diferenciação genética entre as
populações. E a outra abordagem, baseada em estatística Bayesiana foi utilizada para
observar se existe estruturação genética entre as populações.
As distâncias genéticas de Nei (1978) calculadas entre as populações variaram
de 0,0078 a 0,1367. Esta análise revela que as populações estudadas não apresentam
distâncias genéticas significativas. Contudo, os resultados obtidos com o calculo das
distâncias de Nei corroboram com os valores obtidos com as estimativas de FST par-a-
par, e mostram que os valores mais significativos de distâncias foram entre a população
GO-2 e as populações BA-2 e BA-3. Além disso, podemos notar que a distância entre
BA-2 e BA-4 foi alta apesar de ambas estarem próximas geograficamente (Tabela14).
Podemos notar que não houve agrupamento das populações por estado. Uma
vez que as distâncias genéticas entre as populações não são capazes de determinar
essa resolução de agrupamento. Percebe – se que a resolução dos agrupamentos é
baixa, evidenciada pela baixa consistência dos nós obtidos por reamostragem por
bootstrap (Figura 13). Essa baixa resolução indica não haver diferenciações
significativas entre populações. Entretanto apenas podemos observar consistência no
primeiro nó, que agrupou as populações por safra. É importante ressaltar que as
distâncias genéticas são de pequena amplitude e que dessa forma, determinam essa
baixa resolução.

Tabela 14 – Distâncias genéticas de NEI (1978), calculadas entre as populações
BA-2 BA-3 BA-4 GO-1 GO-2 MS-1 MS-2 MT-1 MT-2 MT-3 PB-1
BA-2
BA-3 0,0078 BA-4 0,1367 0,1065
GO-1 0,0101 0,0024 0,1113 GO-2 0,1352 0,1155 0,0236 0,1194
MS-1 0,0257 0,0289 0,0562 0,0180 0,0459 MS-2 0,1026 0,0612 0,0498 0,0653 0,0389 0,0485
MT-1 0,0530 0,0519 0,0732 0,0192 0,0630 0,0370 0,0374 MT-2 0,1100 0,0590 0,0922 0,0828 0,0834 0,0580 0,0224 0,0749
MT-3 0,0884 0,0810 0,0272 0,0793 0,0206 0,0305 0,0232 0,0323 0,0367 PB-1 0,1058 0,0702 0,0168 0,0658 0,0263 0,0329 0,0159 0,0356 0,0737 0,0216

99
Figura 13 – Padrão de divergência genética entre as populações de A. argillacea, definido pelo agrupamento UPGMA, a partir das distâncias genéticas de Nei (1978). Correlação cofenética igual a 0,99%. Para testar a consistência dos nós foram realizadas 1000 reamostragens por bootstrap
Através da análise de estrutura populacional, realizada pelo programa Structure
2.2 (PRITCHARD et al., 2002) utilizando o modelo admixture, que considera que as
populações a priori mantém certa coesão genética; e determinando haver correlação
entre as populações (o parâmetro correleted) indicou não haver estruturação espacial
da variabilidade genética entre as populações estudadas.
Dentre as possibilidades para o valor de K, que variaram de 1 a 16, o valor obtido
através do método proposto por Evanno (2005) foi de dois grupos (Figura 14). A partir
desse valor de K, foi então obtida à melhor saída de dados para esse parâmetro, entre
as cinco repetições de permutações. Pode se observar através da análise bayesiana
que não existe estruturação geográfica da variabilidade genética, entretanto podemos
observar uma tendência à estruturação temporal dessa variabilidade (Figura 15).

100
Figura 14 – Resultado da análise obtida para a determinação do valor de agrupamento que melhor representa a variabilidade genética das populações de A. argillacea estudadas: K variando de 1 a 15, com cinco repetições cada K, burn in de 50.000 permutações e seguidas por 500.000 permutações MCMC
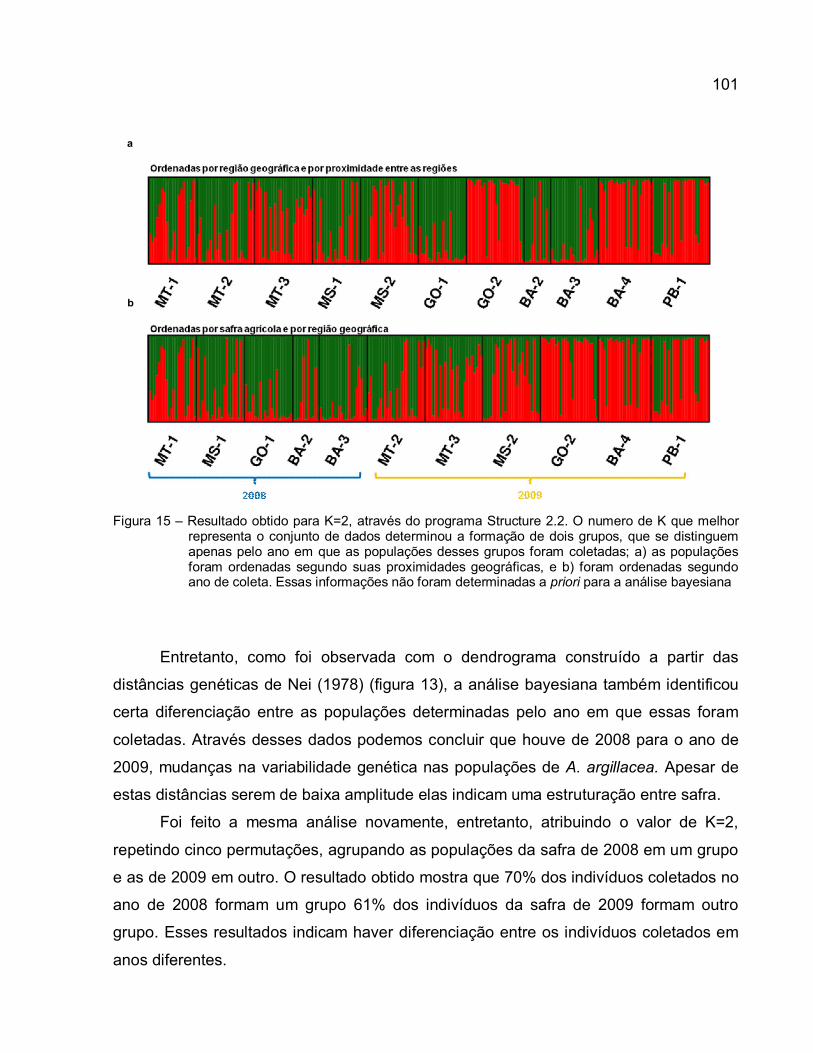
101
Figura 15 – Resultado obtido para K=2, através do programa Structure 2.2. O numero de K que melhor representa o conjunto de dados determinou a formação de dois grupos, que se distinguem apenas pelo ano em que as populações desses grupos foram coletadas; a) as populações foram ordenadas segundo suas proximidades geográficas, e b) foram ordenadas segundo ano de coleta. Essas informações não foram determinadas a priori para a análise bayesiana
Entretanto, como foi observada com o dendrograma construído a partir das
distâncias genéticas de Nei (1978) (figura 13), a análise bayesiana também identificou
certa diferenciação entre as populações determinadas pelo ano em que essas foram
coletadas. Através desses dados podemos concluir que houve de 2008 para o ano de
2009, mudanças na variabilidade genética nas populações de A. argillacea. Apesar de
estas distâncias serem de baixa amplitude elas indicam uma estruturação entre safra.
Foi feito a mesma análise novamente, entretanto, atribuindo o valor de K=2,
repetindo cinco permutações, agrupando as populações da safra de 2008 em um grupo
e as de 2009 em outro. O resultado obtido mostra que 70% dos indivíduos coletados no
ano de 2008 formam um grupo 61% dos indivíduos da safra de 2009 formam outro
grupo. Esses resultados indicam haver diferenciação entre os indivíduos coletados em
anos diferentes.
a
b

102
Para verificar se existe diferenciação dentro de cada safra. Foi então feita análise
separando as populações coletadas em anos diferentes. Foram atribuídos o mesmo
modelo e os mesmos parâmetros, definidos para a análise anterior, apenas mudando a
amplitude de variação de K, que foi de um a dez agrupamentos. Contudo através dessa
análise não foi possível observar estruturação genética dentro de cada ano.
Por fim foi realizada a estimativa de θ, reunindo as populações de 2008 em um
grupo e as populações de 2009 em outro. Foram estimados o θ dentro de cada grupo, e
entre grupos. Os valores do parâmetro θ estimados dentro de cada grupo foram de:
0,010, com IC 95% de -0,001 a 0,026, para o grupo um, composto pelas populações
BA-2, BA-3, GO-1, MS-1 e MT-2 (Safra 2008); e 0,024, com IC 95% de 0,005 a 0,044
para o grupo dois, formado pelas populações BA-4, GO-2, MS-2, MT-2, MT-3 e PB-1
(Safra 2009). Já o valor obtido entre grupos foi de 0,039, e IC 95% de 0.003 a 0.090,
sendo essas estimativas diferentes de zero, evidenciando haver, apesar de baixa, certo
grau de diferenciação entre safras.
Os resultados mostram que não existe estruturação genética significativa entre
as populações das diferentes regiões geográficas. Contudo, foi possível observar certa
diferenciação entre populações coletadas em anos diferentes. As mudanças na
variabilidade genética observadas na safra de 2009 em relação à de 2008, podem estar
ligadas a eventos estocásticos que alteram a aleatoriamente, ou a eventos
determinísticos, como a seleção natural. Entretanto, apenas podemos inferir sobre
ocorrência de mudanças aleatórias, uma vez que não é possível saber sobre a
ocorrência de seleção natural nos locos estudados, pois a princípio os locos
microssatélites neutros, ou seja, pois não é possível saber a ligação deles com genes
funcionais.
A espécie se trata de um inseto herbívoro com alta capacidade de dispersão.
Através de estudos com marcação e recaptura, e monitoramento da ocorrência da
espécie na cultura do algodão, realizados anteriormente, foi possível determinar o
estágio fenológico da planta que inicia a infestação da praga e em que época do ano
essas infestações ocorrem. No Brasil, A. argillacea inicia sua dispersão na região
nordeste, e segue sentido Sul, para a região Centro-Oeste, passando pelos estados de
Mato-Grosso, Mato-Grosso do Sul, Goiás, até atingir o estado de São Paulo e Minas

103
gerais. Por fim, retorna, seguindo o plantio de algodão, a região de onde se iniciou a
sua dispersão. Podemos inferir que esse comportamento esteja por si só, influenciando
as mudanças nas freqüências e na diversidade de alelos encontradas nos locos
estudados, uma vez que a cada evento de dispersão um número reduzido de migrantes
partem para colonizar outra região, determinando dessa forma, eventos de gargalho de
garrafa, seguido de eventos de colonização com rápido crescimento populacional. Além
disso, pelo fato de a cultura do algodão não ocorrer o ano todo, o período em que a
planta não é cultivada, determina uma drástica redução populacional que
conseqüentemente altera a variabilidade genética através de eventos de deriva. Dessa
forma, a estruturação da variabilidade genética temporal pode ocorrer.
Todavia, não se pode descartar a possibilidade das práticas de manejo estar
influenciando a diversidade genética de A. argillacea, uma vez que a mortalidade
causada pelo uso de inseticidas, ou outra forma de controle que seja capaz de reduzir
drasticamente a densidade populacional da praga, pode estar determinando também
eventos de gargalho de garrafa.
Recentemente, muitos estudos examinando a diferenciação genética entre
populações de lepidópteros - pragas, que são alvos do controle de plantas GM, têm
sido feitos, com o objetivo de avaliar e entender processos que estejam determinando
possíveis diferenciações genéticas entre as suas populações e quais seriam as
implicações dessa diferenciação no manejo da resistência a toxina expressa por essas
plantas. Muitos desses estudos tem se concentrado nas espécies de lepidópteros que
causam os maiores prejuízos na agricultura e que apresentam maior risco para
evolução da resistência. Uma das espécies mais estudadas nos Estados Unidos e
Europa é Ostrinia nubilalis (Hübner), que causa grandes prejuízos na cultura do milho e
é a principal praga alvo do controle do milho transgênico. Estudos populacionais com
essa espécie já foram realizados com marcadores Alozimáticos até marcadores
baseados em polimorfismo de DNA (BOURGUET et al., 2000; KRUM et al., 2008;
COATES et al., 2004; 2005). Entretanto, todos esses trabalhos mostraram não haver
estruturação da variabilidade genética mesmo entre populações isoladas
geograficamente. Contudo, muitos desses trabalhos erram na escolha dos marcadores
moleculares, que não foram suficientemente robustos em acessar o polimormismo

104 existente entre as populações, levando assim a resultados errôneos (MARÇON et al.,
1999; COATES et al., 2004). Já outros trabalhos, fora inconclusivos, pois os autores
não tomaram o cuidado em separar nas análises, insetos de O. nubilalis pertencente a
raça feromônio “Z” da “E”, e insetos que possuem padrão de voltinismo diferentes
(univoltina, bivoltina etc), e assim os resultados obtidos levaram a conclusões erradas a
cerca da diferenciação genética. Bourguet e colaboradores, estudando a estruturação
da variabilidade genética dessa espécie na França, não encontraram estruturação
significativa entre populações distantes, mas encontrou entre populações próximas. Já
Krum et al. (2009) estudando a variabilidade genética de O. nubilalis nos Estados
Unidos, utilizando marcador AFLP e RAPD, não identificou diferenciação genética
significativas entre a maior parte das populações, entretanto identificou uma
estruturação moderada entre as populações do Norte em relação as do Sul, devido aos
padrões de voltinismo encontrados entre essas regiões. Já Kim e colaboradores,
(2009), estudando O. nubilalis nos Estados Unidos, tomando os devidos cuidados em
não amostrar populações de raças ferômonio diferentes e de mesmo padrão de
voltinismo, e amostrando um grande número de insetos, dentro de um transecto Norte –
Sul de 270 Km, que cruzou com outro transecto Leste – Oeste também de 270 Km,
também não identificaram estruturação genética espacial significativa. Através das
análises de FST par – a – par, e estudando a taxa de migração entre cada população
puderam concluir que a dimensão geográfica da espécie é grande, e que mesmo ter
havido uma expansão geográfica seguindo a expansão da agricultura, essa não
determinou eventos de gargalos significativos. Sendo assim, os autores corroboram
com a realização do monitoramento da resistência em escala geográfica, e que da
mesma forma que o fluxo gênico pode retardar a evolução da resistência ele pode
espalhar os alelos que conferem resistência, quando a resistência já estiver
desenvolvida.
Outra espécie de inseto que também causa grandes prejuízos na agricultura
mundial e também é alvo de plantas transgênicas é Helicoverpa armigera (Hübner). Os
trabalhos de genética de populações mais importantes realizados com essa espécie
são provenientes da Austrália, nestes foram utilizados marcadores microssatélites
(SCOTT et al., 2006; ENDERSBY et al., 2006). Scott e colaboradores, estudando

105
populações de H. armigera coletadas na reigão do Vale Murrumbidge, Austrália,
durante três safras agrícolas, identificou uma variação nos níveis de estruturação da
variabilidade genética entre as safras. Durante a safra de 2001/2002, não foram
identificadas diferenciações significativas entre as populações. Na safra 2002/2003,
foram identificadas diferenciação significativas entre as populações, e, além disso,
foram identificados três principais “tipos” genéticos: os primeiros tipos foram
constituídos das primeiras duas gerações da espécie naquele, de indivíduos já
presentes nos locais que estavam saindo da diapausa, e a progênies desses indivíduos;
o segundo tipo foi constituído da terceira geração, além dos imigrantes externos; e o
terceiro tipo foi determinado por outros imigrantes de outras regiões. E por fim, foi
identificada alta estruturação genética na safra de 2002/2003 por conta da expansão da
agricultura do algodão e pelo uso mais intenso de inseticidas sintéticos, que
determinaram mudanças na dinâmica populacional da espécie na região. Outro
trabalho, entretanto, não identificou estruturação genética entre populações de H.
armigera, próximas e distantes. Esse trabalho foi realizado em duas regiões, uma
próxima do Vale Murrumbidge, Austrália, outra região mais distante, em Queensland,
Austrália. Foram coletadas 14 populações dentro dessas duas regiões, e sobre
diferentes plantas hospedeiras. Apesar de essas variáveis poderem atuar determinando
a distribuição da variabilidade genética e a direção do fluxo gênico, nenhuma
estruturação foi evidenciada (ENDERSBY et al., 2006).
No Brasil, os trabalhos de variabilidade genética tem se concentrado em
espécies pragas que são controladas pelas principais variedades de plantas
transgênicas plantadas no país. Dentre essas espécies, Spodoptera frugiperda se
destaca como sendo a de maior importância para o manejo da resistência, pois além de
atacar plantas de milho, essa espécie também se alimenta de algodão, estando
portanto exposta continuamente a toxinas do tipo Cry. Sendo assim, Martinelli et al.
(2006) realizaram um estudo de variabilidade genética dessa praga visando responder
se existe diferenciação genética entre populações de S. frugiperda na cultura do milho e
na cultura do algodão, e se essa diferenciação está relacionada com a planta
hospedeira, ou com a proximidade geográfica. Para tanto, foram utilizados marcadores
RAPD, que apesar de ser um marcador dominante, em desuso atualmente, foi capaz de

106 respoder essa pergunta. As populações de S. frugiperda estudadas formam grupos
devido a proximidade espacial, ao invés da preferência pela planta hospedeira. Para
confirmar esses resultados, foram utilizados em outro estudo, marcadores AFLP, que
confirmaram os resultados anteriores (MARTINELLI et al., 2007)
Dentro do contexto do manejo da resistência de insetos a plantas GM, pode –
se perceber a importância de estudos que buscam avaliar a variabilidade genética e
entender quais processos determina a estruturação espacial dessa variabilidade. Sendo
assim, os conhecimentos da área de genética de populações, cada vez mais estão
conseguindo responder questões e auxiliar o direcionamento de pesquisas e a tomada
de decisões para se delinear melhores práticas de manejo da resistência de insetos à
táticas de controle populacional de pragas.
No presente estudo foi possível observar variabilidade na resposta das
populações naturais de A. argillacea a toxina Cry1Ac. Entretanto essa variabilidade não
está ligada a mudanças na suscetibilidade das populações.
Já o estudo de genética de populações, conseguiu identificar que não existem
unidades evolutivas que da espécie, que possam estar passando por processos de
diferenciação genética isoladamente, uma vez que não foi identificada estruturação da
variabilidade genética da espécie no Brasil. Contudo foi identificada indícios de
estruturação genética temporal, uma vez que as populações coletadas em duas safras
apresentaram certa diferenciação genética.
Sendo assim, práticas de manejo da resistência de A. argillacea à toxina
Cry1Ac necessitam ser realizadas uma vez que a espécie possui variabilidade natural
na resposta à toxina, dessa forma a espécie mostrou ter potencial para a evolução da
resistência. Além disso, práticas de manejo da resistência não devem ser decididas
localmente ou regionalmente, mas em escala nacional, uma vez que não existe
diferenciação genética entre as populações da espécie. Outro aspecto importante a ser
considerado é o alto fluxo gênicos encontrado entre as populações de A. argillacea no
Brasil, e os eventos de gargalho genéticos seguidos de eventos fundadores. A alta
coesão genética entre as populações, determinada pelo intenso fluxo gênico influência
dois aspectos importantes para a evolução da resistência: se por um lado, para os
alelos que conferem resistência poderem se fixar em uma determinada população

107
demorará mais tempo, por outro, quando a resistência se tornar problema em um local,
a dispersando desse alelos através do fluxo gênica, será rápida, atingindo rapidamente
todas as populações da espécie no Brasil. Já os eventos de gargalho podem determinar
a rápida fixação de alelos que conferem resistência de uma ano para outro.

108

109
3 CONCLUSÕES
Há variabilidade natural na resposta das populações de A. argillacea à toxina
Cry1Ac;
Os locos microssatélites foram eficientes para detectar variabilidade genética
dentro e entre as populações de A. argillacea no Brasil;
Existe pequena estruturação da variabilidade genética entre as populações nas
diferentes safras, porém não existe estruturação entre populações de diferentes
regiões;
Ocorre alta taxa de migração entre as populações de A. argillacea no Brasil.

110

111
REFERÊNCIAS
ALFENAS, A.C. Eletroforese de isoenzimas e proteínas afins: fundamentos e
aplicações em plantas e microrganismos. Viçosa: UFV, 1998. 574 p.
ALI, M.I.; LUTTRELL, R.G.; YOUNG, S.I. III. Susceptibilities of Helicoverpa zea e
Heliothis virescens (Lepidoptera: Noctuidae) populations to Cry1Ac insecticidal protein.
Jounal of Economic Entomology, Lanham, v. 99, p. 164-175, 2006.
ALSTAD, D.;ANDOW,D.A. Managing the evolution of insect resistance to transgenic
plantas. Science, Washington, v. 268, p. 1894-1896, 1995.
ANDOW, D.A.; HUTCHINSON, W.D. Bt – corn resistance management. In: MELLON,
M.; RISSLER, J. (Ed.). Now or never: serious new plants to save a natural pest control.
Washington: Union of Concerned Scientist, 1998. p. 19-66.
AZEVEDO, F.R.; MATTOS, K.O.; VIEIRA, F.V. Comportamento alimentar de Alabama
argillacea Hubner (Lepidoptera: Noctuidae) em algodoeiro. Ciência Agronômica,
Fortaleza, v. 33, p. 5-9, 2002.
BATES, S.R; ZHAO, J.H.; ROUSH, R.T.; SHELTON. A.M. Insect resistance
management in GM crop: Past, present and future. Nature Biotechnology, London,
v. 23, p. 57-62, 2005.
BETZ, F.S.; HAMMOND, B.G.; FUCHS, R.L. Safety and advantages of Bacillus
thuringiensis protected plants to control insect pests. Regulatory Toxicology and
Pharmacology, Columbia, v. 32, p. 156–173, 2000.
BILLOTTE, N.; LAGODA, P.J.L.; RISTERUCCI, A.; BAURENS, F. Microsatellite-
enriched libraries: applied methodology for the development of SSR markers in tropical
crops. Fruits, Paris, v. 54, p. 277-287, 1999.
BORÉM, A.; CAIXETA, E.T. Marcadores moleculares. Viçosa: UFV, 2006. 374 p.
BOTELHO, P.S.M.; SILVEIRA NETO, S.; LARA, F.M. Flutuação populacional de
curuquerê - do – algodoeiro (Alabama argillacea Hübner), em 4 municípios do Estado
de São Paulo. Anais da Sociedade Entomológica do Brasil, Jaboticabal, v. 5, p. 181-
193, 1976.

112 BOURGUET, D.; BETHENOD, M.T.; PASTEUR, N.; VIARD, F. Gene flow in the
European corn borer ostrinia nubilalis: implications for the sustainability of transgenic
insecticidal maize. Proceedings of the Royal Society of London B, London, v. 267,
p. 117-122, 1999.
BROOKES, A.J. The essence of SNPs. Gene, Amsterdam, v. 234, n. 2, p. 177-186,
1999.
CALCAGNOLO, C. Principais pragas do algodoeiro. In: NEVES, O.S.; CAVALERI, P.A.;
VERDADE, F.C.; JUNQUEIRA, A.A.B.; GRIDI-PAPP, I.L.; ORTOLANI, A.A.; SILVA,
N.M. da; RIGHI, N.R.; FERRAZ, C.A.M.; CORRÊA, D.M.; CALCAGNOLO, G.;
SILVEIRA, A.P.; COSTA, A.S.; CARVALHO, A.M.B.; MENDES, H.C.; FUZATTO, M.G.;
CORRÊA, F.; BERZAGHI, M.N. (Ed.). Cultura e adubação do algodoeiro. São Paulo:
Instituto Brasileiro de Potassa, 1965. p. 319-389.
CAPRIO, M.A.; TABASHNIK, B.E. Gene flow accelerates local adaptation among finite
populations: simulating the evolution of insecticide resistance. Journal of Economic
Entomology, Lanham, v. 85, p. 611-620, 1992.
CAPRIO, M.A.; SUMERFORD, D.V.; SIMS, S.R. Evaluating transgenic plants for
suitability in pest and resistance management programs. In: LACEY, L.A.; KAYA, H.K.
(Ed.). Field manual of techniques in invertebrate pathology. Dordrecht: Kluwer
Academic, 2000. p. 805-825.
COATES, B.S.; HELLMICH, R.L.; LEWIS, L.C. Polymmorphic CA/GT and GA/CT
microsatellite loci for Ostrinia nubilalis (Lepidoptera: Crambidae). Molecular Ecology
Notes, London, v. 5, p. 10-12, 2005.
COATES, B.S.; SUMERFOLD, D.V.; HELLMICH, R.L. Geographic and voltinism,
differentiation among North American Ostrinia nubilalis (European corn borer)
mitochondrial cytochrome c oxidase haplotypes. Journal of Insect Science, Madison,
v. 4, p. 1-9, 2004. Disponível em: <http://www.insectscience.org/4.35>. Acesso em: 10
set. 2008.
CREST, S.; TULMANN NETO, A.; FIGUEIRA, A. Detection of single repeat
polymorphism in denaturing polyacrylamide sequencing gels by silver staining. Plant
Molecular Biology Reporter, Athens, v. 19, p. 299-306, 2001.
CROFT, B.A. Developing a philosophy and program of pesticide resistance
management. In: ROUSH, R.T; TABASHNIK, B.E. (Ed.). Pesticide resistance in
arthropods. New York: Chapman and Hall, 1990. p. 277-296.

113
DENNEHEY, T.J. Decision-making for managing pest resistance to pesticides. In FORD,
M.G.; HOLLOMAN, D.W.; KHANBAY, B.P.S.; SAWICKI, R.M. (Ed.). Combating
resistance to xenobiotics: biological and chemical approaches. Chichester:
EllisHorwood, 1987. p. 118-126.
DOBZHANSKY, T. Genetics and origins of species. 3rd ed. New York: Columbia
University Press, 1951. 364 p.
DOYLE, J.J.; DOYLE, .J.L. Isolation of plant DNA from fresh tissue. Focus, Rockville,
v. 12, p. 13-15, 1990.
ENDERSBY, N.M.; HOFFMAN, A.A.; McKECHNIE, S.W.; WEEKS, A.R. Is there genetic
structure in populations of Helicoverpa armigera from Australia? Entomologia
Experimentalis et Applicata, Dordrecht, v. 122: p. 253-263, 2007.
ESTADOS UNIDOS. Environmental Protection Agency. Biopesticides registration
action document: Bacillus thuringiensis plant incorporated protectants. 2001.
Disponível em: <http://www.epa.gov/pestisides/ biopesticides /reds/brad_bt_pip2.htm>.
Acesso em 18 dez. 2009.
EVANNO, G.; REGNAUT, S.; GOUDET, J. Detecting the number of clusters of
individuals using the software STRUCTURE: a simulation study. Molecular Ecology,
London, v. 14, p. 2611-2620, 2005.
FALCONER, D.S.; MACKAY, T.F.C. Introduction to quantitative genetics. New York:
Longmam Scientific & Technicals, 1989. 438 p.
FENGXIA, M.; SHEN, J.; ZHOU, W.; CEN, H. Long-term selection for resistance to
transgenic cotton expressing Bacillus thuringiensis toxin Helicoverpa armigera (Hübner)
(Lepidoptera: Noctuidae). Pest Management Science, Sussex, v. 60, p. 167-172, 2003.
FERRÉ, J.; VAN RIE J. Biochemistry and genetics of insect resistance to Bacillus
thuringiensis. Annual Review of Entomology, Palo Alto, v. 47, p. 501–533, 2002.
FERREIRA, M.E., GRATTAPAGLIA, D. Introdução ao uso de marcadores
moleculares em análises genéticas. Brasilia: EMBRAPA, CENARGEN, 1998. 220 p.
FFRENCH-CONSTANT, R.H.; ROUSH, R.T. Resistance detection and documentation:
the relative roles of pesticide and biochemical assays. In: ROUSH, R.T.; TABASHNIK,
B.E. (Ed.). Pesticide resistance in arthropods. New York: Chapman and Hall, 1990.
p. 4-38.

114 FIELD, D.; WILLS, C. Long, polymorphic microsatellites in simple organisms.
Proceedings of The Royal Society B: Biological Sciences, London, v. 263, n. 1367,
p. 209-215, 1996.
FITT, G.P.; OMOTO, C.; MAIA, A.H.; WAQUIL, J.M.; CAPRIO, M.; OKECH, M.A.; IA, E.;
HUAN, N.H.; ANDOW, D.A. Resistance risks of bt cotton and their management in
Brazil. In: HILBECK, A.; ANDOW, D.A.; FONTES, E.M.G. (Ed.). Environmental risk
assessment of genetically modified organisms: methodologies for assessing bt
cotton in Brazil. Cambridge: Cabi Publ., 2006. p. 300-345.
GALLO, D.; NAKANO, O.; SILVEIRA NETO, S.; CARVALHO, R.P.L.; BAPTISTA, G.C.;
BERTI FILHO, E.; PARRA, J.R.P. PARRA; ZUCCHI, R.A.; ALVES, S.B.; VENDRAMIM,
J.D.; MARCHINI, L.C.; LOPES, J.R.S.; OMOTO, C. Entomologia agrícola. Piracicaba:
FEALQ, 2002. 920 p. (Biblioteca de Ciências Agrárias Luiz de Queiroz, 10).
GILL, S.S.; COWLES, E.A.; PIETRANTONIO, P.V. The mode of action of Bacillus
thuringiensis endotoxins. Annual Review of Entomology, Palo Alto, v. 37, p. 615–636,
1992.
GONZALEZ-NUNEZ, M.; ORTEGO, F.; CASTANERA, P. Susceptibility of Spanish
populations of the corn borers Sesamia nonagrioides (Lepidoptera: Noctuidae) and
Ostrinia nubilalis (Lepidoptera:Crambidae) to a Bacillus thuringiensis endotoxin. Journal
of Economic Entomology, Lanham, v. 93, p. 459–463, 2000.
GOUDET, J. FSTAT 2.9,3: a program to estimate and test gene diversities and fixation
indices 2001. Disponível em: <http://www.unil.chlizea/softwares/fstat. html>. Acesso em:
15 mar. 2009.
GOULD, F. Sustainability of transgenic insecticidal cultivars: Integrating pest genetics
and ecology. Annual Review of Entomology, Palo Alto, v. 43, p. 701–726, 1998.
GRAVENA, S.; CUNHA, H.F. Artrópodes predadores na cultura Algodoeira.
Jaboticabal: FUNEP, 1991. 120 p.
HALLIDAY, W.R.; BURNHAM; K.P. Choosing the optimal diagnostic dose for monitoring
insecticide resistance. Journal of Economic Entomology, Lanham, v. 83, p. 1151-
1159, 1990.
HARTL, D.L. Princípios de genéticas de população. Tradução I.F. Afonso. 3. ed.
Ribeirão Preto: FUNPEC, 2008. 1 v.

115
HELENTJARIS, T.; SLOCUM, M.; WRIGHT, S.; SCHAEFER, A.; NIENHUIS, J. Construction of genetic linkage maps in maize and tomato using restriction fragment length polymorphisms. Theoretical and Applied Genetics, Berlin, v. 72, p. 761-769, 1986.
HENDRCKS, D.E.; LINGREN, P.D.; HOLLINGSWORTH, J.P. Number of bollworms,
tobacco budworms and cotton leafworms caught in traps equipped with fluorescent
lamps of five colours. Journal of Economic Entomology, College Park, v. 68, p. 645-
649, 1975.
HOFFMANN, C.; VANDERBRUGGEN, H.; HOFTE, H.; VAN RIE, J.; JANSENS, S.;
VAN MELLAERT, H. Specificity of Bacillus thuringiensis to brush-border membrane
vesicles of the cabbage butterfly (Pieris brassica). European Journal of Biochemistry,
Oxford, v. 173, p. 85–91, 1988.
HUANG, F.; LEONARD, R.; MOORE, S., YUE, B.; PARKER, R.; REAGAN, T.; STOUT,
M.; COOK, D.; AKBAR, W.; CHILCUTT, C.; WHITE, W.; LEE, D.; BILES, S.
Geographical susceptibility of Louisiana and Texas populations of the sugarcane borer,
Diatraea saccharalis (F.) (Lepidoptera: Crambidae) to Bacillus thuringiensis Cry1Ab
protein. Crop Protection, Guildford, v.27, p. 799-806, 2008.
INTERNATIONAL SERVICE FOR THE ACQUISITION OF AGRI-BIOTECH
APPLICATIONS. Disponível em: <http://www.isaaa.org>. Acesso em: 15 dez. 2009.
JÁCOME, A.G.; SOARES, J.J.; OLIVEIRA, R.H.; CORDÃO SOBRINHO, F.P. Efeito da
remoção das folhas no desenvolvimento vegetativo e na produção do algodoeiro.
Pesquisa Agropecuária. Brasileira, Brasília, v. 36, p. 751-755, 2001.
JARNE, P.; LAGODA, P.J.L. Microsatellites, from molecules to populations and back.
Trends in Ecology and Evolution, Cambridge, v. 11, p. 424–429, 1996.
JI, Y-J. Polymorphic microsatellite loci for the cotton bollworm Helicoverpa armigera
(Lepidoptera: Noctuidae) and some remarks on their isolation. Molecular Ecology
Notes, London, v. 3, p. 102-104, 2003.
JI, Y-J.; ZHANG, D.X. Characteristic of microsatellite DNA in lepidopteran genomes and
implications for their isolation. Acta Zoologica Sinica, Beijing, v. 50, p. 608-614, 2004.
KEYGENE NV (Nederland). Marc Zabeau; Pieter Vos. Selective restriction fragment
amplification: a general method for DNA fingerprinting EP 0534858. 24 set, 1992.
24 abr. 2005.

116 KINSINGER, R.A.; McGAUGHEY, W.H. Susceptibility of populations of Indianmeal moth
and almond moth to Bacillus thuringiensis. Journal of Economic Entomology,
Lanham, v. 72, p. 347-349, 1979.
KNOWLES, B.H.; ELLAR, D.J. Colloid-osmotic lysis is a general feature of the
mechanism of action of delta-endotoxin with different insect specificity. Biochimica et
Biophysica Acta, Amsterdam, v. 924, p. 509–518, 1987.
KRUMM, J.T.; HUNT, T.E.; SKODA, S.R.;HEIN, G.L.;LEE, D.J.; CLARK, P.L.; FOSTER,
J.E. Genetic variability of the European corn borer, Ostrinia nubilalis, suggests gene flow
between populations in the Midwestern United States. Journal of Insect Science,
Madison, v. 8, p. 1-12, 2008. Disponível em: <http://www.insectscience.org/8.72>.
Acesso em: 19 jan. 2009.
KWOK, P.Y. Methods for genotyping single nucleotide polymorphisms. Annual Review
of Genomics and Human Genetics, Palo Alto, v. 2, p. 235-258, 2001.
LADEIRA, J.S. O aparecimento de curuquerê nos algodoais. Boletim de Agricultura,
Belo Horizonte, v. 6, p. 57-59, 1957.
LEORA SOFTWARE. PoloPLUS: a user´s guide to Probit or Logit analysis. Berkeley,
2002. 20 p.
LEWIS, P.; ZAYKIN, D. Genetic data analysis: computer program for the analysis of
allelic data. Disponível em: <http://alleyn.eeb.uconn.edu/gda/>. Acesso em: 15 mar.
2009.
LU, M.-G.; RUI, C.-H.; ZHAO, J.-Z.; JIAN, G.-L.; FAN, X.-L.; GAO, X.-W. Selection and
heritability of resistance to Bacillus thuringiensis subsp. Kurstaki and transgenic cotton in
Helicoverpa armigera (Lepidoptera: Noctuidae). Pest Management Science, Sussex,
v. 60, p. 887-893, 2003.
LUTTRELL, R.G.; WAN, L.; KNIGHTEN, K. Variation in susceptibility of noctuid
(Lepidoptera) larvae attacking cotton and soybean to purified endotoxin proteins and
commercial formulations of Bacillus thuringiensis. Journal of Economic Entomology,
Lanham, v. 92, p. 21-32, 1999.
MARCHINI, L.C. Avaliação de dano do curuquerê - do – algodoeiro, Alabama
argillacea (Hübner, 1818) em condições simuladas e redução de sua população
através de isca tóxica. 1976. 72 p. Dissertação (Mestrado em Entomologia) – Escola
Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 1976.

117
MARÇON, P.C.R.G.; YOUNG, L.J. STEFFEY, K.L.; SIEGFRIED, B.D. Baseline
susceptibility of European corn borer (Lepidoptera: Crambidae) to Bacillus thuringiensis
toxins. Journal of Economic Entomology, Lanham, v. 92, p. 279-285, 1999.
MARÇON. P.C.R.G.; SIEGFRIED, B.D.; SPENCER, T.; HUTCHINSON, W.D.. Development of diagnostic concentrations for monitoring Bacillus thuringiensis resistance in European corn borer (Lepidoptera: Crambidae). Journal of Economic Entomology, Lanham, v. 93, p. 925-930, 2000.
MARTINELLI, S.; MONTRAZI, R.B.; ZUCHI, M.I.; SILVA-FILHO, M.C.; OMOTO, C.
Molecular variability of Spodoptera frugiperda (Lepidoptera: Noctuidae) populations
associated to maize and cotton crops in Brazil. Journal of Economic Entomology,
Lanham, v. 99, p. 519-526, 2006.
MARTINELLI, S.; CLARK, P.L.; ZUCHI, M.I.; SILVA-FILHO, M.C.; FOSTER, J.E.;
OMOTO, C. Genetic structure and molecular variability of Spodoptera frugiperda
(Lepidoptera: Noctuidae) collected in maize and cotton fields in Brazil. Bulletin of
Entomological Research, London, v. 97, p. 225-231, 2007.
MARTINS, G.L.M.; MARUYAMA, L.C.T.; MARUYAMA, W.I. Agentes microbianos no
controle de Alabama argillacea (Hüebner, 1918) (Lepidoptera: Noctuidae) em
algodoeiro. Arquivos do Instituto Biológico, São Paulo, v. 74, p. 23-27, 2007.
MILLER, M. TOOLS FOR POPULATION GENETIC ANALYSIS. Disponível em:
<http://herb.bio.nau.edu/~miller/tfpga>. Acesso em: 20 fev. 2009.
MULLIS, K.; FALOONA, F. Specific synthesis of DNA in vitro via a polymerase catalyzed chain reaction. Methods in Enzymology, New York, v. 55, p. 335-350, 1987.
NEI, M. F-statistics and analysis of gene diversity in subdivided population. Annals of
Human Genetics, London, v. 41, p. 225-33, 1977.
______. Estimation of average heterozygosity and genetic distance from a small number
of individuals. Genetics, Bethesda, v .89, n. 3, p.583-590, 1978.
NÈVE, G.; MEGLÉCZ, E. Microsatellite frequencies in different taxa. Trends in
Evolution and Ecology, Amsterdam, v. 15, p. 376-377, 2000.
O’CONNELL, M.; WRIGHT, J.M. Microsatellite DNA in fishes. Reviews in Fish Biology
and Fisheries, Dordrecht, v. 7, p. 331-363, 1997.
OLIVEIRA, E.J.; PÁDUA, J.G.; ZUCCHI, M.I.; VENCOVSKY, R.; VIEIRA, M.L.C. Origin,
evolution and genome distribution of microsatellites. Genetics and Molecular Biology,
Ribeirão Preto, v. 29, p. 294-307, 2006.

118 PRITCHARD, J.K.; STEPHENS, M.; DONNELLY, P. Inference of population structure
using multilocus genotype data. Genetics, Bethesda, v. 155, p. 945-959, 2000.
QUIRINO, E.S.; SOARES, J.J. Efeito do ataque de Alabama argillacea no crescimento
vegetativo e sua relação com a fenologia do algodoeiro. Pesquisa Agropecuária
Brasileira, Brasília, v. 36, p. 1005-1010, 2001.
RAMALHO, F.S. Cotton pest management: Part 4. A Brazilian perspective. Annual
Review of Entomology, Palo Alto, v. 39, p. 463-578, 1994.
REED, J.P.; HALLIDAY, W.R. Establishment of Cry9C susceptibility baselines for
European corn borer and southwestern corn borer (Lepidoptera:Crambidae). Journal of
Economic Entomology, Lanham, v. 94, p. 397–402, 2001.
ROBERTSON, J.L.; PREISLER, H.K. Pesticide bioassays with arthropods. Boca
Raton: CRC Press, 1992. 127 p.
ROHLF, J. NTSYSpc 2.1. New York: Applied Biostatistics, 2000. 1 CD-ROM.
ROSENBERG, N.A. DISTRUCT: a program for the graphical display of population
structure. Molecular Ecology Notes, London, v. 4, p. 137-138, 2004.
ROSSITER, M.; YENDOL, W.G.; DUBOIS, N.R. Resistance to Bacillus thuringiensis in
gypsy moth (Lepidoptera: Lymantriidae): genetic and environmental causes. Journal of
Economic Entomology, Lanham, v. 83, p. 2211–2218, 1990.
ROUSH, R.T.; McKENZIE, J.A. Ecological genetics of insecticide and acaricide
resistance. Annual Review of Entomology, Palo Alto, v. 32, p. 361-380, 1987.
ROUSH, R.T.; MILLER, G.L. Considerations for the design of insecticide monitoring
programs. Journal of Economic Entomology, Lanham, v. 79, p. 293-298, 1986.
SAEGLITZ, S.; BARTSCH, D.; EBER, S.; GATHMANN, A.; PRIESNITZ, K.U.;
SCHUPHAN, I. Monitoring the Cry1Ab susceptibility of European corn borer in Germany.
Journal of Economic Entomology, Lanham, v. 99, p. 1768–1773, 2006.
SANTOS, T.M.; BOIÇA JUNIOR, A.L. Resistência de genótipos de algodoeiro
(Gossypium hirsutum L.) a Alabama argillacea (Hüebner) (Lepidoptera: Noctuidae).
Neotropical Entomology, Londrina, v. 30, p. 297-303, 2001.
SANTOS, W.J. Manejo das pragas do algodão no Estado do Paraná. Revista
Agropecuária, São Paulo, v. 21, p. 22-27, 1980.

119
SCHLÖTTERER, C. The evolution of molecular markers – Just a matter of fashion?
Nature Reviews: Genetics, London, v. 5, p. 63-69, 2004.
SCHLÖTTERER, C.; TAUTZ, D. Slippage synthesis of simple sequence DNA. Nucleic
Acids Research, London, v. 20, p. 211-215, 1992.
SCHLÖTTERER, C.; RITTER, R.; HARR, B.; BREM, G. High mutation rate of a long
microssatellite allele in Drosophila melanogaster provides evidence for allele-specific
mutation rates. Molecular and Biology Evolution, Oxford, v. 15, n. 10, p. 1269-1274,
1998.
SCHNEPF, E.; CRICKMORE, N.; VAN RIE, J.; LERECLUS, D.; BAUM, J.; FEITELSON,
J.; ZEIGLER, D.R.; DEAN, D.H. Bacillus thuringiensis and its pesticidal crystal proteins.
Microbiology and Molecular Biology Review, Washington, v. 62, p. 775–806, 1998.
SCOTT, L.J.; LAWRENCE, N. LANGE, C.L.; GRAHAM, G.C.; HARDWICK, S.;
ROSITER, L.; DILLON, M.L.; SCOTT, K.D. Populations dynamics and gene flow pf
Helicoverpa armigera (Lepidoptera: Noctuidae) on cotton and grain crops in the
Marrumbidgee Valley, Australia. Journal of Economic Entomology, Lanham, v. 99,
p. 155-163, 2006.
SEÁRA, H.S. Perdas causadas pelo curuquerê, Alabama argillacea (Hübner, 1818) e
pelo “Ácaro Bronzeado”, Heterotergum gossupii Kiefer, na cultura do algodão Mocó.
Pesquisa Agropecuária Nordestina, Recife, v. 2, p. 5-11, 1970.
SIA, E.A.; BUTLER, C.A.; DOMINSKA, M.; GREENWELL, P.; FOX, T.D.; PETES, T.D.
Analyses of microsatellite mutations in the mitochondrial DNA of Saccharomyces
cerevisae. Proceedings of the National Academy of Sciences of the USA,
Washington, v. 97, p. 250-255, 2000.
SIEGFRIED, B.D.; SPENCER, T.; NEARMAN, J.; Baseline susceptibility of the corn
earworm (Lepidoptera: Noctuidae) to the Cry1Ab toxin from Bacillus thuringiensis.
Journal of Economic Entomology, Lanham, v. 93, p. 1265–1268, 2000.
SIEGFRIED, B.D.; VAUGHN, T.T.; SPENCER, T. Baseline susceptibility of western corn
rootworm (Coleoptera: Chrysomelidae) to Cry3Bb1 Bacillus thuringiensis toxin. Journal
of Economic Entomology, Lanham, v. 98, p. 1320–1324, 2005.
SILVA, A.L.; NETO PRADO, P.C.; CUNHA, H.F. Avaliação da produtividade,
segundo efeito da desfolha e eliminação de estruturas frutíferas nos diferentes
estágios do algodoeiro. Goiânia: Embrapa, 1980. 125 p. Relatório Técnico da
Embrapa-GO.

120 SILVEIRA NETO, S. Levantamento de insetos e flutuação da população de pragas
da ordem Lepidoptera, com o uso de armadilhas luminosas, em diversas regiões
do Estado de São Paulo. 1972. 183 p. Tese (Livre Docência) – Escola Superior de
Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 1972.
SIMS, S.R.; BERBERICH, S.A. Bacillus thuringiensis Cry1Ac Protein levels in raw and
processed seed of transgenic cotton determination using insect bioassay and ELISA.
Journal of Economic Entomology, Lanham, v. 89, p. 247–251, 1996.
SLATKIN, M. A measure of population subdivision based on microsatellite allele
frequencies. Genetics, Bethesda, v.130, p.457-462, 1995.
SOLTIS, D.E.; SOLTIS, P.S. Isozymes in plant biology. Portland: Dioscorides, 1989.
268 p.
SOUZA, A.P. Biologia molecular aplicada ao melhoramento. In: NASS, L.L.; VALOIS,
A.C.C.; MELO, I.S.; VALADARES-INGLIS, M.C. (Ed.). Recursos genéticos &
melhoramento de plantas. Rondonópolis: Fundação MT, 2001. p. 939-966.
STONE, T.B.; SIMS, S.R. Geographic susceptibility of Heliothis virescens and
Helicoverpa zea (Lepidoptera: Noctuidae) to Bacillus thuringiensis. Journal of
Economic Entomology, Lanham, v. 86, p. 989-994, 1993.
STRAND, M.; PROLLA, T.A.; LISKAY, R.M.; PETES, T.D. Destabilization of racts of
simple repetitive DNA in yeast by mutations affecting DNA mismatch repair. Nature,
London, v. 365, p. 274-276, 1993.
TABASHNIK, B.E. Determining the mode of inheritance of pesticide resistance with
backcross experiments. Journal of Economic Entomology, Lanham, v. 38, p. 325-
326, 1991.
______. Evolution of resistance to Bacillus thuringiensis. Annual Reviews of
Entomology, Palo Alto, v. 39, p. 47-79, 1994.
TABASHNIK, B.E.; GASSMANN, A.J.; CROWDER, D.W. CARRIÈRE, Y. Insect
resistance to Bt crops: evidence versus theory. Nature Biotechnology, London, v. 26,
p. 199-202, 2008.
TAUTZ, D.; TRICK, M.; DOVER, G.A. Cryptic simplicity in DNA is a major source of
genetic variation. Nature, London, v. 322, p. 652-656, 1986.
TOTH, G.; GASPARI, Z.; JURKA, J. Microssatellites in different eukaryotic genomes:
survey and analysis. Genome Research, New York, v. 10, p. 967-981, 2000.

121
VAN OOSTERHOUT, C.; HUTCHINSON, W.F. WILLS, D.P.M.; SHIPLEY, P. Micro-
Checker: software for identifying and correcting genotyping error in microsatellite data.
Molecular Ecology Notes, London, v. 4, p. 535-538, 2004.
VAN RIE, J.; JANSENS, S.; HOFTE, H.; DEGHEELE, D.; VAN MALLAERT, H.
Specificity of Bacillus thuringiensis β-endotoxin: importance of specific receptors on the
brush border membrane of the midgut of target insects. European Journal of
Biochemistry, Oxford, v. 186, p. 239–247, 1989.
VANTOAL, T. T.; PENG, J.; MARTINS, S. S. Using AFLP markers to determinate the
contribution of parental genomes during recurrent selection. Soybean Genetics
Newsletter, Iowa, v. 23, p. 214-216, 1996.
VOS, P.; HOGERS, R.; BLEEKER, M.; REIJANS, M.; VAN DE LEE, T.; HORNES, M.;
FRITERS, A.; POT, J.; PALEMAN, J.; KUIPER, M.; ZABEAU, M. AFLP: A new
technique for DNA fingerprinting. Nucleic Acids Research, London, v. 23, p. 4407-
4414. 1995.
WEIR, B.S. Genetics data analysis II: methods for discrete population genetic data.
Suderland: Sinauer Associates, 1996. 455 p.
WEIR, B.S.; COCKERHAM, S.S. Estimating F-statistics for the analyses of population
structure. Evolution, Lawrence, v. 38, p. 1358-1370, 1984.
WILLIAMS, J.G.; KUBELIK, A.R.; LIVAK, K.J.; RAFALSKI, L.A.; TINGEY, S.V. DNA
polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic
Acids Research, London, v. 18, p. 6531-6535, 1990.
WRIGHT, J.M.; BENTZEN, P. Microsatellites: Genetic markers of the future. Reviews in
Fish Biology and Fisheries, Dordrecht, v. 4, p. 384-388, 1994.
WU, K.; GUO, Y.; LV., N.; GREENPLATE, J. T.; DEATON, R. Resistance Monitoring of Helicoverpa armigera (Lepidoptera: Noctuidae) to Bacillus thuringiensis Insecticidal Protein in China. Journal of Economic Entomology, Lanham v. 95, p. 826-831, 2002.
ZANE, L.; BARGRLLONI, L.; PATARNELLO, T. Strategies for microssatellites isolation:
a review. Molecular Ecology, London, v. 11, p. 1-6, 2002.
ZHANG, D.X. Isolation, characterization and cross-species amplification of eight
microsatellite DNA loci in the migratory locust (Locusta migratoria). Molecular Ecology
Notes, London, v. 3, p. 483-486, 2003.
______. Lepidopteran microsatellite DNA: redundant but promising. Trends in Ecology
and Evolution, Amsterdam, v. 19, p. 507-509, 2004.