universidade de sÃo paulo escola de engenharia de lorena - sistema de...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO ESCOLA DE ENGENHARIA DE LORENA
CARLOS ALBERTO SANCHES PEREIRA
Avaliação do efeito de microrganismos probióticos sobre Eimeria spp em Rattus norvegicus
Lorena – SP 2007

CARLOS ALBERTO SANCHES PEREIRA
Avaliação do efeito de microrganismos probióticos sobre Eimeria spp em Rattus norvegicus
Tese apresentada a Escola de Engenharia de Lorena da Universidade de São Paulo para a obtenção do título de Doutor em Biotecnologia Industrial.
Área de Concentração: Microbiologia Aplicada Orientação: Prof. Dr. Ismael Maciel de Mancilha
Lorena – SP 2007

AUTORIZO A REPRODUÇÃO E DIVULGAÇÃO TOTAL OU PARCIAL DESTE TRABALHO, POR QUALQUER MEIO CONVENCIONAL OU ELETRÔNICO, PARA FINS DE ESTUDO E PESQUISA, DESDE QUE CITADA A FONTE.
Catalogação na Publicação Biblioteca Universitária
Escola de Engenharia de Lorena da Universidade de São Paulo
Pereira, Carlos Alberto Sanches
Avaliação do efeito de microrganismos probióticos sobre Eimeria spp. em Rattus norvegicus / Carlos Alberto Sanches Pereira ; orientador Ismael Maciel de Mancilha. -- 2007
75 f. : fig.
Tese (Doutorado – Programa de Pós-Graduação em Biotecnologia Industrial. Área de Concentração: Microbiologia Aplicada) – Escola de Engenharia de Lorena da Universidade de São Paulo, 2007
1. Microbiologia Aplicada 2. Preparado probiótico 3. Eimeria spp. I. Título.
579.6 - CDU

DEDICATÓRIA
A minha mãe... Minha Rainha... Com Amor, admiração e gratidão por seu incentivo, carinho, presença incansável para a obtenção deste título – Mãe, este título é em sua homenagem...

ADRADECIMENTOS
Em primeiro lugar, gostaria de agradecer a DEUS, por mais uma conquista na minha vida. Sem a SUA presença em meu coração, nada disso teria acontecido. A Nossa Senhora de Aparecida, por ter ouvido minhas preces. Ao Professor Dr. Ismael Maciel de Mancilha, pela amizade, atenção e apoio durante toda minha estadia na EEL. À Professora Dra. Maria da Graças de Almeida Felipe, por ter lido meu E-mail em 2003 e ter participado desta etapa da minha vida. Ao meu Grande Amigo Mauro Souza Lima, pelo incentivo nas horas difíceis, e pelos ensinamentos de estatística. Aos Amigos Ronie Cabral, Priscila Filgueiras e Patrícia Lacerda, pelo mês de Janeiro que vocês se dedicaram ao experimento. O trabalho não sairia sem o auxílio de vocês. Ao amigo Renato Teixeira pelo apoio nas referências bibliográficas. À Débora pelas dificuldades vencidas juntos. Ao Centro Universitário Geraldo Di Biase por ter cedido o laboratório no período de férias para a realização dos experimentos.

“Se um homem tem um talento e não tem capacidade de usa-lo, ele fracassou.
Se ele tem um talento e usa somente a metade deste, ele fracassou
parcialmente. Se ele tem um talento e de certa forma aprende a usa-lo em sua
totalidade, ele triunfou gloriosamente e obteve uma satisfação e um triunfo que
poucos homens conhecerão”.
Thomas Wolfe

RESUMO
PEREIRA, C. A. S. Avaliação do efeito de microrganismos probióticos sobre Eimeria spp em Rattus norvegicus. 2006. 74 f. Tese (Doutorado em Biotecnologia Industrial) - Escola de Engenharia Química de Lorena, Universidade de São Paulo, 2007. O presente trabalho teve como objetivo avaliar o efeito antiparasitário em Rattus norvegicus de um preparado probiótico constituído de 4 espécies de lactobacilos, L. casei, L. plantarum, L. fermentum e L. acidophilus. Para tanto foram utilizados 25 animais, divididos em cinco grupos, onde os animais do grupo A receberam ração e água diariamente (controle negativo); grupo B além de ração e água os animais foram infectados com oocistos de Eimeria spp. no sexto dia do experimento (controle positivo da Eimeriose). Os animais do grupo C receberam diariamente 1 ml do preparado probiótico, além de água e ração (controle positivo do efeito probiótico); grupo D, os animais receberam 1 ml do preparado probiótico durante cinco dias e no sexto dia foram infectados com oocistos de Eimeria spp, e continuaram a receber 1 ml do preparado probiótico até o final do experimento. Quanto aos animais do grupo E, até o quinto dia receberam água e ração à vontade, no sexto dia foram infectados com aproximadamente 5000 oocistos de Eimeria spp e a partir do terceiro dia de infecção, passaram a receber diariamente 1 ml do preparado probiótico. Após o 9° dia do experimento, os animais dos cinco grupos, tiveram suas fezes coletadas diariamente, pesadas e submetidas ao exame parasitológico para se proceder a contagem de oocistos de Eimeria spp por grama de fezes (OoPG) e para contagem de lactobacilos na fezes UFC/g fezes. O desempenho zootécnico dos animais foi determinado por meio do consumo de ração, ganho de peso e conversão alimentar dos animais. Para tanto, foi oferecido diariamente aos animais 30 g de ração e após 24 horas a ração residual no comedouro foi devidamente pesada, e por diferença determinou-se o consumo diário de ração de cada animal. Os animais foram pesados todos os dias para se determinar o ganho de peso e, por conseguinte calcular a taxa de conversão alimentar. Para obtenção do leucograma, um animal de cada grupo foi submetido a punção cardíaca no 3º, 6º, 9º, 12º e 15º dia após infecção por Eimeria spp. O sangue foi acondicionado em tubo de ensaio contendo EDTA como anticoagulante. A contagem total e diferencial dos leucócitos foi realizada por meio de contador automático Cell-Dyn 3500, da marca Abbott. As contagens diferenciais foram confirmadas, analisando-se os filmes sanguíneos, com auxílio do microscópio óptico. Os resultados demonstraram que o preparado probiótico ministrado nos animais do grupo D e E foi eficaz no sentido de promover a interrupção da eliminação de oocistos de Eimeira spp nas fezes dos animais no 12o dia de infecção. Estes animais permaneceram com as fezes sob aspecto normal, que favoreceu a higidez dos animais. Esta interrupção pode ser devido a diminuição de sítios disponíveis na membrana dos enterócitos e pelo aumento das células fagocitários (neutrófilos segmentados). O Teste t (p < 0,05) para a eliminação de oocistos nas fezes e o teste ANOVA (p < 0,05) para as contagens de leucócitos não apresentaram diferenças significativas entre os tratamentos D e E. Os animais dos grupos C, D e E obtiveram maior ganho de peso e melhor conversão alimentar quando comparados com os animais dos grupos A e B, o que demonstra a eficácia do preparado probiótico tanto no tratamento quanto na prevenção de Eimeriose, no ganho de peso e na conversão alimentar de Rattus norvegicus. Palavras-chave: Probióticos , Lactobacillus, Eimeria spp, Rattus norvegicus

ABSTRACT
PEREIRA, C. A. S. Evaluation of probiotic microorganismos on Eimeria spp in Rattus norvegicus. 2006. 74 f. Thesis (Doctoral in Industrial Biotechnology) – Escola de Engenharia de Lorena, Universidade de São Paulo, São Paulo, 2007. The present work aimed to evaluate the antiparasitary effect of a probiotic mixture constituted of 4 Lactobacillus species (L. casei, L. plantarum, L. fermentum and L. acidophilus) on Rattus norvegicus. For this purpose, 25 animals were used, divided in five groups, where the animals from the group A received daily ration and water (negative control). In the group B, besides ration and water, the animals were infected with Eimeria spp oocysts at the sixth day of the experiment (positive control of Eimeriosis). The animals from the group C daily received 1 ml of the probiotic mixture, besides water and ration (positive control of the probiotic effect). In the group D, the animals received 1 ml of the probiotic mixture for five days and at the sixth day, they were infected with Eimeria spp oocysts, and continued receiving 1 ml of the probiotic mixture until the end of the experiment. For the animals from the group E, until the fifth day they comfortably received water and ration; at the sixth day, they were infected with ca oocysts of Eimeria spp and from the third day of infection, they started to receive 1 ml of the probiotic mixture daily. After the 9th day of the experiment, the animals from the five groups had their feces collected daily, weighed and submitted to parasitologic exam in order to proceed the counting of Eimeria spp. oocysts per gram of feces (OoPG) and the Lactobacillus counting in the feces (UFC/g feces). The zootecnic performance of the animals was determined by the ration consumption, weight earnings and their alimentary conversion. For this purpose, 30g ration were offered to the animals daily and after 24 hours the residual ration in the feed box was properly weighed and the daily consumption of ration of each animal was determined by the difference. The animals were weighed everyday in order to determine the weight earnings and, consequently to calculate the tax of alimentary conversion. For the leucogram obtainment, one animal of each group was submitted to the heart puncture on the 3rd, 6th, 9th, 12th and 15th days after infection by Eimeria spp. The blood was kept in test tube containing EDTA as anticoagulant. The total and differential counting of the leukocytes was accomplished through automatic accountant Cell-Dyn 3500, mark Abbott. The differential countings were confirmed by analyzing the sanguine films, with aid of optical microscope. The results demonstrated that the probiotic mixture for the animals from the groups D and E was effective for promoting the interruption of Eimeira spp oocysts elimination. in the feces of the animals at the 12th day of infection. These animals remained with the feces under normal aspect, what favored the animals health. This interruption could be due to the decrease of sites available on the enterocytes membrane and to the increase of fagocitary cells (segmented neutrophils). The Test t (p <0,05) for the oocysts elimination from the feces and the test ANOVA (p <0,05) for the leukocytes counting did not present significant differences in relation to the treatments for the groups D and E. The animals from the groups C, D and E obtained larger weight earnings and better alimentary conversion when compared to the animals from the groups A and B, what demonstrates the effectiveness of the probiotic mixture in the treatment and prevention of Eimeriosis, in the weight earnings and in the alimentary conversion of Rattus norvegicus. Key-words: Probiotics, Lactobacillus, Eimeria spp, Rattus norvegicus

LISTA DE ILUSTRAÇÕES
Figura 1 – Ciclo Evolutivo de Eimeria spp .................................................................. 33
Figura 2 – Acomodação dos ratos em gaiolas individuais ........................................... 38
Figura 3 – Administração de preparado probiótico aos animais .................................. 39
Figura 4 – Administração de oocistos de Eimeria nos animais.................................... 40
Figura 5 – Pesagem dos ratos ....................................................................................... 41
Figura 6 – Punção cardíaca........................................................................................... 43

LISTA DE TABELAS
Tabela 1 – Contagem de lactobacilos nas fezes dos animais (Log UFC/g fezes) ........ 44
Tabela 2 – Contagem de oocistos nas fezes dos animais ............................................. 46
Tabela 3 – Avaliação dos resultados (Teste t) de OoPG entre os grupos de animais
tratados e não tratados com probióticos ..................................................... 47
Tabela 4 – Leucograma dos animais ............................................................................ 48
Tabela 5 – Desempenho zootécnico dos animais ......................................................... 52

SUMÁRIO
1 INTRODUÇÃO ....................................................................................................... 12
2 REVISÃO BIBLIOGRÁFICA ............................................................................... 14
2.1 PROBIÓTICOS...................................................................................................... 14
2.2 MICROBIOTA INTESTINAL............................................................................... 15
2.3 ESPÉCIES DE MICRORGANISMOS QUE APRESENTAM PROPRIEDADES
PROBIÓTICAS...................................................................................................... 16
2.3.1 Características do Gênero Lactobacillus ......................................................... 16
2.3.1.1 Lactobacillus acidophilus ................................................................................ 17
2.3.1.2 Lactobacillus casei........................................................................................... 17
2.3.1.3 Lactobacillus fermentum................................................................................. 17
2.3.1.4 Lactobacillus plantarum.................................................................................. 18
2.4 MECANISMO DE AÇÃO ..................................................................................... 18
2.4.1 Adesão, Agregação e Colonização .................................................................... 18
2.4.2 Resistência a Sais Biliares ................................................................................. 20
2.4.3 Produção de bacteriocinas ................................................................................ 21
2.4.4 Produção de Ácidos Orgânicos e Peróxido de Hidrogênio ............................ 23
2.4.5 Atividade anticarcinogênica ............................................................................. 24
2.4.6 Estimulação da Imunidade ............................................................................... 25
2.4.7 Efeito hipocolesterolêmico ................................................................................ 27
2.5 PROBIÓTICOS NA SAÚDE ANIMAL................................................................ 27
2.6 AÇÕES ANTIPARASITÁRIAS DOS PROBIÓTICOS........................................ 30
2.7 EIMERIA E EIMERIOSE....................................................................................... 32
2.7.1 Ciclo de vida ...................................................................................................... 34
3 MATERIAL E MÉTODOS .................................................................................... 36
3.1 MICRORGANISMOS .......................................................................................... 36
3.1.1 Probióticos .......................................................................................................... 36
3.1.2 Eimeria ............................................................................................................... 37
3.2 AVALIAÇÃO DOS MICRORGANISMOS PROBIÓTICOS SOBRE EIMERIA 37
3.3 MÉTODOS ANALÍTICOS ................................................................................... 40
3.3.1 Consumo de ração ............................................................................................. 40
3.3.2 Contagem de Lactobacillus nas fezes .............................................................. 41
3.3.3 Exame Parasitológico ....................................................................................... 41

3.3.4 Exame do Tecido Hematopoético (Leucograma) ........................................... 42
3.4 ANÁLISE DOS RESULTADOS .......................................................................... 43
4 RESULTADOS E DISCUSSÃO ............................................................................ 44
4.1 AVALIAÇÃO DA PREPARAÇÃO PROBIÓTICA NA COLONIZAÇÃO DO
INTESTINO DOS ANIMAIS ................................................................................. 44
4.2 AVALIAÇÃO DOS MICRORGANISMOS PROBIÓTICOS SOBRE EIMERIA 45
4.3 AVALIAÇÃO DO TECIDO HEMATOPOÉTICO (LEUCOGRAMA) .............. 48
4.4 DESEMPENHO ZOOTÉCNICO DOS ANIMAIS................................................ 51
5 CONCLUSÕES ....................................................................................................... 54
6 SUGESTÕES PARA TRABALHOS FUTUROS ................................................. 55
REFERÊNCIAS ........................................................................................................ 56
APÊNDICES ............................................................................................................... 70

12 1. INTRODUÇÃO
Os animais saudáveis caracterizam-se em geral por apresentarem um bom
funcionamento do trato intestinal, o que garante o equilíbrio da microbiota presente. Este fator
é fundamental na utilização de nutrientes para um bom desenvolvimento da produção. No
entanto, o aparente equilíbrio pode ser quebrado, como em situações em que o animal fica
estressado devido a agressões decorrentes do manejo, variações climáticas ou alimentares. A
alteração deste equilíbrio oferece condições para que bactérias patogênicas e alguns
protozoários proliferem e provoquem distúrbios entre os quais a diarréia. É neste momento
que os probióticos podem auxiliar através de sua ação, promovendo o reequilíbrio da
microbiota intestinal.
Os efeitos benéficos dos probióticos podem ser mediados por um efeito antagônico
direto contra grupos específicos de organismos, resultando na redução de suas células viáveis,
através da produção de compostos antibacterianos, competição por nutrientes ou sítios de
adesão no intestino. Seus efeitos ainda podem se notar sobre o metabolismo microbiano,
aumentando ou diminuindo a atividade de enzimas carcinogênicas, ou ainda pela estimulação
da imunidade do hospedeiro, aumentando os níveis de anticorpos ou atividade dos
macrófagos.
A utilização de probióticos em animais, com efeito, antiparasitário, teve início na
década de 70, com aves infectadas por Eimeria tenella, onde os probióticos usados
melhoraram o quadro clínico destas aves. Em bovinos e suínos os trabalhos foram realizados
com Cryptosporidium parvum, também coccídio, causador de diarréias graves em animais
jovens. Em alguns trabalhos observa-se a utilização de ratos como modelo experimental para
se avaliar a ação antiparasitária dos probióticos e os resultados têm sido promissores.
Segundo informações da EMBRAPA (2004), o Brasil detém o maior rebanho bovino
comercial do planeta, representando 15% do total mundial. É o segundo maior produtor de
carne bovina, com 6,9 milhões de toneladas, obtidas a baixo custo de produção, revelando-se
o país mais competitivo internacionalmente. Esses números demonstram os avanços da
pesquisa no setor, colocando o Brasil numa confortável posição em termos de produtividade e
qualidade do produto. Para que estes números sejam mantidos, especial atenção deve ser
direcionada ao manejo dos animais jovens, que apresentam maior sensibilidade aos agentes
patogênicos em geral.
A falta de condições sanitárias elevam os custos do controle de doenças causadas por
endoparasitos, colocando em risco os lucros do produtor. Os animais acometidos por

13 endoparasitos, como espécies Eimeria representam relevantes prejuízos econômicos, devido à
síndrome de má-absorção naqueles que sobrevivem. Este quadro pode ser revertido atuando-
se na prevenção com uso de probióticos promovendo equilíbrio na microbiota do rumem e ou
intestino destes animais.
Com a atual preocupação dos consumidores pela aquisição de alimentos saudáveis,
vários setores como a pecuária, têm procurado atender a essas exigências criando animais
“orgânicos”, os quais são mantidos em pastos sem agrotóxicos e sem adubação química,
tratados apenas com medicamentos homeopáticos e probióticos.
Os animais tratados com probióticos podem ter suas condições sanitárias (ou de saúde)
preservadas, e quando infectados por endoparasitos a evolução para a cura poderá ser mais
rápida quando comparados àqueles que receberam apenas tratamento homeopático, reduzindo
custos para o produtor e melhorando a qualidade de vida do animal.
Desta forma, o presente trabalho teve por objetivo contribuir para o desenvolvimento
de método de controle parasitário por meio da avaliação do efeito de espécies de
Lactobacillus, que apresentam propriedades probióticas, como agente antiparasitário para
Eimeria spp em ratos (Rattus norvegicus) como modelo experimental para bovinos.

14 2 REVISÃO BIBLIOGRÁFICA
2.1 PROBIÓTICOS
O termo probiótico, utilizado em várias áreas da biologia, deriva do grego pro biosis,
ou seja, em favor da vida (O’SULLIVAN et al., 1992).
Fuller (1995) define probiótico como sendo uma preparação constituída de
microrganismos vivos não patogênicos ou estimulantes microbianos que afetam a microbiota
do animal.
O termo probiótico foi utilizado pela primeira vez em 1965 por Lilly e Stillwell para
descrever substâncias secretadas por um microrganismo, as quais estimulavam o crescimento
de um outro, opondo-se dessa forma ao significado de antibiótico. Entretanto, este conceito
foi alterado posteriormente por Sperti, em 1971, para descrever extratos de tecidos que
estimulam o crescimento microbiano (KUNITACHI, 1996).
Em 1974, Parker definiu probióticos como organismos e substâncias que contribuem
para o equilíbrio de microrganismos intestinais e esta definição relaciona o uso de probiótico
com a microbiota intestinal (FULLER, 1989). Na tentativa de melhorar esta definição, Fuller
(1992) redefiniu probióticos como “suplemento alimentar contendo microrganismos vivos que
afeta beneficamente a microbiota do animal pelo estabelecimento do equilíbrio microbiano
intestinal”. Entretanto, a definição aceita internacionalmente é de que os probióticos são
microrganismos vivos, administrados em quantidades adequadas, que conferem benefícios a
saúde do hospedeiro (FOOD AND AGRICULTURE ORGANIZATION OF UNITED NATIONS,
WORLD HEALTH ORGANIZATION, 2001; SANDERS, 2003).
A palavra probiótico relacionada com alimento suplementar é referida apenas a partir
de 1974, enquanto que, suplementos alimentares com microrganismos vivos datam de
milhares de anos, como leites fermentados, que provavelmente foram os primeiros alimentos
contendo microrganismos vivos, documentados no Velho Testamento (KROGER,
KURMANN; RASIC 1989; FULLER, 1992).
Segundo Kroger; Kurmann; Rasic (1989), Hosse; Sozzi (1991) e Fuller (1992) Elie
Metchnikoff, trabalhando no Instituto Pasteur, em Paris, descobriu a chave do processo. Este
pesquisador inferiu que a ingestão de bactérias lácticas exercia, em sua visão, uma influência
positiva na microbiota normal do trato gastrintestinal por prevenir a putrefação, prolongando a
vida. O iogurte pode ter sido visto como uma preparação probiótica e é impossível saber com
certeza quais espécies ele e seus contemporâneos estudaram, mas foram provavelmente

15 misturas não intencionadas de lactobacilos que foram algumas vezes usadas. Qualquer que
fosse a sua intenção, seu trabalho pode ser considerado como o nascimento do probiótico, isto
é, ingestão de microrganismos com o objetivo de promoção da saúde.
2.2 MICROBIOTA INTESTINAL
O feto in útero é estéril, mas na passagem através da vagina durante o nascimento
adquire microrganismos. Estes são rapidamente adquiridos pós-nascimento e o animal recém-
nascido adquire uma microbiota intestinal característica de sua espécie. No estado selvagem o
animal obtém sua microbiota intestinal de seu ambiente imediato o qual encontra-se
contaminado com bactérias da mãe (FULLER, 1989).
Conforme Tannock et al. (1988), a microbiota intestinal indígena final que está
estabilizada nos intestinos compreende uma “coleção” complexa de aproximadamente 1014
microrganismos consistindo de aproximadamente 400 espécies de bactérias. Dentro de um
sistema tão complexo observam-se vários inter-relacionamentos entre diferentes
microrganismos e o hospedeiro. A despeito de todo este escopo por variabilidade, a
microbiota rapidamente se estabelece em uma população estável.
A composição da microbiota é determinada pelo hospedeiro e apesar de haver uma
grande quantidade de bactérias que podem sobreviver e crescer no trato intestinal, algumas
não podem fazê-lo. As bem sucedidas não têm apenas que superar os produtos químicos
antimicrobianos presentes no intestino, mas também evitar os efeitos do peristaltismo que
tende a expeli-las com o bolo fecal. A sobrevivência de microrganismos probióticos no
intestino depende dos fatores de colonização que eles possuem, permitindo que resistam aos
mecanismos antibacterianos (químicos e físicos) que operam no intestino (FULLER, 1988).
A microbiota estável que se desenvolve no intestino auxilia o animal a resistir à
infecções, particularmente no trato gastrintestinal. O fenômeno tem sido descrito por vários
autores como: antagonismo bacteriano, interferência bacteriana, efeito barreira, resistência à
colonização e exclusão competitiva. A melhor evidência para este efeito protetor da
microbiota intestinal origina-se da observação de que animais livres de germes são mais
suscetíveis a doenças do que os animais normais com uma microbiota intestinal completa
(WALLACE; NEWBOLD, 1992)

16 2.3 ESPÉCIES DE MICRORGANISMOS QUE APRESENTAM PROPRIEDADES
PROBIÓTICAS
De acordo com Ferreira (2003), os microrganismos probióticos mais utilizados
pertencem ao grupo das bactérias lácticas, que são representadas por 15 gêneros: Aerococcus,
Atopobium, Bifidobacterium, Brochothrix, Carnobacterium, Enterococcus, Lactobacillus,
Lactococcus, Leuconostoc, Oenococcus, Pediococcus, Streptococcus, Tetrogenococcus,
Vagococcus, Weissella. As bactérias lácticas são Gram-positivas, anaeróbias, microaerófilas
ou aerotolerantes, catalase negativas, apresentando-se na forma de cocos ou bacilos.
As leveduras não patogênicas possuem algumas características probióticas que as
diferenciam das bactérias. A resistência natural que têm aos antibióticos tem-se constituído
numa base interessante para sua aplicação como probióticos. Neste particular, a levedura
Sacharomyces boulardi é a que mais amplamente foi testada em ensaios experimentais e
clínicos, revelando resultados promissores (PENNA et al., 2000).
Segundo Fuller (1989), os efeitos benéficos dos probióticos podem ser mediados por
um efeito antagônico direto contra grupos específicos de organismos, resultando em uma
diminuição do número de patógenos, por efeito do seu metabolismo (produção de
bacteriocinas, peróxido de hidrogênio, efeito barreira) ou por estimulação de imunidade do
hospedeiro (aumento da fagocitose pelos macrófagos e ou produção de anticorpos e citocinas
específicas).
2.3.1 Características do gênero Lactobacillus
O gênero Lactobacillus é constituído de células que variam de longas e delgadas,
algumas vezes bastões curtos e curvos, freqüentemente cocobacilos. Normalmente são
imóveis; quando são móveis, os flagelos são peritríqueos. Não produzem esporos, são Gram-
positivos, catalase-negativos, não reduzem nitrato a nitrito. Utilizam a glicose para
fermentação. Podem ser homofermentativos, produzindo cerca de 85 % de ácido láctico a
partir da glicose ou heterofermentativos, produzindo ácido láctico, CO2, Etanol (KANDLER;
WEISS, 1986; HAMMES, WEISS; HOLZAPFEL, 1992).
O crescimento na superfície em meio sólido é freqüentemente aumentado por
anaerobiose a 5-10 % CO2, a faixa de temperatura para o crescimento está entre 5-53ºC, sendo
a temperatura ótima de 30-40ºC. São acidúricos e o pH ótimo está entre 5,5-5,8. Requerem

17 um complexo nutricional derivados de ácidos nucléicos, vitaminas, sais, ácidos graxos ou
ésteres de ácidos graxos e carboidratos fermentescíveis (DE MAN, et al., 1960).
São caracteristicamente sacarolíticos, atuando notadamente sobre carboidratos
gerando como produto final o lactato. São produtores de acetato, formitato, succinato, gás
carbônico e etanol (BUCHANAN; GIBBONS, 1974).
Os Lactobacillus geralmente não são patogênicos sendo encontrados em intestino de
animais, produtos de plantas onde carboidratos estão disponíveis, também encontrados na
boca, vagina e trato intestinal de vários animais homotérmicos incluindo o homem (FULLER,
1995).
2.3.1.1 Lactobacillus acidophilus
São bacilos com as extremidades arredondadas, variando em largura 0,6-0,9 µm por
1,5-6 µm de comprimento, ocorrendo isolados, em pares ou em pequenas cadeias. Não
crescem bem a 45ºC. Suas colônias são geralmente lisas, convexas, de bordos regulares,
freqüentemente translúcidas e não são pigmentadas. Necessitam de riboflavina, ácido fólico,
pantotenato de cálcio e niacina para seu crescimento. Pertencem ao grupo de lactobacilos
homofermentativos obrigatórios (KANDLER; WEISS, 1986; HAMMES, WEISS;
HOLZAPFEL, 1992).
2.3.1.2 Lactobacillus casei
São bacilos com as extremidades ligeiramente quadradas, variando em largura 0,7-1,1
µm por 2,0-4,0 µm de comprimento, normalmente se apresentam em cadeias. Não crescem a
45ºC, suas colônias são geralmente lisas, com bordos irregulares, freqüentemente translúcidas.
Necessitam de riboflavina, ácido fólico, pantotenato de cálcio e niacina para seu crescimento.
Estão presentes em produtos lácteos, esterco de vaca, silagem, trato intestinal humano, boca e
vagina. Pertencem ao grupo dos lactobacilos heterofermentativos facultativos (KANDLER;
WEISS, 1986; HAMMES, WEISS; HOLZAPFEL, 1992).
2.3.1.3 Lactobacillus fermentum
São bacilos curtos, variando em tamanho 0,5-1,0 µm de largura por 3,0 µm ou mais de
comprimento, ocorrendo isolados, em pares ou cadeias, imóveis e as suas colônias são

18 geralmente lisas ou algumas vezes rugosas, convexas, de bordos regulares ou ligeiramente
irregulares, freqüentemente translúcidas, não são pigmentadas, mas algumas espécies
produzem pigmento alaranjado. Pertence ao grupo de lactobacilos heterofermentativos
obrigatórios com produção de ácidos e gás pela fermentação da glicose, fermentam lactose,
melibiose, algumas vezes rafinose, xilose, arabinose e manose (KANDLER; WEISS, 1986;
HAMMES, WEISS; HOLZAPFEL, 1992).
De acordo com Naito et al. (1995) L. fermentum é a espécie predominante de
Lactobacillus no trato intestinal de suínos, nos primeiros 40 dias de vida e especificamente
sob condições de desmame aos 7 e 35 dias de vida.
2.3.1.4 Lactobacillus plantarum
São bastões com pontas arredondadas, retos geralmente 0,9-1,2 µm de largura por 3-8
µm de comprimento, ocorrem isolados, em pares, ou em curtas cadeias. A mobilidade e a
presença de flagelos são usualmente ausentes e as colônias formadas são lisas e convexas.
Não produz amônia a partir da arginina e os testes para esculina, hidrolisado hipurato e
acetoína são usualmente positivos. São tolerantes a temperatura de 15ºC, mas sensíveis a
45ºC, sendo a temperatura ótima de crescimento usualmente entre 30-35ºC. Fermentam
manitol, sorbitol, salicina, ramnose, lactose e manose, freqüentemente rafinose, xilose,
arabinose e melibiose (DAVIS, 1955).
Pertence ao grupo de lactobacilos heterofermentativos facultativos que são
freqüentemente isolados de material vegetal e alimentos fermentados. Este é também
encontrado como habitante comum do trato intestinal dos animais e o homem, com contagens
médias de 107 a 109 células por grama de fezes em indivíduos colonizados (ADLERBERTH
et al., 1996; GARCIA, 1999).
2.4 MECANISMOS DE AÇÃO
2.4.1 Adesão, Agregação e Colonização
De acordo com Redondo-Lopez et al. (1990) a proteção da microbiota na vagina
exercida por lactobacilos pode ser acompanhada por dois mecanismos: (1) produção de
substâncias antimicrobianas, tais como ácido láctico e bacteriocinas e (2) aderência à mucosa

19 e coagregação, que pode formar uma barreira a qual previne colonização por microrganismos
patogênicos.
Conforme Servin; Coconnier (2003) os lactobacilos dispõem de vários determinantes
na superfície celular (interações eletrostáticas, hidrofóbicas, forças estéricas, ácidos
lipoteicóicos, entre outros), que são envolvidos por suas interações com as células epiteliais
do intestino.
Adlerberth et al. (1996), observaram que a adesão de L. plantarum à células humanas
HT-29 ocorreu por mecanismo de aderência sensível a manose, sendo esta interação
provocada por proteínas do tipo lectinas. Como a parede celular da levedura S. cerevisiae é
constituída de polissacarídeos contendo manose (mananas), Escherichia coli e outras
enterobactérias contendo adesinas para receptores manose específicos, aglutinam células de
levedura (MIRELMAN; ALTMANN; ESHDAT, 1980). A capacidade de aglutinar células de
levedura pode, portanto, ser uma indicação de atividade manose específica.
Em Enterococcus faecalis, a capacidade de promover agregação é devida à secreção
de pequenos peptídeos hidrofóbicos chamados feromônios sexuais, com conseqüente aumento
da freqüência de conjugação (EHRENFELD et al., 1986; MORI et al., 1986). Os feromônios
parecem induzir a síntese de proteínas superficiais codificadas por plasmídeo, as quais
mediam contato célula-célula (BORIS et al., 1997).
A agregação celular pode desempenhar um papel decisivo na patogenicidade de um
organismo (FIRON et al., 1987).
Constituintes da microbiota normal e algumas bactérias patogênicas possuem a
habilidade de colonizar a superfície da mucosa intestinal (GOLDIN; GORBACH; SAXELIN,
1992). Uma vez que alguns microrganismos parecem capazes de se fixar firmemente ao
epitélio intestinal (KLEEMAN; KLAENHAMMER, 1982). Acredita-se ser este um pré-
requisito importante em probióticos para uma sobrevivência a longo prazo no intestino
durante a competição contra outros microrganismos por nichos específicos e subseqüente
multiplicação (O’ SULLIVAN et al., 1992; SALOFF-COSTE, 1997).
A proteção contra a colonização do trato intestinal por microrganismos
potencialmente patogênicos foi denominada exclusão competitiva (SANDERS, 1993), cuja
evidência pioneira fora obtida por NURMI; RANTALA (1973) em aves. Quando estas, logo
após o nascimento, foram inoculadas com conteúdo fecal de uma ave adulta, a frequência de
infecções por espécies de Salmonella foi reduzida significativamente.
Os principais componentes envolvidos no mecanismo de exclusão competitiva são:
competição por receptores na parede intestinal, produção de ácidos graxos voláteis e/ou outras

20 substâncias antibacterianas produzidas pela microbiota anaeróbia e a competição por
nutrientes (FULLER, 1989).
Segundo Fuller; Barrow; Brooker (1978), E. coli presente no intestino de ratos recém-
nascidos teve seu crescimento suprimido por Lactobacillus salivarius, que se encontrava
associado à superfície epitelial do intestino delgado.
De acordo com Costerton; Arrie; Cheng (1985), é importante a existência de
microrganismos benéficos na região do íleo, pois essa região é considerada como o principal
ponto de infecção por linhagens de E. coli enterotoxigênicas no homem e em outros animais,
sendo, portanto a região alvo dos probióticos.
Diversos critérios de seleção para microrganismos probióticos têm sido sugeridos,
porém seria interessante se focar em determinados aspectos uma vez que o preenchimento de
todos os requisitos seria impossível para uma única linhagem (HAVENAAR; BRINK; HUIS
IN’TVELD, 1992; NOUSIAINEN; SETALA, 1998).
2.4.2 Resistência a Sais Biliares
A bile é uma solução salina constituída de sais biliares, colesterol, lecitina, pigmentos
biliares e outros produtos finais do metabolismo orgânico e alguns traços de metais
(NEUMANN; FERREIRA, 1995).
Para ser considerado um bom probiótico e ser usado como adjunto dietético o
microrganismo deverá ser capaz de crescer em concentrações de 0,15 a 0,3% de meio oxgall
contendo bile bovina, pois, o intestino delgado e o cólon apresentam altas concentrações de
ácidos biliares, que permitem inibir ou inativar os microrganismos (GOLDIN; GORBACH,
SAXELIN, 1992; NEUMANN; FERREIRA, 1995; GARCIA, 1999).
Os trabalhos conduzidos por Rowland (1992) demonstram que os principais ácidos
biliares primários, cólico e chenodeoxicólico, são sintetizados no fígado e conjugados com
taurina e glicina, sendo que a desconjugação desses ácidos ocorre durante a circulação
enterohepática e algumas bactérias presentes na microbiota intestinal são capazes de realizar a
desconjugação desses ácidos.
Em estudo com bezerros Gilliland; Staley; Bush (1984) identificaram uma maior
incidência de lactobacilos bile-resistentes no jejuno, região esta que Tannock et al. (1994)
consideraram a mais importante para uma eventual inoculação do bolo alimentar e
disseminação dos microrganismos probióticos ao longo de todo o trato digestivo.

21 De acordo com Gilliland; Speck (1977) algumas linhagens de L. fermentum foram
testadas quanto à resistência aos sais de bile em meio sólido, onde pôde-se observar um ótimo
resultado para as linhagens selvagens e recém cultivadas e resultados de bom a razoável para
as linhagens comerciais. Os autores relatam que o desempenho inferior apresentado pelas
linhagens comerciais pode estar relacionado à perda de viabilidade devido ao processo de
armazenamento.
2.4.3 Produção de Bacteriocinas
As bacteriocinas são substâncias de natureza protéica de origem bacteriana, com ação
antagonista sobre outras bactérias, normalmente linhagens relacionadas. Apresentam
características diferentes dos antibióticos, pela própria estrutura química e por não serem
metabólitos secundários uma vez que a maioria é sintetizada durante a fase exponencial de
crescimento. Seu efeito é bactericida ou em alguns casos bacteriostático. As bacteriocinas são
produzidas por bactérias gram-positivas e gram-negativas, patogênicas ou não-patogênicas
(ÁVILA et al., 2005).
A capacidade de sintetizar uma bacteriocina por uma bactéria é conferida geralmente
pela presença de plasmídeo (TAGG; DAJANI; WANNA MAKER, 1976).
As bactérias lácticas produzem quatro classes de bacteriocinas: I – Lantibióticos, II –
Peptídeos pequenos estáveis ao calor, III – Proteínas grandes termosensíveis e IV – Proteínas
complexas que requerem adicionalmente carboidratos ou lipídeos na molécula para
expressarem a atividade de bacteriocina (KLAENHAMMER, 1993).
O espectro de ação das bacteriocinas normalmente está relacionado com contra
microrganismos intimamente relacionados. Assim, bacteriocinas de bactérias gram-positivas
possuem amplo espectro de inibição contra bactérias gram-positivas, sendo ativas também
contra algumas espécies gram-negativas. Dentre as bactérias gram-positivas, cujas
bacteriocinas têm mostrado inibição a bactérias gram-negativas estão L. acidophilus, Bacillus
cereus, Streptococcus, Staphylococcus e Corynebacterium (TAGG; DAJANI; WANNA
MAKER, 1976).
De acordo com os trabalhos realizados por Klerk; Smit (1967) e Tagg; Dajani; Wanna
Maker (1976) L. fermentum, cepa 466, produz uma única bacteriocina, a qual é uma proteína
termoestável resistente à temperatura de 96ºC por 30 minutos e sensível à tripsina e pepsina,
mas não à lisozima. É uma proteína de natureza lipopolissacarídica, composta de 16
aminoácidos, 4 açúcares, hexosamina e fósforo.

22 Fuller (1992) descreve a atividade antimicrobiana de várias espécies de Lactobacillus
baseada em suas bacteriocinas produzidas.
De acordo com o trabalho de Chateau; Castellanos; Deschamps (1993) várias espécies
de Lactobacillus produtores de bacteriocinas, isoladas de probióticos comerciais identificadas
como L. fermentum, L. rhamnosus, L. plantarum, L. acidophilus e L. pseudoplantarum, foram
testadas contra bactérias patogênicas. Os autores observaram que todos os patógenos testados
foram inibidos por uma ou mais espécies de Lactobacillus, sendo que o maior efeito de
inibição foi observado contra Listeria monocytogenes, e considerado satisfatório contra
Escherichia coli, Salmonella typhimurium e Salmonella enteritidis.
Os mecanismos de ação das bacteriocinas ainda não estão esclarecidos, de acordo com
de Martinis; Alves; Franco (2002). Segundo estes autores, supõem-se que a ação letal das
bacteriocinas se processe em três estágios: (1) ligação da bacteriocina ao receptor de
membrana, que garantirá sua entrada na célula; (2) transporte através da membrana, de
mecanismo desconhecido e (3) morte celular, pela formação de poros na membrana alterando
o seu potencial de ação.
O papel das bacteriocinas in vivo é discutível, uma vez que seriam hidrolisadas por
enzimas proteolíticas. Sua ação permanece, portanto, não demonstrada e a aplicação in vivo
ainda desconhecida (BARROW, 1992 e DE MARTINIS; ALVES; FRANCO, 2002). Existe,
também, a possibilidade de transferência de genes por conjugação, que codificam
bacteriocinas, para outras bactérias intestinais que efetivamente sobrevivem e se implantam
nesse ambiente produzindo bacteriocinas, que as auxiliam na colonização e sobrevivência
(SANDERS, 1993).
Segundo Dicks; Mellett; Hoffman (2004) bacteriocinas e cepas de bactérias lácticas
produtoras de bacteriocinas tem sido foco de extensivas pesquisas nos últimos anos,
favorecendo seu uso como bioprotetores de alimentos.
Estudos iniciais sobre a produção de bacteriocinas por bactérias lácticas estão
enfocados em cepas associadas com produtos da indústria de lacticínios. Recentemente, a
atividade bacteriocinogênica tem sido observada em cepas presentes em carnes, silagem,
vegetais fermentados e vinho vermelho (HEMME; FOUCAUD-SCHEUNEMANN, 2004).
O uso de bacteriocinas pode ser considerado como uma biotecnologia com aplicação
potencial em sistemas alimentares. As bacteriocinas contrastam com os antibióticos,
considerados ilegais como conservadores. Os antibióticos são formados por reações de
condensação enzimática de aminoácidos, enquanto as bacteriocinas são peptídeos ou proteínas

23 com atividade antibacteriana sintetizados ribossomicamente, não sendo letais para as células
que as produzem (DE MARTINIS; ALVES; FRANCO, 2002).
2.4.4 Produção de Ácidos Orgânicos e Peróxido de Hidrogênio
O modo de ação dos ácidos orgânicos (acido acético, ácido fórmico e ácido láctico) é
atribuído pela redução direta do pH intracelular por ionização dos ácidos não dissociados, ou
interrupção do transporte de substrato pela alteração da permeabilidade da membrana celular
(BARBOSA; TORRES, 1999).
Na forma não dissociada os ácidos orgânicos são facilmente solúveis na membrana
celular, interferem com sua permeabilidade e impedem a fosforilação oxidativa a partir do
sistema de transporte de elétrons. Promovem acidificação do conteúdo da célula, causando
sua morte (CLIVER; MARTH, 1990). Enquanto pH baixo por si só é um fator na inibição de
microrganismos, ele também potencializa a atividade de ácidos orgânicos, que em baixos
valores de pH encontram-se na forma não dissociada, que é muito mais bactericida (JUVEN,
MEINERSMANN; STERN, 1991).
A ação intracelular do ácido acético, assim como fórmico, deve-se à diminuição do pH
intracelular, desnaturando as proteínas do protoplasma (LÜCK, 1981).
Parte da ação do ácido fórmico, especialmente sobre as bactérias, deve-se ao
abaixamento do pH intracelular. Ao contrário dos ácidos orgânicos de massa molecular
elevada, o ácido fórmico, assim como os ácidos acético e láctico atuam em concentrações
relativamente altas, que fazem diminuir sensivelmente o pH intracelular. Além disso, o ácido
fórmico tem ação antimicrobiana independente do pH e inibe as descarboxilases e as enzimas
porfirínicas, especialmente a catalase (LÜCK, 1981).
A ação antimicrobiana exercida pelo ácido láctico é relativamente pequena quando
comparada aos ácidos acético e fórmico, porém, é mais eficiente que o ácido cítrico. Atua
principalmente contra as bactérias, sobretudo as anaeróbias. Grande parte de sua atividade
deve-se a diminuição do pH intracelular (LÜCK, 1981).
Os lactobacilos apresentam a capacidade de gerar peróxido de hidrogênio durante o
crescimento por diferentes mecanismos (BEUCHAT; GOLDEN, 1989; DAESCHEL, 1989):
pelo fato dos lactobacilos não possuírem a enzima catalase, ocorre um acúmulo de peróxido
de hidrogênio no meio de cultivo, podendo reagir com outros compostos como o superóxido e
formar o radical (OH-), que é muito mais reativo e poderoso oxidante de biomoléculas
(BUCHANAN; GIBBONS, 1974).

24 Em ambientes naturais como leite e saliva a ação antagonista do peróxido pode
também ser potencializada. Concentrações inócuas podem tornar-se inibitórias na presença de
lactoperoxidase e tiocianatos, onde peróxido pode gerar o radical tóxico hipotiocianato. Tal
sistema é conhecido como sistema Lactoperoxidase (REITER; HARNULV, 1984).
Subramanian; Olson (1968) verificaram que a produção de peróxido pode ser auto
inibitória para culturas láticas. Todas as estirpes de Lactococcus lactis estudadas sofreram
pequenas inibições, expressa em termos de percentagem de acidez titulável. Lactococcus
lactis cremoris foi a estirpe mais sensível, enquanto Lactococcus lactis lactis e subsp.
diacetilactis foram as mais resistentes, fato constatado pelo aumento de acidez titulável entre
3,5 e 6 horas de incubação em meio contendo 40 e 80 ppm de peróxido.
A produção de peróxido foi avaliada em várias espécies de Lactobacillus (PREMI;
BOTTAZZI, 1972). Foi observado que houve variação entre as diferentes espécies e entre
estirpes da mesma espécie. Após 6 dias estocadas a 5oC, quatro estirpes de Lactobacillus
delbrueckii lactis produziram as maiores quantidades de peróxido (8,5; 9,0; 10 e 8,4 ppm)
menor produção foi detectada em estirpes de Lactobacillus helveticus (1,2; 2,9; 1,9; 2,6 e 0,0
ppm) e Lactobacillus delbrueckii bulgaricus (2,4; 0,0; 3,8 e 0,0 ppm).
Segundo Gilliland; Speck (1975) o número de células de Pseudomonas fragi teve um
decréscimo acentuado durante estocagem por 10 dias a 5oC, na presença de Lactobacillus
delbrueckii bulgaricus, enquanto que na ausência, houve um aumento de cerca de 100 vezes
mais. A adição de catalase evidenciou que o peróxido de hidrogênio foi o responsável pelo
efeito inibidor observado.
2.4.5 Atividade Anticarcinogênica
As propriedades anticarcinogênicas dos lactobacilos estão representadas em três
categorias de acordo com Fuller, (1992): destacando-se a) inibição das células dos tumores; b)
supressão das bactérias que produzem enzimas tais como β-glucosidase, β-glucuronidase e
azoredutase as quais são responsáveis pela liberação de carcinógenos a partir de complexos
inócuos e c) destruição dos carcinógenos tais como as nitrosaminas e a supressão da
nitroredutase a qual está envolvida na síntese das nitrosaminas.
Perdigón et al. (1986), mostraram que a administração oral de L. casei inibe o
crescimento de fibrossarcoma após inoculação em camundongos e que esse efeito está
relacionado à dose e ao momento do tratamento.

25 Goldin; Gorbach; Saxelin (1992), mostraram que a introdução de L. acidophilus na
dieta reduziu a incidência de tumores de cólon induzidos quimicamente em ratos. Segundo
estes autores o possível mecanismo de ação consiste na modificação da atividade metabólica
da microbiota colônica reduzindo dessa maneira a formação de carcinogênicos no intestino
grosso.
Tomita et al. (1994), comprovaram o efeito anticarcinogênico do L. casei biovariedade
Shirota em ratos Wistar machos com tumores na bexiga, observando que após o tratamento o
volume do tumor foi consideravelmente reduzido, quando comparado ao controle.
2.4.6 Estimulação da Imunidade
O trato intestinal não somente é responsável pela digestão de alimentos e absorção de
nutrientes, mas é também um órgão do sistema imune mais importante em mamíferos
(JANEWAY et al., 2002). Existem muitas células imunocompetentes na mucosa intestinal,
como linfócitos intraepiteliais e macrófagos, assim como linfócitos T e B na mucosa mais
profunda. No lúmem das placas de Peyer, células M com atividade fagocítica estão presentes
além das células epiteliais intestinais. Acredita-se que as células M forneçam uma barreira
contra a entrada de microrganismos e antígenos dietéticos e mostrem uma resposta imune a
antígenos macromoleculares exógenos no intestino (ELIA; DE SOUZA, 2001). Em animais, a
colonização bacteriana do intestino geralmente começa logo após o nascimento e a microbiota
é mantida durante a vida, e apresenta algumas alterações conforme a condição fisiológica ou
idade do hospedeiro. Provavelmente existe uma relação íntima entre a função imunológica do
intestino e a microbiota intestinal. Como evidência disto, observa-se uma grande diferença na
atividade imunológica entre camundongos isentos de germes e normais, indicando que a
microbiota intestinal desempenha papel importante na manutenção do sistema imune (ELIA;
DE SOUZA, 2001). Portanto, parece razoável que a administração de lactobacilos com uma
potente atividade imunopotenciadora no intestino, que é rico em células imunocompetentes,
possa ter uma influência marcante sobre a imunidade do hospedeiro (PENNA et al., 2000).
Nos últimos anos vários estudos têm investigado a interação entre a presença de
bactérias lácticas na dieta e imunocompetência. Modulando certos parâmetros do sistema
imune específico e não específico essas bactérias poderiam proteger o hospedeiro contra
infecções por patógenos entéricos e desenvolvimento de tumores (GIBSON; ROBERFROID,
1995).

26 Os fatores responsáveis pelo estímulo do sistema imune parecem estar relacionadas a
ativação de macrófagos (PERDIGÓN et al., 1995) e linfócitos (ISOLAURI et al., 1995), e o
aumento nos níveis de imunoglobulinas A (IgA), G (IgG), além de citocinas e Th1 que
produzem gama-interferon (INFγ) (O’SULLIVAN et al., 1992), do fator de necrose tumoral-
TNF (PERDIGÓN et al., 1995) e interleucina 2 (IL-2) que aumentam a imunidade mediada
por células específicas (MATSUZAKI et al., 1997).
Segundo Perdigón et al. (1995), em ratos desnutridos ocorrem alterações do epitélio
intestinal que induzem a translocação de bactérias desse local a outros. Por isso, para
obtenção de efeitos no sistema imune, estes autores sugerem que a administração do L. casei
deva ser feita após o restabelecimento da condição de nutrição adequada para garantir a
permanência do probiótico no trato intestinal.
A ingestão de leite fermentado com várias bactérias produtoras de ácido lático,
incluindo L. casei, L. acidophilus e Bifidobacterium longum, aumenta o número de células
que produzem IgA, a atividade de macrófagos, o nível de IgG sérica, e anticorpos específicos
contra determinados patógenos (SAUCIER et al., 1992).
Nader de Macias et al. (1992), relataram que ratos infectados com Shigella sonnei e
alimentados com leite fermentado com L. acidophilus sobreviviam por mais tempo e
apresentavam títulos de anticorpos anti-Shigella mais elevados no soro e no conteúdo
intestinal quando comparado aos controles.
Estudando o comportamento dos leucócitos de sangue humano de voluntários adultos
alimentados com Lactobacillus acidophilus, Schiffrin et al. (1997) observaram que o número
de granulócitos neutrófilos e a atividade fagocitária dos mesmos foi mais elevada no grupo
tratado após seis semanas de tratamento. Os autores concluíram que estes resultados
alcançados poderiam ter sido em função dos lactobacilos terem estimulado o organismo a
produzir citocinas intimamente ligadas ao aumento destas células e sua atividade fagocitária.
L. rhamnosus reduziu a morbidade de E. coli O157:H7 em camundongos infectados
experimentalmente, estimulando o aumento da atividade fagocitária dos neutrófilos e dos
títulos de anticorpos específicos IgA (SHU; GILL, 2002).
Outras bactérias ácido lácticas também apresentam atividade imuno-moduladoras.
Yasui (1999) comprovou que a administração de Bifidobacterium breve estimulou o sistema
imune humoral em camundongos provocando aumento da produção de IgA anti-Rotavírus e
de IgG antivírus da Influenza, protegendo-os contra essas duas infecções. Shu; Qu; Gill
(2001) relataram que leitões tratados com B. lactis HN109 apresentaram diminuição da
diarréia associada com Rotavírus e E. coli, concomitante ao aumento dos títulos de anticorpos

27 contra patógenos específicos no trato gastrintestinal, da concentração de neutrófilos
sanguíneos e da resposta proliferativa de linfócitos T.
Após duas semanas de tratamento com L. gasseri CECT 5714 e L. coryniformis CECT
5711 o sangue humano de adultos voluntários tratados com ambos lactobacilos apresentou
aumento sérico de neutrófilos e de sua atividade fagocitária (OLIVARES et al., 2006).
2.4.7 Efeito Hipocolesterolêmico
O efeito de redução do colesterol pela ingestão de leite fermentado com bactérias
produtoras de ácido lático, foi sugerido pela primeira vez por Mann; Spoerry (1974), que
observaram uma inesperada diminuição do nível de colesterol sanguíneo em homens jovens
após ingestão diária de uma média de 2,52 l de leite fermentado com espécies de
Lactobacillus durante 21 dias.
Utilizando leite fermentado com S. thermophilus Rao et al. (1998), mostraram que o
probiótico estudado foi capaz de reduzir o nível sanguíneo de colesterol e inibir a síntese
hepática de colesterol em ratos.
Gilliland; Nelson; Maxwell (1985), relataram uma diminuição do nível sanguíneo de
colesterol em suínos alimentados com leite fermentado por uma cepa de L. acidophilus que
foi isolada das fezes dos animais. Esta cepa apresentava resistência aos ácidos biliares e era
capaz de assimilar colesterol. Os autores concluíram que a atividade de assimilação de
colesterol se deve a capacidade da cepa em hidrolisar ácidos biliares conjugados e separar
colesterol das micelas de bile. Se os ácidos biliares conjugados forem hidrolisados no
intestino delgado superior, o colesterol torna-se insolúvel e sua absorção intestinal é
suprimida. Muitas cepas de L. acidophilus e espécies relacionadas de bactérias produtoras de
ácido lático apresentam a capacidade de hidrolisar ácidos biliares conjugados (KLAVER;
MEER, 1993).
2.5 Probióticos na Saúde Animal
Aspectos relevantes sobre a utilização de probióticos nas rações de aves foram
publicados por Miles; Jermingan (1985), Jin et al. (1997), Fernandes et al. (2000), Ferreira
(2000) e Leedle (2000). Microrganismos probióticos quando oferecidos às aves recém
eclodidas favorecem a colonização no trato intestinal, e a ocupação preventiva com
microrganismos probióticos diminui a probabilidade de ocupação por microrganismos

28 indesejáveis. A aplicação de preparações probióticas pode ser realizada através de
pulverização direta sobre as aves, via água de bebida, ou misturando-se nas rações
(ROSTAGNO et al., 2003).
Independente da via de aplicação, para boa atuação do probiótico, a administração
deve ser feita o mais precocemente possível, para que as bactérias benéficas colonizem o trato
intestinal do hospedeiro, antes dos patógenos. Para que o probiótico seja eficiente, os
microrganismos devem ser específicos do hospedeiro, sendo assim, um bom probiótico para
perus pode não alcançar os mesmo resultados em frangos de corte (FULLER, 1992).
A presença de 0,10% de cultura de Lactobacillus nas rações de frangos de 1 a 42 dias,
proporcionou melhora no ganho de peso e na conversão alimentar dos animais (JIN et al.,
1998).
Line (1998) estudaram o efeito da adição de 1g e de 100g de S. boulardii por kg de
ração, em frangos desafiados com S. typhimurium e com Campylobacter jejuni, onde foi
observado que o número de Salmonella diminuiu para os dois tratamentos em relação ao
controle, ao passo que para Campylobacter os tratamentos não tiveram efeito na diminuição
da população deste microrganismo.
Ao avaliar a inclusão de probiótico a base de B. subitilis sobre o desempenho de
frangos de corte, Campos et al. (2002) observaram que a inclusão de probiótico na ração
melhorou a viabilidade e o ganho de peso dos animais criados até os 42 dias de idade, sem
prejudicar as demais variáveis estudadas.
Lan; Rinh; Benno (2003), trabalhando com L. agilis JCM 1048 e L. salivarius
salivarius JCM 1230 isoladas do intestino de frangos, evidenciaram que estas cepas
apresentavam propriedades probióticas fazendo com que os animais apresentassem ganho de
peso 10,7% superior ao controle após 40 dias tratamento. Por outro lado, L. crispatus foi
capaz de inibir a adesão da E. coli O78 no trato intestinal de frangos infectados
experimentalmente (EDELMAN et al., 2003).
Abe et al. (1995) estudaram os efeitos da administração de uma preparação probiótica
contendo Bifidobacterium thermophilum, Enterococcus faecium e L. acidophilus no ganho de
peso e frequência de diarréias em leitões e bezerros recém-nascidos. A frequência de diarréia
em leitões e bezerros foi menor nos grupos tratados com o probiótico. A média do ganho de
peso dos bezerros, após 90 dias de experimento, foi 40% maior no grupo tratado com o
probiótico quando comparado ao grupo controle.

29
A adição de microrganismos probióticos em rações para leitões na fase inicial de
crescimento (10 a 30 Kg) proporcionou aumento significativo no ganho de peso e controle
eficaz na incidência de diarréia (VASSALO et al., 1997).
Shu; Qu; Gill (2001) avaliando Bifidobacterium lactis HN019 em leitões
demonstraram que seu uso foi efetivo no controle de diarréias provocadas por Rotavírus e E.
coli e que esta efetividade foi provocada pelo aumento da atividade fagocitária dos leucócitos,
da proliferação dos linfócitos T e aumento no título de anticorpos contra os agentes
causadores da diarréia.
Avaliando alimentos fermentados por bactérias láticas em suínos em fase de
crescimento, Canibe; Jensen (2003) mostraram que esses animais apresentaram aumento no
ganho de peso e diminuição na concentração de enterobactérias no intestino.
De acordo com Vanbelle; Teller; Focant (1990) e Ruppert; McCoy; Hutjens (1994), as
indicações de produtos probióticos na profilaxia de infecções bacterianas de animais recém-
nascidos e recém-desmamados consistem em ministrar doses de 106 a 109 UFC/g de ração,
por três a cinco dias consecutivos, e, como promotor de crescimento, empregam-se doses
diárias de 106 a 107 UFC/g de ração, no mínimo durante um a dois meses. Os resultados
reportados na literatura sobre o uso de probióticos para fins terapêuticos ou como promotor de
crescimento em bezerros são heterogêneos, irregulares e nem sempre positivos. A resposta do
animal ao uso de probióticos é influenciada pelo tipo de probiótico, pela dose utilizada, idade
e raça do animal, tipo de exploração e de manejo, uso concomitante de antibióticos e do
ambiente de criação.
Segundo Vanbelle; Teller; Focant (1990) o uso de probióticos em dietas de bezerros
lactentes resulta em aumentos médios de 4,5% no ganho de peso vivo e 2,5% na conversão
alimentar, em sistemas de criação nos quais os animais são submetidos à condições de
estresse. Alves et al. (2000) trabalhando com bezerros alimentados com leite adicionado de
microrganismos probióticos encontraram aumentos médios de 6,2% no peso vivo ao abate, de
5,6% no ganho médio diário de peso vivo e de 8,5% na conversão alimentar, em relação aos
animais alimentados com dietas sem a adição do probiótico.
Além de prevenir a ocorrência de diarréia, probióticos bacterianos também são usados
para melhorar o desenvolvimento e manutenção de uma fermentação estável no rúmen.
Lactobacillus e Enterococcus faecium têm demonstrado melhorar a ingestão de alimentos e o
ganho de peso em bovinos jovens (WREN, 1987; LEE; BOTTS, 1988). Umberger et al.
(1989) relataram que Lactobacillus estimulou a ingestão de alimentos e ganho de peso em
cordeiros recém desmamados. Ozawa et al. (1983) constataram que E. faecalis estabilizou a

30 microbiota intestinal e estimulou o ganho de peso em bezerros após tratamento com
antibiótico.
Chaves et al. (1999) trabalhando com L. acidophilus (LT 516) em bezerros mostraram
que este microrganismo foi capaz de reduzir a ocorrência de diarréias, sem alterar o odor e a
consistência das fezes. Os resultados revelaram também que animais que receberam
probiótico nos primeiros dez dias de vida apresentaram maior número de lactobacilos
facultativos no intestino delgado. O número de coliformes totais no intestino delgado foi
maior nos animais controle e menor nos que receberam probióticos nos primeiros 56 dias.
Siuta (2001) avaliou por um período de 137 dias o ganho de peso de 36 novilhos
Hereford, tratados diariamente, ad libitum, com dietas de concentrados à base de silagens de
cevada e de milho, adicionados com 0,5 ml de probiótico “Cytozyme” por cabeça. O
probiótico promoveu aumento no ganho de peso diário em cerca de 10-14% e melhorou a
eficiência da conversão alimentar em cerca de 6%. Esses efeitos foram melhor observados
com silagem de milho.
Utilizando vacina combinada de probióticos (L. acidophilus) com pili K99 e A14 de E.
coli, Avila et al. (1995) obtiveram resultados satisfatórios no tratamento de diarréia em vacas
prenhas e bezerros.
Agarwal et al. (2002) trabalhando com bezerros recém-nascidos, observaram que os
animais tratados com probióticos apresentavam menor tempo de duração da diarréia, quando
comparado aos do grupo controle. Em experimentos com E. coli enterohemorrágica em
bezerros, Zhao et al. (2003) demonstraram que os animais tratados com probióticos
apresentaram menores episódios de diarréia.
O papel do probiótico como biorregulador microbiano reside em manter o equilíbrio
da microbiota intestinal e ruminal, tendo como principal função impedir a colonização
intestinal por bactérias patogênicas como E. coli enterotoxigênica (ÁVILA et al., 1988) e
promover aumento de ganho do peso dos animais (Abe et al., 1995).
2.6 AÇÃO ANTIPARASITÁRIA DOS PROBIÓTICOS
Trabalhando com frangos infectados por Eimeria tenella, Kimura et al. (1976)
mostraram que Lactobacillus spp e Bifidobacterium spp, promoveram diminuição no número
de oocistos após o quinto dia após de infecção, quando surgiram esquizontes e gametócitos.

31
Em presença de número elevado de lactobacilos, foi observado que a Trichomonas
vaginalis não tem bom crescimento e os lactobacilos não crescem bem quando em presença
de taxa elevada de T. vaginalis (SOSZKA; KUCZYNSKA, 1977).
Mcgrory et al. (1994), trabalhando com T. vaginalis e L. acidophilus in vitro,
demonstraram que uma concentração elevada de T. vaginalis foi capaz de reduzir o número de
lactobacilos, e que este efeito poderia ter sido em função da fagocitose das bactérias pelos
protozoários ou por substâncias secretadas que apresentam efeito deletério.
Alak et al. (1997) administraram L. reuteri como probiótico em camundongos
infectados com Cryptosporidium parvum e demonstraram que os animais que não receberam
dietas contendo L. reuteri desenvolveram criptosporidiose persistente e eliminaram grandes
quantidades de oocistos nas fezes. Por outro lado os animais suplementados com L. reuteri
tiveram a colonização do trato intestinal aumentada o qual foi inversamente relacionado à
eliminação de oocistos nas fezes.
Trabalhando com L. reuteri e L. acidophilus, Foster et al., (1997) observaram in vitro
uma redução de 81% da viabilidade do oocisto de C. parvum. Os autores sugeriram que esta
redução pode ter sido devido à presença de substâncias antimicrobianas contra os vários
estágios do ciclo de vida do protozoário.
Utilizando camundongos imunodeficientes e infectados por C. parvum, Alak et al.
(1999) demonstraram redução na eliminação de oocistos nos animais suplementados com
Lactobacillus em sua dieta e sugeriram que L. reuteri e L. acidophilus podem reduzir a
população do parasita C. parvum no epitélio intestinal, sendo que, L. acidophilus foi mais
eficaz que L. reuteri.
Waters (1999) observaram que a colonização de L. reuteri em ratos gnotobióticos
TCR-α-deficientes limitou a infecção por C. parvum e deprimiu o processo inflamatório,
mostrando assim que L. reuteri teve papel importante na limitação da resposta inflamatória.
Avaliando suínos infectados com Cryptosporidium parvum, e alimentados com uma
mistura de L. acidophilus e Bifidobacterium spp em sua dieta, Rotkiewiez et al. (2001)
demonstraram que houve uma redução significativa na eliminação de oocistos de
Cryptosporidium.
Pérez et al. (2001) usando o sobrenadante de uma suspensão de Lactobacillus
johnsonii observaram que houve diminuição na proliferação de trofozoítos de Giardia
intestinalis in vitro após 96 horas de incubação, e que esta diminuição poderia estar
relacionada a substâncias antagônicas produzidas pelos lactobacilos durante o crescimento.

32 Dalloul et al. (2003a) estudando frangos de corte suplementados com probióticos a
base de lactobacilos, demonstraram que estes microrganismos promoveram aumento do
número de linfócitos intraperitoniais, tornando os animais mais resistentes ao parasitismo por
E. acervulina.
A utilização de probióticos a base de lactobacilos em frangos infectados por E.
acervulina, diminuiu a geração de oocistos, diminuindo também a condição de invasão e o
desenvolvimento da E. acervulina. Essa condição pode ser devido à alta produção de
Interleucina-2 (IL-2), que foi detectada no sangue dos animais (DALLOUL et al., 2003b).
Tierney et al. (2004), examinando o efeito de três espécies de lactobacilos isoladas de
fezes de aves, constataram que as mesmas inibiram significativamente a invasão parasitária de
Eimeria tenella por exclusão competitiva.
Glass et al. (2004), mostraram que células livres do sobrenadante de L. acidophilus e
L. reuterii, reduziram significativamente a cultura de células infectadas por C. parvum e C.
hominis isolados de bovinos. A redução da viabilidade e infectividade do C. parvum e do C.
hominis pode estar relacionada hipoteticamente com a produção de compostos
antimicrobianos.
Frangos contaminados com Eimeria acervulina e alimentados com probióticos a base
de lactobacilos, apresentaram redução de 14% na eliminação de oocistos nas fezes. Segundo
os autores esta redução está relacionada ao aumento dos níveis plasmáticos de Interferon-
gama (INF-γ) e IL-2 e aumento de INF-γ na mucosa intestinal, o que estimulou a imunidade
celular, que afetou o ciclo do parasito (DALLOUL, et al., 2005).
2.7 EIMERIA E EIMERIOSE
A eimeriose é uma doença causada por protozoários coccídios do gênero Eimeria, que
acomete principalmente animais jovens (SLAPETA, et al., 2001). É uma parasitose de
distribuição mundial, atingindo rebanhos submetidos aos mais diferentes sistemas de manejo,
embora seja mais grave e mais freqüente em animais criados em sistemas intensivos, daí a sua
importância em rebanhos leiteiros (ROSE, 1987).
Os animais infectados apresentam baixo aproveitamento do alimento consumido,
apresentando assim baixo desempenho com alto consumo de alimentos e maior permanência
no pasto ou em confinamento (SHI et al., 2001).

33 A propagação do parasita se dá através das fezes dos animais infectados, que
contaminam o pasto, outros alimentos e água. O parasita sobrevive por longos períodos na
pastagem e nos confinamentos. Animais sadios expostos ao meio ambiente contaminado são
infectados, se tornando elementos da cadeia infectante (ZHAO; DUSZYNSKI; LOKER,
2001).
Conforme Frandesen (1985) as espécies do gênero Eimeria possuem um hospedeiro
específico. Raramente uma espécie de Eimeria completa o ciclo de infecção em mais de um
hospedeiro. O mecanismo fundamental de especificidade de hospedeiro não está bem
esclarecido, mas a maioria provavelmente inclui fatores genéticos, nutricionais/bioquímicos e
imunológicos.
A enfermidade causada por Eimeria afeta principalmente a parede intestinal do
hospedeiro, além do fígado e rins em alguns casos, em cujos epitélios e endotélios esses
parasitas exercem ação destruidora (LEITÃO, 1983).
De acordo com Faber et al. (2002), aqueles parasitas que penetraram nas células da
parede intestinal de seus novos hospedeiros, inicialmente destruirão essas células, para em
seguida lesarem novas células vizinhas, dando então ao aparecimento de uma lesão na parede
intestinal (úlcera), que passa também a sangrar, agravando o quadro parasitário com a perda
de sangue decorrente e também com uma infecção secundária causada por germes de
associação existentes no próprio local. Com reiteradas infecções e parasitismo concomitante
evolui o quadro para o aparecimento de inflamações catarrais de início dos órgãos digestivos,
seguida de aparecimento de sangue e pus nas fezes para uma enterite mais grave
(hemorrágica). A própria ulceração da parede intestinal pode evoluir para camadas mais
profundas dos intestinos, e ao atingirem a camada muscular e mucosa ou mesmo a serosa,
podem provocar a perfuração dos intestinos e daí uma peritonite generalizada.
Simultaneamente, com o parasitismo, são secretadas toxinas pelas eimerias,
determinantes pelas vias hemáticas ou linfáticas dos chamados fenômenos complementares de
intoxicação ou sensibilidade do animal, que se traduzem por coceiras ou sintomas mais
severos e graves. A existência de uma substância liberada pela E. tenella, quando injetada por
via venosa em coelhos, dá lugar a manifestações de intoxicação de curso mortal (WARUIRU
et al., 2000).
Os órgãos digestivos dos animais apresentam, quando intensamente parasitados pelo
protozoário, uma coloração vermelha (congestão), além de pontos hemorrágicos ou mesmo
placas, úlceras, tumefações e engrossamento da mucosa parasitada. O conteúdo intestinal

34 apresenta-se fluido, espumoso e com estrias de sangue. Pode ocorrer despregamento da
própria mucosa que então terá aspecto caseoso (MARSHALL, 1998).
2.7.1 Ciclo de Vida
O ciclo de vida desta espécie conforme ilustrado na Figura 1, divide-se em três fases:
esporulação; esquizogonia; gametogonia e formação de oocisto (YUN; LILLEHOJ;
LILLEHOJ, 2000).
Oocistos não esporulados, constituídos por uma massa nucleada de protoplasma
circundada por uma parede resistente, são eliminados nas fezes. Em condições adequadas de
oxigenação, alta umidade e temperatura ao redor de 27ºC, o núcleo se divide duas vezes e a
massa protoplasmática forma quatro corpos cônicos, que se irradiam a partir de uma massa
central. Cada um destes cones nucleados torna-se arredondado e forma um esporoblasto,
enquanto em algumas espécies o protoplasma restante forma o corpo residual do oocisto.
Cada esporoblasto secreta uma parede de material refrátil, passando a ser conhecido
como esporocisto, enquanto o protoplasma em seu interior se divide em dois esporozoítos em
forma de banana. Em algumas espécies, o protoplasma restante dentro do esporocisto forma
um corpo residual do esporocisto. O tempo necessário para estas alterações varia de acordo
com a temperatura, mas em condições ideais geralmente requer dois a quatro dias. O oocisto,
composto agora de uma parede externa envolvendo quatro esporocistos, cada um deles com
dois esporozoítos, é conhecido como oocisto esporulado e constitui a forma infectante.
O hospedeiro se infecta ingerindo o oocisto esporulado. Os esporocistos são então
liberados mecanicamente ou por gás carbônico, e os esporozoítos, ativados por ação da
tripsina e da bile, deixam o esporocisto. Na maioria das espécies, cada esporozoíto penetra
numa célula epitelial, tornando-se arredondada, sendo, então, conhecido como trofozoíto.
Após alguns dias, cada trofozoíto se divide por fissão múltipla e forma um esquizonte,
uma estrutura constituída por um grande número de organismos nucleados alongados,
conhecidos como merozoítos. Quando a divisão está completa e o esquizonte maduro, a célula
hospedeira e o esquizonte se rompem e os merozoítos livres, invadem células vizinhas. A
esquizogonia pode repetir-se, sendo o número de gerações de esquizontes dependente da
espécie.
Os esquizontes maduros podem, em alguns casos, ser identificados histologicamente
por sua localização, tamanho e número de merozoítos que contêm. Os merozoítos são
organizados como uma série de organismos de formato em crescente, ao contrário, no

35 microgametócito maduro, os microgametas ficam dispostos ao redor da periferia da célula. O
macrogametóctio possui um grande núcleo central, com pequenos grânulos localizados ao
redor da periferia da célula.
A esquizogonia termina quando os merozoítos dão origem a gametócitos masculinos e
femininos. Os fatores responsáveis por esta mudança para gametogonia não são totalmente
conhecidos. Os macrogametócitos são femininos e permanecem unicelulares, mas aumentam
de tamanho preenchendo a célula parasitada. Podem ser distinguidos de trofozoítos ou
esquizontes em desenvolvimento pelo fato de possuírem um único núcleo avantajado. Cada
microgametócito masculino sofre divisão repetida e forma um grande número de organismos
uninucleados flagelados, os microgametas. É apenas durante este curto estágio que os
coccídios possuem órgãos de locomoção. Os microgametas são liberados por ruptura da célula
hospedeira, cada qual penetrando em um macrogameta, ocorrendo, então, fusão dos núcleos
do micro e do macrogameta. Forma-se uma parede cística ao redor do zigoto resultante,
identificado agora como um oocisto, em geral não se verificando qualquer desenvolvimento
posterior, até que este oocisto não esporulado seja eliminado nas fezes.
O período pré-patente varia consideravelmente, podendo ser tão curto quanto cinco
dias em aves domésticas, ou de três a quatro semanas em algumas espécies de ruminantes.
Figura 1 – Ciclo evolutivo de Eimeria spp
Fonte: ZOETECNOCAMPO, 2005

36 3 MATERIAL E MÉTODOS
O presente trabalho foi desenvolvido no Laboratório de Microbiologia do Centro
Universitário Geraldo Di Biase (UGB) em Volta Redonda/RJ.
3.1 MICRORGANISMOS
3.1.1 Probióticos
A preparação probiótica avaliada no presente trabalho foi constituída de um “pool” de
lactobacilos composto pelas seguintes espécies: Lactobacillus casei, L. plantarum ATCC
8014, L. fermentum ATCC 9338 e L. acidophilus ATCC 4536 da coleção de culturas do
Laboratório de Microbiologia do Centro Universitário Geraldo Di Biase, Volta Redonda, RJ.
As culturas foram mantidas sob congelamento a -20ºC em Caldo MRS (De Mann,
Rogosa e Sharpe) constituído de glicerol 40%. Para ativação, as culturas foram descongeladas
a temperatura ambiente e repicadas três vezes consecutivas em caldo MRS esterilizado a
121ºC/15 min. e incubadas a 37ºC por 24 à 48 horas (BORIS et al., 1997).
Para confirmação da identidade destes microrganismos, as culturas puras foram
submetidas à coloração de Gram e aos testes de fermentação de açúcares e catalase. Para o
teste de fermentação dos principais açúcares (galactose, lactose, maltose, sorbitol, rafinose e
manitol), foi utilizado caldo MRS adicionado de 2% do açúcar-teste e 0,004% do indicador
púrpura de bromocresol, com incubação a 37ºC/48 horas (ARICIA et al., 2004).
Para a formação do referido “pool” de Lactobacillus as culturas foram
individualmente ativadas por três vezes em caldo MRS, sendo do último repique retirada uma
alíquota de 1 ml de cada cultura, a qual foi transferida para frascos Erlenmyer esterilizados
contendo 50 ml de leite desnatado a 10%, esterilizado seguido de incubação a 37ºC por 18
horas. Após este período juntou-se o conteúdo dos 4 frascos formando-se assim o “pool” de
lactobacilos, contendo aproximadamente 14 Log UFC/ml (MACHADO, 2001). Este “pool”
de microrganismos foi preparado a cada três dias e armazenado a 5° C até o momento da
administração aos animais.
A contagem de células foi realizada pela técnica de “pour-plate” em ágar MRS, das
diluições 10-13 10-14 e 10-15 após incubação a 37ºC por 48 horas

37 3.1.2 Eimeria
Os oocistos das espécies de Eimeria foram obtidos de ratos selvagens da espécie
Rattus norvegicus, naturalmente infectados, que foram capturados com auxílio de armadilhas
e iscas. Após a captura, estes foram mantidos em gaiolas, a fim de se obter suas fezes. Os
animais receberam água e ração balanceada, sem coccidiostático, ad libitum.
As fezes colhidas foram examinadas para verificar a presença de oocistos pela técnica
de centrifugo-flutuação, conforme descrito por (MENEZES; LOPES, 1995).
Aos oocistos em suspensão foram adicionados solução de dicromato de potássio a
2,5% na relação de 1:2, para provocar a esporulação dos mesmos. Os oocistos foram mantidos
em solução de dicromato de potássio a 2,5% por uma semana sob refrigeração
(STOCKDALE, 1985).
Antes de infectar os ratos, os oocistos foram lavados com água destilada e
centrifugados a 400 x g por 10 minutos para eliminação do dicromato de potássio
(STOCKDALE, 1985; BILLETER, et al., 2006).
3.2 AVALIAÇÃO DOS MICRORGANISMOS PROBIÓTICOS SOBRE EIMERIA
Neste experimento, com duração de 21 dias, foram utilizados vinte e cinco animais da
espécie Rattus norvegicus Wistar machos, com quatro semanas de idade, cedidos pelo
Biotério da USP-SP. Os animais foram mantidos individualmente em gaiolas plásticas (Figura
2) com serragem esterilizada, a temperatura ambiente, recebendo água e ração balanceada,
sem coccidiostático ad libitum. Foram realizados exames parasitológicos nas fezes dos
animais antes do início do experimento, para confirmar a ausência de oocistos.
Os animais foram divididos em cinco grupos (A, B, C, D e E) com cinco ratos em
cada grupo, sendo que todos os grupos receberam ração sem coccidiostático e água e foram
tratados durante 21 dias da seguinte forma:
Grupo A: Somente ração e água diariamente (controle negativo).
Grupo B: Infecção com oocistos de Eimeria spp. no sexto dia do experimento (controle
positivo da Eimeriose), recebendo ração e água diariamente.
Grupo C: 1 ml da preparação probiótica diariamente (controle positivo do efeito probiótico).
Grupo D: 1 ml da preparação probiótica durante cinco dias e no sexto dia do experimento
foram infectados com oocistos de Eimeria spp.

38 Grupo E: Infecção com oocistos de Eimeria spp no sexto dia do experimento e a partir do
terceiro dia de infecção, administração diária de 1 ml da preparação probiótica.
Figura 2 – Acomodação dos ratos em gaiolas individuais
A preparação probiótica (1 ml) foi administrada aos animais por via oral, sem auxílio
de anestésico com auxílio de seringa descartável de 5 ml, introduzida diretamente na boca dos
animais (Figura 3). Os animais permaneciam 2 horas sem alimentação para receber a referida
preparação.

39
Figura 3 – Administração de preparado probiótico aos animais
Os animais dos grupos B, D e E foram submetidos a valores de éter por 5 minutos,
sendo desta forma anestesiados e logo em seguida infectados com aproximadamente 5000
oocistos de Eimeria spp, por meio de sonda esofágica de plástico (SHI et al., 2001), conforme
mostrado na Figura 4.
Após o 9° dia do experimento, os animais dos grupos B, D e E, tiveram suas fezes
coletadas diariamente, pesadas e submetidas ao exame parasitológico para se proceder a
contagem de oocistos de Eimeria spp por grama de fezes (OoPG), seguindo a técnica descrita
por MENEZES; LOPES, (1995).

40
Figura 4 – Administração de oocistos de Eimeria aos animais.
3.3 MÉTODOS ANALÍTICOS
3.3.1 Consumo de Ração
Os animais receberam, individualmente, 30 g diária de ração. Após 24 horas pesou-se
a ração residual no comedouro, e por diferença determinou-se o consumo diário de ração de
cada animal. Os animais foram pesados (Figura 5) todos os dias para se determinar o ganho de
peso e, por conseguinte calcular a taxa de conversão alimentar.

41
Figura 5 – Pesagem dos ratos
3.3.2 Contagem de Lactobacillus nas fezes
A contagem dos lactobacilos foi feita por grupo no dia 0, 6o, 9o, 12o, 15o, 18o e 21o dia
de experimento (ALAK et al., 1997) .
Em tubo de ensaio esterilizado, com capacidade para 20 ml foram pesados 1,56 g de
fezes e acrescentando-se 14,06 ml tampão fosfato, seguido de homogeneização, com auxílio
de um homogenizador de tubo tipo Vortex (marca Drake). A partir desta suspensão foram
realizadas as respectivas diluições decimais e as células de Lactobacillus foram contadas pela
técnica de “Pour Plate” em Ágar MRS, sendo que as placas receberam 1 ml das respectivas
diluições e foram incubadas a 37ºC por 48 horas.
3.3.3 Exame Parasitológico
As fezes foram processadas pela técnica da centrífugo-flutuação, onde 2 g de fezes de
cada animal, foram maceradas com 100 ml de água destilada em Becker com capacidade de
200 ml sendo filtrado em um tamis. Em seguida transferiu-se 10 ml desta suspensão para tubo

42 de centrífuga, centrifugando-se por 10 minutos a 400 x g, sendo o sobrenadante desprezado.
Ao sedimento adicionou-se solução saturada de sacarose (densidade 1.20-1.25) até a borda do
tubo, homogeneizou-se e centrifugou-se novamente por 10 minutos a 400 x g. Após esta etapa
colocou-se uma lamínula em contato com o sabrenadante no topo do tubo por 5 minutos,
tempo necessário para que os oocistos aderissem à lamínula. Em seguida colocou-se a
lamínula sobre uma lâmina limpa e seca, procedendo-se a contagem dos oocistos com auxílio
do microscópio óptico (Nikon) em objetiva de 40x. O valor encontrado foi multiplicado pelo
fator cinco, para a obtenção do OoPG/g fezes por animal, conforme descrito por MENEZES;
LOPES, 1995.
3.3.4 Exame do Tecido Hematopoético (Leucograma)
Para obtenção do leucograma, um animal de cada grupo foi submetido a punção
cardíaca após o 3º, 6º, 9º, 12º e 15º dia da infecção por Eimeria spp. Para tanto os animais
foram anestesiados com vapores de éter antes da coleta do sangue em conformidade com a
Resolução Nº 714 de 20/07/2002 do Conselho Federal de Medicina Veterinária (COBEA 1996).
O sangue (5 mL) foi colhido com auxílio de seringa e agulha (Figura 6) e imediatamente
transferido para tubo de ensaio contendo EDTA como anticoagulante, sendo este
homogeneizado em seguida.
Imediatamente após a coleta, com auxílio de um esfregaço em lâmina com as bordas
cortadas, confeccionou-se o filme sanguíneo que em seguida foi corado pelo método de
Leishman (BAIN, 1998).
A contagem total e diferencial dos leucócitos foi realizada por meio de contador
automático Cell-Dyn 3500, da marca Abbott. As contagens diferenciais foram confirmadas,
analisando-se os filmes sanguíneos, com auxílio do microscópio óptico em objetiva de 100x
(Nikon).

43
Figura 6 – Punção cardíaca
3.4 ANÁLISE DOS RESULTADOS
Os resultados foram analisados estatisticamente por meio de ANOVA e Teste t de
Student ao nível de significância de 0,05 e (n-1) graus de liberdade.

44 4 RESULTADOS E DISCUSSÃO
4.1 AVALIAÇÃO DA PREPARAÇÃO PROBIÓTICA NA COLONIZAÇÃO DO
INTESTINO DOS ANIMAIS
O fenômeno de adesão nas células epiteliais do intestino é um importante pré-requisito
para a colonização de cepas probióticas, prevenindo sua eliminação imediata pela peristalse,
favorecendo a competitividade neste ecossistema (KOS, et al. 2003).
No presente estudo foi avaliada a colonização do intestino por parte dos
microrganismos que compuseram a preparação probiótica administrada aos animais,
conforme demonstrado na Tabela 1. Para tanto, os animais dos grupos C, D e E receberam 1
ml da preparação probiótica, ao passo que os animais dos grupos A (controle negativo) e B
(controle para Eimeriose) não receberam a referida preparação. Os resultados representados
na Tabela 1 permitem observar que os animais do grupo E passaram a liberar maior
quantidade de lactobacilos nas fezes a partir do 9o dia de experimento, enquanto os animais
dos grupos C e D passaram a liberar a partir do 15o dia, quando comparados com os animais
dos grupos A e B. Estas observações permitem sugerir que para a permanência dos
lactobacilos no intestino torna-se necessária a ingestão diária de pequenas doses de produtos
probióticos, uma vez que espécies de lactobacilos são liberadas constantemente nas fezes, por
meio da peristalse, dificultando assim a colonização no intestino.
Tabela 1 – Contagem de lactobacilos nas fezes dos animais (Log UFC/g fezes)*
Grupos Dia 0 6o dia 9o dia 12o dia 15o dia 18o dia 21o dia
A 8.31 ± 0.12 9.16 ± 0.13 9.42 ± 0.04 9.01 ± 0.08 9.52 ± 0.20 8.71 ± 0.07 9.21± 0.03
B 8.10 ± 0.21 8.52 ± 0.15 9.31 ± 0.20 8.64 ± 0.17 8.93 ± 0.11 9.72 ± 0.03 9.52 ± 0.09
C 9.01 ± 0.15 9.53 ± 0.16 9.25 ± 0.09 9.55 ± 0.13 9.93 ± 0.15 10.3 ± 0.23 10.7 ± 0.17
D 8.43 ± 0.05 8.45 ± 0.07 8.79 ± 0.18 8.78 ± 0.10 9.45 ± 0.06 9.67 ± 0.17 10.1 ± 0.14
E 8.92 ± 0.09 8.67 ± 0.12 9.26 ± 0.10 9.20 ± 0.05 9.48 ± 0.03 9.7 ± 0.12 9.91 ± 0.15
* Valores médios de 3 repetições
O aumento do número de lactobacilos nas fezes dos animais dos grupos C e D a partir
do 15o dia permite inferir que até este momento a preparação probiótica recebida pelos
animais demonstrou ser efetiva, na colonização do intestino dos animais e que a partir de
então cerca de 30% dos 14 Log UFC/ml dos lactobacilos presentes na preparação, ingerida

45 diariamente, foram eliminados nas fezes. Esta situação provavelmente envolve as barreiras
naturais do organismo dos animais que compreendem pH estomacal, a presença de sais
biliares, movimentos de peristalse e o número de sítios disponíveis nas células intestinais
(FULLER, 1989).
Observou-se ainda que os animais do grupo E passaram a eliminar maior quantidade
de lactobacilos a partir do 9o dia possivelmente em função de terem sido infectados 6 dias
antes com os oocistos de Eimeria spp., tendo em vista que este protozoário também necessita
de sítios disponíveis na membrana das células intestinais para penetrar e dar prosseguimento
ao seu ciclo de vida. Desta forma, aproximadamente 35% dos 14 Log UFC/ml de lactobacilos
estariam sendo eliminados juntamente com as fezes. Resultados semelhantes foram obtidos
por Alak et al (1997) que infectaram camundongos imunodeficientes com Cryptosporidium
parvum e os trataram com L. reuteri. Estes autores observaram que os animais eliminaram
maiores quantidades de lactobacilos nas fezes a partir do 10o dia da infecção permanecendo
até o 25o dia.
4.2 AVALIAÇÃO DOS MICRORGANISMOS PROBIÓTICOS SOBRE EIMEIRA
A ação dos microrganismos probióticos estudados no presente trabalho sobre espécies
de Eimeria foi avaliada entre os animais dos grupos D e E (infectados e tratados com
probióticos) comparados com os animais do grupo B (infectados e não tratados), onde os
grupos receberam aproximadamente 5000 oocistos de Eimeria spp, cujos resultados
encontram-se apresentados na Tabela 2. Estes resultados demonstram que os animais do
grupo B, os quais foram infectados e não receberam a referida preparação probiótica, após 3
dias de infecção começaram apresentar quadro de morbidade e fezes com aspecto pastoso,
que durou até o final do experimento. Por outro lado, os animais dos grupos D e E, que foram
infectados e receberam a preparação probiótica, apresentaram-se com aparência física
saudável e fezes com aspecto normal, que expressa a higidez dos animais, o que demonstra a
eficácia das cepas de lactobacilos sobre o ciclo da Eimeria spp no trato intestinal dos animais.
Vale ressaltar que os animais do grupo B apresentam apenas lactobacilos na
composição natural de sua microbiota intestinal, o que não foi suficiente para controlar o
efeito da infecção por Eimeria spp.

46 Tabela 2 – Contagem de oocistos nas fezes dos animais (*)
Dias após a infecção Grupos Dia
0 6o 7o 8o 9o 10o 11o 12o 13o 14o 15o
A 0 0 0 0 0 0 0 0 0 0 0
B 0 50 1175 2710 2260 1165 1355 1500 540 910 600
C 0 0 0 0 0 0 0 0 0 0 0
D 0 30 10 90 185 65 5 0 0 0 0
E 0 10 0 2880 2225 680 25 0 0 0 0
(*) Número de oocistos liberados por g de fezes – OoPG/grupo de animais
Observa-se ainda que os animais dos grupos A e C não eliminaram oocistos, pois não
foram contaminados. Por outro lado, os animais dos grupos B, D e E começaram a eliminar
oocistos nas fezes no sexto dia após a infecção, sendo que os animais dos grupos D e E
interromperam a eliminação no décimo segundo dia da infecção e os do grupo B
permaneceram até o décimo quinto dia. Estes resultados permitem verificar a eficácia da
preparação probiótica avaliada sobre a colonização de Eimeria spp no trato intestinal dos
animais.
Os resultados apresentados na Tabela 1, demonstram que ocorreu aumento do número
de lactobacilos nas fezes dos animais dos grupos D e E, o que permite sugerir que houve uma
colonização positiva por estas espécies no intestino destes animais resultando em um efeito
positivo sobre a ação da Eimeria spp.
Desta forma, avaliou-se a influência da colonização de lactobacilos no intestino dos
animais em relação a infecção por Eimeria spp. Este efeito foi verificado por meio da
contagem de OoPG nas fezes dos animais dos grupos B, D e E. A análise estatística (Teste t)
dos resultados revelou que houve diferença significativa (p < 0,05) na contagem de OoPG nas
fezes dos animais dos grupos B e D, bem como dos grupos B e E. Nota-se que tanto os
animais do grupo D quanto do grupo E interromperam a eliminação de oocistos no décimo
segundo dia de infecção, sendo que o mesmo não ocorreu com os animais do grupo B (Tabela
2).
A análise dos resultados (Tabela 3) pelo Teste t demonstra que as diferenças do
número de lactobacilos entre o grupo B (controle Eimeriose) e os grupos D e E (infectados)
foram significativas (p < 0,05). Sendo assim, foram avaliadas as diferenças do número de
OoPG nas fezes dos animais dos grupos D e E, uma vez que os animais do grupo D
receberam tratamento antecipado (5 dias) e os animais do grupo E (3 dias) após a infecção.

47
Tabela 3 – Avaliação dos resultados (Teste t) de OoPG entre os grupos de animais tratados e não tratados com probióticos
Grupos Teste t
B x D* 0,0062
B x E* 0,0082
D x Ens 0,2637
* significativo p < 0,05 ns não significativo
Verificou-se que os animais do grupo E eliminaram maior quantidade de oocistos em
relação ao grupo D, o que pode ser devido à infecção dos animais do grupo E ter sido anterior
à administração da preparação probiótica. A avaliação destes resultados pelo Teste t revela
que não houve diferença significativa (p < 0,05) entre os valores de OoPG observados nestes
dois grupos de animais (D e E). O intervalo de tempo entre a infecção e os tratamentos dos
animais, permitiu que o protozoário tivesse tempo diferenciado para se instalar e invadir as
células do trato gastrointestinal dando início ao ciclo de infecção mais rapidamente. Quando
estes animais passaram a receber o preparado probiótico três dias após a infecção, os
Lactobacillus presente no preparado passaram a colonizar o intestino dos animais, o que
possivelmente dificultou a perpetuação da Eimeria. Resultado semelhante foi obtido por
Dalloul et al. (2003a) que avaliando a imunidade da mucosa intestinal de frangos alimentados
com probióticos a base de lactobacilos, observaram redução significativa na eliminação de
oocistos nas fezes dos animais do grupo tratado, mostrando resistência a infecção. O
mecanismo exato de proteção provocado pelos lactobacilos ainda não está claro, sendo que,
em parte poderia ser atribuído a estimulação do sistema imune por elementos presentes no
intestino, como por exemplo bactérias probióticas, que estimulariam a resposta contra Eimeria
por meio dos linfócitos intraperitoniais.
Eimeria é um parasita intracelular, que invade as células vizinhas para se reproduzir.
Primeiramente este coccídeo adere à superfície do endotélio, sendo que as bactérias
probióticas podem se adaptar e competir por sítios de adesão, ocupando os receptores das
células epiteliais (TIERNEY et al. 2004). Desta forma, este mecanismo pode ter retardado a
penetração e infiltração dos oocistos nas células epiteliais do intestino dos animais do grupo D
e consequentemente provocado redução de sua proliferação assexuada que resultaria em
redução na eliminação de oocistos nas fezes.
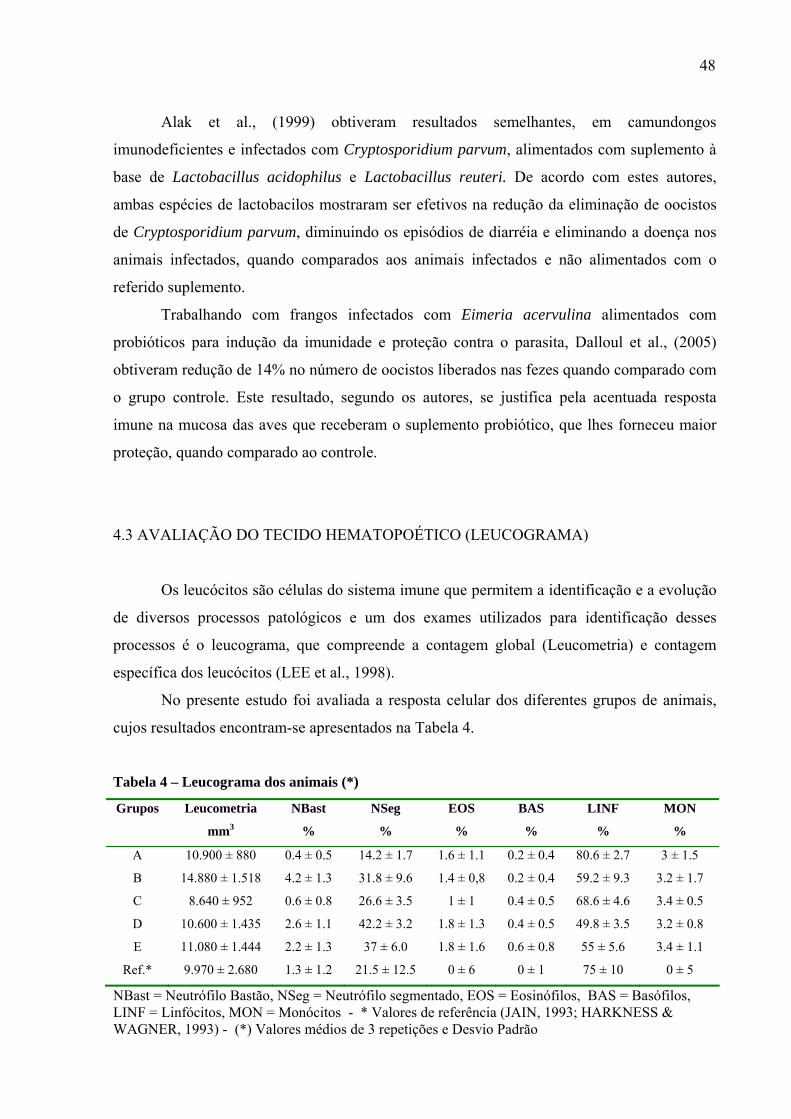
48 Alak et al., (1999) obtiveram resultados semelhantes, em camundongos
imunodeficientes e infectados com Cryptosporidium parvum, alimentados com suplemento à
base de Lactobacillus acidophilus e Lactobacillus reuteri. De acordo com estes autores,
ambas espécies de lactobacilos mostraram ser efetivos na redução da eliminação de oocistos
de Cryptosporidium parvum, diminuindo os episódios de diarréia e eliminando a doença nos
animais infectados, quando comparados aos animais infectados e não alimentados com o
referido suplemento.
Trabalhando com frangos infectados com Eimeria acervulina alimentados com
probióticos para indução da imunidade e proteção contra o parasita, Dalloul et al., (2005)
obtiveram redução de 14% no número de oocistos liberados nas fezes quando comparado com
o grupo controle. Este resultado, segundo os autores, se justifica pela acentuada resposta
imune na mucosa das aves que receberam o suplemento probiótico, que lhes forneceu maior
proteção, quando comparado ao controle.
4.3 AVALIAÇÃO DO TECIDO HEMATOPOÉTICO (LEUCOGRAMA)
Os leucócitos são células do sistema imune que permitem a identificação e a evolução
de diversos processos patológicos e um dos exames utilizados para identificação desses
processos é o leucograma, que compreende a contagem global (Leucometria) e contagem
específica dos leucócitos (LEE et al., 1998).
No presente estudo foi avaliada a resposta celular dos diferentes grupos de animais,
cujos resultados encontram-se apresentados na Tabela 4.
Tabela 4 – Leucograma dos animais (*)
Grupos Leucometria
mm3
NBast
%
NSeg
%
EOS
%
BAS
%
LINF
%
MON
%
A 10.900 ± 880 0.4 ± 0.5 14.2 ± 1.7 1.6 ± 1.1 0.2 ± 0.4 80.6 ± 2.7 3 ± 1.5
B 14.880 ± 1.518 4.2 ± 1.3 31.8 ± 9.6 1.4 ± 0,8 0.2 ± 0.4 59.2 ± 9.3 3.2 ± 1.7
C 8.640 ± 952 0.6 ± 0.8 26.6 ± 3.5 1 ± 1 0.4 ± 0.5 68.6 ± 4.6 3.4 ± 0.5
D 10.600 ± 1.435 2.6 ± 1.1 42.2 ± 3.2 1.8 ± 1.3 0.4 ± 0.5 49.8 ± 3.5 3.2 ± 0.8
E 11.080 ± 1.444 2.2 ± 1.3 37 ± 6.0 1.8 ± 1.6 0.6 ± 0.8 55 ± 5.6 3.4 ± 1.1
Ref.* 9.970 ± 2.680 1.3 ± 1.2 21.5 ± 12.5 0 ± 6 0 ± 1 75 ± 10 0 ± 5
NBast = Neutrófilo Bastão, NSeg = Neutrófilo segmentado, EOS = Eosinófilos, BAS = Basófilos, LINF = Linfócitos, MON = Monócitos - * Valores de referência (JAIN, 1993; HARKNESS & WAGNER, 1993) - (*) Valores médios de 3 repetições e Desvio Padrão

49 Observa-se que os animais do grupo A (controle) mantiveram as contagens de
leucócitos dentro dos valores de referência. Os animais do grupo B, que foram infectados por
oocistos de Eimeria spp apresentaram alteração na leucometria que se mostrou elevada, bem
como no número dos neutrófilos bastão e segmentados, que são células envolvidas com a
imunidade inata dotadas de componentes que permitem realizar intensa atividade fagocitária
contra agentes estranhos (JANEWAY et al., 2002). Nota-se ainda na Tabela 4 que a
porcentagem de linfócitos apresenta-se diminuída e as demais células não expressaram
nenhuma alteração.
Os animais do grupo C, que receberam o preparado probiótico diariamente tiveram sua
leucometria dentro dos valores de referência, entretanto, os valores de neutrófilos
segmentados apresentaram-se levemente aumentados, porém ainda dentro da referência sendo
que os valores das demais células não apresentaram alterações. O grupo D constituído de
animais que receberam preparado probiótico por seis dias antes de serem infectados com
oocistos de Eimeria spp apresentaram os resultados da leucometria dentro dos valores
normais, entretanto o número de neutrófilos segmentados apresentou-se aumentado, sendo os
valores de linfócitos diminuídos e as demais células sem alterações. Os animais do grupo E
que foram infectados com oocistos de Eimeria spp e passaram a receber a preparação
probiótica somente a partir do terceiro dia de infecção, não apresentaram alterações na
leucometria, mas o número de neutrófilos segmentados também apresentou-se aumentado, à
semelhança dos animais do grupo D que tiveram o número de linfócitos reduzidos e as demais
células não expressaram alterações.
Considerando-se que o número de leucócitos nos animais dos diferentes grupos
apresentou valores diferenciados (Tabela 4), o teste ANOVA, demonstrou que as diferenças
encontradas foram significativas (p < 0,05) entre os grupos que receberam o preparado
probiótico (grupos C, D e E) e os que não receberam (A e B). Portanto, conclui-se que os
lactobacilos presentes na preparação probiótica foram capazes de promover variações
leucocitária no sangue dos animais do grupo C, D e E.
Comparando os resultados relativos ao leucograma dos animais dos grupos A e B o
teste ANOVA expressou diferença significativa (p < 0,05) entre os valores observados,
mostrando que os animais do grupo B ao responderem ao parasitismo, apresentaram
alterações compatíveis com o quadro clínico dos animais.
Avaliando-se os resultados dos animais dos grupos A e C, verificou-se que os animais
do grupo C apresentaram leucometria normal, porém com valores menores que o grupo A,

50 demonstrando que houve um achatamento entre os valores mínimo e máximo. O teste
ANOVA destes resultados revelou diferença significativa (p < 0,05) entre estes grupos. Estes
resultados sugerem que as bactérias lácticas avaliadas podem ter estimulado a produção de
citocinas provocando uma redução de número de leucócitos na corrente sanguínea
(JANEWAY et al., 2002).
Quando se compara os resultados da leucometria dos animais dos grupos B e D e
grupos B e E, o teste ANOVA, também expressou diferença significativa (p < 0,05) entre
estes, Esta diferença pode estar relacionada com a produção de citocinas, induzida pelos
lactobacilos quando presentes na mucosa do intestino, que poderiam estar interferindo com as
células da corrente sanguínea. Estas estariam promovendo o deslocamento do “pool” central
de neutrófilos para o “pool” marginal, favorecendo desta maneira a diapedese e em seguida a
fagocitose das formas de Eimeria pelos mesmos, uma vez que estas são as primeiras células a
participarem de processos inflamatórios juntamente com os macrófagos (JANEWAY et al.,
2002).
Os resultados relativos aos grupos D e E submetidos ao teste de ANOVA, não
expressaram diferenças significativas entre os valores de leucometria apresentados (p > 0,05),
o que possivelmente pode estar relacionado ao intervalo de tempo entre a contaminação com
oocistos e a administração do preparado probiótico. Se este intervalo fosse maior
possivelmente seriam observarias diferenças significativas entre os valores obtidos, uma vez
que o parasito teria mais tempo para perpetuar seu ciclo e a resposta do sistema imune seria
mais intensa. A utilização do “pool” de Lactobacillus pode ter contribuído para obtenção
destes resultados, pois a ação sinergética favorece a resposta imune, tornando-a mais rápida e
mais eficiente (TIMMERMAN et al., 2004).
Quando se compara os resultados da contagem específica dos leucócitos entre os
animais dos grupos C, D e E, em relação ao número de neutrófilos segmentados, o teste
ANOVA demonstrou diferença significativa (p < 0,05) entre os valores obtidos. Estes
resultados mostram que o preparado probiótico exerceu efeito imunomodulador estimulando,
possivelmente por meio de citocinas, o aumento de células fagocitárias (neutrófilos
segmentados), que provavelmente auxiliaram na interrupção do ciclo da Eimeria nos animais
dos grupos D e E. Por outro lado, paralelamente houve queda no número dos linfócitos
circulantes que também pode estar relacionada com a presença de citocinas específicas que
promoveram este efeito.
Microrganismos que apresentam propriedades probióticas podem apresentar ação na
imunomodulação do hospedeiro, devido a produção de glicopeptídeos ou outros metabólitos

51 (CHESSON, 1994). Alguns gêneros de bactérias intestinais como Lactobacillus e
Bifidobacterium estão diretamente relacionados com o aumento da resposta imune
correspondendo a produção de anticorpos, ativação de macrófagos, proliferação de células T,
aumento do número de neutrófilos e monócitos, produção de Interferon, Interleucina-2 (IL-2)
entre outros metabólitos (DALLOUL, et al., 2005; BUDINO, et al., 2004; SCHIFFRIN, et al.,
1995; SCHIFFRIN, et al., 1997).
Shu; Qu; Gill (2001) relataram que leitões tratados com B. lactis HN019 apresentaram
diminuição na incidência de diarréia associada com Rotavírus e E. coli, concomitante ao
aumento dos títulos de anticorpos contra patógenos específicos no trato gastrintestinal,
destacando-se a concentração de neutrófilos sanguíneos e da resposta proliferativa dos
linfócitos T.
Avaliando Lactobacillus gasseri CECT 5714 e Lactobacillus coryniformis CECT
5711, Olivares et al., (2006) produziram leite fermentado que foi administrado em humanos
sadios, com a finalidade de se avaliar o seu efeito sobre o sistema imune. Após duas semanas
de tratamento o sangue dos indivíduos que receberam o tratamento constituído por ambas
espécies de Lactobacillus apresentaram aumento significativo no número de neutrófilos,
monócitos e na atividade fagocitária, sendo observado que o aumento dos neutrófilos foi
mantido até o final do experimento. Os autores concluíram que este tipo de estímulo favorece
o organismo diante de microrganismos patogênicos.
Lactobacillus rhamnosus ministrados em camundongos infectados experimentalmente
com E. coli O157:H7, reduziram a morbidade, estimulando o aumento da atividade fagocitária
dos neutrófilos e macrófagos e o aumento de anticorpos específicos na mucosa (IgA)
intestinal (SHU; GILL, 2002).
4.4 DESEMPENHO ZOOTÉCNICO DOS ANIMAIS
Uma característica importante do bom funcionamento do trato intestinal dos animais é
determinada pelo equilíbrio de sua microbiota bacteriana. Esse equilíbrio é muito importante,
mas difícil de ser verificado. Tal equilíbrio pode ser alterado por fenômenos agressivos que
modificam as secreções intestinais e a peristalse (MARUTA, 1993) sendo que nesse caso,
microrganismos patogênicos bem como, a Eimeria podem se proliferar.

52
Tendo em vista a avaliação do efeito da preparação probiótica sobre o desempenho
zootécnico dos animais mediu-se o consumo de ração, ganho de peso e determinou-se a
conversão alimentar, cujos resultados encontram-se apresentados na Tabela 5.
Tabela 5 – Desempenho Zootécnico dos Animais
Grupos Consumo de ração
g/dia/animal
Ganho de peso
g/dia/animal
Conversão alimentar
g/g
A 19,16 5,18 3,72
B 15,98 3,47 4,82
C 18,92 5,36 3,60
D 15,50 4,12 3,76
E 14,42 3,79 3,87
No que se refere ao consumo de ração observou-se que os animais dos grupos B, D e
E apresentaram valores menores comparados aos demais grupos. Esta observação se deve
provavelmente ao fato desses animais estarem infectados por Eimeria spp tendo como
conseqüência a perda de apetite, menor ganho de peso e conseqüente deficiência na conversão
alimentar.
Relacionando-se os parâmetros consumo de ração e ganho de peso, estabeleceu-se o
parâmetro conversão alimentar. Observa-se que os animais do grupo B (controle para
Eimeriose) apresentaram menor ganho de peso, menor conversão alimentar e maior consumo
de ração, quando comparados ao grupo A (controle negativo) e grupo C (animais que
receberam a preparação probiótica). Estas alterações provavelmente foram provocadas por
lesões no epitélio intestinal que são típicas neste quadro de parasitose. Segundo Rose (1987)
estas lesões são responsáveis pelo maior consumo de ração, deficiência no ganho de peso e na
conversão alimentar dos animais
Os animais do grupo C consumiram menos ração que os animais do grupo A, porém
apresentaram melhor conversão alimentar tendo como conseqüência maior ganho de peso.
Esta situação se deve ao fato dos animais do grupo C terem recebido diariamente a preparação
probiótica a base de lactobacilos, o que favoreceu o melhor desempenho zootécnico dos
animais. Resultados semelhantes foram obtidos por Alves et al., (2000) quando utilizaram
suplemento probiótico composto por L. acidophilus, Eenterococcus faecium e Sacharomyces
cerevisae na dieta de vitelos, onde os animais que receberam o probiótico apresentaram maior
ganho de peso diário e melhor conversão alimentar, no período de 92 aos 119 dias de idade.

53
Notou-se ainda que os animais do grupo D consumiram menos ração que os animais
do grupo B e C e obtiveram maior ganho de peso e melhor conversão alimentar que os
animais dos grupos A, B e E. Estas observações se devem possivelmente ao fato que os
lactobacilos presentes no preparado probiótico, favoreceram esta performance. Entretanto, os
animais do grupo E consumiram menos ração que os demais grupos, obtiveram ganho de peso
e conversão alimentar superiores aos animais do grupo B. Esta situação mostra que o
preparado probiótico ingerido antes ou após a infecção exerce efeito no trato intestinal dos
animais, favorecendo o ganho de peso e conversão alimentar.
Corrêa (2003) avaliou um preparado poliprobiótico constituído por L. acidohilus, L.
casei, Streptococcus salivarium E. faecium, B. subtilis, B. toyoi e S. cerevisae no desempenho
zootécnico e rendimento de carcaça de frangos de corte. Os resultados revelaram que as aves
tratadas com o preparado poliprobiótico apresentaram melhor conversão alimentar quando
comparadas ao grupo controle, permitindo desta forma demonstrar o efeito do poliprobiótico
como promotor de crescimento.

54 5 CONCLUSÕES
Os resultados obtidos no presente trabalho permitem concluir que:
- O preparado probiótico mostrou ser efetivo no tratamento de Eimeriose, interrompendo a
eliminação de oocistos de Eimeria spp nos ratos que foram contaminados e
posteriormente tratados com probióticos e vice-versa.
- O “pool” de microrganismos probióticos promoveu aumento do número de neutrófilos no
sangue dos animais que representaram o controle do probiótico, dos animais tratados com
probiótico e depois infectados com oocistos de Eimeria e dos animais infectados com
oocistos de Eimeria e tratados depois com probióticos.
- Os animais que receberam probiótico apresentaram melhor desempenho zootécnico no
que se refere ao ganho de peso e taxa de conversão alimentar.

55 6 SUGESTÕES PARA TRABALHOS FUTUROS
Visando um melhor entendimento sobre a ação de espécies de microrganismos
probióticos sobre Eimeria spp, os resultados do presente trabalho permitem sugerir a
realização de novos experimentos, sob as mesmas condições experimentais:
- Realizar estudos histológicos, com objetivo de identificar lesões provocadas pela
Eimeria spp e a possível fase do ciclo evolutivo que está sendo afetada, bem como a
identificação da espécie de Lactobacillus presentes no preparado probiótico aderidos às
células intestinais.
- Criar um grupo F, onde os animais seriam infectados com oocistos de Eimeria spp,
desenvolveriam o quadro agudo da doença e a partir deste ponto, os animais passariam a
receber o preparado probiótico, com a intensão de se avaliar o efeito do referido preparado na
fase aguda da infecção, onde os animais estão mais debilitados.
- Estudar a resposta Imune compreendendo a Imunidade humoral, dosando citocinas e
anticorpos específicos e a Imunidade celular avaliando-se a população e sub-população de
linfócitos, bem como a atividade fagocitária pelos neutrófilos e macrófagos.

56 REFERÊNCIAS ABE, F. et al. Effect of administration of a probiotic containing bifidobacteria and lactic acid bacteria on newborn piglets and calves. Nutrition Research, v. 8, p. 141-146, 1995. ADLERBERTH, I. et al. A. manose-specific adherence mecanism in Lactobacillus plantarum confering binding to the human colonic cell line HT-29. Applied and Environmental Microbiology, v. 62, n. 7, p. 2244-2251, 1996. AGARWAL, N. et al. Microbial status and rumen enzyme profile of crossbred calves feed on different microbial feed additives. Letters Applied Microbioliology, v. 34, n. 5, p. 329-336, 2002. ALAK, J. I. et al. Effect of Lactobacillus reuteri on intestinal resistence to Cryptosporidium parvum infection in a murine model of acquired immunodeficiency syndrome. Journal Infections Diseases, v. 175, n. 1, p. 218-221, 1997. ALAK, J. I. et al. Supplementation with Lactobacillus reuteri or L. acidophilus reduced intestinal shedding of Cryptosporidium parvum oocysts in immunodeficient C57BL/6 mice. Cellular Mollecular Biology, v. 45, n. 6, p. 855-863, 1999. ALVES, P. A. de P. M. et al. Uso de probiótico composto por Lactobacillus acidophilus, Streptococcus faecium e Sacharomyces cerevisae na dieta de vitelos bovinos: efeitos sobre o desempenho e a qualidade da carne, Brazilian Journal of Veterinary Research and Animal Science , v. 37 n. 5, p.25-34, 2000. ARICIA, M. et al. Some characteristics of Lactobacillus isolates from infant faeces. Food Microbiology, v. 21, p. 19-24, 2004. ÁVILA, F. A. et al. Escherichia coli isolated from calves with diarrhea in the northern region of state of São Paulo, Brazil. Arquivos de Veterinária, v. 4, p. 285-289, 1988. ÁVILA, F. A. et al. A comparative study of the efficiency of a probiotic and the ati-K99 and anti-A14 vaccines in the control of diarrea in calves in Brazil, Revista do Instituto de Medicina Tropical de São Paulo, v. 48, n. 3, p. 239-243, 1995. AVILA, M. et al. Influence of a bacteriocin-producing lactic culture on proteolysis and texture of Hispánico cheese. International Dairy Journal, v. 15, n. 2, p.145-153, 2005.

57 BAIN, B. J. Células sanguíneas. Porto Alegre: Artmed, 1998. 118 p. BARBOSA, H. R.; TORRES, B. B. Microbiologia Básica. Rio de Janeiro: Atheneu, 1999. 196 p. BARROW, P. A. Probiotic fo chickens. In: Probiotics: the scientific basis. FULLER, R. 1992. p. 225-257. BILLETER, S. A. et al. Ponazuril inhibits the development of Eimeria vermiformes in experimentally infected outbred swiss mice. Disponível em <http://www.parasitology.informatik.uni-wuerzbug.de/login/n/h/j_004-1263-9.html.linked> Acesso: 28.out.2006. BEUCHAT, L. R.; GOLDEN, D. A. Antimicrobials occuring naturally in foods. Food Technology, v. 1, p. 134-142, 1989. BORIS, S. et al. Characterization of the aggregation promoting factor from Lactobacillus gasseri, a vaginal isolate. Journal of Applied Microbiology, v. 83, p. 413-420, 1997. BUCHANAN, R. E.; GIBBONS, N. E. Bergey’s manual of determinative bacteriology. 8. ed. Baltimore: Williams & Wilkins, 1974. 1268 p. BUDINO, F. E. L. et al. Influência da adição de probiótico e/ou prebiótico em dietas de leitões desmamados sobre as atividades das enzimas digestivas e parâmetros sanguíneos. Acta Scientiarum Animal Sciences, v. 26, n. 4, p. 529-536, 2004. CAMPOS, D. M. B. et al. Níveis de inclusão de probiótico (Bacillus subtilis) sobre o desempenho de frangos de corte. Revista Brasileira de Ciência Avícola, v. 4, suplemento, p. 36, 2002. CANIBE, N.; JENSEN, B. B. Fermented and nonfermented liquid feed to growing pigs: effect on aspects of gastrointestinal ecology and growth performance. Journal animal Science, v. 81, n. 8, p. 2019-2031, 2003. CHATEAU, N.; CASTELLANOS, I.; DESCHAMPS, A. M. Distribuition of pathogen inhibition in the Lactobacillus isolates of a commercial probiotic consortium. Journal of Applied Bacteriology, v. 74, p. 36-40, 1993.

58 CHAVES, A. H. et al. Efeito da Estirpe LT 516 de Lactobacillus acidophilus como probiótico para bezerros. Revista Brasileira de Zootecnia, v. 5, p. 1075-1085, 1999. CHESSON, A. Probiotics and other intestinal mediators. In: COLE, D. J. A. et al. Principles of pig science. 1994. p. 197-214. CLIVER, D. O.; MARTH, E. H. Preservation, sanitation, and microbiological specifications for food. In: CLIVER, D. O. Foodborne Diseases. 1. ed. San Diego: Academic Press, 1990. p. 45-63. COLÉGIO BRASILEIRO DE EXPERIMENTAÇÃO ANIMAL – COBEA. Manual para técnicos em bioterismo. São Paulo: Winner Graph. 1996. 259 p. CORRÊA, G. S. S. Efeito de antibiótico e probióticos sobre o desempenho e rendimento de carcaça de frangos de corte. Arquivo Brasileiro Medicina Veterinária e Zootecnia, v. 55, n. 4, p. 1-9, 2003. COSTERTON, J. W.; ARRIE, T. J. E.; CHENG, K. J. Phenomena of bacterial adhesion. In Bacterial Adhesion. ed. Savage D. C. E Fletcher M. London: Plenum Press, 1985. p. 3-43. DAESCHEL, M. A. Antimicrobial substances from lactic acid bacteria for use as food preservatives, Food Technology, v. 36, p. 164-167, 1989. DALLOUL, R. A. et al. Enhanced mucosal immunity against Eimeria acervulina in broilers fed a Lactobacillus-based probiotic. Poultry Science. v. 82, n. 1, p. 62-66, 2003a. DALLOUL, R. A. et al. Intestinal immunomodulation by vitamin A deficiency and Lactobacillus-based probiotic in Eimeria acervulina-infected broiler chickens. Avian Disease, v. 47, n. 4, p. 1313-1320, 2003b. DALLOUL, R. A. et al. Induction of local protective immunity to Eimeria acervulina by Lactobacillus-based probiotic. Comparative Immunology Microbiology Infectious Diseases, v. 28, p. 351-361, 2005. DAVIS, G. H. G. The classification of Lactobacilli from the human mouth. Journal General Microbiology, v. 13, p. 481-493, 1955.
DE MAN, J. C. et al. A medium for the cultivation of Lactobacilli. Journal of Applied Bacteriology, v. 23, p. 130-135, 1960.

59 DE MARTINIS, E. C. P.; ALVES, V. E.; FRANCO, B. D. G. M. Fundamentals and perspectives for the use of bacteriocins produced by lacticacid bacteria in meat products. Food Reviews International, v. 18, n. 2, p.191-208, 2002. DICKS, L. M. T.; MELLETT, F. D.; HOFFMAN, L. C. Use of bacteriocin-producing starter cultures of Lactobacillus plantarum and Lactobacillus curvatus in production of ostrich meat salami. Meat Science, v. 66, n. 3, p. 703-708, 2004. EDELMAN, S. et al. In vitro adhesion of an avian pathogenic Escherchia coli O78 strain to surface of the chicken intestinal tract and to ileal mucus. Veterinary Microbiology, v. 91, n. 1, p. 41-56, 2003. EHRENFELD, E. E. et al. Identification of pheromone-induced surface proteins in Streptococcus faecalis and evidence of a role for lipoteichoic acid in formation of mating aggregates. Journal of Bacteriology, v. 168, p. 6-12, 1986. ELIA, C. C. S.; de SOUZA, H. S. P. Imunologia da mucosa intestinal: da bancada ao leito. São Paulo: Atheneu, 2001. 187 p. EMBRAPA. Produção Animal. Disponível em: <http://www.embrapa.br/linhas_acao/alimentos/prod_animal/index.html.> Acesso: 25.mar.2004. FABER, J. E. et al. Eimeria infections in cows in the periparturient phase and their calves: oocyst excretion and levels of specific serum and colostrum antibodies. Veterinary Parasitology, v. 104, n. 1, p. 1-17, 2002. FERNANDES, P. C. C. et al. Viabilidade do uso de probiótico na alimentação de monogástricos. Caderno Técnico de Veterinária e Zootecnia, v. 31, p. 53-71, 2000. FERREIRA, A. J. P. Exclusão competitiva na avicultura. In: SIMPÓSIO SOBRE ADITIVOS ALTERNATIVOS NA NUTRIÇÃO ANIMAL, 2000, Campinas. Anais... Campinas: Colégio Brasileiro de Nutrição Animal, 2000. p. 133-140. FERREIRA, C. L. L. F. Grupo de Bactérias Lácticas: caracterização e aplicação tecnológica de bactérias probióticas. In: FERREIRA, C. L. F. Prebióticos e Probióticos: atualização e Prospecção. Viçosa: Universidade Federal de Viçosa, 2003. p. 7-34.

60 FIRON, N. et al. Aromatic alpha-glycosides of mannose are powerful inhibitors of the adherence of type 1 fimbriated Escherichia coli to yeast and intestinal ephitelial cells. Infection Immunology, v. 55, n. 2, p. 472-476, 1987. FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS, WORLD HEALTH ORGANIZATION. Evaluation of health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. Córdoba, 2001. 34 p. Disponível em: <ftp://ftp.fao.org/es/esn/food/probioreport_ en.pdf>. Acesso em: 10 out. 2006. FOSTER, F. A. et al. Viability of Cryptosporidium parvum during ensilage of perennial ryegrass. Journal Applied Microbiology, v. 82, n. 1, p. 115-120, 1997. FRANDSEN, J. C. Effects of restrictions in dietary protein and vitamin a on the responses of rats to infections by Nippostrongylus brasiliensis (Nematoda) and N. brasiliensis plus Eimeria nieschulzi (Coccidia). International Journal for Parasitology, v. 15, n. 4. p. 523-528, 1985. FULLER, R.; BARROW, A.; BROOKER, B. E. Bacteria associated with the gastric epithelium of neonatal pigs. Applied and Environmental Microbiology, v. 35, n. 3, p. 582-591, 1978. FULLER, R. Basis and efficacy of probiotics. World’s Poultry Science Journal, v. 44, p.69-70, 1988. FULLER, R. Probiotics in man and animals. Journal of Applied Bacteriology, v. 66, p. 365-378, 1989. FULLER, R. History and development of probiotics. Probiotics: the scientific basis. Ed. By Fuller, R. London: Chapaman & Hall, 1992. p. 1-8. FULLER, R. Probiotics, therir development and use: In: Old Herborn University Seminar Monograph 8 (VANN der WAAII. D., HEIDT, P. J. and RUSCH, V. C., eds.). Herborn-Dill: Institute for Microbiology and Biochemistry, 1995. p. 1-8. GARCIA, S. Isolamento e caracterização de bactérias láticas para uso como probiótico. 1999. 157 f. Tese (Doutorado) - Universidade Estadual de Londrina-PR, 1999. GIBSON, G. R.; ROBERFROID, M. B. Dietary modification of the human colonic microbiota: introducing the concept of prebiotics. Journal Nutrition, v. 125, p. 1401-1412, 1995.

61 GILLILAND, S. E.; SPECK, M. L. Inibition of psychrotrophic bacteria by Lactobacilli and pedicocci in nonfermented refrigerated foods. Journal Food Science, v. 40, p. 903-935, 1975. GILLILAND, S. E.; SPECK, M. L. Enumeration and identity of Lactobacilli in dietary produtcs. Journal of Food Protection, v. 40, n. 11, p. 760-762, 1977. GILLILAND, S. E.; STALEY, T. E.; BUSH, L. J. Importance of bile tolerance of Lactobacillus acidophilus used as a dietary adjunt. Journal Dairy Science, v. 67, p. 3045-3051, 1984. GILLILAND, S. E.; NELSON, C. R.; MAXWELL, C. Assimilation of cholesterol by Lactobacillus acidophilus. Applied Microbiology Enviromental, v. 33, p. 15-18, 1985. GLASS, M. D. et al. Effects of Lactobacillus acidophilus and Lactobacillus reuteri cell free supernatants on Cryptosporidium viability and infectivity in vitro. Food Microbiology, n. 12, p. 423-429, 2004. GOLDIN. B. R.; GORBACH S. L.; SAXELIN, M. Survival of Lactobacillus species (strain GG) in human gastrointestinal tract. Digestive Disease Scienci, v. 37, n. 1, p. 121-128, 1992. HAMMES, W. P.; WEISS, N.; HOLZAPFEL, W. The genera Lactobacillus and Carnobacterium. In: BALOWS, A. et al. The prokaryotes: a handbook on the biology of bacteria: ecophysiology, isolation, identification, applications. New York: Springer-Verlag, 1992. v. 2. p. 1535-1578. HARKNESS, J. E.; WAGNER, J. E. Biologia e clínica de coelhos e roedores. 3. ed. São Paulo: Livraria Roca, 1993. 238 p. HAVENAAR, R.; BRINK, B. T.; HUIS IN’TVELD, J. H. J. Selection of strains for probiotic use. In: Probiotics. The scientific basis. Ed. By Fuller, R. London: Chapaman & Hall, 1992. p. 209-224, HEMME, D.; FOUCAUD-SCHEUNEMANN, C. Leuconostoc, characteristics, use in dairy technology and prospects in functional foods. International Dairy Journal, v. 14, n. 6, p. 467-494, 2004. HOSSE, H.; SOZZI, T. Biotechnology group meeting probiotics: fact ou fiction?. Journal of Chemical Technology and Biotechnology, v. 51, p. 539-570, 1991.

62 ISOLAURI, E. et al. T. Improved immunogenicity of oral D x RRV reassortant rotavirus vaccine by Lactobacillus casei GG. Vaccine, v. 13, p. 310-12, 1995. JAIN, N. C. Essentials of veterinary hematology. Philadelphia: Lea & Febiger, 1993. 417 p. JANEWAY, C. A. et al. Imunobiologia: o sistema imune na saúde e na doença. 5. ed. Porto Alegre: Artmed, 2002. 767 p. JIN, L. Z. et al. Probiotics in poultry: modes of action. Word’s poultry Science Journal, v. 53, n. 4, p. 351-368, 1997. JIN L. Z. et al. Growth performance, intestinal microbial population, and serum cholesterol of broilers fed diets containing Lactobacillus cultures. Poultry Science, v. 77, n. 9, p. 1259-1265, 1998. JUVEN, B. J.; MEINERSMANN, R. J.; STERN, N. J. Antagonistic effect of lactobacili and Pediococci to control intestinal colonization by human enteropathogens in live poultry. Journal Applied Bacteriology, v. 70, p. 95-103, 1991. KANDLER, O.; WEISS, N. Genus Lactobacillus Beijerinck 1901, 212AL. In: SNEATH, P. H. A. et al. Bergey’s manual of systematic bacteriology. Baltimore: Willians & Wilkins, 1986. v. 2. p. 1209-1234. KIMURA, N. et al. Studies on the relationship between intestinal flora and cecal coccidiosis in chicken. Poultry Science, v. 55, n. 4, p. 1375-1383, 1976. KLAENHAMMER, T. R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiology Review, v. 12, p. 39-86, 1993. KLAVER, F. A. M.; MEER, R. V. D. The assumed assimilation of cholesterol by Lactobacilli and Bifidobacterium bifidum is due to their salt desconjugation activity. Applied and Environmental Microbiology, v. 59, p. 1120-1124, 1993. KLEEMAN, E. G.; KLAENHAMMER, T. R. Adherence of Lactobacillus species to human fetal intestinal cells. Journal Dairy Science, v. 65, n. 11, p. 2063-2069, 1982. KLERK, H. C.; SMIT, J. A. Properties of a Lactobacillus fermentum bacteriocin. Journal General Microbiology, v. 48, p. 309-316, 1967.

63 KOS, B. et al. Adhesion and aggregation ability of probiótica strain Lactobacillus acidophilus M92. Journal of Applied Microbiology, v. 94, p.981-987, 2003. KROGER, M.; KURMANN, J. A.; RASIC, J. L. Fermented milks: past, present and future. Food Technology, v. 43, n. 1, p. 92-99, 1989. KUNITACHI, Y. Yakult central institute for microbiological research, Tokio, 1996. LAN, P. T.; RINH, le T.; BENNO, Y. Impact of two probiotic Lactobacillus strains feeding on fecal lactobacilli and weight gains in chicken. Journal General Applied Microbiology, v. 49, n. 1, p. 29-36, 2003. LEE, G. R. et al. Wintrobe: hematologia clínica., 9. ed. São Paulo: Manole, 1998. vol. 1, 1424 p. LEE, R. W.; BOTTS, R. L. Evaluation of single oral dosing and continuous feeding of Streptococcus faecium M74 (Suntabac) on performance of incoming feedlot cattle. Journal Animal Science, n. 66, sup. 1, p. 460, 1988. LEEDLE, J. Probiotics and DFMs: mode of action in the gastrointestinal tract. In: SIMPÓSIO SOBRE ADITIVOS ALTERNATIVOS NA NUTRIÇÃO ANIMAL, 2000, Campinas. Anais… Campinas: Colégio Brasileiro de Nutrição Animal, 2000. p. 23-32. LEITÃO, J. da S. Parasitologia Veterinária. Lisboa: Fundação Caloutre, 1983. 529 p. LINE, J. E. Effect of yeast-suplemented fee don Salmonella and Campylobacter populations in broilers. Poultry Science, v. 77, n. 3, p. 405-410, 1998. LÜCK, E. Conservacion quimica de los alimentos. Zaragoza: Acribia, 1981. 348 p. MACHADO, D. F. Efeito probiótico na modulação dos níveis de colesterol sérico e na taxa de translocação de Lactobacillus spp. em ratos. 2001. 111f. Dissertação (Mestrado) - Universidade Federal de Viçosa. Viçosa-MG, 2001. MANN, G. V.; SPOERRY, A. Studies of a surfactant and cholesterelemia in the Massai. American Journal of Clinical Nutrition, v. 27, p. 464-469, 1974.

64 MARSHALL, R. N. Bovine coccidiosis in calves following turnout. Veterinary Immunology and Immunopathology, v. 143, n. 13, p. 366-367, 1998. MATSUZAKI, T. et al. Effect of oral administration of Lactobacillus casei on alloxan-induced diabetes in mice. APMIS, v. 105, p. 637-642, 1997. MARUTA, K. Probióticos e seus benefícios. In: CONFERÊNCIA DE CIÊNCIA E TENCOLOGIA AVÍCOLAS, 1993, Santos. Anais... Santos: APINCO, 1993. p. 203-219. MCGRORY, T. et al. The interactions of Lactobacillus acidophilus and Trichomonas vaginalis in vitro. Jouranl Parasitology, v. 80, n. 1, p. 50-54, 1994. MENEZES, R. de C. A. A. de.; LOPES, C. W. G. Epizootiologia de Eimeria arloingi em caprinos na Microrregião Serrana Fluminense, Rio de Janeiro, Brasil. Revista da Universidade Rural, v. 17, n. 2, p. 5-12, 1995. MILES, R. D.; JERNINGAN, M. A. Probiotics in poutry nutrition: a review. World’s Poultry Science Journal, v. 41, n.2, p. 99-107, 1985. MIRELMAN, D; ALTMANN, G; ESHDAT, Y. Screening of bacterial isolates fo mannose-specific lectin activity by agglutination of yeast. Journal of Clinical Microbiology, v. 11, p. 328-331,1980. MORI, M. et al. Isolation and structure of the Streptococcus faecalis sex pheromone, CAM 373. FEBS Letters, v. 206, p. 69-72, 1986. NADER De MACIAS, M. F. et al. Inhibition of Shigella sonnei by Lactobacillus casei and Lactobacillus acidophilus. Journal of Applied Bacteriology, v. 73, p. 4078-4111, 1992. NAITO, S. et al. Development of intestinal Lactobacilli in normal piglets. Journal of Applied Bacteriology, v. 79, n. 1. p. 230-236, 1995. NEUMANN, E.; FERREIRA, C. L. L. F. Lactobacillus acidophilus as dietary adjunt: in vitro susceptibility to gastric juice, bile salts, lysozyme and chemotherapeutic agents. Revista Brasileira de Microbiologia, v. 26, n. 1, p. 59-65, 1995. NOUSIAINEN, J.; SETALA, J. Lactis acid bacterial as animal probiotics. In: Lactic Acid Bacteria – Microbiology and Functional Aspects Ed. Salminen S. e Wright A. Von . 2nd ed. New York: Marcel Dekker, Inc., 1998. p. 437-473,

65 NURMI, E.; RANTALA, M. New aspects of Salmonella infection in broiler prodution. Nature, v. 241, n.19, p. 210-211, 1973. OLIVARES, M. et al. The consumption of two new probiótica strains, Lactobacillus gasseri CECT 5714 and Lactobacillus coryniformis CECT 5711, Boosts the Immune system of healthy human. International Microbiology, v. 9, p. 47-52, 2006. O’SULLIVAN, M. G. et al. Probiotic bacteria: myth or reality? Trends in Food Science & Technology, v. 3, p. 309-314, 1992. OZAWA, K. et al. Effect of Streptococcus faecalis BIO-4R on intestinal flora of weanling piglets and calves. Applied Environmental Microbiology, v. 45, p. 1513-1518, 1983. PENNA, F. J. et al. Bases experimentais e clínicas atuais para o emprego dos probióticos. Journal of Pediatria, v. 76, supl. 2, p. 209-217, 2000. PERDIGÓN, G. et al. Lactobacilli administered orally induce release of enzymes from peritoneal macrophages in mice. Milchwissenschaft, v. 41, n. 6, p. 344, 1986. PERDIGÓN, G. et al. Immune system stimulation by probiotics. Journal of Dairy Science, v. 78, n.7, p.1597-1606, 1995. PÉREZ, P. F. et al. Inhibition of Giardia intestinalis by extracellular factor from Lactobacilli; an in vitro study. Applied and Environmental Microbiology, v. 67, n. 11, p.5037-5042, 2001. PREMI, L.; BOTTAZZI, V. Hydrogen peroxide formation and hydrogen peroxide splitting activity in lacti and bacteria. Milchwissenschaft, v. 27, n. 12, p. 762-765, 1972. RAO, L. M. et al. Bile salt desconjugation and cholesterol removal from media by Lactobacillus casei. Journal Dairy Scienci, v. 81, p. 2103-2110, 1998. REDONDO-LOPEZ, V. et al. Emerging role of Lactobacilli in the control and maintenance of the vaginal bacterial microflora. Reviews of Infectious Diseases, v. 12, p. 856-872, 1990. REITER, B.; HARNULV, G. Lactoperoxidase antibacterial system: natural occurrence, biological functions and pratical applications. Journal Food Protection, v. 47, n. 9, p. 724-732, 1984.

66 ROSE, M. E. Immunity to Eimeria infections. Veterinary Immunology and immunopathology, v. 17, p. 333-343, 1987. ROSTAGNO, H. S. et al. Utilização de probióticos e prebióticos em aves. In: FERREIRA, C. L. L. F. Prébióticos e probióticos: atualização e prospecção. Viçosa: Universidade Federal de Viçosa, 2003. p.181-202. ROTKIEWIEZ, T. et al. Effect of Lactobacillus acidophilus and Bifidobacterium sp. on the course of Cryptosporidium parvum invasion in new-born piglets. Bulletin Veterinary Pulawy, v. 45, p. 187-195, 2001. ROWLAND, I. R. Metabolic interactions in gut. In: Probiotics: the scientific basis. Ed. By Fuller, R. London: Chapman & Hall, 1992. p.29-53.
RUPPERT, L. D. G.; McCOY, G. C.; HUTJENS, M. F. Feedings of probiotics to calves. Journal of Dairy Science, v. 77, Sup. 1, p.296, 1994. SALLOFF-COSTE, C. J. Lactobacillus acidophilus. Danone Word Newsletter, v. 13, p. 1-7, 1997. SANDERS, M. E. Effect of consumption of lactic cultures on human health. In: Advances in food and nutrition research, Ed. By Kinsella, J. E. 1993. v. 37, p. 67-129. SANDERS, M. E. Probiotics: considerations for human health. Nutrition, v. 61, n. 3, p. 91-99, 2003. SAUCIER, L. et al. Effect of feeding lactic acid bacteria and fermented milk on specific and nonspecific immune responses of mice infected with Klebsiella pneumoniae AD-I: Journal Food Protection, v. 55, p. 595-600, 1992. SCHIFFRIN, E. J. et al. Immunomodulation of human blood cells following the ingestion of lactic acid bacteria. Journal Dairy Scienci, v. 78, p. 491-497, 1995. SCHIFFRIN, E. J. et al. Immune modulation of blood leukocytes in humans by lactic acid bacteria: criteria for strain selection. American Journal Clinical Nutrition, v. 66, p. 515S-520S, 1997.

67 SERVIN, A. L.; COCONNIER, M. H. Adhesion of probiotic strains to the intestinal mucosa and interaction with pathogens. Best practice & Research clinical Gastroenterolgy, v. 17, n. 5, p. 741-754, 2003. SHI, M. et al. Lynphocyte subpopulations in the caecum mucosa of rats after infections with Eimeria separata: early responses in naïve and immune animals to primary and challenge infections. International Journal for Parasitology, v. 31, p. 49-55, 2001. SHU, Q.; QU, F.; GILL, H. S. Probiotic treatement using Bifidobacterium lactis HN019 reduces weanling diarrhea associated with rotavirus and Escherichia coli infection in a piglet model. Journal Pediatric Gastroenterological Nutrition, v. 33, n. 2, p. 171-177, 2001. SHU, Q.; GILL, H. S. Immune protection mediate by the probiotic Lactobacillus rhamnosus HN001 (DR20a) against E. coli O157:H7 infection in mice. FEMS Immunology and Medical Microbiology, v. 34, n. 1, p. 59-64, 2002. SIUTA, A. Effectiveness of the activity of probiotic product "Cytozyme" in feeding young beef cattle. Part. 2. Evaluation of the usefulness of "Cytozyme" in feeding of young beff cattle. Acta Agraria Silvestria, v. 29, p. 135-145, 2001. SLAPETA, J. R. et al. Eimeria telekii n. sp. (Aplicomplexa: Coccidia) from Lemmiscomys striatus (Rodentia: Muridae): morphology, pathology and phylogeny. Parasitology, v. 122, p. 133-143, 2001. SOSZKA, S.; KUCZYNSKA, K. Influence of Thichomonas vaginalis on the physiological flora of the vagina. Wiadomosci Parazytologiczne, v. 23, p. 519-523, 1977. STOCKDALE, P. G. H. Mouse strain variation and effects of oocyst dose in infection of mice with Eimeria falciformes, a coccidian parasite of the large intestine. International Journal for Parasitology, v.15, n. 4, p. 447-452, 1985. SUBRAMANIAN, C. S.; OLSON, N. F. Effect of hydrogen peroxide on activity of lactic cultures in milk. Journal Dairy Science, v. 51, n. 4, p. 517-519, 1968. TAGG, J. R.; DAJANI, A. S.; WANNA MAKER, L. W. Bacteriocins of gram-positive bacteria. Bacteriological Reviews, v. 40, n. 3, p. 722-756, 1976. TANNOCK, G. W. et al. Reconstitution of the gastrointestinal microflora of Lactobacillus-free mice. Applied Environmental Microbiology, v. 53, p. 2971-2975, 1988.

68 TANNOCK, G. W. et al. Deconjugation of bile acids by lactobacilli in the mouse small bowel. Applied Environmental Microbiology, v. 60, n. 9, p. 3419-3420, 1994. TIERNEY, J. et al. In vitro inhibition of Eimeria tenella invasion by indigenous chicken Lactobacillus species. Veterinary Parasitology, v. 122, p. 171-182, 2004. TIMMERMAN, H. M. et al. Monostrain, multistrain and multispecies probiotics – comparasion of functionality and efficacy. International Journal of Food Microbiology, v. 96, p. 219-233, 2004. TOMITA, K. et al. Influence of Lactobacillus casei on rat bladder carcinogenesis: jpn. Journal Urology, v. 85, p.655-663, 1994. UMBERGER, S. H. et al. Evaluation of a Lactobacillus inoculant on feedlot lamb performance. Animal Science Research, v. 8, p. 40-45, 1989. VANBELLE, M.; TELLER, E.; FOCANT, M. Probiotics in animal nutrition: a review. Archives of Animal Nutrition, v. 40, n. 7, p. 543-67, 1990. VASSALO, M. et al. Probióticos para leitões dos 10 aos 30 kg de peso vivo. Revista Brasileira de Zootecnia, v. 1, p. 131-138, 1997. WALLACE, R. J.; NEWBOLD, C. J. Influence of Yucca shidigera extract on ruminal ammonia concentrations and ruminal microorganisms. Applied Environmental Microbiology, v. 60, n. 6, p. 1762-1767, 1992. WARUIRU, R. M. et al. The prevalence and intensity of helminth and coccidial infections in dairy cattle in central Kenya. Veterinary Research, v. 24, n. 1, p. 39-53, 2000. WATERS, W. R. Effects of Lactobacillus reuteri on Cryptosporidium parvum infection of Gnotobiotic TCR-α-deficient mice. Journal Eukaryotic Microbiology, v. 46, n. 5, 1999. WREN, B. Probiotics: fact or fiction. Large Animal Veterinary, p. 28-30, 1987. YASUI, H. Immunomodulatory function of lactic acid bacteria. Antonie Van Leeuwenhoek International Journal of General and Molecular Microbiology. v. 76, n. 1. p. 383-389, 1999.

69 YUN, C. H.; LILLEHOJ, H. S.; LILLEHOJ, E. P. Intestinal immune responses to coccidiosis. Developmental and Comparative Immunology, v. 24, p. 303-324, 2000. ZHAO, T. et al. Pathogenicity of enterohemorrhagic Escherichia coli in neonatal calves and evaluation of fecal shedding by treatment with probiotic Escherichia coli. Journal Food Protection, v. 66, n. 6, p. 924-930, 2003. ZHAO, X.; DUSZYNSKI, D. W.; LOKER, E. S. A simple method of DNA extraction for Eimeria species. Journal of Microbiological Methods, v. 44, p. 131-137, 2001. ZOETECNOCAMPO. Ciclo evolutivo da Eimeria spp. Disponível em <http://www.zoetecnocampo.com/documentos/eimeria/ciclo.jpg>. Acesso: 10.Maio.2005.

70 APÊNDICES

71 APÊNDICE A – BIOGRAFIA
CARLOS ALBERTO SANCHES PEREIRA, filho de Carlos dos Santos Pereira e
Maria Emília Sanches Pereira, nasceu em 05 de novembro de 1968 na cidade do Rio de
Janeiro.
Pai de Ivina Sanches Pereira e Ana Clara Firmino Sanches.
Em 1992 concluiu o curso de graduação em Ciências Biológicas pela Fundação
Educacional Rosemar Pimentel (FERP/UGB).
Em 1993 foi contratado pela Prefeitura Municipal de Barra Mansa, para o Cargo de
Responsável Técnico do Laboratório Municipal (SIRENE I) de Análises Clínicas.
Em 1998 foi contratado pela UGB (Centro Universitário Geraldo Di Biase) para o
cargo de professor de Microbiologia e Imunologia, onde permanece até a presente data.
Em 2000 Concluiu o curso de Pós-Graduação “Latu Sensu” em Hematologia pela
Universidade Federal do Rio de Janeiro.
Em 2001 concluiu o Mestrado em Ciência e Tecnologia de Alimentos na Universidade
Federal Rural do Rio de Janeiro, sob orientação da Profa. Dra. Rosa Helena Luchese, cuja
dissertação versou sobre Propriedades probióticas de Lactobacillus plantarum e Lactobacillus
fermentum e antagonismo frente a linhagens de Escherichia coli Enteropatogênica (EPEC) e
Enterotoxigênica (ETEC).
Em 2003 iniciou o curso de Doutorado em Biotecnologia Industrial, na Escola de
Engenharia de Lorena - USP, sob a orientação do Prof. Dr. Ismael Maciel de Mancilha.
No período de 2004 à 2006 coordenou o curso de Pós-Graduação “Latu Sensu” em
Análises Clínicas junto ao Centro Universitário Geraldo Di Biase.
Em 2005 foi contratado pelo CONMEDH (Convênios Médicos Hospitalares) em
Volta Redonda – RJ, para Gerenciar sua rede de Laboratórios Clínicos, onde permanece até a
presente data.

APÊNDICE B – Contagem de oocistos nas fezes dos animais
Valores expressos em OoPG/g de fezes Grupos A e C não foram infectados X – animal abatido dia anterior
Dias de Infecção Animais 00 01 02 03 04 05 06 07 08 09 10 11 12 13 14 15
A1 00 00 00 00 00 00 00 00 00 00 00 00 00 00 00 00 A2 00 00 00 00 00 00 00 00 00 00 X A3 00 00 00 00 X A4 00 00 00 00 00 00 00 X A5 00 00 00 00 00 00 00 00 00 00 00 00 00 X B1 00 00 00 00 00 00 40 X B2 00 00 00 00 00 00 10 1175 515 520 635 835 900 X B3 00 00 00 00 00 00 00 00 945 960 530 520 600 540 910 600 B4 00 00 00 00 X B5 00 00 00 00 00 00 00 00 1250 780 X C1 00 00 00 00 00 00 00 00 00 00 00 00 00 00 00 00 C2 00 00 00 00 X C3 00 00 00 00 00 00 00 00 00 00 X C4 00 00 00 00 00 00 00 X C5 00 00 00 00 00 00 00 00 00 00 00 00 00 X D1 00 00 00 00 00 00 05 X D2 00 00 00 00 00 00 00 10 10 50 40 05 00 00 00 00 D3 00 00 00 00 X D4 00 00 00 00 00 00 00 00 00 60 25 00 00 X D5 00 00 00 00 00 00 25 00 80 75 X E1 00 00 00 00 00 00 00 00 650 820 500 00 00 X E2 00 00 00 00 00 00 00 00 800 905 X E3 00 00 00 00 00 00 10 X E4 00 00 00 00 X E5 00 00 00 00 00 00 00 00 1430 500 180 25 00 00 00 00
72

APÊNDICE C – Resultados dos Leucogramas
Valores médios de 3 repetições LEUC – Leucometria mm3 BST – Bastões % SEG – Segmetnados % EOS – Eosinóafilos % BAS – Basófilos % LINF – Linfócitos % MONO – Monócitos %
Animais/Grupos LEUC BST SEG EOS BAS LINF MONOA1 11.700 0 13 2 0 84 1 A2 10.900 1 17 0 0 77 5 A3 10.800 0 15 2 0 79 4 A4 11.600 0 13 3 0 81 3 A5 9.500 1 13 1 1 82 2 B1 14.700 3 30 2 0 60 5 B2 15.400 4 38 2 0 55 1 B3 17.100 5 40 0 1 52 2 B4 13.000 3 16 1 0 75 5 B5 14.200 6 35 2 0 54 3 C1 8.300 0 30 2 0 64 4 C2 8.200 2 24 0 0 71 3 C3 9.300 0 27 2 1 66 4 C4 9.900 0 22 0 0 75 3 C5 7.500 1 30 1 1 65 3 D1 10.500 2 41 1 1 53 4 D2 12.900 3 47 1 0 45 3 D3 9.300 1 39 4 0 52 4 D4 10.800 3 44 2 1 47 3 D5 9.500 4 40 1 0 52 3 E1 10.200 1 40 1 0 56 2 E2 11.500 1 38 3 2 52 4 E3 11.800 2 30 4 0 61 3 E4 9.100 4 32 0 0 59 5 E5 12.800 3 45 1 1 47 3
73

APÊNDICE D – Consumo individual de ração (g) pelos animais
Dias de Experimento Animais 02 03 04 05 06 07 08 09 10 11 12 13 14 15 16 17 18 19 20 21
A1 15,10 15,89 15,99 16,45 17,17 13,25 15,93 15,47 13,85 17,10 17,10 14,17 16,16 17,03 17,20 17,20 17.90 20,99 23,48 21,70 A2 16,22 16,88 17,00 18,87 22,22 22,93 21,19 28,76 19,16 22,93 22,93 26,24 24,94 25,41 X A3 16,59 15,98 16,00 16,89 15,29 14,78 18,87 17,92 X A4 14,45 15,09 15,89 15,97 17,17 18,78 23,28 21,24 17,34 19,18 19,18 X A5 13,70 13,40 14,89 15,78 16,97 18,40 20,14 17,95 15,67 22,07 22,07 18,09 18,23 18,53 17,98 17,03 17.88 18.99 X B1 12,47 12,90 12,99 13,90 16,34 15,46 16,39 12,83 11,84 17,02 17,02 X B2 11,10 12,98 13,87 14,82 18,80 15,90 20,28 23,89 16,64 22,23 22,23 22,02 16,75 17,99 17,19 15,52 16,44 17,98 X B3 13,17 13,09 13,89 14,10 16,54 17,43 16,59 18,41 15,21 14,96 14,96 13,50 12,76 18,17 19,81 13,81 13,88 15,88 16,99 16,96 B4 13,49 13,90 14,89 15,89 16,97 14,70 20,99 22,15 X B5 10,12 12,87 11,89 13,18 16,44 17,89 18,58 18,62 17,16 18,00 18,00 18,70 16,13 19,09 X C1 11,19 12,98 12,76 13,28 13,45 14,61 17,32 17,65 13,86 19,46 19,40 17,47 16,80 18,16 17,51 16,32 16,40 18,99 24,84 23,72 C2 11,39 12,09 13,00 12,96 14,40 13,72 21,00 20,73 X C3 12,39 13,98 14,90 16,90 19,32 15,90 23,80 22,08 19,33 22,48 22,48 20,81 19,71 21,50 X C4 13,45 14,88 15,09 15,29 15,23 10,26 19,36 21,16 15,39 19,81 19,81 X C5 11,17 12,87 13,67 12,89 11,16 11,13 19,35 20,33 18,13 23,93 23,93 21,63 15,00 21,42 21,30 20,52 20,55 20,01 X D1 12,18 12,09 12,65 11,99 11,63 16,73 16,29 16,00 14,71 18,27 18,27 X D2 12,49 13,67 13,87 14,87 13,75 12,83 17,01 16,65 16,24 17,47 17,47 18,12 18,18 18,71 19,88 17,23 17,99 18,99 19,87 16,63 D3 11,90 12,90 13,83 14,17 14,55 12,54 18,90 17,50 X D4 12,09 12,87 12,90 11,90 12,87 17,88 13,54 11,88 14,95 17,18 17,18 13,96 15,55 14,52 16,29 14,83 15,00 14,9 X D5 11,09 12,09 13,87 12,78 12,84 14,59 16,32 14,65 11,37 13,85 13,55 13,29 17,01 11,71 X E1 10,23 12,76 12,67 13,98 13,92 16,82 16,36 18,28 16,04 14,66 14,66 15,09 12,04 16,67 15,38 13,67 13,77 14,08 X E2 10,32 11,09 11,99 12,80 10,11 10,79 13,37 15,06 13,76 12,83 12,83 14,30 16,24 19,74 X E3 11,98 12,90 13,10 13,89 14,30 9,46 17,64 17,72 17,61 19,26 19,26 X E4 12,98 12,87 13,78 15,97 16,92 14,65 19,32 14,65 X E5 11,09 12,19 12,45 15,98 16,08 18,91 21,55 18,22 14,96 18,13 18,13 15,56 17,22 17,54 16,43 12,96 13,00 13,88 14,64 14,83 X – animal abatido dia anterior
74

2
APÊNDICE E – GANHO DE PESO (g) PELOS ANIMAIS
Dias de Experimento Animais 01 02 03 04 05 06 07 08 09 10 11 12 13 14 15 16 17 18 19 20 21
A1 143 148 156 160 165 172 176 181 187 198 201 209 211 216 223 230 240 245 254 260 265 A2 205 211 219 224 233 239 237 241 247 263 266 267 277 282 288 X A3 166 173 181 185 192 194 200 205 211 X A4 155 164 171 184 195 199 208 215 223 228 232 240 X A5 159 166 170 172 177 178 186 194 201 209 215 219 225 232 240 247 255 257 X B1 167 173 180 189 197 200 196 194 191 196 200 205 X B2 188 184 195 200 204 209 214 219 225 228 230 238 239 246 247 250 254 258 X B3 145 151 156 159 165 168 171 179 184 189 193 189 194 196 201 207 217 219 210 211 215 B4 168 172 180 184 192 194 199 205 208 X B5 167 171 179 180 184 185 187 192 199 211 215 213 219 220 225 X C1 137 140 146 155 162 165 169 177 182 190 195 203 207 210 216 219 225 236 240 246 257 C2 146 159 167 179 184 186 192 200 205 X C3 188 194 203 209 213 216 224 230 239 250 254 260 267 270 274 X C4 176 184 191 195 199 197 203 212 217 230 236 250 X C5 180 189 197 193 197 190 203 211 216 229 237 244 249 257 265 270 280 290 X D1 152 158 161 168 172 169 171 172 199 187 201 210 X D2 154 158 167 171 175 174 175 180 186 194 199 205 209 216 219 222 226 228 238 245 250 D3 151 159 168 175 183 183 188 192 193 X D4 146 145 150 154 162 164 169 167 166 177 180 197 194 199 205 209 211 217 X D5 155 159 163 169 177 175 181 180 181 189 194 199 196 200 206 X E1 176 179 188 195 200 202 198 199 204 212 216 206 206 210 208 209 212 216 X E2 135 135 140 146 159 159 156 160 163 173 180 170 172 173 175 X E3 146 149 158 162 166 167 167 175 181 184 190 202 X E4 164 167 172 179 188 192 195 203 204 X E5 151 165 171 177 182 181 185 195 199 205 210 213 214 218 220 223 228 231 238 241 247
X – animal abatido dia anterior
75