universidade de pernambuco · ii antonio terto da silva junior evolução cromossômica e...
TRANSCRIPT

i
Universidade Federal de Pernambuco
Centro de Ciências Biológicas
Programa de Pós-Graduacão em Genética
Antonio Terto da Silva Junior
Evolução cromossômica e mapeamento de genes no cromossomo X de Drosophila sturtevanti (grupo saltans).
Recife
2014

ii
Antonio Terto da Silva Junior
Evolução cromossômica e mapeamento de genes no cromossomo X de Drosophila sturtevanti (grupo saltans).
Dissertação apresentada ao Programa de Pós-
Graduação em Genética da Universidade Federal
de Pernambuco como parte dos requisitos
exigidos para obtenção do título de Mestre em
Genética.
Orientadora: Profa. Dra. Rita de Cassia de Moura
Coorientadora: Profa. Dra. Tania Tassinari Rieger
Recife
2014

iii
Catalogação na fonte Elaine Barroso
CRB 1728
Silva Junior, Antonio Terto da Evolução cromossômica e mapeamento de genes no cromossomo X de Drosophila sturtevanti (grupo saltans)/ Recife: O Autor, 2014. 49 folhas : il., fig., tab.
Orientadora: Rita de Cássia de Moura Coorientadora: Tania Tassinari Rieger Dissertação (mestrado) – Universidade Federal de Pernambuco,
Centro de Ciências Biológicas, Genética, 2014.
Inclui bibliografia
1. Mosca- genética 2. Mapeamento cromossômico I. Moura, Rita de
Cássia de (orientadora) II. Rieger, Tania Tassinari (coorientadora) III. Título
595.774 CDD (22.ed.) UFPE/CCB- 2014- 171

iv
Antonio Terto da Silva Junior
Evolução cromossômica e mapeamento de genes no cromossomo X de Drosophila sturtevanti (grupo saltans).
Aprovado em 10/03/2014
____________________________________________
Profa. Dra. Rita de Cássia de Moura
Departamento de Biologia – UPE (Orientadora)
Banca Examinadora:
____________________________________________
Profa. Dra. Neide Santos
Departamento de Genética - UFPE
____________________________________________
Profa. Dra. Cláudia Rohde
Núcleo de Biologia - CAV - UFPE
____________________________________________
Profa. Dra. Vilma Loreto da Silva
Departamento de Genética - UFPE
Recife
2014

v
Dedico aos meus pais, pelo carinho e
incentivo desde o inicio da minha
formação.

vi
Agradecimentos
Agradeço a Dra. Tania Tassinari Rieger (minha coorientadora) e ao Dr.
José Ferreira dos Santos por me receberem no LED como aluno no ano de 2009
e participarem desde então do meu desenvolvimento acadêmico e cientifico.
Agradeço ainda pela orientação e apoio para o desenvolvimento da minha
dissertação.
Agradeço a Dra. Rita de Cassia de Moura por me acolher como
orientando de mestrado e principalmente por abrir meus horizontes e ampliar
minha visão com relação ao mundo científico.
Agradeço a Dra. Vilma Loreto da Silva e a Dra. Neide Santos por
oferecerem ajuda quanto à estrutura necessária para realização do projeto e a
Dra Ana Cristina Lauer Garcia e sua aluna Geórgia Oliveira pela ajuda na
identificação das espécies.
Agradeço a todos os docentes que contribuíram com minha formação
acadêmica durante a realização do mestrado em genética da UFPE. Agradeço
ainda aos funcionários do Departamento de Genética Romildo, Zizi, Nara e
Henrique que sempre foram solícitos.
Agradeço ao CNPq pela bolsa de Mestrado.
Agradeço aos meus companheiros de laboratório Adiles, Morse, Tyago,
Milena e Darlene que me ajudaram infinitamente, tanto no meu trabalho como na
minha vida, dando apoio, conselhos e tornando todos os dias no LED divertidos.
Além disso, agradeço a Evilis, Diego, Augusto, Luciana, Nicole, Rafaela,
Fernanda Ito, Júlio, Italo e vários outros que fizeram parte da historia do LED e
que mesmo depois de irem, sabem que são “pra sempre LED”.

vii
Agradeço a Jéssica, a mulher da minha vida, pelo apoio, carinho e ajuda
desde o inicio do meu mestrado, com o qual sem ela não teria nem conseguido
ingressar.
Agradeço a minha família e a todos que contribuíram nessa etapa da
minha vida.

vii
Resumo
Drosophila sturtevanti é a espécie de moscas com maior dispersão no grupo
saltans, ocorrendo desde a América central até o sul do Brasil. O grupo saltans é
um dos quatro maiores grupos do subgênero Sophophora e consiste de 21
espécies. O cariótipo de D. sturtevanti apresenta seis cromossomos, com o
primeiro par metacêntrico, correspondendo aos cromossomos sexuais, o segundo
par também metacêntrico, menor que o primeiro, e o terceiro par acrocêntrico.
Porém, não há na literatura fotomapa descrito para as espécies do grupo, sendo
os dados citogenéticos escassos. Este trabalho visa ampliar os estudos evolutivos
no subgênero Sophophora através da comparação dos marcadores encontrados
nos braços XL e XR de D. sturtevanti, D. prosaltans e D. willistoni e do
mapeamento, por hibridização in situ, dos genes Ubi e E71 em D. sturtevanti, com
a posterior comparação dos resultados encontrados com dados relatados para
espécies próximas. A relação dos braços XL e XR de D. sturtevanti com os de D.
willistoni e os de D. prosaltans demonstrou a presença de padrões conservados
entre essas espécies e possibilitou inferir que os braços XL e XR de D. sturtevanti
correspondem aos elementos A e D de Müller, respectivamente. A localização
dos genes Ubi e E71 no braço XR de D. sturtevanti e sua conservação de
localização também no braço XR de D. willistoni deu maior embasamento para se
afirmar que esse braço politênico corresponde ao elemento D de Müller.
Palavras chave: Subgênero Sophophora; Relações evolutivas; Elementos de
Müller; Mapeamento cromossômico.

viii
Abstract
Drosophila sturtevanti is a fruit fly species with greater dispersion in saltans group,
occurring from Central America to Southern Brazil. The saltans group is one of the
four major groups of the Sophophora subgenus and consists of 21 species. From
cytogenetic point of view, the karyotype of D. sturtevanti has six chromosomes,
including two metacentric, two acrocentric and two other smaller metacentric.
However, in the literature does not exist a photo map described for the species of
the group, being the cytogenetic data scarce. This work aims to extend
evolutionary studies in the Sophophora subgenus by comparing the makers found
in the XL and XR arms of D. sturtevanti, D. prosaltans and D. willistoni and the
mapping, by in situ hybridization, the Ubi and E71genes in D. sturtevanti with
subsequent comparison of the results with data reported for closely related
species. The relation of XL and XR arms of D. sturtevanti with D. willistoni and D.
prosaltans showed the presence of conserved patterns between these species
and allowed to infer that the XL and XR arms of D. sturtevanti correspond to A and
D Müller elements, respectively. The location of Ubi and E71 genes in XR arm of
D. sturtevanti and their location conserved also in XR arm of D. willistoni gave
better basis for claiming that this polytene arm corresponds to Müller D element.
Key words: Sophophora subgenus; evolutionary relations; Müller elements;
chromosomal mapping.

ix
Sumário
Resumo vii
Abstract viii
Lista de Figuras x
Lista de Abreviaturas xi
1. Introdução 1
2. Revisão da Literatura 3
2.1. Características gerais e importância do gênero Drosophila 3
2.2. Cromossomos politênicos 5
2.3. Comparação de elementos politênicos
2.4. A espécie Drosophila sturtevanti Duda, 1927
2.5. Genes de resposta ao estresse e desenvolvimento
2.6. Mapeamento gênico por hibridização in situ em drosofilídeos
6
9
12
14
3. Objetivos 17
4. Material e métodos 18
4.1. Linhagem utilizada, cultivo e manutenção dos estoques
4.2. Tratamento das laminas e lamínulas
4.2.1. Convencional
4.2.2. Hibridização
4.2.3. Lamínulas
4.3. Preparação citológica
4.4. Preparação da Sonda
4.4.1. Amplificação e marcação de plasmídeos
4.4.2. Hibridização in situ Permanente (PISH)
4.5. Análise Microscópica
5. Resultados
5.1. Fotomapa de D. sturtevanti
5.2. Comparação dos braços XL de D. sturtevanti com outras espécies de Drosophila
5.3. Confecção do braço XR de D. prosaltans e comparação dos braços XR de D. sturtevanti, D. willistoni e D. prosaltans
18
18
18
19
19
19
20
20
20
21
22
22
23
26
5.4. Localização dos genes Ubi e E71 28
6. Discussão 35
7. Conclusões 40
8. Referências Bibliográficas 41
9. Currículo Lattes 47

x
Lista de Figuras
Figura 1
Imagem da fêmea à esquerda (A) e do macho à direita (B) de Drosophila sturtevanti.
10
Figura 2
Fotomapa dos cromossomos politênicos de Drosophila sturtevanti, linhagem DIR. Os
centrômeros estão voltados à esquerda. Setas indicam regiões de pufes.
29
Figura 2
Continuação do Fotomapa dos cromossomos politênicos de Drosophila sturtevanti, linhagem
DIR. Os centrômeros estão voltados à esquerda. Setas indicam regiões de pufes.
30
Figura 3
Correlação entre o braço XL de D. sturtevanti (central) com os braços XL de D. prosaltans
(acima) e D. willistoni (abaixo). Mostrando em azul a correlação de pufes importantes.
31
Figura 4
Correlação entre o braço XR de D. prosaltans publicado por Cavalcanti (acima) e o braço XR
confeccionado no presente trabalho (abaixo). As linhas azuis indicam as relações entre as
seções nos dois braços politênicos. As setas do braço descrito por Cavalcanti (acima) indicam
regiões de contrição em que o cromossomo quebra facilmente.
32
Figura 5
Braço XR de D. prosaltans confeccionado no presente trabalho, com subseções.
32
Figura 6
Correlação entre o braço XR de D. sturtevanti (central) com os braços XR de D. prosaltans
(acima) e D. willistoni (abaixo). Mostrando diversos pontos de similaridade. Em azul escuro são
mostrados os pontos de correspondência mais importantes.
33
Figura 7
Braços cromossômicos XR de D. sturtevanti, mostrando a localização dos genes Ubi (A) e E71
(B) através de hibridização in situ permanente. As barras em azul indicam a correspondência
da marcação nas seções do braço XR. As setas indicam a localização do gene hibridizado.
34
Figura 8
Esquema comparativo da correspondência dos elementos de Müller em Drosophila
melanogaster, Drosophila willistoni e Drosophila sturtevanti considerando a localização de
genes de copia única e a sequência da localização desses genes no elemento D.
39

xi
Lista de Abreviaturas
BCIP – Bromo-cloro indolil fosfato
CAV – Centro Acadêmico de Vitória
Dpp – Decapentaplegic
E71 – Hormonio de Resposta ao hormônio ecdisona
hsp – heat shock protein
HSR – Heat Shock Response
LED – Laboratório de Experimentação em Drosophila
NBT – Azul de nitro tetrazólio
ORFs – Open Reading Frame
PISH – Hibridização in situ Permanente
SAP – Estreptoavidina/ Fosfatase alcalina
Ubi – Ubiquitina
UFPE – Universidade Federal de Pernambuco
UPE – Universidade de Pernambuco

1
1. Introdução
A espécie Drosophila sturtevanti é membro do grupo saltans de Drosophila,
apresentando ampla distribuição geográfica na região neotropical, sendo
encontrada desde o México, ilhas do Caribe, até o Sul do Brasil. O cariótipo de D.
sturtevanti é constituído por seis cromossomos, com o primeiro par metacêntrico,
correspondendo aos cromossomos sexuais, o segundo par também metacêntrico,
menor que o primeiro, e o terceiro par acrocêntrico. Drosophila sturtevanti é pouco
caracterizada do ponto de vista citogenético, não possuindo um fotomapa de
referência publicado para a espécie, uma vez que a maior parte dos estudos
relacionados a esta espécie são restritos a elementos de transposição.
Com a descrição de um mapa de alta resolução para D. sturtevanti foi
possível estabelecer uma comparação direta entre o genoma dessa espécie e o
de espécies próximas. Nesse sentido a escolha de D. prosaltans para
comparação, deve-se ao fato dessa espécie ser a única do grupo saltans que
possui uma maior caracterização citogenética, com um desenho mapa publicado,
o que possibilita uma comparação inicial dentro do grupo saltans. Estando na
base do subgênero Sophophora junto ao grupo saltans, o grupo willistoni
(representado por D. willistoni) se mostra de grande importância para
comparações dentro deste subgênero. Diferente de D. sturtevanti, D. willistoni é
uma espécie amplamente conhecida e estudada, tanto do ponto de vista
citogenético como evolutivo.
A comparação entre os braços do cromossomo X de D. sturtevanti (grupo
saltans), D. prosaltans (grupo saltans) e D. willistoni (grupo willistoni) possibilita
ampliar o entendimento sobre as relações evolutivas existentes dentro do
subgênero Sophophora. Para tal, utilizamos como marcadores o padrão de

2
bandas, interbandas e pufes e a localização dos genes Ubi e E71 por hibridização
in situ permanente.

3
2. Revisão da Literatura
2.1 . Características gerais e importância do gênero Drosophila
A ordem Diptera é composta por insetos que possuem um par de asas,
representados por moscas e mosquitos, que são grupos numerosos e
pertencentes à classe Insecta. Dentro desta ordem destaca-se a família
Drosophilidae constituída por cerca de 3952 espécies distribuídas em 73 gêneros
(Gottschalk, 2008), tendo ampla importância, pois seus representantes estão
envolvidos em aspectos agronômicos ou de saúde humana (Grimaldi e Engel,
2005).
A família Drosophilidae caracteriza-se por possuir dois pares de asas,
sendo o primeiro par membranoso e funcional e o segundo, chamado de halteres,
modificado como órgão de equilíbrio. Possui aparelho bucal do tipo sugador, em
forma de probóscide, apresenta tórax segmentado em três regiões, sendo o
protórax e o metatórax pequenos e fusionados ao mesotórax. Apresenta pernas
compostas por cinco segmentos tarsais que possuem grande importância
sistemática e nas fêmeas o abdome é originalmente segmentado, enquanto que
os machos apresentam os últimos segmentos fusionados, o que denota o
dimorfismo sexual das espécies dessa família. Adicionalmente, é observada
metamorfose completa com larvas ápodas e vermiformes, cabeça reduzida e
retrátil com pupas imóveis (Buzzi, 2002).
A maioria dos drosofilídeos é caracterizada por colonizar frutos caídos e se
alimentarem dos fungos que se desenvolvem nesses frutos. No entanto, algumas
espécies colonizam o fruto ainda na árvore, sendo consideradas insetos pragas e
causando danos a diversos cultivares (Santos et al., 2003; Fernandes e Araújo,
2011). A família tem distribuição em regiões temperadas e tropicais, habitando os

4
mais diversos ecossistemas, de forma que algumas espécies são consideradas
cosmopolitas e outras estão restritas a determinadas partes do planeta. Na
América são encontrados diversos gêneros de drosofilideos devido à complexa
fauna associada a alterações climáticas que promovem a especiação. Nesse
sentido destacam-se os grupos willistoni e saltans com ampla distribuição na
região neotropical (Gao et al., 2011).
Dentre as espécies de moscas a mais estudada tem sido Drosophila
melanogaster, que embora sem importância agronômica é um valioso modelo em
ensino e pesquisa nas áreas de genética, desenvolvimento e evolução. Portanto,
devido a sua frequente utilização como organismo modelo para pesquisa desde o
início do Século XX, a espécie tornou-se a melhor caracterizada e de maior
interesse nas áreas citadas acima (Da Lage et al., 2007; Van der Linde et al.,
2010). O amplo conhecimento gerado nestas áreas permitiu a crescente aplicação
de D. melanogaster também como modelo experimental para o estudo de
diversas condições clínicas humanas. Desta forma, análises comparativas entre
diferentes genomas de espécies do gênero Drosophila apontam de forma
definitiva D. melanogaster como modelo de estudos do sistema imune inato e seu
combate à infecções fúngicas (Hoffmann, 2007; Huang et al., 2010), bacterianas
(Valanne et al., 2011; Buchon et al., 2013) ou virais (Kawai e Akina, 2006; Sabin
et al., 2010), de desordens humanas relacionadas à perda de homeostase,
incluindo o metabolismo de açúcares e lipídios (Leopold e Perrimon, 2007;
Pospisilik et al., 2010) e dos mecanismos de funcionamento cerebral (Fortini e
Bonini, 2000; Vosshall, 2007; Chiang et al., 2011).
Embora seja o grupo de organismos eucariontes superior com o maior
número de genomas sequenciados, o gênero Drosophila ainda desperta diversas

5
dúvidas e controvérsias, tanto sobre a relação interna entre os subgêneros
Sophophora e Drosophila, quanto sobre as relações externas com outros grupos,
como o gênero Zaprionus. Neste sentido, têm-se acumulado evidências de que o
subgênero Drosophila seja um grupo parafilético, necessitando de ampla revisão
taxonômica para refletir a real relação filogenética entre os grupos de espécies
(Robe et al., 2005; Da Lage et al., 2007; Katoh et al., 2007; Yang et al., 2012). A
ampliação da análise genômica e evolutiva destas espécies, bem como a inclusão
de outras espécies, trará novas hipóteses fundamentais e novas informações que
contribuirão para o esclarecimento das relações filogenéticas no subgênero
Sophophora.
2.2 . Cromossomos politênicos
Os cromossomos politênicos são cromossomos gigantes formados a partir
de uma série de ciclos endomitóticos que ocorrem durante a interfase de células
específicas. Esses cromossomos são distribuídos em ordens distintas de insetos
(Diptera, Collembola, Lepidoptera e Orthoptera) e mais raramente em
protozoários, tecidos de mamíferos e células de angiospermas (Zhimulev, 1996;
Zhimulev e Koryakov, 2009).
Cromossomos politênicos se formam a partir do pareamento dos filamentos
homólogos de DNA, cromômero a cromômero, e duplicação do seu material a
cada ciclo endomitótico. As várias cromátides idênticas alinhadas lado a lado,
formam um bandeamento natural, produzido pelo pareamento dos cromômeros
paralelos das cromátides. Este bandeamento representa marcadores estruturais
que possibilitam mapeá-los e estabelecer blocos sintênicos com espécies
próximas. O estudo destes marcadores passou a ser mais informativo a partir do
sequenciamento do genoma de D. melanogaster (Adams et al., 2000),

6
contribuindo para a comparação e o estabelecimento das relações evolutivas dos
elementos cromossômicos entre diversas espécies (Schaeffer et al., 2008).
As bandas e interbandas estão diretamente relacionadas com a
condensação da cromatina e a espessura do politênico, a depender do número de
endomitoses que o núcleo sofreu, de forma que no núcleo politênico de D.
melanogaster são encontrados 1024 filamentos formando o cromossomo
politênico. As regiões de alargamento do cromossomo politênico são
denominadas de pufes e estão intimamente ligados a atividade gênica. Os pufes
são regiões descondensadas em sítios específicos em que ocorre a transcrição
de genes localizados naquela região. O padrão de pufes difere em tecidos
distintos do mesmo indivíduo e em diferentes estágios do desenvolvimento, mas é
constante em um tecido no mesmo estágio de desenvolvimento. Muitos pufes
podem ser induzidos em laboratório por diversos agentes, como o hormônio
ecdisona, modificações de temperatura e etanol (Zhimulev, 1996; Mitrofanov,
2007). Inicialmente acreditava-se que cada região de pufe representava a
atividade de um único gene, atualmente sabe-se que ocorrem vários genes que
são transcritos juntos (Kolesnikova et al., 2011).
2.3 . Comparação de elementos politênicos
A nomenclatura dos cromossomos politênicos é baseada em padrões
distintos, algumas vezes sendo usados números romanos, e em outras, arábicos.
Muitos autores dividem todo o complemento cromossômico em 100 seções, mas
isso não ocorre sempre. Entretanto, todos usam as letras L (left, esquerdo) e R
(right, direito) para identificar os dois braços do mesmo cromossomo. Em 1940,
Müller propôs um método para comparar os cromossomos politênicos. Na sua

7
proposta ele utilizou os braços cromossômicos de D. melanogaster como padrão
para comparação de elementos politênicos entre as espécies e determinou uma
nomenclatura de A-F para esses braços que passaram a ser conhecidos como
elementos de Müller (Müller, 1940). Nesse sistema o X de D. melanogaster
representa o elemento A; IIL, B; IIR, C; IIIL, D; IIIR, E e o IV corresponde ao
elemento F. Assim, Müller propôs que estes elementos estariam presentes em
todas as espécies de drosofilídeos, podendo ocorrer reorganizações (inversões,
translocações, fusões) durante a diversificação das espécies. Com a criação e
padronização do método, para determinar as relações entre os cromossomos
politênicos, foi possível comparar mapas citogenéticos, utilizando a localização de
marcadores físicos e genéticos localizados nos braços politênicos, demonstrando
assim blocos de sintênia que possibilitam inferir sobre a reorganização dos
elementos de Müller. Desta forma, foi reunido desde a década de 40 um grande
conjunto de informações sobre a evolução cromossômica na família Drosophilidae
(Campos et al., 2007; Clark et al., 2007; Schaeffer et al., 2008).
As primeiras análises genéticas no gênero Drosophila foram realizadas a
partir do desenvolvimento dos mapas de cromossomos politênicos de D.
melanogaster (Bridges, 1935). Este autor propôs uma nomenclatura alfanumérica
para a divisão de cada braço em 20 unidades (seções) e para cada uma delas
seis subunidades de A-F (sub-seções), no intuito de atribuir uma designação para
cada uma das bandas presentes. Esta codificação do mapa cromossômico
permitiu a identificação de alterações cromossômicas, pontos de quebra e a
localização de genes associados. Estes mapas, inicialmente produzidos como
desenhos realizados em câmara clara, tiveram a sua resolução ampliada pela

8
produção de mapas fotográficos de mosaico por Lefevre (1976) e aperfeiçoados
por mapas de microscopia eletrônica por Sorsa (1988a, b, c).
O subgênero Sophophora apresenta uma variedade de rearranjos que
alteraram a organização dos elementos de Müller (Schaeffer et al., 2008). Apesar
da conservação dos marcadores encontrados nos braços politênicos homólogos,
os cariótipos das espécies desse subgênero têm mudado através de eventos de
fusão de braços cromossômicos seguidos por eventos de rearranjos do tipo
inversão. Em D. ananassae, por exemplo, o cromossomo X (elemento A)
geralmente acrocêntrico foi modificado em um cromossomo metacêntrico a partir
de uma inversão pericêntrica. Também em D. ananassae o elemento F, que
geralmente é um cromossomo pontual, se apresenta como um metacêntrico de
tamanho equivalente a seu elemento A (Schaeffer et al., 2008).
O cromossomo X de D. willistoni (grupo willistoni), metacêntrico, é
resultado de uma fusão dos elementos A e D de Müller. Essa correspondência
também pode ser vista em D. pseudoobscura e D. persimillis, pertencentes ao
grupo obscura (Schaeffer et al., 2008). É provável que a distribuição dos
elementos politênicos em D. willistoni (AD, BC, EF) e D. melanogaster (A, BC,
DE, F) sejam derivadas da organização que é encontrada em D. pseudoobscura
(AD, B, C, E, F) de forma que teria ocorrido a união dos elementos B e C para dar
origem a um arranjo hipotético AD, BC, E, F que com a união dos elementos E e
F deu origem ao arranjo de D. willistoni, enquanto uma translocação entre AD e E
deu origem a D. melanogaster (Sturtevant e Novitski, 1941). No entanto, a
ausência da fusão entre os elementos A e D em D. subobscura, uma espécie
próxima a D. pseudoobscura e D. persimilis, sugere que as fusões nos grupos
obscura e willistoni não são homólogas (Schaeffer et al., 2008). O cromossomo

9
pontual de D. melanogaster que representa o elemento F de Müller está
intimamente ligado ao cromossomo 3 de D. willistoni de forma que o elemento E e
F sofreram uma fusão (Papaceit e Juan, 1998).
Além de estabelecer relações dentro do gênero Drosophila, as inferências
sobre os elementos de Müller possibilitaram também estabelecer relações entre
os gêneros Drosophila e Zaprionus. Com a utilização do mapa produzido por
Campos et al. (2007) e a localização de seis genes cópia única foi possível
estabelecer a relação dos elementos cromossômicos entre D. melanogaster e Z.
indianus. Desta maneira, os elementos de Müller A, B, C, D, E e F correspondem
aos cromossomos politênicos X, II, IV, III, V e VI de Z. indianus, respectivamente
(Campos et al., 2007). Portanto, embora morfologicamente distinto de Drosophila,
o gênero Zaprionus também conserva a homologia de elementos cromossômicos
proposta por Müller (1940).
2.4 . A espécie Drosophila sturtevanti Duda, 1927
Drosophila sturtevanti pertence ao subgrupo sturtevanti do grupo saltans de
Drosophila. Esta espécie foi descrita por Sturtevant (1942) como membro do
subgênero Sophophora. O subgrupo sturtevanti foi estabelecido por Magalhães
(1962), utilizando caracteres morfológicos, e inclui as espécies D. rectangularis,
D. milleri e D. sturtevanti. As espécies D. magalhaesi e D. dacunhai, foram
incluídas posteriormente neste subgrupo (Mourão e Bicudo, 1967; Bicudo, 1979).
Estas cinco espécies são morfologicamente similares, o que torna o grupo
potencialmente informativo para estudos de especiação.
No que diz respeito à distribuição geográfica, D. sturtevanti apresenta
grande dispersão, sendo encontrada desde o México e ilhas do Caribe até o Sul

10
do Brasil. Esta ampla distribuição torna D. sturtevanti importante para estudos
com a finalidade de esclarecer as relações existentes entre o grupo saltans e
outros grupos de Drosofilídeos.
Os estudos citogenéticos para esta espécie são escassos, sendo a maioria
dos trabalhos relacionados a estudos de evolução mediados por elementos P de
transposição (De Setta et al., 2007). Entre os trabalhos, pode-se citar Dobzhansky
e Pavan (1943) que descreveram o complemento mitótico de D. sturtevanti como
2n=6, com o primeiro par metacêntrico, correspondendo aos cromossomos
sexuais, o segundo par também metacêntrico, um pouco menor que o primeiro
par, e o terceiro par acrocêntrico. No mesmo trabalho os autores apresentam um
desenho parcial (apenas pontas e bases) dos cromossomos politênicos de D.
prosaltans (outro representante do grupo saltans), onde sugerem que D.
sturtevanti apresenta pontas e bases similares, facilitando o reconhecimento dos
elementos. Contudo, segundo Dobzhansky e Pavan (1943), D. prosaltans
apresenta uma translocação entre o cromossomo sexual (metacêntrico maior) e o
Figura 1 - Imagem da fêmea à esquerda (A) e do macho à direita (B) de D. sturtevanti.
B
♂
A
♀

11
autossomo acrocêntrico, de forma que um dos metacêntricos dessa espécie não
corresponde ao encontrado em D. sturtevanti. Além disso, estes autores afirmam
ainda que o complemento cromossômico de D. sturtevanti é ancestral em relação
ao encontrado em D. prosaltans, ou seja, a distribuição dos elementos
cromossômicos dessa espécie teria surgido a partir de um ancestral com a
distribuição semelhante à encontrada em D. sturtevanti.
Posteriormente, Cavalcanti (1948) publicou um desenho mapa de
referência para os cromossomos politênicos de D. prosaltans. O desenho mapa
da espécie, mesmo não oferecendo total resolução dos seus cromossomos,
possibilitou uma comparação do padrão de bandas, interbandas e pufes dessa
espécie, permitindo relacionar este padrão com o de espécies próximas. Desta
forma, D. prosaltans tornou-se uma espécie de interesse para comparações
dentro do grupo saltans, uma vez que é a única espécie do grupo que possui uma
maior caracterização citogenética (Cavalcanti, 1948). D. prosaltans é bastante
dispersa, apresentando distribuição geográfica que vai desde o norte do México,
passando pela América central, sul do Brasil e Paraguai. A espécie possui
número cromossômico 2n=6, com dois pares de cromossomos metacêntricos e
um par acrocêntrico assim como D. sturtevanti. Em contrapartida, assim como em
D. sturtevanti, a maior parte dos estudos relacionados a D. prosaltans estão
restritos a estudos de evolução mediados por elementos de transposição (Castro
e Carareto, 2004).
Quando os cromossomos mitóticos de D. prosaltans e D. sturtevanti são
comparados se verifica que um dos metacêntricos dessa espécie é claramente
mais longo que o outro e o acrocêntrico é muito menor do que qualquer um dos
metacêntricos (Dobzhansky e Pavan 1943). No entanto, não há na literatura um

12
mapa de alta resolução dos cromossomos politênicos de D. sturtevanti ou D.
prosaltans, assim como nenhum dentro do grupo saltans, disponível para uma
melhor caracterização citogenética e genômica.
2.5 . Genes de resposta ao estresse e desenvolvimento
As proteínas de resposta ao estresse conhecidas como hsps (heat shock
proteins) foram descritas inicialmente como proteínas induzidas pelo aumento
brusco de temperatura, no entanto, respondem também à presença de metais
pesados e agentes como arsênicos e etanol (Zhimulev, 1996). A resposta ao
choque térmico e os genes Hsps foram descobertos em Drosophila pelo padrão
de ativação de pufes nos cromossomos politênicos em resposta ao calor e
estresse oxidativo (Ritossa, 1962; 1996). O estresse sofrido pela célula causa
uma resposta chamada de Heat Shock Response (HSR) ou resposta ao choque
térmico que resulta em um aumento extremamente rápido na expressão de hsps.
Estas proteínas estão agrupadas em famílias definidas pelo seu peso
molecular denominadas HSP100, HSP90, HSP70, HSP60, HSP40 e HSP20, que
inclui a Ubiquitina, todas bastante conservadas e encontradas desde procariotos
até o homem (Jolly e Morimoto 2000). Muitas destas proteínas têm função de
chaperonas, uma classe de proteínas caracterizadas pela capacidade de modular
a estrutura e dobramento de outras proteínas, processo muito importante, pois
converte cadeias lineares de polipeptídeos em estruturas tridimensionais que
possibilitam que as proteínas exerçam todas as suas atividades vitais (Kim et al.,
2007). Essas proteínas são abundantemente expressas em múltiplos
compartimentos da célula e compreendem uma significativa fração da maquinaria
celular (Bukau et al., 2006; Ron e Walter 2007).

13
A Ubiquitina pertence à família das menores proteínas Hsps, sendo
constituída por 76 aminoácidos e encontrada em todos os animais, leveduras,
bactérias e plantas superiores, sendo altamente conservada (Jolly e Morimoto
2000). Atua como marcador de proteínas, para degradá-las, e está envolvida em
processos de reparo de DNA e na sinalização para endocitose (Mukhopadhyay e
Riezman, 2007). Em D. melanogaster o gene da Ubiquitina está localizado na
seção 63F do braço 3L (Izquierdo, 1994) e caracteriza-se por não ser diretamente
estimulado pelo aumento da temperatura, mas pelos pulsos do hormônio
ecdisona. Em D. willistoni a ubiquitina está localizada na seção 27A do braço XR
(Rieger, 1999; Schaeffer et al., 2008).
O hormônio ecdisona aciona e coordena muitas das transições de
desenvolvimento do ciclo de vida da Drosophila. Pulsos de ecdisona ocorrem
durante toda a fase embrionária, na fase larval, pré-pupa, pupa e adulto,
regulando numerosos processos fisiológicos, incluindo mudanças celulares,
diferenciação e morte (Enright et al., 2004). Assim como os demais hormônios
esteróides, a ecdisona penetra nas células e liga-se com alta afinidade a
receptores que são fatores de transcrição. Estes receptores se ligam a elementos
de resposta e estimulam a ativação de genes específicos que levam ao
desenvolvimento adequado do organismo (Beckstead et al., 2001). Esta cascata
de ativação gênica favorece o aparecimento de um padrão de pufes
característicos ativos. Dentre estes genes de resposta ao hormônio ecdisona esta
o gene E71.
Em D. melanogaster o gene E71 esta localizado na seção 71E do braço 3L
(Restifo e Gregory, 1986), em D. willistoni está localizado na seção 29A do braço
XR (Rieger, 1999; Schaeffer et al., 2008). Com a localização de genes

14
conservados evolutivamente, como é o caso da Ubiquitina e o E71, é possível
inferir sobre como esses marcadores se comportaram durante a evolução.
2.6 . Mapeamento gênico por hibridização in situ em drosofilídeos
A técnica de hibridização in situ tem sido aplicada em cromossomos de
diversos organismos, principalmente para localização de sequências repetitivas.
No entanto, combinada aos cromossomos politênicos de dípteros ela é
especialmente informativa, tendo sido utilizada em diversos gêneros, tais como
Bactrocera (Drosopoulou et al., 2012), Aedes (Timoshevskiy et al., 2013),
Anopheles (Hall et al., 2013), Zaprionus (Campos et al., 2007), entre outros,
também para localização de genes cópia única. A metodologia de hibridização in
situ tem-se mostrado extremamente útil para estudos evolutivos em Drosophila,
pois, através da comparação da localização dos genes em D. melanogaster e em
outras espécies do gênero é possível identificar blocos de sintenia gênica e de
rearranjos cromossômicos entre as espécies, como nos grupos willistoni, obscura
e repleta (Schaeffer et al., 2008, Rohde e Valente, 2012). O posicionamento dos
fragmentos do arcabouço genômico das 12 espécies de Drosophila foi bastante
facilitado pelos dados gerados através de hibridização in situ disponíveis na
literatura (Bhutkar et al., 2008).
O grupo willistoni inclui vários níveis de divergência evolutiva, incluindo
espécies em status nascendi, subespécies e espécies crípticas (Garcia et al.,
2006). Devido à sua importância como modelo evolutivo, D. willistoni foi incluída
como única espécie Neotropical no projeto que resultou no sequenciamento
completo do genoma de 11 espécies de Drosophila (Clark et al., 2007), além de
D. melanogaster (Adams et al., 2000). A análise de 31 fragmentos do arcabouço

15
cromossômico correspondentes a 65% do genoma de D. willistoni (total 235,5 Mb)
revelou-a como a mais diferente entre as 12 espécies, com o maior genoma e as
maiores ORFs, sendo também a única a não possuir genes para selenoproteínas
(Clark et al., 2007; Bhutkar et al., 2008). Com o objetivo de ampliar o estudo
evolutivo entre as espécies estudadas pelo Consórcio Genoma de Drosophila,
foram publicados novos fotomapas para D. willistoni, onde, com a localização de
nove genes estruturais, foi apontado o posicionamento dos fragmentos do
arcabouço cromossômico de D. melanogaster nos cromossomos de D. willistoni
(Schaeffer et al., 2008).
Dados relatados recentemente identificaram a localização dos genes
Hsp70 e Hsp83 em D. sturtevanti (Monte, 2013), sendo verificada uma
conservação na localização desses genes em relação a D. willistoni. Em ambas
as espécies os genes Hsp70 e Hsp83 estão localizados respectivamente nos
braços III e XR, correspondendo aos elementos E e D de Muller, fortalecendo as
relações existentes entre os grupos saltans e willistoni.
O gene Period foi localizado em D. willistoni apresentando pelo menos dois
sítios principais, localizados nos braços XL e III (Monte, 2009). Pela conservação
dos elementos politênicos, estes genes devem apresentar a mesma localização
em D. sturtevanti. Os genes E71 e Ubi foram localizados no braço XR de D.
willistoni (Rieger, 1999; Schaeffer et al., 2008) que corresponde ao elemento D de
Müller. Por inferência, os genes E71 e Ubi estariam localizados no braço XR de D.
sturtevanti de modo que a localização desses genes preencherá as lacunas
existentes na correlação dos elementos cromossômicos.
Os dados relatados sobre localização gênica em drosofilídeos revelam a
importância e eficácia na identificação de blocos sintênicos entre espécies

16
próximas e possibilita uma maior correlação entre Drosophila willistoni e D.
sturtevanti, ampliando assim as informações sobre as relações evolutivas do
subgênero Sophophora (Schaeffer et al., 2008).

17
3. Objetivos
3.1. Objetivo Geral
Comparar os braços XL e XR e localizar genes nos cromossomos
politênicos de Drosophila sturtevanti, através de hibridização in situ permanente
no intuito de contribuir para o entendimento da evolução cromossômica no
subgênero Sophophora.
3.2. Objetivos Específicos
1. Aprimorar o fotomapa de referência para os cromossomos politênicos de D.
sturtevanti;
2. Confeccionar o fotomapa do braço XR de D. prosaltans;
3. Comparar os braços XL e XR de D. sturtevanti com os de D. prosaltans e D.
willistoni;
4. Localizar por hibridização in situ permanente os genes Ubi (gene de resposta
ao estresse) e E71 (gene de resposta ao hormônio ecdisona);
5. Comparar o posicionamento dos genes Ubi e E71 em D. sturtevanti, D.
willistoni e D. melanogaster, gerando novos dados sobre as reorganizações
cromossômicas sofridas pelas espécies no subgênero Sophophora.

18
4. Material e Métodos
4.1. Linhagem utilizada, cultivo e manutenção dos estoques
As linhagens DIR e DIR 2 (Dois Irmãos Recife) foram coletadas no campus
da UFRPE por nossa equipe em 2009 e 2013, respectivamente e estabelecidas
no Laboratório de Experimentação em Drosophila (LED). A linhagem DIR é de D.
sturtevanti, utilizada para construção do fotomapa e localização dos genes. A
linhagem DIR 2 é de D. prosaltans utilizada para construção do fotomapa do
braço XR. As linhagens foram mantidas em recipientes de 60 mL tampados com
rolhas de espuma. Em cada um destes foi colocado o meio de cultura, onde
ocorreu a postura dos ovos e o posterior desenvolvimento das larvas utilizadas
nas preparações citológicas. As larvas foram mantidas sob luz e temperatura
constantes (22-24 °C) e alimentadas com fermento biológico todos os dias a fim
de se obter um desenvolvimento satisfatório das larvas para posterior obtenção
dos cromossomos politênicos.
O preparo do meio de cultura foi realizado com farinha de milho (150 g),
aveia (50 g) e centeio (25 g), fermento biológico (10 g), mel (30 g), banana (100 g
diluídas em 100 ml de água), antifúngico (Metil parabeno, 1,5 g em 30 ml de
etanol) e os ácidos fosfórico e propiônico (6 ml cada diluídos em 12 ml de água
destilada). O meio foi preparado no micro-ondas durante 15 min sendo
homogeneizado a cada 1 min.
4.2 . Tratamento das lâminas e lamínulas
4.2.1. Convencional
As lâminas foram lavadas com detergente e deixadas para secar por 24 h
protegidas de poeira. Após a secagem foram imersas em uma solução de gelatina

19
comestível dissolvida em água destilada na proporção 7,5 g/L. Após secarem
foram armazenadas em caixas a 4°C.
4.2.2. Hibridização
Após a lavagem inicial as lâminas foram imersas em álcool 70% a
temperatura ambiente por 1h, depois em solução SSC/Denhardt’s a 65° C por 2 h
e 30 min e fixadas em etanol-acético (3:1) por 30 min. Depois de secas foram
armazenadas a 4º C.
4.2.3. Lamínulas
As lavagens das lamínulas foram feitas, inicialmente, com detergente
comercial, etanol e ácido clorídrico. Após a secagem foram imersas em solução
de silicone/clorofórmio na proporção de 1:50.
4.3. Preparação citológica
Larvas de 3º estágio são ideais para a preparação de cromossomos
politênicos, pois eles se apresentam de forma mais desenvolvida. Com o auxílio
de pinças segurou-se os lados anterior e posterior do corpo da larva (dentículos
do aparelho bucal e seção posterior) e puxou-se até retirada da porção anterior
com as glândulas salivares. A glândula foi colocada na lâmina e adicionou-se o
fixador (3 ác. propiônico:2 água destilada:1 ác. lático). Após 3 min o excesso de
fixador foi retirado com papel filtro e para preparação de lâminas pela coloração
convencional foi adicionado o corante orceína-lacto-acética (2%) por 3 min. O
material foi esmagado e espalhado sob a lamínula. Retirou-se o excesso de
corante e para evitar o ressecamento do material as bordas da lamínula foram
seladas com esmalte.
Na preparação das lâminas para hibridização foi seguida a mesma
metodologia das preparações coradas. Entretanto, foi adicionada uma gota de

20
ácido propiônico a 45% e o fixador (3 ác. propiônico:2 água destilada:1 ác. lático)
por 3 min, sem a adição do corante. Após a preparação as lâminas ficaram na
horizontal por no mínimo 18 h na geladeira (4° C). Após este período as lamínulas
foram retiradas com nitrogênio líquido e as lâminas passaram por banhos de
estringência, onde foram colocadas por 2 h em álcool 70%, passaram por uma
solução de SSC 2% em banho-maria (65° C) por 30 min, um banho em álcool
70% (65° C) por 5 min, um banho em álcool 95% (65° C) por 5 min e outros dois
banhos em álcool 70% e 95% de 10 min cada, conforme descrito em Campos et
al. (2007).
4.4. Preparação da Sonda
4.4.1. Amplificação e marcação dos plasmídeos
Os fragmentos dos genes Ubi e E71 foram clonados do genoma de
Drosophila melanogaster. Os fragmentos dos genes foram inseridos em
plasmídeos e os clones utilizados foram amplificados em Escherichia coli (DH5
alfa) no meio LB com o antibiótico ampicilina (100 µg/mL) (Ausubel et al., 2002).
Para a utilização como sondas, os plasmídeos completos foram marcados com
biotina (dUTP), utilizando o sistema BioNick DNA (Invitrogen).
4.4.2. Hibridização in situ permanente (PISH)
A hibridização in situ permanente foi realizada em tampão contendo 50%
de formamida a 37° C por 40 h, utilizando 500 ng de DNA marcado para cada
lâmina. A revelação foi realizada com estreptoavidina/fosfatase alcalina (SAP),
azul de nitro tetrazólio (NBT) e bromo-cloro indolil fosfato (BCIP), em condições
de estringência de 50% (2X SSC à 37o C) (Silva Valenzuela et al., 2006). As
lâminas foram contra coradas com orceína-lacto-acética (2%) e montadas em
Entellan (Merck) ®.

21
A técnica de PISH exige uma frequência de núcleos marcados superior a
30% para se considerar a marca consistente.
4.5 . Análise Microscópica
O material foi analisado em microscópio sob contraste de fase para
determinar o local e o número de marcações nos cromossomos politênicos das
lâminas hibridizadas. Os cromossomos marcados por hibridização in situ foram
fotografados utilizando o fotomicroscópio (Leica) na objetiva de 100X sob
contraste de fase no Departamento de Genética da UFPE.

22
5. Resultados
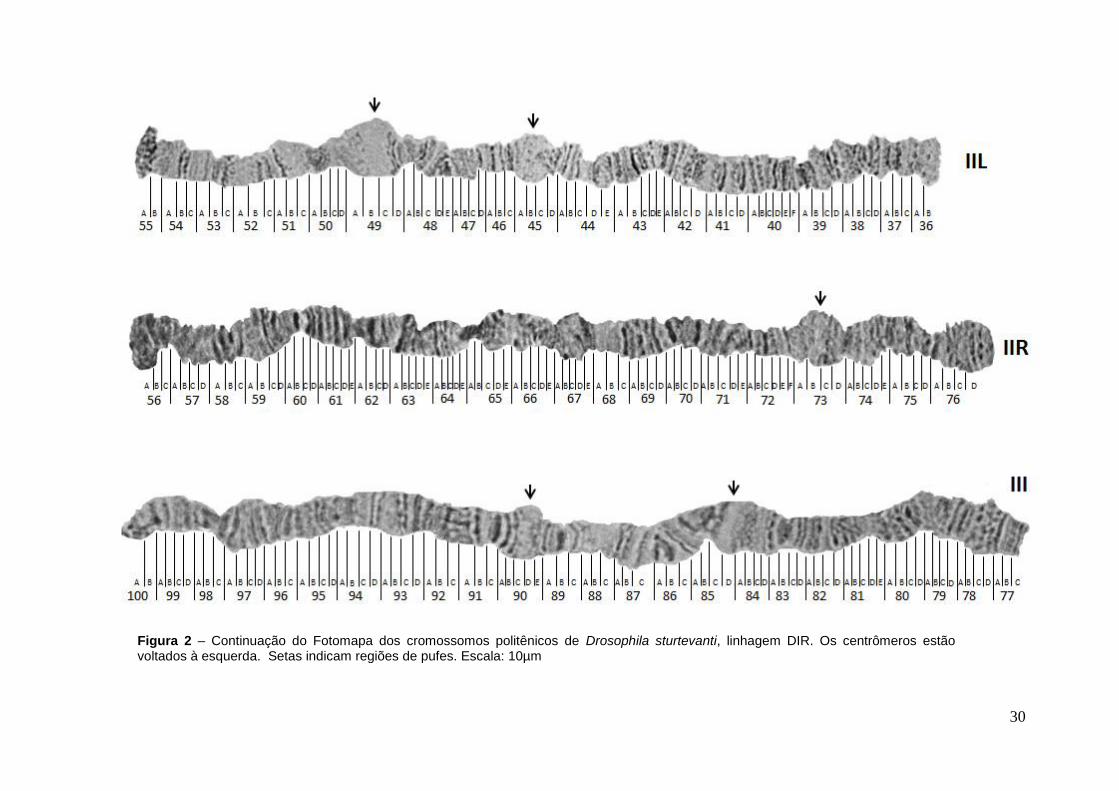
5.1. Fotomapa de D. sturtevanti
A confecção do fotomapa de D. sturtevanti foi realizada por Monte (2013) e
a inserção de subseções foram feitas neste trabalho. No fotomapa de referência
de D. sturtevanti os cromossomos politênicos da espécie foram divididos em 100
seções, tendo cada braço de 15 a 24 seções e de 3 a 6 subseções em cada
seção. Todos os braços foram orientados com as regiões centroméricas para o
lado esquerdo e as regiões teloméricas para o lado direito (Fig.2). A identificação
dos braços foi baseada na tendência a permanecerem conectados e na
comparação e reconhecimento dos padrões de bandas, interbandas e pufes
homólogos aos de D. prosaltans e outras espécies.
O par I de D. sturtevanti, correspondente ao cromossomo X, apresenta
morfologia submetacêntrica, pois o braço XL é menor que o XR, sendo também o
menor de todos os braços politênicos. O braço XL foi subdividido em 15 seções,
da seção 1 a 15, apresentando como principais marcas de reconhecimento a
ponta aberta e constrições evidentes na subseção C da seção 10, subseção A da
seção 11 e subseção A da seção 13, bem como as regiões de pufes bem
evidentes na seção 3, subseções B a E da seção 6, seção 9, subseções A a C da
seção 10 e seção 13. Por sua vez, o braço XR foi subdividido em 20 seções, da
seção 16 a 35 apresentando marcas bem caracterizadas e de fácil identificação.
Apresentou ainda ponta em forma de leque seguida de três bandas de forte
marcação (subseções C e D da seção 35), além de grandes pufes nas seções 18
e 27 e constrições na subseção E da seção 17 e subseção B da seção 21. É
importante destacar uma sequência na seção 25 que pode ser visualizada como
bandas sobressalentes na posição horizontal.

23
O cromossomo II apresenta-se metacêntrico, embora o braço IIL seja
levemente menor que o braço IIR. O braço IIL foi subdividido em 20 seções, de 36
a 55 cujas principais marcas de identificação são o grande pufe na seção 49 e
outro menor na seção 45. Para este braço foram observadas constrições na
subseção A da seção 39, subseção B da seção 44 e subseção A da seção 50. O
braço IIR foi dividido em 21 seções, da 56 a 76. Apresenta ponta em forma de
bulbo com duas bandas evidentes pontilhadas (subseção D da seção 76),
finalizando em uma constrição após uma banda bem evidente. Entretanto, a
constrição mais evidente está na subseção A da seção 65. O maior pufe do braço
está localizado na seção 73.
O cromossomo III representa o único par acrocêntrico, sendo o maior dos
braços politênicos da espécie e foi subdividido em 24 seções, da 77 a 100 e
mostra ponta fusiforme (seções 77 e 78). Os maiores pufes deste cromossomo
são visualizados nas subseções C e D da seção 85 e seção 90. A banda da
subseção A da seção 83 tem aspecto pontilhado. A única constrição evidente
ocorre na subseção B da seção 98.
5.2. Comparação dos braços XL de D. sturtevanti com D. prosaltans e D.
willistoni
Após a confecção do mapa de referência para D. sturtevanti, e posterior
inserção de subseções, foi possível comparar o braço XL desta espécie com os
braços XL das espécies D. willistoni descrito em fotomapa por Schaeffer et al.,
(2008) e D. prosaltans descrito em desenho mapa por Cavalcanti (1948),
observando suas relações de padrão de bandas, interbandas e pufes. Estes
marcadores refletem o padrão de ativação gênica no 3° estágio larval e

24
possibilitam uma relação direta entre os elementos cromossômicos de espécies
que possuem mapas politênicos descritos (Fig.3).
Baseado nos padrões de bandas, interbandas e pufes presentes nos
cromossomos politênicos das espécies analisadas, os braços politênicos
apresentaram um padrão conservado (Fig.3). A ponta do braço XL de D.
prosaltans se apresenta fechada, enquanto que D. sturtevanti e D. willistoni
apresentam ponta aberta. Na comparação do padrão de bandas, interbandas e
pufes entre D. sturtevanti e D. prosaltans podem ser visualizados blocos de
sintênia que se repetem por todo o cromossomo, com especial destaque para
regiões de constrição que ocorrem na subseção C da seção 10 e subseção A da
seção 11 de D. sturtevanti às quais são correspondentes às seções 8 e 6 de D.
prosaltans, respectivamente.
Em D. sturtevanti cinco regiões de pufes se destacam: seção 3, subseções
B a E da seção 6, seção 9, subseções A a C da seção 10 e seção 13 (Fig. 3),
sendo observada correspondência destes pufes em D. prosaltans: o pufe na
seção 3 de D. sturtevanti corresponde a seção 15 de D. prosaltans; o pufe das
subseções B a E da seção 6 de D. sturtevanti corresponde a seção 13 de D.
prosaltans; o pufe da seção 9 tem correspondência com a seção 8 de D.
prosaltans; o pufe das subseções A a C da seção 10 encontra correspondência
com o da seção 7 de D. prosaltans e o pufe da seção 13 de D. sturtevanti tem
correspondência com o da seção 4 de D. prosaltans demonstrando um padrão
conservado de ativação gênica que reflete na expressão desses pufes.
Por outro lado, D. sturtevanti aparentemente apresenta uma duplicação
entre as subseções A e B da seção 6 e a subseção A da seção 7 do braço XL.
Essa característica não foi visualizada no braço descrito de D. prosaltans, não

25
sendo encontrada uma similaridade clara para essas regiões. Porém, é
importante destacar que o braço XL descrito para D. prosaltans é um desenho
que, mesmo possibilitando uma primeira comparação para a espécie, não oferece
total resolução do padrão de bandas, interbandas e pufes da espécie. De maneira
geral, os padrões encontrados entre D. prosaltans e D. sturtevanti apresentam
uma similaridade que reflete a relação existente entre as duas espécies.
Na comparação entre os padrões de bandas encontrados nos braços XL
de D. sturtevanti e D. willistoni foram localizadas diversas regiões de similaridade.
Entre elas destacam-se a constrição encontrada na subseção A da seção 11 de
D. sturtevanti e sua correspondência na subseção A da seção 5 de D. willistoni,
além da correspondência encontrada entre os pufes das subseções B a E da
seção 6, seção 9, subseções A a C da seção 10 e seção 13 de D. sturtevanti
respectivamente na subseção A da seção 9; nas subseções D e E da seção 6;
nas subseções C e D da seção 5 e subseção B da seção 3 de D. willistoni. Esta
comparação revelou um padrão conservado de pufes ativos em ambas as
espécies.
Outro pufe que pode ser destacado, presente na seção 3 de D. sturtevanti,
foi relacionado com a seção 14 de D. willistoni, porém, esta mesma região de D.
willistoni parece estar duplicada na seção 13 desta espécie, característica não
encontrada para esta seção em D. sturtevanti. Da mesma forma, a duplicação
visualizada entre as subseções A e B da seção 6 e a subseção A da seção 7 do
braço XL de D. sturtevanti, não foi visualizada em D. willistoni não sendo
encontrada uma correspondência clara para essas regiões. De maneira geral
esses dois elementos cromossômicos apresentam blocos de similaridade.

26
Na comparação do padrão de bandas, interbandas e pufes do braço XL de
D. sturtevanti, D. prosaltans e D. willistoni foi observado um padrão conservado
entre as espécies do presente estudo.
5.3. Confecção do braço XR de D. prosaltans e comparação dos braços XR
de D. sturtevanti, D. willistoni e D. prosaltans.
Estendendo a comparação dos padrões de bandas, interbandas e pufes,
foi confeccionado o braço politênico XR de D. prosaltans. A confecção de um
mapa parcial de alta resolução torna ainda mais claro o padrão de marcas
encontradas nesse braço politênico e possibilita uma comparação direta e
especifica desses marcadores com os de espécies próximas. Nesse sentido,
primeiro foi utilizada a descrição de Cavalcanti (1948) para identificação do braço
XR de D. prosaltans, então, foi confeccionado o braço XR e estabelecida à
relação entre as seções do braço produzido por Cavalcanti e o braço
confeccionado no presente trabalho (Fig. 4).
A relação entre os braços XR foi estabelecida conservando as 18 seções
em que o cromossomo havia sido dividido anteriormente, que correspondem da
seção 18 a 35 com posterior inserção de subseções estabelecidas de A a F. A
seção 23 chamou atenção por ser bem maior que todas as outras seções
encontradas nesse braço politênico, além do grande pufe encontrado nas
subseções A e B da seção 29 e a ponta em leque encontrada na seção 35 (Fig.
5).
Após a confecção e relação das seções do braço XR de D. prosaltans, foi
possível comparar os padrões encontrados no braço politênico dessa espécie
com os encontrados em D. sturtevanti e D. willistoni. Nesse sentido destaca-se a

27
ponta dos braços politênicos das três espécies que se apresentam em forma de
leque, sendo esta uma característica marcante desse braço politênico e que ajuda
na sua identificação. Esta ponta esta localizada nas subseções C e D da seção 35
de D. sturtevanti, seção 35 de D. prosaltans e subseção B da seção 36 de D.
willistoni (Fig. 6).
Na comparação entre os braços XR de D. sturtevanti e D. prosaltans são
visualizados diversos pontos de similaridade no decorrer do cromossomo, com
especial atenção para os grandes pufes nas seções 18 e 27 do braço XR de D.
sturtevanti que possui correspondência na seção 20 e subseções A e B da seção
29 de D. prosaltans, sendo o pufe da seção 20 de D. prosaltans bem menor que o
encontrado em D. sturtevanti. Duas constrições marcantes são encontradas na
subseção E da seção 17 e subseção B da seção 21 de D. sturtevanti, tendo
relação com a subseção A da seção 20 e subseção B da seção 23 de D.
prosaltans, sendo as constrições de D. sturtevanti bem mais acentuadas que as
encontradas em D. prosaltans. Na seção 25 do braço XR de D. sturtevanti é
encontrada uma sequência de bandas sobressalentes na horizontal, não sendo
encontrada uma correspondência dessas bandas em D. prosaltans. A seção 23
de D. prosaltans, estabelecida pela comparação com o mapa descrito por
Cavalcanti (1948), é relativamente grande e são encontrados poucos pontos de
similaridade na correlação dessa região com o braço XR de D. sturtevanti.
Comparando o braço XR de D. sturtevanti com o braço XR de D. willistoni
foi observado diversos pontos de similaridade. Os maiores pufes desse braço
politênico, nas seções 18 e 27, de D. sturtevanti encontram correspondência nas
subseções A e B da seção 19 e subseção A da seção 29 respectivamente de D.
willistoni. As constrições na subseção E da seção 17 e subseção B da seção 21

28
de D. sturtevanti também estão presentes na subseção B da seção 18 e subseção
C da seção 23 de D. willistoni respectivamente. Chama atenção o pufe das
subseções A e B da seção 20 de D. willistoni que não encontra correspondência
em D. sturtevanti e a sequência de bandas sobressalentes na horizontal
encontradas na seção 25 de D. sturtevanti que não encontra correspondência em
D. willistoni.
Assim como na comparação do braço XL de D. sturtevanti, D. prosaltans e
D. willistoni o braço XR também demonstrou a presença de padrões de bandas,
interbandas e pufes conservados entre essas espécies.
5.4. Localização cromossômica dos genes Ubi e E71
Os genes Ubi e E71 foram hibridizados nos cromossomos politênicos de D.
sturtevanti para estabelecer a relação da localização desses genes com D.
willistoni e D. melanogaster para obter mais informações que possibilitem inferir
sobre a sintênia dos elementos de Müller em D. sturtevanti.
O gene Ubi foi mapeado em D. sturtevanti na subseção B da seção 26 do
braço cromossômico XR e apresentou 62% de frequência de hibridização (Fig.
7a). Na hibridização do gene E71 foi evidenciada a localização do mesmo na
subseção B da seção 27 do braço XR, com 40% de frequência de hibridização
(Fig. 7b). Na hibridização de ambos os genes não foram encontradas outras
marcações significativas.

29
Figura 2 – Fotomapa dos cromossomos politênicos de Drosophila sturtevanti, linhagem DIR. Os centrômeros estão voltados à esquerda. Setas indicam regiões de pufes. Escala: 10µm
30
Figura 2 – Continuação do Fotomapa dos cromossomos politênicos de Drosophila sturtevanti, linhagem DIR. Os centrômeros estão voltados à esquerda. Setas indicam regiões de pufes. Escala: 10µm

31
XL D. prosaltans Cavalcanti, 1948
XL D. willistoni Schaeffer et al., 2008
XL D. sturtevanti Monte, 2013; Presente
Trabalho.
Figura 3 – Relação entre o braço XL de D. sturtevanti (central) com os braços XL de D. prosaltans (acima) e D. willistoni (abaixo). Mostrando em azul a correlação de pufes importantes.

32
Figura 4 – Relação entre o braço XR de D. prosaltans publicado por Cavalcanti (acima) e o braço XR confeccionado no presente trabalho (abaixo). As linhas azuis indicam as relações entre as seções nos dois braços politênicos. As setas do braço descrito por Cavalcanti (acima) indicam regiões de contrição em que o cromossomo quebra facilmente.
D. prosaltans
Figura 5 – Braço XR de D. prosaltans confeccionado no presente trabalho, com subseções.
XR
XR
XR

33
Figura 6 – Relação entre o braço XR de D. sturtevanti (central) com os braços XR de D. prosaltans (acima) e D. willistoni (abaixo). Mostrando diversos pontos de similaridade. Em azul escuro são mostrados os pontos de correspondência mais importantes.
XR D. prosaltans Presente Trabalho.
XR D. sturtevanti Monte, 2013;
Presente Trabalho.
XR D. willistoni Schaeffer et al.,
2008

34
Figura 7 – Braços cromossômicos XR de D. sturtevanti, mostrando a localização dos genes Ubi (A) e E71 (B) através de hibridização in situ permanente. As barras em azul indicam a correspondência da marcação nas seções do braço XR. As setas indicam a localização do gene hibridizado.
XR
A B

35
6. Discussão
A confecção do fotomapa de referência para os cromossomos politênicos
de D. sturtevanti permite localizar marcadores físicos que possibilitam a relação
com espécies que possuem mapas descritos, de modo que é possível fazer
inferências de como esses marcadores se comportaram durante a evolução,
esclarecendo assim divergências dentro do grupo saltans. A comparação dos
marcadores existentes nos braços XL e XR de D. sturtevanti com os braços XL e
XR de D. prosaltans (em especial o braço XR aqui descrito) e D. willistoni,
possibilitou uma análise inicial das relações existentes dentro do grupo saltans e a
comparação com o grupo willistoni, ambos pertencentes ao subgênero
Sophophora.
A comparação entre braços do cromossomo X de D. sturtevanti e D.
prosaltans apresentada neste trabalho evidenciou que existe a conservação do
braço XR. Contudo o braço XL e o cromossomo III de D. prosaltans
correspondem respectivamente ao cromossomo III (acrocêntrico) e ao braço XL
de D. sturtevanti quando se observa os cromossomos mitóticos dessas espécies.
Estas evidências corroboram o trabalho de Dobzhansky e Pavan (1943), o qual
indica que o complemento mitótico de D. sturtevanti é ancestral em relação ao de
D. prosaltans, pois o cariótipo desta ultima é oriundo de um evento de
translocação. Neste trabalho também foi possível observar que o padrão de
bandas, interbandas e pufes é conservado com relação aos cromossomos XL e
XR, mesmo com a quebra cromossômica presente em D. prosaltans.
Os braços XR e XL de D. sturtevanti e D. willistoni ao serem comparados
permitiram inferir sobre a sua correspondência quanto aos elementos de Müller.
Dessa forma, considerando a sintênia estabelecida entre D. willistoni e D.

36
melanogaster por Schaeffer et al. (2008), onde foi visto que o metacêntrico X de
D. willistoni (grupo willistoni) foi gerado a partir da fusão dos elementos A e D de
Müller (X e o braço autossômico 3L de D. melanogaster), evento também
encontrado em D. pseudoobscura e D. persimillis ambas do grupo obscura
(Schaeffer et al., 2008), propomos que os braços XL e XR de D. sturtevanti
correspondem respectivamente aos elementos A e D de Müller. Da mesma forma
pode-se inferir a partir dessa comparação que os braços XL e XR de D.
prosaltans correspondem aos elementos A e D de Müller, ao passo que mesmo
com a translocação presente nessa espécie, os elementos de Müller
permaneceram conservados.
Apesar da conservação dos elementos politênicos nas espécies do
presente estudo, duplicações nos braços XL de D. sturtevanti e D. willistoni foram
encontradas. Estas duplicações são possivelmente resultado da atividade de
elementos de transposição presentes no genoma dessas espécies, os quais
também podem ser responsáveis pela ocorrência de inversões observadas
(Kobayashi e Bicudo 1997; Rohde e Valente, 2012; Monte, 2013). Elementos de
transposição, particularmente elementos P, são encontrados nos principais
grupos de espécies de Drosophila (melanogaster, obscura, saltans e willistoni)
tendo uma grande importância nas alterações que causam variabilidade dentro
das espécies (Castro e Carareto, 2004). Portanto, é sugerido que estes elementos
são provavelmente os responsáveis pelo evento que deu origem ao cariótipo de
D. prosaltans.
A localização do gene Ubi na subseção B da seção 26 do cromossomo XR
de D. sturtevanti possibilitou inferir sobre a correspondência deste braço politênico
com o elemento D de Müller (Fig. 8). Esta correspondência também foi observada

37
em D. willistoni para o gene Ubi (Schaeffer et al., 2008), assim como em D.
melanogaster onde este gene está localizado no braço 3L (Izquierdo, 1994).
Embora tenha sido observada a conservação quanto a localização do gene Ubi no
braço cromossômico XR de D. sturtevanti e D. willistoni, a localização na
subseção B da seção 26 difere da localização esperada (seção 24) pela
comparação do padrão de bandas. Esta localização diferente é possivelmente
oriunda de uma pequena inversão.
A localização do gene E71 em D. sturtevanti na subseção B da seção 27
do braço XR corrobora com a inferência de que esse braço politênico corresponde
ao elemento D de Müller (Fig. 8). Esse gene também se encontra localizado no
braço XR de D. willistoni (Schaeffer et al., 2008) e no 3L de D. melanogaster
(Restifo e Gregory, 1986), correspondendo para ambas às espécies ao elemento
D de Müller.
O gene Hsp83 localizado por Monte (2013) no cromossomo XR de D.
sturtevanti fortalece ainda mais a hipótese de que este braço politênico
corresponde ao elemento D de Müller, pela corespondente localização desse
gene no braço XR de D. willistoni (Schaeffer et al., 2008) e no braço 3L de D.
melanogaster (Holmgren et al., 1981). No braço cromossômico III de D.
sturtevanti, Monte (2013) localizou o gene Hsp70, indicando correspondência com
o elemento E de Muller, pois este gene apresenta conservação da localização no
cromossomo lll de D. willistoni (Bonorino et al., 1993) e no braço 3R de D.
melanogaster (Livak et al., 1978). A relação dos elementos politênicos utilizando
os genes mapeados no presente trabalho e a comparação do padrão de bandas,
interbandas e pufes possibilitou inferir que os elementos de Müller permanecem
conservados em D. sturtevanti (Fig. 8).

38
A localização dos genes Ubi, E71 e Hsp83 no mesmo elemento politênico
de D. melanogaster, D. willistoni e D. sturtevanti possibilitou relacionar o
posicionamento desses genes nas três espécies (Fig. 8). Dessa forma, D.
willistoni e D. sturtevanti conservam o posicionamento dos genes da base à ponta
do cromossomo, sendo a sequência: Ubi – E71 – Hsp83 (Fig. 8). No entanto,
quando comparados com D. melanogaster, foi possível verificar uma alteração do
posicionamento dos genes Ubi e E71 provavelmente oriundo de uma inversão
entre esses genes, que deu origem a sequência de posicionamento da base à
ponta: E71 – Ubi – Hsp83 em D. melanogaster (Izquierdo, 1994; Restifo e
Gregory, 1986; Holmgren et al., 1981) (Fig. 8). Enquanto o gene Hsp83 mantém
sua localização conservada nas três espécies analisadas, sendo mais próximo a
ponta do elemento politênico.
Ainda que os dados apresentados no presente trabalho sejam de grande
contribuição para o estudo evolutivo das espécies em pauta, é importante a
realização de novos estudos de comparação entre os elementos cromossômicos
de D. sturtevanti e espécies próximas, no intuito de revelar novos blocos de
sintênia que possam elucidar as relações existentes entre os grupos saltans e
willistoni, bem como com outras espécies de drosofilídeos. Além disso, é
importante a identificação de novos marcadores a partir da localização de genes
em D. sturtevanti, em especial os genes Dpp e Period, os quais foram mapeados
em D. willistoni e D. melanogaster. Estes estudos poderão contribuir para futuras
inferências sobre rearranjos existentes durante a evolução e diversificação das
espécies.

39
E71
D. melanogaster
X 2L 2R 3L 3R 4
D. willistoni
XL XR IIR III IIL
D. sturtevanti
XL XR IIR IIL III
Elemento A Elemento B Elemento C Elemento D Elemento E Elemento F Figura 8 - Esquema comparativo da correspondência dos elementos de Müller em Drosophila melanogaster, Drosophila willistoni e Drosophila sturtevanti considerando a localização de genes de copia única e a sequência da localização desses genes no elemento D.
Hsp70 Hsp83
Hsp70 Hsp83
Hsp70 Hsp83
E71
Ubi
E71
Ubi
(Monte, 2013; Presente Trabalho)
(Bonorino et al., 1993; Rieger, 1999; Schaeffer et al., 2008)
(Holmgren et al., 1981; Restifo e Gregory, 1986; Izquierdo, 1994)
Ubi

40
7. Conclusões
1. Na comparação do braço XL e XR entre D. sturtevanti, D. prosaltans
e D. willistoni foram encontradas correspondências de bandas,
Interbandas e pufes ativos que refletem as relações existentes entre
essas espécies e possibilitaram inferir, por sintênia estabelecida
entre D. willistoni e D. melanogaster, que estes braços politênicos
correspondem aos elementos A e D de Müller.
2. A confecção do braço XR de D. prosaltans possibilitou uma melhor
visualização do padrão de bandas, interbandas e pufes encontrados
nesse elemento politênico e tornou a comparação com D. sturtevanti
ainda mais informativa.
3. A localização dos genes Ubi e E71 no braço XR de D. sturtevanti
possibilitou relacionar esta localização com D. willistoni e D.
melanogaster de modo que a conservação da localização destes
genes nessas espécies fortaleceu ainda mais a hipótese de que
esse braço politênico corresponde ao elemento D de Müller.
4. A localização dos genes Ubi, E71 e Hsp83 mostrou a conservação
da posição em D. willistoni e D. sturtevanti, enquanto que em D.
melanogaster a posição dos genes Ubi e E71 está invertida. Já o
gene Hsp83 tem seu posicionamento conservado nas três espécies
analisadas.

41
8. Referências Bibliográficas
Adams MD, Celniker SE, Holt RA, Evans CA, Gocayne JD et al., (2000) The
genome sequence of Drosophila melanogaster. Science 287: 2185–2195.
Ausubel F, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA and Struhl K
(2002) Short Protocols in Molecular Biology, 1512 pages, 2º Vol., John Wiley
& Sons.
Beckstead R, Ortiz JA, Sanchez C, Prokopenko SN, Chambon P et al., (2001)
Bonus, a Drosophila Homolog of TIF1 Proteins, Interacts with Nuclear
Receptors and Can Inhibit bFTZ-F1-Dependent Transcription. Mol. Cell. 7:
753-765.
Bhutkar A, Schaeffer SW, Russo S, Xu M, Smith TF and Gelbart WM (2008)
Chromosomal rearrangement inferred from comparisons of 12 Drosophila
genomes. Genetics 179:1657-1680.
Bicudo HEMC (1979) Reprodutive isolations in the saltans group of Drosophila. IV.
The sturtevanti subgroup. Rev. Brasil. Genet, 4: 247-258.
Bonorino CBC, Pereira M, Alonso CEV, Valente VL and Abdelhay E (1993) In situ
mapping of the hsp70 locus in seven species of the willistoni group of
Drosophila. Revista Brasileira de Genetica. 16: 561-561.
Bridges CB (1935) Salivary chromosome maps with a key to thebanding of the
chromosomes of Drosophila melanogaster. J. Hered. 26: 60–64.
Buchon N, Broderick NA and Lemaitre B (2013) Gut homeostasis in a microbial
world: insights from Drosophila melanogaster. Nat. Rev. Microbiol. 16: 615–
26.
Bukau B, Weissman J and Horwich A (2006) Molecular Chaperones and Protein
Quality Control. Cell, 125: 443-451.
Buzzi ZJ (2002) Entomologia didática. 4. ed. Curitiba: UFPR, 347 pp.
Campos SRSLC, Rieger TT and Santos JF (2007) Homology of polytene elements
between Drosophila and Zaprionus determined by in situ hybridization in Z.
indianus. Genet Mol Res 6 (2): 162 -176.
Castro JP and Carareto CMA (2004). Characterization of two full-sized P elements
from Drosophila sturtevanti and Drosophila prosaltans. Genetics and
Molecular Biology. 27( 3): 373-377.

42
Cavalcanti AGL (1948) Geographic variation of chromosome structure
in Drosophila prosaltans. Genetics 33: 529-536.
Chiang AS, Lin CY, Chuang CC, Chang HM, Hsieh CH et al., (2011) Three-
dimensional reconstruction of brain-wide wiring networks in Drosophila at
single-cell resolution. Curr Biol.
Clark AG, Garcia ACL, Rieger TT, Rohde C, Valente VLS et al., (2007) Evolution
of genes and genomes on the Drosophila phylogeny. Nature, 450: 203-215.
Da Lage JL, Kergoat GJ, Maczkowiak F, Silvain JF, Cariou ML and Lachaise D
(2007) A phylogeny of Drosophilidae using the Amyrel gene: questioning the
Drosophila melanogaster species group boundaries. Journal of Zoological
Systematics and Evolutionary Research, 45(1): 47–63.
De Setta N, Costa AP, Lopes FR, Van Sluys MA and Carareto CM (2007)
Transposon display supports transpositional activity of P elements in species
of the saltans group of Drosophila. J. Genet. 86(1): 37--43.
Dobzhansky TH and Pavan C (1943) Chromosome complements of some south-
brazilian species of Drosophilia. Proc. N. A. S. 29: 368-375.
Drosopoulou E, Nakou I, Šíchová J, Kubíčková S, Marec F and Mavragani-
Tsipidou P (2012) Sex chromosomes and associated rDNA form a
heterochromatic network in the polytene nuclei of Bactrocera oleae (Diptera:
Tephritidae). Genetica 140 (4-6): 169–180.
Enright AJ, John B, Gaul U, Tuschl T, Sander C and Marks DS (2004) MicroRNA
targets in Drosophila. Genome biology, 5(1).
Fernandes DRR and Araújo EL (2011) Ocorrência de Zaprionus indianus gupta
(Diptera: Drosophilidae) em frutos de juazeiro Ziziphus joazeiro mart.
(rhamnaceae) no estado do Rio Grande do Norte. Revista Brasileira de
Fruticultura, 33(4): 1356-1358.
Fortini ME and Bonini NM (2000). Modeling human neurodegenerative diseases in
Drosophila. Trends Genet 16: 161–167.
Garcia ACL, Rohde C, Audino GF, Valente VLS and Valiati VH (2006)
Identification of the sibling species of the Drosophila willistoni subgroup
through the electrophoretical mobility of acid phosphatase-1. Journal of
Zoological Systematics and Evolutionary Research, 44 (3): 212–216.
Gao JJ, Hu YG, Toda MJ, Katoh T and Tamura K (2011) Phylogenetic
relationships between Sophophora and Lordiphosa, with proposition of a

43
hypothesis on the vicariant divergences of tropical lineages between the Old
and New Worlds in the family Drosophilidae. Molec. Phylog. Evol. 60(1): 98--
107.
Gottschalk MS (2008) Utilização de recursos tróficos por espécies Neotropicais de
Drosophilidae (Diptera). Ph.D. Thesis, Universidade Federal do Rio Grande
do Sul, Porto Alegre, Brazil.
Grimaldi D and Engel MS (2005) Evolution of the Insects. Cambridge Univ Press,
New York.
Hall BH, Qi Y, Timoshevskiy V, Sharakhova MV, Sharakhov IV et al., (2013) Six
novel Y chromosome genes in Anopheles mosquitoes discovered by
independently sequencing males and females. BMC Genomics.
Hoffmann J (2007) Antifungal defense in Drosophila. Nature Immunology 8: 543-
545.
Holmgren R, Corces V, Morimoto R., Blackman R and Meselson M (1981)
Sequence homologies in the 5'regions of four Drosophila heat-shock
genes. Proceedings of the National Academy of Sciences, 78(6): 3775-3778.
Huang HR, Chen ZJ, Kunes S, Chang GD and Maniatis T (2010) Endocytic
pathway is required for Drosophila Toll innate immune signaling. Proc. Natl.
Acad. Sci. USA 107: 8322–8327.
Izquierdo M (1994) Ubiquitin genes and ubiquitin protein location in polytene
chromosomes of Drosophila. Chromosoma, 103(3):193-197.
Jolly C and Morimoto RI (2000) Role of the heat shock response and molecular
chaperones in oncogenesis and cell death. Journal of the National Cancer
Institute. 92(19): 1564-1572.
Katoh T, Nakaya D, Tamura K and Aotsuka T (2007) Phylogeny of the Drosophila
immigrans species group (Diptera: Drosophilidae) based on Adh and Gpdh
sequences. Zoological Science, 24: 913-921.
Kawai T and Akina S (2006) Innate immune recognition of viral infection. Nature
Immunology 7: 131-137.
Kim HJ, Hwang NR and Lee KJ (2007) Heat shock responses for understanding
diseases of protein denaturation. Mol. Cells 23: 123–131.
Kobayashi MK and Bicudo HE (1997). Inversion polymorphism in laboratory
strains and natural samples of Drosophila sturtevanti (saltans group,
sturtevanti subgroup). Cytobios, 89:7-20.

44
Kolesnikova TD, Semeshin VF, Andreyeva EN, Zykov IA, Kokoza EB,
Kalashnikova DA and Zhimulev IF (2011) Induced decondensation of
heterochromatin in Drosophila melanogaster polytene chromosomes under
condition of ectopic expression of the Supressor of underreplication
gene. Fly, 5(3): 181-190.
Lefevre G (1976) A photographic representation and interpretation of the polytene
chromosomes of Drosophila melanogaster salivary glands. The Genetics and
Biology of Drosophila, edited by M. Ashburner and E. Novitski. Academic
Press, New York. pp. 31–66.
Leopold P and Perrimon N (2007) Drosophila and the genetics of the internal
milieu. Nature, 450: 186-188.
Livak KJ, Freund R, Schweber M, Wensink PC and Meselson M (1978) Sequence
organization and transcription at two heat shock loci in
Drosophila. Proceedings of the National Academy of Sciences, 75(11): 5613-
5617.
Magalhaes LE (1962) Notes on the taxonomy, morphology and distribution of the
saltans group of Drosophila with description of four new species. Univ.
Texas Publs Stud. Genet. 2(6205): 135--154.
Mitrofanov VG (2007) Molecular-genetic mechanisms of the effect of
developmental hormones in insects. Ontogenez 38(5): 330—344.
Monte ES and Rieger TT (2009) Mapeamento por Hibridização in situ do gene
period em Drosophila willistoni. XVII Congresso de Iniciação Científica I
Congresso de Iniciação em Desenvolvimento Tecnológico e Inovação.
Monte ES (2013) Cartografia Genômica e Evolução em Drosophila sturtevanti
Duda, 1927 (Grupo saltans). Dissertação de mestrado do Programa de Pós-
Graduação em Genética da Universidade Federal de Pernambuco.
Mourão CA and Bicudo HEMC (1967) Duas novas espécies de Drosophila do
grupo “saltans” (Drosophilidae, Diptera). Papeis Avulsos de Zoologia, 20:
123-134.
Mukhopadhyay D and Riezman H (2007) Proteasome-independent functions of
ubiquitin in endocytosis and signaling. Science, 315(5809): 201-205.
Muller HJ (1940) Bearings of the 'Drosophila' work on systematics. In: Huxley j
(ed) The New Systematics. Clarendon Press, Oxford, pp 185--268.

45
Papaceit M and Juan E (1998) Fate of dot chromosome genes in Drosophila
willistoni and Scaptodrosophila lebanonensis determined by in situ
hybridization. Chromosome Res. 6: 49–54.
Pospisilik JA, Schramek D, Schnidar H, Cronin SJ, Nehme NT et al., (2010)
Drosophila genome-wide obesity screen reveals hedgehog as a determinant
of brown versus white adipose cell fate. Cell 140: 148–160.
Restifo LL and Gregory MG (1986) Poly (A) shortening of coregulated transcripts
in Drosophila. Developmental biology 115.2: 507-510.
Rieger TT (1999) Mapeamento por hibridação in situ e expressão no
desenvolvimento de genes de reposta a estresses e ao hormônio ecdisona
em espécies do grupo willistoni de Drosophila. Ph.D. Thesis, Universidade
Federal do Rio Grande do Sul, Porto Alegre, Brazil.
Ritossa F (1962) A new puffing pattern induced by temperature shock and DNP
in Drosophila. Experientia;18:571–573.
Ritossa F (1996) Discovery of the heat shock response. Cell Stress &
Chaperones;1:97–98.
Robe LJ, Valente VLS, Budnik M and Loreto ELS (2005) Molecular phylogeny of
the subgenus Drosophila (Diptera, Drosophilidae) with an emphasis on
Neotropical species and groups: a nuclear versus mitochondrial gene
approach. Molecular Phylogenetics and Evolution 36: 623–640.
Rohde C and Valente VLS (2012) Three decades of studies on chromosomal
polymorphism of Drosophila willistoni and description of fifty different
rearrangements. Genetics and molecular biology, 35(4): 966-979.
Ron D and Walter P (2007) Signal integration in the endoplasmic reticulum
unfolded protein response. Nat. Rev. Mol. Cell Biol. 8: 519–529.
Sabin LR, Hanna SL and Cherry S (2010) Innate antiviral immunity in Drosophila.
Curr Opin Immunol, 22(1):4-9.
Santos JF, Rieger TT, Campos SRC, Nascimento ACC, Félix PT, Silva SVO and
Freitas FMR (2003) Colonization of Northeast Region of Brazil by the
drosophilid flies Drosophila malerkotliana and Zaprionus indianus, a new
potential insect pest for Brazilian fruit culture. Drosophila Information Service,
Oklahoma, 86: 92-95.
Schaeffer SW, Rohde C, Valente VLS, Garcia ACL, Rieger TT et al., (2008)
Polytene chromosomal maps of 11 Drosophila species: the order of genomic

46
scaffolds inferred from genetic and physical maps. Genetics, 179: 1601-
1655.
Silva Valenzuela MDGD, Almeida FCS, Antonio LFM, Libório TN, Acquafreda T,
Cazal C, Ferraz A and Nunes FD (2006). In situ hybridization with non-
radioactive riboprobes: principles and applications in pathology. Jornal
Brasileiro de Patologia e Medicina Laboratorial, 42(3): 207-213.
Sorsa V (1988a) Chromosome Maps of Drosophila, Vol. I. CRC Press, Boca
Raton, FL.
Sorsa V (1988b) Chromosome Maps of Drosophila, Vol. II. CRC Press, Boca
Raton, FL.
Sorsa V (1988c) Polytene Chromosomes in Genetic Research. John Wiley &
Sons, New York.
Sturtevant AH and Novitski E (1941) The homologies of the chromosome
elements in the genus Drosophila. Genetics 26: 517–541.
Sturtevant AH (1942) The classification of the genus Drosophila, with description
of nine new species. University of Texas Publication, Austin. 4213: 5-51.
Timoshevskiy VA, Severson DW, Debruyn BS, Black WC, Sharakhov IV and
Sharakhova MV (2013) An integrated linkage, chromosome, and genome
map for the yellow fever mosquito Aedes aegypti. PLoS Negl Trop Dis. 7:
2052.
Valanne S, Wang JH and Ramet M (2011) The Drosophila Toll signaling pathway.
J. Immunol. 186: 649–56.
Van Der Linde K, Houle D, Spicer GS and Steppan SJ (2010) A supermatrix
based molecular phylogeny of the family Drosophilidae.- Genetics Research,
92: 25-38.
Vosshall LB (2007) Into the mind of a fly. Nature, 480: 193-197.
Yang Y, Hou ZC, Qian YH, Kang H and Zeng QT (2012) Increasing the data size
to accurately reconstruct the phylogenetic relationships between nine
subgroups of the Drosophila melanogaster species group (Drosophilidae,
Diptera). Mol Phylogenet Evol. 62: 214–223.
Zhimulev IF (1996) Morphology and structure of polytene chromosomes. Adv.
Genet. 34: 1–497.
Zhimulev IF and Koryakov DE (2009) Polytene chromosomes. eLS.

47
9. Curriculum vitae (Lattes)
Antonio Terto da Silva Junior
Curriculum Vitae
_________________________________________________________________
Dados pessoais
Nome Antonio Terto da Silva Junior
Nascimento 03/10/1990 - recife/PE - Brasil
CPF 089.731.354-22
_________________________________________________________________
Formação acadêmica/titulação
2012 Mestrado em Programa de Pós-Graduação em Genética.
Universidade Federal de Pernambuco, UFPE, Recife, Brasil
Título: Evolução cromossômica e mapeamento de genes no
cromossomo X de Drosophila sturtevanti (grupo saltans).
Orientador: Rita de Cassia de Moura
Co-orientador: Tania Tassinari Rieger
Bolsista do (a): Conselho Nacional de Desenvolvimento Científico
e Tecnológico
2008 - 2011 Graduação em licenciatura plena em ciências biológicas.
Universidade Federal de Pernambuco, UFPE, Recife, Brasil
Bolsista do (a): UNIVERSIDADE FEDERAL DE
PERNAMBUCO/PROACAD
_________________________________________________________________
Formação complementar

48
2009 - 2010 GN006-INTRODUÇAO A GENÉTICA.
UNIVERSIDADE FEDERAL DE PERNAMBUCO/PROACAD,
UFPE, Brasil.
Bolsista do (a): UNIVERSIDADE FEDERAL DE
PERNAMBUCO/PROACAD
_________________________________________________________________
Atuação profissional
1. Universidade Federal de Pernambuco - UFPE
Vínculo institucional
2010 – 2011
Vínculo: Outro (bolsista pibic), Enquadramento funcional: SEMANAL, Regime:
Dedicação exclusiva.
_________________________________________________________________
Projetos
Projetos de pesquisa
2012 – Atual
Evolução cromossômica e mapeamento de genes no cromossomo X de
Drosophila sturtevanti (grupo saltans).
Descrição: Este projeto visa ampliar os estudos evolutivos no gênero Drosophila,
através do mapeamento de genes evolutivamente conservados, nos
cromossomos politênicos de Drosophila sturtevanti e da comparação de
marcadores convencionais dos braços XL e XR de Drosophila sturtevanti com os
de Drosophila prosaltans e Drosophila willistoni.
Situação: Em andamento Natureza: Projetos de pesquisa

49
Alunos envolvidos: Mestrado acadêmico (3);
Integrantes: Antonio Terto da Silva Junior (Responsável); Tânia Tassinari Rieger;
Rita de Cassia de moura.
2011 - 2012
Subprojeto: “Localização cromossômica do gene HSP70 por hibridização in situ
em Drosophila sturtevanti Duda, 1927 (grupo saltans).”
Situação: Concluído Natureza: Projetos de pesquisa
Integrantes: Antonio Terto da Silva Junior (Responsável); Fernanda Ito dos
Santos; Tânia Tassinari Rieger.
2010 - 2011
Análise do complemento mitótico em Drosophila Sturtevanti Duda, 1927 (grupo
saltans).
Descrição: O projeto tem como objetivo ampliar estudos evolutivos no gênero
Drosophila, descrever o complemento mitótico da linhagem de trabalho e
comparar os resultados com dados relatados para a espécies próximas buscando
novos dados sobre as relações filogenéticas no subgênero Sophophora.
Situação: Concluído Natureza: Projetos de pesquisa
Integrantes: Antonio Terto da Silva Junior (Responsável); Tânia Tassinari Rieger.