ultra-estrutura de leucÓcitos, perfil ... - uenf.br · bacelar pela disponibilização de seus...
TRANSCRIPT

ULTRA-ESTRUTURA DE LEUCÓCITOS, PERFIL HEMATOLÓGICO E
BIOQUÍMICO SÉRICO DE AVESTRUZES (Struthio camelus, Linnaeus
1758) CRIADAS NO ESTADO DO RIO DE JANEIRO
SÉRGIO FERNANDES BONADIMAN
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE
CAMPOS DOS GOYTACAZES - RJ
MAIO/2008

II

III
ULTRA-ESTRUTURA DE LEUCÓCITOS, PERFIL HEMATOLÓGICO E
BIOQUÍMICO SÉRICO DE AVESTRUZES (Struthio camelus, Linnaeus
1758) CRIADAS NO ESTADO DO RIO DE JANEIRO
SÉRGIO FERNANDES BONADIMAN
“Dissertação apresentada ao Centro de
Biociências e Biotecnologia da
Universidade Estadual do Norte
Fluminense ”Darcy Ribeiro”, como parte
das exigências para obtenção do título de
Mestre em Biociências e Biotecnologia”
CAMPOS DOS GOYTACAZES - RJ
MAIO/2008

IV
ULTRA-ESTRUTURA DE LEUCÓCITOS, PERFIL HEMATOLÓGICO E
BIOQUÍMICO SÉRICO DE AVESTRUZES (Struthio camelus, Linnaeus
1758) CRIADAS NO ESTADO DO RIO DE JANEIRO
SÉRGIO FERNANDES BONADIMAN
“Dissertação apresentada ao Centro de
Biociências e Biotecnologia da Universidade
Estadual do Norte Fluminense “Darcy Ribeiro”,
como parte das exigências para obtenção do
título de Mestre em Biociências e Biotecnologia
(Biologia Celular)”
Aprovada em 20 de maio de 2008
Comissão Examinadora:
___________________________________________________________________
Profa. Nádia Regina Pereira Almosny (Dra. Medicina Veterinária) - UFF
___________________________________________________________________
Prof. Arthur Giraldi Guimarães (Dr. Biofísica) – UENF
___________________________________________________________________
Prof. Milton Masahiko Kanashiro (Dr. Biociências) – UENF
___________________________________________________________________
Prof. Renato Augusto DaMatta (Dr. Biofísica) – UENF
(Orientador)

V
“Não diga que as estrelas estão mortas
Só porque o céu está nublado.
Não se iluda, o pé que dá fruta é o que mais leva pedra.
A raiz é uma flor, que despreza a fama...”
Wado

VI
Agradecimentos
Ao Prof. Renato DaMatta, não só pela amizade e companheirismo mas pela
orientação, incentivo e troca de idéias.
Ao amigo Luis Fernando Vieira pelo altruísmo na ajuda com as coletas de campo.
Aos meus avós (Honório, Rosa, José e Morcina) e pais (Honory e Rosa) pelos
ensinamentos e exemplos de caráter, honestidade e seriedade.
À minha linda, Roberta, pela ajuda com as diagramações, paciência com a distância
e todo amor dado.
Ao Prof. Clóvis Santos pela amizade, oportunidades de trabalho e revisão desta
dissertação.
Ao Prof. Flávio Miguens pelo auxílio prestado nos experimentos de citoquímica.
Aos Profs. Carlos Veiga, Edilene Silva e Humberto Couto, pelos ensinamentos e
confiança nas colaborações que muito acrescentaram à minha formação.
Aos técnicos do LBCT (Adriana Martins, Artur Rodrigues, Beatriz Ribeiro, Darli
Grativol, Márcia Adriana Dutra, Rosemary Maciel e Sérgio Rangel Braga) pela
prestatividade e ótimo convívio.
Aos Profs. Antônio Albernaz e Francisco de Oliveira, e ao biólogo Josias Machado
pela troca de experiências e auxílio na mensuração da hemoglobina e captura de
imagens.
Ao Dr. José Morgado e ao Msc Daniel Tanaka, INCA/RJ, pela hospitalidade e
disponibilização do microscópio eletrônico em tempos de dificuldade.

VII
As fazendas parceiras, Campos Avestruz e Avestrucenter, nas pessoas do Sr.
Oswaldo e do veterinário Paulo Castro respectivamente, além dos funcionários, pela
disponibilidade e cooperação.
Aos biólogos Carlos Bacelar e Rodrigo Dias e a técnica Layra, do Laboratório Plíno
Bacelar pela disponibilização de seus equipamentos para realização dos ensaios
bioquímicos.
Aos alunos Gustavo Stratievsky, Diego Medina e Nicole Ederly pela ajuda e
aprendizado.
Aos colegas do programa de pós-graduação em biociências e biotecnologia, em
especial ao Guilherme Rabelo, Joseph Evaristo, Emílio Castro, Daniel Zandonadi,
Denise Klein, Cristiane Carvalho, Elaine Roiffé, Luiz Renato Maciel e Jorge André.
Aos meus irmãos (Júnior, Marco Antônio e Marcus César) por toda amizade e apoio
ao longo desses 10 anos em Campos.
Aos grandes amigos que fiz nesse longo período em Campos (República da Mãe
Joana, Bonde da Cirrose e banda Dumêiprufim). Grandes irmãos que deixarão
saudades.
Ao CAPES, CNPq, FAPERJ, TECNORTE e UENF pelo apoio financeiro e logístico.

VIII
Sumário Página
Agradecimentos VI
Sumário VIII
Lista de figuras X
Lista de tabelas XII
Lista de abreviações XIII
Resumo XIV
Abstract XV
Introdução 1
Avestruz 1
Hematologia aviária 3
Heterófilos 5
Eosinófilos 6
Basófilos 6
Linfócitos 7
Monócitos 7
Trombócitos 8
Bioquímica sérica 8
Aspartato aminotransferase 9
Creatina quinase 9
Ácido úrico 10
Glicose 10
Colesterol 10
Proteínas plasmáticas 11
Eletrólitos 11
Considerações finais 12
Objetivos gerais 13
Objetivos específicos 13
Materiais e métodos 14
Animais 14
Coleta de sangue e obtenção dos hemogramas 15
Microscopia óptica 17

IX
Ultra-estrutura 17
Rotina 17
Peroxidase 18
Bioquímica 18
Aspartato aminotransferase 18
Creatina quinase 19
Ácido úrico 19
Glicose 19
Colesterol 20
Proteínas plasmáticas 20
Eletrólitos 20
Resultados 22
Leucócitos 22
Hematologia 29
Bioquímica sérica 31
Discussão 33
Conclusão 43
Referências Bibliográficas 44

X
LISTA DE FIGURAS
Página
Figura 1 Ratitas encontradas no mundo de hoje. A: emú. B: ema,
representante sul americana do grupo. C: avestruzes macho
(esquerda) e fêmea (direita). D: cassoar. E: kiwi.
2
Figura 2 Contenção e coleta de sangue em avestruzes. A: contenção
usando capuz preto. B: coleta de sangue de uma avestruz com
acesso à veia ulnar e utilizando sistema de coleta a vácuo.
15
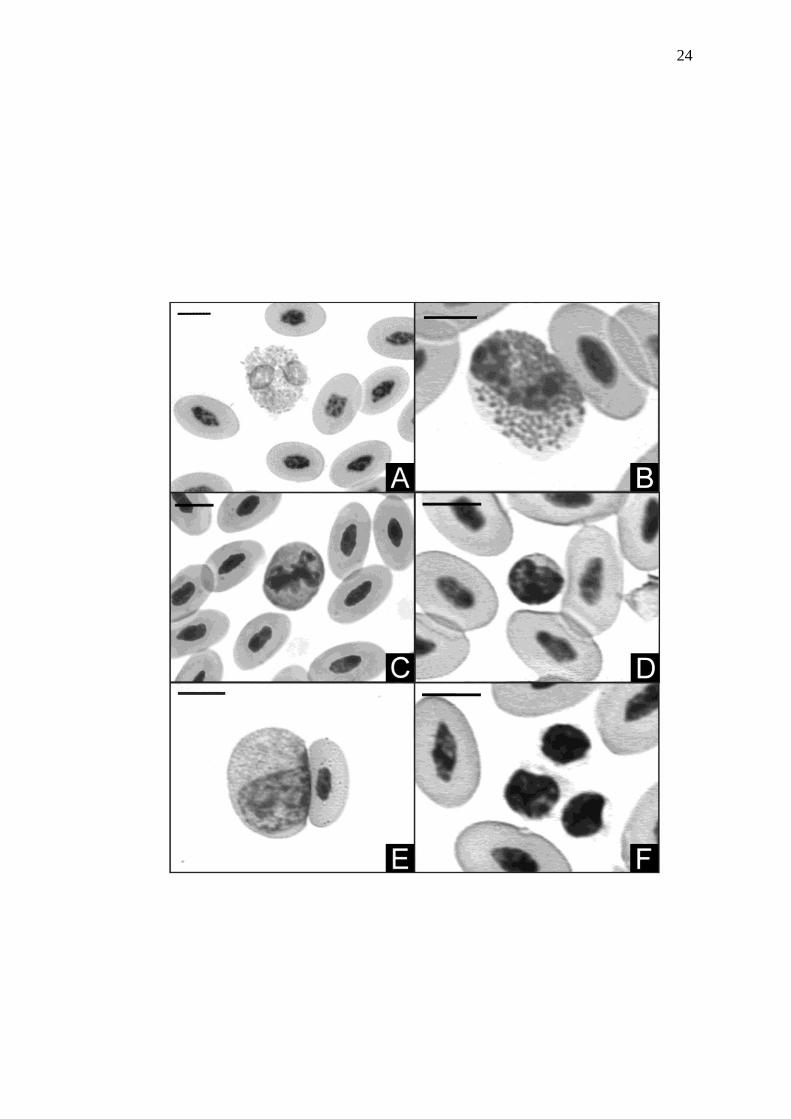
Figura 3 Leucócitos de avestruz, corados com Giemsa, observados em
microscopia óptica de campo claro. A: heterófilo apresentando
núcleo lobulado e grânulos fusiformes; B: eosinófilo; com núcleo
riniforme e citoplasma repleto de grânulos arredondados; e C:
basófilo, notar grânulos grandes e redondos dispostos sobre o
núcleo. D: linfócito; notar alta relação núcleo citoplasma, no qual
este aparece como uma linha tênue ao redor do núcleo e aquele
apresenta grande quantidade de heterocromatina. E: monócito;
notar baixa relação núcleo citoplasma e presença de vesículas
citoplasmáticas. F: agregado de trombócitos, notar núcleo
condensado e citoplasma hialino. Barra = 7 µm
24
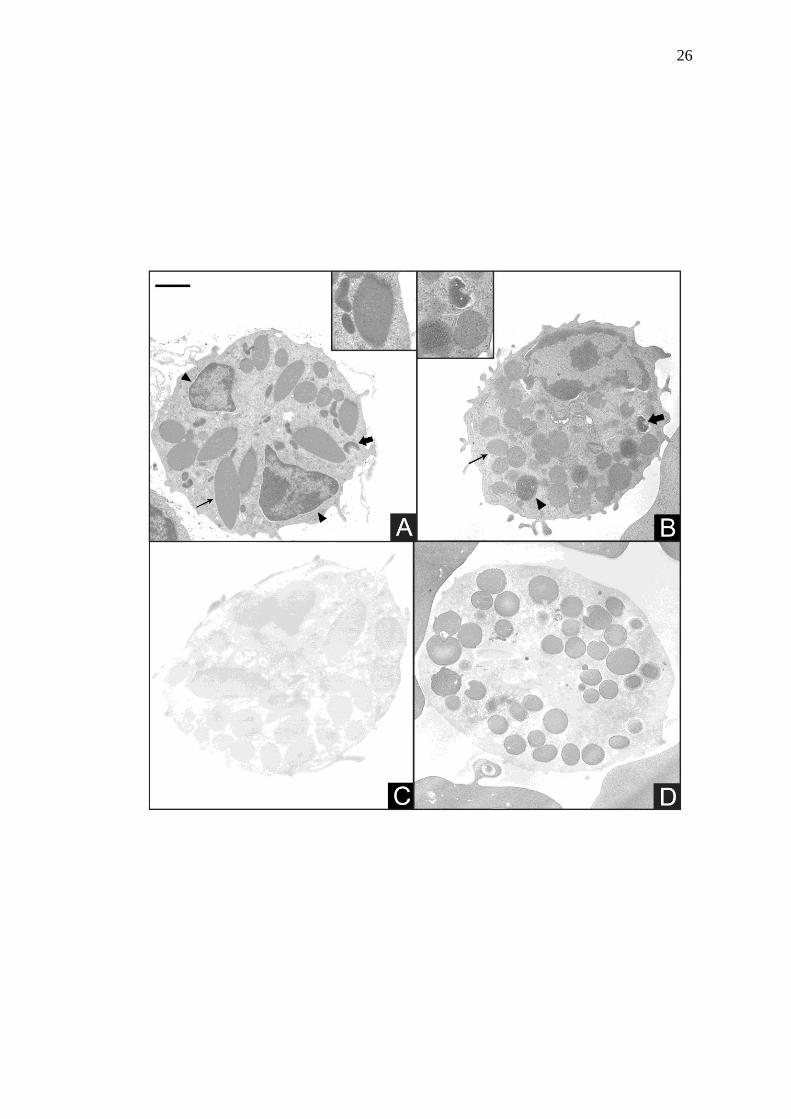
Figura 4 Leucócitos granulocíticos de avestruz, preparados para rotina (A
e B) e para detecção da enzima peroxidase (C e D), observados
por microscopia eletrônica de transmissão. A: heterófilo; notar
núcleo lobulado (cabeça de seta) e dois tipos de grânulos: I) em
forma de bastão e menos eletrondensos (seta) e II) menores, de
forma heterogênea e mais elétrondensos (seta larga). Inset
mostrando diferença entre os grânulos. B: eosinófilo; observar
núcleo riniforme e periférico. No citoplasma, destaque para os
três padrões de grânulos, tipo I (cabeça de seta), II (seta) e III
(seta larga). Inset mostrando diferença entre os grânulos. C:
heterófilo apresentando ausência de atividade dessa enzima em
seus grânulos. D: eosinófilo com grânulos eletrondensos
confirmando atividade da enzima em seu interior. Barra = 1,1
µm.
26

XI
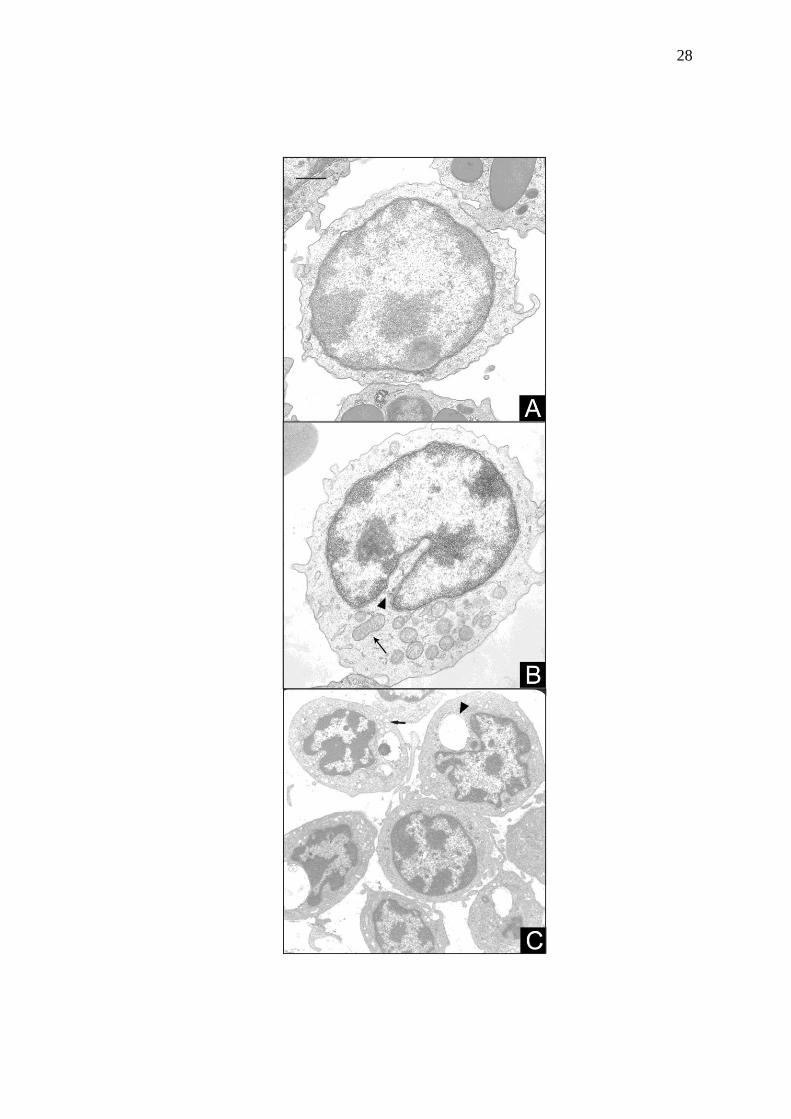
Figura 5 Leucócitos agranulocíticos de avestruz preparados para rotina
em microscopia eletrônica de transmissão. A: linfócito
apresentando núcleo repleto de heterocromatina, e citoplasma
escasso e homogêneo. B: monócito com evidentes
mitocôndrias (seta). Notar núcleo sem lobulações e presença de
sulco na face citoplasmática do núcleo (cabeça de seta). C:
trombócito apresentando núcleo repleto de heterocromatina,
vacúolos citoplasmáticos (cabeça de seta) e sistema canalicular
interno (seta). Barra = 1,1 µm.
28

XII
LISTA DE TABELAS
Página
Tabela 1 Níveis de garantia contidos na ração utilizada na
alimentação das avestruzes. 14
Tabela 2 Valores para série vermelha, contagem de trombócitos e
leucócitos totais de avestruzes. Sangue de machos (n =
50) e fêmeas (n = 50) foram avaliados pelos métodos
descritos e os resultados apresentados em médias com
desvios padrões.
30
Tabela 3 Contagens diferenciais dos leucócitos de avestruz.
Esfregaços de machos (n = 50) e fêmeas (n = 50) foram
analisados e os resultados apresentados em médias com
seus desvios padrões.
30
Tabela 4 Parâmetros bioquímicos séricos de avestruzes. Soro de
machos (n = 25) e fêmeas (n = 25), foi analisado e os
resultados apresentados em média com seus desvios
padrões.
32
Tabela 5 Valores hematológicos encontrados neste trabalho
comparados aos observados em outros estudos. 39
Tabela 6 Parâmetros bioquímicos encontrados neste trabalho
comparados aos observados em outros estudos. 42

XIII
LISTA DE ABREVIAÇÕES
A:G: albumina/globulina
AST: aspartato aminotransferase
BSA: albumina de soro bovina
CHCM: concentração de hemoglobina corpuscular média
CK: creatina quinase
CO2: dióxido de carbono
Cu+2: íon cúprico
ET: hematimetria
GA: glutaraldeído
H2O2: peróxido de hidrogênio
Hb: hemoglobina
K+: potássio
LG: leucometria
Na+: sódio
OsO4: tetróxido de ósmio
PA: formaldeído recém preparado
PT: proteínas totais
TB: trombócitos
VCM: volume corpuscular médio
VG: volume globular

XIV
RESUMO
Avestruzes foram introduzidas no Brasil no início da década passada. Hoje, o
país possui um rebanho de 250 mil aves criadas majoritariamente na região sudeste.
Entretanto, a falta de dados sobre morfologia de leucócitos, perfis hematológico e
bioquímico cria dificuldades no diagnóstico de patologias. Portanto, o objetivo deste
estudo foi estabelecer tais parâmetros nessas aves. Para isto, sangue foi coletado
da veia ulnar de avestruzes adultas (acima de 72 meses) criadas no Estado do Rio
de Janeiro e submetido a técnicas hematológicas (contagem de trombócitos,
leucócitos, hemácias, diferencial dos leucócitos e mensuração da hemoglobina) e
caracterização ultra-estrutural dos leucócitos. Esfregaços sanguíneos foram corados
com Giemsa e observados em microscopia de campo claro. Leucócitos foram
separados das hemácias, fixados, preparados, para rotina e localização da peroxidase,
e observados no microscópio eletrônico de transmissão. Outra amostra sanguínea foi
coagulada e o soro coletado e conservado a -70 oC para posterior análise por
bioquímica seca. Heterófilos apresentaram núcleo lobulado e dois tipos de grânulos
citoplasmáticos. Um grande, elíptico e menos elétrodenso que o outro, menor de
morfologia heterogênea. Ambos grânulos não apresentaram atividade peroxidásica.
Eosinófilos tinham núcleo riniforme, às vezes lobulado, excêntrico e dois padrões de
grânulo com mesma morfologia, mas de diferente elétrodensidade. Estes grânulos
apresentaram reação positiva para peroxidase. Linfócitos, monócitos e trombócitos
mostraram as mesmas características de outras aves. Parâmetros hematológicos e
bioquímicos não tiveram relação com gênero e foram estabelecidos como padrões de
normalidade para avestruzes criadas no Brasil. Essas informações auxiliarão no
diagnóstico de patologias específicas de avestruzes e servirão como conhecimentos
básicos no estudo de imunologia e patologia comparada nessa espécie.
Palavras-chaves: Struthio camelus, leucócitos, hematologia, bioquímica sérica

XV
ABSTRACT
Ostriches were introduced in Brazil in began of the past decade. Nowadays
the country has 250 thousand animals, most of them raised in the southeastern
region. However, the lack of information about the leukocytes morphology,
hematological e biochemistry profiles creates difficulties to diagnose pathologies.
Thus, the aim of this study was the established these parameters in ostriches. Blood
was collected from the ulnar vein of adults (up to 72 months) ostrich of a farm localized
in Rio de Janeiro State. Blood were collected for the hemogram process (thrombocytes,
erythrocytes, leukocytes, differential leukocyte count and hemoglobin measurement) and
preparation for ultra-structural analysis. Blood smears were stained by the Giemsa
method and were observed by light microscopy. Leukocytes were separated from red
cells, fixed and prepared, for routine and peroxidase localization, and examined in a
transmission electron microscope. Another blood sample were clothed, serum was
collected and stored at -70 oC for posterior dry biochemical analysis. Heterophil
presented lobulated nucleus and two kinds of citoplasmatic granules. One big, rod
shaped and less electrondense than the other, small with heterogenic morphology. Both
granules lacked peroxidase activity. Eosinophils had kidney shaped, sometimes
lobulated, eccentric nucleus and two kinds of granules with the same morphology, but
different electrondensity. These granules showed positive reaction for peroxidase.
Lymphocytes, monocytes and thrombocytes reveled the same characteristics of others
avian species. Hematological and serum biochemical profiles had no gender influence,
and were established as default of normality for ostriches raised in Brazil. These
information will help in diagnoses specific ostriches pathologies and serve as basics
knowledges for studies in immunology and comparative pathology in this specie.
Keywords: Struthio camelus, leukocytes, hematology, serum biochemistry

1
1- Introdução
A estrutiocultura, cirção racional de avestruzes, foi introduzida no Brasil no
início da década de 90. Porém, os primeiros investidores não obtiveram sucesso na
criação, devido a falta de conhecimento sobre as necessidades e características
dessa espécie. Mesmo assim, a partir do século XXI, seu potencial econômico é
verificado pelo crescimento do rebanho brasileiro que progride em escala
geométrica. Principalmente após a Portaria no 36 do Ministério do Meio Ambiente em
2002 e a Instrução Normativa Conjunta no 2 em 2003. Ambas determinando que a
regulamentação das criações passa a ser, a partir de então, de responsabilidade do
Ministério da Agricultura Pecuária e Abastecimento. Mudança que desburocratizou o
processo de implantação da atividade nas propriedades. E o interesse de
profissionais brasileiros que, ao buscar conhecimentos em países onde avestruz é
criada com sucesso, possibilitou a estruturação da estrutiocultura no país.
No Brasil, apesar de hoje termos número considerável de criadores
organizados em associações, existe carência de profissionais habilitados na área.
Ademais, os conhecimentos a respeito da fisiologia básica desta ave são escassos,
incluindo os necessários para o desenvolvimento de técnicas diagnósticas, tais
como morfologia de leucócitos, hemograma e bioquímica sérica.
Portanto, este estudo teve como objetivo a caracterização morfológica dos
leucócitos, determinação do perfil hematológico e bioquímico desta ave no Brasil;
subsidiando conhecimentos para o avanço da medicina diagnóstica aviária.
1.1- Avestruz
Avestruzes, Struthio camelus, emas, Rhea americana, kiwis, Apterix sp.,
cassoares, Dromaius casuarius, Casuarius bennetti e C. unappendiculatus, e emús,
D. novaehollandiae, pertencem a Superordem Paleognathae e formam o grupo das
Ratitas (Figura 1). Mesmo com antecessores voadores, as ratitas são incapazes de
voar. A ausência da quilha do externo e o atrofiamento dos músculos peitorais são
fatores determinantes para tal incapacidade. Em compensação, tanto os cinturões
torácico e pélvico quanto os membros anteriores e posteriores sofreram alterações
que permitiram excelente adaptação à vida terrestre, podendo alcançar velocidade
de 60 km/h em sua corrida (Fowler, 1991).

2
Fonte: http://en.wikipedia.org/wiki/Ratites
Figura 1: Ratitas encontradas no mundo de hoje. A: emú. B: ema, representante sul
americana do grupo. C: avestruzes macho (esquerda) e fêmea (direita). D: cassoar. E: kiwi.

3
Originárias de clima semi-árido, a adaptação de avestruzes ao clima brasileiro
ocorreu sem maiores problemas. Estas aves podem atingir uma altura máxima de 3
metros, e peso de cerca de 150 kg. Os machos adultos são pretos e brancos,
enquanto que as fêmeas são castanhas pardas. As fêmeas põem, em média, 60
ovos por ano, gerando até 20 filhotes. Os animais estão prontos para o abate com
18 meses de idade, quando pesam aproximadamente 100 kg e geram 30 kg de
carne limpa, 1,4 m2 de couro, além das plumas. A vida produtiva de uma fêmea é de
30 a 35 anos e a postura inicia aos dois anos de idade. Já o macho inicia a vida
reprodutiva aos 2,5 anos de idade. A expectativa de vida das avestruzes gira em
torno de 65 e 70 anos (ACAB, 2007).
Os principais produtos oriundos da avestruz são as plumas, o couro, os ovos
e a carne. A exploração racional da avestruz no Brasil teve seu início em 1995.
Mesmo com as dificuldades que surgiram decorrente da falta de conhecimento
técnico, a estrutiocultura tornou-se um mercado promissor sendo alvo de grandes
investidores. Entre os principais motivos desta escolha estão o baixo custo de
implantação e a viabilidade em pequenas extensões de terra. Além da alta
reprodutividade, uma fêmea pode gerar 20 pintos por ano, o animal ao ser abatido
possui grande versatilidade de subprodutos (REVISTA BRASILEIRA DE
AGROPECUÁRIA, 1999).
O Brasil conta com um rebanho de aproximadamente 250 mil aves, e o
Estado do Rio de Janeiro tem aproximadamente 40 investidores criando por volta de
3000 avestruzes (ACAB, 2007). A estrutiocultura no Estado cresce à medida que
novos, pequenos e grandes, produtores procuram esta prática, estimulada pela
criação de um abatedouro especializado no município de Quissamã. Hoje, a
associação de criadores do Estado já conta com doze investidores (ACAERJ, 2007).
Seis empresários apostam nessa atividade na região Norte Fluminense onde
compõem um rebanho de, aproximadamente, 300 aves.
1.2- Hematologia aviária
O sangue é um tecido de notória importância na fisiologia dos animais.
Portanto, sua análise, através do hemograma, é uma ferramenta complementar
importante na clínica veterinária. O hemograma fornece acesso a saúde geral do
animal, permite avaliar a capacidade de combate a infecções, além de auxiliar no
diagnóstico, e no progresso ou regresso de doenças (Jain, 1993).

4
A hematologia clínica aviária é relativamente recente. Apenas no início da
década de 60 foi publicado o primeiro trabalho completo sobre as células
sanguíneas de galinhas (Lucas e Jamroz, 1961). Desde então, o estudo dos
leucócitos circulantes ganhou impulso. Sergent et al. (2000) e Bowerman et al.
(2004) utilizaram o hemograma como ferramenta de avaliação do impacto de fatores
estressantes na saúde de pingüins-anões e águias-carecas, respectivamente.
Porém ressaltam a importância de uma boa coleta e de mais estudos visando o
isolamento de variáveis. Manejo, idade e enfermidades são preponderantes na
variação dos índices hematológicos.
Apesar da avestruz possuir importância econômica significativa, a falta de
parâmetros hematológicos traz grandes dificuldades na interpretação de seu
hemograma (Levi et al., 1989). Poucos trabalhos foram realizados, entre os quais,
em Botswana (Mushi et al., 1999a), Israel (Spinu et al., 1999), Turquia (Yüksek et al.,
2002) e no Brasil (Vilar et al., 2003). Ademais, nenhum estudo foi realizado acerca
da morfologia ultra-estrutural de leucócitos desta espécie. Até hoje, a classificação
destes tipos celulares nas avestruzes segue parâmetros descritos para galinhas.
Portanto, se faz importante discutir tais parâmetros.
As avestruzes, assim como todas as aves, possuem peculiaridades nas
células do sangue. Além das hemácias, os trombócitos, homólogos as plaquetas dos
mamíferos, são nucleados. Entre leucócitos, destaque aos heterófilos de galinhas e
gansos que possuem grânulos elípticos e conteúdo enzimático diferente do
encontrado nas células homólogas de mamíferos, os neutrófilos (Rausch e Moore,
1975). As características morfológicas e o perfil enzimático dos grânulos são de
grande valia nos estudos morfo-funcionais dessas células.
Enzimas como a peroxidase, fosfatase ácida e alcalina são importantes na
diferenciação de leucócitos, principalmente quando observados sob microscopia
eletrônica de transmissão. A peroxidase está relacionada com a capacidade
microbicida de neutrófilos humanos, pois possibilita a produção de espécimes
oxidantes (Babior et al., 1973), e modulação dos efeitos vasculares do óxido nítrico
em camundongos (Eiserich, et al., 2002). Essa enzima está ausente nos heterófilos
de diversas aves e roedores, diferente de alguns mamíferos (Jain, 1967; Jain, 1969;
Maxwell, 1984). Brune et al. em 1972 demonstraram que, mesmo com a ausência de
peroxidase no interior de seus grânulos, os heterófilos de galinhas são capazes de
fagocitar e destruir patógenos oportunistas que lesam humanos portadores de falha

5
na atividade desta enzima. Já a atividade oxidativa e microbicida da fosfatase ácida
está presente tanto em eosinófilos de patos (Anas platyrhynchos), gansos (Anser
anser), galinhas d’angola (Numida meleagris), codornas (Coturnix coturnix japonica),
pombos (Columbia lívia) e perus (Meleagris gallopavo) (Maxwell e Siller, 1972) como
nos dos humanos (Ghidoni e Goldberg, 1966). Entretanto, estudos sobre atividade
da fosfatase alcalina em leucócitos são escassos e controversos (Jain, 1969).
1.2.1- Heterófilos
Os heterófilos são os leucócitos circulantes predominantes de avestruzes
(Levi et al., 1989; Mushi et al., 1999a; Spinu et al., 1999; Yüksek et al., 2002; et al.,
2003). Este leucócito está envolvido na resposta inflamatória primária de aves
(Maxwell e Robertson, 1998). Quando sob restrição alimentar ou frio intenso o
número de heterófilos circulantes na galinha pode cair ou aumentar respectivamente
(Maxwell, 1993; Maxwell e Robertson, 1998). Alterações na morfologia desta célula
são encontradas em situações de doença sendo denominadas “tóxicas”. Nesta
situação a célula apresenta vacuolização citoplasmática, degranulação, rompimento
dos grânulos, e degeneração nuclear (Campbell, 2004a). Topp e Carlson (1972)
demonstraram capacidade fagocítica desses leucócitos de perus.
Redondos, os heterófilos são maiores que as hemácias e raramente variam
de tamanho. O citoplasma é transparente e pode estar levemente acidofílico caso
haja degranulação (Campbell, 1995). O núcleo é corado basofilicamente com
regiões mais escuras representando cromatina condensada. Quando maduros
apresentam lobulações nucleares. No citoplasma, estão presentes grânulos
acidofílicos em forma de bastão (Lucas e Jamroz, 1961). Dhingra et al. (1969)
observaram por microscopia eletrônica de transmissão três tipos distintos de
grânulos em heterófilos de galinhas. Um redondo e grande, mais osmiofílico que os
demais, outro elíptico com matriz densa e longitudinal, e o menor menos osmiofílico.
Ericsson e Nair (1973) também descreveram três tipos de grânulos em heterófilos de
galinhas, porém atribuíram ao maior e mais osmiofílico forma elíptica e presença de
matriz densa.

6
1.2.2- Eosinófilos
Os eosinófilos de mamíferos exercem proteção contra infecções helmínticas,
e estão freqüentemente associados a alergias, porém sua função permanece uma
icógnita nas aves (Maxwell, 1995).
Diferente dos heterófilos, os eosinófilos possuem grande variação de
tamanho, porém não de sua forma arredondada. O citoplasma possui coloração azul
claro e o núcleo, riniforme, é basofílico com regiões mais escuras de
heterocromatina (Lucas e Jamroz, 1961). No citoplasma estão contidos grânulos
eosinofílicos redondos (Campbell, 2004a). Maxwell e Siller em 1972, estudando
eosinófilos de diferentes aves, observaram variações nos grânulos de eosinófilos
nas espécies abordadas. Patos e gansos apresentaram em seus grânulos matriz
cristalina como em alguns mamíferos. Eosinófilos de codornas estimulados com soro
eqüino ou albumina bovina apresentam aumento na atividade da fosfatase ácida nos
grânulos, bem como alterações morfológicas destas organelas (Maxwell, 1986).
1.2.3- Basófilos
Os basófilos são normalmente maiores que os heterófilos e eosinófilos.
Embora pertençam a população minoritária de leucócitos circulantes, os basófilo
possuem função importante nos eventos inflamatórios, secreção de aminas
vasoativas, e na hipersensibilidade cutânea (Maxwell e Robertson, 1995). Aves sob
situação de restrição alimentar, calor intenso e medo podem apresentar basofilia,
aumento dos basófilos circulantes (Maxwell, 1993; Maxwell e Robertson, 1995).
O formato dessas células é redondo as quais possuem grande variação de
tamanho entre as espécies de aves. O citoplasma possui coloração basofílica,
porém muitas vezes escondida pela grande quantidade de grânulos citoplasmáticos.
Já o núcleo raramente apresenta lobulações, cora-se basofilicamente e
freqüentemente encontra-se sob os grânulos. Estes grânulos, redondos ou ovais,
são fortemente basofílicos (Campbell, 1995). A ultra-estrutura revela variados graus
de eletrondensidade, mais ou menos fibrilar, entre grânulos de uma mesma célula
(Dhingra et al., 1969).

7
1.2.4- Linfócitos
Os linfócitos possuem as mesmas funções das células homônimas de
mamíferos e representam a população majoritária de leucócitos de algumas aves
(Coke et al., 2004). O número de linfócitos circulantes pode estar elevado ou
reduzido em situações de restrição alimentar e medo, respectivamente (Maxwell,
1993).
Linfócitos podem variar de tamanho, mas são menores do que as hemácias.
O citoplasma é escasso e cora-se em azul vítreo, formando uma linha tênue ao
redor do núcleo, entretanto, pode aumentar de volume em linfócitos maiores.
Projeções citoplasmáticas e pseudópodos podem ser observados com freqüência. O
núcleo é redondo e centralizado, com sulcos raros, e cora-se acidofilicamente
(Lucas e Jamroz, 1961). A cromatina nuclear é, em grande parte, condensada
deixando o núcleo com diversas regiões escuras sob microscopia óptica ou
eletrondensas sob microscopia eletrônica de transmissão. Outra característica
peculiar dessas células é a alta razão núcleo:citoplasma (Campbell, 1995).
1.2.5- Monócitos
Os monócitos das aves são morfologicamente similares aos dos mamíferos.
Precursores de macrófagos, eles são as principais células fagocíticas do sistema
imune das aves. Qualidade anteriormente atribuída aos trombócitos (Chang e
Hamilton, 1979), mas que realizam a fagocitose em baixa intensidade (DaMatta et
al., 1998; Qureshi, 2003).
Normalmente maiores que as hemácias, os monócitos têm formato redondo e
citoplasma azul vítreo com borda hialina, quando observados com coloração tipo
Romanowsky. O núcleo excêntrico é curvo na face membranar, com sulco na face
citoplasmática. As principais características que diferenciam os monócitos dos
linfócitos são a menor quantidade de heterocromatina e menor razão
núcleo:citoplasma (Lucas e Jamroz, 1961). Grânulos azurofílicos podem estar
presentes no citoplasma destas células em algumas espécies de aves e répteis
(Montali, 1988).

8
1.2.6- Trombócitos
Como dito, os trombócitos são equivalentes as plaquetas dos mamíferos. São
caracterizados pela sua forma elíptica ou redonda, tamanho pequeno, presença de
núcleo, sistema canalicular interno e vacúolos citoplasmáticos (Claver, 2005). A
principal função dessas células está na participação dos eventos hemostáticos.
Embora alguns autores (Daimon et al., 1985; Taffarel e Oliveira, 1993) atribuam suas
características citoquímicas a eventos fagocíticos. A presença de proteínas
relacionadas a citotoxidade e da fosfatase ácida em seus vacúolos reforçam esta
tese. Porém, estudos posteriores (DaMatta et al., 1998) revelaram que, comparado
aos macrófagos, sua atividade fagocítica é reduzida quando bacterias interagem
com estas células e não ocorre com os protozoários Toxoplasma gondii e
Trypanosoma cruzi. Portanto, o perfil citoquímico dessas células pode estar
relacionado à remoção de debris celulares e fibrinólise na coagulação.
No esfregaço sanguíneo, os trombócitos são identificados por seu tamanho,
forma e por estarem freqüentemente agregados. Ademais, seu núcleo apresentar
grande quantidade de heterocromatina, quase picnótico, produzindo coloração
intensamente basofílica. O citoplasma é quase translúcido e pode conter grânulos
acidofílicos e vacúolos (Campbell, 1995).
1.3- Bioquímica sérica
O perfil bioquímico sérico é uma abordagem, de avaliação fisiológica,
complementar ao hemograma. Ambos formam o principal pilar do diagnóstico
veterinário (Harr, 2002). O monitoramento das funções renais, hepáticas e
musculares, do metabolismo da glicose e da concentração de íons plasmáticos
possibilita o entendimento do real efeito do manejo e doenças sobre a fisiologia das
aves.
Este dado é de grande importância na avaliação do status nutricional dessas
aves. Por exemplo, avestruzes alimentadas com elevado teor de proteína bruta
apresentam aumento na concentração sérica de ácido úrico (Polat et al., 2003).
A impactação gástrica é uma patologia comum em avestruzes que recebem
manejo nutricional inadequado. O trânsito gastro-intestinal é interrompido e o animal,
se não receber tratamento adequado, vai a óbito. Yüksek et al. (2002) e Komnenou
et al. (2003) encontraram aumento dos valores plasmáticos de enzimas hepáticas,
proteínas totais e ácido úrico em avestruzes com essa patologia.

9
Já em frangos de corte, rações contaminadas com aflatoxinas, ou até mesmo
desbalanceadas, podem alterar a funcionalidade do fígado e rins, e acarretar
mudança das concentrações das proteínas plasmáticas (Quezada et al., 2000;
Oguz, 2002).
1.3.1- Aspartato aminotransferase (AST)
A AST, também chamada de TGO, transaminase glutâmica oxalacética, é
responsável pela transaminação reversível do aspartato e α-cetoglutarato para
glutamato e oxalacetato. Duas isoformas estão presentes nos tecidos de diversos
animais, a mitocondrial e a citosólica, onde aquela é a isoenzima mais abundante
(Kaneko, 1989). Fígado, rins, coração e músculo esquelético são órgãos que
possuem concentração significante dessa enzima (Medway et al., 1973).
Conseqüentemente, em hepatopatias a concentração plasmática da AST pode estar
elevada, porém sem especificidade (Coles, 1984). Entretanto, em degeneração
hepática crônica o nível pode estar baixo indicando perda tecidual importante (Meyer
et al., 1992). Na rotina clínica de aves a AST pode ser utilizada como enzima de
escolha para diagnosticar hepatopatias, melhor ainda quando utilizada com a
creatina quinase para descartar possibilidade de lesão muscular (Dabbert e Powell,
1993 ; Jaensch et al., 2000; Campbell, 2004b).
1.3.2- Creatina quinase (CK)
A creatina quinase, ou fosfocreatina quinase, tem como principais funções
criar reservatório energético e de prótons prontamente disponível; prevenir aumento
do ADP livre intracelular e permitir sinalização para início da glicogenólise no
exercício (Stryer,1995). Três isoformas da CK estão presentes nas aves, todas
envolvidas em metabolismo de alta energia (Kerr, 2003). Essa enzima é encontrada
no citoplasma de células musculares estriadas, lisas e cardíacas (Lassen, 2004). E
por isso é considerada a melhor abordagem na avaliação de danos musculares.
O aumento desta enzima no soro ou plasma pode indicar infarto do miocárdio,
degeneração muscular ou apenas um esforço físico marcante. A CK é uma enzima
muscular específica nas aves (Campbell, 2004b). Portanto, deve ser utilizada para
avaliar danos nesse tecido.

10
1.3.3- Ácido úrico
O ácido úrico é o produto majoritário do catabolismo de proteínas nas aves
(Harr, 2002). Sua síntese ocorre no fígado e quando acumulado acarreta aumento
de sua concentração no plasma. Esta alta concentração pode levar a uma patologia
denominada gota que se caracteriza pela precipitação de cristais de ácido úrico,
principalmente, nas articulações e superfícies de órgãos. Casos crônicos desta
patologia podem culminar na morte do animal (Sturkie, 1965). O aumento de sua
concentração plasmática também pode estar associado a necrose tecidual e jejum
prolongado, com catabolismo de compostos nitrogenados (Campbell, 2004b).
Portanto, o ácido úrico pode ser utilizado como indicador de alterações no
metabolismo renal.
1.3.4- Glicose
A glicose é a molécula de pronta obtenção de energia das células em geral.
Sua concentração no sangue, a glicemia, é influenciada pela ação de hormônios,
tais como glucagon, insulina e corticóides (Sturkie, 1965). Existem grandes
variações nas concentrações plasmáticas entre as espécies de aves granívoras e
carnívoras (Harr, 2002). A hiperglicemia pode ser causada por estresse, diabetes
mellitus e corticóides endógenos ou exógenos, já hipoglicemia está relacionada ao
jejum e insulinomas, neoplasia das células de Langerhans (Kerr, 2003).
1.3.5- Colesterol
O metabolismo do colesterol em aves é idêntico ao de mamíferos (Harr,
2002). Porém, possui participação importante na vitelogênese de algumas espécies
desses ovíparos (Sturkie, 1965).
Aumento no colesterol plasmático pode estar associado à fibrose hepática ou
obstrução dos ductos biliares. Hipotiroidismo, lipemia e dietas com altos níveis de
gordura também podem elevar a concentração de colesterol. Já a diminuição pode
indicar resolução de doença hepática, má-digestão, má-absorção ou jejum
prolongado (Campbell, 2004b).

11
1.3.6- Proteínas plasmáticas
Proteínas plasmáticas podem ser divididas em albumina, globulinas, proteínas
de transporte, enzimas, hormônios e proteínas de coagulação. Exceto pelas
globulinas, produzidas pelas células plasmáticas derivadas dos linfócitos B, essas
moléculas são formadas no fígado. Uma das principais funções dessas moléculas é
a garantia da osmolaridade intra, extra-celular e no interior vascular. Porém existem
as globulinas, fibrinogênio e proteínas de transporte com funções específicas na
imunidade, coagulação e sinalização endócrina, respectivamente (Sturkie, 1965).
Entretanto, apenas a primeira fração, composta pela albumina e imunoglobulinas,
bem como a razão entre as partes (A:G), possui valor de diagnóstico (Campbell,
2004b). Esses parâmetros comumente estão alterados em processos inflamatórios
agudos e crônicos, falência renal, hepatopatias e desidratação (Harr, 2002).
1.3.7- Eletrólitos
Os eletrólitos de maior importância na fisiologia das aves são sódio (Na+) e
potássio (K+), onde este está em menor concentração plasmática do que aquele
(Sturkie, 1965). O Na+ consumido pelas aves é absorvido nos intestinos e excretado
na urina. Sua principal função fisiológica está relacionada ao balanço hídrico.
Avestruzes possuem glândulas de sal, localizadas logo abaixo das órbitas, de onde
derivam ductos até a extremidade do bico. Estas glândulas podem ser responsáveis
por até 80% da excreção de sódio em aves sob alta temperatura (Campbell, 2004b).
Patologias que afetam rins, intestinos e até glândulas de sal conseqüentemente
podem levar a perda excessiva de Na+. Já a hipernatremia pode ser causada por
severa desidratação ou perda de líquido pobre em Na+, como vômito e diarréia (Kerr,
2003).
O K+ é o cátion intracelular predominante (Campbell, 2004b). E compõe, junto
ao Na+, dentre outros exemplos, o sistema de co-transporte celular responsável pela
captação da glicose. Distúrbios na concentração deste elemento acarreta diminuição
da atividade elétrica de células, e normalmente podem estar associados a distúrbios
das glândulas adrenais (Kerr, 2003).

12
1.4- Considerações finais
Com a base de dados gerada a partir desse estudo compreendendo:
morfologia de leucócitos, perfil hematológico e bioquímico das avestruzes, espera-se
contribuir para o aprimoramento do diagnóstico, hematológico e bioquímico, de
patologias que acometem essas aves. Estes dados também servirão como
conhecimentos básicos, e conseqüentemente necessários, para estudos em
patologia comparada e imunobiologia de aves.

13
2- Objetivos
2.1- Objetivo geral
- Caracterizar os leucócitos circulantes de S. camelus, através da citoquímica
sob microscopia óptica e eletrônica de transmissão, e determinar o perfil
hematológico e bioquímico sérico destas aves.
2.2- Objetivos específicos
- Comparar a ultra-estrutura e a presença da atividade da enzima peroxidase
nos leucócitos de S. camelus com de outras aves previamente estudadas.
- Verificar se há diferenças entre machos e fêmeas, quanto aos valores
hematológicos e bioquímicos.
- Determinar os perfis hematológicos e bioquímicos para avestruzes adultas
no Brasil.

14
3- Materiais e métodos
3.1- Animais
As aves examinadas pertencem a fazenda Avestrucenter, localizada no
município de Cabo Frio (S 22° 46’ 31’’; N 42° 01’ 33’’), estado do Rio de Janeiro.
Cem avestruzes, 50 machos e 50 fêmeas, maiores de 30 meses, foram utilizados
para determinar o perfil hematológico. Já para o perfil bioquímico foram 25 aves de
cada sexo do mesmo rebanho. A dieta oferecida era composta de água, gramínea à
vontade e ração específica com níveis de garantia expostos na tabela 1.
Anualmente, entre os períodos de postura, que vai de março a julho, as aves
da propriedade são submetidas a controle parasitológico, com febendazol (30
mg/kg) e ivermectina (0,4 mg/kg). Mesmo assim, as avestruzes passaram por
avaliação clínica, na qual apenas as aparentemente saudáveis e de jejum tiveram o
sangue coletado. A coleta foi realizada entre os meses de maio e junho de 2006 no
período matutino.
Tabela1: Níveis de garantia contidos na ração comercial utilizada na alimentação das avestruzes.
Composto Quantidade (%)
Umidade (máx.) 13
Proteína Bruta (mín.) 12
Extrato Etéreo (mín.) 2
Matéria Fibrosa (máx.) 20
Matéria Mineral (máx.) 20
Cálcio (máx.) 2
Fósforo (mín.) 0,6
Fonte: http://www.guabi.com.br/rc/avestruzes/produtos.asp?marca=9&codigo=197

15
3.2- Coleta de sangue e obtenção dos hemogramas
Depois da contenção física, utilizando gancho e capuz, o sangue das aves foi
coletado através da punção da veia ulnar com tubos a vácuo (Vacutainer BD)
contendo heparina lítica equipados com agulhas 25 x 7 mm (BD) (Figura 2).
Esfregaços sanguíneos foram feitos imediatamente e os frascos, contendo sangue,
acondicionados a 10oC até a chegada ao laboratório.
Figura 2: Contenção e coleta de sangue em avestruzes. A: contenção usando capuz
preto. B: coleta de sangue de uma avestruz com acesso à veia ulnar e utilizando sistema de
coleta a vácuo.

16
Para a série vermelha foram obtidos: i) volume globular (VG) pelo
microhematrócrito; ii) hematimetria (ET) pela contagem direta do número de
hemácias na câmara de Neubauer; e iii) dosagem da concentração de hemoglobina
pelo método colorimétrico.
Para a obtenção do VG pelo microhematócrito, um tubo capilar de vidro foi
preenchido com sangue total e centrifugado a 20000 g por 5 minutos. Passado o
tempo o tubo foi colocado sobre régua específica e o volume de células em relação
ao volume total foi obtido em porcentagem. Para a ET, o sangue foi adicionado ao
diluente de Dacie modificado (1 l de água destilada; 10 ml de formaldeido 40% e
31,3 mg de citrato trissódico) facilitando a visualização e contagem das células na
câmara de Neubauer (Campbell, 2004a). Para contagem, 20 μl de sangue foi
adicionado a 2 ml do diluente acrescido de 20 μl do corante azul cresil brilhante. A
hemoglobina foi mensurada utilizando reação enzimática da cianometemoglobina
(Labtest Ind. Bras.), e posterior leitura por espectrofotometria em 540 nm. A reação
foi realizada adicionando 20 μl de sangue a 5 ml de reagente do kit, seguido de
homogeneização. Após dez minutos, antes da mensuração da hemoglobina, a
solução foi centrifugada a 1600 g por 5 minutos para remover partículas nucleares
das hemácias evitando interferência na leitura.
Com os dados de microhematócrito, hematimetria e concentração de
hemoglobina foram calculados o Volume Corpuscular Médio (VCM) e a
Concentração de Hemoglobina Corpuscular Média (CHCM) (Jain, 1993). Onde:
VCM = VG x 10 / hematimetria (hemácias x 106/mm3)
CHCM = hemoglobina / VG x 100
Para série branca, a leucometria (LG) foi obtida através da contagem dos
leucócitos totais na câmara de Neubauer de sangue heparinizado. Como a heparina
causa aglomeração de leucócitos no sangue de avestruzes, especialmente
heterófilos (Green e Blue-McLendon, 2000), a leucometria também foi obtida através
de contagem das células nos esfregaços feitos com sangue livre de anticoagulante.
Os leucócitos foram contabilizados ao longo do esfregaço em paralelo as hemácias.
Quando duas mil hemácias foram registradas, o LG foi calculado integrando o
numero de leucócitos contados no esfregaço com o ET conforme fórmula proposta
por Tavares-Dias e Moraes, 2006:
LG (leucócitos/mm3) = leucócitos contados no esfregaço x ET (hemácias/mm3) /
2000

17
A contagem diferencial foi feita em extensões sanguíneas coradas pelo
método do Panótico rápido. As células foram avaliadas e contadas utilizando
microscópio Ziess Axioplan.
Os trombócitos e hemácias foram contabilizados simultaneamente na câmara
de Neubaer.
3.3- Microscopia óptica
Extensões sanguíneas recém feitas foram fixadas em metanol por 5 minutos e
coradas com Giemsa por 40 minutos. Depois de lavadas e secas, foram observadas,
em campo claro, em microscópio óptico Ziess Axioplan. As células foram
documentadas digitalmente pelo programa AnaliSys acoplado ao microscópio.
3.4- Ultra-estrutura
Um colchão leucocitário foi obtido, segundo Silva et al., (2004). Para obtenção
do colchão, um tubo plástico foi preenchido com sangue total, vedado com duas
esferas metálicas, colocado em um tubo de 15 ml e centrifugado a 700 g por 15
minutos a 10oC. Após a centrifugação, que separa os leucócitos das hemácias, o
tubo foi cortado, sob lupa, com uma lâmina de bisturi na intersecção de leucócitos e
hemácias. Os leucócitos foram aspirados com auxílio de pipeta Pasteur e
preparados de acordo com os tratamentos descritos a seguir. As imagens foram
obtidas no microscópio eletrônico de transmissão Zeiss 900 operado em 80 kv para
rotina e 50 kv para citoquímica.
3.4.1- Rotina
As células foram fixadas com 2% glutaraldeído (GA), 4% formaldeído recém
preparado (PA) em tampão cacodilato 0,1 M, pH 7,2, durante 1 hora. Em seguida as
células foram lavadas com o mesmo tampão por três vezes. A pós-fixação foi com
tetróxido de ósmio (OsO4) a 1 % em tampão cacodilato contendo 0,8% de
ferricianeto de potássio por 1 hora. Depois de lavadas três vezes com tampão
cacodilato, a amostra foi submetida à desidratação em série crescente de acetona,
50 a 100%, com 30 minutos em cada etapa. A inclusão foi feita em resina epóxi
polimerizada por 48 horas a 60oC. Cortes ultra-finos de 80 nm foram feitos no
ultramicrótomo para posterior montagem em grades de cobre de 200 mesh. A

18
contrastação foi feita com acetato de uranila (5%), por vinte minutos, e citrato de
chumbo (2%), por cinco minutos, na ausência de luz.
3.4.2- Peroxidase (Roels et al., 1975)
As células foram fixadas em GA 0,5% em tampão cacodilato 0,1 M, pH 7,2,
por trinta minutos. Em seguida, foram lavadas em tampão cacodilato com 5% de
sacarose. E pré-incubadas com 0,5 mg/ml de diaminobenzidina por 30 minutos em
tampão cacodilato 0,1 M, pH 7,0 com 5% de sacarose. Após este tempo outra
incubação foi realizada na mesma solução contendo peróxido de hidrogênio (H2O2) a
0,02 %, por mais trinta minutos. As células foram lavadas em tampão cacodilato com
5% de sacarose por 2 vezes. Depois o material foi incluído e cortado como descrito
no item 4.4.1. A contrastação não foi realizada. No controle foi omitido o H2O2.
3.5- Bioquímica
O soro foi obtido através da punção da veia ulnar, com tubos a vácuo
(Vacutainer BD) sem anticoagulante com gel separador. Logo depois da
coagulação, ainda no campo, o tubo foi centrifugado por 5 minutos a 2600g e os
tubos acondicionados a 10 oC. No laboratório o soro foi aliquotado em tubos
plásticos de 1,5 ml e, em seguida, congelados à –70oC. Para análise bioquímica, as
enzimas mensuradas foram AST e CK, além de ácido úrico, glicose, colesterol,
proteínas plasmáticas e eletrólitos. Todos os ensaios enzimáticos foram feitos
utilizando método de bioquímica seca no equipamento Vitros 950 (Johnson &
Johnson’s®) conforme descrito em seguida.
3.5.1- AST
A AST foi mensurada pela taxa de pontos múltiplos, onde 11 μl de soro foi
depositado sobre o reativo seco e difundido para as camadas subjacentes. O reativo
seco é constituído por um elemento analítico seco de múltiplas camadas envolvido
em suporte transparente de poliéster. Neste teste o grupamento amino do L-
aspartato é transferido para α-cetoglutarato, na presença do piridoxal-5-fosfato,
produzindo glutamato e oxaloacetato. Este é convertido em piruvato e dióxido de
carbono (CO2) pela oxaloacetato descarboxilase. Então o piruvato é oxidado a
acetilfosfato e H2O2 pela piruvato oxidase. A última etapa da reação é catalisada
pela peroxidase oxidando um corante branco, produzindo um corante colorido. O

19
índice de oxidação do corante branco é monitorado por espectrofotometria a 37 oC
em 670 nm por 5 minutos. Um filtro de corte de baixo comprimento de onda no
suporte do reativo evita interferências da luz incidente durante a produção do
corante.
3.5.2- CK
A CK foi mensurada pela mesma técnica utilizada para a AST. Onze μl de
soro foi depositado, e difundido, no reativo seco que continha N – acetilcisteína para
ativar a enzima. Assim, a CK catalisa a conversão do fosfato de creatina e ADP em
creatina e ATP. Este possibilita a fosforilação do glicerol, pela glicerol quinase, que
resulta no L- α-glicerofosfato. Em seguida, este é oxidado pela L- α-glicerofosfato
oxidase, gerando fosfato de diidróxiacetona e H2O2. O final desta reação ocorre
assim como no item 4.5.1, onde a transformação do corante branco em colorido é
monitorada em 670 nm a 37 oC por 5 minutos. Para aumentar a precisão no
resultado o soro foi diluído, 1:4, em albumina de soro bovina (BSA) a 7% Vitros.
3.5.3- Ácido úrico
Este metabólito é mensurado pelo teste colorimétrico em reativo seco. Dez μl
de soro foi depositado e uniformemente distribuído pelas camadas do reativo seco.
Então, o ácido úrico, ao migrar para camada do reagente, é oxidado pela uricase
formando alantoína e H2O2. A partir daí a análise ocorre como descrito nos itens
anteriores. O monitoramento da transformação do corante é dado em 670 nm a 37
oC por 5 minutos.
3.5.4- Glicose
A glicose foi mensurada pelo teste colorimétrico. Após difusão de 10 μl de
soro sobre o reagente seco, a glicose foi oxidada pela glicose oxidase para formar
gluconato e H2O2. Este catalisa a oxidação de precursores do corante vermelho
utilizado para leitura da luz refletida. A análise foi realizada em comprimento de onda
de 540 nm a 37 oC por 5 minutos.
3.5.5- Colesterol
O teste colorimétrico também foi escolhido para mensuração do colesterol. Ao
depositar 10 μl de soro, o colesterol, assim como os ésteres de colesterol, foi

20
dissociado de lipoproteínas pela ação do surfactante Triton X-100 ao ser difundido
no reagente seco. Os ésteres de colesterol foram hidrolisados pela éster hidrolase.
Assim, o colesterol livre foi oxidado pela colesterol oxidase para gerar colestenona e
H2O2. Finalmente o H2O2 oxidou o corante branco gerando um corante colorido. A
densidade de corante formado foi proporcional a concentração do colesterol na
amostra. A mensuração foi realizada por espectrofotometria em 540 nm a 37 oC por
5 minutos.
3.5.6- Proteínas plasmáticas
O método de análise das proteínas totais foi baseado na reação do biureto
que produz um complexo violeta após a reação das proteínas com o íon cúprico
(Cu+2) em pH alcalino. Assim, o montante de 10 μl foi depositado e distribuído pelo
reativo. Ao entrarem em contato com o reagente, tartarato de cobre, as proteínas
possibilitam a formação do complexo colorido. A quantidade de proteínas totais foi
dada por espectrofotometria em 540 nm a 37 oC por 5 minutos.
A albumina também foi mensurada pelo método colorimétrico. Ao depositar e
difundir 10 μl de soro sobre o reativo, a albumina reagiu com verde de bromocresol
presente na camada de dispersão. O complexo formado foi medido por
espectrofotometria em 540 nm a 37 oC por 5 minutos.
As globulinas foram obtidas pela diferença entre as proteínas totais e
albumina. Já a relação albumina/globulina (A:G) foi obtida pela divisão entre os
respectivos valores.
3.5.7- Eletrólitos
O Na+ foi mensurado pelo teste potenciométrico. Neste teste, o reativo seco
consistia em dois eletrodos, referência e indicador, de íons seletivos contendo três
camadas cada: i) membrana de íons seletivos com metil monensina, ionóforo para
sódio; ii) camada de referência, com cloreto de sódio e pH 5,6 e iii) outra membrana
de íons seletivos contendo prata e cloreto de prata. Então, 10 μl de soro foram
aplicados sobre o reativo que junto a 10 μl fluido de referência Vitros formaram uma
solução estável. Assim, cada eletrodo produziu um potencial elétrico em resposta à
concentração de sódio aplicada. A diferença de potencial existente entre os
eletrodos foi proporcional à concentração do íon na amostra. O teste durou
aproximadamente 2 minutos a 37 oC.

21
O método potenciométrico também foi utilizado para o K+. Neste, o reativo
seco continha dois eletrodos, referência e indicador, selecionadores de íons
contendo: i) valinomicina, ionóforo para potássio; ii) camada de referência com
cloreto de potássio e iii) prata e cloreto de prata. A partir daí os eventos são os
mesmos descritos para o sódio.
3.6- Análise estatística
Os dados foram tabulados em planilhas do Microsoft Excel e tiveram suas
respectivas médias e desvios padrões calculados. As diferenças entre as médias de
machos e fêmeas foram avaliadas através do teste t de Student.

22
4- Resultados 4.1- Leucócitos
Sob a luz da microscopia óptica os heterófilos (Fig. 3A) apresentaram núcleo
basofílico com até três lóbulos. O citoplasma destas células apresentou coloração
levemente acidofílica, com grânulos em forma de bastão e corados acidofilicamente.
Outra população de granulócitos, os eosinófilos (Fig. 3B), apresentou núcleo
basofílico, excêntrico, riniforme e raramente lobulado. O citoplasma apresentou cor
azulada, semi-translúcido, contendo grânulos eosinofílicos e arredondados em
abundância. Os basófilos (Fig. 3C) apresentaram núcleo basofílico e bastante
lobulado. O citoplasma apresentou coloração basofílica, contendo grânulos grandes
e esféricos, fortemente basofílicos, dispersos sobre e próximo ao núcleo. Os
linfócitos (Fig. 3D) e monócitos (Fig. 3E) apresentaram características tintoriais
semelhantes. Estas células apresentaram núcleo basofílico e sem lobulações. O
citoplasma destas células apresentou coloração levemente azulada e sem grânulos.
Entretanto, foram encontrados eventualmente grânulos citoplasmáticos acidofílicos
em monócitos. A diferença entre essas células foi a relação núcleo:citoplasma, que
foi menor nos monócitos. Ademais o núcleo dos linfócitos apresentou coloração
fortemente heterocromática. Os trombócitos (Fig. 3F) apresentaram forma variada,
desde redondo a elíptico. Normalmente estavam agregados demonstrando núcleo
fortemente basofílico, citoplasma escasso e hialino sem grânulos. Freqüentemente
foram observados vacúolos citoplasmáticos.
Na observação em microscopia eletrônica de transmissão os heterófilos (Fig.
4A) apresentaram núcleo lobulado com heterocromatina distribuída em sua periferia.
Dois tipos de grânulos foram observados. Os fusiformes de diferentes tamanhos
foram chamados de tipo I, e os menores, de forma heterogênea, mais elétrondensos,
foram denominados tipo II. Os eosinófilos (Fig. 4B) apresentaram núcleo excêntrico
com heterocromatina na periferia, e nucléolo. No citoplasma foram observados três
padrões de grânulos com diferença de eletrondensidade. Os grânulos tipo I e II eram
mais comuns e redondos, sendo este mais elétrondenso que aquele. Já o tipo III não
foi observado em todos eosinófilos. Este tipo era redondo como os outros, porém
com material elétrondenso depositado excêntricamente. Infelizmente não foi possível
observar basófilos.

23
Figura 3: Leucócitos de avestruz, corados com Giemsa, observados em
microscopia óptica de campo claro, notar hemácias elípticas e nucleadas. A:
heterófilo apresentando núcleo lobulado e grânulos fusiformes; B: eosinófilo; com
núcleo riniforme e citoplasma repleto de grânulos arredondados; e C: basófilo, notar
grânulos grandes e redondos dispostos sobre o núcleo. D: linfócito; notar alta
relação núcleo citoplasma, no qual este aparece como uma linha tênue ao redor do
núcleo e aquele apresenta grande quantidade de heterocromatina. E: monócito;
notar baixa relação núcleo citoplasma e presença de vesículas citoplasmáticas. F:
agregado de trombócitos, notar núcleo condensado e citoplasma hialino. Barra = 7
µm
24

25
Figura 4: Leucócitos granulocíticos de avestruz, preparados para rotina (A e B) e
para detecção da enzima peroxidase (C e D), observados por microscopia
eletrônica de transmissão. A: heterófilo; notar núcleo lobulado (cabeça de seta) e
dois tipos de grânulos: I) em forma de bastão e menos eletrondenson (seta) e II)
menores, de forma heterogênea e mais eletrondensos (seta larga). Inset mostrando
diferença entre os dois grânulos. B: eosinófilo; observar núcleo riniforme e
periférico. No citoplasma, destaque para os três padrões de grânulos, tipo I (cabeça
de seta), II (seta) e III (seta larga). Inset mostrando os três tipos de grânulos
encontrados. C: heterófilo apresentando ausência de atividade dessa enzima em
seus grânulos. D: eosinófilo com grânulos eletrondensos confirmando atividade da
enzima em seu interior. Barra = 1,1 µm.
26

27
Figura 5: Leucócitos agranulocíticos de avestruz preparados para rotina em
microscopia eletrônica de transmissão. A: linfócito apresentando núcleo repleto de
heterocromatina, e citoplasma escasso e homogêneo. B: monócito com
mitocôndrias evidentes (seta). Notar núcleo sem lobulações e presença de sulco na
face citoplasmática do núcleo (cabeça de seta). C: trombócito apresentando núcleo
repleto de heterocromatina, vacúolos citoplasmáticos (cabeça de seta) e sistema
canalicular interno (seta). Barra = 1,1 µm.
28

29
Enquanto heterófilos (Fig. 4C) e monócitos apresentaram reação positiva
apenas no retículo endoplasmático (não mostrado) para peroxidase. Os eosinófilos
apresentaram reação positiva para esta enzima no interior de seus grânulos (Fig.
4D). Nos controles não houve marcação (não mostrado).
Os linfócitos (Fig. 5A) apresentaram heterocromatina distribuída na periferia
do núcleo, porém em maior quantidade que nos monócitos (Fig. 5B). O citoplasma
apresentou pouco volume e homogeneidade. Os monócitos apresentaram núcleo
excêntrico com heterocromatina distribuída nas bordas, presença de nucléolo e
sulco na face citoplasmática do núcleo. Baixa relação núcleo:citoplasma foi
verificada nestas células, além de mitocôndrias e vacúolos típicos observados com
freqüência. Os trombócitos, freqüentemente agregados, continham núcleo repleto de
heterocromatina, citoplasma com vacúolos e sistema canalicular interno (Fig. 5C).
4.2- Hematologia
Não houve diferenças estatísticas entre os parâmetros hematológicos de
machos e fêmeas. O perfil hematológico das avestruzes avaliadas está exposto nas
tabela 2 e 3. O VG teve a mesma média para ambos sexos, variando entre 39,3 e
46,7 % nos machos de 39,2 a 43,8 % nas fêmeas. A média do ET foi levemente
superior nos machos. Nestes o valor flutuou entre 1,4 e 2,2 x 106/mm3, já nas
fêmeas os valores variaram entre 1,4 e 2 x 106/mm3. A concentração de
hemoglobina apresentou variação entre 10,4 e 12,6 g/dL nos machos e 9,7 e 12,4
g/dL nas fêmeas. O VCM médio das fêmeas foi maior que dos machos. No qual o
das fêmeas variou entre 206,6 e 300,8 fL e dos machos entre 201,6 e 301,4 fL. Já o
CHCM foi maior nos machos, variando entre 24,1 e 29,1 %. Nas fêmeas os valores
variaram entre 22,1 e 29,5 %. A população de trombócitos nos machos variou entre
35,7 e 63.7 x 103/mm3. Esta média foi maior nas fêmeas, a qual variou entre 38,7 e
66,5 x 103/mm3.
A LG apresentou maiores médias em machos, que em fêmeas. As médias
variaram entre 11,4 e 18,2 x 106/mm3 nestas e entre 11,1 e 19,5 naqueles. Ademais
foi confirmada a nocividade da heparina à leucometria verificada por Green e Blue-
McLendon (2000). Já que, com sangue heparinizado, foram obtidas médias de 2,5 e
2,3 x 103/mm3 para machos e fêmeas respectivamente. Portanto houve déficit de
500 % na população de leucócitos ao serem contabilizados com sangue conservado
com heparina lítica.

30
As médias obtidas na contagem diferencial foram bastante próximas entre
machos e fêmeas (tabela 3). Os heterófilos representaram a maior parcela dos
leucócitos circulantes, por volta de 70%, seguido pelos linfócitos. Monócitos,
eosinófilos e basófilos foram visualizados raramente.
Tabela 2: Valores para série vermelha, contagem de trombócitos e leucócitos totais de avestruzes. Sangue de machos (n = 50) e fêmeas (n = 50) foram avaliados pelos métodos descritos e os resultados apresentados em médias com desvios padrões.
Parâmetro
Machos Fêmeas
Média ± DP* Média ± DP
VG (%) 43,0 ± 3,7 43,0 ± 3,8
ET (.106/mm3) 1,8 ± 0,4 1,7 ± 0,3
Hb (g/dL) 11,5 ± 1,1 11,0 ± 1,3
VCM (fL) 251,5 ± 49,9 253,7 ± 47,1
CHCM (%) 26,6 ± 2,5 25,8 ± 3,7
TB (.103/mm3) 49,7 ± 14,0 52,6 ± 13,9
LG (.103/mm3) 15,3 ± 4,2 14,8 ± 3,4
VG: volume globular, ET: hematimetria, Hb: hemoglobina, VCM: volume corpuscular médio, CHCM: concentração de hemoglobina corpuscular média, TB: trombócitos, LG: leucócitos totais.
* Desvio padrão
Tabela 3: Contagens diferenciais dos leucócitos de avestruz. Esfregaços de machos (n = 50) e fêmeas (n = 50) foram analisados e os resultados apresentados em médias com seus desvios padrões.
Leucócito
Machos Fêmeas
Média ± DP*
Média ± DP
Heterófilo (%) 73,5 ± 5,3 73,6 ± 4,5
Heterófilo (.103/mm3) 11,3 ± 3,4 10,7 ± 2,9
Eosinófilo (%) 1,8 ± 1,5 1,8 ± 1,6
Eosinófilo (.103/mm3) 0,3 ± 0,3 0,3 ± 0,2
Basófilo (%) 0 ± 0,1 0,1 ± 0,3
Basófilo (.103/mm3) 0 ± 0 0 ± 0,1
Linfócito (%) 22,1 ± 5,8 22,4 ± 4,3
Linfócito (.103/mm3) 3,3 ± 1,2 3,5 ± 1,0
Monócito (%) 2,6 ± 1,6 2,1 ± 1,5
Monócito (.103/mm3) 0,4 ± 0,3 0,3 ± 0,2 *
Desvio padrão

31
4.3- Bioquímica sérica
Não houve diferença entre as médias de machos e fêmeas nos parâmetros
bioquímicos. As médias da concentração dos elementos avaliados estão exibidas na
tabela 4. A enzima AST apresentou maior média nas fêmeas, que variou entre 304,2
e 481,6 U/l. Já nos machos a atividade desta enzima variou entre 287,1 e 489,2 U/l.
Para CK não foi diferente. A média das fêmeas foi maior que a dos machos. A
variação entre as fêmeas ficou entre 1394,9 e 7644,9 U/l, e nos machos entre
2471,5 e 6307,5 U/l. O ácido úrico obteve a mesma média para os dois sexos. A
glicose apresentou maior concentração nos machos, variando entre 170,2 e 218,2
mg/dl. Já nas fêmeas, variou entre 151,1 e 204,7 mg/dl. O colesterol também foi
maior nos machos que nas fêmeas. Os valores variaram nestas entre 40,1 e 71,5
mg/dl, e entre 44 e 68,8 mg/dl naqueles. Diferença insignificante foi observada para
os valores de PT entre machos e fêmeas. Machos e fêmeas apresentaram mesma
média para albumina. Assim como as proteínas totais (PT), as médias de globulinas
foram bastante próximas entre machos e fêmeas, com variações entre 2,5 e 3,3 g/dl
e 2,4 e 3,6 g/dl respectivamente. Machos apresentaram maior A:G que as fêmeas.
Sódio e potássio tiveram maiores médias nos machos que nas fêmeas. O Na+ variou
nos machos entre 140 e 149,6 mEq/l, e nas fêmeas entre 135,8 e 152 mEq/l. Já o K+
flutuou entre 3,1 e 4,3 mEq/l nos machos e entre 2,9 e 4,1 mEq/l nas fêmeas.
Tabela 4: Parâmetros bioquímicos séricos de avestruzes. Soro de machos (n = 25) e fêmeas (n = 25) foi analisado e os resultados apresentados em média com seus desvios padrões.
AST: aspartato aminotransferase, CK: creatina quinase, PT: proteínas totais, A:G: albumina/globulina.
* Desvio padrão
Parâmetro Machos Fêmeas
Média ± DP*
Média ± DP
AST (U/l) 388,5 ± 101,4 392,9 ± 88,7
CK (U/l) 4389,5 ± 1918 4519,9 ± 3125
Ácido úrico (mg/dl) 4,7 ± 1,2 4,7 ± 1,6
Glicose (mg/dl) 194,2 ± 24,0 177,9 ± 26,8
Colesterol (mg/dl) 56,4 ± 12,4 55,8 ± 15,7
PT (g/dl) 4,6 ± 0,5 4,7 ± 0,8
Albumina (g/dl) 1,7 ± 0,1 1,7 ± 0,2
Globulina (g/dl) 2,9 ± 0,4 3 ± 0,6
A:G 0,6 ± 0,07 0,56 ± 0,04
Na+ (mEq/l) 144,8 ± 4,8 143,9 ± 8,1
K+ (mEq/l) 3,7 ± 0,6 3,5 ± 0,6

32
5- Discussão
A caracterização dos leucócitos de avestruz foi realizada por microscopia
óptica, eletrônica de transmissão e localização ultra-estrutural da atividade da
enzima peroxidase nos granulócitos. Esfregaços sanguíneos, corados com Giemsa e
observados por microscopia óptica de campo claro, mostraram leucócitos com
morfologia similar aos já descritos para outras aves (Campbel, 1995; Canfield, 1998).
Já a ultra-estrutura destas células foi similar as de galinhas (Dhingra et al., 1969)
com algumas considerações feitas a seguir. Ademais, os perfis hematológico e
bioquímico foram estabelecidos para avestruzes criadas no estado do Rio de
Janeiro, formando uma base de dados sólida para auxiliar o diagnostico de
patologias nessa espécie. Como esperado, os valores obtidos diferem dos
estabelecidos em alguns estudos prévios, já que coleta, idade, região, manejo e
metodologia são fatores que influenciam os resultados e serão discutidos neste item.
Os heterófilos de avestruzes apresentaram núcleo lobulado e dois tipos de
grânulos citoplasmáticos. Dhingra et al. (1969) e Damion & Caxton-Martins (1977)
identificaram três tipos de grânulos em heterófilos de galinha. O maior e elíptico foi
descrito como mais eletrodenso dos três, diferindo do observado em S. camelus no
qual este era menos eletrondenso. Outro tipo de grânulo, ausente nas avestruzes,
foi descrito apresentando matriz eletrodensa em seu interior. O terceiro tipo era
menor e menos eletrondenso que os outros dois. Já Topp & Carlson (1972),
estudando leucócitos de perus, observaram apenas um tipo de grânulo nos
heterófilos de forma circular a oval, com variação de afinidade pelo ósmio. Assim
como Santos et al. (2003), que também observaram apenas um tipo de grânulo em
Buteo magnirostris, gavião-carijó, elíptico e bastante eletrodenso. Dhingra et al.,
1969, estudando galinhas, comentam que o grânulo elíptico pode ser encontrado
com matriz elétrodensa em seu interior, dependendo do estagio de maturação do
heterófilo. Durante as observações neste trabalho um heterófilo com estas
características foi encontrado. O ponto comum entre os autores citados e o presente
estudo foi a lobulação característica do núcleo dos heterófilo. A ausência da
atividade da enzima peroxidase no interior dos grânulos dos heterófilos também foi
verificada em outras aves (Rausch & Moore 1975; Damion & Caxton-Martins 1977;
Santos et al., 2003). Desta forma, podemos concluir que os dados apresentados
neste trabalho descrevem de forma clara as características morfológicas dos
heterófilos de avestruz. Também pode-se concluir que não existe um padrão

33
característico de heterófilos de aves. Esta heterogeneidade deve ser levada em
conta em trabalhos que objetivam a caracterização de leucócitos. Para tal se faz
importante a localização da enzima peroxidase como provável não marcadora deste
tipo celular.
Os eosinófilos de avestruz apresentaram núcleo riniforme, às vezes
bilobulado, e dois tipos de grânulos redondos comumente encontrados. Um terceiro
tipo de grânulo, também redondo e com material osmiofílico depositado, foi
observado raramente. Os eosinófilos de galinhas estudados por Dhingra et al.,
(1969) e Damion & Caxton-Martins (1977), apresentaram as mesmas características
de núcleo, porém apenas um tipo de granulo foi encontrado. Este era redondo,
eletrondenso e com conteúdo homogêneo. Maxwell & Siller (1972) estudaram
eosinófilos de cinco espécies de aves diferentes. Patos e gansos demonstraram
grânulos contendo matriz cristalina eletrondensa e retangular, ocupando quase toda
organela, quando contrastados apenas com citrato de chumbo. A principal diferença
encontrada foi que os grânulos do primeiro apresentavam forma alongada,
entretanto, no segundo, a forma variou entre redonda ou oval. Já galinhas d’angola
demonstraram características similares as de galinhas, com dois padrões de
tamanho e eletrondensidades, permitindo classificar dois tipos diferentes de
grânulos. Ou seja, um maior e mais osmiofílico que outro. Codornas também
apresentaram dois padrões distintos de grânulos. Um oval, mais comum, maior e
com periferia mais osmiofílica que o centro. Enquanto o segundo tipo era menor,
mais osmiofílico e de forma variada. Perus apresentaram núcleo sem lobulações, na
maioria das vezes, e grânulos citoplasmáticos escassos e adjacentes ao núcleo que
eram bastante eletrondensos com morfologia irregular. Pombos demonstraram
núcleo lobulado característico, assim como das outras espécies estudadas. Seus
grânulos revelaram três padrões distintos. Um grande, oval ou redondo,
intensamente osmiofílico e homogêneo, outro de morfologia similar, porém menos
eletrondenso e um terceiro tipo, característico de metamielócitos, com forma e
tamanho similares aos demais, porém com aparência granular e espaços vazios no
interior (Maxwell & Siller, 1972). Estas características são similares aos descrito nas
avestruzes neste trabalho. Ademais, os eosinófilos de avestruz apresentaram reação
positiva intensa para peroxidase, assim como em galinhas, patos, codornas e
gaviões-carijós (Damion & Caxton-Martins, 1977; Maxwell, 1986a; Maxwell, 1986b;

34
Santos et al., 2003). Esta característica é um ótimo marcador deste tipo celular
diferindo claramente do heterófilo.
Monócitos e linfócitos observados apresentaram características típicas, entre
as quais diferenças na relação núcleo/citoplasma, quantidade de heterocromatina,
presença marcante de organelas e sulco nuclear na face citoplasmática, como já
descrito anteriormente (DaMatta et al., 1998b; DaMatta et al., 2000). Trombócitos de
avestruzes também foram similares aos observados em anfíbios, répteis e outras
aves, com presença de núcleo, vacúolos citoplasmáticos, sistema canalicular
interno, além da forma variando entre oval ou redonda (Damion et al., 1985; Taffarel
& Oliveira, 1993; DaMatta et al., 1998b; Santos et al. 2003b). Portanto, estes três
tipos celulares são de fácil caracterização morfológica na microscopia eletrônica de
transmissão, e não existe necessidade de localização enzimática para auxiliar sua
diferenciação na microscopia eletrônica de transmissão. Porém, na microscopia
óptica a diferenciação destas requer atenção principalmente às carcterísticas
tintoriais do citoplasma e núcleo.
Embora não houvesse diferença estatística nos parâmetros hematológicos
machos apresentaram médias levemente maiores em ET, CHCM e LG. Enquanto
fêmeas tiveram médias de VCM e TB pouco maiores que dos machos. Dados
similares foram verificados por Levi, et al. (1989). Já que não houve diferenças entre
as médias de machos e fêmeas, elas foram fundidas e estão ao lado das propostas
por outros autores na tabela 5.
O VG obtido foi menor que o encontrado por Palomeque et al. (1991) e igual
ao reportado por Mushi et al. (1999a). Porém foi superior aos encontrados por Levi
et al. (1989), Green (1999), Vilar et al. (2003) e Raukar e Simpraga (2005). Estes
últimos estudando avestruzes de um dia. A ET apresentou médias menores que as
observadas por Palomeque et al. (1991), Mushi et al. (1999a) e Vilar et al. (2003). Já
Levi et al. (1989), Green (1999) e Raukar e Simpraga (2005) obtiveram valores de
ET inferiores que os encontrados no presente estudo. A concentração de
hemoglobina apresentou valor similar ao encontrado por Green (1999). Já Levi et al.
(1989), Palomeque et al. (1991), Mushi et al. (1999a) e Vilar et al. (2003) obtiveram
maiores médias para Hb. Entretanto Raukar e Simpraga (2005), avaliando pintos de
um dia, obtiveram média bastante inferior. Fato explicado pela menor concentração
de hemoglobina em avestruzes mais jovens (Palomeque et al., 1991; Vilar et al.,
2003). Levi et al. (1989), Palomeque et al. (1991), Green (1999), Mushi et al.

35
(1999a), Vilar et al. (2003) e Raukar e Simpraga (2005) observaram menores médias
para VCM. Condição invertida para CHCM, com exceção da apresentada por Raukar
e Simpraga (2005). Mesmo com as diferenças apresentadas, nenhum dos valores
estabelecidos pelos autores revelou grande discrepância em comparação aos
obtidos nesse estudo. Exceto pelo reportado por Raukar e Simpraga (2005), onde
avestruzes mais jovens tendem a apresentar valores menores para parâmetros
eritrocíticos que os adultos. Porém, pode haver controvérsias já que avestruzes
entre quatro e 10 meses podem apresentar ET maior que adultos (Levi et al., 1989;
Green, 1999). Ademais, os maiores valores de hemoglobina encontrados por Mushi
et al. (1999a) e Vilar et al. (2003) podem ser conseqüência da não centrifugação da
amostra, feita após a reação. Esta centrifugação tem a finalidade de remover
partículas nucleares dos eritrócitos que alteram a leitura (Green e Blue-McLendon,
2000; Campbell, 2004a).
A contagem de trombócitos também não apresentou diferenças significativas,
embora tenha sido maior nas fêmeas. Este parâmetro não é amplamente empregado
nos artigos publicados até o momento. É possível que tal fato seja devido a
insegurança existente na identificação destas células na câmara de Neubauer, já
que existe similaridade com linfócitos. Ademais, a contagem automática não é
possível de ser realizada pela mesma razão da manual, ou seja, semelhança com
linfócitos. Mesmo assim, as médias obtidas foram similares as encontradas por Vilar
et al. (2003). Entretanto, média bastante inferior foi proposta por Mushi et al.
(1999a), fruto de um provável equívoco na identificação dos trombócitos. Neste
trabalho, a utilização do diluente de Dacie modificado, com azul cresil brilhante,
proporcionou uma melhor diferenciação entre trombócitos e leucócitos
agranulocíticos. Portanto, para otimizar as contagens de trombócitos e leucócitos na
câmara de Neubauer é fundamental a utilização de uma corante na solução afim de
prevenir erros.
A LG proposta foi similar as encontradas por Palomeque et al. (1991), Green
(1999) e Vilar et al. (2003). Médias muito menores foram observadas por Levi et al.
(1989), Mushi et al. (1999a) e Raukar e Simpraga (2005). Esta discrepância está
provavelmente relacionada ao anticoagulante utilizado. A utilização do EDTA (ácido
etilenodiamino tetra-acético), ou a estimativa do LG através da contagem das células
de esfregaço sanguíneo pela utilização da fórmula proposta por Tavares-Dias e
Moraes, 2006, não influenciou a obtenção deste parâmetro. A comparação dos

36
trabalhos já publicados e do aqui apresentado mostra, claramente, que a LG obtida
diretamente de amostras sanguíneas utilizando a heparina como anticoagulante leva
a um decréscimo na população de linfócitos, (Levi et al., 1989; Mushi et al., 1999a;
Raukar e Simpraga 2005) como já mencionado por Green e Blue-McLendon (2000)
e Campbell (2004a).
As contagens diferenciais dos leucócitos foram similares entre os sexos.
Heterófilos apresentaram maiores médias quando avaliados por Raukar e Simpraga
(2005) em avestruzes de um dia, utilizando citrato de sódio como anticoagulante. Já
Green (1999) e Vilar et al. (2003) observaram médias similares às encontradas pelo
presente estudo. Entretanto, Levi et al. (1989) e Mushi et al. (1999a) propuseram
médias inferiores às demais, provavelmente alteradas pelo uso da heparina como
anticoagulante. Os eosinófilos tiveram médias similares às propostas por todos os
trabalhos consultados, inclusive no realizado em avestruzes de um dia (Levi et al.,
1989; Green, 1999; Mushi et al., 1999a; Vilar et al., 2003; Raukar e Simpraga, 2005).
Entretanto, os basófilos obtiveram média inferior a todos estes artigos, exceto a
apresentada por Levi et al., 1989. Entre os agranulócitos, os linfócitos apresentaram
média inferior à proposta por Mushi et al., (1999a), similar às de Levi et al. (1989) e
Green (1999) e superior às apresentadas por Vilar et al. (2003) e Raukar e Simpraga
(2005). Já a média observada para os monócitos, assim como nos eosinófilos, foi
similar a todos os trabalhos (Levi et al., 1989; Green, 1999; Mushi et al., 1999a; Vilar
et al., 2003; Raukar e Simpraga, 2005). Spinu, et al. (1999) concluíram que
avestruzes domésticas sofrem mais fatores estressantes que as selvagens, já que
aquelas apresentaram eosinofilia, basofilia e menor índice fagocítico quando
comparadas à estas. Em complementação, Kamau et al. (2002) verificaram que
mudanças de ambiente, alimentação e manipulação podem acarretar aumento da
população de heterófilos e diminuição de linfócitos, simulando os efeitos de
corticosteróides. Portanto, o efeito do estresse e a origem das aves são explicações
plausíveis para as diferenças encontradas na contagem diferencial.
Durante as coletas de campo e realização dos primeiros hemogramas em
uma fazenda localizada no município de Campos dos Goytacazes, RJ, foi observada
intensa leucocitose acompanhada de eosinofilia, chegando a 14%, nas amostras
avaliadas. A interpretação clássica desta alteração reportada em mamíferos indica
possível infestação por parasitas do trato gastrointestinal. Portanto, foi realizado
exame coproparasitológico no rebanho desta determinada fazenda. Como resultado

37
obtivemos o diagnóstico e a identificação do parasito do gênero Libyostrongylus sp.
(Bonadiman et al. 2006), acarretando o descarte da propriedade para
estabelecimento de parâmetros hematológico e bioquímicos de avestruzes sadias.
Machos e fêmeas não apresentaram diferenças estatísticas entre seus
parâmetros bioquímicos nesse trabalho, assim como no de Khazraiinia et al., (2006).
Contrário a idade, o sexo exerce pouca influência nos valores bioquímicos,
principalmente devido a produção hormonal, maior nos adultos, e manejo nutricional
dado às diferentes faixas etárias na cadeia produtiva destas aves (Levy et al., 1989;
Palomeque et al., 1991; Okotie-Eboh et al., 1992; Quintavalla et al., 2001).
Entretanto, nem todos os trabalhos evidenciaram esta diferença (Khazraiinia et al.,
2006). Uma única média para machos e fêmeas foi obtida e está apresentada lado a
lado com as estabelecidas por outros estudos (tabela 6).
As médias obtidas para AST e CK foram superiores as apresentadas por Levy
et al. (1989), Palomeque et al. (1991) e Quintavalla et al. (2001), porém foram
similares às reportadas por Verstappen et al. (2002). Okotie-Eboh et al. (1992)
obtiveram médias inferiores para AST, e similares para CK. Este mesmo autor
encontrou maiores médias destas enzimas em fêmeas jovens quando comparadas
às adultas. Já Khazraiinia et al. (2006) observaram média semelhante para AST em
avestruzes no Irã. Os altos valores obtidos para essas enzimas pode ser resultado
do estresse induzido pela captura (Levy et al., 1989; Palomeque et al., 1991;
Verstappen et al., 2002), já que as aves eram maiores de três anos e destinadas à
reprodução. Esta condição aguça o territorialismo e conseqüentemente a
agressividade, resultando em maior estresse durante a coleta de sangue, mesmo
com a utilização do capuz escuro na contenção. Assim, a AST pode ser usada tanto
para avaliação hepática, quanto na avaliação de estresse muscular. A metodologia
empregada também pode explicar diferença entre esses valores, pois os mais
próximos também utilizaram o método da bioquímica seca, ao contrário dos
inferiores que utilizaram kits comercias, onde utilizam os ensaios bioquímicos
clássicos. Outro ponto importante a ser levado em conta é que o presente estudo foi
o único a conservar o soro a – 70 oC, preservando de forma melhor a atividade
enzimática. Ademais, Quintavalla et al. (2001) sugerem que aves criadas em clima
tropical ou sub-tropical apresentam maiores níveis de AST, assim como as
avestruzes alimentadas com maiores níveis de proteínas (Bovera et al., 2007).
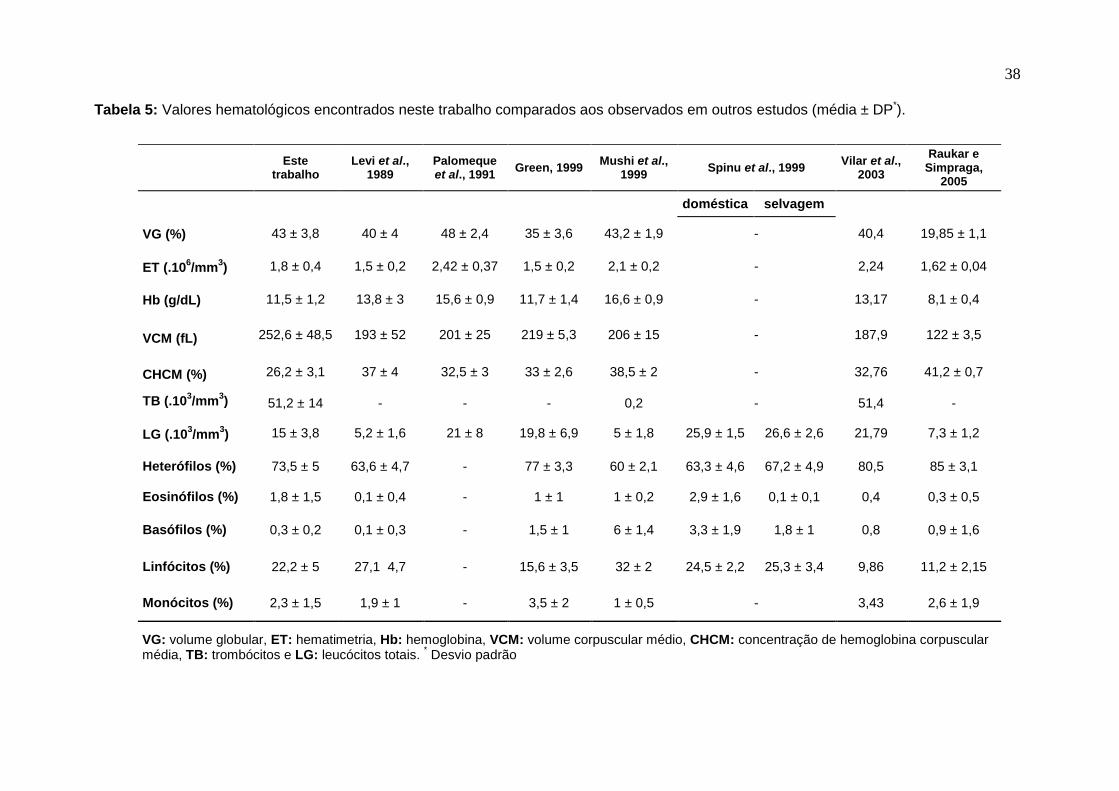
38
Tabela 5: Valores hematológicos encontrados neste trabalho comparados aos observados em outros estudos (média ± DP*).
VG: volume globular, ET: hematimetria, Hb: hemoglobina, VCM: volume corpuscular médio, CHCM: concentração de hemoglobina corpuscular média, TB: trombócitos e LG: leucócitos totais.
* Desvio padrão
Este
trabalho Levi et al.,
1989 Palomeque et al., 1991
Green, 1999 Mushi et al.,
1999 Spinu et al., 1999
Vilar et al., 2003
Raukar e Simpraga,
2005
doméstica selvagem
VG (%) 43 ± 3,8 40 ± 4 48 ± 2,4 35 ± 3,6 43,2 ± 1,9 - 40,4 19,85 ± 1,1
ET (.106/mm
3) 1,8 ± 0,4 1,5 ± 0,2 2,42 ± 0,37 1,5 ± 0,2 2,1 ± 0,2 - 2,24 1,62 ± 0,04
Hb (g/dL) 11,5 ± 1,2 13,8 ± 3 15,6 ± 0,9 11,7 ± 1,4 16,6 ± 0,9 - 13,17 8,1 ± 0,4
VCM (fL) 252,6 ± 48,5 193 ± 52 201 ± 25 219 ± 5,3 206 ± 15 - 187,9 122 ± 3,5
CHCM (%) 26,2 ± 3,1 37 ± 4 32,5 ± 3 33 ± 2,6 38,5 ± 2 - 32,76 41,2 ± 0,7
TB (.103/mm
3) 51,2 ± 14 - - - 0,2 - 51,4 -
LG (.103/mm
3) 15 ± 3,8 5,2 ± 1,6 21 ± 8 19,8 ± 6,9 5 ± 1,8 25,9 ± 1,5 26,6 ± 2,6 21,79 7,3 ± 1,2
Heterófilos (%) 73,5 ± 5 63,6 ± 4,7 - 77 ± 3,3 60 ± 2,1 63,3 ± 4,6 67,2 ± 4,9 80,5 85 ± 3,1
Eosinófilos (%) 1,8 ± 1,5 0,1 ± 0,4 - 1 ± 1 1 ± 0,2 2,9 ± 1,6 0,1 ± 0,1 0,4 0,3 ± 0,5
Basófilos (%) 0,3 ± 0,2 0,1 ± 0,3 - 1,5 ± 1 6 ± 1,4 3,3 ± 1,9 1,8 ± 1 0,8 0,9 ± 1,6
Linfócitos (%) 22,2 ± 5 27,1 4,7 - 15,6 ± 3,5 32 ± 2 24,5 ± 2,2 25,3 ± 3,4 9,86 11,2 ± 2,15
Monócitos (%) 2,3 ± 1,5 1,9 ± 1 - 3,5 ± 2 1 ± 0,5 - 3,43 2,6 ± 1,9

39
Avaliando o metabolismo energético, Palomeque et al. (1991) e Okotie-Eboh
et al. (1992) reportam maiores médias para glicose e colesterol. Já Verstappen et al.
(2002) e Khazraiinia et al. (2006) encontraram valores similares aos apresentados
neste estudo. Apesar da tênue diferença, o tempo de espera necessário para
coagulação da amostra pode ter acarretado na queda da concentração sérica de
glicose (Palomeque et al., 1991). Levy et al. (1989) e Palomeque et al. (1991)
observaram maior concentração de glicose em avestruzes jovens, em contraste com
os dados relatados por Okotie-Eboh et al. (1992). Ademais, em outro estudo,
Moniello et al. (2006) justificam a inexistência da diferença na concentração de
glicose entre os gêneros com a heterogeneidade da faixa etária das aves por eles
estudadas. Justificativa que descartamos com este estudo, pois todas as aves
utilizadas pertenciam ao rebanho destinado a reprodução. Já Polat et al. (2003)
evidenciaram aumento da glicemia nas avestruzes alimentadas com níveis de
proteína bruta de 20%, quando comparadas àquelas alimentadas com dieta
contendo 23% de proteína bruta. Quanto ao colesterol, Levy et al. (1989)
observaram queda da concentração à medida que a ave envelhece. Fenômeno
esperado devido a reabsorção total do saco vitelínico, rica fonte de colesterol para
os pintinhos. Talvez esse decréscimo não ocorresse na natureza, pois Norambuena
e Parada (2005), avaliando flamingos, notaram que aves alimentadas com a dieta
natural apresentam maiores concentrações séricas de colesterol quando
comparadas as que receberam dieta industrializada.
Os níveis de ácido úrico das avestruzes deste estudo apresentaram médias
similares as observadas por Quintavalla et al. (2001). Porém, valores mais altos
foram observados nos estudos de Levy et al. (1989), Palomeque et al. (1991),
Okotie-Eboh et al. (1992), Verstappen et al. (2002) e Khazraiinia et al. (2006). Levy
et al. (1989) e Okotie-Eboh et al. (1992) observaram diminuição da concentração
deste metabólito em avestruzes jovens até seis meses, após esta idade a
concentração tende a estabilizar. Como esperado, Polat et al. (2003) e Bovera et al.
(2007) encontraram maior concentração de ácido úrico em avestruzes alimentadas
com dietas contendo maior concentração de proteínas.
A concentração de PT teve pouca variação, às já reportadas (Levy et al.,
1989; Palomeque et al., 1991; Okotie-Eboh et al., 1992; Quintavalla et al., 2001;
Verstappen et al., 2002). Exceto por Khazraiinia et al. (2006) que obtiveram menor
média para este parâmetro. Os demais índices, proteínas plasmáticas, albumina,

40
globulinas e A:G, apresentaram médias semelhantes aos demais estudos (Okotie-
Eboh et al., 1992; Quintavalla et al., 2001; Khazraiinia et al., 2006). Levy et al.
(1989), Palomeque et al. (1991), Okotie-Eboh et al. (1992) e Moniello et al. (2006)
encontraram, em fêmeas adultas, maior concentração de PT, conseqüência do
aumento de globulinas que é resultante do aumento da produção de estrógenos
nessas avestruzes. Norambuena e Parada (2005), no mesmo estudo citado
anteriormente observaram aumento nas concentrações de PT e globulinas nos
flamingo alimentados com dieta industrializada. Já Polat et al. (2003) obtiveram o
mesmo resultado avaliando avestruzes suplementadas com alfafa substituindo a
silagem de milho. Assim, a baixa variação entre os valores de proteínas plasmáticas
de machos e fêmeas está relacionada a mesma fase fisiológica em que se
encontram. Já variação entre os estudos pode estar relacionada a concentração de
proteínas encontrada na alimentação oferecida.
Finalmente, entre os eletrólitos, Levy et al. (1989), Palomeque et al. (1991) e
Verstappen et al. (2002) observaram médias semelhantes para Na+. Entretanto, para
o K+, Palomeque et al. (1991) obtiveram valor bastante superior, enquanto Levy et
al. (1989) encontraram média semelhante. Embora os eletrólitos apresentem pouca
variabilidade em avestruzes independente das condições fisiológicas (Moniello et al.,
2006), Lumeij (1985) e Verstappen et al., (2002) avaliando a influência do binômio
tempo de estocagem/temperatura nas concentrações plasmáticas de K+ em aves,
concluíram que caso a estocagem do sangue total dure mais que duas horas em
temperaturas de 0 oC ou 20 oC, os valores da concentração deste íon podem variar
15% para menos ou para mais, respectivamente. Portanto, ao comparar os
resultados do presente estudo, levando em conta estas considerações, foi
estabelecido que caso o sangue seja centrifugado logo após a coagulação e o soro
congelado a – 70 oC esta amostra não apresentará alterações decorrentes da
armazenagem.
Os parâmetros apresentados para avestruzes estão de acordo com os valores
gerais indicados para a classe das Aves (Campbell, 2004c). Dentro deste contexto, o
perfil bioquímico obtido neste trabalho, bem como o aprofundamento em outros
parâmetros aqui não estudados e o entendimento a respeito dos fatores que
influenciam esses dados serão de grande importância, especialmente para o
diagnóstico de doenças que acometem essas aves, como a impactação gástrica
(Yüksek et al., 2002) e deformidades no cinturão pélvico (Mushi et al., 1999b).
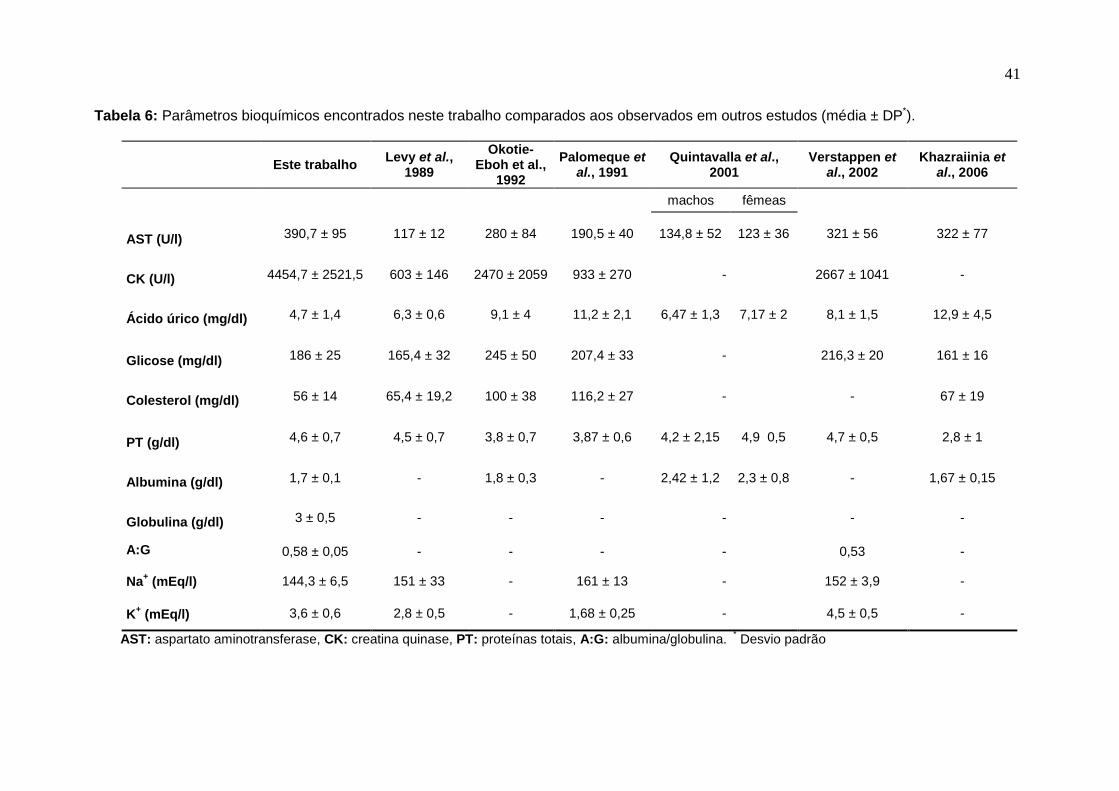
41
Tabela 6: Parâmetros bioquímicos encontrados neste trabalho comparados aos observados em outros estudos (média ± DP*).
Este trabalho Levy et al.,
1989
Okotie-Eboh et al.,
1992
Palomeque et al., 1991
Quintavalla et al., 2001
Verstappen et al., 2002
Khazraiinia et al., 2006
machos fêmeas
AST (U/l) 390,7 ± 95 117 ± 12 280 ± 84 190,5 ± 40 134,8 ± 52 123 ± 36 321 ± 56 322 ± 77
CK (U/l) 4454,7 ± 2521,5 603 ± 146 2470 ± 2059 933 ± 270 - 2667 ± 1041 -
Ácido úrico (mg/dl) 4,7 ± 1,4 6,3 ± 0,6 9,1 ± 4 11,2 ± 2,1 6,47 ± 1,3 7,17 ± 2 8,1 ± 1,5 12,9 ± 4,5
Glicose (mg/dl) 186 ± 25 165,4 ± 32 245 ± 50 207,4 ± 33 - 216,3 ± 20 161 ± 16
Colesterol (mg/dl) 56 ± 14 65,4 ± 19,2 100 ± 38 116,2 ± 27 - - 67 ± 19
PT (g/dl) 4,6 ± 0,7 4,5 ± 0,7 3,8 ± 0,7 3,87 ± 0,6 4,2 ± 2,15 4,9 0,5 4,7 ± 0,5 2,8 ± 1
Albumina (g/dl) 1,7 ± 0,1 - 1,8 ± 0,3 - 2,42 ± 1,2 2,3 ± 0,8 - 1,67 ± 0,15
Globulina (g/dl) 3 ± 0,5 - - - - - -
A:G 0,58 ± 0,05 - - - - 0,53 -
Na+ (mEq/l) 144,3 ± 6,5 151 ± 33 - 161 ± 13 - 152 ± 3,9 -
K+ (mEq/l) 3,6 ± 0,6 2,8 ± 0,5 - 1,68 ± 0,25 - 4,5 ± 0,5 -
AST: aspartato aminotransferase, CK: creatina quinase, PT: proteínas totais, A:G: albumina/globulina. * Desvio padrão

42
6- Conclusões
A morfologia dos leucócitos de S. camelus foi caracterizada. Heterófilos e
eosinófilos apresentaram dois padrões de grânulos característico, cada. As
principais características que os diferencia é a presença da atividade da enzima
peroxidase nos grânulos do eosinófilo e os grandes grânulos elípticos dos
heterófilos.
O perfil hematológico foi estabelecido, sem diferenças significativas entre os
gêneros. A heparina lítica mostrou-se ineficiente para a obtenção dos hemogramas.
Caso seja utilizada, um esfregaço, com sangue livre de anticoagulante, deve ser
feito imediatamente após a coleta evitando alterações no leucograma.
Assim como o perfil hematológico, o bioquímico foi definido mostrando que a
imediata separação do soro e posterior congelamento a – 70 oC minimizam
alterações na amostra, possibilitando o armazenamento prolongado.

43
7- Referências bibliográficas
ACAB (2007): Associação dos Criadores de Avestruz do Brasil. Disponível em:
<http: www.acab.org.br.> acesso em setembro de 2007.
ACAERJ (2007): Associação dos Criadores de Avestruz do Estado do Rio de
Janeiro. Disponível em: <http: www.acaerj.org.br.> acesso em setembro de 2007.
ANGERMULLER, S. e FAHIMI, H. (1984) A new cerium-based method for
cytochemical localization of thiamine pyrophosphatase in the Golgi complex of rat
hepatocytes, comparison with the lead technique. Hystochemistry, 80:107.
BABIOR, B.M.; KIPNES, R.S. e CURNUTTE, J.T. (1973) Biological defense
mechanisms: the production by leukocytes of superoxide, a potential bactericidal
agent. The Journal of Clinical Investigation, 52: 741-744.
BONADIMAN, S.F.; EDERLI, N.B.; SOARES, A.K.P.; MORAES NETO, A.H.A.;
SANTOS, C.P.; DAMATTA, R.A. (2006) Occurrence of Libyostrongylus sp.
(NEMATODA) in ostriches (Struthio camelus Linnaeus, 1758) from the north region
of the state of Rio de Janeiro, Brazil. Veterinary Parasitology, 137: 175-179.
BOWERMAN, W.W.; STICKLE, J.E.; SIKARSKIE, J.G.; GIESY, J.P. (2000)
Hematology and serum chemistries of nestling bald eagles (Haliaeetus
leucocephalus) in the lower peninsula of MI, USA. Chemosphere 41: 1575-1579.
BOVERA, F.; MONIELLO, G.; RIU, N.; MEO, C.; PINNA, W.; NIZZA, A. (2007) Effect
of diet on the metabolic profile of ostriches (Struthio camelus var. domesticus),
Tropical Animal Health and Production, 39: 265-270.
BRASIL. (2003) Instrução Normativa Conjunta no 2. Regulamento Técnico para
Registro, Fiscalização e Controle Sanitário dos Estabelecimentos de Incubação de
Ovos, de Criação e Alojamento de Ratitas. 21 de fevereiro de 2003. MAPA/DAS.
BRASIL. (2002) Portaria no 36 do Ministério do Meio Ambiente em 15 de março de
2002. MMA/IBAMA.
BRUNE, K.; LEFFELL, M. S. e SPITZNAGEL, J. K. (1972) Microbicidal activity of
peroxidaseless chicken heterophile leukocytes. Infection and Immunity, 3: 283-287.
CAMPBELL, T.W. (1995) Avian Hematology and Cytology. Ames: Iowa State
University Press, p. 12-13.
CAMPBELL, T.W. (2004a) Hematology of birds. In: Thrall, M.A. (org.) Veterinary
Hematology and Clinical Chemistry. Lippincott Williams & Wilkins, Philadelphia, USA,
p. 225 – 258.

44
CAMPBELL, T.W. (2004b) Clinical chemistry of birds. In: Thrall, M.A. (org.)
Veterinary Hematology and Clinical Chemistry. Lippincott Williams & Wilkins,
Philadelphia, USA, p. 479 – 492.
CAMPBELL, T.W. (2004c) Blood biochemistry of lower vertebrates. In: 55th Annual
Meeting of the American College of Veterinary Pathologists (ACVP) & 39th Annual
Meeting of the American Society of Clinical Pathology (ASVCP), 6p.
CANFIELD, P.J. (1998) Comparative cell morphology in the peripheral blood film
from exotic and native animals. Australian Veterinary Journal, 76: 793-800.
CHANG, C. e HAMILTON, P.B. (1979) The thrombocyte as the primary circulating
phagocyte in chickens. Journal of Reticuloendothelial Society, 25: 585-590.
COKE, R.L.; WEST, G.D. e HOOVER, J.P. (2004) Hematology and Plasma
Biochemistry of Captive Puna Ibis (Plegadis ridgewayi). Journal of Wildlife Diseases,
40: 141–144.
COLES, E.H. (1984) Patologia Clínica Veterinária. São Paulo, Brasil. Ed. Marele, 566
p.
DABBERT, C.B. e POWELL, K.C. (1993) Serum enzymes as indicators of capture
myopathy in mallards (Anas platyrhynchos). Journal of Wildlife Diseases, 29: 304-
309.
DAIMON, T. e CAXTON-MARTINS, A. (1977) Electron microscopic and enzyme
cytochemical studies on granules of mature chicken granular leucocytes. Journal of
Anatomy, 123: 553-562.
DAIMON, T.; GOTOH, Y.; KAWAI, K.; UCHIDA, K. (1985) Ultrastructural distribution
of peroxidase in thrombocytes of mammals and submammals. Histochemistry, 82:
345-350.
DAMATTA, R.A., SEABRA, S.H., DE SOUZA, W. (1998a) Further studies on the
phagocytic capacity of thrombocytes. Journal of Submicroscopic Cytology &
Pathology, 30: 271-277.
DAMATTA, R.A., MANHÃS, L.; SEABRA, S.H., DE SOUZA, W. (1998b) Coculture of
chicken thrombocytes and monocytes: morphological changes and lectin binding.
Biocell, 22: 45-52.
DAMATTA, R.A., SEABRA, S.H., MANHÃS, L.; DE SOUZA, W. (2000) Nitric oxide is
not involved in the killing of Trypanosoma cruzi by chicken macrophages.
Parasitology Research, 86: 239-243.

45
DHINGRA, L.D.; PARRISH, W.B.; VENZKE, W.G. (1969) Electron microscopy of
granular leukocytes of chicken (Gallus domesticus). American Journal of Veterinary
Research, 4: 637-642.
EISERICH, J.P.; BALDUS, S.; BRENNAN, M.L.; MA, W.; ZHANG, C.; TOUSSON, A.;
CASTRO, L.; LUSIS, A.J.; NAUSEEF, W.M.; WHITE, C.R.; FREEMAN, B.A. (2002)
Myeloperoxidase, a leukocyte-derived vascular NO oxidase. Science 296: 2391-
2394.
ERICSSON, J.L.E. e NAIR, M.K. (1973) Electron microscopy demonstration of acid
phophatase activity in the development and mature heterophils of the chicken.
Histochemistry 37: 97-105.
FOWLER, M.E. (1991) Comparative clinical anatomy of ratites. Journal of Zoo and
Wildlife Medicine, 22: 204.
GHIDONI, J.J. e GOLDBERG, A.F. (1966) Light and electron microscopic localization
of acid phosphatase activity in human eosinophils. The American Journal of Clinical
Pathology, 4: 402-405.
GREEN, R.A. (1999) Reference range for normal ostriches & rheas. Veterinary
clinical pathology laboratory. Texas A & M University Teaching Hospital, Apud:
GREEN, R.A. e BLUE-MCLENDON, A. (2000) Ratite hematology. In: FELDMAN,
B.V.; ZINKL, J.G.; JAIN, N.C. (eds.), Schalm's veterinary hematology, 5. ed,
Baltimore, Maryland, USA: Lippincott Williams & Wilkins, p. 1201-1206.
GREEN, R.A. e BLUE-MCLENDON, A. (2000) Ratite hematology. In: FELDMAN,
B.V.; ZINKL, J.G.; JAIN, N.C. (eds.), Schalm's veterinary hematology, 5. ed,
Baltimore, Maryland, USA: Lippincott Williams & Wilkins, p. 1201-1206.
HARR, K.E. (2002) Clinical chemistry of companion avian species: A Review.
Veterinary Clinical Pathology, 3: 140-149.
JAIN, N.C. (1967) Peroxidase activity in leukocytes of some animal species. Folia
Haematologica, 88: 297-304.
JAIN, N.C. (1969) A comparative cytochemical study of leukocytes of some animal
species. Folia Haematologica, 94: 49-63.
JAIN, N.C. (1993) Essentials of hematology. Lea & Febiger, Philadelphia, USA.

46
JAENSCH, M.J.; CULLEN, L. e RAIDAL S.R. (2000) Assessment of liver function in
galahs/cockatoos (Eolophus roseicapillus) after partial hepatectomy: a comparison of
plasma enzyme concentrations, serum bile acid levels, and galactose clearance
tests. Journal of Avian Medical Surgery, 14: 164-171. Citado por: HARR, K.E. (2002)
Clinical chemistry of companion avian species: A Review. Veterinary Clinical
Pathology, 3: 140-149.
KANEKO, J. (1989) Clinical biochemistry of Domestic Animals. 4a Ed. New York,
U.S.A. Academic Press, 725 p.
KHAZRAIINIA, P.; SAEI, S.; MOHRI, M.; HADDADZADEH, H.R.; DARVISIHHA, H.R.;
KHAKI, Z. (2006) Serum biochemistry of ostrich (Struthio camelus) in Iran.
Comparative Clinical Pathology, 15: 87-89.
KOMNENOU, A.T.; GEORGIADES, G.K.; SAVVAS, I.; DESSIRIS, A. (2003) Surgical
treatment of gastric impaction in farmed ostriches. Journal of Veterinary Medicine
Association, 50: 474-477.
LEVI, A.; PERELMAN, B.; WANER, T.; VAN GREVENBROEK, M.; VAN CREVELD,
C.; YAGIL, R. (1989) Haematological parameters of the ostrich. Avian Pathology, 18:
321-327.
LEVY, A.; PERELMAN, B.; WANER, T.; VAN GREVENBROEK, M.; VAN CREVELD,
C.; YAGIL, R. (1989) Reference blood chemical values in ostrich (Struthio camelus).
American Journal of Veterinary Research, 50: 1548-1550.
LUCAS, A.M. e JAMROZ, C. (1961) Atlas of avian hematology. Agricultural
Monograph. 25, United States Department of Agriculture, Washington D.C.
LUMEIJ, J.T. (1985) The influence of blood sample treatment on plasma potassium
concentrations in avian blood. Avian Pathology, 14: 257-260.
MAXWELL, M.H. (1984) Histochemical identification of tissue eosinophils in the
inflammatory response of the fowl (Gallus domesticus). Research in Veterinary
Science, 37: 7-11.
MAXWELL, M.H. (1986a) Fine structural and cytochemical studies of eosinophils
from fowls and ducks with eosinophilia. Research in Veterinary Science, 41: 135-148.
MAXWELL, M.H. (1986b) Ultrastructural and cytochemical studies in normal
Japanese quail (Coturnix coturnix japonica) eosinophils and in those from birds with
experimentally induced eosinophilia. Research in Veterinary Science, 41: 149-161.
MAXWELL, M.H. (1987) The avian eosinophil: a review. World’s Poultry Science
Journal, 3: 190-207.

47
MAXWELL, M.H. e ROBERTSON, G.W. (1995) The avian basophilic leukocyte: a
review. World’s Poultry Science Journal, 3: 307-325.
MAXWELL, M.H. e ROBERTSON, G.W. (1998) The avian heterophil leukocyte: a
review. World’s Poultry Science Journal, 2: 155-178.
MAXWELL, M.H. e SILLER, W.G. (1972) The ultraesturctural characteristics of the
eosinophil granules in six species of domestic bird. Journal of Anatomy, 2: 289-303.
MEDWAY, W.; PRIER, J.E.; WILKINSON, J.S. (1973) Patologia Clínica Veterinária.
Cidade do México, México. Union Tipográfica Editorial Hispano-americana. 532 p.
MEYER, D.J.; COLES, E.H. e RICH, L.J. (1992) Veterinary Laboratory-Medicine
Interpretation and Diagnosis. Philadelphia, U.S.A. W. B. Saunders Company 349 p.
MONTALI, R.J. (1988) Comparative pathology of inflammation in the higher
vertebrates (reptiles, birds and mammals). Journal of Comparative Pathology, 99:1-
26.
MONIELLO, G.; BOVERA, F.; DE RIU, N.; PICCOLO, G.; PINNA, W.; DI MEO, C.
(2006) Gender effect on the metabolic profile of ostriches (Struthio camelus), Italian
Journal of Animal Science, 5: 229-235.
MUSHI, E.Z.; BINTA, M.G.; CHABO, R.G.; ISA J.F.; KAPAATA, R.W. (1999a)
Selected hematologic values of farmed ostriches (Struthio camelus) in Botswana.
Journal of Veteterinary Diagnostic Investigation, 4: 372-374.
MUSHI, E.Z.; BINTA, M.G.; CHABO, R.G.; ISA, J.F.; PHUTI, M.S. (1999b) Limb
deformities of farmed ostrich (Struthio camelus) chicks in Botswana, Tropical Animal
Health and Production, 31: 397-404.
NORAMBUENA, M.C. e PARADA, M. (2005) Serum biochemistry in andean
flamingos (Phoenicoparrus andinus) natural versus artificial diet. Journal of Zoo and
Wildlife Medicine, 36: 434-439.
OGUZ, H.; KURTOGLU, F.; KURTOGLU, V.; BIRDANE, Y.O. (2002) Evaluation of
biochemical characters of broiler chickens during dietary aflatoxin (50 and 100 ppb)
and clinoptilolite exposure. Research in Veterinary Science, 73: 101-103.
OKOTIE-EBOH, G.; BAILEY, C.A.; HICKS, K.D.; KUBENA, L.F. (1992) Reference
serum biochemical values for emus and ostriches. American Journal of Veterinary
Research, 53: 1765-1768.
PALOMEQUE, J.; PINTO, D. e VISCOR, G. (1991) Hematologic and blood chemistry
values of the Masai ostrich (Struthio camelus). Journal of Wildlife Diseases, 27: 34-
40.

48
POLAT, U.; CETIN, M.; TURKYILMAZ, O.; AK, I. (2003) Effects of different dietary
protein levels on the biochemical and production parameters of ostriches (Struthio
camelus). Veterinarski arhiv 73 : 73-80.
QUEZADA, T.; CUÉLLAR, H.; JARAMILLO-JUÁREZ, F.; VALDIVIA, A.G.;.REYES,
J.L. (2000) Effects of aflatoxin B1 on the liver and kidney of broiler chickens during
development. Comparative Biochemistry and Physiology, Part C 125: 265–272.
QUINTAVALLA, F.; BIGLIARDI, E.; BERTONI, P. (2001) Blood biochemical baseline
values in the ostrich (Struthio camelus). Annali della Facoltà di Medicina Veterinária,
Università degli Studi di Parma, 21: 61-71.
QURESHI, M.A. (2003) Avian macrophage and immune response: an overview.
Poultry Science, 82: 691-698.
RAUKAR, J. e SIMPRAGA, M. (2005) Haematological parameters in the blood of
onde day old ostriches. Israel Journal of Veterinary Medicine, 60: 112-114.
RAUSCH, P.G. e MOORE, T.G. (1975) Granule enzymes of polymorphonuclear
neutrophils: a philogenetic comparison. Blood, 46: 913-919.
REVISTA BRASILEIRA DE AGROPECUÁRIA (1999), ano I, no 02.
ROBINSON, J.M. (1985) Proved localization of intracellular sites of phosphatases
using cerium and cell permeabilzation. Journal of Histochemistry and Cytochemistry,
33: 749.
ROELS, F.; WISSE, E.; DE PREST, B.; VAN DEN MEULEN, J. (1975) Cytochemical
discrimination between catalases and peroxidases using diaminobensidine.
Histochemistry, 41: 281-312.
SANTOS, A.A.; DA SILVA, A.M.; SILVA, M.R.; SEGRETO, H.R.; EGAMI, M.I.
(2003a) Structural, cytochemical, immunocytochemical and ultrastructural
characterization of blood granulocytes of the roadside hawk Buteo magnirostris
(Gmelin, 1788) (Avian, Falconiform). Journal of Submicroscopy Cytology and
Pathology, 35: 351-357.
SANTOS, A.A.; DA SILVA, A.M.; DE CARVALHO, V.A.L.; EGAMI, M.I. (2003b)
Morpho-cytochemical and ultrastructural characterization of peripheral thrombocytes
of roadside hawk Buteo magnirostris (Gmelin, 1788) (avian, falconiform).
International Journal of Morphology, disponivel em:
http://findarticles.com/p/articles/mi_m5EOM/is_4_21/ ai_n18617265.

49
SERGENT, N.; ROGERS, T. E CUNNINGHAM, M. (2004) Influence of biological and
ecological factors on hematological values in wild Little Penguins, Eudyptula minor.
Comparative Biochemistry and Physiology, 138: 333– 339.
SILVA, E.O.; DINIZ, J.P.; ALBERIO, S.; LAINSON, R.; DE SOUZA, W.; DAMATTA,
R.A. (2004) Blood monocyte alteration caused by a hematozoan infection in the lizard
Ameiva ameiva (Reptilia: Teiidae). Parasitology Research, 93: 448-456.
SPINU, M.; SPINU, O. e DEGEN, A.A. (1999) Haematological and immunological
variables in a domesticated and wild subspecies of ostrich (Struthio camelus). British
Poultry Science, 40: 613 – 618.
STRYER, L. (1995) Bioquímica. 4a. Ed. São Paulo, Brasil. Guanabara Koogan.
STURKIE, P.D. (1965) Avian Physiology. 2nd Ed. New York, U.S.A. Cornell University
Press, p. 41-69.
TAVARES-DIAS, M. e MORAES, F.R. (2006) Hematological parameters for the
Brycon orbignyanus Valenciennes, 1850 (Osteichthyes: Characidae) intensively bred.
Hidrobiológica, 16: 271-274.
TOKUMITSU, S.I. e FISHMAN, W.H. (1983) Alkaline phosphatase biosynthesis in the
endoplasmic reticulum and its transport through the Golgi apparatus to the plasma
membrane. Cytochemical evidence. Journal of Histochemistry and Cytochemistry,
31: 647.
TOPP, R.C. e CARLSON, H.C. (1972) Studies on avian heterophils III: phagocytic
proprieties. Avian Diseases, 1: 374-380.
VILAR, T.D.; VOLINO, W.; STELLING, W.; RIBEIRO, I..C.; LEMOS, M.;
NASCIMENTO, M.D. (2003) Determinação dos parâmetros hematológicos em
avestruzes (STRUTHIONIDAE:AVES) Struthio camelus no Estado do Rio de Janeiro,
Brasil. Revista Universidade Rural – Série Ciências da Vida, 1: 301-302.
YUKSEK, N.; AGAOGLU, Z.; KAYA, A.; ASLAN, L.; ERDOGAN, H.M.; AKGUL, Y.
(2002) Stomach impaction in ostriches (Struthio camelus): blood chemistry,
hematology, and treatment. Avian Diseases, 46: 757-760.
VERSTAPPEN, F.A.L.M.; LUMEIJ, J.T.; BRONNEBERG, G.G. (2002) Plasma
chemistry reference values in ostrich. Journal of Wildlife Diseases, 38: 154-159.