transformaÇÃo e mutagÊnese insercional em …livros01.livrosgratis.com.br/cp033914.pdf · 3.2...
TRANSCRIPT

MARCOS ANTÔNIO SOARES
TRANSFORMAÇÃO E MUTAGÊNESE
INSERCIONAL EM Penicillium griseoroseum POR REMI
(INTEGRAÇÃO MEDIADA POR ENZIMA DE
RESTRIÇÃO)
Tese apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Microbiologia Agrícola, para obtenção do título de “Magister Scientiae”
VIÇOSA MINAS GERAIS – BRASIL
2002

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

MARCOS ANTÔNIO SOARES
TRANSFORMAÇÃO E MUTAGÊNESE
INSERCIONAL EM Penicillium griseoroseum POR REMI
(INTEGRAÇÃO MEDIADA POR ENZIMA DE
RESTRIÇÃO)
Tese apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Microbiologia Agrícola, para obtenção do título de “Magister Scientiae”
APROVADA: 26 de Março de 2002
Profª. Célia Alencar de Morais Prof. Sérgio Hermínio Brommonschenkel
Profª. Elza Fernades de Araújo (Conselheira)
Prof. Everaldo Gonçalves de Barros (Conselheiro)
Profª. Marisa Vieira de Queiroz
(Orientadora)

A DEUS,
Aos meus pais Marly e Antônio

AGRADECIMENTOS
A DEUS, pela força inestimável que me mantem todos os momentos de
minha vida;
Aos meus queridos pais, Marly e Antônio, que sempre estiveram ao meu
lado e pelo amor, carinho e atenção;
À minha querida avó, “Marinha”, pelo carinho e preocupação com meu
bem-estar;
Aos meus irmãos José Geraldo, Maria Aparecida, Maria Cabral, Maria da
Consolação e Edilene, pela atenção e pelo compartilhamento de afeto;
À Universidade Federal de Viçosa, à Coordenação de Aperfeiçoamento de
Pessoal de Nível Superior (CAPES) e ao Departamento de Microbiologia pela
oportunidade de realização deste trabalho;
À professora e amiga Marisa Vieira de Queiroz, pela orientação precisa,
pela atenção, carinho, disposição, humildade que fez deste trabalho um grande
prazer, e pelo estímulo constante em todos os momentos desse curso;
À professora Elza Fernandes de Araújo, pelo exemplo de dinamismo na
condução de sua vida profissional;
À professora Célia Alencar de Moraes, aos professores Sérgio Hermínio
Brommonschenkel e Everaldo Gonçalves de Barros pela participação na banca de
defesa e pelas valiosas sugestões que contribuíram para a finalização deste
trabalho;
Ao professore Lúcio A. de O. Campos e à professora Marisia Cyreti Forte
Pontes, pelos ensinamentos, pelo apoio e incentivo durante o meu curso de
graduação, e pelo exemplo de ética e moral profissional;

À professora Maria Catarina Megumi Kasuya e ao professor Marcos
Rogério Tótola, pela amizade, carinho, e principalmente, pela minha iniciação em
pesquisa e treinamento;
Aos professores do Departamento de Microbiologia que contribuíram para
minha formação profissional;
Aos meus grandes amigos Dyana, William, Cláudio, Aliçandro, Bruno,
Klédna e Maurílio pela convivência diária, pelos cuidados e pelos inúmeros
momentos felizes;
Às minhas queridas amigas de república Francismar, Poliana e Wanessa,
pela convivência sempre divertida e que fizeram da minha estadia em Viçosa
muito aconchegante;
Ao meu amigo Maurício, pelo incentivo constante e grande amizade;
Aos amigos de laboratório de Associações Micorrizas e de Genética
Molecular e de Microrganismos, pela ajuda na condução deste trabalho e pelos
momentos de descontração no trabalho;
Aos meus colegas de curso, pelo convívio agradável e amizade;
Aos meus amigos de João Monlevade, que mesmo afastados pela
distância, transmitiram amizade que tanto ajudou-me em Viçosa;
Aos funcionários do Departamento de Microbiologia, representados pelas
secretárias Laura, Nilcéia e Dona Aparecida, pelo empenho e pela amizade;
A todos que, de alguma forma, contribuíram para a realização deste
trabalho e para o meu crescimento pessoal e profissional.

BIOGRAFIA
MARCOS ANTÔNIO SOARES, filho de Antônio Soares Cabral e Marly
Soares Cabral, nasceu em São Domingos do Prata, Minas Gerais, no dia 16 de
junho de 1977.
Em outubro de 2000, graduou-se em Ciências Biológicas pela
Universidade Federal de Viçosa – MG.
Em agosto de 2000, iniciou como estudante do programa de mestrado em
Microbiologia Agrícola na Universidade Federal de Viçosa, concluindo em abril
de 2001.

CONTEÚDO
Resumo........................................................................................................................ vii Abstract...................................................................................................................... ix 1. Introdução.............................................................................................................. 01 2. Revisão de literatura.............................................................................................. 03 3. Material e Métodos................................................................................................ 17
3.1 Microrganismos, plasmídeos e meios de cultura....................................... 17 3.2 Extração de DNA plasmidial..................................................................... 17 3.3 Transformação mediada por REMI........................................................... 19 3.4 Isolamento de mutantes.............................................................................. 19 3.5 – Determinação da atividade de poligalacturonase (PG) e pectina liase
(PL) em meio líquido................................................................................................... 20
3.6 - Análise da estabilidade dos mutantes...................................................... 21 3.7 - Análise do padrão de integração.............................................................. 21
4. Resultados e Discussão.......................................................................................... 23 4.1 – Efeito da enzima de restrição Xba I na eficiência de transformação...... 23 4.2 – Caracterização das integrações nos transformantes................................ 26 4.3 – Isolamentos de mutantes morfológicos e com atividade de pectinases
alterada......................................................................................................................... 31
4.3.1) Morfológicos...................................................................................... 31 4.3.2) Mutantes com atividade de pectinases alterada.................................. 34
4.4 – Isolamentos de mutantes niaD- a partir dos transformantes................... 38 5. Conclusões.............................................................................................................. 42 6. Referências Bibliográficas..................................................................................... 43

RESUMO
SOARES, Marcos Antônio, M.S., Universidade Federal de Viçosa, março de
2002. Transformação e mutagênese insercional em Penicillium griseoroseum por REMI (integração mediada por enzima de restrição). Orientadora: Marisa Vieira de Queiroz. Conselheiros: Elza Fernandes de Araújo e Everaldo Gonçalves de Barros.
A aplicação da mutagênese insercional para etiquetagem gênica,
utilizando-se plasmídeos, requer um eficiente sistema de transformação e
ocorrência de integrações únicas no genoma. O presente trabalho demonstrou que
o uso da enzima de restrição XbaI, concomitantemente com o plasmídeo
linearizado, durante a transformação de Penicillium griseoroseum, resultou num
aumento de 6,77 vezes na eficiência de transformação em comparação com o
tratamento com plasmídeo circular. Análises de hibridização mostraram uma forte
tendência para integrações únicas (80% dos transformantes analisados). A
integração foi direcionada para o sítio de restrição, caracterizando eventos REMI.
Integrações REMI e não-REMI foram estáveis. Entre os transformantes, mutantes
morfológicos, bem como mutantes com redução na atividade total de
poligalacturonases, foram isolados. A porcentagem de mutantes foi de 0,95%.
Esse valor poderia ter sido maior considerando que mutantes auxotróficos não
puderam ser recuperados da população de transformantes. A reversão do fenótipo
nos transformantes foram mais freqüentes (até 90%) do que mutações no gene
niaD de P. griseoroseum (aproximadamente 10%). A perda das duas cópias do
vetor em um dos transformantes, que apresentou alteração morfológica, restaurou
o fenótipo original. Uma dessas cópias estava, provavelmente, etiquetando um
gene importante para a determinação da morfologia do tipo selvagem. Esses

resultados demonstram que o uso de REMI na transformação de P. griseoroseum
permite a recuperação de maior número de transformantes quando comparado a
outros métodos. Linhagens com inserções únicas originadas de eventos REMI e
não-REMI podem ser facilmente isoladas, permitindo e facilitando tanto a
etiquetagem quanto a clonagem de genes.

ABSTRACT
SOARES, Marcos Antônio, M.S., Universidade Federal de Viçosa, March, 2002. Transformation and insertional mutagenesis in Penicillium griseoroseum by REMI (restriction enzyme mediated integration). Major advisor: Marisa Vieira de Queiroz. Committee members: Elza Fernandes de Araújo and Everaldo Gonçalves de Barros.
The application of insertional mutagenesis for gene tagging with plasmids
requires an efficient transformation system and the occurrence of single insertions
in the genome. The present work demonstrates that the use of the restriction
enzyme Xba I, concomitantly with plasmid linearization, during transformation of
Penicillium griseoroseum, resulted in 6.77-fold increase in the transformation
efficiency in comparison to the treatment with a circular plasmid. Hybridization
analyses showed a strong trend to single insertions (80% of the transformants).
Integration was directed to the restriction sites of Xba I, characterizing the
occurrence of REMI events. REMI and non-REMI integrations were stable.
Among the transformants, morphological as well as mutants with reduced
polygaturonase total activity were isolated. The percentage of mutants
corresponded to 0.95%. This value might have been actually higher since
auxotrophic mutants could not be recovered from the population of transformants.
Mutations in the Fusarium oxysporum nia D gene, used as an auxotrophy marker,
were obtained at higher frequencies (up to 90%) than those in the P. griseoroseum
nia D (approximately 10%). The loss of the two vector copies by one of the
transformants presenting morphological alterations restored the original
phenotype. One of these copies was probably tagging the gene necessary for the

usual morphology in the wild-type. These results demonstrate that the use of
REMI for the transformation of P. griseoroseum allows the recovery of a higher
number of transformants. Strains with single insertions resulting from REMI and
non-REMI events can be easily isolated, allowing gene tagging and facilitating the
gene cloning.

1 – INTRODUÇÃO
Fungos filamentosos possuem grande importância ecológica e econômica.
Atuam eficientemente na decomposição da matéria orgânica do solo, o que
possibilita a ciclagem de nutrientes; outras espécies especializaram-se no
parasitismo de plantas e animais; e existem aqueles que chamam a atenção por
colonizar e produzir toxinas em alimentos utilizados na dieta animal, resultando
em sérias intoxicações ou no desenvolvimento de doenças; diversas enzimas e
compostos produzidos por determinadas espécies são amplamente utilizados nas
indústrias têxteis e farmacêuticas.
Assim como os organismos superiores, os fungos são eucariotos. Sua
manutenção em laboratório, na maioria das vezes, é fácil, pois não requerem
grande espaço físico, seu ciclo celular é relativamente rápido e seus requerimentos
nutricionais são simples. Todas as características citadas, tornam esse grupo de
organismos excelentes modelos biológicos nas áreas da fisiologia e genética.
Conseqüentemente, a obtenção de mutantes é primordial para que tais estudos
possam ser realizados.
Embora linhagens mutantes possam ser obtidos espontaneamente numa
população, a freqüência de tais mutações é baixa. O número de mutantes pode ser
aumentado utilizando-se agentes mutagênicos físicos, químicos ou biológicos.
Agentes biológicos, como plasmídeos, transposons ou mesmo T-DNA
(Agrobacterium tumefaciens), são utilizados em procedimentos de transformação
para obtenção de mutantes. Quando a molécula de DNA integra no genoma da
célula receptora ocorre uma interrupção na seqüência original de nucleotídeos,
resultando numa mutação dita insercional.
Mutantes obtidos via agentes biológicos possuem certas vantagens em
relação àqueles originados utilizando-se compostos químicos ou agentes físicos. A
capacidade de relacionar o fenótipo mutante com a seqüência interrompida, por
meio de análise de segregação, fornece um indício da função do gene.
Conseqüentemente, a clonagem da seqüência mutada torna-se menos trabalhosa,
considerando que a mesma está etiquetada com a molécula do agente biológico. O
não emprego de reagentes químicos mutagênicos é outra conveniência da

mutagênese insercional; resíduos de tais compostos poderiam poluir o ambiente,
ou mesmo o manipulador, caso não fosse utilizado corretamente.
Em contrapartida aos benefícios citados anteriormente, alguns
requerimentos são necessários para uma eficiente mutagênese insercional. É
preferível a utilização de vetores que não possuam seqüências homologas ao
genoma da célula hospedeira, propiciando-se integrações aleatórias. Eficiência na
transformação é necessária quando esse método é utilizado na mutagênese
insercional e preferencialmente que as integrações dos agentes biológicos sejam
únicas para facilitar analises de segregação e clonagem da seqüência mutada.
A transformação em fungos filamentosos tem o inconveniente de
apresentar uma baixa eficiência, variando de 0 a 10 transformantes/μg de DNA
(Redman et al., 1994). Diversas variáveis interferem nesse valor, e diversos
trabalhos têm relatado mecanismos para otimizar esse processo.
Schiestl e Petes (1991) constataram que a adição de enzima de restrição,
no momento da transformação, ocasionava um aumento de sete vezes na obtenção
de transformantes em Saccharomyces cerevisiae. Ao mesmo tempo, esse
procedimento propiciou integrações únicas do plasmídeo no genoma hospedeiro.
Essa técnica foi denominada REMI (Integração Mediada por Enzima de
Restrição) e vem sendo usada em um número crescente de fungos, onde a maioria
dos trabalhos alcançou resultados muito similares aos obtidos originalmente.
Diversos genes de fungos foram clonados por meio do emprego dessa técnica,
principalmente aqueles relacionados com patogenicidade em espécies
fitopatogênicas.
Penicillium griseoroseum é uma das espécies estuda pelo Departamento de
Microbiologia/UFV. Esse fungo tem se destacado pelo seu potencial na produção
de enzimas pectinolíticas que podem ser aplicadas nas indústrias de alimento e
farmacêutica. Também pode ser utilizado como modelo biológico para estudos de
regulação da expressão gênica, diferenciação celular, dentre outros.
Esse trabalho tem como objetivo verificar se o emprego da técnica REMI
leva a um aumento da eficiência de transformação de Penicillium griseoroseum e
avaliar o potencial mutagênico desta técnica por meio do isolamento de mutantes.

2 – REVISÃO DE LITERATURA
Os fungos constituem um grupo de organismos que se destacam pela sua
importância científica, econômica e ecológica. Esses organismos são utilizados
como modelo de estudo da célula eucariótica e em vários estudos de diferentes
processos celulares, justamente por apresentarem ciclo sexual; possuírem
diferenciação celular resultando em células especializadas; grande flexibilidade de
suas vias metabólicas, além da razoável facilidade de manutenção em laboratório
(Griffin,1994).
Juntamente com importância científica, diferentes ramos industriais têm
utilizado fungos para a fabricação de produtos alimentícios, tais como cogumelos
e produtos lácteos. A capacidade de produção de enzimas, como pectinases para a
produção e clarificação de sucos, é outro ramo bastante lucrativo, assim como a
fabricação de produtos farmacêuticos e compostos químicos como aminoácidos,
aspartame e etanol. Dois exemplos marcantes da aplicação industrial desses
organismos são Penicillium chrysogenum e Acremonium chrysogenum; essas duas
espécies são utilizadas com grande sucesso na indústria farmacêutica para a
produção dos antibióticos penicilina e cefalosporina, respectivamente (Brakhage,
1998).
O fungo Penicillium griseoroseum vem sendo estudado há mais de dez
anos pelo departamento de Microbiologia da UFV (D’Angelo, 1998; Brito, 1998;
Abreu, 2001). Esse organismo é um excelente produtor de pectinases e diferentes
trabalhos foram desenvolvidos com o objetivo de conhecer os genes que
compõem o complexo pectinolítico e isolar linhagens mutantes hiperprodutoras.
Mutação é uma modificação na seqüência de nucleotídeos do material
genético de um organismo. Conseqüentemente, é transferida de uma geração à
outra, ou seja, tem caráter herdável. Nem toda mutação origina alteração no
fenótipo da célula. Dessa forma, pesquisas com mutantes que resultam em
mudanças fenotípicas são mais freqüentes (Singer et al., 1982).
As mudanças numa seqüência de DNA variam em extensão. Podem ser
observadas modificações no número e/ou estrutura dos cromossomos. Como
conseqüência a célula mutante pode conter cromossomos a mais ou a menos; ou
mesmo cromossomos de tamanho diferente por perda/ganho de fragmentos ou

inversão de regiões. Essas alterações são bem mais extensas e, portanto, acabam
afetando diversos genes ao mesmo tempo. Outras alterações são fisicamente
menos evidente na estrutura do cromossomo, pois afetam pequenas seqüências de
nucleotídeos. Dessa forma acabam restringindo-se a um único gene.
A mutação é o fenômeno primário que gera variabilidade genética dentro
de uma população de indivíduos, sobre a qual a seleção natural atua em
conseqüência da pressão ambiental. Por conseguinte, contribuem para a
adaptabilidade dos organismos às mudanças ambientais.
Descobertas e avanços nas áreas de bioquímica, fisiologia e genética só
foram possíveis graças à obtenção, caracterização e análise de mutantes. Eles são
utilizados em mapeamento de genes com cruzamentos entre linhagens contendo
diferentes mutações; na análise de rotas metabólicas; dentre outras aplicações.
Mutantes podem ser obtidos espontaneamente numa população em baixa
freqüência, pois os organismos estão sujeitos a uma taxa de mutação basal. As
modificações na seqüência de nucleotídeos são conseqüências do próprio
metabolismo celular e/ou compostos químicos ou agentes físicos presentes no
ambiente.
O número de mutantes numa população pode aumentar utilizando-se
agentes mutagênicos, que podem ser físicos, químicos ou biológicos. Moléculas
de DNA como transposons, plasmídeos e T-DNA são freqüentemente utilizadas
como agentes biológicos na mutagênese (Casqueiro, et al., 1999; Souza et al.,
2000; Ramachandran et al., 2001). A mutação, resultante da utilização dessas
moléculas, ocorre por inserção das mesmas no genoma, ocasionando a interrupção
da seqüência original de nucleotídeos no DNA. Esse processo é chamado de
mutagênese insercional. Comprovando-se que o fenótipo mutante segrega com a
molécula de DNA utilizada na mutagênese, é demonstrado que o gene mutado
está marcado pelo vetor, processo conhecido como etiquetagem gênica. A
clonagem de genes etiquetados é mais fácil, justamente por eles conterem
seqüências conhecidas do vetor utilizado na mutagênese (Brown et al., 1998;
Souza et al., 2000).
A mutagênese insercional tem auxiliado na elucidação de genes, que até
então tinham função desconhecida. Isso é feito direcionando-se a inserção para a
seqüência de interesse e observando o fenótipo do indivíduo mutante
(Ramachandran et al., 2001; Vidan et al., 2001).

Dentre os agentes biológicos, os plasmídeos são os mais empregados para
a transformação e mutagênese insercional de fungos filamentosos. Essas
moléculas de DNA podem ocorrer naturalmente em fungos e estão
freqüentemente presentes em populações isoladas do ambiente. Os plasmídeos
que ocorrem naturalmente nesses organismos são citoplasmáticos e localizados,
especificamente, nas mitocôndrias.
A utilização dos plasmídeos isolados de fungos filamentosos na biologia
molecular ainda é limitada (Griffiths, 1995). Por isso, os que são empregados na
transformação desses microrganismos são geralmente isolados de bactérias. Para
sua utilização como vetores de transformação, necessitam conter um gene
marcador para seleção das linhagens transformadas em bactérias, e outro
marcador para a utilização em fungos. Normalmente, os plasmídeos utilizados na
transformação não possuem capacidade de replicação no fungo e, portanto, para
que se mantenha na célula transformada de maneira estável, é necessária a sua
integração no genoma.
Apesar da transformação de bactérias ter sido descrita pela primeira vez
em 1928, por Griffith, foi somente em 1973 que Tatum e colaboradores relataram
pela primeira vez a transformação de um fungo filamentoso. O procedimento
desses pesquisadores foi adicionar o DNA de um isolado selvagem de Neurospora
crassa ao meio de cultivo de um mutante, deficiente na síntese de inositol (inos-),
juntamente com cálcio. A partir da cultura do mutante, foram isoladas colônias
capazes de sintetizar inositol. Cultivando os transformantes, observou-se que a
capacidade de síntese do composto foi estável durante o crescimento vegetativo.
Um dos problemas encontrados pelos pesquisadores durante os experimentos de
transformação foi a taxa de reversão espontânea do mutante inos-. O que forneceu
suporte para a hipótese de transformação foi a taxa de reversão ser 30 vezes
superior após o tratamento, em relação ao controle onde não adicionou-se o DNA
da linhagem selvagem. Desde este trabalho até os dias de hoje, diversas espécies
de fungos foram transformadas em condições de laboratório (Fincham, 1989).
Diversos tratamentos têm sido utilizados para transformação de fungos,
como aplicação de um campo elétrico (eletroporação); bombardeamento de
partículas cobertas com o DNA (biobalística); polietilenoglicol (PEG); alta
concentração de LiCl e utilização do plasmídeo Ti de Agrobacterium tumefaciens

(Chakraborty et al., 1991; Fungaro et al.,1995; Queiroz et al., 1998; Chen et al.,
2000).
O processo de biobalística foi proposto por Sanford et al. (1987) com o
objetivo de introduzir material genético no núcleo de células vegetais.
Atualmente, tal processo tem sido aplicado na transformação de diferentes
organismos como protozoários, fungos, animais e plantas (Klein et al., 1992;
Bhairi et al., 1992; Vainstein et al., 1994; Fungaro et al., 1995).
Basicamente, a introdução do DNA na célula se dá por força mecânica.
Portanto, nessa técnica emprega-se microprojéteis encapados com o ácido
nucléico a ser introduzido na célula ou tecido. O aparelho utilizado é capaz de
acelerar os microprojéteis (0,2 a 4 μm de diâmetro) a velocidades superiores a
1500 Km h-1. Essas partículas perfuram a parede e membrana celular distribuindo-
se aleatoriamente no interior da célula. O emprego da biobalística evita o trauma
químico e elétrico sofrido pelas células em outras metodologias de transformação,
pois as partículas de ouro ou de tungstênio, utilizadas para a confecção dos
microprojéteis, possuem alta densidade e baixa reatividade química (Watts et al.,
1990).
Em contrapartida algumas variáveis interferem com a eficiência dessa
técnica. Molnar et al. (1997), otimizando o bombardeamento de Pisum sativum,
verificaram que parâmetros como pressão, tamanho dos microprojéteis e a
distância entre o material a ser transformado e os microprojéteis precisam ser
testados para obtenção de uma maior eficiência na transformação, e
conseqüentemente redução nos danos celulares. Uma vez no interior da célula, o
ácido nucléico pode se dissociar do microprojétil e, eventualmente, integrar-se no
genoma do organismo a ser transformado. O mecanismo de integração do DNA
no genoma hospedeiro é pouco conhecido. Acredita-se que primeiro ocorra um
estágio de pré-integração e, posteriormente, a integração propriamente dita
(Pawlowski et al. citado por Pawlowski, 1998).
A utilização da biobalística é muito vantajosa em organismos para os quais
a obtenção e regeneração de protoplastos não são possíveis ou não estão
otimizados, ou mesmo quando se quer evitar o uso dos mesmos. Diferentes tipos
celulares de fungos podem ser transformados por essa técnica, como micélio e
esporos. Urediniósporos de Uromyces appendiculatus (Bhairi et al., 1992) e

conídios de Aspergillus nidulans (Fungaro et al., 1995) são exemplos de esporos
utilizados na transformação utilizando-se partículas de tungstênio.
A eletroporação é outra técnica de transformação, que se baseia na
aplicação de um pulso elétrico sobre uma suspensão de células na presença de
moléculas de DNA (Watts et al., 1990). O mecanismo de entrada do DNA na
célula ainda não foi completamente elucidado. Alguns autores sugerem que o
pulso aplicado contribui para a ruptura da membrana plasmática, originando
pequenos poros, tornado-a permeável a pequenas moléculas. O DNA difundiria-se
através destes pequenos poros de aproximadamente 5nm de diâmetro
(Zimmermann e Vienken citados por Watts et al., 1990). Outros pesquisadores
propõem que, pela carga residual da membrana plasmática ser negativa, após o
pulso elétrico, ocorreria uma inversão desta carga. Essa inversão de cargas
facilitaria a agregação e transporte de moléculas carregadas negativamente, como
o DNA (Watts et al., 1987, citados por Watts et al., 1990).
Uma forte barreira à entrada do DNA durante a eletroporação é a parede
celular de bactérias, fungos e plantas. Chakraborty et al. (1991) puderam verificar
esse efeito a partir da eletroporação de conídios de N. crassa germinados na
presença de β-glucoronidase. Esse pré-tratamento pode ser de extrema utilidade,
pois enfraquece a parede celular sem comprometer a integridade da célula. A
eficiência de transformação foi 10 vezes superior quando os conídios foram
tratados com β-glucoronidase. A eletroporação necessita de condições
experimentais precisas, como tempo e quantidade do pulso aplicado, para cada
espécie ou isolado a ser transformado. Outros fatores como idade do conídio,
estágio da germinação, natureza da enzima micolítica usada, escolha dos
marcadores e o meio de seleção são críticos para o sucesso de tal metodologia.
Agrobacterium tumefaciens e A. rhizogenes são bactérias gram- presentes
no solo, capazes de transformar células vegetais, induzindo a proliferação celular
no tecido vegetal.
Esse fenômeno é induzido pelo plasmídeo presente em Agrobacterium
chamado de plasmídeo Ti (“tumour inducing”). Ti é um plasmídeo circular de
aproximadamente 180 Kb. Possui uma região chamada de T-DNA, que é
transferida da bactéria para a célula vegetal durante o processo de infecção e que é
capaz de inserir-se no genoma por recombinação não homóloga. Esse plasmídeo

foi modificado para ser utilizado na biotecnologia com o objetivo de
transformação de vegetais em laboratório.
Em 1995, foi relatada a transformação de S. cerevisiae utilizando A.
tumefaciens(Bundock et al, 1995). As eficiências de transformação relatadas
variaram de 10-6 (Bundock et al., 1995) até 10-3 / receptor recuperado (Piers at al.,
1996). Esses autores concluíram que a indução dos genes vir é necessária para a
transformação de S. cerevisiae, assim como os braços do T-DNA e o
processamento do T-DNA pela proteína VirD1/D2. Desse modo, o mecanismo é,
em parte, similar à transferência observada em plantas. No entanto os genes
necessários para a adsorção da bactéria nas células vegetais não são essenciais
para a transformação de levedura.
A. tumefaciens é capaz de transformar, além de leveduras, os seguintes
fungos filamentosos: Aspergillus awamori, A. niger, Fusarium venenatun,
Trichoderma reesei, Colletotrichum gloeosporioides, N. crassa e Agaricus
bisporus (Groot et al., 1998; Gouka, et al., 1999; Chen et al., 2000). A
transformação é efetiva com conídios, micélio e protoplasto de fungos, além de
fragmentos do basidiocarpo de A. bisporus (Chen et al., 2000).
A integração normalmente ocorre de maneira heteróloga, mas quando o T-
DNA possui homologia com o genoma do fungo transformado a integração
também ocorre nas regiões homólogas (Gouka et al., 1999).
A. tumefaciens também foi capaz de transformar diferentes linhagens de
células humanas (Kunik, et al., 2001), assim como na transformação de células
vegetais, a integração foi heteróloga e genes vir e chv foram essenciais; apesar de
que a indução dos genes vir pela adição de acetoseringona não foi necessária para
a detecção de células transformantes, possivelmente pelo fato do meio de cultura
ou exsudatos das células conterem compostos fenólicos capazes de realizar a
mesma função da acetoseringona.
A obtenção de células, de S. cereviseae, sem parede celular (Hutchison et
al., 1967) possibilitou o aperfeiçoamento da utilização de PEG para a
transformação de fungos. Esses autores submeteram as células de S. cereviseae ao
tratamento com glusulase, uma preparação do suco gástrico de caracol rica em
1,3-glucanase e quitinase. Essa preparação foi capaz de lisar a parede celular da
levedura. Para manter a integridade dos protoplastos, utilizaram-se um
estabilizador osmótico composto de sorbitol 1M. A partir deste trabalho de

obtenção de protoplastos abriu-se a possibilidade de utilização dessa metodologia
no processo de transformação.
Hinnen et al. (1978), utilizando protoplastos preparados a partir de
leveduras deficientes na biosíntese de leucina, leu2-, obtiveram células com o
fenótipo selvagem após o tratamento dos protoplastos com o DNA de células do
tipo selvagem na presença de cloreto de cálcio. Nesse processo de transformação,
os protoplastos, juntamente com o vetor, são tratados com uma solução de PEG e
CaCl2 que auxiliam na transformação.
O mecanismo de entrada do DNA no protoplasto ainda não foi elucidado.
Um dos papéis dos policátions, utilizados na infecção de protoplastos de plantas
com ácido nucléico viral, seria modificar a carga elétrica do ácido nucléico e da
membrana celular para propiciar uma aproximação destes elementos (Motoyoshi
et al., 1974; citados por Watts et al, 1991). Hebert (1963) supôs que o PEG teria
uma função similar, agregando e precipitando o DNA sobre a membrana
plasmática dos protoplastos.
Os fungos Ustilago maydis, Colletotrichum sp, A. bisporus, Botrytis
cinerea, Penicillium chrysogenum, P. griseoroseum e Trichoderma virens são
alguns exemplos de fungos filamentosos cujos protoplastos foram transformados
utilizando PEG (Wang et al., 1988; Redman et al., 1994; van de Rhee et al., 1996;
Baek et al., 1997; Graessle et al., 1997; Levis et al., 1997; Queiroz et al., 1998).
Uma das limitações desta técnica é a necessidade de obtenção e regeneração de
protoplastos da espécie de interesse.
A eficiência de transformação para fungos filamentosos é muito variável,
sendo que algumas espécies apresentam valores baixos, variando de 0 a 10
transformantes/μg de DNA (Redman et al., 1994). Fatores como espécie,
metodologia, adição de inibidores de nucleases ou de enzima de restrição,
quantidade e características do vetor (tamanho, grau de homologia, estado físico,
marca de seleção, presença de seqüências de replicação autônoma, modificações
causadas pela bactéria utilizada na amplificação), influenciam a eficiência e até
mesmo a estabilidade da transformação (Schiestl et al., 1991; Picard et al., 1987,
Diolez et al., 1993; Redman et al., 1994; van de Rhee, et al., 1996; Levis, et al.,
1997; Manczinger et al., 1997).
A integração do plasmídeo no genoma pode ocorrer basicamente por
integração homóloga ou heteróloga. Fatores como grau de homologia entre vetor e

o genoma e os sistemas de recombinação da célula a ser transformada são
determinantes no direcionamento de cada mecanismo de integração (Picard et al.,
1987; Baek et al., 1997; Hogan et al, 1997; Levis et al., 1997; Woods et al., 1998;
Casqueiro at al., 1999; Cormack et al., 1999; Kiechle et al., 2000; Lin et al., 2001;
Perreira et al., 2001).
A integração homóloga, aditiva ou troca gênica, pode ocorrer quando o
vetor possui regiões de homologia com o genoma a ser transformado;
normalmente possuindo algum gene do próprio organismo sendo utilizado como
marca de seleção das células transformadas, ou contendo apenas fragmento de
DNA do organismo. A integração homóloga aditiva é caracterizada por um único
evento de recombinação entre as duas moléculas de DNA, e a troca gênica por
dois eventos de recombinação.
Foram obtidos transformantes numa eficiência de 1,6%, de P.
chrysogenum, deficientes na síntese de lisina por direcionamento da integração do
plasmídeo contendo um fragmento de DNA de 4,9 Kb do gene lys2 desse mesmo
fungo, mas a utilização de vetores com fragmentos menores que o original
resultou na ausência de transformantes, demonstrando que o tamanho da região de
homologia influencia no padrão de integração. Quando esses mesmos autores
utilizaram o gene pyrG como marcador e sua seqüência flanqueada por regiões
homólogas ao gene lys2 de 4,3 e 3 Kb, pode-se recuperar transformantes que
sofreram troca gênica numa eficiência de 0,14%, mas nenhuma troca gênica pôde
ser observada quando seqüências menores do gene lys2 foram utilizadas. Um
hipótese que explica esse evento pode estar relacionada com o próprio locus do
gene lys2 (Casquerio et al. 1999).
Pereira et al. (2001) obtiveram 100 % de integrações homólogas, aditivas
ou troca gênica, quando transformaram P. griseoroseum com o plasmídeo pNPG1
que contém um fragmento de 6,1 Kb que possui o gene nia D desse mesmo fungo.
Em Candida glabrata regiões de homologia de aproximadamente 100 pb
são suficientes para dirigir a integração homóloga (Cormack et al., 1999) e em S.
cerevisiae regiões de 30 pb, de cada lado de um fragmento de DNA não
homólogo, já são suficientes para conseguir uma recombinação eficiente (Hua et
al., 1997).
As diferenças na freqüência de integrações homológas entre diferentes
organismos parecem depender, principalmente, da existência um sistema de

recombinação homólogo eficiente nos organismos. Quando o vetor não apresenta
homologia ou esta é baixa, ele se integra aleatoriamente no genoma hospedeiro
podendo inativar qualquer gene. Esse tipo de integração é chamada de ectópica ou
heteróloga. Mesmo quando um vetor apresenta regiões de homologia com
algumas seqüências do genoma receptor, a integração heteróloga pode ocorrer
devido a um sistema de recombinação homóloga pouco eficiente ou mesmo
ausência desse sistema e/ou por apresentar um sistema de recombinação
heteróloga mais eficiente. Esse sistema de integração é um método eficiente para a
mutagênese insercional, quando o objetivo é obter diferentes tipos de mutantes, já
que sua integração pode ser definida como aleatória.
Na transformação de um mutante deficiente na assimilação de nitrogênio
de B. cinerea com o gene niaD de F. oxysporum, foram obtidos 10
transformantes/μg de DNA, sendo que a eficiência de transformação foi o dobro
quando utilizou-se o gene do próprio B. cinerea (Levis et al., 1997). Penicillium
expansum, quando transformado com o gene nia homólogo, produziu 15-16
transformantes/μg de DNA (Torres, 2001), mas quando transformado com o gene
de F. oxysporum foram obtidos apenas 8 transformantes/μg de DNA (Ribeiro,
1998). A eficiência de transformação com um gene homólogo tende ser maior do
que a obtida com um gene heterólogo.
Estratégias como a utilização de vetores com seqüências teloméricas e/ou
seqüências de replicação autônoma acarretam num aumento acentuado na
eficiência de transformação. Redman et al (1994), ao transformar Colletotrichum
lindemuthianum com o vetor pHA1.3, obtiveram uma eficiência de transformação
de 100 a 10.000 transformantes/μg de DNA. Essa alta eficiência foi
correlacionada com o fato desse vetor possuir seqüências teloméricas de F.
oxysporum e seqüência de replicação autônoma.
Schiestl et al. (1991), transformando S. cerevisiae, propuseram que a
adição de enzima de restrição à mistura de transformação aumentava a eficiência
de transformação em sete vezes. O evento de integração mediado pela enzima de
restrição foi denominado de REMI (Integração Mediada por Enzima de
Restrição). Esses autores sugeriram que a enzima de restrição, utilizada durante a
transformação, induziria quebras duplas no DNA genômico nos seus sítios de

clivagem gerando extremidades coesivas com o vetor linear quando o plasmídeo é
digerido com a mesma enzima.
A clivagem do genoma pela enzima de restrição, durante a transformação,
resulta basicamente em três eventos em relação ao sítio de clivagem da enzima
utilizada: deleções (Figura 1-IA), reparo (Figura 1-IB) e reparo com integração do
vetor (Figura 1-IC). Por meio de sistemas de reparo de quebra dupla, o vetor é
integrado no genoma, podendo regenerar o sítio de clivagem. Dessa forma quando
o DNA total dos transformantes é clivado com a mesma enzima utilizada na
transformação, pode-se detectar um fragmento de DNA de mesmo tamanho que o
vetor. Isso evidência que os dois sítios de clivagem, presentes nas extremidades
do vetor, foram perfeitamente regenerados durante a integração (Figura 1 – IIA).
O sistema de reparo de quebra dupla pode não regenerar um dos sítios de
clivagem, e até mesmo os dois, resultando num evento de mutação não REMI
(Figura 1 – IIB).
Mutantes gerados a partir da transformação utilizando enzimas de restrição
podem ser gerados por outro mecanismo diferente da mutagênese insercional. As
quebras geradas no DNA genômico podem ser corrigidas erroneamente (Figura 1
– IIC), sendo que em alguns casos, deleções e rearranjos já foram observados
(Linnemannstöns et al., 1999).

I
II
Figura 1: Efeito da enzima de restrição utilizada na transformação e integrações do vetor. I- Após a
atuação da enzima no genoma pode haver deleções(A), reparo(B) ou reparo com integração do
vetor no sítio de clivagem da enzima(C). II- Mecanismos de integração do vetor no sitio de
clivagem presente no genoma podem regenerar os dois sítios resultando em evento REMI (A),
regenerar um sítio ou nenhum (B) ou simplesmente regenerar o sítio inicial, mas a integração do
vetor ocorrer em outra região (C).

Quebras de fita dupla podem surgir como conseqüência da ação
de enzimas de restrição, além da ação de topoisomerases, radiações e
agentes químicos. O reparo de quebras duplas no DNA de leveduras
pode ocorrer via um processo reconhecido como reparo por ligação de
terminais não homólogos (Schiestl e Petes, 1991; Moore et al., 1996;
Zhu et al., 1996; Manivasakam et al., 1998; Lewis et al, 1999).
A integração de vetores lineares, sem a presença de enzima, ocorreria por
outro mecanismo de recombinação não homólogo no qual regiões de
microhomologia dos terminais fita simples do vetor atuariam na integração.
Enzimas, como topoisomerase I (Topo I), podem estar envolvidas nesse tipo de
recombinação em S. cerevisiae (Zhu et al., 1996)
Além da diferença no padrão de integrações, existem também diferenças
na estabilidade e no número das mesmas. Integrações homólogas aditivas tendem
a ser mais instáveis por gerar regiões próximas com alta homologia. Essas regiões
podem ser perdidas por um processo de recombinação homóloga. Casqueiro et al.
(1999) observou baixa estabilidade em transformantes que apresentavam esse
padrão de integração.
A presença de cópias únicas e/ou em tandem do plasmídeo podem ocorrer
em qualquer tipo de integração. Provavelmente integrações em tandem surgiriam
a partir de integrações homólogas entre os vetores presentes no núcleo; ocorrendo
antes da integração no genoma, formando concatâmeros ou por meio de ciclos de
recombinação com vetores já integrados no genoma (Fincham, 1989).
Um outro modelo de integração em tandem foi recentemente proposto em
Dictyostelium discoideum (Barth et al., 1998). Os autores do trabalho sugerem que
múltiplas cópias poderiam ser inseridas por meio da replicação circulo rolante a
partir de concatâmeros ou de moléculas simples de plasmídeo, mas
necessariamente circular.
A aplicação da técnica REMI não se limita apenas ao aumento da
eficiência de transformação, podendo ser utilizada na mutagênese insercional,
propiciando um sistema eficiente de etiquetagem gênica, geração de aberrações
cromossômicas e estimulação de recombinação mitótica em sítios específicos
(Schiestl et al., 1991)

Kuspa e Loomis (1992) obtiveram 20 a 60 vezes mais transformantes de
D. discoideum na presença da enzima de restrição utilizada na clivagem do vetor.
A integração do vetor no genoma preservou, em mais de 70% dos transformantes,
o sítio de restrição da enzima utilizada; sugerindo que os terminais compatíveis
entre o vetor e o genoma, gerados pela mesma enzima, são eventualmente
reparados pela maquinaria de reparo da célula. Isso está de acordo com os
resultados obtidos quando a enzima utilizada para a clivagem do vetor foi
diferente da utilizada durante o procedimento de transformação. Enzimas que
geram terminais coesivos, por exemplo, BamH1 e Sau3A1, acarretaram aumento
na transformação, ao contrário do que ocorreu entre BamH1 e EcoR1.
A clonagem do gene Tox1 de Cochliobolus heterostrophus (Lu et al.,
1994) foi realizada utilizando-se o REMI, sendo que de um total de 1310
transformantes, dois eram deficientes na síntese da toxina T e a mutação estava
ligada ao marcador de seleção contido no vetor usado na transformação. Foi
verificado que a forma ativa ou inativa da enzima de restrição adicionada à
suspensão de protoplastos influenciava a eficiência de transformação. A enzima
contida na reação de clivagem foi capaz de aumentar em cerca de 20 vezes a
eficiência de transformação, e a adição posterior de enzima acabou por decrescer
esse valor. Os dois mutantes Tox1- foram gerados pela integração do vetor em
diferentes posições nesse locus, o sítio de restrição da enzima utilizada durante a
transformação foi regenerado, sendo, portanto um evento REMI verdadeiro.
Em Magnaporthe grisea foi relatado um aumento de aproximadamente 10
vezes na eficiência de transformação utilizando-se REMI (Shi et al., 1995). A
eficiência variou com a concentração e a enzima utilizada, onde geralmente
concentrações relativamente altas acabaram por diminuir a eficiência de
transformação. Esse decréscimo não está relacionado com possíveis danos ao
genoma, já que a viabilidade dos protoplastos não foi alterada. A utilização de
enzimas, para linearizar e/ou transformar, que geram terminais abruptos causam
um leve efeito na eficiência de transformação, indicando que terminais coesivos
entre o vetor e o genoma são regenerados com maior eficiência. Após a análise de
600 transformantes foram isolados dois mutantes morfológicos, um auxotrófico e
dois relacionados com a patogenicidade.
Já em U. maydis, não se observou diferença na eficiência de transformação
utilizando enzima de restrição no momento da transformação com o vetor circular

(Bölker, 1995). Após a análise do padrão de integração, pode-se concluir que a
presença da enzima de restrição propiciou, em 90% dos transformantes,
integrações únicas; ao contrário dos transformantes obtidos sem adição de enzima,
onde o vetor apresentou integrações em tandem. Os resultados obtidos por esses
autores demonstram que o REMI é uma técnica eficiente para a etiquetagem
gênica em U. maydis, sendo que de 1000 transformantes analisados, 1 a 2% foram
incapazes de produzir os sintomas de patogenicidade nos testes realizados.
REMI foi aplicado para diversos fungos como Alternaria alternata,
Coprinus cinereus, Mycosphaerella zeae-maydis, Glomerella graminicola, D.
discideum, Colletotrichum magna, Paecilomyces fumosoroseus, A. nidulans, A.
oryzae e A. fumigatus (Akamatsu et al., 1997; Granado et al., 1997; Sánchez, et
al., 1998; Yun et al., 1998; Epstein et al., 1998; Barth et al., 1998; Redman et al.,
1999; Tanaka et al., 1999; Cantone et al., 1999; Yaver et al., 2000; Brookman et
al., 2000; Thon et al., 2000; Zeng et al., 2000; Souza et al., 2000) com os
objetivos de aumentar a eficiência de transformação; isolar mutantes,
principalmente para deficiência na fitopatogenicidade, e etiquetar genes para sua
subseqüente clonagem.
Esses trabalhos corroboram a observação de que REMI é uma técnica
muito eficiente para a etiquetagem de genes porque, em geral, leva a um aumento
da eficiência de transformação, propicia integrações aleatórias no genoma da
espécie a ser transformada e favorece integrações únicas do vetor, diminuindo
integrações em tandem, o que facilita a futura clonagem do gene mutado.
Alguns requerimentos são essenciais para a utilização eficiente do REMI.
É necessário que o vetor possua capacidade de replicação e seleção em E. coli,
para facilitar a recuperação do plasmídeo de um possível mutante de interesse;
para selecionar as células transformantes deve-se utilizar um marcador dominante
ou um marcador que complemente uma mutação auxotrófica, nesse caso, é
preferível que a seqüência do marcador não seja homóloga para propiciar
integrações heterólogas; o vetor precisa ter sítios únicos de restrição; é necessário
padronizar a melhor concentração de enzima que resulte na máxima eficiência.
A partir da aplicação de REMI pode-se obter diferentes mutantes. No
entanto, muitas vezes o mutante pode ser isolado sem a ocorrência de integração
do plasmídeo no gene em estudo. Para comprovar que o gene foi realmente
etiquetado, pode-se utilizar diferentes estratégias, como: análise de segregação

pelo ciclo sexual ou parassexual, transformação da linhagem selvagem com o
plasmídeo recuperado, e pela complementação do mutante com um cosmídeo ou
plasmídeo carregando a região do gene da linhagem selvagem.

3 - MATERIAL E MÉTODOS
3.1 – Microrganismos, plasmídeos e meios de cultura
O organismo utilizado foi Penicillium griseoroseum (CCT 6421), isolado
de sementes florestais pelo Dr. J. J. Muchovej, do Departamento de Fitopatologia
da Universidade Federal de Viçosa. Os experimentos de transformação foram
realizados com uma linhagem mutante, obtida por Pereira (1999) denominada
PG63. Esse isolado possui conídios verdes e apresenta uma deleção de 211 pb no
gene niaD, tornando-o incapaz de assimilar nitrogênio na forma nítrica. A
linhagem auxotrófica foi cultivada em meio completo (Pontecorvo et al., 1953)
modificado por Azevedo e Costa (1973). Utilizou-se meio mínimo (Pontecorvo et
al., 1953) para a seleção e manutenção dos transformantes a 25°C. Para a seleção
e regeneração dos protoplastos adicionou-se sacarose 0,56 M em ambos os meios.
O plasmídeo pNH24 (Diolez et al., 1993), que possui o gene niaD de F.
oxysporum e sítios únicos de restrição para Xba I e Eco RV, foi utilizado nos
experimentos de transformação. O isolamento de mutantes niaD- foi realizado
segundo UnKles et al. (1989), utilizando meio mínimo contendo clorato de sódio
e glutamina.
3.2 – Extração de DNA plasmidial
O plasmídeo pNH24 foi isolado de E. coli DH5α pelo método de extração
alcalina e purificado por cloreto de lítio, como descrito em Sambrook et al.
(1989). O plasmídeo foi linearizado por digestão de 5 μg do DNA com 15
unidades de Xba I num volume de reação de 30 μL, por 2 horas, segundo
instruções do fabricante. Nos experimentos de transformação foram utilizadas
diferentes quantidades da enzima Xba I (Quadro 1).

Quadro 1: Diferentes tratamentos empregados nos experimentos de
transformação de P. griseoroseum.
TRATAMENTO
CONDIÇÃO DO
VETOR USADO NA
TRANSFORMAÇÃO*
CONDIÇAO DA
ENZIMA
ADICIONADA A
REAÇÃO DE
CLIVAGEM
UNIDADES DE
ENZIMA
ADICIONADAS
DURANTE A
TRANSFORMAÇÃO
T1 Ausência do vetor - 0
T2 circular - 0
T3 Linear inativa
(15min/70°C) 0
T4 Linear Ativa 0
T5 Linear Ativa 1
T6 Linear Ativa 5
T7 Linear Ativa 10
T8 Linear Ativa 15
T9 Linear Ativa 20
* O vetor foi linearizado com a enzima Xba I

3.3 – Transformação mediada por REMI
Uma suspensão de esporos, obtida da linhagem PG63, foi plaqueada sobre
meio completo, coberto com papel celofane e incubada por 20 horas a 25°C. Após
a incubação, aproximadamente 800 mg de micélio foram misturados a 5 mL de
tampão de digestão (KCl 0,8 M, pH 5,8; tampão fosfato 10 mM; 15 mg da enzima
GLUCANEX® - Novo Nordisk). Essa mistura foi mantida a 30°C, 80 rpm por 3
horas. Logo após a protoplastização, a suspensão foi filtrada, e lavada por 3 vezes
em STC (Sorbitol 1M; CaCl2 0,05 M e Tris-HCl 0,1 M) com centrifugação de
3000g por 15 min; ao final obteve-se uma suspensão de protoplastos na
concentração de 108 protoplastos/mL.
A transformação seguiu os procedimentos de Yelton et. al (1984) e
Ballance e Turner (1985) com modificações. Num tubo Eppendorff foram
adicionados: 100 μL da suspensão de protoplastos, 5 μg de DNA plasmidial,
quantidades variáveis da Xba I (Quadro 1) e 50μL de polietilenoglicol 6000 25%.
As misturas foram incubadas em banho de gelo por 30 minutos, seguindo-se a
adição de 500 μL da mesma solução de polietilenoglicol. Após 20 minutos a 25
°C, os protoplastos foram plaqueados em meio mínimo contendo nitrogênio
nítrico como única forma de nitrogênio e sacarose 0,56 M, pelo método “pour
plate”, e mantidos a 250C por 5 dias. A eficiência de transformação foi dada pela
contagem dos transformantes que cresceram no meio seletivo.
O controle negativo da transformação foi realizado nas mesmas condições,
exceto pela ausência do plasmídeo, assim como os controles de regeneração dos
protoplastos.
3.4 - Isolamento de mutantes
Mutantes morfológicos que apresentaram modificações na cor, na
esporulação ou no crescimento foram selecionados pela observação direta das
colônias. Para esses isolados o número de conídios foi estimado por transferência
de discos das colônias para 4 mL de Tween 80 0,1%. A contagem foi feita em
câmara de Neubauer.

A produção diferenciada de pectinases dos transformantes foi estimada
pela medição do halo de degradação de pectina em placa de Petri. Cerca de 500
transformantes purificados foram crescidos em meio mineral tamponado (MMT) a
pH 7,2 (2,0 g de KH2PO4, 7,0 g de K2HPO4, 1,0 g de (NH4)2SO4, 1,0 g de
MgSO4.7H2O, 3,0 g de pectina cítrica, 0,6 g de extrato de levedura, 13 g de ágar –
água q.s.p 1000 mL) por 5 dias a 25°C. Após esse período, discos de 7 mm do
micélio foram transferidos para placas de Petri contendo o meio tamponado Mac
Ilvaine (17,95 g de NaHPO4, 7,74 g de ácido cítrico, 2,5 g de pectina cítrica, 13 g
de agar – água q.s.p. 1000 mL) e incubados por 48 h a 40 °C. Para revelação do
halo de degradação foi utilizada uma solução de iodo/iodeto (5,0 g de KI, 1,0 g de
iodo – água q.s.p. 330 mL). Os transformantes que apresentaram medidas do halo
de degradação menor que o selvagem foram selecionados para posterior dosagem
da atividade enzimática em meio líquido.
3.5 – Determinação da atividade de poligalacturonase (PG) e pectina
liase (PL) em meio líquido
As dosagens enzimáticas foram realizadas no sobrenadante das
culturas crescidas em meio líquido. Para dosagem de poligalacturonase
o meio utilizado foi o meio mineral não tamponado (0,6 g de KH2PO4, 2
g de K2HPO4, 1 g de (NH4)2SO4, 5,5 g de MgSO4.7H2O, 3 g de pectina,
3 g de extrato de levedura, pH 6,3 – água q.s.p. 1000 mL) e para
dosagens de pectina liase utilizou-se o meio mineral tamponado (3,8 g
de KH2PO4, 6,8 g de K2HPO4, 1 g de (NH4)2SO4, 5,5 g de MgSO4.7H2O,
3 g de pectina, 3 g de extrato de levedura – água q.s.p. 1000 mL). Uma
suspensão de esporos foi preparada a partir das linhagens de P.
griseoroseum crescidas por 7 dias em ágar-aveia. Frascos Erlenmeyer
contendo 50 mL de meio foram inoculados com tal suspensão numa
concentração final de 1 x 106 esporos/mL. As culturas foram incubadas

a 25 °C, a 150 rpm por 36 horas para dosagens de PG e por 120 horas
para PL.
A dosagem de poligalacturonase baseou-se no método do açúcar
redutor do DNS (Miller, 1959). A mistura de reação constou de 1 mL
do sobrenadante da cultura e 1 mL de solução do substrato (ácido
poligaraturônico 0,6 %). Imediatamente após a mistura retirou-se uma
alíquota de 0,25 mL a qual foi adicionada a um tubo contendo 1 mL de
DNS e 0,25 mL de água. A mistura de reação foi mantida por 20 min a
40 °C. Após esse período, retirou-se 0,25 mL da mistura e adicionou-se
1 mL de DNS e 0,25 mL de água. Os tubos foram mantidos a 95-100 °C
por 5 min e logo em seguida, 10 mL de água foram adicionados a cada
tudo. A absorbância foi medida a 540 nm.
Para a dosagem de PL 1 mL do substrato (pectina cítrica 2,5 %) e
1,5 mL do sobrenadante da cultura foram misturados. Uma alíquota de
0,5 mL foi retirada e adicionada num tubo de ensaio contendo 4,5 mL
de HCl 10 mM para interromper a reação. Após 30 min, foram
retiradas mais duas alíquotas realizando-se o mesmo procedimento. A
absorbância foi lida a 235 nm, como descrito por Albersheim e Killias
(1962).
3.6 - Análise da estabilidade dos mutantes
Os mutantes selecionados foram transferidos para meio completo e
mantidos por 7 dias, a 25°C. Após esse período foram novamente transferidos
para meio completo, sendo esse processo repetido por 5 vezes. Após essas
transferências testou-se a capacidade de crescimento em meio mínimo com nitrato
na forma nítrica como a única fonte de nitrogênio. Mutantes estáveis foram
capazes de manter sua capacidade de assimilar o nitrogênio do meio mínimo.

3.7 - Análise do padrão de integração
Para extração de DNA, as linhagens transformantes foram crescidas em
meio mínimo coberto com papel celofane. O isolado PG63 foi cultivado em meio
completo.
Realizou-se a maceração do micélio congelado em nitrogênio líquido com
auxílio de almofariz e pistilo. Pequena quantidade do micélio em pó foi
transferida para tubos Eppendorfs e a extração foi realizada segundo metodologia
de Speacht et al. (1982). Cerca de 3 μg de DNA foi clivado num volume 50 μL de
reação de clivagem utilizando Eco RV ou Xba I, de acordo com recomendações
do fabricante. Os fragmentos de DNA da reação de clivagem foram separados por
eletroforese em gel de agarose 0,8 % e transferidos para membrana de nylon
(Stratagene®) de acordo com Sambrook et al. (1989).
O plasmídeo pNH24, empregado como sonda, foi marcado com dUTP –
fluoresceína, utilizando-se flúor-12-dUTP ( Dig hight primer DNA labeling and
detection starter Kit II, Roche) segundo instruções do fabricante, ou com α-32P-
ATP (Sambrook et al., 1998). As condições de hibridização e lavagens da
membrana, quando hibridizadas com sonda marcada por dUTP – fluoresceína,
seguiram as recomendações do fabricante para sondas homólogas, assim como a
detecção da hibridização.
A sonda marcada com α-32P-dATP foi fervida por 10 minutos e mantida
em banho de gelo até sua utilização. A membrana foi incubada por 16 horas a
68°C, sob agitação lenta.
Seguiram-se quatro lavagens, na mesma temperatura de hibridização: uma
vez em SSC 2x, SDS 0,1%; duas vezes em SSC 1X, SDS 0,1%, e uma vez em
SSC 0,1x, SDS 0,1%, cada uma com duração de 20 minutos.
O resultado da hibridização foi detectado após exposição de um filme
radiográfico à membrana por 15 min, quando utilizou-se marcação com dUTP –
fluoresceína, e por 24 horas a –80°C para marcação com α-32P-ATP.

4 – RESULTADOS E DISCUSSÃO
4.1 – Efeito da enzima de restrição Xba I na eficiência de
transformação de P. griseoroseum
O vetor linearizado mostrou-se mais eficiente que a forma
circular para a obtenção de transformantes (Figura 2, T2 e T3). As
extremidades do vetor linear podem ser utilizadas de forma
semelhante ao que ocorre em S. cerevisiae, onde os terminais livres de
DNA são usados pelo sistema de integração ilegítima.
De uma forma geral, o estado físico do vetor influenciou a
transformação de diversos outros fungos (Wang et al., 1988; Shi et al.,
1995; van de Rhee et al, 1996; Manivasakam et al., 1998). No entanto,
em determinadas espécies, essa diferença não foi observada (Granado
et al., 1997; Thon et al., 2000). Na maioria dos trabalhos com REMI
citados anteriormente, a linearização do vetor resulta num efeito
positivo na eficiência de transformação.
Em Ustilago maydis a linearização do plasmídeo pHL1 acarretou
um aumento de 20 vezes na eficiência de transformação (Wang et al.,
1988), assim como trabalhos com Agaricus bisporus, Magnaporthe
grisea e S. cerevisiae têm mostrado o mesmo resultado (Shi et al.,
1995; van de Rhee et al, 1996; Manivasakam et al., 1998). No caso de
Cochliobolus heterostrophus, a utilização do plasmídeo pUCATPH
circular inviabilizou a obtenção de transformantes, o que não ocorreu
quando esse plasmídeo foi linearizado, apesar da taxa de
transformação ter sido baixa (Lu et al., 1994)
Espécies como Colletotrichum graminicola (Thon et al., 2000) e
Coprinus cinereus (Granado et al., 1997) não apresentaram diferenças

na eficiência de transformação quando foi utilizada a forma linear ou
circular do plasmídeo.
A diferença entre o tratamento T3 e o tratamento T4 está na presença da
enzima de restrição utilizada na clivagem do plasmídeo, inativa ou ativa,
respectivamente; portanto, Xba I foi capaz de aumentar em 3,24 vezes a eficiência
transformação de P. griseoroseum (Figura 2).
Após o trabalho inicial de Schiestl e Petes (1991), diversos outros têm
demonstrado que enzimas de restrição são capazes de influenciar a transformação
de diversos fungos.
A influência na eficiência de transformação varia entre os tipos e
concentrações das enzimas utilizadas. Asp718, EcoRV, HindIII, SalI, SmaI, HpaI,
MscI e SnaBI não influenciaram na eficiência da transformação de S. cerevisiae
(Manivasakam et al., 1991) e BamHI em Ustilago maydis (Bölker et al., 1995).
Inversamente, HindIII e SalI atuaram positivamente na eficiência de
transformação de A. oryzae (Yaver et al., 2000).
Devido à variação de respostas em relação ao tipo de enzima e ao
organismo estudado, acredita-se que a incapacidade de uma enzima de restrição
em afetar a taxa de transformação não se relaciona com suas propriedades, mas
sim com o ambiente celular, habilidade de entrada na célula e condições de
transformação (Manivasakam et al., 1998).
A quantidade que resulte numa maior eficiência de
transformação precisa ser otimizada empiricamente. A eficiência de
transformação parece ser inversamente proporcional às concentrações
da enzima testada. Os resultados obtidos nesse trabalho indicam que a
adição de 1U é suficiente para a obtenção da maior eficiência de
transformação de P. griseoroseum (Figura 2, T5). Quantidades maiores
resultaram num decréscimo da eficiência, apesar dos números de
transformantes serem superiores ao tratamento no qual a enzima foi
inativada (Figura 2, T3 e T6 a T9).

Granado (1997) verificou que a enzima de restrição não tem
efeito mutagênico que leve a uma menor viabilidade, pelo menos que
seja mensurável, em protoplastos de C. cinereus.
A enzima EcoRI, quando utilizada na transformação de S.
cerevisiae, também não diminuiu a viabilidade das células (Manivasakam
e Schiestl, 1998), mas quando expressa neste organismo resultou em
morte celular provavelmente por induzir inúmeras quebras duplas na
molécula de DNA que não puderam ser corrigidas, eficientemente, pelo
sistema de reparo (Lewis et al., 1999).
Possivelmente as concentrações utilizadas nos procedimentos de
transformação não são nocivas o suficiente para causar inviabilidade
celular. Quantidades maiores da enzima de restrição podem atuar no
sítio de reparo, decrescendo a freqüência de inserção e refletindo uma
menor eficiência de transformação.

Figura 2: Influência do estado físico do plasmídeo e da quantidade de enzima de
restrição na eficiência de transformação de P. griseoroseum. Os tratamentos
correspondem aos mesmos do Quadro 1. Médias seguidas pela mesma letra
não diferem entre si pelo teste de Scott-Knott a 5 % de probabilidade. T1-
ausência de plasmídeo; T2-plasmídeo circular; T3-plasmídeo linearizado, e
com inativação da enzima utilizada na sua clivagem; T4 a T9-plasmídeo

linearizado, com a enzima ativa utilizada na reação de clivagem, e com adição
de 0, 1, 5, 10, 15 e 20U, respectivamente, de enzima durante a
transformação.

4.2 – Caracterização das integrações dos plasmídeos nos transformantes
Para determinar o padrão de integração do plasmídeo nos transformantes,
o DNA total foi digerido com a enzima Xba I e Eco RV.
O plasmídeo pNH24 possui um único sítio de clivagem para a
enzima Eco RV, conseqüentemente o número de cópias e a
aleatoriedade da integração podem ser inferidos baseados no número
de fragmentos de DNA gerados quando o DNA dos transformantes é
clivado com essa enzima e hibridizado com o plasmídeo.
Se o transformante possuir apenas uma cópia do vetor no seu
genoma, a clivagem do seu DNA produzirá dois fragmentos de DNA.
Isso é resultado da clivagem da enzima uma única vez na seqüência do
plasmídeo, gerando dois fragmentos, sendo que cada um está ligado ao
DNA do fungo. Quando dois transformantes apresentarem diferentes
padrões de bandas, pode-se inferir que a integração ocorreu em
diferentes sítios no genoma. Integrações em tandem são visualizadas
pelo aparecimento de três bandas, sendo uma delas correspondente ao
tamanho do vetor e outras duas de tamanhos variáveis (Figura 3,
canaleta 13). Algumas plasmídeos integrados podem sofrer rearranjos
resultando em pequenas deleções do vetor, nesse caso a hibridização
gera apenas uma banda (Figuras 3, canaleta 10).
A transformação utilizando Xba I favoreceu integrações únicas
em 80 % dos casos analisados, nos casos restantes as integrações
ocorreram em tandem ou foram produzidos rearranjos (Figuras 3 e 4).
Apesar de dois transformantes obtidos com o plasmídeo pNH24 na forma
circular também terem resultado em integrações únicas, pode-se observar que esse
padrão ocorreu em baixa freqüência, como já verificado por Queiroz et al.(1998)
que transformaram pela primeira vez P. griseoroseum utilizando o plasmídeo
pNE24 (Figura 3, canaletas 2 e 3).

Integrações únicas e aleatórias são os principais requerimentos para a
mutagênese insercional, pois facilita a verificação da integração no gene
responsável pelo fenótipo do mutante e a futura clonagem do gene interrompido.
Bölker et al. (1995) apesar de não ter verificado aumento na eficiência de
transformação de U. maydis via REMI, concluíram que essa técnica é eficiente
para a mutagênese insercional, pois favorece justamente esse tipo de integração.
Alguns autores questionam a aleatoriedade fornecida pelo REMI por
isolarem transformantes com o mesmo fenótipo e/ou genes idênticos
interrompidos (Lu et al., 1994). Isso pode estar relacionado com o tamanho do
gene em questão, o número de sítios de clivagem presentes no mesmo para a
enzima utilizada na transformação, ou mesmo o número de genes relacionados
com um determinado fenótipo.
Quando a integração do vetor é direcionada pela enzima de restrição
utilizada durante a transformação, os dois sítios de restrição nos seus terminais
podem ser regenerados. Então, a hibridização gera um fragmento de mesmo
tamanho que o plasmídeo quando o DNA genômico do transformante for clivado
com a mesma enzima.
Três tipos de integração ocorreram entre os transformantes analisados. O
primeiro tipo mostrou um fragmento de tamanho idêntico ao do plasmídeo
pNH24, indicando que nesse transformante os dois sítios foram regenerados
(Figura 4, canaleta 6). Um outro tipo ocorreu quando nenhum dos sítios foi
regenerado ou a regeneração ocorreu em apenas um deles, esse caso é evidenciado
por um fragmento maior que o fragmento do vetor (Figura 4, canaletas 3 e 4). E
por fim, o terceiro tipo surgiu quando o plasmídeo circular integrou mantendo o
sítio de XbaI na sua seqüência, dessa forma fragmentos menores que o do vetor
foram evidenciados pela hibridização (Figura 4 canaleta 2).

A
B
Figura 3 – Análise dos transformantes de P. griseoroseum. (A) Clivagem do DNA total com Eco
RV: M – Marcador de tamanho λ Hind III; P – plasmídeo pNH24, 1 e 17 – P.
griseoroseum PG63; 2 a 16 transformantes niaD+. (B) Auto-radiografia da hibridização
utilizando como sonda o plasmídeo pNH24 marcado com α -32P- ATP.

A B
Figura 4 – Análise dos transformantes com produção de poligalacturonase alterada. (A) Clivagem
do DNA total com Eco RV M – Marcador de tamanho λ Hind III; P – plasmídeo pNH24,
1 – P. griseoroseum PG63; 2 - Tr7 e 3 - Tr8. (B) Auto-radiografia da hibridização
utilizando como sonda o plasmídeo pNH24 marcado com dUTP – fluoresceína.
Transformantes apresentam integrações únicas pela presença de duas bandas.

A B
Figura 5 – Analise dos transformantes de P.griseoroseum. (A) Clivagem do DNA total
com Xba I: M – Marcador de tamanho λ Hind III; P – plasmídeo pNH24, 1 – P.
griseoroseum PG63; 2 – transformantes obtidos com plasmídeo circular; 3 a 6
– transformantes obtidos com plasmídeo linear (B) Auto-radiografia da
hibridização utilizando como sonda o plasmídeo pNH24 marcado com dUTP –
fluoresceína. A seta indica um fragmento de DNA do mesmo tamanho que o
plasmídeo.

4.3 – Isolamentos de mutantes morfológicos e com atividade enzimática de
pectinases alterada
4.3.1 - Mutantes Morfológicos
Diferentes tipos de mutantes podem ser isolados de uma população. O
isolamento restringe-se àqueles que possuem alterações fenotípicas capazes de
serem visualizadas por alguma metodologia. Nessa categoria podemos isolar
indivíduos com alterações morfológicas, auxotróficas, resistentes e/ou sensíveis a
drogas, relacionados com patogenicidade e produção alterada de metabólitos e/ou
enzimas.
Mutantes morfológicos caracterizam-se por apresentar
diferentes fenótipos como alterações na taxa de crescimento linear,
no padrão de ramificação, no grau de esporulação, na pigmentação dos
esporos e na degeneração celular.
De um total de 843 transformantes obtidos foram isolados 6 mutantes
morfológicos (Figura 6). Esses mutantes foram originados somente dos
tratamentos onde houve adição da enzima de restrição. Essa origem deve-se ao
fato de ter sido analisado maior número de transformantes originados desses
tratamentos.
Os mutantes Tr 1, Tr 2 e Tr 3 possuem tonalidade verde mais claro em
relação ao selvagem. Alterações na cor podem ser causadas pela redução na
quantidade de pigmentos, atraso no desenvolvimento dos conídios ou mesmo
redução no número deles. Nesses mutantes esse efeito parece relacionado com a
redução na esporulação, verificado após a contagem de esporos.
O mutante Tr4 apresentou esporos de coloração marron. Esse isolado pode
apresentar um gene da via de produção do pigmento verde mutado ou outro gene
envolvido na deposição do pigmento na parede celular. Em A. nidulans diversos
genes contribuem para os nuances da cor do esporo, mas os dois principais são
wA e yA responsáveis pelas tonalidades branca e amarela, respectivamente. O
produto desses dois genes atuam diretamente na via de formação do pigmento.
Outros genes como chA, fwA, bwA ou dilA e dilB podem alterar a quantidade e a

qualidade do pigmento ou em algum aspecto da parede celular (Adams et al.,
1998).
O mutante Tr5 apresentou redução na esporulação e alterações no padrão
de crescimento. Depois de alguns dias de crescimento, a borda da colônia
começava a sofrer alterações da morfologia, podendo ser resultado de
modificações no padrão de ramificação, o que foi observado por meio de
microscopia.
O mutante Tr6 mostrou áreas do micélio deficientes na esporulação,
provavelmente acarretadas por mutação em um gene envolvido no controle da
esporulação. Fenótipos semelhantes foram obtidos em A. nidulans (Adams et al.,
1998).
Os padrões de integração do vetor nesses mutantes podem ser reunidos em
três grupos: o primeiro apresentando integrações únicas no genoma como os Tr1,
Tr5 e Tr6 (Figura 3, canaletas 12, 8, 11, respectivamente); o segundo
apresentando integrações múltiplas, representado pelos transformantes Tr4 e Tr2
(Figura 3, canaletas 13 e 15, respectivamente), e por fim transformantes que
apresentaram rearranjos no vetor como no Tr3 (Figura 3, canaleta 10).
No mutante Tr4, as integrações ocorreram em tandem e no Tr2
possivelmente houve dois eventos de integração em diferentes regiões do genoma.
Nos transformantes Tr1 e Tr5, os dois sítios de clivagem para Xba I foram
regenerados durante a inserção do vetor (dados não mostrados)
Para os demais mutantes não foi verificado se a mutação responsável pelo
fenótipo dos transformantes ocorreu devido a inserção do plasmídeo ou a outras
mutações causadas pela enzima de restrição utilizada durante a transformação.
Essa estudo poderá ser realizado por meio da análise de segregação entre o
fenótipo e o vetor pelo ciclo parassexual ou mesmo transformando a linhagem
PG63 com o vetor recuperado a partir do mutante.
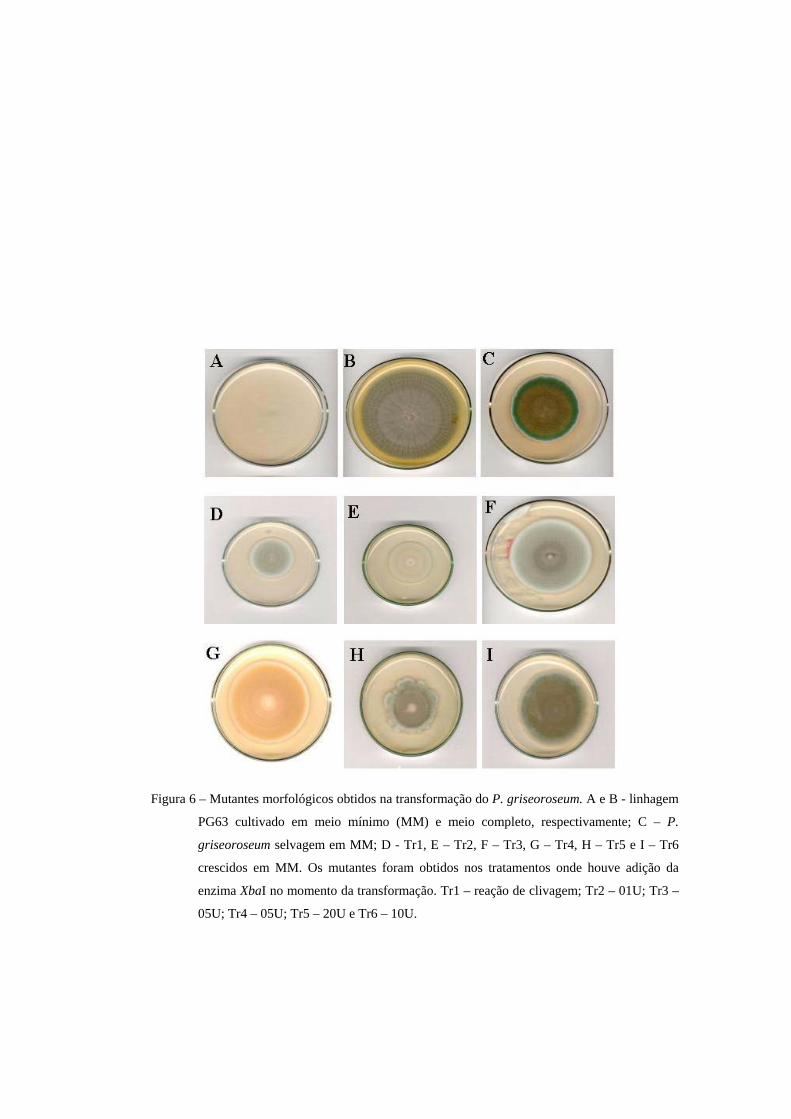
Figura 6 – Mutantes morfológicos obtidos na transformação do P. griseoroseum. A e B - linhagem
PG63 cultivado em meio mínimo (MM) e meio completo, respectivamente; C – P.
griseoroseum selvagem em MM; D - Tr1, E – Tr2, F – Tr3, G – Tr4, H – Tr5 e I – Tr6
crescidos em MM. Os mutantes foram obtidos nos tratamentos onde houve adição da
enzima XbaI no momento da transformação. Tr1 – reação de clivagem; Tr2 – 01U; Tr3 –
05U; Tr4 – 05U; Tr5 – 20U e Tr6 – 10U.

4.3.2 - Mutantes com atividade de pectinases alterada
Penicillium griseoroseum tem demonstrado ser um eficiente produtor de
enzimas pectinolíticas, tais como pectina liase e poligalacturonase. Nesse fungo já
foram isolados dois genes que codificam para pectina liase (Bazzolli,
comunicação pessoal) e dois genes que codificam endopoligalacturonases
(endoPG) (Ribon, 2001). A obtenção de mutantes para produção dessas enzimas é
importante para o isolamento de novos genes, que ainda não tenham sido
encontrados.
A atividade total de pectinases foi analisada em 500 transformantes pelo
método da medida do halo de degradação em placa de Petri. Esse método não é
muito sensível, mas tem-se prestado para uma triagem inicial quando é necessário
analisar um grande número de isolados.
A maioria dos transformantes apresentou valores muito similares ao
isolado PG63 utilizado como controle. Dessa primeira análise foram selecionados
três transformantes que apresentaram halos de degradação menores que o controle
(Figura 7), sendo que um deles, o Tr5,também apresentou alterações
morfológicas.
Dosagens da atividade enzimática em meio líquido foram realizadas para
confirmação dos resultados obtidos pela medida do halo de degradação em meio
sólido. As dosagens foram realizadas para poligalacturonases e pectina liases para
verificar qual das duas enzimas tinha sido afetada.
Os transformantes Tr7 e Tr8 apresentam menor atividade de
poligalacturonase (PG); não diferindo na atividade de pectina liase (PL) em
relação à linhagem transformada, P. griseoroseum PG63 (Figura 8). O
transformante Tr5 destacou-se pela sua maior atividade de PL.
Esses resultados indicam uma possível mutação em genes relacionados
com a síntese de PG. O sequenciamento das regiões que flanqueiam o vetor
poderá elucidar se o gene mutado corresponde aos genes de endoPG já
caracterizados. Caso a mutação não seja especificamente nos genes de PG, genes
de regulação ou genes com efeitos pleiotrópicos na expressão ou secreção de PG
podem ter sido afetados pela mutação. No caso do transformante Tr5, uma
mutação pode ter ocorrido em algum gene que codifica uma proteína repressora,
ou em outro gene de efeito pleiotrópico.

A obtenção de dois mutantes com redução da atividade de PG e ao mesmo
tempo apresentado alterações morfológicas indica uma possível mutação num
gene de efeito pleiotrópico.
A obtenção de mutantes deficientes na produção de PG, com mutações
especificamente no gene que codifica essa enzima, pode ser dificultada quando
existem diferentes cópias do gene no genoma. Mesmo que se tenha uma cópia
mutada, a segunda cópia funcional poderá mascarar o efeito da mutação
resultando num fenótipo muito discreto e de difícil identificação. Quando se
trabalha com genes representados uma única vez no genoma, a seleção de
mutantes, resultantes de mutagênese insercional, pode ser mais evidente e precisa.
Os três mutantes isolados possuem apenas uma integração do vetor (Figura 3 e 4).
O potencial mutagênico da técnica de REMI foi demonstrado pela
obtenção de 8 mutantes, entre morfológicos e com produção de pectinases
alterada, num total de 843 transformantes. A eficiência de mutação foi de 0,95%
por meio de REMI, e 0,63 % para mutagênese utilizando UV (Brito, 1998).
Mutantes auxotróficos não puderam ser isolados já que a linhagem
transformada, PG63, possui uma marca auxotrófica e a seleção dos transformantes
é realizada em meio mínimo.
A técnica de REMI tem demonstrado ser eficaz pela etiquetagem do gene
que possivelmente está determinando o fenótipo. Outros fatores que determinam
sua eficácia são a aleatoriedade das integrações no genoma e aumento da
eficiência de transformação.
Para a futura clonagem desses genes é necessário verificar se a mutação
resultou da inserção do vetor ou originou-se por outro mecanismo, como a ação da
endonuclease adicionada na transformação.

Figura 7 – Estimativa da atividade de pectinases em meio sólido. I - Transformantes
crescidos em meio mineral tamponado. A: P. griseoroseum PG63, B: Tr 5; C: Tr
7 e D: Tr 8. II – Medidas dos halos de degradação em meio tampão Mac
Ilvaine. A: Tr 5 (2,3 cm) e S (2,9 cm); B: Tr 7 (2,3 cm) e S (2,6 cm); C: Tr 8
(2,4 cm) e S (2,7 cm).

Figura 8 – Dosagem da atividade de pectinases em meio líquido. A – Atividade total de pectina
liase (PL). B – Atividade total de poligalacturonase (PG). Médias seguidas pela mesma
letra não diferem entre si pelo teste de Scott-Knott a 5 % de probabilidade.
A
B

4.4 – Isolamento de revertentes niaD- a partir dos transformantes niaD+
A obtenção de mutantes incapazes de assimilar nitrogênio na
forma nítrica pode ser realizada facilmente por obtenção de colônias
resistentes ao clorato.
A enzima nitrato redutase pode converter o clorato em clorito,
que é um composto tóxico à célula, portanto colônias que não assimilam
nitrato são capazes de crescerem no meio contendo clorato. As
mutações podem ocorrer em diferentes genes responsáveis pela
assimilação de nitrogênio nítrico, tais como os genes que codificam
permeases (crn), relacionados com a síntese do cofator (cnxA),
reguladores positivos (nirA e niiA) e finalmente o gene estrutural da
nitrato redutase (niaD).
Após a seleção, é necessário realizar a caracterização da
mutação em diferentes fontes de nitrogênio. Mutantes
especificamente no gene niaD são incapazes de crescer em meio com
NO3-, mas apresentam crescimento em meio contendo NO2
-,
hipoxantina e NH4+.
Os transformantes possuem o gene niaD de F. oxysporum.
Revertentes niaD- obtidos a partir desses transformantes podem
surgir por mutação ou por perda do plasmídeo. Para analisar a
estabilidade dos transformantes obtidos, foram realizadas tentativas
de isolamento de linhagens revertentes, em meio com clorato, de
alguns transformantes, para correlacionar a obtenção de colônias niaD-
com a perda das integrações do plasmídeo pNH24.
Foram utilizadas as linhagens de transformantes que
apresentaram alterações morfológicas e foram observadas freqüências

variações no número de revertentes (Quadro 3). Dos 6 transformantes
testados, 4 apresentaram freqüências maiores que 40%; um
apresentou uma freqüência de 10%, semelhante a obtida por Brito
(1998) para P. griseoroseum e o Tr6 não originou nenhuma colônia nia
D-. Em P. expansum a obtenção de mutantes niaD- por meio da
resistência ao clorato é próxima de 24 % das colônias resistentes ao
clorato (Pio, 2001).
Diversas seqüências levam a rearranjos nos cromossomos em
fungos filamentosos. Seqüências repetidas no genoma são substrato
para a recombinação podendo ocasionar rearranjos e até mesmo
deleções (Fierro et al., 1999). Integrações em tandem foram instáveis
em transformantes de P. chrysogenum (Casqueiro et al., 1998)
justamente por serem regiões homólogas e muito próximas.
Entre os mutantes morfológicos, o Tr2 e o Tr4 apresentaram
integrações múltiplas. O primeiro apresentou integrações em tandem e
o segundo apresentou duas integrações em diferentes locais do genoma
(Figura 3, canaletas 13 e 15, respectivamente). As freqüências de
reversão foram discrepantes entre esses dois isolados.
O Tr4 resultou numa reversão ao fenótipo niaD- de
aproximadamente 90%, correlacionado-se com possível rearranjo ou
deleção na região de integração do plasmídeo. Pelo padrão de
hibridização, de um dos isolados nia D-, é provável que tenha ocorrido
uma recombinação entre as cópias da inserção resultando numa única
cópia com deleção de parte do plasmídeo contendo o gene niaD (Figura
3, canaleta 14). O revertente analisado parece reter uma atividade
alterada da nitrato redutase, pois seu crescimento em meio mínimo
com nitrato, apesar de ser residual, freqüentemente origina regiões de
crescimento compacto e inclusive com esporulação (Figura 10).

A porcentagem de revertentes niaD- do transformante Tr2 foi
relativamente baixa quando comparado aos mutantes Tr1, Tr3, Tr4 e
Tr5 (Tabela 4). Por apresentar duas integrações, seria necessário
mutações ou perdas das duas cópias para resultar em linhagens
resistentes ao clorato, e a recombinação entre essas cópias deve ser
menos freqüente por aparentemente estarem inseridas em diferentes
regiões do genoma. Um fato interessante é que entre os mutantes
niaD- recuperados do mutante Tr2, três mutantes reverteram o
fenótipo (Figura 9). A reversão foi causada por perda das duas cópias
dos vetores, observadas pela falta de sinal de hibridização (Figura 3,
canaleta 16). Isso leva a crer que a inserção de uma das cópias resultou
na mutação.
Os mutantes Tr1, Tr3 e o Tr5, que possuem uma única cópia do
gene niaD de F. oxysporum, apresentaram porcentagens de mutação no
gene niaD- maiores em relação ao gene endógeno. Essa diferença pode
ser conseqüência de integrações em regiões que favoreçam a mutação
nesse gene ou mesmo a perda do vetor. O que pode ser o contrário
para o Tr6, que não apresentou mutantes niaD- quando colocado em
meio seletivo.

Quadro 3: Porcentagem de colônias nia D- resistentes ao clorato
obtidas a partir do plaqueamento de esporos dos transformantes com
alterações morfológicas.
Transformante Revertentes (%)
Tr1 40,62
Tr2 09,68
Tr3 80,00
Tr4 89,75
Tr5 57,14
Tr6 00,00

Figura 9– Mutantes nia D- obtidos a partir do transformante Tr2 isolados em meio seletivo. I e II
são placas replicas contendo meio completo e meio mínimo com nitrato, respectivamente.
A – P. griseoroseum PG63 nia D-; B – transformante Tr2, niaD+, do P. griseoroseum
PG63, que apresenta alteração morfológica; C – três mutantes nia D- obtidos a partir do
Tr2 em meio com clorato e que reverteram ao fenótipo do P. griseoroseum PG63.
Figura 10 – Crescimento do mutante niaD-, em meio mínimo contendo nitrato, obtido a partir do
Tr4. A – plaqueamento de esporos e B – inoculação pontual. Mutantes niaD- possuem
crescimento residual nesse tipo de meio, mas esse mutante apresenta regiões onde o
crescimento é mais vigoroso chegando a esporular (seta).

5 – CONCLUSÕES
A linearização do plasmídeo pNH24 para transformação de Penicillium
griseoroseum resultou num aumento da eficiência de transformação, assim como a
utilização de enzima de restrição XbaI.
A adição de apenas 1U da enzima XbaI, concomitantemente com a
linearização do plasmídeo, foi suficiente para aumentar 6,77 vezes a eficiência de
transformação em relação ao tratamento que utilizou o plasmídeo circular.
Quantidades maiores resultaram num decréscimo da transformação.
A maioria dos transformantes apresentou integrações únicas e aleatórias no
genoma. Nos transformantes obtidos na presença da enzima de restrição, a
integração do vetor restituiu os sítios de clivagem, indicando que a ocorrência de
REMI nesse fungo.
Mutantes com alterações morfológicas, com atividade reduzida de
poligalacturonases (PGs) e atividade aumentada de pectina liase (PL) foram
isolados dos transformantes obtidos nos tratamentos onde utilizou-se a enzima
XbaI.
A freqüência de reversão do fenótipo Nia- nos transformantes foi maior do
que a freqüência de mutação no gene niaD do próprio P. griseoroseum. A
obtenção de uma reversão do fenótipo de um dos mutantes morfológicos, pela
perda do plasmídeo, indicou que o gene está etiquetado por uma das duas cópias
do vetor contida nesse mutante.
A utilização de REMI em P. griseoroseum é indicada para a obtenção de
maiores freqüências de transformação, obtenção de transformantes com
integrações únicas e para a etiquetagem de genes.

6 - REFERÊNCIAS BIBLIOGRÁFICAS
Abreu, L. M. (2001) Análise genética de isolados de fungos ectomicorrízicos e
de Penicillium spp. Viçosa, UFV. Imprensa Universitária (Tese de Mestrado), 66p.
Adams, T. H.; Wieser, J. K.; Yu, J. (1998) Asexual sporulation in Aspergillus
nidulans. Microbiology and Molecular Biology Reviews, 62 (1): 35 – 54. Akamatsu, H.; Kodama, Y. I.; Otani, H.; Kohmoto, K. (1997) Aal- toxin-
deficient mutants of Alternaria alternata tomato pathotype by restriction enzyme mediated integration. Phytopathology, 87 (9): 967 – 972.
Albersheim, P.; Killias, U. (1966) Pectin lyase from fungi. Methods in
Enzymology, 8: 628 – 635. Baek, J; Kenerley, C. M. (1998) The arg2 gene of Trichoderma virens: cloning
and development of a homologous transformation system. Fungal Genetics and Biology, 23: 34 - 44.
Barth, C.; Fraser, D. J.; Fisher, P. R. (1998) Co-insertional replication is
responsible for tandem multimer formation during plasmid integration into the Dictyostelium genome. Plasmid, 39: 141 – 153.
Bhairi, S. M.; Staples, R. C. (1992) Transient expression of the β-glucuronidase
gene introduced into Uromyces appendiculatus uredosporos by particle bombardment. Phytopathology, 82 (9): 986 - 989.
Black, M.; Seeber, F; Soldati, D.; Kim, K; Boothroyd, J. C. (1995) Restriction
enzyme-mediated integration integration elevates tranformation frequency and enables co-transfection of Toxoplasma gondii. Molecular and Biochemical Parasitology, 74: 55 – 63.
Bölker, M.; Böhnert, H. U.; Braun, K. H.; Görl, J.; Kahmann, R. (1995) Tagging
pathogenicity genes in Ustilago maydis by restriction enzyme-mediated integration (REMI). Molecular General Genetics, 248: 257 – 552.
Brakhage, A. (1998) Molecular Regulation of β-Lactam Biosynthesis in
Filamentous Fungi. Microbiology and Molecular Biology Reviews, 62 (3): 547 - 585.
Brito, A. R. T. (1998) Isolamento e caracterização de mutantes de Penicillium
griseoroseum. Dissertação (Mestrado em Microbiologia Agrícola), Universidade Federal de Viçosa, 68p.
Brookman, J.; Denning, D. W. (2000) Molecular genetics in Aspergillus
fumigatus. Current Opinion in Microbiology, 3: 468-474. Brown, J. S.; Holden, D. (1998) Insertional mutagenesis of pathogenic fungi.
Current Opinion in Microbiology, 1:390 – 394.

Bundock, P.; den Dulk-Ras, A.; Beijersbergen, A.; Hooykaas, P. J. J. (1995)
Trans-kingdom T-DNA transfer from Agrobacterium tumefaciens to Saccharomyces cerevisiae. The EMBO Journal, 14: 3206 - 3214.
Bundock, P.; Hooykaas, P. J. J. (1996) Integration of Agrobacterium tumefaciens
T – DNA in the Saccharomyces cerevisiae genome by illegitimate recombination. Proceedings of the National Academy of Sciences U.S.A., 93: 15272 – 15275.
Cantone, F. A.; Vandenberg, J. D. (1999) Genetic transformation and
mutagenesis of the entomopathogenic fungus Paecilomyces fumosoroseus. Journal of Invertebrate Pathology, 74: 281 – 288.
Casqueiro, J.; Gutiérrez, S.; Bañuelos, O.; Hijarrubia, M. J.; Martín, J. F. (1999)
Gene targeting in Penicillium chrysogenum: disruption of the lys2 gene leads to penicillin overproduction. Journal of Bacteriology, 181 (4): 1181 – 1188.
Chakraborty, B. N.; Patterson, N. A.; Kapoor, M. (1991) An electroporation-
based system for high-efficiency transformation of germinated conidia of filamentous fungi. Canadian Journal of Microbiology, 37: 858 - 863.
Chen, X.; Stone, M.; Schlagnhaufer, C.; Romaine, C. P. (2000) A fruiting body
tissue method for efficient Agrobacterium-mediated transformation of Agaricus bisporus. Applied and Environmental Microbiology, 66 (10): 4510 - 4513.
Cormack, B. P.; Falkow, S. (1999) Efficient homologous and illiegitimate
recombination in the opportunistic yeast pathogen Candida glabrata. Genetics, 151: 979 – 987.
D’Angelo (1998) Purificação parcial e caracterização de poligalacturonases de
Penicillium griseoroseum. Tema de monografia, Universidade Federal de Viçosa, 53p.
Diolez, A.; Langin, T.; Gerlinger, C.; Brygoo, Y. & Daboussi, M.J. (1993) The
nia gene of Fusarium oxysporum: isolation, sequence and development of a homologous transformation system. Gene, 131: 61 – 67.
Epstein, L.; Lusnak, K.; Kaur, S. (1998) Transformation mediated
developmental mutants of Glomerella graminicola (Colletotrichum graminicola). Fungal Genetics and Biology, 23: 189 – 203.
Fierro, F.; Martín, J. F. (1999) Molecular mecanisms of chromossomal
rearrangement in fungi. Critical reviews in microbiology, 25(1): 1-17. Fincham, J. R. S. (1989) Transformation in fungi. Microbiological reviews,
53(1): 148-170.

Fungaro, M. H.; Rech, E.; Muhlen, G. S.; Vainstein, M. H.; Pascon, R. C.; Queiroz, M. V.; Kleiner, A. A. P.; Azevedo, J. L. (1995) Transformation of Aspergillus nidulans by microprojectile bombardment on intact conidia. FEBS Microbiology Letters, 125: 293 – 298.
Gouka, R. J.; Gerk, C.; Hooykaas, P. J. J.; Bundock, P.; Musters, W.; Verrips, C. T.; Groot, M. J. A. (1999) Transformation of Aspergillus awamori by Agrobacterium tumefaciens-mediated homologous recombination. Nature Biotechnology, 17:598 - 601.
Granado, J. D.; Chaloupková, K. K.; Aebi, M.; Kües, U. (1997) Restriction
enzyme mediated DNA integration in Coprinus cinereus. Molecular General Genetics, 256: 28 – 36.
Griffin, D. H. (1994) Fungal physiology. 2ºed. Wiley-Liss, New York,. 458 p. Griffiths, A. J. F. (1995) Natural plasmids of filamentous fungi. Microbiological
Reviews, 59 (4): 673 - 685. Graessle, S.; Haas, H.; Friedlin, E.; Kürnsteiner, H.; Stöffler, G.; Redl, B. (1997)
Regulated system for heterologous gene expression in Penicillium chysogenum. Applied and Environmental Microbiology, 63 (2):753-756.
Groot, M. J. A.; Bundock, P.; Hooykaas, P. J. J.; Beijersbergen, A. G. M. (1998)
Agrobacterium tumefaciens mediated transformation of filamentous fungi. Nature Biotechnology, 16: 839 – 842.
Hebert, T. T. (1963) Precipitation of plant viruses by polyethylene glycol.
Phytopathology, 53: 362-366. Hinnen, A.; Hicks, J. B.; Fink, G. R. (1978) Transformation of yeast chimaeric
ColE1 plasmid carrying LEU2. Proceedings of the National Academy of Sciences USA, 75: 1929 - 1933.
Hogan, L. H.; Klein, B. S. (1997) Transforming DNA integrates at multiple sites
in the dimorphic fungal pathogen Blastomyces dermatitidis. Gene, 186: 219-226.
Hutchison, H. T.; Hartwell, L. H. (1967) Macromolecule synthesis in yeast
spheroplasts. Journal of Bacteriology, 94(5):1697-1705. Hua, S.; Qiu, M.; Chan, E.; Zhu, L.; Luo, Y. (1997) Minimum length of
sequence homology required for in vivo cloning by homologous recombination in yeast. Plasmid, 38: 91 – 96.
Kahmann, R.; Basse, C. (1999) REMI (restriction enzyme mediated integration)
and its impact on the isolation of pathogenicity in fungi attacking plants. European Journal of Plant Pathology, 105: 221 – 229.

Klein, T.M., Arentzen, R.; Lewis, P.A.; Fitzpatrick-McElligott, S. (1992) Transformation of microbes, plants and animals by particle bombardment. Bio/Technology, 10:286 - 291.
Kuspa, A.; Loomis, W. F. (1992) Tagging developmental genes in Dictyostelium
by restriction enzyme-mediated integration of plasmid DNA. Proceedings of the National Academy of Sciences U.S.A., 88: 8803 – 8807.
Kunik, T.; Tzfira, T.; Kapulnik, Y; Gafni, Y.; Dingwall, C.; Citovsky, V. (2001)
Genetic transformation of HeLa cells by Agrobacterium. . Proceedings of the National Academy of Sciences U.S.A., 98 (4): 1871 – 1876.
Lewin, B. Genes VII, 1ª ed. Oxford University Press, New York, 990 p Lewis, L. K., Westmoreland, J. W.; Resnick, M. A. (1999) Repair of
endonuclease-induced double-strand breaks in Saccharomyces cerevisiae: essencial role for genes associated with nonhomologous end-joining. Genetics, 152: 1513 – 1529.
Levis, C.; Fortini, D.; Brygoo, Y. (1997) Transformation of Botrytis cinerea
with the nitrate reductase gene (niaD) shows a high frequency of homologous recombination. Current Genetics, 32: 157 - 162.
Lin, Y.; Waldman, A.S. (2001) Capture of DNA sequences at double-strand
breaks in mammalian chromosomes. Genetics, 158: 1665-1674. Linnemannstöns, P.; Vob, T.; Hedden, P; Gaskin, P.; Tudzynski, B. (1999)
Deletions in the gibberellin biosynthesis gene cluster of Gibberella fujikuroi by restriction enzyme mediated integration and conventional transformation mediated mutagenesis. Applied and Environmental Microbiology, 65 (6): 2558 – 2564.
Lu, S.; Lyngholm, L.; Yang, G.; Bronson, C.; Yoder, O. C.; Turgeon, B. G.
(1994) Tagged mutations at the Tox 1 locus of Cochliobolus heterostrophus by restriction enzyme-mediated integration. . Proceedings of the National Academy of Sciences U.S.A., 91: 12649 – 12653.
Manivasakam, P.; Schiestl, R. H. (1998) Nonhomologous end joining during
restriction enzyme-mediated DNA integration in Saccharomyces cerevisiae. Molecular and Celular Biology, 18 (3): 1736 – 1745.
Manczinger, L.; Komonyi, O.; Antal, Z.; Ferenczy, L. (1997) A method for high-
frequency transformation of Trichoderma viride. Journal of Microbiological Methods, 29: 207 – 210.
Miller, G. L. (1959) Use of dinitrosalicylic acid reagent for determination of
reducing sugar. Analitical Chemical, 31: 426-428.

Mishra, N. C.; Tatum, E. L. (1973) Non-Mendelian inheritance of DNA-induced inositol independence in Neurospora. Proceedings of the National Academy of Sciences U.S.A., 70 (12): 3875 – 3879.
Molnar, Z.; Jenes, B; Ördög, V (1997)
http://www.movar.pate.hu/~plantph/tatra99.html Moore, J. K.; Haber, J. E. (1996) Cell cycle and genetic requeriments of two
pathways nonhomologous end-joining repair of double-strand breaks in Saccharomyces cerevisiae. Molecular and Cellular Biology, 16 (5): 2164 – 2173.
Morton, J. B. (1999) Fungi. In: Sylvia, D. M.; Fuhrmann, J. J.; Hartel, P. G.;
Zuberer, D. A. Principles and applications of soil microbiology, 1ª ed. Prentice Hall, Upper Saddle River, New Jersey, p. 72 - 92.
Pawlowski, W.; Somers, D. A. (1998) Transgenic DNA integrated into the oat
genome is frequently interspersed by host DNA. Proccedings of the National Academy of Sciences U.S.A., 95 (21): 12106 – 12110.
Pereira, J. F. (1999) Isolamento e caracterização do gene da nitrato redutase de
Penicillium griseoroseum. Tema de monografia, Universidade Federal de Viçosa, 36p.
Picard, M.; Debuchy, R.; Julien, J.; Brygoo, Y. (1987) Transformation by
integration in Podospora anserine. Molecular General Genetics, 210: 129-134. Piers, K. L.; Heath, J. D.; Liang, X.; Stephens, K. M.; Nester, E. W. (1996)
Agrobacterium tumefaciens-mediated transformation of yeast. Proceedings of the National Academy of Sciences USA, 93: 1613-1618.
Pontecorvo, G., Roper, J. A., Hemmons, L. M., Macdonald, K. D. & Bufton, A.
W. J. (1953) The genetics of Aspergillus nidulans. Advances in Genetics, 5: 141 - 238.
Queiroz, M. V.; Barros, A. O.; Barros, E. G.; Guimarães, W. V.; Araújo, E. F.
(1998) Transformation of Penicillium griseoroseum nitrate reductase mutant with the nia gene from Fusarium oxysporum. Canadian Journal of Microbiology., 44: 487 - 489.
Ramachandran, S.; Sundaresan, V. (2001) Transposons as tools for functional
genomics. Plant Physiology Biochemical, 39: 243 – 252. Redman, R. S.; Rodriguez, R. J. (1994) Factors affecting the efficient
transformation of Colletotrichum species. Experimental Mycology, 18: 230 - 246.
Redman, R. S.; Ranson; J. C.; Rodriguez, R. J. (1999) Conversion of the
pathogenic fungus Colletotrichum magna to a nonpathogenic, endophytic

mutualist by gene disruption. Molecular Plant-Microbe Interactions, 12 (11): 969 – 975.
Ribon, A. O. B. (2001) Organização e regulação de genes que codificam
poligalacturonases em Penicillium griseoroseum. Dissertação (Doutorado em Microbiologia Agrícola), Universidade Federal de Viçosa, 68p.
Ribeiro, J. B. (2001) Isolamento e caracterização de genes que codificam
poligalacturonases e transformação de Penicillium expansum. Dissertação (Mestrado em Microbiologia Agrícola), Universidade Federal de Viçosa, 68p.
Roos, D. S.; Sullivan, W. J.; Striepen, B.; Bohne, W.; Donald, R. G. K. (1997)
Tagging genes and trapping promoters in Toxoplasma gondii by insertional mutagenesis. Methods: a Companion to Methods in Enzymlogy, 13:112 - 122.
Sambrook, J.; FRITSCH, E.F.; MANIATS, T. (1989) Molecular cloning. A laboratory manual. 2 ed. Cold Spring Harbor, New York. Cold Spring Harbor Laboratory.
Sánchez, O.; Navarro, R. E.; Aguirre, J. (1998) Increased transformation
frequency and tagging of developmental genes in Aspergillus nidulans by restriction enzyme-mediated integration (REMI). Molecular General Genetic, 258: 89 – 94.
Sanford, J. C.; Klein, T. M.; Wolf, E. D.; Allen, N. (1987) Delivery of
substances into cells tissues using particle bombardment process. Part. Sci. Techn. 5:27-37.
Schiestl, R. H.; Petes, T. D. (1991) Integration of DNA fragments by illegitimate
recombination in Saccharomyces cereviseae. Proceedings of the National Academy of Sciences USA, 88: 7585-7589.
Shi, Z.; Christian, D.; Leung, H. (1995) Enhanced transformation in
Magnaporthe grisea by restriction enzyme mediated integration of plasmid DNA. Phytopathology, 85 (3): 329 – 333.
Singer, B.; Kusmierek, J. T. (1982) Chemical mutagenesis. Annual Review
Biochemical, 52: 655 - 693. Southern, E.M. (1975) Detection of specific sequences among DNA fragments
separated by gel electrophoresis. Journal of Molecular Biology, v. 98, p. 503-517.
Souza, C. C.; Goldman, M. H. S.; Goldman; G. H. (2000) Tagging of genes
involved in multidrug resistance in Aspergillus nidulans. Molecular General Genetic, 263: 702-711.
Speacht, C. A.; Dirusso, C.C.; Novotny, C.P.; Ullrich, R. C. (1982) A method
for extracting high-molecular-weight deoxyribonucleic acid from fungi. Analytic Biochemistry, 119: 158 - 163.

Tanaka, A; Shiotani; H.; Yamamoto, M.; Tsuge, T. (1999) Insertional mutagenesis and cloning of the genes required for biosynthesis of the host-specific AK-toxin in the japanese pear pathotype of Alternaria alternata. Molecular Plant-Microbe Interactions, 12 (8): 691 – 702.
Thon, M. R.; Nuckles, E. M.; Vaillancourt, L. J. (2000) Restriction enzyme-
mediated integration used to produce pathogenicity mutants of Colletotrichum graminicola. Molecular Plant-Microbe Interactions, 12 (13): 1356 – 1365.
Torres, R. T. (2001) Isolamento e caracterização do gene que codifica nitrato
redutase em Penicillium expansum. Dissertação (Mestrado em Microbiologia Agrícola), Universidade Federal de Viçosa, 47 p.
Unkles, S. E.; Campbell, E. I.; Rtuiter-Jacobs, Y. M. J. T.; Brockhuijsen, M.;
Macro, J. A.; Carrez, D.; Contreras, R.; Van den Hondel, C. A. M. J.J.; Kinghorn, J. R. (1989) The development of homologous transformation system for Aspergillus oryzae based on the nitrate reductase assimilation pathway: a covenient and general selection system for filamentous fungal transformation. Molecular and General Genetics, 218: 99-104.
Wang, J.; Holden, D. W.; Leong, S. A. (1988) Gene transfer system for the
phytopathogenic fungus Ustilago maydis. Proceedings of the National Academy of Sciences USA, 85: 865 - 869.
Watts, J. W.; Stacey, N. J. (1991) Novel methods of DNA transfer. In: Peberdy,
J. F.; Caten, C. E.; Ogden, J. E.; Bennet, J. W. Applied Molecular Genetics of fungi, 1ª ed.,. University Press, Cambridge, p. 45-65.
Van de Rhee, M. D.; Graça, P. A. M.; Huizing, H. J.; Mooibroek, H. (1996)
Transformation of the cultivated mushroom, Agaricus bisporus, to hygromycin B resistance. Molecular and General Genetics, 250: 252-258.
Vainstein, M. H.; Alves, S. A.; Lima, B. D.; Aragão, F. J. L.; Rech, E. L. (1994)
Stable DNA transfection in a flagellate trypanosomatid by microparticle bombardment. Nucleic Acids Research, 22 (15): 3263 – 3264.
Vidan, S.; Snyder, M. (2001) Large-scale mutagenesis: yeast genetics in the
genome era. Current Opinion in Biotechnology, 12:28-34. Yaver, D. S.; Lamsa, M.; Munds, R.; Brown, S. H.; Otani, S.; Franssen, L.;
Johnstone, J. A.; Brody, H.; (2000) Using DNA-tagged mutagenesis to improve heterologous protein production in Aspergillus oryzae. Fungal Genetics and Biology, 29: 28 – 37.
Yelton, M. M.; Hamer, J. R.; Timberlake, W. E. (1984) Transformation of
Aspergilus niger by using a trpC plasmid. Proceedings of the National Academy of Sciences U.S.A., 81:1470-1474.
Yun, S. H.; Turgeon, B. G.; Yoder, O. C. (1998) REMI-induced mutants of
Mycosphaerella zeae-maydis lacking the polyketide PM-toxin are deficient in

pathogenesis to corn. Physiological and Molecular Plant Pathology, 52: 53 – 66.
Zhu, J.; Schiestl, R. H. (1996) Topoisomerase I involvement in illegitimate
recombination in Saccharomyces cerevisiae. Molecular and Cellular Biology, 16 (4): 1805 – 1812.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo