sinais envolvidos na comunicaÇÃo de a sahlberg, … · revisÃo de literatura ... anatomia do...
TRANSCRIPT

SINAIS ENVOLVIDOS NA COMUNICAÇÃO DE AMERRHINUS YNCA SAHLBERG, 1823 (COLEOPTERA: CURCULIONIDAE)
RODOLFO MOLINÁRIO DE SOUZA
"Dissertação apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense Darcy Ribeiro, como parte das exigências para obtenção do título de Mestre em Produção Vegetal".
Orientadora: Ana Maria Matoso Viana-Bailez
CAMPOS DOS GOYTACAZES - RJ JULHO – 2006

SINAIS ENVOLVIDOS NA COMUNICAÇÃO DE Amerrhinus ynca
SAHLBERG, 1823 (COLEOPTERA: CURCULIONIDAE)
RODOLFO MOLINÁRIO DE SOUZA
"Dissertação apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense Darcy Ribeiro, como parte das exigências para obtenção do título de Mestre em Produção Vegetal".
Aprovada em 21 de julho de 2006
Comissão Examinadora:
_________________________________________________________________ Profa. Terezinha M. C. Della Lucia (D. Sc., Fitotecnia) – UFV
_________________________________________________________________ Prof. Omar Eduardo Bailez (D. Sc., Biologia do Comportamento) – UENF
_________________________________________________________________ Prof. Carlos Ruiz-Miranda (Ph.D., Comportamento Animal) – UENF
_________________________________________________________________ Profa. Ana Maria M. Viana-Bailez (D. Sc., Biologia do Comportamento) – UENF
Orientadora

ii
Dedico esta dissertação à minha mãe, Dona Cida, que dedicou sua vida à minha
formação; e ao meu irmão, Marcondes.

iii
AGRADECIMENTOS
Agradeço primeiramente a Deus por ter me dado mais esta oportunidade
e por ter colocado em minha vida pessoas maravilhosas que contribuíram
imensamente para o meu sucesso.
À CAPES pelo fomento da bolsa e à Prefeitura de Quissamã, na pessoa
de Olavo Pimentel, que tornou possível nossas várias idas ao campo.
À UENF, ao Centro de Ciências e Tecnologias Agropecuárias e ao
Laboratório de Entomologia e Fitopatologia, pelo uso de suas instalações e
equipamentos e pela oportunidade de realizar este trabalho.
Ao Laboratório de Ciência Ambientais da UENF, setor de Ecologia por ter
cedido os equipamentos para as análises acústicas.
A minha orientadora Profª. Ana Maria, pela paciência, carinho e amizade.
Aos professores Carlos Ruiz, Omar Bailez e Terezinha Della-Lucia por
aceitarem participar da banca e pelas críticas e correções que enriqueceram o
trabalho.
Ao grande parceiro e amigo Nilson pelos momentos de força e por ter
estado sempre ao meu lado, ajudando-me com as traduções, coletas, correções,
etc.
À Profa. Maria Luisa Lopes Alvarez pelas imagens feitas em microscopia
eletrônica na Universidade do Chile.

iv
A Thiago, Silvio e Alexandre por terem me ajudado nas coletas de campo
e atividades dentro do laboratório.
A Denise e Arli pelos momentos de descontração, carinho e amizade.
Aos colegas de laboratório, Gilson, Carol e Vinicius pelos momentos de
discussões os quais foram importantes para a execução deste trabalho.
À Camila que foi meu porto seguro durante todo o período em que fiquei
em Campos e que sempre me ouviu e acolheu nos momentos mais difíceis.
Aos colegas de Pós-graduação Marcinha, Eliane, Jathinder, Adriano,
Josimar e Laerciana pela amizade e convivência. Em especial a Jô e a Lala pelas
conversas, por estarem sempre dispostas a ajudar e pelo que fizeram por mim.
À Dona Rosa que sempre me tratou como um filho durante o tempo em
que fui seu inquilino.
À minha eterna amiga Nathalie por sempre ter estado ao meu lado e
sempre ter elevado meu astral.
E finalmente à minha mãe e ao meu irmão, sempre interessado e
prestativo, por acreditarem em mim, pelo carinho e amor. A toda minha família,
especialmente às minhas tias Lu e Bidu que sempre me acolheram com tanto
carinho.

v
SUMÁRIO
RESUMO ...............................................................................................................vii 1. INTRODUÇÃO .................................................................................................... 1
2. REVISÃO DE LITERATURA............................................................................... 4
2.1. A cultura do Coqueiro .................................................................................4
2.2. Broca-da-ráquis-foliar do coqueiro - Amerrhinus ynca................................5
2.3. Comportamento Reprodutivo......................................................................9
2.3.1. O Acasalamento ............................................................................9
2.3.2. A "guerra dos sexos" ...................................................................12
2.4. Comunicação química em insetos ............................................................14
2.4.1. Comunicação química em Curculionídeos ..................................15
2.5. Comunicação sonora em insetos..............................................................21
2.5.1. Produção de Som em Coleoptera ............................................24
3. TRABALHOS .................................................................................................... 31
DIMORFISMO SEXUAL DE Amerrhinus ynca SAHLBERG, 1823 (COLEOPTERA: CURCULIONIDAE).......................................................................................... 31
COMPORTAMENTO DE CORTE E ACASALAMENTO DE Amerrhinus ynca SAHLBERG, 1823 (COLEOPTERA: CURCULIONIDAE)................................ 42
BIOENSAIOS COM ODORES DE COESPECÍFICOS E DA PLANTA HOSPEDEIRA DE Amerrhinus ynca SAHLBERG, 1823 (COLEOPTERA: CURCULIONIDAE). ......................................................................................... 62

vi
ANATOMIA DO APARATO ESTRIDULATÓRIO, ANÁLISE DO SOM E PAPEL DA ESTRIDULAÇÃO NO COMPORTAMENTO DE CORTE E ACASALAMENTO. DE Amerrhinus ynca SAHLBERG, 1823 (COLEOPTERA: CURCULIONIDAE)82
4. RESUMOS E CONCLUSÕES......................................................................... 108
5. REFERÊNCIAS BIBLIOGRÁFICAS................................................................ 111

vii
RESUMO
SOUZA, RODOLFO MOLINÁRIO; Engenheiro Florestal, Msc.; Universidade Estadual do Norte Fluminense Darcy Ribeiro; Junho – 2006, Sinais envolvidos no comportamento de Amerrhinus ynca Sahlberg, 1823 (Coleoptera: Curculionidae). Profª. Orientadora Ana Maria Matoso Viana-Bailez, Prfs. Conselheiros: Omar Eduardo Bailez e Carlos Miranda Ruiz.
Amerrhinus ynca é uma coleobroca cujas larvas se alimentam dos tecidos
internos da ráquis foliar do coqueiro. Até pouco tempo era considerada praga
secundária, mas devido à periodicidade e intensidade dos surtos tem atingido o
status de praga primária em algumas regiões. Pouco se conhece sobre sua
biologia, por isso este trabalho tem por objetivo: encontrar e descrever caracteres
morfológicos que permitam a distinção dos sexos; descrever a seqüência
comportamental associada à corte e acasalamento; identificar possíveis
semioquímicos envolvidos nas interações intra e interespecíficas; identificar e
descrever as estruturas associadas à estridulação bem como qualificar e
quantificar os parâmetros temporais e espectrais dos sons produzidos por A.
ynca. Adultos de A. ynca utilizados nos estudos foram coletados em um plantio
comercial de coco e levados para o laboratório. Após a confirmação dos sexos
procurou-se identificar, através de observações da morfologia externa,
caracteristicas distintas entre machos e fêmeas. O comprimento e a largura do
corpo foram medidos. O comportamento de corte e acasalamento foi analisado

viii
através de observação direta e por filmagem. Registraram-se a freqüência e a
duração dos principais atos comportamentais. Os bioensaios para verificar a
presença de semioquímicos foram feitos utilizando-se três tipos de olfatômetros,
nos quais os estímulos utilizados foram insetos coespecíficos e partes do
coqueiro. Os sons foram gravados com gravador digital e analisados com ajuda
do software Canary. Para se verificar o papel da estridulação no comportamento
de acasalamento foram realizadas observações de quatro tipos de casais (com ou
sem partes dos élitros). Foram encontrados três caracteres morfológicos distintos
e as fêmeas são maiores que os machos. Os acasalamentos ocorreram tanto em
fotofase quanto em escotofase. A monta iniciou-se aos 119 ± 75 min após os
casais terem sido colocados em observação. A corte e a cópula duraram em
média 3 ± 2,5 min e 51 ± 49 min, respectivamente. A produção de som durante a
corte, através da estridulação, é o evento mais freqüente entre os machos. As
fêmeas de A. ynca acasalaram múltiplas vezes e em geral com vários machos.
Após a primeira cópula, os machos apresentaram comportamento de guarda da
fêmea. Nos testes de atratividade de odores a maioria dos indivíduos não
respondeu aos estímulos testados em três tipos de olfatômetros. Machos e
fêmeas foram mais estimulados nos testes com partes vegetais. Verificou-se que
machos e fêmeas emitem sons por estridulação. Existem duas estruturas distintas
envolvidas na produção de som: a pars stridens e o plectrum. A pars stridens está
localizada na região ventral da extremidade posterior dos élitros direito e
esquerdo e o plectrum está localizado no sétimo esternito abdominal. O som
produzido pelos machos, por estridulação, durante o comportamento de corte, é
diferente daquele que produzem quando são perturbados. Observous-se, durante
a corte, que as fêmeas movimentam o abdômen de forma similar ao movimento
que os machos desempenham ao estridularem, mas só foi possível obter um
padrão espectral do som específico quando elas eram pertubadas. Tal som difere
daqueles produzido pelos machos. No tratamento: fêmeas que tiveram sua pars
stridens removida, a porcentagem de casais que acasalaram (montas e cópulas)
reduziu. Já os machos gastam mais tempo tentando cortejar as fêmeas, quando
suas partes stridens são retiradas.

ix
ABSTRACT
SOUZA, RODOLFO MOLINÁRIO; Forest Engineer, Msc.; Universidade Estadual do Norte Fluminense Darcy Ribeiro; June – 2006, Signs involved in behavior of Amerrhinus ynca Sahlberg, 1823 (Coleoptera: Curculionidae). Advisor Ana Maria Matoso Viana-Bailez, Committee members: Omar Eduardo Bailez and Carlos Miranda Ruiz.
Amerrhinus ynca is a woodborer whose larvae feed on the internal tissues of the
leaf-stalk of coconut palm. Until recently it was considered a secondary pest, but
due to the periodicity and intensity of the attacks it has been reaching the status of
primary pest in some areas. The Amerrhinus ynca`s biology is very little known,
therefore this work has the following objectives: to find and to describe
morphological characters to differentiate the sexes; to describe the behavioral
sequence associated to the courtship and mating behaviors; to identify possible
semiochemicals involved in the intra and inter-specific interactions; to identify and
to describe the structures associated to stridulation; to analyze the temporary and
spectral parameters of the produced sounds, and to verify the role of stridulation
on the reproductive behavior. Adults of A. ynca used in the studies were collected
in a commercial plantation of coconut palms and taken to laboratory. After
confirmation of the sexes, different characters between males and females were
sought through observations of the external morphology. The length and the width
of the body were measured. Mating and courtship behaviors were monitored

x
through direct observation and filming. In both cases frequency and duration of
behavioral acts were registered. In order to verify the presence of semiochemicals,
bioassays were conducted by using three types of olfactometers in which the
stimuli were cospecifics and vegetable parts. The sounds were recorded by a
digital tape recorder and analyzed by the Canary software. To verify the role of
stridulation on the mating behavior, observations of four types of couples were
accomplished, couples with or without parts of the elytra. Three different
morphological characters were found, besides the females being larger than the
males. The matings happened in photophase as much as in scotophase. Mount
began at 119 ± 75 min later. The courtship and copulation lasted 3 ± 2,5 min and
51 ± 49 min average, respectively. The sound production during courtship, through
stridulation, was the most frequent event among the males. The females of A.
ynca mated multiple times and in general with several males. After the first
copulation the males guarded the females. In the three types of olfactometer, no
response to coespecifics tested as stimulus was observed. Males and females
were most stimulated when vegetable parts were tested in the olfactometers. It
was verified that males and females produce sounds by stridulation. There are two
different structures involved in the sound production: the pars stridens and the
plectrum. The pars stridens is located in the ventral area of the posterior extremity
of the right and left elytra, and the plectrum this located in the seventh abdominal
sternite. The stridulation of males during courtship behavior was different than that
produced when they were disturbed. Females presented movement similar to
stridulatory one during the courtship and they stridulated differently than males
when were disturbed. When females had their partes stridentes removed, the
percentage of couples that mated (mount and copulation) was reduced. Males
spent more time trying to court the females when their partes stridentes were
removed.

1
1. INTRODUÇÃO
O coqueiro constitui uma importante cultura para os trópicos, sendo
amplamente explorado devido aos seus múltiplos usos e finalidades e por ser
uma grande fonte geradora de divisas (Ferreira et al. 1997). Considerada a mais
importante das culturas perenes, é capaz de gerar um sistema auto-sustentável
de exploração (Cuenca 1997), fornecendo alimento para consumo in natura e
matéria-prima para o processamento agro-industrial (Mirizola-Filho 2002).
Para Mirizola-Filho (2002), a incidência de pragas e de doenças nos
coqueirais do Brasil constitui um problema limitante, respondendo sobremaneira
pelos baixos índices de produtividade e pelo aumento no custo de implantação.
Segundo o autor, a ação nociva das pragas pode ser observada desde a
implantação da cultura, ocasionando grande número de replantes, atraso no
desenvolvimento e, conseqüentemente, queda na produtividade. Nesta fase, é
intensa a ação das coleobrocas, cujas larvas se alimentam dos tecidos da planta
e afetam a produção, reduzindo-a drasticamente (Ferreira et al. 1997).
Dentre as coleobrocas, destaca-se a broca-da-ráquis foliar do coqueiro,
Amerrhinus ynca Sahlberg, 1823 (Coleoptera: Curculionidae). Suas larvas se
alimentam dos tecidos internos da ráquis foliar, fazendo galerias longitudinais que
se estendem tanto em direção ao tronco, como em direção à extremidade da folha
causando o seu amarelamento e enfraquecimento (Ferreira et al. 1997). Segundo
Mirizola-Filho (2002), com o crescimento populacional do inseto, a ráquis quebra-
se facilmente sob a ação do vento, provocando atraso no desenvolvimento da
planta e conseqüente queda da produção. O autor menciona ainda que, até pouco

2
tempo A. ynca era considerada praga secundária, mas devido a altos índices de
infestação em coqueirais do Estado da Bahia, Sergipe, Espírito Santo e Rio de
Janeiro seu status, nessas regiões, tem se elevado ao de praga primária.
O controle efetivo de seus níveis populacionais utilizando-se produtos
químicos só é possível quando o inseto atinge o estádio adulto (Gomes 1992).
Entretanto, com o crescimento dos coqueiros torna-se difícil e oneroso combatê-
lo. O correto monitoramento desta praga permitiria, além do controle eficiente do
inseto, a redução do número de aplicações de inseticidas. Isto contribuiria para o
aumento dos rendimentos, melhora qualitativa dos frutos, e reduções na poluição
ambiental.
Visando desenvolver novas ferramentas capazes de auxiliar os
produtores rurais no controle dos níveis populacionais de insetos-praga, várias
técnicas de monitoramento ou até mesmo de controle massal, baseadas em
mecanismos de comunicação, têm sido propostas. Por exemplo, a utilização de
armadilhas iscadas com substâncias químicas atrativas (semioquímicos) tem se
constituído em uma alternativa valiosa no Manejo Integrado de Pragas (MIP),
podendo levar a uma redução significativa do uso de inseticidas convencionais
(Vilela & Mafra-Neto 2001). Além disso, até o presente, ainda não foi constatado
nenhum perigo dessas substâncias ao meio ambiente e o seu uso não tem
resultado na resistência de insetos, por serem substâncias que regulam
comportamentos essenciais à sobrevivência da praga (Ferreira & Zarbin 1998).
Outras tecnologias utilizando-se da comunicação acústica também têm sido
aplicadas como ferramenta para detectar as densidades populacionais de insetos-
alvo, principalmente as infestações de insetos que ficam escondidos, tais como os
escarabeídeos do gênero Phyllophaga (Zhang et al. 2003). Adultos de A. ynca em
suas relações intraespecíficas produzem um som, através da estridulação. A
investigação deste mecanismo sonoro pode abrir novos caminhos na busca por
técnicas possíveis de serem utilizadas no Manejo Integrado desta praga.
Portanto, o objetivo deste trabalho é, através do conhecimento do
comportamento reprodutivo de A. ynca, identificar e descrever componentes
comportamentais, químicos e acústicos, que possam ser manipulados e utilizados
em técnicas de controle comportamental que auxiliem os produtores de coco no
monitoramento e/ou redução dos níveis populacionais deste inseto-praga. Para
isso, como ponto de partida, pretende-se: (i) encontrar e descrever caracteres

3
morfológicos que permitam a distinção dos sexos no campo e em laboratório, e (ii)
descrever a seqüência comportamental associada aos comportamentos de corte
e acasalamento, elucidando o papel dos componentes acústicos e químicos
dentro do comportamento reprodutivo de A. ynca. Os próximos passos,
entretanto, serão: (iii) identificar possíveis semioquímicos envolvidos tanto nas
interações intraespecíficas quanto nas interações com seu hospedeiro e; (iv)
identificar e descrever as estruturas associadas à estridulação bem como
qualificar e quantificar os parâmetros temporais e espectrais dos sons produzidos
por A. ynca.

4
2. REVISÃO DE LITERATURA
2.1. A cultura do Coqueiro O coqueiro (Cocus nucifera Linnaeus) é uma monocotiledônea
pertencente à família Palmae e à subfamília Cocoideae (Passos 1998b). É uma
espécie monotípica, arbórea, de caule ereto (do tipo estipe) e freqüentemente
sem ramificações e com folhas terminais (do tipo penada) (Mirizola-Filho 2002).
Esta palmeira, assim como toda monocotiledônea, não possui uma raiz principal,
mas um sistema radicular fasciculado, sendo as raízes primárias (mais grossas)
responsáveis pela sustentação e produzidas continuamente (Passos 1998a). O
fruto do coqueiro é do tipo drupa, constituído de uma epiderme, seguida por um
epicarpo, mesocarpo e endocarpo onde se localiza a semente (Mirizola-Filho
2002).
O coqueiro é considerado a “árvore da vida”, pois exerce papel importante
para a subsistência de populações das regiões tropicais quentes e úmidas do
globo (Ferreira et al. 1997). A cultura do coqueiro, graças à capacidade de gerar
um sistema auto-sustentável de exploração, pela geração de divisas e
fornecimento de alimento (fonte de proteínas e calorias), é considerada, segundo
Cuencas (1997), a mais importante das culturas perenes. Os frutos do coqueiro,
suas folhas, inflorescência, madeira etc. proporcionam a milhares de pequenos
proprietários alimentos, bebidas, combustíveis e abrigo (Mirizola-Filho 2002).

5
No Brasil, grande parte da produção nacional de coco, segundo Cuencas
(1997), se destina à fabricação de alimentos industrializados, que são
transportados da Região Nordeste até a Região Sul, para serem processados sob
a forma de sorvetes, doces, produtos à base de chocolate e iogurtes. Para
Mirizola-Filho (2002), o aumento da procura e do consumo do coco verde “in
natura”, tem sido responsável, nos últimos anos, pela expansão da cocoicultura
para os Estados das regiões Sudeste, Centro-Oeste e Sul, não só ao longo do
litoral, mas também no interior dessas regiões.
Vários são os fatores que limitam o desenvolvimento da cultura do
coqueiro no país. Segundo Passos (1998a), os fatores climáticos, tais como as
elevadas taxas de evapotranspiração associadas à irregularidade das chuvas na
região Nordeste; a elevada umidade relativa do ar no Norte; e as baixas
temperaturas e umidade atmosférica durante os meses de menor pluviometria no
Sudeste e Centro-Oeste, são os principais problemas enfrentados pelos
produtores destas regiões. Para Fontes et al. (1998), as características dos solos
da região do litoral nordestino, tais como: baixa fertilidade natural, excessiva
permeabilidade, baixos teores de matéria orgânica e pouca capacidade de
retenção de água, têm sido responsáveis por grandes perdas de produtividade.
Acrescenta-se ainda a este cenário a ação de pragas e doenças.
As injúrias causadas pelos insetos podem ser observadas desde a
implantação do coqueiral até o estabelecimento da planta adulta, quando é
intensa a ação nociva de larvas de coleobrocas (Ferreira et al. 1997). Por
exemplo, as larvas de Homalinotus coriaceus Gillenhal (Coleoptera:
Curculionidae) alimentam-se do pedúnculo floral e da bainha da folha do coqueiro,
impedindo a formação de frutos (Moura & Vilela 1998). Outras coleobrocas
importantes, segundo Ferreira et al. (1997), são: Rhinostomus barbirostris
Fabricius, 1775, Rhynchophorus palmarum Linnaeus, 1764 e Amerrhinus ynca
Salhberg, 1823, todas pertencentes à família Curculionidae.
2.2. Broca-da-ráquis-foliar do coqueiro - Amerrhinus ynca A broca-da-ráquis-foliar do coqueiro ou broca-do-pecíolo, Amerrhinus
ynca (Figura 1D), é um coleóptero originário do Brasil (Gomes 1992) e

6
pertencente à família Curculionidae. Segundo Ferreira et al. (1998), o adulto de A.
ynca é um gorgulho cujo comprimento varia de 14 a 22 mm, com um rostro que
pode chegar a seis milímetros nos maiores exemplares. Ainda segundo esses
autores, a coloração do corpo é tipicamente amarelada com pontos pretos
brilhantes e salientes, além de manchas escuras irregulares que se formam pela
variação na quantidade de escamas presentes nos élitros. De hábito diurno, o
adulto é encontrado com maior freqüência durante o verão e se alimentando da
inflorescência do coqueiro (Renard et al. 1987). Para a oviposição as fêmeas
fazem com o rostro pequenos orifícios no pecíolo foliar e em seguida introduzem
de três a quatro ovos, que são arredondados e de coloração esbranquiçada
(Figura 1A). As pupas (Figura 1C) se formam dentro das galerias construídas
pelas larvas (Figura 1B) ao longo do pecíolo, de onde emergem os adultos (Figura
1D) (Costa et al. 1973) após 20 dias de incubação (Bondar 1940).
Figura 1 – Fases do ciclo de vida de Amerrhinus ynca. (A) ovo, (B) larva de último ínstar, (C) pupa e (D) adulto se alimentando da inflorescência.
As larvas de A. ynca em seu último estágio chegam a atingir trinta
milímetros de comprimento e são ápodas e de coloração branca (Mirizola-Filho

7
2002). Elas se alimentam dos tecidos internos da ráquis foliar do coqueiro (Moura
& Vilela 1998), construindo galerias longitudinais que podem atingir oito
milímetros de diâmetro por 40 cm de comprimento (Ferreira et al. 1998), tanto em
direção ao tronco quanto em direção à extremidade da folha (Gomes 1992). Os
restos alimentares são expelidos pelo orifício de postura juntamente com a seiva
que exsuda da palmeira, o que caracteriza os danos iniciais da praga (Bondar
1940).
Esse inseto utiliza, além do coqueiro, outras plantas hospedeiras como a
carnaúba (Copernicia cerifera Martius), o dendezeiro (Elaeis guineensis Jacq.), o
licurizeiro (Syagrus coronata (Martius)), a macaubeira (Platymiscium duckei
Huber) e algumas palmeiras ornamentais como o pati (Syagrus botryophora
Martius) (Ferreira et al. 1997). Amerrhinus ynca já foi observado nos Estados de
Alagoas, Amazonas, Bahia, Espírito Santo, Pará, Paraná, Rio de Janeiro, São
Paulo e Sergipe (Ferreira et al. 1998). Até há alguns anos este gorgulho era
considerado uma praga secundária (Mirizola-Filho, 2002), mas dada à
periodicidade e intensidade de ataque este quadro vem se revertendo em alguns
coqueirais do país, como os da região Norte Fluminense, onde esta coleobroca já
atingiu o status de praga primária.
Ao se alimentarem dos tecidos internos do pecíolo, as larvas causam o
amarelamento das folhas e, conseqüentemente, o enfraquecimento da ráquis que
se quebra facilmente com a ação dos ventos (Figura 2C) (Moura & Vilela 1998).
Segundo Gomes (1992), a perda de seiva bruta e elaborada da folha atinge
diretamente as funções fotossintéticas da planta, principalmente se há a queda
das folhas, o que repercute negativamente sobre a produtividade do coqueiro.
Além disso, as galerias formadas pelas larvas de A. ynca se tornam uma porta de
entrada para patógenos, como o fungo Phyllachora torendiella, causador da lixa
do coqueiro, doença que promove o envelhecimento precoce das folhas (Moura e
Vilela 1998).

8
Figura 2 - Coqueiro atacado por Amerrhinus ynca. Em (A) restos alimentares e seiva que exsuda pelos orifícios de postura (em destaque). (B) orifícios por onde emergem os adultos (em destaque) e (C) queda e amarelamento das folhas atacadas por Amerrhinus ynca.
A incidência desse besouro em coqueirais pode comprometer mais de
50% das folhas de uma planta (Ferreira et al. 1998). Em 1987 foi relatado no
município de Una, Estado da Bahia, um forte ataque de A. ynca em um coqueiral
da variedade anão-amarelo (Renard et al. 1987). No município de Quissamã,
Norte do Estado do Rio de Janeiro, numa rápida visita a vários coqueirais, é
possível verificar em uma única planta, que a maioria das folhas foram atacadas
por esta coleobroca (observações de campo 2005). Além disso, o controle dos
níveis populacionais abaixo do nível de dano econômico é limitado pelo fato de as
larvas se encontrarem alojadas dentro do pecíolo (Gomes 1992), o que muitas
vezes eleva o custo do controle em áreas extensas da cultura. No entanto, o
controle cultural através da poda é o mais recomendado (Mirizola-Filho 2002;
Ferreira et al. 1998; Moura & Vilela 1998; Moura 1993; Ferreira 1987 e Bondar
1940). Por exemplo, em um estudo sobre a poda das folhas atacadas por A. ynca
no sul da Bahia, Moura (1993) constatou que, após um ano de controle, a técnica
conseguiu reduzir em 65% os níveis populacionais da praga. Moura e Vilela
(1998) recomendam ainda o controle preventivo e o biológico, sendo que neste
último se utiliza o díptero Paratheresia menezesi Townsend (Diptera: Tachinidae)
que parasita as larvas e as pupas de A. ynca. A pulverização com produtos
químicos também é recomenda por Moura e Vilela (1998), que ainda sugerem
aplicações localizadas de inseticidas através de injeções nos canais construídos
pelas larvas. A manipulação do comportamento através do uso de semioquímicos
ainda não é aplicada como técnica de manejo integrado desta praga, mas o uso

9
de feromônios associados a cairomônios tem sido feito para capturar adultos da
broca-do-olho-do-coqueiro, Rhynchophorus palmarum (Linneaus)
(Coleoptera:Curculionidae), apresentando bons resultados no campo (Navarro et
al. 2002).
2.3. Comportamento Reprodutivo
2.3.1. O Acasalamento Através do comportamento de acasalamento é possível compreender
atividades e eventos como a procura, identificação, atração, avaliação e por fim a
aceitação de parceiros para a reprodução (Lloyd 1979). O acasalamento,
portanto, pode ser dividido em etapas ou processos. A primeira etapa é o
reconhecimento do parceiro, seguida pela corte, cópula e pós-cópula. Dentro de
cada etapa pode ser usada uma série de atos esteriotipados.
Para o reconhecimento e localização do parceiro, os insetos podem
dispor de estímulos visuais, químicos e auditivos (Alcock 1998). Alguns estímulos
químicos envolvidos na localização de parceiros são os feromônios de agregação
e sexuais liberados principalmente por coleópteros e lepidópteros. Por exemplo, o
feromônio de agregação liberado por Rhynchophorus palmarum atrai machos e
fêmeas para o acasalamento (Moura & Vilela 2001).
A utilização de sinais acústicos para a localização e reconhecimento dos
parceiros é amplamente utilizada por insetos (Bailey 2003). Para este fim, é
importante que eles possam analisar as características estruturais destes sinais
tais como seu espectro e padrão de modulação de amplitude para determinar seu
conteúdo e sua localização (Pollak 2000). Entre os ortópteros, principalmente
aqueles da subordem Ensifera, os machos produzem um som espécie-específico
que pode ser: (i) utilizado pelas fêmeas para a localização de parceiros
potenciais, (ii) utilizado durante o comportamento de corte, (iii) na marcação de
território e (iv) para mediar interações agressivas entre machos (Mason 1996). Em
muitos sistemas de sinalização acústica de insetos é rara a produção de um canto
de chamamento pelas fêmeas, mas quando isso acontece freqüentemente está
associado a um dueto entre os sexos (Bailey 1991). Segundo Bailey (2003), este

10
dueto sonoro exerce, entre outras funções, o papel de reconhecimento de
coespecíficos para o acasalamento. Entre os insetos também podem ocorrer
duetos através de sinalização por bioluminescência, nos quais um dos sexos
responde às mudanças nos sinais luminosos do outro sexo (Bailey 2003).
Em relação aos estímulos visuais, os machos de borboletas são
geralmente o sexo mais ativo em procurar por oportunidades de acasalamento
(Kemp 2002). Segundo Ziemba & Rutowski (2000), borboletas que utilizam os
sinais visuais para localização de parceiros podem se comportar de duas
maneiras, em pouso, enquanto aguardam, ou em vôo, procurando ativamente por
oportunidades enquanto voam. Em espécies de Papilionoidea e Hesperioidea as
fêmeas utilizam a tática de pouso para localizar os machos para o acasalamento
e os machos geralmente localizam as fêmeas patrulhando (Scott 1974).
A corte consiste basicamente de uma sucessão de atos comportamentais
prévios ao acasalamento, efetuados na maioria dos casos pelos machos. Nesta
etapa, os insetos emitem e recebem uma sucessão de estímulos para os quais
são elaborados padrões apropriados de respostas (Viana & Vilela 1996). Exemplo
de estímulos envolvidos na corte podem ser movimentos, danças, estimulação
tátil, odores, vibração das asas e produção de sons. Em Drosophila montana
Stone, Griffen & Patter, 1942 (Diptera: Drosophilidae) por exemplo, os sons
produzidos pela vibração das asas dos machos são essenciais para estimular as
fêmeas ao acasalamento, e em certas circunstâncias estes sons de corte
influenciam a escolha de parceiros pelas fêmeas (Suvanto et al. 2000). Em muitos
gafanhotos, machos e fêmeas se comunicam acusticamente durante a corte,
sendo que os machos exibem uma série de sinais estridulatórios e as fêmeas
respondem a seus coespecíficos com estridulação similar, e complexa exibição
visual (Bailey 2003). Já em outros insetos, como em Dectes texanus texanus
LeConte, 1862 (Coleoptera: Cerambycidae), toque de antenas entre os sexos
estimulam os machos a montar nas fêmeas imediatamente após tocá-las (Crook
et al. 2004). No lepidóptero Phyllocnistis citrella Staiton, 1856 (Lepidoptera:
Gracillariidae), as fêmeas chamam a atenção do macho levantando as asas e
estendendo as antenas para trás do corpo enquanto o macho se movimenta
rapidamente em várias direções pulando e exibindo vigoroso movimento antenal
(Parra-Pedrazzoli et al. 2006). Em Bephratelloides pomorum (Fabricius, 1908)
(Hymenoptera: Eurytomidae), após a monta o macho expõem o edeago, toca a

11
fêmea com as antenas, curva a extremidade do abdome vibrando as asas e
antenas em movimentos rítmicos e então efetua a cópula (Pereira et al. 1998). A cópula, por sua vez, consiste na transferência direta de espermatóforo
para a espermateca da fêmea. Durante a cópula o macho pode transferir
substâncias que irão contribuir para a oviposição ou repressão da receptividade
pela fêmea. Neste ponto eles podem executar certos padrões motores de
comportamento, conhecidos como comportamento de corte copulatória, que
estimulam as fêmeas a permanecerem em cópula e pós-cópula, garantindo assim
que seus gametas sejam transferidos para a próxima geração (Eberhard 1991).
No besouro Tribolium castaneum (Herbst, 1797) (Coleoptera: Tenebrionidae),
Edvardsson & Arqnqvist (2000) verificaram que táticas apropriadas de
estimulação da fêmea durante a cópula pelos machos induzem as fêmeas a
ajudar a transportar o esperma do local de deposição de ejaculados para o local
de estocagem do esperma, aumentando o sucesso de paternidade pós-
acasalamento.
Já os comportamentos associados à pós-cópula consistem basicamente
em guardar a fêmea impedindo que ela copule com outros machos, seja através
da guarda da fêmea (permanecendo sobre o dorso da fêmea por longos períodos,
por exemplo) ou pela liberação de substâncias que irão inibir a aproximação de
outros machos. A esta fase também podem estar associados tanto
comportamentos esteriotipados, como a estimulação tátil e sinalização sonora,
quanto comportamentos passivos. No curculionídeo Cosmopolites sordidus
(Germar, 1824) (Coleoptera: Curculionidae), os machos na presença de outros
machos permanecem sobre o dorso da fêmea por um determinado período após
o acasalamento (Viana & Vilela 1996). Também no heteróptero Gerris lateralis
Schummel, 1832 (Heteroptera: Gerridae) os machos permanecem passivamente
sobre o dorso da fêmea por um longo período de tempo sem contato genital
(Arnqvist 1988) e em alguns casos, como em Gerris remigis Say, 1832, os
machos que guardam suas parceiras estridulam para reduzir as tentativas de
intromissão de outros machos (Wilcox & Stefano 1991). Já em outras espécies
desta família, os machos, após o término da cópula, descem da fêmea e
permanecem próximos a ela impedindo agressivamente que outros machos se
aproximem enquanto ela oviposita (Arnqvist 1997). Já os machos do coccinelídeo
Menochilus sexmaculatus (Fabricius, 1781) (Coleoptera: Coccinellidae), após a

12
cópula, permanecem alguns segundos sobre o dorso da fêmea girando e
lambendo seu élitro e pronoto, tornando-a, provavelmente, não atrativa a outros
machos (Maisin et al. 1997).
2.3.2. A "guerra dos sexos" O comportamento sexual é a chave para o entendimento da biologia das
espécies, sua origem e a maneira como elas vivem hoje. Além disso, a biologia
das espécies é fundamentada no seu sucesso sexual. Por exemplo, machos
aumentam seu sucesso reprodutivo acasalando com muitas parceiras em altas
taxas de acasalamento enquanto que aumentam seu sucesso reprodutivo
maximizando o número de ovos viáveis, embora um ou poucos acasalamentos
sejam suficientes (Arnqvist & Nilsson 2000).
Neste ponto atuam os custos de acasalamento, uma vez que a
reprodução sexual requer uma sincronização das atividades de dois organismos,
a qual gera um gasto de energia e tempo, além do fato de que os interesses dos
parceiros não são iguais (Daly 1978) criando um "conflito sexual". Isso quer dizer
que a interação entre machos e fêmeas freqüentemente não termina com a
cópula; os machos devem persuadir as fêmeas para que elas usem seu esperma
e não o de outros machos (Wiley 1997) e, por outro lado, as fêmeas podem fazer
escolhas crípticas variando seu investimento reprodutivo dependendo da
atratividade de seu parceiro (Edvardsson & Arqnqvist 2005). Dentro deste conflito,
o “gene egoísta” exerce papel fundamental desde o início da procura até o final da
fertilização, não produzindo nada sem um lucro reprodutivo líquido (Lloyd 1979).
Para Daly (1978), os custos caem principalmente sobre os machos, mas são
substanciais nas fêmeas e podem, também, variar de acordo com o sistema de
acasalamento e com o cuidado parental.
Para Arqnvist & Rowe (2002), a coevolução entre os sexos é reconhecida
como o processo central da evolução, com potencial para modelar várias
interações entre os sexos e seus gametas, bem como taxas de diversificação,
especiação e excitação. Dentro desta "competição armada", gerada pelas
interações coevolutivas, os sexos utilizam estratégias que aumentam o seu
sucesso reprodutivo variando o sucesso reprodutivo do outro sexo (Stockley
1997). A competição por esperma é uma destas estratégias utilizadas por machos
e afeta o sucesso reprodutivo das fêmeas. Carbone e Rivera (1998) definem a

13
competição por esperma como a competição que ocorre dentro da fêmea, entre o
esperma de dois ou mais machos pela fertilização dos ovos. Para Stockley
(1997), os machos podem adotar uma série de estratégias (como, por exemplo, o
prolongamento do acasalamento) para impedir que as fêmeas acasalem com
outros machos, evitando a competição por esperma e aumentando assim seu
sucesso reprodutivo. A guarda do parceiro através do prolongamento do
acasalamento é um fenômeno bem descrito em insetos (Arnqvist 1997),
entretanto, para que os machos evitem a competição por esperma e garantam
seu sucesso reprodutivo eles dispõem de tempo e energia que poderiam ser
utilizados para encontrar e acasalar com fêmeas adicionais (Harari et al. 2003).
Esta estratégia utilizada por machos para impedir ou suceder em competição por
esperma pode influenciar negativamente o sucesso reprodutivo das fêmeas,
reduzindo sua eficiência de forrageamento, sua fertilidade e as chances que elas
teriam de lucrar com os benefícios de múltiplos acasalamentos (Stockley 1997).
Dentro do contexto da competição por esperma, as fêmeas podem fazer
escolhas crípticas como uma possível estratégia para aumentar o seu sucesso
reprodutivo. Neste caso, segundo Wedell (1996), as fêmeas fazem uma escolha
pós-copulatória priorizando espermas de machos escolhidos para fertilização dos
ovos ou aumentando seu sucesso reprodutivo após acasalar com um macho
escolhido. Segundo Birkhead (1998) o termo críptico é utilizado porque tal
processo ocorre dentro do corpo da fêmea e não pode ser observado
diretamente, sendo que sua compreensão requer um entendimento de
mecanismos associados com a inseminação, armazenagem de esperma e
fertilização. Entretanto, o autor salienta que, apesar de existirem evidências
fisiológicas e estruturais presentes em fêmeas de muitos organismos (diferentes
locais de armazenagem de esperma, por exemplo) que possam sustentar a
hipótese da escolha críptica de fêmeas como um processo ligado à seleção
sexual, ainda não foi comprovada em nenhum organismo vantagem reprodutiva
para fêmeas.

14
2.4. Comunicação química em insetos A comunicação é um componente comportamental de extrema
importância para a adaptação dos animais, destacando-se por permitir a interação
entre as espécies. Lewis & Gower (1980) definem comunicação como a
transmissão de sinal ou sinais entre dois indivíduos, favorecendo a seleção tanto
da produção quanto da recepção do sinal ou sinais.
Ao longo da evolução, alguns seres vivos, diante da necessidade de
interagir com indivíduos da mesma espécie ou de espécies diferentes,
desenvolveram uma comunicação química característica. Os sinais químicos,
envolvidos neste tipo de comunicação, são os responsáveis pela transmissão de
informações biologicamente importantes (Paiva & Pedrosa-Macedo 1985), o que
assegura a tais indivíduos oportunidades de sobrevivência e de preservação da
espécie.
A comunicação química estabelecida entre os organismos é feita com o
auxílio de substâncias voláteis, as quais são biossintetizadas e liberadas pelo
emissor e captado e assimilado pelo receptor que, por sua vez, pode sofrer uma
resposta imediata ou em longo prazo, promovendo alterações em seu
comportamento ou em sua fisiologia (Paiva & Pedrosa-Macedo, 1985). Aos sinais
químicos envolvidos na comunicação entre os seres vivos dá-se o nome de
semioquímicos.
Quando os compostos químicos são utilizados para mediar interações
entre indivíduos de espécies diferentes, atuando em dois ou mais níveis da cadeia
trófica, diz-se tratar de substâncias aleloquímicas (Vilela & Della Lucia 2001). De
acordo com os benefícios e prejuízos provocados por essas substâncias
químicas, os aleloquímicos podem ser classificados como cairomônios, alomônios
e sinomônios. Já os feromônios são compostos químicos que promovem uma
modificação no comportamento e na fisiologia de indivíduos receptores da mesma
espécie (Tegoni et al. 2004), ou seja, possuem uma ação intraespecífica.
Os insetos se comunicam basicamente através de substâncias químicas
e, segundo Vilela & Della Lucia (2001), os odores liberados por eles são
fundamentais para o bom desempenho de suas atividades durante toda a sua
existência, contribuindo para a localização e seleção do alimento e do parceiro
sexual, e para a orientação na busca por abrigo, presas e plantas hospedeiras.

15
No caso de insetos sociais, os odores contribuem para a organização das
atividades da colônia, especialmente os processos de defesa e de marcação de
território.
Os odores percebidos pelos insetos são moléculas orgânicas pequenas
geralmente hidrofóbicas e voláteis (Pelosi 1996) detectadas por sensilas
especializadas presentes nas antenas e ligadas a neurônios sensoriais (Tegoni et
al. 2004). Segundo Dickens & Mori (1989), a perfeita orientação dos insetos até
seus coespecíficos, seja para agregação ou acasalamento, depende da
sensitividade e especificidade de detectar e traduzir a mensagem feromonal.
Dessa forma, a presença de quiralidade e a composição precisa de uma mistura
feromonal são de extrema importância para a uma perfeita comunicação química
(Ferreira 2001).
Para Landolt & Phillips (1997), muitos insetos fitófagos utilizam as suas
plantas hospedeiras não somente para se alimentarem, mas também como sítio
de reprodução. Fisiologicamente, os compostos químicos adquiridos através do
consumo, absorção e inalação de materiais da planta hospedeira podem agir no
sistema endócrino do inseto, estimulando a liberação de hormônios responsáveis
por redimensionar os metabólitos das rotas primárias do seu metabolismo no
sentido da biossíntese de compostos feromonais (Tillman et al. 1999). Além disso,
a liberação e a recepção de feromônio também sofrem as influências dos
compostos liberados pela planta hospedeira, aumentando a produção e liberação
de feromônio e causando mudanças na percepção quantitativa do receptor
(Landolt & Phillips 1997). Tais compostos químicos podem, ainda, atuar como
sinergistas, aumentando a atração de insetos para as armadilhas iscadas com
feromônio no campo (Giblin-Davis et al. 1996), uma técnica de controle e
monitoramento utilizada no manejo integrado de pragas.
2.4.1. Comunicação química em Curculionídeos
De acordo com El-Sayed (2005), já foi identificada, em quase 40 famílias
da ordem Coleoptera, a presença de substâncias químicas mediando interações
intra e inter específicas. Dentro da família Curculionidae, 43 espécies distribuídas
em 20 gêneros apresentam algum tipo de atraente químico, sendo que a grande
maioria é representada por feromônios de agregação (El-Sayed 2005).

16
Na subfamília Curculioninae trabalhos de campo e laboratório já
evidenciaram a presença de feromônio de agregação produzido por machos em
três espécies do gênero Anthonomus (A. grandis Boheman, 1843; A. eugenni
Cano, 1894 e A. rubi (Herbst, 1795)) e uma espécie do gênero Curculio. Estudos
sobre o comportamento de acasalamento de A. grandis, o bicudo do algodoeiro,
revelaram que, logo após os adultos machos encontrarem sua planta hospedeira,
estes passam a liberar substâncias que são atrativas (Cross & Mitchell, 1966) a
seus coespecíficos (Hardee et al. 1969 e Tumlinson et al. 1971). Com isso, no
final dos anos 60, pesquisadores identificaram, isolaram e sintetizaram a partir de
machos do bicudo quatro compostos terpenóides, entre eles o grandisol
(Tumlinson et al. 1969), que ficaram conhecidos como grandlure, um feromônio
de agregação produzido por A. grandis. Segundo Eller et al. (1994), o feromônio
de agregação do gorgulho da pimenta, A. eugenii, é constituído por uma mistura
de seis compostos, sendo que três deles são comuns ao feromônio de agregação
produzido pelos machos de A. grandis. Já para o gorgulho da flor do morango,
Anthonomus rubi, o seu feromônio de agregação é constituído de três compostos,
sendo o grandisol comum à mistura feromonal do bicudo do algodoeiro (Innocenzi
et al. 2001). O feromônio de agregação produzido pelo gorgulho da noz, C. caryae
Horn, 1873, também pertencente à subfamília Curculioninae, é constituído pelos
mesmos compostos atrativos produzidos pelos machos de A. grandis, a diferença
está apenas na proporção dos compostos utilizados para promover uma resposta
eficiente nas fêmeas (Hedin et al. 1997).
Na subfamília Pissodinae, três espécies também produzem grandisol
como seu feromônio de agregação, além do seu aldeído correspondente
(grandisal). Booth et al. (1983) isolaram e identificaram, a partir do abdômen e
intestino posterior de machos de Pissodes strobi (Peck, 1817) e de P.
approximatus Hopkins, 1911, estes dois compostos como sendo componentes de
seus feromônios de agregação. Em 1984 foi demonstrado através de testes de
campo e laboratório que o gorgulho da casca do pinus e cedro, P. nemorensis
Germar, 1824, também utiliza o grandisol e grandisal como um feromônio de
agregação produzido por machos para atrair ambos os sexos (Phillips et al. 1984).
Outras substâncias atraentes são relatadas em outras espécies de Pissodes,
como lineatin para Pissodes gyllenhali (Sahlberg, 1834) e Pissodes pini

17
(Linnaeus, 1758) (Martikainen 2001) e α-pineno para Pissodes affinis Randall,
1838 e Pissodes fasciatus LeConte, 1876 (Miller & Heppner 1999).
Na subfamília Cryptorhynchinae, o gorgulho da ameixa, Conotrachelus
nenuphar (Herbst, 1797), teve seu feromônio de agregação isolado e sintetizado,
a partir de voláteis de machos, com o nome de ácido grandisóico que atrae
ambos os sexos (Eller & Bartelt 1996). Este feromônio de agregação sofre um
efeito sinergista muito forte quando na presença de voláteis liberados pela planta
hospedeira, como o benzaldeido (Piñero & Prokopy 2003). Na família
Curculionidae, vários são os exemplos de feromônios de agregação produzidos
por machos que sofrem um aprimoramento na presença de voláteis liberados
pelas suas plantas hospedeiras (Leskey & Prokopy 2001). Por exemplo, o
gorgulho do arroz, Sitophilus oryzae (Linnaeus, 1763), só libera seu feromônio de
agregação produzido por machos na presença de grãos de arroz, na ausência do
alimento não há liberação (Landolt & Phillips 1997).
Os gorgulhos do gênero Sitophilus, pertencentes à subfamília
Rhynchophorinae, são conhecidos como importantes pragas de grãos
armazenados. A primeira evidência de um feromônio de agregação produzido por
machos deste gênero foi registrada em 1981, por Phillips & Burkholder, para o
gorgulho do arroz Sitophilus oryzae (Philips & Burkholder 1981). Em seguida
demonstrou-se a presença em Sitophilus granarius (Linnaeus, 1758), o gorgulho
do celeiro (Faustini et al. 1982) e Sitophilus zeamais Motschulsky, 1855, o
gorgulho do milho (Walgenbach et al. 1983). Em 1984 foi identificado o composto
sitofilure como um feromônio de agregação produzido por machos de S. oryzae e
S. zeamais (Schmuff et al. 1984).
2.4.1.1. Semioquímicos produzidos por gorgulhos das Palmáceas
Dentro da subfamília Rhynchophorinae, encontram-se quatro gêneros de
pragas de importância econômica para a cultura de palmáceas nos trópicos, o
gênero Dynamis, Metamasius, Rhabdoscelus e Rhynchophorus, apesar de
algumas espécies também utilizarem outras plantas hospedeiras, para
alimentarem, ovipositarem e desenvolverem suas larvas. Os compostos
majoritários e minoritários do feromônio de agregação produzido por machos
desta subfamília, são constituídos principalmente por moléculas de 8, 9, ou 10

18
carbonos, ramificações de metil e álcoois secundários (Tabela 1) (Giblin-Davis et
al. 1996).
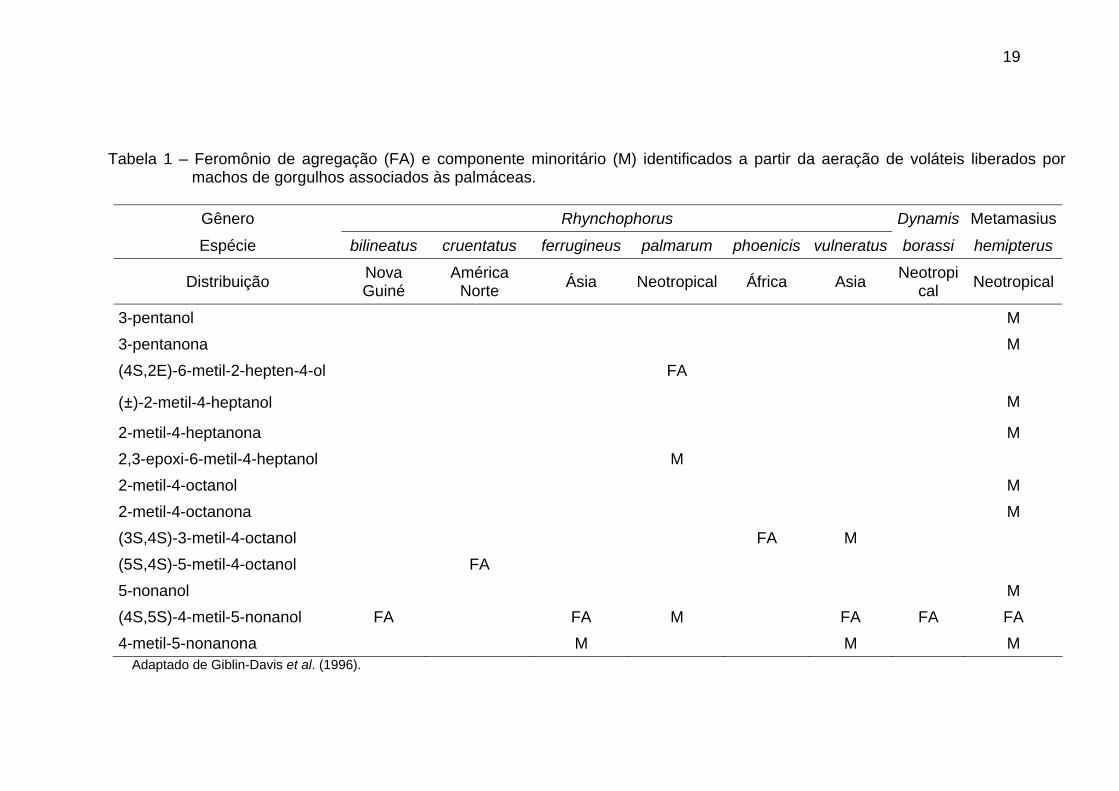
19
Tabela 1 – Feromônio de agregação (FA) e componente minoritário (M) identificados a partir da aeração de voláteis liberados por machos de gorgulhos associados às palmáceas.
Gênero Rhynchophorus Dynamis Metamasius
Espécie bilineatus cruentatus ferrugineus palmarum phoenicis vulneratus borassi hemipterus
Distribuição Nova Guiné
América Norte Ásia Neotropical África Asia Neotropi
cal Neotropical
3-pentanol M 3-pentanona M (4S,2E)-6-metil-2-hepten-4-ol FA
(±)-2-metil-4-heptanol M
2-metil-4-heptanona M 2,3-epoxi-6-metil-4-heptanol M 2-metil-4-octanol M 2-metil-4-octanona M (3S,4S)-3-metil-4-octanol FA M (5S,4S)-5-metil-4-octanol FA 5-nonanol M (4S,5S)-4-metil-5-nonanol FA FA M FA FA FA 4-metil-5-nonanona M M M
Adaptado de Giblin-Davis et al. (1996).

20
A primeira evidência da produção de um composto químico de função
atrativa nesta subfamília foi demonstrada para o gorgulho das palmeiras
ornamentais, Rhabdoscelus obscurus (Boisduval, 1835) (Chang & Curtis 1972).
Este gorgulho produz um feromônio liberado pelos machos, e o composto
principal da mistura feromonal foi identificado como 2-metil-4-octanol, mas eles
também produzem (E2)-6-metil-2-heptan-4-ol (rincoforol) e 2-metil-4-heptanol
(Giblin-Davis et al. 2000).
O rincoforol é o composto principal da mistura feromonal produzida por
machos da broca-do-olho-do-coqueiro, Rhynchophorus palmarum (Jaffé et al.
1993). A presença deste feromônio de agregação foi demonstrada em 1991 em
trabalhos de campo e laboratório com machos vivos de R. palmarum (Rochat et
al. 1991) e logo em seguida a atividade do enantiômero S do composto 6-metil-2-
heptan-4-ol foi confirmada por Oehlschlager et al. (1992). Alguns compostos
liberados pelos tecidos danificados da planta hospedeira, como o acetato de etila
e o etanol, também exercem uma função de orientação nesses gorgulhos,
provavelmente a curtas distâncias, além de potencializar a atração do feromônio
de agregação (Jaffé et al. 1993).
A partir destes trabalhos tornaram-se intensos, durante a década de
noventa, os estudos com feromônios de agregação produzidos por machos dentro
do gênero Rhynchophorus (que causam sérios prejuízos às palmáceas). Em
1993, estudos de campo e laboratório demonstraram a presença de feromônio de
agregação em Rhynchophorus cruentatus (Fabricius, 1775), o gorgulho do palmito
(Weissling et al. 1993), Rhynchophorus phoenicis Fabricius 1801, o gorgulho da
palmeira africana (Gries et al. 1993), Rhynchophorus ferrugineus (Olivier, 1790), o
gorgulho vermelho e em Rhynchophorus vulneratus Panzer, 1798 (Hallett et al.
1993). O composto 5-metil-4-octanol (cruentol) (Weissling et al. 1994) e o 3-metil-
octan-4-ol (fonicol) foram identificados, através da extração de voláteis produzidos
por machos, como os componentes principais do feromônio de agregação de R.
cruentaus e R. phoenicis, respectivamente. Estes dois compostos possuem
quatro estereoisômeros, mas apenas o (4S,5S) – cruentol e o (3S,4S) – fonicol
são produzidos pelos machos e capazes de provocar uma resposta
comportamental (Perez et al. 1994). Outros estudos feitos utilizando-se
eletroantenograma revelaram a atividade do acetato de etila, propranato de etila,
propranato de isobutila, butirato de etila e isobutirato de etila (todos liberados pela

21
planta hospedeira) em machos e fêmeas de R. phoenicis, mas apenas o
proprianato de etila aumentou significativamente a captura deste gorgulho em
armadilhas iscadas com o feromônio no campo (Gries et al. 1994).
Para R. ferrugineus foram identificados os compostos 4-metil-5-nonanol
(ferruginol) e a sua cetona correspondente, 4-metil-5-nonanona, como sendo
constituintes do seu feromônio de agregação (Hallett et al. 1993). O ferruginol
também é o componente principal dos feromônios de agregação de R. vulneratus
(Hallett et al. 1993) e Rhynchophorus bilineatus Montrouzier, 1857 (Oehlschlager
et al. 1995), além de estar presente na mistura feromonal produzida por machos
de R. palmarum (Hallett et al. 1993).
Além disso, o ferruginol também é o componente principal do feromônio
de agregação de outras duas espécies de gorgulhos da subfamília
Rhynchophorinae, Dynamis borassai (Giblin-Davis et al. 1997) e Metamasius
hemipterus sericeus (Olivier, 1807) (Perez et al. 1997). O gorgulho da cana-de-
açúcar do oeste da Índia, M. hemipterus sericeus, que também utiliza palmáceas
como hospedeiro, produz além do ferruginol mais sete compostos macho-
específicos (Perez et al. 1997). Segundo Giblin-Davis et al. (1996) este gorgulho
pode representar uma forma mais primitiva do que outras espécies de
Rhynchophorus, uma vez que os machos produzem um grande número de
álcoois e cetonas em seu feromônio de agregação em contra partida com os
feromônios de agregação de espécies de Rhynchophorus, que produzem,
tipicamente, um ou dois compostos.
2.5. Comunicação sonora em insetos A comunicação, como visto anteriormente, é baseada na transmissão de
uma mensagem, através de sinais, por uma fonte emissora até um receptor.
Estes sinais podem ser químicos, visuais, táteis ou auditivos. Os sistemas de
comunicação que utilizam a audição como um canal para a transmissão e a
recepção de sinais acústicos têm como princípio a análise dos parâmetros
temporais e de freqüência destes sinais (Landa 2000). Segundo Bailey (1991),
estes sistemas possuem certas vantagens sobre outros processos de sinalização,
uma vez que não há a necessidade de condições do ambiente, como o vento para

22
a efetiva transferência de sinais, como ocorre com as trilhas de feromônio e, além
disso, eles podem ser utilizados por espécies diurnas e noturnas e agir muito
além das próprias dimensões do corpo.
Muitos sons e sinais produzidos por insetos são espécie-específicos e são
utilizados principalmente para a localização, reconhecimento e formação de pares
(Cokl & Virant-Doberlet 2003). Em alguns coleópteros, a função da produção de
sons, muitas vezes, está associada à liberação de feromônio, desempenhando
um papel mais complexo no comportamento de acasalamento (Michael &
Rudinsky 1972). Esta sinalização sonora também pode agir além do
comportamento intrasexual, sendo útil, por exemplo, em mecanismos de defesa
presentes em alguns coleópteros e heterópteros (Virant-Doberlet & Cokl 2004), na
localização de hospedeiros por insetos parasitas (Haynes & Yeargan 1999), no
cuidado parental como em Membracidae (Cocroft 1999), na marcação de território
como nas lagartas Depranidae (Yack et al. 2001) e na regulação de interações
sociais complexas como em algumas abelhas domésticas (Lewis et al. 2002).
Além disso, alguns insetos podem imitar o som de outros insetos, como as
largatas Maculinea (Lepidoptera: Lycaenidae) que imitam o som de algumas
formigas operárias do gênero Myrmica (Hymenoptera: Formicidae) (Devries et al.
1993). O som, enquanto sinalizador de uma informação, é representado pelas
suas características estruturais, tais como o seu espectro e padrão de modulação
de amplitude, o que vai permitir ao receptor analisar e localizar a mensagem
mesmo quando vários sinais chegam simultaneamente de diferentes fontes
(Pollack 2000). Segundo Machens et al. (2001), o reconhecimento do sinal não é
baseado exclusivamente na aceitação de padrões sonoros espécie-específicos
corretos, mas também envolve a atividade de rejeição de sinais que contêm
componentes fortes ou duvidosos, como aqueles que ocorrem entre as cigarras,
grilos e gafanhotos. Segundo Bailey (1991), os insetos podem apresentar, em seu sistema
auditivo, receptores localizados em diferentes partes do corpo que permitem a
percepção das variações físicas que ocorrem com a transmissão de sons. Dessa
forma, Bailey (1991) divide a produção de som em: (i) som de campo próximo, (ii)
som de origem aérea e (iii) som através do substrato. Vale ressaltar que, apesar

23
desta classificação, os insetos podem utilizar vários mecanismos para a produção
de sons e sinais dificultando uma classificação segura (Haskell 1974).
As vibrações de campo próximo são geradas pelo deslocamento de
partículas de ar ou água próximas da fonte emissora e são captadas por
receptores do tipo pêlos mecanoreceptores, mas este tipo de produção de som é
pouco comum em insetos (Bailey 1991). Já a comunicação originária do substrato
está relacionada com uma variedade de interações e exerce diversas funções
comportamentais (Virant-Doberlet & Cokl 2004). Tais sons são produzidos pelos
insetos através da fricção de partes de seu corpo e são transmitidos pelo solo ou
pelos materiais nos quais eles vivem, servindo, dessa forma, como substrato para
a transferência da energia sonora até o receptor da mensagem (Haskell 1974).
Para a produção de sons de origem aérea os insetos utilizam a cutícula como um
sistema de sinalização sonora, através da fricção, puxão e toques sobre sua
superfície dura, os quais permitirão que áreas percutidas ao redor da cutícula, ou
pela deformação causada pela contração dos músculos gerados pela cutícula
sozinha, vibrem com alguma liberdade ou deformação (Bailey 1991).
Para a recepção de um sinal acústico o inseto necessita de órgãos
mecanoreceptores apropriados (Hoy & Robert 1996). Estes órgãos são receptores
auditivos que respondem às oscilações do meio ao redor, sendo que cada
receptor consiste de um ou mais neurônios sensoriais, um grupo de células
acessórias e estruturas cuticulares do tipo sensilas e pêlos (Bailey 1991). As
sensilas campaniformes, localizadas próximas das juntas intersegmentais das
pernas do inseto, e os órgãos escolopidiais ou órgão subgenal, localizados na
tíbia de todas as seis pernas, são os melhores receptores para as vibrações
originárias do substrato (Virant-Doberlet & Cokl 2004). Já os órgãos cercais de
baratas, o órgão de Johnston de mosquitos e as arestas de drosófilas são bons
exemplos de receptores de sons de campo-próximo (Hoy & Robert 1996). Os
órgãos de Johnston, que se localizam na base da antena dos insetos, constituem
um mecanoreceptor mais complexo funcionando como um proprioreceptor e que
também pode ser utilizado, entre outras funções, para perceber os sons de
origem aérea (Bailey 1991).
Além disso, outras estruturas mais complexas também exercem a função
de receptores de sons como os ouvidos timpanais (Hoy & Robert 1996). Segundo
Forrest et al. (1997), estas estruturas possuem três características: (i) uma área

24
afinalada pela cutícula, conhecida como membrana timpânica; (ii) esta membrana
está associada a um saco de ar, formado por uma expansão do sistema traqueal;
e (iii) a tradução da vibração pela membrana dentro dos sinais neurais é realizada
pelas células sensóriais bipolar no órgão sensorial cordotonal.
A utilização de ouvidos timpanais é a maneira mais usual de detecção de
sons de origem aérea e evoluiu tanto em vertebrados quanto em várias ordens de
insetos como em Orthoptera, Lepidoptera, Mantodea, Hemiptera (Bailey 1991),
bem como em coleópteros das famílias Cicindelidae e Scarabaeidae (Hoy &
Robert 1996). Segundo Forrest et al. (1997), a função primária dos ouvidos de
insetos é perceber e localizar os sinais acústicos emitidos por seus coespecíficos
ou para detectar os sons gerados por seus predadores e presas.
2.5.1. Produção de Som em Coleoptera
A produção de som entre os coleópteros já foi verificada em várias
famílias (Carabidae, Hygrobiidae, Dytiscidae, Hydrophilidae, Silphidae, Lucanidae,
Trogidae, Acanthoceridae, Passalidae, Geotrupidae, Buprestidae, Anobiidae,
Tenebrionidae, Cerambycidae, Chrysomelidae, Attelabidae, Curculionidae,
Scolytidae, Scarabeidae e Bostrichidae) (Lobanov 2002). Nos coleópteros, os
mecanismos de estridulação evoluíram e se diversificaram entre as larvas, pupas
e adultos que podem utilizar várias partes do corpo como a cabeça, tórax,
abdômen, asas, pernas e o próprio élitro para produzir sons (Haskell 1974).
A estridulação, muito comum entre os besouros, pode ser definida como o
processo por meio do qual o som ou vibração é produzido pela fricção de duas
partes do corpo que se movem uma contra a outra (Cokl & Virant-Doberlet 2003).
Segundo Lobanov (2002), uma das superfícies de atrito se assemelha a uma
estrutura em forma de múltiplas cristas paralelas (tipo “lima” ou pars stridens) e a
outra se assemelha a um dentículo (“raspador” ou plectrum) e ambas se movem
uma em relação à outra para gerar um som.
Os órgãos estridulatórios de besouros podem estar localizados em
posições amplamente diferentes. Adultos de Lilioceris lilii (Scopoli, 1763)
(Coleptera: Chrysomelidae) produzem um trinado através de seu aparato
estridulatório localizado entre os élitros e o abdômen (Smith 2003). No gênero
Pelobius Hermanni (Coleoptera: Dytiscidae), a produção de um barulho
estridulatório se dá quando os sulcos paralelos (pars stridens) localizados

25
próximos da sutura marginal dos élitros são raspados contra o plectrum situado
na margem extrema do abdômen (Darwin 1871). Em machos do gênero
Laccobius Erichon, 1837 (Coleoptera: Hydrophilidae) a pars stridens está situada
no terceiro pleurito abdominal e possui uma estrutura eulamelada e o plectrum
está localizado no interior dos élitros (Pirisinu et al. 1988).
O aparato estridulatório de Eupsophulus castaneus Horn, 1870
(Coleoptera: Tenebrionidae) consiste em um plectrum, localizado na face interior
do fêmur posterior e uma pars stridens, localizada em uma placa lateral entre o
tergito e esternito do primeiro segmento abdominal (Slobodchikoff & Spangler
1979). Segundo Serrano et al. (2003), o mecanismo de estridulação em Oxychelia
tristis (Coleoptera: Carabidae) é produzido tanto por machos quanto por fêmeas
ao friccionarem a margem interna do fêmur (raspador ou plectrum) sobre a
epipleura elitral (lima ou pars stridens), sendo que os movimentos do abdômen
parecem funcionar com um modulador de amplitude e as características
temporais do som são fortemente diferentes entre os sexos, mas os espectros de
freqüência são similares. Os autores mencionam ainda que estas estruturas de
estridulação também são encontradas em outras espécies de Oxycheila, bem
como no grupo próximo Cheiloxya binotata longipennis Horn, 1891. Também no
gênero Heterocercus Fabricius, 1972 (Coleoptera: Heteroceridae) as raspadeiras
são localizadas no fêmur e a estridulação é produzida pela sua fricção sobre as
limas localizadas nas laterais do primeiro segmento abdominal (Darwin 1871). Em
alguns Cerambycidae a área de estridulação está situada na parte anterior do
mesonoto que fica coberto pelo pronoto e o som é produzido quando o protórax
se move em direção ao mesotórax, deslizando a margem posterior e afiada do
mesonoto contra a área de estridulação (Lobanov 2002).
Os órgãos estridulatórios também são amplamente distribuídos entre as
larvas de Coleoptera. Em algumas larvas de Melolonthinae, Rutelinae, Dynastinae
e Cetoninae, espinhos sobre a maxila são esfregados contra uma placa estriada
situada sobre a mandíbula e em larvas de Lucanidae, Passalidae e Geotrupidae,
uma série de sulcos (pars stridens) sobre a coxa das pernas medianas é
friccionada sobre os raspadores (plectrum) localizados sobre o trocânter das
pernas posteriores (Haskell 1974). O aparato de estridulação das larvas muitas
vezes não apresenta uma função muito clara, como nas larvas do passalídio,
Lucanu cervus, no qual os sons possivelmente direcionam as larvas para o

26
propósito de se manterem juntas ou para reivindicarem um espaço, sendo as
funções de defesa, proteção e orientação no substrato descartáveis (Sprecher
2003). Segundo Verdú & Galante (2001) algumas larvas de espécies do gênero
Pedaridium Harold (Coleoptera: Scarabaeidae) podem ser classificadas pela
presença ou ausência de áreas de estridulação na mandíbula e a presença deste
aparato de estridulação impediria o contato entre larvas, evitando o canibalismo
pela produção de um som de sinalização.
Na ordem Coleoptera, a emissão de sinais acústicos pode estar
relacionada tanto aos comportamentos de reprodução quanto aos de ataque e
ambos podem estar associados à liberação de feromônios. Por exemplo, o
escolitídio Dendroctonus pseudotsugae Hopkins, 1905 possui um comportamento
antiagregação no qual a ocorrência de estridulação e a liberação de feromônio
antiagregação exercem um papel importante, pois juntos criam uma zona
territorial, dentro da qual nenhum ataque é permitido e além da qual os ataques
são permitidos (Prenzel et al. 1999).
Dentro do contexto do comportamento reprodutivo, a estridulação pode
ser utilizada por outros escolitídeos do gênero Ips para estabelecer uma
comunicação no início da formação das câmaras nupciais, e este som geralmente
é produzido pelas fêmeas (López & Oliveira 2002). Os besouros platipodídios,
Platypus quercivorus (Murayama, 1925) (Coleoptera: Platypodidae) também
produzem vários sons pré-acasalamento. Segundo Ohya & Kinuura (2001), as
fêmeas de P. quercivorus produzem um “trinado de aproximação” para chamar a
atenção dos machos e em seguida produzem um zunido de pré-acasalmento que
induz os machos a empurrá-las para dentro da galeria onde vai ocorrer o
acasalamento; por fim os machos produzem um cricrido enquanto entram na
galeria (Figura 3).

27
Figura 3 - Três comportamentos seqüenciais de pré-acasalmento acompanhado de sons em Platypus quercivorus. (A) uma fêmea caminhando próximo ao buraco da galeria com um “trinado de aproximação”. (B) uma fêmea produzindo um zunido de pré-acasalamento o qual direciona um macho a ajudá-la a entrar na galeria. (C) um macho produzindo um cricrido na galeria com sua extremidade posterior permanecendo fora do buraco após a introdução da fêmea.
Em besouros do gênero Aphodius (Coleoptera: Scarabaeidae), são os
machos que cantam durante as interações macho-fêmea e a cópula só ocorre nos
intervalos entre um canto e outro (Kasper & Hirschberger 2005). Segundo
Breidbach (1986), o mecanismo de estridulação de Hylotrupes bajulus (Linnaeus,
1758) (Coleoptera: Cerambycidae) também está envolvido no comportamento de
acasalamento, exercendo um papel na estimulação tátil das fêmeas pelos machos
coespecíficos. Breidbach (1986) menciona ainda a participação da estridulação no
comportamento agressivo destes besouros. No tenebrionídio, Eupsophulus
castaneus, além da estridulação que pode estar envolvida no comportamento de
defesa, os machos desta espécie ainda produzem um som típico, ao baterem o
abdômen contra o substrato, e que possivelmente esteja envolvido de alguma
forma no comportamento de acasalamento (Slobodchikoff & Spangler 1979).
A produção de som em coleópteras também pode não estar associada a
nenhum destes comportamentos. Por exemplo, machos e fêmeas do cerambicídio
Dectes texanus texanus só estridulam quando são estressados (Crook et al.

28
2004). Isso também ocorre em P. quercivorus, que estridulam também
espontaneamente (Ohya & Kinuura 2001). Já em Odontotaenius disjunctus (Illiger,
1800) (Coleoptera: Passalidae), a estridulação por distúrbio possui capacidade
deterrente, favorecendo o ataque constante do predador, o corvo Corvus
brachyrhynchos Brehm, 1822 (Buchler et al. 1981).
2.5.1.1. Aparato estridulatório e produção de som em Curculionidae
A produção de sons em diversos Curculionidae se dá pelo método de
estridulação élitro-abdominal, descrito por Durmortier, em 1963, o qual consiste
na fricção do plectrum, presente no abdômen, sobre a pars stridens, localizada
nos élitros (Durmortier 1963). Este mecanismo pode ser encontrado em várias
espécies do gênero Conotrachelus (Gibson 1967), Pissodes (Harman & Kranzler
1969), Smicronyx (Hyder & Oseto 1989), Phrydiuchus (Wilson et al. 1993), entre
outros (Durmortier 1963).
Segundo Harman & Harman (1972), tanto o élitro direito quanto o
esquerdo de Pissodes strobi contêm a pars stridens, sendo que esta estrutura
difere, quanto ao número e espaçamento dos sulcos, entre machos e fêmeas. Os
autores também observaram diferenças nas raspadeiras, localizadas no sétimo
tergito abdominal, de machos e fêmeas, quanto ao número de dentes. A
estridulação em P. strobi e P. approximatus, para ambos os sexos, é composta de
chiados diplossilábicos de duas partes com duração de 50 e 80 ms e com
intervalo entre chiados variando de 5 a 25 ms; apenas as taxas de repetição de
chiados dos machos são mais rápidas do que as das fêmeas para ambas as
espécies (Harman & Kranzler 1969).
Gibson (1967) observou que a pars stridens de três espécies do gênero
Conotrachelus (C. naso LeConte, 1876; C. posticatus Boheman, 1837; e C.
carinifer Casey, 1892) está situada na borda do élitro esquerdo, prolongando-se
debaixo do direito; que os sulcos ou stridulitra são maiores nos machos do que
nas fêmeas, e que o comprimento e a largura da pars stridens e o seu número de
dentes são variáveis entre os sexos e entre as espécies. De acordo com suas
análises, Gibson observou que, nas três espécies de Conotrachelus, ambos os
sexos possuem plectra semelhantes, compostos de dois pares de tubérculos e
localizados no sexto tergito abdominal. Em C. naso e C. carinifer, a estridulação

29
dos machos difere daquela das fêmeas quanto ao tom (agudo e grave), mas
também foram observadas diferenças em velocidade e freqüência.
Para Hyder & Oseto (1989), o aparato estridulatório de Smicronyx fulvus
LeConte, 1876 e Smicronyx sordidus LeConte, 1876 é constituído pela pars
stridens localizada sob a superfície apical dos élitros com sulcos individuais,
formando um arco oblíquo em direção à margem sulural, e por dois plectra
localizados no sexto tergito abdominal. A análise do som, segundo o estudo dos
autores, revelou que machos das duas espécies produzem freqüências mais altas
do que as fêmeas coespecíficas, mas as freqüências entre esses machos são
diferentes. Os autores observaram também que o alcance da freqüência é
diferente entre os grupos e a distribuição das densidades de freqüências são
diferentes entre os sexos.
Em Phrydiuchus tau Warner, 1969 o plectrum é formado por estruturas
emparelhadas, dorso-laterais presentes no sétimo tergito abdominal e a pars
stridens é constituída de uma série de fendas descontínuas, transversais,
situadas ao longo da margem da sutura apical de cada élitro (Wilson et al. 1993).
Duas categorias de sons são relatadas para Curculionidae, bem como
para outros artrópodes, segundo Alexander (1967): a estridulação por distúrbio e
a estridulação de corte. A primeira está relacionada ao som produzido por ambos
os sexos em resposta às interações interespecíficas e intraespecíficas,
associadas aos comportamentos de defesa e alarme. Já a estridulação de corte é
utilizada na comunicação intraespecífica, relacionada com os comportamentos de
acasalamento.
Em P. tau foi observada a presença destes dois tipos de som, sendo que
a estridulação de corte consiste em dois pulsos curtos a intervalos de 40 ms
separados por aproximadamente 500 ms e os sons de distúrbio, também de pulso
duplo, são repetidos de forma irregular (Wilson et al. 1993). Em S. fulvus e S.
sordidus, a produção de som também está relacionada ao comportamento de
corte, sendo que o segundo plectrum é usado para produzir sons antes e durante
a cópula (Hyder & Oseto 1989). Em C. nenuphar, Manpe & Neuzig (1966)
observaram que gorgulhos adultos de ambos os sexos podem atrair o sexo
oposto pela estridulação, entretanto, a cópula nesta espécie aconteceu muitas
vezes sem ser precedida pela estridulação. Em outras espécies do gênero, não
foi observada nem registrada a produção de som quando os gorgulhos não eram

30
induzidos a isto (Gibson 1967), o que pode caracterizar neste gênero uma
estridulação por distúrbio. Segundo Harman & Kranzler (1969), diferenças nas
taxas de chiado entre os sexos de Pissode strobi e P. aproximatus podem ser
uma característica sexo-específica utilizada dentro do comportamento de
acasalamento.

31
3. TRABALHOS
DIMORFISMO SEXUAL DE AMERRHINUS YNCA SAHLBERG, 1823 (COLEOPTERA: CURCULIONIDAE)
RESUMO
Amerrhinus ynca é um coleóptero da família Curculionidea, originário do Brasil.
Este inseto é encontrado danificando a ráquis foliar de coqueiros. Este trabalho foi
realizado para estabelecer um método de sexagem de adultos de Amerrhinus
ynca que permita a realização de estudos comportamentais. Adultos de A. ynca
capturados em um plantio comercial de coco foram levados para o laboratório e
com base em características citadas na literatura para sexar outros
Curculionidaes, formaram-se casais que foram observados até a ocorrência de
cópulas. Em seguida, machos e fêmeas (n=60) foram sacrificados e sob lupa
estereoscópica (40 X) observou-se a sua morfologia externa e com um
paquímetro digital mediu-se o comprimento e a largura corporal. Foram
encontradas três características morfológicas distintas entre os sexos de adultos
de A. ynca: (i) os dois primeiros esternitos abdominais nas fêmeas apresentam
um formato convexo enquanto nos machos os mesmos esternitos são
ligeiramente côncavos, (ii) a margem posterior do 5o esternito nas fêmeas

32
apresenta um entalhe mediano, enquanto nos machos é totalmente arredondada
e (iii) o oitavo tergito abdominal nas fêmeas é recoberto pelo sétimo e, nos
machos, é visível. Médias de comprimento e largura do corpo foram maiores nas
fêmeas que nos machos.
ABSTRACT
Sexual dimorphism of the Amerrhinus ynca Sahlberg, 1823 (Coleoptera: Curculionidae)
Amerrhinus ynca is a beetle of the family Curculionidae, native of Brazil. This
insect damages the petiole of coconut trees. The aim of this work is to determine a
sexing method of A. ynca adult to be usede in behavioral studies. Adult insects of
A. ynca captured in a commercial plantation of coconut were taken to the
laboratory, individualized and fed. One hundred and twenty adults of A. ynca were
observed (60 couples) and the sexual identity of the insects was obtained by
observing the characteristics mentioned in the literature for sexing Homalinotus
coriaceus (Coleoptera; Curculionidae), and confirmed by observing the courtship.
The search for external morphological characters was accomplished under
magnifying stereoscopic microscope (40 X). The the distance between the
extremities: from the head to the posterior part of the elytra was used to measure
body length, for the width, the largest distance among the sides of the body. Three
morphological characters can be used to differentiate sexes of adults of A. ynca in
the field and in the laboratory: (i) the configuration of the 1st and of the 2nd
abdominal sternite, (ii) the shape of the posterior margin of the 5th sternite and (iii)
the design and the shape of the last abdominal tergite. The length and width
averages differed statistically for both sexes, being the females larger than the
males.

33
INTRODUÇÃO A broca-do-pecíolo ou broca-da-ráquis-foliar do coqueiro, Amerrhinus
ynca Sahlberg 1823, é um coleóptero Curculionidea, originário do Brasil (Gomes,
1992). As populações deste inseto são encontradas danificando a ráquis foliar de
coqueiros adultos (Ferreira et al. 1998) e também podem ser encontradas
atacando os talos foliares do licurizeiro (Bondar 1940).
O adulto de A. ynca é uma broca de coloração amarela a esbranquiçada
(Moura & Vilela 1998), com manchas pretas espalhadas pelo corpo (Gomes
1992), cujo comprimento varia de 14 mm a 22 mm de comprimento, além de um
rostro que pode variar de 4 mm, nos menores exemplares, até 6 mm, nos maiores
(Ferreira et al. 1998). É um besouro de hábitos diurnos que pode ser observado
alimentando-se das flores masculinas do coqueiro (Moura & Vilela, 1998).
Em algumas fases dos estudos de biologia, comportamento e ecologia,
faz-se necessário a determinação dos sexos de insetos adultos (Duan et al.
1999), como, por exemplo, em estudos de comportamento que envolvam
respostas a sinais químicos (feromônios), nos quais há a necessidade de
determinar a natureza do feromônio e as diferenças sazonais entre machos e
fêmeas (Innocenzi et al. 2002).
Pouco se conhece sobre a biologia de A. ynca (Mirizola-Filho 2002),
portanto, visando desenvolver estudos sobre sua biologia, ecologia e
comportamento, este trabalho pretende descrever diferenças morfológicas que
sejam distintas entre os sexos e que sirvam na diferenciação sexual durante as
atividades de campo e laboratório.
MATERIAIS E MÉTODOS Insetos adultos de A. ynca foram coletados no município de Quissamã,
Estado do Rio de Janeiro, no período de março a setembro de 2004. Os insetos
coletados foram mortos em solução de álcool 70%, em laboratório.
Os estudos foram realizados no Laboratório de Entomologia e
Fitopatologia do Centro de Ciência e Tecnologia Agropecuária da Universidade

34
Estadual do Norte Fluminense, Rio de Janeiro. Adultos de A. ynca coletados no
campo foram acondicionados dentro de gaiolas de acrílico (30 X 30 X 30 cm) com
dieta artificial de cana-de-açúcar. Assim que se formava um casal, possíveis
machos e fêmeas foram separados em recipientes diferentes até a confirmação
dos sexos. Esta, por sua vez, foi feita da seguinte maneira: após separar 60
individuos de cada sexo, os possíveis machos e fêmeas tiveram seus sexos
confirmados ao se observar a presença ou ausência do oitavo tergito abdominal,
conforme característica utilizada para diferenciar outros Curculionidae (Sarro et al.
2004).
A avaliação das características morfológicas externas foi realizada sob
lupa estereoscópica com capacidade de aumento de até 40 vezes. Procuraram-se
características que servissem na diferenciação dos sexos dos indivíduos dessa
espécie, tais como o tamanho do corpo, sua coloração, o comprimento do rostro e
da cabeça, a forma das antenas e das pernas, e variações nos segmentos
torácicos e abdominais (Duan et al. 1999). Para a medida do comprimento,
considerou-se a distância entre a extremidade anterior da cabeça e a posterior
dos élitros, e para a largura, a maior distância entre os lados do corpo (Anjos
1992) (Figura 1). As medidas de comprimento e largura dos insetos foram
coletadas com o auxilio de um paquímetro digital e os dados submetidos à análise
de variância (ANOVA).
Figura 1 – Esquema mostrando como foram obtidas as medidas corporais de Amerrhinus ynca.

35
RESULTADOS Três caracteres morfológicos podem ser utilizados para diferenciar os
sexos de adultos de A. ynca no campo e no laboratório: (i) a conformação do 1o e
do 2o esternitos abdominais, (ii) a forma da margem posterior do 5o esternito e (iii)
a disposição e forma do último tergito abdominal.
Adultos de A. ynca apresentam, na parte ventral, cinco esternitos
abdominais visíveis. Os dois primeiros não apresentam mobilidade e são maiores
que os demais. Entretanto, nas fêmeas deste gorgulho, os dois primeiros
esternitos abdominais apresentam um formato convexo enquanto nos machos os
mesmos esternitos são de ligeiramente côncavos a planos (Figura 2).
Figura 2 - Esquema da vista lateral do abdômen de ambos os sexos de Amerrhinus ynca, mostrando a diferença anatômica existente entre os dois primeiros segmentos abdominais.
Constatou-se que a margem da extremidade posterior do 5o esternito das
fêmeas de A. ynca apresenta um entalhe mediano, enquanto nos machos esta
característica não foi observada, ou seja, neles a extremidade se apresenta de
forma totalmente arredondada (Figura 3).

36
Figura 3 – Esquema da vista ventral do final do abdômen de ambos os sexos de Amerrhinus ynca, mostrando a diferença existente na margem posterior do quinto esternito.
A terceira característica observada se encontra na parte dorsal do
abdômen (Figura 4). Observou-se que em fêmeas adultas de A. ynca o oitavo
tergito abdominal é recoberto pelo sétimo, ao passo que nos machos o oitavo
tergito é visível. Sarro et al. (2004), ao estudarem a morfologia da broca do
pedúnculo floral do coqueiro, Homalinotus coriaceus (Gyllenhal, 1836), obtiveram
os mesmos resultados, e suas observações serviram de base no momento de se
decidir entre os sexos dos adultos de A. ynca. Além disso, o oitavo tergito dos
machos de A. ynca é mais curto do que o sétimo tergito. Observou-se ainda que a
extremidade final do abdômen, formada pelo oitavo tergito nos machos e pelo
pigídio nas fêmeas, também apresenta diferenças, sendo que nas fêmeas esta
extremidade é mais afinalada quando comparada com a dos machos (Figura 4).

37
Figura 4 – Esquema da vista doral do abdômen, mostrando a diferença entre machos e fêmeas de Amerrhinus ynca quanto à disposição dos tergitos abdominais.
As médias do comprimento e largura do corpo de adultos de A. ynca, bem
como as suas amplitudes, encontram-se na tabela 1. As médias de comprimento
e largura diferem estatisticamente para ambos os sexos, sendo as fêmeas
maiores que os machos. Entretanto, as medidas corporais não podem ser
utilizadas para separar os adultos, uma vez que as amplitudes se sobrepõem,
tanto em relação à largura, quanto em relação ao comprimento do corpo.
Tabela 1 – Morfometria corporal de adultos, machos e fêmeas, de Amerrhinus ynca.
Adultos Comprimento do corpo1 (amplitude) (mm)
largura do corpo2 (amplitude) (mm)
Número de exemplares
Macho 17,49 ± 0,20a (13,44 - 20,14) 7,29 ± 0,73a (5,33 - 8,43) 60
Fêmea 18,70 ± 0,12b (16,35 - 20,85) 8,00 ± 0,41b (6,76 - 8,74) 60 1, 2, Médias dos comprimentos e larguras seguidas de mesma letra, na mesma coluna, não
diferem entre si (ANOVA; P = 0,05).

38
DISCUSSÃO A diferença morfológica entre os dois primeiros esternitos abdominais de
machos e fêmeas de A. ynca também foi observada no gênero Conotrachelus
(Coleoptera: Curculionidae) nas espécies C. nenuphar (Herbst, 1797) (Thomson
1932), C. neomexicanus Fall, 1913 (Bodenham et al. 1976) e C. schoofi Papp,
1978 (Tedders & Payne 1986). Já para a tribo Molytini (Coleoptera:
Curculionidae), Craw (1999) também observou diferenças sexuais na margem do
quinto esternito abdominal, que é menor e mais transversal nos machos e maior,
mais comprido e mais arredondado, nas fêmeas. Em fêmeas de Ganglionus
mitigatus Franz, 2001 (Coleoptera: Curculionidae), Franz & O’Brien (2001)
observaram que a margem posterior do oitavo esternito abdominal é que é
distintamente emarginada.
O recobrimento do oitavo tergito abdominal em fêmeas parece ser uma
característica comum entre alguns besouros da família Curculionidae, tendo sido
relatada em C. neomexicanus Fall, (Bodenham et al. 1976), Phrydiuchus tau
Warner, 1969 (Wilson et al. 1993) e Pissodes strobi (Peck, 1817) (Harman &
Harman 1972).
Estudos sobre a morfometria corporal de adultos de Conotrachelus
schoofi, desenvolvidos por Tedders & Payne (1986) mostraram que as fêmeas
são em média ligeiramente maiores que os machos e que as variações no
tamanho do corpo se sobrepõem, tornando-se uma característica distintiva não
confiável. Já para H. coriaceus, Sarro et al. (2004) não encontraram diferenças
corporais entre os adultos (machos e fêmeas) trazidos do campo. Os autores
observaram apenas diferenças na largura do corpo de indivíduos vindos do
campo em relação àqueles criados em laboratório.
Não foram encontradas, para A. ynca, diferenças na coloração nem na
forma das pernas e antenas. Duan et al. (1999), após examinarem centenas de
adultos de Anthonomus pomorum Linneaus, 1758 (Coleoptera: Curculionidae),
também não observaram diferenças discriminatórias entre machos e fêmeas em
relação a cor e morfologia das pernas e antenas. Para Anthonomus rubi (Herbst,
1795), Innocenzi et al. (2002) observaram algumas vezes diferenças entre os
sexos pela posição das antenas e pela curvatura das tíbias, mas concluíram que
a diferença intra-sexual destas características não permite uma discriminação

39
confiável. Já em Mauritinus seferi Bondar, 1960 (Coleoptera: Curculionidae),
Barbosa & Valente (2003) observaram que machos adultos deste gorgulho
apresentam no centro distal da face interna da protíbia uma franja de longas setas
castanho-amareladas, que é ausente nas fêmeas e que serve, portanto, como
característica discriminatória dos sexos. Existem outras diferenças morfológicas
associadas ao aparato responsável pela produção de som em A. ynca que serão
tratadas com maiores detalhes no quarto trabalho desta dissertação.
RESUMOS E CONCLUSÕES Este trabalho foi realizado com o objetivo de estabelecer um método de
sexagem de adultos de Amerrhinus ynca que permita a realização de estudo
sobre sua biologia, ecologia e comportamento. Para isso, adultos de A. ynca
capturados em um plantio comercial de coco foram levados para o laboratório e,
após observação de casais em cópula, possíveis machos e fêmeas foram
separados e a confirmação de sua identidade sexual se deu ao observar a
presença ou ausência do oitavo tergito abdominal. em cópula. Em seguida,
machos e fêmeas (n=60) foram sacrificados e a sua morfologia externa foi
observada sob lupa estereoscópica com aumento de até 40 vezes e com um
paquímetro digital foram medidos o comprimento e a largura corporal. Foram
encontradas três características morfológicas distintas entre os sexos de adultos
de A. ynca: (i) os dois primeiros esternitos abdominais nas fêmeas apresentam
um formato convexo enquanto que nos machos os mesmos esternitos são
ligeiramente côncavos, (ii) a margem posterior do 5o esternito nas fêmeas
apresenta um entalhe mediano, enquanto que nos machos é totalmente
arredondada e (iii) o oitavo tergito abdominal nas fêmeas é recoberto pelo sétimo
e, nos machos, é visível. Médias de comprimento e largura do corpo foram
maiores nas fêmeas que nos machos.
Portanto, os sexos dos adultos de A. ynca podem ser facilmente
diferenciados pela verificação da morfologia dos dois primeiros esternitos
abdominais, forma da margem posterior do quinto esternito, tamanho e forma do
penúltimo tergito abdominal. Dentre estes, os dois primeiros caracteres são os
mais adequados por apresentarem confiabilidade e praticidade, não provocando

40
danos nem alterando o comportamento dos insetos durante a verificação das
características. As medidas morfométricas desses coleópteros, por sua vez, não
servem para a diferenciação dos sexos, uma vez que suas amplitudes se
sobrepõem.
REFERÊNCIAS BIBLIOGRÁFICAS
Anjos, N. (1992) Taxonomia, ciclo de vida e dinâmica populacional de Costalimaita ferruginea (Fabr., 1801) (Coleoptera: Chrysomelidae), praga de Eucalyptus spp. (Myrtaceae). Tese (Doutorado em Entomologia). Piracicaba.-SP, Escola superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 165 p.
Barbosa, M. L. L. & Valente, R. M. (2003) Mauritinus seferi Bondar, 1960: bionomy, description of immature stages and redescription of adult (Coleoptera: Curculionidae). Revista Brasileira de Entomologia, 47 (1):81-88.
Bodenham, J.; Stevens, R. E. & Thatcher, T. O. (1976) A cone weevil, Conotrachelus neomexicanus, on Ponderosa Pine in Colorado: life history, habits, and ecological relationships (Coleoptera: Curculionidae). The Canadian Entomologist, 108:693 – 699.
Bondar, G. (1940) Insetos Nocivos e Moléstias do Coqueiro (Cocos nucifera) no Brasil. Salvador:Tipografia Naval, 156 p.
Craw, R. C. (1999) Molytini (Insecta: Coleoptera: Curculionidae: Molytinae). Fauna of New Zealand 39:68p. Disponível em: < http://www.landcareresearch.co.nz/research/biodiversity/invertebratesprog/faunaofnz/extracts/FNZ39/fnz39mor.asp>, acesso em: março de 2005.
Duan, J. J.; Weber, D. C.; Hirs, B. A. & Dorn, S. (1999) A new character for sex differentiation of adults of Anthonomus pomorum L. (Col., Curculionidae). Journal Applied Entomology. 123:319-320.
Ferreira, J. M. S.; Lima, M. F.; Santana, D. L. Q. & Moura, J. I. L. (1998) Pragas do coqueiro. In: Sobrinho, R. B.; Cardoso, J. E. & Freire, F. C. (eds.). Pragas de Fruteiras Tropicais de Importância Agroindustrial. Brasília: Embrapa-SPI, p. 81-118.
Franz, N. M. & O’Brien, C. W. (2001) Ganglionus, a new genus of Derelomini (Coleoptera: Curculionidae) associated with Carludovica (Cyclanthaceae). Annals of the Entomological Society of America, 94 (6):835-850.
Gomes, R. P. (1992) O Coqueiro da Bahia. 6. ed. São Paulo: Nobel, 111 p.

41
Harman, D. M. & Harman, A. L. (1972) Stridulatory mechanisms in the White Pine Weevil, Pissodes strobi. Annals of the Entomological Society of America, 65:1076 – 1079.
Innocenzi, P. J.; Hall, D. R.; Cross, J. V. & Green, S. V. (2002) Sexing adults of the strawberry blossom weevil, Anthonomus rubi (Col., Curculionidae). Journal Applied Entomology,126:159-160.
Mirizola-Filho, L. A. (2002) Cultivo de coco anão. 1. ed. Viçosa: Aprenda Fácil, 322p.
Moura, J. I. L. & Vilela, E. F. (1998) Pragas do Coqueiro e Dendezeiro. 2. ed. Viçosa: Aprenda Fácil, 124 p.
Sarro, F. B.; Crocomo, W. B. & Ferreira, J. M. S. (2004) Aspectos da biologia e morfologia da broca do pedúnculo floral do coqueiro, Homalinotus coriaceus (Gyllenhal) (Coleoptera: Curculionidae). Neotropical Entomology, 33 (1):7-12.
Tedders, W. L. & Payne, J. A. (1986) Biology, life history, and control of Conotrachelus schoofi (Coleoptera: Curculionidae) on Pecans. Journal of Economic Entomology, 79:490-496.
Thomson, J. R. (1932) Sex differentiation of adults of Conotrachelus nenuphar. Journal of Economic Entomology, 25:807-810.
Wilson, L. M.; Henry, C. S.; Johnson, J. B. & McCaffrey, J. P. (1993) Sound production in Phrydiuchus tau (Coleoptera: Curculionidae). Annals of the Entomological Society of America, 86:621 – 630.

42
COMPORTAMENTO DE CORTE E ACASALAMENTO DE AMERRHINUS YNCA SAHLBERG, 1823 (COLEOPTERA: CURCULIONIDAE)
RESUMO
A chave para se entender a biologia de uma espécie é conhecer o seu
comportamento reprodutivo. Assim é possível identificar e manipular pontos
vulneráveis dentro deste processo os quais permitirão que medidas de controle e
monitoramento sejam desenvolvidas com o propósito de reduzir a ação de
organismos competidores, como os insetos-praga. Amerrhinus ynca é um
curculionídio de importância econômica para algumas regiões do Brasil, e pouco
se conhece sobre sua biologia, por isso o objetivo deste trabalho foi investigar e
descrever seu comportamento de corte e acasalamento. Casais de A. ynca foram
observados de forma direta e através de filmagem. Registrou-se a freqüência dos
atos comportamentais envolvidos no comportamento de acasalamento e a
duração da cópula, corte e pós-cópula. Acasalamentos ocorreram tanto em
fotofase quanto em escotofase. Os machos iniciaram a monta 119 ± 75 min após
a formação dos grupos, permanecendo em corte durante 3 ± 2,5 min. A cópula
durou 51 ± 49 min. Os atos mais freqüentes dos machos durante a pré-cópula
foram a estridulação e a estimulação do corpo da fêmea com as pernas
anteriores. Durante a cópula o ato mais observado foi a estimulação do protórax
da fêmea com o primeiro par de pernas e a raspagem com o rostro no pronoto
das fêmeas. As fêmeas de A. ynca acasalaram múltiplas vezes e em geral com
vários machos. Após a primeira cópula os machos apresentaram comportamento

43
de guarda da fêmea, permanecendo sobre seu dorso por períodos que variaram
de 40 min a 33 horas. A. ynca é uma espécie poliândrica que apresenta uma
seqüência comportamental de pré-cópula e cópula similar à de outras espécies da
família Curculionidae. Os machos permanecem sobre o dorso da fêmea evitando
que elas sejam copuladas por outros machos.
ABSTRACT
Courtship and mating behavior of the Amerrhinus ynca Sahlberg, 1823 (Coleoptera: Curculionidae)
Understanding the biology of species and knowing their sexual behavior
one can identify and manipulate vulnerable points that make possible controling
and monitoring in order to reduce the action of competitor organisms, as the
insect-pests. The objective of this work was to investigate and describe the
behavioral acts involved in the precopulation, copulation and poscopulation as well
as the system mating of this insect. Couples of A. ynca were analyzed through
direct observation and images were also obtained by filming. In both cases
frequency and duration of acts involved in mating behavior were registered. Mating
happened under photophase as well as under scotophase. The males began
mounting the females 119 ± 75 min after the formation of the couples and they
stayed courting them during 3 ± 2,5 min. The copulation lasted 51 ± 49 min. Most
frequent behavior acts among the males during the precopulation were stridulation
and stimulation of the female's body with the front legs. During copulation the most
observed behavioral act was stimulation of the protorax of the female with the first
pair of legs and the scratching of the pronotum of the female with the rostrum. The
females of A. ynca mated multiple times and in general with several males. After
the first copulation the males presented guarding behavior of the female, staying
on its back for periods that varied from 40 min to 33 h probably to avoid the sperm
competition. A. ynca is a polyandrous species that presented a behavioral
sequence of precopulation and copulation similar to those of other curculionidae
species.

44
INTRODUÇÃO A chave para se entender a biologia de uma espécie é conhecer o seu
comportamento reprodutivo. Assim é possível identificar e manipular pontos
vulneráveis dentro deste processo os quais permitirão que medidas de controle e
monitoramento sejam desenvolvidas com o propósito de reduzir a ação de
organismos competidores (Lloyd 1979), como os insetos-praga. Por exemplo, os
estudos sobre o comportamento de acasalamento são fundamentais para o
desenvolvimento de técnicas de atração e esterilização utilizadas para reduzir
níveis de infestação de insetos-praga (Wojcik 1969).
Associados ao comportamento de acasalamento, principalmente durante
o processo de corte, estão a emissão e a recepção de sucessivos estímulos que
irão gerar no receptor um padrão elaborado e apropriado de respostas (Viana &
Vilela 1996). Todo este processo é moldado e controlado pela relação de custos e
benefícios intimamente ligada à competição intra e intersexual (Daly 1978) e que
varia de acordo com o sistema de acasalamento (Suzuki et al. 2005). Por
exemplo, fêmeas de muitas espécies de animais geralmente acasalam com vários
machos (Olsson et al. 1994). A poliandria provavelmente evoluiu porque os
benefícios de múltiplos acasalamentos em fêmeas poliândricas excedem os
custos ou porque os machos manipulam a interação no sentido de seu sucesso
reprodutivo (Campbell 2005).
As interações entre machos e fêmeas geralmente não terminam com a
cópula (Wiley 1997). Neste ponto, tem-se o conflito sexual que ocorre quando
características que aumentam o sucesso reprodutivo de um dos sexos levam à
redução no sucesso do outro sexo (Stockley 1997). Assim, fêmeas podem se
beneficiar de diversas maneiras pela discriminação entre machos potenciais,
antes, durante e depois do acasalamento (Wedell 1996). Os machos devem
"persuadir" as fêmeas para que elas usem o seu esperma melhor do que o de
qualquer outro macho (Wiley 1997).
Estudos sobre a biologia e o comportamento reprodutivo em
Curculionidae já foram realizados em diversas espécies, como Anthonomus
grandis Boheman, 1843 (Bartlett et al. 1968), Conotrachelus nenuphar (Herbst,
1797) (Johnson & Hays 1969), Sitophilus zeamais Motschulsky, 1855
(Walgenbach & Burkholder 1987), Cosmopolites sordidus (Germar,1824) (Viana &

45
Vilela 1996), Gonipterus scutellatus Gyllenhal, 1833 (Carbone & Rivera 1998),
Pissodes strobi (Peck) (Lewis et al. 2002), Sitophilus oryzae Lineaus (Campbell
2005) e Homalinotus coriaceus (Gyllenhal, 1836) (Duarte et al. 2002). Em
Amerrhinus ynca é a primeira vez que se estuda o comportamento de corte e
acasalamento, por isso o objetivo deste trabalho foi investigar e descrever os atos
comportamentais envolvidos na pré-cópula, cópula e pós-cópula deste inseto.
MATERIAIS E MÉTODOS Adultos de Amerrhinus ynca coletados num plantio comercial de coco
localizado no município de Quissamã, RJ, foram transportados para o Laboratório
de Entomologia e Fitopatologia do Centro de Ciências e Tecnologias
Agropecuárias da Universidade Estadual do Norte Fluminense. No laboratório, os
insetos foram separados por sexo e mantidos até a realização dos experimentos
em caixas de acrílico (30 x 30 cm) com alimento (cana-de-açúcar). Os insetos
foram mantidos a 25 ± 2 °C, fotoperíodo de (12:12h) luz-escuro, com escotofase
iniciada às 20 horas. O comportamento de acasalamento foi analisado através de
observações diretas e por imagens obtidas através de filmagem.
As imagens utilizadas na observação do comportamento de acasalamento
foram obtidas com a filmagem focal contínua dos insetos, utilizando-se micro-
câmeras monocromáticas com luz infravermelha, para filmar durante escotofase.
Estas micro-câmeras foram acopladas a um computador para digitalizar as
imagens e seu funcionamento foi gerenciado através do programa de
monitoramento de imagens “Geovision GV 600”. Foram observados cinco casais
mantidos juntos dentro de uma caixa, cujas paredes inferiores foram constituídas
de vidro (30 X 20 X 10 cm), e as paredes superiores de isopor (25 cm de altura
com uma inclinação de 45º) com uma tampa de vidro (30 X 20 cm) (Figura 1). Os
insetos foram identificados com tinta à base de água no ventre e no dorso e
filmados continuamente durante 36 horas. As imagens foram analisadas
utilizando-se o programa Windows Media Player© v. 10.0 em um computador
Windows XP©. Durante as análises foram observados os atos envolvidos no
comportamento de acasalamento e algumas variavés associadas às interações
entre os indivíduos (Tabela 1).

46
Figura 1 – Esquema da arena utilizada para filmar os casais de Amerrhinus ynca.
A observação direta do comportamento de corte e acasalamento de A.
ynca foi realizada com 30 casais, mantidos em caixas Gerbox® (10 x 10 x 5 cm)
durante 5 horas. Durante este período foram registrados os atos comportamentais
associados ao processo de acasalamento. Os insetos foram observados em seis
módulos com cinco casais por observação, durante três dias. As observações
foram feitas em sala climatizada com temperatura de 25º C, alternando manhã e
tarde. Os atos comportamentais (Tabela 1) foram anotados em planilhas
confeccionadas para este fim, nas quais foi registrada a frequência de cada ato
comportamental, além do início e fim da corte, cópula e pós-cópula.
Foi elaborado um etograma dos atos comportamentais envolvidos no
processo de acasalamento, e apresentada a freqüência de cada ato em 30 casais
observados. Optou-se pela observação direta porque através das imagens foi
impossível determinar o momento exato do início e término da cópula, pois,
dependendo da posição da fêmea na arena de filmagem, não era possível
quantificar alguns atos comportamentais.

47
Tabela 1 – Descrição dos atos comportamentais e variáveis envolvidas no comportamento de acasalamento de Amerrhinus ynca.
Ato comportamental Descrição
Monta normal O macho sobe na fêmea tocando a cabeça e o pronoto com as antenas.
Monta em giro O macho sobe na fêmea tocando a parte final do abdômen com as antenas.
Antenação Movimento intenso das antenas.
Estridulação Produção de som ao friccionar os élitros contra o abdômen.
Estimular o protórax da fêmea com as pernas anteriores
O macho esfrega, utilizando um espinho localizado na tíbia das pernas anteriores, as laterais do protórax da fêmea.
Estimular o abdômen da fêmea com as pernas medianas e posteriores.
O macho esfrega as laterais do abdômen da fêmea utilizando as pernas medianas e posteriores para estimulá-la.
Tentativas de cópula O macho expõe a genitália para tentar penetrar a fêmea.
Raspar com o rostro. O macho raspa o rostro sobre o pronoto da fêmea em movimentos de vai-e-vem.
Variáveis Descrição
Tempo de disponibilidade da fêmea
Tempo desde o momento em que um macho desce do dorso da fêmea até o momento em que outro ou o mesmo macho inicia uma nova monta.
Monta de pós-cópula Tempo que o macho permanece sobre o dorso da fêmea, após a primeira cópula, até a sua descida.
Latência Tempo desde o ínicio das observações até o início da monta do macho sobre o dorso da fêmea.
Duração da corte Tempo desde o início da monta até o início da cópula.
Duração da cópula Tempo desde o início da penteração até a retirada do edeago do macho.
Duração da pós-cópula Tempo desde o fim da cópula até o momento em que o macho desce da fêmea ou inicia uma nova seqüência comportamental de corte.
RESULTADOS
Os acasalamentos de Amerrhinus ynca ocorreram tanto em fotofase
quanto em escotofase. Dos 30 casais, 90 % iniciaram o acasalamento e foram
bem sucedidos. Os machos iniciaram a monta 119,19 ± 75,91 min após o inicio

48
das observações e permaneceram 2,67 ± 2,59 min cortejando as fêmeas até
iniciarem a cópula, que teve uma duração de 50,98 ± 49,45 min (Tabela 2).
Tabela 2 – Média e amplitude dos tempos de início da monta, duração da corte e da cópula em Amerrhinus ynca
Média ± desvp (min) Amplitude (min)
Latência 119,19 ± 75,91 10,00 – 288,00
Duração da corte 2,67 ± 2,59 0,50 – 11,00
Duração da cópula 50,98 ± 49,45 4,00 – 163,00 Após a cópula, 81% dos machos permaneceram em cima das fêmeas,
8% abandonaram a fêmea e não acasalaram novamente e 11% se afastaram da
fêmea, mas voltaram a acasalar novamente. Daqueles que permaneceram em
cima da fêmea após a primeira cópula, 36 % chegaram a copular mais uma vez,
36% copularam mais duas vezes, 5% copularam mais quatro vezes e 23% não
copularam novamente, apesar de terem sido observados novos cortejos (Figura
2).
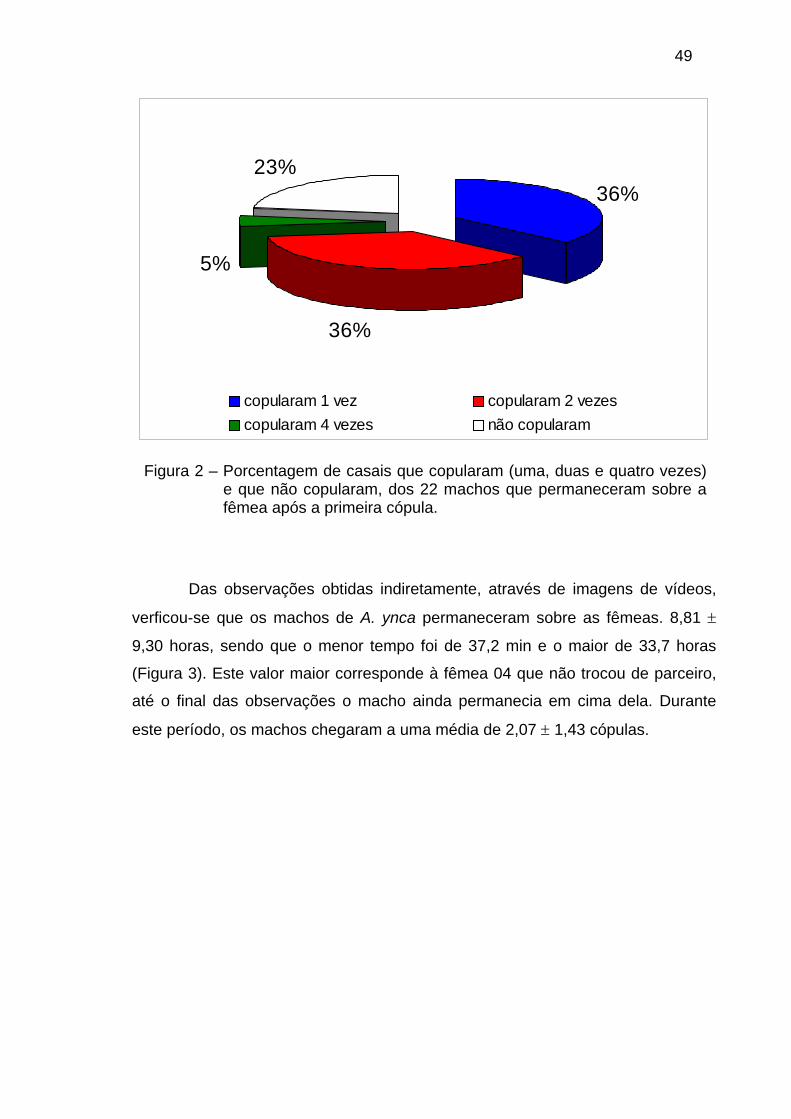
49
36%
36%
5%
23%
copularam 1 vez copularam 2 vezescopularam 4 vezes não copularam
Figura 2 – Porcentagem de casais que copularam (uma, duas e quatro vezes) e que não copularam, dos 22 machos que permaneceram sobre a fêmea após a primeira cópula.
Das observações obtidas indiretamente, através de imagens de vídeos,
verficou-se que os machos de A. ynca permaneceram sobre as fêmeas. 8,81 ±
9,30 horas, sendo que o menor tempo foi de 37,2 min e o maior de 33,7 horas
(Figura 3). Este valor maior corresponde à fêmea 04 que não trocou de parceiro,
até o final das observações o macho ainda permanecia em cima dela. Durante
este período, os machos chegaram a uma média de 2,07 ± 1,43 cópulas.

50
05
10152025303540
primeiro segundo terceiro quarto quintoAcasalamentos
Tem
po d
e pe
rman
ênci
a (h
oras
)
fêmea 01 fêmea 02 fêmea 03 fêmea 04 fêmea 05
Figura 3 – Tempo de permanência dos machos de Amerrhinus ynca sobre o dorso das fêmeas após a primeira cópula.
Quatro das cinco fêmeas de A. ynca trocaram de parceiro mais de uma
vez ou voltaram a ser montadas pelo mesmo macho mais de uma vez (Tabela 3).
O tempo médio que a fêmea ficou disponível a uma nova monta variou entre 0,27
e 14,77 horas (Tabela 4).
Tabela 3 – Quantidade de vezes que as fêmeas de Amerrhinus ynca trocaram de parceiros e quantas vezes ela voltou a ser montada pelo mesmo macho. A letra X representa os parceiros e a quantidade de monta
MACHO FÊMEA 1 2 3 4 5
TOTAL DE MONTAS
1 X X X 3 2 X X X X 4 3 X X 2 4 X 1 5 X X X X X 5

51
Tabela 4 – Tempo de disponibilidade das fêmeas de Amerrhinus ynca para um novo acasalamento
Tempo de predisposição (horas)
Fêmea 1a a 2a monta 2a a 3a monta 3a a 4a monta 4a a 5a monta
1 1,83 0,85 - - 2 0,07 1,89 - - 3 14,77 - - - 4 - - - - 5 0,27 0,06 1,25 1,65 A seqüência dos atos comportamentais envolvidos na pré-cópula, cópula,
e pós-cópula está representada no etograma da figura 4. Assim que o macho
encontra a fêmea, ele a toca com as antenas e com os tarsos das pernas
anteriores. Este primeiro contato pode se dar por qualquer lado do corpo da
fêmea, mas a monta ocorre em maior frequência pelas laterais. Há duas
possibilidades de monta, uma em que o macho sobe na fêmea tocando sua
cabeça com o rostro e as antenas, e outra em giro, na qual o macho monta no
sentido contrário a cabeça da fêmea tocando primeiro o final do abdômen. Logo
após e independentemente do tipo de abordagem, o macho dá início à corte,
durante o qual ele desenvolve vários atos comportamentais não sucessivos como
a produção de som ao friccionar os élitros contra o abdômen; estimular as laterais
do protórax da fêmea com as pernas anteriores e as laterais do abdômen com as
pernas medianas e posteriores, usando um espinho localizado na extremidade
final da tíbia; expor sua genitália e fazer de uma a várias tentativas de cópula.
Algumas fêmeas são receptivas e aceitam a monta e a corte passivamente, mas
outras se movem erguendo o corpo enquanto caminham. Quando isso ocorre, os
machos seguram as fêmeas com suas pernas medianas e posteriores, ao mesmo
tempo em que estridulam e esfregam as laterais do protórax da fêmea com as
pernas anteriores tentando penetrá-las com seu edeago.
Durante a cópula os machos seguram as fêmeas com os três pares de
pernas, flexionando seu abdômen de forma rítmica. Durante este período outros
atos comportamentais podem se exibidos pelos machos, como raspar o rostro no
pronoto da fêmea e estimular as laterais do protórax e do abdômen da fêmea. Os

52
dois últimos atos comportamentais ocorrem frequentemente após a introdução da
genitália do macho, mas podem ocorrer em espaços de tempo maiores ou
associados com o ato de raspar o rostro sobre o pronoto da fêmea. Este ato, por
sua vez, pode ocorrer de uma a três vezes, em uma seqüência de três a cinco
raspadas por vez, durante o período de copulação.
Assim que o macho finaliza a cópula ele pode novamente estridular e
esfregar as pernas anteriores nas laterais do protórax da fêmea, possivelmente
sinalizando o final da cópula. Alguns machos descem em seguida da fêmea e
outros permanecem sobre seu dorso por um longo período.
Figura 4 – Etograma dos atos comportamentais envolvidos na monta, pré-
cópula, cópula e no final da cópula dos machos de Amerrhinus ynca. Os valores indicados na figura representam a freqüência com que os atos ocorreram nos 30 casais.
Os atos comportamentais que os machos mais desempenharam durante
o processo de cortejamento foram a estridulação, o ato de estimular a fêmea com
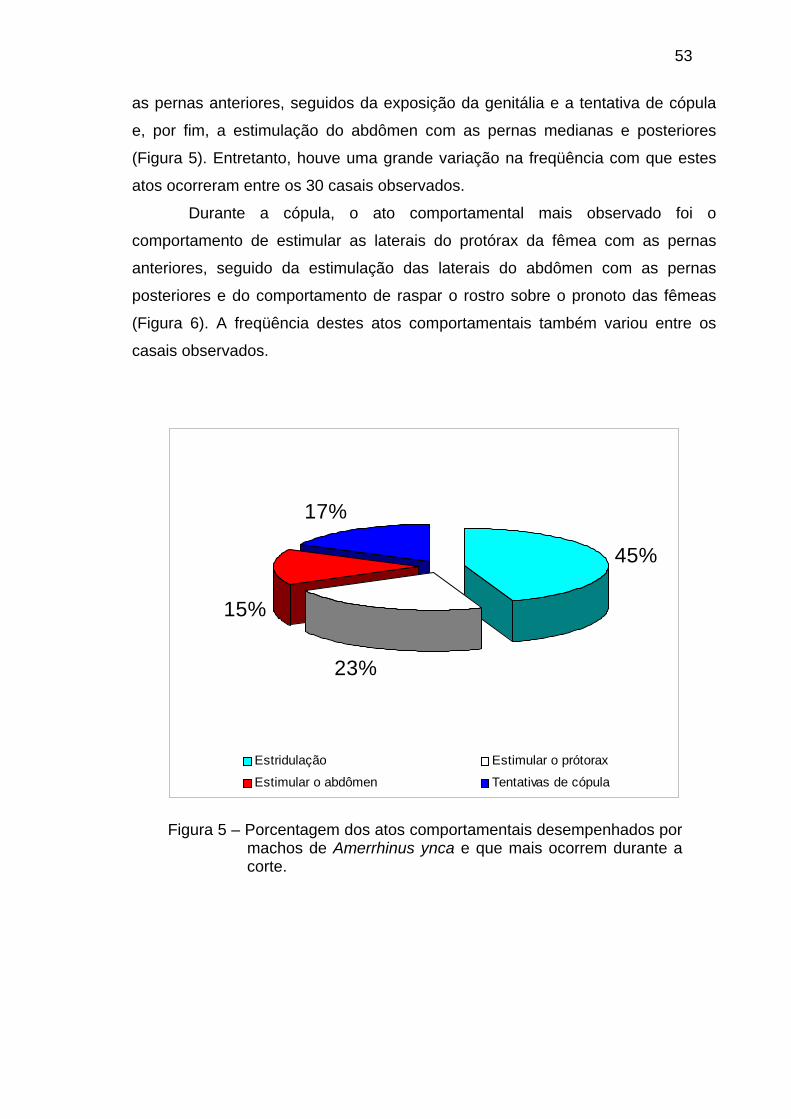
53
as pernas anteriores, seguidos da exposição da genitália e a tentativa de cópula
e, por fim, a estimulação do abdômen com as pernas medianas e posteriores
(Figura 5). Entretanto, houve uma grande variação na freqüência com que estes
atos ocorreram entre os 30 casais observados.
Durante a cópula, o ato comportamental mais observado foi o
comportamento de estimular as laterais do protórax da fêmea com as pernas
anteriores, seguido da estimulação das laterais do abdômen com as pernas
posteriores e do comportamento de raspar o rostro sobre o pronoto das fêmeas
(Figura 6). A freqüência destes atos comportamentais também variou entre os
casais observados.
45%
23%
15%
17%
Estridulação Estimular o prótorax
Estimular o abdômen Tentativas de cópula
Figura 5 – Porcentagem dos atos comportamentais desempenhados por machos de Amerrhinus ynca e que mais ocorrem durante a corte.

54
3%
71%
26%
Raspar com o rostro Estimular o protórax Estimular o abdômen
Figura 6 – Porcentagem dos atos comportamentais desempenhados por machos de Amerrhinus ynca e durante a cópula.
DISCUSSÃO Amerrhinus ynca apresentou uma grande amplitude quanto à duração da
cópula. Essa variação corrobora o observado em outros Curculionidaes como
Sitophilus zeamais (Walgenbach & Burkholder 1987); Sitophilus granarius
Linneaus, 1758 (Wojcik 1969); Homalinotus coriaceus (Gyllenhal, 1836) (Duarte et
al. 2002); Cosmopolites sordidus (Viana & Vilela 1996) e Anthonomus grandis
(Bartlett et al. 1968). Esta grande variação na duração da cópula em A. ynca
possivelmente está relacionada com a heterogeneidade dos indivíduos, uma vez
que estes foram coletados no campo sem se conhecer as idades nem seu estado
fisiológico e de maturação.
Os comportamentos de pré-cópula exibidos por A. ynca são similares aos
observados em outros gêneros da família Curculionidae. Em S. granarius (Wojcik
1969) e S. zeamais (Walgenbach & Burkholder 1987) os machos abordam as
fêmeas a partir de qualquer posição e, em contato, eles a montam imediatamente.
Carbone & Rivera (1998) observaram que, durante a tentativa de cópula, os
machos de Gonipterus scutellatus (Coleoptera: Curculionidae) estimulam as

55
laterais do tórax da fêmea com as pernas anteriores expondo sua genitália.
Entretanto, Cortero et al. (2004) observaram que machos de Trichobaris
championi Barber (Coleoptera: Curculionidae), após o contato frontal ou lateral,
mudam rapidamente de posição para ter contato por trás e então sobem na fêmea
segurando-a com as pernas anteriores e posteriores ao mesmo tempo em que
esfregam com as pernas medianas as laterais do abdômen da fêmea.
Durante a cópula o posicionamento dos machos de S. granarius é similar
àquele de machos de A. ynca e eles também raspam o rostro sobre o pronoto das
fêmeas (Wojcik 1969). Em S. zeamais o comportamento dos machos é similar aos
descritos anteriormente (Walgenbach & Burkholder 1987). Entretanto, no
curculionídeo, Rhynchophorus cruentatus (Fabricius, 1775) (Coleoptera:
Curculionidae), o fato de raspar a fêmea utilizando a ponta distal do pronoto foi
observado em machos desta espécie como um ato que antecede a monta, e ao
invés de raparem o pronoto eles rapam o élitro da fêmea (Vanderbilt et al. 1998).
Os autores argumentam que este comportamento pode ser utilizado para
apaziguar a fêmea ou para discriminar entre os hidrocarbonetos cuticulares
heteroespecíficos liberados pela cutícula da fêmea.
Os movimentos rítmicos do abdômen dos machos de A. ynca durante a
cópula também foram observados por Walgenbach & Burkholder (1987) em S.
zeamais. Segundo os autores este movimento pode estar associado à
transferência de espermatóforos para a espermateca da fêmea.
Os machos de A. ynca, logo após iniciarem a cópula e durante a cópula,
executam atos comportamentais similares aos observados durante a corte
(estimulação do protórax e do abdômen utilizando os três pares de pernas).
Segundo Rodriguez & Eberhad (1994) este tipo de comportamento, conhecido
como corte copulatória, é comum e provavelmente está associado ao sucesso
reprodutivo dos machos, uma vez que eles, através destes atos, estimulam as
fêmeas a usarem seu esperma de preferência àquele de outros machos para a
fertilização dos ovos. Em Tribolium castaneum (Herbst, 1797) (Coleoptera:
Tenebrionidae) machos com mais atividade de corte copulatória conseguem
maior sucesso de paternidade enquanto acasalam com fêmeas já acasaladas
(Edvardsson & Arnqvist 2005).
As fêmeas de A. ynca são altamente poliândricas, e podem copular com
outro parceiro imediatamente após uma cópula. A ocorrência de acasalamentos

56
múltiplos e de poliandria também foi observada em outros Curculionidaes. Em S.
zeamais (Walgenbach & Burkholder 1987) e Conotrachelus nenuphar
(Coleoptera: Curculionidae) (Johnson & Hays 1969), as fêmeas foram observadas
acasalando várias vezes com o mesmo macho e com machos diferentes.
Campbell (2005) verificou que, apesar de múltiplos acasalamentos serem
freqüentemente algo custoso para várias fêmeas, em S. oryzae elas lucram com
os benefícios de múltiplos acasalamentos, por aumentarem o tempo de
oviposição. Duarte et al. (2002) também observaram múltiplos acasalamentos em
fêmeas de H. coriaceus. Segundo Arnqvist & Nilsson (2000), a ocorrência de
múltiplos acasalamentos afeta o sucesso reprodutivo das fêmeas, seja pelo ato
em si (estimulando a produção de ovos), pela presença de esperma (estimulando
a produção de ovos e aumentando a fertilidade) ou por substâncias acessórias
que são transferidas juntamente com a ejaculação tornando-as férteis. Entretanto,
múltiplos acasalamentos também podem atuar negativamente no sucesso
reprodutivo e a manutenção evolutiva deste sistema está associada ao equilíbrio
entre os custos e os benefícios de múltiplos acasalamentos para as fêmeas
(Arnqvist 1989). Segundo Birkhead (1998), quando uma fêmea é inseminada por
mais de um macho, o resultado de paternidade pode ser conseqüência da
combinação de qualquer um dos seguintes processos: competição por esperma,
aborto diferencial ou escolha críptica de fêmeas.
O prolongamento do acasalamento, através da permanência do macho
sobre o dorso da fêmea pode ser conseqüência de uma disputa inter ou intra-
sexual ligada à seleção sexual. Carbone & Rivera (1998) observaram que os
machos de G. scutellatus permanecem sobre a fêmea por um tempo muito longo,
que pode variar de 42 minutos a 55 horas. Este comportamento também foi
observado em H. coriaceus (Duarte et al. 2002), C. sordidus (Viana e Vilela 1996),
R. cruentatus (Vanderbilt et al. 1998), Ectopis ferrugalis Broun, 1881 (Coleoptera:
Curculionidae) (Hutcheson 1991), S. granarius (Wojcik 1969) e Diaprepes
abbreviatus (Linneaus, 1758) (Coleoptera: Curculionidae) (Harari et al. 2003).
Este comportamento pós-copulatório poderia ser justificado pela tentativa
do macho de prevenir novas cópulas da fêmea com machos competidores antes
de ovipositar, prevenindo ou reduzindo o risco de competição por esperma, ou
mesmo permitiria ao macho preencher a espermateca com esperma (Carbone &
Rivera 1998). Segundo Viana & Vilela (1996), os machos de C. sordidus guardam

57
as fêmeas após a cópula em resposta à presença de competidores, uma vez que
a freqüência com que os machos permanecem sobre as fêmeas, quando na
presença de outros machos, era maior do que quando os machos estavam
sozinhos. Em A. ynca, não se fez esta comparação, mas foi possível observar que
tanto machos isolados quanto na presença de outros machos exibem este tipo de
comportamento. Entretanto, deve-se fazer novos testes para procurar
compreender este tipo de comportamento com observações de campo e
laboratório. Esse comportamento pós-cópula, exibido pelos machos, segundo
Arnqvist (1989) pode ser prejudicial à fêmea, aumentando seu gasto com tempo e
energia e reduzindo, por exemplo, sua eficiência de forrageamento. Para Arnqvist
(1988), a manutenção desse tipo de comportamento ao longo da evolução
provavelmente esteve associada a um benefício contrabalanceado, uma vez que
os machos, enquanto guardam as fêmeas, gastam tempo e energia que poderiam
ser usados para encontrar e acasalar com outras fêmeas. Em D. abbreviatus,
Harari et al. (2003) verificaram que os machos enquanto guardam as fêmeas,
prolongando o acasalamento, incorrem em custos de oportunidade, acasalando
com um número reduzido de fêmeas, mas eles fertilizam aproximadamente 75%
dos ovos e reduzem suas chances de um novo acasalamento.
A produção de som através da estridulação também foi observada
fazendo parte do comportamento de corte de alguns Curculionidae. Em
Phrydiuchus tau Warner, 1969 (Coleoptera: Curculionidae), Wilson et al. (1993)
observaram que os machos estridulam ao se aproximarem das fêmeas e, durante
a pré-cópula, voltam a estridular enquanto acariciam a cabeça da fêmea com os
tarsos das pernas anteriores. Os autores não observaram estridulação de corte
nas fêmeas. Em A. ynca, durante as observações do comportamento de
acasalamento, também não seu ouviu qualquer emissão de som por parte das
fêmeas, entretanto, foram observados movimentos do abdômen da fêmea (similar
ao produzido por machos ao friccionarem o élitro contra o abdômen) antes da
monta e durante o cortejamento do macho. Provavelmente estes movimentos de
abdômen estejam associados à produção de um som não audível. Segundo
Hyder & Oseto (1989), em Smicronyx fulvus LeConte, 1876 e Smicronyx sordidus
LeConte, 1876 (ambos Coleoptera: Curculionidae) a estridulação de corte
produzida por machos pode estar associada à atração da fêmea e a estridulação
de corte da fêmea provoca uma resposta para o acasalamento. Em Euscepes

58
postfasciatus (Fairmaire, 1849) (Coleoptera: Curculionidae) a produção de som
pelos machos parece ser um passo da seqüência comportamental que levará à
cópula, e as fêmeas respondem a este sinal permanecendo estacionárias
(Yasuda & Tokuzato 1999).
RESUMOS E CONCLUSÕES A chave para se entender a biologia de uma espécie é conhecer o seu
comportamento reprodutivo. Assim é possível identificar e manipular pontos
vulneráveis dentro deste processo os quais permitirão que medidas de controle e
monitoramento sejam desenvolvidas com o propósito de reduzir a ação de
organismos competidores, como os insetos-praga.
Amerrhinus ynca é um curculionídio de importância econômica para
algumas regiões do país, e pouco se conhece de sua biologia, por isso o objetivo
deste trabalho foi investigar e descrever seu comportamento de corte e
acasalamento. Para isso, casais de A. ynca foram observados de forma direta,
quantificando-se a freqüência de atos comportamentais e variáveis temporais
como a duração da cópula, pré-cópula e pós-cópula, também foram observados
através de filmagem, a qual permitiu avaliar as interações entre os indíviduos.
A partir dos resultdos verificou-se que: os acasalamentos ocorreram tanto
em fotofase quanto em escotofase; os machos iniciaram a monta 119 ± 75 min
após a formação dos grupos, permanecendo em corte durante 3 ± 2,5 min; a
cópula durou 51 ± 49 min; os atos mais freqüentes dos machos durante a pré-
cópula foram a estridulação e a estimulação do corpo da fêmea com as pernas
anteriores; durante a cópula o ato mais observado foi a estimulação do protórax
da fêmea com o primeiro par de pernas e a raspagem com o rostro no pronoto
das fêmeas; as fêmeas acasalaram múltiplas vezes e em geral com vários
machos e após a primeira cópula os machos apresentaram comportamento de
guarda da fêmea, permanecendo sobre seu dorso por períodos que variaram de
40 min a 33 h.
Amerrhinus ynca apresentou, portanto, uma seqüência comportamental
de pré-cópula, cópula e pós-cópula similar àquela de outras espécies da família
Curculionidae. As fêmeas acasalam com mais de um macho, e várias vezes com

59
o mesmo macho, caracterizando um sistema poliândrico de acasalamento. Os
machos produzem um som durante a fase de pré-cópula e cópula e permanecem
sobre a fêmea por períodos longos.
REFERÊNCIAS BIBLIOGRÁFICAS
Arnqvist, G. & Nilsson, T. (2000) The evolution of polyandry: multiple mating and female fitness in insects. Animal Behaviour, 60:145 – 164.
Arnqvist, G. (1988) Mate guarding and sperm displacement in the water strider Gerris lateralis Schumm (Heteroptera: Gerridae). Freshwater Biology, 19:269 – 274.
Arnqvist, G. (1989) Multiple mating in water strider: mutual benefits or intersexual conflict? Animal Behaviour, 38:749 – 756.
Birkhead, T. R. (1998) Cryptic female choice: criteria for establishing female sperm choice. Evolution, 52(4):1212 – 1218.
Bartlett, A. C.; Mattix, E. B. & Wilson, N. M. (1968) Multiple matings and use of sperm in the boll weevil, Anthonomus grandis. Annals of the Entomological Society of America, 61:1148 – 1155.
Campbell, J. F. (2005) Fitness consequences of multiple mating on female Sitophilus oryzae L. (Coleoptera: Curculionidae). Behavior, 34:833 – 843.
Carbone, S. S. & Rivera, A. C. (1998) Sperm competition, cryptic female choice and prolonged mating in the Eucalyptus Snout Beetle, Gonipterus scutellatus (Coleoptera: Curculionidae). Etologia, 6:33 – 40.
Cortero, H. G. C.; Tovar, J. C.; Martinez, N. B. & Collado, J. L. (2004) Comportamento de alimentación, cortejo, cópula y oviposición de Trichobaris championi Barber (Coleoptera: Curculionidae). Agrociência, 38:365 – 373.
Daly, M. (1978) The cost of mating. The American Naturalist, 112:771 – 774.
Duarte, A. G.; Benvenuto, I. D.; Amorim, S. C. R.; Lima, I. S. & Ferreira, J. M. S. (2002) Comportamento de acasalamento de Homalinotus coriaceus, Gyll. (Coleoptera: Curculionidae) em laboratório. In.: XVII Congresso Brasileiro de Fruticultura 2002 Belém-PA. Disponível em: <tche.br/sbfruti/modules.php?name=Content&pa=showpage&pid=14> acesso em: abril de 2006.
Edvardsson, M. & Arnqvist, G. (2005) The effects of copulatory courtship on differential allocation in the Red Flour Beetle Tribolium castaneum. Journal of Insect Behavior, 18:313 – 322.

60
Harari, A. R.; Landolt, P. J.; O'Brien, C. W. & Brockmann, H. J. (2003) Prolonged mate guarding and sperm competition in the weevil Diaprepes abbeviatus (L.). Behavioral Ecology, 14(1):89 – 96.
Hutcheson, J. (1991) Notes of the ecology of Ectopsis ferrugalis (Curculionidae). New Zealand Entomologist, 14:41 – 44.
Hyder, D. E. & Oseto, C. Y. (1989) Structure of the stridulatory apparatus and analysis of the sound produced by Smicronyx fulvus and Smicronyx sordius (Coleoptera, Curculionidae, Erirrhininae, Smicronychini). Journal of Morphology, 201:69 – 84.
Johnson, A. W. & Hays, S. B. (1969) Laboratory mating behavior of the Plum Curculio. Journal of Economic Entomology, 62:438 – 440.
Lewis, K. G.; Liewlaksaneeyanawin, C.; Alfaro, R. I.; Ritland, C.; Ritland, K. & El-Kassaby, Y. A. (2002) Sexual reproduction in the pine weevil (Pissodes strobi [Peck] [Coleoptera: Curculionidae]): implications for population genetic diversity. The Journal of Heredity, 93:165 – 169.
Lloyd, J. E. (1979) Mating behavior and natural selection. The Florida Entomologist, 62:17 – 34 .
Olsson, M.; Gullberg, A.; Tegelström, H.; Madsen, T. & Shine, R. (1994) Can female adders multiply? Nature, 369:528.
Rodriguez, R. L. & Eberhard, W. G. (1994) Male courtship before and during copulation in two species of Xyonysius Bugs (Hemiptera: Lygaeidae). Journal of the Kansas Entomological Society, 67:37 – 45.
Stockley, P. (1997) Sexual conflict resulting from adaptations to sperm competition. Trends Ecology Evolution, 12:154 – 159.
Suzuki, S.; Nagano, M. & Trumbo, S. T. (2005) Intrasexul competition and mating behavior in Ptomascopus morio (Coleoptera: Silphidae Nicrophotine). Journal of Insect Behavior, 18:233 – 242.
Vanderbilt, C. F.; Giblin-Davis, R. M. & Weissling, T. J. (1998) Mating behavior and sexual response to aggregation pheromone of Rhynchophorus cruentatus (Coleoptera: curculionidae). The Florida Entomologist, 81:351 – 360.
Viana, A. M. M. & Vilela, E. F. (1996) Comportamento de corte e acasalamento de Cosmopolites sordidus Germar (Coleoptera: Curculionidae). Anais da Sociedade Entomológica do Brasil, 25:347 – 350.
Walgenbach, C. A. & Burkholder, W. E. (1987) Mating behavior of the maize weevil, Sitophilus zeamais (Coleoptera: Curculionidae). Annals of the Entomological Society of America, 80:578 – 583.
Wedell, N. (1996) Mate quality affects reproductive effort in a paternally investing species. The American Naturalist, 148:1075 – 1088.
Wiley, R. H. (1997) Coevolution of the sexes. Science, 275:1075 – 1076.

61
Wilson, L. M.; Henry, C. S.; Johnson, J. B. & Mccaffrey, J. P. (1993) Sound production in Phrydiuchus tau (Coleoptera: Curculionidae). Annals of the Entomological Society of America, 86:621 – 630.
Wojcik, D. P. (1969) Mating behavior of 8 stored-product beetles (Coleoptera: Dermestidae, Tenebrionidae, Cucujidae, and Curculionidae). The Florida Entomologist, 52:171 – 197.
Yasuda, K. & Tokuzato, M. (1999) Sound production during mating and disturbance in the West Indian Sweet Potato weevil, Euscepes postfasciatus (Fairmaire) (Coleoptera: Curculionidae). Applied Entomology and Zoology, 34:443 –447.

62
BIOENSAIOS COM ODORES DE COESPECÍFICOS E DA PLANTA HOSPEDEIRA DE AMERRHINUS YNCA SAHLBERG, 1823 (COLEOPTERA:
CURCULIONIDAE).
RESUMO
A comunicação é de extrema importância para a adaptação dos animais,
destacando-se por permitir a interação entre as espécies. Aos sinais químicos
envolvidos na comunicação entre os seres vivos dá-se o nome de semioquímicos. Este trabalho foi realizado com a finalidade de se verificar a existência de
semioquímicos atraentes, que atuem nas interações intra e interespecíficas de
Amerrhinus ynca. Os bioensaios foram feitos utilizando-se três tipos de
olfatômetros: um olfatômetro em Y conectado a um fluxo de ar e outros dois
olfatômetros tipo arena com dupla escolha sem fluxo de ar. As fontes de estímulo
utilizadas foram: (i) machos de A. ynca, (ii) fêmeas de A. ynca, (iii) cana-de-
açúcar, (iv) inflorescência jovem de coqueiro (v) partes do coco e (vi)
combinações de alguns deles. Foram realizados testes até se obter a resposta de
20 insetos, nos três tipos de olfatômetros; não se repetiram os insetos em testes
com um mesmo estímulo. Os insetos machos e fêmeas, testados individualmente
ou em grupo não reponderam aos estímulos em nenhum dos oltatômetros
utilizados. A cana-de-açúcar foi o único estímulo que provocou uma atividade
maior dos insetos nos olfatômetros, mas as respostas a odores não foram
significativas. Ambos os sexos também responderam mais ao estímulo e ao
controle quando se utilizou a inflorescência como estímulo nos três tipos de

63
olfatômetro. As combinações que mais estimularam a locomoção foram a cana-
de-açúcar + machos e inflorescência + machos.
ABSTRACT
Bioassays with cospecfics and host plants odor of Amerrhinus ynca Sahlberg, 1823 (Coleoptera: Curculionidae)
Communication has extreme importance to the adaptation of animals; it allows the
interaction among species. Chemical signs involved in the communication among
alive beings are named semiochemical. This work was accomplished with the
purpose of verifying the existence of attractive semiochemicals that act in the intra
and interespecific interactions of Amerrhinus ynca. The bioassays were conducted
using three types of olfactometers, an olfactometer in "Y" connected to a flow of air
and two other arena olfactometers with double choice chambers but without air
flow. The sources of stimuli were: (i) males of A. ynca, (ii) females of A. ynca, (iii)
sugar cane, (iv) young inflorescence, (v) fruit and (vi) combinations of some of
them. Not all the stimuli were tested in the three types of olfactometers. Tests
were accomplished until obtaining the behavioral response of 20 insects, in the
three types of olfactometers. The same insects were never reused with a same
stimulus. When both males and females were tested together or separated,
insects did not give any responses, staying still in the three types of olfactometers.
The sugar cane, used in the bioassays in the olfactometers type I and type II, was
the stimulus that provoked the highest activity of insects, but failed to present a
choice for this stimulus. Both sexes also responded to the stimulus and to the
control mostly when inflorescence was used in the three types of olfactometers.
The combinations of stimuli that mostly stimulated males and females of A. ynca
were sugar cane + males, and inflorescence + males. The data presented here
were not sufficient to confirm the presence of chemical medators of interactions
intraspecific due to the high percentile of non-response. The results with vegetable
tissues (sugar cane, inflorescence and fruit) evidenced the greatest stimulation,
suggesting a possible attractiveness to volatiles liberated by the host plant.

64
INTRODUÇÃO
A comunicação é um dos componentes comportamentais de grande
importância para a adaptação dos animais, por permitir a interação entre as
espécies. Lewis & Gower (1980) definem comunicação como a transmissão de
sinal ou sinais entre dois indivíduos, favorecendo a seleção tanto da produção
quanto da recepção do sinal ou sinais. Aos sinais químicos envolvidos na
comunicação entre os seres vivos dá-se o nome de semioquímicos, os quais são
responsáveis pela transmissão de informações biologicamente importantes que
asseguram a tais indivíduos oportunidades de sobrevivência e de preservação da
espécie (Paiva & Pedrosa-Macedo, 1985).
Em curculionídeos, a presença de sinais químicos envolvidos na
comunicação, sua identificação e isolamento, foram bastante estudados em
diversas espécies como em Anthonomus eugenii Cano (Eller et al. 1994), Curculio
caryae Horn, 1873 (Hedin et al., 1997), Pissodes strobi (Peck, 1817) (Booth et al.
1983), Conotrachelus nenuphar (Herbst, 1797) (Eller & Bartelt, 1996), Sitophilus
granarius Linneaus, 1758 (Phillips et al. 1987), Rhynchophorus palmarum
Linnaeus, 1764 (Jaffé et al. 1993), entre outros. A utilização de armadilhas
iscadas com feromônio e cairomônio, como técnica de monitoramento e controle
dos níveis populacionais de algumas destas espécies, tem demonstrado
resultados satisfatórios e essas armadilhas já são empregadas em campo com
sucesso.
Em Amerrhinus ynca, o uso de semioquímicos em armadilhas de captura
permitiria monitorá-lo na cultura, possibilitando, além de seu controle mais
eficiente, a redução no número de aplicações de inseticidas, contribuindo assim
para a melhoria na rentabilidade do produtor e para a redução da poluição
ambiental. Dentro deste contexto, este trabalho foi realizado com a finalidade de
verificar a existência de semioquímicos que atuem nas interações intra e
interespecíficas de A. ynca.

65
MATERIAIS E MÉTODOS
Adultos de A. ynca coletados num plantio comercial de coco localizado no
município de Quissamã, RJ, nos meses de outubro de 2005 a janeiro de 2006,
foram levados ao Laboratório de Entomologia e Fitopatologia do Centro de
Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte
Fluminense. No laboratório, os insetos foram separados por sexo e mantidos até
a realização dos experimentos em caixas de acrílico (30 x 30 cm) com alimento
(cana-de-açúcar). Os insetos foram mantidos a 25 ± 2 °C, e fotoperíodo de
12:12h.
Para se verificar a presença de sinais químicos envolvidos no
comportamento de A. ynca, foram utilizados três tipos de olfatômetro:
I) Olfatômetro em "Y" olfatômetro de duas vias similar ao utilizado em testes
olfativos com outros curculionídeos (Weissling et al. 1994; Ruiz-Montiel et al.
2003; Tafoya et al. 2003). O olfatômetro foi constituído de um tubo de vidro em “Y”
de 1,5 cm de diâmetro, possuía um braço principal de 20 cm de comprimento e
dois braços laterais de 15 cm. O arraste de odores no olfatômetro foi provocado
por um compressor. Um filtro de carvão ativado foi responsável pela purificação
do ar que entrava no sistema. Do filtro o ar era conduzido até o lavador de gases
(1.000 ml) contendo 500 ml de água destilada onde, por meio do borbulhamento,
o ar era novamente purificado e umedecido. O fluxo de ar utilizado nos testes foi
regulado, com ajuda de um fluxômetro, em 30 ml/s. As extremidades dos braços
secundários do olfatômetro foram conectadas a câmaras de vidro (2l) contendo o
estímulo olfativo ou ar puro como controle (Figura 1).

66
Figura 1 - Esquema do olfatômetro de duas vias, modelo em “Y”, (Esq.). Foto do dispositivo olfatométrico (dir.).
II) Olfatômetro tipo arena constituído por duas câmaras-testes contendo o
estímulo ou o ar puro como controle, interligadas à câmara principal e sem fluxo
de ar. Cada câmara-teste era constituída por duas caixas Gerbox (10 X 10 X 5
cm) coladas e ligadas à câmara principal por braços de borracha transparente (6
cm de comprimento por 2 cm de diâmetro). Quando o estímulo utilizado foi inseto
(macho ou fêmea ou os dois juntos) foi colocada no meio das duas caixas uma
tela de filó, para impedir que os insetos entrassem na câmara principal. A câmara
principal era constituída por um copo de plástico transparente invertido com oito
cm de comprimento X dois cm de espessura com maior diâmetro igual a 10,5 cm
e menor diâmetro igual a sete cm (Figura 2).
III) Olfatômetro de câmara múltipla constituído por um recipiente de vidro (30
X 15 X 15 cm) dividido em três partes (câmaras) de 10 cm de comprimento cada
uma. As divisórias eram de plástico duro de cor amarela, coladas à parede de
vidro. Cada parede de plástico tinha uma abertura na parte mediana superior de
3,5 cm de comprimento X 1,5 cm de altura, por onde os insetos passavam. Os
insetos eram colocados no meio da câmara central e os estímulos ou controle nas
câmaras laterais. O recipiente era tampado por uma tampa de vidro e não havia
fluxo de ar (Figura 3).
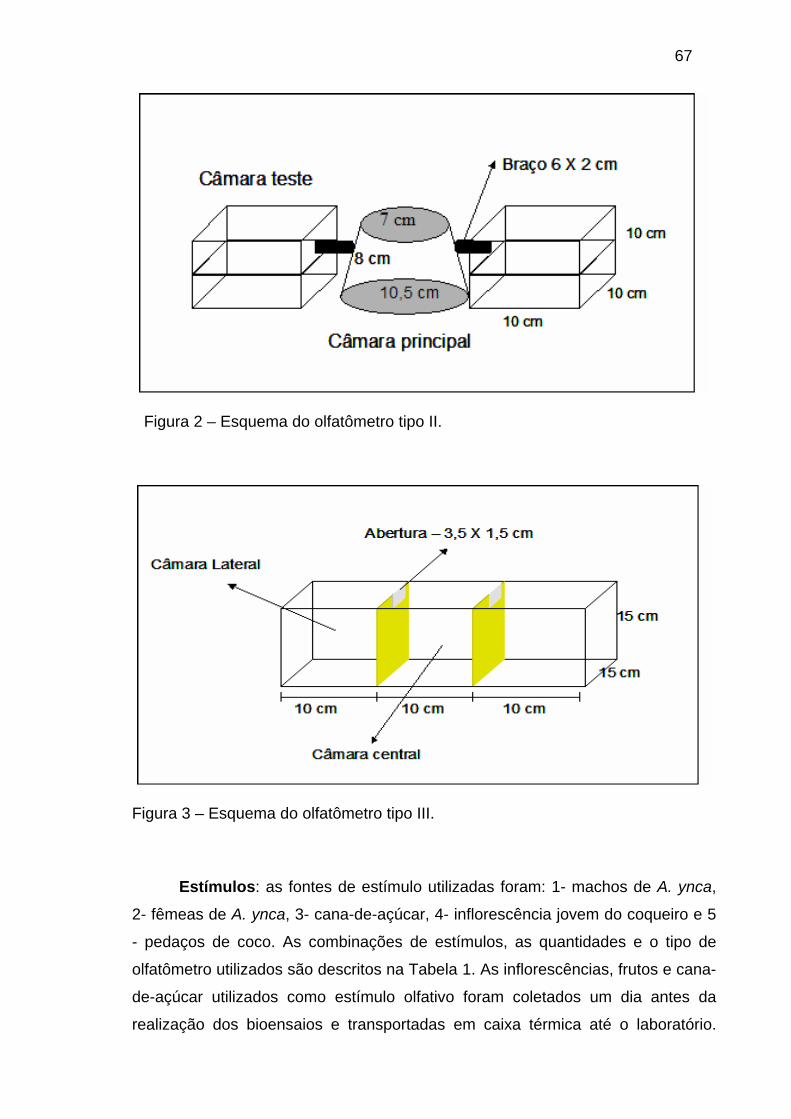
67
Figura 2 – Esquema do olfatômetro tipo II.
Figura 3 – Esquema do olfatômetro tipo III. Estímulos: as fontes de estímulo utilizadas foram: 1- machos de A. ynca,
2- fêmeas de A. ynca, 3- cana-de-açúcar, 4- inflorescência jovem do coqueiro e 5
- pedaços de coco. As combinações de estímulos, as quantidades e o tipo de
olfatômetro utilizados são descritos na Tabela 1. As inflorescências, frutos e cana-
de-açúcar utilizados como estímulo olfativo foram coletados um dia antes da
realização dos bioensaios e transportadas em caixa térmica até o laboratório.
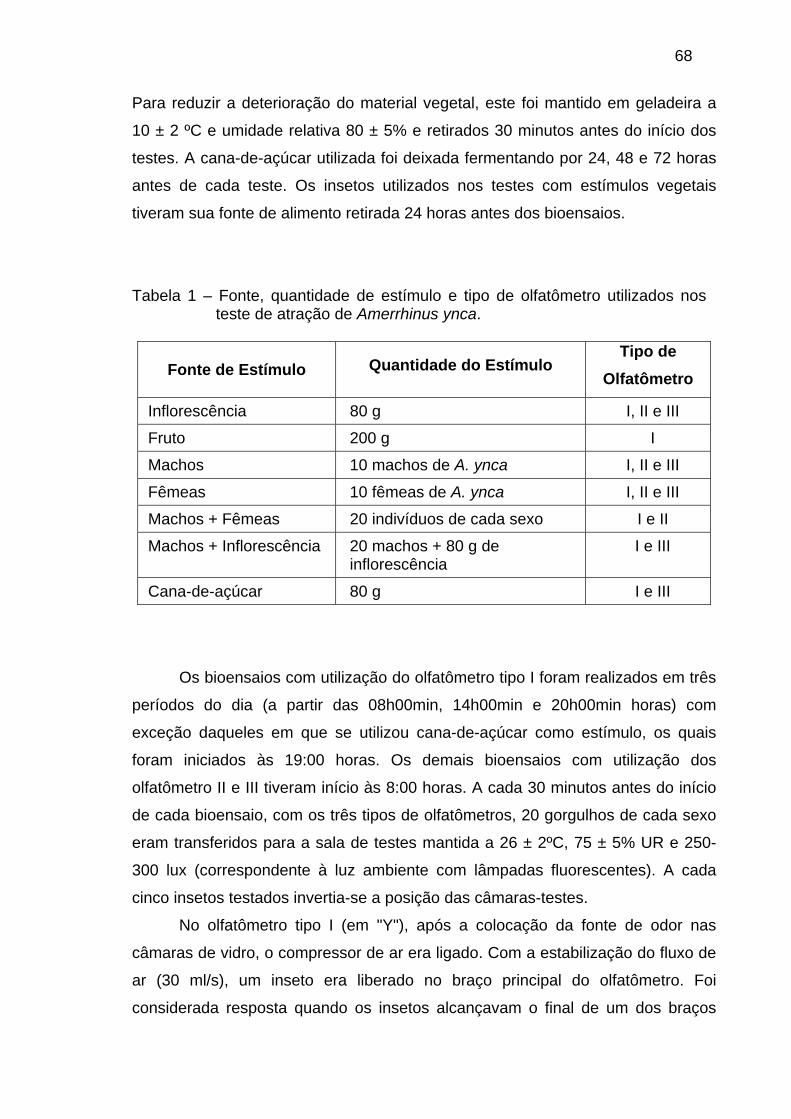
68
Para reduzir a deterioração do material vegetal, este foi mantido em geladeira a
10 ± 2 ºC e umidade relativa 80 ± 5% e retirados 30 minutos antes do início dos
testes. A cana-de-açúcar utilizada foi deixada fermentando por 24, 48 e 72 horas
antes de cada teste. Os insetos utilizados nos testes com estímulos vegetais
tiveram sua fonte de alimento retirada 24 horas antes dos bioensaios.
Tabela 1 – Fonte, quantidade de estímulo e tipo de olfatômetro utilizados nos teste de atração de Amerrhinus ynca.
Fonte de Estímulo Quantidade do Estímulo Tipo de
Olfatômetro
Inflorescência 80 g I, II e III
Fruto 200 g I
Machos 10 machos de A. ynca I, II e III
Fêmeas 10 fêmeas de A. ynca I, II e III
Machos + Fêmeas 20 indivíduos de cada sexo I e II
Machos + Inflorescência 20 machos + 80 g de inflorescência
I e III
Cana-de-açúcar 80 g I e III Os bioensaios com utilização do olfatômetro tipo I foram realizados em três
períodos do dia (a partir das 08h00min, 14h00min e 20h00min horas) com
exceção daqueles em que se utilizou cana-de-açúcar como estímulo, os quais
foram iniciados às 19:00 horas. Os demais bioensaios com utilização dos
olfatômetro II e III tiveram início às 8:00 horas. A cada 30 minutos antes do início
de cada bioensaio, com os três tipos de olfatômetros, 20 gorgulhos de cada sexo
eram transferidos para a sala de testes mantida a 26 ± 2ºC, 75 ± 5% UR e 250-
300 lux (correspondente à luz ambiente com lâmpadas fluorescentes). A cada
cinco insetos testados invertia-se a posição das câmaras-testes.
No olfatômetro tipo I (em "Y"), após a colocação da fonte de odor nas
câmaras de vidro, o compressor de ar era ligado. Com a estabilização do fluxo de
ar (30 ml/s), um inseto era liberado no braço principal do olfatômetro. Foi
considerada resposta quando os insetos alcançavam o final de um dos braços

69
secundários. A escolha e o tempo de teste foram registrados. O tempo máximo de
duração dos testes, baseado em experimentos similares com outros
curculionídeos (Rhynchophorus cruentatus (Fabricius, 1775), (Weissling et al.
1994) e Scyphophorus acupuncatatus Gyllenhal (Ruiz-Montiel et al. 2003)) foi
estabelecido em 5 minutos.
Com os olfatômetros tipo II e III o tempo máximo foi de 30 minutos e os
insetos eram colocados no centro da câmara principal, um por vez. A cada cinco
insetos testados a posição das câmaras-testes foi invertida.
No olfatômetro tipo III para os testes com insetos como fonte de estímulo,
estes eram colocados dentro de uma caixa Gerbox (10 X 10 X 5cm) tampada com
tecido de filó, a qual era colocada em uma das câmaras laterais. Este
procedimento evitou que os insetos circulassem pelo olfatômetro.
Foram realizados testes até se obter a resposta de 20 insetos, nos três
tipos de olfatômetros, e não se repetiram os insetos em testes com um mesmo
estímulo. Após cada sessão de testes, todo o material foi colocado em solução
Extran® por duas horas, depois lavado em água corrente, para se evitar qualquer
resíduo de odor.
Verificou-se em teste preliminar de tendenciosidade que os insetos são
foto trópicos. Para minimizar este efeito, nos testes feitos durante a fotofase com
o olfatômetro tipo I, foi colocada uma lâmpada de 40 Watts em frente à bifurcação
do olfatômetro a uma distância de 50 cm. Nos demais olfatômetros os testes
foram realizados dentro do túnel de vento, o que impedia a penetração direta da
luz.
RESULTADOS
a) Olfatômetro em "Y"
Através dos testes realizados com o olfatômetro em Y não foi possível
verificar atração de nenhum dos estímulos testados. Os insetos não responderam
aos odores dos próprios insetos. Tanto odores de machos quanto de fêmeas,
separadamente (Figura 4 e 5) ou colocados juntos (Figura 6), não provocaram o
comportamento de caminhamento no olfatômetro. Quando o estímulo foi macho, o
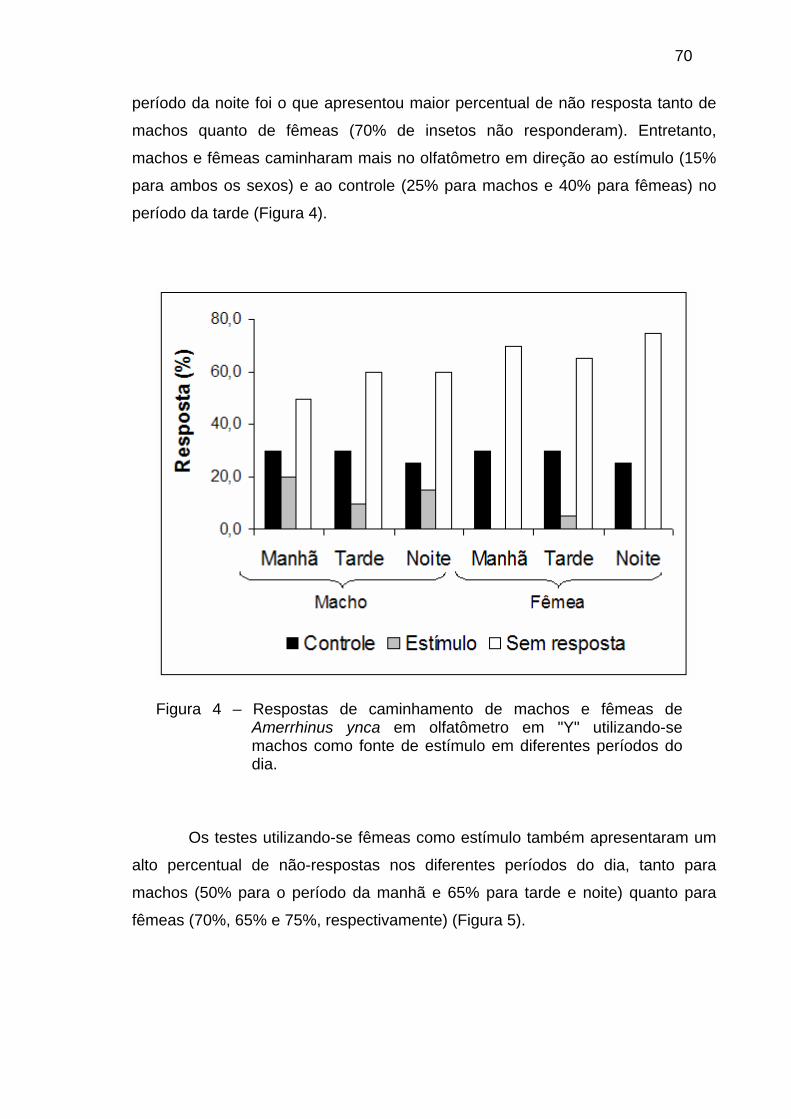
70
período da noite foi o que apresentou maior percentual de não resposta tanto de
machos quanto de fêmeas (70% de insetos não responderam). Entretanto,
machos e fêmeas caminharam mais no olfatômetro em direção ao estímulo (15%
para ambos os sexos) e ao controle (25% para machos e 40% para fêmeas) no
período da tarde (Figura 4).
Figura 4 – Respostas de caminhamento de machos e fêmeas de Amerrhinus ynca em olfatômetro em "Y" utilizando-se machos como fonte de estímulo em diferentes períodos do dia.
Os testes utilizando-se fêmeas como estímulo também apresentaram um
alto percentual de não-respostas nos diferentes períodos do dia, tanto para
machos (50% para o período da manhã e 65% para tarde e noite) quanto para
fêmeas (70%, 65% e 75%, respectivamente) (Figura 5).

71
Figura 5 – Respostas de caminhamento de machos e fêmeas de Amerrhinus ynca em olfatômetro em "Y" utilizando-se fêmeas como fonte de estímulo em diferentes períodos do dia.
Figura 6 – Respostas de caminhamento de machos e fêmeas de Amerrhinus ynca em olfatômetro em "Y" utilizando-se como fonte de estímulo machos e fêmeas juntos.

72
Já nos testes com cana-de-açúcar, houve maior atividade dos insetos
dentro do olfatômetro. A porcentagem de insetos (machos e fêmea) que
responderam ao controle foi maior do que nos demais tratamentos. O estímulo
que atraiu um número maior de insetos foi cana-de-açúcar com 24 horas de
fermentação (Figura 7).
Figura 7 – Respostas de caminhamento de machos e fêmeas de Amerrhinus ynca em olfatômetro em "Y" utilizando-se diferentes estágios de fermentação da cana-de-açúcar como estímulo.
Quando se testou a inflorescência em diferentes períodos do dia o
percentual de não resposta também foi o maior (Figura 8). As fêmeas
responderam mais durante a noite e os machos à tarde.
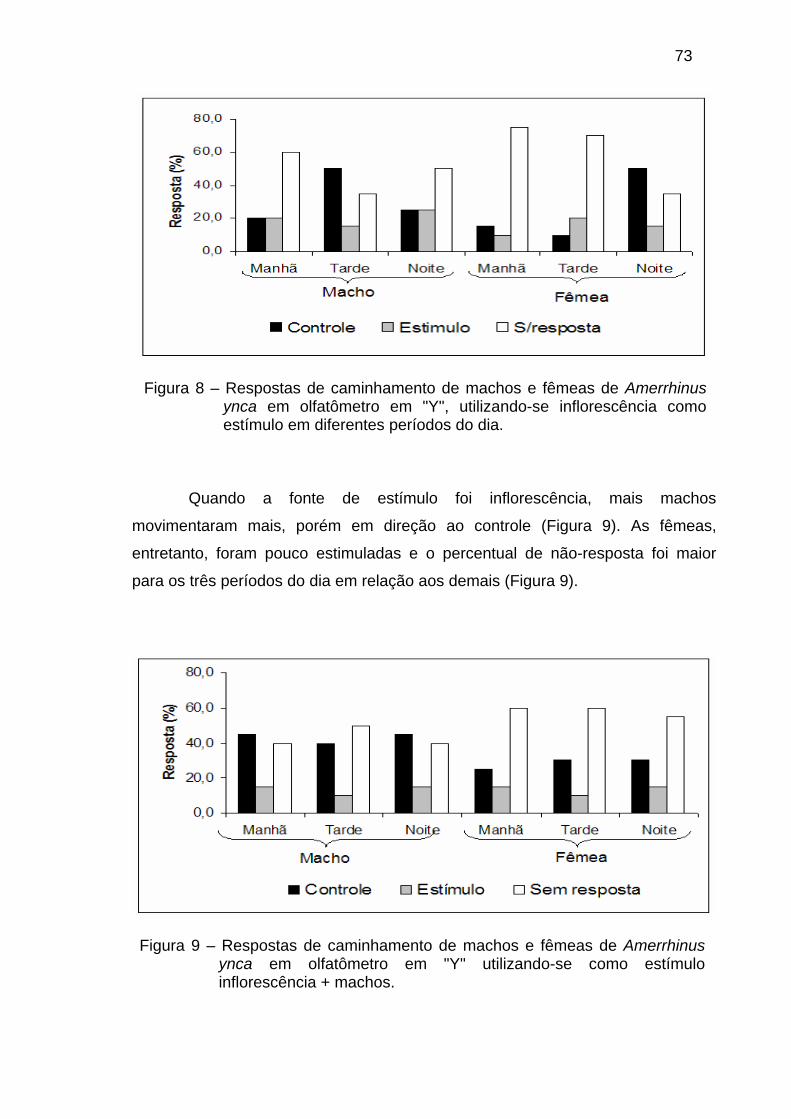
73
Figura 8 – Respostas de caminhamento de machos e fêmeas de Amerrhinus ynca em olfatômetro em "Y", utilizando-se inflorescência como estímulo em diferentes períodos do dia.
Quando a fonte de estímulo foi inflorescência, mais machos
movimentaram mais, porém em direção ao controle (Figura 9). As fêmeas,
entretanto, foram pouco estimuladas e o percentual de não-resposta foi maior
para os três períodos do dia em relação aos demais (Figura 9).
Figura 9 – Respostas de caminhamento de machos e fêmeas de Amerrhinus ynca em olfatômetro em "Y" utilizando-se como estímulo inflorescência + machos.
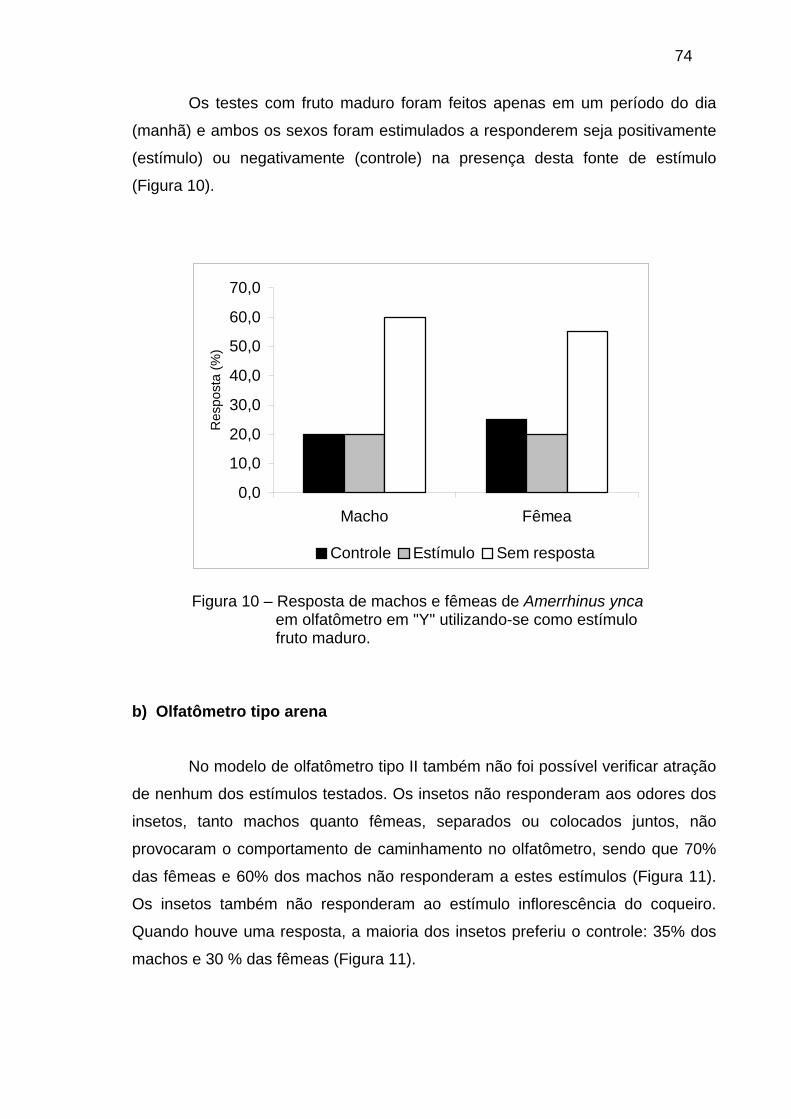
74
Os testes com fruto maduro foram feitos apenas em um período do dia
(manhã) e ambos os sexos foram estimulados a responderem seja positivamente
(estímulo) ou negativamente (controle) na presença desta fonte de estímulo
(Figura 10).
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
Macho Fêmea
Res
post
a (%
)
Controle Estímulo Sem resposta
Figura 10 – Resposta de machos e fêmeas de Amerrhinus ynca em olfatômetro em "Y" utilizando-se como estímulo fruto maduro.
b) Olfatômetro tipo arena
No modelo de olfatômetro tipo II também não foi possível verificar atração
de nenhum dos estímulos testados. Os insetos não responderam aos odores dos
insetos, tanto machos quanto fêmeas, separados ou colocados juntos, não
provocaram o comportamento de caminhamento no olfatômetro, sendo que 70%
das fêmeas e 60% dos machos não responderam a estes estímulos (Figura 11).
Os insetos também não responderam ao estímulo inflorescência do coqueiro.
Quando houve uma resposta, a maioria dos insetos preferiu o controle: 35% dos
machos e 30 % das fêmeas (Figura 11).

75
Figura 11 – Respostas de adultos de Amerrhinus ynca aos testes em olfatômetro tipo arena aos diferentes tipos de estímulos. Acima, respostas de fêmeas e, abaixo, respostas de machos.
c) Olfatômetro de câmara múltipla
Nos testes com o olfatômetro tipo III novamente se constatou o mesmo
comportamento. Os insetos não responderam aos odores de machos ou fêmeas
coespecíficos (Figura 12).
Apesar de 45% dos insetos não terem dado nenhum tipo de resposta,
machos e fêmeas mostraram um pouco mais de atividade quando se utilizou
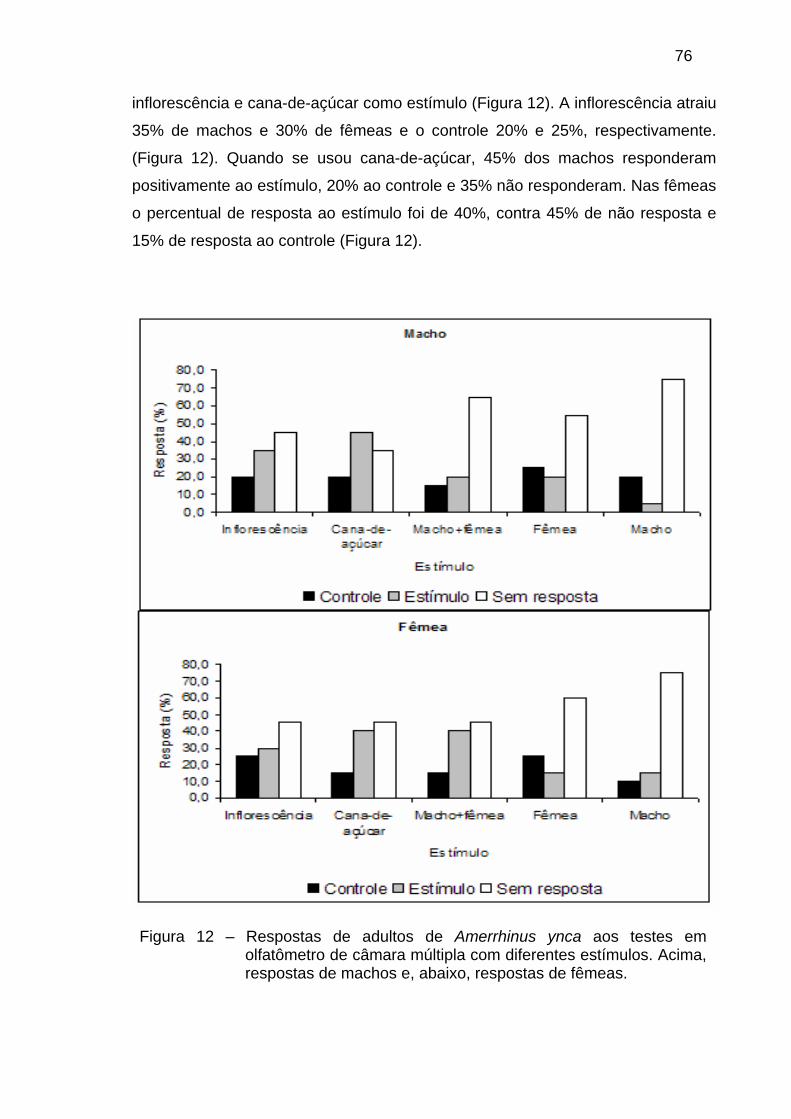
76
inflorescência e cana-de-açúcar como estímulo (Figura 12). A inflorescência atraiu
35% de machos e 30% de fêmeas e o controle 20% e 25%, respectivamente.
(Figura 12). Quando se usou cana-de-açúcar, 45% dos machos responderam
positivamente ao estímulo, 20% ao controle e 35% não responderam. Nas fêmeas
o percentual de resposta ao estímulo foi de 40%, contra 45% de não resposta e
15% de resposta ao controle (Figura 12).
Figura 12 – Respostas de adultos de Amerrhinus ynca aos testes em
olfatômetro de câmara múltipla com diferentes estímulos. Acima, respostas de machos e, abaixo, respostas de fêmeas.

77
DISCUSSÃO
Neste trabalho foram utilizados três tipos de dispositivos olfatométricos na
tentativa de se verificar a presença de alguma substância química mediadora de
interações dentro da espécie (feromônio sexual ou de agregação) ou entre
espécies, principalmente voláteis liberados pela planta hospedeira (cairomônios).
Entretanto, os dados aqui obtidos não foram suficientes para confirmar a
presença de substâncias químicas mediadoras de interações interespecíficas. O
baixo percentual de resposta demonstra que os dispositivos olfatométricos
utilizados não foram capazes de evidenciar algum tipo de resposta olfativa desta
espécie. Vários fatores podem ter contribuído, como a metodologia utilizada, as
condições físicas do ambiente de teste, como fluxo de ar, luminosidade,
temperatura e umidade relativa, que podem não ter sido bem ajustadas às
necessidades da espécie. Ou, talvez, esses insetos utilizem outros mecanismos
associados à sinalização acústica e/ou visual em sua comunicação.
De fato, nos dispositivos olfatométricos II e III, o percentual de
estimulação (respostas positivas + negativas) foi maior quando os sexos foram
utilizados como fonte de estímulo do que nos bioensaios com o olfatômetro tipo I.
Possivelmente isto ocorreu por causa da produção de som que pode ter atraído
seus coespecíficos mais do que uma possível substância química, ou ambos
agem em sinergismo. Outros testes devem ser feitos excluindo-se este sinal
acústico em ambos os sexos.
Leskey & Prokopy (2001) verificaram após colarem o élitro das fêmeas de
C. nenuphar, eliminando a produção de som, que este não influenciou na
atratividade de possíveis fontes de odores. Entretanto, no cerambicídeo Dectes
texanus texanus, Crook et al. (2004), ao estudarem seu comportamento de
acasalamento, verificaram que o feromônio sexual (material lipossolúvel), liberado
pelas fêmeas e presente na sua cutícula, provavelmente não atua em distâncias
maiores do que poucos centímetros, sendo que outros estímulos, visuais ou
acústicos, são utilizados para a localização dos parceiros. Alguns homópteros
movem-se entre as plantas e utilizam a comunicação através de vibrações para
encontrar seus coespecíficos, entretanto, em certos casos, a comunicação
química através de feromônio exerce uma função de atração à longa distância

78
(Cokl & Virant-Doberlet, 2003). Já em algumas mariposas é comum a utilização
de um sistema de sinalização acústica em associação com a liberação de
feromônios para atração de coespecíficos (Conner 1999).
Os odores das partes vegetais (cana-de-açúcar, inflorescência e fruto)
provocaram uma maior atividade dos insetos, sugerindo um possível efeito sobre
sua locomoção, apesar da cana-de-açúcar e do fruto maduro não serem fontes
naturais de alimento para adultos de A. ynca. Compostos voláteis liberados pela
planta hospedeira e que exercem atratividade já foram documentados em várias
espécies de curculionídeos. Segundo Gunawardena et al. (1998), ambos os sexos
de Rhynchophorus ferrugineus são atraídos por compostos liberados pelos
tecidos de sua planta hospedeira. Alguns desses, liberados pelos tecidos
danificados da planta hospedeira de R. palmarum, como o acetato de etil e o
etanol, também exercem uma função de orientação nesses gorgulhos,
provavelmente a curtas distâncias, além de potencializar a atração do feromônio
de agregação (Jaffé et al. 1993). Em Anthonomus grandis, α-pineno, limoneno, β-
cariofileno e alguns sesquiterpenos presentes nos botões florais e nas folhas do
algodão, atraem, segundo Minyard et al. (1969), os gorgulhos em uma mistura
complexa. Em C. nenuphar, Butkewich & Prokopy (1993) sugerem que tais
gorgulhos são capazes de detectar a grandes distâncias os odores liberados por
vários frutos de suas plantas hospedeiras e que estes estímulos químicos podem
ser utilizados em armadilhas para monitorar sua atividade em pomares.
Possivelmente voláteis liberados pelas flores masculinas (provavelmente
presentes na cana-de-açúcar e no fruto maduro do coqueiro) possam exercer em
adultos de A. ynca atração até o sítio de alimentação. Outros testes devem ser
realizados tentando se eliminar possíveis interferências físicas do ambiente e
procurando se obter maior suporte na biologia, ecologia e comportamento de A.
ynca, na tentativa de desvendar a presença de substâncias voláteis atrativas
liberadas pela planta hospedeira.
RESUMOS E CONCLUSÕES A comunicação é de extrema importância para a adaptação dos animais,
destacando-se por permitir a interação entre as espécies e dentro da espécie.

79
Este trabalho foi realizado com a finalidade de se verificar, através de testes em
laboratório, a existência de substâncias químicas liberadas por coespecíficos e
pela planta hospedeira que atraem adultos de A. ynca.
Os bioensaios foram feitos utilizando-se três tipos de olfatômetros: um
olfatômetro em Y conectado a um fluxo de ar e outros dois olfatômetros tipo arena
com dupla escolha sem fluxo de ar. As fontes de estímulo utilizadas foram: (i)
machos de A. ynca, (ii) fêmeas de A. ynca, (iii) cana-de-açúcar, (iv) inflorescência
jovem de coqueiro (v) partes do coco e (vi) combinações de alguns deles. Foram
realizados testes até se obter a resposta de 20 insetos, nos três tipos de
olfatômetros; não se repetiram os insetos em testes com um mesmo estímulo.
Através desse trabalho foi possível verificar que: os insetos machos e
fêmeas, testados individualmente ou em grupo, não responderam aos estímulos
em nenhum dos olfatômetros utilizados; a cana-de-açúcar foi o único estimulo que
provocou uma atividade maior dos insetos nos olfatômetros, mas as respostas a
odores não foram significativas; ambos os sexos também responderam mais ao
estímulo e ao controle quando se utilizou a inflorescência como estímulo nos três
tipos de olfatômetro e; as combinações que mais estimularam a locomoção foram
a cana-de-açúcar + machos e inflorescência + machos.
Portanto, neste trabalho foram utilizados três tipos de dispositivos
olfatométricos na tentativa de evidenciar possíveis odores de coespecíficos e de
tecidos vegetais que promovessem alguma resposta comportamental em A. ynca.
Entretanto, os dados aqui apresentados não foram suficientes para confirmar a
presença de substâncias químicas mediadoras de interações intra-especificas
devido ao alto percentual de não-resposta. Já os resultados com partes vegetais
(cana-de-açúcar, inflorescência e fruto) estimularam uma maior locomoção dos
insetos, sugerindo uma possível atratividade a voláteis liberados pela planta
hospedeira.
REFERÊNCIAS BIBLIOGRÁFICAS
Booth, D. C.; Phillips, T. W.; Claesson, A.; Silverstein, R. M.; Lanier, G. N. & West, J. R. (1983) Aggregation pheromone components of two species of Pissodes weevils (Coleoptera: Curculionidae): isolation, identification, and field activity. Journal of Chemical Ecology, 9:1 – 12.

80
Butkewich, S. L. & Prokopy, R. J. (1993) The effect of short-range host odor stimuli on host fruit finding and feeding behavior of Plum Curculio adult (Coleoptera: Curculionidae). Journal of Chemical Ecology, 19:825 – 835.
Cokl, A. & Virant-Doberlet, M. (2003) Communication with substrate-borne signals in small plant-dwelling insects. Annual Review of Entomology, 48:29 – 50.
Conner, W. E. (1999) ‘Un chant d’appel amoureux’: acoustic communication in moths. The Journal of Experimental Biology, 202:1711 – 1723.
Crook, D. J.; Hopper, J. A.; Ramaswamy, S. B. & Higgins, R. A. (2004) Courtship behavior of the Soybean Stem Borer Dectes texanus texanus (Coleoptera: Cerambycidae): evidence for a female contact sex pheromone. Annals of the Entomological Society of America, 97:600 – 604.
Eller, F. J. & Bartelt, R. J. (1996) Grandisoic acid, a male-produced aggregation pheromone from the plum curculio, Conotrachelus nenuphar. Journal of Natural Products, 59:451 –453.
Eller, F. J.; Bartelt, R. J.; Shasha, B. S.; Schuster, D. J.; Riley, D. G.; Stansly, P. A.; Mueller, T. F.; Shuler, K. D.; Johnson, B.; Davis, J. H. & Sutherland, C. A. (1994) Aggregation pheromone for the pepper weevil, Anthonomus eugenii Cano (Coleoptera: Curculionidae): identification and field activity. Journal of Chemical Ecology, 20:1537 – 1555.
Gunawardena, N. E.; Kern, F.; Janssen, E.; Meegoda, C.; Schafer, D.; Vostrowsky, O. & Bestmann, H. J. (1998) Host attractants for red weevil, Rhynchophorus ferrugineus: identification, electrophysiological activity, and laboratory bioassay. Journal of Chemical Ecology, 24:425 – 437.
Hedin, P. A.; Dollar, D. A.; Collins, J. K.; Dubois, J. G.; Mulder, P. G.; Hedger, G. H.; Smith, M. W. & Eikenbary, R. D. (1997) Identification of male pecan weevil pheromone. Journal of Chemical Ecology, 23:965 – 977.
Jaffé, K.; Sanchez, P.; Cerda, H.; Hernandez, J. V.; Jaffé, R.; Urdaneta, N.; Guerra, G.; Martinez, R. & Miras, B. (1993) Chemical ecology of the palm weevil Rhynchophorus palmarum (L.) (Coleoptera: Curculionidae): attraction to host plants and to a male-produced aggregation pheromone. Journal of Chemical Ecology, 19:1703 – 1720.
Leskey, T. C. & Prokopy, R. J. (2001) Adult Plum Curculio (Coleoptera: Curculionidae) attraction to fruit and conspecific odors. Annals of the Entomological Society of America, 94:275 – 288.
Lewis, B. & Gower, M. (1980) Biology of communication. London: Blackie, 239 p.
Minyard, J. P.; Hardee, D. D.; Gueldner, R. C.; Thompson, A. C.; Wiygul, G. & Hedin, P. A. (1969) Constituents of the Cotton Bud: compounds attractive to the Boll Weevil. Journal Agriculture Food Chemical, 17:1093 – 1097.
Paiva, M. R. & Pedrosa-Macedo, J. H. P. (1985) Feromonas de Insetos. Curitiba: GTZ, 94 p.

81
Phillips, J. K.; Miller, S. P. F.; Andersen, J. F.; Fales, H. M.; & Burkholder, W. E. (1987) The chemical identification of the granary weevil aggregation pheromone. Tetrahedron Letters, 28:6145 – 6146.
Ruiz-Montiel, C.; González-Hernández, H.; Leyva, J.; Llanderal-Cazares, C.; Cruz-Lopez, L. & Rojas, J. C. (2003) Evidence for a male-produced aggregation pheromone in Scyphophorus acupunctatus Gyllenhal (Coleoptera: Curculionidae). Journal of Economic Entomology, 96:1126 – 1131.
Tafoya, F.; Lopez-Collado, J.; Stanley, D.; Rojas, J. C. & Cibrian-Tovar, J. (2003) Evidence of an aggregation pheromone in males of Metamasius spinolae (Coleoptera: Curculionidae). Environmental Entomology, 32:484 – 487.
Weissling, T. J.; Giblin Davis, R. M.; Gries, G.; Gries, R.; Perez, A. L.; Pierce, H. D. Jr.; & Oehlschlager, A. C. (1994) Aggregation pheromone of palmetto weevil, Rhynchophorus cruentatus (F.) (Coleoptera: Curculionidae). Journal of Chemical Ecology, 20:505 – 515.

82
ANATOMIA DO APARATO ESTRIDULATÓRIO, ANÁLISE DO SOM E PAPEL DA ESTRIDULAÇÃO NO COMPORTAMENTO DE CORTE E ACASALAMENTO.
DE AMERRHINUS YNCA SAHLBERG, 1823 (COLEOPTERA: CURCULIONIDAE)
RESUMO Os insetos que utilizam a comunicação acústica nas relações interindividuais
podem produzir suas emissões sonoras através de distintos meios, a estridulação
é o mais difundido. Em Curculionidae a estridulação se dá através da fricção do
plectrum do abdômen com a pars stridens localizada nos élitros. Este trabalho
descreve a morfologia do aparato estridulatório de Amerrhinus ynca, analisa as
suas emissões sonoras e investiga o papel da estridulação no seu
comportamento de corte. Os sons foram gravados com gravador digital e
analisados com ajuda do software Canary. Para verificar o papel da estridulação
no comportamento de acasalamento de A. ynca foram realizados quatro
tratamentos: (i) casais não manipulados, (ii) casais em que ambos os sexos
tiveram parte de seus élitros direito e esquerdo removidos, (iii) casais nos quais
apenas o macho teve parte de seus élitros removidos e (iv) casais nos quais
apenas as fêmeas tiveram seus élitros direito e esquerdo removidos. A parte dos
élitros que foi removida corresponde ao local onde se encontram as partes
stridentes. Os casais foram observados durante cinco horas. Foram analisadas a
freqüência de monta e cópula, a freqüência dos atos comportamentais, a duração
do período de latência, pré-cópula, cópula e pós-cópula. A pars stridens de A.

83
ynca está localizada na região ventral da extremidade posterior dos élitros direito
e esquerdo. Nos machos ela é constituída de fileiras de cristas de altura e
espaçamentos variáveis. Nas fêmeas as cristas são menos acentuadas e estão
distribuídas de forma irregular. Machos e fêmeas apresentaram dois plectra
localizados no sétimo esternito abdominal, debaixo de cada pars stridens.
Verificou-se que machos e fêmeas emitem sons por estridulação. Os machos
estridularam durante o comportamento de corte. As fêmeas também
apresentaram movimento similar ao estridulatório, mas um espectro específico
não foi registrado. Os machos apresentaram estridulações quando foram
perturbados através de pressões nas laterais do abdômen. A característica
temporal e espectral destes sons foi distinta da observada durante a corte. As
fêmeas também estridularam quando perturbadas e o espectro de emissão do
som foi diferente do observado nos machos. Nos tratamentos nos quais as
fêmeas tiveram sua pars stridens removida a porcentagem de casais que
acasalaram (montas e cópulas) reduziu. Já os machos gastaram mais tempo
tentando cortejar as fêmeas, quando sua pars stridens foi retirada. Evidenciou-se
a existência de aparato estridulatório em A.ynca, morfologicamente diferente nos
dois sexos. Nos machos foi possível diferenciar dois tipos de sons, uma
estridulação associada possivelmente a encontros com outros machos e uma
estridulação de corte. A remoção da par stridens interfere no acasalamento
aumentando o número de montas e reduzindo o número de cópula, além de
alterar a duração da pré-cópula, cópula e pós-cópula.
ABSTRACT
Anatomy of the stridulatory apparatus, analysis of the sound and role of stridulation in courtship and mating behavior of Amerrhinus ynca Sahlberg, 1823
(Coleoptera: Curculionidae)
Insects that use the acoustic communication in the inter-specific relationships can
produce their sound emissions through different means. Stridulation is the most
spread one. In Curculionidae stridulation happens through the friction of the

84
plectrum of the abdomen with the pars stridens located in the elytra. This work
describes the morphology of the stridulatory apparatus of Amerrhinus ynca; it
analyzes the sound emissions and investigates the role of stridulation in courtship
behavior. The sounds were recorded with digital tape recorder and analyzed by
means of the Canary software. In order to verify the role of stridulation on the
mating behavior of A. ynca, four treatments were accomplished: (i) manipulated
couples, (ii) couples in which both sexes had part of their right and left elytra
removed, (iii) couples in which only the male had part of its elytra removed (iv)
couples in which only the female had part of its elytra removed. The part of the
elytra that was removed corresponded to the place of pars stridens location. The
couples were observed for five hours. The frequency of mount and copulation; the
frequency of the behavioral acts; the duration of the latency period, pre-copulation,
copulation and post-copulation, were analyzed. The pars stridens of A. ynca is
located in the ventral area of the posterior extremity of the right and left elytra. In
males it is constituted of arrays of ridges of variable height and spacing. In the
females, the ridges are less prominent and are distributed in an irregular way.
Males and females presented two plectra located in the seventh abdominal
sternite, under each pars stridens. It was verified that males and females emit
sounds by stridulation. Males stridulated during the courtship behavior. Females
also presented a movement similar to the stridulatory one, but a specific spectrum
was not registered. Males presented stridulation when they were disturbed by
pressures on the lateral parts of the abdomen. The temporary and spectral
characteristic of these sounds was different from that observed during courtship.
Females also stridulated when they were disturbed and the spectrum of sound
emission was different from that observed in males. In the treatments in which
females had their partes stridentes removed the percentage of couples that
coupled (mount and copulation) was reduced. Males spent more time and energy
trying to court the females, when their partes stridentes were removed. It was
evidenced the existence of stridulatory apparatus in A. ynca, morphologically
different in both sexes. In males it was possible to differentiate two types of
sounds, a disturbance stridulation and a courtship stridulation. The removal of the
pars stridens interferes in the mating behavior by increasing the number of mounts
and reducing the number of copulation, besides altering the duration of the pre-
copulation, copulation and post-copulation.

85
INTRODUÇÃO A comunicação é baseada na transmissão de uma mensagem, através de
sinais, de uma fonte emissora até um receptor. Estes sinais podem ser químicos,
visuais, táteis ou auditivos. Os sistemas de comunicação que utilizam o canal
acústico para a transmissão e recepção de sinais, têm como princípio a análise
dos parâmetros temporais e de freqüência destes sinais (Landa 2000).
Muitos sons e sinais produzidos por insetos são espécie - específicos e
utilizados principalmente para a localização, reconhecimento e formação de pares
(Cokl & Virant-Doberlet, 2003). O som, enquanto sinalizador de uma informação,
é representado pelas suas características estruturais, tais como o seu espectro e
padrão de modulação de amplitude, o que vai permitir ao receptor analisar e
localizar a mensagem mesmo quando vários sinais chegam simultaneamente de
diferentes fontes (Pollack 2000).
A estridulação, comum entre os besouros, pode ser definida como o
processo por meio do qual o som ou vibração é produzido pela fricção de duas
partes do corpo que se movem uma contra a outra (Cokl & Virant-Doberlet 2003).
Em Curculionidae a produção de som se dá pelo método de estridulação élitro-
abdominal, descrita por Durmortier, em 1963, e que consiste na fricção do
plectrum presente no abdômen sobre a pars stridens localizada nos élitros
(Durmortier 1963). Este mecanismo pode ser encontrado em várias espécies do
gênero Conotrachelus (Gibson 1967), Pissodes (Harman & Kranzler 1969),
Smicronyx (Hyder & Oseto 1989), Phrydiuchus (Wilson et al. 1993), entre outros
(Durmortier 1963).
A produção de som através da estridulação pode estar associada tanto a
interações inter e intra-específicas quanto a interações intra-sexuais. Segundo
Haskell (1974), para as duas primeiras interações, ambos os sexos produzem
uma estridulação específica e envolvida primariamente no comportamento de
defesa e alarme, que pode ser denominada estridulação de distúrbio. Já a
estridulação de corte é utilizada na comunicação intra-sexual, relacionada com os
comportamentos de corte e acasalamento.
O objetivo deste trabalho foi descrever a anatomia das estruturas
associadas à produção de som, analisar o som produzido e investigar o seu papel
no comportamento de acasalamento de Amerrhinus ynca.

86
MATERIAIS E MÉTODOS Os insetos foram coletados em coqueirais comerciais no município de
Quissamã, localizado na região Norte do Estado do Rio de Janeiro, e
transportados para o Laboratório de Entomologia e Fitopatologia. No laboratório,
os insetos foram mantidos dentro de potes de vidros (capacidade 3.250 ml) com
tampas teladas contendo fragmentos de cana-de-açúcar como alimento. A criação
foi mantida em sala com temperatura de 25 ± 2 °C e fotoperíodo de (12:12h) luz-
escuro.
Anatomia do aparato estridulatório - Para descrever a anatomia do aparato
estridulatório de A. ynca foram feitas observações em microscópio eletrônico e em
lupa estereoscópica com aumento de 40 vezes. A presença da pars stridens nos
élitros direito e esquerdo e do plectrum no abdômen de machos e fêmeas foi
confirmada pelas observações sob lupa estereoscópica utilizando-se cinco
indivíduos de cada sexo.
O detalhamento da estrutura localizada nos élitros direito e esquerdo de
ambos os sexos foi feito sob microscopia eletrônica de alta resolução. Para tal fim
se utilizou um indivíduo de cada sexo, o qual teve seus élitros retirados e
mantidos em dessecador por 48 horas, e submetidos a uma fina aplicação de
ouro. Já as observações do plectrum nos tergitos do abdômen foram feitas
diretamente com o uso de lupa estereoscópica. Tanto nas observações em
microscopia eletrônica quanto naquelas com lupa estereoscópica foram utilizados
insetos que morreram logo após o acondicionamento no laboratório.
Análise do som produzido - Uma imagem de vídeo, produzida em uma câmera
digital Sony® capaz de sincronizar som e imagem, foi utilizada para verificar de
que forma os insetos produziam o som. Nessa observação, um único indivíduo
macho foi perturbado, pressionando-se as laterais de seu abdômen com o polegar
e o dedo indicador, o tempo suficiente para que se conseguissem várias imagens
de sua estridulação.
Posteriormente foram registrados os sons de distúrbio produzidos por
machos e fêmeas e o de corte produzido pelos machos durante o processo de

87
acasalamento. O som foi gravado utilizando-se um gravador digital Sony® MZ-
R70 e um microfone condensador Sennheiser (MM6/K6P).
A estridulação de distúrbio foi registrada ao se pressionar a lateral do
abdômen de sete indivíduos de cada sexo (n=7) selecionados aleatoriamente da
população vinda do campo. Cada registro individual durou 60 segundos. Já a
estridulação de corte produzida por machos de A. ynca durante o acasalamento
foi gravada a partir de sete casais (n=7) mantidos individualizados dentro de
caixas Gerbox® (10X10X5 cm). A gravação teve início assim que o macho
entrava em contato com a fêmea. Caso ocorresse a monta, a gravação
continuava até o início da cópula, caso contrário era interrompida até uma nova
tentativa de monta. Os sons de corte e de distúrbio foram gravados nos finais de
semana para se evitar ao máximo a interferência de sons externos, uma vez que
não havia dentro da Universidade uma sala acusticamente apropriada.
No Laboratório de Ciências Ambientais, setor de bioacústica da
Universidade Estadual do Norte Fluminense, as gravações foram digitalizadas em
um computador Mac Os9, utilizando-se o software de pesquisa bioacústica Canary
(versão 1.2). A digitalização foi feita a uma taxa amostral de 44100 Hz e
transferida em formato de 16 bits por amostra. O gravador utilizado na
digitalização foi o mesmo utilizado nas gravações, garantindo-se assim igualdade
de velocidade e reprodução entre coleta e processamento de sons. Após a
digitalização dos sons gravados (14 sons de distúrbio e sete de corte) seus
respectivos oscilogramas e espectrogramas foram criados utilizando-se o Canary
(Figuras 1 e 2).
As análises das variáveis acústicas (Tabela 1) foram feitas da seguinte
maneira: para os sons produzidos por machos (distúrbio e corte) foram escolhidos
aleatoriamente, e sempre que possível, cinco cantos dentro do intervalo de tempo
total da gravação (para o som de distúrbio o tempo total de gravação foi de 60
segundos). Foram analisados o primeiro pulso destes cinco cantos e, sempre que
possível, mais cinco pulsos escolhidos aleatoriamente. Nem sempre foi possível
selecionar cinco pulsos, porque alguns cantos eram compostos por três ou quatro
pulsos e em alguns casos havia menos de cinco cantos por tempo de gravação.
A grande dificuldade encontrada no momento de analisar os sons
produzidos pelas fêmeas foi o fato de que elas estridulam praticamente dentro da
faixa de ruído ambiental (± 2 kHz). Assim, ao se construir o espectrograma das

88
sete gravações feitas para as fêmeas, foi muito difícil separar o que era ruído do
que era estridulação. Apenas em duas fêmeas foi possível fazer nitidamente esta
distinção e proceder às análises. Dessa forma, foram analisados 35 pulsos
consecutivos dentro do intervalo de 60 segundos para cada uma das duas
gravações.
As variáveis temporais e espectrais dos sons produzidos por machos e
fêmeas foram medidas a partir de gráficos do espectrograma (Resolução de
freqüência = 86,13 Hz, Resolução de tempo = 1,45 ms e FFT Size = 512 pts).
Tabela 1 – Definições das variáveis acústicas medidas para as estridulações de corte (macho) e distúrbio (macho e fêmea) de Amerrhinus ynca.
Variáveis Definição Sexo analisado
Duração do pulso (ms) Tempo total de duração do pulso. Macho e fêmea
Duração total do canto (s) Tempo total do canto, medido a partir do início do primeiro pulso até o final do último pulso.
Macho
Duração interpulso (ms) Tempo de duração entre dois pulsos consecutivos.
Macho e fêmea
Freqüência máxima (kHz) Maior freqüência do pulso. Macho e fêmea
Freqüência mínima (kHz) Menor freqüência do pulso (padronizada em 1kHz).
Macho e fêmea
Intensidade média (dB) Intensidade média na porção selecionada do espectrograma.
Macho e fêmea
Taxa de repetição do canto Freqüência de cantos por unidade de tempo. Macho
Quantidade de pulsos Quantidade de pulsos do canto. Macho
Quantidade de cantos Quantidade de cantos presentes no tempo total de gravação. Macho
A comparação dos parâmetros dos sons de distúrbio e de corte
produzidos pelos machos foi feita utilizando-se 7 cantos e cinco pulsos por canto
(excluindo-se o primeiro pulso), totalizando 35 pulsos por indivíduo. Já para se
comparar o primeiro pulso com os demais foram utilizados os primeiros pulsos de
cada canto mais cinco pulsos distintos do primeiro, escolhidos aleatoriamente,
totalizando cinco pulsos por indivíduo para cada tipo de pulso (primeiro e demais

89
pulsos). Para comparar a estridulação de distúrbio de machos e de fêmeas, foram
selecionados aleatoriamente dois machos. Destes, 25 pulsos foram escolhidos
aleatoriamente entre os cinco cantos dos dois machos para se comparar os
parâmetros espectrais (freqüência máxima e intensidade média) e temporais
(duração do pulso).
As comparações dos dados observados foram feitas através da análise
de variância utilizando-se o software de análise estatística Statistica v. 7.0.
Papel do som no acasalamento - Para se verificar o papel da estridulação no
comportamento de acasalamento de A. ynca foram realizados quatro tratamentos:
(i) casais não manipulados, (ii) casais em que ambos os sexos tiveram parte de
seus élitros direito e esquerdo removida, (iii) casais dos quais apenas o macho
teve parte de seus élitros removida e (iv) casais dos quais apenas as fêmeas
tiveram seus élitros direito e esquerdo removidos. A parte dos élitros que foi
removida correspondia ao local onde se encontra a pars stridens.
Os casais, mantidos individualizados dentro de caixas Gerbox® (10 x 10 x
5 cm) foram observados durante cinco horas. Cada tratamento foi dividido em seis
módulos com cinco casais por tempo de observação (n=30). Eles foram
observados em sala climatizada (temperatura de 25º C) e durante a fotofase,
alternando manhã e tarde.
As variáveis registradas neste experimento foram: a freqüência de monta
e cópula, a freqüência dos atos comportamentais, a duração do período de
latência, pré-cópula, cópula e pós-cópula (Tabela 2). Sendo que, para a
freqüência de monta se considerou toda a monta ocorrida durante as cinco horas
de observação e para a freqüência de cópula, o número de cópulas que
ocorreram desde o início da primeira monta até a descida do macho do dorso da
fêmea. Para os dados de freqüência dos atos comportamentais foram
considerados nas análises aqueles que mais ocorreram durante a primeira pré-
cópula (Tabela 2).
Como os machos precisam se fixar na fêmea, agarrando com as pernas
posteriores as laterais de seu abdômen, o efeito mecânico da retirada de parte
dos élitros pode interferir nos resultados, impedindo-se a permanência dos
machos sobre o dorso das fêmeas e dificultando o acasalamento. Por isso, um
novo tratamento foi realizado colando-se os élitros das fêmeas no abdômen,

90
utilizando-se corretivo líquido a base d’água, e impedindo-se que elas
produzissem algum tipo de som. Neste caso foram utilizados 30 casais, nos quais
foi analisado se houve ou não monta, durante as cinco horas de observação.
As comparações entre os tratamentos foram feitas através do software de
análise estatística Statistica© v.7.0, utilizando-se a análise de variância.
Tabela 2 – Descrição dos atos comportamentais de Amerrhinus ynca e das variáveis utilizados nas comparações entre os tratamentos.
Variáveis Descrição
Freqüência de monta Todas as montas que ocorreram durante as cinco horas de observações.
Freqüência de cópula
Todas as cópulas que ocorreram durante todo o primeiro tempo em que os machos permaneceram sobre o dorso das fêmeas, ou seja, desde o momento da primeira monta até o momento em que os machos descem do dorso da fêmea.
Freqüência dos atos comportamentais
Todos os atos comportamentais que ocorreram durante a primeira pré-cópula.
Latência Tempo decorrido desde o início das observações até o início da monta.
Duração da pré-cópula Tempo decorrido desde a monta até o início da cópula.
Duração da cópula Período que os machos permanecem com seu edeago introduzido dentro da genitália da fêmea.
Duração da pós-cópula Tempo decorrido desde a retirada do edeago da genitália da fêmea até a descida do macho de seu dorso.
Ato Comportamental Descrição
Estridulação Produção de som pela fricção dos élitros contra o abdômen.
Estimular o protórax Esfregar o espinho localizado nas tíbias das pernas anteriores do macho nas laterais do protórax da fêmea.
Estimular o abdômen Esfregar o espinho localizado na tíbia das pernas posteriores do macho nas laterais do abdômen da fêmea.
Tentativas de cópula O macho expõe a genitália e faz tentativas de cópula.
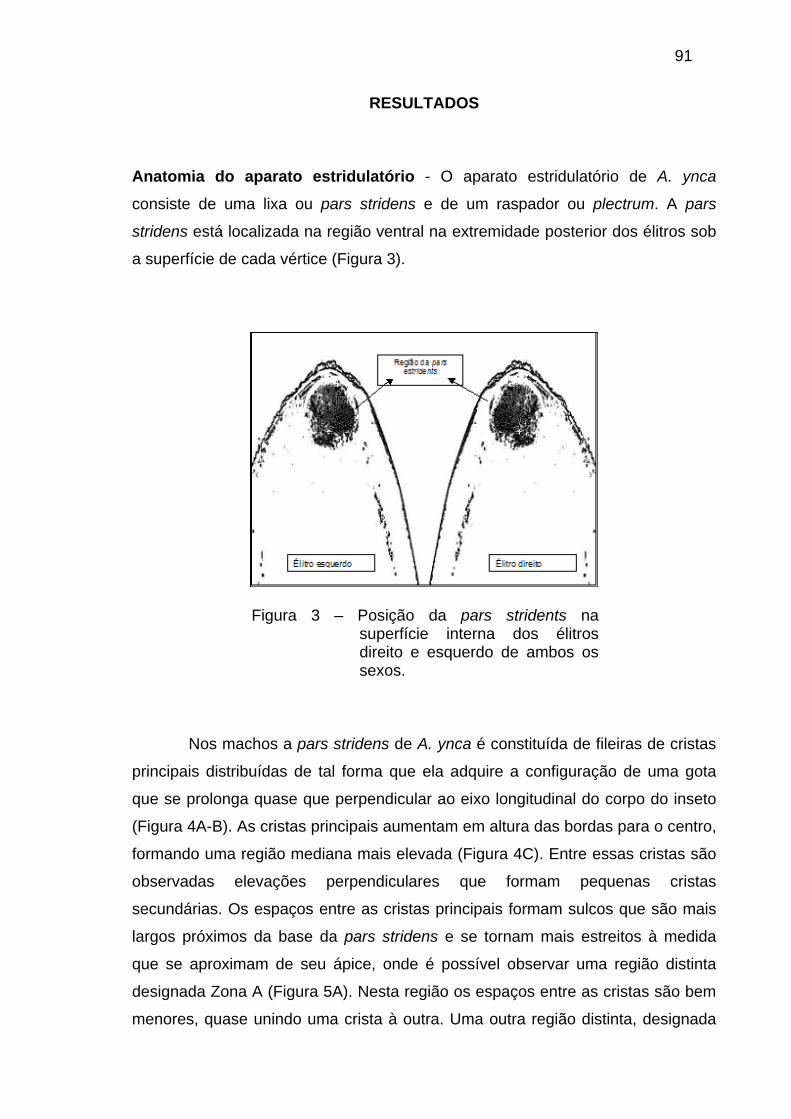
91
RESULTADOS
Anatomia do aparato estridulatório - O aparato estridulatório de A. ynca
consiste de uma lixa ou pars stridens e de um raspador ou plectrum. A pars
stridens está localizada na região ventral na extremidade posterior dos élitros sob
a superfície de cada vértice (Figura 3).
Figura 3 – Posição da pars stridents na superfície interna dos élitros direito e esquerdo de ambos os sexos.
Nos machos a pars stridens de A. ynca é constituída de fileiras de cristas
principais distribuídas de tal forma que ela adquire a configuração de uma gota
que se prolonga quase que perpendicular ao eixo longitudinal do corpo do inseto
(Figura 4A-B). As cristas principais aumentam em altura das bordas para o centro,
formando uma região mediana mais elevada (Figura 4C). Entre essas cristas são
observadas elevações perpendiculares que formam pequenas cristas
secundárias. Os espaços entre as cristas principais formam sulcos que são mais
largos próximos da base da pars stridens e se tornam mais estreitos à medida
que se aproximam de seu ápice, onde é possível observar uma região distinta
designada Zona A (Figura 5A). Nesta região os espaços entre as cristas são bem
menores, quase unindo uma crista à outra. Uma outra região distinta, designada
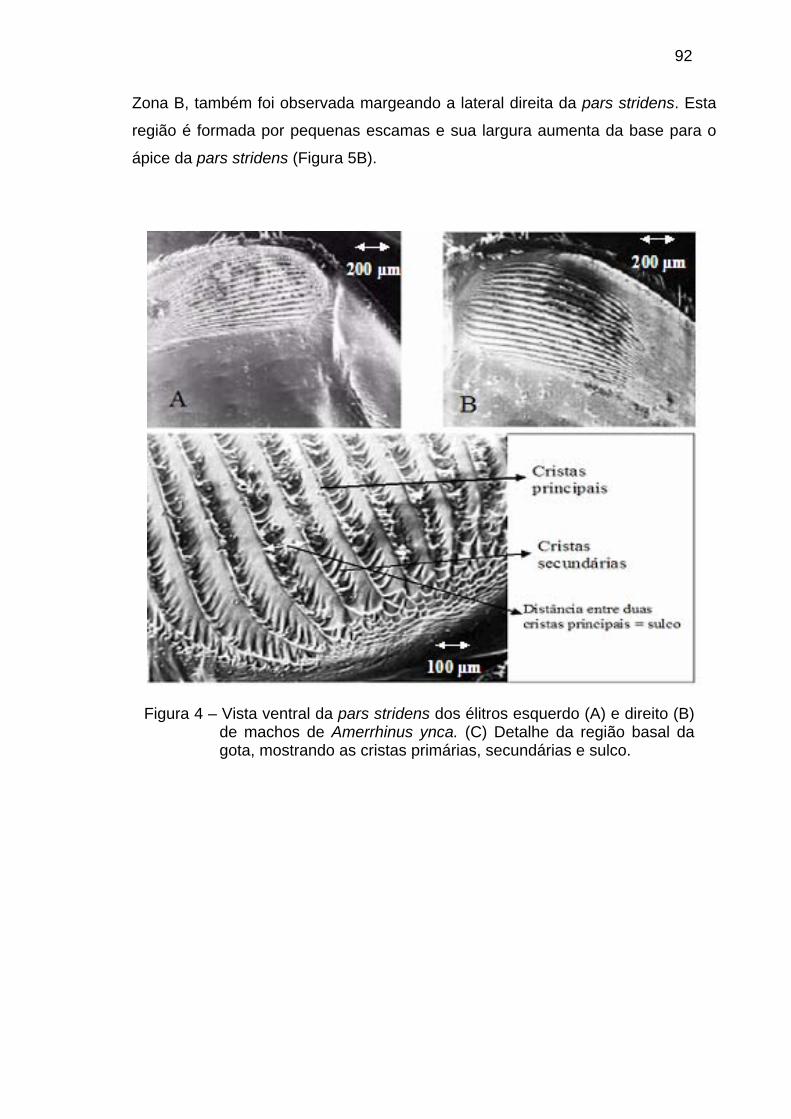
92
Zona B, também foi observada margeando a lateral direita da pars stridens. Esta
região é formada por pequenas escamas e sua largura aumenta da base para o
ápice da pars stridens (Figura 5B).
Figura 4 – Vista ventral da pars stridens dos élitros esquerdo (A) e direito (B) de machos de Amerrhinus ynca. (C) Detalhe da região basal da gota, mostrando as cristas primárias, secundárias e sulco.

93
Figura 5 – Vista da pars stridens (PS) do élitro direito de machos de Amerrhinus ynca mostrando as duas regiões distintas (Zona A e B).
Nas fêmeas de A. ynca a pars stridens é constituída de fileiras de cristas,
mas estas estão distribuídas de forma irregular, são similares em altura e
apresentam sulcos de largura homogênea (Figura 6B). Na pars stridens das
fêmeas não foram observadas cristas secundárias.
Figura 6 – Vista ventral dos élitros direitos de macho (A) e fêmea (B), mostrando
as diferenças estruturais na pars stridens de Amerrhinus ynca.

94
Tanto os machos quanto as fêmeas de A. ynca apresentam dois plectra,
eqüidistantes, localizados na margem anterior do sétimo esternito abdominal,
abaixo da pars stridens dos élitros esquerdo e direito (Figura 7A). Os plectra são
constituídos por dentes esclerotizadas que se prolongam lateralmente (Figura
7B). Sob a superficie do sétimo tergito abdominal é possível verificar a presença
de pontuações que são mais evidentes nos machos do que nas fêmeas (Figura
7A). Estas pontuações também são esclerotizadas, mas não foi observado se
elas atuam na produção de som.
Figura 7 - Desenho esquemático do abdômen de macho e fêmea de Amerrhinus ynca. (A) vista ventral mostrando os dois dentes plectrais e as pontuações sobre o penúltimo tergito abdominal do macho e no pigídio da fêmea. (B) vista lateral mostrando o prolongamento dorsolateral do dente plectral.
Análise da produção do som - Através da imagem de vídeo foi possível
observar que a produção de som em machos de A. ynca coincide com a fricção
do plectrum contra a pars stridens, sendo que o som é produzido ao se
movimentar o abdômen em um golpe para frente e outra para trás (Figura 8).
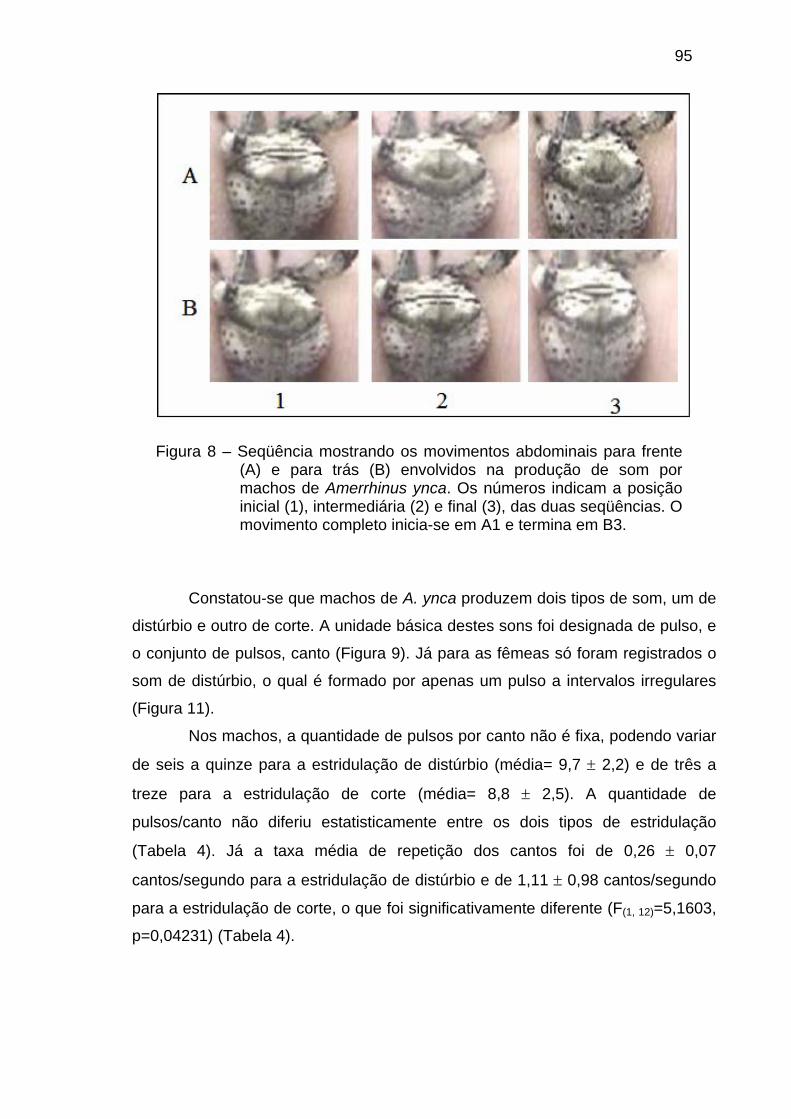
95
Figura 8 – Seqüência mostrando os movimentos abdominais para frente (A) e para trás (B) envolvidos na produção de som por machos de Amerrhinus ynca. Os números indicam a posição inicial (1), intermediária (2) e final (3), das duas seqüências. O movimento completo inicia-se em A1 e termina em B3.
Constatou-se que machos de A. ynca produzem dois tipos de som, um de
distúrbio e outro de corte. A unidade básica destes sons foi designada de pulso, e
o conjunto de pulsos, canto (Figura 9). Já para as fêmeas só foram registrados o
som de distúrbio, o qual é formado por apenas um pulso a intervalos irregulares
(Figura 11).
Nos machos, a quantidade de pulsos por canto não é fixa, podendo variar
de seis a quinze para a estridulação de distúrbio (média= 9,7 ± 2,2) e de três a
treze para a estridulação de corte (média= 8,8 ± 2,5). A quantidade de
pulsos/canto não diferiu estatisticamente entre os dois tipos de estridulação
(Tabela 4). Já a taxa média de repetição dos cantos foi de 0,26 ± 0,07
cantos/segundo para a estridulação de distúrbio e de 1,11 ± 0,98 cantos/segundo
para a estridulação de corte, o que foi significativamente diferente (F(1, 12)=5,1603,
p=0,04231) (Tabela 4).

96
Figura 9 - Oscilograma mostrando a forma das ondas dos cantos de estridulação de distúrbio (A) e de corte (B) produzidos por machos de Amerrhinus ynca.
Tabela 4 – Pulsos por canto e taxa de repetição dos cantos de machos de Amerrhinus ynca.
Pulsos/canto (n=35) Cantos/segundo (n=7)
Estridulação Média1 ± sd amplitude Média2 ± sd* Amplitude
Distúrbio 9,7 ± 2,2 6,0 – 15,0 0,26 ± 0,07* 0,13 – 0,33
Corte 8,8 ± 2,5 3,0 – 13,0 1,11 ± 0,98* 0,30 – 3,03 * Diferença significativa (ANOVA P < 0,001).
Outra diferença constatada foi que a duração do pulso (F(1, 316)=6,1592,
p=0,01) e a intensidade média (F(1, 316)=17,658, p=0,001) são maiores na
estridulação de distúrbio, enquanto que o alcance máximo da freqüência (F(1,
316)=74,652, p=0,00000), em média, foi maior na estridulação de corte (Tabela 5 e
Figura 10). A duração do intervalo entre pulsos não difere entre os dois tipos de
estridulação.
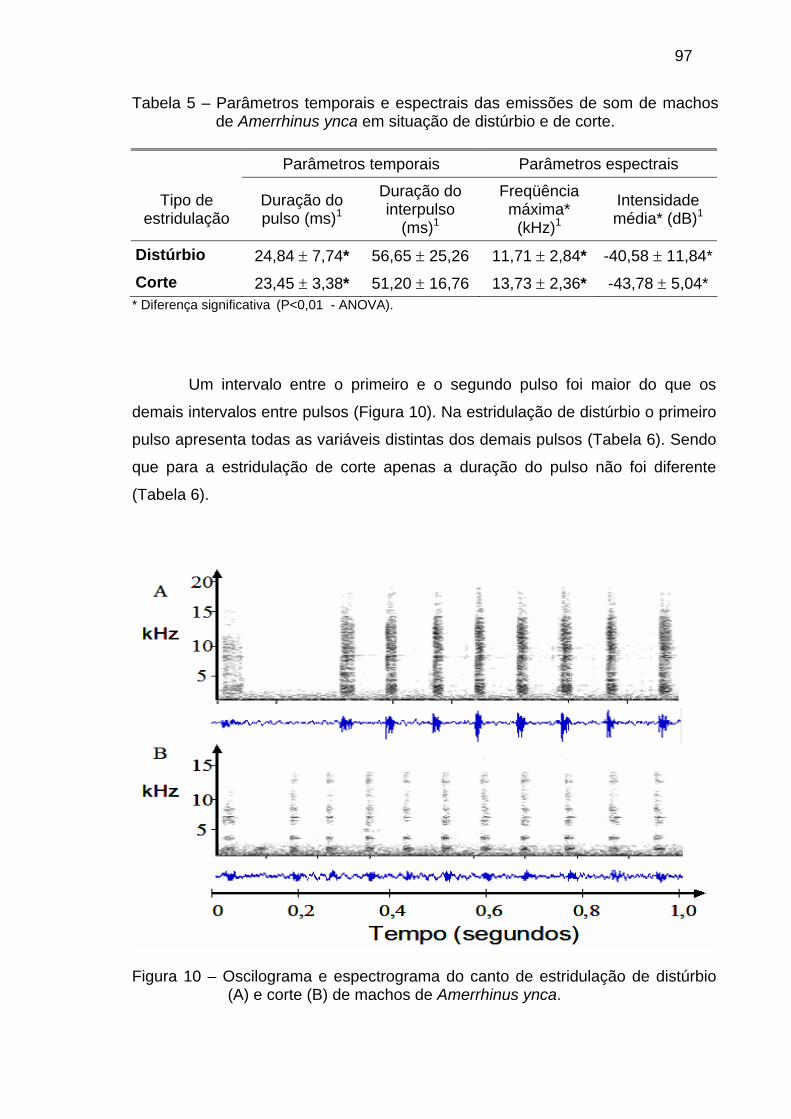
97
Tabela 5 – Parâmetros temporais e espectrais das emissões de som de machos de Amerrhinus ynca em situação de distúrbio e de corte.
Parâmetros temporais Parâmetros espectrais
Tipo de estridulação
Duração do pulso (ms)1
Duração do interpulso
(ms)1
Freqüência máxima*
(kHz)1
Intensidade média* (dB)1
Distúrbio 24,84 ± 7,74* 56,65 ± 25,26 11,71 ± 2,84* -40,58 ± 11,84*Corte 23,45 ± 3,38* 51,20 ± 16,76 13,73 ± 2,36* -43,78 ± 5,04* * Diferença significativa (P<0,01 - ANOVA).
Um intervalo entre o primeiro e o segundo pulso foi maior do que os
demais intervalos entre pulsos (Figura 10). Na estridulação de distúrbio o primeiro
pulso apresenta todas as variáveis distintas dos demais pulsos (Tabela 6). Sendo
que para a estridulação de corte apenas a duração do pulso não foi diferente
(Tabela 6).
Figura 10 – Oscilograma e espectrograma do canto de estridulação de distúrbio (A) e corte (B) de machos de Amerrhinus ynca.

98
Tabela 6 – Média dos parâmetros temporais e espectrais do primeiro pulso e interpulso em relação aos demais pulsos e interpulsos, para a estridulação de corte e de distúrbio em machos de Amerrhinus ynca.
Tipo de pulso/interpulso Tipo de estridulação Parâmetros
Primeiro Demais
Duração do pulso (ms) 36,34 ± 11,83* 24,36 ± 5,67*
Duração interpulso (ms) 151,56 ± 64,27* 59,42 ± 23,30*
Freqüência máxima (kHz) 8,71 ± 4,26* 11,69 ± 2,84* Distúrbio
Intensidade média (dB) -45,68 ± 4,80* -41,41 ± 6,93*
Duração do pulso (ms) 25,93 ± 7,17 NS 23,67 ± 3,35 NS
Duração do interpulso (ms) 96,93 ± 46,83* 50,21 ± 21,13*
Freqüência máxima (kHz) 11,74 ± 2,99* 13,63 ± 2,30* Corte
Intensidade média (dB) -48,26 ± 4,65* -44,58 ± 5,10* * Diferença significativa (P< 0,001, ANOVA).
Para as fêmeas de A. ynca só foi possível descrever o som de distúrbio
produzido por duas fêmeas. Para as duas fêmeas analisadas apenas a
intensidade média não diferiu significativamente (Tabela 7).
Tabela 7 – Média ± desvio padrão dos parâmetros temporais e espectrais do canto produzido pela estridulação de distúrbio de fêmeas de Amerrhinus ynca.
Média1 ± desvio padrão
Parâmetros Fêmea 01 Fêmea 02
Duração do pulso (ms) 39,4 ± 7,8* 83,3 ± 11,7*
Intervalo entre cantos (s) 0,8 ± 0,7 * 0,6 ± 0,3*
Freqüência máxima (kHz) 7,8 ± 3,5* 5,6 ± 4,0*
Intensidade média (dB) -42,3 ± 2,3 -41,7 ± 3,1 * Diferença significativa (P<0,05 – ANOVA).
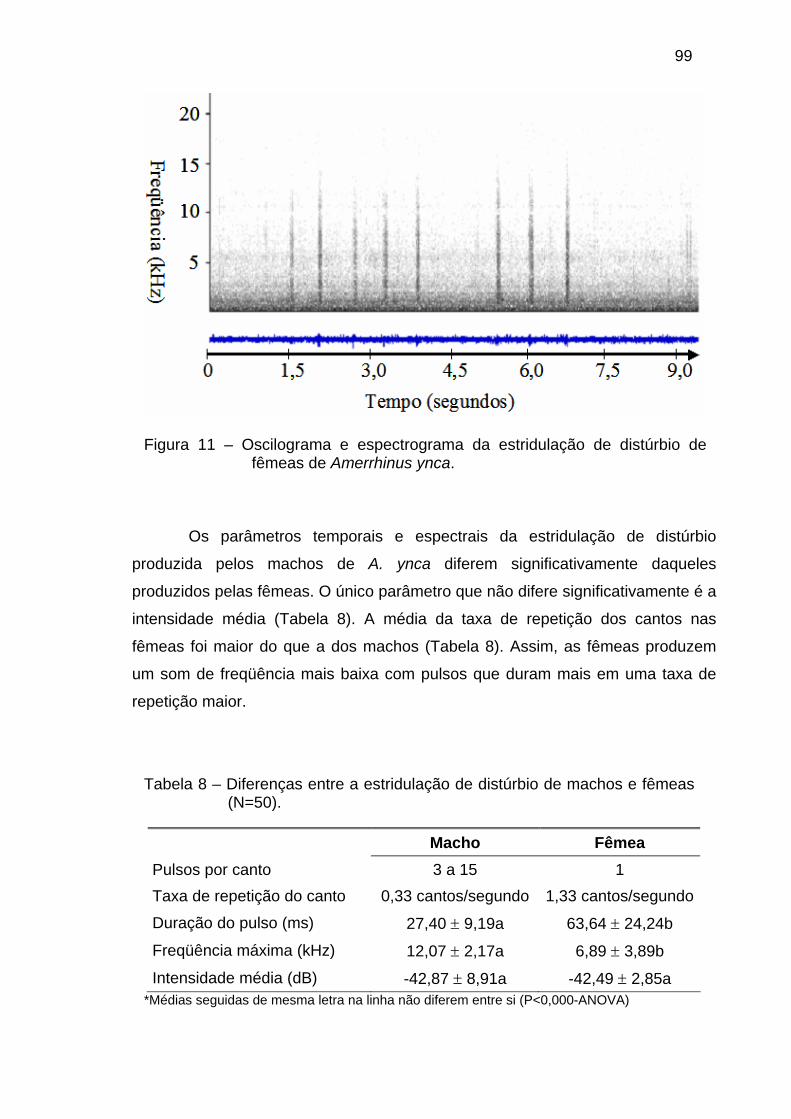
99
Figura 11 – Oscilograma e espectrograma da estridulação de distúrbio de fêmeas de Amerrhinus ynca.
Os parâmetros temporais e espectrais da estridulação de distúrbio
produzida pelos machos de A. ynca diferem significativamente daqueles
produzidos pelas fêmeas. O único parâmetro que não difere significativamente é a
intensidade média (Tabela 8). A média da taxa de repetição dos cantos nas
fêmeas foi maior do que a dos machos (Tabela 8). Assim, as fêmeas produzem
um som de freqüência mais baixa com pulsos que duram mais em uma taxa de
repetição maior.
Tabela 8 – Diferenças entre a estridulação de distúrbio de machos e fêmeas (N=50).
Macho Fêmea Pulsos por canto 3 a 15 1 Taxa de repetição do canto 0,33 cantos/segundo 1,33 cantos/segundo
Duração do pulso (ms) 27,40 ± 9,19a 63,64 ± 24,24b
Freqüência máxima (kHz) 12,07 ± 2,17a 6,89 ± 3,89b
Intensidade média (dB) -42,87 ± 8,91a -42,49 ± 2,85a *Médias seguidas de mesma letra na linha não diferem entre si (P<0,000-ANOVA)

100
Papel do som no acasalamento - Houve redução no número de casais que
copularam quando a pars stridens dos machos foi removida. Entretanto, quando a
pars stridens das fêmeas foi removida tanto o número de montas quanto os de
cópulas sofreram reduções (Figura 12). O mesmo ocorreu quando as partes
stridentes de ambos os sexos foram removidas. Já quando os élitros das fêmeas
foram colados, apenas 43% dos machos chegaram a montar pelo menos uma
vez.
Figura 13 – Quantidade de casais que montaram e copularam pelo menos uma vez (n=30).
As freqüências de monta (F3,116=3,2579, p=0,024) e de cópula
(F3,116=9,2087, p=0,000) foram significativamente diferentes para os quatro
tratamentos (Figura 14).
O período de latência e a duração do cortejo nos casais não manipulados
diferiram dos demias tratamentos. A duração da cópula foi maior (F3, 110=8,0667,
p=0,00007) e o tempo de guarda das fêmeas também foi maior (F3, 107=5,1276,
p=,002) (Figura 15D) nos casais não manipulados.

101
Quanto aos atos comportamentais envolvidos no comportamento de pré-
cópula (estridulação1, tentativa e esfregar com as pernas anteriores e
posteriores), não houve diferença significativa entre os tratamentos. Entretanto, as
médias de freqüência destes atos comportamentais foram maiores no tratamento
3 do que nos demais tratamentos (Figura 14).
Figura 14 – Freqüência média dos atos comportamentais envolvidos no comportamento de pré-cópula dos machos de Amerrhinus ynca para os quatro tratamentos.
DISCUSSÃO
Aparato estridulatório - Como em outros curculionídeos, a pars stridens de A.
ynca também se localiza na superfície interna dos élitros. Harman & Harman
(1972), ao estudarem o aparato estridulatório de Pissodes strobi (Peck, 1817)
(Coleoptera: Curculionidae), observaram que tanto o élitro direito quanto o
esquerdo de ambos os sexos contém a pars stridens e que esta estrutura está
1 Os movimentos do abdômen dos machos nos tratamentos 2 e 3 foram considerados como estridulatórios, apesar de os machos não terem produzido som.

102
localizada em sua superfície interna, próxima à sutura apical do vértice, junto com
outras áreas distintas que a circundam. Em Smicronyx fulvus LeConte, 1876 e
Smicronyx sordidus LeConte, 1876, Hyder & Oseto (1989) também descreveram
a localização da pars stridens de ambas as espécies como sendo sob a superfície
de cada ápice elitral. O mesmo também ocorre em Phrydiuchus tau Warner , 1969
(Coleoptera: Curculionidae) (Wilson et al. 1993). Já em quatro espécies do gênero
Conotrachelus (C. naso LeConte, 1876; C. posticatus Boheman, 1837; C. carinifer
Casey, 1892; e C. nenuphar (Herbst, 1797)), a pars stridens está localizada na
borda do élitro esquerdo, estendendo-se sob o direito, e freqüentemente pode
ocorrer nos machos uma pars stridens menor, sobre a borda do élitro direito
(Gibson 1967).
Diferenças entre a pars stridens de machos e de fêmeas com relação ao
espaçamento e à altura das cristas também foram observadas em outros
curculionídeos. Hyder & Oseto (1989) observaram diferenças na altura das cristas
de machos e fêmeas de ambas as espécies do gênero Smicronyx, sendo que nos
machos as cristas são mais altas do que nas fêmeas. Gibson (1967) observou
que em C. posticatus a pars stridens dos machos varia quanto à altura e ao
espaçamento das cristas, enquanto nas fêmeas esta variação é aparentemente
uniforme. O autor menciona ainda que a variação em altura e espaçamento das
cristas da pars stridens dos machos aumenta a partir da porção anterior para a
posterior. Hyder & Oseto (1989) mencionam uma área mediana da pars stridens
formada pela variação na altura das cristas nas duas espécies do gênero
Smicronyx, que ficou caracterizada como a região ótima de produção de som. A
mesma variação em altura mencionada por estes autores ocorre na pars stridens
dos machos de Amerrhinus ynca, criando uma região central onde as cristas são
mais altas. Provavelmente esta região central também esteja ligada à produção
ótima de som em A. ynca.
Em curculionídeos o plectrum pode estar localizado no sexto ou no sétimo
tergito abdominal. Assim como em machos e fêmeas de A. ynca o plectrum de
Phrydiuchus tau está localizado, segundo Wilson et al. (1993), na margem
anterior do sétimo tergito abdominal e consiste de duas estruturas pareadas que
se estendem lateralmente. O plectrum de ambos os sexos de P. strobi também
está localizado no sétimo tergito abdominal, entretanto, segundo Harman &
Harman (1972), o plectrum nesta espécie consiste de duas fileiras de dentes, uma

103
em cada lado do tergito, sendo que o número de dentes por fileiras varia entre os
sexos. Já para S. fulvus e S. sordidus, existem dois plectra, um localizado no
sexto tergito abdominal e outro situado na margem anterior do sétimo tergito
abdominal de ambos os sexos (Hyder & Oseto 1989). Nas espécies do gênero
Conotrachelus estudado por Gibson (1967), o plectrum está localizado em uma
porção mediana do sexto tergito abdominal e é constituído de pares de
tubérculos. Segundo esse autor, C. naso, C. posticatus e C. carinifer possuem
dois pares de tubérculos e o plectrum de machos e fêmeas são similares, ao
passo que em C. nenuphar o plectrum do macho é constituído de apenas um par
de tubérculos e as fêmeas são desprovidas desta estrutura estridulatória.
Produção de som - A produção de som em A. ynca se dá pela fricção do
plectrum localizado no sétimo tergito abdominal contra a pars stridens do élitro,
através de dois movimentos abdominais: um golpe para frente e outro para trás.
Em P. tau (Wilson et al. 1993) e em Conotrachelus sp. (Gibson 1967), cada dois
pulsos produzidos pela estridulação são resultados de um golpe para frente e
outro para trás. Isto pode ser verdadeiro para A. ynca, se o número de pulsos por
canto for sempre um número par, a não ser que o último pulso produzido seja de
intensidade e freqüência abaixo da faixa de ruído e não tenha sido detectado
neste estudo.
A presença de dois sons distintos também foi observada por Wilson et. al
(1993) em machos de P. tau. Gibson (1967) ao estudar a produção de som em
Conotrachelus só faz menção à estridulação de distúrbio produzida por C. naso,
C. carinifer e C. posticatus. Mas, segundo o autor, as estridulações individuais
produzidas por estas três espécies variam fortemente quanto à velocidade e à
freqüência do som. Hyder & Oseto (1989) também encontraram grande
variabilidade no alcance de freqüência nas estridulações produzidas por machos
de Smicronyx sordidus e Smicronyx fulvus. Segundo os autores, esta variação
está associada à anatomia do aparato estridulatório, principalmente à variação
estrutural da pars stridens. Outras espécies de coleópteros, como o besouro
Aphodius ater (Degeer, 1774) (Coleoptera: Scarabaeidae), também exibem uma
alta variabilidade individual ao estridular (Hirschberger, 2001).
Diferenças entre o som produzido por machos e fêmeas também foram
observadas dentro dos curculionídeos. Em S. sordidus e S. fulvus os machos

104
produzem sons com alcance de freqüência mais alta do que o das fêmeas (Hyder
& Oseto 1989). Em P. strobi e P. approximatus Hopkins, 1911, além do alcance
de freqüência, a taxa de repetição do canto também variou entre os sexos, sendo
mais rápida nos machos do que nas fêmeas (Harman & Kranzler, 1969). Entre os
sexos de C. naso e C. posticatus, Gibson (1967) observou diferenças quanto à
tonalidade dos pulsos produzidos por machos e fêmeas.
Portanto, este trabalho corrobora os demais estudos da comunicação
acústica em coleópteros evidenciando que os dois sexos de A. ynca produzem
sons. Estes sons são diferentes e são produzidos por aparatos estridulatórios
morfologicamente distintos, sendo que os machos possuem um aparato com
maior capacidade de produção de som que o das fêmeas. Nos machos foi
possível diferenciar dois tipos de sons, uma estridulação de distúrbio associada a
encontros ou disputas com outros machos e uma estridulação de corte que foi
registrada durante o acasalamento.
Papel da estridulação no comportamento de corte e acasalamento - A
remoção da pars stridens de machos e fêmeas de A. ynca afetou o
comportamento de corte e acasalamento. Através dos resultados é possível
perceber que nos tratamentos em que as fêmeas tiveram sua pars stridens
removida a porcentagem de casais que acasalaram (montas e cópulas) reduziu.
Mesmo quando as fêmeas tiveram seus élitros colados no abdômen a
porcentagem de acasalamentos (montas) também sofreu redução. Provavelmente
as fêmeas utilizam uma estridulação específica para sinalizar ao macho que
aceitam sua corte e, conseqüentemente, a cópula. Entretanto, outros sinais
liberados pelas fêmeas, como odores, também podem servir como sinalizadores e
agir sinergisticamente com os sinais acústicos. Segundo Haskell (1974), em um
padrão de comportamento de corte no qual o inseto utiliza a sinalização sonora
para estimulação e discriminação específica, outros estímulos (visuais, olfativos e
táteis) também são utilizados, mais como uma regra do que como uma exceção.
Os machos, por sua vez, gastam mais tempo tentando cortejar as fêmeas,
quando sua pars stridens é retirada. Entretanto, a ausência de som não os
impede de copular, mesmo porque outros atos comportamentais podem ser
utilizados no cortejamento, como esfregar as pernas anteriores e posteriores nas
laterais do corpo da fêmea. De qualquer forma, o fato de a fêmea não ouvir os

105
sons produzidos pelos machos faz com que eles gastem mais tempo na pré-
cópula e menos tempo em cópula e pós-cópula.
RESUMOS E CONCLUSÕES Os insetos que utilizam a comunicação acústica nas relações
interindividuais podem produzir suas emissões sonoras através de distintos
meios, a estridulação é o mais difundido. Em Curculionidae a estridulação se dá
através da fricção do plectrum do abdômen com a pars stridens localizada nos
élitros. Este trabalho descreveu a morfologia do aparato estridulatório de
Amerrhinus ynca, analisou as suas emissões sonoras e investigou o papel da
estridulação no seu comportamento de corte.
Os sons foram gravados com gravador digital e analisados com ajuda do
software Canary. Para verificar o papel da estridulação no comportamento de
acasalamento de A. ynca foram realizados quatro tratamentos: (i) casais não
manipulados, (ii) casais em que ambos os sexos tiveram parte de seus élitros
direito e esquerdo removidos, (iii) casais nos quais apenas o macho teve parte de
seus élitros removidos e (iv) casais nos quais apenas as fêmeas tiveram seus
élitros direito e esquerdo removidos. A parte dos élitros que foi removida
corresponde ao local onde se encontram as pars stridens. Os casais foram
observados durante cinco horas. Analisou-se a freqüência de monta e cópula, a
freqüência dos atos comportamentais, a duração do período de latência, pré-
cópula, cópula e pós-cópula.
A pars stridens de A. ynca está localizada na região ventral da
extremidade posterior dos élitros direito e esquerdo. Nos machos ela é constituída
de fileiras de cristas de altura e espaçamentos variáveis. Nas fêmeas as cristas
são menos acentuadas e estão distribuídas de forma irregular. Machos e fêmeas
apresentaram dois plectra localizados no sétimo esternito abdominal, debaixo de
cada pars stridens. Verificou-se que machos e fêmeas emitem sons por
estridulação e que os machos estridulam durante o comportamento de corte e
quando são pertudados através de pressões nas laterais do abdômen. A
característica temporal e espectral destes sons foi distinta. As fêmeas também
apresentaram movimento similar ao estridulatório durante o acasalamento, mas

106
um espectro específico não foi registrado, apenas foi possível analisar o som
produzido quando as fêmeas eram pertubadas e o espectro de emissão do som
foi diferente do observado nos machos. Evidenciou-se a existência de aparato
estridulatório em A.ynca, morfologicamente diferente nos dois sexos. Nos machos
foi possível diferenciar dois tipos de sons, uma estridulação associada
possivelmente a encontros com outros machos e uma estridulação de corte.
Nos tratamentos nos quais as fêmeas tiveram sua pars stridens removida
a porcentagem de casais que acasalaram (montas e cópulas) reduziu. Já os
machos gastaram mais tempo tentando cortejar as fêmeas, quando sua pars
stridens foi retirada. A remoção da par stridens interfere no acasalamento,
aumentando o número de montas e reduzindo o número de cópula, além de
alterar a duração da pré-cópula, cópula e pós-cópula.
Este trabalho corrobora, portanto, os demais estudos da comunicação
acústica em coleópteros, evidenciando que os dois sexos de A. ynca produzem
sons e apresentam aparatos estridulatórios morfologicamente distintos. Nos
machos foi possível diferenciar dois tipos de sons, uma estridulação de distúrbio
associada a encontros ou disputas com outros machos e uma estridulação de
corte que foi registrada durante o acasalamento.
A remoção da pars stridens interfere no acasalamento, aumentando o
número de montas e reduzindo o número de cópulas, além de alterar a duração
da pré-cópula, cópula e pós-cópula.
REFERÊNCIAS BIBLIOGRÁFICAS
Cokl, A. & Virant-Doberlet, M. (2003) Communication with substrate-borne signals in small plant-dwelling insects. Annual Review of Entomology, 48:29 – 50.
Durmortier, B. (1963) Morphology of sound emission apparatus in Arthropod. In: R. G. Busnel (ed.). Acoustic Behavior of Animals. New York: Elsevier Publishing Co., 933 p.
Gibson, L. P. (1967) Stridulatory mechanisms and sound production in Conotrachelus (Coleoptera: Curculionidae). Annals of the Entomological Society of America, 60:43 – 54.

107
Harman, D. M. & Harman, A. L. (1972) Stridulatory mechanisms in the White Pine Weevil, Pissodes strobi. Annals of the Entomological Society of America, 65:1076 – 1079.
Harman, D. M. & Kranzler, G. A. (1969) Sound production in the White-Pine Weevil, Pissodes strobi, and the Northern Pine Weevil, P. aproximatus. Annals of the Entomological Society of America, 62:134 – 136.
Haskell, P. T. (1974) Sound production. In.: Morris, R. (ed.) The physiology of insecta. New York: Academic Press, p. 353 – 409.
Hirschberger, P. (2001) Stridulation in Aphodius dung beetles: behavioral context and intraspecific variability of song patterns in Aphodius ater (Scarabaeidae). Journal of Insect Behavior, 14:69 – 88.
Hyder, D. E. & Oseto, C. Y. (1989) Structure of the stridulatory apparatus and analysis of the sound produced by Smicronyx fulvus and Smicronyx sordius (Coleoptera, Curculionidae, Erirrhininae, Smicronychini). Journal of Morphology, 201:69 – 84.
Landa, S. (2000) Communication. Saint-Petersburg: Zoological Institute RAS. Beetles (Coleoptera) and Coleopterologists. Disponível em:< http://www.zin.ru/Animalia/Coleoptera/eng/biol41.htm>. Acesso em: 15 de setembro de 2005.
Pollack, G. (2000) Who, what, where? Recognition and localization of acoustic signals by insects. Neurobiology of Behaviour, 10:763 – 767.
Wilson, L. M.; Henry, C. S.; Johnson, J. B. & Mccaffrey, J. P. (1993) Sound production in Phrydiuchus tau (Coleoptera: Curculionidae). Annals of the Entomological Society of America, 86:621 – 630.

108
4. RESUMOS E CONCLUSÕES
Amerrhinus ynca é uma coleobroca cujas larvas se alimentam dos tecidos
internos da ráquis foliar do coqueiro. Até pouco tempo era considerada praga
secundária, mas devido à periodicidade e intensidade dos surtos tem atingido o
status de praga primária em algumas regiões. Pouco se conhece de sua biologia,
por isso este trabalho teve por objetivo: encontrar e descrever caracteres
morfológicos que diferenciem os sexos; descrever a seqüência comportamental
associada à corte e ao acasalamento; identificar possíveis semioquímicos
envolvidos nas interações intra e interespecíficas; identificar e descrever as
estruturas associadas à estridulação; analisar os parâmetros temporais e
espectrais dos sons produzidos e verificar o seu papel no comportamento
reprodutivo.
Os adultos de A. ynca utilizados nos estudos foram coletados em um
plantio comercial de coco e levados para o laboratório. Após a confirmação dos
sexos procurou-se identificar, através de observações da morfologia externa,
caracteres distintos entre machos e fêmeas. O comprimento e a largura do corpo
foram medidos. O comportamento de acasalamento e corte foi analisado através
de observação direta e por filmagem. Registraram-se em ambos os casos a
freqüência e a duração dos atos comportamentais. Os bioensaios para verificar a
presença de semioquímicos foram feitos utilizando-se três tipos de olfatômetros,
nos quais se utilizou como estímulo insetos coespecíficos e partes do coqueiro.
Os sons foram gravados com gravador digital e analisados com ajuda do software
Canary. Para verificar o papel da estridulação no comportamento de

109
acasalamento foram realizadas observações de quatro tipos de casais (com ou
sem partes dos élitros).
Foram encontrados três caracteres morfológicos distintos; as fêmeas são
maiores que os machos. Os acasalamentos ocorreram tanto em fotofase quanto
em escotofase. A monta iniciou-se aos 119 ± 75 min após os casais terem sido
colocados em observação. A corte e a cópula duraram em média 3 ± 2,5 min e 51
± 49 min, respectivamente. A produção de som durante a corte, através da
estridulação, é o evento mais freqüente entre os machos. As fêmeas de A. ynca
acasalaram múltiplas vezes e em geral com vários machos. Após a primeira
cópula os machos apresentaram comportamento de guarda da fêmea.
Nos testes de atratividade de odores, a maioria dos indivíduos não
responderam aos estímulos testados em três tipos de olfatômetros. Machos e
fêmeas foram mais estimulados quando se testaram partes vegetais.
Verificou-se que machos e fêmeas emitem sons por estridulação através
da fricção de duas estruturas: a pars stridens e o plectrum. A pars stridens está
localizada na região ventral da extremidade posterior dos élitros direito e
esquerdo e o plectrum está localizado no sétimo esternito abdominal de ambos os
sexos. O som produzido pelos machos, por estridulação, durante o
comportamento de corte, é diferente daquele que produzem quando são
perturbados. Observous-se, durante a corte, que as fêmeas movimentam o
abdômen de forma similar ao movimento que os machos desempenham ao
estridularem. Elas também estridulam quando são perturbadas, mas o som
produzido difere daquele dos machos.
No tratamento: fêmeas que tiveram sua pars stridens removida, a
porcentagem de casais que acasalaram (montas e cópulas) reduziu. Já os
machos gastam mais tempo e energia tentando cortejar as fêmeas, quando sua
pars stridens é retirada.
Assim pode-se concluir que:
- existem diferenças morfológicas entre machos e fêmeas que podem ser
utilizadas com segurança para diferenciá-los no campo e no laboratório sem que
haja a necessidade de feri-los.
- os acasalamentos ocorreram tanto em fotofase quanto em escotofase e
os machos durante a pré-copula estridulam e estimulam as fêmeas com as

110
pernas anteriores, e durante a cópula a estimulação do protórax e do abdômen da
fêmea com o primeiro par de pernas e a raspagem do pronoto da fêmea com o
rostro foram os atos comportamentai mais frequentes. As fêmeas de A. ynca
acasalaram múltiplas vezes e em geral com vários machos e estes apresentaram
o comportamento de guarda da fêmea, permanecendo sobre seu dorso por
longos períodos.
- não foi possível verificar a atratividade de machos e fêmeas aos odores
testados no três tipos de dispositivos olfatométricos, ou seja, os dados aqui
apresentados não foram suficientes para confirmar a presença de substâncias
químicas mediadoras de interações intraespecificas e interespecíficas devido ao
alto percentual de não resposta.
- são duas as estruturas associadas à produção de som nesta espécie: a
pars stridens e o plectrum. A primeira estrutura está localizada na região ventral
da extremidade posterior dos élitros direito e esquerdo. Nos machos elas são
constituídas de fileiras de cristas de altura e espaçamentos variáveis. Nas fêmeas
as cristas são menos acentuadas e estão distribuídas de forma irregular. Machos
e fêmeas apresentaram dois plectra localizados no sétimo esternito abdominal,
debaixo de cada pars stridens.
- machos e fêmeas emitem sons por estridulação. Os machos
estridularam durante o comportamento de corte. As fêmeas também
apresentaram movimento similar ao estridulatório, mas um espectro específico
não foi registrado. Os machos apresentaram estridulações quando foram
perturbados através de pressões nas laterais do abdômen. A característica
temporal e espectral destes sons foi distinta da observada durante a corte. As
fêmeas também estridularam quando perturbadas e o espectro de emissão do
som foi diferente do observado nos machos.
- nos tratamentos nos quais as fêmeas tiveram sua pars stridens removida
a porcentagem de casais que acasalaram (montas e cópulas) reduziu. Já os
machos gastam mais tempo tentando cortejar as fêmeas, quando sua pars
stridens é retirada.

111
5. REFERÊNCIAS BIBLIOGRÁFICAS
Alcock, J. (1998) Animal behavior: In: evolutionary approach. Editora Sinauer Associates. 6ª ed. p.185-189.
Alexander, R. D. (1967) Acoustical communication in arthropods. Annual Review of Entomology, 12:495 – 526.
Arnqvist, G. & Nilsson, T. (2000) The evolution of polyandry: multiple mating and female fitness in insects. Animal Behaviour, 60:145 – 164.
Arnqvist, G. & Rowe, L. (2002) Antagonistic coevolution between the sexes in a group of insects. Nature, 415:787 – 789.
Arnqvist, G. (1988) Mate guarding and sperm displacemente in the water strider Gerris lateralis Schumm (Heteroptera: Gerridae). Freshwater Biology, 19:269 – 274.
Arnqvist, G. (1997) The evolution of water strider mating system: causes and consequences of sexual conflicts. In: Choe, J. C. & Crespi, B. J. (eds.). The evolution of mating systems in insects and arachnids. Cambridge: Cambridge University Press, p. 146 – 163.
Bailey, W. J. (1991) Acoustic behaviour of insects: an evolutionary perspective. 1.ed. London: Chapman and Hall. 225 p.
Bailey, W. J. (2003) Insect duets: underlying mechanisms and their evolution. Physiological Entomology, 28:157 – 174.
Birkhead, T. R. (1998) Cryptic female choice: criteria for establishing female sperm choice. Evolution, 52(4):1212 – 1218.
Bondar, G. (1940) Insetos Nocivos e Moléstias do Coqueiro (Cocos nucifera) no Brasil. Salvador:Tipografia Naval, 156 p.

112
Booth, D. C.; Phillips, T. W.; Claesson, A.; Silverstein, R. M.; Lanier, G. N. & West, J. R. (1983) Aggregation pheromone components of two species of Pisssodes weevils (Coleoptera: Curculionidae): isolation, identification, and field activity. Journal of Chemical Ecology, 9:1 – 12.
Breidbach, O. (1986) Studies on the stridulation of Hylotrupes bajulus (Coleoptera: Cerambycidae) communication through support vibration morphology and mechanics of the signal. Behavioural Processes, 12:169 – 186.
Buchler, E. R.; Wright, T. B. & Brown, E. D. (1981) On the functions of stridulation by the passalid beetle Odontotaenius disjunctus (Coleoptera: Passalidae). Animal Behavior, 29:493 – 486.
Carbone, S. S. & Rivera, A. C. (1998) Sperm competition, cryptic female choice and prolonged mating in the Eucalyptus Snout Beetle, Gonipterus scutellatus (Coleoptera: Curculionidae). Etologia, 6:33 – 40.
Chang, V. C. S. & G. A. Curtis. (1972) Pheromone production by the New Guinea sugarcane weevil. Environmental Entomology, 1:476 – 481.
Cocroft, R. B. (1999) Parent-offspring communication in response to predators in a subsocial treehopper (Hemiptera: Membracidae: Umbonia crassicornis). Ethology, 105:553 – 568.
Cokl, A. & Virant-Doberlet, M. (2003) Communication with substrate-borne signals in small plant-dwelling insects. Annual Review of Entomology. 48:29 – 50.
Costa, J. M. ; Barreto, A. C. & Filho, J. L. (1973) Pragas das palmáceas. Cruz das Almas, Bahia: Instituto de Pesquisas Agropecuárias do Leste,
Crook, D. J.; Hopper, J. A.; Ramaswamy; S. B. & Higgins, R. A. (2004) Courtship behavior of the Soybean Stem Borer Dectes texanus texanus (Coleoptera: Cerambycidae): evidence for a female contact sex pheromone. Annals of the Entomological Society of America, 97:600 – 604.
Cross, W. H. & Mitchell, H. C. (1966) Mating behavior of the boll weevil. Journal of Economic Entomology, 59:1503 – 1057.
Cuenca, M. A. G. (1997). Importância econômica do coqueiro. In.: Ferreira, J. M. S.; Warwick, D. R. N. & Siqueira, L. A. (eds.). A Cultura do coqueiro no Brasil. 2. ed. Brasília: Embrapa-SPI; Aracaju: Embrapa-CPATC, p. 17 – 56.
Daly, M. (1978) The cost of mating. The American Naturalist, 112:771 – 774.
Darwin, C. (1871) The Descent of Man: Chapter 10 - Secondary sexual characters of insects. In: The Online Literature Library. Desenvolvido por Knowledge Matters Ltda. Disponível em: <http://www.literature.org/authors/darwin-charles/the-descent-of-man/chapter-10.html> Acesso em: agosto de 2005.
Devries, P. J.; Cocroft, R. B. & Thomas, J. (1993) Comparison of acoustical signals in Maculinea butterfly caterpillars and their obligate host Myrmica ants. Biol. J. Linn. Soc., 49:229 – 238.

113
Dickens, J. C. & Mori, K. (1989) Receptor chirality and behavioral specificity of the boll weevil, Anthonomus grandis Boh. (Coleoptera: Curculionidae), for its pheromone, (+)-grandisol. Journal of Chemical Ecology, 15:517 – 528.
Durmortier, B. (1963) Morphology of sound emission apparatus in Arthropod. In: R. G. Busnel (ed.). Acoustic Behavior of Animals. New York: Elsevier Publishing Co., 933 p.
Eberhard, W. G. (1991) Copulatory courtship and cryptic female choice in insects. Biological Reviews, 66:1 – 31.
Edvardsson, M. & Arnqvist, G. (2000) Copulatory courtship and cryptic female choice in red flour beetles Tribolium castaneum. Proc. R. Soc. Lond., 267:559 – 563.
Edvardsson, M. & Arnqvist, G. (2005) The effects of copulatory courtship on Differential Allocation in the Red Flour Beetle Tribolium castaneum. Journal of Insect Behavior, 18:313 – 322.
Eller, F. J. & Bartelt, R. J. (1996) Grandisoic acid, a male-produced aggregation pheromone from the plum curculio, Conotrachelus nenuphar. Journal of Natural Products, 59:451 –453.
Eller, F. J.; Bartelt, R. J.; Shasha, B. S.; Schuster, D. J.; Riley, D. G.; Stansly, P. A.; Mueller, T. F.; Shuler, K. D.; Johnson, B.; Davis, J. H. & Sutherland, C. A. (1994) Aggregation pheromone for the pepper weevil, Anthonomus eugenii Cano (Coleoptera: Curculionidae): identification and field activity. Journal of Chemical Ecology, 20:1537 – 1555.
El-Sayed, A. M. (2005) The Pherobase: Database of Insect Pheromones and Semiochemicals. Disponível em:<http://www.pherobase.net> Acesso em outubro de 2005.
Faustini, D. L.; Giese, W. L.; Phillips, J. K.; & Burkholder, W. E. (1982) Aggregation pheromone of the male granary weevil, Sitophilus granaries (L.). Journal of Chemical Ecology, 8:679 – 687.
Ferreira, J. M. S. (1987) Proteção fitossanitária do coqueiral: III. Controle de pragas no campo. Aracaju: Embrapa-CNPCo, 23 p. (Circular Técnico, 7).
Ferreira, J. M. S.; Lima, M. F.; Santana, D. L. Q. & Moura, J. I. L. (1998) Pragas do coqueiro. In: Sobrinho, R. B.; Cardoso, J. E. & Freire, F. C. (eds.). Pragas de Fruteiras Tropicais de Importância Agroindustrial. Brasília: Embrapa-SPI, p. 81 – 118.
Ferreira, J. M. S.; Lima, M. F.; Santana, D. L. Q.; Moura, J. I. L. & Souza, L. A. (1997). Pragas do coqueiro. In.: Ferreira, J. M. S.; Warwick, D. R. N. & Siqueira, L. A. (edts.) (1997). A cultura do coqueiro no Brasil. 2 ed., Brasília: Embrapa-SPI, p. 189 – 221.
Ferreira, J. T. B. & Zarbin, P. H. G. (1998) Amor ao primeiro odor: a comunicação química entre os insetos. Química Nova na Escola, 7:3 – 6.

114
Ferreira, J. T. B. (2001) A estrutura química: código de comunicação. In.: Vilela, E. F. & Della Lucia, T. M.C. (eds.) Feromônios de insetos: biologia, química e emprego no manejo de pragas. 2. ed. Ribeirão Preto: Holos, p. 41 – 44.
Fontes, H. R.; Cintra, F. L. D. & Filho, O. M. C. (1998) Implantação e manejo da cultura do coqueiro. In: Ferreira, J. M. S.; Warwick, D. R. N. & Siqueira, L. A. (editores). A Cultura do coqueiro no Brasil. 2. ed. Brasília: Embrapa-SPI; Aracaju: Embrapa-CPATC, p. 99 – 128.
Forrest, T. G.; Read, M. P.; Farris, H. E. & Hoy, R. R. (1997) A tympanal hearing organ in scarab beetles. The Journal of Experimental Biology, 200:601 – 606.
Giblin Davis, R. M.; Gries, R.; Crespi, B.; Robertson, L. N.; Hara, A. H.; Gries, G.; O' Brien, C.W. & Pierce, H. D., Jr. (2000) Aggregation pheromones of two geographical isolates of the New Guinea sugarcane weevil, Rhabdoscelus obscurus. Journal of Chemical Ecology, 26:2763 – 2780.
Giblin-Davis, R. M.; Gries, R.; Gries, G.; Peña-Rojas, E.; Pinzon, I. L.; Pena, J. E.; Perez, A. L.; Pierce, H. D., Jr. & Oehlschlager, A. C. (1997) Aggregation pheromone of palm weevil, Dynamis borassi. Journal of Chemical Ecology, 23:2287 – 2297.
Giblin-Davis, R. M.; Peña, J. E.; Oehlschlager, A. C. & Perez, A. L. (1996) Optimization of semiochimical-based trapping of Metamasius hemipterus sericeus (Oliver) (Coleoptera: Curculionidae). Journal of Chemical Ecology, 22:1389 – 1410.
Gibson, L. P. (1967) Stridulatory mechanisms and sound production in Conotrachelus (Coleoptera: Curculionidae). Annals of the Entomological Society of America, 60:43 – 54.
Gomes, R. P. (1992) O Coqueiro da Bahia. 6. ed. São Paulo: Nobel, 111 p.
Gries, G.; Gries, R.; Perez, A. L.; Gonzales, L. M.; Pierce, H. D. Jr.; Oehlschlager, A. C.; Rhainds, M.; Zebeyou, M. & Kouame, B. (1994) Ethyl propionate: synergistic kairomone for African Palm weevil, Rhynchophorus phoenicis L. (Coleoptera: Curculionidae). Journal of Chemical Ecology, 20:889 – 897.
Gries, G.; Gries, R.; Perez, A. L.; Oehlschlager, A. C.; Gonzales, L. M.; Pierce, H. D.; Kouda, M.; Zebeyou, M. & Nanou, N. (1993) Aggregation pheromone of the African Palm weevil, Rhynchophorus phoenicis F. Naturwissenschaften, 80:90 – 91.
Hallett, R. H.; Gries, G.; Gries, R.; Borden, J. H.; Czyzewska, E.; Oehlschlager, A. C.; Pierce, H. D., Jr.; Angerilli, N. P. D. & Rauf, A. (1993) Aggregation pheromones of two Asian Palm Weevils, Rhynchophorus ferrugineus and R. vulneratus. Naturwissenschaften, 80:328 – 331.
Harari, A. R.; Landolt, P. J.; O'Brien, C. W. & Brockmann, H. J. (2003) Prolonged mate guarding and sperm competition in the weevil Diaprepes abbeviatus (L.). Behavioral Ecology, 14(1):89 – 96.

115
Hardee, D. D.; Cross, W. H. & Mitchell, E. B. (1969) Male boll weevils are more attractive than cotton plants to other boll weevils. Journal Economic Entomology, 62:165 – 169.
Harman, D. M. & Harman, A. L. (1972) Stridulatory mechanisms in the White Pine Weevil, Pissodes strobi. Annals of the Entomological Society of America, 65:1076 – 1079.
Harman, D. M. & Kranzler, G. A. (1969) Sound production in the White-Pine Weevil, Pissodes strobi, and the Northern Pine Weevil, P. aproximatus. Annals of the Entomological Society of America, 62:134 – 136.
Haskell, P. T. (1974). Sound production. In: Rockstein, M. (ed.) The Physiology of Insect. New York: Academic Press. p. 354 – 410.
Haynes, K. F. & Yeargan, K. V. (1999) Exploitation of intraspecific communication systems: elicit signalers and receivers. Annals of the Entomological Society of America, 92:960 – 970.
Hedin, P. A.; Dollar, D. A.; Collins, J. K.; Dubois, J. G.; Mulder, P. G.; Hedger, G. H.; Smith, M. W. & Eikenbary, R. D. (1997) Identification of male Pecan Weevil pheromone. Journal of Chemical Ecology, 23:965 – 977.
Hoy, R. R. & Robert, D. (1996) Tympanal hearing in insects. Annual Reviews Entomology, 41:433 – 450.
Hyder, D. E. & Oseto, C. Y. (1989) Structure of the stridulatory apparatus and analysis of the sound produced by Smicronyx fulvus and Smicronyx sordius (Coleoptera, Curculionidae, Erirrhininae, Smicronychini). Journal of Morphology, 201:69 – 84.
Innocenzi, P. J.; Hall, D. R. & Cross, J. V. (2001) Components of male aggregation pheromone of Strawberry Blossom Weevil, Anthonomus rubi Herbst. (Coleoptera: Curculionidae). Journal of Chemical Ecology, 27:1203 – 1218.
Jaffé, K.; Sanchez, P.; Cerda, H.; Hernandez, J. V.; Jaffé, R.; Urdaneta, N.; Guerra, G.; Martinez, R. & Miras, B. (1993) Chemical ecology of the Palm Weevil Rhynchophorus palmarum (L.) (Coleoptera: Curculionidae): attraction to host plants and to a male-produced aggregation pheromone. Journal of Chemical Ecology, 19:1703 – 1720.
Kasper, J. & Hirschberger, P. (2005) Stridulation in Aphodius dung beetles: songs and morphology of stridulatory organs in North American Aphodius species (Scarabaeidae). Journal of Natural History, 39:91 – 99.
Kemp, D. J. (2002) Visual mate-searching behaviour in the evening brown butterfly, Melanitis leda (L.) (Lepidoptera: Nymphalidae). Australian Journal of Entomology, 41:300 – 305.
Landa, S. (2000) Communication. Beetles (Coleoptera) and Coleopterologists. Saint-Petersburg: Zoological Institute RAS. Disponível em:< http://www.zin.ru/Animalia/Coleoptera/eng/biol41.htm>. Acesso em: setembro de 2005.

116
Landolt, P. J. & Phillips, T. W. (1997) Host plant influences on sex pheromone behavior of phytophagous insects. Annual Review of Entomology, 42:371-391.
Leskey, T. C. & Prokopy, R. J. (2001) Adult plum curculio (Coleoptera: Curculionidae) attraction to fruit and conspecific odors. Annals of the Entomological Society of America, 94:275-288.
Lewis, B. & Gower, M. (1980) Biology of communication. London: Blackie, 239 p.
Lewis, L. A.; Schineider, S. S & Degrandi-Hoffman, G. (2002) Factors influencing the selection of recipients by workers performing vibration signals in colonies of the honeybee, Apis mellifera. Animal Behaviour, 63:361 – 367.
Lloyd, J. E. (1979) Mating behavior and natural selection. The Florida Entomologist, 62:17 – 34 .
Lobanov, A. L. (2002) Sound. Saint-Petersburg: Zoological Institute RAS. Beetles (Coleoptera) and Coleopterologists. Disponível em: < http://www.zin.ru/Animalia/Coleoptera/eng/biol412.htm>. Acesso em: setembro de 2005.
López, A. B. & Oliveira, M. R. V. (2002) Praga quarentenária 7: Ips spp (Coleoptera: Scolytidae). Comunicado Técnico 62, 1 º ed. Brasília: EMBRAPA Distrito Federal.
Machens, C. K.; Stemmler, M. B.; Prinz, P.; Krahe, R.; Ronacher, B. & Herz, A. V. M. (2001) Representation of acoustic communication signals by insect auditory receptor neurons. The Journal of Neuroscience, 21:3215–3227.
Maisin, N.; Hassan, S. T. S.; Hussein, M. Y. & Sajap, A .S. (1997) Mating behaviour of Menochilus sexmaculatus Fabr. (Coleoptera: Coccinellidae). Malaysian Applied Biology, 26:57 – 61.
Manpe, C. D. & Neuzing, H. H. (1966) Function of the stridulating organs of Conotrachelus (Coleoptera: Curculionidae). Annals of the Entomological Society of America, 59:614 – 615.
Martikainen, P. (2001) Non-target beetles (Coleoptera) in Trypodendron pheromone traps in Finland. Anz. Schädlingskd, 74:150 – 154.
Mason, A. C. (1996) Territoriality and the function of song in the primitive acoustic insect Cyphoderris monstrosa (Orthoptera: Haglidae). Animal Behaviour, 51:211 – 224.
Michael, R. R. & Rudinsky, J. A. (1972) Sound production in Scolytidae: sound specificity in male Dendroctonus beetles. Journal of Insect Physiology, 18:2189 – 2201.
Miller, D. R. & Heppner, D. (1999) Attraction of Pissodes affinis and P. fasciatus (Coleoptera: Curculionidae) to pityol and alpha -pinene in a coastal stand of western white pine and douglas-fir. Journal of Entomology Society. B.C., 96:73 – 76.

117
Mirizola-Filho, L. A. (2002). Cultivo de côco Anão. Viçosa: Aprenda Fácil, 322 p.
Moura, J. I. L. & Vilela, E. F. (1998) Pragas do Coqueiro e Dendezeiro. 2 .ed. Viçosa: Aprenda Fácil, 124 p.
Moura, J. I. L. & Vilela, E. F. (2001) Feromônios no controle de Rhynchophorus palmarum (l.) (Coleoptera: Curculionidae). In: Vilela, E. F. & Della-Lucia, T. M. C. (eds.). Feromônios de insetos: biologia, química e emprego no manejo de pragas 2. ed. Riberão Preto: Holos. p. 145 – 146.
Moura, J. I. L. (1993) Táticas para o controle de Amerrhinus ynca, broca-da-ráquis foliar do coqueiro. Anais do Congresso Brasileiro de Entomologia, 14, Piracicaba, SP: SEB, p.611.
Navarro, D. M. A. F.; Murta, M. M.; Duarte, A. G.; Lima, I. S.; Nascimento, R. R. & Sant’ana, A. E. G. (2002) Aspectos práticos relacionados ao uso do Rincoforol, o feromônio de agregação da broca-do-olho-do-coqueiro Rhynchophorus palmarum L. (Coleóptera: Curculionidae) no controle de pragas do coqueiro. Análise de sua eficiência em campo. Química Nova, 25:32-36.
Oehlschlager, A. C.; Pierce, H. D., Jr.; Morgan, B.; Wimalaratne, P.D.C.; Slessor, K. N.; King, G. G. S.; Gries, G.; Gries, R.; Borden, J. H.; Jiron, L. F.; Chinchilla, C. M. & Mexzan, R. G. (1992) Chirality and field activity of rhynchophorol, the aggregation pheromone of the American Palm Weevil. Naturwissenschaften, 79:134 – 135.
Oehlschlager, A. C.; Prior, R. N. B.; Perez, A. L.; Gries, R., Gries, G., Pierce, H. D., Jr. & Laup, S. (1995). Structure, chirality, and field testing of a male-produced aggregation pheromone of Asian Palm Weevil Rhynchophorus bilineatus (Montr.) (Coleoptera: Curculionidae). Journal of Chemical Ecology, 21:1619 – 1629.
Ohya, E. & Kinuura, H. (2001) Close range sound communications of the oak platypodid beetle Platypus quercivorus (Murayama) (Coleoptera: Platypodidae). Applied Entomology and Zoology, 36:317 – 321.
Paiva, M. R. & Pedrosa-Macedo, J. H. P. (1985) Feromonas de Insetos. Curitiba: GTZ, 94 p.
Parra-Pedrazzoli, A. L.; Cossé, A.; Murata, Y.; Bento, J. M. S.; Vilela, E. F. & Leal, W. S. (2006) Towards the identification and synthesis of the sex pheromone of the citrus leafminer, Phyllocnistis citrella Stainton (Lepidoptera: Gracillariidae). Neotropical Entomology, 35:12 – 18.
Passos, E. E. M. (1998a) Ecofisiologia do coqueiro. In: Ferreira, J. M. S.; Warwick, D. R. N. & Siqueira, L. A. (eds.). A Cultura do coqueiro no Brasil. 2. ed. Brasília: Embrapa-SPI; Aracaju: Embrapa-CPATC, p. 65-72.
Passos, E. E. M. (1998b) Morfologia do Coqueiro. In: Ferreira, J. M. S.; Warwick, D. R. N. & Siqueira, L. A. (eds.). A Cultura do coqueiro no Brasil. 2. ed. Brasília: Embrapa-SPI; Aracaju: Embrapa-CPATC, p. 57-64.

118
Pelosi, P. (1996) Perireceptor events in olfaction. Journal of Neurobiology, 30:3 – 19.
Pereira, M. J. B.; Anjos, N. & Eiras, A. E. (1998) Cortejo y cópula en la broca de la semilla de guanábana Bephratelloides pomorum (Hymenoptera: Eurytomidae). Revista de Biología Tropical, 46:105 – 108.
Perez, A. L.; Campos, Y.; Chinchilla, C. M.; Oehlschlager, A. C.; Gries, G.; Gries, R.; Giblin-Davis, R. M.; Castrillo, G.; Pena, J. E.; Duncan, R. E.; Gonzalez, L. M.; Pierce, H. D., Jr.; Mcdonald, R. & Andrade, R. (1997) Agregation pheromones and host kairomones of West Indian Sugarcane Weevil, Metamasius hemipterus sericeus. Journal of Chemical Ecology, 23:869 – 888.
Perez, A. L.; Gries, G.; Gries, R.; Giblin Davis, R. M. & Oehlschlager, A. C. (1994) Pheromone chirality of African Palm Weevil, Rhynchophorus phoenicis (F.) and Palmetto Weevil, Rhynchophorus cruentatus (F.) (Coleoptera: Curculionidae). Journal of Chemical Ecology, 20:2653 – 2671.
Phillips, J. K. & Burkholder, W. E. (1981) Evidence for a male-produced aggregation pheromone in the rice weevil. Journal of Chemical Ecology, 74:539 – 542.
Phillips, T. W.; West, J. R.; Foltz, J. L.; Silverstein, R. M. & Lanier, G. N. (1984) Aggregation pheromone of the deodar weevil, Pissodes nemeorensis (Coleoptera: Curculionidae): isolation and activity of grandisol and grandisal. Journal of Chemical Ecology, 10:1417 – 1423.
Piñero, J. C. & Prokopy, R. J. (2003) Field evaluation of plant odor and pheromonal combinations for attracting plum curculios. Journal of chemical Ecology. 29:2735 – 2748.
Pirisinu, Q.; Spinelli, G. & Bicchierai, M. C. (1988) Stridulatory apparatus in the Italian species of the genus Laccobius Erichson (Coleoptera: Hydrophilidae). International Journal of Insect Morphology and Embryology, 17:95 – 101.
Pollack, G. (2000) who, what, where? Recognition and localization of acoustic signals by insects. Neurobiology of Behaviour, 10:763 – 767.
Prenzel, B. G.; Laidlaw, W. G. & Wieser, H. (1999) Whitin-tree dynamics of mass attack by Dendroctonus pseudotsugae (Coleoptera: Scolytidae) on its host. The Canadian Entomologist, 131:635 – 643.
Renard, J. L.; Mariau, D. & Dollet, M. (1987) Rapport de mission defense dês cultures au Brésil. Paris: Le cocotier. : IRHO, 48 p.
Rochat, D.; Gonzalez, A. V.; Mariau, D., Villanueva, A. G. & Zagatti, P. (1991) Evidence for male-produced aggregation pheromone in American palm weevil, Rhynchophorus palmarum (L.) (Coleoptera: Curculionidae). Journal of Chemical Ecology, 17, p.1221 – 1230.
Schmuff, N. R.; Phillips, J. K.; Burkholder, W. E., Fales, H. M.; Chen, C. W.; Roller, P. P. & Ma, M. (1984) The chemical identification of the rice weevil and maize weevil aggregation pheromone. Tetrahedron Letter, 25:1533 – 1534.

119
Scott, J. A. (1974) Mate-locating behavior of butterflies. American Midland Naturalist, 91:103 – 117.
Serrano, A. R.; Diogo, A. C.; Viçoso, E. & Fonseca, P. J. (2003) New stridulatory structures in a Tiger Beetle (Coleoptera: Carabidae: Cicindelinae): morphology and sound characterization. The Coleopterists Bulletin, 57:161 – 166.
Slobodchikoff, C. N. & Spangler, H. G. (1979) Two types of sound production in Eupsophulus castaneus (Coleoptera: Tenebrionidae). The Coleopterists Bulletin, 33:239 – 244.
Smith, I. (2003) The lily leaf beetle (Coleoptera: Chrysomelidae). Ontario: Laboratory Services Division, University of Guelph. Disponível em: < http://www.uoguelph.ca/pdc/Factsheets/PDFs/056LilyLeafBeetle.pdf>. Acesso em: setembro de 2005.
Sprecher, E. (2003) The status of Lucanus cervus in Switzerland. Annals of the Pan-European Conference on Saproxylic Beetles, 2, London: People’s Trust for Endangered Species, London, 2003. p. 6-8.
Stockley, P. (1997) Sexual conflict resulting from adaptations to sperm competition. Trends Ecology Evolution, 12:154 – 159.
Suvanto, L.; Liimatainen, J. O.; Tregenza, T. & Hoikkala, A. (2000) Courtship signals and mate choice of the flies of inbred Drosophila montana strains. Journal of the Evolutionary Biology, 13:583 – 592.
Tegoni, M.; Campanacci, V. & Cambillau, C. (2004) Structural aspects of sexual attraction and chemical communication in insects. Trend in Biochemical Sciences, 29 (5):257-264.
Tillman, J. A.; Seybold, S. J.; Jurenka, R. A. & Blomquist, G. J. (1999) Insect pheromones - an overview of biosynthesis and endocrine regulation. Insect Biochemistry and Molecular Biology, 29:481–514.
Tumlinson, J. H.; Gueldner, R. C.; Hardee, D. D.; Thompson, A. C.; Hedin, P. A. & Minyard, J. P. (1971) Identification and synthesis of the four compounds comprising the boll weevil sex attractant. Journal Organic Chemistry, 36:2616 – 2621.
Tumlinson, J. H.; Hardee, D. D.; Gueldner, R. C.; Thompson, A. C.; Hedin, P. A. & Minyard, J. P. (1969) Sex pheromones produced by male boll weevil: isolation, identification, and synthesis. Science, 166:1010 – 1012.
Verdú, J. R. & Galante, E. (2001) Larval morphology and breeding behavior of the genus Pedaridium Harold (Coleoptera: Scarabaeidae). Annals of the Entomological Society of America, 94:596 – 604.
Viana, A. M. M. & Vilela, E. F. (1996) Comportamento de corte e acasalamento de Cosmopolites sordidus Germar (Coleoptera: Curculionidae). Anais da Sociedade Entomológica do Brasil, 25:347 – 350.

120
Vilela, E. F. & Della Lucia, T. M. C. (2001) Introdução aos semioquímicos e terminologia. In: Vilela, E. F. & Della Lucia, T. M. C. (eds.) Feromônios de insetos: biologia, química e emprego no manejo de pragas. 2. ed. Riberão Preto: Holos, p. 9 – 12.
Vilela, E. F. & Mafra-Neto, A. (2001) Registro de Feromônios Comerciais e Legislação. In: Vilela, E. F. & Della Lucia, T. M. C. (eds.) Feromônios de insetos: biologia, química e emprego no manejo de pragas. 2. ed. Riberão Preto: Holos, p. 151 – 159.
Virant-Doberlet, M. & Cokl, A. (2004) Vibrational communication in insects. Neotropical Entomology, 33:121 – 134.
Walgenbach, C. A.; Phillips, J. K.; Faustini, D. L. & Burkholder, W. E. (1983) Male-produced aggregation pheromone of the maize weevil, Sitophilus zeamais, and interespecific attraction between three Sitophilus species. Journal of Chemical Ecology, 9:831 – 841.
Wedell, N. (1996) Mate quality affects reproductive effort in a paternally investing species. The American Naturalist, 148:1075 – 1088.
Weissling, T. J.; Giblin Davis, R. M. & Scheffrain, R. H. (1993) Laboratory and field evidence for male-produced aggregation pheromone in Rhynchophorus cruentatus (F.) (Coleoptera: Curculionidae). Journal of Chemical Ecology, 19:1195 – 1203.
Weissling, T. J.; Giblin Davis, R. M.; Gries, G.; Gries, R.; Perez, A. L.; Pierce, H. D. Jr.; & Oehlschlager, A. C. (1994) Aggregation pheromone of palmetto weevil, Rhynchophorus cruentatus (F.) (Coleoptera: Curculionidae). Journal of Chemical Ecology, 20:505 – 515.
Wilcox, R. S. & Stefano, J. D. (1991) Vibratory signals enhance mate-guarding in a water strider (Hemiptera: Gerridae). Journal of Insect Behavior, 4:43 – 50.
Wiley, R. H. (1997) Coevolution of the sexes. Science, 275:1075 – 1076.
Wilson, L. M.; Henry, C. S.; Johnson, J. B. & Mccaffrey, J. P. (1993) Sound production in Phrydiuchus tau (Coleoptera: Curculionidae). Annals of the Entomological Society of America, 86:621 – 630.
Yack, J. E.; Smith, M. L. & Weatherhead, P. J. (2001) Caterpillar talk: acostically mediated territoriality in larval Lepidoptera. Proceeding of Natural Academy of Sciences of USA, 98:11371 – 11375.
Zhang, M.; Crocker, R. L.; Mankin, R. W.; Flanders, K. L. & Brandhorst-Hubbard, J. (2003) Acoustic identification and measurement of activity patterns of white grubs in soil. Journal Economic Entomology, 96:1704 – 1710.
Ziemba, K. & Rutowski, R. (2000) Sexual dimorphism in eye morphology in a butterfly (Asterocampa leilia; Lepidoptera: Nymphalidae). Psyche, 103:25 – 36.