salom menezes atributos de macroinvertebrados … · 2013-09-11 · material publicado). n o h...
TRANSCRIPT

3
Universidade de Aveiro 2010
Departamento de Biologia
SALOMÉ MENEZES LACERDA NEVES
ATRIBUTOS DE MACROINVERTEBRADOS PARA BIOMONITORIZAÇÃO EM CENÁRIOS AGRÍCOLAS MACROINVERTEBRATE TRAITS AS BIOMONITORING TOOLS IN AGRICULTURAL SCENARIOS
Dissertação apresentada à Universidade de Aveiro para cumprimento dos requisitos necessários à obtenção do grau de Doutor em Biologia, realizada sob a orientação científica do Professor Doutor Amadeu Mortágua Velho da Maia Soares, Professor Catedrático do Departamento de Biologia da Universidade de Aveiro, e co-orientação científica do Professor Doutor Donald J. Baird, Research Professor do Department of Biology da University of New Brunswick.
Apoio financeiro da FCT e do FSE no âmbito do III Quadro Comunitário de Apoio através da Bolsa de Doutoramento SFRH/BD/18514/2004


5
¡Pura Vida!


7
o júri
presidente Prof. Doutor Joaquim Borges Gouveia professor catedrático do Departamento de Economia, Gestão e Engenharia da Universidade de Aveiro
Prof. Doutora Lúcia Maria das Candeias Guilhermino professora catedrática do Instituto de Ciências Biomédicas Abel Salazar da Universidade do Porto
Prof. Doutor Amadeu Mortágua Velho da Maia Soares professor catedrático do Departamento de Biologia da Universidade de Aveiro
Prof. Doutor Rui Godinho Lobo Girão Ribeiro professor associado com agregação do Departamento de Ciências da Vida da Universidade de Coimbra
Prof. Doutor António José Arsénia Nogueira professor associado com agregação do Departamento de Biologia da Universidade de Aveiro
Prof. Doutor Fernando Manuel Raposo Morgado professor auxiliar com agregação do Departamento de Biologia da Universidade de Aveiro
Prof. Doutor José Vitor Sousa Vingada professor auxiliar do Departamento de Biologia da Universidade do Minho
Prof. Doutor Donald John Baird professor e investigador do Department of Biology da University of New Brunswick (Fredericton)
Prof. Doutor Carlos Barata Martí investigador principal do Consejo Superior de Investigaciones Científicas (Barcelona)


9
agradecimentos
Aos meus orientadores, Amadeu Soares, Donald Baird e Luisa Castillo, por terem aceite supervisionar, de forma muito paciente, este que foi, sem dúvida, um dos maiores desafios da minha vida. Às instituições e laboratórios que acolheram o meu projecto e cederam todas as condições para que se pudesse concretizar: DeBio e CESAM (Universidade de Aveiro), IRET e LARNAVISI (Universidad Nacional) e CRI (University of New Brunswick). À Área de Conservación Cordillera Volcánica Central do Ministerio de Ambiente y Energía da Costa Rica e à Consellería de Medio Ambiente e Desenvolvemento Sostible da Xunta de Galicia pelas autorizações para levar a cabo colheitas de material biológico nas áreas de estudo. À Fundação para a Ciência e Tecnologia pelo apoio financeiro. Às tão internacionais e profissionais equipas de campo com quem tive a honra de trabalhar: Carla e Ramiro na Galiza; Alice, Sílvia, Colin, Meyer e Mauricio na Costa Rica; Joseph, Eric, Dave, Daryl e Rick no Canadá. Aos técnicos Abel, Margaret e Kristie. Aos investigadores que contribuíram com valiosas sugestões e contribuições científicas: António Nogueira do DeBio (pelo apoio e discussões sobre traits e análise multivariada), Maria Luisa Fournier do IRET (pelo apoio na Costa Rica), Maria José Cerejeira do ISA (pelas sugestões e saídas a potenciais áreas de estudo), José Tarazona do INIA (pelas sugestões referentes à Galiza), Marc Cadotte e dois revisores anónimos do JAE (pelas revisões ao material publicado). Não há parte melhor de um trabalho do que apercebermo-nos de que fizemos bons amigos entre os colegas. Às !labecas"! Carla, Marta, Joanne, Sandrina, Guida, Maria e Fabianne. E Matilde!! Muita mulher junta até pode resultar... À Carla, amiga fora e dentro do laboratório, em Portugal e no estrangeiro, todos os dias e a todas as horas!:) Aos !letais"! São muitos mas não posso deixar de destacar Sara, Zé, Raquel, Henrique, Marco, Jeamylle, Vanessa e Pestana. A todos os que me acolheram durante o tempo passado no Ecotox e na UNB. Aos amigos que fizeram com que estar longe de casa fosse quase tão bom como estar perto, Dennis, Colin, Eva, Alice, Leo, Silvia e Claudia. E àqueles que me fazem sempre ter vontade de regressar, Nelson, Dora, Mix, Ana Margarida, Fernando, Nuno, Inês, Eduardo, Cat, Ana Chorão, Bernardo, toda a Biomalta e a família das danças, Norma, Belinda e Renato. À minha família, pelo seu apoio incondicional: à minha mãe Rosário, ao meu irmão Jaime, ao meu padrinho Filipe, à minha tia Ângela, aos meus primos Bernardo, Simão, Rita e Carlota e à minha avó Alice. Aos meus avós Miguel e Vita, por terem feito parte da minha vida de uma forma tão única e inesquecível. Obrigada, thank you e gracias!


11
palavras-chave
atributos de macroinvertebrados; ecossistemas de água doce; biomonitorização; agricultura; ribeiros; comunidades biológicas; regiões biogeográficas; gradientes de impacto
resumo
Os ecossistemas de água doce – responsáveis por funções ambientais importantes e pelo fornecimento de bens e serviços insubstituíveis – têm vindo a ser severamente afectados por perturbações antropogénicas. A conversão de floresta em terreno agrícola afecta os sistemas aquáticos através de uma série de mecanismos: sedimentação; excesso de nutrientes; contaminação; alterações hidrológicas; e remoção de vegetação ripícola. As comunidades de macroinvertebrados de água doce – devido à sua diversidade, ubiquidade e sensibilidade às perturbações ambientais – revelam-se como particularmente adequadas para estudos de avaliação da integridade ecológica destes sistemas expostos simultaneamente a múltiplos factores de impacto.
O uso sistemático de respostas biológicas para avaliação de mudanças ambientais – ou biomonitorização – pode ser levado a cabo através de diversas metodologias, que, de uma forma geral, não consideram aspectos funcionais das comunidades biológicas e têm aplicabilidade geograficamente restrita. A biomonitorização através de atributos biológicos (características que reflectem a adaptação das espécies ao seu meio ambiente) revela-se como uma ferramenta promissora na resolução dos problemas referidos, apresentando vantagens adicionais: relações causa-efeito directas; melhoria na diferenciação de impactos; e integração da variabilidade natural.
O presente estudo apresenta uma revisão critica do estado-da-arte actual na área do uso de atributos biológicos em biomonitorização. Até à data de publicação, não estava disponível nenhum outro trabalho com a base conceptual do uso de atributos de macroinvertebrados enquanto descritores de comunidades e para efeitos de biomonitorização e gestão de sistemas de água doce. Descrevem-se as teorias ecológicas de suporte destas metodologias (conceitos de habitat-molde e de filtros paisagísticos) e os estudos que aplicaram estas teorias em cenários reais, tendo-se chamado a atenção para questões técnicas e possíveis soluções. As necessidades futuras nesta área englobam: o desenvolvimento de uma só ferramenta de biomonitorização de aplicação alargada; uma maior compreensão da variabilidade natural nas comunidades biológicas; diminuição dos efeitos de soluções de compromisso biológico e sindromas; realização de estudos autoecológicos adicionais; e detecção de impactos específicos em cenários de impacto complexos.
Um dos objectivos deste estudo foi contribuir para a melhoria das técnicas de biomonitorização através de atributos, focalizando em comunidades de macroinvertebrados ribeirinhas em diferentes regiões biogeográficas (as bacias hidrográficas dos rios: Little e Salmon em New Brunswick, Canadá; Anllóns na Galiza, Espanha; Reventazón em Cartago, Costa Rica). Em cada região, foram estudados gradientes de uso agrícola de solo, incluindo desde bacias hidrográficas quase exclusivamente cobertas por floresta até bacias sob a influência maioritária de actividades agrícolas intensivas.
Em cada gradiente de uso de solo, a caracterização da comunidade biológica (por amostragem de macroinvertebrados em troços de rápidos) foi acompanhada pela caracterização do habitat circundante (incluindo propriedades da bacia hidrográfica, análise química das águas e outras propriedades à escala local). A comunidade de macroinvertebrados foi caracterizada através de informação taxonómica, métricas estruturais, índices

12
de diversidade, métricas de tolerância, índices bióticos e através da compilação de atributos biológicos e fisiológicos gerais, de história de vida e de resistência a perturbações. Análises estatísticas univariadas e multivariadas foram usadas para evidenciar os gradientes biológicos e físico-químicos, confirmar a sua co-variação, testar a significância da discriminação de níveis de impacto e estabelecer comparações inter-regionais.
A estrutura de comunidades revelou os complexos gradientes de impacto, que por sua vez co-variaram significativamente com os gradientes de uso de solo. Os gradientes de impacto relacionaram-se sobretudo com entrada de nutrientes e sedimentação. Os gradientes biológicos definidos pelas medidas estruturais seleccionadas co-variaram com os gradientes de impacto estudados, muito embora apenas algumas variáveis estruturais tenham individualmente discriminado as categorias de uso de solo definidas a priori. Não foi detectada consistência nas respostas das medidas estruturais entre regiões biogeográficas, tendo-se confirmadado que as interpretações puramente taxonómicas de impactos são difíceis de extrapolar entre regiões.
Os gradientes biológicos definidos através dos atributos seleccionados também co-variaram com os gradientes de perturbação, tendo sido possível obter uma melhor discriminação de categorias de uso de solo. Nas diferentes regiões, a discriminação de locais mais impactados foi feita com base num conjunto similar de atributos, que inclui tamanho, voltinismo, técnicas reproductivas, microhabitat, preferências de corrente e substrato, hábitos alimentares e formas de resistência. Este conjunto poderá vir a ser usado para avaliar de forma predictiva os efeitos das modificações severas de uso de solo impostas pela actividade agrícola. Quando analisadas simultaneamente através dos atributos, as comunidades das três regiões permitiram uma moderada mas significativa discriminação de níveis de impacto. Estas análises corroboram as evidências de que as mudanças nas comunidades de macroinvertebrados aquáticos em locais sob a influência de agricultura intensiva podem seguir uma trajectória convergente no espaço multidimensional, independentemente de factores geográficos. Foram fornecidas pistas para a identificação de parâmetros específicos que deverão ser tidos em conta no planeamento de novos programas de biomonitorização com comunidades de macroinvertebrados bentónicos, para aplicação numa gestão fluvial verdadeiramente ecológica, nestas e noutras regiões. Foram ainda sugeridas possíveis linhas futuras de investigação.

13
keywords
macroinvertebrate traits; freshwater ecosystems; biomonitoring; agriculture; streams; biological communities; biogeographic regions; impact gradients
abstract
Freshwater ecosystems - engineers of environmental functions and
important providers of irreplaceable goods and services - are being severely affected by anthropogenic perturbations. The conversion of forested land to agriculture is affecting these aquatic systems through a series of mechanisms: sedimentation; nutrient enrichment; contaminant input; hydrologic alteration; and riparian clearing. Freshwater macroinvertebrate communities, being diverse, widespread, and sensitive to environmental perturbation, are particularly interesting for the ecological integrity assessment of those aquatic systems affected by such a multitude of stressors.
Biomonitoring - the systematic use of biological responses to evaluate environmental changes - can be undertaken through diverse methodologies. But these do not usually consider the functional intricacies of biological communities and are geographically constrained. Biomonitoring through sets of biological traits (characteristics that reflect species adaptation to their environment) appears as a promising tool to overcome these issues, offering a series of other advantages: direct causal relationships with stressors; better differentiation of impacts; and integration of natural fluctuations.
In this study, the current state-of-the-art of the usage of biological traits in biomonitoring is presented in a critical review of the existing published literature. Until publication date, no such work was available to provide freshwater ecologists with the conceptual underpinning for the use of traits as community descriptors and for freshwater biomonitoring and management. The support from ecological theory (the habitat templet concept and the landscape filtering hypothesis) was reviewed and studies applying this knowledge under real scenarios were presented. Technical issues were addressed and solutions proposed. Specific future needs are: a broader unified trait biomonitoring tool; more accurate understanding of the natural variation of community patterns; approaches to deal with trait trade-offs and syndromes; additional life history and ecological requirement studies; and the detection of specific impacts under multiple stressor scenarios.
The aim was to address the improvement of biomonitoring through traits, focusing on freshwater macroinvertebrate communities from streams of different biogeographic regions (in the Little and Salmon River, New Brunswick, Canada; Anllóns River, Galicia, Spain; Upper Reventazón River, Cartago, Costa Rica) spanning comparable gradients from low (watersheds with percentages of forest cover >75%) to high agricultural land use intensity.
In each land use gradient, the characterization of the biological community (by macroinvertebrate kick sampling in riffle areas) was accompanied by the characterization of the surrounding habitat (watershed scale properties, water chemistry and other reach scale properties). The macroinvertebrate community was characterized through taxonomic information, structural metrics, diversity indices, tolerance metrics, biotic indices and through the attribution of traits reflecting general biological and physiological features, life history and resistance to disturbance. Univariate and multivariate statistical data analyses were used to highlight biological and physico-chemical gradients, confirm their co-variation, test the significance of impact level discrimination and establish interregional comparisons. Community structure was used to reveal complex impact gradients, that significantly covaried with watershed agricultural land use gradients. These

14
impact gradients were mostly related with nutrient input and sedimentation. The biological gradients defined by selected structural measures co-varied with the disturbance gradients, although few structural variables individually discriminated the a priori defined land use categories. No consistency in the responses of the structural measures across biogeographic regions was detected. It was therefore confirmed that pure taxonomic interpretations of potential impacts are difficult to extrapolate between regions.
The biological gradients defined by the selected traits also co-varied with the disturbance gradients and an improved discrimination of land use categories was obtained. Across regions, a similar set of traits discriminated higher impact sites, including size, voltinism, reproductive techniques, microhabitat, current and substrate preferences, feeding habits and resistance forms. This set could be further studied and used to predictably assess the effects of severe land uses changes posed by agricultural scenarios. When analyzed simultaneously using traits, the communities of the three regions allowed a moderate but significant discrimination of impact levels. These analyses support the evidence that freshwater macroinvertebrate community shifts in sites impacted by intensive agriculture may follow convergent trajectories in multi-dimensional space, regardless of geography. Overall, clues were given to identify specific features that should be considered in the design of future freshwater biomonitoring programs using benthic macroinvertebrate communities for application in true ecologically oriented river management in these and other regions. Future research needs were also suggested.

15
contents
Chapter 1. General Introduction 5
1.1. Streams: importance and threats to integrity 7
1.2. Stream integrity monitoring using benthic macroinvertebrates 10
1.2.1. Biomonitoring through sets of biological traits 17
1.3. Environmental impacts of agriculture: the case of the potato crop 17
1.3.1. Potato production practices 18
1.3.2. Environmental concerns raised by potato crops 19
1.4. Conceptual framework of the study 22
1.5. References 24
Chapter 2. Beyond taxonomy: a review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring 31
2.1. Introduction 33
2.2. Biological traits and ecological theory: the habitat templet concept 34
2.2.3. The River Habitat Templet 35
2.3. Biological traits and ecological theory: the habitat filtering hypothesis 37
2.4. Progress in the development of traits as biomonitoring tools 39
2.5. “Reference” state studies in freshwater ecosystems 40
2.6. Technical aspects of biomonitoring 43
2.7. Statistical tools 44
2.8. Trade-offs and trait syndromes 45
2.9. Applications in human impact scenarios 46
2.10. Conclusions 49
2.11. References 51
2.12. Supporting information: databases offering information on freshwater macroinvertebrate traits 59
Chapter 3. Study areas and methods 61
3.1. Choice of study regions 63
3.1.1. Little River and Salmon River watersheds (Canada) 65
3.1.2. Anllóns River watershed (Spain) 66
3.1.3. Reventado, Birrís and Turrialba watersheds (Costa Rica) 67
3.2. Materials and Methods 71
3.2.1. Habitat characterization 71
3.2.2. Biological community taxonomic characterization 73
3.2.3. Biological community characterization through traits 74
3.3. References 77

16
Chapter 4. Freshwater macroinvertebrate community gradients in three agricultural regions: a non-functional approach 81
4.1. Introduction 83
4.2. Methods of data analysis 85
4.2.1. Unresponsive variables 85
4.2.2. Impact gradients 85
4.2.3. Biological gradients 86
4.2.4. Relationships between the biological and environmental gradients 88
4.3. Results 88
4.3.1. Unresponsive variables 88
4.3.2. Impact gradients 90
4.3.3. Biological gradients 94
4.4. Discussion 103
4.5. References 111
Chapter 5. Macroinvertebrate traits in watersheds from different biogeographic regions and their application in biological monitoring 117
5.1. Introduction 119
5.2. Methods of data analysis 121
5.2.1. Trait gradients 121
5.2.2. Discrimination of land use categories (taxa and traits) 122
5.2.3. Co-structure of environmental and biological variables 122
5.2.4. Interregional comparisons 123
5.3. Results 124
5.3.1. Discrimination of land use categories 124
5.3.2. Co-structure of environmental and biological gradients 129
5.3.3. Interregional comparisons 130
5.4. Discussion 134
5.5. References 139
Chapter 6. General Discussion and Conclusions 143
6.1. References 149

1
abbreviations
ANNA - Assessment by Nearest Neighbor Analysis
ANOVA – one-way ANalysis Of Variance
AQEM - Assessment system for the ecological Quality of streams and rivers
throughout Europe using benthic Macroinvertebrates
AUSRIVAS - AUStralian RIVer Assessment Scheme
BEAST - BEnthic Assessment SedimenT
EPT – Ephemeroptera, Plecoptera and Trichoptera
FPCA - Fuzzy Principal Components Analysis
PCA – Principal Components Analysis
RIVPACS - River InVertebrate Prediction And Classification System
RLQ – R-array (environmental variable-site table), Q-array (taxon-trait table)
and L-link (taxon-site table) between R and Q
SPEAR - SPEcies At Risk
USEPA – United States Environmental Protection Agency


3
figures and tables
Fig. 1.1 - Multiple stressor scenario posed by intensive agriculture on freshwater systems. Fig. 3.1 - World map with the location of the three regions studied: eastern Canada (Little River and Salmon River in New Brunswick); northwestern Spain (Anllóns River); and central Costa Rica (Upper Reventazón River). Country/province outlines and hydrological maps of each river. Fig. 3.2 – Sampling site location in Canada, Spain and Costa Rica. Photographs show streams belonging to each land use category (Foley Brook, Outlet Brook, Dead Brook, A Regueira, Rego da Balsa, Río Archeiro, Río Retes, Río Coliblanco, Quebrada Pacayas). Fig. 3.3 – Decision tree employed to fill the macroinvertebrate taxa x traits matrix. Examples of the affinity score assignment of three categories of a generic trait y for a generic macroinvertebrate genus x. Fig. 4.1 - Mean elevation of Costa Rican reference, medium impact and high impact field sites. Fig. 4.2 – Biplot of PCA of impact variables at Canadian reference, medium impact and high impact field sites. Fig. 4.3 - Mean values of the environmental variables used to characterize the land use impact gradient in Canadian reference, medium impact and high impact field sites. Fig. 4.4 – Biplot of PCA of impact variables at Spanish reference, medium impact and high impact field sites. Fig. 4.5 - Mean values of the environmental variables used to characterize the land use impact gradient in Spanish reference, medium impact and high impact field sites. Fig. 4.6 – Biplot of PCA of impact variables at Costa Rican reference, medium impact and high impact field sites. Fig. 4.7 - Mean values of the environmental variables used to characterize the land use impact gradient in Costa Rican reference, medium impact and high impact field sites. Fig. 4.8 - Mean values of derived biological variables for Canadian reference, medium impact and high impact field sites. Fig. 4.9 – Biplots of PCA using macroinvertebrate taxa abundances at reference, medium impact and high impact field sites in three biogeographic regions. Mean values of the axes scores of each PCA.
22
64
70
76
88
90
91
92
93
94
95
97
98

4
Fig. 4.10 - Mean values of derived biological variables for Spanish reference, medium impact and high impact field sites. Fig. 4.11 - Mean values of derived biological variables for Costa Rican reference, medium impact and high impact field sites. Fig. 5.1 – Biplot of FPCA of macroinvertebrate trait category abundances at Canadian reference, medium impact and high impact field sites. Mean values of the axes scores of the PCA. Fig. 5.2 – Frequency distribution of 100 simulated values (for 72 randomly selected taxa) and observed values of between land use category variances for taxa and trait composition of macroinvertebrate communities at Canadian field sites. Fig. 5.3 – Biplot of FPCA of macroinvertebrate trait category abundances at Spanish reference, medium impact and high impact field sites. Mean values of the axes scores of the PCA. Fig. 5.4 – Frequency distribution of 100 simulated values (for 72 randomly selected taxa) and observed values of between land use category variances for taxa and trait composition of macroinvertebrate communities at Spanish field sites. Fig. 5.5 – Biplot of FPCA of macroinvertebrate trait category abundances at Costa Rican reference, medium impact and high impact field sites. Mean values of the axes scores of the PCA. Fig. 5.6 – Frequency distribution of 100 simulated values (for 72 randomly selected taxa) and observed values of between land use category variances for taxa and trait composition of macroinvertebrate communities at Costa Rican field sites. Fig. 5.7 – Total number of macroinvertebrate taxa found in stream field sites of three biogeographic regions. The number of taxa found in at least one other region and the number of taxa exclusively found in each region. Fig. 5.8 – Biplots of PCA and FPCA of macroinvertebrate taxa and trait abundances at reference, medium impact and high impact field sites in Canada, Spain and Costa Rica. Fig. 5.9 – Mean values of the PCA axes scores for taxa abundances and of the FPCA axes scores for trait category abundances considering land use categories and regions. Fig. 5.10 – Frequency distribution of 100 simulated values (for 72 randomly selected taxa) and observed values of between land use category variances and between region variances for taxa and trait composition of macroinvertebrate communities in three different biogeographic regions. Table 1.1 - Community-level measures used in freshwater macroinvertebrate biomonitoring accompanied by examples. Table 3.1 – Macroinvertebrate traits and trait categories. Table 4.1 - Variables (measured and derived) used in the analyses and respective codes used in the text and figures. Table 4.2 - Mean values of environmental variables measured in reference, medium impact and high impact field sites. Table 5.1 – First axis and RV-coefficient results of co-inertia analyses relating environmental variables associated with land use gradients and biological variables (taxa and trait category abundances).
100
102
124
125
126
127
128
128
131
132
133
134
13
75
87
89
129

Chapter 1. General Introduction


General Introduction
7
1.1. Streams: importance and threats to integrity
“Unfortunately, the frequency of unpleasant environmental perturbations and
the extent of the areas affected as well as the duration of effect have increased
markedly during the past few years. […] Industrial societies invariably have
operated on the assumption that natural ecosystems have a certain capacity for
assimilating societal wastes without themselves being significantly degraded.”
(Cairns Jr, 1980)
“One of the nonecologists present plaintively but humorously asked, "Why
can't you environmental toxicologists just give us a freeze dried, talking fish on a
stick? This fish could be inserted in any aquatic ecosystem where pollution
problems were suspected and the fish would immediately expand, determine the
biological condition of the water, and give the person holding the stick the answer
verbally." […] those two simple sentences contain a beautiful description of the
kinds of methods the rest of the world wants from ecologists and environmental
toxicologists.”
(Cairns Jr, 1985)
These two citations define a problem - environmental pollution - that, over the
last decades, has led to a scientific search for new tools for ecosystem integrity
assessment. Pollution (either urban, agricultural or industrial) is a growing global
issue and aquatic systems are particularly at risk, since they are the receiving
environment for the myriad contaminants released as a result of human activities.
Currently, aquatic ecosystems are being severely altered or destroyed at a greater
rate than at any other time in human history, and much more rapidly than they are
being restored (Baron et al., 2002).
The importance of freshwater ecosystems (rivers, lakes, groundwater, and
wetlands) as providers of ecosystem goods and services is undeniable, in terms of
natural productivity, energy flow and the cycling of matter. They constitute

Chapter 1
8
important habitats for a wide variety of organisms, supporting and regulating
ecosystem productivity, their ability to receive materials generated from the
landscape (allowing the creation of physical habitat structure and nutrient supply
and storage), the definition (by the flow regime) of rates and pathways by which
precipitation enters and circulates within the ecosystems, the ability to regulate
climate, the storing and cycling of soil and sediments (Baron et al., 2002, Hooper
et al., 2005). Their dynamic, relatively scarce and insular nature aggravates the
consequences of the destruction of these supporting systems.
In addition, humans are particularly dependent on these systems and their
surroundings as sources of drinking and irrigation water, sources of food,
navigation pathways, sources of hydroelectric power, waste disposal and
purification systems, recreational spaces, as well as for the establishment of
agricultural land and many other types of activities. These activities that are
connected to societal well-being have long taken precedence over other goods
and services provided by freshwater ecosystems (Baron et al., 2002), although
they constitute expensive and sometimes irreplaceable benefits.
The crucial importance of freshwater systems to mankind has in some way
defined the magnitude of the threats that affect them, as human communities and
activities are spatially focused around these systems. Climate change, the
construction of dams and the modifications of channels, the introduction of alien
species and land use alterations are major threats affecting stream ecosystems
(Cushing & Allan, 2001; Allan, 2004). Freshwater habitats are embedded within
the terrestrial world, deriving much of their character from their drainage basins;
human activities in the drainage basin often have strong effects on freshwater
habitats (Strayer, 2006). One major driver of freshwater – and in particular stream
- ecosystem degradation is in fact the conversion of watershed land cover from
forest to human-altered forms, mainly for agricultural activities or for urban uses,
but also for forestry, mining, and recreation.
Agricultural land, for example, occupies the largest fraction of land area in
many developed catchments (Allan, 2004), and its presence leads to:
- decreased stream bank stability, large woody debris presence and

General Introduction
9
energy inputs (as leaf litter) and increased water temperatures and light
exposure due to the removal of riparian forest (Broadmeadow & Nisbet,
2004);
- increased nonpoint inputs of pollutants and nutrients (phosphorus
and nitrogen) by pesticide and fertilizer applications (Carpenter et al.,
1998);
- stream flow alterations by extraction of water for irrigation,
increased soil compaction, the presence of drainage ditches, alterations of
vegetation evapotranspiration rates (crop vs. natural vegetation) and loss of
wetland areas (Nilsson & Renöfält, 2008);
- increased sediment inputs and deposition due to intensive tillage
practices and removal of riparian vegetation (Henley et al., 2000).
The main mechanisms of stream ecosystem impairment by land use
changes are therefore diverse and can be summarized in the following categories:
sedimentation; nutrient enrichment; contaminant input; hydrologic alteration;
riparian clearing/canopy opening; loss of large woody debris. These mechanisms
may trigger a series of chemical, physical and biological effects in freshwater
ecosystems, generating marked deviations from natural patterns. These effects
include decreased habitat quality, physical damage of living organisms, alterations
in algal biomass, ecosystem productivity and food quality (consequently, food web
disruption), shifts in species composition and abundance, species extinction,
chemical toxicity (by heavy metals, organic compounds, etc.), changes in
invertebrate drift and emergence, depression of fish growth, reproduction and
survival, changes in flood magnitude and frequency, and so on (see Allan, 2004
for a review).
Given the importance of freshwater ecosystems and the deleterious effects
human activities can have in those systems, the assessment, protection and
restoration of their integrity is a priority. The concept of stream health or integrity
implies the preservation of a “balanced, integrated, adaptive system having the full

Chapter 1
10
range of elements (genes, species, assemblages) and processes (mutation,
demography, biotic interactions, nutrient and energy dynamics, metapopulation
processes) expected in areas with minimal influence from modern human society”
(Karr, 1999). In order to maintain the “full range” mentioned by Karr, there is an
urgent need to restore balance in the costly ecological trade-offs associated with
human exploitation of freshwater ecosystems.
1.2. Stream integrity monitoring using benthic macroinvertebrates
The development of new assessment and monitoring methodologies for the
aquatic environment (the search for the “talking fish on a stick”) was particularly
stimulated by the need to assess compliance with a multitude of laws and
regulations concerning and regulating aquatic systems and the human activities
associated with them. There was a growing recognition that functionally intact and
biologically complex aquatic ecosystems provide many economically valuable
services and long-term benefits to society (Baron et al., 2002). Therefore, following
the assumption that measurement of the condition of the biota can be used to
assess the condition of an ecosystem (Herricks & Cairns, 1982), environmental
decision-making is now highly dependent on the systematic use of biological
responses to evaluate changes in the environment with the intent to use this
information in a quality control program – or biomonitoring (Matthews, 1982).
Diverse methodologies for biological monitoring have been developed, since
the first references to the concept of biological indicator of environmental
conditions at the beginning of the 20th century (Cairns Jr & Pratt, 1993; Bonada et
al., 2006). The diversification of methodologies was justified by the informative
potential of different methods concerning different types of disturbances, the
complexity of freshwater systems, the different types of assessment and the
different precisions those types require, the appearance of new laws and
regulations and even the variation of priorities among freshwater ecologists
(Bonada et al., 2006).

General Introduction
11
Different biological response types have been used to quantify human
impact, for example: bacterial luminescence, biomarkers, concentrations of
nutrients or pollutants in plant or animal tissues, invertebrate behavior, fluctuating
asymmetry or community structure and function (Statzner et al., 2001). In fact, it
has been long accepted by ecologists and environmental scientists that biological
communities reflect the environmental conditions they find themselves in and
respond to disturbances in a predictable manner (Cranston, 1990). Lower levels of
biological organization elicit mechanisms of contaminant effects in an often more
obvious way, whereas ecological significance is generally more apparent at higher
levels. As a result, biomonitoring of freshwater benthic communities became a
valuable tool in the assessment of stream integrity.
The communities of freshwater invertebrates are particularly interesting and
offer a series of generic advantages that motivate the use of this group in
freshwater biomonitoring and ecological integrity assessment. While defining the
criteria for an “ideal biomonitoring tool” – and taking into account that aquatic
invertebrates do not constitute perfect tools – aquatic ecologists highlight some of
the advantages they bring to biomonitoring (Rosenberg & Resh, 1996; Bonada et
al., 2006): the widespread distribution, in many different aquatic habitats; the
usually high abundances; the high species richness, offering a spectrum of
environmental responses, specifically to stress; their sedentariness (e.g. in
comparison to fish), facilitating spatial analysis of pollution effects (they are good
indicators of localized conditions); the potential to use drift ability to indicate
pollutant presence; the possibility of tracing effects over longer periods due to long
life cycles of certain species; the inexpensive sampling gear needed and ease of
use; the well-described taxonomy for genera and families (with exceptions for
tropical taxa); the differential sensitivities of common species to types of pollution;
and the opportunity to use many of the invertebrate species in experimental
studies of pollution effects, allowing the association of these studies with the
outcomes of aquatic monitoring programs. Furthermore, combinations of
invertebrate taxa can be categorized so as to permit ecological questions to be
addressed at the functional level (Merritt & Cummins, 1996). In particular, the

Chapter 1
12
functional importance of aquatic macroinvertebrates in freshwater ecosystems is
undeniable and mostly related to their diverse array of feeding habits (Wallace &
Webster, 1996): they constitute important links between their food and higher
trophic level vertebrates and they contribute immensely to nutrient cycling and
turnover of organic material from inside the system or entering through the riparian
zone.
The weaknesses of using this biological group in biomonitoring are related
with their inability to respond directly to all types of impacts, the fact that their
abundance and distribution can be affected by factors other than water quality
(e.g. seasonal changes, type of substrate), the fact that their dispersal abilities
may carry them to areas where they naturally do not occur and the need for deep
taxonomic knowledge for correct identification (Rosenberg & Resh, 1996).
Methods developed over the past decades (for an historical review refer to
Cairns Jr & Pratt, 1993) for biomonitoring of stream integrity using aquatic
invertebrate communities are diverse and span from the usage of relatively simple
biotic indices, to multimetric approaches that combine several of those indices or
even to relatively complex multivariate approaches that aim to recognize and
predict patterns of disturbance. Although community-level freshwater
biomonitoring has received more attention, tools developed for other levels of
biological organization are worthy of mention; these include (Rosenberg & Resh,
1996): sub-individual monitoring (e.g. measurements of changes in enzyme
activities or respiratory metabolism); individual level monitoring (e.g. morphological
deformities or bioaccumulation studies); population monitoring (e.g. some biotic
indices using populations as indicator taxa or fluctuating asymmetry
measurements); and ecosystem monitoring (e.g. monitoring effects on the
structure of a food web or alterations in productivity).

General Introduction
13
For community-level assessments, the different approaches typically include
one or several measures (usually called metrics) that in someway describe
community parameters (Table 1.1). Taxa richness, the number of taxa in a
community, is a commonly used structural metric. It is based on the premise that
the number of taxa decreases as water quality decreases (Rosenberg & Resh,
1996). Another common richness measure is EPT richness (the number of
Ephemeroptera, Plecoptera and Trichoptera taxa in a sample), working under the
assumption that most taxa in these orders are pollution sensitive. If abundances
are taken into consideration, several composition measures can be calculated:
total number of individuals, percent of EPT, ratio of EPT abundance to
Chironomidae, percent of dominant taxon and so on (Barbour et al., 1999). In
these cases the assumption is that certain stresses cause variations in total
numbers of individuals and that a healthy and stable assemblage will be relatively
consistent in its proportional representation, though individual abundances may
vary in magnitude. When both the richness of the sample and the number of
individuals of each species (evenness) are combined in a unique metric, we get a
diversity index (e.g. the Shannon"s Index), and comparisons are made assuming
Type of measure Examples
Richness measures Taxa richness; EPT richness
Composition measuresTotal number of individuals; Percent of EPT; Ratio of EPT; Abundance to
Chironomidae; Percent of dominant taxon
Diversity indices Shannon’s Index
Similarity indices Coefficient of Community Loss; Pinkham-Pearson Index
Tolerance/intolerance measures Number of intolerant taxa; Percent of tolerant organisms
Biotic indicesSaprobic Index; Belgian Biotic Index; Biological Monitoring Working Party
Score; Biological Condition Index; Hilsenhoff Biotic Index
Multimetric indices Index of Biotic Integrity; AQEM assessment system
Multivariate approaches RIVPACS; AUSRIVAS; BEAST; ANNA
Functional feeding group measures Proportion of shredders in a leaf pack; Ratio of scrapers to collectors
Sets of biological traits
Table 1.1 - Community-level measures used in freshwater macroinvertebrate biomonitoring accompanied by examples (descriptions and bibliographic references in the text).

Chapter 1
14
that high diversity would associate with a balanced and stable community. But the
application of this approach in biomonitoring has been highly criticized, and
suggestions have been made not to use diversity indices except when in
combination with other indices (Boyle et al., 1990; Lydy et al., 2000). Another type
of index compares community structure between two sites or at the same site at
different times and is based on the rationale that communities of disturbed and
undisturbed sites will become more dissimilar as stress increases. Examples of
similarity indices include the Coefficient of Community Loss or the Pinkham-
Pearson Index (Rosenberg & Resh, 1996).
When pollution tolerances are assigned to different taxa,
tolerance/intolerance measures can be calculated in order to get a
representation of relative sensitivity to perturbation. Examples of these metrics are
the number of intolerant taxa or the percent of tolerant organisms, but also the
popular biotic indices - coded numerical expressions representing the combined
tolerances or intolerances of the organisms in a sample, ideally to specific types of
pollution. Examples include the Saprobic Index (dealing with organic pollution;
Rolauffs et al., 2004), the Belgian Biotic Index (Pauw & Vanhooren, 1983), the
Biological Monitoring Working Party Score (Armitage et al., 1983) and the
Hilsenhoff Biotic Index (Hilsenhoff, 1988). The application of these methodologies
will depend on the accurate assessment of taxa tolerance used to calculate the
index and, as tolerance values are usually specific for the geographic region for
which they were developed, the indices are to a large extent geographically
constrained.
Multimetric indices use a combination of several of these individual metrics
in order to assess stream integrity (e.g. the Index of Biotic Integrity or the AQEM
assessment system). In this type of approach, the first step is the selection and
calibration of metrics and subsequent aggregation of these metrics into a
combination index that is applicable to homogeneous sites (this phase involves
characterization of reference conditions that will form the basis for assessment); in
a second stage, the biological condition at a particular site is assessed, and levels
of impairment are determined (Barbour et al., 1999). These indices use various

General Introduction
15
measures of richness and composition, tolerance values, functional feeding group
designations, etc. and this results in the diminishing of the risk of making incorrect
assessments. Once more, one limitation of this approach is the lack of large-scale
applicability across ecoregions (Bonada et al., 2006). Another way of assessing
stressor impacts by comparing community patterns from impacted and non-
impacted sites is the application of multivariate approaches (e.g. RIVPACS,
AUSRIVAS, BEAST, ANNA). In this case, reference sites are classified using the
biological assemblage to establish the variance expected to occur (as opposed to
classifying reference sites based on geographic and physical attributes as done in
multimetric approaches) and the species-site matrix is used to determine
deviations from these references (metric scores are not computed until after this
analysis, to determine the types of stressors that may motivate those deviations)
(Lücke & Johnson, 2008). Multivariate approaches assess consequences of
human impacts in community composition and are currently not assessing
ecological functions (Bonada et al., 2006).
The biomonitoring methods described previously usually do not consider
measures that somehow relate to ecological functions of the freshwater
ecosystems. The use of functional feeding group measures (e.g. proportion of
shredders in a leaf pack or ratio of scrapers to collectors) is one example of
methodologies that consider the functional roles played by the biological
communities on the systems under evaluation, instead of just using measures of
the structure of macroinvertebrate communities. This methodology is based on the
assumption that organisms have evolved certain morphological-behavioral food-
gathering mechanisms or locomotion-attachment adaptations and can be placed
into particular groups that relate with particular food source availability or specific
habitat types (Vannote et al., 1980; Rosenberg & Resh, 1996). Perturbations of the
communities could be detected through changes in the expected abundances
inside these functional groups. In biomonitoring, simple associations between
abundances and the classification of the communities according to feeding groups
can be used; but functional feeding groups have also been associated with
multimetric approaches or approaches using sets of biological traits (cf. next

Chapter 1
16
section). One advantage of using such a methodology is that the proper
assignment of feeding groups across regions would enable the application of this
approach across ecoregions, although only impacts that change food availability
can be detected (Bonada et al., 2006).
More recently, biomonitoring approaches that use sets of biological traits
have been developed and these will be addressed in the following section and in
Chapter 2.
Although a great diversity of biomonitoring methodologies has been
developed over the years, not all these methods can be considered totally
adequate for all environmental assessment situations. The choice will always
depend on the objectives of the study (e.g. baseline monitoring, trends analysis or
detecting unknown sources of pollution). It is important that the macroinvertebrate
biomonitoring method chosen can actually detect change that has occurred at an
impacted site, yet can also point towards the underlying cause(s) of the observed
changes, whether natural or human induced. A predictive behavior of these
methodologies would also be useful, that is, the ability to predict potential changes
in a biological community based on ecological theory alone. Furthermore, ideal
biomonitoring tools should be applicable across large spatial scales, so that
knowledge developed for one particular region can be easily applied in other
regions. This is particularly important considering that environmental law and
regulations are generally defined for large geographic areas that comprise different
ecoregions (e.g. the European Union Water Framework Directive or the Canada
Water Act). Previous works have highlighted these but also some other criteria that
define the “ideal biomonitoring tool using aquatic invertebrates” (Bonada et al.,
2006): potential to assess ecological functions, important in valuing environmental
services; low costs, reproducibility and simplicity of sampling, sorting, taxonomic
identifications and standardized experimentation; reliable identification of changes
of overall and specific human impacts without interference by natural variability
patterns in reference conditions; and the indication of impact on a linear scale so
that freshwater management costs and ecological improvements can be linearly
related.

General Introduction
17
1.2.1. Biomonitoring through sets of biological traits
One of the most promising biomonitoring tools using aquatic invertebrates
currently under development is the use of sets of traits of these organisms, which
are characteristics related mostly with their biology and ecology. The use of traits
allows a view of the community that goes beyond mere structure (taxa richness
and abundance), including predictors of functional roles and adaptation skills to a
changing environment. The ecological theory behind this tool states that
organisms have inherent traits for survival - for example optima for natural
conditions, habitat preferences and life-history characteristics - and hypothesis
that certain traits are selectively removed by anthropogenic environmental
changes can be posed (based on the concept of habitat templet - Southwood,
1977).
With this “trait approach” the assessment of impaired conditions is possible,
but there is a great potential for inference of specific causes of impairment, and
this would be particularly interesting in multiple stressor scenarios. As mentioned
before, on freshwater systems, the tools currently used for the assessment of the
negative effects of human impacts are generally not applicable across large
geographic units because they were developed and adapted to national or regional
characteristics of the aquatic biota (Gayraud et al., 2003). This is another point
where the application of the trait approach can be advantageous, as a functional
image of a community will be more comparable among different ecoregions than a
structural, taxonomy-based image. These and other issues are further developed
in Chapter 2.
1.3. Environmental impacts of agriculture: the case of the potato crop
As mentioned in Section 1.1, agriculture - as a form of land-use alteration -
can impair aquatic ecosystems through a variety of mechanisms. This multiple
stressor scenario posed by agricultural activities appears as an interesting setup to
test hypothesis raised from the attempts to apply the “trait approach” to aquatic

Chapter 1
18
macroinvertebrate biomonitoring in a more diagnostic and widely applicable way.
One example of a highly environmental aggressive crop is the potato crop,
due to a series of production practices that origin a multitude of alterations to
natural environmental conditions. The uses of the potato are diverse and range
from human consumption (fresh or processed) to re-usage as seed tubers for
growing the next season"s potato crop; it is the world"s fourth most important food
crop, after maize, wheat and rice (FAO, 2008). Potatoes also serve as food to farm
animals and are used for several purposes in the pharmaceutical, textile, wood, oil
drilling and paper industries, and even in the production of fuel-grade ethanol.
Potato is therefore an economically important and widespread crop, potatoes
being grown in more than 100 countries, under temperate, subtropical and tropical
conditions (FAO, 2008).
1.3.1. Potato production practices
Growing potatoes involves extensive ground preparation (ploughing,
harrowing and rolling) so that the soil reaches a suitable condition (FAO, 2008).
”Seed potatoes” are then sown, and during the four weeks of potato canopy
development, weeds have to be controlled my mechanical removal or herbicide
application. Chemical fertilizer application will depend on the available soil
nutrients, but requirements are usually relatively high. As an example, in volcanic
soils a deficiency in phosphorus is typically detected (FAO, 2008) and
compensated by heavy fertilizer application. The predominantly used fertilizers
contain three macronutrients (nitrogen, potassium, and phosphorus) and have
been shown to improve yield and quality of potato tubers where native supplies are
limiting (Davenport et al., 2005). Animal manures, sludge, slurries, and plant
material incorporated into the soil have also been used as a partial or total
replacement for soluble mineral fertilizers in potato production (Muñoz et al.,
2005). The soil moisture content must also be maintained at a relatively high level,
and frequent irrigation is frequently needed in order to maximize crop yield.
In terms of crop protection, some precautions are usually taken to avoid crop

General Introduction
19
loss due to diseases and pests, namely selection of tolerant varieties, use of
certified seed tubers, and crop rotation (Muñoz et al., 2005) with other, dissimilar
crops, such as maize, beans and alfalfa. In the field, potato cultures may be
affected by fungi, viruses, viroids, bacteria, nematodes, insects and weeds (Struik
et al., 2007). Chemical control is a widespread methodology used in their
elimination. It includes soil fumigant treatments, fungicide, insecticide, nematicide
and herbicide applications at different moments of potato production.
When the crop is reaching maturity, tuber maturation is artificially induced by
killing the potato vine before harvesting using mechanical killing, chemical killing
(using compounds such as diquat, paraquat, sulfuric acid or glyphosinate) or a
combination of both methods (Kempenaar & Struik, 2007). Potatoes are then
harvested, using once again processes associated with intensive soil tillage (FAO,
2008).
1.3.2. Environmental concerns raised by potato crops
As shown, from all the pests that farmers potentially have to deal with when
maintaining their crops – namely weeds, animals (insects, mites, nematodes,
rodents, slugs, snails or birds) and plant pathogens (viruses, bacteria, fungi) –
many have to be simultaneously minimized when maintaining a potato crop. All of
the mentioned pest groups are of high economic importance because vegetative
propagation predominates in potato production (Oerke, 2006) and therefore lack of
chemical control applications can result in major economic losses to the producer
(Noronha et al., 2008): in the absence of the different forms of crop protection
almost 75% of attainable potato production would be lost. Therefore the potato
industry has experienced the rise of chemical crop protectants, including
fungicides, insecticides, nematicides, herbicides, growth regulators and vine killing
agents (Struik et al., 1997). For aquatic organisms such as macroinvertebrates,
the main route of exposure to these compounds is usually via the water and only
to a lesser extent through sediment or food (Liess et al., 2005). The routes of
pesticide exposure include uptake from pore water and overlying water across

Chapter 1
20
body walls and across respiratory surfaces, and ingestion of contaminated
sediment. Agricultural insecticide and herbicide runoff as been shown to increase
aquatic invertebrate deformities and mortality rates, shift their abundance, drift and
emergence patterns, disrupt endocrine systems and cause physical avoidance
(Allan, 2004).
The protection of the potato crop also includes other types of intervention,
besides chemical pest control, including frequent soil disturbance and intensive
fertilizer application. As mentioned previously, potato cultivation usually involves
intensive soil tillage throughout the cropping period (during soil preparation,
mechanical weeding and mechanized harvesting), which often leads to soil
degradation, erosion and leaching of nitrates (Fiener & Auerswald, 2007; FAO,
2008). High and sustained levels of sediment entering streams may cause
permanent alterations in community structure, diversity, density, biomass, growth,
and rates of reproduction and mortality by affecting food webs and habitats
(Henley et al., 2000). Sediment in transport can have an abrasive quality and
increases turbidity, it can reduce the quantity of periphyton that grows on stream
substrata, limit light penetration and therefore reduce phytoplankton production,
reduce macrophyte biomass, growth, and diversity and coat invertebrate gills and
respiratory surfaces; as sediment settles, interstitial spaces between coarse
substrata are filled, which reduces available habitat for macroinvertebrates and
insect community structure may change with this alterations, favoring burrowing
insects tolerant of low oxygen levels; also, insect escape through drift has been
shown to increase (Ryan, 1991; Henley et al., 2000).
As for fertilizer utilization, efforts have focused on phosphorus as a
potential contaminant of surface waters and nitrate as a potential groundwater
contaminant (Davenport et al., 2005). Agriculture has in fact been identified as a
potential major contributor of nitrogen and phosphorus to surface waters
(Carpenter et al., 1998; Randall & Mulla, 2001). The negative consequences of
this nutrient surplus to aquatic life are related with eutrophication, increases in
autotrophic biomass and production, loss of aquatic plant beds as habitats,
acceleration of litter breakdown rates, decrease in dissolved oxygen, and resulting

General Introduction
21
shifts from sensitive to more tolerant species (Carpenter et al., 1998; Allan, 2004).
Other important factors to consider are changes in stream hydrology
caused by the need to irrigate the crops and build proper drainage systems, as
well as by the changes in evapotranspiration rates from natural to crop vegetation.
These changes increase flood magnitude and frequency, often lowering base flow,
and there is a substantial body of evidence indicating that both high and low flow
disturbances play a central role in structuring stream communities (Poff & Ward,
1989). Due to hydrological alterations, in-stream habitats can become more
degraded due to more efficient transport of nutrients, sediments and contaminants
from the surrounding fields; habitat characteristics like water temperature, oxygen
content, water chemistry and substrate particle sizes can be altered (Richter et al.,
1996).
The potato field can also extend to the stream margin, by means of the
removal of natural riparian vegetation. As a consequence, water temperatures,
light penetration and plant growth tend to increase due to reduced shading; bank
stability decreases and channel erosion increases; there are less inputs of litter
and wood and less retention of nutrients and trophic structure shifts occur (Allan,
2004). Loss of large woody debris in stream ecosystems induces a series of
hydrologic and hydraulic consequences, resulting in reduced availability of feeding,
attachment and cover microhabitats and losses of sediment and organic material
storage and transfer (Gurnell et al., 1995). These changes can ultimately result in
reductions on macroinvertebrate diversity, density and/or biomass.
As shown, the negative impact of the potato crop on the aquatic environment
can be significant. For freshwater systems in the vicinity of potato plantations there
are multiple and complex ecological consequences of the agricultural interventions
associated with this crop (Fig. 1.1). Aquatic systems will ultimately constitute sinks
for the contaminants (nonpoint inputs of crop protectants and fertilizers) and
abnormal soil amounts released from the nearby fields; but they will also
potentially be more exposed to sun radiation, have less large woody debris
presence and have lower leaf litter inputs due to riparian vegetation clearing; and

Chapter 1
22
flow pattern alterations will be induced due to irrigation and drainage practices
(among other detrimental factors). This myriad of environmental stressors can be
further complicated by the interactions between different stressors, which are
mostly unknown.
Fig. 1.1 - Multiple stressor scenario posed by intensive agriculture on freshwater systems (agriculture-related activities are underlined and resulting stressors are in italic). T – temperature; CDW – coarse woody debris; P – phosphorus; N – Nitrogen.
1.4. Conceptual framework of the study
Many studies have focused on evaluating how species characteristics vary
across complex landscapes. Trait-based approaches offer significant advantages
over traditional taxonomic descriptors of ecological communities, by virtue of a
generic set of community descriptors that are not constrained by biogeography.
The potential value of this approach in aquatic biomonitoring of anthropogenic
stressors is reviewed by examining if unique trait patterns can be associated with
specific gradients of habitat disturbance and whether these tools aid in the
diagnose of causal agents under multiple stressor scenarios.
P N
! Crop protectants application
Fertilizer application
Soil tillage
Riparian clearing
Precipitation
Stream
Water extraction
Nutrient
runoff
Pollutant runoff
Sedimentation
Leaf litter and CWD inputs
! !
Flow pattern alterations
Light, T
Increased water T and light exposure

General Introduction
23
In this study, focus was on freshwater benthic macroinvertebrate
communities from streams of different biogeographic regions of the world and on
the potential impacts of intensive agricultural land use (intensive potato farming).
Each of the three study areas included streams spanning a gradient from low to
high agricultural land use intensity and the null hypotheses tested were:
- that the interdependence of watershed land use gradients and stream
environmental gradients cannot be detected;
- that the disturbance gradients studied do not affect the biological gradients
defined by the macroinvertebrate communities of the three biogeographic regions;
- that the trait pattern response to disturbance of the macroinvertebrate
communities in the three biogeographic regions does not differ from the structural
community response.
The stressor regime gradient was chosen to include nutrient enrichment,
chemical protectant input, sedimentation, riparian clearing and hydrological
alteration, and traditional taxonomy-based biomonitoring methods were compared
to more recent trait-based methodologies. Moreover, the potential of the trait
approach to highlight macroinvertebrate sensitivity to the multiple stressor
scenario was assessed. The implications of the results in the further development
of a diagnostic trait approach are discussed.
In order to assess the current state-of-the-art of the usage of biological traits
in monitoring and disturbance diagnostic methodologies, a critical review of the
existing published literature was undertaken. Until publication date, and despite
the decades of research in the area, no such work was available to provide
freshwater ecologists with the conceptual underpinning for the use of traits as
community descriptors and for freshwater biomonitoring and management. This
review work is presented in chapter two: “Beyond taxonomy: a review of trait-
based community descriptors as tools for biomonitoring”.
In chapter three, “Study areas and methods”, a detailed description of the
selected study sites, selection criteria, and general methodology is given. In this

Chapter 1
24
chapter, the procedures that were carried out are substantiated, methodological
problems are discussed and information on the study areas, which is usually not
easily available through the scientific publication system, is systematized.
Before the attempt to further develop methods that associate specific trait
patterns with gradients of disturbance, it is important to detect if disturbance and
biological response gradients actually exist. In chapter four, “Freshwater
macroinvertebrate community gradients in three agricultural regions: a non-
functional approach”, the disturbance and biological response gradients are
defined for the three regions using known community-level evaluation tools. The
differences in response gradients among regions are highlighted.
In chapter five, “Macroinvertebrate traits in watersheds from different
biogeographic regions and their application in biological monitoring”, species traits
are included in the community level analysis in order to evaluate if response
gradients match between different biogeographic regions where similar
disturbance gradients occur. The advantages of this more generic biomonitoring
tool are explored and a comparison with more traditional approaches is presented.
In chapter six, the results are discussed altogether, general conclusions are
drawn and future research needs addressed.
1.5. References
Allan, J. D. (2004) Landscape and Riverscapes: The Influence of Land Use on
Stream Ecosystems. Annual Review of Ecology, Evolution, and
Systematics, 35 (1), 257-284.
Armitage, P. D., Moss, D., Wright, J. F. & Furse, M. T. (1983) The performance of
a new biological water quality score system based on macroinvertebrates

General Introduction
25
over a wide range of unpolluted running-water sites. Water Research, 17
(3), 333-347.
Barbour, M. T., Gerritsen, J., Snyder, B. & Stribling, J. (1999) Rapid
Bioassessment Protocols For Use in Streams and Wadeable Rivers:
Periphyton, Benthic Macroinvertebrates and Fish, Second Edition.
Washington, D.C., U.S. Environmental Protection Agency. 339 pp.
Baron, J. S., Poff, N. L., Angermeier, P. L., Dahm, C. N., Gleick, P. H., Hairston, J.,
Jackson, R. B., Johnston, C. A., Richter, B. D. & Steinmanj, A. D. (2002)
Meeting Ecological and Societal Needs for Freshwater. Ecological
Applications, 12 (5), 1247-1260.
Bonada, N., Prat, N., Resh, V. H. & Statzner, B. (2006) Developments in Aquatic
Insect Biomonitoring: A Comparative Analysis of Recent Approaches.
Annual Review of Entomology, 51 (1), 495-523.
Boyle, T. P., Smillie, G. M., Anderson, J. C. & Beeson, D. R. (1990) A Sensitivity
Analysis of Nine Diversity and Seven Similarity Indices. Research Journal of
the Water Pollution Control Federation, 62 (6), 749-762.
Broadmeadow, S. & Nisbet, T. R. (2004) The effects of riparian forest
management on the freshwater environment: a literature review of best
management practice. Hydrology and Earth System Sciences, 8 (3), 286-
305.
Cairns Jr, J. (1980) Biological monitoring part I—Early warning systems. Water
Research, 14 (9), 1179-1196.
Cairns Jr, J. (1985) Just give me a freeze dried, talking fish on a stick. Journal
Water Pollution Control Federation, 57 (10), 980.
Cairns Jr, J. & Pratt, J. R. (1993) A history of biological monitoring using benthic
macroinvertebrate. In: D. M. Rosenberg & V. H. Resh (Ed.). Freshwater
biomonitoring and benthic macroinvertebrates. London, Chapman & Hall,
10-27.
Carpenter, S. R., Caraco, N. F., Correll, D. L., Howarth, R. W., Sharpley, A. N. &
Smith, V. H. (1998) Nonpoint pollution of surface waters with phosphorus
and nitrogen. Ecological Applications. 8, 559-568.

Chapter 1
26
Cranston, P. S. (1990) Biomonitoring and Invertebrate Taxonomy. Environmental
Monitoring and Assessment, 14 (2-3), 265-273.
Cushing, C. E. & Allan, J. D. (2001) Streams - Their Ecology and Life. San Diego,
California, Academic Press. 366 pp.
Davenport, J. R., Milburn, P. H., Rosen, C. J. & Thornton, R. E. (2005)
Environmental impacts of potato nutrient management. American Journal of
Potato Research. 82, 321-328.
FAO (2008) International Year of the Potato 2008: New light on a hidden treasure.
Rome, Food and Agriculture Organization of the United Nations. 148 pp.
Fiener, P. & Auerswald, K. (2007) Rotation Effects of Potato, Maize, and Winter
Wheat on Soil Erosion by Water. Soil Science Society of America Journal,
71, 1919-1925.
Gayraud, S., Statzner, B., Bady, P., Haybach, A., Scholl, F., Usseglio Polatera, P.
& Bacchi, M. (2003) Invertebrate traits for the biomonitoring of large
European rivers: an initial assessment of alternative metrics. Freshwater
Biology, 48 (11), 2045-2064.
Gurnell, A. M., Gregory, K. J. & Petts, G. E. (1995) The role of coarse woody
debris in forest aquatic habitats: implications for management. Aquatic
Conservation, 5 (2), 143-166.
Henley, W. F., Patterson, M. A., Neves, R. J. & Lemly, A. D. (2000) Effects of
Sedimentation and Turbidity on Lotic Food Webs: A Concise Review for
Natural Resource Managers. Reviews in Fisheries Science, 8 (2), 125 —
139.
Herricks, E. E. & Cairns, J. (1982) Biological Monitoring. Part III. Receiving System
Methodology Based on Community Structure. Water Research, 16 (2), 141-
153.
Hilsenhoff, W. L. (1988) Rapid Field Assessment of Organic Pollution with a
Family-Level Biotic Index. Journal of the North American Benthological
Society, 7 (1), 65-68.
Hooper, D. U., Chapin, I., Ewel, J. J., Hector, A., Inchausti, P., Lavorel, S., Lawton,
J. H., Lodge, D. M., Loreau, M., Naeem, S., Schmid, B., Setala, H.,

General Introduction
27
Symstad, A. J., Vandermeer, J. & Wardle, D. A. (2005) Effects Of
Biodiversity On Ecosystem Functioning: A Consensus Of Current
Knowledge. Ecological Monographs, 75 (1), 3-35.
Karr, J. R. (1999) Defining and measuring river health. Freshwater Biology, 41 (2),
221-234.
Kempenaar, C. & Struik, P. C. (2007) The Canon of Potato Science: 33. Haulm
Killing. Potato Research, 50, 341–345
Liess, M., Brown, C. D., Dohmen, P., Duquesne, S., Hart, A., Heimbach, F.,
Kreuger, J., Lagadic, L., Maund, S. J., Reinert, W., Streloke, M. &
Tarazona, J. V. (2005) Effects of Pesticides in the Field. Pensacola, SETAC
Press. 138 pp.
Lücke, J. D. & Johnson, R. K. (2008) Detection of ecological change in stream
macroinvertebrate assemblages using single metric, multimetric or
multivariate approaches. Ecological Indicators, 9, 659-669.
Lydy, M. J., Crawford, C. G. & Frey, J. W. (2000) A comparison of selected
diversity, similarity, and biotic indices for detecting changes in benthic-
invertebrate community structure and stream quality. Archives of
Environmental Contamination and Toxicology. 39, 469-479.
Matthews, R., Buikema Jr, A L, Cairns Jr, J, Rodgers Jr, J H (1982) Biological
monitoring Part IIA—receiving system functional methods, relationships and
indices. Water Research, 16 (2), 129-139.
Merritt, R. W. & Cummins, K. W. (1996) An Introduction to the Aquatic Insects of
North America. Dubuque, IA, Kendal/Hunt Publishing Company. 441 pp.
Muñoz, F., Mylavarapu, R. S. & Hutchinson, C. M. (2005) Environmentally
responsible potato production systems: A review. Journal of Plant Nutrition.
28, 1287-1309.
Nilsson, C. & Renöfält, B. M. (2008) Linking flow regime and water quality in rivers:
a challenge to adaptive catchment management. Ecology and Society, 13
(2), 18.

Chapter 1
28
Noronha, C., Vernon, R. S. & Vincent, C. (2008) Les insectes ravageurs
importants de la pomme de terre au Canada. Cahiers Agricultures, 17 (4),
375-381.
Oerke, E. C. (2006) Crop losses to pests. Journal of Agricultural Science, 144, 31-
43.
Pauw, N. & Vanhooren, G. (1983) Method for biological quality assessment of
watercourses in Belgium. Hydrobiologia, 100 (1), 153-168.
Poff, N. L. & Ward, J. V. (1989) Implications of Streamflow Variability and
Predictability for Lotic Community Structure: a Regional Analysis of
Streamflow Patterns. Canadian Journal of Fisheries and Aquatic Sciences,
46 (10), 1805-1818.
Randall, G. W. & Mulla, D. J. (2001) Nitrate nitrogen in surface waters as
influenced by climatic conditions and agricultural practices. Journal of
Environmental Quality. 30, 337-344.
Richter, B. D., Baumgartner, J. V., Powell, J. & Braun, D. P. (1996) A method for
assessing hydrologic alteration within ecosystems. Conservation Biology.
10, 1163-1174.
Rolauffs, P., Stubauer, I., Zahrádková, S., Brabec, K. & Moog, O. (2004)
Integration of the saprobic system into the European Union Water
Framework Directive – Case studies in Austria, Germany and Czech
Republic. Hydrobiologia, 516 (1), 285-298.
Rosenberg, D. M. & Resh, V. H. (1996) Use of aquatic Insects in Biomonitoring. In:
R. W. Merritt & K. W. Cummins (Ed.). An Introduction to the Aquatic Insects
of North America. Dubuque, IA, Kendal/Hunt Publishing Company, 87-97.
Ryan, P. A. (1991) Environmental effects of sediment on New-Zealand streams - A
review. New Zealand Journal of Marine and Freshwater Research, 25 (2),
207-221.
Southwood, T. R. E. (1977) Habitat, the Templet for Ecological Strategies? Journal
of Animal Ecology, 46 (2), 336-365.
Statzner, B., Bis, B., Dolédec, S. & Usseglio Polatera, P. (2001) Perspectives for
biomonitoring at large spatial scales: a unified measure for the functional

General Introduction
29
composition of invertebrate communities in European running waters. Basic
and Applied Ecology, 2 (1), 73-85.
Strayer, D. L. (2006) Challenges for freshwater invertebrate conservation. Journal
of the North American Benthological Society, 25 (2), 271-287.
Struik, P. C., Askew, M. F., Sonnino, A., MacKerron, D. K. L., Bang, U., Ritter, E.,
Statham, O. J. H., Kirkman, M. A. & Umaerus, V. (1997) Forty years of
potato research: highlights, achievements and prospects. Potato Research,
40 (1), 5-18.
Struik, P. C., Lommen, W. J. M., Haverkort, A. J. & Storey, R. M. J. (2007) The
Canon of Potato Science. Potato Research, 50 (3-4).
Vannote, R. L., Minshall, G. W., Cummins, K. W., Sedell, J. R. & Cushing, C. E.
(1980) The River Continuum concept. Canadian Journal of Fisheries and
Aquatic Sciences, 37, 130-137.
Wallace, J. B. & Webster, J. R. (1996) The Role of Macroinvertebrates in Stream
Ecosystem Function. Annual Review of Entomology, 41, 115-139.


Chapter 2. Beyond taxonomy: a review of macroinvertebrate trait-
based community descriptors as tools for freshwater
biomonitoring


Trait-based community descriptors
33
Note: the contents of this chapter have been published by Salomé Menezes, Donald J. Baird and
Amadeu M. V. M. Soares in volume 47 of the Journal of Applied Ecology (2010, pp: 711-719).
2.1. Introduction
There has been a long publication history examining the relationships
between species biological traits and environmental constraints (see Statzner et
al., 2001b for an historical review since the 19th century). While this interest in the
association between habitat affinities and the traits of an organism started with
early ecologists, the perception of its significance has changed over time (Statzner
et al., 2001b). A trait is defined as a characteristic that reflects a species
adaptation to its environment. Traits are usually divided in two categories:
biological traits (e.g. life cycle, physiological and behavioural characteristics, such
as maximum body size, lifespan, feeding and reproductive strategies, mobility,
etc.) and ecological traits (related to habitat preferences, like pH and temperature
tolerances, tolerance to organic pollution, biogeographic distribution, etc.). In fact,
an organism!s life history is a set of co-adapted traits arising from natural selection
to solve particular ecological problems (Stearns, 1976), such as limited food
resources or the presence of predators. During the 1970s, there was a focus on
life-history tactics in general and mechanisms for their evolution in particular
(Stearns, 1976) in the absence of a global theoretical ecological framework linking
habitat pressures and species traits.
Certain traits, by influencing organismal performance, can affect ecosystem
functioning (functional traits; McGill et al., 2006). Therefore, besides the potential
to define biological communities and predict changes in those communities due to
disturbance, species traits can also be used as measures of community functional
diversity (Petchey & Gaston, 2006). Both of these aspects can produce tools to
predict the functional consequences of biological change caused by human
activities.

Chapter 2
34
2.2. Biological traits and ecological theory: the habitat templet concept
Research into the predictive potential of using species traits to define
biological communities (the trait-based approach sensu Baird et al., 2008) has
increasingly appeared in the scientific literature in the last three decades. The first
works addressing this issue were in plant ecology and hypothesized that
competition, stress and disturbance are interactive determinants of herbaceous
vegetation by invoking different strategies on plants (Grime, 1974). The value of
this approach to plant management was presented by Grime as the ability to
detect competition and stress and to predict the intensity of disturbance at
particular sites. These ideas and theoretical models concerning plant
attributes/traits were the basis for the development of new plant community
descriptors and for the functional interpretation of vegetation data.
In 1977, Thomas Southwood formulated a general theoretical model for the
classification of ecological strategies in his presidential address to the British
Ecological Society (Southwood, 1977). Southwood showed that, in theory, the
ecological strategies of a species have evolved in response to a non-rigid 'habitat
templet' – the characteristics of the habitat are said to select and favour certain
sets of biological characters in the individual. This and other theoretical models for
life-history strategies (including Grime!s model) were later compared in their ability
to detect patterns and predict the evolution of strategies from the combination of
traits under the action of habitat selective forces (Southwood, 1988), and
similarities were found among the predictions resulting from the application of the
different models.
The underlying hypothesis in Southwood's work has inspired several
successful applications of the habitat templet perspective to different areas of
ecological knowledge, particularly studies on freshwater systems (including the
River Continuum Concept; Vannote et al., 1980), macroinvertebrate communities
and biomonitoring.

Trait-based community descriptors
35
2.2.3. The River Habitat Templet
Using the river habitat templet, stream ecologists have made a priori
predictions about stream community changes (Townsend & Hildrew, 1994) by
predicting trends in species traits along gradients of disturbances (i.e. temporal
variability) and refugia (i.e. spatial variability). These types of predictions were
tested by applying the trait approach to stream communities taxonomically well
characterized (e.g. Resh et al., 1994; Townsend et al., 1997).
Many examples of this application come from studies in one of the major
rivers in Europe, the Rhône. One initial approach linked lists of aquatic coleopteran
taxa to environmental and biological variables using correspondence analysis, in
order to organize them along a current-substrate gradient (Bournard et al., 1992).
In 1994, an entire issue of the Journal of Freshwater Biology (volume 31, issue 3)
was devoted to the long-term ecological research done on the Upper Rhône and to
the attempt to relate theoretical habitat templets, freshwater species traits, and
species richness. This was the first major test of the river habitat templet concept,
by means of a linkage of species traits to a set of temporal and spatial variables,
using a fuzzy coding approach of the data (Chevenet et al., 1994). This approach
codes biological and environmental information on a scale that describes the
affinity between two items, from 0 (when no affinity exists between those items, for
example a species and a trait modality) to x (strong affinity between the two items).
The advantages of this coding methodology are: the standardized coding of
information coming from diverse sources or concerning very distinct taxa; the
inclusion of trait variation found within a species; and the potential for statistical
analysis by ordination methods like correspondence analysis (Chevenet et al.,
1994).
The overall design of the research strategy of the “Rhône team” was to
develop a general tool for ecologically-oriented river management, where a priori
predictions were derived from available ecological theories (Statzner et al., 1994).
A physical habitat templet for the Rhône River was scaled using available sets of
physico-chemical data and trait data was structured through fuzzy coding.

Chapter 2
36
Alternative mathematical tools allowing the simultaneous analysis of species and
environment data as well as their relationships arose, namely a two-table
ordination method - co-inertia analysis (Dolédec & Chessel, 1994). This allowed
the testing of relationships between species traits and habitat utilization, and trait
trends were related with the river habitat templet (Townsend & Hildrew, 1994).
Based on Southwood!s theory (Southwood, 1977), Townsend & Hildrew
(1994) predicted that populations in more spatially heterogeneous habitats will be
less disturbed by certain temporal variations, stating that the likelihood of an
organism removal by disturbance will be influenced by spatial heterogeneity. The
authors made autecological predictions on life-history, morphological and
physiological traits, predicting how they would vary with disturbance.
The “Rhône team” analyzed thirteen animal and plant taxonomic groups of
the Upper Rhône River and concluded that spatio-temporal variability did not serve
as a templet for the species traits (Resh et al., 1994). The matching between
species traits and habitat characteristics appeared to be more complex than
initially predicted: alternative trade-offs between trait combinations could have
been neglected (Resh et al., 1994) or the set of taxonomic groups was too
heterogeneous.
In a New Zealand river, researchers tested the habitat templet with only
benthic insects and redefining the templet axes specifically for this taxonomic
group, by using frequency of disturbance as the temporal axis and variation of
features associated with refugia as the spatial axis (Townsend et al., 1997). The
result was that many of the initial habitat templet predictions proved to be correct.
This work has supported the idea that the habitat templet analysis had to take into
consideration the groups of organisms of interest and their possible relationships
with stream disturbance.
Benthic macroinvertebrates constitute a taxonomically well-described,
widespread, highly abundant and diverse group of aquatic organisms that respond
to a wide range of stressors. They can be good indicators of localized conditions
and long-term effects through the application of low cost and easy-to-use sampling
methodologies. Their undeniable functional importance in freshwater ecosystems

Trait-based community descriptors
37
is mostly related to their diverse array of feeding habits (Wallace & Webster,
1996).
Macroinvertebrate traits have been used to predict biotic interactions with
their predators (e.g. fish). A conceptual model and procedure for ranking
invertebrates based on functional relationships with their fish predators has been
proposed (Rader, 1997) as an important step in predicting disturbance effects on
fish populations. Invertebrates were ranked according to traits for drift propensity
and availability as food and significant correlations between the predicted ranks
and the actual ranks of invertebrates in fish guts were obtained (Rader, 1997).
Others (Statzner et al., 1997) have focused on reproductive traits of aquatic
insects to establish a relationship with the habitat characteristics at different
scales, following the approach of the “Rhône team” but using bibliographic
searches to extract worldwide insect data. This has introduced bias toward
abundant and widely distributed species, which may have reduced the trait range
to that of r-strategist species: small-sized species with short generation times, high
fecundity and high levels of dispersal, as opposed to the k-strategists (Southwood,
1977). A co-structure of the reproductive and habitat traits was detected and the
authors positively tested Southwood!s habitat templet theory for reproductive traits
of aquatic insects. The agreement with the habitat templet predictions was not true
to all traits studied though. The authors conclude that “one should not expect that
a given habitat acts as a templet in an uniform way for all traits of all its species”
(Statzner et al., 1997), and there seems to exist a certain effect of the habitat
scale.
2.3. Biological traits and ecological theory: the habitat filtering
hypothesis
The concept of habitats working as environmental trait filters was further
developed by several authors due to a growing recognition that there is a need for
general predictive models that are able to deal with an increasing number of

Chapter 2
38
environmental issues. Resetting the community-level research goal from
description to prediction has led to attempts to establish assembly rules that
forecast presence/absence of species as well as their abundance according to the
available set of environmental conditions, assuming these conditions work as
filters (Keddy, 1992). Essentially, the least suited sets of biological traits are
eliminated in a given environment, and only taxa possessing traits which pass
through the habitat filter will be present in the community.
Addressing the interplay of regional and local biotic and abiotic factors that
governs freshwater community composition, Poff (1997) presented one possible
general framework for understanding and predicting the distribution and
abundance of species in those communities, where species are described in terms
of their functional relationships to the habitat selective forces (landscape filters).
These filters are said to constrain expression of local selective forces or biotic
potential, so that species in a regional pool must possess appropriate functional
attributes (species traits) to "pass! through these filters and join a local community.
Once more, this means that species distribution and abundance and community
composition could be predicted if environmental constrains imposed at different
scales are considered (habitat data). The habitat filter scales specified by Poff
were basin or watershed, stream valley bottom or stream reach, channel unit
(riffle, pool) and microhabitat; the importance of hierarchically integrating different
spatial and temporal scales as factors affecting species distribution and
abundance is therefore highlighted by Poff!s model. In contrast with the regression
techniques used for example in the Rhône River data, this mechanistic approach
to community prediction implies explicit, quantitative assumptions about the
filtering of species by habitat factors (Poff, 1997). Highly detailed, a priori
knowledge on both traits and filters is needed in order to establish this predictive
model, and thus it is sensitive to data availability.
This importance of habitat properties at different scales in influencing species
traits was addressed in a study where reach- and catchment-scale characteristics
were used to predict trait patterns of freshwater insects of east-central Michigan
agricultural catchments (Richards et al., 1997). The authors successfully predicted

Trait-based community descriptors
39
life history and behavioural attributes using the reach-scale characteristics, but
found some difficulties in observing species trait patterns at smaller scales.
Following these observations, attempts to separate the respective influence of
various habitat filters were made (Lamouroux et al., 2004). The complexity of the
environmental filter theory development is undeniable (Statzner et al., 2004), in the
sense that multiple filters, both biotic and abiotic and working at very different
spatial and temporal scales, are expected to affect biological traits, often in
antagonistic ways.
2.4. Progress in the development of traits as biomonitoring tools
Charvet and co-workers (Charvet et al., 1998a; Charvet et al., 1998b) have
directly addressed stream biomonitoring using macroinvertebrate traits, and the
need to develop more generic biomonitoring tools. Traditionally used diversity
measures, biotic indices and community structure analysis – taxonomy-based
tools – do not allow the establishment of causal relationships with stressors and do
not integrate natural fluctuations. The authors hypothesized that, based on the
habitat templet concept and using a species-abundance table as well as a
species-traits table, it was possible to obtain a functional image of the study
system and detect pollution impact (in a simple upstream-downstream study
design). While comparing the traditional approaches that use physico-chemical or
taxonomic data (the latter using the calculations of eight biological indices – e.g.
Margalef Index, Shannon Index – and community structure analysis) with the
"functional approach! (using biological traits weighted by abundances), a better
separation of the upstream and downstream sites was obtained when including
trait information. This worked as an initial assessment of the potential of traits to
overcome some of the issues raised by traditional biomonitoring methodologies.
Dolédec et al. (1999) evidenced that, at the time, much work had still to be done in
the adaptation of the trait approach to a biomonitoring technique.

Chapter 2
40
In a search for a conceptual framework for trait freshwater biomonitoring,
Dolédec et al. (1999) highlighted that an ideal biomonitoring tool – generic in terms
of geographic application, specific in terms of stressor identification, reliable and
derived from sound theoretical ecological concepts – is possible to obtain using
benthic macroinvertebrate ecological and biological traits, as an alternative to the
traditional taxonomy-based approaches. These taxonomic approaches (metrics
such as abundance, taxa richness of species or families) are said to fail when
generalizations to different types of freshwater systems are needed (regional
constraints) and result in losses of ecological information. The authors again
tested the river habitat templet, using multivariate analysis to evaluate how
patterns of species traits in macroinvertebrate communities of a large river could
discriminate differences in overall human impact, by comparison with taxonomy-
based approaches (species abundance-by-sites). All tools tested revealed human
impact effects; the analysis based on biological traits was less confounded by
natural spatial gradients and was the best indicator of stressor impact.
Meanwhile, following the French trait research studies, a subset of the
“Rhône team” continued to fill the biological and ecological trait database and
published a revised version for the macroinvertebrate fauna of French rivers
(Tachet et al., 2000), later complemented with information for other European
areas and taxa (Statzner et al., 2007). They have also revised the
macroinvertebrate trait applications to running water biomonitoring, highlighting
that a broader unified trait biomonitoring tool covering not only France but also
larger geographical scales was needed, in order to meet current European policies
(Statzner et al., 2001a).
2.5. “Reference” state studies in freshwater ecosystems
Charvet et al. (2000) tested whether biological and ecological traits used as
biomonitoring tools would yield the same results for semi-natural (non-impacted)
streams of different ecoregions. They found that trait-based community structure

Trait-based community descriptors
41
was in fact stable across environmental gradients, in contrast with the taxonomy-
based community structure, which varied significantly with geologic and altitudinal
differences. Other authors tested both the spatial stability of the reference state
(referring to the example of the “reference condition approach”, Wright et al., 2000)
and the potential for indication of specific human impacts using data at the
European scale (Statzner et al., 2001a). A high spatial and temporal stability of the
functional composition of non-impacted freshwater communities at the European
scale was detected and this was related with a landscape filter (sensu Poff, 1997)
limiting taxa abundance at the local scale. Although differences in the trait patterns
of reference and impacted locations were detected, the human impact examples
chosen (dams and sewage input) were not sufficient to highlight clear mechanistic
explanations for the patterns found.
Archaimbault et al. (2005) have assessed the influence of geology on trait
profiles of macroinvertebrate communities of reference sites belonging to the same
biogeographic area, confirming that the functional structure of reference
communities was constant across geology types, even when taxonomic variability
was observed. Statzner et al. (2005) have shown that the expected variation
across “reference” rivers can be predicted by trait patterns, more or less accurately
depending on the chosen models of analysis. Bonada et al. (2007b) concluded
that stream flow permanence constrains the invertebrate community both
structurally and functionally, relating various trait profiles with the Mediterranean
climatic characteristics.
Over the last two decades, the majority of the macroinvertebrate trait
applications to freshwater systems have been focused on the European continent
but the interest in developing a trait biomonitoring tool applicable also to North
American freshwater invertebrates has lead to the publication of a trait database
with more than 14000 records (Vieira et al., 2006). North-American studies have
also started to examine the relationships between physical variables and benthic
trait patterns, particularly under “reference state" conditions. The benthic
community structure and functioning of high-gradient mountain streams was
studied and the influence of longitudinal and reach-scale variables on those

Chapter 2
42
communities was assessed (Finn & Poff, 2005). For this particular small spatial
extent scenario, defining assemblages functionally provided no greater
understanding of community patterns than taxonomic definitions, given several
known environmental variables (see also the study of Finnish headwater streams
by Heino et al., 2007), and the issue of trait trade-offs and alternative adaptive
solutions for the same habitat was once more raised. The results did not agree
with previous studies which observed a functional stability of unimpacted
communities across environmental gradients (Charvet et al., 2000), but the
authors hypothesize that, although human impact was low, the natural stressors
associated with more extreme zones (alpine zone, harsher environment, limited
human influence) could explain the functional instability detected.
Another recent geographical expansion of the freshwater trait approach was
to the neotropical benthic communities (Tomanova & Usseglio-Polatera, 2007).
Taxonomic and functional structures of reference benthic communities were
compared and, after overcoming the difficulty of limited availability of neotropical
invertebrate trait information, the authors found that communities that differed
taxonomically were functionally similar. They have also found that some of the
traits related to the environmental variables, but not sufficiently matching the trait-
habitat relationships detected for temperate environments (not enough to confirm
the existence of general benthic trait rules that would be applicable over different
climatic regions).
As for natural temporal variations, several studies (Bêche et al., 2006; Bêche
& Resh, 2007) highlight the importance of an accurate understanding of the natural
variation of community patterns in “reference” systems (natural ecological filters),
so that the biological trait approach can be properly used to detect differences
between these systems and the ones affected by human stressors (anthropogenic
filters).

Trait-based community descriptors
43
2.6. Technical aspects of biomonitoring
The way in which taxonomic and spatial resolution influence trait-based
predictability is a substantial issue. Dolédec et al. (2000) combined abundance
data obtained at different scales and trait data, and investigated the accuracy of
community descriptions expressed at different levels of taxonomic and spatial
resolution. They concluded that scale significantly affected the accuracy of
community descriptions based on abundance but that while using biological traits
accurate descriptions were achieved by species, genus or family identifications
(Dolédec et al., 2000). This suggests that species identification may not always be
necessary in future stream biomonitoring studies, a contribution for the
simplification of the tools under development as the taxonomic expertise required
would be significantly decreased.
Under this premise, data of benthic macroinvertebrate traits from French
freshwaters were used, at the genus level, to explore issues that limited the
successful testing of the river habitat templet, like trait trade-offs (Resh et al.,
1994) - variable combinations of traits that organisms may possess that confound
the establishment of matches between habitat and trait patterns. Non-taxonomic
aggregations of taxa - functional groups of organisms with high relationship
similarities among their biological and ecological traits – were defined in order to
investigate the mechanisms affecting species distributions (Usseglio-Polatera et
al., 2000). The application of these groupings for human impact differentiation was
later attempted (Usseglio-Polatera et al., 2001), and functional trait diversity
(Shannon diversity index calculated using the non-taxonomic groupings) seemed a
better indicator of human impact than taxonomic diversity, with more impacted
sites exhibiting higher proportions of organisms with traits assumed to adapt them
to frequent disturbances or reduce the impact of certain stressors.
Gayraud et al. (2003) suggest once more (cf. Dolédec et al., 2000) that
descriptions of the functional structure of communities more or less affected by
human activities at the species level may not be needed. Attention should also be
given to the choices of weighting of species or excluding alien species. The

Chapter 2
44
authors suggest the development of a low cost community functional description
tool using presence-absence of genera or families of native freshwater taxa, that
could be used to discriminate strong human impact gradients. Other tests
(Haybach et al., 2004) have also detected high robustness of the trait approach
with decreased taxonomic resolution as well as against faunal changes, such as
the seasonal ones. In this study the authors were also able to demonstrate the
functional quality of the approach by detecting associations between the trait
pattern responses to disturbance of a certain site with the predominating rK-
strategies.
Another technical aspect to consider in a trait biomonitoring programme is
the effect of different sampling efforts, as these effects on taxonomic variables are
well known (Kerans et al., 1992). It is therefore important to test how the sampling
effort would affect the functional diversity measures based on biological traits.
While confirming the influence of sampling effort on taxonomy-based measures,
Bady et al. (2005) concluded that functional diversity indices (using traits) show
greater accuracy with less sampling effort and higher precision across season and
location gradients.
2.7. Statistical tools
The inclusion of species traits in ecological studies brings a variety of
advantages but also great challenges for the simultaneous analysis of three data
matrices: environmental, trait and species composition tables. The RLQ analysis,
a three-table ordination method, was presented as a solution for this problem
(Dolédec et al., 1996). It was followed by the testing procedures proposed by
Legendre and coworkers with the intent of establishing a link between an
environmental variable and a species trait through a table containing presence–
absence data, presented as the fourth-corner problem (Legendre et al., 1997).
Some limitations of the proposed methodologies were highlighted subsequently,
namely the lack of consideration for evolutionary linkages among traits, the

Trait-based community descriptors
45
analysis of a single trait and a single environmental variable at a time, and the
impossibility of using abundance data, since both methods imply the use of binary
data (Nygaard & Ejrnaes, 2004; Poff et al., 2006). To address at least part of these
difficulties, an improved fourth-corner method was proposed (Dray & Legendre,
2008).
2.8. Trade-offs and trait syndromes
Since the first attempts to test the river habitat templet, the issue of
alternative trade-offs between trait combinations has been discussed (e.g. Resh et
al., 1994; Usseglio-Polatera et al., 2000). It was noted that different possible
combinations of traits in an organism, working as adaptive solutions, may
confound the establishment of matches between habitat and trait patterns. In fact,
trade-offs in species performances of different ecological functions is one of the
most common explanations for coexistence in communities (Kneitel & Chase,
2004). They are defined as negative functional interactions between traits, with
investments in one trait leaving fewer resources available for investments in
another (Verberk et al., 2008b). The need arises for a formal analysis of trade-offs,
accounting for phylogenetic relationships and potential confounding effects on trait
measures.
Poff et al. (2006) have highlighted that, while the developments of the trait
approach in biomonitoring are promising, there is a lack of adequate
understanding of how individual traits are intercorrelated and how this dependence
among traits reflects phylogenetic constraints. The fact that traits are often linked
together by evolution, forming the so-called trait syndromes (Nylin & Gotthard,
1998), hinders their treatment as independent entities. The authors have explored
this issue for lotic insects of North America by creating a trait database, by
demonstrating the importance of trait state linkage within a taxon, by examining
trait correlations and using this to provide information on future trait selection for
biomonitoring purposes. They have also examined phylogenetic associations of

Chapter 2
46
certain traits and suggested that robust (unlinked, uncorrelated) traits would
provide better insights to predictive community ecology and multiple trait response
along environmental gradients (Poff et al., 2006).
As species traits cannot be viewed independently, one possible solution can
be to see them as part of complex adaptations, combinations of co-evolved
attributes based on known functional relationships among them, i.e. life-history
strategies (Verberk et al., 2008b). These strategies enable the organism to deal
with a range of ecological problems. Verberk and co-workers present these
strategies in four groups (dispersal, synchronization, reproduction and
developmental trade-off), based on species traits and their interrelations known
from life-history theory. Once more biological knowledge on the different species is
available, these strategies should provide the connections between traits and
environmental conditions, pursued by those that have been testing the habitat
templet theory over the last decades (particularly since Resh et al., 1994). This
approach has already been tested for lentic macroinvertebrate species (Verberk et
al., 2008a), where different species are assigned to the same life-history strategy,
according to their ability to solve similar ecological problems by employing
combination of species traits. Differences in "strategy composition! of freshwater
systems were related with known environmental conditions and good prospects for
environmental quality programmes were presented. This approach illustrates the
importance of considering comparative phylogenetic methods as employed in
evolutionary ecology (e.g. Freckleton et al., 2002) where convergent phenotypic
solutions to ecological problems across phylogenies can be understood and
placed in their correct context, with the potential to contribute towards global
analyses of species in relation to their environment (Westoby, 2006).
2.9. Applications in human impact scenarios
Initial studies dealing with the ability of the biological trait approach to detect
the effects of deleterious human impacts on freshwater ecosystems have already

Trait-based community descriptors
47
been mentioned (e.g. for sewage pollution Charvet et al., 1998a; for overall human
impact Dolédec et al., 1999; Statzner et al., 2001a; for agricultural land use
Richards et al., 1997). More specific studies under various anthropogenic impact
scenarios followed, attempting to address the issue of human impact
differentiation.
In New Zealand streams affected by agricultural development, it was shown
that both taxonomy- and trait-based methods were able to discriminate land use
practices, but the trait approach worked better for this purpose, accounting for
more between-land use variance (Dolédec et al., 2006). Shifts in
macroinvertebrate trait patterns with increasing land use were described, proving
once more the great potential of this approach for the biomonitoring of stream
communities, particularly the ones affected by agricultural stressors. Díaz et al.
(2008) have suggested a habitat templet for SE Spain streams, including a
disturbance axis (natural climatic variation) and an adversity axis (intensive
agriculture pressure), and a RLQ analysis was used for identifying species traits
that respond to impacts of land use change at different scales. Although
confounding effects of geology, altitude and climate were detected, the authors
were able to link macroinvertebrate community ecological organization with
disturbance and human pressure. In the case of this particular type of human
impact, it would be important to distinguish, for example, pesticide effects from
other particular stressors associated with agriculture or from natural stressors.
With this purpose in mind, the concept of classifying species according to their
vulnerability towards pesticides (Species At Risk, or SPEAR) defined by certain
ecological and physiological traits was tested in Northern Germany streams (Liess
& Von Der Ohe, 2005) and across European biogeographic regions (Schaefer et
al., 2007). This indicator system successfully discriminated reference and
pesticide contaminated sites, and it was demonstrated that these pollutants altered
both the community structure and function of the studied lotic systems (Schaefer et
al., 2007).
Dealing with hydrological alteration at a continental scale (across Canada),
Horrigan & Baird (2008) proposed a set of flow-sensitive traits to be used in the

Chapter 2
48
development of biomonitoring through flow metrics, addressing the possible effects
of trait syndromes. Additionally, they suggest the preliminary establishment of
typical ranges of flow-sensitive traits for different types of reference streams
(differing in hydrological regime, size, order and slope) and their use as
biomonitoring metrics with potential global applicability.
Attempts to use trait profiles to differentiate types of stressors heavily
affecting large European rivers (namely heavy metal pollution and cargo-ship
traffic), did not result in an adequate discrimination of impacts, highlighting that the
detection of specific impacts under multiple stressor scenarios is very complex
(Dolédec & Statzner, 2007). In order to tackle this problem, individual and
combined effects of the principal stressors affecting New Zealand agricultural
streams were investigated (Townsend et al., 2008) by using both taxonomy-based
and trait-based measures. A change from management solutions based on
individual stressor analysis to solutions based on multiple stressor analysis is
recommended. Another multiple stressor scenario is the one imposed by wildfires,
and the comparison of successional patterns in burned and reference streams has
highlighted interactions between functional traits of local taxa and the postfire
environmental conditions (Vieira et al., 2004).
Other types of studies dealt with the implications of future climate change
scenarios by comparing ecological (trait and taxonomic) differences between
Mediterranean and temperate European streams (Bonada et al., 2007a). It was
highlighted that climate change may produce large changes in the taxonomic
composition without substantially altering the pattern of functional traits.
One example of another potential of the trait approach is the prediction of the
efficiency of restoration management activities by using macroinvertebrate
communities to assess the effects of restoration measures (working as trait filters).
There are studies recommending phased restoration procedures in order to ensure
the survival of species not possessing the traits that would allow them to survive to
the future application of such measures (van Kleef et al., 2006). Other studies
(Tullos et al., 2009) report that both the taxonomic and the functional-trait
approach showed the effects of restoration measures. Both these works (and also

Trait-based community descriptors
49
e.g. Reckendorfer et al., 2006; Paillex et al., 2009) prove that there is an
interesting potential for the applications of trait metrics in the establishment of
informed management resolutions by proper understanding the functional
implications of such measures.
2.10. Conclusions
This review highlights much of the progress towards the development of a
theoretical framework for ecologically oriented stream biomonitoring and
management. There is still a long way to go before scientists will be able to use
community level data to accurately diagnose causes of stream impairment
(stressor biodiagnosis), besides the mere identification of degradation presence.
Scientists have to be able to convey the importance of protecting freshwater
ecosystem functions, particularly to managers and legislators. In the last decades
the functional trait approach has been identified as one of the tools with more
potential for biomonitoring and management of stream ecosystems. This potential
focuses mainly around the concept of the ideal biomonitoring tool – generic in
terms of geographic application, specific in terms of stressor identification, reliable
and derived from sound theoretical ecological concepts – and in the fact that
traditional taxonomy-based approaches many times fail to work in these ideal
ways
By looking at species trait patterns, significantly affected by human impacts,
we can make mechanistic interpretations of the effects of anthropogenic activities
by examining how traits co-vary with specific environmental pressures and drivers.
This is possible both in freshwater environments and in other ecosystems where a
suitable knowledge-base exists, and environmental management can be
substantially improved. The fact that issues with natural spatial and temporal
variability can be overcome is a huge advantage for those trying to assess and
maintain ecosystem quality over broad spatial scales or in situations of limited

Chapter 2
50
availability of taxonomic information (e.g. in less-developed countries, the tropics
or the polar regions).
Together, the trait studies reviewed here attest to the ability of the functional
trait approach to form the basis of a new generation of biomonitoring tools.
However, they also highlight the need for further research in the following areas: (i)
a broader, unified trait biomonitoring tool covering larger geographical scales (this
implies the need for a confirmation of the existence of general benthic trait rules
that would be applicable over different climatic regions); (ii) an accurate
understanding of the natural variation of community trait patterns in unimpacted
systems; (iii) new trait analysis protocols that diminish the effects of trait trade-offs
and trait syndromes; (iv) undertaking additional life-history and ecological niche
studies, promoting open access trait databases where the compiled results of
these studies can be found (cf. Section 2.12 for a list of available databases); (v)
improved understanding of which traits are functionally important and how to use
them as relevant measures of functional diversity at the community level,
especially to guide restoration programmes; and (vi) increased effort to apply trait-
based approaches to the detection of specific impacts under multiple stressor
scenarios.

Trait-based community descriptors
51
2.11. References
Archaimbault, V., Usseglio Polatera, P. & Bossche, J. (2005) Functional
Differences Among Benthic Macroinvertebrate Communities in Reference
Streams of Same Order in a Given Biogeographic Area. Hydrobiologia, 551,
171-182.
Bady, P., Dolédec, S., Fesl, C., Gayraud, S. & Bacchi, M. (2005) Use of
invertebrate traits for the biomonitoring of European large rivers: the effects
of sampling effort on genus richness and functional diversity. Freshwater
Biology, 50, 169-173.
Baird, D. J., Rubach, M. N. & Van den Brink, P. J. (2008) Trait-based ecological
risk assessment (TERA): the new frontier? Integrated environmental
assessment and management, 4, 2-3.
Bêche, L. A., McElravy, E. P. & Resh, V. H. (2006) Long-term seasonal variation in
the biological traits of benthic-macroinvertebrates in two Mediterranean-
climate streams in California, U.S.A. Freshwater Biology, 51, 56-75.
Bêche, L. A. & Resh, V. H. (2007) Biological traits of benthic macroinvertebrates in
California mediterranean-climate streams: long-term annual variability and
trait diversity patterns. Fundamental and Applied Limnology, 169, 1-23.
Bonada, N., Dolédec, S. & Statzner, B. (2007a) Taxonomic and biological trait
differences of stream macroinvertebrate communities between
mediterranean and temperate regions: implications for future climatic
scenarios. Global Change Biology, 13, 1658-1671.
Bonada, N., Rieradevall, M. & Prat, N. (2007b) Macroinvertebrate community
structure and biological traits related to flow permanence in a
Mediterranean river network. Hydrobiologia, 589, 91-106.
Bournard, M., Richoux, P. & Usseglio Polatera, P. (1992) An approach to the
synthesis of qualitative ecological information from aquatic coleoptera
communities. Regulated Rivers: Research & Management, 7, 165-180.

Chapter 2
52
Charvet, S., Kosmala, A. & Statzner, B. (1998a) Biomonitoring through biological
traits of benthic macroinvertebrates: perspectives for a general tool in
stream management. Archiv für Hydrobiologie, 142, 415-432.
Charvet, S., Roger, M. C., Faessel, B. & Lafont, M. (1998b) Biomonitoring of
freshwater ecosystems by the use of biological traits. Annales de
Limnologie–International Journal of Limnology, 34, 455–464.
Charvet, S., Statzner, B., Usseglio Polatera, P. & Dumont, B. (2000) Traits of
benthic macroinvertebrates in semi-natural French streams: an initial
application to biomonitoring in Europe. Freshwater Biology, 43, 277-296.
Chevenet, F., Dolédec, S. & Chessel, D. (1994) A fuzzy coding approach for the
analysis of long-term ecological data. Freshwater Biology, 31, 295-309.
Díaz, A. M., Alonso, M. L. S. & Gutierrez, M. R. V. A. (2008) Biological traits of
stream macroinvertebrates from a semi-arid catchment: patterns along
complex environmental gradients. Freshwater Biology, 53, 1-21.
Dolédec, S. & Chessel, D. (1994) Co-inertia analysis: an alternative method for
studying species-environment relationships. Freshwater Biology, 31, 277-
294.
Dolédec, S., Chessel, D., Ter Braak, C. J. F. & Champely, S. (1996) Matching
species traits to environmental variables: a new three-table ordination
method. Environmental and Ecological Statistics, 3, 143-166.
Dolédec, S., Olivier, J. M. & Statzner, B. (2000) Accurate description of the
abundance of taxa and their biological traits in stream invertebrate
communities: effects of taxonomic and spatial resolution. Archiv fuer
Hydrobiologie, 148 25-43.
Dolédec, S., Phillips, N., Scarsbrook, M., Riley, R. H. & Townsend, C. R. (2006)
Comparison of structural and functional approaches to determining landuse
effects on grassland stream invertebrate communities. Journal of the North
American Benthological Society, 25, 44-60.
Dolédec, S. & Statzner, B. (2007) Invertebrate traits for the biomonitoring of large
European rivers: an assessment of specific types of human impact.
Freshwater Biology, 53, 617-634.

Trait-based community descriptors
53
Dolédec, S., Statzner, B. & Bournard, M. (1999) Species traits for future
biomonitoring across ecoregions: patterns along a human-impacted river.
Freshwater Biology, 42, 737-758.
Dray, S. & Legendre, P. (2008) Testing the Species Traits-Environment
Relationships: The Fourth-Corner Problem Revisited. Ecology, 89, 3400-
3412.
Finn, D. S. & Poff, N. L. (2005) Variability and convergence in benthic communities
along the longitudinal gradients of four physically similar Rocky Mountain
streams. Freshwater Biology, 50, 243-261.
Gayraud, S., Statzner, B., Bady, P., Haybach, A., Scholl, F., Usseglio Polatera, P.
& Bacchi, M. (2003) Invertebrate traits for the biomonitoring of large
European rivers: an initial assessment of alternative metrics. Freshwater
Biology, 48, 2045-2064.
Grime, J. P. (1974) Vegetation Classification by Reference to Strategies. Nature,
250, 26-31.
Freckleton, R. P., Harvey, P. H. and Pagel, M. (2002) Phylogenetic analysis and
comparative data: a test and review of evidence. American Naturalist, 160,
712-726.
Haybach, A., Scholl, F., Konig, B. & Kohmann, F. (2004) Use of biological traits for
interpreting functional relationships in large rivers. Limnologica, 34, 451-
459.
Heino, J., Mykra, H., Kotanen, J. & Muotka, T. (2007) Ecological filters and
variability in stream macroinvertebrate communities: do taxonomic and
functional structure follow the same path? Ecography, 30, 217-230.
Horrigan, N. & Baird, D. J. (2008) Trait patterns of aquatic insects across gradients
of flow-related factors: a multivariate analysis of Canadian national data.
Canadian Journal of Fisheries and Aquatic Sciences, 65, 670–680.
Keddy, P. A. (1992) Assembly and Response Rules: Two Goals for Predictive
Community Ecology Journal of Vegetation Science, 3, 157-164.
Kerans, B. L., Karr, J. R. & Ahlstedt, S. A. (1992) Aquatic Invertebrate
Assemblages - Spatial And Temporal Differences Among Sampling

Chapter 2
54
Protocols. Journal of the North American Benthological Society, 11, 377-
390.
Kneitel, J. M. & Chase, J. M. (2004) Trade-offs in community ecology: linking
spatial scales and species coexistence. Ecology Letters, 7, 69-80.
Lamouroux, N., Dolédec, S. & Gayraud, S. (2004) Biological traits of stream
macroinvertebrate communities: effects of microhabitat, reach, and basin
filters. Journal of the North American Benthological Society, 23, 449-466.
Legendre, P., Galzin, R. & Harmelin-Vivien, M. L. (1997) Relating behavior to
habitat: Solutions to the fourth-corner problem. Ecology, 78, 547-562.
Liess, M. & Von Der Ohe, P. C. (2005) Analyzing effects of pesticides on
invertebrate communities in streams. Environmental Toxicology and
Chemistry, 24, 954-965.
McGill, B. J., Enquist, B. J., Weiher, E. & Westoby, M. (2006) Rebuilding
community ecology from functional traits. Trends in Ecology & Evolution,
21, 178-185.
Nygaard, B. & Ejrnaes, R. (2004) A new approach to functional interpretation of
vegetation data. Journal of Vegetation Science, 15, 49-56.
Nylin, S. & Gotthard, K. (1998) Plasticity in Life-History Traits. Annual Review of
Entomology, 43, 63-83.
Paillex, A., Dolédec, S., Castella, E. & Merigoux, S. (2009) Large river floodplain
restoration: predicting species richness and trait responses to the
restoration of hydrological connectivity. Journal of Applied Ecology, 46, 250-
258.
Petchey, O. L. & Gaston, K. J. (2006) Functional diversity: back to basics and
looking forward. Ecology Letters, 9, 741-758.
Poff, N. L. (1997) Landscape filters and species traits: towards mechanistic
understanding and prediction in stream ecology. Journal of the North
American Benthological Society, 16, 391-409.
Poff, N. L., Olden, J. D., Vieira, N. K. M., Finn, D. S., Simmons, M. P. &
Kondratieff, B. C. (2006) Functional Trait Niches of North American Lotic
Insects: Trait-based Ecological Applications in Light of Phylogenetic

Trait-based community descriptors
55
Relationships. Journal of the North American Benthological Society, 25,
730-755.
Rader, R. B. (1997) A functional classification of the drift: traits that influence
invertebrate availability to salmonids. Canadian Journal of Fisheries and
Aquatic Sciences, 54, 1211-1234
Reckendorfer, W., Baranyi, C., Funk, A. & Schiemer, F. (2006) Floodplain
restoration by reinforcing hydrological connectivity: expected effects on
aquatic mollusc communities. Journal of Applied Ecology, 43, 474-484.
Resh, V. H., Hildrew, A. G., Statzner, B. & Townsend, C. R. (1994) Theoretical
habitat templets, species traits, and species richness - a synthesis of long-
term ecological research on the Upper Rhone river in the context of
concurrently developed ecological theory. Freshwater Biology, 31, 539-554.
Richards, C., Haro, R. J., Johnson, L. & Host, G. E. (1997) Catchment and reach-
scale properties as indicators of macroinvertebrate species traits.
Freshwater Biology, 37, 219-230.
Schaefer, R. B., Caquet, T., Siimes, K., Mueller, R., Lagadic, L. & Liess, M. (2007)
Effects of pesticides on community structure and ecosystem functions in
agricultural streams of three biogeographical regions in Europe. Science of
the Total Environment, 382, 272-285.
Southwood, T. R. E. (1977) Habitat, the Templet for Ecological Strategies? .
Journal of Animal Ecology, 46, 336-365.
Southwood, T. R. E. (1988) Tactics, Strategies and Templets. Oikos, 52, 3-18.
Statzner, B., Bady, P., Dolédec, S. & Scholl, F. (2005) Invertebrate traits for the
biomonitoring of large European rivers: an initial assessment of trait
patterns in least impacted river reaches. Freshwater Biology, 50, 2136-
2161.
Statzner, B., Bis, B., Dolédec, S. & Usseglio Polatera, P. (2001a) Perspectives for
biomonitoring at large spatial scales: a unified measure for the functional
composition of invertebrate communities in European running waters. Basic
and Applied Ecology, 2, 73-85.

Chapter 2
56
Statzner, B., Bonada, N. & Dolédec, S. (2007) Conservation of taxonomic and
biological trait diversity of European stream macroinvertebrate communities:
a case for a collective public database. Biodiversity and Conservation, 16,
3609-3632.
Statzner, B., Dolédec, S. & Hugueny, B. (2004) Biological trait composition of
European stream invertebrate communities: assessing the effects of various
trait filter types. Ecography, 27, 470-488.
Statzner, B., Hildrew, A. G. & Resh, V. H. (2001b) Species Traits And
Environmental Constraints: Entomological Research and the History of
Ecological Theory. Annual Review of Entomology, 46, 291-316.
Statzner, B., Hoppenhaus, K., Arens, M. & Richoux, P. (1997) Reproductive traits,
habitat use and templet theory: a synthesis of world-wide data on aquatic
insects. Freshwater Biology, 38, 109-135.
Statzner, B., Resh, V. H. & Roux, A. L. (1994) The synthesis of long-term
ecological research in the context of concurrently developed ecological
theory - design of a research strategy for the Upper Rhône River and its
floodplain. Freshwater Biology, 31, 253-263.
Stearns, S. C. (1976) Life-History Tactics - Review Of Ideas. The Quarterly Review
of Biology, 51, 3-47.
Tachet, H., Richoux, P., Bournaud, M. & Usseglio-Polatera, P. (2000) Invertébrés
d'eau douce: systématique, biologie, écologie. CNRS Editions, Paris. 588
pp.
Tomanova, S. & Usseglio-Polatera, P. (2007) Patterns of benthic community traits
in neotropical streams: relationship to mesoscale spatial variability.
Fundamental and Applied Limnology, 170, 243-255.
Townsend, C. R. & Hildrew, A. G. (1994) Species Traits in Relation to a Habitat
Templet for River Systems. Freshwater Biology, 31, 265-275.
Townsend, C. R., Scarsbrook, M. R. & Dolédec, S. (1997) Quantifying disturbance
in streams: alternative measures of disturbance in relation to
macroinvertebrate species traits and species richness. Journal of the North
American Benthological Society, 16, 531-544.

Trait-based community descriptors
57
Townsend, C. R., Uhlmann, S. S. & Matthaei, C. D. (2008) Individual and
combined responses of stream ecosystems to multiple stressors. Journal of
Applied Ecology, 45, 1810-1819.
Tullos, D. D., Penrose, D. L., Jennings, G. D. & Cope, W. G. (2009) Analysis of
functional traits in reconfigured channels: implications for the
bioassessment and disturbance of river restoration. Journal of the North
American Benthological Society, 28, 80-92.
Usseglio-Polatera, P., Bournard, M., Richoux, P. & Tachet, H. (2000) Biological
and ecological traits of benthic freshwater macroinvertebrates: relationships
and definition of groups with similar traits. Freshwater Biology, 43, 175-205.
Usseglio-Polatera, P., Richoux, P., Bournard, M. & Tachet, H. (2001) A functional
classification of benthic macroinvertebrates based on biological and
ecological traits: application to river condition assessment and stream
management. Archiv fuer Hydrobiologie, Suppl., 139, 53-83.
van Kleef, H. H., Verberk, W. C. E. P., Leuven, R. S. E. W., Esselink, H., van der
Velde, G. & van Duinen, G. A. (2006) Biological traits successfully predict
the effects of restoration management on macroinvertebrates in shallow
softwater lakes. Hydrobiologia, 565, 201-216.
Vannote, R. L., Minshall, G. W., Cummins, K. W., Sedell, J. R. & Cushing, C. E.
(1980) The River Continuum concept. Canadian Journal of Fisheries and
Aquatic Sciences, 37, 130-137.
Verberk, W. C. E. P., Siepel, H. & Esselink, H. (2008a) Applying life-history
strategies for freshwater macroinvertebrates to lentic waters. Freshwater
Biology, 53, 1739-1753.
Verberk, W. C. E. P., Siepel, H. & Esselink, H. (2008b) Life-history strategies in
freshwater macroinvertebrates. Freshwater Biology, 53, 1722-1738.
Vieira, N. K. M., Clements, W. H., Guevara, L. S. & Jacobs, B. F. (2004)
Resistance and resilience of stream insect communities to repeated
hydrologic disturbances after a wildfire. Freshwater Biology, 49, 1243-1259.

Chapter 2
58
Vieira, N. K. M., Poff, N. L., Carlisle, D. M., Moulton, S. R., Koski, M. L. &
Kondratieff, B. C. (2006) A database of lotic invertebrate traits for North
America. U.S. Geological Survey Data Series 187.
Wallace, J. B. & Webster, J. R. (1996) The Role of Macroinvertebrates in Stream
Ecosystem Function. Annual Review of Entomology, 41, 115-139.
Westoby, M. (2006) Phylogenetic ecology at world scale, a new fusion between
ecology and evolution. Ecology 87 (Supplement), S163-S165.
Wright, J. F., Sutcliffe, D. W. & Furse, M. T. (2000) Assessing the biological quality
of fresh waters - RIVPACS and other techniques. Freshwater Biological
Association, Ambleside, UK.

Trait-based community descriptors
59
2.12. Supporting information: databases offering information on
freshwater macroinvertebrate traits
A list of published (paper and online) references that provide lists of
freshwater macroinvertebrate taxa associated with arrays of species traits.
Heneghan, P.A., Biggs, J., Jepson, P.C., Kedwards, T., Maund, S.J., Sherratt, T.N., Shillabeer, N.,
Stickland, T.R. & Williams, P. (1999) Pond-FX: ecotoxicology from pH to population
recovery [online]. 1st edition. Oregon State University: Department of Entomology.
http://www.ent.orst.edu/PondFX.
Merritt, R. W., Cummins & K. W., Berg, M. B. (2008) An introduction to the aquatic insects of North
America. Kendall/Hunt, Dubuque, Iowa, 4th edition.
Moog, O. (2002) Fauna aquatica Austriaca. 2nd edition. Wasserwirtschaftskataster,
Bunderministerium für Land- und Forstwirtschaft, Umwelt und Wasserwirtschaft, Wien.
Multiple articles/authors (the “Rhône team”) (1994) Special Issue of Freshwater Biology, number 3,
volume 31.
Schmedtje, U. & Colling M. (1996) Ökologische Typisierung der aquatischen Makrofauna.
Informationsberichte des Bayerischen Landesamtes für Wasserwirtschaft, Munich. 543 pp.
Schmidt-Kloiber, A. & Vogl R. - database administrators (2009) Freshwaterecology.info – The taxa
and Autoecological Database for Freshwater Organisms [online]. Version 4.0.
http://www.freshwaterecology.info/index.php.
Statzner, B., Hoppenhaus, K., Arens, M. & Richoux, P. (1997) Reproductive traits, habitat use and
templet theory: a synthesis of world-wide data on aquatic insects. Freshwater Biology, 38,
109-135.
Tachet, H., Richoux, P., Bournaud, M. & Usseglio-Polatera, P. (2000) Invertébrés d'eau douce:
systématique, biologie, écologie. CNRS Editions, Paris. 588 pp.
Tomanova, S. & Usseglio-Polatera, P. (2007) Patterns of benthic community traits in neotropical
streams: relationship to mesoscale spatial variability. Fundamental and Applied Limnology,
170, 243-255.
Verberk, W. C. E. P., Siepel, H. & Esselink, H. (2008) Applying life-history strategies for freshwater
macroinvertebrates to lentic waters. Freshwater Biology, 53, 1739-1753.
Vieira, N. K. M., Poff, N. L., Carlisle, D. M., Moulton, S. R., Koski, M. L. & Kondratieff, B. C. (2006)
A database of lotic invertebrate traits for North America. U.S. Geological Survey Data
Series 187. http://pubs.usgs.gov/ds/ds187/.


Chapter 3. Study areas and methods


Study areas and methods
63
3.1. Choice of study regions
In order to answer the hypotheses raised in the Conceptual Framework
(Section 1.4), a sampling methodology was defined and applied equally in three
study areas of three different biogeographic regions. The overall similarity between
land use gradients of very different biogeographic regions was an initial generic
criterion for site selection. The preference was for gradients of agricultural land
use, particularly crops known to have severe impact on the aquatic systems. Thus,
the study regions were chosen according to the presence of river watersheds
under the influence of intensive potato farming activities. More specific criteria for
study region selection included:
- presence of sufficiently replicated small streams (order 1-3 considering the
Strahler stream order system at 1:50000);
- substantial/predominant presence of cobble substrate (riffle/run) habitats in
the stream so that a sampling approach focused only on these habitats provides a
representative sample of the stream reach;
- permanent character of these small streams;
- absence of major human alteration along the river course (i.e. dams or
channelization);
- possibility to establish a land use gradient, spanning from low to high
agricultural land use under potato crops;
- distance from municipalities, in order to avoid intensive human presence
and its consequences in terms of environmental impacts;
- existence of previous scientific studies done in that particular region that
could provide useful information to the project.
The verification of these criteria was made by: analysis of geographical
information available online or made available by local research groups; direct
contact with local researchers working in the areas of expertise that this project
relates to; field observations; and review of published and unpublished literature.

Chapter 3
64
At the end of this process, the three study regions were selected and are
here defined by the major watercourses including all the study streams (Fig. 3.1):
- the Little River and Salmon River watersheds, in northwestern
New Brunswick, Canada;
- the Anllóns River watershed, in northern Galicia, Spain;
- the Upper Reventazón River watershed, in Cartago, Costa Rica.
In each region, and using topographic features, soil conditions, and most
importantly crop and management practices, sub-watersheds were identified and
selected in a way that allowed studying the impacts of agriculture on stream
communities. Study watersheds were constrained to those with 1st-3rd order
streams to avoid or minimize the presence of confounding effects from non-
agricultural stressors, as they drain areas of a single geological type and possess
more simplified land use patterns.
Fig. 3.1 - World map with the location of the three regions studied: eastern Canada (Little River and Salmon River in New Brunswick; A); northwestern Spain (Anllóns River; B); and central Costa Rica (Upper Reventazón River; C). Boxes enclose country/province outlines and hydrological maps of each river. Scale corresponds to the hydrological maps.
C
A B
A
B
C

Study areas and methods
65
3.1.1. Little River and Salmon River watersheds (Canada)
The Canadian field sites were located in the northwestern part of the
maritime province of New Brunswick (eastern Canada, Fig. 3.1A) between
latitudes 46º 59! and 47º 12! N and longitudes 67º 31! and 67º 44! W. The field
sites were distributed along two watersheds, the Little River and the Salmon River
watersheds, both draining into the upper Saint John River near the town of Grand
Falls.
In terms of bedrock geology, the area is underlain by Ordovician and/or
Silurian calcareous and argillaceous sedimentary rocks interbedded with igneous
volcanic rocks; surficial geology consists of compact till, some of which has been
reworked, with or without a surficial capping of ablational till or residual,
glaciofluvial and alluvial deposits (Fahmy et al., 1986). Soils within the studied
watersheds were primarily Orthic Humo-Ferric Podzols - acid soils with a
subsurface accumulation of iron-aluminum-compounds - and Podzolic Luvisols -
soils with subsurface accumulation of high activity clays and high base saturation
(Langmaid et al., 1976; FAO, 1998).
The topography, ranging from about 100 to 300 m above sea level, was
characterized by predominantly undulating to rolling surface expressions with
average slopes of 5-15%. The climate is moderately cool boreal with a humid to
prehumid soil moisture regime (Langmaid et al., 1976). The upper Saint John
watershed has an average summer temperature between 16 and 18°C, while
average winter temperatures range from -8 to -12°C; it receives about 1100 mm of
precipitation annually (Department of Environment, 2007).
In terms of land use characteristics, forest, mainly mixed woods (yellow birch,
white spruce and sugar maple on the upland areas and balsam fir and white and
black spruce in the valleys), occupied more than 80% of the region. The main crop
on the agricultural land was potato, in rotation with grains, peas, and hay; some
land was maintained under pasture.
The Little River is a 5th order watercourse draining an area of 383 Km2 (NB
Aquatic Data Warehouse, 2008). Its mean annual discharge was estimated at
approximately 3.2 x 107 m3 (Gray, 2003). The Salmon River is a 6th order

Chapter 3
66
watercourse, and its watershed occupies an area of 573 Km2 (NB Aquatic Data
Warehouse, 2008).
Focus on these catchments was due to their location within New Brunswick!s
intense potato producing region. Potatoes are the most important cash crop in the
province, with approximately 230 Km2 in production, most of it in the St. John River
valley manly between Grand Falls and Woodstock (Cunjak & Newbury, 2005). The
uppermost region of both study catchments is predominantly forested while
agricultural fields (mainly devoted to potato farming) dominate the landscape in the
lower regions (Gray, 2003), so land use gradients can easily be established.
3.1.2. Anllóns River watershed (Spain)
The Anllóns River can be found in the northern part of the province of Galicia
(district of Bergantiños), in northwestern Spain (Fig. 3.1B). Traditionally, Galician
agriculture is characterized by small landholdings, and has been based on multi-
crop and intensive farming (Xunta de Galicia, 2004). Attention to Galicia, and the
region of Bergantiños in particular, was drawn by its similarity with the Atlantic
provinces of Canada (mentioned in the previous section), namely in terms of
adequacy for potato cultivation (Nogueira, 2004).
The Spanish field sites are situated between latitudes 43º 08! and 43º 15! N
and longitudes 8º 30! and 8º 54! W. The Anllóns River drains a rural coastal area
of 516 Km2, being one of the few rivers in Galicia whose natural regime has not
been altered by dams (Devesa-Rey et al., 2008). This 5th order watercourse runs
from East to West through the district of Bergantiños, and drains to the Atlantic
Ocean in the town of Laxe. Its average annual flow is 10.77 m3s-1 (Rubinos et al.,
2003).
The climate of the region is atlantic humid; precipitation is abundant, with a
winter maximum typical of oceanic climates, and an annual average of 1200 mm.
Temperature varies from 8-9ºC in winter time to never more than 20ºC in the
summer (Devesa-Rey et al., 2008).
The bedrock geology of the watershed consists of metamorphic rocks

Study areas and methods
67
(schists) in its upper area and igneous basic rocks (gabbros and amphibolites) in
the middle area; the lower stretch of the river runs over granite of two micas
(igneous rock), followed by biotite gneiss (metamorphic) at the mouth (Devesa-
Rey et al., 2008). Dominant soil types in the area are Umbrisols - acid soils with a
thick, dark topsoil rich in organic matter - and Cambisols - weakly to moderately
developed soils (FAO, 1998; Vázquez & Anta, 2005).
In terms of topography, in the upper part of the river high slopes can be
found, mostly in the first 7 Km, where altitudes of 400-500 m are detected. The
Anllóns River turns into a smooth profile in its middle area, with an average slope
of 4%, descending from around 100 m until sea level. In the final 13 Km, slopes
are below 3%. Overall, topographically, the area could be described as having a
gentle undulating to rolling surface (slopes from 2 to 25%).
In terms of land use, the area includes mixed forest of Eucaliptus globulus,
Eucaliptus alba and Pinus pinaster (about 60% of the total land cover), cultivated
lands (30%), pastures (10%) and urban uses (1%) (Consellería do Medio Rural,
2004). The most important crops of the region are cereals and potatoes, in rotation
with several types of vegetables. Forage crops also occupy a significant
percentage of the watersheds! cultivated area.
Land-use gradients are relatively easy to establish within the Anllóns River
watershed, as sub-watersheds with different levels of agricultural land cover can
be found. Sub-watersheds draining areas located almost entirely in forested areas
can be used as representatives of reference conditions.
3.1.3. Reventado, Birrís and Turrialba watersheds (Costa Rica)
The Costa Rican field sites (Fig. 3.1C) are situated in the vicinity of the town
of Pacayas, in the province of Cartago (Atlantic slope of the central valley of Costa
Rica). They are located between latitudes 9º 56! and 9º 58! N and longitudes 83º
47! and 83º 53! W.
Sampling was focused in three 4th order tributaries of the Upper Reventazón
River: the Reventado River, the Birrís River and the Turrialba River. In total, the

Chapter 3
68
three sub-watersheds occupy an area of about 200 Km2, of the total 1500 Km2
drained by the Upper Reventazón River.
This area is known for its intensive agricultural activity, mostly based in the
production of potatoes in rotation with carrots and cabbages, and providing more
than half of the potatoes consumed nationwide (Marchamalo, 2004). The proximity
of a protected area (Irazú Volcano National Park), allows the establishment of a
reference condition for the region.
The bedrock geology of the area consists of igneous volcanic rocks
(pyroclastic material like agglomerates, lava and tuff) and sedimentary rocks (like
conglomerates, limonite, sandstone and limestone) (Escoto, 1993). In terms of soil
typology, the area is dominated by Andosols - young soils mostly resulting from
volcanic deposits (FAO, 1998; Marchamalo, 2004).
The studied rivers are characterized by abrupt slope changes along their
course – the altitudes varying from around 3000 to 600 m above sea level in a
distance of 12 Km (van Westen, 2000). In these watersheds, high slopes can
therefore be detected, especially in the upper areas (in the surroundings of the
Irazú and Turrialba volcanoes), and rolling to hilly surfaces along with steeply
dissected to mountainous relieves characterize the topography of the region
(slopes range from around 3% to more than 90% - Peréz & van Es, 2005).
As the study area is located in the Atlantic slope of Costa Rica, it is under the
influence of the Caribbean Sea. The climate favors heavy rains that can increase
the annual average precipitation to 2289 mm (Villanueva, 2001). The dry season
usually goes from January to April. The temperature regime presents two seasons;
the warmer months are March through May and the coolest are November through
January. The annual average temperature is 21.7ºC, this average being
considerably decreased in the upper areas of the Reventazón watershed.
Focus on this particular area was due to the fact that, in Costa Rica, the
vegetable production was mostly concentrated in these regions that surround the
Irazú and Turrialba volcanoes (Peréz & van Es, 2005). The upper part of the study
watersheds is mostly covered with secondary forest, belonging to the Prusia sector
of the Irazú Volcano National Park; the areas outside the Park are used for

Study areas and methods
69
agriculture (mostly annual crops) and cattle farming (Müller et al., 1998).
Agricultural land (more than 30% of the watershed area) is mostly devoted to
potato and other vegetable crops (cabbage, carrot, broccoli), but part of the land is
devoted to pasture (30%); forested areas (around 30%) are covered with Andean
alder, cypress, cedar, oak and, more recently, eucalyptus; urban areas occupy
less than 2% of the watersheds (Marchamalo, 2004).

Chapter 3
70
! "
#
#
!
"
!
!
"
"
"
#
#
#
!
#
!
#
#
" " "
! "
#
Fig. 3.2 – Sampling site location in the Little River and Salmon River (Canada; A), in the Anllóns River (Spain; B); and in the Upper Reventazón River (Costa Rica; C). Sampled streams were categorized according to the level of watershed land use change: no or minimal human impact (!); medium impact (26-50% of watershed devoted to agriculture, "); high impact (more than 50% of watershed devoted to agriculture, #). Scale corresponds to the hydrological maps. Photographs show streams belonging to each land use category (from top to bottom, left to right: A - Foley Brook, Outlet Brook, Dead Brook; B - A Regueira, Rego da Balsa, Río Archeiro; C - Río Retes, Río Coliblanco, Quebrada Pacayas).
! "
#
!
"
#
!
!
!
" "
#
#

Study areas and methods
71
3.2. Materials and Methods
In all three study areas, the same sampling methodology was followed using
a gradient study design based on the Canadian Aquatic Biomonitoring Network
protocols (Reynoldson et al., 2003). With the sampling design adopted here,
impact is inferred from the spatial pattern alone in a situation where there is no
knowledge of when specifically the impact occurred. Sampling was undertaken in
August 2005 in Canada, May 2006 in Costa Rica and in November 2006 in Spain.
The characterization of the biological community (by macroinvertebrate sampling)
was accompanied by the characterization of the surrounding habitat in each
sampling point. In each region, 10 sampling points were chosen (Fig. 3.2),
corresponding to 10 different streams with watersheds that spanned a gradient
from no or minimal human impacts (watersheds with percentages of forest cover
higher than 75%, working as references) to streams where intensive agricultural
land use was clearly apparent (mainly row-crop farming). Watershed area land use
was determined from field surveys and land use map analysis. In each stream, a
flowing reach (riffle) representative of the stream characteristics was chosen.
Sampled reaches were always located at least 50 meters upstream from any road
or bridge crossing to minimize their effects on stream velocity, depth, and overall
habitat quality; also, care was taken to ensure that the sample reaches had no
major tributaries discharging immediately upstream or downstream (Barbour et al.,
1999).
3.2.1. Habitat characterization
Several watershed-scale properties were obtained from geographical
information available from various sources and compiled into the project database.
These properties included catchment area (CatcArea), elevation, Strahler system
stream order at 1:50000 (StrOrder) and forest cover as percentage of watershed
(VegPerc).
Surface water samples were collected (one per reach) and kept refrigerated

Chapter 3
72
for subsequent analyses in external certified laboratories. Parameters analyzed
include total suspended solids (TSS), alkalinity (Alk), major ion concentrations
(potassium (K), sodium (Na), calcium (Ca), magnesium (Mg), fluoride (F), chloride
(Cl) and sulphate (SO4)), and major nutrient concentrations (organic nitrogen
(Norg), ammonia (Namm), nitrites and nitrates (NO3NO2), total (TotP) and soluble
(SolP) phosphorus; analyses to carbon contents were not possible in both Costa
Rica and Galicia and had to be excluded from the analysis). Pesticide residue
analysis was also undertaken. The lists of pesticides analyzed by each laboratory
were based on pesticides most commonly applied in each of the study areas.
Since most of the outcomes were no-detect results, these results were not further
considered.
The remaining environmental parameters were measured in situ at the time
of macroinvertebrate sampling. Geographical coordinates and altitude of each
sampling point were determined using a portable GPS device. In each sampled
reach, water pH, dissolved oxygen (expressed both as concentration - DOmgL -
and percent saturation - DOperc), specific conductance (SpeCond), and
temperature (T) were measured using field water quality meters (WTW, Germany
and ISY, USA handheld meters). Embeddedness estimates (as percentage of
substrate area covered by fine sediment, Embedd) were obtained by visual
assessment of 3 square replicate areas of 0.25 m2 along the reach. Channel
wetted width (ChanWidth) was determined in a representative cross section of
each stream, using a measuring tape. Surface water velocity (WatVeloc) was
determined using a portable electromagnetic flow meter (when available - Marsh-
McBirney Model 2000 Flo-Mate) or a floating object travelling over a measured
distance (3 replicates). Measurements of canopy cover density (as percentage of
area covered by overhanging vegetation - CanoDens) were obtained using a
spherical densiometer (Robert E. Lemmon, USA) and 3 replicates were taken.
Due to the influence on substratum stability and refuge availability,
microhabitat heterogeneity in each stream reach was visually assessed
considering the presence of the following habitat types: rootwads, coarse woody
debris, leaf packs, bedrock, fine sediment, pebbles, cobbles and boulders. Due to

Study areas and methods
73
its influence on bank stability and allocthonous food input, riparian vegetation in
each stream reach was assessed considering the categories: grasses, shrubs,
coniferous trees and deciduous trees.
Chlorophyll a is widely used to estimate photosynthetically capable biomass
in the periphyton community and the concentration of this chlorophyll has been
shown to relate to primary productivity (Vadeboncoeur et al., 2006). Estimates of
chlorophyll a concentration (Chla, expressed as µg/cm2) were obtained from
periphyton samples scraped off from known surface areas. Five replicate 4 cm2
scrapings were obtained from five small boulders randomly collected from the
bottom of each riffle. The samples were maintained in water from the sampling
location and refrigerated until laboratory delivery. In the laboratory, samples were
frozen for posterior determination of chlorophyll a concentrations by a
spectrophotometric method following the USEPA protocol (Arar, 1997).
3.2.2. Biological community taxonomic characterization
According to Barbour and co-workers (Barbour et al., 1999),
macroinvertebrate diversity and abundance are usually higher in cobble substrate
(riffle) habitats, and a single-habitat approach allows standardized assessments
among streams having those habitats. A single-habitat approach was therefore
selected to obtain an estimate of macroinvertebrate presence in each reach:
benthic macroinvertebrate sampling was undertaken in the bottom of riffle areas,
using a hand-operated D-frame kick net (0.5 mm mesh aperture). In each reach,
bottom material was kicked (and animals dislodged and caught by the net) for 3
minutes, with the operator adopting a zigzag trajectory moving upstream in order
to sample different locations of the reach. Large debris was inspected and
discarded and samples were preserved with 10% formalin and then transferred to
and stored in 70% ethanol. This process is advantageous since it kills specimens
quickly with minimum preservative, provides tissue fixation without dissolving
calcareous deposits in the exoskeletons of some taxa, preserves colour, replaces
most of the water in organisms with alcohol, and reduces the amount of formalin in

Chapter 3
74
the sample making sorting more comfortable (Reynoldson et al., 2003).
In the laboratory, samples were sorted under a stereoscopic microscope. In
order to reduce the effort required for sorting and identification, subsampling was
carried out when necessary. The method followed was the fixed-count method,
targeting for a count of 300 organisms (as described in Reynoldson et al., 2003)
and based on the Marchant sub-sampling device. Sorted organisms were
identified to the lowest possible taxonomic level (except for Oligochaeta and
Chironomidae) and counted under a stereoscopic microscope. As a form of
taxonomic adjustment between regions, a comparative analysis of taxa lists was
performed and equivalent taxonomic levels were used for similar groups found
simultaneously in the different regions; e.g., if this comparison resulted in the
decision of usage of genus level, the abundances of all species within a genus
were summed.
3.2.3. Biological community characterization through traits
For each collected taxon, 72 categories or 'modalities' of 16 biological traits
were documented, adapted from the strategies of the "Rhône team! (Tachet et al.,
2000 and related studies). The selected traits reflected general biological and
physiological features, life history and potential resistance to disturbance (Table
3.1).
Affinity of each taxon for the trait categories was fuzzy coded (Chevenet et
al., 1994) due to the multiplicity of information sources and to include the variability
within each taxon (since a taxon might have affinity for different categories of the
same trait e.g. at different life stages). A score from 0 (no affinity or absence of
information) to n (high affinity) was assigned to each category, reflecting the
relative strength of association of each taxon for that category. For example, a
taxon that predominantly uses gills for respiration but might also depend on
cutaneous respiration to satisfy some portion of its oxygen uptake is assigned an
affinity of 3 for the category gills and an affinity of 1 for the category cutaneous of

Study areas and methods
75
the trait respiration (being assigned 0 for
the remaining categories of the trait).
Traits scored 0 for all categories if no
information was available.
Information was collected from
available databases (major sources of
information: Tachet et al., 2000 and
related studies; Tomanová, 2007; Vieira
et al., 2006, by fuzzy coding their
information; additional information from
the databases already listed in Section
2.12). For taxa for which information
was unavailable in the databases,
coding was done by using additional
published information, personal
communications from experts, or by
calculating means for taxa within the
same coarser taxonomic level (e.g.
means for species within the same
genus in order to obtain genera affinity
scores; Fig. 3.3).
The sequences of scores per taxon
were then transformed into sequences
of frequencies per trait (affinity scores
were standardized so that their sum for
a given taxon and a given trait was 1).
This procedure yielded a [taxon x trait
category] matrix that was multiplied by
the [site x taxon abundance] matrix to
produce the [site x trait category
abundance] matrix containing the
Table 3.1 – Macroinvertebrate traits and trait categories
Trait Categories0-2.5 2.5-5 5-10 10-2020-40 >40 less than 1yearmore than 1yearsemivoltineunivoltinemultivoltineegglarvapupaadultovoviviparity+parental carefree egg/clutchescemented egg/clutchesterrestrial egg/clutchesasexualstrong dispersalmedium dispersalweak dispersalresistance eggs, statoblastscocoonsdiapause or dormancyno resistance formfine sediment+microorganismsdetritus <1mmplant detritus !1mmmicrophytesmacrophytesdead animals !1mmmicroinvertebratesmacroinvertebratesdeposit feedershredderscraperfiltererpiercer (plants or animals)predatorparasitecutaneousgillsplastronspiraclestenothermal psychrophilic (<15ºC)stenothermal thermophilic (>15ºC)eurythermallowlands (<1000 m)mountain level (1000-2000 m)alpine level (>2000 m)rock/bouldersgravelsandsiltmacrophytesalgaeLWDdetritusswimmercrawler /sprawlerburrowerattachedquietslowfast laminarfast turbulentstreamlined/fusiformflattenedcylindricalspherical
Microhabitat
Attachement to substrate
Current velocity preference
Body Shape
Feeding habits
Respiration
Temperature preference
Altitude preference
Reproduction and oviposition
Dissemination potential
Resistance form
Diet preferences
Maximum size (mm)
Life Cycle duration
No. reproductive cycles/year
Aquatic stage

Chapter 3
76
distribution of abundances per trait categories in each site, and used to describe
the functional composition of the macroinvertebrate communities. For statistical
purposes, this matrix was log-transformed to approximate a normal distribution.
Fig. 3.3 – Decision tree employed to fill the macroinvertebrate taxa x traits matrix. Examples show how the affinity score assignment of three categories (tr) of a generic trait y could occur for a generic macroinvertebrate genus x.
Therefore, the described methods yielded, for each study region, three data
matrices: the site x environmental variable matrix; the site x taxon matrix; the site x
trait category matrix. Choices in terms of specific data analysis will be described in
each of the following chapters.
Available in published databases?
Add to taxon x trait matrix.
Fuzzy code.
Available from experts?
Leave blank.
yes
yes
no
yes
no
no
yes no
no yes
tr1
? Genus x
tr3
?
tr2
?
Trait y
tr1 3 Genus x
tr3 1
tr2
0
Trait y tr1
0 Genus x
tr3 0
tr2
0
Trait y
Available in other publications?
Fuzzy coded?
Calculate mean of species information.
Available information on species within genus x?

Study areas and methods
77
3.3. References
Arar, E. J. (1997) Method 446.0, In Vitro Determination of Chlorophylls a, b, c1+c2
and Pheopigments in Marine And Freshwater Algae by Visible
Spectrophotometry. Cincinnati, Ohio, U. S. Environmental Protection
Agency. 26 pp.
Barbour, M. T., Gerritsen, J., Snyder, B. & Stribling, J. (1999) Rapid
Bioassessment Protocols For Use in Streams and Wadeable Rivers:
Periphyton, Benthic Macroinvertebrates and Fish, Second Edition.
Washington, D.C., U.S. Environmental Protection Agency. 339 pp.
Chevenet, F., Dolédec, S. & Chessel, D. (1994) A fuzzy coding approach for the
analysis of long-term ecological data. Freshwater Biology, 31, 295-309.
Consellería do Medio Rural (2004) Anuario de Estatística Agraria 2004. Xunta de
Galicia. 257 pp.
Cunjak, R. A. & Newbury, R. (2005) Atlantic Coast Rivers of Canada. In: A. C.
Benke & C. E. Cushing (Ed.). Rivers of North America, Elsevier, 939-980.
Department of Environment, N. B. (2007) New Brunswick watersheds - St. John
River. Env. Reporting Series, http://www.gnb.ca/0009/0371/0013/index-
e.asp.
Devesa-Rey, R., Moldes, A. B., Diaz-Fierros, F. & Barral, M. T. (2008) Toxicity of
Anllóns River sediment extracts using microtox and the Zucconi
phytotoxicity test. Bulletin of Environmental Contamination and Toxicology,
80 (3), 225-230.
Escoto, O. A. S. (1993) Determinación de áreas críticas mediante sistemas de
información geográfica, cuenca del río Reventado, Costa Rica. CATIE,
Turrialba, Costa Rica. MSc thesis. 131 pp.
Fahmy, S. H., Rees, H. W. & MacMillan, J. K. (1986) Soils of New Brunswick - A
First Approximation. New Brunswick Department of Agriculture. 109 pp.
FAO (1998) World reference base for soil resources. Rome, FAO - International
Society of Soil Science. 88 pp.

Chapter 3
78
Gray, M. A. (2003) Assessing non-point source pollution in agricultural regions of
the upper St. John River basin using the slimy sculpin (Cottus cognatus).
University of New Brunswick, PhD thesis. 184 pp.
Langmaid, K. K., MacMillan, J. K. & Losier, J. G. (1976) Soils of Northern Victoria
County New Brunswick. New Brunswick Department of Agriculture. 152 pp.
Marchamalo, M. (2004) Ordenación del territorio para la producción de servicios
ambientales hídricos. Aplicación a la cuenca del Río Birrís (Costa Rica).
Universidad Politecnica de Madrid, PhD thesis. 400 pp.
Müller, S., Ramírez, S. M. J. N. L., Núñez, J. & Ramírez, L. (1998) Indicadores
para el uso de la tierra: El caso de la Cuenca del Río Reventado, Costa
Rica. Instituto Interamericano de Cooperación para la Agricultura. 58 pp.
NB Aquatic Data Warehouse, C. R. I. (2008) Saint John River Atlas: Release 2.
Retrieved 29/08/2008, http://cri.nbwaters.unb.ca/sjratlas/site/index.castle.
Nogueira, J. F. (2004) La patata - Breve introducción a su historia y descripción.
Retrieved 18/09/2008, from http://www.elgrelo.com/52/52_03.htm.
Peréz, Y. & van Es, J. (2005) Plan de manejo de la microcuenca de las quebradas
Pacayas y Plantón, Cartago, Costa Rica. Heredia, Universidad Nacional -
Facultad de Tierra y Mar. 68 pp.
Reynoldson, T. B., Logan, C., Pascoe, T. & Thompson, S. P. (2003) CABIN
(Canadian Aquatic Biomonitoring Network) Invertebrate Biomonitoring Field
and Laboratory Manual. National Water Research Institute - Environment
Canada. 47 pp.
Rubinos, D., Barral, M. T., Ruiz, B., Ruiz, M., Rial, M. E., Álvarez, M. & Diaz-
Fierros, F. (2003) Phosphate and arsenate retention in sediments of the
Anllóns river (northwest Spain). Water Science & Technology, 48 (10), 159-
166.
Tachet, H., Richoux, P., Bournaud, M. & Usseglio-Polatera, P. (2000) Invertébrés
d'eau douce: systématique, biologie, écologie #. Paris, CNRS Editions. 588
pp.

Study areas and methods
79
Tomanová, S. (2007) Functional aspect of macroinvertebrate communities in
tropical and temperate running waters. Masaryk University, PhD thesis. 158
pp.
Vadeboncoeur, Y., Kalff, J., Christoffersen, K. & Jeppesen, E. (2006) Substratum
as a driver of variation in periphyton chlorophyll and productivity in lakes.
Journal of the North American Benthological Society, 25 (2), 379-392.
van Westen, C. (2000) Estudio preliminar de amenazas naturales en la cuenca del
Rio Turrialba, Canton Turrialba, Costa Rica. Capacity Building For Natural
Disaster Reduction & Regional Action Program For Central America. 88 pp.
Vázquez, F. M. & Anta, R. C. (2005) Mapa de solos de Galicia - 1:50.000. Xunta
de Galicia, 43, 45, 69.
Vieira, N. K. M., Poff, N. L., Carlisle, D. M., Moulton, S. R., Koski, M. L. &
Kondratieff, B. C. (2006) A database of lotic invertebrate traits for North
America. U.S. Geological Survey Data Series 187.
Villanueva, E. S. M. (2001) Análisis espacial del tipo de uso de la tierra en la
cuenca del río Turrialba, Costa Rica. CATIE, Turrialba, Costa Rica. MSc
thesis. 84 pp.
Xunta de Galicia (2004) Galicia 2004. Retrieved 18/09/2008, from
http://www.xunta.es/galicia2004/index.htm.


Chapter 4. Freshwater macroinvertebrate community gradients in
three agricultural regions: a non-functional approach


Non-functional approach to community gradients
83
4.1. Introduction
The pernicious effects that land use changes can have on stream
ecosystems have been a matter of long and well documented debate. One
particular form of change – the conversion of forest to agricultural land – raises
serious concerns due to the multiplicity of mechanisms by which it influences the
stream environment: sedimentation, nutrient enrichment, chemical pollution,
hydrologic alteration, riparian clearing and loss of large woody debris (Allan, 2004).
Several practices associated with row-crop agriculture, in the absence of riparian
buffer strips, can lead to bank erosion, streambed disturbance, and increased
inputs of fine sediment, nutrients and chemical contaminants to nearby streams
(Townsend & Riley, 1999). This leads to impacts on aquatic habitats by increasing
deposition of fine sediments and habitat instability. Such physico-chemical
responses to land use changes may act as stressors of stream macroinvertebrate
communities, producing increasingly deleterious consequences for biodiversity
and ecosystem processes with increasing physical changes to the streambed
(Riley et al., 2003).
Macroinvertebrates are exceptional signals of environmental conditions
affecting streams running through agricultural catchments since their responses at
the community level can be integrated to assess land-use influences at multiple
spatial scales through properties of both physical habitat and water chemistry
(Weigel et al., 2003). Their assemblage structure reacts predictably to
environmental change, in terms of gross community composition, indicating stream
integrity (Barbour et al., 1999), and they are frequently used in freshwater
biomonitoring programs. As mentioned in Section 1.2, different approaches for
macroinvertebrate community assessment were developed over the years, and
they typically include one or several measures describing community parameters,
such as richness and composition measures, tolerance measures, indices of
diversity and biotic indices (Bonada et al., 2006). A proliferation of indices,
especially for pollution assessment, reflects the value of using the biological

Chapter 4
84
components of stream ecosystems (Norris & Georges, 1993), but they are not
intended to replace proper statistical analysis of data, nor are they absolute
measures of a community, and they are subject to sampling error and, more
importantly, to natural community variation (Reynoldson et al., 2003). It is
recognized that indices are useful in condensing data down to single numbers and
that they must be interpreted cautiously, in combination with other indices as well
as other data collected at the sites.
A fundamental objective of most biomonitoring programs is the assessment
of the ecological integrity of an ecosystem, which requires confidence that the
methods used can detect existing changes effectively (Lücke & Johnson, 2008).
Comparing the reliability of different biological methods of detection is therefore
crucial. But variations in sampling and processing of macroinvertebrates among
sites and studies can add to the complication of assessing stream impairment and
comparing among different assessment methodologies (Clarke et al., 2002). When
different sampling methods are used, sampling intensity is inconsistent and
taxonomic resolutions inevitably differ among studies (Claret et al., 1999).
Consistency of the various aspects of biomonitoring is therefore imperative.
The objective of this study was, after the selection of catchments in settings
that were similar except for their degree of agricultural land use, to evaluate the
ability of stream community structure (taxonomic information, structural metrics
and diversity indices) and physiological responses to pollution (tolerance metrics
and biotic indices), commonly used in biomonitoring programs, to reveal impact
gradients with appropriate discriminatory power of different levels of impact.
Providing a starting point for water quality managers to think about not only how to
partition naturally occurring variation in the aquatic communities but also how
differences between regions may influence how stressors affect biota, is
considered fundamental (Hawkins et al., 2000). Consistent taxonomic
measurements of macroinvertebrate communities of different biogeographic
regions withstanding the conditions of similar types of impacts were compared in
order to highlight possible differences and common points, which might contribute
for future water quality management decisions regarding the study areas.

Non-functional approach to community gradients
85
4.2. Methods of data analysis
For all statistical tests the significance level used was 0.05. One-way
Analysis of Variance (ANOVA) or its nonparametric equivalent, Kruskal-Wallis
rank-sum test (when normality or homoscedasticity was not achieved) were used
for univariate statistical testing. When ANOVA/Kruskal-Wallis found significant
differences, Tukey/Dunn!s tests were performed for post-hoc comparisons. All
multivariate analyses were run with the ade4 library version 1.4-14 (Dray & Dufour,
2007) implemented in the R freeware version 2.11.1 (R Development Core Team,
2010). The remaining analyses were run with SigmaStat version 3.5 (Systat
Software, Inc., 2006).
4.2.1. Unresponsive variables
Principal components analyses (PCA) of environmental variables considered
to be unresponsive to the land use gradient were carried out to assess how
comparable the sites were if land use change was not considered. The variables
(refer to Table 4.1 for variable identification) considered were StrOrder, CatcArea,
ChanWidth and Elevation. One-way ANOVAs were used to compare these
potentially unresponsive variables of 3 land use categories: reference sites
(representing watersheds with over 75% vegetated area, less than 25% devoted to
agricultural activities); medium impact (26-50% of watershed devoted to
agriculture); high impact (more than 50% of watershed devoted to agriculture).
4.2.2. Impact gradients
PCA on centered and standardized variables was used to correlatively
assess the impact-associated habitat characteristics in each region and establish
the structure of the environmental gradients. For the PCA the following variables
were used: TotP, SolP, NO3NO2, Namm, SO4 (as measures of chemical input),
SpeCond, Embedd (sediment input), CanoDens (riparian vegetation clearing) and

Chapter 4
86
WatVeloc (hydrologic alterations). One-way ANOVAs were used to compare
impact-associated environmental variables of the 3 land use categories.
4.2.3. Biological gradients
Several measures of stream integrity using macroinvertebrates were
determined. Single richness, composition and tolerance metrics commonly used in
biomonitoring programs were selected (Barbour et al., 1999): taxa richness
(TxRich); total and relative abundance (TotAbd and RelAbd); number and percent
of Ephemeroptera, Plecoptera and Trichoptera (E, P, Tri, EPT, PercE and
PercEPT); percent of Chironomidae (PercChi); percent of Oligochaeta (PercOli);
percent of dominant taxon (PercDTx). Traditional biotic and diversity indices were
also determined: the Shannon-Wiener diversity (ShanDiv), the Pielou!s evenness
(PielEve) and the Simpson!s Diversity Indices (SimDiv) (Legendre & Legendre,
1998); the Family Biotic Index (FBI) (Mandaville, 2002); the Biological Monitoring
Working Party (BMWP) adapted to the Iberian Peninsula (Alba-Tercedor &
Sánchez-Ortega, 1988; Alba-Tercedor, 1996), Costa Rica (Ministerio del Ambiente
y Energia, 2006) and North America (Mandaville, 2002); and the Average Score
Per Taxon (ASPT), calculated by dividing the BMWP by the number of families
represented in the sample.
PCA on transformed (log[x+1]) abundance data was used as a multivariate
tool to obtain synthetic multivariate scores for sites based on taxa composition
(summarizing macroinvertebrate community variation). In order to compare among
analyses, axis ordination scores were standardized and scaled to values between
0 and 1 using the formula:
yi=(xi-min)/(max-min) (Equation 1)
where xi is the value of score for site i, min is the minimum site score and max is
the maximum site score.
One-way ANOVA/Kruskal-Wallis rank-sum tests were used to compare all
biological variables among the 3 land use categories (in order to assess
discriminatory power).

Non-functional approach to community gradients
87
Table 4.1 - Variables (measured and derived) used in the analyses and respective codes used in the text and figures.
Type Variable Code
Water alkalinity (mg/L) Alk
Water calcium concentration (mg/L) Ca
Canopy cover density (%) CanoDens
Catchment area (Km2) CatcArea
Channel maximum depth (m) ChanDepth
Channel wetted width (m) ChanWidth
Substrate benthic chl a ($g/cm%) Chla
Water chloride concentration (mg/L) Cl Water dissolved oxygen (mg/L) DOmgL
Water dissolved oxygen (%) DOPerc
Site elevation (m) Elevation
Substrate embeddedness in fines (%) Embedd
Water fluoride concentration (mg/L) F
Water potassium concentration (mg/L) K
Site latitude (º) Latit Site longitude (º) Longit Water magnesium concentration (mg/L) Mg
Water sodium concentration (mg/L) Na
Water ammonia concentration (mg/L) Namm
Water nitrites and nitrates concentration (mg/L) NO3NO2
Water organic nitrogen concentration (mg/L) Norg
Water pH pH
Water sulphate concentration (mg/L) SO4
Water soluble phosphorous concentration (mg/L) SolP
Water specific conductance ($S/cm) SpeCond
Strahler system stream order at 1:50000 StrOrder Water temperature (ºC) T
Water total phosphorous concentration (mg/L) TotP
Water total suspended solids (mg/L) TSS
Watershed forest cover (%) VegPerc
Env
iron
me
nta
l
Surface water velocity (m/s) WatVeloc
Average Score Per Taxon ASPT
Biological Monitoring Working Party BMWP
Average Score Per Taxon (Costa Rica) CR-ASPT
Biological Monitoring Working Party (Costa Rica) CR-BMWP
Ephemeroptera taxa richness (taxa) E
Ephemeroptera, Plecoptera and Trichoptera taxa richness (taxa) EPT
Family Biotic Index FBI Average Score Per Taxon (Iberian Peninsula) I-ASPT
Biological Monitoring Working Party (Iberian Peninsula) I-BMWP
Average Score Per Taxon (North America) NA-ASPT
Biological Monitoring Working Party (North America) NA-BMWP
Plecoptera taxa richness (taxa) P
Percent of Chironomidae (%) PercChi Percent of dominant taxon (%) PercDTx
Percent of Ephemeroptera (%) PercE
Percent of Ephemeroptera, Plecoptera and Trichoptera (%) PercEPT
Percent of Oligochaeta (%) PercOli Pielou!s evenness Index PielEve
Relative macroinvertebrate abundance RelAbd
Shannon-Wiener diversity index ShanDiv
Simpson!s Diversity Index SimDiv
Total macroinvertebrate abundance (individuals) TotAbd
Trichoptera taxa richness (taxa) Tri
Bio
log
ical
Macroinvertebrate taxa richness (taxa) TxRich

Chapter 4
88
4.2.4. Relationships between the biological and environmental
gradients
To address the relationships (co-variation) between the biological information
and the environmental variables that better define the impact gradient, correlations
between pairs of arrays (biological and environmental measures) were calculated
(Spearman correlation coefficient).
4.3. Results
4.3.1. Unresponsive variables
The three land use categories could not be distinguished in the unresponsive
variables PCAs (data not shown). StrOrder, CatcArea, ChanWidth and Elevation
(see exception below) did not significantly differ between the three land use
categories in the three regions. This supports the assumption that, for each region,
the selected watersheds were reasonably similar in terms of their estimated
unresponsive characteristics. The mean values of
the environmental characteristics are shown in
Table 4.2.
The exception was Elevation in Costa Rica,
which was significantly higher (ANOVA F=7.187,
p=0.026) in reference sites (ranging from 2500 to
2850m) when compared to medium impact sites
(ranging from 1634 to 1733m; Fig. 4.1). Despite
the intention of a selection of watersheds with
equivalent elevations, there was an unavailability
of non-impacted areas in the lower elevation
region of the Costa Rican study area (lower areas
were fully devoted to agricultural practices, higher
areas were protected by the Irazú Volcano
National Park).
Fig. 4.1 - Mean elevation (± 1 standard error) of Costa Rican reference (ref), medium impact (med) and high impact (high) field sites. Different letters indicate significant differences between impact levels at p&0.05.

Non-functional approach to community gradients
89
Veg
Perc
(%
)97.6±1.28
66.3±4.99
24.2±4.57
85.3±3.04
50.3±1.45
22.7±1.45
97.0±2.00
63.3±3.33
27.6±5.64
Lati
t (º
)47.1±0.05
47.1±0.04
47.0±0.03
43.2±0.02
43.2±0.01
43.2±0.01
10.0±0.00
10.0±0.00
9.9±0.00
Lo
ng
it (
º)-67.6±0.05
-67.6±0.04
-67.7±0.04
-8.8±0.02
-8.8±0.07
-8.5±0.02
-83.9±0.01
-83.8±0.00
-83.8±0.01
Ele
vati
on
(m
)175.0±19.35
169.5±13.88
140.0±19.92
152.0±61.55
111.0±16.00
319.0±89.09
2675.0±175.001677.7±29.17
2238.5±194.97
Catc
Are
a (
Km
2)
25.8±4.36
19.2±3.31
19.2±2.32
8.0±2.18
22.5±1.08
9.7±3.01
4.9±0.10
6.1±0.58
6.3±1.39
Str
Ord
er
2.7±0.33
2.5±0.29
2.7±0.33
2.0±0.41
2.3±0.33
2.0±0.00
3.0±0.00
2.0±0.00
2.5±0.29
Ch
an
Wid
th (
m)
2.45±0.236
1.73±0.445
1.90±0.854
1.83±0.346
2.17±0.268
3.13±0.371
2.18±0.075
1.53±0.260
1.83±0.554
Ch
an
Dep
th (
m)
0.12±0.000
0.17±0.026
0.14±0.027
0.31±0.063
0.58±0.220
0.33±0.083
0.21±0.090
0.12±0.009
0.21±0.035
T (
ºC)
12.78±0.188
14.67±1.237
14.92±0.616
12.41±0.385
12.50±0.202
11.62±0.303
13.01±0.890
17.09±0.382
12.97±0.142
DO
Perc
(%
)94.63±4.049
95.76±6.277
89.72±4.880
98.38±0.800
94.67±2.522
95.00±1.041
80.95±8.550
76.70±2.358
77.90±0.508
DO
mg
L (
mg
/L)
10.25±0.248
9.53±0.568
9.27±0.671
10.26±0.130
10.03±0.228
10.03±0.093
7.13±0.633
7.39±0.184
8.21±0.054
pH
7.82±0.058
8.19±0.185
8.04±0.045
6.45±0.208
6.99±0.152
6.53±0.250
7.25±0.087
6.96±0.214
6.79±0.773
TS
S (m
g/L
)1.47±0.485
1.92±0.790
1.21±0.474
8.25±3.065
8.33±0.882
13.00±4.726
12.48±0.525
0.27±0.167
21.96±16.126
F (m
g/L
)0.03±0.003
0.06±0.010
0.07±0.012
0.05±0.005
0.09±0.010
0.10±0.006
1.45±0.260
0.16±0.028
0.70±0.350
K (m
g/L
)0.40±0.071
0.67±0.108
1.49±0.220
1.10±0.134
2.33±0.333
2.73±0.145
5.24±0.880
3.58±0.415
6.99±0.362
Na (m
g/L
)1.99±0.431
3.38±1.247
5.88±1.400
10.54±1.679
9.43±1.269
8.52±0.745
9.38±1.540
5.33±0.903
8.99±1.521
Ca (m
g/L
)27.57±8.306
46.43±4.185
82.97±1.462
2.56±0.117
4.23±1.718
2.50±0.159
25.50±2.900
7.11±1.135
22.12±6.550
Mg
(m
g/L
)2.57±0.614
4.18±0.630
7.33±0.622
1.60±0.167
4.76±1.687
1.58±0.295
13.13±1.680
3.82±0.596
9.39±1.963
Alk
(m
g/L
)79.50±15.803114.48±11.057184.67±4.055
11.84±1.335
21.70±6.151
10.74±0.743
44.96±3.360
42.73±7.176
45.35±14.852
Cl (
mg
/L)
0.76±0.055
8.09±2.079
20.93±4.106
9.27±2.481
9.89±2.461
10.91±0.910
3.34±0.260
2.23±0.170
6.30±1.647
6.14±4.742
9.87±6.407
9.13±2.959
8.09±2.245
44.13±19.914
23.54±0.000
22.06±22.057
21.24±6.075
60.26±26.793
ref
med
hig
hV
ari
ab
le
Can
ad
aS
pain
Co
sta
Ric
a
ref
med
hig
hre
fm
ed
hig
h
Ta
ble
4.2
- M
ean
val
ues
(± 1
sta
ndar
d e
rror
) of
env
iron
men
tal v
aria
bles
mea
sure
d in
re
fere
nce
(re
f), m
edi
um im
pact
(m
ed
) a
nd h
igh
impa
ct (
hig
h)
field
site
s o
f thr
ee b
ioge
ogra
phic
reg
ions
. Ref
er to
Tab
le 4
.1 fo
r va
riabl
e id
entif
icat
ion.

Chapter 4
90
Correlation analysis of the reference and medium impact sites showed that
no significant correlations exist between Elevation and the remaining
environmental and biological variables, with the exception of Embedd (r=0.975,
p=0.017).
4.3.2. Impact gradients
In Canada, the PCA on the impact-associated variables showed that
SpeCond, SO4, SolP, TotP and NO3NO2 defined the first ordination axis (this axis
explained 62% of the total variance; Fig. 4.2). From the 9 impact-associated
variables, these were the most important ones in defining the complex nutrient and
sediment enrichment gradient of the Canadian field sites assigned to the different
land use categories. Sorted axis 1 PCA sample scores indicated that there was no
site overlap between land use categories.
Fig. 4.2 – Biplot of principal components analysis of impact variables at Canadian reference (ref), medium impact (med) and high impact (high) field sites. First (horizontal) and second (vertical) axes explain 62% and 14% of the variance, respectively. Rectangles indicate each land use category centroid and ellipses give and idea of the dispersion of the points in each category, with d indicating the scale of the plot. Arrows show the position of the impact variables. Refer to Table 4.1 for variable identification.
d = 2
high
med
ref
SpeCond SO4
SolP NO3NO2
TotP Namm
WatVeloc CanoDens
Embedd

Non-functional approach to community gradients
91
For all 5 variables and also Embedd significant differences were detected
among land use categories: the average values of SpeCond, SO4, SolP, TotP,
NO3NO2 and Embedd were significantly higher in high impact sites (ANOVAs,
SpeCond F=26.025, p<0.001; SO4 F=45.734, p<0.001; SolP F=6.486, p=0.025;
TotP F=10.622, p=0.008; NO3NO2 F=9.903, p=0.009; Embedd F=7.522, P=0.018;
Fig. 4.3).
0.000
0.005
0.010
0.015
0.020
0.025
ref med high
Sol
P (
mg
/L)
a
b
ab
0.000
0.005
0.010
0.015
0.020
0.025
ref med high
Tot
P (
mg/
L)
a
b
ab
0
100
200
300
400
500
600
ref med high
Spe
Co
nd (
µS
/cm
)
*
*
*
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
ref med high
NO
3NO
2 (m
g/L
)
*
0
5
10
15
20
25
ref med high
SO
4 (m
g/L)
*
*
*
Fig. 4.3 - Mean values (± 1 standard error) of the environmental variables used to characterize the land use impact gradient in Canadian reference (ref), medium impact (med) and high impact (high) field sites. Symbol (*) and different letters (a, b) denote significant differences at p&0.05. Refer to Table 4.1 for variable identification.
0
5
10
15
20
25
30
35
ref med high
Em
bedd
(%
)
*

Chapter 4
92
In Spain, the PCA on the impact-associated variables showed that Embedd,
SpeCond, TotP and SolP better defined the first ordination axis (this axis explained
63% of the total variance) and that SO4, TotP and SolP better defined the second
ordination axis (this axis explained additional 29% of the total variance; Fig. 4.4). A
complex nutrient and sediment enrichment gradient of the Spanish field sites was
defined by these variables. The PCA showed that site groupings according to land
use categories do not overlap.
Significant differences were detected among land use categories for SO4,
TotP, SolP and Namm. Reference sites had significantly lower average values of
three variables (ANOVAs, SO4 F=63.599, p<0.001; TotP F=108.900, p<0.001;
SolP F=126.776, p<0.001; Fig. 4.5) but a higher concentration of Namm (ANOVA,
F=6.677, p=0.024).
Fig. 4.4 – Biplot of principal components analysis of impact variables at Spanish reference (ref), medium impact (med) and high impact (high) field sites. First (horizontal) and second (vertical) axes explain 63% and 29% of the variance, respectively. Rectangles indicate each land use category centroid and ellipses give and idea of the dispersion of the points in each category, with d indicating the scale of the plot. Arrows show the position of the impact variables. Refer to Table 4.1 for variable identification.
SpeCond
SO4
SolP
NO3NO2
TotP
Namm
WatVeloc CanoDens
Embedd
d = 2
ref
med
high

Non-functional approach to community gradients
93
In Costa Rica, the PCA on the impact-associated variables showed that
TotP, SO4 and SpeCond defined the first ordination axis (this axis explained 38%
of the total variance) and that NO3NO2 and Namm defined the second ordination
axis (this axis explained additional 23% of the total variance; Fig. 4.6). These
variables defined the complex nutrient and sediment enrichment gradient of the
Costa Rican field sites assigned to the different land use categories. Significant
differences were detected among land use categories for TotP, SpeCond,
Embedd, Namm and SO4. There was a significant decline of the average values in
the medium impact sites when compared with both reference and high impact sites
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
ref med high
Nam
m (
mg/
L)
a
b
ab
0
2
4
6
8
10
12
14
ref med high
SO
4 (m
g/L)
*
*
*
0.000
0.050
0.100
0.150
0.200
0.250
0.300
0.350
ref med high
Tot
P (
mg/
L)
*
0
10
20
30
40
50
60
70
ref med high
Em
bed
d (%
)
0.000
0.050
0.100
0.150
0.200
0.250
0.300
0.350
ref med high
Sol
P (
mg
/L)
*
0
20
40
60
80
100
120
ref med high
Spe
Co
nd (
µS
/cm
)
Fig. 4.5 - Mean values (± 1 standard error) of the environmental variables used to characterize the land use impact gradient in Spanish reference (ref), medium impact (med) and high impact (high) field sites. Symbol (*) and different letters (a, b) denote significant differences at p&0.05. Refer to Table 4.1 for variable identification.

Chapter 4
94
(ANOVAs, TotP F=5.355, p=0.046; SpeCond F=6.780, p=0.029; Embedd F=5.776,
p=0.040; SO4 F=7.035, p=0.027; Fig. 4.7). For Namm, reference sites had
significantly lower average values (ANOVA F=11.922, p=0.008).
4.3.3. Biological gradients
A total of 166 different taxa (belonging to 98 families) were collected and
identified in the sampling sites of the three biogeographic regions: 75 in the
Canadian field sites, 71 in Spain and 64 in Costa Rica.
In Canada (Fig. 4.8), reference sites had significantly higher TxRich and E
than high impact sites (ANOVAs, TxRich F=6.454, p=0.026; Kruskal-Wallis, E
H=7.594, p=0.004). Reference sites had significantly lower FBI and PercOli than
high impact sites (ANOVAs, FBI F=4.961, p=0.046; PercOli F=6.564, p=0.025).
Impact sites had higher RelAbd than the remaining sites (Kruskal-Wallis, H=6.409,
Fig. 4.6 – Biplot of principal components analysis of impact variables at Costa Rican reference (ref), medium impact (med) and high impact (high) field sites. First (horizontal) and second (vertical) axes explain 38% and 23% of the variance, respectively. Rectangles indicate each land use category centroid and ellipses give and idea of the dispersion of the points in each category, with d indicating the scale of the plot. Arrows show the position of the impact variables. Refer to Table 4.1 for variable identification.
SpeCond
SO4
SolP
NO3NO2
TotP
Namm
WatVeloc
CanoDens Embedd
d = 1
med
ref
high

Non-functional approach to community gradients
95
p=0.034). As for the remaining metrics and indices, decreasing trends along the
impact gradient were observed for PercEPT, EPT, P, Tri, SimDiv, ShanDiv, NA-
BMWP and NA-ASPT. An increasing trend along the impact gradient was
observed for TotAbd and PercDTx.
It is worth noting that the average values for the FBI for reference sites were
generally indicative of excellent water quality, the values for medium impact sites
indicated good water quality and the values for high impact sites indicated fair
water quality (Mandaville, 2002). As for the NA-ASPT, reference and medium
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
ref med high
Nam
m (
mg/
L)
*
0
20
40
60
80
100
120
ref med high
SO
4 (m
g/L)
*
0
1
2
3
4
5
6
7
ref med high
NO
3NO
2 (m
g/L
)
0
50
100
150
200
250
300
350
ref med high
Spe
Co
nd (
µS
/cm
)
*
0.000
0.050
0.100
0.150
0.200
0.250
0.300
0.350
0.400
0.450
0.500
ref med high
Tot
P (
mg/
L)
*
0
10
20
30
40
50
60
70
80
90
100
ref med high
Em
bed
d (%
)
*
0.000
0.050
0.100
0.150
0.200
0.250
0.300
ref med high
Sol
P (
mg
/L)
Fig. 4.7 - Mean values (± 1 standard error) of the environmental variables used to characterize the land use impact gradient in Costa Rican reference (ref), medium impact (med) and high impact (high) field sites. Symbol (*) denotes significant differences at p&0.05. Refer to Table 4.1 for variable identification.

Chapter 4
96
impact sites had values that indicate clean water, and high impact sites had
probable moderate pollution (Mandaville, 2002). NA-BMWP indicated high impact
stream waters has moderately contaminated and reference and medium impact
streams as very clean.
TxRich and PercOli significantly correlated with SpeCond (TxRich, r=-0.828,
p=0.000; PercOli, r=0.939, p=0.000) and SO4 (TxRich, r=-0.905, p=0.000; PercOli,
r=0.939, p=0.000). E was significantly correlated with SpeCond (r=-0.926,
p=0.000), SO4 (r=-0.951, p=0.000), TotP (r=-0.840, p=0.000), SolP (r=-0.815,
p=0.002) and NO3NO2 (r=-0.957, p=0.000). The FBI was significantly correlated
with SpeCond (r=0.830, p=0.000), SO4 (r=0.879, p=0.000) and SolP (r=0.806,
p=0.003). Significant correlations between the majority of the remaining single
metrics and indices and the impact gradient variables were detected, with the
exceptions of RelAbd, SimDiv, PielEve, PercE, PercDTx, PercChi and P, that did
not significantly correlate with any of the variables that better define the impact
gradient.
The first two axis of the PCA done on the Canadian abundance data
explained 36.9% (axis 1: 20.0%; axis 2: 16.9%) of the total variance of the
assemblage composition among streams. Differences among land use categories
were not significant for both axis 1 and axis 2 scores, although a good
discrimination of reference sites and a moderate discrimination of the group of high
impact sites can be observed in the PCA, with the exception of one medium
impact site, that clearly separates from all the others (Fig. 4.9-A). Analysis of the
species fit showed that Lepidostoma, Hygrobates, Oligochaeta and Nematoda had
the highest contributions in defining macroinvertebrate community ordination. Axis
1 scores significantly correlated with SpeCond (r=0.842, p=0.000), SO4 (r=0.770,
p=0.007), SolP (r=0.648, p=0.038) and NO3NO2 (r=0.927, p=0.000). Axis 2 scores
significantly correlated with SpeCond (r=0.636, p=0.043) and SO4 (r=0.648,
p=0.034).

Non-functional approach to community gradients
97
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
Sim
Div
0
0.5
1
1.5
2
2.5
3
ref med high
ShanD
iv
0.000
0.020
0.040
0.060
0.080
0.100
0.120
ref med high
RelA
bd
*
Fig. 4.8 - Mean values (± 1 standard error) of derived biological variables for Canadian reference (ref), medium impact (med) and high impact (high) field sites. Symbol (*) and different letters (a, b) denote significant differences at p&0.05. Refer to Table 4.1 for variable identification.

Chapter 4
98
Fig. 4.9 – Biplots of principal components analyses using macroinvertebrate taxa abundances at reference (ref), medium impact (med) and high impact (high) field sites in three biogeographic regions: A – Canada (first and second axes explain 20% and 17% of the variance respectively); B – Spain (first and second axes explain 31% and 14% of the variance respectively); C – Costa Rica (first and second axes explain 29% and 17% of the variance respectively). Stars connect sites to the corresponding land use category centroid and ellipses give and idea of the dispersion of the points in each category, with d indicating the scale of the plot. Mean values (± 1 standard error) of the transformed axes scores of each PCA are shown on the right. Different letters (a, b) denote significant differences at p&0.05.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
2nd a
xis
score
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
1st
axis
score
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
1st
axis
score
b
a
ab
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
2nd a
xis
score
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
1st
axis
score
b
a
ab
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
2nd a
xis
score
b
a
ab
A d = 5
high
med
ref
d = 5
high
med
ref
B
C d = 5
high
med
ref

Non-functional approach to community gradients
99
In Spain (Fig. 4.10), medium sites had significantly higher TxRich when
compared with reference sites (ANOVA, F=5.655, p=0.035). In reference sites, E
was significantly lower (Kruskal-Wallis, H=6.144, p=0.017) but PercEPT and
RelAbd were significantly higher (ANOVAs, PercEPT F=6.760, p=0.023; RelAbd
F=5.772, p=0.033). Trends for decreased values along the impact gradient were
observed for PercChi, PielEve and I-BMWP. A trend for increasing values along
the impact gradient was observed for P, PercE, FBI, PercDTx, PercOli and I-
ASPT. FBI for reference sites and medium impact sites indicated good water
quality and the values for high impact sites indicated fair water quality (Mandaville,
2002). As for the I-ASPT, its values indicated clean waters and I-BMWP (Alba-
Tercedor, 1996) indicated not contaminated waters for all land use categories.
PercDTx was significantly correlated with SO4 (r=0.636, p=0.043), TotP
(r=0.742, p=0.011) and SolP (r=0.693, p=0.022) and SimDiv was significantly
correlated with TotP (r=-0.744, p=0.011) and SolP (r=-0.699, p=0.022). None of
the remaining single metrics and indices was significantly correlated with the
variables that better define the Spanish impact gradient.
The first two axis of the PCA done on the abundance data explained 44.8%
(axis 1: 30.7%; axis 2: 14.1%) of the total variance of the assemblage composition
among streams. Differences among land use categories were significant for axis 1
scores (ANOVA, F=5.894, p=0.032) and the groups of sites could be moderately
discriminated in the PCA, although the high impact sites appear closer to
reference sites than to medium impact sites (Fig. 4.9-B). Analysis of the species fit
showed that Sericostoma, Oulimnius and Atherix had the highest contributions in
defining macroinvertebrate community ordination, mainly differentiating medium
impact sites. Axis 1 scores did not significantly correlate with the variables that
better define the impact gradient. Axis 2 scores significantly correlated with
NO3NO2 (r=0.673, p=0.029)
In Costa Rica (Fig. 4.11), PielEve was significantly higher in medium impact
sites when compared to high impact sites (ANOVA, F=6.528, p=0.031). TotAbd
was significantly lower in medium impact sites (ANOVA, F=5.990, p=0.037). The

Chapter 4
100
metrics and indices that exhibited decreasing trends from reference sites to
impacted sites were TxRich, E, Tri, EPT, PercEPT, SimDiv, ShanDiv, CR-BMWP
0
1
2
3
4
5
6
ref med high
FBI
Fig. 4.10 - Mean values (± 1 standard error) of derived biological variables for Spanish reference (ref), medium impact (med) and high impact (high) field sites. Symbol (*) and different letters (a, b) denote significant differences at p&0.05. Refer to Table 4.1 for variable identification.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
ref med high
Pie
lEve
0.000
0.010
0.020
0.030
0.040
0.050
0.060
ref med high
RelA
bd
a
b
ab

Non-functional approach to community gradients
101
and CR-ASPT. Increasing trends along the gradient were observed for FBI,
PercDTx, PercChi and PercOli. FBI for reference sites was indicative of fairly poor
water quality, the values for medium impact sites indicated fair water quality and
the values for high impact sites indicated poor water quality. As for the CR-ASPT,
reference sites had values that indicate doubtful quality, medium impact sites
classified as probably moderately polluted and high impact sites as probably
severely polluted. CR-BMWP indicated high impact stream waters were
contaminated, medium impact streams were very contaminated and reference
streams were acceptable (Ministerio del Ambiente y Energia, 2006).
PielEve was significantly correlated with TotP (r=-0.833, p=0.002) and SolP
(r=-0.720, p=0.025). TotAbd was significantly correlated with SpeCond (r=0.750,
p=0.016) and TotP (r=0.767, p=0.012). From the remaining single metrics and
indices, only TxRich, FBI, CR-BMWP, CR-ASPT and PercDTx significantly
correlated with at least one of the variables that better define the impact gradient.
The first two axis of the PCA done on the abundance data explained 45.7%
(axis 1: 28.8%; axis 2: 16.9%) of the total variance of the assemblage composition
among Costa Rican streams. Differences among land use categories were
significant for axis 1 scores (Kruskal-Wallis, H=6.144, p=0.014) and axis 2 scores
(Kruskal-Wallis, H=5.800, p=0.030). The groups of sites belonging to each land
use category can be discriminated with no overlapping in the PCA (Fig. 4.9-C),
although one of the reference sites appears to highly separate from all the others.
Analysis of the species fit showed that Lampyridae, Contulma and Tipula had the
highest contributions in separating the macroinvertebrate communities of high
impact and medium impact sites. Axis 1 scores significantly correlated with
SpeCond (r=0.817, p=0.004), Embedd (r=0.740, p=0.020) and SO4 (r=0.783,
p=0.009). Axis 2 scores significantly correlated with SpeCond (r=-0.700, p=0.030),
SO4 (r=-0.750, p=0.016), CanoDens (r=-0.683, p=0.036) and TotP (r=-0.783,
p=0.009).

Chapter 4
102
Fig. 4.11 - Mean values (± 1 standard error) of derived biological variables for Costa Rican reference (ref), medium impact (med) and high impact (high) field sites. Symbol (*) and different letters (a, b) denote significant differences at p&0.05. Refer to Table 4.1 for variable identification.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
ref med high
Sim
Div
0
0.5
1
1.5
2
2.5
ref med high
ShanD
iv
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
Pie
lEve
a
b
ab
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
ref med high
Tri (
No.
taxa)

Non-functional approach to community gradients
103
4.4. Discussion
An attempt was made to select catchments in settings that were similar
except for their degree of agricultural land use. By assessing how comparable the
sites were if land use change was not considered inside each ecoregion, it was
shown that the catchments were indistinguishable in variables that do not respond
to land use, with the exception that medium impact Costa Rican catchments were
located lower in elevation than reference catchments. Analysis of only the medium
and reference sites showed no significant relationships between elevation and
most of the environmental and biological variables. Only Embedd significantly
correlated with Elevation, therefore responses of biological variables attributed to
differences in sediment input have to be interpreted with caution.
The three land use gradients studied combined mostly features of nutrient
input and sedimentation intensities, as expected when considering the agricultural
practices used in row-crop agriculture (e.g. ploughing, harrowing and rolling, that
cause frequent soil disturbances; the chemical inputs due to fertilizer application;
irrigation practices that alter stream hydrology; Richter et al., 1996; Randall &
Mulla, 2001; Fiener & Auerswald, 2007).
In Canada, more intensely farmed sites had, on average, higher nutrient
(phosphorus and nitrogen) and sulphate concentrations and higher deposition of
fine particles than lower impact sites. Soil erosion is a serious problem in the
potato-growing regions of Atlantic Canada either by water or tillage (Tiessen et al.,
2007), with consequences for the water chemistry of nearby streams (such as
sedimentation and nutrient leaching). In Spain, least impacted sites had, on
average, lower sulphate and nutrient (phosphorus) concentrations than higher
impact sites (fine sediment input also tended to increase). These patterns match
the literature on the effects of agricultural land use, particularly row-crop
agriculture, on environmental variables (Richards et al., 1996; Riley et al., 2003;
Davenport et al., 2005). Some authors suggest that conversion of forest areas to
agriculture can have significant influences on stream temperatures and light
penetration (with consequences on primary productivity) due to natural riparian

Chapter 4
104
vegetation removal (Allan, 2004), though no such effect on environmental
variables could be detected from the present study design.
It should be noted that Spanish reference sites may have been affected by a
non-identified source of ammonia, since this form of nitrogen appears with
elevated concentrations when compared to medium impact sites. This fact or
others associated with it, although not altering the overall impact (physico-
chemical) gradient, could have altered the biological gradient, as responses of the
macroinvertebrate community do not discriminate reference from high impact sites,
as initially predicted.
The Costa Rican gradient exhibited a pattern marked mostly by non
significant differences among reference and high impact sites in terms of impact-
associated variables, with the exception of ammonia concentration (although a
separation without overlaps of the land use categories was obtained in the PCA),
and by an inverted pattern from reference to medium impact sites when compared
with the expected trends (higher phosphorus, sulphate and sediment levels were
detected in the references, instead of the opposite, only observed for nitrogen
concentrations). An analysis of the remaining non-biological variables highlighted
the fact that the chosen reference sites exhibited very particular characteristics
(mostly related to bedrock geology and altitude) that might mask the detection of
human-associated effects. Reference streams, although draining catchments with
an average 97% of vegetation cover (protected by a National Park), are located in
higher elevations (as mentioned before) and had high non-agricultural inputs of
sediment and sulphates. The volcanic nature of the area (Alvarado et al., 2006)
and the steeper slopes usually associated with higher elevations – and frequently
correlated with higher topsoil erosion and sediment loads (Krishnaswamy et al.,
2001) – might have contributed to these sediment and sulphate loads observed in
reference sites. Other water quality studies have found this same pattern for this
area (Osorio, 1995) and studies in this and other areas in Costa Rica also highlight
that streams under the influence of volcanic processes are more soluble-rich
(including higher sulphate, magnesium and even phosphorus concentrations) than

Non-functional approach to community gradients
105
co-occurring (soluble-poor) streams that do not receive volcanic inputs (Pringle,
1991; Marchamalo, 2004).
Nevertheless, an impact increase from medium to high impact sites could still
be detected by analyzing environmental variables. High impact streams, while with
significantly higher percentages of their watersheds devoted to agriculture, had
higher nutrient and sulphate concentrations and higher inputs of sediment, when
compared to medium impact streams. These increases could be expected in areas
under the effects of row-crop agricultural practices, particularly in tropical
watersheds, where high rainfall erosivity is associated with land uses that provide
inadequate soil protection (Krishnaswamy et al., 2001).
With the above-mentioned exceptions, the complex physico-chemical
gradients observed in Canada, Spain and Costa Rica tend to support the
classification of sites which was defined a priori based on the observed degree of
agricultural impact. This indicates the presence of impact gradients that could be
used to compare with any observed biological gradients. Having confirmed the
existence of impact variable gradients associated with land use changes, it is
important to evaluate if and how the biological components of stream ecosystems
respond to these impacts, which is a fundamental objective of most biomonitoring
programs (Lücke & Johnson, 2008). The correlative analysis of the relations
between a number of single-metrics and indices commonly used in biomonitoring
programs and a number of impact variables revealed that only some biological
metrics/indices can give impact related information, and it was confirmed that
appropriate separation of sites according to levels of impact can be given by these
biological variables.
In Canada, richness and tolerance measures were able to separate heavily
impacted sites, and showed significant relationships with the variables that better
defined the impact gradient, namely those associated with sediment, sulphate and
nutrient inputs. High impact sites had, on average, lower taxa variety and less taxa
belonging to the order Ephemeroptera, but higher relative abundance, percentage
of Oligochaeta and a higher FBI. The trends for increased relative abundance

Chapter 4
106
(relative contribution of individuals to the total fauna) and decreased taxa and
Ephemeroptera richness with decreasing water quality are in agreement with the
expected for these measures (Barbour et al., 1999). For relative abundance, it is
predicted that a healthy and stable assemblage will be relatively consistent in its
proportional representation, though individual abundances may vary in magnitude
(Barbour et al., 1999). Overall richness indicates the health of the community
through its! diversity, increasing with increases of habitat diversity, suitability and
water quality (Barbour et al., 1999). Ephemeropterans are an insect group
generally considered to be sensitive to pollution (along with Plecopterans and
Trichopterans), and their diversity should therefore increase with increasing water
quality (Barbour et al., 1999). On the other hand, Oligochaetes are generally
considered as pollution tolerant (supported by these results), although there are
come caveats concerning this generalization for all types of chemical pollution
(Chapman, 2001).
The FBI appropriately classified the streams according to the levels of
agricultural impact, but both the NA-ASPT and the NA-BMWP performed poorly in
differentiating the three levels of impact. The FBI works as a low-cost and rapid
summary of multiple tolerances for the benthic arthropod community, and the
results obtained in Canada are in agreement with the expected increases in family
tolerances with high levels of impact (Mandaville, 2002). Consistency of these FBI
results with studies examining forested versus agricultural stream sites of Atlantic
Canada has been noted (Brisbois et al., 2008), but the authors did not find taxa
richness to be a useful indicator.
Abundance data (macroinvertebrate community variation) did not completely
differentiate all the impact categories, although there was a good separation of
reference streams and a moderate separation of the other two categories (with the
exception of one site). The separation of reference sites was better explained by
Lepidostoma and Hygrobates abundances and high impact sites were separated
by the abundances of oligochaetes (generally considered as tolerant to pollution)
and nematods. Lepidostoma has been categorized as sensitive to phosphorus and
sulphate concentrations (Yuan, 2004), impact variables that have been in fact

Non-functional approach to community gradients
107
higher in Canadian high impact sites when compared to references. Hygrobates
(Smith et al., 2001) and Nematoda (Poinar Jr., 2001) have also been described as
a potentially good indicators of environmental quality, particularly chemical
pollution or physical disturbance.
In Spain, several structural metrics were able to separate reference from
medium impact sites, but no correlations with the impact variables were significant.
Reference sites (lower sulphate and phosphate concentrations but higher levels of
ammonia) had lower taxa richness and less taxa belonging to the Ephemeroptera
order but a higher percentage of EPT and higher relative abundance, when
compared to medium impact sites. Lower taxa (and Ephemeroptera) richness and
higher relative abundance in a macroinvertebrate community might happen as a
response to increasing perturbation (Barbour et al., 1999). Unexpected nitrogen
inputs to our a priori selected reference sites could potentially have worked as this
perturbation, masking the overall separation of sites according to the increasing
impact categories (correspondent to increasing inputs of phosphates, sulphates
and sediment, detected in the impact gradient analysis). A confirmation of this
hypothesis could not be obtained with the present study design. By verifying the
correlation matrices of biological variables and environmental variables not directly
used for impact gradient assessment, it can be detected that the concentration of
organic nitrogen negatively and significantly correlated with taxa richness.
Generally, the soluble, inorganic forms of nitrogen are the most biologically
available; particulate and dissolved organic fractions of nitrogen are generally not
immediately available (USEPA, 2000). However, they can be converted to
ammonium by bacteria and fungi. This supports the hypothesis of unexpected
nitrogen excess in reference sites having had an negative impact on the biological
components of these streams, through deregulations of the nutrient cycling
patterns (Vanni, 2002). The source of this nitrogen input remains obscure,
although it could be related to cattle farming known to exist in other areas of the
Anllóns basin (Consellería do Medio Rural, 2004).

Chapter 4
108
As for classification-based indices (FBI, I-ASPT and I-BMWP), these failed to
distinguish water quality differences among categories of impact. Abundance data
(macroinvertebrate community variation) was not able to completely differentiate
the impact categories, although there was a good separation of reference and high
impact streams from medium impact streams, supporting once more the notion
that reference streams might have been affected by unpredicted perturbations
(possibly nitrogen contamination) that altered the macroinvertebrate community
patterns. The separation of medium impact sites was better explained by the
abundances of Sericostoma – categorized as a sensitive taxon in several
biomonitoring methods (e.g. Alba-Tercedor, 1996) -, Oulimnius – mentioned as
sensitive to phosphorus and sulphate concentrations (Yuan, 2004) - and Atherix –
categorized as relatively sensitive to nutrient inputs (Carlisle et al., 2007). Other
studies of northern Spain streams (González et al., 2003) also found responses of
taxa richness, Oulimnius and Atherix abundances along nutrient input gradients.
In Costa Rica, two structural measures were able to discriminate medium
impact from high impact sites, evidencing significant relationships with variables
associated with sediment and phosphate inputs. When compared to medium
impact sites, high impact streams had, on average, higher taxa abundances and
the organisms were less evenly distributed along the taxa. Evenness is a
component of diversity expected to be high in more healthy communities, with the
effects of stress (sediment and phosphate inputs in this study) often reducing
numbers of intermediate taxa and increasing numbers of one or two (more
tolerant) taxa (Reynoldson et al., 2003). In another study of Costa Rican streams
(Pringle & Ramirez, 1998), it was observed that benthic invertebrate communities
were also dominated by relatively fewer taxa in sites disturbed by agricultural
activities, although lower densities and taxa richness were also observed (these
were not observed in the present study). One study using impact sites common
with the ones of the present study has also detected important decreases in taxa
evenness, with correlations with phosphate concentrations, and increases in
abundances, as well as similar trends for other metrics (Marchamalo, 2004). The

Non-functional approach to community gradients
109
variability and unpredictability of responses to impact of the measures of overall
abundances alone have been discussed (Barbour et al., 1992). The use of
combined information of more than one measure can give a better picture of the
biological change. In the present study (as well as in Marchamalo!s study), it
seems that higher levels of land use change are related with significant increases
of macroinvertebrate abundances with strong dominance of certain taxa
(translating in lower evenness values).
Classification-based indices (CR-ASPT and FBI) indicated reference sites as
fairly poor or doubtful in terms of water quality. This fact points towards the
hypothesis that (probably natural) stressors could be affecting the
macroinvertebrate communities of the a priori selected reference sites. High
impact sites were always correctly classified as having poor water quality. Overall
trends of the remaining metrics also support the probable poorer water quality of
high impact sites. Abundance data evidenced the separation of categories with no
overlaps (Lampyridae, Contulma and Tipula had the highest contributions and
separated high impact sites). Tipula has been categorized as tolerant to
phosphorus concentrations (Yuan, 2004), that were higher in high impact sites
when compared to medium impact sites, possibly promoting their higher
abundances.
The distinction of reference sites, also detected previously while assessing
the impact gradient and the CR-ASPT and FBI water quality categories, was
highlighted by the taxa abundance gradient, but attention should given to one
particular reference sample (collected inside the Irazú National Park): the
macroinvertebrate assemblage responded to the particular natural characteristics
of this stream in ways that distinguish it from all other assemblages. Taxa found in
high abundances in this site have been referred as relatively tolerant to the percent
of fines (Carlisle et al., 2007), fact that would agree with a response to the
elevated percentage of embeddedness of this stream, due to potential volcanic
inputs. The idiosyncrasies of the Costa Rican reference sites have already been
discussed. A higher number of sampled reference streams would have helped
solving this particular problem, but the difficulties of finding real "reference

Chapter 4
110
conditions! in these heavily humanized scenarios have to be bared in mind, and
solutions have been highly discussed among researchers (Reynoldson et al.,
1997). One possible immediate solution for the problem of the Costa Rican study
region might be in focusing on medium impact sites (that have moderately
separated from high impact sites in both impact and biological gradients) as "lower
impact systems! for comparison with high impact sites.
Low stressor sensitivity or error prone biomonitoring approaches could impair
the main objective of protecting freshwater ecosystems, by promoting disputes
among those responsible for decision-making. Despite the existence of impact
gradients associated with agricultural land use, based on several impact-
associated variables, only some of the biological variables significantly covaried
with these impact variables, and discriminated the a priori defined land use
categories. Nevertheless, studies like the one presented, may give clues to identify
specific features that should be considered in the design of future biomonitoring
programs for each of the studied regions.
The biological variables that better discriminated land use categories were
different in each of the three ecoregions. Taxa richness and relative abundance
had consistent discriminatory power in Canada and Spain, but they did not prove
useful for the Costa Rican sites. A variable useful in one region might not highlight
changes in a different community gradient, due to, for example, natural stressors.
This problem could be partially solved with the use of higher numbers of reference
samples, which could highlight natural variability. But finding adequate references
is many times difficult (Nijboer et al., 2004). Another interesting option could be the
use of functional images of the communities from the different regions, instead of
mere structural/taxonomic measures. Structural metrics lack the ability of
assessing ecological functions (Bonada et al., 2006), and are therefore limited
predictors of changes in ecosystem function. Since the biological communities
along geographically constrained gradients of land use were different in terms of
structure (richness, composition metrics and diversity indices) and physiological
responses to pollution (tolerance metrics and biotic indices), it can be

Non-functional approach to community gradients
111
hypothesized that, even though the structure of the community adapts differently in
different areas, due to the different taxonomic compositions, the functional
response to stress will be similar, and more global biomonitoring approaches could
be developed.
One important objective of the present study was to identify parameters and
metrics that could be useful for classifying the impacts of agricultural activity on
stream health in three biogeographic areas. Although interesting in terms of data
exploration and detection of true (statistically significant) trends in stream ecology,
univariate analyses - such as the correlation analyses and ANOVAs performed -
can give limited insight when dealing with biological measures, due to subtle
biological trend masking (since there is a need to summarize the characteristics of
a site in a single value). Multivariate techniques that simultaneously compare sets
of environmental and biological variables can be used to complement univariate
comparisons for better detection and understanding of spatial trends in benthic
fauna (Norris & Georges, 1993), suggesting in addition causal relationships
between environmental and biological gradients (that could only be confirmed
through proper experimentation). This type of analysis of the data set will be dealt
with in the following chapter. Also, a functional enrichment of taxonomic
approaches used to detect land use impacts on aquatic systems will be explored,
with the introduction of species traits in the analysis.
4.5. References
Alba-Tercedor, J. (1996) Macroinvertebrados acuáticos y calidad de las aguas de
los ríos. IV Simposio del Agua en Andalucía, Almeria, 203-213.
Alba-Tercedor, J. & Sánchez-Ortega, A. (1988) Un método rápido y simple para
evaluar la calidad biológica de las aguas corrientes basado en el de
Hellawell (1978). Limnetica, 4, 51-56.

Chapter 4
112
Allan, J. D. (2004) Landscape and Riverscapes: The Influence of Land Use on
Stream Ecosystems. Annual Review of Ecology, Evolution, and
Systematics, 35 (1), 257-284.
Alvarado, G. E., Carr, M. J., Turrin, B. D., Swisher III, C. C., Schmincke, H. &
Hudnut, K. W. (2006) Recent volcanic history of lrazú volcano, Costa Rica:
Alternation and mixing of two magma batches, and pervasive mixing.
Geological Society of America, Special Paper 412, 259-276.
Barbour, M. T., Gerritsen, J., Snyder, B. & Stribling, J. (1999) Rapid
Bioassessment Protocols For Use in Streams and Wadeable Rivers:
Periphyton, Benthic Macroinvertebrates and Fish, Second Edition.
Washington, D.C., U.S. Environmental Protection Agency. 339 pp.
Barbour, M. T., Plafkin, J. L., Bradley, B. P., Graves, C. G. & Wisseman, R. W.
(1992) Evaluation of EPAs Rapid Bioassessment Benthic Metrics - Metric
Redundancy and Variability among Reference Stream Sites. Environmental
Toxicology and Chemistry, 11 (4), 437-449.
Bonada, N., Prat, N., Resh, V. H. & Statzner, B. (2006) Developments in Aquatic
Insect Biomonitoring: A Comparative Analysis of Recent Approaches.
Annual Review of Entomology, 51 (1), 495-523.
Brisbois, M. C., Jamieson, R., Gordon, R., Stratton, G. & Madani, A. (2008) Stream
ecosystem health in rural mixed land-use watersheds. Journal of
Environmental Engineering and Science, 7, 439-452.
Carlisle, D. M., Meador, M. R., Moulton, S. R. & Ruhl, P. M. (2007) Estimation and
application of indicator values for common macroinvertebrate genera and
families of the United States. Ecological Indicators, 7 (1), 22-33.
Chapman, P. M. (2001) Utility and relevance of aquatic oligochaetes in Ecological
Risk Assessment. Hydrobiologia, 463, 149-169.
Claret, C., Marmonier, P., Dole-Olivier, M. J., Des Chatelliers, M. C., Boulton, A. J.
& Castella, E. (1999) A functional classification of interstitial invertebrates:
supplementing measures of biodiversity using species traits and habitat
affinities. Archiv fuer Hydrobiologie, 145 (4), 385-403.

Non-functional approach to community gradients
113
Clarke, R. T., Furse, M. T., Gunn, R. J. M., Winder, J. M. & Wright, J. F. (2002)
Sampling variation in macroinvertebrate data and implications for river
quality indices. Freshwater Biology, 47, 1735-1751.
Consellería do Medio Rural (2004) Anuario de Estatística Agraria 2004. Xunta de
Galicia. 257 pp.
Davenport, J. R., Milburn, P. H., Rosen, C. J. & Thornton, R. E. (2005)
Environmental impacts of potato nutrient management. American Journal of
Potato Research. 82, 321-328.
Dray, S. & Dufour, A. B. (2007) The ade4 Package: Implementing the Duality
Diagram for Ecologists. Journal of Statistical Software, 22 (4), 1-20.
Fiener, P. & Auerswald, K. (2007) Rotation Effects of Potato, Maize, and Winter
Wheat on Soil Erosion by Water. Soil Science Society of America Journal,
71, 1919-1925.
González, J. M., Basaguren, A. & Pozo, J. (2003) Macroinvertebrate communities
along a third-order Iberian stream. Annals of Limnology, 39 (4), 287-296.
Hawkins, C. P., Norris, R. H., Gerritsen, J., Hughes, R. M., Jackson, S. K.,
Johnson, R. K. & Stevenson, R. J. (2000) Evaluation of the Use of
Landscape Classifications for the Prediction of Freshwater Biota: Synthesis
and Recommendations. Journal of the North American Benthological
Society, 19, 541-556.
Krishnaswamy, J., Richter, D. D., Halpin, P. N. & Hofmockel, M. S. (2001) Spatial
patterns of suspended sediment yields in a humid tropical watershed in
Costa Rica. Hydrological Processes, 15, 2237-2257.
Legendre, P. & Legendre, L. (1998) Numerical Ecology. Amsterdam, Elsevier. 870
pp.
Lücke, J. D. & Johnson, R. K. (2008) Detection of ecological change in stream
macroinvertebrate assemblages using single metric, multimetric or
multivariate approaches. Ecological Indicators, 9, 659-669.
Mandaville, S. M. (2002) Benthic Macroinvertebrates in Freshwaters: Taxa
Tolerance Values, Metrics, and Protocols. Soil & Water Conservation
Society of Metro Halifax. 128 pp.

Chapter 4
114
Marchamalo, M. (2004) Ordenación del territorio para la producción de servicios
ambientales hídricos. Aplicación a la cuenca del Río Birrís (Costa Rica).
Universidad Politecnica de Madrid, PhD thesis. 400 pp.
Ministerio del Ambiente y Energia (2006) Reglamento para la evaluación y
clasificación de la calidad de cuerpos de aguas superficiales. Ministerio del
Ambiente y Energía, Costa Rica, 14.585, 1-18.
Nijboer, R. C., Johnson, R. K., Verdonschot, P. F. M., Sommerhauser, M. &
Buffagni, A. (2004) Establishing reference conditions for European streams.
Hydrobiologia, 516, 91-105.
Norris, R. H. & Georges, A. (1993) Analysis and Interpretation of Benthic
Macroinvertebrate Surveys. In: D. M. Rosenberg & V. H. Resh (Ed.).
Freshwater Biomonitoring and Benthic Macroinvertebrates. New York,
Chapman & Hall, 234-286.
Osorio, J. G. V. (1995) Evaluación de la calidad del agua en la cuenca del río
Reventado, Cartago, Costa Rica, bajo el enfoque de indicadores de
sostenibilidad. CATIE, Turrialba, Costa Rica. MSc thesis. 158 pp.
Poinar Jr., G. O. (2001) Nematoda and Nematomorpha. In: J. Thorp & A. Covich
(Ed.). Ecology And Classification Of North American Freshwater
Invertebrates, Academic Press, 255-295.
Pringle, C. M. (1991) Geothermally Modified Waters Surface at La Selva Biological
Station, Costa-Rica - Volcanic Processes Introduce Chemical
Discontinuities into Lowland Tropical Streams. Biotropica, 23, 523-529.
Pringle, C. M. & Ramirez, A. (1998) Use of both benthic and drift sampling
techniques to assess tropical stream invertebrate communities along an
altitudinal gradient, Costa Rica. Freshwater Biology, 39, 359-372.
R Development Core Team (2010) R: A language and environment for statistical
computing. Vienna, Austria, R Foundation for Statistical Computing.
http://www.R-project.org.
Randall, G. W. & Mulla, D. J. (2001) Nitrate nitrogen in surface waters as
influenced by climatic conditions and agricultural practices. Journal of
Environmental Quality. 30, 337-344.

Non-functional approach to community gradients
115
Reynoldson, T. B., Logan, C., Pascoe, T. & Thompson, S. P. (2003) CABIN
(Canadian Aquatic Biomonitoring Network) Invertebrate Biomonitoring Field
and Laboratory Manual. National Water Research Institute - Environment
Canada. 47 pp.
Reynoldson, T. B., Norris, R. H., Resh, V. H., Day, K. E. & Rosenberg, D. M.
(1997) The reference condition: a comparison of multimetric and
multivariate approaches to assess water-quality impairment using benthic
macroinvertebrates. Journal of the North American Benthological Society,
16 (4), 833-852.
Richards, C., Johnson, L. B. & Host, G. E. (1996) Landscape-scale influences on
stream habitats and biota. Canadian Journal of Fisheries and Aquatic
Sciences, 53, 295-311.
Richter, B. D., Baumgartner, J. V., Powell, J. & Braun, D. P. (1996) A method for
assessing hydrologic alteration within ecosystems. Conservation Biology.
10, 1163-1174.
Riley, R., Townsend, C., Niyogi, D., Arbuckle, C. & Peacock, K. (2003) Headwater
stream response to grassland agricultural development in New Zealand.
New Zealand Journal of Marine and Freshwater Research, 37 (2), 389-403.
Smith, I. M., Cook, D. R. & Smith, B. P. (2001) Water Mites (Hydrachnida) And
Other Arachnids. In: James Thorp & Alan Covich (Ed.). Ecology And
Classification Of North American Freshwater Invertebrates, Academic
Press, 551-659.
Tiessen, K. H. D., Lobb, D. A., Mehuys, G. R. & Rees, H. W. (2007) Tillage erosion
within potato production in Atlantic Canada: II - Erosivity of primary and
secondary tillage operations. Soil & Tillage Research, 95 (1-2), 320-331.
Townsend, C. R. & Riley, R. H. (1999) Assessment of river health: accounting for
perturbation pathways in physical and ecological space. Freshwater
Biology, 41 (2), 393-405.
USEPA (2000) Nutrient Criteria Technical Guidance Manual - Rivers and Streams.
EPA-822-B-00-002. Washington. 253 pp.

Chapter 4
116
Vanni, M. J. (2002) Nutrient Cycling by Animals in Freshwater Ecosystems.
Annual Review of Ecology and Systematics, 33 (1), 341-370.
Weigel, B. M., Wang, L., Rasmussen, P. W., Butcher, J. T., Stewart, P. M., Simon,
T. P. & Wiley, M. J. (2003) Relative influence of variables at multiple spatial
scales on stream macroinvertebrates in the Northern Lakes and Forest
ecoregion, U.S.A. Freshwater Biology, 48 (8), 1440-1461.
Yuan, L. L. (2004) Assigning macroinvertebrate tolerance classifications using
generalised additive models. Freshwater Biology, 49 (5), 662-677.

Chapter 5. Macroinvertebrate traits in watersheds from different
biogeographic regions and their application in biological
monitoring


Macroinvertebrate traits and biomonitoring
119
5.1. Introduction
When freshwater macroinvertebrate community composition is described
only by the abundances of the different species present, the detection of changes
due to anthropogenic stressors can be done only by looking at species loss or
change in abundance (Dolédec et al., 2006). Describing those communities using
taxa attributes or traits - characteristics that reflect species adaptation to the
environment -, and looking at changes in trait category frequencies, may increase
the discrimination of impact levels (e.g. Charvet et al., 1998; Dolédec et al., 1999)
and give indications of shifts in community function (McGill et al., 2006). Another
advantage of the trait approach is its potential for wider geographic application
(Charvet et al., 2000; Gayraud et al., 2003), since it is known that biomonitoring
tools relying only on taxonomic measures risk being limited to the spatial scale for
which they are developed (Dolédec et al., 1999). The !functional image" of
communities could potentially be used as a generic tool for the ecologically
oriented management of streams and other ecosystems (Charvet et al., 2000).
Community ecology is often applied across highly variable temporal and
spatial scales, yet generalizing phenomena across biogeographic regions remains
challenging. Relating species traits to habitat characteristics in the light of habitat
templet theory (Southwood, 1977; Southwood, 1988; Poff & Ward, 1990;
Townsend & Hildrew, 1994) can give biogeographically unconstrained models for
the study of the interactions of environment and biota.
According to Poff"s theoretical model of landscape filters (Poff, 1997), it is
important to integrate different spatial and temporal scales hierarchically when
developing predictive models of community composition. Geographical constraints
reflecting climatic, topographic and geological gradients, in addition to human
impact levels, can influence the biological trait structure of the communities. By
comparing the same stressor gradient in very different geographical scenarios, the
opportunity to test if broad scale comparisons and biomonitoring methodology
extrapolations are actually possible arises.

Chapter 5
120
This study examines the possibility of developing globally-applicable
biomonitoring tools for freshwaters of different continents, by using traits not
simply to replace taxonomic descriptions but as a functional enrichment of such
approaches. As the backdrop for trait-based community analysis, the habitat
templet concept (e.g. Townsend et al., 1997), presents the hypothesis that
present-day habitat conditions are matched by present-day traits in the organisms.
Here, an application of the habitat templet approach focuses on the discrimination
of environmental impacts by including measures of the multiple stressors arising
from intensive agricultural activities. One can predict (Southwood, 1977;
Southwood, 1988; Townsend & Hildrew, 1994) that the main traits involved in the
differences in macroinvertebrate communities under the influence of agricultural
stressors are those that favor resistance and/or resilience (e.g. multivoltinism,
asexual reproduction, short life cycles, small size, resistance forms), being
predominant in highly impacted environments. Traits negatively related to
environmental harshness (e.g. semi-voltinism, long life cycles, larger sizes,
unprotected eggs) should be predominant in least impacted sites. Intermediate
impact streams should exhibit a mixture of the mentioned characteristics. Although
different biogeographic regions exhibit different characteristics in terms of geology,
vegetation, land use and topography, it is expected that the functional response of
the biological communities to similar impact gradients will be convergent, that is to
say, the trait profiles of very different taxonomic communities are expected to shift
in similar ways when exposed to similar anthropogenic disturbances.
In order to test the potential of using traits for geographically-unconstrained
biomonitoring of agricultural streams, data was collected using similar
methodologies across different biogeographic regions (cf. Chapter 3), removing
the problem of inconsistent observation (i.e. sampling) methods (e.g. Gayraud et
al., 2003; Statzner et al., 2004 compile data collected in diverse ways into a unique
data set).

Macroinvertebrate traits and biomonitoring
121
5.2. Methods of data analysis
For all statistical tests the significance level used was p=0.05. One-way
Analysis of Variance (ANOVA) or its nonparametric equivalent, the Kruskal-Wallis
rank-sum test (when normality or homoscedasticity was not achieved) were used
for univariate statistical testing. When either ANOVA or Kruskal-Wallis yielded
significant differences, Tukey/Dunn"s tests were performed for post-hoc
comparisons. All multivariate analyses were run with the ade4 library version 1.4-
14 (Dray & Dufour, 2007) implemented in the R freeware version 2.11.1 (R
Development Core Team, 2010). The remaining analyses were run with SigmaStat
version 3.5 (Systat Software, Inc., 2006).
5.2.1. Trait gradients
Fuzzy principal components analysis (FPCA; the linear equivalent to the
fuzzy correspondence analysis described by Chevenet et al., 1994) of trait
category abundance data was used as a multivariate tool for two purposes: (i) to
obtain synthetic multivariate scores for sites based on trait composition, and (ii) to
describe functional variation in the macroinvertebrate community. PCA was
chosen over correspondence analysis because it discriminates sites based on
absolute rather than relative changes (Van den Brink et al., 2003). FPCA in
particular provides a joint ordination of sites and traits, considering the latter as
sets of fuzzy coded categories. In order to compare among analyses, axis
ordination scores were standardized and scaled to values between 0 and 1 using
the formula:
yi=(xi-min)/(max-min) (Equation 1)
where xi is the value of the score for site i, min is the minimum site score and max
is the maximum site score.
To quantify the relationship (co-variation) between the trait information and
the environmental variables that better define the impact gradients, correlations
between pairs of arrays (FPCA axis scores and environmental measures) were

Chapter 5
122
calculated (Spearman correlation coefficient).
5.2.2. Discrimination of land use categories (taxa and traits)
One-way ANOVA or Kruskal-Wallis rank-sum tests were used to compare
FPCA site scores (axis 1 and 2) among the three land use categories, in order to
assess their discriminatory power. The degree of discrimination among land use
categories by structural (i.e. taxon-abundance matrices) and functional (trait-
abundance matrices) community information was assessed using between-class
analysis (with land use category as a categorical variable), a type of PCA with
respect to instrumental variables (Lebreton et al., 1991). This analysis ordinates
sites under the constraint that the ordination maximally separates sites in the
various classes of the categorical variable (in this case, land use categories). The
significance of the overall difference between categories (between-class inertia)
was tested against simulated values obtained after 10000 permutations of the
rows of the taxonomic and trait-composition matrices (Romesburg, 1985).
Explained variance of the between-class PCA (taxa) and FPCA (traits) was also
used for direct comparisons of discrimination degrees of the taxonomic and trait
approaches. However, since the number of variables differed between taxonomic-
and trait-composition tables (166 taxa and 72 trait categories) and given that the
amount of variability within a table is related to the number of variables, explained
between-category variance was recalculated from simulated taxonomic-
composition matrices with 72 randomly selected taxa.
5.2.3. Co-structure of environmental and biological variables
The relationships between the biological measures (taxa and traits) and the
set of environmental variables expected to respond to land use were analyzed
through co-inertia analyses (Dray et al., 2003), in order to better understand the
drivers of biological change in the studied streams. This ordination method aims to
study the common structure of pairs of data tables by quantifying the variability in
taxon/trait composition explained by the environmental gradient. However, when

Macroinvertebrate traits and biomonitoring
123
compared with other constrained analyses, co-inertia analysis is less sensitive to
the inclusion of relatively high numbers of collinear variables, which is the case
when many biotic and environmental variables are measured in only a few sites
(Dolédec & Chessel, 1994). The RV-coefficient, a multivariate counterpart of
univariate correlation coefficients (Robert & Escoufier, 1976), gave a measure of
correlation between the pairs of matrices (taxon-environmental and trait-
environmental) and the resulting observed total covariance was thus an
expression of the match of the pairs of matrices. The statistical significance of this
co-structure between the matrices was tested against simulated match values
obtained after 10000 permutations of the matrices" rows (Romesburg, 1985).
5.2.4. Interregional comparisons
PCA performed on taxa abundance and FPCA performed on trait category
abundance, considering all three regions, were used to summarize
macroinvertebrate community variation, and to visualize the discrimination of both
land use categories and regions. The degrees of discrimination among land use
categories and regions by structural (taxa abundances) and functional (trait
abundances) community information were assessed using between-class
analyses of data from the three regions combined (one between-class analysis
with land use category as the categorical variable and a second one with region as
the categorical variable). The significance of the overall differences between
categories/regions was tested against simulated values obtained after 10000
permutations of the rows of the taxonomic and trait-composition matrices
(Romesburg, 1985). Once more, since the number of variables differed between
taxonomic- and trait-composition tables, for direct comparisons between the two
approaches the total explained between-class variance could not be used directly.
The analyses were repeated with 100 simulated taxonomic-composition matrices
with 72 randomly selected taxa, in order to obtain a frequency distribution of
between-class variance to compare with the observed values.

Chapter 5
124
5.3. Results
5.3.1. Discrimination of land use categories
In Canada, the first 2 axes of the FPCA performed on trait abundances
explained 85.3% of the total variance, (75.0% and 10.3% for axes 1 and 2
respectively, Fig. 5.1). The discrimination of land use categories along the factorial
axes can be depicted from the visual analysis of the plot. High impact sites had
higher variability and were more distributed along the axes than references or
medium impact sites. Focusing on each of the two first axes, sample scores did
not significantly discriminate land-use categories (Fig. 5.1), as had already
happened with the taxa composition PCA scores. The first axis sample scores
significantly correlated with variables that defined the impact gradient: SpeCond
(r=0.770, p=0.007), SO4 (r=0.697, p=0.022) and NO3NO2 (r=0.648, p=0.038). As
for the second axis scores, significant correlations were detected with SO4
(r=0.600, p=0.050) and NO3NO2 (r=0.685, p=0.025).
Fig. 5.1 – Biplot of fuzzy principal components analysis of macroinvertebrate trait category
abundances at Canadian reference (ref), medium impact (med) and high impact (high)
field sites. First (horizontal) and second (vertical) axes explain 75% and 10% of the
variance respectively. Stars connect sites to the corresponding land use category centroid
and ellipses give and idea of the dispersion of the points in each category, with d
indicating the scale of the plot. Mean values (± 1 standard error) of the transformed axes
scores of the PCA are shown on the right.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
1st
axis
score
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0,.9
1
ref med high
2nd a
xis
score
d = 5
high
med
ref

Macroinvertebrate traits and biomonitoring
125
The between-class analyses showed that overall between land use
categories variance was significant for both taxa (between-class variance
explained was 26.7%; p=0.024) and trait abundances (between-class variance
explained was 30.5%; p=0.045). The results of simulated between-category
variances with 72 randomly selected taxa (identical to the number of trait
categories) statistically confirmed (p=0.01; Fig. 5.2) that the degree of
discrimination of land use categories was higher for the trait approach when
compared with the taxonomic approach.
Analysis of the trait category scores, considering their relative influence on
the macroinvertebrate community ordination, showed that the typical organisms of
high impact sites were large (>40 mm), multivoltine, asexual, burrowers and
deposit feeders, with preferences for detritus as microhabitat and quiet water
flows, and using cocoons as resistance form. Medium impact sites were separated
by small (2.5-5 mm), streamlined, spiracle and/or plastron breathers, swimmers,
parasites and organisms preferring algae as their microhabitat. In references,
macroinvertebrates were mostly ovoviviparous and offering parental care of eggs
or producing free eggs, crawlers, predators, medium sized (10-40 mm), with
preferences for rocks, boulders or large woody debris as microhabitats.
Fig. 5.2 – Frequency distribution of 100
simulated values (for 72 randomly selected
taxa) and observed values (dashed lines) of
between land use category variances for taxa
and trait composition of macroinvertebrate
communities at Canadian field sites. To
improve visualization, x-axis scale was focused
(origin is not 0).

Chapter 5
126
In Spain, the first two axes of the FPCA explained 59.9% of the variability
(axis 1: 35.6%; axis 2: 24.3%; Fig. 5.3), therefore they were maintained for further
analyses. There is a good discrimination of land use categories in the plot,
although reference sites appear to distribute with higher variability than medium
and high impact sites and are closer to high impact sites than medium impact
sites. As for each of the two first axes viewed separately, axis 2 scores
significantly discriminated reference sites (ANOVA, F=13.416, p=0.004; Fig. 5.3).
These scores significantly correlated with SO4 (r=0.673, p=0.05). Axis 1 scores
did not significantly correlate with the variables that defined the impact gradient.
The between-class analysis showed that overall between land use categories
variance was significant when considering trait abundances (between-class
variance explained was 38.3%; p=0.013). The same was true for taxa abundances
(between-class variance explained was 30.9%; p=0.013). The variance values can
only be compared among approaches through the results of simulated between-
category variances with 72 randomly selected taxa (identical to the number of trait
Fig. 5.3 – Biplot of fuzzy principal components analysis of macroinvertebrate trait category
abundances at Spanish reference (ref), medium impact (med) and high impact (high) field
sites. First (horizontal) and second (vertical) axes explain 36% and 24% of the variance
respectively. Stars connect sites to the corresponding land use category centroid and
ellipses give and idea of the dispersion of the points in each category, with d indicating the
scale of the plot. Mean values (± 1 standard error) of the transformed axes scores of the
PCA are shown on the right. Symbol (*) denotes significant difference at p#0.05.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
1st
axis
score
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
2nd a
xis
score
*
d = 5
high
ref
med
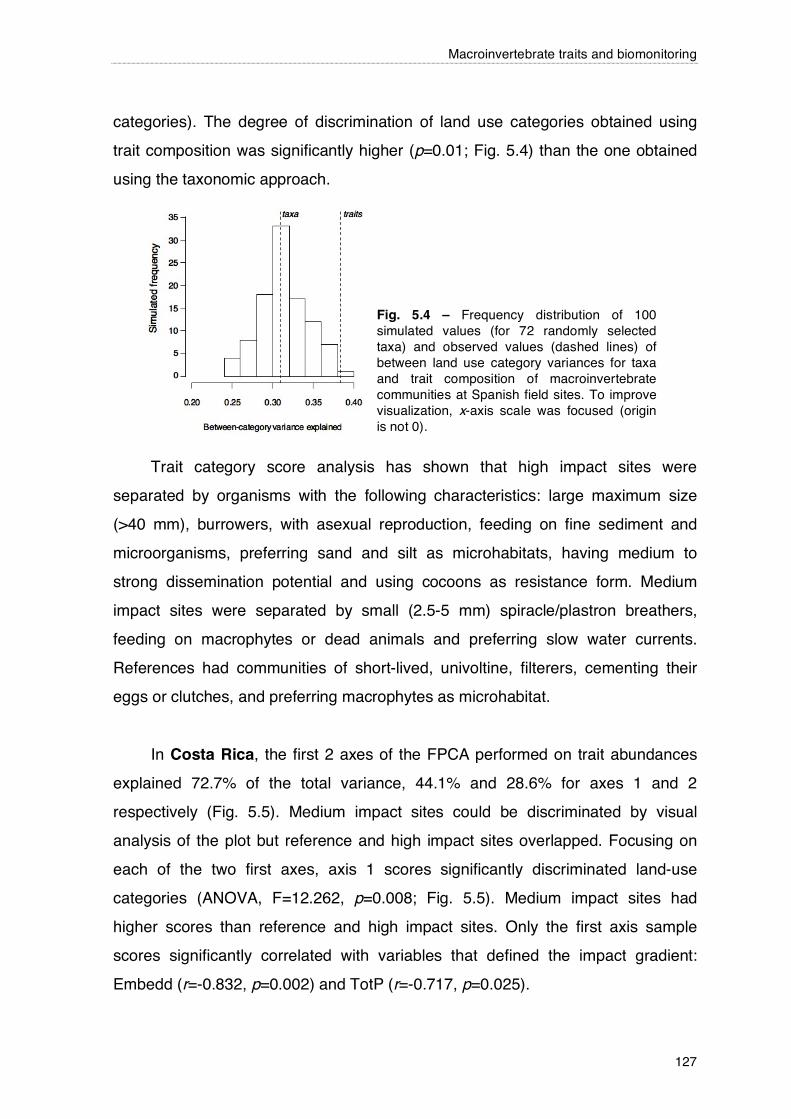
Macroinvertebrate traits and biomonitoring
127
categories). The degree of discrimination of land use categories obtained using
trait composition was significantly higher (p=0.01; Fig. 5.4) than the one obtained
using the taxonomic approach.
Trait category score analysis has shown that high impact sites were
separated by organisms with the following characteristics: large maximum size
(>40 mm), burrowers, with asexual reproduction, feeding on fine sediment and
microorganisms, preferring sand and silt as microhabitats, having medium to
strong dissemination potential and using cocoons as resistance form. Medium
impact sites were separated by small (2.5-5 mm) spiracle/plastron breathers,
feeding on macrophytes or dead animals and preferring slow water currents.
References had communities of short-lived, univoltine, filterers, cementing their
eggs or clutches, and preferring macrophytes as microhabitat.
In Costa Rica, the first 2 axes of the FPCA performed on trait abundances
explained 72.7% of the total variance, 44.1% and 28.6% for axes 1 and 2
respectively (Fig. 5.5). Medium impact sites could be discriminated by visual
analysis of the plot but reference and high impact sites overlapped. Focusing on
each of the two first axes, axis 1 scores significantly discriminated land-use
categories (ANOVA, F=12.262, p=0.008; Fig. 5.5). Medium impact sites had
higher scores than reference and high impact sites. Only the first axis sample
scores significantly correlated with variables that defined the impact gradient:
Embedd (r=-0.832, p=0.002) and TotP (r=-0.717, p=0.025).
Fig. 5.4 – Frequency distribution of 100
simulated values (for 72 randomly selected
taxa) and observed values (dashed lines) of
between land use category variances for taxa
and trait composition of macroinvertebrate
communities at Spanish field sites. To improve
visualization, x-axis scale was focused (origin
is not 0).

Chapter 5
128
Between-category analyses showed that overall between land use categories
variance was significant for both taxa (between-class variance explained: 35.1%;
p=0.001) and trait abundances (between-class variance explained: 48.0%;
p=0.009). The results of simulated between-category variances with 72 randomly
selected taxa statistically confirmed (p<0.01; Fig. 5.6) that the degree of
discrimination of land use categories was higher for the trait approach when
compared with the taxonomic approach.
Fig. 5.6 – Frequency distribution of 100
simulated values (for 72 randomly selected
taxa) and observed values (dashed lines) of
between land use category variances for taxa
and trait composition of macroinvertebrate
communities at Costa Rican field sites. To
improve visualization, x-axis scale was focused
(origin is not 0).
Fig. 5.5 – Biplot of fuzzy principal components analysis of macroinvertebrate trait category
abundances at Costa Rican reference (ref), medium impact (med) and high impact (high)
field sites. First (horizontal) and second (vertical) axes explain 44% and 29% of the
variance respectively. Stars connect sites to the corresponding land use category centroid
and ellipses give and idea of the dispersion of the points in each category, with d
indicating the scale of the plot. Mean values (± 1 standard error) of the transformed axes
scores of the PCA are shown on the right. Symbol (*) denotes significant difference at
p#0.05.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
1st
axis
score
*
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
ref med high
2nd a
xis
score
d = 5
med
high
ref

Macroinvertebrate traits and biomonitoring
129
Analysis of the trait category scores shows that the separation of the high
impact sites in the PCA was mainly due to large (>40 mm), multivoltine, asexual,
burrowers and deposit feeders, with preferences for detritus or silt as microhabitat
and quiet water flows, and using cocoons as resistance form. Medium impact sites
had higher abundances of small organisms (2.5-5 mm), plastron breathers,
scrapers, small detritus (<1 mm) feeders, and organisms that cement their
eggs/clutches. References had higher abundances of flattened, ovoviviparous
organisms, feeding on dead animals.
5.3.2. Co-structure of environmental and biological gradients
In Canada, the random permutation tests indicated that the selected
environmental variables were significantly related to the biological components,
whether dealing with taxa or traits (taxa-environment co-inertia analysis: RV-
coefficient=0.64, simulated p=0.002; trait-environment co-inertia analysis: RV-
coefficient=0.76, simulated p=0.001; Table 5.1). Focus was on the first axis of the
co-inertia analysis since it represented a prominent part of the total variability
(70.3% for the taxonomic analysis; 82.7% for the trait analysis). Along the first
axis, the proportion of variance extracted from the environmental and biological
datasets was higher than 72% on both co-inertia analyses (Table 5.1). The scores
of the environmental variables for the first co-inertia axis were similar in the two
approaches and NO3NO2, SpeCond, SO4, TotP and SolP had the highest scores.
Site ordination, based on biological information, was therefore linked to the
gradient described by the selected environmental variables, i.e., taxa and trait
abundances were not randomly distributed along the environmental gradient.
Taxa Traits Taxa Traits Taxa Traits
Proportion of covariance (%) 70.3 82.7 54.9 59.9 50.2 76.1
Variance extracted from environmental matrix (%) 99.9 99.6 73.0 98.7 95.6 95.7
Variance extracted from biological matrix (%) 72.8 93.9 95.0 43.6 60.9 91.6
RV-coefficient 0.64 0.76 0.68 0.62 0.72 0.63
Simulated p 0.002 0.001 0.005 0.042 0.009 0.029
Canada Spain Costa Rica
Table 5.1 – First axis and RV-coefficient results of co-inertia analyses relating environmental
variables associated with land use gradients and biological variables (taxa and trait category
abundances) of three biogeographic regions. p denotes probability based on 10000
permutations (see text for details).

Chapter 5
130
The first two axes of the co-inertia analysis done on the Spanish data
explained 71.5% of the total variability when using taxa abundances and 85.2%
when using traits. Along the first axis, the proportion of variance extracted from the
environmental and biological datasets was higher than 43% on both co-inertia
analyses (Table 5.1). The highest environmental variable scores for the first co-
inertia axis were Namm, CanoDens, TotP and SolP in the taxa co-inertia analysis
and TotP, SolP, SO4 and Namm in the trait co-inertia analysis. Random
permutation tests revealed significant relationships between the environmental
variables and taxonomic information (RV-coefficient=0.68, simulated p=0.005) and
between the environmental variables and trait abundances (RV-coefficient=0.62,
simulated p=0.042). The biological gradients defined by two different approaches
were related to the impact gradient.
In Costa Rica, the first two axes of the co-inertia analysis represented 77.8%
of the total variance for the taxonomic analysis and 90.9% of the total variance for
the trait analysis. Along the first axis, the proportion of variance extracted from the
environmental and biological datasets was higher than 60% on both co-inertia
analyses (Table 5.1). The scores of the environmental variables for the first co-
inertia axis were similar in the two approaches and SpeCond, SO4, TotP and
Embedd had the highest scores. Permutation tests have shown that environmental
and biological components were significantly related (taxa-environment co-inertia
analysis: RV-coefficient=0.72, simulated p=0.009; trait-environment co-inertia
analysis: RV-coefficient=0.63, simulated p=0.029; Table 5.1). Site ordination,
based on biological information, was therefore linked to the gradient described by
the environmental variables included in the analyses.
5.3.3. Interregional comparisons
As expected, given their biogeographic separation, low taxa overlap was
observed among different regions, with the majority of taxa occurring in only one

Macroinvertebrate traits and biomonitoring
131
region (Fig. 5.7), and merely 11 of the total 166
different taxa collected in the three study areas were
present in all three regions. The axis of the PCA
performed on the abundance data (all sites, all
regions) explained 98.8% of the variance in
assemblage composition among streams (97.7% for
axis 1 and 1.1% for axis 2). Different land use
categories were not clearly separated in the PCA
(Fig. 5.8A1) but a good discrimination of sites could
be observed in the PCA if only regions were
considered (Fig. 5.8A2). Differences among land
use categories were not significant for both axis 1
and axis 2 scores but both axis scores were
significantly different when comparing regions
(ANOVA, axis 1: F=85.381, p<0.001; axis 2:
F=22.947, p<0.001; Fig. 5.9A 1-4).
The first 2 axes of the FPCA performed on trait
abundances of the three regions explained 55.1% of the total variance (32.9% and
22.2% for axes 1 and 2 respectively). The discrimination of land use categories
and regions along the factorial axes could be moderately discerned from the visual
analysis of the plot (Fig. 5.8B1), although more overlaps can be observed between
different land use groupings than between regional groupings (Fig. 5.8B2).
Nevertheless, when comparing the trait FPCA with the taxa PCA, the regional
groupings appear less discriminated and the land use groupings appear more
discriminated in the FPCA. Significance tests on FPCA axis scores showed that
only axis 2 scores were significantly different among impact levels (ANOVA,
F=5.805, p=0.008; Fig. 5.9B2) and axis 1 scores were significantly different among
regions (ANOVA, F=23.835, p<0.001; Fig. 5.9B3).
When considering all three regions simultaneously, the between-land use
category analyses showed that overall between categories variance was
significant when considering traits abundance (between-class variance explained
Fig. 5.7 – Total number of
macroinvertebrate taxa
found in stream field sites of
three biogeographic regions.
The number of taxa found in
at least one other region
(shared) and the number of
taxa exclusively found in
each region (exclusive) are shown.

Chapter 5
132
was 13.0%; p=0.025) but the same was not true for taxa abundance (between-
class variance explained was 7.0%; p=0.4034). It was statistically confirmed that
the degree of discrimination of land use categories obtained using trait
composition was significantly higher (p<0.01; Fig. 5.10A) than the one obtained
using the taxonomic approach (considering the simulated between-category
variances with 72 randomly selected taxa).
d = 5
r e f
m e d
h i g h
d = 5
c a
c r
s p
d = 5
r e f
m e d
h i g h
Fig. 5.8 – Biplots of principal components analyses and fuzzy principal components
analysis of macroinvertebrate taxa (A) and trait (B) abundances at reference (ref), medium
impact (med) and high impact (high) field sites in Canada (ca), Spain (sp) and Costa Rica
(cr). The variance explained by first (horizontal) and second (vertical) axes is: A – 98%
and 1%; B – 33% and 22%. Stars connect sites to the corresponding land use category
centroid (A1 and B1) or region centroid (A2 and B2) and d indicates the scale of the plots.
A1 A2
B1 B2 d = 5
c a
c r
s p

Macroinvertebrate traits and biomonitoring
133
As for the between-class analyses considering regions as instrumental
variables, overall between region variance was significant when considering trait
abundance (between-class variance explained was 31.4%; p<0.001) or taxa
abundance (between-class variance explained was 40.3%; p<0.001). As expected,
the degree of discrimination of regions obtained using trait composition was
significantly lower (p=0.02; Fig. 5.10B) than the one obtained using the taxonomic
approach (considering the simulated between-category variances with 72
randomly selected taxa).
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
ref med high
1st
axis
score
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
ref med high
2nd a
xis
score
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
cr sp ca
2nd a
xis
score
*
*
*
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
cr sp ca
1st
axis
score
*
A1 A2 A3 A4
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
ref med high
1st
axis
score
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
cr sp ca
2nd a
xis
score
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
ref med high
2nd a
xis
score
*
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
cr sp ca
1st
axis
score
*
*
* B1 B2 B3 B4
Fig. 5.9 – Mean values (± 1 standard error) of the principal components analysis
transformed axes scores for taxa abundances (A) and of the fuzzy principal components
analyses transformed axes scores for trait category abundances (B) considering land use
categories (A1, A2, B1, B2) and regions (A3, A4, B3, B4). Codes denote reference (ref), medium impact (med) and high impact (high) field sites in Canada (ca), Spain (sp) and
Costa Rica (cr). Symbol (*) denotes significant differences at p#0.05.

Chapter 5
134
5.4. Discussion
By using trait composition in ordination analyses rather than the more
traditional approach of taxa composition, it was shown that an improved
discrimination of land use categories could be obtained across the different
biogeographic regions. The relationship of the trait gradient with the impact
gradient variables could be established, and even the reference state problems
detected earlier in the taxonomic analysis of Costa Rican and Spanish study areas
were highlighted. Co-inertia analyses complemented the analysis by showing how
both taxonomic and functional information significantly related to environmental
variables that integrate the land use gradients (as proven in the previous chapter).
A large proportion (always higher than 50%) of covariance between biological and
environmental variables was explained by both the structural and the trait data and
the biological datasets were strongly correlated with the environmental ones.
Biological information was therefore strongly linked to the impact gradients, and
traits and taxa gave an equally good match with the patterns of sediment and
Fig. 5.10 – Frequency distribution of 100 simulated values (for 72 randomly selected taxa) and
observed values (dashed lines) of between land use category variances (A) and between
region variances (B) for taxa and trait composition of macroinvertebrate communities in three
different biogeographic regions. To improve visualization, x-axis scale was focused (origin is
not 0).
A B

Macroinvertebrate traits and biomonitoring
135
nutrient inputs of all three regions viewed independently, while using a similar
study design. Similar results were reported for streams in intensive agriculture
areas in southeaster Spain (Díaz et al., 2008) and a gradient of agricultural
development in New Zealand (Dolédec et al., 2006).
Inconsistency of study design, relating to differences in sampling technique
and taxonomic resolution, is a constant problem in comparative trait-related
studies across different biogeographic areas which use data collected by different
researchers in different regions. Indeed it could be argued that in such cases,
detected trait patterns could be strongly influenced by methodological differences -
a factor eliminated by the study design employed here.
It was shown that trait composition could discriminate sites along land use
gradients across different biogeographic regions, irrespective of their taxonomic
composition. The discrimination of high impact sites in particular was reflected in a
similar set of trait categories for the different regions: >40 mm (maximum size),
multivoltine (number of reproductive cycles per year), asexual (reproduction),
burrower (attachment to substrate), deposit feeder or fine sediment (feeding
habits), detritus or sand and silt (microhabitat), quiet (current velocity preference)
and cocoons (resistance form). In medium impact sites, common distinguishing
trait categories included 2.5-5 mm (maximum size), spiracle and plastron
(respiration mode). These results indicate a potential set of macroinvertebrate
traits that could be used to predictably assess the effects of severe land uses
changes posed by agricultural scenarios. Consistencies were found with the
predictions of selection of traits related to higher resistance and resilience in sites
under the influence of !impact filetrs". The number of reproductive cycles per year,
reproductive mode and feeding habit have been reported has respondent to
human impacts (Gayraud et al., 2003). In agricultural scenarios, one study has
also found similar significant responses to disturbance of the number of
reproductive cycles, feeding habit and respiration mode (Dolédec et al., 2006) and
another has found similar responses of voltinism, feeding modes and resistance
form (Díaz et al., 2008). Nevertheless, large body sizes, predicted for more stable

Chapter 5
136
and benign environments, were abundant in heavily impacted sites (a similar
pattern was observed by Díaz et al., 2008). This could be due to the presence of
large (coded with affinity for >40 mm in the trait maximum size), fast-growing,
resistant organisms i.e., Oligochaetes, which were detected with high abundances
in the more impacted sites of all three regions (cf. previous chapter). The fact is
that maximum size was distributed as predicted in the remaining land use
categories in Canada: reference sites had high abundances of larger organisms
(10-40 mm) while in medium impacted sites smaller (2.5-5 mm) organisms
prevailed (this size class was also predominant in medium impact sites of the
remaining regions). Actual measurements of the body size (Basset et al., 2004) of
the organisms found in these communities could work as a more appropriate
descriptor of this trait for biomonitoring purposes.
Following habitat templet theory (Southwood, 1977; Southwood, 1988; Poff &
Ward, 1990; Townsend & Hildrew, 1994), other expected trait-environment
relationships were observed: in Canadian reference communities, the high
abundances of organisms producing free eggs (less investment in parental care),
with preference for rocks/boulders (and probably averse to high sediment inputs)
and large woody debris (probably averse to riparian cleared sites), as well as the
high presence of medium sized organisms and predators could be associated with
least impacted and more stable stream ecosystems. Reference communities were
not truly comparable among regions, since both in Costa Rica and Spain the
presence of other stressors (natural and anthropogenic) has led to impact gradient
shifts. But as a possible response to that, in Spain, reference sites had
communities where characteristics usually associated with impact were present in
higher abundances (short life cycles, egg/clutch cementing), supporting the
hypothesis of a possible impairment leading to macroinvertebrate communities
that invest more in egg care and reproduction and less in growth (Verberk et al.,
2008). In Costa Rican references, the high abundances of flattened body shapes,
ovoviviparity and more investment in parental care, and dead animal feeders could
be an indicator of a stressed/less-stable community. A study of neotropical species

Macroinvertebrate traits and biomonitoring
137
traits has found streamlined bodies as an adaptative advantage in hydraulically
rough environments (Tomanova & Usseglio-Polatera, 2007).
Overall, in the three study regions, there was no taxonomic correspondence,
and very different taxa were found in each of the ecoregions (less than 7% overlap
between the three regions). In interregional comparisons, considering a matrix with
taxa abundances from all regions, strong discrimination across the three regions
was obtained but the discrimination of land use categories was no longer
statistically significant. The biological communities of the three regions were very
different in terms of their taxonomy, not allowing an overall discrimination of land
use intensities. This confirms that biomonitoring methods based exclusively on
taxonomic measures are necessarily limited to the spatial scale for which they
were developed and pure taxonomic interpretations of potential impacts may be
difficult to extrapolate from one region to the other. Traits yielded a better
(significant) overall discrimination of levels of impact (land use categories),
exhibiting less variability among regions than taxa abundances. Some biological
traits are geographically constrained and can have limitations similar to the ones
presented by taxonomic approaches: e.g., groups having shorter life cycles in the
equator when compared to northern latitudes (Pinder, 1986). But although still
significantly different in terms of their trait structure, the communities of the three
regions in fact allowed a moderate but significant discrimination of impact levels,
indicating once more that the freshwater macroinvertebrate community shifts in
sites impacted by the results of intensive agricultural practices may follow
convergent trajectories in multi-dimensional space, regardless of geography. The
discrimination of land use levels would probably be increased if more references
and actual unimpacted sites could be used to represent Spanish and Costa Rican
stream communities. Alternative approaches, considering the difficulties of finding
appropriate reference sites, suggest that references for bioassessment purposes
could be established from modeling knowledge of regional taxa pools (and taxa
environmental preferences and tolerances) and key environmental features, but
this “filters approach” still needs further development (Chessman & Royal, 2004).

Chapter 5
138
The results highlight the potential importance of the trait approach in
between-region extrapolation of biomonitoring criteria and adaptation of
methodologies usually developed for certain regions and completely inexistent in
others. The enrichment of identity-based information with the corresponding
species characteristics seems highly advantageous for freshwater biomonitoring
programs that intend to have: better discrimination of human impacts, more global
applicability (Bonada et al., 2006) and - if true functional traits are considered
(Violle et al., 2007) - ecological relevance.
Some methodological considerations regarding the trait approach still require
to be addressed, however. While quantification of species traits began to be
addressed more than a decade ago (Poff, 1997), consistent ways of achieving this
remain to be developed, particularly for tropical species (Tomanova & Usseglio-
Polatera, 2007). The fact is that most trait information collected and published by
stream ecologists is related to common, widespread species of
macroinvertebrates, which often conform the 'r-strategist' life-history type,
potentially biasing any of the performed analyses (Statzner et al., 1997).
Also important is the fact that the expression “taxonomic differences” among
macroinvertebrate communities from different biogeographic regions could relate
not only to the different taxa found in each community but also to differences in our
ability to identify those taxa with similar accuracy. Trait attribution in these taxa is
difficult since, particularly in the tropics, ecological descriptions which form the
basis for trait analysis are scarce, even for common species (Tomanova &
Usseglio-Polatera, 2007). In Costa Rica, for example, taxonomic keys are not
available for all groups and some species" autecology is completely unknown.
Higher taxonomic levels (mostly genus) had to be used many times. And although
it has been proven that the usage of coarse taxonomic levels in the functional
description of macroinvertebrate communities for biomonitoring purposes might
not constitute a problem (Gayraud et al., 2003), difficulties in the process of trait
attribution for those species where less autecological information is available still
exist. It is a fact that, when genus level is used, genus trait profiles are many times

Macroinvertebrate traits and biomonitoring
139
described from species profiles. The trait approach therefore still suffers from a
basic lack of autecological studies of less common species, and species from less-
visited regions.
5.5. References
Basset, A., Sangiorgio, F. & Pinna, M. (2004) Monitoring with benthic
macroinvertebrates: advantages and disadvantages of body size
descriptors. Aquatic Conservation, 14, S43-S58.
Bonada, N., Prat, N., Resh, V. H. & Statzner, B. (2006) Developments in Aquatic
Insect Biomonitoring: A Comparative Analysis of Recent Approaches.
Annual Review of Entomology, 51 (1), 495-523.
Charvet, S., Roger, M. C., Faessel, B. & Lafont, M. (1998) Biomonitoring of
freshwater ecosystems by the use of biological traits. Annales de
Limnologie–International Journal of Limnology, 34, 455–464.
Charvet, S., Statzner, B., Usseglio Polatera, P. & Dumont, B. (2000) Traits of
benthic macroinvertebrates in semi-natural French streams: an initial
application to biomonitoring in Europe. Freshwater Biology, 43 (2), 277-296.
Chessman, B. & Royal, M. (2004) Bioassessment without reference sites: use of
environmental filters to predict natural assemblages of river
macroinvertebrates. Journal of the North American Benthological Society,
23 (3), 599-615.
Chevenet, F., Dolédec, S. & Chessel, D. (1994) A fuzzy coding approach for the
analysis of long-term ecological data. Freshwater Biology, 31, 295-309.
Díaz, A. M., Alonso, M. L. S. & Gutierrez, M. R. V. A. (2008) Biological traits of
stream macroinvertebrates from a semi-arid catchment: patterns along
complex environmental gradients. Freshwater Biology, 53, 1-21.

Chapter 5
140
Dolédec, S. & Chessel, D. (1994) Co-inertia analysis: an alternative method for
studying species-environment relationships. Freshwater Biology, 31 (3),
277-294.
Dolédec, S., Phillips, N., Scarsbrook, M., Riley, R. H. & Townsend, C. R. (2006)
Comparison of structural and functional approaches to determining landuse
effects on grassland stream invertebrate communities. Journal of the North
American Benthological Society, 25 (1), 44-60.
Dolédec, S., Statzner, B. & Bournard, M. (1999) Species traits for future
biomonitoring across ecoregions: patterns along a human-impacted river.
Freshwater Biology, 42 (4), 737-758.
Dray, S., Chessel, D. & Thioulouse, J. (2003) Co-Inertia Analysis and the Linking
of Ecological Data Tables. Ecology, 84 (11), 3078-3089.
Dray, S. & Dufour, A. B. (2007) The ade4 Package: Implementing the Duality
Diagram for Ecologists. Journal of Statistical Software, 22 (4), 1-20.
Gayraud, S., Statzner, B., Bady, P., Haybach, A., Scholl, F., Usseglio Polatera, P.
& Bacchi, M. (2003) Invertebrate traits for the biomonitoring of large
European rivers: an initial assessment of alternative metrics. Freshwater
Biology, 48 (11), 2045-2064.
Lebreton, J. D., Sabatier, R., Banco, G. & Bacou, A. M. (1991) Principal
component and correspondence analysis with respect to instrumental
variables : an overview of their role in studies of structure-activity and
species-environment relationships. In: J. Devillers & W. Karcher (Ed.).
Applied multivariate analysis in SAR and environmental studies. Dordrecht,
The Netherlands, Kluwer Academic Publishers, 85-114.
McGill, B. J., Enquist, B. J., Weiher, E. & Westoby, M. (2006) Rebuilding
community ecology from functional traits. Trends in Ecology & Evolution, 21
(4), 178-185.
Pinder, L. C. V. (1986) Biology of Freshwater Chironomidae. Annual Review of
Entomology, 31 (1), 1-23.

Macroinvertebrate traits and biomonitoring
141
Poff, N. & Ward, J. (1990) Physical habitat template of lotic systems - recovery in
the context of historical pattern of spatiotemporal heterogeneity.
Environmental Management, 14 (5), 629-645.
Poff, N. L. (1997) Landscape filters and species traits: towards mechanistic
understanding and prediction in stream ecology. Journal of the North
American Benthological Society, 16 (2), 391-409.
R Development Core Team (2010) R: A language and environment for statistical
computing. Vienna, Austria, R Foundation for Statistical Computing.
http://www.R-project.org.
Robert, P. & Escoufier, Y. (1976) A unifying tool for linear multivariate statistical
methods: the RV-coefficient. Journal of Applied Statistics, 25, 257–265.
Romesburg, H. C. (1985) Exploring, confirming, and randomization tests.
Computers & Geosciences, 11 (1), 19-37.
Southwood, T. R. E. (1977) Habitat, the Templet for Ecological Strategies? Journal
of Animal Ecology, 46 (2), 336-365.
Southwood, T. R. E. (1988) Tactics, Strategies and Templets. Oikos, 52 (1), 3-18.
Statzner, B., Dolédec, S. & Hugueny, B. (2004) Biological trait composition of
European stream invertebrate communities: assessing the effects of
various trait filter types. Ecography, 27 (4), 470-488.
Statzner, B., Hoppenhaus, K., Arens, M. & Richoux, P. (1997) Reproductive traits,
habitat use and templet theory: a synthesis of world-wide data on aquatic
insects. Freshwater Biology, 38 (1), 109-135.
Tomanova, S. & Usseglio-Polatera, P. (2007) Patterns of benthic community traits
in neotropical streams: relationship to mesoscale spatial variability.
Fundamental and Applied Limnology, 170, 243-255.
Townsend, C. R. & Hildrew, A. G. (1994) Species Traits in Relation to a Habitat
Templet for River Systems. Freshwater Biology, 31, 265-275.
Townsend, C. R., Scarsbrook, M. R. & Dolédec, S. (1997) Quantifying disturbance
in streams: alternative measures of disturbance in relation to
macroinvertebrate species traits and species richness. Journal of the North
American Benthological Society, 16, 531-544.

Chapter 5
142
Van den Brink, P. J., van den Brink, N. W. & Ter Braak, C. J. F. (2003) Multivariate
analysis of ecotoxicological data using ordination: demonstrations of utility
on the basis of various examples. Australasian Journal Ecotoxicology, 9,
141-156.
Verberk, W. C. E. P., Siepel, H. & Esselink, H. (2008) Life-history strategies in
freshwater macroinvertebrates. Freshwater Biology, 53 (9), 1722-1738.
Violle, C., Navas, M., Vile, D., Kazakou, E., Fortunel, C., Hummel, I. & Garnier, E.
(2007) Let the concept of trait be functional! Oikos, 116 (5), 882-892.

Chapter 6. General Discussion and Conclusions


Discussion and Conclusions
145
Macroinvertebrates do not depend on the names we give them. They do
depend on the characteristics selected through evolution, by the imposition of
habitat filters (Keddy, 1992) that allow them to withstand not only naturally variable
environments, but also the “artificial filters” imposed by the harsh conditions of
heavily humanized ecosystems. So one could question the reasons why identity-
based analysis is still the bulk technique used in ecosystem integrity assessment.
If the intention is to have true ecologically-oriented river management, with
recognition of the dependence of human society on naturally functioning
ecosystems (Baron et al., 2002), it is demanding that the protection of natural
ecosystems and the recovery of heavily impacted ones consider the functional
intricacies of biological communities. A shift in paradigm, from a pure structural
view of those communities to a more functionally enriched one, is crucial if one
wants to detect the true consequences of anthropogenic stressors to ecosystem
functions. The development of biomonitoring through functional parameters of the
communities has been carried out over the last decades (Bonada et al., 2006),
although the !ideal biomonitoring tool" is still not fully developed. Traits, as
characteristics that reflect species adaptation to their environment, may work as
an interesting improvement of traditionally used biomonitoring methods. The
present work intended to be a contribution for this improvement.
An assessment of the biological integrity of the different study ecosystems
was undertaken, in a consistent manner across different biogeographic regions.
Stream community structure was used to reveal complex impact gradients
(Chapter 4), defined by impact-associated variables that significantly covaried with
watershed agricultural land use gradients (supporting the a priori defined
classification of sites in each region). These gradients were mostly related with
nutrient input and sedimentation – two of the main stressors affecting freshwater
biological communities in scenarios of severe land use change due to intensive
agriculture (Allan, 2004).
It was proven that the biological gradients defined by selected structural
measures of the macroinvertebrate communities co-varied with the disturbance

Chapter 6
146
gradients studied, although few structural variables individually discriminated the a
priori defined land use categories. Also, there was no consistency in the
responses of the structural measures across biogeographic regions, confirming
their lack of global applicability for biomonitoring purposes using
macroinvertebrates. When analyzed simultaneously, the taxonomically defined
communities of one region clearly separate from the ones in other regions. This
confirms that pure taxonomic interpretations of potential impacts may be difficult to
extrapolate from one region to the other.
By reviewing the current state-of-the-art on macroinvertebrate biomonitoring
through traits (Chapter 2), it was highlighted that when compared with taxonomy-
based approaches the use of traits as a functional enrichment of such approaches
has the following advantages:
- the possibility of establishment of causal relationships with stressors;
- a better detection/differentiation of gradients of impact;
- the integration of natural fluctuations;
- the definition of functional images of the communities, that allow the
detection of functional consequences at the ecosystem level;
- the geographic unconstrained application, with the possibility of
generalizations to different types of (less studied) freshwater systems.
Problems in the definition of reference states in both the impact and the
biological gradients were evident for both Spanish and Costa Rican study areas,
highlighting the difficulties of both including natural fluctuations in result
interpretation and finding real !reference conditions" in heavily humanized
scenarios. One solution for these types of problems would reside in higher
replication of reference samples, which could highlight natural variability, but this is
not always a viable solution. Additionally, trait patterns, when compared with taxa
patterns, could be easier and more specifically associated with unexpected
sources of pollution (the possibility raised for Spanish references) or ranges of
natural variation (the possible explanation for Costa Rican reference state issues).

Discussion and Conclusions
147
It was proven that the biological gradients defined by selected traits of the
macroinvertebrate communities co-varied with the disturbance gradients studied
(Chapter 5). Additionally, when the taxonomic approach was enriched with the use
of taxa traits, an improved discrimination of land use categories was obtained in
each of the different biogeographic regions.
Across regions, a similar set of traits discriminated higher impact sites,
including size, voltinism, reproductive techniques, microhabitat, current and
substrate preferences, feeding habits and resistance forms. This set could be
further studied and used to predictably assess the effects of severe land uses
changes posed by agricultural scenarios in freshwater biomonitoring programs.
When analyzed simultaneously using traits, and contrary to what happened with
the overall taxonomic patterns, the communities of the three regions allowed a
moderate but significant discrimination of impact levels. These analyses support
the evidence that freshwater macroinvertebrate community shifts in sites impacted
by intensive agriculture may follow convergent trajectories in multi-dimensional
space, regardless of geography – with obvious implications in the development of
geographically unconstrained freshwater biomonitoring tools.
In future research studies, it would be interesting to see focus on:
- how trait pattern responses are altered with time and natural fluctuations;
- pairing trait studies with measures of community and ecosystem functions
(like leaf litter decay, e.g. Piscart et al., 2009) in order to confirm direct functional
relationships;
- producing life-history and ecological studies on species that are less
frequent or from less studied regions in order to populate taxon-trait databases;
this is imperative in the improvement of trait attribution and analysis since highly
detailed, a priori knowledge on traits is needed to establish predictive models (thus
they are sensitive to data availability; Poff, 1997);
- promoting open and easy access to the populated trait databases, with the
results of collaborations of different research groups working in the area (and with

Chapter 6
148
investment in common trait category definitions, unified coding of information,
etc.);
- relating biomonitoring results with experimental studies on invertebrate
species and communities, so that cause-effect relationships can be properly
established; these studies should assess individual anthropogenic stressor effects,
effects of multiple simultaneous stressors and effects of natural stressors.
Mesocosms (Caquet et al., 2000; Wong et al., 2004) in particular are a type of
experimental setting that could allow a better understanding of cause-effect
relationships between agricultural stressors, natural stressors and biological
consequences. The detection of specific impacts under multiple stressor scenarios
is very complex (Dolédec & Statzner, 2007) and experimentation would probably
facilitate the process of selection of the traits that better relate with each specific
impact;
- further developing the use of the trait approach for the analysis of
ecosystem recovery after restoration measures, considering those measures as
trait filters (like e.g. van Kleef et al., 2006; Paillex et al., 2009).
The effects of taxa loss in a community due to anthropogenic activities
cannot be diminished, but that is many times the case. There are questions many
times left unanswered in biomonitoring programs that contribute to this: What does
the loss of a particular taxon mean? How does this loss affect the overall
ecosystem? If we address the problem by showing how actual ecosystem
functions are partially or completely lost due to anthropogenic impacts, the effects
can no longer be ignored and ecosystem protection needs will be better conveyed
to managers and legislators, and to the general public. Clues were given to identify
specific features that should be considered in the design of future biomonitoring
programs for each of the studied regions. The advantageous use of traits for
interregional extrapolations was also highlighted. Hopefully, this study will
contribute for the implementation of a new generation of biomonitoring tools, closer
to a “polyglot” “ecofunction-friendly” “freeze dried, talking fish on a stick”.

Discussion and Conclusions
149
6.1. References
Allan, J. D. (2004) Landscape and Riverscapes: The Influence of Land Use on
Stream Ecosystems. Annual Review of Ecology, Evolution, and
Systematics, 35 (1), 257-284.
Baron, J. S., Poff, N. L., Angermeier, P. L., Dahm, C. N., Gleick, P. H., Hairston, J.,
Jackson, R. B., Johnston, C. A., Richter, B. D. & Steinmanj, A. D. (2002)
Meeting Ecological and Societal Needs for Freshwater. Ecological
Applications, 12 (5), 1247-1260.
Bonada, N., Prat, N., Resh, V. H. & Statzner, B. (2006) Developments in Aquatic
Insect Biomonitoring: A Comparative Analysis of Recent Approaches.
Annual Review of Entomology, 51 (1), 495-523.
Caquet, T., Lagadic, L., Sheffield, S. R., Ware, G. W., Albert, L. A., Hutzinger, O.,
Knaak, J. B., Mayer, F. L., Morgan, D. P., Park, D. L., Tjeerdema, R. S.,
Voogt, P. D., Whitacre, D. M., Yang, R. S. H. & Gerba, C. P. (2000)
Mesocosms in ecotoxicology (1): Outdoor aquatic systems. Reviews of
Environmental Contamination and Toxicology, 165, 1-38.
Dolédec, S. & Statzner, B. (2007) Invertebrate traits for the biomonitoring of large
European rivers: an assessment of specific types of human impact.
Freshwater Biology, 53, 617-634.
Keddy, P. A. (1992) Assembly and Response Rules: Two Goals for Predictive
Community Ecology Journal of Vegetation Science, 3 (2), 157-164.
Paillex, A., Dolédec, S., Castella, E. & Merigoux, S. (2009) Large river floodplain
restoration: predicting species richness and trait responses to the
restoration of hydrological connectivity. Journal of Applied Ecology, 46 (1),
250-258.
Piscart, C., Genoel, R., Dolédec, S., Chauvet, E. & Marmonier, P. (2009) Effects of
intense agricultural practices on heterotrophic processes in streams.
Environmental Pollution. 157, 1011-1018.

Chapter 6
150
Poff, N. L. (1997) Landscape filters and species traits: towards mechanistic
understanding and prediction in stream ecology. Journal of the North
American Benthological Society, 16 (2), 391-409.
van Kleef, H. H., Verberk, W. C. E. P., Leuven, R. S. E. W., Esselink, H., van der
Velde, G. & van Duinen, G. A. (2006) Biological traits successfully predict
the effects of restoration management on macroinvertebrates in shallow
softwater lakes. Hydrobiologia, 565, 201-216.
Wong, D. C. L., Maltby, L., Whittle, D., Warren, P. & Dorn, P. (2004) Spatial and
temporal variability in the structure of invertebrate assemblages in control
stream mesocosms. Water Research, 38 (1), 128-138.