respostas fisiológicas de plantas jovens variantes de pau-brasil a
TRANSCRIPT

UNIVERSIDADE ESTADUAL DE SANTA CRUZ
PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO VEGETAL
VIVIANE MARIA BARAZETTI
RESPOSTAS FISIOLÓGICAS DE PLANTAS JOVENS VARIANTES DE PAU-
BRASIL A DIFERENTES NÍVEIS DE IRRADIÂNCIA
ILHÉUS - BAHIA
2013

2
VIVIANE MARIA BARAZETTI
RESPOSTAS FISIOLÓGICAS DE PLANTAS JOVENS VARIANTES DE PAU-
BRASIL A DIFERENTES NÍVEIS DE IRRADIÂNCIA
Dissertação apresentada ao Programa de Pós-Graduação em
Produção Vegetal, da Universidade Estadual de Santa Cruz -
UESC, como parte dos requisitos para a obtenção do título
de Mestre em Produção Vegetal.
Área de Concentração: Cultivos em Ambiente Tropical
Úmido.
Orientador: Prof. Dr. Alex-Alan Furtado de Almeida.
Co-orientador: Prof. Dr. Marcelo Schramm Mielke
ILHÉUS - BAHIA
2013
ii

3
VIVIANE MARIA BARAZETTI
RESPOSTAS FISIOLÓGICAS DE PLANTAS JOVENS VARIANTES DE PAU-
BRASIL A DIFERENTES NÍVEIS DE IRRADIÂNCIA
Ilhéus, BA, 19/04/2013
________________________________________________________
Alex-Alan Furtado de Almeida – DSc
UESC/DCB
(Orientador)
________________________________________________________
Marcelo Schramm Mielke – DSc
UESC/DCB
________________________________________________________
Fábio Pinto Gomes – DSc
UESC/DCB
________________________________________________________
Rogério Ferreira Ribas – DSc
UFRB
iii

4
DEDICATÓRIA
Dedico este trabalho à minha filha Stephany Arícia, pelo amor incondicional.
Desejo que você...
Não tenha medo da vida, tenha medo de não vivê-la.
Não há céu sem tempestades, nem caminhos sem acidentes.
Só é digno do pódio quem usa as derrotas para alcançá-lo.
Só é digno da sabedoria quem usa as lágrimas para irrigá-la.
Os frágeis usam a força; os fortes, a inteligência.
Seja um sonhador, mas una seus sonhos com disciplina,
Pois sonhos sem disciplina produzem pessoas frustradas.
Seja um debatedor de ideias. Lute pelo que você ama.
(Augusto Cury)
iv

5
AGRADECIMENTOS
Inicio meus agradecimentos com a frase célebre de Roger Von Oech, que em seus
momentos de criação escreveu que “... descobrir consiste em olhar para o que todo mundo
está vendo e pensar uma coisa diferente”. Assim, descobri também que descobrir é agradecer
à dádiva do Grande Arquiteto do Universo pela possibilidade de estar presente na vida
daqueles que compartilharam de seu conhecimento, seu convívio, seus afazeres e suas
amizades, no desenvolvimento desta pequena obra, oportuna para a transmissão de
conhecimentos e levar a verdade científica à tona dos dados da academia.
A família é fonte de prosperidade. Assim, agradeço aos meus pais, Ivanete e Aldo
Barazetti, uma vez que a melhor herança que puderam deixar foi a firmeza em aspirar triunfos
e a capacidade de assimilar derrotas. Pessoas simples que dedicaram parte do seu valioso
tempo em me preparar para o futuro. Aos meus irmãos Geferson e Erivaldo, pela paciência e
incentivo, mesmo à distância.
Nunca estamos sós, é verdade! Mesmo longe de meus pais, jamais estive só.
Agradeço ao meu marido, Ivan Montenegro, por tudo, pelos momentos em que chorei, pelos
momentos em que me desesperei, pelos momentos mais maravilhosos da minha vida, pelo
socorro nos momentos angustiantes, pelos carinhos e beijos que me fizeram sorrir e despertar
e pelos momentos de alegria que fez questão de dividir comigo e, finalmente, pelos momentos
que com muita esperança pensou junto comigo no nosso futuro.
O amor de mãe é diferente de qualquer outra coisa no mundo. Não obedece lei ou
piedade, ele ousa todas as coisas imagináveis e inimagináveis. Assim, agradeço a uma
“pessoinha” muito especial, minha filha Stephany Arícia que com sua graça, simpatia e
“carinha mais linda do mundo” me deu forças para continuar nessa árdua batalha.
Agradeço ao meu orientador, Alex-Alan Furtado de Almeida, com uma singela frase
de um autor completamente desconhecido que diz que “... ser professor é acreditar que um
outro mundo é possível... é aquele que caminha com o tempo, propondo paz e despertando
sabedoria, estendendo a mão, iniciando o diálogo”. Meu orientador foi aquele que não
ensinou fórmulas, regras ou raciocínios, mas sim aquele que me despertou para a realidade,
compreendendo, estimulando, me enriquecendo com seu saber e sua presença. Ensinou-me
que sempre serei um discípulo na escola da vida.
Aos professores Raúl R. Valle, Marcelo S. Mielke e Fábio P. Gomes, meus
agradecimentos. Faço minhas as palavras de Carlos Drumond de Andrade que diz que “...
professor não disserta sobre o ponto difícil do programa. Um aluno dorme, cansado das
v

6
canseiras dessa vida. O professor vai sacudi-lo? Vai repreendê-lo? Não. O professor baixa a
voz com medo de acordá-lo”. Foi assim que aprendi a admirar os valores destes amigos
professores. Suas contribuições foram essenciais para o desenvolvimento desse trabalho.
Universidade diferente, curso novo, colegas novos. Era tudo muito novo,
interessante, pessoas diferentes, salas diferentes, cursos diferentes, e a estranha sensação de
muitos desafios que haveria pelo caminho. Os mestres aplicavam suas matérias e nós
tentávamos nos adaptar ao estilo de ensino acadêmico. Muitas dúvidas começaram a surgir,
dúvidas essas que fortaleceriam ainda mais a nossa curiosidade sobre os assuntos. Então
começaram os trabalhos, as pesquisas, as leituras, e foi aí que se formaram os grupos e os
laços de amizades, na faculdade ou fora dela. Assim, fortalecidos pelos laços da amizade,
quero agradecer a todos os colegas que partilharam seu tempo em todas as etapas do
mestrado, lembrando sempre de Martielly, Leandro, Flávia, Priscila, Alayana, Cristiane,
Eliane, Téssio, Romária, Joelson e Felipe. Esse álbum de amigos será levado para todo o
sempre.
Laboratórios, centros de pesquisa, instrutores e analistas. Um mundo novo a ser
descoberto com pessoas que fazem parte do dia-a-dia de nosso aperfeiçoamento. Agradeço,
assim, aos amigos Heriberto, Rubens, Cleide, Dan Lobão, Robélio, Paulo Marrocos, Gerson,
Adelino e Marcelo, funcionários da Comissão Executiva do Plano da Lavoura Cacaueira e
Universidade Estadual de Santa Cruz, pelo esforço e ajuda no desenvolvimento das coletas e
análises dos experimentos. Assim, agradeço com uma frase de Roberto Shinyashiki dizendo a
vocês que “... tudo o que um sonho precisa para ser realizado é alguém que acredite que ele
possa ser realizado”.
Agradeço à FAPESB, pela concessão da bolsa de mestrado, a CEPLAC, pelo espaço
e pela oportunidade da realização das análises do meu experimento e a UESC, pela
oportunidade de ingresso no curso de Pós-Graduação em Produção Vegetal. Agradeço,
também, àqueles que indiretamente contribuíram para a finalização de meus objetivos. Para
estes, e para todos aqueles que acreditaram em mim, encerro os agradecimentos citando e
focando a notória frase de Albert Einstein, onde “Nenhuma mente que se abre para uma nova
ideia voltará a ter o tamanho original”.
vi

7
RESPOSTAS FISIOLÓGICAS DE PLANTAS JOVENS VARIANTES DE PAU-
BRASIL AO SOMBREAMENTO
RESUMO
Caesalpinia echinata Lam. é uma espécie arbórea nativa da Mata Atlântica em risco de
extinção. As informações oriundas de estudos com a espécie são contraditórias, dificultando o
seu manejo e a sua conservação. Objetivou-se, no presente trabalho, avaliar os efeitos do
sombreamento no crescimento, na assimilação do carbono e na composição química de
plantas jovens variantes de pau-brasil (folha pequena - SV; folha média - MV e folha grande -
LV), por meio da análise de crescimento, medições de trocas gasosas foliares, pigmentos
fotossintéticos e carboidratos. Os variantes foram cultivados a pleno sol, no interior de viveiro
e no interior de casas de sombra com telas sombreadoras, que proporcionaram condições
ambientais com percentuais de irradiância de 100, 60, 35, 25 e 15% durante 244 dias.
Observou-se, durante este período, no ambiente de 100% de irradiância, que o variante SV
apresentou maior número de folhas, maiores biomassas secas de raiz, caule e total, maior
razão de massa de raiz, maior taxa de assimilação líquida (TAL) e maior taxa fotossintética
bruta máxima em irradiância de saturação (Amax, 8,92 µmol m-2
s-1
). Nesta mesma irradiância,
o variante MV apresentou maior diâmetro do caule, maior razão de massa de caule (RMC),
maior teor de amido (AM) na raiz e de açúcares solúveis totais (AST) na folha e maior massa
foliar específica; ao passo que o variante LV apresentou maior taxa de crescimento relativo
(TCR), maior TCR em altura e em diâmetro do caule, maior razão entre o teor de carotenóides
(Car) e o teor de clorofila total (Chl T) (Car/Chl T), maiores taxas de fotossíntese líquida (A)
e transpiratória (E), maior condutância estomática (gs) e maior razão entre a concentração
intercelular e ambiente de CO2. Entretanto, em condições de 60% de irradiância, o variante
LV apresentou maior teor de AM na folha e maior área foliar; ao passo que o variante MV
apresentou maior valor de rendimento quântico aparente (α, 0,073 µmol CO2 µmol fótons-1
) e
maior teor de Car. No ambiente de 35% de irradiância, o variante LV apresentou maior teor
de AST na raiz e maiores eficiências instantânea e intrínseca no uso da água; enquanto que o
variante SV apresentou maior altura e maior biomassa seca foliar. Em 25% de irradiância, o
variante SV apresentou maior razão de área foliar; maior teor de AM no caule e maior teor de
clorofila b (Chl b); ao passo que o variante LV mostrou maior razão de massa de folha e
maior razão entre os teores de clorofila a (Chl a) e Chl b (Chl a/b); e o variante MV
apresentou maiores teores de Chl a e Chl T. Entretanto, em 15% de irradiância, o variante SV
apresentou maior teor de AST no caule, enquanto que o variante MV apresentou maior valor
de α e menor taxa respiratória na ausência de luz (Rd, 0,16 µmol m-2
s-1
). Em suma, os
variantes morfológicos de pau-brasil ajustaram seu maquinário fotossintético, no sentido de
maximizar os ganhos de energia, por meio de plasticidade fisiológica diferenciada, durante a
aclimatação aos diferentes ambientes de radiação luminosa. A variação da irradiância
influenciou positivamente a assimilação do CO2, quando relacionada com gs e E, dando
condições aos variantes morfológicos de atingirem o seu crescimento potencial, demonstrado
pelo aumento de TCR com o incremento da intensidade de luz. As mudanças nos padrões
biométricos dos variantes de pau-brasil foram afetadas pela irradiância, em consequência das
variações nos ganhos de biomassa determinados por TAL e TCR. Houve uma diferenciação
dos variantes morfológicos em relação à assimilação do carbono e a variação da irradiância. O
variante LV se aclimatou melhor em condição de 100% de irradiância; ao passo que os
vii

8
variantes SV e MV se aclimataram melhor em condições de 35 e 15% de irradiância,
respectivamente.
Palavras-chave: C. echinata, estresse por irradiância, produção de biomassa, morfotipo de
pau-brasil, trocas gasosas foliares, pigmentos cloroplastídicos, carboidratos.
viii

9
PHYSIOLOGICAL RESPONSES OF YOUNG PLANT VARIANTS OF
BRAZILWOOD TO SHADE
ABSTRACT
Caesalpinia echinata Lam. is a native tree species of the Atlantic Forest at risk of extinction.
Information from studies with this species are contradictory, making difficult its management
and conservation. The aim of the present study was to evaluate the effects of shading on
growth, carbon assimilation and chemical composition of seedlings of brazilwood variants
(small leaf - SV; medium leaf - MV and large leaf - LV) through growth analysis,
measurements of leaf gas exchange, photosynthetic pigments and carbohydrates. The variants
were grown in full sun, inside the nursery and inside shade houses shaded with screens, which
provided environmental conditions with irradiance percentage of 100, 60, 35, 25 and 15%
over 244 days. It was observed during this period, at the 100% irradiance environment that the
SV variant had the greatest number of leaves, higher root, shoot and total dry biomass, higher
root mass ratio, higher net assimilation rate (NAR) and higher maximum gross photosynthetic
rate at saturation irradiance (Agmax, 8.92 µmol m-2
s-1
). At the same irradiance, the MV variant
showed greater stem diameter, greater stem mass ratio (CMR), higher starch (AM)
concentration in roots, total soluble sugars content (AST) in leaves and higher specific leaf
mass, while the LV variant showed higher relative growth rate (RGR), higher RGRs of height
and stem diameter, higher ratio of carotenoids and total chlorophyll (Car/Chl T), higher rates
of net photosynthesis (A) and transpiration (E) rates, high stomatal conductance (gs) and
higher internal to external CO2 concentration. However, under conditions of 60% of
irradiance, the variant LV showed higher leaf AM content and greater leaf area, whereas the
variant MV showed higher apparent quantum yield value (α, 0.073 µmol CO2 µmol photons-1
)
and higher content of Car. In the 35% irradiance environment, the LV variant showed higher
levels of AST in roots and greater instantaneous and intrinsic water use efficiencies, while the
SV variant showed greater height and leaf dry biomass. In 25% of irradiance, the SV variant
showed higher leaf area ratio, higher levels of stem AM and higher chlorophyll b (Chl b)
concentration, while the variant LV showed high leaf mass ratio and greater ratio of
chlorophyll a (Chl a) to Chl b (Chl a/b). The variant MV showed higher contents of Chl a and
Chl T. However, at 15% irradiance, the SV variant showed higher content of AST in the stem,
while the MV variant showed higher α and lower respiration rate (Rd, 0,16 µmol m-2
s-1
). In
summary, the morphological variants of brazilwood adjusted their photosynthetic apparatus,
in order to maximize energy gains, through differential physiological plasticity, during
acclimation to the light radiation environments. The variation of irradiance positively
influenced the assimilation of CO2, when related to gs and E, giving conditions to the
morphological variants to achieve their growth potential, demonstrated by the increased RGR
with increasing light intensity. Changes in biometric patterns of brazilwood variants were
affected by irradiance, demonstrated by variations in biomass gain determined by NAR and
RGR. There was a differentiation of the morphological variants in relation to carbon
ix

10
assimilation and the change in irradiance. The LV variant is better acclimated in 100%
irradiance; while VS and MV variants better acclimatized under conditions of 35% and 15%
of irradiance, respectively.
Keywords: C. echinata, stress irradiance, biomass production, morphotypes of Brazil wood,
leaf gas exchange, chloroplastidic pigments, carbohydrates.
x

11
LISTA DE TABELAS
Tabela 1 Análise físico-química do solo utilizado no experimento
31
Tabela 2 Equações utilizadas na determinação das concentrações de clorofilas a (Chl
a), b (Chl b) e total (Chl T) e de carotenóides (Car).
33
Tabela 3 Valores médios, máximos e mínimos, e porcentagem em relação ao pleno
sol, para os diferentes ambientes de sombreamento. Dados coletados em
Ilhéus, BA, Brasil, entre agosto de 2011 e abril 2012.
35
Tabela 4 Parâmetros derivados das curvas de saturação de luz para plantas jovens de
três variantes morfológicos de C. echinata crescidas em diferentes níveis de
irradiância. Médias intra e intermorfotípicas, seguidas pelas mesmas letras
maiúsculas e minúsculas, respectivamente, não diferem entre si pelo teste de
Duncan (p≤0,05). Valores médios de cinco repetições (± EP).
39
Tabela 5 Taxa fotossintética líquida por unidade de área foliar (A), condutância
estomática ao vapor de água (gs), taxa transpiratória (E) e razão entre a
concentração intercelular e ambiente de CO2 (Ci/Ca) de plantas jovens de
três variantes morfológicos de C. echinata crescidas em diferentes níveis de
irradiância, aos 24 meses após a emergência, em RFA ≥600 μmol fótons m-2
s-1
. Médias intra e intermorfotípicas, seguidas pelas mesmas letras
maiúsculas e minúsculas, respectivamente, não diferem entre si pelo teste de
Duncan (p≤0,05). Valores médios de cinco repetições (± EP).
40
xi

12
LISTA DE FIGURAS
Figura 1 Áreas de concentração de Caesalpinia echinata no Brasil
23
Figura 2 Comparação das folhas entre os três variantes morfológicos da espécie C.
echinata variante folha pequena (SV), variante folha média (MV) e variante
folha grande (LV) nos cinco níveis de irradiância.
24
Figura 3 Radiação fotossinteticamente ativa (RFA) medida a pleno sol (A), no interior
do viveiro (B) e no interior das casas sombreadas com telas sombreadoras de
25% (C), 50% (D) e 75% (E).
36
Figura 4 Curvas de resposta da taxa fotossintética líquida (A) à radiação
fotossinteticamente ativa (RFA) para plantas jovens dos variantes morfológicos
folha pequena (SV) [A], folha média (MV) [B] e folha grande (LV) [C] de C.
echinata crescidas em diferentes níveis de irradiância [100% (◊), 60% (□),
35% (Δ), 25% (○) e 15% (×) de irradiância], aos 24 meses após o plantio.
Pontos são médias de cinco repetições (± EP). Equação: A = Amax (1- exp (-α
RFA/Amax)) - Rd.
38
Figura 5 Eficiências instantânea (A/E) (A) e intrínseca (A/gs) do uso da água (B) em
plantas jovens de três variantes morfológicos de C. echinata (folha pequena
(SV); folha média (MV) e folha grande (LV)) crescidas em diferentes níveis de
irradiância, aos 24 meses após a emergência. Médias intra e intermorfotípicas,
seguidas pelas mesmas letras maiúsculas e minúsculas, respectivamente, não
diferem entre si pelo teste de Duncan (p≤0,05). Valores médios de cinco
repetições (± EP).
41
Figura 6 Teores de clorofila a (Chl a) (A), clorofila b (Chl b) (B) e razão entre os teores
de Chl a e Chl b (Chl a/b) (C) em folhas de plantas jovens de três variantes
morfológicos de C. echinata (folha pequena (SV); folha média (MV) e folha
grande (LV)) crescidas em diferentes níveis de irradiância aos 24 meses após a
emergência. Médias intra e intermorfotípicas, seguidas pelas mesmas letras
maiúsculas e minúsculas, respectivamente, não diferem entre si pelo teste de
Duncan (p≤0,05). Valores médios de cinco repetições (± EP).
43
Figura 7 Teores de carotenóides (Car), (A) clorofila total (Chl T) (B) e razão entre os
teores de carotenóides e clorofila total (Car/Chl T) (C) em folhas de plantas
jovens de três variantes morfológicos de C. echinata (folha pequena (SV);
folha média (MV) e folha grande (LV)) crescidas em diferentes níveis de
irradiância, aos 24 meses após a emergência. Médias intra e intermorfotípicas,
seguidas pelas mesmas letras maiúsculas e minúsculas, respectivamente, não
diferem entre si pelo teste de Duncan (p≤0,05). Valores médios de cinco
repetições (± EP).
44
xii

13
Figura 8 Número de folhas (A), altura total (B) e diâmetro do caule (C) de plantas
jovens de três variantes morfológicos de C. echinata (folha pequena (SV);
folha média (MV) e folha grande (LV)) crescidas em diferentes níveis de
irradiância aos 24 meses após a emergência. Médias intra e intermorfotípicas,
seguidas pelas mesmas letras maiúsculas e minúsculas, respectivamente, não
diferem entre si pelo teste de Duncan (p≤0,05). Valores médios de cinco
repetições (± EP).
46
Figura 9 Massas secas de raiz (MSR) (A), folha (MSF) (B), caule (MSC) (C) e total
(MST) (D) de plantas jovens de três variantes morfológicos de C. echinata
(folha pequena (SV); folha média (MV) e folha grande (LV)) crescidas em
diferentes níveis de irradiância, aos 24 meses após a emergência. Médias intra e
intermorfotípicas, seguidas pelas mesmas letras maiúsculas e minúsculas,
respectivamente, não diferem entre si pelo teste de Duncan (p≤0,05). Valores
médios de cinco repetições (± EP).
47
Figura 10 Razões de massa de raiz (A), caule (B) e folha (C) de plantas jovens de três
variantes morfológicos de C. echinata crescidas em diferentes níveis de
irradiância. Médias intra e intermorfotípicas, seguidas pelas mesmas letras
maiúsculas e minúsculas, respectivamente, não diferem entre si pelo teste de
Duncan (p≤0,05). Valores médios de dez repetições (± EP)
48
Figura 11 Área foliar específica (AFE) (A), razão de área foliar (RAF) (B) e área foliar
total (AF) (C) de plantas jovens de três variantes morfológicos de C. echinata
crescidas em diferentes níveis de irradiância. Médias intra e intermorfotípicas,
seguidas pelas mesmas letras maiúsculas e minúsculas, respectivamente, não
diferem entre si pelo teste de Duncan (p≤0,05). Valores médios de dez
repetições (± EP).
50
Figura 12 Massa foliar especifica (MFE) (A) e taxa assimilatória liquida (TAL) (B) de
plantas jovens de três variantes morfológicos de C. echinata crescidas em
diferentes níveis de irradiância, aos 24 meses após a emergência. Médias intra e
intermorfotípicas, seguidas pelas mesmas letras maiúsculas e minúsculas,
respectivamente, não diferem entre si pelo teste de Duncan (p≤0,05). Valores
médios de dez repetições (± EP).
51
Figura 13 Taxa de crescimento relativo (TCR) (A), taxa de crescimento relativo em altura
(TCRA) (B) e taxa de crescimento relativo em diâmetro (TCRD) (C) de plantas
jovens de três variantes morfológicos de C. echinata crescidas em diferentes
níveis de irradiância, aos 24 meses após a emergência. Médias intra e
intermorfotípicas, seguidas pelas mesmas letras maiúsculas e minúsculas,
respectivamente, não diferem entre si pelo teste de Duncan (p≤0,05). Valores
médios de dez repetições (± EP).
52
xiii

14
Figura 14 Teor de amido na raiz (A), no caule (B) e na folha (C) de plantas jovens de três
variantes morfológicos de C. echinata (folha pequena (SV); folha média (MV)
e folha grande (LV)) crescidas em diferentes níveis de irradiância aos 24 meses
após a emergência. Médias intra e intermorfotípicas, seguidas pelas mesmas
letras maiúsculas e minúsculas, respectivamente, não diferem entre si pelo teste
de Duncan (p≤0,05). Valores médios de cinco repetições (± EP).
54
Figura 15 Teor de açúcares solúveis totais (AST) na raiz (A), no caule (B) e na folha (C)
de plantas jovens de três variantes morfológicos de C. echinata (folha pequena
(SV); folha média (MV) e folha grande (LV)) crescidas em diferentes níveis de
irradiância aos 24 meses após a emergência. Médias intra e intermorfotípicas,
seguidas pelas mesmas letras maiúsculas e minúsculas, respectivamente, não
diferem entre si pelo teste de Duncan (p≤0,05). Valores médios de cinco
repetições (± EP).
55
xiv

15
LISTA DE ABREVIATURAS E SIGLAS
α Rendimento quântico aparente
A Taxa de fotossíntese líquida por unidade de área foliar
Amax Taxa fotossintética bruta máxima em irradiância de saturação
Rd Taxa respiratória na ausência de luz
E Taxa transpiratória foliar
gs Condutância estomática ao vapor de água
Ci/Ca Razão entre a concentração intercelular e ambiente de CO2
RFA Radiação fotossinteticamente ativa
NF Número de folhas
AF Área foliar
DC Diâmetro do caule
ALT Altura de planta
MSC Massa seca de caule
MSF Massa seca de folha
MSR Massa seca de raiz
MST Massa seca total
MFE Massa foliar especifica
RAF Razão de área foliar
RMC Razão massa de caule
RMF Razão massa de folha
RMR Razão massa de raiz
TCR Taxa de crescimento relativo
TCRA Taxa de crescimento relativo em altura
TCRD Taxa de crescimento relativo em diâmetro
TAL Taxa assimilação liquida
AST Açúcar solúvel total
SV Variante morfológico folha pequena
MV Variante morfológico folha média
LM Variante morfológico folha grande
xv
iii

16
SUMÁRIO
RESUMO .......................................................................................................................... vii
ABSTRACT ....................................................................................................................... ix
1. INTRODUÇÃO................................................................................................................. 17
2. OBJETIVOS ...................................................................................................................... 20
2.1 Geral .................................................................................................................................. 20
2.2 Específico .......................................................................................................................... 20
3. REVISÃO DE LITERATURA ......................................................................................... 21
3.1 História e descrição da espécie .......................................................................................... 21
3.2 Respostas fisiológicas das plantas à variação da irradiância ............................................. 25
4. MATERIAL E MÉTODOS............................................................................................... 31
4.1 Espécies e condições de cultivo ........................................................................................ 31
4.2 Medições de trocas gasosas foliares .................................................................................. 32
4.3 Determinação dos teores de clorofila e carotenóides ........................................................ 33
4.4 Avaliação de crescimento .................................................................................................. 33
4.5 Determinação dos teores de carboidratos .......................................................................... 34
4.6 Análise estatística .............................................................................................................. 34
5. RESULTADOS ................................................................................................................. 35
6. DISCUSSÃO ..................................................................................................................... 56
7. CONCLUSÕES ................................................................................................................. 65
8. REFERÊNCIAS ................................................................................................................ 66
xvi

17
1. INTRODUÇÃO
O efeito do sombreamento sobre os processos fisiológicos de diversas culturas é
complexo, envolvendo, além dos fatores bióticos fundamentais, a variação da intensidade e
qualidade da luz, a temperatura, o movimento do ar, a umidade relativa do ar e a umidade do
solo. Desta forma, observados os fatores dinâmicos do desenvolvimento, plantas que crescem
sob a sombra desenvolvem estrutura e aparência diferentes daquelas que crescem sob plena
luz (COSTA et al., 1998; ZHANG et al., 2003; DAMATTA, 2004).
A radiação luminosa influencia o crescimento e a distribuição das espécies vegetais
em diversos ecossistemas (CHAZDON et al., 1996; BOARDMAN, 1977; LAMBERS et al.,
1998). Os valores de radiação fotossinteticamente ativa (RFA) variam temporal e
espacialmente, induzindo as plantas a desenvolverem mecanismos de aclimatação e
plasticidade à variação dos regimes luminosos (ZHANG et al., 2003). Níveis altos ou baixos
de luz podem ocasionar decréscimo na produção de carboidratos, diminuição no crescimento
e desenvolvimento, além de danificar o maquinário fotossintético (LAMBERS et al., 1998).
Decorrente destes fatores, as plantas desenvolveram várias estratégias para enfrentar a
diversidade de influências bióticas e abióticas, como a plasticidade de aclimatação às
condições de sol e sombra (DEMMIG-ADAMS; ADAMS, 1996).
Respostas às variações da irradiância estão relacionadas à aclimatação do sistema
fotossintético, como resultado das mudanças dos padrões de crescimento e distribuição de
fotoassimilados (CHAZDON et al., 1996). As plantas possuem elevada capacidade de
aclimatação às diferentes intensidades de radiação luminosa. Isso ocorre em função da
regulação morfológica, da composição e da estrutura fisiológica, determinando alterações nas
vias metabólicas primárias e secundárias (DAMATTA, 2004). A planta busca, por meio
destas variações, uma resposta fisiológica, visando alcançar um estado funcional máximo
(NOBEL, 1991; CHAZDON et al., 1996).

18
Diferenças na morfologia, anatomia e fisiologia têm sido documentadas para
espécies aclimatadas a ambientes sombreados ou a pleno sol (BOARDMAN, 1977;
BJÖRKMAN, 1981; GIVNISH, 1988; LARCHER, 1995). Diversos atributos fisiológicos
podem estar associados à aclimatação ou à adaptação a irradiância, como: (i) alterações nas
taxas fotossintéticas e respiratórias (BJÖRKMAN, 1981; GIVNISH, 1988; CHAZDON et al.,
1996); (ii) aumento na concentração e atividade da ribulose-1,5-bisfosfato carboxilase-
oxigenase - Rubisco (KANECHI et al., 1996; RAMALHO et al., 1999); (iii); recuperação
relativamente rápida da fotoinibição (DAMATTA; MAESTRI, 1997); dentre outros.
As plantas desenvolveram diversos mecanismos para maximizar a aquisição de luz e
evitar condições ambientais desfavoráveis. Esses mecanismos são empregados na
coordenação de mudanças no crescimento e desenvolvimento, resultando em plasticidade
fenotípica, otimização de recursos e capacidade para evitar ou tolerar condições menos
favoráveis (ALVES et al., 2002). Para estes mecanismos, a luz participa diretamente da
fotossíntese, influenciando a abertura estomática e a síntese de clorofila (KOZLOWSKI et al.,
1991; FELFILI et al., 1999), provocando alterações morfofisiológicas, com grau de
aclimatação relacionado às características genéticas e condições edafoclimáticas (ALVES et
al., 2002).
A luz fornece energia para a fotossíntese da planta e envia sinais que regulam seu
desenvolvimento por meio de receptores de luz sensíveis a diferentes intensidades, variações
na qualidade espectral, quantidade, presença ou ausência e estado de polarização (POGGIANI
et al., 1992). Modificações nos níveis de irradiância condicionam diferentes respostas
fisiológicas em suas características bioquímicas, anatômicas e de crescimento (POGGIANI et
al., 1992; ATROCH et al., 2001).
A radiação solar fornece para a atmosfera 1367 W m-² de energia ano
-1. Cerca de
metade desta energia é recebida como luz visível na parte de frequência mais alta do espectro
eletromagnético e o restante na do infravermelho e como radiação ultravioleta
(GUIMARÃES, 2003). Somente a energia luminosa, dentro de uma faixa reduzida de
comprimento de onda, é captada pelas plantas, alterando as reações químicas ou induzindo
correntes elétricas nos tecidos e células. Dessa forma, o processo fotossintético, em que essa
energia é capturada, é usado na produção de carboidratos, participando da morfogênese e das
respostas de crescimento diferenciadas em função das irradiância recebida (REID et al.,
1991). O desenvolvimento das plantas exterioriza a capacidade de aclimatação das espécies às
condições de irradiância do ambiente em que estão se desenvolvendo, no sentido de
maximizar o ganho total de carbono (ALMEIDA et al., 2005).

19
No presente estudo, avaliaram-se as respostas de variantes morfológicos de plantas
jovens de Caesalpinia echinata (pau-brasil) ao sombreamento. Esta espécie é endêmica da
floresta atlântica brasileira, ocupa o estrato médio da floresta. Sua longevidade chega a 300
anos, necessitando de sombreamento moderado na sua fase juvenil. Apresenta crescimento
lento e irregular, brotações após o corte, e não são recomendados plantios a pleno sol
(RIZZINI, 1971; CARVALHO, 2003). É encontrada em regiões semiáridas, típica da floresta
primária densa, próximas do litoral e raramente nas formações secundarias (CARVALHO,
2003). Ocorre naturalmente desde o estado do Rio Grande do Norte até o Rio de Janeiro
(LEWIS, 1998). Na região sul da Bahia, C. echinata pode ser encontrada em condições de
Cabruca, um sistema agroecológico de cultivo onde a espécie Theobroma cacao é cultivada
na floresta atlântica raleada, sob a sombra de espécies arbóreas de grande porte, responsável
pela manutenção dos maiores e bem mais distribuídos remanescentes de pau-brasil (LOBÃO
et al., 2007).

20
2. OBJETIVOS
2.1 Geral
Avaliar o crescimento, a assimilação do carbono e a composição química de três
variantes morfológicos de pau-brasil submetidos a diferentes níveis de irradiância.
2.2 Específico
Realizar medições de trocas gasosas foliares; determinar os teores foliares de
clorofilas e carotenóides, bem como os de carboidratos em folhas, caules e raízes ; e avaliar o
crescimento de três variantes de pau-brasil submetidos a diferentes níveis de irradiância.

21
3. REVISÃO DE LITERATURA
3.1 História e descrição da espécie
O Brasil foi descoberto na região sul da Bahia e, por se tratar de uma região muito
rica em espécies florestais, como o pau-brasil, o interesse da coroa portuguesa voltou-se
imediatamente à sua exploração econômica (LOBÃO, 2007). O pau-brasil passou a ser o
centro das atenções durante boa parte da colonização brasileira, tornando-se a primeira grande
riqueza, procurada e disputada por varias nações (CASTRO, 2002). A razão principal foi a
brasilina, o corante vermelho extraído do pau-brasil que servia para tingir tecidos, pintar ovos
de páscoa e era utilizado como base para cosméticos, como o atual rouge. A madeira do pau-
brasil, sendo de alta qualidade, era empregada em construções civis, fabricação de móveis de
luxo e arcos de violino. Os indígenas fabricavam armas que lhes serviam na guerra, caça e
pesca (BARBEDO et al., 2002). Em 1789 o naturalista francês Jean Baptiste Lamarck (1744 a
1829) estudou e descreveu a espécie cientificamente, denominando-a por Caesalpinia
echinata. O nome do gênero Caesalpinia foi dado em homenagem ao botânico e médico
Andreas Caesalpinus que viveu entre 1519 a 1603, e o nome específico echinata por ser uma
árvore que possui acúleos, semelhantes aos encontrados nos troncos e galhos de roseiras
(CARVALHO, 2003).
No início do século XVII, o pau-brasil estava quase erradicado de suas áreas naturais
por causa da extensa exploração (LOBÃO et al., 1997; BARBEDO et al., 2002; CASTRO,
2002). Hoje está relacionado na lista oficial de espécies da flora brasileira ameaçadas de
extinção (IBAMA, 2013). Na área de ocorrência natural, ainda existem pequenos fragmentos
da espécie de pau-brasil, como em alguns pontos do sul da Bahia, a exemplo da Reserva
Ecológica do Pau-Brasil, em Porto Seguro, BA, em Pernambuco e no Rio de Janeiro (Figura
1) (CARVALHO, 2003). Recentemente, foi encontrado um núcleo populacional
completamente fora da área conhecida de ocorrência natural, na Chapada da Conquista, BA
(CARVALHO, 2003).

22
Caesalpinia echinata é uma espécie lenhosa pertencente à família Fabaceae,
semidecídua, podendo alcançar 30 m de altura e 40 cm de diâmetro, possui tronco reto, com
casca cor cinza escura, coberta por acúleos, especialmente nos ramos mais jovens. As folhas
são perenes, compostas e alternadas, na grande maioria, bipinadas e apresentam coloração
verde-escura brilhante, são ovais e subdivididas em pinas e estas em folíolos (LEWIS, 1998).
Suas flores são aromáticas, de coloração amarelo-ouro, formando um cacho cônico
(CARVALHO, 2003). Quando jovem, o alburno é espesso e apresenta coloração branco-
amarelada, diminuindo com a idade, cedendo lugar ao cerne, de cor vermelha intensa, mais
resistente à umidade. O cerne é a parte mais valiosa, fonte do corante brasilina (CARVALHO,
2003).
A espécie C. echinata possui três variantes morfológicos, popularmente conhecidos
como folha-de-arruda, folha-de-café e folha-de-laranja (Figura 2), porém, ainda não
classificados botanicamente. Doravante passaremos a identificá-los como pau-brasil variante
pequeno, médio e grande, baseados no tamanho da folha, de acordo com Juchum et al. (2008).
Estes autores designaram os variantes morfológicos de pau-brasil como variante folha
pequena (small variant - SV), variante folha média (medium variant - MV) e variante folha
grande (large variant - LV), respectivamente (JUCHUM et al., 2008).
O variante morfológico SV, encontrado com maior frequência e maior distribuição
geográfica por toda a costa brasileira, do Rio de Janeiro ao Rio Grande do Norte, tem como
característica folhas pequenas, alternas, compostas bipinadas, constituída normalmente de sete
a 11 pinas e de nove a 22 folíolos. O variante morfológico MV, mais comumente encontrado
nos estados do Espírito Santo e Bahia, possui folha de tamanho médio, constituída de três a
seis pinas e de sete a 12 folíolos. O variante morfológico LV, espécie de ocorrência restrita na
região cacaueira do sudeste da Bahia, possui folha grande, com duas a quatro pinas e quatro a
seis folíolos. Isoladamente, esta árvore atinge a altura de oito a 12 m. Na mata pode alcançar o
dossel florestal de até 30 m (CURVELO et al., 2008).
Na natureza, a disponibilidade de luz influencia diretamente o crescimento e o porte da
C. echinata. Nestas condições, competindo por luminosidade, a espécie apresenta crescimento
lento, com caule retilíneo e diâmetro reduzido. Em contrapartida, na ausência de competição
por luz, há formação mais rápida da copa, seguida de maior diâmetro do caule, além do
aumento do número de ramificações (LIMA et al., 2002).

23
Figura 1 - Áreas de concentração de Caesalpinia echinata no Brasil Fonte: LOBÃO (2007), com modificações.

24
Figura 2 – Comparação das folhas entre os três variantes morfológicos da espécie C. echinata
variante folha pequena (SV), variante folha média (MV) e variante folha grande
(LV) nos cinco níveis de irradiância.

25
3.2 Respostas fisiológicas das plantas à variação da irradiância
O desenvolvimento das plantas é influenciado por fatores ambientais e sua
modificação traz como consequência diferentes respostas. A irradiância intervém em
numerosos processos fisiológicos, dos quais o mais importante é a assimilação de carbono
(fotossíntese), atribuindo valores e escalas de importância na produtividade dos ecossistemas
(CARVALHO, 2003). A aclimatação do maquinário fotossintético e as alterações anatômicas
proporcionam variações nos padrões de crescimento e alocação de biomassa como respostas à
variação da irradiância (CHAZDON et al., 1996).
A radiação solar torna possível a realização da fotossíntese e o aquecimento do ar e do
solo, favorecendo os processos vitais da planta. É necessário que a planta receba de 1 a 2% de
radiação solar para que possa se manter. Na natureza, a luta pela sobrevivência exige que a
planta obtenha o máximo de irradiância para acentuar os ganhos fotossintéticos. Assim, por
meio do processo fotossintético, a energia radiante é fixada em energia química potencial,
interferindo diretamente na produtividade (FLORES et al., 2004).
A atividade fotossintética é afetada diretamente pela intensidade de radiação luminosa,
temperatura, concentração de CO2, teor de nitrogênio da folha e a umidade do solo, estando o
funcionamento dos estômatos, abertura e fechamento, diretamente relacionado à intensidade
luminosa e ao estatus hídrico da folha (MARENCO e LOPES, 2005). Costa e Marenco (2007)
demonstraram que a relação entre funcionalidade dos estômatos e a área foliar influenciam a
produtividade vegetal, com estômatos controlando a absorção de CO2 e a área foliar a
captação e interceptação de luz. Para Larcher (2000), as trocas gasosas variam durante o
desenvolvimento e são diretamente proporcionais ao curso diário e anual das flutuações
abióticas ambientais ao nicho onde a planta está localizada.
A parte aérea das plantas recebe diversos tipos de radiação luminosa durante o dia.
Larcher (2000) classifica essa radiação como sendo a radiação solar direta, a radiação que
sofre espalhamento na atmosfera, a radiação difusa em dias nublados e a radiação refletida da
superfície do solo. A radiação na copa das árvores também sofre variação em função da forma
de crescimento, do tipo de ramificação e da posição da folha, permitindo que a planta
condicione o melhor ordenamento da superfície de assimilação no sentido de permitir melhor
arranjo espacial para que as folhas recebam radiação solar direta de forma permanente
(ZHANG et al., 2003). As plantas normalmente se aclimatam, por meio de plasticidade
fenotípica, às condições de radiação ambiente. A diferenciação fenotípica de órgãos e tecidos

26
geralmente não é reversível. Se as condições de radiação mudam, novos ramos são produzidos
e as folhas dos ramos originais não adaptadas senescem e sofrem abscisão (COSTA et al.,
1998; ZHANG et al., 2003; DAMATTA, 2004).
A fotossíntese de plantas sombreadas requer maximização da quantidade de radiação
absorvida, relacionada a pequenas taxas de perda de carbono por meio da respiração e
fotorrespiração (ZHANG, et al., 2003). A aclimatação das plantas ao ambiente de radiação
depende do ajuste de seu maquinário fotossintético, de modo que a radiação seja utilizada da
maneira mais eficiente possível (ENGEL, 1989). As respostas a essa adaptação são refletidas
no crescimento global da planta. Bunce et al. (1977) e Kamaluddin; Grace (1992) observaram
que o crescimento está relacionado com a capacidade de adaptação das plantas às variações de
intensidade de radiação luminosa do ambiente, tanto para espécies arbóreas da floresta
tropical como para espécies arbustivas.
Boardman (1977), Kamaluddin e Grace, (1992), Müller et al., (1992) constataram que
a taxa fotossintética, respiração, eficiência quântica do fotossistema 2 (PS2), anatomia,
estrutura foliar e características fenológicas, como longevidade da folha, podem ser alteradas
pelo efeito da intensidade da radiação durante a ontogenia foliar. Sims e Pearcy (1991) e Chen
e Klinka (1997) observaram que plantas expostas ao sombreamento apresentam valores
extremamente reduzidos de irradiância de compensação, fundamentado, principalmente, pelos
baixos valores da taxa respiratória na ausência de luz (Rd).
Folhas de sombra possuem maior área foliar, menor massa foliar específica e menor
quantidade de cloroplastos se comparadas com as de sol, devido à redução da espessura do
mesofilo, a existência de grana mais largas e maior teor de clorofila; ao passo que as folhas de
sol alcançam taxa fotossintética máxima e são mais espessas, devido à formação de grandes
células paliçádicas e, ou aumento no número de camadas dessas células (LAMBERS et al.,
1998).
De acordo com Björkman (1981), Anderson (1986), Anderson e Osmond (1987),
Terashima e Hikosaka (1995) e Noguchi et al. (1996), folhas a pleno sol e sombreadas
apresentam diferenças estruturais, tendo quantidades diferentes de Rubisco, citocromos,
centros de reação dos fotossistemas 1 (PS1) e PS2 e atividades de enzimas respiratórias. Para
Evans (1989), Hikosaka; Terashima (1995) em irradiâncias elevadas a quantidade de Rubisco
e o centro de reação de PS2 aumentam com a expansão do complexo coletor de luz de PS2.
A folha é especializada na absorção de radiação luminosa e as propriedades
morfológicas e estruturais entre os parênquimas paliçádico e lacunoso resultam em uma
absorção de radiação de forma mais uniforme (TAIZ; ZEIGER 2009). Baixos níveis de

27
radiação luminosa podem ocasionar estresses nas plantas em decorrência da diminuição da
taxa de assimilação de CO2, do decréscimo na produção de carboidratos e da diminuição no
crescimento e no desenvolvimento. Em contrapartida, níveis elevados de intensidade
luminosa podem danificar o maquinário fotossintético (LAMBERS et al., 1998), de forma, às
vezes, irreversível. De acordo com Souza et al. (2009), em uma mesma copa, as folhas
sujeitas a diferentes regimes de radiação apresentam diferenças anatômicas, como a redução
no comprimento das células do parênquima paliçádico e na espessura do mesofilo nas folhas
sombreadas. A arquitetura da vegetação afeta não somente as trocas de matéria e energia entre
a planta e o ambiente, como também pode revelar a estratégia da planta para se ajustar aos
processos evolutivos, como adaptações aos fatores físicos, químicos e biológicos (SOUZA et
al., 2009). Para Demmig-Adams e Adams (1996), estes processos de ajuste tratam-se de
plasticidade e aclimatação às condições de sol e sombra, bem como mecanismos de
dissipação do excesso de energia.
Diversos atributos morfofisiológicos estão associados à aclimatação em irradiâncias
elevadas, como (i) o aumento na densidade estomática (CAI et al., 2005); (ii) as mudanças na
fotossíntese e respiração (GIVNISH, 1988; CHAZDON et al., 1996); (iii) as mudanças nos
padrões de crescimento ou de alocação de biomassa (POPMA; BONGERS, 1991; POORTER
et al., 2003); (iv) a diminuição da área foliar específica; (v) a presença de cutícula mais
espessa (VOLTAN et al., 1992; FAHL et al., 1994), com maior quantidade de cera
epicuticular (AKUNDA et al., 1979); (vi) os cloroplastos com poucos grana e menor
quantidade de tilacóides por granum (FAHL et al., 1994); (vii) os aumentos na concentração e
atividade da Rubisco (KANECHI et al., 1996); e (viii) a recuperação relativamente rápida da
fotoinibição (DAMATTA; MAESTRI, 1997).
Ainda assim, as alterações na fisiologia da folha e a produção de novas folhas com
morfologia e fisiologia apropriadas ao ambiente luminoso são componentes de respostas à
aclimatação. Além das mudanças físicas, ocorrem alterações na relação pigmentar de
clorofilas e pigmentos acessórios (CHAVES, 2009). Os pigmentos envolvidos na fotossíntese
são as clorofilas a e b, os carotenóides e as ficobilinas. A clorofila b (Chl b), os carotenóides e
as ficobilinas constituem os chamados pigmentos acessórios. As plantas utilizam a radiação
fotossinteticamente ativa (RFA), compreendida entre os comprimentos de onda de 390 e 760
nm, denominada espectro visível (STREIT et al., 2005). A energia absorvida é transferida
para os centros de reação, nas membranas tilacóidais. Existem dois centros de reação: (i) um
absorvendo energia em 680 nm e (ii) outro a 700 nm, interagindo por meio de transportadores
de elétrons (STREIT et al., 2005).

28
A clorofila é um pigmento fotossintético presente nas plantas superiores e a sua
abundância varia de acordo com a espécie. A clorofila a (Chl a) está em presente em todos os
organismos que realizam fotossíntese, o primeiro estágio do processo fotossintético; enquanto
que os pigmentos acessórios auxiliam na absorção de luz e na transferência da energia
radiante para os centros de reação (TAIZ; ZIEGER, 2009). A concentração e a proporção de
pigmentos fotossintéticos das folhas variam com a espécie, o meio e a idade da folha
(KRAMER; KOZLOWSKI, 1979; POPMA; BONGERS, 1991).
As Chl a e b absorvem radiação visível principalmente na faixa do azul e do vermelho
e os carotenóides entre o azul e o ultravioleta (STREIT et al., 2005). A partir da molécula de
Chl a, cujo pico de absorção de irradiância é no comprimento de onda de 680 nm do espectro
visível, os elétrons oriundos da oxidação da molécula de água são transferidos para a cadeia
transportadora de elétrons da fotossíntese. Conforme Engel e Poggiani (1991), a combinação
entre as Chl a e Chl b e os pigmentos acessórios permite maior captação da energia luminosa,
conferindo às plantas maior eficiência fotossintética, crescimento, plasticidade e
adaptabilidade.
Os carotenóides, pigmentos amarelos e laranjas, desempenham duas funções distintas:
(i) absorção de luz nos complexos de captação e (ii) ação fotoprotetora do maquinário
fotoquímico, prevenindo danos foto-oxidativos às moléculas de clorofila. Absorvem a luz
azul e repassam energia para a clorofila para uso na fotossíntese (KERBAUY, 2004). De
acordo com Oliveira et al. (2008), os carotenóides dissipam, na forma de calor, a energia
química armazenada pelas clorofilas.
Logo, clorofilas e carotenóides absorvem radiação visível, desencadeando as reações
fotoquímicas da fotossíntese responsáveis pelo metabolismo primário (SEIFERMAN-
HARMS, 1987). A qualidade e a quantidade da radiação são responsáveis pelo tipo,
quantidade e incorporação de carotenóides no maquinário fotossintético (MACMAHON et
al., 1991). A faixa espectral absorvida necessária para ativar o processo fotossintético está
relacionada diretamente ao tipo de pigmento existente, variando essa absorção energética
entre o azul e o vermelho.
A aclimatação das plantas a uma determinada condição de irradiância está associada
aos teores de clorofilas e nitrogênio foliar (ALVARENGA et al., 2003). A molécula de
clorofila encontra-se em constante processo de síntese e degradação (foto-oxidação)
(BRAND, 1997). Sob alta irradiância, a foto-oxidação ocorre de forma pronunciada, enquanto
sob condições de sombreamento, as concentrações foliares de clorofilas tendem a aumentar
(BRAND, 1997; ALVARENGA et al., 2003).

29
De acordo com Larcher (2000), plantas de sombra realizam um ganho fotossintético
igual à metade ou até um terço em relação às plantas de sol; ao passo que espécies que
desenvolvem pequena área de assimilação (acículas ou folhas pequenas) interceptam pouca
radiação e apresentam um modesto ganho fotossintético. Segundo Souza et al. (2009), os
maiores valores de absorbância são encontrados nas folhas de sombra em comparação às de
sol, indicando maior concentração de pigmentos naquelas folhas. Tais alterações possibilitam
a maximização da captura de radiação (NAKAZONO et al., 2001).
Os teores de clorofilas são mais elevados em folhas sombreadas do que as expostas à
radiação solar plena (SOUZA et al., 2009; Araújo, 2009). Em plantas de sol, ambiente
intensamente iluminado, é menor a concentração de moléculas de clorofila por cloroplasto,
principalmente de Chl b, uma vez que essas plantas não necessitam investir na produção de
pigmentos coletores de energia luminosa (SALISBURY; ROSS, 1992). A maior proporção
relativa de Chl b, em plantas sombreadas, permite a captação de energia de comprimentos de
onda diferentes, com a correspondente transferência energética para uma molécula específica
de Chl a, participante das reações fotoquímicas da fotossíntese (WHATLEY; WHATLEY,
1982; TAIZ; ZEIGER, 2009). Segundo Thornber (1975), o aumento relativo do teor de
clorofila pode estar ligado ao incremento da proporção do complexo coletor clorofila a/b,
proteína associada ao PS1 nos grana, em relação ao complexo P680 (que contém somente Chl
a – proteína ) do PS2 nos tilacóides, facilmente foto-oxidado, assim como abordou Mitchell
(1979), ao indicar a existência de maior proporção de grana em cloroplastos de folhas de
sombra.
Quando folhas sombreadas são expostas a alta irradiância ocorre fotoinibição da
fotossíntese (OBERBAUER et al., 1985) e alterações anatômicas qualitativas de seus
constituintes (WALLER, 1986; LAMBERS; POORTER 1992). Para Bjorkman et al. (1972),
folhas de sombra investem maior energia na produção de pigmentos fotossintetizantes,
procurando otimizar a captura de luz incidente. De acordo com Pezzopane et al. (2003), Lunz
(2006) e Costa et al. (2011), as alterações microclimáticas, provocadas pela utilização de
sistemas de sombreamento, que modificam a temperatura e umidade relativa do ar e a
radiação solar, influenciam no desenvolvimento e crescimento vegetal, mostram alterações
fisiológicas e são dependentes da intensidade, qualidade e duração da radiação solar.
Independentemente das alterações que ocorrem nos pigmentos, a variação da intensidade de
irradiância, conforme a capacidade de captação e absorção da planta, afeta a quantidade de
matéria da planta, sua biomassa e sua biometria, evidenciando aspectos fisiológicos distintos
para indivíduos expostos à grande variação de incidência luminosa.

30
A análise de crescimento permite avaliar o desempenho da planta e a contribuição dos
diferentes órgãos no crescimento total. Com base em dados amostrais de crescimento é
possível estimar as causas de variações de crescimento entre plantas geneticamente diferentes
ou entre plantas crescendo em ambientes diferentes (BENINCASA, 1988). A capacidade de
uma determinada espécie em aclimatar-se à condição de sol ou de sombra pode ser
evidenciada pela avaliação do crescimento inicial das plantas em diferentes condições de
disponibilidade luz. Espécies sombreadas, que dependem de maior irradiância, possuem a
capacidade de crescer em altura mais rapidamente (ENGEL, 1989; VALLADARES et al.,
2000; ZANELLA et al., 2006). Logo, altura e diâmetro de caule são os parâmetros usados
com maior frequência na avaliação do crescimento das plantas em relação ao sombreamento
(VERDIAL et al., 2000; ZANELLA et al., 2006). O crescimento em diâmetro depende da
atividade cambial, que é estimulada por carboidratos produzidos pela fotossíntese e por
hormônios transportados das regiões apicais (SCALON et al., 2001). O diâmetro é indicador
da assimilação líquida de carbono, logo depende diretamente da fotossíntese (ENGEL, 1989),
e garante maior sustentação a planta (SCALON et al., 2001). Adicionalmente, determinações
de área foliar, produção de matéria seca e relações de biomassa entre as partes aérea e
radicular também fazem parte da análise de crescimento (FARIAS et al., 1997).
A determinação do ponto de crescimento satisfatório, ou ótimo, de algumas espécies,
crescidas em diferentes percentuais de luminosidade, pode ser atribuída à capacidade de
ajustes no mecanismo fotossintético, no sentido de maximizar a aquisição de energia (DIAS-
FILHO, 1997; ALVES et al., 2002), e à concentração de carboidratos (açúcares e amido), cuja
relação está diretamente relacionada ao desenvolvimento da planta. Durante o processo
fotossintético, a maioria das espécies vegetais produz sacarose e acumula quantidade
suficiente de amido, durante o período de luz, utilizado principalmente para suprir a demanda
de energia no período escuro (ZEEMAN et al., 2004; AMARAL 2007), e a sacarose
translocada para sustentar o crescimento (MACHADO et al., 1989). Tecidos
fotossinteticamente ativos possuem maior capacidade de produção de carboidratos do que
necessitam, para a manutenção do metabolismo e crescimento. O excedente, na forma de
sacarose, é transportado para tecidos menos fotoativos ou inativos, como folhas jovens, ramos
e raízes, que, após o florescimento, são direcionados para os frutos, tubérculos e raízes de
reserva (DANTAS et al., 2007). Além de fonte de reserva, a formação e a utilização dos
carboidratos não estruturais desempenham papéis importantes na distribuição de matéria seca
entre raiz e parte aérea (MACHADO et al., 1989).

31
4. MATERIAL E MÉTODOS
4.1 Espécies e condições de cultivo
O experimento foi conduzido entre agosto de 2011 a abril de 2012, no Campus Soane
Nazaré de Andrade, Universidade Estadual de Santa Cruz (UESC), Rodovia Jorge Amado,
km 16, Bairro Salobrinho, localizado no município de Ilhéus, BA, nas coordenadas
39°13’59’’ de longitude oeste e 14°45’15’’ de latitude sul.
Foram utilizadas plantas jovens, com dois anos de idade, de três variantes
morfológicos de pau-brasil, nativos da Mata Atlântica Brasileira, obtidos de sementes,
coletadas de árvores da floresta atlântica do sul da Bahia, na Estação Ecológica Pau-Brasil –
ESPAB/CEPLAC, localizada no município de Porto Seguro, BA. As sementes foram
germinadas em tubetes plásticos pretos cônicos de 235 cm3, contendo composto orgânico
(casca de Pinus e fibra de coco, na proporção de 1:1) e adubo peletizado (Osmocote®).
Posteriormente, as mudas, com um ano de idade, foram transplantadas para vasos plásticos
com capacidade de 20 L, contendo solo como substrato, e transportadas para o viveiro da
UESC, onde permaneceram por 60 dias. Posteriormente, parte das plantas jovens dos
diferentes morfotipos foi mantida no viveiro e parte transferida para casas de sombra com
telas sombreadoras de 25% (C), 50% (D) e 75% (E), e para a condição de pleno sol
(tratamento controle). Durante o experimento foram realizadas adubações com fosfato mono-
amônio (MAP) purificado, uréia e KNO3, na dosagem de 4,21, 3,07 e 2,93 g L-1
,
respectivamente, a cada 15 dias, por um período de 244 dias, de acordo com a análise físico-
química do solo (Tabela 1).
Tabela 1 – Análise fisico-química do solo utilizado no experimento.
pH Al Ca Mg Ca + Mg K P
[CaCl2] [cmolc dm-3
] [mg dm-3
]
4,7 0,4 7 3,5 10,4 0,16 63

32
Os valores de radiação fotossinteticamente ativa (RFA) medidos na extremidade
superior das plantas, submetidas aos diversos tratamentos de irradiância, foram obtidos com
sensor de radiação luminosa S-LIA-M003, acoplado a uma estação climatológica Hobo Micro
Station Data Logger (Onset, EUA).
4.2 Medições de trocas gasosas foliares
A avaliação das trocas gasosas foi realizada no final do experimento, sempre entre
8:00 e 12:30 h, em folha completamente expandida e madura de cinco plantas de cada
variante morfológico por tratamento. A curva de saturação de radiação luminosa foi obtida
utilizando-se sistema portátil para medições de fotossíntese LI-6400 (Li-Cor, Nebraska,
USA), equipado com uma fonte de luz artificial 6400-02B RedBlue. Foram realizadas
medições em nove níveis de RFA (0, 25, 50, 100, 200, 400, 600, 800 e 1000 µmol fótons m-2
s-1
), iniciando-se a sequência sempre em ordem decrescente.
O tempo mínimo fornecido para a estabilização das leituras, em cada nível de RFA,
foi de 60 s e o máximo, para salvar cada leitura, de 120 s. O coeficiente de variação (CV)
máximo admitido para salvar cada leitura foi de 0,3%. Além da RFA, foram mantidos
constantes o CO2 atmosférico no interior da câmara foliar e a temperatura do bloco a 26°C. O
fluxo de CO2 foi ajustado, mantendo-se uma concentração de 380 μmol mol-1
no interior da
câmara. As taxas de fotossíntese líquida (A), de transpiração (E) por unidade de área foliar, a
condutância estomática ao vapor de água (gs) e a razão entre a concentração interna e
atmosférica de CO2 (Ci/Ca) foi estimada a partir dos valores da variação de CO2 e da umidade
no interior da câmara, determinados pelo analisador de gases por infravermelho do aparelho,
em RFA ≥ 600 μmol fótons m-2
s-1
. Ajustes para as curvas de saturação da radiação luminosa
foram realizados por modelo de regressão não linear para equações exponenciais. Utilizou-se
a equação A = Amax (1- exp (-α RFA/Amax)) - Rd (IQBAL et al., 1997) para a estimar a curva de
resposta da taxa fotossintética líquida (A) em função da radiação fotossinteticamente ativa,
onde Amax é a taxa fotossintética bruta máxima em irradiância de saturação, α o rendimento
quântico aparente e Rd a taxa respiratória quando RFA = 0 μmol fótons m-2
s-1
. Foi também
calculada a eficiência instantânea (A/E) e intrínseca (A/gs) do uso da água

33
4.3 Determinação dos teores de clorofila e carotenóides
Os teores de clorofila e carotenóides foram determinados em base de área em extratos
de dimetilsulfóxido (DMSO) de discos foliares (HISCOX; ISRAELSTAM, 1979), com
algumas modificações. Após incubação de três discos foliares (0,5 cm2) com 2 mL de DMSO
saturado com CaCO3 por 24 h a 65ºC, a absorbância dos extratos foi lida em
espectrofotômetro de microplacas (VERSAmax) nos comprimentos de onda de 645, 663 e
480 nm, para a determinação das concentrações de clorofila a (Chl a),clorofila b (Chl b) e de
carotenóides (car), respectivamente (Tabela 2). A concentração de clorofila total (Chl T) foi
estimada pela soma dos teores das Chl a e b.
Tabela 2 – Equações utilizadas na determinação das concentrações de clorofilas a (Chl a), b
(Chl b) e total (Chl T) e de carotenóides (Car).
Comprimento de onda (nm) Teor de pigmento (µg/cm-2
)
480 Car = ((1000 x A480) - (1,63 x Chl a) - (53,78 x Chl b))/220
645 Chl b = (25,06 x A649) - (6,5 x A665)
663 Chl a = (12,47 x A665) - (3,62 x A649)
4.4 Avaliação de crescimento
O crescimento foi avaliado medindo 10 plantas de cada variante morfológico, em cada
tratamento, no inicio e final do período experimental. Os dados coletados foram altura (ALT)
e diâmetro do coleto (DC), usando régua milimetrada e paquímetro, respectivamente. Foi
contado o número de folhas (NF) e, em seguida, procedeu a divisão das plantas em partes
(raiz, caule e folha) que, após a medição da área foliar total por planta, foram armazenadas em
sacos de papel e colocadas para secar em estufa de circulação forçada de ar a 75ºC, até massa
constante. A área foliar foi determinada utilizando um medidor de área foliar Li-3100 (Li-Cor,
Nebraska, USA).
A partir dos dados de massa seca, das diferentes partes da planta, e de área foliar
foram calculadas as seguintes variáveis de crescimento (HUNT, 1990):
a) Massa foliar específica: MFE = MSF/AF (g cm-2
);
b) Razão de área foliar: RAF = AF/MST (cm² g-1
);

34
c) Razão de massa de raiz: RMR = MSR/MST (g g-1
);
d) Razão de massa de caule: RMC = MSC/MST (g g-1
);
e) Razão de massa de folha: RMF = MSF/MST (g g-1
);
f) Taxa de crescimento relativo: TCR = [ln(MST T2)-ln(MST T1)]/(T2-T1) (g g-1
dia-1
);
g) Taxa de crescimento relativo em altura TCRA=lnALT2-lnALT1/(T2-T1) (cm cm-1
dia-1
);
h) Taxa de crescimento relativo em diâmetro TCRD = lnDC2-lnDC1/(T2 - T1) (mm mm-1
dia-1
);
i) Taxa de assimilação líquida TAL=(MST2-MST1)/(T2-T1) (lnAF2-lnAF1)(AF2- F1) (g cm-2
dia-1
)
Onde: AF representa área foliar, T o tempo, ALT a altura, DC o diâmetro do coleto,
MSF a massa seca foliar, MSC a massa seca de caule, MSR a massa seca de raiz e MST a
massa seca total.
4.5 Determinação dos teores de carboidratos
4.5.1 Teor de açúcares solúveis totais
Nas diferentes partes da planta, o teor de açúcares solúveis totais (AST) foi
determinado pelo método de antrona (CLEGG, 1956).
4.5.2 Teor de Amido
A determinação do teor de amido, nas diversas partes da planta, foi realizada de
acordo com o método descrito por McCready et al. (1950).
4.6 Análise estatística
Empregou-se o delineamento experimental inteiramente casualizado, com 15
tratamentos, em esquema fatorial 5 x 3 [4 níveis de sombreamento + tratamento controle
(pleno sol) e 3 três variantes morfológicos do pau-brasil], 5 repetições e 10 plantas jovens por
unidade experimental. Foram realizadas análises de variância (ANOVA) e teste F.
Posteriormente, fez-se comparação de médias dos tratamentos por meio do teste de Duncan
(p≤0,05).

35
5. RESULTADOS
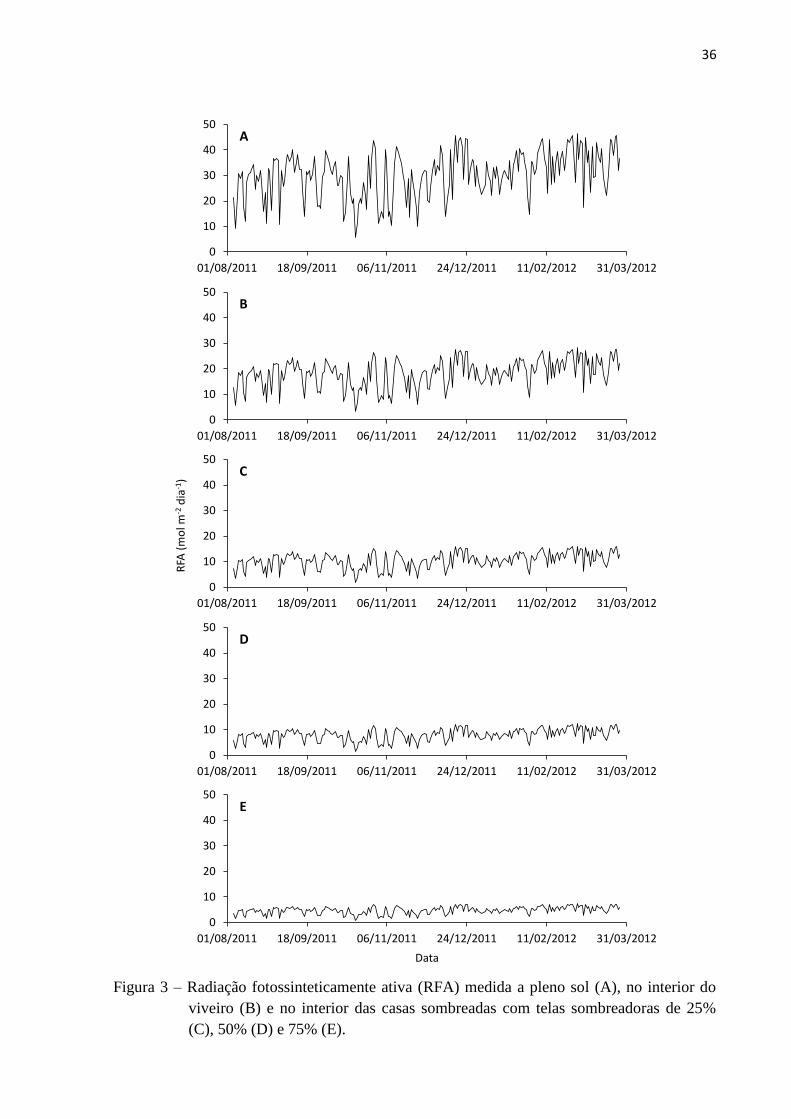
A radiação fotossinteticamente ativa (RFA) medida nos tratamentos a pleno sol, no
interior do viveiro e no interior das casas sombreadas com telas sombreadoras de 25%, 50% e
75%, logo acima da superfície das plantas, corresponderam a 100%, 60%, 35%, 25%, e 15%
de irradiância, respectivamente (Tabela 3 e Figura 3). Os dados, coletados entre agosto de
2011 e abril de 2012 apontaram variações de RFA (Tabela 3). Para os tratamentos 100%,
60%, 35%, 25%, e 15% de irradiância, as variações mínima, máxima e média diárias foram de
5,7; 46,6 e 30,3 mol m-2
dia-1
, de 3,5; 28,2 e 18,4 mol m-2
dia-1
, de 2,0; 16,1 e 10,5 mol m-2
dia-1
, de 1,5; 12,3 e 8,0 mol m-2
dia-1
e de 0,9; 7,3 e 4,8 mol m-2
dia-1
, respectivamente.
Tabela 3 – Valores médios, máximos e mínimos, e porcentagem em relação ao pleno sol, para
os diferentes ambientes de sombreamento. Dados coletados em Ilhéus, BA, Brasil,
entre agosto de 2011 e abril 2012.
Pleno sol Viveiro 25% 50% 75%
[mol m-2
dia-1
]
Média 30,3 18,4 10,5 8,0 4,8
Máximo 46,6 28,2 16,1 12,3 7,3
Mínimo 5,7 3,5 2,0 1,5 0,9
% Pleno sol 100% 60% 35% 25% 15%
36
0
10
20
30
40
50
01/08/2011 18/09/2011 06/11/2011 24/12/2011 11/02/2012 31/03/2012
Data
A
0
10
20
30
40
50
01/08/2011 18/09/2011 06/11/2011 24/12/2011 11/02/2012 31/03/2012
Data
B
0
10
20
30
40
50
01/08/2011 18/09/2011 06/11/2011 24/12/2011 11/02/2012 31/03/2012
RFA
(m
ol m
-2d
ia-1
)
Data
C
0
10
20
30
40
50
01/08/2011 18/09/2011 06/11/2011 24/12/2011 11/02/2012 31/03/2012
Data
D
0
10
20
30
40
50
01/08/2011 18/09/2011 06/11/2011 24/12/2011 11/02/2012 31/03/2012
Data
E
Figura 3 – Radiação fotossinteticamente ativa (RFA) medida a pleno sol (A), no interior do
viveiro (B) e no interior das casas sombreadas com telas sombreadoras de 25%
(C), 50% (D) e 75% (E).

37
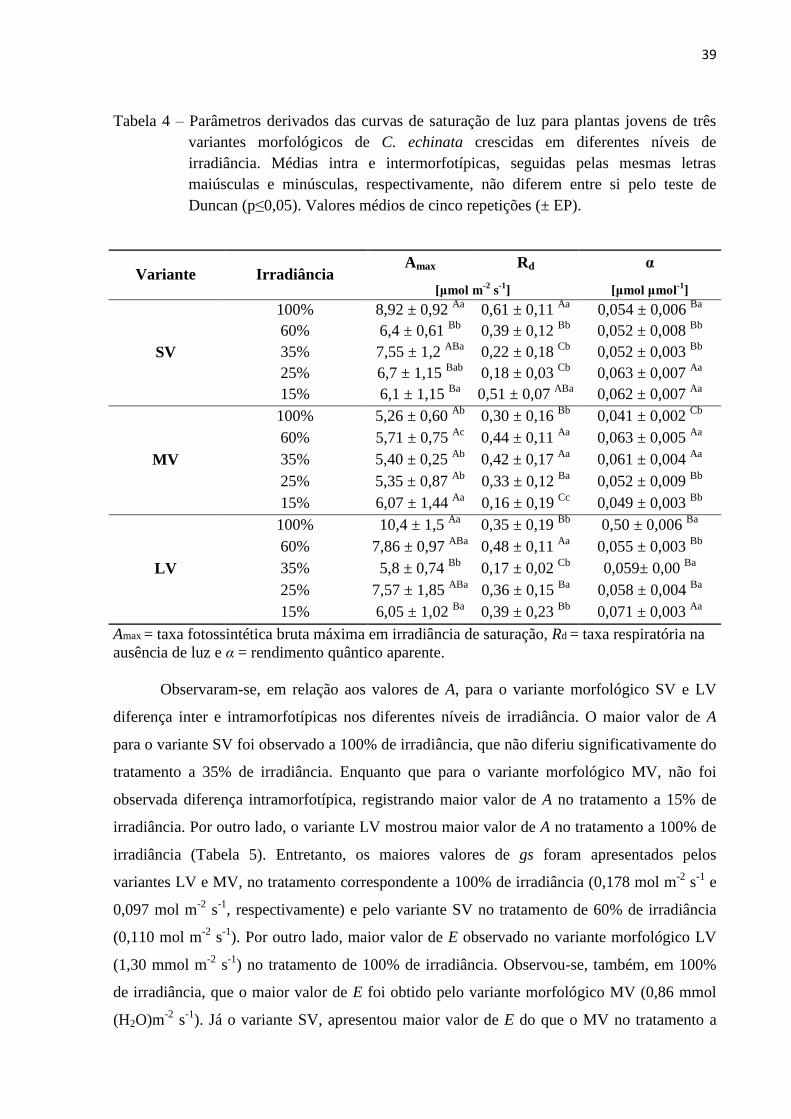
Embora tenha sido observadas diferenças inter e intramorfotípicas para os variantes
de pau-brasil, em relação aos parâmetros derivados das curvas de respostas da fotossíntese à
radiação fotossinteticamente ativa (Figura 4, Tabela 4), não foi possível diferenciá-los com
precisão em relação a todos os níveis de irradiâncias avaliados Constatou-se, a partir destes
parâmetros, que o variante morfológico SV de C. echinata apresentou maior valor de Amax no
tratamento com 100% de irradiância. Valores máximos de Rd a 100% e α a 25% de irradiância
também não diferindo do tratamento a 15%. O variante morfológico MV não mostrou
diferença significativa (p≤ 0,05) para os valores de Amax nos diferentes níveis de irradiância. O
maior valor de Amax foi observado a 15% de irradiância. Entretanto, o valor máximo de Rd no
tratamento de 60% de irradiância não diferiu dos tratamentos com 35% de irradiância. O
maior valor de α, para este variante morfológico, foi observado no tratamento com 60% de
irradiância, não diferindo do tratamento com 35%. Por outro lado, o variante morfológico LV
registrou o maior valor de Amax no tratamento 100% de irradiância. Em relação a Rd,, o maior
valor foi encontrado para este morfotipo no tratamento de 60% de irradiância. O maior valor
de α foi encontrado no tratamento de 15% de irradiância.

38
Figura 4 – Curvas de resposta da taxa fotossintética líquida (A) à radiação fotossinteticamente
ativa (RFA) para plantas jovens dos variantes morfológicos folha pequena (SV)
[A], folha média (MV) [B] e folha grande (LV) [C] de C. echinata crescidas em
diferentes níveis de irradiância [100% (◊), 60% (□), 35% (Δ), 25% (○) e 15% (×)
de irradiância], aos 24 meses após o plantio. Pontos são médias de cinco repetições
(± EP). Equação: A = Amax (1- exp (-α RFA/Amax)) - Rd.
39
Tabela 4 – Parâmetros derivados das curvas de saturação de luz para plantas jovens de três
variantes morfológicos de C. echinata crescidas em diferentes níveis de
irradiância. Médias intra e intermorfotípicas, seguidas pelas mesmas letras
maiúsculas e minúsculas, respectivamente, não diferem entre si pelo teste de
Duncan (p≤0,05). Valores médios de cinco repetições (± EP).
Variante Irradiância Amax Rd α
[μmol m-2
s-1
] [μmol μmol-1
]
SV
100% 8,92 ± 0,92 Aa
0,61 ± 0,11 Aa
0,054 ± 0,006 Ba
60% 6,4 ± 0,61 Bb
0,39 ± 0,12 Bb
0,052 ± 0,008 Bb
35% 7,55 ± 1,2 ABa
0,22 ± 0,18 Cb
0,052 ± 0,003 Bb
25% 6,7 ± 1,15 Bab
0,18 ± 0,03 Cb
0,063 ± 0,007 Aa
15% 6,1 ± 1,15 Ba
0,51 ± 0,07 ABa
0,062 ± 0,007 Aa
MV
100% 5,26 ± 0,60 Ab
0,30 ± 0,16 Bb
0,041 ± 0,002 Cb
60% 5,71 ± 0,75 Ac
0,44 ± 0,11 Aa
0,063 ± 0,005 Aa
35% 5,40 ± 0,25 Ab
0,42 ± 0,17 Aa
0,061 ± 0,004 Aa
25% 5,35 ± 0,87 Ab
0,33 ± 0,12 Ba
0,052 ± 0,009 Bb
15% 6,07 ± 1,44 Aa
0,16 ± 0,19 Cc
0,049 ± 0,003 Bb
LV
100% 10,4 ± 1,5 Aa
0,35 ± 0,19 Bb
0,50 ± 0,006 Ba
60% 7,86 ± 0,97 ABa
0,48 ± 0,11 Aa
0,055 ± 0,003 Bb
35% 5,8 ± 0,74 Bb
0,17 ± 0,02 Cb
0,059± 0,00 Ba
25% 7,57 ± 1,85 ABa
0,36 ± 0,15 Ba
0,058 ± 0,004 Ba
15% 6,05 ± 1,02 Ba
0,39 ± 0,23 Bb
0,071 ± 0,003 Aa
Amax = taxa fotossintética bruta máxima em irradiância de saturação, Rd = taxa respiratória na
ausência de luz e α = rendimento quântico aparente.
Observaram-se, em relação aos valores de A, para o variante morfológico SV e LV
diferença inter e intramorfotípicas nos diferentes níveis de irradiância. O maior valor de A
para o variante SV foi observado a 100% de irradiância, que não diferiu significativamente do
tratamento a 35% de irradiância. Enquanto que para o variante morfológico MV, não foi
observada diferença intramorfotípica, registrando maior valor de A no tratamento a 15% de
irradiância. Por outro lado, o variante LV mostrou maior valor de A no tratamento a 100% de
irradiância (Tabela 5). Entretanto, os maiores valores de gs foram apresentados pelos
variantes LV e MV, no tratamento correspondente a 100% de irradiância (0,178 mol m-2
s-1
e
0,097 mol m-2
s-1
, respectivamente) e pelo variante SV no tratamento de 60% de irradiância
(0,110 mol m-2
s-1
). Por outro lado, maior valor de E observado no variante morfológico LV
(1,30 mmol m-2
s-1
) no tratamento de 100% de irradiância. Observou-se, também, em 100%
de irradiância, que o maior valor de E foi obtido pelo variante morfológico MV (0,86 mmol
(H2O)m-2
s-1
). Já o variante SV, apresentou maior valor de E do que o MV no tratamento a

40
60% de irradiância (1,02 mmol m-2
s-1
), não deferindo estatisticamente com o tratamento a
35% de irradiância (Tabela 5). Além disso, verificou-se, para os variantes morfológicos de
pau-brasil, diferença significativa, inter e intramorfotípicas em relação aos valores de (Ci/Ca).
O variante SV mostrou maior valor Ci/Ca no tratamento a 60% de irradiância. Entretanto,
para o variante LV, o maior valor foi encontrado no tratamento de 100% de irradiância, ao
passo que para o variante MV, o maior valor foi verificado em 100% de irradiância, que não
diferiu significativamente de 15% de irradiância.
Tabela 5 – Taxa fotossintética líquida por unidade de área foliar (A), condutância estomática
ao vapor de água (gs), taxa transpiratória (E) e razão entre a concentração
intercelular e ambiente de CO2 (Ci/Ca) de plantas jovens de três variantes
morfológicos de C. echinata crescidas em diferentes níveis de irradiância, aos 24
meses após a emergência, em RFA ≥600 μmol fótons m-2
s-1
. Médias intra e
intermorfotípicas, seguidas pelas mesmas letras maiúsculas e minúsculas,
respectivamente, não diferem entre si pelo teste de Duncan (p≤0,05). Valores
médios de cinco repetições (± EP).
Variante Irradiância A gs E
Ci/Ca
[μmol (CO2) m-2
s-1
] [mol (H2O) m-2
s-1
] [mmol (H2O) m-2
s-1
]
SV
100% 7,55 ± 0,50 Aa
0,053 ± 0,002 Bb
0,63 ± 0,04 Bb
0,40 ± 0,04 Bb
60% 6,58 ± 0,40 ABa
0,110 ± 0,012 Aa
1,02 ± 0,08 Aa
0,50 ± 0,04 Aa
35% 7,48 ± 0,55 Aa
0,062 ± 0,004 Ba
0,90 ± 0,06 Aa
0,39 ± 0,04 Ba
25% 6,41 ± 0,47 ABab
0,054 ± 0,004 Bb
0,71 ± 0,04 Bb
0,43 ± 0,04 Ba
15% 5,72 ± 0,44 Bab
0,052 ± 0,006 Bb
0,71 ± 0,07 Ba
0,36 ± 0,05 Bb
MV
100% 5,56 ± 0,39 Ab
0,097 ± 0,012 Aab
0,86 ± 0,09 Aab
0,50 ± 0,05 Aa
60% 4,68 ± 0,31 Bb
0,051 ± 0,005 Bb
0,63 ± 0,05 Bb
0,47 ± 0,04 ABa
35% 4,97 ± 0,42 Bb
0,033 ± 0,002 Cb
0,41 ± 0,03 Cb
0,36 ± 0,04 Ca
25% 5,18 ± 0,38 ABb
0,043 ± 0,003 Cc
0,60 ± 0,04 Bb
0,43 ± 0,04 Ba
15% 6,04 ± 0,51 Aa
0,06 ± 0,004 Ba
0,76 ± 0,05 ABa
0,53 ± 0,03 Aa
LV
100% 8,93 ± 0,62 Aa
0,178 ± 0,021 Aa
1,30 ± 0,13 Aa
0,50 ± 0,05 Aa
60% 6,57 ± 0,48 Ba
0,047 ± 0,004 Cb
0,57 ± 0,04 Cb
0,37 ± 0,05 Bb
35% 5,03 ± 0,34 Cb
0,033 ± 0,002 Db
0,46 ± 0,03 Cb
0,32 ± 0,04 Ba
25% 7,23 ± 0,57 ABa
0,083 ± 0,011 Ba
1,02 ± 0,11 Ba
0,45 ± 0,04 ABa
15% 5,09 ± 0,41 Cb
0,042 ± 0,003 Cc
0,57 ± 0,04 Cb
0,44 ± 0,04 ABab
Não houve diferença intermorfotípica para A/E, com a variação dos níveis de
irradiância, exceto para o variante SV em 35% de irradiância (Figura 5A). As diferenças
significativas (p≤ 0,05) foram apenas intramorfotípica, cujos maiores valores de A/E foram
apresentados pelo variante SV em 100% de irradiância, seguido pelo MV em 100%, 60% e
35%, que não diferiram significativamente (p≤ 0,05) entre si, e pelo LV em 60% e 35% de

41
irradiância. Em contrapartida, verificaram-se diferenças intra e intermorfotípicas para A/gs
(Figura 5B). Os maiores valores de A/gs foram observados pelo variante SV em 15% de
irradiância, pelo LV em 60%, que não diferiu significativamente (p≤ 0,05) de 35% de
irradiância, e pelo MV em 100% de irradiância, que também não diferiu significativamente
(p≤ 0,05) de 60%, 35% e 25% de irradiância.
Figura 5 – Eficiências instantânea (A/E) (A) e intrínseca (A/gs) do uso da água (B) em plantas
jovens de três variantes morfológicos de C. echinata (folha pequena (SV); folha
média (MV) e folha grande (LV)) crescidas em diferentes níveis de irradiância,
aos 24 meses após a emergência. Médias intra e intermorfotípicas, seguidas pelas
mesmas letras maiúsculas e minúsculas, respectivamente, não diferem entre si
pelo teste de Duncan (p≤0,05). Valores médios de cinco repetições (± EP).
Observaram-se variações intra e intermorfotípicas significativas (p≤0,05) para os
teores de pigmentos cloroplastídicos, com as mudanças dos níveis de irradiância. Verificou-se
que os teores de Chl a, Chl b e Chl T foram consideravelmente maiores nas plantas mais
sombreadas em relação às que foram cultivadas a 100% de irradiância, cujos valores médios
diferiram significativamente (p≤ 0,05) para os três variantes morfológicos de pau-brasil

42
(Figuras 6 e 7). Os maiores teores de Chl a encontrados para os variantes morfológicos MV,
LV e SV foram de 7,97; 6,25 e 5,55 mg dm-2
, respectivamente, quando expostos ao
tratamento de 25% de irradiância, embora para SV este valor não diferiu significativamente
(p≤ 0,05) do tratamento de 35% de irradiância (Figura 6A). Por outro lado, os maiores teores
de Chl b, foram apresentados pelo variante morfológicos SV, no tratamento com 35% de
irradiância, e pelos variantes MV e LV, ambos no tratamento com 25% de irradiância, cujos
teores foram de 4,95, 4,92 e 3,74 mg dm-2
, respectivamente (Figura 6B). Além disso, os
maiores teores de Chl T foram apresentados pelo variante morfológico MV (12,9 mg dm-2
) no
tratamento com 25% de irradiância; seguido pelo SV (10,4 mg dm-2
) no tratamento com 35%
de irradiância, que não deferiu significativamente (p≤0,05) do tratamento a 15% de
irradiância; e pelo LV (9,99 mg dm-2
) exposto ao tratamento com 25% de irradiância (Figura
7B). Houve uma variação na razão de Chl a/b em relação aos tratamentos, as maiores razões
foram encontradas em condições de baixa irradiância, cujos valores foram de 1,57 1,62 e 1,67
mg dm-2
para os variantes morfológicos SV, MV e LV, respectivamente, no tratamento
correspondente a 25% de irradiância (Figura 6C).
Em relação aos carotenóides (Figura 7A), os maiores teores foram apresentados
pelos variantes morfológicos MV (1,60 mg dm-2
), no tratamento de 60% de irradiância; LV
(1,33 mg dm-2
), no tratamento de 35% de irradiância, que não diferiu significativamente (p≤
0,05) dos tratamentos de 25 e 100% de irradiância; e SV (1,31 mg dm-2
) no tratamento a
100% de irradiância. Por outro lado, as maiores razões Car/Chl T (Figura 7C), ao contrário da
razão Chl a/b, foram encontradas nos maiores níveis de irradiância, cujos maiores valores
foram apresentados pelos variantes morfológicos MV, SV e LV, nas irradiâncias
correspondentes a 35, 60 e 100% respectivamente.

43
Figura 6 – Teores de clorofila a (Chl a) (A), clorofila b (Chl b) (B) e razão entre os teores de
Chl a e Chl b (Chl a/b) (C) em folhas de plantas jovens de três variantes
morfológicos de C. echinata (folha pequena (SV); folha média (MV) e folha
grande (LV)) crescidas em diferentes níveis de irradiância aos 24 meses após a
emergência. Médias intra e intermorfotípicas, seguidas pelas mesmas letras
maiúsculas e minúsculas, respectivamente, não diferem entre si pelo teste de
Duncan (p≤0,05). Valores médios de cinco repetições (± EP).

44
Figura 7 – Teores de carotenóides (Car), (A) clorofila total (Chl T) (B) e razão entre os teores
de carotenóides e clorofila total (Car/Chl T) (C) em folhas de plantas jovens de
três variantes morfológicos de C. echinata (folha pequena (SV); folha média
(MV) e folha grande (LV)) crescidas em diferentes níveis de irradiância, aos 24
meses após a emergência. Médias intra e intermorfotípicas, seguidas pelas
mesmas letras maiúsculas e minúsculas, respectivamente, não diferem entre si
pelo teste de Duncan (p≤0,05). Valores médios de cinco repetições (± EP).

45
Observou-se, para os variantes morfológicos de C. echinata, que os valores de NF
diferiram apenas intramorfotipicamente, com a variação da irradiância (Figura 8A). Os
maiores valores de NF foram encontrados para os variantes SV, MV e LV em 100%, 100% e
60% de irradiância, respectivamente.
Houve variações intra e intermorfotípicas significativas (p≤ 0,05) para ALT e DC para
as variantes morfológicos de C. echinata, em função dos níveis de irradiância (Figura 8B e
8C). Os maiores valores de ALT foram encontrados para os variantes SV e LV em 35% e
100% de irradiância, respectivamente, ao passo que para o variante MV não se verificou
diferenças significativas para ALT em relação à variação da irradiância (Figura 8B).
Entretanto, para DC os maiores valores foram apresentados, para todos variantes
morfológicos avaliados, em 100% de irradiância (Figura 8C).
Verificaram-se diferenças intra e intermorfotípicas significativas (p≤ 0,05) para MSR,
MSF, MSC e MST dos variantes de C. echinata, com a variação dos níveis de irradiância
(Figura 9A-D). Os maiores valores de MSR, MSF, MSC e MST foram apresentados pelos
variantes morfológicos SV, MV e LV em 100% de irradiância (Figura 9A-D), exceto para o
variante SV, cujo maior valor de MSF foi encontrado em 35% de irradiância.
Para RMR, RMC e RMF, observaram-se variações intra e intermorfotípicas, para os
variantes morfológicos de C. echinata, com as mudanças dos níveis de irradiância (Figura
10). Os maiores valores de RMR foram alcançados pelos variantes MV e LV no tratamento de
100% de irradiância. Para o variante SV os maiores valores foram obtidos em 100% e 60% de
irradiância, que não diferiram significativamente (p≤ 0,05) entre si. Entretanto, para RMC, os
maiores valores foram apresentados pelo variante LV em 100% de irradiância e pelo MV em
100% e 60% de irradiância, que também não diferiram significativamente entre si. Para o
variante morfológico SV não houve diferença intramorfotípica para RMC, com a variação da
irradiância. Em contrapartida, para RMF, os maiores valores foram apresentados pelos
variantes MV e LV em 35, 25 e 15% de irradiância, que não diferiram intramorfotipicamente
(p≤ 0,05); ao passo que para SV, o maior valor de RMF foi encontrado em 25% de
irradiância.

46
Figura 8 – Número de folhas (A), altura total (B) e diâmetro do caule (C) de plantas jovens
de três variantes morfológicos de C. echinata (folha pequena (SV); folha média
(MV) e folha grande (LV)) crescidas em diferentes níveis de irradiância aos 24
meses após a emergência. Médias intra e intermorfotípicas, seguidas pelas
mesmas letras maiúsculas e minúsculas, respectivamente, não diferem entre si
pelo teste de Duncan (p≤0,05). Valores médios de cinco repetições (± EP).

47
Figura 9 – Biomassas secas de raiz (MSR) (A), folha (MSF) (B), caule (MSC) (C) e total
(MST) (D) de plantas jovens de três variantes morfológicos de C. echinata (folha
pequena (SV); folha média (MV) e folha grande (LV)) crescidas em diferentes
níveis de irradiância, aos 24 meses após a emergência. Médias intra e
intermorfotípicas, seguidas pelas mesmas letras maiúsculas e minúsculas,
respectivamente, não diferem entre si pelo teste de Duncan (p≤0,05). Valores
médios de cinco repetições (± EP).

48
Figura 10 – Razões de massa de raiz (A), caule (B) e folha (C) de plantas jovens de três
variantes morfológicos de C. echinata crescidas em diferentes níveis de
irradiância. Médias intra e intermorfotípicas, seguidas pelas mesmas letras
maiúsculas e minúsculas, respectivamente, não diferem entre si pelo teste de
Duncan (p≤0,05). Valores médios de dez repetições (± EP).

49
Para AF os variantes SV e MV não diferiram intramorfotípicamente, no entanto, os
variantes SV e LV não diferiram intermorfotípicamente, exceto o variante SV em 60% de
irradiância (Figura 11). Houve variações intramorfotípicas significativas (p≤0,05) para AFE
entre os variantes morfológicos de C. echinata. Entretanto, os variantes MV e LV não
diferiram intermorfotipicamente. Os maiores valores de AFE foram alcançados pelos
variantes SV no tratamento de 25% de irradiância, ao passo que para o variante MV os
maiores valores foram obtidos em 35 e 25% de irradiância, que não diferiram
significativamente (p≤ 0,05) entre si. Já para o variante LV o maior valor de AFE foi
encontrado no tratamento a 35% de irradiância, que não diferiu significativamente (p≤0,05)
do tratamento a 25 e 15% de irradiância.
Em relação a RAF, observou-se variação intra e intermorfotípicas, para os variantes
morfológicos com as mudanças dos níveis de irradiância. Os maiores valores de RAF foram
obtidos por SV e MV em 25% de irradiância, enquanto que para LV os maiores valores foram
alcançados em 35, 25 e 15% de irradiância, uma vez que não diferiram significativamente (p≤
0,05) entre si (Figura 11).
Evidenciou-se, com a variação da irradiância, a existência de diferenças intra e
intermorfotípicas em relação às variáveis MFE, TCR, TCRA, TCRD e TAL para os diversos
variantes morfológicos de C. echinata avaliados (Figura 12 e 13). Os maiores valores de MFE
foram observados para os variantes SV, MV e LV em 100% de irradiância, embora para SV o
maior valor de MFE não tenha diferido significativamente de 60% de irradiância. Entretanto,
para TCR, os três variantes morfológicos avaliados alcançaram os maiores valores também
em 100% de irradiância, a exceção do variante LV, cujos valores de TCR não diferiram
significativamente (p≤ 0,05) entre si em 100% e 60% de irradiância. Em relação a TCRA, os
maiores valores foram obtidos pelo variante SV em 35%, pelo MV em 100% e 35% e pelo
LV em 100% de irradiância. Em contrapartida, para TCRD, os maiores valores foram
atingidos, pelos três variantes morfológicos, em 100% de irradiância, a exceção do variante
SV, cujos maiores valores não diferiram significativamente entre si, em 100% e 60% de
irradiância. Por outro lado, os maiores valores de TAL foram alcançados pelos três variantes
de C. echinata em 100% de irradiância, a exceção do variante morfológico LV, cujos maiores
valores de TCR não diferiram significativamente (p≤ 0,05) em 100% e 60% de irradiância.

50
Figura 11 – Área foliar específica (AFE) (A), razão de área foliar (RAF) (B) e área foliar total
(AF) (C) de plantas jovens de três variantes morfológicos de C. echinata crescidas
em diferentes níveis de irradiância. Médias intra e intermorfotípicas, seguidas
pelas mesmas letras maiúsculas e minúsculas, respectivamente, não diferem entre
si pelo teste de Duncan (p≤0,05). Valores médios de dez repetições (± EP).

51
Figura 12 – Massa foliar especifica (MFE) (A) e taxa assimilatória liquida (TAL) (B) de
plantas jovens de três variantes morfológicos de C. echinata crescidas em
diferentes níveis de irradiância, aos 24 meses após a emergência. Médias intra e
intermorfotípicas, seguidas pelas mesmas letras maiúsculas e minúsculas,
respectivamente, não diferem entre si pelo teste de Duncan (p≤0,05). Valores
médios de dez repetições (± EP).
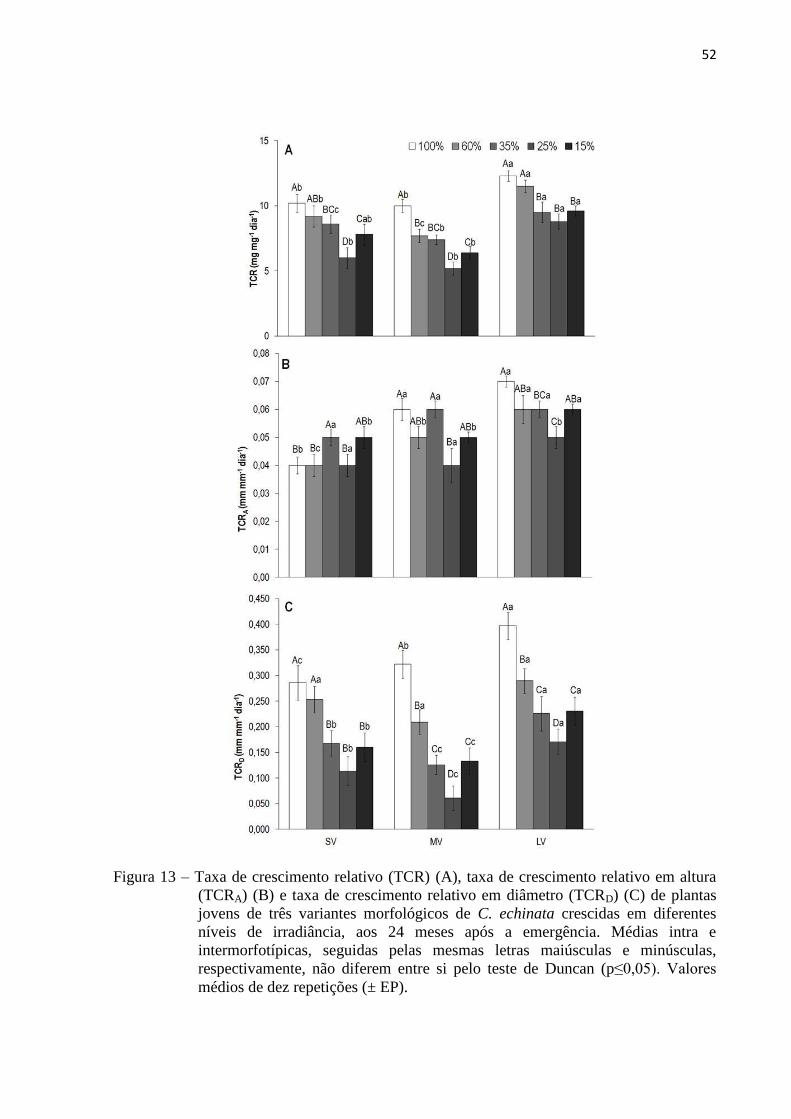
52
Figura 13 – Taxa de crescimento relativo (TCR) (A), taxa de crescimento relativo em altura
(TCRA) (B) e taxa de crescimento relativo em diâmetro (TCRD) (C) de plantas
jovens de três variantes morfológicos de C. echinata crescidas em diferentes
níveis de irradiância, aos 24 meses após a emergência. Médias intra e
intermorfotípicas, seguidas pelas mesmas letras maiúsculas e minúsculas,
respectivamente, não diferem entre si pelo teste de Duncan (p≤0,05). Valores
médios de dez repetições (± EP).

53
Houve variação intra e intermorfotípica, para o teor de amido, somente em nível foliar
(Figura 14C). Os maiores teores de amido foliar foram obtidos pelo variante SV, em 60% de
irradiância, que não diferiu significativamente (p≤ 0,05) de 100% e 25% de irradiância, e
pelos variantes MV e LV em 60% de irradiância. Em contrapartida, o teor de amido no caule
manteve-se relativamente constante, independentemente de variantes morfológicos e da
variação dos níveis de irradiância, a exceção dos variantes SV e LV que obtiveram o maior e
o menor teor de amido no caule em 25% e 15% de irradiância, respectivamente (Figura 14B).
O mesmo fato se observou para o teor de amido na raiz, a exceção do morfotipo MV, que
apresentou o menor teor em 60% de irradiância (Figura 14A).
Observou-se, nos diferentes órgãos das plantas dos diversos variantes morfológicos de
C. echinata, que as variações no teor AST se comportaram de maneira similar às variações
dos teores de amido (Figura 15). Basicamente, as variações intra e intermorfotípicas, em
relação aos teores AST, ocorreram principalmente em nível foliar, cujos maiores teores foram
encontrados para o variante SV em 60 e 35% de irradiância e para os variantes MV e LV em
100% de irradiância (Figura 15C). Os teores de AST no caule, para os variantes morfológicos
MV e LV, não diferiram significativamente (p≤ 0,05) em relação aos níveis de irradiância, a
exceção do variante SV que apresentou um aumento no teor de AST no caule em 15% de
irradiância (Figura 15B). Em contrapartida, os maiores teores de AST na raiz foram
encontrados para os variantes MV e LV em 35% de irradiância, ao passo que para o variante
SV não se observou alterações significativas (p≤ 0,05) nos teores de AST com a variação da
irradiância (Figura 15A).

54
Figura 14 – Teor de amido na raiz (A), no caule (B) e na folha (C) de plantas jovens de três
variantes morfológicos de C. echinata (folha pequena (SV); folha média (MV) e
folha grande (LV)) crescidas em diferentes níveis de irradiância aos 24 meses
após a emergência. Médias intra e intermorfotípicas, seguidas pelas mesmas
letras maiúsculas e minúsculas, respectivamente, não diferem entre si pelo teste
de Duncan (p≤0,05). Valores médios de cinco repetições (± EP).
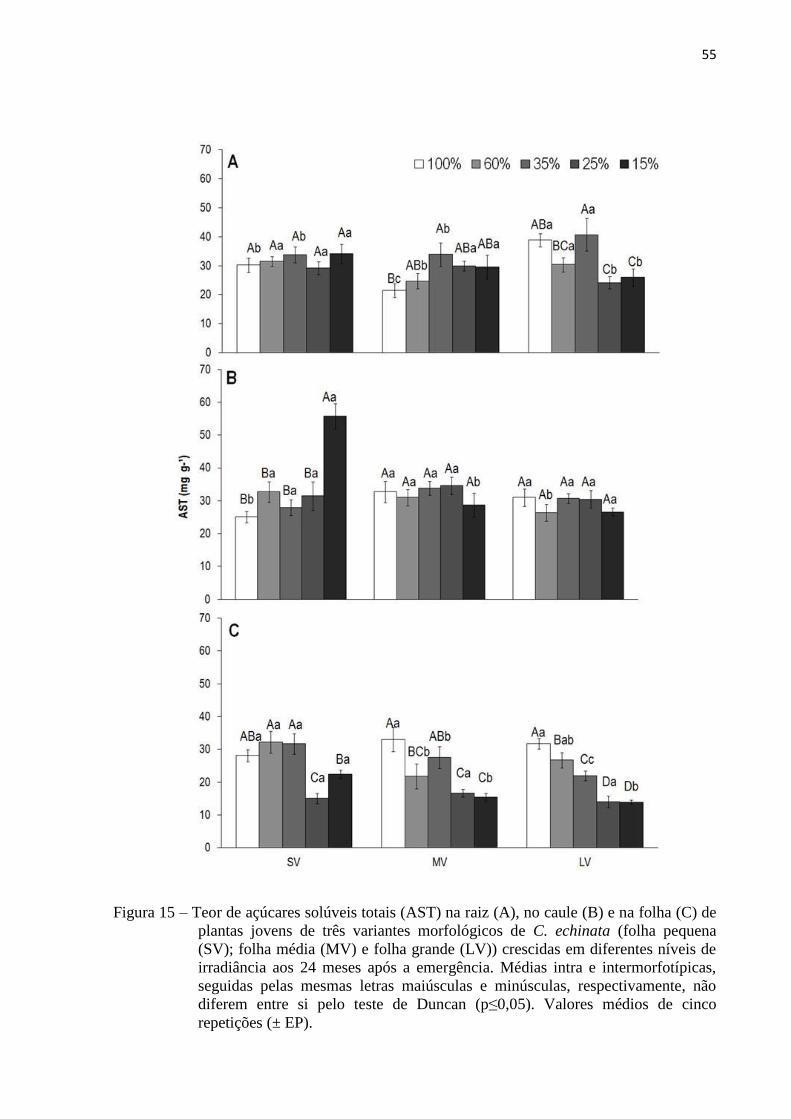
55
Figura 15 – Teor de açúcares solúveis totais (AST) na raiz (A), no caule (B) e na folha (C) de
plantas jovens de três variantes morfológicos de C. echinata (folha pequena
(SV); folha média (MV) e folha grande (LV)) crescidas em diferentes níveis de
irradiância aos 24 meses após a emergência. Médias intra e intermorfotípicas,
seguidas pelas mesmas letras maiúsculas e minúsculas, respectivamente, não
diferem entre si pelo teste de Duncan (p≤0,05). Valores médios de cinco
repetições (± EP).

56
6. DISCUSSÃO
Os maiores valores de Amax (Tabela 4) dos variantes morfológicos LV e SV foram
encontrados no tratamento a 100% de irradiância, com decréscimos nos níveis subsequentes
de sombreamento, demonstrando a capacidade de aclimatação dessa espécie a ambientes com
elevada RFA. Constatou-se que os maiores valores para Rd e α foram registrados para o
variante SV a 100% de irradiância. Segundo SACCO (1980) são características de espécies
heliófitas encontradas nos limites externos das matas preservadas, florestas secundárias e em
áreas de restingas litorâneas. Por outro lado, se observou uma inversão nos valores de Amax
para o variante morfológico MV (Tabela 4). Plantas adaptadas a ambientes sombreados são
mais eficientes fotossinteticamente, perdendo esta eficiência em níveis elevados de RFA
(LEVERENZ, 1995). Zhang et al., (2003) afirmaram que o valor elevado de Amax para
plântulas crescendo sob sombreamento é um indicativo da plasticidade fisiológica ao adaptar-
se a ambientes com reduzida RFA.
Os vegetais são perfeitamente capazes de adaptação, em maior ou menor grau, em
função dos diferentes gradientes de luz (GIVNISH, 1988), denotando que a plasticidade
fenotípica, independente das condições de sol e sombra, é de elevada importância, tanto para a
própria sobrevivência como seu crescimento em ambientes heterodinâmicos, como as
florestas tropicais úmidas (VALLADARES et al., 2000; ROZENDAAL et al., 2006). Essa
capacidade determina o potencial da espécie em adaptar-se às condições de sol ou sombra,
podendo ser avaliada pelo crescimento inicial sob diferentes condições de disponibilidade de
luz (MIELKE; SCHAFFER, 2010).
A variação decrescente dos valores de Rd (Tabela 4) e de TCR (Figura 13) com a
diminuição de RFA, indica que quanto menor for Rd, menor também será a taxa de
crescimento. Os dados discordam da existência de uma correlação inversa da Rd e TCR, onde
o menor consumo de carbono na respiração representa maior proporção de carbono disponível
para o crescimento (HOPKINS, 2000).
Notou-se um aumento dos valores de gs dos variantes LV e MV no tratamento a
100% de irradiância e para o SV no tratamento a 60% de irradiância (Tabela 5). O

57
relacionamento destes dados com os valores de A para os variantes SV, MV e LV indicam
alterações na abertura dos estômatos pela variação da RFA. Mesmo não havendo uma
diferença significativa (p≤0,05) de gs e A para os tratamentos a 100%, 60%, 35% e 15% de
irradiância, observou-se similaridade na concentração de Ci/Ca dos variantes LV, SV e MV
nos tratamentos de 100%, 60% e 15% de irradiância, respectivamente. Essa variação também
foi descrita por Wong et al. (1978), Daley et al. (1989), Santos (2009) e Lavinscky (2009),
indicando que uma menor A pode estar associada a uma menor gs, implicando em elevação da
eficiência intrínseca do uso da água (A/gs). Entretanto, Costa (1996), observando tais
variações, determinou que na relação de RFA com A e gs, a energia capturada não utilizada
no sistema pode ser prejudicial ao maquinário fotossintético, causando fotodestruição das
moléculas de clorofila.
Segundo Silva et al. (2004), Reis e Campostrini (2011), a estimativa da resposta de A
às variações ambientais assume que a razão Ci/Ca influencia a taxa de assimilação de CO2,
que é calculada sob condições de saturação de luz, regulada pela regeneração da ribulose-1,5-
bisfosfato (RuBP), ou pela atividade catalítica da Rubisco, quando a concentração de RuBP
no cloroplasto está próxima a saturação. Nesse sentido, somente os variantes morfológicos
LV e SV obtiveram maior taxa assimilatória de CO2, quando expostos ao tratamento com
100% de irradiância e o MV ao tratamento com 15% de irradiância. O interessante é o fato de
dois variantes reagirem de forma completamente opostas quanto à intensidade de RFA e Rd. O
variante LV atingiu melhor desempenho em assimilação de CO2 com taxa respiratória
elevada, quando em 100% de irradiância (Tabela 2), ao passo que o variante MV obteve
maior A e Rd elevado, quando a 15% de irradiância. Os dados confirmam as informações de
Siebke et al. (2002) e de Reis e Campostrini (2011) que apontam que o balanço de CO2 está
nitidamente relacionado a Rd e a condutância de CO2 no mesofilo, sendo esta última a variável
chave para a determinação de Ci/Ca. Ao contrário de Lloyd e Farqhar (1994), o que se pôde
comprovar, exceto para os variantes LV e SV, é que os valores de Ci/Ca variam de acordo
com o ambiente, com tendência a serem mais elevados em espécies mesomórficas do que em
espécies xeromórficas, onde o variante MV, mais tolerante ao sombreamento, apresentou
valores mais elevados de Ci/Ca do que os demais variantes, consideradas plantas de pleno sol.
Verificou-se, em relação A/E, que os maiores valores foram registrados para os
variantes morfológicos de pau-brasil em ambientes de maior irradiância, que corresponderam
aos menores valores de gs e E, o que reduziu a perda de água por transpiração, exceto para o
variante MV no tratamento com 100% de irradiância, acordando com as ideias de Liberato et
al. (2006) e Gonçalves et al. (2009), pois a pleno sol pode ter havido deficiência hídrica, que

58
por sua vez, pode ter afetado o potencial hídrico foliar, o status nutricional e as trocas gasosas
foliares, influenciando diretamente o desenvolvimento da planta.
Os valores de A/E traduzem, também, a capacidade demonstrada pelos variantes
morfológicos quanto a A/gs, registrando padrões semelhantes durante a aclimatação de cada
variante morfológico (Figura 5). Ferreira et al. (2012), trabalhando com plantas jovens de
Bertholletia excelsa, detectaram que a manutenção de elevadas taxas fotossintéticas
associadas a menores valores de gs e E são características de plantas tolerantes a menor
quantidade de água disponível no solo. Esta característica se reflete diretamente nos elevados
valores de A/E e A/gs, cujos patamares servem como indicadores para monitoramento do
estabelecimento de plantas e demonstração da plasticidade fisiológica aos diversos
tratamentos e complementos abióticos ambientais.
As maiores concentrações de CO2 assimilado foram registradas para os variantes LV
quando exposto ao tratamento com 35% de irradiância, com valores de A/E na ordem de 10,3
µmol CO2 mmol-1
H2O. Isso representa a eficiência com que os variantes morfológicos
utilizam a água ao mesmo tempo em que realizam a fotossíntese, sendo o movimento
estomático o principal mecanismo de controle das trocas gasosas em nível foliar (TATAGIBA
et al., 2007).
Entretanto, Tenhunem et al. (1987); Tatagiba et al. (2007) estabeleceram que o
funcionamento dos estômatos constitui um comprometimento fisiológico, pois, quando
abertos, permitem a assimilação de CO2 e, quando fechados, promovem uma maior eficiência
do uso da água. Logo, a eficiência no uso da água muda quando a difusão do CO2 ou da água
são alteradas. Segundo Larcher (2000), quando os estômatos estão totalmente abertos, a
absorção de CO2 é mais limitada pela resistência de transferência do que a perda de água por
transpiração, sendo que a melhor relação entre absorção de CO2 e perda de água é alcançada
quando os estômatos estão parcialmente fechados. Machado et al. (2005) estudando Citrus,
também detectou os menores valores de A/E com a elevação dos valores de A, gs e E.
A eficiência fotossintética das plantas está fortemente relacionada aos teores de
clorofila em nível foliar (ALMEIDA et al., 2004), uma vez que esses pigmentos
cloroplastídicos são responsáveis pela captura da radiação eletromagnética usada na
fotossíntese, sendo, portanto, essenciais na conversão da radiação luminosa em energia
química, na forma de ATP e NADPH + H+ (JESUS; MARENCO, 2008). O maior acúmulo de
clorofila em níveis mais sombreados pode ser devido ao efeito compensatório da espécie à
menor quantidade de radiação luminosa disponível (ALMEIDA et al., 2004; MARTINAZZO
et al., 2007). Os dados corroboram as ideias de Fermino-Jr (2004) que aponta que a redução

59
no teor de clorofila, em ambientes de maiores irradiações, pode atuar como um mecanismo de
redução da fotoinibição. De acordo com Tan et al. (2000), há uma maior taxa de degradação
de clorofila em ambientes sob alta irradiância, tratando-se de um ajuste fisiológico mediante
baixa concentração de clorofila.
Rego e Possamai (2006) verificaram que plantas de Cariniana legalis respondem ao
aumento da luminosidade com a redução dos teores de clorofila a, b e total. Morais et al.
(2007), por sua vez, observaram que espécies arbóreas da Amazônia, que crescem sob
sombreamento, apresentam maior teor foliar de clorofila. Almeida et al. (2004) e Fermino-Jr
(2004), além de observar maior teor de clorofila em folhas sombreadas, verificaram que, com
o aumento da irradiância, as plantas apresentam redução da razão Chl a/b. A clorofila b e os
carotenóides são considerados pigmentos acessórios do processo fotossintético, pois, embora
não estejam relacionados à captura da energia luminosa nos centros de reação (papel
desenvolvido pela clorofila a), promovem fotoproteção ao sistema (MARENCO; LOPES,
2009). Desta forma, a relação Chl a/b e Car/Chl T são indicadores da plasticidade fisiológica
em relação à alteração nas condições de luminosidade (SCALON et al., 2003; KITAJIMA;
HOGAN, 2003). Almeida et al. (2004) também observaram redução da razão Chl a/b em
plantas de Cryptocaria aschersoriana cultivadas em maiores irradiância, quando comparado
ao cultivo sombreado (30, 50 e 70% sombreamento). Enquanto maiores níveis de clorofila b
podem ser encontrados em condições de baixa luminosidade, atuando no aumento da captação
de luz (captação de energia em comprimentos de onda diferentes que a clorofila a) (SCALON
et al. 2003), a clorofila a é mais sensível ao aumento da radiação do que a clorofila b, sendo
degradada mais facilmente (TAN et al., 2000; MAGALHÃES et al. 2009). Assim, a razão
clorofila a/b tende a diminuir tanto em plantas que se desenvolvem em sombreamento
intenso, quando em plantas submetidas à elevada irradiância.
A relação entre Car/Chl T foi menor nas plantas em pleno sol, fato também
observado por Magalhães et al. (2009) em plantas de Minquartia guianensis. O aumento do
teor de carotenóides pode atuar na aclimatação à alta irradiância por desempenharem um
papel relevante na dissipação de energia sob a forma de calor (DEMMIG-ADAMS; ADAMS,
2006; MARENCO; LOPES, 2009).
Verificou-se que o sombreamento influenciou o crescimento, de maneira
significativa, dos três variantes morfológicos de C. echinata. O aumento do NF teve um
comportamento diferenciado para os diversos tratamentos de irradiância (Figura 8A), sendo o
variante SV o que apresentou maior NF no tratamento a 100% de irradiância. No geral,
quanto maior o número de folhas, maior será o número de estômatos e maior a transpiração.

60
Isso permite também uma maior absorção de CO2 e um aumento da atividade fotossintética
em resposta ao sombreamento (PASSOS, 1997).
Em relação ao crescimento em ALT, os maiores valores foram atingidos pelos
variantes SV e LV em 35% e 100% de irradiância, respectivamente; ao passo que não houve
resposta significativa (p≤ 0,05) do variante MV à variação da irradiância. Segundo Osunkoya
et al. (1994) e Claussen (1996), o crescimento em altura de muitas plantas é limitado pela
quantidade de luz disponível, e muitas desenvolvem estratégias para sobreviverem e se
estabelecerem em ambiente de pouca luz. Por outro lado, o aumento de DC foi diretamente
proporcional à intensidade de radiação luminosa. Todos os variantes morfológicos analisados
apresentaram maior incremento de DC principalmente em 100% de irradiância. Estes
resultados diferem daqueles obtidos por Aguiar et al. (2005), que encontraram maiores
valores de DC para mudas de pau-brasil cultivadas em 20% e 40% de sombreamento.
O estudo da massa seca revelou aspectos interessantes quanto ao comportamento das
raízes dos variantes morfológicos de pau-brasil. Os resultados mostraram a existência de um
decréscimo significativo (p≤0,05) de MSR, com a redução da intensidade de radiação
luminosa (Figura 9A). Este decréscimo se deveu, provavelmente, à menor disponibilidade de
carboidratos para planta em condições de baixa irradiância, uma vez que o maior acúmulo de
MSR foi registrado para todos os variantes morfológicos crescendo em 100% de irradiância.
Em contrapartida, o variante SV respondeu contrariamente aos efeitos da diminuição da
luminosidade no tratamento de 15% de irradiância. Isto ocorreu, provavelmente, devido à
diminuição do período de recepção de assimilados, sob baixos níveis de radiação, em razão da
maior capacidade mobilizadora (força do dreno) do sistema radicular. Fanti et al. (2002),
trabalhando com Adenanthera pavonina, também encontraram resultados semelhantes, em
relação a diminuição de MSR com a redução da irradiância. Além disso, estes autores
verificaram que os maiores valores de MSR foram obtidos por plantas crescendo em alta
irradiância e em solo adubado, seguido por plantas crescendo também em alta luminosidade,
mas em solo sem adubação. Por outro lado, observou-se, para os três variantes de C. echinata,
que o acúmulo de MSF não foi diretamente proporcional ao aumento da irradiância. Os
resultados revelaram que MSF apresentou maiores valores para o variante SV, quando
submetido a 35%, 100% e 15% de irradiância, respectivamente (Figura 8B). Em
contrapartida, Lima et al. (2010), trabalhando com três espécies arbóreas nativas (C. echinata,
Cariniana legalis e Genipa americana) apontaram acúmulo de MSF proporcionalmente ao
aumento da disponibilidade de luz.

61
Ao contrário de MSF, verificou-se, no presente estudo, que houve maior acúmulo de
MSC com o incremento da irradiância. Fato também observado por Illenseer e Paulino (2002)
trabalhando com Euterpe edulis e por Lima et al. (2010). Entretanto, para todos os variantes
morfológicos de C. echinata avaliados, o acúmulo de MST foi diretamente proporcional ao
aumento da irradiância. De acordo com Mengarda (2009), o crescimento e desenvolvimento
das plantas são afetados por diversos fatores ambientais, sendo a irradiância o parâmetro
diretamente ligado à produção de biomassa, regulando o ganho de energia e o metabolismo de
carboidratos por meio da fotossíntese. Desta forma, a luz é um dos fatores abióticos mais
importantes no estabelecimento da vegetação. Mudas cultivadas a altas irradiâncias
apresentaram RMF inferior àquelas cultivadas sob condições de menores intensidades de
irradiância, demonstrando maior intensidade de exportação de fotoassimilados da folha sob
alta luminosidade.
Alteração da taxa de crescimento relativo (TCR) é dependente de alteração na
fisiologia da planta, medida pela capacidade fotossintética em relação à área fotossintetizante,
ou seja, na TAL e, ou na morfologia, medida pela razão de área foliar (RAF) (HUNT, 1990).
Sob baixa irradiância, as plantas tendem a aumentar a RAF ou a RMF como uma estratégia de
adaptação ao sombreamento para ganho de carbono (ALMEIDA et al., 2005; SOUZA;
VALIO, 2003).
Os variantes LV, SV e MV apresentaram maior valores de AF quando submetidos
aos tratamentos com 60%, 35% e 35% de irradiância, respectivamente (Figura 11). Por outro
lado, estes variantes morfológicos apresentaram maior MFE e menor AF, quando cultivados
nos tratamentos a 100% e 35% de irradiância, respectivamente, quando comparado com os
tratamentos sombreados. Em estudos realizados por Mengarda (2009), com a mesma espécie,
avaliando o crescimento das plantas em diferentes regimes de luz, pôde-se observar também
que o aumento da irradiância afetou diretamente AF das plantas, cujos melhores resultados
para AF foram obtidos nos tratamentos sombreados. Voltan et al. (1992) e Fahl et al. (1994),
ao avaliarem o crescimento de plantas de Coffea arabica, encontraram maiores valores de
MFE e menor de AF em plantas submetidas a pleno sol, sugerindo que a radiação excessiva
foi limitante para o crescimento foliar. A expansão foliar em condições de baixa irradiância é
frequentemente relatada e indica uma maneira da planta compensar a diminuição da luz, com
melhor utilização deste recurso, aumentando área de superfície (CAMPOS; UCHIDA, 2002).
Normalmente, AF e sua taxa de expansão aumentam com a diminuição da densidade do fluxo
radiante; consequentemente, há um decréscimo na densidade de estômatos, o que indica um
mecanismo de aclimatação da planta a baixos níveis de luz. Assim, as plantas tendem a

62
promover a expansão máxima da superfície foliar, captando, com maior eficiência, a luz
disponível.
RAF representa o tamanho da superfície assimilatória em relação à biomassa seca
total. Essa característica de crescimento apresentou valores mais elevados, quanto os diversos
variantes morfológicos de C. echinata foram submetidos aos menores níveis de irradiância.
Segundo Fanti et al. (2002), um menor valor de RAF, em plantas submetidas a altas
intensidades de radiação luminosa, é considerado como uma resultante da capacidade da
planta em se aclimatar a diferentes condições de luz, dentro de certos limites. Por outro lado,
TAL, que descreve a eficiência da produção líquida do aparelho assimilatório, também
aumentou nos maiores níveis de irradiância, principalmente para o variante morfológico SV.
Fato também observado por Mesquita et al. (2002) em relação ao aumento de TAL. Ademais,
TCR, que representa o incremento de biomassa em relação à biomassa pré-existente, também
aumentou com o incremento de luz, uma vez que TCR resulta de TAL x RAF. Logo,
comparativamente, os resultados do presente trabalho mostraram haver similaridade de TCR e
TAL, cujos maiores valores foram encontrados para os variantes morfológicos crescidos a
pleno sol. Estes resultados contradizem os de Mengarda (2009) que demonstrou maiores
valores de TCR e TAL para plantas jovens de C. echinata crescidas em 50% de irradiância.
Interessante, entretanto, é a relação existente entre as variáveis RAF, AF, TCR e TAL.
Apesar de os maiores valores de TCR e TAL terem sido obtidos em 100% de irradiância e os
de RAF em 25 e 35% de irradiância, ficou demonstrado que tais incrementos condizem com
plantas crescendo sob baixa intensidade de radiação luminosa, indicando alterações
fisiológicas no sentido de maximizar ganhos fotossintéticos em função da diminuição de
RAF. A relação existente entre aumento de AF e RAF, diminuição de TCR e TAL e redução
da fotossíntese em baixos níveis de luz, representa mais uma estratégia da planta para
sobreviver do que para crescer. Fato também evidenciado por Almeida et al. (2005) e Souza e
Valio (2003), que alegaram que as plantas tendem a aumentar a AF e RAF, quando colocadas
sob condições de baixa irradiância, como sendo uma estratégia de adaptação ao
sombreamento para ganho de carbono. Logo, pôde-se observar, para o variante morfológico
LV, que o maior valor da A foi obtido em 100% de irradiância (8,93 μmol m-2
s-1
). Nestas
condições, a TAL conferiu melhores resultados àquele variante, indicando melhor
aclimatação e adaptabilidade do aparato fotossintético. Entretanto, se levarmos em conta a
relação de A, gs, E, Ci/Ca com TAL, verificou-se que os melhores resultados foram obtidos
com os variantes morfológicos localizados nos tratamentos intermediários (35 e 15% de
irradiância), concordando com os resultados obtidos por Mergarda (2009). De acordo com

63
Encinas et al. (2005), existem árvores dominantes e dominadas. O tamanho e a forma das
árvores são claramente dependentes das características locais e das condições edafoclimáticas
em que estão se desenvolvendo. Desta forma, os variantes morfológicos de pau-brasil, ou seja,
árvores da mesma espécie, podem apresentar diferenças marcantes nas suas variáveis
dendrométricas, na tentativa de otimizar o crescimento por meio de aclimatação aos efeitos do
local e, principalmente, da intensidade de luz disponível para o crescimento correspondente.
Para os variantes morfológicos SV, MV e LV, o tratamento com 100% de irradiância
favoreceu o crescimento, quando comparados aos demais tratamentos. Os menores valores de
MFE, TCR, TCRA e TCRD, apresentados nos tratamentos 60%, 35%, 25% e 15% irradiância
(Figura 12 e 13), foram fatores limitantes para o desenvolvimento destes variantes
morfológicos nestas condições. Os resultados obtidos contradizem as informações relatadas
por Poggiani et al. (1992) e Demuner et al. (2004) que, estudando Piptadenia rigida,
Schizolobium parahyba e Albizzia lebbeck, demonstraram que tais espécies apresentaram
melhor rendimento em tratamentos sombreados, principalmente a 80% de sombreamento.
Logo, as respostas de crescimento dos variantes morfológicos do C. echinata à irradiância
podem indicar a capacidade de aclimatação destes variantes para maximizar o uso eficiente de
nutrientes e da água em condições de variação da radiação luminosa. Assim, o variante
morfológico LV está melhor aparelhado para se desenvolver e crescer a pleno sol.
Os variantes morfológicos SV, MV e LV, em resposta aos diferentes níveis de
irradiância, mostraram aclimatações fisiológicas e bioquímicas na tentativa de maximizar os
ganhos fotossintéticos. Fato também observado por Soriani et al. (2012) em trabalhos
realizados com as espécies Pterogyne nitens, Hymenaea courbaril e Copaifera langsdorffii,
avaliando teores de carboidratos e crescimento destas espécies aclimatadas em três condições
de irradiância. O amido, um dos carboidratos de armazenamento mais abundante encontrado
nas plantas, que suporta o metabolismo e crescimento no escuro, normalmente não é
transportado do cloroplasto para os drenos metabólicos até que seja totalmente hidrolisado em
moléculas de glicose, sendo degradado dentro dos plastídeos onde foi sintetizado (ZEEMAN
et al., 2004; TAIZ; ZEIGER, 2009). Desta forma, os padrões de armazenamento do amido
podem estar diferenciados em vários tecidos, sofrendo, com isso, os efeitos imediatos das
variações de irradiância. No presente trabalho, o aumento da irradiância provocou elevação
dos teores de AST e de amido nas folhas para todos os variantes morfológicos avaliados, ao
passo que para o variante morfológico SV os teores de AST e de amido foram maiores no
caule, a 15% e 25% de irradiância, respectivamente. Entretanto, os teores de amido na raiz
não sofreram influência direta da radiação luminosa, ao passo que os maiores teores de AST

64
para os variantes MV e LV foram encontrados a 35% de irradiância. Estes resultados
contradizem com os encontrados por Casagrande Jr. et al. (1999), avaliando outras espécies,
ao afirmaram que aumento do sombreamento eleva os teores de amido nas folhas. Por outro
lado, Soriani et al. (2012), trabalhando com três espécies arbóreas, não observaram diferenças
significativas na concentração de amido em folhas, com a variação da irradiância. Segundo
estes autores, a espécie Hymenaea courbaril apresentou maior teor de amido em condições de
sombra do que a pleno sol.

65
7. CONCLUSÕES
Os variantes morfológicos de pau-brasil ajustaram seu aparato fotossintético, no
sentido de maximizar os ganhos de energia, por meio de plasticidade fisiológica diferenciada,
durante a aclimatação aos diferentes ambientes de radiação luminosa.
A variação da irradiância influenciou positivamente a assimilação do CO2, quando
relacionada com gs e E, dando condições aos variantes morfológicos de atingirem o seu
crescimento potencial, demonstrado pelo aumento de TCR com o incremento da intensidade
de luz.
As mudanças nos padrões biométricos dos variantes de pau-brasil foram afetadas
pela irradiância, em consequência das variações nos ganhos de biomassa determinados por
TAL e TCR.
Houve uma diferenciação dos variantes morfológicos em relação à assimilação do
carbono e a variação da irradiância. O variante LV se aclimatou melhor em condição de 100%
de irradiância; ao passo que os variantes SV e MV se aclimataram melhor em condições de 35
e 15%, respectivamente.

66
8. REFERÊNCIAS
AGUIAR, F. F. A.; KANASHIRO, S.; TAVARES, A. R.; PINTO, M. M.; STANCATO, G.
C.; AGUIAR, J.; NASCIMENTO, T. D. R. Germinação de sementes e formação de mudas de
Caesalpinia echinata Lam. (PAU-BRASIL): efeito de sombreamento. Revista Árvore,
Viçosa-MG, v.29, n.6, p.871-875, 2005
AKUNDA, E.M.W.; IMBAMBA, S.K.; KUMAR, D. High density planting of coffee. II.
Adaptive changes in some plant characteristics. East African Agricultural and Forestry
Journal, v. 45, p. 133-136, 1979.
ALMEIDA, L. P.; ALVARENGA, A. A.; CASTRO, E. M.; ZANELA, S. M.; VIEIRA, C. V.;
Crescimento inicial de plantas de Cryptocaria aschersoniana Mez. submetidas a níveis de
radiação solar. Ciência Rural, v. 34, n.1, jan-fev, 2004.
ALMEIDA, S.M.Z.; SOARES, A.M.; CASTRO, E.M.; VIEIRA, C.V.; GAJEGO, E.B.
Alterações morfológicas e alocação de biomassa em plantas jovens de espécies florestais sob
diferentes condições de sombreamento. Ciência Rural, v. 35, n. 1, p. 62-68, 2005.
ALVARENGA, A.A.; CASTRO, E.M.; LIMA JUNIOR, E.C.; MAGALHAES, M.M. Effects
of different light levels on the initial growth and photosynthesis of Croton urucurana Baill. In
southeastern Brazil. Revista Árvore, v.27, n.1, p.53-57, 2003.
ALVES, P.L.; MAGALHÃES, A.C.N.; BARJA, P.R. The phenomenon of photoinhibition of
photosynthesis and its importance in reforestation. The Botanical Review, v. 68, n. 2, p. 193-
208, 2002.
AMARAL, L. I. V.; GASPAR, M.; COSTA, P. M. F.; AIDAR, M. P. M.;
BUCKERIDGE, M. S. Novo método enzimático rápido e sensível de extração e dosagem de
amido em materiais vegetais. Hoehnea 34(4): 425-431, 2007.
ANDERSON, J.M. Photoregulation of the composition, function, and structure of thylakoid
membranes. Annual Review of Plant Physiology, v. 37, p. 93-136, 1986.

67
ANDERSON, J.M.; OSMOND, C.B. Shade-sun responses: compromises between
acclimation and photoinhibition. In: KYLE, D.J.; OSMOND C.B.; ARNTZEN, C.J. (Eds.)
Photoinhibition. Amsterdam: Elsevier, 1987. p 1-38.
ATROCH, E. M. A. C.; SOARES, Â. M.; ALVARENGA, A. A.; CASTRO, E. M.
Crescimento, teor de clorofilas, distribuição de Biomassa e características anatômicas de
plantas Jovens de bauhinia forficata link submetidas à diferentes condições de sombreamento.
Ciências agrotécnica, Lavras, v.25, n.4, p.853-862 , jul./ago., 2001
ARAÚJO, D. D. Crescimento de mudas de jatobá (Hymenaea courbaril L.) sob quatro
níveis de sombreamento. 2009. 65 f. Dissertação (Mestrado em Botânica ). Universidade
Federal Rural da Amazônia – Belém, 2009.
BARBEDO, C.J.; BILIA, D.A.C.; RIBEIRO, R.C.L.F. Tolerância à dessecação e
armazenamento de sementes de Caesalpinia echinata Lam. (Pau-brasil), espécie da Mata
Atlântica. Rev. Bras. Bot., v. 25, n. 4, p. 431-439, Dez. 2002.
BENINCASA, M. M. P. Análise de crescimento de plantas (noções básicas).
Jaboticabal: FUNEP, 1988. 42 p.
BJÖRKMAN, O. Responses to different quantum flux densities. In: Encyclopedia of Plant
Physiology. Vol. I. New Series. LANGE, O.L.; NOBEL, P.S.; OSMOND, C.B.; ZIEGLER,
H. (Eds.) Berlin: Springer-Verlag, 1981. p. 57-107.
BJÖRKMAN, O.; BOARDMAN, N.K.; ANDERSON, J.M.; THORNE, S.W.;
GOODCHILD, D.J.; PYLIOTIS, N.A. Effect of light intensity during growth of Atriplex
patula on the capacity of photosynthetic reactions, chloroplast components and structure.
Carnegie Institute Washington Year Book, v. 71, p. 115-135, 1972.
BOARDMAN, N.K. Comparative photosynthesis of sun and shade plants. Annual Review of
Plant Physiology, v. 28, p. 355-377, 1977.
BRAND, M.H. Shade influences plant growth, leaf color and chlorophyll content of Kalmia
latifolia L . cultivars. Hort Science , v.32, n.2, p.206-208, 1997
BUNCE, J.A.; PATTERSON, D.T.; PEET, M.M.; ALBERTE, R.S. Light acclimation during
and after leaf expansion in soybean. Plant Physiology, v. 60, p. 255-258, 1977.

68
CAI, Z.Q.; RIJKERS, T.; BONGERS, F. Photosynthetic acclimation to light changes in
tropical monsoon forest woody species differing in adult stature. Tree Physiol., 25: 1023–
1031, 2005.
CAMPOS, M.A.; UCHIDA, T. Influência do sombreamento no crescimento de mudas de três
espécies amazônicas. Pesquisa Agropecuaria Brasileira, 37: 281-288, 2002.
CARVALHO, P. E. R. Espécies arbóreas brasileiras. Brasília: Embrapa, 2003. 1039 p.
CASAGRANDE JUNIOR, J. G.; BIANCHI,V. J.; STRELOW, E. Z.; BACARIN, M. A.;
FACHINELLO, J. C. Influência do sombreamento sobre os teores de carboidratos e
fenóis em estacas semilenhosas de araçazeiro. Pesq. agropec. bras., Brasília, v.34, n.12,
p.2219-2223, dez. 1999.
CASTRO, C. F. A. Gestão florestal no Brasil Colônia. 2002. 199 f. Tese (Doutorado em
Desenvolvimento Sustentável) - Centro de Desenvolvimento Sustentável, Universidade
Nacional de Brasília, Brasília, 2002.
COSTA, L.C.B.; ALMEIDA, A-A.F.; VALLE, R.R. Crescimento, teor de clorofila e estrutura
anatômica em plântulas de Theobroma cacao submetidas a diferentes irradiâncias e doses de
nitrogênio. Agrotrópica, v. 10, n. 1, p. 21-30, 1998.
COSTA, G. F.; MARENCO, R. A.. Fotossíntese, condutância estomática e potencial hídrico
foliar em árvores jovens de andiroba (Carapa guianensis). Acta Amaz. [online]. vol.37. n.2,
2007.
COSTA, E.; LEAL, P. A. M. Medidas radiométricas em casas de vegetação com cobertura
plástica na região de Campinas – SP. Engenharia Agrícola, Vol 31, Nº3, Jaboticabal, SP.
2011
COSTA, L.C.B. Trocas gasosas, atividade da redutase do nitrato, composição química e
estrutura anatômica em plântulas de Theobroma cacao submetidas a diferentes
irradiâncias e doses de nitrogênio. 1996. 88 f. Dissertação (Mestrado) – Universidade
Federal da Bahia, Salvador, 1996.
CHAVES, A. R. M. Aspectos fisiológicos do crescimento e da produção de cafeeiro. 2009.
79f. Tese (Doutorado em Fisiologia Vegetal). Universidade Federal de Viçosa. MG. 2009.
CHAZDON, R.L.; PEARCY, R.W.; LEE, D.W.; FETCHER, N. Photosynthetic responses of
tropical forest plants to contrasting light environments. In: MULKEY, S.S.; CHAZDON,

69
R.L.; SMITH, A.P. (Eds.) Tropical Forest Plant Ecophysiology. New York: Chapman and
Hall, 1996. p. 5-55.
CHEN, H.Y.H.; KLINKA, K. Light availability and photosynthesis of Pseudotsuga menziesii
seedlings grown in the open and in the forest understory. Tree Physiology, v. 17, p. 23-29,
1997.
CLAUSSEN, J.W. Acclimation abilities of three tropical rainforest seedlings to an increase in
light intensity. Forest Ecology and Management 80:245-255. 1996.
CLEGG, K.M. The application of the anthrone reagent to the estimation of starch in cereals. –
Journal Science Food Agriculture. 7: 40-44, 1956.
CURVELO, K. B. et al. Manual de plantio e indicações técnicas para manutenção de
mudas de Pau-brasil (Caesalpinia echinata) no campo. Ilhéus: CEPLAC, 2008. 26 p.
DAMATTA, F.M. Ecophysiological constraints on the production of shaded and unshaded
coffee: a review. Field Crops Research, v. 86, p. 99-114, 2004.
DAMATTA, F.M.; MAESTRI, M. Photoinhibition and recovery of photosynthesis in Coffea
arabica and C. canephora. Photosynthetica, v. 34, p. 439-446, 1997.
DANTAS, B. F.; RIBEIRO, L. S.; PEREIRA, M. S.. Teor de açúcares solúveis e insolúveis
em folhas de videiras, cv. syrah, em diferentes posições no ramo e épocas do ano. Rev. Bras.
Fruticultura, Jaboticabal - SP, v. 29, n. 1, p. 042-047, Abril 2007.
DALEY, P.F.; RASCHKE, K.; BALL, J.Y.; BERRY, J.A. Topography of photosynthetic
activity of leaves obtained from video images of chlorophyll fluorescence. Plant Physiology,
v. 90, p. 1233-1238, 1989.
DEMMIG-ADAMS, B.; ADAMS, W.W. The role of xanthophyll cycle carotenoids in the
protection of photosynthesis. Trends in Plant Science, v. 1, p. 21-26, 1996.
DEMMIG-ADAMS, B; ADAMS, W. W. emmig-Adams B, Adams W.W. Photoprotection in
an ecological context: the remarkable complexity of thermal energy dissipation. New Physiol
172:11-21. 2006
DEMUNER, V.G.; HEBLING, A.S.; DAGUSTINHO, D.M. Efeito do sombreamento no
crescimento inicial de Gallesia integrifólia (spreg)Harms. Bol. Mus. Biol. Mello Leitão. 2004

70
DIAS-FILHO, M.B. Physiological response of Solanum crinitum Lam. to contrasting light
environments. Pesquisa Agropecuária Brasileira, v. 32, n. 8, p. 789-796, 1997.
ENCINAS, J. M.;SILVA, G.F.;PINTO, J. R. R., Idade e crescimento das
árvores.Comunicações Técnicas Florestais. Vol. 7, n1, Brasília. 2005.
ENGEL, V.L. Influência do sombreamento sobre o crescimento de mudas de essências
nativas, concentração de clorofila nas folhas e aspectos de anatomia. Piracicaba, 1989. 202p.
(M. Sc-ESALQ/USP).
ENGEL, V.L.; POGGIANI, F. Estudo da concentração de clorofila nas folhas e seu espectro
de absorção de luz em função do sombreamento em mudas de quatro espécies florestais
nativas. Revista Brasileira de Fisiologia Vegetal, v. 3, n. 1, p. 39-45, 1991.
EVANS, J.R. Photosynthetic and nitrogen relationships in leaves of C3 plants. Oecologia, v.
78, p. 9-19, 1989.
FAHL, J.I.; CARELLI, M.L.C.; VEGA, J.; MAGALHÃES, A.C. Nitrogen and irradiance
levels affecting net photosynthesis and growth of young coffee plants (Coffea arabica L.).
Journal of Horticultural Science, v. 69, p. 161-169, 1994.
FANTI, S.C.; PEREZ, S.C. J.G.A. Influência do sombreamento artificial e da adubação
química na produção de mudas de Adenanthera pavonina L. Ciência Florestal, Santa Maria,
v. 13, n. 1, p. 49-56. 2002
FARIAS, V.C.; SILVA, D.E.; ALMEIDA, A.J. Análise de crescimento de mudas de
cedrorana (Cedrelinga catenaeformis (Ducke) Ducke) cultivadas em condições de viveiro.
Revista Brasileira de Sementes, v. 19, n. 2, p. 193-200, 1997.
FELFILI, J.M.; ALVEZ, C.V.; COSTA, D.R. Comportamento de plântulas de Sclerolobium
paniculatum Vog. var. rubiginosum (Tul.) Benth. sob diferentes níveis de sombreamento em
viveiro. Revista Brasileira de Botânica, v. 22, n. 2, p. 297-301, 1999.
FERREIRA, M. J.; GONÇALVES, J. F. C.; FERRAZ, J. B. S. Crescimento e eficiência do
uso da água de plantas jovens de castanheira-da-amazônia em área degradada e submetidas à
adubação. Ciência Florestal, Santa Maria, v. 22, n. 2, p. 393-401, abr.-jun., 2012.

71
FERMINO-JR, P.C.P. Anatomia ecológica comparada de folhas de Guapira opposita
(Vell.) Reitz (Nyctaginaceae) na vegetação de restinga e na Floresta Ombrófila Densa.
Dissertação, Universidade Federal de Santa Catarina. 2004
FLORES, O.; GOURLET-FLEURY, S.; PICARD, N.; Local disturbance, forest structure and
dispersal effects on sapling distribution of light-demanding and shade-tolerant species in a
French Guianian forest. France. 2004
GIVNISH, T. J. Adaptation to sun and shade: a whole-plant perspective. Australian Journal
of Plant Physiology, v. 15, p. 63-92, 1988.
GONÇALVES, J. F. C; SILVA, E. M.; GUIMARÃES, D. G. Características fotossintéticas e
potencial hídrico foliar de plantas jovens de andiroba submetidas à deficiência hídrica e à
reidratação. Pesquisa Agropecuária Brasileira, Brasília, v. 44, n. 1, p. 8-14, jan. 2009.
GUIMARÃES, A. P. C. Estimativa de parâmetros da camada atmosférica para
Cálculo da irradiação solar incidente na superfície .2003.185f. Tese (Doutor em ciências
em engenharia mecânica). Universidade Federal do Rio de Janeiro, RJ, 2003.
HIKOSAKA, K.; TERASHIMA, I. A model of the acclimation of photosynthesis in the
leaves of C3 plants to sun and shade with respect to nitrogen use. Plant, Cell and
Environment, v. 18, p. 605-618, 1995.
HISCOX, J.D.; ISRAELSTAM, G.F. A method for the extraction of chlorophyll from leaf
tissue without maceration. Canadian journal of botany 57: 1332-1334. 1979.
HOPKINS, W. G. Introduction to Plant Physiology. 2ed. New York: John Wiley &
Sons,Inc., 512p, 2000,.
HUNT, R. Basic growth analysis. Londres: Unwin Hyman, 1990. 112p.
IBAMA. Instituto Brasileiro de Amparo ao Meio Ambiente. Lista Oficial de flora
ameaçada de Extinção. Disponível em: ≤http://www.ibama.gov.br/flora/extincao.htm>.
Acesso em: 15/02/2013
IQBAL, R.M.;RAO, AUR-R.; RASUL, E.;WAHID, A. Mathematical models and response
functions in photosynthesis: an exponential model. In: PESSARAKLI, M. (Ed.) Handbook of
Photosynthesis. NewYork: Marcel Dekker Inc., 1997. p. 803-810.

72
ILLENSEER, R.; PAULILO, M.T.S. Crescimento e eficiência na utilização de nutrientes em
plantas jovens de Euterpe edulis mart. sob dois níveis de irradiância, nitrogênio e fósforo.
Acta Botanica Brasílica, v.16, n.4, 2002.
JESUS, S. V.; MARENCO, R. A.; O SPAD-502 como alternativa para a determinação dos
teores de clorofila em espécies frutíferas. Acta Amazônica, v. 38, n. 4, p. 815 – 818, 2008.
JUCHUM, F. S.; COSTA, M. A.; AMORIM, A. M.; CORRÊA, R. X. Phylogenetic
relationships among morphotypes of Caesalpinia echinata Lam. (Caesalpinioideae:
Leguminosae) evidenced by trnL intron sequences. Naturwissenschaften. 95:1085–1091.
2008
KAMALUDDIN, M.; GRACE, J. Photoinhibition and light acclimation in seedlings of
Bischofia javanica, a tropical forest tree from Asia. Annals of Botany, v. 69, p. 47-52, 1992.
KANECHI, M.; UCHIDA, N.U.; YASUDA, T.; YAMAGUCHI, T. Nonstomatal inhibition
associated with inactivation of Rubisco in dehydrated coffee leaves under unshaded and
shaded conditions. Plant and Cell Physiology, v. 37, p. 455-460, 1996.
KERBAUY, G.B.Fisiologia Vegetal. Guanabara Koogan, Rio de Janeiro, RJ, Brasil. 452pp.
2004
KITAJIMA, K.; HOGAN, K. P. Increases of chlorophyll a/b ratios during acclimation of
tropical woody seedlings to nitrogen limitation and high light. Plant, Cell and Environment,
v. 26, p. 857-865, 2003.
KOZLOWSKI, T.T.; KRAMER, P.J.; PALTARDY, S.G. The physiological ecology of
woody plants. San Diego: Academic Press, 1991. 657 p.
KRAMER, T.; KOZLOWSKI, T. Physiology of woody plants. New York: Academic Press,
1979. 811 p.
LAMBERS, H.; CHAPIN III, F.S.; PONS, T.L. Plant physiological ecology. New York:
Springer Verlag, 1998.
LAMBERS, H.; POORTER, H. Inherent variation in growth rate between higher plants: a
search for physiological causes and ecological consequences. Advances in Ecological
Research, v. 23, p. 187-261, 1992.

73
LARCHER, W. Ecofisiologia Vegetal, São Carlos: Rima, 2000. 531p.
LARCHER, W. Physiological Plant Ecology: Ecophysiology and stress physiology of
functional groups. 3 ed. Berlin: Springer-Verlag Berlin Heidelberg, 1995. 506 p.
LAVINSCKY, A.O. Características fotossintéticas e crescimento inicial de mudas
de Euterpe edulis em ambientes de ‘Cabruca’. Ilhéus: UESC. Dissertação de
Mestrado. Ilhéus-BA, 74 p., 2009.
LEVERENZ, J.W. Shade shoot structure of conifers and the photosynthetic response to light
at two CO2 partial pressures. Functional Ecology, v. 9, p. 413-421. 1995
LEWIS, G. P. C. A Revision of the Poincianella – Erytrostemon Group. Royal Botanic
Gardens, Kew. Ed. J.M; Lock. 1998.
LIBERATO, M.A.R.; GONÇALVES, J.F.C.; CHEVREUIL, L.R.; NINA JUNIOR, A.R.;
FERNANDES, A.V.; SANTOS JUNIOR, U.M. Leaf water potential, gas exchange and
chlorophyll a fl uorescence in acariquara seedlings (Minquartia guianensis Aubl.) under water
stress and recovery. Brazilian Journal of Plant Physiology, v.18, p.315-323, 2006.
LIMA, H.C.; LEWIS, G.P. & BUENO, E. Pau-Brasil. In: BUENO, E. et al. (eds) Pau-brasil:
uma biografia. São Paulo: Axis Mundi, 2002. p. 39-76.
LIMA, M. A. O.; MIELKE, M. S.; LAVINSKY, A. O.; FRANÇA, S.; ALMEIDA, A.A.
F.; GOMES, F. P. Crescimento e plasticidade fenotípica de três espécies arbóreas com uso
potencial em sistemas agroflorestais. Scientia Forestalis, Piracicaba, v. 38, n. 87, p. 527-
534, set. 2010.
LLOYD, J.; FARQUHAR, G.D. 13C discrimination during CO2 assimilation by the
terrestrial biosphere. Oecologia, v. 99, p. 201-215, 1994.
LOBÃO, D. E. V.P. Agroecossistema cacaueiro da Bahia: Cacau-cabruca e fragmentos
florestais na conservação de espécies arbóreas. 2007. 109 f. Tese (Doutorado em Produção
Vegetal) - Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista
“Júlio de Mesquita Filho”, Jaboticabal, 2007.
LOBÃO D. E.; CARVALHO A. M.; CARVALHO D. L. Ecossistemas e agroecossistemas do
Sudeste da Bahia - Bioma Mata Atlântica. In: Universidade Federal da Bahia. Revista dos
Mestrandos em Direito Econômico da UFBA: edição especial, 5 - Direito Ambiental.
Salvador, p. 32-45. 1997.

74
LOBÃO, D. E. et al. Cacau Cabruca – sistema agrossilvicultural tropical. In: VALLE, R. R.
(Ed.). Ciência, Tecnologia e Manejo do Cacaueiro. Itabuna: Gráfica e Editora Vital, 2007.
p. 290-323.
LUNZ, A. M. P. Crescimento e produtividade do cafeeiro sombreado e a pleno sol. 2006.
94f. Tese (doutorado). Escola Superior de Agricultura Luiz de Queiroz. Piracicaba, 2006
MACHADO, R. C. R.; MÜLLER, M. W.; BARRETO,E W. S. Variações diurnas no teor de
amido e de açúcar solúveis em Folhas de cacau (Theobroma cacao L.). Rev. Bras. Fisiol.
Vegetal 1(2) : 163-167, 1989.
MACHADO, E. C.; SCHMIDT, P. T.; MADINA, C. L.; RIBEIRO, R. V. Respostas da
fotossintese de três espécies de citros a fatores ambientais. Pesq. Agropec. Bras., Brasília, v.
40, n. 12, p. 1161-1170, 2005.
MACMAHON, M.J.; KELLY, J.W.; DECOTEAU, D.R.; YOUNG, RE.; POLLOCK, R.K.
Growth of Dendranthema x Grandiflorum (Ramat.) Kitamura under various spectral filters.
Journal of American Society of Horticultural Science, v. 116, p. 950-954, 1991.
MAGALHÃES, N.S, MARENCO, R. A, MENDES, K. R. Aclimatação de mudas de
acariquara à alta irradiância. Pesquisa Agropecuária Brasileira 44(7):687-694. 2009
MARENCO, R. A.; LOPES, N. F. Fisiologia vegetal: fotossíntese, respiração, relações
hídricas e nutrição mineral. Viçosa-MG, Editora UFV. 451 p. 2005.
MARENCO, R. A.; LOPES, N.F. Fisiologia Vegetal, Viçosa: UFV, 2009.
MARTINAZZO,E.G.; ANESE,S.; WANDSCHEER,A.C.D.; PASTORINI, L.H. Efeito do
Sombreamento sobre o Crescimento Inicial e Teor de Clorofila Foliar de Eugenia uniflora
Linn (Pitanga) – Família Myrtaceae. Rev. Bras. Bioc., Porto Alegre, v. 5, supl. 2, p. 162-164,
jul. 2007
McCREADY, R.M., GUGGOLZ, J., SILVEIRA, V., OWENS, H.S. Determination of starch
and amylose in vegetables. Application to peas. – Analytical Chemistry. 22: 1156-1158,
1950.

75
MENGARDA, L. H. G.; SOUZA, R. L. F.; CAMPOSTRINI, E.; REIS, F. O.; VENDRAME,
W. A.; CUZZUOL, G. R. F.Light as an indicator of ecological succession in brazilwood
(Caesalpinia echinata Lam.). Brazilian society of plant physiology. 21(1): 55-64, 2009.
MESQUITA, A.C.; FREITAS, R.B.; OLIVEIRA,L.E.M. Análise de crescimento de plantas.
Universidade federal de lavras. 2002.
MITCHELL, R.L. Crop Growth and Culture. Ames: The Iowa State University Press,
1979, 349 p.
MORAIS, R.R. et al. Chloroplastid pigment contents and chlorophyll a fluorescence in
Amazon tropical three species. Árvore 31(5):959-966. 2007
MÜLLER, M.W.; SERRANO, P.; BIEHL, B. Photosynthetic characteristics during
development of leaves from Theobroma cacao L. Physiologia Plantarum, v. 85, n. 3, p. 599-
609, 1992.
NAKAZONO, E. M.; COSTA, M. C.; FUTATSUGI ,K.; PAULILO, M. T. S. Crescimento
inicial de Euterpe edulis Mart. em diferentes regimes de luz. Revista brasil. Bot., São Paulo,
V.24, n.2, p.173-179, jun. 2001.
NOBEL, P.S. Physicochemical and Environmental Plant Physiology. New York:
Academic Press, 1991.
NOGUCHI, K., SONOIKE, K.; TERASHIMA, I. Acclimation of respiratory properties of
leaves of Spinacia oleracea L., a sun species, and of Alocasia macrorrhiza (L.) G. Don., a
shade species, to changes in growth irradiance. Plant and Cell Physiology, v. 37, p. 377-384,
1996.
OBERBAUER, S. F.; STRAIN, B. R.; FETCHER, N. Effect of CO2 enrichment on seedling
physiology and growth of two tropical tree species. Phisiologia Plantarum, 65, 352-356.
1985.
OLIVEIRA, J. R.; DUARTE, N. F.; FASSIO, P. O. Análise dos teores de clorofila e
carotenóides como Indicadores de fitotoxicidade de herbicidas em Toona ciliata
var. australis. I Jornada Científica e VI FIPA do CEFET Bambuí Bambuí/MG – 2008
OSUNKOYA, O. O. et al. Influence of seed size and seedling ecological attributes on shade-
tolerance in northen Queensland. Journal of Ecology, v. 82, p. 149-163. 1994

76
PASSOS, E.E.M. Ecofisiologia do Coqueiro. In: WARWICK; D.R.N.; SIQUEIRA, L.A.
(Ed.) Cultura do coqueiro no Brasil. Aracaju: Embrapa-SPI, 1997. p. 65-72.
PEZZOPANE, J.R.M; GALLO, P.B.; PEDRO JÚNIOR, M.J.; ORTOLANI, A.A.
Caracterização microclimática em cultivos consarciados café/coqueiro anão verde. Revista
brasileira de Agrometeorologia, Santa Maria, v11, n. 2, p. 293-302, 2003.
POORTER, L.; BONGERS, F.; STERCK, F.J.; WÖLL; H. Architecture of 53 rainforest tree
species differing in adult stature and shade tolerance. Ecology, v. 84, p. 602-608, 2003.
POPMA, J.; BONGERS, F. Acclimation of seedlings of three Mexican tropical rainforest tree
species to a change in light availability. Journal of Tropical Ecology, v. 7, p. 85-97, 1991.
POGGIANI, F.; BRUNI, S.; BARBOSA, E.S.Q. Efeito do sombreamento sobre o
crescimento das mudas de três espécies florestais. Revista do Instituto Florestal de São
Paulo, v.4, n.2, p.564-569, 1992.
RAMALHO, J.D.C., CAMPOS, P.S., QUARTIN, V.L., SILVA, M.J., NUNES, M.A. High
irradiance impairments on photosynthetic electron transport, ribulose-1,5-bisphosphate
carboxylase/oxygenase and N assimilation as a function of N availability in Coffea arabica L.
plants. J. Plant Physiology, v. 154, p. 319-326, 1999.
REID, D.M.; BEALL, F.D.; PHARIS, R.P. Environmental Cues in Plant Growth and
Development. In: STEWARD, F.C. (Ed.). Plant Physiology. San Diego: Academic Press
Inc.. Volume X: Growth and Development. p. 65-181. 1991
REIS, F. O.; CAMPOSTRINI, E. Microaspersão de água sobre a copa: um estudo relacionado
às trocas gasosas e à eficiência fotoquímica em plantas de mamoeiro. R. Brasileira
agrociência, Pelotas, v.17, n.1-4, p.66-77, jan-mar, 2011.
REGO, G.M,; POSSAMAI, E. Efeito do sombreamento sobre o teor de clorofila e
crescimento inicial de jequitibárosa. Bol Pesq Flo 53:179-194. 2006.
RIZZINI, Carlos Toledo. Árvores e madeiras úteis do Brasil: Manual de Dendrologia
Brasileira. Ed 1. São Paulo: Edgard Blücher, 1971. 294 p.
SACCO, J.C. Flora Ilustrada Caibutarinense: Passifloráceas. Itajaí: R. Reitz, 1980. 132 p.

77
SALISBURY, F.B.; ROSS, C.W. Plant Physiology. Belmont: Wadsworth, 1992.
SANTOS, M. L. Estabelecimento e crescimento de mudas de euterpe edulis em três
ambientes florestais. Dissertação (Mestrado em Produção Vegetal). Universidade Estadual
de Santa Cruz – UESC. ILHÉUS. 2009
SEIFERMAN-HARMS, D. The light-harvesting and protective functions of carotenoids in
photosynthetic membranes. Physiology Plantarum, v. 69, p. 561-568, 1987.
SIEBKE, K.; GHANNOUM, O.; CONROY, J. P.; VON CAEMMERER, S. Elevated CO2
increases the leaf temperature of two glasshouse-grows C-4 grasses. Funcional Plant
Biology, v.29, n.12, p.1377-1385, 2002.
SILVA, E.C.; NOGUEIRA, R.J.M.C.; NETO, A.D.A.; BRITO, J.Z.; CABRAL, E.L.
Aspectos ecofisiológicos de dez espécies em uma área de caatinga no município de
Cabaceiras, Paraíba, Brasil. Iheringia, Série Botânica, 59(2): 201-205. 2004
SIMS, D.A.; PEARCY, R.W. Photosynthesis and respiration in Alocasia macrorrhiza
following transfers to high and low light. Oecologia, v. 86, p. 447-453, 1991.
SOUZA, G. M.et al. Estratégias de utilização de luz e estabilidade do desenvolvimento de
plântulas de Cordia superba Cham. (Boraginaceae) crescidas em diferentes ambientes
luminosos. Acta Bot. Bras. [online]. 2009, vol.23, n.2, pp. 474-485.
SOUZA, R.P.; VALIO, I.F.M. Seeedling growth of fifteen Brazilian tropical tree species
differing in successional status. Revista Brasileira de Botânica, v.26, n.1, p.35-47, 2003.
SORIANI, H. H.; CONTIN, D. R.; FAVARETTO, V.F.; MARTINEZ, C. A. Teor de
carboidratos e crescimento em espécies arbóreas Aclimatadas em três condições de
luminosidade. Trabalho de Pesquisa, parte da tese de doutorado do primeiro autor – USP.
2012
SCALON, S.P.Q.; FILHO, H.S.; RIGONI, M.R.; VERALDO, F. Germinação e crescimento
de mudas de pitangueira (Eugenia uniflora L.) sob condições de sombreamento. Revista
Brasileira de Fruticultura, v. 23, n. 3, p. 652-655, 2001.
SCALON, S.P.Q. et al.Crescimento inicial de mudas de Bombacopisis glabra (Pasq.) A.
Robyns sob condição de sombreamento. Árvore 27(6):753-758. 2003.

78
STREIT, N. M.; CANTERLE,L. P.; CANTO,M. W.; HECKTHEUER, L. H. H. As clorofilas.
Ciência Rural, v.35, n.3, mai-jun, 2005.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 4 ed. São Paulo: Artmed, 2009. 720 p.
TAN Y. et al. (2000) Resolution of kinetic system of simultaneous degradation of chlorofill a
and b by Parafac. Analytic Chem Acta 412: 195-202
TATAGIBA, S. D.; PEZZOPANE, J. E. M.; REIS, E. F.; DARDENGO, M. C. J. D.;
EFFGEN, T. A. M. Comportamento fisiológico de dois clones de Eucalyptus na época seca e
chuvosa. Cerne, Lavras, v. 13, n. 2, p. 149-159, 2007.
TENHUNEN, J. D.; PEARCY, R. W.; LARANCE, O. L. Diurnal variation in leaf
conductance gas exchange in natural environments. In: ZEIGER, E.; FARQUHAR, G.;
COWAN, I. (Eds.). Stomatal function. Stanford: Stanford University, 1987. p. 323-351.
TERASHIMA, I.; HIKOSAKA, K. Comparative ecophysiology of leaf and canopy
photosynthesis. Plant, Cell and Environment, v. 18, p. 1111-1128, 1995.
THORNBER, J.P. Chlorophyll-proteins: light-harvesting and reaction center components of
plants. Annual Review of Plant Physiology, v. 26, p. 127-58, 1975.
VALLADARES, F.; WRIGHT, S.J.; LASSO, E.; KITAJIMA, K.; PEARCY, R.W. Plastic
phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology,
v. 81, n. 7, p. 1925-1936, 2000.
VERDIAL, M.F.; LIMA, M.S.; NETO, J.T.; DIAS, C.T.; BARBANO, M.T. Métodos de
formação de mudas de maracujazeiro amarelo. Scientia Agricola, v. 57, n.4, p. 795-798,
2000.
VOLTAN, R.B.Q.; FAHL, J.I.; CARELLI, M.L.C. Variação na anatomia foliar de cafeeiros
submetidos a diferentes intensidades luminosas. Revista Brasileira de Fisiologia Vegetal, v.
4, p. 99-105, 1992.
WALLER, D.M. The dynamics of growth and form. In Plant Ecology, M.J.CRAWLEY,
ed.Oxford, London.496 p.1986.

79
WHATLEY, J.M.; WHATLEY, F.R. A luz e a vida das plantas. São Paulo: EPU-EDUSP,
1982. 101 p.
WONG, S.C.; COWAN, I.R.; FARQUHAR, G.D. Leaf conductance in relation to
assimilation in Eucalyptus pauciflora Sieb. ex. Spreng. influence of irradiance and partial
pressure of carbon dioxide. Plant Physiology, v. 62, p. 670-674, 1978.
ZANELLA, F.; SONCELA, R.; LIMA, A.L.S. Formação de mudas de maracujazeiro
“amarelo” sob níveis de sombreamento em Ji-Paraná/RO. Ciência Agrotécnica, v. 30, n. 5, p.
880-884, 2006.
ZEEMAN, S.C.; SMITH, S.M.; SMITH, A.M. The breakdown of starch in leaves. New
Phytologist 163: 247-261. 2004
ZHANG, S.; MA, K.; CHEN, L. Response of photosynthetic plasticity of Paeonia
suffruticosa to changed light environments. Environmental and Experimental Botany, v.
49, p. 121-133, 2003.