respostas fisiológicas de jeju (hoplerythrinus ... · piruvato aumentou após 6 horas de...
TRANSCRIPT

00257
2009Rev. Bras. Saúde Prod. An., v.l0, n.l, p.210-223, jan/mar, 2009ISSN 15199940 SP-00257
Respostas fisiológicas de jeju (Hoplerythrinus unitaeniatus) expostos ao aratmosférico
Physiological responses injeju ("Hoplerythrinus unitaeniatus ") to atmospheric airexposure
MARIANO, Wagner dos Santos'; OBA, Eliane Tie2, SANTOS, Laila Romagueira
Bichara ', FERNADES Marisa Narciso4
IFaculdades Anhanguera, Curso de Medicina Veterinária, Dourados, Mato Grosso do Sul, Brasil.2Empresa Brasileira de Pesquisa Agropecuária, Ministério da Agricultura, Pecuária e Abastecimento,
Macapá, Amapá, Brasil.3Universidade de São Paulo, Instituto de Biociência, Departamento Fisiologia Humana, São Paulo, Brasil.4Universidade Federal de São Carlos, Centro de Ciências Biológicas, São Carlos, São Paulo, Brasil.*Endereço para correspondência: [email protected]
RESUMO
As respostas fisiológicas de jeju (Hoplerythrinusunitaeniatusy à exposição ao ar foram analisadasapós I, 6 e 12 horas de exposição ao ar e I e 6horas após o retomo ao ambiente aquático. Asconcentrações plasmáticas de cortisol e glicoseforam significativamente maiores que as do grupocontrole em I, 6 e 12 horas de exposição ao ar,respectivamente. Desequilíbrio iõnico e ácido-basedurante exposição aérea foram evidenciados pelaredução de Na+ e K+ plasmáticos e do pH comsubsequente restauração do equilíbrio ácido-basequando o animal retomou ao meio aquático.Acúmulo de amônia foi observado em todos osgrupos experimentais e aumentou progressivamentedurante a exposição ao ar. Durante recuperação emmeio aquático, a amônia foi significativamentemaior que a do grupo controle. A concentração delactato foi significativamente maior apenas após 1hora de recuperação em meio aquático, enquanto opiruvato aumentou após 6 horas de recuperação nomeio aquático. O hematócrito, o número deeritrócitos, a concentração de hemoglobina e aconcentração de hemoglobina corpuscular médiaaumentaram significativamente nos animaisexpostos ao ar atmosférico. A exposição ao aratmosférico pode ser caracterizada como um agenteestressor para H. unitaeniatus e implica ajustesfisiológicos para manter a transferência do O2
atmosférico para os tecidos e a homeostase emobilização de energia.
Palavras-chave: cortisol, estresse, glicose,intermediários metabólicos, variáveishematológicas
SUMMARY
Physiological responses to air exposure wereanalyzed in jeju, Hoplerythrinus unitaeniatus after1, 6 and 12 hours of air exposure and I and 6hours after retuming to water environment. Plasmacortisol and glucose were significantly higher thanthose of control group after I, 6 and 12 hours of airexposure, respectively. Acid-base and ionicunbalances during air exposure were evidenced bylow plasma pH and low plasma concentration ofNa + and K+ followed by pH and acid-baseequilibrium recovery as fish retumed to waterenvironment. Ammonia accumulation was foundin all experimental groups increasing continuouslyduring air exposure. During the recovery period inwater ammonia concentration was significantlyhigher than the control. Plasma concentration oflactate was significantly higher than the controlsonly after I hour of recovery in water whilepiruvate increased after 6 hours of rccovery inwater. Hematocrit, red blood cells, hemoglobinconcentration and mean corpuscular hemoglobinconcentration increased significantly during airexposure. Air exposure can be characterized as astressor agent to H. unitaeniatus and imply inphysiological adjustments to keep the O2 transferfrom air to tissue and homeostasis as well asenergy mobilization.
Keywords: cortisol, glucose, haematologicalvariables, metabolic intermediaries, stress
Respostas fisiológicas de jeju2009 SP - 00257
1111111111111111111111111111111111111111111111111111111111I111111111111111114461-1
210

Rev. Bras. Saúde Prod. An., v.l0, n.l, p.210-223, jan/mar, 2009ISSN 1519 9940
http://www.rbspa.utba.br
INTRODUÇÃO
o ciclo anual de cheia e seca é ofenômeno ecológico mais importante deum no.A área de abrangência de inundação e otempo de permanência das águas noscampos determinam a disponibilidadede habitats e alimentação para peixes,crustáceos, aves, répteis, mamíferos,plantas aquáticas e semiaquáticas. Comessa dinâmica, o Pantanal sul-mato-grossense (Brasil), que possuicaracterísticas climáticas e topográficaspeculiares, apresenta períodos deestiagem com duração média de quatromeses no ano (VAL VERDE, 1972;CADAVID GARCIA, 1984),provocando a diminuição do nível daságuas dos rios, dando origem aambientes conhecidos popularmentecomo baías, vazantes e corixos(AMARAL FILHO, 1986; MARIANOet aI., 2007). Durante esse período, osorganismos aquáticos ficam confinadosnessas lagoas ou corixos, queapresentam condições estressantes,como hipóxia, exposição ao ar e falta dealimento (MARIANO et a!., 2007).Os corixos são caracterizados porcorpos de águas rasas, muitas vezesestagnadas, com altas temperaturas epobres em oxigênio, nos quais espéciesque possuem respiração bimodal, queutilizam as brânquias para obtenção deoxigênio dissolvido no meio aquático eoutro órgão modificado para arespiração aérea, têm maiorpossibilidade de sobreviver aos períodosde estiagem e mesmo à completaemersão durante curtas migrações entrecorpos d'água (CADAVID GARCIA,1984; MARIANO et aI., 2007).Animais que vivem nessas condiçõesmantêm diferentes estratégias para lidarcom a pouca disponibilidade deoxigênio e o alimento. Quando o
oxigemo e o alimento diminuem, ospeixes normalmente escapam paraoutras regiões. Diferentes estratégias,como a hiperventilação, bradicardia,sincronismo cardio-respiratório,constrição periférica vascular econstrição do trato digestório, sãoalgumas das respostas fisiológicas emorfológicas de peixes que vivemnestas condições (MORAES et aI.,2002; MARIANO et aI., 2007).A exposição ao ar durante as migraçõesentre os ambientes alagados impõe aopeixe uma situação de estresse, emdecorrência do risco de predação poranimais aéreos, dessecação ehipertermia e, ainda, pode causardisfunção branquial, uma vez que asbrânquias, além da respiração, têmpapel importante na regulação osmóticae iônica (PIlPER, 1989; PERRY, 1997),no equílibrio ácido-base (GILMOUR etaI., 1997) e na excreção de produtosnitrogenados (RANDALL et aI., 2004).As Alterações hematológicas(DETHLOFF et aI., 1999), hormonais,metabólicas (BARTON & IW AMA,1991; CARRAGHER & REES, 1994) eno equilíbrio hidromineral (ACERETEet aI., 2004) refletem as respostas a esseestresse e à dinâmica dos ajustesfisiológicos durante a exposição ao ar.Na América do Sul, existem apenas trêsgêneros da família Erythrinidae:Erythrinus; Hoplias e Hoplerythrinus(GODOY, 1975). Os peixes do gêneroHoplias apresentam respiraçãoexclusivamente aquática e aqueles dosoutros dois gêneros possuem respiraçãoaérea facultativa. A distribuiçãogeográfica do gênero Hoplerythrinusabrange o Peru, Bolívia, Venezuela,Guiana, Paraguai e Brasil (GODOY,1979; PIlPER, 1989). Vulgarmenteconhecido por jeju (SMITH, 1979), oHoplerythrinus unitaeniatus é a únicaespécie citada desse gênero; é ativa,carnívora, de crescimento rápido e
211

Rev. Bras. Saúde Prod. An., v.l0, n.l, p.210-223, jan/rnar, 2009ISSN 15199940
ocorre em águas de inundaçãotemporária (LOWE-MCCONNEL,1987). Durante os períodos de pesca noPantanal, é muito utilizada como iscapara peixes como o pintado(Ps eudop Iatys toma corruscans) edourado (Salminus maxillosus) e podeser encontrado com facilidade nas lojasde iscas-vivas da região (STEVENS &HOLETON, 1978; JUCÁ-CHAGAS,2004).No Pantanal, o jeju é comumenteencontrado em corpos d' águatemporários formados durante o períodode estiagem (FINK & FINK, 1978). Aespécie apresenta respiração aéreafacultativa e utiliza a bexiga natatóriaricamente vascularizada como o órgãopara a respiração aérea (GODOY, 1975;KRAMER, 1978; FERNANDES &MORON, 1996; GRAHAM, 1997).Durante severa hipóxia e anóxia, ooxigênio é absorvido via bexiganatatória, o que permite suasobrevivência nos períodos de estiagem.Quando os corixos são formados noPantanal, jejus são frequentementeencontrados migrando entre essescorpos alagados e, consequentemente,ficam expostos ao ar atmosféricos(KRAMER, 1978; JUCÁ-CHAGAS,2004).Muito pouco se conhecia sobre asrespostas fisiológicas desta espcciedurante o estresse de exposição aérea eo retorno à água. Desta maneira,realizou-se este trabalho com o objetivode avaliar as respostas fisiológicas aoestresse induzido pela exposição ao aratmosférico e após o retomo ao meioaquático em jeju, H. unitaeniatus, pormeio de marcadores hormonais,bioquímicos e hematológicos.
http://www.rbspa.utba.br
MATERIAIS E MÉTODOS
Foram coletados cerca de 100exemplares de jeju (Hoplerythrinusunitaeniatusi, adultos de ambos ossexos, com peso de 187,8 ± 26,08 g. Ospeixes foram coletados na bacia do RioParaná, município de Bataguaçu, MatoGrosso do Sul, e mantidos em tanquesde 1.000 L com circulação contínua deágua sem cloro, termostatizada(24 ± 2°C), aeração constante efotoperíodo natural (~ 12h: 12h). Osparâmetros fisico-químicos da águaforam monitorados e se mantiveramaproximadamente constantes:pH = 6,7 a 7,5; D02 = 6,0 a 7,5 mg/L;dureza = 25 a 30 mg/L (como CaC03);a1calinidade = 9,5 a 10,2 mg/L (comoCaC03); e condutividade = 65 a 72~S/cm. Semanalmente os tanques eramhigienizados e sifonados paramanutenção do bem-estar e da sanidadedos peixes. Os animais foramalimentados com peixes de pequenoporte três vezes por semana até 24 horasantes dos experimentos, quando aalimentação foi suspensa.Depois de um período de 90 dias para ocondicionamento ao manejolaboratorial, os peixes foram divididosem scis grupos de oito peixes. O grupocontrole (TO) foi mantido submerso nasmesmas condições de aclimatação; trêsgrupos foram expostos ao meio aéreodurante 1 hora (T 1); 6 horas (T6) e 12horas (T 12) e dois grupos foramexpostos ao meio aéreo durante 12horas antes de serem lançados ao meioaquático durante 1 hora (TR1) e 6 horas(TR6) para recuperação.Um tanque experimental abrigou ospeixes por 24 horas antes do inicio dosexperimentos. A água do tanque foilentamente removida e reintroduzidapor meio de um sifão para não causardistúrbio aos animais. Um fino filme de
212

Rev. Bras. Saúde Prod. An., v.IO, n.I, p.2IO-223, jan/mar, 2009ISSN 15199940
http://www.rbspa.ufba.br
água foi deixado no fundo dos tanquespara manter a umidade do ambiente eevitar a dessecação, porém sem permitira respiração aquática durante todo operíodo de exposição ao meio aéreo.Imediatamente após o período deexposição ao meio aéreo ou período derecuperação em meio aquático,amostras de sangue foram coletadas porpunção caudal com seringa heparinizadapara determinação dos parâmetroshematológicos. O pH sanguíneo foideterminado utilizando-se umpeagâmetro Micronal B375. Ohematócrito (Hct=%) foi determinado,em duplicata, em tubo capilarheparinizado utilizando-se centrífugapara microhematócrito Fanen mod.211.A concentração de hemoglobina(Hb=g/dL) foi determinada de acordocom o método da formação decianometahemoglobina utilizando-seespectrofotômetro e leitura em 540 nrn.A contagem do número de eritrócitos(RBC= x 106/mrrr') foi feita em câmarade Neubauer após diluição do sangueem formol citrato a 4%. A partir dosdados de Hct, concentração de Hb eRBC, foram determinados os índiceshematimétricos: VCM (volumecorpuscular médio um'), HCM(hemoglobina corpuscular média =pg/célula), CHCM (concentração dehemoglobina corpuscular média = %)(VERRASTRO et aI., 1998).O sangue remanescente foi centrifugadoe o plasma foi separado em alíquotaspara determinação das concentrações decortisol, glicose, piruvato, lactato,amoma, osmolalidade e íons. Aconcentração da glicose foi determinadapor colorimetria, de acordo com ométodo descrito por Duboie et al.(1956), e a leitura realizada emespectrofotômetro Spectronic Genesysem 480 nrn; a concentração de piruvatofoi determinada pelo método descritopor Lu (1939) e leitura em 440 nrn; e a
de lactato, pelo método descrito porHarrower & Brown (1972) e leitura em570 nrn. A determinação da amônia foifeita pelo método de Gentzkon &Masen (1942) e leitura emespectrofotômetro em 420 nrn. Aosmolalidade foi determinada porcongelamento utilizando um micro-osmômetro 1l0SMETTE Precision. Asconcentrações de Na+ e K+ foramdeterminadas por fotômetro de chamaDigimed DM-61, após diluição 1:100 eo cr foi quantificado utilizando-se kitreagente comercial (!dt LabtestDiagnóstica). O cortisol foi medido pormeio do kit COAT -A-COUNT Cortisol(COAT-A-COUNT® Cortisol,Diagnostic Products Corporation, LosAngeles, USA), que permite quantificaro nível do cortisol (hidrocortisona,Composto F) por radioimunoensaioutilizando o marcado r 1251 e umcontador Gamma-Cl Z.A partir dos resultados dos parâmetrosavaliados, foi utilizado o teste deBartlett para determinar ahomogeneidade dos dados e definir aaplicação de testes paramétrico ou nãoparamétrico. Em dependência ao tipo deanálise, foi utilizado o teste nãoparamétrico de Kruskal-Wallis e o pós-teste Dunn, nas variáveis paramétricasfoi utilizada a análise de variância e oteste Dunnett, a 5% de significância. Oprograma estatístico utilizado foi oGraphPad Instat versão 2.01 (GraphPadsoftware, San Diego, CA - USA).
RESULT ADOS E DISCUSSÃO
Logo após a remoção da água do tanque,os peixes se movimentaram muito, mas,no decorrer da exposição aérea,diminuíram gradualmente amovimentação até permanecerempraticamente imóveis, com raros
213

Rev. Bras. Saúde Prod. An., v.l0, n.l, p.210-223, jan/mar, 2009ISSN 15199940
movimentos. Durante a respiração aérea,foi possível perceber aumento do mucocutâneo dos peixes. Após o retomo aomeio aquático, os peixes voltaram aapresentar os movimentos aleatórioscaracterísticos da espécie. Não houvemortalidade durante o experimento.A concentração de lactato foisignificativamente maior em uma horaapós o retomo ao meio aquático. Apósseis horas de recuperação, aconcentração de lactato plasmático foiestatisticamente igual ao grupo controle.A concentração de piruvato foisignificativamente maior no grupo comtempo de recuperação de seis horas.A concentração de amônia em todos osgrupos experimentais foisignificativamente mais elevada que nogrupo controle (Tabela 1).Nas primeiras seis horas de exposição aoar atmosférico, os níveis de cortisolaumentaram significativamente, porém,após 12 horas, baixaram para níveissemelhantes aos do grupo controle. Aconcentração de glicose aumentou até 12horas de exposição ao meio aéreo,entretanto, uma hora após o retomo aomeio aquático, já havia regredido avalores próximos ao do grupo controle(Tabela 2).As variáveis hematológicas: hematócrito(Hct), concentração de hemoglobina(Hb) e contagem de eritrócitos (RBC)aumentaram significativamente durante aexposição ao ar, entretanto, durante operíodo de recuperação na água, nãodiferiram das observadas no grupocontrole. Não houve diferençasignificativa nos valores do VolumeCorpuscular Médio (VCM) e daHemoglobina Corpuscular Média(HCM) em nenhum dos gruposexperimentais. A Concentração deHemoglobina Corpuscular Média(CHCM) aumentou significativamenteem todos os grupos de exposição aérea(1 hora, 6 horas e 12 horas), inclusive no
http://www.rbspa.ufba.br
grupo que ficou uma hora emrecuperação em ambiente aquático. Apósseis horas de recuperação na água, aConcentração de HemoglobinaCorpuscular Média retomou a níveissemelhantes ao do grupo controle(Tabela3).A osmolaridade plasmática aumentousignificativamente seis horas após aexposição ao ar. A concentraçãoplasmática de íons Na + diminuiu emtodos os grupos experimentais, enquantoa de K+ foi mais baixa nos gruposexpostos ao ar por uma e seis horas. Em12 horas de exposição ao ar e durante operíodo de recuperação em meioaquático, a concentração plasmática deK+ foi semelhante à do grupo controle. Oíon Cl não foi alterado durante o períodode exposição ao ar e recuperação emmeio aquático. O pH sanguíneo diminuiusignificativamente em todos os gruposde exposição aérea, quando comparadoao grupo controle. O grupo que serecuperava por 1 hora em meio aquáticonão apresentou diferença no pH emcomparação ao grupo controle,entretanto, no grupo que se recuperavapor 6 horas em meio aquático, houveaumentou significativo (Tabela 4).
DISCUSSÃO
Embora o JCJu tHoplerythrinusunitaeniatus) possa sobreviver 24 horasem meio aéreo (STEVENS &HOLETON, 1978), os dados obtidosneste estudo evidenciam que a exposiçãoao ar atmosférico, situação que o animalenfrenta em seu habitat durante operíodo de seca, confirmam asdenominadas respostas primanas esecundárias ao estresse concomitantecom disfunção branquial associada àexcreção da amônia.
214
Rev. Bras. Saúde Prod. An., v.tO, n.1, p.210-223,jan/mar, 2009ISSN 15199940
http://www.rbspa.ufba.br
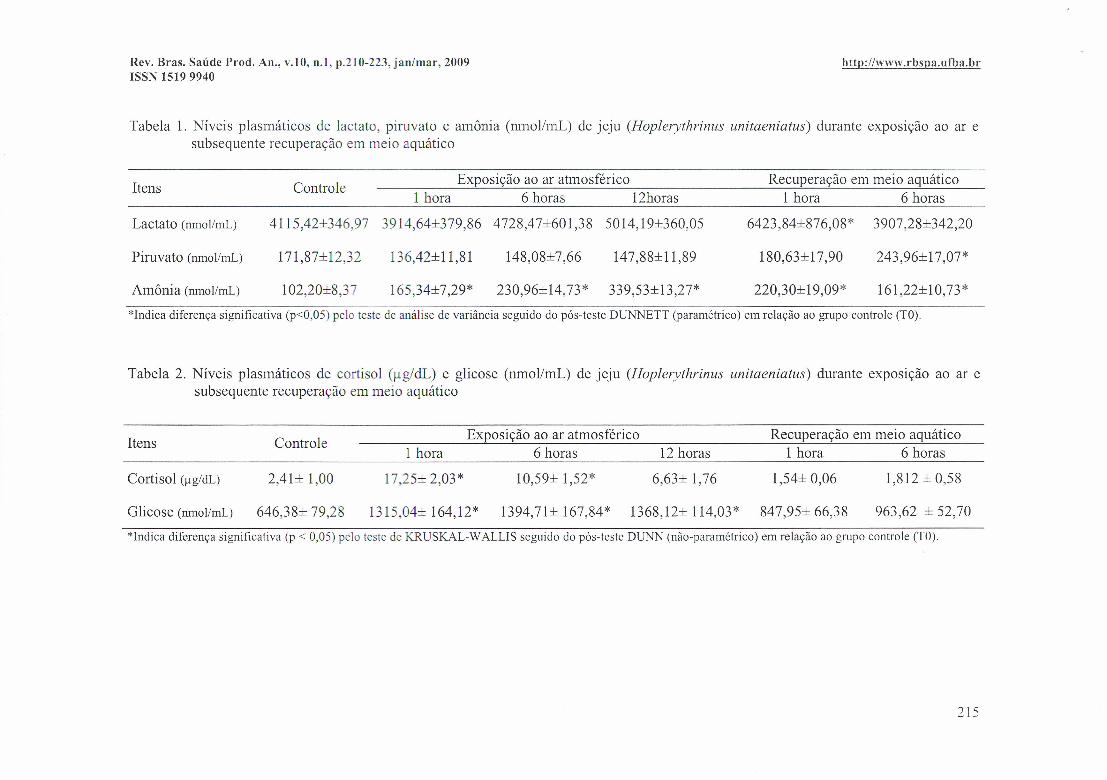
Tabela 1. Níveis plasmáticos de lactato, piruvato e amônia (nmol/mL) de jeju (Hoplerythrinus unitaeniatus) durante exposição ao ar esubsequente recuperação em meio aquático
Itens ControleExposição ao ar atmosférico Recuperação em meio aquático
1 hora 6 horas 12horas 1 hora 6 horas
Lactato (nmoIlmL) 4115,42±346,97 3914,64±379,86 4728,47±601,38 5014,19±360,05 6423,84±876,08* 3907,28±342,20
Piruvato (nmoIlmL) 171,87±12,32 136,42±11,81 148,08± 7,66 147,88±11,89 180,63±17,90 243,96±17,07*
Amônia (nmoIlmL) 102,20±8,37 165,34±7,29* 230,96±14,73* 339,53±13,27* 220,30±19,09* 161,22±10,73*
*lndica diferença significativa (p<0,05) pelo teste de análise de variância seguido do pós-teste DUNNETT (paramétrico) em relação ao grupo controle (TO).
Tabela 2. Níveis plasmáticos de cortisol (ug/dl.) e glicose (nmol/mL) de jeju (Hoplerythrinus unitaeniatus) durante exposição ao ar esubsequente recuperação em meio aquático
Itens ControleExposição ao ar atmosférico Recuperação em meio aquático
1 hora 6 horas 12 horas 1 hora 6 horas
Cortisol (llg/dL) 2,41± 1,00 17,25± 2,03* 10,59± 1,52* 6,63± 1,76 1,54± 0,06
1315,04± 164,12* 1394,71± 167,84* 1368,12± 114,03* 847,95± 66,38
1,812 ± 0,58
Glicose (nmoIlmL) 646,38± 79,28 963,62 ± 52,70
*lndica diferença significativa (p < 0,05) pelo teste de KRUSKAL- WALLIS seguido do pós-teste DUNN (não-paramétrico) em relação ao grupo controle (TO).
215
Rev. Bras. Saúde Prod. An., v.l0, n.l, p.210-223, jan/mar, 2009ISSN 15199940
http://www.rbspa.ufba.br
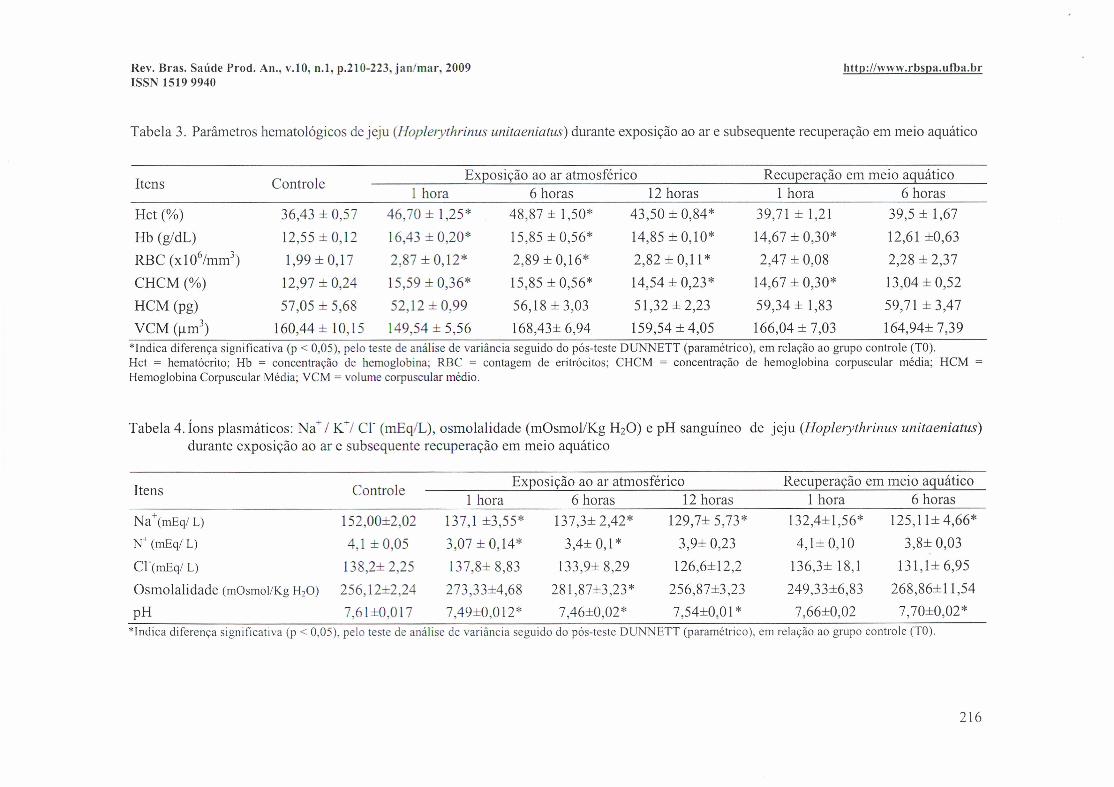
Tabela 3. Parâmetros hematológicos de jeju (Hoplerythrinus unitaeniatus) durante exposição ao ar e subsequente recuperação em meio aquático
Itens ControleExposição ao ar atmosférico Recuperação em meio aquático
1 hora 6 horas 12 horas 1 hora 6 horasHct (%) 36,43 ± 0,57 46,70 ± 1,25* 48,87 ± 1,50* 43,50 ± 0,84* 39,71 ± 1,21 39,5 ± 1,67
Hb (g/dL) 12,55 ± 0,12 16,43 ± 0,20* 15,85 ± 0,56* 14,85 ± 0,10* 14,67 ± 0,30* 12,61 ±0,63
RBC (xl06hnm3) 1,99 ± 0,17 2,87 ± 0,12* 2,89 ± 0,16* 2,82 ± 0,11 * 2,47 ± 0,08 2,28 ± 2,37
CHCM (%) 12,97 ± 0,24 15,59 ± 0,36* 15,85 ± 0,56* 14,54 ± 0,23* 14,67 ± 0,30* 13,04 ± 0,52
HCM (pg) 57,05 ± 5,68 52,12 ± 0,99 56,18 ± 3,03 51,32 ± 2,23 59,34 ± 1,83 59,71 ± 3,47
VCM (/lm3) 160,44 ± 10,15 149,54 ± 5,56 168,43± 6,94 159,54 ± 4,05 166,04 ± 7,03 164,94± 7,39*lndicadiferençasignificativa(p < 0,05), pelo teste de análisede variânciaseguidodo pós-testeDUNNETT(paramétrico),em relaçãoao grupocontrole(TO).Hct = hematócrito;Hb = concentração de hemoglobina; RBC = contagem de eritrócitos; CHCM = concentraçãode hemoglobina corpuscular média; HCM =HemoglobinaCorpuscularMédia;VCM= volumecorpuscularmédio.
Tabela 4. Íons plasmáticos: Na +/ K+/ Cl (mEq/L), osmolalidade (mOsmol/Kg H20) e pH sanguíneo de jeju (Hoplerythrinus unitaeniatus)durante exposição ao ar e subsequente recuperação em meio aquático
Itens ControleExposição ao ar atmosférico Recuperação em meio aquático
1 hora 6 horas 12 horas 1 hora 6 horasNa+(mEq/ L) 152,00±2,02 137,1 ±3,55* 137,3± 2,42* 129,7± 5,73* 132,4±1,56* 125,11± 4,66*
N+(mEq/L) 4,l±0,05 3,07±0,14* 3,4±0,1* 3,9±0,23 4,1±0,10 3,8±0,03
Cr(mEq/ L) 138,2± 2,25 137,8± 8,83 133,9± 8,29 126,6±12,2 136,3± 18,1 131,1± 6,95
Osmolalidade (mOsmol/KgH20) 256,12±2,24 273,33±4,68 281 ,87±3,23 * 256,87±3,23 249,33±6,83 268,86±11,54
pH 7,61±0,017 7,49±0,012* 7,46±0,02* 7,54±0,01 * 7,66±0,02 7,70±0,02**lndica diferençasignificativa(p < 0,05), pelo teste de análisede variânciaseguidodo pós-testeDUNNETT(paramétrico),em relaçãoao grupocontrole(TO).
216

Rev. Bras. Saúde Prod. An., v.10, n.1, p.210-223, jan/mar, 2009ISSN 15199940
http://www.rbspa.ulba.br
o estresse em peixes é caracterizadoprincipalmente pela elevação daconcentração plasmática de cortisol(MAZEAUD et al., 1977; BARTON &IWAMA, 1991; WENDELAARBONGA, 1997). O aumento deaproximadamente 6 vezes naconcentração do cortisol plasmático(2,90 a 17,24 u g/dl.) após a primeirahora de exposição ao ar seguido dediminuição gradual e o retomo aosníveis semelhantes ao controle(mantidos em meio aquático) sugereque a exposição ao ar pode sercaracterizada como estresse agudo parajejuo Pickering & Pottinger (1989)ressaltaram que, em situações deestresse agudo, o nível de cortisol empeixes pode aumentar até 10 e 100vezes o valor do nível basa1. Aconcentração de cortisol plasmático emSparus aurata exposto ao aratmosférico (3 minutos) foi, 1 hora apóso estímulo, 23 vezes maior que o nívelbasal, enquanto o confinamento (24horas) provocou aumento de 8 vezes(ARENDS et al., 1999). EmOncorhynchus mykiss, 30 segundos deexposição ao ar resultou em aumento de60 vezes na eoncentração basal decortisol (SLOMAN et a\., 2001).O aumento da glicose plasmática é umaresposta comum ao estresse (MORAESet aI., 2002; ACERETE et al., 2004) econ titui fonte extra de energia quepossibilita ao animal superar osdistúrbios causados pelo agenteestressor (WENDELAAR BONGA,1997). Em geral, o aumento da glicoseplasmática tem sido considerado oresultado da ação das catecolaminas nofigado (JANSSENS & WATERMAN,1988; RANDALL & PERRY, 1992;WENDELAAR BONGA, 1997),entretanto também está associado aoaumento da glicose plasmática,principalmente durante estresse
prolongado, via gliconeogênese(WENDELAAR BONGA, 1997).O aumento da glicose e a ausência deaumento na concentração de lactato epiruvato sugerem que o O2 absorvidopela bexiga natatória, mediado pelosajustes hematológicos, manteve osuprimento de O2 aos tecidos e ometabolismo aeróbico durante aexposição aérea. A absorção de O2 pelabexiga natatória por jeju é maior que 30% em condições de hipóxia ambiental elivre acesso ao ar atmosférico e aumentaà medida que a concentração de O2
diminui na água (STEVENS &HOLETON, 1978), extraindoaproximadamente 7 ml.Os/respiraçãoaérealkg quando em severa hipóxia(JUCÁ-CHAGAS, 2004). Emcondições de exposição ao ar, 100% doO2 tem que ser obtido via bexiganatatória.O aumento dos parâmetroshematológicos, do hematócrito, daconcentração de hemoglobina e contagemde eritrócitos, como observado em jeju,aumenta a capacidade de transporte do O2
e provavelmente ocorreu para reduzir osefeitos do agente cstrcssor e manter ahorneostasia interna preservando amanutenção do suprimento de O2 aostecidos. O hematócrito depende donúmero de eritrócitos presentes e podeser influenciado por sua forma e seutamanho. Como o volume corpuscularmédio e a osrnolalidadc (exceto em T6)não foram alterados, pode-se sugerirque o aumento no Hct em jeju ocorreucomo resultado da liberação de novoseritrócitos pelos centroshematopoiéticos durante a exposição aoar (AFFONSO et al., 2002; MORAES,et al., 2002; MARIANO et al., 2007).Após 12 horas de exposição ao ar, ohematócrito continuava apresentandodiferença estatística, porém indicandoredução numérica em comparação aogrupo mantido por seis horas de
217

Rev. Bras. Saúde Prod. An., v.IO, n.I, p.2IO-223, jan/mar, 2009ISSN 15199940
http://www.rbspa.ufba.br
exposição ao ar. Essa redução numéricapode indicar que, após 12 horas deexposição aérea, os centroshematopoiéticos diminuem a liberaçãode eritrócitos, porém essa afirmativanão pôde ser confirmada, pois não foiquantificado tempo superior a 12 horasde exposição aérea.O aumento do lactato nos peixes naprimeira hora após o retomo ao meioaquático, que possui menordisponibilidade de O2 que o aratmosférico, pode ter sido ocasionadopelo aumento brusco da atividadeanimal, uma vez que, segundo Stevens& Holeton (1978), o consumo de O2 domeio aquático em jeju, após 24 deexposição ao ar, não é muito alto e nãoevidencia débito de O2 muito alto.Arends et al. (1999) tambémconsideraram que o aumento do nível delactato plasmático durante a exposiçãoaérea (3 minutos) de Sparus auratapossivelmente foi ocasionado peloaumento da atividade muscular.Peixes que possuem respiração aérea,como jeju, mantêm as brânquias comoos órgãos primários para a excreção deCO2, amônia e equivalentes ácidos(T-JYDE & PERRY, 1987; RANDALLet al., 2004). A ausência do fluxo deágua sobre as brânquias, em decorrênciada supressão da ventilação branquialdurante a exposição aérea, elimina aação deste órgão como potencial sítio deregulação iônica c ácido-base cexcreção de amônia. O balançohidromineral em peixes, embora possaser modificado por muitos agentesestressores, está associado à atividadeda enzima Na+/K+(NH/)-ATPase naregião basolateral das células de cloretoe à ação de trocadores iônieos Na +/H+(NH/) e Cr/OR(HC03-) na regiãoapical dessas células que compensam asperdas iônicas por difusão. Além disso,a absorção de Na +, via canais de Na +,também ocorre durante a excreção ativa
de H+ (via V-ATPase) na região apicaldas células pavimentosas (RANDALLet al., 2004). Processo semelhantetambém ocorre nos túbulos renais para areabsorção de íons (CAMERON &WOOD 1978).A diminuição na concentração dos íonsplasmáticos (Na + e K+) durante aexposição aérea em jeju pode terocorrido via excreção renal. SegundoCameron e Wood (1978), em condiçõesde repouso e em meio aquático, o fluxourinário de íons e a excreção de amôniapelos rins de jeju são comparáveis aosde outros teleósteos e ligeiramentemaiores que em traira, Hopliasmalabaricus, espécie da famíliaErythinidae com respiraçãoexclusivamente aquática.Um pH corpóreo estável requer que aprodução de ácido seja equiparada a suaexcreção. Em animais aquáticos, assuperfícies externas têm a capacidade deexpelir ácido de maneira semelhante àobservada no dueto coletor do rim demamíferos. As brânquias dos peixes deágua doce têm H+-ATPase que excretaprótons na sua superfície apical doepitélio. Outro mecanismo de troca é oHCO-3/Cr, que também ocorrem nasbrânquias (RANDALL et. aI., 2000).No entanto, para que estes mecanismosejam utilizados, a presença da águatorna-se indispensável. Durante aexposição ao ar, o pH em jeju reduziu,comprovando que a falta d'água altera aregulação ácido-base.A redução do pH e o acúmulo deamônia durante a exposição aéreaindicam que jeju tem uma capacidadelimitada para a regulação ácido-base.Nessas condições, a redução do pHsanguíneo durante a exposição aéreapode estar relacionada também àretenção de CO2 que é excretado pelasbrânquias por difusão e hidratado,participando da regulação ácido-base,uma vez que uma hora após o retorno ao
218

Rev. Bras. Saúde Prod. An., v.10, n.1, p.210-223, jan/mar, 2009ISSN 15199940
http://www.rbspa.ufba.br
meio aquático, o pH sanguíneo foirestaurado (CAMERON & WOOD,1978; HYDE & PERRY, 1987;PERRY, 1997).Em jeju a participação de NH4+ naacidificação da urina pelos rins é de87% em condições de repouso(CAMERON & WOOD, 1978). EmAnguilla rostrata exposta ao ar, ocorrepequeno aumento na excreção de H+pelos rins e a recuperação do pH, após oretomo ao meio aquático, deve-seprimariamente ao trocadorNa+/H\NH/) nas brânquias. HYDE ePERRY (1987) enfatizaram que oprincipal papel dos rins durante aexposição aérea é reduzir O possívelrisco de desidratação, pois regulará aperda hídrica. De acordo com Randall etaI. (2000), os rins de teleósteos de águadoce, como é o caso do jeju, têmgrandes glomérulos, a maioria delesmaiores que dos teleósteos marinhos.Uma vez que seus corpos sãohipertônicos em relação ao meio e aágua se difunde para seus corpos, osteleósteos de água doce mantêm oequilíbrio de água produzindo grandesvolumes de urina diluída (RANDALLet a!., 2000).O aumento da concentração de amôniaplasmática em todos o gruposexperimentais pode estar associado àdificuldade de excreção via brânquias,pois houve aumento gradativo em 1, 6 e12 horas de exposição aérea e, com oretorno da água, o nível tambémdiminuiu gradativamente, mas seishoras de recuperação não foramsuficientes para que a amoruaplasmática retomasse ao nonna1. Polezet aI. (2003) também verificaram quejeju exposto ao ar por duas horasapresentou aumento significativo daamônia plasmática. Nesse mesmoestudo, esses autores demonstraramainda que o jeju aumenta a atividade daenzima glutamina sintetase, uma das
enzimas do ciclo da uréia que converteamônia em glutamina, reduzindo osefeitos da toxicidade da amônia(MCKENZIE et aI. 1999; MOMMSEN& WALSH, 1992; MOMMSEN &W ALSH, 1989). Além disso, essaconversão pode auxiliar no controle dopH, uma vez que, para cada molécula deglutamina metabolizada, surgem doisíons amônio (NH4+) que poderão serexcretados pelo rim e dois íonsbicarbonatos (HCO} +) que serãolançados na corrente sanguínea(SCHWARTZ, 1995). Entretanto, osdados obtidos neste estudo mostraramque, embora possa estar ocorrendo 12horas de exposição ao ar, essa estratégianão impediu o aumento da amôniaplasmática.A exposição aérea em jeju
(Hoplerythrinus unitaeniatus) atuacomo estressor agudo, caracterizadopelo aumento na concentração dohormônio cortisol e da glicoseplasmática. Seis horas em meioaquático após a exposiçao aérea sãosuficientes para a recuperação.Considerando os resultados de lactato epiruvato plasrnático, durante arespiração a' rea, o metabolismoaeróbico é possivelmente mantido,provavelmente pela eficiência da bex iganatatória na obtenção d O2, associadoaos ajustes hematológicos mostradosdurante a exposição aérea.O jeju apresenta, durante 12 horas deexposição ao ar, desequilíbrio iônico,caracterizado pela diminuição dos íonsNa + e K+, e 6 horas de recuperação emmeio aquático não são suficientes para arecuperação. A exposição aérea dificultaainda a excreção de amônia,evidenciada pelo acúmulo crescente deamônia durante todos os grupos deexposição aérea. Seis horas derecuperação não são suficientes paraque o nível de amônia retome aononnaJ.
219

Rev. Bras. Saúde Prod. An., v.IO, n.I, p.2IO-223, jan/mar, 2009ISSN 15199940
Durante a exposição aérea, há síntese eliberação de eritrócitos, evidenciadaspelos resultados de hematócrito,concentração de hemoglobina,contagem de eritrócitos e volumecorpuscular médio. A exposição aéreaprovoca desequilíbrio ácido-base emjeju, caracterizado pela redução do pHem todos os grupos expostos ao ar.
REFERÊNCIAS
ACERETE, L.; BALASCH, r.c.,ESPINOSA, E.; JOSA, A.; TORT, L.Physiological responses in Eurasianperch (Percafluviatilis, L.) subjected tostress by transport and handling.Aquaculture, n.237, 167-178,2004.[ Links]
AFFONSO, E.G.; POLEZ, V.L.P.;CORRÊA, C.F.; MAZON, A.F.;ARAÚJO, M.R.; MORAES, G.;RANTIN, F.T. Blood parameters andmetabolites in the teleost fishColossoma macropomum exposed tosulfide or hypoxia. ComparativeBiochernistry and Physiology C,v.133, p.375-382, 2002. [ Links ]
AMARAL FILHO, Z.P. Solos doPantanal Mato-grossense. In:SIMPÓSIO SOBRE RECURSOSNATURAIS E SÓCIO-ECON6MICOSDO PANTANAL, 1986, Corumbá.Anais ... Corumbá: EMBRAPAlDDT,1986. p.29-42. [ Links ]
ARENDS, RJ.; MANCERA, J.M.;MUNÕZ, J.L.; WENDELAARBONGA, S.E. The stress response ofthe gi1thead sea bream (Sparus aurataL.) to air exposure and confinement.J ournal of Endocrinology, n.163,p.149-157, 1999. [Links]
http://www.rbspa.ulba.br
BARTON, B.; IW AMA, G.Physiological changes in fish fromstress in aquaculture with emphasis onthe response and effects ofcorticosteroids. Annual Review of FishDieases, 1, p.3-26, 1991. [Links]
CADA VID GARCIA, E.A. O clima doPantanal Mato-grossense. Corumbá:EMBRAPA Pantanal, 1984. 39p.(Circular Técnica, 14). [ Links ] .
CAMERON, J.N.; WOOD, C.M. Renalfunction and acid-base regulation in twoAmazonian erythrinid fishes: Hopliasmalabaricus, a water breather, andHoplerythrinus unitaeniatus, afacultative air breather. CanadianJournal of Zoology, n.56, p.9l7-930,1978. [ Links ]
CARRAGHER, J.F.; RESS, C.M.Primary and secondary stress responsesin golden perch, Macquaria ambigua.Comparative Biochemistry andPhysiology A, 107, pA9-56, 1994.[Links]
DETHLOFF, G.M.; SCHLENK, D.;KHAN, S.; BAILEY, TT.C.The effcctsof copper on blood and biochemicalparameters of rainbow trout(Oncorhynchus mykiss). Archives ofEnvironmntal Contamination andToxicology, v.36, nA, pA15-423, 1999.[ Links ]
DUBOLE, M.G., GILLES, K.A.;HAMILTON, J.K.; ROBERTS, P.A.;SMITH, F. Colorimetric method fordetermination of suggars and relatedsubstances. Analytical Chemistry,v.28, p.350-358, 1956. [ Links ]
220

Rev. Bras. Saúde Prod. An., v.IO, n.I, p.2IO-223, jan/mar, 2009ISSN 15199940
http://www.rbspa.ufba.br
FERNANDES, M.N.; MORON, S.E.Respiratory organs in erythrinid fishes.ln VAL, A.L., RANDALL, D.I;MACKINLEY, D. (Eds.). Proceedingof Physiology of Tropical fishes. SanFrancisco: San Francisco University,1996. p.141. [ Links ]
FINK, W.I.; FINK, S.V. A Amazôniacentral e seus peixes. Acta Amazônica,v.8, n.4, p.19-42, 1978. [Links]
GENTZKON, C.J.; MASEN, IM. Anaccurate method for the determinationofblood urea nitrogen by directnesslerization. Journal of BiologicalChemistry, n.l43, p.531-544, 1942.[ Links ]
GILMOUR, K.M., HENRY, R.P.,WOOD, CM., PERRY, S.F.,Extracellular carbonic anhydrase and anacid-base disequilibrium in the blood ofthe dogfish Squalus acanthias. Journalof Experimental Biology, n.200, p.173-183,1997. [Links]
GODOY, M.P. Família erythrinidae. In:Peixes do Brasil, subordemCharacoidei: bacia do rio MogíGuassú. Piracicaba: Franciscana, 1975.p.440-444. [ Links ]
GODOY, M.P. Rio Iguaçu, Paraná,Brasil: reconhecimento da ictiofauna,modificações ambientais e usosmúltiplos dos reservatórios. Rio deJaneiro: Eletrosul, 1979. 33p.[ Links ]
GRAHAM, IB. Air-breathing fishes:evolution, diversity and adaptation.San Diego: Academic Press, 1997.299p. [ Links ]
HARROWER, J.R.; BROWN, CH.Blood lactic acid. micromethod adaptedto field collection of microliter samples.Journal of Applied Physiology, v.32,n.5, p.224-228, 1972. [ Links ]
HYDE, D.A.; PERRY, S.F. Acid-baseand ionic regulation in the American ell(Anguilla rostrata) during and afterprolonged aerial exposure: branchialand renal adjustments. Journal ofExperimental Biology, n.133, p.429-447,1987. [Links]
JANSSENS, P.A.; WATERMAN, I;Hormonal regulation of glucogenesisand glycogenolysis in carp (Cyprinuscarpio) liver pieces cultured in vitro.Comparative Biochemistry andPhysiology A, v.91, 451-455,1988.[ Links ]
JUCÁ-CHAGAS, R. Air breathing ofthe neotropical fishes Lepidosirenparadoxa, Hoplerythrinus unitaeniatusand Hoplosternum littorale duringaquatic hypoxia. ComparativeBiochemistry and Physiology A,v.139, 49-53,2004. [ Links ]
KRAMER, D.L. Ventilation oftherespiratory gas bladder inHoplerythrinus unitaeniatus (pisces,Characoidei, Erythrinidae). CanadianJournal of Zoology, n.56, p.931-938,1978. [ Links ]
LOWE-MCCONNEL, R.H. Ecologicalstudies in tropical fish communities.Cambridge: Cambridge UniversityPress, 1987. 382p. [ Links ]
LU, G.D. The mctabolism of pyruvicaeid in normal and vitamin B- deficientstate. A rapid specific and sensitivemethod for the estimation of bloodpyruvate. Biochemical Journal, n.33,p.249-254, 1939. [Links]
221

Rev. Bras. Saúde Prod. An., v.IO, n.I, p.2IO-223, jan/mar, 2009ISSN 15199940
MARIANO, W.S.; WEBSTER, V.;TOLEDO, 1.R.S.; GARCIA, R.G.Aspectos Fisiológicos e morfométricosde tuvira, Gymnotus carapo(LINNAEUS, 1758), submetidas àprivação alimentar. Faces daAcademia, v.2, n.2, p.51-61, 2007.[ Links ]
MAZEAUD, M.M.; MAZEAUD, F.;DONALDSON, E.M. Primary andsecondary effects of stress in fish: somenew data with a general review.Transactions of the AmericanFisheries Society, v.106, n.1, 201-212,1977. [ Links ]
MCKENZIE, D.J.; PIRACCINI, G.;FELSKlE, A.; ROMANO, P.; BRONZI,P.; BOLIS, c.L. Effects ofplasma totalammonia content and pH on ureaexcretion in Nile tilapia. Physiologicaland Biochemical Zoology, v.72, p.1l6-125,1999. [Links]
MOMMSEN, T.P.E.; WALSH, PJ.Evolution ofurea synthesis invertebrate: the piscine connection.Science, v.243, p.72-75, 1989. [Links]
MOMMSEN, T.P.E.; WALSH, PJ.Biochemical and environmentalperspectives on nitrogen metabolismoExperientia, v.48, 583-593, 1992.r Links ]
M01~ES, G.; AVILEZ, I.M.;ALTRAN, A.E.; BARBOSA, C.C.Biochemical and hematologicalresponses of the banded knife fishGymnotus carapo (Linnaeus, 1758)exposed to environrnental hypoxia.Brazilian Journal of Biology, v.62,n.4, p.633-640, 2002. [ Links ]
http://www.rbspa.ufba.br
PERRY, S.F. The chloride cell:structure and function in the gills offreshwater fishes. Annual Review ofPhysiology, v.59, p.325-347, 1997.[Links]
PICKERING, A.D.; POTTINGER, T.G.Stress responses and disease resistancein salmonid fish: effects of chronicelevation of plasma cortisol. FishPhysiology and Biochemistry, v.7, n.1-4, p.253-258, 1989. [ Links ]
PIIPER,1. Factors affecting gas transferin respiratory organs of vertebrates.Canadian Journal of Zoology, n.67,p.2956-2960, 1989. [ Links ]
POLEZ, V.L.P.; MORAES, G.;SANTOS NETO, C. Differentbiochemical strategies of twoNeotropical fish to cope with theimpairment of nitrogen excretion duringair exposure. Brazilian Journal ofMedical and Biological Research,v.36, n.2, p.279-285, 2003. [ Links ]
RANDALL, DJ.; PERRY, S.F.Catecolamines. In: HOAR, W.S.;RANDALL, DJ.; FARRELL, A.P.,Fish Physiology. San Diego: AcademicPress, p.255-300, 1992. [ Links ]
RANDALL, D.; BURGGREN, W.;FRENCH, K. Fisiologia Animal:mecanismos e adaptações. 4.ed. Rio deJaneiro: Guanabara Koogan, 2000.729p. [ Links ]
RANDALL, D.J.; IP, Y.K.; CHEW,S.F.; WILSON, J.M. Air breathing andammonia excretion in the giantmudskipper, Periophthalmusschlosseri. Physiological andBiochemical Zoology, V. 77, p.783-788, 2004. [ Links ]
222

Rev. Bras. Saúde Prod. An., v.IO, n.I, p.2IO-223, jan/mar, 2009ISSN 15199940
http://www.rbspa.ufba.br
SCHWARTZ, lH. Renal acid-basetransport: the regulatory role of theinner medullary collectin duct. KidneyInternational, 47, 333,1995. [Links]
SLOMAN, K.A.; DESFORGES, P.R.;GILMOUR, K.M. Evidence for amineralocorticóide-like receptor linkedto branchial chloride cell proliferationin freshwater rainbow trout,Oncorhynchus mykiss. Journal ofExperimental Biology, n.204, p.3953-3961,2001. [Links]
SMITH, NJ.H. A Pesca no rioAmazonas. Manaus: IMPA, 1979.154p. [ Links ]
STEVENS, E.D.; HOLETON, G.F. Thepartitioning of oxygen uptake from airand from water by erythrinids.Canadian Journal of Zoology, n.56,p.965-969, 1978. [ Links ]
VALVERDE, 1Fundamentosgeográficos do planejamento rural nomunicípio de Conunbá. RevistaBrasileira de Geografia, v.34, n.l,p.149-144, 1972. [Links]
VERRASTRO, T.; LORENZI, T.F.;NETO, S.W. Hematologia eHemoterapia. Rio de Janeiro: Atheneu,1998. 303p. [Links]
WENDELAAR BONGA, S.E. Thestress response in fish. PhysiologicalReviews, v.77, p.591-625, 1997.[Links]
Data de recebimento: 17/06/2008Data de aprovação: 26/01/2009
223